/

























































































































































































































































































Текст
АКАДЕМИЯ НАУК СССР
ИНСТИТУТ БИОЛОГИИ РАЗВИТИЯ
им. Н. К. КОЛЬЦОВА
ТЕРМОДИНАМИКА
БИОЛОГИЧЕСКИХ
ПРОЦЕССОВ
ОБЩИЕ ПОЛОЖЕНИЯ,
КОНСТИТУТИВНЫЕ ПРОЦЕССЫ,
ДИССИПАТИВНЫЕ СТРУКТУРЫ,
НЕКОТОРЫЕ ПРОБЛЕМЫ ЭВОЛЮЦИИ
ИЗДАТЕЛЬСТВО «НАУКА»
МОСКВА 1976
УДК 557.2; 577.95; 536.7
Термодинамика биологических процессов. М., «Наука», 1976, 280 с.
В коллективной монографии рассматриваются современные проб-
лемы термодинамики нелинейных необратимых процессов и вопросы
использования термодинамики в биологии. Обсуждаются принци-
пиальные вопросы, связанные с теоретическим и экспериментальным
обоснованием качественной и количественной феноменологической тео-
рии развития, роста и старения организмов. Приводятся эксперимен-
тальные данные по теплопродукции и энергетическому обмену при
росте культуры микроорганизмов, в процессе оогенеза, во время раз-
вития и роста животных. В заключительных главах монографии рас-
сматриваются современные теории диссипативных структур, проблемы
прогрессивной эволюции и классификации животных.
Монография рассчитана на биофизиков, физиологов, эмбриоло-
гов, а также физиков и математиков, интересующихся проблемами
современной термодинамики и ее использованием в биологии.
Табл. 17, илт. 70, список лит.—712 назв.
Ответственный редактор
доктор биологических наук
А. И. ЗОТИН
Коллектив авторов:
И, Г, Владимирова, В. А. Васильев, С. С. Васильев, В, А. Грудницкий,
А. А. Зотин, А. И. Зотин, Р. С. Зотина, В, А. Коноплев,
И. С. Никольская, Н. Д. Озернюк, Е. В. Преснов, Е. А. Прокофьев,
Л, И, Радзинская, В, Е. Соколов, Н. Ш. Шагиморданов —
СССР, Москва.
Р. Бретель, Р. Вальтер, И. Лампрехт, К, Д. Лёр,
П. Сайади, Б. Шааршмидт — Западный Берлин.
© Издательство «Наука», 1976 г.
21004—423
Т 055(02^-76 436'76’ ки-2
ПРЕДИСЛОВИЕ
Попытки использования термодинамики для описания процес-
сов, происходящих в живых организмах, имеют длительную исто-
рию, однако до сих пор не отпали сомнения в возможности построе-
ния термодинамики биологических процессов (Блюменфельд,
1974, Чернавский, 1975). Основная трудность, как нам кажется,
заключается в том, что сама термодинамика как общая феноменоло-
гическая теория процессов, сопровождающихся диссипацией
энергии, еще далека от завершения. Если два больших раздела
термодинамики: теория равновесных и теория линейных неравно-
весных процессов, в основном завершены к настоящему времени,
то теория нелинейных необратимых процессов, к которым отно-
сятся и многие процессы, протекающие в живых системах, еще на-
ходится в стадии своего становления. Основные поиски идут в на-
правлении создания вариационных принципов термодинамики
(Biot, 1955, 1970; Ono, 1961; Дярматы, 1965, Дьярматы, 1974;
Циглер, 1966; Finlayson, Scriven, 1967; Vojta, 1967; Бахарева,
1968, 1971; Филюков, Митрофанов, 1968; Farkas, 1968; Gyarmati,
1968, 1969, 1974; Lesse, 1968; Farkas, Noszticzius, 1972, Lavenda,
1972, 1973, 1974; Vincze, 1972; Lebon, Lambermonl, 1973; Lebon,
Casas-Varquez, 1974), распространения положений локальной тер-
модинамической теории Пригожина на нелинейные процессы
(Glansdorff, Prigogine, 1954, 1964; Glansdorff, 1956, 1960; Приго-
жин, 1959, 1960; Prigogine, 1966; Nicolis, 1971, 1972; Глансдорф,
Пригожин, 1973), использования теории графов (Oster, Desoer,
1971; Oster et al., 1971; Katchalsky et al., 1973), использования
обобщенных соотношений взаимности (Стратонович, 1967, 1969,
1971) и глобальных характеристик (Edelen, 1972,. 1973а, Ь, 1974а,
Ь, с, 1975; Преснов, 1973, Преснов и др., 1973; Van Kampen, 1973;
Зотин, 1974; Зотин, Коноплев, Преснов, 1975), вероятностного рас-
смотрения термодинамических процессов (Бахарева, Бирюков, 1969,
1970, 1974; Коноплев, Зотин, 19756; Зотин, Коноплев, Преснов,
1975; Преснов, Зотина, 1975) и т. д. Какое из этих направлений
окажется справедливым и будет положено в основу термодинами-
ки нелинейных необратимых процессов, сказать сейчас трудно.
Однако необходимость этой работы для понимания явлений, про-
исходящих в системах, далеких от равновесия и особенно в био-
логических системах, очевидна. Дело в том, что в биологии имеют-
3
ся данные, показывающие, что термодинамика необратимых про-
цессов может быть использована для решения важнейших проблем
физиологии, биохимии и биологии развития (Katchalsky, Curran,
1965; Morowitz, 1968; Зотин, 1974). Именно факты всегда явля-
лись главным стимулом, который приводил к построению или со-
вершенствованию теории.
Следует также отметить, что внедрение термодинамики в био-
логию долгое время сдерживалось несовершенством методов изме-
рения слабых тепловых потоков, характерных для внутриклеточ-
ных биохимических процессов. Лишь в последние десятилетия
были сконструированы и получили распространение достаточно
чувствительные микрокалориметры (см. Кальве, Прат, 1963; Са1-
vet, Prat, 1963; Boivinet, 1964; Boivinet, Rybak, 1969; Brown,
1969; Wadso, 1970; Lamprecht et al., 1971; Forrest, 1972; Spink,
Wadso, 1975). Можно надеяться, что широкое распространение
и использование этих приборов позволит создать достаточно на-
дежную экспериментальную базу для термодинамики биологичес-
ких процессов.
Данная монография является результатом сотрудничества
двух лабораторий: Лаборатории биофизики развития Института
биологии развития Академии наук СССР и Лаборатории биофизи-
ки Центрального института биохимии и биофизики Западно-
берлинского университета. Все авторы, принимавшие уча-
стие в написании этой монографии, так или иначе связаны с ука-
занными лабораториями. Естественно, что мы не смогли охватить
все разделы биологии, в которых используется термодинамика
необратимых процессов, и ограничились только теми вопросами,
которые разрабатываются в наших лабораториях. В связи с этим
следует подчеркнуть, что основные теории, изложенные в книге,
строятся на базе глобального подхода к нелинейным необратимым
процессам, которого придерживается большинство авторов этой
монографии. Этот подход подробно изложен и обоснован в первых
главах книги.
Материалы, представленные в книге, обсуждались на семинаре
по теоретической биофизике в Физическом институте АН СССР,
на различных семинарах и коллоквиумах в Институте биологии
развития АН СССР, в Центральном институте биохимии и биофи-
зики Западноберлинского университета, в Обществе математи-
ческой и физической биологии ГДР. Всем участникам этих об-
суждений авторы приносят свою искреннюю благодарность.
А. И. Зотин, И. Лампрехт
I
ОБЩИЕ ПРОБЛЕМЫ ТЕРМОДИНАМИКИ
БИОЛОГИЧЕСКИХ ПРОЦЕССОВ
Ни один из разделов теоретической физики не связан с биоло-
гией так тесно и не дает столь эффективного описания процессов
жизни, как термодинамика. Хотя механика, гидродинамика, элек-
тродинамика или оптика находят большое применение в биологии,
они описывают явления, которые не столь характерны для живых
систем, как теплопродукция и связанные с нею процессы метабо-
лизма.
Современной термодинамике присущи и другие чрезвычайно
важные для биологии понятия, такие, как возникшие в 60-х годах
нашего века понятие «стрелы времени» (см. Popper, 1965, 1967а, Ь)
или различные критерии эволюции, характеризующие направление
изменений термодинамических систем (Glansdorff, Prigogine,
1964; Prigogine, 1966; Nicolis, 1971; Zolin, 1972; Преснов, 1973;
Зотин, 1974). Несмотря на это, вопрос об использовании тех или
иных разделов термодинамики в биологии во многом еще остается
дискуссионным (Лазарев, 1945; Ubbelohde, 1947; Шредингер, 1947;
Brillouin, 1949, 1955; Raymond, 1950; Пасынский, 1953, 1963;
Touloukian, 1956; Волькенштейн, 1958, 1965, 1973; Wilkie, 1960;
Bernhard, 1964; Тринчер, 1965; Riegel, 1965; Calloway, 1966; Мол-
чанов, 1967; Blum, 1967; Антонов, 1968; Зотин, 1971, 1974; Кобо-
зев, 1971; Николаев, 1971; Ребане, 1972; Блюменфельд, 1974; Приц,
1974; , Чернавский, 1975 и др.).
Ниже приводятся обзоры материалов о применимости понятий
классической термодинамики и термодинамики линейных необра-
тимых процессов к некоторьш биологическим явлениям, а также
основные положения так называемой глобальной теории нелиней-
ных необратимых процессов, на которую опираются многие теоре-
тические и экспериментальные работы, представленные в этой
книге.
5
ИСПОЛЬЗОВАНИЕ ПОНЯТИЙ
КЛАССИЧЕСКОЙ ТЕРМОДИНАМИКИ В БИОЛОГИИ
И. Лампрехт
Термодинамика возникла около 200 лет тому назад, когда в ре-
зультате экспериментов Лавуазье была разрушена теория флоги-
стона Шталя *. С самого начала своего развития термодинамика
использовала данные о теплопродукции мелких животных, полу-
ченные с помощью калориметров типа Лавуазье — Лапласа. Эта
тесная связь термодинамики и биологии никогда не прекращалась.
Так два врача сделали первые шаги к современной теории: Майер
и Гельмгольц сформулировали первый закон термодинамики.
Позднее в Берлине Рубнер продемонстрировал применимость пер-
вого начала к живым системам в экспериментах по метаболизму
микроорганизмов. С тех пор живая материя является любимой
темой классической термодинамики, а в настоящее время и термо-
динамики необратимых процессов. Нет сомнений, что термодина-
мика ведет к более глубокому пониманию живой материи и пока-
зывает пути решения проблем, которые не могут быть решены дру-
гими способами. В данной работе мы рассмотрим характерные для
термодинамики величины и обсудим их связь с биологическими
проблемами.
Внутренняя энергия U
Хотя существует математический, аксиоматический путь по-
строения термодинамики, в котором функции состояния термодина-
мической системы вводятся как следствия различных законов,
мы используем более эмпирический путь, определяя различные
величины и рассматривая первое и второе начала термодинамики
как обобщение человеческого опыта 2.
Если в системе, отделенной фиксированной или подвижной
стенкой от среды, происходит изменение ее состояния, то это может
происходить только в результате прохождения тепла из системы
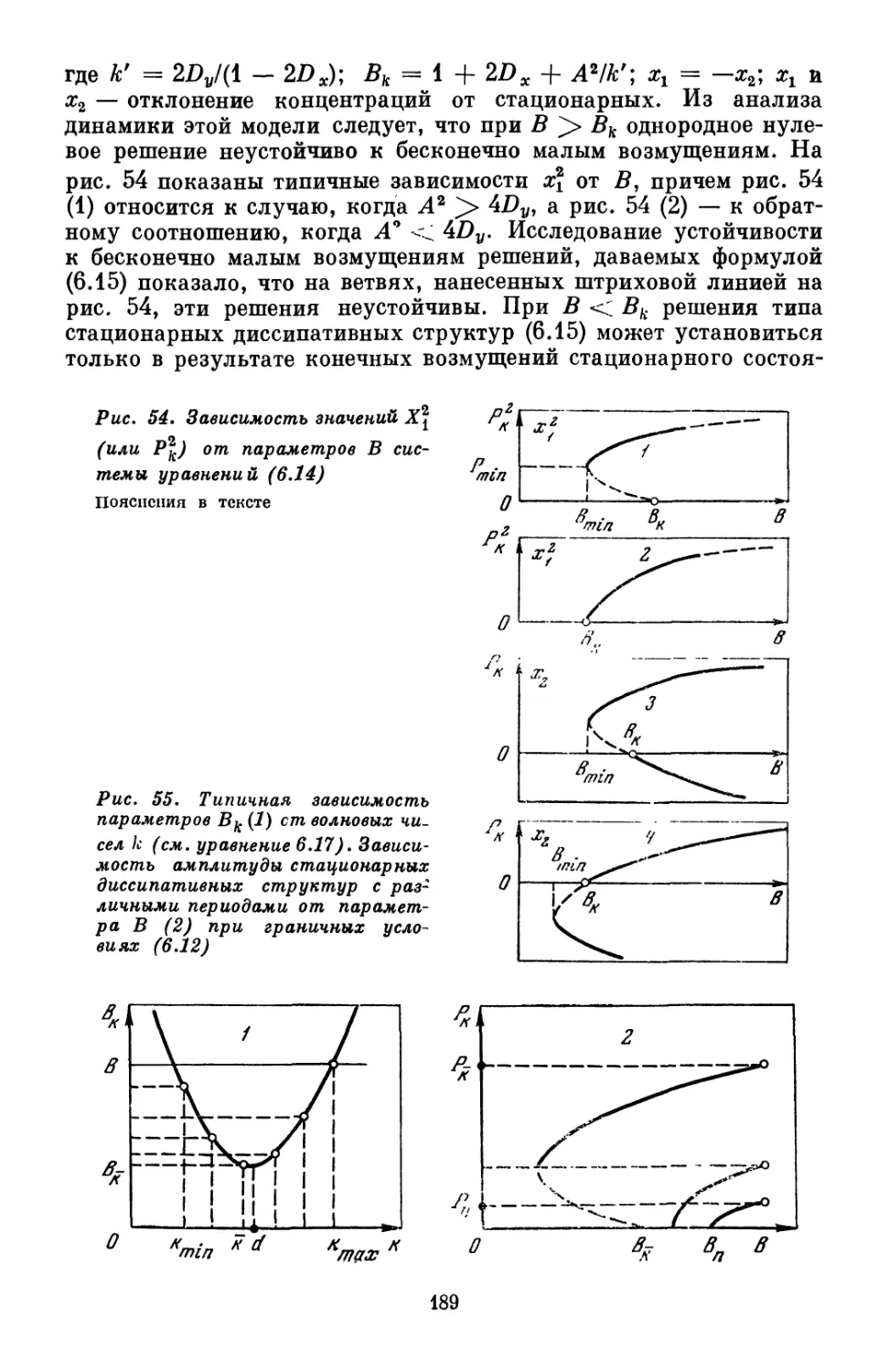
или в систему и совершения работы над системой или самой систе-
мой. Обозначая тепло, входящее в систему, через (?, а работу, со-
вершенную системой, через /1, мы можем записать изменение функ-
ции состояния системы, внутренней энергии U:
Ли = Лф + ЛА. (1.1)
© J. Lamprecht, 1976 г.
1 Подробнее с историей возникновения термодинамики можно познако-
миться в монографии Гельфера (1969). Прим. ред.
2 Планк сформулировал идею, согласно которой невозможность изобретения
вечного двигателя является самым строгим доказательством первого на-
чала термодинамики.
6
Очень часто работа связана с расширением газа внутри системы,
на которую действует внешнее давление. Приписывая отрицатель-
ный знак работе, совершенной системой, мы можем, следуя закону
Бойля—Мариотта, написать: АЛ = — p/W и, подставляя это
значение.в (1.1), получим:
ДС7 = \Q - pbV. (1.2}
Интегрируя (1.2), можно получить абсолютную величину внут-
ренней энергии CZ, однако в большинстве случаев нас будет инте-
ресовать только изменение внутренней энергии АС/, когда система
переходит из одного состояния в другое. Величина \Q может быть
измерена калориметрически, тогда как остальные параметры мож-
но получить только в манометрических экспериментах или путем
расчетов известных реакций. Мы различаем три различных слу-
чая:
а) калориметрический сосуд сообщается с окружающей сре-
дой, так что в нем не происходит изменения давления Др = 0;
б) калориметрический сосуд закрыт, и давление в нем изме-
няется (Др 0), но объем остается константным: ДУ = 0;
в) хотя один из случаев [а) или б)] выполняется, мы принимаем
ДУ = Др = 0, если не возникает газовой фазы в исследуемой
реакции.
Рассмотрим эти различия на простом примере, который имеет
большое значение в биологии. В процессе гликолиза глюкоза рас-
падается в цепи энзиматических реакций до спирта или лактата.
В первом случае этот процесс можно записать в виде уравнения:
F С6Н12Ов -> 2С2Н5ОН + 2СО2.
Для простоты мы возьмем один моль сахара, растворенного в од-
ном литре воды, который помещен в двухлитровый сосуд. Если
сосуд открыт для воздуха (Др = 0), мы можем рассчитать ДУ
для двух молей газа: 2 X 22,4 л = 44800 см3 СО2. Для нормаль-
ного давления в 760 мм рт. ст. имеем: — рДУ = 1,013 кр/см2*
•44800 см3 = 454 мкр ~ 4450 дж ~ 1060 кал.
Изменение тепла в этом процессе равно &Q = —17 ккал/моль.
Поэтому общее изменение внутренней энергии равно: At/ = &Q —
— рДУ = 67 кдж/моль = 16 ккал/моль. В этих расчетах мы молча-
ливо допускаем, что вода насыщена углекислотой и в системе отсут-
ствуют процессы растворения. В большинстве биологических экс-
периментов этими явлениями можно пренебречь.
Когда в процессе гликолиза глюкоза превращается в лактат:
СвН12Ов -> 2СН3СНОНСООН,
образования газа не происходит, и изменение внутренней энер-
гии соответствует продукции тепла в системе, которое равно
151 кдж/моль = 36 ккал/моль. Обычно отклонение от этой вели-
чины тем больше, чем больше образуется газа в процессе метабо-
лизма глюкозы. Так, в случае дыхания на каждый моль глюкозы
7
образуется 6 молей углекислого газа и эффект работы расширения
следует утроить. Однако по сравнению с изменением внутренней
энергии, равным At/ = 678 ккал/моль, этим эффектом также мож-
но пренебречь.
Уравнение (1.1) является одной из формулировок первого нача-
ла термодинамики: вся энергия, получаемая системой, превра-
щается во внутреннюю энергию системы (или более выразительно —
энергия никогда не создается и не исчезает). Этот закон, сформу-
лированный для физических газовых систем, был с успехом про-
верен на живых объектах. В начале этого столетия в серии работ
с микробными культурами Рубнер показал, что энергия, содер-
жащаяся в пище, разделяется в процессе потребления организма-
ми на три части, сумма которых постоянна: 1) переходящая в теп-
ло и выделяющаяся в среду; 2) запасаемая в клеточном материале
(эта энергия выявляется с помощью сжигания объектов в калори-
метрической бомбе); 3) связанная с метаболитами, продуктами
жизнедеятельности организмов.
Позднее Седлачек (Sedlaczek, 1964) повторил эти измерения
с высокой степенью точности на различных бактериях (Е. coli,
Р. vulgaris, S. aureus, Р. fluoresceus и др.). Он сравнивал умень-
шение теплоты сгорания культуральной среды в начале и в кон-
це опыта с суммой теплопродукции и теплосодержания бактериаль-
ной культуры. Рассмотрим один пример из этой серии работ, ка-
сающийся роста Е. coli:
Теплота сгорания (в кал) среды в начале опыта 17 338
То же в конце опыта 16 264
Разница 1074
Теплота сгорания бактериальной культуры 528
Теплопродукция (в кал) во время роста культуры 468
Сумма 996
Разница 1074 — 996 = 78 кал составляет 7,3%, однако если
принять во внимание, что уменьшение энергии в среде получено
как разность двух очень больших величин, эта разница будет нич-
тожно малой (78 кал от 17338 составляет 0,4%). Эти опыты показа-
ли хорошее согласие с первым началом термодинамики.
Энтальпия Н
Как показано выше, изменение внутренней энергии может быть
обусловлено потоком тепла и увеличением объема. Для многих
химических реакций, связанных с образованием газа, существен-
на функция состояния, зависящая от давления, определенная сле-
дующим образом:
Н = U + pV, (1.3)
которую называют энтальпией системы. В дифференциальном виде
8
мы можем написать:,
ДЯ = \U + Д (р7) - \U + Vkp + рДИ,
или, учитывая (1.2):
ДЯ = Д(? - p\V + V\p + рД7 = Д(? + W- (14)
В том случае, когда в течение эксперимента не происходит из-
менения давления (например, если калориметрический сосуд со-
общается со средой) Др = 0, изменение энтальпии равно погло-
щению или образованию тепла в изучаемой реакции. Так как
в большинстве биохимических реакций давление остается постоян-
ным, то увеличение энтальпии является теплотой реакций изучае-
мого процесса.
Мы уже отмечали выше, что вклад в изменение внутренней
энергии, связанный с совершением работы, мал по сравнению с
вкладом тепловых явлений. Поэтому данные о приросте внутрен-
ней энергии можно приравнивать энтальпии и оценивать послед-
нюю с помощью калориметрических измерений.
Температура Т
Для того чтобы перейти к другим термодинамическим функ-
циям состояния и, в частности, к энтропии, следует ввести поня-
тие температуры — понятия, хорошо известного в повседневной
жизни. Температура в противоположность внутренней энергии,
энтальпии и энтропии является интенсивной величиной. Мы при-
нимаем, что два тела, находящихся в контакте, имеют одинаковую
температуру, если отсутствует тепловой обмен между этими телами.
Обычно используют две шкалы температур: шкалу Цельсия,
в которой температурный интервал между точкой замерзания
(принятой за нуль) и точкой кипения воды делят на 100 частей,
и шкалу Фаренгейта, где этот же интервал делят на 180 частей,
причем точке замерзания приписано значение +32. В физике обыч-
но используется шкала Кельвина, где температурный интервал,
как и в шкале Цельсия, делят на 100 частей, но принимается, что
точка замерзания соответствует +273° К. Недавно рекомендовано
использовать только эту шкалу температур.
Температура является величиной, очень характерной для жи-
вых систем. В отличие от внутренней энергии и энтальпии можно
говорить не только об изменении температуры, но и об ее абсолют-
ной величине.
В норме жизнь связана с очень узкой областью температур;
она лежит, примерно, между 10 и 40° С. Высшие организмы имеют
специальные системы адаптаций к экстремальным температурам
в пределах, примерно, от —100 до + 55° С. Известны также тер-
мофильные организмы, в особенности бактерии и водоросли, кото-
рые так хорошо адаптированы к своим биотопам, что могут расти
даже при температурах, близких к +100° С. Можно поэтому ду-
9
мать, что жизнь может существовать до тех пор, пока существует
вода в жидком состоянии. С этой точки зрения температура замер-
зания воды является нижней границей существования жизни,
хотя точка замерзания воды может быть сдвинутой при высоких
концентрациях солей или органических веществ.
Энтропия S
Очень существенной, но часто неверно истолковываемой функ-
цией состояния системы является энтропия — экстенсивная ве-
личина подобно внутренней энергии, энтальпии и объему. При
данной температуре Т и тепловом обмене со средой Д(>, осущест-
вляемом обратимым путем, изменение энтропии системы равно:
Согласно второму началу термодинамики, в изолированной сис-
теме общая энтропия не может убывать. В идеализированных об-
ратимых экспериментах она может оставаться константной (буду-
чи при этом функцией состояния!), но во всех реальных процессах,
происходящих в изолированных системах, энтропия стремится
к максимуму. В этом смысле энтропия тесно связана с изменением
времени и поэтому ее увеличение иногда называют «стрелой вре-
мени».
Больцман смог объяснить эту тенденцию энтропии к достиже-
нию максимального значения в своей статистической теории. Он
показал, что энтропия связана с вероятностью состояния системы
Рт уравнением:
S = к\пРт, (1.5)
где Рт — термодинамическая вероятность состояния системы, к —
константа Больцмана х.
Так как система всегда стремится к более вероятному состоя-
нию, то из (1.5) следует вывод об увеличении энтропии в реальных
процессах. И наоборот, увеличение энтропии, вытекающее из вто-
рого начала термодинамики, соответствует (1.5) увеличению ве-
роятности, т. е. уменьшению порядка, в системе. Так как кажется,
что растущие организмы противоречат этому утверждению, то час-
то этим доказывают неприменимость второго начала в живой ма-
терии. Однако второй закон сформулирован только для изолиро-
ванных систем, а организмы не являются системами, изолирован-
ными от среды. Если мы хотим использовать понятие энтропии
классическим путем, мы должны рассмотреть изолированную сис-
1 Шеннон (Shannon, 1948) вывел идентичное по виду уравнение для коли-
чества информации. Это явилось основанием для введения термина энтро-
пия в теории информации. Хотя имеется подобие между отрицательной
энтропией и информацией, их связь не следует переоценивать: выражение
информации через джоули по меньшей мере бесполезно.
10
тему, включающую в себя организм вместе с окружающей его сре-
дой. В этой новой системе относящаяся к организму часть энтропии
может уменьшаться при одновременном увеличении ее в большой
(неживой) части системы. Несмотря на уменьшение в одной из
частей, общее изменение энтропии системы в целом при этом всегда
положительно.
Согласно Шредингеру (1947), в живой материи имеет место
устойчивый поток отрицательной энтропии (негэнтропии) из окру-
жающей среды, компенсирующий непрерывную продукцию энтро-
пии в системе: организмы питаются негэнтропией! Ту же мысль
в очень современной форме высказал 90 лет тому назад австрийский
физик Больцман (см. Broda, 1970): «Всеобщая борьба за существо-
вание живых существ не является таким образом борьбой за ве-
щества или энергию, которая в форме тепла, к сожалению, не мо-
гущего быть использованным, имеется в достаточном количестве
в каждом теле, а является борьбой за энтропию (точнее, негэнтро-
пию г), возможность использования которой появляется в резуль-
тате перехода энергии от горячего солнца к холодной земле. Чтобы
возможно полнее использовать этот переход, растения простирают
неизмеримую поверхность своих листьев и заставляют солнечную
энергию, прежде чем она дойдет до уровня температуры поверхно-
сти земли, осуществить химические синтезы, о которых мы в на-
ших лабораториях не имеем никакого представления».
Поясним приложимость второго начала в биологии на двух при-
мерах.
а. Животные живут за счет растений или других животных
и получают негэнтропию из них. Но откуда в этом случае получают
негэнтропию сами растения? В первую очередь в результате об-
разования макромолекул из углекислоты и воды в присутствии
света и уменьшения энтропии веществ. Согласно закону радиации,
максимум интенсивности излучаемого спектра соответствует тем-
пературе источника. Температура Солнца Т$ (5500° К) приблизи-
тельно в 20 раз больше температуры поверхности Земли Те (290° К).
Если количество тепла Q, связанного с солнечной радиацией, аб-
сорбируется Землей и частично реизлучается ею, то изменение
энтропии будет:
Величину Д5 можно рассчитать: она равна Д5 = 150 п кал/
/град, моль, где п число квант, необходимых для образования од-
ной молекулы сахара в процессе фотосинтеза:
6СО2 + 6Н2О -> С6Н12О6 + 6О2
с уменьшением энтропии, равном Д5 = —40 кал/град. моль.
Примерно 3 кванта используется в этой реакции, т. е. + 450 кал/
1 Замечание Broda, (1970).
11
/град-моль для компенсации —40 кал/град* моль. Эффектив-
ность в 10 % достаточна для выполнения требований второго начала
(Britten, Gamow, 1961).
Можно также рассчитать максимальную плотность размещения
растений на всей поверхности Земли так, чтобы изменение энтро-
пии, связанное с ростом растений, равнялось потоку энтропии от
Солнца. Эта плотность может быть равна 2-104 грамм растений
на см2 по сравнению с приблизительно 1 г/см2 реально наблюдае-
мым на Земле (Stein, 1960).
б. Другой пример показывает, что пределы, поставленные вто-
рым началом, никогда не достигаются. Размножение микроорга-
низмов может быть представлено в виде (Morowitz, 1960):
1 клетка + субстрат -> 2 клетки + метаболиты.
Можно различать различные стадии этого процесса: поступле-
ние пищи в клетку; встраивание молекул субстрата в клеточную
поверхность; увеличение энтропии в процессе метаболизма суб-
страта; тепловой поток в среду. Разрушение субстрата порядка
0,03 грамма сырого материала на грамм новых клеток достаточно
для соответствия размножения клеток второму началу. Мы же на-
шли для Е. coli эту величину, равную 4,95. Продукция энтро-
пии в этом случае в 150 раз больше необходимой.
В калориметрических опытах с дрожжами мы нашли, что в про-
цессе ферментации AS = + 128 кал/град. моль, тогда как негэн-
тропия клеток равна —7 кал/град. моль. Даже если реакционный
сосуд передает всю теплоту, образовавшуюся в процессе ме-
таболизма, в окружающую среду, поток энтропии равняется
—85 кал/град. моль, и сохраняется положительный баланс, необ-
ходимый для соответствия со вторым началом.
Свободная энергия Р
Согласно первому началу термодинамики, максимальная рабо-
та, которую можно получить, равна:
ДА = AU - AQ
или для обратимых процессов:
ДА = AU - TAS = AF. (1.6)
Свободная энергия F вводится как изменение внутренней энер-
гии системы минус энтропийный член. Если AU не зависит от
температуры, дифференцирование ДА дает:
= - Д5
al
И
&F = ДС7 - Т-^- ,
al
(1-7)
12
где (1.7) — уравнение Гиббса — Гельмгольца. Только при Т = О
или очень малых температурных коэффициентах максимальной
работы имеет место прямой переход внутренней энергии в работу.
Свободная энтальпия
(свободная энергия Гиббса) G
Тем же путем, что и свободная энергия, вводится понятие сво-
бодной энергии Гиббса:
G = H—TS (1.8)
или в дифференциальной форме (при ДТ = 0):
\G = ДЯ- T\S. (1.9)
Изменение свободной энергии Гиббса или свободной энтальпии
существенно для направления химических реакций. Химическая
система может спонтанно изменять свое состояние только в том
случае, если это изменение сопровождается отрицательной Дбг,
т. е. если обратимые реакции совершают работу. Отрицательный
знак Дбг может быть обусловлен экзотермическим эффектом (ДЯ А
<ч0) независимо от знака Д5, строго положительным изменением
энтропии или их комбинацией. Рассмотрим по одному примеру
для каждого случая.
Связывание Я-ацетил-й-глюкозоамина как ингибитора фермен-
та лизоцима в фермент-ингибиторный комплекс. Термодинамичес-
кие показатели имеют следующие величины: ДЯ= —5,6 ккал/моль;
Д5 = 11,4 кал/град. моль; Дб? = —5,6—298 (—0,0114) =
= —2,2 ккал/моль (Bjurulf et al., 1970).
Образование комплекса между а-хемотрипсином и трипсином
поджелудочной железы при pH = 8,0 и 25° С дают следующие тер-
модинамические данные: ДЯ = +3 ккал/моль; А5 = 47 кал/град.
моль; ДО = 3—298 (+0,048) = —11 ккал/моль. Они показывают,
что сильное увеличение энтропии компенсируется для эндотерми-
ческих реакций (Vincent, Ladenski, 1973).
Для гидролиза АТФ при pH = 7,0 и 37° С найдено ДЯ =
= —4,8 ккал/моль; Д5 = 9,45кал/град. моль; AG = —4,8—310*
•(+0,00945) = — 7,7 ккал/моль (Benzinger, Hems, 1956).
Мы видим на этих примерах, что хотя потребления тепла нет
(ДЯ < 0), но свободная энтальпия (ДО < 0) существенна для
спонтанного протекания реакций.
Удельные теплоемкости ср и cv
Менее важными для биологической термодинамики являются по-
нятия теплоемкости при постоянном давлении ср или объеме cv.
Они определяются как температурные коэффициенты внутренней
13
энергии или энтальпии:
/ dU \ . / дН \
Су ~ \ дТ )v’ Ср~ \ дТ /р'
Поэтому теплота, передаваемая системе, приводит к увеличению
энергии U и энтальпии Н:
dU = cvdT; dH = cpdT.
С помощью значений ср и су можно делать выводы относи-
тельно состояния системы, например формы воды в живой клетке
или в растворе альбумина [в смеси 0,3 г воды на 1 г альбумина
содержится «калорически связанная» форма, тогда как остальная
вода является свободной (Hast, Pauly, 1971)1.
Термодинамические измерения
Для того чтобы новые теории, представленные в этой книге,
можно было сопоставлять с экспериментальными результатами,
имеет смысл привести здесь данные, касающиеся техники экспери-
ментальной работы. При этом следует отметить, что основное зна-
чение для обсуждаемых проблем имеет измерение теплопродукции
живых систем. Поэтому прямая калориметрия играет существен-
ную роль в доказательствах теории. Кроме прямой калориметрии,
мы имеем дело с непрямой калориметрией, которая связана с ма-
нометрическими измерениями, измерением концентрации субстра-
тов, pH, напряжения кислорода и т. д.
Различают два типа аппаратуры в прямой калориметрии —
изотермическую и адиабатическую. За исключением некоторых
специальных приложений (разложение навоза или силоса) адиа-
батическая калориметрия менее употребима в биологических экс-
периментах, так как теплота, выделяемая организмами в таких
калориметрах, увеличивает температуру среды, в то время как
при нормальных условиях температура сохраняется постоянной
или периодически меняется с известным временным ритмом. В от-
личие от этого в изотермических калориметрах сохраняется посто-
янная температура, и измеряется поток тепла между реакционным
сосудом и большим термостатируемым блоком. В термопарах (сей-
час чаще всего конструируются так называемые батареи Пельтье)
тепловой поток преобразуется в электродвижущую силу, которая
регистрируется с помощью потенциометров в виде «термограмм».
Эти термогрдммы показывают теплопродукцию за единицу времени
как функцию времени. Суммирование данных дает возможность
судить об общей теплопродукции до известного момента или в те-
чение всего эксперимента. Все другие данные рассчитываются из
двух величин теплопродукции Q = dQldt и общего тепла Q, кото-
рое, как показано выше, приблизительно равно изменению энталь-
пии системы Н.
14
Современные инструменты имеют чувствительность порядка
70—100 мкв/мвт. С помощью некоторых технических усовершен-
ствований, особенно дающих прямую и константную нулевую ли-
нию, можно достичь чувствительности 1 мкв или даже меньше.
Однако превысить этот предел невозможно, так как каждая термо-
пара и усилитель имеют собственный термический шум, который
соответствует типичному напряжению 50 пв. Этот шум может быть
снижен уменьшением температуры, но в биологических калори-
метрах мы ограничены определенной оптимальной для организма
величиной температуры. Чувствительность в 1 мкв соответствует
измерению теплового потока, равного 10 мквт. Растущая культура
дрожжей в обычной среде высвобождает 3,7 «10-12 вт/клетка, рав-
ное 3,2-10“9 кал/час. «клетка. Необходимо использовать прибли-
зительно 3 • 105 клеток дрожжей, чтобы достигнуть наименьшего
предела чувствительности аппаратуры в 10 мквт. Из этого видно,
что мы очень далеки от возможности измерения теплопродукции
единичными микроорганизмами.
Главным инструментом в непрямой калориметрии являются
аппараты Варбурга, которые измеряют продукцию или потребле-
ние газов в процессе метаболизма. Для примера мы рассмотрим
окисление глюкозы дрожжами. Изменение свободной энтальпии
будет, как известно, равно:
СвН12О6 + 6О2 -+ 6СО2 + 6Н2О AG = —686 ккал/моль.
Если углекислый газ поглощать КОН, то можно измерить пот-
ребление кислорода по уменьшению давления в реакционнохм сосу-
де. Эти измерения можно перевести в микролитры потребленного
кислорода, которые соответствуют количеству свободной энталь-
пии, образующейся за это время. В некоторых случаях более под-
ходящим или может быть единственно возможным способом ис-
следования реакций является измерение уменьшения субстратов
или увеличения продуктов реакции с помощью специальных био-
химических тестов или аппаратуры. Если реакции известны, то
можно рассчитать количество свободной энергии, как показано
для манометрических экспериментов.
Все термодинамические понятия, введенные выше, относятся
к классической теории, однако многие из них сохранены и в совре-
менной теории необратимых процессов. В то же время следует от-
метить, что некоторые понятия классической теории трудно ис-
пользовать для описания жизненных процессов и их применение
для этой цели может приводить к ошибкам. Что же это за понятия?
Прежде всего классическая термодинамика имеет дело с за-
крытыми, или изолированными, системами. В то же время нет
организмов, на долгое время изолированных от окружающей среды,
они нуждаются для своего существования в непрерывном потоке
энергии и вещества.
Обратимые процессы классической термодинамики являются
бесконечно медленными — все процессы в живой материи проис-
15
ходят с реальной скоростью и сопровождаются поэтому продук-
цией энтропии в каждый данный момент времени.
Для применения термодинамики необходимо четко определить
фазы системы. Для живых систем это сделать трудно, так как они
содержат множество органелл и небольших изолированных кле-
точных участков, содержащих иногда малое количество макромо-
лекул. Более того, микроорганизмы и отдельные изолированные
клеточные участки и число молекул в них могут быть настолько
малы, что вероятность флуктуаций становится значительной, так
как согласно статистической термодинамике флуктуации концен-
трации пропорциональны квадратному корню из общего числа
молекул.
ВТОРОЕ НАЧАЛО, НЕГЭНТРОПИЯ,
ТЕРМОДИНАМИКА ЛИНЕЙНЫХ
НЕОБРАТИМЫХ ПРОЦЕССОВ
А, И. Зотин
Если по поводу применимости первого начала термодинамики
в биологии давно все уже стало ясным, то относительно возмож-
ности использования второго начала до сих пор продолжаются дис-
куссии. Этот вопрос не раз поднимался на протяжении истории
термодинамики (см. Мейергоф, 1928; Оппенгеймер, 1934; Лазарев,
1945; Пасынский, 1953; Гельфер, 1973 и др.). Например, один из
основоположников термодинамики Герман Гельмгольц, как и та-
кие крупные ученые, как Льюис (1929) и Вернадский (1967), счи-
тали, что второе начало в некоторых случаях не применимо к био-
логическим явлениям. Гельмгольц и Льюис, в частности, думали,
что второе начало неприменимо к вирусам и бактериям, так как,
по их мнению, эти мельчайшие организмы способны избирательно
использовать тепловые флуктуации и нужные молекулы, подобно
«демонам Максвелла». Однако, как показали специальные подсче-
ты, тепловые флуктуации не могут быть использованы живыми
организмами вне зависимости от их размеров (см. Тарусов, 1960).
Еще менее правдоподобно предположение о наличии в живых сис-
темах механизма, подобного «демонам Максвелла» (Мейергоф,
1928).
Сильным аргументом против приложимости второго начала
к живым организмам одно время считали данные расчетов, показы-
вающие, что коэффициент полезного действия животных выше,
чем КПД, вычисленный по формуле ц = 1 — Тг1Тъ где ц —
КПД машины Карно, 7\ и Г2 — температура нагревателя и хо-
16
лодильника. Этот аргумент, однако, потерял всякий смысл как
только стало очевидным, что живые организмы не являются тепло-
выми машинами (Оппенгеймер, 1934; Лазарев, 1945).
К настоящему времени большинство авторов не сомневается
в приложимости основных положений второго начала к живым ор-
ганизмам (Мейергоф, 1928; Оппенгеймер, 1934; Лазарев, 1945;
Butler, 1946; Пасынский, 1953, 1963; Touloukian, 1956; Wilkie,
1960; Тарусов, 1960; Eyringet al., 1960; Morowitz, 1960, 1968; Воль-
кенштейн, 1965; Антонов, 1968; Glaser, 1971; Зотин, 1971; Ребане,
1972; Блюменфельд, 1974 и др.). Однако даже в учебных руковод-
ствах иногда еще встречаются сомнения по этому поводу. Напри-
мер, в известном курсе биофизики Байера (1962) сказано, что вто-
рое начало термодинамики не применимо к живым организмам,
так как, по мнению автора, второе начало справедливо только для
изолированных систем, а живые организмы являются открытыми
системами.
По-видимому, подобного рода заключения связаны с не совсем
точным толкованием смысла второго начала. В основе второго на-
чала термодин мики лежит утверждение, что любые неравновесные
процессы сопровождаются образованием теплоты диссипации. Вот
как, например, формулирует второе начало Пригожин (1960, стр.
стр. 33): «... в любом макроскопическом участке системы прира-
щение энтропии, обусловленное течением необратимых процессов,
является положительным». Эта формулировка эквивалентна фор-
мулировке Планка (1936): «образование тепла путем трения необра-
тимо» и всем остальным формулировкам второго начала. Следова-
тельно, независимо от того имеем ли мы дело с изолированной, за-
крытой или открытой системами, в соответствии со вторым нача-
лом, в результате протекания необратимых процессов в системе
всегда происходит возникновение энтропии диссипации. В откры-
той системе (см. Пригожин, 1960; де Гроот и Мазур, 1964; Popper,
1965 и др.) изменение энтропии системы равно:
dS = deS + dtS, (1.10)
где deS — поток энтропии из среды, d{S — продукция энтропии
внутри системы в результате протекания необратимых процессов.
Согласно второму началу в формулировке Пригожина:
diS 0. (1.11)
Из (1.10) и (1.11) вытекает, что для адиабатически изолированной
системы (deS — 0) имеем:
dS > 0.
Таким образом, справедливость этого неравенства для изолиро-
ванной системы, которое многие считают вторым началом термо-
динамики, является частным случаем соотношений (1.10) и (1.11),
т. е. второе начало применимо к открытым системам, а, следова-
тельно, и к живым организмам.
17
Тринчер (1965) пришел к выводу, что второе начало тер-
модинамики не применимо к живым системам. Все свои рассужде-
ния Тринчер строит на том, что не существует эксперименталь-
ного доказательства применимости второго начала к живым си-
стемам. Легко, однако, показать, что это неверно. И дело не только
в том, что многими авторами косвенными методами показана воз-
можность использования второго начала при описании некоторых
процессов, происходящих в живых системах (Лазарев, 1927; 1945;
Мейергоф, 1928; Оппенгеймер, 1934; Butter, 1946; Пасынский, 1953,
1963; Тарусов, 1960; Eyringet al., 1960; Morowitz, 1960; и др.). Весь
многолетний опыт изучения энергетики животных и растений по-
казывает, что и в состоянии покоя и при выполнении работы про-
исходит непрерывное выделение тепла живыми системами. Теплота,
выделяемая живыми организмами, является теплотой диссипации,
так как в конечном счете она представляет собой продукт дегра-
дации энергии, заключенной в пище. Это означает, что физиоло-
гические процессы в живых организмах, включая различные реак-
ции метаболизма, проницаемость, осмотическую, электрическую
и механическую работы, сопровождаются образованием теплоты
диссипации. Но это и составляет основное содержание второго
начала термодинамики. Таким образом, применимость второго
начала термодинамики к живым системам давно уже доказана
многочисленными опытами по сопоставлению дыхания и тепло-
продукции организмов с теплосодержанием пищи. Более того,
эти наблюдения показали, что степень необратимости процессов,
происходящих в организмах, чрезвычайно велика, т.е. живые
системы далеко отстоят от состояния равновесия. Так, по под-
счетам Кальве и Прата (1963), человек выделяет на 1 г веса 1 ка-
лорию за час, в то время как Солнце — 2-10“4. Другими словами,
1 г человеческого тела выделяет тепла в 10 000 раз больше, чем 1г
Солнца. Человек на бегу выделяет такое же относительное коли-
чество тепла, как большой океанский пароход, дрозофила в по-
лете — как автомобиль на полной скорости, бактерия — как реак-
тивный самолет. Хотя в данных расчетах не учтены соотношения
поверхности к объему у этих объектов (что несомненно не пра-
вильно при сопоставлении таких разных по размерам объектов,
как человек и Солнце), они все же показывают высокую степень
необратимости процессов, протекающих в живых системах.
Большая путаница существует относительно использования
понятия энтропии в биологии. Одни авторы (например, Тринчер,
1965; Blum, 1967) предлагают вообще исключить это понятие из
обихода биологов, другие хотели бы охарактеризовать такие
сложные явления, как развитие зародышей (Быховский, 1965),
старение (Нагорный, 1938; Sacher, 1967), эволюцию организмов
(Быховский, 1965; Campbell, 1967) и биосферы (Ребане, 1972),
используя представления об изменении энтропии. Эта путаница,
как нам кажется, возникла главным образом в результате несколь-
ко произвольного толкования работ Шредингера (1947) и Брил-
18
люэна (Brillouin, 1949, 1955; Бриллюэн, 1960, 1962), в которых
сделана попытка использовать понятие энтропии для характери-
стики упорядоченности и организованности биологических про-
цессов.
По Шредингеру (1947), живые организмы «питаются» отрица-
тельной энтропией и таким путем компенсируют увеличение энтро-
пии в системе, происходящее в результате процессов жизнедея-
тельности. Само по себе это утверждение, если его не понимать
слишком буквально, не вызывает особых возражений с точки
зрения термодинамики открытых систем. Действительно, в ней
принимается, что в стационарном состоянии открытая система
характеризуется равенством:
dS dS d.S
dt dt ' dt
(1.12)
системе,
потоком
и следовательно:
d{S dS
dt dt
Другими словами, продукция энтропии в открытой
находящейся в стационарном состоянии, компенсируется
энтропии, идущем из среды. В этом смысле можно говорить, что
открытые системы (в том числе и живые организмы) «питаются»
отрицательной энтропией (Bakke et al., 1955; Хаазе, 1967). Не-
смотря на близость этих понятий, последнее из них менее удачно,
так как создает иллюзию, что процессы, происходящие в живых
организмах, можно охарактеризовать понятиями увеличения или
уменьшения энтропии системы. Между тем, давно уже стало оче-
видным, что открытую систему, в отличие от изолированной или
закрытой, нельзя продуктивно описывать, используя понятие
увеличение или уменьшение энтропии системы, так как энтропия
открытой системы может увеличиваться, уменьшаться или оста-
ваться неизменной в зависимости от того, будет ли поток энтропии
больше продукции энтропии, меньше или равным ей (Волькен-
штейн, 1965).
Путаница у последователей Шредингера возникла также в
результате его попытки связать понятие отрицательной энтропии
с понятием порядка и организованности живых систем. По Шре-
дингеру (1947), «энтропия, взятая с обратным знаком, есть сама
по себе мера упорядоченности» и организм поддерживает себя на
высоком уровне упорядоченности благодаря тому, что он «не-
прерывно извлекает упорядоченность из окружающей среды».
Однако, как было сказано раньше, поток энтропии (отрицатель-
ная энтропия) необходим для компенсации возникновения энтро-
пии в организмах, а не для поддержания высокого уровня упоря-
доченности этих процессов. По сути дела в этом случае, как отме-
чает ряд авторов (Кобозев, 1948; Пасынский, 1957, 1963; Wilkie,
1960; Тарусов, 1960 и др.), речь идет о потоке свободной энергии,
19
идущей на компенсацию убыли свободной энергии в организмах.
Это можно показать для любой открытой системы, включая жи-
вые системы, в которых осуществляются тепловые изменения,
совершается работа и протекают химические реакции (см. Tol-
man, Fine, 1948; Eyring et al., 1960). Приведем доказательство для
закрытой системы, которое легко распространить и на открытые
системы. Первое начало для системы, в которой протекают хими-
ческие реакции, можно записать:
dU = TdS — pdV - У, Akdt,k,
k=l
где dU — изменение внутренней энергии системы, р — давление,
V — объем, А — сродство, — степень полноты реакции к. Тер-
модинамический потенциал Гиббса (свободная энергия при по-
стоянном объеме) из (1.8) и (1.3) равен:
G = U + pV - TS.
Учитывая это соотношение, получим:
dG = —SdT + Vdp — '^\Akdt,k.
fc=l
При условии Т = const; р = const:
m
dG=-^Akd^k.
fc=l
Известно (см. Пригожин, 1960; Хаазе, 1967 и др.), что для хи-
мических реакций выполняется равенство:
dis _ 1 V л
at ~ т dt *
Поэтому имеем:
dis 1 dG
dt ~~ T~~dT'
Следовательно, скорость продукции энтропии в системе равна
убыли свободной энергии. Учитывая (1.12), мы можем сказать,
что в открытой системе, находящейся в стационарном состоянии,
поток энтропии равен потоку свободной энергии, идущей из сре-
ды в систему:
deS _ 1 dG
dt Т dt '
Отсюда следует, что поток энтропии (отрицательная энтропия)
20
связан не с поддержанием упорядоченности системы, а с уменьше-
нием свободной энергии в живых организмах, происходящим в
результате протекания в системе необратимых процессов.
Судя по всему, Шредингер (1947) предполагал, что вся отри-
цательная энтропия, поступающая в живые организмы, идет на
построение порядка или, как теперь иногда говорят, на увеличе-
ние информационного содержания системы. Более осторожно к
этому вопросу подошел Бриллюэн (1960, 1962), который ввел
специальный термин — негэнтропия, аналогичный, но не иден-
тичный термину отрицательная энтропия. Он сделал попытку
связать термодинамическое понятие энтропии (в статистическом
истолковании — мера беспорядка) с понятием информации (мера
порядка). Для этой цели Бриллюэн использует понятие связанной
информации, как информации, которой можно поставить в соот-
ветствие некоторое микросостояние физической системы. Таким
путем можно связать понятие информации с некоторыми чисто
физическими изменениями, происходящими в системе. Бриллюэн
(1960) показал, что связанная информация равна убыли энтропии
или увеличению негэнтропии. Это, так называемый, «негэнтро-
пийный принцип информации», согласно которому информация
может быть превращена в негэнтропию и обратно. Связь между
информацией и энтропией в этом случае определяется тем, что
для получения, хранения и использования информации необхо-
димо совершить некоторые физические действия, которые неиз-
бежно сопровождаются изменением энтропии системы. Энергия,
необходимая для чтения информации, как считает Бриллюэн,
доставляется негэнтропиёй. Подсчет показал, что количество тре-
буемой негэнтропии на каждый бит информации равно 10“16 CGS
(Бриллюэн, 1960). Это показывает, что реальная доля изменения
энтропии системы, связанная с чтением информации, ничтожно
мала по сравнению с обычными изменениями энтропии, происхо-
дящими в физических системах. Это же показано и другими авто-
рами (Morrison, 1964; Сырников, 1965).
К сожалению, даже для такой ограниченной задачи, о которой
говорит Бриллюэн, понятие негэнтропии не необходимо и вполне
перекрывается понятием энтропии (Woolhause, 1967а, b; Wilson,
1968 а, Ь). Для этих целей вполне достаточно понятие потока энт-
ропии, часть которого используется для компенсации продукции
энтропии в системе, связанной с осуществлением информационных
процессов.
Попытка связать термодинамику с теорией информации и та-
ким путем получить возможность судить о степени упорядоченно-
сти живых систем, естественно, вызывала большой интерес у био-
логов и физиков, интересующихся проблемами биологии. Выска-
заны различные точки зрения и в том числе такие экстравагантные,
по выражению Вулхауза (Woolhause, 1967а), как утверждение,
что поток негэнтропии в живых системах равен продукции энтро-
пии (Buchel, 1967; Тринчер, 1965; Быховский, 1965, 1966). Экстра-
21
вагантность этой точки зрения состоит в том, что в данном слу-
чае вся теплопродукция организма приравнивается негэнтропии,
т.е. увеличению содержания информации в организме.
Вообще вопрос об изменении количества информации в орга-
низме в процессе его жизни остается во многом неясным. Так,
Бриллюэн (Brillouin, 1955) пришел к выводу, что колйчество
связанной информации не должно возрастать во время развития
животных. Равен (1964), наоборот, считает, что количество инфор-
мации во время развития и роста организмов возрастает в огром-
ных количествах. Эптер и Вольперт (Apter, Wolpert, 1965) пришли
к выводу, что проблемы изменения содержания информации в
живых системах в настоящее время вообще не существует, так как
количество информации в организмах не поддается измерению.
С нашей точки зрения, Эптер и Вольперт правы и еще рано гово-
рить о количестве, ценности или измерениях информации в живых
организмах. Единственно, о чем можно твердо говорить, это то,
что если даже в процессе жизни организмов происходит увеличение
содержания информации, для оценки ее количества нельзя исполь-
зовать понятие негэнтропии или отрицательной энтропии. В ор-
ганизмах, как и в любой другой открытой системе, поток энтро-
пии из среды необходим для компенсации продукции энтропии
в системе, возникающей в результате протекания необратимых
процессов.
Следует особо подчеркнуть, что хотя для живых организмов
несомненно справедливо второе начало термодинамики и понятие
энтропии, представление об увеличении или уменьшении энтро-
пии системы мало что дает при характеристике процессов, проте-
кающих в живых системах. Гораздо большее значение имеют
такие понятия, как продукция энтропии в системе и поток энтро-
пии из среды. Здесь особенно отчетливо проявляется различие
в подходах классической термодинамики и термодинамики необ-
ратимых процессов: классическая термодинамика имеет дело с
изменением величин (энтропия, объем, масса и т.д.), термодинамика
необратимых процессов — со скоростями изменения величин (про-
дукция энтропии, поток энтропии и др.).
Как известно, понятие времени не входит в соотношения клас-
сической термодинамики. Поэтому только с созданием термоди-
намики линейных необратимых процессов, только с введением
времени и понятия скоростей процессов стало возможным исполь-
зование термодинамики для описания биологических явлений.
Теория, которая легла в основу современной термодинамики ли-
нейных необратимых процессов, была создана сравнительно давно
(Onsager, 1931а, Ь), однако ее быстрое развитие и широкое при-
менение в различных областях химии, физико-химии и физики
началось лишь в последние 25—30 лет (Prigogine, 1947, 1955;
Денбиг, 1954; де Гроот, 1956; Пригожин, 1960; Лыков, Михайлов,
1963; де Гроот, Мазур, 1964; Хаазе, 1967; Glaser, 1971; Залевски,
1973; Зотин, 1974 и др.). В основе термодинамики линейных необ-
22
ратимых процессов, кроме уравнений и законов классической тер-
модинамики, лежат следующие положения.
1. Линейные законы, согласно которым вблизи от равновесия
для необратимых процессов выполняется соотношение:
п
(i = 1,..., n), (1.13)
где Ji — удельные термодинамические потоки, Х7- — термодина-
мические силы, Lu — феноменологические коэффициенты.
2. Соотношение взаимности Онзагера, согласно которому пе-
рекрестные феноменологические коэффициенты в уравнении (1.13)
равны между собой:
= (1.14)
Это положение доказывается методами статистической физики
(см. Onsager, 1931а, Ь; де Гроот, 1956; де Гроот, Мазур, 1964).
Иногда соотношение взаимности Онзагера принимают без доказа-
тельств в качестве нового начала термодинамики (Miller, 1956,
1960; Хаазе, 1967).
3. Утверждение, согласно которому сумма произведений по-
токов на силы равна скорости возникновения энтропии в системе:
>=1
Уравнение (1.15) можно получить исходя из различных термоди-
намических соображений. Оно показывает, что протекание необ-
ратимых процессов в системе всегда сопровождается возникно-
вением энтропии в результате диссипации энергии.
Экспериментальной основой термодинамики линейных необ-
ратимых процессов являются многочисленные феноменологические
законы, устанавливающие наличие пропорциональности между
скоростью того или иного процесса и силами, вызывающими этот
процесс. Например, закон Фурье связывает тепловой поток с
градиентом температуры
Q------grad Т.
Закон Фика — поток диффундирующего вещества с градиентом
концентраций:
~ grad cm,.
Закон Ома — силу тока с разностью электрического потенциала:
ё — grad v
и т.д.
23
Применимость соотношений термодинамики линейных необ-
ратимых процессов ограничена областью изменений системы неда-
леко отстоящей от состояния равновесия, когда в изучаемом про-
цессе выполняется условие линейности между потоками и силами
(1.13). Как показывает опыт, эта область для некоторых процес-
сов (диффузия, теплопередача и др.) довольно велика, в то время
как для других процессов (химические реакции и др.) она ограни-
чена узкой зоной вблизи состояния равновесия.
Важнейшим понятием термодинамики линейных необратимых
процессов является понятие стационарного состояния, которое
позволяет значительно расширить область применимости этого
раздела термодинамики. В отличие от изолированных систем,
которые по мере протекания в пей необратимых процессов при-
ближаются к состоянию равновесия, открытые системы могут до-
стигать не равновесного, а стационарного состояния. Отличие
стационарного состояния от равновесного заключается в том,
что в этом состоянии макроскопические процессы не прекращаются,
как в случае равновесного состояния, а идут с постоянной
скоростью. Основным вопросом теории стационарного состояния
системы является вопрос о критериях такого состояния. В тер-
модинамике линейных необратимых процессов принимается, что
стационарное состояние может быть охарактеризовано экстремаль-
ным принципом. В частности, согласно теореме или принципу
Пригожина: в станционарном состоянии при фиксированных внеш-
них параметрах, скорость продукции энтропии в системе постоян-
на по времени и минимальна по величине. Из этой теоремы следует,
что если система по той или иной причине выведена из стационар-
ного состояния, то она будет изменяться до тех пор, пока удель-
ная скорость продукции энтропии не примет наименьшего значе-
ния. Другими словами, эволюция открытой системы к стационар-
ному или равновесному состоянию описывается неравенством:
(1Л6)
где
Т d,S
* = ТТГ. О-*’)
ф — удельная диссипативная функция системы, V — объем си-
стемы.
Мы не будем останавливаться на доказательствах этой теоремы
(их можно найти в монографиях Prigogine, 1947, 1955; Пригожин,
1960; де Гроота, 1956). Отметим только, что в этом доказательстве
используются линейные законы и соотношение взаимности Онза-
гера, т.е. теорема Пригожина справедлива для области линейных
необратимых процессов, а не для любых необратимых процессов.
Как подчеркивается в теореме Пригожина, стационарное состоя-
ние наступает при неизменных внешних параметрах системы.
Это утверждение не является абсолютно необходимым условием
24
появления стационарного состояния: оно может наступить и при
изменяющихся внешних параметрах системы в том случае, если
их изменение происходит значительно медленнее, чем изменение
параметров самой системы. Понятие стационарного состояния
поэтому предполагает существование двух масштабов времени.
Когда по тем или иным причинам в системе и в среде имеется двой-
ной масштаб времени протекания тех или иных процессов, то пере-
ход системы к состоянию равновесия сам собой распадается на
два этапа: появление стационарного состояния и эволюция ста-
ционарного состояния к равновесию.
Хотя применимость термодинамики линейных необратимых
процессов ограничена областью систем, недалеко отстоящих от
равновесия или стационарного состояния, она с интересом была
встречена биологами и использована для описания таких явлений,
как проницаемость и активный транспорт, мышечное сокращение
и процессы возбуждения, развитие и рост животных (см. Kat-
chalsky, Curran, 1965; Kedem, 1965; Caplan, 1966; Wilkie, Wo-
ledge, 1967; Антонов, 1968; Rappaport, 1968; Glaser, 1971; Нико-
лаев, 1971; Van Uden, 1971; Зотин, 1974 и др.). Одновременно
выяснились большие трудности ее использования в биологии и
насущная необходимость в построении термодинамики нелиней-
ных необратимых процессов. В данной книге большинство попы-
ток описания биологических явлений опирается на так называе-
мую глобальную теорию нелинейных необратимых процессов,
которая подробно излагается ниже.
ФОРМАЛИЗМ НЕРАВНОВЕСНОЙ
ФЕНОМЕНОЛОГИЧЕСКОЙ ТЕРМОДИНАМИКИ
Е. В. Преснов
В основе термодинамического описания биологических про-
цессов должна лежать нелинейная теория необратимых процессов,
так как живые системы являются системами, далекими от равно-
весия. Поэтому мы должны уделить особое внимание рассмотре-
нию современного состояния термодинамики и выбрать или разра-
ботать нелинейную теорию, которую можно было бы положить
в основу рассмотрения процессов развития, роста и старения ор-
ганизмов. Здесь неизбежен некоторый субъективизм, который
может быть оправдан только успешным применением полученных
общих результатов.
25
Общие положения
Современная термодинамика необратимых процессов ведет свое
начало с классических работ Онзагера (Onsager, 1931а, Ь). Даль-
нейшее ее развитие мы находим в работах де Донде (de Donder,
1927; de Donder, Rysselberghe, 1936), Казимира (Casimir, 1945)
и некоторых других ученых. К середине 60-х годов термодинамика
необратимых процессов превращается в достаточно обширную
и содержательную теорию, имеющую большое прикладное зна-
чение (см. Денбиг, 1954; Meixner, 1954; де Гроот, 1956; Пригожин,
1960; Лыков, Михайлов, 1963; де Гроот, Мазур, 1964; Хаазе, 1967).
Однако в отличие от классической термодинамики, имеющей хо-
рошую аксиоматическую базу (см. Boyling, 1972, 1973), термоди-
намика необратимых процессов пе имеет еще аксиоматической
основы, что лишает ее необходимого обоснования. В связи с этим
следует уделить внимание прежде всего математическому форма-
лизму неравновесной феноменологической термодинамики, явля-
ющемуся основной составной частью любой аксиоматической мо-
дели.
В свою очередь ясно, что для построения аксиоматической ос-
новы термодинамики следует либо формализовать понятие энтро-
пии физической системы, либо отказаться от такового (Мейкснер,
1970). К сожалению, корректное физическое определение энтро-
пии существует сейчас только для равновесных систем. Несмотря
на большое количество попыток определения неравновесной энтро-
пии (Prigogine et al., 1973; Hosokawa, 1973; Landauer, 1973; Пе-
летминский, Соколовский, 1974 и др.), единого мнения в этом
вопросе пока нет. Обычно ограничиваются рассмотрением случая
таких неравновесных систем, когда выполняется принцип локаль-
ного равновесия, предложенный Пригожиным (Prigogine, 1947).
Пусть дана термодинамическая система, разделенная на мик-
роскопически большие и макроскопически малые подсистемы.
Предполагается, что, хотя в целом система неравновесна, каждая
из подсистем находится в состоянии равновесия (принцип локаль-
ного равновесия). Это предположение дает возможность определе-
ния для неравновесной системы понятия энтропии в тех же са-
мых независимых переменных, что и в случае равновесной систе-
мы. Соответственно, для такой системы в локальной форме будет
справедливо уравнение Гиббса:
п
Т ds = du 4- pdv -—
i=i
где v = p"1 — удельный объем (p — плотность массы), p — дав-
ление, rii — массовые концентрации, рг- — химический потенциал
(па единицу массы), Т — температура, s, и — плотности энтропии
26
и энергии (на единицу массы), причем выполняется функциональ-
ная зависимость:
s = s (и, v, rtf).
Как оказалось, предположение локального равновесия охва-
тывает довольно большой круг задач, соответствующих системам,
далеким от термодинамического равновесия (Nicolis et al., 1969;
Глансдорф, Пригожин, 1973).
Согласно Пригожину (1960), изменение энтропии открытой
системы можно представить в виде (1.10), а второе начало термо-
динамики — в виде (1.11). Важнейшим понятием в локальной
термодинамической теории Пригожина является локальная ско-
рость продукции энтропии о:
d-S (•
^ = -2-= Udr, (1.18)
ttt J
V
где V — объем, занимаемый системой. Предположение локального
равновесия дает:
о > 0. (1.19)
Локальное производство энтропии о является основным понятием
феноменологической термодинамики и вычисляется, исходя из
уравнения Гиббса по формуле (Пригожин, 1960; де Гротт 1956;
Nicolis, 1971):
п
б = (1.20)
г=1
Данная формула представляет термодинамическую величину а
в виде билинейной формы соответствующих потоков связанных
с необратимыми процессами, и обобщенных сил вызывающих
эти потоки. Тензорный ранг потоков и сил, сопряженных между
собой, должен быть один и тот же, однако из формулы (1.20)
следует, что тензорным характером сил и потоков можно прене-
бречь, сводя их к простой совокупности скалярных величин (Седов,
1973; Глансдорф, Пригожин, 1973; Дярматы, 1974; Nicolis, 1971;
Edelen, 1974а, b).
Заметим, что потоки и силы в тепловом равновесии отсутствуют.
Продукция энтропии в равновесии также равна нулю.
Феноменологические уравнения
В феноменологической термодинамике предполагается суще-
ствование неких феноменологических законов, связывающих тер-
модинамические потоки и силы:
7i = 7i (•£].» • • • , #п)« (1.21)
27
В линейной аппроксимации законов (1.21):
п
ji — У1) (1.22)
/С=1
Уравнение (1.22) справедливо для необратимых процессов, про-
текающих вблизи равновесия. Второй закон термодинамики по-
зволяет утверждать, что диагональные коэффициенты 1ц неотри-
цательны (1и >0). Более того, показано (Onsager, 1931а, b; Casimir,
1945; Meixner, 1973), что всегда можно выбрать потоки и силы
такими, что матрица (lik) будет симметрична (соотношения взаим-
ности Онзагера— Казимира):
I* = hi (1 < (1.23)
Соотношения (1.23) широко обсуждаются в литературе и, в
частности, в полемических работах Трусделла (1970а, Ь) и Мейк-
снера (1970, Meixner, 1973). Особо следует подчеркнуть такое важ-
ное обстоятельство, что при доказательстве соотношений (1.23)
учитывалась только лишь квадратичная часть величины Д5,
выражающей отклонение энтропии от равновесия в зависимости
от внутренних параметров системы. Холмс и Мортимер (Holmes,
Mortimer, 1967), учитывая все члены в тейлоровском разложении
величины Д5 по степеням внутренних параметров, получают
«почти» симметричную матрицу (Zifc), т. е. соотношения взаимности
верны с точностью до бесконечно малых величин (см. также По-
пов, 1955). Последнее заставляло многих авторов искать иные
формулировки соотношений взаимности, обобщающих соотноше-
ния (1.23) в нелинейной теории необратимой термодинамики.
Прежде всего остановимся на математическом формализме,
который используется в нелинейных теориях. Будем интерпрети-
ровать потоки Д (х), . . ., jn (х), как компоненты некоторого век-
торного поля G в n-мерном евклидовом пространстве Rn с коор-
динатным репером . . ., хп. Тогда мы можем воспользоваться
известной теоремой (см., например, Картан, 1971). Соотношения:
д/. di\.
4 = 4 (1.24)
являются необходимым условием потенциальности векторного
поля G, а в случае односвязности области определения этого
векторного поля и достаточным условием его потенциальности.
Потенциальность векторного поля G можно записать компактно
таким образом:
grad I (х) = G, (1.25)
где I (х) скалярная функция.
Легко видеть, что нелинейные соотношения (1.24) являются
обобщением соотношений (1.23). Более того, в указанном форма-
28
лизме содержится не только формализм линейной теории Онза-
гера (Onsager, 1931а, Ь), но также и формализм нелинейных
теорий Дярматы (Gyarmati, 1969). Впервые соотношения (1.24)
были записаны Дярматы (Gyarmati, 1961), а доказаны для элект-
рических цепей Брайтоном и Мозером (Brayton, Moser, 1964).
В последнее время эти соотношения широко обсуждаются в лите-
ратуре (Edelen, 1972, 1973а, b; Lavenda, 1972; Oster et al., 1973;
Van Kampen, 1973; Farkas, Noszticzius, 1972; Lambermont, Lebon,
1972).
Как уже говорилось, потенциальность векторного поля и со-
отношения взаимности, по^существу, являются эквивалентными
положениями, поэтому при построении термодинамической тео-
рии обычно пытаются из каких-либо физических соображений,
обосновать либо то, либо другое. Приведем два различных при-
мера, иллюстрирующих подобные построения:
1-й пример. Формальное доказательство соотношений (1.24)
возможно на основаниях постулата Кельвина о независимости
процессов (Li, 1958; Pitzer, 1961; Oster et al., 1973). Так, если
постулировать существование невзаимодействующих «кинетиче-
ски независимых» процессов:
j'k = ёк С4) (1 < к < т), (1.26)
а также, что любой произвольный поток ft есть линейная комбина-
ция независимых потоков
7И
7i = (1.27)
/£=1
тогда из (1.27) и (1.20) получим, что силы, сопряженные потокам
4, даются выражением:
а-; = ^01^. (1.28)
i=l
Отсюда следует равенство:
А = У, Ягкёк ( У, , (1.29)
/С=1 1 = 1
а значит и соотношения:
Впрочем, ограниченность постулата Кельвина влечет частный
характер приведенных рассуждений (Oster et al., 1973).
29
2-й пример. Потенциальный характер (1.25) носит также и
квазиоднородная теория Циглера (1966). Она заключается в том,
что рассматриваются функции а (х1? . . . , хп), удовлетворяющие
функциональному уравнению:
(1.31)
К=1 к
где / произвольная функция. Далее Циглер предлагает записы-
вать феноменологические законы (1.21) в таком виде:
<5 дз
7(<з) 9
(1.32)
Эти законы выводятся у него из некоего принципа ортогонально-
сти, который имеет и физическое обоснование (Ziegler, 1970).
Законы (1.32) приводят к потенциалу:
ф = С1-33)
и значит удовлетворяются соотношения (1.24). Полученный по-
тенциал ср удовлетворяет дополнительному условию, вытекающему
из (1.20);
= (г34)
г=1 г
Функции а, удовлетворяющие уравнению (1.31), Циглер назы-
вает квазиоднородными, поскольку соотношение (1.31) является
обобщением условия однородности:
= (1.35)
/с=1 *
(здесь а суть однородная функция степени г). Автор также гово-
рит о том, что квазиоднородные функции могут служить для ап-
проксимации нерегулярных функций а с определенной степенью
точности.
Полная потенциальная теория построена в работах Эделена
(Edelen, 1974а) и наших (Преснов и др., 1973; Преснов, 1973).
Кроме того, Остер и др. (Oster et al., 1973), интепретируя термо-
динамическую систему посредством графа связей, также приходят
к соотношениям (1.24). Они предлагают абстрактную топологи-
ческую модель термодинамики необратимых процессов, в которой
соотношения взаимности (1.24) выступают в роли лагранжевого
подмногообразия 2п-мерного евклидового пространства. Интересно
отметить, что взаимность в этой модели не является внутренним
свойством системы, а зависит от выбора координатной системы
30
для лагранжевого подмногообразия (ср. с аналогичными сооб-
ражениями для соотношений взаимности Онзагера — Казимира
(Трусделл, 1970а, б; Мейкснер, 1970; Meixner, 1973). Необхо-
димо, впрочем, отметить и то, что при всей привлекательности
модели Остера, она еще не имеет своих оригинальных резуль-
татов, хотя в ее пределах находятся все идеи термодинамической
теории Глансдорфа — Пригожина.
Наконец, важно заметить, что потенциальный характер разоб-
ранных теорий лишает возможности рассмотрения систем, нахо-
дящихся в магнитном поле. По-видимому, в подобных случаях
следует воспользоваться общей термодинамической теорией, раз-
рабатываемой Эделеном (Edelen, 1972, 1973а, Ь, 1974а — d),
которая будет сформулирована несколько ниже.
Вариационная техника
и экстремальные принципы
необратимой термодинамики
Обсуждавшиеся выше феноменологические законы, а также
соотношения взаимности позволяют изучать большое число не-
обратимых явлений. Однако современная термодинамика разви-
вается и в ином направлении: в направлении поиска вариацион-
ных принципов. Здесь главный вопрос заключается в следующем:
существует ли общий принцип, характеризующий неравновесные
состояния независимо от деталей явления, происходящих в сис-
теме? Остановимся более подробно на этом вопросе. В изолирован-
ной системе deS — 0 и согласно второму закону энтропия системы
возрастает, пока не достигнет своего максимального значения.
Система таким образом стремится к состоянию термодинамиче-
ского равновесия. Если же мы будем рассматривать открытую
систему, которая может обмениваться с внешним миром массой
и энергией (или даже замкнутую систему), то при условии достаточ-
но больших внешних резервуаров, неизменяющихся со временем,
система может стремиться к постоянному режиму, отличному от
равновесия. Пригожин называет такое состояние стационарным
неравновесным состоянием. Но этот режим уже больше не харак-
теризуется максимумом энтропии (так как deS Ф 0) или минимумом
свободной энергии. Другими словами, вариационные принципы,
справедливые в тепловом равновесии, не применимы за пределами
этого состояния. Поэтому поставленный Пригожиным вопрос,
сформулированный выше, заключается в том, чтобы найти новые
принципы, которые обобщают понятие термодинамического потен-
циала для стационарных (или медленно меняющихся со временем)
неравновесных состояний.
Первые попытки поиска подобных вариационных принципов
предпринимались для области линейной термодинамики (Onsa-
ger, 1931а, Ь; Пригожин, 1960; Дярматы, 1965; Бахарева, 1968;
Biot, 1970; Lambermont, Lebon, 1972), а впоследствии и для не-
31
линейной термодинамики (Циглер, 1966; Ziegler, 1970; Глансдорф,
Пригожин, 1973). Кроме того имеются попытки обобщения прин-
ципов линейной теории на нелинейную область (Vojta, 1967; Ба-
харева, Крылов, 1971; Farkas, Noszticzius, 1972; Lambermont,
Lebon, 1972; Дярматы, 1974). Подчеркнем, что последние из
перечисленных работ не имеют не только доказательств своей
правомочности (в первую очередь, конечно, физических), но и
четко определенного физического смысла. Авторы ограничились
лишь косвенными аргументами в пользу своих принципов. Здесь
уместно вспомнить несколько резкую, но справедливую критику
Трусделла (1970а, б).
Дискуссионны, с нашей точки зрения, и попытки обобщения
линейных принципов и автоматический перенос их на нелинейную
теорию. Поскольку экстремальные принципы имеют немалую
популярность, приведем аналитические выражения этих прин-
ципов.
Для необратимых процессов вблизи равновесия и для систем,
находящихся в механическом равновесии, при постоянных тер-
модинамических силах и варьируемых макроскопических потоках
справедливо равенство (Onsager, 1931а):
6 [б (7, X) — е (/, 7)]ж = о, (1.36)
где
п
(1-37)
г, К=1
причем матрица" (Ь^) обратна матрице (Zif£) системы (1.22) и сим-
метрична. Ясно, что принцип наименьшей диссипации энергии
(1.36) эквивалентен (что легко проверить варьируя (1.36)) линей-
ным законам:
п
(1.38)
/1=1
с соотношением взаимности:
bik = bki, (1.39)
которые в свою очередь тождественны законам (1.22) и (1.23).
Законы же (1.22) и (1.23) эквивалентны условию потенциальности
потоков. Принцип наименьшей диссипации таким образом экви-
валентен предположению о потенциальности потоков.
Дярматы (1965, 1974) дал дальнейшее развитие принципа Он-
загера (1.36), основываясь на указанных выше эквивалентностях.
Он воспользовался двойственной (1.37) функцией:
п
<р=4" У,
г, к—1
32
и предложил следующий вариационный принцип:
б[о (/, х) — 0 (/, /) — ср (х, z)] = 0, (1.41)
где варьирование происходит как по силам, так и по потокам,
но каждый раз варьируются либо только потоки, либо только си-
лы [нелинейное обобщение этого принципа дал Эделен (Edelen,
1974а)]. Таким образом, принцип (1.41) состоит из принципа Он-
загера и вариационного принципа, записанного в виде:
б[о (/, х) — ф (ж, z)]; = 0. (1-42)
Ясно, что эквивалентности, имеющие место для принципа Онза-
гера, справедливы и в случае принципа Дярматы. Заметим также
и то, что впервые варьировал функцию [о (/, х) — 0 (/, /) — ф (х,х)]
Оно (Опо, 1961).
Дярматы, впрочем, не ограничился локальной формулировкой
принципов (1.36), (1.41) и (1.42), а сформулировал вариационный
принцип в глобальной форме для континуума:
б (б — 0 — ф) dv = 0. (1.43)
v
Остальные принципы (Biot, 1955; 1970; Циглер, 1966; Баха-
рева, 1968) основаны так же, как и принципы Онзагера и Дярма-
ты, на механических аналогиях в термодинамике и эквивалент-
ны соотношениям взаимности для нее. Возникающий вопрос об
эквивалентности этих принципов между собой был подробно ис-
следован (Бахарева, 1971). Результат, полученный Бахаревой,
которая провела детальную аналогию линейной термодинамики
с аналитической механикой, заключается в том, что принципы
Онзагера, Дярматы, Био, Циглера эквивалентны принципу:
п
-6(Д«) +£/^ = 0, (1.44)
1=1
A 1 V Р №
где As ж----5 gi^XiX^ — параметры системы и
г
Xi = ii их потоки. И более того, в основе этих принципов лежит
принцип:
п
У, - = 0, (1.45)
1=1 х 1 7
где Xi = d&s/dxi — термодинамические силы.
Таким же образом в аналитической механике все многообразие
принципов (Эйлера, Лагранжа, Якоби, Гамильтона) является
всего лишь различными формулировками принципа Даламбера,
который дает полное решение задач механики (Ланцош, 1965).
33
Вопрос об эквивалентности этих принципов в линейной термоди-
намике обсуждался также в работе Кумар и Гупта (Kumar, Gupta,
1973). Известна работа, в которой доказывается, что не сущест-
вует общей вариационной теоремы для стационарных состояний
диссипативных систем в координатах физического пространства
(Gage et al., 1966). Это утверждение, однако, не лишает возмож-
ности формулировать специальные вариационные теоремы для
частных систем (например, линейных) и не запрещает образования
экстремумов над другим множеством координат (теория Приго-
жина — Глансдорфа).
Как видно из изложенного выше, существует довольно большое
число различных экстремальных принципов. Мы уже отмечали
(см. также Oster et al., 1973), что подобные экстремальные форму-
лировки зависят от соответствующих соотношений взаимности.
И хотя по свидетельству Остера и др. (Oster et al., 1973) нет новой
информации от таких принципов, они часто упрощают числовой
анализ и имеют определенную эстетическую привлекательность.
Обобщение этих принципов на нелинейные термодинамические
теории нуждается в особом обосновании, выходящем за рамки
формальных рассуждений. В настоящее время, как нам кажется,
заслуживают детального обсуждения уже упоминавшийся прин-
цип ортогональности Циглера и критерий эволюции Глансдор-
фа — Пригожина (Глансдорф, Пригожин, 1973).
В 1970 г. Циглер (Ziegler, 1970) предложил доказательство
экстремального принципа, названного им принципом ортогональ-
ности. Этот принцип, эквивалентный в линейном случае соотно-
шениям Онзагера — Казимира, сохранил важное свойство линей-
ной теории: двойственность между представлением через силы и
через потоки, которая является некоторым обобщением преобра-
зования Лежандра. Наконец, само доказательство принципа ор-
тогональности предполагает справедливой статистическую меха-
нику Гиббса для необратимых квазистатических процессов. Ана-
литическое выражение принципа Циглера в силовом представлении
дается выражением:
л=(Ё^Жл) (1-46)
Таким образом, принцип Циглера заключается в определенном
выборе нелинейных феноменологических законов, так что имеет
место формула (1.20).
Анализ нелинейной теории Циглера показывает, что эта теория
не имеет потенциального характера, хотя важный частный случай
этой теории — квазиоднородная теория потенциальна.
Прежде чем переходить к изложению теории Глансдорфа —
Пригожина следует заметить, что последняя отлична от теории
Циглера. Этот вывод следует из нашей потенциальной модели
нелинейной теории Глансдорфа — Пригожина, изложенной ниже.
34
Теория Глансдорфа — Пригожина для процессов, далеких от
термодинамического равновесия, включает три существенных ас-
пекта (Глансдорф, Пригожин, 1973; Nicolis, 1971): 1) установление
общего критерия эволюции для стационарных состояний, далеких
от равновесия, 2) построение термодинамических потенциалов,
характеризующих эти состояния, 3) изучение устойчивости этих
состояний.
Критерий эволюции утверждает возможность построения такой
линейной дифференциальной формы со, зависящей от переменных со-
стояния, что
(1.47)
где знак равенства относится к стационарному состоянию. Для
диссипативных процессовdx^ dv= dx9\ т. е. в этом случае
форма со является вариацией продукции энтропии в единицу вре-
мени, обусловленной изменением обобщенных сил. В пределах
линейных явлений, для которых выполняются соотношения вза-
имности Онзагера — Казимира, со = 1/2 т.е. в данном случае
критерий эволюции сводится к теореме о минимуме продукции
энтропии. Следует отметить также, что условием, при котором
справедлив критерий (1.47), является неизменность граничных
условий в термодинамической системе. Критерий эволюции под-
вергся и подвергается широкому обсуждению различными авто-
рами (см. Mel, 1954; Meijer, Edwards, 1970; Зубарев, 1971; Рубин,
Фохт, 1972; Oster et al., 1973; Преснов, 1973).
Тот факт, что дифференциальная форма ю не является полным
дифференциалом, привело Глансдорфа и Пригожина к понятию
так называемого локального потенциала, который разделяет до
некоторой степени свойства потенциалов классической термоди-
намики. Строгое определение локального потенциала, универсаль-
ное для всех физических систем, и исследование вопроса о его
существовании можно найти в работе Родэ (Rohde, 1972).
Пусть Q область в n-мерном евклидовом пространстве Rn,
граница этой области dQ есть гладкая (п — 1) — мерная гипер-
поверхность в Rn, и замыкание нашей области Q ограничено
в Rn.
Пусть теперь {Li \i = 1, . . ., N [} есть семейство интегро-диф-
ференциальных операторов от п независимых пространственных
переменных и пусть система уравнений:
[А (х, 0] = 0 (1 < i < TV) (1.48)
с граничными и начальными условиями:
Gi [А] = 0 (1 < i < TV), (1.49)
где
А (х, i) = (Ах (х, t), . . ., Ап (х, £)) (1.50)
имеет решение Ао.
35
Определение. Мы скажем, что функционал:
Ф [И, [X, Н, К, DH, DK] dQ + $ Ф ddtl,
где L некий лагранжиан, a D оператор дифференцирования, есть
локальный потенциал тогда и только тогда, когда
Ф [Л, Ло] (0 > Ф [40, Ло] (i) > - оо (1.51)
для всех достаточно гладких Л, удовлетворяющих граничным ус-
ловиям (1.49) в каждый момент времени, где знак равенства имеет
место только, если Л ~ Ло,
б) неизменность вектор-функции Н и оптимизация этого функ-
ционала по вектор-функции К ведет к системе уравнений Эйле-
ра-Лагранжа, которые в случае тождественного совпадения функ-
ции Н и функции К превращаются в систему (1.48).
В связи с этим определением Родэ (Rohde, 1972) обсуждает
важный вопрос о существовании локального потенциала в рам-
ках данного определения. Автор получает некоторые достаточ-
ные условия для его существования (которые не отличаются,
впрочем, общностью) и указывает на неединственность локального
потенциала в пределах приведенного определения.
Физический смысл, который вкладывают в это определение его
создатели, следующий: локальный потенциал Ф понимается как
функционал от двух множеств функций, от усредненных функ-
ций, соответствующих квазистационарным решениям макроско-
пических уравнений, и от флуктуационных величин. Расширен-
ный вариационный принцип понимается в терминах теории
флуктуаций. Уравнение Эйлера — Лагранжа, соответствующее ло-
кальному потенциалу Ф, редуцируются тогда к уравнениям мак-
роскопической физики. Эта расширенная вариационная проце-
дура дополняется свойством минимальности, выражающим, что
избыток локального потенциала положительно определен вблизи
нефлуктуационного состояния.
Свойство дифференциальной формы ю не быть общим дифферен-
циалом и отсутствие точного вариационного принципа приводит
к тому, что стационарные состояния больше не характеризуются
в общем случае экстремумом термодинамического потенциала.
Таким образом, устойчивость этих состояний не всегда обеспечена.
Это разделение между эволюцией и устойчивостью привело ис-
следователей к задаче отыскания независимого критерия устой-
чивости для состояний, далеких от равновесия. Была разработана
полная теория инфинитезимальной устойчивости для неравновес-
ных состояний (Глансдорф, Пригожин, 1973). Главным резуль-
татом этой теории является утверждение о возможности построе-
ния отрицательно определенной квадратичной формы:
6г2-б2«-в<0, (1.52)
Зв
где v — средняя гидродинамическая скорость, 6 — обозначает ва-
риацию соответствующих величин, как результат флуктуаций.
Кроме того, в пределах малых флуктуаций:
^-5^>0 (1.53)
и значит стационарное состояние устойчиво по Ляпунову. Впро-
чем в работах Шлегля (Schldgl, 1967, 1971 а, Ь; см. также Janssen,
1972) получен обобщенный критерий устойчивости справедли-
вый не только для инфинитезимальных отклонений. Интересна
также и дискуссия о необходимости и достаточности критерия
устойчивости Глансдорфа —Пригожина для устойчивости тер-
модинамических систем (Keizer, Fox, 1974; Glansdorff et al., 1974).
Канонический формализм
неравновесной феноменологической термодинамики
Выше были кратко изложены различные теории термодина-
мики необратимых процессов: Онзагера, Дярматы, Циглера, При-
гожина и некоторых других авторов. В этом разделе будет по-
строен некий математический формализм, связывающий и обоб-
щающий рассмотренные теории. Следует заметить, что фактически
все теории неравновесной феноменологической термодинамики
основаны на предположении локального равновесия, или другими
словами выводятся из локального уравнения Гиббса. Рассмот-
ренные ниже локальная и глобальная теории требуют для своего
обоснования менее сильное утверждение, чем уравнение Гиббса.
Предлагаемый формализм опирается на следующую лемму
(см., например, Милнор, 1965; Арнольд, 1971).
Лемма Адамара. Пусть е — дифференцируемая (класса Ст)
функция, заданная на n-мерном евклидовом пространстве Кп.
И пусть она равна в точке (хг, . . ., хп) = (0, . . ., 0) нулю вместе
со всеми своими частными производными. Тогда:
п
е = У, h ...,xn)xit (1.54)
2=1
где it — дифференцируемые / класса Ст~г в окрестности точки
(0, . . ., 0) функции, причем:
1
(* дв
П=у^-<,хх1,---,ххп)ах (г = 1,..., п). (1.55)
о г
Заметим, что лемма Адамара ни в коей мере не утверждает
единственности разложения (1.54). В действительности, как легко
видеть, выбор функций ц осуществляется с точностью до функций
37
yi (хъ . . xn) таких, что:
п
^yiXi = Q. (1.56)
1=1
Общий случай (jj + yi) служит предметом систематического изу-
чения в очень интересных работах Эделена (Edelen, 1973а, 1974а).
В этих работах был исследован также случай yt = 0, рассмот-
ренный нами в ряде работ (Преснов и др., 1973; Преснов, 1973).
Назовем теперь, для определенности, независимые перемен-
ные Xi силами, а функции Л ЕЭ С1 — каноническими потоками
и сформулируем основные утверждения канонического формализ-
ма (см. Циглер, 1966; Картан, 1971; Преснов и др., 1973; Преснов,
1973; Edelen, 1974а).
Утверждение 1. Канонические потоки (1.55) удовлетворяют
соотношениям:
Э/. dju
-±- = -± (Л А: = 1, ..п). (1.57)
Утверждение 2. Канонические потоки (1.55) допускают пред-
ставление:
п
= хп)хк = (1.58)
К=1
так что имеет место соотношение:
Цг) = hiW (г,п), (1.59)
где
11
lik № = т э/Х ’ • • • ’ хкс^ dt dK- (* -60>
0 0 г к
Утверждение 3. Соотношения (1.57) в общем случае не тож-
дественны соотношениям (1.59), но в линейной теории (/{ =
= SZ^ft) оба эти соотношения редуцируются к виду lik =
= lki = const.
Утверждение 4. Канонические потоки (1.55) потенциальны:
Л = Д- (i = l,..., и) (1.61)
с потенциалом:
1
/ (х) = \ — е (тах,. .., таЛ) dx.
о
(1.62)
38
Утверждение 5. Потенциал (1.62) является единственной,
с точностью до произвольной постоянной, функцией, удовлетво-
ряющей условиям (1.54) и (1.61).
Утверждение 6. Имеет место формула:
x) = f(x) +g(j), (1.63)
где
i
g(j) = Tjn] dr (1.64)
О
и этот формализм преобразований Лежандра (1.61) — (1.64)
определяет двойственность между представлением через силы и
представлением через потоки.
Следует отметить, что построенный канонический формализм
включает в себя не только формализм линейной теории термоди-
намики необратимых процессов, но также формализм квазиодно-
родной теории Циглера (1966) и формализм нелинейных теорий
Дярматы (Gyarmati, 1969).
Локальная каноническая теория
Канонический формализм, приведенный выше, обобщает ряд
известных формализмов феноменологической термодинамики. Рас-
смотрим взаимосвязь этого формализма с формализмом локальной
термодинамики Пригожина.
Пусть дана термодинамическая система, занимающая в про-
странственно-временном континууме SR область D CZ SR с прост-
ранственным объемом V = V (i). Основным видом параметров,
характеризующих состояние термодинамической системы, явля-
ется множество интенсивных величин {xt : D R (i = 1, ...
..., п)}, называемых локально определенными термодинамическими
силами (или просто локальными силами). Из предположения ло-
кального равновесия и уравнения Гиббса следует, что специаль-
ная интенсивная величина: локальное производство энтропии
о функционально зависима от локальных сил (о = е . . .
. . ., я:п)). Если е ее С2, то, опираясь на лемму Адамара, можно
построить локальную каноническую теорию феноменологической
термодинамики. Основой этой теории являются соотношения
(1.57), а основным положением, требующим обоснования — проб-
лема канонического выбора локальных потоков. Мы покажем,
что канонические потоки (1.55) термодинамически уместны, но
предварительно подчеркнем, что канонический выбор локальных
потоков (1.55) = 0) является своего рода принципом термо-
динамики необратимых процессов (другой принцип см. в работах
Циглера (1966; Ziegler, 1970).
Итак, пусть j° суть «истинные» потоки для локальных сил
Xi ~ ’ аit ~ канонические потоки для тех же локальных
39
сил. Легко видеть, что преобразование (xh fy —► (хь ]t) оставляет
инвариантным локальное производство энтропии:
п п
^7°^ = Mi (1-65)
1=1 1=1
и кроме того, по определению канонического выбора (1.55), всег-
да верно неравенство:
п
5^Mi>0. (1.66)
г=1
Таким образом, условие инвариантности (Глансдорф, Пригожин,
1973) не нарушено.
Обсудим в связи с этим критерий эволюции Глансдорфа — При-
гожина (Глансдорф, Пригожин, 1973; Nicolis, 1971).
Для диссипативных термодинамических систем при постоянных
граничных условиях имеет место неравенство:
(!«>
1=1
т. е. изменение локальных сил протекает всегда так, чтобы умень-
шить величину производства энтропии в системе. По свидетель-
ству Глансдорфа и Пригожина (1973), критерий (1.67) не зависит
ни от каких предположений о феноменологических соотношениях
между потоками и силами. Значит можно сделать вывод, что этот
критерий справедлив и для канонических потоков (1.55).
Утверждение 7. (Преснов, 1973). Критерий эволюции Гланс-
дорфа — Пригожина (1.67) имеет потенциальный характер:
Т=4г<0’ (1-68)
где
11
R = е (ххи .., тхп) dx dv. (1.69)
о о
Ясно, что утверждение 7 решает проблему устойчивости стацио-
нарных состояний, так как знак равенства в неравенствах (1.67)
и (1.68) относится только к ним.
Утверждение 7 показывает также, что критерий эволюции
Глансдорфа — Пригожина не описывает колебательных режимов
в стационарных состояниях открытых систем [так как потенциал
(1.69) постоянен в стационарном состоянии]. Но этот критерий да-
ет некую информацию о затухающих колебательных процессах,
которые могут иметь место при приближении к стационарному
состоянию.
40
Интересно отметить, что хотя критерий эволюции Глансдорфа—
Пригожина формулируется в терминах локальных сил, он отно-
сится к полному производству энтропии в системе и значит, по
существу, является глобальным утверждением, как и критерий
эволюции, сформулированный Зотиным и др. (Зотин, 1974; Зотин,
Коноплев, Преснов, 1975)
Глобальная каноническая теория
Зотиным и др. (Зотин, 1974; Зотин, Коноплев, Преснов, 1975)
предложен критерий эволюции, отличный от критерия эволюций
Глансдорфа — Пригожина. Он сводится к утверждению, что
при приближении термодинамических систем, далеких от равно-
весия, к стационарному или равновесному состоянию выполняет-
ся неравенство (2.22). Так как этот критерий не равнозначен кри-
терию (1.68), то встает вопрос об отношении введенного Зотиным
критерия эволюции к локальной теории. На самом деле вопрос этот
имеет более общий характер, так как критерий Зотина фактически
отражает другой подход к термодинамике необратимых процес-
сов, который можно назвать глобальным подходом. При таком гло-
бальном подходе используются только глобальные характеристи-
ки термодинамических систем, такие, как общая продукция эн-
тропии, объем системы, масса, время и т. д., поэтому эта глобаль-
ная теория допускает непосредственную экспериментальную про-
верку с большей легкостью, чем локальная теория. Глобальная
теория, по-видимому, не может быть сведена к локальной теории,
хотя в основе обеих теорий лежит один и тот же канонический ма-
тематический формализм, обсуждавшийся выше.
В отличие от критерия эволюции Глансдорфа — Пригожина,
критерий эволюции Зотина (2.22) не имеет аналитического выра-
жения. Это не позволяет провести детальное сравнение двух кри-
териев. Тем не менее мы можем предложить все же некий подход
к проблеме поиска аналитического выражения для критерия
(2.22). Он заключается в построении теории, оперирующей толь-
ко глобальными характеристиками термодинамической системы,
в частности, такой важной глобальной характеристикой системы,
как ее удельная диссипативная функция ф = ф (0 = T/V ёР.
Подчеркнем, что функция ф может быть определена, например,
и для локальной термодинамики Пригожина (1960) и для линейной
термодинамики Онзагера (Onsager, 1931а, Ь).
Глобальная теория основана на следующих положениях.
Пусть дана термодинамическая система, занимающая в простран-
ственно-временном континууме SR область D CZ 9R. И пусть {xt :
: D i? (i — 1, . . ., п)} есть множество интенсивных величин,
таких, что для любых точек (р; t), (р'\ t) ЕЕ D имеют место равен-
ства xt (р, t) — Xi (pr; £) (i = 1, . . ., ri). Таким образом, интен-
сивные величины Xi : D R редуцируются к функционалам
41
Xi : R+ R (здесь R+ = [0, oo], t. e. Xf = Xf (t)), которые яв-
ляются глобальными характеристиками данной системы.
Утверждение 8. Существует отображение Е : Rn R такое,
что
E(Xlt ...,Хп)е=я|>. (1.70)
Если же существуют однозначные отображения Х^1 : R
R+ класса С”1 игр = ф (0 GE Ст, то отображение Е также будет
однозначным отображением класса Ст. Назовем теперь функцио-
налы Xi глобальными силами, если Е ЕЕ С2. Тогда опять, как и
при построении локальной канонической теории, опираясь на
лемму Адамара, можно построить глобальную каноническую тео-
рию.
В связи с построением глобальной теории возникает вопрос,
который касается взаимосвязи локальной и глобальной теорий.
По-видимому, в общем случае глобальная теория не имеет интер-
претации в терминах локальной теории, хотя, например, для
термодинамических систем с увеличивающимся объемом (dV/dt >
> 0) можно определить глобальные силы таким образом:
= = (1.71)
что отображение Xi1, по крайней мере, однозначно в силу стро-
гой монотонности функции Xi (t). Следовательно, если Xf1 ЕЕ С2
и ф GE С2, то на основании утверждения 8 существует глобальная
теория, в которой силы заданы формулой (1.71).
В заключение сформулируем кратко основные положения, на
которых базируется глобальная теория: 1) лемма Адамара, 2) ут-
верждение 8,3) принцип наименьшей диссипации энергии (стр. 97)
и критерий эволюции (2.22).
Исходя из этих положений имеют место следующие главные
результаты глобальной теории:
п
x|, = £jJi(X)Xi (1.72)
i=l
п
К=1
Lin (X) = Lki (X)
^-<0.
(i == 1,..., n)
(г, к = 1,..n)
(1.73)
(1.74)
(1-75)
II
КАЧЕСТВЕННАЯ
ФЕНОМЕНОЛОГИЧЕСКАЯ ТЕОРИЯ
РАЗВИТИЯ ОРГАНИЗМОВ
Живые организмы являются открытыми системами, так как они
обмениваются с окружающей средой веществом и энергией. В свя-
зи с этим давно уже стало ясным, что хотя во взрослом состоянии
основные параметры, которыми можно описывать животных и
растения, остаются неизменными во времени, живые организмы
находятся при этом не в равновесном, а в стационарном состоянии
(Hill, 1931; Bertalanffy, 1932, 1950). Осознание этой истины поз-
воляло надеяться, что термодинамика необратимых процессов
и теория открытых систем могут быть распространены на живые
системы. Одной из попыток распространить основные положения
термодинамики линейных необратимых процессов на явления раз-
вития, роста и старения организмов, является теория Пригожина
и Виам (Prigogine, Wiame, 1946; Пригожин, 1960).
Согласно этой теории, к процессам развития и роста живот-
ных приложимо соотношение (1.17) и критерий эволюции термо-
динамики линейных необратимых процессов (1.16). Предполагает-
ся, что развитие, рост и старение живых организмов представляет
собой процесс непрерывного приближения живой системы к ко-
нечному стационарному состоянию, сопровождающийся умень-
шением удельной скорости продукции энтропии, уменьшением
удельной диссипативной функции ф. При этом стационарное со-
стояние организмов, как и неживых систем, характеризуют ми-
нимальной и константной скоростью продукции энтропии.
Таким образом, согласно термодинамической теории Приго-
жина — Виам, во время развития, роста и последующих возра-
стных изменений организмов происходит непрерывный процесс
старения системы, выражающийся в уменьшении удельной дисси-
пативной функции системы. Так как удельную диссипативную
функцию с известным приближением можно приравнять интен-
сивности теплопродукции (д) и дыхания «о,) организмов (см. Зо-
гин, Грудницкий, 1970):
(2.1)
то это положение теории Пригожина — Виам можно свести к ут-
верждению, что во время развития, роста и старения организ-
мов происходит непрерывное снижение интенсивности теплопро-
дукции или дыхания. Имеющиеся в физиологии данные о скоро-
43
Рие. 1. Схематическая модель уклонения и приближения живой системы
к стационарному состоянию
Объяснения в тексте
сти теплопродукции и дыхания животных и человека хорошо под-
тверждают этот вывод (Зотин, 1974).
В обычных условиях покоя животные и человек, как известно,
находятся в стационарном состоянии с минимальной интенсив-
ностью теплопродукции, соответствующей основному обмену.
Под воздействием внешних или внутренних факторов организмы
могут уклониться от стационарного состояния, а после прекра-
щения действия этих факторов снова вернуться к прежнему со-
стоянию. Одновременно, в соответствии с теорией Пригожина —
Виам, в процессе развития, роста и старения происходит непре-
рывное приближение организмов к конечному стационарному
состоянию. Следовательно, организмы характеризуются двумя
типами стационарных состояний: одно из них — стационарное
состояние в каждый данный момент времени, другое — конечное
стационарное состояние, к которому эволюционирует живая си-
стема в процессе жизни. Было предложено поэтому уклонение
и приближение живой системы к конечному стационарному состоя-
нию назвать конститутивным процессом, а временное уклонение
или приближение системы к текущему стационарному состоя-
нию — индуцибильным (Озернюк и др., 1971).
Теорию Пригожина — Виам и различия в конститутивных и
индуцибильных процессах можно иллюстрировать схемой
(рис. 1), в которой шарик катится вниз по наклонному желобу.
В каждый данный момент времени под действием внешней силы
44
шарик может уклониться от стационарного состояния (б), а затем
вновь вернуться на дно желоба (в) и продолжать движение к ко-
нечному стационарному состоянию (г). Движение шарика по дну
желоба (а, г) соответствует конститутивному приближению ор-
ганизмов к конечному стационарному состоянию. Уклонение
и возвращение шарика к текущему стационарному состоянию
(б, в) соответствует индуцибильному уклонению живой системы
(Зотин, 1974).
Индуцибильное уклонение от стационарного состояния мо-
жет быть кратковременным в связи с импульсным воздействием
каких-либо факторов на живую систему. В этом случае процесс
уклонения и возвращения системы к стационарному состоя-
нию можно назвать индуцибильно импульсным (Зотин, 1974).
Исходя из (1.15) и (1.17), изменение удельной диссипативной
функции системы, связанное с индуцибильно импульсными про-
цессами, будет описываться уравнением:
= 4- const,
(2.2)
где J i — удельные потоки, связанные с индуцибильными явле-
ниями; Xi — силы, а ф(о — удельная диссипативная функция,
связанные с этими потоками.
Примером индуцибильно импульсного процесса является лю-
бое временное усиление теплопродукции или дыхания при кратко-
временном воздействии факторов среды или под влиянием
внутренних причин.
Индуцибильное уклонение организмов от текущего стационар-
ного состояния под воздействием внешних или внутренних при-
чин может сопровождаться переходом системы в новое стационар-
ное состояние без возвращения на каком-то достаточно продол-
жительном отрезке времени к исходному состоянию. Подобный
процесс можно назвать индуцибильно адаптивным (Зотин, 1974)
и он может быть описан уравнением:
т
^(а)=X ^аХа+const’ (2‘3)
а=1
где Ja — удельные потоки, связанные с явлением адаптации;
Ха и ф(а) — связанные с этими потоками силы и удельная дис-
сипативная функция.
Примером индуцибильно адаптивного процесса является пе-
реход организма к новому уровню теплопродукции и дыхания
при изменении внешней среды (температуры, газового и солевого
составов, давления и т. д.) или при длительных изменениях внут-
ренних параметров системы (паталогических, гормональных,
нервных и т. д.)
45
Соответственно для конститутивных процессов, к которым
относятся все явления развития (оогенез, эмбриональное разви-
тие, рост, старение, регенерация, злокачественный рост и т. д.),
можно написать:
п
*Ф(/О = У1, J+ 'Ф(О), (2.4)
fc=l
где J а — удельные потоки, связанные с конститутивным процес-
сом; Хк и — связанные с этими потоками силы и удельная
диссипативная функция,,*^)—диссипативная функция основно-
го обмена.
Индуцибильно импульсные, индуцибильно адаптивные и кон-
ститутивные процессы отличаются друг от друга временем ре-
лаксации, так как импульсные процессы длятся обычно минуты и
часы, адаптивные — дни и недели, конститутивные — месяцы
и годы. Поэтому к ним можно применить «принцип простоты»
(см. Романовский и др., 1975), согласно которому несколько яв-
лений, значительно отличающихся друг от друга по времени ре-
лаксации, можно рассматривать независимо друг от друга, что
и записано в уравнениях (2.2), (2.3), (2.4).
Как видно из изложенного, в наиболее общей форме можно
выделить четыре типа явлений, определяющих удельную дисси-
пативную функцию живых систем: индуцибильно импульсные,
индуцибильно адаптивные, конститутивные процессы и основной
обмен. Другими словами, в самом общем виде уравнение для дис-
сипативной функции, которое включает в себя все основные про-
цессы жизни организма, должно иметь вид:
= 'Ф(О) + 'Ф(г) + 'ф(а) + 'Ф(К)- (2.5)
В этом уравнении фактически представлена вся термодинамика
жизненных процессов, так как оно включает в себя реакции ор-
ганизма на внешние воздействия, проведение различного рода
работ, адаптацию организма к меняющимся внешним и внутрен-
ним условиям, паталогические процессы, а также процессы разви-
тия, роста и старения. Последние (2.4) и относятся собственно
к тем явлениям, которые описывает термодинамическая теория
развития организмов Пригожина — Виам.
ЭКСПЕРИМЕНТАЛЬНАЯ И ТЕОРЕТИЧЕСКАЯ ОСНОВЫ
КАЧЕСТВЕННОЙ ФЕНОМЕНОЛОГИЧЕСКОЙ
ТЕОРИИ РАЗВИТИЯ
А, И,Зотин, Р. С. Зотина
Качественная феноменологическая теория развития организ-
мов основывается на предположении Пригожина и Виам
(Prigogine, Wiame, 1946), согласно которому для описания биологи-
ческих процессов можно использовать критерий эволюции термо-
динамики линейных необратимых процессов (1.16). Из этого кри-
терия следует, что если система не находится в стационарном
состоянии, то она будет изменяться до тех пор, пока скорость про-
дукции энтропии не примет наименьшего из всех возможных зна-
чений. В соответствии с предположением Пригожина — Виам
критерий эволюции (1.16) может быть использован, в частности,
для описания процессов развития и роста животных. Многочис-
ленные экспериментальные наблюдения хорошо согласуются
с этим предположением (см. Zotin, 1972; Зотин, 1974). Кратко
остановимся на этих данных.
Развитие, рост и старение организмов
Приложимость теории Пригожина — Виам к эмбриональному
развитию животных вызывала большие сомнения и горячие дис-
куссии (Тринчер, 1965; Беркинблит, 1965; Быховский, 1965; Сыр-
ников, 1965; Волькенштейн и Сырников, 1966; Зотин, 1966; Ни-
колаев, 1966). Мы не будем подробно останавливаться на различ-
ных мнениях, но должны отметить, что тщательное и корректное
рассмотрение экспериментальных данных дает основание ут-
верждать, что эта теория применима к эмбриональному разви-
тию животных (Zotin, 1972; Зотин, 1974). Главное здесь четко раз-
личать понятия конститутивного и индуцибильного процессов.
Так, давно показано, что в первое время после оплодотворения
происходит заметное усиление дыхания яиц морских ежей, мор-
ских звезд, асцидий и др. (Ротшильд, 1958; Monroy, 1965), причем
это усиление состоит из двух фаз: быстрого усиления в первые
минуты после оплодотворения и последующего более медленного
увеличения. Согласно теории Пригожина — Виам, во время раз-
вития должно происходить непрерывное уменьшение интенсив-
ности дыхания и теплопродукции, а эти наблюдения как бы
противоречат этому утверждению. Однако при оплодотворении про-
исходит импульсное воздействие сперматозоида на яйцо и его акти-
вация, поэтому первый пик подъема интенсивности дыхания мы
можем отнести к индуцибильно импульсному процессу. В то же
время в момент оплодотворения яйца попадают из тела самки
47
в окружающую среду. При этом меняются внешние параметры
системы, что вызывает переход системы к новому стационарному
состоянию. Поэтому второе более медленное усиление интенсив-
ности дыхания мы можем отнести к категории индуцибильно адап-
тивного процесса. Теория Пригожина — Виам описывает только
конститутивные процессы, поэтому данные об усилении дыхания
яиц после оплодотворения не могут ей противоречить (Зотин,
1974).
К индуцибильно адаптивному уклонению живой системы от
стационарного состояния можно отнести также хорошо известные
наблюдения, о том, что после рождения млекопитающих проис-
ходит устойчивое усиление интенсивности теплопродукции и ды-
хания, которое продолжается разное время у разных видов.
Например, через 3—4 дня после рождения у мышат начинает увели-
чиваться интенсивность теплопродукции, которая достигает мак-
симума к 10—12-му дню, а затем снижается (Walker, 1967). При
рождении млекопитающих происходит резкое изменение внешних
и внутренних параметров системы: внешних, так как организм
переходит к свободному существованию, внутренних, так как
после рождения меняется температура тела, и возникает хими-
ческий механизм терморегуляции животного. Характерно, на-
пример, что у мышат, максимальная интенсивность теплопродук-
ции у которых достигается на 10—12-й день, становление хими-
ческой терморегуляции происходит в первые 10 дней (Джелинео,
1959). Здесь особенно ясно выступает тот факт, что уклонение си-
стемы от стационарного состояния при рождении млекопитающих
происходит не только в результате изменения внешних парамет-
ров системы, но и внутренних, связанных со становлением хими-
ческой терморегуляции и изменением температуры тела.
Очевидно, что если причина временного усиления интенсив-
ности теплопродукции и дыхания у млекопитающих после рожде-
ния заключается в том, что в это время происходит изменение
внешних и внутренних параметров системы, то такое же явление
должно наблюдаться и у других животных: при выклеве птенцов
птиц, линьке насекомых и т. д. И действительно, такого рода
факты хорошо известны в литературе. Так, у насекомых наблю-
даются периодические падения и подъемы интенсивности дыха-
ния, совпадающие по времени с периодом линек личинок. Для
примера, на рис. 2 приведены данные об изменении интенсивности
дыхания личинок американской белой бабочки. В период личиноч-
ного развития насекомых на фоне непрерывного снижения интен-
сивности дыхания, предполагаемого теорией Пригожина — Ви-
ам, происходят периодические падения и подъемы, совпадающие
по времени с линькой личинок. Эти подъемы и падения мы можем
отнести к индуцибильно адаптивным уклонениям, происходящим
в периоды, когда меняются внешние и внутренние параметры си-
стемы. Все эти изменения происходят на фоне непрерывного кон-
ститутивного снижения интенсивности теплопродукции и дыха-
48
ния. Подобные данные получены и на других объектах. На
рис. 3, например, показано уменьшение интенсивности дыхания
зародышей кур и уток в период инкубации, а на рис. 4 — умень-
шение интенсивности дыхания коров и рыб в период зародышевого
развития.
Все сказанное выше позволяет представить процессы, проис-
ходящие в период эмбрионального и постэмбрионального разви-
тия с точки зрения термодинамической теории Пригожина — Виам
в виде схемы (рис. 5). Согласно этой схеме, на фоне непрерыв-
ного конститутивного приближения живых организмов к конеч-
ному стационарному состоянию в отдельные периоды эмбриональ-
ного и постэмбрионального развития происходят индуцибильно
импульсные и адаптивные уклонения. Индуцибильные адаптив-
ные уклонения происходят вслед за изменением внешних и внут-
ренних параметров, индуцибильно импульсные — при раздра-
жении системы в момент оплодотворения, при рождении, выклеве,
вылуплении, а также во время линек личинок насекомых. В тех
случаях, когда индуцибильно адаптивному уклонению предше-
ствует изменение внешних параметров системы, ему предшествует
и индуцибильно импульсный процесс. После окончания инду-
цибильного процесса живая система вновь возвращается к состоя-
нию конститутивного приближения к конечному стационарному
состоянию.
Если период эмбрионального развития животных представляет
известные трудности при рассмотрении его с точки зрения теории
Пригожина — Виам, то рост и старение организмов являются
той базой, на которую с самого начала опиралась эта теория.
Действительно, хорошо известно, что во время роста животных
и человека происходит уменьшение интенсивности дыхания и уров-
ня основного обмена. Это утверждение следует из многочислен-
ных исследований, в которых показано, что между скоростью
дыхания и весом животных существует степенная зависимость
вида:
<?о2 = aW\ (2.6)
где Qot — скорость дыхания, W — вес животного, а и к констан-
ты. Справедливость уравнения (2.6) установлена для большин-
ства групп животных от простейших до млекопитающих (см. Zeut-
hen, 1953; Винберг, 1956; Ивлев, 1959, 1963; Hemmingsen, 1960;
Бызова, 1965, 1972; Дольник, 1968; Сущеня, 1972; Зотин, 1974
и др.). Можно сказать, что зависимость (2.6) является эмпириче-
ским законом, справедливым для всех животных. Однако из
(2.6) следует, что интенсивность дыхания животных (скорость пот-
ребления кислорода на единицу веса животного) уменьшается во
время роста животного. Действительно, деля правую и левую
части уравнения (2.6) на вес, имеем:
$0. = аЖ*-1, (2.7)
49
Рис. 2. Интенсивность дыха-
ния личинок и куколок амери-
канской белой бабочки на раз-
ных стадиях развития (Иван-
нин, 1969)
Рис. 2
Рис. 3
Рис. 3. Интенсивность тепло-
продукции зародышей кур (из
Зотина, 1966) и уток (Хас-
кин, 1961) на разных стадиях
инкубации яиц
1 — утки; 2 — куры
Рис. 4. Интенсивность тепло-
продукции на разных стадиях
развития форели (1) (из Зо-
тина, 1966) и интенсивность
дыхания во время зародышевого
развития коров джерсейской
породы (2) (из Нагорного и
др., 1963)
200 Сдщни
ipp
Рис. 4
Рис, 5, Схема индуцибилъных и конститу-
тивных процессов во время развития и
роста различных животных
j — птицы и млекопитающие; 2 — рыбы и амфи-
бии; з — насекомые; л — линька, р — рождение
Рис. 6. Зависимость интенсивности дыха-
ния от веса тела у Rana pip tens (1),
R. sylvatica (2), Xenopus versicolar (3)
(из Davison' 1955)
а так как константа k меньше единицы, то к — 1 = 6<0и (2.7)
можно записать:
?о2 = aW-b (2.8)
ИЛИ
log <7о2 = log а — 6 log W. (2.9)
Следовательно, чем больше вес животного, тем меньше интен-
сивность дыхания. Для примера на рис. 6 приведены данные, по-
лученные при изучении дыхания лягушек разного веса: в логариф-
мической системе координат имеет место линейная зависимость
между интенсивностью дыхания и весом животных, т. е. зависи-
мость (2.9). Тот факт, что зависимость (2.6) справедлива для всех
животных, означает, что во время роста, когда происходит увели-
чение массы тела животного, интенсивность дыхания уменьшает-
ся, как это и следует из теории Пригожина — Виам.
Непосредственное измерение основного обмена и интенсивно-
сти дыхания у разных видов животных в зависимости от возраста
также показывает, что во время роста и старения происходит па-
дение удельной скорости теплопродукции. Для примера на рис. 7
приведены наблюдения об изменении основного обмена во время
роста птиц и млекопитающих. Так же как и в случае эмбриональ-
ного развития, в процессе роста и старения происходит уменьше-
ние интенсивности дыхания не только в организме в целом, но
и в большинстве органов и тканей животных. В табл. 1 приведены
51
наблюдения разных авторов о дыхании различных органов и тка-
ней белых крыс в зависимости от возраста животных.
Во всех случаях наблюдается заметное уменьшение интен-
сивности дыхания во время роста крыс. Сходные данные получе-
ны и для других видов животных (птицы, морские свинки, кроли-
ки, телята и др.) (см. Нагорный и др., 1963).
Таблица 1
Возрастные изменения интенсивности дыхания (Oz мл/час/г сухого веса)
различных органов и тканей белых крыс (из Нагорного и др., 1963)
Объект Возраст, месяцы Снижение дыхания, %
1 3 6 12 18
Весь организм 6,4 5,6 4,9 3,4 56,9
Почка 20,6 17,76 19,73 18,85 16,67 19,0
Селезенка 9,1 7,9 8,1 — 6,9 24,2
Мышцы 8,4 3,7 3,7 3,6 3,2 60,8
Печень 7,31 6,55 — 5,52 5,45 26,9
Мозг 5,91 5,19 — 4,86 4/75 18,0
Семенные железы 5,31 3,55 3,98 3,53 3,04 42,8
Кишечник 4,74 3,67 3,35 2,24 1,66 65,0
Кожа 2,3 0,9 0,5 0,3 — 85,2
Хрящ 1,8 0,6 0,3 0,2 0,2 74,6
Таким образом, многочисленные данные, полученные при
изучении дыхания животных, показывают, что во время роста
происходит уменьшение удельной скорости теплопродукции
(удельной скорости продукции энтропии), причем это относится
не только к организму в целом, но и к отдельным органам и тка-
ням. Как и в случае зародышевого развития, во время роста этот
процесс может нарушаться индуцибильно импульсными и адап-
тивными уклонениями, которые возникают при различных раз-
дражениях, при совершении работы, при длительных или крат-
ковременных изменениях внутренних или внешних параметров
системы.
Несколько слов о возможном механизме конститутивного
приближения живых организмов к конечному стационарному сос-
тоянию. С точки зрения феноменологической теории развития,
конститутивное приближение живых систем к конечному стацио-
нарному состоянию является процессом старения живых систем.
Отличие периода эмбрионального развития, роста и старения,
с этой точки зрения, заключается в скорости процесса старения:
она наибольшая на ранних стадиях развития, меньше во время
роста и наименьшая на тех стадиях, которые мы называем процес-
52
сом старения. Представление о старении, как о процессе умень-
шения диссипативной функции системы, имеет большие преиму-
щества перед другими определениями старения, так как оно поз-
воляет легко перейти к изучению конкретных механизмов старе-
ния. Действительно, скорость продукции энтропии пропорцио-
нальна скорости теплопродукции организма, которая в свою
очередь пропорциональна скорости дыхания организма. Таким об-
разом, изучение механизмов уменьшения интенсивности дыхания
во время зародышевого развития и роста является, с точки зре-
Рис. 7. Изменение основного обмена во время роста птиц и млекопитающих
(из Зотина, 1974)
1 — куры; 2 — утки; 3 — крысы; 4 — овцы; 5 — свиньи; 6 — лошади; 7 — коровы
Рис, 8. Схематическое изображение оогенеза, зародышевого развития, роста,
старения, заживления ран, регенерации и злокачественного роста с точки
зрения качественной феноменологической теории развития (Зотин, Зотина,
53
ния термодинамической теории развития, изучением механизма
старения. В настоящее время еще трудно представить себе в пол-
ном объеме механизм снижения интенсивности дыхания в про-
цессе роста и старения организмов. Однако несколько упростить
эту проблему все же можно, так как, отвлекаясь от различного
рода механизмов регуляции дыхания, можно считать, что базис-
ный уровень дыхания в клетках и тканях (основной обмен) опре-
деляется главным образом концентрацией митохондрий в них
(Зотин, 1974). Тогда механизм конститутивного процесса, меха-
низм старения организмов мы можем свести к механизму умень-
шения концентрации митохондрий в клетках и тканях организ-
ма во время развития. То, что это утверждение возможно, пока-
зывают некоторые данные, имеющиеся в литературе: несмотря на
некоторые противоречия в данных разных авторов, можно считать
вероятным, что именно возрастное уменьшение митохондрий
определяет возрастное уменьшение интенсивности дыхания жи-
вотных и отдельных органов и тканей (Зотин, 1974). В этом слу-
чае мы должны сказать, что конститутивный процесс представля-
ет собой главным образом процесс изменения концентрации
митохондрий в клетках и механизм конститутивного процесса в зна-
чительной степени может быть связан с механизмом размножения
митохондрий и изменения числа митохондрий в клетках и тка-
нях животных в процессе развития.
Основные следствия теории Пригожина — Виам
Как следует из теории Пригожина — Виам и эксперименталь-
ных данных, приведенных выше, во время развития, роста и ста-
рения животных происходит непрерывное уменьшение удельной
диссипативной функции, нарушаемое лишь индуцибильно им-
пульсными и адаптивными процессами. Естественным следствием
этой теории является утверждение, что в жизни организмов дол-
жен существовать период конститутивного уклонения от стацио-
нарного состояния, сопровождающийся не уменьшением, а увели-
чением удельной диссипативной функции. Явления, которые про-
исходят в это время, имеют большой интерес для теоретической
биологии, так как в эти периоды, с точки зрения термодинами-
ческой теории развития, происходит омоложение живых систем.
Можно выделить три таких периода в жизни организмов (Zotin,
Zotina, 1967; Зотин и Зотина, 1969; Зотин, 1970, 1974; Zotin,
1972): период оогенеза, начальные стадии заживления ран и ре-
генерации органов и тканей, начальные стадии злокачественного
роста (рис. 8).
В процессе оогенеза происходит увеличение удельной дисси-
пативной функции и она достигает максимального значения, до-
статочного для того, чтобы живая система вступила на путь раз-
вития, на путь конститутивного приближения к конечному ста-
ционарному состоянию. С точки зрения феноменологической
54
теории развития, оогенез представляет собой процесс естествен-
ного омоложения живой системы, тогда как все остальные периоды
жизни — только процессы старения. Начальные стадии регене-
рации и злокачественного роста также представляют собой про-
цесс конститутивного уклонения живой системы от стационарного
состояния с последующим возвращением к нему (см. рис. 8). В слу-
чае злокачественного роста уклонение системы от стационарного
состояния происходит таким неудачным путем, что живая система
не может вернуться к прежнему стационарному состоянию. Зло-
качественные клетки — это клетки, омолодившиеся таким неу-
дачным путем, что они не могут вступить на путь старения и оста-
ются молодыми до тех пор, пока не убьют живую систему, частью
которой они являются.
Данные, полученные при изучении интенсивности дыхания и
гликолиза в процессе оогенеза, регенерации и злокачественного
роста, достаточно хорошо подтверждают указанные выше след-
ствия теории Пригожина — Виам (см. Зотин, 1974). Так, при
изучении интенсивности дыхания и гликолиза ооцитов в процессе
оогенеза вьюна было найдено, что интенсивность дыхания, рассчи-
танная на 1 мг белка ооцита, быстро увеличивается в период ма-
лого роста, достигает максимума, а затем снижается (см. рис. 43)
(Озернюк, 1970). Интенсивность анаэробного гликолиза в ооге-
незе вьюна изменяется сходным образом (Озернюк и др., 1971).
Отличие заключается в том, что максимум интенсивности глико-
лиза приходится па более позднюю фазу оогенеза по сравнению
с максимумом интенсивности дыхания: максимум интенсивно-
сти гликолиза соответствует стадии начала вителлогенеза, а ин-
тенсивности дыхания — началу вакуолизации цитоплазмы. Сни-
жение интенсивности дыхания со стадии, соответствующей нача-
лу вакуолизации цитоплазмы зарегистрировано также в оогенезе
шпорцевой лягушки (рис. 33) (Зотин, 19756). Исходя из приве-
денных наблюдений о дыхании ооцитов вьюна и лягушки, можно
считать, что интенсивность теплопродукции ооцитов увеличивает-
ся на стадиях, предшествующих вителлогенезу, а затем уменьшает-
ся. Таким образом, в этих работах установлено, что конститутив-
ное уклонение, естественное омоложение живых систем, проис-
ходит на узком участке онтогенеза животных, в период малого ро-
ста ооцитов.
Вторым пунктом, в котором согласно схеме, показанной на
рис. 8, возможен процесс конститутивного уклонения живой сис-
темы от стационарного состояния, являются процессы регенера-
ции и заживления ран. Действительно, если процессы развития
и роста животных сопровождаются конститутивным приближением
системы к конечному стационарному состоянию, то это должно
быть справедливым и для таких явлений, как регенерация, так
как у большинства животных в процессе регенерации имеет место
рост и дифференцировка регенерирующих органов и тканей.
Более спорным является вопрос о заживлении ран, так как этот
55
процесс трудно сравнивать с явлениями развития и роста. Кроме
того при нанесении раны происходит нарушение внешних и внут-
ренних параметров системы и можно было бы толковать явления,
которые разыгрываются при ранении как индуцибильно импульс-
ное или индуцибильно адаптивное уклонение, подобное процессу
возбуждения.
Мы остановимся лишь на процессах регенерации. Процесс ре-
генерации начинается с повреждения или ранения органа или тка-
ни. Поэтому на самых ранних стадиях этого явления может иметь
место индуцибильно импульсное уклонение системы, характерное
для процесса возбуждения. Однако в дальнейшем развивается
сначала процесс конститутивного уклонения, а затем приближе-
ния к стационарному состоянию, характерному для данной
стадии развития животного. Сразу после ампутации хвоста у аксо-
лотля происходит кратковременное усиление дыхания и гликоли-
за, связанное с нанесением рапы и возбуждением окружающих
рану тканей (рис. 46). Максимум интенсивности анаэробного гли-
колиза у половозрелых аксолотлей наблюдается через 2 часа,
а интенсивности дыхания — через 16 час. К 22-му часу после ампу-
тации тот и другой процессы снижаются до нормального уровня,
до уровня дыхания и гликолиза в контроле, а затем развивается
новый процесс, процесс их контитутивного усиления. Владимиро-
ва (1974, 1975) обнаружила, что у взрослых аксолотлей в процессе
регенерации хвоста имеют место два максимума интенсивности ды-
хания: первый на стадии бластемы, второй на стадии начала диф-
ференцировки (см. рис. 46). При изменении интенсивности гли-
колиза имеется один максимум к середине процесса регенерации
хвоста (см. рис. 46). У молодых, трехнедельных аксолотлей имеет
место один максимум изменения интенсивности дыхания при реге-
нерации хвоста, который приходится на 5-й день после ампутации
и соответствует стадии бластемы (Владимирова, 1975). Следова-
тельно, имеются определенные возрастные различия в процессе
регенерации хвоста у аксолотлей. Они касаются не только продол-
жительности этого процесса, но и особенностей дыхания и глико-
лиза. Усиление интенсивности дыхания при регенерации обнару-
жено, кроме аксолотля, у земляного червя, планарий, у млеко-
питающих при регенерации печени и у некоторых других живот-
ных (O’Brien, 1957; Needham, 1960; Сидорова и др., 1966).
Таким образом, процесс регенерации начинается с кратко-
временного, индуцибильно импульсного усиления интенсивности
дыхания и гликолиза, связанного с раздражением и возбуждением
поврежденных тканей, а затем развивается процесс конститутив-
ного усиления и дыхания и гликолиза. Интенсивность этих про-
цессов достигает максимума примерно к середине регенерации,
а затем постепенно, по мере завершения регенерации, снижается
до нормы. В случае регенерации хвоста у аксолотлей имеют место
два максимума интенсивности дыхания.
Еще в начале века Варбург сделал важное открытие, согласно
56
которому злокачественное перерождение тканей сопровождается
значительным усилением аэробного и анаэробного гликолиза
(Warburg, 1930). Варбург считал, что для раковых клеток столь
же характерно нарушение нормального дыхания. Последующие
исследования показали, что по наличию и активности окислитель-
ных ферментов, по способности к окислительному фосфорилирова-
нию, по уровню дыхания раковые клетки не отличаются сущест-
венным образом от нормальных (Вейнхаус, 1957; Ельцина, 1960;
Хореккер, Хайят, 1961; Aisenberg, 1961; Шапот, 1968; Зотин,
1974). В своих последних работах Варбург (Warburg, 1956а, Ь;
Warburg et al., 1958), хотя не отказался от идеи дефектного дыха-
ния, все же допускал, что в количественном отношении дыхание
раковых клеток мало отличается .от нормальных.
Исследования Варбурга вызвали большой интерес и стимули-
ровали исследования по метаболизму злокачественных образова-
ний. В подавляющем числе случаев было подтверждено утвержде-
ние о том, что злокачественные образования обладают повышен-
ным аэробным и анаэробным гликолизом по сравнению с нормаль-
ными тканями. Заметного отличия в дыхании опухолевых и нор-
мальных клеток не наблюдается (см., например, Aisenberg, 1961).
В более поздней работе Айзенберг и Морис (Aisenberg, Morris,
1961) пришли к выводу, что в некоторых штаммах крысиных ге-
патом Морриса интенсивность гликолиза не выше, чем в нормаль-
ной печени. Однако в работе Барка и др. (Burk et al., 1967) пока-
зано, что повышенный гликолиз характерен и для указанных штам-
мов гепатом. Особенно отчетливо выявилась связь величины ана-
эробного гликолиза со степенью злокачественности тканей при
изучении гепатом Морриса с разной степенью злокачественности
и разной скоростью роста. Эти данные показаны в табл. 2. Как
Таблица 2
Гликолиз в нормальной печени крыс и в гепатомах Морриса разной скорости
роста (Burk et al., 1967)
Ткани Qqq2 ~~ гликолиз (0,1—0.5% глюкозы в среде) Число изучен* ных случаев
минимум максимум .среднее
Нормальная печень 0,18 0,50 0,32 6
Наиболее медленно растущие гепато- мы 8624, 7794В, 7787, 7793 0,40 1,06 0,69 9
Медленно растущие гепатомы 5123В-Д, Н-35 0,58 1,80 1,03 13
Средняя по скорости роста гепатома 7288С 2,00 6,40 4,10 15
Быстро растущие гепатомы 3924А 4,5 8,0 6,7 3
Н-35ТС2, 3683 11,8 17,5 13,7 6
57
Рис. 9. Интенсивность дыхания в
присутствии глюкозы (1), в
отсутствие глюкозы (2) и аэроб-
ный гликолиз (3) в процессе роста
асцитного рака Эрлиха (Ач,
Штрауб, 1954)
видно, чем больше скорость роста
гепатом, тем выше уровень ана-
эробного гликолиза в них. Таким
образом, к настоящему времени
твердо доказано, что злокачест-
венное перерождение тканей со-
провождается резким усилением
аэробного и анаэробного гликоли-
за, причем степень увеличения
интенсивности гликолиза тем боль-
ше, чем больше степень злокачест-
венности тканей.
Существенным отличием злока-
чественного роста от процессов,
которые происходят при нормаль-
ном развитии или при регенера-
ции, является то, что в процессе
злокачественного перерождения
тканей, по-видимому, не происхо-
дит перехода живой системы от
процесса конститутивного (уклоне-
ния к конститутивному прибли-
жению системы, к конечному стационарному состоянию. Например,
на рис. 9 приведены данные Ач и Штрауб (1954) по дыханию и гли-
колизу при возникновении асцитного рака Эрлиха. Из этих дан-
ных следует, что после начального увеличения интенсивности гли-
колиза и значительно меньшего увеличения интенсивности дыха-
ния эти процессы сохраняются на постоянном уровне не переходя
в фазу уменьшения, как это имеет место во время оогенеза или
при регенерации.
При возникновении злокачественных образований происходит
усиление интенсивности аэробного и анаэробного гликолиза,
но не происходит существенного изменения дыхания. Исследование
механизмов конститутивного уклонения живых систем в процессе
оогенеза, регенерации и злокачественного роста может таким об-
разом вскрыть не только механизмы естественного омоложения
организмов, но и пролить некоторый свет на такие важнейшие
проблемы, как механизм регенерации и механизм малигнизации.
В этом плане мы пока имеем немного данных, позволяющих су-
дить о механизме конститутивного уклонения живых систем от
стационарного состояния. Однако и в этом случае, по-видимому,
он во многом связан с механизмом изменения концентрации и строе-
ния митохондрий в клетках. Так, например, Озернюк (1972) обна-
ружил, что в период малого роста ооцитов вьюна происходит уве-
личение содержания митохондриального белка в ооцитах парал-
лельно увеличению интенсивности дыхания, а в период большого
роста — снижение концентрации митохондрий и интенсивности
дыхания (см. рис. 43). Оказалось также, что в период малого роста
58
ооцитов вьюна происходит интенсивный синтез митохондриаль-
ных белков: включение С14-валина в белки митохондрий на ука-
занной стадии в 7 раз выше, чем на стадии конца большого роста
(Озернюк, 1974). Наконец, прямые определения содержания ми-
тохондрий, проведенные путем подсчета числа митохондрий в ооци-
тах на электронномикроскопических фотографиях, показали (см.
табл. 7), что концентрация митохондрий увеличивается от ста-
дии малого роста к стадии начала вакуолизации цитоплазмы
и снижается на последующих стадиях оогенеза (Озернюк, Пальм-
бах, 1975). Следовательно, можно считать установленным, что
увеличение интенсивности дыхания и конститутивное уклонение
живой системы в период оогенеза определяется изменением кон-
центрации митохондрий в ооцитах.
Относительно изменения концентрации митохондрий при ма-
лигнизации клеток известно немногое. Согласно наблюдениям
Клода и др. (Claude et al., 1952), число митохондрий в печени крыс
равно 33,0-1010, а в гепатоме 39,4-1010 шт/г сырого веса ткани.
Если эти данные подтвердятся для других опухолей, то можно
будет считать, что конститутивное уклонение живой системы
в процессе злокачественного перерождения сопровождается уве-
личением концентрации митохондрий в клетках. Это, однако, как
говорилось, не приводит к увеличению интенсивности дыхания
клеток, что, возможно, связано с нарушением структуры и функ-
ции митохондрий раковых клеток. Существует большое число ра-
бот (см. обзор Виленчик, 1973), в которых показано, что молеку-
лярный механизм злокачественного перерождения клеток в зна-
чительной степени определяется появлением в них мутантных ми-
тохондрий. Это позволило Виленчику высказать предположение,
что одним из механизмов канцерогенеза является селективное раз-
множение определенного типа митохондрий, содержащих специ-
фически измененную митохондриальную ДНК.
Критерий эволюции и его использование
в качестве теоретической базы
качественной феноменологической теории развития
Как видно из изложенного, феноменологическая теория разви-
тия организмов достаточно хорошо согласуется с эксперименталь-
ными данными, а ее следствия имеют большое значение для пони-
мания и выяснения механизма таких существенных явлений, как
старение и омоложение, регенерация и злокачественный рост.
Вместе с тем теоретическая основа, на которой базировалась теория
Пригожина — Виам, слаба. Она исходит из предположения, что для
описания процессов развития и роста организмов можно использо-
вать критерий эволюции термодинамики линейных необратимых про-
цессов (1.16). Имеются, однако, серьезные теоретические возраже-
ния против использования критерия (1.16) для описания этих про-
цессов. Применимость уравнений термодинамики линейных не-
59
обратимых процессов и4 соответственно, критерия (1.16) ограничены
областью необратимых процессов, протекающих в системах, неда-
леко уклонившихся от состояния равновесия, в которых выпол-
няются линейные законы и соотношение взаимности Онзагера.
В то же время большинство реальных необратимых процессов,
протекающих в физических, химических или биологических си-
стемах, существенно нелинейны. Поэтому необходимо специальное
доказательство возможности применения критерия (1.16) или по-
добного ему критерия для описания нелинейных процессов.
Попытки распространить методы феноменологической термоди-
намики на нелинейные необратимые процессы известны довольно
давно (см. Пригожин, 1960), но в последние годы в этом направле-
нии сделано особенно много. Одна из таких попыток связана с ис-
пользованием в термодинамике необратимых процессов вариаци-
онных принципов, впервые предложенных Онзагером (Onsager,
1931а, Ь). Неясно, однако, может ли принцип наименьшей дисси-
пации энергии, сформулированный Онзагером, быть распростра-
нен на системы, в которых протекают нелинейные необратимые про-
цессы. Многие авторы считают, что вариационные принципы приме-
нимы для описания нелинейных необратимых процессов (Vojta, 1967;
Gyarmati, 1969; Бахарева, 1971; Бахарева, Крылов, 1971; Дярматы,
1974). С другой стороны, доказана теорема, согласно которой не
существует общего вариационного принципа для диссипативных
систем в координатах физического пространства (см. Gage et al.,
1966; Richardson, 1969). Из этой теоремы следует, что вариацион-
ные принципы, предложенные Онзагером, Дярматы, Бахаревой,
и неравенство (1.16) не могут быть распространены на все виды
нелинейных процессов. Подобный же вывод для неравенства (1.16)
можно сделать исходя из критерия эволюции, предложенного Гланс-
дорфом и Пригожиным (Glansdorff, Prigogine, 1954, 1964; Glans-
dorff, 1956, 1960; Prigogine, 1955, 1966; Nicolis, 1971; Глансдорф,
Пригожин, 1973). Авторы показали, что в наиболее общем случае
эволюция термодинамической системы к равновесному или стаци-
онарному состоянию может быть описана неравенством:
т<0’ (2-1°)
где
п
(2-11)
5=1
И
п п
dip = d^ip 4- djip = У, J} dXj + У Xj dJj. (2.12)
J=1 5=1
В отличие от dtp линейная дифференциальная форма dxф не во
всех случаях является полным дифференциалом некоторой функ-
60
ции состояния системы. В системе, где выполняются линейные за-
коны и соотношение взаимности Онзагера, неравенство (2.10) пе-
реходит в (1.16) (см. Пригожин, I960). Таким образом, из теоремы
Глансдорфа — Пригожина следует, что неравенство (1.16) спра-
ведливо только для линейных необратимых процессов. В случае
нелинейных необратимых процессов диссипативная функция
не всегда будет уменьшаться и в стационарном состоянии не обя-
зательно примет минимальное значение, т. е. для нелинейных необ-
ратимых процессов будет выполняться следующее неравенство:
4г <2ЛЗ>
при условии выполнения неравенства (2.10). Действительно, было
показано, что в отдельных случаях неравенство (1.16) не выпол-
няется, и скорость продукции энтропии в стационарном состоянии
системы не минимальна (Петров, 1966; Richardson, 1969; Nicolis,
1971; Рубин, Фохт, 1972). Следовательно, из теории Глансдорфа —
Пригожина следует, что неравенство (1.16) не может характеризо-
вать процессы развития и роста организмов на тех стадиях, когда
они описываются нелинейными феноменологическими уравнения-
ми: в этом случае следует использовать критерий эволюции (2.10).
Однако критерий эволюции (2.10) не имеет того ясного физическо-
го смысла, который имеет критерий (1.16). Действительно, удель-
ную диссипативную функцию системы можно с известным приб-
лижением приравнять интенсивности теплопродукции (2.1) и поэ-
тому критерий эволюции (1.16) для линейной области можно на-
писать в виде:
4г < °- (2-14)
Очевидно, что критерий (2.14) не следует из критерия эволюции
Глансдорфа — Пригожина (2.10).
Таким образом, качественная феноменологическая теория раз-
вития, с одной стороны, хорошо обоснована экспериментальными
данными, с другой стороны,— в нелинейном варианте не имеет
твердого теоретического фундамента, так как из критерия (2.10)
не следует критерий (2.14). Учитывая, что теория Пригожина —
Виам имеет чрезвычайно важное значение для биологии развития,
необходимо было в какой-то форме восстановить теоретическую ба-
зу этой теории. Мы предприняли такую попытку, опираясь на
принцип, аналогичный принципу наименьшей диссипации энер-
гии Онзагера (Зотин, 1974; Зотин, Коноплев, Преснов, 1975).
Согласно второму началу термодинамики, любые необратимые
процессы сопровождаются образованием теплоты диссипации. В
изолированной системе образование теплоты диссипации приво-
дит к увеличению энтропии системы в целом. Если система адиаба-
тически не изолирована, теплота диссипации может покинуть пре-
делы системы, и общая энтропия системы может не изменяться или
61
даже убывать. Если открытая система находится вблизи от равно-
весия, и необратимые процессы протекают достаточно медленно,
теплота диссипации может полностью покинуть пределы системы.
Если система далека от равновесия и скорость образования тепло-
ты диссипации достаточно велика, не вся теплота диссипации будет
покидать пределы системы: часть ее может использоваться для не-
которых необратимых процессов. Другими словами, не вся тепло-
та, образующаяся как результат протекания необратимых процес-
сов, будет в этом случае теплотой диссипации в прямом смысле это-
го термина. Следовательно, для термодинамических систем далеких
от равновесия мы можем разбить удельную диссипативную функ-
цию на две части (Зотин, 1974):
(2.15)
где
Т d S т d-S,
фа — функция связанной диссипации; — функция внешней
диссипации. Сходное разбиение предложено и некоторыми други-
ми авторами (Landauer, 1973).
Принципиальное отличие разбиения (2.15) от разбиения Гланс-
дорфа — Пригожина (2.12) заключается в том, что в разбиении
(2.12) речь идет о механизме (об источниках) возникновения тепло-
ты диссипации в системе, в то время как в разбиении (2.15) —
о судьбе образовавшейся теплоты диссипации. Как видно из раз-
биения (2.12), источник возникновения теплоты диссипации двой-
ствен: частично она образуется за счет изменения потоков, частич-
но — за счет изменения сил. Двойственна и судьба образовавшей-
ся теплоты диссипации (2.15): часть ее используется внутри систе-
мы, другая часть покидает пределы системы. Так как только та
часть теплоты диссипации, которая покидает пределы системы,
может характеризовать степень диссипации энергии в системе, мы
сформулировали принцип наименьшей внешней диссипации энер-
гии (Зотин, 1974; Зотин, Коноплев, Преснов, 1975) следующим об-
разом: в стационарном состоянии любой термодинамической систе-
мы функция внешней диссипации принимает наименьшее из всех
возможных значений.
Логически возможны четыре случая.
1. Функция внешней диссипации системы равна удельной дис-
сипативной функции и равна нулю:
фа = ф = 0; ф^ = min. (2.17)
Этот случай относится к области классической термодинамики
и соответствует состоянию равновесия системы (энтропия системы
максимальна и скорость ее изменения равна нулю). Если система
не находится в состоянии равновесия и адиабатически изолирова-
на, то:
“Фа = 0; яр = > 0
62
(2-18)
и эволюция системы к равновесному состоянию определяется не-
равенством:
^->0.
dt
2. Функция внешней диссипации системы равна удельной дис-
сипативной функции, минимальна и константна:
фй = ф = const; фй = min. (2.19)
Это соответствует теореме Пригожина в термодинамике линейных
необратимых процессов, для которых выполняются линейные за-
коны и соотношение взаимности Онзагера. Если система является
открытой, то:
п
и эволюция системы к стационарному состоянию или равновесию
определяется:
< о
dt dt
3. Функция внешней диссипации системы не равна удельной
диссипативной функции, минимальна и константна:
фй =#Ф; фс/ = const; фй = min. (2.21)
В этом случае справедливо неравенство:
dt <
(2.22)
и эволюция системы к стационарному состоянию определяется:
dt
4. Функция внешней диссипации системы не равна удельной
диссипативной функции, не константна, но минимальна:
Ф(/ =АФ; Фа =# const; = min. (2.23)
Этот случай, как и предыдущий, относится к области термодина-
мики нелинейных необратимых процессов. Принцип наименьшей
внешней диссипации в этом случае может означать только то, что
величина функции внешней диссипации в стационарном состоянии
совершает колебания около некоторого усредненного значения.
При этом
ТЙ»
и эволюция системы к стационарному состоянию определяется
63
выражением:
><0, (2.24)
где
t
1 г
ЧЧ (0 = —^d [X(T)]dr
6
средняя величина функции внешней диссипации.
В первых трех случаях стационарное состояние системы может
быть представлено точкой в пространстве состояний. В четвертом
случае стационарное состояние представляет собой цикл (замкну-
тую траекторию) в пространстве состояний, т. е. в этом случае
необходимо некоторое уточнение понятия стационарного состоя-
ния.
Реальная удельная скорость теплопродукции в живых систе-
мах соответствует функции внешней диссипации. Поэтому прин-
цип наименьшей внешней диссипации энергии может служить ба-
зой для качественной феноменологической теории развития. Эво-
люция системы к конечному стационарному состоянию определяет-
ся в этом случае неравенствами (2.22) или (2.24). Эти критерии
эволюции хорошо соответствуют экспериментальным данным, по-
лученным при изучении основного обмена растущих животных
и развивающихся зародышей.
Разбиение (2.15), лежащее в основе критерия эволюции (2.22),
также может быть проверено экспериментально. Действительно,
в живых системах все диссипативные процессы определяются в ко-
нечном счете процессами дыхания и гликолиза. Другими словами,
диссипативная функция системы ф пропорциональна интенсивно-
сти дыхания и гликолиза организмов (Schaarschmidt et al., 1975;
Зотин, Шааршмидт, Лампрехт, 1975) или, если аэробный гликолиз
в организмах незначителен, пропорциональна интенсивности ды-
хания (Зотин, 19756). С другой стороны, функция внешней дисси-
пации фй по определению должна быть пропорциональна интенсив-
ности теплопродукции системы. Следовательно, для живых систем
мы можем вычислять функцию связанной диссипации фи по разно-
сти между данными непрямой и прямой калориметрии. Такие дан-
ные получены как для растений (см. Семихатова, 1967; Жолкевич,
1968) и микроорганизмов (Schaarschmidt et al., 1975; Зотин, Ша-
аршмидт, Лампрехт, 1975), так и для животных (Грудницкий,
19726, 1975; Зотин, 19756; Грудницкий, Никольская, 1976).
Более подробно этот вопрос будет рассмотрен в главе 4. Здесь мы
лишь упомянем о данных Студенского (см. Чередниченко, 1965),
который еще в конце прошлого века показал, что при беременно-
сти собак наблюдается расхождение данных прямой и непрямой
калориметрии, при этом расчет теплопродукции по дыханию на
14,3% превышал данные прямого измерения теплопродукции.
64
В опытах с проростками пшеницы доля рассеянной энергии
составляла от 38 до 95% от ф (Doyer, 1915), а в растущих листь-
ях — 20% (Жолкевич и др., 1964; Жолкевич, 1968). Таким обра-
зом, уже в этих опытах была доказана реальность существования
функции связанной диссипации.
СТАТИСТИЧЕСКАЯ ИНТЕРПРЕТАЦИЯ ФУНКЦИИ
ВНЕШНЕЙ ДИССИПАЦИИ И КРИТЕРИЯ ЭВОЛЮЦИИ
В. А, Коноплев, А, Н. Зотин
В термодинамике линейных необратимых процессов степень
необратимости процессов, протекающих в системе, определяется
величиной удельной диссипативной функции. Очевидно, что чем
больше степень необратимости процессов, тем дальше от равнове-
сия находится система, тем меньше вероятность состояния такой
системы. Следовательно, величина удельной диссипативной функ-
ции связана с вероятностью состояния открытой системы (Зотин,
Коноплев, Преснов, 1975). Чтобы это стало ясным, рассмотрим
вопрос о вероятности флуктуации в изолированной системе, близ-
кой к состоянию равновесия. Согласно Эйнштейну (см. Глансдорф
и Пригожин, 1973), для такой системы:
Р = Р0 expAL, (2.25)
где PQ — вероятность состояния системы в равновесии;
Д5 = S (а) — 50; So — значение энтропии системы в состоя-
нии равновесия; к — константа Больцмана.
Преобразуя (2.25) в:’
AS = Л: In
*0
и беря производную по времени, получим:
dS _ к dP
dt ~ Р dt ‘
(2.26)
В открытой системе общее изменение энтропии происходит за
счет возникновения энтропии (продукция энтропии) внутри систе-
мы и в результате обмена вещества и энергии со средой (поток эн-
тропии). Формула Эйнштейна (2.25) получена для адиабатически
изолированных систем. Поток энтропии в таких системах отсут-
ствует и величина dSIdt характеризует интенсивность продукции
энтропии в системе. Следовательно, уравнение (2.26) относится
только к продукции энтропии внутри системы, но не к потоку
65
энтропии. Учитывая, что в формуле (2.25) рассматривается изме-
нение энтропии в единице объема системы, можно написать:
_ 1 diS __ к dP
V dt ~ Р dt
или
_ Т d.S _ кТ dP
— V dt ~ Р dt
(2.27)
(2.28)
где а — удельная скорость продукции энтропии в системе.
Легко показать, что для случая систем, в которых протекают
линейные необратимые процессы, скорость изменения вероятност-
ного состояния системы описывается уравнением:
-^- = ?(Р0-Р), (2.29)
где р — константа.’
Действительно, решая уравнение (2.29) с граничным условием
Р<х> = Pq, имеем:
Р = Ро [1 - С exp (-pi)],
где С — произвольная положительная константа.
Подставляя это выражение в (2.27), получим:
_ кР^С exp (— 30 _ г ехр(—3Q
Ро [1-0 exp (-301 ‘ [1-Сехр(-301 ‘
Учитывая, что предположение (2.29) сделано для систем, на-
ходящихся вблизи от состояния равновесия, мы можем пренебречь
в знаменателе членом С ехр (—pi), который значительно меньше 1.
Тогда получаем:
а = к PC exp (—-Pi).
Это уравнение хорошо согласуется с зависимостью а = а (i),
полученной в термодинамике линейных необратимых процессов
на основании совершенно иных соображений (Onsager, 1931а, Ь;
де Гроот, Мазур 1964).
Теперь подставляя (2.29) в (2.27), получаем:
где а = &р.
Из уравнения (2.30) следует, что удельная скорость продукции
энтропии в системе, не слишком далеко уклонившейся от равнове-
сия, тем больше, чем меньше вероятность состояния данной си-
стемы.
Если открытая система далека от состояния равновесия, то
уравнение (2.29) может не выполняться, и изменение вероятност-
ного состояния системы должно описываться нелинейным уравне-
66
нием. В общем виде это нелинейное уравнение можно записать:
= W)L (2.31)
Требование, которое мы должны предъявить к виду функции / (Р)
состоит в следующем: при Р -> Ро, / (Р) 1, так как вблизи от
равновесия должно выполняться уравнение (2.29). Подставляя
(2.31) в (2.27), получим зависимость между удельной скоростью
продукции энтропии и вероятностью состояния для нелинейной
открытой системы:
б = аЛ_а/(/>). (2.32)
Для открытых систем, далеких от равновесия, должно быть спра-
ведливым разбиение (2.15):
cr = + ad, (2.33)
где аи — удельная скорость связывания энтропии диссипации
в системе, od — удельная скорость выведения энтропии диссипации
из системы.
Согласно принципу наименьшей внешней диссипации энергии
(Зотин, 1974; Зотин, Коноплев, Преснов, 1975), функция внешней
диссипации или удельная скорость выведения энтропии диссипации
в (2.15) и (2.33) может только уменьшаться. Так как вид функции
/ (Р) не определен и второй член в правой части уравнения (2.32)
может не уменьшаться по мере приближения системы к равновес-
ному или стационарному состоянию, то мы можем приписать сле-
дующие значения ои и ad:
ou = af(P) (2.34)
od:=a^. (2.35)
Для функции внешней диссипации из (2.35), получаем:
(2.36)
В наиболее общем случае, когда система характеризуется усред-
ненным значением функции внешней диссипации (система очень
далека от равновесия), связь функции внешней диссипации с ве-
роятностью состояния системы принимает вид:
to to to
Yd = ^d dt = -j-dt = aTP0-^\-^dt. (2.37)
0 0 0
Следовательно, функция внешней диссипации открытой системы
обратно пропорциональна вероятности состояния системы. Прин-
цип наименьшей внешней диссипации энергии, согласно которому
в стационарном состоянии функция внешней диссипации принима-
67
ет наименьшее из всех возможных значений, утверждает таким об-
разом, что стационарное состояние является наиболее вероятным
состоянием системы.
Отсюда ясен и физический смысл критерия эволюции в гло-
бальной теории нелинейных необратимых процессов. Так, беря
производную по времени от (2.36)
__ „ TPQ dP
dt & Р2 dt
(2.38)
и учитывая, что вероятность состояния системы по; мере проте-
кания в ней необратимых процессов не может убывать, т. е.
dPIdt^O, получаем критерий эволюции (2.22):
Следовательно, эволюция системы к равновесию или стационарно-
му состоянию всегда сопровождается переходоли системы в бо-
лее вероятное состояние.
В свою очередь вероятность самих стационарных состояний
зависит от того, насколько данная система находится далеко от
равновесия. В равновесном состоянии о = 0, / (Р) = 1 и из (2.32)
следует, что:
а - а = 0; Р = Ро = max. (2.39)
Вероятность состояния системы в равновесии максимальна.
В линейной области для системы, находящейся в стационарном
состоянии, имеем а = а = const, / (Р) = 1 и из (2.32) следует,
что:
(2.40)
где Р<?> — вероятность стационарного состояния термодинамиче-
ской системы в линейной области. Так как а 0, то (а + а) а
иа/(а + а) < 1. Отсюда следует, что:
р(л) п
1 ст \ о.
(2-41)
Вероятность стационарного состояния открытой системы, в кото-
рой протекают линейные необратимые процессы, меньше, чем ве-
роятность равновесного состояния.
Для нелинейной области имеем из (2.35):
азЙг = 6’ = (2-42)
^ст
где Ре? — вероятность стационарного состояния; Ь —константа.
68
Подставляя йз (2.40) значение Ро в (2.42), получаем:
Р™ = ±+±Р£\ (2.43)
Так как а < би а 1 (а = Л0, где к константа Больцмана, рав-
ная 1.38-10"16 эрг-град"1), то из (2.43) следует, что:
Р^<Р^<Р0. (2М)
Стационарное состояние термодинамических систем, далеких
от равновесия, имеет меньшую вероятность по сравнению со ста-
ционарным состоянием систем, близких к равновесию, и тем более
систем, находящихся в состоянии равновесия.
СТОХАСТИЧЕСКОЕ РАССМОТРЕНИЕ
КОНСТИТУТИВНЫХ ПРОЦЕССОВ
И КРИТЕРИЯ ЭВОЛЮЦИИ
JE, В. Преснов, Р. С, Зотипа
Не раз высказывалось предположение, что развитие и рост ор-
ганизмов представляют собой стохастический процесс (Medawar,
1942; 1945; Zotin, 1972; Зотипа, Зотин, 1973; Parks, 1973; Гончаров
и др., 1975; Преснов, Зотина, 1975). В феноменологической теории
развития, например, предполагается, что в процессе конститутив-
ного приближения организмов к конечному стационарному состоя-
нию происходит переход системы из менее вероятного состояния
к более вероятному. До настоящего времени, однако, вопрос о раз-
витии и росте организмов, как о стохастическом процессе, оставал-
ся на уровне чисто интуитивных соображений. Если бы удалось
связать удельную скорость теплопродукции с вероятностью состо-
яния живой системы, то критерий эволюции (2.14), который вы-
полняется в процессе развития и роста, можно было бы толковать
на языке стохастических представлений.
Ранее представлена такая попытка, опирающаяся на теорию
флуктуаций Эйнштейна и соотношения термодинамики необрати-
мых процессов. Мы попытались подойти к этой проблеме с других
позиций, исходя из представлений о развитии и росте, как о мар-
ковском процессе (Преснов, Зотина, 1975).
Введем следующие предположения относительно термодинами-
ческой системы.
1. Термодинамическая система описывается конечным набо-
ром независимых сил {XJ.
69
2. Процесс изменения сил является стохастическим с плотно-
стью вероятности р (Хц . . ., Хп, t)
\p(Xlt...,Xn, t)dXt- dXn = i Vt (2.45)
Плотность вероятности p (Xx, . . Xn, t) должна удовлетво-
рять следующим условиям, которые очевидны из ее физического
смысла:
а) р (Xi, . . ., Хп, t) -> 0 при Xi -> ±оо (i = 1,2, . . ., п);
б) р(0, . . О, Хп, t),
т. е. в любой момент времени плотность вероятности имеет абсо-
лютный максимум в начале координат;
в) равновесное состояние термодинамической системы (Xt = О,
г = 1,2, . . ., п) имеет в качестве плотности вероятности 6-функ-
цию;
г) в устойчивых стационарных состояниях плотность р (Х1?
Х2, . . ., Хп, t) имеет локальные максимумы, а в неустойчивых
стационарных состояниях — локальные минимумы.
Для живых систем, являющихся частным случаем термодина-
мической системы, введем следующие дополнительные условия.
3. Стохастический процесс {Xt (Z)} является марковским про-
цессом.
4. Поведение живой системы (процесс развития и роста) соот-
ветствует принципу наименьшего спуска (Преснов, 1973), т. е.
описывается динамической системой в ее силовом пространстве:
dX. л \г
= ° (2.46)
Здесь удельная теплопродукция организмов является функцией
сил Xi, т. е. с = М (Х1? . . ., Хп). Из этих предположений сле-
дует, что к развивающимся организмам применимо уравнение
Фоккера — Планка — Колмогорова:
’ (p“V (2.47)
dt । dAj dXi I v 7
i=i
Представляется интересным и более общее толкование уси-
ленного критерия эволюции в виде динамической системы:
+ = : (i = l,..., п), (2.48)
где — случайные функции времени типа белых гаус-
совских шумов с матрицей спектральных плотностей || || =
= const) и корреляционной матрицей || Ищ (т) | — в (т) ||
(6 (т) — дельта функция).
В этом случае уравнение Фоккера — Планка — Колмогорова
70
будет иметь вид (см., например, Красовский, 1974):
п п
“ 4 1 ' г“1 1 *
Таким образом, уравнение (2.47) или уравнение (2.49) в слу-
чае воздействия на систему гауссовских белых шумов связывает
удельную скорость теплопродукции организма, а, следовательно,
и функцию внешней диссипации системы, с плотностью вероятно-
сти, которая характеризует распределение случайных величин
(/), . . ., Хп (0, являющихся глобальными термодинамически-
ми силами. Эти уравнения, учитывающие в общем случае как инду-
цибильные, так и конститутивные процессы, еще не имеют в на-
стоящее время общей теории их решения в аналитическом виде.
Ниже будут приведены два частных случая аналитического реше-
ния уравнений (2.47) и (2.49).
Пусть живой организм находится в квазистационарном режи-
ме, при котором удельная скорость теплопродукции убывает доста-
точно медленно. Более того, предположим, что живая система
находится в установившемся режиме, т. е. плотность вероятности
неизменна во времени dpldt = 0. Тогда уравнение (2.47) принимает
следующий вид:
1 V д ° дм 1 V д2м л /о
w-+L^f=0- (2-50>
i—1 г-=1 г
Для нахождения точного решения этого уравнения необходимы
следующие вполне естественные предположения:
р (Хх, . . Хп) = Р1 (XJ-. . .-рп (Хп) (2.51)
М (Х1г . . ., Хп) = Мх (Хх).. . .-Мп (Хп). (2.52)
Эти предположения сводят уравнение (2.50) к уравнению:
yi Г 1 1 dM{ i
dX. М. дХ. + м. J *
Ясно, что полученное уравнение распадается на систему уравне-
ний:
1 rfn. 1 dM. 1 d2M.
Умножив i-тое уравнение этой системы соответственно на Mi9
получим:
1 dp. dM. d2M.
+ = (* = !.....») (2.53)
;i
Решая каждое из уравнений системы (2.53) относительно Mt (Xf),
получим п интегралов:
М;(Х;) = сД —+ (г
о г г
Для определения постоянных зададим начальные условия:
М (0, . . 0) = Л/^О)-. . .-71/п(0), так как теплопродукция в со-
стоянии равновесия равна нулю. Тогда С i = 0 и решение получим
в виде:
п
М = П G ( -^dXi
i=1 о 1
или
М = С ^-ydX, (2.54)
п
где С = JJ Ci, а В есть тг-мерная область силового фазового про-
1=1
странства В = [0, XJ X . . . X[0, Хп\. Константа С может быть
определена из условия нормировки для плотности вероятности
(2.45).
Наконец, уравнение (2.49) для установившегося режима в слу-
чае некоррелированных белых шумов, спектральная плотность
которых St есть:
St = 1/а, а = const (2.55)
может быть приведено к виду:
ЕдМ д In р 1 уч о Г д21пр /5 In ”\21 V д2М
ЬХ. dXi 2 dXf +(^i ‘
i^l г=1 u г 4 ' J 1=1 1
(2.56)
Это уравнение имеет следующее решение:
In р = - 2аМ + С, (2.57)
где константа С, как и в решении уравнения (2.47), определяется
из условия нормировки (2.45) (см. Красовский, 1974).
Приведенное исследование, основанное на усиленном крите-
рии эволюции, подтверждается многочисленными эксперименталь-
ными данными (Зотин, 1974) и позволяет по-новому взглянуть на
понятие удельной скорости теплопродукции живых систем. Это
понятие, по-видимому, можно связать с вероятностью состояния
живого организма, которую следует понимать, как вероятность
того, что глобальные термодинамические силы принимают опре-
деленные значения. Эта связь особенно отчетливо видна из реше-
72
ния (2.57) уравнения (2.56) и состоит в том, что минимум удельной
скорости теплопродукции соответствует максимуму плотности ве-
роятности и, наоборот. В этом смысле можно говорить об «обрат-
ной пропорциональности» между этими величинами. Отсюда сле-
дует, что в процессе развития живых организмов происходит
(имеется в виду конститутивный процесс) переход живой системы
из менее вероятного состояния в более вероятное.
УСИЛЕННЫЙ КРИТЕРИЙ ЭВОЛЮЦИИ
В БИОЛОГИИ РАЗВИТИЯ
Е. В. Преснов
Пригожин и Виам (Prigogine, Wiame, 1946) рассмотрели про-
цессы развития и роста организмов, опираясь на критерий эволю-
ции термодинамики линейных необратимых процессов. Они пред-
полагали при этом, что удельную диссипативную функцию живой
системы можно приравнять интенсивности теплопродукции. Одна-
ко теоретические соображения, изложенные выше, показали, что
интенсивность теплопродукции живой системы следует приравни-
вать не диссипативной функции, а удельной функции внешней дис-
сипации:
4>d = Я (2.58)
и использовать критерий эволюции (2.22). Этот критерий в форме
(2.14) достаточно хорошо подтверждается экспериментальными
данными, но носит чисто качественный характер.
В связи с этим отметим, что в настоящее время существует до-
статочно много прямых и косвенных указаний на то, что разви-
тие живой природы, на каком бы уровне мы его ни рассматрива-
ли (филогенез, онтогенез и т. д.), а также функционирование живой
системы на этих уровнях происходит оптимальным образом (Ро-
зен, 1969). По-видимому, процесс развития, роста и старения с тер-
модинамической точки зрения имеет свой принцип оптимальности
(см. Преснов, 1972, 1973). Этот принцип (усиленный критерий
эволюции) формулируется таким образом: в процессе развития,
роста и старения основной обмен организмов убывает наискорей-
шим возможным способом. Согласно соответствия (2.58) и зависи-
мости (2.46), аналитическое выражение сформулированного прин-
ципа оптимальности имеет следующий вид в силовом термодина-
мическом представлении:
Xi = - (X) (1 < i < п). (2.59)
73
Человек (из Thompson, 1942; Brody, 1945)
Возраст, годы 3 4 5 6 7 8 9 10 И 12 13 14 15
dr) —2,9 —2,9 —2,3 —2,7 —2,1 —1,8 —1,7 —1,7 —1,8 —1,9 —1,7 —1,4 —1,0 dM —— —о,86 —0,57 —0,36 —0,74 —0,64 —0,44 —0,48 —0,33 —0,91 —1,45 —1,74 —1,88 —2,18 at ’ ’ ’ ’ ’ ’
Возраст, годы 16 17 18 19 20 25 30 35 40 50 60 70
-4т- —0,7 —0,6 —0,6 —0,4 —0,05 —0,05 —0,05 —0,05 —0,05 —0,05 —0,05 —0,05 at ’ ’ ’ ’ ’ ’ ’ ’ ’ ’ —0,63 —0,54 —0,18 —0,13 —0,062 —0,050 —0,050 —0,050 —0,050 —0,051 —0,052 —0,051 at 1 ’ ’
Относительная средняя квадратическая ошибка ± 0,297.
Свиньи (из Brody, 1945)
Возраст, месяцы 4 6 8 10 12 14 16 18 20 22 24 26 28 30
—45,6 —30,7 —20,2 —13,0 —8,4 —5,5 —3,7 —2,5 —1,7 —1,0 —0,5 —0,2 —0,3 —1,4
at » » ’ 1
—22,2 —8,9 —5,0 —3,2 —2,2 —1,6 —1,2 0,9 0,7 0,6 —0,5 —0,4 —0,39 —0,405
dt ’ 1
Относительная средняя квадратическая ошибка ± 0,39.
Кролики (из Томме и Ким, 1938)
Возраст, месяцы 1/3 2/3 1 1,5 2 3 4 5 6 7
—1- —13 6 —8,0 —4,4 —1,5 —0,39 —0,35 —0.39 —0,02 —0,05 —0,40 dM —т— —12,4 —7,1 —4,3 —2,1 —1,08 —0,33 —0,20 —0,23 —0,27 —0,22 at 1 1 1 1 1 1 Относительная средняя квадратическая ошибка ± 9,8. Коровы (из Brody, 1945)
Возраст, месяцы 2 4 6 8 10 12 14 16 18 20 22 24
м -4т- —2,37 —1,93 —1,57 —1,28 —1,08 —0,84 —0,69 —0,56 —0,46 —0,37 —0,29 —0,21 сл at 1 1 ’ ’ dM —— —3,34 —1,77 —1,23 —0,96 —0,80 —0,68 —0,59 —0,52 —0,46 —0,40 —0,36 —0,31 at ’ ’ ’ ’ ’ ’ ’ Относительная средняя квадратическая ошибка ±0,038
Куры (из Barott, Pringle, 1946)
Возраст, недели 1 2 5 8 12 18 23 28* 33 40
dq dt —1,380 —1,213 —0,801 —0,503 —0,245 —0,062 —0,017 —9,012 —0,013 —9,001
dM --гг- —1,118 —1,187 —0,676 —0,387 at 1 1 1 ’ Добавлено нами при аппроксимации. Отнозительная средняя квадратическая ошибка ± 0,061. —0,192 —9,066 —0,026 —0,012 —0,008 —9,007
В дальнейшем для простоты положим к2(Х) = 1, но заметим, что
этот множитель может иметь нетривиальный биологический смысл
(Rosen, 1973).
Автономная динамическая система (2.59) позволяет выписать
формулу:
п п
(2.60)
Ясно, что критерии эволюции (2.59) и (2.60) сильнее критерия
(2.22).
В свою очередь, подставляя в формулу (2.60) выражение сил
роста Xg = rg (WnlW'1* — 1), дифференцировки Xd = rd (tm — t)
и формообразования Xf — rf (Wm — Ж1/з) (см. (3.39), (3.40),
(3.43)), можно провести то дополнительное исследование критерия
эволюции (2.22), о котором было сказано выше. Это исследование
заключается в установлении соответствия:
dq dM
dt dt
В качестве критерия для соответствия (2.61) была выбрана функ-
ция:
N
Q {rg, rd, rf) = У,
(2.62)
где (d(Jdt)i — экспериментальные значения скорости изменения
основного обмена животных в процессе роста. Значения констант
rd, г/ были вычислены исходя из условия минимума функции
Q (см., подробнее Преснов, Зотина, 1975). Численные расчеты,
выполненные для ряда животных и человека на ЭВМ М-222, дали
результаты, приведенные в таблице.
Эти материалы позволяют сделать вывод, что теоретические
построения, приведенные выше, подтверждаются эксперименталь-
ными наблюдениями. Следовательно, усиленный критерий эволю-
ции, в котором использованы силы роста, дифференцировки и фор-
мообразования, может достаточно хорошо описывать развитие
животных и человека.
Ill
КОЛИЧЕСТВЕННАЯ
ФЕНОМЕНОЛОГИЧЕСКАЯ ТЕОРИЯ
РАЗВИТИЯ ОРГАНИЗМОВ
Качественная феноменологическая теория развития организ-
мов базируется на критерии эволюции (2.22) и (2.24). В основе ее
лежит положение, вытекающее из статистической интерпретации
функции внешней диссипации и критерия эволюции, согласно
которому развитие, рост и старение организмов представляет со-
бой процесс перехода живой системы из менее вероятного состоя-
ния к более вероятному. Эта теория имеет чрезвычайно важные
следствия, касающиеся путей исследования и познания механиз-
мов основных явлений биологии развития, включая проблему вос-
становления органов и тканей, злокачественного роста, старения
и омоложения. Однако эта теория не содержит предпосылок, поз-
воляющих построить количественную теорию развития, хотя сама
термодинамика необратимых процессов содержит такие предпо-
сылки. Речь идет о построении системы дифференциальных урав-
нений, аналогичных уравнениям математической физики, которая
позволила бы описать явления роста, дифференцировки и формо-
образования, наиболее характерных моментов всякого развития.
К сожалению, здесь мы сталкиваемся с той же проблемой, что
и при рассмотрении критерия эволюции термодинамических сис-
тем к равновесному или стационарному состоянию: в настоящее
время законченная термодинамическая теория относится только
к системам, недалеко отстоящим от положения равновесия.
Как уже отмечалось, существуют различные попытки постро-
ить термодинамику нелинейных необратимых процессов. Наибо-
лее обоснованной является попытка группы Пригожина, в основу
которой положен принцип локального равновесия. Этот принцип,
однако, резко ограничивает область применимости локальной те-
ории необратимых процессов, так как допущение локального рав-
новесия в каждой точке неравновесной системы возможно только
для систем, не слишком далеко уклонившихся от равновесия. Сле-
довательно, локальная теория не способна описывать системы, да-
лекие от равновесия, когда интенсивность необратимых процес-
сов, протекающих в системе, настолько велика, что говорить о ло-
кальном равновесии невозможно.
Живые организмы являются системами, далекими от равнове-
сия, поэтому локальная теория не может служить базой для по-
строения термодинамики биологических процессов. Здесь нужна
77
другая термодинамическая теория, свободная от допущений
локальной теории. Первый вариант такой теории под названием
«глобальная теория нелинейных необратимых процессов» пред-
ставлен в статье Преснова. Она опирается на принцип наименьшей
внешней диссипации энергии, лемму Адамара и глобальные соот-
ношения термодинамики линейных необратимых процессов Онза-
гера (Преснов и др., 1973; Зотин, 1974; Зотин, Коноплев, Зотин,
1975). Можно надеяться, что на базе глобальной теории в ближай-
шем будущем будет построена количественная феноменологиче-
ская теория процессов развития, роста и старения организмов,
как и феноменологическая теория многих других биологических
явлений.
НЕЛИНЕЙНЫЕ
ФЕНОМЕНОЛОГИЧЕСКИЕ УРАВНЕНИЯ
A. If. Зотин, В. А. Коноплев, Е. В. Преснов
В глобальной теории нелинейные феноменологические урав-
нения (1.73) записываются в следующем виде:
п
= (г = 1,..., п), (3.1)
7=1
где
г dJ.
..., Хп) = ВД
б J
д7.
^> (°) =-5^7 (°),
а обобщенное соотношение взаимности:
Ltj (Хи . . ., Xn) = Ln (X1; . . Xn) (i, 7 = 1,..., n).
(3.3)
Система уравнений (3.1) является наиболее общим видом не-
линейных феноменологических уравнений. Нелинейность такой
системы максимальна, так как феноменологические коэффициенты
зависят от всех сил, участвующих в процессе: Ьц = (А\, . . .
. . ., Хп). При рассмотрении той или иной реальной термодинами-
ческой системы нет гарантий, что необратимые процессы, проте-
кающие в системе, должны описываться уравнениями с максималь-
ной степенью нелинейности. Этот вопрос можно решить только
опытным путем. Но для этого следует установить некоторые
78
уровни аппроксимации, которые соответствовали бы разной степе-
ни нелинейности системы.
При рассмотрении уровней аппроксимации естественно следо-
вать традициям, установившимся в термодинамике, поэтапного рас-
смотрения термодинамических систем с разной степенью необрати-
мости процессов, протекающих в них: вначале строится термоди-
намика равновесных процессов, затем термодинамика неравновес-
ных процессов близких к равновесным (линейные необратимые
процессы) и наконец термодинамика нелинейных необратимых про-
цессов, протекающих в системах, далеких от равновесия. Мы бу-
дем рассматривать уровни аппроксимации необратимых процессов
соответствующие послойному удалению от состояния равновесия
системы: линейная область, квазилинейная и нелинейная (Зотин,
Преснов, Коноплев, 1975).
Первый уровень — линейная теория. Выполняются линейные
законы:
п
(z = (3.4)
7=1
и соотношение взаимности Онзагера:
Li} = Ьц (z, ; = п). (3.5)
Второй уровень — квазилинейная теория. Нелинейные фено-
менологические уравнения имеют вид:
п
7; = МХ;)Х1 (/ = 1,...,п) (3.6)
7 = 1
i г^З
и выполняется соотношение взаимности Онзагера (3.5).
Третий уровень — нелинейная теория. Нелинейные феномено-
логические уравнения имеют вид:
п
Л = 7»(Х0Хг+^7;з(Х„Х7)Х3- (i = l............п) (3.7)
7=1
1^3
и выполняется обобщенное соотношение взаимности в форме:
Ltj (Xh Xj) = Ln (Xit Xj). (3.8)
Четвертый уровень — нелинейная теория. Выполняются не-
линейные феноменологические уравнения (3.1) и обобщенное соот-
ношение взаимности (3.3).
Основное допущение для 2-го и 3-го уровней аппроксимации со-
стоит в предположении, что вблизи от области линейности феноме-
нологические коэффициенты в первую очередь зависят от сил, оп-
ределяющих данный поток. Появление добавочной зависимости
для перекрестных коэффициентов от сил, сопряженных с этими ко-
79
эффициентами, 3-го уровня аппроксимации определяется обобщен-
ным соотношением взаимности в виде (3.8).
Такой многоуровневый подход в нелинейной феноменологиче-
ской термодинамике позволяет переходить от линейных систем,
близких к равновесному или стационарному состоянию, к систе-
мам, далеким от равновесия, с все более увеличивающейся сте-
пенью нелинейности. В процессе моделирования того или иного
явления в зависимости от степени нарушения линейности между
потоками и силами в изучаемом процессе можно переходить к все
более высокому уровню аппроксимации, пока не будут получены
уравнения, соответствующие реально наблюдаемым эксперимен-
тальным данным.
Очевидно, однако, что для того, чтобы перейти к более конкрет-
ному виду феноменологических уравнений разных уровней аппрок-
симации, необходимо сделать некоторые предположения относи-
тельно вида функциональной зависимости феноменологических
коэффициентов от сил. Мы рассмотрим только третий уровень
аппроксимации, на базе которого будут построены нелинейные
феноменологические уравнения роста, дифференцировки и формо-
образования.
При высказывании предположения о виде функциональной за-
висимости феноменологических коэффициентов от сил следует учи-
тывать, что они должны зависеть от сил таким образом, чтобы по
мере приближения системы к равновесному или стационарному
состоянию феноменологические коэффициенты превращались в
константы. Этому требованию удовлетворяют зависимости:
La = Lit exp (— aj?) (Z, j = 1, . . ., n) (3.9)
(Xh Xj) = Ltj exp [- (aj? + <*^)]. (3.10)
Подставляя эти выражения в уравнения (3.7), получим для треть-
его уровня аппроксимации:
п
Ji = aiLiiXi+'^aiajLijXj (г = 1,..., п), (3.11)
7=1
jVi
где
ai = exp (— алХ*) (3.12)
aj = exp (— ayXj).
Возможен и другой подход к построению общего вида нелиней-
ных феноменологических уравнений роста, дифференцировки и
формообразования (Коноплев, Зотин, 19756), опирающийся на сто-
хастическую теорию нелинейных неравновесных процессов, раз-
работка которой начата сравнительно недавно Бахаревой и Бирю-
ковым (1969, 1970, 1974). На основании стохастических соображе-
ний эти авторы получили нелинейное кинетическое уравнение
80
общего вида:
rfa zi /' \ Г а (°0 dSS (7)
— = С (a) exp
dt ' 1 r I к ду
(3.13)
где а — макроскопический параметр системы, С (а) и а (а) — ки-
нетические коэффициенты, к — константа. Авторы ограничились
случаем, когда С (а) = const и а (а) = const и использовали в сво-
ей работе уравнение вида:
det „ Га
— = Сехр^
ЭД5(«)
da J
(3.14)
Однако это ограничение сильно сужает область применимости
уравнения (3.13) и возможность его использования в биологии
развития или в физиологии. Для того, чтобы придать уравнению
(3.13) более общий вид по сравнению с (3.14), мы попытались (Ко-
ноплев, Зотип, 19756) раскрыть конкретный вид функциональной
зависимости С (а) и а (а). Напомним, что в термодинамике линей-
ных необратимых процессов имеют место следующие соотношения
(см. де Гроот, 1956):
J = ; X = 9Af(ct) (3.15)
dt ’ det v '
m
n) (3.16)
1=1
или в случае одной переменной:
=------~gaa. (3.17)
Теперь воспользуемся соотношением, полученным Бахаревой и
Бирюковым (1969) методами теории случайных процессов:
-Д5(«) = а(а)^1, (3.18)
где А5 (а) — отклонение энтропии системы от равновесного или
стационарного состояния.
Комбинируя (3.17) и (3.18), имеем:
, ч дДУ (а) 1 /о л п\
а —эГ2" = Т gaa“ (3-19)
Учитывая, что в стационарном состоянии и вблизи от него спра-
ведливо равенство:
получим, учитывая (3.19):
а (а) = 4 Г1 (3-20)
81
В простейшем случае, когда в процессе участвует один поток и
одна сила, уравнение (3.13), принимая во внимание (3.15), можно
записать:
J = С (а) ехр • (3.21)
Вид функции а (а) для этого случая вытекает из равенства (3.20).
Для нахождения функции С (а) воспользуемся очевидным утвер-
ждением: при приближении нелинейной системы к равновесному
или стационарному состоянию феноменологическое уравнение
(3.21) должно принять вид:
J = LX. (3.22)
Перепишем (3.21), используя (3.20), в виде:
J = C(a)exp(4-g’1X2) (3.23)
Разложив (3.23) в ряд:
J = C(a)[l (3.24)
и, пренебрегая вторым членом по сравнению с первым, получим:
J = С (а). (3.25)
Из сопоставления (3.22) с (3.25) следует, что:
С (a) = LX. (3.26)
Очевидно, что в более общем случае С (а) будет некоторой функци-
ей С (X) стремящейся при X 0 к LX.
Подставляя значение функции С (а) и о (а) соответственно из
(3.20) и (3.26) в (3.21), получаем нелинейное кинетическое урав-
нение для случая одного потока и силы:
J = LX ехр (4- Г1*2) (3.27)
или для случая п числа сил:
п
= £^3Х,ехр (4-gy%?) (*' = 1, • • «)• (3.28)
3=1
Таким образом, опираясь на стохастическую теорию нелинейных
необратимых процессов, развитую Бахаревой и Бирюковым (1969,
1970, 1974), мы получили нелинейные феноменологические урав-
нения, сходные с уравнениями (3.11), если принять что:
(3.29)
Это сопоставление вполне возможно, так как при выводе уравне-
ния (3.28) мы использовали соотношения линейной теории (3.15)
82
и (3.17) и приближение (3.25). Поэтому уравнение (3.28) относится
нр ко всей нелинейной области, а только к нелинейным системам,
недалеко отстоящим от линейной области (третий уровень аппрок-
симации). Очевидно, что в уравнении (3.28) не учтено обобщенное
соотношение взаимности, поэтому в полном виде его следует
писать:
п
+ У, ЗДехр [- (а;Х? + (3.30)
7=1
Jr-i
ДИФФЕРЕНЦИАЛЬНЫЕ УРАВНЕНИЯ
БИОЛОГИИ РАЗВИТИЯ
Р. С, Зотина, А. И. Зотин
Развитие, рост и старение организмов включают в себя три рода
явлений: изменение массы (рост), появление различий в разных
частях системы (дифференцировка) и изменение формы тела (мор-
фогенез). В количественной феноменологической теории развития
(Zotina, Zolin, 1972; Зотина, Зотин, 1973а; Zolin, 1972; Зотин,
1973, 1974) принимают, что взаимодействие этих трех явлений во
время жизни организмов аналогично взаимодействию потоков и
сил в термодинамике линейных необратимых процессов и может
быть описано уравнениями вида (1.13). Обозначая удельный поток
изменения веса животных через:
удельный поток дифференцировки через:
А.=-^ (3.32)
и удельный поток формообразования через:
= (3.33)
мы можем записать в линейном приближении (1.13) взаимодейст-
вие процессов изменения веса, дифференцировки и формообразо-
вания во время роста в виде следующей системы уравнений:
= kggXg + kgdXd + kgfXh
= kdgXg + kddXd + kdiXh (3.34)
jp = kfgXg + kfdXd + knXh
83
где Xg — сила, вызывающая поток изменения веса, Xd — сила,
вызывающая поток дифференцировки, X; — сила, вызывающая
поток формообразования, кц — константы (Z = g, d, f; j = g, d, /).
Трудным вопросом для количественной феноменологической
теории развития и роста организмов является вопрос о конкретном
значении сил, входящих в уравнение (3.34). В термодинамике не-
обратимых процессов в качестве феноменологических уравнений
обычно используются эмпирические законы, установленные опыт-
ным путем. Например, при рассмотрении процессов диффузии
используют закон Фика, при рассмотрении процессов теплопереда-
чи — закон Фурье и т. д. По этому же пути мы пошли при опреде-
лении сил Xg и Xd в уравнении (3.34), воспользовавшись уравне-
ниями роста животных, которые достаточно хорошо подтверждены
экспериментальными данными (Zotin, Zotina, 1967; Зотин, Зотина,
1969, 1975; Зотин, Грудницкий, 1970; Грудницкий и др., 1971;
Zotin et al., 1971; Зотина, Зотин, 1973а).
К настоящему времени предложено большое число различных
эмпирических и полуэмпирических уравнений и формул для описа-
ния процессов роста животных и человека (см. обзоры Гофман,
1938; Brody, 1945; Medawar, 1945; Зотина, Зотин, 1973а; Мина,
1973; Зотин, 1974, 1975а). Для того чтобы решить, какие из этих
уравнений можно принять в качестве феноменологических законов,
характеризующих развитие и рост организмов, следует напомнить
те требования, которые предъявляет термодинамика необрати-
мых процессов к силам. Таких требований два: 1) параметры, оп-
ределяющие термодинамическую силу, должны быть интенсивными
величинами, 2) эти интенсивные величины должны создавать
некоторый градиент. Этим требованиям не удовлетворяют многие
эмпирические уравнения роста, например, уравнение Робертсона.
Не останавливаясь подробнее на этом вопросе, можно сказать, что
этому условию удовлетворяет популярное в настоящее время урав-
нение роста Пюттера — Берталанфи, которое после некоторых
преобразований можно записать в следующем виде:
* ^- = к1), (3.35)
И7 dt 8 \ jyb у ' 7
где Wm — максимальный вес животного, kg и b — константы.
Многие другие теории роста, например, теория Броди — Найде-
нова, Дэвидсона — Лэйрд, Шмальгаузена, могут быть при извест-
ных условиях идентифицированы с теорией Пюттера — Берта-
ланфи (Зотин, 1972, 1974, 1975а). Другое уравнение, которое удов-
летворяет требованиям термодинамики и не сводится к уравнению
Берталанфи (3.35), предложено нами (Зотина, Зотин, 1967) и имеет
вид:
= (з.зб)
84
где tm — время достижения максимального веса животного,
kd — константа.
Из всей массы известных уравнений роста, только эти два урав-
нения пригодны для наших целей. Встает вопрос, какое из них
мы должны выбрать в качестве феноменологического закона рос-
та? Оба эти уравнения достаточно хорошо описывают рост живот-
ных. Оба эти уравнения применены для описания роста не во всех
случаях. Так, уравнение Берталанфи мало пригодно для описания
веса птиц и млекопитающих на поздних этапах онтогенеза. Это
видно хотя бы из следующих рассуждений. Решение уравнения
Берталанфи (3.35) имеет вид:
1
w = Wm (1 - e’V')b , (3.37)
из которого следует, что вес животного непрерывно увеличивается
и максимальное значение веса может быть достигнуто лишь при
t = оо. В то же время хорошо известно, что максимальный вес
птиц и млекопитающих достигается в первой трети жизни. В свою
очередь уравнение (3.36) описывает достаточно хорошо лишь пе-
риод быстрого увеличения веса животного, так как его решение
имеет вид:
k .
W = Wme 2 \ (3.38)
Из формулы (3.38) видно, что максимальный вес животного до-
стигается при t = tm, при t > tm вес животного должен умень-
шаться, причем по кривой, симметричной кривой предшествую-
щего увеличения веса. Этого, естественно, не бывает. Поэтому
возникла идея: что оба уравнения и (3.35), и (3.36) справедливы,
но отражают разные явления, происходящие во время роста. Было
высказано предположение (Zolin, Zolina, 1967; Зотин, Зотина,
1969; 1975; Зотин, Грудницкий, 1970; Грудницкий и др., 1971;
Zotin el al., 1971; Зотина, Зотин, 1973а; Зотин, 1974), что одно из
них уравнение (3.35) является феноменологическим уравнением
роста, а другое (3.36) — феноменологическим уравнением диффе-
ренцировки. Тогда можно записать для силы, определяющей рост:
[ wb \
xg = 1), (3.39)
а для силы, определяющей дифференцировку:
xd = rd (tm - t), (3.40)
где rg и rd — константы размерности, tm в этом случае время до-
стижения максимального уровня дифференцировки. Подставляя эти
значения сил в уравнение (3.34), получим уравнение роста в виде:
1 dW у / W-m j \ . г /, /Q /4\
(Ц “ ^gg y—jyb / 3’
85
и уравнение дифференцировки:
/ \
(М2>
учитывающее взаимодействие этих процессов (здесь = к^).
Для проверки правильности уравнения (3.41), как уравнения
роста животных, были использованы данные по росту кур, приво-
димые в работе Баротта и Прингль (Barott, Pringle, 1946). Как
видно из рис. 10, это уравнение удовлетворительно описывает
рост кур. Более прямое доказательство преимущества уравнения
(3.41) было получено при рассмотрении кривых изменения веса
животных за весь период онтогенеза (Грудницкий и др., 1971).
Обычно изменение веса животных рассматривают только от момен-
та рождения до достижения максимального веса животного. И в
этой области многие уравнения роста хорошо описывают кривые
изменения веса. Однако изменение веса многих животных продол-
жается и на последующих этапах жизни: после достижения мак-
симального веса он сохраняется некоторое время на стационарном
уровне, а затем постепенно снижается. Насколько нам известно,
только уравнение (3.41) может в некоторых случаях описывать
всю кривую изменения веса животных, включая рост, стационар-
ную фазу и уменьшение веса к концу жизни. Это было показано
на примере изменения веса золотистого хомячка (рис. И). Теорети-
ческая кривая, основанная на уравнении (3.41), хорошо согласует-
ся с экспериментальными данными, заимствованными из справоч-
ника Альтмана и Диттмера (Altman, Dittmer, 1964). Характерно,
что константа Lgd при описании роста хомячка (рис. 11) почти на
5 порядков меньше константы Lgg. Это показывает, что при описа-
нии изменения веса животных в процессе роста, сила, определяю-
щая дифференцировку, оказывает значительно меньшее влияние
на изменение веса, по сравнению с силой, определяющей рост.
Другими словами, выбор сил (3.39) и (3.40) оправдывается при прак-
тическом использовании уравнения (3.41). Можно привести также
следующее соображение в пользу того, что выбор уравнения (3.36)
в качестве феноменологического закона дифференцировки являет-
ся разумным. Общий объем дифференцировок в процессе развития
и роста организмов не велик на ранних стадиях развития и роста
по сравнению с более поздними стадиями, затем он достигает из-
вестного максимума и уменьшается к концу жизни, т. е. процесс
изменения общего объема дифференцировок описывается кривой
с максимумом, типа гауссовской кривой. Решение уравнения (3.36)
представляет собой гауссовскую кривую нормального распреде-
ления (3.38). Все сказанное, как нам кажется, оправдывает те
предположения, которые мы сделали при получении уравнений
(3.41) и (3.42).
Сложным оказался вопрос о силе, вызывающей формообразо-
вание. Рассуждения, аналогичные приведенным выше, показали,
что нельзя подыскать какое-либо уравнение роста, отличное
86
Рис. 10. Сравнение экспериментальных данных (1) по росту кур (Barott,
Pringle, 1946) с кривой (2), рассчитанной по уравнению (3.41) (Грудниц-
кий и др., 1971)
Константы: = 0,035, Lgd = 0,000008, tm — 1 год, Wm = 2,5 кг
Рис. 11. Сравнение экспериментальных данных (1), полученных при изучении
изменения веса золотистых хомячков на протяжении всей жизни (Altman,
Dittmer, 1964) с кривой (2), рассчитанной по уравнению (3.41) (Грудницкий
и др., 1971)
Константы Lgg = 0,126, Lgd 0,000006, tm = 180 дн, Wm = 112 г
Рис. 12. Сравнение эксперимен-
тальных данных (1), полученных
при изучении изменения веса на
протяжении жизни (а) бобров
(Бородина, 1970), белоногих хо-
мячков (б) (Layne, 1966) и золо-
тистых хомячков (в) (Altman,
Dittmer, 1964) с кривой (2), рас-
считанной по уравнению (3.45)
Константы:
а — Lgg = 18,0, Lgd = 0,010, Lgf =
= —4,7, tm = 12,5 лет = 100%
= 19,8 кг = 100%;
6 - Lgg = 21,1, L d = 0,006, Lgf =
= —6,1» tm = 55 недель 100%,
Wm =- 27,1 г = 100%;
e-L^7,16;5’ L«d~ 0’0,1> Lgf =
- —4,3, tm = 62 недели = loO%,
IVm = 114 г = 100%
80 120 160 i,°/>
от (3.35) и (3.36), которое тем или иным путем можно было бы
использовать в качестве уравнения формообразования. Оказалось,
однако, что есть другой путь, позволяющий подойти к решению
этого вопроса — путь рассмотрения размерностей различных по-
казателей, входящих в уравнение (3.34) (Зотин, Зотина, 1972,
1975; Зотин, 1973, 1974). Не останавливаясь подробнее на различ-
ных соображениях и допущениях, которые мы использовали в
этих работах, приведем окончательное выражение для силы, опре-
деляющей формообразование:
Xf = rf (W?n - И^). (3.43)
Эта сила соответствует уравнению роста:
_L^ = /C/(<_Wb). (3.44)
Расчеты на ЭВМ показали, что уравнение (3.44) достаточно хоро-
шо описывает рост собак и других млекопитающих (Гинис и др.
1974; Зотин, 1974). Следовательно, предположения, положенные
в основу вывода уравнений (3.43) и (3.44) не беспочвенны.
Теперь подставляя (3.43) в (3.34), получим после соответствую-
щих преобразований, систему уравнений:
V Т = Л -
W = Л - + bd/) + Ldd (tm - t) (3.45)
= (^ — И">)(ф- + Llt} + Lfd (tn - t),
которая является системой линейных феноменологических урав-
нений роста, дифференцировки и формообразования, учитываю-
щей взаимодействие этих процессов друг с другом.
Для проверки правильности первого уравнения в системе (3.45),
как уравнения, описывающего изменение веса животных на про-
тяжении всей жизни, были использованы данные, полученные
при изучении изменения веса белоногих и золотистых хомячков
и бобров. Расчеты, проведенные В. А. Коноплевым с помощью
ЭВМ «Мир-2», показали, что уравнение (3.45) достаточно хорошо
описывает изменение веса этих животных на протяжении жизни
(рис. 12). Следовательно, система линейных относительно сил фе-
номенологических уравнений (3.45) может быть использована при
рассмотрении роста и старения некоторых животных.
До сих пор мы исходили из допущения, что развитие и рост
организмов может описываться линейными феноменологическими
уравнениями. С точки зрения термодинамики необратимых про-
цессов это равноценно утверждению, что процессы развития и
роста осуществляются в системе не слишком далеко уклонившейся
от стационарного состояния. Очевидно, что такое предположение
88
может быть справедливым только для определенного участка жиз-
ненного пути организма, когда он приближается к конечному
стационарному состоянию. Так, не удалось получить хорошего
совпадения эмпирических данных (Грудницкий и др., 1971), по-
лученных при изучении роста людей в процессе всей жизни, с тео-
ретической кривой рассчитанной по уравнению (3.41). Рост людей
значительно отличается от роста многих млекопитающих. Это свя-
зано, по-видимому, с тем, что дети после рождения еще долго
сохраняют некоторые эмбриональные черты. Еще в большей сте-
пени это, вероятно, относится к сумчатым животным. Неприло-
жимость уравнения (3.41) для описания роста людей можно
объяснить тем, что уравнение (3.41), как и (3.45), линейно и хоро-
шо описывает рост только на сравнительно поздних стадиях раз-
вития. Можно думать, что в период зародышевого развития и на
начальных стадиях роста организмы далеки от конечного стацио-
нарного состояния и феноменологические уравнения роста, диффе-
ренцировки и формообразования должны иметь нелинейный вид.
В экспериментальном плане, критерием правильности таких нели-
нейных уравнений может быть то, что они лучше чем линейные
уравнения (3.45) описывают рост людей. Другими словами, они
должны описывать не только рост животных после рождения, но
и зародышевый рост.
В настоящее время можно попытаться построить нелинейную
количественную теорию развития и роста животных, опираясь на
полученные в предыдущей статье нелинейные феноменологические
уравнения вида (3.30). В наиболее общем виде нелинейные урав-
нения роста, дифференцировки и формообразования, учитывая
(3.1) и (3.34), можно записать (Преснов и др., 1973; Зотин, 1974):
Jg = kgg (X) Xg + kgd (X) Xd + kgf (X) xf
Jd = kdg (X) Xg + kdd (X) Xd + kdf (X) Xf (3.46)
J, = kfg (X) Xg + kfd (X) Xd + kff (X) X„
где X = (Xg, Xd, Xf).
Система (3.46) является наиболее общим видом нелинейных фе-
номенологических уравнений биологии развития и соответствует
наивысшей степени нелинейности живой системы, максимальной
степени удаления от конечного стационарного состояния. Мы мо-
жем, однако, опираясь на уровни аппроксимации, ограничиться
пока третьим уровнем (3.7) и, исходя из уравнений (3.31), (3.32),
(3.33), написать систему (3.46) в виде:
-Т = (Xg) + kgd (Xg, Xd) Xd + kgl (Xg, Xf) Xf
(Xd, Xg) Xg 4- kdd (Xd) Xd + kdf (Xd, Xf) Xf (3.47)
V X^ X* + k* X“> xd + kff (Xf) Xf.
89
На основании некоторых приближений (Зотин, 1974; Зотин,
Зотина, 1975) и стохастического подхода в теории нелинейных
необратимых процессов (Коноплев, Зотин, 19756) было получено
феноменологическое нелинейное уравнение (3.30). Подставляя
в это уравнение значение сил из (3.39), (3.40), и (3.43), получим
дифференциальные уравнения биологии развития в следующем
более конкретном виде:
1 dW
W dt
/ \
ai Lgg ---1azLgd {tm — t) + (И7™ — Wb)
1 dD
[V dt
— &2
/ wb
„ т I m
— 1) + Ldd (tm — t) 4“
+ a3Ldf(Wbm~W4 (3.48)
1 dF
TF dt
+ Lff(Wbm-Wb)
— 1 ) + a2^/d (Jm — t) +
где
=- exp
Wb
r m
a2 = exp[— ad(tm — t)2\
(3.49)
a3 = exp [— az (W\bn — Wb)2].
Очевидно, что по мере приближения организма в процессе раз-
вития и роста к конечному стационарному состоянию нелинейные
уравнения роста, дифференцировки и формообразования (3.48)
переходят в линейные уравнения (3.45).
При получении уравнений (3.48) мы предполагали, что констан-
та Ъ остается неизменной на всех стадиях роста организма. Это
предположение допустимо, если рассматривать только постэмбри-
ональный рост животных: не исключено, что во время зародышево-
го развития и на ранних этапах постэмбрионального роста констан-
та Ъ зависит от времени. Мы предположили, что эта зависимость
выражается уравнением: b (t) = b [1 + ехр (— 0£)] (Зотина, Зо-
тин, 19736; Зотин, 1974; Зотин, Зотина, 1975). Однако, практиче-
ское использование уравнения (3.48) для описания изменения
веса людей на протяжении жизни показало, что лучше использо-
вать зависимость вида: Ь (t) = b [1 + yt ехр (— 0£)] (Зотина, Зо-
тина и др., 1976). Так как общая феноменологическая теория раз-
вития должна охватывать все этапы жизни организма, включая
зародышевый и ранний постэмбриональный периоды, в фено-
менологических уравнениях биологии развития следует учесть
90
зависимость константы b от времени в уравнениях (3.48):
1 d,W _ „ \ т / Пт
W dt, '
1 I + a^gd (tm — 0 +
1 dD
IV dt
+ a3Lg/(^-JVb(O)
= a2
— 1 ) + ^dd (Jm — 0 +
+ «Mwbm-ww)
1 dF
W dt
ar = exp
/ yy1^ \
dlLfg ^(,0) — 1J + «2^/d (fm — t) +
+ L„(Wbm- W»W)
где
/ irb x2
- °U—тт- 1
s \ iv^ ) /
a2 = exp [— ad(tm — Z)2]
a3 = exp[-*f(Wbm-WWy]
b(t) = b[i + exp (— 3£)].
(3.50)
(3.51)
Из (3.51) очевидно, что с некоторого момента времени константа
b практически становится независимой от времени. Детальный эк-
спериментальный анализ уравнения роста, входящего в систему
(3.48) и (3.50), будет дан в следующей статье. Из материалов, при-
водимых в этой работе, видно, что уравнение роста удовлетвори-
тельно описывает изменение веса людей в процессе жизни (см.
рис. 13 и 16).
Как видно из изложенного, количественная феноменологиче-
ская теория развития организмов имеет в некотором отношении
завершенный характер и получила первые экспериментальные под-
тверждения. Несмотря на это многие вопросы, связанные с этой
теорией, остаются еще не ясными и спорными. Неясны вопросы,
связанные с такими понятиями как поток дифференцировки, поток
формообразования, силы, вызывающей рост, дифференцировку и
формообразования. Эти вопросы, как мы надеемся, станут в конце
концов более ясными при рассмотрении молекулярных механиз-
мов роста, дифференцировки и формообразования. Система диф-
ференциальных уравнений (3.50) описывает изменение веса людей
на протяжении всей жизни, начиная с момента рождения. Это не
означает, что система (3.50) может быть использована для описа-
ния роста зародышей и, тем более роста ооцитов животных и чело-
века. Возможно, что использование четвертого уровня аппрокси-
91
мации, т. е. феноменологических уравнений вида (3.46), позволит
построить такую систему дифференциальных уравнений, которая
описывала бы изменение веса живой системы от периода оогенеза
до самых поздних стадий онтогенеза животного. Возможно также,
что дальнейшее развитие феноменологической теории развития
организмов приведет к получению иных уравнений, чем те, кото-
рые приведены в данной работе. Важно, однако, как нам кажется,
не то, что уравнения вида (3.45), (3.48) или (3.50) могут оказаться
неправильными, а то, что путь и логика их получения открывает
возможность построения количественной теории развития орга-
низмов. Вместе с тем, совпадение теоретических расчетов с экспери-
ментальными данными (рис. 13, 16) вселяет надежду, что дифферен-
циальные уравнения вида (3.45), (3.48) и (3.50) окажутся справед
ливыми.
компьютерный анализ
НЕЛИНЕЙНЫХ УРАВНЕНИЙ РОСТА
В. А. Коноплев, А. И. Зотин, Е. А. Прокофьев,
Н. Ш. Шаги мор дано в
Исходя из термодинамической теории развития организмов,
были предложены линейные феноменологические уравнения, учи-
тывающие взаимодействие процессов роста и дифференцировки
(Зотин, Зотина, 1969; Грудницкий и др., 1971). Проверка этих
уравнений с помощью ЭВМ показала, что они удовлетворительно
описывают рост кур и изменение веса хомячков на протяжении
всей жизни (Грудницкий и др., 1971; Зотин, 1974). Вместе с тем
оказалось, что полученное линейное уравнение не может быть
применено для описания изменения веса людей в процессе жизни.
Поэтому в дальнейшем была сделана попытка учесть в уравнении
роста не только процессы дифференцировки, но и формообразова-
ние, а также получить нелинейные относительно сил уравнения
роста (Зотин, Зотина, 1972, 1975; Зотина, Зотин, 1973а; Зотин,
1973; 1974; Гинис и др., 1974; Коноплев, Зотин, 19756). Целью
этих работ было получение уравнения роста, удовлетворительно
описывающего изменение веса людей на протяжении всей жизни,
так как возможность описания изменения веса людей в экспери-
ментальном плане может служить критерием правильности того
или иного нелинейного уравнения роста животных (Зотин, 1974).
Оказалось, что нелинейное уравнение роста (3.48), приведенное
в работе Зотина и Зотиной (1975), в общем удовлетворяет этому
критерию (Коноплев, Зотин и др., 1975).
92
Данные об изменении веса людей (мужчин) на протяжении
всей жизни были взяты из книги Кэтлэ (1913). Расчет кон-
стант в уравнениях и сопоставление этих уравнений с кривыми
изменения веса людей проводили с помощью аналоговой ЭВМ
типа МН-14 методом подбора коэффициентов и на ЭВЦМ «Мир-2»—
методом Рунге — Кутта.
Нелинейное феноменологическое уравнение роста организмов,
учитывающее взаимодействие роста, дифференцировки и формооб-
разования, было получено опираясь па некоторые термодинами-
ческие соображения (Зотин, 1974; Зотин, Зотина, 1975; Зотин,
Преснов, Коноплев, 1975) и стохастический подход к нелинейным
термодинамическим системам (Коноплев, Зотин, 19756). Из си-
стемы (3.48) видно, что это уравнение можно записать в следую-
щем виде:
1 dW _
W dt —ai
/ wb \
Lgg -------1 j + (tm — 0 +
+ a3Lgf(Whm-W»)
(3.52)
где
ai = exp
/ Wb
— a J—£
g\ Wb
a2 = exp[—ad (tm — t)2]
a3 = exp [— a/ (Wbm — TVb)2].
Здесь W — вес организма, t — время, Wm — максимальный вес,
tm — время достижения максимального уровня дифференциров-
ки; Lu, aj, b — константы.
В уравнении (3.52) учтено обобщенное соотношение взаимности
(см. Преснов й др., 1973). Без учета этого соотношения уравнение
(3.52) принимает вид:
1 dW т ( Wm Д , „ т tt ,
W dt Ч “г a2^gdVm Ч "г
+ a3Lgf(Whm-Wb).
(3.53)
Мы проводили проверку следующих вариантов уравнения (3.52):
1) члены а± = а2 = а3 = 1 — линейный вариант уравнения
(3.52);
2) коэффициенты Lgg, Lgd, Lgf попарно полагали равными
нулю, что соответствует присутствию в уравнении (3.52) или
(3.53) только одной из сил;
3) коэффициенты Lgg, Lgd, Lgf поочередно полагали равными
нулю, что соответствует присутствию в уравнении (3.52) и (3.53)
двух сил;
93
4) в уравнениях (3.52) и (3.53) присутствуют все коэффициенты
и нелинейные члены, что позволяет выяснить справедливость обоб-
щенного соотношения взаимности, которое отсутствует в уравне-
нии (3.53).
Все полученные результаты представлены на рис. 13—15. Как
видно, из этих наблюдений, линейный вариант плохо соответствует
кривой изменения веса людей (рис. 13, а). В нелинейном уравне-
нии роста (3.52) и (3.53) должны присутствовать все силы для того,
чтобы оно соответствовало экспериментальной кривой изменения
веса людей на протяжении всей жизни (рис. 13, б, в). Так, если
нелинейные уравнения (3.52) и (3.53) включают в себя только одну
силу (только один из коэффициентов Lgg1 Lgd, Lgf не равен нулю),
их решение не соответствует кривой изменения веса людей
(рис. 14, а—в). То же самое получается, если в уравнения (3.52) и
(3.53) входят только две из обобщенных сил (один из коэффициен-
тов Lgg, Lgd, Lgf равен нулю) (рис. 14, г—ё) или в уравнениях от-
сутствуют нелинейные члены (рис. 15). Из рис. 13—15 следует так-
же, что наличие члена, определяющего соотношение взаимности
в уравнении (3.52), не сказывается существенным образом на опи-
сании роста людей; кривые, полученные на основании уравнения
(3.52), практически не отличаются от кривых, полученных на осно-
вании (3.53) рис. 146, в.
Различия сводятся только к различиям в величине коэффи-
циентов.
Таким образом, расчеты на ЭВМ показали, что нелинейное урав-
нение роста (3.52), так же, как уравнение (3.53), удовлетворитель-
но описывает изменение веса людей на протяжении всей жизни.
В то же время линейное уравнение типа (3.45) и нелинейные урав-
нения (3.52) или (3.53), в которых отсутствует хотя бы одна из
обобщенных сил, не могут правильно описать кривые изменения
веса людей. Это показывает, что изменение веса людей может быть
описано только нелинейным уравнением роста и что в процессе
роста людей происходит взаимодействие всех трех основных яв-
лений, связанных с развитием организмов: роста, дифференциров-
ки и формообразования.
Вместе с тем более детальный анализ применимости уравнения
(3.52) к описанию изменения веса людей на протяжении жизни
показал, что на ранних стадиях роста теоретическая и эксперимен-
тальная кривая совпадают недостаточно хорошо (рис. 16, 6). Мы
предположили, что это связано с тем, что в уравнении (3.52)
Ь = const, в то время как имеются данные об изменении этого
коэффициента в процессах эмбрионального и раннего постэмбрио-
нального развития животных и человека. К сожалению в настоя-
щее время мы можем определить зависимость Ъ (t) только эмпири-
ческим путем. Единственное ограничение, которое накладывается
на эту зависимость, состоит в том, что константа b должна за-
висеть от времени только на ранних стадиях роста. В отличие от
более ранних работ (Зотина, Зотин, 19736; Зотин, 1974) мы пришли
94
Рис. 13. Сравнение экспери-
ментальных данных (1), по-
лученных при изучении изме-
нения веса людей на протяже-
нии жизни (Кэтлэ, 1913) с
кривыми (2), рассчитанны-
ми по: а — уравнению (3.45);
б — по уравнению (3.52);
в — по уравнению (3.53)
Рис. 14. Сравнение экспери-
ментальных данных (1), по-
лученных при изучении изме-
нения веса людей на протя-
жении жизни, с кривыми (2) у
рассчитанными с помощью
уравнения (3.52)
Варианты уравнения:
= Lgd = 0;
6 “ fed - Lgf = 0:
в - L.gg - hf -0:
г Lgd — О»
О __ rgg - о-
rgd “а0,
e~Lgf-°
a~Lgg~
95
Рис. 15. Сравнение экспери-
ментальных данных (1), по-
лученных при изучении изме-
нения веса людей на протя-
жении жизни, с кривыми (2),
рассчитанными с помощью
уравнения (3.52)
Варианты уравнения: а — а 0 =
= аа = 0;
б — aj = = 0;
в — ag = af — 0
Рис. 16. Сравнение экспериментальных данных (1), полученных при изу-
чении роста людей (Кэтлэ, 1913), с кривыми, рассчитанными: по уравне-
нию (3.52) (2) (Lgg = 1,7; Lgd = 0,0006; Lg< = —0,3; ag = 7,1; ad -*
мало; af = 2,0; Wm = 69.1; tm ~ 51) и no уравнению (3.55) (3) (Lga = 0,93;
Lad = 0,0007; Laf = 0,0001; ag = 0,3; ad ~ 0; af = 1,2; у = 0,761; p =
= 0,503; Wm = 69,1; lm = 31)
к выводу, что эта зависимость выражается уравнением:
b (t) = b [1 + у t exp (- Р0], (3.54)
где у и р — константы.
Уравнение (3.54) несомненно требует еще теоретического обо-
снования, но оказалось, что с его помощью можно улучшить приме-
нимость уравнения (3.52) для описания изменения веса людей.
Для того чтобы показать это, мы записали уравнение (3.52) в виде:
1 dw _ Г г (Д . т .. .. ,
IF“dt----611 VFw)— 1/ — +
+ a3Lg/(Wl-HW))
(3.55)
где
wb
m
2
- I ’' m л I
a, = exp — I —r-rr — 1 I
a2 = exp [— (tm — t)2]
6l3 = exp[-a/(^-Wi’('))2J
5(Z) = Ml +vexp(— ^)]
и с помощью ЭВМ «Мир-2» проверили возможность его использо-
вания для описания изменения веса людей (Зотин, Зотина и др.,
1976). Оказалось, что в этом случае теоретическая кривая на ран-
них стадиях роста лучше соответствует росту людей (рис. 16).
Это показывает, что для описания развития и роста животных и че-
ловека следует использовать уравнения биологии развития вида
(3.55), если мы хотим исследовать эти процессы с достаточно ран-
них стадий.
Проведенный анализ показывает, что положения, на которых
строится количественная феноменологическая теория развития,
не беспочвенны. В то же время следует отметить, что соответствие
уравнений (3.52) или (3.55) кривым изменения веса людей являет-
ся необходимым, но недостаточным критерием правильности этих
уравнений. Вероятно, можно построить и другие уравнения, ко-
торые будут достаточно хорошо описывать кривые изменения веса
людей и животных на протяжении жизни. Для доказательства пра-
вильности уравнений (3.52) и (3.55) и всей феноменологической
теории развития организмов в целом требуются и иные экспери-
ментальные подтверждения, связанные с проверкой различных
следствий, вытекающих из этой теории.
97
СОВРЕМЕННЫЕ ТЕОРИИ И УРАВНЕНИЯ РОСТА
Р. Вальтер, И. Лампрехт
В каждой науке первым шагом является простое описание яв-
ления, наблюдаемого в природе или в эксперименте. Однако очень
быстро такой чисто качественный подход оказывается недостаточ-
ным для объяснения наблюдаемых эффектов и установления их
связи с другими явлениями. Поэтому появляется потребность в ко-
личественной оценке и математическом обосновании и часто оказы-
вается, что научная проблема становится понятней, если мы спо-
собны перевести ее на язык математического формализма.
Достигли ли мы этой цели при изучении процессов развития и
роста организмов и каковы могут быть практические результаты
подобной работы?
Начнем со второго вопроса. Рост является сложным процессом,
связанным с различными внутренними и внешними параметрами.
Эти параметры могут быть совершенно разными для единичных
клеток, органов, организмов, культуры микробов или популяции
животных. Если мы хотим использовать эти параметры для полу-
чения некоторых общих соотношений и предсказаний, имеется
только одна возможность — математизация результатов измерений.
Например, почти во всех случаях роста можно наблюдать длитель-
ную или кратковременную экспоненциальную фазу со специфиче-
ской константой роста. Последняя дает возможность сравнивать
рост различных организмов или различные ситуации при
росте.
Что касается первого вопроса, то следует сказать следующее.
Имеется много попыток построить уравнения роста, которые в от-
дельных случаях хорошо приспособлены к решению специальных
проблем, но мало удачны в других случаях. Одна из таких попыток,
основанная на термодинамическом подходе к явлениям развития
и роста организмов, подробно описана выше. Она ограничена,
однако, только рассмотрением самих процессов роста без учета
влияния среды; в ней не рассматриваются механизмы роста и она
не касается роста популяций. Следовательно, в настоящее время
нет всеобъемлющей теории и математической формулировки роста,
а имеются лишь различные подходы (см. ниже).
v Описать рост, по-видимому, можно было бы довольно легко,
если бы не было влияния окружающей среды, которая замедляет
и ограничивает развитие организмов. В результате экспоненциаль-
ная кривая роста, например микроорганизмов, ограничена не-
сколькими генерациями, а затем наступает стационарное состояние
© R. Walter, I. Lamprecht, 1976 г.
98
(которое характерно не только для микробных культур, но и для
высших организмов). Замедление роста может быть связано: с огра-
ниченными источниками питания; с ограниченным пространством,
что ведет к негативным изменениям окружающей среды в резуль-
тате роста популяции (например, уменьшение pH среды в фермен-
терах в результате накопления углекислоты или молочной кисло-
ты); с воздействием хищников в ограниченной системе (взаимодей-
ствие типа хищник — жертва); с внутренними часами организма
и т. д. Так как эти условия по своему характеру и действию раз-
личны, они нуждаются в различных выражениях при математиче-
ском описании.
В качестве переменных для характеристики роста можно ис-
пользовать различные параметры. Если рассматривать культуру
микроорганизмов, рост которой начинается с иноклюации в среду
небольшого объема культуры, мы можем в качестве переменных
взять число клеток в одном мл или во всем культуральном сосу-
де и оценивать рост по числу отдельных клеток. Число клеток с не-
которым приближением можно считать пропорциональным вели-
чине экстинкции светового потока, пропускаемого через культуру
и эта оптическая величина может быть использована для характе-
ристики роста микробной культуры. Для этой же цели можно ис-
пользовать гематокрит, в котором общий объем осажденной куль-
туры микроорганизмов может служить мерой ее роста, а также из-
мерения сырого и сухого веса, теплот сгорания или содержания
азота в популяции микробов. Для характеристики популяций,
например, в проблеме Лотка — Вольтерра, в качестве наиболее
подходящей величины выбирают число животных. Подобно мик-
робным культурам различные параметры используют и при описа-
нии роста высших организмов. Рассматривая отдельные индиви-
дуумы (животных или человека) выбирают вес, длину или величи-
ну поверхности организма.
После этого краткого биологического введения мы перейдем
к математической стороне дела.
Конечно, всегда можно аппроксимировать какую-либо стацио-
нарную (т. е. бесконечно-дифференцируемую) функцию полино-
мом n-степени. Но такое описание лишено содержания и биологи-
ческого значения. В основном мы будем различать три класса
математических моделей роста биологических систем: детермини-
рованные, стохастические и параметрические.
Детерминированные модели. Подобные модели исходят из на-
личия причинно-следственных связей при явлениях роста. Обычно
изменение во времени начальной популяции клеток приравнивают
сумме членов, которые отражают влияние специфических процес-
сов на развитие и рост. Этот набор членов приводит к системе диф-
ференциальных уравнений, которые в основном решаются с по-
мощью ЭВМ. Для простоты мы начнем со случая одного или двух
членов, а затем распространим эту систему на более сложную
модель.
99
Стохастические модели. Если система переменных становится
очень сложной или если имеется лишь один параметр, но его изме-
нения во времени прослеживается недостаточно четко, то для кон-
струирования модели используются вероятностные допущения.
Это можно сделать, например, когда мы имеем дело с небольшим
числом клеток и время деления отдельных клеток неизвестно, хотя
и ограничено некоторыми пределами. С переходом от малого
числа клеток к большому средние величины становятся независи-
мыми от стохастических моментов и описание становится детерми-
нированным.
Параметрические модели. В некоторых случаях для описания
роста недостаточны ни детерминированные, ни стохастические мо-
дели. Тогда вводят новые параметры (очень часто в виде экспо-
нент), что приводит к лучшему соответствию с экспериментальны-
ми данными. Часто эти параметры являются чисто математи-
ческим приемом и лишены биологического значения или смысла.
Эти модели поэтому имеют меньший интерес в контексте данного
обзора.
Детерминированные модели роста
Принимая, что изменение числа клеток dN за бесконечно ма-
лую величину времени dt пропорционально числу клеток N (£),
получаем дифференциальное уравнение:
d V
4r=«yw (3-56)
и, интегрируя, получаем уравнение Мальтуса:
N (t) = Noeat. (3.57)
Здесь а так называемая константа роста, которая связана с вре-
менем жизни одного положения т соотношением:
а=1п2/т (3.58)
a Nq — число клеток к началу роста.
Так как экспонента еа/ является ускоренно-возрастающей
функцией, то, очевидно, что закон (3.57) применим только в огра-
ниченных масштабах. Поэтому, когда этот закон используется
для описания роста, то молчаливо подразумевается, что он отно-
сится к небольшому интервалу индивидуальной жизни организмов
(см. Kostizin, 1939). В то же время этот закон является составной
частью всех детерминированных моделей, описывающих «логариф-
мическую» или «экспоненциальную» фазу развития. Например, на
рис. 17, а показано соответствие уравнения (3.57) данным, полу-
ченным при изучении изменения сырого и сухого веса рыжего
таракана (Blattella germanica L.).
100
Рис. 17. Сравнение экспериментальных данных по росту таракана (Blattella,
germanica L.) с кривыми, рассчитанными по уравнениям: а — Мальтуса
(3.57); б — Ферхлюста — Пирля (3.63); в — Найденова (3.67); г — Лэйрд
(3.70); д — Шмальгаузена (3.73); е — Зотиной и Зотина (3.75); ж —
Яношека (3.90); з — Гомпертца (3.92); и — Пюттера — Берталанфи
w
2
3
Рис» 18» Схема трех фаз роста
организмов
— скрытая или лаг-фаза;
— экспоненциальная (логарифмиче-
ская) или лог-фаза;
— постоянная или стационарная
фаза
линейную зависимость между
t
Логарифмирование (3.57) дает
числом клеток N (I) и временем:
In N (0 = In No + at.
В этой форме очень часто проводят математическое описание
экспериментальных результатов.
Если мы рассматриваем нормальный рост организмов или куль-
туры, мы ясно различаем три главные фазы роста: лаг-фазу, свя-
занную с адаптацией к новым условиям среды; лог-фазу, или экс-
поненциальную фазу, соответствующую максимальной скорости
роста, и стационарную фазу, когда число клеток, вес или объем
сохраняются константными (рис. 18). Если рост не происходит и
показатели роста падают, то константа а становится отрицатель-
ной. При постоянном числе клеток константа а равна нулю. Это
наводит на мысль, что величина а не является неизменной, но яв-
ляется функцией времени или таких параметров, как концентра-
ция субстратов или величины pH.
Броди (Brody, 1945) разделил кривую увеличения веса организ-
мов на отдельные участки, каждый из которых имеет специфиче-
скую константу в уравнении:
И
W (t) = Woifi', (3.60)
где W — вес организма ко времени t, Woi — начальный вес, с ко-
торого ведется исследование роста по уравнению (3.59).
Логистическая функция.
Если принять, что в уравнении (3.56) а не является константой
и зависит от числа клеток по формуле:
а = р _ yN (j), (3.61)
то мы получим логистическую функцию или закон Ферхюльста —
Пирля (Pearl, 1924):
где Р и у — константы.
(3.59)
(3.62)
102
Решение этого уравнения имеет вид:
N (/) =---. (3.63)
3 + ^о(е₽,-1)
Исходя из уравнения (3.63), обсудим некоторые особенности
логистической функции: 1) при очень малых t значение величин
в скобке близко нулю и мы получаем закон Мальтуса; 2) при очень
больших t единица в скобках и константа Р в знаменателе (3.63)
могут быть опущены и получаем: N (/) = Р/у = 7Vmax стационарная
фаза роста; 3) рост останавливается, когда в (3.62) р — yN (t^) =
= 0, т. е. при у = P/7Vmax- Следовательно, во время роста р —
— y7V0 0; 4) максимальная скорость роста достигается в сред-
ней части кривой, когда число клеток достигает половины конеч-
ной величины; 5) кривая симметрична относительно точки пере-
гиба. Кривые роста, лишенные этого свойства, не могут быть опи-
саны с помощью формулы (3.63).
Соответствие формулы (3.63) кривой роста таракана (по данным
об изменении сырого веса) показано на рис. 17, б, а кривой роста
культуры дрожжей (по калориметрическим наблюдениям) — на
рис. 19. Мы видим, что соответствие вполне удовлетворительное,
однако значение констант р и у в том и другом случае несколько
различно. Константа р является константой скорости для лог-фа-
зы. Константа у у дрожжей в отличие от тараканов зависит от про-
цессов диффузии питательных веществ из среды.
Уравнения роста Робертсона, Оствальда и Найденова. Как
показано выше, константа у связана с конечным числом клеток
в культуре. Робертсон (Robertson, 1908) и Оствальд (Ostwald,
1908) также ограничили рост максимальным весом Wm к концу
развития и утверждением, что скорость роста пропорциональна
разности между максимальным весом и весом организма в данный
момент, а также пропорциональна данному весу:
J^==lW(t)(Wn-W(t)) (3.64)
или
-^Р = (:з_т^(О)^(О, (3.65)
где yWm = р.
Уравнение (3.65) соответствует логистической функции (3.62).
Если мы примем, что скорость роста пропорциональна только
разности (Wm — W), то (3.64) переходит в:
-^ = а(Жт-Ж(<)). (3.66)
Это уравнение Найденова (1928), интегрируя которое получим:
W (0 = Wm [1 - ехр (- at) 1 (3.67)
103
Рис, 19. Кривая роста дрожжей (Saccharomyces cerevisiae L.), полученная с
помощью калориметрических измерений
1 — скорость роста; 2 — рост: квадратики — опытные данные, линия — математические
расчеты
Если (3,66) подставить в (3.67), то получим:
^-=аТГтое-“'. (3.68)
Очевидно, что dW/dt уменьшается во времени и уравнение (3.67)
может описать только часть кривой роста, после точки перегиба,
т. е. переходный период от экспоненциальной к стационарной фазе
роста (рис. 17, в). Зотин (Zotin, 1972) указал, что только интенсив-
ные, но не экстенсивные величины могут быть использованы в
качестве термодинамических сил. Исходя из этого разность
(lVm — W) нельзя использовать в качестве движущей силы роста.
Уравнение роста Дэвидсона и Лэйрд. Дэвидсон (Davidson,
1928) и Лэйрд (Laird, 1964, 1966) приняли, что константа в уравне-
нии (3.56) является функцией времени. Исходя из двух дифферен-
циальных уравнений:
1 dW _
W dt
1 da ___
a dt
(у — константа) получается после интегрирования:
W (0 = exp (1 - е-п] (3-70)
при граничных условиях t = 0, W = Wo и a = a0. При малых
значениях t уравнение (3.70) по мере протекания процесса перехо-
дит в закон Мальтуса (3.57). При больших значениях t член e~yt
становится много меньше единицы и (3.70) переходит в:
W (0 = Woeao^ = const, (3.71)
т. е. W (£) асимптотически приближается к Wm = WQ exp a0/y.
Поэтому уравнение (3.70) дает сигмоидную кривую, необходимую
104
для описания роста. Но это уравнение, в отличие от (3.63) не сим-
метрично относительно точки перегиба (рис. 17, г и 20, б).
Уравнение роста Шмальгаузена. Шмальгаузен (1935) предпо-
ложил, что удельная скорость изменения веса обратно пропорцио-
нальна времени:
1 dW 1
W dt "" а t ’
(3.72)
что дает:
Рис. 20. Сравнение экспериментальных данных по росту дрожжей (черные
квадратики) с кривыми, рассчитанными: а — по уравнению Найденова (3.67);
б — по уравнению Лэйрд (ЗЛО)
Рис. 21. Сравнение экспериментальных данных по скорости роста дрожжей
(черные квадратики и кружочки) с кривыми, рассчитанными: а — по урав-
нению Шмальгаузена (3.73); б — по уравнению Лампрехта (3.81)
105
Это параболическое уравнение не дает Сигмоидной кривой ро-
ста, но может описать, как видно из рис. 17, д и 21, а, лаг- и лог-
фазы роста.
Уравнение роста Зотиной и Зотина. В отличие от Робертсона,
Оствальда и других авторов Зотина и Зотин (1967, 1973а) норми-
ровали рост не максимальным весом, а общим временем роста tm.
Принимается, что удельная скорость изменения веса пропорцио-
нальна времени оставшегося развития (tm — t):
= ₽(<„-«) (3.74)
или после интегрирования:
W(t) = (3.75)
для t tm (3.75) переходит в:
IV (/) = Wme~^tm ~ .
Принимая Wm ехр (—(3/™) = Wm и 2$tm = а, получим закон
Мальтуса (3.57).
Уравнение (3.75) представляет собой гауссовское распределе-
ние. Это становится особенно ясным, если придать этому уравне-
нию следующий вид (Клименко, 1971, 1975):
1 -JLfJsLZL?
= и 0 ’> (з-76)
где р = 1/2<т2, Р' ~ б
Впоследствии авторы (Zotin, 1972; Zotina, Zotin, 1972; Зоти-
на, Зотин, 1973а) предположили, что константа (3 (3.74) зависит
от времени:
р = ре“7(,"“0’ (3.77)
и преобразовали уравнение (3.74) в:
=4(tm- 0(3.78)
При t -> tm уравнение (3.78) переходит в (3.74). Интегрирование
(3.78) дает:
W (0 = к ехр ехр (- у (tn — 02)] . (3.79)
Уравнение (3.79) хорошо соответствует экспериментальным дан-
ным (рис. 17, е). Приведенные рассуждения, однако, переводят
всю проблему из чисто детерминированных моделей и перемещают
внимание на стохастические элементы, как видно из уравнения
(3.76).
106
Уравнение роста Лампрехта и Меггере. Совершенно иной под-
ход использован Лампрехтом и Меггере (Lamprecht, Meggers,
1971) при изучении с помощью микрокалориметрии растущей куль-
туры дрожжей. В неперемешиваемой культуре клетки опускаются
на дно сосуда и снабжение их питательными веществами опреде-
ляется только диффузией. В таких экспериментах термограммы,
связанные с изменением числа клеток или их сухого веса, хорошо
описываются логистической функцией (см. рис. 19). Одновремен-
ное наблюдение за концентрацией питательных веществ показало,
что перегиб зависит не от количества субстратов, а только от вре-
мени их диффузии. Эта ситуация полностью изменяется, если со-
держание сосуда хорошо перемешивается. Для этого случая полу-
чена система из двух дифференциальных уравнений следующим
путем. В начале экспоненциальной фазы увеличение числа клеток
в культуре описывается уравнением (3.56):
4r = ajVW’
где а является функцией концентрации субстратов в среде (с).
В этом специальном случае а соответствует гиперболической
функции
8С
Л = С I
(3.80)
где е и 6 — константы.
В то же время потребление клетками субстратов приводит к
изменению концентрации питательных веществ в среде пропорцио-
нально скорости изменения числа клеток:
4г = - ф 4г = - Фа w N W- <3-81 >
Невозможно дать точное решение этих трех уравнений, поэтому
результат был получен приближенными методами и сравнен с тер-
мограммами и с нормальной кривой роста (см. рис. 21).
Полученные уравнения справедливы только для гомогенной
культуры микроорганизмов, но их легко использовать и в других
случаях, если известна зависимость между константой роста и
концентрацией субстратов или можно сделать некоторые предпо-
ложения относительно этой зависимости. Кроме концентрации пи-
тательных веществ, можно брать и другие параметры, такие, как
величина pH, пространство (см. ниже) и др. Если а описывается
линейной функцией от числа клеток a = (3 — yN (/) или другими
параметрами, пропорциональными числу клеток, то мы прихо-
дим снова к логистической функции (3.62).
107
Стохастические модели роста
Вероятность Pg, что число клеток N (t) увеличится на одну
клетку за интервал времени dt, пропорциональна N (0 и ft:
Pg = N (ОМ + v (ft), (3.82)
где v (ft) — корректирующая функция, X — константа. Таким же
образом вероятность смерти одной клетки равна:
Pd = N(t)bp + v' (ft). (3.83)
В общем случае мы принимаем, что увеличение числа клеток
О = N (0 м + V (Ф) - (N (t) + V' (О)). (3.84)
Для большого числа клеток N (0 и короткого периода ft v (ft) =
= v' (ft) и поэтому:
-^ = МГ(0-ИЛГ(0 = аЛГ(0, (3.85)
где а = X — р,.
При этих условиях стохастическая модель переходит в детер-
минированную .
Уравнение роста Яношека. Рост организмов можно рассмат-
ривать с точки зрения «теории мишени» (см. Janoschek, 1957; Но-
ерре, 1959). Одно столкновение молекулы питательного вещества
с клеткой приводит к реакции, которая проявляется в росте. Идея
состоит в том, что из т, биологически существенных мишеней толь-
ко г (г < т) этих мишеней реагируют в интервале времени t. Мож-
но подсчитать вероятность этих событий и число клеток, которые
реагируют на субстрат, т. е. растут. Мы не будем здесь приводить
полностью математические выкладки, связанные с этой пробле-
мой — их легко найти в литературе (Timofeeff-Ressovsky, Zim-
mer, 1947). Рост осуществляется с вероятностью Pg, если перед
этим происходят р одновременных реакций g с вероятностью:
Pg = 1 - (3.86)
где к вероятность одного столкновения. Тогда:
Pg = (1 - e'kt)p. (3.87)
Если только одно из этих р событий достаточно для начала роста,
(3.87) переходит в:
Pg = 1 - (e~ki)p (3.88)
Окончательно^получаем:
Pg = [1 - (e*')p]w, (3.89)
если принимаются во внимание т мишеней. Уравнение (3.89),
полученное из теории мишеней, является приближенной формой
108
закона Яношека. Однако имеется некоторое различие между урав-
нением (3.89) и законом Яношека:
W (0 = Wm [1 - e-(ft0P], (3.90)
которое дано автором без доказательств. По этой причине уравне-
ние (3.90) может быть отнесено к третьей группе параметрических
моделей, р — согласно Яношеку «внутренний фактор», связанный
с цитоплазмой клеток. Результат выглядит простым при р = 1,
что справедливо согласно Яношеку для рыб (сельди) и млекопи-
тающих (люди, свиньи). Для большей части организмов, изученных
Яношеком и др., р больше 1 (см. рис. 17, ж).
В литературе имеются и другие стохастические модели (Collins,
Richmond, 1961; Szekely et al., 1972), но они настолько специфич-
ны, что не представляют интереса в контексте данной книги.
Функция Гомпретца. Несколько авторов использовали в ка-
честве уравнения роста функцию Гомпертца, которая может быть
записана:
или в интегральной форме:
w = Wo exp |J- (1 - exp (- 30)] , (3.92)
где аир — константы.
При небольших t (3.92) переходит в закон Мальтуса (3.57):
W = Жо exp (1 - (1 - .))] ~ Woext.
Уравнение (3.91) и (3.92), которое было использовано Гом-
пертцом (Gompertz, 1825), для описания кривых смертности
людей, было применено также! для описания роста коров
(Davidson, 1928), моллюсков (Weymouth, McMillin, 1930), роста
опухолей (Laird, 1964) и зародышей (Laird, 1966). Это широко ис-
пользуемое уравнение показывает, почему на соответствующих ста-
диях роста увеличение веса следует экспоненциальному закону
(см. рис. 17, з).
Параметрические модели роста
Уравнение роста Ферхюльста. В 1838 г. Ферхюльст (см. Lol-
ka, 1925) попытался объяснить сигмоидную кривую роста популя-
ции организмов, принимая, что скорость роста пропорциональна
оставшемуся свободному пространству, занимаемому популяцией.
Исходя из этого можно написать:
<3-93>
где R — общий объем пространства (или площади), который мо-
109
жет занять популяция, N (t) — число организмов, пропорциональ-
ное занятому ими пространству в R. При у = 0//? (3.93) переходит
в логистическую функцию (3.62):
-^- = [Р-тЛГ(0]ЛГ(О.
Уравнение роста Пюттера. Это уравнение опирается на пред-
положение, что синтез массы микроорганизма пропорционален
поверхности, а деление — объему (Putter, 1920). Так как синтез
(пропорциональный квадрату длины Z2) ведет к увеличению веса
организмов, а деление (пропорциональное кубу длины Z3) — к
торможению роста, то изменение веса, в частности микробов опре-
деляется уравнением:
= kJ? - kJ*. (3.94)
Можно нормировать длину в определенный момёнт времени I (/) =
= 1 длиной I в середине развития, выражением:
I' = 111 (3.95)
при kJ2 — kJ? = v и I = кг1кг (3.94) переходит в:
4^ = 4-Z'2(l — I'). (3.96)
В этом случае dWIdt становится отрицательной, когда Z' > 1,
т. е. имеется точка перегиба на кривой роста.
Применимость этой модели ограничена одноклеточными орга-
низмами, так как трудно определять размеры тела животных че-
рез длину и использовать в этом случае понятие деления. Но
формально этот путь дает хорошую возможность описывать сиг-
моидные кривые роста организмов.
Уравнение роста Берталанфи. Берталанфи (Bertalanffy, 1934,
1960) видоизменил уравнение роста Пюттера таким путем, чтобы
в нем шла речь не о длине организма, а о его весе, который пропор-
ционален объему:
4^ = kjrf^ - k2W, (3.97)
где п и т — целые числа.
Интегрирование (3.97) приводит к следующему уравнению
роста.]
1
п
wit) = (3-98)
L К2 \ К2 f J
Принимая, что конечный вес Wm много больше начального ТУ0,
НО
получим из (3.98):
W) = Wm[l (3.99)
Берталанфи различает три типа роста животных: при п/т = 1
уравнение (3.97) ведет к закону Мальтуса (3.57); при nhn = 2/3
— к уравнению Пюттера (3.94); при 2/3 <inhn < 1 — к уравне-
нию Берталанфи (3.97). Хотя эти уравнения получены для описа-
ния роста отдельных организмов (рис. 17, и), они могут быть при-
менены к культуре микроорганизмов, так как число отдельных кле-
ток здесь достаточно велико, а качественное состояние их остается
константным за время развития.
Уравнение роста Винберга. Винберг (1968) попытался проана-
лизировать уравнение Берталанфи (3.97) с энергетической точки
зрения. По его мнению, одна часть А свободной энергии, заключен-
ной в пище, используется для образования новой клеточной мас-
сы, а другая — для поддержания метаболизма. При некоторых спе-
циальных условиях выполняется соотношение:
= *• <З.Ю0>
Величина В может быть измерена по скорости потребления кисло-
рода, которое пропорционально Wn т, тогда как А прямо связа-
на со скоростью увеличения веса dW/dt:
A = = = -L-в^
dt 1 —к 1 — к 1
ИЛИ
= (3.101)
где Bl9 Z, к, к' — константы.
Если вес организма ограничен максимальной величиной Wm
необходимо ввести лимитирующий фактор С:
х-А х-21
1-—
W т
гг т
так что (3.101) трансформируется в:
(3.102)
dt
и мы получаем
Берталанфи и
п
w1 т
ТП
уравнение Берталанфи (3.97). Так как уравнения
Винберга требуют пропорциональности между
111
скоростью роста и весом тела, которое не выполняется у многих
организмов, эти уравнения ограничены специальными периодами
жизни организма.
Таким образом, в данной работе мы попытались рассмотреть
некоторые уравнения роста, которые имеются в литературе, и об-
судить их особенности. Очень трудно оценить, какое из этих урав-
нений является более правильным, так как разные авторы исполь-
зуют их для описания роста различных видов организмов и в не-
сравнимых условиях. Поэтому мы провели проверку всех этих
уравнений только па двух видах организмов: на насекомом Blat-
tella germanica и на культуре дрожжей Saccharomyces cerevisiae.
Более полные кривые роста таракана при различных условиях
разведения даны на рис. 34, а для дрожжей в калориметрических
экспериментах — на рис. 27, 29, 30.
Если в одних случаях математическое описание дает хорошее
совпадение с экспериментальными данными (рис. 17, б, е, з, 19),
то в других — только ограниченный период роста может быть опи-
сан предложенными уравнениями (рис. 17, а, в, д, 20, а, 21, а). Сейчас
еще трудно предложить универсальное уравнение роста, пригод-
ное для всех организмов и случаев, однако некоторые полезные
шаги в этом направлении уже сделаны.
IV
ТЕПЛОПРОДУКЦИЯ
В ЖИВЫХ СИСТЕМАХ
В литературе имеется сравнительно мало работ, в которых теп-
лопродукцию измеряли прямыми методами. Это объясняется слож-
ностью методов прямой калориметрии, их дороговизной и трудоем-
костью. В начале нашего века техника калориметрических иссле-
дований начала быстро развиваться (см. Чередниченко, 1965) и
достигла известного совершенства, однако она вскоре была вытес-
нена методами непрямой калориметрии, так как оказалось, что ме-
тоды измерения дыхания значительно чувствительнее, проще и
дешевле методов, связанных с измерением теплопродукции орга-
низмов. Широкое распространение методов непрямой калоримет-
рии определялось также тем, что была высказана и доказана для
взрослых нерастущих животных идея о том, что дыхание организ-
мов прямо пропорционально теплопродукции. При этом исследо-
ватели опирались на закон сохранения энергии, т. е. на представ-
ление о применимости первого начала термодинамики к живым ор-
ганизмам (Гинецинский, Лебединский, 1956; Durnin, Passmor§,
1967). Хотя нет сомнений в применимости первого начала термо-
динамики к организмам, постепенно стало очевидным, что в про-
цессах развития и роста, а также при некоторых функциональных
нагрузках на организм, имеет место расхождение данных прямой
и непрямой калориметрии (Чередниченко, 1965; Семихатова, 1967;
Зотин, Грудницкий, 1970). Это вытекает и из соображений, разви-
тых во второй главе настоящей книги. Как было показано, для си-
стем, далеких от равновесия, должно выполняться разбиение (2.15),
согласно которому не вся теплота, образовавшаяся в результате
протекания необратимых процессов, выделяется в среду в виде
теплоты диссийации: часть ее может использоваться внутри си-
стемы. Это утверждение должно быть справедливым для любой
открытой системы, но оно может быть легко проверено только на
живых системах. Действительно, в живых организмах все дисси-
пативные процессы определяются процессами дыхания и гликоли-
за, так как энергетика организмов в конечном счете определяется
этими двумя процессами. Другими словами, интенсивность дыха-
ния и гликолиза организмов пропорциональна диссипативной
функции системы:
п
Я5 — JjXj — + (.гл = CjM, (4-1)
;=i
ИЗ
где (м — удельная скорость метаболизма, (02 — интенсивность
дыхания, (гл —интенсивность гликолиза. С другой стороны,
функция внешней диссипации по определению (Зотин, 1974)
пропорциональна интенсивности теплопродукции:
фа = (4.2)
где q — интенсивность теплопродукции организмов.
Следовательно, для живой системы имеем:
Фи = 9м —<• (4.3)
Во время развития и на начальных стадиях роста организмы
находятся в состоянии, далеком от стационарного, или равновес-
ного, поэтому в таких системах должно наблюдаться расхождение
данных прямой и непрямой калориметрии. Часть энергии обра-
зованной внутри организма за счет дыхания и гликолиза, должна
фиксироваться внутри живой системы в виде функции связанной
диссипации в соответствии с уравнением (4.3). Из уравнения (4.3)
следует также, что во время развития и роста организмов, когда
живая система находится далеко от конечного стационарного со-
стояния, об эволюции системы нельзя судить по данным об изме-
нении дыхания — требуются измерения теплопродукции систе-
мы прямыми методами. К сожалению, таких данных немного в
литературе и лишь в последние годы стали быстро развиваться
исследования, в которых теплопродукцию организмов измеряют
прямыми, калориметрическими методами.
ТЕПЛОПРОДУКЦИЯ
ПРИ ПРОТЕКАНИИ ЖИЗНЕННЫХ ПРОЦЕССОВ
Б* Шааршмидт, Лампрехт
Каждое событие, происходящее в живой системе, сопровожда-
ется выделением или поглощением тепла, так как любые химиче-
ские или физические процессы связаны с изменением энтальпии.
Следовательно, непрерывная регистрация термогенеза с помощью
термограмм должна точно отражать все изменения жизненных
функций как в субклеточных биохимических системах, так и в изо-
лированных органах или в целых организмах.
Тепловой обмен между системой и окружающей средой можно
легко измерить с помощью калориметрических методов, которые
давно уже используются в термохимических исследованиях. Раз-
витие техники калориметрии в настоящее время достигло такой
стадии, когда даже очень слабые тепловые процессы, происходя-
© В. Schaarschmidt, I. Lamprecht, 1976 г.
114
щие в биологических системах, могут быть успешно измерены с
помощью микрокалориметров. Поэтому измерение теплопродук-
ции является в принципе очень чувствительным индикатором из-
менения энергии в биологической системе независимо от того, на-
сколько сложны протекающие в ней лроцессы.
С другой стороны этот метод неспецифичен и приносит инфор-
мацию только об интегральных эффектах. На практике, однако,
часто имеет место преобладание нескольких простых реакций, так
что можно выявить достаточно точную корреляцию между измеря-
емой теплотой и основными продуктами реакции (Forrest, 1972;
Spink, Wadso, 1975). Кроме того, путем применения специальных
калориметров и устройств (Wadso, 1970), а также путем математи-
ческой обработки термограмм и результатов экспериментов, мож-
но получить сведения о многих количественных параметрах (Wil-
kie, 1960; Wendlandt, 1964; Garn, 1965), таких как внутренняя
энергия, энтальпия, энтропия, теплоемкость, скорость реакций,
константа Михаэлиса — Ментона, тип реакций, число реакций,
переход фаз и т. д.
Таким образом, термодинамический подход может широко при-
меняться в биохимии, биологии и многих других родственных об-
ластях науки (Wilkie, 1960; Brown, 1969). Ниже дается лишь срав-
нительно короткий обзор работ в широком спектре исследований
теплопродукции при протекании жизненных процессов. Естест-
венно, что здесь использована далеко не вся литература по данной
проблеме и предпочтение отдано, насколько это возможно, обзор-
ным статьям.
Одиночные биохимические реакции
Описано почти необозримое количество экспериментов с био-
химически хорошо определенными макромолекулами, целью ко-
торых являлось определение теплоемкостей, констант равновесия,
а также теплот сгорания, гидролиза, растворения, сольватации,
ионизации, разбавления, испарения, связывания и денатурации.
Большой интерес вызвали ферментативные реакции (см. об-
зор Brown, 1969) и особенно различного типа реакции гидролиза
АТФ (Ohlmeyer, 1946; Kitzinger, Benzinger, 1955; Benzinger,
Hems, 1956) и других фосфатных соединений (Ohlmeyer, Sha-
tas, 1952), так как они имеют большое значение как поставщики
и переносчики энергии в промежуточных реакциях метаболизма
(Sturtevant, 1962). Изучены также гидролазные реакции, связанные
с внутриклеточным синтезом^ распадом белков (Sturtevant,г 1953;
Benzinger, Kitzinger, 1963). Хорошие результаты дает измерение
теплопродукции во время реакции ингибирования ферментов. Так,
определение теплот реакций позволило высказать предположение
об образовании в’ процессе’ ингибирования^ фермент-ингибитор-
ного комплекса и определить число центров связей (Ross, Gins-
burg, 1969; Shiao, 1970; Bolen et al., 1971; Bjurulf, Wadso, 1972).
115
Субклеточные системы
В литературе имеется немного данных о комплексных биологи-
ческих подсистемах и их теплопродукции. Причиной этого, по-
видимому, является слишком сложная процедура изоляции орга-
нелл в достаточном количестве. Поэтому измерения проводили
главным образом с целыми клетками или организмами.
При изучении транспорта веществ через мембраны очень прос-
тые опыты провели Браун и др. (Brown et al., 1965). Разделяя ка-
лориметрический сосуд с помощью кожицы лука на две части (со-
держащую глюкозу или глюкозоксидазу), они измеряли теплоту
окисления глюкозы, связанную с переносом ее через мембрану из
одной части системы в другую. Эти результаты были сопоставлены
с переносом глюкозы через другие естественные и искусственные
мембраны. Сходные опыты проведены с искусственными липопро-
теидными мембранами (Steim et al., 1969; Melchior et al., 1970)
и при изучении связывания кальция в фосфолипидном комплексе
(Joos, Carr, 1969).
Недавние исследования, касающиеся более сложных реакций,
таких, как реакция антиген — антитело (Steiner, Kitzinger, 1956;
Barisas et al., 1972), связывания белков с поверхностно активны-
ми веществами (Jones et al., 1973), синтез белка в бесклеточных си-
стемах (Lamprecht et al., 1973), показали, насколько полезны дан-
ные по теплопродукции для анализа биологических процессов.
Следует отметить большое число калориметрических эксперимен-
тов с белками и нуклеиновыми кислотами, в которых исследова-
лась роль воды (см. Каюшин, 1967), реакции разворачивания
(Barisas et al., 1972) и других структурных переходов (см. обзор
Привалов, 1970).
Исследовали также изменение энтальпии в митохондриях при
протекании в них процессов, катализируемых другими субклеточ-
ными частицами (Рое et al., 1967), при этом особенно интенсивно
исследовали энергетическое сопряжение обмена калия и каль-
ция между средой и митохондриями (Рое, Estabrook, 1968; Рое,
1969).
Микроорганизмы
Вероятно, наибольшее число калориметрических измерений
в биологии проведено с микроорганизмами. Это связано, по-ви-
димому, с тем, что микроорганизмы легко культивируются, как
правило, хорошо охарактеризованы генетически, а также пред-
ставляют большой интерес^для микробиологической промышлен-
ности и медицины.
^Одиночные) микроорганизмы слишком малы для того, чтобы
получить данные о теплопродукции отдельных особей. Для про-
ведения одного измерения обычно необходимо более 106 индиви-
дуальных клеток. Хотя в этом случае тепловой поток обусловлен
116
Рис. 22. Пальцеобразные термо-
граммы шести различных видов
бактерий (Boling et al., 1973)
а — Enterobacter aerogenes;
б — Klebsiella;
в — Proteus vulgaris;
г — Enterobacter cloacae;
d — Escherichia coli;
e — Proteus rettgeri
суммой многих биохимических реакций измерение теплопродук-
ции очень полезно, так как термогенез микробных культур
воспроизводим и хорошо характеризует особенности орга-
низмов.
Микроорганизмы сильно отличаются по метаболизму, по по-
требности в питании, по их способности адаптации к среде. Это
открывает пути для того, чтобы различать организмы, сравнивая
только их тепловую эволюцию (Prat, 1953; Кальве, Прат, 1963;
Jaworski et al., 1968). Например, Прат обнаружил три различные
термограммы для трех типов тифозных бактерий (Prat, 1953).
Для семнадцати видов Enterobacteriaceae калориметрические экс-
перименты (см. рис. 22), продолжающиеся от 8 до 14 часов, показа-
ли возможность быстрой и специфической характеристики бакте-
рий с помощью «пальцеобразных» термограмм (Boling et al.,
1973). Эти исследования были распространены на некоторые дру-
гие организмы, такие, как Pseudomonas и другие грамотрицатель-
ные палочки и грамположительные кокки (Staples et al., 1973).
Кроме того, у дрожжей удается различать нормальные линии и
линии с дефицитным дыханием (Lamprecht et al., 1971), дикие ра-
сы и радиационно чувствительные линии (Schaarschmidt, Lamp-
recht, 1974), а также линии разной плоидности (Lamprecht et al.,
1975).
Систематическое изучение роста и метаболизма бактерий и
дрожжей (Boivinet, 1964, 1971; Belaich et al., 1968; Belaich, Sa-
ri, 1969; Brown, 1969; Lamprecht et al., 1971; Lamprecht, Meg-
gers, 1972; Forrest, 1972; Lamprecht et al., 1973) показали, что в
микробной микрокалориметрии имеются широкие возможности
для проведения экспериментов, их интерпретации и истолкования.
Эти работы касаются влияния различных субстратов и их кон-
центраций; эффекта перемешивания культуры и различного газо-
вого состава, с разным атмосферным давлением; различий в обмене
метаболитов в процессе роста культуры; химических и физиче-
ских воздействий, а также влияния температуры, pH среды и ан-
117
тибиотиков. Общим результатом этих исследований явилось поло-
жение, согласно которому теплопродукция во время метаболизма
связана, главным образом, с процессами катаболизма, анаболити-
ческие процессы могут несколько модифицировать тепловой по-
ток, но в очень слабой степени (Morowitz, 1960; Boivinet, 1964;
Brown, 1969; Forrest, 1972).
Большой интерес для микробиологии и биохимии представляют
непрерывные культуры и культуры синхронизированных клеток.
С конструированием проточных калориметров (Wadsd, 1970) ста-
ло возможным изучение теплопродукции таких культур. Комби-
нация проточного калориметра с ферментером позволила воздей-
ствовать на культуру извне без нарушения теплового режима внут-
ри калориметра (Eriksson, Wadsd, 1971; Tillberg et al., 1971; Poo-
le et al., 1973), а также исследовать регуляторные процессы при
переходе культуры в различные стационарные состояния под вли-
янием различных воздействий (Brettel et al., 1972).
Измерение теплового потока может быть с успехом исполь-
зовано при изучении сложных смесей микробов в природе. Экспе-
рименты касались таких вопросов, как активность микробов в
почве и влияние на этот процесс нагревания и радиации (Morten-
sen et al., 1973); развитие бактерий в молоке при хранении (Cliffe
et al., 1973), активность термофильных бактерий при самовозго-
рании сырого сена (Hussain, 1973), а также микробной фермента-
ции в рубце крупного рогатого скота (Houpt, 1968) и овец (Forrest,
Walker, 1964). Результаты последнего эксперимента показали, что
несмотря на сложность системы, реакция идет по типу реакции
первого порядка. Было показано также, что при силосовании тра-
вы наблюдается концентрационный эффект, равный примерно 10%
от обычной величины энергии в 4,4 ккал/г, заключенной в траве
(McDonald et al., 1973).
Среди фотосинтезирующих организмов изучался в основном
метаболизм водорослей, теплопродукция которых хорошо корре-
лирует с темносветовым периодом и с фазой роста культуры (Till-
berg et al., 1971).
Высшие организмы
За последние 20 лет в литературе описано большое число экс-
периментов, в которых измерялся тепловой поток у животных и
растений (см. обзор Prat, 1969). У животных исследовалась тепло-
продукция в большом диапазоне весов от очень маленьких, толь-
ко что родившихся мышей (рис. 23) и летающих насекомых (Са1-
vet, Prat 1963; Prat, 1969) до шимпанзе и крупных домашних
животных (Dale et al., 1967). У растений исследовали'теплопродук-
цию во время прорастания семян (Calvet, Prat, 1963), роста кор-
ней (Prat, 1948) и цветения (Prat, 1952). Тепловой обмен был из-
мерен у зимующих млекопитающих (Calvet, Prat, 1963), у”мигри-
рующих птиц во время безостановочного полета над водой (Odum
et al., 1965), у птенцовых птиц (Myrcha et al., 1973), во время мета-
118
Рис, 23. Рост и теплопродукция мышей от рождения до 35 дней (Prat, 1969)
Q — теплопродукция, W — вес, q — удельная теплопродукция. Двойная линия в на-
чале кривых Q и q соответствует двум опытам, проведенным при разных температурах
(согласно этим данным гомойотермия устанавливается у мышей приблизительно па
пятый день)
морфоза насекомых (Calvet, Prat, 1963; Prat, 1969), в процессе
эмбрионального развития рыб и кур (Зотин, 1966; Зотин, Зотина,
1969). Последние эксперименты были связаны с попыткой построе-
ния феноменологической теории развития, роста и старения, опи-
рающейся на термодинамику необратимых процессов (Zotin, 1972;
Зотин, 1974). Эта теория изложена во 2 главе настоящей книги.
Хорошо известно, что органы могут сохранять свою биологи-
ческую активность при отделении их от организма и помещении
в соответствующие условия. Теплопродукция таких органов явля-
ется чувствительным индикатором сохранения жизненных функ-
ций. Поэтому было сконструировано сложное экспериментальное
устройство для получения данных о термогенезе мышц и изоли-
рованных нервов (Gibbs et al., 1967; Hill, 1965; Wilkie, 1968) и
даже целого сердца лягушки при различных условиях оксигена-
ции, механических сокращений и отведения электрокардиограмм
(Boivinet, Rybak, 1969). Буавине (Boivinet et al., 1968) продемон-
стрировал возможность использования данных о теплопродукции
для определения активности гормонов и ответа тканей на действие
гормонов (рис. 24).
Обзор почти всех работ, опубликованных до 1964 г., связанных
с тепловыми измерениями в мышцах, дан Хиллом (Hill, 1965),
а обзор работ, проведенных на нервах, мышцах и электрическом
органе, опубликованных до 1973 г., имеется в статье Эббот и Хоу-
эрт (Abbott, Howarth, 1973).
Человек
Один из принципиальных вопросов физиологии касается
основного обмена людей. Обычно основной обмен измеряется непря-
мым методом (путем измерения дыхания). Прямой метод использо-
вали редко, причем измерения проводили или в специально скон-
119
Рис. 24. Термограмма жировой
ткани, показывающая увеличение
теплопродукции при добавлении
инсулина и уменьшение при до-
бавлении NaF (Boivinet et al.,
1968)
струированных больших калориметрах (Benzinger et al., 1958;
Carlson et al., 1964; Tiemann, 1969; Snellen et al., 1972), или рассчи-
тывали теплопродукцию всего организма по данным, полученным
при измерении теплопродукции конечностей (Eldblom, 1935; Rich-
ter, 1959). Приблизительная величина теплопродукции, получен-
ная прямыми методами, равна 2200 ккал/день. Эта величина из-
меняется с возрастом людей.
Важное поле деятельности открывается при изучении адапта-
ции людей к меняющимся условиям окружающей среды. Бензин-
гер (Benzinger et al., 1958) исследовал в калориметре почти мгно-
венную реакцию людей на изменение температуры или инъекцию
метахолина, который, как известно, стимулирует мышечное напря-
жение и кровообращение в коже (рис. 25). Благодаря механизму
потоотделения скорость возвращения к исходному уровню тепло-
продукции после температурного сдвига различалась на порядок
в зависимости от того, проводилась ли адаптация к более низкой
или к более высокой температуре. В связи с изменениями среды
могут наблюдаться сезонные или суточные ритмы метаболитиче-
ской активности (Carlson et al., 1964). Теплопродукция человека
Рис. 25. Теплопродукция после внутримышечной инъекции метахолина
После инъекции (стрелка — начало отсчета времени) теплопродукция возрастает до
40 кал/сек, но через 20 мин. уменьшается до величины 21,2 кал/сек, которая ниже на-
чального уровня в 23,8 кал/сек (Benzinger et al., 1958)
120
после питья воды хорошо коррелирует с изменением температуры
тела и потовыделением (Snellen et al., 1972). В серии точных экс-
периментов была показана возможность предсказывать теоретиче-
ски частоту сердечных сокращений и скорости потовыделения при
работе в тепле (Givoni, 1969). При изучении теплопродукции лю-
дей в больших калориметрах был показан индуцируемый пищей
термогенез, который является специфически динамическим дейст-
вием пищи и не зависит от общего содержания энергии в пище
(Pittet el al., 1974).
Очень интересной является область клинических приложений
тепловых измерений. Например, были исследованы с помощью
специальных устройств нарушения периферического кровообра-
щения (Richter, 1959) и такие клинические болезни, как ишиас
(Eldblom, 1935). Широкие исследования проведены также при
изучении метаболизма, аггрегации и действия ингибиторов на
эритроцитах (Levin, Воуо, 1971), лейкоцитах (Levin, 1971) и тром-
боцитах (Monti, Wadso, 1973; Ross et al., 1973) здоровых и боль-
ных людей. Было найдено увеличение скорости теплопродукции
эритроцитов пациентов с различного рода анемией (Воуо, Ikomi-
Kumm, 1972; Levin,1973). С системой лейкоцитов и тромбоцитов
исследовали эффект фагоцитоза (Prat, 1962).
Терморегуляция
Обычно тепло, образующееся в результате жизненных процес-
сов, покидает пределы организма и переходит в окружающую сре-
ду. Если концентрация реагентов, участвующих в процессах дос-
таточно велика, как например, в микробных культурах хорошо
снабженных питанием, то в термически изолированных материалах
температура может увеличиться, что ведет в конечном счете к
самовозгоранию (сена, шерсти, навоза, алкоголя при фермента-
ции и др.). В очень точных измерениях на разных последователь-
ных фазах разогрева сырого сена было установлено, что быстрое
самовозгорание начинается примерно при 265° (Hussain, 1973).
Если отсутствует регуляция тепла, как в приведенных выше
примерах, биологическая система погибает. У одиночных микро-
организмов такая катастрофа предотвращается в результате огра-
ничения размеров клетки: все тепло, возникающее внутри клетки,
может выделиться наружу (Morowitz, 1960). У высших организ-
мов для этой цели вырабатываются специальные механизмы тепло-
выделения, например, кровообращение, которое у гомойотермных
организмов осуществляет’главный контроль за постоянством тем-
пературы тела*при варьирующей температуре среды. Однако теп-
лопродукция не всегда является только ненужным результатом
метаболизма организмов — в некоторых случаях она использу-
ется для самосогревания животных.
В некоторых работах, цитированных выше (Benzinger et.al.,
1958; Calve!, Prat, 1963; Carlson et al., 1964; Dale et al., 1967; Prat,
121
1969; Snellen et al., 1972), приводятся данные о регуляции темпе-
ратуры тела. Мы рассмотрим лишь некоторые из этих работ.
Измерение теплопродукции у новорожденных мышей (Calvet,
Prat, 1963) дали следующие цифры теплового потока: при 25°
2,7 кал/г-час, при 37° — 7,5 кал/г-час. Подобная реакция на из-
менение внешних условий (резкое усиление термогенеза при по-
вышении температуры) типична для пойкилотермных организ-
мов. Реакция гомойотермных животных как раз обратна. Этот
результатат поэтому показывает, что температура тела у новорож-
денных млекопитающих не регулируется. У взрослых мышей ме-
ханизмы, контролирующие температуру, могут быть нарушены с
помощью спазмолитических агентов. После их введения термоге-
нез животных уменьшается в течение 1 часа до половины нормаль-
ного уровня, что приводит животное в неактивное состояние, по-
добное зимней спячке.
Другой очень интересный контроль температуры наблюдается
у насекомых (Prat, 1969). Если измерять теплопродукцию группы
насекомых одного вида, то оказывается, что средний термогенез
особей ниже, чем термогенез при индивидуальном исследовании.
Этот групповой эффект является следствием хорошо известной кол-
лективной терморегуляции общественных насекомых, например,
в ульях пчел, где температура поддерживается постоянной и летом
и зимой.
Принимая во внимание тот факт, что теплопродукция микро-
организмов намного выше, чем других организмов (табл. 3), Прат
(Prat, 1962) поставил следующий интересный вопрос: дает ли что-
либо повышенная теплопродукция паразитическим бактериям,
живущим в тканях человека и животных? Возможно, что клетки
хозяина повреждаются не только в результате токсичности бакте-
рий, но и в результате «термокаутеризации» их паразитами.
В заключение отметим, что удельная теплопродукция различ-
ных организмов по величине отличается на несколько порядков
(см. табл. 3). Некоторые причины такого расхождения вполне
объяснимы. У микроорганизмов большая часть теплопродукции
связана с очень интенсивными клеточными делениями. У высших
животных теплопродукция определяется главным образом работой
мышц и активностью различных желез, контролируемых нервной
системой. Кроме того, микроорганизмы, как известно, далеки от
термодинамического равновесия и должны иметь активный мета-
болизм в связи с высокой теплопродукцией и потоком энтропии.
В частности, закон поверхности Рубнера, согласно которому теп-
лопродукция является функцией поверхности, не выполняется
для гомойотермных животных (Kleiber, 1961).
Так как физическая размерность времени (минута или час)
очень небиологична, то в качестве стандартного времени для теп-
лопродукции было выбрано время генерации. Однако и при этом
данные для различных организмов значительно отличаются друг
от друга.
122
Таблица 3
Сравнение теплопродукции различных организмов
Темпера- тура, °C Вес, г Время генера- ции, мин. Теплопродукция одного организма, (* кал \ Удельная теплопро- дукция. ( кал \ К час-г / Теплопродукция за одну генерацию ( кал \ Автор
час-организм/ \ г. гене рация /
Бактерии 25 4.10-12 40 1,8-Ю-з 450 300 Prat, 1969
со Дрожжи 30 2-10-п 102 3,5-Ю-з 180 300 Lamprect et al., 1971
Дрозофилы 31 1,2-10-8 1,4-10^ 35-10-8 30 700 Кальве, Прат, 1963
Мыши 25 5 105 40 8 14-103 Prat, 1969
Люди 20 70-Юз 107 100 1,4 250-103 Кальве, Прат, 1963
Сердце лягушки 27 47-10-3 5,3-Ю5 47.10-8 1 8,9-Юз Boivinet, Rybak, 1969
Эритроциты человека 37 2-10-и 1,7-105 30-10-12 о,з 850 Monti, Wadso, 1973
Зерна пшеницы 30 6-10-2 2,6-105 15-Ю-з 0,25 НО Кальве, Прат, 1963
Солнце 5000 Юзз — 1,6-1029 1,610-4 —
Тем не менее теплопродукция организмов отражает характер-
ные черты живой материи и позволяет не только проводить анализ
простых процессов, но и открывает новые возможности для экс-
периментальных исследований и построения новых гипотез в об-
ласти сравнительной физиологии.
ИЗМЕНЕНИЕ %-ФУНКЦИИ
ВО ВРЕМЯ РОСТА КУЛЬТУР МИКРООРГАНИЗМОВ
Б. Шааршмидт, Р. Бретель
В главе 2 настоящей книги было предложено разделять дисси-
пативную функцию гр системы на две части (2.15), одна из которых
представляет собой функцию внешней диссипации (ф^-функцию),
другая — функцию связанной диссипации (фи-функцию). В живых
организмах все диссипативные процессы связаны с деградацией
пищи в процессе энергетического метаболизма (гликолиза и дыха-
ния), скорость которого может быть измерена методом Варбурга.
В то же время внешняя диссипация (фа-функция) в организмах
представляет собой теплопродукцию, возникающую в процессе
метаболизма и выделяемую в окружающую среду. Ее можно из-
мерить с помощью калориметров. Используя указанные два мето-
да прямой и непрямой калориметрии, можно определять функцию
связанной диссипации в жизненных процессах по уравнению (4.3).
Эти рассуждения справедливы в отношении процессов развития
и роста животных. Что же касается микроорганизмов, то они
слишком малы, чтобы можно было измерять теплопродукцию или
метаболизм отдельных особей. Поэтому для работы с микроорга-
низмами приходится брать всю культуру, содержащую миллио-
ны клеток. Такая культура не равнозначна отдельному орга-
низму, но все же некоторое подобие между ними очевидно.
В данной работе приводятся эксперименты, с помощью кото-
рых можно оценить величину фи-функции во время роста культу-
ры дрожжей. Были использованы три различных метода для вы-
ращивания культур. В двух случаях культуры выращивались
с самого начала до конца в определенном объеме жидкой среды
без замены питательной среды или клеток. Одна из этих культур
развивалась почти без доступа кислорода, другая — в условиях
снабжения кислородом, близким к оптимальному. Третий метод
выращивания представлял собой метод непрерывного культиви-
рования, когда дрожжи росли в хемостате при нескольких раз-
личных скоростях обмена среды.
© В. Schaarschmidt, R. Brettel, 197G г.
124
Относительно анаэробные культуры
В этой серии экспериментов культура дрожжей (Saccharomy-
ces cerevisiae) была приготовлена на соответствующей ростовой
среде и затем инокулирована в калориметрический сосуд объемом
100 мл. Теплопродукцию растущей культуры регистрировали не-
прерывно в виде термограмм. Через определенные интервалы
времени рост культуры прерывали и измеряли скорость метабо-
лизма клеток монометрическим или полярографическим мето-
дами. Детали описания условий культивирования, методы изме-
рения теплопродукции и метаболизма описаны раньше (Lamp-
recht et al., 1971, 1973; Schaarschmidt et al., 1975).
Для улучшения условий роста культура непрерывно переме-
шивалась. Однако из-за сравнительно небольшой площади по-
верхности сосуда, в котором росла культура (~ 10 см2), и малой
скорости перемешивания снабжение культуры кислородом было
недостаточным: клеткам было доступно только 0,8% от макси-
мально растворимого при данных условиях количества кислоро-
да. Поэтому глюкоза, потребляемая в процессе роста культуры,
расщеплялась главным образом через гликолитический путь.
Это следует из рис. 26, на котором показаны теплопродукция,
убыль глюкозы из среды и появление этанола в среде во время
роста культуры дрожжей.
На рис. 27 показаны увеличение сухого веса (ТУ) и числа кле-
ток (7V) во время роста культуры дрожжей. Очевидно, что кривая
изменения числа клеток не совпадает с кривой изменения сухого
веса: вначале роста культуры число клеток увеличивается мед-
леннее веса, а затем — быстрее. Это не является следствием оши-
бок в счете клеток, которые в отношении дрожжей легко сделать
из-за процесса почкования клеток, а результатом изменения объе-
ма клеток: анализ изменения клеточного объема на различных фа-
зах роста культуры показал, что максимальный объем имеет ме-
сто в начале экспоненциальной фазы роста и минимальный —
в конце. Поэтому коэффициент VK/7V, представляющий собой ус-
редненный сухой вес отдельной клетки, не остается постоянным
в процессе роста (рис. 27), и расчет теплопродукции на 1 мг су-
хого веса или на одну клетку дает совершенно разную картину
изменения теплопродукции культуры. Между лаг-фазой и экс-
поненциальной фазой роста культуры образуется много поч-
кующихся клеток. Поэтому значительная доля теплопродукции,
по-видимому, определяется процессом деления. Это утверждение
важно учитывать при рассмотрении кривой изменения фи-функ-
ции, показанной на рис. 28.
Кривая изменения фи-функции имеет колоколообразный вид
с максимумом в начале экспоненциальной фазы роста культуры
(рис. 28). После достижения максимума она уменьшается до очень
низкой величины и долгое время остается константной. Такой
характер изменения фи-функции во время роста дрожжей коррели-
125
рует с кривой исчезновения глюкозы из среды и увеличением кле-
точной массы культуры (рис. 29): кривая изменения удельной
скорости образования клеток 1/W• AN/At имеет такой же харак-
тер, как и изменение фи-функции и совпадает с последней лучше,
чем кривая изменения удельной скорости сухого веса культуры
i/W • AW/At. Отсюда следует, что изменение функции связанной
диссипации лучше коррелирует с образованием клеток, чем с из-
менением их массы.
Большая скорость связывания диссипации в начале экспонен-
циальной фазы роста культуры дрожжей легко объяснима, так
Рис. 26. Теплопродукция (1), потребление глюкозы (2) и продукция этано-
ла (3) культурой дрожжей, растущей в анаэробных условиях
Рис. 27. Изменение концентрации клеток (N), сухого веса (W) и удельной
концентрации клеток (N/W) во время роста культуры дрожжей
126
как в это время многие биохимические процессы, протекающие
в клетках, также достигают максимальной скорости (например,
синтез различных ферментов, связанных с удвоением ДНК,
поступление пуринов в клетки, синтез компонентов клеточной
стенки и т. д.). Высокая скорость различных процессов в свою
очередь сопровождается высокой скоростью диссипации энергии
и соответственно высокой скоростью связывания диссипации,
что указывает на то, что живая система далека от конечного ста-
ционарного состояния. В последующей экспоненциальной фазе
роста микробной культуры все процессы, связанные с ростом,
по-видимому, хорошо сбалансированы и удельная скорость}[роста
культуры остается постоянной.
Рис. 28. Изменение функции связанной диссипации (1) и удельной ско-
1 bN
росши изменения числа клеток (2) в культуре дрожжей, растущей
в анаэробных условиях
Рис. 29. Рост культуры дрожжей в анаэробных условиях
1 Д¥7
1 — удельная скорость изменения сухого веса —(мг/мг/час); 2 — удельная ско-
рость изменения числа клеток ~ <10’ клеток/мг/час) и 3 -удельная скорость потребле-
1
НИЯ ГЛЮКОЗЫ (мг/мг/час)
W At
127
Относительно аэробные культуры
В этой серии экспериментов культура дрожжей росла в фер-
ментере объемом 1 л и в результате хорошего перемешивания
клетки находились в оптимальных условиях снабжения кислоро-
дом и питанием. Небольшая часть объема культуры непрерывно
прогонялась через проточный калориметр, где определялась
скорость теплопродукции культуры. Благодаря тому, что сосуд
с культурой был отделен от калориметра, в нем можно было по-
местить кислородный и СО2-электроды, а также брать пробы для
энзиматического определения глюкозы и этанола. Кроме тепло-
продукции, гликолиза и дыхания можно было также определять
Рис. 30. Увеличение сухого веса (W), изменение концентрации глюкозы (cg)
и этанола (са) в культуре дрожжей, растущей в аэробных условиях
Рис. 31. Изменение удельной скорости теплопродукции (q), удельной ско-
рости метаболизма (q^) и ^и-функции в культуре дрожжей, растущей в
аэробных условиях
Первый максимум связан с изменением концентрации глюкозы в среде, второй -- с изме-
нением концентрации этанола (см. рис. 30)
128
сухой вес культуры, используя простое фотоэлектрическое уст-
ройство, измеряющее оптическую плотность культуры.
Исчезновение глюкозы из среды, продукция и последующее ис-
пользование этанола, а также увеличение сухого веса культуры
показаны на рис. 30. Из двувершинного характера кривой роста
культуры следует, что после распада глюкозы в результате про-
цесса гликолиза весь образующийся этанол полностью и быстро
окисляется благодаря оптимальному снабжению культуры кис-
лородом. В соответствии с этим скорость метаболизма см, ско-
рость теплопродукции q и фи-функция имеют два максимума
(рис. 31), каждый из которых соответствует одной из двух стадий
роста.
В этих экспериментах, если не учитывать двувершинного ха-
рактера кривой роста культуры, изменение фи-функции не отли-
чалось существенным образом от ее изменений в относительно
анаэробных культурах. В обоих случаях, максимум фи-функции
приходился на переходный период между лаг-фазой и экспонен-
циальной фазой. В период экспоненциальной фазы роста культу-
ры фи-функция уменьшалась почти до нуля. В этот период рост
культуры сбалансирован и имеет постоянную скорость. В отли-
чие от анаэробных у аэробных культур появляется новый источ-
ник энергии в виде этанола. Поэтому после короткого периода
адаптации с несколькими переходными колебаниями метаболизма
начинается новый рост культуры с таким же изменением фи-
функции, как и в первом периоде роста.
Непрерывное культивирование
Непрерывное культивирование проводилось в системе, где
ферментер был скомбинирован с проточным микрокалориметром
с помощью переливной трубки, сохраняющей объем культуры
константным. Приток свежей среды, содержащей глюкозу, как
вещества, лимитирующего рост, и обезвоженный экстракт дрож-
жей, как источник азота, был константным в стационарных ус-
ловиях. В этих условиях культура дрожжей росла экспонен-
циально, и удельная скорость роста р была равна скорости регули-
руемого притока свежей среды. Эта скорость изменялась от
одного дня к другому, но учитывалась только такая скорость, при
которой глюкоза — источник углерода — полностью окислялась
(Brettel et al., 1972; Brettel, 1974).
Результаты опытов, представленные на рис. 32, показывают,
что и удельная скорость теплопродукции q, и удельная скорость
метаболизма увеличиваются почти линейно относительно удель-
ной скорости роста р. Соответственно, фи — функция близка ну-
лю при всех изменениях р, что совпадает с результатами, получен-
ными в предыдущих экспериментах для экспоненциальной фазы
роста при непроточном культивировании. Небольшие изменения
фи-функции лежат в пределах ошибки измерения. Возможно так-
129
Рис. 32. У дельная скорость тепло-
продукции (q), удельная скорость
метаболизма (q^) и функция
в зависимости от скорости роста
дрожжей при непрерывном куль-
тивировании
же, что они связаны с колебаниями состава дрожжевого экст-
ракта, используемого в культуральной среде, которые с
трудом поддаются учету.
Таким образом, при непрерывном культивировании (с одина-
ковой скоростью роста культуры дрожжей в течение нескольких
недель) не обнаружено изменения фи-функции.
Рассматривая кинетику роста микробных культур, можно вы-
делить три главные фазы роста. Первая — лаг-фаза кажется
очень статичным периодом без каких-либо внешних изменений
культуры. Однако это очень активное состояние, сопровождаю-
щееся образованием ферментов, обусловливающих адаптацию
клеток к питательным веществам и регуляцию процессов, пред-
шествующих клеточным делениям. Вторая фаза, длящаяся весь
период экспоненциального роста, является очень эффективной
фазой с точки зрения энергетики. С другой стороны, это хорошо
сбалансированное состояние культуры, так как в это время не
происходит развития или изменения культуры в целом. При не-
прерывном культивировании эта фаза может продолжаться не-
сколько недель или даже больше без заметного изменения ско-
рости роста, состава ферментов или скорости метаболизма. Это
состояние при постоянных внешних параметрах, таких, как тем-
пература или pH среды, изменяется только в том случае, когда
меняются условия питания: истощаются источники питания
в случае замкнутых культур или меняется скорость притока све-
жей среды при непрерывном культивировании. Наконец, третья
фаза роста культуры — стационарная — является малоактивной
фазой, так как из-за отсутствия субстратов и других неблаго-
приятных условий организмы впадают в состояние покоя с мини-
мальной скоростью метаболизма и прекращают рост.
Исходя из определения фи-функции, как выражения для свя-
яанной продукции энтропии в процессах развития и роста, ста-
130
циопарная фаза кажется неестественным состоянием, сохраняю-
щимся ограниченное время и лежащим вне нормального цикла
жизни. Поэтому она не интересна с точки зрения изучения фи-
функции. Во время фазы экспоненциального роста культура хо-
рош о адаптирована к условиям среды и жизненные процессы
в ней протекают с постоянной скоростью. Это означает, что она
достигла стационарного состояния, сопоставимого со стационар-
ным состоянием многоклеточных организмов. Поэтому биохими-
ческие процессы в такой системе протекают как бы в системе,
не очень далекой от равновесия. Соответственно, фи-функция,
как и следует из ее определения, константна и близка к нулю.
Переходный период между лаг-фазой и экспоненциальной фа-
зой является наиболее интересным периодом роста культуры
дрожжей. В это время происходит развитие и ускорение внутри-
клеточных процессов и их изменение в сторону более экономич-
ных процессов экспоненциальной фазы. Как результат этих из-
менений фи-функция уменьшается от своего максимального зна-
чения до минимальной величины. Переходный период между лаг-
фазой и экспоненциальной фазой является такой стадией роста
культуры, во время которой на фоне медленного роста происходят
многие изменения типа старения. В связи с этим только этот пе-
риод можно считать той частью жизни микробной культуры, ко-
торая сопоставима с ростом многоклеточного организма.
Как видно из приведенных опытов, поведение фи-функции не
зависит от субстратов, вида метаболизма и типа культивирова-
ния. Для дальнейшего прояснения проблем, связанных с изуче-
нием фи-функции, полезно было бы исследовать синхронные куль-
туры делящихся клеток, а также воздействие на культуры ра-
диации и химических веществ, активно изменяющих клеточный
метаболизм.
ИЗМЕНЕНИЕ i|>d И ^-ФУНКЦИЙ
ВО ВРЕМЯ ООГЕНЕЗА ШПОРЦЕВОЙ ЛЯГУШКИ
А. И. Зотин
С точки зрения феноменологической теории развития орга-
низмов оогенез представляет особый интерес, так как в этот пе-
риод происходит омоложение живой системы и вступление ее на
путь развития и роста (Zotin, Zotina, 1967; Зотин, Зотина, 1969;
Озернюк и др., 1971; Zotin, 1972; Зотин, 1974). В то же время
имеется очень мало экспериментальных данных об интенсивности
дыхания и теплопродукции в этот период. По существу, надеж-
131
ные данные по интенсивности дыхания ооцитов разных ста-
дий оогенеза получены только для вьюна (Озернюк, 1970);
а интенсивности дыхания и теплопродукции для ооцитов шпор-
цевой лягушки (Зотин, 1975). Между тем подобного рода
данные могут иметь интерес не только для термодинамической
теории развития организмов, но и для более широкого круга про-
блемы термодинамики нелинейных необратимых процессов. Дело
в том, что при изучении теплопродукции и дыхания ооцитов легче
всего проверить одно из основных положений, составляющих ос-
нову критерия эволюции открытых термодинамических систем,
далеких от равновесия (2.22). Этот критерий основан на разбие-
нии (2.15), в котором функция фи пропорциональна теплоте дис-
сипации, связываемой в единице объема за единицу времени, а
функция — теплоте диссипации, покидающей пределы систе-
мы. Когда система находится вблизи от равновесия или стационар-
ного состояния, вся теплота диссипации, возникающая внутри
системы, покидает ее пределы. Если система далека от равнове-
сия, то только часть теплоты диссипации выделяется в окружа-
ющую среду: другая часть связывается внутри системы в виде
фи-функции. Чем выше степень нелинейности системы, чем даль-
ше она находится от равновесия, тем большая часть теплоты дис-
сипации должна фиксироваться внутри системы.
В живых системах все диссипативные процессы так или иначе
связаны с дыханием и гликолизом, и диссипативную функцию
можно экспериментально определять по уравнению (4,1). Так как
для ооцитов можно пренебречь интенсивностью аэробного гли-
колиза (см. Озернюк и др., 1971), то зависимость (4.1) перехо-
дит в:
= <о2. (4.4)
Функция внешней диссипации пропорциональна интенсивности
теплопродукции системы (4.2), поэтому для ооцитов с учетом (4.3)
можно написать:
tu = <0, — G- (4.5)
Как и в случае роста культуры дрожжей (Schaarschmidt et al.,
1975; Зотин, Шааршмидт, Лампрехт, 1975), на ооцитах можно про-
верить правильность разбиения (2.15): на ранних стадиях раз-
вития и роста организмы находятся в состоянии, далеком от ко-
нечного стационарного состояния, а в период оогенеза удаленность
от этого состояния особенно велика. Очевидно, что чем дальше
открытая система находится от равновесного или стационарного
состояния, тем больше должна быть величина функции связан-
ной диссипации, поэтому период оогенеза особенно удобен для
выявления фи-функции.
Измеряя интенсивность теплопродукции на разных стадиях
оогенеза, можно проверить также правильность утверждения фе-
132
номепологической теории развития, что именно в период оогенеза
происходит конститутивное уклонение живых систем от стацио-
нарного состояния и переход системы на путь конститутивного
приближения к конечному стационарному состоянию. Хотя этот
вопрос стал более или менее ясным после работы Озернюка (1970),
проведенной на ооцитах вьюна (см. рис. 43), однако требовалось
подтверждение этих данных на других объектах. Необходимо
было также получить прямые данные об интенсивности теплопро-
дукции ооцитов. Эти данные были получены для ооцитов шпор-
цевой лягушки Xenopus laevis (Зотин, 1975).
В соответствии с периодизацией Дьюмон (Dumont, 1972),
исследовали 6 стадий оогенеза шпорцевой лягушки: I — пре-
вителлогенез (диаметр ооцитов 50—100 мк), II — стадия конца
превителлогенеза (диаметр ооцитов до 450 мк), III — начало ви-
теллогенеза (начало синтеза пигмента и желтка, диаметр ооцитов
до 600 мк), IV — вителлогенез (диаметр ооцитов 600—1000 мк),
V — конец вителлогенеза (диаметр ооцитов 1000—1200 мк), VI —
зрелые ооциты (диаметр 1200—1300 мк). Выделение ооцитов из
яичников проводили по методу, разработанному Озернюком
(1970), с небольшим видоизменением: ооциты конца большого ро-
ста и частично середины вителлогенеза получали путем механи-
ческого выдавливания из яичника. Этим достигалась большая
сохранность крупных ооцитов по сравнению с ооцитами, получен-
ными после обработки яичника трипсином.
Дыхание ооцитов измеряли методом Варбурга. В сосудики
Варбурга в 3 мл раствора Рингера помещали следующее число
ооцитов: на стадии превителлогенеза — 2000—2500, на стадии на-
чала вителлогенеза — 1000, на стадии вителлогенеза — 600—
800, на стадии конца вителлогенеза и зрелых ооцитов — SOO-
SOO. Измерение дыхания и теплопродукции проводили при 20°.
Теплопродукцию измеряли в микрокалориметре Кальве (тип
MS-70, Сетрам, Франция) с рабочим объемом ячеек 15 мл и чув-
ствительностью 70 мкв/мвт [детали работы с микрокалориметром
описаны в работе Лампрехта и Шааршмидта (Lamprecht et al.,
1971, 1973)]. После измерения дыхания ооциты переносили из
сосудиков Варбурга в калориметрические стаканчики в 1 мл
раствора Рингера. В течение первых 2—3 часов устанавливался
стационарный режим, а затем к ооцитам добавляли 1 млО,ЗМТХУ
для определения нулевой точки отсчета теплопродукции ооцитов.
Стационарный режим для нулевой точки (недышащие ооциты)
достигался через 2—3 часа. Теплопродукцию ооцитов измеряли
в мкал/час, дыхание — в мкл/час. Для сравнения данные по ды-
ханию пересчитывали на мкал/час путем умножения их на коэф-
фициент 5,05 (см. Гинецинский, Лебединский, 1965). Для опре-
деления интенсивности дыхания и теплопродукции измеряли
сухой вес ооцитов. Для этого их помещали на мембранный фильтр
с диаметром пор 0,4 мк, отмывали от среды и высушивали под ва-
куумом при 100° в течение 3—4 часов.
133
В табл. 4 представлены данные по дЫхайию и теплопродукции
ооцитов шпорцевой лягушки разных стадий оогенеза. Как видно,
в процессе оогенеза происходит значительное увеличение скорости
дыхания и теплопродукции. В расчете на один ооцит дыхание уси-
ливается почти в 4 раза от стадии превителлогенеза до стадии зре-
лого яйца, а теплопродукция — в 7 раз. Очевидно, однако, что
интенсивность этих процессов уменьшается во время оогенеза,
так как сухой вес ооцитов на этих же стадиях увеличивается
Таблица 4
Дыхание и теплопродукция ооцитов Xenopus laevis
Стадия оогенеза Сухой вес ооцита, мг Дыхание ооцита Теплопродукция ооцита
мкл|час п мкал|час п
Превителлогенез 0,0078 0,0081 10 0,013 7
Вител- логенез начало середина 0,031 0,224 0,0146 0,0194 10 4 0,016 0,048 5 4
, конец 0,376 0,0283 6 0,058 3
Зрелое яйцо 0,420 0,0314 3 0,094 1
в 53 раза. Данные об изменении интенсивности дыхания и тепло-
продукции ооцитов шпорцевой лягушки в сопоставимых едини-
цах (мкал/час/мг сухого веса ооцита) в зависимости от стадий
оогенеза и размеров ооцитов приведены на рис. 33. Происходит
значительное уменьшение интенсивности дыхания и теплопродук-
ции в процессе оогенеза, особенно быстрое в период, предшест-
вующий началу вителлогенеза. Для ооцитов вьюна показано, что
интенсивность дыхания возрастает в период малого роста и бы-
стро снижается, начиная со стадии вакуолизации цитоплазмы,
т. е. с началом периода большого роста (Озернюк, 1970). Период
превителлогенеза в оогенезе лягушек, по-видимому, соответствует
стадии вакуолизации цитоплазмы ооцитов костистых рыб, так как
в это время у амфибий происходит возникновение кортикальных
гранул (Гинзбург, 1968). Поэтому можно считать, что изменение
интенсивности дыхания ооцитов шпорцевой лягушки со стадии
превителлогенеза сходно с изменением интенсивности дыхания
ооцитов костистых рыб. Это подтверждают и расчеты константы Ь
в уравнении зависимости интенсивности дыхания ооцитов в период
превителлогенеза и начала вителлогенеза по формуле (2.9). Рас-
чет по данным, приведенным на рис. 33, показал, что для ооцитов
шпорцевой лягушки b = 0,70. Эта величина близка по величине
константе b для сходных стадий оогенеза вьюна, где Ь = 0,67
(Озернюк, Зотин, 1974).
Одновременное изучение интенсивности дыхания и теплопро-
дукции ооцитов шпорцевой лягушки позволило рассчитать вели-
134
Рис. 33. Изменение интенсивности дыхания (1) ,^и-функции (2) и ^-функ-
ции (3) в процессе оогенеза Xenopus laevis L. (Зотин, 1975)
чину и характер изменения г|)и-функции в процессе оогенеза.
Как видно из рис. 33, величина фи-функции наибольшая в пе-
риод превителлогенеза и наименьшая у зрелых яиц. Более дроб-
ные определения показали, что у ооцитов шпорцевой лягушки
весом 0,0038 мг (сухого веса) фи-функция равна 12,10, у ооцитов
весом 0,0110—2,35, весом 0,0236—1,41, весом 0,224—0,242, ве-
сом 0,403—0,179 и весом 0,430—0,146 мкал/час/мг. Эти наблю-
дения являются экспериментальным доказательством правиль-
ности разбиения (2.15) и утверждения (2.22), согласно которому
в процессе приближения открытой системы к конечному стацио-
нарному состоянию величина функции связанной диссипации
уменьшается. Это же одновременно показано нами для растущей
культуры дрожжей (Schaarschmidt etal., 1975b; Зотин, Шаарш-
мидт, Лампрехт, 1975).
Физический смысл функции связанной диссипации пока ос-
тается неясным. Здесь возможны различного рода предположения.
По моему мнению, величина фи-функции должна коррелировать
со скоростями химических реакций, протекающих в открытой
системе. Функция связанной диссипации представляет собой часть
теплоты, возникающей в системе, но не покидающей ее пределы.
Эта теплота может использоваться, главным образом, для ускоре-
ния химических реакций. Поэтому чем больше величина фи-
функции системы, тем больше должна быть скорость всех химичес-
ких процессов, протекающих в системе. Этот вывод может быть
легко проверен экспериментально: при одной и той же темпера-
туре среды скорости химических реакций на ранних стадиях ооге-
неза должны быть выше, чем на поздних.
135
ТЕПЛОПРОДУКЦИЯ И ДЫХАНИЕ
ВО ВРЕМЯ РАЗВИТИЯ И РОСТА
ДВУХ ВИДОВ НАСЕКОМЫХ
К. %. Лёр, П, Сайади, И. Лампрехт
Существует очень мало данных о теплопродукции насекомых,
полученных прямыми методами. Еще меньше работ посвящено
сопоставлению теплопродукции и дыхания во время развития
насекомых. Нам известна лишь одна работа (Fourche, Lemaitre,
1974), в которой проведено такое сопоставление для стадии мета-
морфоза мухи — Ceratitus capitata. Между тем насекомые яв-
ляются очень удобным объектом для изучения возрастных изме-
нений метаболизма и для получения данных об изменении и
ф^-функций во время развития и роста, так как продолжитель-
ность жизни многих насекомых невелика.
Для изучения теплопродукции и дыхания были выбраны два
вида насекомых: мучной хрущак (Tenebrio molitor L.) и рыжий
таракан (Blattella germanica L.).
Tenebrio molitor L. (Coleoptera) содержали в термостатируе-
мой комнате с температурой 30° и относительной влажностью 45—
50%. Развитие насекомых проходило в цилиндрических стеклян-
ных сосудах, которые были помещены в закрытый контейнер,
выстланный изнутри мокрой тканью. В контейнере находился
открытый сосуд с пересыщенным раствором хлористого натрия.
Относительная влажность в контейнере была выше 75%. Из ос-
новной культуры, хранящейся в контейнере, было выделено не-
сколько групп насекомых (приблизительно по 20 животных
в каждой группе), которые содержались вместе в течение всего
экспериментального периода. В процессе роста число особей по-
степенно уменьшалось, поэтому сырой вес группы в целом, как
правило, оставался постоянным. Насекомых кормили пшенич-
ными отрубями и три раза в неделю ломтиками моркови. Личиноч-
ное развитие продолжалось приблизительно 90 дней (с несколь-
кими линьками см. рис. 34), стадия куколки 4—5 дней.
Blattella germanica L. (Blattoptera) содержали в тех же усло-
виях или в специальном инкубаторе при 25°. Насекомых кормили
собачьими брикетами, содержащими (в % от сырого веса) 21,6%
белков, 12,8% жиров и 22,3% углеводов. Иногда в пищу добав-
ляли ломтики свежего яблока. У этого вида насекомых самка вы-
нашивает оотеку до момента выхода из нее молоди. Из одной ооте-
ки рождается 40 или 50 личинок. Насекомых содержали группа-
ми по 20 животных в каждой, но за время роста число их умень-
шалось до одного на группу.
Для определения теплопродукции был использован дифферен-
циальный микрокалориметр Кальве (Сетрам/Лион) с объемом со-
© К..- Г). Loehr, Р. Sayyadi, I. Lamprecht, 1976 г.
136
судов 15 мл и чувствительностью 60,0 мкв/мвт. Регистрацию теп-
лопродукции проводили с помощью самописца Киппа — Зенена
с регулировкой шкалы от 50 до 500 мкв. Нулевая линия — глав-
ная проблема подобного рода калориметрических измерений —
была определена в опытах с пустыми сосудами. В этих контроль-
ных измерениях, после небольшого периода времени, необходи-
мого для установления равновесия, термограммы показывали
константный результат. Как показывают термограммы опытов,
в отличие от контрольных термограмм, на них имеются флуктуа-
ции, связанные, по-видимому, с быстрыми изменениями двигатель-
ной активности насекомых. Такие колебания метаболизма не
улавливаются при монометрических исследованиях, так как
в последнем случае регистрация идет не непрерывно, а усреднен-
ие за достаточно продолжительные отрезки времени. Исследова-
ние теплопродукции насекомых проводили как в закрытых, так
и в открытых для воздуха сосудах. Расчет показал, что количест-
ва воздуха в закрытых сосудах было достаточно и за время опыта
содержание кислорода и углекислого газа в них изменялось мало.
Манометрические исследования проводили с помощью аппа-
рата Варбурга (фирма Браун/Мелсунген) с сосудиками объе-
мом 14 и 20 мл. Все измерения, как манометрические, так и кало-
риметрические, проводили при 25° — температуре, наиболее бла-
гоприятной для изученных насекомых.
На рис. 34 показаны данные об изменении сырого веса двух
изученных видов насекомых во время развития и роста.
Изменение веса Т. molitor происходило по сигмоидной кривой
с лаг-фазой в течение первых 10 дней и экспоненциальной фазой
до 50 дня (см. рис. 34). Показатель степени при экспоненциальном
росте равен 0,131/день, что соответствует удвоению сырого веса
личинок за каждые 5,3 дня. После 85-го дня развития, на стадии
куколки, сырой вес насекомого резко уменьшается (на рис. 34
этот момент показан треугольником). Это уменьшение веса свя-
зано, по-видимому, с прекращением питания и потерей воды ку-
колками. Стрелки на рис. 34 указывают дни линек в период личи-
ночного развития Т. molitor. Сырой вес взрослых насекомых прак-
тически не изменяется.
У В. germanica экспоненциальный характер кривой роста не
так хорошо выражен, как у Т. molitor (см. рис. 34). Однако и для
них можно вычислить показатель степени, который для самок
равен 0,072/сутки (время удвоения веса — 9,7 дня) и для самцов—
0,062/сутки (время удвоения веса — 11,2 дня). Колебания в из-
менении веса рыжего таракана во время роста трудно объяснимы.
Возможно, что они связаны с ритмами питания таракана.
Потребление кислорода изученными видами насекомых, как
функция времени, показано на рис. 35. Сопоставление данных,
полученных на трех группах Т. molitor, показывают, что во время
личиночного развития мучного хрущака происходит непрерывное
уменьшение интенсивности дыхания и некоторое усиление мета-
137
болизма после образования куколки. Сходные данные получены
и для В. germanica (рис. 35). Во время роста происходит снижение
интенсивности дыхания насекомых. В период формирования ооте-
ки интенсивность дыхания самок сохраняется на одном уровне,
тогда как интенсивность дыхания самцов возрастает.
Как было отмечено выше, теплопродукция изученных живот-
ных не представляет собой прямой линии, что связано с флуктуа-
циями метаболизма и подвижностью насекомых во время опытов.
Иногда во время дневной активной фазы термограммы показыва-
ли сильные усиления теплопродукции. Внезапные флуктуации
теплопродукции иногда наблюдались и в ночное время. Эти рез-
кие усиления теплопродукции продолжались около 5 минут и до-
стигали 70% от средней теплопродукции. Они повторялись с час-
тотой от 1 до 3 часов (рис. 36). По-видимому, эти флуктуации
можно объяснять тем, что тараканы закрывают свои трахеи на
довольно длительное время, используя для дыхания запасенный
там кислород, а затем внезапно освобождаются от углекислого
газа и забирают новый запас воздуха (спонтанный пароксизм).
Средние величины интенсивности теплопродукции для двух
изученных видов насекомых показаны на рис. 37. В обоих слу-
Рис. 34. Изменение сырого веса мучнйго хрущака и рыжего таракана во время
роста
а — мучной хрущак, стрелки указывают даты линек, треугольник — стадию куколки;
б — рыжий таракан; 1 — самцы, 2 — самки
138
чаях она уменьшается во время развития и роста животных. С та-
раканами измерения проводили только на самцах. Как видно из
рис. 37, после 80 дня, как и в случае манометрических измерений,
наблюдается усиление теплопродукции этих насекомых.
Интенсивность дыхания, как постулировано соотношением
(4.4), соответствует ty-функции, а интенсивность теплопродукции
(4.2) \|)d-функции. Поэтому из данных, приводимых на рис. 35 и
37, можно рассчитать изменение фи-функции во время развития
и роста Т. molitor и В. germanica. У тараканов происходит непре-
рывное уменьшение этой величины во времени (рис. 38), тогда как
у мучного хрущака она сильно уменьшается в первые тридцать
Ч К
но во <20 го но во во юо
Сутки
Рис. 35. Интенсивность дыхания трех групп хрущака (а) и одной группы
тараканов (б)
На рис. а‘ внутренние графики расположены таким образом, чтобы время совпадало
с указанным на оси абсцисс. На рис. 6 1 — самцы, 2 — самки; треугольником отме-
чено время образования оотек у самок
139
i i
Сутки
Рис. 37. Интенсивность теплопродукции (q) двух групп мучных хруща-
ков (а) и самцов тараканов (б)
Треугольником показана стадия куколки
4
1
ZZh
ЧО
80 /20
Сутки
ZO
40
60
80
8
Ч
О
Рис. 38. Изменение функции свя-
занной диссипации (хри) во время
роста двух групп хрущаков (а) и
самцов таракана (б)
Рис. 39. Изменение диссипативной
функции функции внешней дис-
сипации (tya) и функции связанной
диссипации (tyu) во время роста
самцов рыжего таракана
20 40 60 Сутки
дней развития, а затем сохраняется на более или менее постоян-
ном уровне (рис. 38). Увеличение фи в период перехода от кукол-
ки к имаго не противоречит теории яри-функции.
На рис. 39 сопоставлены данные об изменении ф, ф^ и фи-
функций во время роста В. germanica. Следует отметить, что чис-
ло проведенных измерений недостаточно для математического
описания кривых изменения этих функций. Тем не менее резуль-
таты, полученные в данной работе, показывают, что феномено-
логическая теория развития (глава 2) может быть использована
в случае роста насекомых. Дальнейшие эксперименты могут про-
лить свет не только на эту проблему, но и на природу флук-
туаций теплопродукции и дыхания насекомых.
ТЕПЛОПРОДУКЦИЯ И ДЫХАНИЕ
НА РАННИХ СТАДИЯХ РОСТА АКСОЛОТЛЕЙ
В. А. Г рудницкий, И. С, Никольская
Измерение теплопродукции организмов в калориметрических
установках при изучении энергетического обмена до сих пор не
нашло широкого распространения в исследовательской практике.
Сложность методики и значительные затраты времени при экспе-
риментах являются причиной предпочтения и широкого распро-
странения более простого и удобного метода косвенной оценки об-
мена по скорости потребления кислорода организмом. Вместе
с тем эти методы, основанные на различных физических явлениях,
по разному характеризуют исследуемый процесс, что находит
отражение в известных случаях расхождения данных, получаемых
обоими методами. Такие расхождения получены при некоторых
патологических состояниях организма, при гипоксии, откармли-
вании после голодания и т. д. (Лихачев, 1893; Авроров, 1899; Ве-
селкин, 1960; Чередниченко, 1965; Граменицкая, 1967; Полуш-
кина, 1967). Особый интерес для исследования энергетики орга-
низма представляют опыты, в которых было найдено расхождение
рассчитанной по потреблению кислорода теплопродукции над фак-
тически измеренной в калориметре у беременных собак (Студен-
ский, 1897). Автор связывал причину такого расхождения с по-
вышенной величиной процессов синтеза при беременности.
Исходя из этого, следует ожидать расхождения в данных пря-
мой и непрямой калориметрии во время роста, когда синтез
веществ в организме велик. Теоретически было получено анали-
тическое выражение для расхождения данных прямой и непря-
мой калориметрии при росте (Зотин, Грудницкий, 1970), и за-
тем экспериментально показано существование этого явления
(Грудницкий, 1972а, б, 1975) во время роста меченосцев (Xip-
141
кал/мс[г
Рис. 40. Из менение интенсивности
дыхания (Яо2) и интенс ивности те-
плопродукции (q) в зависимости от
веса у личинок аксолотля
hophorus helleri L.), при росте ооцитов и микроорганизмов (Зо-
тин, 1975; Зотин и др., 1975). Исследованию найденной особенно-
сти энергетики растущих животных посвящена и данная работа.
Опыты проводили на аксолотлях начиная от момента вылуп-
ления зародышей из икры. Параметром, относительно которого
систематизировали данные по дыханию и теплопродукции, являл-
ся сырой вес тела животных. На начальных этапах, когда личинка
имеет запасы питания, вес тела определяли без желточного меш-
ка. Исследовали энергетический обмен во время роста аксолотлей
в диапазоне весов 0,007—2,7 г. Измерение теплопродукции про-
водили спустя двое суток после кормления и, затем, на том же
материале измеряли дыхание. До веса 0,1 г измерение проводили
на партиях численностью 30—35 штук, более крупных бра-
лись по 1 экземпляру в опыт.
Для определения теплопродукции использовали адиабатичес-
кий калориметр, описанный ранее (Грудницкий, 1972а, б). Дли-
тельность опыта 12 часов. Дыхание измеряли на аппарате Вар-
бурга. Температура в обоих случаях равнялась 22,7°. Было про-
ведено 80 замеров, равномерно распределенных по исследованному
весовому диапазону.
Приводимые кривые зависимости теплопродукции и дыхания
от веса тела получены при помощи двойного линейного усреднения
по трем точкам. Оксикалорийный коэффициент принят 5 кал/мл О2:
Как можно видеть на рис. 40, изменение интенсивности дыха-
ния и теплопродукции имеет сложный характер. В момент вылуп-
ления из икры, который представлен на графике минимальным
исследованным весом, уровень обмена, рассчитанный по дыханию,
в 3 раза превышает значение, полученное прямой калориметрией.
После вылупления интенсивность дыхания аксолотлей резко
уменьшается, а теплопродукция растет и становится выше уров-
ня, рассчитанного по дыханию. В последующем величина тепло-
продукции снижается, и у животных весом более 0,6 г тепла вы-
деляется меньше, чем можно ожидать из данных по скорости пот-
ребления кислорода. Постепенное замедление расхождения между
данными непрямой и прямой калориметрии при возрастании веса
112
тела, по-видимому, указывает йа то, что эта величина стремится
к постоянному значению на данном участке роста аксолотлей.
Изменения интенсивности дыхания и теплопродукции сразу
после вылупления имеют вид переходного процесса между состоя-
ниями энергетического обмена до и после вылупления. Можно
полагать, что данное явление связано с переходом организма
к активному образу жизни, однако выяснение конкретных меха-
низмов этой перестройки требует экспериментальной разработки.
Изучение энергетического обмена растущих животных мето-
дами прямой и непрямой калориметрии и сопоставление получен-
ных результатов ограничивается немногими примерами, упомя-
нутыми выше. Во всех исследованных случаях наблюдается мень-
ший уровень теплопродукции, чем уровень обмена, рассчитанный
по скорости дыхания. Этот же факт подтверждают данные, полу-
ченные в настоящей работе. Отличительной особенностью прове-
денной работы является исследование периода роста, характери-
зующегося резкой сменой условий развития при вылуплении и
сменой типа питания. В этот интересный момент роста между пот-
реблением кислорода и теплопродукцией организма не сущест-
вует жесткого, однозначного соотношения и возможны колебания,
в которых теплопродукция превышает уровень обмена, рассчи-
танный по дыханию. Не исключено, что это обстоятельство будет
иметь значение при выяснении механизма, определяющего соот-
ношение между теплопродукцией и скоростью дыхания в расту-
щем организме.
ЗАВИСИМОСТЬ ТЕПЛОПРОДУКЦИИ ОТ ВЕСА ТЕЛА
У РАСТУЩИХ ОРГАНИЗМОВ
В. А. Грудницкий, Б. Шааршмидт
Хорошо известно, что интенсивность дыхания животных с уве-
личением веса тела выражается логарифмической зависимостью
(2.9). Эту зависимость распространяют и на теплопродукцию, так
как существует представление о том, что потребление кислорода
и теплопродукция организма являются тождественными при
оценке уровня энергетического обмена. Однако теоретический
анализ, проведенный с использованием соотношений термодина-
мики необратимых процессов и уравнения роста (3.36), показал,
что теплопродукция и величина веса тела находятся в полуло-
гарифмической зависимости (Зотин, Зотина, 1969; Зотин, Груд-
ницкий, 1970; Зотин, 1974).
q = с — d\v%W. (4.6)
© В. А. Грудницкий, 1976 г. © В. Schaarschmidt, 1976 г.
143
кал/час/г
Рис. 41. Зависимость удельной скорости теплопродукции от логарифма веса
тела у меченосцев
Рис. 42. Зависимость удельной скорости теплопродукции от логарифма
веса у S. cerevisiae при 30° (1), S. cerevisiae при 40° (2), В. stearothermophilus
(3) (Schaarschmidt et al., 1976)
Согласно уравнению (4.6) графики зависимости q = / (log W)
в полулогарифмической шкале координат должны быть представ-
лены прямыми линиями.
На рис. 41 показан график зависимости q — / (log TV), най-
денный для меченосцев (рыбы сем. Poeciliidae) (Грудницкий,
1972а). Исследование теплопродукции меченосцев проводили на
изготовленном в Лаборатории биофизики развития ИБР АН
СССР адиабатическом калориметре (Грудницкий, 1972а). Тепло-
продукцию измеряли у рыб в диапазоне сырого веса тела, равном
0,15—2,5 г. Так как рыбки живородящие и это могло бы затруд-
нить толкование результатов, получаемых на самках, в опыт бра-
ли только самцов. Как было выяснено (Грудницкий, 1972а, б,
1975), рост самцов меченосцев происходит непрерывно, хотя и
значительно замедляется после начального периода. Это обстоя-
тельство позволяло определять величину теплопродукции у рыб
без учета возраста, ориентируясь только на вес тела. Однако ос-
новная масса замеров относится к возрасту 3,5—14 месяцев, и
лишь несколько наиболее крупных экземпляров взяты неизвест-
ного возраста. Измерение теплопродукции проводили на третий
день после кормления. Температура в ходе опытов 22,7°.
На рис. 42 представлены результаты, полученные при измере-
нии теплопродукции культур микроорганизмов. Эти опыты про-
водили на микрокалориметрах типа Кальве (Seteram/France)
с объемом калориметрических сосудов 100 и 10 мл (Schaarsch-
midt et al., 1975а). Детальное описание экспериментальных усло-
144
вий и микрокалориметрической методики Дано ранее (Lamprecht
et al., 1973). Для большинства экспериментов использованы
дрожжи диплоидной линии 211 (Saccharamyces cerevisiae). Часть
экспериментов проводили на соответствующих гаплоидах и гекса-
плоидах. Также измерена теплопродукция у термофильной бакте-
рии Bacillus stearothermophilis. Ростовая^среда состояла из дрож-
жевого экстракта (10 г/л), пептона (5 г/л) и глюкозы (5—20 г/л).
Для измерения сухого веса было применено два метода. При
одном из них рост культуры в калориметрическом сосуде преры-
вался в разное время, и растущая масса высушивалась и взве-
шивалась. При другом одновременно с измерением теплопродук-
ции в калориметрическом сосуде измерялась оптическая плот-
ность культуры, и затем рассчитывалась величина сухого веса.
Измерение теплопродукции во всех случаях проводили на фазе
экспоненциального роста культуры. Так как хорошо известно,
что объем и сухой вес дрожжевых клеток увеличивается со сте-
пенью плоидности (Laskowski, 1962), серия культур разной пло-
идности может рассматриваться как последовательные фазы роста
одного организма. Исходя из этого, для расчетов были использо-
ваны также соответствующие данные по теплопродукции и сухо-
му весу разноплоидных культур.
Как видно из рис. 41 и 42, экспериментальные данные под-
тверждают справедливость зависимости (4.6) как для высших
животных, так и для микроорганизмов. Этот вывод следует так же
из анализа данных разных авторов для некоторых других живот-
ных (Зотин, 1974). Таким образом, можно считать, что во время
роста организма удельная теплопродукция меняется в зависимо-
сти от величины веса иным образом, чем интенсивность дыхания.
В общем случае зависимость между теплопродукцией и весом тела
при росте животных является полулогарифмической, выражае-
мой уравнением (4.6). Однако данное уравнение может быть рас-
пространено только на периоды роста, которые соответствуют
закономерности уравнения роста, применявшегося для его вы-
вода. Вне границ применимости уравнения (4.6) зависимость меж-
ду теплопродукцией и весом тела, вероятно, имеет иной характер.
К таким случаям, очевидно, относится изменение теплопродукции
на ранних этапах роста аксолотля (см. стр. 142). Полученные для
аксолотля данные не показывают полулогарифмической зависи-
мости теплопродукция — вес тела, что, по-видимому, объясняется
особенностью роста аксолотлей в период после вылупления из
икры. Не удается обнаружить также полулогарифмическую за-
висимость при анализе данных о теплопродукции В. mirabilis
(Jaworski et al., 1968).
Данные примеры еще раз подчеркивают необходимость даль-
нейшей экспериментальной и теоретической разработки вопроса
о границах применимости уравнения (4.6) и характере зависимо-
сти теплопродукция —вес тела на этапах роста, где уравнение
(4.6) не соответствует действительности.
V
НЕКОТОРЫЕ ПРОБЛЕМЫ
ЭНЕРГЕГИКИ ПРОЦЕССОВ РАЗВИТИЯ
Согласно качественной феноменологической теории развития,,
во время эмбрионального развития, роста и старения организмов
происходит уменьшение, а в процессе оогенеза, на начальных
стадиях регенерации и злокачественного роста — увеличение
функции внешней диссипации (2.16), измеряемой по интенсивности
теплопродукции (4.2). Как указывалось, изменение интенсивно-
сти теплопродукции в организмах определяется изменением ин-
тенсивности процессов дыхания и гликолиза. Следовательно, из
качественной феноменологической теории развития вытекает, что-
механизм конститутивных процессов связан с изменением интен-
сивности этих процессов. Так как конститутивное приближение
организмов к конечному стационарному состоянию с точки зрения
этой теории представляет собой процесс старения организмов,
а конститутивное уклонение — процесс омоложения, то следствие,
указанное выше, фактически указывает пути исследования меха-
низмов старения и омоложения живых систем.
В настоящее время еще трудно себе представить в полном
объеме механизм снижения интенсивности дыхания и гликолиза
в процессе развития и роста животных и механизм увеличения
интенсивности этих процессов в период оогенеза, на начальных-
стадиях регенерации и злокачественного роста. Однако несколько1
упростить эту проблему все же можно. Во-первых, у большин-
ства организмов вклад дыхания в энергетику намного больше,,
чем вклад гликолиза, и поэтому для начала можно ограничить-
ся изучением механизмов изменения интенсивности дыхания..
Во-вторых, отвлекаясь от различного рода частных механизмов:
регуляции дыхания, можно считать, что базисный уровень дыха-
ния в клетках и тканях определяется концентрацией митохонд-
рий. Если это так, то механизм старения организмов можно связы-
вать с механизмом уменьшения концентрации митохондрий, а ме-
ханизм естественного омоложения —- с механизмом увеличения:
концентрации митохондрий. То, что такое утверждение возможно,,
показывают данные, полученные при изучении энергетики разви-
тия животных (Зотин, 1974). Однако для окончательного доказа-
тельства этого положения требуется еще большая работа, так как
таких данных в литературе немного и они в значительной степени
противоречивы.
146
ИЗМЕНЕНИЕ МИТОХОНДРИЙ
ВО ВРЕМЯ РАЗВИТИЯ И РОСТА ЖИВОТНЫХ
И. С, Никольская, JL. И. Радзинская
С тех пор, как интактные митохондрии были выделены из кле-
ток (Hogeboom et al., 1948) и получены первые электронно-микро-
скопические фотографии их структуры (Palade, 1956; Sjostrand,
1956), накопились многочисленные данные о морфологии и функ-
циональной специфике митохондрий.
Выяснено, что размер и форма митохондрий могут широко
варьировать в разных клетках. Обычна удлиненная или сфериче-
ская форма митохондрий, встречается кольцевидная, ветвистая,
звездчатая или спиральная (Ленинджер, 1966). Средний размер ми-
тохондрий в клетках печени равен 3 X 0,5—1,0 мк, эндокрин-
ных клеток поджелудочной железы — около 10 мк, летательной
мышцы стрекоз — 8 X 8 мк. Недавно Гир и Беднерик (Gear,
Bednarek, 1972) разработали метод прямого измерения размеров
митохондрий в растворе и показали, что средний диаметр мито-
хондрий печени равен 0,93 мк, а средний объем одной митохондрии
печени составляет 0,42 мк3, сердца — 0,60 мк3, коры почек —
0,23 мк3. Морфологическая гетерогенность митохондрий различ-
ных тканей была найдена и другими авторами (Bondi et al., 1972;
Bulos et al., 1972) и др.
Показано также, что существуют определенные различия
в строении митохондрий клеток печени, почек, мозга, мышц и
других тканей. Эти различия касаются расположения и количества
крист, формы крист и матрикса, формы и размеров митохондрий.
Все это свидетельствует об определенной тканевой специфичности
митохондрий. Однако часто в одной и той же ткани встречаются
митохондрии различного размера и строения. Например, при
исследовании митохондрий мышц (Edington, McCafferty, 1973),
было найдено, что срезы отдельных митохондрий колеблются от
1-7.10-2 до 109-10"2 мк2, т. е. почти в 70 раз.
Особенно наглядно гетерогенность митохондрий тканей была
продемонстрирована в работах с выделением различных субфрак-
ций митохондрий. Так, в сердце кролика обнаружено несколько
субфракций митохондрий, которые отличаются по ультраструк-
туре и свойствам (Монахов, 1964). Интересно, что по данным
этого автора, количественные соотношения субфракций в ткани
оказались постоянными. Из тканей мозга выделено 60 субфракций
митохондрий, отличающихся по размеру (Blokhins, Veldstra,
1970). Обнаружена также гетерогенность популяции митохонд-
рий, выделенных из печени (Swick et al., 1967). В настоящее время
таких работ довольно много. В частности, Сатев и др. (Satav et al.,
1973) выделили из печени митохондрии, которые они обозначили
147
как легкие, тяжелые и пенистые. Авторы показали, что ДНК
всех фракций не различается по количеству и плотности, РНК
содержится в легкой фракции в 3 раза больше, чем в тяжелой,
биосинтез белка происходит интенсивнее в легкой фракции. Фрак-
ции отличаются по ультраструктуре: в тяжелых митохондриях
хорошо развиты кристы, мембраны двойные; в легких митохон-
дриях мембраны также двойные, но крист мало. Пенистые мито-
хондрии состоят из хорошо развитых, но деградирующих ми-
тохондрий. Авторы считают, что легкие митохондрии являются
предшественниками тяжелых.
По-видимому, гетерогенность митохондрий можно связать
с их биогенезом — с образованием, увеличением их числа, услож-
нением организации, а в такой ткани, как мозг, и с определенной
специализацией. Качественные изменения митохондрий могут
происходить при их деградации.
В последние годы биогенез митохондрий привлекает присталь-
ное внимание исследователей. Имеются подробные сводки работ,
выполненных по биогенезу митохондрий (Рудин, Уилки, 1970;
и др.). Многое в биогенезе митохондрий еще не ясно, так как
этот вопрос в настоящее время находится в стадии разработки.
Известно, что митохондрии способны к самовоспроизведению
частично за счет собственного генетического аппарата, а частично
за счет генетического аппарата ядра (Нейфах, 1973, 1974а, б).
ДНК митохондрий несколько отличается от ядерной ДНК по со-
ставу нуклеотидов и плавучей плотности. Митохондрии могут
самостоятельно синтезировать РНК и некоторые белки. Часть
белков (например, цитохром с) синтезируется вне митохондрий,
некоторые другие — например, АТФ-за и цитохромоксидаза —
синтезируются частично в митохондриях, частично вне их (Tza-
goloff et al., 1973). Киселев и Гайцхоки (1972) считают, что в ми-
тохондрии может проникать ядерная РНК и играть роль матрицы
при биосинтезе белка при использовании рибосом митохондрий.
Свансон (Swanson, 1971) также показал, что в митохондрии Хе-
nopus laevis могут проникать полинуклеотиды и служить матри-
цами для синтеза полипептидов на рибосомах.
Считается, что биогенез митохондрий — многоэтапный про-
цесс, включающий постепенное усложнение самих митохондрий
и их мембран (Нейфах, 1967, 1974а, б). Мембраны митохондрий
представляют собой очень сложные образования, в них входят
около 30 белков, 10 липидов, нуклеиновые кислоты, сиаловые
кислоты. Показано, что ферменты наружной и внутренней мем-
бран митохондрий печени синтезируются асинхронно, сборка этих
мембран также происходит не одновременно (Gear, 1970).
Вопрос о длительности существования отдельной митохондрии
до сих пор неясен. Время полусуществования цитохрома С и
гема а в митохондриях печени и сердца составляет 5,5—5,9 дня,
белка — 5—7 дней (Aschenbrenner et al., 1970). В то же время
имеются указания на больший срок существования митохондрий.
148
В мозге молодых крыс белки митохондрий имеют период полусу-
ществования около 27 суток (Menzies, Gold, 1972).
В ходе индивидуального развития животных можно условно
выделить следующие периоды, когда происходят изменения био-
генеза митохондрий: 1) оогенез, 2) зародышевое развитие, 3) ткани
сформировавшегося организма в обычных условиях существова-
ния, 4) ткани животных после различных воздействий (при гипо-
ксии, усиленной физической нагрузке, при беременности, при
действии температуры и т. д.). По-видимому, общий план биоге-
неза митохондрий для каждого этапа сходен, однако, возможно
имеется и определенная специфика этого процесса, касающаяся
темпа увеличения массы митохондрий, их дифференцировки и др.
Можно думать, что в зависимости от потребностей организма на
одних этапах преобладает процесс образования митохондрий,
их сборки или деления, на других — качественные изменения и
дифференцировка.
Вопрос об образовании митохондрий в течение оогенеза рас-
смотрен в сообщении Озернюка (стр. 232). Автор показал, что в те-
чение оогенеза происходит многократное увеличение массы мито-
хондрий и их дифференцировка.
Сведений о воспроизводстве митохондрий в течение зароды-
шевого развития крайне мало. На основании исследования био-
синтеза рибосомальной ДНК в митохондриях Xenopus laevis
высказано предположение, что образование новых митохондрий
начинается на поздних стадиях эмбриогенеза, а до этого функцио-
нируют митохондрии ооцитов (Chase, Dawid, 1972). Однако морфо-
логическая и биохимическая дифференцировка митохондрий все
же происходит в течение эмбрионального развития. Например,
митохондрии тритона мало дифференцированы на ранних стадиях
эмбрионального развития (стадия ранней гаструлы) и полная их
дифференцировка достигается лишь на стадии нейрулы (Buchner,
1966).
Биогенез митохондрий млекопитающих изучен, главным обра-
зом, на тканях взрослых животных. В течение зародышевого раз-
вития большинство работ посвящено изучению изменения струк-
туры и функции митохондрий различных тканей. Из имеющихся
отрывочных данных можно заключить, что в ходе эмбриогенеза
и ранних стадий постнатального развития изменяется как размер
и форма митохондрий, так и содержание в них ферментов. Так,
в сердце эмбриона крысы на 10-й день развития митохондрии мел-
кие, имеют слабо выраженные кристы, количество крист невелико.
К 14-му дню развития митохондрии уже имеют вид, характерный
для молодых животных, они становятся крупнее, с хорошо выра-
женными кристами. За это же время (с 10-го по 14-й день) актив-
ность АТФ-азы, НАДН-оксидазы и цитохромоксидазы митохонд-
рий возрастает почти в 3 раза (Mackler et al., 1971). В то же время
в митохондриях печени эмбрионов крысы активность дыхатель-
ных ферментов (цитохромоксидазы, сукцинатдегидрогеназы, сук-
149
цинат — цитохром С — редуктазы и НАДН — цитохром С —
редуктазы) очень низка и составляет х/2—х/8 часть от активности
этих ферментов у взрослых животных. Увеличение активности
этих ферментов наблюдается только на 2—3-й день постэмбрио-
нального развития, в это же время отмечается возрастание числа
митохондрий (Jakovcic et al., 1971). Эти данные подтвердили ре-
зультаты Леви и Тоири (Levy, Тонгу, 1970), которые также по-
казали, что активность ферментов внутренней и наружной мем-
бран митохондрий печени крыс низка перед рождением и только
к 5—8-му дню после рождения их активность возрастает, а фер-
менты цикла трикарбоновых кислот в матриксе митохондрий пе-
чени достигают активности, наблюдаемой у взрослых животных
только непосредственно перед рождением.
На основании этих данных можно предположить наличие связи
между степенью функциональной активности и дифференцировки
органа к моменту рождения у млекопитающих и степенью морфо-
логической и биохимической зрелости митохондрий. Вероятно,
сердце эмбриона крыс является более «зрелым» органом, в то
время как печень начинает функционировать с большой нагрузкой
после рождения, а морфологическое и функциональное созрева-
ние мозга завершается в первые недели после рождения крысы,
в это же время обнаружено изменение митохондрий (Samson et al.,
1960).
Можно представить себе, что после завершения дифференци-
ровки тканей, которая, по-видимому, сопровождается структур-
ной и функциональной дифференцировкой митохондрий, в тканях
устанавливается определенное равновесие между образованием
и разрушением митохондрий в соответствии с длительностью
существования митохондрий, характерной для данной ткани.
В тканях поддерживается определенный уровень концентрации
митохондрий с определенным соотношением субфракций. Так,
есть работы, в которых показано, что митохондрии взрослых
годовалых крыс качественно не отличаются от митохондрий
2-х годовалых крыс (Cold et al., 1968), а соотношение субфракций
митохондрий в клетках печени взрослых животных постоянно
(Монахов, 1964).
Какие факторы поддерживают этот постоянный уровень мито-
хондрий, что вызывает их образование, деление или дифферен-
цировку, пока не известно.
Возможно, что в дальнейшем при изучении механизмов, управ-
ляющих биогенезом митохондрий окажутся полезными данные
по изменению образования и функциональной активности мито-
хондрий при различных воздействиях, при усилении и уменьше-
нии функциональной активности тканей.
В последние годы появились работы, связывающие образование
и дифференцировку митохондрий с функциональной активностью
ткани. Так, в митохондриях молочной железы мышей такие пара-
метры, как количество митохондриального белка, активность
150
Цитохромоксидазы, сукцинатоксидазы и ферментов окислитель-
ного фосфорилирования не изменялись на протяжении беремен-
ности, но резко возрастали при наступлении периода лактации
(Jones, Rosano, 1972).
Авторы считают, что при этом происходит образование новых
митохондрий.
В мышцах крыс при тренировке было обнаружено увеличе-
ние мелких митохондрий, которые являются предшественниками
крупных, более дифференцированных митохондрий (Edington,
McCaferli, 1973). Получены данные, свидетельствующие, что при
отсутствии функциональной нагрузки митохондрии мышц пре-
терпевают ряд изменений (Мах, 1972). Уже через сутки после обез-
движивания крыс дыхательный контроль в митохондриях мышц
уменьшается в 2 раза. К 6-му дню дыхательный контроль
полностью утрачивается.
Еще одним примером влияния функциональной нагрузки
органа на митохондрии может служить наследственная дистрофия
мышц. При этом заболевании скорость дыхания и окислительное
фосфорилирование в митохондриях мышц и печени снижены
(Jakobson et al., 1970; Howland, Challberg, 1973). Локализация
нарушений пока не ясна. Дыхательная цепь, по-видимому, не
нарушена, так как НАДН2 и янтарная кислота окисляются с вы-
сокой скоростью и количество переносчиков в митохондриях
не изменяется по сравнению с нормой. Авторы высказывают пред-
положение, или повреждены каким-то образом ферменты ЦТК и
механизмы сопряжения дыхательной цепи (Jakobson et al.,
1970), или может быть нарушена проницаемость мембран мито-
хондрий для ионов Н+ (Howland, Challberg, 1973).
Показано также, что нарушение функциональной активности
тканей при старении сопровождается изменениями митохондрий.
Количественные и качественные изменения в митохондриях мышц
домашней мухи при старении отмечают ряд авторов (Clark, Rock-
stein, 1964; Rockstein, Bhatnager, 1965; Bulos et al., 1972). При
старении у мухи Phormia regina изменяются структура и функ-
циональные возможности митохондрий летательных мышц, что
коррелирует с уменьшением способности к полету (Bulos et al.,
1972). Другие авторы считают, что ультраструктура митохонд-
рий летательных мышц старых мух не изменяется (исследованы
мухи 7—15 и 65—95 дневного возраста), однако функциональные
нарушения имеются: синтез белка в митохондриях снижен и ды-
хательный контроль при окислении а-глицерофосфата в 2 раза
ниже у старых мух. Скорость окисления экзогенного НАДН
митохондриями старых мух в 2 раза выше, чем у молодых, что мо-
жет быть связано с нарушением проницаемости мембран мито-
хондрий (Tribe, Ashnurst, 1972).
Имеются данные об изменении митохондрий и их биогенеза при
изменении среды инкубации или при воздействии гипоксии на
культуру дрожжей (см. Рудин, Уилки, 1970).
151
Несмотря на то, что как в онтогенезе, так и при различных воз-
действиях на организм наблюдаются значительные не только ко-
личественные, но и качественные изменения митохондрий, нам
представляется возможным попытаться провести сопоставление
между потреблением тканями (или организмом) кислорода и со-
держанием (концентрацией) в них митохондрий или митохондри-
ального белка.
Имеется большое число работ, в которых показано, что в те-
чение развития и роста различных животных происходит увели-
чение скорости дыхания (см. Needham, 1931; Bracket, 1950; Tuit,
1953; Boell, 1955; Коржуев и др., 1960; Браше, 1961; Lovtrup,
1963; Никольская, 1965; Gustafson, 1965; Радзинская, Николь-
ская, 1972; Зотин, 1974 и др.). Характер изменения скорости
дыхания и стадии, на которых происходит подъем дыхания, у раз-
личных животных различны. Так, у морского ежа увеличение скоро-
сти дыхания наблюдается на стадии ранней бластулы (Gustafson,
1965), у амфибий и некоторых рыб — начиная со стадии средней
бластулы — гаструлы и особенно резко на стадии вылупления
(Фаустов и др., 1968), у насекомых (в частности, у тутового
шелкопряда) — неуклонно увеличивается, начиная с первых
часов после откладки яиц (Радзинская, Никольская, 1972).
Известно также, что скорость дыхания связана с весом жи-
вотного. В общем виде зависимость между скоростью дыхания и
весом выражается уравнением вида (2.6).
Во многих работах, связанных с изучением энергетического
обмена, кроме скорости дыхания определяется интенсивность
дыхания — количество кислорода, потребляемого в единицу вре-
мени, отнесенное к единице веса. На основании большого факти-
ческого материала в настоящее время можно считать установлен-
ным, что в течение зародышевого развития, роста и при старении
животных интенсивность дыхания снижается. Кроме того, установ-
лено, что в это же время происходит уменьшение интенсивности
дыхания не только всего организма, но и отдельных органов и
тканей (Нагорный и др., 1963; Romanolf, 1967; Зотин, 1974).
Казалось весьма перспективным связать изменение скорости
или интенсивности дыхания в течение зародышевого развития,
роста и старения с изменением митохондрий — их структуры,
функционального состояния и количества.
Попытки установить зависимость между потреблением кисло-
рода и количеством митохондрий в тканях предпринимались
давно.
Наиболее четко идея зависимости интенсивности дыхания от
числа митохондрий была выражена Смитом (Smith, 1956). Автор
показал, что потребление кислорода тканями прямо пропорцио-
нально числу митохондрий в них. У животных разного веса была
показана зависимость между интенсивностью дыхания тканей
печени и числом митохондрий. Таким образом, согласно Смиту,
зависимость между дыханием и весом тела (2.8) у животных раз-
152
ного размера определяется количеством митохондрий в тканях.
Имеются работы, в которых удается проследить косвенную
связь между содержанием митохондрий и интенсивностью дыха-
ния тканей. Это работы, в которых авторы проводили определе-
ние содержания цитохромов или активности некоторых ферментов
в тканях и интенсивностью дыхания. Показано, что коэффициенты
в уравнении, выражающем зависимость между весом органов и
содержанием в них цитохрома С или активности некоторых окси-
даз, близки по значению для коэффициентов зависимости дыхания
от веса животного (Rosenthal, Drabkin, 1943; Lin et al., 1959).
Подобные данные были получены также в отношении зависимо-
сти активности цитохромоксидазы в печени у животных разного
веса (Kunkel, Campbell, 1952).
Перечисленные работы касаются связи между потреблением ки-
слорода тканями взрослых животных и содержанием митохонд-
рий или митохондриальных ферментов в них. Работ, показываю-
щих подобную зависимость в течение развития и роста, очень
мало и они довольно разноречивы. Поэтому авторы связывали
изменение числа митохондрий или митохондриальных ферментов
как с изменением скорости, так и с изменением интенсивности
дыхания в течение развития и роста.
По данным Густафсона (Gustafson, 1965), при развитии мор-
ского ежа наблюдается увеличение числа митохондрий, начиная
со стадии ранней гаструлы, что совпадает с увеличением в этот
момент скорости дыхания. Другие исследователи не подтвердили
этих наблюдений. Было показано, что число митохондрий на ста-
дии ранней гаструлы действительно увеличивается, но потом
происходит снижение их числа, несмотря на продолжающееся
увеличение скорости дыхания (Shaver, 1956).
Имеются работы по определению цитохромоксидазной актив-
ности в тканях в течение развития. Однако полученные резуль-
таты также были весьма противоречивы. Так, по данным одних
авторов цитохромоксидазная активность возрастает в течение за-
родышевого развития амфибий (Boell, 1955; Boell, Weber, 1955;
Boell et al., 1971) и рыб (Hishida, Nakano, 1954; Донцова,1965).
Другие авторы указывают на то, что она не изменяется (Spiegel-
man, Steinbach, 1945; Petrucci, 1957; Lang, Grant, 1961; Браше,
1961).
В последние годы в этот вопрос была внесена некоторая ясность.
Было показано (Абрамова, Васильева, 1973), что имеющиеся в ли-
тературе данные, указывающие на увеличение цитохромоксидаз-
ной активности в митохондриях и гомогенатах тканей в течение
развития рыб и амфибий (Boell, Weber, 1955; Estes, 1960; Дон-
цова, 1965), являются результатом того, что в них не было учтено
изменение проницаемости митохондрий для экзогенного цитохрома
С в течение развития. В недавно вышедшей работе было показано
(Kistler, Weber, 1974), что активность цитохромоксидазы в мито-
хондриях в течение развития амфибий не изменяется и ранее полу-
153
ченные одним из авторов данные, говорящие об увеличении цито-
хромоксидазной активности митохондрий (Weber, Boell, 1962),
являются результатом загрязнения митохондриального материала
желточными гранулами.
В литературе имеются указания на то, что масса митохондри-
ального белка в течение развития зародышей остается постоянной
(Абрамова, Васильева, 1973), несмотря на значительное увеличе-
ние массы зародыша, однако и интенсивность их дыхания в этот
период не претерпевает существенных изменений (Зотин, 1974).
Соответствие уровня интенсивности дыхания и концентрации
митохондрий было установлено в ооцитах вьюна (Озернюк, 1972
и стр. 155 наст. сб). Увеличение интенсивности дыхания и соот-
ветственно увеличение массы митохондрий было продемонстри-
ровано при регенерации у аксолотля (Владимирова, стр. 165).
Связь изменения интенсивности дыхания и числа митохондрий
с возрастом можно наблюдать также в опытах на крысах. Было
показано, что количество митохондрий мозга крысы возрастает
с 3-го до 33-го дня постэмбрионального развития (Samson et al.,
1960), что коррелирует с увеличением интенсивности дыхания
ткани мозга в этот период. Активность сукцинатоксидазы и
лактатдегидрогеназы почек и скелетных мышц крыс падает с воз-
растом (Barrows et al., 1960; Barrows, 1966; Schmukler, Barrows,
1966), что согласуется co снижением интенсивности дыхания в
тканях этих органов (Нагорный и др., 1963).
Удается проследить определенную связь между снижением
интенсивности дыхания животных при старении и числом мито-
хондрий в тканях. Так, уменьшение концентрации митохондрий
ряд авторов наблюдали в мышцах мух при старении (Clark et
al., 1964; Rockstein, Bhatnager, 1965). Снижение интенсивности
дыхания с возрастом у человека сопровождается четкими количе-
ственными и качественными изменениями митохондрий. В мио-
карде (Левкова, Трунов, 1970) с возрастом число митохондрий
в клетке уменьшается, увеличиваются их размеры, уменьшается
также количество крист. Происходит набухание митохондрий,
просветление матрикса, деформация крист и разрежение про-
странства между ними. Увеличение объема митохондрий в стар-
ческом возрасте автор рассматривает как компенсаторную реак-
цию на уменьшение их количества. Количество митохондрий
в миокарде крыс раннего возраста при расчете на единицу пло-
щади также уменьшается. При электронномикроскопическом под-
счете числа митохондрий на срезах печени получены подобные
данные (Tauchi et al., 1964; Гришко, Левкова, 1966; Левкова,
Трунов, 1970).
Уменьшение числа митохондрий в печени с возрастом было
показано также у людей (Tauchi, Sato, 1968). Так, в срезе печени
размером 5000 мк2 число митохондрий составляло: у людей до
49 лет - 1893, 50-59 лет - 1829, 60-64 лет - 1434, 65-69 лет-.
1223 и свыше 70 лет — 1200.
154
Таким образом, усиленное образование митохондрий в оогёнё-
зе сменяется, по-видимому, более или менее стабильным уровнем
митохондрий в эмбриогенезе, а в ходе органогенеза, роста и диффе-
ренциации органов происходит усложнение организации мито-
хондрий, их дифференцировка (возможно, в соответствии со сте-
пенью дифференцировки тканей к моменту рождения у млекопи-
тающих или выклеву у рыб и амфибий). Далее, по-видимому,
устанавливается какой-то баланс между образованием и разру-
шением митохондрий. Функционирует в каждый данный момент,
по сути, определенная, стационарная масса митохондрий.
Уровень концентрации митохондрий в тканях, по предполо-
жению Зотина (1974), определяет уровень основного обмена.
Удается проследить определенную зависимость между интенсив-
ностью дыхания животных и отдельных тканей и концентрацией
митохондрий. Снижение интенсивности дыхания в течение онто-
генеза, по-видимому, связано со снижением концентрации мито-
хондрий в тканях.
Механизмы, управляющие делением митохондрий, их биоге-
незом, в настоящее время не ясны, и только активная разработка
этих вопросов, накопление и сопоставление экспериментальных
данных поможет разобраться в этом сложном вопросе.
РОЛЬ МИТОХОНДРИЙ
В РЕГУЛЯЦИИ ДЫХАНИЯ ВО ВРЕМЯ ООГЕНЕЗА
Н, Д, Озернюк
В процессе оогенеза происходит существенное изменение мета-
болизма и ультраструктуры ооцитов направленное главным обра-
зом на накопление запасных питательных веществ и обеспечение
энергетики последующего зародышевого развития. В связи с этим
в настоящее время тщательно изучены ультраструктурные особен-
ности накопления ооцитами запасных питательных веществ (Кага-
saki, Komoda, 1958; Ward, 1962; Айзенштадт, 1965; Srivastawa,
1965; Wischnitzer, 1967). Показана высокая активность энергети-
ческого обмена ооцитов (Мещерская, 1935; Платова, 1946, 1948;
Gonse, 1955, 1957; Petrucci, 1960). Детально исследован механизм
накопления гликогена в ооцитах (Юровицкий и др., 1972; Юро-
вицкий, 1973). Подробно исследованы вопросы, связанные с из-
менением интенсивности дыхания, гликолиза и количества мито-
хондрий в процессе оогенеза (Озернюк, 1970; 1972; 1974; Озернюк
и др., 1971; Озернюк, Пальмбах, 1975). Все это позволяет в нас-
тоящее время обсудить более детально роль изменения ультра-
155
структуры и количества митохондрий в энергетическом метабо-
лизме ооцитов, а также такую общебиологическую проблему,
как механизм воспроизведения митохондрий.
Дыхание ооцитов
Манометрическим методом было измерено дыхание ооцитов
вьюна стадии малого роста, вакуолизации цитоплазмы, начала
вителлогенеза, окончания большого роста и зрелых яиц (Озер-
нюк, 1970). Потребление кислорода ооцитами значительно уве-
личивается в течение роста ооцитов. Ооциты стадии малого роста
потребляют 0,07 мкл/час/100 шт., а в закончивших рост ооцитах
потребление кислорода составляет 4,93 мкл/час/100 шт. Этот ре-
зультат не удивителен, так как усиление дыхания происходит
параллельно росту ооцитов. Принципиально иная зависимость
получается при пересчете дыхания на единицу массы ооцита,
в данном случае на мг общего белка ооцита (интенсивность дыха-
ния) (рис. 43). На стадии малого роста интенсивность дыхания
равна 1,00 мкл/час/мг общего белка ооцита. К стадии вакуоли-
зации цитоплазмы (начало большого роста) эта величина возра-
стает до 2,21 мкл/час/мг белка, а к началу вителлогенеза снижается
до 1,12 мкл/час/мг белка и незначительно падает на последующих
стадиях оогенеза. В расчете на мг общего азота ооцитов интен-
сивность дыхания меняется сходным образом.
При рассмотрении возможного механизма изменения ин-
следует учитывать два теорети-
чески возможных способа регу-
ляции дыхания. Во-первых, оно
может регулироваться изосте-
рическими или аллостерически-
ми механизмами, включающими
изменения активности дыха-
тельных энзимов, изменение
количества или доступности
субстратов или кофакторов ре-
акций, конформационные изме-
нения молекул фермента, выз-
ванных лигандом (Скулачев,
1969). Одним из таких регуля-
торов дыхания является «ды-
хательный контроль» — регуля-
ция скорости потребления кис-
лорода соотношением АДФ -f-Ф
неорг./АТФ.
Второй способ увеличения
интенсивности дыхания связан
с ростом количества митохон-
дрий.
мн л / час/мг белна
тенсивности дыхания в оогенезе
Рис. 43. Изменение интенсивности
дыхания (1) и концентрации мито-
хондриального белка (2) в процессе
оогенеза вьюна
156
Концентрация митохондриального белка в оОцитах
Была проведена проверка второго способа регуляции дыхания
в ооцитах. Для этой цели было изучено изменение концентрации
митохондрий в ооцитах разных стадий оогенеза. Измеряли кон-
центрацию белка митохондрий, выделенных из ооцитов разных
стадий, а также подсчитывали число митохондрий под электрон-
ным микроскопом.
Количество митохондриального белка в расчете на ооцит,
так же как и потребление кислорода ооцитами, существенно уве-
личивается в процессе оогенеза. Такое совпадение является вполне
закономерным, учитывая тот факт, что эти процессы протекают
на фоне интенсивного роста ооцитов. Концентрация митохондри-
ального белка (количество белка митохондрий/количество общего
белка ооцита), так же как и интенсивность дыхания, значительно
увеличивается на ранних стадиях оогенеза и снижается на более
поздних стадиях (см. рис. 43). Так, концентрация белка мито-
хондрий в ооцитах малого роста равна 0,061 мкг митохондриаль-
ного белка/мкг общего белка ооцита. К стадии вакуолизации
цитоплазмы она возрастает до 0,156, к стадии начала вителлоге-
неза снижается до 0,134 мкг белка митохондрий/мкг общего белка
и незначительно падает к окончанию большого роста ооцитов.
Таким образом, интенсивность дыхания и концентрация мито-
хондриального белка в ооцитах изменяется сходным образом.
Следовательно, можно сделать заключение, что интенсивность
дыхания во время оогенеза определяется концентрацией мито-
хондрий в ооцитах.
Синтез митохондриальных белков в ооцитах
Увеличение концентрации митохондрий в ооцитах ранних
стадий может быть связано с усилением синтеза митохондриаль-
ных белков, что приводит к росту мембран митохондрий и их по-
следующему делению. При этом нужно учитывать тот факт, что
формирование структуры митохондрий обеспечивается цитоплаз-
матической и митохондриальной белоксинтезирующими системами
(Рудин, Уилки, 1970). Для проверки этого предположения была
измерена интенсивность включения меченой аминокислоты в белки
митохондрий ооцитов стадии конца малого — начала большого
роста, когда концентрация митохондриального белка максималь-
ная и включение в митохондриальные белки ооцитов начала малого
роста, а также начала вителлогенеза, когда концентрация белка
митохондрий минимальная. Учитывая двойственный контроль над
биогенезом митохондрий как со стороны генома ядра, так и генома
самой митохондрии, интенсивность включения меченого предшест-
венника в митохондриальные белки измерялась in vivo и in vitro.
В табл. 5 представлена интенсивность включения С14-валина
in vivo в белки митохондрий ооцитов стадии начала малого роста,
157
Таблица б
Включение Си-валина (10 мккюри 1мл) in vivo в белки митохондрий ооцитов
(имп/мин/ме белка)
Номер опыта Начало малого роста Конец малого — начало большого роста Конец большого роста Номер опыта Начало малого роста Конец малого — начало большого роста Конец большого роста
1-й 1342 3 263 689 5-й 2289 458
2-й 1760 5 426 437 6-й — 3671 1597
3-й 3416 11773 1266 7-й — 8059 —
4-й — 10 044 999 С реднее 2173 6359 908
окончания малого — начала большого роста и стадии окончания
большого роста ооцитов.
Максимальная интенсивность включения меченого предшест-
венника наблюдается в митохондрии ооцитов окончания малого—
начала большого роста, когда концентрация митохондриального
белка наиболее высокая. Включение С14-валина в митохондриаль-
ные белки ооцитов более ранних стадий, а также более поздних
стадий значительно ниже, чем на стадии конца малого — начала
большого роста ооцитов. Из этих данных следует, что увеличение
концентрации митохондриального белка на ранних стадиях оогене-
за связано с усилением интенсивности синтеза митохондриальных
белков. Учитывая тот факт, что эти опыты проводились in vivo,
можно заключить, что речь идет об усилении синтеза митохон-
дриальных белков как на цитоплазматических, так и на мито-
хондриальных рибосомах. Последняя возможность, а именно
синтез белка самими митохондриями, была проверена в опытах
in vitro.
Было измерено включение С14-валина в белки изолированных
митохондрий ооцитов стадии малого — начала большого роста,
Таблица 6
Включение Сп-валина (1 мккюри/мл) in vitro в белки изолированных
митохондрий ооцитов (имп/мин/ме белка)
Номер опыта Стадия малого — начала большого роста Стадия окончания большого роста Номер опыта Стадия малого — начала большого роста Стадия окончания большого роста
1-й 1032 1615 5-й 1005 996
2-й 810 806 6-й 951 —
3-й 597 504 С реднее 1012 1121
4-й 1680 1685
158
когда включение меченого предшественника в белки митохонд-
рий in vivo максимальное, и включение в митохондриальные белки
ооцитов конца большого роста, когда включение предшественника
in vivo минимальное. Из табл. 6 следует, что интенсивность вклю-
чения С14-валина в белки митохондрий этих стадий практически
не отличается. Эти результаты можно объяснить или отсутствием
усиления синтеза белка в изолированных митохондриях ооцитов
малого—начала большого роста, или исчезновением факторов,
вызывающих такое усиление, при выделении митохондрий.
Рост митохондриальных мембран
во время оогенеза
Очевидно, что концентрация митохондриального белка в ооци-
тах может увеличиваться или за счет возрастания концентрации
митохондрий (число митохондрий в единице объема ооцита), или
за счет роста размеров ранее существующего количества мито-
хондрий. Для того чтобы выяснить, какая из этих возможностей
реализуется во время оогенеза, было изучено изменение числа,
распределения и ультраструктуры митохондрий в ооцитах раз-
ных стадий оогенеза.
Измерение объема митохондрий в ооцитах различных стадий
показало, что этот параметр возрастает во время оогенеза (табл. 7),
и, следовательно, существует вклад роста размеров митохондрий
в усиление интенсивности дыхания на ранних стадиях оогенеза.
Рост митохондриальных мембран в ооцитах имеет ряд особен-
ностей. Так, в ооцитах определенных стадий оогенеза митохонд-
рии находятся в тесном контакте с гранулярным эндоплазмати-
Таблица 7
Количество и размеры митохондрий, в ооцитах вьюна
Показатель Стадии оогенеза
начало конец вакуолизация начало малого роста малого роста цитоплазмы вителлогенеза
Диаметр ооцитов, мк 75 150 250 400 Число митохондрий в 0,05XЮб 0,6x10е 4,1 ХЮ6 12,9X 10е ооците, шт. Концентрация митохонд- о,26 0,36 0,50 0,39 рий в мк3, шт. Объем митохондрий, мк3 0,05±0,006 0,10±0,003 0,11±0,003 0,18±0,006 Протяженность крист 16,2±2,7 31,7^2,1 79,0^4,3 83,4±6,4 в 1 митохондрии (безраз- мерные ед.) Протяженность крист 324,2 317,3 718,7 463,6 в 1 мк3 объема митохонд- рии
159
ческим ретикулумом. Речь идет о таких ситуациях, когда мито-
хондрии на 25—100% своего периметра находятся в контакте
с гранулярным эндоплазматическим ретикулумом. На самых
ранних стадиях оогенеза (начало малого роста) эти контакты
отсутствуют и появляются только к концу малого роста. На этой
стадии 14% митохондрий находятся в контакте с гранулярным
эндоплазматическим ретикулумом. К стадии вакуолизации цито-
плазмы эта цифра возрастает до 17%, а к началу вителлогенеза —
до 19%. На более поздних стадиях оогенеза и во время зароды-
шевого развития (от стадии 2 бластомеров до начала гаструляции)
контакты митохондрий с мембранами эндоплазматического ретику-
лума не обнаружены. Необходимо отметить, что на участках
эндоплазматического ретикулума, контактирующего с митохон-
дриями, количество рибосом в 3 раза больше по сравнению с дру-
гими участками мембраны ретикулума. Роль этих контактов
становится более ясной, если учесть, что значительная часть мито-
хондриальных белков синтезируется на цитоплазматических рибо-
сомах и переносится затем в митохондрии (Рудин, Уилки, 1970).
Прямой перенос белков микросомной фракции в митохондрии
in vitro был показан Каденбахом (Kadenbach, 1967). Близость
цитоплазматических рибосом, сидящих на мембранах эндоплазма-
тического ретикулума, к митохондриям в ооцитах необходима,
вероятно, для более эффективного транспорта в митохондрии бел-
ков, синтезированных на цитоплазматических рибосомах. Для
проверки этого предположения были проведены опыты по инги-
бированию синтеза белка в митохондриях ооцитов разных стадий
антибиотиком циклогексимидом. Последний, как известно, инги-
бирует синтез белка в цитоплазме на 80S рибосомах. Однако
в экспериментах in vivo этот ингибитор подавляет биосинтез
белка не только в цитоплазме, но и в митохондриях, что является
одним из доказательств контроля биогенеза митохондрий со сто-
роны цитоплазматических факторов. Опыты с влиянием цикло-
гексимида на синтез белка в митохондриях in vivo были проведены
на ооцитах двух стадий: на ооцитах начала малого роста, где кон-
Таблица 8
Влияние циклогексимида на включение С1*-валина (10 мккюри/мл) в белки
митохондрий ооцитов (имп/мин1мг белка)
Номер опыта Контроль (без ингиби- тора) Циклогек- симид (0.5 мкг|мл) Ингибиро- вание (в % от контроля) Номер опыта Контроль (без ингиби- тора) Циклогек- симид (0,5 мкг/мл) Ингибиро- вание (в % от контроля)
Начало малого роста Вакуолизация цитоплазмы ООЦИТОВ
1-й 1342 1139 14,5 1-й 5426 1413 74,0
2-й 1760 1760 0 2-й 2289 582 74,6
3-й 4058 2992 26,3 3-п 1326 257 80, Г)
160
такты митохондрий с эндоплазматическим ретикулумом отсут-
ствуют, и на ооцитах стадии вакуолизации цитоплазмы, где 17 %
митохондрий контактируют с эндоплазматическим ретикулумом.
Включение С14 валина в митохондриальные белки ооцитов стадии
вакуолизации цитоплазмы существенно ингибируется циклогек-
симидом (в среднем на 76% по сравнению с контролем), тогда как
на стадии начала малого роста ооцитов степень ингибирования
включения значительно меньше и составляет в среднем 14%
(табл. 8).
Таким образом, предположение о роли контактов между мито-
хондриями и эндоплазматическим ретикулумом в транспорте бел-
ков из цитоплазмы в митохондрии, по-видимому, справедливо.
Концентрация митохондрий в ооцитах
Число митохондрий в ооцитах разных стадий оогенеза значи-
тельно увеличивается и повторяет в общих чертах изменение ко-
личества митохондриального белка на этих стадиях оогенеза (см.
табл. 7). В начале малого роста в одном ооците содержится
0,05 X 106 митохондрий, а к началу вителлогенеза их число
увеличивается до 12,9 X 10е. Ни одна из изученных в этом отно-
шении клеток, даже более крупных, чем ооцит, не содержит такого
большого количества митохондрий (табл. 9).
Концентрация митохондрий в единице объема ооцита увели-
чивается'от стадии малого роста до стадии вакуолизации цито-
плазмы и снижается на более поздних этапах оогенеза (см. табл. 7),
т. е. концентрация митохондрий, митохондриального белка и ин-
тенсивность дыхания изменяются сходным образом. Из этих дан-
ных следует, что изменение интенсивности дыхания в процессе
оогенеза определяется концентрацией митохондрий в ооцитах.
Если сравнить концентрацию митохондрий в ооцитах стадии ва-
куолизации цитоплазмы и в других типах клеток различных орга-
низмов, оказывается, что в ооцитах этот показатель наиболее
высокий (см. табл. 9).
Многократное увеличение числа митохондрий во время ооге-
неза свидетельствует об их интенсивном воспроизведении в ооци-
тах. О механизме увеличения количества митохондрий в оогенезе
можно судить, изучая распределение митохондрий в радиальном
направлении от центра ооцита к краю (рис. 44). В ооцитах малого
роста" число митохондрий увеличивается от центра ооцита к его
периферии. При переходе к большому росту (стадия вакуолизации
цитоплазмы) характер распределения митохондрий существенно
меняется. Так, в околоядерной зоне ооцитов этой стадии наблю-
дается резкое увеличение числа митохондрий, которое снижается
в‘средней зоне ооцита и возрастает к его периферии. Если сравнить
характер распределения митохондрий в ооцитах малого роста и
вакуолизации цитоплазмы, то видно, что увеличение числа мито-
хондрий осуществляется, главным образом, за счет митохондрий
161
Таблица 9
Количество митохондрий в различных типах клеток
Тип клеток Число мито- хондрий в 1 клетке, шт. Число мито- хондрий в 1 мк2 (шт.) Автор
Средний лимфоцит крысы 6-8 0,03 Ток ин 1074
Клетки эпителия 12-перстной 100—200 — Тот же
кишки крысы Клетки эпителия ворсинок 100—200 »
12-перстной кишки крысы Клетки печени крысы 1200-1500 0,10
» » » 1500—3000 0,23 Loud, 1962
» » » 2550 0,19 Allard et. al., 1952
Клетки гепатомы крысы 1390 0,10 Тот же
Клетки регенерир. печени 1800 0,13 »
крысы (1 день) Клетки регенерир. печени 2500 0,15 »
крысы (20 день) Амеба 500 000 0,03 Ленинджер, 1966
Яйцеклетка мор. ежа 14 000 0,06 Гот же
» » » 200 000 0,32 Piko et al., 1967
Ооциты вьюна, стадии: начало малого роста 50 000 0 26 Озернюк, ^альм®ах’
конец малого роста 600000 0,39 Те же
вакуолизация цитоплазмы 4100 000 0,55 »
начало вителлогенеза 12 900 000 0,40
околоядерной зоны. Митохондрии околоядерной и перифериче-
ской части ооцита существенно отличаются по некоторым ультра-
структурным характеристикам. Так, объем митохондрий и про-
тяженность их крист в околоядерной зоне значительно ниже, чем
в периферической части ооцита (рис. 45). Таким образом, попу-
ляцию митохондрий в ооцитах стадии вакуолизации цитоплазмы
можно разделить на две части: крупные периферические части
ооцита, содержащие много крист, и мелкие — околоядерной
зоны, за счет которых и происходит, в основном, увеличение числа
митохондрий во время оогенеза. Можно предположить, что глав-
ным образом в околоядерной зоне ооцита происходит воспроизве-
дение митохондрий, так как увеличение их числа происходит
в этой части ооцита и митохондрии этой зоны имеют мелкие раз-
меры, возможно, вследствие деления,
162
Рис. 44. Распределение числа мито-
хондрий в ооцитах вьюна разных
стадий
1 — начало малого роста; 2 — конец
малого роста; 3 — начало вакуолизации
цитоплазмы; 4 — конец вакуолизации
цитоплазмы; 5 — начало вителлогенеза
Рис. 45. Распределение числа мито-
хондрий (1), их объема (2) и про-
тяженности крист (3) в ооцитах
вьюна на стадии вакуолизации ци-
топлазмы
Механизм воспроизведения митохондрий
Наиболее популярной точкой зрения на механизм увеличения
числа митохондрий в клетке (или поддержания на постоянном
уровне) является предположение о делении митохондрий. Эта
гипотеза получила серьезное обоснование в экспериментах Лака
(Luck, 1963) и других исследователей (Droz, Bergeron, 1965;
Parsons, Rustad, 1968), а также после обнаружения в митохонд-
риях собственной ДНК (Nass, Nass, 1963). Основным доказатель-
ством деления митохондрий является обнаружение ДНК в мито-
хондриях всех типов клеток, тканей органов и организмов и оди-
наковая кодирующая способность митохондриальной ДНК во
всех перечисленных объектах (Нейфах, 1974а). Очевидно, что та-
кая ситуация возможна только при образовании новых мито-
хондрий из ранее существующих и попадании реплицировавшейся
митохондриальной ДНК при делении митохондрии в каждую до-
чернюю органеллу. Задолго до открытия митохондриальной ДНК
и экспериментов Лака на нейроспоре, авторы многочисленных
работ приходили к выводу о делении митохондрий на основании
обнаружения в клетке гантелевидных органелл (см. обзоры Novi-
koff, 1961; Schatz, 1970), являющихся, как принято считать, ста-
диями деления митохондрий.
Гантелевидные митохондрии обнаружены и в ооцитах разных
стадий оогенеза. В ооцитах вьюна обнаружено 8% таких мито-
хондрий. Столь небольшое количество делящихся митохондрий
может свидетельствовать о том, что акт деления происходит до-
вольно быстро.
Неясным остается вопрос о механизмах, запускающих деление
митохондрий. Логично было бы предположить, что митохондрии
делятся, достигнув максимального размера. Однако размеры ми-
тохондрий, по-видимому, не являются фактором, запускающим де-
ление митохондрий, так как на электронно-микроскопических фо-
тографиях обнаруживается много гигантских неделящихся мито-
хондрий, в то время как гантелевидные митохондрии, являющие-
ся стадиями деления, имеют средние размеры. Роль инициатора
деления митохондрий приписывалась митохондриальной ДНК
(Hawley, Wagner, 1967). Известно, что в быстроделящихся клет-
ках содержание митохондриальной ДНК в расчете на митохон-
дриальный белок в несколько раз выше по сравнению с покоя-
щимися клетками, например клетками печени. Этот факт показан
для митохондрий зародышевых, регенерирующих и опухолевых
тканей (Nass, 1969). Однако связь между репликацией митохон-
дриальной ДНК и делением митохондрий, вероятно, отсутствует,
т. к. во время зародышевого развития происходит многократное
усиление биосинтеза митохондриальной ДНК (Михайлов, Гаузе,
1974), а количество митохондриального белка в этот период не
меняется (Chase, Dawid, 1972; Абрамова, Васильева, 1973; Kist-
ler, Weber, 1974).
164
Отмеченная в данной работе локализация митохондрий вблизи
ядра, а также увеличение числа этих органелл в клетке за счет
их деления в околоядерной зоне может служить косвенным аргу-
ментом в пользу участия ядра в регуляции концентрации мито-
хондрий в клетке.
Сокращение митохондриальной мембраны при формировании
перетяжки может быть вызвано локальным действием АТФ на со-
кратительные белки митохондрий (Озернюк, Пальмбах, 1975).
ЭНЕРГЕТИКА ПРОЦЕССА РЕГЕНЕРАЦИИ
И. Г. Владимирова
Усиление интенсивности дыхания регенерирующих тканей об-
наружено более тридцати лет назад. Вольский (Wolsky, 1941), ис-
следуя регенерацию хвоста аксолотлей, показал, что на стадии
бластемы дыхание регенератов примерно в 1,7 раза выше, чем
в контрольных тканях. Рывкина (1942), изучая регенерацию ко-
нечности аксолотлей, обнаружила подъем дыхания не только на
стадии бластемы, но и в первую половину стадии дифференцировки.
Наивысший уровень дыхания отмечен в конце стадии внутрен-
ней дифференцировки, когда потребление кислорода увеличивает-
ся в 2 раза.
По данным других авторов (Фаустов, Карлсон, 1967; Faustov,
1969), интенсивность дыхания подэпителиальной части регенерата
конечности на стадии дифференцировки превышает контрольный
уровень в 4 раза. Снижение интенсивности дыхания до контроль-
ных значений происходит в конце стадии дифференцировки (Рыв-
кина, 1942; Фаустов, Карлсон, 1967; Faustov, 1969).
Дальнейшие исследования (Владимирова, 1974, 1975) показа-
ли, что в регенерирующих тканях хвоста аксолотлей интенсив-
ность дыхания возрастает в первые часы после операции. У поло-
возрелых аксолотлей через 16 часов после операции потребление
кислорода тканями, прилегающими к раневой поверхности, уве-
личивается в 1,5 раза. К концу первых суток интенсивность дыха-
ния уменьшается до контрольного уровня (рис. 46). На стадии
разрушения и дедифференцировки вновь наблюдается подъем ды-
хания. В конце стадии разрушения и де дифференцировки интен-
сивность дыхания тканей культи хвоста выше контрольного уров-
ня у половозрелых аксолотлей на 80%. Дальнейшее усиление ды-
хания наблюдается в регенератах хвоста в начале стадии бластемы.
В конце стадии бластемы, в начале дифференцировки хряща пот-
ребление кислорода регенератами хвоста половозрелых аксолотлей
165
уменьшается, но тем не менее превышает контрольный уровень
на 40—50%. В начале дифференцировки мышц отмечен новый
подъем дыхания. Интенсивность дыхания в это время в расчете
на мг сырого веса имеет примерно тот же уровень, что и на стадии
бластемы. При расчете на мг сухого веса потребление кислорода
на стадии дифференцировки (1,18 и 1,63 мкл/час/мг) несколько
выше, чем на стадии бластемы (1,00 и 1,47 мкл/час/мг). Уменьше-
ние интенсивности дыхания до контрольного уровня происходит
в конце стадии дифференцировки (рис. 46).
У неполовозрелых аксолотлей также наблюдается некоторое
усиление интенсивности дыхания тканей в первые сутки после
операции. Наиболее высокий уровень дыхания отмечен на 5-е
сутки регенерации на стадии бластемы. В течение стадии диффе-
ренцировки интенсивность дыхания постепенно уменьшается
(рис. 47).
Исследования разных возрастных групп аксолотлей показали,
что интенсивность дыхания регенератов хвоста зависит от началь-
ного уровня обмена и в целом уменьшается с возрастом аксолот-
лей (Владимирова, Никольская, 1971; Владимирова, 1975). Наи-
более высокая интенсивность дыхания регенерирующих тканей —
0,280 мкл/час/мг сырого веса — отмечена у одной из партий трех-
недельных аксолотлей, наиболее низкая —0,073 мкл/час/мг сырого
веса — у одиннадцатимесячных аксолотлей. По сравнению с на-
чальным уровнем потребление кислорода увеличивается при реге-
нерации хвоста у разных партий неполовозрелых аксолотлей в 1,4—
2,1 раза, у половозрелых аксолотлей — в 2,4—3,2 раза. Вместе
с тем абсолютная величина подъема дыхания регенерирующих тка-
ней одинакова и составляет 0,05—0,07 мкл/час/мг сырого веса у
всех исследованных возрастных групп животных (Владимирова,
Никольская, 1971; Владимирова, 1975).
Для того чтобы выяснить, является ли усиление интенсивно-
сти дыхания признаком, характерным только для регенерации,
было измерено дыхание тканей у животных, утративших способ-
ность к регенерации после местного рентгеновского облучения тка-
ней хвоста (Владимирова, 1975). Оказалось, что у облученных
аксолотлей интенсивность дыхания увеличивается в тканях куль-
ти хвоста, прилегающих к ампутационной поверхности, в первые
часы после операции и на стадии разрушения, через 5—6 дней.
В последующие две недели заживления раны потребление кисло-
рода тканями хвоста облученных аксолотлей не изменяется (Вла-
димирова, 1975). Полученные результаты приводят к выводу, что
усиление дыхания тканей культи хвоста происходит и в процессе
регенерации, и в процессе гладкого заживления ампутационной
раны. Увеличение интенсивности дыхания во время роста и диф-
ференцировки регенерата характерно только для регенерирую-
щих тканей.
За счет каких же механизмов происходит увеличение интенсив-
ности дыхания в процессе регенерации?
166
Данные, имеющиеся к настоящему времени в литературе, не-
достаточны, чтобы ответить на этот вопрос. Поэтому в нашей рабо-
те поставлены специальные опыты по измерению потенциальной
мощности ферментов дыхания и концентрации митохондриального
белка.
Для выяснения потенциальной мощности дыхательной системы
было определено максимальное дыхание срезов тканей в присут-
ствии субстрата дыхания (сукцината натрия) и разобщителя (ди-
нитрофенола).
Рис. 46. Интенсивность дыхания (I),
анаэробного (2) и аэробного (3)
гликолиза в регенерирующих тканях
хвоста половозрелых аксолотлей
Рис. 47. Интенсивность дыхания
(1), анаэробного гликолиза (2) и
концентрация АТФ (3) в регене-
рирующих тканях хвоста неполово-
зрелых аксолотлей (в % к контроль-
ному уровню)
Вверху: 1-й квадрат—дедифференцировка
и эпителизация
167
Максимальное дыхание регенератов хвоста неполовозрелых
аксолотлей на стадиях бластемы и дифференцировки на 50% выше
максимального дыхания контрольных тканей. На стадии бласте-
мы максимальное дыхание регенератов имеет примерно тот же
уровень, что и дыхание на эндогенных субстратах. На стадии диф-
ференцировки стимулирующее действие сукцината натрия и ди-
нитрофенола выражено сильнее, чем на стадии бластемы: макси-
мальное дыхание регенератов значительно выше дыхания на эндо-
генных субстратах.
У половозрелых аксолотлей максимальное дыхание увеличи-
вается в процессе регенерации хвоста на 70—100%. Наивысшие
значения отмечены в регенератах на стадии бластемы и в начале
дифференцировки мышц, одновременно с пиком дыхания на эндо-
генных субстратах (рис. 48).
Соотношение между максимальным дыханием и дыханием на эн-
догенных субстратах у половозрелых аксолотлей, так же как у не-
половозрелых аксолотлей, изменяется в течение регенерации.
В начальные периоды подъема дыхания добавление сукцината нат-
рия и динитрофенола к срезам регенерирующих тканей половозре-
лых аксолотлей усиливает потребление кислорода на 50—70%.
На стадии бластемы и в начале дифференцировки мышц, когда
Рис. 48. Интенсивность дыхания на эндогенных субстратах (1) и макси-
мального дыхания (2) регенератов хвоста тринадцатимесячных аксолотлей
1к и 2к соответствующие показатели у контрольных животных.
Вверху: 1-й квадрат—дедифференцировка, 2-й квадрат—бластома
168
эндогенное дыхание регенератов достигает наивысшего уровня,
сукцинат натрия и динитрофенол увеличивают потребление кис-
лорода на 10—20%. Уменьшение стимулирующего эффекта не свя-
зано с недостаточным количеством субстрата, так как специальные
опыты показали, что применяемые концентрации сукцината нат-
рия являются оптимальными и для контрольных, и для регене-
рирующих тканей.
Полученные результаты свидетельствуют, что изменение макси-
мального дыхания регенератов в течение стадии бластемы происхо-
дит одинаково у неполовозрелых и половозрелых аксолотлей. Это
позволяет предположить, что механизм усиления интенсивности
дыхания регенератов хвоста на стадии бластемы у исследованных
возрастных групп аксолотлей одинаков.
Таким образом, в процессе регенерации происходит увеличе-
ние потенциальной мощности ферментов дыхания. Эффективность
использования потенциальной мощности ферментов изменяется
в течение регенерации. Наиболее высокая эффективность отмечена
у неполовозрелых аксолотлей на стадии бластемы, у половозрелых
аксолотлей — на стадии бластемы и в начале дифференцировки
мышц. На основании полученных данных можно сделать вывод,
что в регенерирующих тканях хвоста аксолотлей происходит уве-
личение концентрации и активности ферментов дыхания.
Усиление активности ферментов дыхания отмечено в исследова-
ниях ряда авторов. Чайльд (Child, 1948, 1951) показал, что повы-
шенная активность цитохромоксидазы присуща регенерирующим
тканям тритонов и планарий с момента операции до стадии наруж-
ной дифференцировки. По данным Нивелинской (Niwelinski, 1960,
1964), увеличение уровня цитохромоксидазы происходит на стадии
дифференцировки. На ранних стадиях регенерации высокая ак-
тивность цитохромоксидазы наблюдается только в регенерацион-
ном эпителии.
Помимо цитохромоксидазы в регенерирующих тканях амфибий
измерена активность сукцинат-, малат- и изоцитрат дегидрогена-
зы. В «прогрессивной фазе» регенерации (стадия бластемы и диф-
ференцировки) активность сукцинатдегидрогеназы в 1,5 раза пре-
вышает контрольный уровень. Активность малатдегидрогеназы
возрастает в меньшей степени, хотя гистохимические исследования
(Wolfe, Cohen, 1963; Schmidt, Weidman, 1964) выявили высокую
активность малат- и изоцитратдегидрогеназы. Сукцинатдегидро-
геназа имеет высокий уровень в регенерационном эпителии (Niwe-
linski, 1960, 1964; Wolfe, Cohen, 1964; Schmidt, Weidman, 1964;
Johnson, Singer, 1964). Клетки подэпителиальной части регенерата
на стадии бластемы, по данным Нивелинской (Niwelinski, 1960;
1964), обладают вполне отчетливой реакцией на сукцинатдегидро-
геназу, но другие исследователи (Schmidt, Weidman, 1964; Johnson,
Singer, 1964) отмечают, что активность фермента или незначитель-
на, или вообще отсутствует. К концу стадии бластемы происхо-
дит уменьшение активности сукцинатдегидрогеназы в регенера-
169
ционном эпителии. В клетках подэпйтелиальной части регенерата
активность дегидрогеназ повышается в процессе дифференциров-
ки. В раннем прохондральном сгущении обнаружена умеренная
активность сукцинатдегидрогеназы, более активными являются
малат- и изоцитратдегидрогеназы (Niwelinski, 1960, 1964; Wolfe,
Cohen, 1963; Schmidt, Weidman, 1964; Johnson,’ Singer, 1964).
Высокий уровень дегидрогеназ отмечен в дифференцирующихся
мышцах (Niwelinski, 1960, 1964; Wolfe, Cohen, 1963; Schmidt,
Weidman, 1964).
Изучение активности сукцинат де гидрогеназы в регенерирую-
щих тканях земляного червя Eisenia foetida показало, что на про-
тяжении регенерации происходит увеличение активности фермен-
та. Наиболее высокий уровень обнаружен в регенератах на стадии
дифференцировки (O’Brien, 1957). У планарий повышенная по
сравнению с контролем активность сукцинатдегидрогеназы наб-
людается в регенератах на стадии дифференцировки (Ортабаева,
1973).
Таким образом, к настоящему времени установлено, что высо-
кая активность ферментов дыхания характерна для регенерирую-
щих тканей амфибий и червей на стадиях бластемы и дифференци-
ровки. Некоторое противоречие вносят результаты гистохимичес-
кого определения сукцинатдегидрогеназы (Schmidt, Weidman,
1964; Johnson, Singer, 1964), которые свидетельствуют о том, что
на стадии бластемы активность фермента в подэпителиальной
части регенерата незначительна.
Благодаря электронно-микроскопическим исследованиям поя-
вилась возможность изучить тонкую структуру клеток регенери-
рующего органа. В регенерационном эпителии взрослых тритонов
изменения тонкой структуры клеток не обнаружено (Norman,
Schmidt, 1967). Существенные изменения отмечены в клетках под-
эпителиальной части регенерата, но митохондрии, по данным всех
исследователей (Нау, 1958, 1959; Salpeter, Singer, 1960; Norman,
Schmidt, 1967), изменяются меньше, чем другие органеллы. На ста-
дии бластемы митохондрии имеют небольшие размеры (диаметрО,3—
0,5 мк у неполовозрелых амфибий), немногочисленные и нерегуляр-
но расположенные кристы (Нау, 1958, 1959; Salpeter, Singer, 1960;
Norman, Schmidt, 1967). Характерной особенностью стадии бла-
стемы является тесная связь митохондрий и эндоплазматического
ретикулума (Нау, 1958; Salpeter, Singer, 1960). Особенно ярко вы-
ражена эта особенность на стадии ранней бластемы у неполовозре-
лых амфибий (Нау, 1958).
При дифференцировке хряща и мышц утрачивается койтакт
митохондрий и эндоплазматического ретикулума. Митохондрии
по-прежнему имеют немногочисленные кристы. Размеры митохон-
дрий существенно не изменяются (Norman, Schmidt, 1967).
В нашей работе проведено измерение количества митохондри-
ального белка в регенератах хвоста половозрелых аксолотлей.
Митохондрии выделяли в среде, содержащей 0,25М сахарозы,
170
0,05М триса, 0,025М КС1, pH 7,4. Ядра удаляли двукратным
центрифугированием при 1000 g в течение 10 минут. Митохондрии
осаждали центрифугированием при 20 000 g в течение 30 минут.
Результаты показали, что содержание митохондриального белка в
регенератах увеличивается с 32,9 мкг на стадии ранней бластемы до
44,2 мкг в начале дифференцировки хряща и до 234,2 мкг на реге-
нерат в конце стадии дифференцировки. Рост регенерата, следова-
тельно, сопровождается увеличением количества митохондрий.
Пересчет митохондриального белка на мг белка регенерата
показал, что на протяжении большей части процесса регенерации
в регенерирующих тканях концентрация митохондриального белка
выше, чем в контрольных тканях (рис. 49).
Эти данные в некоторой степени подтверждаются тем, что в ре-
генерирующих тканях в начале стадии дифференцировки отмечено
повышенное включение меченых аминокислот в белки митохондрий
(Теплиц, 1974). Кроме того, в процессе регенерации увеличивается
количество изозима лактатдегидрогеназы, связанного с мембрана-
ми митохондрий, что, по мнению авторов (Chan, Balek, 1973), сви-
детельствует об увеличении концентрации митохондрий.
Таким образом, в наших исследованиях показано, что во вре-
мя роста и дифференцировки регенерата хвоста половозрелых
аксолотлей происходит увеличение интенсивности дыхания на эн-
догенных субстратах, максимального дыхания и концентрации
митохондриального белка. Аналогичные изменения обнаружены
в ооцитах животных на стадии малого роста (Платова, 1948; Gon-
171
se, 1957; Озернюк, 1970, 1972). Электронно-микроскопические ис-
следования показали, что усиление процессов вызвано увеличени-
ем концентрации митохондрий (Озернюк, Пальмбах, 1975).
Изучение процесса регенерации планарий выявило увеличение
количества митохондрий в необластах на стадии бластемы и на ста-
дии дифференцировки (Sauzin, 1966, 1967; Ортабаева, 1973).
На основании имеющихся данных можно предположить, что
усиление интенсивности дыхания регенерирующих тканей обу-
словлено увеличением концентрации митохондрий.
Помимо увеличения концентрации митохондрий подъем дыха-
ния может происходить за счет эффективного использования ды-
хательной системы (Зотин, 1974). Известно, что аллостерическим
регулятором активности некоторых ключевых ферментов цикла
трикарбоновых кислот является АТФ и другие адениловые ну-
клеотиды (см. Гайцхоки, 1967;Скулачев,р1969). Адениловые нуклео-
тиды играют большую роль в регуляции скорости тока электронов
в дыхательной цепи: АТФ тормозит, АДФ ускоряет перенос элек-
тронов (Чанс, Хаджихара, 1962). Поэтому содержанию аденило-
вых нуклеотидов и их соотношению приписывают ведущую роль
в механизме изменения дыхания (см. Зотин, 1974).
При регенерации конечности и хвоста половозрелых аксолот-
лей на стадии дифференцировки выявлена обратная зависимость
между изменением содержания АТФ и дыхания (Фаустов, Карл-
сон, 1967; Faustov, 1969; Владимирова, 1975). Авторы высказали
предположение, что АТФ принимает участие в регуляции дыхания
регенерирующих тканей на стадии дифференцировки.
Известно, что на активность цикла трикарбоновых кислот мо-
гут влиять конкурентные процессы, такие, как гликолиз, гексо-
зомонофосфатный цикл. В связи с этим интересно проследить, как
изменяется интенсивность этих процессов во время регенерации.
Наиболее полные данные получены для процесса гликолиза.
У половозрелых аксолотлей кратковременный подъем анаэробного
гликолиза наблюдается в тканях культи хвоста, прилегающих
к раневой поверхности в первые часы после операции (Владимиро-
ва, 1975). Через 1—16 часов после операции анаэробная продукция
молочной кислоты примерно в 1,5 раза выше контрольного уровня
(рис. 46). К концу первых суток интенсивность гликолиза умень-
шается и на стадии разрушения и дедифференцировки существен-
но не отличается от контрольных значений. Увеличение продук-
ции молочной кислоты отмечено в регенератах хвоста половозре-
лых аксолотлей на стадии бластемы. В конце стадии бластемы,
в начале дифференцировки хряща достигается наивысший уровень,
по сравнению с контролем анаэробный гликолиз увеличивается
в 2 раза. В начале дифференцировки мышц интенсивность ана-
эробного гликолиза в регенератах хвоста уменьшается (рис. 46).
Усиление интенсивности анаэробного гликолиза па стадии
бластемы обнаружено также при исследовании регенератов конеч-
ности шести — девятимесячных аксолотлей (Рывкина. 1942, 1945).
172
Наиболее высокий уровень отмечен в регенератах на стадии кону-
са, когда в регенератах начинается дифференцировка хряща. Интен-
сивность анаэробного гликолиза на этой стадии регенерации в 2
раза выше, чем в контрольных тканях (Рывкина, 1942).
В регенерирующих тканях хвоста половозрелых аксолотлей
образование молочной кислоты наблюдается также в аэробных
условиях (Владимирова, 1975). Подъем аэробного гликолиза про-
исходит одновременно с подъемом анаэробного гликолиза. Наи-
более высокие значения аэробного гликолиза отмечены в тканях
культи через 2—16 часов после операции и в регенератах в конце
стадии бластемы и в начале дифференцировки хряща (см. рис. 46).
В конце стадии дифференцировки образование молочной кислоты
в аэробных условиях прекращается.
Об усилении аэробного гликолиза свидетельствует увеличение
содержания молочной кислоты в регенератах, обнаруженное в ра-
ботах Окунева (Okuneff, 1933) и Владимировой (1935). Содержа-
ние молочной кислоты увеличивается в регенератах на стадии поч-
ки и достигает наивысшего уровня на стадии конуса, т. е. одновре-
менно с подъемом аэробного гликолиза. По Окуневу (Okuneff,
1933), содержание молочной кислоты увеличивается в 2,2 раза, по
Владимировой (1935) — в 1,75 раза. В течение стадии дифферен-
цировки содержание молочной кислоты в регенератах снижается.
Изучение разных возрастных групп аксолотлей показывает
(Владимирова, 1975), что усиление аэробного гликолиза харак-
терно только для регенерирующих тканей половозрелых аксолот-
лей. У неполовозрелых аксолотлей образования молочной кисло-
ты в аэробных условиях не обнаружено. При регенерации хвоста
у неполовозрелых аксолотлей происходит увеличение интенсив-
ности анаэробного гликолиза (см. рис. 47). Наиболее высокий
уровень анаэробного гликолиза отмечен в регенератах хвоста че-
рез 5 дней после операции, на стадии бластемы. В течение стадии
дифференцировки интенсивность анаэробного гликолиза умень-
шается до контрольной величины.
Сопоставление изменения гликолиза и дыхания регенерирую-
щих тканей показывает, что у неполовозрелых аксолотлей интен-
сивность дыхания и гликолиза изменяется параллельно (рис. 47).
Такое же положение наблюдается и в течение большей части про-
цесса регенерации хвоста половозрелых аксолотлей. Лишь в кон-
це стадии бластемы и в начале стадии дифференцировки наблю-
дается обратная зависимость между изменением дыхания и глико-
лиза (см. рис. 46). На этих стадиях интенсивность гликолиза до-
стигает наивысших значений, а интенсивность дыхания падает.
Усиление гликолиза в регенерирующих тканях половозрелых
амфибий сопровождается подъемом активности лактатдегидроге-
назы. Увеличение уровня лактатдегидрогеназы отмечено в тканях
культи, прилегающих к раневой поверхности, к концу первых су-
ток после операции (Geczik, Wolsky, 1959). По-видимому, усиле-
ние активности фермента, так же как увеличение интенсивности
173
анаэробного и аэробного гликолиза, вызвано повреждением тка-
ней при операции. Известно (Браун и др., 1967), что повреждение
тканей вызывает наряду с увеличением дыхания увеличение про-
дукции молочной кислоты в анаэробных и аэробных условиях.
В «прогрессивной фазе» регенерации активность лактатдегидро-
геназы более чем в 1,5 раза выше контрольного уровня (Geczik,
Wolsky, 1959). Высокая активность фермента характерна и для
регенерационного эпителия, и для подэпителиальной части регене-
рата (Wolfe, Cohen, 1963; Johnson, Singer, 1964; Schmidt, Weidman,
1964). Однако активность альдолазы, другого фермента гликолиза,
в регенератах на стадии бластемы незначительна. Высокая актив-
ность альдолазы наблюдается в клетках дифференцирующегося
хряща (Schmidt, Weidman, 1964).
Низкая активность альдолазы дала возможность авторам
(Schmidt, Weidman, 1964; Schmidt, 1966,1968) сомневаться в реаль-
ности подъема гликолиза на этой стадии регенерации. По мнению
этих исследователей, гликолитическое окисление углеводов в ре-
генератах на стадии бластемы невелико и основную роль в энерге-
тическом обеспечении клеток играет окисление липидов и пенто-
зомонофосфатный цикл (Schmidt, 1966, 1968).
Глюкозо-6-фосфатдегидрогеназа является одним из ключевых
ферментов гексозомонофосфатного цикла. При регенерации конеч-
ности амфибий его повышенная активность отмечена в тканях куль-
ти на стадии разрушения и дедифференцировки, в регенератах на
стадии бластемы, в дифференцирующихся хряще, костях, мышцах
(Wolfe, Cohen, 1963; Schmidt, Weidman, 1964).
У червей Gineus sanguineux обнаружен подъем активности
глюкозо-6-фосфатдегидрогеназы в регенератах переднего конца
тела (Reutter, Sasse, 1970). Наиболее высокий уровень характерен
для регенератов на стадии бластемы и для закладок некоторых
органов на стадии дифференцировки. Авторы предполагают, что
в регенератах на стадии бластемы и дифференцировки происходит
активация гексозомонофосфатного цикла.
Таким образом, в регенератах на стадии бластемы наблюдается
увеличение интенсивности дыхания, гликолиза, пентозомонофосфат-
ного цикла. Одновременно с усилением этих процессов в регене-
рирующих тканях резко (в 4,7 раза у неполовозрелых аксолотлей
и в 6,4 раза у половозрелых аксолотлей) возрастает концентрация
АТФ (Владимирова, 1975). Вероятно в регенератах на стадии бла-
стемы включаются механизмы, направленные на повышение уров-
ня энергетического обмена. Подъем интенсивности дыхания в про-
цессе регенерации обусловлен в значительной степени увеличе-
нием концентрации митохондрий. Механизмы усиления гликолиза
в регенерирующих тканях пока не выяснены. Постепенное при-
ближение интенсивности энергетического обмена регенерирую-
щих тканей к контрольному уровню наблюдаются на стадии диф-
ференцировки у неполовозрелых аксолотлей, и в конце стадии
дифференцировки и на стадии роста у половозрелых аксолотлей.
VI
ДИССИПАТИВНЫЕ СТРУКТУРЫ
Теория диссипативных структур, развитая Пригожиным и его
сотрудниками (Prigogine, 1969, 1972; Nicolis, 1971; Prigogine et
aL, 1972a, b; Babloyantz, Nicolis, 1972; Goldbeter, Nicolis, 1972;
Глансдорф, Пригожин, 1973; Пригожин, Николис, 1973; Nicolis,
Auchmuty, 1974), получила в настоящее время широкое признание
(Tondeur, 1970; Martinez, 1972; Кариг, 1972; Волькенштейн, 1973;
Эйген, 1973а, b; Hanson, 1974; Романовский и др., 1975), хотя мно-
гое в этой теории остается неясным и недоказанным. Тем не менее
существование в системах далеких от равновесия диссипативных
структур (структур в широком понимании этого слова, т. е. не
только некоторых морфологически выраженных образований, но
и временных стабильных структур, колебательных процессов
и т. д.) сомнения не вызывает. Примеров подобных структур можно
привести очень много, как из области химии, так и биологии.
Сложнее обстоит дело с теоретическим обоснованием возможности
их возникновения в системе. Можно надеяться, что теорию дисси-
пативных структур удастся связать с представлениями, развиты-
ми в главе второй настоящей книги, касающимися функции свя-
занной диссипации. В этом случае станут ясными энергетические
источники поддержания и существования диссипативных струк-
тур. Во всяком случае можно будет рассматривать диссипативные
структуры в рамках глобальной теории нелинейных необратимых
процессов.
ДИССИПАТИВНЫЕ СТРУКТУРЫ В ФИЗИКЕ,
ХИМИИ И БИОЛОГИИ
И. Лампрехт
Обычно, когда говорят о структурах, представляют себе меха-
нические образования, такие, как кристалл, здание, лист и т. д.
Но в нашей жизни наряду с пространственными встречаются и вре-
менные структуры. К последним относятся все периодические или
квази-периодические процессы.
© I. Lamprecht, 1976 г.
175
Что имеют в виду, Когда говорят о диссипатиВйых структурах?
Известно, что равновесные структуры остаются неизменными и не-
зависимыми от окружающей среды, пока эта среда не изме-
няется. Эти структуры могут иметь чисто механическую природу,
как кристалл, спущенная пружина, шарик на дне ямы или пред-
ставлять собой некое распределение: градиент плотности в жидко-
сти или газе, распределение системы по фазам и т. д. В отличие от
этого в природе имеются структуры, которые самым тесным обра-
зом связаны со средой. Для существования таких структур необ-
ходим постоянный обмен энергии, а иногда и вещества и они ста-
бильны только до тех пор, пока связаны с окружающей средой.
Так как при этом происходит диссипация энергии, то их назвали
диссипативными структурами (Prigogine, Nicolis, 1967). Эти струк-
туры широко представлены в физике, химии и биологии.
Стабильное и нестабильное состояние систем
Как известно из классической термодинамики, в изолированной
системе при протекании необратимых процессов происходит увели-
чение энтропии, которая достигает максимума в состоянии равно-
весия. Эволюция системы определяется неравенством (2.18) и в
равновесном состоянии, когда изменение энтропии системы равно
нулю и она принимает максимальное значение, система устойчива
и стабильна.
В открытой системе, если внешние параметры системы остаются
неизменными, может наступать не равновесное, а стационарное
состояние. Если такая система находится недалеко от равнове-
сия и относится к так называемой линейной области термодинами-
ки необратимых процессов, то ее эволюция описывается неравен-
ством (1.16). Неравенство (1.16) соответствует теореме о минимуме
продукции энтропии в стационарном состоянии (см. Пригожин,
1960). С помощью теоремы Ляпунова можно показать, что стацио-
нарное состояние в этом случае устойчиво, так как всегда можно
найти положительно определенную квадратичную форму, которая
принимает минимальное значение в некоторой точке пространства
переменных (Глансдорф, Пригожин, 1973).
Для открытых систем, далеких от равновесия, относящихся
к области нелинейных необратимых процессов, показано, что эво-
люция системы определяется неравенством (2.10), а не (1.16) (При-
гожин, 1960; Глансдорф, Пригожин, 1973). Соотношение (2.10)
является распространением неравенства (1.16) на область нели-
нейных процессов и в этом смысле (2.10) можно считать неким уни-
версальным критерием эволюции систем, не связанным с линейны-
ми законами.
Для неравенства (1.16) было показано, что состояние с мини-
мальной продукцией энтропии устойчиво во времени. В противо-
положность (1.16) соотношение (2.10) не является дифференциалом
функции состояния системы и (2.10) не включает в себя представ-
176
Ление об устойчивости системы в стационарном состоянии. Однако
если потоки и силы в стационарном состоянии подчиняются требо-
ванию:
п
(6.1)
г=1
то это состояние стабильно во времени (Пригожин, Николис, 1973).
Неравенство (6.1) справедливо как вблизи от равновесия, так и в
системах далеких от равновесия, если в этих системах отсутствуют
процессы обратной связи автокаталитического типа.
Диссипативные структуры
Пригожин и Лефеве (Prigogine, Lefever, 1973) показали, что
в зависимости от степени удаленности от равновесия термодинами-
ческие системы ведут себя двумя различными способами: 1) вблизи
от термодинамического равновесия система стремится к максималь-
ной энтропии, т. е. к состоянию максимального беспорядка (вбли-
зи от равновесия происходит нарушение порядка); 2) вдали от
равновесия, за пределами состояния с нормальным термодинами-
ческим поведением, т. е. в области, где не выполняется критерий
стабильности, происходит возникновение структур (вдали от рав-
новесия имеет место процесс возникновения упорядоченности). Эта
ситуация хорошо известна в гидродинамике, где выше определен-
ного порога неустойчивости под влиянием сил гравитации и кон-
векционных потоков возникают гексагональные структуры — фе-
номен Бенара (Benard, 1900). На этом примере ясно, что внешняя
среда имеет существенное значение для возникновения подобного
рода структур.
Имеется много чисто диссипативных систем, где нелинейность
феноменологических законов достигается различными путями.
Большое внимание в последнее время привлечено к системе регу-
ляций различных метаболических путей в биохимической сети
реакций (Hess, 1963; Hess, Boiteux, 1968). Нелинейность возникает,
например, в результате автокаталитических процессов, при ингиби-
ровании и активации, при перекрестном катализе и т. д. Эти явле-
ния приводят к пространственно упорядоченному распределению
веществ или к колебаниям концентрации веществ во времени и про-
странстве.
Во всех этих случаях возникновение порядка связано с нали-
чием флуктуаций и поэтому получило название «порядок через
флуктуации» (Prigogine, 1969; 1973; Prigogine, Lefever, 1973). По-
казано, что возможность существования нарушающих симметрию
нестабильностей приводит к спонтанной самоорганизации, кото-
рая имеет особенно большое значение в биологии (Hess, 1963; Hess,
Boiteux, 1968; Plesset, Winet, 1974). Тюринг (Turing, 1952), по-
видимому, первым показал, что в химических системах как след-
177
ствие химических реакций и процессов диффузии могут возникать
структуры типа диссипативных. Подобные временные и пространст-
венные структуры, возможно, являются первыми ступенями воз-
никновения биологической упорядоченности.
Неравенство (6.1) является условием устойчивости стационар-
ных состояний. Избыточные потоки (6//) и силы (6Хг) относитель-
но средних значений этих величин можно связать с избыточной
энтропией, возникшей вследствие флуктуации следующим путем.
Разлагая в ряд значение энтропии S около состояния So имеем:
5 = 50 + 6S + V2625 + . . . (6.2)
Так как б2 5 является квадратической формой, получим неравен-
ство:
б2 5 < 0. (6.3)
Из (6.2), (6.1) и (1.15) следует, что:
44‘га=Ёв-'-«=- <6-4>
г=1
Поэтому для стационарного состояния:
4^>0. (6.5)
Это означает, что избыточная энтропия всегда отрицательна (6.3),
но увеличивается со временем (6.4) (Prigogine, Lefever, 1973).
Избыточная энтропия б2 5 определяет вероятность малых флук-
туаций около исходного состояния. Распространяя уравнение Эйн-
штейна на неравновесные условия (Глансдорф, Пригожин, 1973;
Пригожин, Николис, 1973) эту вероятность можно записать в виде:
(6.6)
где к — константа Больцмана.
Выражение (6.6) показывает, что всегда имеется небольшая,
но положительная вероятность перехода за порог нестабильности
в область, где исходно малые флуктуации могут усилиться, а затем
стабилизироваться благодаря потоку вещества и (или) энергии
проходящих через систему. Эти вновь возникшие структуры несут
на себе отпечаток исходной флуктуации, т. е. обладают неким ви-
дом примитивной памяти (Nicolis, Babloyantz, 1969; Prigogine,
Lefever, 1973).
Диссипативные структуры
в физических системах
Как указывалось выше, диссипативные структуры могут воз-
никать только за пределами определенного порога устойчивости
в системах далеких от термодинамического равновесия. В самом
простом случае энергия системы (возникающая внутри или посту-
178
пающая в систему из среды в виде теплового потока) не тратится
целиком на поддержание случайного, теплового движения моле-
кул, но частично используется для создания макроскопических
конвекционных структур, т. е. для возникновения пространствен-
ной организации.
Неустойчивость Бенара. Наиболее известным примером дис-
сипативной структуры является феномен Бенара (Benard, 1900).
Тонкий слой вязкой жидкости подогревают снизу так, что возни-
кает температурный градиент противоположный градиенту плот-
ности, возникающему в результате действия гравитационных сил.
Выше некоторого числа Рэлея флуктуации в жидкости усиливают-
ся и в слое возникают гексагональные структуры. Эти эксперимен-
ты подробно обсуждаются в известной книге Чандрасекара (Chan-
drasekhar, 1961). Появление структур является следствием
перехода от случайного теплового движения в макроскопический,
пространственно упорядоченный поток вязкой жидкости, который
для своего поддержания нуждается в поступлении энергии из сре-
ды. Эффект обусловлен взаимодействием двух диссипативных про-
цессов; движением вязкой жидкости и тепловым потоком.
В соответствии с принципом Больцмана структуры возникают,
когда энергия преобладает над энтропией и появлению упорядо-
ченных структур способствует понижение температуры. В отличие
от этого способа образования структур в неустойчивости Бенара
и в других подобных случаях мы имеем дело с другим принципом,
принципом «порядка через флуктуации», который связан с усиле-
нием малых флуктуаций стабилизацией их на новом уровне (дис-
сипативные структуры).
Полимерные структуры. Гамбале и Глиоци (Gambale, Gliozzi,
1971) описали явление, аналогичное феномену Бенара. Они нано-
сили тонкий слой полимера на специальную насыщенную водой
мембрану. Осмотическая разность в этих двух фазах приводила
к высвобождению воды из мембраны в результате чего возникал
концентрационный градиент в растворе полимера. Выше опреде-
ленного порога начиналась конвекция и возникали диссипативные
структуры. Они представляли собой полигональные ячейки, раз-
мерами в несколько миллиметров, которые были лучше видны,
если вода в мембране была слегка подкрашена.
Вендкер и др. (Wendker et al., 1976) наблюдали появление дис-
сипативных структур при полимеризации акриламида, раствор
которого был подкрашен метиленовым синим. Структуры появля-
лись через 10 минут после начала опыта, по-видимому, в результа-
те автокаталитической реакции, происходящей в системе. Так как
полимер выделяет воду и раствор через полчаса после начала опы-
та становится плотным, эти структуры как бы фиксируются в рас-
творе полимера. Этот процесс является интересным примером пе-
рехода диссипативной структуры в равновесную.
Временные диссипативные структуры. Известно очень много
интересных физических примеров временных диссипативных струк-
179
тур. Здесь мы коротко упомянем лишь об одном несколько необыч-
ном явлении.
Лампрехт и Шааршмидт (Lamprecht, Schaarschmidt, 1972) по-
казали, что яркость горящего фитиля при использовании разных
горючих смесей испытывает колебания, подобно колебаниям при
неустойчивом горении свечи. Эти колебания проявляются только
в узкой зоне по длине фитиля и только ниже некоторого порога
яркости. Фактически в этом случае мы имеем дело с диссипатив-
ной структурой, появление которой можно объяснить авторегуля-
ционными эффектами, связанными с поглощением горючей смеси,
испаряющейся с фитиля и диффузией горючей смеси в пламя (Lam-
precht, Schaarschmidt, 1976). По небольшой продолжительности
этих колебаний и по малому числу циклов (если фитиль смочен,
а не погружен в резервуар с горючим) это явление напоминает
хорошо известную реакцию Жаботинского.
Диссипативные структуры
в химических системах
Вблизи от термодинамического равновесия или в области ли-
нейных необратимых процессов решения уравнений для химичес-
ких реакций устойчивы, поэтому, если нас интересуют диссипатив-
ные структуры, мы должны рассматривать кинетику химических
реакций, протекающих вдали от равновесия.
Стационарное состояние в химических системах возникает в
результате взаимодействия двух противодействующих процес-
сов: химических реакций, ведущих к возникновению пространст-
венной неоднородности и диффузии продуктов реакций, ведущей
к однородности в системе. Такое стационарное состояние поддержи-
вается непрерывным потоком энергии из среды. При истощении
источников энергии стационарное состояние становится нестабиль-
ным и система стремится к равновесию. При решении нелинейных
кинетических уравнений выявляется целая иерархия стационар-
ных состояний (Seshadri, 1974).
Теоретические модели диссипативных структур. Лефеве (Le-
fever, 1968) предложил простую модель, включающую два реаген-
та А, В, два продукта реакций D, Е и два промежуточных продук-
та х, у для того, чтобы показать разные типы решений кинетичес-
ких уравнений для системы реакций:
А^В,
2х + У Зя,
В + х у + D,
я £*.
Наиболее важной в этой системе является вторая, тримолекуляр-
ная, автокаталитическая реакция. Для простоты принимается,
что концентрация веществ Л, В, D, Е постоянны во времени и из-
180
меняются только х и у, В зависимости от выбора начальных усло-
вий можно получить два типа решений для стационарного состоя-
ния: однородное и неоднородное. Однородное решение появляется
вблизи от термодинамического равновесия. Другие решения пред-
ставляют собой пространственные и временные диссипативные
структуры. Они могут рассматриваться в качестве прототипа пе-
редачи информации на макроскопические расстояния с помощью
химических сигналов (Prigogine, Lefever, 1973).
Сходную модель рассматривали Глансдорф и Пригожин (1973)
для выяснения зависимости решений кинетических уравнений от
констант диффузии и начальных условий, а также для выяснения
поведения предельного цикла данной системы.
Мы не будем больше углубляться в эту проблему, покажем
только некоторые результаты расчетов, проведенных с помощью
ЭВМ, для изменений концентрации х во времени, как функции
начальных условий (рис. 50). Подобные же колебания характерны
для плоскости yt и плоскости ху.
Реакция Белоусова — Жаботинского. Наиболее известной из
химических реакций, в которых обнаруживаются диссипативные
структуры, является реакция впервые описанная Белоусовььм
и интенсивно исследованная Жаботинским (Zaikin, Zhabotinsky,
1970; Жаботинский, 1974) и некоторыми другими авторами (Field,
Noyer, 19^2; Field et al., 1972; Koros, Burger, 1973; Koros et al.,
1973b; Thoenes, 1973; Janjic et al., 1974; Stroot, Janjic, 1975).
Эта реакция состоит в окислении малоновой кислоты броматом
калия при участии ионов церия в качестве катализатора. Механизм
реакций довольно сложен и содержит ряд промежуточных продук-
тов. Следить за реакцией можно с помощью потенциометрических
или фотометрических измерений концентрации Се4+ и Се3+. Най-
дено несколько вариантов реакций, в которых вместо малоновой
кислоты используются яблочная кислота или ацетилацетон, а в
качестве катализаторов — марганец или комплексные ионы желе-
за. Общим для всех этих реакций является период индукции и вне-
запное возникновение колебаний, которые могут продолжаться дол-
гое время. В наших калориметрических исследованиях с системой,
включающую яблочную кислоту, колебания при 19° наблюдались
более 48 часов. Амплитуда убывала со временем и колебания по-
степенно ослаблялись, но вновь восстанавливались при добавле-
нии свежего раствора бромата, который является лимитирующим
фактором реакции, и таким образом источником потока энергии
для временной диссипативной структуры. На рис. 51 приведены
данные об изменении теплового потока в реакции Жаботинского,
измеренного с помощью микрокалориметра.
Если использовать в качестве реакционного сосуда колонку,
временная структура переходит в пространственную или в про-
странственно-временную. После появления небольшой негомо-
генности в неперемешивающемся растворе пространственная струк-
тура распространяется по всей колонке в форме горизонтальных
181
Рис. 50. Флуктуация концен-
трации промежуточного веще-
ства х по времени как функ-
ция начальных условий
Для всех кривых А = 1,0: (1)
В = 1,0; (2) В = 1,5; (3)
В = 1,9; (4) В = 2,0; (5) В = 2,5
Рис. 51. Изменение теплового
потока в реакции Жаб о мин-
ского (3 мл 0,8 М яблочной
кислоты; 3 мл 0,2 М бромата
калия; 0,4 мл 0,046 М суль-
фата магния) при 25°. Общая
теплопродукция равна 35,6 дж
Рис. 52. Изменение теплового потока в реакции Жаботинского (малоновая
кислота, бромат калия и т. д.)
а — запись термограммы реакции; б — термограмма на фоне желоба (объяснения
в тексте)
йоЛос, которые могу! под влиянием вертикального температурно-
го градиента перемещаться по всему сосуду (Thoenes, 1973). Тот
факт, что структуры появляются в небольшой области начальных
концентраций, исчезают со временем в закрытой системе и хорошо
восстанавливаются, является убедительным доказательством их
обусловленности диссипативными процессами, возникающими за
пределами нарушающих симметрию нестабильностей. Реакция
Жаботинского совершенно отличается от хорошо известного фено-
мена колец Лизиганга, которые являются статичными, равновес-
ными структурами, не поддерживаемыми потоком энергии в си-
стеме.
В главе 2 настоящей книги было показано, что развитие жи-
вотных происходит в направлении минимума функции внешней
диссипации %. С теоретической точки зрения развитие может быть
описано механической аналогией: шариком, катящимся вниз по
наклонному желобу (рис. 1). Этот шарик не обязательно все время
остается точно на дне желоба: он может испытывать небольшие от-
клонения от линии дна в горизонтальной плоскости. Подобное же
поведение наблюдается при снятии калориметрических термограмм
в реакции Жаботинского (рис. 52). Такое сопоставление, опираю-
щееся только на сходство процессов истощения источников энер-
гии, поддерживающих скорости реакции, и уменьшение теплопро-
дукции системы, разумеется, условно, но оно дает представление
об идее наклонного желоба и уклонений от средней линии дна
желоба.
Диссипативные структуры
в биологических системах
Диссипативные структуры имели бы чисто академический ин-
терес, если бы их нельзя было использовать для объяснения воз-
никновения биологических структур. Как показано выше, в тер-
модинамических системах наблюдается два рода упорядоченности:
одна возникает в результате уменьшения энтропии при низких
температурах, другая — в результате усиления небольших флук-
туаций и их стабилизации потоком энергии. Этот последний тип
упорядоченности наиболее привлекателен для термодинамики
биологических процессов, так как живые организмы являются
открытыми системами, далекими от равновесия, и они существуют
при относительно высоких температурах. Поэтому в заключение
мы рассмотрим появление диссипативных структур (или структур
сходных с диссипативными) в биохимических и биологических
системах. Они могут наблюдаться на различных уровнях сложно-
сти: 1) на молекулярном уровне — колебание концентраций суб-
стратов при реакциях, катализируемых ферментами; 2) на кле-
точном уровне — колебания типа индукции и репрессии генома,
описанные Жакобом и Моно; 3) на надклеточном уровне — циркад-
ные ритмы в организмах; 4) на популяционном уровне — коле-
бания численности организмов в системе хищник — жертва.
183
Гесс и др. (Hess, 1963; Hess, Boiteux, 1968, см. также Сельков,
1975) провели много исследований, касающихся первого уровня —
молекулярных колебаний. Наибольший интерес представляют ре-
гуляции по типу обратных связей некоторых ступеней гликолиза.
В этом процессе, по-видимому, важную роль играет фосфофрукто-
киназа, активируемая АДФ — продуктом этой реакции. Другие
исследования касались колебаний, происходящих при фотосинте-
зе в цикле Кельвина.
Образование структур в клеточных агрегатах. Гериш (Gerisch,
1971) показал, что в сосуде со слизневиком Dictyostelium discoi-
deum под влиянием стимулирующего агента, называемого акра-
зином, вначале раздельные амебоидные организмы агрегируют и
образуют структуры. Эти структуры формируются в течение минут
или часов и напоминают наборы концентрических колец. Они воз-
никают в различных точках сосуда и взаимно уничтожаются в
местах контакта. Иногда, когда центры агрегации работают в раз-
ных фазах, образуются спиральные линии. Могут возникнуть
также структуры типа волн, если многие независимо колеблющи-
еся группы клеток постепенно попадают под влияние одной из
групп, являющейся «генератором» колебаний. В этом случае они
утрачивают свой авторитм и превращаются в структуры, передаю-
щие возбуждение.
Стимулирующий агент «акразин» был идентифицирован с цик-
лической АМФ. Он разрушается в среде создавая таким путем
условия для периодического возникновения хемотакса в системе.
Авторитмическая активация агрегационных центров характери-
зуется периодом от 3 до 5 минут.
Биоконвекционные структуры. В некоторых культурах бакте-
рий, жгутиковых и ресничатых можно наблюдать биоконвекцион-
ные структуры, которые по внешнему виду, на первый взгляд
напоминают фигуры Бенара, но на деле совершенно отличны от
этого феномена, так как температурный градиент не принимает
никакого участия в установлении биоконвекционных структур.
Плессе и Вине (Plesset, Winet, 1974) изучали поведение культу-
ры ресничатой инфузории (Tetrahymena pyriformis). Эти организ-
мы тяжелей воды, но стремятся передвинуться к поверхности жид-
кости и скапливаются вместе в поверхностном слое. В результате
они образуют обратный градиент плотности с однородным слоем
повышенной плотности у поверхности. Из этого слоя формируют-
ся структуры, содержащие живые организмы, типа «пальцев»,
которые вырастают со скоростью, вдвое превышающей обычную
скорость плавания этих организмов. Эта физическая ситуация гра-
витационной нестабильности хорошо известна в гидродинамике,
под названием нестабильность Рэлея — Тэйлора. Возникает лишь
небольшая полоса шириной 1 см, внутри которой образуются «па-
дающие пальцы», что хорошо соответствует теории. В случае смер-
ти организмов нестабильная ситуация обратного градиента плот-
ности нарушается и система стремится к механическому равнове-
184
Рис. 53. Изменение соотноше-
ния жертва—хищник во вре-
мени в системе Didinium па-
зи turn в качестве хищника (X)
и Paramaecium cauda turn в ка-
честве жертвы (Ж)
МП
о--------1--------1--------1--------1-----
5 10 15 Z0 Сутки
сию, так как больше нет
потока энергии, т. е. диссипативная
структура исчезает.
Система хищник — жертва. На популяционном уровне можно
наблюдать, например, колебания численности двух видов орга-
низмов, живущих вместе в ограниченной области. Один из них
является хищником, питающимся другим — жертвой. Увеличе-
ние числа хищников автоматически вызывает уменьшение чис-
ленности жертв, так как шанс избежать хищника у жертвы стано-
вится меньше. Математически эта проблема исследована Лотка и
Волтерра (Lotka, 1925; Volterra, 1926). Временная колебательная
структура популяций была изучена в самых различных биологи-
ческих системах таких, как смесь популяций Paramaecium bursaria
с Schizosaccharomyces pombe; Bacterium proteus и Eheyletus eru-
ditus c Aleuroglyphus agilis; Acarus siro и Phytoseiulus persimilis
c Tetranychus urticae. Первые были хищниками, вторые — жерт-
вами. На рис. 53 приводятся временные относительные изменения
концентрации жертвы в системе Didinium nasutum и Paramaeci-
um caudatum.
Седжел и Джексон (Segel, Jackson, 1972) показали, что в систе-
ме эксплуататор — жертва (как они предпочитают называть си-
стему хищник — жертва) при особых условиях таких, как взаимо-
действие между жертвами, борьба между эксплуататорами и др.
происходит случайный разброс обоих диффузионных нестабиль-
ностей. Это интересный факт поскольку обычно диффузия приво-
дит к гомогенности системы и стабилизации равновесия, тогда как
реакции ведут к негомогенности. В данном случае мы имеем эко-
логически важный пример диссипативной структуры, которая в
то же время является диффузионной структурой. Как и в случае
отмеченной выше биоконвекционной структуры, данная структу-
ра может поддерживаться только до тех пор пока система живая,
т. е. пока через нее проходит постоянный поток энергии.
СТАЦИОНАРНЫЕ ДИССИПАТИВНЫЕ СТРУКТУРЫ
В. А. Васильев
Пространственно-периодические структуры широко распрост-
ранены в природе. Одним из примеров таких структур являются
разнообразные кристаллы. В биологических системах, которые в
отличие от кристаллов являются открытыми системами, также
имеются пространственно-периодические структуры. Поскольку
эти структуры в открытых системах возникают и поддерживаются
за счет диссипативных процессов, связанных с обменом между
средой и системой, то их называют диссипативными структурами
(Глансдорф, Пригожин, 1973).
Диссипативные структуры целесообразно подразделить на три
вида: 1) стационарные, например, стоящие страты в плазме разря-
да, структуры Тюринга в гомогенных «автоколебательных» хи-
мических средах и др. (Жаботинский, 1974; Романовский и др.,
1975); 2) локализованные, примеры которых приведены в ряде
работ (Глансдорф, Пригожин, 1973; Пригожин, Николис, 1973;
Васильев, Романовский, 1976); 3) динамические, например, фено-
мен Бернара в гидродинамике (Глансдорф, Пригожин, 1973),
концентрационные стоящие волны в химической кинетике (Василь-
ев, Романовский, 1976) и т. д.
Темой данной работы является кинетическая теория стацио-
нарных диссипативных структур. Здесь возникают две основные
проблемы: получение стационарных решений системы уравнений
исследуемой модели и исследование устойчивости этих решений
к различным возмущениям. Основное внимание в данной работе
уделяется первой задаче и приводятся только некоторые резуль-
таты, относящиеся ко второй.
Постановка задачи. Система уравнений, описывающая кине-
тику распределенной химической или биологической системы мо-
жет быть записана в виде:
71
^Xi = Фх(хх...хп) 4- (Dij-^Xj) (i = 1, и), (6.7)
где п — число компонент системы; Ф$ (хи . . ., хп) — нелинейные
функции, описывающие кинетику реакций; последний член в пра-
вой части уравнения описывает процессы масса — переноса (ра-
ди простоты в данной работе предполагается, что матрица
диагональна и ее элементы не зависят от х^ т. е. в рассмотрение
не включены процессы активного и «сопряженного» транспорта).
Алгебраическая система уравнений . . ., хп) = 0 име-
ет хотя бы одно положительное решение, в противоположном слу-
чае уравнения (6.7) при достаточно’ больших t имели бы только
неограниченные решения, что не имеет смысла для реальных хими-
186
ческих или биологических систем. Компоненты этого решения обоз-
начим Xt, и назовем это решение стационарным состоянием. В даль-
нейшем будут исследоваться системы с единственным стационарным
состоянием.
Исследуем устойчивость однородного стационарного состоя-
ния — тривиального решения системы уравнений (6.7). Для это-
го удобно перейти к другим переменным по формуле: zt — Xi — Xi,
где Zi представляет собой отклонение от однородного стацио-
нарного состояния. В этих переменных уравнения (6.7) прини-
мают вид:
= zJ + Du-^Zi, (г, 7 = 1, п), (6.8)
где Мц — матрица п X п с постоянными коэффициентами; Ф[ —
нелинейные функции, не содержащие переменных первой степени.
Исследование устойчивости тривиального решения уравнения
(6.7) эквивалентно исследованию устойчивости нулевого решения
уравнения (6.8). Представим возмущения нулевого решения в
виде:
Zi (t, г) = zQi ехр [XfcZ + jjtkr] (i, к = 1, . . ., n). (6.9)
Подставив (6.9) в (6.8) и оставив только линейные члены, получим
алгебраическую линейную однородную систему уравнений и, ис-
пользуя условия существования ее нетривиальных решений, при-
ходим к характеристическому уравнению вида:
X* + gn-i (. • . + go (• • : *8) = 0.
(к = 1, . . п). (6.10)
Тривиальное решение неустойчиво к бесконечно малым возмуще-
ниям с длиной волны 2/к, если реальная часть хотя бы одного кор-
ня уравнения (6.10) больше нуля (см. Романовский и др., 1975).
Можно показать, что в случае неустойчивости (не колебательного
типа), приводящей к стационарным диссипативным структурам
периода 2/к, уравнение (6.10) имеет нечетное количество корней
с положительными реальными частями. Для этих к выполняется
условие g0 <; 0. Последнее возможно в рассматриваемых системах
(go (к = 0) > 0) лишь при наличии положительной обратной свя-i
зи в цепи реакций, что предполагает авто- или кросс-каталитиче-
ские реакции в моделируемой системе.
Решения типа стационарных диссипативных структур должны
обеспечивать равенство нулю правых частей уравнений (6.8) и
выполнение граничных условий. В работе будут использованы гра-
ничные условия двух типов: 1) постоянные стационарные концент-
рации на «стенках»:
xt (/, 0) = Xi (t, 1) = Xi (i = 1, . . ., n). (6.11)
187
2) непроницаемые «стенки» на границе:
0) = -^а:{(«, 1) (г = 1, .... п). (6.12)
Иначе говоря, стационарные диссипативные структуры соответ-
ствуют решениям краевой задачи, состоящей из уравнений:
Du -^Zi + MijZj 4- Ф^!, ..., zn) = 0 (6.13)
и граничных условий (6.11) или (6.12).
Ниже рассмотрен один способ решения нелинейных краевых
задач такого типа и его использование на примере известной мо-
дельной системы (Глансдорф, Пригожин, 1973, формулы (15,1;
15,2). Отметим, что большая часть полученных результатов спра-
ведлива и для других моделей. Уравнения этой модельной систе-
мы следующие:
-^х = (В — i) х 4- А2у + Ф (х, y) + Dx^x
~^У = — Вх— А*у—Ф(х,у)+ Dy^y (6.14)
Ф (*> У) = 4^ + + х2У’
где х и у — отклонения значений концентраций от стационарного
состояния ($= А; у = В/А).
Подчеркнем, что присутствие квадратичных членов х2 в (6.14)
принципиально необходимо для того, чтобы концентрации не при-
нимали отрицательных значений. В уравнениях (6.14) при В >
Вк В-^ (выражения для Вк и В^ приведены ниже, см. (6.18)
и (6.19)) стационарное состояние неустойчиво при возмущениях
с длиной волны 2/к.
Точные решения уравнения (6.14) получить не удается, поэто-
му ниже будет показан приближенный метод получения одного
класса нетривиальных решений — решения типа стационарных
диссипативных структур. Чтобы наглядно себе представить роль
различных членов в (6.14), вначале кратко рассмотрим простей-
шие конечно-разностные модели краевой задачи.
Конечно-разностные модели задачи
Простейшей конечно-разностной аппроксимацией системы урав-
нений (6.14) при граничных условиях (6.12) является модель двух
связанных посредством диффузии объемов, в которых протекают
моделируемые реакции. Сделав замену переменных по формулам,
приводимым ранее (Васильев, Романовский, 1975), легко получить
точное нетривиальное решение этой задачи:
х* - (242 - 2DV - Вк')х2 - к'А2 (В - ВК) = 0, (6.15)
188
где kf = 2DV/(1 — 2DX); Bk = 1 + 2Z>X + A2lkf\ xr = xr и
x2 — отклонение концентраций от стационарных. Из анализа
динамики этой модели следует, что при В > Вк однородное нуле-
вое решение неустойчиво к бесконечно малым возмущениям. На
рис. 54 показаны типичные зависимости х* от В, причем рис. 54
(1) относится к случаю, когда А2 4Dy, а рис. 54 (2) — к обрат-
ному соотношению, когда 4° < 4Dy. Исследование устойчивости
к бесконечно малым возмущениям решений, даваемых формулой
(6.15) показало, что на ветвях, нанесенных штриховой линией на
рис. 54, эти решения неустойчивы. При В <; Вк решения типа
стационарных диссипативных структур (6.15) может установиться
только в результате конечных возмущений стационарного состоя-
Рис. 54. Зависимость значений X2
(или Р2) от параметров В сис-
темы уравнений (6.14)
Пояснения в тексте
Рис. 55. Типичная зависимость
параметров Вк(1) ст волновых чи-
сел к (см. уравнение 6.17). Зависи-
мость амплитуды стационарных
диссипативных структур с раз-
личными периодами от парамет-
ра В (2) при граничных усло-
виях (6.12)
189
ния. Формулы, описывающие стационарные дйссийативпые струк-
туры при большом числе диффузионно-связанных объемов более
громоздки, хотя качественно дают такие же результаты.
Для моделирования распределенной системы при граничных
условиях (6.11) рассмотрим три объема, связанных диффузией.
В двух крайних объемах отклонения от стационарного состояния
будем считать равными нулю. Для отклонения в среднем объеме
получим уравнение:
= (6.16)
На рис. 54 (3, 4) показаны зависимости х2 от В, причем Вк = Sinin
только, если Л2 = 47)у. Анализ устойчивости стационарных дис-
сипативных структур дает такие же результаты, что и в предыду-
щем случае. Для возникновения стационарных диссипативных
структур при В < Вк необходимы конечные возмущения стацио-
нарного состояния. При В Вк возможны две такие структуры,
отличающиеся знаком отклонения переменной от стационарного
состояния. Вопрос о том, какие из этих решений осуществятся,
решается выбором начальных условий задачи.
Если несколько усложнить модель и между крайними объема-
ми поместить не один, а два объема, то наряду с нулевыми и типа
(6,16) появятся и другие: график одной из зависимостей (х2 —
— х3)2/2 от В имеет тот же вид, что и на рис. 54(7, 2), и при В >
Вк (здесь kf — 3Dy/(l + 3D J) может быть только одна устой-
чивая ветвь. В данной работе не приводится уравнения, определяю-
щего это решение, так как оно громоздко и не имеет «наглядного»
аналитического решения.
Хотя системы малого числа связанных объемов являются гру-
быми моделями соответствующей краевой задачи, они позволяют
сделать ряд выводов, которые следует учесть при исследовании
стационарных диссипативных структур в распределенной систе-
ме: 1) уравнения (6.15) и (6.16) показывают, что появление дисси-
пативных структур, не предсказываемых линейными теориями и,
в частности, термодинамикой, связано с квадратичными членами
в нелинейных операторах; 2) ограниченность решений типа дис-
сипативных структур обусловлено кубическими членами, т. е.
необходимо пользоваться только такими приближениями, которые
позволяют одновременно учитывать квадратичные и кубичные
члены в нелинейных операторах; 3) при граничных условиях (6.11)
поведение решений различной формы с четным и нечетным числом
полуволн качественно различно.
О методе получения решений
типа стационарных диссипативных структур
Задача нахождения периодических по пространственной ко-
ординате решений, обращающих правую часть уравнений (6.14)
в ноль (т. е. получение решений системы уравнений типа (6.13))
190
в некоторых отношениях близка к традиционным задачам теории
колебаний. Из теории колебаний известно, что период колебаний
нелинейной системы зависит от амплитуды. Так как в нашем слу-
чае (краевая задача на отрезке от 0 до 1) период должен быть стро-
го равен 2/Л, то необходимо найти такую амплитуду для стацио-
нарной диссипативной структуры, при которой это равенство
выполняется. Отсюда вытекает следующая схема метода: 1) вы-
делить из уравнений линейный оператор, который для собствен-
ного значения ноль имеет собственную функцию периода 2/А;
2) найти зависимость нелинейных поправок к этому периоду от
амплитуды; 3) найти амплитуду для периода 2/Л, при которой эти
поправки обращаются в ноль.
Рассмотрим одну из возможных реализаций этой схемы. Для
этого перепишем уравнение (6.13) для исследуемой модели сле-
дующим образом:
Л2
#х * + (Вк — 1) X + А2у = — Фк (х, у),
& (6.17)
DV У - в*х - А2у = ф* У^
где Фк = (В — Вк)х + Ф (х, у); г изменяется от 0 до 1; х (t, г)
и У г) должны удовлетворять граничным условиям (6.11) или
(6.12); Вк определяется соотношением:
Вк = 1 + rfk2Dx + А2/кк, (6.18)
где кк = n2k2Dyl(i + n2k2Dх); к — целое число. Это соотноше-
ние получено из условия, что оператор, стоящий в левой части
(6.17), имеет единственную не нулевую собственную функцию
периода 2/к, соответствующую собственному значению 0. На рис.
55 (7) приведена типичная зависимость Вк от к (штриховые ли-
нии показывают, что к принимает только целые значения). При
всех к ее [Amin; &maxl соответствующие характеристики урав-
нения (6.10) имеют один положительный корень и в этом случае
однородное стационарное состояние системы (6.14) неустойчиво
к возмущениям с этими длинами волн. Рассматриваемая зависи-
мость имеет минимум при:
1
7c~d = -L(_f_) \ (6.19)
Л \ &Х™у f '
_ 1
где к — ближайшее к d целое число и Вк [1 + A (px/Dv) 2 ]2.
Выбрав к ищем решение системы уравнений (6.17) в виде:
У (г) = РкУк + pkyi + Ркуп + - (6.20)
Аналогичное выражение будет для х (г). В случае граничных ус-
ловий (6.11) ук = sin (лЛг), в случае граничных условий (6.12)
ук = cos (лАт). Соответствующим граничным условием удовлет-
191
воряют также yi и уц. Подставив (6.20) в (6.17) и собрав члены
при одинаковых степенях Рк, получим систему линейных диффе-
ренциальных уравнений для нахождения yi (в правой части стоят
функции от Рк)9 уц (в правой части стоят функции от ук и уц)
и т. д. Чтобы быть последовательными при получении этих урав-
нений следует считать, что:
В - Вк = К&2 + . . . (6.21)
Полученная система уравнений имеет нетривиальное решение,
удовлетворяющее граничным условиям не при любом а тогда
и только тогда, когда правая часть системы (6.17) ортогональна
к решению системы, сопряженной левой части (6.17). Конкрет-
ный вид условий ортогональности для системы (6.17) следующий:
1
&к (Ркхк + Ркх1 + •••*> РкУк + В"кУ1 + •••) Ук (6.22)
о
Легко видеть, что условие (6.22) при граничных условиях
(6.12) есть приравненная нулю поправка на период колебаний,
вычисляемая методами Пуанкаре — Ляпунова или Боголюбо-
ва — Митропольского (Моисеев, 1966; Боголюбов, Митрополь-
ский, 1963). Специфика рассматриваемой задачи в неоднознач-
ности выделения линейной части, т. е. возможны решения с^раз-
личными периодами.
Вычислив интеграл (6.22), получим уравнение для нахожде-
ния Рк, которое, как правило, можно легко решить. Подставив
это Рк в (6.20), получим приближенное решение соответствующей
краевой задачи. Выбор числа членов в ряде (6.20) для построе-
ния первого приближения определяется видом нелинейностей в
уравнениях и граничными условиями. Например, при (6.12) и
наличии квадратичных и кубичных членов необходимо брать и
Ут, так как ху ~ х2~ркутук; х2у ~ ркук одного порядка по Рк.
Примеры решений
типа стационарных диссипативных структур
Вначале рассмотрим случай граничных условий (6.12) — не-
проницаемые «стенки» на границе рассматриваемого пространст-
ва. В этом случае х (г) и у (г) представим в виде следующих рядов:
я(г) = Pm cos (лтг); у (г) = Pmcos (лтг). (6.23)
?п=0 т—о
Такая форма записи позволяет конкретизировать вид z/i, z/ц, . . .
в (6.20). Из (6.17) получим следующее соотношение:
Рт = —n2m2DyPm/(l + n2m2Dx) = —ккРт. (6.24)
Подставив (6.20) в (6.22) и использовав (6.24), найдем у^ и уц.
192
В результате вид выражения (6.20) в рассматриваемом случае
следующий:
у (г) = Рк cos (лкг) + Рк
X
cos (2лкг) 1
Вк'2к — 47)^2^ __ Л2 ““ I2
/Вкк — 2 А2^ , з / к\ __cos (ЗлЛт)
Х \ 2Л ) к\4 вк^ — АЪ — Щ^к*
Действуя по вышеописанной схеме, получим уравнение для
нахождения Рк, одно из которых Рк = 0, а другое:
к
92Pl + 9iPl + -f(Bk-B) = 0
(6.26)
позволяет найти амплитуду нетривиального решения задачи. При
выводе уравнения (6.26) отбрасывались все члены со степенями
выше, чем Р^ (включительно). Выражения для q± и q2 громоздки,
поэтому они не выписаны полностью. Укажем, что всегда q2
0, а при В = Вк:
qx = -1- Л'Я 3 - (Л« - л2^Р„) (•
/с 4 \
вп-в* + А.
(6.27)
Из (6.27) следует, что, если А2 4> n2k2Dy, то возможно, что qt <
<Z 0. Это означает, что и при В <С Вк решение, имеющее период
2/к существует. Характерная для этого случая зависимость рк
от В показана на рис. 54 (7). Если qr < 0 при В = Вк, то зави-
симость имеет вид, приведенный на рис. 54 (2) — стационарные
диссипативные структуры с периодом 2/к не существуют, если
В <г Вк)
Теперь рассмотрим краевую задачу при граничных условиях
(6.11) — на границе фиксированы концентрации, равные стацио-
нарному состоянию. Для х и у в этом случае следует использовать
представление:
у = у1| Рт sin (лтпг); х = У*, Рт sin (лтг). (6.28)
7П=1 ТП=1
Связь между Рт и Рт дается соотношением (6.24). Подставив
(6.20) в (6.17), находим, что:
yi = У, fm, н sin (тяг), (6.29)
где т принимает только нечетные значения; fm,k — дробно-ра-
циональная функция от иг2, к2 и параметров уравнения (6.17).
Вычисления для получения уравнений, определяющих Рк
различны в случаях, когда к — четное и когда к — нечетное.
193
В последнем случае г/i дает вклад при вычислении интеграла
(6.22) только в коэффициенты при Р* и больших степеней Рк.
С точностью до этих величин уравнение для нахождения ампли-
туд нетривиальных решений имеет следующий вид:
<В - ВД - -Sta (В - Р> + + - = °- <6-30)
Наличие ненулевого коэффициента при Рк в (6.30) обусловлено
присутствием квадратичных членов в ФЛ (см. (6.18)). Знак этого
коэффициента зависит от соотношения между А2 и 3i2k2Dy, т. е.
от периода исследуемой стационарной диссипативной структуры.
На рис. 54 показаны типичные для нечетных к зависимости Р к
от В: рис. 54 (3) — А2 л2к21)у; рис. 54 (4) — А2 < n2k2Dy. Из
графиков и приведенных соотношений видно, что практически
с любым периодом решения для стационарных диссипативных
структур существуют и при В С Вк. При В > Вк существует
два решения с одним и тем же периодом — 2/7с. Этому факту мож-
но дать наглядную интерпретацию. Пусть для простоты к = 1,
тогда распределение концентраций имеет колоколообразную фор-
му — в рассматриваемом приближении yr = Pr sin (лг). Ясно,
что из-за квадратичной нелинейности величины > 0 и Р1>2 <С
< 0 неодинаковы и по абсолютной величине; также очевидно, что,
если к четное или граничные условия (6.12), то этой «неодинако-
вости» быть не может — это отразится на виде уравнений для на-
хождения амплитуд, в них входит Рк только в четных степенях.
Если к — четное, то при подстановке (6.20) в (6.22), вследствие
наличия квадратичных членов, нельзя ограничиться только пер-
вым слагаемым, следует оставить которое является бесконеч-
ным рядом (6.29). Поэтому Рк придется искать из уравнения,
в коэффициенты которого входят эти ряды. Так как просуммиро-
вать их в общем виде не удалось, то были проведены расчеты для
ряда значений параметров А, В, Z)x, Dy — в численных примерах,
как правило, легко получить оценки сумм и величины коэффици-
ентов искомого уравнения. В результате такого подхода было ус-
тановлено, что при достаточно больших А2 (по сравнению с n2k2Dy)
возможны стационарные диссипативные структуры периода 2/Л
и при В < Вк, а при В > Вк существует единственная струк-
тура с этим периодом; т. е. качественно зависимость Рк от В (на-
помним, что к — четное) имеет такой же вид, как и показанная
на рис. 54 (7) или 54 (2).
При выделении линейного оператора в системе уравнений
(6.17) выбор к был в достаточной степени произволен, но из выше-
изложенного следует, что не при любом к (6.17) имеет решение
для стационарных диссипативных структур. На рис. 55 (2) пока-
зан график зависимости Рк от В при граничных условиях (6.12).
Заметим, что при (6.12) не монотонная зависимость Рк (В), как
правило, имеет место лишь для Р^ [В- — наименьшее из всех
194
Вк, см. рис. 55 (7)] и иногда близких к нему значений к. Пересе-
чения вертикальной линии, соответствующей значению парамет-
ра В с кривыми Рк и определяют те Л, для которых существует
решение для стационарных диссипативных структур соответст-
вующего периода.
Устойчивость стационарных диссипативных структур.
Некоторые результаты численных экспериментов
В предыдущем разделе получены неоднородные решения си-
стемы уравнений (6.17) с различными периодами. Ясно, что все
решения системы (6.17) будут стационарными решениями исход-
ной динамической системы уравнений (6.14). Но вопрос об устой-
чивости этих решений и начальных условиях, к ним приводящим,
остался открытым. Частичный ответ на этот вопрос дается ниже.
Устойчивость стационарных диссипативных структур к ма-
лым возмущениям, удовлетворяющим соответствующим гранич-
ным условиям, исследовали по следующей схеме. В уравнения
(6.14) подставляли у (/, г), равное исследуемой стационарной дис-
сипативной структуре плюс у (t, г), где у (t, г) — малое возму-
щение, для х (Z, г) использовали аналогичное выражение. В силу
предположения о малости возмущений, оставлялись только ли-
нейные по у и х члены. Вопрос о поведении у и х во времени опре-
деляется спектрами полученных задач на собственные значения.
Опустим примеры реализации этой схемы и укажем основные
факты, которые удалось установить с ее помощью: 1) при В В^
устойчива только одна стационарная диссипативная структура
периода 2/Л, т. е. часть ветви, идущая из Вк влево, всегда неустой-
чива (см. рис. 54); 2) при В несколько больших, чем 1 + А2
и граничных условиях (6.12) стационарные состояния становятся
неустойчивыми — в системе возникают автоколебательные режимы
(напомним, что при В > 1 + А2 «точечная» кинетика системы явля-
ется автоколебательной Глансдорф, Пригожин, 1973; Романовский
и др., 1975); 3) для ряда частных примеров удалось показать, что
стационарная диссипативная структура с периодами, близкими
к 2//с1Пах или 2/Amin, неустойчивы; в некоторых случаях устой-
чивой оказывалась только одна диссипативная структура — ее
период 2/к.
Дальнейшее аналитическое исследование устойчивости на-
толкнулось на определенные технические трудности, преодолеть
которые пока не удалось. Но следующее утверждение, пока не
доказанное, представляется весьма правдоподобным: в системах
устойчивыми являются либо одна стационарная диссипативная
структура с периодом 2/Л, либо немногие с периодами, близки-
ми к нему — другими словами, устойчивые диссипативные струк-
туры с наибольшими амплитудами (см. рис. 55 (2)). Результаты
численного интегрирования системы уравнений (6.14), приводи-
195
Рис, 56. Стационарные решения
системы уравнений (6.14), полу-
ченные с помощью численных рас-
четов на ЭВМ при
а — граничных условиях (6.12). D =
= 0,1; Dy = 0,5; А = 4; В, = 8,4;
— В = 0,1; п = 1; б — граничных
условиях (6.11). Dx = 0,038; D =
= 0,5; А = 7; В2 = 8,7; В2 — b =
= 0,1; п = 2
мне в работах Полякова (1974), где использовались граничные
условия (6.12), и численные расчеты автора хорошо согласуются
с этим утверждением. Его также, по-видимому, подтверждают
экспериментальные работы, в которых исследовались стацио-
нарные диссипативные структуры, возникающие при протекании
реакции Белоусова — Жаботинского (Жаботинский, 1974; Zai-
kin, Zhabotinsky, 1970).
Численный эксперимент проводился в целях исследования
динамики возникновения стационарных диссипативных структур
и проверки точности решений, полученных в предыдущем
разделе. Начальные условия задавались в виде одного не-
большого «перепада». С этими начальными условиями система
решалась на ЭВМ при В Вк. Установившееся распределение
бралось в качестве начальных данных для интегрирования системы
при В < Вк -— при том же В система интегрировалась с пре-
дыдущими начальными условиями. В результате было установле-
но, что при В < Вк возможен только жесткий режим возникнове-
ния стационарных диссипативных структур, т. е. однородное ста-
ционарное состояние при В < Вк может быть неустойчивым к
конечным возмущениям, тогда как при В > Вк оно неустойчиво
к произвольным возмущениям. Стационарные распределения кон-
центраций, полученные на ЭВМ, представлены на рис. 56.
Сравнение результатов численного интегрирования и резуль-
татов, полученных аналитически, показало, что последние каче-
ственно правильно описывают решения для стационарных дис-
сипативных структур. При В близких к В к (при вычислении амп-
литуд распределений) точность равна 5—10%. Несколько хуже,
но качественно верно в рассмотренных приближениях описы-
вается форма решения для стационарных диссипативных структур.
На примере рассмотренной в данной работе модельной систе-
мы (6.14) можно увидеть, что свойства возникающих в ней ста-
ционарных диссипативных структур определяются соотношения-
ми между скоростями химических реакций и переноса вещества.
Форма этих структур зависит и от граничных условий, но дина-
196
мика процессов становления (в частности, наличие жесткого ре-
жима возникновения) качественно одинакова при различных гра-
ничных условиях. Системы рассмотренного тина в некотором
смысле «грубые». Так, например, простейшие конечно-разностные
модели, учитывающие лишь нелинейные свойства системы и в
известном смысле диффузий веществ в системе, имеют качествен-
но те же режимы, что и распределенная система. Эта же «грубость»
систем наблюдается и при образовании динамических диссипа-
тивных структур — стоящих концентрационных волн (Василь-
ев, Романовский, 1975).
Подчеркнем то обстоятельство, что возможность жесткой ди-
намики стационарной диссипативной структуры, типична для
концентрационных систем. Действительно, ее наличие связано
с квадратичными членами (6.14), а члены этого типа всегда при-
сутствуют в обсуждаемых системах — это связано с условием
положительности концентрационных переменных. Анализ трех-
компонентных моделей, приведенных в некоторых работах, по-
казал, что и для них характерно наличие жесткой динамики (Жа-
ботинский, 1974; Field, Noyes, 1974).
Граничные условия (6.12), по-видимому, характерны лишь
для случаев, когда моделируются реакции, проводимые в «про-
бирке». Ситуации, в которых следовало бы использовать условие
(6.11), как правило, выглядят весьма искусственными. В то же
время для реальных биологических объектов типичны ситуации,
в которых следует использовать граничные условия смешанного
типа. В работе этот случай не рассматривался, но, поскольку
оказалось, что режимы стационарных диссипативных структур
качественно похожи в случаях (6.11) и (6.12), то и при граничных
условиях смешанного типа будут наблюдаться такие же явления.
В теоретической биологии представления о диссипативных
структурах играют значительную роль при рассмотрении процес-
сов морфогенеза, дифференцировки тканей ит. д. (Глансдорф, При-
гожин, 1973; Романовский и др., 1975, Zhabotinsky, Zaikin, 1973).
Во всех этих процессах жесткая динамика стационарных дисси-
пативных структур может играть важную роль. В самом деле,
в мягком режиме система может управляться только изменениями
параметров и форма возникающих стационарных диссипативных
структур может зависеть от начальных условий, которые в дос-
таточной степени случайны. Другая ситуация при жесткой ди-
намике: сигнал начала построения диссипативной структуры
может быть подан при помощи изменения концентрации специфи-
ческого вещества (х или у в рассмотренной модели) на краю за-
нятого системой пространства — при этом в стационарной дис-
сипативной структуре будет строиться период за периодом, т. е.
диссипативная структура как бы достраивает себя. Такие режимы
возникновения стационарных диссипативных структур, по-ви-
димому, более реальны для биологии.
ДИНАМИЧЕСКИЕ ДИССИПАТИВНЫЕ СТРУКТУРЫ
В, А, Васильев
В биохимических или химических гомогенных системах, имею-
щих авто- или кросскаталитические реакции возможно периоди-
ческое во времени протекание реакций. Например, в работе Гланс-
дорфа и Пригожина (1973) рассмотрены автоколебания в «точеч-
ной» системе, соответствующей распределенной системе (6.14).
«Точечной» — называется система уравнений, получаемая из
распределенной, если в последней формально положить все коэффи-
циенты диффузии равными нулю; подробно физический смысл
этого термина обсуждается в работе Романовского и др. (1975).
Большое количество примеров биохимических точечных автоко-
лебательных систем и их математических моделей содержится в
работах Жаботинского (1974) и Селькова (1975).
С другой стороны, исследования точечных систем хотя и не-
обходимы, но не достаточны для понимания процессов, происхо-
дящих в биологических объектах, имеющих относительно большую
протяженность — например, большую, чем ]ZZ)Zxap, где D —
коэффициент диффузии, ТХаР —- характерное для системы вре-
мя (в частности, период колебаний) (Романовский и др., 1975).
В данной работе, не учитывая явлений сопряженного и активного
транспорта, будет показано, что в распределенных системах с
простой диффузией могут возникать пространственно неоднород-
ные режимы автоколебаний, например, концентрационные стоя-
чие волны. Такие волны будем называть динамическими дисси-
пативными структурами, подчеркивая, что они возможны лишь в
термодинамически открытых системах и их пространственная пе-
риодичность аналогична периодичности стационарных диссипа-
тивных структур. Ниже в основном обсуждаются квазигармони-
ческие автоколебательные режимы, т. е. рассматриваются систе-
мы с «малой» нелинейностью, а некоторые результаты, относящие-
ся к релаксационным системам приводятся с целью подчеркнуть
тот факт, что пространственно неоднородные автоколебательные
режимы типичны для диффузионных распределенных систем.
Анализ квазигармонических автоколебаний в (6.7) удобно на-
чать с определения координат особой точки соответствующей то-
чечной системы, т. е. получения однородного стационарного ре-
шения системы (6.7), а затем исследовать его устойчивость. Как
указывалось в предыдущей статье, однородное стационарное со-
стояние неустойчиво к бесконечно малым возмущениям, если
среди корней характеристического уравнения (6.10) имеются
корни с положительными действительными частями. Автоколеба-
ния возникнут в случае четного числа таких корней и будут ква-
зигармоническими, если отношение ReKJJn^. мало.
198
Рис. 57. Типичная длч двухком-
понентной системы зависимость
Re от k (1) в случае gQ (к) > 0.
Одни из возможных типов зависи-
мости Re kfc от к (2) для трехкомп-
онентной системы go (к) > О
Коэффициенты характеристического уравнения (6.10) в слу-
чае двухкомпонентных моделей имеют следующий вид:
qx ( • к) = -Ми - М22 + kW (Dn + Z>22) (6.31)
q0 (. ..,&) = det (M) — к2 л1 (DnM22 + Z>22MU — Z>uZ>22n2A:2),
где использованы те же обозначения, что и при написании урав-
нений (6.8). Из (6.7) следует, что, если точечная система уравне-
ний имеет устойчивую особую точку (Мп + М22 < 0), то в соот-
ветствующей распределенной системе автоколебания не могут
возникнуть в результате малых возмущений. Если же М1г +
+ Л722 > 0, то в распределенной системе могут возбуждаться
моды с различными к, меньшими чем к (см. рис. 57 (1).
Нелинейные члены в уравнениях (6.8) обусловливают взаимо-
действие мод и их устойчивость (неустойчивость). Если получать
приближенные решения в виде одной моды, то нельзя будет вы-
яснить вопрос о ее устойчивости к возмущениям с произвольной
длиной волны. Исходя из этого, решение системы уравнений (6.8)
в случае граничных условий (6.12) (непроницаемые «стенки» на
границах линии) следует искать в виде:
со
Zi = ак (0 cos (лАт) = 1, • , и)- (6.32)
fc=0
Подставляя (6.32) в (6.8) и используя метод гармонического ба-
ланса по пространственному аргументу, получим бесконечную
систему уравнений для аК (/) в обыкновенных производных. Ес-
тественно, получение точного решения этой системы нисколько
не проще решения исходной системы уравнений. Но при таком
подходе легко ограничить задачу и исследовать взаимодействия
малого числа мод, например, парные взаимодействия между мо-
дами с к = 0 и всеми остальными с к ф 0. Более того, обычно
199
представляют интерес только к к, а это уже достаточно простая
задача. Полученные системы четырех уравнений точно такие же,
как и в случае модели исследуемой реакции, протекающей в двух
связанных посредством диффузии объемах (для численного сов-
падения коэффициентов следует формально считать,что л2 =2).
Мода (стоящая волна) с к 0 устойчива только тогда, когда в
двух связанных объемах возможны устойчивые несинфазные ко-
лебания. В работе Васильева и Романовского (1975) было пока-
зано, что в случае квазигармонических режимов в этих объемах
для двухкомпонентных систем, устойчивыми являются только
синфазные колебания. Так что в распределенных двухкомпонент-
ных системах одномодовые автоколебания с к 0 неустойчивы,
устойчив синфазный режим колебаний, т. е. стоящие волны не
возможны.
В случае граничных условий (6.11) (стационарные концентра-
ции на границах линии) решение следует искать в виде следую-
щего ряда:
Zi(t, г) = (t) sin(лАт) (Z = 1, п). (6.33)
к=1
Из (6.33) и (6.7) следует, что (поскольку к не меньше 1) при до-
статочно больших коэффициентах диффузии стационарное состояние
устойчиво, хотя и в точечной модели были возможны автокоде’
бания, т. е. диффузия в случае граничных условий (6.11) стабили-
зирует однородное стационарное состояние. Как правило, квази-
гармонические автоколебания возникают с к — 1, а эту моду едва
ли целесообразно считать динамической диссипативной структу-
рой. Поэтому можно утверждать, что в двухкомпонентных систе-
мах таких структур быть не может.
Перейдем к рассмотрению трехкомпонентных систем. Для та-
ких систем коэффициенты характеристического уравнения (6.10)
имеют вид:
g0(..., к) = — del Л? + л2А:2[У|£>;.У;! — л2/г2 4-
4“ ^11^33^22 4“ ^22^33^11) 4“ J (6.34)
gi (..., к) = — л2А2 [-0ц (Л^22 4“ М33) 4- Р22(МП 4- М33) 4“
4- O33 (Мц 4" М22) — я? к2 (ОцО22 4'ОцОзз4-£)22Озз)];
к) = У, — Мц + n2k2Du;
где Мц — минор Мн-го элемента матрицы М (6.8). Если g0 <
<С 0, то характеристическое уравнение имеет нечетное число кор-
ней с положительными действительными частями — в этом слу-
чае (6.8) может иметь решения типа стационарных диссипатив-
200
ных структур с периодом 2/Л, рассмотренных выше. Поэтому мы
будем предполагать, что g0 0. В этом случае уравнение (6.10)
всегда имеет хотя бы одип отрицательный корень. Если выполня-
ется условие (Демидович, 1967):
go (• gi (• • •> *)g2 (• ..,*)> 0, (6.35)
то остальные два корня имеют положительные действительные
части; однородное стационарное состояние неустойчиво к возму-
щениям с длинами волн, для которых выполняется условие (6.35).
Сопоставляя (6.34) и (6.35) можно заметить, что возможны трех-
компонентные системы, характеристические уравнения которых
имеют корни С Rekk ДЛЯ к GE &maxL причем /cmin > 0,
см. рис. 57 (2), что является важным различием между клас-
сами двух и трехкомпонентных систем. Как правило, для этих к
Rehk\Imh \ << 1 и | Imh | > | |- Таким образом, в распреде-
ленных трехкомпонентных системах диффузия может приводить
к возникновению автоколебаний даже, если в точечной системе их
быть не может.
Рассмотрим пример одной модельной системы, построенной на
основе широкоизвестной двухкомпонентной системы (Глансдорф,
Пригожин, 1973 уравнения 15.1 и 15.2). Дополним эту систему
третьим уравнением и для удобства химической интерпретации
запишем в концентрационных переменных:
X = А + х2у — (В 4- 1) х + Dxx х
^у = В — х2у + DVy-fr2y (6.36)
Л Я2
А А = W + Рх - RA + Daa А,
где В, W, Р, R — постоянные коэффициенты, a A (f, г) описыва-
ет поведение концентрации вещества «кросскатализатора»; А
и х в этой модели образуют дополнительную цепь автокатализа.
При достаточно больших по сравнению с Dxx и Dyy величинах
Daa график зависимости от к для системы (6.36) имеет вид, пока-
занный на рис. 576.
Для получения квазигармонических автоколебательных ре-
шений систем типа (6.36) с граничными условиями (6.11) или (6.12)
можно использовать метод, коротко изложенный выше. Некото-
рые детали в его применении при граничных условиях (6.12) бу-
дут приведены в работе Васильева и Романовского (1976), в кото-
рой третья компонента системы является ингибитором. В настоя-
щей работе приведем только результаты, полученные из анализа
ряда трехкомпонентных систем:
1) В случае устойчивой точечной системы в распределенной
системе могут возбуждаться стоячие волны с к ей [Amin; &max]
201
(рис. 57 (2)). Естественно, присутствие нелинейных членов при-
водит к тому, что невозможны строго одномодовые режимы авто-
колебаний, но, как правило, амплитуды колебаний в одной из мод
значительно (в 10 и более раз) превышают остальные. В качест-
ве примера на рис. 58 показана зависимость амплитуды колебаний
от пространственной переменной г, полученная численным ин-
тегрированием системы уравнений (6.36) на ЭВМ. При сравнении
рис. 58 и 56 (на последнем дана стационарная диссипативная струк-
тура) становится очевидной правомерность термина; «динамиче-
ская диссипативная структура»; 2) Структуры этого типа могут
Рис. 58. Зависимость амплитуды
автоколебаний А от простран-
ственной координаты для сис-
темы уравнений (6.36)
Dxx = Dyy = °: DAA = 0.03; В =
= 3,9; W = 0,342; Р = 1,8; К — 2
(при этих параметрах точечная система
устойчива)
возникать и в случае автоколебательной точечной системы, если
отношение 7?еЛ0/7тоЛ0 мало. При этом устойчивыми могут быть
только те моды, частоты колебаний в которых превышают частоту
автоколебаний точечной системы; 3) Как правило, в системах мо-
жет возбуждаться ряд мод, большинство из которых устойчиво к
бесконечно малым возмущениям. Но динамические диссипативные
структуры с более низкими частотами колебаний относительно
менее устойчивы — обычно достаточно сравнительно небольшого
возмущения, чтобы низкочастотная перешла в более высокочас-
тотную, поэтому в системах может существовать лишь малое чис-
ло мод; 4) Низкочастотные моды могут возникнуть как результат
специфических начальных условий.
Отметим, что все вышеизложенные результаты получены ква-
зигармоническими методами, которые дают удовлетворительные
результаты, если для любого к выполняются: 1.
К сожалению, в настоящее время не существуют надежные методы
анализа распределенных релаксационных систем. Для этих си-
стем основным методом исследования моделей является численный
эксперимент, дополненный качественным анализом моделей. Как
оказалось, в случае релаксационных систем «одномодовые» стоя-
щие волны невозможны. Но и в этом случае имеются неоднород-
ные по пространству режимы автоколебаний, например: режим
ведущих центров описан в работах Жаботинского (1974), Василь-
ева и Заикина (1976), где также приводится пример решения двух-
компонентной системы типа «зыби», существуют и другие про-
странственно неоднородные режимы. Как правило, частоты коле-
баний в пространственно неоднородных режимах выше частот
колебаний соответствующих точечных систем.
Динамические диссипативные структуры и другие простран-
ственно неоднородные режимы автоколебаний, по-видимому, мо-
гут играть роль в понимании пространственной и временной ор-
ганизации биологических объектов, в известной степени, объеди-
няя эти понятия, давая четкую связь между пространственной
и временной ритмикой.
ДИССИПАТИВНЫЕ СТРУКТУРЫ И фи-ФУНКЦИЯ
А, И, Зотин
При обсуждении проблем, связанных с критерием эволюции
нелинейных термодинамических систем, было предложено раз-
биение (2.15), согласно которому в открытых системах далеких
от равновесия не вся образующаяся в результате протекания не-
обратимых процессов теплота диссипации покидает пределы систе-
мы—часть ее используется внутри системы в виде функции связан-
ной диссипации (Зотин, 1974; Зотин, Коноплев, Преснов, 1975).
Экспериментальные исследования, проведенные на дрожжах,
ооцитах лягушки и растущих рыбах (Schaarschmidt et aL, 1975;
Зотин, Шааршмидт, Лампрехт, 1975; Зотин, 19756; Грудницкий,
1975; Грудницкий, Никольская, 1976) показали реальность су-
ществования фи-функции и возможность ее изучения на биологи-
ческих объектах (см. главу 4). Главный вопрос, который возни-
кает при обсуждении проблемы фи-функции, сводится к вопросу о
том, что собой реально представляет процесс связывания теплоты
диссипации в нелинейных системах.
По моему мнению, возможны три пути фиксации теплоты дис-
сипации, образующейся при протекании нелинейных необрати-
мых процессов внутри системы: 1) локальное увеличение темпера-
туры в системе и, как следствие, локальное ускорение химических
процессов; 2) образование диссипативных структур; 3) фиксация
теплового излучения путем циклизации энергии на внутрикле-
точных структурах.
1. Быстрое образование теплоты диссипации при протекании
нелинейных необратимых процессов может приводить к локаль-
ному увеличению температуры и самоускорению самого процесса
или соседних химических реакций. Фактически в этом случае
речь идет не о реальном нагреве локальных областей системы,
а о снижении энергии активации химических реакций. Результат
связывания части теплоты диссипации может привести к увеличе-
нию скорости всех процессов в системе без регистрируемого уве-
личения температуры. Этот вывод легко проверить на живых объек-
203
тах. Известно, что чем меньше величина организма, тем выше у
пего интенсивность дыхания (2.8) и теплопродукции (4.6). Сле-
довательно, организмы небольшого размера дальше отстоят от
состояния равновесия, чем более крупные, и скорости всех про-
цессов, протекающих в них при одинаковой температуре среды,
должны быть выше. Это же самое относится к разным стадиям
развития растений и животных: в период оогенеза или зароды-
шевого развития скорости всех химических реакций в организме
должны быть больше, чем во время роста и тем более старения
организма.
2. Как показали Пригожин и др. (Prigogine, 1969, 1972; Ni-
colis, 1971; Глансдорф, Пригожин, 1973; Пригожин, Николис,
1973), в нелинейной термодинамической системе возможно обра-
зование нестабильных структур. Эти структуры, названные
Пригожиным «диссипативными структурами», представляют со-
бой некую пространственную упорядоченность системы (напри-
мер, нестабильность Бенара, реакции Жаботинского), периоди-
ческий процесс (например, гликолиз) и др. Для всех диссипатив-
ных структур характерно, ч*го они возникают только в системах
далеких от равновесия за счет дополнительных потоков энергии,
существующих внутри системы. По моему мнению, таким допол-
нительным источником энергии может служить та часть теплоты
диссипации, которая связывается внутри системы при протека-
нии в ней нелинейных необратимых процессов, т. е. фи-функция.
Таким образом, теорию диссипативных структур, базирующую-
ся на локальной термодинамической теории Пригожина, можно,
по-видимому, связать через фи-функцию с глобальной термоди-
намической теорией, развитой в настоящей книге.
3. Очевидно, что образование теплоты диссипации в системе
должно сопровождаться возникновением теплового, инфракрас-
ного излучения. Это излучение может поглощаться в соседних
областях системы, но может и использоваться непосредственно
в виде энергии возбуждения макромолекул и внутриклеточных
структур.
Так, согласно теории Васильева (1955, 1956, 1959), подробно
изложенной ниже (см. стр. 205), взаимодействие электромагнит-
ного излучения с макромолекулами или внутриклеточными обра-
зованиями, например, хлоропластами, может приводить к цикли-
зации энергии в виде энергии возбуждения. Предполагается, что
молекулярные, структурные ансамбли могут играть роль электро-
магнитных колебательных контуров — «молекулярных колеба-
тельных контуров» (Васильев, 1956).
При прохождении электромагнитной волны резонансной часто-
ты через такой молекулярный колебательный контур происходит
циклизация свободной энергии и переход ее в энергию возбуж-
дения структурного ансамбля. Например, можно рассматривать
гранулы хлорофилла в качестве «молекулярных колебательных
контуров», в которых при длине волны фотохимически активного
204
света возникает резонанс между колебаниями электромагнитного
поля в падающей волне и электромагнитными колебаниями в гра-
нуле, что ведет к возбуждению хлорофилла (Васильев, 1959).
Если эта теория окажется справедливой, то можно себе пред-
ставить еще один механизм связывания теплоты диссипации внут-
ри живых систем. Например, можно считать, что такие внутри-
клеточные структуры, как митохондрии, являются «молеку-
лярными колебательными контурами», способными связывать
энергию электромагнитного теплового излучения. Тепловое излуче-
ние, возникающее в результате диссипации энергии при проте-
кании необратимых процессов, будет улавливаться в этом случае
митохондриями и создавать или усиливать электрическое поле на
мембранах митохондрий. Это может привести к усилению скоро-
сти синтеза АТФ и всех других энергетических процессов в клетке.
Так как указанное усиление электрического поля митохондрий
может осуществляться только за счет процессов диссипации, его
можно относить к категории диссипативных структур. Очевидно,
что можно придумать различные варианты опытов, позволяющих
проверить возможность преобразования энергии инфракрасного
излучения в энергию электрического поля митохондрий или дру-
гих внутриклеточных или надклеточных образований.
ЗНАЧЕНИЕ ЦИКЛИЗАЦИИ СВОБОДНОЙ ЭНЕРГИИ
В БИО-ФИЗИКО-ХИМИЧЕСКИХ ПРОЦЕССАХ
С. С. Васильев
В термодинамике необратимых процессов живые организмы
относятся к типу открытых систем, т. е. систем, способных обмени-
ваться с окружающей средой энергией и веществом. При этом вво-
дится представление о стационарных процессах, протекающих в
условиях сохранения локального равновесия между соответст-
вующими промежуточными стадиями преобразования энергии и
вещества. Поэтому используемое в данном случае понятие «энтро-
пия» имеет характер «энтропии по Клаузису»,т. е. некоторой функ-
ции состояния. Но это значит, что применение в биологии термо-
динамических понятий в их строгом смысле может иметь лишь
ограниченное значение.
В самом деле, если, например, рассматривать две заряженные
мембраны каких-либо клеточных органелл как пластины плоского
конденсатора, то запас свободной энергии электрического поля
между мембранами будет равен:
Z = ^Lq, (6.37)
205
где е0 — «электрическая постоянная» системы единиц СИ; е —
относительный диэлектрический коэффициент и Q - объем среды
между мембранами; Е — напряженность электрического поля в
рассматриваемом объеме. При этом здесь Z является «свободной
энергией» в термодинамическом смысле этого слова (Абрагам-Бек-
кер, 1936), поскольку коэффициент 8 в общем случае является
функцией температуры, давления и химического состава рассмат-
риваемой среды. Но напряженность Е может меняться независи-
мо от 8 (например, при 8 — const; Т — const) в результате не-
посредственного контакта данных мембран с какими-либо источ-
никами электрического напряжения. Также заряд мембран мо-
жет изменяться под воздействием переменного электрического
поля потока излучения, исходящего от внешнего источника. При
этом, если рассматриваемый конденсатор является частью неко-
торого колебательного контура или является частью эквивалент-
ного такому контуру волновода, то способность восприятия энер-
гии излучения такими системами посредством резонанса зависит
от их геометрической структуры, определяющей присущие им
«собственные частоты» или «моды колебаний». Необходимым ус-
ловием подобного накопления свободной энергии является ус-
тойчивость структуры рассматриваемых систем по отношению к
тепловым воздействиям. Отсюда следует, что употребляемое в
данном случае понятие «система» не имеет того смысла, который
этот термин имеет в термодинамике. Поэтому подобного рода сис-
темы следует называть структурными ансамблями, накопление
же свободной энергии в таких ансамблях, сопровождаемое пра-
вильной сменой фаз колебания, можно назвать циклизацией сво-
бодной энергии, а колеблющуюся безватную энергию — структурной
энергией.
Однако только формальное сопоставление живых организмов
с макроскопическими резонаторами еще недостаточно для объясне-
ния роли циклизации свободной энергии в кинетике био-физико-
химических процессов. В связи с этим следует отметить, что Гур-
вич, развивая теорию биологического поля, ввел представление
о «циркуляции фотонов между неравновесными констелляциями»
молекул (Гурвич, 1944).
В наших же исследованиях (Васильев, 1955, 1956, 1959) мы
стремились выяснить биофизический смысл устойчивых структур-
ных элементов клеток и их органелл, имеющих специфическую
морфологию. При этом в качестве первого примера были избраны
гранулы хлоропластов, наличие которых, как известно, необхо-
димо для осуществления процесса фотосинтеза. Результаты этих
работ излагаются ниже в несколько упрощенном, но зато и в бо-
лее наглядном виде, причем приводятся соответствующие допол-
нения и обобщения.
206
Цйклйзация свободной энергии
в хлоропластах
Известйо, что гранулы хлоропластов состоят из набора дис-
кообразных пластин, диаметры которых имеют порядок 0,2—2 мк.
Пластины состоят из липидов. Гидрофобная цепь молекулы хло-
рофилла (фитол) погружена в эти пластины, а гидрофильное
кольцо молекулы (хлорофиллин) располагается между ними. Зна-
чение относительного диэлектрического коэффициента между пла-
стинами полагается равным е = 1,58. На ранних стадиях раз-
вития гранул электронный микроскоп обнаруживает их сетчатую
структуру (Рабинович, 1951).
Представляется естественным допустить, что подобная пластин-
чатая структура, свойственная многим органеллам живых клеток,
в частности — митохондриям — должна обеспечивать какие-то
особые биофизические функции. В соответствии с этим мы пред-
положили, что пластины гранул выполняют роль плоских конден-
саторов, соединенных в грануле «последовательно». Тогда, если
обозначить через ха долю поверхности пластины, способной не-
сти электрические заряды, и через ра — радиус пластин, емкость
единичного конденсатора рассматриваемого типа в единицах
системы СИ будет равна:
а~ Ла
(6.38)
Здесь 80 — электронная постоянная системы СИ; е — относи-
тельный диэлектрический коэффициент; яра — геометрическая
площадь пластин; Ла — расстояние между ними. Индекс «а»
означает, что мы рассматриваем величины, характеризующие
«структурные ансамбли». Для того, чтобы такой ансамбль мог
являться электромагнитным колебательным контуром, к емкости
Са должна быть присоединена индуктивность La. Значение этой
индуктивности можно получить на основании следующих рас-
суждений. Представим себе две пластины конденсатора в виде
цилиндра радиуса ра и пусть эти пластины заряжаются за счет
энергии поля электромагнитной волны, создающей «ток смещения»
величиною ia между пластинами. Такой ток создаст магнитное
поле, напряженность которого на расстоянии
стин, согласно системе СИ, будет равна: Н?
ра от центра пла-
= W -О^ввдно,
что на оси рассматриваемого цилиндра Н = 0; поэтому на рассто
янии г от этой оси соответствующая напряженность будет равна
#г==^—;Г. Объем рассматриваемого цилиндра
радиуса г равен
Q — яг2Ла; тогда c?Q — 2лгЛайг. Таким образом, вся магнитная
207
энергия Wm тока ia в объеме £2 при р, = 1 будет равна:
Wm= У“2-йй = п&г = — •
О i
Отсюда получается:
La~ 8л •
(6.39)
(6.40)
И тогда из (6.38) и (6.40) имеем:
(6.41)
где Ха — длина электромагнитной волны в вакууме, соответствую-
щая круговой частоте соа, и с0 == l/F^^oHo — скорость света в
вакууме.
Аналогичное выражение получается и в том случае, если
рассматривать гранулу как объемный цилиндрический резона-
тор СВЧ-волн (Сарбахер, Эдсон, 1947). Поэтому, если в (6.41)
положить ]Леха/2 1, то для диаметра 2раа = 0,2—2,0 мк длина
волны Ха окажется имеющей порядок 0,3—3,0 мк, чему соот-
ветствуют волны, активные в фотосинтезе, волны митогенетиче-
ского излучения и волны инфракрасного излучения, соответствую-
щие колебаниях многих функциональных групп молекул, прини-
мающих участие в био-физико-химических процессах.
Следует при этом указать, что при последовательном соеди-
нении к конденсаторов емкостью Са их общая емкость равна
Ск = Са/к; при последовательном же соединении к индуктивно-
стей La их общая индуктивность равна Lk = kLa. Поэтому соб-
ственная частота набора пластинчатых гранул = l/|Ac\Lfc
будет равна частоте соа = 1/|ЛСаЬа.
Таким образом, гранулы хлоропластов могут рассматриваться
как колебательные макроскопические системы, способные резо-
нировать на электромагнитные волны той же длины, которые яв-
ляются активными для возбуждения молекул хлорофилла.
Заметим, что для стабилизации генераторов электромагнит-
ных волн (клистронов) в современной СВЧ-технике применяется
наполнение полости их волноводов аммиаком (Smithet al., 1947),
ротационные переходы в молекулах которого сопровождаются
поглощением и излучением электромагнитных волн в том же диа-
пазоне частот, на который настраивается клистрон. Такого рода
системы, к которым могут быть отнесены и рассматриваемые нами
гранулы хлоропластов, а также и любые органеллы клеток, мо-
гут быть названы элементарными структурными ансамблями с
согласованными частотами. При таком согласовании не только
208
молекулы могут стабилизировать макроскопические колебания
в ансамбле, но и концентрация возбужденных молекул, включен-
ных в ансамбль, будет поддерживаться на уровне, отличном от
термодинамического равновесия.
Коэффициент фокусировки свободной энергии
Было показано (Васильев, 1956), что для учета влияния
резонансного накопления энергии в структурном ансамбле на про-
цессы возбуждения включенных в ансамбль молекул удобно моле-
кулы представлять моделью «молекулярного колебательного
контура», состоящего из индуктивности Lnm, емкости Спт и сопро-
тивления Rnm, соединяемых последовательно. При этом введен-
ные здесь индексы пт означают, что возбуждение молекул рас-
сматривается как квантовый процесс п т, т. е. как переход
молекулы с квантового уровня т на квантовый уровень п. Соот-
ветственно, собственная круговая частота рассматриваемого моле-
кулярного колебательного контура может быть обозначена че-
рез (Опт.
Введение величины Lnm может быть мотивировано тем, что
во всех системах единиц отношение е2/те, где е — заряд электрона
и те — его масса, имеет ту же размерность, что и отношение
l2/Lnm, где I — условное расстояние между зарядами в класси-
ческом осциллирующем диполе. При этом квантовый характер
рассматриваемого процесса может быть учтен тем, что вместо
массы те вводится величина me/fnm, где fnm — «сила осцилля-
тора», определяемая соотношением
2(0 т
/,im = -^-e|rnm|2, (6.42)
причем | гпт | — «матричный элемент», выражающий вероят-
ность рассматриваемого квантового перехода п т. Таким об-
разом, величина Lnm может быть определена равенством:
/2 тп
= (6.43)
' пт
Тогда можно положить:
^пт пт
здесь утп — «коэффициент затухания», имеющий размерность
[сек-1], который в простейшем случае может быть приравнен
коэффициенту спонтанного излучения Эйнштейна Атп; в более
же общем случае он может быть представлен как показатель ско-
рости преобразования энергии поглощенных квантов hvnm в дру-
гие формы энергии. В силу этого, величина Rnm может рассмат-
риваться как «сопротивление излучения» или вообще как «сопро-
209
Рис, 59, Объяснения в тексте
тивление активной нагрузки» процесса квантового возбуждения,
рассматриваемого молекулярного контура.
Включение молекулярных контуров в структурный ансамбль
может быть представлено следующими последовательно усложня-
ющимися схемами.
На рис. 59, а изображен конденсатор См между обкладками
которого находится N молекул, находящихся на квантовом уров-
не возбуждения тп, причем для простоты здесь принято, что
N = 2. Этот конденсатор сначала мы считаем «ориентированным»
относительно внешнего поля, т. е. полагаем, что нормаль п к
пластинам конденсатора совпадает с направлением вектора Е
напряженности внешнего поля, которое колеблется с круговой
частотой со. Тогда параллельное пластинам активное сопротивле-
ние будет равно Rn = Rnm/N и выделяемая в конденсаторе См
на этом сопротивлении активная мощность Wm может рассматри-
ваться как мощность, расходуемая на квантовое возбуждение
молекул п <— т.
Электрическое поле электромагнитной волны напряженностью
Е может зарядить пластину конденсатора, сообщая им действую-
щее напряжение Um в соответствии с чем величина Wm будет оп-
ределяться равенством:
t/2
WM = . (6.45)
Теперь представим себе, что к конденсатору См присоединя-
ется последовательно индуктивность LM в силу чего конденсатор
См преобразуется в конденсатор колебательного контура, кото-
рый мы обозначим как «структурный ансамбль а». При этом рас-
чет показывает, что емкость такого контура Са ~ См» Под индук-
тивностью же контура мы будем подразумевать вычисляемую по
формуле (6.40) индуктивность La = LM- Кроме того, для просто-
210
ты, будем считать, что входящая в (6.43) величина I = 1а ~ Ла.
Таким образом, ансамбль а может быть представлен следующей
схемой на рис. 59, б. Пусть собственная частота <о0 такого ансамб-
ля будет удовлетворять условию соа = conm ~ со. Тогда рассмат-
риваемая электромагнитная волна сможет сообщить пластинам
конденсатора Са напряжение Uа UMi так как в этом случае
возникает явление «резонанс напряжений». И тогда выделяемая
на сопротивлении 7?# активная мощность будет:
и*
wa=^->wM.
Схема рис. 59, в эквивалентна схеме рис. 59, б но в этом
случае сопротивление 7?# между пластинами конденсатора Са за-
менено сопротивлением 7?а, включенным последовательно конден-
сатору Са и индуктивности La, При таком преобразовании соблю-
дается соотношение (Гинкин, 1948):
1
(6.46)
(6.47)
Л" * л„см •
(6.48)
(6.49)
При резонансе напряжений подводимое извне к контуру напря-
жение U (в нашем случае U = UM) равно напряжению на сопро-
тивлении Ra, а напряжение Ua между пластинами конденсатора
больше этого напряжения во столько раз, во сколько раз ем-
костное сопротивление конденсатора Хс = 1]Саыа больше сопро-
тивления 7?а. Это отношение называется «добротностью контура»
и может быть обозначено через Qa. Таким образом, с учетом (6.47)
получается:
и Хг 1
<2а = -(Ц = fl" = Я С <0 RNCa6ia.
М а а а а
Тогда из (6.45), (6.46), (6.48) найдем, что отношение мощностей
Wa, Wm будет равно:
“ Qa &a
Здесь величина d обозначает «логарифмический декремент зату-
хания» рассматриваемого контура а; величина d/л показывает
относительное уменьшение напряжения Uа за один цикл колеба-
ния, обусловленное преобразованием безваттной структурной
энергии контура в ваттную энергию, расходуемую на активном
сопротивлении контура.
Следует отметить, что обозначения Qa, да, аа в кратной за-
писи означают величины Qnm,a> $пт,а> °пт,^ так как все они от-
носятся к процессу п <— т, протекающему в ансамбле а.
На схемах рис. 59, г, д показано, что к «ориентированному»
ансамблю а присоединяется антенна, состоящая из индуктивности
211
Ца), емкости C(a) и сопротивления 7?(a). В этом случае образо-
ванный из ансамбля а и указанной антенны ансамбль А будет
иметь индуктивность La, емкость С а и сопротивление Ra, причем
будут иметь место соотношения:
С С
La == La 4- Z/(a) La\ С А = ТГ-пг-ТтД- <с Ra — Ra + ^(a)*
а "Г ° (a)
(6.50)
Ввиду условия LA>La и С а < Са может оказаться, что соб-
ственная частота со а ансамбля А не будет отличаться от соа, т. е.
будет удовлетворяться условие соа — Поэтому мож-
но считать, что отличие со а от соа будет изменять напряжение Uа
между пластинами конденсатора Са до значения kaUa, где коэф-
фициент ка может рассматриваться как коэффициент электриче-
ской связи ансамбля с питающим его электромагнитным полем;
также можно представлять его как «коэффициент модуляции»
напряженности поля в ансамбле.
Далее, можно считать, что антенна как бы увеличивает длину
отрезка 1а до некоторой длины 1а, на котором внешнее поле соз-
дает электрическое напряжение. С другой же стороны, такой про-
вод, будучи жестко скреплен с конденсатором Са, при поворотах
может изменять угол Фа между нормалью к пластинам конденса-
тора и направлением электрического вектора внешнего поля.
Поэтому можно сказать, что величина Ia cos Фа является «дейст-
вующей величиной антенны», изменяющей напряжение kaUa rq
lA
значения cos Фа- В соответствии с этим, с учетом неравен-
Л
ства Са Ф С а, определим «коэффициент» настройки ансамбля на
внешнее поле равенством:
к А = Ло-у--^-С031|>А. (6.51)
а а
Тогда найдем, что напряжение Uа между пластинами конденсато-
ра Са при рассматриваемых условиях окажется равным:
UA = kAUa. (6.52)
Наконец, собственное активное сопротивление 7?(а) антенны мо-
жет рассматриваться как «сопротивление утечки» энергии ансамб-
ля. Наличие этой утечки изменит выделяемую на сопротивлении
мощность vRa/RA раз. Таким образом, с учетом (6.47), (6.48),
(6.51), (6.52), найдем, что на сопротивлении R^ в активном
ансамбле А будет выделяться мощность:
А—
А *N*A «X Ло)
1+ %Qa
(6.53)
1 + RNQa
212
И тогда, с учетом (6.45) отношение мощностей Wa, Wm будет
равно:
_ _ _ кА@а __ Гр __ Гр _ я2 /R кд\
Опт, А - <5д - ------/Г" - Qa - Qnm, А - ~£— • (6.54)
м 1 । cP ”пт,А
1 + HN Ча
Теперь учтем, что скорость возбуждения молекул 7Vm, испыты-
вающих квантовой переход п ч- т равна (Шпольский, 1950):
d\N 1
= пт [^mL (6.55)
где [7VnJ, [7Vn] — концентрации возбужденных молекул, находя-
щихся на квантовых уровнях тп, тг; uVt пт— спектральная плот-
ность излучения частоты v = vnm; и коэффициент Bvm возбужде-
ния Эйнштейна, который с учетом системы единиц СИ, имеет вид:
Bnm=-ggr|rnm|2. (6.56)
Если во внешнем электрическом поле спектральная плотность
излучения пт и при резонансном поглощении этой энергии
структурным ансамблем А ее плотность возрастает до значения
птвпт, а, то скорость процесса возбуждения молекул Nm в
ансамбле А соответственно возрастает до значения:
------ = Впти», птРпт, А [^тп]а- (6.57)
При этом будет иметь место равенство:
В пт ^пт, А = ge^21 rnm Р ^пт, А = I г пт, А |2* (6.58)
т. е. указанное увеличение спектральной плотности можно ин-
терпретировать как увеличение вероятности квантового перехода,
определяемого величиной | гП7П |2 до значения:
| ГпТп, А Р = | Г пт Р бдти, А* (6.59)
Это значит, что коэффициент бпт,А может быть определен как
коэффициент структурного усиления квантового перехода п <— ш
в элементарном структурном ансамбле А с согласованными час-
тотами. Структурный ансамбль А может также рассматриваться
и как структурная линза, фокусирующая свободную энергию
внешнего электромагнитного поля на квантовых переходах п ч—
т. Поэтому коэффициент оП7П> д может также называться коэф-
фициентом фокусировки свободной энергии.
Известно, также, что матричные элементы | гпт | являются
элементами соответствующих матриц (| г„т |), которые определя-
ют характер распределения квантовых переходов в соответствую-
щих атомах или молекулах.
213
Поэтому и величины | гппчд | должны рассматриваться как
элементы некоторой матрицы ( | rnzn>A[ ), которая может характе-
ризовать распределение’ или фокусировку свободной энергии в
органеллах клеток. Путем сочетания отдельных матриц (| |)
для органелл клеток можно перейти к клеткам и органам, подобно
тому, как схему сложной радиотехнической системы можно рас-
сматривать состоящей из некоторой совокупности «четырехполюс-
ников», сочетающихся между собою по законам перемножения
матриц. Другими словами, подобная совокупность матриц (| гП7П) а|)
может рассматриваться как принципиальная схема системы управ-
ления радиостанции живого организма.
Приведенные выше рассуждения вполне применимы и для слу-
чая акустических волн. Это может иметь весьма важное значение,
так как, согласно Шредингеру, хромосомы могут быть сравнимы
с апериодическими твердыми кристаллами (Шредингер, 1947).
Поэтому весьма возможно, что хромосомы и другие органеллы
клеток могут функционировать не только как чисто электромаг-
нитные, но и как электроакустические структурные ансамбли.
При этом надо учесть, что согласно современным представлениям,
электроны, ионы и «дырки» в твердых телах образуют плазму во
многом аналогичную плазме газовых разрядов. В такой плазме
обнаруживаются различные типы электромагнитных волн, спо-
собные взаимодействовать с акустическими колебаниями кри-
сталлической решетки (Канер, Яковенко, 1975). К этому надо
добавить, что различие типов электронно-дырочной проводимости
в смежных слоях биологических структур может обеспечивать
этим структурам свойства транзисторов, т. е. самовозбуждающих-
ся генераторов колебаний релаксационного типа. Таким образом,
указанные эффекты могут обеспечивать участие органелл клеток
в процессах генерирования, усиления и преобразования различ-
ных ритмов электромагнитных и электроакустических колебаний,
начиная от низкочастотного радиодиапазона до оптической и
ближней ультрафиолетовой области.
В соответствии с этим, в настоящее время установлено, что,
например, многие клетки центральной нервной системы способны
генерировать ритмические импульсы даже в отсутствие внешних
раздражений. С помощью внутриклеточных микроэлектродов в
таких клетках обнаруживаются постоянные медленные колеба-
ния мембранного потенциала, сопровождаемые одиночными или
множественными разрядами «потенциалов действия» (Бабский и
др., 1966).
Кинетика эндотермического синтеза
в структурном биологическом ансамбле
Нам представляется целесообразным попытаться представить
в наиболее простой форме возможную энергетическую схему ки-
нетики какого-либо эндотермического синтеза, протекающего в
214
структурном биологическом ансамбле без участия тепловой акти-
вации веществ, вступающих в реакцию.
Для определенности будем считать, что рассматриваемый син-
тез является матричным синтезом белка в рибосомах, причем ог-
раничимся рассмотрением кинетики начального звена такого
синтеза, протекающего при участии фермента Ф, макроэргиче-
ских молекул М и молекул субстрата 7V, превращающихся в пер-
вичный продукт синтеза X*. Активные формы молекул будем
обозначать знаками: Ф(1), Л/(2), А(3), где нижние индексы^ 1, 2,
3 будут означать номера сортов рассматриваемых активных час-
тиц, концентрации которых в рибосомах мы обозначим соответ-
ственно через тгх, дг2, п3. При составлении кинетических урав-
нений будем пользоваться способом обозначений, который при-
менялся нами в кинетическом анализе цепных реакций (Васильев,
1959).
Инициирующей стадией реакции будем считать активацию
фермента Ф за счет поглощения им кванта энергии hv. Этот про-
цесс (1) изобразим так:
ф (6.60)
Источником квантов hv может являться какая-либо хемолюминес-
центная реакция, протекающая, например, в некоторой области
клетки, из которой поток излучения может распространяться по
эндоплазматической сети а-цитомембран, несущей на всем своем
протяжении многочисленные рибосомы и могущей выполнять роль
волновода для электромагнитных волн длиною волны порядка
0,2 мк, так как диаметр каналов этой сети имеет порядок 0,1 мк.
Также источником распространяемых по эндоплазматической
сети электромагнитных колебаний могут быть процессы зарядки
и разрядки клеточных мембран, питаемых каким-либо ионофор-
ным механизмом. Важно то, что плотность свободной энергии,
доставляемой в виде квантов hv к рибосомам, может усиливаться
или ослабляться в зависимости от резонансной настроенности на
кванты hv рассматриваемого волновода, а также подобной же
настроенностью сцепляющихся в полисомные структуры рибосом.
Таким образом, усиление или ослабление процесса (6.60) за счет
изменения такой настроенности будет вполне эквивалентно пере-
ключению контактов на электростанции, питающей свободной
энергией процесс (6.60).
Возможные физико-химические процессы (2), ведущие к дезак-
тивации активных частиц Ф(1) кратно представим так:
Ф(\) Ф. (6.61)
Далее допустим, что активная частица Ф*Г) взаимодействует с
макроэргической молекулой М, приводя ее в активное состояние
в соответствии со схемой процесса (3):
Ф(П + м М(‘2). (6.62)
215
Процесс (4) дезактивации частиц М(2), по аналогии с (6.61), пред-
ставим так:
М(2) -> М. (6.63)
Но учтем, что макроэргическая молекула, взаимодействуя с q
молекулами фермента Ф, может приводить их в активное состоя-
ние. Этот процесс (5) представим так:
М(2) + дФ -> дФ*(у). (6.64)
Другими словами, мы допускаем, что инициирующий процесс
(6.60) может иметь весьма малую интенсивность, хотя величина
кванта hv или соответствующая амплитуда электрического на-
пряжения, «включающие» процесс (6.60), должны быть достаточно
большими. Концентрация же частиц Ф*± может быть значитель-
но увеличена за счет совокупности процессов (6.62), (6.64), пред-
ставляющих простейшую цепную реакцию, в которой размноже-
ние частиц осуществляется за счет энергии распада М(2).
Частица M(2) может, конечно, активировать и молекулы
субстрата N в соответствии со схемой процесса (6):
+7V^2V(*3) (6.65)
Частицы 7V(3) могут дезактивироваться по схеме процесса (7):
Л\*з) N, (6.66)
а также реагировать с другими молекулами субстрата L, давая
первичный продукт синтеза X*, который, для общности, можпо
считать также находящимся в активном состоянии. Эту реакцию (8)
запишем так:
7V(3) + L -> X* (6.67)
Обозначим концентрации молекул Ф, М, N, L, X*, а также
и квантов hv соответствующими знаками, заключенными в квад-
ратные скобки, и будем отмечать константы скоростей рассмот-
ренных процессов (6.60) — (6.67) индексами этих процессов.
Тогда систему кинетических уравнений, определяющих концен-
трации пх, тг2, n3, [X*] рассматриваемых активных частиц, запи-
шется в такой форме:
= kl [ф] + 9*5 [ф] «2 — *2«1 — *3 [М] П1
— = к3 [М] — fc4n2 — к3 [Ф] п2 — к3 [N]п2
= *в [Л/J п2 — Л7«3 — к3 [L] п3 (6.68)
di — и3.
216
Введем обозначения:
ЛДЛуЦФ] = иг к3 [£] = гх3
дЛ5[Ф] = г12 к2 + Л3[М] = Si
к3[М1 = г21 к4 + Л8[Ф] + к3 [АГ] = $8
к3 [7V] = г32 к, + к3 [£] = $3.
С учетом (6.69) система (6.68) представится так:
dn-t ,
= Ui 4- Г12п2 — Silli
dn<>
= r2lW! — S27Z2
Удобно также ввести обозначения:
Г21 _ . Г12 _ . Г32 __ м . ГХЗ _
— — со21, — — ®12, — — со32, — — сох3,
о2 л2 Лд
а также положить:
^12^21 =
(6.69)
(6.70)
(6.71)
(6.72)
Здесь величины £/, Sj означают обобщенные константы скорости
исчезновения активных частиц сорта f, /; величины ri7- означают
обобщенные константы скорости появления частиц сорта i, в
результате реагирования частиц сорта / по тому или иному нап-
равлению. Соответственно этому величины coi7- означают матема-
тические ожидания появления частиц сорта i в результате реаги-
рования частиц сорта /. При этом из сопоставления (6.69), (6.71)
видно, что co2i < 1’, 0032 < 1» но, поскольку число 7, образующих-
ся по схеме активных частиц Ф(1) может быть q> 1, возможен
случай, когда со12> 1. Тогда может оказаться, что и определяе-
мая равенством (6.72) величина со > 1. В нашей работе (Василь-
ев, 1964) было показано, что значение величины со определяет
характер сопряженно-цепных реакций. Если со’> 1, реакция
протекает е самоускорением вплоть до момента ее торможения, нас-
тупающему в результате расхода исходных веществ; если же
со < 1, то концентрации активных частиц, принимающих участие
в развитии цепной реакции, нарастает вначале по линейному от
времени закону, который в конечном итоге сменяется стационар-
ной стадией, когда:
dn.
W = (6.73)
Здесь мы ограничимся рассмотрением именно этого случая, т. е,
217
co <1. Тогда из (6.70) — (6.73) найдем:
= ^зСОзг^гх- (6-74)
При условии со < 1 величина 1/1 — со означает «длину цепи»
рассматриваемого цепного процесса. Скорость же инициирующего
процесса (6.60) представим так:
U1 = = *1 [0][/zv] = Вп1иъп1ап1чА [Ф], (6.75)
где кг = Вп1 — коэффициент Эйнштейна фотовозбуждения моле-
кул Ф, находящихся в нормальном квантовом состоянии т = 1
и переходящих при процессе (6.60) в состояние «п»; [Av] = иъп1
ал1, а — спектральная плотность излучения в структурном ан-
самбле А, причем и^п1 — спектральная плотность энергии в по-
токе излучения, приносящего кванты Av, а &п1,а — «коэффициент
структурного усиления», определяемый в общем виде формулой
(6.54).
Величину
ХЛ = (6.76)
можно назвать «коэффициентом цепного сопряжения» рассматри-
ваемого первичного процесса фотовозбуждения (6.60), результа-
том которого является синтез продукта X* (ставя индекс А при
мы, хотим отметить, что структура ансамбля А может также
оказывать влияние на величину сог-7- и со). Таким образом, скоро-
сть нарастания концентрации продукта на основании (6.74),
(6.75) и (6.76) может быть представлена в виде:
J — Вп1 [Ф] ni^m, аХа* (6.77)
Конечно, для разных типов био-физико-химических процессов
величины, входящие в формулу (6.77), могут иметь разное значе-
ние. Но, по-видимому, в большинстве процессов, протекающих
в биологических структурных ансамблях, инициирование реакций
осуществляется «инженцией» некоторой порции свободной энер-
гии, т. е. процессом, аналогичным процессу фотовозбуждения:
всегда плотность инжентируемой энергии может быть увеличина
или уменьшена за счет «фокусирующего» действия ансамбля, в
котором протекают реакции, степень вовлечения которых в синтез
продукта X* будет определяться тем или иным выражением, эк-
вивалентным коэффициенту %а-
Биологические структурные ансамбли
и диссипативные структуры
В известной книге Глансдорфа и Пригожина (1973) рассмат-
риваются различные схемы циклических процессов ферментных
реакций, протекающих при участии, так называемых, «диссипа-
218
тйвных структур». В качестве одного из возможных примеров та-
ких структур приводят фермент фосфофруктокиназу. Здесь гово-
рится, что указанный фермент «может находиться в двух конфор-
мациях: неактивной и активной, имеющей большее сродство к
субстрату. Конформационное равновесие — кооперативный эф-
фект; это означает, что, если одна из единиц, образующих данную
молекулу, изменяет свою конформацию, то одновременно с ней
это делают и остальные» (Глансдорф, Пригожин, 1973, стр. 241).
При этом смещение продуктов реакции конформационного равно-
весия в сторону активной формы дает «дестабилизирующий вклад
в производство избыточной энтропии». Как всегда, здесь имеется
ввиду «энтропия по Клаузиусу». Но это значит, что конформаци-
онные процессы рассматриваются как зависимые от температур-
ных флуктуаций. Именно это обстоятельство и соответствует наз-
ванию таких систем — диссипативные структуры.
С нашей точки зрения, такие структуры можно рассматривать
как структурные ансамбли, обладающие столь малой добротно-
стью, что в них не может осуществляться резонансное накопление
свободной энергии. И в этом случае, конечно, процесс диссипации
энергии может рассматриваться как «производство энтропии»,
а задержку этого рассеяния — как «дестабилизация диссипации».
Но, во-видимому (и это самое главное), биологические структур-
ные ансамбли — не «диссипативные структуры», а «резонансные
системы с большей добротностью». Поэтому вопрос о регулирова-
нии биологических систем выходит за пределы методов, применяе-
мых в настоящее время в термодинамике необратимых процессов.
В заключение отметим, что для вопроса о регуляции распре-
деления свободной энергии в клетках и их органеллах особое
значение могут иметь процессы, связанные с возможными сверх-
проводящими свойствами входящих в их состав молекул. Так,
известно, что явление сверхпроводимости основано на электрон-
фононном взаимодействии и что сверхпроводящее состояние ха-
рактеризуется некоторой «длиной когерентности £» (Харрисон,
1972). Смысл величины состоит в том, что любые возмущения в
какой-либо точке сверхпроводника обязательно сказываются на
свойствах сверхтекучих электронов, находящихся на расстоянии
порядка или меньше g от этой точки. При этом весьма интересно,
что величина g имеет порядок ~2000 А, т. е. порядок длины
именно тех эклектромагнитных волн, которые оказываются ак-
тивными при фотосинтезе, при восприятии зрительных впечат-
лений и при митозах (Quickenden, QueHee, 1974). Это и указывает
на возможную роль явлений сверхпроводимости и когерентности
в протекании и регулировании ряда био-физико-химических про-
цессов в живых организмах.
VII
ВЕРОЯТНОСТЬ состояния
И УПОРЯДОЧЕННОСТЬ
БИОЛОГИЧЕСКИХ СИСТЕМ
В биологии давно уже используются такие термины, как вы-
сота организации и упорядоченности биологических систем
(Бауэр, 1935; Гурвич, 1944; Опарин, 1957; Шмальгаузен, 1972).
Несмотря на это, до сих пор они носят большей частью интуитив-
ный характер, точных критериев упорядоченности живых систем
пока нет (см. Блюменфельд, 1974). Вместе с тем вопрос об упоря-
доченности, о высоте организации животных имеет большое зна-
чение для некоторых проблем индивидуального развития орга-
низмов и биологической эволюции животных и особенно для ре-
шения вопросов о направлении эволюционного прогресса, о пу-
тях биологического совершенствования и закономерностей макро-
эволюции (Шмальгаузен, 1972). Понятие упорядоченности живых
систем важно и для многих других проблем общей биологии. Осо-
бенно остро эти вопросы встают, когда исследователи пытаются
определить понятие жизни — выяснить те отличия, которые су-
ществуют между живыми и неживыми системами (Опарин, 1966).
Обычно утверждается, что живые системы являются организо-
ванными системами, где все процессы упорядочены. Однако когда
дело доходит до конкретных определений жизни, то со времени
Клода Бернара так и не удалось ввести точное определение жизни.
Не менее важные проблемы возникают в биологии развития, ког-
да речь заходит о проблемах морфогенеза (Гурвич, 1944; Белоусов,
1971). Происходит ли в процессе формообразования и дифферен-
цировки увеличение степени упорядоченности живых систем или
развивающиеся организмы непрерывно приближаются к наибо-
лее вероятному состоянию? Интуитивно кажется очевидным, что
в процессе дифференцировки происходит увеличение упорядочен-
ности живой системы, но тогда как согласовать это утверждение
с более или менее доказанным положением, что во время развития
и роста организмов происходит непрерывное приближение живой
системы к более вероятному состоянию (глава 2)?
Все эти вопросы трудно разрешить при отсутствии точных
критериев понятий организации и упорядоченности. Между тем,
исходя из статистического истолкования диссипативной функции
и функции внешней диссипации (2.36), т. е. исходя из самых об-
щих понятий современной науки, удается выработать такой кри-
терий (Коноплев, Зотин, 19756). Естественно, что этот критерий
220
применим не только к живым системам, но и к неживым. Поэтому,
для использования функции внешней диссипации в качестве кри-
терия упорядоченности живых организмов, приходится вводить в
этот критерий чисто биологические закономерности, свойственные
только живым организмам (Коноплев, Зотин, 19756; Зотин,
Озернюк и др., 1975).
Зависимость (2.36) получена двумя путями: в одном случае,
исходя из теории флуктуаций Эйнштейна (стр. 65), в другом —
рассматривая развитие и рост организмов, как стохастический,
марковский процесс (стр. 69). Это позволяет думать, что критерий
упорядоченности, о котором будет речь идти ниже, теоретически
достаточно хорошо обоснован. Как же можно использовать соот-
ношение (2.36) для характеристики степени упорядоченности
термодинамической системы?
С точки зрения термодинамики, наиболее вероятным состоя-
нием системы являются состояния равновесия (полный молекуляр-
ный хаос в системе). Если система уклонится от равновесного
состояния, то вероятность ее состояния уменьшится и увеличится
степень упорядоченности системы. Чем дальше находится термо-
динамическая система от состояния равновесия, тем более упоря-
доченной она должна быть в стабильном состоянии. С другой
стороны, уклонение от равновесия сопровождается увеличением
величины функции внешней диссипации и уменьшением вероят-
ностного состояния системы. Поэтому, вероятность состояния и
величина функции внешней диссипации характеризуют степень
упорядоченности процессов, протекающих в открытой системе.
Мы можем, следовательно, использовать понятие вероятности
состояния для характеристики упорядоченности живой системы,
вычисляя относительную вероятность состояния по формуле
(2.36).
Ниже приводятся попытки использования соотношения (2.30)
и (2.36) для решения некоторых вопросов, связанных с происхож-
дением прокариот, для характеристики прогрессивной эволюции,
классификации животных и изменения упорядоченности живых
систем в процессе биологической эволюции и индивидуального
развития организмов.
ВОЗМОЖНЫЙ МЕХАНИЗМ
ПРОИСХОЖДЕНИЯ БАКТЕРИЙ
Н. Д. Озернюк, А. И, Зотин, В, А. Коноплев,
А, А. Зотин
Согласно теории Опарина (1957, 1966), точка зрения которого
принимается большинством современных исследователей, живые
организмы возникли из первичного бульона, насыщенного угле-
водами, аминокислотами, нуклеотидами и др. Первичные орга-
низмы (протобионты), по мысли Опарина, представляли собой ко-
ацерватные системы, эволюция которых постепенно привела к
возникновению всего разнообразия жизни на Земле. С этой точ-
ки зрения, эволюция первичных форм жизни шла монофилити-
ческим путем. В последние годы, однако, получили широкое
распространение представления о симбиотическом происхожде-
нии клеток и организмов, т. е. о полифилитическом происхож-
дении организмов. Особенно убедительные данные представлены
относительно возможности возникновения митохондрий и хлоро-
пластов в животных и растительных клетках в результате симбио-
за соответственно бактерий и синезеленых водорослей с амебоид-
ными первичными организмами (Nass, 1969; Пинус, Метлицкая,
1970; Рудин, Уилки, 1970; Margulis, 1970; Schnepf, Brown, 1971;
Нейфах, 1973, 1974а; Остроумов, 1973; Тахтаджян, 1973; Meyer,
1973). Предполагается, что из первичного бульона независимо
друг от друга возникли и микроорганизмы, и значительно более
крупные по размеру амебоидные клетки. Вопрос о том, какая
точка зрения является более правильной, можно решить, опи-
раясь на рассмотрение вероятности состояния живых систем (Зо-
тин, Озернюк и др., 1975). Поставим этот вопрос следующим
образом: какова вероятность возникновения из первичного буль-
она микроскопических живых систем по сравнению с более круп-
ными организмами. Чтобы решить этот вопрос, напомним, что
первичный бульон, согласно Опарину, представлял собой равно-
весную среду, в то время как живые организмы являются сильно
неравновесными открытыми системами. Очевидно, что вероят-
ность возникновения из равновесной среды открытой, неравно-
весной системы тем меньше, чем больше степень неравновесности
процессов, протекающих в открытой системе. В термодинамике
степень неравновесности системы пропорциональна величине
удельной диссипативной функции для линейных необратимых
процессов и величине функции внешней диссипации — для нели-
нейных процессов. Как говорилось, первичный бульон представ-
лял собой равновесную среду. Поэтому, возникающие на первых
этапах эволюции живые системы (протобионты) должны были
представлять собой системы близкие к равновесию. К таким
системам применимы соотношения термодинамики линейных не-
обратимых процессов, для которых справедливо соотношение
222
(2.30) или, переходя к удельной диссипативной функции, соот-
ношение вида:
(-£ -1) . (7.1)
Из уравнения (7.1) следует, что величина удельной диссипатив-
ной функции обратно пропорциональна вероятности состояния
системы: чем больше диссипативная функция, тем меньше вероят-
ность состояния данной системы и наоборот. Следовательно, веро-
ятность возникновения живой системы, удельная диссипативная
функция которой велика, из равновесного первичного бульона
значительно меньше, чем системы с малой величиной диссипативной
функции.
Удельную диссипативную функцию организмов, с известным
приближением (см. Зотин, Грудницкий, 1970), можно прирав-
нять интенсивности теплопродукции или дыхания (2.1). Исходя
из этого мы можем оценить возможность появления микроорга-
низмов в качестве первых живых систем на Земле по сравнению
с более крупными организмами. Для этого используем хорошо
обоснованную в физиологии зависимость между интенсивностью
дыхания и весом тела животных (2.8). Из зависимости (2.8) сле-
дует, что чем меньше размеры (вес) организма, тем больше интен-
сивность дыхания и, следовательно, тем больше величина удель-
ной диссипативной функции (2.1), тем меньше вероятность состоя-
ния организма (7.1). Бактерии представляют собой наименьшие
по размерам живые организмы, способные к самостоятельному
существованию, поэтому, вероятность их или аналогичных им
по размерам живых систем возникновения из первичного бульона
наименьшая по сравнению с более крупными организмами.
Исходя из уравнения (7.1), можно оценить в относительных едини-
цах эту вероятность. Для этого примем, что вероятность состоя-
ния микроорганизмов равна Ри а вероятность всех остальных
организмов — Р2. Тогда:
_ ^1^2 (^0 — ^1) П
^2 “ ^PjPo-Л,) ‘
Так как вероятность равновесного
чем вероятность состояния живой,
можно считать, что (Ро — Р) ~ Р.
(2.8) и (7.2):
Р2 _ «,^2
Pi a2W\'T\'
состояния намного больше,
неравновесной системы, то
Получаем, используя (2.1),
(7-3)
Принимая значение констант для микроорганизмов равными
а = 0,084 кал/час, b = 0,25 (см. Hemmingsen, 1960) и Тг = Т2 —
= const, получаем формулу:
Pl _ /И^ОДЗ
Pl ~ иг,/ ’
(7.4)
223
по которой можно оценить вероятность возникновения живых си-
стем разного веса из равновесной среды. Пусть вероятность состоя-
ния условного организма весом 1 г равна 1, тогда, используя
(7.4), получаем:
Pw = И/0,25, (7.5)
где Pw — относительная вероятность состояния, зависящая только
от веса организма; W — вес организма.
Легко подсчитать по формуле (7.5), что относительная вероят-
ность возникновения микроорганизма весом 10“12 г равна 0,001,
условная вероятность возникновения живой системы размерами
10"6 г равна 0,03, размерами 10"3 г — 0,2, размерами 1 г — 1,
размерами 103 г — 6, размерами 106 г — 32. Отсюда следует, что
первые организмы, появившиеся на Земле, вряд ли имели микроско-
пические размеры. Скорее всего, это были огромные скопления жи-
вого вещества порядка нескольких тонн. Вероятность возник-
новения таких систем из первичного бульона в 104—10б раз
больше, чем живых систем микроскопических размеров (Зотин,
Озернюк и др., 1975). Эти образования были намного проще
современных организмов и характеризовались низкой величиной
константы а. Малейшее нарушение их размеров в результате
механических или иных причин приводило к уменьшению вероят-
ности существования таких систем и они исчезали. Однако внутри
этих гигантских протобионтов могли возникнуть меньшие по раз-
мерам живые системы, обладающие большой диссипативной функ-
цией. Вероятность их возникновения внутри первичных прото-
бионтов значительно выше, чем прямо из первичного бульона.
Подобные системы могли покидать пределы материнского организ-
ма при механических разрушениях последних. Они имели несом-
ненное преимущество перед первичными протобионтами, поэто-
му эволюционный процесс, постулированный Опариным (1957,
1966), для таких систем должен был привести к вытеснению ги-
гантских протобионтов системами меньшего размера с более высо-
ким значением диссипативной функции. Этот процесс мог много-
кратно повторяться вплоть до появления микроскопических
организмов, обладающих достаточно сложной организацией и ма-
лой вероятностью существования. Таким образом, монофилитиче-
ская теория Опарина лучше согласуется с выводами, вытекающи-
ми из термодинамикинеобратимыхпроцессов, по сравнению с тео-
рией симбиотического происхождения первичных организмов.
В связи со сказанным маловероятной кажется гипотеза о про-
исхождении митохондрий животных и растительных клеток из
бактериоподобных организмов, а фотосинтезирующих органелл
растительных клеток — из синезеленых водорослей. Факты, кото-
рые обычно приводят для доказательства этой гипотезы, касаю-
щиеся сходства митохондрий и бактерий, допускают и обратное
толкование: на их основе вполне допустимо предположить, что не
митохондрии произошли из прокариот, а прокариоты из мито-
224
хондрий. Как было показано выше, эта последняя гипотеза лучше
согласуется с термодинамикой необратимых процессов, так как
она не требует, чтобы бактерии и синезеленые водоросли появля-
лись на Земле раньше первичных организмов.
Сходство митохондрий и бактерий
Исследования последних 10—15 лет показали, что митохондрии
являются в значительной степени автономными структурами, спо-
собными к саморазмножению. Этот вывод был сделан после обна-
ружения в митохондриях собственной ДНК и системы ее репли-
кации, а также автономного аппарата транскрипции и трансляции
(Roodyn et al., 1961; Nass, Nass, 1963; Nass, 1969; Пинус, Метлиц-
кая, 1970; Рудин, Уилки, 1970; Хансон, Иванова, 1971 и др.).
Учитывая, что митохондрии имеют размеры и форму, близкую
к бактериям, а также много других свойств, близких к бактериаль-
ным, все большее распространение получила гипотеза происхож-
дения митохондрий из бактерий (Казакова, 1967; Скулачев, 1969;
Рудин, Уилки, 1970; Тахтаджян, 1973; Нейфах, 1974а; Зотин,
Озернюк и др., 1975 в).
Важнейшее сходство митохондрий и бактерий касается строе-
ния и свойств мембран: считается, что внутренняя мембрана ми-
тохондрий аналогична клеточной мембране бактерий. Показано,
например, что существует большое сходство между дыхательными
комплексами внутренней мембраны митохондрий и мембраны бак-
териальной клетки, а также сходство в липидном составе этих
мембран (Рудин, Уилки, 1970). Большое сходство между мито-
хондриями и бактериями найдено в отношении строения и функ-
ционирования ДНК. Известно, что ДНК бактерий имеет кольце-
вое строение. Оказалось, что мт-ДНК также имеет кольцевое
строение (Nass, 1969; Пинус, Метлицкая, 1970; Хансон, Иванова,
1971). В митохондриях содержится собственная ДНК—полиме-
раза, отличающаяся от ядерной, а репликация мт-ДНК происходит
независимо от репликации яДНК. Сходство между митохондриаль-
ной и бактериальной ДНК было обнаружено и при определении
частоты встречаемости динуклеотидов. В митохондриях обнару-
жены транспортная и рибосомальная РНК, а недавно было пока-
зано существование собственной информационной РНК (Нейфах,
1973, 1974а).
Значительную автономность и сходство с бактериями обнару-
живает белоксинтезирующая система митохондрий (Roodyn et al.,
1961; Рудин, Уилки, 1970; Пинус, Метлицкая, 1970). Так, напри-
мер, константа седиментации бактериальных рибосом, равная
70S, близка константе седиментации митохондриальных рибосом,
равной 55—65S, и обе они отличаются от константы седиментации
цитоплазматических рибосом, равной 80S (Borst, Grivell, 1971).
Большое сходство обнаружено в отношении влияния антибиоти-
ков на синтез белка в митохондриях и в бактериях. Так, антибио-
225
Таблица 10
Сходство между митохондриями и бактериями (Зетин, Озернюк и др., 1975)
Признак Клетка эукариот Митохондрии Бактерии
Строение Сходные размеры и форма Внутренняя мембрана митохондрии аналогична мембране бактерий: сход- ство дыхательных ансамблей мембран; сходство состава липидов в мембра- нах; сходная система пермеаз
Размножение Деление
Днк В составе оформ- Нить ДНК вместо оформленного ядра ленного ядра Нитчатая Кольцевая Связана с гистона- Связана с мвхмбранами ми
Аппарат реплика- ции ДНК, транс- крипция и транс- ляция Независимый аппарат репликации ДНК (ДНК-полимераза), транскрип- ции (РНК-полпмераза) и трансляции (т-РНК, рибосомы, активирующие ферменты) м РНК стабильна м-РНК нестабильна в инициации в инициации транс- трансляции участвует формилметго- ляции участвует ни л-т-РНК метионил-т-РНК Одинаковая чувствительность РНК- полимеразы к этидиумбромиду и ан- тибиотикам
Рибосомы 806* 55—656* 706* Не синтезируют бе- Синтезируют белок с фаговой м-РНК лок с фаговой м-РНК
Чувствитель- ность к: хлорамфени- колу циклогексп- миду Не чувствительны Чувствительны Чувствительны Не чувствительны
тик хлорамфеникол подавляет включение меченых аминокислот
в митохондрии и в бактерии и не действует на включение в опытах
с цитоплазматическими рибосомами (Borst, Grivell, 1971). Полу-
чены и другие данные о действии антибиотиков на синтез белков
в митохондриях и бактериях. Все эти данные, как и другие наблю-
дения о сходстве между митохондриями и бактериями, полученные
разными авторами (см. Казакова, 1967; Nass, 1969; Скулачев,
1969; Рудин, Уилки, 1970; Meyer, 1973; Нейфах, 1973, 1974а)
суммированы в табл. 10. Как видно, имеется много сходных черт
как в строении, так и в биохимии митохондрий и бактерий.
226
Возмсжный механи; м сбразований митохондрий
п возникновения бактерий в процессе эволюции
В гипотезе происхождения митохондрий из бактерий механизм
возникновения митохондрий более или менее ясен: сначала симби-
оз микроорганизма с первичной амебоидной клеткой, затем внед-
рение части генома бактерии в геном ядра, потеря полной автоном-
ности симбиотического организма и превращение его в клеточную
органеллу. Трудный для понимания пункт этой гипотезы состоял
в том, что не вся бактериальная ДНК должна была ассоцииро-
ваться с геномом хозяина при возникновении митохондрий —
часть ее оставалась в органелле в виде мт-ДНК. Однако не-
давно Мейер (Meyer, 1973) показал, что мт-ДНК правильней со-
поставлять не с ДНК нуклеоида бактерий, а с кольцевой плазмид-
ной ДНК бактериальной клетки. Исходя из факта наличия двух
видов ДНК в бактериальной клетке, легко себе представить, что
в процессе превращения бактерий в митохондрии происходило
внедрение нуклеотидной ДНК в геном хозяина, и образование
мт-ДНК из плазмоидпой ДНК бактерии.
Принимая обратную гипотезу, гипотезу возникновения про-
кариот из митохондрий, естественно было бы также рассмотреть
возможный механизм этого процесса. И здесь, конечно, трудно
обойтись без предположений; важно только, чтобы они были до-
статочно правдоподобными. Первое и главное предположение, ко-
торое мы используем, состоит в утверждении, что в процессе эво-
люции митохондрии возникали из ядерной оболочки первичных
клеток. Это предположение, ввиду наличия в митохондриях коль-
цевой ДНК, равноценно предположению, что к внутренней мемб-
ране ядра клеток примыкает (или примыкала в период появления
первичных митохондрий) кольцевая ДНК, в которой закодиро-
ван синтез структурных белков и аппарат синтеза этих белков.
Принимая это допущение, легко объяснить появление двух видов
ДНК при возникновении бактерий из митохондрий.
Предположение о том, что митохондрии могут образовываться
из ядерной оболочки, высказывалось неоднократно (см. Ленин-
джер, 1966; Казакова 1967; Рудин, Уилки, 1970). В последние
годы в связи с данными, полученными при изучении строения и
биохимии ядерных мембран, многие другие авторы пришли к вы-
воду, что цитоплазматические мембраны образуются из ядерной
(Збарский, 1969, 1972; Пинус, 1973). Возможность образования
митохондрий из ядерной мембраны подтверждается следующими
фактами. Митохондрии часто расположены на поверхности или
в складках ядерной оболочки (Hartman, 1954; Ornstein, 1956;
Band, 1959; Дмитриева, 1963; Stang-Voss, Straubesand, 1970). Ядер-
ная оболочка иногда образует изгибы и выпячивания, от которых
отрываются и переходят в цитоплазму пузырьки (см. Збарский,
1969). Образование такого рода пузырьков из ядерной мембраны
наблюдали в ооцитах (Baud, 1959), а также при патологических и
227
фармакологических воздействиях (David, 1964). Учитывая, что
ядерная оболочка состоит из двух мембран толщиной 70—80 А,
легко себе представить, что при образовании выпячиваний или пу-
зырьков из ядерной оболочки могут образовываться митохондрио-
подобные структуры. Это особенно очевидно, если принять во
внимание полученные в последние годы в лаборатории Збарского
данные о ферментном составе и биохимии ядерных оболочек
(Збарский и др., 1968; Збарский, 1969, 1972; Кузьмина и др., 1970;
Бульдяева и др., 1972; Бухвалов, 1973). Этими авторами показано,
что изолированные из печени крыс ядерные мембраны содержат
все основные ферменты дыхания и дыхательной цепи. Мембраны
способны к полноценному дыханию, окислительному фосфорили-
рованию и активному транспорту, т. е. обладают теми же свойст-
вами, что и мембраны митохондрий или бактерий. Следовательно,
образующиеся из ядерной мембраны пузырьки по своему строению
и по своим функциям могут быть вполне аналогичными митохонд-
риям. Кроме этих косвенных данных существуют и прямые дока-
зательства образования митохондрий из ядерной мембраны. Наи-
более убедительными в этом отношении являются результаты, по-
лученные Брантом и Паппсом (Brandt, Pappas, 1959). Эти авторы
показали, что из выпячиваний ядерной оболочки амебы форми-
руются митохондрии (рис. 60). Следует отметить, что отшнуровав-
шиеся структуры являются не просто пузырьками ядерной мемб-
раны, а полноценными митохондриями, содержащими кристы.
Последние появляются еще до отшнурования митохондрий от ядер-
ной мембраны. Наконец, в ряде работ митохондрии были обнару-
жены внутри ядра. Так Гофман и Григ (Hoffman, Grigg, 1958)
показали наличие митохондрий в ядрах лимфоцитов крыс и мы-
шей, а также в сперматогониях и сперматоцитах крыс. Было по-
казано также присутствие митохондрий в ядре сперматид (рис. 61)
(Stang-Voss, Straubesand, 1970).
Таким образом, гипотеза об образовании митохондрий из ядер-
ной оболочки имеет достаточное экспериментальное обоснование.
Несмотря на это, требуется еще одно допущение, которое нуждает-
ся в экспериментальном обосновании: следует предположить, что
к внутренней мембране ядра прикреплены кольцевые ДНК типа
мт-ДНК, а также примыкают рибосомы митохондриального типа.
Тогда образующиеся из ядерной оболочки пузырьки могут при
некоторых условиях становиться митохондриями. Таких данных
пока нет, хотя сведения о наличии в изолированных ядерных
мембранах ДНК имеются в литературе (Перевощикова и др.,
1968; Franke et al., 1970; Збарский, 1972). Необходимо еще дока-
зать, что эта ДНК не захвачена из ядра при выделении ядерных
мембран и что она является нуклеиновой кислотой типа мт-ДНК.
Во всяком случае, следует подчеркнуть, что гипотеза об образова-
нии митохондрий из ядерной мембраны не может существовать без
объяснения механизма появления в митохондриях автономного
кодирования и синтеза части структурных белков мембран.
228
Рис, 60. Образование митохондрий из ядерной оболочки у амебы (Brandt,
Pappas, 1959)
М — митохондрии, N — ядро
Рис. 61. Митохондрии внутри ядра сперматиды Eisenia foetida (S ang-
Voss, Staubesand, 1970)
Рис. 62. Две стадии возможного механизма образования бактериоподобных
структур из митохондрий
I — стадия перехода части генома, кодирующего синтез митохондриальных белков,
* вырост ядерной мембраны; i I — стадия отшнуровки бактериоподобного организма от
ядерной мембраны и его размножение в цитоплазме клетки. Обозначения: м — мито-
хондрии, Я — ядро, Д — ДНК ядра, В — вырост ядерной мембраны, Я — кусок ДНК.
содержащий гены, необходимые для синтеза белков митохондрий, R — рентгеновский
квант, б — бактериоподобный организм
Описанный механизм образования митохондрий не исключает,
конечно, и другие способы размножения митохондрий, например
путем деления, но этот способ может лежать в основе размноже-
ния митохондрий в периоды особенно быстрого их увеличения
в клетках (в период оогенеза, после клеточного деления и т. д.).
Если описанный механизм образования митохондрий сущест-
вовал в тот исторический период, когда только возникали первые
клеточные органеллы, то легко себе представить, каким образом
в процессе формирования митохондрий могли возникнуть полно-
стью автономные бактериоподобные органеллы, способные к само-
воспроизведению (рис. 62). Для этого достаточно представить
себе, что тем или иным путем в образующиеся выросты ядерной
оболочки клетки попадает та часть ядерной ДНК, в которой за-
кодированы синтезы всех белков митохондрий, отсутствующих
в мт-ДНК. В эти выросты, конечно, могут случайно попадать и
другие части ядерной ДНК, образующейся в ядре в результате по-
ломки хромосом, например под действием радиации. Однако такая
новая органелла может существовать в клетке только в том случае
если в выросты ядерной мембраны попадет часть ядерного генома,
кодирующего вместе с мт-ДНК все митохондриальные белки.
Пузырек, окруженный двумя мембранами и содержащий полный
код и аппарат синтеза белка, превращается при этом в автономный
организм, способный размножаться внутри материнского орга-
229
низма. Очень быстро такая авторазмножающаяся органелла обра-
зует внутри клетки колонию бактериообразных организмов (см.
рис. 62). Такая клетка, несомненно, должна погибнуть и попав-
шие в среду первичного бульона новообразованные бактерии лег-
ко могут существовать самостоятельно или внедряться в другие
организмы.
НАПРАВЛЕНИЕ
ЭВОЛЮЦИОННОГО ПРОГРЕССА ОРГАНИЗМОВ
А. 2Г. Зотин, В. А. Коноплев
В настоящее время считают, что нет объективных критериев
эволюционного прогресса и поэтому расчленяют это понятие на
ряд представлений: о биологическом или адаптационном прогрес-
се, о биотехническом, групповом, неограниченном и биогеоценоти-
ческом прогрессе (Тимофеев-Ресовский и др., 1969). Вместе с тем
подчеркивается, что палеонтологические данные с несомненностью
свидетельствуют о появлении в процессе эволюции все более со-
вершенных форм животных и растений (Тимофеев-Ресовский и др.,
1969). Следовательно, речь идет не о том,что в процессе эволюции
не происходит совершенствования организмов, а об отсутствии
точных критериев этого прогресса. Такой критерий, по-видимому,
может быть разработан, опираясь на статистическую интерпрета-
цию функции внешней диссипации и использование специфиче-
ской для живых организмов зависимости (2.6) и (2.8) (Коноплев,
Зотин, 1975а; Зотин, Озернюк и др., 1975).
Как видно из соотношения (2.36), функция внешней диссипа-
ции обратно пропорциональна вероятности состояния термодина-
мической системы. Так - как функцию внешней диссипации с
известным приближением можно приравнять интенсивности дыха-
ния, то для живых организмов мы можем связать вероятность
состояния с величиной интенсивности дыхания и на этой основе ввес-
ти критерий упорядоченности биологических систем. На рис. 63
приведены данные, суммированные Хеммингсеном (Hemmingsen,
1960), о зависимости скорости теплопродукции, измеренной не-
прямым методом, от веса организмов разных групп. Константы
в уравнении (2.6), согласно данным Хеммингсена (рис. 63), равны:
для одноклеточных 0,084 кал/час, для пойкилотермных животных
0,69 кал/час, для гомойотермных животных 19,68 кал/час. Как
видно из рис. 63, эти три группы животных, которые заведомо от-
личаются друг от друга по высоте организации, четко отличаются
и по уровню дыхания, а, следовательно, по вероятности состояния.
Мы можем охарактеризовать относительную вероятность сущест-
230
Рис. 63. Зависимость скорости дыхания от веса тела у одноклеточных, по
килотермных и гомойотермных (Hemmingsen, 1960)
вования каждой из этих групп, если, например, вероятность со-
стояния одноклеточных весом 1 грамм примем за единицу. Пусть
в уравнении (7.3) вероятность состояния одноклеточных Рх = 1,
вес Wi = 1, температура 293° К, константа а = 0,084 кал/час.
Тогда относительная вероятность состояния других организмов
(Ротн) будет равна: |
Ротн = 2,86-10-“ X- И*’. (7.6)
Пользоваться формулой (7.6) для определения степени упоря-
доченности организмов неудобно, так как в эту формулу входят
размеры организма W и вероятность состояния сильно зависит
от веса. Чтобы выявить более четко зависимость величины относи-
тельной вероятности от степени упорядоченности живых систем ус-
ловимся вычислять ее только для организмов весом 1 г. Фактически,
в этом случае мы рассматриваем вероятность состояния организмов
одного веса и тем самым избавляемся от зависимости вероятности
состояния от веса организмов. В этом случае из (7.6) получаем:
Ptl = 2,86-16-“ X (7.7)
и введя обозначение:
Сг = 4-, (7.8)
а
231
приходим к критерию упорядоченности следующего вида:
С? = 3,49«103-^-, (7.9)
где Ра — относительная вероятность состояния организма весом
1 г; Сг — критерий упорядоченности.
Из критерия (7.9) следует, что степень упорядоченности и вы-
соты организации живых систем прямо пропорциональна констан-
те а в уравнении (2.8), связывающего интенсивность дыхания и
веса организмов, т. е. пропорциональна основному обмену. Сход-
ный подход к вопросу о характеристике высоты организации жи-
вых предложили Ивлев (1959, 1963) и Дольник (1968), опи-
раясь на совершенно иные соображения. Применяемые ими кри-
терии отличаются от нашего, но главное отличие состоит не в этом,
а в том, что критерий (7.9) выведен исходя из термодинамических
и вероятностных соображений, в то время как критерии Ивлева
и Дольника являются чисто эмпирическими.
Возвращаясь к вопросу о путях эволюционного прогресса, сле-
дует отметить, что критерий (7.9) может служить критерием совер-
шенствования форм жизни в процессе эволюции. Очевидно, что
живые организмы с малой вероятностью состояния и высокой
степенью упорядоченности не могли возникнуть из первичного,
равновесного бульона на ранних этапах происхождения жизни на
Земле. Только шаг за шагом в процессе биологической эволюции
происходило появление организмов с все более высокой степенью
упорядоченности. Следовательно, критерий упорядоченности (7.9)
позволяет решать вопрос об одном из важнейших направлений
биологического прогресса, который приводил к все большему уда-
лению живых систем от равновесного состояния, появлению в свя-
зи с этим высокой степени упорядоченности процессов, протекаю-
щих в организмах, и освобождению от зависимости организмов от
окружающей среды. В связи с этим следует сказать несколько
слов об ароморфозах. Согласно А. Н. Северцову, ароморфоз
представляет собой приспособительное изменение организмов,
в результате которого общая энергия жизнедеятельности взрослых
потомков повышается (см. Шмальгаузен, 1969). Ароморфоз озна-
чает усложнение организации организмов, поднятие ее на более
высокий уровень, что дает возможность организмам расширить
использование внешней среды, расширить ареал распространения
вида. Ароморфоз в первую очередь связан с повышением энергии
жизнедеятельности организмов — критерий упорядоченности (7.9)
также в первую очередь характеризует этот показатель, так как
он характеризует относительный уровень основного обмена орга-
низмов. Чтобы пояснить возможность использования критерия
(7.9), рассмотрим данные, приводимые на рис. 63. Подсчет вели-
чины критерия упорядоченности по этим данным показывает, что
одноклеточные, холоднокровные и теплокровные имеют величину
критерия соответственно 1, 8 и 222. Следовательно, мы можем выде-
232
лить два крупнейших ароморфоза, происшедших в процессе эво-
люции: при переходе от одноклеточных к многоклеточным, когда
степень упорядоченности увеличилась от 1 до 8, и при появлении
теплокровных животных, когда упорядоченность увеличилась
с 8 до 222. Следует учитывать, что данные, приводимые на рис. 63,
сильно усреднены и внутри каждой группы для животных раз-
ных типов и классов степень упорядоченности может сильно отли-
чаться. Подсчет критерия упорядоченности по данным, получен-
ным разными авторами для различных типов и классов животных^
показал (табл. 11, 12), что имеются значительные различия в ор-
Таблица 11
Критерий упорядоченности беспозвоночных животных
Тип и класс а. кал |час ь Ст Автор
Protozoa (Простейшие) 0,084 0,25 1,0 Hemmingsen, 1960
Porifera (Губки) 0,120 0,45 1,4 Ивлев, 1963
Scyphozoa (Сцифоидные) 0,055 0,15 0,6 Яковлева, 19G4
Anthozoa (Кораллы) 0,167 0,14 2,0 Ивлева, 1964
Oligochaeta (Олигохеты) 0,440 0,25 5,2 Бызова, 1965
Merostomata (Мечехвосты) 0,404 0,19 4,8 Сущеня, 1972
Crustacea (Ракообразные) 0,825 0,19 8,8 Дольник, 1968
0,625 0,24 7,4 Сущеня, 1972
Mollusca (Моллюски) 0,770 0,25 9,2 Ивлев, 1963
Insecta (Насекомые) 2,710 0,24 32,1 Wigglesworth, 1953
Таблица 12
Критерий упорядоченности позвоночных животных
Класс а. кал] час ь Ст Автор
Amphioxi (Ланцетники) 1,37 0,09 16,3 Ивлев, 19(1
Cyclostomata (Круглоротые) 1,42 0,19 16,9 Винберг, 1956
Teleostei (Рыбы) 1 ,43 0,19 17,0 Винберг, 1956
1,15 0,23 13,7 Дольник, 1968
Amphibia (Амфибии) 1,25 0,23 14,9 Brown, 1964
1,11 0,25 13,2 Hutchison, et al., 1968
Reptilia (Рептилии) 3,94 0,23 47,1 Brown, 1964
2,12 0,24 25,2 Дольник, 1968
Aves (Птицы) 18,1 0,26 215 King, Earner, 1961
35,2 0,34 410 Дольник, 1968
Mammalia (Млекопитающие) 18,4 0,27 219 Brody, 1945
15,5 0,25 185 Kleiber, 1961
233
ганизации разных животных. Так, например, кораллы и губки
по степени упорядоченности только в полтора — два раза выше,
чем простейшие, а медузы по своей организации оказались
даже ниже, чем простейшие. Позвоночные, естественно, по высоте
организации оказались выше, чем беспозвоночные, исключая, мо-
жет быть, насекомых, высота организации которых по этому кри-
терию сопоставима с рептилиями (табл. 11). Внутри типа позво-
ночных наибольший критерий упорядоченности характерен для
птиц и млекопитающих, наименьший — для бесчерепных, кругло-
ротых и рыб. Критерий упорядоченности для рептилий выше, чем
Та блица 13
Критерии упорядоченности у некоторых реликтовых животных
Животные а, кал|час ь Сг
Ракообразные (Сущеня, 1972)
Artemia salina 0,39 0,30 4,5
Apus concriformis — щитни 2,21 0,19 26,0
Общее для ракообразных 0,63 0,24 7,5
Рыбы (Винберг, 1956)
Cyprinidae (карповые) 1,19 0,19 14,2
Acipenseridae (осетровые) 1,92 0,19 23,0
Общее для рыб 1,43 0,19 17,0
для амфибий, но почти не отличается у амфибий по сравнению с
рыбами. Это согласуется, как отметил Дольник (1968), с точкой
зрения Северцова, что ароморфоз у позвоночных произошел не
тогда, когда они перешли от водного образа жизни к наземному, а
при появлении класса рептилий.
В заключение остановимся еще на одном вопросе, который
может представлять интерес для проблем, связанных с изучением
вопроса о причинах длительного сохранения в процессе эволюции
отдельных реликтовых форм животных. Как следует из табл. 13.
критерий высоты организации у щитней из подкласса жаброногих
ракообразных отличается очень высоким значением. Он равен 26
против 4,5 у Artemia salina из того же подкласса и 7,5 для всего
класса ракообразных в целом. Возможно, что ароморфоз, который
привел к появлению щитней более 200 миллионов лет тому назад,
оказался настолько важным для этой группы животных, что они
продолжают существовать и в настоящее время несмотря на все
превратности судьбы. То же самое можно сказать о такой древней
группе рыб, как осетровые. Критерий упорядоченности у этих
234
животных заметно выше, чем, например у карповых рыб и всего
класса рыб в целом. Вопрос этот, несомненно, требует дополни-
тельного рассмотрения и доказательств, однако, не исключено,
что большая величина критерия упорядоченности современных
реликтовых форм является одной из причин их успеха в борьбе за
жизнь на протяжении многих миллионов лет.
КРИТЕРИЙ УПОРЯДОЧЕННОСТИ
И ПРОБЛЕМЫ КЛАССИФИКАЦИИ ЖИВОТНЫХ
В. А. Коноплев, В. Е, Соколов, А, И. Зотин
Как было показано выше, критерий упорядоченности (7.9) мо-
жет быть использован для характеристики ароморфозов и прогрес-
сивной эволюции организмов. Естественно было поэтому предпо-
ложить, что критерий упорядоченности может быть использован
для построения классификации животных, в которой отражена
степень высоты организации и сложности организмов (Коноплев,
Зотин, 1975а; Коноплев, Соколов, Зотин, 1975).
Существуют различные подходы к классификации животных
(Майр, 1971; Антонов, 1974; Юдин, 1974). Одни из них преследуют
чисто диагностические цели, связанные с определением таксоно-
мической принадлежности, другие направлены на построение есте-
ственной системы животных, учитывающей происхождение и
эволюцию тех или иных групп животных. Возможны и другие
классификации, например, по среде обитания (водные, наземные,
почвенные животные), по способам размножения (яйцекладущие,
живородящие, яйцеживородящие), по терморегуляции (пойкило-
термпые, гомойотермные) и т. д. Все эти классификации полезны,
если только правильно очертить круг их задач и цели, которые они
преследуют.
Критерий упорядоченности (7.9) дает нам в руки достаточно
объективный критерий для классификации животных по высоте
их организации, показывая пути прогрессивной эволюции в той
или иной группе животных. Действительно, как указывалось,
критерий упорядоченности связан с вероятностью состояния ор-
ганизмов (7.8): чем больше критерия упорядоченности, тем меньше
вероятность состояния системы. Очевидно, что в момент происхож-
дения жизни на Земле возникали живые системы с большой веро-
ятностью состояния и, соответственно, с малым критерием упоря-
доченности. По мере прогрессивной эволюции организмов они все
дальше удалялись от наиболее вероятного состояния и критерий
упорядоченности, как и высота организации, их должны были воз-
растать. Это видно из табл. И, 12: критерий упорядоченности на-
235
именыпий у простейших и медуз, возрастает с повышением уровня
организации у других беспозвоночных и достигает наибольшей:
величины у птиц и млекопитающих.
Критерий упорядоченности основан на очень общих положе-
ниях науки и может быть достаточно точно измерен по интенсив-
ности дыхания взрослых организмов. Следует, однако, учитывать,
что опыты по определению интенсивности дыхания часто проводят
в несравнимых условиях и разными методами, плохо сопоставимы-
ми друг с другом. Разброс данных по стандартному обмену живот-
ных настолько велик у разных авторов и число измерений настоль-
ко мало, что чаще всего невозможно определить достоверность
полученных значений интенсивности дыхания, а, следовательно,
и достоверность различий между разными группами животных.
Мы считаем, однако, что это не может послужить препятствием
для использования критерия упорядоченности для классифика-
ции животных, так как в настоящее время важно решить принци-
пиальные вопросы, связанные с этим критерием, а уточнение и уве-
личение числа данных по дыханию организмов на современном
этапе не представляет особых трудностей.
В качестве примера использования критерия упорядоченности
рассмотрим класс млекопитающих, для многих отрядов которого
удалось подобрать более или менее достаточное число данных по
основному (стандартному) обмену (Коноплев, Соколов, Зотин,
1975). Рассмотрим различные отряды млекопитающих в том по-
рядке, в котором они расположены в общепринятой классификации
(Simpson, 1945; Соколов, 1973).
1. Отряд однопроходных (Monotheria). Представлен всего дву-
мя семействами: утконосами (Orhithorhynchidae) и ехиднами
(Tachyglossidae), из которых семейство утконосов представлено
всего одним видом. В связи с этим данных по метаболизму этого
отряда не много. Все они представлены на рис. 64 в логарифмиче-
ской системе координат, удобной для определения констант а и Ь
(см. уравнение (2.8). На этом рисунке, как и на последующих,
пунктирная прямая соответствует уравнению (2.8) для всего клас-
са млекопитающих в целом (Kleiber, 1961). Так как температура
тела млекопитающих в среднем равна 37° С или 310° К, то крите-
рий упорядоченности (7.9) для класса млекопитающих равен:
Сг = 11,2 а. (7.10)
Вычисляя константу а методом наименьших квадратов по данным,
приводимым на рис. 64, и подставляя ее значение в уравнение
(7.10), получим, что критерий упорядоченности для отряда одно-
проходных Сг = 69.
2. Отряд сумчатых (Marsupialia). Сравнительно хорошо изу-
чен в физиологическом отношении и поэтому имеется достаточно
данных по метаболизму. Все они показаны на рис. 64. Вычисление
критерия эволюции показывает, что для отряда сумчатых Сг —
= 142.
236
3. Отряд насекохмоядных (Insectivora). К этому отряду отно-
сятся самые маленькие по размерам млекопитающие. Так, среди
семейства землероек (Soricidae) имеются звери весом 4—5 (Sorex
vagrane vagranus) и даже 3,5 г (Sorex cinereus). Уровень стан-
дартного обмена этих животных оказался экстремально вы-
сок: он составляет 300—400% от уровня, соответствующе-
го кривой Клайбера для млекопитающих сходного веса (рис. 65).
Считается, что причинами высокого уровня стандартного об-
мена у землероек являются повышенная подвижность и пи-
щеварительная активность этих животных: мелкие землеройки
в течение суток съедают количество пищи, равное весу тела (Mor-
rison, Pearsen, 1946; Morrison et al., 1959; Слоним, 1961). Землерой-
ки весом более 5 г и меньше 25—30 г также имеют аномаль-
но высокий уровень стандартного обмена (рис. 65). Возможное
объяснение причин чрезмерно высокого метаболизма экстремаль-
но маленьких животных будет дано ниже. Расчет критерия упоря-
доченности для представителей отряда насекомоядных крупнее,
чем 25—30 г, показал, что Сг = 214.
Рис, 64, Зависимость стандартного обмена от веса тела у однопроходных
(1), сумчатых (2), неполнозубых (3) и зайцеобразных (4)
Пунктирная прямая соответствует уравнению Клайбера для всех млекопитающих
237
4. Отряд рукокрылых (Chiroptera). Данные по стандартному
обмену рукокрылых представлены на рис. 65. Расчет критерия
упорядоченности показал, что для рукокрылых Сг = 181.
5. Отряд приматов (Primates). Для характеристики этого от-
ряда можно использовать только данные, полученные для подот-
Рис. 65, Зависимость стандартного обмена от веса тела у насекомоядных
(1), рукокрылых (2) и грызунов (3)
238
ряда человекообразных обезьян, или высших приматов. По низ-
шим приматам данных о стандартном обмене в литературе мы не
нашли. Следует отметить, что у приматов, как правило, измеряет-
ся не стандартный, как у других млекопитающих, а основной об-
мен. Как известно, основной обмен примерно на 25% ниже стан-
дартного (Brody, 1945), поэтому, чтобы сопоставить метаболизм
приматов с другими млекопитающими, пришлось ввести соответ-
ствующую поправку. Данные по стандартному обмену высших
приматов представлены на рис. 66. Критерий упорядоченности,
вычисленный по этим данным, для высших приматов Сг = 374.
6. Отряд неполнозубых (Edentata). Данные об интенсивности
дыхания неполнозубых (броненосцы, муравьеды, ленивцы) пред-
ставлены на рис. 64. Как видно из этих данных, имеет место ано-
мально низкий уровень метаболизма у этих животных, по сравне-
нию с метаболизмом других млекопитающих сходного веса. При-
чиной этого, возможно, является то, что у неполнозубых вес тка-
ней, активно участвующих в метаболизме, составляет 40—50%
по сравнению с 70—75% у остальных млекопитающих (Goffart,
1971). Если учесть это обстоятельство и вести расчет интенсивно-
сти дыхания на единицу активнодышащей массы, равной остальным
млекопитающим, то критерий упорядоченности для неполнозубых
Сг = 146.
7. Отряд зайцеобразных (Lagomorpha). Зайцеобразные пред-
ставлены сравнительно немногочисленными наблюдениями (рис.
64). Критерий упорядоченности, вычисленный по этим данным,
Сг - 236.
8. Отряд грызунов (Rodentia). Отряд грызунов представлен
наибольшим количеством данных (рис. 65). Критерий упорядочен-
ности для всего отряда в целом равен Сг — 248. Для этого отряда
получено такое количество данных, что можно определить крите-
рий упорядоченности и для отдельных семейств. Соответствующие
расчеты показали, что по этому показателю грызуны довольно
однородны и критерий упорядоченности для разных семейств ко-
леблется в пределах 220—270.
9. Отряд хищных (Carnivora). Данные по стандартному мета-
болизму хищных представлены на рис. 66. Расчет критерия упо-
рядоченности дает значение Ст = 299.
10. Отряд непарнокопытных (Perissodactyla) и парнокопыт-
ных (Arliodactyla). Так как данных по метаболизму парнокопыт-
ных и непарнокопытных не очень много и они мало отличаются
друг от друга, мы их объединили (рис. 67). Критерий упорядочен-
ности для этих двух отрядов Сг = 344.
Все полученные данные о критерии упорядоченности разных от-
рядов класса млекопитающих суммированы в табл. 14. В ней раз-
личные отряды расположены в соответствии с увеличением крите-
рия упорядоченности. Как видно, в целом критерий упорядочен-
ности хорошо отражает увеличивающуюся сложность организации
млекопитающих от однопроходных до высших приматов.
239
Можно привести и другие примеры характеристики отрядов или
семейств с помощью критерия упорядоченности (Коноплев, Зотин,
1975а). Так, в табл. 15 представлены данные о критерии упорядо-
ченности для разных семейств отряда бесхвостых земноводных
класса амфибий. Как видно, критерий упорядоченности для жаб
в 6 раз выше, чем для чесночницы и в 2 раза выше, чем для лягу-
шек. Критерий (7.9) позволяет расположить четыре семейства от-
ряда бесхвостых амфибий по высоте организации в следующей
Рис, 66, Зависимость стандартного обмена от веса тела у высших прима-
тов (1), даманов (2) и хищных (3)
Рис, 67, Зависимость стандартного обмена от веса тела у парнокопытных
и непарнокопытных
240
Таблица 14
Критерий упорядоченности для различных отрядов класса млекопитающих
(Коноплев, Сколов, Зотин, 1975)
Классификация в соответствии с критерием (7.10) а, кал/час ъ Ст Классификация в соответствии с критерием (7-10) кал/час ъ Сг
Однопроходные 5,8 0,21 69 Грызуны 20,8 0,31 248
Сумчатые 12,0 0,26 142 Хищные 25,1 0,27 299
Неполнозубые 12,2 0,23 146 Непарнокопытные 28,9 0,27 344
Рукокрылые 15,7 0,27 181 Парнокопытные 28,9 0,27 344
Насекомоядные 17,9 0,27 214 Приматы (высшие) 31,4 0,27 374
Зайцеобразные 19,8 0,28 236
Таблица 15
Критерий упорядоченности для разных семейств бесхвостых земноводных
при 15° (расчеты по данным Hutchison et al,, 1968)
Семейство а, кал/час ь Сг
Чесночницы (Pelobatidae) 0,35 0,06 4
Квакши (Hylidae) 0,64 0,18 8
Лягушки (Ranidae) 1,17 0,36 14
Жабы (Bufonidae) 2,26 0,41 27
Класс амфибий в целом 1,11 0,25 13
Таблица 16
Критерий упорядоченности для разных отрядов класса ракообразных
(расчеты по данным Сущени, 1972)
Отряд а, кал/час ь Сг
Phyllopoda (листоногие) 0,39 0,30 4,5
Amphipoda (бокоплавы) 0,68 0,21 8,1
Cladocera (ветвистоусые) 0,69 0,19 8,2
Jsopoda (равноногие) 0,69 0,18 8,2
Decapoda (раки, омары, крабы) 0,70 0,15 8,5
Copepoda (веслоногие) 0,96 0,23 11,4
Для всего класса ракообразных 0,63 0,24 7,5
Таблица 17
Критерий упорядоченности для различных родов класса ракообразных
(расчеты по данным Сущени, 1972)
Род и вид а, кал/час ъ Сг
Раки, омары, крабы
Xantbo hudrophilus 0,39 0,22 4,6
Eriphia spiniforons 0,46 0,17 5,6
Uca pugnax 0,59 0,18 7,0
Pachygrapsus crassipes 0,59 0,35 7,0
Carcinus maenas 0,62 0,20 7,4
Hemigrapsus nudus 0,62 0,34 7,4
Pachygrapsus marmoratus 0,69 0,25 8,3
Pugettia producta 1,15 0,18 13,6
Menippe mercenaria 1,57 0,31 18,7
Равноногие
Armadillidium vulgare 0,19 0,39 2,3
Asellus meridianus 0,26 0,35 3,1
Porcellio laevis 0,29 0,28 3,5
Spbaeroma hookeri 0,48 0,39 3,7
Asellus aquaticus 0,53 0,22 6,5
Mesidothea entomon 0,58 0,15 6,9
Ligia oceanica 0,85 0,27 9,9
Idotea baltica 0,88 0,18 10,1
Oniscus asellus 0,87 0,28 10,3
Porcellio laevis 1,02 0,17 12,1
Campecapea hirsuta 1,14 0,07 13,6
Веслоногие
Temora longicornis 0,15 0,37 1,8
Acartia clausi 0,39 0,14 4,5
Rbincalanus cornutus 0,39 0,19 4,5
Macrocyclops alb id us 0,48 0,37 3,7
Acartia latistosa 0,66 0,22 7,8
Calanus finmarchicus 0,72 0,22 8,6
Centropages hamatus 1,57 0,20 18,7
последовательности: чесночницы, квакши, лягушки, жабы. Такое
расположение может быть не совсем обычно для систематики, но
его, по-видимому, следует учитывать при рассмотрении вопросов
прогрессивной эволюции в классе амфибий.
То же самое относится к классификации отрядов в классе ра-
кообразных, для которых собраны достаточно надежные данные
в монографии Сущени (1972). Здесь по высоте организации выде-
242
ляются веслоногие (табл. 16). Мы не будем сопоставлять классифи-
кацию, приведенную в табл. 16, с общепринятым подразделением
отрядов в классе ракообразных. Это просто еще одна иллюстрация
того, что критерий упорядоченности может быть использован для
выводов, касающихся прогрессивной эволюции внутри того или
иного класса животных.
Наконец, критерий упорядоченности, по-видимому, может
быть использован и в классификации родов, а может быть и видов.
Для примера мы покажем величины этого критерия, рассчитанные
по данным разных авторов (Сущеня, 1972) для разных родов неко-
торых отрядов ракообразных (табл. 17). Как видно из этих наблю-
дений, имеются заметные различия по критерию упорядоченности
между разными родами отрядов ракообразных.
Сейчас еще трудно судить о том, являются ли эти различия от-
ражением последовательности происхождения различных родов
или они связаны с различиями экологии и поведения животных.
Не исключено, однако, и то, что значительные различия в критерии
упорядоченности связаны с внутрисемейным, внутриотрядным или
внутриклассовым ароморфозом, ведущим к возникновению новой
группы животных более высокой организации. Эти группы могут
образовывать тупиковые ветви, как, например, щитни или осетро-
вые рыбы (см. табл. 13), которые все же благодаря более высокой
степени упорядоченности оказываются чрезвычайно живучими и
образуют группу реликтовых форм. Другие дают начало все более
прогрессирующим формам животных, быстро завоевывающим ши-
рокие ареалы и образующим обширные таксономические подраз-
деления. Например, не исключено, что крупный ароморфоз среди
млекопитающих, связанный с возникновением отряда насекомояд-
ных, привел к появлению приматов и в конце концов к высшей
форме приматов — человеку, обладающему наивысшим критерием
высоты организации в животном царстве. Все подобного рода пред-
положения, однако, получат право на существование только пос-
ле того, как мы сможем более точно сравнивать величину критерия
упорядоченности у разных групп животных, а для этого прежде
всего необходима тщательная систематизация и оценка имеющих-
ся экспериментальных данных об интенсивности дыхания живот-
ных, а также целенаправленное исследование по этому признаку
наиболее интересных групп животных.
ВОПРОСЫ НЕЛИНЕЙНОСТИ
ПРИ ИСПОЛЬЗОВАНИИ КРИТЕРИЯ
УПОРЯДОЧЕННОСТИ
А. И, Зотин, В. А. Коноплев, В. А. Грудницкий
При выводе критерия упорядоченности (7.9) и (7.10) мы пред-
полагали, что интенсивность дыхания организмов можно прирав-
нять интенсивности теплопродукции. Это, казалось бы, вполне оп-
равдано, так как давно уже показано, что у взрослых животных
наблюдается полное соответствие данных прямой и непрямой кало-
риметрии (см. Гинецинский, Лебединский, 1956; Чередниченко,
1965; Durnin, Passmore, 1967). Однако, как следует из теоретиче-
ских построений (Зотин, Грудницкий, 1970; Зотин, 1974) и экспе-
риментальных данных, полученных при изучении теплопродукции
и дыхания растущих культур микроорганизмов (Зотин, Шаарш-
мидт, Лампрехт, 1975; Schaarschmidt et al., 1975), ооцитов ля-
гушки (Зотин, 19756) и листьев растений (Жолкевич и др., 1964;
Жолкевич, 1968), во время роста имеет место несовпадение тепло-
продукции, измеренной прямыми и непрямыми методами.
В настоящее время мы можем подвести под явление расхожде-
ния данных прямой и непрямой калориметрии во время роста жи-
вотных и растений более глубокий теоретический фундамент,
чем это было сделано раньше (Зотин, Грудницкий, 1970). Для
этого воспользуемся разбиением (2.15) и теми положениями, на
которых базируется это разбиение. Согласно этим положениям
(Зотин, 1974; Зотин, Коноплев, Преснов, 1975), чем дальше от
равновесия или стационарного состояния находится термодина-
мическая система, тем больше должна быть величина функции
связанной диссипации фи. В случае живых систем мы можем при-
равнять диссипативную функцию интенсивности дыхания организ-
мов (2.1), а функцию внешней диссипации — интенсивности тепло-
продукции (4.2). Отсюда следует, что функция связанной дисси-
пации равна разности между данными непрямой и прямой кало-
риметрии (4.5). Эта разность должна быть тем больше, чем дальше
от конечного стационарного состояния находится живая система.
Последнее положение достаточно хорошо подтверждается экспе-
риментальными данными (см. рис. 33, 39). Таким образом, вызы-
вает большое сомнение возможность использования критерия
(7.9) для характеристики степени упорядоченности живых систем
во время развития и роста: в этом случае интенсивность дыхания
не пропорциональна функции внешней диссипации.
Подобного рода сомнений, казалось бы, не должно быть, когда
мы сравниваем между собой животных, достигших стационарного
веса. У таких животных функция связанной диссипации должна
быть равна нулю и не должно наблюдаться расхождения данных
244
прямой и непрямой калориметрии. Многочисленные наблюдения,
как говорилось, хорошо подтверждают этот вывод: у взрослых
животных и людей теплопродукция, рассчитанная по дыханию,
совпадает с теплопродукцией, измеренной прямыми методами
(Студенский, 1897; Лихачев, Авроров, 1902; Atwater, 1904; Lusk,
1915, 1921; Murlin, Lusk, 1915; Benedict, 1938; Wohl, James,1942;
Чередниченко, 1965; Семихатова, 1967; Durnin, Passmore, 1967;
и др.). Однако в связи с исследованием вопроса о применимости
критерия упорядоченности для характеристики различных групп
животных (Коноплев, Соколов, Зотин, 1975) встал вопрос о том,
справедливо ли это утверждение для животных любого размера,
включая экстремально малых животных.
Дело в том, что стандартный метаболизм экстремально малых
по размеру млекопитающих и птиц оказался на 300—400% выше
метаболизма других животных этих классов. Например, на рис. 68
представлены данные, взятые из работ различных авторов, об ин-
тенсивности дыхания млекопитающих (представлены данные толь-
ко для плацентарных млекопитающих) в зависимости от веса тела.
Как видно из этих наблюдений, в классе млекопитающих в целом,
как это отмечали многие авторы (Brody, 1945; Kleiber, 1961; Сло-
ним, 1961 и др.), выполняется степенная зависимость между ин-
тенсивностью дыхания и весом тела (2.8): экспериментальные точ-
ки лежат вокруг так называемой прямой Клайбера (рис. 68). Од-
нако данные, полученные для очень маленьких по размерам мле-
копитающих (в основном для землероек из отряда насекомоядных),
лежат выше прямой Клайбера, причем чем мельче животные этой
группы, тем больше уклоняются они от клайберовской прямой.
Рис. 68, Зависимость стандартного обмена от веса тела в классе млеко-
питающих (кроме однопроходных и сумчатых)
Пунктирная прямая соответствует уравнению Клайбера
245
Аналогичная ситуация имеет место в классе птиц, данные по
стандартному метаболизму которых представлены на рис. 69.
И в этом случае животные очень малых размеров имеют уровень
метаболизма значительно выше, чем уровень метаболизма, характер-
ный для всего класса в целом. Как и в случае млекопитающих,
уклонение от среднего уровня метаболизма, представленного на
рис. 69 пунктирной прямой, происходит у животных, размеры ко-
торых меньше 30—50 г. В связи с этим представляется вероятным
предположение, что у животных очень малых размеров, как и у ра-
Рис. 69. Зависимость стандартного обмена от веса тела в классе птиц
(из King, Farner, 1961; Dawson, Hudson, 1970)
Пунктирная прямая соответствует уравнению Кинга и Фарнера (King, Farner, 1961)
Рис. 70. Расхождение данных непрямой и прямой калориметрии ($и~ФУнк~
ция) у различных организмов от бактерий до человека в зависимости от веса
организмов
246
стущих животных, имеет место расхождение данных прямой и не-
прямой калориметрии.
Легко себе представить, что уровень теплопродукции у живот-
ных экстремально малых размеров в связи с очень большим отно-
шением поверхности к объему настолько велик, что даже в стацио-
нарном состоянии у них функция связанной диссипации не равна
нулю. Эти системы столь далеко отстоят от равновесия, что для
них и в стационарном состоянии справедливо разбиение (2.15).
Если это так, то у животных малых размеров, в отличие от более
крупных, должно экспериментально наблюдаться расхождение
данных прямой и непрямой калориметрии.
К сожалению, в литературе имеется пока немного наблюдений
о теплопродукции животных, полученных прямыми методами.
Поэтому мы попытались собрать данные, касающиеся не какой-либо
отдельной группы животных, а всех организмов от бактерий до
человека. Эти данные, взятые из работ различных авторов (Кальве,
Прат, 1963; Dale et al., 1967; Prat, 1969; Lamprecht et al., 1971;
Hudson, Deavers, 1973; Грудницкий, 1975; Schaarschmidt et al.,
1976), суммированы на рис. 70. Как видно, фи-функция, вычислен-
ная по формуле (4.5), равна нулю только у организмов размерами
больше, чем 50—100 г. При этом чем меньше размеры организмов,
тем больше величина функции связанной диссипации.
Следовательно, у животных, размеры которых меньше 50—
100 г, наблюдаются расхождения данных прямой и непрямой кало-
риметрии и использовать критерий упорядоченности (7.9) для
таких групп организмов можно лишь с большими оговорками.
Здесь требуется иной критерий, основанный на данных прямого
измерения теплопродукции организмов. К сожалению, в на-
стоящее время таких данных слишком мало, чтобы попытаться ис-
пользовать этот новый критерий для целей классификации орга-
низмов.
ЗАКЛЮЧЕНИЕ
Термодинамика биологических процессов, некоторым пробле-
мам которой посвящена настоящая книга, в принципе должна бы
охватить с единой точки зрения все основные биологические
явления. Только в этом случае она могла бы послужить надежным
фундаментом для теоретической биологии. Нет, конечно, никаких
гарантий, что в настоящее время можно построить такой фун-
дамент для теоретической биологии. Наоборот, история биологии
учит нас, что все прежние попытки создать на базе термодина-
мики основы теоретической биологии оказались неудачными.
На данном этапе развития науки основная трудность заключается
в отсутствии общепринятой теории нелинейных необратимых про-
цессов. Но даже после завершения термодинамики нелинейных
необратимых процессов, как отметил, в частности, Николаев
(1971), мы окажемся, по существу, в самой начальной фазе
построения термодинамики жизненных процессов, так как в
живых системах все процессы осуществляются под контролем
управляющих систем. Следовательно, для описания этих про-
цессов необходима такая термодинамическая теория, которая
учитывала бы процессы регуляции и управления, происходящие в
системе. Дело несколько облегчается тем фактом, что одновремен-
но с развитием термодинамики нелинейных необратимых процес-
сов происходит быстрое развитие наук, изучающих процессы регу-
ляции и управления в живых системах и в автоматических устрой-
ствах, таких, как теория информации, автоматического регулиро-
вания, оптимального управления и т. д. По-видимому, смыкание
этих теорий с термодинамикой и даст в конце концов основу для
построения общей феноменологической теории, которая явится
базой для термодинамики биологических процессов.
Исходя из сказанного, на вопрос, который часто обсуждают и
решают отрицательно (например, Волькенштейн, 1973; Эйген,
1973а, б; Блюменфельд, 1974), нужна ли новая физика при рассмот-
рении биологических явлений или достаточно старой физики, но
необходимо ее умелое использование, следует ответить положи-
тельно, если речь идет об использовании термодинамики. Да,
нужна новая термодинамика, которая описывала бы не только про-
цессы, происходящие в равновесии или вблизи от равновесия, но
и широкий класс явлений, протекающих в системах, далеких от
248
равновесия. Нужна новая термодинамика, способная описывать
процессы, протекающие в организованных системах, т. е. в систе-
мах, в которых осуществляются процессы регуляции и управле-
ния.
Несмотря на то, что построение такой теории является еще
делом будущего, уже сейчас существует потребность рассмотреть и
обобщить первые попытки построения термодинамики жизненных
явлений с тем, чтобы в какой-то степени подвести теоретическую
базу под те исследования, которые стимулированы термодинами-
кой, несмотря на ее незавершенный вид. Дело в том, что, как вид-
но из материалов, приводимых в настоящей книге, термодинамика
необратимых процессов может служить базой для совершенно но-
вых подходов к таким важнейшим биологическим проблемам, как
теория эволюции, индивидуального развития организмов, реакции
живых систем на внешние воздействия и т. д. Уже сейчас термоди-
намический подход к явлениям развития, роста и старения позво-
ляет наметить пути изучения таких важнейших проблем, как ме-
ханизм старения, заживления ран и регенерации, злокачественно-
го роста и др. Такие многообещающие, практически важные
задачи, которые на данном этапе своего развития может ставить тер-
модинамика жизненных процессов, заставляет предпринимать рис-
кованные попытки обобщения и подведения итогов, что и сделано
частично в данной книге. Мы опирались при этом на самые послед-
ние достижения в области термодинамики нелинейных необрати-
мых процессов. К сожалению, в этой области многое все еще оста-
ется неясным и ни одна нелинейная термодинамическая теория
пока не касается систем, в которых возможны процессы регуля-
ции и управления.
Таким образом, термодинамика биологических процессов мо-
жет рассматриваться в настоящее время скорее как заявка на бу-
дущее, чем как завершенная или достаточно развитая область зна-
ний. Целью этой книги было, главным образом, постановка задач,
выявление основных проблем и подведение первых итогов, а также
привлечение внимания к биологическим проблемам, которые воз-
никают в биологии под влиянием термодинамики, и к проблемам,
которые возникают в термодинамике под влиянием биологии.
ЛИТЕРАТУРА
А брагам-Беккер. 1936. Теория элек-
тричества. М.—Л., ОНТИ.
Абрамова Н. Б., Васильева М. В.
1973. Некоторые свойства эмбри-
ональных митохондрий вьюна.—
Онтогенез, 4, 288—293.
Авроров П. П. 1899. Прямая и не-
прямая калориметрия у живот-
ных в состоянии азотистого равно-
весия, голодания и откармливания
их. СПб., Изд. К. Л. Риккер.
А йзенштадт Т. Б. 1965. Некото-
рые особенности ультраструктуры
ооцитов в связи с синтезом желт-
ка.— Ж. общ. биол., 26, 230—236.
Андреев В. М. 1962. Критические
явления в открытых системах.
Ж. физ. химии, 36, 42—52.
Антонов А. С. 1974. Геносистема-
тика. Достижения, проблемы и
перспективы. — Усп. совр. биол.,
77, № 2, 31—47.
Антонов В. Ф. 1968. Термодинами-
ка биологических систем. В сб.
«Биофизика». М., «Высшая школа»,
13-47.
Арнольд В. И. 1971. Обыкновенные
дифференциальные уравнения. М.,
«Наука».
Ач Г., Штрауб Ф. Б. 1954. Об
обмене веществ асцитических ра-
ковых клеток.— Докл. АН СССР,
95, 1021-1024.
Бабский Е. Б., Зубков А. А., Ко-
сицкий Г. И., Ходоров Б. И.
1966. Физиология человека. М.,
«Медицина».
Байер В. 1962. Биофизика. М., ИЛ.
Бауэр Э. С. 1935. Теоретическая био-
логия. М.—Л., Изд. ВИЭМ.
Бахарева И. Ф. 1968. Вариацион-
ные принципы неравновесной тер-
модинамики.— Ж. физ. химии, 42,
2394-2398.
Бахарева И. Ф.. 1971. О вариацион-
ных принципах неравновесной тер-
350
модинамики.— Инж.-физ. ж., 2(Г,
1105-1110.
Бахарева И. Ф. 1974. Экстремальные
свойства энтропии стационарного
состояния линейных систем.—
Ж. физ. химии, 48, 2727 — 2731.
Бахарева И. Ф., Бирюков А. А.
1969. Некоторые вопросы нелиней-
ной неравновесной термодинами-
ки.— Теорет. эксп. химия, 5,
340-346.
Бахарева И. Ф., Бирюков А. А.
1970. К стохастической теории
нелинейных неравновесных про-
цессов.— Изв. высш, учебн. за-
вед., Физика, 9, 58—63.
Бахарева И. Ф., Бирюков А. А.
1974. Стохастическая интерпрета-
ция уравнений нелинейной не-
равновесной термодинамики.—
Ж. физ. химии, 48, 1959—1964.
Бахарева И. Ф., Крылов А. Ф.г
1972. Вариационный метод описа-
ния нелинейных неравновесных
процессов.— Инж.-физ. ж., 21,
161-164.
Белоусов Л. В. 1971. Проблемы эм-
брионального формообразования.
М., Изд. МГУ.
Беркинблит М. Б. 1965. О приме-
нимости теоремы Пригожина к про-
цессу эмбриогенеза и к эволю'-
ции.— Биофизика, 10, 1103—
1105.
Блюменфельд Л. А. 1974. Пробле-
мы биологической физики. М.,
«Наука».
Боголюбов Н. Н., Митрополь-
ский Ю. А. 1963. Асимптотические
методы в теории нелинейных коле-
баний. М., Физматгиз.
Бородина М. Н. 1970. Возрастная
изменчивость некоторых морфоло-
гических признаков бобров мок-
шанской популяции. —Труды Мор-
довского рос, зад., 5, 91—130.
Браше Ж. 1961. Биохимическая эм-
бриология. М., ИЛ.
Браун А. Д., Булычев А. Г., Гане-
лина Л. Ш. 1967. Изменение ме-
таболизма клетки при поврежде-
нии.— Цитология, 9, 1225—1247.
Бриллюэн Л. 1960. Наука и теория
информации. М., Физматгиз.
Бриллюэн Л. 1962. Термодинами-
ка, статистика и информация.—
Усп. физ. наук, 77, 337—352.
Булъдяева Т. В., Кузьмина С. Н.,
Збарский И. Б. 1972. Окислитель-
ное фосфорилирование в изоли-
рованных ядерных оболочках и яд-
рах клеток печени крысы. —
Докл. АН СССР, 202, 467—470.
Бухвалов И. Б. 1973. Ультраструк-
турная локализация ферментных
систем клеточного ядра.— Усп.
совр. биол., 76, 382—394.
Бызова Ю. Б, 1965. Зависимость
потребления кислорода от образа
жизни и размера тела на примере
дождевых червей (Lumbricidae,
Oligochaeta).— Ж. общ. биол., 26,
555-562.
Бызова Ю. Б. 1972. Дыхание поч-
венных беспозвоночных. Сб. эко-
логия почвенных беспозвоночных.
М., «Наука», 3—39.
Быховский А. И. 1965. Еще к вопро-
су о применимости теоремы Приго-
жина в биологии (о повышенном
термогенезе на начальных ста-
диях развития живых организ-
мов).— Биофизика, 10, 1105 —
1108.
Быховский А. И. 1966. О тепловых
эффектах и надежности процес-
сов синтеза белков и нуклеиновых
кислот. Докл. АН СССР, 171,
992-995.
Васильев В. А., Заикин А. М. 1976.
Волновые режимы в реакции окис-
ления броммалоновой кислоты
броматом, катализируемой комп-
лексными ионами железа. В сб.
«Кинетика и катализ». М., «Нау-
ка».
Васильев В. А., Романовский Ю. М,
1975. Связанные автоколебатель-
ные системы как модели ведущего
центра. Теорет. эксп. биофизика.
Калининград, 5, 52—60.
Васильев В. А,, Романовский Ю. М.
1976. О роли диффузии в системах
с автокатализом.— Теорет. эксп.
биофизика. Калининград, 6.
Васильеве. С. 1955. Кинетика возбуж-
дения молекул электромагнитными
и механическими волнами. I. Зна-
чение циклизации свободной энер-
гии при возбуждении молекул
в структурных ансамблях.— Ж.
физ. химии, 29, 2018—2030.
Васильев С. С. 1956. Кинетика воз-
буждения молекул электромаг-
нитными и механическими вол-
нами. II. Расчет коэффициента
структурного усиления процес-
сов возбуждения электромагнит-
ными волнами молекул, входящих
в состав элементарных структур-
ных ансамблей.— Ж. физ. химии,
30, 2456-2468.
Васильев С. С. 1959. Кинетика воз-
буждения молекул электромаг-
нитными и механическими волна-
ми. III. О роли структурных ан-
самблей в первичном активацион-
ном акте процесса фотосинтеза.—
Ж. физ. химии, 33, 2053—2062.
Васильев С. С. 1964. Кинетический
анализ цепных реакций. 7. Преоб-
разование форм решений основных
уравнений кинетики цепных реак-
ций методом структурной алгебры
для любого числа сортов актив-
ных частиц.— Ж. физ. химии, 38,
2214-2222.
Вейнхаус С. 1957. Окислительный
обмен в опухолевых тканях. В сб.
«Успехи в изучении рака». М.,
ИЛ, 3, 185-248.
Веселкин П. Н. 1960. Об изменениях
энергетического обмена при лихо-
радочных состояниях. В сб. «Фос-
форилирование и функция». Л.,
Изд. ИЭМ, 327-335.
Вернадский В. И. 1967. Биосфера.
М., «Мысль».
Виленчик М. М. 1973. Изменения
цитоплазматической ДНК при
малигнизации клетки. — Усп. совр.
биол., 75, 388—405.
Винберг Г. Г. 1956. Интенсивность
обмена и пищевые потребности
рыб. Минск, Изд. Белорусок, гос.
ун-та.
Винберг Г. Г. 1968. Взаимозависи-
мость интенсивности обмена и ско-
рости роста у животных. В сб.
«Биология моря», 15, 5—15. Киев,
«Наукова думка».
Владимирова Е. А. 1935. Содержа-
ние молочной кислоты в регене-
рирующей конечности аксолотля
на ранних стадиях регенерации. —
251
Труды Лаб. эксп. зоол. и морфол.
живот., 4, 119—133.
Владимирова И. Г. 1974. Рост п ды-
хание регенерирующих тканей
хвоста аксолотлей.— Онтогенез,
5, 96-99.
Владимирова И. Г. 1975. Дыхание
и гликолиз в регенерирующих
тканях хвоста аксолотля. — Ж. общ.
биол., № 6, 920—928.
Владимирова И. Г., Никольская
И. С. 1971. Изменение ин-
тенсивности дыхания при регене-
рации хвоста у аксолотлей раз-
личного возраста.— Онтогенез, 2,
502-506.
Волькенштейн М. В. 1958. Макро-
молекулы и биология. — Изв. АН
СССР, сер. биол., 1, 3—25.
Волькенштейн М. В. 1965. Молеку-
лы и жизнь. Введение в молеку-
лярную биофизику. М., «Наука».
Волькенштейн М. В. 1973. Биоло-
гия и физика.— Усп. физ. наук,
109, 499-515.
Волькенштейн М. В., Сырников
Ю. П. 1966. Еще раз об «Эле-
ментах биологической термодина-
мики» К. С. Тринчера.— Биофи-
зика, И, 379—380.
Гайцхоки В. С. 1967. Структурная
организация биологического окис-
ления и сопряженных процессов.
В сб. «Механизмы интеграции кле-
точного обмена». Л., «Наука»,
153-176.
Гельфер Я. М. 1969. История и ме-
тодология термодинамики и ста-
тистической физики, 1. М.,
«Высшая школа».
Гельфер Я. М. 1973. История и ме-
тодология термодинамики и ста-
тистической физики, 2. М., «Выс-
шая школа».
Гинецинский А. Г., Лебединский
А. В. 1956. Курс нормальной фи-
зиологии. М., Медгиз.
Гинзбург А. С, 1968. Оплодотворение
у рыб и проблема полиспермии.
М., «Наука».
Гинис Б.Ю., Шагимординов Н. Ш.,
Зотин А. И. 1974. Видоизменен-
ное уравнение роста животных
Робертсона.— Онтогенез, 5, 284—
286.
Гинкин Г. Г. 1948. Справочник по
радиотехнике. М.—Л., Госэнерго-
издат.
Глансдорф 11., Пригожин И. 1973.
Термодинамическая теория струк-
туры, устойчивости и флуктуа-
ций. М., «Мир».
Гончаров А. И., Поддубный А. Г.,
Сметанин М. М. 1975. Опыт ве-
роятностного анализа роста рыб.
Сб. «Количественные аспекты рос-
та организмов». М., «Наука».
Гофман П. Б, 1938. Количествен-
ные закономерности роста живот-
ных.— Усп. совр. биол., 9, 39—
67.
Граменицкая Е. С. 1967. О несо-
ответствии данных прямой и не-
прямой калориметрии после крат-
ковременного пребывания живот-
ных в условиях пониженного и
повышенного парциального давле-
ния кислорода. Всес. конф, по
теплообмену и теплорегуляции.
Л., 19-20.
Гришко Ф,И., Левкова Н. А. 1966.
О метаболизме мышечной ткани
у старых крыс.— Физиол. ж.,
52, 1455-1459.
Гроот С. Р. де. 1956. Термодинами-
ка необратимых процессов. М.,
Гос. изд. техн.-теорет. литер.
Гроот С. Р. де, Мазур Г. 1964. Не-
равновесная термодинамика. М.,
«Мир».
Грудницкий В. А. 1972а. Зависи-
мость теплопродукции от веса во
время роста меченосцев (Xipho-
phorus helleri Н.).— Докл. АН
СССР, 202, 1435-1437.
Грудницкий В. А. 19726. Энерге-
тические закономерности во вре-
мя роста меченосцев (Xiphopho-
rus helleri Н.). В сб. «Энергети-
ческие аспекты роста и обмена
водных животных». Киев, «Нау-
кова думка», 40—41.
Грудницкий В. А. 1975. Энергетиче-
ские закономерности при росте рыб.
В сб. «Количественные аспекты
роста организмов». М., «Наука»,
48-53.
Грудницкий В. А., Никольская И. С.
1976. Теплопродукция на ран-
них стадиях роста аксолотля по
данным прямой и непрямой кало-
риметрии (в печати).
Грудницкий В. А., Зотин А. И.,
Терентьева Н. В., Шагиморда-
нов Н. Н1. 1971. Уравнение роста,
учитывающее взаимодействие рос-
та и дифференцировки.—Онтоге-
нез, 2, 451—454.
252
Гурвич А. Г. 1944. Теория биоло-
гического поля. М., «Советская
наука».
Демидович Б. П. 1967. Лекции по
математической теории устойчиво-
сти. М., «Наука».
Денбиг К. 1954. Термодинамика ста-
ционарных необратимых процес-
сов. М., ИЛ.
Джелинео С. 1959. Развитие гомо-
термии у млекопитающих.— Усп.
совр. биол., 47, 108—120.
Дмитриева Н. П. 1963. О тесных
топографических контактах мем-
бранных структур в клетках опу-
холей (электронно-микроскопи-
ческое исследование).— Архив па-
тологии, 25, № 10, 16—24.
Дольник В. Р. 1968. Энергетический
обмен и эволюция животных.—
Усп. совр. биол., 66, 276—293.
Донцова Г. В, 1965. Изменение ци-
тохромоксидазной активности в
раннем развитии вьюна и иссле-
дование роли ядер в этом процес-
се.— Ж. общ. биол., 26, 585 —
596.
Дярматы И. 1965. Об общем ва-
риационном принципе неравновес-
ной термодинамики.— Ж. физ. хи-
мии, 39, 1489—1493.
Дярматы И. 1974. Неравновесная
термодинамика. Теория поля и ва-
риационные принципы. М., «Мир».
Ельцина Н. В. 1960. Энергетиче-
ский обмен раковых клеток.— Био-
химия, 25, 135—142.
Жаботинский А. М. 1974. Концент-
рационные автоколебания. М.,
«Наука».
Жолкевич В. Н. 1968. Энергетика ды-
хания высших растений в условиях
водного дефицита. М., «Наука».
Жолкевич В. Н., Холлер В, А.,
Рогачева А. Я. 1964. О соотно-
шении между дыханием и тепло-
отдачей в растущих листьях.—
Докл. АН СССР, 158, 1213-1216.
Залевски К, 1973. Феноменологи-
ческая и статистическая термоди-
намика. М., «Мир».
Збарский И. Б. 1969. Структура
и функции ядерной оболочки.—
Усп. совр. биол., 67, 323—341.
Збарский И. Б, 1972. Связь ядер-
ной оболочки со структурами ци-
топлазмы и механизмы ядерно-
цитоплазматических отношений.—
Усп. совр. биол., 73, 3—25.
Збарский И, Б,, Покровский А. А.,
Перевощикова К, А., Гаппа-
ров М. М., Делекторская Л. Н.,
Лашнева Н. В. 1968. Содержание
некоторых окислительных фер-
ментов в ядерных оболочках пе-
чени крысы.— Докл. АН СССР,
181, 993-996.
Зотин А, И. 1966. Изменение ско-
рости продукции энтропии во вре-
мя эмбрионального развития и
роста.— Биофизика, И, 554—
557.
Зотин А. И. 1970. Старение и омо-
ложение с точки зрения термо-
динамики необратимых процес-
сов.— Природа, 9, 49—55.
Зотин А. И. 1971. По поводу ис-
пользования понятий энтропия
и негэнтропия в биологии.— Ж.
общ. биол., 32, 27—36.
Зотин А. И. 1972. Идентификация
уравнения роста животных Бер-
тал анфи с функцией Гомпертца.—
Онтогенез, 3, 616—618.
Зотин А. И. 1973. Феноменологи-
ческая теория развития организ-
мов.— Онтогенез, 4, 3—10.
Зотин А. И. 1974. Термодинами-
ческий подход к проблемам раз-
вития, роста и старения. М., «На-
ука».
Зотин А. И. 1975а. Количественные
теории роста (история, современное
состояние и перспективы). В сб.
«Количественные аспекты роста ор-
ганизмов». М., «Наука», 267—272.
Зотин А. И. 19756. Теплопродук-
ция и дыхание ооцитов Xenopus
laevis L. на разных стадиях ооге-
неза.— Онтогенез, 6, 623—626.
Зотин А. Я., Грудницкий В. А.
1970. Соотношение теплопродук-
ции и дыхания во время роста
животных.— Онтогенез, 1, 437—
444.
Зотин А. Я., Зотина Р. С. 1969.
Термодинамический подход к про-
блемам развития, роста и старе-
ния.— Ж., общ. биол., 30, 94—110.
Зотин А. Я., Зотина Р. С. 1972.
Размерность потоков, сил и дру-
гих показателей в термодинамиче-
ской теории развития.— Теорет.
эксп. биофизика. Калининград, 3,
95-101.
Зотин А. Я., Зотина Р. С. 1975.
Феноменологическая теория рос-
та. В сб. «Количественные аспекты
роста организмов». М., «Наука»,
57-68.
253
Зотин А. И., Зотина Р. С., Про-
кофьев Е. А., Коноплев В, Л. 1976.
Использование уравнений роста
для определения продолжитель-
ности жизни млекопитающих и че-
ловека.— Изв. АН СССР, сер.
биол. (в печати).
Зотин А. И., Коноплев В. А., Прес-
нов Е. В. 1975. Физический смысл
и статистическая интерпретация
критериев эволюции.— Ж. физ.
химии, 49, 2901—2905.
Зотин А. И., Озернюк Н. Д.ч Зо-
тин А. А., Коноплев В. А. 1975.
Возможный путь происхождения
прокариот.— Ж. общ. биол., 36,
163-172.
Зотин А. И., Преснов Е. В., Ко-
ноплев В. А. 1975. Уровни аппрок-
симации для нелинейных фено-
менологических уравнений биоло-
гии развития. В сб. «Количествен-
ные аспекты роста организмов».
М., «Наука».
Зотин А. И., Шааршмидт Б.,
Лампрехт И. 1975. Функция свя-
занной диссипации и ее изменения
во время роста культуры дрож-
жей.—Изв. АН СССР, сер. биол.
4, 593—598.
Зотина Р. С., Зотин А. И. 1967.
Количественные соотношения меж-
ду весом, длиной, возрастом, раз-
мерами яиц и плодовитостью у жи-
вотных.— Ж. общ. биол., 28, 82—
92.
Зотина Р. С., Зотин А. И. 1973а.
Объединенные уравнения роста.—
Ж. общ. биол., 34, 606—616.
Зотина Р. С., Зотин А. И, 19736.
Количественная феноменологиче-
ская теория роста животных. В сб.
«Механизмы управления размно-
жением и дифференцировкой кле-
ток животных тканей». Красно-
ярск, 283—284.
Зубарев Д. И., 1971. Неравновесная
статистическая термодинамика. М.,
«Наука».
Иванчик Е. П. 1969. Динамика га-
зообмена у гусениц и куколок аме-
риканской белой бабочки (Hyphan-
tria cunea Drury). В сб. «Перио-
дичность индивидуального разви-
тия насекомых». М., «Наука»,
224-236.
Ивлев В. С. 1959. Опыт оценки эво-
люционного значения уровней
энергетического обмена.—Ж. общ.
биол., 20, 94—103. И
Ивлев В. С. 1961. Уровень энерге
тического обмена ланцетника.—
Докл. АН СССР, 140, 1217-1219.
Ивлев В. С. 1963. О параметрах,
характеризующих уровни энерге-
тического обмена животных. В сб.
«Применение математических ме-
тодов в биологии», 2, 146—151.
Л., Изд. ЛГУ.
Ивлева И. В. 1964. Элементы энер-
гетического баланса актиний.—
Труды Севаст. биол. ст., 15, 410 —
428.
Казакова Т. Б. 1967. Генетический
контроль над синтезом структур-
ных элементов дыхательной цепи.
В сб. «Механизмы интеграции кле-
точного обмена». Л., «Наука»,
157-204.
Кальве Э., Прат А. 1963. Микро-
калориметрия. М., ИЛ.
Канер Э, А., Яковенко В, М. 1975.
Гидродинамические неустойчивости
в твердотельной плазме.— Усп.
физ. наук, 115, 41—72.
Карие Е. 1972. Некоторые примене-
ния диссипативных структур в фи-
зических, физико-химических и
биологических системах. В сб.
«Термодинамика необратимых про-
цессов и ее применение». Чернови-
цы, 59—61.
Картан А. 1971. Дифференциаль-
ное исчисление. Дифференциаль-
ные формы. М., «Мир».
Кашкаров Д. Н., Станчинский В. В.
1940. Курс зоологии позвоноч-
ных животных. М.—Л., Изд. АН
СССР.
Каюшин Л. П. (ред.) 1967. Состоя-
ние и роль воды в биологических
объектах. М., «Наука».
Киселев О. И., Гайцхоки В. С. 1972.
Перенос ядерной РНК в изолиро-
ванные митохондрии.— Биохи-
мия, 37, 1224—1231.
Клименко В. В. 1971. Уравнение
роста шелковичного червя Вот-
byx mori L.— Онтогенез, 2, 617 —
625.
Клименко В. В. 1975. Анализ урав-
нений роста на примере шелкович-
ного червя. В сб. «Количественные
аспекты роста организмов». М.,
«Наука», 36—41.
Кобозев Н. И. 1948. Несколько кри-
тических замечаний о книге
Э. Шредингера «Что такое жизнь
с точки зрения физики».— Бюлл.
МОИП, отд. биол., 53, 31—35.
254
Кобозев Н. И. 1971. Исследование
в области термодинамики процес-
сов информации и мышления. М.,
Изд. МГУ.
Коноплев В. А., Зотин А. И. 1975а.
Критерий упорядоченности и его
применение к некоторым эволю-
ционным проблемам.— Теорет.
эксп. биофизика. Калининград, 5,
98-108.
Коноплев В. Л., Зотин А. И. 19756.
Нелинейное феноменологическое
уравнение роста, основанное на
стохастических представлениях.
В сб. «Количественные аспекты
роста организмов». М., «Наука».
Коноплев В. Л., Зотин А. И., Ша-
гиморданов Н. Ш., Скачкова А. И.
1975. Анализ нелинейных уравне-
ний роста на аналоговой ЭВМ.—
Онтогенез, 6, 527—529.
Коноплев В. Л., Соколов В. Е., Зо-
тин А. И. 1975. Классификация
животных по энергетическому ме-
таболизму,— Зоол. Ж., 54, 1429—
1441.
Коржуев П. Л., Никольская И, С.,
Радзинская Л. И. 1960. Дыхание
икры осетровых рыб в период ин-
кубации.— Вопр. ихтиол., 14,
113-118.
Красовский А. А. 1974. Фазовое
пространство и статистическая тео-
рия динамических систем. М.,
«Наука».
Кузьмина С. Н.^ Монахов Н. К.,
Гайцхоки В. С., Нейфах С. А.,
Збарский И. Б. 1970. Дыхатель-
ная активность изолированных
ядерных оболочек и ядер печени
крыс.— Докл. АН СССР, 191,
215-217.
Кэтлэ А. 1913. Социальная физика
или опыт исследования о развитии
человеческих способностей, 2,
кн. 3. Киев.
Лазарев П, П. 1927. О возможности
приложения физико-химических
методов к живой материи.— Докл.
АН СССР, сер. А, 3, 31—34.
Лазарев П. П. 1945. Современные
проблемы биофизики. М.—Л.,
Изд-во АН СССР.
Ланцош К. 1965. Вариационные
принципы механики. М., «Мир».
Левкова Н. А., Трунов В. И. 1970.
Особенности строения митохонд-
рий старческого миокарда.— Кар-
диология, 10, № 9, 94—97.
Ленинджер А. 1966. Митохондрия.
Молекулярные основы структуры
и функции. М., «Мир».
Лихачев Л. Л. 1893. Теплопроиз-
водство здорового человека при
относительном покое. Докт. дисс.
№ 14. СПб.
Лихачев Л. Л., Авроров Н. Н. 1902.
Исследование газового и теплово-
го обмена при лихорадке. Изв.
Импер. Военно-мед. акад., 5,
239-281.
Лыков А. В., Михайлов Ю. А. 1963.
Теория тепло- и массопереноса.
М.— Л., Госэнергоиздат.
Льюис Дж. Н. 1929. Анатомия нау-
ки. М.— Л., Госиздат.
Майр Э. 1971. Принципы зоологи-
ческой систематики. М., «Мир».
Мейергоф О. 1928. Термодинамика
жизненных процессов. М,— Л.,
Госиздат.
Мейкснер И. 1970. Процессы в про-
стых термодинамических системах.
В сб. «Механика», 3, 129—148.
Мещерская К. А. 1935. О некото-
рых физиологических свойствах
ооцитов низших позвоночных на
различных стадиях роста.— Ар'
хив анат., гистол., эмбриол., 14,
656—667.
Милнор Дж. 1965. Теория Морса.
М., «Мир».
Мина М. В. 1973. Рост рыб. Методы
исследования в природных попу-
ляциях. В сб. «Итоги науки и тех-
ники. Зоология позвоночных», 4,
68-115.
Михайлов В. С., Гаузе Г. Г. 1974.
Активация биосинтеза митохонд-
риальной ДНК в эмбриогенезе
вьюна.— Онтогенез, 5, 501—504.
Моисеев Н. Н. 1966. Асимптотиче-
ские методы нелинейной механики.
М., «Наука».
Молчанов А. М. 1967. Термодинами-
ка и эволюция. В сб. «Колебатель-
ные процессы, в биологических
и химических системах». М.,
«Наука», 292—308.
Молчанов А. М. 1971. Колебатель-
ные режимы как частный случай
критических режимов. В сб. «Ко-
лебательные процессы в биологи-
ческих и химических системах»,
т. 2, 327—331. Пущино на Оке.
Монахов Н. К. 1964. О биохими-
ческих функциях сердечных мито-
хондрии различного типа.—Био-
химия, 29, 955—963.
Нагорный А. В. 1938. О закономер-
855
ностях возрастных изменении
и втором принципе энергетики.—
Труды научно-иссл. зоол.-биол.
ин-та Харьковск. гос. ун-та, сек-
тор общей физиол., 5, 19—20.
Нагорный А. В., Никитин В. Н.,
Буланкин И, Н. 1963. Проблема
старения и долголетия. М., Гос.
изд. мед. лит.
Найденов Н. В. 1928. Закономерно-
сти в росте молодняка крупного
рогатого скота и свиней.— Зап.
Белорусск. гос. акад, сельск.
хоз-ва, 7, 1—68.
Нейфах А, А. 1960. Радиационная
пнактивация клеточных ядер как
метод исследования их роли в
развитии дыхания у зародышей
рыб.— Биохимия, 25, 658—668.
Нейфах С, А. 1967. Динамические,
структурные и генетические фак-
торы интеграции обмена. В сб.
«Механизмы интеграции клеточ-
ного обмена». Л., «Наука», 9—65.
Нейфах С. А. 1973. Генетические
функции митохондрий.— Вести.
Акад. мед. наук СССР, 1, 8—18.
Нейфах С. А. 1974а. Проблемы ци-
топлазматической наследствен-
ности. В сб. «Генетические функ-
ции органоидов цитоплазмы».
Л., «Наука», 7—19.
Нейфах С. А. 19746. Механизмы
клеточной интеграции в процессе
биогенеза митохондрий. В сб. «Ге-
нетические функции органоидов
цитоплазмы». Л., «Наука», 58—70.
Николаев А. Я. 1966. О термодина-
мике живых систем,— Биофизика,
И, 551-554.
Николаев Л. А. 1962. Термодинами-
ка необратимых процессов и про-
блемы биогенеза.— Ж. фпз. хи-
мии, 36, 3—14.
Николаев Л. А. 1971. Основы физи-
ческой химии биологических про-
цессов. М., «Высшая школа».
Никольская И. С. 1965. Потребле-
ние кислорода развивающимися
зародышами невского лосося
Salmo salar L. в процессе разви-
тия.— Вопр. ихтиол., 5, 742—747.
Озернюк Н. Д. 1970. Интенсивность
дыхания и содержание АТФ в
оогенезе вьюна.— Докл. АН
СССР, 192, 242—245.
Озернюк Н. Д. 1972. Изменение ко-
личества митохондрий в процессе
оогенеза вьюна.— Докл. АН
СССР, 207, 974-977.
Озернюк Н. Д. 1974. Биосинтез бел-
ков митохондрий в ооцитах вьюна
Misgurnus fossilis L.— Онтогенез,
5, 294-296.
Озернюк Н. Д., Зотин"А. И. 1974.
Зависимость между скоростью ды-
хания и весом ооцитов вьюна.—
Онтогенез, 5, 70—72.
Озернюк Н. Д., Пальмбах Л. Р.
1974. Связь между митохондриями
и эндоплазматическим ретикулу-
мом в ооцитах вьюна Misgurnus
fossilis L.— Онтогенез, 5, 404—
407.
Озернюк Н. Д., Пальмбах Л. Р.
1975. Рост и воспроизведение ми-
тохондрий в ооцитах вьюна.—
Онтогенез, 6,442—449.
Озернюк Н. Д., Зотин А. И., Юрови-
цкийЮ. Г. 1971. Оогенез как модель
процесса уклонения живой систе-
мы от стационарного состояния.—
Онтогенез, 2, 565—571.
Опарин А. И. 1957. Возникновение
жизни на Земле. М., Изд-во АН
СССР.
Опарин А, И. 1966. Возникновение
и начальное развитие жизни. М.,
Медицина.
Оппенгеймер К. 1934. Химические
основы жизненных процессов. М.—
Л., Биомедгиз.
Ортабаева Л. М. 1973. Изучение ди-
намики активности сукцинатде-
гидрогеназы в необластах в про-
цессе регенерации молочно-белых
планарий,— Цитология, 15,1476—
1480.
Остроумов С. А. 1973. О чем гово-
рит сходство митохондрий, хло-
ропластов и прокариотов.— При-
рода, 3, 21—29.
Пасынский А. Г. 1953. Второе на-
чало термодинамики в биологии.—
Биохимия, 18, 644—653.
Насынский А. Г. 1957. Теория от-
крытых систем и ее значение для
биохимии.—Усп. совр. биол., 43,
263—279.
Пасынский А. Г. 1963. Биофизиче-
ская химия. М., «Высшая школа».
Пелетминский С. В., Соколов-
ский А. И. 1974. К вопросу о по-
строении неравновесной энтро-
пии.— Теорет. мат. физика, 20,
85-99.
Перевощикова К, А., Делектор-
ская Л. Н., Делекторский В. В.,
Збарский И. Б. 1968. Выделение
и некоторые свойства ядерных
256
оболочек животных клеток. — Ци-
тология, 10, 573—580.
Петров Ю. П. 1966. К вопросу о
принципе минимума скорости воз-
растания энтропии в стационар-
ном режиме.— Биофизика, 11,
926-928.
Пинус Е. А. 1973. Об автономии
митохондрий. В сб. «Проблемы
возникновения и сущности жизни».
М., «Наука», 246—250.
Пинус Е. А., Метлицкая А. 3. 1970.
Синтез митохондриальных бел-
ков,— Усп. совр. биол., 70, 3—25.
Планк М. 1936. Об основании второ-
го закона термодинамики. В кн.
Ван-дер-Ваальс и Констамм «Курс
термостатики», 1, 438—452.
Платова Т. П. 1946. О значении
ядра и цитоплазмы в окислитель-
ном обмене овоцитов тритона.—
Докл. АН СССР, 53, 259—262.
Платова Т. П. 1948. Окислитель-
ный обмен овоцитов тритона в свя-
зи с процессами синтеза плазмы
и желтка.— Докл. АН СССР, 60,
1397-1400.
Полушкина А. Ф. 1967. О соотноше-
нии данных прямой и непрямой
калориметрии у кроликов, под-
вергнутых противотуберкулезной
вакцинации. В сб. «Всес. конф,
по теплообмену и теплопродукции».
Л., 84-85.
Полякова М. С. 1974. О диссипатив-
ных структурах в одной модели
химической реакции с диффу-
зией.— Вести. МГУ, 6, 643—648.
Попов К. 1955. Математические ос-
новы теории необратимых термо-
динамических процессов.— Ж.
эксп. теорет. физики, 28, 257 —
282.
Преснов Е. В. 1972. Математиче-
ская модель термодинамической
теории развития.— Труды IV
межд. биофиз. конгресса. М., 2,
6-7.
Преснов Е. В, 1973. Потенциальный
характер критерия эволюции тер-
модинамики необратимых процес-
сов.—Ж. физ. химии, 47, 2902—
2904.
Преснов Е. В., Зотина Р. С. 1975.
Развитие, основной обмен и веро-
ятность состояния живой системы.
Сб. теорет. эксп. биофизики. Ка-
лининград, 5, 48—51.
Преснов Е. В., Зотина Р. С., Зо-
тин А, И. 1973. Общий вид фено-
менологических уравнений роста,
дифференцировки и формообразо-
вания. Сб. Теорет. эксп. биофизи-
ка. Калининград, 4, 42—45.
Привалов П. Л, 1970. Тепловые ис-
следования внутримолекулярных
превращений биополимеров. Пер-
спективы сканирующей микрокало-
риметрии.— Биофизика, 15, 206—
214.
Пригожин И. 1959. Проблема эво-
люции в термодинамике необрати-
мых явлений. В сб. «Возникнове-
ние жизни на Земле». М., Изд-во
АН СССР, 408—416.
Пригожин И. 1960. Введение в тер-
модинамику необратимых процес-
сов. М., ИЛ.
Пригожин И., Николис Ж. 1973.
Биологический порядок, струк-
тура неустойчивости.— Усп. физ.
наук, 109, 517—544.
Приц А. К. 1974. Принцип стационар-
ных состояний открытых систем
и динамика популяций. Изд. Ка-
лининград. гос. ун-та.
Рабинович Е, 1951. Фотосинтез. М.,
ИЛ.
Рабинович П. Г. 1974. Неединст-
венность прямоугольных решений
задачи Бенара. В сб. «Теория ветв-
ления и нелинейные задачи на
собственные значения». М., «Мир»,
209-227.
Равен X. 1964. Оогенез. Накопление
морфогенетической информации.
М., «Мир».
Радзинская Л.И.У Никольская И. С,
1972. Энергетика развития туто-
вого шелкопряда. 1. Дыхание
и уровень АТФ в зародышевом
развитии.— Онтогенез, 3, 602—
608.
Ребане К. 1972. К вопросу исполь-
зования понятия энтропии в био-
логии и проблемах защиты при-
роды.— Изв. АН Эст.ССР, 21,
282-288.
Розен Р. 1969. Принцип оптималь-
ности в биологии. М., «Мир».
Романовский Ю. М., Степано-
ва Н. В., Чернавский Д. С. 1975.
Математическое моделирование
в биофизике. М., «Наука».
Ротшильд. 1958. Оплодотворение.
М., ИЛ.
Рубин A. Б., Фохт А. С. 1972.
Об определении скорости прироста
энтропии для систем, далеких от
257
термодинамического равновесия.—
Биофизика 17, 96—102.
Рудин Д., Уилки Д. 1970. Биоге-
нез митохондрий. М., «Мир».
Рывкина Д. Е. 1942. Дыхание тка-
ней при регенерации.— Изв. АН
СССР, сер. биол., 1—2, 17—29.
Рывкина Д. Е. 1945. Дыхательный
коэффициент тканей при регенера-
ции.— Докл. АН СССР, 49, 469—
471.
Сарбахер Ф., Эдсон В. 1947. Техни-
ка сверхвысоких частот. М., Гос-
издат связи и радио.
Седов Л. М. 1973. Механика сплош-
ной среды, т. 1. М., «Наука».
Селъков Е. Е. 1975. Регуляция поли-
ферментных систем. М., «Наука».
Семихатова О. А. 1967. Методы
оценки энергетической эффектив-
ности дыхания растений Л.,
«Наука».
Сидорова В. Ф., Рябинина 3. А.,
Лейкина Е. М. 1966. Регенерация
печени у млекопитающих. Л., «Ме-
дицина».
Скулачев В. П, 1969. Аккумуляция
энергии в клетке. М., «Наука».
Слоним Л. Д. 1961. Основы общей
экологической физиологии млеко-
питающих. М.—Л., Изд-во АН
СССР.
Соколов В. Е. 1973. Систематика
млекопитающих. М., «Высшая
школа».
Стратонович Р. Л, 1960. Флуктуа-
ционная термодинамика неравно-
весных процессов.— Ж. эксп. теор.
физики, 39, 1647—1659.
Стратонович Р. Л, 1967. О важней-
ших соотношениях нелинейной
термодинамики необратимых про-
цессов.— Вести. МГУ, сер. 3, 4,
84-89.
Стратонович Р. Л. 1969. К нелиней-
ной термодинамике неравновесных
процессов.— Вести. МГУ, сер. 3,
1, 40-46.
Стратонович Р. Л. 1971. Примене-
ние линейной и нелинейной флук-
туационно-диссипативной термо-
динамики к механике.— Вести.
МГУ, сер. 3, 1, 3—10.
Студенский А. А. 1897. Опыт со-
поставления количеств теплоты,
вычисленных (на основании дан-
ных Рубнера) по обмену, с коли-
чествами её, определяемыми кало-
риметром у животных (собак)
в норме, лихорадке и беременности.
Докторская дисс. № 61. СПб.,
изд-во М. Гутзац.
Сущеня Л. М, 1972. Интенсивность
дыхания ракообразных. Киев,
«Наукова думка».
Сырников Ю. П. 1965. О примени-
мости второго начала термодина-
мики к живой материи.— Биофи-
зика, 10, 1102-1103.
Тарусов Б. Н. 1960. Основы биофи-
зики и биофизической химии, 1.
М., «Высшая школа».
Тахтаджян А. Л. 1973. Четыре
царства органического мира.—
Природа, 2, 22—32.
Теплиц Н.А. 1974. Включение ме-
ченых аминокислот в белки кле-
точных органелл мышечной ткани
при восстановлении регенерацион-
ной способности конечностей аксо-
лотлей, подавленной рентгенов-
ским облучением.— Онтогенез, 5,
391—397.
Тимофеев-Ресовский Н. В., Ворон-
цов Н, Н., Яблоков А. В. 1969.
Краткий очерк теории эволюции.
М., «Наука».
Токин И, Б. 1974. Проблемы радиа-
ционной биологии. Л., «Медицина».
Томме М. Ф., Ким М. М. 1938. Га-
зообмен, обмен веществ и энергии
у растущих кроликов.— Усп. зо-
отехн. наук, 5, 100—115.
Тринчер К. С. 1965. Биология и ин-
формация. Элементы биологиче-
ской термодинамики. М., «Нау-
ка».
Трусделл К. 1970а. Термодинамика
для начинающих. В сб. «Механи-
ка», 3, 116-128.
Трусделл К. 19706. Главы из книги
«Шесть лекций по современной
натурфилософии». В сб. «Механи-
ка», 4, 99—136.
Фаустов В. С,, Карлсон Б, М. 1967.
Изменение уровня АТФ и дыхания
в регенерирующих конечностях ак-
солотля Ambystoma mexicanum.—
Докл. АН СССР, 176, 728—730.
Фаустов В. С., Зотин А. И., Рад-
зинская Л. И., Никольская И. С.
1968. Аденозинтрифосфат в раз-
витии зародышей иглокожих, рыб
и амфибий.— Ж. эвол. биох. и
физиол., 4, 224—230.
Филюков А. А., Митрофанов В, Б.
1968. Об особенностях вариацион-
ной формулировки общего крите-
рия эволюции в методе локального
258
потенциала.— Инж.-физ. ж., 14,
658—665.
Хаазе Р. 1967. Термодинамика не-
обратимых процессов. М., «Мир».
Хансон К. П., Иванова Л. В., 1971.
Митохондриальная дезоксирибо-
нуклеиновая кислота животных.—
Усп. совр. биол., 72, 3—23.
Харрисон У. 1972. Теория твердого
тела. М., «Мир».
Хаскин В. В. 1961. Теплообмен
птичьих яиц при инкубации.—
Биофизика, 6, 91—99.
Хореккер Б. Л., Хяйят X. X. 1961.
Пути углеводного обмена в здоро-
вых и опухолевых клетках. В сб.
«Современные проблемы биохимии».
М., ИЛ, 362-396.
Циггеп Г. 1966. Экстремальные прин-
ципы термодинамики необрати-
мых процессов и механики сплош-
ной среды. М., «Мир».
Чанс Б., Хаджихара Б. 1962. Пря-
мые спектрофотометрические изме-
рения взаимодействия компонен-
тов дыхательной цепи с АТФ, АДФ,
фосфатом и разобщающими веще-
ствами.— Труды V Междунар.
биох. конгресса, симп. 5, 10—43.
Чередниченко Л. К. 1965. Физиоло-
гическая калориметрия. М.— Л.,
«Наука».
Чернявский Д. С. 1975. Термоди-
намика и биология. Сб. Теорет.
эксп. биофизика. Калининград, 5,
3-27.
Чернявский Ф. Б. 1962. О размноже-
нии и росте снежного барана.—
Зоол. ж., 41, 1556—1566.
Шапот В. С. 1968. Злокачественный
рост глазами биохимика.— Приро-
да, 3, 29—39.
Шмальгаузен И. И. 1935. Определе-
ние основных понятий и методики
исследования роста. В сб. «Рост
животных». М.— Л., Госиздат
биол. мед. литер., 8—60.
Шмальгаузен И. И. 1969. Пробле-
мы дарвинизма. Л., «Наука».
Шмальгаузен И, И. 1972. Факторы
прогрессивной (ароморфной) эво-
люции^ снижение энтропии. В сб.
«Закономерности прогрессивной
эволюции». Л., 5—24.
Шпольский Э. В. 1949. Атомная фи-
зика. М., Гос. изд. техн. теор.
литер, т. I.
Шполъский Э. В, 1950. Атомная
физика. М., Гос. изд. техн. теор.
литер, т., II.
Шредингер Э. 1947. Что такое жизнь
с точки зрения физики? М., ИЛ.
Эйген М. 1973а. Молекулярная са-
моорганизация и ранние стадии
эволюции.— Усп. физ. наук, 109,
545-589.
Эйген М. 19736. Самоорганизация
материи и эволюция биологических
макромолекул. М., «Мир».
Юдин К. А. 1974. О понятии «приз-
нак» и уровнях развития система-
тики животных. В сб. «Теоретиче-
ские вопросы систематики и фи-
логении животных». Л., «Наука»,
5-29.
Юровицкий Ю. Г., Милъман Л. С.,
Озернюк Н. Д. 1972. Некоторые
особенности регуляции гликолиза
в ооцитах вьюна.— Онтогенез, 3,
405-414.
Юровицкий Ю, Г. 1973. Особенности
регуляции углеводного обмена
в оогенезе при созревании ооцитов
и в раннем эмбриогенезе.— Ж.
общ. биол., 34, 97—109.
Яковлева К. К. 1964. Интенсивность
дыхания у некоторых черномор-
ских медуз.— Труды Севаст. биол.
ст., 17, 364—367.
Abbott В. С., Howarth J. V. 1973.
Heat studies in exitable tissues.—
Physiol. Rev., 53, 120—158.
Aisenberg A. C. 1961. The Glyco-
lysis and Respiration of Tumours.
N. Y., Acad. Press.
Aisenberg A. C., Morris H. P. 1961.
Energy pathways of hepatoma.—
Nature, 191, N 5123, 1314—1315.
Allard C., Lamirande J, de, Caufe-
ro A. 1952. Mitochondrial popula-
tion in mammalian cells. IV.
Preliminary results on the variation
in the mitochondrial population of
the average rat liver cell during
azodyescarcinogenesis.— Canad. J.
Med. Sci., 30, 542—543.
Altman P. L,, Dittmer D. S. 1964.
Biology Data Book. Washington,
Federat. Amer. Soc. Exper. Biol.
Apter M, J., Wolpert L. 1965.
Cybernetics and development.
I. Information theory.— J. Theo-
rem Biol., 8, 244-257.
A schenbrenner V., DruyanR., A IbinR.,
Rabinowitz M. 1970. Haem a, cy-
tochrome c and total protein tur-
nover in mitochondria from rat
heart and liver. — Biochem. J.,
119, 157-160.
Atwater W, O, 1904. Ncue Vcrsuche
259
fiber Stoff- und Kraftwechsel im
menschlichen Korper.— Ergebn.
Physiol., 3, 1-497.
Babloyantz A., Nicolis G. 1972.
Chemical instabilities and multi-
ple steady state transitions in
Monod-Jacob type models. — J. The-
oret. Biol., 34, 185—192.
Bakke F., Olsen H., Wergeland H.
1955. On Schrodinger’s negative
entropy.— Kongel. norske vid. sels-
kabs forhandl., 28, 21—24.
Barisas B. G., Singer S. J., Stur-
tevant J. M. 1972. Thermodyna-
mics of the binding of 2,4-dini-
trophenol and 2,4,6-trinitrophenol
haptens to the homologous and
heterologous rabbit antibodies.—
Biochemistry, 11, 2741—2744.
Barott H. G., Pringle E. M. 1946.
Energy and gaseous metabolism
of the chicken from hatch to matu-
rity as affected by temperature.—
J. Nutrit., 31, 35-50.
Barrows С. H. 1966. Enzymes in the
study of biological aging. In: Per-
spective in Experimental Geron-
tology. Illinois, Thomas.
Barrows С. H., Falzone J. A.,
Shock N. W. 1960. Age differences
in the succinoxidase activity of
homogenates and mitochondria
from the livers and kidneys of
rats.— J. Gerontol., 15, 130.
Baud C. A. 1959. Ultrastructure et
function de la membrane nucleai-
re. En: Problemes d’ultrastructu-
res et la fonctions nucleaires. Pa-
ris, p. 1—34.
Belaich J. P., Sari J. C. 1969. Micro-
calorimetric studies on the forma-
tion of magnesium complexes of
adenine nucleotides.— Proc. Nat.
Acad.Sci. USA, 64, 763—770.
Belaich J. P., SenezJ. C., Murgier M.
1968. Microcalorimetric study of
glucose permeation in microbial
cells.— J. Bacteriol., 95, 1750 —
1757.
Benard H. 1900. Les tourbiLons cel-
lulaires dans une nappe liquide.
I. Description general des pneno-
menes. II. Proc6des mecaniques
et optiques d’examen lois numeri-
ques des phenomenes.— Rev. g£n.
sci. pure et appl., 11, 1261—1271,
1309-1328.
Benedict F. G. 1938. Vital Energe-
tics. Washington, Carnegie Inst.
Benzinger T. H., Hems R. 1956.
Reversibility and equilibrium of
the glutaminase reaction observed
calorimetrically to find the free
energy of ATP hydrolysis.— Proc.
Nat. Acad. Sci. USA, 42, 896-900.
Benzinger T. H., Huebscher R. G.,
Minard D., Kitzinger C. 1958.
Human calorimetry by means of
the gradient principle.— J. Appl.
Physiol., 12, Suppl. 1, 1—24.
Benzinger T. H., Kitzinger C. 1963.
Microcalorimetry, new methods
and objectives.— In: Temperatu-
re — its Measurement and Control
in Science and Industry, v. 3,
pt 3, N. Y., Reinhold, p. 43-60.
Bernhard R. 1964. Survey of some
biological aspects of irreversible
thermodynamics.— J. Theoret.
Biol., 7, 532-557.
Bertalanffy L. von. 1932. Theoretische
Biologic. Berlin.
Bertalanjfy L. von. 1934. Untersuchun-
gen uber die Gesetzlichkeit des
Wachstums. I. Allgemeine Grund-
lagen der Theorie: Mathematische
und physiologische Gesetzlichkei-
ten des Wachstums bei Wassertie-
ren.— W. Roux’ Arch., 131, 613—
653.
Bertalanjfy L. von. 1950. The theory
of open systems in physics and
biology.— Science, 111, 23—29.
Bertalanjfy L. von. 1960. Principles
and theory of growth.— In: Fun-
damental Aspects of Normal and
Malignant Growth. Amsterdam,
Elsevier, p. 137—259.
Biot M. A. 1955. Variational prin-
ciples in i reversible thermodyna-
mics with application to viscoelas-
ticity.— Phys. Rev., 97, 1463 —
1469.
Biot M. A. 1970. Variational prin-
ciples in heat transfer. Oxford,
Clarendon Press.
Bjurulf C., Laynez J., Wadsd J.,
1970. Thermochemistry of lysozy-
me-inhibitor binding.— Europ.
J. Biochem., 14, 47—52.
Bjurulf C., Wadsd I. 1972. Thermo-
chemistry of lysozyme-inhibitor
binding.— Europ. J. Biochem., 31,
95-102.
Blokhius G. G. D., Veldstra H. 1970.
Heterogenity of mitochondria in
rat brain.— FEBS Lett., 11,197 —
199.
Blum H. F. 1967. The concepts of
entropy and energy in biology.
260
Ann. N. Y., Acad. Sci., 138, 894 -
895.
Boell E. J. 1955. Energy exchange
and enzyme development during
embryogenesis.— In: Analysis of
Development, Philadelphia, Saun-
ders. p. 520—555.
Boell E. J., D'Anna T., Greehfield P.,
Petrucci D. 1971. Cytochrome oxi-
dase activity during amphibian
development.— J. Exper. Zool.,
178, 151-163.
Boell E. J., Weber R. 1955. Cyto-
chrome oxidase activity in mitochon-
dria during amphibian develop-
ment.— Exper. Cell Res., 9, 559 —
567.
Boivinet P. 1964. Perfectionnement
aux methodes d’etalonnage et
d’emploi du microcalorimetre E. Ca-
Ivet. Utilisation en microbiologie.
Theses, Aix-Marseille.
Boivinet P. 1971. Contributions de la
calorimetric aux recherches biolo-
giques actuelles.— Proc. First
Europ. Biophys. Congr., 4, 277 —
288.
Boivinet P., Garrigues J. C., Gran-
getto T. 1968. Dosage microcalori-
metrique de 1’insuline.— Compt.
rend. Soc. biol. France, 162,
1770-1774.
Boivinet P., Rybak B. 1969. Etude
microcalorimetrique du fonction-
nement electro-mecanique du coe-
ur de grenouille.— Life Sci., 8,
11-20.
Bolen D. W., Fibgel M.t Biltonen R.
1971. Calorimetric studies of pro-
tein-inhibitor interaction. I. Bin-
ding of 3’-cytidine monophosphate
to ribonuclease A at pH 5,5.—
Biochemistry, 10, 4136—4140.
Boling E. A., Blanchard G. C.,
Russell W. J. 1973. Bacterial
identification by microcalorime
try.— Nature, 241, 472—473.
Bondi E. E., Devlin T. M., Ch'ihJ. J.
1972. Distribution of two mito-
chondrial populations in rabbit
kidney cortex and medulla.—
Biochem. and Biophys. Res. Com-
mune., 47, 574—580.
Bornmann L,, Busse H., Hess B.
1973. Oscillatory oxidation of
malonic acid by bromate 1.
Thin-layer chromatography.—
Z. Naturforsch., 28b, 93—97.
Bornmann L., Busse H., Hess В. 1973a.
Oscillatory oxidation of malonic
acid by bromate 2. Analysis of
products by gas chromatography
and mass spectrometry. — Z. Na-
turforsch., 28b, 824—827.
Bornmann L., Busse H., Hess В. 1973b.
Oscillatory oxidation of malonic-
acid by bromate 3. CO2 and BrO3-
titration.— Z. Naturforsch., 28c,
514-516.
Borst P., Griiell L, A, 1971. Mito-
chondrial ribosomes. — FEBS
Lett., 13, 73—88.
Boyling J. B. 1972. An axiomatic
approach to classical thermody-
namics.— Proc. Roy. Soc. London,
Ser. A, 329, 35—70.
Boyling J. B. 1973. Carnot engines
and the principle of increase of
entropy.— Internet. J. Theoret.
Phys., 7, 291-299.
Воуо A. E., Ikomi-Kumm J. A. 1972.
Increase metabolic heat produc-
tion by erythrocytes in sickle cell
disease.— Lancet, 1, 1215—1216.
Bracket J. 1950. Chemical Embryolo-
gy. N. Y., Hafner.
Brandt P. W., Pan pas G. I). 1959.
Mitochondria. II. The nuclear-
mitochondrial relationship in
Pelomyxa caroeinensis Wilson
(Chaos chaos L.).— J. Biophys.
and Biochem. Cytol., 6, 91—96.
Brayton R., Moser J. 1964. A theory
of non-linear networks. Quart.
Appl. Math., 22, 1-33.
Brettel R. 1974. Microcalorimetric
measurements of the energy utili-
zation in chemostat cultures of
Saccharomyceb cerevisiae.— Proc.
4 Internet. Sympos. on Yeasts,
p. 87—88, Inst. Food Technol.,
Vienna.
Brettel R., Corti L., Lamprecht I.,
Schaarschmidt B. 1972. Combina-
tion of a continuous culture with
a flow microcalorimeter.— Studio
Biophys., 34, 71-76.
Brillouin L. 1949. Life, thermodyna-
mics, and cybernetics.— Amer.
Scientist, 37, 554—568.
Brillouin L. 1955. Entropy and
growth of an organism.— Ann.
N. Y. Acad. Sci., 63, 454-455.
Britten G., Gamow G. 1961. Negative
entropy and photosynthesis.—
Proc. Nat. Acad. Sci. USA, 47,
724-727.
Broda E. 1970. The evolution of
bioenergetic processes. Progress in
261
Biophysics and Molecular Biology,
21.
Brody S. 1945. Bioenergetics and
Growth. N. Y., Reinhold.
Brown H. D. 1969. Calorimetry of
enzyme-catalyzed reactions.— In:
Biochemical Microcalorimetry.
N. Y., Acad. Press, p. 149—165.
Brown H. D., Evans W. J., Altschul
A. M. 1965. Application of calori-
metry. Scheme for the continuous
observation of the movement of
glucose across a biological memb-
rane.— Biochim. et biophys. acta,
94, 302-304.
Brown L. 1964. Physiology of the
Amphibia. N. Y.
Biichel W. 1967. Entropy and infor-
mation in the universe.— Nature,
213 319_____320.
Buchner F. 1966. DNS, RNS und
Protein Stoffwechsel im normalen
und im atmungsgestorben Wizbel-
tierkeim (nach historadiographi-
scher und electronmicroscopischen
Untersuchungen.)— Bull. Schweiz.
Acad. med. Wiss., 22, 56—79.
Bulos B., Shukla S., Sacktor B. 1972.
Bioenergetic properties of mito-
chondria from flight muscle of
aging blowflies.— Arch. Bio-
chem. and Biophys., 149, 461 —
469.
Burk D., Woods M., Hunter J. 1967.
On the significance of: glucolisis
for cancer growth, with special
reference to Morris rat hepatomas.—
J. Nat. Cancer Inst., 38, 839 —
867.
Butler J. A. V. 1946. Life and the
second law of thermodynamics. —
Nature, 158, 153—154.
Calloway N. 0. 1966. The role of
entropy in biologic senescence.—
J. Amer. Geriatr. Soc., 14, 342—
349.
Calvet E., Prafr H. 1963. Recent
Progress in Microcalorimetry. Ox-
ford, Pergamon Press.
Campbell B. 1967. Biological entropy
pump.— Nature, 215, 1308.
Caplan S, R. 1966. A characteristic
of selfregulated linear energy con-
verters. The Hill force — veloxcity
relation for muscle.— J. Theoret.
Biol., 11, 63-86.
Carlson L. D., Honda N., Sasaki T.t
Jvdy W. W. 1964. A human calo-
rimeter.— Proc. Soc. Exper. Biol.
Med., 117, 327-338.
Casimir H. B. G, 1945. On Onsager’s
principle of microscopie reversibi-
lity.— Rev. Mod. Phys., 17, 343 —
350.
Chan T. H. C., Balek R. W. 1973.
Changes in lactate dehydrogenase
zymograms from forelimb and tail
regenerates of two species of
Salamander.— Compar. Biochem.
Physiol., Ser. A, 44, 1093-1100.
Chandrasekhar S. 1961. Hydrodyna-
mic and Hydromagnetic Stabi-
lity. Oxford, Clarendon Press.
Chase J. W., Dawid I. B. 1972.
Biogenesis of mitochondria during
Xenopus laevis development.—
Develop. Biol., 24, 504—518.
Child С. M. 1948. Differential oxi-
dation and reduction of indicators
in reconstitution of Hydra and a
planarian.— Physiol. Zool., 21,
327—351.
Child С. M. 1951. Oxidation-reducti-
on indicator patterns in regenera-
tion of the foreleg and tail of Tri-
turus torosus larvae.— Biol. Bull.,
101, 121-130.
Clark A. M., Rockstein M. 1964. Aging
in insects.— In: Physiology of In-
sects, v. 1. N. Y., Acad. Press,
p. 227-281.
Claude A., Mathieu R., Lamirande G.
de, Cantero A. 1952. Mitochondrial
population in mammalian cells.
I. Description of a counting tech-
nique and preliminary results on
rat liver in different physiologi-
cal and pathological condition.—
Cancer Res., 12, 407—412.
Cliffe A. J., Me Kinnon С. H.,
Berridge N. J. 1973. Microcalo-
rimetric estimation of bacteria in
milk.— J. Soc. Dairy Technol.,
26, 209—210.
Cold P. H„ Gee M. V.,Stpehler B. L.
1968. Effect of age on oxidative
phosphorylation in the rat.— J. Ge-
rontol., 23, 509—512.
Collins J. F., Richmond M. H. 1961.
Rate of growth of Bacillus cereus.—
J. gen. Microbiol., 28, 15—35.
Dale H. E., Shanklin M. D., John-
son H. D., Brown W. H. 1967.
Energy metabolism of the chim-
panzee: a comparaison of direct
and indirect calorimetry.— J. Appl.
Physiol., 22, 850-853.
David H. 1964. Physiologische und
pathologische Modifikationen der
submicroskopischen Kernstructur.
262
11. Die Kernmembranen.— Z. mik-
rosk.-anat. Forsch., 71, 526—550.
Davidson F. A. 1928. Growth and
senescence in purebred Jersey cows.
Bull. Illinois agric. Exper. Stat.,
302, 183—235.
Davison J. 1955. Body weight, cell
surface, and metabolic rate in
anuran amphibia.— Biol. Bull.,
109, 407-419.
Dawes G. 5., Mott J. C. 1959. The
increase in oxygen consumption of
the lamb after birth.— J. Physiol.,
146, 295—315.
Dawson W. R., Hudson J. W. 1970.
Birds. In Comparative Physiology
of Thermoregulation, v. I. N. Y.,
Acad. Press, p. 223—310.
Deen H. 1967. Effect of bromine
derivatives of malonic acid in the
oscillating reaction of malonic acid,
cerium ions and bromate.— Nature,
213, 589-590.
Donder Th. de. 1927. L’affinite.
Paris, Gauthier — Villars.
Donder Th. de, Rysselberghe P. van.
1936. Affinity. Stanford Univ.
Press.
Doyer L. C. 1915. Energie-Umgetzun-
gen wahrend der Keimung von
Weizenkornern.— Rec. Trav. bot.
neerl., 12, 369-423.
Droz B., Bergeron M. C. 1965.
Defection radioautographique de
proteines nouvellement synthezisees
a pariir de leucine-Зн dans les
mitochondries du foil et du rein
chez le rat.— C. r. Acad. sci. Pa-
ris, 261, 2757-2760.
Dumont J. N. 1972. Oogenesis in
Xenopus laevis (Dandier.)—
J. Morphol., 136, 153—180.
Durnin J. V. G. A., Passmore R.
1967. Energy, work and leisure.
London, Heinemann.
Edelen D. G. B. 1972. A nonlinear
Onsager theory of irreversibility.—
Internal. J. Engng Sci., 10, 481 —
490.
Edelen D. G. B. 1973a. Asymptotic
stability, Onsager fluxes and rea-
ction kinetics.— Internal. J. Engng
Sci., 11, 819-839.
Edelen D. G. B. 1973b. On the existen-
ce of symmetry relations and dis-
sipation potentials.— Arch. Ra-
tion. Meeh. Analyses, 51, 218—227.
Edelen D. G. B. 1974a. Generalized
Onsager fluxes and forces: a non-
linear phenomenological theory.—
Z. phys. Chem. Nene Folge, 88,
27__53
Edelen D. G. B. 1974b.| Primitive
thermodynamics: a new look at
the Clausius — Duhem inequali-
ty.— Internat. J. Engng Sci., 12,
121—141.
Edelen D. G. B. 1974c. On the chara-
cterization of fluxes in nonlinear
irreversible thermodynamics.—
Internat. J. Engng Sci., 12, 397 —
411.
Edelen D. G. B. 1974d. Irreversible
thermodynamics of nonlocal sys-
tems.— Internat. J. Engng Sci.,
12, 607-631.
Edington D. W., McCafferty PF. B.
1973. Mitochondrial size distri-
bution analysis in the soleus muscle
of trained and aged rats.— Experi-
entia, 29, 692—693.
Eldblom E. 1935. Hauttemperatur-
essungen bei Ischias.— Klin.
Wschr., 14, 639-641.
Eriksson R., Wadsd I. 1971. Design
and testing of a flow microcalori-
meter for studies of aerobic bacteri-
al growth. — Proc. First Europ.
Biophys. Congr., 4, 319—327.
Estes Z. E. 1960. Cytochrome oxidase
activity of intact and digitonin-
treated chick liver mitochondria
during development.— Anat. Rec.,
138, 346.
Eyring H., Boyce R. P., Spikes J. D.
1960. Thermodynamics of living
systems.— In: Comparative Bioche-
mistry, v. 1. N. Y., Acad. Press,
p. 15-73.
Farkas H. 1968. The reformulation of
the Gyarmati principle in a gene-
ralized «Г» picture.— Z. Phys.
Chem., 239, 124-132.
Farkas H., Noszticzius Z. 1972. On
the non-linear generalization of
the Gyarmati principle and theo-
rem.— Ann. Phys., 27, 341—348.
Faustov V. S. 1969'. Changes in ATP
levels and respiration in regenra-
ting axolotl limbs.— Anat. Rec.,
163, 301.
Field R. J., Noyes R. M. 1972.
Explanation of spatial band pro-
pagation in the Belousov reaction.—
Nature, 237, 390—392.
Field R. J., Noyes R. M. 1974.
Oscillations in chemical systems.
IV. Limit, cycle behaviour in a
model of real chemical reaction.—
J. Chem. Phys., 60, 1877—1884.
263
Field R. J., Кiiros В., Noyes В. M.
1972. Oscillations in chemical
systems. II. Thorough analysis of
temporal oscillation in the broma-
te — cerium-malonic acid system. —
J. Amer. Chem. Soc., 94, 8649 —
8664.
Finlayson B. A., Scriven L. E. 1967.
On the search for variational prin-
ciples.— Internal. J. Heat Tran-
sfer, 10, 799—821.
Forrest W. W. 1972. Microcalorimet-
ry.— In: Methods in Microbiology,
v. 6B, London, Acad. Press,
p. 285-318.
Forrest W. W., Walker D. J. 1964.
Change in entropy during bacte-
rial metabolism.— Nature, 201,
49-52.
Fourche J., Lemaitre C. 1974. Le
metabolisme energetique au cours
des metamorphoses de Ceratitis
capitata — relations thermogenese—
respiration.— Bull. biol. France
et Belgique, 108, 97—102.
Franke W. W., Deumling B., Er men
B.t Jarasch E. Kleining H,
1970. Nuclear membranes from
mammalian liver. I. Isolation pro-
cedure and general characteriza-
tion.—J. Cell. Biol., 46, 379 —
395 •
Gage D. H., Schiffer M., Kline S.
Reynolds W. C. 1966. The non —
existence of a general thermokinetic
variational principle.— In: Non-
equilibrium Thermodynamics, Va-
riational Techniques and Stability.
Univ. Chicago Press, p. 283 —
286.
Gambale F.t Gliozzi A. 1971. Dynamic
pattern formation of convective
Hows. Proc. First Europ. Biophys.
Congr., 4, 167—171.
Garn P. D. 1965. Thermoanalytical
Methods of Investigation. N. Y.,
Acad. Press.
Gear A. B. L. 1970. Inner- and outer-
membrane enzymes of mitochond-
ria during liver regenration.—
Biochem. J., 120, 577—587.
Gear A, B. L.t Bednarek J. M. 1972.
Direct counting and sizing of mito-
chondria in solution.— J. Cell.
Biol., 54, 325—345.
Geczik R., Wolsky A. 1959. Dehydro-
genase activity during regeneration
in the tail of Tri turns viridescens:
a histological and enzymological
study.— Anat. Rec., 134, 568.
Gerisch G. 1971. Periodische Signale
steuern die Musterbildung in
Zellverbanden. — Naturwissenschaf
ten, 58, 430—438.
Gibbs C. L., Mommaerts W. F.t Ric-
chiuti N. V. 1967. Energetics of
cardiac contractions.— J. Physiol.,
191, 25-46.
Givoni B. 1969. Man, climate and
architecture. London, Elsevier.
Glansdorff P, 1956.J Sur une loi de
moderation des transformations
chimiques irreveribles.— Bull. Cl.
sci. Acad. roy. Belg., 42, 628—630.
Glansdorff P. 1960. On a non-linear
/law of the irreversible phenomena
with stationary constraints.— Mo-
lec. Phys., 3, 277-283.
Glansdorff P., Nicolis G.f Prigogine I.
1974. The thermodynamic stabi-
lity theory of non-equilibrium sta-
tes.— Proc. Nat. Acad. Sci. USA,
71, 197—199.
Glansdorff P,, Prigogine I. 1954. Sur
|les proprietes dilierentielles de la
production d’entropie.— Physica,
Z0, 773-780.
Glansdorff P., Prigogine I. 1964. On
a general evolution criterion in
macroscopic physics.— Physica,
30, 351—374.
Glansdorff P., Prigogine I. 1971.
Thermodynamic Theory of Stru-
cture, Stability and Fluctuations.
London, Wiley and Sons, Ltd.
Glaser R. 10/1. Einfuhrung id die
Biophysik. Jena, VEB Gustav
Fiscner Verlag.
Goff art M. 1971. Function and Form
ш the Sloth. N. Y., Acad. Press.
Goldbeter A., Nicolis G. 1972. Far from
equilibrium synthesis of small poly-
mer chains^ and cnemical evoluti-
on.— Biophysik, 8, 212.
Gompertz B. 1825.1 On the nature of
the function expressive of the law
ol human mortality, and on a new
mode oi determining the value of
life contingencies.— Philos. Trans.
/Roy. Soc. London, 115, 513—565.
Gonse P, H, 1955. Reciieiches sur la
physiologic de I’ovogenese chez
Phascolosoma vulgare.^ Exper.
Cell Res., 8, 550—1>53.
Gonse P. H. 1957. L’ovogenese chez
Phascolosoma vulgare. Ill. Res-
piration exogene et endogene de
Г ovocyte efiet de 1’eau de mer.-^
Bioclum. et biophys. acta, 24,
267—278.
264
Gorski F. 1966. Plant growth and
entropy production. Krakow,
Zaklad Fiziol. Poglin Pan.
Gustafson T. 1965. Morphogenetic
significance of biochemical patterns
in sea urchin embryos.— in: The
Biochemistry of Animal Develop-
ment, v. 1, N. Y., Acad. Press,
p. 139—202.
Gyarmati 1. 1961. On the phenome-
nological basis of irreversible ther-
modynamics.— Periodica poly-
techn., Budapest, 5, 219—243,
321-339.
Gyarmati I. 1968. On the most gene-
ral form of the thermodynamic in-
tegral principle. — Z. phys. Ghem.,
2з9, 133—13/.
Gyarmati I. 1969. On the «Governing
principle of dissipative processes»
and its extension to non-linear
problems.— Ann. Phys., 23, 353 —
o78.
Gyarmati I, 1970. Nonequilibrium
Thermodynamics. Berlin, Sprin-
ger.
Gyarmati I. 1974. Generalization of
the governing principle of dissipa-
tive processes to complex scalar
iields.— Ann. Phys., 31, 18—32.
Hanson 11. P. 1974. Spatial structu-
res in dissipative systems.—J.
Chem. Phys., 60, 321U—3214.
Hartman J. F. 1954. Electron mic-
roscopy of motor nerve cells follo-
wing section of axones.— Anat.
Rec., 118, 19—34.
Hast G., Pauly H. 1971. Kalorische
Eigenscnaften des gebundenen Was-
sers in Proteinlosungen.—Biophy-
sik, 7, 283—294.
Hawley E. 5’., Wagner R. P. 1967.
Synchronous mitochondrial divi-
sion in Neurospora crassa.—J.
Cell Biol., 35, 489-499.
Hay E. D. 1958. The fine structure
oi blastema cells and differentia-
ting cartilage cells in regenerating
limbs oi Amblystoma larvae.—
J. Biophys. and Biochem. Gytol.,
4, 583—592.
Hay E. D. 1959. Electron microsco-
pic observations of muscle dedif-
ierentiation in regenerating Amb-
lystoma limbs.— Develop. Biol.,
1, 555—585.
Hemmingsen A. H. 1960. Energy me-
tabolism as related to body size
and respiratory surface, and iis
evolution.— Repts Steno memor.
Hospital Nordisk Insulinlabor., 9,
7—110.
Hess В. 1963. Funktionelle und morp-
hologische Organization der Zelle.
Berlin, Springer Verlag.
Hess B., Boiteux A. 1968. Regulatory
Functions of Biological Membranes.
Amsterdam, Elsevier.
Hill A. V. 1931. Adventures in Biop-
hysics. Univ Pennsylvania Press
Philadelphia.
Hill A. V. 1965. Trails and Trials in
Physiology. London, Arnold.
Hishida T., Nakano E. 1954. Respira-
tory metabolism during fish deve-
lopment.— Embryologia, Z, 67 —
79.
Hoeppe K. 1959. Das reaktionskine-
tische Grundgesetz auf das Wachs-
tum bezogen. Dissertation, Gieben.
Hoffman H.t Grigg G. W. 1958. An
electron microscopic study of
mitochondria formation.— ьхрег.
Cell. Res., 15, 118—131.
Hogeboom G. H.t Schneider W. C.,
Palade G. E. 1948. Cytochemical
studies of mammalian tissues. 1.
Isolation of intact mitocnondna
from rat liver; some biochemical
properties oi mitochondria and
submicroscopic particulate mate-
rial.—J. Biol. Chem., 172, 619—
635.
Holmes C. F.t Mortimer R. G. 1967.
Derivation oi higher order recip-
rocal relations.— Industr. Engin
Cnem. Fundament, 6, 321—325.
Hosokawa J. 1973. Entropy produc-
tion in the lunction randomwalk
model.— J. Math. Buys., 14, 13/4—
1377.
Houpt T. R. 1968. Suction divice for
measurement of rumen exchange
in vivo.—Amer. J. Veterin. Res.,
Z9, 201—204.
Howland J. L.t Challberg M. D. 1973.
Altered respiration and proton per-
meability in liver mitochondria
from genetically dystrophic mice.—
Biochem. and Biophys. Res. Com-
muns, 50, 574—580.
Hudson J. W.t Deavers D. R. 1973.
Metabolism, pulmocutaneous wa-
ter loss and respiration of light spe-
cies of ground squirrels from dif-
ferent environments.— Compar.
Biochem. Physiol., 45, 69—1UU.
Hulbert A. J., Dawson T. J. 1974.
Standard metabolism and body
temperature ol penameloid man-
265
supials from different environ
ments.— Compar. Biochem. Phy-
siol., 47, 583—590.
Hussain H. M. 1973. Okologische
Untersuchungen uber die Bedeu-
tung thermophiler Mikroorganis-
men fur die Selbsterhitzung von
Heu.— Z. allg. Mikrobiol., 13,
323____334.
Hutchison V. H., Whitford W. G.,
Kohl M. 1968. Relation of body
size and surface area to gas exchan-
ge in anurans.— Physiol. Zool.,
41, 65-85.
Jakobson В. E., Blanchaer M. C.,
Wrogemann K. 1970. Defective
respiration and oxidative phospho-
rilation in muscle mitochondria of
hamsters in the late stages of he-
reditary muscular dystrophy.—
Canad. J. Biochem., 48, 1037—
1042.
Jakovcic S., Haddock J., Getz G. S.,
Rabinowitz M., Swift H. 1971.
Mitochondrial development in li-
ver of foetal and new-born rats.—
Biochem. J., 121, 341—347.
Janjic D., Stroot P.t Burger U. 1974.
Oscillations chimiques du systeme
BrOs“ (Ce4+) acetylacetone.—
Helv. chim. acta 56, 266—271.
Janoschek A, 1957. Das reaktionski-
netische Grundgesetz.—Statist.
Vierteljahresschrift, 10, 26—37.
Janssen H. K. 1972. Stability of
transport.— Z. Phys., 253, 176—
182.
Jaworski A., Sedlazek L., Czerniaw-
ski F., ZablockiB. 1968. Thermo-
genesis of the stable L-forms of
Proteus mirabilis and their parent
bacteria.— Acta microbiol., 17,
219-230.
Johnson E. A., Singer M. 1964.
A hictochemical study of succinic
and lactic dehydrogenases in the
regenerating forelimb of the adult
newt, Triturus.—Proc. Soc. Exper.
Biol, and Med., 117, 27—31.
Jones D. H., Rosano T, G. 1972. Stu-
dies ofj mitochondrial development
prior to lactogenesis in the mouse
mammary gland.— Arch. Biochem.
and Biophys., 153, 130—138.
Jones M. N.f Skinner H. A., Tipping
E., Wilkinson A. 1973. The interac-
tion between ribonuclease A and
surfactants.— Biochem. J., 135,
231—236.
Joos R. W., Carr C. W. 1969. The
binding of calcium to phospholipid-
protein complexes.— Proc. Soc.
Exper. Biol, and Med., 132, 865 —
870.
Kadenbach B. 1967. Synthesis of mi-
tochondrial proteins: demonstra-
tions of a transfer of proteins from
microsoms into mitochondria.—
Biochim. et biophys. acta, 134,
430-442.
Karasaki 5., Komoda T. 1958. Elect-
ron micrographs of a crystalline
lattice structure in yolk platelets
of the amphibian embryo.— Natu-
re, 181, 407-408.
Katchalsky A., Curran P. F.
1965. Nonequilibrium Thermodyna-
mics in Biophysics. Massachusetts,
Harvard Univ. Press.
Katchalsky A., Oster G. F., Per el-
son A. S. 1973. Network thermody-
namics: dynamic modelling of
biophysical systems.— Quart.
Rev. Biophys., 6, 1—134.
Kedem O. 1965. Water flow in the
presence of active transport.—Sym-
pos. Soc. Exp. Biol., 19, 61—73.
Keizer J., Fox R. F. 1974. Qualms
regarding the range of validity of
Glansdorff—Prigogine criterion
for stability of non-equilibrium
states.— Proc. Nat. Acad. Sci.
USA, 71, 192—196.
King J. K., Farner D. S. 1961. Ener-
gy metabolism, thermoregulation
and body temperature.— In: Bio-
logy and Comparative Physiology
of Birds, v. 2. N. Y., Acad. Press,
p. 215-288.
Kistler A., Weber R. 1974. Enzyme
patterns in mitochondria of eggs,
liver and skeletal muscle during
larval development of Xenopus.—
Develop. Biol., 37, 236—247.
Kitzinger C. Benzinger T. 1955. War-
metonung der Adenosintriphosphor-
saure-Spaltung.— Z. Naturforsch.,
10b, 375-382.
Kleiber M. 1961. The Fire of Life.
N. Y., Wiley.
К bros E. 1974. Monomolecular treat-
ment of chemical oscillation.—
Nature, 251, 703—704.
Korbs E., Burger M. 1973. Bromide-
selective electrode for use in fol-
lowing the Zhabotinsky-type oscil-
lating chemical reaction. Buda-
pest, Akad. Kiado, p. 191—203.
Korbs E., Orbdn M., Nagy Zs, 1973a.
Periodic heat evolution during
266
temporal chemical oscillations.—
Nature Phys. Sci., 242, 30—31.
Koros E., Orb&n M., Nagy Zs. 1973b.
Periodicity in the rate of heat evo-
lution during the temporal oscilla-
tion in the 2,4-pentanedione-bro-
mate-catalyst system.— J. phys.
Chem., 77, 3122-3123.
Kostizin У. A. 1939. Biologic Mathe-
matique 1937 (Mathematical Bio-
logy) London.
Kumar I. J., Gupta L. N. 1973. On
some relations between variational
principles in irreversible thermody-
namics.— Acta chim. Acad. Sci.
Hung., 76, 345-357.
Kunkel H. О., Campbell J. E. 1952.
Tissue cytochrom oxidase activity
and body weight.— J. Biol. Chem.,
198, 229—236.
Laird A. K. 1964. Dynamics of tumo-
ur growth.— Brit. J. Cancer, 18,
490—502.
Laird A. K. 1966. Dynamics of emb-
ryonic growth.— Growth, 30, 263—
275.
Lambermont J., Lebon G. 1972. A rat-
her general variational principle
for purely dissipative non-statio-
nary processes.— Ann. Phys., 28,
15-30.
Lamprecht I., Lochmann E. R.,
Pietsch I., Uttech K. 1973. Micro-
calorimetric experiments on a cell-
free protein synthesizing system
of Saccharomyces.— In: Proteides
of the Biological Fluids, v. 20,
Oxford, Pergamon Press, p. 551 —
553.
Lamprecht I., Meggers C. 1971. Mat-
hematische Beschreibung mikro-
kalorimetrisch ermittelter Wachs-
tumskurven von Saccharomyces.—
Proc. First Europ. Biophys. Congr.,
4, 335-339.
Lamprecht I., Meggers C. 1972. Mik-
rokalorimetrische Untersuchungen
zum Stoffwechsel von Hefen. II.
Wachs turn auf festen Nahrboden.—
Biophysik, 8, 316—325.
Lamprecht I., Meggers C., Stein W.
1971. Mikrokalorimetrische Unter-
suchungen zum Stoffwechsel von
Hefen. I. Wachstum in fliissigen
Medien.— Biophysik, 8, 42—52.
Lamprecht I., Schaarschmidt B. 1972.
The flickering of a dying flame.—
Nature, 240, 445-446.
Lamprecht I., Schaarschmidt B. 1976.
Dissipative oscillations of a candle
flame, (to be published).
Lamprecht I., Schaarschmidt B.,
Stein ИЛ 1973. Mikrokalorimetri-
sche Untersuchungen zum Stoff-
wechsel von Hefen. III. Betriebs-
stoffwechsel in Glucosepuffer.—
Biophysik, 10, 177-180.
Lamprecht I., Schaarschmidt B.,
Welge G. 1975. Microcalorimetric
investigations of the metabolism
of yeast. V. Influence of ploidy on
growth and metabolism.— Rad.
Environm. Biophys., 12.
Landauer R. 1973. Entropy changes
for steady-state fluctuations.— J.
Stat. Phys., 9, 351—371.
Lang C, A., Grant Ph. 1961. Respi-
ratory enzyme changes during from
embryogenesis. — Proc. Nat. Acad.
Sci. USA, 47, 1236—1244.
Laskowski W. 1962. Inaktivierungs-
versuche mit homozygoten Hefes-
tammen verschiedenen Ploidiegra-
des.— Z. Naturforsch., 17b, 93—
108.
Lavenda В. H. 1972. Concepts of
stability and symmetry in irrever-
sible thermodynamics.— Foundat.
Phys., 2, 161-179.
Lavenda В. H. 1973. Thermodynamics
of nonlinear interacting irrever-
sible processes.— Foundat. Phys.,
3, 53-88.
Lavenda В. H. 1974. Principles and
representations of nonequilibrium
thermodynamics.— Phys. Rev., 9,
929—942.
Layne J. N. 1966. Postnatal develop-
ment and growth of Peromyscus
floridanus.— Growth, 30, 23—45.
Lebon G. J., Casas-Varquez J. 1974.
Lagrangian formulation of unsteady
non-linear heat transfer. — J. Engin.
Math., 8, 31-44.
Lebon G. J., Lambermont J. K. 1973.
Generalization of Hamiltons prin-
ciple to continuous dissipative
systems.— J. Chem. Phys. 59,
2929-2936.
Lefever R. 1968. Dissipative structures
in chemical systems.— J. Chem.
Phys., 49, 4977-4978.
Lesse P. 1968. Non-equilibrium ther-
modynamics. II. Contribution to
the theory of a macroscopic evolu-
tion criterion.— Collect. CzechosL
Chem. Commons, 33, 801—807.
Levin K. 1971. Heat production by
leucocytes and thrombocytes mea.
267
sured with a flow microcalorimeter
in normal man and during thyroid
disfunction. — Clin. chim. Acta,
32, 87—94.
Levin K. 1973. Determination of
heat production from erythrocytes
in normal man and in anemie
patients with flow microcalori-
metry.— Scand. J. Clin. Lab. In-
vest., 32, 55—65.
Levin К., Boyo A. E. 1971. Heat
production from erythrocytes.—
Scand. J. Clin. Lab. Invest., 27,
Suppl. 118, 55.,
Levy M., Toury R. 1970. Etude de
1’evolution des activites des enzy-
mes mitochondriaux de Г hepato-
cyte au cours du developpement du
rat.— Biochim. et biophys. acta,
216, 318-327.
Li J. С. M. 1958. Thermodynamics
for nonisothermal systems. The
classical formulation.— J. Chem.
Phys., 29, 747-754.
Lin E. C., Rivlin R. S., Knox W. E.
1959. Effect of body weight and
sex on activity of enzymes invol-
ved in amino acid metabolism.—
Amer. J. Physiol., 196, 2, 303—
306.
Lotka A. J. 1925. Elements of Phy-
sical Biology. Baltimore, Willi-
ams a. Wilkins Co.
Loud A. V. 1962. A method for quan-
titative estimation of cytoplasma-
tic structures.— J. Cell. Biol., 15,
481.
L^vtrup S. 1963. On the oxygen upta-
ke during early amphibian embryo-
genesis.— J. Exper. ZooL, 152,
307-314.
Lusk G. 1915. Animal calorimetry.
II. An investigation into the cau-
ses of the specific dynamic action
of the foodstruffs.— J. Biol. Chem.,
20, 555—717.
Lusk G. 1921. Animal calorimetry. 17.
The behaviour of various inter-
mediary metabolites upon the heat
production.— J. Biol. Chem., 49,
453-478.
Luck D. J. L. 1963. Genetic of mi-
tochondria in Neurospora crassa.—
Proc. Nat. Acad. Sci. USA, 49,
233-240.
Muckier B., Grace R., Duncan H. M.
1971. Studies of mitochondrial
development during embryogenesis
in the rat.— Arch. Biochem. and
Biophys., 144, 603-610.
Margulis L. 1970. Origin of Eucaryo-
tic Cells. New Haven, Yale Univ.
Press.
Martinez H. M. 1972. Morphogenesis
and chemical'dissipative structures.
A computer simulated case study.
— J. Theoret. Biol., 36, 479—501.
Max S. R. 1972. Disuse atrophy of
sceletal muscle: loss of functional
activity of mitochondria.— Bioc-
hem. and Biophys. Res. Communs,
46, 1394-1398.
McDonald P., Henderson A. R., Rat-
ion J. 1973. Energy changes during
ensilage.— J. Sea. Fd. Agric., 24,
827-834.
Medawar P. В. 1942. Discussion of
growth and new growth.— Proc.
Roy. Soc. Med., 35, 500.
Medawar P. B. 1945. Size, shape and
age. In: Essays on Growth and
Form. Oxford Univ. Press,
p. 157-187.
Meijer P. H. E., Edwards J. C. 1970.
Modification of the minimum ent-
ropy-production principle.— Phys.
Rev., set. A, 1, 1527-1536.
Meixner J. 1954. Thermodynamik der
Irreversible Prozesse. Aachen.
Meixner J. 1973. Consistency of the
Onsager-Casimir reciprocal rela-
tions.— Adv. Mol. Relax. Proces-
ses, 5, 319—331.
Mel H. C. 1954. Entropy production
and the stationary state of chemi-
cal reactions.— Bull. Cl. sci. Acad,
roy. Belg., Ser. 5, 40, 834—845.
Melchior D. L., Morowitz H. J.4
Strutevant J. M., Tsong T. У.
1970. Characterization of the plas-
ma membrane of Mycoplasma laid-
lawii.— Biochem. et biophys. acta,
219, 114-122.
Menzies R. A., Gold P. H. 1972. The
apparent turnover of mitochondria,
ribosomes and s-RNA of the brain
in young, adult and aged rats.—
J. Neurochem., 19, 1671—1683.
Meyer R. R. 1973. On the evolutiona-
ry origin of mitochondrial DNA.—
J. Theoret. Biol., 38, 647—663.
Miller D. G. 1956. Thermodynamic
theory of irreversible processes.
I. The basic macroscopic concepts.—
Amer. J. Phys., 24, 433—444.
Miller D. G. 1960. Thermodynamics
of irreversible processes. The expe-
rimental verification of the Onsa-
ger reciprocal relations.— Chem.
Rev., 60, 15-37.
268
Monroy A. 1965. Chemistry and Phy-
siology of Fertilization. N. Y.,
Holt, Rinehart and Winston.
Monti M., Wadsd I. 1973. Microcalo-
rimetric measurements of heat
production in human erythrocytes.—
Scand. J. Clin. Lab. Invest.,
32, 47—54.
Morowitz H. J. 1960. Some consequ-
ences of the application of the se-
cond law of thermodynamics to
cellular systems.— Biochem. et
biophys. acta, 40, 340—345.
Morowtiz H. J. 1968. Energy Flow
in Biology. N. Y., Acad. Press.
Morrison P. 1964. A thermodynamic
characterization of self-reproduc-
tion.— Rev. Mod. Phys., 36, 517—
524.
Morrison P. R., Pearsen О. P. 1946.
The metabolism of a very small
mammals.— Science, 104, 287 —
289.
Morrison P. R., Ryser F., Dawe A. R.
1959. Studies on the physiology of
the masked shnew Sorex cinereus.—
Physiol. ZooL, 32, 256-271.
Mortensen K., Noren B., Wadsd J.
1973. Microcalorimetry in the study
of the activity of microorganisms.—
Bull. Ecol. Res. Comm. Stock-
holm, 17, 189-197.
Mur Lin J. R., Lusk G. 1915. Animal
calorimetry.— J. Biol. Chem., 22,
15—41.
Myrcha A., Pinowski J., Тошек T.
1973. Variations of the water and
ash contents in the caloric values
of nestling starlings (Sturnus vulga-
ris L.) during their development.—
Bull. Acad, polon. sci., Ser.
sci. biol., 21, 649—655.
Nass M. M. K. 1969. Mitochondrial
DNA: advances, problems and
goals. Studies of size and structure
of mitochondrial DNA relate to
biogenesis and function of this
organelle.— Science, 165, 25—35.
Nass M. M. K., Nass S. 1963. Intra-
mitochondrial fibers with DNA
characteristics. I. Fixation and
electron detaining reactions.— J.
Cell Biol., 19, 539—611.
Needham J. 1931. Chemical Embryo-
logy, v. 3, Cambridge Univ. Press.
Needham A. E. 1960. Regeneration
and growth. In: Fundamental As-
pects of Normal and Malignant
Growth. Amsterdam, Elsevier, p.
588—663.
Nicolis G. 1971. Stability and dissi-
pative structures in open systems
far from equilibrium.— Adv. Chem.
Phys., 19, 209-324.
Nicolis G. 1972. Fluctuations around
non-equilibrium states in open non-
linear systems.— J. Stat. Phys.,
6, 195-222.
Nicolis G., Auchmuty J. F. G. 1974.
Dissipative structure catastrophes,
and pattern formation: a bifurca-
tion analysis.— Proc. Nat. Acad.
Sci. USA, 71, 2748-2751.
Nicolis G., Babloyantz A. 1969. Fluc-
tuations in open systems. — J.
Chem. Phys., 51, 2632—2637.
Nicolis G., Wallenborn J,, Velar-
de M. G. 1969. On the validity of
Gibbs entropy law in strongly
coupled systems.— Physica, 43,
263—276.
Niwelinski J. 1960. An enzymologie
study of the regenerating forelimb
of the newt Triturus vulgaris.—
Folia Biol., 8, 1—9.
Niwelinski J. 1964. Activity of int-
racellular enzymes in regenerating
newt limbs.— Sympos. Biol. Hung.
Budapest, Akad. Kiado, 3, 47—53.
Norman W. P., Schmidt A. J. 1967.
The fine structure of tissues in the
amputated-regenerating limb of the
adult newt, Diemictylus virides-
cens.— J. MorphoL, 123, 271—312.
Novikoff A.B. 1961. Mitochondria
(chondriosomes). In: The Cell, v.
2. N. Y., Acad. Press, p. 299-422.
O'Brien B. R. A. 1957. Tissue meta-
bolism during posterior regenera-
tion in the earthworm.— Avstral.
J. Exper. Biol, and Med. Sci., 35,
373—380.
Odum E. P., Marshall S, G., Marp-
les T. G. 1965. The caloric content
of migrating birds.— Ecology, 46,
901—904.
Ohlmeyer P. 1946. Warmemessungen
bei Fermentreaktionen. — Z. Na-
turforscb., 1, 30—35.
Ohlmeyer P., Shat as R. 1952. Heat of
hydrolysis of some phosphate com-
pounds.— Arch. Biophys. and Bio-
chem., 36, 411—420.
Okuneff N. 1933. Uber einige physiko-
chemische Erscheinungen wahrend
der Regeneration. — Biochem. Z.,
257, 242-244.
Ono S. 1961. Variational principles
in thermodynamics and statistical
269
mechanics of irreversible processes. —
Adv. Chem. Phys., 3, 267—321.
Onsager L. 1931a. Reciprocal relations
in irreversible processes. I.— Phys.
Rev., 37, 405—426.
Onsager L. 1931b. Reciprocal rela-
tions in irreversible processes. II.—
Phys. Rev., 38, 2265-2279.
Ornstein L. 1956. Mitochondrial and
nuclear interaction.— J. Biophys.
and Biochem. Cytol., 2, 351—352,
Suppl.
Oster G. F., Desoer C. A. 1971. Telle-
gen’s theorem and thermodynamic
inequalities.— J. Theoret. Biol.,
32, 219-241.
Oster G.,Perelson A., Katchalsky A.
1971. Network thermodynamics.—
Nature, 234, 393-399.
Oster G. F., Perelson A. S., Katchals-
ky A. 1973. Network thermodyna-
mics: dynamic modelling of biophy-
sical systems.— Quart. Rev, Biop-
hys., 6, 1—134.
Ostwald W. 1908. Uber die zeitlichen
Eigenschaften der Entwicklungsvor-
gange.— Vortr. Aufs. Entwicklung-
mech. Org., 5, 1—71.
Palade G. E. 1956. Electron micro-
scopy of mitachondria and other
cytoplasmic structures. In: Enzy-
mes: Units of Biological Structure
and Function. Acad. Press, chap.
9, p. 185—215.
Parks J, R. 1973. A stochastic model
of animal growth.— J. Theoret.
Biol., 42, 505—518.
Parsons J. A., Rustad R. C. 1968.
The distribution of DNA among di-
viding mitochondria of Tetrahy-
mena pyriformis.— J. Cell Biol.,
37, 683-693.
Pearl R. 1924. The curve of popula-
tion growth.— Trans. Amer. Phi-
los. Soc. 63, 10—17.
Petrucci D. 1957. Citocromo-ossidasi
e respirazione rell’embriogenesi deg-
li anfibi Anuri. I. Contenuto di
citocromo-ossidase e velocita di
consumo di O2 nella embriogenesi
di В ufa bufa.— Acta embryol.
morphol. exper., 1, 105—117.
Petrucci D. 1960. Respiration et cy-
tochrom oxydase au cours de l’o-
ogenese chez les Amphibiens.—
Arch, internal, physiol, biochem.,
68, 225—226.
Piko L., Tyler A., Vinograd J. 1967.
Amount location, priming capacity,
circularity and other properties of
cytoplasmic DNA in sea urchin
eggs.— Biol. Bull., 132, 68—90.
Pittet P., Gygax P. H., Jequier E.
1974. Thermic effect of glucose and
amino acids ?in man studied by
direct and indirect calorimetry.—
Brit. J. Nutr., 31, 343—349.
Pitzer K. S. 1961. Irreversible ther-
modynamics.— Pure Appl. Chem.,
2, 207-210.
Plesset M. S., Winet H. 1974. Bio-
convection patterns in swimming
microorganism cultures as an
example of Ryaleigh—Taylor ins-
tability.— Nature, 248, 441—443.
Poe M. 1969. Kinetic studies of tem-
perature changes and oxygen up-
take in a differential calorimeter:
energy balance during calcium ac-
cumulation by mitochondria.—
Arch. Biochem. and Biophys., 132,
377-387.
Poe M., Estabrook R. V. 1968. Kine-
tic studies of temperature changes,
oxygen utilization, pH and potas-
sium concentration in a differen-
tial calorimeter: the enthalpy of
potassium accumulation by rat
liver mitochondria.— Arch. Bio-
chem. and Biophys., 128, 725—733.
Poe M., Gutfreund H., Estabrook R. W.
1967. Kinetic studies of tempera-
ture change and oxygen uptake in
a differential calorimeter: the heat
of oxidation of NADH and succi-
nate.— Arch. Biochem. and Biop-
hys., 122, 204-211.
Poole R. K., Lloyd Kemp R. B.
1973. Respiratory oscillations and
heat evolution in synchronously
dividing cultures of the fission
yeast Schizosaccharomyces pombe
972h.— J. Gen. Microbiol., 77,
209-220.
Popper K. R. 1965. Time’s arrow and
entropy.— Nature, 207, 233—234.
Popper K. R. 1967a. Time’s arrow
and feeding on negentropy.— Natu-
re, 213, 320.
Popper K. R. 1967b. Structural in-
formation and the arrow of time.—
Nature, 214, 322.
Prat H. 1948. Histo-physiological
gradient and plant organogenesis.
— Bot. Rev., 14, 603-643.
Prat H. 1952. Microcalorimetric stu-
dies on dermination of cereals.—
Canad. J. Bot., 30, 379-394.
Prat H. 1953. Observations sur la
270
thermogenese bacterienne.— Rev.
canad. biol., 12, 19—34.
Prat H. 1962. Thermogenese bacteri-
enne et evolution des foyers infec-
tieux.— Compt. rend. Soc. biol.
France, 156, 1135—1137.
Prat H. 1969. Calorimetry of higher
organisms. In: Biochemical Mic-
rocalorimetry. N. Y., Acad. Press,
p. 181—198.
Prigogine I, 1947. Etude thermody-
namique des phenomenes irrever-
sibles. Lifcge, Desoer.
Prigogine I, 1955. Introduction to
Thermodynamics of Irreversible
Processes. Chicago, Thournes.
Prigogine I. 1966. Evolution criteria,
variational properties and fluctua-
tions. In: Non-equilibrium Thermo-
dynamics, Variational Techniques
and Stability. Univ. Chicago Press,
p. 3—16.
Prigogine I. 1969. Structure, dissipa-
tion and life.— In: Theoretical
Physics and Biology. North-Hol-
land Publ. Co., p. 23—52.
Prigogine I. 1972. La thermodynami-
que de la vie.— Recherche, 3,
547-562.
Prigogine I. 1973a. Time, irreversi-
bility and structure In: The Phy-
sicist’s Conception of Nature. Dor-
drecht, D. Reidel Publ. Co.
Prigogine I. 1973b. Irreversibility as
a symmetry-breaking process.—
Nature, 246, 67—71.
Prigogine I., George G., Henin F.
1973. A unified formulation of
dynamics and thermodynamics
with special reference to nonequi-
librium statistical thermodyna-
mics.— Chem. Seripta (Sweden), 4,
5—32.
Prigogine I., Lefever R. 1973. Theory
of dissipative structures. In: Sy-
nergetics — cooperative Phenomena
in Multi-component Systems. Stut-
tgart, B. G. Teubner.
Prigogine I,, Lefever R., Goldbeter A.,
H er schkowitz-Kaufman M. 1969.
Symmetry breaking instabilities
in biological systems.— Nature,
223, 913—916.
Prigogine I,, Nicolis G. 1967. On sym-
metry-breaking instabilities in
dissipative systems.— J. Chem.
Phys., 46, 3542-3549.
Prigogine I., Nicolis G. 1971. Fluctu-
ations and the mechanism of ins-
tabilities.— Proc. 3rd Internal.
Conf. From Theoret. Phys, to Biol.,
Versailles, 89—109.
Prigogine I., Nicolis G. 1971. Biolo-
gical order, structure and instabi-
lities.— Quart. Rev. Biophys.,
4, 107—148.
Prigogine I., Nicolis G., BabloyantzA.
1972a. Thermodynamics of evolu-
tion. I. Physics today, 11, 23—28.
Prigogine INicolis G., Babloyantz A.
1972b. Thermodynamics of evolu-
tion. II. Physics today, 12, 38—
44.
Prigogine Z., Wiame J. M. 1946.
Biologie et thermodynamique des
phenomenes irreversibles.— Expe-
rientia, 2, 451—453.
Putter A. 1920. Studien fiber physio-
logische Ahnlichkeit. VI. Wachstu-
msahnlichkeiten.— Pfliiger’s Arch,
ges. Physiol., 180, 298—340.
Quickenden T. QueHee Sh. S. 1974.
Weak luminescence from yeast Sac-
charomyces cerevisiae and the exis-
tence of mitogenetic radiation.—
Biochem. and Biophys. Res. Com-
muns, 60, 764—770.
Rappavort S. I. 1968. Active trans-
port in isotropic and anisotropic sys-
tems. A critical analysis.— J. The-
oret. Biol., 19, 247—274.
Rawitscher M., Sturtevant J. M. 1961.
The heats of ionization of deoxy-
nucleotides and related compounds.
— J. Amer. Chem., 82, 3739—3740.
Raymond R. C. 1950. Communication,
entropy, and life.— Amer. Sci-
entist, 38, 273—278.
Reutter K., Sasse D. 1970. Chemomor-
phologische Befunde (Glycogen,
G—6—P—DH, RNS) am Regene-
rationsblastem von Lineus eangui-
neus Rathke (Nemertini).— W.
Roux’ Arch., 165, 285—295.
Richardson I. W. 1969. On the prin-
ciple of minimum entropy produc-
tion.— Biophys. J., 9, 265—267.
Richter A. 1959. Die Stromungskalo-
rimetrie und ihre Verwendbarkeit
bei der Diagnostik peripher Dur-
chblutungsstorungen. Dissertation,
Leipzig.
Riegel J. 4. 1965. Energy, Life and
Animal Organization. London.
Robertson T. В. 1908. On the normal
rate of growth of an individual and
its biochemical significance.— W.
Roux’ Arch., 25, 581—614.
Rockstein M., Bhatnagar P. J. 1965.
Age change in size and number of
271
the giant mitochondria in the flight
muscle of the common housefly
(Musca domestica).— Insect Phy-
siol., 11, 481.
Rohde S. M. 1972. Minimally proper-
ties of the local potential.— Phy-
sica, 61, 53—66.
Romanoff A. L. 1967. Biochemistry
of Avian Embryo. — N. Y., Wiley.
Roodyn D. ВReis P. J., Work T. S.
1961. Protein synthesis in mitot-
hondria.— Biochem. J., 80, 9—21.
Rosen R. 1973. On the generation of
metabolic novelties in evolution.—
In: Biogenesis, Evolution, Home-
ostasis., N. Y., Springer, p. 113—
118.
Rosenthal 0., Drabkin D. L. 1943.
The cytochrome C content of nor-
mal and neoplastic mammalian epi-
thelium and its correlation with
body mass.— J. Biol. Chem., 150,
131-141.
Ross P. D., 1 letcher A. P., Jamie-
son G. A, 1973. Microcalorimetric
study of isolated blood plattelts
in the presence of thrombin and
other aggregating agents.— Bio-
chim. et biophys. acta, 313, 116—
118.
Ross P. D.j Ginsburg A. 1969. A calo-
rimetric study ot the binding of
two feedback inhibitors to the glu-
tamine synthetase from E. coli.—
Biochemistry, 8, 4690—4695.
Sacher G. 4. 1967. The complementa-
rity of entropy terms for the tem-
perature — dependens of develop-
ment and aging.— Ann. N.Y. Acad.
Sci., 138, 680—712.
Salpeter M. M., Singer M. 1960. The
fine structure of mesenchymatous
cells in the regenerating iorelimb
of the adult newt Triturus.— De-
velop. Biol., 2, 516—534.
Samson F. E., Balfour W. M., Ja-
cobs R. J, 1960. Mitochondrial
changes in developing rat brain.—
Amer. J. Physiol., 199, 693—696.
Satav J. G., Raj wade M.S., Katya-
re S. S., JVetrawali M. 5., Fatter-
paker P., Sreenivasan A. 1973. The
significance of promitochondrial
structures in rat liver for mitochon-
drial biogenesis.— Biochem. J.,
134, 687—b95.
Sauzin M. J. 1966. Etude au micros-
cope electronique du neoblaste de
la planaire Dugesia gonocephala
(Turnellarie, Triclade) et de ses
changements ultrastructuraux au
cours des premiers stades de la re-
generation.— C. r. Acad. sci. Pa-
ris, D263, 605-e-608.
Sauzin M. J. 1967. Etude ultrastruc-
turale de la differenciation du ne-
oblaste au cours de la regeneration
de la planaire Dugesia gonocephala.
I. Differenciation en cellule ner-
veuse.— Bull. Soc. zool. France,
92, 313-318.
Schaarschmidt B., Lamprecht I. 1974.
Calorimetric measurement of the
glucose metabolism of U. V.-sensi-
tive mutants.— Internat. J. Radi-
at. Biol., 25, 167—173.
Schaarschmidt B., Lamprecht I., Zo-
tin A. I. 1976. Quantitative rela-
tion between heat production and
weight during growth of microbial
cultures.— Exper. Cell Res.
Schaarschmidt B., Zotin A. I., Bret-
tel R., Lamprecht I. 1975. Expe-
rimental investigation of the bound
dissipation function. Change of the
w-function during the growth of
yeast.— Arch. Microbiol., 105,
13-16.
Schatz G. 1970. Biogenesis of mito-
chondria. In: Membranes of Mito-
chondria and Chloroplasts. N. Y.,
Van Nosirand, Reinhold, p. 251 —
314.
Schldgl F. 1967. On the statistical
foundation of the thermodynamic
evolution criterion of Glansdorff
and Prigogine.— Ann. Phys., 45,
155____163.
Schldgl F. 1971a. Stability of steady
state.— Z. Phys., 243, 303—309.
Schldgl F. 1971b. On thermodynamics
near a steady state. Z. Phys., 248,
446—458.
Schmidt A. J. 1966. The molecular
basis of regeneration: enzymes. Il-
linois Univ. Press, 4, N 4.
Schmidt A. J. 1968. Cellular Biology
of Vertebrate Regeneration and Re-
pair. Univ. Chicago Press.
Schmidt A. J., Weidman T. 1964.
Dehydrogenases and aldolase in
regenerating forelimb of the adult
newt Diemictylus viridescens.— J.
Exper. Zool., 155, 303—316.
Schmukler M., Barrows С. Я. 1966.
Age differences in lactic and malic
dehydrogenase in the rat.— J. Ge-
rontol., 21, 109—111.
Schnepf E., Brown R. M. 1971. On
relationships between endosymbio-
272
sis and the origin of plastides and
mitochondria.— In: Origin and Con-
tinuity of Cell Organells. N. Y.,
Pringer, p. 299—322.
Sedlaczek L. 1964. Heat balance of
growth of some bacterial cultures.—
Acta microbiol. polon., 13, 101 —
112.
Segel L. A,, Jackson J. L. 1972. Dis-
sipative structure: An explanation
and an ecological example.— J. The-
oret. Biol., 37, 545—559.
Seshadri M. S. 1974. A reaction-dif-
fusion coupled system with multi-
ple steady states.— J. Theoret.
Biol. 47, 351-365.
Shannon С. E. 1948. A mathematical
theory of communication.— Bell
Syst. Techn. J., 27, 379—423.
Shaver J. R. 1956. Mitochondrial po-
pulations during development of
the sea urchin.— Exper. Cell Res.,
11, 548-559.
Schiao D. D. F. 1970. Calorimetric
investigations of the binding of in-
hibitors to a-chymotrypsin 2. A
systematic comparison of the ther-
modynamic functions of binding of
a variety of inhibitors to a-chymo-
trypsin.— Biochemistry, 9, 1083—
1090.
Simpson G. G. 1945. The principles
of classification and a classifica-
tion of mammals.— Bull. Amer.
Museum Natur. Hist., 85, 1—350.
Sjostrand F. S. 1956. Ultrastructure of
cell as revealed by the electron mic-
roscope.— Internal. Rev. CytoL,
5, 455.
Smith R. E. 1956. Quantiative rela-
tions between liver mitochondria
metabolism and total body weight
in mammals.— Ann. N. Y. Acad.
Sci., 62, 403-422.
Smith W. F., Quevedo J. L. G. de,
Carter R. L., Bennett FK. S. 1947.
Frequency stabilization of micro-
wave oscillators by spectrum li-
nes.— J. Appl. Phys., 18, 1112—
1115.
Snellen J. ТУ., Mitchell D., Busan-
sky M. 1972. Calorimetric analysis
of the effect of drinking saline so-
lution on whole body sweating.—
Pfliigers Arch. ges. Physiol., 333,
124-144.
Spangler R. A., Snell F. M. 1967.
Transfer function analysis of an
oscillatory model chemical system. —
J. Theoret. Biol., 16, 381—405.
Spiegelman S., Steinbach H. B. 1945.
Substrate — enzyme orientation
during embryonic development.—
Biol. Bull., 88, 254-268.
Spink E., Wadsd J. 1975. Calorimetry
as an analytic tool in biochemistry
and biology.— In: Methods in
Biochemical Analysis, v. 23, N. Y.
Wiley.
Srivastawa M. D. L. 1965. Cytoplas-
mic inclusions in oogenesis.— In-
ternal. Rev. CytoL, 18, 73—98.
Stang-Voss C., Straubesand J. 1970.
Uber die Neubildung von Mitochon-
drion: Electronenmikroskopische
Untersuchungen an Spermatiden
von Eisenia foetida (Annelidae).—
Z. Zellforsch., Ill, 127—142.
Staples B. R., Prosen E. J., Gold-
berg R. N. 1973. National Bureau
of Standards Interim Report,
NBSIR A73-181.
Steim J. M., Tourtellotte M. E.,
Reinert J. C., McElhaney R. N.,
Rader R. L. 1969. Calorimetric evi-
dence for the liquidcrystalline
state of lipids in a biomembrane.—
Proc. Nat. Acad. Sci. USA, 63,
104-109.
Stein W. 1960. Zur Entropiebalanz
organischer Strukturen.— Naturwis-
senschaften, 47, 374.
Steiner R. F., Kitzinger C. 1956. A
ealorimetric determination of the
heat of an antigen — antibody reac-
tion.— J. Biol. Chem., 222, 271 —
284.
Stroot P., Janjic D. 1975. Oscillati-
ons chimiques lors de 1’oxidation
de dicetones par 1c bromate en pre-
sence d’un catalyseur (Reaction
du type Belousov — Zhabatinsky).
Helv. chim. acta, 58, 116—120.
Sturtevant J. M. 1953. Heats of hydro-
lysis of amide and peptide bonds.—
J. Amer. Chem. Soc., 75, 2016—
2017.
Sturtevant J, M. 1962. Heats of bio-
chemical reactions.— In: Experi-
mental Thermochemistry, v. 2,
N.Y., Wiley, p. 427-442.
Swanson R. F. 1971. Incorporation
of high molecular weight polynu-
cleotidea by isolated mitochond-
ria.— Nature, 231, 31—34.
Swick R.W.,Stange J. L., Nance S. L.,
Thomson J. F. 1967. The hetero-
geneous distribution of mitochon-
dria enzymes in normal rat liver. —
Biochemistry, 6, 737—744.
273
Szekely J. G.' Kennedy G. G., Hojf-
manJ.G. 1972. Monte Carlo simu-
lation of clone growth.— Cell Tis-
sue Kinetics, 5, 203—213.
Tauchi H., Sato T. 1968. Age chan-
ges in size and number of mito-
chondria of human hepatic cells.—
J. Gerontol., 23, 454—461.
Tauchi H., Sato T., Hoshino M., Ko-
bayashi H., Adochi F., Aoki J.,
Masuko T. 1964. Studies on corre-
lation between ultrastruction and
enzymatic activities of parenchy-
mal cells in senescence.— In: Age
with a Future. Copenhagen, Munk-
sgaard, p. 203—211.
Thoenes D. 1973. «Spatial oscillati-
ons» in the Zhabotinskii reaction.
Nature Phys. Sci., 243, 18—20.
Thompson W. D'Arcy. 1942. Growth
and Form. Cambridge Univ. Press.
Tillberg J. E., Kylin A., Sundberg J.
1971. Microcalori metric measure-
ments of the heat evolution and its
sensitivity to DNP during different
steps of synchronous cultures of
Scenedesmus.— Plant Physiol., 48,
779—782.
Tiemann J. 1969. Ein Respirationska-
lorimeter zur Bestimmung des Trans-
formationsgrades der Verbrennung
im menschlichen Organismus.—
Elektromedizm., 14, 135—145.
T imofeeff-Ressovsky N. W.f Zim-
mer K. G. 1947. Das Trefferprin-
zip in der Biologie. Leipzig, Sit-
tirzel.
Tolman R. C., Fine P. C. 1948. On
the irreversible production of en-
tropy.— Rev. Med. Phys., 20, 51 —
77.
Tondeur D. 1970. Les structures dis-
sipatives. Un concept recent et
thermodynamique des processus
irreversibles.— Entropie, 34—35,
3-12.
Touloukian Y. S. 1956. The Concept
of Entropy in Communication,
Living Organisms, and Thermody-
namics. Lufayette, Purdue Univ.
Tribe M. A., Ashhurst D. E. 1972.
Biochemical and structural vari-
ations in the flight muscle mito-
chondria of aging blowflies Calli-
phoraerythrocephala.— J. Cell Sci.,
10, 443—469.
Tuft P. 1953. Energy change in de-
velopment. Arch, neerl., zool., 10,
Suppl. 1, 59-75.
Turing A. M. 1952. The chemical ba-
sis of morphogenesis.— Philos.
Trans. Roy. Soc. London, Ser. B,
237, 37—72.
TzagoloffA .,Rubin M.S.'S ierra M. T.
1973. Biosynthesis of mitochondrial
enzymes.— Biochim. et biophys.
acta, 301, 71—104.
Ubbelohde A. R. 1947. Time and Ther-
modynamics. Oxford Univ. Press.
Van Kampen N. G. 1973. Nonlinear
irreversible processes.— Physica,
67, 1-22.
Van Uden N. 1971. Thermodynamics
of the chemostat. — Z. allg. Mi-
krobiol., 11, 541-550.
Vincent J. P.' Ladenski M. 1973. The
interaction between a-chymotryp-
sin and pancreatic trypsin inhibi-
tor (Kunutz inhibitor).— Europ.
J. Biochem., 38, 365—372.
Vincze Gy. 1972. Deduction of the
Reynolds equations of turbulence
from the Gyarmati principle.— Ac-
ta chim. Acad. sci. Hung., 75, 33 —
43.
Vojta G. 1967. Hamiltonian formalisms
in the thermodynamic theory of
irreversible processes in continuous
systems.— Acta chim. Acad. sci.
Hung., 54, 55-64.
Volterra V. 1926. Variazioni e flut-
tuazzioni del numere individui in
speci animali conviventi.— Mem.
Accad. Lincei, set II, 6, 31.
Wadso I. 1970. Microcalorimeters.—
Quart. Rev. Biophys., 3, 383—427.
Walker M. G. 1967. Heat production
of the albino mouse during
growth.— Experientia, 23, 541.
Warburg O. 1930. The Metabolism
of Tumours. London, Constable.
Warburg O. 1956a. On the origin of
cancer cells.— Science, 123, 309—
314.
Warburg O. 1956b. On respiratory
impairment in cancer cells. — Sci-
ence, 124, 269—270.
Warburg O.' Gawehn К., Geissler A. W.
1958. Stoffwechsel der weipen Blut-
zellen.— Z. Naturforsch., 13b, 515—
516.
Ward R. 1962. The origin of protein
and fatty yolk in Rana pipiens.
II. Electron microscopical and cy-
tochemical observation of young and
matural cocytes.— J. Cell Biol.,
14, 309-341.
Weber R.' Boell E. J.1962. Enzyme
patterns in isolated mitochondria
274
from embryonic and larval tissues
of Xenopus.— Develop. Biol., 4,
452-472.
Wendker U., Lamprecht I., Schaar-
schmidt B. 1976. Occurrence of dis-
sipative structures during polyme-
rization of acrylamid and transi-
tion to an equilibrium structure,
(in press).
Wend land t W. W. 1964. Thermal Me-
thods of Analysis. N. Y., Wiley.
Weymouth F. W., McMillin H. C.
1930. The relative growth and mor-
tality of the Pacific razor cham (Si-
liqua patula Dixon), and their bea-
ring on the commercial fishery.—
Bull. Bur. Fisch. Wash., 46, 543 —
567.
Wigglesworth V. B. 1953. The Princi-
ples of Insect Physiology. London,
Methuen.
Wilkie D. R. 1960. Thermodynamics
and the interpretation of biological
heat measurements. — Progr. Biop-
hys. Biophys. Chem., 10, 259—
298.
Wilkie D. R. 1968. Heat work and
phosphorylcreatine breakdown in
muscle.— J. Physiol., 195, 157 —
183.
Wilkie D. 7?., Woledge R. C. 1967.
The application of irreversible
thermodynamics to muscular con-
traction. Comments on a recent
theory by S. R. Caplan.— Proc.
Roy. Soc., Ser. B, 169, 17—29.
Wilmanski K, 1972. On thermodyna-
mics and functions of state of non-
isolated systems.— Arch. Ration.
Meeh. Analyses, 45, 251—281.
Wilson J. A. 1968a. Increasing entro-
py of biological systems. — Nature,
219, 534—535.
Wilson J. A. 1968b. Entropy, not
negentropy.— Nature, 219, 535—
536.
Winfree A. T, 1972. Spiral waves of
chemical activity.— Science, 175,
634-636.
Wischnitzer 5. 1967. The cytoplasmic
inclusions of the salamander oocy-
te. II. Yolk plattelts.— Acta em-
bryol. et morphol. exper., 10, 15—
30.
Wohl K., James W. O. 1942. The ener-
gy changes associated with plant
respiration. — New Phytologist, 41,
230-256.
Wolfe H. J., Cohen R. B. 1963. His-
tochemical studies on regenerating
urodele limb. I. Oxidative and gly-
cogenic enzymes.— Develop. Bi-
ol., 8, 48—66.
Wolsky A. 1941. Contributions to the
physiology of regeneration. Allat-
tani kozl., Hung., 38, 9—16.
Woolhouse H. W. 1967a. Negentropy,
information and feeding of orga-
nisms.— Nature, 213, 952.
Woolhouse H. W. 1967b. Entropy and
evolution. — Nature, 216, 200.
Zaikin A. N., Zhabotinsky A. M.
1970. Concentration wave propaga-
tion in two-dimensional liquid-pha-
se self-osciHating system. — Nature,
225, 535-537.
Zeuthen E. 1953. Oxygen uptake as
related to body size in organisms.—
Quart. Rev. Biol., 28, 1—12.
Zhabotinsky A. MZaikin A. N. 1973.
Autowave processes in a distribu-
ted chemical system.— J. Theoret.
Biol., 40, 45-61.
Ziegler H. 1970. Proof an orthoganali-
ty principle in irreversible thermo-
dynamics.— Z. angew. Math. Phys.,
21, 853—863.
Zotin A. I. 1972. Thermodynamic As-
pects of Developmental Biology.
Basel, S. Karger A. G.
Zotin A. Faustov V. £., Radzinska-
ja L. I., Ozernyuk N. D. 1967. ATP
level and respiration of embryos.—
J. Embryol. Exper. Morphol., 18,
1-12.
Zotin A. I., Grudnitzky V. A., Shagi-
mordanov N. Sh., Terentieva N. B.
1971. Growth equation with al-
lowances made for interaction of
growth and differentiation.— W.
Roux’ Arch., 168, 169—173.
Zotin A. I., Zolina R.' S., 1967. Ther-
modynamic aspects of developmen-
tal biology.— J. Theoret. Biol.,
17, 57-75.
Zotina R. S., Zotin A. I. 1972. To-
wards a phenomenological theory
of growth.— J. Theoret. Biol., 35,
213-225.
ОГЛАВЛЕНИЕ
Предисловие............................................... 3
I
Общие проблемы термодинамики биологических про-
цессов ................................................... 5
И. Лампрехт
Использование понятий классической термодинамики в био-
логии .................................................... 6
А. И. Зотин
Второе начало, негэнтропии, термодинамика линейных необра-
тимых процессов.......................................... 16
Е. В. Преснов
Формализм неравновесной феноменологической термодинамики 25
II
Качественная феноменологическая теория развития
организмов............................................... 43
А. И. Зотин, Р. С, Зотина
Экспериментальная и теоретическая основы качественной фено-
менологической теории развития........................... 47
В, Л. Коноплев, А. И. Зотин
Статистическая интерпретация функции внешней диссипации и
критерия эволюции........................................ 65
Е. В. Преснов, Р. С. Зотина
Стохастическое рассмотрение конститутивных процессов и кри-
терия эволюции........................................... 69
Е. В. Преснов
Усиленный критерий эволюции в биологии развития........ 73
III
Количественная феноменологическая теория развития
организмов............................................... 77
А. И. Зотин, В. А. Коноплев, Е. В, Преснов
Нелинейные феноменологические уравнения.................. 78
276
ТЕРМОДИНАМИКА
БИОЛОГИЧЕСКИХ ПРОЦЕССОВ
Общие положения,
конститутивные процессы,
диссипативные структуры,
некоторые проблемы эволюции
Утверждено к печати
Институтом биологии развития
им. Н. К. Кольцова
Академии наук СССР
Редактор И. С. Никольская
Редактор издательства А. М. Гидагевич
Художник О. Л. Орман
Художественный редактор Н. Н. Власик
Технический редактор Р. Г. Грузинова
Корректоры Г. Н. Джиоева, Л. Ф. Кривенко,
Л. II. Харитонова
Сдано в набор 18/V 1976 г.
Подписано к печати 7/Х 1976 г.
Формат 60 x 90Vie Бумага типографская № 1.
Усл. печ. л. 17,62. Уч.-изд. л. 18,5
Тираж 3100. Т-16055. Тип. зак. 738.
Цена 1 р. 41 к.
Издательство «Наука»
103717 ГСП, Москва К-62, Подсосенский пер. ,21
2-я типография издательства «Наука».
121099, Москва, Г-99, Шубинский пер., 10