/









































































































































Текст
INSTITUT ZOOLOGIQUE DE L’ACADfiMIE DES SCIENCES DE L’URSS
NOUVELLE SERIE n° 6
FAUNE DE L’URSS
Sous la direction de S. A. SERNOV, Membre de ГAcademie Redige par A. A. STACKELBERG
CRUSTACEES
Vol. VII, n° 3
ISOPODES DES MERS ORIENTALES
par E. F. GURJANOVA
EDITION DE I’ACADEMIE DES SCIENCES DE I’URSS MOSCOU - 1936 • LENINGRAD
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ АКАДЕМИИ НАУК СССР
НОВАЯ СЕРИЯ № 6
ФАУНА СССР
Главный редактор акад. С. А. ЗЕРНОВ Редактор А. А. ШТАКЕАЬБЕРГ
РАКООБРАЗНЫЕ
Том VII, вып. 3
РАВНОНОГИЕ ДАЛЬНЕВОСТОЧНЫХ МОРЕЙ
Составила
Е. Ф. ГУРЬЯНОВА
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР МОСКВА • 1936 • ЛЕНИНГРАД
Напечатано по распоряжению Академии Наук СССР Июнь 1936 г.
Непременный секретарь академик Н. Горбунов
Редактор издания А. А. Штакельберг
Технический редактор М. И. Стеблин-Каменский. — Ученый корректор А. М. Налетов
Сдано в набор 10 февраля 1936 г. — Подписано к печати 25 июня 1936 г.
280-ь XII стр.
Формат бум. 72 X 1Ю см,—18 V4 печ. л.—48 550 тип. зн. в л.—22,00 уч.-авт. л.—Тираж 1200
Ленгорлит № 16655. — АНИ № 713. — Заказ № 647
Типографии Академии Наук СССР. Ленинград, В. О., 9 линии 12
ПРЕДИСЛОВИЕ
В связи с интенсивными исследованиями за последние десять лет наших дальневосточных морей, связанными с выдвинутыми социалистическим строительством проблемами освоения Дальнего Востока и использования его природных богатств, выяснилось, как богата и разнообразна в частности его морская фауна и как ничтожно мало мы о ней знаем. Вместе с тем именно на Дальнем Востоке морская фауна беспозвоночных имеет особенно важное значение. Очень многие представители этой фауны сами являются объектами промысла, другие служат пищей промысловых животных, третьи тесно связаны с ними разного рода биоцено-тическими связями; многие формы, сами по себе не могущие быть объектом промысла, важны, однако, как биологические показатели экологических условий и возможности присутствия здесь промысловых рыб и беспозвоночных и т. д. Особенно важно разобраться в тех сложных и чрезвычайно богатых биоценозах, которые развиваются в Беринговом, Охотском и Японском морях. Для этого совершенно необходимо точное определение систематического положения отдельных компонентов этих биоценозов. Отсюда ясно, какое важное значение имеет появление сейчас, в период больших работ, развертываемых на Дальнем Востоке, сводок и определителей по морской фауне Дальнего Востока. Вот почему, ясно отдавая себе отчет в том, что наши знания по фауне дальневосточных морей еще крайне скудны, и что списки животных, зарегистрированных в этих морях, будут быстро увеличиваться, я тем не менее считаю нужным уже сейчас дать такую сводку с определителем по фауне Isopoda Берингова, Охотского и Японского морей. Благодаря богатейшему материалу по группе Isopoda, собранному русскими экспедициями за период с 1900 г. до наших дней и охватывающему глубины от 0 до 3500 м, во всех трех дальневосточных морях мы можем указать уже 136 видов морских Isopoda. На основании этих материалов, хранящихся в коллекциях Зоологического института Академии Наук, а также и литературных данных (главным образом работы Ричардсон и Бенедикта) мною составлена настоящая сводка и определитель. Виды и формы, уже зарегистрированные в Японском, Охотском и Беринговом морях (кроме побережья южной части Японии) отмечены звездочкой, стоящей перед порядковым номером вида. Все новые нахождения по нашим материалам, впервые публикуемые, даются
VI
ПРЕДИСЛОВИЕ
после короткого описания вида под рубрикой „распространение". В тех случаях, когда нахождение в том или другом из дальневосточных морей дается на основании литературных данных, после указания мест нахождения в скобках указывается литературный источник. В сводку вошли также и те виды, которые пока еще не зарегистрированы в пределах рассматриваемых морей, но присутствие которых в их фауне возможно.
Краткие диагнозы видов и подвидов составлены на основании имеющегося в коллекциях Института материала; в тех случаях, когда данный вид, зарегистрированный в фауне Дальнего Востока, в наших коллекциях не был представлен, диагноз дается по работе автора, который описал этот вид или форму.
Работа моя преследует задачу дать возможность исследователю морской фауны Дальнего Востока самому ориентироваться в группе Iso-poda, поэтому я даю краткий морфологический очерк этой группы с указанием наиболее важных для определения систематических признаков. Таблицы для определения подотрядов, семейств, родов и видов и полный список с краткими диагнозами всех видов Isopoda, найденных до сих пор в Японском, Охотском и Беринговом морях, составляют содержание предлагаемой работы. Для каждого вида даются схематические рисунки, краткое. описание и общее географическое распространение по материалам, обработанным мною, и по литературным данным. Не считая нужным в общедоступном определителе приводить всю синонимику каждого вида, я ограничиваюсь лишь приведением главнейшей синонимики и указанием на работы, где данный вид описан впервые, и на работы, заключающие в себе лучшие описания и лучшие рисунки данной формы.
Большая часть рисунков, помещенных здесь, оригинальна и сделана по экземплярам, хранящимся в коллекциях Зоологического института Академии Наук; часть заимствована из работ Ричардсон, Сарса, Ганзеиа„ Бенедикта и др. (в таких случаях указывается, источник).
В данном выпуске приводится краткий морфологический очерк и: самые общие замечания по экологии группы, необходимые для пользования определительными таблицами. В конце морфологического очерка дан краткий зоогеографический очерк фауны Isopoda Берингового, Охотского и Японского морей, а также список литературы, заключающий в себе наиболее важные работы, посвященные фауне изопод Дальнего-Востока и Тихого океана.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
Отряд Isopoda
I. Подотряд Asellota
1. Сем. Janiridae
1. Род Janira Leach
Стр
1. J. alascensis (Benedict)...............*.................... . 41
2. J. erostrata (Rich.)...................................... 41
3. J. soldatovi Gurjan......................................... 42
-4. J. holmesi (Rich.).......................................... 42
5. J. sarsi (Rich.).......................................... 43
2. Род Janiropsis G. Sars
1. J. kincaidi Rich. .......................................... 44
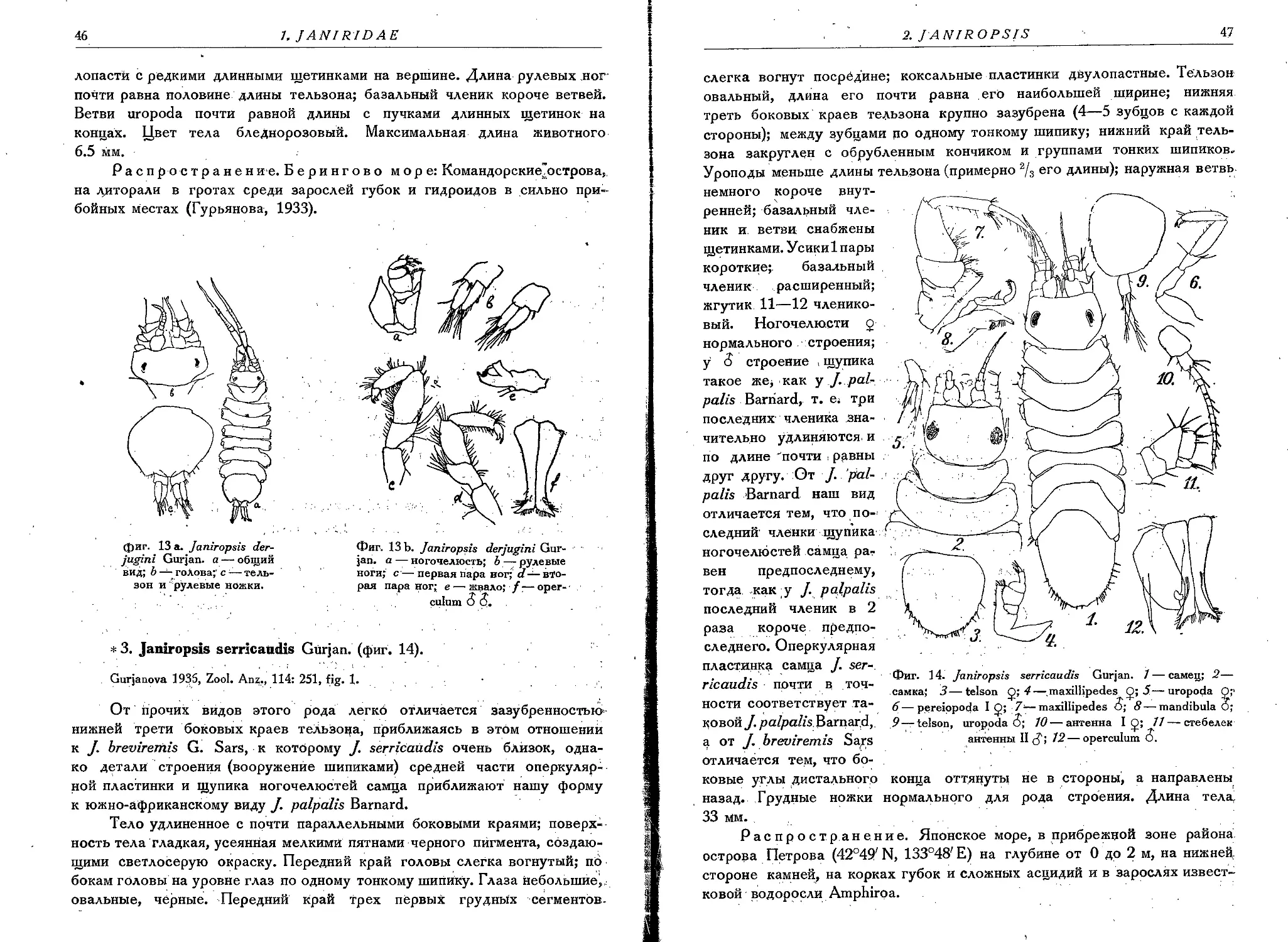
2. J. derjugini Gurjan ......................................... 45
3. J. serricaudis Gurjan.................................. . 46
3. Род Microprotus Richardson
1. M. coecus Rich................................... ............48
2. Сем. Munnidae
1. Род Munna Kroyer
1. M. stephenseni Gurjan. ...................................... 50
2. M. arnholdi Gurjan........................................... 51
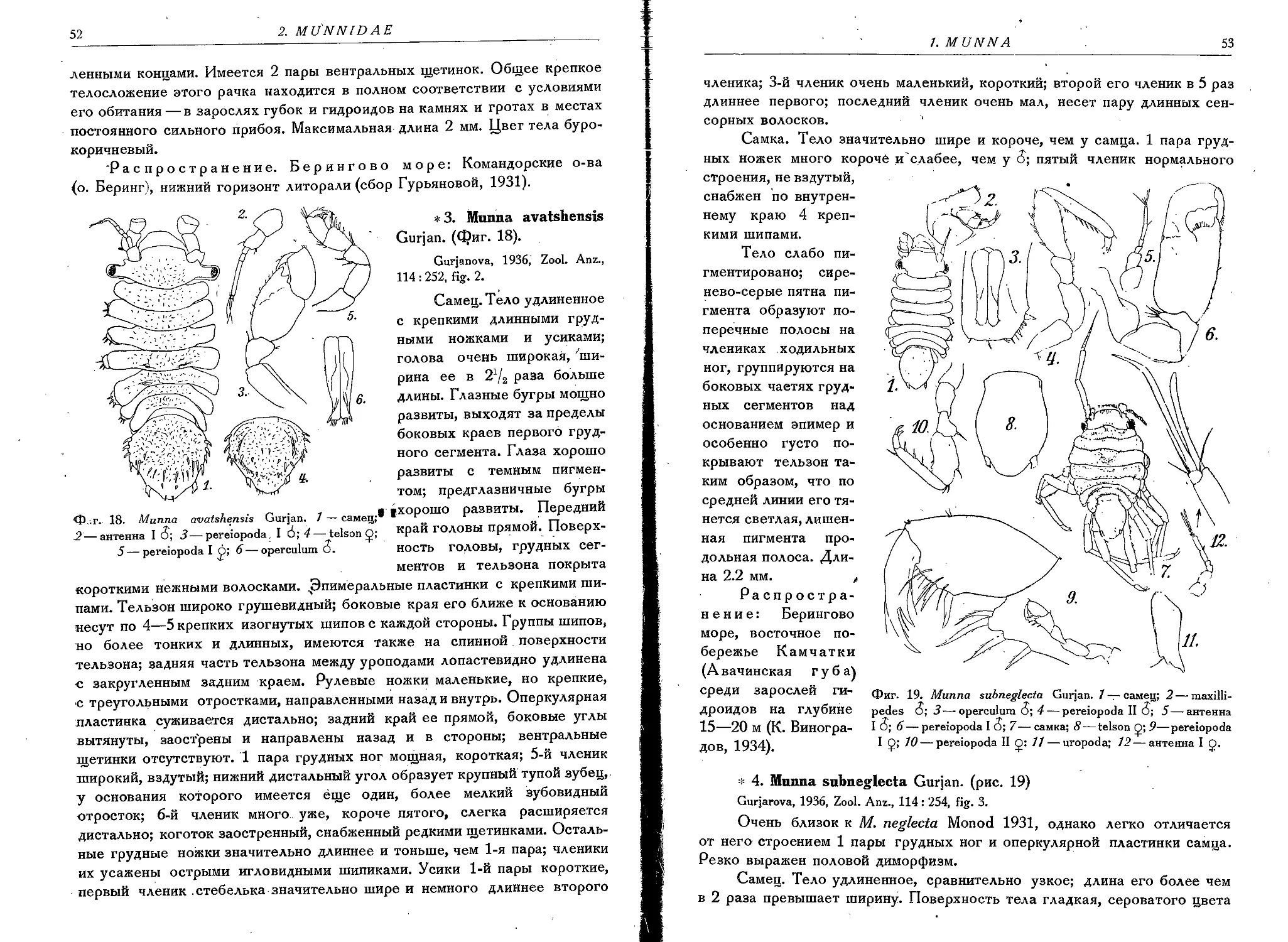
3. M. avatshensis Gurjan....................................... 52
4. M. subneglecta Gurjan........................................53
3. Сем. Mnnnopsidae
1. Род Ilyarachna G. Sars.
1. I. zachsi Gurjan......................................... . 56
2. 1. starokadomskii Gurjan. ................................... 57
,2. Род Munnppsurus Richardson
1. M. giganteus (G. Sars)....................... .............. 58
la. M. giganteus var.'ochotensis Gurjan. ................... . 59
2. M. laevis (Rich.)...................................... 60
.3. M. minutus Gurjan......................... ..............- . 61
VIII
РАВНОНОГИЕ РАКИ
3. Род Eurycope G. Sars
Стр.
1. Е. spinifrons Gurjan................................................... 62
2. Е. pavlenkoi Gurjan..................................................... 63
4. Род Munnopsis М. Sars
1. М. latifrons Beddard.................................................... 64
II. Подотряд Flabellifera
4. Сем. Cymothoidae
1. Подсем. C i г о 1 a n i n a e
1. Род Cirolana. Leach
1. C. japonensis Rich.................................,........... • 67
2. Род Bathynomus M.-Edw.
1. B. doderleini Ortmann.......................................‘ . . . 68
2. Подсем. A e g i n a e
3. Род Aega Leach
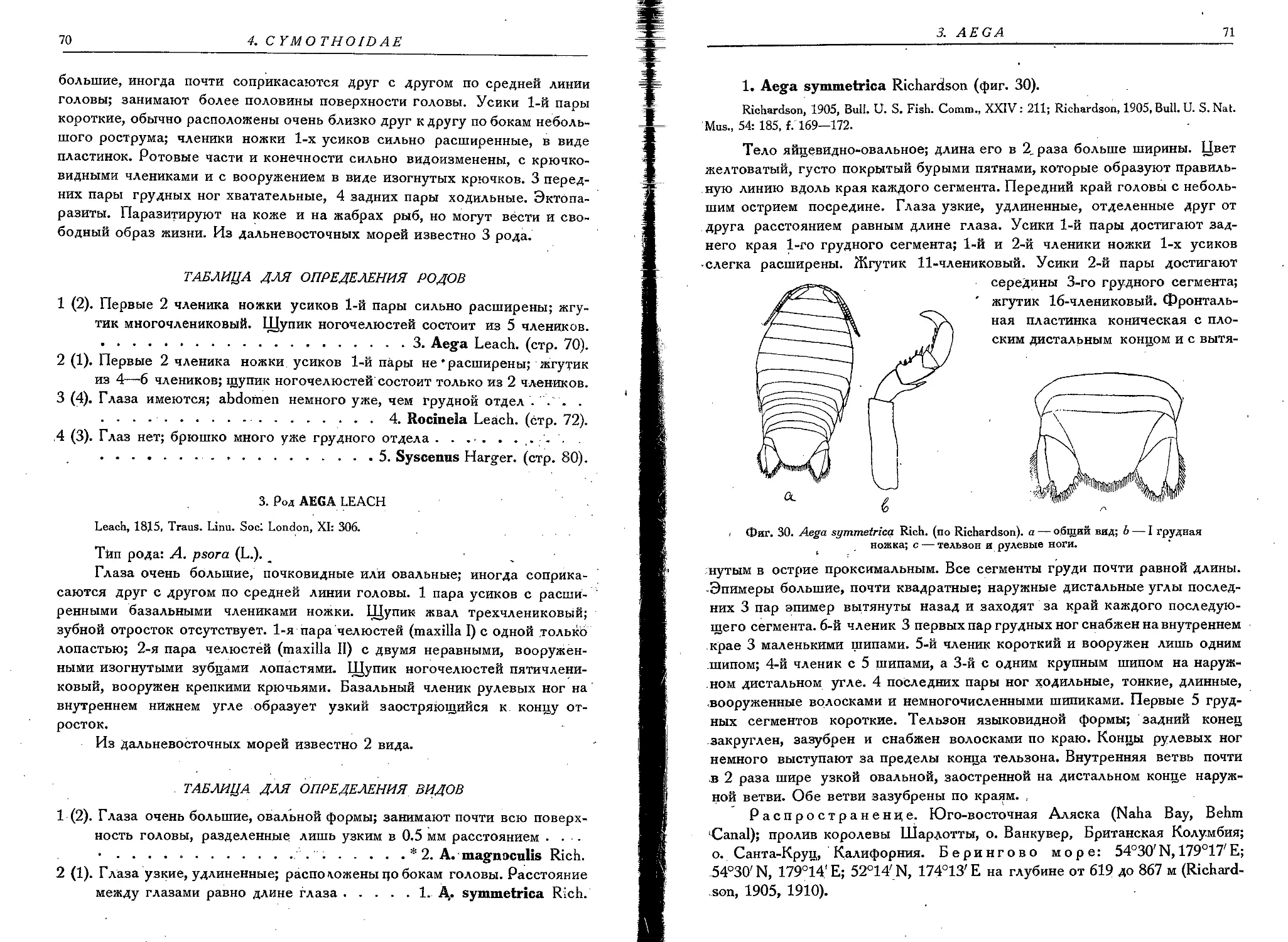
1. A. symmetrica Rich. . . .................................... 71.
2. A. magnoculis Rich......................................... 72
4. Род Rocinela Leach
1. R. cornuta Rich............................................. 73
2. R. maculata Sch. a. Meinert................................. 74
3. R. belliceps (Stimpson)................................... 75
4. R. angustata Rich. ..;...............,............. 77
5. R. japonica Rich............................................ 78
6. R. niponia Rich............................................. 79
5. Род Syscenus Harger
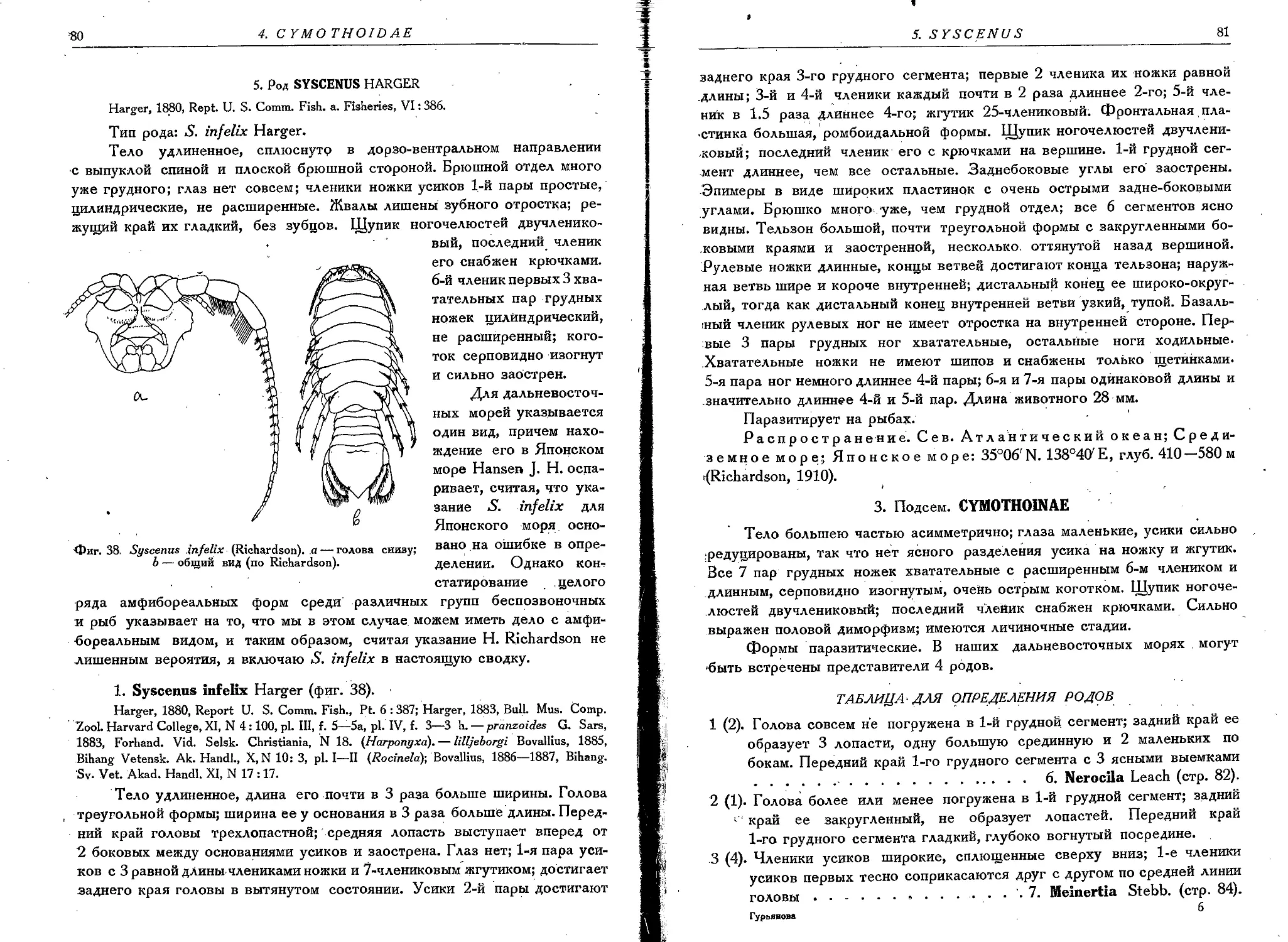
1. S. infelix Harger........................................... 80
3. Подсем. Cymothoinae
6. Род Nerocila Leach
1. N. japonica Sch. a. Meinert........................................... 82
2. N. californica Sch. a. Mein. ........................................ 83
7. Род Meinertia. Stebbing
1. M. oxyrrhynchaena Koelbel............................................. 84
8. Род Livoneca Leach
1. L. amurensis Gerstf.................................................. 87
2. L. raynaudii M.-Edw................................................ 88-
3. L. propinqua Rich.................................................... 89-
‘СИСТЕМАТИЧЁСКИЙ УКАЗАТЕЛЬ IX
Стр.
4. L. sacciger Rich.......................................... . 90
5. L. epimerias Rich.............................................. 91
6. L. vulgaris Stimpson . . . '................................. 92
7. L. californica Sch. a. Mein.................................... 93
8. L. caudata Sch. a. Mein....................................... 93
9. Род Irona Schioedte et Meinert
1. I. melanostica Sch. a. Mein.................................... 94
5. Сем. Sphaeromidae
1. Подсем. Limnorinae
1. Род Limnoria Leach.
1. L. lignorum (Rathke).......................................... 97
2. L. japonica Rich........................ . . .......... 98
2. Подсем. Sphaerominae
2. Род Tecticeps Richardson
1. T. renoculis Rich..............................................102
la. T. renoculis..var. laevis Gurjan. ... .......................103
lb. T. renoculis var. tnarginalis Gurjan. ....................... 105
lc. T. renoculis var. carinatus Gurjan............................105
Id. T. renoculis var. nodulosus Gurjan............................106
le. T. renoculis var. serratus Gurjan. ...........................106
2. T. alascensis Rich...................'.........................107
3. T. glaber Gurjan............................................. 110
4. T. convexus Rich............................................. Ill
5. T. leucophthalmus Gurjan.......................................Ill
3. Род Holotelson Richardson
1. H. tuberculatus Rich..................................... . . 112
4. Род Dynamene Leach
1. D. glabra Rich..... ........ ........... '. . . . 114
5. Род Cilicaea Leach
1. C. cordata Rich. .......................................... 115
6. Род Cymodoce Leach
1. C. japonica Rich........................................... 117
2. C. acuta Rich................................................ 119
7. Род Sphaeroma Latreille
1. S. sieboldi Dollfus........................................... 120
8. Род Exosphaeroma Stebbing
1. E. amplicauda (Stimpson) . . .............................. 122
X
РАВНОНОГИЕ РАКИ
9. Род Neosphaeroma Baker
Стр.
1. N. oregonensis (Dana)............................................123
2. N. ovata (Gurjan.)...............................................124
б. Сем. Anthuridae
1. Род Cyathura Norm. a. Stebb.
1. C. carinata (Kroyer).............................................126
2. Род Paranthura Bate a. Westw.
1. P. japonica Rich.................................................128
3. Род Calathura Norm. a. Stebb.
1. C. brachiata (Stimpson) . .......................................129
III. noAOTpaflValvifera
7. Сем. Idotheidae
1. Род Idothea Fabricius
1. I. metallica Bose.............................................. 134
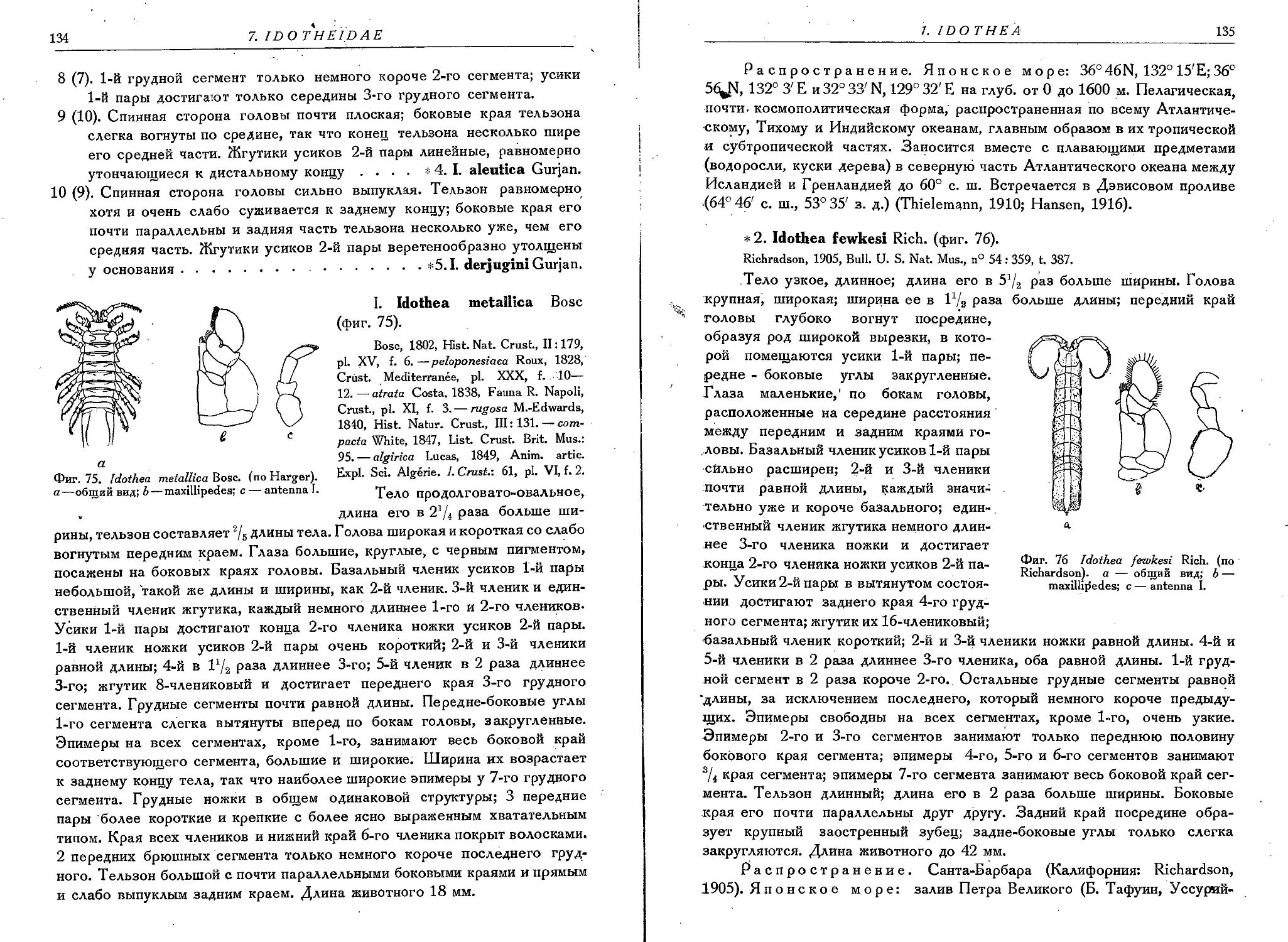
2. I. fewkesi Rich..................................................135
3. I. ochotensis Brandt........................................... 136
4. I. aleutica Gurjan............................................. 137
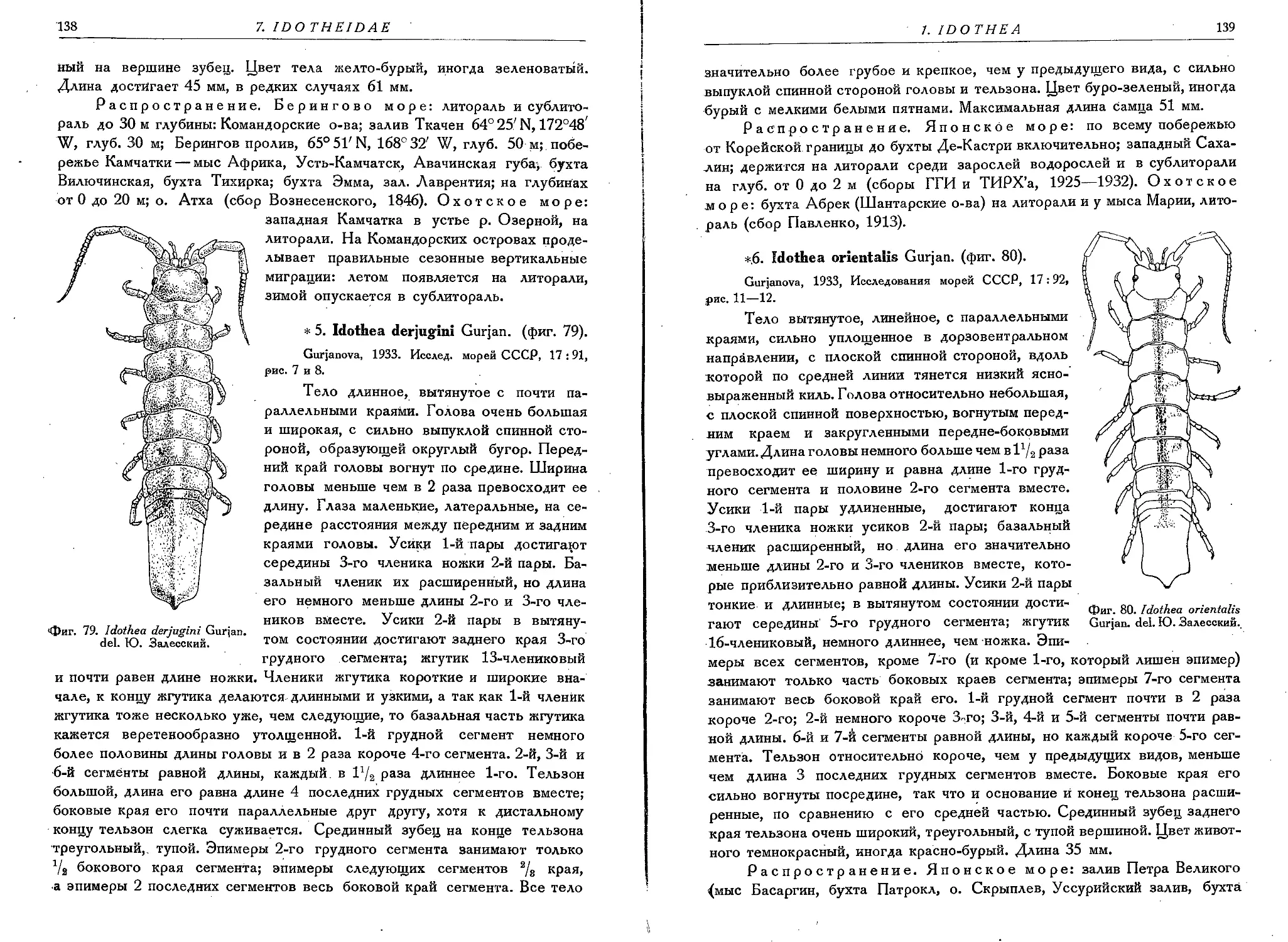
5. I. derjugini Gurjan.......................................... 138
6. I. orientals Gurjan...-......................................... 139
2. Род Pentidothea Richardson
1. P. japonica Rich.................................................140
2. P. rotundata Rich............................................... 141
3. P. wosnesenskii (Brandt).........................................142
4. P. whitei (Stimpson) ............................................144
3. Род Mesidothea. Richardson
1. M. entomon (L.) subsp. orientalis Gurjan.......... . .........145
4. Род Peniias Richardson
1. P.hayi Rich................................................... 147
5. Род Symmius Richardson
1. S. caudatus Rich. . . . *...................................... 149
6. Род Synidothea Harger
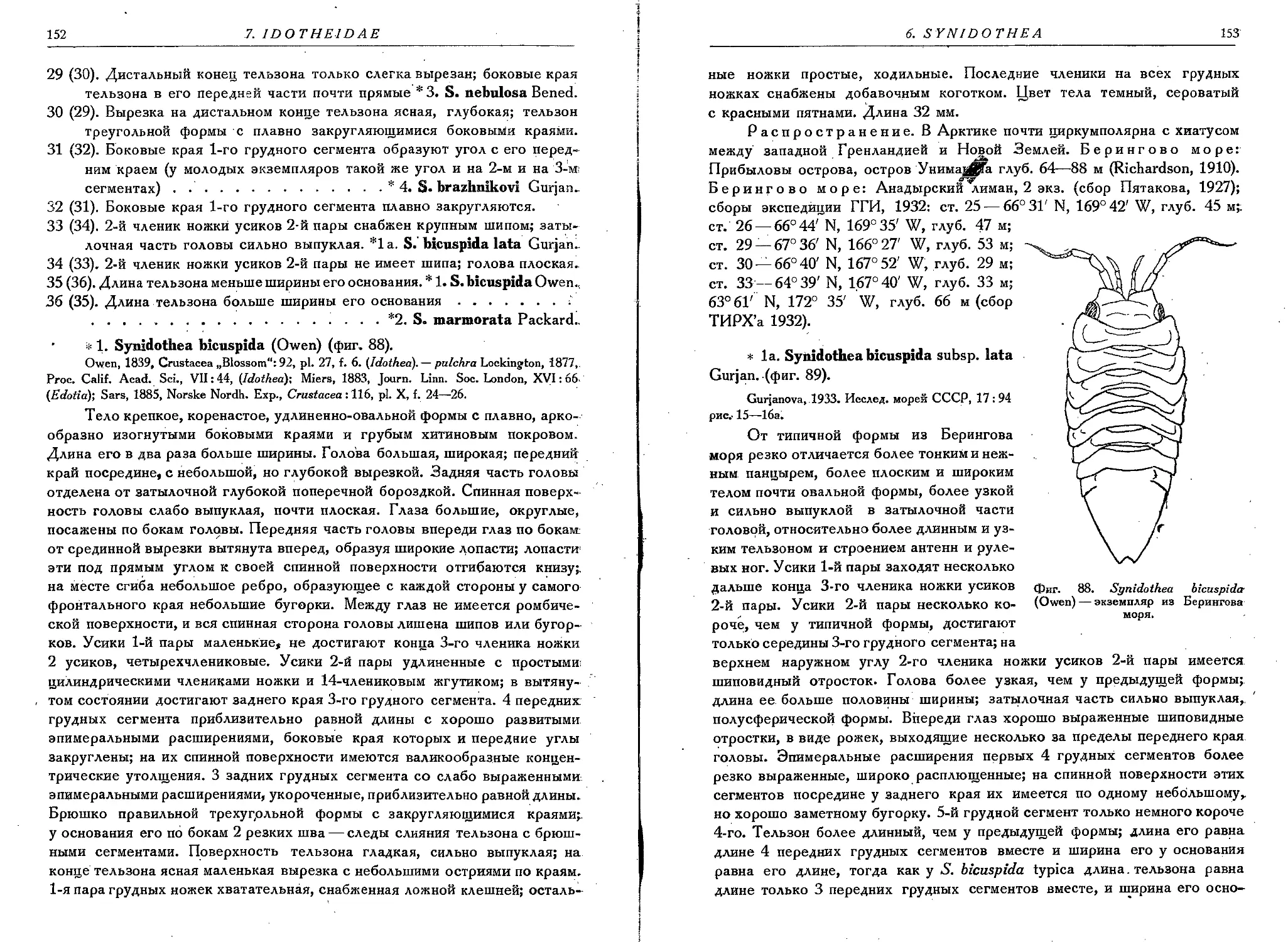
1. S. bicuspida (Owen) . . ........... . ..........................152
la. S. bicuspida subsp. lata Gurjan................................153
2. S. marmorata (Packard)..........................................154
3. S. nebulosa Bened...............................................155
4. S. brazhnikovi Gurjan.......................................... 156
СИСТЕМАТИЧЕСКИМ УКАЗ АТЕЛ& XI
Стр.
5. S. erosa Bened...........................................................157
6. S. pallida Bened.........................................................158
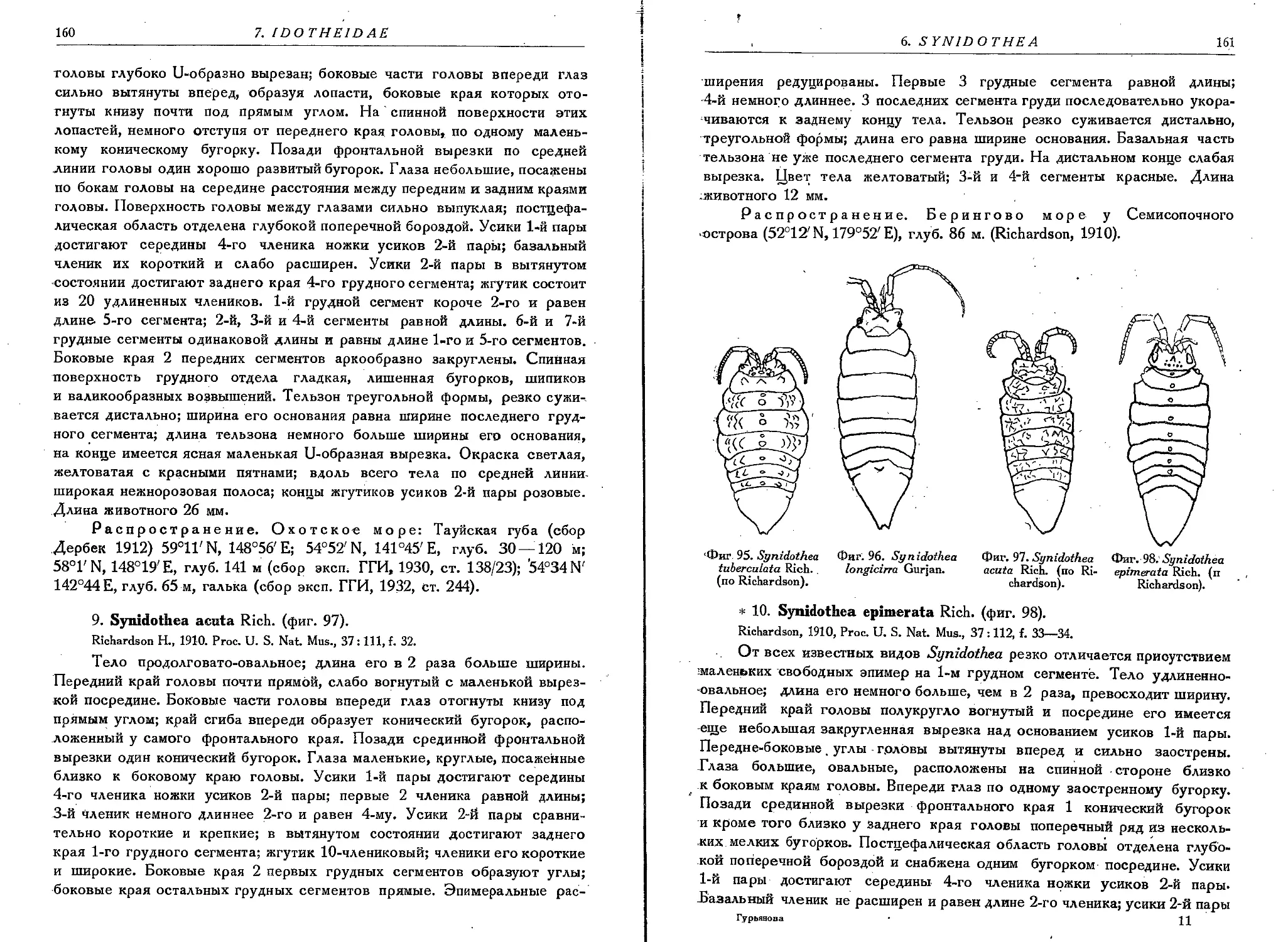
7. S. tuberculata Rich......................................................159
8. S. longicirra Gurjan.....................................................159
9. S. acuta Rich. .........................................................160'
10. S', epimerata Rich. ....’...............................................161
11. S. excavata Gurjan.......................................................162
12. S. harfordi Bened........................................................163
13. S. nodulosa (Kroyer) .................................................. 164
14. S. laevis Bened. '.......................................................165
15. S. muricata (Harford)....................................................166
16. S. spinosa Gurjan........................................................167
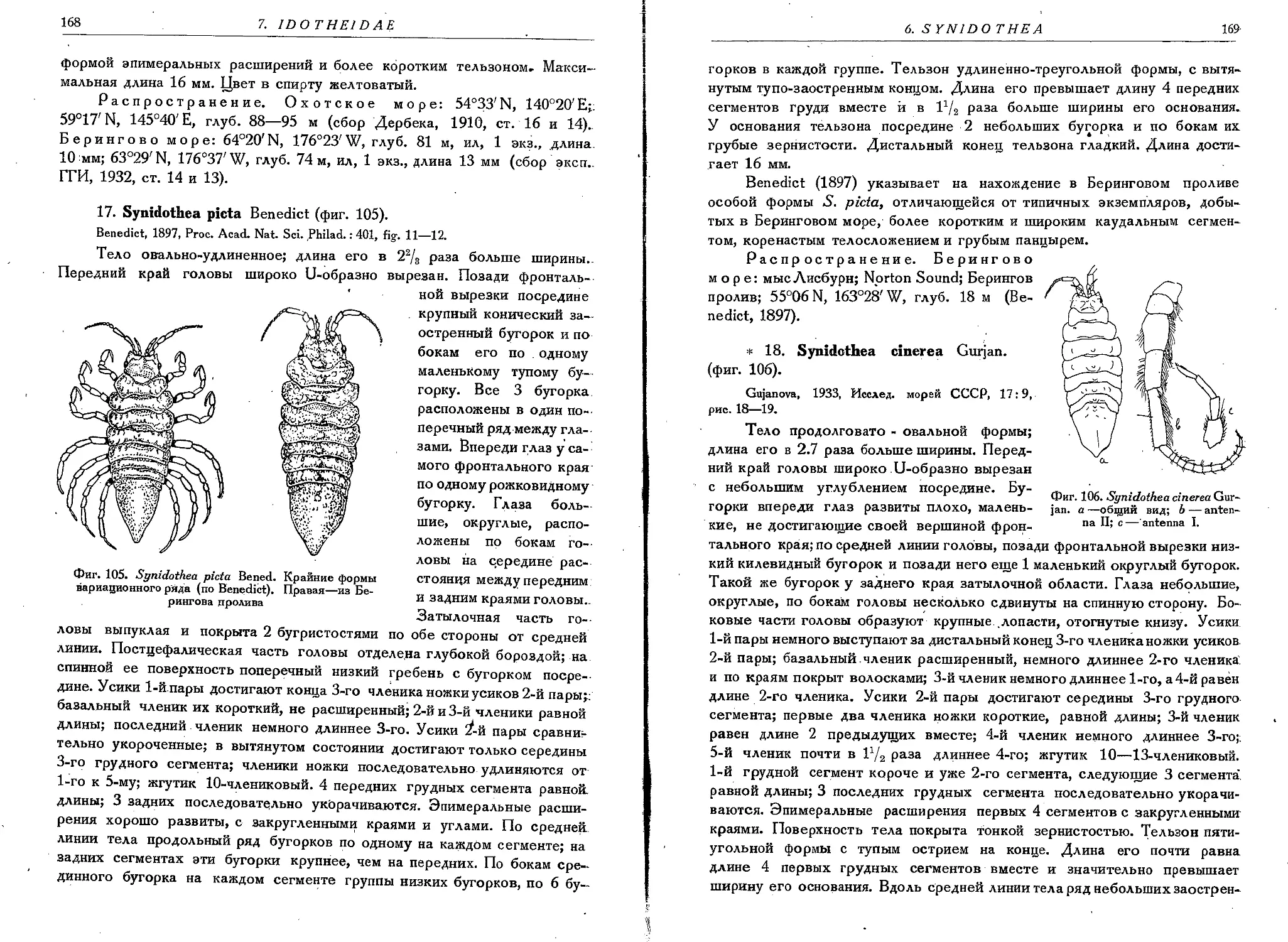
17. S. picta Bened......................................................... 168
18. S. cinerea Gurjan.......................................................169"
7. Род Cleantis Dana
1. C. planicauda Bened......................................................170-
2. C. isopus Grube in Miers................................................ 172:
8. Сем. Arcturidae
1. Род Arcturus Latreille
1. A. baffini (Sab.)...................................................... 175
2. A. ulbani Gurjan..........................-..............................177
3. A. hastiger Rich..................................................... 178-
4. A. longispinis Bened. . ................................................180-
5. A. setosus Gurjan.................................................... 181
5a. A. setosus var. seminudus Gurjan........................................183-
5b. A. setosus var. acuticaudalis Gurjan....................................183
6. A. diversispinis Rich................................................... 184
7. A. magnispinis Rich.................................................. 185-
8. A. crassispinis Rich.....................................................186
9. A. brevispinis Rich................................................. 188-
10. A. granulatus Rich..................................................... 189
11. A. beringanus Bened. ................................................. 191.
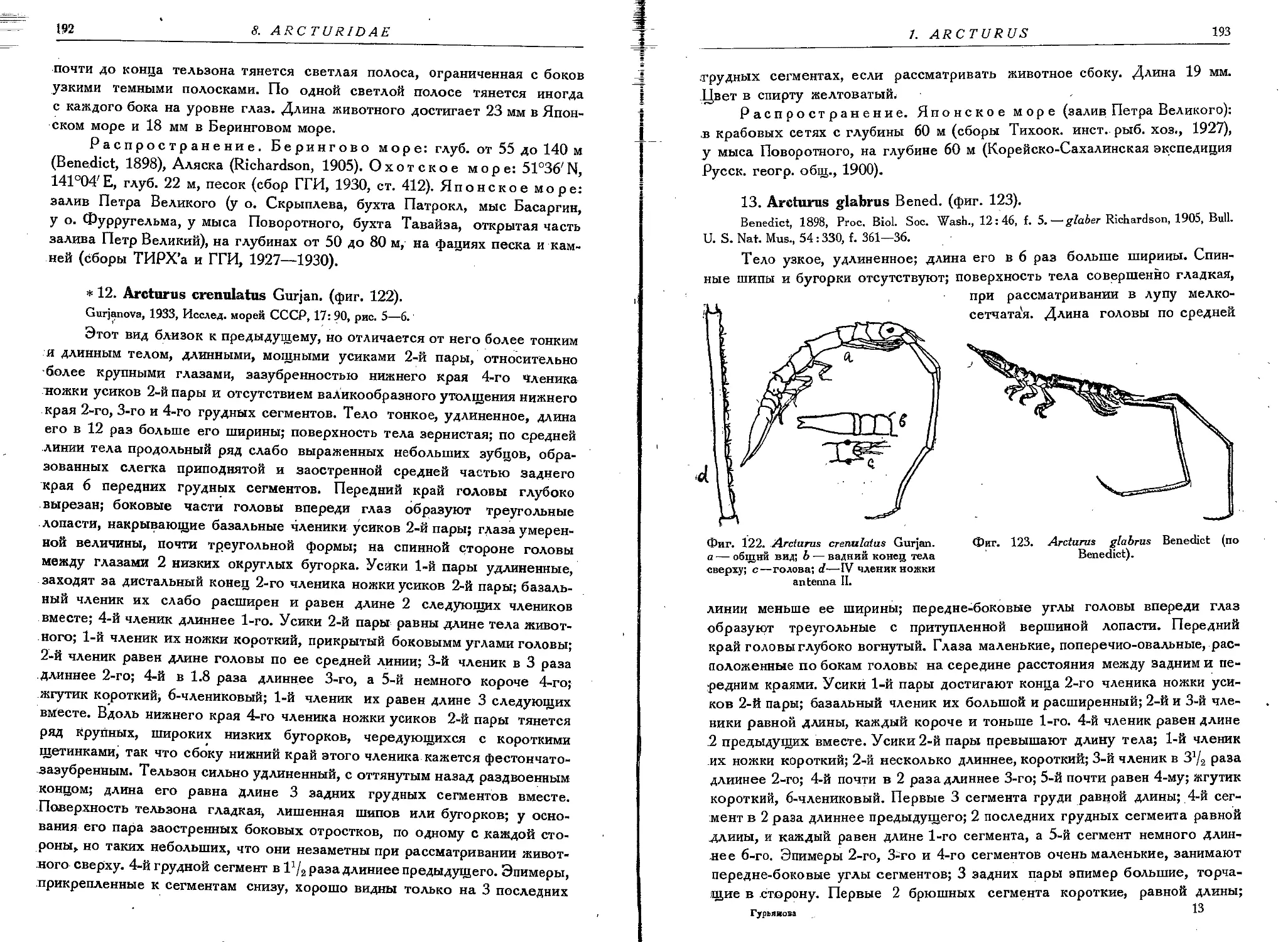
12. A. crenulatus Gurjan.................................................. 192
13. A. glabrus Bened....................................................... 193-
14. A. tritaeniatus Rich.....................................................194
15. A. hirsutus Rich.........................................................194
16. A. beddardi Gurjan................................... • .............196'
2. Род Pleuroprion zur Strassen
1. P. murdochi (Benedict).................................................. 198
2. P. hystrix (G. Sars).................................................... 200'
3. P. intermedium (Rich.)...................................................901
4. P. tarasovi Gurjan.......................................................201.
3. Род Astacilla Cordiner
1. A. dilatata Rich....................................................... 203-
2. A. polita Gurjan.................................................... 204
XII
РАВНОНОГИЕ РАКИ
IV. Подотряд Oniscoidea
9. Сем. Ligiidae
1. Род Ligia (Fabricius) Стр
1. L. cinerascens Bpdde-Lund.................................... 208
2. L. pallasi Brandt..............................................210
2. Род Ligidium Brandt
1. L. tenue Budde-Lund........................................ 211
1О.Сем. Trichoniscidae
1. Род Trichoniscus Brandt
1. T. papillicornis Rich. . ......................................213
11. Сем. Oniscidae
1. Род Porcellio Latreille
1. P. scaber Latr................. . . ...........................215
V. Подотряд Epicar idea
12. Сем. Bopyridae
1. Род Phryxus Rathke
1. P. abdominalis (Kroyer)..................................... 218
2. Род Argeia Dana
1. A. pugettensis Dana ................................. ........ 219
3. Род Bopyroides Stimpson
1. B. hippolytes (Kroyer) . . ................................ 221
13. Сем. Dajidae
1. Род Holophryxus Richardson
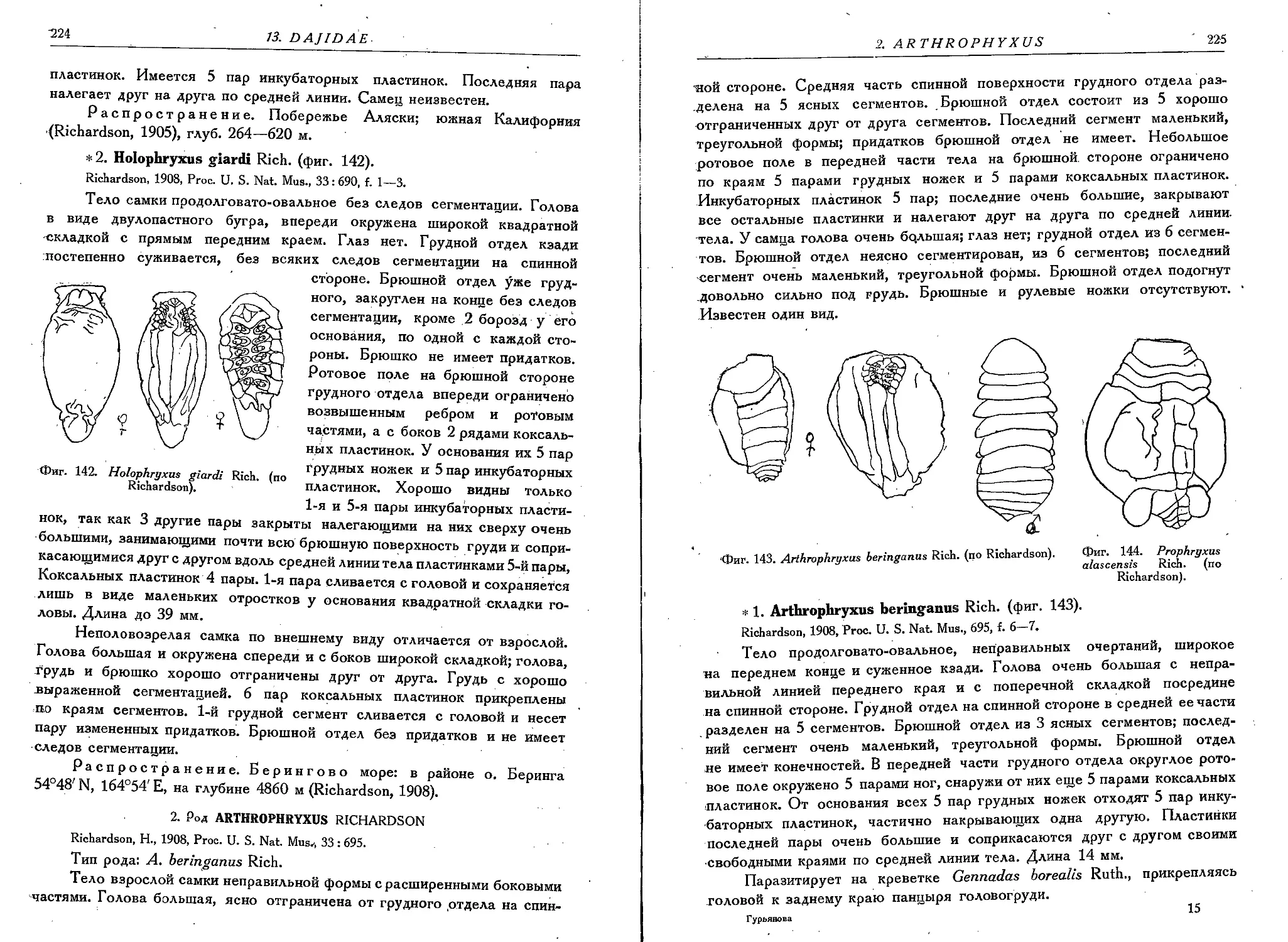
1. H. alascensis Rich.............................................223
2. H. giardi Rich...........................................• • • 224
2. Род Arthrophryxas Richardson
1. A. beringanus Rich. . .'..................................... 225
3. Род Prophryxus Richardson
1. P. alascensis Rich. ........................................ 226
VI. Подотряд Gnathiidea
14. Сем. Gnathiidae
1. Род Gnathia Leach
1. G. tuberculata Rich. .....'.................................. . 229
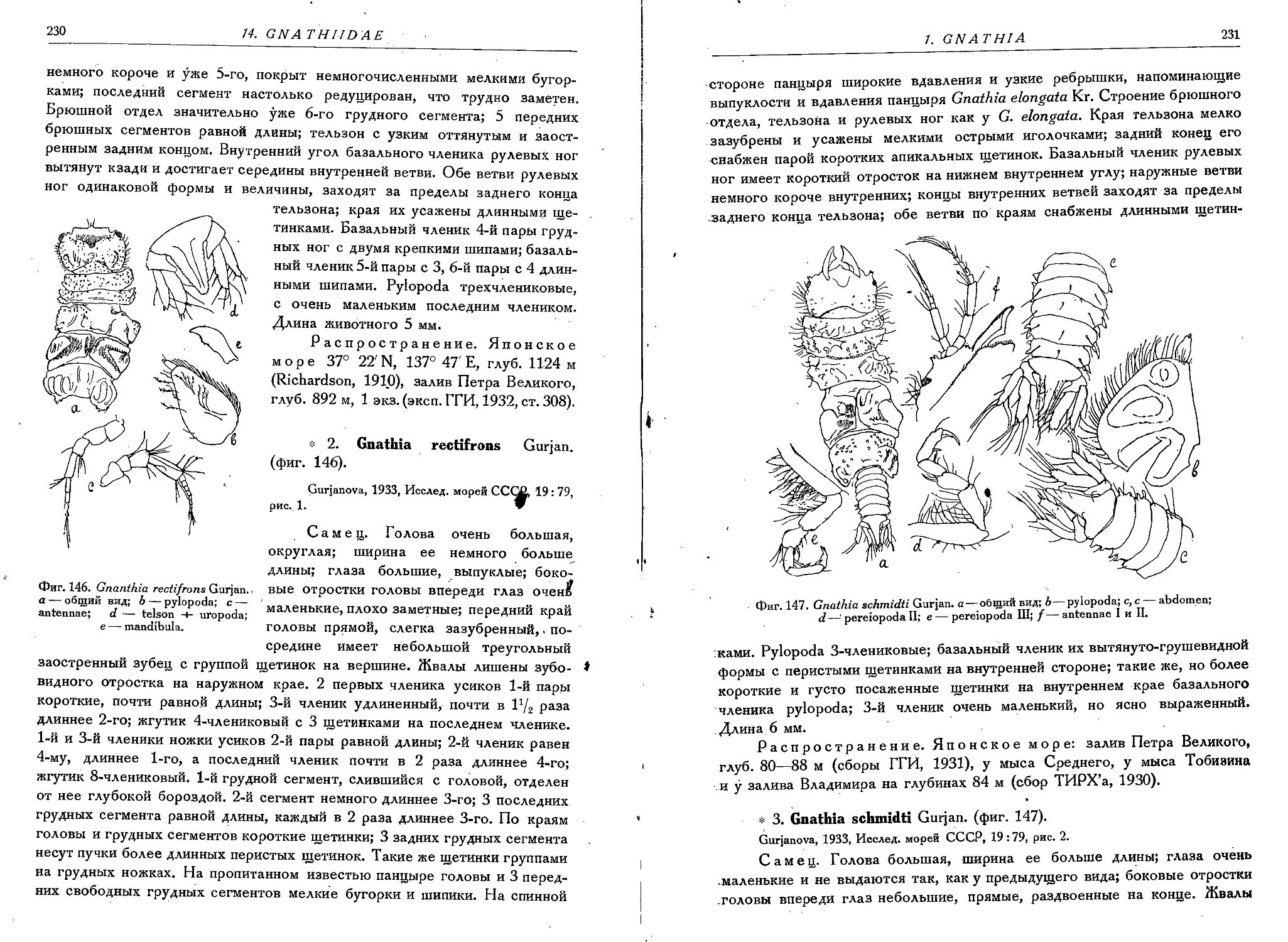
2. G. rectifrons Gurjan..........................................: 230
3. G. schmidti Gurjan.............................................231
A. G. derzhavini Gurjan.......................................... 232
5. G; elongata (Kroyer) ........ . .. ....................... . 233
6. G. (Elaphognathia) monodi Gurjan....... . .............. . . . 236
ВВЕДЕНИЕ МОРФОЛОГИЧЕСКИЙ ОЧЕРК
Отряд равноногих раков (Isopoda} принадлежит к группе высших ракообразных (Malacostraca) и характеризуется следующйми основными признаками: 1-й, а иногда и 2-й, грудные сегменты, как правило, сливаются с головой; тельзон или хвостовая пластинка, за исключением сем. Anthu-ridae, срастается с последним брюшным сегментом, образуя каудальный сегмент; глаза сидячие, т. е. лишены стебельков; грудные ножки одноветвистые, так как экзоподит отсутствует; 1-я пара грудных ножек, срастаясь по средней линии у основания, образует ногочелюсти. Брюшные ножки двуветвистые, пластинчатые и приспособлены главным образом для дыхания в водной или воздушной среде, но во многих случаях несут и локомоторную функцию; ветви брюшных ног в большинстве случаев одночлениковые; сердце целиком или отчасти лежит в брюшной области.
Большей частью это маленькие животные, величиною от нескольких миллиметров до нескольких сантиметров; наиболее, крупный рачок из этой группы, Bathynomus giganteus, достигает 27 см длины. Очертания тела чаще всего вытянутоовальные, иногда линейной формы с параллельными краями или же вытянутое цилиндрическое тело почти червеобразной формы. Окраска весьма разнообразна. Сухопутные и прибрежные формы обычно окрашены в темный, бурый или серый цвет, иногда с пестрым рисунком. Наоборот, формы глубоководные и живущие в местах с осла- ' Пленным освещением или пещерные — бесцветны (лишены пигмента). В некоторых случаях окраска является защитной; например, формы, живущие на песчаном грунте (различные виды Syn.idoth.ed), окрашены под цвет его: серовато-желтоватые с темными и красными пятнами; формы, населяющие заросли бурых водорослей (различные виды Idothea), окрашены в бурый и коричневый цвета; виды, населяющие заросли красных водорослей, например, Idothea orientalis, окрашены в яркокрасные тона и т. д. У некоторых видов окраска сильно вариирует (Idothea baltica и др.). Часто пигментирована только верхняя поверхность тела, а нижняя — бесцветна; у многих пигментные пятна образуют сложный узор и т. д.
Тело большей частью равномерно расчленено на сегменты; иногда отдельные сегменты значительно превосходят все остальные по размерам (Astacilld) или же, наоборот, некоторые сегменты редуцируются и очень
Гурьянова 1
2
РАВНОНОГИЕ РАКИ
коротки по сравнению с остальными (Anthuridae, Gnathiidae) и даже совсем исчезают. У форм паразитических тело самки асимметричное,, часто бесформенное, округлое, в виде простого нерасчлененного мешка, набитого яйцами, со слабыми следами сегментации или вовсе без намека, на расчленение (Epicaridae). Иногда животное способно свертываться в шар. У многих форм (особенно у Asellota) отдельные сегменты тела,, брюшные или грудные, сливаются друг с другом, чаще всего на спинной стороне, но иногда по всей длине пограничного края и даже без следов-слияния в виде швов или борозд. Чаще всего наблюдается слияние брюш- -ных сегментов друг с другом и с тельзоном. Обычно в этих случаях по бокам крупного сегмента, получившегося в результате слияния нескольких сегментов, сохраняются 2-3 хорошо выраженных шва в виде линий,, указывающих на число слившихся сегментов. Обычно тело сплюснуто' в дорзо-вентральном направлении и спинная сторона его сводчатовыпуклая, а брюшная — плоская. В некоторых случаях животное приобретает внешний вид, похожий на черепаху (Tecticeps).' В других случаях тело веретенообразно вытянуто, почти цилиндрическое (Arcturidae) или же: сильно вытянутое цилиндрическое, червеобразное (Anthuridae). Самцы подотряда Gnathiidea, благодаря сильному развитию жвал, выступающих, впереди головы и образующих настоящие щипцы или клещи, по внешнему’ виду напоминают насекомых.
Поверхность тела покрыта хитиновой оболочкой или скелетом. Хитин обычно довольно крепкий, в некоторых случаях (Ghathiidea) инкрустируется известью. У паразитических форм хитин тонкий, мягкий или вовсе-отсутствует. Поверхность тела в большинстве случаев гладкая, но иногда покрыта особой скульптурой, гранулированная, или сетчатая, или покрыта: точечными вдавлениями. На теле часто имеются отростки, шипы, бугорки,, гребни, кили, волоски, щетинки и пр.
Нормально тело Isopoda распадается на 3 отдела: голову, грудной отдел и брюшко. Голова имеет 2 пары усиков, верхние и нижние, а на. брюшной стороне снабжена ротовыми частями, парными и непарными,, состоящими из отдельных члеников. Грудной отдел нормально состоит из 7 сегментов; каждый сегмент несет пару грудных нбжек; брюшной отдел, состоит из различного числа сегментов у разных групп; наибольшее число свободных сегментов брюшка 6. Брюшной отдел нормально несет 5 пар брюшных ног (pleopoda) и 1 пару рулевых ног (uropoda). Брюшные и рулевые ножки нормально двуветвистые. По бокам грудных сегментов, имеются расширенные пластинки, подвижно сочлененные с ними или же срастающиеся неподвижно — эпимеры. В некоторых случаях слияние эпи- ' мер с боковыми частями полное, без следов шва, и тогда получающееся расширение боковых частей сегмента носит название эпимерального-или плеврального расширения. Нормально эпимеры 1-го грудного сегмента нацело сливаются с сегментом, но иногда и 1-я пара эпимер свободна. (Synidothea epimerata и др.). -Свободные эпимеры могут занимать или весь.
МОРФОЛОГИЧЕСКИЙ ОЧЕРК
боковой край сегмента, или же только его часть, обычно переднюю (фиг. 1). Эпимеральные пластинки представляют собой расширенный 1-й членик ножки — коксальный членик или соха. Часто эпимеральные расширения имеются и на брюшных сегментах.
Голова. Как правило, голова срастается без всяких следов шва с 1-м грудным сегментом, иногда и 2-й грудной сегмент сливается с головой (Gnathiidea}. Передний край головы закругленный или прямой, вогнутый или выпуклый, цельный или посредине с вырезкой; иногда средняя часть переднего края головы вытянута вперед в виде клюва или широ-
Фиг. 1. Limnoria lignorum L; общий вид сбоку; 2—7—плевральные или коксальные пластинки (эпимеры).
кой лопасти, образующей так называемое лобное поле (area frontalis). Глаза сидячие, фасеточные, хорошо развиты, иногда рудиментарные или вовсе отсутствуют; расположены они или на спинной стороне головы (дорзальные глаза), или по бокам головы (латеральные), или иногда на брюшной стороне (вентральные). В некоторых случаях глаза помещаются на выдающихся по бокам головы крупных буграх (Munnidae}. Придатков на голове 2 пары, а именно верхняя и нижняя пары усиков или антенн. С нижней стороны головы находятся ротовые части или буккальная масса. Первая или верхняя пара усиков или антенн (antennulae или antennae I) обычно короткая и состоит из трехчленикового стержня или ножки (pars basalis) и жгутика (flagellum), слагающегося из отдельных члеников. Жгутик .иногда многочлениковый (Asellota}, иногда — одночлениковый (Idotheidae}-, в редких случаях все членики жгутика сливаются вместе со следами, или без следов слияния, образуя один крупный членик (род Cleantis}. Добавочный жгутик обычно отсутствует; иногда наблюдается его рудимент (у Baihynomus, лачияск. Cryptoniscus}. У некоторых форм, особенно у наземных Oniscoidea, усики 1-й пары сильно редуцированы и настолько коротки, что не видны сверху, со спинной сто-1*
'4
РАВНОНОГИЕ РАКИ
роны Вторая или нижняя пара усиков или а н т е н н (antennae, antennae II или antennae inferiores) обычно длиннее 1~й пары, часто значительно длиннее тела животного, мощно развиты (Arcturidae); состоят из пятичлениковаго стержня или ножки и многочленикового жгутика. Иногда присутствует еще добавочный жгутик (flagellum accessorium). У многих форм Asellota на 3-м членике ножки сохраняется короткий экзоподит, в виде чешуйки (squama); у некоторых форм (сем. Anthuridae)' на 4-м членике ножки многочлениковый, но короткий, добавочный жгутик. Место прикрепления усиков к голове может быть различным; так, они могут быть расположены или на самом краю передней части головы -(терминальные усики) или же сдвинуты несколько на спинную (дорзальные усики) или нижнюю, брюшную сторону головы (вентральные усики). В редких случаях усики помещаются на конце особых боковых выростов ' головы (род Dendrotiori). У некоторых форм (некоторые представители сем. Cymothoidae) обе пары усиков редуцированы, и нет ясного разделения их на ножку и жгутик, а имеются просто членистые придатки.
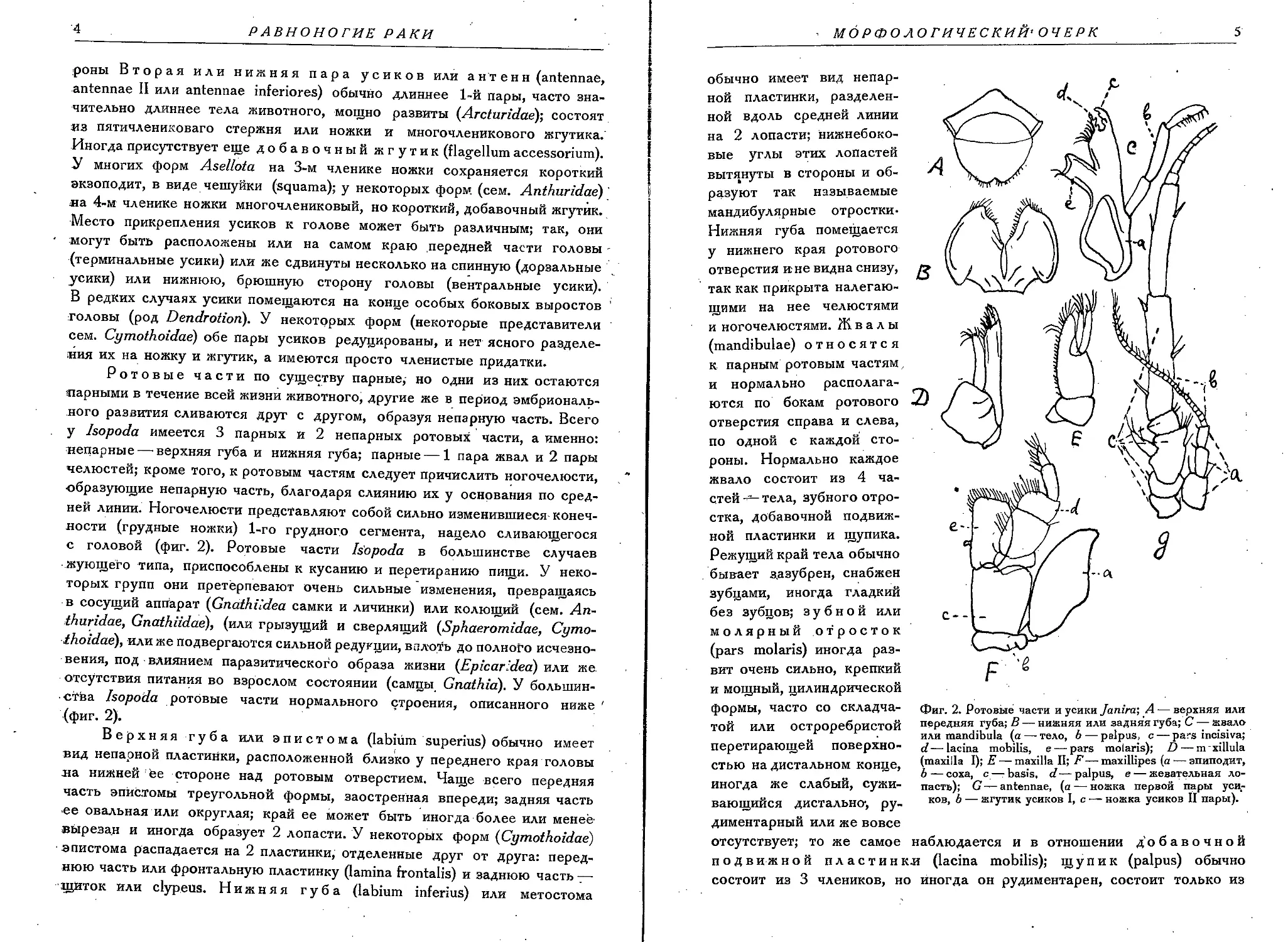
Ротовые части по существу парные,- но одни из них остаются парными в течение всей жизни животного, другие же в период эмбрионального развития сливаются друг с другом, образуя непарную часть. Всего у Isopoda имеется 3 парных и 2 непарных ротовых части, а именно: непарные — верхняя губа и нижняя губа; парные — 1 пара жвал и 2 пары челюстей; кроме того, к ротовым частям следует причислить ногочелюсти, образующие непарную часть, благодаря слиянию их у основания по средней линии. Ногочелюсти представляют собой сильно изменившиеся конечности (грудные ножки) 1-го грудного сегмента, нацело сливающегося с головой (фиг. 2). Ротовые части Isopoda в большинстве случаев • жующего типа, приспособлены к кусанию и перетиранию пищи. У некоторых групп они претерпевают очень сильные изменения, превращаясь в сосущий аппарат (Gnathiidea самки и личинки) или колющий (сем. Anthuridae, Gnathiidae), (или грызущий и сверлящий (Sphaeromidae, Cymothoidae}, или же подвергаются сильной редукции, вплоть до полного исчезновения, под влиянием паразитического образа жизни (Epicaridea) тилм же отсутствия питания во взрослом состоянии (самцы Gnathiai). У большинства Isopoda ротовые части нормального строения, описанного ниже ' (фиг. 2).
Верхняя губа или эпистома (labium superius) обычно имеет вид непарной пластинки, расположенной близко у переднего края головы ла нижней be стороне над ротовым отверстием. Чаще всего передняя часть эпйс.томы треугольной формы, заостренная впереди; задняя часть ее овальная или округлая; край ее может быть иногда более или менее-вырезан и иногда образует 2 лопасти. У некоторых форм (Cymothoidae) эпистома распадается на 2 пластинки, отделенные друг от друга: переднюю часть или фронтальную пластинку (lamina frontalis) и заднюю часть — щиток или clypeus. Нижняя губа (labium inferius) или метостома
- МОРФОЛОГИЧЕСКИЙ’ 04ЕРК
5
обычно имеет вид непарной пластинки, разделенной вдоль средней линии на 2 лопасти; нижнебоковые углы этих лопастей вытянуты в стороны и образуют так называемые мандибулярные отростки-Нижняя губа помещается у нижнего края ротового отверстия ине видна снизу, так как прикрыта налегающими на нее челюстями и ногочелюстями. Ж в а л ы (mandibulae) относятся к парным ротовым частям и нормально располагаются по бокам ротового отверстия справа и слева, по одной с каждой стороны. Нормально каждое жвало состоит из 4 частей тела, зубного отростка, добавочной подвижной пластинки и щупика. Режущий край тела обычно бывает з.азубрен, снабжен зубцами, иногда гладкий без зубцов; зубной или молярный отросток (pars molaris) иногда развит очень сильно, крепкий и мощный, цилиндрической
Фиг. 2. Ротовые части и усики Janira', А — верхняя или передняя губа; В — нижняя или задняя губа; С — жвало или mandibula (а — тело, 6 — palpus, с — pars incisiva; d~ lacina mobilis, e — pars molaris); D — m xillula (maxilla I); E— maxilla П; Г—• maxillipes (a •— эпиподит, b — coxa, c—- basis, d — palpus, e — жевательная лопасть); G — antennae, (a — ножка первой пары усиков, b — жгутик усиков I, с —• ножка усиков II пары).
формы, часто со складчатой или остроребристой перетирающей поверхностью на дистальном конце, иногда же слабый, суживающийся дистально-, рудиментарный или же вовсе
отсутствует; то же самое наблюдается и в отношении добавочной
подвижной пластинка (lacina mobilis); щупик (palpus) обычно состоит из 3 члеников, но иногда он рудиментарен, состоит только из
б
РАВНОНОГИЕ РАКИ
2 или даже 1 членика или же отсутствует вовсе. У самцов рода Gnathia жвалы, прикрепляясь по бокам ротового отверстия, вытянуты вперед далеко за пределы переднего края головы и образуют настоящие клещи, смыкаясь по средней линии. Зубной отросток у них иногда имеется в виде небольшого бугра на наружном крае. Щупик отсутствует; режущий край или гладкий, или иногда слегка зазубрен. Первая пара челюстей (maxillulae). Каждая из них состоит из трехчленикового основания, так называемого протоподита (protopoditus), и 2 лопастей, наружной и внутренней; наружная ветвь или лопасть представляет собой сохранившийся эндоподит; а внутренняя лопасть — просто расширенный вырост членика протоподита; экзоподит отсутствует. Обе лопасти обычно снабжены' на внутреннем и верхнем краях щетинками или более или менее крепкими, часто зазубренными, шипами. Вторая пара челюстей (maxillae, maxillae И). Каждая из них состоит из трехчленикового основания или базальной части, протоподита и 3 лопастей. Самая внутренняя лопасть представляет собой вырост последнего членика протоподита; обе наружные лопасти образованы вторичным расщеплением на 2 части эндоподита; наружная ветвь или экзоподит отсутствует вовсе. Все 3 лопасти обычно снабжены простыми или перистыми щетинками.
Ногочелюсти (maxillipes, maxillipedes) образуют непарную ротовую часть, прикрывающую снизу все другие ротовые части. Ногочелюсти образованы 1-й парой грудных ножек? слившихся у основания вместе и образовавших непарную добавочную ротовую часть. Ногочелюсти состоят из 3 основных частей — базальной части, щупика и эпиподита. Базальная часть нормально состоит из 3 члеников (Н. Hansen считает, что 1-й членик— это ргаесоха, 2-й членик образовался в результате слияния в один соха и epipoditus, а 1-й членик — результат слияния basis с endopo-ditus); последний членик базальной части на внутреннем верхнем угле образует большую жевательную лопасть (lobus masticatorius); щупик (palpus) хорошо развит и состоит из различного числа члеников; максимальное число члеников щупика ногочелюстей 5. У основания базальной части, прикрепленной к ее коксальному (2-му) членику, обычно сохраняется хорошо развитый эпиподит (epipoditus) в виде плоской закругленной, треугольной, или неправильной формы пластинки, которая прикрывает собою наружные части всех подлежащих ротовых частей. У подотряда Gnathiidea, благодаря полному слиянию с головой и 2-го грудного сегмента, очень сильно изменена также 2-я пара грудных ножек, приобретая сходство с ногочелюстями. Ножки эти, носящие названия pylopoda, с сильно расширенным 1-м члеником, как крышечкой прикрывают снизу все ротовые части и ногочелюсти. Обычно (у рода Gnathia) pylopoda состоят из 3 члеников — сильно расширенного 1-го членика, образованного слиянием базального и ишиального члеников, маленького морального и очень маленького конечного членика; иногда конечный членик сильно 6
МОРФОЛОГИЧЕСКИЙ ОЧЕРК
7
редуцирован и отсутствует вовсе или представлен в виде простого бугорка на 2-м членике. Внутренний край 1-го членика аркообразно изогнут и усажен перистыми щетинками; наружный край его обычно прямой. У рода Paragnathia pylopoda состоят из 5 расширенных члеников (базального, ишиального, морального, карпального и проподиаль-ного), снабженных щетинками.
У некоторых групп Isopoda все ротовые части очень сильно уклоняются от нормального жующего типа, образуя сосущий и колющий аппарат; у многих отдельные ротовые части или все они целиком редуцируются. Так, у самОк и личинок Gnathiidea и у сем. Anthuri-dae все ротовые части сильно вытягиваются вперед, стилетообразны, с острыми колющими кончиками и пиль-чато зазубренными краями; сложенные вместе они образуют ротовой конус, приспособленный для сосания; на внутренней стороне ротовых частей имеются желоба, по которым стекает высасываемая жидкость. У взрослых самцов Gnath.idae, которые совсем не питаются, жвалы образуют клещи впереди головы; верхняя губа отсутствует, челюсти 1-й пары срастаются вместе; челюсти 2-й пары сохраняются лишь в виде маленьких рудиментов. У самок Gna-thiidea жвалы также срастаются вместе, челюсти 1-й пары отсутствуют, а челюсти 2-й пары рудиментарны. У некоторых Gnathiidea (Paragnathia) редуцируются ногочелюсти (фиг. 3). У Ер'са-ridea верхняя и нижняя губы срастаются -вместе, образуя бугорок или конус
е
Фиг. 3. Видоизмененные ротовые части; А — Calathura (а — общий вид с нижней стороны ногочелюстей, жвал й челюстей, 6 — жвало, с — maxilla 1, d — нижняя губа); В— Gnathia (а — жвалы самца; Ъ — самки); С — No-tophryxus с нижней стороны, ротовая область.
с отверстием на конце. Из этого отверстия торчат концы жвал, сильно зазубренные по краям. Челюсти 1-й пары отсутствуют, челюсти 2гй пары рудиментарны или тоже исчезают. Ногочелюсти рудиментарны или отсутствуют. Таким образом, у паразитических Isopoda редукция ротовых частей
доходит до того, что сохраняются лишь эпи- и метастома.
8
РАВНОНОГИЕ РАКИ
Грудной отдел. Грудь или мезозома (thorax) нормально состоит и® 7 свободных подвижно-сочлененных друг с другом сегментов; 1-й грудной сегмент, как правило, сливается с головой у всех Isopoda, поэтому счет сегментам ведется со следующего (2-го) сегмента. Таким образом 1-м сег- . ментом называют 1-й свободный сегмент; хотя иногда и этот сегмент (2-й) сливается с головой (сем. Gnathiidae и Ischnomesidae), но он сохра- , няет наименование „первого грудного сегмента". Очень часто отдельные сегменты или группы сегментов грудного отдела срастаются вместе; слия- 1 ние сегментов друг с другом наблюдается или только посредине на спинной стороне, или же по всему краю сегмента, со следами шва на месте слияния или вовсе без всяких видимых следов слияния. У многих форм1 (Asellota) грудной отдел распадается на 2 участка: передний участок обычно образован 4 передними, сходными по форме сегментами; задний участок в таком случае состоит из 3 последних грудных сегментов, имеющих резко отличную от передних сегментов форму. Например, у сем. Мип-nidae передний отдел мезозомы образован 4 широкими передними сегмен- ' тами, имеющими вытянутые вперед боковые участки, а задний отдел состоит из 3 узких и коротких маленьких последних сегментов груди. У рода Ilyarachna, наоборот, 4 передние сегмента груди более слабые и короткие, чем 3 задних грудных сегмента. Иногда последний (7-й грудной), сегмент рудиментарен (сем. Anthuridae, Gnathiidae)', часто наблюдается ' слияние последнего торакального сегмента с брюшным отделом. Каждый грудной сегмент несет пару одноветвистых конечностей — грудных ножек. Нормально 1-й членик грудной конечности (соха) превращен в широкую плоскую пластинку (эпимер ы; боковые, коксальные или плевральные пластинки). Эпимеры подвижно сочленяются с сегментом, занимая весь, боковой край сегмента или только его часть. 1-я пара эпимер, как правило, срастается с 1-м сегментом без следов шва, но в некоторых случаях, например, у Synidothea epim.era.ta, свободна. У многих форм (род Synidothea и др.) все эпимеры сливаются со своими сегментами без видимых следов слияния, образуя так называемые „плевральные расширения" грудных сегментов.
Г рудные ножки (thoracopoda, peraeopoda, pereiopoda). Нормально грудных ножек, соответственно числу свободных грудных сегментов» 7 пар. У подотряда Gnathiidea только 5 пар, так как передняя пара превращается в pylopoda и присоединена к голове вследствие слияния с головой и 2-го грудного сегмента, а последняя пара, вследствие редукции последнего грудного сегмента, отсутствует вовсе. Все грудные ножки одноветвистые, так как экзоподит не развивается. Каждая ножка состоит из 7 отдельных члеников; 1-й членик (соха), уплощаясь, превращается, как было указано выше, в эпимеральную или плевральную пластинку, или же целиком сливается с телом сегмента, образуя его плевральное расширение.
Только у очень немногих Isopoda (Phreotoicidae и некоторые Asellota) соха существует как свободный членик ножки. Каждая грудная ножка,
МОРФОЛОГИЧЕСКИЙ. ОЧЕРК.
9 >
таким образом, нормально состоит из 6 дистальных члеников; из которых каждый или просто нумеруется II, III, IV и т. д., или же носит свое название (basipodit или basis, ischium, merus, carpus, propodus и dactylus или endopodit). Последний членик часто называется просто коготком и часто • снабжен еще добавочным когтем. У самок у основания грудных ножек прикрепляется еще особая, усаженная по краям волосками и щетинками, . пластинка, оостегит или инкубаторная пластинка. Число оосте-гитов у разных форм Isopoda различное, от 7 до 3 пар; складываясь вместе, оостегиты образуют инкубаторную или выводковую камеру (marsupium), куда самка откладывает яйца и где первое время остается вылупившаяся из них молодь.
В зависимости от образа жизни у разных групп Isopoda грудные ножки имеют различное строение; так бывают ноги ходильные, с удлиненными в общем цилиндрическими или слегка уплощенными члениками; , бывают ноги хватательные с сильно расширенным предпоследним члеником, к которому пригибается серповидно изогнутый коготок (dactylus), так что вместе они образуют нечто в роде клешни или ложную клешню (subchela); может быть и настоящая клешня (chela у рода Katianira). У некоторых форм (Cymotholdae и Epicar idea) ноги снабжены крепкими шипами.. и последний членик превращается в мощный изогнутый крючок; при. помощи таких ног рачок цепляется за кожу животного, на котором он паразитирует. У настоящих паразитов (Epicaridea) ноги сильно редуцированы, превращены в крючкообразные придатки или отсутствуют вовсе. У форм пелагических ноги плавательного типа, с сильно расширенными в виде пластинки последними члениками, которые по краям усажены длинными перистыми плавательными щетинками (сем. Munnopsidae). У некоторых форм грудные ножки (4 передних пары) усикообразные, . с удлиненными узкими члениками, усаженные длинными щетинками (сем. Arcturidae). Иногда у одной и той же формы одновременно грудные ножки разных типов; например, у Munnopsis 4 передние пары грудных ног ходильные, а 3 задние пары плавательные; часто передняя пара ног хватательная и снабжена ложной или настоящей клешней, а остальные простые,... ходильные; у Arcturidae первые 4 пары ног усиковидные, а 3 последних пары ходильные и т. п. (фиг. 4). Нормально членики грудных ножек снабжены всякого рода шипами, отростками, волосками и т. п. вооруже- нием; почти всегда задние пары ног бывают несколько длиннее передних, * но иногда только одна из пар грудных ножек отличается заметно по своей длине и крепости от других.
Брюшной отдел. Брюхо, абдомен или метазома (abdomen, metasoma) • короткое; нормально состоит из 6 сегментов; обычно передний абдоми--нальный сегмент срастается с задним грудным сегментом. Хвостовая, пластинка или т е л ь з он (telson) только в редких случаях (сем. Anthu-ridae) остается свободной, в подавляющем же большинстве случаев она . срастается с последним абдоминальным сегментом, образуя общий, так.
10
РАВНОНОГИЕ РАКИ
называемый каудальный сегмент. Отдельные брюшные сегменты обычно сливаются друг с другом и число их редуцируется. Иногда все брюшные сегменты и тельзон сливаются вместе в один каудальный сег-мент (например, у родов Synidothea, Pentias) и только по бокам этого ;каудального сегмента у его основания сохраняются швы, указывающие на слияние сегментов друг с другом. Боковые части брюшных сегментов часто снабжены расширениями или лопастями. Нормально брюшко несет € пар конечностей — 5 пар брюшных или плавательных ног и 1 пару рулевых ног.
Фиг. 4. Типы грудных ног: 7 — цепляющаяся нога (Aega psira); 2 — ходильная ножка (Aega psora) (а — basis, Ъ — ishium, с — merus, d— carpus, e — propodus, f—-dactylus); 3—хватательная нога (pleurogonium). снабженная ложной клешней; 4 — плавательная нога (Ilyarachna); 5— видоизмененная 1-я пара грудных ног Gnathia (левая pylopoda).
Брюшные ножки (pleopoda) нормально в числе 5 пар, но у различных групп число их вариирует; у паразитических форм (Epicaridea) брюшные ножки большей частью исчезают нацело. Каждая брюшная ножка состоит из стержневой части или ножки (sympodit) и 2 в е т-в е й —н аружной(экзоподит) и внутренней (эндо подит). Стержневая часть состоит обычно из 2 члеников, но у некоторых форм, например у Bathynomus, имеется praecoxa (по Hansen’y) и ножка трех-члениковая. Ветви (rami) обычно одночлениковые, но иногда одна из них двучлениковая, в редких случаях многочлениковая (фиг. 5). Брюшные ножки служат, главным образом, для дыхания, редко для плавания. В тех случаях, когда брюшные ножки несут локомоторную функцию, по краям ветвей имеются так называемые маргинальные перистые щетинки. В тех случаях, когда брюшные ножки несут функцию респираторную, они жаберного типа или же снабжены воздушными камерами. Обычно ветви имеют пластинчатую форму, и одна ветвь налегает сверху на другую. При жаберном дыхании по краям ветвей, несущих эту функцию, нет маргинальных щетинок, консистенция жаберной пластинки
МОРФОЛОГИЧЕСКИЙ ОЧЕРК
11
мясистая и стенки часто снабжены кровеносными лакунами и поперечными складками на верхней поверхности;» иногда часть ветви расщепляется, образуя пучки жаберных нитей у своего основания (Bathynomus). При воздушном дыхании ветви брюшных ножек снабжены так называемыми белыми телами или псевдотрахеями. В одних случаях все брюшные ножки жаберные (Cymothoidae), в других наблюдается специализация не только
Фиг. 5. Плавательные или брюшные ножки (pleopoda): 7 — вторая пара pleopoda Idothea $ (а — мужской придаток); 2 — пятая пара pleopoda Idothea-, 3 — первая пара pleopoda Janita $ (operculum и сложный копулятивный аппарат); 4 — Operculum Janira у ; 5— pleopoda с жаберными отростками (Bathynomus),
различных пар брюшных ножек, но и различных ветвей одной и той же пары. Так, у Sphaeromidae обычно 4-я и 5-я пара брюшных ног респираторные, а первые 3 пары локомоторные. У многих внутренняя ветвь мясистая и несет функции дыхания, а наружная ветвь или плавательная, снабженная маргинальными щетинками, или же служит крышечкой, защищающей лежащие под ней дыхательные ветви (Sphaeromidae, Onis-coidea). У некоторых форм (Anihuridae, Gnathiided) обе ветви всех брюшных ног несут как респираторную, так и локомоторную функции. Часто одна из пар брюшных ног, срастаясь по средней линии, образует тонкую широкую пластинку или крышечку (operculum), при
12
РАВНОНОГИЕ РАКИ
крывающую снизу остальные брюшные ножки (Asellota). Строение и форма: крышечки у самца и самки различны; у самки она обычно в виде простой цельной более или менее выпуклой округлой пластинки, у самца — узкая,, вытянутая, частично расщепленная вдоль по средней линии, вооруженная щетинками, отростками и пр. По Hansen’y operculum самки Asellota образована слиянием 2-й пары брюшных ног, a operculum самца — слиянием 1-й пары брюшных ножек. Обычно 2-я пара брюшных ног несет мужской придаток (у самцов) в виде длинного узкого отростка или щетинки (appendix masculina). У Asellota ветви второй пары брюшных, ног образуют сложный копулятивный аппарат.
Рулевые ноги (uropoda) имеются у равноногих раков в числе только одной пары; у многих форм они сильно редуцированы’ или отсутствуют. Нормально каждая из рулевых ног состоит из основного членика (basis) и 2 ветвей. Иногда сохраняется только одна ветвь, или их нет совсем; сами ветви могут быть одно- или двучлениковыми и претерпевают различную степень редукции, иногда сливаясь с основным члеником.. У некоторых групп (Sphaeromidae) одна ветвь неподвижно прикреплена и только одна свободно-подвижна и может заходить под неподвижно' укрепленную ветвь. Сами ветви или расширенные в виде пластинок,, или же узкие цилиндрические, палочкообразные. Существует два основных типа., прикрепления рулевых ног к последнему абдоминальному сегменту — боковой или латеральный и концевой или терминальный. Кроме того, они могут быть расположены или на брюшной стороне (брюшные или вентральные рулевые ножки) или на спинной (спинные или дорзальные). При боковом, (латеральном) типе прикрепления место сочленения основного членика рулевых ног с последним абдоминальным (каудальным) сегментом находится по бокам этого последнего сегмента близ его основания. В таком: случае они образуют вместе с последним сегментом брюшка хвостовой веер (подотряд Flabellifera и подотряд Gnathiidea) или в виде двух створок прикрывают снизу брюшные ножки (подотряд Valviferd). У некоторых, (сем. Anthurldae) прикрепленные по бокам свободного тельзона рулевые ноги образуют чашечку, так как наружная ветвь uropoda сильно сдвинута на спинную сторону и нависает над основанием тельзона и внутренними: ветвями (фиг. 6). При концевом (терминальном) типе прикрепления рулевые ноги располагаются на заднем крае последнего абдоминального сегмента-
Хвостовая пластинка или тельзон (telson) у всех Isopoda,. за исключением сем. Antharidae, целиком срастается с последним брюшным сегментом, образуя вместе с ним очень характерный терминальный или каудальный сегмент. Края этого сегмента или тельзона часто бывают зазубрены, с зубцами, вырезками, отростками, шипами и пр.
КРАТКИЙ ОЧЕРК ЭКОЛОГИИ И БИОЛОГИИ ISOPODA
Почти все Isopoda животные водные, и подавляющее большинство’ их морские. По своей экологии они сильно отличаются друг от друга и,
ОЧЕРК ЭКОЛОГИИ И БИОЛОГИИ
13
•обладая широкой приспособляемостью, образуют ряд экологических групп. Так, среди них есть и свободно живущие, и паразитические формы, и бентонические, ползающие по дну, и пелагические, плавающие в толще воды или на поверхности; среди них встречаются и формы, зарывающиеся глубоко в грунт, и формы, сверлящие дерево, и т. д. Многие виды держатся среди зарослей водорослей, под камнями на отливе; иногда живут в пустых раковинах, в трубках червей, или строят свои домики.
Кроме чисто морских и типично пресноводных форм, существуют формы, населяющие переходную область — эстуарии рек, прибрежные опреснённые места и т. п. Многие из них способны переносить значитель-
"Фиг. 6. Типы прикрепления рулевых ног: 7 — латеральный тип (Aega psora) — рулевые ноги и тельзон образуют хвостовый веер; 2 — латеральный тип (Idothea) — рулевые ноги прикрывают pteopoda снизу в виде двустворчатой крышечки (вид снизу; левая uropoda снята, чтобы показать прикрытые ею плавательные ножки. 3— тоже у Arcturus (вид сбоку) -4— терминальный тип прикрепления (у Janira); 5 — латеральный тип—рулевые ноги и тельзон образуют чашечку (Anthuridae).
ные и резкие колебания солености, как, например,/него albifrons, которая в большом количестве встречается в устьях рек, где приливо-отливные колебания солености очень велики (от 33°/00 до 0) и, следовательно, амплитуда суточных колебаний солености достигает 33°/00, причем колебания эти отличаются крайней резкостью, почти мгновенностью.
Равноногие раки заселяют самые разнообразные глубины, начиная от глубин, исчисляемых километрами, й кончая приливо-отливной зоной. Некоторые формы, например Mesidothea entomon, заселяя эстуарии крупных •рек, могут, однако, далеко подниматься вверх по реке (главным образом, цепляясь за кожу рыб и передвигаясь вместе с ними), заходить в притоки этой реки и встречаться на расстоянии более тысячи километров от устья.
14
РАВНОНОГИЕ РАКИ
Многие живут в области переходной к наземной, например, род Llgia, которая массами держится в зоне прибоя выше линии максимального прилива; наконец, имеется ряд форм чисто сухопутных, так называемые мокрицы (Oniscoidea), которые (РогсеШо и др.) живут большей частью на земле во влажной среде под камнями, листьями, досками и пр., а также в жилых домах, но некоторые из них (Hemilepistus) живут в сухих пустынях.
В подавляющем большинстве случаев Isopoda раздельнополы, но есть и гермафродиты (сем. Cymothoidae)-, половой диморфизм, за исключением подотрядов Gnathiidea, Epicaridea и некоторых представителей сем. Sphae-romidae, выражен очень слабо. Самцы, однако, значительно крупнее самок; у форм паразитических, наоборот, огромная бесформенная самка, совершенно утратившая внешнее сходство с раками, и маленький сегментированный, сохранивший общие черты ракообразного, самец. Самки вынашивают яйца и молодь в выводковых сумках (инкубаторных камерах).
У всех Isopoda, за исключением Gnathiidea и Epicaridea, все развитие происходит внутри яйца, из которого вылупляется уже почти готовое, сформировавшееся животное, с полной сегментацией тела и конечностями; вылупившаяся молодь живет еще некоторое время (у различньтх форм продолжительность жизни различная), в выводковой камере матери. У подотряда Gnathiidea наблюдается метаморфоз с несколькими личиночными стадиями; у подотряда Epicaridea метаморфоз регрессивный.
КРАТКИЙ ЗООГЕОГРАФИЧЕСКИЙ ОЧЕРК ISOPODA БЕРИНГОВА, ОХОТСКОГО И ЯПОНСКОГО МОРЕЙ
Весь материал по дальневосточным морям по группе равноногих раков, хранящийся в коллекциях Зоологического института Академии Наук, обработанный мною и легший в основу предлагаемой сводки, охватывает богатые сборы и коллекции, собранные как различными экспедициями, так и отдельными лицами. Важнейшие из этих коллекций представлены сборами И. Вознесенского 1845—1846 гг.; д-ра Безе 1866 г.; д-ра Полякова 1881—1883 гг.; д-ра Слюнина 1886—1903 гг.; д-ра Зандера 1890 г.; д-ра Бунге 1896 г.; Янковского 1899 г.; Б. Гейнемана 1907 г.; Бражникова 1907—4908 гг.; Пальчевского 1908 г.; Н. Смирнова и Бегака 1907 г.; П. Ю. Шмидта 1900—1901 гг. и 1929 г.; д-ра Холодного 1909 г.; д-ра Арнгольда и д-ра Старокадомского во время экспедиции Вилькицкого 1910—1913 гг.; Ф. Дербека 1909—1912 гг.; Белоусова 1913 г.; В. Солдатова и Павленко 1910—-1912 гг.; Павленко 1913—1915 гг.; Гидрографической экспедиции Восточного океана 1912—1917 гг.; Рожковского в 1917 г.; Тихоокеанского института рыбного хозяйства за период 1925—1932 гг.; экспедиций Государственного гидрологического института (ГГИ) в течение 1925—1933 гг.; Е. Гурьяновой за 1930—1932 гг.; сборы Камчатской станции Государственного гидрологического института
ЗООГЕОГР АФИЧЕСКИЙ ОЧЕРК-
15
в 1933 г. и некоторые другие. В результате обработки этих обширных материалов число зарегистрированных ранее для Берингова, Охотского и Японского морей видов Isopoda резко увеличилось ив настоящее время, насчитывает уже 127 видов. Поотдельным морям эти формы распределены таким образом: в Беринговом море имеется 54 вида,,в Охотском 47,, в Японском море 62 формы. Прежде чем дать общую зоогеографическую характеристику фауны морских Isopoda Дальнего Востока, рассмотрим ее по отдельным морям. Следует помнить, что это лишь первая попытка подвести итоги тому немногому, что мы знаем о морской фауне равноногих раков Дальнего Востока и что провести углубленный зоогеографический анализ ее в настоящий момент невозможно: слишком мало мы еще знаем условия жизни в наших дальневосточных морях, слишком малы и скудньп наши сборы по их фауне. Наименее изучено до сих пор Охотское море, более других известна фауна моря Японского. Однако, экспедиции ГГИ за последние годы произвели исследования максимальных глубин Охотского и Японского морей, а в Беринговом море сборы бентоса производились до глубины почти 4000 м. Таким образом, в наших коллекциях, были сборы как береговые, литоральные, мелководные, так и сборы с глубин 3500—4000 м.
Берингова море. В этом море в настоящее время зарегистрировано 55 следующих видов Isopoda:
Подотряд ASELLOTA
Сем. Janiridae 1. Janira erostrata (Rich.) 2. J. holmesi (Rich.) 3. /. sarsi (Rich.) 4. Janiropsis kincaidi Rich. 5. J. derjugini Gurjan. 6. Microprotus coecus Rich. Сем. Cymothoidae. 12. Aega symmetrica Rich. 13. A. magnoculis Rich. 14. Rocinella cornuta Rich. 15. R. belliceps (Stimpson). 16. R. angustata Rich. Сем. Sphaeromidae 17. Limnoria lignorum (Rathke). Сем. Idotheidae 24. Idothea ochotensis Brandt 25. Z. aleutica Gurjan. Сем. Munnidae 7. Munna stephenseni Giirjan. 8. M. arnholdi Gurjan. 9. M. avatshensisGurjan Сем. Munnopsidae 10. Munnopsurus giganteus giganteus (G-, Sars) 11. M. laevis (Rich.) Подотряд FLABELLIFERA 18. Tecticeps renoculis Rich. var. . marginalis . Gurjan. 19. Г. alascensis Rich. , 20. C.licaea cordata Rich. 21. Exosphaeroma amplicauda (Stimps.) 22. Neosphaeroma oregonensis (Dana) Сем. Anthuridae.. 23. Calathara brachiata (Stimps) Подотряд VALVIFERA 26. Pentidothea wosnesenskii (Brandt) 27. P. whitei (Stimps. ) 28. Mestldothea entomon (L.) subsp. orientals Gurjan..
16
РАВНОНОГИЕ РАКИ
29. Synidothea bicuspida (Owen) . 30. Л marmorata (Packard)
31. б', nebulosa Bened.
32. б', acuta Rich.
. 33. б', nodulosa (Kr.)
34. 6". laevis Bened.
35. б', spinosa Gurjan.
36. б'. picta Benedict
Сем. Arcturidae
.. 37. Arcturus longispinis Benedict
38. A, diversispinis Rich.
39. A. magnispinis Rich.
40. A, brevispinis Rich.
41. A. beringanus Bened.
42. A. glabrus Bened.
43. A. tritaeniatus Rich.
44. A. hirsutus Rich.
45. Pleuroprion murdochi (Bened.)
46. P. intermedium (Rich.)
Подотряд ONISCOIDEA
Сем. Ligiidae Сем. Trichoniscidae
47. Ligia pallasi Brandt 49. Trichoniscus papillicarnis Rich.
- 48. Ligidium tenue Budde-Lund. Сем. Oniscidae
50. Porcellio scaber Latreille
'Подотряд EPICARIDEA
Сем. Bopyridae Сем. Dajidae
. 51. Ph'yxus abdominalis (Kr.) 54. Holophryxus giart/i Rich.
.. 52. Argeia pugettensis Dana 55. Arthrophryxus beringanus Rich.
. 53. Bopyroides hippolytes (Kr.)
Рассматривая этот список с точки зрения зоогеографической, можно легко видеть, что состав фауны Isopoda Берингова моря довольно пестр и в общих чертах состоит из различных групп. Основу фауны Isopoda составляют виды, известные до сих пор только из Берингова моря; условно назовем эту группу эндемиками или аутохтонами Берингова моря. „Эндемиков" в Беринговом море 19, а именно: Janira holmesi, Janiropsis derju-gini, Microprotus coecus, Munna arnholdi, M. stephenseni, Tecticeps reno-culis marginalis, Synidothea acuta, S. laevis, S. picta, Arcturus longispinis, A. diversispinis, A. magnispinis, A. brevispinis, A. glabrus, A. tritaeniatus, A. hirsutus, Pleuroprion intermedium, Ligidium tenue и Holophryxus giardi. Все эти формы, кроме Arcturus hirsutus и Holophryxus giardi, более или менее мёлководные, могли выработаться в самостоятельные виды в особых физико-географических условиях Берингова моря и их аутохтонный характер весьма вероятен. Две же другие формы, A. hirsutus и Н giardi, принадлежат к глубоководной фауне, добыты на глубинах свыше 2000 м и, вероятно, являются представителями абиссальной фауны северной части Тихого океана, так как Н. giardi паразитирует на абиссальной тихоокеанской креветке, а Н. hirsutus . по строению тела и тельзона относится «к группе видов Arcturus, добытых экспедицией „Challenger" в абиссальной зоне Тихого океана.
Вторую группу образуют виды, встреченные лишь в Беринговом ..и Охотском морях; их всего 5.форм, а именно: Janira sarsi, Tecticeps «alascensis, Idothea aleutica, Synidothea nebulosa, S. spinosa, Pentidothea
ЗООГЕОГ Р АФИЧЕСКИИ ОЧЕРК
17
’uiosnesenskii. Характерно, что все это также формы прибрежные, главным образом мелководные и, вероятно, должны быть отнесены к аутохтонам мелководий азиатского побережья северной части Тихого океана.
В третью группу входят виды, встречающиеся во всех трех дальневосточных морях; таких видов всего 6: Janira erostrata, Rocinela angustata, R. belliceps, Neosphaeroma oregonensis, Idothea ochotensis, Arcturus beringanus. Почти все они—. представители мелководной фауны и очевидно вообще характерны для западного побережья северной части Тихого океана, а оба, вида Rocinela и Neosphaeroma вообще широко распространены в этой части океана по обоим его берегам.
Имеются в Беринговом море также формы, общие с Японским морем, не найденные пока в холодном суровом Охотском море -- Janiropsis kin-caidi и Trichoniscus papillicornis. Возможно, однако, что отсутствие их в списках фауны Охотского моря объясняется лишь крайне плохой изученностью побережья этого моря, и в частности литоральной и супрали-торальной зон.
Особую группу образуют виды арктические, встречающиеся в Северном Ледовитом океане и в Беринговом море; таких видов 7 — Munnopsurus giganteus giganteus, Calathura brachiata, Synidothea bicuspida, S. marmo-rata, S. nodulosa, Pleuroprion murdochi и Mesidothea entomon, образующая в Тихом океане особый подвид М. entomon subsp. orientalis. Что касается Munnopsurus, Calathura и Mesidothea, то эти формы арктические по происхождению, тогда как Synidothea и Pleuroprion, повидимому, выходцы из Тихого океана, расселившиеся в Арктике (см. об этом ниже). Все они встречены или в северном районе Берингова моря, или же у его западных берегов. Кроме того, здесь имеются широко распространенные арктическо-бореальные формы {Bopyroides hippolytes) и формы с наклонностью тс космополитизму — Limnoria lignorum и Phryxus abdominalis.
Особый интерес представляет присутствие в Беринговом море группы типично-абиссальных видов северной части открытого Тихого океана: Munnopsurus laevis, Aega magnoculis, Arcturus hirsutus, Holophryxus giardi и Arthrophryxus beringanus. Присутствие этих форм в Беринговом море на глубинах свыше 1000—2000 м, указывает на открытую связь абиссальной фауны Берингова моря с абиссальной открытой частью Тихого океана.
Хорошо выражены в фауне Isopoda Берингова моря также и элементы бореальной Тихоокеанской фауны; таких видов в Беринговом море насчитывается 6: Pentidothea whitei, Exosphaeroma amplicauda, Calicaea cordata, Munnopsurus laevis, Janiropsis kincaidi, Neosphaeroma oregonensis, причем все они сконцентрированы в юговосточной части моря у Алеутской цепи и отсутствуют у Камчатки. Имеется одна форма типично субарктическая тихоокеанская, Argeia pugettensis, широко распространенная по обоим берегам Тихого океана, и ряд субарктических тихоокеанских 4»орм, преимущественно американского побережья — Rocinela cornuta, Гурьянова 2
18
РАВНОНОГИЕ РАКИ
Idothea aleutica, Pentidothea wosnesenskii, Aega symmetrica и Ligia pallasi.
Уже этот грубый анализ состава фауны Isopoda Берингова моря дает возможность заключить о существовании широкой связи абиссали: этого моря с абиссальной фауной Тихого океана и о существовании обмена фаунами между Беринговым морем и Арктикой, с одной стороны^ и между Беринговым и Охотским морем, с другой. Связь с глубинами Тихого океана осуществляется главным образом в районе Командор и так называемого Камчатского моря й через проливы между Алеутскими островами, так как все 5 абиссальных тихоокеанских форм добыты в этом районе Берингова моря (см. распространение этих форм в систематической части определителя).
Некоторые мелководные и прибрежные формы юго-восточной части Берингова моря (Алеутской цепи и побережья Аляски) проникают на западное побережье Камчатки в Охотское море через Первый Курильский пролив у мыса Лопатки. Idothea aleutica, Synidothea nebulosa — формы западно-американские, встречены вдоль побережья Алеутских островов,, на Командорах, в Авачинской губе на Восточной Камчатке и наконец у Большерецка на западном берегу Камчатки в Охотском море, и больше в Охотском море нигде не найдены.1 О том, что проникновение это идет: через Первый Курильский пролив, указывает нахождение Н. Смирновым, и Бегаком западноамериканской формы Janira sarsi как раз в самом проливе у Лопатки. Влияние фауны Охотского моря на фауну Берингова моря также имеет место; укажем хотя бы на существование в Беринговом море типично охотского вида Tecticeps renoculis, образовавшего, правда,, в условиях Берингова моря особукэ форму—-var. marginalis. Центр происхождения этого вида лежит несомненно в Охотском море, где имеется. 4 формы этого вида и другие близкие к нему виды Tecticeps.
Двусторонний обмен фаунами существует также и между Северным. Ледовитым океаном и Беринговым морем. Наличие в фауне Берингова моря типично арктических форм (Munnopsurus giganteus, Calathura hra-chiata, Mesidothea entomon) указывает на проникновение в Тихий океан' арктической фауны. С другой стороны, Тихий океан оказывает большое влияние на фауну Арктической области; это влияние выражается в проникновении через’Берингов пролив в Чукотское море и море Бофора некоторых видов Synidothea и Pleuroprion. Центр происхождения обоих этих родов, повидимому, лежит в северной части Тихого океана, так как из всех известных до сих пор видов рода Synidothea большинство (около 25) видов распространены в Беринговом, Охотском и Японском морях и в прилегающих частях Тихого океана, 2 вида известны из западного района северной части Атлантического океана и 3 вида из Арктики. Все три
1 На литорали Тауйской губы и Шантарских островов обычна Idothea ochotensis, и ни П. Ушаков, ни И. Закс, ни другие исследователи этих районов Охотского моря ни разу не добывали там Id. aleutica.
ЗООГЕОГРАФИЧЕСКИЙ ОЧЕРК
19
арктических вида Synidothea и оба- атлантических распространены, кроме того, и в Тихом океане.
Месторождения этих видов заставляют предполагать следующий путь проникновения тихоокеанских видов в Арктику. Пройдя Берингов пролив, тихоокеанские формы затем распространяются в Арктике по двум направлениям; одни идут вдоль Сибирского побережья на запад (Syn. bicuspida? Syn. nodulosa), проходят, распространяясь по мелководьям Восточносибирского, Лаптевых и Карского морей, до Новой Земли и Вайгача и проникают через проливы Югорский Шар и Карские Ворота в Печорский район Варенцова моря. Так, «S', bicuspida доходит до меридиана ст. Хар-лавки на восточном Мурмане и заходит в горло Белого моря, a «S', nodulosa поднимается вдоль побережья Новой Земли на север, становится циркумполярной, образуя ряд географических рас. Другие формы {Pleuroprion? S. marmorata, Arcturus baffini) идут на восток в море Бофора, проходят вдоль северного побережья Аляски и Канады и появляются у берегов Лабрадора, Баффиновой Земли, западной Гренландии, Исландии и Фарерских островов. Существованием этих двух путей распространения тихоокеанских форм в Арктике объясняется существование хиатуса в ареале распространения некоторых почти циркумполярных мелководных форм в районе между Гренландией, Исландией, Скандинавией и Шпицбергеном; обе ветви путей распространения тихоокеанских форм в Арктике, направляясь на восток и на запад, тем не менее не образуют замкнутого кольца, нарушая этим циркумполярность распространения в частности арктических видов Synidothea и Pleuroprion. '
Распространение в пределах Берингова моря форм арктических и форм тихоокеанских бореальных, заставляет выделить два обособленных друг от друга района: один район охватывает мелководную северную часть Берингова моря и восточное побережье Камчатки; здесь наблюдается арктический тип фауны Isopoda (присутствие настоящих арктических видов — Munnopsurus giganteus, Calathura brachiata, Mesidothea entomon и отсутствие типично бореальных тихоокеанских видов — Calicaea, Neo-sphaeroma, Exosphaeroma, Janiropsis, и др.); другой район—более тепловодный, охватывает Командорские острова, юговосточную часть Берингова моря и Алеутскую гряду; он характеризуется отсутствием форм арктических и присутствием типично бореальных видов Isopoda. Еще W. Dall в 1886 г. и 1899 г.1 на основании распределения Mollusca намечал такое районирование Берингова моря, а в 1933 г., А. Иванов,2 на основании распределения морского бентоса приходит к такому же выводу. Распределение Isopoda, таким образом, подтверждает выводы W. Dall и А. Ива
1 W. Dall. 1886. Report on Benns' Island Mollusca... Proc. U. S. Nat. Mus., 9.; W. Dall. 1899. The Mollusk Fauna of the Pribilof Islands. Report of Fur-Seal Investigations 1896—1897. part. 3.
' 2 Доклад А. Иванова в 1933 г. на научном совещании Гидробиологического совета
ГГИ о результатах работ экспедиции ГГИ в Беринговом море в 1932 г.
2
20
РАВНОНОГИЕ РАКИ
нова о существовании в Беринговом море двух зоогеографических про-винций — арктической Камчатской провинции и тепловодной Алеутской провинции, куда проникают как тепловодные абиссальные формы Тихого океана (Aega magnoculis, Arthrophryxus, Holophryxus, Munnopsurus laevis) особенно в районе Командор и Атту, так и ряд прибрежных американских форм, спускающихся на юг до южной Калифорнии (Cilicaea cordaia, Exosph. amplicauda, Pentidothea whitei, Ligia pallasi, Aega symmetrica, Janiropsis, виды Arcturus и пр.). Гидрологический режим этих двух районов Берингова моря совершенно различный, а распределение глубин, течений и льдов позволяют ясно нанести на карту границы этих двух провинций — более или менее мелководной замерзающей холодноводной Камчатской и глубоководной, никогда не замерзающей, отепленной глубинными теплыми тихоокеанскими водами Алеутской провинции.
Охотское море. В настоящее время в этом море зарегистрировано 47 форм Isopoda', формы эти следующие:
Подотряд ASELLOTA
Сем. Janiridae Сем. Munnopsidae
1. Janira erostrata (Rich.) 4. Munnopsurus giganteus giganteus (G.
2. Janira sarsi (Rich.) Sars)
3. Janira soldatovi Gurjan. 5. M. giganteus (G. Sars) ochotensis Gurjan-
6. Ilyarachna starokadomskii Gurjan.
Подотряд FLABELLIFERA
Сем. Anthuridae
7. Paranthura japonica Rich.
8. Cyathura carinata (Kr.)
9. Calathura brachiata (Stimps.)
Сем. Cymothoidae
10. Aega magnoculis Rich.
11. Rocinela maculata Sch. a. Mein.
12. R. belliceps (Stimps)
13. R. angustata Rich.
14. Livoneca raynaudii M. Edw.
15. L. amurensis (Gerstfeldt)
Сем. Sphaeromidae
16. Tecticeps alascepsis Rich.
17. T. renoculis Rich, renoculis Rich.
18. T. renoculis Rich, laevis Gurjan.
19. T. renoculis Rich, nodulosus Gurjan.
20. T. renoculis Rich, carinatus Gurjan.
21. T. leucophthalmus Gurjan.
Подотряд VALVIFERA
Сем. Idotheidae Сем. Arcturidae.
22. Mesidothea entomon (L.) orientalis Gurjan.
23. Idothea ochotensis Brandt.
24. /. aleutica Gurjan.
25. I. derjugini Gurjan.
26. I. fewkesi Rich.
27. I. orientalis Gurjan.
28. Pentidothea wosnesenskii (Brandt).
29. Synidothea bicuspida (Owen) lata Gurjan.
30. X nebulosa Bened.
31. X longicirra Gurjan.
32. Л tuberculata Rich.
33. 5L muricata (Harford).
34. A. spinosa Gurjan.
35. «5. cinerea Gurjan.
36. Arcturus baffini (Sab.).
37. A. ulbani Gurjan.
38. A. setosus Gurjan.
39. A. setosus Gurjan. acuticaudalis Gurjan..
40. A. setosus Gurjan. seminudus Gurjan.
41. A. crassispinis Rich.
42. A.’hastlger Rich.
43. A. beringanus Bened.
44. A. granulatus Rich.
45. A. beddardi Gurjan.
46. Pleuroprion hystrix G. Sars.
47. P. tarasovi Gurjan.
ЗООГЕОГ Р АФИЧЕСКИЙ ОЧЕРК
21
Отсутствие представителей 3 других подотрядов, а именно Gnathiidea, Epicaridea и Oniscoidea объясняется исключительно слабой изученностью фауны Охотского моря и особенно берегового населения и паразитов раков.
Рассматривая этот список, мы видим, что видовой состав фауны изопод Охотскойо моря отличается еще большей пестротою, чем в Беринговом море. Кроме тех групп, с которыми мы имели уже дело в Беринговом море; здесь появляются еще элементы иного зоогеографического характера. Прежде всего, следуя порядку, принятому для Берингова моря, укажу на существование в Охотском море аутохтонных эндемичных форм.1 Таких форм в настоящее время насчитывается 16, а именно: Munnopsurus giganteus var. ochotensis, Tecticeps renoculis renoculis, T. renoculis laevis, T. renoculis nodulosus, T. renoculis carinatus, T. leucophthalmus, Synidothea longicirra, S. tuberculata, S. cinerea, Arcturus ulbani, A. granulates, A. setosus, A. setosus seminudus, A. setosus acuticaudalis, A. beddardi, Pleuroprion tarasovi. To, что среди аутохтонов здесь имеются ряды подвидов й вариететов, указывает на сравнительную молодость фауны Охотского моря и на существование интенсивных процессов видообразования, сказывающихся на расщеплении местных видов на подвиды и морфы и на образований особых форм от видов, проникающих в Охотское море из других районов мирового океана (напр. Munnopsurus giganteus ochotensis)
Вторую группу составляют виды общие для Охотского и Берингова морей —15 видов, а именно: Janira sarsi, J. erostrata, Munnopsurus giganteus giganteus, Calathura brachiata, Rocinela belliceps, R. angustata, Aega magnoculis, Tecticeps alascensis, Mesidothea entomon orientalis, Idothea ochotensis, I. aleutica, Pentidothea wosnesenskii, Synidothea spinosa, S. nebu-lasa, Arcturus beringanus. Из них часть, как указывалось выше, проникла в Охотское море из Берингова моря (/. aleutica, Р. wosnesenskii, S. nebu-losan J. sarsi) по побережью Камчатки и дошла до Болыперецка по ее западному берегу через Первый Курильский пролив; часть вероятно прошла через другие, более глубокие проливы Курильской гряды {Calathura, Munnopsurus), а часть (Aega magnoculis, R. angustata) является глубоководными формами Тихого океана и через глубокие проливы проходит как в Берингово, так и в Охотское море.
Третью группу составляют виды общие для Охотского и Японского морей; таких форм 18: Janira soldatovi, J. erostrata, Ilyarachna starokadom-skii, Paranthura japonica, Rocinela maculata, R. belliceps, R. angustata, Tecticeps renoculis laevis, Idothea fewkesi, I. ochotensis, I. derjugini, I. orientalis, Synidothea bicuspida lata, Arcturus setosus seminudus, A. setosus acuticaudalis, A. beringanus, A. hastiger и A. crassispinis. Существование этих форм в обоих морях и их распределение в пределах того и другого
1 Эндемизм этот, конечно, следует понимать условно, так как весьма вероятно, что при дальнейшем изучении фауны дальневосточных морей часть этих видов окажется и в других районах Тихого океана.
22
РАВНОНОГИЕ РАКИ
моря с определенностью указывает на существование двустороннего обмена фаунами между Охотским и Японским морями;, мало того, анализ этого списка указывает, что имеются 2 пути, по которым происходит этот обмен. С одной стороны, он энергично совершается при помощи Лаперузова пролива, с другой же стороны—через пролив Невельского и Амурский лиман.
Так, Rocinela maculata, II. starokadomskii, Idothea fewkesi, I. orientalis, I. derjugini, S. bicuspida lata и A. crassispinis, будучи распространены широко в Японском море, являясь там банальными мелководными формами, проходят в Татарский пролив (см. их распространение в систематической части работы) и оттуда попадают в залив Анива и югозападную часть Охотского моря. В других районах Охотского моря эти формы, кроме I. derjugini и I. fewkesi, не были встречены. Ясно, что путь их проникновения из Японского в Охотское море лежит через Лаперузов пролив. С другой стороны, типично охотские формы, в изобилии заселяющие мелководья Охотского моря и в частности добытые в заливе Анива, напр. Tecticeps renoculis laevis, A. setosus seminudus и acuticaudalis и A. hastiger, встречаются в северной части Японского моря, главным образом в Татарском проливе. Надо думать, что формы эти проникли из Охотского моря в Японское через Лаперузов пролив.
Однако, несмотря на совершенно исключительные условия пролива Невельского, создаваемые эстуарными условиями Амурского лимана,3 и здесь наблюдается частичный обмен формами между Охотским и Японским морями. Так, Paranthura japonica, встречающаяся в эгтуариях рек, южного происхождения, добытая на западном берегу Охотского моря в бухте Нагайя и в Амурском лимане, несомненно проходит из Японского моря в Охотское через пролив Невельского. Точно также Idothea fewkesi» южная тепловодная форма, добытая кроме Японского моря также на Шантарских островах и у острова Лангр в Амурском лимане, несомненно проникла в Охотское море из Японского через пролив Невельского. С другой стороны, Idothea ochotensis, переносящая значительные колебания солености и сильное опреснение, широко распространенная в Охотском море, где она является банальнейшей литоральной формой (напр., на Шантарах), встречается также и в северной части Японского моря в Татарском проливе. Так как она добыта также и на острове Лангр в Амурском лимане, то естественно предположить, что I. ochotensis проникает из Охотского моря в северную часть Японского моря через пролив Невельского. Следовательно, имеется, хотя и слабый, но взаимный обмен фаунами между Японским и Охотским морями не только через цролив Лаперуза, но и через пролив Невельского. Затем имеется группа видов,
1 По мнению К. Дерюгина, Амурский лимаи создает гидрологическую пробку, непроходимую для морской фауны, и обмен формами между Охотским и Японским морями через пролив Невельского невозможен.
ЗООГЕОГРАФИЧЕСКИЙ ОЧЕРК 23
общих для всех трех дальневосточных морей (6 форм, см. выше в разборе фауны Берингова моря).
Особую группу составляют в Охотском море арктические виды; их 5, а именно: Munnospurus giganteus giganteus, который попадая в условия Охотского моря отщепляет еще особую местную форму (М. giganteus ochotensis), Calathura brachiata, Mesidothea entomon, образующая подвид orientalis, Synidothea muricata и Arcturus baffini. Первые 2 вида, очевидно, проникают, из Арктики в Охотское море через Берингово море а 2 последние, S- muricata и A. baffini, наоборот, из ТихоЬо океана попадают в море Бофора («S', muricata) и, распространяясь на запад, доходят до Карского моря (.S’. muricata), а распространяясь на восток вдоль берегов полярной Канады, появляются у Западной Гренландии, Исландии и Фарерских островов (A. baffini). Следующая группа — тихоокеанские абиссальные, виды, Aega magnoculis и вероятно Arcturus beddardi (пока отнесенный к группе эндемиков), добыты в южной части Охотского моря и проникли сюда, по всей вероятности, через глубокие проливы между южными островами Курильской гряды из открытой части Тихого океана. i В Охотское море проникают также и тепловодные почти тропические формы с космополитическим распространением—Cyathura carinata, найденная в устьи реки в бухте Нагаево. Имеется здесь и форма антарктическая— Livoneca raynaudii, добытая Вознесенским в 1847 г. у порта Аяна. Наконец, в Охотском море имеется форма и субтропического типа — Livoneca amurensis. Эта паразитическая изопода, распространенная в бассейне реки Амура, в бассейне Лефу и на Гималаях, спускается по Амуру в Амурский лиман. Особую группу представляет собою Pleuroprion hystrix, добытый у порта Аяна и известный ранее из северной части Атлантического океана. Следуя терминологии Л. С. Берга (1934), мы можем назвать ее формой амфибореальной.
Таким образом, фауна изопод Охотского моря, несмотря на ее весьма малую изученность, оказывается богатой и разнообразной. Особые, весьма суровые условия прибрежной и мелководной частей Охотского моря вырабатывают специфическую охотскую фауну; широкая связь через Курильские проливы с глубинами Тихого океана, обусловливает существование в абиссальной зоне Охотского моря глубоководных тихо-: океанских изопод. Обмен Охотского моря с фауной моря Японского приводит к обогащению Охотского моря тепловодными и япономорскими видами. Разнообразие физико-географических условий в пределах самого Охотского моря как в горизонтальном (в смысле районов), так и вертикальном (по глубинам) направлении вызывает расщепление охотских аутохтонов на мелкие таксономические единицы, подвиды, вариетёты, морфы. Расщепление на ряд форм Tecticeps renoculis (Rich.) и Arcturus setosus Gurjan. является ярким примером этого явления. Двойственная природа Охотского моря, в смысле гидрологического режима его верхних и глубоких частей, позволяет уживаться рядом формам типично арктиче-
24
РАВНОНОГИЕ РАКИ
сжим, холодноводным с формами более тепловодными, даже субтропичес^ кого типа.
Японское море. До сих пор в Японском море, за исключением субтропической части его у берегов Японии, зарегистрировано 66 форм, Isopoda, принадлежащие к 64 видам. Формы эти следующие:
Сем. Janiridae
1. Janira soldatovi Gurjan.
2. J. erostrata (Rich.).
3. Janiropsis kincaidi Rich.
4. /. serricaudis Gurjan.
5. Manna subneglecta Gurjan.
Подотряд ASELLOTA
Сем. Munnopsidae
6. Ilyarachna starokadomskii Gurjan.
7. I. zachsi Gurjan.
8. Munnopsurus minutus Gurjan.
9. Eurycope spinifrons Gurjan.
10. E. pavlenkoi Gurjan.
Подотряд FLABELLIFERA
Сем. Anthuridae
11. Paranthura japonica Rich.
Сем. Cymothoidae
12. Rocinela maculata Sch. a Mein.
13. R. belliceps (Stimps).
14. R. angustata Rich.
15. R. japonica Rich.
16. R. niponia Rich.
17. Nerocila japonica Sch. a Mein.
18. Meinertia oxyrrhynchaena (Koelbel)
19. Livoneca amurensis (Gerstfeldt)
10. L. epimerias Rich.
21. L. sacciger Rich.
Сем. Sphaeromidae
22. Limnoria .lignorum (Rathke)
23. L. japonica Rich.
24. Tecticeps • renoculis Rich. var. laeyis Gurjan.
25. T. renoculis Rich. var. serratus Gurjan.
26. T. glaber Gurjan. .
27. Cymodoce japonica Rich.
28. Dynamene glabra Rich.
29. Holotelson tuberculatus Rich.
20. Sphaeroma sieboldi Dollfus
31. Neosphaeroma oregonensis (Dana)
32. N. ovata Gurjan.
Подотряд
Сем. Idotheidae
33. Idothea metallica (Bose.)
, 34. I. derjugini Gurjan.
35. I. ochotensis Brandt
36. I. orientalis Gurjan.
37. I. fewkesi Rich.
38. Pentidothea rotundata Rich.
39. P. japonica Rich.
40. Pentias hayi Rich.
41. Synidothea brazhnikovi Gurjan.
42. X epimerata Rich.
43. Л. excavata Gurjan.
44. S. harfordi Benedict.
Подотряд
Сем. Ligiidae
56. Ligia cinerascens Budde-Lund,
VALVIFERA
45. ȣ. bicuspida (Owen) lata .Gurjan.
46. Cleantis planicauda Benedict.
47. C. isopus Grube a. Miers.
Сем. Arcturidae
48. Astacilla dilatata Rich.
49. A. polita Gurjan
50. Arcturus hastiger Rich.
51. A. crassispinis Rich.
52.. A. setosus Gurjan. var. seminudus Gurjan.
53. A. setosus Gurjan. var. acuticaudalisx Gurjan.
54. A. crenulatus Gurjan.
55. A. beringanus Benedict.
ONISCOIDEA
Сем. Trichoniscidae
57. Trichoniscus papillicomis Rich.
ЗООГЕОГРАФИЧЕСКИЙ ОЧЕРК
25
Подотряд EPICARIDEA
Сем. Bopyridae 59. Bopyroides hippolytes (Кг.)
58. Phryxus abdominalis (Кг.) 60. Argeia pugettensis Dana.
Подотряд GNATHIIDEA
63. G. schmidti Gurjan.
Сем. Gnathiidae 64. G. derzhavini Gurjan.
61. Gnathia tuberculata Rich. - 65. G. elongata (Kr.)
62/G. rectifrons Gurjan. 66. G. (Elaphognathia) monodi Gurjan.
Японское море, благодаря почти десятилетним работам Тихоокеанского института рыбного хозяйства и Гос. Гидрологического института/ изучено значительно лучше, чем Берингово и Охотское море и особенно район залива Петра Великого. Неудивительно поэтому, что число зарегистрированных для Японского моря видов Isopoda значительно превышает списки Охотского и Берингова морей; выяснен лучше не только состав, но и распределение фауны в Японском море; и здесь можно уже наметить некоторые закономерности не касаясь больше форм общих для Японского и Охотского морей и форм общих для всех 3 наших дальневосточных морей, о которых уже говорилось выше, посмотрим, что представляет собою с точки зрения зоогеографической фауна Японского моря. Анализ ее выяснил, что зоогеографический состав фауны Isopoda Японского моря очень пестр и состоит из различных зоогеографических групп.
1-ю группу образуют эндемики Японского моря, 23 вида: Janiropsis serricaudis, Миппа subneglecta, Munnopsurus minutus, Ilyarachna zachsi, Eurycope spinifrons, E. pavlenkoi, Limnoria japonica, Tecticeps glaber* T. renoculis serratus, Sphaeroma sieboldi, Neosphaeroma ovata, Rocinela niponia, R. japonica, Livoneca epimerias, L. sacciger, Synidothea epimerata* S. brazhnikovi, S. excavata, Arcturus crenulatus, Gnathia tuberculata, G. derzhavini, G. schmidti и G. rectifrons. Большинство их добыто в заливе Петра Великого на малых глубинах, часть в районе Хакодате у Японских берегов, и 3 формы — Eurycope spinifrons, Gnathia tuberculata и G. schmidti пока что добыты на глубинах около 1000 м в заливе Петра. Великого (первая из них встречалась на глубине 663 м).
2-я группа — арктические формы, здесь представлена только одним видом Gnathia elongata, обладающим в Японском море очень широким диапозоном вертикального распределения и встречающимся на глубинах от 76 до 2900 м.
3-я группа — арктическо-бореальные виды, широко распространенные в Арктике и спускающиеся далеко к югу в Атлантическом и Тихом океанах, представлена также только одним видом — Bopyroides hippolytes.
4-ю группу составляют субарктические тихоокеанские формы Азиатского побережья, т. е. виды,^’которые распространены главным образом
26
РАВНОНОГИЕ РАКИ
у наших берегов, в западной части Северного Тихого океана. В Японском море таких форм 10: *Janira soldatovi, J. erostrata, Ilyarachna starokadom-skii, Paranthura japonica, Rocinela maculata, *Idothea ochotensis, *Syni-dothea bicuspida lata, Arcturus beringanus, *A. hastiger, A. crassispinis. Большинство из них встречается по всему морю, а часть только в северной его части в Татарском проливе (4 формы, общие с Охотским морем, отмечены звездочкой).
5-ю группу образуют субарктические тихоокеанские формы, широко распространенные по обоим берегам северной части Тихого океана,, числом 4: Trichoniscus papillicornis, Rocinela angustata, R. belliceps, Argeia pugettensis. Интересно отметить, что 3 последние являются эктопаразит тами рыб (Rocinela) и ракообразных (Argeia)) обеспечивая этим себе возможность широкого распространения,
В состав 6-й группы входят 6 форм тихоокеанско-бореальных: Janiropsis kincaidi, Neosphaeroma oregonensis, Idothea orientalis, I. derjugini, Pentidothea japonica и Astacilla dilatata. Все эти формы прибрежные, мелководные, кроме Astacilla) встреченной у берегов Японии.
7-ю группу образуют формы субтропические — 10 видов: Nerocild japonica, Meinertia oxyrrhynchaena, Livoneca amurensis, Holotelson tuber-culatus, Cymodoce japonica, Dynamene glabra, Pentidothea rotundata, Cleantis isopus, Pentias hayi и Ligia cinerascens. Весьма характерно, что все это или литоральные, или очень мелководные прибрежные формы. Livoneca amurensis встречена в устье р. Лефу.
8-ю группу образует одна амфибореальная форма Cleantis planicauda (вест-индская форма).
9-ю группу образуют 2 калифорнийские тепловодные мелководные формы — Idothea fewkesi и Synidothea harfordi.
Наконец, 10-ю группу образуют формы с наклонностью к космополитизму— Idothea metallica, Phryxas abdominalis и Limnoria lignorum.
Таким образом, по сравнению с Охотским и Беринговым морями, Японское море обладает более тепловодной фауной равноногих раков. Из общего числа 66 форм около 30% составляют эндемики (условные; 23 формы), около 25% (16 форм) — умеренные субарктические, около 30% (21 форма) — тепловодные и менее 2% — арктические формы (1 вид).
Настоящие субтропические формы, прибрежные, мелководные, однако ограничиваются лишь заливом Петра Великого и дальше на север не идут, за исключением Ligia cinerascens и Cymodoce japonica, которые распространяются до бухты Преображенья. Весьма знаменательно полное отсутствие в Японском море абиссальных тихоокеанских форм, указывающее на изоляцию глубинной фауны Японского моря от абиссали Тихого океана^ что вполне согласуется с общим рельефом дна моря и мелководностью проливов между островами, отделяющими Японское море от океана. Если обмена между глубинами Японского моря и Тихого океана не существует, то точно также не существует и специфических глубоководных
ЗООГЕОГРАФИЧЕСКИМ ОЧЕРК 27
форм Японского моря. Абиссальная зона заселена теми же самыми формами Isopoda, которые встречаются в батиали и сублиторали, что указывает на относительную молодость глубин Японского моря. Как на характерную черту Японского моря следует указать, что сублиториальные изоподы здесь имеют очень широкий диапазон вертикального распределения, встречаясь с глубины 50—60 м и достигая максимальных глубин Японского моря около 3000 м. Если глубины Японского моря, благодаря очень мелким проливам оказываются изолированными от других соседних участков мирового океана, то между фауной более мелководной, литоральной и сублиторальной, существует взаимный обмен с окружающими морями. Так об обмене между Японским и Охотским морем через проливы Лаперуза и Невельского уже говорилось выше (см. в разделе, посвященном фауне Охотского моря). Из Желтого моря вместе с ветвью теплого Цусимского течения через Цусимский пролив устремляется к заливу Петра Великого целый поток субтропических форм: по берегам Японии в Японское море частично проходят тропические формы {Ligia exotica, Idothea metallica, представители родов Tylos, Cirolana, Sphaeromidae и пр.), не вошедшие в настоящую сводку, как элементы отсутствующие у берегов СССР. * !
Характерно также и то, что более тепловодные формы заселяют сильно прогреваемые летом поверхностные горизонты моря, тогда как формы умеренного типа и более холодноводные держатся на глубинах, начиная от 60—70 м и до 3000 м, т. е. обратно тому, что мы наблюдаем в Охотском и Беринговом морях, где тепловодная фауна приурочена к отепленным глубинным слоям, а более холодноводные формы держатся в охлажденных поверхностных горизонтах моря.
Рассмотрев вкратце фауну изопод каждого из дальневосточных морей в отдельности, попробуем теперь дать общую характеристику морской фауны Isopoda Дальнего Востока. В настоящий момент во всех трех морях вместе зарегистрировано 128 форм Isopoda, относящихся к 119 видам, 44 родам, 14 семействам и 6 подотрядам. Слабая изученность паразитических форм Японского моря (Cymothoidae и Epicaridea) и. малая исследованность Охотского и Берингова морей не дает возможности полнее охватить нашу дальневосточную морскую фауну. В дальнейшем, конечно, по мере изучения ее списки видов будут сильно увеличены и, весьма вероятно, в связи с этим изменятся и наши взгляды на зоогеографический характер фауны. В настоящий момент всех известных для Берингова, Охотского и Японского морей изопод можно разделить с точки зрения зоогеографической на 11 групп (распределение этих групп по морям представлено на табл. 1).
1-ю группу образуют наиболее богато представленные аутохтонные формы, эндемичные при современном уровне наших знаний для одного из трех морей. Их насчитывается 59 форм, т. е. 46.0°/0 числа всех известных морских дальневосточных форм (списки см. по отдельным
28
РАВНОНОГИЕ РАКИ
Таблица 3
РАСПРЕДЕЛЕНИЕ ОСНОВНЫХ ЗООГЕОГРАФИЧЕСКИХ ГРУПП ISOPODA В ДАЛЬНЕВОСТОЧНЫХ МОРЯХ
VERBREITUNG DER WICHTIGSTEN GRUPPEN DER ISOPODEN IN DEN NORDOSTASIAT1SCHEN MEEREN
Берингово море Berings-See Охотское море Ochotski-Меег Японское море Japanisches Meer. Во всех трех морях In alien drei Meeren
Название группы эщему tizahl § -с flj » s t s s 3 0) er:
Bezeichnung der Gruppe s s о о S 5 s ° . I s ° , i
о . . . Lz^ и я ° И Nfc. м Я ° g Sb о 30 g 0 - Гг, s N“*
Число форм Anzahl der % отношен: числу форм Prozentsatz allgemeinen Число форм Anzahl der % отношен числу форм Prozentsatz allgemeinen Число форм Anzahl der % отношен числу форм Prozentsatz allgemeinen Число форм Anzahl derJ % отиошен числу форм Prozentsatz ^allgemeinen
Широкораспростра-
й Й £ нениые. — Weit ver-
И e breitete Formen . . 6 10.9 4 8.6 4 6.0 6 4.6
Формы азиатского по-
® 1 s бережья. — Formen
der asiatischen К.Ц-
8 IS sten 3 5.5 10 21.2 13 19.7 10 7.9
S о У S X ttl Формы американского
Субаркт анские <] tische p побережья. — Formen der amerikani-schen Kiisten . . . 7 12.7 4 8.6 • 7 5.4
Всего. — Im ganzen . 16 29.1 18 38.4 17 25.7 23 17.9
Эндемики. — Endemische For-
men . 18 31.7 16 34.0 25 38.0 59 46.0-
Тихоокеанские бореальные
формы. — Pacifische boreale Formen 6 11.0 2 4.3 6 9.0 10 7.9
Арктические формы.—Arktische Formen Субтропические формы. — Sub- 7 12.7 5 10.7 1 10 1.5 10 10 7.9
. 2.1 15.1
tropische Formen Абцссальные формы Тихого —* — 1 7£
океана. — Tiefseeformen des Stillen Ozeans 5 9.1 1 2.1 5 3.9
Космополиты.—KosmopoliPsche 4.5 3.9
Formen . 2 3.6 1 2.1 3 5
Субтропические калифорнийские
формы. —Subtropische kali-fornische Formen — i 2.1 2 3.0 2 1.5
Антарктические формы. — Ant-arktische Formen Амфибореальные формы. — — — 1 2.1 — 1.5 1 0.8
2.1 1.5
Amphiboreale Formen .... Аркгаческо-бореальн. формы.— —- 1.8 1 1 2
1.5 0.8
' Arktisch-boreale Formen . . . 1 — — 1 1
Всего 55 100% 47 100% 66 100% 128 I 100%
ЗООГЕОГР АФИЧЕСКИЙ ОЧЕРК
29
морям). Распределены они таким образом, что в Беринговом море их 18 форм, в Охотском 16, в Японском 25 форм. Все они, за исключением глубоководных охотских форм Arcturus beddardi и Tecticeps leacoph-thalmus, литоральные формы или же представители фауны верхнего отдела сублиторали.
2-ю группу составляют тихоокеанские субарктические формы; их 23 формы, т. е. около 18°/0, и распадаются они на три группы; одни из них, напр. Rocinela angustata, R. belliceps, Argeia pugettensis, широко распространены как по азиатскому побережью, так и по американскому берегу и Алеутской гряде (6 форм), другие, напр., Arcturus beringanus, Janira soldatovi, Idothea ochotensis, образуют группу субарктических форм азиатского побережья (10 форм), третьи же, как напр., Idothea aleutica, Pentidothea wosnesenskii, Aega symmetrica, могут быть названы субарктическими тихоокеанскими формами американского побережья (7 форм).
В 3-ю группу входят тихоокеанские бореальные формы, как напр. Cilicaea cordata, Pentidothea japonica, Neosphaeroma oregonensis, Idothea derjugini (10 форм). Бореальные формы хорошо проникают в Берингово и Японское моря и слабо в Охотское море.
4-ю группу образуют арктические виды; их 10 видов, т. е. около 8%; одни из них, напр. Mesidothea entomon, Calathura brachiata, Munnopsurus giganteus, Gnathia elongata—спускаются в Тихий океан из Северного Ледовитого океана; другие же виды рода Synidothea, Pleuroprion и Arcturus, наоборот из Тихого океана проникли в Арктику и там распространились. Характерно, что элементы, общие с Арктической областью (Северного Ледовитого моря) имеются во всех трех морях и чем дальше к югу, тем их меньше и тем глубже они опускаются (напр., в Японском море только один вид—Gnathia elongata на глубинах от 76 до 900 м).
К 5-й группе относятся тихоокеанские субтропические виды (10 форм), как напр. Holotelson, Dynamene, Cleantis, Ligia, Cymodoce и др. Все они встречены только в Японском море и не заходят ни в Охотское, ни в Берингово моря. Исключение представляет лишь пресноводная, паразитирующая на рыбах Livoneca amurensis (гималайская форма), спускающаяся по Амуру в Амурский лиман. Субтропические формы все без исключения прибрежные, мелководные или литоральные и заходят в наши воды, очевидно, вместе с теплыми водами Цусимского течения, дающего небольшую ветвь по направлению к Владивостоку.
6-ю группу составляют настоящие абиссальные формы Тихого океана; их 5 форм — Munnopsurus laevis, Aega magnoculis, Arcturus hir-sutus, Holophryxus giardi. Они заходят в Желтое, Охотское и Берингово моря, но совершенно отсутствуют в Японском море. Выше уже указывалось, что причина этого явления (отрицательная черта фауны Японского моря) заключается в изоляции глубин Японского моря от Тихого океана при помощи мелководных (не глубже 200 м) проливов. Наоборот, Охотское море соединяется с Тихим океаном глубокими (до 2000 м) проливами,
30
РАВНОНОГИЕ РАКИ
и в его абиссали встречена Aega magnoculis; охотские абиссальные эндемики Arcturus beddardi и Tecticeps leucop hthalmus, вероятно, являются пришельцами из Тихого океана и просто пока еще не были добыты в Тихом океане.
7-ю группу составляют виды с тенденцией к космополитическому распространению, напр. Cyathura carinata, Idothea metallica, Phryxus abdominalis, Limnoria (5 видов).
8-ю группу образуют субтропические виды, известные только из южной Калифорнии (2 формы) — Idothea fewkesi и Synidothea harf ordi; обе они добыты в Японском море, причем S. harfordi не встречается севернее залива Петра Великого, a I. fewkesi проходит в Татарский пролив и оттуда через пролив Невельского проходит в Охотское море (этот путь кажется более вероятным, чем путь через Лаперузов пролив, так как на восточном берегу Сахалина она не обнаружена, но имеется у о. Лангра в Амурском лимане в сильно опресненном районе и на Шантарских островах). »
В 9-ю группу приходится выделить единственную антарктическую форму Livoneca raynaudii из Охотского моря.
10-ю группу составляют формы арктическо-бореальные, т. е. циркумполярные в Арктике, и спускающиеся в бореальные воды в Атлантике и в Тихом океане; в дальневосточных морях в эту группу Isopoda входит лишь один вид—Bopyroides hippolytes, встречающийся в Беринговом и Японском морях.
Наконец-, последнюю, 11-ю группу составляют так называемые амфи-бореальные1 формы, обладающие прерывистым распространением, известные из северных частей Атлантики и Тихого океана с хиатусом посредине в Арктике и отсутствующие также в тропиках; к ним относятся Pleuroprion hystrix и Cleantis planicauda, а также Syscenus infelix, встреченный Richardson у восточного побережья Японии.
Как видим, состав фауны дальневосточных морей весьма пестрый,, и пестрота эта зависит от наложения друг на друга элементов различных фаун вследствие большой сложности гидрологического режима, различий в физико-географических условиях во всех трех морях и связи их между собой и с прилегающими участками Северного Ледовитого и Тихого океанов. Распределение течений и рельеф дна, как указывалось уже выше,, в значительной мере определяют собою состав фауны Isopoda дальневосточных морей. Однако уже просто фактор широты оказывает влияние на различие фаун Берингова, Охотского и Японского морей. Так, наиболее северное по географическому положению Берингово море, хотя и обладает богатой и разнообразной фауной, но в основном мы имеем там представителей холодноводных арктических групп, семейств, родов;;
1 Л. С. Берг. 1934. Об амфибореальном (прерывистом) распространении морской фауны в северном полушарии. Изв. Геогр. общ., 66, вып. I.
ЗООГЕОГРАФИЧЕСКИЙ ОЧЕРК
31
напр., здесь богаче, чем в Японском море, представлены Asellota и почти отсутствуют Sphaeromidae; Asellota представлены арктическими родами Janira, Munnopsurus, Миппа. Наоборот, в южном Японском плохо развиты Asellota и богато развиваются представители южного тепловодного субтропической природы семейства Sphaeromidae (роды Sphaeroma, Exosphae-roma, Holotelson, Dynamene, Cymodoce и др.). Южный характер фауны Японского моря и умеренный характер фаун Берингова и Охотского морей подчеркивается также разнообразием состава фауны Isopoda. Обычно для теплых субтропических и особенно тропических групп характерно, на ряду с обилием видов, также и обилие родов; если с этой точки зрения рассмотреть фауну наших морей, то Японское море обладает значительно большим ^разнообразием видов и родов, чем Берингово и Охотское моря;, так, на каждый род, известный из Японского моря, приходится только 2 формы, в Беринговом море, включая его' тепловодную часть, также 2 формы, в Охотском море около 3 форм на род. Этим подчеркивается также ярко выраженная тенденция охотских форм к расщеплению и образованию новых форм. Фактор широты сказывается и на родовом составе фауны Isopoda по всем трем морям; так, в Японском море имеется 11 типично тепловодных родов, в Беринговом уже только 3, а в Охотском нет ни одного тепловодного рода. Различия в физико-географических условиях этих трех морей приводят к чрезвычайно любопытному явлению развертывания как бы вееров видов одного и того же рода в каждом из морей; роды Tecticeps, Arcturus и Synidothea дают прекрасный пример этого явления. Так, в Беринговом море имеется 2 эндемичных вида Tecticeps, 6 форм Arcturus и 6 форм Synidothea; в Охотском море 5 эндемичных форм Tecticeps, 6 Arcturus и 4 Synidothea; в Японском море 2 эндемичных вида Tecticeps, 1 Arcturus и 4 Synidothea.
Если посмотреть, как распределяются представители Isopocla в дальневосточных морях по глубинам, то оказывается, что богатая разнообразная тепловодная фауна верхних отделов сублиторали и литорали в Японском море сменяется более бедной, однообразной умеренной фауной на глубинах; в Беринговом море фауна изопод богата и разнообразна как на мелководьях, так и на глубинах, а в Охотском море богатая своеобразная фауна мелководий сменяется своеобразной же, но более бедной фауной глубоководной.
О распределении изопод по различным биоценозам почти ничего неизвестно. Для Японского моря по заливу Петра Великого можно отметить ряд характерных форм Isopoda для некоторых сублиториальных биоценозов; так, для зарослей красных водорослей на глубинах в 20—25 м характерна Idothea orientalis, для тех же глубин на фациях скал и камней Synidothea epimerata; для биоценоза Zostera на глубинах от 0.5—10 м характерны Janiropsis kincaidi, Synidothea brazhnikovi, Pentias hayi, Pentidothea rotundaia, Holotelson tuberculatus; для устричников и банок мидий — Cymodoce japonica. Для супралиторали на фации камней харак
32
РАВНОНОГИЕ РАКИ
терно присутствие Ligia cinerascens и Trichoniscus papillicornis, а для глубоководного ила на глубинах свыше 1000 м — Eurycope spinifrons и Gnaihia elongaia.
ЛИТЕРАТУРА
Литература по морским Isopoda дальневосточных морей довольно обширна, но все это, главным образом, списки видов, добытых в различных районах северной части Тихого океана разными экспедициями и отдельными лицами. Наиболее важны для каждого работающего над дальневосточными Isopoda следующие труды:
Baker W. 1908. Notes on Some Species of the Isopod family Sphaeromidae, from the South Australian Coast. Part I. Trans. Proc. Rept. Roy. Soc. South Australia, XXXII.
---- 1910. Idem, part II. Ibid., XXXIV.
---- 1926. Species of the Isopod Family Sphaeromidae from the Eastern, Southern and Western Coasts of Australia. Ibid., L.
---- 1928. Australian Species of the Isopod Family Sphaeromidae. Ibid., LIL
Barnard K. 1914. Contributions of the Crustacean Fauna of South Africa, I. Additions to the
Marine Isopoda. Ann. South African Museum, X, part XI.
---- 1914. Idem,''III. Additions to the Marine Isopoda. Ibid., X, part 16.
—— 1920, Idem, VI. Further Additions to the List of Marine Isopoda. Ibid., XVII, part 5.
---- 1925 a. Idem, IX. Further Additions to the List of Isopoda. Ibid., XX, part 5.
---- 1925 b. A Revision of the Family Anthuridae (Crustacea-Isopoda), with Remarks on Cer-
tain Morphological Peculiarities. Journ. Linn. Soc., 36, № 241.
В a г г о w A. 1919. The Occurrence of a Rockboring Isopod along the Shore of San Francisco Bay. California, Univ. Calif. Publ., Zool., 19.
BeddardF. 1885. Preliminary Notice of the Isopoda Collected during the Voyage of H. M. S.
Challenger, part II. Munnopsidae. Proc. Zool. Soc. London.
---- 1886. Isopoda. „Challenger". Reports on the Scientific Results of the Exploring Voyage of H. M. S. Challenger. 1873-1876. Zoology, XVII.
Benedict J. 1897. A Revision of the Genus Synidothea. Proc. Acad. Nat. Sci. Phila., 53,. part I.
---- 1898 a Two New Isopods of the Genus Idotea from the Coast of California. Proc. Bio). Soc. Washington, 12.
---- 1898 b. The Arcturidae in the U. S. Nat. Museum. Proc. Biol. Soc. Washington, 12.
Brandt F. 1833. Conspectus Monographiae Crustacearum Oniscoidorum Latreillei. Bull. Soc.
Imper. Natur. Moscou, 6.
---- 1851. Krebse. Middendorff’s Reise in den Aussersten Norden und Osten Sibiriens.
Zoologie, Bd. II. , .
Boone P. 1919. Description of Ten New Isopods. Proc. U. S. Nat. Mus., 54.
--- 1920. Isopoda. Report of the Canadian Arctic Exped. 1913—1918. Crustacea, II, part D.
Bonnier. J. 1900. Contributions a 1’etude des Epicarides. Traveaux Station Zoologique Wimereux, VIII, Paris.
Budde-Lund G. 1885. Crustacea Isopoda Terrestria per familias et genera et species descripta. Hauniae.
•Chilton Ch. 1891. On the Changes in Form of a Parasitic Isopod (Nerocila). Trans. N. Z. Inst., (6), 23.
---- 1914. Revision of Species of Genus Limnoria. Ann. Mag. Nat. Hist., (8), 13,
С о 11 i n g e W. 1916 a. Description of a New Species of Marine Isopoda of the genus Pentias Rich. Journ. Zool. Research, I, Ns 1,
---- 1916 b. On the Marine Isopoda Idotea ochotensis Brandt. Jonrn. Zool. Research, I, № 2.
ЛИТЕРА ТУРА
33
С о 11 i n g е W. 1916 с. On the Structure of the Marine Isopod Mesidothea sibirica (Birula) with Some Remarks upon Allied Genera. Ibid.
---- 1917 a. Description of a New Species of Isopoda of the Genus Synidotea from the Golf of Manaar. Rec. Indian Mus., XIII, part I, № 1.
---- 1917 b. A Revision of the British Idoteidae, a Family of Marine Isopoda. Trans. Roy. Soc. Edinburgh, 57, part 3,
---- 1918. Some Observations upon two Rare Marine Isopods. Journ. Zool. Research, III.
D a n a J. 1849. Conspectus Crustaceorum. Amer. Jour. Sci. Arts, ser. 2, 8.
---- 1852. United States Exploring Expedition during the Years 1838—1842 under the Command of Charles Wilkes, U. S. N. XIII, Crustacea, part I, Philadelphia.
---- 1853. Idem., XIV, part II. .
---- 1855. Catalogue and Descriptions of Crustacea by Dr. John L. Le-Conte. Proc. Acad. Nat. Sci. Philadelphia, 7.
D o 11 f u з A. 1889. Sur.quelques Isopodes du Musee de Leyde, Notes Leyd. Mus., II.
Fee A. R. 1926. The Isopoda of Departure Bay and Vicinity, with Descriptions of New Species, Variations and Colour Notes. Contr. Canadian Biology. Toronto, n. s., 3, № ?.
Gerstfeldt G. 1858. Uber einige. zum Teil neue Arten Platoden, Anneliden, Myriapoden und Crustaceen Sibiriens, namentlich seines ostlichen Teils und des Amurgebietes. Memoires savants etrangers. Acad. Sci. St. Petersbourg, t. VIII.
Liard et В о n n i e r. 1893. Contributions, a I’etude des Epicarides. Bull. Scient. France Belgique, ser. 4, 25.
;----- 1895. Memoire.de 1’Epicarides. Bull. Soc. Biol. France Belgique, 25.
JGurjanova, E.] Гурьянова E. 1933 а. К^фауне равноногих раков (Isopoda) Тихого океана. I. Новые виды Valvifera и Flabellij'era. Исследов. морей СССР, вып. 17.
---- 1933 Ь. Idem, II. Новые виды Gnathiidea и Asellota. Ibid., вып. 19.
—-— 1934. Idem, III. Новые виды сборов экспедиций ГГИ в 1932 г. Ibid., вып. 22.
---- 1936. Zur Fauna der Isopoda des Stillen Ozeans. Zool. Anzeiger, 114.
Haan de W. 1850. Fauna Japonica auctorePh.Fr.de Siebold. Crustacea Lugduni Bata-vorum, 50.
Hale H. 1925. Review of Australian Isoppds of the Cymothoid Group, part I. Trans. Roy. Soc. South Australia, XLIX.
---- 1926. Idem, part 11. Ibid., L.
Hansen H. 1890. Cirolanidae et Familiae nonnulae propinquae Musei Hauniensis. Vidensk, Selsk. Skr., 6, Afd. V, 3. . ’
----,1897. Reports on'the Dredging Operations of the West Coast of Central America to the Galapagos, to the West Coast of Mexico and in the Gulf of California... (Albatross 1881), XXII. The Isopoda. Bull. Mus. Compar. Zool. Harvard College, 31, № 5, Cambridge.
Hansen H. 1905 a. On the Morphology and Classification of the Asellota group of Crustaceans ... Proc. Zool. Soc., London, 19Q4, II.
—:— 1905 b. Revision of the European Marine Forms of the Cirolaninae . .. Journ. Linn. Soc. London, 29. f
----1905c. On the Propagation, Structure and Classification of the family Sphaeromidae. Quart. Jour. Microsc. Sci., XLIX, part I, new ser. .
---- 1916. Isopoda. The Danish Ingolf. Exp., Ill, part 5.
Harger O. 1879. Notes on New England Isopoda. Proc. U. S. Nat. Mus., II.
---- 1880. Report on the Marine Isopoda of New England and Adjacent Waters. Rep. U. S. Corn. Fish__________and Fisheries for 1878.
---- 1883. Reports on the Results of Dredging, under the Supervision of Alex. Agassiz, XXIII. Report on the Isopoda. Bull. Mus. Compar. Zool. Harvard College, XI, № 4. Cambridge.
Hilgendorf F. 1893- Bemerkungen iiber 2 Isopoden die Japanische Siisswasser-Assel (Asselus hilgendorfi Bov.) und eine neue Munna-Art. Sitzungsb. Geselsch. Naturf. Freunde zu Berlin.
о Г урьявова
34
РАВНОНОГИЕ РАКИ
Hodgson Т. 1910. Crustacea. IX. Isopoda, Nat. Antarctic-Exped. 1901—1904. Nat. Hist.. IV, № 3.
Holmes S. 1904 a. Remarks on the Sexes of Sphaeromids with a Description of a New Species of Dynamene. Proc. Calif. Acad. Sci., 3 series, Zoology, III.
----- 1904 b. On Some New or Imperfectly Known Species of West American Crustacea. Proc.. Calif. Acad. Sci., 3 series, Zoology, III.
I wasa Masao. 1934. Description of a New Isopqd Crustacean (Sphaeromidae) irom Hokkaido.. Proc. Imp. Acad., Tokyo, X, n° 6.
Jackson H. 1922. A Revision of the Isopod Cenus Ligia (Fab.). Proc. Zool. Soc., London, part II.
К о e 1 b e 1 C. 1879. Ober einige neue Cymothoiden. Sitzungsb. math. nat. Klasse Kais. Akad. Wissensch., Wien, Band 78, Abt. I.
Leach W. 1814. Article „Crustaceology". Brewster Edinburgh Encycl., VII.
---- 1815. A Tabular View of the External Characters of Four Classes of Animals, which Linne Arranged under Insecta, with the Description of the Genera Composing three of' these Classes in Orders, and Descriptions of Several new Genera and Species; Trans.
Linn. Soc. London, XI, pt. 2. - '
----- 1818. Article ,,Cymothoidees“. Diet. Sci. Nat., XII.
Lockington W. 1877. Description of Seventeen-New Species of Crustacea Proc. Calif-, Acad. Sci;, VII. 1876. :
M i e r s E. 1877. On a Collection of Crustacea Decapoda and Isopoda Chiefly from South-America with Description of New Genera and Species. Proc. Zool. Soc. London. . - -
---- 1883. Revision of the Idotheidae, a-Family of Sessile-Eyed Crustacea. Jourh. Linn. Soc..
Zool., 16. ...
Miller M. 1933. A-New Blind Isopod Asellus californicus and a Revision of the Subterranean Asellids. Publi Zool. Union California, 39, № 4.
Monod T.1926. Les Gnathiidae. Essai monographique (Morphologie, biblogie, systematique)., Mem. Soc. Sci. Nat. Maroc. № 13. '
---- 1930. Contribution a1I’etiide des Cirolanidae. Ann. Sci. Nat. Zool. (10), ХШ. " ‘ —- 1925—1931. Tanaidaces et isopodes aquatiques de 1’Afrique occidentale ef septenfrionale
parts I, II, III. Mem. Soc. Sc. Nat. Maroc., V, № 3, 6, 29.
Murdo ch. 1885. Marine Invertebrates. Report of the International Polar Expedition to Point* Barrow. Alasca, Washington.
N i e r’s t r as z H. 1917. Die Isopoden Sammlung in Naturhistorischen Refchs-Museum zuLeiden.. Cymothoidae, Sphaeromidae, Serolidae, Idotheidae, Asellidae, Janiridae, Munnopsidae..
Zool. M£dedeel’s Rijks Mus. Nat. Hist., Leiden, Deel III, Afl.-2—3.
---- 1918. Alte und neue Isopoden. Ibid, Deel-IV, Afl. 2. ' .............1 " - *
---- 1931. Die Isopoden der Siboga-Expedition III. Isopoda Genuina II. Flabellifera. Siboga-Exp-Monographie. XXXIIC. 1
Nordenstam A. 1933. Marine Isopoda of the Families Serolidae, Idotheidae, Pseiudidothei-dae, Arcturidae, Parassellidae and Stenetriidae, Mainly froth the South Atlantic. Further Zool. Results Swedisch Antarc. Exped. 1901—1903, III, № 1. .
Norman A. 1904. British Isopoda of the Families Aegidae, Cirolanidae, Idoteidae and Arcta-ridae. Ann. Mag. Nat. Hist. (7), 14. 1
О hl in A. 1901. Isopoda from Tierra del Fuego and Patagonia. I Valvifera. SvenskaExp. Magellansland., Bd. II, № 11.
Ortmann A. 1895. A New Species of the Isopod Genus Bathynomus. Proc. A₽ad. Nat. Sci. Phil., 1894.
Owen R. 1835. Appendix to Ross’s Second Voyage in Search of a North West Passage London. ;
---- 1839. The Zoology of Captain Beechey’s Voyage to the Pacific Ocean and Bering’s Straits, performed in the H. M. S. Blossom in the Years 1825—1828, London.
ЛИТЕРАТУРА
.35
R а с о v it z а Е. 1907. Isopodes terrestres, I ser. Arch. Zool. ser. IV, VII.
---- 1908. Idem, 2 ser., ibid., IX.
----- 1910. Sphaeromiens. 5 ser., ibid., IV.
—— 1912. Cirolanides, 5 ser., ibid., X.
—1919 a. Notes sur les Isopodes, 1, 2, ibid., 53.
----1919 b. Idem, 3, 4. 5, ibid., 58.
---- 1919 c. Idem, 6, 7, ibid., 58.
------ 1920. Idem, 8, 9, ibid., 59.
---- 1923. Idem, 10, 11, ibid., 61.
---- 1924. Idem, 12, ibid., 62.
------ 1925. Idem, 13, ibid., 63. J
Richardson H. 1897. A New Genus and Species of Sphaeromidae from Alaskan waters-Proc. Biol. Soc., Washington, XL
----1898. Description of Four New Species of Rocinela, .with a Synopsis of.the Genus. Proc, Amer. Philos. Soc., 37.
-----1 1899. Key to the Isopods of the Pacific Coast of North America, with Description of 22’ New Species. Proc. U. S. Nat. Mus., 21.
----- 1900 a. Synopsis of Nor th American Invertebrates. VIII. Isopoda. ..American Naturalist 34, Boston. . ;. .
---- 1900 b. Description of a New Species of Idotea from Hakodate Bay, Japan. Proc. U. S. Nat Mus., 22.
---- 1904 a. Contributions to the Natural History of the Isopoda. I. Isopoda Collected in Japan» in the Year 1900 by the U. S. Fish. Commission Steamer Albatross and the Year 1881 by the U. S. S. Palos. Proc. U. S. Nat. Mus., 27.
---- 1904 b. Idem. II. Isopoda collected in Japan by Jordan and Snyder. Ibid., 27.
---- 1905 a. Isopods from the Alaska Salmon Investigations, Bull. Bureau Fish., 24. 1904.
---- 1905 b. Monograph of the Isopods of North America. Bull. U. S. Nat. Mus., № 54.
---- 1906. Description of new Isopod of the family Sphaeromidae. Proc. U. S. Nat-Mus., 31.
---- 1908 a. On some Isopods of the Fam. Dajidae from the North West Pacific Ocean with, Description of One'New Genus and Two New Species. Ibid., 33.
---- 1908 b. Description of a New Isopod Genus Family Dajidae. Ibid., 34.
---- 1908 c. Some new fsopoda Aselloidea from North America. Ibid., 35.
---- 1910. Isopods Collected in the North-West Pacific by the U. S. Bureau from the Fisheries Steamer Albatross in 1906. Ibid., 37.
---- 1911. Description of a New Genus and Species of Janiridae from the North-West Pacific-Ibid., 40.
S a r s G. 1885. Crustacea. Den norske Nordhavs Exped., VI.
---- 1899. Isopoda. An Account of the Crustacea of Norway, II, Bergen.-
Sheppard E. 1933. Isopod Crustacea. Part I. The Family Serolidae. Discovery Reports VIL
Schioedte J. et Meinert F. 1879. Symbolae and monographiam Cymothoarum crusta-ceorum isopodum familiae. I. Aegidae. Naturh. Tidsskr. (3), Bd. 12.
---- 1883. Idem, II, III. Anilocridae, Saophridae. Ibid., Bd. 13.
---- 1884. Idem. IV. Cymothoidae. Ibid., Bd. 14.
Stebbing T. 1900. Crustacea from the Falkland Islands Collected by Mr. Rupert Vallentin. Part I. Proc. Zool. Soc., London.
----1910a. Isopoda from the Indian Ocean and British East Africa, Percy Sladen Exped-Trans. Linn. Soc., London, ser. 2, Zool. XIV, part I.
---- 1910 b. General Catalogue of South African Crustacea. Ann. S. African Mus., VI, part, IV, London.
---- 1914. Crustacea from the Falkland Island Collected by Mr Rupert Vallentin. Part. II. Proc-Zool. Soc. London.
----1919. Idem, part, III, ibid.
3*
36
РАВНОНОГИЕ РАКИ
Stephensen К. 1927. Papers from Dr. Mortensen’s Pacific Expedition 1914—1916. 40. Crustacea from Auckland and Campbell Islands. Vidensk. Midd. Naturh. Foren., 83.
Stimpson W. 1857. The Crustacea and Echinodermata of the Pacific Shores of North America. Boston Journ. Nat. Hist., 6.
---- 1863. On an Oceanic Isopod found near the South-Eastern Shores of Massachusetts Proc. Acad. Nat. Sci. Phil., 14.
---- 1864. Descriptions of New Marine Invertebrates from Puget Sound — Ibid., 16.
Strassen zur. 1902. Uber die Gattung Arcturus und die Arcturiden der deutschen Ti^fsee-Expedition. Zool. Anz., 25.
---- 1903. Zusatz zu meinem Artikel fiber die Arcturiden. Ibid., 26.
Tattersail W. 1921. Tanaidacea and Isopoda. British Antarctic (Terra Nova) Exp, .1910.
Nat. Hist. Report, Zoology, III, № 8.
Th i e 1 e m a n n M. 1910. Beitrage zur Kentniss der Isopoden Fauna Ostasiens F. Doffein. Betrage zur Naturgeschichte Ostasiens. Abhandl. Math. Phys. Klasse K. Bayer Akad. Wiss., II. Suppl. Bd. 3 Abt.
Ueno Masuzo. 1927. Notes on Some Subterranean Isopods and Amphipods of Japan. Mem. College Sc. Kyoto Imp. Univer., ser. B., Ill, № 3.
V a n h о f f e n J. 1914. Die Isopoden der Deutschen Siidpolar-Exped'. 1910—1903. Deutsche Siidpolarr Expedition, 15, Zoology, 7. •
СПЕЦИАЛЬНАЯ ЧАСТЬ
Отряд равноногих раков — Isopoda, принадлежит к группе высших раков (Malacostraca) и характеризуется следующими основными признаками: тело большей частью сплюснуто в дорзо-вентральном направлении (иногда цилиндрическое, вытянутое червеобразно) и расчленено на сегменты более или менее равномерно; все тело распадается на 3 отдела — голову, грудной отдел и брюшко. Первый грудной сегмент всегда нацело сливаетея с головой, и 1-я пара грудных ножек превращается в ногочелюсти; иногда с головой срастается и 2-й грудной сегмент. Тельзон или хвостовая пластинка срастается с последним брюшным сегментом. Глаза сидячие, т. е. лишены стебельков. Грудные ножки одноветвистые, так как экзоподит отсутствует. Брюшные ножки двуветвистые ,с очень коротким базальным члеником й широкими пластинчатыми ветвями, приспособленными главным образом для дыхания; ветви брюшных ножек большей частью одночлениковые. Сердце целиком или отчасти лежит в области брюшка.
Отряд Isopoda распадается на 7 подотрядов. В дальневосточных морях известны представители лишь 6 подотрядов, так как представители тропической группы Phreotoicidea не заходят в умеренные воды. Наиболее важными признаками для крупных систематических единиц, подотрядов, служат число свободных грудных ножек, строение и характер прикрепления рулевых ног (uropoda) и строение брюшных ножек (pleopoda).
Для семейств наиболее важными признаками являются строение ротовых частей, характер сегментации тела и строение торакальных конечностей; при определении рода следует обращать главное внимание на строение жвал (mandibulae), ногочелюстей (maxillipedes), грудных ножек (thoracopoda) и рулевых ног (uropoda), а также на характер сегментации брюшного отдела; видовые отличия выражаются в деталях строения и вооружения сегментов, в форме конечностей, глаз, усиков, в присутствии или отсутствии на теле всевозможных выростов, шипов, отростков бугров, волосков и пр.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДОТРЯДОВ
1 (6). Рулевые ножки латеральные, т. е. прикреплены по бокам тельзона у его основания или на его боковом крае,. образуя или каудальный
38
РАВНОНОГИЕ РАКИ
веер или чашечку, или же створки, открывающиеся на брюшной стороне.
2 (5). Рулевые ножки вместе с тельзоном образуют каудальный веер или чашечку.
3 (4). Свободных грудных ножек 7 пар .... II. Flabellifera (стр. 65). 4 (3). Свободных грудных ножек только 5 пар, так как передняя пара превращается в pylopoda и в виде крышечки прикрывает снизу ротовые части, а последняя пара нацело редуцирована .......
............................. . . . VI. Gnathiidea (стр. 227);
5 (2). Рулевые ноги в виде створок залегают на брюшной стороне тель-зона, накрывая собой все брюшные ножки . III. Valvifera (стр.' 131).
6 (1). Рулевые ножки терминальные, т. е. прикреплены к заднему краю тельзона, большей частью палочкообразные.
7 (8). Все брюшные ножки прикрыты снизу тонкой цельной пластинкой в виде крышечки . . ;................;..........I. Asellota (стр. 38).
8 (7). Брюшные ножки лежат свободно и не прикрыты сверху тонкой цельной крышечкой.
9 (10). Формы свободноживущие с нормальным расчленением тела на сегменты и нормальными конечностями. Брюшные ножки служат для воздушного дыхания. . . . . ................IV. Oniscoidea (стр. 205).
10 (9). Формы паразитические; тело самки обычно асимметричное, лишь со следами сегментации или вовсе без всяких швов. Брюшные ножки 1 ' приспособлены для жаберного дыхания....................
- ;........................' . . . . . . V. Epicaridea (стр, 215).
I. Подотряд ASELLOTA
• Все сегменты метазомы сливаются друг с другом и с хвостовой пластинкой, образуя общий хвостовой сегмент или тельзон. Pleopoda служат исключительно для дыхания; самцы имеют 5 пар брюшных ножек; pleopoda 1-й пары, сливаясь друг с другом, образуют крышечку (operculum) в виде тонкой пластинки, накрывающей сверху остальные брюшные ножки; у самок 4 пары брюшных ножек,. причем 1-я пара отсутствует, а крышечка образуется слиянием 2-й пары брюшных ножек. Uropoda терминальные, т. е. прикреплены на конце тельзона, одноветвистые или двуветвистые; в некоторых случаях (Munnidae) вовсе не имеют ветвей и: представлены лишь маленьким видоизмененным базальным члеником. Грудные сегменты часто сливаются друг с другом со следами шва или вовсе без швов. Очень часто 4 передних сегмента груди свободны, а 3 последних срастаются друг с другом, и таким образом весь thorax образует 2 отдела — передний и задний. Грудные ножки ходильные, иногда 3 последних пары превращаются в плавательные ножки с расширенными в виде пластинок члениками, снабженными длинными щетинками по краям (сем. Munnopsidae). Коксальные пластинки (эпимеры) маленькие
ASELLOTA
39
или отсутствуют вовсе. Голова свободна или сливается с 1-м грудным сегментом. Антенны 1-й йары обычно маленькие; антенны 2-й пары часто очень длинные,, значительно превышающие длину тела. Ротовые части нормально развиты. Этот подотряд сохраняет , много примитивных черт; у многих представителей имеются ясный экзоподит и 3 членика сим-подита антенн, а также часто сохраняется чешуйка (squama) на антеннах. - .................
Наиболее характерными чертами этого подотряда является присутствие крышечки (operculum), прикрывающей брюшные ножки, и терминальный тип прикрепления рулевых ног (uropoda).
В. наших дальневосточных морях известны 22 формы, принадлежащие к 3 семействам — Janiridae, Munnidae и Munnopsidae. . -
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ
1 (2). Зубной отросток на жвалах нацело редуцирован или же короткий, суживающийся дистально и направлен назад. 3 задних пары грудных ножек плавательные..................... . 3. Munnopsidae (стр. 54).
2,(1). Зубной отросток на жвалах хорошо развит, почти цилиндрический и направлен вперед или вертикально. 3 задних пары грудных ног ходильные. . . .
3 (4). Все грудные сегменты вследствие равномерного расчленения,тела более или менее одинаковой формы и размеров и все свободно подвижны . . .... . , , . . ... • . .1. Janiridae (стр. 39).
4(3). Грудной отдел вследствие, неравномерного расчленения тела ясно распадается на 2 участка, передний и задний; передний участок из 3 сегментов значительно больше и шире, чем задний, состоящий из 3 сегментов . . . . . . ,. .... .........2. Munnidae (стр. 49).
I. Сем. JANIRIDAE
Тело удлиненное, равномерно расчлененное, сплюснутое в дррзо-вецтральном направлении; голова и все.грудные сегменты свободно сочленены друг с другом. Глаза, если они имеются, — на спинной стороне головы; передне-боковые углы головы обычно пластинчато расширены и треугольно заострены. Ротовые части хорошо развиты; жвалы нормаль-ные с хорошо развитым зубным отростком и щупиком. Щупик ногоче-люстей хорошо развит, с расширенным 2-м члеником и нормально развитыми2 концевыми члениками. Все грудные ножки ходильные; 1-я пара хватательная. Рулевые. ноги больщей частью хорошо развиты; одно- или .двуветвистые, но иногда сильно редуцированы. , ,
В дальневосточных морях известно 3 рода.
40
7. JANIRIDAE
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Глаза отсутствуют; рострум короткий с прямым передним краем.. Грудные ножки 1-й пары короткие, слабые и значительно меньше,, чем остальные 6 пар. 1-й (базальный) членик верхних усиков большой, широкий, с вытянутым в виде лопасти передне-боковым углом
................................3. Microprotus Rich. (стр. 48)..
2 (1). Глаза имеются; рострум заострен или его нет вовсе; 1-я пара грудных ног мало отличается по размерам и строению от остальных* пар грудных ножек. Базальный членик верхней пары усиков нормальный., 3 (4). 2-й членик щупика ногочелюстей много шире лопасти 2-го членика
базальной части............... . 2. Janiropsis G. Sars (стр. 44).
4 (3). 2-й членик щупика ногочелюстей не шире лопасти 2-го членика
базальной части..................... 1. Janira Leach, (стр. 40).
1. Род JANIRA LEACH
Leach, 1814, Edinbourgh Encycl., VII: 434.
Тип рода: I. maculosa Leach.
Тело нормально расчленено на сегменты, сплюснуто в дорзо-вен-тральном направлении; ротовые части нормального строения; зубной отросток жвал почти цилиндрический, направлен вперед и внутрь. 2-й членик щупика ногочелюстей такой же ширины, как лопасть 2-гоь членика базальной части. Рулевые ноги (uropoda) хорошо развиты, двуветвистые. Грудные ножки 1-й пары немного отличаются по величине от остальных. Глаза обычно хорошо развиты, расположены на дорзальной стороне головы. Голова часто снабжена заостренным рострумом: и боковыми заостренными лопастями. Базальный членик усиков 1-й пары нормального строения, т. е. лишь немного крупнее остальных, цилиндрической формы. В дальневосточных морях известно 5 видов этого рода.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Голова снабжена длинным, достигающим 2/з Длины головы рос-трумом....................................... 1. J. alascensis Bened.
2 (1). Рострума нет вовсе, или он еле заметен.
3 (4). Боковые углы головы впереди глаз закруглены; поверхность тела густо покрыта короткими жёсткими волосками . . * 5. J. sarsi Rich.
4 (3). Боковые углы головы впереди глаз вытянуты в хорошо выраженные заостренные лойасти; поверхность тела гладкая, лишенная волосков.
5 (6). Боковые отростки головы впереди глаз оканчиваются 2-мя шипи-ками; передний край головы почти прямой . . . 4. J. hohnesi Rich.
6 (5). Боковые отростки головы впереди глаз просто заострены; передний, край головы выпуклый и слегка заострен посредине.
1. JANIR A
41
Фиг. 7. Janira alascensis Rich, (по Н. Richardson).
7 (8). 1-й членик ножки 1-й пары усиков узкий, не шире 2-го членика;: спинная сторона тельзона гладкая..............* 2. J. erostrata Rich.
8 (7). 1-й членик ножки 1-й пары усиков широкий, много шире 2-го членика. На спинной стороне тельзона у его дистального конца труппы веерообразно расположенных тонких иголочек. . * 3. J. soldatovi Gurjan.
* 1. Janira alascensis (Benedict) (фиг. 7).
Benedict in Richardson, 1905, Bull. U. S. Nat. Mus., 54:- 464, f. 519 (Jolella).
Голова снабжена длинным равным длине головы, суживающимся дистально тупым рострумом. Ширина головы в 2 раза больше ее длины. Боковые углы головы впереди глаз образуют расширенные лопасти, заостренные на концах. Задний край 1-го грудного сегмента сильно выпуклый. Боковые части 4 передних грудных сегментов расщеплены на 2 лопасти. Эпимеры двулопастные, маленькие, ясно видные между боковыми лопастями грудных сегментов. Боковые части 3 задних грудных сегментов
образуют по одной расширенной лопасти. Тельзон образует 3 продольных закругленных на конце лопасти; боковые лопасти тельзона больше и выдаются за пределы срединной. Проподит 1-й пары грудных ног расширенный с тонкозазубренным хватательным краем.
Распространение. Берингово море: 71°02'N, 157°46'W, глубина 34 м (Richardson, 1905).
Фиг. 8. Janira erOstrata
Rich.
* 2. Janira erostrata (Rich.) (фиг. 8).
Richardson, 1899, Proc. U. S. Nat. Mus., XXI, 858 (Janthe); Richardson, 1905, Bull. U. S. Nat. Mus, 54: 465, f. 520 (Jolella).
Ширина головы в 2J/2 раза больше ее длины; вместо рострума имеется маленькое заострение посредине переднего края головы. Боковые лопасти головы впереди глаз хорошо развиты, заостряющиеся дистально. Глаза на спинной стороне головы. Усики 1-й пары маленькие, короткие; длина их меньше ширины головы. 1-й членик их ножки ненормально узкий и маленький. 4 передних грудных сегмента с расщепленными на 2 лопасти боковыми частями. Эпимеры маленькие, в виде лопасти, расположенные между боковыми лопастями сегментов. Задний край тельзона образует 3 тупые трехугольные лопасти; срединная лопасть меньше боковых, вершины которых выдаются за пределы вершины срединной лопасти. Uropoda почти равны длине тельзона^.
ветви uropoda почти равной длины. 1-я пара грудных ног хватательная. Длина 6 мм.
42
7. JAttIRIDAE
i • Распространение. Берингово мор е: о. Атту, бухта Чичагова, Аляска (Richardson, 1905). Японское море: бухта Ольги 43°38'N, 135°8'Е, глуб. 65 м, 6 экз., сбор ГГИ, 1930; Бражников, 1907, ст. 10; залив Петра Великого, мыс Астафьева (полуостров Краббе), глуб. 16 м, 14 экз. Охотское море: ст. 202 и 203: 45°40'N, 141°59/Е, глуб. 56 м 9 экз., галька (сборы ГГИ 1932).
* 3. Janira soldatovi Gurjanova (фиг. 9).
Gurjanova, 1933. Исслед. Морей СССР, 19.: 81, рис. 4.
; Голова широкая, но ширина ее немного меньше, чем в 2 раза превышает длину. Переднебоковые углы головы впереди глаз вытянуты в небольшие треугольно заостренные лопасти; передний,край головы слегка выпуклый, закругленный посредине; рострум отсутствует. Глаза большие Дорзально расположенные; жгутик усиков 1-й пары восьмичлениковый; членики узкие, удлиненные. Эпимеры хорошо развиты на всех грудных сегментах и снабжены острыми шипиками:. Спинная сторона мезозрмы гладкая, лишенная шипов, отростков или волосков. Заднебоковые углы тельзона оттянуты книзу в виде крупных широких треугольно-заостренных лопастей. С]рединная лопасть между ними закругленная и меньших размеров, На спинной стороне тельзона по бокам его над основанием боковых лопастей группы веерообразно расположенных тонких щетинок. Uropoda хорошо развиты, снабжены щетинками и шипиками. Наружные их . ветви значительно короче внутренних. Ближе всего стоит к виду J. erostrata, отличаясь от него закругленным фронтальным краем, слабыми переднебоковыми отрезками головы и строением тельзона, рулевых ног и усиков. .Максимальная длина тела 5.5 мм,
Р а с п р о ст р а н е н и е. О хо т с к о е море: у мыса Укоп на глубине 50 м. 1 экз. 53°37z N, lSe^l'E (Павленко, 1911, ст. 44). Японское море: Татарский пролив.(против р. Нельмы) на глубинах от 31 до 96 м (ТИРХ, 1931).
4. Janira holmesi (Richardson) (фиг. 10).
Richardson, 1905, Bull. U. S. Fish. Comm., XXIV: 216 (Joie); Richardson, 1905, Bull.’ ’U. S. Nat. Mus., №54: 465, f. 521 (Jolella). !
Голова уже, чем у предыдущих видов; длина ее лишь немного меньше .ширины; рострум отсутствует. Передний край головы почти 1пр'ямо$ с небольшим слабо выраженным заострением посредине. По бокам головы .впереди глаз по одному небольшому отростку, заканчивающемуся 2 шипиками. Глаза большие, дорзальные, расположенные близко к переднему краю головы у основания боковых отростков головы. Усики 1-й пары достигают конца 4-го членика ножки у усиков 2-й пары. Базальный .членик усиков I большой и расширенный, жгутик 20-члениковый. Усики ,2-й пары длиннее тела животного. Эпимеры хорошо развитые, расщепленг ные на 2 заостряющиеся лопасти. Боковые части 2-го и 3-го грудных
1. JANIRA
43
сегментов двулопастные, тогда как 1-й;и 4 задних грудных сегмента по бокам имеют по одной небольшой лопасти. Тельзон на конце между рулевыми ножками образует широкую закругленную лопасть; заднебоковые углы тельзона вытянуты в треугольные заостренные отростки. Рулевые ноги равны длине тельзона; наружные ветви немного короче внутренних. Базальный членик рулевых ног вытянутый, равен длине наружной ветви. Грудные ножки снабжены добавочным когтем. 1-я пара хватательная с большим, вооруженным шипиками по внутреннему краю
Фи^. 9. Janira soldatovi Gurjan.
Фиг. 10. Janira holmesi Rich, (no Richardson).
Фиг. 11. Janira sarsi Rich, (no Richardson).
карпальным члеником; проподит зазубрен вдоль внутреннего края в проксимальной части.
Распространение. Берингово море: юго-восточная Аляска, пролив Стефенс и бухта Нага в Behm Canal,, на глубине от 80 до 360 м (Richardson, 1905).
*5. Janira sarsi (Richardson) (фиг. 11).
Richardson, 1905, Bull. U. S. Nat. Mus., 54: 467, f. 523 (Jolella). ' .
Ширина головы почти в 2 раза больше ее длины; рострум отсутствует. Передний край головы почти прямой с очень маленьким притупленным бугорком посредине. Передне-боковые углы головы закруглены и; не имеют вытянутых впереди глаз лопастей, а лишь небольшие выпуклости, снабженные 4 тонкими шипиками. Глаза небольшие, округлые> расположенные близко у краев головы. Базальный членик усиков 1-й пары большой и расширенный; жгутик многочлениковый; членики
44
1. J AN I RID AE
короткие и широкие. Эпимеры хорошо развиты, с 2 вытянутыми узкими лопастями. Боковые части 3 передних грудных сегментов образуют по 2 узких удлиненных отростка, между которыми видны лопасти эпимер. 4-й грудной сегмент имеет по бокам только по одному отростку в передней части. 3 задних грудных сегмента с расщепленными на 2 неровных лопасти боковыми частями. Боковые отростки сегментов и лопасти эпимер усажены шипиками. Тельзон очень широкий; задний край его образует 2 небольших заостренных отростка на задне-боковых углах и 3 небольшие закругленные лопасти между ними. Все тело животного густо покрыто короткими жесткими волосками. Длина животного 10.5 мм.
Распространение. Берингово море: о. Амчитка у Аляски (Richardson, 1905); 1-й Курильский пролив южной Лопаткинской подводной гряды (Смирнов и Бегак, 1907, ст. XIX, № 101), на глубине от 15 до 25 мм. р
2. Род JANIROPSIS G. SARS
G. Sars, 1899, Account of the Crustacea of Norway. II. Isopoda: 102.
Тип рода: /. breviremis G. Sars.
Тело равномерно расчленено, все сегменты и голова свободны. Боковые части грудных сегментов слегка расширены. Глаза хорошо развиты. Рострум и боковые лопасти головы впереди глаз отсутствуют. Усики 1-й пары маленькие, жгут с ограниченным числом члеников. Усики 2-й пары длинные, у самцов развиты более мощно, чем у самок. Чешуйка (squama) маленькая, но хорошо развитая у обоих полов. 2-й и 3-й членики щупика ногочелюстей пластинчатые, сильно расширенные, так что ширина их превосходит ширину лопасти 2-го членика базальной части ногочелюстей. Ноги сравнительно короткие и толстые с добавочным коготком. 1-я пара грудных ножек у самца хватательная и сильно превосходит по длине остальные ножки; карпальный членик ее веретенообразно расширен. Средняя часть оперкулярной пластинки самца сильно расширяется у вершины. В остальном сходен с родом Janira.
В дальневосточных морях известно 3 вида. ' I
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Боковые края тельзона пильчатозазубрены. * 3.J. serricaudis Gurjan. 2 (1). Боковые края тельзона гладкие.
3 (4). Длина тельзона больше его ширцны . . . .. . * 1. J. kincaidi Rich. 4 (3). Длина тельзона меньше или равна его ширийе ................
.......................................* 2. J. derjugini Gurjan.
* 1. Janiropsis kincaidi Richardson (фиг. 12).
Richardson, 1904, Harriman Alaska Expedition. Crustacea, X; Richardson, 1905, Bull. U. S. Nat. Mus. № 54: 456, f. 509—514.
Длина головы больше ее ширины. Передний край головы прямой с закругленными боковыми углами. Глаза большие, черные, дорзальн»
2. JANIROPSIS
45
Фиг. 12. Janiropsis kincaidi Rich. ("экз. из Берингова моря, Командоры , а-т-общий вид; Ь — голова; с — I грудная ножка; d—II грудная ножка; е — рулевая ножка.
расположенные. Усики 1-й пары короткие; жгутик 8-члениковый у самки и 10-члениковый у самца. Усики 2-й пары длинные, достигают в вытянутом состоянии заднего края тельзона. 1-й грудной сегмент с прямым передним краем и прямыми боковыми краями. Эпимеры хорошо развиты, простые. Боковые части грудных сегментов не образуют лопастей. Тельзон удлиненный, слегка суживающийся дистально. Длина его больше ширины. Задний край тельзона образует небольшие заостренные лопасти по бокам широкой закругленной срединной лопасти. Uropoda короткие, немного меньше половины длины тельзона. Базальный членик их короче ветвей. Наружные ветви рулевых ног немного короче внутренних.
Цвет тела оливково-коричневый с густо расположенными черными точками. Длина животного 5.5 мм.
Распространение. Берингово море: Якутат, Аляска (Richardson, 1904); Командорские острова на литорали в массовом количестве под камнями (Гурьянова, 1930— 1931); Японское море: залив Петра Великого в зарослях Zostera папа на глубине 1 —3 м (Шурин, 1925—1930).
* 2. Janiropsis derjugini Gurjanova (фиг. 13).
Gurjanova, 1933, Иссл. морей СССР, 19: 82, рис. 5—6.
От предыдущего вида отличается более широким и коротким телом, коренастого сложения и крепкими более короткими грудными ножками, 6-й членик которых снабжен тупым отростком, нависающим над последним члени
ком. Голова очень короткая и широкая, ширина ее в 2 раза больше длины. Передний край головы образует волнистую линию с округлыми слабыми возвышениями по бокам и между основаниями первых антенн и слабыми вогнутостями у основания антенн. Передне-боковые углы головы слегка оттянуты вперед; глаза хорошо развиты, дорзальные. Базальные членики усиков 1-й пары расширенные, значительно превосходящие по ширине ширину вторых члеников ножки. Жгутик 11—12-чле-никовый. Усики 2-й пары в вытянутом состоянии достигают заднего края последнего грудного сегмента. Передний край 1-го грудного сегмента вогнут посредине с оттянутыми вперед боковыми частями. Эпимеры хорошо развиты на всех сегментах. Поверхность тела гладкая; тельзон короткий и широкий; ширина его значительно превосходит длину. Заднебоковые углы тельзона образуют небольшие треугольно-заостренные
46
7, JANIRIDAE
лопасти с редкими длинными щетинками на вершине. Длина рулевых ног почти равна половине длины тельзона; базальный членик короче ветвей. Ветви uropoda почти равной длины с пучками длинных щетинок на концах. Цвет тела бледнорозовый. Максимальная длина животного 6.5 мм.
Распространен не. Берингово море: Командорские^острова, на диторали в гротах среди зарослей губок и гидроидов в сильно прибойных местах (Гурьянова, 1933).
фиг. 13 a. Janiropsis derjugini Gurjan. а — общий вид; Ъ голова;' с — тель-зон И рулевые ножки.
Фиг. 13 b. Janiropsis derjugini Gurjan. а — ногочелюсть; Ь — рулевые ноги; с — первая пара ног; d — вторая пара ног; е — жвало; f ,— operculum S $.
* 3. Janiropsis serricaudis Gurjan. (фиг. 14).
Gurjanova 1935, Zool. Anz,, 114: 251, fig. 1. •
От прочих видов этого рода легко отличается зазубренностью» нижней трети боковых краев тельзона, приближаясь в этом отношений к J. breviremis G. Sars, к которому J. serricaudis очень близок, однако детали строения (вооружение шипиками) средней части оперкулярной пластинки и щупика ногочелюстей самца приближают нашу форму к южно-африканскому виду J. palpalis Barnard.
Тело удлиненное с почти параллельными боковыми краями; поверхность тела гладкая, усеянная мелкими пятнами черного пигмента, создающими светлосерую окраску. Передний край головы слегка вогнутый; по бокам головы на уровне глаз по одному тонкому шипйку. Глаза небольшие^, овальные, черные. Передний край трех первых грудных сегментов-
2. JANIROPSIS
47
слегка вогнут посредине; коксальные пластинки двулопастные. Тельзон овальный, длина его почти равна его наибольшей ширине; нижняя треть боковых краев тельзона крупно зазубрена (4—5 зубцов с каждой стороны); между зубцами по одному тонкому шипику; нижний край тельзона закруглен с обрубленным кончиком и группами тонких шипиков» Уроподы меньше длины тельзона (примерно 2/з его длины); наружная ветвь
немного короче внутренней; базальный членик и. ветви снабжены щетинками. Усики 1 пары короткие; базальный членик расширенный; жгутик 11—12 членико-вый. Ногочелюсти 9 нормального строения; у S строение , щупика такое же^ как у J. palpalis Barnard, т. е. три последних членика значительно удлиняются, и по длине "почти > равны друг другу. От J. palpalis Barnard наш вид отличается тем, что последний' членки щупика Г ногочелюстей самца ра-вен предпоследнему, тогда как у J. palpalis последний членик в 2 раза короче предпоследнего. Оперкулярная пластинка самца J. ser-. ricaudis почти в точности соответствует таковой J. palpalis Barnard, а от J. breviremis Sars отличается тем, что боковые углы дистального
12.
serricaudis Gurjan. 7 — самец; 2—
14. Janiropsis
3—telson g; 4—maxillipedes J; 5—uropoda
Фиг.
самка;
6—pereippoda I 9, 7‘—maxillipedes S; 8—mandibula (J; 9 — telson, uropoda $; 10 — антенна I 9; 11 — стебелек антенны II Л; 12—operculum $.
10.
II 12— operculum $.
ковые углы дистального конца оттянуты назад. Грудные ножки нормального для 33 мм. .
Распространение. Японское море, в прибрежной зоне района острова Петрова (42°49/N, 133°48'Е) на глубине от 0 до 2 м, на нижней стороне камней, на корках губок и сложных асцидий и в зарослях известковой водоросли Amphiroa.
не в стороны, а направлены рода строения. Длина тела,
48
1. JANIRIDAE
3. Род MICROPROTUS RICH.
Richardson, 1910, Proc. U. S. Nat. Mus., 37:116.
Тип рода: M. coecus Rich.
Голова несет короткий, широкий с прямым передним краем рострум. Глаз нет. Базальный членик усиков 1-й пары расширенный с вытянутым в виде лопасти передним наружным углом; жгутик многочлениковый. Усики 2-й пары в 2’/2 раза превышают длину тела животного. Молярный отросток жвал хорошо развит; щупик трех-/ \ члениковый. -Все тело животного явственно \ / распадается на 2 отдела — передний из 4 пе-
\ / редких грудных сегментов и головы, и зад-
\ I ний, состоящий из 3 последних грудных сег-
I I ментов и тельзона. Все сегменты тела и эпи-
I / меры снабжены длинными шиповидными
| I отростками. Все грудные ножки без исклю-
I чения ходильные; 1-я пара ног значительнЬ
н короче и слабее остальных. Uropoda хорошо
11 развиты, двуветвистые, длина их больше noil J ловины длины тельзона. Тельзон снабжен
I \ til шиповидными выростами. Тупой короткий
11 I 1 рострум, отсутствие глаз, расширенный ба-
111 I зальный членик усиков 1-й пары, и распадение
Ц I | тела на 2 отдела сближают этот вид с сем.
ЛЦга, Munnopsidae, однако строение жвал и руле-
JSweZ вых ножек и отсутствие плавательных ног
типично для семейства Janiridae. Известен ТТлЖТ только один вид.
1. Microprotus coecus Richardson
J: (фиг- 15)-
Richardson, 1910, Proc. U. S. Nat. Mus., 37: ТИС 16—118, f. 38-42.
« ** Голова широкая и короткая. Глаза от-
Фиг. 15. Microprotus coecus Rich, сутствуют. Усики 1-й пары достигают конца (по Richardson). 5_го членика ножки усиков 2-й пары; базаль-
ный членик их несет расширенную лопасть на переднем наружном крае; жгутик многочлениковый. Усики 2-й пары очень длинные, в 2J/2 раза длиннее тела; 2-й и 3-й членики ножки их несут по одному длинному шипу; жгутик многочлениковый. Эпимеры 2-го, 3-го и 4-го сегментов снабжены парой длинных шиповидных отростков. 4 передних грудных сегмента несут каждый по 3 длинных шипа — 1 посередине и по одному с каждой стороны у основания эпимер. Эпимеры 3 задних сегментов простые, округлые. 3 задних грудных сегмента несут по 3 срединных острых шипа у заднего края и по одному длинному шиповидному отростку с каждой стороны сегмента на передне-боковом угле.
3. MICROPROTUS 49
Тельзон округлой формы с 2 крепкими ‘ шиповидными отростками на дистальном конце между рулевыми ножками и 2 парами боковых отростков. На спинной стороне тельзона 2 коротких конических шипика — один у основания, другой на середине расстояния между основанием и концом тельзона. Базальный членик uropoda длинный и тонкий, достигает конца терминальных шипов тельзона, ветви uropoda равной длины. Грудные ноги 1-й пары ходильные, короткие и слабые; остальные ножки такого же строения, но значительно крепче и длиннее. Все членики ног длинные и тонкие, особенно 5-й и 6-й членики. Имеется добавочный коготок.
Распространение. Берингово море: 52°14/ N, 174°13' Е на глубине 600 м в чистом сером песке (Richardson, 1910). »
2. Сем. MUNNIDAE
Зубной отросток жвал хорошо развит, почти цилиндрический и направлен вперед или вертикально. Голова очень широкая и короткая, снабженная крупными, иногда очень сильно выступающими боковыми буграми. Тело вследствие неравномерного расчленения явственно распадается на 2 отдела, так как 4 передних грудных сегмента и голова значительно шире, чем 3 задних сегмента и тельзон. Глаза, если они имеются, расположены на хорошо развитых боковых буграх головы; у слепых форм эти бугры также имеются и часто заостряются на концах. Жвалы лишены щупика вовсе или же он сильно редуцирован. 1-я пара грудных ног хватательная с сильно развитым 5-м члеником. Все остальные ноги ходильные. Рулевые ножки очень маленькие, одноветвистые.
В дальневосточных морях известен один род Миппа с 4 видами.
1 . Род MUNNA KROYER
Kroyer, 1839, Naturh. Tidsskr., II: 612.
Тип рода: N. boecki Кг.
Тело короткое, грушевидное, распадается на 2 отдела — передний состоит из головы и 4 первых грудных сегментов и задний отдел — из 3 последних грудных сегментов и тельзона. Г олова широкая и короткая с резко выраженными боковыми буграми. Глаза обычно хорошо развиты и сидят на вершине этих бугров. Жвалы снабжены хорошо развитым щупиком. Рулевые ноги в виде коротких одноветвистых придатков на конце тельзона. Тельзон обычно грушевидной формы. Эпимеры маленькие, но ясно заметные, иногда снабжены шипами. Фронтальная часть головы между основаниями усиков вытянута в широкую тупую лопасть. Усики 1-й пары маленькие и слабые; усики 2-й пары крепкие и длинные, иногда превышающие длину тела животного.
В дальневосточных морях известно 4 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (6). Впереди глаз имеются небольшие предглазничные бугры, иногда с группой щетинок на вершине, все эпимеры несут крепкие шипы.
г- 4
1урьянова ’
50
2. MUNNIDAE
Фиг. 16. Munna stephenseni Gurjan. а — общий вид; Ъ — усики I пары; с — I пара ног; d— конец тельзона и рулевые ножки; е — II пара ног; /—operculum <5.
2 . (3). Края тельзона гладкие, лишенные шипов. Поверхность тела и тельзона гладкая, лишенная шипиков и волосков. *4. М. subneglecta Gurjan.
3 (2). Края тельзона несут от 2 до 4 шипов с каждой стороны. Поверхность тела гладкая или покрыта нежными волосками.
4 (5). Поверхность тела гладкая; по бокам тельзона только по 2 шипа с каждой стороны.................... • • • * 1. М. stephenseni Gurjan.
5 (4). Тело покрыто короткими нежными волосками. По бокам тельзона по 4 шип.! с каждой стороны................* 3. М. avatshensis Gurjan.
6 (1). Предглазничных бугров нет. Только 4 и 5 пары эпимер несут по 2 шипика ................................ . . * 2. М. arnholdi Gurjan.
* 1. Munna stephenseni Gurjanova (фиг. 16).
Gurjanova, 1933, Ис-след. морей СССР, 19 : 88, рис. 15.
Тело короткое и очень широкое; ширина его (у 3-го грудного сегмента) равна длине головы -+- ре-гаеоп. Голова большая и очень широкая; ширина ее почти в 3 раза больше длины. Глаза большие, прекрасно развитые и расположены на сильно развитых боковых буграх головы,
которые выходят за пределы боковых краев 1-го грудного сегмента. Впереди глаз по одному крупному предглазничному бугру, несущему группу щетинок на притупленной вершине. Усики 1-й пары с двухчлениковым жгутиком; последний членик длиннее 1-го и снабжен 4 длинными перистыми щетинками на конце. Усики 2-й пары относительно короткие и крепкие, также как и грудные ножки. Эпимеры сильно развиты на всех сегментах и снабжены длинными крепкими шипами по наружному краю. Тельзон восьмиугольной формы с прямым задним краем и несет с каждой стороны по паре очень крепких толстых, изогнутых боковых шипов, примерно на середине расстояния между основанием тельзона и его задним краем. На конце тельзона 2 пучка длинных простых щетинок, расположенных на задне-боковых его углах. Рулевые ноги маленькие, но крепкие; как и у М. kroyeri снабжены зубовидными отростками, направленными внутрь и назад; operculum самца с широким основанием, постепенно суживается
1. MUNNA
51
дистально; вентральные щетинки отсутствуют. Поверхность тела гладкая, без волосков или шипиков. Цвет животного белесовато-желтоватый с бурыми пятнами; максимальная длина 3 мм.
Распространение. В большем количестве держится под камнями, в нижнем горизонте литорали на о. Беринга; Командорские о-ва, Берингово море (Гурьянова, 1931).
* 2. Munna arnholdi Gurjanova (фиг. 17).
Gurjanova, 1933, Исслед. морей СССР., 19: 87, рис. 13—14.
Тело удлиненное с крепкими, относительно короткими усиками и трудными ножками. Ширина головы почти в 3 раза превышает ее длину.
. Фиг. 17а. Munna arnholdi Gurjan. а — общий вид; b — голова; с — задний край тельзона.
Фиг. 17 Ь. То же. а -1- рулевая нзжка; 6 — operculum (5; с — antenna I; d — peraeopoda I; е — pe-raeopoda II; f— peraeopoda III.
Глаза большие, округлые, посаженные на короткие, но мощные боковые бугры головы. Обе пары усиков короткие и крепкие. На спинной поверхности грудных сегментов редкие жесткие короткие волоски и посаженные правильными поперечными рядами короткие, толстые с тупыми вершинами шипики. На боковых расширенйях грудных сегментов по 2 более длинных и тонких шипа. Толстые короткие шипики более многочисленны на 3 последних грудных сегментах (по 5-6 шипиков в ряду на каждом сегменте). Тельзон удлиненный с округлыми очертаниями, слегка суживается у основания; на заднем конце между уроподами широкая закругленная лопасть. На спинной стороне тельзона короткие’ толстые притупленные шипики, расположенные правильными поперечными и продольными рядами таким образом, что получается 3 поперечных ряда (2 ряда по 4 шипика и один ряд из 2 более тонких шипов) и 4 продольных ряда (2 медиальных из 3 шипиков в каждом ряду и 2 латеральных по 2 шипика в каждом). На конце тельзона 2 пары тонких шипов. Рулевые ножки короткие, крепкие, с направленными внутрь и назад треугольно-заостренными отростками. Operculum самца широкий у основания, суживается дистально с закруг-4*
52
2. MU'NNIDAE
ленными концами. Имеется 2 пары вентральных щетинок. Общее крепкое телосложение этого рачка находится в полном соответствии с условиями его обитания—в зарослях губок и гидроидов на камнях и гротах в местах постоянного сильного прибоя. Максимальная длина 2 мм. Цвет тела бурокоричневый.
"Распространение. Берингово море: Командорские о-ва (о. Беринг), нижний горизонт литорали (сбор Гурьяновой, 1931).
*3. Munna avatshensis
Gurjan. (фиг. 18).
Gurjanova, 1936, Zool. Anz., 114 : 252, fig. 2.
Самец. Тело удлиненное с крепкими длинными грудными ножками и усиками; голова очень широкая, 'ширина ее в 22/2 раза больше длины. Глазные бугры мощно развиты, выходят за пределы боковых краев первого грудного сегмента. Глаза хорошо развиты с темным пигментом; предглазничные бугры
Ф...Г. 18. Munna avatshensis Gurjan. 1 - самец;’ Хорошо развиты. Передний 2—антенна I 5; 3—pereiopoda. I 6; 4 — telson край ГОЛОВЫ Прямой. Поверх-5—pereiopoda I j; б—operculum S. НОСТЬ ГОЛОВЫ, груДНЫХ СвГ-
ментов и тельзона покрыта короткими нежными волосками, лЭпимеральные пластинки с крепкими шипами. Тельзон широко грушевидный; боковые края его ближе к основанию несут по 4—5 крепких изогнутых шипов с каждой стороны. Группы шипов, но более тонких и длинных, имеются также на спинной поверхности тельзона; задняя часть тельзона между уроподами лопастевидно удлинена с закругленным задним краем. Рулевые ножки маленькие, но крепкие, с треугольными отростками, направленными назад и внутрь. Оперкулярная пластинка суживается дистально; задний край ее прямой, боковые углы вытянуты, заострены и направлены назад и в стороны; вентральные щетинки отсутствуют. 1 пара грудных ног мощная, короткая; 5-й членик широкий, вздутый; нижний дистальный угол образует крупный тупой зубец, у основания которого имеется еще один, более мелкий зубовидный отросток; 6-й членик много уже, короче пятого, слегка расширяется дистально; коготок заостренный, снабженный редкими щетинками. Остальные грудные ножки значительно длиннее и тоньше, чем 1-я пара; членики их усажены острыми игловидными шипиками. Усики 1-й пары короткие, первый членик .стебелька значительно шире и немного длиннее второго
Z. MUNNA
53
членика; 3-й членик очень маленький, короткий; второй его членик в 5 раз длиннее первого; последний членик очень мал, несет пару длинных сен
сорных волосков.
Самка. Тело значительно шире и короче, чем у самца. 1 пара грудных ножек много короче и'слабее, чем у 5; пятый членик нормального
строения, не вздутый, снабжен по внутреннему краю 4 крепкими шипами.
Тело слабо пигментировано; сиренево-серые пятна пигмента образуют поперечные полосы на члениках ходильных ног, группируются на боковых чаетях грудных сегментов над основанием эпимер и особенно густо покрывают тельзон таким образом, что по средней линии его тянется светлая, лишенная пигмента продольная полоса. Длина 2.2 мм. f
Распространение: Берингово море, восточное побережье Камчатки (А вачинская губа) среди зарослей гидроидов на глубине 15—20 м (К. Виноградов, 1934).
Фиг. 19. Munna subneglecta Gurjan. 7 — самец; 2 — maxillipedes 3; 3 — operculum 3; 4— pereiopoda II 3; 5—антенна I 3; 6— pereiopoda I 3; 7— самка; 8 — telson J; 9—pereiopoda I J; 10— pereiopoda II J: 77 — uropoda; 12—антенна I J.
* 4. Munna subneglecta Gurjan. (рис. 19)
Gurjarova, 1936, Zool. Anz., 114: 254, fig. 3.
Очень близок к M. neglecta Monod 1931, однако легко отличается от него строением 1 пары грудных ног и оперкулярной пластинки самца. Резко выражен половой диморфизм.
Самец. Тело удлиненное, сравнительно узкое; длина его более чем в 2 раза превышает ширину. Поверхность тела гладкая, сероватого цвета
54
3. MU NNOPSID A Е
с мелкими темными пятнами. Боковые края грудных сегментов закруглены. Эпимеры закругленные, снабженные парными шипами на 4 и 5 сегментах. Передний край головы прямой; глазные бугры большие, широкие; пред-глазничные бугры хорошо развиты. Глаза большие, черные. Антеннальные вырезы глубокие. Усики 1-й пары маленькие; жгутик 5-члениковый; базальный членик ножки расширенный, равен по длине второму; третий членик маленький и узкий, равный первому членику жгутика. Последний членик жгутика несет тонкие длинные щетинки. 1-я пара грудных ног мощная; второй членик удлиненный; третий короче второго, а четвертый короче третьего членика. 5-й членик мощно развит, много длиннее и шире второго членика с хорошо развитым внутренним дистальным отростком. Шестой членик крючкообразно изогнут с двумя бугорками по внутреннему 1фаю. Последний членик узкий, саблевидный. Средняя часть оперкулярной пластинки сильно напоминает 7И. neglecta Monod, но дистально суживается, и концы лопастей ее плавно закруглены.
Самка обладает более коротким и широким телом, слабее развитыми глазными буграми. Усикц второй пары короткие, короче тела. Грудные, ножки нормального строения, крепкие и сравнительно короткие. Длина.2 мм.
Распространение: Японское море, район острова Петрова (42°49' N, 133°48z Е); литоральная зона на нижней стороне камней, на корках губок и асцидий; спускается и ниже, до глубины 0.5 м в самый верхний горизонт сублиторали.
3. Сем. MUNNOPSIDAE
Глаз нет. Зубной отросток жвал короткий, суживающийся дистально или нацело редуцирован. Тело распадается на 2 отдела. Голова и 4 передних грудных сегмента образуют передний отдел, а 3 задних грудных сегмента и тельзон — задний отдел. Базальный членик усиков 1-й пары сильно расширен в виде пластинки; 1-я пара грудных ножек простая, меньше и слабее, чем остальные. 3 следующих пары ног ходильные, а 3 задних пары грудных ножек плавательные, с расширенными в виде пластинок и снабженными плавательными щетинками 2 дистальными члениками. Рулевые ноги маленькие, одно- или двуветвистые. Распадается на 3 группы или подсемейства — подсем. Eurycopinae с характерным родом Еигусоре, подсем. Ilyarachninae с основным родом Ilyarachna и подсем. Munnopsi-пае с типичным родом Munnopsis. Так как на Дальнем Востоке известно еще очень мало представителей этого семейства, мы рассмотрим его по родам. Из дальневосточных морей известно 4 рода.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Усики терминальные; тельзон узко-треугольной формы, суживается и заостряется к дистальному концу . .1. Ilyarachna G. Sars (стр. 55).
/. IL YARACHNA 55
2 (1). Усики дорзальные; тельзон овальной формы с закругленным задним концом.
3 (4). Жвалы лишены зубного отростка; 7-й членик 3 пар задних (плавательных) грудных ножей отсутствует. Все тело удлиненное; ’ задний отдел тела много уже переднего . . 4. Munnopsis М. Sars (стр. 64).
4 (3). Зубной отросток жвал хорошо выражен или же образует крупный широкий бугор. 7-й членик 3 задних (плавательных) грудных ножек имеется; все тело укороченное; задний отдел тела не уже переднего.
5 (6). Жвалы имеют lacina mobile; зубной отросток хорошо развит, цилиндрический; 7-й членик плавательных ножек хорошо развит и длиннее, чем плавательные щетинки.................3. Еигусоре G. Sars (стр. 62).
6 (5). Жвалы лишены lacina mobile; зубной отросток в виде широкого отростка; 7-й членик плавательных ног маленький, короче плавательных щетинок. - ........................2. Munnopsurus Rich. (стр. 57).
1. Род ILYARACHNA G. SARS.
G. Sars, 1869, Christ. Vid. Selsk. ForhandL: 167. — Mesostenus G. Sars, 1863, ibid.: 8 (nom. praeoccup.).
Тип рода: 1. longicornis G. Sars.
Tело суживается к заднему концу; голова очень большая и широкая, часто шире переднего грудного сегмента. Глаз нет; фронтальная лопасть отсутствует; 2-й членик щупика ногочелюстей длинный и широкий; 2 дистальных членика хорошо развиты. Жвалы имеют трехчлениковый щупик, хорошо развитый зубной отросток, но не имеют lacina mobile. 4 передних грудных сегмента узкие, сильно сближены друг с другом; передние края их часто зазубрены. 3 задних грудных сегмента большие и широкие. 5-й сегмент глубоко вырезан по средине заднего края, так что край этот аркообразно вогнут. 2-й членик 3-й и 4-й пар ног короткий, а 3-й членик более, чем в 2 раза длиннее 4-го. Тельзон треугольной формы, тупозаостряющийся на конце. Усики терминальные. Рулевые ножки одноветвистые, очень маленькие, вентрально расположенные.
На Дальнем Востоке известно 2 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Передний край четырех первых грудных сегментов крупно пильчато зазубрен. На голове 6 крупных шипов . . . . * 1. I. zachsi Gurjan.
2 (1). Передние края 4 первых грудных сегментов несут по 4 крупных шиповидных отростка. На голове 4 крупных шипа и 2 более мелких по бокам их.............................*2. I. starokadomskii Gurjan.
56
3. MUNNOPSIDAE
* 1. Ilyarachna zachsi Gurjan. (фиг. 20).
Gurjanova, 1933, Исслед. морей СССР, 19: 83, рис. 7.1
Фиг. 20. Ilyarachna zachsi Gurjan. а — общий вид; b — antenna I; с — регаео-poda I; d—uropoda.
Голова очень широкая с очень сильно развитыми лопастями и выпуклой затылочной частью. Ширина головы больше, чем ширина 1-го и 2-го сегментов. груди. -На выпуклой затылочной части, покрытой ячеистой скульптурой, 6 крупных шипов, по 3 шипа в ряду с каждой стороны от средней линии головы; передние края 4 передних грудных сегментов очень крупно пильчато зазубрены, как у видов. Echinozone, однако присутствие щупика на жвалах, одноветвистые рулевые ножки, присутствие продольного срединного киля на operculum самки и строение усиков заставляют с несомненностью отнести эту форму к роду Ilyarachna.1 2 Очень-характерно строение усиков 1-й пары. Базальный членик ее более узкий, чем у других известных видов этого рода и, суженный у основания, расширяется дистально; наружный край его усажен крепкими изогнутыми шипами; особенно характерен очень длинный 3-й членик (1-й членик жгутика),, который почти в 2 раза длиннее 2-го членика и немного длиннее даже 1-го, базального членика, и снабжен тонкими длинными щетинками. Рулевые ноги маленькие, одноветвистые; базальный членик их длинный, узкий, усажен по краям длинными перистыми щетинками, так что единственная ветвь, скрытая этими щетинками, плохо видна. В остальном /. zachsi близка к I. hirticeps G. Sars.. Максимальная длина животного 9 мм.
Распространение. Японское море: залив Петра Великого, открытая часть на глубинах от 121 до 780 м; у мыса Гамова (район Посьета) на глубине 105 м, песок; в районе залива Владимира на глубине 1002 м; Татарский пролив на глубине 250 м (сборы ТИРХ’а и ГГИ, 192& и 1930—1931).
1 В моей работе 1933 г., по недосмотру технического редактора, на стр. 83 перепутаны места рисунков. Левый рисунок (под номером 7) относится к виду I. starokadomskii' правый рисунок, поставленный под номером 8, — к I. zachsi. Е. Г.
2 Наоборот, Ilyarachna quadrispinosa Beddard должна быть отнесена к роду Echinozone G. Sars, так как у этого вида,, по описанию и рисункам (Beddard, 1886), жвалы. лишены щупика, рулевые ноги имеют 2 ветви, передний край 5-го грудного сегмента -несет зубцы, как и предыдущие сегменты, и на конце тельзона над основанием рулевых ног-имеются небольшие вырезки, характерные для рода Echinozone.
1. IL YARACHNA
57
Фиг. 21. Ilyarachna siarokadomskii Gurjan. a —"общий вид; b — peraeopoda I; c — ножки antenna II; d—antenna L_i,
* 2. Ilyarachna starokadomskii Gurjan. (фиг. 21). Gurjanova, 1933, Исслед. морей СССР, 19: 84, рис. 8.
Голова значительно уже, чем у предыдущего вида, не шире 1-го грудного сегмента. Передний край ее прямой; затылочная часть уплощенная и несет 4 крупных шипа в среднем поперечном ряду и по 2 более мелких шипа по бокам этого ряда, немного отступя кзади. Передний край 3 первых грудных сегментов несет 4 очень крупных, направленных вверх и вперед шипа. 4-й сегмент с гладким, лишенным шипов, передним краем, но его задний край снабжен многочисленными мелкими шипиками. Эпимеры заостренные. Над эпимерами на боковых частях 4 передних грудных сегментов имеется по 2-3 шипа с каждой стороны. Базальный членик усиков 1-й пары такого же строения, как и у предыдущего вида; 3-й членик (т. е. 1-й членик жгутика) крепкий,
длинный, равный длине базального членика, лишен щетинок, но имеет редкие шипы. Весь жгутик очень длинный, превосходит длину базального членика в 4 раза и состоит из 19 (не считая 1-го длинного) удлиненных, узких члеников. Верхние края 3 члеников ножки усиков 2-й пары усажены длинным и тонкими шипами. Все тело более короткое и крепкое, чем у II. zachsi. Максимальная длина животного 6.5 мм.
Распространение. Японское море: залив Петра Великого на глубине 780—730 м; Татарский пролив на глубине 250 м; у о. Фурру-гельма на 130 м (сбор ТИРХ’а и ГГИ 1930—1931). Охотское море: ст. 216 —51°90' N, 154°17/ Е, 591 м; ст. 262 — 52°34' N, 154o06z Е на 100 м (сборы ГГИ, 1932).
2. Род MUNNOPSURUS RICH.
Richardson H., 1912, Bull. Inst. Ocean. Monaco, 227.
Тип рода: M. giganteus (G. Sars).
В основном очень близок к роду Еигусоре, отличаясь от него строением головы, жвал и грудных ножек. Жвалы лишены подвижной пластинки
58
3. MUNNOPSIDAE
(lacina mobile); режущий край их простой, иногда только с одним зубцом. Зубной отросток в виде большего вытянутого бугра с закругленной вершиной и пучком жестких щетинок. Предпоследний членик щупика ного-челюстей с вытянутым в виде узкой лопасти верхним внутренним углом. 7-й членик 3 задних (плавательных) ножек очень маленький и слабый, короче, чем плавательные щетинки на краях расширенных 5-го и 6-го члеников. 2-й членик й-ой пары ног длинный, длиннее 3-го членика. Все тело распадается на 2 участка: 4 передних грудных сегмента с не-сколько расширенными боковыми частями, направленными вперед; 3 задних сегмента с аркообразно-изогнутыми краями и боковыми частями, направленными назад. Тельзон широкий с закругляющимся задним краем. Рулевые ноги терминальные, маленькие, двуветвистые. Эпимеры маленькие, закругленные, имеются на всех сегментах.
Из дальневосточных морей известно 3 вида и 1 вариетет.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ И ФОРМ
1 (4). Наружный край 2-го и 3-го члеников щупика ногочелюстей голый. 2 (3). На спинной стороне 3 последних грудных сегментов имеется борозда по средней линии и по бокам ее по одному низкому широкому бугорку
........................................*3. М. minutus Gurjan.
3 (2). Спинная сторона 3 последних грудных сегментов гладкая, лишенная бугорков и продольной борозды .....................2. М. laevis Rich.
4 (1). Наружный край 2-го и 3-го члеников щупика ногочелюстей усажен короткими щетинками.
5 (6). По средней линии 3 последних грудных сегментов имеется глубокая борозда и по одному бугорку с каждой стороны ее....................
.....................................*1. М. giganteus G. Sars.
6 (5). На 3 последних грудных сегментах нет ни борозды ни бугорков .
......................*1а. М. giganteus var. ochotensis Gurjan.
* 1. Munnopsurus giganteus (G. Sars.) (фиг. 22).
G. Sars, Arch. f. Math, og Naturv., II: 353 (Eurycope)-, G. Sars, 1885, Den Norske Nord-, havs Exp.: 130, pl. XI, f. 1—25 (Eurycope). — arctica Richardson, 1912, Bull. Inst. Ocean. Monaco, 227.
Тело продолговатое, ясно распадающееся на 2 отдела. Передний отдел (голова и 4 передних грудных сегмента) значительно короче, чем задний и отделен от него ясной перетяжкой. Все сегменты груди и голова свободны. Голова не имеет ясно выраженной фронтальной лопасти, и все ротовые части (буккальная масса) вытянуты вперед. 1-й сегмент груди немного шире головы; 5 следующих сегментов с ясными маленькими округлыми эпимерами; эпимеры 1-го сегмента не видны сверху; 3 задних грудных сегмента одинаковых размеров, длиннее передних сегментов и снабжены глубокой бороздой по средней линии. По бокам этой борозды
2. MUNNOPSURUS
59
по одному округлому бугорку. Тельзон очень большой' и широкий со слегка оттянутым посредине закругленным задним концом. Длина его почти равна длине 3 последних грудных сегментов вместе. Базальный членик усиков 1-й пары в виде расширенной плоской пластинки. Длина его почти равна длине головы. 2-й и 3-й членики очень маленькие; жгутик многочлениковый/ в 2 раза длиннее базального членика. Усики 2-й пары в 4 раза длиннее базального членика. Усики 2-й пары в 4 раза длиннее тела; 2 последних членика ножки их очень длинные; длина каждого из них почти равна длине тела; жгутик многочлениковый в 2 раза длиннее, чем последний членик ножки. Глаз нет. Режущий край жвалы гладкий, без зубцов; зубной отросток в виде широкого округлого бугра с пучком щетинок на вершине; щупик хорошо развит. 2-й и 3-й членики щупика ногочелюстей расширены и густо усажены по внутреннему краю щетинками. Коготок 1-й пары грудных ног рудиментарный. Все 4 передних пары ног ходильные и очень длинные; 3 задних пары ног плавательные с расширенными в виде пластинок 5 и 6 члениками, по краям усаженными перистыми щетинками. 7-й членик их рудиментарный и составляет всего !/8 Длины 6-го членика. Рулевые ножки очень маленькие, двуветвистые, прикреплены на брюшной стороне заднего конца тельзона; наружная ветвь их значительно короче внутренней. Длина до 33 мм.
Распространение. Берингово море: 55°25' N, 165°46'Е, глуб. 136—250 м, 2 экз., длина 13 мм (ст. 5, сбор ГГИ, 1932). Охотское море: 49°13' N, 146°06' Е, глуб. 645 м, 1 экз., длина 13 мм (сбор ГГИ, 1932, ст. 212).Сев. Ледовитый океан от Гренландии до моря Лап- тевых на глубинах от 200 до 1100 м.
*la. Munnopsurus giganteus var. ochotensis Gurjan. (фиг. 23).
Gurjanova, 1933, Исслед. морей СССР, 19: 84, рис. 9.1
От типичной формы отличается меньшими размерами, более удлиненным и узким телом, строением плавательных ног и тельзона и структурой 3 последних грудных сегментов, на спинной стороне которых нет ни срединной борозды ни бугорков по бокам ее. Длина тельзона заметно превосходит его ширину и задний конец его сильнее оттянут, чем у преды-
1 Места рисунков в моей работе 1933 перепутаны. Правый рисунок под № 10 должен быть перенесен влево под № 9. Е. Г.
60
3. M UNNO PS IDAS
дущего. 5-й и 6*-й членики плавательных ног относительно более длинные и узкие, равной длины; 7-й членик очень узкий и длинный, более половины длины 6-го членика. В остальном не отличается от типичной арктической формы. Длина 15 мм.
Распространение. Охотское море: 56°45' N, 145°50'jE, глуб. 218 м, 6 экз. (сб. ГГИ, 1930, ст. 135/20);"57°52/ N, 143°11/Е, глуб.80 м, 1 экз. (сб. Гейнеман, 1910, ст. 18); 49°13 N, 146°06' Е, глуб. 645 м, 3 экз. (сб. ГГИ, 1932, ст. 212).
Фиг. 24. Munnopsurus laevis (Rich.) (no Richardson); a — общий вид; Ъ — peraeopoda 5.1
Фиг. 23., Munnopsurus giganteus (G. Sars) var ochotensis Gurjan. a — общий вид; b — тельзон; с — uropoda; d — peraeopoda 5; e — mandibula; /— maxillipedes; к—*antennae I и'II.
2 . Munnopsurus laevis (Richardson) (фиг. 24).
Richardson H., 1910, Proc. U. S. Nat. Mus., 37: 120, f. 45—46 (Еигусоре).
Хотя Richardson и не дает описания ротовых частей этой формы, однако, судя по общему строению тела, присутствию маленьких закругленных эпимер на всех грудных сегментах и отсутствию ясно выраженной, столь характерной для видов рода Еигусоре фронтальной области (area frontalis), этот вид должен быть отнесен к роду Munnopsurus.
Тело продолговато-овальное, длина его в 272 раза больше ширины. Поверхность тела совершенно гладкая. Ширина головы равна ширине 1-го грудного сегмента; 3 следующих все одинаковой ширины и немного шире первого. 3 задних сегмента груди Постепенно увеличиваются в длине таким образом, что 5-й в 1х/2 раза, 6-й в 2 раза, а 7-й в 2т/2 раза длиннее 4-го сегмента, хотя ширина их остается той же самой. Эпимеры в виде маленьких закругленных пластинок на всех грудных сегментах. Спинная
2. MUNNOPSURUS
61
Фиг. 25. Munnopsurus minutus Gurjan. а — общий вид; Ь — antenna II; с — antenna I; d—• maxillipedes; е — peraeopoda 5;/—cephalon; к—telson, uropoda.
сторона 3 последних грудных сегментов гладкая, лишенная срединной борозды и бугорков. Тельзон яйцевидно удлиненный с оттянуто-закругленным задним концом. Рулевые ножки очень маленькие, двуветвистые; наружная ветвь немного короче внутренней. 1-я пара грудных ног короче и слабее, чем 3 следующих очень длинных пары. Расширенные 5 и 6 членики плавательных ножек (3 задних пар) равной длины со щетинками только на внутреннем крае. 7-й членик очень тоненький, палочкообразный и составляет 1/3 длины 6-го членика. Дл. 10 мм. '
Распространение. Японское море: 31°39' N, 129°24' Е, глуб. 800 м. Берингово море: 54°33' N, 178Р45' Е, глуб. 1100 м. Северная часть Тихого океана 42°02/ N, 142°33' Е, глуб. 680 м (Richardson, 1910).
* 3. Munnopsurus minutus Gurjan. |фиг. 25).
Gurjanova, 1933, Исслад. морей СССР, 19: 84, рис. 10.1
Голова очень широкая, шире, чем 1-й грудной сегмент. Глаз нет; базальные членики 1-й пары усиков сильно расширены у основания; жгутик 9-членико-вый, членики тонкие, длинные, снабженные на их проксимальном конце 3—4 длинными щетинками. 2-й и 3-й членики щупика ногочелюстей с голым, лишенным щетинок внутренним краем, только на внутреннем дистальном угле 3-го членика группа щетинок. Эпимеры малень
кие, округлые, ясно выраженные на первых 4 грудных сегментах. На спинной стороне каждого из 3 последних сегментов ясная продольная срединная борозда и по бокам ее по одному резкому бугорку. Тельзон в общем пятиугольной формы со сглаженными углами. Ширина его у основания немного больше длины. Рулевые ножки относительно крупнее, чем у других видов, двуветвистые. 6-й и 5-й членики плавательных ног (3 задних пар грудных ног) не так сильно расширены, как у предыдущих видов. 6-й членик почти в 2 раза меньше 5-го; 7-й членик длинный, червеобразный, достигает 2/з длины 6-го членика. Длина животного 7 мм.
1 В моей работе 1933 г. по недосмотру технического редактора места рисунков 9 и 10 на стр. 85 перепутаны. Левый рисунок осносится к М. minutus, правый — к М. giganteus var. ochotensis.
62
3. MUNNOPSIDAE
Распространение. Японское море, залив Петра Великого, к югу от острова Русского на глуб. от 230 до 440 м; к югу от мыса Гамова на глуб. 190 м и на 42'43' N, 135°59' Е на глуб. от 197 до 202 м (сб. ТИРХ, 1930).
. 3. Род EURYCOPE G. SARS
G. Sars, 1863, Chr. Vid. Selsk. Forhandl.: 5.
Тип рода: E. cornuta G. Sars.
.Тело слабо-вытянутое, овальное, сводчато-выпуклое на спинной стороне и плоское на брюшной стороне; распадается на 2 отдела — передний состоит из головы и 4 передних коротких и широких грудных сегментов и задний отдел из больших и широких 3 последних грудных сегментов и тельзона. Грудные сегменты обнаруживают тенденцию к слиянию друг с другом. Голова с хорошо выраженной, ясно отграниченной фронтальной лопастью, благодаря глубоким вырезкам переднего края у основания антенн. Глаз нет. Усики 1-й пары дорзальные с расширенными в виде чешуйки базальными члениками. Усики 2-й пары тонкие й длинные, обычно значительно длиннее тела. Жвалы снабжены lacina mobile; режущий край с зубцами; зубной отросток хорошо развит; щупик с изогнутым пластинчатым последним члеником. Предпоследний членик щупика ногочелюстей с вытянутым в виде узкой допасти внутренним верхним углом. 4 передних грудных сегмента снабжены заостренными, направленными вперед эпимерами. Тельзон большой, полукруглой или полуовальной формы с закругленным задним краем. Рулевые ножки маленькие, двуветвистые, прикрепленные на брюшной стороне тельзона. 4 пары передних грудных ножек ходильные, причем 1-я пара короче и слабее следующих. 3 задних пары грудных ног плавательные, с расширенными в виде пластинок 5-м и 6-м члениками, густо усаженными по краям перистыми щетинками; коготок нормально развит. Большое значение при определении видов этого рода имеет форма брюшной крышечки (operculum) самца и самки, а также форма фронтальной лопасти.
Из дальневосточных морей известно два вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Фронтальная лопасть копьевидно заострена на конце..........
......................................* 1. Е. spinifrons Gurjan.
2 (1). Фронтальная лопасть широкая с прямым передним краем ....
......................................* 2. Е. pavlenkoi Gurjan.
51! 1. Eurycope spinifrons Gurjan. (фиг. 26).
Gurjanova, 1933, Исслед. морей СССР, 19: 85, рис. И а, рис. 12 а—д.
Все сегменты груди свободны; тело вытянуто-овальной формы. Длина сегментов кзади постепенно увеличивается; предпоследний и последний
3. EUR YCOPE
63
Фиг. 26. Eurycope spinifrons Gurjan.
их сегментов.
грудные сегменты равной длины и превышают длину 5-го грудного сегмента почти в 2 раза. Фронтальная лопасть головы копьевидно заостряется и несет у своего дистального конца 2 пары шипов. Внутренний дистальный угол расширенного базального членика усиков первой пары образует очень слабо развитую лопасть с 3—4 щетинками на вершине. Тельзон полукруглой формы с небольшими вырезками над основанием рулевых ног;, ширина его почти в 1.5 раза превышает длину. Рулевые ноги двуветвистые, внутренняя ветвь их в 1.5 раза длиннее наружной; на концах обоих ветвей пучки щетинок. Макс. дл. 5 мм.
Глубоководная форма Японского моря.
Распространение. Япон-ское море: залив Петра Великого, на глуб. от 730 до 3000 м; бухта Владимира 43°55' N, 135°56' Е на глуб. от 308 до 1002 м; 42°38' N, 134°1/ Е на глуб. 669 м (сб. ГГИ, 1931).
*2. Eurycope pavlenkoi Gurjan. (фиг. 27).
Gurjanova, 1933, Исслед. морей СССР, 19: 85, рис. 11-6, 12-т.
Все сегменты тела свободны; тело короткое, удлиненно-овальное. Длина передних четырех грудных сегментов одинаковая, меньше, чем длина более крупных 3 послед
равной длины с сильно выпуклым передним и вогнутым задним краями. Фронтальная лопасть головы образует широкую закругленную, с прямым и валикообразно утолщенным передним краем лопасть между основаниями первых усиков. Базальный членик усиков 1-й пары сильно расширен у основания, так что ширина его основания в 1.5 раза превышает длину всего членика; жгутик 8-члениковый; членики вытянутые, снабжены длинными щетинками. Тельзон сравнительно короткий, правильной полукруглой формы; ширина его у основания почти в 2 раза превышает длину (1.7 раза). Внутренняя лопасть предпоследнего (4-го) членика щупика ногочелюстей равна или немного длиннее последнего членика. Operculum самки с хорошо выраженным срединным продольным килем. Ножки и грудные и рулевые обломаны, так что строение их неизвестно. Длина 4 мм.
Распространение. Японское море: залив Петра Великого к югу от мыса Тобизина на глубине 60 м, 2 экз., длина 7 мм.
Фиг. 27. Eurycope pavlenkoi Gurjan.
Все 3 задних сегмента
64
3. MUNNOPSIDAE
Фиг. 28. Munnopsis latifrons Beddard; a — общий вид (по Beddard); b—telson (no Richardson).
4. Род MUNNOPSIS M. SARS.
M. Sars, 1860. Chr. Vid. Selsk. Forhandl.: 84.
Тип рода: M. fу pica M. Sars.
Тело вытянутое и ясно распадается на 2 отдела. Голова и 4 передних грудных сегмента образуют передний широкий отдел; 3 задних грудных сегмента и тельзон узкие, длинные и образуют задний отдел, значительно более узкий, чем передний. Голова умеренной величины; передний край ее у основания антенн глубоко вырезан; между усиками образуется длинная, вытянутая вперед язычковидная лопасть. Боковые части 4 передних грудных сегментов слегка вздуты. 3 задних сегмента тесно соприкасаются друг с другом с сильно выпуклой спинной поверхностью. Тельзон вытянуто овальный. Усики 1-й пары с очень большим расширенным базальным члеником. Усики 1-й пары очень длинные, во много раз превышают длину тела. 4 передних пары грудных ножек ходильные; 1-я и 2-я пары много короче и слабее последующих, которые очень длинные, в несколько раз длиннее тела. Три задних пары грудных ножек плавательные с расширенными листовидно 5-м и 6-м члениками. 7-й членик редуцирован нацело. Рулевые, ноги терминальные, одноветвистые. Глаз нет; жвалы лишены зубного отростка; режущий край с зубцами, щупик хорошо развит; lacina mobile имеввся. Предпоследний членик щупика ногочелюстей простой, не имеет лопа-стевидного отростка на верхнем внутреннем угле.
Из дальневосточных морей известен только один вид.
1. Munnopsis latifrons Beddard (фиг. 28).
Beddard, 1885, Proc. Zool. Soc., London, 917, pt. 4; Beddard, 1886, Report voyage Challenger, XVII: 56, pl. X., f. 1—4.
Голова очень большая и широкая; длина ее равна длине 3 передних грудных сегментов вместе; ширина равна ширине 1-го сегмента. Расстояние между основаниями маленьких усиков 1-й пары очень большое, рав-
ное 7з ширины головы. Эпимеры маленькие, плохо заметные; 3 задних грудных сегмента сильно разнятся по величине друг от друга; 1-й из них (5-й грудной сегмент) самый короткий, длина его равна всего 1/3 длины каждого из 2 следующих сегментов; 6-й и 7-й сегменты почти равной длины. Спинная поверхность их покрыта неглубокими поперечными бороздками, отделяющими друг of друга небольшие выпуклости. Тельзон большой, овальный; длина .его равна длине 4 последних грудных сегментов
4. MUNNOPSIS
65
вместе; задний край закруглен. Боковые части тельзона над основанием рулевых ног образуют по одному с каждой стороны крепкому отростку, направленному назад. Рулевые ножки стилетовидные, одноветвистые; длина их почти равна длине тельзона; снабжены длинными простыми щетинками. Усики второй пары очень длинные, в 6 раз превышают длину тела. 5-й и 6-й членики ножки очень длинные, усажены крепкими шипами; жгутик короткий и не разбит на отдельные членики. 1-я пара грудных ног короткая, хватательная; внутренний край 3-го и 4-го члеников ее усажен крепкими шипами, гребневидно расположенными. 3 задних пары ног плавательные с листовидно расширенными 5-м и 6-м члениками; 7-й членик их отсутствует. Длина 15 мм.
Распространение. Восточное побережье Японии 35°11' N, 139°28' Е, глуб. 621 м (Beddard, 1886), 34°5Z N, 137°59' Е.;Жел-т о е м о р е: 30°34' N, 129°19z Е, глуб. от 792 до 1190 м (Richardson, 1910).
И. Подотряд FLABELLIFERA
В состав подотряда входят очень разнообразные группы Isopoda, часто с сильно измененными конечностями и ротовыми частями под влия-янием паразитического и полупаразитического образа жизни. Единственными, правда, очень характерными, чертами, объединяющими представителей этого подотряда, будут, во-первых, присутствие 7 пар грудных ножек и, во-вторых, строение рулевых ножек, которые прикреплены по бокам тельзона у его основания и вместе с тельзоном образуют хвостовой веер, подобный хвостовому вееру Decapoda и Schizopoda. Брюшные ножки -имеют характер настоящих плавательных органов, но в то же время они служат также для дыхания. У большинства грудные ножки хватательного или цепляющегося типа; ротовые части специально модифицированы, часто превращены в сосательные органы. Тельзон не срастается с брюшными сегментами и образует свободную хвостовую пластинку. Глаза часто очень большие, занимающие иногда почти всю поверхность головы, но иногда отсутствуют. »
В дальневосточных морях представлен 3 семействами Anthuridae, Sphaeromidae и Cymothoidae.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ
1 (2). Тело вытянутое, червеобразное, узкое, почти цилиндрическое; эпимеры отсутствуют; тельзон и рулевые ножки вместе образуют хвостовой веер в виде чашечки...............6. Anthuridae (стр. 125).
2 (1). Тело широкое, овальных очертаний, сплюснуто в дорзо-вентральном направлении. Эпимеры хорошо развиты или, сливаясь с сегментами, образуют плевральные расширения; тельзон вместе с рулевыми ногами образует плоский хвостовой веер.
Гурьянова 5
66
4. CYMO THOIDAE
3 (4). Ветви рулевых ног не одинаковые; одна из ветвей или неподвижно прикреплена, или же рудиментарна, значительно короче другой или совсем отсутствует.....................5. Sphaeromidae (стр. 95).
4 (3). Ветви рулевых ног более или менее одинаковые в виде расширенных, пластинок, большей частью равной или почти равной длины, обе подвижные ..............................4. Cymothoidae (стр. 66).
4. Сем. CYM0TH01DAE
Глаза хорошо развиты; тело сплющено в дорзо-вентральном направлении й более или менее равномерно расчленено; нередко наблюдается асимметрия. Эпимеры все хорошо развиты и не сливаются с сегментами. Рулевые ноги двуветвистые; ветви обычно расширены в виде пластинки,, равной длины. Рулевые ноги вместе с тельзоном образуют плоский веер,' так как прикреплены они по бокам тельзона у его основания и ветви лежат в одной плоскости с тельзоном. Ротовые части й конечности несколько-видоизменены, часто с крючковидными изогнутыми члениками и вооружены крючками. Эпистома разделена на фронтальную пластинку и clypeus.
Большей частью эктопаразиты. В дальневосточных морях представлены 3 подсемействами и 9 родами.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ
1 (2). 2 последних членика щупика ногочелюстей усажены щетинками, но не крючками; щупик лежит свободно . . . .1. Cirolaninae (стр. 66).: 2 (1). Щупик ногочелюстей окружен конусом, образованным дистальными концами ротовых частей; членики щупика вооружены не щетйнками,. а крючками
3 (4). Обе пары усиков явственно состоят из ножки и жгутиков. Членики щупика жвал узкие; щупик ногочелюстей состоит из S члеников (в редких случаях из 2) ........... 2. Aeginae (стр. 69) 4 (3). Обе пары усиков редуцированы с почти незаметным разделением » их на ножку и жгутик; членики щупика жвал расширены; щупик ногочелюстей всегда двухчлениковый ... 3. Cymothoinae (стр. 81).
1. Подсем. CIROLANINAE
Тело широкое, вытянутое, сильно выпуклое со спинной стороны;, брюшной отдел состоит из 6 сегментов. Глаза обычно хорошо развиты, небольшие, расположены по бокам головы или на нижней стороне. Усики 1-й пары очень маленькие; усики 2-й пары длинные и тонкие; 2 последних членика щупика ногочелюстей лишены крючков и усажены щетинками. Только 3 первых пары грудных ножек хватательные, остальные ножки ходильные. Рулевые ноги хорошо развиты, большие. Из дальневосточных морей известно 2 рода.
1. CIROLANA
67
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Брюшные ножки нормального строения, без пучков жаберных нитей у основания их ветвей. Глаза расположены по бокам головы и лишь частично, заходят на нижнюю сторону . 1. Cirolana Leach, (стр. 67).
2 (1). У основания ветвей брюшных ножек густые пучки жаберных нитей; глаза расположены по бокам головы, но на нижней (брюшной) ее стороне . ...................... 2. Bathynomus M.-Edw. (стр. 68).
1. Род CIROLANA LEACH
Leach, 1818, Diet. Sc. Nat., XII: 347.
Тип рода: С. стanchii Leach.
Тело более или менее удлиненное с гладкой, выпуклой спинной поверхностью. Задний край головы вогнутый. 1-й грудной сегмент образует боковые расширения, охватывающие голову с боков. Ножка усиков 2-й пары пятичлени-ковая. Зубной отро
сток жвал вытянут фиг. 29. Cirolana japonensis Rich, (по Richardson), а — усики В виде пластинки, уса- С нижней стороны; Ъ — брюшяо; с — I грудная ножка; d — II гр.
„ ножка; е — IV гр. ножка; /—VII гр. ножка.
женной по переднему ..........
краю рядом зубчиков. Рулевые ноги с оттянутым внутрь отростком базального членика; наружная ветвь уже внутренней. Брюшные ножки лишены жаберных нитей.
В дальневосточных морях известен один вид.
1. Cirolana japonensis Richardson (фиг. 29).
Richardson, 1904, Proc. U. S. Nat. Mus., 27: 35, f. 3—5.
Длина тела в 3 раза больше его ширины; глаза очень маленькие, округлые, расположенные на переднебоковых углах головы, еветлокорич-невые. Перёдний край головы с маленьким острием посредине и небольшими вдавлениями у основания усиков. Усики 1-й пары очень короткие, достигают конца 4-го членика ножки усиков 2-й пары; жгутик из 10 очень коротких слабо отграниченных друг от друга члеников. Усики 2-й пары достигают переднего края 4-го грудного сегмента; жгутик из 24 члеников.
5*
68
4. CYMO THOIDAE
Фронтальная (интерантеннальная) пластинка узкая и удлиненная; clypeus не вооружен, совершенно гладкий. 1-й грудной сегмент немного длиннее 2-го. 4-й, 5-й и б-й сегменты равной длины и равны по длине 1-му. Эпимеры 2-го и 3-го сегмента простые; эпимеры следующих 4 сегментов с оттянутыми назад задними углами. Острия эпимер 7-го сегмента достигают заднего края 2-го брюшного сегмента. Все эпимеры широкие и гладкие со слабыми следами килей. Первые 4 брюшных сегмента равной ширины и длины. Задне-боковые углы 3-го и 4-го сегментов заострены и оттянуты назад, так что боковые части 5-го сегмента накрыты этими заострениями 4-го сегмента. Тельзон треугольной формы с притупленной вершиной и зазубренным задним краем, усаженным 10 шипами и волосками. Внутренняя ветвь рулевых ног в 2 раза шире наружной. Обе ветви равной длины с зазубренными и вооруженными шипиками и волосками краями. 3-й и 4-й членики 1-й пары ног с вытянутыми дистально отростками. Отросток 4-го сегмента достигает середины 6-го членика. У 2-й и 3-й пары ног отросток 4-го сегмента достигает конца 5-го членика. Базальный членик б-й и 7-й пары ног сильно расширен, образуя высокий гребень. Все ножки снабжены длинными перистыми волосками и шипиками по краям.
Распространение. Добыт у маяка Jokkaichi, Япония (Richardson, 1904).
2. Род BATHYNOMUS M.-EDW.
A. Milne-Edwards, Comptes rendus Acad. Sc. Paris, 88:172.
Тип рода: В. giganteus A. M.-Edw.
Ножка усиков второй пары состоит из 5 члеников; базальный членик рулевых ног не имеет отростка с внутренней стороны. Брюшные ножки снабжены пучками жаберных нитей у основания ветвей. Глаза расположены по бокам головы на ее нижней (брюшной) поверхности. В остальном очень близок к роду Cirolana. Из дальневосточных морей известен 1 вид.
1. Bathynomns doederleini Ortmann.
Ortmanh, 1894, Proc. Acad. Nat. Sci. Philad., 191.
От гигантской изоподы Bathynomns giganteus, описанной A. Milne-Edwards в 1879 г., В. doederleini отличается более узким телом, длина которого в 3 раза больше ширины; тельзон снабжен 7 шипами на заднем крае, средний из которых наибольший по величине; ширина тельзона лишь немного больше его длины; по средней линии верхней поверхности проходит ясный продольный киль; обе ветви рулевых ног заострены на концах. Эти признаки позволяют легко отличить В. doederleini от В. giganteus
Вся верхняя поверхность тела тонко зернистая и покрыта точечными вдавлениями. Передний край головы посредине слабо вогнут с коротким отростком, отогнутым книзу. Глаза расположены на нижней поверхности
2. BATH YNOMUS
69
головы около фронтального края головы, который ясно приподнят. Фронтальная пластинка треугольная, с закругленными краями, впереди соприкасается с фронтальным отростком. Clypeus вытянут вперед в виде тупого треугольного отростка, выходит за пределы переднего края головы, так что виден сверху; его длина больше, чем длина губы. Жгутик усиков 1-й пары равен длине их трехчлениковой ножки и состоит из 30 члеников; 1-й членик ножки усиков 2-й пары короткий и не виден сверху; жгутик вариирует в длине, 25-члениковый, достигает заднего края 1-го грудного сегмента. 1-й грудной сегмент равен длине головы, остальные значительно короче, укорачиваясь последовательно к заднему концу тела. 2 передние пары эпимер почти одинаковые, не вытягиваются кзади; 4 следующих пары с вытянутыми заостренными задними углами, особенно у 5-го, 6-го и 7-го сегментов. 3 передние пары грудных ног крепкие; 2-й членик их не расширен, 3-й удлиненный; внутренний край 4-го членика шиповатый; отросток на его наружном крае достигает середины 6-го членика (только на 1-й паре ног этот-отросток очень короткий); 6-й членик удлиненный и изогнутый. 4 следующих пары грудных ног одинаковой структуры, последовательно увеличивающиеся по длине к заднему концу тела. 2-й членик их более или менее нормальный, не утолщенный очень сильно. 3-й членик расширяется дистально, на переднем крае его острые шипы, такие же, как на переднем крае 4-го и 5-го члеников. 6-й членик узкий, немного длиннее 5-го. Брюшные сегменты сводчато-выпуклые, аркообразно изогнутые; боковые углы их от 2-го до 5-го сегмента оттянуты назад и снабжены продольным ребром; это ребро наиболее длинное на 3-м сегменте. Ветви брюшных ног закругленные, почти равные; на нижнем крае их основания пучки жаберных нитей, характерные для рода. Тельзон, так же как и все остальные сегменты, покрыт точечными вдавлениями и мелкой зернистостью; ширина его основания немного больше длины; боковые края немного сходятся кзади, а не параллельны друг другу. Задний край прямой и снабжен 7 шипами. По средней линии тельзона на спинной стороне продольный киль, тянущийся до конца срединного, наиболее длинного шипа заднего края. Шипы заднего края неравной длины; срединный шип немного длиннее остальных. Базальный членик рулевых ног с вытянутым внутренним задним углом; наружная ветвь удлиненная, с почти параллельными краями, заостряется на конце. Внутренняя ветвь почти треугольной формы длиннее чем наружная, заостряется на конце и выходит за пределы заднего края тельзона. По краям обе ветви несут шипики. Ноги и тельзон покрыты волосками.
Распространение. Японское побережье (около EnoshimaВ бухте Сагами) (Ortmann, 1894), 35с05' N, 138°4Г Е, глуб. 527 м (Richardson, 1910).
2. Подсем. AEGINAE
Тело широкое, овальнре, с плоской брюшной и сводчато-выпукло спинной стороной. Поверхность тела совершенно гладкая; глаза очень
70
4. C YMOTHOIDAE
большие, иногда почти соприкасаются друг с другом по средней линии головы; занимают более половины поверхности головы. Усики 1-й пары короткие, обычно расположены очень близко друг к другу по бокам небольшого рострума; членики ножки 1-х усиков сильно расширенные, в виде пластинок. Ротовые части и конечности сильно видоизменены, с крючковидными члениками и с вооружением в виде изогнутых крючков. 3 передних пары грудных ног хватательные, 4 задних пары ходильные. Эктопаразиты. Паразитируют на коже и на жабрах рыб, но могут вести и свободный образ жизни. Из дальневосточных морей известно 3 рода.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Первые 2 членика ножки усиков 1-й пары сильно расширены; жгутик многочлениковый. Щупик ногочелюстей состоит из 5 члеников.
......................................3. Aega Leach, (стр. 70).
2 (1). Первые 2 членика ножки усиков 1-й пары не • расширены; жгутик из 4—6 члеников; щупик ногочелюстей состоит только из 2 члеников.
3 (4). Глаза имеются; abdomen немного уже, чем Грудной отдел . . . .
............................... 4. Rocinela Leach, (стр. 72).
,4 (3). Глаз нет; брюшко много уже грудного отдела . . . . . .....
..................................5. Syscenus Harger, (стр. 80).
3. род AEGA LEACH
Leach, 1815, Trans. Linn. Sod London, XI: 306.
Тйп рода: A. psora (L.). _ , •
Глаза очень большие, почковидные или овальные; иногда соприкасаются друг с другом по средней линии головы. 1 пара усиков с расширенными базальными члениками ножки. Щупик жвал трехчлениковый; зубной отросток отсутствует. 1-я пара челюстей (maxilla I) с одной только лопастью; 2-я пара челюстей (maxilla II) с двумя неравными, вооруженными изогнутыми зубцами лопастями. Щупик ногочелюстей пятичлени-ковый, вооружен крепкими крючьями. Базальный членик рулевых ног на внутреннем нижнем угле образует узкий заостряющийся к концу отросток.
Из Дальневосточных морей известно 2 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Глаза очень большие, овальной формы; занимают почти всю поверхность головы, разделенные лишь узким в 0.5 мм расстоянием . . .
• ................................. * 2. A. magnoculis Rich.
2 (1). Г лаза узкие, удлиненные; распо \ожены цо бокам головы. Расстояние между глазами равно длине глаза.............1. Д. symmetrica Rich.
3. AEGA
71
1. Aega symmetrica Richardson (фиг. 30).
Richardson, 1905, Bull. U. S. Fish. Comm., XXIV : 211; Richardson, 1905, Bull. U. S. Nat. Mus., 54:185, f.'169—172.
Тело яйцевидно-овальное; длина его в 2. раза больше ширины. Цвет желтоватый, густо покрытый бурыми пятнами, которые образуют правильную линию вдоль края каждого сегмента. Передний край головы с небольшим острием посредине. Глаза узкие, удлиненные, отделенные друг от друга расстоянием равным длине глаза. Усики 1-й пары достигают заднего края 1-го грудного сегмента; 1-й и 2-й членики ножки 1-х усиков слегка расширены. Жгутик 11-члениковый. Усики 2-й пары достигают середины 3-го грудного сегмента;
' жгутик 16-члениковый. Фронтальная пластинка коническая с плоским дистальным концом и с вытя-
I Фиг. 30. Aega symmetrica Rich, (по Richardson), а — общий вид; Ь — I грудная ножка; с — тельзон и рулевые ноги.
нутым в острие проксимальным. Все сегменты груди почти равной длины. -Эпимеры большие, почти квадратные; наружные дистальные углы последних 3 пар эпимер вытянуты назад и заходят за край каждого последующего сегмента. 6-й членик 3 первых пар грудных ног снабжен на внутреннем крае 3 маленькими шипами. 5-й членик короткий и вооружен лишь одним шипом; 4-й членик с 5 шипами, а 3-й с одним крупным шипом на наружном дистальном угле. 4 последних пары ног ходильные, тонкие, длинные, вооруженные волосками и немногочисленными шипиками. Первые 5 грудных сегментов короткие. Тельзон языковидной формы; задний конец закруглен, зазубрен и снабжен волосками по краю. Концы рулевых ног немного выступают за пределы конца тельзона. Внутренняя ветвь почти .в 2 раза шире узкой овальной, заостренной на дистальном конце наружной ветви. Обе ветви зазубрены по краям. ,
Распространение. Юго-восточная Аляска (Naha Bay, Behm Canal); пролив королевы Шарлотты, о. Ванкувер, Британская Колумбия; о. Санта-Круц, Калифорния. Берингово море: 54°30'N, 179°17zЕ; 54°30' N, 179°14' Е; 52°14' N, 174°13' Е на глубине от 619 до 867 м (Richardson, 1905, 1910).
72
4. C YMOTHO1DAE
* 2. Aega magnoculis Richardson (фиг. 31).
Richardson, 1910, Proc. U. S. Nat. Mus., 37: 80, f. 7.
Тело овальное, длина его почти в 2 раза больше ширины. Поверх' ность тела гладкая. Ширина головы больше ее длины; передний край головы посредине слегка заострен. Глаза большие, овальные, занимают почти всю поверхность головы и отделены друг от друга коротким в 0.5 мм
расстоянием. 2 первых членика ножки усиков 1-й пары равной длины, не
расширенные. 3-й
Фиг. 31. Aega magno-’ calls Rich. — экз. яз
Охотского моря.
членик их ножки узкий, удлиненный, равный длине 2 первых члеников вместе; жгутик 18-члениковый. Усики I достигают заднего края 1-го торакального сегмента." Усики 2-й пары достигают заднего края 3-го грудного сегмента; жгутик 21-члениковый. Фронтальная пластинка коническая с плоским овальным дистальным и вытянутым в острие проксимальным концами. Все сегменты груди равной длины, только-последний немного короче остальных. Эпимеры большие, почти квадратные; у последних 3 пар заднебоковые углы вытянуты и заострены. 1-й брюшной сегмент почти целиком закрыт последним грудным' сегментом. Следующие 3 сегмента равной длины-Тельзон округлой формы с вытянутым в маленькое острие задним концом, по бокам которого края тельзона зазубрены. Рулевые ноги немного длиннее тель-
зона. Внутренняя ветвь немного длиннее и шире наружной; задний край ее косо срезан, а наружный угол заострен. Наружная ветвь овальная с заострен-
ным дистальным концом. Края обеих ветвей зазубрены. Первые 3 пары грудных ног хватательные. На 6-м членике на
дистальном его конце маленький шип; на 5-м членике 1 большой шип.
Последние 3 пары ног ходильные и вооружены волосками и шипиками. Длина животного 21 мм, из Охотского моря 25 мм.
Распространение. Беринговоморе: 54°30' N Е, 179°14z Е; 50°30' Е, 179°17' N; 52°14' Е, 174°13 N, глуб. 680-950 м. Китайское море: 31°39/N, 129°20' Е наглуб. 730 м (?) (Richardson 1910). Охотское море: 49°23' N, 148°4б'Е наглуб. 1076 м, 2 экз. (Сбор ГГИ, 1932, ст. 214)-
4. Род ROCINELA LEACH
Leach, 1815, Diet. Sc. Nat., XII: 346.
Тип рода: J?, danmoniensis Leach.
Тело широкое, овальное с плоской нижней и слабо выпуклой спинной поверхностью. Abdomen немного уже, чем грудной отдел. Тельзон закруглен на конце; глаза большие, иногда соприкасаются друг с другом:
4. ROCINELA
73
по средней линии. Первые 2 членика ножки усиков 1-й пары не расширены; фронтальная пластинка маленькая и узкая. Зубной отросток жвал в виде языковидной пластинки, щупик ногочелюстей двучлениковый; последний членик его вооружен изогнутыми крючками. 6-й членик 3 первых пар грудных ножек расширенный и вооружен шипами; коготок их образует очень большой, изогнутый крючок. 6-й членик остальных 4 пар ног короткий (ножки ходильные).
Из дальневосточных морей известно 5 видов.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Переднебоковые углы 1-го грудного сегмента вытянуты по бокам головы в виде маленьких рожек . . . . . . . . 1. R. cornuta Rich.
2 (1). Переднебоковые углы 1-го грудного сегмента не образуют роговидных отростков.
3 (4). На 6-м членике 3 первых пар грудных ножек более 4 шипов . . .
...............................................6. R. niponia Rich. 4 (3). 6-й членик 3 первых пар грудных ножек несет от 2 до 4 шипов. 5 (6). На поверхности тела нет темных пигментных пятен............
.......................................* 4. R. angustata Rich.
6 (5). На поверхности тела имеются темные пигментные пятна.
7 (8). Темные пятна по бокам 4-го грудного сегмента и у основания тельзона ................................. * 2. R. maculata Sch. a. Mein.
8 (7). Пятна темного пигмента только по бокам брюшных сегментов и у основания тельзона.
9 (10). Передний край головы плавно закруглен и не имеет вырезок по бокам средней линии..........................* 3. R. belliceps Stimps.
10 (9). Передний край головы с лопастью посредине и по бокам этой лопасти вырезки, так что фронтальный край трехвершинный . . .
........................................ 5. R. japonica Rich.
1. Rocinela cornuta Richardson (фиг. 32).
Richardson, 1898, Proc. Amer. Philos. Soc., 37: 12, f. t. 2; Richardson, 1905, Bull. U. S. Nat. Mus., 54: 192, f. 176—178.
Длина тела немного больше, чем в 2 раза превосходит его ширину; очертания тела овальные; поверхность гладкая с рассеянными по ней точечными вдавлениями. Голова почти, треугольной формы, с вытянутой вперед в виде носика передней частью. Глаза большие, расположенные в заднебоковых частях головы. Усики 1-й пары достигают переднего края 1-го сегмента; жгутик 6-члениковый; усики 2-й пары достигают в вытянутом состоянии заднего края 2-го грудного сегмента; жгутик 16-члениковый. Грудные сегменты почти равной длины. Переднебоковые углы 1-го сегмента сильно вытянуты,< вперед и образуют по бокам головы рожки, которые достигают средины головы. Этот признак отличает R. cornuta.
74
4. CYMOTHOIDAE
от всех прочих известных видов этого рода. Эпимеры всех сегментов заостряются кзади; острия направлены назад. 1-й брюшной сегмент почти .целиком закрыт последним грудным сегментом. Тельзон вытянутый, языковидный с закругленным задним краем, тонко зазубренным. Ветви рулевых ног одинаковой формы и величины, внутренняя ветвь немного более длинная, чем наружная, достигает конца .тельзона. И тельзон и рулевые ножки усажены волосками. 6-й членик хватательных ног с 3 шипами; на 5-м членике 3 тупых шипа. Ходильные ноги длинные, тонкие, покрытые
шипами.
Фиг. 32. Rocinela сог-nuta Rich, (по Richardson).
Фиг. 33. Rocinela macalata Sch. & Meinert. (no Richardson), a — усики с брюшной стороны; Ь —-I грудная ножка; с — общий вид.
Распространение. Берингово море: Шумагинские о-ва, Аляска, глуб. 1200 м (Richardson, 1905), 54°30' N, 179°14' Е на глуб. 621 м (Richardson, 1910).
*2. Rocinela maculata Sch. &. Mein. (фиг. 33).
Schioedte & Meinert, 1879, Naturh. Tidsskrift, (3), XII: 393, pl. XII, f. 10—12.
Тело широкое, овальной формы; поверхность тела гладкая, усеянная мелкими пятнышками; на спинной поверхности 4-го грудного сегмента по бокам его по одному округлому темному пятну в виде глазков. У основания тельзона по бокам также по темному пятну. Фронтальная часть головы треугольной формы с притупленным концом. Фронтальная пластинка маленькая, ромбоидальной формы. Глаза маленькие, пентагональ-
4. ROCINELA
75
ной формы. Расстояние между глазами равно г/8 ширины головы. Усики 1-й пары маленькие, достигают до средины 5-го членика. усиков 2-й пары; жгутик 6-члениковый. Усики 2-й пары достигают заднего края 2-го сегмента. Передний край 1-го грудного сегмента двувыемчатый и поэтому образует волнистую линию с вогнутостью посредине и по бокам, позади глаз. Эпимеры большие и широкие. Заднебоковые углы задних эпимер оттянуты назад и заострены. Острие эпимеры 7-го грудного сегмента достигает середины 2-го брюшного сегмента. 3 передних пары грудных ножек хватательные, короткие; 4-й членик их короткий и вооружен 3 или 4 крепкими тупыми шипами; 6-й членик с 4 длинными
острыми шипами вдоль нижнего края; коготок длинный, тонкий, крючковидно изогнут :и несет 4 киля. Ходильные ноги длинные, крепкйе, вооруженные короткими шипами. 1-й брюшной сегмент почти нацело закрыт последним грудным сегментом; боковые части остальных брюшных сегментов вытянуты назад и заострены. Тельзон короткий, широкий, в общем округлой формы с гладким краем; поверхность гладкая, имеются
2 вдавления по бокам тельзона у его основания. Рулевые ноги большие, длинные; внутренняя ветвь немного длиннее и шире, чем наружная ветвь. Края ветвей зазубрены. Длина животного 35 мм.
Распространение. Зап. Гренландия (Bovallius, 1885). Япон-ское море: По всему морю от Корейской границы до бухты де-Кастри включительно на глубинах от 0 до 220 м. На поверхности моря собирается на свет фонаря (сборы ТИРХ’а и ГГИ, 1925—1932; Павленко и Солдатова, 1911—‘1915; Дербека, 1909; СтароКадомского, 1912; Гидрогр. эксп. Вост, океана, 1915—1920; Бражникова, 1907, 1908). Охотское море: залив Анива, глуб. 20 м (сбор Шмидта, 1901); 48°58/ N, 140°35/ Е, гл. 75 м; 45°40'N, 141°59'Е, гл. 56 м (ГГИ, 1930, 1932).
* 3. Rocinela belliceps (Stimpson) (фиг. 34).
Stimpson, 1864, Proc. Acad. Nat. Sci. Phil., XVII: 155 (Aega); Richardson, 1905, Bull. U. S. Nat. Mus., № 54 :199, f. 187—-192. —• alascensis Lockington, 1877, Proc. Cal. Acad. Sci. VII: 46 (Aega); Richardson, 1898, Proc. Amer. Philos. Soc., 37 :11.
Тело продолговато-овальное, длина его немного больше, чем в 2 раза превышает ширину. Голова треугольной формы, ширина ее больше длины-
76
4. CYMO THOIDAE
Глаза большие, овальные, разделенные друг от друга расстоянием равным ширины основания головы. 1-й членик ножки усиков 1-й пары почти целиком прикрыт сверху вытянутой передней частью головы; 2-й членик короткий; 3-й в 11/2 раза длиннее 2-го; жгутик из 4 члеников, достигает заднего края головы. Усики 2-й пары в вытянутом состоянии достигают заднего края 2-го грудного сегмента; жгутик 14-члениковый. Фронтальная пластинка очень маленькая, ромбоидальной формы у основания. 1-й грудной сегмент немного длиннее, чем остальные, со слегка выпуклым передним краем. 2-й, 3-й и 4-й — равной длины; 5-й, 6-й и 7-й — прогрессивно укорачиваются, так что 7-й сегмент почти в два раза короче 6-го. Первые 3 пары эпимер закругленные. Последние 4 пары с заостренными концами и легкими диагональными килями. 1-й брюшной сегмент закрыт последним грудным сегментом. Тельзон, широкий, закругленный, с тонко зазубренным задним краем. Ветви рулевых ног одинаковой ширины с закругленными концами; внутренняя ветвь немного длиннее наружной, но не достигает конца тельзона. Базальный членик с отростком на внутренней стороне, достигающим конца наружной ветви. Первые 3 пары грудных ног хватательные, остальные ходильные. На 4-м членике хватательных ног 3 шипа и 3 шипа на 6-м членике. Боковые части 2-го, 3-го, 4-го и 5-го брюшных сегментов и основание тельзона покрыты темным пигментом. Максимальная длина животного 37 мм.
Распространение. Северная часть Тихого океана от Калифорнии до Аляски, Алеутские острова; Берингово море; Японское море, глуб. от 100 до 450 м —(52°55'N, 173°27'Е; 52°55'N, 173°26'Е; 46°42z N, 151°45/ Е; 46°42z N, 151°47' Е; 38°33' N, 138°40' Е; 36°18' N. 129°44z Е; 52°11ZN, 179°57'W (Richardson, 1910). Японское море: Дербек, 1912, ст. 5, 46°57',N 138°41ZE, глуб. 60м, 2 экз.; сборы ТИРХ 1928—1931: залив Петра Великого у о. Фурругельма и у м. Поворотного, глуб.. 167—340 м, 1 экз., длина 35 мм; Татарский пролив у мыса Олимпиады, 46°12z N, 138°15'Е, глуб. 62—100 м, 1 экз.; у м. Пещерного, глуб. 260 м, 3 экз.,. длиной 38 мм; в бухте Силантьева, глуб. 177—-192 м; бухта Владимира 43°55ZN, 135°46'Е, глуб. 152 м; сбор Павленко, 1913, ст. 342,—42°17/ N, 139°04z Е, глуб. 72 м; сборы ГГИ 1930—1931: 50°36' N, 140°33' Е, глуб. 70 м; у мыса Кекурного, глуб. 82 м, 1 экз., 43°51' N, 135°35' Е,- глуб. 80 м. О х о т-ское море: 56°3'N, 143°52ZE (Гидрогр. эксп. Вост, океана, ст. 8, 1916); сборы ГГИ 1930, ст. 118/3,—43°46/N, 135°23ZE, глуб. 100 м; ст. 134/19,— 56°08'N, 144°55'Е, глуб. 287 м, 2 экз.; 47°04/N, 138°56'Е, глуб. 73, экз. 1; сборы ГГИ, 1932: ст. 248—56°10z N, 143°15' Е, глуб. 182 м, 2 экз.; ст. 249— 56°24'N, 143°18'Е, глуб. 100, м, 1 экз.; ст. 252—55°32'N, 149°14'Е, глуб. 335 м, 2-экз.; ст. 254—56°00z N, 150°06z Е, глуб. 235 м, 1 экз. Б е р и н-говоморе: о. Кадьяк, 12 экз. (сборы Вознесенского 1846 и Петелена), Уналашка (Вознесенский, 1846), Командорские о-ва у о. Медного и о. Беринг (сборы ТИРХ'а 1929 и 1930, сбор Гурьяновой, 1931). Побережье Японии (сбор Рожковского, 1917).
4. ROCINELA
Ti
* 4. Rocinela angustata Rich. (фиг. 35).
Richardson, 1899, Proc. U. S. Nat. Mus.,' 21 :828; Richardson, 1905, Bull. U. S. Nat. Mus., 54 : 206, f. 203—208. — laticaufy. Hansen, Richardson, 1898, Proc. Amer. Philos. Soo., 37 :14, f. 5—6.
Тело удлиненно-овальное; передняя часть головы вытянута вперед в виде короткого широкого тупого отростка. Глаза большие, овальные. Усики 1-й пары достигают заднего края головы; жгутик 6-члениковый. Усики 2-й пары достигают середины 2-го грудного сегмента; жгутик 15-члениковый. Грудные сегменты неравной длины: 1-й сегмент, самый боль
Фиг. 35. Rocinela angustata Rich, (по Richardson), а — голова; telson; d—uropoda.
Ь — pereiopoda I; с —
шой с глубоко вогнутым передним краем, так что боковые его части охватывают голову с боков. Первые 3 пары эпимер с закругленными углами; 3 задних пары с вытянутыми заостренными углами. 1-й брюшной сегмент почти целиком закрыт последним грудным сегментом. 5-й сегмент уже, чем основание тельзона. Тельзон широкий, полуовальный с закругленным зазубренным задним краем, покрыт грубыми волосками. У основания тельзон вдавлен по бокам приподнимающейся середины. Наружная ветвь рулевых ног почти в 2 раза шире внутренней ветви; обе ветви одинаковой длины, с неясно зазубренными, покрытыми велосками краями. Хватательные ноги короткие и крепкие; на 6-м их членике 4 шипа; на 4-м — 6 шипов и волоски. Ходильные ножки (4 пары последних грудных ног) относительно короткие и крепкие, вооруженные шипами и волосками. Длина животного достигает 44 мм.
Распространение. Сев. часть Тихого океана от Калифорнии до Аляски; Puget Sound; о. Унимак (Аляска); Япония (Манацура). Я п о н с к о е
78
4. CYMOTHOIDAE
море: 41°58/ N, 142°30' Е; 42O11'N, 142°12'Е; глуб. от 120 у Аляски до 920 м в Японском море (Richardson, 1905, 1910). Берингово море’: у острова Беринга (Командорские, о-ва), 1 экз., длина 44 мм (сборСлюнина). Охотское море: ст. 211—49°12/N, 145°21ZE, глуб. 443 м, 1 экз.; ст. 216 — 51°10'N, 154°17/Е, глуб. 591 м, длина 42 мм (сбор ГГИ, 1932).
=;= 5. Rocinela japonica Rich. (фиг. 36).
Richardson, 1898, Proc. Amer. Phil. Soc., 37, Ks 157: 15, f. 7—9.
Поверхность тела покрыта точечными вдавлениями и мелкими черными или коричневыми пятнами. У основания каудаЛьного сегмента (тельзона) 2 небольших темнокоричневых пятна, отделенных друг от друга расстоянием, равным половине ширины сегмента. Голова в общем треугольной формы с сильно вытянутой вперед фронтальной частью. Передний край головы образует трехвершинную кривую, так как средняя часть его образует крупную трапециевидную лопасть, у основания этой лопасти по бокам ее с каждой стороны по одной небольшой вырезке
Фиг. 36. Rocinela japonica Rich, а — голова; Ь — pereiopoda I; с-pereiopoda II; d- pereiopoda И Передне-боковые уГЛЫ ГОЛОВЫ III; е — uropoda. впереди глаз треугольной фор-
мы. Края всех 3 лобных вершин впереди глаз приподняты. Глаза большие, расположены по. бокам головы на ее задне-боковых углах; расстояние между глаз равно половине ширины головы. Усики 1-й пары достигают заднего края головы, жгутик их 6—7-члениковый. Усики 2-й пары в вытянутом состоянии достигают заднего края 2-го грудного сегмента; жгутик 15-членико-вый. Эпимеры на всех сегментах (кроме 1-го, который не имеет свободных эпимер), заостренные; более закругленные задние углы у первых 2 пар эпимер, у остальных они очень острые. Грудные сегменты почти одинаковой длины. 1-й брюшной сегмент почти нацело закрыт налегающим на него последним грудным сегментом. Задние углы этого сегмента сильно заострены и вытянуты в стороны. Тельзон широкий с закругленными краями и тупым острием на конце. Задний край его неясно зазубрен и снабжен волосками. Ветви рулевых ног выходят несколько за пределы конца тельзона. Внутренняя ветвь немного длиннее наружной, но обе ветви равной ширины; наружные края обеих ветвей вооружены шипиками и волосками. 3 передние хватательные пары грудных ног сравнительно тонкие; на предпоследнем членике на внутреннем крае, образующем не-
4 ROCINELA
19
большую лопасть, 3—4 изогнутых тонких шипа; на меральном членике по 3—5 крупных тупых зубца. От R. maculafa этот вид легко отличается формой головы и отсутствием темных, в виде глазков, пятен на 4-м сегменте, а от R. belliceps формой фронтального края и строением ног. Максимальная длина 37 мм.
Распространение. Бухта Хакодате (на о. Хоккайдо) на глубине 27 м (Richardson, 1898). Японское море: 51°26/ N, 140°49/ Е, глуб. 64 м, 1 экз.; 50°06'N, 140°33/Е, глуб. 20 м (сбор ГГИ 1931, ст. 425/13 и 436/24);. 43°39'N, 135°16'Е, глуб. 54 м (Сбор. ГГИ 1930, ст. 375/42).
6. Rocinela niponia Rich. (фиг. 37).
Richardson, 1910, Proc. U. S. Nat. Mus., 37: 83, f. 9—10.
Тело вытянуто-овальное; длина его немного более, чем в 2 раза превышает ширину. Голова треугольной формы с вытянутой в виде широкого треугольного, закругленного на конце отростка передней- частью. Глаза большие, отделенные друг от друга расстоянием, равным длине глаза. Усики 1-й пары достигают заднего края головы и почти доходят до конца ножки усиков 2-й пары; жгутик 6-члениковый; 1-й членик его в 2 раза
длиннее 2-го. Усики 2-й пары достигают заднего края 2-го грудного сегмента; жгутик из 16 члеников. 1-й, 2-й и 7-й грудные сегменты равной, длины; 3-й, 4-й, 5-й и 6-й немного длиннее. 2 первых пары эпимер закругленные, остальные с заостренными задними углами. 1-й брюшной сегмент закрыт последним грудным сегментом, но боковые его части видны сверху. Тельзон треугольный на конце, вооруженный по краю короткими ширами и волосками; рулевые ножки не достигают конца тельзона; наружная ветвь немного короче и уже, чем внутренняя; обе ветви с шипиками • и волосками по. краям. 6-й членик 1-й (хватательной) пары грудных ножек снабжен широким ладонным (пальмарным) отростком, покрытым по краю рядом из 10 изогнутых шипов; у 2 следующих пар хватательных ног 6-й членик с 8 шипами. 5-й членик снабжен 1 длинным шипом; 4-й членик с 5 длинными шипами, причем передний шип почти в 2 раза длиннее остальных. Ходильные ноги также вооружены многочисленными шипами. Длина животного 18 мм.
Распространение. Японское море: 38°16/N, 138°52' Е, глуб. 140 м (Richardson, 1910).
£
Фиг. 37. Rocinela niponia Rich. (по Richardson).. 1— голова; 2—pereiopoda I.
so
4. C YMO THOIDAE
5. Род SYSCENUS HARGER
Harger, 1880, Rept. U. S. Comm. Fish. a. Fisheries, VI: 386.
Тип рода: 5. infelix Harger.
Тело удлиненное, сплюснутр в дорзо-вентральном направлении с выпуклой спиной и плоской брюшной стороной. Брюшной отдел много уже грудного; глаз нет совсем; членики ножки усиков 1-й пары простые, цилиндрические, не расширенные. Жвалы лишены зубного отростка; режущий край их гладкий, без зубцов. Щупик ногочелюстей двучленико-вый, последний членик его снабжен крючками.
7—6-й членик первых 3 хва-\ \ /г””" 'Л тательных пар грудных
( ножек цилиндрический,
л Сг~не расширенный; кого-
ЗА ___-Jyh ток серповидно изогнут
тЗ и сильно заострен.
Ou Для дальневосточ-
J И ных морей указывается
И U )Х у один вид, причем нахо-
fl ' f /г 7 ' ждение его в Японском
// (Л лН море Hansen J. Н. оспа-н ривает, считая, что ука-
. # W зание б', infelix для
® Японского моря осно-
фиг. 38. Syscenus infelix (Richardson), а — голова снизу; вано на ошибке в опре-Ъ — общий вид (по Richardson). Делении. ОднаКО КОНт
статирование целого ряда амфибореальных форм среди различных групп беспозвоночных и рыб указывает на то, что мы в этом случае можем иметь дело с амфи-бореальным видом, и таким образом, считая указание Н. Richardson не лишенным вероятия, я включаю «S', infelix в настоящую сводку.
1. Syscenus infelix Harger (фиг. 38).
Harger, 1880, Report U. S. Comm. Fish., Pt. 6 :387; Harger, 1883, Bull. Mus. Comp. Zool. Harvard College, XI, N 4 :100, pl. Ill, f. 5—5a, pl. IV, f. 3—3 h. — pranzoides G. Sars, 1883, Forhand. Vid. Selsk. Christiania, N 18. (Harponyxa). — lilljeborgi Bovallius, 1885, Bihang Vetensk. Ak. Handl., X, N 10: 3, pl. I—II (Rocinela}-, Bovallius, 1886—1887, Bihang. Sv. Vet. Akad. Handl. XI, N 17 :17.
Тело удлиненное, длина его почти в 3 раза больше ширины. Голова треугольной формы; ширина ее у основания в 3 раза больше длины. Передний край головы трехлопастной; средняя лопасть выступает вперед от 2 боковых между основаниями усиков и заострена. Глаз нет; 1-я пара усиков с 3 равной длины члениками ножки и 7-члениковым жгутиком; достигает заднего края головы в вытянутом состоянии. Усики 2-й пары достигают
5. SYSCENUS
81
заднего края 3-го грудного сегмента; первые 2 членика их ножки равной .длины; 3-й и 4-й членики каждый почти в 2 раза длиннее 2-го; 5-й членик в 1.5 раза длиннее 4-го; жгутик 25-члениковый. Фронтальная пластинка большая, ромбоидальной формы. Щупик ногочелюстей двучлени-ковый; последний членик его с крючками на вершине. 1-й грудной сегмент длиннее, чем все остальные. Заднебоковые углы его заострены. Эпимеры в виде широких пластинок с очень острыми задне-боковыми углами. Брюшко много уже, чем грудной отдел; все 6 сегментов ясно видны. Тельзон большой, почти треугольной формы с закругленными боковыми краями и заостренной, несколько, оттянутой назад вершиной. Рулевые ножки длинные, концы ветвей достигают конца тельзона; наружная ветвь шире и короче внутренней; дистальный конец ее широко-округлый, тогда как дистальный конец внутренней ветви узкий, тупой. Базаль-1ный членик рулевых ног не имеет отростка на внутренней стороне. Первые 3 пары грудных ног хватательные, остальные ноги ходильные. .Хватательные ножки не имеют шипов и снабжены только щетинками. 5-я пара ног немного длиннее 4-й пары; 6-я и 7-я пары одинаковой длины и значительно длиннее 4-й и 5-й пар. Длина животного 28 мм.
Паразитирует на рыбах.
Распространение. Сев. Атлантический океан; Средиземное море; Японское море: 35°06'N. 138°4(УЕ, глуб. 410 —580 м ^Richardson, 1910).
3. Подсем. CYMOTHOINAE
Тело большею частью асимметрично; глаза маленькие, усики сильно редуцированы, так что нет ясного разделения усика на ножку и жгутик. Все 7 пар грудных ножек хватательные с расширенным 6-м члеником и длинным, серповидно изогнутым, очейь острым коготком. Щупик ногочелюстей двучлениковый; последний члейик снабжен крючками. Сильно выражен половой диморфизм; имеются личиночные стадии.
Формы паразитические. В наших дальневосточных морях . могут быть встречены представители 4 родов.
ТАБЛИЦА- ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Голова совсем не погружена в 1-й грудной сегмент; задний край ее образует 3 лопасти, одну большую срединную и 2 маленьких по бокам. Передний край 1-го грудного сегмента с 3 ясными выемками ...................................... 6. Nerocila Leach (стр. 82).
2 (1). Голова более или менее погружена в 1-й грудной сегмент; задний с' край ее закругленный, не образует лопастей. Передний край 1-го грудного сегмента гладкий, глубоко вогнутый посредине.
3 (4). Членики усиков широкие, сплющенные сверху вниз; 1-е членики усиков первых тесно соприкасаются друг с другом по средней линии головы......................... 7. Meinertia Stebb. (стр. 84).
Гурьявова
82
4. CYMO THOIDAB
4 (3). Членики усиков не расширены и сжаты с боков; 1-е членики усиков первых отставлены друг от друга на значительное расстояние.
5 (6). Передняя часть брюшного отдела очень мало, только слегка погружена в заднюю часть грудного отдела. Все сегменты груди равной длины или же 1-й сегмент длиннее, а последний сегмент короче, чем все остальные .............................8. Livoneca Leach (стр. 86).
6 (5). Брюшной сегмент глубоко погружен в грудной отдел; 1-й сегмент груди значительно длиннее 2-го; б остальных грудных сегментов-постепенно делаются все короче и короче...............................
........................................9. Irona Sch. a. Mein. (стр. 94).
6. Род NEROCILA LEACH
Leach, 1818, Diet. Sc. Nat., XII: 351.
Тип рода: N. bivitiaia (Risso).
Тело уплощенное; задняя часть головы образует 3 лопасти, но совсем не погружена в грудной отдел. Усики 1-й пары почти соприкасаются: своими базальными члениками. Передний край 1-го грудного сегмента образует 3 глубоких выемки, соответственно 3 задним лопастям головы. Заднебоковые углы сегментов, начиная со 2-го, вытянуты и заострены, причем, чем дальше к заднему концу тела, тем больше, так что на последних сегментах образуются длинные отростки. Эпимеры, особенно на задних сегментах, с вытянутыми и заостренными задне-боковыми углами. Брюшко свободное, не погруженное: в .заднюю часть грудного отдела; тельзон; большой, щитовидный. Брюшко заметно уже грудного отдела:
В наших водах можно ожидать встретить 2 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Глаза не видны; заднебоковые углы всех грудных сегментов, вытянуты в заостренные, направленные назад отростки.................... . .:
...............< . .. . . . . ... . . . . 1 N. japonica Sch. a. Mein. 2 (1). Глаза хорошо выраженные; заднебоковые углы только 2 последних грудных сегментов образуют заостренные отростки .......
....................................2. N. californica Sch. a. Mein..
1, Nerocila japonica Sch. a. Meinert (фиг. 39).
. 1881, Schioedte a. Meinert, Naturh, Tidsskrift, Ser. Ill, XIII: 20, tab. II, f. 1—2
Tело овальное; длина его в 2 раза больше ширины; голова большая? почти квадратная; передний край ее широкозакругленный, длина ее больше длины 1-го сегмента; глаза почти совершенно редуцированы; усики 1-й и 2-й пары почти равной длины, 8—9-члениковые. Передний, край 1-го грудного сегмента имеет 3 выемки, соответственные 3 лопастям задней части головы. Задние углы всех грудных сегментов оттянуты и заострены; начиная со 2-го Сегмента эти заострения превращаются уже:
6. NEROCILA
83
Фиг. 39. Nerocila japnica Sch. & Mein, (no Schioedte и Meinert).
в отростки, величина которых последовательно увеличивается к заднему концу, так что конец боковых отростков последнего грудного сегмента достигает основания тельзона. Эпимеры на всех сегментах ясно отделены. 4-я, 5-я, 6-я и 7-я пара эпимер с сильно ©ттянутыми назад задними углами, образующими тупые отростки. Все грудные ножкй хватательные; длина их постепенно увеличивается к заднему концу тела, так что последняя пара самая длинная. Коготки на всех ножках тонкие, очень острые, серповидно-изогнутые. Тельзон широкий, слегка суживается дистально; края его вакруглены. Посредине заднего края тупой треугольный зубец, с закругленной вершиной. Боковые части брюшных сегментов вытянуты в длинные, направленные в стороны, заостряющиеся отростки. Рулевые ножки длиннее тельзона; наружная ветвь длиннее внутренней. Наружная ветвь уже, чем основание внутренней. Длина животного 21 мм. Цвет желтоватый.
Распространение. Японское' море (Schi^dte a. Meinert), без точного местонахождения.
2. Nerocila californica Sch. a. Meinert (фиг. 40).
Schioedte a. Meinert, 1881—1883, Naturh. Tidsskrift (3), XIII: 72—76, pl. V, 12—13,
pi. VI, f. 1—2.
Фиг. 40. Nerocila californica Sch. & Mein, (до Schioedte et Meinert).
Тело продолговато-овальное; длина его в 2.5 раза больше ширины; ширина головы больше ее длины; передний край головы широко закругленный; задний край образует 3 лопасти, из которых срединная много больше боковых. Глаза неясные и почти целиком исчезают; усики 1 пары 7-члениковые, достигают конца 7-го членика
усиков 2-й пары; усики 2-й пары 10-члениковые и достигают середины 1-гО грудного сегмента. Щупик ногочелюстей 2-члениковый, щупик жвал
6*
84
4. CYMO THOIDAE
3-члениковый. 1-й грудной сегмент длиннее, чем 3 следующих сегмента; 3 задних сегмента длиннее чем 3 предыдущих и приблизительно равны длине 1-го. Только у 2 последних грудных сегментов задние углы вытянуты в заостренные отростки. Эпимеры всех грудных сегментов, за исключением 1-го, ясно отделены от сегментов; у первых 3 пар эпимер задние углы закруглены; у последних 3 пар оттянуты назад и заострены. Все сегменты брюшка ясно видны. Тельзон широкий, ширина его равна длине, почти квадратной формы, но с закругленными краями; задний край посредине образует тупой зубец. Рулевые ножки длиннее тельзона; их наружная ветвь длиннее внутренней; обе ветви вытянутые, узкие; наружная ветвь уже, чем основание внутренней. Все грудные ножки хватательные, постепенно увеличиваются в длине к заднему концу тела. Коготки длинные, острые, серповидно-изогнутые. Вдоль тела по спинной стороне 2 продольных светлокоричневых полосы на общем темном фоне окраски тела. Длина животного 19 мм.
Паразитирует на рыбах, прицепляясь к их плавникам.
Распространение. Сан-Диего> Калифорния; Панамская бухта; у мыса Шур и у National City, Калифорния. На плавниках рыб из рода Promicrops, Gyropleurodus, Paralabrax, Scorpaena, Triakis, . Myliobatis (Richardson, 1905).
7. Род MEINERTIA STEBBING
Stebbing, 1893, Hist. Crust.: 354.
Тип рода: M. imbricata (Fabr.).
Тело вытянутое, с почти параллельными краями. Голова слегка погружена в 1-й грудной сегмент. Глаза маленькие. Первая пара усиков с расширенными члениками; основания усиков соприкасаются по средней линии. Членики усиков 2-й пары сдавлены с боков. Передний край 1-го грудного сегмента закруглен. Эпимеры 1-го грудного сегмента у самки с направленным вперед гребнем, образующим нечто вроде , ложечки или бугра. Эпимеры передних сегментов не достигают ’ задних углов сегментов на большое расстояние; эпимеры задних сегментов достигают конца сегмента и даже грудного отдела. Тельзон поперечный. Грудные ножки, за исключением 2 первых пар, длинные; коготки длинные, острые, серповидно изогнутые; у самок коготок 3-й пары ног заметно длиннее остальных. На базальном (втором) членике 4 задних пар грудных ножек имеется’ высокий киль. Самец меньше самки.
Из дальневосточных морей известен один вид.
1., Meinertia oxyrrhynchaena (Koelbel) (фиг. 41).
Koelbel, 1879, Sitzungsb. mathem.-natur. KI Kais. Akad. Wiss., Wien,.78, I Abt: 1, tab. 1, f. la — le (Ceratothoa); Thielemann, 1910, Abh. nat. phys. Klasse Bayer. Akad. Wiss., II. Suppl., Bd. 3, Abh. 1 :1, tab. f. I. la— le.
Тело вытянутое с выпуклой спинной и плоской брюшной стороной. Голова глубоко погружена в 1-й грудной сегмент, трехугольной формы
7. ME IN ER TIA
85
с треугольно заостренным передним краем. Глаза хорошо развиты, овальной формы, посажёны по бокам головы на середине расстояния между передним и задним краем головы. Членики обеих пар усиков расширенные, сплюснутые. Усики 1-й пары 7-члениковые, достигают переднего края 1-го грудного сегмента. Усики 2-й пары заходят за передние углы 1-го грудного сегмента, 9-члениковые; последние 2 членика очень маленькие. Передний край 1-го грудного сегмента глубоко вырезан спереди, и передние боковые углы его охватывают голову с боков. Длина 2^о грудного сегмента немного больше длины головы; 2-й и 3-й сегменты равной длины, и каждый из них в 1.5 раза длиннее 1-го; 4-й грудной сегмент немного длиннее предыдущих сегментов. 3 задних грудных сегмента постепенно
Фиг 41b. Meinertia oxyrrhynchaena Sch. & Mei-nert (no Schioedte & Meinert).
самый короткий сегмент последний. Эпимеры
Фиг. 41a. Meinertia oxyrrhynchaena Sch. & Mein, (no Thielemann).
укорачиваются, так что
хорошо развиты на всех сегментах. Эпимеры 1-й пары не достигают перед
него угла 1-го грудного сегмента; эпимеры 2-й, 3-й, 4-й и 5-й пар не достигают заднего угла сегментов. Эпимеры 6-го й 7-го сегментов занимают боковые стороны сегментов по всей их длине. Задние углы 3 передних пар эпимер закруглены, а передние углы их тупые. Задние углы 4 последних пар эпимер слегка крючковидно изогнуты и заострены, а передние их углы широко закруглены. Абдомен погружен передней своей частью в грудной отдел. 5 передних брюшных сегментов очень короткие, но немного уже грудного отдела. Боковые части 1-го сегмента закрыты сверху последним грудным сегментом. Тельзон поперечный; ширина его почти в 2 раза больше длины; задний край с вырезкой посредине; у основания спинная сторона тельзона вдавлена, а посредине проходит продольная борозда до срединной вырезки. Рулевые ножки длиннее тельзона, наружные ветви немного длиннее внутренних; на базальных члениках 4 задних пар грудных ножек высокие и широкие кили. Коготки всех грудных ножек острые, длинные, серповидно изогнутые. Длина животного 26 мм. Цвет желтый.
86
4. CYMOTHOIDAE
Распространение. Японское море (Schioedte а. ' Meinert, 1884); бухта Сагами, 120—110 м, (Thielemann, 1910); восточное.побережье Японии.
8. Род LIVONECA LEACH
' Leach, 1818, Diet. Sc. Nat., XII: 351.
Тип родас L. redmanni Leach.
Тело широкое, овальное, часто более или менее асимметричное; голова глубоко погружена в передний грудной сегмент; боковые углы этого сегмента выдаются вперед, охватывая заднюю часть головы; задний край головы закругленный, не образует лопастей; усики 1-й пары широко расставлены, так что их основания удалены друг от друга на некоторое расстояние. Передний край грудного сегмента обычно глубоко вырезан посредине, соответственно погруженной в нее голове, а иногда вырезки имеются и по бокам его. Брюшко слабо погружено в грудной отдел и обычно не уже, чем грудной отдел, хотя у некоторых (L. amurensis)* оно резко уже, чем грудь. Глаза маленькие, часто неясные; эпимеры на всех задних 6 сегментах свободные. Щупик ногочелюстей 2-члениковый; щупик жвал трехчлениковый. Все грудные ноги одинаковой скульптуры, хватательные с серповидно-изогнутыми острыми коготками. Тельзон обычно, широкий, закругленный,
На Дальнем Востоке известно 8 видов.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (12). Тело симметричное.
2 (7). Брюшной отдел уже, чем предпоследний грудной сегмент.
3 .(6). Глаза хорошо развиты.
4 (5). Глаза большие, выпуклые, округлые; передняя часть головы вытянута вперед и образует впереди глаз широкую лопасть с прямым передним краем. Брюшко немного уже предпоследнего грудного сегмента . •
......................................... 6. L. vulgaris Stimps.
5 (4). Глаза небольшие, плоские, удлиненно овальные; передний край головы плавно закругляется; брюшко много уже предпоследнего грудного сегмента...................................* 1. L. amurensis Gerstf.
6 (3). Глаза почти нацело редуцированы; имеются лишь следы глаз по бокам головы. .............................. 4. L. sacciger Rich.
7 (2). Брюшной отдел такой же ширины, как предпоследний грудной сегмент и шире последнего сегмента.
8 (11). Ветви рулевых ног неравной длины.
9 (10). Основание тельзона такой же ширины, как передние брюшные сегменты; последняя парк эпимер в виде маленьких закругленных пластинок ........................... 8. L. caudata Sch. a. Mein.
8. LIVONECA
87
10 (9). Основание тельзона уже, чем передние брюшные сегменты; последняя пара эпимер в виде больших широких пластинок.....................
.......................................... 5. L. epimerias Rich.
11 (8). Ветви рулевых ног равной длины. . . . * 2. L. raynaudii M.-Edw.
12 (1). Тело несимметричное, изогнутое направо или налево.
13 (14). Ширина основания тельзона в 2 раза больше его длины ....
....................................... 3. L. propinqua Rich.
14 (13). Ширина основания тельзона меньше его длины.................
...................................7. L. californica Sch. a. Mein.
* 1. Livoneca amurensis (Gerstfeldt) (фиг. 42).
Gerstfeldt, 1858, Mem. de savants etrangers de 1’Academie -des Sciences de St. Petersbourg, VIII: 18 (Cymothoa). — daurica Miers, 1877, Proc. Zool. Soc., London: 676, pl. 69, f. 4. (Lironeca). — .mohtanus Schioedte a. Meinert, 1884, Naturh. Tidskr. (3): 303, tab. XI, f. 10—11 (Ichtyoxenus).
Этот вид был описан в 1858 г. Gerstfeldt по материалам экспедиции Маака 1855 г. в бассейн Амура. Экземпляры, описанные им из реки Онон, хранятся в коллекциях Зоологического института Академии Наук и подвергнуты мною изучению. Тщательно сравнив их с описанием Livoneca daurica^ данным в 1877 г. Miers’oM по одному экземпляру из реки Онон, я могла убедиться в идентичности этих видов. Повидимому, Ichtyoxenus montanus (из района Гималаев), описанный Schioedte a. Meinert’oM, также
Фиг. 42. Livoneca amurensis (Gerstfeldt) (оригинальный экземпляр).
должен быть отнесен к этому же виду. Судя по тем материалам, которые имеются в коллекциях Академии Наук в Ленинграде, взрослые экземпляры
сильно вариируют по форме тела и особенно в отношении ширины брюшка.
Тело широкое, овальное, с наибольшей шириной на 4-м грудном сегменте. Голова маленькая, овальная; ширина ее немного больше длины;
передний край головы плавно закругляется; глаза маленькие, овальные, с черным пигментом, расположены по бокам передней части головы. Усики 1-й пары очень короткие, достигают только середины глаза, 7—8-члени-ковые; усики 2-й пары достигают заднего края головы, 1—8-члениковые. 1-й грудной сегмент очень длинный и значительно уже 2-го и почти в 2 раза .длиннее его. Передний край 1-го грудного сегмента глубоко вырезан по
средине, соответственно погруженной части головы, так что передне-боковые широко-закругленные части сегмента охватывают голову до уровня, глаз. Следующие 5 грудных сегментов все значительно шире, но короче, чем 1-й сегмент; ширина сегментов увеличивается постепенно, так что наиболее широкими являются 3-й и 4-й сегменты, а затем тело суживается. Последний грудной сегмент сильно изогнут, почти полулунной формы, охватывая с боков погруженное в него брюшко. Таким образом, грудной
88
4. CYMO THOIDAE
отдел имеет форму широкого овала, иногда почти круглый. Эпимеры узкие,, на всех сегментах занимают только переднюю часть боковых краёв сегмента;: задние края эпимер закруглены. Брюшной отдел погруженный; боковые части, первых 2, а иногда и 3 брюшных сегментов прикрыты боковыми частями полулунно-изогнутого последнего грудного сегмента. Все брюшко-значительно уже грудного отдела; тельзон широкий с закругленным задним краем; ширина его основания немного больше длины. Спинная поверхность плоская или слабо выпуклая; иногда по средней линии тельзона слабо-выраженный продольный киль. Рулевые ноги короткие, не достигают заднего края тельзона. Ветви узкие, равной длины, суживаются дистально. Все грудные ножки крепкие, одинаковой структуры, с короткими толстыми члениками и короткими острыми изогнутыми крючками. Передний край; базального членика 3 первых пар грудных ножек снабжен в своей проксимальной части небольшим килем; такой же невысокой киль, несколько более резко выраженный у проксимального конца, на заднем крае базального членика 4 последних пар грудных ножек. Вся поверхность тела покрыта мелкими точечными пятнами Черного пигмента, хорошо видными под лупой; особенно густо, эти пятнышки расположены на голове и брюшных сегментах. Длина взрослой формы достигает 32 мм.
Распространение. Бассейн реки Амура, район Гималаев (?)-(Gerstfeldt, Miers, Schioedte a. Meinert). Река Онон, 5 экз. на теле Cypri-nus amurensis (сбор экспедиции Маак, 1855), максим, длина 22 мм. У дер. Яковлевки Спасского уезда из полости тела чебака; река Даубихе,. 2 экз., максим, длина 32 мм (сбор А. Дьяконова и Н. Филипьева, 1927). Река Шилка около Базыкиной, 1 экз., в полости рта чебака, длина 26 мм (сбор В. Подгорбунского). Река Амур: 1 экз., длина 27 мм (Большов, 1876), с молодью в сумке. Амурский залив против Малышевского-протока, 4 экз., максм. длина 14 мм (сбор Солдатова, 1910). . То же,. 1 экз., длиною 13 мм из полости тела Idus walceki. Нижнее течение Амура, в 18 км от Хабаровска, в полости тела чебака, 2 экз., максим, длина 23 мм (сбор Дмитриева, 1916). Пальгинская протока р. Амура, из полости тела, сигов (Idus walceki), 3 экз., длина 10 мм (сбор Солдатова, 1910).
* 2. Livoneca raynaudii M.-Edw. (фиг. 43).
Milne-Edwards, 18^0, Hist. nat. Crust., Ill; Schioedte a. Meinert, 1884, Naturh. Tidsskr.,. (3), 14: 367, tab. 15. f. 9—10. — novae-zelandiae Miers, 1876, Ann. Mag;. Nat. Hist., (4)k 17 :228.
Тело широкое, овальное, симметричное; ширина его 18 мм при 31 мм длины; голова в общем треугольной формы с вытянутой вперед трапецевидно передней частью; глаза небольшие в задне-боковых углах головы; затылочная часть закругленная, выдается несколько назад; вся голова погружена в 1-й грудной сегмент, передне-боковые углы которого охватывают голову с боков на уровне глаз. Усики 1-й пары 8-члениковые, достигают заднего края головы; усики 2-й пары 9-члениковые, достигают середины 1-го сегмента. 1-й грудной сегмент в 2 раза длиннее 2-го; остальные сег-
^LIVONECA
89
Фиг. 43. Livoneca гаупаи~ dii M.-Edw. (экз. из Охотского моря).
менты последовательно укорачиваются к заднему концу тела. Эпимеры сравнительно узкие, занимают весь боковой край сегмента; 2 последние Пары эпимер немного заходят за пределы заднего края своего сегмента. Задние углы эпимер тупо заострены, почти закруглены. Брюшной отдел немного погружен в грудной отдел, так что боковые части 1-го брюшного сегмента накрыты боковыми частями последнего торакального сегмента. Все брюшные сегменты короткие и широкие, почти равной длины; ширина брюшных сегментов несколько больше ширины последнего грудного. Тельзон почти полукруглой формы, ширина его больше длины (длина тельсона 7 мм при ширине его у основания в 9.5 мм). Задний край тельзона широко закруглен, гладкий, без зазубрин и волосков. Спинная поверхность тельзона совершенно плоская, вдавленная у осно. вания. Рулевые ноги достигают заднего края тельзона; обе ветви одинаковой длины и ширины; внутренняя ветвь на конце закруглена, наружная слегка заострена; ветви длиннее базального членика, нижний внутренний угол которого образует короткий тупой отросток; края ветвей гладкие. Все грудные ножки одинаковой структуры, крепкие, с короткими толстыми члениками и длинными острыми серповидно изогнутыми коготками. Базальный членик 4 задних пар грудных ног с низким килем вдоль заднего края; киль этот образует тупой, конический, короткий отросток в прокси
мальной своей части. Экземпляр из Охотского моря отличается от кали-форнских экземпляров более выпуклой спинной стороной тела, более длинным тельзоном (у калифорнских ширина тельзона у основания ровно в 2 раза больше его длины) и более длинными и широкими ветвями рулевых ног. От описания и рисунков Schioedte et Meinert охотский и кали-форнские экземпляры отличаются более длинными рулевыми нОгами, ветви которых достигают заднего края тельзона, тогда как рулевые ноги австралийских экземпляров достигают только середины длины тельзона.
Распространение. Южная часть Тихого океана (Австралия, Новая Зеландия); мыс Доброй Надежды, Земля Вандимена; Иокогама (Schioedte a. Meinert, 1884); Калифорния, 2 экз. сбор Вознесенского, 1846; 1 экз., снятый с акулы у берегов Росса (Вознесенский); Охотское море: порт Аян, 1 экз. (сбор Вознесенского, 1847).
3. Livoneca propinqua Richardson (фиг. 44).
Richardson, 1904, Proc. U. S. Nat. Mus., 27: 37, f. 6—7.
Тело широкое, асимметричное, изогнутое направо или налево почти параллельными краями; брюшной отдел не уже грудного; голова маленькая,.
90
4. CYMO THOIDAE
треугольной формы; передний край головы посредине образует притупленное острие; задний край головы прямой; глаза большие, овальные, расположенные в задне-боковых углах головы. Усики 1-й пары 7-члениковые,
достигают передне-боковых углов 1-го грудного сегмента; усики 2-й пары 13-члениковые, заходят в вытянутом состоянии несколько дальше заднего края головы. 1-й грудной сегмент значительно длиннее всех остальных; передние углы его охватывают голову с боков; задние углы широко закругленные. Остальные грудные сегменты почти равной длины, только последний немного короче, чем остальные. Эпимеры 2-го и 3-го сегмента длинные, узкие, занимают всю боковую поверхность сегмента; эпимеры 4-го и 5-го сегментов короткие, заостряющиеся сзади и занимают только половину боковой части сегмента. Эпимеры б-го сегмента также сзади заострены и занимают 3/4 длины боковой части сег-
Фиг. 45. Livoneca sae-ciger Rich, (no Richardson).
Фиг. 44. Livoneca pro- мента; эпимеры 7-ой пары сзади закругляются и pinqua Rich, (по Ri- занимают всю боковую часть сегмента. Брюшные chardson): сегменты короткие, но ширина их равна ширине грудных. Тельзон поперечный, ширина его в 2 раза, больше длины, с закругленным задним краем. Рулевые ножки в виде узких уховидных придатков с закругленными концами; наружная ветвь немного длиннее внутренней и почти достигает заднего края тельзона. Грудные ножки все одинаковой структуры, хватательные, с крючковидными коготками. Базальные членики 4 последних пар ног снабжены высоким килем.
Длина животного 18 мм.
Распространение. Берега Японии (порт Хеда);Японское море:35°0б'N, 138/О40Е;33°25'N» 135°37/ Е, глубина от 380 до 480 м. Паразитируют в ротовой полости Chalinura (Richardson, 1904).
4. Livoneca sacciger Richardson (фиг. 45).
Richardson, 1910, Proc. U. S. Nat. Mus., 87: 37, f. 12.
Тело продолговато овальное, совершенно симметричное; брюшной отдел уже, чем грудной. Ширина головы почти равна ее длине; передний край головы закруглен с маленьким острием посредине; задний край головы закругленный; глаза почти на-- цело редуцированы, остались лишь слабые следы
их. Усики 1-й пары 8-члениковые достигают передне-боковых углов 1-го грудного сегмента. Усики 2-й пары 12-члениковые, достигают средины 1-го грудного сегмента. 1-й грудной сегмент немного длиннее .2тгр;
г
t
F 8r^I V О NEC A 91
Г 3-й, 4-й, 5-й и 6-й сегменты длиннее 1-го, а 7-й сегмент короче не только 1-го, но и 2-го сегмента. Передне-боковые углы 1-го сегмента заострены и слегка оттянуты вперед. Эпимеры на всех сегментах, кроме 1-го, занимают всю длину боковой части сегмента; представлены в виде удлиненных пластинок, постепенно от 1-го до последнего делаются все шире и шире; задние края всех пар эпимер мешковидные, закругленные. Брюшко погру-L жено в 7-й грудной сегмент; первые 4 сегмента равной длины; 5-й сегмент немного длинее предыдущих. Ширина тельзона немного больше его длины; задний край тельзона широко-закругленный; к заднему концу тельзон слегка суживается. Ветвй рулевых ног равной длины, почти достигают конца тельзона; внутренняя ветвь немного шире наружной; концы ветвей закруглены. Все грудные ноги хватательные; на базальном членике 4 последних пар ног очень низкие гребни. Длина животного 34 мм. Самец отличается от самки присутствием глаз, более длинными усиками, которые достигают заднего края * 1-го сегмента, более узким телом и меньшими размерами.
; Распространение. Японское море:
42°10' N, 142О14' Е, на 618 м глубины, в ротовой < полости Synaphobranchus (Richardson, 1910). Жел-
; тое море: 32O36< N, 132°23/ Е, на глубине Фиг. 46. Livoneca epi-
осп .. merias Rich, (no Ri-
860 M- chardson).
5. Livoneca epimerias Richardson (фиг. 46).
Richardson, 1910, Proc. U. S. Nat. Mus., 37:88, f, 13.
Тело симметричное, удлиненное, длина его почти в 2 раза больше ширины, а боковые края почти параллельны друг другу. Длина головы равна ее ширине; передний край головы широко-закругленный; глаза маленькие, но хорошо выраженные, посаженные в боковых углах головы; задний край головы закруглен.. Усики 1-й пары 8-члениковые, достигают передне-боковых углов 1-го сегмента; усики 2-й пары 8—12-чле-никовые, достигают до конца первой трети длины бокового края 1-го сегмента. 1-й грудной сегмент длиннее всех остальных; 7-й сегмент груди самый короткий. Ясные эпимеры на всех сегментах, кроме 1-го. Эпимеры в виде узких пластинок, только 7-я пара эпимер в виде очень широких закругленных пластинок. Эпимеры всех сегментов, кроме 4-го и 5-го, достигают заднего угла сегментов, занимая всю длину боковой части его. Брюшко глубоко погружено в последний грудной сегмент, но не уже грудного отдела. Первые 4 брюшных сегмента очень ; короткие, равной длины; 5-й сегмент в 1.5 раза длиннее предыдущих. Тель ? зон поперечный, ширина его' почти в 2 раза больше длицы (10:6,5). Задний край его закруглен. Наружная ветвь рулевых ног намного шире
92
4. CYMO THOIDAE
и длиннее внутренней ветви;, рулевые ноги короткие, достигают лишь середины длины тельзона. Все грудные ноги хватательные; базальные членики 4 последних пар ног вооружены высоким гребнем. Длина животного 29 мм..
Распространение: Японское море — порт Хакодате (Richardson, 1910); сборы Albrecht, 2 экз., дл. 32 мм.
6. Livoneca vulgaris Stimpson (фиг. 47).
Stimpson, 1857, Bost. Journ. Nat. Hist. VI: 508, pl. XXII, f. 9. — occidenialis Richardson, 1899, Proc. U. S. Nat. Mus., 21:830 (Anilocra).
Тело симметричное, широкое, овальное; голова треугольной формы, наибольшая ширина ее больше длины, погружена в 1-й грудной сегмент.
Фиг. 47. Livoneca vulgaris Sch. & Mein, (no Schioedte-a. Meinert). / -—личинка; 2—3— взрослые <J.
Глаза большие выпуклые, овальные, расположены в задних углах головя, близко к переднему краю головы; задний край головы закруглен. Усики 1-й пары 8-члениковые, достигают конца 6-го членика 2-й пары усиков; базальные членики на расстоянии 1 мм друг от друга. Усики 2-й пары 10-члениковые, достигают середины 1-го грудного сегмента; 1-й- и 4-й грудные сегменты равной длины, длиннее всех прочих; 3-й и 5-й сегменты равной длины, немного короче, чем 1-й и 4-й; 2-й и 6-й сегменты равной длины, каждый немного короче 3-го и 5-го сегментов. 7-й сегмент самый короткий, в 2 раза короче, чем 1-й. Эпимеры в виде узких пластинок, занимающих весь боковой край соответствующего сегмента. Брюшко такой же ширины, как последний грудной сегмент; 1-й брюшной сегмент по бокам закрыт последним грудным сегментом. Тельзон широкий с широко закругленным задним краем; ширина его в 2 раза больше длины. Рулевые ноги такой же длины, как тельзон; ветви их одинаковой формы, равной длины и ширины; дистальные концы ветвей закруглены. Все грудные ноги хвата
8. LIVONECA
93
тельные; коготки длинные; узкие; базальный членик последних 4 пар снабжен низким килем. Длина животного 28 мм.
Распространение. Калифорния; паразитирует на жабрах Нурег-prosopon, Steindachneria, Ophidon (Richardson, 1905)
Фиг. 48. Livoneca californica Sch. & Mein, (no’ Schioedte et Meinert).
7. Livoneca californica Sch. a. Meinert (фиг. 48).
Schioedte a. Meinert, 1883—1884. Naturh. Tidsskrift, (3), XIV : 372, pl. XVI, f. 1—2.
Тело удлиненно-овальное, несимметричное, слегка изогнутое направо. Голова треугольной формы, широкая у основания, с вытянутой закругленной узкой передней частью. Глаза большие, овальные, расположенные в задне-боковых углах головы, отделены друг от друга расстоянием, равным длине глаза. Задний край головы закруглен. Усики 1-й пары 8-члениковые; основания их почти соприкасаются друг с другом. Усики 2-й пары 8-члениковые, достигают конца 7-го членика 1-й пары, очень слабые и тонкие. 1-й сегмент груди самый длинный; 2-й и 3-й сегменты равной длины, каждый короче 1-го. 4-й, 5-й и 6-й сегменты короткие, равной длины; 7-й сегмент самый короткий, в 2 раза короче 1-го. Передний край 1-го сегмента глубоко вогнут, так что боковые его части охватывают погруженную в нем голову с боков. Эпимеры в виде узких пластинок; 2-я и 3-я пары эпимер не доходят до конца бокового края соответствующих сегментов; 4 задних пары эпимер занимают весь боковой край своего сегмента. Брюшко такой, же ширины, как последний грудной сегмент; 1-й брюшной сегмент по бокам прикрыт последним грудным сегментом. Тельзон удлиненно-языковидный, ширина его немного больше длины; задний край его закруглен. Рулевые ноги достигают конца тельзона; ветви равной длины и ширины и одинаковой формы с закругленными дистальными концами. Все грудные ножки хватательные с длинными тонкими изогнутыми коготками. Базальный членик лишен гребней на всех грудных ножках. Длина животного 16 мм.
Распространение. Калифорния; паразитирует на акулах (Richardson, 1905). .
8. Livoneca caudata Sch. a. Meinert (фиг. 49).
Schioedte a. Meinert, 1883—1884, Naturh. Tidsskr., (3), XIV: 360, pl. XV, f. 1—2.
Тело широкое, удлиненное, симметричное, с почти параллельными краями; длина его больше чем в 2 раза больше ширины. Голова треугольной формы, погруженная; передний край ее двувыемчатый. Глаза небольшие, овальные. Усики 1-й пары у основания широко расставлены. Усики
94
4. CYMO THOIDAE
2-й пары 12-члециковые, достигают передне-бокового угла 1-го грудного сегмента. 1-й грудной сегмент равен по длине 4-му и 5-му сегментам; 2-й, 3-й и 6-й сегменты равной длины, короче 1-го; седьмой сегмент короче всех остальных. Эпимеры в виде изогнутых пластинок; на всех сегментах занимают весь боковой край своего сегмента. Брюшко широкое;
ширина его несколько больше, чем ширина последнего грудного сегмента;
передний край
Фиг. 49. Livoneca caudata Sch. & Mein, (no Schioedte et Meinert).
1-го брюшного сегмента прикрыт последним грудным сегментом. Тельзон широкий, полукруглый; ширина его почти в 2 раза больше длины. Задний край широкозакругленный; рулевые ноги короткие, не достигают середины длины тельзона; внутренняя ветвь немного уже и длиннее, чем наружная. Длина животного 41 мм; цвет серый.
Распространение. Япония ('Schioedte a. Meinert), побережье; бухта Сагами и Мисаки (Thielemann^ 1910).
9. Род IRONA SCHIOEDTE a. MEINERT
Schioedte a. Meinert, 1884, Naturh. Tidsskr. (3), XIV : 381.
i '
Тип рода. I. renardi Blecker.
Тело овальное, у самки асимметричное; голова глубоко погружена. Усики 1-й пары широко расставлены, так что Между их основаниями значительное расстояние; передний край грудного сегмента глубоко вогнут и боковые края его охватывают голову с боков. 1-й грудной сегмент значительно длиннее 2-го; остальные сегменты груди последовательно укорачиваются к'Заднему концу тела, так что последний грудной сегмент самый короткий. Передняя пара эпимер узкая й
' длинная, постепенно расширяющаяся кзади; остальные эпимеры короткие и широкие. Брюшко глубоко погружено в грудной отдел, но ширина его равна ширине грудного отдела. Все грудные ножки
хватательные; базальные членики их лишены гребня. Из дальневосточных морей известен один вид.
1. Irona melanostica Sch. a. Meinert (фиг. 50).
Schioedte a. Meinert, 1883—1884, Naturh. Tidsskr. (3), XIV : 388, pl. XVII, f. 3—5.
Тело овальное, изогнутое влево, несимметричное; длина его почти в 2 раза больше ширины; голова полукруглая, глубоко погруженная. Глаза большие, пентагональной формы. Усики 1-й пары 8-члениковые; достигают концом своего 6-го членика передних углов 1-го сегмента, груди; у основания усики широко разделены друг от друга. Усики 2-й пары 10-члениковые, тонкие, достигают своим 8-м члеником передних
9. IRONA
95
углов 1-го сегмента. 1-й грудной сегмент почти в 1.5 раза длиннее 2-го;
3-й сегмент Немного короче 2-го; 4 задних сегмента почти равной длины; каждый из них в 2 раза короче 2-го сегмента. Эпимеры 1-й и 2-й пары узкие, удлиненные с закругленными задними краями; эпимеры 3-й пары удлиненные и расширенные; 4-й и 5-й пары — короче и шире; 6-я пара — широкие с прямым задним краем. Брюшко глубоко погружено в послед-
ний грудной сегмент, так что боковые части 1-го брюшного сегмента, 2-го и частично 3-го сегмента закрыты последним грудным сегментом. Тельзон широкий с плавно закругляющимся задним краем. Ширина его в 2 раза больше лины. Рулевые ножки немного заходят за конец тельзона; наружная ветвь немного длиннее внутренней. Длина тела самки 20 мм. Тело коричневатое с мелкими темными пятнышками.
Распространение. Японское море и моря восточной Азии; Сандвичевы острова (Schioedte et Meinert). Побережье Японии — Окаяма и Иокогама (Thielemann, 1910).
Фиг. 50. Irona melanostica (по Thielemann).
5. Сем. SPHAEROMIDAE
\
Тело способно свертываться в шарик. Все 7 грудных сегментов свободны. БрЮшные сегменты обычно сливаются друг с другом или же только тельзон срастается с последним сегментом и сильно развит. Ветви рулевых ног неодинаковые; они или очень неравной длины и одна ветвь (наружная) рудиментарна, очень короткая, или совсем исчезает (иногда. исчезают обе ветви), или же одна ветвь (внутренняя) неподвижно укреплена, срастаясь с базальным члеником. Эпистома хорошо развита, не разделяется на фронтальную пластинку и clypeus. Ножка усиков 1-й пары 3-члениковая; ножка усиков 2-й пары 5-члениковая. Ротовые части кусающие или жующие, У $ J часто сильно видоизменены и не служат для питания. Инкубаторные пластинки самок только на 2-м, 3-м и 4-м сегментах. Брюшные ножки пластинчатые; внутренние ветви всех брюшных, ножек и наружные ветви 1-й и 2-й пары одночлениковые. Обе ветви 1-й и 2-й пары с длинйыми перистыми щетинками по краям. Обе ветви следующих пар не имеют таких щетинок и приспособлены для дыхания.
Распадается на 3 подсемейства, из которых на Дальнем Востоке известны только два.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ
1 (2). Abdomen состоит из двух сегментов; одна ветвь рулевых ног прикреплена неподвижно; тело способно свертываться в шар ....
.............................. . 2. Sphaerominae (стр. 98)..
*96 5. SPHAEROMIDAE
2 (1). Abdomen большей частью состоит из б сегментов. Обе ветви рулевых ног подвижные. Тело большей частью неспособно свертываться в шар. Ветви рулевых ног узкие, цилиндрические, одна ветвь отсутствует вовсе или рудиментарна . . . .1. Limnorinae (стр. 96).
1. Подсем. LIMNORINAE
1-й грудной сегмент не имеет эпимер; на всех остальных грудных сегментах эпимеры свободные. Брюшко состоит из 6 свободных сегментов. Жвалы крепкие, но с рудиментарной lacina mobile, лишенные пластинки на левой стороне. Зубной отросток редуцирован; щупик жвал 3-членико-вый. Челюсти 1-й пары с хорошо развитой внутренней лопастью. Челюсти 2-й пары с 3 маленькими, но хорошо развитыми лопастями. Ногочелюсти на лопасти 2-го членика имеют 1 крючок; эпиподит ногочелюстей большой, длина его больше ширины. Обе ветви брюшных ножек 1-й, 2-й, 3-й и 4-й пар с длинными маргинальными щетинками. Ветви 5-й пары брюшных ног не имеют таких щетинок и лишены квадратных полей или бугорков. Имеется всего один род.
1. Род LIMNORIA LEACH
Leach, 1814, Edinb. Encycl., VII: 433.
Тип рода L. lignorum (Rathke).
Тело продолговатое co сводчато-выпуклой спинной стороной. Все сегменты тела резко отделены друг от друга; голова короткая, круглая; ротовые части оттянуты книзу, 1-й грудной сегмент длиннее остальных и накрывает заднюю часть головы. Эпимеральные пластинки все свободно сочленены с сегментами, кроме 1-го. грудного сегмента, лишенного эпимер совершенно. Абдомен состоит из 6 сегментов; передние 5 брюшных сегментов снабжены эпимеральными расширениями; тельзон очень широкий, приплюснутый. Антенны короткие, с 3—4 члениками жгутика. Глаза круглые, расположены на боковых сторонах головы. Жвалы лишены зубного отростка, снабжены 3-члениковым щупиком; режущий край оканчивается коническим долотообразным острием. Челюсти 2-й пары заканчиваются 3 лопастями с пучками щетинок на концах. Жевательная лопасть челюстей 1-й пары с конической верхушкой, усаженной грубыми шипами, внутренняя лопасть узкая, с 3 перистыми щетинками на вершине. Щупик ногочелюстей 5-члениковый; членики его расширенные; базальная часть ногочелюстей узкая, вытянутая. Грудные ножки первых 3 пар короче остальных, снабжены добавочным когтем и короткими грубыми шипиками. Рулевые ноги маленькие; ветви очень неравной длины; наружная ветвь крючковидная, много короче внутренней.
Сверлящая форма; проделывает сложные ходы в деревянных частях морских сооружений.
Из дальневосточных морей известно 2 вида.
1. LIMNORIA
97
Фиг. 51. Limnoria lignorum (Rathke) (по G. Sars). a — передний конец тела; b — telson; с—uropoda.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Поверхность 5-го брющного сегмента и тельзона гладкая ....
.....................................* 1. L. lignorum Rathke.
2 (1). На спинной поверхности 5-го брюшного сегмента 2 низких бугорка; по средней линии тельзона 3 бугорка и 2 параллельных друг другу продольных ребрышка............................2. L. japonica Rich.
* 1. Limnoria lignorum (Rathke) (фиг. 51).
Rathke, 1799, Skrivt. Natur. Selsk., V: 101, pl. Ill, f. 14 (Cymothoa); G. Sars, 1899, An •account..., II, Isopoda: 76, pl. XXXI. — terebrans Leach, 1814, Edinb. Encyclop., VII: 433.
Тело полуцилиндрической формы,вытянутое; длина его почти в Зраза больше ширины. Эпимеры по краям усажены волосками; 2> передние пары эпимер овально-четырехугольные. 4 задних пары значительно -больше, и их заостренные задние углы оттянуты кзади. Длина тельзона почти равна длине всех 5 передних брюшных сегментов вместе; ширина его в 1.5 раза больше длины; форма -тельзона овальная, задний край плавно закругляется и усажен волосками, спинная поверхность его уплощена, гладкая. Глаза маленькие, круглые, черные с небольшим числом крупных линз. Верхние усики немного более крепкие, чем нижние, жгутик их 3-члениковый с длинными сензорными щетинками. Жгутик усиков 2-й пары 5-члениковый. 1-я пара грудных ножек немного больше, чем следующие, но такой же структуры; 4-й и 5-й/ членики грудных ножек снабжены короткими шипиками и бугорками; последняя пара ног более длинная и тонкая и лишена бугорков и шипиков на членйках. Все ножки -снабжены раздвоенным на конце добавочным когтем. Базальный членик рулевых ног сильно грубо зазубрен по наружному. краю; внутренняя ветвь палочковидная с пучком длинных щетинок на конце; наружная ветвь короткая, изогнутая крючком. Длина животного 5 мм.
Сверлит деревянные части судов, пристаней, доков и пр.
Распространение. Средиземное и Черное моря; побережье Западной Европы; Немецкое море; Норвежское побережье; Баренцово море; Белое море; по восточному побережью Сев. Америки от залива Лаврентия до Флориды; в Тихом океане от Берингова моря до Калифорнии (Richardson, 1905); Японское море: залив Петра Великого (сборы ТИРХ’а); Татарский пролив (сборы Булатова).
Г урьяиова 7
98
5. SPH AEROMIDAE
2. Limnoria japonica Rich. (фиг. 52).
Richardson, 1910, Proc. U. S. Nat. Mus., 37: 95, f. 21.
Тело удлиненно-овальное; голова вздутая, округлая с вдавленной: лобной частью; от ее переднего края отходит округлый вырост, оттянутый вверх. Глаза большие, круглые, посажены по бокам головы. Усики 1-й пары 4-члениковые;'2 первых членика равной длины, 2-й немного длиннее предыдущих; 4-й членик маленький с пучком длинных волосков на вершине. Усики 2-й пары с 5-члениковой ножкой и 5-члениковым жгутиком. 1-й грудной сегмент в 2 раза длиннее головы; остальные грудные сегменты равной длины, каждый короче 1-го в 2 раза. Эпимеры подвижно сочлененные с сегментами, образуют широкие пластинки, которые становятся, более широкими на задних сегментах. Первые 4 сегмента брюшка равной длины; 5-й сегмент в 2 раза длиннее каждого из пре-1 / \ дыдущих и снабжен 2 низкими бугорками, образующими
\ продольный ряд по средней линии сегмента. Тельзон
.. к,'j 1. большой, почти круглый по очертаниям, вогнутый на Мк О Л дорзальной стороне. У его основания посредине имеется; «С''4---т . большой резко выраженный бугорок и 2 более малень-
_ _ ких, близко посаженных друг к другу бугорка; эти бу-
Фиг. 52. Limnoria п •
iaponica Rich, (по горки продолжаются в 2 низких продольных киля, не Richardson). доходящих до заднего края тельзона. Рулевые ноги; почти такой же длины, как тельзон; внутренняя ветвь, короче базального членика с пучком щетинок на вершине; наружная ветвь-очень короткая. Длина животного 5 мм; цвет тела желтоватый, 3 последних грудных сегмента красноватобурые.
Распространение. Японское море. 37° 23z N, 137° 36" Е, на куске плавающего в воде бамбука (Richardson, 1910).
2. Подсем. SPHAEROMINAE
1-й грудной сегмент лишен эпимер; на всех остальных грудных сегментах эпимеры неподвижно срастаются с боковыми частями сегментов, со следами или без всяких следов шва. 5 передних брюшных сегментов-сливаются вместе в один сегмент с 3 или 2 поперечными бороздками по. его бокам, указывающими на следы слияния сегментов. Тельзон срастается с последним сегментом; таким образом имеются только 2 свободных брюшных сегмента. Жвалы крепкие, lacina mobile хорошо развита, с пластинкой на левой стороне; зубной отросток хорошо развит; щупик 3-чле-никовый. Челюсти 1-й пары с хорошо развитой внутренней лопастью;, челюсти 2-й пары с 3 удлиненными лопастями. На 2-м членике ногочелюстей на лопасти только один крючок; эпиподит маленький, ширина его-больше длины. Ветви 5-й пары брюшных ног не имеют перистых маргинальных щетинок; внутренняя ветвь 4-й пары обычно без щетинок; вау-
Z. LIMNORIA
99
тренние ветви 4-й и 5-й пары дыхательные. Наружная ветвь 5-й пары обычно с 3 утолщениями или выступами, густо усеянными чешуевидными мелкими шипиками. Внутренняя ветвь рулевых ног неподвижно срастается с базальным члеником или отсутствует.
На Дальнем Востоке известно 8 родов.
Н. Hansen (1906) на основании строения брюшных ножек разделяет подсем. Sphaerominae на 3 основных группы: 1) Hemibranchiatae-, 2) Eubran-chiatae', 3) Platybranchiatae. Первая группа характеризуется тем, что внутренняя ветвь 4-й и 5-й пар брюшных ног толстая, мясистая, с глубокими поперечными складками; наружная ветвь тонкая, перепончатая, 2-чле-никовая, прозрачная и обе ветви этих пар ног лишены перистых маргинальных щетинок; наружная ветвь 5-й пары имеет высокий чешуйчатый субапикальный вырост на нижней поверхности. Обе ветви 3-й пары брюшных ног густо усажены длинными перистыми щетинками, по крайней мере на дистальном крае. На Дальнем Востоке группа Hemibranchiatae представлена родами Sphaeroma, Exosphaeroma, Neosphaeroma, Cymodoce и Cilicaea.
2-я группа, Eubranchiatae, имеет обе ветви 4-й и 5-й пар брюшных ног почти одинаковые, часто мясистые, с глубокими поперечными складками, также без маргинальных щетинок; наружная ветвь 5-й пары дву-члениковая с очень высоким субапикальным чешуйчатым выростом. Обе ветви 3-й пары ног с длинными перистыми щетинками, по крайней, мере на дистальном конце. На Дальнем Востоке эта группа представлена родами Dynamene и Holotelson.
3-я группа, Platybranchiatae, характеризуется тем, что обе ветви 4-й и 5-й пар брюшных ног совершенно не имеют поперечных складок и их наружная ветвь одночлениковая. Внутренняя ветвь 4-й пары брюшных ног с немногими короткими терминальными перистыми щетинками; у рода Tecticeps наружная ветвь с многочисленными длинными маргинальными перистыми щетинками. Обе ветви 5-й пары брюшных ног без маргинальных щетинок; наружная ветвь со слабым чешуйчатым отростком, или совсем лишена его. 3-я пара брюшных ног иногда имеет перистые маргинальные щетинки на обеих ветвях, как и на ветвях 2-й пары, иногда одна или обе ветви голые. На Дальнем Востоке представлена одним родом Tecticeps.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). 1-я, а у самца и 2-я, пара грудных ножек хватательные, снабженные ложной клешней..........................2. Tecticeps Rich. (стр. 100).
2 (1). Первые 2 пары грудных ног, так же как и остальные, простые, ходильного типа, без ложной клешни.
3 (4). Наружная ветвь рулевых ног не может заходить под внутреннюю ветвь; ветви сильно отличаются друг от друга по форме и размерам
...................................5. Cilicaea Leach (стр. 115).
7*
100
5. SPHAEROMIDAE
4 (3). Наружная ветвь рулевых ног может заходить под внутреннюю; обе ветви более или менее одинаковой формы и размеров.
5(12). Задний край тельзона цельный, лишенный вырезки посредине.
6(11). Задний край тельзона плавно закругляется.
7 (8). Членики щупика ногочелюстей простые, без лопастевидных отростков на верхних внутренних углах . . 7. Sphaeroma Latr. (стр. 120).
8 (7). 2-й, 3-й и 4-й членики щупика ногочелюстей имеют внутренние верхние углы, оттянутые в виде лопастей с пучками щетинок на их вершине.
9 (10). Эндоподит 3-Й пары брюшных ног с тремя бранхиальными складками .............................9. Neosphaeroma Back. (стр. 122).
10(9). Эндоподит 3-й пары брюшных ног лишен бранхиальных складок.
...............................8. Exosphaeroma Stebb. (стр. 121).
11 (6). Задний край тельзона образует 3 зубца; один крупный с закругленной вершиной посредине и два маленьких, заостренных по бокам, у основания этого срединного зубца . 3. Holotelson Rich. (стр. 112).
12 (5). Задний край тельзона снабжен глубокой вырезкой посредине.
13 (14). Внутри вырезки на заднем крае тельзона имеется лопастевидный отросток..............................6. Cymodoce Leach, (стр. 117).
14 (13). Вырезка на заднем крае тельзона лишена внутреннего отростка.
.................................4. Dynamene Leach, (стр. 114).
2. Род TECTICEPS RICHARDSON
Richardson, 1897, Proc. Biol, Soc. Washington, XI: 180.
Тип рода: T. alascensis Rich.
Тело щитовидное* овальное с выпуклой спинной и плоской брюшной поверхностью. Голова очень широкая, в общем треугольной формы; передний край ее в виде щита накрывает собою усики, которые залегают под краями головы и грудных сегментов и не видны сверху. Глаза большие, почковидные или овальные, расположенные на спинной стороне головы. 1-я пара грудных ног, а у самца и 2-я пара, хватательные, снабженные ложной клешней; остальные грудные ножки ходильные, одинаковой структуры. Тельзон широкий, цельнокрайний, закругленный на конце или слегка заостряющийся. Эпимеры полностью сливаются с сегментами без следов шва. Абдомен из 2 сегментов; передний сегмент образован слиянием первых брюшных сегментов, со следами швов по бокам. Рулевые ножки двуветвистые; ветви неравной длины; внутренняя ветвь укреплена неподвижно. 2-й, 3-й и 4-й членики щупика ногочелюстей с оттянутыми в виде лопастей верхними внутренними углами.
В дальне-восточных морях известно 5 видов и 5 форм этого рода.
2. TECTICEPS
101
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ И ВАРИЕТЕТОВ
1 (12). Глаза узкие, почковидные., .
2 (7). Поверхность тела совершенно гладкая; нет ни зернистости, ни бугорков, ни килей на спинной поверхности сегментов.
Фиг. 53. Tecticeps renoculis renoculis Rich, (no Richardson).
Фиг. 54. Tecticeps renoculis laevis Gurjan. del. Ю. Залесский.
Фиг. 55. T. renoculis var. marginalis Gurjan.
Фиг. 56. T. renoculis var. catinatus Gurjan.
Фиг. 57. T. renoculis var. nodulosus Gurjan.
Фиг. 58. T. renoculis var. serratus Gurjan.
3 (6). Края эпимеральных расширений гладкие, без шипиков и зазубрин. 4 (5). Глаза широко-почковидные; голова плоская с уплощенной фронтальной частью. Контуры тела образованы плавно закругляющимися линиями..................... * la. Т. renoculis var. laevis Gurjan.
5 (4). Глаза узко-почковидные; край головы валикообразно утолщен, так что посредине фронтального края ясновыраженный небольшой
102
5. SPHAEROMIDAE
низкий, широкий бугорок. Контуры тела образованы ступенчатой
линией............... . * lb. Т. renoculis var. marginaiis Gurjan.
6 (3). Верхние края 4-го и 3~го грудных сегментов ^и нижние края эпиме-ральных расширений 5-го, 6-го и 7-го сегментов несут маленькие зубчики и шипики..............* le. Т. renoculis var. serratus Gurjan.
7 (2). Спинная сторона грудных сегментов несет бугорки, поперечные кили или же вся поверхность тела мелкозернистая и покрыта неправильной бугристостью.
8 (9). Наружная ветвь рулевых ног короче внутренней; по средней линии тела тянется продольный ряд бугорков, по одному на каждом сегменте ..............‘ . . . . * Id. Т. renoculis var. nodulosus Gurjan.
9 (8). Наружная ветвь рулевых ног длиннее внутренней. Имеется или зернистость и бугристость или же поперечные кили на спинной поверхности тела.
10 (11). Поверхность тела мелкозернистая; кроме зернистости на спинной поверхности сегментов неправильная бугристость......................
................................... 1. Т. renoculis renoculis Rich.
11 (10). На спинной стороне каждого, грудного сегмента у его заднего края невысокий поперечный киль с зазубренным верхним краем . .
..........................* 1с. Т. renoculis var. carinatus Gurjan.
12 (1). Глаза округлой формы.
13 (14). Глаза молочно-белого цвета . . . * 5. Т. leucophthalmus Gurjan.
14 (13). Глаза с темным почти черным пигментом.
15 (16). Боковые углы 5-го грудного сегмента вытянуты назад, образуя большие заостренные лопасти; 2-й членик ножки усиков 1-й пары снабжен длинным заостренным отростком . . * 3. Т. glaber Gurjan.
16 (15). Задние боковые углы 5-го грудного сегмента образуют небольшие закругленные лопасти; 2-й членик ножки усиков 1-й пары не имеет отростка.
17 (18). Глаза расположены в задне-боковых углах головы, округлые; конец тельзона заострен..............................* 2. Т. alascensis Rich.
18 (17). Глаза расположены на средине расстояния между передним и задним краями головы; задний конец тельзона закруглен ....
............................................4. Т. convexus Rich.
1. Tecticeps renoculis Richardson (фиг. 53).
Richardson, 1910, Proc. U. S. Nat. Mus., 37 : 89, f. 14—16.
Тело продолговато-овальное, очень широкое. Поверхность тела мелкозернистая, бугорчатая,'цвет розовый, края тела беловатые. Ширина головы почти в 2 раза превышает ее длину; передняя ее часть (впереди глаз) шире, чем затылочная (позади глаз). Глаза большие, широко почковидные, с черным или бурым пигментом, расположенным на спинной стороне в задне-боковых углах головы. Фронтальный край вытянут вперед и закруглен. Базальные членики усиков 1-й пары большие и рас
2. TEC TICEPS
103
ширенные; 2-й членик почти в 2 раза короче 4-го базального; 3-й удлиненный и тонкий; жгутик 11-члениковый и достигает заднего края 1-го грудного сегмента. Усики 2-й пары достигают заднего края 3-го грудного сегмента. Передне-боковые углы 1-го грудного сегмента вытянуты вперед, закруглены и охватывают с боков- голову. Все грудные сегменты почти •одинаковой длины. Эпимеры целиком сливаются с сегментами, образуя на 3 задних сегментах широкие, с закругленными краями, лопасти, направленные назад. Брюшной отдел из двух сегментов — переднего, получившегося от слияния первых брюшных сегментов с 3 линиями швов, и заднего или тельзона. Ширина тельзона в 2 раза больше его длины; задний край его более или менее закруглен. У основания тельзона имеются кили, по одному с каждой стороны, идущие по диагонали над основаниями рулевых ног. Ветви рулевых ног неравной длины; более короткая и широкая, неподвижно укрепленная, внутренняя ветвь немного не достигает заднего края тельзона; более узкая заостренная подвижная наружная ветвь заходит дальше конца тельзона у самца, а у самки такой же длины, как и внутренняя. 1-я пара грудных ног хватательная, снабженная ложной клешней; 6-й членик их сильно расширен, яйцевидно-овальный, по нижнему краю вооруженный грубыми жесткими короткими волосками или щетинками; 7-й членик образует длинный, изогнутый, прижимающийся ж внутреннему краю проподита коготь, У самца и 2-я пара грудных ног несет ложную клешню; 6-й членик ее слабо расширенный, почти такой же ширины, как карпальный членик, с тупым выростом у проксимального конца на внутреннем крае; 7-й членик длиннее, чем 6-й, и достигает в ело-' женном состоянии середины 5-го сегмента ножки. Следующие 5 пар грудных ног ходильные, постепенно возрастающие по длине.
Длина животного 20.5 мм,
Распространение. Охотское море: у восточного берега Сахалина около мыса Терпения (48°43'N, 145°13/Е) и на 48°43/N, 144°53'Е (Richardson, 1910).
В наших дальневосточных водах Т. renoculis распадается на <6 отдельных форм; их отличительные признаки приведены в таблице (стр. 108—109).
* la. Tecticeps renoculis var. laevis Gurjan. (фиг. 54).
Gurjanova, 1935, Исслед. морей СССР, 22 : 26, рис. З.1
Эта форма, в большом количестве добытая на мелководьях Охотского моря, значительно отличается от типичной формы, описанной Н. Richardson в 1910 г. из Охотского моря с восточной стороны Сахалина, относительно более широким телом и .более короткими и широкими тельзоном и головой.
1 В моей работе (1935 : 26) рисунки Т. renoculis marginaiis и Т. renoculis laevis оказались перевернутыми, почему подписи к ним следует поменять местами.
104
5. SPHAEROMIDAE
Поверхность тела Т. renoculis laevis совершенно гладкая, покрытая лишь мелкими точечными вдавлениями, тогда как у Т. renoculis Rich.— бугорчатая и покрыта мелкой зернистостью. Ширина тела Т. renoculis laevis составляет 75% его длины (у Т. renoculis Rich, только 63.4%); ширина головы больше чем в 2 раза превышает ее длину (у Т. renoculis: Rich, ширина головы немного меньше чем в 2 раза превышает ее длину, так как фронтальная часть сильно вытянута вперед). Глаза у Т. renoculis Rich, помещены в задне-боковых углах головы у самого заднего края ее, а у Т. renoculis laevis глаза отодвинуты от заднего края вперед и расположены на равный расстояниях от переднего и заднего краев головы. Ширина головы впереди глаз у Т. renoculis Rich, больше, чем ее ширина позади глаз, а у Т. renoculis laevis ширина головы впереди глаз и позади глаз одинаковая, так что боковые края головы параллельны друг другу. Ширина тельзона у Т. renoculis laevis больше чем в 2 раза превосходит его длину, тогда как у Т. renoculis Rich, ширина тельзона немного меньше: чем в 2 раза превышает его длину. Конец тельзона Т. renoculis laevis треугольной формы, заостряющийся (у типичной формы конец тельзона трапециевидный с слабо закругленным задним краем). 2-й членик ножки усиков 1-й пары у Т. renoculis laevis в 1.5 раза короче базального членика; жгутик 12-члениковый. Цвет тела розовато-серый с светлобурыми пятнами и бурыми полосками на эпимеральных расширениях сегментов. 3-й грудной сегмент заметно длиннее всех остальных. Проподйальный членик 2-й пары грудных ножек самца снабжен пучком длинных щетинок на наружном дистальном угле. Усики 1-й пары достигают заднего края 1-го грудного сегмента, а усики 2-й пары заднего края 3-го сегмента. В остальном признаки сходны с описанием и рисунками Н. Richardson для типичной формы. Длина животного 22 мм.
Распространение. Японское море: сбор Павленко и Солдатова, 1911, ст. 65 — 50°00'N, 141°51'Е, глуб. 110 м, 1 экз.; 1913, ст. 431 — Татарский пролив, 48°08' N, 140°08'Е, глуб. 121 м, I экз.; сбор ГГИ, 1931, ст. 438/26 — 50°06'N, 141 °25'Е, глуб. 150 м, 3 экз. Охотское море: сбор Дербека, 1910, ст. 9. — у мыса Мария 54°20' N, 142°12' Е, глуб. 60 м, 5 экз.; ст. 14 — 54 33' N, 140°20' Е, глуб. 84 м, 2 экз,; ст. 8 — 53°45' N, 141° 11'Е, глуб. 25 м, 18 зкз.; Дербек, 1912, ст. 10 —54°52'N, 141°15'Е, глуб. 120 м, 6 экз.;’ 54°22' N, 141 °26' Е, глуб. 50 м, 3 экз.; сборы Солдатова и Павленко, 1911: ст. 44—-53O37'N, 136°54'Е, глуб. 50 м, 32 экз.; ст. 53 — 54°21'N, 141°33'Е, глуб. 90 м, 5 экз.; ст. 65 —50°00'N, 141О51'Е, глуб. 110 м, 1 экз.; 1912, ст. 168 — 55°57'N, 138°13'Е, глуб. 85 м, 1 экз.; 1913, ст. 374, 54°17'N, 140°4б', 2 экз.; ст. 394 —55°34'N, 141°30'Е, глуб. 40 м, 2 экз.; 1915, ст. 477 — бухта Надежда (Сахалин), глуб. 35 м, 2 экз.; ст. 478 — 54О35” N, 142°16'Е, 9 экз.; ,сборы Гйдрогр. эксп. Тихого океана, 1914—1918: 53° 01'N, 143°35'Е, 7 экз.; 55°32'N, 137°37'Е, I экз.; 52°19'N, 144°17' Е, 5 экз. Сборы Н. Смирнова и Бегака, 1907, ст. 26—53°20' N, 154°52'Е, глуб. 160 м, 2 экз.; ст. 94, 52°06'N, 155°35'Е, глуб. 100 м, 8 экз..
2. TECTICEPS
105
Сборы ГГИ, 1930: ст. 130/15 — 53°38' N, 141°35' Е, глуб. 25 м, 14 экз.; ст. 131/16 — 54°00'N, 141°53/Е, глуб. 47 м, 9 экз.; ст. 133/18 — 54°33'N, 142°40'Е, глуб. 67 м, 2 экз.; ст. 134/19 — 56°08'N, 144° 55'Е, глуб. 287 м, 2 экз.; ст. 136/21 — 57°30'N, 147°10'Е, глуб. 146 м, 1 экз.; ст. 137/22 — 57°40' N, 147°50'Е, глуб. 144 м, 10 .экз.; ст. 138/23, 58°01'N, 148°19'Е, глуб. 141 м, 9 экз.; сборы ГГИ, 1932: ст. 237 —54°18'N, 142°00'Е, глуб. 53 м, 1 экз.; ст. 238—53°56' N, 142°14' Е, глуб. 28 м, 4 экз.; ст. 236—53°47' N, 143°22'Е, глуб. 84 м, 4 экз.; ст. 239 —54°19'N, 140°18'Е, глуб, 60 м,2 экз.; ст. 241—54°19'N, 140°18'Е, глуб. 60 м, 1 экз.; ст. 244—54°34'N, 142°44'Е, глуб. 65 м, 7 экз.; ст. 246 — 55°04' N, 142°55' Е, глуб. 128 м, 1 экз.; ст. 257 — 55°41' N, 154°35' Е, глуб. 132—150 м, 1 экз.
Форма, характерная для фауны песка на мелководьях Охотского моря; проникает в северную часть Японского моря (Татарский пролив).
* lb. Tecticeps renoculis var. marginalis Gurjan. (фиг. 55).
Gurjanova, 1935, Исслед. морей СССР, 22:26, рис. 2.1
От Т. renoculis laevis сильно отличается утолщенными краями головы, эпимер и тельзона и ступенчатой линией очертания тела. Поверхность тела совершенно гладкая; все тело более крепкое и массивное, чем у Т. renoculis laevis. Длина тела 23.01 мм при ширине в 14 мм; длина головы 4.5 мм, ширина 7 мм; расстояние между глазами 4.2 мм. Усики 1-й пары в вытянутом состоянии достигают заднего края 1-го грудного сегмента, жгутик 11-члениковый; усики 2-й пары достигают середины 3-го грудного сегмента, жгутик 11-члениковый. Строение ротовых частей и ножек такое же, как и у основной формы. Лобная часть плоская, но передний край головы посредине утолщен и образует широкий расплывчатый бугорок. Цвет тела темносерый; край головы, утолщенные эпимеры и края тельзона беловатые. Глаза более узкие, чем у двух предыдущих форм, почковидные. Голова относительно более широкая, а рулевые ноги несколько короче, чем у Т. renoculis laevis.
Распространение. Берингово море: 61°22'N, 178°02'Е, глуб. 360 м, грунт — песок. Добыты 17 экз. экспедицией ГГИ, 1932, ст. 53.
*lc. Tecticeps renoculis var. carinatus Gurjan. (фиг. 56).
Gurjanova, 1933, Исслед. морей СССР, 17:99, рис. 26—27.
От типичной формы отличаемся коротким и широким телом и сильно уплощенной спинной стороной. Поверхность тела покрыта мелкой зернистостью. По средней линии тела продольный ясно выраженный киль, доходящий до конца тельзона. На каждом грудном сегменте у его заднего края имеется валикообразный поперечный низкий киль с неправильно зазубренным краем. Такой же киль на голове между глазами, иногда. очень резко выраженный. Усики 1-й пары достигают заднего края
1 См. сноску на странице 113.
106
5. SPHAEROMIDAE
2-го сегмента; жгутик 12-члениковый; усики 2-й пары достигают заднего края 3-го, а иногда середины 4-го грудного сегмента; жгутик 11-членико-вый. Конец тельзона треугольный, заостренный. Наружная ветвь рулевых ножек заходит дальше конца тельзона. Эпимеры уплощенные с поперечными зазубренными килями. 1-й грудной сегмент равен по длине 3-му, остальные сегменты груди короче и тем короче, чем ближе к заднему концу тела. Длина животного 17 мм. Цвет розовый.
Распростр.анение. 1932, Охотское море: ст. 217—51°35'N, 154°46'Е, глуб. 418 м, песок, 4 экз.; 1930, ст. 135/20—56°45'N, 145°50'Е, глуб. 218 м, 7 экз. Явинская банка, 53°20' N и 154°52/ Е, глуб. 144 м., песок, 2 экз. (Сбор Н. Смирнова и Бегак, 1907).
* Id. Tecticeps renoculis var. nodulosus Gurjan. (фиг. 57).
Gurjanova, 1935, Исслед. морей СССР, 22: 27, рис. 4.
От типичной формы отличается плоской спинной поверхностью, присутствием мелких шипиков на эпимеральных расширениях и присутствием продольного ряда бугорков по средней линии тела. Длина тела 13 мм при ширине его в 9 мм; голова широкая, полукруглая, с сильно вогнутым задним краем; длина головы 3 мм при ширине в 4 мм; расстояние между, глазами 3 мм; глаза узко-почковидные. Посредине вогнутого заднего края головы маленький бугорок. По такому же бугорку посредине каждого грудного сегмента у его заднего края. Усики 1-й пары достигают заднего края 2-го грудного сегмента; жгутик 12-члениковый; усики 2-й пары достигают середины 4-го грудного сегмента; жгутик 11-члениковый. Конец тельзона закруглен. Ветви рулевых ног равной длины или наружная ветвь немного короче внутренней; наружная ветвь достигает конца тельзона. Поверхность тела мелкозернистая; на эпимеральных расширениях имеются слабые кили и грубая зернистость. 1-й грудной сегмент в 1.5 раза длиннее 2-го; остальные сегменты тем короче, чем ближе к заднему концу тела.
Распространение. Охотское море, 55°32'N, 149°14'Е, >глуб. 335 м, грунт — ил и галька (эксп. ГГИ, 1932, ст. 252), 2 экз.
* le. Tecticeps renoculis v. serratus Gurjan. (фиг. 58).
Gurjanova, 1935, Исслед. морей СССР, 22 : 27, .рис. 5.
От основной формы отличается более коротким телом и гладкой поверхностью тела, присутствием шиповатой зазубренности на эпимерах и формой головы. Спинная сторона тела сильно выпуклая; длина животного 16 мм при ширине в 10.5 мм. Голова короткая, широкая; длина ее 3.5 мм при ширине в 5 мм; расстояние между узкопочковидными глазами 3 мм. Линии тела по бокам плавно закругляются. Вдоль верхнего края эпимеральных расширений 3-го и 4-го грудных сегментов, а на 5-м, 6-м и 7-м сегментах, вдоль нижнего края, небольшие шипики и зубчики; кроме того, поверхность эпимеральных расширений мелко зернистая. Усики
2. TECTICEPS
107
1-й пары достигают заднего края 1-го сегмента; жгутик 11-члениковый. 'Усики 2-й пары достигают середины 3-го грудного сегмента; жгутик 11-члениковый. Конец тельзона тупой, в общем треугольный со слабым продольным килем на конце. Наружная ветвь рулевых ног немного длиннее внутренней и едва достигает конца тельзона. Первые 3 грудных сегмента одинаковой длины, остальные немного короче. Цвет зеленовато-желтоватый с неясными серыми пятнами.
Распространение. Японское море: Татарский пролив, глуб. 80—223 м (Сборы ТИРХ’а, 1931).
* 2. Tecticeps alascensis Richardson (фиг. ,59). •
Richardson, 1897, Proc. Biol. Soc. Washington, XI : 181; Richardson, 1905, Bull. U. S. Nat.
Mus., n° 54: 276, f. 286—289.
Тело овальное, с плавно закругляющимися боковым’и линиями; поверхность тела гладкая, покрытая точечными вдавлениями. Голова
большая, в два раза длиннее каждого из грудных сегментов; передний край вытянут вперед, тупой, с легким вдавлением посредине. Глаза черные, округлые, расположенные в задне-боковых углах головы. Усики 1-й пары достигают заднего угла 1-го грудного сегмента; жгутик 10-члениковый. Усики 2-й пары достигают средины 2-го сегмента; жгутик 12-члениковый. Грудные сегменты почти равной длины. Спинная сторона сводчато-выпуклая. Ширина головы впереди глаз больше, чем ее ширина позади глаз. Эпимеральные расширения 5-го грудного сегмента почти квадратной формы. Задний край 1-го брюшного сегмента образует 2 острых уголка по бокам над основанием тельзона. Тельзон треугольной
Фиг. 59. Т. alascensis Rich. -
формы со сводчато-выпуклой спинной поверхностью и заостряющимся концом. Внутренняя ветвь рулевых ног короче и шире наружной, не
достигает конца тельзона, с тупым дистальным концом; наружная подвиж
ная ветвь узкая, заостренная на конце, заходит дальше конца тельзона. Цвет тела вариирует от коричневого до желтоватого с темными пятнышками. Измерения наиболее крупного из экземпляров, добытых у Алеутских островов Вознесенским в 1846 г., дали следующие результаты: длина тела 15 мм; ширина 10 мм; длина головы 3 мм; ширина головы впереди глаз 5 мм, ширина головы позади глаз 4.5 мм. Длина тельзона 4 мм; ширина его у основания 8 мм; расстояние между глазами 3 мм.
Распространение. Берингово море: Прибыловы о-ва; Алеутские о-ва; берега Аляски и Камчатки на глубине от 18 до 200 м-Охотское море: бО^Чб7 N, 167°41/ W; у Курильских островов; северная часть Тихого океана: Бристольская бухта; о-в Якутан, Унимак, .Амак на глубине 16 м (Richardson, 1905, 1907). У Алеутских островов, внутренности кита, 4 экз. (сбор Вознесенского, 1846).
108
5. SPHAEROMIDAE
i
2. TECTICEPS
109
СРАВНИТЕЛЬНАЯ ТАБЛИЦА ФОРМ I TECTICEPS RENOCULIS RICH.
Т. г. renoculis Т, г. carmaius 1 Т, г. nodulosus Т. г. laevis Т. г. marginaiis Т. г. serratus
Длина тела 20.5 17.5 13 22 23 16
Наибольшая ширина тела в мм . 13 12 9 14 14 10.5
Отношение ширины тела к длине в°/о 63.4 68.5 69.2 63.6 60.8 65.6
Длина головы в мм 4 4 3 4.5 4 3.5
Ширина головы в мм 7 6 4 7 7 5
Отношение длины головы к ее ширине в % 57.1 57.1 75.0 64.3 57.1 7010
Длина тельзона в мм 5.5 4.5 3.5 5 6 4.5
Ширина тельзона в мм .... 11 10 7 11 И 9
Отношение длины тельзона к его ширине в % 50 45.0 50 45.4 54.5 50
Поверхность тела Мелкозернистая, бугорчатая Мелкозернистая. Поперечные зазубренные кили на голове и грудных сегментах Мелкозернистая. По средней линии тела продольный ряд бугорков Гладкая; точечные вдавления по всей поверхности Гладкая. Точечные вдавления по всей поверхности Гладкая; на эпимерах мелкая, зернистость
Цвет Розовый; края тела беловатые Розовый Розовато-серый с темными пятнами на эпимерах Серовато-розоватый с бурыми пятнами Темносерые края тела беловатые Зеленовато-желтый с серыми пятнами.
Конец тельзона Закруглен Треугольно-заостренный Закруглен Треугольно-заострен Треугольно заострен с бугорком на конце Треугольный конец тупой
Спинная сторона тела .... Выпуклая Плоская Плоская Выпуклая Слабо выпуклая Очень сильно свод-чато-вы пуклая
Усики 1-й пары Достигают заднего края 1-го сегмента; жгутик 11-членико-вый Достигают заднего края 1-го сегмента. Жгутик 12-членико-вый Достигают заднего края 2-го сегмента. Жгутик 12-членико-вый Достигают заднего угла 1-го сегмента. Жгутик 12-членико-вый Достигают середины 2-го сегмента. Жгутик 11-члениковый Достигают заднего края 1-го сегмента. Жгутик 11-члениковый
Усики 2-й пары Достигают заднего края 3-го сегмента; жгутик 11-членико-вый Достигают заднего края 4-го сегмента. Жгутик 11-членико-вый Достигают середины 4-го сегмента. Жгутик 11-члениковый Достигают заднего 'края 3-го сегмента. Жтутик 11-члениковый Достигают заднего края 3-го сегмента Жгутик 11-члениковый. Достигают середины 3-го сегмента. Жгутик 11-члениковый
Длина сегментов тела Первый сегмент самый длинный, немного длиннее 2-го сегмента; остальные становятся все короче и короче, по мере продвижения к заднему концу тела Первый сегмент равен длине 3-го сегмента, остальные грудн. сегменты более короткие 1-й сегмент в 1.5 раза длинее 2-го. Остальные грудные сегменты еще короче Самый длинный 4-й сегмент, равный длине 5-го и 6-го сегментов вместе 1-й сегмент немного длиннее 2-го; остальные сегменты постепенно становятся короче к заднему концу тела Первые 3 сегмента одинаковой длины; остальные немного короче.
110
5. SPHAEROMIDAE
* 3. Tecticeps glaber Gurjan. (фиг. 60).
Gurjanova, 1933, Исслед. морей СССР, 17 : 98, рис. 24—25. —japonicus Iwasa Masaoj. 1934, Proc. Imp. Acad. Tokyo, 10, n° 6 :370, fig. 1—2.
Тело яйцевидно-овальной формы с широкой выпуклой спинной поверхностью. Передний край головы закругленный; задний край прямой^ Впереди глаз голова резко расширяется, образуя боковые лопасти, прикрывающие сверху усики. Глаза черные, округлые, слегка сдавленные на медиальной стороне (намек на почковидность); расположены в заднебоковых углах головы, немного отступя от заднего края. Усики 1-й пары, достигают заднего края 2-го грудного сегмента; второй членик их ножки снабжен длинным, почти достигающим конца 3-го членика заостренным отростком на наружном дистальном угле; жгутик 13-члени-ковый. Усики 2-й пары достигают заднего края 3-го грудного сегмента; жгутик 14-чле-никовый. Эпимеральные расширения 5-го грудного сегмента сильно оттянуты назад, образуя’ длинную, резко заостряющуюся лопасть на задне-боковом угле сегмента; острие этой лопасти достигает почти середины эпи-мерального расширения последнего 7-го грудного сегмента. 1-й брюшной сегмент, образованный слиянием всех передних брюшных
Фиг. 60. Т. glaber Gurjan. а — сегментов, длинный; длина его равна длине общий вид; Ь — antenna I.
двух последних грудных сегментов, вместе-Задний край его по бокам у основания тельзона образует 2 маленьких заостренных отростка. Тельзон треугольной формы с заостряющимся дистальным концом; длина его в 2 раза больше длины 1-го грудного сегмента (5 мм) при ширине основания 8 мм. Внутренние ветви рулевых ног не достигают конца тельзона; наружные ветви узкие, резко заостренные, длинные, заходят далеко за пределы конца тельзона. Длина тела 24 мм при максимальной ширине у 4-го сегмента в 15 мм.. Длина головы 3 мм, ширина головы позади глаз 4 мм; ширина головы, впереди глаз 5.5 мм. Цвет тела серый с желтоватыми краями и темными пятнами вдоль задних краев грудных сегментов и на швах 1-го брюшного сегмента.
Распространение. Японское море: залив Петра Великого; бухта Венсан на глубине 14 м; залив Броутона, глуб. 28 м (сборы П. Шмидта,. 1900); бухта Патрокл, на глуб. 30 м (сбор ТИРХ’а); бухта де-Кастри на глубинах от 2 до 20 м на песчаном грунте. Бухта Аккеши, Хоккайдо, гл. 10 м (Iwasa 1934).
2. TECTICEPS
111
4. Tecticeps convexus Richardson (фиг. 61).
Richardson, 1899, Proc. U. S. Nat. Mus., 21 : 837; Richardson, 1905, Bull. U. S. Nat. Mus., n° 54 :278, f. 290—291.
между передним и
Фиг. 61. Т. convexus Rich, (по RichardsonJ. а — голова; Ь — telson;
краем. Обе ветви
Тело овальное, более или менее плоское; поверхность тела гладкая.. Как у предыдущего вида, голова впереди глаз сильно расширяется, образуя щитовидные лопасти, накрывающие сверху обе пары усиков. Передний край головы закруглен и посредине несколько утолщен, образуя 2 маленьких выпуклых возвышения. Базальные членики усиков 1-й пары слегка выступают за пределы накрывающего их головного щита. Задний край головы прямой. Глаза неправильно округлой формы; расположены на спинной стороне головы на середине расстояния задними углами головы. Усики 1-й пары достигают заднего угла 3-го грудного сегмента; жгутик 16-чле-никовый. Усики 2-й пары достигают середины 4-го грудного сегмента; жгутик 13-члениковый. Грудные сегменты равной длины. Передне-боковые углы 1-го сегмента охватывают с боков заднюю часть головы, доходя до середины глаза, образуя широкие пластинки по бокам сегмента. Эпимеральные расширения грудных сегментов широкие, ширина их почти в 2 раза больше длины. На 5-м грудном сегменте эпимеральное расширение почти квадратной формы с прямым передним краем. 1-й брюшной сегмент длинный с 3 линиями швов по бокам, получившимися в результате слияния сегментов. Задний край его по бокам у основания тельзона образует 2 маленьких заостренных отростка. Тельзон в общем треугольной формы с широко закругленным задним
рулевых ног почти равной длины и не достигают вершины тельзона; наружная ветвь узкая и сильно заостренная на конце; внутренняя ветвь-широкая, закругленная на вершине. Цвет светложелтый с бурыми пятнами.
Распростране ни е. Калифорния, на глубине 10 м, песчаный, грунт (Richardson, 1905).
* 5. Tecticeps leucophthalmus Gurjan. (фиг. 62).
Gurjanova, 1935, Исслед. морей СССР, 22 : 25, рис. 1.
От прочих видов этого рода легко отличается молочнобелым цветом, глаз, присутствием волосков на эпимерах й конце тельзона. Тело удлиненно-овальной формы с плавно-закругляющимися на обоих концах тела контурами. Поверхность тела гладкая., Длина 11 мм при ширине 3.5 мм.. Расстояние между глазами 2 мм. Глаза молочно-белого цвета, удлиненно овальной формы, посажены по бокам головы на ее спинной стороне,, непосредственно, у заднего края. Передний край головы несколько вытянут
112
5. SPHAEROMIDAE
вперед, закругленный. Впереди глаз голова слегка расширяется, образуя маленькие лопасти. Усики 1-й пары достигают середины 3-го грудного сегмента; жгутик 10-члениковый. Усики 2-й пары достигают середины 5-го грудного сегмента; жгутик 10-члениковый. Все грудные сегменты и 1-й брюшной сегмент почти равной длины. Эпимеры 5-го сегмента почти квадратные. Тельзон трехугольной формы с узким задним закругленным концом, с небольшим бугорком на самом конце. Рулевые ножки с широкими закругленными на концах неподвижными внутренними ветвями и более
длинными узкими, заостряющимися наружными ветвями, выходящими за пределы конца тельзона. Цвет тела розовато-желтоватый с белесыми краями головы, эпимер и тельзона. Ротовые части и грудные ножки нормального для рода строения.
Распространение. Охотское море. 49°29' N, 152°00' Е, глуб. 1366 м, в жидком илу, 1 экз. (экспедиция ГГИ, 1932, ст. 215),
Фиг. 62. Т. leucoph-thalmus Gurjan.
3. Род HOLOTELSON RICHARDSON
Richardson, 1910, Proc. U. S. Nat. Mus., 37: 93.
Тип рода: H. tuberculatus Rich.
Все грудные ножки ходильные, лишенные ложной клешни. Голова нормальной величины; базальный членик усиков 1-й пары не имеет острого отростка на дистальном конце. У самца задний край 7-го грудного сегмента образует пару маленьких
треугольных отростков по одному с каждой стороны от средней линии тела; у самки эти отростки очень малы, но ясно выражены. Задний край 1-го брюшного сегмента образует 3 конических бугорка или отростка, один посредине и по одному с каждой стороны около бокового края. Задний край тельзона снабжен 3 зубцами; срединный зубец очень большой, узкий, вытянутый назад с закругленной вершиной; 2 другие зубца расположены по бокам у основания срединного зубца и значительно меньшей величины. У самки все 3 зубца на заднем крае тельзона развиты слабее, чем у самца. Ветви рулевых ног одинаковых размеров и формы у самца, у самки наружная ветвь немного меньше чем внутренняя. 2-я пара брюшных ножек самца снабжена стилетом. Наружная ветвь 3-й пары брюшных ног двучлениковая. Обе ветви 4-й и 5-й пар брюшных ног мясистые, с поперечными складками и не имеют краевых щетинок. Наружная ветвь 5-й пары двучлениковая.
Известен только один вид этого рода.
* 1. Holotelson tuberculatus Richardson (фиг. 63).
Richardson, 1910, Proc. U. S. Nat. Mus., 37: 93, f. 17—20.
Тело удлиненное с закругленным передним краем и почти параллельными боковыми краями; длина его в 2 раза больше ширины. Спинная
3. HOLOTELSON
113
сторона сильно выпуклая, сводчатая. Голова очень широкая, полукруглой формы, с широко закругленным передним и почти прямым задним краем, передний край валикообразно утолщен. Задне-боковые углы головы слегка оттянуты назад, образуют маленькие закругленные лопасти, на которых помещаются глаза. Базальный членик усиков 1-й пары большой и расширенный; длина его в 2 раза больше ширины; 2-й членик короткий и широкий, длина его почти равна ширине; 3-й членик узкий и длинный, в 2 раза длиннее 2-го членика; жгутик 14-члениковый, в вытянутом состоянии достигает середины 1-го грудного сегмента. Усики 2-й пары достигают середины 2-го грудного сегмента, жгутик их 1б-члениковый. 1-й грудной сегмент в 1.5 раза длиннее 5 следующих, которые почти одинаковой длины; эпимеральные расширения этих сегментов короткие и широкие, задне-боковые углы их слегка оттянуты и образуют тупые острия. Последний грудной сегмент по средней линии длиннее предыдущих; средняя часть его заднего края оттянута назад и образует 2 коротких закругленных отростка, по одному, с каждой стороны от средней линии, которые нависают над 1-м брюшным сегментом, прикрывая его спереди. 1-й брюшной сегмент по бокам имеет по 2 поперечных шва с каждой стороны (следы слияния брюшных сегментов); задний его край образует 3 ко-, ротких треугольных отростка, 1 посредине и 2 по бокам; вершины этих трех отростков утолщены в виде бугорков. Спинная сторона тельзона выпуклая и посредине несет один поперечный ряд из 3 бугорков; расши-
ренный у основания тельзон суживается дистально, в общем треугольной; формы и на заднем конце снабжен тремя отростками; один отросток, срединный, очень большой, сильно вытянут назад с закругленной вершиной 2 другие расположены по бокам от него у самого его основания, маленькие, притупленные. Рулевые ноги короткие, достигают лишь середины длины тельзона; ветви их уховидные, закругленные, одинаковой формы и размеров. Самки сильно отличаются от самцов почти прямым задним краем 7-го грудного сегмента, лишенным срединных отростков, слабо выраженными бугорками на заднем крае 1-го абдоминального сегмента и треугольно заостренной вершиной тельзона, не имеющей срединного и боковых отростков на его конце; если эти отростки имеются, то выражены они очень слабо, по сравнению с самцом. Длина животного достигает 11 мм; цвет розоватый, часто голова и тельзон темно окрашены. Свертывается в неполный шар.
Распространение. Японское море: Мороран (Richardson» 1910); залив Петра Великого (Посьет, бухта Патрокл, б. Соболь, б. Тихая в Уссурийском заливе, б. Валентин, б. Тафуин в заливе Восток, мыс Басаргин); на север доходит до бухты Преображение (эксп. ЗИНАН 1934).
Гурьянова 8
114
5. SPHAEROMIDAE
Характерная форма биоценоза Zostera-, держится массами в ее корнях.; встречается в зарослях Corallina, на мидиях, на глубинах от 0 до 10 м< и на литорали среди красных и бурых водорослей и на сваях пристаней (Сборы ТИРХ'а, 1926—1929).
4. Род DYNAMEME LEACH
Leach, 1814, Brewster Edinb. Encycl., VII: 433.
Тип рода: D. bidentata (Adams).
Брюшной отдел состоит из 2 сегментов; первый из них образован1 слиянием нескольких сегментов вместе, и следы этого слияния сохраняются в виде швов по бокам сегмента; тельзон дистально суживается и на конце, имеет вырезку; иногда вместо вырезки округлое отверстие или задний конец свертывается в трубочку, незамыкающуюся на брюшной стороне.. Ветви рулевых ног обычно одинаковой формы, у самок одинаковых размеров, но у самцов наружная ветвь обычно значительно, иногда в 2 раза,, длиннее внутренней^ Внутренняя неподвижная ветвь прикреплена к боковому краю последнего брюшного сегмента. Наружная ветвь свободная, и способна подгибаться под внутреннюю ветвь. 2-й, 3-й и '4-й членики щупика ногочелюстей имеют с внутренней стороны вытянутые лопасти. Все грудные ножки ходильные, лишенные ложной клешни. Наружная ветвь-3-й пары брюшных ног одночлениковая. 2-я пара брюшных ног самца, не имеет стилета. У самца задний край 6-го грудного сегмента имеет пару небольших отростков, у самки этот край прямой. Базальный членик усиков. 1-й пары нормальный, без особых расширений или отростков.
На Дальнем Востоке известен 1 вид.
* 1. Dynamene glabra Richardson (фиг. 64).
Richardson, 1899, Proc. U. S. Nat. Mus., 21:834; Holmes, 1904, Proc. Cal. Acadi Sci.. (3),. Ill, N 11:304; Richardson, 1905, Bull. U. S. Nat. Mus., n° 54:301, f. 321—322.
Тело овальной формы; поверхность его совершенно гладкая; голова маленькая с закругленным передним и прямым задним краями. Глаза расположены в задне-боковых углах головы. Усики 1-й пары в вытянутом, состоянии достигают передне-бокового угла 1-го грудного сегмента; базальный членик их удлиненный. 2-й членик короткий, в 2 раза короче 1-го; жгутик 6-члениковый. Усики 2-й пары достигают. заднего края 1-го грудного сегмента; жгутик из 10 члеников. 1-й грудной сегмент-.немного длиннее каждого из остальных сегментов груди, которые равной, длины. Тельзон треугольной формы с сильно выпуклой спинной стороной; задний узкий конец его на конце завертывается в трубочку, открытую на брюшной стороне; сверху на конце тельзона видна лишь полукруглая вырезка. Внутренняя ветвь рулевых ног большая, закругленная на. конце и достигает конца тельзона; наружная ветвь маленькая, такой же; формы, но достигает только половины длины внутренней ветви. Цвет-
4. DYNAMENE, 5. CILICAEA
115
животного темносерый, иногда с розоватыми краями; конец тельзона обычно более светло окрашен, чем остальная его часть. Длина 2 мм. Половой диморфизм почти не выражен, кроме того, что-самец крупнее самки.
Распространенно. Калифорния — бухта Монтерей и Сан-Диего Richardson, 1899). Японское море: залив Петра Великого; держится среди зарослей Zostera на глубинах от 0 до 5 м, на литорали среди красных водорослей; ночью приплывают на свет фонаря и могут быть пойманы на поверхности моря. В большом количестве собраны в бухтах Патрокл, Соболь, у мыса Басаргина, в заливе Владимира, у о. Чихачева на литорали и в Zostera до 5 м глубины (сборы ТИРХ’а и экспедиции ГГИ, 1927).
5. Род CILICAEA LEACH
Leach, 1818, Diet. Sc. Nat, XII: 342. ' \
Тип рода: C. latreillei Leach.
Два последних сегмента груди равной длины, каждый из них длиннее каждого из $ предыдущих. Брюшной отдел из 2 сегментов; на 1-м брюшном сегменте (по крайней мере у самца) имеется ДЛИННЫЙ отросток, напра- Фиг. 64 Dynamene glabra Rich, вленный назад. Тельзон на конце имеет (по Richardson). a — telson, Ь — pleopoda.
простую или фигурную вырезку; иногда внутри этой вырезки имеется еще длинный, иногда очень большой, отросток. 2-й, 3-й и 4-й членики щупика ногочелюстей имеют лопасти на дистальном внутреннем угле. Наружная ветвь 3 пары брюшных ног всегда двучлениковая. У самца ветви рулевых ног очень неравные; наружная неспособна подгибаться под внутреннюю, обычно больших размеров; внутренняя ветвь очень маленькая или совсем редуцируется. У самки обе ветви рулевых ног одинаковой формы и размеров. Половой диморфизм выражен резко, особенно в строении рулевых ног и тельзона.
На Дальнем Востоке известен один вид.
* 1. Cilicaea cordata Richardson (фиг. 65).
Richardson, 1899, Proc. U. S. Nat. Mus., 21 : 839, (<5), f. 16; Richardson, 1905, Bull. U. S. Nat. Mus., n° 54 : 310, f. 336—340. — tuberculosa Richardson, 1899, Proc. U. S. Nat. Mus.,. 21 : 833, f. 9, (<J) (Dynamene). — cordata Holmes, 1904, Proc. Cal. Acad. Sci. (3), III: 303. (Dynamene).
Самец. Тело удлиненно-овальное, суживается к переднему концу. Передний край головы закруглен, валикообразно утолщен, имеет небольшое тупое острие по середине. Задне-боковые углы головы образуют небольшие закругленные лопасти, на которых расположены глаза; задний край головы вогнутый. Усики 1-й пары достигают заднего края головы; базальный чле-8*
116
5. SPHAEROMIDAE
ник их удлиненный; 2-й членик очень короткий; жгутик 8—9-члениковый.
Усики 2-й пары в вытянутом состоянии достигают заднего угла 3-го груд-
Фиг. 65 а. СШ-саеа cordala Rich. $ (no Richardson).
состоящую из узким каналом,
ного сегмента; жгутик 15-члениковый. 1-й грудной, сегмент немного длиннее 2-го; следующие 5 сегментов одинаковой длины; 7-й сегмент немного длиннее каждого из 5 предыдущих. На заднем крае последнего грудного сегмента поперечный ряд бугорков, группами по 2 бугорка; иногда такие же бугорки и на заднем крае б-го грудного сегмента. Эпимеры очень широкие с закругленными вершинами, оттянуты вниз и вбок и не видны поэтому сверху. 1-й брюшной сегмент с 2 парами швов слияния по бокам; на его заднем крае имеется 5 раздвоенных на вершине бугорков. Тельзон очень большой, больше половины длины грудного отдела и головы вместе; у основания тельзона 3 крупных двойных бугорка, образующих один поперечный ряд. Задний край тельзойа имеет фигурную вырезку, 3, синусов; ’ 2 вер>хних синуса, соединенные друг с другом сердцевидной формы, 3-й последний синус образует ши-;
рокую, в общем полукруглую вырезку на самом конце тельзона, сообщающуюся с обоими глубже лежащими сердцевидными синусами (см. фиг. 65). Внутренняя ветвь рулевых ног неподвижно укреплена, широкая, короткая, заходит только немного дальше половины длины тельзона; наружная ветвь длинная и более тонкая, широкая и плоская у основания, суживается дистально, несколько изогнутая и заходит немного дальше конца тельзона; ее наружный край зубчатый и ближе к дистальному концу вооружен 4 крупными зубовидными шипами. Все грудные ноги тонкие, длинные, ходильные”; снабжены добавочным коготком.
Самка отличается от самца тем, что усики 2-й пары немного короче и достигают лишь заднего угла 1-го грудного сегмента. Задние края 2 последних грудных сегментов гладкие, лишенные бугорков; у заднего 1-го брюшного сегмента 3 простых округлых бугорка, так же как и на базальной, сильно выпуклой части тельзона. Тельзон треугольной формы с простой широкой каналовидной вырезкой на конце и небольшим бугорком над нею. Обе ветви рулевых ног одинаковой формы и величины, закругленные на конце; наружная немного короче внутренней; обе ветви не достигают конца тельзона. Самка меньше самца. Цвет желтовато-беловатый с розовыми нежными пятнами. Длина животного 7 мм.
Фиг. 65b. Cilicaea cordata Rich, g (no Richardson).
Распространение. Берингово море: Попов остров (Алеутская гряда); Каталина — остров и Гуалала (Калифорния). Живет в ниж-
6. CYMODOCE
117
нем горизонте литоральной зоны в зарослях известковых водорослей Corallina (Richardson, 1899).
6. Род CYMODOCE LEACH
Leach, 1814, Brewster-Edinb. Encycl., VII: 433.
Тип рода: C. trunctata (Mont.).
Все грудные ножки простые, ходильные, лишенные ложной клешни. На конце тельзона у обоих полов имеется вырезка, иногда полукруглая, большей частью двухлопастная, разделенная вдоль пополам длинным срединным выростом. У самки ротовые части сильно видоизменяются; на 4-м, 5-м и б-м члениках щупика ногочелюстей вытянутые лопасти на внутреннем их крае; эпистома без свободного отростка впереди. Наружная ветвь 3-й пары брюшных ног двучлениковая; инкубаторные пластинки налегают друг на друга по средней линии; ; наружная ветвь рулевых ног хорошо развита; внутренняя ветвь умеренной величины, никогда не бывает рудиментарной. У самца на спинной стороне 1-го брюшного сегмента нет длинного, направленного назад отростка. Резко выражен половой диморфизм.
На Дальнем Востоке известно 2 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Тело покрыто волосками 1. С. japonica Rich. 3.
2 (1). Поверхность тела гладкая, без волосков.
3 (4). Задний край 1-го брюшного сегмента по бокам имеет по одному маленькому треугольному отростку, вогнут по средине и имеет по бокам этой вогнутости еще по одному маленькому треугольному выросту.......................................... 2. С. acuta Rich.
4 (3). Задний край 1-го брюшного сегмента между двумя зубцами прямой, а не вогнутый ................................ * 1. С. japonica Rich. g.
*1. Cymodoce japonica Richardson (фиг. 66).
Richardson, 1907, Proc. U. S. Nat. Mus., 31:7, f. 11 (<5). — affinis Richardson, 1907, Proc. U. S. Nat. Mus., 31: 11, f. 15 (g).
Самец. Тело вытянутое, уплощенное, способно свертываться в шар. В расправленном состоянии боковые края грудного отдела параллельны друг другу. Длина тела в 2 раза больше его ширины. Боковые части сегментов (эпимеральные расширения) направлены вниз, отгибаясь почти под прямым углом к плоскости спины. Голова очень широкая; передний край ее закруглен, с валикообразным утолщением и небольшим бугорком посредине; задний край головы прямой и также валикообразно утолщен. Задне-боковые углы головы образуют небольшие, закругленные лопасти, выдающиеся за пределы заднего края; на этих лопастях распо-
118
5. SPHAEROMIDAE
Фиг. 66. Cymodoce japonica Rich.
ложены глаза. 1-й грудной сегмент в 2 раза длиннее 2-го; остальные б грудных сегментов равной длины. Задние углы 1-го сегмента отогнуты книзу под прямым углом и образуют направленные назад узкие треугольные, заостренные отростки, достигающие заднего края следующего сегмента. Боковые части (эпимеральные расширения) всех остальных грудных сегментов образуют такие же отростки, направленные вниз и назад, но только более толстые и тупые. Усики тонкие, слабые; 1-я пара усиков в вытянутом состоянии достигает середины 1-го грудного сегмента жгутик 15—19-члениковый; базальный членик ножки большой и удлиненный; 2-й членик в 2 раза короче 1-го, но такой же ширины; 3-й тонкий, немного длиннее 2-го членика. Усики 2-й пары достигают заднего края 3-го сегмента; жгутик 18—24-члениковый, каждый членик расширяется дистально. Спинная поверхность грудного отдела зернистая; на задних грудных сегментах более крупные зерна концентрируются в поперечные ряды, по 2 на каждом сегменте. Кроме зернистости на задних сегментах появляются пучки очень коротких волосков. Эпимеральные расширения, особенно задние пары, покрыты, короткими жесткими темнокоричневыми волосками. Боковые части 1-го брюшного сегмента сильно оттянуты назад и книзу и несут пару швов. Благодаря оттянутым боковым частям задний край 1-го брюшного сегмента (если животное в расправленном виде) оказывается изогнутым дугой, образуя почти полуокружность. Посредине заднего края пара довольно крупных конических бугров, направленных назад и нависающих над основанием тельзона. Вся поверх
ность брюшного отдела (1-го сегмента и тельзона) покрыта мелкими бугорками и зернами и очень короткими, жесткими, темнокоричневыми волосками. Тельзон в общем треугольной формы, суживается дистально. Спинная поверхность его выпуклая; вдоль средней линии по спинной стороне тельзона тянется широкое желобовидное вдавление. Края этого желоба образованы двумя парами бугорков; задняя пара значительно крупнее передней пары. Конец этого желоба упирается в широкое округлое возвышение над основанием срединного отростка на конце тельзона. На конце тельзона вилка из 3 толстых обрубленных отростков. Рулевые ноги достигают конца тельзона; ветви их равной длины с прямо обрезанными задними краями. Жесткие коричневые волоски, покрывающие всю поверхность ветвей становятся несколько длиннее и гуще на краях ветвей. Длина самца достигает 22 мм.
Самка меньших размеров; отличается от самца сильно выпуклой спинной поверхностью грудного отдела, отсутствием зернистости, мелких
6. CYMODOCE
119
0-
Фиг. 67. Cymodoce acuta Rich, (по Richardson), а — общий вид; Ь— голова снизу; с — антенна I.
бугорков и волосков не только на грудном, но и на брюшном отделах. Задний край 1-го грудного сегмента по бокам основания тельзона имеет по одному маленькому треугольному отростку; между ними край сегмента гладкий, ровный, не имеет никаких выростов или бугорков. Базальная часть тельзона шаровидно-выпуклая, гладкая; посредине ее едва намечается пара маленьких бугорков и небольшое продольное желобовидное вдавление между ними. Тельзон оканчивается 3 небольшими отростками, из которых срединный несколько крупнее, чем 2 боковых. Концы рулевых ног не достигают конца тельзона.
Распространение. Хакодате; Отару; Нанао, Мороран (Япония); в зоне прилива, на поверхности моря и на глубинах до 106 м; Японское море: 34°17'N, 130°15'Е; 34°20'N, 130°1ГЕ; 36°46'N, 132°15'Е; 34°20z N, 130°10' Е, на глубинах от 106 до 1520 м (Richardson, 1907, 1910).
Одна из самых обычных форм в заливе Петра Великого. Встречается в большом количестве в корнях Zostera (от 1.5 до 19 м глубины), на устричниках (от 0.5 до 24 м), в зарослях Corallina, Lithophyllum и красных водорослей до 18—25 м; в пароходных шлаках (Золотой Рог, от 1 до 15 м глуб.), в песке до 3 м глуб. (б. Холуай), в зарослях Sargassum. (б. Та-фуин, Уссурийский залив, бухты Парис, Улисс, Патрокл, Тавайза, Новик, Стрелок, Назимова, Посьет, Сидими, Преображенье, Гайдамак, у о. Скрыплев, мыс Басаргин, м. Гамова). Заходит в Соленое озеро в бухте Назимова, в р. Усхеза (приток р. Лефу, сбор Полякова). На
поверхности моря приплывает на свет фонаря (сборы ТИРХ и ГГИ, 1925— 1932). Побережье Япония (Хакодате, Нагасаки, Иокогама по сборам Полякова 1883 г.). Желтое море: Порт Артур (сборы Бунге, 1898). Вдоль берега Азии идет на север до б. Преображения (эксп. ЗИНАН, 1934).
_ 2. Cymodoce acuta Richardson (фиг. 67).
Richardson, 1904, Proe. U. S. Nat. Mus., 27: 38, f. 8,9.
Известна только самка. Поверхность тела гладкая, лишенная волосков. Голова большая и широкая, с небольшим срединным острием, на закругленном переднем крае. Глаза большие, расположены на небольших закругленных лопастях головы в ее задних боковых углах. Усики 1-й пары достигают заднего края 1-го грудного сегмента; 1-й членик их ножки большой и широкий; 2-й такой же ширины, но в несколько раз короче 1-го; 3-й членик тонкий и удлиненный; жгутик 17-члениковый; усики 2-й пары достигают заднего края 2-го грудного сегмента; жгутик состоит из 20 члеников. 1-й грудной сегмент в 2 раза длиннее каждого
120
5. SPHAEROMIDAE
из следующих, которые равной длины. Эпимеральные расширения узкие,, заостренные, отделенные от сегментов тонким швом. 1.-й брюшной сегмент в 2 раза длиннее последнего грудного сегмента, е 3 швами по бокам (следы слияния сегментов). Задний край его образует по бокам 2 небольших треугольных отростка; край сегмента между этими боковыми отростками вогнутый и имеет еще пару срединных небольших треугольных отростков. Тельзон широкий у основания, суживается дистально, треугольной формы. Вершина тельзона трехзубчатая; срединный зубец крупнее боковых. На середине выпуклой спинной стороны тельзона 2 возвышения, по 1 с каждой стороны от средней линии. Ветви рулевых ног равной длины, далеко не достигают конца тельзона; обе ветви заострены на дистальном конце. Грудные ножки все одинаковой структуры, ходильные, с добавочным коготком. Цвет беловатый с многочисленными черными пятнами. Длина 8 мм.
Распространение. Берег Японии у Уено-ури на поверхности моря (Richardson, 1904).
7. Род SPHAEROMA LATREILLE IN BOSC
Latreille In Bose, 1802, Hist. nat. Crustac., 1:182.
Тип рода: Д. serratum (Fabr.).
Тело свертывается в плотный шар; брюшной отдел из 2 сегментов; тельзон с закругленным задним Краем, никогда не имеет вырезки, широкий у основания, слегка суживается дистально. 4-й, 5-й и б-й членики щупика ногочелюстей не имеют вытянутых лопастей на внутреннем крае, но снабжены пучками очень длинных щетинок. Жвалы нормальные; конец режущего отростка тупой или с маленькими зубцами; 3 передних пары грудных ног с густыми длинными жесткими перистыми волосками на наружном крае 3-го и 4-го члеников. Все грудные ножки ходильные, лишенные ложной клешни. Наружная ветвь 3-й пары брюшных ног одно-члениковая. Инкубаторные пластинки самки перекрывают друг друга по-средней линии. Основание тельзона не шире последнего грудного сегмента. Ветви рулевых ног одинаковых размеров и формы; наружная зазубрина вдоль внешнего края; внутренняя ветвь неподвижно укреплена, наружная ветвь подвижная и может подгибаться под внутреннюю ветвь.
На Дальнем Востоке известен один вид.
1. Sphaeroma sieboldi Dollfus (фиг. 68).
Dollfus, 1889, Notes Leyden Museum, 11:93, pl. 5, f. 3a, b.
Поверхность тела покрыта мелкими бугорками и гранулами, наиболее резко выраженными на переднем конце тела и на тельзоне. Голова маленькая и короткая. Передний край головы с небольшим острием по
7. SPHAEROMA, 8. EXOSPHAEROMA
121
средине и 2 вдавлениями и 2 выпуклостями по бокам его. Передне-боковые углы головы образуют маленькие треугольные заостренные лопасти. Глаза большие, овальные, помещаются в задне-боковых углах головы.
2
Усики 1-й пары достигают у длины усиков 2-й пары; 2 первых членика
ножки усиков 1-й пары расширенные; 3-й членик узкий, удлиненный; жгутик 8-члениковый. Жгутик усиков 2-й пары 10-члениковый. Передне-
боковые части 1-го грудного сегмента вытянуты вперед, образуя большие треугольной формы лопасти по бокам головы; концы этих лопастей почти достигают переднего края головы. Благодаря грануляции швы эпимер
слабо заметны. Тельзон сильно гранулирован; бугорки равномерно распределены по всей его поверхности; по средней линии образуются 2 про-
дольных, расходящихся кзади ряда более крупных бугорков. Конец тельзона закруглен и поверхность его вогнутая. Рулевые ножки выходят немного за пределы конца тельзона; обе ветви почти одинаковые; внутренняя ветвь ланцетовидная, усажена короткими щетинками по наружному краю. Внутренний край наружной ветви почти прямой усажен щетинками, наружный край снабжен семью зубцами со щетинками. Длина животного 7 мм.
В коллекциях Зоологического института Академии .Наук вид не представлен.
Распространение. Япония (Dollfus, 1889); Хакодате на литорали (Richardson, 1910).
Фиг. 68. Sphaeroma sie-boldi Dollfus (no Dollfus),. голова и тельзон.
8. Род EXOSPHAEROMA STEBBING •
Stebbing, 1900, Proc. Zool. Soc. London: 553.
Тип рода: E. gigas Stebb.
От предыдущего рода отличается тем, что 2-й, 3-й и 4-й членики-щупика ногочелюстей имеют длинные лопасти на внутреннем крае;, наружная ветвь рулевых ног с гладким незазубренным внешним краем; 3 передние пары грудных ног не имеют длинных жестких плавательных', щетинок на наружном крае 3-го и 4-го члеников. Наружная ветвь 3-й пары брюшных ног 2-члениковая; у самки инкубаторные пластинки маленькие,, далеко не достигают свободными краями средней линии тела. Конец, тельзона слегка оттянут, но не заострен. Наружная ветвь всех 5 пар брюшных ног никогда не несет дыхательной функции; внутренняя ветвь-1-й пары широкая и короткая; внутренняя ветвь 3-й пары не несет дыхательных функций; ветви 4-й пары на дистальном конце снабжены: перистыми маргинальными щетинками.
На Дальнем Востоке известен один вид.
122
5. S PH AERO MID АЕ
1. Exosphaeroma amplicauda (Stimpson) (фиг. 69).
Stimpson, 1857, Bost. Journ. Nat. Hist-, VI: 510 (Sphaeroma)', Richardson 1905, Bull. U. S. Nat. Mus., n° 54: 288, f. 301—302.
Тело очень широкое сзади, суживается на переднем конце; наибольшая его ширина на последнем грудном сегменте в 1.5 раза меньше длины тела. Голова большая, широкая; передний край закруглен и снабжен маленьким срединным зубцом. Глаза небольшие, круглые, расположены в задне-боковых углах головы. Базальный членик усиков 1-й пары длинный, равен длине 3-го членика; 2-й членик в 2 раза короче 1-го; жгутик
• Фиг. 69. Exosphaeromaa amplicauda - (Stimps.) (по Richardson), а — общий вид; Ь —maxillipedes.
5-члениковый и достигает в вытянутом состоянии заднего края головы. 2 первые членика усиков 2-й пары очень короткие, равной длины; 3-й членик в 2 раза длиннее 2-го и равен длине 4-го; 5-й в 1.5 раза длиннее 4-го; жгутик 9-члениковый и достигает заднего края 1-го грудного сегмента. 1-й грудной сегмент немного длиннее других. Боковые части всех грудных сегментов отогнуты книзу под почти прямым углом к спинной поверхности. Эпимеры неясно отграничены от сегментов легким швом. 1-й брюшной сегмент с 3 линиями швов с каждой стороны на боковых
частях сегмента; тельзон треугольной формы с закругленной тупой вершиной. Рулевые ноги очень большие, обе ветви достигают вершины тельсона и равной ширины. Наружная подвижная ветвь закруглена сзади;
внутренняя неподвижная ветвь с закругленным наружным краем, но внутренний край образует оттянутую небольшую лопасть на конце. Последние 3 сегмента груди и 1-й сегмент брюшного отдела на заднем крае несут рудиментарные бугорки; иногда эти бугорки хорошо развиты, конические. На спинной стороне тельзона 2 пары бугорков, образующих 2 продольных ряда по бокам от средней линии. Длина животного * 2.5 мм.
Распространение. Побережье Аляски (Kyska-Harbor и северный берег Амчитка); Калифорния (Richardson, 1905). На песчаном пляже
на литорали.
9. Род NEOSPHAEROMA BAKER
Baker, 1926, Trans. Proc. Roy. Soc. South Australia, 50 : 252.
Тип рода: N. laticauda Baker.
От предыдущего рода отличается тем, что наружные ветви 4-й и 5-й шар брюшных ног более или менее дыхательные; внутренняя ветвь 1-й
8. EXOSPH AEROMA, 9. NEOSPHAEROMA 123
пары брюшных ног узкая; внутренняя ветвь 3-й пары несет дыхательную функцию, а наружная ветвь двучлениковая; на дистальных концах ветвей 4-й napj>i перистые щетинки.
На Дальнем Востоке 2 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Усики 1-й пары в вытянутом состоянии доходят до заднего края 2-го, а усики 2-й пары до заднего края 4-го грудного сегмента.........
. ..................................... * 2. N. ovata Gurjan.
2 (1). Усики 1-й пары в вытянутом состоянии достигают только заднего края головы, а усики 2-й пары — заднего края 2-го грудного сегмента
.................................. * 1. N. oregonensis Dana.
Фиг. 70. Neosphaeroma oregonensis (Dana) (по Tielle-mann).
*1. Neosphaeroma oregonensis (Dana) (фиг. 70).
Dana,' 1851, Pros. Acad. Nat. Sci. Phila., VII (Sphaeroma); Kichardson, 1905, Bull. U. S. Nat. Mus., n° 54: 246, f. 315—316 (Exo-sphaeroma); Monod, 1931, Mem. Soc. Sci. Nab Maroc., n° 29:76, f. 74. — olivacea Lockington, 1877, Proc. Calif. Acad. Sc., VII: 177 (Sphaeroma).
Тело удлиненное с закругленными задними и передним концами и с параллельными боковыми краями; способно свертываться в плотный шар. Длина тела немного больше чем в 2 раза превосходят его ширину. Голова широкая, передний край ее закруглен, с небольшим тупым не видным сверху, острием по средине. Глаза маленькие, круглые, расположены в заднебоковых углах головы; усики 1-й пары достигают сере
дины 1-го грудного сегмента; базальный членик их удлиненный, 2-й членик в 2 раза меньше 1-го, 3-й в 1т/2 раза длиннее 2-го; жгутик 8—13-чле-никовый. Усики 2-й пары достигают заднего края 2-го грудного сегмента. 1-й членик их ножки очень короткий, 2-й в 3 раза длиннее 1-го, 3-й равен длине 2-го, 4-й в 2 раза длиннее 3-го и равен длине 5-го членика; жгутик 13-члениковый. 1-й грудной сегмент немного длиннее каждого из следующих, которые равной длины. Эпимеральные расширения с закругленными краями,слегка оттянуты назад, со следами швов на месте слияния эпимер с сегментами. Задние углы их тупо заострены. 1-й брюшной сегмент с 3 швами по бокам с каждой стороны. Тельзон широкий, у осно вания слегка суживается дистально; задний край его закруглен. Ветви рулевых ног неравной длины; внутренняя неподвижно укрепленная ветвь достигает конца тельзона, слегка суживается дистально и закруглена на вершине; наружная подвижная ветвь составляет только 2/3 длины внутренней, также закруглена на вершине. Наружная ветвь 3-й, 4-й
124 5. SPHAEROMIDAE
Фиг. 71. Neosphaeroma ovata Gurjan. а — общий вид; Ъ — антенна I.
и 5-й пар брюшных .ног двучлениковая; внутренняя ветвь 3-й и 4-й пар-брюшных ног с несколькими щетинками на дистальном конце. Внутренняя ветвь 2-й пары брюшных ног самца снабжена стилетом. Цвет животного-темносерый, иногда розоватый. Длина достигает 8 мм.
Распространение. Побережье Тихого Океана Северной Америки от Калифорнии и Британской Колумбии до Аляски включительно; Алеутские о-ва, Уналяска, Командоры, на глубине от 0 до 12 м и на литорали (Richardson, 1905, 1910). Японское море: все побережье от Корейской границы до зал. Де-Кастри включительно на литорали и в сублиторали на глубине от 0 до 15 м; на песке, гальке, под камнями,, в зарослях водорослей (бурых, зеленых, красных и в известковых Corallina и Lithothamnion), на сваях пристаней. Заходит в устья рек (р. Нянь-няку в Советской гавани, р. Датта в Татарском проливе, р. Холуай в бухте Владимир); Лебяжья лагуна у Сидими. В Беринговом море: на Командорах в устье реки Гаванской.
* 2. Neosphaeroma ovata (Gurjan.) (фиг. 71).
Gurjanova, 1933, Исслед. морей СССР, 17 :99, рис. 28-(Exosphaeroma).
Отличается от предыдущего вида более широким овальной формы телом, сильно выпуклой спинной стороной, длинными усиками и формой тельзона. Тело широкое, овальных очертаний;, длина его немного меньше, чем в 2 раза превосходит его ширину. Поверхность тела гладкая, свод
чато-выпуклая. Эпимеральные расширения по форме напоминают язычки, оттянуты в стороны и отделены от сегментов заметными швами слияния. Голова полукруглой формы, широкая, с закругленным передним краем и небольшим тупым острием посредине. Глаза большие, выпуклые, круглые; расположены в задне-боковых углах головы. Усики 1-й пары в вытянутом состоянии достигают заднего края 2-го грудного сегмента; базальный членик сильно расширяется к дистальному концу, удлиненный; 2-й членик в 2 раза короче 1-го, 3-й немного длиннее 2-го; жгутик 8—-9-чле-никовый. Усики 2-й пары в вытянутом состоянии достигают заднего края 4-го грудного сегмента; 1-й членик их очень короткий, 2-й удлиненный; 3-й в 2 раза короче 2-го; 4-й немного длиннее 3-го, а 5-й равен 4-му; жгутик 10—12-члениковый. Тельзон полукруглой формы с плавно закругленным задним краем; ширина его у основания больше длины; ветви рулевых ножек закруглены на концах; наружная ветвь такой же ширины, но составляет только 2/з длины внутренней неподвижной ветви. Брюшные ножки совершенно такого же строения, как и у N. oregonensis. Цвет тела оранжево-желтый с розовыми пятнами или же темносерый. Максимальная длина 9 мм.
9. NEOSPHAEROMA 125
Распространение. Японское море: м. Басаргин в супралиторали; Советская гавань, Де-Кастри, Б. Новый Джигит, на литорали; устье р. Суйфун, устье реки в бухте Ольга на литорали на песке и среди .водорослей; Лебяжья лагуна у Сидими в зарослях Zostera на глубине 0.5 м (сборы ТИРХ’а и ГГИ, 1925—1929).
6. Сем. ANTHURIDAE
Тело очень длинное, цилиндрическое, с узкими, равными ширине головы вытянутыми сегментами. Голова маленькая; обе пары усиков короткие, почти равной длины с ясным разделением на ножку и жгутик; .жгутики обеих пар усиков с малым числом члеников, иногда одночленико-вые; у рода Xenanthura имеется добавочный жгутик у 2-й пары; глаза большей частью отсутствуют или сильно редуцированные. РотовЫе части обычно сильно видоизменены, сосущие или колющие, редко нормальные. 1-я пара грудных ног значительно крепче и массивнее, чем остальные, снабжена ложной клешней. 2 следующих пары значительно более слабые, с несовершенной ложной клешней. 4 задние пары грудных ног ходильные. Брюшные ложки служат исключительно для дыхания; 1-я пара большая, накрывает сверху все остальные. Эпимеры отсутствуют. На спинной стороне 4-го, 5-гб и б-го грудных ^дегментов имеются небольшие желобовидные вдавления. Брюшной отдел состоит из различного числа сегментов, так как часто они сливаются друг с другом со следами или без следов шва. Наиболее характерно для этого семейства строение хвостового веера; тельзон свободный, л виде язычка, закругленный на вершине и вогнутый на спинной стороне; рулевые ножки двуветвистые с очень коротким базальным члеником; ветви неравной длины и расположены таким образом, что широкая лопастевидная .наружная ветвь сдвинута на спинную сторону и нависает сверху над вогнутым тельзоном. Вследствие этого, рулевые ножки и тельзон вместе образуют не плоский хвостовой веер, а нечто в роде чашечки, нижняя стенка которой образована тельзоном, а боковые и верхняя стенки — изогнутыми поверхностями ветвей рулевых ног. У основания тельзона имеется парный или непарный статоцист. Громадное значение в классификации Anthuridae имеет строение тельзона и ротовых частей; Barnard, 1925, разбивает все -семейство на 2 секции—-с кусающими и колющими ротовыми частями; из встреченных на Дальнем Востоке 3 родов, одйн (род Cyathurd) относится к секции А, характеризующейся нормальными кусающими или жующими ротовыми частями, а 2 рода—Calathura и Paranthura — к секции В с видоизмененными колющими ротовыми частями.
ТАБЛИЦА ДЛЯ- ОПРЕДЕЛЕНИЯ РОДОВ
1 (4). Щупик ногочелюстей состоит из 3 члеников.
2 (3). Жгутик усиков 2-й пары нормальный, многочлениковый.
.................. 3. Calathura Norm. a. Stebb. (стр. 129).
126
6. ANTHURIDAE
3 (2). Жгутик усиков 2-й пары рудиментарный, одночлениковый ....
....................... . 2. Paranthura Bate a. Westw. (стр. 127)..
4 (1). Щупик ногочелюстей состоит из 2 члеников..................
.........................1. Cyathura Norm. a. Stebb. (стр. 126).
1. Род CYATHURA NORM. a. STEBB.
Norman a. Stebbing, 1886 Trans. Zool. Soc. London, 2:152.
Тип рода: C. carinata (Кг.).
Глаза обычно имеются, иногда отсутствуют. На грудных сегментах, имеются боковые кили и дорзальные желобовидные вдавления. Брюшные сегменты сливаются друг с другом, сохраняя следы слияния в виде швов, по бокам брюшка. Форма тельзона на поперечном разрезе чечевицеобразная. Усики 1-й пары с 1—3-члениковым жгутиком, который у самца иногда покрыт длинными щетин-
. _— 1 . ками, образующими щетку. Усики
/jV 2-й пары с одночлениковым редуциро-
ванным жгутиком. Щупик жвал 3-чле-\ никовый, последний членик обычно Jkjf/ больше 1-го, с пучком щетинок на.
что вершине. Ногочелюсти состоят из 4 4 члеников. На 6-м членике 1-й пары.
Фиг. 72. Cyathura carinata Кг. (экз. из Амур- грудных ног, образующем вместе ского лимана) с длинным когтем ложную клешню,,
имеется’ крупный ладонный зуб.
6-й членик 2 следующих пар грудных ножек простой, цилиндрический. 1-я пара брюшных ножек, рулевые ноги и тельзон не затвердевшие, нормальной консистенции. Наружная ветвь рулевых ног охватывает тельзон, нависая над ним. У самок 3 пары инкубаторных пластинок.
На Дальнем Востоке известен лишь один вид.
* 1. Cyathura carinata (Kroyer) (фиг. 72).
Kroyer, 1846, Naturh. Tidskr. (2), II: 402 (Anthura)-, Kroyer, 1849, Voyag. Scand., pl.. 27, f. 3 (Anthura). — estuaria Barnard, 1914, Ann. S. Afr. Museum, X, 11:334a.
Тело очень узкое, удлиненное, длина его более, чем в 7 раз превосходит ширину. Голова короткая, длина ее в l1^ раза меньше ширины; глаза, очень маленькие, но хорошо видные; щупик жвал очень велик, по сравнению с телом жвал; усики 1-й пары с 3-члениковым жгутиком; первые 2 членика ножки равной длины, 3-й немного короче 2-го; конец жгутика, достигает конца 4-го членика ножки усиков 2-й пары. Усики 2-й пары, немного длиннее 1-й пары; базальные членики их короткие; 2-й в 2 раза длиннее 1-го, 3-й в 2 раза короче 2-го, 4-й немного короче 3-го, 5-й чле
1. CYATHURA, 2. PARANTHURA 127
ник в 1г/2 раза длиннее 4-го; жгутик одночлениковый, очень короткий, равен половине длины 5-го членика ножки. 1-й, 4-й и 5-й грудные сегменты равной длины, каждый в 2 раза длиннее головы. 2-й, 3-й и 2 последних грудных сегмента равной длины, каждый в 1г/2 раза длиннее головы. 5-й членик 1-й пары грудных ножек с вытянутой и тупозаостренной вершиной; пальмарный край с крепким зубцом у основания, б-й членик 2. следующих пар грудных ножек цилиндрический. Все 6 передних брюшных сегментов сливаются вместе без всяких следов, образуя 1 сегмент, рав- ный длине 2-го грудного сегмента. Тельзон овальной формы с прямыми боковыми краями, постепенно переходящими в закругленный задний край; . спинная сторона тельзона слабо вогнута. Внутренняя ветвь рулевых ног овальной формы, ширина ее почти равна длине; конец ее закруглен; наружная ветвь овально-треугольная, слегка нависает над основанием тельзона, почти налегая на его спинную поверхность. Цвет тела беловатый с бурыми пятнами; длина 13—27 мм.
Распространение. Северная часть Атлантического океана: побережье Северной Америки и Европы, Немецкое море, Датские воды, . Балтийское море (Кильская бухта); южная Африка (Barnard, 1914), Китай (W. Tattersail, 1920); часто в солоноватых водах, эстуариях и в пресной воде (Barnard, 1925). Охотское море: бухта Нагайя в 3 км от устья реки в пресной воде (сбор Державина, 1927).
2. Род PARANTHURA BATE a. WESTW., 1868
Bate a. Westwood, 1868, Hist. Brit, sessile-eyed Crustac., II: 163.
Глаза хорошо развиты, иногда редуцированы или отсутствуют. Спинно-боковые кили на грудных сегментах слабо развиты; небольшое -желобовидное вдайление пересекает передний край заднего сегмента; . последний грудной сегмент очень короткий. Все брюшные сегменты сливаются вместе, сохраняя следы слияния по бокам сегмента в виде -швов.' Тельзон плоский или вогнут на спинной стороне, иногда выпуклый, тонкий, не затвердевший. Статоцисты отсутствуют. Жгутик усиков первых 4—б-члениковый у J и щетковидный у 3. Усики 2-й пары с одночлениковым жгутиком. Щупик жвал трехчлениковый со щеткой щетинок на последнем членике. Ногочелюсти состоят из 3 члеников; 2-й и 3-й членики их равной длины; 2-й членик не имеет отростка на вершине.... Ложная клешня 1-й пары грудных ног с цельным пальмарным краем,.. небольшим зубцом у его основания и коротким когтем. 5-й членик 4 задних грудных ножек цилиндрический, не образует отростка, прилегающего к б-му членнку. 1-я пара брюшных ног не затвердевшая. Наружные ветви рулевых ног узкие, нависают над тельзоном. Инкубаторных пласти- -нок 3—4 пары.
Из дальневосточных морей известен один вид.
:128
б. ANTHVRIDAE
* 2. Paranthura japonica Rich. (фиг. 73).
Richardson, 1910, Proc. U. S. Nat. Mus., 37: 77, f. 4—5.
Тело узкое, удлиненное; длина его в 8 раз больше ширины. Голова короткая, длина ее равна ширине. Передний край головы вогнутый, с небольшим срединным острием. Глаза большие, круглые, расположенные у основания передне-боковых углов головы. 1-й членик усиков 1-й пары удлиненный; 2-й и 3-й членики короткие, равной длины;
жгутик 4-члениковый. Усики 2-й пары немного длин- f I
..нее предыдущих; 2-й членик их ножки удлинен- I/ ( ный; следующие 2 очень короткие, равной длины; 'а
А
“Фиг. 73. Paranthura japonica Rich, а—общий вид; Ъ— задний конец тела; с—uropoda; d— pereiopoda I; е — cephalon; f— abdomen.
5-й в iy2 раза длиннее 4-го; жгутик из 1 членика, снабженного волосками. Первые 5 грудных сегментов равной длины, каждый в 1 1/2 раза длиннее головы, б-й сегмент более короткий, а 7-й грудной сегмент в 2 раза, короче 6-го. • 5 передних сегментов брюшного отдела сливаются вместе на спинной стороне, оставляя следы слияния в виде швов по бокам 1-го свободного брюшного сегмента, получившегося от слияния. 2-й брюшной сегмент равен половине длины предыдущего. Тельзон удлиненный с закругленной вершиной. Базальный членик рулевых ног достигает '8/i Длины терминального сегмента. Внутренняя ветвь укороченная, •с закругленным концом, не достигает конца тельзона. Наружная ветвь
2. PARANTHURA, 3.,C ALATHURA 129
аркообразно нависает над тельзоном и равна длине базального членика. 3 передние пары грудных ног хватательные; 1-я пара снабжена ложной клешней. Последние 4 пары грудных ног ходильные. Длина животного 8—9 мм. Цвет желтоватый с неправильным!} темными пятнами.
Распространение. Японское море: Мороран, на литорали (Richardson, 1910); залив Петра Великого (бухта Рында, б. Соболь, б. Патрокл), на глуб. 3—5 м (сборы Тихоокеанск. инет. рыб. хоз., 1928). Охотское море: бухта Нагайя в Амурском лимане, в 3 км от устья реки (Державин, 1927).
3. Род CALATHURA NORM. a. STEBB.
Norman a. Stebbing, 1886. Trans. Zool. Soc. London, 2:131.
Тип рода: C. brachiata (Stimps.).
Глаза отсутствуют вовсе или имеются их слабые следы. Дорзолате-ральные кили на грудных сегментах развиты слабо, но спинные желоба имеются. Последний, 7-й, грудной сегмент короткий. Все б брюшных сегментов отделены друг от друга ясными швами поперек всей спинной поверхности брюшного отдела; тельзон с вогнутой дорзальной поверхностью, не имеет статоцистов. Жгутик обеих пар усиков 8—12-членико-вый. 2-й членик щупика немного длиннее 2 других, равных друг Другу; на конце 3-го членика его пучок щетинок. Ногочелюсти состоят из 5 члеников; 3-й членик очень короткий; 2-й с вытянутым дистальным концом. Пальмарный край ложной клешни 1-й пары грудных ног цельный с зубцом у основания и коротким когтем. 5-й членик 3 последних пар грудных ног треугольной формы. 1-я пара брюшных ног не затвердевшая. Наружная ветвь рулевых ног широкая, нависает над основанием тельзона. У самки 3 пары инкубаторных пластинок.
Известен только один вид рода.
* 1. Calathura brachiata (Stimpson) (фиг. 74).
Stimpson, 1853, Smiths. Contr. knowl., VI: 43 (Anthura); G. Sars, 1899, Account, II: 46, pl. XIX, f. 2. — norvegica Sars, 1899, Account, II: 45, pl. XIX, f. 1.
Тело очень узкое, длинное, цилиндрическое; длина его почти в 15 раз больше ширины. Голова небольшая, короткая, ширина ее немного больше длины; передний край головы вогнутый с маленьким острием посредине. Глаз нет. Усики 1-р пары с 3-члениковой ножкой и 12-члени-ковым жгутиком; 1-й членик ножки удлиненный, 2-й членик немного короче 1-го, а 3-й равен половине длины 1-го; членики жгутика короткие, толстые, 2-й членик в 2 раза длиннее 1-го. Усики 2-й пары с 14-члени-ковым жгутиком. 1-й грудной сегмент в 2 раза длиннее головы; 2-й сегмент в 1г/2 раза длиннее 1-го, суживается кзади; 3-й сегмент немного длиннее 2-го с небольшой перемычкой у заднего края; 4-й сегмент Гурьянова 9
130
6. ANTHURIDAE
Фиг. 74. Calathura brachiata (Stimpson) (по G. Sars).
в 2 раза длиннее 1-го и на спинной стороне своей узкой передней части посредине имеет резко выраженное углубление; такие же углубления у переднего края посредине спинной поверхности имеют 5-й и б-й сегменты; 5-й сегмент равен длине 3-го; б-й сегмент равен длине 1-го; 7-й сегмент совсем короткий, равен половине длины 1-го. Боковые кили слабо выражены лишь по обе стороны 1-го грудного сегмента, тогда как на всех остальных сегментах, в противоположность арктическим экземплярам, эти кили отсутствуют. Первые 3 пары грудных ног снабжены ложной клешней. 4 задних пары ног ходильные, тонкие, слабые. 1-я пара ног резко отличается от других своей мощностью, б-й членик более вытянут, чем у арктических представителей этого рода; пальмарный край не имеет зубца у основания и густо покрыт тонкими волосками. Все 5 первых брюшных сегментов с ясно-выраженными границами их, не только по бокам, но и на спинной стороне, б-й сегмент короткий, расширяющийся посредине. Тельзон узкий, вытянутый, узко-овальной формы, с почти заостренным концом й сильно выгнутой спинной поверхностью. Наружная ветвь рулевых ног широкая с закругленным нижним краем, сильно сдвинута на спинную сторону и нависает над тельзоном. Тихоокеанские экземпляры отличаются от арктических и североатлантических представителей этого вида значительно более длинным телом (длина тела атлантических экземпляров только" в 10 раз больше его ширины), удлиненным б-м члеником 1-й пары грудных ног,, более узким, почти заостренным на конце тельзоном и отсутствием боковых килей на грудных сегментах.. Максимальная длина животного из Охотского моря 45 мм..
Распространение. Северная часть Атлантического побережья, вдоль берегов Северной Америки от зал. Лаврентия до мыса Sabi; вдоль европейского побережья от Финмаркена до Бискайского залива включительно; Арктическая область: западная и восточная Гренландия, Ян-Майен, Медвежий остров, Шпицберген, Земля Франца Иосифа, Баренцево и Карское моря, северная часть моря Лаптевых. На Дальнем Востоке найден экспедициями Гос. гидрологического института в 1932 г. Охотское море: ст. 211 — 49°12'N, 145°21/Е, глуб. 443 м, ил и галька, 1 экз.; ст. 212—49°13'’ N, 14б°0б' Е, глуб. 645 м, серо-зелен, ил, б экз.; ст. 214—: 49°23' N, 148°4б' Е, глуб. 1076 м, серо-зелен, ил, 2 экз.; ст. 216—51 °10' N, 154°17' Е, глуб. 591 м, ил, 6 экз.; ст. 217—51°35' N, 154°4б' Е, глуб. 418 м, илистый песок, 20 экз.; ст. 228—53°13'N, 152°35'Е, глуб. 664 м, ил, 5 экз.; ст. 252—55°32'N, 149°14'Е, глуб. 335 м, ил и галька, 1 экз.; ст. 255— 55°36' N, 153°02' Е, глуб. 503 м, 14 экз. Беринговоморе: ст. 53—61°22' N, 178°02'Е, глуб. 360 м, песок, 16 экз.; 54°47/N, 167O2'E, глуб. 108 м, галька, 5 экз.
3. C ALA THUR А
131
* III. Подотряд VALVIFERA
. Все 7 пар грудных ножек хорошо развиты; по строению грудные ножки дифференцированы, так что передние 3 или 4 пары ног иного строения, чем задние пары. Брюшные ножки служат как для дыхания, так иногда и для плавания. Рулевые ножки латеральные, прикреплены по бокам тельзона у его основания, но не образуют каудального веера, а расположены на брюшной стороне брюшного отдела и имеют форму створок, накрывающих сверху брюшные ножки. Тельзон срастается целиком с последним брюшным сегментом и образует верхнюю стенку камеры, в которой залегают брюшные ножки; рулевые ноги служат нижней, брюшной стенкой этой камеры. Тело обычно более или менее равномерно расчленено; иногда 4-й грудной сегмент значительно превышает по размерам все остальные сегменты. Тело большей частью сплющено в дорзо-вентральном направлении, но иногда сильно вытянутое, цилиндрическое. Глаза обычно хорошо развиты; ротовые части нормальные.
На Дальнем Востоке представлен 2 семействами и 10 родами.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ
1 (2). Тело более или менее широкое, сплюснутое' в дорзо-вентральном направлении; расчленение на сегменты равномерное. Усики 2-й пары всегда короче тела. 3 передние пары ног часто снабжены ложной клешней хватательного типа..............7. Idotheidae. (стр. 131).
2 (1). Тело длинное, узкое, почти цилиндрическое; некоторые сегменты тела, чаще всего 4-й грудной сегмент, сильно отличаются по форме и размерам от остальных. 4 передних пары грудных ног усиковидные, направлены вперед; членики их узкие, удлиненные, снабженные длинными щетинками. 3 задних пары ног ходильные. Усики 2-й пары очень длинные, часто длиннее тела .... 8. Arcturidae. (стр. 173).
7. Сем. IDOTHEIDAE
Тело более или менее сплюснуто в дорзо-вентральном направлении, удлиненной, овальной или линейной формы. Грудной отдел расчленен на однородные по форме и величине сегменты. Эпимеры хорошо развиты в виде широких пластинок, свободно сочлененных с сегментами, или же сливаются с сегментами со следами или без следов швов, образуя плевральные расширения. Сегменты брюшка обычно сливаются друг с другом и с тельзоном, образуя один цельный большой каудальный сегмент. 3 передние пары ног обычно хватательного типа, снабженные ложной клешней; 4 задние пары грудных ног ходильные. Брюшные ножки двуветвистые, 9*
132 7. IDOTHEIDAE
ветви их пластинчато расширены; передние пары с ветвями густо усаженными по краям щетинками; у самцов обычно 2-я пара снабжена стилетами. Рулевые ножки очень большие, в виде створок, закрывающих на брюшной стороне камеру, в которой помещаются брюшные жаберные ножки. Усики < 1-й пары маленькие с одночлениковым жгутиком; усики 2-й пары длинные, но всегда короче тела; жгутик их многочлениковый, но иногда все членики жгутика сливаются вместе, образуя 1 крупный членик. Ротовые части нормальные. Жвалы всегда лишены щупика. Щупик ногочелюстей с различным числом члеников от 3 до 5.
В наших дальневосточных морях известно 7 родов.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (10). Щупик ногочелюстей 4-или 5-члениковый.
2 (3). Боковые части головы впереди глаз вытянуты вперед и образуют расщепленные вдоль на 2 закругляющиеся лопасти отростки; брюшко состоит из 5 сегментов.................3. Mesidothea Rich. (стр. 145).
3 (2). Боковые части головы впереди глаз не образуют расщепленных-на-двое лопастей; брюшной отдел меньше чем из 5 сегментов.
4 (5). Членики жгутика усиков 2-й пары слиты вместе, образуя один цельный кусок или же их всего 2—3, неотличимых по форме от члеников ножки . .........................7. Cleantis Dana. (стр. 170).
5 (4). Жгутик усиков 2-й пары нормальный, многочлениковый.
6 (7). Щупик ногочелюстей из 4 члеников . . 1. Idothea Fabr. (стр. 132).
7 (6). Щупик ногочелюстей 5-члениковый
8 (9). Брюшной отдел из 3 хорошо развитых сегментов, 2 передних сегмента и тельзона.................... 2. Pentidothea Rich. (стр. 140).
9 (8). Передние сегменты брюшка слиты в одно целое с тельзоном, образуя только один большой каудальный сегмент со следами слияния в виде швов по бокам сегмента ... 4. Pentias Rich. (стр. 147).
10 (1). Щупик ногочелюстей 3-члениковый.
11 (12). Эпимеры всех 7 грудных сегментов слиты с сегментами, образуя плевральные расширения; брюшко состоит из 1 сегмента; жгутик усиков 2-й пары нормальный, многочлениковый . . ....................
.............................. 6. Synidothea Harg. (стр. 150).
12 (11). Последние 3 грудных сегмента имеют хорошо развитые свободные эпимеры. Брюшной отдел из 3 сегментов; членики жгутика усиков 2-й пары слиты вместе, образуя один цельный кусок . . . ..........................................5. Symmius Rich. (стр. 148).
1. Род IDOTHEA FABRICIUS
Fabricius, 1798, Entom. Syst. Suppl.: 303.
Тип рода: I. emarginata Fabr.
Тело овальное или вытянутое с почти параллельными краями, сплюснуто в дорзо-вентральном направлении. Эпимеры за исключением
/. IDO THEA
133
1-го сегмента, свободные в виде узких пластинок, занимающих весь боковой край сегмента или только его^5асть. Голова почти квадратная с закругленными передне-боковыми углами и слабо вогнутым передним краем. Глаза хорошо развиты, посажены по бокам головы. Жгутик усиков 1-й пары короткий, одночлениковый; жгутик усиков 2-й пары удлиненный, многочлениковый. Жвалы очень крепкие, сильные с широким большим зубным отростком; режущий край с 2 зубцами. Щупик отсутствует. Челюсти нормального строения; щупик ногочелюстей из 4 члеников; последний членик пластинчато расширенный. Ноги однообразного строения с сильным раздвоенным на конце коготком; передние 3 пары обычно несколько короче и сильнее, чем 4 задние пары и снабжены несовершенной ложной клешней. У самца ноги обычно крепче и сильнее, чем у самки. Брюшной отдел состоит из 3 сегментов; передние 2 хорошо развиты; 3-й и 4-й брюшные сегменты сливаются друг с другом и с тельзоном, образуя один каудальный сегмент с ясным швом по бокам. Рулевые ножки только с одной ветвью, вместо второй ветви имеется длинная перистая щетинка. Наиболее характерные признаки для различения видов — строение усиков, тельзона и относительные размеры тела.
Виды этого рода обычно селятся на мелководьях, чаще всего в зоне прилива, среди камней и водорослей.
В дальневосточных морях известно б видов этого рода.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Задний край тельзона прямой или слабо выпуклый; тело относительно короткое и крепкое.......................1. I. metallica Bose.
2 (1). Задний край тельзона имеет срединный зубец; тело сильно вытянуто, линейное.
3 (4). Задний край тельзона трехзубчатый; 1, более крупный, зубец посредине и 2 более коротких по бокам от него, образованные заостренными и слегка оттянутыми назад задне-боковыми углами тельзона .
...........................................* 2. 1. fewkesi Rich.
4 (3). Задне-боковые углы тельзона закругленные; имеется только 1 срединный тупой зубец.
5 (б). Усики 2-й пары тонкие, длинные, в вытянутом состоянии заходят дальше заднего края 4-го грудного сегмента, достигая середины 5-го сегмента. Боковые края тельзона ясно вогнутые, так что задний конец его ясно шире его срединной части и даже базальной части.......................................* б. I. orientaiis Gurjan.
б (5). Усики 2-й пары более короткие и толстые, не достигают заднего края 4-го грудного сегмента. Задний конец тельзона не шире основания.
7 (8). 1-й грудной сегмент в 2 раза короче 2-го, усики 1-й пары достигают середины 4-го грудного сегмента..............*3.1. ochotensis Brandt.
134
7. [DO THEIDAE
8 (7). 1-й грудной сегмент только немного короче 2-го сегмента; усики 1-й пары достигают только середины 3-го грудного сегмента.
9 (10). Спинная сторона головы почти плоская; боковые края тельзона слегка вогнуты по средине, так что конец тельзона несколько шире его средней части. Жгутики усиков 2-й пары линейные, равномерно утончающиеся к дистальному концу .... * 4. I. aleutica Gurjan.
10 (9). Спинная сторона головы сильно выпуклая. Тельзон равномерно хотя и очень слабо суживается к заднему концу; боковые края его почти параллельны и задняя часть тельзона несколько уже, чем его средняя часть. Жгутики усиков 2-й пары веретенообразно утолщены у основания....................................*5.1. derjugini Gurjan.
а
Фиг. 75. Idothea metallica Bose, (по Harger). а—общий вид; Ь — maxillipedes; с — antenna I.
I. Idothea metaiHca Bose (фиг. 75).
Bose, 1802, Hist. Nat. Crust., II: 179, pl. XV, f. 6. — peloponesiaea Roux, 1828, Crust. Mediterranee, pl. XXX, f. 10— 12. — atrata Costa, 1838, Fauna R. Napoli, Crust., pl. XI, f. 3. — rugosa M.-Edwards, 1840, Hist. Natur. Crust., Ill: 131. — com-pacta White, 1847, List. Crust. Brit. Mus.: 95. — algirica Lucas, 1849, Anim. artic. Expl. Sci. Algerie. I. Crust.: 61, pl. VI, f. 2.
Тело продолговато-овальное, длина его в 21/4 раза больше ши-
рины, тельзон составляет 2/5 Длины тела. Г олова широкая и короткая со слабо вогнутым передним краем. Глаза большие, круглые, с черным пигментом, посажены на боковых краях головы. Базальный членик усиков 1-й пары небольшой, такой же длины и ширины, как 2-й членик. 3-й членик и единственный членик жгутика, каждый немного длиннее 1-го и 2-го члеников. Усики 1-й пары достигают конца 2-го членика ножки усиков 2-й пары. 1-й членик ножки усиков 2-й пары очень короткий; 2-й и 3-й членики равной длины; 4-й в 1г/2 раза длиннее 3-го; 5-й членик в 2 раза длиннее 3-го; жгутик 8-члениковый и достигает переднего края 3-го грудного сегмента. Грудные сегменты почти равной длины. Передне-боковые углы 1-го сегмента слегка вытянуты вперед по бокам головы, закругленные. Эпимеры на всех сегментах, кроме 1-го, занимают весь боковой край соответствующего сегмента, большие и широкие. Ширина их возрастает к заднему концу тела, так что наиболее широкие эпимеры у 7-го грудного сегмента. Грудные ножки в общем одинаковой структуры; 3 передние пары более короткие и крепкие с более ясно выраженным хватательным типом. Края всех члеников и нижний край б-го членика покрыт волосками. 2 передних брюшных сегмента только немного короче последнего грудного. Тельзон большой с почти параллельными боковыми краями и прямым и слабо выпуклым задним краем. Длина животного 18 мм.
7. IDOTHEA
135
Распространение. Японское море: 36° 46N, 132° 15'Е; 36° 5£ЦЧ, 132° З'Е и 32° 33’ N, 129° 32' Е на глуб. от 0 до 1600 м. Пелагическая, почти, космополитическая форма/ распространенная по всему Атлантическому, Тихому и Индийскому океанам, главным образом в их тропической и субтропической частях. Заносится вместе с плавающими предметами (водоросли, куски дерева) в северную часть Атлантического океана между Исландией и Гренландией до 60° с. ш. Встречается в Дэвисовом проливе (64° 46' с. ш., 53° 35' з. д.) (Thielemann, 1910; Hansen, 1916).
больше длины; передний край
а
Фиг. 76 Idothea fewkesi Rich, (по Richardson), а — общий вид; Ъ — maxillipedes; с — antenna I.
* 2. Idothea fewkesi Rich. (фиг. 76).
Richradson, 1905, Bull. U. S. Nat. Mus., n° 54:359, t. 387.
Тело узкое, длинное; длина его в 5]/2 раз больше ширины. Голова крупная, широкая; ширина ее в 173 раза головы глубоко вогнут посредине, образуя род широкой вырезки, в которой помещаются усики 1-й пары; передне - боковые углы закругленные. Глаза маленькие,1 по бокам головы, расположенные на середине расстояния между передним и задним краями головы. Базальный членик усиков 1-й пары сильно расширен; 2-й и 3-й членики почти равной длины, каждый значительно уже и короче базального; един-. ственный членик жгутика немного длиннее 3-го членика ножки и достигает конца 2-го членика ножки усиков 2-й пары. Усики 2-й пары в вытянутом состоянии достигают заднего края 4-го грудного сегмента; жгутик их 16-члениковый;
-базальный членик короткий; 2-й и 3-й членики ножки равной длины. 4-й и 5-й членики в 2 раза длиннее 3-го членика, оба равной длины. 1-й грудной сегмент в 2 раза короче 2-го. Остальные грудные сегменты равной длины, за исключением последнего, который немного короче предыдущих. Эпимеры свободны на всех сегментах, кроме 1-го, очень узкие. Эпимеры 2-го и 3-го сегментов занимают только переднюю половину бокового края сегмента; эпимеры 4-го, 5-го и б-го сегментов занимают 3/4 края сегмента; эпимеры 7-го сегмента занимают весь боковой край сегмента. Тельзон длинный; длина его в 2 раза больше ширины. Боковые края его почти параллельны друг другу. Задний край посредине образует крупный заостренный зубец; задне-боковые углы только слегка закругляются. Длина животного до 42 мм.
Распространение. Санта-Барбара (Калифорния: Richardson, 1905). Японское море: залив Петра Великого (Б. Тафуин, Уссурий
136
7. IDO THEIDAE
ский залив, б. Патрокл, б. Тихая, залив Восток, Сидими, б. Посьет)г в корнях Zostera и на фации песка на глубинах от 0 до 8 м; Татарский пролив 52° 3' N, 141° 30' Е, глуб. 18 м; бухта Де-Кастри; бухта Ольга; Западный Сахалин (бухта Мауки)(сборы ТИРХ’а и ГГИ, 1925—1932). Охотское море: Шантарские *о-ва (бухта Абрек, бухта Нагаева; о. Лангр на г 1913, и ГГИ 1930).
Укома), бухта от 0 до 2 мина литорали (сборы Павленко,.
ине
Фиг. 77. Idothea ochotensis Brandt del. Ю. Залесский (оригинальный экземпляр).
* 3. Idothea ochotensis Brandt фиг. 77).
Brandt, 1851, Middendorff’s Sibirische Reise, II,. Crust.: 145, pl. VI, f. 33.
Тело сильно вытянутое, узкое, с почти параллельными боковыми краями. Голова имеет округлые очертания со слабо вогнутым передним краем; кзади слегка расширяется. Глаза маленькие, посажены по бокам головы на середине расстояния между передним и задним краями головы. Спинная поверхность головы слегка выпуклая. Базальный членик усиков 2-й пары расширенный; 2 следующих более узкие и длинные; последний членик достигает дистального конца 2-го членика ножки усиков 2-й пары. Усики 2-й пары сравнительно короткие; в вытянутом состоянии достигают середины 4-го грудного сегмента; жгутик 10—12-члениковый, в l1^ раза короче ножки. Членики тонкие, длинные,, удлиняющиеся постепенно к дистальному концу антенны. 1-й грудной сегмент в 2 раза,
короче 2-го сегмента; остальные почти равной длины, только последний, 7-й грудной сегмент немного короче предыдущих. Эпимеры 2-го, 3-го, 4-го и 5-гб сегментов занимают только переднюю часть бокового края сегмента. Эпимеры 2 последних сегментов занимают всю поверхность бокового” края сегмента, расширяются кзади. Тельзон удлиненный с почти параллельными боковыми краями; спинная поверхность его сводчато-выпуклая; по средней линии тянется слабо выраженный продольный киль, более ясно видный на самом конце тельзона, образующем крупный тупой зубец. Цвет тела желтовато-зеленый. Длина животного достигает в редких случаях 53 мм, чаще 38—40 мм.
Распространение. Тихоокеанское побережье Северной Америки от о.Ванкувер до Аляски включительно,Калифорния (?), Курильские острова, Авачинская бухта (Берингово море) литораль (Richardson, 1905, 1910). Берингово море: Авачинская губа, на литорали-
1. IDO THEA
137
и в сублиторали в зарослях Zostera на глубине от 0 до 16 м; сильно опресненная лагуна р. Анатаванной (у Петропавловска); о. Карагинский, литораль. Охотское море: порт Аян; Шантарские острова (б. Абрек, губа Якшина на литорали), Тауйская губа, бухта Нагаева; о. Ольский; залив Байкал; о. Удд (Залив Счастье), о. Феоклистов 54° 53' N, 136° 56' Е), Тугурский залив; Еринейская губа; о. Лангр на литорали и в зарослях водорослей до 2 м. Японское море: бухта Гайдамак в зал. Рында
(Татарский пролив); бухта Де-Кастри, литораль под 1929 и ТИРХ’а, 1931).
* 4. Idothea aleutica Gurjan. (фиг. 78).
Gurjanova, 1933, Исслед. морей СССР, вып. 17: 91, рис. 9, 10. — ochotensis Richardson, 1900, Proc. U. S. Nat. Mus., 22 : 732, f. 2, 3a, 4a, 5b, 6b, (non Brandt); Richardson, 1899, Proc. U. S. Nat. Mus., 21:846, f. 26.
Этот вид, принимаемый Richardson, а также Miers и Benedict, за 1. ochotensis Brandt, должен быть выделен в самостоятельный вид, I. aleutica. От I. ochotensis Brandt, оригинальные экземпляры которого, хранящиеся в Зоологическом институте Академии Наук СССР, я имела в руках для сравнения, эта форма отличается более широким и коротким телом, почти -четырехугольной плоской головой, более крупной, чем у /. ochotensis, короткими усиками 2-й пары, формой ^тельзона и деталями строения ногочелюстей, грудных ножек и рулевых ног.
Голова плоская, широкая с почти прямым передним и вогнутым посредине задним краем. Глаза небольшие по бокам головы на середине растояния между передним и задним краями головы. Усики 1-й пары достигают конца 2-го членика ножки усиков 2-й пары; базальный членик
камнями (сборы ГГИ,..
Фиг. 78. Idothea aleutica Gurjan. del. Ю. Залесский.
их расширенный, длина его равна длине 2-го и
3-го члеников вместе;
последний членик немного длиннее предпоследнего. Усики 2-й пары в вытянутом состоянии достигают только середины 3-го грудного сегмента; жгутик 11-члениковый. 1-й грудной сегмент в 2 раза короче головы; 2-й, 3-й и 4-й грудные сегменты равной длины, немного длиннее 1-го. Последние 3 грудных сегмента равной длины, немного короче предыдущего, но длиннее 1-го сегмента. Эпимеры 2 последних пар занимают всю боковую поверхность сегмента. Все грудные сегменты короткие и широкие, ширина их больше чем в 3 раза больше длины. Брюшной отдел длинный: при длине тела в 35 мм равен 12.5 мм. Тельзон с вогнутыми посредине боковыми краями, так что расширяется и у основания и у дистального конца.' Задний конец оттянут в тупой крупный, закруглен-
138
7. IDO THEIDAE
от 0 до 20 м; о. Атха
•Фиг. 79. Idothea derjugini Gurjan. del. Ю. Залесский.
ный на вершине зубец. Цвет тела желто-бурый, иногда зеленоватый. Длина достигает 45 мм, в редких случаях 61 мм.
Распространение. Берингово море: литораль и сублитораль до 30 м глубины: Командорские о-ва; залив Ткачен 64° 25'N, 172°48 W, глуб. 30 м; Берингов пролив, 65O51'N, 168° 32' W, глуб. 50 м;. побережье Камчатки — мыс Африка, Усть-Камчатск, Авачинская губа; бухта Вилючинская, бухта Тихирка; бухта Эмма, зал. Лаврентия; на глубинах Вознесенского, 1846). Охотское море: западная Камчатка в устье р. Озерной, на литорали. На Командорских островах проделывает правильные сезонные вертикальные миграции: летом появляется на литорали, зимой опускается в сублитораль.
* 5. Idothea derjugrni Gurjan. (фиг. 79).
Gurjanova, 1933. Исслед. морей СССР, 17 :91, рис. 7 и 8.
Тело длинное, вытянутое с почти параллельными краями. Голова очень большая и широкая, с сильно выпуклой спинной стороной, образующей округлый бугор. Передний край головы вогнут по средине. Ширина головы меньше чем в 2 раза превосходит ее длину. Глаза маленькие, латеральные, на середине расстояния между передним и задним краями головы. Усики 1-й пары достигают середины 3-го членика ножки 2-й пары. Базальный членик их расширенный, но длина его немного меньше длины 2-го и 3-го члеников вместе. Усики 2-й пары в вытянутом состоянии достигают заднего края 3-го грудного сегмента; жгутик 13-члениковый
и почти равен длине ножки. Членики жгутика короткие и широкие вначале, к концу жгутика делаются длинными и узкими, а так как 1-й членик жгутика тоже несколько уже, чем следующие, то базальная часть жгутика кажется веретенообразно утолщенной. 1-й грудной сегмент немного более половины длины головы и в 2 раза короче 4-го сегмента. 2-й, 3-й и 6-й сегменты равной длины, каждый, в 1*/2 раза длиннее 1-го. Тельзон большой, длина его равна длине 4 последних грудных сегментов вместе; боковые края его почти параллельные друг другу, хотя к дистальному концу тельзон слегка суживается. Срединный зубец на конце тельзона треугольный,, тупой. Эпимеры 2-го грудного сегмента занимают только */2 бокового края сегмента; эпимеры следующих сегментов 2/8 края, а эпимеры 2 последних сегментов весь боковой край сегмента. Все тело
1. IDOTHEA
139
значительно более грубое и крепкое, чем у предыдущего вида, с сильно выпуклой спинной стороной головы и тельзона. Цвет буро-зеленый, иногда бурый с мелкими белыми пятнами. Максимальная длина самца 51 мм.
Распространение. Японское море: по всему побережью от Корейской границы до бухты Де-Кастри включительно; западный Сахалин; держится на литорали среди зарослей водорослей и в сублиторали на глуб. от 0 до 2 м (сборы ГГИ и ТИРХ’а, 1925—1932). Охотское море: бухта Абрек (Шантарские о-ва) на литорали . раль (сбор Павленко, 1913).
*6. Idothea orientalis Gurjan. (фиг. 80).
Gurjanova, 1933, Исследования морей СССР, 17:92, рис. 11—12.
Тело вытянутое, линейное, с параллельными краями, сильно уплощенное в дорзовентральном направлении, с плоской спинной стороной, вдоль которой по средней линии тянется низкий ясно-выраженный киль. Г олова относительно небольшая, с плоской спинной поверхностью, вогнутым передним краем и закругленными передне-боковыми углами. Длина головы немного больше чем в 1т/2 раза превосходит ее ширину и равна длине 1-го грудного сегмента и половине 2-го сегмента вместе. Усики 1-й пары удлиненные, достигают конца 3-го членика ножки усиков 2-й пары; базальный членик расширенный, но длина его значительно меньше длины 2-го и 3-го члеников вместе, которые приблизительно равной длины. Усики 2-й пары тонкие и длинные; в вытянутом состоянии достигают середины 5-го грудного сегмента; жгутик 1б-члениковый, немного длиннее, чем ножка. Эпи
меры всех сегментов, кроме 7-го (и кроме 1-го, который лишен эпимер) занимают только часть боковых краев сегмента; эпимеры 7-го сегмента занимают весь боковой край его. 1-й грудной сегмент почти в 2 раза короче 2-го; 2-й немного короче 3-го; 3-й, 4-й и 5-й сегменты почти равной длины, б-й и 7-й сегменты равной длины, но каждый короче 5-го сегмента. Тельзон относительно короче, чем у предыдущих видов, меньше чем длина 3 последних грудных сегментов вместе. Боковые края его сильно вогнуты посредине, так что и основание й конец тельзона расширенные, по сравнению с его средней частью. Срединный зубец заднего края тельзона очень широкий, треугольный, с тупой вершиной. Цвет животного темнокрасный, иногда красно-бурый. Длина 35 мм.
Распространение. Японское море: залив Петра Великого {мыс Басаргин, бухта Патрокл, о. Скрыплев, Уссурийский залив, бухта
и у мыса Марии, лито-
Фиг. 80. Idothea orientalis Gurjan. del. Ю. Залесский.
140
7. IDO THEIDAE
Посьет, мыс Тобизина, о. Фурругельма, м. Гамова, о-ва Унковского),, бухта Ольга, бухта Владимир; среди зарослей красных водорослей и на. ламинариях, иногда на песке и гальке, на глуб. от 10 до 20—25 м (сборы ГГИ и ТИРХ’а, 1926—1932). Охотское море: залив Анива, глуб. 20— 30 м (сбор П. Шмидта, 1902); между мысом Красным и м. Палец, на песке,. 14 м глуб. (сбор Павленко и Солдатова, 1912).
2. Род PENTIDOTHEA RICHARDSON
Richardson Н., 1905, Bull. U. S. Nat. Mus., 54:368.
Тип рода: P. resecata (Stimps.)
Тело длинное, вытянутое, иногда с параллельными краями. Брюшной отдел из 3 сегментов; жгутик усиков 2-й пары многочлениковый. Эпимеры: свободно сочленены на всех 6 последних грудных сегментах. Щупик ногочелюстей состоит из 5-ти члеников. В остальном сходен с родом Idothea*
В дальневосточных морях род представлен 4 видами.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (6). Грудные сегменты тела соприкасаются друг с другом по всей их. длине.
2 (3). Эпимеры занимают всю длину бокового края сегментов только у последнего грудного сегмента; тельзон на конце несет крупный, треугольный тупой зубец...........................* 1. Р. japonica Rich-
3 (2) Эпимеры занимают всю длину бокового края у 3 последних грудных сегментов; тельзон закруглен на конце.
4 (5). Тело узкое, вытянутое; длина его почти в 6 раз превышает ширину;: тельзон вытянутый в длину; длина его в 2 раза превышает ширину его основания. Задний конец тельзона плавно закругляется ... .
..........................................* 2. Р. rotunda!a Rich-
5 (4). Тело укороченное, широкое; длина его только в 3 раза превышает ширину; тельзон широкий; длина его около l1^ раза больше ширины основания; конец тельзона широко-закруглен с маленьким тупым зубчиком по средине.........................*3. Р. wosnesenskii Brandt.
6 (1). Грудные сегменты соприкасаются друг с другом только посредине,, так как боковые части их суживаются и оттянуты в стороны . . .
............................................4. Р. whitei Stimps..
* 1. Pentidothea japonica Rich. (фиг. 81).
Richardson, 1900, Proc. U. S. Nat. Mus., 22 :131.
Тело очень длинное, узкое, с параллельными боковыми краями; длина его почти в 6 раз превосходит ширину. Поверхность тела покрыта, мелкой зернистостью. Вдоль спины по средней линии, начиная с заднего края головы, тянется до конца тельзона ясный киль. Голова удлиненная;
2. PENTIDO THEA
141
с сильно вогнутым посредине передним и закругленным выпуклым задним краем; длина головы в l1^ раза меньше ее ширины. Затылочная часть слабо выпуклая. Глаза маленькие, расположены по бокам головы немного ближе к переднему краю, чем к заднему. Усики 1-й пары достигают середины 3-го членика ножки усиков 2-й пары; базальный членик их расширенный, длина его немного меньше длины 2 следующих члеников вместе. Усики 2-й пары тонкие, длинные; достигают заднего края 5-го грудного сегмента; жгутик 20-члениковый, постепенно утончающийся дистально. 1-й грудной сегмент несколько уже и значительно короче 2-го сегмента. >> У
:2-й, б-й и 7-й сегменты равной длины, ff чг\Т1
3-й, 4-й и 5 сегменты равной длины, каждый Я В
немного длиннее 2-го. Эпимеры 2-го сегмента \ /у у\ Н занимают только половину бокового края сегмента; эпимеры 7-го сегмента занимают весь край сегмента. Остальные эпимеры занимают часть края сегмента и тем большую, .
чем ближе к заднему концу тела. Все эпимеры -/Z"
свободные, в виде очень узких пластинок.
Брюшко состоит из 3 сегментов, длина его равна длине 5 передних сегментов груди вместе. 2 передних свободных брюшных сег- Г >
мента очень короткие, длина их вместе I I
меньше длины 1-го грудного сегмента. Тель- 1 ]
вон длинный, узкий, слегка суживается ди- X. /
стально. Задний конец его вытянут в длин- \/
ный широкий у основания и закругленный л „ ., , .
Фиг. ox. Pentidothea japonica на конце треугольный зубец. Задне-боковые Rich.
углы тельзона по бокам основания средин
ного зубца закруглены и незаметно переходят в боковые края тельзона. Длина тельзона равна длине 4 последних грудных сегментов1 вместе и немного больше чем в 2 раза превосходит его ширину у основания. Цвет зеленовато-бурый. Длина животного 42 мм.
Распространение. Побережье Японии (Токио, Сали, Рикуоку, Мороран, Хакодате, на литорали: Richardson, 1900, 1910). Японское море: залив Петра Великого (Б. Тафуин, Посьет, Стрелок, залив Восток, бухта Патрокл, бухта Таваиза, Амурский залив, Сидими, Золотой Рог); бухта Де-Кастри, бухта Найна; на фации песка и среди Zostera и водорослей на глуб. О—б м (сборы ГГИ и ТИРХ’а 1926—1932).
*2. Pentidothea rotundata Rich. (фиг. 82).
Richardson, 1910, Proc. U. S. Nat. Mus., 37:109, f. 30—31.
Тело узкое, удлиненное; длина его в 5т/2 раз больше ширины. Поверхность тела гладкая; цвет светлозеленый с темнозеленым пятнами
142
7. IDOTHE1DAE
Фиг. 82. Pentidothea rotun-data Rich, (по Richardson).
и бурыми продольными полосами, в виде тонких линий. Голова удлиненная с глубоко вырезанным передним краем и трапециевидной задней частью; длина ее немного меньше ширины. Спинная поверхность головы плоская. Глаза очень маленькие, круглые, расположены на самом краю боковой поверхности ближе к заднему краю головы и едва видны сверху. Брюшная поверхность тела плоская; спинная сильно сводчато-выпуклая. Базальные членики усиков 1-й пары расширенные, почти квадратной формы; 3 последних членика короткие, маленькие, равной длины; достигают конца 2-го членика ножки усиков 2-й пары. Усики 2-й пары относительно короткие и толстые; в вытянутом состоянии достигают только середины 2-го сегмента; жгутик 9-члени-ковый, в 1]/2 раза короче ножки. 1-й грудной сегмент такой же ширины, как голова; его передне-боковые углы вытянуты вперед и плотно охватывают заднюю часть головы по бокам.. Длина его в 2 раза меньше длины 2-го сегмента. 2-й, 3-й и 4-й сегменты почти равной длины. 5-й, б-й и 7-й грудные сегменты равной длины, каждый короче 2-го, но длиннеее 1-го сегмента. Эпимеры на всех сегментах, кроме 1-го, свободные, в виде очень узких пластинок, занимающих весь-край сегмента только у последних 3 грудных сегментов. 3 передние пары эпимер не видны при рассматривании животного сверху. Абромен из 3 сегментов; 2 передние очень короткие; тельзон удлиненный, суживающийся дистально-с овально закругленным задним краем. Грудные ножки короткие и слабые. Рулевые ножки не достигают конца тельзона, оставляя непокрытым его закругленный конец на брюшной стороне. Длина животного до 38 мм.
Распространение. Рикуоку, Япония (Richardson,. 1910), Японское море: залив Петра Великого; в больших количествах держится в зарослях Zostera в бухте Соболь, у мыса Басаргин, в бухте Тихой (Уссурийский залив),,
на глуб. О—3 м; преимущественно в корнях и на стеблях Zostera и Phyllospadix; характерная форма биоценоза Zostera (сборы А. Шурин, 1926— 1929, коллекции ТИРХ’а). На север вид доходит до бухты Преображения, (эксп. ЗИН 1934 г.).
* 3. Pentidothea wosnesenskii (Brandt) (фиг. 83).
Brandt, 1851, Middendorff’s Sibir. Reise, Crust: 146 (Idothea); Miers, 1883, Journ.. Linn. Soc. London, XVI: 40 (Idothea); Richardson, 1905, Bull. U. S. Nat. Mus, n° 54:370, f. 402—404. — hirtipes Dana, 1853, U- S. Expl. Exp., Crust., XIV : 704, pl. 46, f. 6 (Idothea).— oregonensis Dana, 1854, Proc. Acad. Nat. Sci. Philad. VII: 175 (Idothea).
Тело удлиненно-овальное с почти параллельными боковыми краями,, длина его в 3 раза больше наибольшей ширины. Длина брюшка равна длине 6 передних грудных сегментов вместе. Голова большая, длина ее
2. PENTIDOTHEA
143
<Х
Фиг. 83. Pentidothea wosnesenskii (Brandt) (по Richardson). а—общий вид; Ъ — maxillipedes; с — antennula.
всю длину бокового коая сегмента и оасши-
в 11j4 раза меньше ширины и равна длине 2 передних сегментов вместе. Передний край головы закругленно вогнутый. Позади глаз голова расширяется. Задняя часть головы по бокам охватывается вытянутыми вперед переднебоковыми закругленными углами 1-го грудного сегмента. Глаза маленькие поперечно-овальные, посажены по бокам головы ближе к ее переднему краю. Базальные членики усиков 1-й пары расширенные; длина всех, члеников ножки равная; одночлениковый жгутик немного длиннее каждого из предыдущих члеников и достигает середины 3-го членика ножки усиков 2-й пары. Усики 2-й пары достигают передне - бокового угла 2-го грудного сегмента; жгутик 16-члениковый. Базальный членик ножки короткий; 2-й и 3-й членики равной длины, каждый длиннее 1-го. Последние 2 членика равной длины, каждый в 1 т/2 раза длиннее 3-го. Все грудные сегменты равной длины. Эпимеры 2-го, 3-го и 4-го сегментов занимают немного больше 3/4 длины бокового края сегмента. Эпимеры 3 последних грудных сегментов занимают
ряются кзади. Все эпимеры большие, хорошо видные сверху; наиболее широкие эпимеры на задних сегментах. Передние края 3 первых грудных сегментов сильно вогнутые. Грудные ножки одинаковой структуры, у самца густо усажены волосками на свободных краях члеников. У самки 5-й членик ног усажен толстыми щетинками вдоль нижнего края. Тельзон большой и широкий. Задний конец его широко закруглен с маленьким острием посредине. На верхней поверхности рулевых нрг косо проходит киль, образованный утолщением передней части. Длина животного 32 мм. Цвет зеленоватый, самцы желтоватые, самки более темно окра
шенные.
Распространение. Охотское море; Камчатское море; западное побережье Северной Америки, Британская Колумбия и Калифорния на юг до Сан-Диего, 0—17 м глубины; Аляска, Уналяска, Попов, остров; Алеутская гряда (Юкатат, Атту, Атха), на литорали (Richardson* 1905, 1910); литораль острова Атха, 1 экз. (сбор Вознесенского* 1846).
144
7. IDO THEIDAE
4. Pentidothea white! (Stimpson) (фиг. 84).
Stimpson, 1864, Proc. Acad. Nat. Sci. Philad. 155 {Idothea); Richardson, 1905, Bull. U. S. Nat. Mus., 54 : 373, f. 405-406.
Тело узкое, удлиненное, с почти параллельными краями. Длина тела в 3 раза больше его ширины. Голова широкая с почти прямым, слегка вогнутым передним краем; ширина ее в 2 раза больше длины. Глаза небольшие, поперечно-овальные, посажены по бокам головы на середине
<Х-
Фиг. 84. Pentidothea whitei (Stimps.) (по Richardson), а — общий вид; b — maxillipedes; с — antennula.
-расстояния между передним и задним ее краями. Усики 1-й пары с расши--ренным базальным члеником, достигают конца 2-го членика ножки усиков 2-й пары; 2-й и 3-й членики ножки одинаковой длины, оба немного короче базального; жгутик одночлениковый. Усики 2-й пары с 17-чле-никовым жгутиком, достигают в вытянутом состоянии заднего угла 2-го грудного сегмента; 1-членик их ножки короткий, в 2 раза короче следующего; 2-й и 3-й членики равной длины; 4-й и 5-й членики равной длины, каждый в 2 раза длиннее 3-го. Все грудные сегменты равной длины; боковые части их несколько суживаются, оттянуты в стороны, у 3 передних сегментов направлены немного вперед, а у 3 задних назад, так что грудные сегменты соприкасаются друг с другом только в средней своей части, а боковые участки их свободны. Эпимеры свободные на всех грудных сегментах, кроме 1-го и занимают всю .поверхность бокового края сегментов. 1-я пара эпимер в виде пластинок, суживающихся к заднему концу; эпимеры 2-й и 3-й пары одинаковой ширины по всей их длине, а эпимеры 3 задних пар с расширенным задним концом. Грудные
3. MESIDOTHEA
145
ножки одинаковой структуры, мощно развитые, крепкие. Внутренний край 6-го сегмента на проксимальном конце снабжен группой твердых щетинок. Брюшной отдел состоит из 3 сегментов; тельзон с сильно выгнутыми краями, расширяющийся и у основания и к заднему концу. Задне-боковые углы его закругленные; задний край посредине образует тупой зубец. Створки рулевых ног снабжены косо пересекающим базальную часть их килем.
Распространение. На банках Mytilus в Калифорнии; Puget Sound; Берингово море (остров Атха) (Richardson, 1910).
• 3. Род MESIDOTHEA RICHARDSON
Richardson Н., 1935, Bull. U. S. Nat. Mus., 54 : 347.
Тип рода: M. eniomon (L).
Грудной отдел равномерно расчленен; тело овально-вытянутое, сплющенное в дорзо-вентральном направлении, с плоской брюшной и выпуклой спинной стороной. Эпимеры на всех сегментах, кроме 1-го, свободны, в виде расширенных пластинок, занимающих всю длину бокового края сегмента. Голова широкая, расплющенная; глаза, если они имеются, маленькие, расположенные на спинной стороне головы. Передний край головы имеет вырезку посредине. Передне-боковые части головы вытянуты впереди глаз и образуют по 2 с каждой стороны маленькие закругленные лопасти. Брюшной отдел состоит из 4 сегментов — 3 свободных брюшных и тель-
Фиг. 85. Mesidothea entomon (L.) subsp. orientalis Gurjan. (экз. из Берингова моря).
зон. Первые 3 пары грудных ног хватательные, снабженные ложной клешней. 4 задних пары ног ходильные. Усики 1-й пары маленькие с одно-члениковым жгутиком. Жгутик усиков 2-й пары многочлениковый. Щупик ногочелюстей состоит из 5 члеников. Крупные формы.
На Дальнем Востоке известен 1 вид, М. entomon (L.), представлен-
ный одним подвидом.
* 1. Mesidothea entomon (L.) subsp. orientalis Gurjan. (фиг. 85).
Gurjanova, 1933, Fauna arctica, VI, Lief. 5 : 439.
Тело удлиненное с плавно аркообразно закругляющимися боковыми линиями и сильно выпуклой спинной поверхностью. Отношение наибольшей ширины тела к его длине от 27.8 до ЗО.7°/о. Голова уплощенная. Глаза маленькие, дорзальные. Впереди глаз по бокам срединной вырезки на переднем крае головы по одному поперечному килю. От переднего края головы по средней линии идет ясно видная продольная бороздка, занимающая первую треть головы. Передняя лопасть головы впереди глаз немного шире, чем задняя. Усики 1-й пары достигают конца 4-го чле-
Гурьянова 10
146
7. IDOTHEIDAE
ника ножки усиков 2-й пары; базальный членик их расширенный;. 2-й членик короткий; 3-й членик удлиненный, тонкий, равный длине базального членика. Усики 2-й пары сравнительно короткие, достигают заднего угла 1-го грудного сегмента; членики ножки сплющенные^ 2-й членик расширенный, базальный членик не виден сверху; 3-й и 4-й членики короткие, 5-й удлиненный, равной длине 2 предыдущих члеников', вместе; жгутик 10—12-члениковый. На первых 4 грудных сегментах по бокам на спинной стороне no 1 продольному килю, особенно резко выраженному на 2-м и 3-м сегментах. Эпимеры широкие в виде пластинок, с оттянутыми назад в виде остриев задне-боковыми углами на 5 последних грудных сегментах. Задние углы эпимеральных расширений 1-го грудного-сегмента закруглены. Спинная поверхность эпимер 2-го и 3-го сегмента гладкая, ровная, ясно вогнутая. Тельзон удлиненно-пятиугольный формы с вытянутым в тупое острие задним концом. Поверхность тела густо покрыта мелкими точечными вдавлениями, хорошо видными на сухих, экземплярах даже невооруженным глазом. Наибольшая ширина тела на 4-м грудном сегменте.
Таблица 2
Mesidothea entomon subsp. orientalis среднее в % М. entomon subsp. glacialis среднее в °/0 М. entomon subsp. entomon среднее в %,
Отношение наибольшей ширины к длине тела 29.1 29.9 28.4
Отношение длины головы к ее наиб, ширине 47.1 48.0 47.1
Отношение ширины основания тельзона к длине тельзона 59.3 48.9 48.9
Отношение длины конца тельзона к длине тельзона 34.8 36.9 36.2
Отношение ширины конца тельзона к ширине основания тельзона 61.0 56.3 62.8
Отношение ширины конца тельзона к длине конца тельзона 104.3 75.0 88.3
Отношение длины головы к длине тела . . 8.7 8.9 8.9
Отношение длины тельзона к длине тела . . 36.7 41.3 40.4
Отношение ширины конца тельзона к длине тельзона » 32.2 27.1 30.9
Дальневосточная форма образует особый подвид, хорошо отличимый.. от северной и балтийской формы. От северного подвида, М. entomon (L.) subsp. glacialis Gurjan., дальневосточный подвид отличается более крепким укороченным и сильно выпуклым на спинной стороне телом; хитиновый покров толстый, грубый; сегменты тела прочно соединены, плотно прилегая друг к другу. Голова шире и более выпуклая, чем у арктической формы; линия затылка большею частью слегка вогнута посредине;
3. MESIDOTHEA. 4. PENT I AS
147
эпимеральные расширения 1-го сегмента плоские и более круто направлены книзу вместе с боковыми частями самого сегмента. Эпимеры 2-го и 3-го сегмента плоские и значительно уже, чем двух других подвидов (М. entomon entomon и М. entomon glacialis). Тельзон резко укорочен и расширен у основания; укорочение тельзона идет за счет укорочения длины его дистального конца; ноги крепче и массивнее, а на эпимерах и конечностях волоски гуще и длиннее, чем у балтийской и арктической форм. Особенно хорошо все эти отличия восточной формы от арктической и балтийской видны из следующей таблицы биометрических признаков.
Эта форма характерна для сильно опресненных районов Берингова и Охотского морей; в изобилии встречается в устьях рек, в протоках" озер (напр., у Усть-Камчатска), но иногда встречается и в прибрежной части в местах с нормальной морской соленостью (в отличие от арктической формы, которая держится только в устьях рек в почти пресной воде с соленостью не выше 1О°/оо).
Распространение. Берингово море: Анадырский лиман, Авачинская губа, устье р. Камчатки, протока оз. Нерпичьего (у Усть-Камчатска), Камчатское побережье, в устьях речек. Охотское море: Шантарские о-ва на литорали и на глуб. О—5 м; устье р. Кутима; Тауйская губа; о, Лангр; Амурский лиман; за мысом Невельского 42° 02' N, 141° 30'Е; 53°59'N, 136° 59'Е; взрослые экземпляры до 75 мм длины; заходя высоко в реки мельчают; самки с инкубаторными пластинками в 30 мм длиною.
4. Род PENTIAS RICHARDSON
Richardson, 1904, Proc. U. S. Nat. Mus., 27 :47.
Тип рода: P. hayi Rich.
Тело очень узкое, удлиненное; 1-й грудной сегмент такой же ширины, как голова. Усики 2-й пары короткие, жгутик состоит из малого числа члеников. Щупик ногочелюстей пятичлениковый. Эпимеры на всех грудных сегментах, кроме 1-го, свободные, но очень маленькие и узкие. Брюшной отдел состоит только из одного сегмента, так как все брюшные сегменты сливаются с тельзоном и сохраняются лишь 3 пары швов по бокам тельзона у. его основания. По внешнему виду очень сходен с представителями рода Idothea.
В дальневосточных морях известен один вид.
* 1. Pentias hayi Richardson (фиг. 86).
Richardson, 1904, Proc. U. S. Nat. Mus., 27:47, f. 24—25.
Тело узкое, сильно вытянутое с параллельными краями. Ширина его почтй такой же ширины, как голова. Длина тела в 4г/а раза больше его ширины. Брюшная поверхность плоская, спинная сильно выпуклая. Поверхность тела совершенно гладкая; голова узкая, ширина ее лишь 10*
148
7. IDOTHEIDAE
немного больше длины (по описанию Richardson ширина головы в 2 раза больше ее длины). Передний край головы слегка вогнутый с очень маленьким, едва заметным срединным зубчиком. Глаза небольшие, черные, округлые, посажены на боковых краях головы ближе к ее заднему краю. Базальные членики усиков 1-й пары очень широкие, почти квадратные. 3 следующих членика удлиненные, равной длины. Конец жгутика (одночлениковый) достигает середины 3-го членика ножки 2-й пары усиков; Усики 2-й пары очень короткие, конически суживающиеся дистально. в вытянутом состоянии едва достигают заднего края 1-го грудного
Сс
Фиг. 86. Pentias hayi Rich, (по Richardson), а — общий вид; Ь — maxilfpedes.
сегмента; жгутик состоит из шести коротких члеников. 1-й грудной сегмент в 2 раза короче 2-го; передне-боковые углы его закруглены, вытянуты вперед и охватывают голову с боков, достигая нижнего края глаза. 3-й сегмент длиннее 2-го и равен длине 4-го сегмента. 3 задних сегмента короче предыдущего, равной длины. Эпимеры 2-го, 3-го и 4-го грудных сегментов занимают только переднюю половину бокового края сегмента; эпимеры 6-го и 7-го сегментов занимают всю длину бокового края сегмента. Тельзон очень узкий и. длинный; длина его равна длине первых 5 грудных сегментов вместе. У основания тельзона по бокам следы слияния брюшных сегментов в виде 3 пар швов; к дистальному концу постепенно суживается и заканчивается тупым треугольным острием.
Ноги очень маленькие и слабые, одинаковой структуры. Коготки крепкие,
изогнутые, раздвоенные на конце, и обычно окрашены в коричневый цвет. Цвет тела беловато-зеленоватый, более интенсивный на каудальном сегменте. Длина животного 25 мм.
Распространение. Побережье Японии (Хакодате, Мисаки, бухта Сагами) (Richardson, 1904, 1910). Японское море: залив Петра Великого в больших количествах в зарослях Zostera (б. Патрокл, б. Соболь, мыс Басаргин, бухта Тихая в Уссурийском заливе), на глуб. О-—-3 м. Вместе с Pentidothea rotundata является одной из самых характерных форм биоценоза Zostera (сборы ТИРХ’а, 1925—1929).
5. Род SYMMIUS RICHARDSON
Richardson, Н., 1904, Proc. U. S. Nat. Mus., 27:40.
Тип рода: S. caudatus. Rich.
Передне-боковые углы головы вытянуты вперед и вбок и образуют широкие, но не расщепленные лопасти, у основания которых расположены
5. SYMMIUS
149
O'-
Фиг. 87. Symmius caudatus Rich, (по Richardson), а — общий вид; b — antenna I; с — antenna II; d — maxillipedes.
очень маленькие глаза. Усики 1-й пары удлиненные, состоят из 4 члеников; последний членик их (жгутик) длиннее всех предыдущих члеников. Усики 2-й пары очень короткие, такой же длины, как усики 1-е, с одночле-никовым жгутиком. Щупик ногочелюстей из 3 члеников. Эпимеры имеются только на 3 последних грудных сегментах; на 4 передних сливаются с сегментами, образуя эпимеральные расширения. Брюшной отдел состоит из 3 сегментов, 2 передние короткие, последний, каудальный сегмент сильно вытянутый, длинный. Створки рулевых ног.одночлени-' ковые, так как 2-й их членик нацело сливается с 1-м даже без следа шва. Известен только один вид этого рода.
1. Symmius cauda-tus Rich. (фиг. 87).
Richardson, Proc. U. S. Nat. Mus., 27 : 40, f. 11—15.
Тело удлиненное, резко суживается к заднему концу; наибольшая ширина его на 3-м грудном сегменте. Голова широкая, ширина ее основания в 2 раза больше длины. Передний край головы между основаниями широких переднебоковых лопастей закругленно-выпуклый; края
Этих лопастей цельные, не расщепленные; глаза очень маленькие, расположенные у основания передне-боковых лопастей головы. Усики 1-й пары удлиненные, четырехчлениковые; последний членик их, единственный членил жгутика, длинный, в 2 раза превышающий длину предпоследнего членика. Усики 2-й пары очень короткие, по длине не превышающие усики 1-й пары; все членики жгутика сливаются вместе, образуя один короткий с закругленной вершиной членик. Первые 4 грудных сегмента равной длины; последние 3 сегмента постепенно укорачиваются и суживаются кзади. Эпимеры 4 передних сегментов сливаются с сегментами, образуя плевральные (эпимеральные) расширения. 3 задних грудных сегмента снабжены широкими свободными эпимерами. Боковые части 2 передних брюшных сегментов оттянуты в стороны, образуя заостренные отростки, по одному
150
7. IDO TH El DAE
с каждой стороны. Тельзон не имеет следов слияния с брюшными сегментами, цельнокрайний; задний конец его вытянут в длинный, узкий отросток, закругленный на конце. У основания этого отростка боковые края тельзона слегка вздуты. 1-я пара грудных ног значительно крупнее и крепче всех остальных; последняя пара ног короткая и слабая. Цвет тела белый с серовато-коричневыми пятнами.
Распространение. Берега Японии (Ose Zaki), на глуб. 120— 140 м (Richardson, 1904).
6. Род SYNIDOTHEA HARGER
Harger, 1878, Amor. Journ. Sc., (3). XV: 374.
Тип рода: S. nodulosa (Kr.).
Тело уллиненно-овальное; передний край головы с вырезкой посредине, иногда глубоко U-образно вырезан. Жгутик усиков 1-й пары много-члениковый; щупик ногочелюстей состоит из 3 члеников; брюшной отдел образован только одним каудальным сегментом, так как все брюшные сегменты сливаются с тельзоном. Эпимеры нацело сливаются с грудными сегментами, образуя на 4 передних сегментах плевральные расширения. В передней части головы между глазами обычно имеется ромбовидная площадка, по углам вооруженная бугорками или шипами. Постцефалическая область головы часто отделена глубокой поперечной бороздкой. Базальный членик створок рулевых ног обычно пересечен диагональным килем. Тельзон заострен на конце или же слегка7раздвоен.
В дальневосточных морях известно 18 видов и одна форма.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (12). Тельзон заостряется на конце.
2 (3). Голова, грудь и тельзон на спинной стороне покрыты крупными шипами.....................................*16. S. spinosa Gurjan.
3 (2). Крупных шипов на поверхности тела нет, иногда имеются лишь низкие бугорки, короткие шипики или валикообразные возвышения. ‘4 (5) Тельзон пятиугольной формы ....... *18. S. cinerea Gurjan.
5 (4). Тельзон вытянутой треугольной формы.
6 (9). Плевральные расширения грудных сегментов лишены бугорков и шипиков.
7 (8). Предглазничные бугорки расположены позади фронтального края головы ..................................*13. S. nodulosa Kroyer.
8 (7). Предглазничные бугорки расположены на самом фронтальном крае головы, образуя его часть . ................*14. S. laevis Bened.
9 (6). Плевральные расширения грудных сегментов несут группы бугорков или шипиков.
6. SYNIDOTHEA
151
10 (11). На спинной стороне головы, позади фронтальной вырезки 4 шиповидных бугорка, расположенных в виде ромба; на плевральных расширениях грудных сегментов группы мелких шипиков..............
. . .................................. *15. S. muricata Harf.
11 (10). На спинной стороне головы позади фронтальной вырезки только 3 клиновидных, сливающихся основаниями, бугорка, расположенных в 1 ряд. На плевральных расширениях грудных сегментов группы низких, неясных бугорков........................*17. S. picta Bened.
12 (1). Тельзон на конце раздвоен или имеет небольшую вырезку.
13 (14). 1-й грудной сегмент снабжен маленькими свободными эпимерами . ...........................................*10. S. epimerata Rich.
14 (13). Эпимеры отсутствуют на всех сегментах без исключения.
.15 (16). Передний край 1-го грудного сегмента по бокам головы имеет ясные полукруглые вырезки................ . *11. S. excavata Gurjan.
16 (15). Передний край 1-го грудного сегмента не имеет вырезок по бокам . головы.
т-17 (18). Передний край головы совершенно прямой, без срединной вырезки ............................................. *12. S. harfordi Bened.
18 (17). Передний край головы U-образной формы или имеет вырезку посредине.
19 (28). Посреди срединной вырезки переднего края головы имеется один или два хорошо развитых бугорка.
20 (23). Позади срединной вырезки только один бугорок.
21 (22). Фронтальная вырезка головы неглубокая; усики 2-й пары в вытянутом состоянии достигают заднего края 1-го грудного сегмента; боковые края 1-го грудного сегмента образуют угол с его передним краем ............................................9. S. acuta Rich.
22 (21). Фронтальная вырезка очень глубокая, U-образная; усики 2-й пары достигают заднего края 4-го грудного сегмента; передне-боковые углы 1-го грудного сегмента плавно закругляются..................
.................................- . . *8. S. longicirra Gurjan.
, 23 (20). Позади срединной фронтальной вырезки 2 хорошо развитых бугорка.
24 (25). Кроме 2 крупных бугорков позади лобной вырезки на спинной стороне головы поперечный ряд из 6 хорошо развитых бугорков.
.......................................*7. S. tuberculata Rich.
25 (24). Кроме 2 бугорков позади лобной вырезки на спинной стороне головы нет никаких бугорков.
26 (27). Оба нависающие над лобной вырезкой бугорка сливаются основаниями друг с другом * 6. S. pallida Bened.
27 (26). Оба нависающие над срединной лобной вырезкой бугорка свободны у основания.................................5. S. erosa Bened.
28 (19). Позади срединной вырезки переднего края головы бугорков нет совсем.
152
7. IDOTHEIDAE
29 (30). Дистальный конец тельзона только слегка вырезан; боковые края тельзона в его передней части почти прямые *3. S. nebulosa Bened.
30 (29). Вырезка на дистальном конце тельзона ясная, глубокая; тельзон треугольной формы с плавно закругляющимися боковыми краями.
31 (32). Боковые края 1-го грудного сегмента образуют угол с его передним краем (у молодых экземпляров такой же угол и на 2-м и на 3-м сегментах).................................* 4. S. brazhnikovi Gurjan.
32 (31). Боковые края 1-го грудного сегмента плавно закругляются.
33 (34). 2-й членик ножки усиков 2-й пары снабжен крупным шипом; заты-
лочная часть головы сильно выпуклая. *1 a. S. bicuspida lata Gurjan.. 34 (33). 2-й членик ножки усиков 2-й пары не имеет шипа; голова плоская.. 35 (36). Длина тельзона меньше ширины его основания. * 1. S. bicuspida Owen., 36 (35). Длина тельзона больше ширины его основания...............?
.....................................*2. S. marmorata Packard..
' * 1. Synidothea bicuspida (Owen) (фиг. 88).
Owen, 1839, Crustacea „Blossom": 92, pl. 27, f. 6. (Idothea). — pulchra Lockington, 1877,. Proc. Calif. Acad. Sci., VII: 44, (Idothea)', Miers, 1883, Journ. Linn. Soc. London, XVI: 66 (Edotia); Sars, 1885, Norske Nordh. Exp., Crustacea : 116, pl. X, f. 24—-26.
Тело крепкое, коренастое, удлиненно-овальной формы с плавно, аркообразно изогнутыми боковыми краями и грубым хитиновым покровом. Длина его в два раза больше ширины. Голова большая, широкая; передний край посредине, с небольшой, но глубокой вырезкой. Задняя часть головы отделена от затылочной глубокой поперечной бороздкой. Спинная поверхность головы слабо выпуклая, почти плоская. Глаза большие, округлые, посажены по бокам головы. Передняя часть головы впереди глаз по бокам от срединной вырезки вытянута вперед, образуя широкие лопасти; лопасти эти под прямым углом к своей спинной поверхности отгибаются книзу;, на месте сгиба небольшое ребро, образующее с каждой стороны у самого фронтального края небольшие бугорки. Между глаз не имеется ромбической поверхности, и вся спинная сторона головы лишена шипов или бугорков. Усики 1-й пары маленькие, не достигают конца 3-го членика ножки 2 усиков, четырехчлениковые. Усики 2-й пары удлиненные с простыми: цилиндрическими члениками ножки и 14-члениковым жгутиком; в вытянутом состоянии достигают заднего края 3-го грудного сегмента. 4 передних грудных сегмента приблизительно равной длины с хорошо развитыми эпимеральными расширениями, боковые края которых и передние углы закруглены; на их спинной поверхности имеются валикообразные концентрические утолщения. 3 задних грудных сегмента со слабо выраженными эпимеральными расширениями, укороченные, приблизительно равной длины. Брюшко правильной трехугрльной формы с закругляющимися краями;, у основания его по бокам 2 резких шва — следы слияния тельзона с брюшными сегментами. Поверхность тельзона гладкая, сильно выпуклая; на конце тельзона ясная маленькая вырезка с небольшими остриями по краям. 1-я пара грудных ножек хватательная, снабженная ложной клешней; осталь
6. SYNIDOTHEA
153
ные ножки простые, ходильные. Последние членики на всех грудных ножках снабжены добавочным коготком. Цвет тела темный, сероватый с красными пятнами. Длина 32 мм.
Распространение. В Арктике почти циркумполярна с хиатусом между западной Гренландией и Новой Землей. Берингово море: Прибыловы острова, остров Унимаиййа глуб. 64—88 м (Richardson, 1910). Берингово море: Анадырский лиман, 2 экз. (сбор Пятакова, 1927); сборы экспедиции ГГИ, 1932: ст. 25 — 66° 31' ст. 26 —66° 44' N, 169° 35' W, глуб. 47 ст. 29 — 67° 36' N, 166° 27' W, глуб. 53 ст. 30 —66° 40' N, 167° 52' W, глуб. 29 ст. 33 —64° 39' N, 167° 40' W, глуб. 33
63° 61' N, 172° 35' W, глуб. 66 м (сбор ТИРХ’а 1932).
м;
м; м; м;
N, 169° 42' W, глуб. 45 м;;
88. Synidothea
bicuspida Берингова
имеется
* la. Synidotheabicuspida subsp. lata Gurjan. (фиг. 89).
Gurjanova, 1933. Иеслед. морей СССР, 17 : 94 рис.-15—16а.
От типичной формы из Берингова моря резко отличается более тонким и нежным панцирем, более плоским и широким телом почти овальной формы, более узкой и сильно выпуклой в затылочной части головой, относительно более длинным и узким тельзоном и строением антенн и рулевых ног. Усики 1-й пары заходят несколько
дальше конца 3-го членика ножки усиков фиг. 2-Й пары. Усики 2-Й пары несколько КО- (Owen) — экземпляр из ' ч моря,
роче, чем у типичной формы, достигают только середины 3-го грудного сегмента; на верхнем наружном углу 2-го членика ножки усиков 2-й пары шиповидный отросток. Голова более узкая, чем у предыдущей формы; длина ее больше половины ширины; затылочная часть сильно выпуклая, полусферической формы. Впереди глаз хорошо выраженные шиповидные отростки, в виде рожек, выходящие несколько за пределы переднего края головы. Эпимеральные расширения первых 4 грудных сегментов более резко выраженные, широко расплющенные; на спинной поверхности этих сегментов посредине у заднего края их имеется по одному небольшому,, но хорошо заметному бугорку. 5-й грудной сегмент только немного короче 4-го. Тельзон более длинный, чем у предыдущей формы; длина его равна длине 4 передних грудных сегментов вместе и ширина его у основания равна его длине, тогда как у S. bicuspida typica длина. тельзона равна длине только 3 передних грудных сегментов вместе, и ширина его осно
154
7. IDOTHEIDAE
вания больше его длины. Дистальный членик рулевых ног крупйый, с широко закругленной вершиной, в 2г/2 раза меньше проксимального {базального) членика, тогда как у типичной формы дистальный членик рулевых ног маленький, и базальный членик в 3*/2 раза длиннее его. Цвет тела серый с густо усеивающими серое поле мелкими черными пятнышками; конец тельзона лишен пигмента; по бокам его у основания также пара бесцветных пятен. Вырезка на заднем конце тельзона несколько грубее .и шире, чем у типичной S. bicuspida. Длина животного 20 мм.
Фиг. 89. Sgnidothea bicuspida var. lata Gurjan. a — общий вид; b — uropoda.
Фиг. 90. Sgnidothea marmorata Packard (no Benedict).
Распространение. Японское море: северная часть Татарского пролива в бухте Де-Кастри, глуб. 5—20 м на песчаном грунте (сбор ГГИ, 1929). Охотское море: бухта Чайво, на северо-восточном побережье о. Сахалина (сбор Дербека, 1910А ст. 10 и 11), глуб. 7 м.
* 2. Synidothea marmorata (Packard) (фиг. 90).
Packard, 1867, Mem. Boston Soc. Nat. Hist., 1:296, pl. VIII, f. 6 (Idotaea); Benedict, 1897, Proc. Acad. Nat. Sci. Philad : 392, f. 2.
Очень близок к «S'. bicuspida (Owen), отличаясь от него боЛее узким и длинным телом, сильно редуцированными эпимеральными расширениями, более резко суживающимся дистально тельзоном и гладкой поверхностью тела. Благодаря значительно более узким редуцированным эпимерам длина тела в 2т/2 раза больше его ширины, тогда как у 5. bicuspida длина тела только в 2 раза больше ширины; постцефалическая лопасть S. marmdratd гладкая, ровная и отделена от затылочной части неясной бороздой, а у S. bicuspida борозда глубокая и На постцефалической области поперечный низкий гребень. Тельзон «S', marmorata относительно более длинный
6. SYNIDOTHEA.
155
Фиг. 91. Synidothea nebalosa Bened. (по Benedict).
и узкий, длина его равна ширине его базиса, тогда как у «S. bicuspida тельзон более широкий и длина его меньше ширины основания. Фронтальный край головы у S. bicuspida посредине имеет небольшую щелевидную вырезку, а у S. marmorata эта вырезка отсутствует,"и фронтальный край головы цельный, слабо вогнутый. Эпимеральные расширения грудных сегментов у «$. bicuspida расплющенные, широкие на всех сегментах, у .S'. marmorata узкие плохо выраженные эпимеральные расширения только на первых 4 сегментах груди. У .S. bicuspida на эпимеральных расширениях концентрические валикообразные утолщения, тогда как у S. marmorata они отсутствуют и узкие эпимеральные расширения грудных сегментов гладкие. На первых 4 сегментах груди у S. bicuspida по 2 поперечных валикообразных ребра, у bicuspida спинная поверхность всех грудных сегментов гладкая. Тельзон S. bicuspida правильный треугольной формы, у .S'. marmorata — конец тельзона слегка оттянут, и вырезка на его заднем крае более глубокая и узкая. Усики 2-й пары 6'. bicuspida достигают заднего края 3-го грудного сегмента и жгутик их 14—16-членико-вый; у 5. marmorata (экземпляры из Берингова моря) усики 2-й пары достигают середины 4-го грудного сегмента и жгутик их 7— 10-члениковый. В остальное сходство этих 2 видов полное.
Распространение. Атлантический океан: Kynetarbuck bay (Лабрадор), глуб. 14 м, песок; 43°00'N, 50° 47z W, гл. 240 м; 43°36ZN, 50°Об' W, глуб. 65 м (Richardson, 1905). Берингово море: ст. 44 — 64° 20' N, 176° 23' W, глуб. 81 м в глинистом илу, 3 экз., длина 14 мм (сборы ГГИ, 1932).
* 3. Synidothea nebulosa Benedict (фиг. 91).
Benedict, 1897, Proc. Acad. Nat. Sei. Philad.: 394, f. 5.
Тело удлиненно-овальное с наибольшей шириной на 4-м грудном сегменте. Фронтальный край головы глубоко U-образно вырезан. Переднебоковые углы головы на концах фронтальной вырезки образуют маленькие тупые острия. Поверхность головы между глазами гладкая; впереди глаз имеются по одному с каждой стороны слабо выраженному бугорку. Поперечная борозда, отделяющая постцефалическую лопасть, глубокая. Глаза большие, латеральные; боковые части головы впереди глаз под прямым углом отогнуты книзу. Базальный. членик усиков 1-й пары не расширен, равен длине 2-го членика; 3-й членик в 1’/2 раза длиннее 2-го и равен последнему; в вытянутом состоянии достигают конца 4-го членика ножки
156
, 7. 1D0THEIDAE
усиков 2-й пары. Усики 2-й пары достигают заднего края 2-го сегмента; жгутик 10—12-члениковый. 4 передних грудных сегмента длиннее последних 3; 4-й несколько длиннее предыдущих. На эпимеральных расширениях имеются валикообраЗные возвышения. Тельзон удлиненный, неправильно языковидной формы; длина его составляет ]/з длины всего тела. На заднем конце широкая и очень слабая вырезка, по бокам которой края тельзона косо срезаны и закруглены. Голова и 4 передних грудных сегмента и тельзон темно окрашены, густо покрытые мелкими черными пятнышками; последние 3 грудных сегмента светлее, светлокоричневые. Максимальная длина животного 19 мм.
Распространение. Уналяска; Кизка Харбор; Унимак; о. Семиди; Б. Константина, Аляска; о. Амак (Аляска); 55° Об' N, 163° 28' Е, глуб. 10— 60 м (Benedict, 1847; Richardson, 1905). Восточный берег Камчатки, бухта Вилючинская, глуб. 13.7 м, песок, 2 экз. (сбор Камчатской станции ГГИ, 1932); западный берег Камчатки, у Болыперецка, мыс Левашева 43 экз. (Сбор ТИРХ’а, 1927).
* 4. Synidothea brazhnikovi Gurjan. (фиг. 92).
Gurjanova, 1933, Исслед. морей СССР, 17 : 97, рис. 22.
По общему строению тела и тельзона ближе всего стоит к У. Ь'си~ spida (Owen). Отличается от него слабым и тонким хитиновым покровом, более тонким расплющенным слабым телом, удлиненными конечностями, слабо развитыми плевральными расширениями, более глубокой и широкой вырезкой на конце тельзона и более длинными и тонкими усиками. Передний край головы почти прямой с глубокой маленькой вырезкой посредине.; боковые части головы впереди глаз под прямым углом отогнуты книзу. Передне-боковые углы головы слегка вытянуты вперед и заострены. Впереди глаз у фронтального края пара заостренных отростков, по одному с каждой стороны. Глаза умеренной величины, с темным пигментом, округлые и посажены не по бокам головы, а несколько сдвинуты на спинную сторону и помещаются на возвышениях, в виде бугров. Никаких бугорков на спинной стороне головы между глазами и на затылке не имеется. Усики 1-й пары достигают середины 4-го членика ножки усиков 2-й пары; базальный членик слабо расширен; 2-й членик длиннее и уже 1-го, 3-й членик длиннее 2-го; членик жгутика несколько короче последнего, 3-го членика ножки. Усики 2-й пары достигают заднего края 4-го грудного сегмента; жгутик 15-члениковый. Эпимеральные расширения 1-го грудного сегмента треугольные; 2-го и 3-го сегментов закругленные, но со следами таких же углов. У молодых экземпляров эпимеральные расширения 4 передних сегментов треугольной формы, заостренные с оттянутыми в стороны задне-боковыми углами. Грудные ножки тонкие, длинные с очень длинными и тонкими коготками, длина которых равна или несколько больше длины предпоследнего, проподиального членика ножки. Тельзон удлиненнотреугольной формы с закругляющимися боковыми краями; длина его равна
6. SYNIDOTHEA
157
длине 4 передних сегментов груди вместе; вырезка на конце тельзона глубокая, полукруглая. Цвет тела желтовато-зеленый с мелкими темными пятнами; более крупные пятна образуют 2 параллельных продольных линии вдоль спины животного, начиная от головы и до конца тельзона; по средней линии тела продольная светлая полоса; боковые края тельзона также более светлые, чем его базальная и средняя часть. Длина животного 20 мм.
Фиг. 92. Synidothea brazhnikovi Gurjan.
Фиг. 93. Synidothea erosa Benedict (no Benedict).
Распространение. Японское море: залив Петра Великого, (бухта Сидими, бухта Холуай, залив Посьет); залив Владимира; бухта Ольга, бухта Преображение, бухта Джигит; зал. Рында Татарского пролива, на глубинах от 5 до 25 м на фации песка и на камнях. Характерная для биоценоза Zostera pacifica форма.
5. Synidothea erosa Benedict (фиг. 93).
Benedict, 1897, Proc. Acad. Nat. Sei. Philad., p. 397, f. 8.
Тело удлиненное, с почти параллельными краями; длина его в 3 раза больше ширины. Голова большая, широкая; ширина ее в 2. раза больше длины. Передний край головы глубоко U-образно вырезан; передне-боковые углы головы тупые. Глаза маленькие, круглые, расположенные на спинной стороне головы, отступя от ее боковых краев. Боковые части головы слегка отогнуты книзу, образуя как бы узкий бордюр по бокам глаз. Позади срединной фронтальной вырезки 2 хорошо выраженных конических бугорка, нависающих над срединной фронтальной вырезкой. Впереди глаз также имеется по одному низкому бугорку. На спинной стороне между глазами имеется 2 выпуклости, разделенные посредине глубокой про
158
7. I DO THE! DAE
дольной бороздой. Позади всех выпуклостей низкий поперечный киль. Усики 1-й пары достигают середины 4-го членика ножки усиков 2-й пары;: базальный членик короткий, равен длине 2-го членика; 3-й членик в 1’/2 раза длиннее 2-го и равен длине последнего членика. Усики 2-й пары, длинные, тонкие; жгутик 17-члениковый. 1-й грудной сегмент немногокороче 2-го; 2-й, 3-й и 4-й сегменты равной длины. Эпимеральные расширения на первых 3 сегментах закругленные; у основания их имеется возвышение; на 1-м грудном сегменте, кроме того, имеется еще 2 низких бугорка. Тельзон удлиненно языкообразной формы, слегка суживающийся и закругляющийся дистально; на заднем его конце имеется слабая, неглубокая вырезка. Длина тельзона больше ширины его основания и равна длине 4 передних- грудных сегментов вместе. Длина животного 22 мм. Поверхность тельзона грубозернистая; у заднего края каждого из 4 передних грудных сегментов имеется по одному срединному неясному бугорку.
Распространение. Остров Саннак (Аляска) 54°00'N, 1б2°40' W, глуб. 869 м (Benedict, 1897).
б. Synidothea pallida Benedict (фиг. 94).
Benedict, 1897, Proc. Acad. Nat. Sci. Philad.: 396, f. 7.
Тело удлиненное, с плавно закругляющимися краями; длина его в 2’/2 раза больше ширины. Передний край глубоко выгнутый без срединной маленькой вырезки. Боковые края головы несколько оттянуты и образуют бордюр по бокам головы с прямыми краями. Над серединой переднего^ края нависают 2 бугорка в виде рожек, основания которых сливаются вместе. По бокам головы впереди глаз по одному маленькому, бугорку. Спинная сторона головы между глазами сильно выпуклая с продольной бороздой посредине. Глаза небольшие, круглые, посажены на спинной стороне головы, отступя от ее краев. Первые 2 членика усиков 1-й пары узкие, равной длины; 3-й членик в П/а раза длиннее 2-го и равен последнему членику; усики 1-й пары достигают конца 4-го членика ножки усиков 2-й пары. Усики 2-й пары тонкие, длинные, достигают заднего края 3-го грудного сегмента; жгутик 10-члениковый; 4 передних грудных сегмента более длинные, чем 3 задних, с закругленными задне-боковыми углами. Вдоль средней линии тела продольный ряд бугорков по одному на каждом сегменте. По бокам срединного бугорка пара небольших шипиков по одному с каждой стороны на всех грудных сегментах. Тельзон уже, чем последний грудной сегмент; удлиненной формы; длина его равна длине 4 передних, грудных сегментов вместе; к заднему концу суживается; на закругленном конце имеет небольшую вырезку, неясно выраженную. Максимальная длина животного 22 мм.
Распространение. Берингово море: у о. Чирикова (55°26'N, 155°26/W), на глуб. 1380 м (Benedict, 1897).
6. SYNIDOTHEA
159'
* 7, Synidothea tuberculata Rich. (фиг. 95).
Richardson H., 1910, Proc. U. S. Nat. Mus., 37:113, f. 35.
Тело продолговато-овальное; длина его в 2 раза больше ширины; передний край гоЛовы широко U-образно вырезан; боковые части головы вперед# глаз пбд прямым углом отогнуты книзу; передняя часть этого сгиба приподнята и образует нависающие над фронтальным краем бугорки по одному с каждой стороны впереди глаз. Позади фронтальной вырезки посредине 2 больших конических бугорка; позади их на спинной стороне головы поперечный ряд из 6 низких бугорков, по 3 с каждой стороны средней линии головы; у заднего края головы 3 бугорка, образующих 2-й поперечный ряд бугорков на голове. Глаза большие, круглые, расположенные на боковых буграх головы. Усики 1-й пары достигают конца 4-го членика ножки усиков 2-й пары. Базальный членик короткий,, равный длине 2-го членика; 3-й и 4-й членики равной длины, каждый немного длиннее 2-го. Усики 2-й пары короткие, крепкие, достигают заднего края 2-го грудного сегмента. 2 первых членика ножки их короткие, равной длины; 3-й в 2 раза длиннее 1-го; 4-й длиннее 3-го и 5-й членик самый длинный; жгутик 13-члениковый. Эпимеральные расширения 4 первых грудных сегментов с закругленными "краями; 1-й сегмент немного короче 2-го; второй сегмент равен длине 4-го, но немного короче 3-го грудного сегмента. 3 задних грудных сегмента значительно короче предыдущих, и длина их уменьшается кзади. На 1-м грудном сегменте имеется крупный бугорок посредине и по одному более вытянутому бугорку по бокам этого срединного бугорка. По средней линии следующих 3 сегментов продольный ряд бугорков, по 2 на каждом сегменте; у основания эпимеральных расширений на 2-м, 3-м и 4-м сегментах группы из 5—6 низких бугорков. По средней линии 3 задних грудных сегментов продольный ряд бугорков, по одному на каждом сегменте; по бокам его по 2 крупных бугорка, окруженных низкими валиками. Тельзон треугольной формы, резко-суживающийся дистально с маленькой U-образной вырезкой на конце. 1-я пара грудных ног хватательная, короче следующих; остальные ноги ходильные. На вторых (базальных) члениках ходильных ног на верхних поверхностях их двулопастной гребень. Последний признак отличает «S', tuberculata от всех остальных видов рода Synidothea. Максимальная длина животного 16.5 мм.
Распространение. Охотское море: 48°32'N, 145°(У7 Е; 48°32'N,145°08/E; 48°43' N, 145°03' Е; 48°43'N, 144°59'Е, глуб. 120—135 м (Richardson, 1910); 46о02' N, 143°36' Е, глуб. 135м, в илу, 2 экз., длина 12 мм (сбор ГГИ, 1932, ст. 204).
* 8. Synidothea longicirra Gurjan. (фиг. 96).
Gurjanova, 1933. Исслед. морей СССР, 17 : 95, рис. 17.
Тело удлиненное с редуцированными плевральными расширениями; длина его почти в 3 раза (в 2.7 раза) больше ширины. Передний край
160
7. IDOTHEIDAE
головы глубоко U-образно вырезан; боковые части головы впереди глаз сильно вытянуты вперед, образуя лопасти, боковые края которых отогнуты книзу почти под прямым углом. На спинной поверхности этих лопастей, немного отступя от переднего края головы, по одному маленькому коническому бугорку. Позади фронтальной вырезки по средней линии головы один хорошо развитый бугорок. Глаза небольшие, посажены по бокам головы на середине расстояния между передним и задним краями головы. Поверхность головы между глазами сильно выпуклая; постцефалическая область отделена глубокой поперечной бороздой. Усики 1-й пары достигают середины 4-го членика ножки усиков 2-й пары; базальный членик их короткий и слабо расширен. Усики 2-й пары в вытянутом состоянии достигают заднего края 4-го грудного сегмента; жгутик состоит из 20 удлиненных члеников. 1-й грудной сегмент короче 2-го и равен длине. 5-го сегмента; 2-й, 3-й и 4-й сегменты равной длины. 6-й и 7-й грудные сегменты одинаковой длины и равны длине 1-го и 5-го сегментов. Боковые края 2 передних сегментов аркообразно закруглены. Спинная поверхность грудного отдела гладкая, лишенная бугорков, шипиков и валикообразных возвышений. Тельзон треугольной формы, резко суживается дистально; ширина его основания равна ширине последнего грудного сегмента; длина тельзона немного больше ширины его основания, на конце имеется ясная маленькая U-образная вырезка. Окраска светлая, желтоватая с красными пятнами; вдоль всего тела по средней линии-широкая нежнорозовая полоса; концы жгутиков усиков 2-й пары розовые. Длина животного 26 мм.
Распространение. Охотское море: Тауйская губа (сбор Дербек 1912) 59°11'N, 148°56'Е; 54°52'N, 141°45'Е, глуб. 30 — 120 м; 58°1'N, 148°19'Е, глуб. 141 м (сбор эксп. ГГИ, 1930, ст. 138/23); ’54°34N' 142°44Е, глуб. 65 м, галька (сбор эксп. ГГИ, 1932, ст. 244).
9. Synidothea acuta Rich. (фиг. 97).
Richardson Н., 1910. Proc. U. S. Nat. Mus., 37: 111, f. 32.
Тело продолговато-овальное; длина его в 2 раза больше ширины. Передний край головы почти прямой, слабо вогнутый с маленькой вырезкой посредине. Боковые части головы впереди глаз отогнуты книзу под прямым углом; край сгиба впереди образует конический бугорок, расположенный у самого фронтального края. Позади срединной фронтальной вырезки один конический бугорок. Глаза маленькие, круглые, посаженные близко к боковому краю головы. Усики 1-й пары достигают середины 4-го членика ножки усиков 2-й пары; первые 2 членика равной длины; 3-й членик немного длиннее 2-го и равен 4-му. Усики 2-й пары сравнительно короткие и крепкие; в вытянутом состоянии достигают заднего края 1-го грудного сегмента; жгутик 10-члениковый; членики его короткие и широкие. Боковые края 2 первых грудных сегментов образуют углы; боковые края остальных грудных сегментов прямые. Эпимеральные рас
6. SYNIDOTHEA
161
ширения редуцированы. Первые 3 грудные сегмента равной длины; 4-й немного длиннее. 3 последних сегмента груди последовательно укорачиваются к заднему концу тела. Тельзон резко суживается дистально, треугольной формы; длина его равна ширине основания. Базальная часть тельзона не уже последнего сегмента груди. На дистальном конце слабая вырезка. Цвет тела желтоватый; 3-й и 4-й сегменты красные. Длина ^животного 12 мм.
Распространение. Берингово море у Семисопочного острова (52O12ZN, 179°52ZE), глуб. 86 м. (Richardson, 1910).
'Фиг 95. Synidothea tuberculata Rich.. (по Richardson).
Фиг. 96. Synidothea longicirra Gurjan.
Фиг. 97. Synidothea acuta Rich, (no Richardson).
Фиг. 98.‘ Synidothea epimerata Rich, (n Richardson).
* 10. Synidothea epimerata Rich. (фиг. 98).
Richardson, 1910, Proc. U. S. Nat. Mus., 37 :112, f. 33—34.
От всех известных видов Synidothea резко отличается присутствием маленьких свободных эпимер на 1-м грудном сегменте. Тело удлиненноовальное; длина его немного больше, чем в 2 раза, превосходит ширину. Передний край головы полукругло вогнутый и посредине его имеется еще небольшая закругленная вырезка над основанием усиков 1-й пары. Передне-боковые. углы головы вытянуты вперед и сильно заострены. Глаза большие, овальные, расположены на спинной стороне близко к боковым краям головы. Впереди глаз по одному заостренному бугорку. Позади срединной вырезки фронтального края 1 конический бугорок и кроме того близко у Заднего края головы поперечный ряд из нескольких мелких бугорков. Постцефалическая область головы отделена глубокой поперечной бороздой и снабжена одним бугорком посредине. Усики 1-й пары достигают середины 4-го членика ножки усиков 2-й пары. Базальный членик не расширен и равен длине 2-го членика; усики 2-й пары
Гурьянова ’ 11
162
7. IDO THEIDAE
длинные, тонкие, достигают заднего края 3-го грудного сегмента; 2 первых членика ножки короткие, равной длины; 2 следующих членика также, равной длины и каждый в 2 раза длиннее 1-го; 5-й членик в V/a раза длиннее 4-го. Жгутик 12-члениковый. 1-й грудной сегмент короче и значительно уже следующих сегментов груди; боковые края его образуют угол. Передние края его вогнутые и снабжены маленькими округлыми эпимерами,, прикрепленными с нижней стороны, но хорошо видные сверху. Боковые, края 2-го сегмента также образуют углы; у 3-го сегмента закруглены, а у остальных прямые. 2-й, 3-й и 4-й сегменты равной длины; 3 задние сегмента последовательно укорачиваются кзади. Вдоль средней линии тела тянется ряд бугорков, по одному на каждом сегменте груди. Тельзон с закругленными краями, суживается дистально, удлиненно-треугольной формы. Длина его равна длине первых 4 грудных сегментов вместе и больше ширины его основания; на дистальном конце ясная полукруглая вырезка. Цвет животного песочно-серый с темными и красными пятнами.. Длина 13 мм.
Распространение. Японское море: залив Петра Великого (Амурский залив, залив Америка, залив Посьет, у о. Фурругельма, Сивучья бухта, открытая часть залива), глуб. 20—80 м. Форма очень характерная для фации песка и для камней; держится обычно массами (сборы ГГИ и ТИРХ’а, 1925—1932).
* 11. Synidothea excavata Gurjan. (фиг. 99).
Gurjanova, 1933, Исслед. морей СССР, 17 : 97, рис. 23.
Тело удлиненное; длина его в 2 раза больше ширины; голова очень-большая, равная длине 2 первых сегментов вместе. Передний край головы вогнутый с небольшой вырезкой посредине. Передне-боковые углы заострены; глаза большие, округлые, посажены по бокам головы; боковые части головы впереди глаз отогнуты книзу под прямым углом; ребро сгиба впереди образует с каждой стороны по одному коническому бугорку. Спинная сторона головы сильно выпуклая, совершенно гладкая; постцефалическая часть отделена поперечной бороздой. Усики 1-й пары достигают конца 4-го членика ножки. Усики 1-й пары достигают конца 4-го членика ножки усиков 2-й пары; базальный членик их слабо расширен и немного длиннее 2-го; 3-й членик равен длине 1-го. Усики 2-й пары достигают середины 3-го грудного сегмента; жгутик 11-члениковый;. 1-й и 2-й членики ножки равной длины; 5-й членик в 2 раза длиннее 1-го; 3-й членик немного длиннее 2-го и равен 4-му. 1-й грудной сегмент короче и уже 2-го; передние края его по бокам головы вогнутые; 2-й, 3-й и 4-й сегменты равной длины; боковые края 2-го сегмента закругленные, у остальных сегментов прямые; на спинной стороне 2-го, 3-го и 4-го сегментов посредине по одному округлому бугорку. Последние 3 сегмента груди последовательно укорачиваются к заднему концу тела. Тельзон очень большой, удлиненно-треугольной формы с закругленными.
6. S YNIDO THEA
163
краями; длина его равна длине первых 4 сегментов и половине 5-го вместе. На дистальном конце глубокая полукруглая вырезка. Все тело плотное, крепкое; цвет буро-серый с более интенсивно окрашенными головой и тельзоном. Длйна 12 мм.
Распространение. Северная часть Японского моря, бухта Жонкьер (сев. Сахалин) в Татарском проливе, 1 экз. (сбор Полякова, 1881); на набережной о. Сахалина (точнее нет указания) 1 экз. (сбор Полякова, 1881).
Фиг. 99. Synidothea ехса- Фиг. 100. Synidothea har-vata Gurjan. fordi Bened. (no Benedict).
* 12. Synidothea harfordi Benedict (фиг. 100).
Фиг. 101. Synidothea nodulosa Kriyer.
B enedict, 1897, Proc. Acad. Nat. Sci. Philad.: 402, f. 1. — marmorata' Harford, 1877, Proc. Acad. Sci. Calif., VII: 117 {Idotea ).
Тело удлиненно-овальное, длина его в 22/8 раза больше ширины; голова большая, округлая; длина ее равна длине 2 первых сегментов груди вместе; ширина головы в 1х/2 раза больше ее длины. Передний край головы прямой, лишенный срединной вырезки. Глаза большие, черные, посажены по бокам головы; спинная сторона головы выпуклая, гладкая, лишенная бугорков и шипиков. Усики 1-й пары достигают конца 3-го членика ножки усиков 2-й пары; базальный членик слабо расширен, короткий; 2-й и 3-й членики равной длины, каждый немного длиннее 1-го; 4-й членик в 2 раза длиннее 3-го. Усики 2-й пары тонкие, длинные, в вытянутом состоянии достигают середины 7-го грудного сегмента; жгутик из 31 членика; 1-й членик ножки короткий, 2-й в 2 раза длиннее 1-го; 3-й немного длиннее 2-го; 4-й в 2 раза длиннее 3-го, а 5-й, самый длинный, в 1% раза длиннее 4-го. 1-й грудной сегмент короткий, в 2 раза короче И*
164
7. IDOTHE1DAE
2-го по средней линии; 2-й, 3-й и 4-й сегменты почти равной длины; 3. задних сегмента короткие, равной длины; эпимеральные расширения слабо выражены. Тельзон удлиненный, слегка суживается дистально; длина его больше ширины его основания и равна длине 5 передних сегментов груди вместе. На заднем конце тельзона неглубокая полукруглая вырезка. Поверхность тела совершенно гладкая без бугорков, шипиков и валикообразных возвышений; середина передней части 2-го, 3-го и 4-го грудных сегментов отделена полукруглой бороздой. Цвет тела буровато-серый, иногда с мраморным рисунком и светлой продольной полосой по средней линии грудного отдела и основания тельзона. Длина животного достигает 18 мм.
Распространение. Калифорния, бухта Магдалена и Сан-Диего (Richardson, 1905). Японское море: залив Петра Великого (Амурский залив, бухта Сидими, у р. Перевозной; залив Посьет, бухта Тавайза), глуб. О—10 м, на песчаном грунте и на устричниках (сборы ГГИ, 1925— 1928). Хакодате у мола на песке, глуб. 8 м (сбор Державина, 1928), в заливе Исе на устричнике глуб. 0.5 м (сбор ТИРХ’а 1928).
* 13. Synidothea nodulosa (Kroyer) (фиг. 101).
Kroyer, 1846, Naturh. Tidsskr. (2), 11: 100 (TdoiAea), Kroyer, 1849, Voy. Scand., Crusty pl. XXV, f. 2 (Idothea); Harger, 1878, Amer. Journ. Sei., 131 XV: 374; Benedict, 1897, Proc. Acad. Nat. Sci. Philad.: 398.
Тело продолговато-овальное, длина его немного больше чем в 2 раза превосходит ширину. Передний край головы слегка вырезан посредине. Глаза небольшие, круглые, расположены по бокам головы. Впереди глаз, немного отступя от переднего края головы по одному коническому бугорку. Между глазами на спинной поверхности ромбическое поле с 4 бугорками по его углам. Голова позади этого ромбического поля сильно выпуклая с легкой продольной бороздой посредине. Постцефалическая область головы отделена глубокой бороздой и несет 1 бугорок посредине. Усики 1-й пары достигают конца 4-го членика ножки усиков 2-й пары; базальный членик короткий, равен длине 2-го членика; 3-й членик в lJ/2 раза длиннее 2-го, а 4-й в 2 раза длиннее 2-го членика. Усики 2-й пары короткие, достигают только середины 2-го грудного сегмента; жгутик 6-члениковый; первые 2 членика ножки короткие, равные-3-й и 4-й равной длины, каждый в 2 раза длиннее 2-го; 5-й членик в 11/2 раза длиннее 4-го. Первые 4 сегмента груди равной длины; 3 задних сегмента Короче и последовательно укорачиваются к заднему концу тела. Эпимеральные расширения на всех грудных сегментах хорошо выражены, особенно на перзых 4; по средней линии тела продольный ряд бугорков, по одному на каждом сегменте; иногда эти бугорки сильно редуцированы и едва заметны. На эпимеральные расширениях группы валикообразных возвышений, чаще всего концентрических или же (в 2 ряда) серповидные. Тельзон в общем треугольной формы, с утончающимся заостренным
6. SYNIDOTHEA
165
дистальным концом. Длина тельзона в l1/^ раза больше ширины его основания. Цвет тела желтовато-сероватый, часто с красными пятнами. Длина тела до 14 мм.
Распространение. Восточный сектор Арктики от Новой Земли до Берингова пролива; вдоль западного берега Северной Америки до Британской Колумбии, глуб. 10—198 м (Richardson, 1899, 1901). У Алеутских островов из желудка кита, 2 экз. (сбор Вознесенского, 1846).
14. Synidothea laevis Benedict (фиг. 102).
Benedict, 1897, PrOc. Acad. Nat. Sci, Philad. : 399.
Тело узкое, удлиненное; длина его немного больше чем в 3 раза превосходит ширину. Передний край головы широко U-образно вырезан
Фиг. 102. Synidothea laevis Bened. (по Benedict).
Фиг. 103. Synidothea muricaia Harf, (no Benedict).
с небольшой щелевидной вырезкой посредине, между основаниями усиков 1-й пары. Позади этой срединной вырезки треугольное поле с 3 бугорками по углам. Боковые части головы впереди глаз отогнуты книзу почти под прямым углом и передний край сгиба образует рожковидные бугорки по одному с каждой стороны, нависающие над фронтальным краем. Глаза небольшие, округлые, посажены по бокам головы на середине расстояния между передним и задним краями головы. Затылочная часть выпуклая, разделенная посредине продольной бороздкой; постцефалическая лопасть отделена глубокой бороздой. Усики 1-й пары достигают середины 3-го членика ножки усиков 2-й пары; базальный членик не расширен, короткий, равен длине 2-го членика. Усики 2-й пары сравнительно короткие, достигают средины 3-го грудного сегмента; жгутик 10-члениковый; 1-й членик ножки очень короткий, невидимый сверху; 3 следующих членика равной длины; 5-й в 1х/2 раза длиннее 4-го. Первые 4 грудных сег
166
7. IDOTHEIDAE
мента равной длины, с резко выраженными широкими закругленными эпимеральными расширениями; боковые края 3 последних, более коротких сегментов, слабо закруглены. Тельзон удлиненно-треугольной формы с узким оттянутым в тупое острие задним концом; длина его равна длине 5 последних сегментов груди- вместе, и в l1^ раза больше ширины его основания. Иногда по средней линии тянется продольный ряд неясных бугорков по одному на каждом сегменте. На эпимеральных расширениях неясные валикообразные возвышения. Цвет серый с темными и красными пятнами. Длина животного 15 мм.
Распространение. Берингово море: между Бристольской бухтой и Прибыловыми островами: 57°22' N, 1б4°24' W, глуб. 52 м; 57°05' N, 164°27'W, глуб. 64 м (Benedict, 1897). Бухта Лаврентия (Мечиг-мен), 1 экз.
* 15. Synidothea muricata (Harford) (фиг. 103).
Harford, 1876, Proc. Acad. Sci.'Calif., VII: 117 (Wotaea); Richardson, 1905, Bull. U. S. Nat. Mus., n° 54:390, f. 433-434.
Тело удлиненно-овальное; длина его в 3 раза больше ширины; передний край головы глубоко U-образно вырезан и передне-боковые углы заострены. Боковые части головы отогнуты книзу почти под прямым углом. Глаза небольшие, округлые, посажены по бокам головы на сере-дийе расстояния между передним и задним краями головы. Позади фронтальной вырезки между глазами, немного впереди них ромбовидное поле с коническими бугорками по углам ромба. Впереди глаз по бокам головы пара нависающих над фронтальным краем рожковидных выростов. Затылочная часть позади ромбовидного поля бисквитообразно выпуклая с бугристой поверхностью. Постцефалическая часть отделена глубокой бороздой и несет 3 бугорка, расположенных в 1 поперечный ряд. Усики 1-й пары достигают конца 4-го членика ножки усиков 2-й пары; базальный членик расширенный, почти в 2 раза шире 2-го членика, но равен его длине; последние 2 членика равной длины, каждый в 2 раза длиннее 2-го. Усики 2-й пары достигают заднего края 3-го грудного сегмента; 1-й членик ножки короткий, невидимый сверху; 3 следующих членика последовательно увеличиваются в длине; 2-й членик несет конический бугорок на наружном дистальном углу; 5-й членик в 2 раза длиннее 4-го; жгутик 13-члениковый. 1-й грудной сегмент немного короче и уже 2-го; 3 следующих сегмента равной длины; последние три сегмента груди последовательно укорачиваются к заднему концу тела. Эпимеральные расширения хорошо развиты на всех сегментах груди; на первых 4 сегментах с закругленными углами. Вдоль средней линии тела ряд бугорков по одному на каждом сегменте тела; по бокам от этого срединного ряда бугорков группы небольших конических шипиков, по 4—5 в каждой группе. Тельзон узкий, вытянуто-треугольной формы, заостряющийся дистально. Длина его равна длине 4 передних сегментов груди вместе и в 2 раза
6. SYNIDOTHEA
167
•больше ширины основания; базальная часть тельзона на спинной стороне покрыта небольшими шипиками и зернистостью; поверхность дистального конца гладкая. Максимальная длина животного 22 мм.
Распространение. Чукотское море и море Бофора, 70°15'N, 162°55'W; 41°03'N,154°15' W, глуб. 45 м (Benedict 1847). Охотское море: экспед. ГГИ, 1932, ст. 249 — 56°24/ N, 143°18л Е, глуб. 100 м, 1 экз., длина 16 мм.
* 16. Synidothea spinosa Gurjan. (фиг. 104).
Gurjanova, 1933, Исслед. морей СССР, 17 : 96, рис. 20—21.
Фиг. 104. Synidothea spinosa Gurjan. а — общий вид; 6 — antenna И; с — antenna I.
Тело вытянутое с почти параллельными боковыми краями; длина его в 3 раза больше ширины; вся спинная поверхность головы, грудных сегментов и тельзона покрыта крупными коническими шипами; боковые части головы отогнуты книзу под прямым углом; глаза небольшие, круглые, латеральные. Передний край головы глубоко U-образно •вырезан. Впереди глаз у самого переднего края головы по одному крупному отростку с каждой стороны, образующих рожки. Эпимеральные расширения грудных сегментов слегка вздуты на конце, конической формы. Шипы на грудных сегментах расположены правильными поперечными рядами; центральные 3 шипа каждого ряда крупнее всех остальных. Усики 1-й пары достигают конца 4-го членика ножки усиков
2-й пары. Базальный членик не расширен, цилиндрический; 2-й членик короче базального; 3-й членик равен длине 1-го и 4-го члеников. Усики 2-й пары в вытянутом состоянии достигают заднего края 3-го грудного сегмента; жгутик 11-члениковый; первые 2 членика ножки короткие, равной длины; 3-й членик равен длине 1-го и 2-го члеников вместе и длине 4-го членика; 5-й в 1х/2 раза длиннее 4-го. 4 передних грудных сегмента равной длины; 3 последних сегмента последовательно укорачиваются к заднему концу тела. Тельзон в общем треугольной формы, резко суживающийся дистально. Задний конец заострен. Длина тельзона равна 3 передним и половине 4-го грудного сегмента вместе и почти в 2 раза больше ширины его основания. Шипы только на спинной стороне проксимального конца тельзона; дистальный конец гладкий. От предыдущего вида отличается резко очень крупными шипами и их расположением конической
168 7. IDOTHEIDAE
формой эпимеральных расширений и более коротким тельзоном- Максимальная длина 16 мм. Цвет в спирту желтоватый.
Распространение. Охотское море: 54°33ZN, 140°20'Е;; 59°17' N, 145°40' Е, глуб. 88—95 м (сбор Дербека, 1910, ст. 16 и 14). Берингово море: 64o20, N, 176°23/W, глуб. 81 м, ил, 1 экз., длина. 10 мм; 63°29'N, 176°37'W, глуб. 74 м, ил, 1 экз., длина 13 мм (сбор эксп.. ГГИ, 1932, ст. 14 и 13).
17. Synidothea picta Benedict (фиг. 105).
Benedict, 1897, Proc. Acad. Nat. Sci. Philad.: 401, fig-. 11—12.
Тело овально-удлиненное; длина его в 2% раза больше ширины.. Передний край головы широко U-образно вырезан. Позади фронталь-' ной вырезки посредине
Фиг. 105. Synidothea picta Bened. Крайние формы вариационного ряда (по Benedict). Правая—ив Берингова пролива
крупный конический заостренный бугорок и по бокам его по одному маленькому тупому бугорку. Все 3 бугорка расположены в один поперечный ряд между глазами. Впереди глаз у самого фронтального края по одному рожковидному бугорку. Глаза большие, округлые, расположены по бокам головы на середине расстояния между передним и задним краями головы.. Затылочная часть го-
ловы выпуклая и покрыта 2 бугристостями по обе стороны от средней линии. Постцефалическая часть головы отделена глубокой бороздой; на
спинной ее поверхность поперечный низкий гребень с бугорком посредине. Усики 1-й.пары достигают конца 3-го членика ножки усиков 2-й пары;: базальный членик их короткий, не расширенный; 2-й и 3-й членики равной длины; последний членик немного длиннее 3-го. Усики 2-й пары сравни
тельно укороченные; в вытянутом состоянии достигают только середины 3-го грудного сегмента; членики ножки последовательно удлиняются от 1-го к 5-му; жгутик 10-члениковый. 4 передних грудных сегмента равной, длины; 3 задних последовательно укорачиваются. Эпимеральные расширения хорошо развиты, с закругленными краями и углами. По средней линии тела продольный ряд бугорков по одному на каждом сегменте; на задних сегментах эти бугорки крупнее, чем на передних. По бокам срединного бугорка на каждом сегменте группы низких бугорков, по 6 бу—
6. S YN1D0 THEA
169
Фиг. 106. Synidothea cinerea Gurjan. а —общий вид; Ь — antenna II: с—antenna I.
горков в каждой группе. Тельзон удлиненно-треугольной формы, с вытянутым тупо-заостренным концом. Длина его превышает длину 4 передних сегментов груди вместе й в l1/^ раза больше ширины его основания. У основания тельзона посредине 2 небольших бугорка и по бокам их грубые зернистости. Дистальный конец тельзона гладкий. Длина достигает 16 мм.
Benedict (1897) указывает на нахождение в Беринговом проливе особой формы S. picta, отличающейся от типичных экземпляров, добытых в Беринговом море, более коротким и широким каудальным сегментом, коренастым телосложением и грубым панцырем.
Распространение. Берингово море: мысЛисбурн; Norton Sound; Берингов пролив; 55°О6 N, 163°28'W, глуб. 18 м (Benedict, 1897).
* 18. Synidothea cinerea Gurjan. (фиг. 106).
Gujanova, 1933, Исслед. морей СССР, 17:9, рис. 18—19.
Тело продолговато - овальной формы; длина его в 2.7 раза больше ширины. Передний край головы широко U-образно вырезан с небольшим углублением посредине. Бугорки впереди глаз развиты плохо, маленькие, не достигающие своей вершиной фрон
тального края; по средней линии головы, позади фронтальной вырезки низкий килевидный бугорок и позади него еще 1 маленький округлый бугорок. Такой же бугорок у заднего края затылочной области. Глаза небольшие, округлые, по бокам головы несколько сдвинуты на спинную сторону. Боковые части головы образуют крупные .лопасти, отогнутые книзу. Усики 1-й пары немного выступают за дистальный конец 3-го членика ножки усиков. 2-й пары; базальный, членик расширенный, немного длиннее 2-го членика, и по краям покрыт волосками; 3-й членик немного длиннее 1-го, а 4-й равен длине 2-го членика. Усики 2-й пары достигают середины 3-го грудного сегмента; первые два членика ножки короткие, равной длины; 3-й членик равен длине 2 предыдущих вместе; 4-й членик немного длиннее 3-го;: 5-й членик почти в 1У2 раза длиннее 4-го; жгутик 10—13-члениковый. 1-й грудной сегмент короче и уже 2-го сегмента, следующие 3 сегмента’ равной длины; 3 последних грудных сегмента последовательно укорачиваются. Эпимеральные расширения первых 4 сегментов с закругленными краями. Поверхность тела покрыта тонкой зернистостью. Тельзон пятиугольной формы с тупым острием на конце. Длина его почти равна длине 4 первых грудных сегментов вместе и значительно превышает ширину его основания. Вдоль средней линии тела ряд небольших заострен
170 7. IDOTE1DAE
ных бугорков, по одному на каждом сегменте у его заднего края; 2 таких же бугорка у основания тельзона. Членики ножки усиков 2-й пары покрыты длинными волосками. Цвет тела розовато-серый с темными и мелкими красными пятнами. Длина тела 26 мм.
Распространение. Охотское море, 53°37'N, 140°30'Е, глуб. 22 м, на песчаном грунте, 14 экз. (сбор. Павленко и Солдатова, 1913, ст. 404); Тауйская губа, бухта Куэгда, глуб. 12—15 м, галька (сбор. ГГИ, 1930).
7. Род CLEANTIS DANA
Dana, 1849, Amer. Journ. Sei. Arts, (2), 8: 427.
Тип рода: C. linearis Dana.
Тело узкое, вытянутое, более или менее равномерно расчлененное, с параллельными боковыми краями. Глаза маленькие, латеральные; щупик ногочелюстей состоит из 5 члеников. Членики усиков 1-й пары очень короткие, редуцированные. Членики жгутика усиков 2-й пары, сливаясь вместе, образуют 1 крупный членик со следами слияния в виде вдавлений и пучков волосков на границах слившихся члеников. Эпимеры на всех сегментах, кроме 1-го, свободны, узкие, но хорошо заметные. Брюшной отдел состоит из нескольких (3—5) сегментов.
В дальневосточных морях известно 2 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Тельзон языковидный, закруглен на конце . *1.С. planicauda Bened. ~2 (1). Тельзон пятиугольной формы с тупым концом . * 2. С. isopus Grube.
* Г. Cleantis planicauda Benedict (фиг. 107).
Benedict in Richardson, 1905, Bull. U. S. Nat. Mus., n° 54: 404, f. 4, 452—454; Richardson, 1905, Bull. U. S. Nat. Mus., 54: 404, f. 452—454.
Тело линейно вытянутое с параллельными боковыми краями. Длина его в 5 раз больше ширины; грудные сегменты не шире головы. Поверхность тела зернистая. Голова почти квадратная, погружена в 1-й грудной сегмент; задний край ее закруглен, передний край слегка вогнут посредине. Глаза расположены по бокам головы близко к переднему краю. По середине лобной части небольшое вдавление, доходящее по средней линии до средины головы. Постокципитальная лопасть ясная. Усики 1-й пары очень короткие, 4-члениковые; базальные членики их короткие и расширенные; следующие членики последовательно укорачиваются и делаются уже; последний членик маленький с пучком коротких волосков на вершине. Усики 2-й пары крепкие, короткие, 6-члениковые. 1-й членик ножки очень короткий, неподвижный; 2-й короткий и крепкий, 3-й равен длине 2-го членика, но более тонкий; 4-й и 5-й членики равной длины, каждый немного
7. CLEANTIS
171
длиннее 2-го. Последний членик, жгутик, равен длине 5-го членика ножки; все членики жгутика слились вместе, образуя только 1 (б-й членик усиков 2-й пары) членик со следами слияния в виде коротких щетинок на границах слившихся члеников. Длина усиков 2-й пары равна длине головы и 2 передних сегментов груди вместе. Усики 1-й пары достигают середины 3-го сегмента усиков 2-й пары. Все сегменты груди почти равной длины и ширины. Эпимеры свободные на всех сегментах, кроме 1-го, но маленькие, узкие и видны только при рассматривании животного сбоку.
Л
Фиг. 107. Cleantis planicauda Rich, (по Richardson). а — общий вид; Ь — maxillipedes; с — antenna I; d — antenna И.
Фиг. 108. Cleantis isopus Grube (no Miers).
Эпимеры последних 3 сегментов значительно крупнее. Задне-боковые углы их оттянуты назад и заострены; выходят за пределы заднего края своего сегмента. Брюшной отдел состоит из 4 сегментов —3 очень коротких передних брюшных сегментов и тельзона, у основания которого по бокам сохранились следы слияния сегментов в виде пары коротких швов. Длина тельзона равна длине 31/2 передних сегментов вместе; боковые края его прямые, параллельные друг другу; задний край правильно закруглен. Спинная сторона тельзона сводчато выпуклая, но задний конец его приплюснут, образуя косо срезанную круглую площадку, ограниченную задним краем тельзона и закругленной линией, идущей через спинную поверхность тельзона полукругом и замыкающей полный круг при встрече с задним краем. 3 передних пары грудных ног хватательные, более крепкие, чем последние 4 пары ходильных ног. Величина ног возрастает от 1-й пары до 3-й, самой крупной; 4-я пара короткая и слабая; 5-я, 6-я и 7-я пары ног снова последовательно возрастают в размерах. Базальный членик рулевых ног, образующих двустворчатую крышку на брюшной стороне тельзона, не имеют диагонального киля.. Цвет тела
172
7. IDO TH El DAE
желтовато-розоватый, co светлым задним краем тельзона и концами: усиков; вдоль спины тянутся 4 тонких черных линии. Кроме того 2 широких черных полосы, по одной с каждой стороны тянутся по бокам тела у самого-его края, захватывая и эпимеры. Длина животного 15 мм.
Распространение. Атлантический океан: Вест-Индия (Пенсакола, Флорида и Porto-Rico). Японское море: бухта Сидими,. глуб. 3—6 м; залив Посьет, бухта Постовая, глуб. 3 м (сборы Тихоок. инет. рыб. хоз., 1927 и 1928), Нагасаки (сбор Полякова, 1883).
Один из ярких примеров амфибореального распространения.
* 2. Cleantis isopus Grube in Miers (фиг. 105).
Grube in Miers, 1883, Journ. Linn. Soc. London, 16: 80, pl. Ill, f. -9—11.
Тело вытянутое с параллельными боковыми краями; длина его-в 3.2 раза больше ширины; по средней линии грудного отдела продольный,, ясно выраженный киль; поверхность тела зернистая. Голова глубоко погружена; задняя половина ее охвачена по бокам выростами 1-го грудного сегмента; передний край головы- глубоко вогнутый; затылочная часть выпуклая; глаза поперечно-овальные, посажены по бокам головы ближе к переднему краю. Усики 1-й пары короткие, достигают середины 3-го членика ножки усиков 2-й пары; базальный членик расширенный и удлиненный; 3 последних членика короткие. Усики 2-й пары достигают заднего края 3-го грудного сегмента; 1-й членик ножки очень короткий, неподвижный, 2-й членик равен длине 3-го и снабжен килем вдоль верхнего-внутреннего края; 4-й членик равен длине 2-го и 3-го члеников вместе;. 5-й равен длине 4-го. Жгутик одночлениковый, образовавшийся слиянием всех члеников жгутика вместе, слегка расширяется дистально с закругленной вершиной; длина жгутика почти в l1^ раза больше длины последнего (5) членика ножки. Сегменты груди шире, чем голова, одинаковой ширины и длины, кроме 1-го сегмента, длина которого по средней линии в 2 раза меньше длины остальных. Передние боковые углы 1-го сегмента вытянуты, вперед по бокам головы и образуют закругленные лопасти, плотно охватывающие голову. Эпимеры на всех сегментах, кроме 1-го, свободны, очень маленькие и узкие, занимают лишь переднюю часть бокового края сегментов; только эпимеры последнего грудного сегмента занимают всю длину бокового края сегмента. Брюшной отдел состоит из 2 сегментов — одного очень короткого, свободного брюшного сегмента и тельзона; у основания тельзона по бокам 2 пары швов, указывающих на слияние с тельзоном еще 2 брюшных сегментов. Тельзон большой, пятиугольной формы, слегка суживающийся дистально; ширина его основания равна ширине последнего грудного сегмента; длина его равна длине 4 передних грудных сегментов вместе. Спинная поверхность тельзона плоская со слабым срединным килем. Базальные членики рулевых ног не имеют диагонального киля.. Трудные ножки слабые, одинаковой структуры. Все тело равномерно окрашено в светлобурый или же в очень темный, почти черный цвет..
7. CLEANTIS
173
Длина животного достигает 18 мм (в заливе Петра Великого) и 26 мм <(из Мисаки).
Распространение. Китайское море: побережье Чифу; 33°12/N, 129°5' Е; литораль Японского побережья — Хакодате, Мороран, Ожика, о. Гото (Richardson, 1910). Японское море: залив Петра Великого (бухта Патрокл, Б. Посьет, Уссурийский залив), между реками Перевозной и Мангугаем, глуб. 2—4-5 м, в зарослях водорослей, на устричниках (сборы ГГИ, 1926—1928); литораль Мисаки (сборы Рожковского, 1917).
8. Сем. ARCTURIDAE
Тело узкое, вытянутое, иногда цилиндрическое. Сегменты тела различной формы и размеров; часто 4-й сегмент значительно превосходит длину всех остальных грудных сегментов. Усики 1-й пары короткие^ с одночлениковым жгутиком; усики 2-й пары длинные, мощные, часто превышают длину тела животного, жгутик многочлениковый, короткий; 2 последних членика ножки очень длинные. 1-й грудной сегмент неподвижно сливается с головой. Ротовые части как у сем. Idotheidae", жвалы лишены щупика. 4 передние пары ног усиковидные, покрыты длинными щетинками и направлены вперед; 3 задние пары грудных ног короткие, крепкие, ходильные. 1-я пара грудных ног очень маленькая, сильно приближена к ротовым частям, по форме и строению похожа на ногочелюсти. 2 передние пары брюшных ног плавательные, остальные служат только для дыхания. Рулевые ноги в виде створок накрывают снизу брюшные ножки. Самец обычно мельче самки, с более вытянутым телом. Брюшной отдел из различного числа сегментов, которые иногда сливаются друг с другом или с тельзоном. На Дальнем Востоке семейство представлено 3 родами.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). 4-й грудной сегмент много длйннее всех остальных грудных сегментов; инкубаторный карман самки образован 3 или 4 парами инкубаторных пластинок..................3. Astacilla Cord. (стр. 202).
2 (1). 4-й грудной сегмент немного длиннее остальных; инкубаторный карман самки образован 5 парами инкубаторных пластинок.
3 (4). Брюшной отдел состоит из 3 ясно ограниченных сегментов — 2 передних и каудального (тельзона).............1. Arcturus Latr. (стр. 173).
.4 (3). Брюшной отдел состоит из 2 сегментов, 1 короткого свободного
брюшного сегмента и тельзона . 2. Pleuroprion zur Strassen (стр. 197).
1. Род ARCTURUS LATREILLE
Latreille, 1829, in Cuvier, Regne Animale, ed. 2. IV: 139.
Тип рода: A. baffini (Sab.).
Тело узкое, длинное, цилиндрическое; грудные сегменты часто снабжены длинными шипами и отростками. Все сегменты груди более или менее
174
8. ARCTURIDAE
равных размеров, 4-й грудной сегмент лишь немного больше остальных-Абдомен состоит из 3 сегментов — 2 свободных передних брюшных сегментов и тельзона. Эпимеры небольшие, свободные на всех сегментах,, кроме 1-го. Инкубаторный карман самки образован 5 парами инкубаторных пластинок. Щупик ногочелюстей 5-члениковый. Усики 1-й пары маленькие? усики 2-й пары очень длинные, часто превышающие длину тела животного;, последние членики ножки очень длинные, цилиндрические; жгутик относительно короткий, многочлениковый.
В дальневосточных морях известно 16 видов и две формы этого рода.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (24). На спинной стороне грудных сегментов имеются крупные парные шипы.
2(15). Конец тельзона раздвоен.
3 (Ю). Парные спинные шипы на всех 7 грудных сегментах и на каждом: брюшном сегменте.
4 (7). На наружном дистальном углу 2-го членика ножки усиков 2-й пары имеется шипик.
5 (6). Усики 2-й пары очень длинные, в 1У2 раза длиннее тела животного . . ..........................................*5. A. setosus Gurjan-
6 (5). Усики 2-й пары лишь немного длиннее тела животного.........
.........................* 5а. A. setosus var. seminudus Gurjan-
7 (4). На наружном дистальном углу 2-го членика усиков 2-й пары нет шипика.
8 (9). Членики усиков 2-й пары гладкие; спинные шипы тела очень длинные, длиннее высоты сегмента.................7. A. magnispinis Rich.
9 (8). Членики усиков 2-й пары покрыты волосками; спинные шипы тела небольшие, короче, чем высота сегмента . . * 8. A. crassispinis Rich-
10 (3). Некоторые грудные или брюшные сегменты лишены парных спинных шипов.
11 (12). На первом грудном сегменте нет спинных шипов..............
. .................................. . 4. A. longispinis Bened-
12 (11). На спинной стороне 1-го грудного сегмента- есть пара крупных шипов.
13 (14). Тело животного и усики покрыты волосками.* 6. A.diversispinis Rich-14 (13). Тело животного и усики лишены волосков, но покрыты грубой
зернистостью...............................* 9. A. brevispinis Rich-
15 (2). Конец тельзона не раздвоен, а заострен, или же слегка закруглен и снабжен длинными шипами.
16 (21). Тельзон заострен на конце.
17 (18). Поверхность тела гладкая, лишенная волосков . ...........
........................................ * 2. A. ulbani Gurjan-
18 (17). Поверхность тела и спинные шипы покрыты волосками.
1. ARCTURUS
175-
19 (20). На передне-боковых углах головы по одному шипику; спинные шипы. тонкие, длинные, превышающие высоту самого сегмента . .
...........*...........*5 b. A. setosus var. acuticaudalis Gurjan.
20 (19). На передне-боковых углах головы нет шипиков; спинные шипы укороченные, меньше, чем высота сегмента . . . *1. A. baffini Sab.
21 (16). Тельзон на конце закруглен и снабжен длинными шипами.
22 (23). Шипы, покрывающие тело, несут пучки волосков в виде розеток на их вершинах; апикальные шипы тельзона покрыты волосками .
......................................... 15. A. hirsutes Rich.
23 (22). Шипы, покрывающие тело, и апикальные шипы тельзона гладкие, без волосков.................................* 16. A. beddard! Gurjan.
24 (1). Спинная сторона грудных сегментов гладкая или несет небольшие низкие бугооки. •
25 (30). Тельзон раздвоен на конце.
26 (29). Поверхность тела гладкая; бугорки на грудных сегментах и на тельзоне отсутствуют.
27 (28). Нижний край 4-го членика ножки усиков 2-й пары несет продольный ряд округлых бугорков, чередующихся с короткими щетинками
....* 12. A. crenulatus Gurjan.
28 (27). Нижний край 4-го членика ножки усиков 2-й пары гладкий . .
* 11. A. beringanus Bened.
29 (26). Поверхность тела грубо-зернистая; на переднем и заднем краях грудных сегментов по одному поперечному ряду тесно *посаженных маленьких бугорков; спинная поверхность тельзона покрыта низкими бугорками............................ 10. A. granulates Rich.
30 (25). Конец тельзона заострен или тупой, закругленный.
31 (34). Спинная поверхность головы и грудных сегментов гладкая, лишенная бугорков.
32* (33). Поверхность тела совершенно гладкая; эпимеры задних грудных сегментов видны сверху; вдоль тела тянутся 3 продольных светлых полосы; конец тельзона тупой, закругленный. 14. A. tritaeniatus Rich.
33 (32). Поверхность тела тонкосетчатая (под лупой); эпимеры задних грудных сегментов не видны сверху; конец тельзона заострен . .
....13. A. glabrus Bened.
34 (31). На голове и грудных сегментах имеются парные спинные более или менее развитые бугорки.......................*3. A. hastiger Rich.
* 1. Arcturus baffini (Sab.) (фиг. 109).
Sabine, 1824, Suppl. Cap. Parry’s First Voyage: 228, pl. I, f. 4—5 (Idothea)', G. Sars,. 1885, Norske Nordhavs Exp., Crust.: 97, pl. 9, f. 1—22.
. Тело длинное, узкое, цилиндрическое; на спинной стороне головы каждого грудного и каждого брюшного сегмента по паре длинных, конических отростков, не превышающих, однако, высоты сегмента. Наиболее крупные по размеру отростки на голове, на 2-м, 3-м и 4-м грудных
176
8. AR-СТ URIDAE
сегментах; наиболее слабо развиты спинные отростки 1-го грудного сегмента; все отростки направлены кверху. Поверхность тела зернистая и покрыта короткими жесткими волосками; более* длинные волоски покрывают поверхность спинных отростков тела. Длина головы примерно равна ее ширине; фронтальный край закругленно вогнутый; глаза большие, выпуклые, посажены по бокам головы. Усики 4-й пары маленькие, немного длиннее головы; 1-й членик их расширенный, пластинчатой формы с грубой зазубренностью по внутреннему (медиальному) краю; 2 следующих членика тонкие, короткие, с бугорками на медиальном крае; жгутик одночлениковый, достигает конца 2-го членика ножки усиков 2-й пары, длина его равна 2 предыдущим членикам вместе и несет на внутреннем крае щетку сензорных щетинок. Усики 2-й пары мощные, очень длинные, равные длине всего тела животного; первые 2 членика ножки короткие и толстые, неправильной формы; 3-й членик длинный, цилиндриче-
baffini (Sab.) — экз. из Охотского скии, длина его больше МОРЯ- чем в 3 раза превышает
длину головы; 4-й членик длинный, более чем в 11/2 раза длиннее 3-го, 5-й чле-узкий, немного короче 4-го. Жгутик сравнительно ко-
Фиг. 109. Arcturas
более узкий и ник еще более роткий, немного более половины последнего членика ножки, 8—10 чле-никовый. Все членики ножки усиков 2-й пары покрыты зернистостьЧо и волосками. Первые 3 сегмента груди приблизительно равной длины, каждый немного длиннее головы; 4-й сегмент немного крупнее предыдущих; 3 задних грудных сегмента последовательно укорачиваются. Эпимеры развиты на всех сегментах кроме 1-го, 3 передние пары очень маленькие, 3 задние пары зубовидные, крупные, с заостренными концами, направленными в стороны. Эпимеры 1-го грудного сегмента нацело сливаются с сегментом и образуют лопасти, в виде языков, направленных вперед и книзу по бокам головы. 2 передних свободных брюшных сегмента короткие, равной длины, несут по паре небольших конических заостренных спинных шипов и по одному заостренному бугорку с каждой стороны у своего нижнего края. Тельзон вытянут, треугольной формы, заостряющийся на конце. По бокам основания тельзона по одному крупному зубовидному отростку, направленному в сторону. Спинная поверхность тель-. зона сильно выпуклая с парой маленьких продольных килей посредине.
Поверхность тельзона зернистая и покрыта волосками. 4 передние пары
1. ARCTURUS
177
трудных ножек усиковидные, слабые, направлены вперед и прижимаясь ж ротовым частям служат как бы добавочными частями к ним, особенно 1-я пара, самая маленькая, которая приобретает облик, близкий к ногоче-люстям; внутренний край члеников всех четырех передних пар грудных ножек усажен длинными щетинками. 3 задних пары грудных ног крепкие, ходильные, последовательно уменьшающиеся в размерах к заднему концу тела; коготки их расщеплены на конце. Цвет тела желтовато-бурый; длина достигает 40 мм. Самки часто вынашивают молодь, вылупившуюся из яиц, внутри инкубаторного кармана, на своих длинных усиках. Самцы обладают -более узким и тонким Телом и удлиненным 4-м грудным •сегментом.
Распространение. Северный Атлантический океан (Фарерские -о-ва); восточная Исландия, глуб. 300 м.; Арктическая область (зап. Гренландия, вост. Гренландия, Баффинов залив, Дэвисов пролив, острова полярного североамериканского архипелага, о. Бичи; Земля Эллис-мера, западный Шпицберген), 9—740 м глубины (Гурьянова, 1932); Охотское море: 55°41' N, 141°25'Е (сбор Гидрогр, эксп. Восточного океана, 1918).
Экземпляры из Охотского моря отличаются от гренландских экземпляров более грубой зернистостью поверхности тела, местами превращающейся в бугристость; спинные отростки грудных сегментов и головы более длинные и грубые, покрытые длинными жесткими волосками; на спинной поверхности тельзона вместо двух маленьких низких продольных килей, пара крупных направленных назад конических выростов.
* 2. Arcturus ulbani Gurjan. (фиг. 110).
Gurjanova, 1933, Исслед. морей СССР, 17 :87, рис. 1.
Тело длинное, узкое, цилиндрическое, несколько более укороченное, по сравнению с предыдущим видом; длина его (у самки) почти в 7 раз больше ширины. Поверхность тела гладкая, лишенная волосков, но покрытая очень тонкой зернистостью и слабо выраженными вдавлениями м морщинками на боковых сторонах первых 5 сегментов груди. На спин-Гурьянова 12
178 8. ARCTURIDAE
ной стороне головы и всех 7 грудных сегментов по паре длинных заостренных, направленных несколько назад шипов или отростков; наиболее крупные из них расположены на 3-м и 4-м грудных сегментах, вместо пары спинных отростков 2 маленьких заостренных бугорка; 2-й брюшной сегмент несет пару дорзальных изогнутых отростков, заостренных на концах. Тельзон на конце заострен, с сильно выпуклой спинной поверхностью, на которой пара параллельных, слабо выраженных коротких продольных килей. У основания тельзона с каждой стороны по одному коническому зубовидному отростку. Г лаза очень большие, выпуклые, слегка почковидные, посажены по бокам головы. Усики 1-й пары едва достигают конца 2-го членика ножки 2-й пары усиков; базальный членик их пластинчатый, расширенный, равный длине 2 следующих члеников вместе. Усики 2-й пары длиннее тела животного; 1-й членик ножки короткий, частично закрыт боковыми краями головы; 2-й членик немного длиннее, толстый,, неправильной формы; 3-й членик в 2х/2 раза длиннее 2-го, цилиндрический; 4-й и 5гй членики цилиндрические, равной длины, каждый равен длине 2-го и 3-го члеников вместе; жгутик 13-члениковый, причем 1-й членик жгутика равен длине 3 следующих члеников. Передний край головы глубока вогнутый с маленьким острием посредине между основаниями базальных члеников усиков 1-й пары. Эпимеры 2-го, 3-го и 4-го грудных сегментов маленькие, зубовидные; эпимеры последних 3 сегментов крупные, почти квадратной формы со слабо оттянутыми книзу нижними задними углами. Поверхность спинных отростков головы и всех сегментов тела гладкая, лишенная волосков. Ротовые части и конечности нормального для рода строения. Самец обладает более узким и тонким телом.
Распространение. Охотское мрре: Ульбанский залив, 54°O3ZN, 138о08'Е, 2 экз.; 54°01'N, 138°20'Е, глуб. 10 м, 16 экз.; 53°30'N, 142°20' Е (залив Байкал), глуб. 8 м, песок, 1 экз. (сбор Павленко и Солдатова, 1911); 54°22'N, 138°15'Е, 1 экз. (Павленко, 1913); 53°56 N, 138°10'Е, 6 экз.; 59°2Г N, 158°35' Е, 20 экз.; 60°13' N, 158°13' Е, (сбор Гидрогр. экспед., 1916); 58°1' N, 148°19z Е, глуб. 141 м, 1 экз. (сбор эксп. ГГИ, 1930» ст. 138/23).
*3. Arcturus hastiger Rich. (фиг. Ill)
Richardson, 1910, Proc. U. S. Nat. Mus., 37 :99, f. 24. •
Тело длинное, узкое, цилиндрическое; длина его в 6 раз больше ширины. Поверхность тела мелкозернистая. На спинной стороне головы и всех 7 грудных сегментов по паре «небольших округлых бугорков. На спинной стороне брюшных сегментов нет ни шипов, ни бугорков. Передний край головы глубоко вогнутый с небольшим острием посредине. Передне-боковые углы головы вытянуты вперед и заострены. Глаза очень большие, выпуклые, поперечно-овальной формы, посажены по бокам головы. Усики 1-й пары достигают дистального конца 2-го членика ножки усиков 2-й пары; базальный .членик большой и расширенный;
Z. ARCTURUS
179
2-й и 3-й членики в 2 раза короче 1-го, узкие, удлиненные, । равной длины; последний членик в ly/s раза длиннее 3-го. Усики 2-й пары очень длинные, в V/2 раза длиннее тела животного. 1-й членик ножки короткий, почтй! скрытый боковыми краями головы; 2-й членик толстый, равный длине головы; 3-й членик цилиндрический, в 3 раза длиннее 2-го; 4-й в 1.7 раза длиннее 3-го, 5-й немного длиннее 4-го; жгутик 10-члениковый, причем 1-й членик жгутика почти такой же длины, как все остальные членики жгутика вместе. 1-й грудной сегмент короче'толовы; 2-й, 3-й и 5-й сегменты равной длины, каждый немного длиннее 1-го; 4-й грудной сегмент в 12/2 раза длиннее 3-го. 3 передние пары эпимер маленькие и прикрепляются к передней части нижнего края соответствующего сегмента; 3 последних пары эпимер большие, зубовидные и Прикреплены по средине нижнего края соответствующего сегмента. 2 передних брюшных сегмента равной длины, каждый почти равен длине 1-го грудного сегмента; спинная поверхность их гладкая. Тельзон длинный, вытянуто треугольной формы, заострен на конце. У основания тельзона по бокам по одному небольшому коническому отростку; спинная сторона тельзона очень сильно выпуклая с парой маленьких коротких продольных килей по средине. Задний заостренный конец тельзона плоский, так что при рас-
Фиг. 111. Arcturus hastiger Rich. (noRichardson).
сматривании тельзона сбоку, круто спускающаяся линия, ограничивающая сильно выпуклую срединную и базальную части тельзона, образует тупой угол с плоской поверхностью конечного острия. Молодые экземпляры и самцы обладают более узким и длинным телом. Самки вынашивают подрастающую молодь на своих антеннах. Вынашивание яиц и вылупление происходит внутри инкубаторного кармана.
12*
180
8. ARCTURIDAE
Распространение. Японское море:. 43°00' N, 140° 10'Е, 43°01', N 140°10/Е, глуб. 702—770 м (Richardson, 1910). Татарский пролив: у мыса Олимпиада, 46°12' N, 138°15' Е, глуб. 62 м; 48° 5 5' N, 140°53' Е, глуб. 120 м; 50°37'N, 141°30'Е, глуб. 92 м; Советская гавань, глуб. 200 м; 54°22'N, 139°54'Е, глуб. 60 м; 54°27'N 140°59'Е, глуб. 58 м (сборы ТИРХ’а и ГГИ, 1931, 1932).
4. Arcturus longispinis Benedict (фиг. 112).
Benedict, 1898, Proc. Biol. Soc. Washington, XII : 44, f. 4; Richardson, 1905, Bull. U. S. Nat. Mus. 54:329, f. 360.
Тело узкое, цилиндрическое, несколько укороченное по сравнению с предыдущими видами; длина его почти в 5 раз больше ширины. На
Фиг. 112. Arcturus longispinis Benedict (по Benedict).
спинной стороне головы, всех сегментов груди, кроме 1-го, и всех брюшных сегментов пара длинных тонких заостряющихся шипов. 1-й грудной сегмент на спинной 9тороне несет пару рудиментарных бугорков. Ширина головы почти в два раза больше ее длины; передней край головы глубоко вырезан; передне-боковые углы вытянуты вперед и тупо заострены. Глаза маленькие, круглые, расположены по бокам головы на середине расстояния между передне-боковыми углами и задним краем. Шипы на голове очень длинные, почти в 2 раза превышающие высоту головы. Усики 1-й пары достигают дистального конца 2-го членика ножки усиков 2-й пары; базальный членик их удлиненный, слабо расширенный; 2-й и 3-й членики равной длины, каждый много короче 1-го; 4-й членик в l1^ раза длиннее 3-го. Усики 2-й пары в 1х/2 раза длиннее тела; 1-й членик ножки короткий, полускрыт боковыми углами головы, 2-й членик немного длиннее, толстый, неправильной формы; 3-й членик в 4 раза длиннее 2-го, цилиндрический; 4-й почти в l1^ раза длиннее 3-го, 5-й немного короче 4-го; жгутик короткий, 8-члениковый; 1-й членик его в несколько раз длиннее 2-го, остальные короткие. На 2-м, 3-м и 4-м грудных сегментах спинные шипы очень длинные, превышающие высоту сегментов, заостренные; спинные шипы 3 последних грудных сегментов более короткие, короче, чем высота сегмента. Эпимеры на всех сегментах груди, кроме 1-го, хорошо развиты, свободные, с закругленными краями; 3 первые пары эпимер маленькие, занимающие переднюю часть нижнего края соответ
/. ARCTURUS
181
ствующих им сегментов; 3 задние пары эпимер более крупные, сдвинуты к середине нижнего края сегментов. Эпимеры 2-го, 3-го и 4-го сегментов прикрыты сверху большими шишковидными отростками бокового края сегмента, по одному с каждой стороны на каждом сегменте. 2 передних брюшных сегмента короткие; на спинной стороне 1-го из них пара рудиментарных шипиков; на спинной стороне 2-го — пара коротких загнутых назад заостренных шипов. Нижний край 1-го брюшного сегмента снабжен треугольным расширением, по одному с каждой стороны. Нижний
Фиг. 113. Arcturus setosus Gurjan. а — общий вид; 6 — голова; с — telson.
край 2-го брюшного сегмента простой, не имеет расширения. Тельзон на конце раздвоен; спинная сторона его образует сдавленный с боков сильно выпуклый бугор с 2 заостренными, направленными назад короткими шипами посредине. По бокам основания тельзона по одному треугольному отростку, направленному назад и в сторону. Строение ротовых частей и конечностей нормально для рода. Волосков нет ни на члениках усиков, ни на всей поверхности тела. Длина животного 35 мм.
Распространение. Берингово море: Алеутские острова, 52°O5'N, 177°40'Е, глуб. 110 м (Benedict, 1898).
* 5. Arcturus setosus Gurjan. (фиг. 113).
Gurjanova, 1933, Исслед. морей СССР, 17 :88, рис. 2.
Тело несколько укороченное, веретенообразной формы; длина его почти в 6 раз больше ширины. Поверхность тела бугристая, покрыта
182
8. ARCTURIDAE
грубой зернистостью и довольно длинными, жесткими, закручивающимися волссками, которые покрывают также проксимальную часть двух усиков, тельзон и спинные шипы. Голова, все сегменты груди, брюшко и тельзон несут по паре длинных, саблевидных, изогнутых, направленных назад спинных шипов. Передний край головы сильно вогнутый; передне-боковые углы головы вытянуты вперед, образуя треугольные лопасти, снабженные небольшим шипиком на вершине. Глаза выпуклые, большие, округлопочковидные, по бокам головы. Усики 1-й пары заходят немного дальше дистального края 2-го членика ножки усиков 2-й пары; базальный членик их слабо расширен; 2-й членик узкий, цилиндрический, равен длине 3-го членика и половине длины 1-го; 4-й членик в 2 раза длиннее 3-го. Усики 2-й пары очень длинные, больше чем в l^s раза длиннее тела животного; 1-й членик их ножки короткий и не виден, скрытый боковыми лопастями ' головы; 2-й членик короткий, толстый и несет острый шип на дистальном боковом углу. 3-й членйк цилиндрический, почти в 4 раза длиннее 2-го; 4-й членйк в 5 раз длиннее 2-го, а 5-й членик немного короче 4-го; жгутик короткий, 14-члениковый, все членики его приблизительно равной длины, эпимеральные расширения. 1-го 'грудного сегмента заостряются книзу; 3 передние пары свободных эпимер маленькие, конические, слегка заостряющиеся книзу; нижние края 2-го, 3-го и 4-го грудных сегментов над основанием эпимер шишковидно вздуты. 3 последние пары эпимер большие, треугольные, с направленным вниз и назад тупым острием. На нижних задних углах 4-го, 5-го, 6-го и 7-го грудных сегментов позади основания эпимер по одному направленному назад небольшому заостренному отростку. Такие же шиповидные отростки, по одному с каждой стороны, на нижних краях 2 первых брюшных сегментов. Тельзон раздвоен на конце; базальная часть тельзона сильно вздута и на вершине несет пару острых, направленных назад шипов; задний раздвоенный конец тельзона плоский. По бокам основания тельзона треугольные заостренные отросткй, по одному с каждой стороны. По такому же, но более мелкому отростку по бокам тельзона, приблизительно на середине его нижнего края. Длина животного 35 мм. Самец отличается более узким и длинным телом. Густота развития волосков на теле, длина спинных шипов и величина шипиков на боковых углах головы и на 2-м членике ножки усиков 2-й пары вариирует. Максимальная длина 37 мм.
Распространение. Охотское море: сборы Павленко, 1911, ст. 24, Ульбанский залив 54°03' N, 138°08' Е, глуб. 40 м, 10 экз.; ст. 33, Шантарские о-ва, 54°46' N, 137°27'Е, глуб. 14 — 20 м, 2 экз.; ст. 36, у о. Феоклистова, 54°53' N, 136°56' Е, глуб. 30 м, 1 экз.; сбор Дербека, 1912, ст. 11, 57°5Q'N, 141°47'Е, глуб. 144 м, 1 экз.; сбор Гидрол. экспедиции, 1915, 1917; 53°56'N, 138°10'Е, 6 экз.; 60°18'N, 158°13'Е, 1 экз.; сбор эксп. ГГИ, 1932, ст. 236 — 53°47'N, 143°22'Е, глуб. 84 м, 2 экз.; 58°15'N, 148°19'Е, глуб. 141 м, 1 экз.; ст. 247—55°38'N, 143°00'Е, глуб. 500 м. 1 экз.; ст. 248 — 56°10'N, 143°15'Е, глуб. 182 м, 3 экз.
1. ARCTURUS
183
* 5 a. Arcturus setosus var. seminudus Gurjan. (фиг. 114).
Gurjanova, 1933, Цсслед. морей СССР, 17:89, рис. 3.
От типичного A. setosus отличается более тонким и длинным телом, •слабым развитием волосков, которые расположены главным образом на задней половине тела, почти прямыми спинными шипами, более острыми и вытянутыми книзу эпимерами, слабым развитием боковых зубцов в средней части боковых краев тельзона и относительно более короткими усиками 2-й пары, которые меньше чем в 1х/2 раза превышают длину тела. Шипы позади эпимер 4-го, 5-го, 6-го и 7-го грудных сегментов, хорошо развиты; эпимеры 3 последних грудных сегментов на- t Ц /
цело сливаются с сегментами, почти без следов швов в месте слияния. Шипы впереди глаз и на 2-м членике ножки усиков IIII П Uj
2-й пары хорошо развиты; жгу- // Ч-’.г \ 4’^
тик усиков 2-й пары 8-членико- М. I l/rw *
вый; 1-й членик его в 4—5 раз длиннее каждого из остальных.
Членики ножки 2-х усиков и Ча спинные шипы тела покрыты \
мелкими зазубринами и ши пиками, НО голые, лишенные ВО- Фиг. 114. Arcturus setosus var. seminudus Gurjan. a — telson. •
aockob. 1ельзон на конце раздвоен. Длина животного 36 мм. •
Распространение. Охотское море: 60°19'N, 158°13'Е (Гижигинская губа), сбор Гидрогр. эксп. Вост, океана, 1915; северная часть Японского моря (Татарский пролив): у мыса Олимпиада, глуб. 30 м, на каменистом грунте (сбор Павленко, 1913).
* 5 b. Arcturus setosus var. acuticaudalis Gurjan. (фиг. 115).
Gurjanova, 1933, Исслед. морей СССР, 17 :89, рис. 4.
От основной формы A. setosus отличается более вытянутым и заостряющимся на конце тельзоном с полным отсутствием боковых отростков в его средней части. На спинных шипах и усиках 2-й пары сохраняются волоски и грубая зернистость, зазубрины и шипики. Шипы на боковых углах головы впереди глаз хорошо развиты, но на 2-м членике ножки усиков 2-й пары вместо шипа (как у A. setosus и A. setosus seminudus) только слабо выраженный бугорок. Тело, как и у предыдущей вариации, более узкое и длинное; усики второй пары несколько укороченные по сравнению с основным видом, хотя и не в такой степени, как у предыдущей формы: лишь в 1г/2 раза длиннее тела животного; жгутик их 10-члениковый, все членики короткие; Шипы позади эпимер на ниж-
184
8. ARCTURIDAE
них краях. 4 последних грудных сегментов отсутствуют. Шишковидные утолщения на нижнем крае 2-го, 3-го и 4-го грудных сегментов над основаниями эпимер резко выражены. Поверхность тела бугристая, грубо зернистая и покрыта редкими жесткими волосками. Длина тела 39 мм.
Распространение. Охотское море: 59°17/N, 145°40/Е, глуб. 88 м (сбор Дербек, 1910), на илистом грунте с галькой; 54°34' N,
Фиг. 115. Arcturus setosus var. acuticaudalis Gurjan. a — telson.
142°44z E, глуб. 6S м, на галечном грунте (сбор П. Ушакова, 1932). Северная часть Японского моря: Татарский пролив, 48°55/N, 140°33z Е, глуб. 124 м (сбор Гидрогр. эксп. Вост, океана, 1914).
* 6. Arcturus diversispinis Rich. (фиг. 116).
Richardson, 1910, Proc. U. S. Nat. Mus., 37 :104, f. 27.
Тело длинное, цилиндрическое; длина его в 6У2 раза больше ширины. На голове, на 6 передних грудных сегментах и на 2-м брюшном сегменте Тара длинных заостренных спинных шипов; на последнем грудном сегменте дара рудиментарных бугорков, а спинная поверхность 1-го брюшного сегмента гладкая, лишенная щипрр или бугорков. Поверхность тела гладкая,, покрытая длинными волосками. Передний край головы глубоко вогнутый; глаза большие, выпуклые, посажены по бокам головы. Боковые части головы впереди глаз .вытянуты в треугольно заостряющиеся лопасти, прикрывающие 1-й членик ножки усиков 2-й пары, Шипы на голове длинные, кони-
J. ARCTURUS
185
ческие, co сближенными основаниями и расходящимися в стороны вершинами. Усики 1-й пары удлиненные, заходят за конец 1-й трети длины 3-го членика ножки усиков 2-й пары; базальные членики их слабо расширены, немного длиннее 2-го членика; 3-й членик равен длине 2-го, а 4-й равен длине 2-го и 3-го члеников вместе. Усики 2-й пары длинные, больше чем в 11/2раза длиннее тела животного; 1-й членик ножки их короткий; 2-й членик толстый, равен длине головы и несет тупой шип на наружном дистальном углу; 3-й членик в 3 раза длиннее 2-го, цилиндрический; 4-й членик больше чем в Р/2 раза длиннее 3-го, 5-й членик немного короче 4-го; жгутик короткий, 8-члениковый (?); 1-й членик их длиннее следующих в 2—3 раза. Все членики ножки усиков 2-й пары покрыты длинными волосками. 3 передние пары эпимер маленькие, шишковидные, закругленные; занимают переднюю часть нижнего края 2-го, 3-го и 4-го грудных сегментов. 3 задние пары эпимер большие, зубовидные, с тупыми закругленными вершинами, направленными в стороны. На нижнем крае 2 первых свободных брюшных сегментов конические тупые отростки. На спинной стороне 1-го брюшного сегмента нет ни шипов, ни бугорков. Пара спинных шипов 2-го брюшного сегмента короткая, заостренная, направленная назад. Тельзон удлиненный, равный длине Зх/2 передних грудных сегментов вместе; конец его суживается и раздвоен. По бокам его основания по одному широкому треугольному отростку; пара маленьких отростков приблизительно на середине бокового края его, по одному с каждой стороны. На спинной поверхности тельзона, посредине, немного ближе к его заднему концу, пара небольших конических шипов, направленных назад. Длина 54 мм.
Фиг. 116. Arcturus dlversispi-nis Rich, (no Richardson).
Распространение. Берингово море: район Командорских, островов, 52°55' N, 173°2б' Е, глуб. 270 м (Richardson, 1910).
7. Arcturus magnispims Rich. (фиг. 117).
Richardson H., 1910, Proc. U. S.'Nat. Mus., 37:103, f. 26.
Тело значительно укороченное по сравнению с прочими видами;; длина его меньше чем в 4 раза превосходит его ширину (в 3.7 раза). На.
186
8. ARCTURIDAE
голове и на всех сегментах грудного и брюшного отдела тела по паре спинных шипов; спинные шипы на голове и на 2-м, 3-м и 4-м грудных сегментах очень длинные, превышающие высоту сегмента; спинные шипы 2-го грудного сегмента и 6 последних сегментов тела, включая и тельзон, более короткие, не превышающие высоту сегментов. Передний край головы глубоко вогнутый; переднебоковые углы головы не имеют шипов на вершине; глаза большие, выпуклые, по бокам головы. Усики 1-й пары достигают конца 2-го членика ножки усиков 2-й пары; базальный членик их не расширенный и равен длине 2 следующих члеников вместе. 4-й членик длиннее 3-го. Усики 2-й пары немного меньше чем в раза длиннее тела животного; 1-й членик их ножки короткий; 2-й членик толстый, равен длине головы по средней линии; 3-й членик в 3 раза длиннее 2-го; ,4-й в Г/2 раза длиннее 3-го, а 5-й немного короче 4-го; жгутик короткий, 8-члениковый (?), 1-й членик его значительно длиннее остальных. Вся поверхность тела и спинные шипы густо покрыты волосками; только на 1-м и 2-м члениках ножки 2 усиков имеются волоски, остальные членики голые. На наружном дистальном углу 2-го членика ножки 2 усиков нет шипа, в противоположность предыдущему виду. Тельзон раздвоен на конце, с укороченным дистальным концом; длина его равна длине 3 первых сегментов груди вместе и в 2 раза больше ширины его основания. По бокам тельзона близко у его базиса пара треугольных заостренных отростков по одному с каждой стороны. Небольшие заостренные отростки имеются также по бокам тельзона над основанием его суженной дистальной части. 3 Передние пары эпимер маленькие, шишковидные, расположены в передней части"нижнего края сегментов. 3 задние пары свободных эпимер большие, зубовидные с закругленными краями. Длина животного 30 мм. От близкого к нему A. setosus этот вид отличается более коротким и толстым телом, голыми члениками ножки усиков 2-й пары, отсутствием шипов на передне-боковых углах головы и дистальном конце 2-го членика ножки 2 усиков и более широким и коротким тельзоном с укороченным дистальным концом.
Распространение. Берингово море, 52°11'N, 179°49/Е, 52°12' N, 179°52' Е, глуб. от 80 до 100 м (Richardson, 1910).
* 8. Arcturus crassispinis Rich. (фиг. 118).
Richardson, 1910, Proc. U. S. Nat. Mus., 37: 106, fig. 29.
Тело длинное, цилиндрическое; длина его немного более, чем в 5 раз . превышает его ширину. Поверхность тела бугорчатая, покрытая крупной зернистостью и короткими жесткими волосками. На голове, на всех грудных и 2 передних брюшных сегментах по паре коротких спинных шипов, приблизительно одинаковых размеров, равных половине высоты сегмента. Передний край головы глубоко вырезан; передне-боковые углы головы образуют треугольные лопасти с притупленной вершиной. Глаза большие выпуклые, по бокам головы. Усики 1-й пары в вытянутом состоянии захо-
1. ARCTURUS
187
Фиг. 118. Arcturus crassispinis Rich (no Richardson).
’Фиг. 117. Arcturus magnispi-nis Rich, (no Richardson).
188
8. ARCTURIDAE
дят несколько за пределы конца 2-го членика ножки усиков 2-й пары;, базальный членик их слабо расширен, удлиненный; 2-й членик узкий, равен длине базального членика; 3-й членик немного короче 2-го; 4-й членик немного длиннее базального. Усики 2-й пары длинные,’ больше чем в 1]/2 раза длиннее тела. Все членики ножки их покрыты волосками; 1-й членик короткий, прикрытый боковыми краями головы; 2-й членик толстый, в 1х/2 раза длиннее головы по средней линии; на наружном дистальном углу его нет шипов; 3-й членик почти в 4 раза длиннее 2-го; 4-й членик равен длине 1-го, 2-го и 3-го члеников вместе; 5-й членик короче 4-го. Жгутик короткий, 6-члениковый; 1-й членик их очень длинный, равный длине всех остальных члеников жгутика вместе. 3 передние пары свободных эпимер маленькие, вздутые, округлые; три задних пары более крупные, зубовидные с прямым нижним краем. На нижних краях 2 первых брюшных сегментов маленькие шиповидные отростки, по одному с каждой стороны. Тельзон длинный, суживающийся дистально, длина его равна длине 4 последних сегментов тела вместе. Задний конец вытянут, узкий, раздвоен на конце; на середине сильно выпуклой спинной стороны его пара маленьких коротких продольных килей. У основания тельзона с каждой стороны по одному небольшому треугольному отростку. Боковые края тельзона при переходе их к узкому дистальному концу образуют, углы. Цвет тела серый. Длина 44 мм. Самцы обладают более узким телом с вытянутым 4-м сегментом; самки вынашивают подросшую молодь на своих антеннах.
Распространение-. Японское море и Желтое море, 40—140 м (Richardsop, 1910). Японское море: залив Петра Великого (залив Посьет, открытая часть залива Петр Великий), глуб. 30—100 м; Татарский, пролив (обычная форма на фации песка, глуб. 80—150 м; залив Владимир, Де-Кастри, Советская гавань (сборы ТИРХ’а и ГГИ, 1925—1932). Охотское море: 58°1'N, 149°19' Е, глуб. 141 м;57°12' N, 142° 18' Е, глуб. 27 м; 58°49' N, 144°2б' Е (сборы ГГИ, 1930), залив Анива, глуб. 32 м (сборы П. Шмидта, 1901).
* 9. Arcturus brevispinis Rich. (фиг. 119).
Richardson Н., 1910, Proc. U. S. Nat. Mus., 37:105, f. 28.
Тело длинное, цилиндрическое, длина его почти в 9 раз больше ширины. Вся поверхность тела покрыта мелкими острыми бугорками. На голове и 4 передних грудных сегментах по паре коротких конических спинных шипов. На 3 последних грудных и 2 первых брюшных сегментах шипы эти или рудиментарны, или отсутствуют вовсе. Передний край головы глубоко вогнутый; боковые углы головы впереди глаз образуют треугольные лопасти. Глаза большие, выпуклые по бокам головы. Усики 1-й пары достигают конца 1-й четверти длины 3-го членика усиков 2-й пары; базальный членик их мало расширен, равен длине 2 следующих члеников вместе; 4-й членик удлиненный, равный длине всех 3 предыду
/. ARCTURUS
189
щих члеников вместе. Усики 2-й пары почти в 1х/2 раза длиннее тела; 1-й членик их ножки не виден, закрытый краями головы; 2-й членик короткий, расширяющийся дистально,, снабженный шипом на наружном дистальном углу; 3-й членик цилиндрический, немного более чем в 3 раза длиннее 2-го; 4-й членик длиннее 3-го, а 5-й немного короче 4-го, но длиннее 3-го; жгутик короткий, 7—8-члениковый; 1-й членик его удлиненный, равен длине всех следующих члеников вместе. Тельзон раздвоен на конце; длина его равна длине 5 последних сегментов тела вместе; у основания тельзона по бокам его крупные треугольные отростки, по одному с каждой стороны. По середине сильно выпуклой спинной поверхности тельзона пара маленьких конических шипов. Боковые края тельзона на уровне этих срединных шипов образуют углы. Длина 32 мм.
Распространение. Берингово море, район Командорских островов, 52°55z N, 173°26z Е, глуб. 243 м (Richardson, 1910).
10. Arcturus granulatus Rich, (фиг. 120).
Richardson, 1910, Proc. U. S. Nat. Mus., , 37 : 9§, f. 23.
Тело удлиненное, веретенообразное; длина его в 4 раза превосходит ширину. Поверхность тела грубозернистая, бугорчатая. Парные спинные шипы отсутствуют; вдоль переднего и заднего краев каждого
трудного сегмента по одному попе- Фиг. 119. Arcturas brevispinis Rich, (по речному ряду мелких, тесно поса- Richardson).
женных бугорков. Спинная поверх-
ность тельзона покрыта низкими округдыми бугорками. Ширина головы немного больше ее длины; передний край головы глубоко вы
190
8. ARCTURIDAE
резан с маленьким острием посредине между основаниями базальных
члеников усиков 1-й пары. Глаза большие, круглые, посаженные близко-к боковым краям головы на середине расстояния между передним и задним, краями головы. На спинной стороне головы между глазами бугристость
Фиг. 120. Arcturus granulatus Rich, (по Richardson).
образует 2 низких возвышения, по одному с каждой стороны от срединной линии. Усики 1-й пары достигают конца 2-го членика ножки усиков 2-й пары; базальные членики их большие и расширенные; следующие 2 членика тонкие, короткие, равной длины; 4-й членик в 2 раза длиннее предыдущего. Усики 2-й пары равны длине тела животного; 1-й членик их ножки очень короткий, закрытый вытянутыми боковыми лопастями головы; 2-й членик, короткий, толстый, равный длине головы; 3-й членик почти в два раза длиннее 2-го; 4-й в 2 раза длиннее 3-го, 5-й почти такой же длины, как 4-й; жгутик короткий, 7-чле-никовый, членики все короткие, приблизительно равной длины. Первые три сегмента груди равной длины, 4-й немногодлиннее 3-го; 3 последних грудных сегмента равной длины, каждый в l1/^ раза короче 1-го,. Передняя часть бокового края 2-го, 3-го и 4-го сегментов образует маленькую лопасть с каждой стороны над основанием эпимер, представленных в виде
узких пластинок, расширяющихся кзади-Эпимеры 3 последних1 грудных сегментов
в^виде больших угловатых отростков; эпимеры 5-го сегмента занимают половину
бокового края сегмента, эпимеры 6-го сегмента — 2/з бокового края,' а последняя
пара эпимер занимает почти всю длину бокового края 7-го грудного сегмента. 2 передних брюшных сегмента короткие; тельзон удлиненный с закругленными краями и раздвоенным концом; длина его равна длине последних 5 сегментов тела
вместе; у основания тельзона пара треугольных отростков, по одному с каждой стороны. Срединные боковые углы отсутствуют. Длина 15.5 мм.
Распространение. Охотское мор е: около острова Симу-шир (Курильская гряда) 4б°42' N, 151°45'Е и 46°42' N, 151°47'Е, глуб. 402 м, в гравии и черном песке (Richardson, 1910).
/. ARCTURUS
191
* 11. Arcturus beringanus Bened. (фиг. 121).
Benedict, 1898, Proc. Biol. Soc. Washington, 12:46, f. 6.
Тело узкое, удлиненное; длина его немного больше чем в 7 раз превосходит ширину; на спинной стороне тела нет ни шипов, ни бугорков, поверхность тела густо покрыта точечными вдавлениями и мелкой зернистостью. Передний край головы глубоко вырезан; передне-боковые углы головы образуют треугольные лопасти с закругленной вершиной; длина головы по средней линии равна ее ширине. Глаза маленькие, круглые, расположены по бокам головы, на середине расстояния между передним и задним краями. Усики 1-й пары достигают конца 2-го членика ножки усиков 2-й пары; базальные членики их расширенные, равные длине 2 следующих члеников вместе; 4-й членик немного длиннее 1-го. Усики 2-й пары, по сравнению с прочйми видами, тонкие, слабые и достигают только
Фиг. 121. Arcturus beringanus Benedict (экз. из Японского моря).
основания тельзона; 1-й членик их ножки короткий, прикрыт боковыми углами головы; 2-й немного длиннее; 3-й членик в 2 раза длиннее 2-го, 4-й в 2 раза длиннее 3-го, а 5-й немного короче 4-го; жгутик короткий, 4-члениковый; 1-й чл'еиик их значительно длиннее следующих; последний членик оканчивается иглой. Первые 3 сегмента груди равной длины; 4-й сегмент в 2 раза длиннее 3-го; 5-й и 6-й сегменты равные и равны длине 1-го сегмента; 7-й грудной сегмент немного короче 6-го. Задние края всех грудных сегментов слегка вогнуты по бокам от средней линии, образуя маленькое направленное назад острие посредине. Задние нижние углы сегментов закругленные. Нижний край 2-го, 3-го и 4-го грудных сегментов валикообразно утолщен; 3 первых пары свободных эпимер маленькие, закругленные, видны только при рассматривании животного сбоку и занимают передний нижний угол соответствующих сегментов;, последние 3 пары эпимер более крупные, зубовидные, с притупленными вершинами и занимают переднюю половину нижнего края последних сегментов груди. Первые 2 брюшных сегмента короткие, равной длины; тельзон удлиненный с закругленными краями и оттянутым узким дистальным концом с U-образной вырезкой на вершине (раздвоен на конце). По бокам основания тельзона небольшие треугольные, плохо выраженные отростки, а на спинной стороне вдавлеиие, в виде поперечной бороздки. Вдавления имеются и на голове как позади, так и впереди глаз с каждой стороны. Цвет тела серовато-бурый, зависящий от густо расположенных мелких серовато-бурых пятен; вдоль средней линии грудного и брюшного отделов,
192
8. ARCTURIDAE
почти до конца тельзона тянется светлая полоса, ограниченная с боков узкими темными полосками. По одной светлой полосе тянется иногда с каждого бока на уровне глаз. Длина животного достигает 23 мм в Японском море и 18 мм в Беринговом море.
Распространение. Берингово море: глуб. от 55 до 140 м (Benedict, 1898), Аляска (Richardson, 1905). Охотское море: 51°36ZN, 141°04/Е, глуб. 22 м, песок (сбор ГГИ, 1930, ст. 412). Японское море: залив Петра Великого (у о. Скрыплева, бухта Патрокл, мыс Басаргин, у о. Фурругельма, у мыса Поворотного, бухта Тавайза, открытая часть залива Петр Великий), на глубинах от 50 до 80 м, на фациях песка и камней (сборы ТИРХ’а и ГГИ, 1927—1930).
* 12. Arcturus crenulatus Gurjan. (фиг. 122).
Gurjanova, 1933, Исслед. морей СССР, 17: 90, рис. 5—6.
Этот вид близок к предыдущему, но отличается от него более тонким и длинным телом, длинными, мощными усиками 2-й пары, относительно более крупными глазами, зазубренностью нижнего края 4-го членика ножки усиков 2-й пары и отсутствием ваЛикообразного утолщения нижнего края 2-го, 3-го и 4-го грудных сегментов. Тело тонкое, удлиненное, длина его в 12 раз больше его ширины; поверхность тела зернистая; по средней линии тела продольный ряд слабо выраженных небольших зубцов, образованных слегка приподнятой и заостренной средней частью заднего края 6 передних грудных сегментов. Передний край головы глубоко вырезан; боковые части головы впереди глаз образуют треугольные лопасти, накрывающие базальные членики усиков 2-й пары; глаза умеренной величины, почти треугольной формы; на спинной стороне головы между глазами 2 низких округлых бугорка. Усики 1-й пары удлиненные, заходят за дистальный конец 2-го членика ножки усиков 2-й пары; базальный членик их слабо расширен и равен длине 2 следующих члеников вместе; 4-й членик длиннее 1-го. Усики 2-й пары равны длине тела животного; 1-й членик их ножки короткий, прикрытый боковыми углами головы; 2-й членик равен длине головы по ее средней линии; 3-й членик в 3 раза длиннее 2-го; 4-й в 1.8 раза длиннее 3-го, а 5-й немного короче 4-го; жгутик короткий, 6-члениковый; 1-й членик их равен длине 3 следующих вместе. Вдоль нижнего края 4-го членика ножки усиков 2-й пары тянется ряд крупных, широких низких бугорков, чередующихся с короткими щетинками, так что сбоку нижний край этого членика кажется фестончатозазубренным. Тельзон сильно удлиненный, с оттянутым назад раздвоенным концом; длина его равна длине 3 задних грудных сегментов вместе. Поверхность тельзона гладкая, лишенная шипов или бугорков; у основания его пара заостренных боковых отростков, по одному с каждой стороны, но таких небольших, что они незаметны при рассматривании животного сверху. 4-й грудной сегмент в I1 /2 раза длиннее предыдущего. Эпимеры, прикрепленные к сегментам снизу, хорошо видны только на 3 последних
1. ARCTURUS
193
.трудных сегментах, если рассматривать животное сбоку. Длина 19 мм. Цвет в спирту желтоватый.
Распространение. Японское море (залив Петра Великого): jb крабовых сетях с глубины 60 м (сборы Тихоок. инет. рыб. хоз., 1927), у мыса Поворотного, на глубине 60 м (Корейско-Сахалинская экспедиция Русск. геогр. общ., 1900).
13. Arcturus glabrus Bened. (фиг. 123).
Benedict, 1898, Proc. Biol. Soc. Wash., 12:46, f. 5.—glaber Richardson, 1905, Bull. U. S. Nat. Mus., 54:330, f. 361—36.
Тело узкое, удлиненное; длина его в 6 раз больше ширины. Спинные шипы и бугорки отсутствуют; поверхность тела совершенно гладкая,
при рассматривании в лупу мелко-
Фиг. 122. Arcturus crenulatus Gurjan. Фиг. 123. Arcturus glabrus Benedict (no a — общий вид; Ъ — вадний конец тела Benedict).
сверху; с—голова; d—IV членик ножки antenna II.
линии меньше ее ширины; передне-боковые углы головы впереди глаз образуют треугольные с притупленной вершиной лопасти. Передний край головы глубоко вогнутый. Глаза маленькие, поперечио-овальные, расположенные по бокам головы на середине расстояния между задним и передним краями. Усики 1-й пары достигают конца 2-го членика ножки усиков 2-й пары; базальный членик их большой и расширенный; 2-й и 3-й членики равной длины, каждый короче и тоньше 1-го. 4-й членик равен длине 2 предыдущих вместе. Усики 2-й пары превышают длину тела; 1-й членик их ножки короткий; 2-й несколько длиннее, короткий; 3-й членик в 31/2 раза длиннее 2-го; 4-й почти в 2 раза длиннее 3-го; 5-й почти равен 4-му; жгутик короткий, 6-члениковый. Первые 3 сегмента груди равной длины; 4-й сегмент в 2 раза длиннее предыдущего; 2 последних грудных сегмента равной длины, и каждый равен длине 1-го сегмента, а 5-й сегмент немного длиннее 6-го. Эпимеры 2-го, 3-го и 4-го сегментов очень маленькие, занимают передне-боковые углы сегментов; 3 задних пары эпимер большие, торчащие в сторону. Первые 2 брюшных сегмента короткие, равной длины;
Гурьянова 13
194
8. ARCT URIDAE
тельзон с сильно оттянутым назад тонким задним концом с тупой вершиной; спинная поверхность тельзона сильно выпуклая; у его основания по бокам по одному небольшому треугольному отростку. Длина 31 мм.
Распространение. Берингово море: 52°05'N, 179°40'W, глуб. 99 -м (Benedict, 1898); 52°55' N, 173°27' Е и между Бристольской-бухтой и Прибыловыми островами, глуб. 64—101 м. (Richardson, 1910).
14. Arcturus tritaeniatus Rich. (фиг. 124).
Richardson H., 1910, Proc. U. S. Nat. Mus., 37:101, f. 25.
Тело узкое, веретенообразной формы; длина его немного более, чем; в 4 раза превышает его ширину. Поверхность тела совершенно гладкая. Цвет в спирту желтоватый с густо усеивающими его бурыми пятнами.-Вдоль тела тянутся 3 продольных светлых полосы, одна по средней линии, 2 другие по бокам от нее. Ширина головы больше ее длины; передний край глубоко вырезан с небольшим острием посредине между базальными члениками усиков 1-й пары. Глаза большие, выпуклые, расположение по бокам головы на середине расстояния между передним и задним краями головы. Усики 1-й пары достигают конца 2-го членика ножки усиков 2-й пары; базальные членики большие и расширенные; 2 следующих узкие, короткие, равной длины; последний членик равен длине 2 предыдущих вместе. Усики 2-й пары мощные, утолщенные, превышающие длину тела. 1-й членик их ножки короткий, прикрытый с боков передними краями головы. 2-й членик почти равен длине головы по средней линии; 3-й в 2 раза длиннее 2-го; 4-й почти в 2 раза длиннее 3-го, а 5-й в 2% раза длиннее 2-го членика; жгутик короткий, 5-члениковый; 1-й членик его равен длине всех остальных вместе. Все сегменты груди равной длины,, за исключением 4-го, который в l1^ раза длиннее каждого из остальных. Передняя часть боковых краев 2-го, 3-го и 4-го сегментов образует-небольшую лопасть, по одной с каждой стороны. Позади этих лопастей расположены эпимеры в виде узких, расширяющихся кзади пластинок. Эпимеры 3 последних грудных сегментов большие, торчащие в стороны; эпимеры 5-го сегмента занимают половину бокового края сегмента, на 6-м, сегменте 2/3 края, и последняя пара эпимер занимает почти всю длину нижнего края сегмента. 2 передних брюшных сегмента короткие; тельзон; вытянутый с плавно закругляющимися краями и тупой вершиной. У основания его по бокам по одному закругленному отростку.. На спинной стороне тельзона между этими отростками небольшое вдавление, пересекающее поперек проксимальный конец тельзона. Длина 19 мм.
Распространение. Берингово море: 52°12'N, 179°52'Е, глуб. 77 м, в мелком черном гравии (Richardson, 1910).
15. Arcturus hirsutus Rich. (фиг. 125).
Richardson, 1904, Proc. U. S. Nat. Mus., 27: 41, f. 16.
Тело густо покрыто иглами или шипами, вершины которых окружены розеткой коротких жестких волосков, расходящихся во все стороны по-
1. ARCTURUS
195
радиусам. Длина тела почти в 4 раза больше его ширины. Передний край головы вогнут посредине. В передней части головы между глазами 2 длинных шипа, превышающих длину всех остальных шипов грудного отдела. В задней части головы между глазами 4 шипа равной длины, расположенных в один поперечный ряд. На передне-боковых углах головы по одному
Фиг. 124. Arcturus tritaeniatus Rich, (по Richardson).
Фиг. 125. Arcturus hirsutus Rich, (no Richardson).
маленькому шипику, а на задне-боковых частях 2 группы мелких Шипиков, по 2—3 шипа в каждой группе. Глаза большие, выпуклые, расположенные по бокам головы. Усики 1-й пары почти достигают конца 3-го членика ножки усиков 2-й пары; базальные членики их несут по одному длинному шипу. Усики 2-й пары сравнительно тонкие и слабые, превышают длину тела животного. На 2-м коротком членике ножки усиков 2-й пары 3 длинных шипа; 3-й членик снабжен 4 шипами, расположенными в один продольный ряд и длинный шип на наружном дистальном углу; 4-й членик, в 2 раза превышающий длину 3-го, снабжен 1 длинным шипом на наружном дистальном углу. 5-й членик невооруженный; все членики ножки 2-х усиков 13*
196
S. ARCTURIDAE
густо покрыты длинными волосками. В передней части 1-го грудного сегмента 2 длинных шипа по каждую сторону от средней линии; в задней его части маленький срединный шипик и по 3 шипа по каждую сторону от него; на нижних краях этого сегмента по 4 мелких шипа с каждой стороны. По бокам от средней линии 2-го грудного сегмента по 2 больших и 2 маленьких шипа с каждой стороны; у заднего его края маленький срединный шип; на 1-й паре эпимер 4 мелких шипа. 3-й грудной сегмент несет по 7 длинных, 3 мелких шипов с каждой стороны от средней линии и 1 длинный шип у середины его заднего края. 4-й сегмент имеет по 8 длинных и 2 мелких шипа по бокам от средней линии и 2 маленьких шипа посредине у его заднего края; эпимеры' 4-го сегмента несут по 2 шипа. 5-й и 6-й сегменты имеют по 5 длинных шипов с каждой стороны от средней линии; эпимеры их снабжены 3 шипами. 7-й сегмент несет по 3 шипа с каждой стороны от средней линии и на эпимерах. Каждый из 2 передних брюшных сегментов снабжен 4 шипами с каждой стороны от средней линии; тельзон закруглен на конце; окаймляя боковой край, вдоль него тянется ряд шипов из 8 или 7 с каждой стороны; спинная поверхность тельзона покрыта многочисленными и короткими шипами. На заднем конце тельзона 2 очень длинных, превышающих длину всех остальных шипов тела, шипа, направленных назад. Между ними 2 более коротких, также направленных назад шипа. Створки рулевых ног покрыты мелкими шипиками. Все членики грудных ног вооружены шипами. Длина тела животного 27 мм.
Распространение. Берингово море: у Крысьих островов 54°30' N, 179°14' Е; 54°31' N, 179°15' Е; 54°30' N, 179°17 Е; 54°30' N, 179°14' Е; 52°0ГЬ1, 174°39'Е; 52°14'N, 174°13/Е; 52°55N, 173°26'Е; 54°5Г N, 167°14'Е; Корейский пролив 34°00z N, 137°49'Е; гл. 243— 1882 (Richardson, 1910).
* 16 Arcturus beddardi Gurjan. (фиг. 126).
Gurjanova, 1935, Исслед. морей СССР, 22:28, рис. 6.
Ближе всего этот вид стоит к A. brunneus Beddard, но отличается короткими усиками и более многочисленными шипами на теле. Тело сравнительно короткое и толстое; брюшной отдел состоит из 3 сегментов; эпимеры ясно ограничены и покрыты мелкими шипами. Все тело покрыто тонкими и длинными, легко обламывающимися заостренными шипами; число этих шипов на каждом сегменте и особенно длина их вариируют у разных экземпляров. Волосков на теле и на шипах нет совсем; поверхность тела и шипов гладкая. Передний край головы вогнутый; глаза небольшие, круглые, выпуклые, посажены по бокам головы; голова короткая, лишь немного длиннее 1-го грудного сегмента; впереди глаз у самого переднего края головы пара длинных заостренных шипов в виде рогов, нависающих над фронтальным краем; у некоторых экземпляров эти шипы очень длинные, достигающие середины. 3-го членика ножки усиков 2-й пары, у других, наоборот, эти шипы совсем короткие. Поперек головы
1. ARCTURUS, 2. PLEUROPRION
197
посредине ряд более мелких шипов по 2 шипа с каждой стороны от средней линии головы. Усики 1-й пары длинные, тонкие, в вытянутом состоянии заходят дальше середины 3-го членика ножки усиков 2-й пары. Усики 2-й пары значительно короче тела; при длине тела в 22 мм длина их всего 18 мм. Членики ножки их вооружены шипами. Каждый из грудных сегментов имеет 2 поперечных ряда шипов, один у переднего, и другой у заднего края сегмента; на 1-м грудном сегменте в переднем ряду только 2 коротких шипа, в заднем ряду 6 шипов, наиболее крупные из них расположены по краям . ряда. На следующих 3 грудных сегментах в 1-м ряду от 6 до 8 шипов и в заднем ряду 8—10 шипов; 3 задних груд-
Фиг. 126. Arcturus beddardi Gurjan. а — telson сверху.
ных сегмента имеют только по одному поперечному ряду шипов, по б—8 шипов в ряду. Первые 2 брюшных сегмента снабжены 2 рядами более мелких шипов, по 3—5 шипов в ряду. Задний конец тельзона закругленный и снабжен 2 очень крупными изогнутыми апикальными шипами; между ними, по середине закругленного конца тельзона, немного отступая от края, 1 значительно более мелкий шип; вся спинная поверхность тельзона выпуклая и усажена многочисленными шипами. Ясно выраженные свободные эпимеры; членики всех грудных ножек, как усовидных, так и ходильных, и створки рулевых ног вооружены многочисленными мелкими шипами. Цвет животного беловатый с буро-красными и розовыми полосами; спинные шипы розовые. Максимальная длина 22 мм. От предыдущего вида резко отличается отсутствием волосков на шипах.
Распространение. Охотское море: 55°13'N, 146°52/Е, глуб. 592 м, в илистом песке; 49°23' N, 148°46' Е, глуб. 1076 м (сборы ГГИ, 1932, ст. 214 и 251).
2. Род PLEUROPRION ZUR-STRASSEN1
Zur-Strassen, 1903. Zool. Ant., 26:00.—Antares zur-Strassen, 1902, Zool. Ant., 25:687.
Тип рода: P. chuni zur-Strassen.
Тело веретенообразно вытянутое, расчленено более или менее равномерно, хотя 4-й грудной сегмент заметно длиннее всех остальных сегментов груди. Глаза, если имеются, латеральные. Усики 1-й пары маленькие, 4-члениковые; усики 2-й пары длинные, но обычно не длиннее
1 В 1902 г. zur-Strassen описал новый род, назвав его Antares, но так как это оказалось nomen praeoccupatum, в 1903 г. автор заменил его названием Pleuroprion.
198
8. ARC TURIDAE
тела. На теле часто имеются шипы или зернистость. Брюшной отдел состоит только из 2 сегментов — 1-го свободного и каудального сегмента, образованного слиянием задних брюшных сегментов с тельзоном. Жвалы лишены щупика; щупик ногочелюстей 5-члениковый. Грудные ножки 2-родов; 4 первых пары усиковидные, слабые, покрытые длинными волосками, направлены вперед; 3 задние пары крепкие, нормального строения, ходильные; коготок раздвоен на конце. Рулевые ножки в виде створок на брюшной стороне, прикреплены по бокам тельзона и накрывают снизу брюшные ножки. У самки 5 пар инкубаторных пластинок. Голова неподвижно срастается на спинной стороне с 1-м грудным сегментом. Эпимеры выражены неясно.
Из дальневосточных морей известно 3 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (6). На спинной стороне грудных сегментов имеются крупные шипы; тельзон раздвоен на конце или снабжен 2 длинными расходящимися шипами.
2 (5). На спинной поверхности тельзона имеются шипы; конец тельзона раздвоен; 3-й членик ножки усиков 2-й пары имеет 1 или 2 шипа у дистального конца.
3 (4). У переднего края головы посредине имеется -1 крупный шип; 3-й членик ножки усиков 2-й пары несет 2 шипа у дистального конца .....................................i . * 1. Р. murdochi Bened.
4 (3). Передний край головы не имеет крупного срединного шипа; 3-й членик ножки усиков 2-й пары несет 1 крупный шип у дистального конца . ...................................... . *2. Р. hystrix G. Sars.
5 (2). Верхняя поверхность тельзона гладкая; 3-й членик ножки усиков 2-й пары гладкий, без шипов ..................3. Р. intermedium Rich.
6 (1). Крупных шипов на спинной стороне тела нет; тельзон заострен на конце; все тело покрыто грубой зернистостью и мелкими острыми бугорками......................................* 4. Р. tarasovi Gurjan.
* 1. Pleuroprion murdochi (Benedict) (фиг. 127).
Benedict, 1898, Proc. Biol. Soc. Washington, 12:49, f. {Arcturus)-, Richardson, 1905, Bull. U. S. Nat. Mtis., 54:342, f. 371—372.
Тело вытянутое, веретенообразной формы; длина его в 3 раза больше ширины. На спинной поверхности всех сегментов тела, включая голову и тельзон, короткие конические шипы, расположенные поперечными рядами. Ширина головы в 2 раза больше ее длины; фронтальный край глубоко вырезан, а передне-боковые углы сильно вытянуты вперед, покрывают основания усиков 2-й пары и несут маленький шипик на вершине. Глаза маленькие, округлые, по бокам головы на середине расстояния между передним и задним краями ее. Между глазами, немного впереди их,
2. PLEUROPRION
199
на спинной стороне головы, поперечный ряд из 3 шипов; срединный, более
крупный шип, как раз посредине фронтальной вырезки; позади глаз 2-й поперечный ряд из 8 шипов. Усики 1-й пары достигают конца 2-го членика ножки усиков 2-й пары; базальный членик их большой и расширенный, 2-й членик почти такой же длины, 3-й в 2 раза короче 2-го, а 4-й равен длине 2-го членика. Усики 2-й пары относительно короткие и слабые, достигают в вытянутом состоянии только заднего края 1-го брюшного
сегмента; базальный членик ножки очень короткий, 2-й членик вооружен 3 шипами; 3-й членик в 1У2 раза длиннее 2-го, с 2 шипами; 4-й в 2У2 раза не имеет шипов; 5-й членик в 2 раза без шипов; членики его
жгутик очень короткий, очень неравной длины.
длиннее 3-го, длиннее 3-го, 3-члениковый,
Фиг. 127. Pleuroprion murdochi Ben. (по Benedict).
Pleuroprion hystrix G. Sars (no Hansen). a—telson; b—antennae I и И.
Фиг. 128.
Фиг. 129. Pleuroprion intermedium Rich, (no Richardson).
На спинной стороне 1-го Трудного сегмента поперечный ряд из 8 коротких конических шипов; 2-й сегмент несет б шипов, 3-Й сегмент с 6 шишами в поперечном ряду и по бокам его, немного сдвинуты вперед еще 2 мелких шипа, по одному с каждой стороны. На 4-м сегменте 2 поперечных ряда шипов и перетяжка тела сегмента между обоими рядами в 1-м ряду 10, во 2-м б шипов и кроме того имеются добавочные шипики. .На 5-м сегменте 18 шипов, расположенных продольными и поперечными рядами; б-й сегмент с поперечным рядом из 8 шипов; последний грудной сегмент тоже с 8, расположенными в поперечный ряд шипами. На 1-м брюшном сегменте 2 поперечных ряда шипов, по 3 шипа в каждом ряду „и по 2 добавочных по бокам сегмента. Тельзон имеет 2 пары боковых отростков, конец его раздвоен; на спинной стороне 2 продольных ряда мелких шипов. Грудные ножки, ротовые части и рулевые ноги нормального для рода строения. Длина 13 мм. Поверхность тела покрыта густыми короткими волосками.
200
8. ARCTURiDAE
Распространение. Море Бофора (мыс Франклин, Benedict,. 1898). Берингово море: Анадырский лиман (сбор М. Пятакова,. 1927).
* 2. Pleuroprion hystrix (G. Sars) (фиг. 128).
Sars, 1877 Arch. Math. Naturw., II: 35Q (Arcturus); Sars, 1885, Norske Nordhavs. Exp., Crust.: 104, pl. IX (Arcturus); Hansen, 1916, Dan. Ingolf Exp., Ill, 5:194, pl. XV, f. 6a— b.
Тело удлиненное, веретенообразной формы; на спинной стороне всех сегментов тела, включая голову и тельзон, имеются длинные шипы. Передний край головы сильно вогнутый; передне-боковые углы головы вытянуты вперед и несут по одному шипику у вершины; глаза маленькие, округлые, по бокам головы, расположенные ближе к переднему краю. Спинные шипы тонкие, большей частью длинные, расположены поперечными рядами, за исключением тельзона. На голове 2 поперечных ряда шипов, передний ряд из 3, задний из 8 длинных шипов. На каждом грудном сегменте по одному поперечному ряду длинных шипов, за исключением 4-го сегмента, который имеет 2 таких ряда шипов и еще добавочные из более мелких шипов. 1-й брюшной сегмент несет 2 пары дорзальных шипов, пару боковых шипов у его основания, по одному с каждой стороны, и цару крупных широких заостренных отростков по бокам заднего края сегмента. Тельзон раздвоен на конце; у его основания пара крупных боковых отростков; на спинной стороне тельзона пара крупных шипов и несколько более мелких. Ус ;ки 1-Й пары достигают конца первой трети 3-го членика ножки усиков 2-й пары; членики их не имеют шипов; усики 2-й пары сравнительно слабые, не достигают в вытянутом состоянии конца грудного отдела; 1-й членик ножки очень короткий, закрытый боковыми частями головы; 2-й членик толстый, почти равен длине 3-го, наружный дистальный угол его снабжен 2 крупными острыми шипами; немного ближе к его проксимальному концу имеется маленький шипик на спинной поверхности членика; 3-й членик, равный длине 2-го, несет 1 заостренный крупный отросток на наружном дистальном углу, 4-й членик тонкий, в 2 раза длиннее 3-го; на его наружном дистальном углу небольшой заостренный отросток. 5-й членик немного короче 4-го, без шипов и отростков; жгутик очень короткий 3-члениковый, 1-й из них очень длинный, 2 последних короткие. Длина 9 мм.
От близкого к нему вида Р. frigidum, известного из Атлантического океана, легко отличается раздвоенным концом тельзона, наличием только 1 пары боковых отростков у основания тельзона. и отсутствием шипа: на базальном членике усиков 1-й пары.
Распространение. Северная часть Атлантического океана между 68°21/ N, 10°40' W и 74°52/ N, 17°1б' W; на юг спускается до 60° 10' N, 6°25' W; глуб. 350—830 м (Hansen, 1916). Охотское море, у порта Аяна (сбор Солдатова и Павленко, 1911) 1 экз., длина 8 мм.
2. PLEUROPRION
201
3. Pleuroprion intermedium (Rich.) (фиг. 129).
Richardson, 1899, Proc. U. S. Nat. Mus., 21:854 (Arcturus); Richardson, 1905, Bull.. U. S. Nat. Mus., 54:346, f. 373.
Тело удлиненное, веретенообразное, покрытое длинными спинными шипами, за исключением тельзона. Передний край головы глубоко вырезан; боковые стороны головы впереди глаз вытянуты вперед, образуя раздвоенные на вершине отростки. У середины фронтального края головы большой шип. Глаза небольшие, округлые, по бокам головы; позади глаз на спинной стороне головы пара длинных шипов. На боковых сторонах головы позади глаз по одному заостренному шипику. Усики 1-й пары маленькие, короткие, не достигают конца 2-го членика ножки усиков 2-й пары; членики их невооруженные. Усики 2-й пары едва достигают в вытянутом состоянии основания абдомена; 2-й членик ножки короткий, скрытый боковыми выростами головы; 2-й членик вооружен 3 шипами; 3-й в 2 раза длиннее 2-го, не вооружен; 4-й и 5-й членики равной длины, каждый в 2 раза длиннее 3-го, не вооруженные; жгутик короткий, 3-членико-вый, с удлиненным 1-м и маленькими последними члениками. Первые 3 грудных сегмента каждый несет поперечный ряд из крупных шипов; 4-й грудной сегмент в 2 раза длиннее каждого из предыдущих, суживается кзади и имеет 2 поперечных ряда длинных шипов, по б шипов в каждом ряду. В поперечном ряду 5-го сегмента 12 шипов; 6-й сегмент с 10 шипами,, а последний грудной с 8 шипами, расположенными в поперечный ряд. 1-й сегмент брюшного отдела короткий, с 12 шипами разной длины, расположенными двумя рядами. Спинная поверхность тельзона гладкая, лишенная шипов; на заднем конце его 2 крупных, расходящихся в стороны шипа; по бокам тельзона 2 пары таких же шипов, 1 пара у основания тельзона и 1 пара на середине бокового края. Длина животного 6.5 мм. Этот вид легко отличить от всех прочих по отсутствию шипов на спинной стороне тельзона.
Распространение. Алеутские острова (Kyska Harbor), глуб.. 18 м (Richardson, 1899).
* 4. Pleuroprion tarasovi Gurjan. (фиг. 130).
Gurjanova, 1935, Исслед. морей СССР, 22 : 29, рис. 7.
Тело вытянутое, тонкое, веретенообразной формы; длина его почти в 9 раз больше ширины; поверхность тела покрыта грубой зернистостью и мелкими острыми бугорками, которые густо покрывают также членики, грудных ножек и усиков. Кроме того, все тело густо покрыто короткими жесткими волосками, кроме заднего конца тельзона. Никаких шипов на спинной поверхности тела и на усиках нет. Голова со слабо вогнутым передним краем; глаза маленькие, округлые, по бокам головы на середине расстояния между задним и передним краями головы. Усики 1-пары достигают дистального конца 3-го членика ножки усиков 2-й пары; базальные
202
8. ARCTURIDAE
членики их не расширены. Усики 2-й пары сравнительно толстые и короткие, крепкие, равные половине длины тела; 1-й членик их ножки закрыт боковыми краями головы; 3-й членик немного длиннее 2-го; 4-й в 2 раза длиннее 3-го, 5-й немного короче 4-го членика; жгутик очень короткий, 4-члениковый, причем 1-й членик длиннее всех остальных вместе. Первые 3 сегмента груди равной длины; 4-й сегмент в 1х/2 раза длиннее 3-го; 3 задних грудных сегмента последовательно укорачиваются к заднему концу тела. 3 передних пары эпимер маленькие, с закругленными краями; 3 задних пары эпимер более крупные, зубовидные, с прямыми нижними краями.
Т ельзон сильно вытянутый, с утончающимся задним концом, заостреннный; длина его равна 4 последним грудным сегментам вместе; спинная сторона
Фиг. 130. Pleuroprion farasovi Gurjan. его В базалЬНОЙ ЧаСТИ СВОД-
чато - выпуклая; боковые края тельзона с закругленными контурами, лишенные отростков. Ноги и ротовые части нормального для рода строения; коготки 3 последних : ходильных пар грудных ножек толстые, короткие. На грудных сегментах и голове неправильные вдавления, особенно ясно заметные на 2-м, 3-м ; и 4-м сегментах. Цвет светлосерый. Длина 10 мм.
Распространение. Охотское море: 55°41' N, 154°35/Е,глуб. 122 м, на илистом грунте (сбор ГГИ, 1932, ст. 257).
3. Род ASTACILLA CORDINER
Cordiner, 1795, Rem. Ruins N. Brit.; Flemming- (ex Cordiner 1795), Quart. Journ. Sci. Mar., 1830:107.
Тип рода: A. longicornis (Sout).
Тело удлиненное, цилиндрическое; 4-й грудной сегмент очень длинный, часто значительно длиннее всех остальных грудных сегментов, взятых вместе. Брюшной отдел состоит только из 2 сегментов — короткого •свободного сегмента и тельзона. Глаза большие, выпуклые, латеральные. Усики 1-й пары короткие, у самца развиты сильнее, чем у самки; на наружном крае последнего членика ряд обонятельных щетинок. Усики 2-й пары длинные, иногда достигают в вытянутом состоянии конца грудного отдела, последние членики ножки сильно удлинены; жгутик очень короткий, 3-чле-никовый. Ногочелюсти с 5-члениковым тупиком. Последние сегменты груди очень подвижно сочленены друг с другом, так что передняя часть тела может изгибаться назад почти под прямым углом к задней части тела. Брюшной отдел состоит из 2 сегментов —короткого переднего и большого заостренного на конце тельзона. Грудные ножки нормального для семейства Arcturidae строения — 3 задних пары ног ходильные, с короткими
3. ASTACILLA
203
раздвоенными на конце коготками, 4 передних пары усиковидные. Рулевые ноги в виде створок накрывают брюшные ножки с нижней стороны.
Поверхность тела иногда зернистая, снабжена бугорками, шипами и пр.
Инкубаторных пластинок 3 или 4 пары; 1-й грудной стороне сливается с головой.
Из дальневосточных морей известно только 2 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Поверхность тела грубо зернистая . . . .
......................1. A. dilatata Rich.
2 (1). Поверхность тела гладкая..............
.................. * 2. A. polita Gurjan.
1. Astacilla dilatata Rich. (фиг. 131).
Richardson H., 1910, Proc. U. S. Nat. Mus., 37: 96, f. 22.
Тело узкое, веретенообразно удлиненное; длина его в 3.8 раза больше ширины; 4-й грудной сегмент не такой длинный, как у прочих видов и равен длине лишь 3 первых сегментов груди вместе. Поверхность тела грубо зернистая и покрыта мелкими бугорками. Передний край головы глубоко вырезан, с небольшим острием посредине, между основаниями усиков 1-й пары; переднебоковые утлы головы вытянуты вперед и раздвоены на конце. Глаза большие, круглые, выпуклые, расположены по бокам головы на середине расстояния между передним и задним краями. На спин-
сегмент на спинной
НОЙ поверхности И голове между глазами пара КОНИ- фиг 131 Astacilla dilatata ческих бугорков, резко отличающихся ОТ мелких Rich, (по Richardson), бугорков, покрывающих всю поверхность головы.
Усики 1-й пары заходят немного за пределы дистального конца 2-го чле
ника ножки усиков 2-й пары; базальный членик их большой и расширенный, покрыт зернистостью; 2-й и 3-й членики короткйе, тонкие, равной длины, вместе равны длине 1-го членика; 4-й членик в 2 раза длиннее 3-го. Усики 2-й пары длинные, равные длине тела; 1-й членик ножки очень короткий, 2-й немного длиннее, покрыт мелкими бугорками; 3-й членик почти в 2 раза длиннее 2 первых вместе; 4-й и 5-Й равной длины, каждый в 2 раза длиннее 3-го. Жгутик 3-члениковый, 1-й из них почти в 3 раза длиннее 2-го, а последний в 2 раза короче 2-го членика. Голова сливается с 1-м грудным сегментом; передне-боковые углы его вытянуты и заострены. 3 первые грудные сегмента равной длины; 4-й сегмент больше чем в 2 раза длиннее 3-го, суживается дистально; 3 задних грудных сегмента (каждый немного
204
8. ARC TURIDAE
короче 1-го) последовательно укорачиваются к концу тела. У заднего края 2-го и 3-го сегментов на спинной стороне по паре крупных конических бугорков; у заднего края 4-го сегмента 2 пары небольших бугорков, расположенных двумя продольными рядами. Передние части бокового края 2-го,, 3-го и 4-го сегментов образуют маленькие лопасти над основанием эпимер, которые имеют каждая форму 2 маленьких треугольных отростков. На спинной поверхности 3 последних грудных сегментов по 2 пары маленьких бугорков, расположенных 2 продольными рядами; эпимеры в виде больших треугольных отростков, направленных в стороны. Брюшной отдел состоит из 2 сегментов, переднего короткого свободного сегмента и каудального сегмента, образовавшегося от слияния брюшных сегментов и тельзона; на 1-м брюшном сегменте поперечный ряд мелких бугорков; основание тельзона имеет резко выраженную перемычку на месте слияния его с остальными брюшными сегментами и несет 2 поперечных ряда мелких бугорков. Боковые края тельзона образуют 2 пары широких треугольных отростков; задний конец его заострен; спинная поверхность тельзона покрыта бугорками. Richardson, описывая этот вид, указывает, что самка имеет 2 пары инкубаторных пластинок, но Hansen (1916), проверявший число их у всех других видов, считает, что для рода Astacilla характерна присутствие 4 пар инкубаторных пластинок. Длина 9.5 мм.
Распространение. Японское море: 38°16ZN, 137°52'Е, глуб. 126 м, на песчаном грунте (Richardson, 1910).
* 2. Astacilla polita Gurjan. (фиг. 132).
Gurjanova, 1936, Zool. Anz., 114: 255, fig. 4.
Тело веретенообразно вытянуто; четвертый грудной сегмент равен длине трех первых грудных сегментов вместе, сильнР расширяется кпереди. Поверхность тела совершенно гладкая, лишенная зернистости, бугорков и волосков; только на спинной стороне головы между глазами два грубозернистых бугорка, покрытые редкими короткими жесткими волосками. Передний край головы глубоко вырезан с небольшим острием посредине между основаниями первых усиков. Передне-боковые углы головы вытянуты вперед и заострены. Глаза небольшие, слабо выпуклые, треугольноовальной формы, черные. Усики 1-й пары не достигают дистального края 2-го членика ножки усиков 2-й пары; базальный членик широкий, плоский, овальной формы; последний членик несет 5 обонятельных щетинок. Усики 2-й пары длинные, но в вытянутом состоянии доходят лишь до конца' грудного отдела; 2-й и 3-й членики почти равной длины, 4-й членик в 2 раза длиннее 3-го и равен длине 5-го членика: жгутик 3-члениковый. Инкубаторных пластинок 4 пары. Брюшко из двух сегментов; коксальные пластинки хорошо развиты; 3 задних пары грудных ног крепкие, с раздвоенными коготками. Тельзон с двумя парами треугольных зубовидных отростков— одна у основаниядругая пара у конца тельзона. Конец тельзона тупо заострен. Цвет тела желтовато-серый. Длина 9 мм. От предыдущего
3. ASTACILLA
205
вида легко отличается гладкой поверхностью тела и тупым, почти закругленным дистальным концом тела.
Распределение. Японское море: район острова Петрова, '(42° 49' N, 133О48/ Е), на глубине 60 м.
•Фиг. 132. Astacilla
polita Gurjan. 7—2 — самка; 3 — maxillipedes g; 4 — антенна I (J; 5—pereiopoda I
IV. Подотряд ONISCOIDEA
Формы наземные; на морском побережья заселяют супралиторальную зону. Тело сплюснуто в дорзовентральном направлении и равномерно расчленено на сегменты; некоторые способны свертываться в шар. Голова маленькая, погружена в 1-й грудной сегмент, глаза обычно хорошо развиты, но имеются и слепые формы. Усики 1-й пары очень маленькие, редуцированные, состоят не больше, чем из 3 члеников и расположены между основаниями усиков 2-й пары. Усики 2-й пары нормального строения,
206 9. LI GJ I DAE
: •
с 5-члениковой ножкой и жгутиком, умеренной длины, обычно не больше половины длины тела, но иногда почти достигают конца тела. Ротовые части нормального строения, жующие. Жвалы мощные, но всегда лишенные щупика. Maxilla II в виде пластинки, без разделения на лопасти. Ь/огоче-люсти небольшие, иногда более или менее редуцированные. Все 7 грудных сегментов свободные, одинаковой формы и размеров с хорошо развитыми эпимеральными расширениями, со следами или без следов шва на месте слияния эпимер с сегментами. Все 7 пар грудных ног ходильные. Брюшной отдел из б свободных сегментов, также снабженных эпимеральными расширениями; тельзон срастается с последним брюшным сегментом. 5 пар брюшных ножек двуветвистые, приспособленные к воздушному дыханию во влажных местах. Внутренние ветви брюшных ножек нежные, губчатой структуры; наружные ветви их более грубые, расширенные в виде крышечек, прикрывают сверху нежные внутренние ветви; 2 передние пары, а иногда и все 5 пар этих крышечек с воздушными полостями, носящими название псевдотрахей. У самцов внутренняя ветвь 2-й пары, а иногда и 1-й пары брюшных ножек, видоизменяется в копулятивный орган. Рулевые ноги терминальные, двуветвистые; ветви всегда одночлениковые.
Из 5 известных для этого подотряда семейств на побережье Берингова, Охотского и Японского морей встречаются представители только 3 семейств — Ligiidae, Trichoniscidae и Oniscidae.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ
1 (6). Рулевые ноги терминальные, не образуют крышечки над брюшными ножками; все 5 пар брюшных ножек хорошо развиты.
2 (3). Жгутик усиков 2-й пары многочлениковый; penis самца парный . . ...............................................9. Ligiidae (стр. 206).
3 (2). Жгутик усиков 2-й пары из малого числа члеников; penis непарный. 4 (5). Жгутик усиков 2-й пары больше чем из 3 члеников; зубной отросток
жвал хорошо развит..................10. Trichoniscidae (стр. 212).
5 (4). Жгутик усиков 2-й пары 2—3-члениковый; зубной отросток жвал редуцирован............................... , 11. Oniscidae (стр. 214).
6 (1). Рулевые ноги образуют крышечку над брюшными ножками; 1-я пара брюшных ножек отсутствует (остается лишь 4 пары брюшных ножек).
7 (8). Все брюшные сегменты свободны; передняя пара (2-я пара) брюшных ножек одноветвистая..................................[Tylidae].
8 (7). Все брюшные сегменты сливаются друг с другом на спинной стороне;, передняя пара (2-я пара) брюшных ножек двуветвистая [Syspastidae]..
9. Сем. LIGIIDAE
Тело овально-удлиненное, с выпуклой спиной и плоской брюшной: стороной. Все грудные и все брюшные сегменты свободны, с хорошо раз
1. LIGIA
207
витыми эпимеральными расширениями. Голова овальной формы, совершенно гладкая; передний край ее плавно закругляется без каких бы то ни было выростов, лопастей или углов. Глаза, если они имеются, обычно большие, овальные, часто занимают большую часть головы. Усики 1-й пары 3-члениковые, последний членик очень маленький, плохо заметный. Усики 2-й пары с длинным многочлениковым жгутиком. Ротовые части сильно выдаются книзу; Maxilla I с 3 перистыми щетинками на внутренней лопасти; жвалы снабжены большим и широким сильным зубным отростком; перетирающая поверхность его густо покрыта мелкими бороздками. На внутреннем крае Maxilla II пара очень толстых перистых щетинок. Щупик ногочелюстей 5-члениковый. Все 5 пар брюшных ножек хорошо развиты, двуветвистые; наружные оперкулярные ветви их без воздушных полостей; внутренние ветви 1-й пары и у самца и у самки одинакового строения; внутренние ветви 2-Й пары брюшных ножек у самца видоизменены, вытягиваются в длинный стилет, слегка расширяющийся дистально, превращаясь в парный. Рулевые ноги терминальные, двуветвистые; ветви заострены на конце, палочковидные.
На Дальнем Востоке известно 2 рода.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Базальный членик рулевых ног простой, цилиндрический; ветви рулевых ног равной длины; задне-боковые углы тельзона образуют маленькие треугольные, заостренные лопасти . 1. Ligia Fabr. (стр. 207).
2 (1).' Базальный членик рулевых ног снабжен отростком на дистальном внутреннем углу; ветви рулевых ног неравной длины; задне-боковые углы тельзона закруглены ................2. Ligidium Brandt (стр. 210).
1. Род LICIA (FABRICIUS)
Fabricius, 1798, Entom. Syst. Suppl.: 301.
Тип рода: L. oceanica (L.).
Тело продолговато-овальное, равномерно расчлененное; брюшко немного уже грудного отдела; поверхность тела зернистая или почти гладкая; эпимеры сливаются с сегментами, часто даже без следов швов; задние углы эпимеральных расширений заострены, а у задних грудных сегментов и у всех брюшных вытянуты назад в короткие острия. Голова выпуклая, овальная; передний край закруглен; усики 1-й пары сильно редуцированы, не выступают за пределы фронтального края и не видны сверху; последний (3-й) членик их очень сильно редуцирован и сохраняется лишь в виде бугорка. Усики 2-й пары длинные, в вытянутом состоянии иногда достигают конца грудного отдела; жгутик многочлениковый. ГлаЗа очень большие, латеральные. Ногочелюсти с короткими закругленными эпигнатами; щупик их 5-члениковый; членики расширенные. 2 первых
208
9. LIGIIDAE
брюшных сегмента с боков прикрыты последним грудным сегментом. Все грудные ноги ходильные, снабжены добавочным коготком. Рулевые ноги с простым удлиненным цилиндрическим базальным члеником и 'равной длины заостренными ветвями. Тельзон широкий и сравнительно короткий; середина заднего края тельзона образует небольшое острие; задние боковые углы его оттянуты в короткие заостренные отростки.
На нашем дальневосточном побережье известно 2 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
.1 (4). Тело удлиненно-овальное; у самца на б-м членике 1-й пары грудных ног имеется отросток в виде бугра, нависающего над основанием
коготка на нижнем крае членика
> Фиг. 133. Ligia cinerascens Budd-Lund. а — telson.
2 (3). Поверхность тела тонко зернистая, почти гладкая; усики 2-й пары в вытянутом состоянии достигают средины б-го грудного сегмента; членики жгутика крепкие, короткие ..................
. . . . * 1. L. cinerascens Budde-Lund.
3 (2). Поверхность тела зернистая; усики 2-й пары значительно длиннее, в вытянутом состоянии достигают заднего края 5-го брюшного сегмента; членики жгутика тонкие, длинные. [L. exotica Roux].
4 (1). Тело широко-овальное; б-й членик 1-й пары грудных ног самца не имеет
отростка у основания коготка ....
...................2. L pallasi Brandt.
* 1. Ligia cinerascens Budde-Lund (фиг. 133).
Budde-Lund, 1885, Crust. Isopoda Terrestria: 265; Gurjanova, 1933, Исслед. морей СССР, 17 :100, рнс. 29—31.
Тело удлиненно-овальное, при максимальной длине в 37 мм, ширина ,его 17 мм, поверхность тела тонкозернистая, почти гладкая. Усики 1-й пары не видны сверху; усики 2-й пары крепкие, длинные, в вытянутом состоянии достигают середины 6-го или заднего края 7-го грудного сегмента; ножка их достигает заднего края 3-го грудного сегмента; жгутик 27-члениковый; членики крепкие, короткие, утолщенные. Первые 2 членика ножки короткие, равной длины, 3-й членик в 2 раза длиннее 2-го. Глаза большие, овальные, по бокам головы. Maxilla II имеет вторую лопасть. Спинная сторона грудных сегментов сводчато-выпуклая. Хорошо видны следы слияния эпимер с сегментами в виде тонких бороздок, особенно . на задних грудных сегментах. У некоторых экземпляров эти следы швов
1. LIGIA
209
видны даже на 1-м грудном сегменте, б-й членик 1-й пары грудных ног самца несет на дистальном крае под основанием коготка крупный бугор, сплюснутый с боков. Брюшной отдел немного подогнут книзу с сильно выпуклой спинной поверхностью. Базальный членик рулевых ног расширяется посредине; ветви длинные, тонкие, заостряются на конце; наружная ветвь немного короче внутренней. Тельзон широкий с оттянутыми в заостренные отростки задними боковыми углами. Середина заднего края с тупым острием; постлатеральные углы его, между срединным острием и задне-боковыми отростками, тупые. Окраска темносерая; концы эпимер, ножек, усиков и Тельзона оранжево-желтого цвета.
Так как этот вид легко смешать с близким видом Ligia exotica Roux, привожу здесь небольшую сравнительную табличку признаков того и другого вида.
L. exotica Roux (Нагасаки) L. cinerascens Budde-Lund (Владивосток)
Длина тела в мм .... 34 31
Ширина тела в мм . . 15 13
Ножка усиков 2-й пары . Достигает заднего края 3-го Достигает середины 3-го
грудного сегмента грудного сегмента
.Конец усиков 2-й пары . Достигает заднего края послед- Достигает заднего края 6-го
'Число члеников жгутика. него брюшного сегмента 30—31 членик; членики грудного сегмента 24 членика; членики креп-
тонкие, удлиненные кие, короткие, утолщенные
Поверхность тела .... Грубозернистая Тонкозернистая, почти
Базальный членик рулевых ног Боковые края его параллельны, гладкая Расширяется посредине
если смотреть сверху
Форма тела ... . . Тело уплощенное, со слабо Тело с сильно выпуклой
Тельзон. выпуклой спинной поверхностью. Брюшко плоское, не подгибается книзу совсем Срединный зубец и отростки на спинной стороной; брюшко сводчато-выпуклое, довольно резко подогнуто книзу Срединный зубец и отростки
Распространение . . постлатеральных углах острые Тропическое и субтропическое на постлатеральных углах тупые Японское море (залив Петра
побережье Тихого, Индийского и Атлантического океанов Великого и Хакодате); Мисаки; Манилла
Распространение. Побережье Японии (порт Хакодате) (Richardson, 1910). Японское море: побережье залива Петра Великого от Посьета и далее на север по крайней мере до бухты Преображения, (остров Петрова). Форма супралиторальная, массами бегает по камням и между ними (сборы экспедиции ГГИ, 1925—1932, ЗИН АН 1934).
Гурьянова 1^
210
9. LIG1IDAE
2. Ligia pallasi Brandt (фиг. 134).
Brandt, 1833, Bull. Soe. Nat. Moscou; Budde-Lund, 1885, Crust. Isop. Terrestria-. 261.— dilatata Stimpson, 1857, Bost. Journ. Nat. Hist., VI: 507, pl. 12, f. 8. —stimpsoni Miers, 1877r Proc. Zool. Soc. London : 671.
Тело широко-овальное; длина его немного меньше, чем в 2 раза превосходит, ширину (20 мм и 11 мм). Поверхность тела зернистая. Голова поперечно-овальная, ширина ее в 21/2 раза больше длины; передний край широко закругленный; глаза большие, круглые, по бокам головы. Усики I-й пары 3-члениковые, не видны сверху; последний членик очень маленький, бугорковидный. Усики 2-й пары в вытянутом состоянии достигают середины 4-го грудного сегмента. 2 первые членика ножки короткие,, равной длины; 3-й почти в 2 раза длиннее 2-го; 4-й в 1% раза длиннее 3-го; 5-й в l1^ раза длиннее 4-го; жгутик 12-члениковый. Первые 4 грудных сегмента равной длины; последние 3 немного короче по средней линий; боковые части их оттянуты кзади. Эпимеральные расширения очень большие, отделенные на всех грудных сегментах ясными линиями швов. Брюшной отдел почти такой же ширины, как грудной. Боковые части 2 первых брюшных сегментов закрыты оттянутыми назад боковыми частями последнего грудного сегмента и не развиты. Эпимеральные расширения 4 последних брюшных сегментов мощно развиты с оттянутыми назад задними углами. На боковых частях 3-го, 4-го и 5-го сегментов ясные кили, косо идущие от середины сегмента к его задним углам. Задний край тельзона закруглен; задне-боковые углы образуют небольшие заостренные отростки. Рулевые ноги сравнительно короткие и толстые;, базальный членик короткий; ветви равной длины, каждая в 2 раза длиннее базального членика. На б-м членике 1-й пары грудных ног самца нет отростка на его дистальном конце под основанием коготка.
Распространение. Аляска, Уналаска, острова Алеутской гряды (Атту, Атха); на юг спускается до Британской Колумбии и Калифорнии (Richardson, 1910); Ново-Архангельск (Thielemann, 1910). Живет в супра-литоральной зоне и на берегу моря.
2. Род LIGIDIUM BRANDT
Brandt, 1833, Consp. Monogr. Onisc.: 12.
Тип рода: L. hypnorum (Cuv.).
Тело удлиненно-овальное; брюшной отдел значительно уже грудного; спинная сторона сводчато-выпуклая. Голова широкая, поперечно-овальная, с закругленным передним краем; глаза большие, латеральные'; усики 1-й пары 3-члениковые, выходят за пределы переднего края головы и хорошо видны сверху; усики 2-й пары нормально развиты, сравнительно короткие, не достигают средины грудного отдела. Щупик ногочелюстей со слабо расширенными члениками; эпигнаты узкие, язычковидные. Эпимеральные расширения грудного отдела не такие большие, как у преды
2. LIGIDIUM
211
дущего рода, и не имеют следов слияния эпимер с сегментами. Грудные ноги тонкие, последовательно делаются длиннее к заднему концу тела. Задний край тельзона закруглен; задне-боковые углы его не образуют заостренных отростков. Рулевые ножки короткие; внутренний дистальный угол базального членика оттянут в конусовидный отросток, к которому прикрепляется внутренняя ветвь рулевых ног- Ветви неравной длины; внутренняя ветвь короче наружной и снабжена 2 длинными тонкими апикальными щетинками.
На Дальнем Востоке известен пока 1 вид.
Фиг. 134. Ligia pallasi Brandt (по Richardson).
Фиг. 135. Ligidium tenue Budde-Lund. (по оригинальному экземпляру).
1. Ligidium tenue Budde-Lund (фиг. 135).
Budde-Lund, 1885, Crust. Isop. terrestria: 258.
Оригинальные экземпляры, хранящиеся в Зоологическом институте Академии Наук, очень плохой сохранности, так что трудно разобраться в деталях строения брюшных ног и усиков.
Тело узкое, удлиненное, суживается к заднему концу; длина его в 3 раза больше ширины. Спинная поверхность сводчато-выпуклая; поверхность тела гладкая. Эпимеральные расширения плохо выражены; задние углы 3 последних грудных сегментов оттянуты назад и заострены. 1-й брюшной сегмент короткий и прикрыт последним грудным сегментом; Задние углы всех брюшных сегментов слегка оттянуты назад и расширены. Голова большая, длина ее больше длины 1-го грудного сегмента; передняя часть головы вытянута немного вперед,, передний край ее закруглен; глаза большие, по бокам Головы, заходят немного на брюшную сторону; спинная поверхность головы между глазами слегка вдавлена посредине. Усики 1-й пары короткие, но хорошо видны сверху, оба их членика 14*
212
10. TRICHONISCIDAE
одинаковой длины и ширины, вооружены щетинками. Усики 2-й пары в вытянутом состоянии достигают почти середины грудного сегмента. Жгутик их 7—8-члениковый (?) (из-за плохой сохранности трудно точно установить число члеников жгутика); последний членик несет пучок щетинок одинаковой длины. Внутренний отросток базального членика рулевых ног по описанию Budde-Lund составляет 1/4 длины наружной ветви. Внутренняя ветвь длинная, заходит далеко за пределы вершины наружной вет.ви и несет 2 коротких терминальных нити, равных половине длины наружной ветви. Цвет тела коричневый, без пятен. Длина животного 9 мм.
Распространение. Остров Ситха (сбор Вознесенского, 1896), 2 экз. (типы Budde-Lund’a).
10. Сем. TRICHONISCIDAE
Тело удлиненное; передне-боковые части головы образуют широкие лопасти; передний край головы между лопастями закруглен. Глаза простые или сложные, маленькие, дорзальные; имеются слепые формы. Внутренняя лопасть Maxilla I снабжена 3 толстыми перистыми щетинками. Щупик ногочелюстей большой, неясно расчлененный; эпигндты короткие и узкие, язычковидные. Зубной отросток жвал хорошо развит. Усики 1-й пары очень маленькие, но не видны сверху; последний членик их хорошо развит и снабжен обонятельными волосками. Усики 2-й пары короткие, но нормального строения — имеется пятичлениковая ножка и короткий жгутик из малого числа члеников. Эпимеральные расширения грудных сегментов небольшие; наружные ветви брюшных ног тонкие и не имеют воздушных камер. Базальный членик рулевых ног расширенный у основания; ветви короткие, равной длины, тонкие, конические, с пучками волосков на вершине. Все брюшные сегменты свободны; penis непарный, конический; внутренние ветви 1-й и 2-й пары брюшных ног превращаются в копулятивный орган. Расширенная базальная часть основного членика рулевых ног сверху прикрыта тельзоном.
На Дальнем Востоке известен 1 род.
1. Род TRICHONISCUS BRANDT
Brandt, 1833, Consp. Monogr. Onisc.: 12.
Тип рода: T. pusillus Brandt.
Тело удлиненное, суживается кзади. Передний край головы закруглен; передне-боковые углы головы образуют широкие лопасти. Эпимеральные расширения 4 первых грудных сегментов развиты слабее, чем 3 последних. Брюшной отдел значительно уже грудного; эпимеральные расширения 2 передних сегментов хорошо видны, не закрыты последним грудным сегментом. Тельзон резко суживается дистально с прямым задним краем. Глаза маленькие, но ясно видные, только с 3 визуальными
/. TRICHONISCUS
213
элементами. Базальный членик усиков 1-й пары большой и изогнутый; последний членик обычно длиннее 2-го. Усики 2-й пары сравнительно короткие, с очень коротким жгутиком, длина которого меньше длины ножки. Ротовые части удлиненные, вытянуты вперед и образуют кбнус. Щупик ногочелюстей одночлениковый, так как все 4 членика его сливаются друг с другом без следов шва; жевательная лопасть почти такой же величины, как щупик, и на внутреннем дистальном конце ее перистый бичик; эпигнаты узкие, языковидные, расширенные в закругленную лопасть
у основания.
На Дальнем Востоке известен один вид.
* 1. Trichomscus
Rich.
1904, Ехр.,
papillicornis (фиг. 136).
Richardson, Harriman Alaska
Crust.: 213; Richardson H., Bull. U. ; S. Nat. Mus., 54:695, f. 734-738.
Тело удлиненное; поверхность его покрыта мелкими бугорками. Боковые ло-
пасти ГОЛОВЫ впереди Фиг- 136‘ Trichoniscus papillicornis Rich, (по Richardson), глаз большие и ши-
рокие; фронтальный край головы образует волнистую линию с выпуклостью посредине и вогнутостями по бокам у основания усиков; срединная выпуклость слегка вырезана посредине. Глаза маленькие, черные, простые; расположены на спинной стороне головы у основания боковых лопастей. Ножка усиков 2-й пары пятичлениковая; на внутреннем крае 3 последних члеников округлые бугорки в виде больших бородавок с пучками коротких волосков на вершине. Наружный дистальный угол последнего членика ножки вытянут в заостренный отросток. Жгутик состоит из 7 члеников и на конце снабжен пучком волосков. Буккальная масса (ротЬвые части) сильно выдается книзу. Грудные сегменты почти равной длины; задне-боковые углы всех грудных сегментов, за исключением 1-го, оттянуты назад особенно сильно у 3 последних сегментов. Брюшной отдел значительно уже грудного; все сегменты примерно одинаковой длины, короткие. Тельзон в общем треугольной формы, короткий и очень широкий у основания, с тупой вершиной. Рулевые ножки короткие с коническими ветвями; базальный членик сильно расширяется дистально; наружная ветвь крепче и толще, чем внутренняя, и немного длиннее ее; на концах обеих ветвей пучки коротких волосков. Цвет бурый, иногда светлорозовый; длина 5 мм.
Распространение. Cook Inlet, Seldovia, на пляже (Richardson, 1904). Японское море: Б. Патрокл в заливе Петра Великого,
214
//. ONISCiDAE
о. Петров у бухты Преображенье; Советская гавань. Под камнями в супра-литоральной зоне вместе с Ligia cinerascens. Берингово море: о. Беринга (Командоры) и коса Култучного озера (Авачинская губа, Петропавловск на Камчатке), под камнями в супралиторальной зоне (сборы ТИРХ’а, 1928 и Гурьяновой, 1930—1932).
11. Сем. ONISCIDAE
Тело овальное или продолговатое, иногда с сильно выпуклой спинной стороной, с трудом свертывается в шар. Усики 2-й пары длинные, но с коротким жгутиком, который состоит только из 2—3 члеников. Глаза хорошо развиты. Зубного отростка на жвалах нет, он заменен пучком щетинок. Наружная лопасть 1-й пары челюстей вооружена шипами на вершине; внутренняя лопасть с 2 толстыми перистыми щетинками. Maxilla II на вершине образует 2 короткие лопасти. Щупик ногочелюстей из 3 члеников, немного длиннее жевательной лопасти; эпигнаты большие. Грудные ножки довольно длинные; рулевые ноги удлиненные, всегда длиннее чем тельзон, выходят за его пределы. Все брюшные сегменты свободны, penis' непарный. Все брюшные ножки развиты, двуветвистые; на оперкулярных (наружных) ветвях их иногда имеются воздушные камеры. Внутренние ветви 1-й и 2-Й пары брюшных ног самца видоизменены в копулятивный орган.
На морском побережье Дальнего Востока известен только один род с одним видом этого семейства.
1. Род PORCELLIO LATREILLE
Latreille, 1804, Hist. Crust. Insect., 7 :45.
Тип рода: P. scaber Latr.
Тело овальное, более или менее сплюснутое дорзовентрально; боковые части грудных сегментов сильно расширены. Передне-боковые углы головы впереди глаз образуют лопасти; передний край головы между лопастями конически вытянут вперед и слегка вырезан посредине. Глаза хорошо развиты, расположены на спинной стороне. Усики 2-й пары длинные; жгутик состоит только из 2 члеников, 1-й обычно длиннее, чем 2-й, иногда оба равной длины. Брюшной отдел почти такой же ширины, как задняя, часть грудного отдела; боковые части 3-го, 4-го и 5-го сегментов хорошо развиты; тельзон- конической формы. Оперкулярные пластинки (наружные ветви) первых 2, а иногда всех 5 пар брюшных ног имеют воздушные камеры. Базальный членик рулевых ног расширен у основания; внутренняя ветвь, значительно более короткая, слабая и тонкая, чем наружная, прикрепляется почти у основания базального членика, тогда как крепкая, длинная наружная ветвь прикрепляется к дистальному концу базального членика.
1. PORCELLIO
215
* 1. Porcellio scaber Latr. (фиг. 137).
Latreille, 1804, Hist. Crust. Ins., VII: 45; Sars, 1899, Accojint, II: 176, pl. 77.
Тело удлиненно-овальное, неспособно свертываться в шар; длина «го немного меньше, чем в 2 раза превосходит ширину. Голова широкая, ширина ее в 2 раза больше длины; лопасти впереди глаз небольшие; передний край головы между этими лопастями треугольно вытянут, с тупой вершиной. Глаза маленькие, круглые, на спинной стороне у основания передне-боковых лопастей головы. Усики 1 3-члениковые, не видны сверху. Усики 2-й пары в вытянутом состоянии достигают середины 3-го грудного сегмента; базальный членик ножки очень короткий; 2-й членик в 12/2 раза длиннее 1-го, 3-й равен длине 2-го; 4-й почти в 2 раза длиннее 3-го, 5-й почти в 2 раза длиннее 4-го. Жгутик из 2 члеников; 1-й его членик короче 2-го. Грудные сегменты почти одинаковой длины; эпимеральные расширения хорошо развиты; следов слияния эпимер с сегментами нет совсем; брюшной, отдел немного уже грудного. Боковые части 2 первых брюшных сегментов закрыты 7-м грудным сегментом; эпимеральные расширения брюшных сегментов боль- j шие, направлены назад и заострены. Тельзон широкий у основания, резко суживается ди
стально, образуя узкую треугольную лопасть, закругленную на вершине. Рулевые ножки длиннее тельзона; наружная ветвь выходит далеко за пределы конца тельзона; концы внутренних ветвей достигают конца тёль-зона. Вся поверхность, тела покрыта мелкими бугорками. Цвет обычно серый, иногда очень темный, почти черный. Края сегментов светло-желтые образуют кайму вокруг всего тела. Длина 10 мм.
Распространение. Живет под листьями, кусками дерева, под камнями на суше, но часто встречается и на морском берегу в супрали-торальной зоне, вместе с Ligia-, почти космополит. Тихоокеанское побережье Северной Америки до Калифорнии и южнее; Атлантическое побережье Северной Америки от залива Лаврентия до Флориды; Гренландия, Исландия, вся Европа, Камчатка, Командорские о-ва; Гавайские о-ва, мыс Доброй Надежды (Thielemann, 1910); о. Ситха (сбор Вознесенского, 1846).
V. Подотряд EPICARIDEA
Все представители этого подотряда, как указывает их название, паразитируют на ракообразных. Только маленький самец сохраняет внеш
и пары рудиментарные,
’. 137. Porcellio scaber (по G. Sars). а — uropoda.
216
12. BOPYRIDAE
ний облик ракообразного, тогда как самка очень сильно деградирована, часто совсем непохожа на рачка и иногда превращается в простой мешок, набитый яйцами и лишенный каких-либо конечностей, с полным отсутствием сегментации. Здесь наблюдается обычно регрессивный метаморфоз и имеются по крайней мере 2 стадии личинки, сильно отличающиеся по внешнему виду от взрослой половозрелой формы. У некоторых имеется еще одна, третья стадия личинки. Самец, по сравнению с самкой, очень маленький, живет на теле самки у ее полового отверстия, сегментирован,, мало отличается от 3 стадии личинки, за исключением присутствия у него генеративных органов. В некоторых случаях животное является эндопаразитом. В случаях эктопаразитизма взрослая самка иногда имеет сегментацию и придатки. Голова в таких случаях обычно несет 2 пары рудиментарных усиков и сильно редуцированные ротовые части. Грудной отдел иногда имеет ножки сильно видоизмененные, снабженные цепляющейся лапкой. Брюшные ножки, если имеются, жаберные, в виде пластинок, у личинок служат для плавания. Рулевые ноги, если они имеются, всегда маленькие, простые.
На Дальнем Востоке известны представители 2 семейств этого, подотряда.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ
1. (2). Тело самки асимметричное, распадается на сегменты; ногочелюсти пластинчатые, двучлениковые. Имеется 7 пар ног, которые, однако, иногда сохраняются только На одной стороне; инкубаторных пластинок 5 пар; у самца все сегменты грудного отдела свободны, жгутик усиков 2-й пары последней личиночной стадии 4-члениковый; паразитируют на Decapoda ................. 12. Bopyridae. (стр. 216)-
2. (1). Тело самки симметрично; иногда сохраняются следы сегментации;
5 пар конечностей. Инкубаторные пластинки маленькие, часто редуцированы. Брюшные ножки отсутствуют или они совсем рудиментарны. Голова самца сливается с 1-м грудным сегментом. У последней личиночной стадии жгутик усиков 2-й пары 5-члениковый. Паразитируют на Schizopoda.........13. Dajidae. (стр. 222)..
12. Сем. BOPYRIDAE
Тело самки ясно сегментировано; асимметричное; спинная сторона приплюснутая. Голова глубоко погружена в грудной отдел. Имеется 2 пары рудиментарных усиков; иногда имеются глаза, расположенные на спинной стороне. Ногочелюсти из 2 плоских члеников; щупик их одночлениковый; имеется пара изогнутых ланцетовидных эпигнат. Имеется обычно 7 пар грудных ножек одинаковой структуры; иногда ноги только на одной стороне. Инкубаторных пластинок 5 пар, нависающих над брюшной поверхностью Груди. Брюшной отдел сегментирован^ имеет. простые пластинЧа
7. PHR YXUS
217
тые ножки; иногда брюшные ножки отсутствуют. Рулевые ноги ланцетовидные, простые. Самец маленький, вполне симметричный с узким удлиненным телом. Голова его закруглена впереди. Все грудные сегменты свободны; брюшные свободны или частично сливаются друг с другом. У личинки 1-й стадии усики 2-й пары с 4-члениковым жгутиком; рулевые ножки с неравными ветвями.
Паразитирует на десятиногих раках. На Дальнем Востоке имеется 3 рода.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Тело самки сильно изогнутое, асимметричное, так что одна сторона ее значительно короче другой. Грудные ножки развиты только на одцой стороне, только передняя представлена в виде пары. Брюшной отдел состоит из 5 сегментов . . 1. Phryxus Rathke (стр. 217).
2 (1). Все 7 грудных ножек парные, т. е. имеются и на той и на другой стороне. Тело более или менее симметричное. Брюшной отдел из 6 сегментов.
3 (4). У самки имеются и брюшные и рулевые ноги, одноветвистые. Заднебоковые углы всех грудных сегментов образуют вытянутые узкие отростки................................. 2. Argeia Dana (стр. 219).
4(3). У самки нет ни брюшных, ни рулевых ног. Заднебоковые углы
грудных сегментов без отростков . 3. Bopyroides Stimpson (стр. 221).
1. Род PHRYXUS RATHKE
Rathke, 1843, Nova Acta Acad. Leopold. CaroL Nat. Curios., 20 : 40.
Тип рода: Ph. abdominalis (Kr.).
Паразитируют на брюшке десятиногих раков- Тело самки совершенно несимметрично; одна его сторона много больше другой. Сегментация грудного отдела ясно выражена на спинной стороне. Эпимеры и хорошо ограниченные боковые части грудных сегментов только на одной, короткой стороне тела. Брюшной отдел из 5 свободных сегментов; последний сегмент маленький, узкий, на конце раздвоен. Ногочелюсти лишены одночленикового щупика. Инкубаторные пластинки на увеличенной стороне тела большие,, пластинчатые и образуют настоящий инкубаторный карман; инкубаторные пластинки противоположной стороны тела маленькие. Только 1-е грудные ноги парные; остальные 6 только по .одной стороне тела, а на другой стороне отсутствуют. 4 пары брюшных ног и виде широких листовидных пластинок. На „короткой стороне тела они меньше, чем на противоположной, более крупной стороне тела. У самца все грудные сегменты свободны, но все брюшные сливаются вместе,, образуя 1 каудальный сегмент. Рулевые и брюшные ножки у самца, отсутствуют. На Дальнем Востоке известен один вид.
"218
12. BOPYRIDAE
* 1. Phryxus abdominalis (Kroyer) (фиг. 138).
Kroyer, 1840, Nat. Tidsskr., Ill: 102, pl. I, II (Bopyrus); Sars, 1899, Account, II: 215, pl. 90—91.
Асимметрия тела самки очень сильная; передний край головы прямой и по бокам имеет 2 маленьких отростка; задний край ее закруглен. Глаз нет. 7-й грудной сегмент на спинной стороне хорошо отграничен по обеим своим сторонам. Инкубаторный карман занимает всю брюшную поверхность самки. На одной стороне тела все 7 грудных ног; на другой стороне только 1 ножка 1-й пары, остальные отсутствуют. Ясные эпимеры на всех сегментах на более длинной стороне тела; на короткой стороне только на 3 последних сегментах. Рулевых ног нет, но имеются 4 пары двуветвистых брюшных ножек; наружные ветви их большие, овальные, листовидные.. Внутренние ветви маленькие. Инкубаторные пластинки на длинной стороне тела очень большие (их — 5), закрывают собою всю брюшную поверхность тела; инкубаторные
Фиг. 138. 'Phryxus abdominalis Кг. (по G. Sars). пластинки на короткой стороне а — самец. тела маленькие, редуцированные
и не принимают участия в образовании инкубаторного кармана. Самец с узким длинным телом, 3 мм длины и 1. мм ширины. Передний край головы его закруглен: глаза имеются. Усики 1-й пары из 3 члеников; усики 2-й пары из 4—5-чле-'ников. Все грудные сегменты свободны. Все сегменты брюшка сливаются вместе, образуя в общем трехугольной формы суживающийся дистально каудальный сегмент. Нет ни рулевых ни брюшных ножек.
Распространение. В Арктике циркумполярен. По Атлантическому побережью Америки доходит до Массачусетса. По Тихоокеанскому до Pyget Sound и Калифорнии. Берингово море: остров Akutan; 52°55ZN, 173°27ZE; мыс Строгонова (Аляска); Шумагины острова..Охотское море: у о. Кадиак (Курильская гряда), глуб. 5—600 м (Richardson, 1905). Японское море: 38°32z N, 138°43z Е; 45°24' N, 140°49? Е; Зб°08 N, 129°49'Е; 48°32Z.N, 145°07zE; 48°32ZN, 145°08zE; 42°11'N, 142°12ZE, глуб. .100—800 м (Richardson, 1910).
Паразитирует на Spirontocaris sp.
2. ARGEIA
219
2. Род ARGEIA DANA
Dana, 1853, U. S. Explor. Exped., 14:804.
Тип рода: A. pugettensis Dana.
Тело самки асимметрично. На брюшной стороне все 7 сегментов груди и все б брюшных ясно отделены друг от друга. Эпимеры имеются на всех сегментах. Задне-боковые углы грудных сегментов образуют узкие заостренные отростки. Брюшной отдел резко суживается дистально; последний, б-й сегмент, на конце образует 2 маленькие лопасти. Брюшные ножки двуветвистые, их 5 пар; наружные ветви в виде узких длинных пластинок, прикрепленных к боковым краям сегментов и образуют бордюр, окружающий весь брюшной, отдел. Внутренние ветви в виде маленьких закругленных пластинок, уменьшающихся в размерах при продвижении к заднему концу тела животного. Рулевые ноги простые в виде 2 узких удлиненных пластинок, похожих на наружные ветви брюшных ног, прикрепленных к последнему сегменту. Имеются все 7 пар грудных ног. У самца все брюшные сегменты сливаются друг с другом; брюшные и рулевые ножки отсутствуют. Паразитирует на жабрах десятиногих раков.
На Дальнем Востоке известен один вид.
* 1. Argeia pugettensis Dana (фиг. 139).
Dana, 1853, U. S. Expl. Exp., Crust., XIV: 804; Richardson, 1905, Bull. U. S. Nat. Mus., 54:544, f. 586—597.
У взрослой самки грудные отростки могут быть совершенно редуцированы, но у некоторых экземпляров они хорошо развиты; особенно сильно развиты они у молодых неполовозрелых самок. Тело взрослой самки асимметрично; длина его 14 мм, ширина 11 мм. Голова широкая, иногда образует 2 слегка намечающиеся лопасти по бокам. Передний край головы закруглен. Глаз нет. Усики 1-й пары из 2 члеников, базальный членик большой, 2-й очень маленький. Усики 2-й пары из 4 члеников. Обе пары усиков не видны со спинной стороны. Все 7 грудных сегментов резко отделены Друг от друга. На 4 передних сегментах бугры от яичников. Эпимеры в виде узких пластинок сбоку от этих бугров, но имеются также и на 3 последних грудных сегментах. Задне-боковые углы грудных сегментов образуют отростки, иногда очень большие; на одной, более длинной стороне тела, они обычно сильнее развиты, чем на другой. Все 6 брюшных сегментов свободны. Последний сегмент маленький с прямым задним краем. Рулевые ножки в виде простых неправильной формы удлиненных пластинок, прикрепленных к последнему сегменту. Имеется 5 пар брюшных ног; все они двуветвистые; наружные ветви в виде удлиненных пластинок такой же формы, как рулевые ножки и прикреплены к краю сегмента на нижней стороне. Внутренние ветви
220
12. BOPYRIDAE
округлые, мешковидные, уменьшаются в размерах к заднему концу тела. Инкубаторных пластинок 5 пар, которые образуют открытый карман, так
как края противоположных пластинок не соприкасаются друг с другом. Все* 7 пар ног хватательные; базальный членик их снабжен высоким килем. / \
Фиг. 139. Argeia pugettensis Dana (по Richardson). Возр. стадии QQ; а—самец; Ь—личинка.
Самец с узким удлиненным симметричным телом; глаза имеются. Передний край головы широко закругленный. Усики 1-й пары 3-члениковые; усики 2-й пары 4-члениковые. Все грудные сегменты свободны; грудные ноги хватательные. Все сегменты брюшка сливаются вместе в 1 сегмент, тупо заостренный на конце.
Распространение. Берингово море: побережье Аляски, Прибыловы острова, побережье Камчатки, Алеутских островов; Калифорния, Ванкувер, Бристольская бухта в северной части Тихого
3. BOPYROIDES
221
океана. Японское море: Хакодате; 47°35'N, 141°43'Е; 47°32'N, 141°45'Е; 36°30'N, .ПЭЧЗ'Е, глуб. 0—160 м (Richardson, 1905, 1910).
Паразитирует в жаберной полости различных Decapoda, на видах рода Cragon, Nectocrangon.
3. Род BOPYROIDES STIMPSON
Stimpson, 1864, Proc. Acad. Nat. Sci. Phila.: 156.
Тип рода В. hippolytes (Kr.).
Тело самки .асимметричное, широкое, расплющенное. Сегментация ясно выражена и на брюшном и на грудном отделах. Голова погружена
Фиг. 140. Bopyroides hippolytes Kr. а—самка со спинной стороны; Ь — самец; с —самка с брюшной стороны (по G. Sars).
в первый грудной сегмент. Усики первый из 3, а второй из 4 члеников. Эпимеры присутствуют, но маленькие и узкие; инкубаторных пластинок 5 пар; полость инкубаторная широко открыта, так как инкубаторные пластинки короткие и прикрывают лишь края полости; имеется 7 пар грудных ног; брюшные ножки отсутствуют и заменены плоскими ребрами. Рулевых ног тоже нет. У самца все сегменты брюшного отдела сливаются вместе. Брюшные и рулевые ножки отсутствуют. Паразитируют в жаберной полости Decapoda.
Известен один вид этого рода на Дальнем Востоке.
х * 1. Bopyroides hippolytes (Kroyer) (фиг. 140).
Kroyer, 1838, Danske Vid. Selsk. Natur. math. Aid., VII: 306, pl. IV, f. 22 (Bopyrus)-, Richardson, 1905, Bull. U. S. Nat. Mus., 54:567, f. 628.
Тело самки асимметричное, овальное; голова глубоко погружена в грудной отдел; передний край ее почти прямой; задний — закругленный. По бокам головы у ее переднего края имеются небольшие закругленные.
TXL 13. DAJIDAE
лопасти. Глаз нет. Усики 1-й пары 3-члениковые; усики 2-й пары 4—5-чле-никовые; обе пары их не видны со спинной стороны головы. Все 7 сегментов груди ясно отделены друг от друга. Эпимеры первых 4 сегментов в виде очень узких пластинрк, сбоку от возвышений яичника. На 3 задних сегментах эпимеры такой же формы и расположены в передней части бокового края сегментов. Все 6 брюшных сегментов ясно выражены, с прямыми боковыми краями. Последний сегмент очень маленький, с прямым задним краем. Брюшные ножки в виде плоских ребер; рулевых ног нет совсем. 5 пар инкубаторных пластинок короткие, так что в центре карман широко открыт и зияет. Грудные ножки (3 пары) очень маленькие; лапки их хватательные; на базальном членике высокий, более или менее закругленный киль. Длина самки 8 мм; самца 3 мм. Тело самца удлиненное; передний край головы закруглен; глаза имеются; усики 1-й пары 3-члениковые; усики 2-й пары 4-члениковые. Боковые края грудных сегментов прямые. Ноги хватательные. Абдоминальный сегмент тупо заострен на конце и лишен конечностей.
Распространение. В Арктике циркумполярен. По атлантическому побережью спускается до Массачусетса. По тихоокеанскому побережью — до Британский Колумбии включительно; Ванкувер; Орегон, Puget Sound. Берингово море: 52°12/N, 179°52'Е; 54°50'N, 167°13'Е; побережье Аляски; Шумагинские о-ва; Прибыловы о-ва. Японское море: 38°09' N, 138°32'Е, глуб. 80—400 м (Richardson, 1905, 1910).
13. Сем. DAJIDAE
Тело самки симметричное; сегментация тела, если она имеется,, заметна лишь на спинной стороне; чаще нет никаких следов сегментации. Ногочелюсти не имеют щупика. Инкубаторные пластинки маленькие, число их уменьшено; инкубаторных полостей как бы две, по бокам тела,, по одной с каждой стороны. Грудных ног только 4 или 5 пар. Все они расположены вокруг ротового отверстия. Брюшные ножки рудиментарные или отсутствуют вовсе. Рулевые ноги также иногда исчезают. У самца голова сливается с 1-м грудным сегментом.
Паразитируют, на Schizopoda. На Дальнем Востоке известно 3 рода.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Брюшной отдел состоит только из 1-го сегмента...... .
............................1. Holophryxus Rich. (стр. 223).
2 (1). Брюшной отдел состоит из 5 сегментов.
3 (4). Голова и 3 передних грудных сегмента хорошо отделены друг от друга ясными границами по всей Их длине .... .................
...................................3. Prophryxus Rich. (стр. 226)-4 (3). Только на средней части спинной стороны тела видны следы шва
между 5 сегментами...........2. Arthrophryxus Rich. (стр. 224).
1. HOLOPHR YXUS
223
Фиг. 141. Holophryxus alascensis Rich, (по Richardson).
1. Род HOLOPHRYXUS RICHARDSON
Richardson H. 1905, Bull. U. S. Fish. Comm., XXIV : 220.
Тип рода: H. alascensis Rich.
Тело самки без всяких следов сегментации. Брюшной отдел значительно уже грудного, цельный, не расчлененный на сегменты и тупо закруглённый на конце. Брюшные и рулевые ножки отсутствуют. Грудных но'жек 5 пар, все они расположены вокруг рта. Имеется 5 пар инкубаторных пластинок.
На Дальнем Востоке известно 2 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Голова самки окружена мягкой складкой с прямым передним краем; по бокам брюшного отдела у его основания по одной небольшой пограничной вырезке . •
. . . * 2. Н. giardi Rich. 2 (1). Голова не имеет складки впереди, образует 2 лопасти; брюшной отдел у своего основания не имеет вырезки, отграничивающей его от грудного отдела. [3 (4). Тело узкое, овальное.
Эпимеральных пластинок
4 пары — Калифорния . .
. . Н. californiensis Rich.].
4 (3). Тело широкое, с грудным отделом почти квадратной формы; эпимер
5 пар.....................................1 Н. alascensis Rich..
1. Holophryxus alascensis Rich. (фиг. 141).
Richardson, 1905, Bull. U. S. Fish. Comm., XXIV: 220.
Тело самки неправильной формы, светложелтого цвета. Голова в виде двулопастного бугра, приподнимающегося над передней, почти квадратной частью тела. Глаз нет. На спинной поверхности грудного отдела никаких следов сегментации. Имеются лишь линии морщин и складок в разных местах тела. Брюшной отдел в виде более узкого конического придатка, также лишен сегментации и не имеет никаких придатков. По бокам грудного отдела в передней его части имеется 5 пар эпимеральных пластинок, которые окружают ротовое поле и не отделены швами от спинной поверхности тела. Ротовое поле на брюшной стороне спереди ограничено головой, а по бокам парой расходящихся рядов эпимеральных пластинок. Обе пары усиков рудиментарны; усики 1-й пары из 2 члеников, усики 2-й пары 3-члениковые. Имеется 5 пар грудных ног, окружающих ротовое поле и прикрепленных как раз у края эпимеральных
224
13. DAJIDAE-
пластинок. Имеется 5 пар инкубаторных пластинок. Последняя пара налегает друг на друга по средней линии. Самец неизвестен.
Распространение. Побережье Аляски; южная Калифорния (Richardson, 1905), глуб. 264—620 м.
*2. Holophryxus giardi Rich. (фиг. 142).
Richardson, 1908, Proc. U. S. Nat. Mus., 33:690, f. 1—3.
Тело самки продолговато-овальное без следов сегментации. Голова в виде двулопастного бугра, впереди окружена широкой квадратной складкой с прямым передним краем. Глаз нет. Грудной отдел кзади постепенно суживается, без всяких следов сегментации на спинной
стороне. Брюшной отдел уже грудного, закруглен на конце без следов сегментации, кроме 2 борозд у его основания, по одной с каждой стороны. Брюшко не имеет придатков. Ротовое поле на брюшной стороне грудного отдела впереди ограничено возвышенным ребром и ротовым частями, а с боков 2 рядами коксальных пластинок. У основания их 5 пар
Фиг. 142. Holophryxus giardi Rich, (по "РУДНЫХ Н°Ж^ И 5 П&Р ИНКУбаТОРНЫХ Richardson). пластинок. Хорошо видны ТОЛЬКО
1-я и 5-я пары инкубаторных пластинок, так как 3 другие пары закрыты налегающими на них сверху очень большими, занимающими почти всю брюшную поверхность груди и соприкасающимися друг с другом вдоль средней линии тела пластинками 5-й пары, Коксальных пластинок 4 пары. 1-я пара сливается с головой и сохраняется
лишь в виде маленьких отростков у основания квадратной складки головы. Длина до 39 мм.
Неполовозрелая самка по внешнему виду отличается от взрослой. Голова большая и окружена спереди и с боков широкой складкой; голова, грудь и брюшко хорошо отграничены друг от друга. Грудь с хорошо выраженной сегментацией. 6 пар коксальных пластинок прикреплены по краям сегментов. 1-й грудной сегмент сливается с головой и несет пару измененных придатков. Брюшной отдел без придатков и не имеет
следов сегментации.
Распространение. Берингово море: в районе о. Беринга 54°48' N, 164°54' Е, на глубине 4860 м (Richardson, 1908).
2. Род ARTHROPHRYXUS RICHARDSON
Richardson, Н., 1908, Proc. U. S. Nat. Mus., 33: 695.
Тип рода: A. beringanus Rich.
Тело взрослой самки неправильной формы с расширенными боковыми частями. Голова большая, ясно отграничена от грудного отдела на спин-
2. ARTHROPHYXUS
225
•ной стороне. Средняя часть спинной поверхности грудного отдела разделена на 5 ясных сегментов. Брюшной отдел состоит из 5 хорошо отграниченных друг от друга сегментов. Последний сегмент маленький, треугольной формы; придатков брюшной отдел не имеет. Небольшое ротовое поле в передней части тела на брюшной, стороне ограничено по краям 5 парами грудных ножек и 5 парами коксальных пластинок. Инкубаторных пластинок 5 пар; последние очень большие, закрывают все остальные пластинки и налегают друг на друга по средней линии, тела. У самца голова очень бо.льшая; глаз нет; грудной отдел из 6 сегментов. Брюшной отдел неясно сегментирован, из б сегментов; последний сегмент очень маленький, треугольной формы. Брюшной отдел подогнут довольно сильно под грудь. Брюшные и рулевые ножки отсутствуют. Известен один вид.
‘Фиг. 143. Arthrophryxus beringanus Rich, (по Richardson).
Фиг. 144. Prophryxus alascensis Rich. (no Richardson).
* 1. Arthrophryxus beringanus Rich. (фиг. 143).
Richardson, 1908, Proc. U. S. Nat. Mus., 695, f. 6—7.
Тело продолговато-овальное, неправильных очертаний, широкое на переднем конце и суженное кзади. Голова очень большая с неправильной линией переднего края и с поперечной складкой посредине на спинной стороне. Грудной отдел на спинной стороне в средней ее части разделен на 5 сегментов. Брюшной отдел из 3 ясных сегментов; последний сегмент очень маленький, треугольной формы. Брюшной отдел не имеет конечностей. В передней части грудного отдела округлое ротовое поле окружено 5 парами ног, снаружи от них еще 5 парами коксальных пластинок. От основания всех 5 пар грудных ножек отходят 5 пар инкубаторных пластинок, частично накрывающих одна другую. Пластинки последней пары очень большие и соприкасаются друг с другом своими свободными краями по средней линии тела. Длина 14 мм.
Паразитирует на креветке Gennadas borealis Ruth., прикрепляясь головой к заднему краю панцыря головогруди.
Гурьянова 15
226- '
13. dajidAe
Тело самца удлиненноё. Голова большая; глаз нет. 1-й грудной сегмент сливается с головой; остальные б сегментов свободны. Имеется 7 пар грудных цожек, все хватательного типа. Брюшной отдел неясно разделен на 6 сегментов; последний сегмент очень мал и треугольно заострен на конце; конечности брюшного отдела отсутствуют. Грудной • отдел изогнут в виде горба, а брюшной отдел подогнут под него.
Распространение. Берингово море: 54°48' N, 164°54/Е. в районе острова Беринг, глуб. 4860 м; (Richardson, 1908). 53°53' N, 144°53'W, глуб. 3960 м. Датская гавань (Аляска), Авачинская бухта, у острова Беринга, глуб. 4860 м; Британская Колумбия (Richardson, 1910)..
3. Род PROPHRYXUS RICHARDSON
Richardson Н., 1910, Proc. U. S. Nat. Mus., 37:124.
Тип рода: P. alascensis Rich.
Очертания тела взрослой самки неправильные. Голова и 3 первых грудных сегмента отграничены друг от друга и от следующей части; грудного отдела на спинной стороне. Последние 4 сегмента отделены друг от друга бороздками только по средней линии тела на спинной стороне. Боковые части грудного отдела расширенные и образуют пару небольших закругленных лопастей, свисающих по бокам брюшного отдела. Брюшной отдел состоит из 5 сегментов; имеются 5 пар грудных ног,, окружающих ротовое поле. Самец неизвестен.
На Дальнем Востоке 1 вид.
1. Prophryxus alascensis Rich. (фиг. 144).
Richardson, 1910, Proc. U. S. Nat. Mus., 37 :124, f. 47—48.
Тело взрослой самки в общем овальное с сдавленный, почти квадратной формы передним концом. Передний край головы прямой. С одной стороны головы у ее переднего края на спинной стороне 2 маленьких черных пятна (глаз?). 3 передних грудных сегмента резко отделены от остальной части грудного отдела и ограничены поперечными бороздами друг от друга на спинной стороне от одного края до другого. Остальные 4 сегмента отделены друг от друга только в средней части спинной стороны. Боковые части этого отдела груди расширены и не имеют следов сегментации. По бокам брюшного отдела задние углы грудного отдела образуют крупные закругленные лопасти, свисающие по бокам брюшка и почти достигающие его дистального конца. Брюшной отдел из 5 сегментов; последний сегмент состоит из двух лопастей. Ротовое поле окружено 5 парами грудных ножек. На брюшке заметны рудименты брюшных ножек.
Распространение. Северная часть Тихого океана: 53°05'N, 138°31'W, глуб. 3600 м (Richardson, 1910).
3. PRO PHR YXUS
227
VI. Подотряд GNATHIIDEA
Представители этого подотряда резко отличаются от прочих групп Isopoda прежде всего тем, что у них имеется метаморфоз, сопровождающийся несколькими личиночными стадиями, и резко выраженный половой диморфизм. Взрослый самец сильно отличается от взрослой самки, которая сохраняет внешний облик личинки. Во взрослом состоянии самец всегда свободноживущий, но с редуцированным, нефункционирующим кишечником, так как пищи взрослый самец не принимает. Самка во взрослом состоянии или свободноживущая, или же является эктопаразитом рыб; 1-я личиночная стадия всегда свободноплавающая; 2-я, иногда и 3-я, стадия личинки — эктопаразиты. У некоторых форм наблюдается живорождение. Для подотряда Gnathiidea характерно, что, кроме 1-го грудного сегмента, с головой сливается и 2-й. грудной сегмент (голомолог 1-го свободного сегмента груди Isopoda других подотрядов) и соответствующая ему пара грудных ножек превращается в особые, сильно специализированные ножки pylopoda, в виде крышечек, накрывающие с брюшной стороны ротовые части. Последняя пара грудных нрг исчезает совсем, а последний (7-й) грудной сегмент сохраняется в виде рудимента. Таким образом для подотряда Gnaihiidea характерно наличие только 5 пар нормальных грудных ног. Тело состоит из 3 отделов — головы, грудного и брюшного отделов. Грудной отдел, как указывалось выше, состоит только из 5 свободных сегментов; брюшной отдел из 6 сегментов (последний сегмент сильно редуцирован) и тельзона. Грудной отдел распадается на 2 части, перетянутые талией; передняя часть образована 2 первыми свободными сегментами, задняя из 3 крупных сегментов, часто сливающихся на спинной стороне. Брюшной отдел очень узкий, состоит из 6 сегментов; тельзон и рулевые ноги образуют каудальный веер, как у Flabellifercl. Усики короткие, слабые, с многочлениковым жгутиком. Ротовые части сильно изменены и редуцированы; у самца очень характерные жвалы, выходящие сильно вперед за пределы переднего края головы и образующие настоящие щипцы. Ротовые части самки и личинки тесно сближены друг с другом, заострены на концах, образуют ротовой конус, который приспособлен для сосания. Глаза, если имеются, большие, выпуклые, по бокам головы. У самца обычно голова и панцырь 2 передних свободных сегментов пропитаны известью.
На Дальнем Востоке представлен одним родом единственного семейства Gnathiidae.
14. Сем. GNATHIIDAE
Характеристика семейства совпадает с характеристикой под отряда.
15*
228
14. GNATHIIDAE
1. Род GNATHIA LEACH
Leach, 1814, Brewster Edinburgh Encyclop., VII: 386. — Anceus Risso (самец), Praniza Latr. (личинка).
Тип рода: G. maxillaris (Mont.).
Тело самца сплющено дорзовентрально; голова очень большая с далеко выступающими за пределы переднего края головы жвалами, образующими щипцы; грудной отдел распадается на 2 части — 2 передних свободных сегмента отделены перетяжкой (талией) от 3 последних брюшных сегментов, несущих ноги. 1-й грудной сегмент сливается с головой; следы этого слияния в виде глубокой поперечной борозды у заднего края головы; 1-я пара грудных ног превращается в pylopoda; pylopoda большие, 2- или 3-члениковые, накрывают снизу ротовые части в виде крышечки. Ногочелюсти не имеют эпигнат; щупик их — 4-члениковый с сильно расширенными, члениками, окаймленными снаружи длинными перистыми щетинками. 5 пар свободных грудных ног небольшие, крепкие, ходильные. Брюшной отдел резко уже, чем грудной, состоит из 5 коротких узких свободных сегментов; брюшные ножки двуветвистые; ветви одночленико-вые, равной длины, усаженные по краям щетинками. Рулевые ноги образуют вместе с тельзоном хвостовой веер; тельзон обычно треугольной формы, суживается дистально.
Тело самки и личинки веретенообразной формы. Голова маленькая с резко выраженным ротовым конусом. .Грудные сегменты иногда срастаются друг с другом неподвижно. Абдомен и хвостовой веер такого же Как у самца строения. Самка иногда, 2-я личиночная стадия всегда, эктопаразиты. Только в 1885 г. Hesse установил, что Praniza Latr. есть не что иное, как самка. и личинка рода Anceus (или Gnathia). Самки известны далеко не для всех видов. Определять приходится главным образом по самцам.
На Дальнем Востоке пока известно б видов.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РИДОВ (55)
1 (10). Передний край головы без вырезки.
2 (3). Передний край головы на брюшной стороне имеет по 4 шипа с каждой стороны у основания усиков . . . * 1. G. tuberculata Rich.
3 (2). Передний край головы на брюшной стороне лишен шипов.
4 (5). Передний край головы прямой, с маленьким треугольным острием
посредине.............................. *2. G. rectifrons Gurjan.
5 (4). Передний край головы иной формы.
б (7). Глаза маленькие, не выдаются за пределы боковых краев головы; боковые части брюшных сегментов несут шиповидные заостренные отростки.......................................* 3 G. schmidti Gurjan.
7 (б). Г лаза большие, сильно выпуклые; боковые части брюшных сегментов лишены острых шиповидных отростков.
Z. GN A THIA
229
Фиг. 145. Gnathia tuberculata Rich, (по Richardson). а — передняя часть тела ; Ь — abdomen; с — передний ' край головы.
8 (9). Боковые части брюшных сегментов вытянуты в виде язычков с пучками длинных щетинок на концах; передний край головы междуоснованиями жвал образует 2 небольшие округлые лопасти; 3-й членик pylopoda сильно редуцирован и плохо заметен.........................
........................................* 4. G. derzhavini Gurjan.
9 (7). Боковые части абдоминальных сегментов нормально'развиты; передний край головы между основаниями жвал образует трехвершинный вырост; 3-й членик pylopoda маленький, но хорошо развитый
* 5. G. elongata Кг. 10(1). Передний край головы имеет глубокую фигурную вырезку * б. G. monodi Gurjan.
* 1. Gnathia tuberculata Rich. (фиг. 145).
Richardson H., 1910, Proc. U. S. Nat. Mus., 37:75, f. 1—3.
Сам e ц. Голова большая, почти четырех-. угольная, очень широкая; передний край головы по середине между основаниями жвал образует закругленную лопасть; передне-боковые углы вытянуты в заостренные от
ростки. Глаза маленькие/ круглые, расположены у основания переднебоковых отростков по бокам головы. Поверхность головы зернистая и покрыта многочисленными мелкими шипами. 2 первых членика усиков 1-й пары короткие, почти равной длины; 3-й в 2 раза длиннее 2-го; жгутик из 5 члеников. 1-й членик усиков 2-й пары короткий; последние 2 членика удлиненные; жгутик 7-члениковый. Жвалы очень большие, вытянуты прямо вперед; внутренние края их, слегка зазубренные посредине, соприкасаются; концы перекрещиваются друг с другом. Передний край головы на брюшной стороне у основания усиков снабжен шипами, по 4 с каждой стороны. 1-й грудной сегмент, слившийся с головой, очень узкий и такой короткий, что почти незаметен. 2-й (1-й свободный) грудной сегмент в 2 раза короче 3-го. По краям оба сегмента несут шипы, а поверхность их покрыта мелкими шипиками и бугорками. 4-й сегмент немного длиннее и уже 3-го, покрыт бугорками; 5-й в 1г/2 раза длиннее 4-го, с очень редкими бугорками на спинной поверхности, б-й сегмент
230
14. GNATHIIDAE
немного короче и уже 5-го, покрыт немногочисленными мелкими бугорками; последний сегмент настолько редуцирован, что трудно заметен. Брюшной отдел значительно уже 6-го грудного сегмента; 5 передних брюшных сегментов равной длины; тельзон с узким оттянутым и заостренным задним концом. Внутренний угол базального членика рулевых ног вытянут кзади и достигает середины внутренней ветви. Обе ветви рулевых ног одинаковой формы и величины, заходят за пределы заднего конца
Фиг. 146. Gnanthia rectifrons Gurjan. а — общий вид; Ъ — pylopoda; с — antennae; d — telson -+- uropoda; е — mandibula.
тельзона; края их усажены длинными ще- . тинками. Базальный членик 4-й пары грудных ног с двумя крепкими шипами; базальный членик 5-й пары с 3, 6-й пары с 4 длинными шипами. Pylopoda трехчлениковые, с очень маленьким последним члеником. Длина животного 5 мм.
Распространение. Японское море 37° 22'N, 137° 47'Е, глуб. 1124 м (Richardson, 1910), залив Петра Великого, глуб. 892 м, 1 экз.(эксп. ГГИ, 1932, ст. 308).
* 2. Gnathia rectifrons Gurjan. (фиг. 146).
Gurjanova, 1933, Исслед. морей ССС£, 19: 79, рис. 1.
Самец. Голова очень большая, округлая; ширина ее немного больше длины; глаза большие, выпуклые; боковые отростки головы впереди глаз очен/ маленькие, плохо заметные; передний край головы прямой, слегка зазубренный,. посредине имеет небольшой треугольный
заостренный зубец с группой щетинок на вершине. Жвалы лишены зубо- ♦ видного отростка на наружном крае. 2 первых членика усиков 1-й пары короткие, почти равной длины; 3-й членик удлиненный, почти в 11/2 раза длиннее 2-го; жгутик 4-члениковый с 3 щетинками на последнем членике. 1-й и 3-й членики ножки усиков 2-й пары равной длины; 2-й членик равен 4-му, длиннее 1-го, а последний членик почти в 2 раза длиннее 4-го; жгутик 8-члениковый. 1-й грудной сегмент, слившийся с головой, отделен от нее глубокой бороздой. 2-й сегмент немного длиннее 3-го; 3 последних грудных сегмента равной длины, каждый в 2 раза длиннее 3-го. По краям головы и грудных сегментов короткие щетинки; 3 задних грудных сегмента несут пучки более длинных перистых щетинок. Такие же щетинки группами на грудных ножках. На пропитанном известью панцыре головы и 3 передних свободных грудных сегментов мелкие бугорки и шипики. На спинной
1. GN AT HI A
231
стороне панциря широкие вдавления и узкие ребрышки, напоминающие выпуклости и вдавления панцыря Gnathia elongata Кг. Строение брюшного отдела, тельзона и рулевых ног как у G. elongata. Края тельзона мелко зазубрены и усажены мелкими острыми иголочками; задний конец его снабжен парой коротких апикальных щетинок. Базальный членик рулевых ног имеет короткий отросток на нижнем внутреннем углу; наружные ветви немного короче внутренних; концы внутренних ветвей заходят за пределы -заднего конца тельзона; обе ветви по краям снабжены длинными щетин-
Фиг. 147. Gnathia schmidti Gurjan. а — общий вид; Ъ—pylopoda; с, с — abdomen; d—' pereiopoda И; е — pereiopoda III; /— antennae I и II.
:ками. Pylopoda 3-члениковые; базальный членик их вытянуто-грушевидной формы с перистыми щетинками на внутренней стороне; такие же, но более короткие и густо посаженные щетинки на внутреннем крае базального членика pylopoda; 3-й членик очень маленький, но ясно выраженный. Длина 6 мм.
Распространение. Японское море: залив Петра Великого, глуб. 80—88 м (сборы ГГИ, 1931), у мыса Среднего, у мыса Тобизина и у залива Владимира на глубинах 84 м (сбор ТИРХ’а, 1930).
* 3. Gnathia schmidti Gurjan. (фиг. 147).
Gurjanova, 1933, Исслед. морей СССР, 19:79, рис. 2.
Самец. Г олова большая, ширина ее больше длины; глаза очень .маленькие и не выдаются так, как у предыдущего вида; боковые отростки .головы впереди глаз небольшие, прямые, раздвоенные на конце. Жвалы
232 14. GN AT HI ID AE.
сравнительно слабые, лишенные зубца на наружном крае; при смыкании концы их не заходят друг за друга. Передний край головы между основаниями жвал выпуклый, с небольшим острием посредине. Усики 1-й пары равной длины с 2 первыми короткими члениками ножки; 3-й членик в 2 раза длиннее 2-го; жгутик 5-члениковый. 2 первых членика ножки усиков 2-й пары короткие, равной длины; 3-й немного длиннее их; 4-й в 2 раза длиннее 3-го, а 5-й в 1% раза длиннее 4-го; жгутик 6—7-членико-вый. На месте слияния 1-го грудного сегмента с головой глубокая борозда. Pylopoda 3-члениковые, но 3-й членик почти нацело редуцирован и плохо заметен; базальный членик очень широкий, округлый, с сильно-аркообразно-изогнутым внутренним краем, усаженным щетинками; наружный край его прямой. 3 первых свободных грудных сегмента одинаковой длины; 2 последних равной длины, каждый более, чем в 2 раза длиннее 3-го. На спинной стороне панцыря головы и грудных сегментов мелкие известковые шипики и бугорки и широкие выпуклости и вдавления. Сегменты брюшного отдела не равной длины, последовательно удлиняются к заднему концу тела; боковые края брюшных сегментов снабжены парными шиповидными отростками. На спинной стороне брюшных сегментов, по 3 коротких толстых щетинки. Грудные ножки и края головы и первых трех свободных грудных сегментов усажены щетинками. Тельзон широкий у основания, резко суживается дистально, образуя удлиненную треугольную лопасть, покрытую на спинной стороне мелкими волосками; кроме волосков на задней вытянутой часЙи тельзона 3 пары дорзальных и 1 пара апикальных коротких щетинок. Вершина тельзона не -заходит дальше концов ветвей рулевых ног. Базальный членик рулевых ног с оттянутым, нижним внутренним углом с длинной щетинкой на его вершине; наружные ветви немного короче внутренних. Длина 5.5 мм.
Распространение. Японское море: залив Владимира,, 43° 56' N, 135°56'Е, глуб. 308—1002 м (сбор А. Державина, 1927), 5 экз.
* 4. Gnathia derzhavini Gurjan? (фиг. 148).
Gurjanova, 1933, Исследов. морей СССР, 19:81, рис. 3.
Голова большая, округлая, ширина ее незначительно превосходит длину. Глаза очень большие, выпуклые. Боковые отростки головы впереди глаз вытянуто-треугольной формы, заостренные или слегка раздвоенные на конце. Жвалы длинные, узкие; при смыкании концы их перекрещиваются;, зубцы на наружном крае жвал отсутствуют. Передний край головы образует волнистую линию с 2 маленькими округлыми лопастями посредине между основаниями жваЛ. На месте слияния 1-го грудного сегмента с головой глубокая борозда. 1-й свободный грудной сегмент короче 2-го;, 3-й немного длиннее 2-го; 4-й в 2 раза длиннее 3-го, а 5-й по средней линии в 2 раза короче 4-го; последний рудиментарный грудной сегмент хорошо виден, такой же, длины, как брюшные сегменты. На спинной, стороне головы и грудных сегментов шипики, бугорки и неправильной;
1. GN A THIA
233
формы вдавления; края головы и грудных сегментов усажены длинными щетинками. Pylopoda 3-члениковые; базальный членик удлиненной формы с прямым наружным и закругленным внутренним краем, усаженным щетинками. На брюшной стороне базального членика имеются перистые щетинки; последний членик pylopoda настолько редуцирован, что со-
храняется только в виде еле заметного бугорка на 2-м коротком членике. Членики ходильных. грудных ног усажены бугорками, шипами и длинными щетинками; на дистальном конце базального и 3-го ^члеников последней пары шпоровидные отростки. Брюшные сегменты одинаковой длины; боковые , части их оттянуты в стороны, образуя языковидные отростки с пучками длинных щетинок на вершине. Тельзон в общем треугольной формы, суживается дистально и несет пару длинных дорзальных и 3 апикальных щетинки. Конец тельзона не выходит за пределы концов ветвей рулевых ног. Базальный
Фиг. 148. Gnathia derzhavini Gurjan. а — общий вид; Ь — pylopoda; с —голова; d—telson -+- uropoda; е, f, I, k— periopoda II, III, IV и V. w
членик рулевых ног с оттянутым кзади внутренним углом, снабженным щетинкой на вершине. Обе ветви по краям имеют длинные щетинки; наружные ветви короче внутренних. Длина 5.1 мм;
Распространение. Японское море: залив Петра Великого., к югу от о. Аскольд на глубине 121 м и к югу от мыса Среднего на глубине 110 м (сбор ТИРХ’а 1930).
* 5. Gnathia elongata (Кг.). (фиг. 149).
Kroyer, 1847, Gaimard. Voy. Scand., Crust.: 388, pl. 30, f. 3a — 3g (Anceus); Sars, 1899, [Account, 11:55, pl. 23, f. 1.
Самец. Голова большая, закругленно-четырехугольная. Боковые отростки головы впереди глаз хорошо выраженные, заостренные; глаза
'234
14. GN A THUD A E
большие, выпуклые; жвалы снабжены зубцом на наружном крае, концы их при смыкании перекрещиваются; передний край головы между основаниями жвал образует слабо выпуклую трехвершинную лопасть. На месте слияния 1-го грудного сегмента с головой глубокая борозда. Первые 2 членика ножки усиков 1-й пары одинаковой длины; 3-й членик почти в 2 раза длиннее 2-го; жгутик 4-члениковый; последний членик ножки усиков 2-й пары в 172 раза длиннее предпоследнего. Жгутик 7-члениковый. Первые 2 свободных грудных сегмента почти равной длины; 3-й в 2 раза длиннее 1-го, 4-й длиннее 3-го, а 5-й по средней линии равен длине 3-го. Поверхность головы и грудных сегментов покрыта мелкими бугорками, шипиками, короткими волосками и широкими вдавлениями. Последний рудиментарный грудной сегмент очень короткий, но ясно заметный. Брюшные сегменты равной длины; тельзон треугольной формы с парой коротких щетинок у заднего края на спинной стороне и с 2 апикальными щетинками; конец тельзона не выходит за пределы концов ветвей рулевых ног; наружные ветви рулевых ног короче внутренних; по краям обеих ветвей длинные щетинки. Внутренний нижний угол базального членика оттянут кзади. Pylopoda 3-члениковые; базальный членик вытянуто-грушевидной формы; передний членик маленький, но хорошо развит; вдоль внутреннего края базального членика перистые щетинки. На заднем брюшном сегменте оттянутые эпимеры. Длина 4 мм.
Взрослая самка. Тело удлиненное, веретенообразной формы; ширина головы больше ее длины; передние углы впереди глаз образуют маленькие отростки; глаза по бокам головы, выпуклые; передняя часть головы между основаниями усиков конусообразная; на вершине конуса * маленькая вырезка или вдавление. Поверхность головы гладкая, без шипов-бугорков или волосков. Грудной отдел вздутый, яйцевидный; 2 первых i грудных сегмента короткие, 3 последних сегмента очень длинные, 1-й из них, наиболее длинный, почти в 6 раз превышает длину 1-го свободного сегмента груди. Последний редуцированный сегмент плохо заметен под вздувшимся краем 5-го сегмента. Брюшной отдел узкий с одинаковой . длины сегментами; хвостовой веер как у самца. Все тело самки гладкое, вздутое, без бугорков и шипов, имеются лишь волоски по краям 2 первых И' последнего грудных сегментов.
Личинки очень похожи на самок, но легко отличимы от них присутствием шейки (голова не просто причленяется к 1-му свободному сегменту, а имеет узкую вытянутую шейку), более узким и удлиненным телом и сильно вытянутым вперед ротовым конусом. Глаза очень большие, ^занимающие весь боковой край головы.
7. GNATHIA
235
Дальневосточные экземпляры отличаются от арктических относительно более длинным и тонким телом (особенно экземпляры с больших глубин), сильно удлиненными жвалами и более узким тельзоном.
Распространение. Арктическая область: Норвежское побережье от Лофотена до Финмаркена; Баренцево море; восточная и западная
Рис. 150. Gnathia (Elap hognat hid) monodi Gurjan. 7—Самец; 2 — telson -+- uropoda О;
-3—голова <5; 4 — maxillipedes 5—pylopoda <5; 6—pereiopoda II; 7 — pereiopoda III; 8—pereiopoda IV; 9— pereiopoda V; 10 — pereiopoda VI.
Гренландия; Дэвисов пролив; Ян-Майен; восточная и северо-восточная Исландия; Шпицберген, Карское море; Северная часть Атлантического океана — Фарерские о-ва (Гурьянова, 1932). Японское море: залив Петра Великого, в открытой его части, глуб. 76—2900 м в илу (сборы ГГИ и ТИРХ’а 1926—1932).
236
14. GNATHIIDAE
* 6. Gnathia (Elaphognathia) monodi Gurjan. (фиг. 150).
Gurjanova, 1936, Zool. Anz., 114 : 256, fig. 5.
Голова большая, почти квадратная, с закругленными углами и глубоким трехугольным вдавлением в лобной части. Передний край головы имеет глубокую сложную фигурную вырезку. Жвалы сильно вытянуты вперед, удлиненные, с крючкообразно изогнутыми концами и неправильно зазубренным внутренним краем. Глаза большие, плоские, светлокоричневые с крупными фасетками. Обе пары усиков короткие. 2-й членик ножки уриков 1 пары короче 1-го; 3-й членик в 3 раза длиннее 2-го и длиннее 1-го; жгутик 5-члениковый; усики II пары немного длиннее головы, жгутик 7-члениковый. Pylopoda из трех члеников; 3-й членик очень маленький; у его основания 2 крупных щетинки. Внутренний’ аркообразный изогнутый край 1-го членика усажен щетинками. Грудные ножки сравнительно слабые, слабо вооруженные щетинками и шипиками. Плавательные брюшные ножки с удлиненными ветвями, нижний конец которых закруглен и снабжен длинными щетинками. Тельзон треугольной формы с тонким оттянутым кончиком, снабженным тремя длинными щетинками. Концы ветвей рулевых ног не выходят за пределы конца тельзона. Обе ветви уроподов равной длины, вооружены щетинками; наружная ветвь немного уже внутренней. Цвет молочно-беловатый; длина 2.5 мм.
Этот вид относится к индо-пацифическому тропическому подроду Ela.phognath.ia, выделенному Th. Monod в 1926 г., так как строение жвал и переднего края головы приближает его к видам Gnathia ferox (Haswell), G. insolita Stebb., Elaphognathia lucanoides Monod и E. rangifer Monod, которые объединяются в один подрод Elaphognathia.
Распространение. Японское море: район острова Петрова. (42° 49 N, 138°48' Е) на глубине от 0 до 2 м в толстых корках Lithothamnion,. на камнях.
ZUSAMMENFASSUNG
KURZE ZOOGEOGRAPHISCHE OBERSICHT DER ISOPODEN DER BERINGS-SEE, DES OCHOTSKISCHEN UND JAPANISCHEN MEERES
Nach der Bearbeitung der Sammlungen des Zoologischen Institutes der Akademie der Wissenscbaften hat sich die Artenzahl der fiir die Berings-See, das Ochotskische und Japanische Meer bekannten Isopoden stark vergrossert. Es sind zur Zeit aus diesen Meeren 128 Arten in 50 Gattungen, 14 Familien und 6 Unterordnungen bekannt. Dabei entfallen fiir die Berings-See 54, fiir das Ochotskische Meer 47 und fiir das Japanische Meer 62 Formen.
Zunachst soli eine Obersicht der Isopoden nach den einzelnen Meeren gegeben werden, doch muss gleich bemerkt werden, dass unsere Kenntnis dieser Gruppe im nordlichen Pazifik zur Zeit noch fragmentarisch ist und diese „Obersicht" nur als Versuch einer provisorischen Einteilung gelten kann. Am wenigsten erforscht ist die Fauna des Ochotskischen Meeres, am besten bekannt diejenige des Japanischen Meeres. Immerhin sind in den letzten Jahren durch Expeditionen des Hydrologischen Institutes, Leningrad, auch die grossten Tiefen des Ochotskischen und Japanischen Meeres erforscht worden, wahrend in der Berings-See der Benthos bis zu Tiefen von 4000 m Icollektiert wurde. Auf diese Weise finden sich in unseren Sammlungen Materiale von der Brandungslinie und dem Litoral bis zu Tiefen von 3500 bis 4000 m.
Berings-See. Aus dieser See sind zur Zeit 55 Arten Isopoden bekannt, welche in 28 Gattungen, 13 Familien und 5 Unterordnungen verteilt sind (siehe Verzeichnis auf S. 15—16).
Beim Betrachten des Verzeichnisses sehen wir, dass es aus Gruppen verschiedener Herkunft zusammengesetzt ist. Den Kern der Isopodenfaune bilden 20 Arten, welche friiher nur aus der Berings-See bekannt waren und deshalb provisorisch als autochthone oder endemische Formen bekannt werden sollen. Zu ihnen gehoren Janira holmesi, Janiropsis derjugini, Microprotus coecus, Munna arnholdi, M. stephenseni, M. avatshensis, Tecticeps renoculis marginaiis, Synidothea acuta, S. laevis, S. picta, Arcturus longispinis, A. diversispinis, A. magnispinis, A. brevispinis, A. glabrus, A. tritaeniatus, A. hirsutus, Pleuroprion intermedium, Ligidium tenue, Holophryxus giardi. Alle diese Arten, ausser Arcturus hirsutus und Holophryxus giardi, sind mehr oder weniger -Flachwasserformen. Ihre Differenzierung in den Lebensverhaltnissen der Berings-See ist deshalb sehr wahrscheinlich, so dass sie mit vollem Recht
238
ISOPODA
als Autochthon fur dieses Meer gelten konnen. Die 2 obengenannten Formen aber, A. hirsutus und H. giardi, gehoren der Tiefsee an. Sie sind erst von 2000 m abgefunden worden und gehoren wahrscheinlich zu den Vertretern der abyssalen Fauna des nordlichen Pazifik, da H. giardi auf einer pazifischen Tiefseegarneele parasitiert und A. hirsutus nach dem Bau seines Telsons zu einer Gruppe von Arten gehort, welche vom „Challenger" im Abyssal des Stillen Ozeans gefunden wurde.
Zur zweiten Gruppe gehoren die Arten, welche nur in der Berings-See und im Ochotskischen Meere gefunden worden sind. Hierher gehoren 5 Formen: Janira sarsi, Tecticeps alascensis, Idothea aleutica, Synidothea nebulosa, S. spinosa, Penthidothea wosnesenskii. Bezeichnenderweise sind das auch Kiisten- und Flachseeformen, die wohl wahrscheinlich als Autochthone der nordasiatischen pazifischen Kiisten angesehen werden miissen.
Die dritte Gruppe wird durch 6 Arten gebildet, die iiber alle 3 nord-ostasiatischen Meere verbreitet sind. Das sind Janira erostrata, Rocinela angustata, Rocinela belliceps, Neosphaeroma oregonensis, Idothea ochotensis, Arcturus beringanus. Fast alle diese Formen gehoren dem Flachwasser an und sind augenscheinlich fur die altweltlichen Kiisten des nordlichen Pazifik charakteristisch, wahrend beide Arten von Rocinela und Neosphaeroma auch langs den amerikanischen Kiisten weit verbreitet sind.
Einige Formen der Berings-See sind wohl im Japanischen Meere gefunden, scheinen aber im kalten Ochotskischen Meere zu fehlen. Das sind Jani-ropsis kincaidi und Trichoniscus papillicornis. Es ist aber moglich, dass. diese Arten nur infolge der sehr ungeniigenden Erforschung des ochotskischen. Litorals und Supralitorals dort noch nicht aufgefunden worden sind.
Eine besondere Gruppe bilden die arktischen Formen, die bis die Berings-See eindringen. Dazu gehoren 7 Arten: Munnopsurus giganteus, Calathura brachiata, Synidothea bicuspida, S. marmorata, S. nodulosa, Pleuroprion murdochi und Mesidothea entomon, welche im Stillen Ozean eine besondere Unterart, M. e. orientalis bildet. Was nun Munnopsurus,. Calathura und Mesidothea betrifft, so sind das arktische Autochthone, wahrend Synidothea und Pleuroprion sich wahrscheinlich aus dem Stillen Ozean. iiber die Arktis verbreitet haben (siehe weiter unten). Alle diese Arten sind entweder nur im nordlichen Teile der Berings-See, oder auch an den asia-tischen Kiisten derselben gefunden worden.
Ausserdem finden sich auch weit verbreitete artkisch-boreale Formen (Bopyroides hippolytes) und Formen, die eine Tendenz zur kosmopolitischen Verbreitung besitzen (Limnoria lignorum und Phryxus abdominalis).
Ein besonderes Interesse beansprucht das Vorhandensein in der Berings-See einer Gruppe von typisch abyssalen Arten des nordlichen Pazifik: Munnopsurus laevis, Aega magnoculis, Arcturus hirsutus, Holophryxus giardi und Arthrophryxus beringanus. Das Vorkommen dieser Formen in der Berings-See in Tiefen von 1000—2000 m weist auf eine Verbindung der beringianischen Tiefsee mit derjenigen des nordlichen Pazifik bin.
ZOOGEOGRAPHISCHE OBERSICH1
239
Gut vertreten sind in der Berings-See auch die Elemente der borealen pazifischen Fauna. Zu ihnen gehoren 5 Formen: Pentidothea white!, Exosphaeroma amplicauda, Cilicaea cordata, Janiropsis kincaidi, Neosphaeroma oregonensis. Diese konzentrieren sich im siidostlichen Teile des Meeres, an der Kette der Aleuten, und fehlen bei Kamtshatka. Eine Form, Argeia pugettensis, ist typisch subarktisch-pazifisch, wahrend eine Reihe von Formen — Rocinela cornuta, Idothea aleutica, Pentidothea wosnesenskii, Aega symmetrica und Ligia pallasi—hauptsachlich langs den subarktischen amerikanischen Kiisten verbreitet ist.
Schon aus dieser allgemeinen Ubersicht der Isopodenfaunen der Berings-See kann man entnehmen, dass eine Verbindung zwischen der Tiefsee-fauna dieses Meeres mit derjenigen des Stillen Ozeans existiert, ferner auf einen Austausch zwischen der Fauna der Berings-See und des Ochotskischen Meeres einerseits, der Berings-See und dfcm Arktischen Meere andrer-seits. Die Verbindung mit der pazifischen Tiefsee findet hauptsachlich in der Gegend der Kommandor-lnseln und in der sog. Kamtshatka-See, sowie durch die Meeresstrassen zwischen der Aleuten statt (siehe die Verbreitung im systematischen Teile).
Einige Flachwasser- und Kiistenformen des siidostlichen Teiles der Berings-See (der Aleutischen Kette und der Kiisten von Alaska) dringen bis zur Westkiiste von Kamtshatka und dem Ochotskischen Meer durch die erste Kurilische Strasse bei Кар Lopatka. Die westamerikanischen Formen Idothea aleutica und Synidothea nebulosa sind langs den Aleuten bei den Kommandor-lnseln, in der Avatshinsk-Bucht auf Kamtshatka und endlich bei Bolsheretzk an der Westkiiste von Kamtshatka gefunden worden, wahrend sie sonst im Ochotskischen Meere nicht vorzukommen scheinen.1 Dass die Verbindung iiber die erste Kurilische Strasse stattfindet, ist durch N. Smirnov uhd Begak nachgewiesen, welche gerade in dieser Strasse die westamerikanische Form Janira sarsi gefunden hatten.
Die Fauna des Ochotskischen Meeres iibt auch einigen Einfluss auf die-jenige der Berings-See aus. So kommt im Berings-Meer die typisch-ochots-kische Tecticeps renoculis, zwar in einer eigenen Form, var. marginalis, vor. Das Zentrum der Verbreitung dieser Art liegt zweifellos im Ochotskischen Meere, wo 4 Formen dieser Art und noch andere Arten der Gruppe vorhan-den sind.
Es existiert auch ein Austausch zwischen den Faunen der Berings-See und dem Eismeere. Das Vorkommen von Munnopsurus giganteus, Mesidothea entomon und Calathura brachiata im Beringsmeere weist deutlich auf das Eindringen der arktischen Fauna in diese Gewasser. Andrerseits iibt der Stille Ozean auch einen starken Einfluss auf die Fauna des Eismeeres aus. Dieser Einfluss aussert sich durch das Eindringen einiger Arten von Syn-
1 Im Litoral des Tauj-Meerbusens und der Shantaren ist Idothea ochotensis .geniein, wahrend /. aleutica weder von Ushakov, noch von Zachs, oder anderen Forschern gefunden worden ist.
240
ISOPODA
idothea und Pleuroprion. durch die Beringstrasse in die Tshuktschen- und Beaufort-See. Das Entstehungszentrum dieser beiden Gattungen liegt augen-scheinlich im nordlichen Teile des Stillen Ozeans, worauf ihre Verbreitung hinweist. So ist der grosste Teil der Arten von Synidothea, etwa 25, eben hier gefunden worden, namlich im nordlichen Pazifik, in der Berings-See, im Ochotskischen und Japanischen Meere. Nur 2 Arten sind aus dem westlichen Teil des nordlichen Atlantik und 3 Arten aus der Arktis bekannt. Diese 5 Arten sind aber auch uber den Stillen Ozean verbreitet. Nach den Fundorten nun kann man sich das Eindringen dieser Gattung und ahnlicher Faunelemente in die Arktis auf folgendem Wege erklaren. Die pazifischen Formen dringen durch die Beringstrasse in das Eismeer und schlagen dann 2 Wege ein. Die einen wenden sich langs der sibirischen Kiiste nach Westen {Synidothea bicuspida, S. nodulosa), dringen durch die Ostsibirische, Laptev-See und’das Karische Meer bis Novaja Semlja vor und verbreiten sich weiter durch die Jugorsche und Karische Strasse in den siidbstlichen Teil der Barents-See. So geht, z. B., «S', bicuspida nach Westen bis zum Meridian von Bucht Charlovka am ostlichen Teile der Murman-Kiiste und bis zum Eingange in das Weisse Meer, wahrend 5. nodulosa langs der Kiiste von Novaja Semlja nach Norden vordringt, zur zirkumpolaren Form wird und eine Reihe geographischer Formen bildet. Andere Formen (Pleuroprion, Synidothea marmorata, Arcturus baffini) gehen nach Osten in die Beaufort-See, weiter langs der Nordkiiste von Alaska und Kanada und erscheinen an den Kiisten von Labrador, Baffinsland, Westgronland, Island und den Faroem. Das Vorhandensein dieser 2 Verbreitungsstrassenerklartauchden Umstand, dass verschiedene fast zircumpolare Formen der Flachsee in ihrer Verbreitung einen Hiatus aufweisen, und zwar in der Gegend zwischen Gron-land, Island, Skandinavien und Spitzbergen. Verschiedene Formen (z. B. einige Arten von Synidothea und Pleuroprion) dringen auf diese Weise in der Arktis nach Osten und Westen vor, ohne dabei vollstandig zircumpolar zu werden.
Die Verbreitung der arktischen und pazifischen Formen in der Berings-See gibt uns Veranlassung diese in 2 Bezirke zu teilen. Der eine umfasst deri nordlichen flachen Teil der Berings-See, sowie die Ostkiiste von Kamtshatka. Hier hat die Isopodenfauna einen deutlichen arktischen Anstrich (Vorhandensein typisch-arktischer Arten, wie Munnopsurus' giganteus, Calathura brachiata, Mesidothea entomon, sowie Fehlen typisch-borealer pazifischer .Arten: Cilicaea, Neosphaeroma, Janiropsis etc.). Der andere, sich durch warmeres Wasser unterscheidende Bezirk, umfasst den siidostlichen Teil des Meeres, die Gegend um die Kommandor-Inseln, sowie die Kette der Aleuten. Er unterscheidet sich von dem Vorigen durch das Fehlen der arktischen und Vorhandensein von borealen pazifischen Formen. Eine solche Einteilung der Berings-See hatte schon W. Dall1 in den Jahren 1886 und 1899 auf Grund
1 W. Dall. 1886. Report on Bering Island Mollusca... Proc. U. S. Nat. Mus., 9. W. Dall. 1899. The Mollusk Fauna of the Pribilof Islands. Report of Fur-seal Investigations 1896—1897, Part 3, 1899.
ZOOGEOGRAPHISCHE (JBERSICHT
241
der Verteilung der Mollusken vorgeschlagen und auch A. Ivanov kam im Jahre 1933 bei der Bearbeitung des beringianischen Benthos zu einem gleichen Resultate.1
Die Verbreitung der Isopoden bestatigt also die von Dall und Ivanov gemachten Schliisse uber das Vorhandensein von 2 Provinzen in der Beririg-See: der arktischen Kamtshadalischen und der warmeren Aleutischen Pro-vinz, in welche sowohl abyssale Warmwasserformen des Stillen Ozeans eindringen (wie Aega magnoculis, Arthrophryxus, Holophryxus, Munnopsu-rus laevis, besonders in der Gegend der Kommandor-Inseln und Attu), ,als auch amerikanische Kiisteiiformen, die nach Siiden bis Siid-Kalifornien verbreitet sind {Cilicaea cordata, Exosphaeroma amplicauda, Penthidothea whi-tei, Ligia pallasi, Aega symmetrica, Janiropsis, einige Arten von Arcturus etc.). Die hydrologischen Eigenheiten dieser 2 Provinzen sind ganz verschieden und auch die Verteilung der Tiefen, Stromungen und des Eises lasst eine sehr deutliche Abgrenzung derselben zu: der flachen, kalten und im Winter «eisbedeckten Kamtshadalischen und der tiefen, nie zufrierenden und durch die pazifischen Tiefenwasser erwarmte AJeutische Provinz.
Ochotskisches Meer. Zur Zeit sind im ganzen 47 Isopodenformen bekannt, die in 41 Arten, 16 Gattungen, 7 Familien und 3 Unterordnungen verteilt sind (siehe Verzeichnis auf S. 20). Das Fehlen der 3 iibrigen Unterordnungen {Gnathiidea, Epicaridea und Oniscoidea) lasst sich leicht durch unsere zur Zeit noch sehr ungeniigende Kenntnis der ochotskischen Fauna, insbesondere der Kiistenzone und der Schmarotzer der Krebstiere.
Das Verzeignis der Isopoden des Ochotskischen Meeres zeigt eine noch weit grossere Verschiedenartigkeit, als wie sie an, der Berings-See bekannt ist. Ausser den zoogeographischen Gruppen, die in der Berings-See vertreten waren, findet man hier noch einige neue. Zuerst wollen wir die Gruppe der endemischen Formen mustern, die im Ochotskischen Meere nicht weniger als 16 betragt:1 2 Munnopsurus giganteus var. ochotensis, Tecticeps renoculis, T. renoculis laevis, T. renoculis nodulosus, T. renoculis cari-natus, T. leucophthalmus, Synidothea longicirra, S. tuberculata, S. cinerea, Arcturus ulbani, A. granulatus, A. setosus, A. setosus seminudus, A. setosus acuticaudalis, A. beddardi, Pleuroprion tarasovi. Wie man sieht, finden sich unter den endemischen Formen verschiedene Unterarten und Varietaten. Dieser Umstand weist wohl darauf hin, dass die Fauna des Ochotskischen Meeres verhaltnismassig jung ist. Andrerseits sehen wir aber hier eine intensive Artenbildung, da verschiedene Arten sich hier in Reihen von Lokalfor-men aufspalten und weitverbreitete, wenig variable Formen besondere, ochot-skische Unterarten bilden (z. B. Munnopsurus giganteus ochotensis).
1 Vortrag von Ivanov im Jahre 1933 des hydrobiologischen Vorstandes des Hydrologischen Institutes iiber die Resultate der Arbeiten in der Berings-See im Jahre 1932.
2 Es ist moglich, dass bei besserer Durchforschung einige dieser Formen auch in den angrenzenden Meeren gefunden werden.
Гурьянова
242
ISOPODA
Zur zweiten Gruppe gehoren 15 Arten, die fiir das Ochotskische Meer und die Berings-See gemeinsam sind. Dazu gehoren Janira sarsi, J. erostrata, Munnopsurus giganteus, Calathura brachiata, Rocinela belliceps, R: angus-tata, Aega magnoculis, Tecticeps alascensis, Mesidothea entomon orientalis, Idothea ochotensis, I. aleutica, Pentidothea wosnesenskii, Synidothea spinosa,. S. nebulosa, Arcturus beringanus. Von ihnen ist ein Teil, wie schon vorher erwahnt war, aus der Berings-See langs der Kiiste von Kamtshatka durch die erste Kurilische Strasse bis Bolsheretsk vorgedrungen (Calathura, Munnopsurus). Ein anderer Teil (Aega magnoculis, Rocinela angustata) gehort der pazifischen Tiefsee an und ist von ihr aus sowohl in das Ochotskischen Meer, als auch in die Berings-See eingedrungen.
Die dritte Gruppe wird aus Arten gebildet, die sowohl im Ochotskischen Meere, als auch im Japanischen Meere vorkommen. Das sind folgende 18 Formen: Janira soldatovi, J. erostrata, Ilyarachna starokadomskii, Paran-thura japonica, Rocinela maculata, R. belliceps, R. angustata, Tecticeps renoculus laevis, Idothea fewkesi, I. ochotensis, I. derjugini, I. orientalis, Synidothea bicuspida lata, Arcturus setosus seminudus^ A. setosus acuticau-dalis, A. beringanus, A. hastiger und A. crassispinis. Das Vorhandensein dieser Formen sowohl im Ochotskischen, als auch im Japanischen Meere, deutet mit Sicherheit auf einen Austausch zwischen den Faunen dieser Meere hin.. Dieser Austausch geht auf 2 Wegen vor sich, zum grossten Teil durch die La Perouse-Strasse, teilweise aber auch durch die Amur-Bai und die Nevelski-Strasse. So gehen, z. B., Rocinela maculata, Ilyarachna starokadomskii, Idothea fewkesi, I. orientalis, I. derjugini, Synidothea bicuspida lata und Arcturus crassispinis, welche im Japanischen Meere weit verbreifet und dort die gewohnlichsten Flachwasserformen sind, durch die Tataren-Strasse und gelangen in die Aniva-Bai und in den sudwestlichen Teil des Ochotskischen Meeres. In anderen Teilen des Ochotskischen Meeres1 sind diese Formen,. ausser Idothea derjugini und I. fewkesi, nicht gefunden worden. Augenschein-lich haben sie sich aus dem Japanischen in das Ochotskischen Meer durch die La Perouse-Strasse verbreitet. Andrerseits sind Formen, wie Tecticeps renoculis laevis,-Arcturus setosus, A. setosus acuticaudalis, und A. hastiger, die im Ochotskischen Flachwasser sehr gemein sind und speziell auch in der Aniva-Bai vorkommen, in den nordlichen Teil des Japanischen Meeres (hauptsachlich in die Tataren-Strasse) eingedrungen. Auch hier ist der Weg durch die La Perouse-Strasse klar genug. Aber auch durch den Nevelski-Sund geht eine Vermittelung zwischen den Faunen des Ochotskischen und Japanischen Meeres vorstatten, trotz der stark verringerten Salzhaltigkeit des Wassers in dieser Strasse.1 So ist, z. B., Paranthura japonica, eine Astuarienform siidlicher Herkunft, welche aber an der Westkiiste des Ochotskischen Meeres
1 Nach der Meinung von Derjugin bildet das Astuarium des Amur eine hydrologische Barriere, die von der marinen Fauna nicht iiberschritten werden kann, so dass der Nevelski-Sund fiir diese unpassierbar ist.
ZOOGEOGRAPHISCHE OBERSICHT 243
in der Nogajev-Bucht, sowie an der Miindung des Amur gefunden ist, zweifel-los aus dem Japanischen Meer in das Ochotskischen Meer auf dem eben erwahnten Wege gelangt. Auch Idothea fewkesi, eine siidliche Form, die aber ausser dem Japanischen Meere noch bei den Shantaren Und der Insel Langr an der Amurmiindung gefunden ist, muss zweifellos aus dem Japanischen in das Ochotskischen Meer durch den Nevelski-Sund gelangt sein. Andrerseits ist Idothea ochotensis, eine Form, die einen sehr verschiedenen Grad des Salzgehaltes vertragt, eine der gewohnlichsten Litoralformen sowohl im Ochotskischen Meere (z. B. bei den Shantaren), als auch in der Tataren-Strasse. Da sie ebenfalls bei der Insel Langr an der Amurmiindung gefunden worden ist, so muss man annehmen, dass auch sie sich durch die Nevelski-Strasse verbreitet hat. So sieht man denn, dass auch durch die Nevelski-Strasse ein wenn auch schwacher, so doch unleugbar beiderseitiger Austausch zwischen den Faunen des Japanischen und des Ochotskischen-Meeres stattfindet.
Alsdann findet sich eine Gruppe von 6 Arten, die in alien 3 Meeren vorkommen (das Verzeichnis ist bei der Beschreibung der Berings-See angegeben).
Eine besondere Gruppe bilden im Ochotskischen Meere die arktischen Arten, von denen im ganzen 5 vorhanden sind: Munnopsurus giganteus (welcher unter der Einwirkung der Eigentiimlichkeiten des Ochotskischen Meeres dort noch eine Lokalform, M. g. ochotensis, bildet), Calathura bra-chiata, Mesidothea entomon (welche die Unterart orientalis bildet), Synidothea muricata und Arcturus bajfini. Die ersten 2 Arten gelangen augenscheinlich aus dem Eismeer durch die Berings-See in das Ochotskischen Meer, wahrend die 2 letzten, Synidothea muricata und Arcturus baffini, urspriinglich aus dem Stillen Ozean stammen und erst spater in die Beaufort-See gelangt sind, von wo «S'. muricata sich nach Westen bis zum Karischen Meer, A. baffini sich nach Osten, langs der Kanadischen Kiiste bis Westgronland, Island und den Faroern verbreitet hat. Zur nachsten Gruppe gehoren die pazifischen Tiefseeformen, zu denen Aega magnoculis und wahrscheinlich Arcturus beddardi (welcher bis jetzt als endemische Form angesehen wird) gehoren. Sie sind im siidlichen Teile des Ochotskischen Meeres gefunden worden und sind hierher wahrscheinlich durch die tiefen Strassen zwischenden siidlichsten der Kurilen aus dem Stillen Ozean eingedrungen.
Im Ochotskischen Meere findet sich auch warmeliebende, fast tropische sehr weit verbreitete Form, Cyathura carinata (in einer Flussmiindung in der Nogajev-Bucht). Weiterhin, ist eine antarktische Form, Livoneca raynaudi von Woznessenski im Jahre 1847 bei Ajan gefunden worden. Zuletzt findet sich im Ochotskischen Meere eine siidliche schmarotzende Siisswasserform, Livoneca amurensis. Diese ist uber den Himalaya und das Gebiet des Amur samt Lefu-Miindung Verbreitet und geht bis in das Astuarium des Amur. Eine eigentiimliche Verbreitungbesitzt Pleuroprion hystrix, welcher bei Ajan erbeutet worden ist, wahrend er friiher nur aus dem nordlichen Teile 16*
244
ISOPODA
des Atlantischen Ozeans bekannt war. Nach der Terminologie von L. Berg (1934) kann er zu den amphiborealen Formen gerechnet werden.
Auf diese Weise muss die Isopodenfauna des Ochotskischen Meeres als zweifelloS reich'und mannigfaltig angesehen werden, trotzdem sie noch sehr ungeniigend erforscht ist. Die eigenartigen Lebensbedingungen der sehr kalten kiisternahen und flachen Teile des Ochotskischen Meeres wirken in hohem Grade modifizierend auf die Fauna ein, so dass hier eigentiimliche, spezifisch ochotskische Formen ausgebildet werden. Die Verbindung der ochotskischen Tiefsee durch die kurilischen Strassen mit dem Stillen Ozean haben zur Folge, dass im Ochotskischen Meere pazifische Tiefsee-Isopoden gefunden werden. Die Nachbarschaft und Verbindung mit dem Japanischen Meere bereichert die Fauna des Ochotskischen Meeres durch japanische Warmwasserformen. Die mannigfaltige Gliederung des Ochotskischen Meeres (sowohl in horizontaler, als auch in senkrechter Richtung) und die Verschie-denheit der Lebensbedingungen in einzelnen Teilen, bedingt eine reiche Gliederung der autochthonen Arten in Lokalformen und Varietaten. Die Einteilung von Tecticeps. renoculis Rich, und Arcturus setosus Gurjan. in zahlreiche Rassen bildet ein gutes Beispiel einer solchen durch verschiedene Lebensbedingungen verursachten Gliederung. Die Verschiedenheit der hydro-logischen Bedingungen zwischen Flachwasser und Tiefsee haben zur Folge, dass im Ochotskischen Meere sowohl arktische und boreale, als auch war-meliebende, fast subtropische Formen vorkommen.
Japanisches Meer. Bis jetzt sind vom Japanischen Meer (ausser dem subtropischen siidlichen Teil an den japanischen Kiisten) 66 Formen Isopor den bekannt, die in 64 Arten, 30 Gattungen, 11 Familien und 6 Unterord-nungen verteilt sind (siehe Verzeichnis auf S. 24—25).
Das Japanische Meer ist im Resultate fast zehnjahriger Forschungs-arbeitendes Pazifischen Fischerei-Institutes in Wladiwostok, sowie des Hydro-logischen Institutes in Leningrad zur Zeit weit besser bekannt als die Berings-See und des Ochotskische Meer. Insbesondere ist die Bai Peters des Grossen gut erforscht. Es ist deshalb nicht zu verwundern, dass die Artenzahl der fur des Japanische Meer bekannten Isopoden die Verzeichnisse der Berings-See und das Ochotskischen Meeres bedeutend iibertrifft. Und zwar ist nicht nur der Artbestand besser bekannt, sondern auch die Verbreitung der Arten innerhalb des Japanischen Meeres schon erforscht, so dass man hier schon von einiger Gesetzmassigkeit derselben reden kann. Von den Formen, die im Ochotskischen und Japanischen Meere, oder in der Berings-See und dem Japanischen Meere vorkommen, oder fiir alle 3 Meere gemeinsam sind, war schon oben die Rede. Aber auch wenn man diese Formen abrechnet, so ist doch die Isopodenfauna des Japanischen Meeres sehr reichhaltig und zusam-mengewiirfelt zu nennen.
Zur ersten Gruppe gehoren die endemischen Formen des Japanischen Meeres, im ganzen 25 Arten: Janiropsis serricaudis, Munna subneglecta, Munnopsurus minutus, Ilyarachna zachsi, Eurycope spinifrons, E. pavlenkoi,
ZOO GEO GRAPHISCHE 0BERS1CHT 245
Limnoria japonica, Tecticeps glaber, T. renoculis serratus, Sphaeroma sieboldi, Exosphaeroma ovata, Rocinela niponia, R. japonica, Livoneca epimerias, L. sacciger, Synidothea epimerata, S. brazhnikovi, S, excavata, Arcturus cre-nulatus, Astacilla polita, Gnathia tuberculata, G. derzhavini, G. schmidti, G. rectifrons und G. (Elap ho gnathia) monodi. Der grosste Teil dieser Formen ist nur in geringen Tiefen der Bai Peters des Grossen gefunden worden, einige an der Japanischen Kiiste bei Hakodate und nur 3 (Eurycope spinifrons, Gnathia tuberculata, G. schmidti) in grosseren Tiefen (etwa 1000 m) der Bai Peters des Grossen, doch muss erwahnt werden, das E. spinifrons schon von 663 m an gefunden worden ist.
Die zweite Gruppe, die der arktischen Formen, ist nur durch 1 Art Gnathia elongata vertreten. Diese Art kommt im Japanischen Meere in sehr verschiedenen Tiefen, von 76 m bis 2900 m vor.
Zur dritten Gruppe gehoren arktisch-boreale Formen, die in der Arktis weit verbreitet sind und sowohl im Atlantischen, als auch im Stillen Ozean weit nach Siiden gehen. Zu ihnen gehort im Japanischen Meere auch nur eine Art — Bopyroides hippolytes.
Zur vierten Gruppe gehoren subarktische pazifisch-asiatische Formen, die hauptsachlich an unseren Kiisten verbreitet sind. Im Japanischen Meere finden sich 10 solche Arten: * Janira soldatoyi, J. erostrata, Ilyarachna sta-rokadomskii, Paranthura japonica, Rocinela maculata, * Idothea ochotensis, * Synidothea bicuspida lata, Arcturus beringanus, * A. hastiger, A. crassispi-nis. Die meisten von ihnen sind iiber das ganze Japanische Meer verbreitet, einige kommen aber nur im nordlichen Teile, im Tataren-Sunde vor (4 Arten die mit Sternen bezeichnet sind; sie sind auch im Ochotskischen Meere gefunden).
Die fiinfte Gruppe bilden die subarktisch-pazifischen Formen, welche sowohl an den altweltlichen, als auch an den amerikanischen Kiisten vorkommen. Hierher gehoren 4 Arten: Trichoniscus papillicornis, Rocinela angus-tata, R. belliceps, Argeia pugettensis. Es muss bemerkt werden, dass die 3 letzten Ektoparasiten an Fischen (Rocinela) und Krebstieren (Argeia) sind, was ihre weite Verbreitung bedeutend erleichtert.
Zur sechsten Gruppe gehoren 6 pazifisch-boreale Formen: Janiropsis kincaidi, Neosphaeroma oregonensis, Idothea orientalis, I. derjugini, Pentidothea japonica und Astacilla dilatata. Das sind alles Flachwasser- und Kiistenformen, ausser Astacilla, welche auf der japanischen Seite des Meeres gefunden wird.
Die siebente Gruppe wird durch 10 subtropische Formen gebildet. Zu ihnen gehoren Nerocila japonica, Meinertia oxyrrhynchaena, Livoneca amu-rensis, Holotelson tuberculatus, Cymodoce japonica, Dynamene galbra, Pentidothea rotundata, Cleantis isopus, Pentias hayi und Ligia cinerascens. Sehr bezeichnend ist es, dass alle diese Formen entweder im Litoral, oder im ganz seichten passer an den Kiisten gefunden werden. Livoneca amuren-sis kommt in der Miindung des Lefu vor.
246
ISOPODA
Zur achten Gruppe gehort eine amphiboreale Form, Cleantisplanicauda । (westindische Form).
Die neunte Gruppe wird durch 2 kalifornische warmeliebenden Flach-wasserformen gebildet — Idothea fewkesi und Synidothea harfordi.
Zur zehnten Gruppe gehoren endlich Formen, die eine Tendenz zur kosmopolitischen Verbreitung aufweisen, nahmlich Idothea metallica, Phryxus abdominalis und Limnoria lignorum.
Man sicht also, dass die Isopodenfauna des Japanischen Meeres im Ver-gleich mit derjenigen der Berings-See und des Ochotskischen Meeres mehr Warmwasserformen aufweist. Aus der Gesamtzahl von 66 Formen sind 21, d. h. 31/%, was indessen nur provisorisch angenommen werden kann. Etwa 25% (16 Formen) sind gemassigt-subarktisch, ungefahr 31 % (21 Formen) Warmwasserformen und weniger als 2% (1 Form) arktische Arten. Eigent-liche subtropische Kiisten- und Flachwasserformen sind aber nur in der Bai Peters des Gross'en gefunden worden und gehen nicht weiter nach Norden, ausser Ligia cinerascens und Cymodpce japonica, welche bis zur Pre-obrazhenje- Bucht verbreitet sind. Sehr bezeichnend fur das Japanische Meer ist das vollstandige Fehlen von eigentlichen pazifischen Tiefseeformen, was auf die vollstandige Angeschiedenheit der Tiefer des Japanischen Meeres von derjenigen des eigentlichen Stillen Ozeans hinweist. Bekanntlich ist das ja auch der Fall, da die Meeresstrassen und Sunde, welche das Japanische Meer mit dem Ozean verbindefi, samtlich seicht sind. Man muss hinzu-fiigen, dass das Japanische Meer iiberhaupt keine echten Tiefseeformen besitzt. Die abyssale Zone des Japanischen Meeres ist von Formen bewohnt, die im Bathyal und Sublitoral weit verbreitet sind, was auf ein geringeS Alter der Tiefsee dieses Meeres hindeutet. Daraus geht noch eine Eigen-tiimlichkeit der Isopodenfauna des Japanischen Meeres hervor: die Sublito-ralformen haben hier eine sehr grosse Vertikalverbreitung, von 50—60 m, bis zu den grossten Tiefen von etwa 3000 m. Wenn nun die Tiefsee des Japanischen Meeres von derjenigen des Ozeans isoliert ist, so findet zwischen der Fauna geringerer Tiefen, der sublitoralen und litoralen, ein sehr bemerk-barer Austausch statt. Uber den Austausch der Faunen zwischen dem Ochotskischen und dem Japanischen Meere durch die La Perouse-Strasse und. den Nevelski-Sund haben wir schon erwahnt (siehe Ubersicht des Ochotskischen Meeres). Aus dem Gelben Meere dringt durch die Zushima-Strasse, zusammen mit einem Zweige der warmen Zushima-Stromung, eine Anzahl subtropischer Formen bis in die Bai Peters des Grossen ein. Langs den Kiisten Japans verbreiten sich in das Japanische Meer einige tropische Formen {Ligia exotica, Idothea metallica, einige Vertreter der Gattungen Tylos, Cirolana, Sphaeromidae etc.), die aber im Verzeichnis nicht ange-fiihrt waren, da sie an den Kiisten von USSR nicht gefunden werden. Sehr bezeichnend fiir das Japanische Meer ist auch der Umstand, das die Warmwasserformen hier nur die obersten, im Sommer stark durchwarmten Schich-ten bewohnen, wahrend von 50—60 m an und weiter abwarts nur boreale
Z О О G Е О G R А Р НIS С Н Е UBERSICHT
247
und Kaltwasserformen vorkommen. Dieses Verhaltnis ist genau entgegenge-setzt demjenigen, welches wir in der Berings-See und dem Ochotskischen Meere finden: dort finden sich die gemassigten und wenigen Warmwasserformen ausschliesslich in den verhaltnismassig warmeren Tiefenschichten, wahrend in den sehr kalten Oberflachenschichten nur kalteliebende Formen vorkommen.
Nach dieser Einzelcharakteristik der Isopodenfauna der Berings-See, der Ochotskischen und Japanischen Meeres wollen wir zu einer allgemeinen zoogeographischen Obersicht iibergehen. Zur Zeit kennen wir von alien 3 Meeren 128 Fprmen von Isopoden, die in 119 Arten, 49 Gattungen, 14 Fa-milien und 6 Unterordnuhgen verteilt sind. Da aber die schmarotzenden .Formen (Cymothoidae und Epicar idae) des Japanischen Meeres noch wenig bekannt sind, das Ochotskische Meer und die Berings-See aber iiberhauj>t * noch mangelhaft erforscht sind, so kann man diese Zahl durchaus nicht als endgiiltig ansehen. Nach einer fortgesetzten Forschungstatigkeit wird nicht nur die Gesamtzahl der Arten mit Sicherheit noch wachsen, sondern es ist auch moglich, dass unsere jetzigen zoogeographischen Ausfiihrungen teil-weise verandert werden miissen. Jetzt aber konneri wir fur die Berings-See, das Ochotskische und Japanische Meer im ganzen 10 zoogeographische Gruppen unterscheiden (die Anordnung dieser Gruppen in den einzelnen Meeren ist im Verzeichnis 3 auf S. 28 angegeben).
Die erste, am zahlreichsten vertretene Gruppe bilden die autochthonen Elemente, die nach dem heutigen Stande unserer Kenntnisse als endemische Formen der einzelnen Meere angesehen werden miissen. Die Zahl dieser Formen betragt 59, d. h. 45.3% aller • bekannten Formen (die Verzeichnisse siehe bei der Besprechung der einzelnen Meere). Wie schon erwahnt, finden sich endemische Formen in der Berings-See 18, im Ochotskischen Meere 16, im Japanischen Meere 25. Von ihnen gehoren nur 2 Arten, Arcturus beddardi und Tecticeps leucophthalmus der Tiefsee des Ochotskischen-Meeres an, die iibrigen sind entweder Litoralformen, oder gehoren der oberen Stufe «des Sublitorals an.
Zur zweiten Gruppe gehoren die pazifischen subarktischen Formen, von denen bis jetzt 23 verzeichnet sind, d. h. 19% der Gesamtzahl. Sie zerfallen in 3 Untergruppen. Die einen von ihnen (z. B. Rocinela angustata, R, belliceps, Argeia pugettensis) sind sowohl an den asiatischen, als auch an den ameri-kanischen Kiisten (bis Alaska) weit verbreitet (6 Formen). Andere Formen (z. B. Arcturus beringanus, Janira soldatovi, Idothea ochotensis, im ganzen 10), sind auf die asiatischen subarktischen Kiisten beschrankt. Die dritten endlich (wie, z. B., Idothea aleutica, Pentidothea wosnesenskii, Aega symmetrica— im ganzen 7) miissen eigentlich als subarktisch-pazifische Formen der amerikanischen Kiisten angesehen werden.
Zur dritten Gruppe gehoren die pazifisch-borealen Formen, wie Cilicaea cordata, Pentidothea japonica, Neosphaeroma oregonensis, Idothea deriuginl (im ganzen 10). Die borealen Formen dringen in grosserer Anzahl
248 ISOPODA
in. die Berings-See und das Japanische Meer ein, sind aber im Ochotskischen’ Meere nur schwach vertreten.
Die vierte Gruppe wird von arktischen Formen gebildet, von denen im ganzen 10 bekannt sind, was 6.2% der Gesamtzahl der Arten ausmacht. Die einen von ihnen, wie Mesidothea entomon, Calathura branchiata, Munnopsurus giganteus, Gnathia elongata, dringen in den Stillen Ozean aus dem Eismeere ein. Andere aber, wie verschiedene Arten von Synidothea, Pleuroprion und Arcturus, sind pazifische Autochthonen, die sich erst spater uber die Arktis verbreitet hatten. Bezeichnend ist es, dass alle 3 Meere gemeinsame Formen mit dem Eismeere aufweisen, doch nimmt ihre Zahl nach Siiden hinab und zugleich senken sie sich in immer grossere Tiefen hinab (so ist, z. B., im Japanischen Meere nur eine arktische Art, Gnathia elpngata, die nur in Tiefen von 76 bis 900 m vorkommt).
Zur fiinften Gruppe gehoren die pazifischen subtropischen Formen, wie z. B., Holotelson, Dynamene, Cleantis, Ligia, Cymodoce (im ganzen 10).. Samtliche Arten dieser Gruppe sind nur im Japanischen Meere gefunden worden und fehlen sowohl im Ochotskischen Meere als auch in der Berings-See. Eine Ausnahme bildet nur Livoneca. amurensis, eine himalayanische SiisswasSerform, die auf Fischen parasitiert und langs dem Amur bis zu des-sen Miindung hinabsteigt, also bis in das Ochotskische Meer vordringt.. Die subtropischen Formen sind alle Kiisten- oder Flachwasserformen, oder gehoren dem Litoral an. Sie sind zweifellos mit dem warmen Wasser der Ziishima-Stromung, welche einen schwachen Zweig bis in die Gegend von Wladiwostok entsendet, in das Japanische Meer gelangt.
Die sechste Gruppe bilden regelrechte Tiefseeformen des Stillen Oze-ans, im ganzen 5 Formen: Munnopsurus laevis, Aega magnoculis, Arcturus, hirsutus, Holophryxus giardi, Arthrophryxus beringanus. Sie gelangen in die Berings-See und das Ochotskische Meer, fehlen aber im Japanischen Meere vollstandig. Dieses Fehlen von pazifischen Tiefseeformen im Japani— schen Meere ist, wie schon oben erwahnt war, durch die vollstandige Isolation der Tiefsee dieses Meeres leicht zu erklaren, da die Strassen zwischen ihm und dem Stillen Ozean eine Tiefe von 200 m nicht iiberschreiten. Das-Ochotskische Meer ist aber mit dem Stillen Ozean durch die Kurilischen Strassen mit Tiefen bis 2000 m verbunden, und dort kommt deshalb auch Aega magnoculis vor. Die abyssalen Formen Arcturus beddardi und Tecti— ceps leucophthalmus, welche zur Zeit als endemische Formen des Ochotskischen Meeres gelten, stammen wahrscheinlich urspriinglich auch aus dem Stillen Ozean und sind dort wohl noch einfach nicht gefunden.
Zur siebenten Gruppe gehoren Formen, die eine Tendenz zu kosmo-politischer Verbreitung bekunden, z. B., Cyathura carinata, Idothea metallica, Phryxus abdominalis, Limnoria lignorum, Livoneca raynaudii — im ganzen 5 Arten.
Die achte Gruppe wird von subtropischen Arten gebildet, die sonst nur an der amerikanischen Kiiste bei Siid-Kalifornien gefunden werden--
ZOOGEOGRAPHISCHE OBERSICHT
249’
Idothea fewkesi und Synidothea harfordi. Beide Formen sind im Japanischen Meere gefunden worden. Wahrend aber «S'. harfordi nordlich der Bai Peters des Grossen nicht getroffen wird, kommt I. fewkesi auch im Tataren-Sunde vor und ist von dort aus durch den Nevelski-Sund bis zum Ochotskischen Meere vorgedrungen (dieser Weg der Verbreitung scheint am wahrschein-lichsteri, da diese Form bei der Insel Langr und den Shantaren gefunden ist, wahrend sie an der Ost-Kiiste von Sachalin fehlt, so dass der Weg uber die La Perouse-Strasse nicht in Frage kommt).
Zur neunten Gruppe gehoren die arktisch-borealen Formen, d. h. solche, die in der Arktis zircumpolar verbreitet sind und im nordlichen Teile sowohl1 des Atlantischen, als auch des Stillen Ozeans getroffen werden. Hierher gehort auch nur eine Art, Bopyroides hippolytes, die in der Berings-See und dem Japanischen Meere vorkommt.
Zur zehnten Gruppe gehoren endlich die amphiborealen Formen1 mit disjunktem Areale, welche im nordlichen Teile des Atlantischen und Stillen Ozeans vorkommen, aber sowohl in der Arktis, als auch in siidlicheren Breiten fehlen. Zu ihnen gehoren Pleuroprion hystrix und Cleantis plani-cauda, aber auch Syscenus infelix, welcher von Richardson an der Ost-Kiiste Japans gefunden worden ist.
Wir sehen also, dass die Isopodenfauna der 3 besprochenen Meere reichhaltig und aus verschiedenen zoogeographischen Gruppen zusammen 7 gewiirfelt ist. Diese Verschiedenartigkeit der Faunenelemente lasst sich durch die Kompliziertheit der hydrologischen und geographischen Bedingungen dieser 3 Meere, sowie durch das Vorhandensein von Verbindungen sowohl zwischen denselben, als auch mit dem Eismeer und dem Stillen Ozean erklaren. Wie schon oben erwahnt war, wird die Verbreitung der Isopodenfauna zum grossen Teil durch die Bodenbeschaffenheit und das Relief, sowie durch die Stromungen bedingt. Nicht minder wichtig ist aber auch die geographische Lage der Meere: der Unterschied in der Entfernung vom Aquator zwischen dem Japanischen, Ochotskischen und Berings-Meere.. So kann man sagen, dass das am weitesten nordlich gelegene Berings-meer, obgleich es eine reiche Isopodenfauna besitzt, im ganzen doch haupt-sachlich Kaltwasserformen aufweist. So sind, z. B., in diesem Meere die Asellota reicher vertreten, als im Japanischen Meere, wahrend die Sphaeromidae im Gegenteil fast fehlen. Die Asellota sind durch arktische Gattungen Janira, Munnopsurus und Munna vertreten. Im Japanischen Meere sind aber die Asellota sehr schwach vertreten, wahrend die siidliche Warmwas-sergruppe Sphaeromidae hier in vielen Formen vorkommt (die Gattungen Exosphaeroma, Holotelson, Dynamene, Cymodoce etc.). Auch im Grade der Mannigfaltigkeit der Formen sehen wir einen Unterschied zwischen dem verhaltnissmassig warmen Japanischen Meere und den beiden anderen, nordlichen Meeren. Gewohnlich findet man zwischen den Formen subtropischer
1 L. Be r g. Ober die amphiboreale (disjunkte) Verbreiterung der Meeresfauna auf der nordlichen Halbkugel. Zoogeographica, II, 3, 1934.
250
ISOPODA
und tropischer Gruppen eine starkere Differenzierung, als in kalten Gewas-sern, weshalb zwischen den Wendekreisen die Zahl der Gattungen auch ungleich grosser zu sein pflegt als weiter polwarts. Es ist deshalb verstand-lich, dass das Japanische Meer eine grossere Mannigfaltigkeit der Arten und Gattungen aufweist, als das Ochotskische Meer und die Berings-See. So kommen auf jede Gattung des Japanischen Meeres nur 2 Formen, 6in Ver-haltnis, das auch in der Berings-See (einschliesslich den verhaltnismassig warmen Teil) beibehalten wird, wahrend im Ochotskischen Meere auf jede Gattung etwa 3 Formen entfallen. Man sieht darin aber auch deutlich die Tendenz der ochotskischen Arten zur Aufteilung in Lokalformen. Auch in der Verteilung der Gattungen spiegelt sich die geographische Lage der Meere wieder. So findet man im Japanischen Meere 11 Warmwassergattun-gen, in der Berings-See nur 3, im Ochotskischen Meere aber keine einzige. Der Unterschied in den Lebensbedingungen dieser 3 Meere hat zur Folge, dass die weitverbreiteten Gattungen in jedem von ihnen eigene Formen Widen. Als Beispiele kann man die Gattungen Tecticeps, Arcturus und Synidothea anfiihren. So findet man im Berings-Meere 2 endemische Formen von Tecticeps, 6 Formen Arcturus und 6 Formen Synidothea, im Ochotskischen Meere 5 endemische Formen Tecticeps, 6 Arcturus und 4 Synidothea, im Japanischen Meere endlich 2 endemische Formen von Tecticeps, 1 Arcturus und 4 Synidothea.
' Die Tiefengliederung der 3 Meere lasst sich in der Isopodenfauna folgendermassen verfolgen. Im Japanischen Meere ist die Warmwasserfauna des oberen Sublitorals sehr reich, wahrend grossere Tiefen von einer an Artenzahl armen gemassigten Fauna bewohnt werden. In der Berings-See ist die Isopodenfauna sowohl in den oberen Schichten, als auch im Abyssal artenreich, wahrend im Ochotskischen Meere die reiche Flachwasserfauna wiederum von der armeren, aber sehr eigentiimlichen Tiefenfauna absticht.
Was die Anordnung der Isopoden in verschiedenen Biozoenosen be-trifft, so muss man sagen, dass bei dem derzeitigen Stande unserer Kennt-nisse iiber die Berings-See und das Japanische Meer kaum was gesagt werden kann. Fur die Bai Peters des Grossen im Japanischen Meere kennen wir die Verteilung der sublitoralen Isopoden in einige Biozoenosen. So wis-sen wir, z. B., dass die Bestande der roten Algen in 20—25 m Tiefe charak-teristische Biotope fur Idothea orientalis sind, wahrend in gleicher Tiefe auf steinigem Grunde Synidothea epimerata vorkommt. In den Bestanden von Zostera von 0.5 bis 10 m Tiefe wohnen Janiropsis kincaidi, Pentias hayi, Pentidothea rotundata, Synidothea brazhnikovi und Holotelson tuberculatus, auf Austerbanken und Banken von Miesmuscheln Cymodoce japonica. Im .Supralitoral kommen auf Steinen als Charakterformen Ligia cinerascens und Trichoniscus papillicornis vor, wahrend fur den Tiefseeschlamm von 1000 m an Eurycope spinifrons und Gnathia elongata typisch sind.
SYSTEMATISCHER TEIL
BESTIMMUNGSTABELLE DER UNTERORDNUNGEN
t
1 (6)f Die Uropoden sind lateral angeordnet, d. h. an den Seiten der Basis des Telson, oder an dessen Seitenrandern befestigt und bilden entweder einen Caudalfacher, eine Napfchen, oder Klappen, die sich an der Ven-tralseite offnen.
2 (5) Die Uropoden bilden zusammen mit dem Telson einen Caudalfacher, oder -ein Napfchen.
3 (4). 7 Paar freie Thoracalfiisse vorhanden ........ II. Flabellifera.
4 (3). Nur 5 Paar freie Thoracalfiisse vorhanden, da das erste Paar in Pylo-poden umgewandelt ist, welche in Form eines Deckels die Mundteile von unten bedecken, wahrend das letzte Paar vollstandig reduziert ist
............................................ VI. Gnathiidea.
5 (2). Die Uropoden bedecken die Ventralseite des Telson in der Form von Klappen, unter denen die Pleopoden vollstandig verdeckt sind..........
. . . .............................................III. Valvifera.
6 (1). Die Uropoden sind terminal, d. h. am hinteren Rand des Telson angeordnet, und gewohnlich stabchenformig.
7 (8). Alle Pleopoden sind von unten mit einem diinnen Plattchen, wie mit einem Deckel bedeckt........................................I. Asellota.
8 (7). Die Pleopoden liegen frei und sind nicht mit einem Plattchen bedeckt.
9 (10). Freilebende Formen mit normaler Segmentierung des Rumpfes und normalen Extremitaten. Die Pleopoden dienen zur Luftatmung ....
. . . •......................................... IV. Oniscoidea.
10 (9). Schmarotzende Formen. Der Rumpf des Weibchens ist gewohnlich assymmetrisch und besitzt nur Spuren einer Segmentation, oft auch nicht einmal solche. Die Pleopoden sind zum Atmen im Wasser specia-lisiert...................................................V. Epicaridea.
I. Subord. ASELLOTA
1 (2)..An den Mandibeln ist der Molar ganz reduziert, oder kurz, dorsalwarts verschmalert und nach hinten gekehrt. Die drei hinteren Paar Thora-calfusse dienen zum Schwimmen . . . . . . . . • . 3. Munnopsidae.
252
ISOPOD A
2 (1). An den Mandibeln ist der Molar gut entwickelt, fast cylinderformig und entweder nach vorher gekehrt, oder befindei sich in senkrechter Lage. Die 3 hinteren Paare der Thoracalfiisse sind Gehfiisse.
3 (4). Alle Brustsegmente sind infolge einer gleichmassigen Gliederung ungefahr von gleicher Form und Grosse und alle beweglich . 1. Janiridae.
4 (3). Der Thorax zerfallt infolge ungleichmassiger Segmentierung in 2 Teile. Der vordere, welcher aus 4 Segmenten besteht, ist bedeutend grosser und breiter, als der hintere, aus 3 Segmenten bestehende . 2. Munnidae.
1. Fam. JANIRIDAE
1 (2). Augen fehlen. Das Rostrum ist kurz, mit geradem Vorderrande. Das erste Paar Thoracalfiisse ist kurz, schwach und bedeutend kleiner, als die iibrigen 6 Paar. Das erste (Basal-) Glied der Antennulen ist gross, breit und mit spatelformig ausgezogenem vorderem Seitenrande. . . .
....................................................3. Microprotus Rich.
2 (1). Augen immer vorhanden. Das Rostrum ist zugespitzt oder fehlt ganz. Das erste Paar Thoracalfiisse unterscheidet sich in Grosse und Batt wenig von den iibrigen.
3 (4). Das zweite Glied des Tasters der Maxillipeden ist viel breiter, als der Lappen des zweiten Gliedes des Basalteiles . . 2. Janiropsis G. Sars.
4 (3). Das zweite Glied des Tasters der Maxillipeden ist nicht breiter, als der Lappen des zweiten Gliedes des Basalteiles ... 1. Janira Leach.
1. Gatt. JANIRA LEACH
1 (2). Am Kopfe befindet sich ein langes Rostrum, das bis 2/s Kopflarige erreicht. — Вег. M............................... 1. J. alascensis Bened.
2 (1). Ein Rostrum fehlt vollstandig, oder ist kaum bemerkbar.
3 (4). Die seitlichen Ecken deS Kopfes vor den Augen sind abgerundet. Der Korper ist mit dichten kurzen und steifen Borsten bedeckt. — Вег. M.
. . ............................................. 5. J. sarsi Rich.
4 (3). Die seitlichen Ecken des Kopfes vor den Augen sind in sp.atelformige zugespitzte Fortsatze ausgezogen. Die Oberflache des Korpers ist glatt, ohne Borsten.
5 (6). Die. seitlichen Fortsatze des Kopfes von den Augen enden in 2 Sta-cheln. Der Vorderrand des Kopfes ist fast gerade. — Вег. M................
..................................................4. J. holmesi Rich.
6 (5). Die seitlichen Fortsatze des Kopfes vor den Augen sind einfach zugespitzt. Der Vorderrand des Kopfes ist konvex und in der Mitte etwas. zugespitzt.
7 (8). Das erste Glied des Stammes des ersten Antennenpaares ist schmal. nicht breiter, als das zweite Glied. Die Dorsalseite des Telson ist glatt. — Вег. M., Och. M., Jap. M. ....... 2. J. erostrata Rich.
1. JANIRIDAE. 2. MUNNIDAE. 3. MUNNOPSIDAE
253
8 (7). Das erste Glied des Stammes des ersten Antennenpaares ist breit, viel breiter, als das zweite Glied. Der Telson tragt auf dem distalen Ende der Dorsalseite Gruppen facherartig angeordneter Nadeln.— Och. M., Jap. M................................3. J. soldatovi Gurjan.
2. Gatt. JANIROPSIS G. SARS
1 (4). Das untere Drittel der Seitenrander des Telsons glatt.
2 (3). Der Telson ist langer, als breit. — Вег. M., Jap. M • 1. J. kincaidi Rich.
3 (2)' Die Lange des Telson ist geringer, als seine Breite, oder gleicht ihr. — Вег. M. . . ... . . . .........................2. J. derjugini Gurjan.
4 (1). Das untere Drittel der Seitenrander der Telsons sageformig— Jap. M. ........................................ 3. J. serricaudis Gurjan.
3. Gatt. MICROPROTUS RICH.
1 (1). Вег. M.......................................1. M. coecus Rich.
2. Fam. MUNNIDAE
1. Gatt. MUNNA KROYER
1 (6). Vor den Augen befinden sich kleine Hocker. Alle Epimerenplatten sind mit starken Stacheln besetzt.
2 (3). Rander des Telson glatt, ohne Stacheln. Oberflache des Korpers des Telson glatt, ohne Stacheln und Borsten. — Jap. M...................
. ................................... 3. M. subneglecta Gurjan.
3 (2). Rander des Telson von jeder Seite mit 2—4 Stacheln besetzt. Oberflache des Korpers des Telson glatt oder mit feinen Harchen besetzt.
4 (5). Oberflache des Korpers glatt; Rander des Telson an jeder Seiten mit nur 2 Stacheln. — Вег. M.....................1. M. stephenseni Gurjan.
5 (4). Oberflache des Korpers mit kurzen feinen Harchen bedeckt; Rander des Telson an jeder Seite mit 4 Stacheln. — Вег. M..................
. . . •............................. 4. M. avatshensis Gurjan.
6 (1). Vor den Augen befinden sich keine Hocker. Nur 4 oder 5 Paare von Epimeralplatten sind mit je 2 Stacheln besetzt. — Вег. M............
............................... 2. M. arnholdi Gurjan.
3. Fam. MUNNOPSIDAE
1 (2). Die Antennen sind terminal angeordnet. Der Telson ist schmal dreieck-ig, nach dem distalen Ende verschmalert und zugespitzt .......
..........................................1. Ilyarachna G. Sars.
2 (1). Die Antennen sind dorsal angeordnet. Der Telson ist oval, hinten abgerundet. ,
254
ISOPODA
3 (4). An den Mandibeln kein Molar vorhanden. Das siebente Glied der 3 hinteren (zum Schwimmen dienenden) Paare der Thoracalfiisse fehlt. Der ganze Korper ist gestreckt, sein hinterer Teil viel schmaler, als der vordere..........................................4. Munnopsis M. Sars.
4 (3). An den Mandibeln ist der Molar gut ausgebildet, oder bildet einen grossen, breiten Hocker. Das siebente Glied an den 3 hinteren (zum Schwimmen dienenden) Paar Thoracalfiisse vorhanden. Der Korper ist verkiirzt. Der hintere Abschnitt des Korpers ist nicht schmaler, als der vordere.
5 (6). Die Mandibeln besitzen eine lacina mobile. Der Molar ist gut ausgebildet, von cylindrischer Form. Das siebente Glied der Schwimmfiisse ist gut entwickelt, langer, als die Schwimmborsten....................
...........................................3. Еигусоре G. Sars.
6 (5). Keine lacina mobile an den Mandibeln vorhanden. Der Molar hat die Form eines breiten Fortsatzes. Das siebente Glied der Schwimmfiisse ist klein, kiirzer, als die Schwimmborsten ...........................
...........•...............................2. Munnopsurus Rich.
1. Gatt. ILYARACHNA G. SARS
4 (2). Der Vorderrand der vier ersten Brustsegmente grob sageformig gezackt. Auf dem Kopfe sind 6 grosse Stacheln vorhanden. — Jap. M..............
............................................1. I. zachsi Gurjan.
2 (1). Der Vorderrand der vier ersten Brustsegmente tragt je zu 4 grossen stachelformigen Fortsatzen. Auf dem Kopfe befinden sich 4 grosse Stacheln und 2 kleine an den Seiten derselben. — Och. M., Jap. M. . .
. . . ...............................2. I. starokadomskii Gurjan.
2. Gatt. MUNNOPSURUS RICHARDSON.
1 (4). Der Vorderrand des zweiten und dritten Gliedes der Maxillipeden ist glatt.
2 (3). Auf der Dorsalseite der 3 letzten Thoracalsegmente befindet sich eine Langsrinne und an den Seiten derselben je zu einem flachen breiten Hocker. — Jap. M............................... 3. M. minutus Gurjan.
3 (2). Die Dorsalseite der 3 letzten Brustsegmente ist glatt, ohne Rinnen und Hocker. — Вег. M., Jap. M., N. St. Oz...............2. M. laevis (Rich.).
4 (1). Der Vorderrand des 2 und 3 Gliedes des Palpus der Maxillipeden ist mit kurzen Borsten besetzt.
5 (6). An der Sagittallinie der 3 letzten Brustsegmente befindet sich eine tiefe Rinne und an deren Seiten je zu einem Hocker.—Arct. Oz., Ber. M., Och. M........................1. M. giganteus giganteus (G. Sars).
6 (5). An den 3 letzten Brustsegmenten ist weder eine Rinne, noch Hocker vorhanden. — Och. M................la. M. giganteus ochotensis Gurjan.
3. MUNNOPSIDAE, 4. CYMOTHOIDAE
255
3. Gatt. EURYCOPE G. SARS
1 (2). Frontallappen lanzettformig, am Ende zugespitzt. — Jap. M....
..........................................1. E. spinifrons Gurjan.
2 (1). Frontallappen breit, mit geradem Vorderrande. — Jap. M.......
..........................................2. E. pavlenkoi Gurjan.
, . 4. Gatt. MUNNOPSIS M. SARS
1 (1). Jap. M., Gelb. M. . . . :.................1. HL latifrons Bedd.
IL Subord. FLABELLIFERA
1 (2). Korper gestreckt, wurmformig, fast cylinderformig. Epimeren fehlen. Telson und Uropoden bilden zusammen den napfformigen Schwan-zfacher..............................................6. Anthuridae.
2 (1). Korper breit, oval, in dorso-ventraler Richtung abgeplattet. Die Epimeren sind gut entwickelt, oder verschmelzen mit den Segmenten und bilden Pleuralverbreiterungen. Der Telson bildet zusammen mit den Uropoden den flachen Schwanzfacher.
3 (4). Der Кёгрег капп grosstenteiis in eine Kugel zusammengerollt werden. Die Aste der Uropoden sind entweder ungefahr gleich lang und dabei einer von ihnen unbeweglich, oder aber beide beweglich, aber von verschiedener Lange, oft auch einer von ihnen rudimentar oder ganz fehlend............................................5. Sphaeromidae.
4 (3). Der Korper kann nicht kugelformig zusammengerollt werden. Beide Aste der Uropoden sind beweglich und von gleicher oder beinahe gleicher Lange ....................................4. Cymothoidae.
4. Fam. CYMOTHOIDAE
1 (2). Die beiden Endglieder des Palpus der Maxillipeden sind mit Borsten besetzt, aber ohne Hackchen. Der Palpus liegt frei . . 1. Cirolaninae.
2 (1). Der Palpus der Maxillipeden ist von einem Kegel umgeben, der von den distalen Mundteilen gebildet ist. Die Glieder des Palpus sind nicht mit Borsten, sondern mit Hakchen bewehrt.
3 (4). Beide Antennenpaare bestehen aus einem Stamm und einer Geisseh. Die Glieder des Palpus der Mandibeln sind schmal. Der Palpus der Maxillipeden besteht aus 5 Gliedern (seiten aus 2) . . , .2. Aeginae.
4 (3). Beide Antennenpaare sind reduziert, so dass ihre Einteilung in Stamm und Geissel kaum sichtbar ist. Die Glieder des Palpus der Mandibeln sind verbreitet. Der Palpus der Maxillipeden besteht immer aus 2 Gliedern...........................................3. Cymothoxnae.
256
ISOPODA
1. Subfam. CIROLANINAE
1 (2). Die Pleopoden sind normal gebaut, ohne Biindeln von Riemenfaden an den Wurzeln ihrer Verzweigungen. Die Augen befinden sich an den Seiten des Kopfes und sind nur teilweise nach unten gerichtet ....
............................................1. Cirolana Leach.
2 (1). Die Pleopoden besitzen an den Wurzeln ihrer Verzweigungen Biischel von Riemenfaden. Die Augen sind seitwarts gerichtet, befinden sich aber an der Abdominalseite des Kopfes.........................
........................................2. Bathynomus M.-Edw,
1. Gatt. CIROLANA LEACH
1 (1). Jap. M............................. . . 1. C. japonensis Rich.
2. Gatt. BATYNOMUS M.-EDW
1 (1). Jap. M................................... B. doderleini Ortm.
2. Subfam. AEG1NAE
1 (2). Die ersten 2 Glieder der Stammes des ersten Antennenpaares sind stark verbreitet. Das Flagellum ist vielgliederig. Der Palpus der Maxil-lipeden besteht aus 5 Gliedern.......................3. Aega Leach.
2 (1). Die ersten 2 Glieder des Stammes des ersten Antennenpaares sind nicht vebreitet. Palpus der Maxillipeden besteht aus 2 Gliedern.
3 (4). Augen sind vorhanden. Der Abdomen ist etwas schmaler, als der Brustteil.........................................4. Rocinela Leach.
4 (3). Keine Augen vorhanden. Der Abdomen ist viel schmaler, als der Brustteil ................. .......... ............... . 5. Syscenus Harger.
3. Gatt AEGA LEACH .
1 (2). Die Augen sind sehr gross, oval. Sie bedecken fast die ganze Ober-flache des Kopfes und sind nur durch einen schmalen Zwischenraum von 0.5 mm von einander getrennt. — Gelb. M., Вег. M., Och. M. . .
...................................... 2. A. magnoculis Rich.
2 (1). Die Augen sind schmal und langlich, befinden sich an den Seiten des Kopfes. Der Zwischenraum zwischen den Augen ist gleich einer Augen-lange. — N. St Oz., Вег. M....................1. A. symmetrica Rich.
4. Gatt. ROCINELA LEACH
1 (2). Die vorderen Seitenecken des ersten Brustsegmentes sind in kleine Hornchen ausgezogen, die sich langs den Kopfseiten hinziehen — Вег. M.
............................................1. R. cornuta Rich.
2 (1). Die vorderen Seitenecken des ersten Brustsegmentes bilden keine hornformigen Fortsatze.
4. C YMOTHOIDAE
257
3 (4). Das sechste died der ersten 3 Paar Thoracalfiisse tragt mehr als
4 Stacheln.— Jap. M. . .......................6. R. niponia Rich.
4 (3). Das sechste died der ersten 3 Paar Thoracalfiisse tragt von 2 bis 4 Stacheln.
5 (6). Auf der Oberflache des Korpers sind keine Pigmentflecken vorhanden.— Вег. M., Och. M...................................4. R. angustata Rich.
6 (5). Auf der Oberflache des Korpers befinden sich Pigmentflecken.
7 (8). Dunkle Flecken firiden sich an den Seiten des vierten Brustsegmentes und an der Basis des Telson. — W. Gron., Och. M., Jap. M...............
................................... 2. R. maculata Sch. u. Mein.
8 (7). Dunkle Flecken finden sich nur an den Seiten der Abdominalsegmente und an der Basis des Telson.
9 (10). Der vordere Rand des Kopfes ist sanft abgerundet, ohne Aussch-nitte. — N. St. Oz., Вег. M. . . ................ 3. R. beiliceps (Stimps.).
10 (9). Der vordere Rand des Kopfes besitzt in der Mitte einen spatelfonni-gen Auswuchs und an seinen Seiten je zu einem Ausschnitt, so dass der Frontalrand ein dreispitziges Aussehen erhalt. — Jap. M............
. . . ..........................................5. R. japonica Rich.
5. Gatt. SYSCENUS HARGER
1 (1). N. St. Oz., Mitt. M., Jap. M. ...............IS. infelix Harger.
3. Subfam. CYMOTHOINAE
1 (2). Der Kopf ist gar nicht im ersten Brustsegment eingelagert; sein Hinterrand bildet drei spatelformige Fortsatze, einen grossen in der Mitte und 2 kleine an den Seiten. Der Vorderrand des ersten Thoracal-segmentes besitzt 3 gut sichtbare Ausschnitte . . . 6. Nerocila Leach..
2 (1). Der Kopf ist mehr oder weniger im ersten Brustsegment eingelagert; sein Hinterrand ist abgerundet, ohne.Fortsatze zu bilden. Der Vorderrand des ersten Brustsegmentes ist glatt, in der Mitte starkausge-buchtet.
3 (4). Die Glieder der Antennen sind breit, von oben und unten abgeplattet. Die ersten Glieder des ersten Antennenpaares beriihren einander an der Mittellinie des Kopfes...........................7. Meinertia Stebb.
4 (3). Die Glieder der Antennen sind nicht verbreitet und seitlich zusam-mengedriickt. Die ersten Glieder des ersten Antennenpaares beriihren einander niemals.
3 (6). Der vordere Teil des Abdomens ist nur wenig in den Thorax eingelagert. Alle Brustsegmente sind von gleicher Lange, oder aber ist das erste Segment langer, das letzte kiirzer, als die iibrigen.............
............................................... 8. Livoneca Leach.
Гурьянова 17
258
ISOPODA
6 (5). Der vordere Teil des Abdomens ist tief in den. Thorax eingelagert. Das erste Brustsegment ist bedeutend langer, als das zweite; die iibrigen nehmen von vorne nach hinten allmahlich an Lange ab......................
............................................9. Irona Sch. u. Mein.
6. Gatt. NEROCILA LEACH
1 (2). Die Augen sind nicht sichtbar. Die hinteren Seitenecken alle Brust-segmente sind in lange, nach hinten gerichtete, spitze Fortsatze ausgezogen— Jap. M............................... 1. N. japonica Sch. u. Mein.
2 (1). Die Augen sind gut ansgebildet. Nur an den beiden hinteren Segmenten sind die hinteren Seitenecken in zugespitzte Fortsatze ausgezogen. — N. St. Oz.................................. 2. N. californica Sch. u. Mein..
7. Gatt. MEINERTIA STEBB.
1 (1). N. St. Oz., Jap. M........... . . 1. M. oxyrrbynchaena (Koelb.J.
8. Gatt. LIVONECA LEACH .
s
1 (12). Der Korper ist symmetrisch.
2 (7). Das Abdomen ist schmaler, als das vorletzte Brustsegment.
3 (6). Die Augen sind gut ausgebildet.
4 (5).- Die Augen sind gross, rund und .gewolbt. Der Vorderteil des Kopfes-ist stark nach vorne ausgezogen und bildet vor den Augen einen breiten spatelformigen Fortsatz mit geradem Vorderrande. Der Abdomen ist wenig schmaler, als das vorletzte Brustsegment. — California ... .
. . .'........................................6. L. vulgaris Stimps.
5 (4). Die Augen sind klein, langlich-oval und flach. Der Vorderteil des Kopfes ist sanft zugerundet. Der. Abdomen ist viel schmaler, als das vorletzte Brustsegment. — Bassin-Amur. . • . 1. L. amurensis Gerstf.
6 (3). Die Augen sind fast vollstandig reduziert; es sind nur Spuren von Augen an den Kopfseiten vorhanden. — Gelb. M., Jap. M....................
................................................ 4. L. sacciger Rich.
7 (2). Der Abdomen ist eben so breit, wie das vorletzte Brustsegment un<J. breiter, als das letzte.
8 (11). Die Aste der Uropoden sind von ungleicher Lange.
9 (10). Die Basis des Telson ist von gleicher Breite, wie die ersten Abdo-minal-Segmente. Die Epimeren des letzten Paares haben die Form von abgerundeten Plattchen. — Jap. M..............8. L. caudata Sch. u. Mein.
10 (9). Die Basis des Telson ist schmaler, als die vorderen Abdominal--Segmente. Die Epimeren des letzten Paares haben die Form von gros-sen, breiten Flatten. — Jap. M......................5. L. epimerias Rich.
11 (8). Die Aste der Uropoden sind von gleicher Lange. — S. St. Or. Och.
. M. California ..............................2. L. raynaudii M.-Edw.
12 (1). Der Korper ist assymmetrisch, rechts oder links gebogen. • -
4. CYMOTHOIDAE F. 5. SPHAEROMIDAE
259
13 (14). Die Breite der Basis des Telson ist doppelt so gross, wie seine Lange. — Jap. M. . . . ..................... . . . 3. L. propiuqua Rich.
14 (13).. Die Breite der Telsonbasis ist geringer, als seine Lange. — California. ............................ . ... . . 7. L. californica Sch. u. Mein.
9. Gatt. IRONA SCH. u. MEIN.
1 (1). N. St. Oz., Sandw. I., Jap. M....1. I. melanostica Sch. u. Mein.
5. Fam. SPHAEROMIDAE
2 (1). Der eine von den Asten der Uropoden ist unbeweglich. Beide Aste haben die Form von fast gleich grossen Plattchen. Der Abdomen . besteht aus 2 Segmenten ...........................2. Sphaerominae..
2 (1). Beide Aste der Uropoden sind beweglich, schmal, fast cylindrisch. Der eine von ihnen ist bedeutend riickgebildet, rudimentar oder fehlt 'zuweilen ganz. Der Abdomen besteht gewohnlich aus 6 Segmenten .
............•.....................................1. Limnorinae.
1. Subfam. LIMNORINAE
1. Gatt. LIMNORIA LEACH
1 (2). Die Oberflache des fiinften Brustsegmentes und des Telson ist glatt.-
Atl. Oz., Arct. Oz., Вег. M., Jap. M. ...... 1. L. lignorum Rich.
2 (1). Auf der Dorsalseite des fiinften Brustsegmentes befinden sich 2 Hocker. Langs der Mittellinie des Telson sind 3 Hocker und 2 einander parallele Leisten angeordnet. — Jap. M. ...... . 2. L. japonica (Rathke).
2. Subfam. SPHAEROMINAE
1 (2). Das erste (beim Mannchen auch das zweite) Paar Thoracalfiisse ist zum Greifen befahigt und mit einer Subchela ausgeriistet .............
.............................................. . 2. Tecticeps Rich.
2(1). Die ersten Paare der Thoracalfiisse sind Gehfiisse und unterscheiden • sich nicht von den iibrigen.
3 (4). Der Aussenast der Uropoden lasst sich nicht unter den Innenast schieben. Die beiden Aste sind in Form und Grosse sehr ungleich . .
................... ...........................5. Cilicaea Leach.
4 (3). Der Aussenast der Uropoden lasst sich unter den Innenast schieben. Die beiden Aste sind ungefahr von gleicher Grosse und Form.
5 (12). Der Hinterrand des Telson ist ganz randig, ohne Ausschnitt in der Mitte.
6 (11). Der Hinterrand des Telson ist sanft zugerundet.
7 (8). Die Glieder des Palpus der Maxillipeden sind eirifach, ohne spatelfor-mige Fortzatze an den distalen inneren Ecken . . 7. Sphaeroma Latr.
8 (7). Das zweite bis vierte Glied des Palpus der Maxillipeden besitzt an der distalen Inneneckeje einen spatelformigen Fortsatz mit Borsten-biischeln an der Spitze.
17
260
ISOPODA
9 (10). Der Endopodit des dritten Paares Pleopoden besitzt 3 Branchialfalten ............................................... 9. Neosphaeroma Baker.
10 (9). Der Endopodit des dritten Paar Pleopoden besitzt keine Branchialfalten ..................................... 8. Exosphaeroma Stebb.
11 (6). Der Hinterrand des Telson bildet 3 Zacken, einen grossen zugerun-deten in der Mitte und zwei kleine, zugespitzte zu beiden Seiten der ersten und an deren Basis...............................3. Holotelson Rich.
12 (5). Der Hinterrand des Telson besitzt in der Mitte einen grossen Ausschnitt.
13 (14). Am Hinterrande des Telson, am Grunde des Ausschnittes, befindet sich ein spatelformiger Fortsatz . ..................6. Cymodoce Leach.
14 (13). Der Ausschnitt am Hinterrande des Telson ist ohhe Fortsatz . . . .....................................................4. Dynamene Loach.
2. Gatt. TECTICEPS RICH.
,1 (12). Die Augen sind schmal, nierenformig.
2 (7). Die Oberflache des Korpers ist vollstandig glatt, die Dorsalseite der Segmente ohne Kornchen, Hocker oder Kiele.
3 (6). Die Rander der Epimeralverbreitungen sind glatt, ohne Stacheln oder Zahnelung.
4 (5). Die Augen sind breit nierenformig. Der Kopf ist flach, mit abge-flachtem Frontalteile. Die Abrisse des Korpers sind sanft abgerun-det,— Och. M., Jap. M...................la. T. renoculis var. laevis Gurjan.
5 (4). Die Augen sind schmal nierenformig. Der Kopfrand ist verdickt, so dass in der Mitte des Frontalrandes sich ein breiter, niedriger, aber gut sichtbarer Wulst vorhanden ist, — Вег. M................. . ... . .
..............................lb. T. renoculis v. marginaiis Gurjan.
6 (3). Die Oberrander des vierten, fiinften und sechsten Brustsegmentes, -sowie die unteren Rander der Epimeralverbreiterungen des fiinften, eechsten und siebenten Segmentes sind mit kleinen Zacken und Stacheln besetzt. — Jap. M.................le. T. renoculis v. serratus Gurjan.
7 (2). Die Dorsalseite der Brustsegmente ist mit Hockern und Querleisten besetzt, oder aber ist der ganze Korper mit Kornern und unregelmas-sigen Wiilsten bedeckt.
S (9). Der aussere Ast der Uropoden ist kiirzer, als der innere. Langs der Sagittallinie des Korpers zieht sich eine Reihe von Hockern je zu einem auf jedem Segment. — Och. M.........................................
..................- . . . . Id. T. renoculis var. nodulosus Gurjan.
9 (8). Der aussere Ast der Uropoden ist langer, als der innere. Die Dorsalseite des Korpers ist gekornelt, mit Hockern oder mit Querleisten bedeckt-*10 (11). Die Oberflache des Korpers ist fein gekornelt. Ausserdem ist der Dorsalteil der Segmente mit unregelmassigen Hockern bedeckt. — Och. M......................................1. T. renoculis renoculis Rich.
5. SPHAEROMIDAE
261
11 (10). Am Hinterrande der Dorsalseite jedes Brustsegmentes befindet sich eine niedrige Querleiste, deren Rand gezahnelt ist.— Och. M. . . .
...............................1c. T. renoculis v. carinatus Gurjan.
12 (1). Die Augen sind rund.
13 (14). Die Augen sind milchweiss. — Och. M . . ....................
...................................5. T. leucophthalmus Gurjan.
14 (13). Die Augen sind dunkel, fast schwarz pigmentiert.
15 (16). Die Seitenecken des fiinften Brustsegmentes sind in grosse spatel-formige zugespitzte Fortsatze ausgezogen, die nach hinten gekehrt sind. Das zweite Glied des Stammes des ersten Antennenpaares ist mit einem langen, zugespitzten Fortsatz versehen. — Jap. M............
..............................................3. T. glaber Gurjan.
16 (15). Die caudalen Seitenecken des fiinften Brustsegmentes bilden kleine zugespitzte spatelformige Fortsatze. Das zweite Glied des Stammes des ersten Antennenpaares ist ohne Fortzatz.
17 (18). Die Augen befinden sich an den hinteren Seitenecken des Kopfes. Das Ende des Telsorjist zugespitzt. — Вег. M. . .2. T. alascensis Rich.
18 (17). Die Augen befinden sich in der Mitte zwischen den vorderen und hinteren Seitenecken des Kopfes. Der hintere Rand des Telson ist zugerundet. — California........................... 4. T. convexus Rich.
3. Gatt. HOLOTELSON RICH.
1 (1). Jap. M................................ . 1. H. tuberculatus Rich.
4. Gatt. DYNAMENE LEACH.
1 (1). Jap. M., California . '.......................1. D. glabra Rich.
5. Gatt CILICAEA LEACH.
1 (1). Вег. M., California..........................1. C. cordata Rich
6. Gatt. CYMODOCE LEACH
1 (2). Der Korper ist mit Borsten bedeckt. — Jap. M.............. .
...................................... . 1. C. japonica Rich. (<?).
2 (1). Der Korper ist glatt, ohne Borsten.
3 (4). Der hintere Rand des ersten Abdominalsegmentes besitzt an den Seiten je zu einem kleinen, dreieckigen Auswuchse; ausserdem ist er in der Mitte mit einer Ausbuchtung versehen, an den Seiten welcher sich wiederum je zu einem kleinen, dreieckigen Auswuchse befindet. — Jap. M. . . .........................................2. C. acuta. Rich.
4 (3). Der hintere Rand des ersten Abdominalsegmentes ist zwischen den zwei Zacken gerade, nicht ausgebuchtet. — Jap. M......................
• • .................................... 1. C. japonica. Rich. (j).
262
ISOPODA
7. Catt. SPHAEROMA LATR.
1 (1)- Jap. M................................1. Sph. sieboldi Doi.
8. Gatt. EXOSPHAEROMA STEBB.
1 (1). Вег. M., California.............1. Ex. amplicauda (Stimps.).
9. Gatt. NEOSPHAEROMA BAKER
1 (2). Die Antennen des ersten Paares erreichen, nach hinten gebogen, den. Hinterrand des zweiten, diejenigen des zweiten Paares den Hinterrand des vierten Brustsegmentes. — Jap. M.........2. N. ovata (Gurjan.).
2 (1). Die Antennen des ersten Paares erreichen, nach hinten gebogen nur ч den Hinterrand des Kopfes, diejenigen des zweiten Paares den Hinterrand des zweiten Brustsegmentes. — N. St. Oz., Вег. M., Jap. M. . .
.................................. 1. N. oregonensis (Dana).
6. Fam. ANTHURIDAE
1 (4). Der Palpus der Maxillipeden besteht aus drei Gliedern.
2 (3). Das Flagellum des zweiten Antennenpaares ist normal, vielgliederig. • ..................................... . 3. Calathura Norm. a. Stebb.
3 (2). Das Flagellum des zweiten Antennenpaares ist rudimentar, ohne Gliederung....................... . 2. Paranthura Bate a. Westw.
4 (1). Der Palpus der Maxillipeden besteht aus zwei Gliedern...
.................. 1. Cyathura Norm. a. Stebb.
1. Gatt. CYATHURA NORM. a. STEBB.
1 (1). Trop. cosmopol.........................1. C. carinata(I^r^.
2. Gatt. PARANTHURA BATE a. WESTW.
1 (1). Jap. M. .............................. 1. P. japonica Rich.
3. Gatt. CALATHURA NORM. a. STEBB.
1 (1). Arct. Oz., N. Atl. Oz., Вег. M., Och. M. . .1. C. brachiata (Stimps.).
III. Subord. VALVIFERA
1 (2). Der Korper ist mehr oder weniger breit, in dorso-ventraler Richtung abgeflacht. Die Gliederung in Segmente ist regelmassig. Die Antennen des zweiten Paares sind immer kiirzer, als der Korper. Die 3 ersten Beinpaare sind oft mit Subchelae zum Greifen bewehrt . 7. Idotheidae.
2 (1). Der Korper ist lang, schmal, fast cylindrisch. Einige Segmente (gewohnlich das vierte Brustsegment) unterscheiden sich in Form und Grosse stark von den iibrigen. Die 4 ersten Beinpaare sind antennen-
6. ANTHURID AE. 7. IDOTHEIDAE
263
formig, nach vorne gerichtet; ihre Glieder sind schmal, lang und mit langen Borsten versehen. Die Fiisse der 3 letzten Paare sind Gehfiisse. Die Antennen des zweiten Paares sind sehr lang, oft danger als der Korper . . . ....... ...............................8. Arcturidae.
7. Fam. IDOTHEIDAE
1 (10). Der Palpus der Maxillipeden besteht aus 4 oder 5 Gliedern.
2 (3). Die Kopfseiten sind vor den Augen in nach vorne gerichtete Fortsatze ausgezogen^ die .langsgespalten sind und je zwei zugerundete Spatein bilden. Der Abdomen besteht aus 5 Segmenten .........................
................................... 3. Mesidothea Rich.
3 (2). Die Kopfseiten sind vor den Augen nicht in doppelte spatelformig^ Fortsatze ausgezogen. Die Zahl der Abdominalsegmente ist unter 5.
4(5). Die Glieder des Flagellums des zweiten Antennenpaares sind vollstandig zusammengewachsen, oder es sind 2—3 Glieder unterscheidbar, r die in der Form den Gliedern des Stammes gleichen..................
................................................7. Cleantis Dana.
5 (4). Die Flagellen des zweiten Antennenpaares sind normal, vielgliederig. 6 (7). Der Palpus der Maxillipeden aus 4 Gliedern .... 1. Idothea Fabr. 7 (6). Der Palpus der Maxillipeden besteht aus 5 Gliedern.
8 (9). Abdominalteil besteht aus 3 gut unterscheidbaren Teilen. 2 Segmenten und dem Telson....................................2. Pentidothea Rich.
9 (8). Die zwei vorderen Abdominalsegmente sind mit dem Telson verwachsen ' und bilden auf diese Weise ein einziges grosses Caudalsegment, das nur an den Seiten Spuren der einstigen Gliederungen in der Form von Querfurchen aufweist................................ 4. Pentias Rich.
10 (11). Der Palpus der Maxillipeden besteht aus 3 Gliedern.
11 (12). Die Epimeren der 7 Brustsegmente sind mit den Segmenten verwachsen und bilden Pleuralerweiterungen. Der Abdomen besteht aus einem Segment. Die Flagellum des . zweiten • Antennenpaares sind normal, vielgliederig..........................6. Synidothea Harger.
12 (11). Die letzten 3 Brustsegmente besitzen gut entwickelte freie Epimeren. Der Abdomen besteht aus 3 Segmenten. Die Glieder der Flagellum des zweiten Antennenpaares sind untereinander vollstandig verwachsen .................................................... 5. Symmius Rich.
1. Gatt. IDOTHEA FABR.
1 (2). Der Hinterrand des Telson ist gerade oder schwach gewdlbt. Der Korper ist relativ kurz und kraftig. — Jap. M., Atl. Oz., St. Oz. . . .
............................................1. I. metallica Bose.
2 (1). Der Hilterrand des Telson besitzt in der Mitte eine Zacke. Der Korper ist sehr gestreckt, linear»
264
ISOPODA
3 (4). Der Hinterrand des Telson ist dreizahnig: eine grossere befindet sich in der Mitte, zwei kiirzere an den Seiten, welch letztere durch ausge-zogene und zugespitzte caudale Seitenecken des Telson gebildet werden. — Jap. M., Och. M., Calif. . . ..............2.1, fewkesi Rich.
4 (3). Die caudalen Seitenecken des Telson sind abgerundet. An dem= Hinterrande des Telson befindet sich nur eine stumpfe Zacke.
5 (6). Die Antennen des zweiten Paares sind lang und reichen diinn und nach hinten gelegt bis uber das vierte Brustsegment hinaus (bis zur Mitte des fiinften). Die Seiten des Telson sind merklich eingebuchtet, so dass der Hinterrand bedeutend breiter ist, als die Mitte, sogar am Basalteile. — Och. M., Jap. M................. . . 6. I. orientalis Gurjan.
6 (5). Die Antennen des zweiten Paares sind nicht sehr lang, so dass sie den Hinterrand des vierten Brustsegmentes nicht erreichen. Der Hinterrand des Telson ist nicht breiter, als dessen Bassalteil.
7 (8). Das erste Brustsegment ist halb so lang, wie das zweite. Die Antennen: des ersten Paares reichen bis zur Mitte des vierten Brustsegmentes. — N. St. Oz., Вег. M., Och. M., Jap. M............3. 1. ochotensis Brandt..
8 (7). Das erste Brustsegment ist nur wenig kiirzer, als das zweite. Die Antennen des ersten Paares reichen bis zur Mitte des dritten Brustsegmentes.
9 (10). Die Dorsalseite des Kopfes ist fast ganz flach. Die Seitenflachen des Telson sind in der Mitte etwas eingebogen, so dass der Hinterrand des Telson einigermassen breiter ist, als die Mitte. Die Flagellen des zweiten Antennenpaares sind linear und verdiinnen sich gleichmassig distalwarts. — Вег. M., Och. M. ........ 4. I. aleutica Gurjan.
10 (9). Die Dorsalseite des Kopfes ist stark gewolbt. Der Telson verengt sich gleichmassig, wenn auch schwach, caudalwarts. Die Seitenrander sind einander fast parallel und das hintere Ende ist etwas schmaler,. als die Mitte. Die Flagellen des zweiten. Antennenpaares sind an der
? Basis spindelformig verdickt. — Jap. M., Och. M.....................
. . . . . . .................5.1. derjugini Gurjan.
2. Gaft. PENTIDOTHEA RICHARDSON
1 (6). Die Brustsegmente beriihren einander in ihrer ganzen Lange.
2 (3). Die Epimeren erstrecken sich nur beim letzten Brustsegment auf die ganzen Seitenrander. Am Hinterrande des Telson befindet sich eine stumpfe, dreieckige Zacke. — Jap. M.................1. P. japonica Rich.
3 (2). Die Epimeren erstrecken sich bei den 3 letzten Brustsegmenten auf den ganzen Innenrand. Der Telson ist hinten abgerundet.
4 (5) Der Korper ist schmal, gestreckt, fast 6 Mai langer als breit; der Telson auch gestreckt, seine Lange 2 Mai grosser, als die Breite an der Basis. Der Hinterrand des Telson ist zugerundet. — Jap. M..............
..............................................2. P. rotundata Rich.
7. IDOTHEIDAE
265
5 (4). Der Korper ist gedrungen und breit, 3 Mai so lang- als breit, der Telson auch breit, 1Mai so lang, als die Breite an der Basis. Das Ende des Telson ist breit abgerundet mit einer kleinen stumpfen Zacke an der Spitze. — Вег M., Och. M., N. St. Oz.....................
........................................3. P. wosnesenskii (Brandt).
6 (1). Die Brustsegmente beriihren einander nur in ihren mittleren Teilen, da die lateralen Teile verschmalert und seitwarts gebogen sind. 4. P. whitei Rich.
3. Gatt. MESIDOTHEA RICH.
1 (1). Вег. M., Och. M.........о ... 1. M. entomon orientalis Gurjan.
4. Gatt. PENTIAS RICH.
1 (1). Jap. M. . . ......................................1. P. hayi Rich,
5. Gatt. SYMMIUS RICH.
1 (1). Jap. M........................................1. S. candatus Rich.
6. Gatt. SYNIDOTHEA HARGER.
1 (12). Der Telson ist hinten zugespitzt.
2 (3). Kopf, Thorax und Telson sind an der Dorsalseite mit grossen Stacheln besetzt. — Вег. M., Och. M...........................6. S. spinosa Gurjan,
3 (2). Auf der Dorsalseite des Korpers sind keine grossen Stacheln vorhan-. den, sondern nur flache Hocker und ganz kurze Stacheln, oder leisten-formige Wiilste. , . ...
4 (5). Der Telson ist fiinfeckig. — Och. M..........18. S. cinerea Gurjan.
5 (4). Der Telson ist gestreckt dreieckig.
6 (9). Die Pleuralverbreiterungen der Brustsegmente sind ohne Hocker und Stacheln. ...
7 (8). Die praeokularen Hocker befinden sich hinter dem Vorderrande des Kopfes. — Arct. Oz., Вег. M., N. St. Oz..............19. S. nodulosa (Kr.).
8 (7). Die praeokularen Hocker befinden sich auf dem Vorderrande des Kopfes und bilden einen Teil desselben. — Вег. M. . .....................
..............................................14. S. laevis Bened.
9 (6). Die Pleuralverbreiterungen der Brustsegmente sind mit Gruppen von Hockern und Stachelchen besetzt.
10 (11). Die Dorsalseite des Kopfes tragt hinter dem Frontalausschnitt .4 stachelahnliche Hocker, in Form eines Rhombus angeordnet. Auf den Pleuralverbreiterungen der Brustsegmente befinden sich Gruppen von kleinen Stachelchen. — Arct. Oz., Och. M. . ...........................
..................... . . .. . . . . . 15. S. muricata (Harford).
11 (10). Die Dorsalseite des Kopfes tragt hinter dem Frontalausschnitt nur 3 keilformige Hocker, die in einer Langsreihe angeordnet sind und
266
ISOPODA
mit ihren Basalteilen zusammenstossen. Auf den Pleuralverbreiterungen der Brustsegmente befinden sich Grupperi niedriger schwachbegrenzter Hocker. — Вег. M............................. 17. S. picta: Bened.
12 (1). Der Telson ist hinter gespalten, oder besitzt einen massigen Ausschnitt.
13 (14). Das erste Brustsegment ist mit kleinen freien Epimeren versehen.— Jap. M..............................................10. S. epimerata Rich.
14 (13). An alien Segmenten fehlen Epimeren.
15 (16). Der Vorderrand des ersten Brustsegmentes hat an den Seiten des Kopfes gut sichtbare halbkiesformige Ausschnitte. — Och. M., Jap. M.
.......................................... 11. S. excavata Gurjan.
16 (15). Der Vorderrand des ersten Brustsegmentes hat keine Ausschnitte an den Seiten des Kopfes.
17 (18). Der Vorderrand des Kopfes ist gerade, ohne Ausschnitt in der Mitte. — California, Jap. M.........................12. S. harfordi Bened.
18 (17). Der Vorderrand des Kopfes ist V-formig, d. h. hat in der Mitte einen Ausschnitt.
19 (28). Hinter dem Ausschnitte am Vorderrande des Kopfes befinden sich ein oder zwei gut sichtbare Hocker.
20 (23). Hinter dem Ausschnitte am Vorderrande des Kopfes befindet sich nur ein Hocker.
21 (22). Der Frontalausschnitt am Kopfe ist nicht tief. Die Antennen des zweiten Paares erreichen nach hinten gebogen den Hinterrand deS ersten Brustsegmentes. Die Seitenrander des ersten Brustsegmentes bilden mit seinem Vorderrande einen Winkel. — Вег. M. ..... .
.............. • . . •...........................9. S. acuta Rich.
22 (21). Der Frontalausschnitt ist sehr tief, V-formig. Die Antennen des zweiten Paares erreichen zuriickgebogen den Hinterrand des vierten Brustsegmentes. Die vorderen Seitenecken des ersten Brustsegmentes sind sanft zugerundet. — Och. M......................8. S. longicirra Gurjan.
23 (20). Hinter dem mittleren Frontalausschnitte befinden sich 2 gut entwic-kelte Hocker. ... - . -
24 (25). Auf dem Kopfe befinden sich hinter dem Frontalausschnitte ausser 2 Hockern noch 6 weitere, die in einer Querreihe angeordnet sind.— Och. M............................................ 7. S. tuberculata Rich.
25 (24). Auf dem Kopfe befinden sich hinter dem Frontalausschnitte nur 2 Hocker.
26 (27) Die beiden Hocker, die am Kopfe uber den Frontalausschnitt geneigt sind, verschmelzen in ihren Basalteilen miteinander. — Вег. M. . . . .
.............................................. 6. S. pallida Bened.
27 (26)., Die 2 . am Kopfe uber dem Frontalausschnitte befindlichen Hocker stehen einzeln. — Вег. M. . . ................ 5. S. erosa Bened.
28 (19). Hinter dem Frontalausschnitte des, Kopfes sind keine Hocker vorhanden. >
7. IDOTHEIDAE. 8. ARCTURIDAE
267
29 (30). Das Distalende des Telson ist nur schwach ausgeschnitten. Die Seitenrander des Telson sind in ihrem vorderen Teile fast gerade.— Вег. M.
........................................ . 3. S. nebulosa Bened.
30 (29). Der Ausschnitt am Distalende des Telson ist tief. Der Telson ist von dreieckiger Form, mit sanft zugerundeten Seitenrandern.
31 (32). Die Seitenrander des ersten Brustsegmentes bilden mit seinem Vorderrande einen Winkel (bei jungen Exemplaren auch am zweiten und dritten Segmente). — Jap. M....................4. S. brazbnikovi Gurjan.
32 (31). Die Seitenrander des ersten Brustsegmentes sind sanft abgerundet.
33 (34). Das Zweite Glied am Stamme des zweiten Antennenpaares ist mit einem grossen Stachel ausgeriistet. Der Hinterkopf ist stark gewolbt. — Och. M., Jap. M........................... 1 a. S. bicuspida lata Gurjan.
34 (33). Das zweite Glied am Stamme des zweiten Antennenpaare^ ist ohne Stachel. Der Kopf ist flach.
35 (36). Die Lange des Telson ist geringer, als seine Breite an der Basis.— Arct. Oz., Вег. M.............................1. S. bicuspida (Owen).
36 (35). Die Lange des Telson ist grosser, als seine Breite an der Basis.— N. Atl. Oz., Вег. M. . . . . . . .. . . . 2. S. marmorata (Packard).
7. Gatt. CLEANTIS DANA * .
1 (2). Der Telson ist zungenformig, am Ende abgerundet. — Atl. Oz;, Jap. M.
...................................... 1. C. planicauda Bened.
2 (1). Der Telson ist funfeckig mit stumpfem Ende. — Gelb. M.,Jap. M. . .
.................................. 2. C. isopus Grube.
8 Fam. ARCTURIDAE
1 (2). Der vierte Brustsegment ist viel langer, als alle iibrigen. Der Brustraum der Weibchen ist aus 3—4 Plattchenpaaren gebildet. . 3. Astacilla Cord.
2 (1). Der vierte Brustsegment ist wenig langer als die iibrigen. Der Brustraum der Weibchen ist aus 5 Plattchenpaaren gebildet.
3 (4). Der Abdomen besteht aus 3 gut voneinander geschiedenen Segmenten, 2 vorderen und 1 Caudalsegment (Telson). ..... 1. Arcturus Latr.
4 (3). Der Abdomen besteht aus 2 Teilen, einem kurzen Abdominalsegment und dem Telson. ......................... 2. Pleuroprion zur-Strassen.
1. Gatt. ARCTURUS LATREILLE
1 (24). Auf der Dorsalseite der Brustsegmente befinden sich grosse paarige Stacheln.
2 (15). Das Ende des Telson ist gespalten.
3 (10). Auf alien Brust- und Abdominalsegmenten sind paarige Riickensta-cheln vorhanden. ,
4 (7). Das zweite Glied des Stammes des zweiten Antennenpaares hat an der distalen Aussenecke einen Dorn. '
268
ISOPODA
5 (6). Die Antennen des zweiten Paares sind sehr lang, l’/2 Mai so lang wie der Korper. — Och. M...............................5. A. setosus Gurjan.
6 (5). Die Antennen des zweiten Paares sind'wenig langer, als der Korper.— Och. M., Jap. M...................5 a. A. setosus var. seminudus Gurjan.
7 (4). Das zweite Glied des Stammes des zweiten Antennenpaares hat an der distalen Aussenecke keinen Dorn.
8 (9). Die Glieder des zweiten Antennenpaares sind glatt. Die dorsalep Stacheln sind sehr lang, langer, als die Segmente.—Вег. M. . . . ; . .
.........................................7. A. magnispinis Rich.
9 (8). Die Glieder des zweiten Antennenpaares sind mit Harchen bedeckt. Die dorsalen Stacheln sind nicht lang, ihre Lange ist geringer, als die Hohe der Segmente. — Och. M., Jap. M., Gelb. M........................
. . .................................... 8. A. crassispinis Rich.
10 (3). Einige der Brust- und Abdominalsegmente sind ohne paarige Riicken-stacheln.
11 (12). Das erste Brustsegment ist ohne Dorsalstacheln.—Вег. M. . . . . ..............................................4. A. longispinis Bened.
12 (11). Das erste Brustsegment besitzt ein Paar Dorsalstacheln.
13 (14).jDer Korper und die Antennen sind mit Harchen bedeckt. —Вег. M.
........................................... 6. A. diversispinis Rich.
14 (13). Der Korper und die Antennen sind ohne Harchen, aber gekornelt.
Вег. M............................... . . . 9. A. brevispinis Rich.
15 (2). Das Ende des Telson ist nicht gespalten, sondern zugespitzt odef zugerundet und mit langen Dornen versehen.
16 (21). Der Telson ist am Ende zugespitzt.
17 (18). Die Oberflache des Korpers ist glatt, ohne Harchen. — Och. M. . ................................................... 2. A. ulbani Gurjan.
18 (17). Die Oberflache des Korpers und die Dorsalstacheln sind mit Harchen besetzt. ‘
19 (20). Die vorderen Seitenecken des Kopfes tragen je zu einem Stachel. Die Dorsalstacheln sind lang und diinn, ihre Lange ist grosser, als die Hohe der Segmente.— Och. M., Jap. M................................
........................ 5 b. A. setosus var. acuticaudalis Gurjan.
20 (19). Die vorderen Seitenecken des Kopfes sind ohne Stacheln. Die Dorsalstacheln sind kurz, ihre Lange ist geringer, als die Hohe der Segmente.— N. Atl. Oz., W. Gron., Och. M...............1. A. baffini (Sab.).
21 (16). Der Telson ist am Ende abgerundet und mit langen Stacheln besetzt.
22 (23). Die den Korper bedeckenden Stacheln tragen an den Spitzen Grup-pen von Harchen, die in der Form von Rosetten angeordnet sind. Die apicalen Stacheln des Telson sind behaart. — N. St. Oz., Вег. M. . .
.............................................15. A. hirsutus Rich.
23 (22). Die den Korper bedeckenden Stacheln, sowie die apicalen Stacheln sind nicht mit Harchen besetzt. — Och. M- * • 16. A. beddardi Gurjan.
8. ARCTURIDAE
269
| 24 (1). Die Dorsalseite der Brustsegmente ist glatt, oder mit kleinen, niedri*
! gen Hockern besetzt.
I 25 (30). Der Telson ist am Ende gespalten.
| 26 (29). Die Oberflache des Korpers ist glatt. Auf den Brustsegmenten und
i auf dem Telson sind keine Hocker vorhanden.
J 27 (28). Der Unterrand des vierten Gliedes des Stamme^ zweiten Antennen-i paares tragt eine Langsreihe von rundlichen Hockern, welche mil
kurzen Borsten abwechseln. — Jap. M. . . . 12. A. crenulatus Gurjan.
28 (27). Der Unterrand des vierten Gliedes des Stammes des zweiten Anten-nenpaares ist glatt. — Вег. M., Och. M.,Jap.M. 11. A. beringanus Bened.
29 (26). Die Oberflache des Korpers ist grob gekornelt. Der Vorder- und Hinterrand der Brustsegmente ist mit einer Querreihe von dicht stehen-den kleinen Hockern bedeckt. Die Dorsalseite ist mit niedrigen Hockern bedeckt. — Och. M. . . . .......................10. A. granulatus Rich.
30 (25). Das Ende des Telson ist zugespitzt, oder abgerundet, stumpf.
31 (34). Die Dorsalseite des Kopfes und der Brustsegmente ist glatt, ohne Hocker.
32 (33). Die Oberflache des Korpers ist ganz glatt. Die Epimeren der hinteren Brustsegmente sind von oben sichtbar. Langs dem Korper ziehen, sich 3 helle Streifen hin. Das Ende des Telson ist stumpf, abgerundet.— Вег. M. . .............................. 14. A. tritaeniatus Rich.
33 (32). Die Oberflache des Korpers ist sehr fein netzformig granuliert (dutch die Lupe gesehen). Die Epimeren der hinteren Brustsegmente. sind von oben nicht sichtbar. Das Ende des Telson ist zugespitzt.— Вег. M., N. St. Oz. ............................. 13. A. glabrus Bened.
34 (31). Auf der Dorsalseite des Kopfes und der Brustsegmente sind paarige mehr oder weniger entwickelte Hocker vorhanden. — Jap. M. . . . .
............................................ 3. A. hastiger Rich.
2. Gatt. PLEUROPRION zur STRASSEN
1 (6). Auf der Dorsalseite der Brustsegmente sind grosse Stacheln vorhanden. Der Telson ist hinten gespalten, oder mit 2 langen divergierenden Stacheln besetzt.
2 (5). Die Dorsalseite des Telson ist mit Stacheln besetzt. Das Caudalende des Telson ist gespalten. Das dritte Glied des Stammes des zweiten Antennenpaares hat einen oder zwei Stacheln am Distalende.
3(4). In der Mitte des Vorderrandes des Kopfes ist ein grosser Stachel vorhanden. Das dritte Glied des Stammes des zweiten Antennenpaares tragt 2 Stacheln am Distalende. — Arct. Oz., Вег. M....................
....................................... 1. P. murdoefai (Bened.).
4 (3). Der Vorderrand des Kopfes ist ohne Stachel. Das dritte Glied des Stammes des zweiten Antennenpaares tragt am Distalende einen grossen
; Stachel. — N. Atl, Oz., Och. M. . ..... . 2. P. hystrix (G. Sars).
270
/SO POD A
-----------.---------------------------------------------------------- j ii
5 (2). Die Dorsalseite des Telson ist glatt. Das dritte Glied des Stammes i
des zweiten Antennenpaares ist glatt, ohne Stacheln. — Вег. M. . . . |
......................................3. P. intermedium (Rich.).
6 (1). Die Dorsalseite des Korpers ist ohne grosse Stacheln. Der Telson ist-hinten zugespitzt. Der ganze Korper ist mit einer groben Kornelung und kleinen spilzen Hockern bedeckt. — Och. M.........................
..........................................4. P. tarasovi Gurjan,
3. Gatt. ASTACILLA CORDINER.
1 (2). Die Oberflache des Korpers ist grob gekomelt. — Jap. M. . . . . . i
............................................ 1. A. dilatata Rich.
2 (1). Die Oberflache des Korpers glatt. — Jap. M. . . . 2. A. polita Gurjan.
IV. Subord.. ONISCOIDEA
1 (6). Die Uropoden sind terminal und bilden liber den Abdominalfiissen keinen Deckel. Alle 5 Paar Abdominalfusse sind gut entwickelt.
2 (3). Die Geisseln des zweiten Antennenpaares sind vielgliederig. Der Penis ist paarig . .......................................9. Ligiidae.
3 (2). Die Geisseln des zweiten Antennenpaares bestehen aus wenigen Gliederh. Der Penis ist nicht paarig.
4 (5). Die Geisseln des zweiten Antennenpaares bestehen aus uber 3 Glie-dern. Der Molar der Mandibeln ist gut entwickelt. . 10. Trichoniscidae.
5 (4). Die Geisseln des zweiten Antennenpaares bestehen aus 2—3 Gliedern.
Der Molar der Mandibeln ist reduziert...............11. Oniscidae.
6 (1). Die Uropoden bilden einen Deckel liber deri Pledpoden. Das erste Paar Pleopoden fehlt (so dass im ganzen 4 Paar ubrig bleiben).
7 (8). Alle Abdominalsegmente sind frei. Das erste (zweite) Paar Pleopoden
ist unverzweigt.........,........................... ; [Tylidae],
8 (7). Die Abdominalsegmente sind an der Dorsalseite vollstandig miteinan-der verschmolzen. Das vordere (zweite) Paar Pleopoden besteht aus zwei Asten............................................ [Syspastidae].
9. Fam. LIGIIDAE
1 (2). Das Basalglied der Uropoden ist einfach, cylindrisch. Die Aste der Uropoden sind von gleicher Lange. Die hinteren Seitenecken des Telson bilden kleine, dreieckige, zugespitzte, spatelformige Fortsatze . .
.............................................. . 1. Ligia Fabr.
2 (1). Das Basalglied der Uropoden ist an der distalen Innenecke mit einem Fortsatze versehen. Die Aste der Uropoden sind von ungleicher Lange.
.............................................2. Ligidium Brandt.
9. LIGIIDAE. 10. TRICHONISCIDAE. 11. ONISCIDAE
271
1. Gatt. LIGIA FABRICIUS
1 (4). Der Korper ist gestreckt oval. Beim Mannchen befindet sich am sechsten Gliede der ersten Thoracalfiisse ein Fortsatz in der Form eines Hockers, der liber die Klaue iiberhangt.
2 (3). Die Oberflache des Korpers ist sehr fein granuliert, fast glatt. Die Antennen reichen zuriickgelegt bis zur Mitte des sechsten Brustsegmentes. Die Glieder der Geissel sind kurz und kraftig. — Jap. M. . . . .
.....................................L. cinerascens Budde-Lund.
3 (2). Die Oberflache des Korpers ist gekornelt. Die Antennen des zweiten Paares sind viel langer, so dass sie nach hinten gebogen den Hinterrand des fiinften Abdominalsegmentes erreichen. Die Glieder der Geissel sind lang und diinn. — Tropisch. St. Oz. . . (L. exotica Roux).
4 (1). Der Korper ist- breit oval. Beim Mannchen ist am ersten Paare Thora-calfusse an der Basis der Kralle (am sechsten Gliede) kein Fortsatz
- - vorhanden. —• N. St. Oz., Вег. M.......7. - 2. L. pailasi Brandt.
2. Gatt. LIGIDIUM BRANDT.
1 (1). Вег. M.............................. 1. L. tenue Budde-Lund.
10. Fam. TRICHONISCIDAE
1. Gatt. TMCHONISCUS BRANDT.
1 (1). Вег M., Jap. M. . .............. . . .1. Tr. papillicornis Rich.
11. Fam. ONISCIDAE.
1. Gatt. PORCELLIO LATR.
1 (1). Kosmopol....................................1. P. scaher Latr.
V. Subord. EPICARIDEA..................
1 (2). Der Korper des Weibchens. ist assymmetrisch segmentiert. Die Maxillipeden sind plattchenformig, zweigliederig. 7 Paar Fiisse, die aber zuweilen nur an einer Seite erhalten bleiberi. 5 Paar Inkubationsplatt-' chen. Beim Mannchen sind alle Brustsegmente beweglich. Im letzten Larvenstadium bestehen die Flagellen des zweiten Antennenpaares aus 4 Gliedern. Schmarotzer auf Decapoden.................12. Bopyridae.
2 (1). Der Korper des Weibchens ist symmetrisch. Zuweilen sind Spuren' einer Segmentation vorhanden. Nur fiinf Paar Fiisse. Die Inkubations-plattchen. sind klein, oft stark reduziert. Abdominalfiissc fehlen oder sind ganz rudimentar. Beim Mannchen verschmilzt der Kopf mit dem ersten Brustsegment. Im letzten Larvenstadium bestehen die Flagellen des zweiten Antennenpaares aus 5 Gliedern. Schmarotzer auf Schizo-poden.................................................. 13. Dajidae.
272
ISOPODA
12. Fam. BOPYRIDAE
1 .(2). Der Korper des Weibchens ist stark assymmetrisch und gekriimmt, so dass die eine Seite bedeutend kiirzer ist, als die andere. Nur das vordere Paar Thoracalfiisse ist beiderseitig entwickelt, die iibrigen sind auf einer Seite ganz reduziert. Der Abdomen besteht aus 5 Segmenten
...........................................1. Phryxus Rathke.
2 (1). Der Korper ist mehr oder weniger symmetrisch. Alle. 7 Paar Tho-racalfiisse sind vollstandig entwickelt Der Abdomen besteht aus 6 Segmenten.
3 (4). Das Weibchen besitzt sowohl Pleopoden, als auch Uropoden (letztere einastig). Die caudalen Seitenecken aller Brustsegmente bilden gestrec-kte, schmale Fortsatze..............................2. Argeia Dana.
4 (3). Das Weibchen besitzt weder Pleopoden, noch Uropoden. Die caudalen Seitenecken der Brustsegmente sind ohne Fortsatze ........
.......................................3. Bopyroides Stimpson. ./
1. Gatt. PHRYXUS RATHKE.
1 (1). N. Alt. Oz., Arct. Oz...............• . 1. Ph. abdominals (Kr.).
2. Gatt. ARGEIA DANA
1 (1). N. St. Oz.............................1. A. pugettensis Dana.
3. Gatt. BOPYROIDES STIMPS.
1 (1). N. Atl. Oz., Arct. Oz., N. St. Oz. ...... 1. B. hippolytes (Kr.).
13. Fam DAJIDAE
1 (2). Der Abdomen besteht aus einem einzigen Segment......... .
...........................................1. Holophryxus Rich.
2 (1). Der Abdomen besteht aus 5 Segmenten.
3 (4). Der Kopf und die ersten 3 Brustsegmente sind vollstandig voneinander geschieden.......................................3. Prophryxus Rich.
4 (3). Die ersten 5 Segmente sind untereinander verwachsen, bloss auf der Dorsalseite sieht man Reste der Einteilung. . . 2. Arthrophryxus Rich.
1. Gatt. HOLOPHRYXUS RICH.
1 (2). Der Kopf des Weibchens ist von einer weichen Falte mit geradem Rande umringt. An den Seiten des Abdomens, an der Grenze der Basis findet sich je ein Ausschnitt. — Вег. M............2. H. giardi Rich.
2 (1). Der Kopf ist vorne ohne Falte und tragt 2 spatelformige Fortsatze. Der Abdomen hat an der Basis und der Grenze zum Thorax keinen Ausschnitt.
12. DAJIDAE. 13. GNATHIIDEA
273
3 (4). Der Korper ist schmal, oval. 4 Paar Epimeralplatten. — California . .
........................................(H. caiiforniensis Rich.).
4 (3). Der Korper ist breit, mit fast quadratformigem Thorax. 5 Paar Epimeren.— N. St. Oz.................................l.H. alascensis Rich.
2. Gatt. ARTHROPHRYXUS RICH.
1 (1). Вег. M., N. St. Oz......................1. A. beringanus Rich.
3. Gatt. PROPHRYXUS RICH.
1 (1). N. St. Oz. ...............................1. P. alascensis Rich.
VI. Subord. GNATHIIDEA
14. Fam. GNATHIIDAE
1. Gatt. GNATHIA LEACH
1 (10). Der Vorderrand des Kopfes besitzt keine Ausschnitte.
2 (3). Am Vorderrande des Kopfes befinden sich an der Ventralseite, von jeder Seite der Antennen, 4 Stacheln. — Jap. M. . . . . ..........
.........................................1. G. tuberculata Rich.
3 (2). Der Vorderrand des Kopfes ist an der Ventralseite ohne Stacheln.
4 (5). Der Vorderrand des Kopfes ist gerade, mit einem kleinen, spitzen Fortsatz in der Mitte.— Jap. M................2. G. rectifrons Gurjan.
5 (4). Der Vorderrand des Kopfes ist von anderer Form.
6 (7). Die Augen sind klein und ragen nicht uber die Seitenecken des Kopfes hinaus. An den Seiten der Abdominalsegmente sind dornartige Fortsatze vorhanden. — Jap. M.................3. G. schmidti Gurjan.
7 (6). Die Augen sind gross, stark gewolbt. An den Seiten der Abdominalsegmente sind keine Stacheln vorhanden.
8 (9). Die Seiten der Abdominalsegmente sind in zungenformige Fortsatze ausgezogen, an den Enden sind Biischel langer Harchen vorhanden. Am Kopfe befinden sich zwischen den Basen der Mandibeln 2 kleine, spa-telformige, zugerundete Fortsatze. Das dritte Glied der Pylopoda ist stark reduziert und kaum sichtbar.—Jap. M. . .4. G. derzhavini Gurjan.
9 (8). Die Seiten der Abdominalsegmente sind normal entwickelt. Am Kopfe befindet sicht zwischen den Basen der Mandibeln ein dreispitziger Auswuchs. Das dritte Glied der Pylopoda ist klein, aber vollstandig entwickelt. — N. Atl., Arct. Oz., Jap. M.........5. G. elongata (Kr.).
10 (1). Der Vorderrand des Kopfes besitzt einen tiefen und komplizierten Ausschitt — Jap. M............... . 6.G. (Elaphognathia) monodi Gurjan.
Гурьянова
18
INDEX
abdominalis, Bopyrus, Phryxus, 16, 17, 25, 26, 30, 217, 218, 238, 246, 248, 272.
acuta, Cymodoce, 117, 119, 261.
acuta, Synidothea, 151, 160, 237, 266.
acuticaudalis, Arcturus setosus, 20, 21, 22, 24, 175,183, 241, 242, 268,
Aega 15,17,18, 20, 21, 23, 29, 30, 70, 256.
Aeginae 66, 69, 256.
affinis, Cymodoce, 117. 1
alascensis, Holophryxus, 223, 273.
alascensis, Janira, 40, 41, 252. -
alascensis, Rocinela, Aega, 75.
alascensis, Prophryxus, 226, 273.
alascensis, Tecticeps, 15, 16, 20, 21, 100, 102, 107, 238, 242, 261.
albifrons, Jaera, 13.
aleutica, Idothea, 15, 16, 18, 20, 21, 29, 134, 137,238,239,242,247, 264.
algirica, Idothea, 134.
amplicauda, Exosphaeroma, Sphaeroma 17, 20, 122, 239, 241, 262.
amurensis, Cymothoa, Livoneca, 20, 23, 24, 26, 29, 86, 87, 243, 245, 248, 208.
Anceus 228.
angustata, Rocinela, 15, 17, 20, 21, 24, 26, 29, 73, 77, 238, 242, 271, 247, 287.
Anilocra 92.
Antares 197.
Anthura 126, 129.
Anthuridae 1, 2, 4, 7, 8, 9, 11, 12, 15, 20, 24, 65, 125, 255, 262.
arctica, Eurycope, Munnopsurus, 58.
Arcturidae 2, 4, 9, 16, 20, 24, 131, 173, 202, 262, 267.
Arcturus 16,17, 19, 20, 21, 22, 23, 24, 25, 26, 29, 30, 31,173, 241, 248, 250, 267.
Argeia 16, 17, 25, 26, 29, 217, 219, 272.
amholdi, Munna, 15, 16, 50, 51, 237, 253.
Arthrophryxus 16, 17, 20, 222, 224, 241, 272, 273.
Asellota 2, 3, 4, 8,12, 15, 20, 24, 31, 38,. 249, 251.
Astacilla 1, 24, 26, 173, 202, 245, 267, 270.
atrata, Idothea, 134.
avatshensis, Munna, 15, 50, 52, 237, 253.
baffini, Arcturus, 19, 20, 23, 173, 175, 240, 243, 268.
baltica, Idothea, 1. '
Bathynomus 1, 3, 10, 67, 68, 256. beddardi, Arcturus, 20, 21, 23, 29; 30, 175, 196, 241, 243, 247, 248, 268.
belliceps, Aega, Ricinela, 15, 17, 20, 21, 24, 26, 29, 73, 75, 79, 238, 242, 245, 247, 257.
beringanus, Arcturus, 16, 17, 20, 21, 24, 26, 29, 175, 191, 238, 242, 245, 247, 269.
beringanus, Arthrophryxus, 16, 17, 224, 225, 238, 248, 273.
bicuspida, Edotia, Idothea, Synidothea, 16, 17„ 19, 22, 152,155, 238, 240, 267.
bidentata, Dynamene, 114.
bivittata, Nerocila, 82.
boecki, Munna, 49.
Bopyridae 16, 25, 216, 271, 272.
Bopyroides 16,17, 30, 217, 221, 272.
brachiata, Calathura, 15, 17, 18, 19, 20, 23, 29, 129, 238, 239, 240, 242, 243, 248, 262.
brazhnikovi, Synidothea, 24, 25, 31, 152, 156, 245, 250, 267.
breviremis, Janiropsis, 44, 47. , brevispinis, Arcturus, 16, 174, 188, 237, 268.
Calathura 15, 17, 18, 19, 20, 23, 29, 125,129,’
238, 262.’
califomica, Livoneca, 87,' 93, 259.
californica, Nerocila, 82, 83, 258. californiensis, Holophryxus, 223, 273.
carinata, Anthura, Cyathura, 20, 23, 30, 126, 243,248,262.
carinatus, Tecticeps renoculis, 20, 21, 102, 105, 108, 241, 261.
caudata, Livoneca, 86, 258.
INDEX
275
caudatus, Symmius, 148, 149, 265.
Ceratothoa 84.
Cilicaea 17, 19, 20, 29, 99, 115, 240, 259, 261.
cinerascens, Ligia, 24, 26, 32, 208, 209, 214, 245, 246, 250, 271.
cinerea, Synidothea, 20, 21, 150, 169, 241, 265.
Cirolana 27, 67, 246, 256.
Cirolaninae 66, 255, 256.
Cleantis 3, 24,26, 29, 30,132,170, 248, 263, 267.
coecus, Microprotus, 15, 16, 48, 237, 253.
compacta, Idothga, 134. '
convexus, Tecticeps, 102, 111, 261.
cordata, Cilicaea, Dynamene, 17, 20, 29, 115, 239, 241, 247, 261.
comuta, Eurycope, 62.
cornuta, Rocinela, 15, 17, 73, 257.
cranchii, Cirolana, 67.
crassispinis, Arcturus, 20, 21, 22, 24, 26, 174, 186,242/245,268.
crenulatus, Arcturus, 24, 25,175, 192, 245, 269.
Cryptoniscus 3.
Cyathura. 20, 23,30,125, 126, 262.
Cymodoce 24, 26, 29,31, 99, 100, 117, 248, 249, 260,26.1.
Cymothoa 87.
Cymothoidae 4, 9, 11, 14, 20, 24, 27, 65, 66, . 247,255,
Cymothoinae 66, 81, 256, 257.
Dajidae 16, 216, 222, 271, 272.
danmoniensis, Rocinela, 72.
daurica, Lironeca, 87.
Dendrotion 4.
perjugini, Idothea, 20, 21, 22, 24, 26, 29, 134, ' 138, 242, 245, 247, 264.
derjugini, Janiropsis, 15, 16, 44, 45, 237, 253. derzhavini, Gnathia, 24, 25, 229, 232, 245, 273. dilatata, Astacilla, 24, 26, 203, 245, 270.
dilatata, Ligia, 210.
diversispinis, Arcturus, 16, 174, 184, 237, 268. doderleini, Bathynomus, 68, 256.
Dynamene 24, 26, 29, 31, 99,100, 104, 248, 249, 260,261. ; .•/
Echinozone 56. ; ' ,
Edotia 152.
emargjnata, Idothea, 132.
entomon, Mesidothea, 13, 15, 17, 18, 19, 20, 23, 29,145, 238, 239, 240, 248, 265.
Elaphognathia 25, 246, 273.
elongata, Anceus, Gnathia, 25, 29, 32, 229, 233, . 245,248,250,273.
Epicaridae 2, 247.
Epicaridea 4, 7, 9, 10, 14, 16, 21, 25, 27, 38, 215, 241, 251, 271.
| epimerata, Synidothea, 2, 8, 24, 25, 31, 151,161» 245, 250, 265.
epimerias, Livoneca, 24, 25, 87, 91, 245, 259.
erosa, Synidothea, 151, 157, 256.
erostrata, Janira, 15, 17 20, 21, 24, 26, 41, 238, 242, 245, 252.
estuaria, Cyathura, 126.
Eubranchiatae 99.
Eurycope 24, 25, 32, 54, 55, 57, 62, 254, 255.
Eurycopinae 54.
excavata, Synidothea, 24, 25, 151, 162, 245, 266. Exosphaeroma 17, 19, 20, 31, 99, 100, 121, 249, 260, 262.
exotica, Ligia, 27, 208, 209, 246, 271.
ferox, Gnathia, 246.
fewkesi, Idothea, 20, 21, 22, 24, 26, 30, 133, 135, 242, 243, 246, 249, 264.
Flabellifera 12,15, 20, 24, 38, 55, 227, 251, 256.
giardi, Holophryxus, 16, 17, 29, 223, 224, 237, 238, 248, 272.
gigantea, Eurycope, 58.
giganteus, Bathynomus, 1, 68.
giganteus, Munnopsurus, 15, 17, 18, 19, 20, 23, 29, 57, 58, 238, 239, 240, 242, 243, 254.
gigas, Exosphaeroma, 121.
glaber, Arcturus, 193.
glaber, Tecticeps, 24, 25, 102, 110, 245, 261.
glabra, Dynamene, 24, 26, 114, 245, 261.
glabrus, Arcturus, 16, 175,193, 237, 269. glacialis, Mesidothea entomon, 146, 147.
Gnathia 4, 6, 25, 29, 32, 228, 273.
Gnathiidae 2, 4, 7, 8, 25, 227, 273.
Gnathiidea 2, 3, 4, 6, 7, 8, 11, 12, 14, 21, 25, ' 38, 227, 241, 251, 273.
.granulatus, Arcturus, 20, 175, 189, 241, 269.
harfordi, Synidothea, Idothea 24, 26, 30, 151, 163, 246, 249, 266.
hastiger, Arcturus, 20, 21, 22, 24, 26, 175, 178, 242, 245, 269.
hayi, Pentias, 24, 26, 31, 147, 245, 250, 265.
Hemibranchiatae 99.
Hemilepistus 14.
hippolytes, Bopyroides, Bopyrus, 16, 17, 30, 221, 238, 245, 249, 272.
hirsutus, Arcturus, 16, 17, 29, 175, 194, 237, 238, 248, 268.
hirticeps, Jlyarachna, 56.
hirtipes, Idothea, 142. . .
holmesi, Janira, 15, 16, 40, 42, 237, 252.
Holophryxus 16,17, 20, 29, 222, 223, 241, 272.. Holotelson 24, 26, 29, 31, 99,100, 112, 248,249, 260, 361.
.hypnorum, Ligidium, 210.
18*
276
РАВНОНОГИЕ РАКИ
hystrix, Pleuroprion, 20, 23, 30, 198, 200, 243, 249, 269.
Ichtyoxenus 87.
Idotaea 154, 166;
Idoteal54, 163, 164.
Idothea 1, 15, 16, 17, 18, 20, 21, 22, 24, 25, 26, 29, 30, 31, 132, 263.
Idotheidae 3, 15, 20, 24, 131, 173, 262, 263
Ilyarachna 8, 22, 24, 25, 26, 54, 55, 253, 254.
Ilyarachninae 54.
imbricata, Meinertia, 84.
infelix, Syscenus, 30, 80, 249, 257.
insolita, Gnathia, 246.
intermedium; Arcturus, Pleuroprion, 16, 198, 201, 237, 270.
Irona 82, 94, 258, 259.
Ischnomesidae 8.
isopus,- Cleantis, 24, 26,170, 171, 172, 245, 267.
Jaera, 13.
Janira, 15, 16,17, 18, 20,21, 24, 26, 29, 31, 40, 249, 251.
Janiridae 15, 20, 24, 30, 251.
Janiropsis 15,16, 17, 19, 24, 25, 26, 31, 40, 44, 240, 241, 251.
Janthe 41. ,
japonensis, Cirolana, 67, 256.
japonica, Cumodoce, 24, 26, 31, 117, 245; 246, 250, 261.
japonica, Idothea, Pentidothea, 24, 26, 29, 140, 245, 247, 264.
japonica, Limnoria, 24, 25, 98, 245, 259.
japonica, Nerocila, 24, 26, 82, 245, 258.
Japonica, Paranthura, 20, 22, 24, 26, 128, 242, 245, 262.
japonica, Rocinela, 24, 25, 73, 78, 245, 257.
japonicus, Tecticeps, 110.
Jolella 41, 42, 43.
Katianira 9.
kincaidi, Janiropsis, 15, 17, 24, 26, 31, 44, 238, 239, 245, 250, 253.
laevis, Eurycope, Munnopsurus, 15, 17, 20, 29, 58, 60, 238, 241, 248, 254.
laevis, Synidothea, 16, 150, 165, 237, 265.
laevis, Tecticeps renoculis, 20, 21, 24, 101,103, 104, 105,109, 241, 242, 260.
lata, Synidothea bicuspida, 20, 21, 22, 24, 96, 152, 153,242,245,267.
laticaudata, Rocinela, 77.
laticauda, Neosphaeroma, 122.
latifrons, Muhno'psis, 64, 255.
latreillei, Cilicaea, 115. '
Ligia 14,16,18, 20, 24, 26, 27,29, 32, 207, 248, 270,271.
Ligidium 16, 207, 210, 270, 271.
Ligiidae 16, 24, 206, 270.
lignorum, Limnoria, Cymothoa, 15, 17, 24, 26,. 96, 97, 238, 246, 248, 259.
lilljeborgi, Rocinela, Syscenus, 80.
Limnoria, 15, 17, 24, 26, -30, 96, 259.
Limnorinae 96, 259.
linearis, Cleantis, 170.
Lironeca 87.
Livoneca 20, 23, 24, 25, 26, 29, 30, 82, 86, 258..
leucophthalmus, Tecticeps, 20, 21, 29, 30, 102, 111, 241, 247, 248, 261.
longicirra, Synidothea, 20, 21,151,159, 241, 266, longicornis, Astacilla, 202.
longicornis, Ilyarachna, 55.
longispinis, Arcturus, 16, 174, 180, 268.
lucanoides, Elaphognathia, 246.
maculata, Rocinela, 20, 21, 22, 24, 26, 74, 75, 79, 242, 245, 257.
maculoSa, Janira, 40.
magnispinis, Arcturus, 16, 174, 185, 237, 268, magnoculis, Aega, 15, 17, 20, 23, 29, 30, 70, 72, 238, 241, 242, 243, 248, 256.
marginalis, Tecticeps renoculis, 15, 16, 18, 102, 105, 109, 237, 239, 260.
marmorata, Idotea, 163.
marmorata, Synidothea, 16, 17,. 19, 152, 154, 155, 238,’240, 267.
maxillaris, Gnathia, 228.
Meinertia 26, 81, 84, 257, 258', melanostica, Irona, 94, 259.
Mesidothea 13, 15, 17,18, 19, 20, 23; 29, Г32;. 145,238,263,265,
Mesosterius 55.
metallica, Idothea, 26, 27, 30, 133, 134, 246,248, 263.
Microprotus 15, 16, 40, 47, 251, 253.
minutus, Munnopsurus,. 24, 25, 29, 58, 61, 244, 254.
monodi, Gnathia, Elaphognathia,, 25, 229,. 245,. 273.
montanus, Ichtyoxenus, 87.
Munna 15,16, 24, 25, 31, 49; 249,-253.
Munnidae 3, 8, 15, 24, 38, 39; 49, 251, 253.
Munnopsidae, 9, 15, 20, 24, 38, 39,54, 251, 253i.
Munnopsinae 54.
Munnopsis 9,54’, 55,, 64, 254, 255.
Munnopsurus 15, 17, 18, 19, 20, 21, 23; 24, 25,. 29, 31, 55, 57, 238, 249, 354.
murdochi, Arcturus, Pleuroprion, 16, 17, 198,. 238, 269.
muricata, Synidothea, Idotaea, 20, 23, 151, 166,. 243, 265.
INDEX
Til
nebulosa, Synidothea, 16, 18, 20, 21, 152, 155, 238, 239, 242, 267.
neglecta, Munna, 53, 54.
Neosphaeroma 15, 17,19, 24,25, 26,29, 99, 100, 122, 240, 260, 262.
Nerocila 24, 26, 81, 257, 258.
niponia, Rocinela, 24, 25, 73, 79, 245, 257.
nodulosa, Synidothea, Idotaea, 16, 17, 19, 150, 164, 238, 240, 265.
nodulosus, Tecticeps renoculis, 20, 21, 102, 106, 109, 241, 260.
norvegica, Calathura, 129.
novae.zelandiae, Livoneca, 88.
occidentalis, Anilocra, 92.
oceanica, Ligia 207.
ochotensis, Idothea, 15, 17, 20, 21, 22, 26, 29, , 133, 136, 238, 239, 242, 243, 245, 247, 264.
ochotensis, Munnopsurus giganteus,.20, 21, 23, 58, 59, 241, 254.
olivacea, Sphaeroma, 123.
Oniscidae 16, 206, 214, 270, 271.
Oniscoidea 3, 11, ,14, 16, 21, 24, 38, 205, 241, 251, 270.
oregonensis, Idotea, 142.
oregonensis, Neosphaeroma, Exosphaeroma, Sphaeroma, 15, 17, 24, 26, 29, 123, 238, 239, 245, 247,262. ' '
orientalis, Idothea, 1, 20, 21, 24, 26, 31, 133, 139, 242, 243, 245, 250, 264.
orientalis, Mesid. entomon, 15, 17, 20, 23, 145, .238, 242, 265.
ovata, Neosphaeroma, Exosph., 24, 25, 123, 124, 245, 262.
oxyrrhynchaena, Meinertia, Ceratothoa, 24, 26, 84, 245, 258.
pallasii, Ligia, 16, 18, 20, 208, 210, 239, 241, 271.
pallida, Synidothea, 151, 158, 266.
palpalis, Janiropsis, 46, 47.
papillicornis, Trichoniscus, 16, 17, 24, 26, 32, 213, 238, 245, 250, 271. ' .
Paragnathia 7.
Paranthura 20, 21, 22, 24, 26, 125, 126, 127, 262.
pavlenkoi, Eurycope, 24, 25, 62, 63, 244, 255.
peloponesiaca, Idothea, 134.
Pentias 10, 24, 26, 31, 132, 147, 263, 265.
Pentidothea 15, 16, 17, 18, 20, 21, 24, 26, 29, 31, 122, 140, 263, 264.
Phreotoicidea 8, 37.
Phryxus 16, 17, 25, 26, 30, 217, 272.
picta, Synidothea, 16, 151, 178, 237, 266.
planicauda, Cleantis, 24, 30, 170, 246, 249, 267.
Platybranchiatae 99.
Pleuroprion, 16, 17, 18,19, 20, 21,23,29,30, 173, 197, 238, 240; 248, 267, 269.
polita, Astacilla, 24, 203, 204, 245, 270.
Porcellio 14, 16, 214, 271.
Praniza 228.
pranzoides, Harponyxa, Syscenus, 80.
Prophryxus 222, 226, 272, 273.
propinqua, Livoneca, 87, 89, 259.
psora, Aega, 70.
pugettensis, Argeia, 16, 17,25, 26, 29, 219, 239, 245, 247, 272.
pulchra, Idothea, 152.
pusillus, Trichoniscus, 212.
quadrispinosa, Ilyarachna, Echinozone, 56.
rangifer, Elaphognathia, 246.
raynaudii, Livoneca, 20, 23, 30, 87, 88,243,,248,. 250.
resecata, Pentidothea, 140.
rectifrons, Gnathia, 25, 228, 230,. 245, 273.
redmarni, Livoneca, 86.
remrdi, Irona, 94.
renoculis, Tecticeps renoculis, 18, 20, 21, 23,. 102, 104, 108, 241, 244, 260.
Rocinela, 15, 17, 20, 21, 22,24, 25, 26, 29, 70, 72, 256, 257.
rotundata, Pentidothea, Idothea, 24, 26, 31, 140, . 141,245,250,264
rugosa, Idothea, 134.
sacciger, Livoneca, 24, 25, 86, 245, 258.
sarsi, Janira, 15, 16,18, 20, 21, 40, 43,238, 239,.
242, 252.
scaber, Porcellio, 214, 215, 271.
schmidti, Gnathia, 25, 228, 231, 245, 273.
seminudus, Arcturus setosus, 20, 21,22, 24,174,.
183, 241, 242, 268.
serratum, Sphaeroma, 120.
serratus, Tecticeps renoculus, 24, 25, 102, 106,.
109, 245, 260.
serricaudis, Janiropsis, 24, 25, 44, 46, 244, 253. setosus, Arcturus, 20, 21,23,174,181, 241, 244, 268.
sieboldi, Sphaeroma, 24, 25, 120, 245, 262.
soldatovi, Janira, 20, 21, 24, 26, 29, 41, 42, 242, 245, 247, 253.
Sphaeroma, 31, 99, 100, 120, 259, 262.
Sphaeromidae 4, 11, 12, 14, 15, 24, 27, 31, 65, 66, 95, 246, 249, 255, 259.
Sphaerominae 95, 98, 99, 259.
spinifrons, Eurycope, 24, 25, 32, 62, 244, 250, 255.
spinosa, Synidothea, 20, 21, 150, 167, 238, 242,. 265.
278
РАВНОНОГИЕ РАКИ
starokadomskii, Ilyarachna, 20, 21, 22, 24, 26, 55, 57, 242, 245, 254,
stephenseni Munna, 15, 16, 50, 237, 253. subneglecta, Munna, 24, 25, 50, 53, 244, 253. stimpsoni, Ligia, 210.
symmetrica, Aega, 18, 20, 29, 70, 71, 239, 241, 247, 256.
Symmius 132, 148, 263, 265.
Synidothea 1, 8,10, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 29, 30, 31,132, 150, 238> 240, 248, 250, 263, 265.
b.' Syscenns 30, 70, 80, 256, 257.
Syspastidae 206, 270.
tarasovi, Pleuroprion, 20,21, 198, 201, 241, 270, Tecticeps 2, 15,16,18, 20, 21, 23, 24, 25, 29, 30, 31, 99, 100, 250, 259, 260.
tenue, Ligidium, 16, 211, 237,271.
Tole 42.
Trichoniscidae 16, 24, 206,212, 270, 271. 'Trichoniscus 16,17, 24, 26, 32, 212, 271.
tritaeniatus, Arturus, 16, 175, 194, 237, 269.
truncata, Cymodoce, 117.
tuberculosa, Dynamene, 115.
tuberculata, Gnathia, 25, 228, 229, 245, 273.
tuberculata, Synidothea, 151, 159, 241, 266.
tuberculatus, Holotelson, 24, 26, 31, 112, 245, 250, 261.
terebrans, Limnoria, 97.
Tylidae 206, 270.
Tylos 27, 246.
typica, Munnopsis, 64.
ulbani, Arcturus, 174, 177, 241, 268.
Valvifera, 12, 15, 20, 24, 38, 131, 251, 262.
vulgaris, Livoneca, 86, 92, 258.
whitei, Pentidothea, 17, 20, 140, 144, 239, 241, 265.
wosnesenskii, Pentidothea,. 17, 18, 20, 21, 29, 140, 142, 238,239, 242, 247, 265. :
zachsi, Hyarachna, 55, 56, 244, 254.
СОДЕРЖАНИЕ
к Стр.
Предисловие ..............................-...................................... V
Систематический указатель видов.......................................... VII _
Введение
Морфологический очерк . ........................................................ 1
Краткий очерк экологии и биологии Isopoda .................. 12
Краткий зоогеографический очерк Isopoda Берингова, Охотского и Японского морей 14 -
Литература..................................................................... 32
Специальная часть
Таблица для определения подотрядов ............................................ 37 '
I. Подотряд Asellota_ .............................• . . 38
1. Сем. Janiridae ........................................................ 39
2. Сем. Munnidae .......... . ............... 49 '
3. Сем. Munnopsidae ............. ......................................... 54
II. Подотряд Flabellifera ....... .................... 65
4. Сем. Cymothoidae ....................................................... 66
5. Сем. Sphaeromidae ............. ........... 95
6. Сем. Anthuridae .... •................................................. 125
III. Подотряд Valvifera....................................................... 131
7. Сем. Idotheidae ....................................................... 131
8. Сем. Arctiiridae . . . •................................................ 173 '
IV. Подотряд Oniscoidea.~...................................................... 205
9. Сем. Ligiidae.......................................................... 206 ‘
10. Сем. Trichoniscidae ......................... 212
11. Сем. Oniscidae ........................................................ 214
V. Подотряд Epicaridea.......................................................... 215 •
12. Сем. Bopyridae ........................................................ 216
13. Сем'. Dajidae................. . ... . ...... 222
VI. Подотряд Gnathiidea ........................... 227
14. Сем. Gnathiidae ....................... . . 227
Резюме (Zusammenfassung)..................... ................................ 247
Алфавитный указатель (Index).................................................. 274