/









































































































































Текст
фауна
ссср
РАКООБРАЗНЫЕ
том.
вып.
3
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА <ЛЕНИНГРАД
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ АКАДЕМИИ НАУК СССР
НОВАЯ СЕРИЯ 35
ФАУНА СССР
Главный редактор акад. Е. Н. ПАВЛОВСКИЙ
Редактор А. А. ШТАКЕЛЬБЕРГ
РАКООБРАЗНЫЕ
Т. III, вып. 3
В. М. РЫЛОВ
CYCLOPOIDA
ПРЕСНЫХ ВОД
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА • 1948 • ЛЕНИНГРАД
Главный редактор
Директор Зоологического Института АН СССР'
акад. Е. Н. Павловский
Редактор издания А. А. Штакельберг
ВЯЧЕСЛАВ МИХАЙЛОВИЧ РЫЛОВ
(1889-1942)
Вячеслав Михайлович Рылов родился 22 марта 1889 г. в г. Крон-
штадте. Детство свое он провел в г; Кологриве Костромской губернии,
где отец В. М. состоял преподавателем Сельскохозяйственного училища.
В 1901 г. отец В. М. переводится в г. Новозыбков Черниговской губ.,
и В. М. поступает в Черниговское реальное училище, шесть классов
которого оканчивает в 1907 г. Седьмой класс В. М. кончил в 1908 г.
уже в Петербургском реальном училище Черняева.
Свое высшее образование В. М. начал в Петербургском политехни-
ческом институте, однако с первого курса института, сдав экзамен
по латинскому языку за курс гимназии, В. М. переходит на физико-
математический факультет Петербургского университета. В 1916 г. В. М.
оканчивает Университет с званием кандидата естественных науки остав-
ляется при Университете для подготовки к профессорскому званию.
Студенческие годы дали возможность В. М. познакомиться с научно-
исследовательской работой и испытать свои силы на педагогическом
поприще — в 1911—1912 гг. он принимает участие в исследовании
пресноводной фауны Нижегородской губернии, в 1913—1915 гг. коман-
дируется на Бородинскую и Мурманскую биологические станции;
в эти же годы В. М. ассистирует на кафедре рыбоводства Петербург-
ского сельскохозяйственного института и ведет летний курс гидробио-
логии на Никольском рыбоводном заводе.
В год окончания университета (1916) В. М. приглашается на долж-
ность сверхштатного младшего зоолога Зоологического музея Академии
Наук, а в 1918 г. получает там место штатного младшего зоолога,
на котором остается до 1929 г., когда получает звание и должность
старшего зоолога. В 1934 г. В. М. присваивается ученая степень доктора
биологических наук без защиты диссертации, а в 1939 г. ученое звание
профессора. За годы работы в Зоологическом музее (в 1932 г. реорга-
низованном в Зоологический институт) В. М. последовательно руково-
дит отделением губок и кишечнополостных (1918—1925), отделением
ракообравных и планктона (1925—1942) и заведует отделом гидробиоло-
гии (1931—1935).
Одновременно В. М. работал ряд лет в Петергофском естественно-
научном институте, Государственвбм Гидрологическом институте и читал
курс планктонологии в Ленинградском Государственном университете.
Два раза В. М. командировался Народным комиссариатом просвещения
за границу для участия в международных лимнологических конгрессах.
В. М. не был только кабинетным ученым, он принимал участие
в экспедициях и совершил поездки во многие места Советского Союза:
на берег Белого моря, на Кавказ, в Среднюю Азию, в среднюю и север-
4
В. И. Ж. АДИН
ную части европейской территории СССР, на водохранилища канала
им. Москвы и на Волгу у Жигулей.
Скончался В. М. Рылов 22 марта 1942 г. после эвакуации из блоки-
рованного Ленинграда. Истощение на почве недоедания и флегмона
руки с последующей ампутацией ее привели к трагическому концу.
Научная деятельность В. М. была в высокой степени плодотворна:
он оставил после себя около 120 печатных работ, средн которых ряд
крупных монографий, несколько научно-популярных книг и множество
статей по вопросам гидробиологии и зоогеографии.
В. М. был общепризнанным специалистом по зоопланктону и весло-
ногим рачкам; известность его в этих областях выходила далеко
за пределы Советского Союза: В. М. неоднократно предлагалось автор-
ство в международных изданиях.
Из руководств и монографий, принадлежащие перу В. М., отметим
его определитель свободноживущих веслоногих ракообразных, изданный
в 1922 г., краткое руководство к исследованию пресноводного планктона,
определитель пресноводных Galanoida СССР, зоопланктон внутренних
вод, напечатанный на немецком языке. В рукописи после смерти В. М.
осталась монография пресноводных циклопид, печатаемая в настоящем
выпуске „Фауны СССР". К глубокому сожалению, неоконченной осталась
работа В. М. по зоопланктону пресных вод СССР.
В. М. не удовлетворяла работа только для сравнительно узкого
круга читателей, которые пользуются большими монографиями и руко-
водствами; он стремился привлечь к своей науке внимание широких
слоев интеллигенции. В этих целях он пишет две популярные книжки —
о свободно плавающих организмах и планктоне, делает обзор низших
ракообразных для „Животного мира СССР" и принимает деятельное
участие в составлении и редактировании „Жизни пресных вод СССР".
Большое внимание В. М. уделял методическим вопросам — он соста-
вил инструкцию для полевого исследования пресноводного планктона
и написал статьи о методике исследования нейстона и о применении
камеры Кольквица для исследования планктона на разных глубинах
в мелких водоемах. В известном методическом руководстве Абдергаль-
дена В. М. принадлежат главы о лимнонейстоне и экскурсионном иссле-
довании лимнопланктона.
Фактический материал для своих обобщающих работ В. М. черпал
из обработки многочисленных сборов фауны из различных концов нашей
обширной родины. Copepoda и Cladocera им были изучены по сборам
из Калининской, Новгородской, Ленинградской и Воронежской областей,
из Лапландии, с Мурманского побережья, из Сибири, с Байкальского
хребта, с Кавказа, Дальнего Востока, Сахалина, из Уссурийского края,
из бассейна Сыр-дарьи, из озер Могильного, Иссык-куля, Камышлы-баш-
ских, Балхаша и т. д. Среди этих сборов В. М. обнаружил и описал
до 15 новых для науки видов и разновидностей (Heterocope soldatovi,
Diaptomus fischeri, Diaptomus amurensis, Diaptomus birulai, Diaptomus
fadeevi, Epischur a chankensis, Bryocamptus der/ugini и др.).
Помимо ракообразных, В. М. уделял внимание и обработке сборов
по гидроидам. Он написал небольшую статью о гидрах Петроградской
губернии, о пресноводном полипе Cordylophora и ряд статей о гидрои-
дах Баренцова моря.
Фаунистические работы давали возможность В. М. делать важные
зоогеографические заключения. Так, он пишет (1920) о ледниковых
реликтах в пресноводной фауне, глубоко анализируя само понятие
ПАМЯТИ В. М. РЫЛОВА
5
о реликтах. „Под реликтом, — пишет В. М.,— мы подразумеваем остаток,
в зоогеографии остаток фауны, в прежние геологические эпохи (или
эпоху) имевшей более обширное распространение и сохранившейся
в данное время лишь в некоторых местах". В работе о Cordylophora
(1922) В. М. говорит: „Мне кажется гораздо более вероятным относить
возникновение Cordylophora к верхнему миоцену, именно к Сарматскому
бассейну".
Большой оригинальностью и свежестью отличаются работы В. М.
по вопросам гидробиологии. Он прокладывает здесь новые пути и ста-
рается критически пересмотреть казалось бы твердо установившиеся
понятия. В статье „Что понимать под планктонным организмом" (1922),
например, В. М. пишет: „В противоположность бентосу как группе
организмов, неразрывно связанной с субстратом как точкой опоры, мне
кажется единственно правильным под планктоном понимать организм,
ведущий свободно плавающий, взвешенный в водкой массе образ жизни,
независимо от субстрата как точки опоры; последней для планктонов
служит только водная масса как таковая. Планктон есть совокупность
планктонных организмов, биоценоз, элементы которого (бионты) харак-
теризуются указанным экологическим признаком".
В другой своей работе „Об одной из основных проблем гидробио-
логии и о некоторых ближайших задачах гидробиологических исследо-
ваний" (1923) В. М. выдвигает важную задачу изучения влияния водных
организмов на водоем, которую он и пытается решать в последующих
своих исследованиях.- В. М. пишет „Изучая водоем как биологическое
целое... мы необходимо сталкиваемся не только с влиянием среды
на водное население, но равным образом и с влиянием последнего
на водную массу"... „Именно в водной среде, объекте исследования
гидробиологии, подобные влияния необычайно значительны, водная масса
является как бы аккумулятором воздействий со стороны происходящих
в ней биологических процессов. В силу своих физических свойств, .
аккумулируя эти воздействия, водная масса непосредственно отражает
их на своем населении".
Здесь же В. М. декларирует и пути решения гидробиологических
проблем: „Метод наблюдения в самой природе и метод эксперименталь-
ный в гидробиологии должны итти рука об руку... Метод сравнитель-
ный (сравнительная гидробиология) должны завоевать... все права
гражданства".
Эти свои положения В. М. более полно развивает в докладе
на Лимнологическом конгрессе (1927). Он различает здесь следующие
виды воздействия планктонных организмов на среду: 1) поглощение
углекислоты водорослями (и накопление углекислоты вследствие жизне-
деятельности животных), 2) влияние планктона на содержание (спектр)
растворенного в воде кислорода (В. М. отмечает, что это влияние еще
мало изучено), 3) влияние на спектр кальция — например, биогенное
декальцинирование, 4) роль диатомей в спектое кремнезема, 5) минера-
лизация органических веществ организмами в процессе самоочищения
воды (В. М. подчеркивает, что особенно благодарной ареной для иссле-
дования этой проблемы может служить эпилимнион эвтрофных озер,
населенный как бы чистой культурой планктона в природе), б) умень-
шение прозрачности воды, 7) посмертное поглощение планктоном рас-
творенного кислорода. В. М. предлагает поставить изучение вопроса
о воздействии планктонных организмов на водную среду на количествен-
ной основе. Далее он вводит в гидробиологию понятие „биодинами-
6 В. И. ЖАДИН
ческий планктонный титр“ и иллюстрирует применимость его на примере
учета декальцинирования воды водорослью Melosira. В заключение
доклада В. М. формулирует понятие „Биодинамический стандарт планк-
тона" и дает формулу для его выражения.
В другой своей работе, посвященной проблеме триптона (1931),
В. М. касается вопросов питания планктонных организмов, т. е. основ-
ных вопросов теории биологической продуктивности водоемов. В. М.
оспаривает мнение Пюттера о малом количестве органического триптона
в водоемах и незначительной роли его в питании водных организмов.
Наоборот, В. М. считает установленной большую роль триптона в пита-
нии зоопланктона. Источниками происхождения триптона в водоемах
В. М. считает планктон, бентос, макрофиты, приток извне, ил (из кото-
рого триптон поднимается с пузырьками газа). В. М. предлагает прин-
ципы количественного изучения триптона — дифференциация по размерам
частиц, подсчет в камерах, сбор осаждением.
Вопросу изучения питания планктонных животных В. М. посвятил
и одну из последних своих работ (1940). В ней он констатировал отри-
цательную роль минерального сестона в питании речных ракообразных
и тем самым пролил свет на нерешенную дотоле проблему преобладания
в речном планктоне коловраток над кладоцерами.
В. М. был одним из немногих гидробиологов, которые изучали
своеобразный биоценоз поверхностной пленки воды — так называемый
нейстон. В. М. не только в совершенстве описал этот биоценоз, изучен-
ный им на водоемах Петергофа (ныне Петродворца), но и предложил
свою методику изучения нейстона. Данные В. М. по нейстону и его
фотографии вошли в русские и зарубежные руководства по гидробио-
логии.
Как подлинно советский ученый, В. М. большое внимание уделял
приложению данных своей науки к практической жизни. Он внес извест-
ный вклад во все отрасли прикладной гидробиологии. Вопросов рыбного
хозяйства касаются его работы по замору рыб в Велетьминском пруду
(1923) и по питанию ладожского сига, пересаженного в оз. Севан (1939).
К проблеме использования сапропелей имеет отношение работа В. М.
о животных остатках в сапропелитах (1923). Биологии личинок малярий-
ного комара касается интересная статья о биотопе Anopheles plumbeus
(1927). Особенно же большую работу В. М. проделал по изучению водо-
хранилищ канала им. Москвы (1941), по исследованию Волги в районе
Жигулей, где должно в будущем возникнуть грандиозное водохранилище
(работа печатается), и р. Невы как источнике водоснабжения Ленинграда
(рукопись).
В. М. Рылов никогда не был одинок в своей научной работе, никогда
не делал секретов из нее. Где бы он ни работал—в Зоологическом
институте Академии Наук, в Государственном Гидрологическом инсти-
туте, в Петергофском институте, в Ленинградском университете или
в экспедициях, — он всегда был окружен товарищами и учениками.
Вячеслав Михайлович погиб в расцвете творческих сил. Многочис-
ленным его планам не суждено было осуществиться. Но то, что им
сделано и написано, будет на долгие годы служить примером беззавет-
ного служения науке. На книгах В. М. будет учиться не одно поколение
советских гидробиологов.
В. Жадин
Ленинград,
15 мая 1947.
ПАМЯТИ В. М. РЫЛОВА
7
СПИСОК ПЕЧАТНЫХ РАБОТ В. М. РЫЛОВА
1. Beitrag-e zur Copepoden-and Cladocerenfauna des Goavernments Twer (Mittelrussland).
Zool. Anz., 43, 8, 1914:339—342.
'2. К фауне Cladocera Воронежской губернии. Tp. СПб. общ. ест., XLV, 1, прот., 3,
1914:64-80.
3. К планктону некоторых озер Витебской губернии. Фауна Copepoda н Cladocera.
Там же, XLV1, 4, 1915:111—163.
-4. К сведению о пресноводной фауне озера Могильного (Мурман, остр. Кильдин)
Там же, XLV1, 1, 7-8, 1915:273-292.
5. Материалы к познанию фауны Copepoda и Cladocera Тверской губернии. Тр. Бород
биол. ст., IV, 1915:1-92.
6. К фауне Copepoda Новгородской губернии. Тр; Петр. обш. ест., XLV1, 1, прот,
5-6, 1915:186—206.
7. К фауне Cladocera Русской Лапландии. Там же, XLV, 4, 1916:109—136.
8. Гидробиологический курс на Никольском рыбоводном заводе в 1915 г. Вести, рыбо-
промышл., 1916:149—164.
9. Обзор пресноводных водоемов Мурманского побережья, исследованных летом 1915 г.
Ежегод. Зоолог, муз. Акад. Наук, XXI, 1916:XV1—XXV11.
10. К планктону озера Бологое Новгородской губернии. Тр. Бород, биол. ст IV
1916:204-218.
11. Diaptomus madag'ascariensis— новый вид пресноводного ракообразного (Eucopepoda,
Calanoida). Ежегод. Зоолог, муз. Росс. Акад. Наук, XXII, 1917:1Х —XI.
12. Заметка о систематическом положении Camptocamptus senilis Lili. (Eucopepoda,
Harpacticoida). Ежегод. Зоолог, муз. Росс. Акад. Наук, ХХ111, 1917:211—217.
13. Hetero соре soldatovi, n. sp., новый вид пресноводного веслоногого ракообразного
(Copepoda, Calanoida). Ежегод. Зоол. муз. Росс. Акад. Наук; ХХ111, 1917:164—178.
14. О новых и малоизвестных видах рода Diaptomus (Copepoda, Calanoida). Ежегод.
Зоол. муз. Росс. Акад. Наук, XXI11, 1917:218—239.
15. Материалы к фауне свободноживущих пресноводных Copepoda Северной России^
Часть 1. Calanoida и Cyclopoida. Ежегод. Зоол. муз. Росс. Акад. Наук, XXII,
1917:247—310.
16. Материалы к фауне свободноживущих пресноводных Copepoda Северной России.
Часть 11, Cyclopoida и Harpacticoida. Ежегод. Зоол. муз. Росс. Акад. Наук,
ХХШ, 1918:43—96.
17. Заметки о Calanipeda aquae-dulcis Kritsb. и о Popella g’uernei Richard. Тр. Петр,
общ. ест., XLV1U, 1, 3—8, 1918:1—13.
18. Отзыв о программе деятельности Биогидрологического отделения Общенаучного
отдела. Отчет о деят. Ком. по нзуч. ест. произв. сил России, 17, 1919:165—166.
19. К сведениям о фауне водоемов г. Петрограда н его окрестностей. 1. Список орга-
низмов, найденных в 1914 г. в водоемах Детскосельских парков. Журн. Петр,
агр. инет., 2, 1920:1—23.
20. О летнем планктоне прудов окрестностей Старого Петергофа. Тр. Петр. общ. ест.,
LI, 1, № 7-8, 1920:175-187, 205—206.
'21 . К биологии Diaptomus amblyodon Marenz (Copepoda, Calanoida)^ Тр. Петр. общ.
ест., LI, 1, прот., 1—4, 1920:2—10.
'22 . К вопросу о ледниковых реликтах в пресноводной фауне. Изв. Росс. Гидр, инет.,
1, 1920:97—113.
23. Пресноводные свободноживущие Copepoda Петроградской губернии. Фауна Петро-
градской губернии, 11, 2, 1921:3—11.
24. Гидры (Coelenterata, Hydrozoa, Hydridae) Петроградской губернии. Фауна Петро-
градской губернии, 11, 3, 1921:3—6.
25. К вопросу о происхождении гидроида Cordylophora lacustris Allman. Докл. Росс.
Акад. Наук, 1922:24—27.
26. Что понимать под „планктонным" организмом. Русс, гидроб. журн., 1, 8, 1922:1—7.
27. Об исследовании планктона прудов окрестностей Старого Петергофа в 1920—1922 гг.
Бюлл. Росс. Гидр, инет., 5, 1922:7—8.
28. Пресноводная фауна Европейской России. Вып. I. Свободноживущие веслоногие
ракообразные (Eucopepoda) Изд. „Природы" н Научрыббюро Главрыбы,
1922:1—126.
29. О новых видах Copepoda Calanoida. Тр. Петр. общ. ест., LII, в. I, №1—8, 1922:1—16.
30. Новая форма североамериканского Diaptomus (Entomostraca, Copepoda) из Восточ-
ной Сибири. Русск. энтом. обозр., XVIII, 1922:1—7.
.31. К фауне Crustacea-Eucopepoda Маньчжурии. Ежегод. Зоол. муз. Росс. Акад. Наук,
XXIV, 1923:52-95, табл. 3.
8
В. И. ЖАДИН
32. Zoologische Ergebnisse der Russischen Expeditionen nach Spitzbergen. Ежегод. Зоол-
муз. Акад. Наук, XXIV, 1923:140—160, табл. I.
33. Жизнь пресных вод. I. Свободно плавающие организмы (планктон). Энцикл. необх.
знай., сер. И, биологии., вып. I, 1923:5—91.
34. О животных остатках сапропелита Толполовского болота. Изв. Сапроп. ком., I,
1923:131-137.
35. О влиянии растворенного в воде кислорода и сероводорода на цикл развитии
и вертикальное распределение инфузории Loxodes rostrum. Тр. I Всеросс. съезда
зоол., анат. и гистол. в Петр. 15—21 XII 1922, 1923:67—68.
36. Об одной из основных проблем гидробиологии и о некоторых ближайших задачах
гидробиологических исследований. Тр. I Всеросс. съезда зоол., анат. и гистол.
в Петр. 15-21 XII 1922, 1923:68-69.
37. (и Перфильев Б. В.) Предварительные результаты ботанического и зоологиче-
ского исследования сапропелита некоторых озер Средней России. Изв. Сапропел.
ком. Росс. Акад. Наук, 1, 1923:65—76, табл. 7.
38. Наблюдения над вертикальным распределением растворенного кислорода в Криста—
теллевом пруде (Петербургской губ.) и некоторые сведения о планктоне послед-
него. Русск. гидроб. журн., II, 1—2, 1923:1—14, ием. рез.
39. Uber den Einfluss des im Wasser gelosten Sauerstofts and Schwefelwasserstoffs auf
den Lebenszyklus und die vertikale Verteilung des Infusors Loxodes rostrum.
O. F. Mull (Aspirotricha, Amphileptina). Int. Rev. f. Hydrobiol., XI, Va>
1923:179—192.
40. Заметка о цветении Anabaena scheremetievi Elenk. и Euglena sanguinea Enrbg-
в прудах окрестностей Старого Петергофа (Петербургской губ.). Русск. гидроб.
журн., II, 5—7, 1923:1—5, нем. рез.
41. Зоопланктон Невской губы. Исследования реки Невы и ее бассейна, вып. 3,.
1923:1—85, табл. 1.
42. О заморе рыбы в Велетьминском пруде Нижегородской губернии. Изв. Росс. Гидроб.
инет., 6, 1923:1—6.
43. Жизнь пресных вод. Планктон. Изд. „Наука н школа", 1924:1—128.
44. Zur Methodik der Untersuchungen des Kammerplanktons in sehr seichten Gewasser-
Sehriften fur Siisswasser- und Meereskunde, 1—4, 1924:3—7.
45. Zuz Biologie des seltenen planktonischen Infusoriums EpistyKs rotans Svec. Int. Revue-
f. Hydrobiol., XII, 3-4, 1924:218-227.
46. Einige Beobachtungen fiber den Einfluss der Schwefelwasserstoffgiing in den Schlamm-
sedimenten kleiner Gewasser auf die Produktion und die vertikale Verteilung- des
Zooplanktons. Verh. d. Int. Ver. f. Limnol., 1924:289—303.
47. Ober zwei wenig bekannte Heterocope-Arten (Copepoda, Calanoida). Zool. Anz., LIX,.
1924:225—232.
48. Ober eine neue Diaptomus-Art aus dem Goktscha-See (Diaptomus faddeevi, sp. n.).-
Zool. Anz., LXII, 1925:224—228.
49. Zur Copepoden Fauna des aussersten Ostens. Zool. Anz., LXIII, 1925:307—318.
50. Ueber das Neuston in einem Teiche der Umgebung von Alt-Peterhof (Gouv. Peters-
burg). Arch. f. Hydrob., XV, 1925:497-508.
51. О биосестонных окрасках воды в водоемах окрестностей Старого Петергофа. Русск..
гидроб. журн., IV, 3—6, 1925:84—95.
52. Beitrage zur Kenntnis der Hochprodiiktionen des Heleoplanktons 1. Ober eine Wasser-
bliite von Anabaena scheremetievi Elenkin im Triton-Teich (Gouv. Petrograd). Int-
Revue f. Hydrobiol., XIII, 3/4, 1925:171—184.
53. Zur Methodik der Neuston-Untersuchungen. Arch. f. Hydrobiol., XVI, 3, 1926:484—493.
54- Ober einige Faile der Neustonfarbung des Sfisswassers. Arch. f. Hydrobiol., XVI, 3,..
1926:484 -489.
55. Планктон озера Ильменя. Мат. иссл. р. Волхова и его бассейна, X, 1926:295—364»..
табл. 3.
56. К познанию фауны Rotatoria некоторых водоемов Олонецкого края (Пудожский'
уезд). Тр. Олонецк. научн. эксп. Росс, гидрол. инет., 1926:3—33.
57. Hydroidea и Alcyonaria, собранные в Баренцевом море Мурманской биологической
станцией во время рейсов по Кольскому меридиану в 1921 г. Ежегод. Зоол. муз..
Акад. Наук СССР, XVIII, 2-3 1926:233-246.
58. Краткое руководство к исследованию пресноводного планктона. Руководства и посо-
бия при изучении жизни пресных вод, 1, 1926:3—80.
59. Hemidiaptomus tarnogradskii, sp. nov., новый вид веслоногого ракообразного (Еисоре-
poda, Calanoida) с Северного Кавказа. Работы Сев.-Кавказск. гидробиол. ст., 1, 2,
1926:19-225, табл. II.
60. Краткое руководство к исследованию пресноводного планктона. Изд. Волжской биол-
ст. (руков. и пособия по изучению жизни пресных вод), в. 1, № 1, 1926:1—81.
ПАМЯТИ В. М. РЫЛОВА
9
61. Alcyonaceae, собранные экспедицией Пловучего морского научного института в 1921 г..
Тр. Плов, морск. научн. инет., II, 1, 1926:63—78.
62. Исследование над планктоном прудов окрестностей Петергофского естественно-науч-
ного института. Тр. Петерг. ест. науч, инет., 4, 1927:5—53, табл. 7.
63. О миозональном распределении камерного планктона в очень мелких водоемах..
Русск. гидроб. журн., VI, 1—2, 1927:3—13.
64. Ein Besteck fur Neustonuntersuehungen. Arch. f. Hydrobiol., XVIII, 1927:57—59.
65. Uber die anmittelbare Verwendung der Kolkwitzschen Planktonkammer. Zur Entnahme
des Planktons aus versebiedenen Tiefen. Arcb. f. Hydrobiol., XVIII, 1927:60—64.
66. Uber das planktische Vorkommen von Anthophysa steini Senn. Int. Rev. Hydrobio'.,
XVIII, 1/2. 1927:75284.
67. Uber die Scblammablagerunden des Ilmen-Sees. Arch. f. Hydrobiol., XVI I, 1927:207—•
223, табл. 1.
68. Einige Gesichtspunkte zur Biodynamik des Limnoplanktons. Verh. d. Int. Ver. d. Lim-
nol, III, 1927:405-423.
69. К биологической характеристике населяемого личинками Anopheles plumbeus Steph,
биотопа по анализу воды из дупла ясеня (Fraxinus excelsior). Докл. Акад. Наук-
СССР, 1927.
70. О включении жира у Diaptomus coeruleus Fisch. Сб. в честь Н. М. Книповича,
1927:123—134.
71. Uber zwei nene Diaptomus-Arten aus dem Kaukasus. Zool. Anz., LXXIII, 3—4,
1927:59-67.
72. К познанию фауны Eucopepoda некоторых водоемов Олонецкого края. Тр. Олон..
научн. эксп. Росс. Гидрол. инет., VI, 1927:3—44.
73. Zur Synonymik von Diaptomus mucronatus Brian 1926 und Diaptomus mucronatus
Rylov 1927. Zool. Anz., LXXVII, 5—6, 1928:49—51.
74. Uber eine neue Episcbure-Art aus dem fernen Osten (Epischura chankensis sp. nov.,.
Copepoda, Calanoida). Zool. Anz., LXXVII, 3—6, 1928:125—137.
75. К фауне Eucopepoda Кавказа, Закавказья и Северной Персии. Работы Сев.-Кавк..
гидроб. ст., П, 2—3, 1928:1—17.
76. Дополнение к статье В. М. Рылова „К фауне Eucopepoda Кавказа, Закавказья,
и Северной Персии". Работы Сев.-Кавк. Гидробиол. ст., II, 2—3, 1928:167, табл. 1..
77. Материалы по фауне пресноводных свободноживущих веслоногих ракообразных
северной Сибири. Тр. Ком. изуч. Якутской АССР, XI, 1928:1—33.
78. Некоторые данные по химизму и биологии вод Силурийского плато Ленинградской,
губернии. Русск. гидроб. журн., VIII, 1—3, 1929:1—13.
79. Некоторые наблюдения над концентрацией водородных ионов в водоемах окрест-
ностей Петергофского есТеств.-науЧн. института. Русск. гидроб. журн, VIII, 4—5,.
1929:115—124.
80. Anleitung zur Untersuchungen des Limnoneustons. Abderhalden’s Handbuch der biolog..
Arbeitsmethoden, Abt. IX, Teil 2/n, 1929:1385—1418.
81. Einige Bemerkungen betreffs des regional-limnologischen Studiens. Verh. d. Int. Ver.
f. Limnol, IV, 1929:538-548.
82. К методике исследования планктона. Тр. II Всес. Гидрол. съезда, 1930:211—218.
83. К лимнологии Кавказа. I. Некоторые данные по гидрологии и химизму озера Кар-
дывач (Зап. Закавказье). Изв. Росе, гидрол. инет, 31, 1930:91—115.
84. Некоторые наблюдения над захватом сестона у Diaptomus coeruleus Fisch. Тр.,
Петер, общ. ест, LX, 2, 1930:149—176.
85. Vorlaufige Mitteilung fiber das Plankton des Kardywatsch-Sees (NW Kaukasus). Докл.
Акад. Наук СССР, 14, 1930:346—380.
86. Данные о зоопланктоне озера Иссык-куль. Иссык. эксп, 1928, 1, 1930:111—135.
87. Uber die typische Form von Diaptomus coeruleus Fischer (Copepoda, Calanoida). Zoo',
Anz., 88, 5—6, 930:111—121.
88. Пресноводные Calanoida СССР. Изд. Инет. рыб. хоз. при Всес. Акад. с.-х. наук
им. Ленина. „Определители орган, пресн. вод", 1, 1930:1—288.
89. Инструкция для полевого исследования пресноводного планктона. Изд. Гос. Гидр,
инет, сер. „Инструкции по биол. иссл. вод", ч. II, 1, 1931:1—81.
90. Uber das Tripton-Problem. Verh. d. Int. Ver. Limnol, V, 1931:540—546.
91. Einige Resultate der limnologischen Untersuchungen an Kardywatsch See. Arch. f. Hyd-
robiol, XXII, 1931:389—409.
92. Alcyonacea сборов экспедиции на землю Франца-Иосифа в 1929 г. Тр. Инет. изуч..
Севера, 49, 1931:166-170.
93. Zur Kenntniss des Copepoda- und Cladocerfauna der Insel Sachalin. Zool. Anz, 99»
3—4, 1932:101—108.
94. Ein neuer Bryocamptus aus dem Kaukasus (Bryocamptus derjugini sp. nov.). Zool. Anz.,.
99, 5—5, 1932: 171—174.
.ю
В. И. ЖАДИН
‘95. Uber das Verkommen von Pseudosida bidentata Herr, im Kaukasus. Arch. f. Hydrob.,
XXV, 1933:133-137.
'95. Научные результаты Дальневосточной гидрофаунистической экспедидии Зоологического
музея в 1927 г. IV. Пресноводные Eucopepoda Уссурийского края. Тр. Зоолог.
Инет. Акад. Наук СССР, 1, 1933:243—280, табл. б. ,
97. Die Exkursionsuntersuchungen des Limnoplanktons. Abderhalden’s Handbuch der
Biolog'. Arbeitsmethoden, Abt. IX, Teil 2Д1, 1933:1729—1774.
98. К сведению о зоопланктоне горных озер окрестностей Теледкого озера (Алтай).
Исследования озер СССР, 3, 1933:121—130, табл. 1.
99. К сведению о планктоне озера Балхаш. Исследования озер СССР, 4, 1933:57—-69.
100. (и Гладков Н. А.). Рыбохозяйственное обследование Камышлы-Башских озер.
Тр. Арал. отд. Всес. Инет. мор. рыб. хоз., III, 1934:81—100.
101. Некоторые качественные и количественный данные о зоопланктоне озера Иссык-
куль. Тр. Киргизск. компл. эксп. Акад. Наук СССР, III, 2, 1935:87—96.
102. Состав и вертикальное распределение зоопланктона некоторых озер Центральной
Якутии. Исследования озер СССР, 8, 1935:85—124.
103. Das Zooplankton der Binnengewasser. Einfiihrung in die Systematik and Okologie
des tierischen Limnoplanktons mit besonderer Berucksichtigung der Gewasser
Mitteleuropas. Die Binnengewasser, herausgegeben von A. Thienemann, XV,
1935:1—262, Taf. 30.
104. Низшие ракообразные. — Crustacea Entomostraca. „Животный мир СССР", изд.
Акад. Наук СССР, 1, 1936:544-554, табл. 1.
105. К биологии Diaptpmus blanci Guerney. Тр. Зоол. инет. Акад. Наук СССР, IV,
1936:149-155.
106. К лимнологии Кавказа. III. Некоторые данные по химизму и зоопланктону водоемов
окр. Хосты и Красной Поляны. Тр. Зоол. инет. Акад. Наук СССР, III, 1936:79—88.
107. Ober das Vorkommen von Acanthodiaptomus tibetanus Daday im Nordfinland nebst
einer Komplettieren dem Beschreibung dieser Art. Ann. Zool. Soc. Vann., II, 2,
1936:3—9.
108. К фауне Eucopepoda озер поймы низовьев р. Амура. Вести. Дальневост. фил.
Акад. Наук СССР, 21, 1936:143—160, табл. 6.
109. Зоопланктон некоторых горных водоемов Байкальского хребта. Тр. Байк. лимн. ст.,
VII, 1937:75-86.
110. О гидробиологической характеристике водоемов Рошпи (воды Силурийского плато
Ленингр. обл.). Уч. зап. Ленинград Госуд. университета, 15, 1937:32—40.
111. Cyclopoida пресных вод. Фауна СССР, Ракообразные, III, 3,1948:1—000,000 фиг.
ПРЕДИСЛОВИЕ
К семейству Cyclopidae принадлежит целый ряд видов, являющихся
промежуточными хозяевами паразитов человека, диких и домашних
млекопитающих и промысловых птиц и рыб. Паразитологу неизбежно
приходится иметь дело с этой группой мелких низших ракообраз-
ных, причем изучение вопросов, связанных с их биологией и эко-
логией, ставится в задачи работ специальных институтов, изучающих
паразитарные болезни. Так, например, уничтожение очага дракункулеза
(ришты) у нас в СССР (Бухара) было связано со специальным
исследованием над циклопами.
В связи с паразитологическими задачами за последние 10 лет
к нам неоднократно обращались с консультацией по систематике и био-
логии (особенно по циклам развития) пресноводных, Cyclopidae, так как
в советской литературе по ним нет никаких сводок монографического
характера, если не считать опубликованной нами в 1922 г. в издании
редакции журнала „Природа" и Научрыббюро Главрыбы краткого опре-
делителя, в настоящее время сильно устаревшего.
Пресноводные Cyclopidae, кроме того, представляют бесспорно весьма
важный пищевой ресурс многих промысловых рыб, значение которого
пока еще едва ли оценено в достаточной степени.
Вследствие указанных обстоятельств автор счел необходимым начать
серию выпусков „Фауны СССР", посвященных веслоногим ракообраз-
ным, не в систематической последовательности, а е семейства Cyclopidae,
отложив обзор других семейств на следующие выпуски этого издания.
В предлагаемом выпуске отводится значительное место биологии
и экологии пресноводных циклопов, что необходимо для выяснения
вопроса о переносе ряда паразитарных болезней и для проведения мер
борьбы с ними, в частности профилактических. В этом отношении
настоящий выпуск сильно отличается от сводок по свободно живущим
веслоногим, имеющихся в западноевропейской литературе, носящих
почти исключительно характер систематического обзора, в котором
биологии и экологии отводится нередко буквально несколько слов.
Предлагаемая автором система семейства отличается от систем,
принимаемых немецкими, английскими и американскими карцинологами
В ее основу положен сравнительно-морфологический критический пере-
смотр фауны пресноводных Cyclopidae всего земного шара.
Выпуск охватывает не только уже известные в СССР виды,
но и вообще известные в Палеарктике. Несмотря на некоторое, впрочем
очень незначительное, увеличение, это было необходимо сделать в виду
того обстоятельства, что если не все, то, по крайней мере, большинство
из еще не показанных для СССР видов будет найдено и в пределах
«нашей страны.
12
ПРЕДИСЛОВИЕ
Рисунки даются преимущественно оригинальные; они сделаны
в большинстве по материалам из СССР. Длина животных везде пока-
зана без фуркальных щетинок. Литература использована до середины
1938 г., когда рукопись была закончена.
По редакционным условиям список литературы представлен в воз-
можно сжатом виде. Приведены лишь главнейшие работы специально
по Cyclopidae. В тексте, особенно в разделе биологии и экологии,
автору пришлось ссылаться на целый ряд планктологических и обще-
лимнобиологических работ; эти ссылки ограничиваются, за редкими
исключениями, только приведением автора и года работы, так как по-
дробный перечень упомянутых работ увеличил бы список литературы
в несколько раз. Не приводятся также названия тех специальных работ
по Cyclopidae, в отношении коих в синонимах указаны издания, в кото-
рых они опубликованы. Специальные работы (главным образом моногра-
фического характера), для которых в синонимах упомянут только год.
издания (с ссылкой на страницы описания и. рисунки), в списке литера-
туры приводятся.
Ряду лиц автор весьма признателен за оказание содействия при
составлении этого выпуска. Особенную благодарность автор должен
высказать акад. Е. Н. Павловскому за просмотр главы о циклопах как
промежуточных хозяевах паразитарных заболеваний и за дополнения:
к этой главе, а также сотруднику ЗИН С. С. Смирнову за ряд указаний
и предоставление ценных препаратов.
систематический указатель видов
L Подсем. Eucyclopinae
1. Род Macrocyclops Claus
Стр.
1. М. fuscus (Jur.)........................................................ . 130
2. М. distinctus (Rich.)....................................................... 132
-3. М. al bid us (Jur.)........................................................ 134
2. Род Eucyclops Claus
1. E. serrulatus (Fisch.)....................................................139
la. E. serrulatus var. proximus Lili..........................................143
lb. E. serrulatus var. speratus Lili.............................................143
1c. E. serrulatus var. nagasakai (Ueno) .................................... 144
2. E. teras (E. Graeter).....................................................145
3. E. macruroides (Lili.)...................................................145
За. E. macruroides var. denticulatus (A. Graeter).............................148
4. E. macrurus (Sars)........................................................148
5. E. graeteri (Chapp.).................................................... 150
3. Род Tropocyclops Kiefer
1. T. prasinus (Fisch.) ........................................................151
У
4. Род Ochridacyclops Kiefer
1. O. arndti Kief............................................................. 154
5. Род Paracyclops Claus
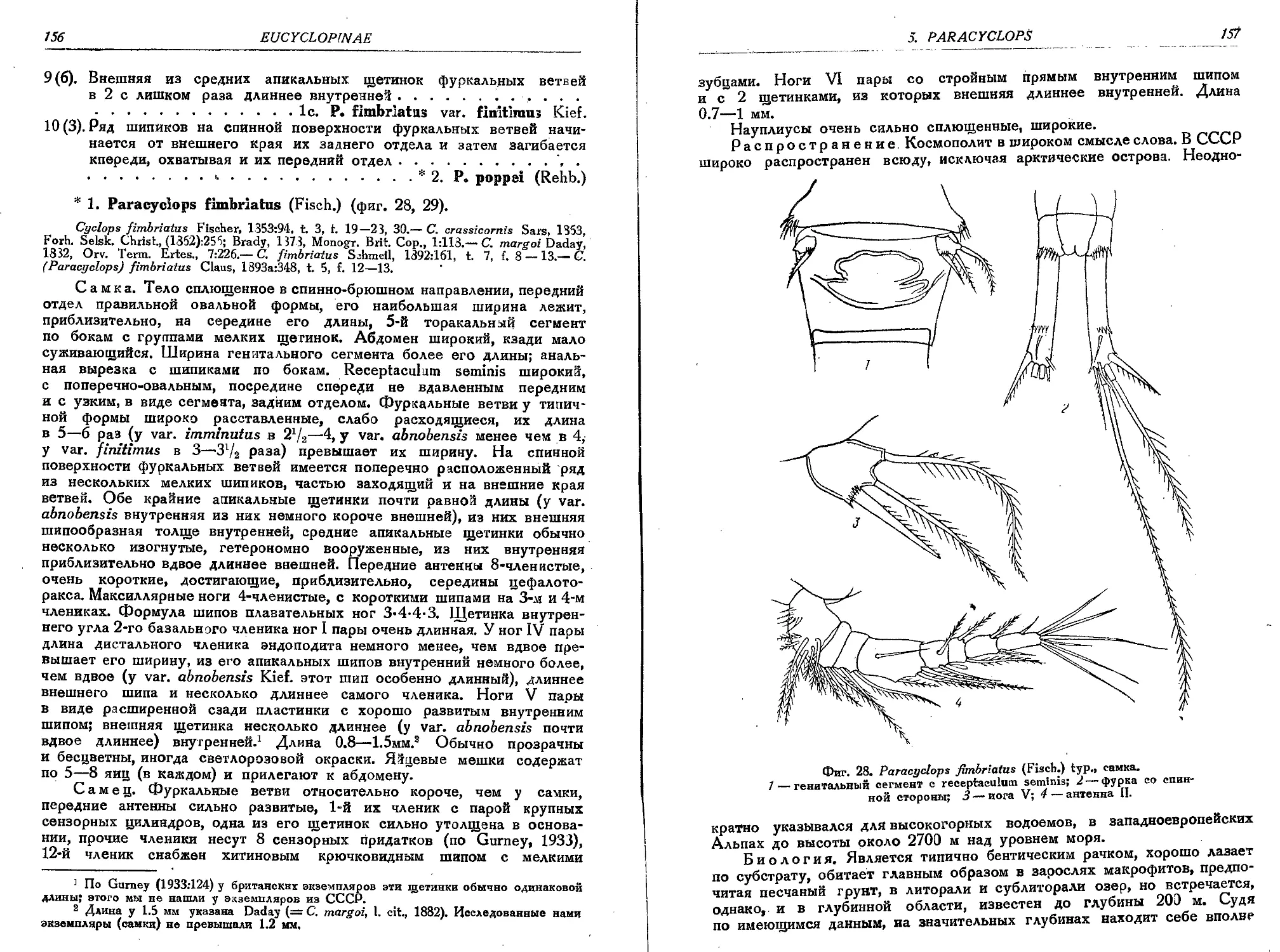
1. P. fimbriatus (Fisch.)................................................... 156
la. P. fimbriatus var. imminutus Kief.........................................158
lb. P. fimbriatus var. abnobensis (Kief.).................................... 159
1c. P. fimbriatus var. finitimus (Kief.)......................................160
'2. P. poppei (Rehb).........................................................160
3. P. affinis Sars..........................................................161
6. Род Ectocyclops Brady
1. E. phaleratus (Koch).................................................... 163
II. Подсем. Cyclopinae
7. Род Cyclops O. F. Muller
1. C. strenuus Fisch......................................................173
la. C. strenuus var. landei Kozm........................................ • 176
lb. C. strenuus var. vranae Kozm...........................................ISO
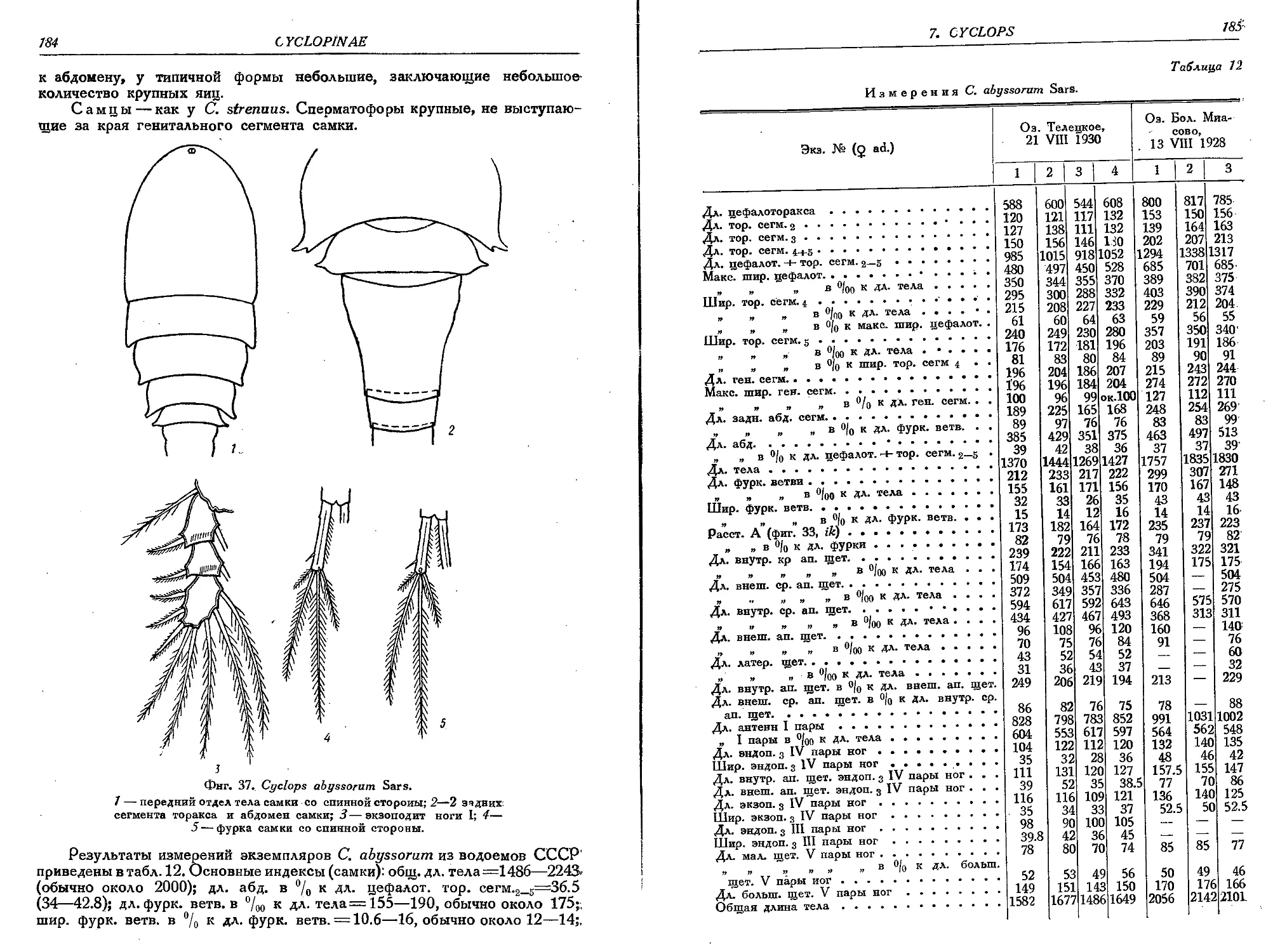
2. C. abyssorum Sars................................................... 183
14
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
Стр.
3. С. bohater Kozrn...................................................................186
4; С. lacustris Sars............................................................... 187
5. С. tatricus Koim...................................................................189
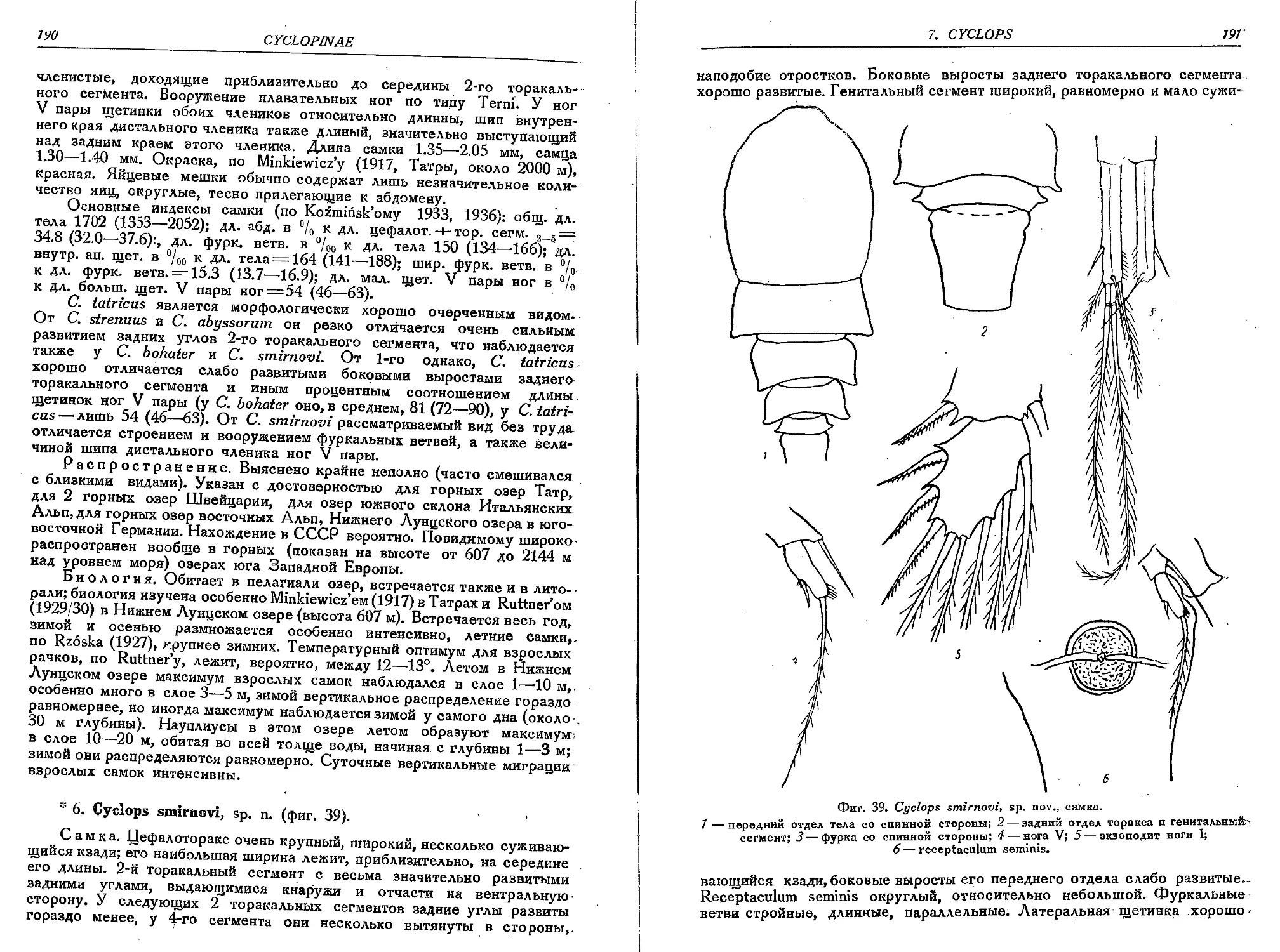
6. C. smirnovi, sp. n.................................................................190
7. C. ochridanus Kief................................................................ 195
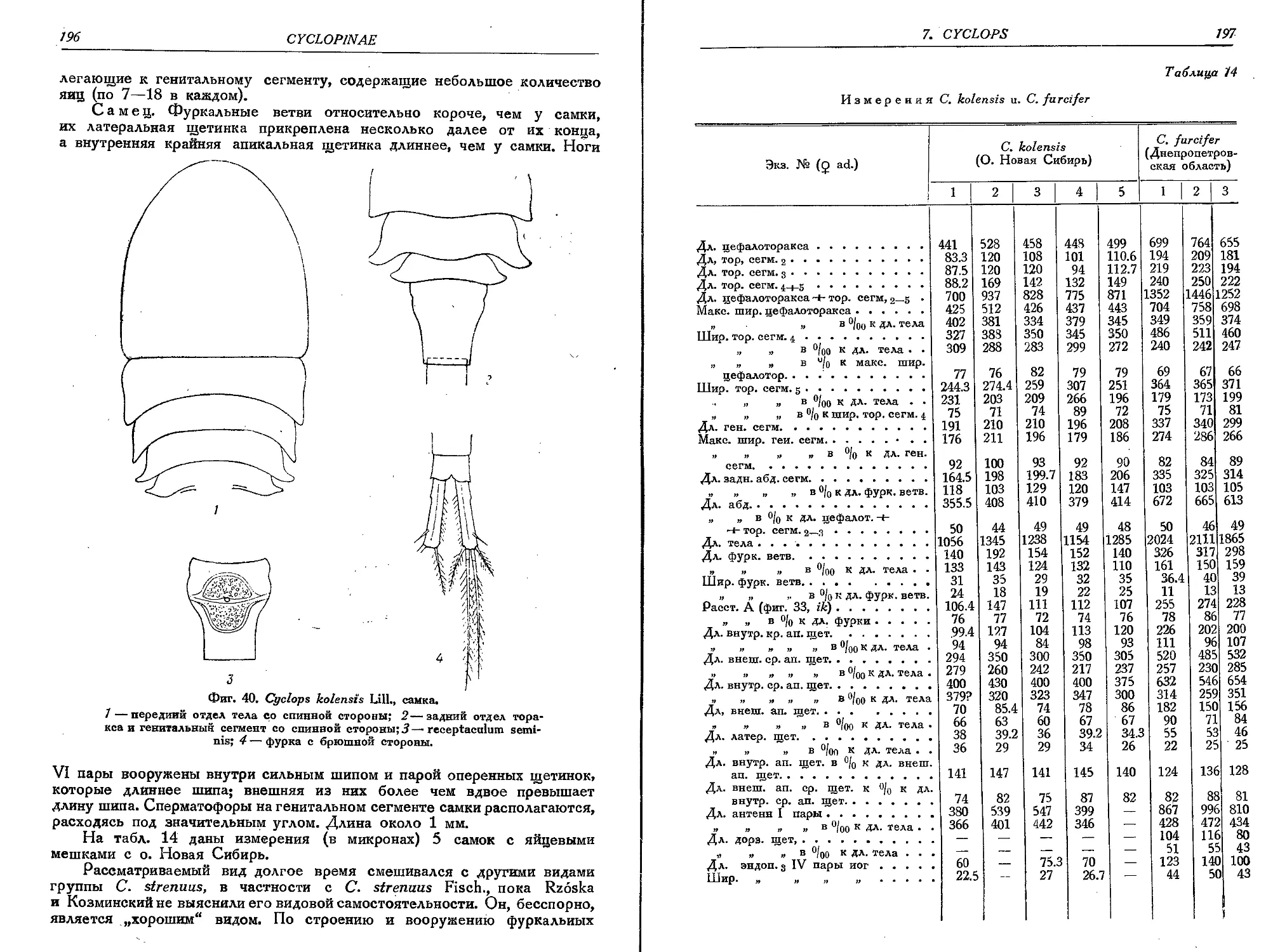
8. C. kolensis Lili....... 195
9. C. furcifer Claus................................................................. 198
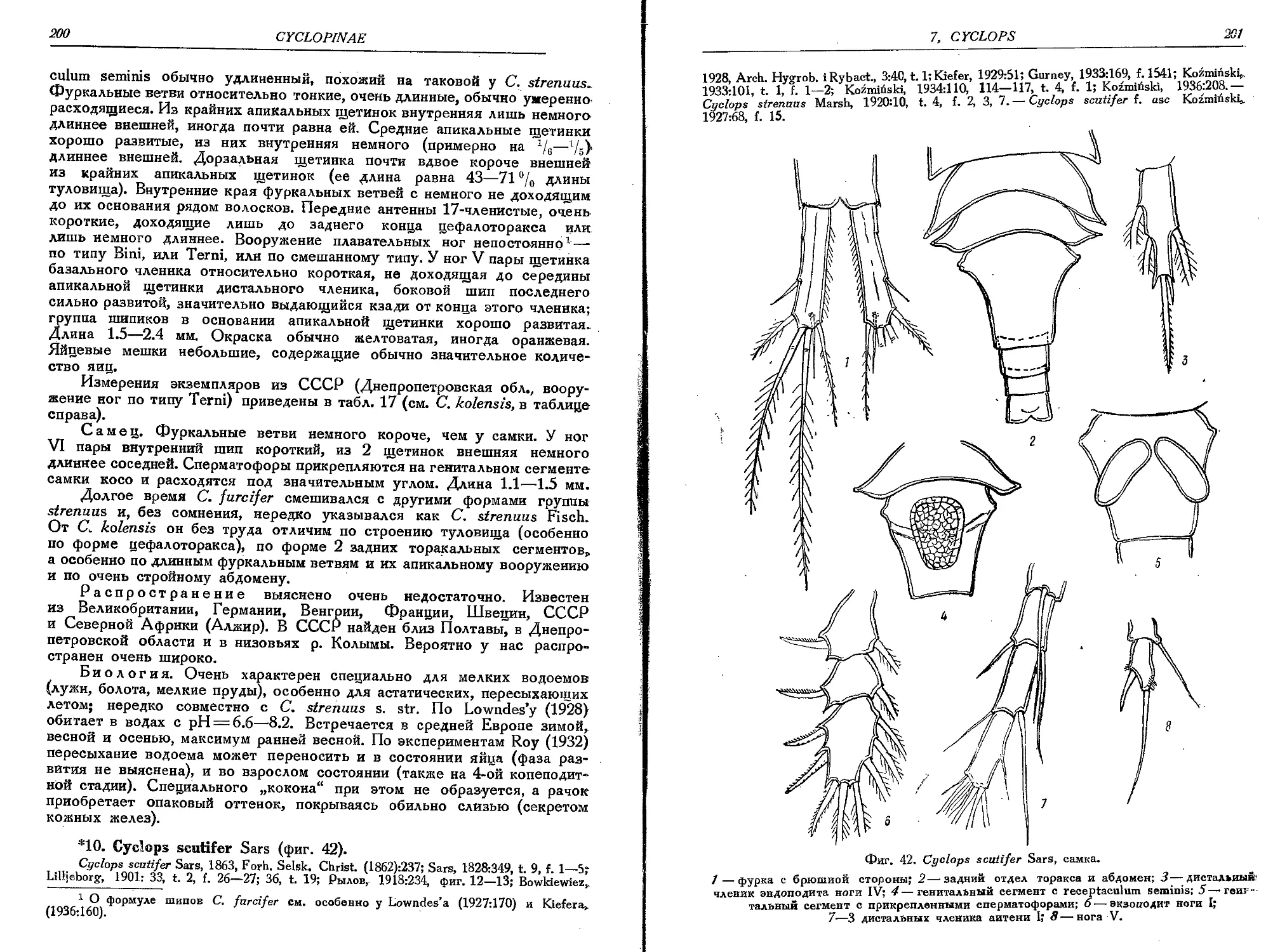
10. C. scutifer Sars........... . •....................................................200
10a. C. scutifer var. wigrensis Kozm...................................................204
11. C. vicinus Uljan...............................................•.................... 204
Ila. C. vicinus var. kikuchii Smirn. .................................................... 207
12. C. insignis Claus.....................................................................208
8. Род Acanthocyclops Kiefer
1. A. viridis (Jur.).............................................................. 215
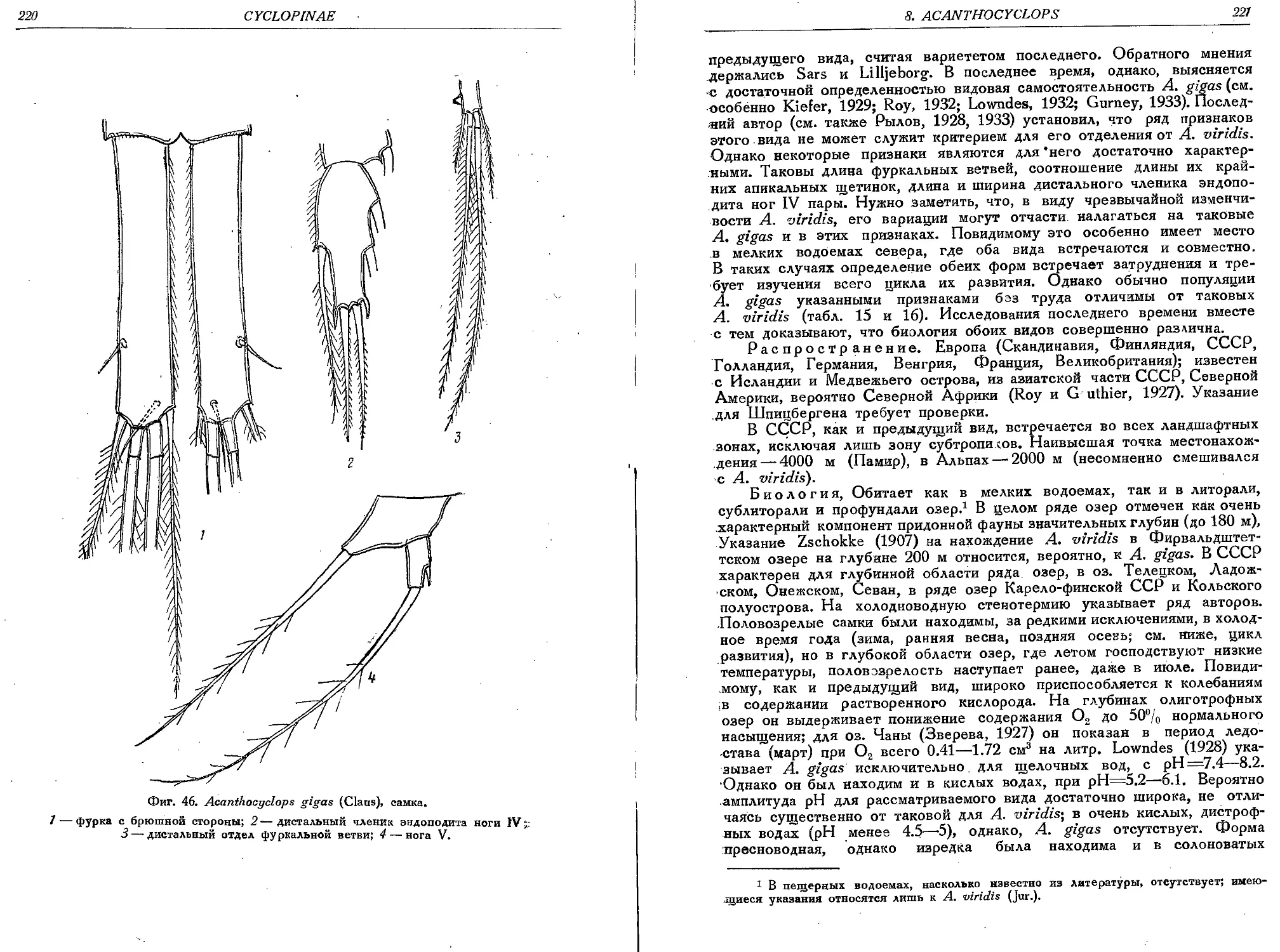
2. A. gigas (Claus)................................................................219
2a. A. gigas var. latipes (Lowndes).................................................223
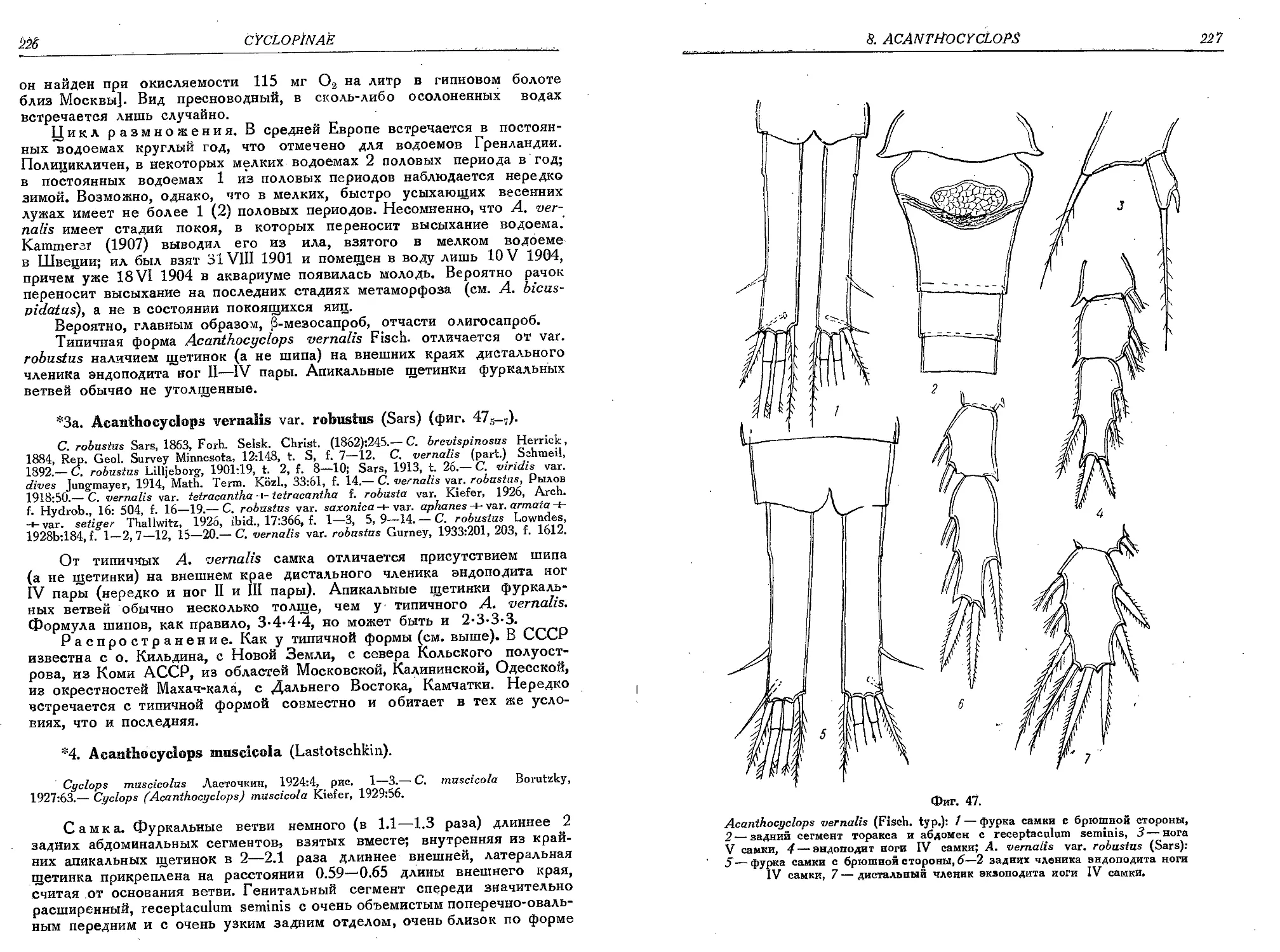
3. A. verualis (Fisch.)........................................................ '. 223
4. A. muscicola (Last.)........................................................... 226
5. A. americanus .Marsh)...........................................................228
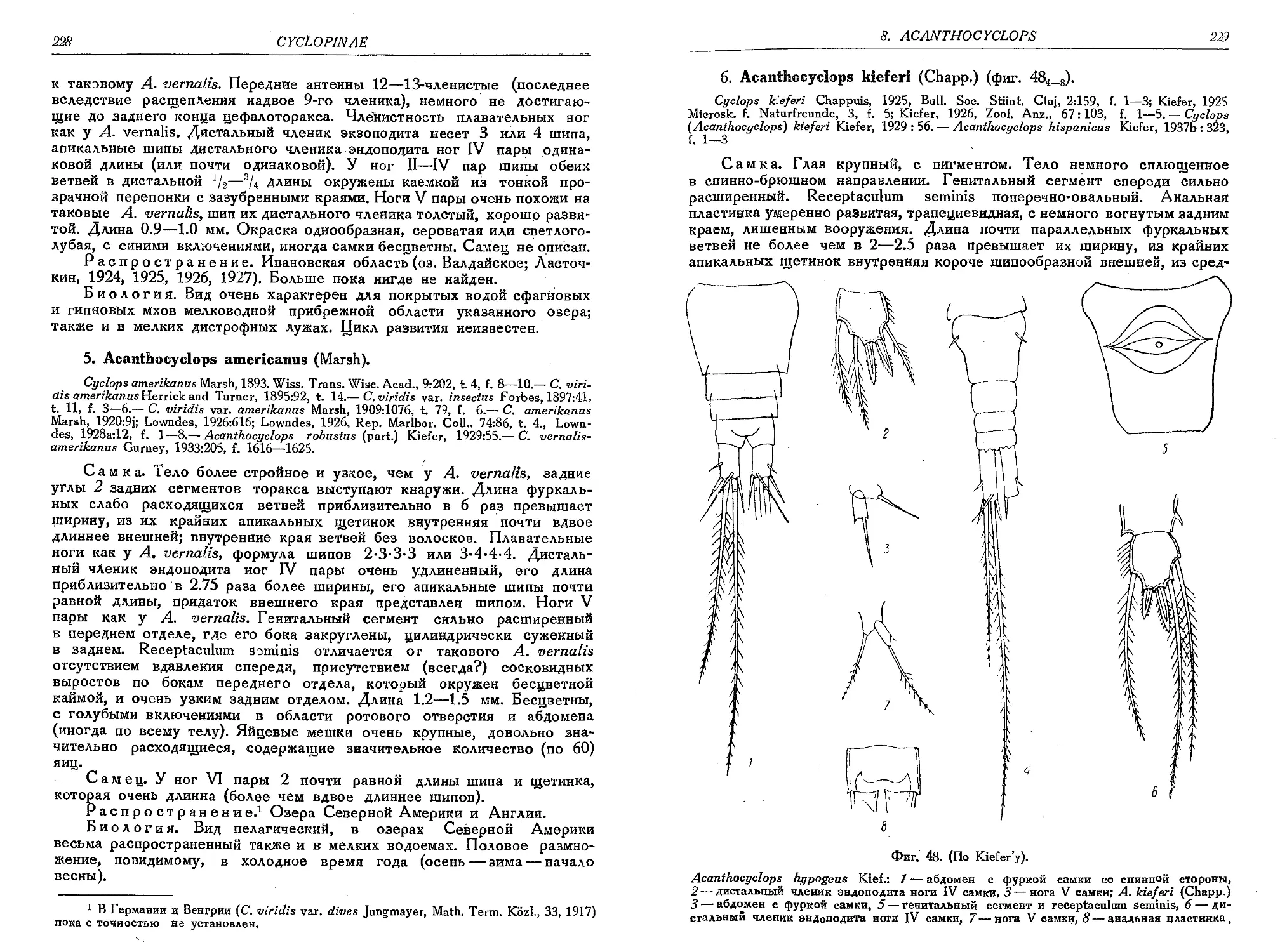
6. A. kieferi (Chapp.).............................................................229
7. A. phraeticus (Chapp.)........................................................ 230
8. A. venustus (Norm, et Scott).................................................. 230
8a. A. venustus var. crinitus (E. Graeter) . . 232
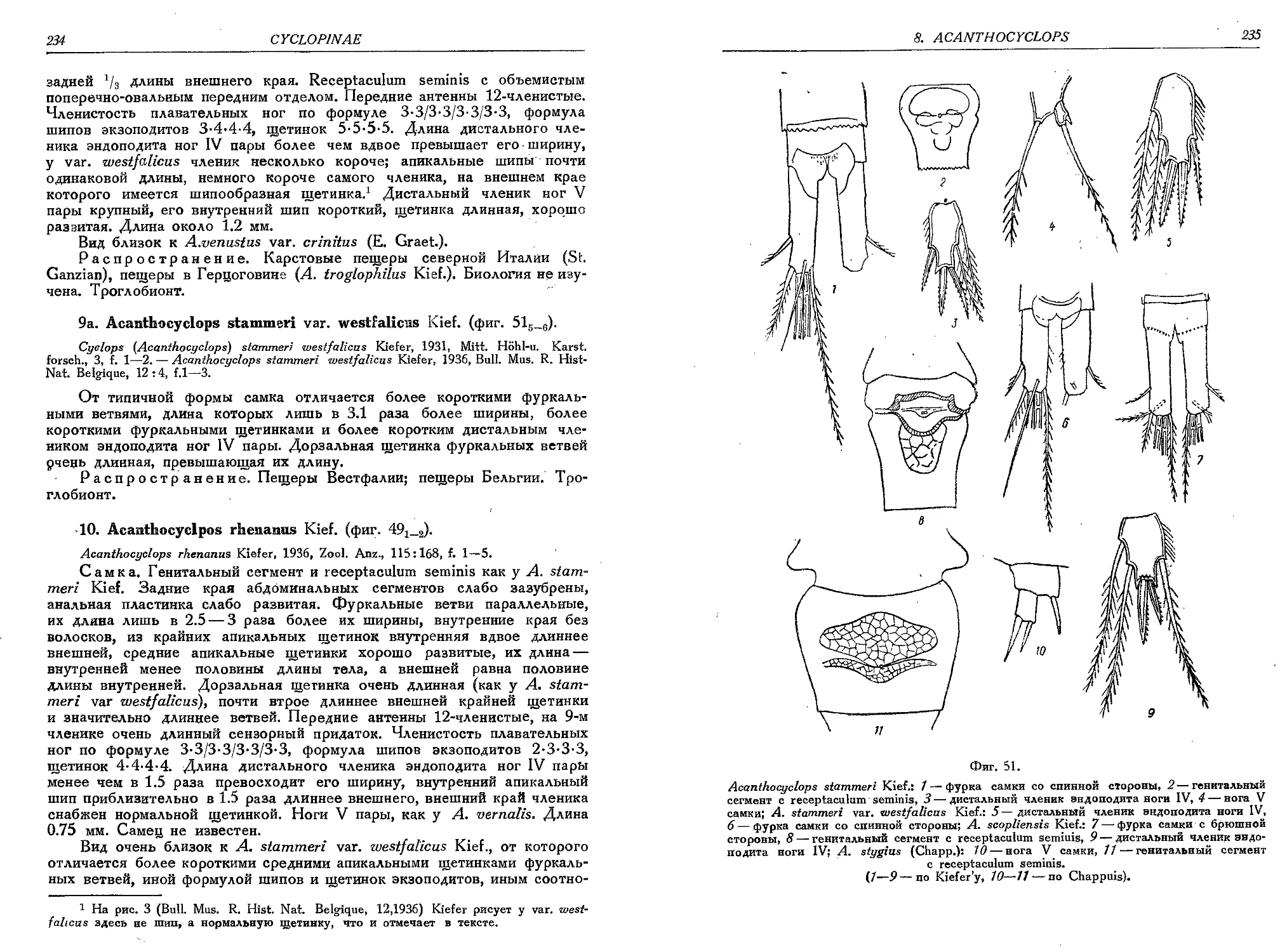
9. A. stammeri Kief.............................................................. 232
9a. A. stammeri var. westfalicus Kief.............................................. 234
10. A. rhenanus Kief...................................................... . . 234
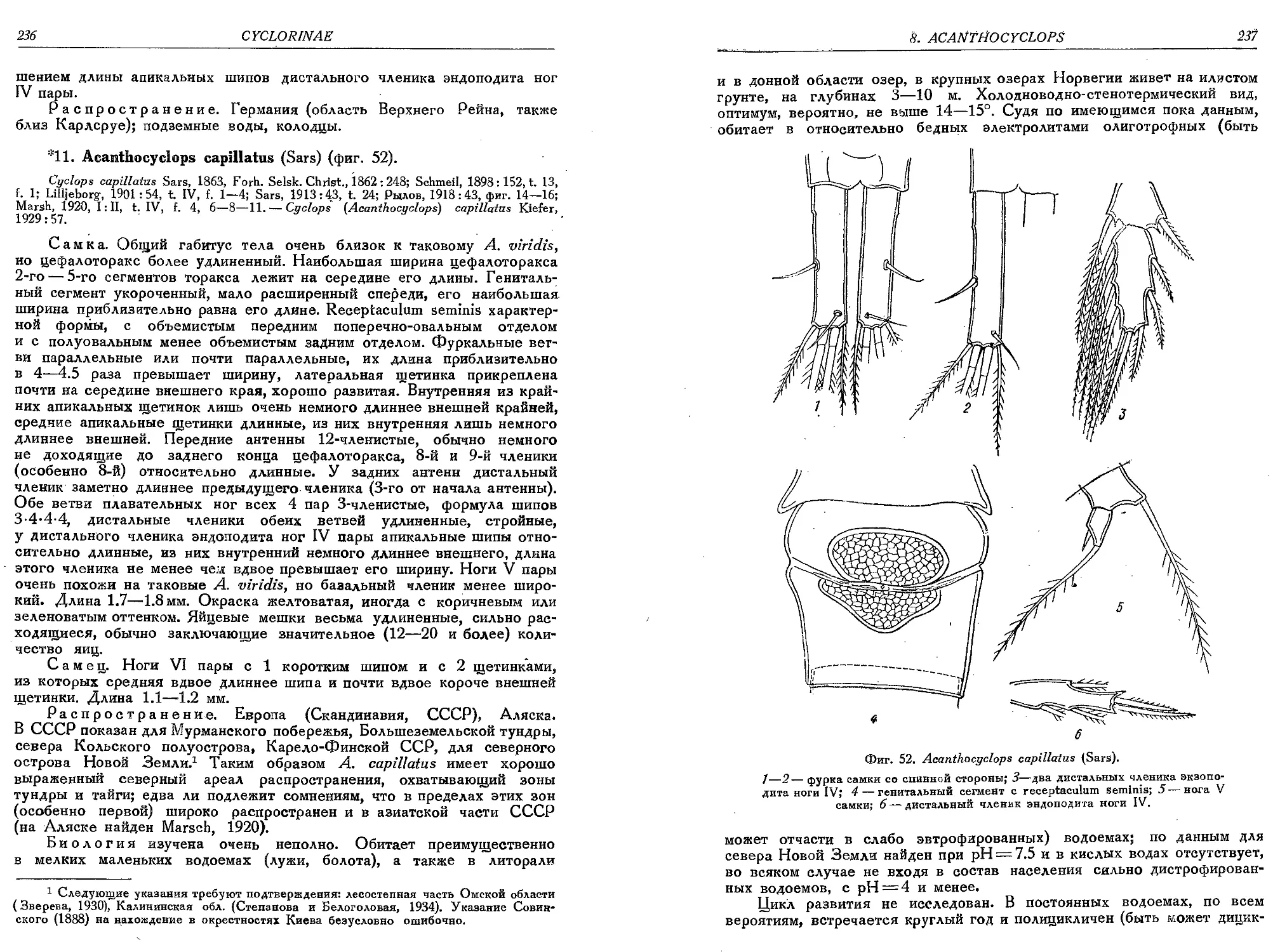
11. A. capillatus (Sars)........................
12. A. sensitivus (Graet. et Chapp.)............
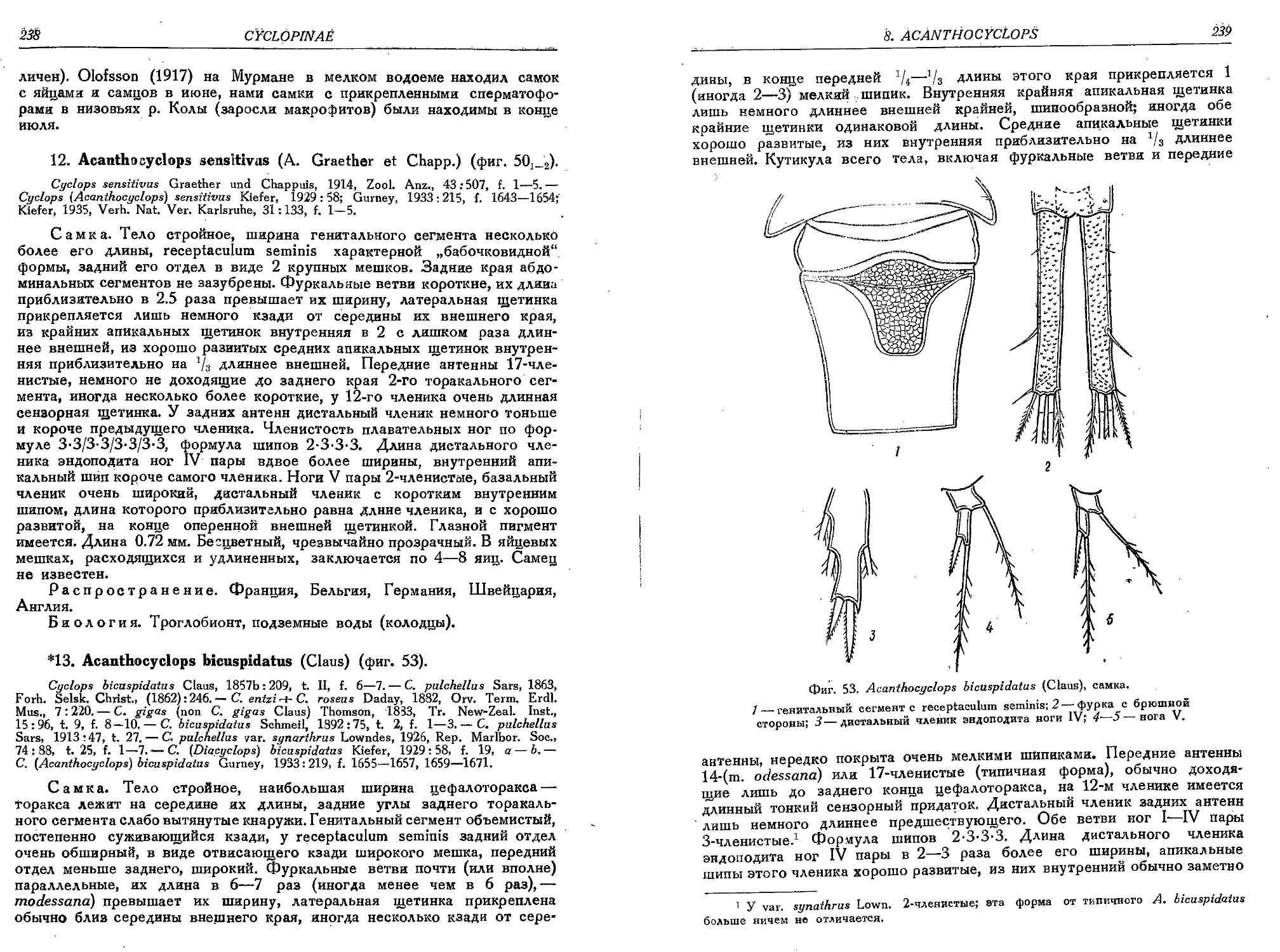
13. A. btcuspidatus (Claus).....................
13a. A. biscupidatus m. odessanus (Schmank.) . •
14. A. karamani Kief.............................
15. A. bisatosus (Rehb.)........................
16. A. crassicaudis (Sars)......................
16a. A. crassicaudis var. brachycercus Kief. . .
16b. A. crassicaudis var. cretensis Kief.........
17. A. scopliensis Kief.........................
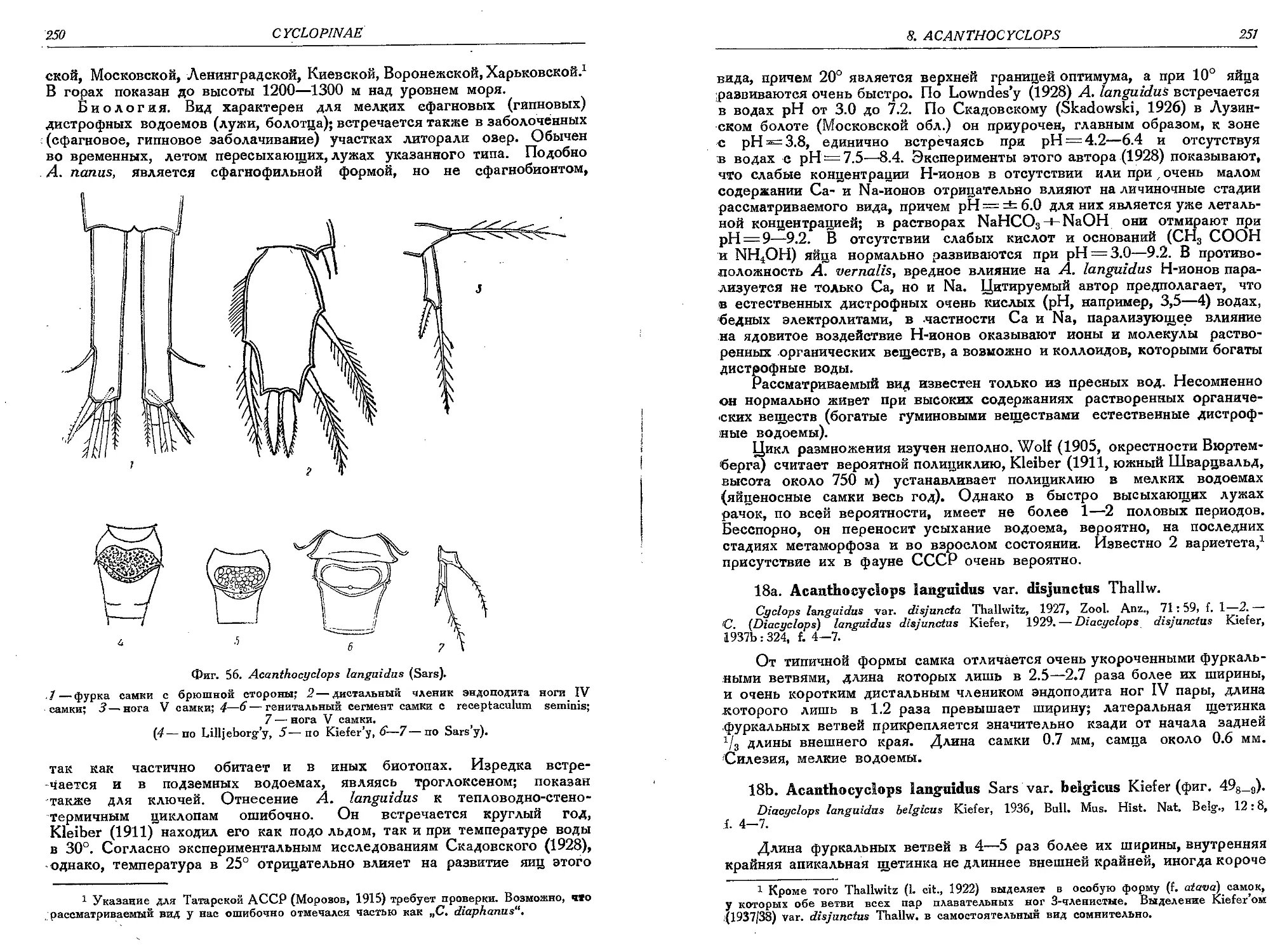
18. A. languidus (Sars).........................
18a. A. languidus var. disjunctus Thall..........
18b. A. languidus var. belgicus Kief...........
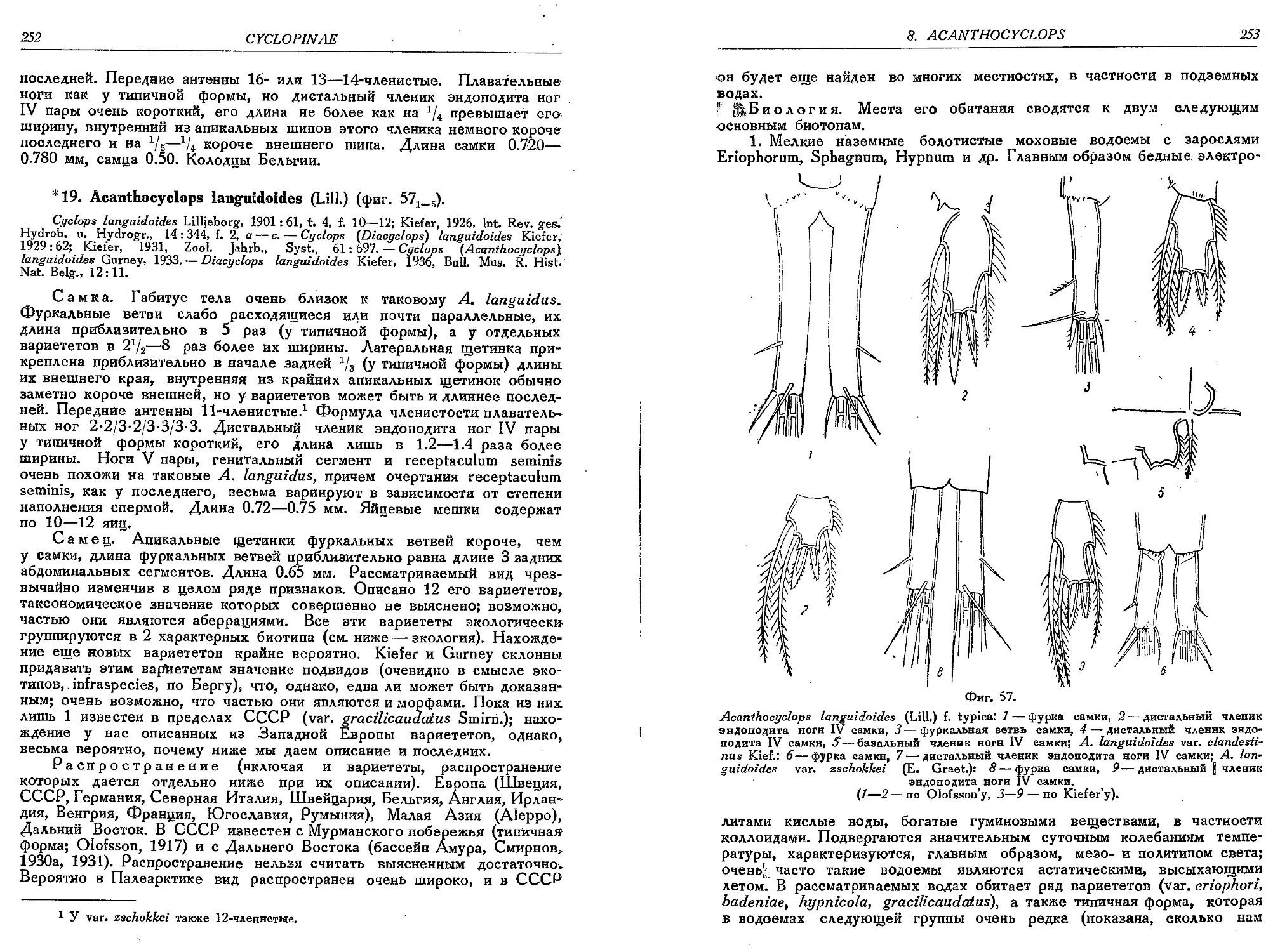
19. A. languidoides (Lili.)................ . .
19a. A. languidoides var. eriophori Gurn. . . . .
19b. A. languidoides var. hypnicola Gurn..........
19c. A. languidoides var. badeniae Kief..........
19d. A. languidoides var. gracilicaudatus Smirn. .
19e. A. languidoides var. hiberniae Gurn.........
19f. A. languidoides var. clandestinus Kief. . .
19g. A. languidoides var. nagysalloensis Kief. • .
19h. A. languidoides var. zschokkei (E. Graeter)
19i. A. languidoides var. putealis Chapp.........
19k. A. languidoides var. italianus Kief. . . . .
191. A. languidoides var. goticus Kief...........
19m. A. languidoides var. tantalus (Kief.) . . . .
20. A. nanus (Sars)..............................
20a. A. nanus var. incertus (Wolf)...............
21. A. stygius (Chapp.).........................
22. A. hypogeus Kief............................
23. A. abyssicola (Lili.).......................
236
- 238
238
241
242
243
245
248
248
. 248
. 249
. 251
. 251
. 252
255
255
255
257
257
258
258
258
259
259
259
259
260
263
263
264
264
9. Род Microcy clops Claus
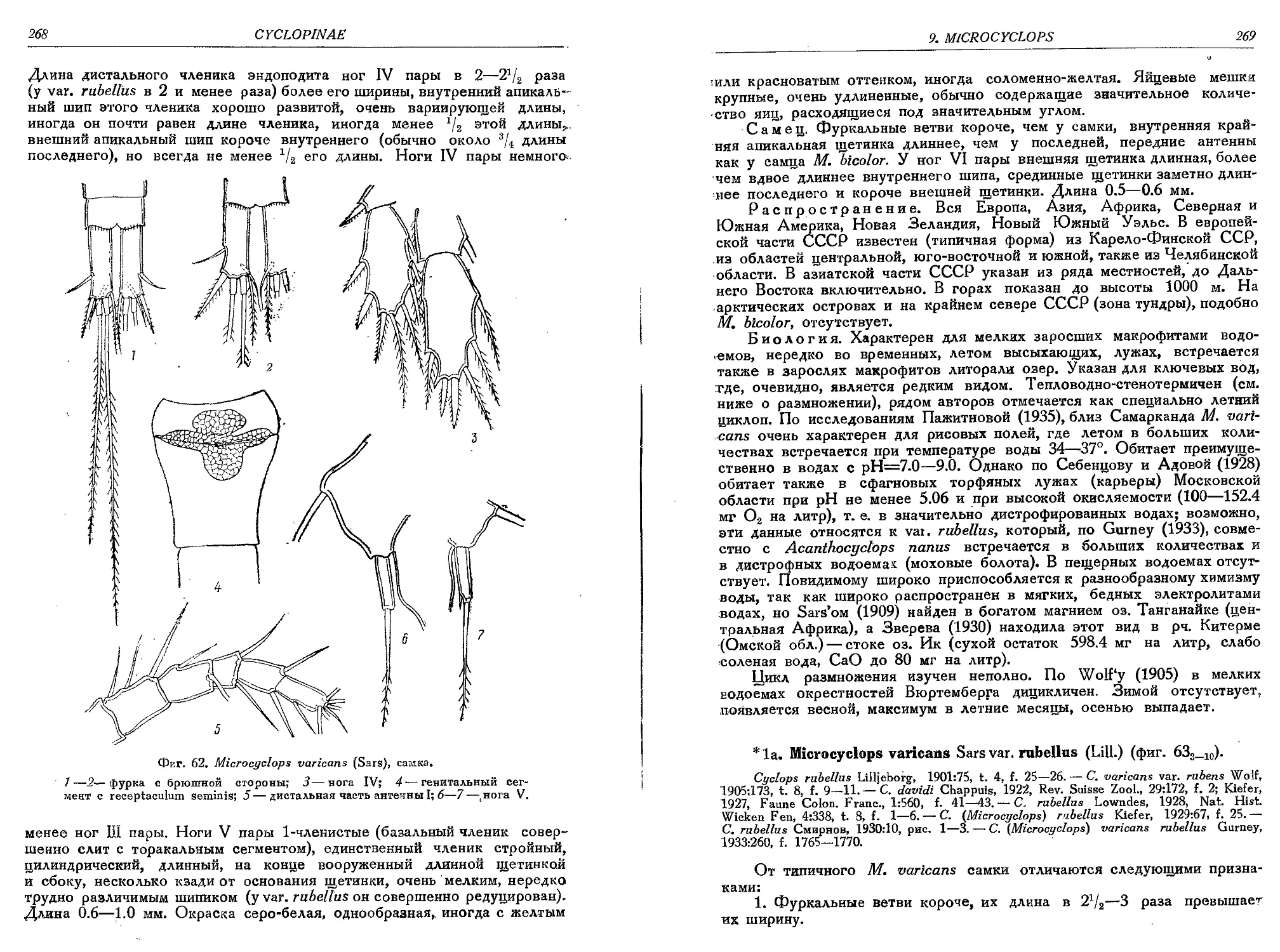
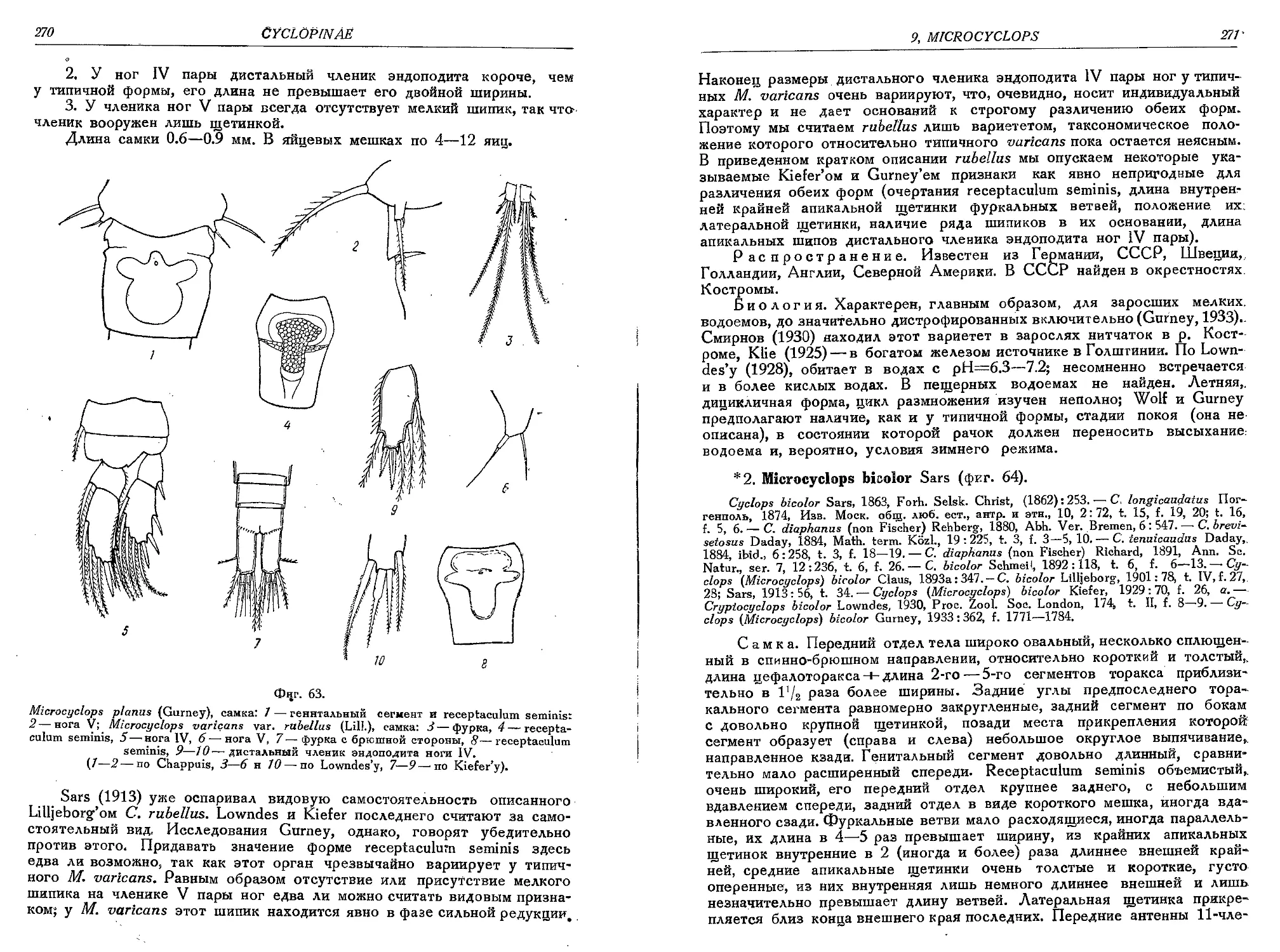
1. M. varicans Sars.....................................................................267
la. M. varicans var. rnbellus (Lili.)....................................................269
2. M. bicolor Sars................................................................... • 271
Invertebrate Zoology
СИСТЕМАТИЧЕСКИЕ УКАЗАТЕЛЬ ВИДОВ (Crustacea) 15"
Стр.
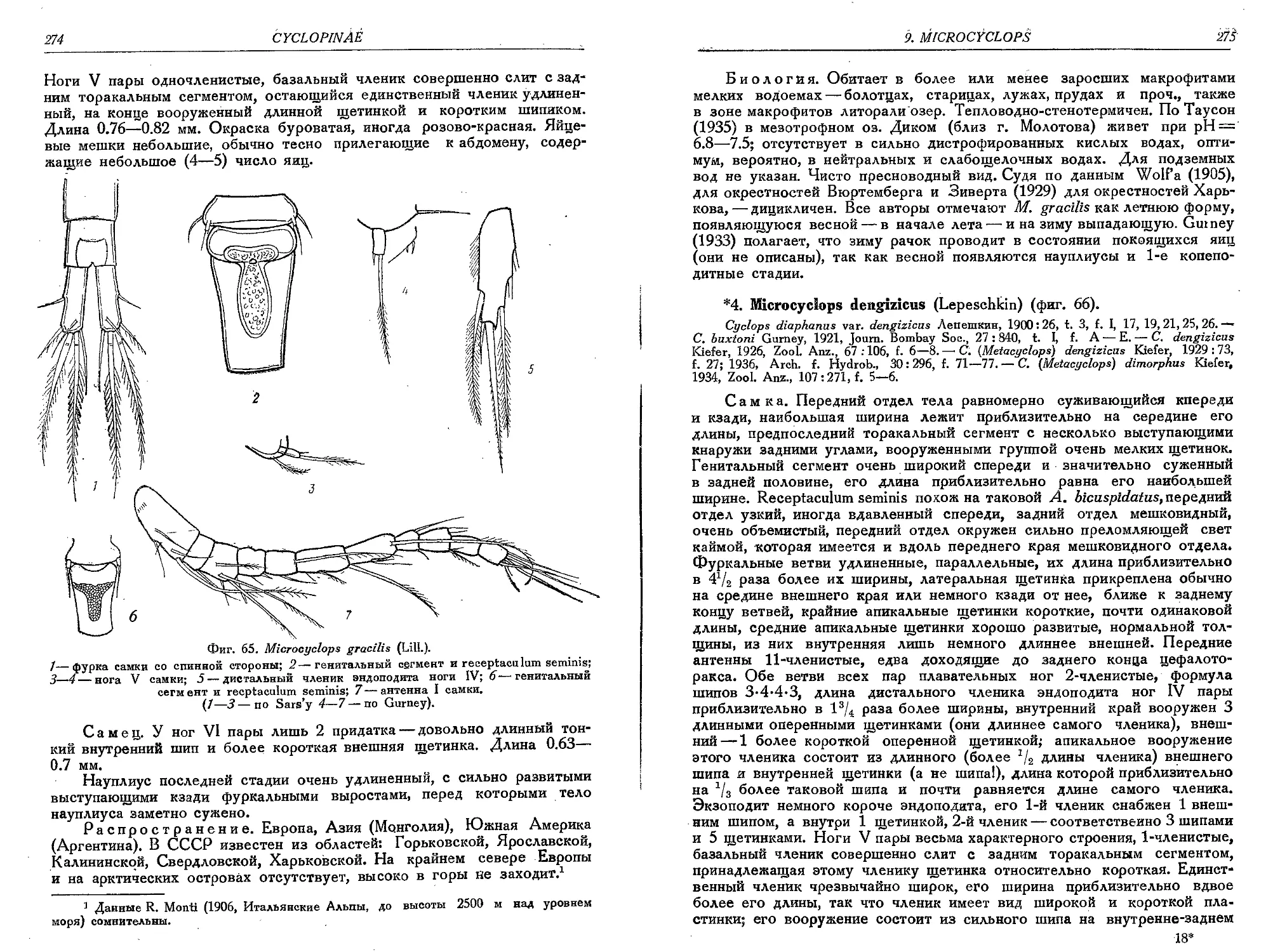
3. М. gracilis (Lili.).............................................................. 273-
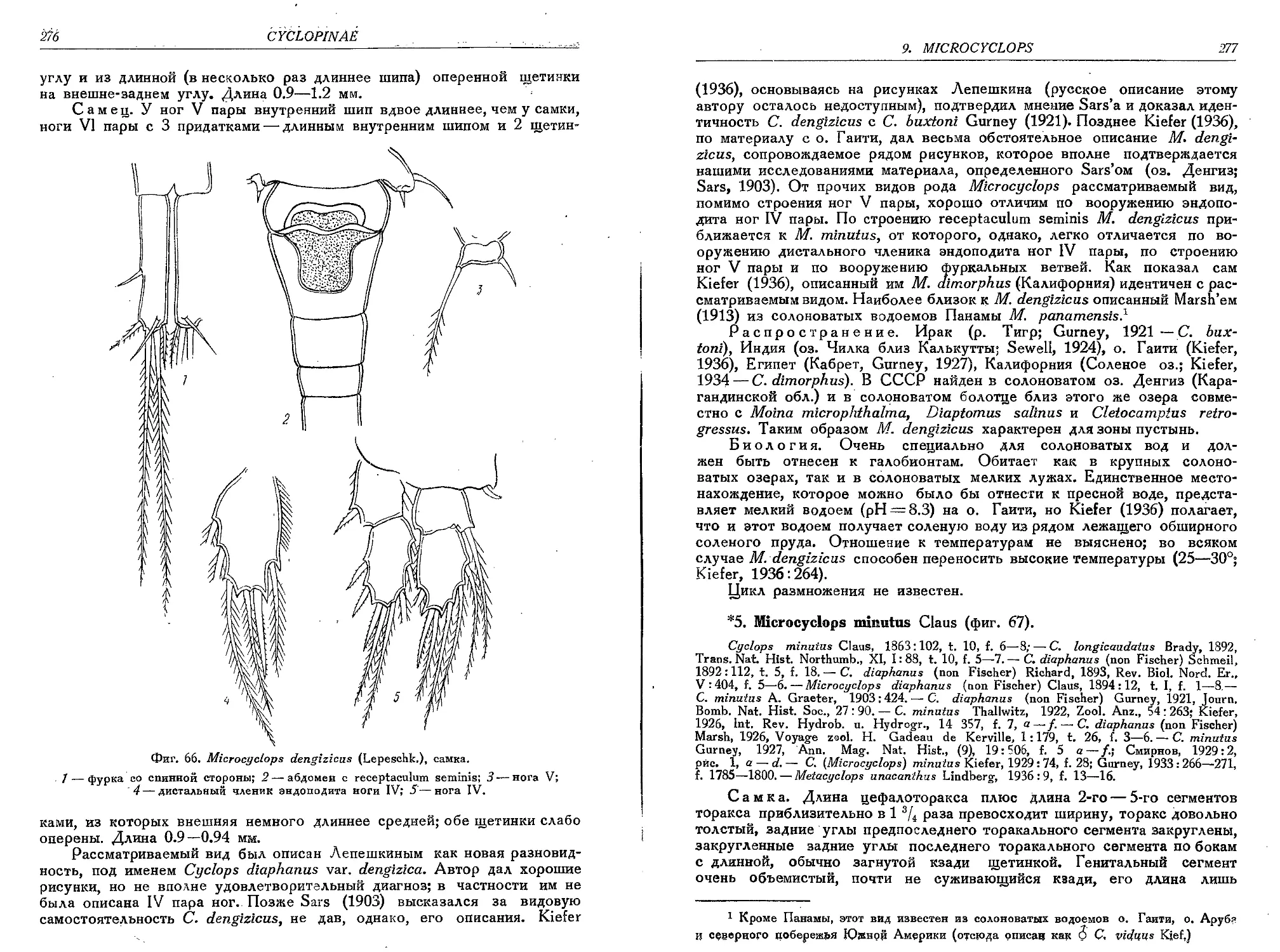
4. М. dengizicus (Lepeschk.)........................................................275
5. М. minutus Claus..................................' . . . .........................277
6. M. planus (Gurn.)................................................................ 280"
10. Род Speocyclops Kiefer
1. S. racovitzai (Chapp.)...........................................................282
2. S. infernus Kief.................................................................282
3. S. sisyphus Kief.................................................................283
4. S. troglodytes (Chapp.)..........................................................285
. 5. S. sebastianus Kief.............................................................285
6. S. plutonis Kief..................,................................................286
7. S. demeliensis (Scourf.)......................................................... 287
8. S. cerberus (Chapp.).............................................................287
9. S. spelaeus Kief.................................................................288
10. S. proserpinae Kief.............................................................. 288
11. S. orcinus Kief................................................................ 288.
12. S. colchidanus (Borutzky)........................................................289
11. Род Graeteriella Brehm
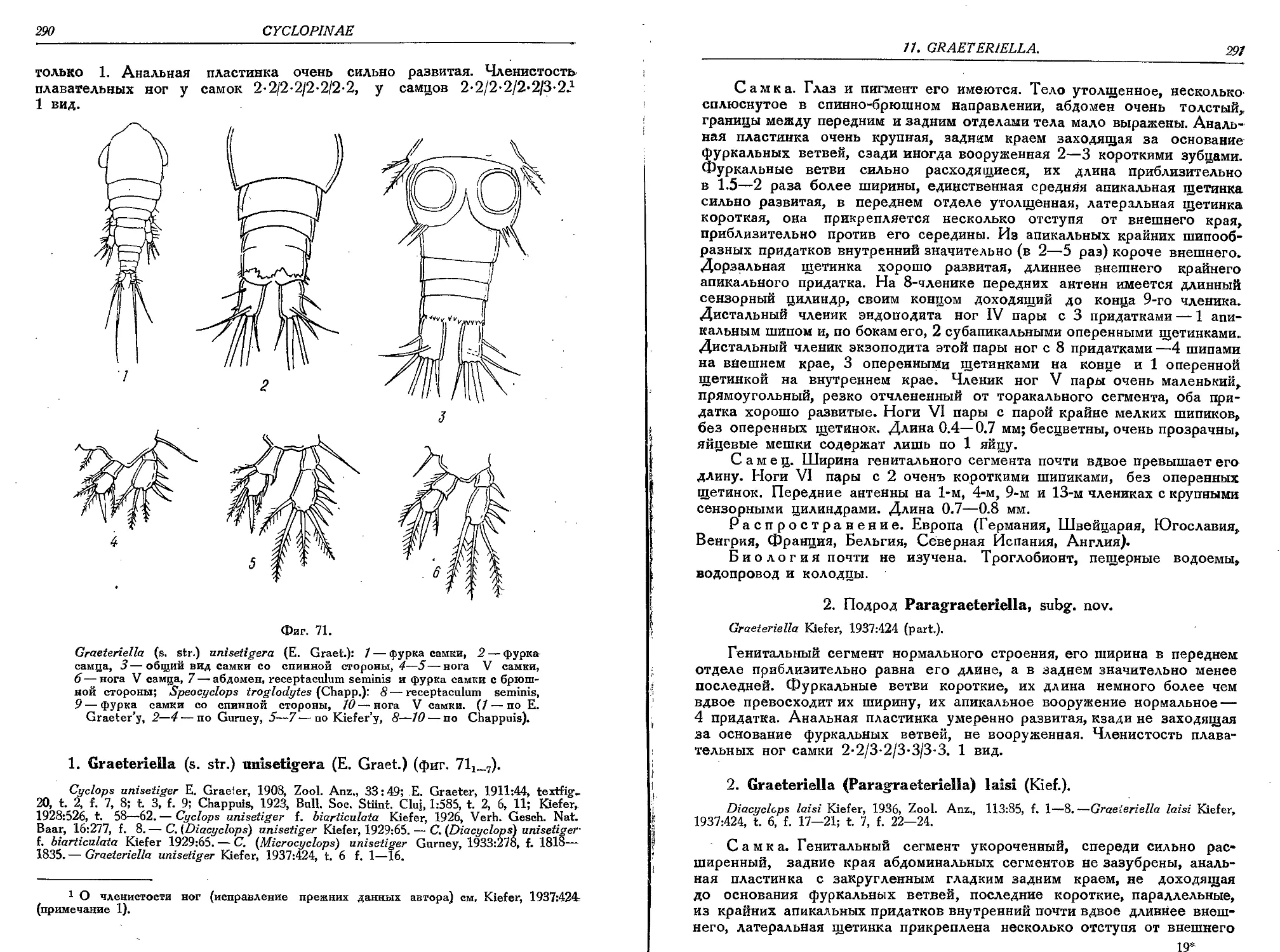
1. G. (s. str.) unisetigera (E. Graeter)............................................290
2. G. (P.) laisi (Kief.)........................................................... 291
12. Род Mesocyclops Sars
Подрод Mesocyclops s. str.
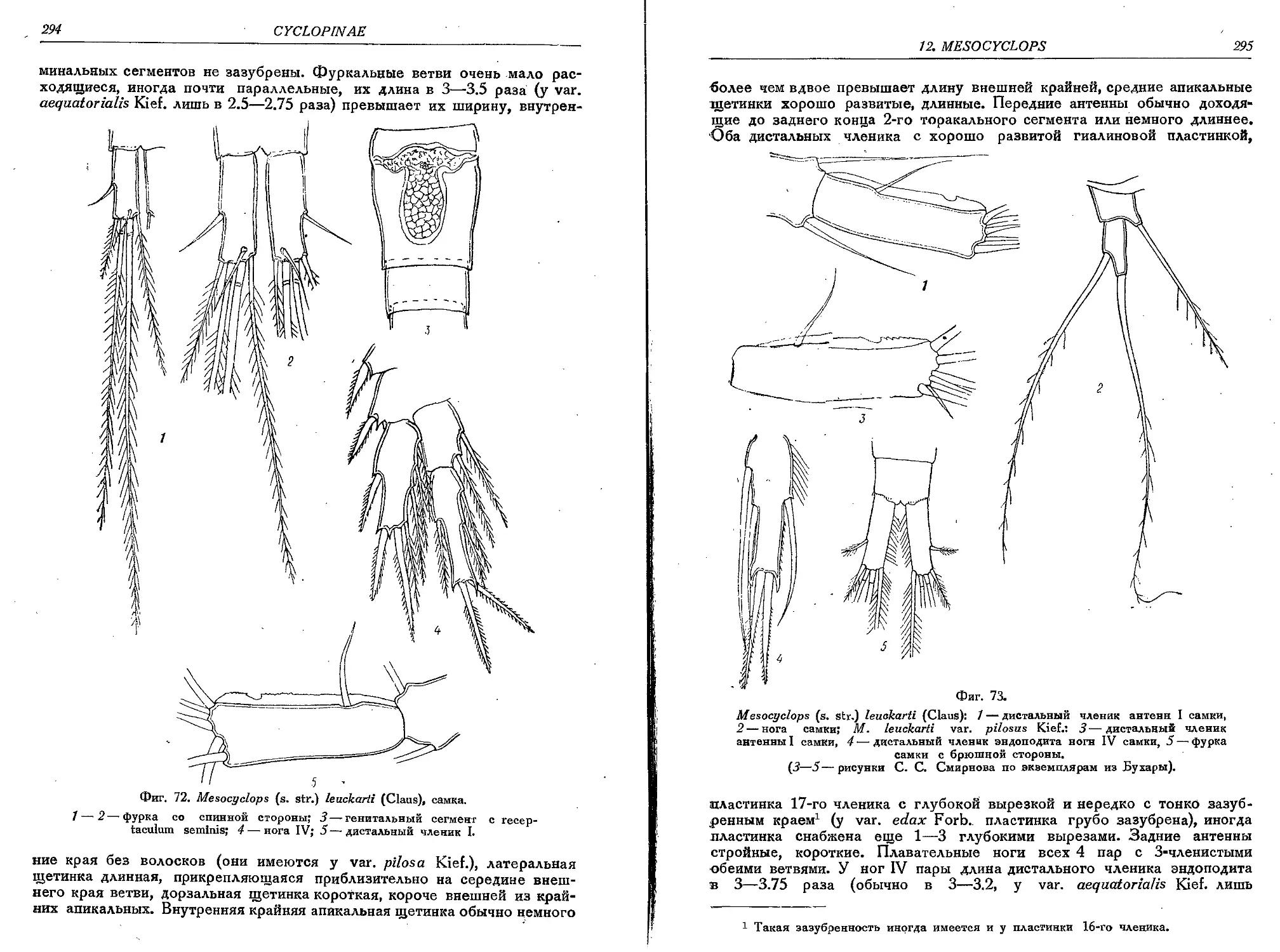
1. M. (s. str.) leuckarti Claus.................................................... 293
la. M. (s. str.) leuckarti var. pilosus Kief...........................................297
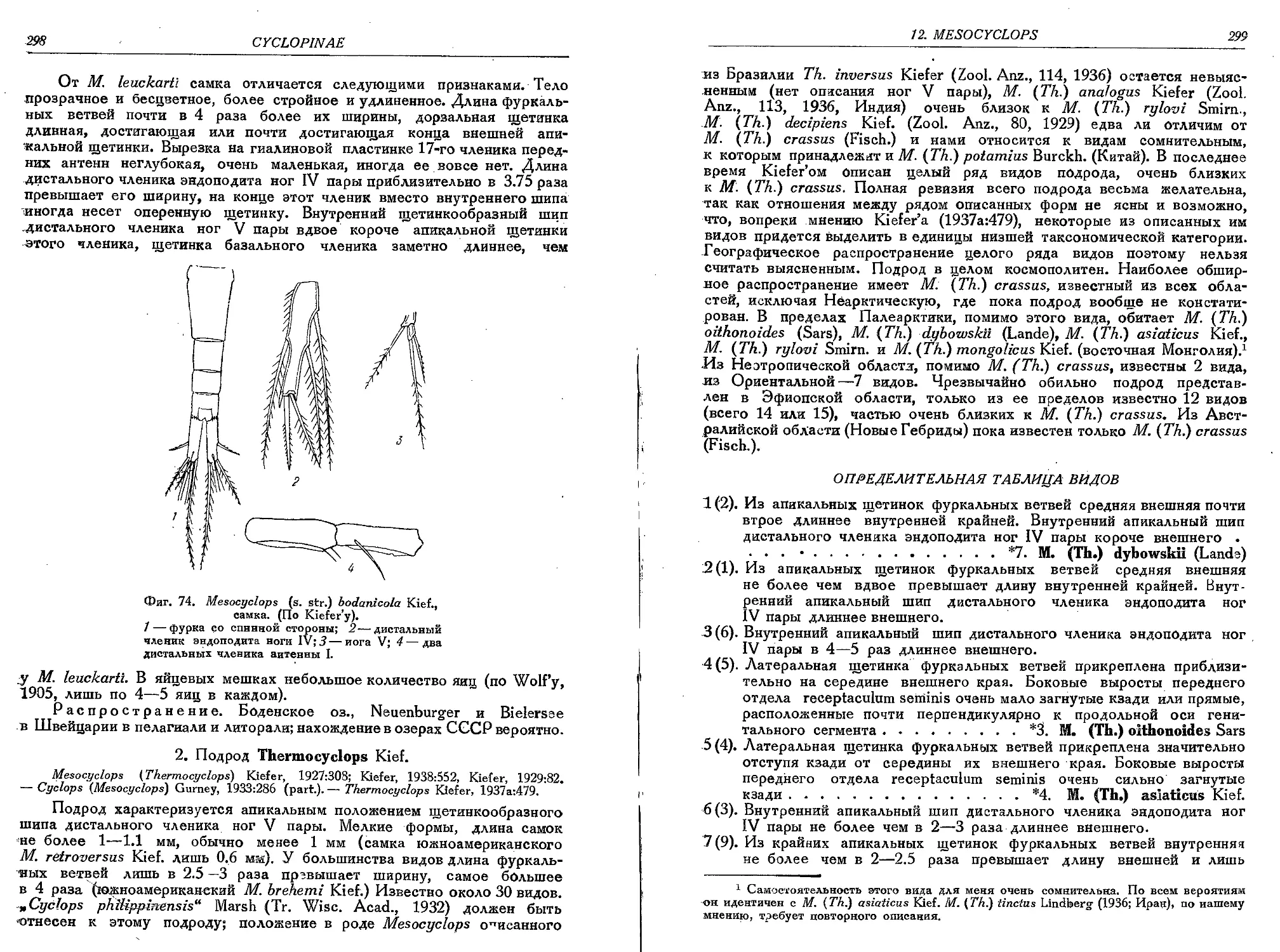
2. M. (s. str.) bodanicola Kief.......................................................297
Подрод Thermocyclops Kiefer
3. M. (T.) oithonoides Sars.........................................................300
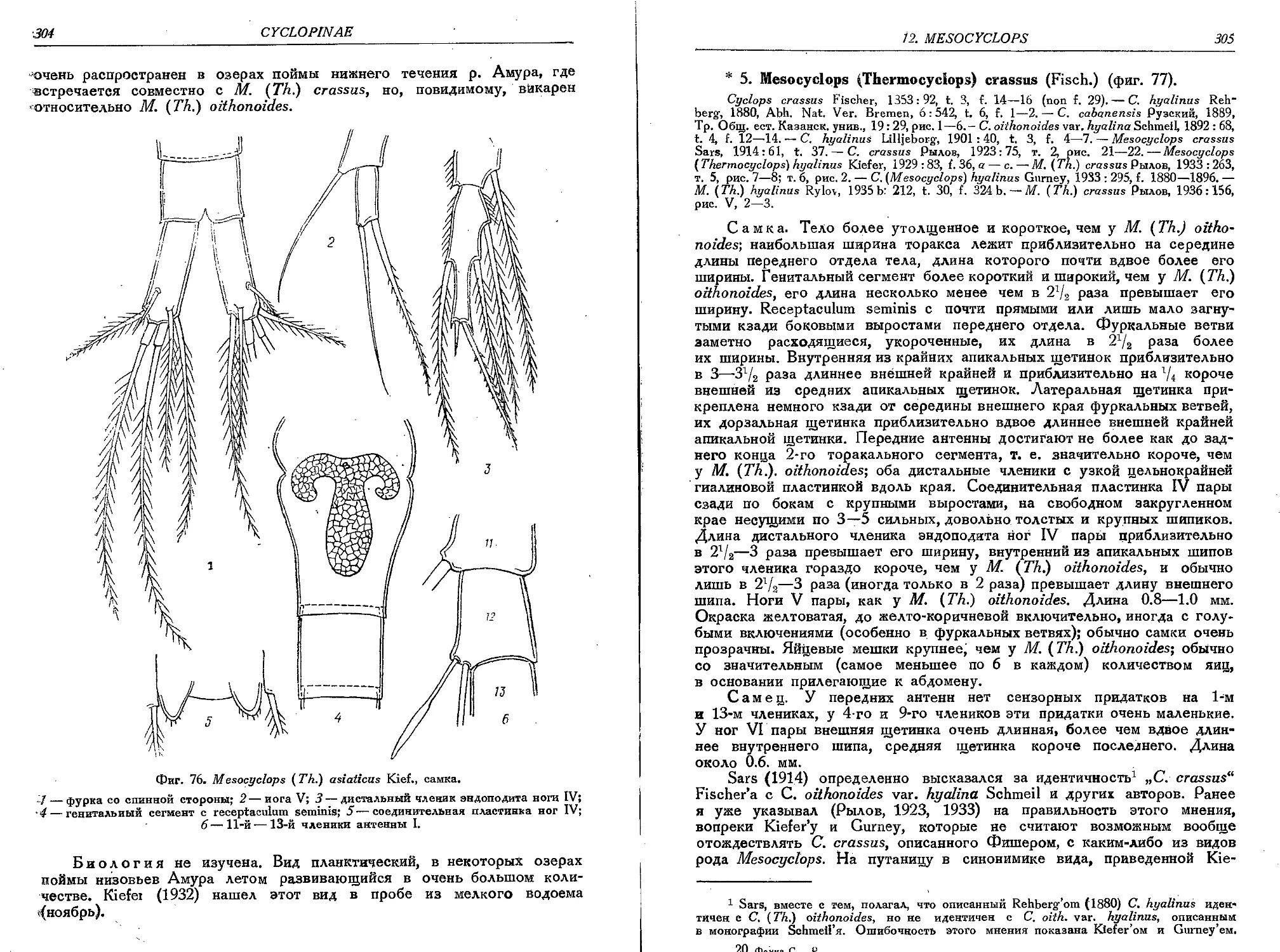
4. M. (T.) asiaticus Kief...........................................................303
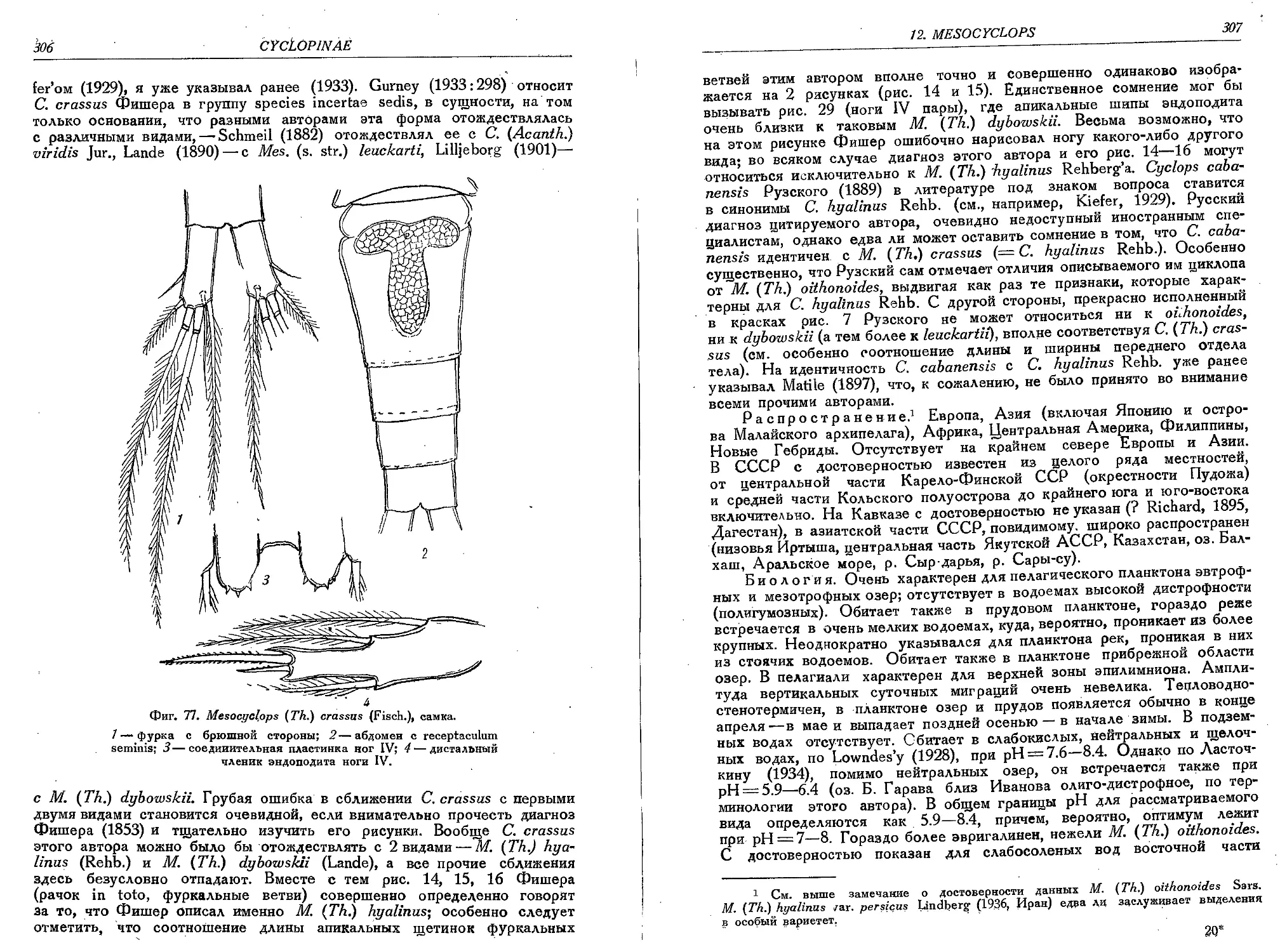
5. M. (T.) crassus (Fisch.).........................................................305
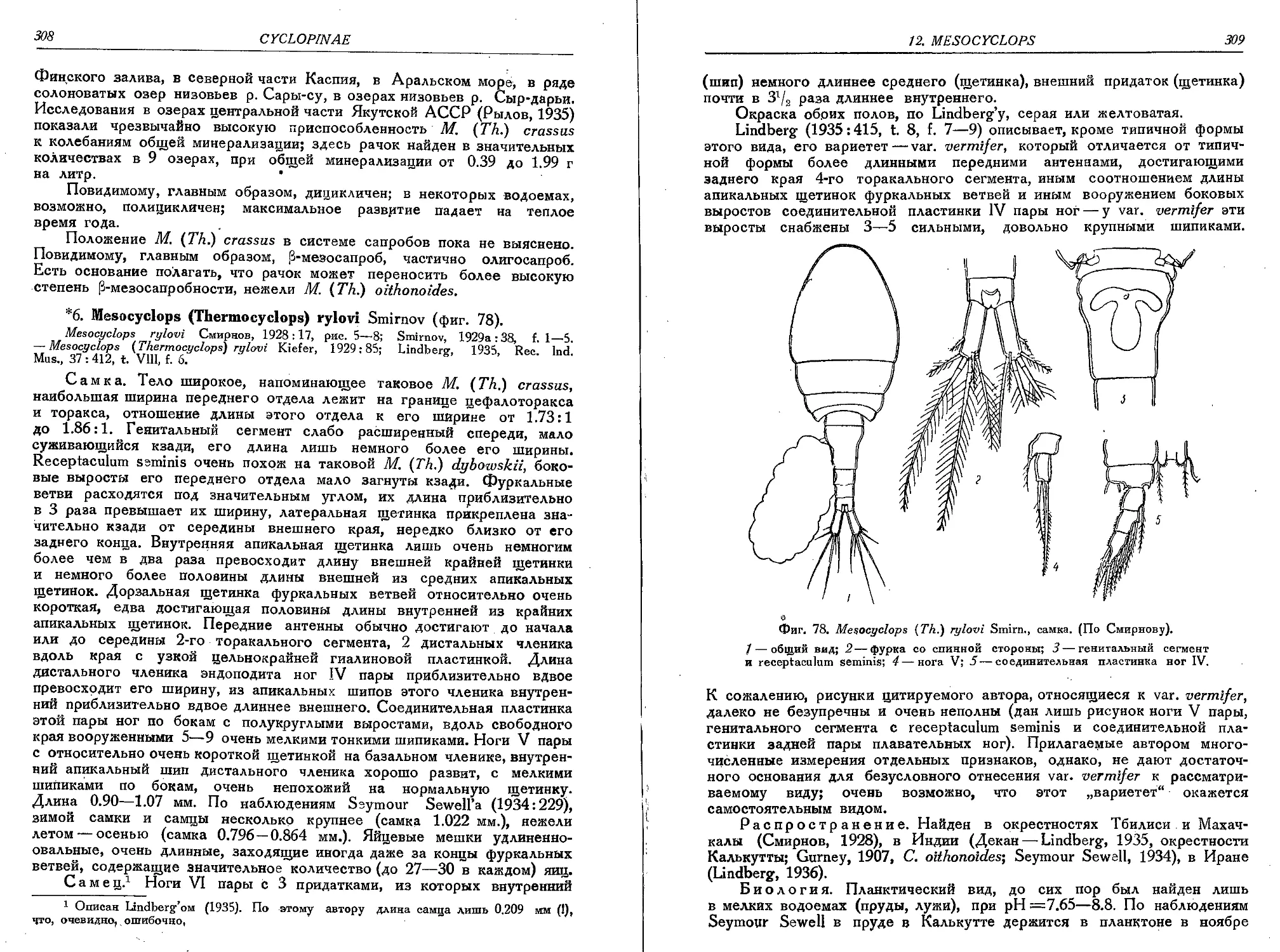
6. M. (T.) rylovi Smirn............................................................ 308
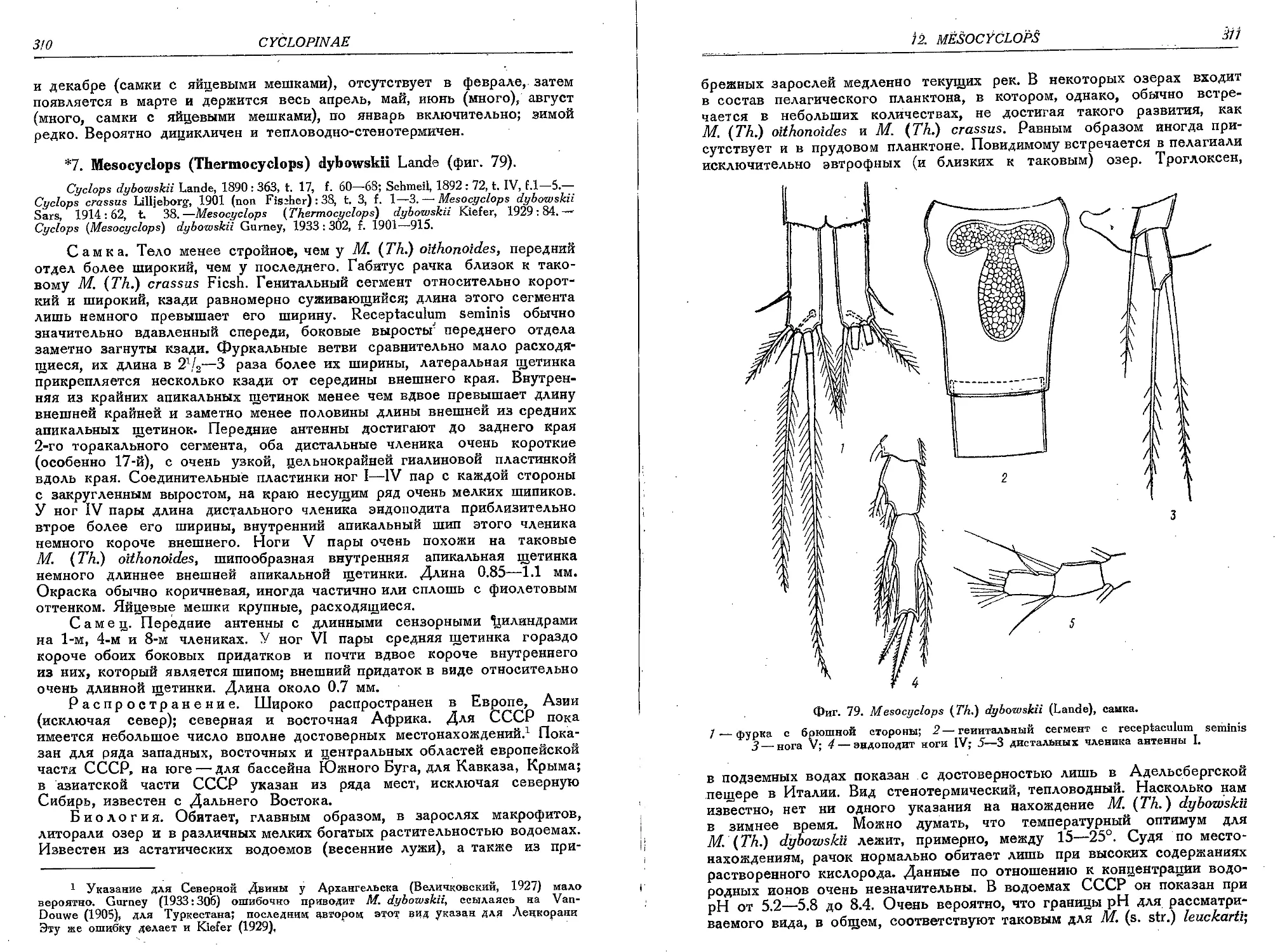
7. M. (T.) dybowskii (Lande).......................................................310'
ВВЕДЕНИЕ
МОРФОЛОГО-АНАТОМИЧЕСКИЙ ОЧЕРК
Внешняя морфология Cyclopidae уже давно стала предметом спе-
циального исследования и в настоящее время изучена достаточно полно.
В ином положении находятся наши сведения по анатомии группы. Не-
смотря на то, что их анатомией начали заниматься уже с 50-х годов
прошлого столетия, в настоящее время отдельные системы органов
Cyclopidae оказываются изученными далеко не одинаково полно. Благо-
даря работам последних 15—20 лет, выполненным методами современной
гистологической техники, наиболее точными сведениями мы располагаем
теперь по органам пищеварения и размножения. Применение методов
прижизненной окраски (Gicklhorn) внесло значительную ясность в строе-
ние некоторых, до последнего времени у Cyclopidae крайне недостаточно
изученных, органов. Ряд органов, однако, остается исследованным далеко
неудовлетворительно, и имеющиеся литературные данные значительно
устарели. Это особенно относится к мускулатуре и нервной системе,
которые еще требуют тщательного изучения.
Тело представителей семейства Cyclopidae (фиг. 1), как и прочих
свободно живущих веслоногих раков, подразделяется на 3 основных
отдела: передний (голова), средний (грудь, торакс) и задний (брюшко,
абдомен), заканчивающийся придатком в виде 2 более или менее рас-
ходящихся наподобие вилки ветвей (фурка, каудальные ветви, фуркаль-
ные ветви). В пределах всего рассматриваемого семейства голова и 1-й
сегмент торакса у обоих полов прочно слиты в один общий отдел —
головогрудь (цефалоторакс), за которой следуют 4 резко дифферен-
цированных сегмента; головогрудь наиболее обширна и может состав-
лять более половины длины остальных четырех грудных сегментов
(особенно -обширна у морского рода Euryte). У некоторых видов 4-й
(предпоследний) торакальный сегмент снабжен сильно вытянутыми
задними углами (например у Cyclops vicinus, С. scutifer), иногда в виде
крылообразных выростов; у С. kolensis эти углы своими окончаниями
загнуты несколько кпереди. Равным образом и 5-й (задний) торакаль-
ный сегмент у некоторых видов (Cyclops vicinus, С. scutifer) образует
по бокам сильно развитые заостренные выросты. В виде редкого исклю-
чения (С. bohater) наблюдается особенно сильное развитие задних углов
2-го торакального сегмента, несколько отвисающих кзади и своими
окончаниями отчасти загибающихся на брюшную сторону торакса. У мно-
гих видов (особенно подсемейства Eucyclopinae) 5-й торакальный сег-
мент с боков имеет вооружение из группы коротких тонких щетинок,
изредка имеющихся и по бокам 4-го сегмента. Головогрудь и следую-
щие за ней 3 торакальных сегмента функционально составляют переднюю
2 Фауна СССР
18
ВВЕДЕНИЕ
часть тела, тогда как последний (5-й) торакальный сегмент вместе
с абдоменом и фуркой составляют его заднюю часть.
Абдомен состоит из 5 сегментов, причем у взрослых самок первые
2 сегмента сливаются в 1 крупный генитальный сегмент, так что весь
абдомен состоит из 4 сегментов. У представителей морского рода Euryte
Фиг. 1. Общий вид циклопа (Eucyclops}', 1 — самка; 2—самец. (По Ульянину,
схематизировано).
а—передние антенны; Ь — яйцевые мешки; с—глаз; d—фурка с фуркаль-
ными щетинками; I—цефалоторакс (~ голове со сращенным с нею 1-м
торакальным сегментом);//—V—-2-й, 3-й, 4-н и 5-й сегменты торакса; 7—5 —
сегменты абдомена, из них 1-й — генитальный сегмент.
слияние передних 2 сегментов неполное и у взрослых самок, причем
граница этих сегментов хорошо различима на спинной поверхности
генитального сегмента. У самцов слияния указанных 2 сегментов не проис-
ходит и во взрослом состоянии, т. е. абдомен состоит у них из 5 сег-
ментов. Генитальный сегмент самок расширен в передней своей
части, где по бокам иногда образует более или менее сильно развитые
выросты (особенно у некоторых пресноводных родов). У обоих полов
(у самцов ближе к брюшной поверхности) по бокам генитального сег-
мента имеются особые шиповидные или щетинковидные придатки —
МОРФОЛОГО-АН АТОМИЧЕСКИМ ОЧЕРК
19
рудиментарная VI пара ног (см. ниже). Задние края абдоминальных
сегментов гладкие или зазубренные. На заднем конце заднего абдоми-
нального сегмента имеется прикрывающая сверху анальное отверстие
особая, так называемая анальная пластинка, у некоторых видов (осо-
бенно у Speocyclops и Graeteriella) довольно значительно развитая,
у большинства же видов очень маленькая (фиг. 2).
Фурка состоит из 2 расположенных симметрично ветвей, в основа-
нии окруженных кольцом из очень мелких шипиков, на брюшной поверх-
ности, однако не доходящих до анального отверстия; эти шипики
принадлежат заднему абдоминальному сегменту. У некоторых видов
(Paracyclops poppei) на брюшной стороне ряды этих шипиков заги-
Фиг. 2. Анальный сегмент (со спинной стороны) с анальной пластинкой.
(По Kiefer’y).
1 — Acanthocyclops 6icaspidatus (Claus); 2—A. abyssicola (Lili.); 3—A.
kieferi (Chapp.); 4 — A. reductus (Chapp.); 5 — A. crassicaudis (Sars); 6—A.
penustus (Norm, et Scott).
баются и доходят до основания анальной пластинки. Длина, толщина
н степень расхождения фуркальных ветвей у разных видов очень раз-
личны. Некоторые виды (особенно Eucyclops macrurus) отличаются
очень тонкими, длинными и параллельно поставленными фуркальными
ветвями, у других видов они очень короткие и толстые. Так у рода
Graeteriella их длина самое большее в 1г/2 раза превышает толщину,
а у некоторых Halicyclops их длина не более толщины. Внутренние
края фуркальных ветвей большей частью без вооружения, гладкие.
У некоторых видов (род Cyclops, Масг осу clops fuscus, Acanthocyclops
viridis) эти края вооружены более или менее сплошным рядом тонких
волосков; у рода Eucy clops внешние края ветвей несут нередко сплош-
ной ряд мелких шипиков (наподобие пилы), причем последние могут
частью заходит на спинную поверхность ветвей. У видов рода Cyclops
на этой поверхности имеется продольная, более или менее явственно
выраженная, кутикулярная складка.
2*
20
ВВЕДЕНИЕ
Вооружение ветвей, помимо указанных волосков и шипиков, состоит
из 6 основных придатков, представленных щетинками и шипами. Внеш-
ний край фуркальных ветвей несет боковую щетинку, прикрепляющуюся
обычно близ заднего конца ветви, сравнительно редко близ середины
или на середине края. На спинной поверхности ветви, очень близко
от ее заднего конца, прикрепляется направленная кзади и обычно
несколько кнаружи тонкая спинная щетинка, длина которой у разных
видов очень различна, но сравнительно редко превышает длину внеш-
ней из апикальных щетинок. Остальные 4 щетинки прикрепляются
к заднему концу ветви: 2 близ внешнего и внутреннего угла ветви
(крайние внешняя и внутренняя апикальные щетинки) и 2 между ними
(средние внешняя и внутренняя апикальные щетинки). Последние 2 щетинки,
особенно внутренняя из них, длиннее крайних апикальных щетинок,
из которых внешняя, как правило, короче внутренней и у некоторых
видов (например у Cyclops insignis) преобразована в шип; редко обе
крайние апикальные щетинки шипообразны (например Graeteriella unise-
tigera). Взаимоотношения длины всех апикальных щетинок могут быть
весьма различными. У Gr. unisetigera из срединных апикальных щетинок
имеется лишь 1 мощно развитая щетинка. Апикальные щетинки оперены
с боков рядом тонких волосков, которые могут быть одинаковыми
(гомономное оперение) или несколько отличающимися друг от друга
(гетерономное оперение).
У некоторых видов (виды рода Cyclops, Acanihocyclops bicuspidatus,
A. bisetosus) на внешнем крае фуркальных ветвей, помимо обычной
боковой щетинки, имеется сильно редуцированная, очень мелкая, иногда
с трудом различимая щетинка недалеко от начала края, приблизительно
в передней 1/4—1/3 его длины.
Сегментация тела представителей семейства Cyclopidae, рассмотрен-
ная выше, кратко резюмируется табл. 1 (по Kiefer’y, 1928).
, Таблиир 1
3 основных Морфологически Физиоло гнческн
отдела 11 основных сегментов 11 основных сегментов при их частичном слиянии 2 основных отдела
I. Голова II. Торакс (грудь) Голова 1-й торакальный сегмент 2-й „ „ 3-и п » 4-й „ „ 5-й „ „ Цефалоторакс (головогрудь) 2-й торакальный сегмент 3-й 4-й „ „ 5-й „ Передний отдел тела
III. Абдомен (брюшко) и Фурка 1-й абдоминальный сегмент 2-й 3-й 4-й „ „ 5-й „ „ Фурка Генитальный сегмент 3-й абдоминальный сегмент 3-й 3-й „ „ Фурка Задний отдел тела
МОРФОЛОГО-АНАТОМИЧЕСКИЙ ОЧЕРК
21
На головном отделе (передний отдел цефалоторакса, исключая первый
сегмент торакса) имеется 6 пар конечностей.
Передние антенны (I антенны, антеннулы; фиг. 3) у обоих
полов одноветвистые. У самок они состоят из ряда цилиндрических
члеников различной длины. Число члеников в пределах семейства
Cyclopidae колеблется от 6 до 21. Передние 21-членистые антенны
свойственны лишь морским формам (род Euryte), тогда как у пресно-
водных и солоноватоводных Cyclopidae имеется от 6 до 18 члеников,
причем 18-членистые антенны (образовались из 17-членистых вследствие
Фиг. 3. Передние антенны самки. (По Gurney).
7 — Acanthocyclops viridis (Jur.); 2—Рат асу clops fimbriatus (Fisch.).
более или менее полного разделения 7-го члеиика надвое) составляют
редкое исключение. Общая длина антенны не всегда стоит в непосред-
ственной связи с числом члеников. Так у морского рода Euryte перед-
ние антенны 21-членистые, но очень короткие, едва достигающие сере-
дины цефалоторакса. Сравнительно с Calanoida, у Cyclopoida мы нахо-
дим вообще гораздо более укороченные антеннулы, никогда не превос-
ходящие длины переднего отдела тела, как правило, достигающие
не далее конца 2-го торакального сегмента, иногда же едва достигаю-
щие середины цефалоторакса. Особенно коротки антеннулы у рода
Halicyclops, где они не достигают середины цефалоторакса.
На табл. 2 (по Schmeil’y, 1892) показано взаимоотношение отдель-
ных члеников передних антенн (с различным числом члеников) приме-
нительно к главнейшим родам пресноводных Cyclopidae. Взаимоотношения
эти установлены на основании данных метаморфоза.
22
ВВЕДЕНИЕ
Таблица 2
Взаимоотношение члеников передних антенн главнейших родов
пресноводных Cyclopidae
Paracyclops fimbriatus (8) 1 1 2 3 4 5 6 7 8
Ectocyclops phaleratas (10) 1-2- 3 4 5 6 7 8 9 10
Microcyclops bicolor (11) . 1-2 3- —4— -5 6 7 8 9 10-11
Eucyclops serrulatus (12) . 1-2- —3—4 -5— -6 7 8 9 10 11-12
Cyclops insignis (14) . . . 1-2 3-4 5 - 6- -7 8 9—10—11 12 13-14
Macrocyclops fuscus (17) . 1-2 3-4 5 6 -7 8-9-10-11—12—13—14 15 16-17
Соотношения с морскими родами Euryte и Halicyclops (оба из под-
семейства Halicyclopinae) показаны в табл. 3 (по Gurney, 1933).
Таблица
Соотношение члеников передних антенн пресноводных и мор-
L. ских родов
Euryte . . . . . 1 2 3 4 5 6 7 8 9 10,11,12,13,14,15,16 17, 18 19, 20, 21
Halicyclops (aequoreus) . . . 1 2 3 4 5 6
Cyclops1 2 . . . 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Euryte . . . , . 1 2 3 4, 5, 6 73 9 пГТГ 12 13 14 15 16 17 18 ll 20 21
Вооружение передних антенн представлено оперенными и неопе-
ренными, обычно короткими, щетинками, сензорными придатками в виде
более или менее длинных щетинок с очень нежным тонкостенным
дистальным прозрачным отделом, нередко расширенным (сензорные
колбы), в виде прозрачных тонкостенных цилиндров (сензорные цилиндры),
а также сензорными придатками в виде узких нежных прозрачных
гиалиновых мембран; кроме того, имеются ряды обычных мелких шипи-
ков, иногда (главным образом у 1-го членика) образующих поперечно-
и косорасположенные ряды. Число оперенных и неоперенных придатков
для каждого отдельного членика постоянно. У форм с 17-членистыми
передними антеннами 10-й и 13-й членики щетинок не имеют. У форм
с меньшим числом члеников нет щетинок' у члеников, гомологичных
2 указанным. Большинство придатков прикреплено ближе к внешней
поверхности члеников, но у 3 дистальных члеников щетинки располо-
жены на внутренней, обращенной к телу животного, поверхности. Апи-
кальный членик антеннулы несет группу, обычно состоящую из 7 щетинок
разной длины.
Сензорные придатки, являющиеся видоизмененными щетинками,
у форм с 17-членистыми антеннулами постоянно имеются по 1 на 12-м, 16-м,
и 17-м члениках. У форм с иным числом члеников эти 3 сензорных при-
датка прикреплены к членикам, гомологичным 3 указанным. Гиалиновые
1 В скобках (за видовым названием) показано число члеников I антенны.
2 С 17-членистыми передними антеннами.
МОРФОЛОГО-АН АТОМИЧЕСКИЙ ОЧЕРК
23
мембраны (например у Macrocyclops, Eucyclops, Mesocyclops), в виде
узких, очень прозрачных и тонких, иногда зазубренных пластинок, свой-
ственны внутреннему краю 2 или 3 дистальных члеников антеннулы;
у некоторых видов вместо них вдоль краев этих члеников имеются
ряды часто расположенных, очень тонких и коротких, прозрачных гиа-
линовых шипиков (например у Eucyclops).
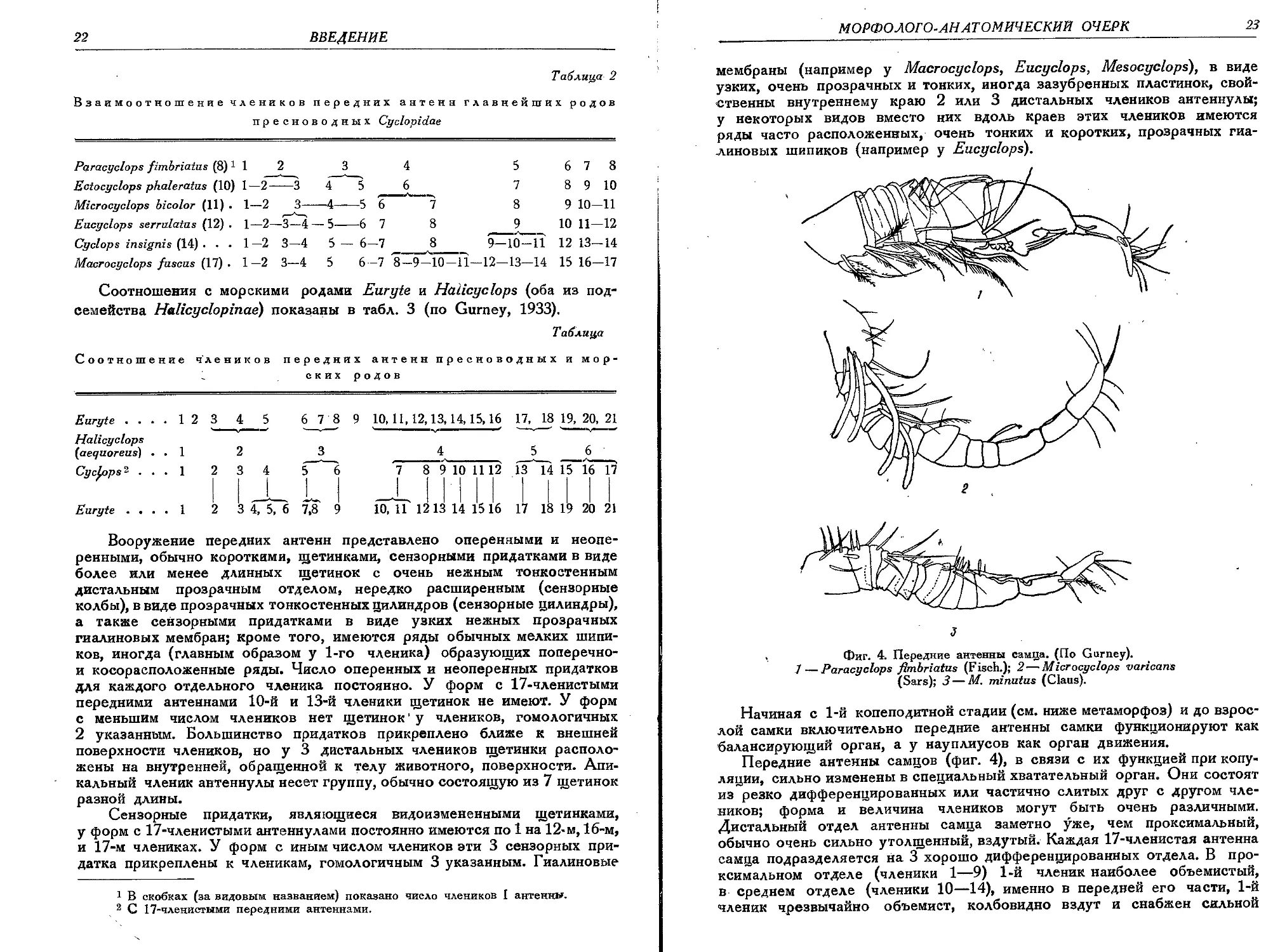
Фиг. 4. Передние антенны самца. (По Gurney).
/ — Paracyclops fimbriatus (F isch.); 2 — Microcyclops varicans
(Sars); 3—M. minutus (Clans).
Начиная с 1-й копеподитной стадии (см. ниже метаморфоз) и до взрос-
лой самки включительно передние антенны самки функционируют как
балансирующий орган, а у науплиусов как орган движения.
Передние антенны самцов (фиг. 4), в связи с их функцией при копу-
ляции, сильно изменены в специальный хватательный орган. Они состоят
из резко дифференцированных или частично слитых друг с другом чле-
ников; форма и величина члеников могут быть очень различными.
Дистальный отдел антенны самца заметно уже, чем проксимальный,
обычно очень сильно утолщенный, вздутый. Каждая 17-членистая антенна
самца подразделяется на 3 хорошо дифференцированных отдела. В про-
ксимальном отделе (членики 1—9) 1-й членик наиболее объемистый,
в среднем отделе (членики 10—14), именно в передней его части, 1-й
членик чрезвычайно объемист, колбовидно вздут и снабжен сильной
24
ВВЕДЕНИЕ
мускулатурой. Дистальный отдел (членики 15—17) антенны состоят
из 3 постепенно суживающихся члеников, из которых передний длин-
нее 2 прочих и имеет сильно хитинизированный утолщенный внешний
край. 2 дистальных членика резко дифференцированы или слиты друг
с другом. Вооружение передних антенн самца состоит из простых щети-
нок различной длины, особенно многочисленных в базальном отделе
антенны, а также из сензорных придатков в виде маленьких коротких
волосков, длинных узких цилиндров и сензорных колб. У самцов под-
семейства Cyclopinae сензорные колбы распределены следующим обра-
зом: 3 на 1-м членике, по 1 на 4-м, 9-м и 15-м члениках. У самцов
подсемейства Eucyclopinae проксимальный отдел антенны вместо сен-
зорных колб несет определенное число длинных сензорных цилиндров.
На табл. 4 показано (по Gurney, 1933) расположение сензорных при-
датков1 на передних антеннах самцов Cyclops, Eucyclops и Macrocy-
cle ps.
Таблица 4
Сегменты
Cyclops
Eucyclops
Macrocyclops
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
А, А, А
А
А
А
А
F
С, С, F
С
С
С, F
С
С, С, F
С
С
С, F
С
С
С
С, F
А
А
F
А
А
F
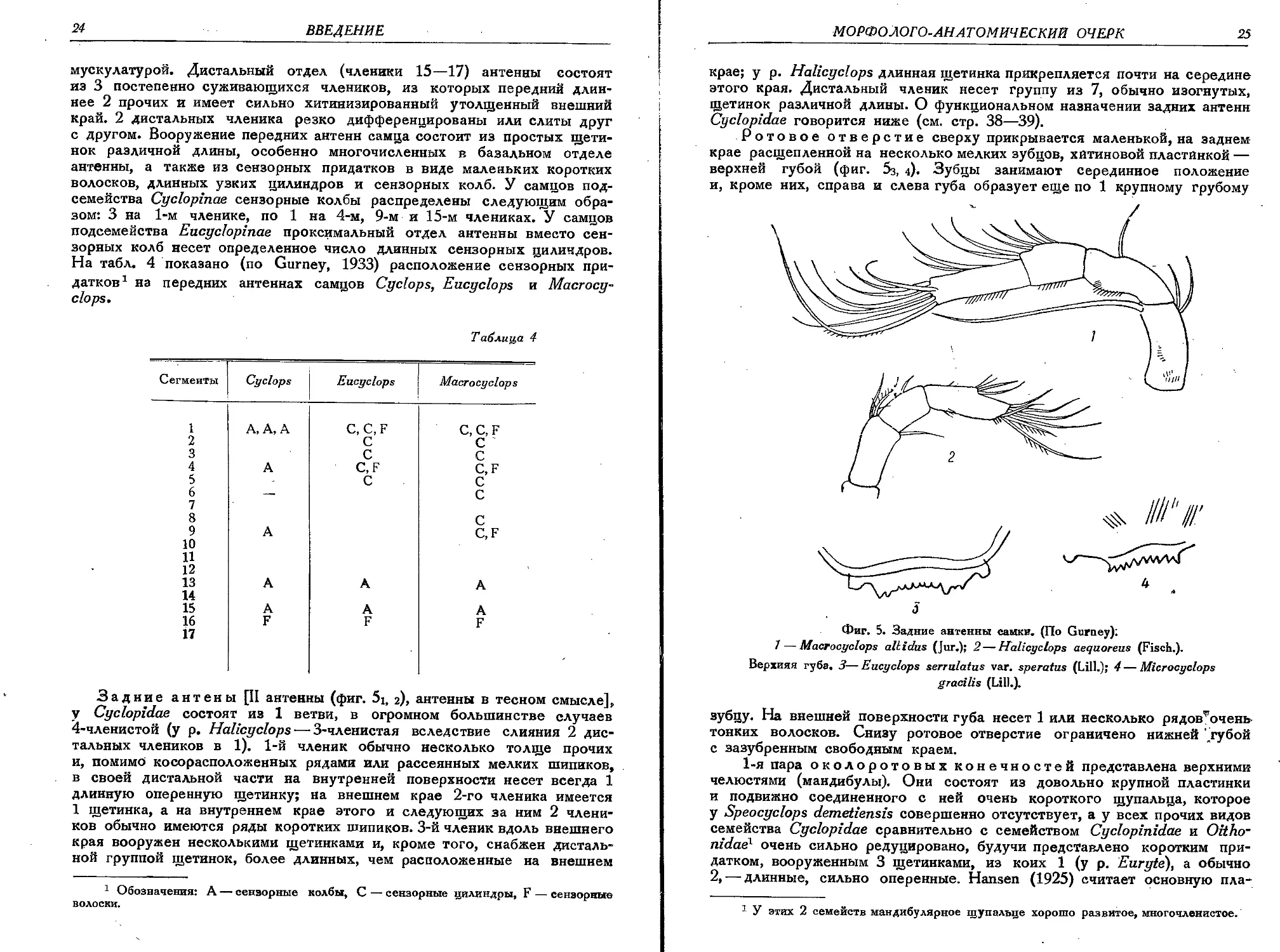
Задние антены [II антенны (фиг. 5j, 2), антенны в тесном смысле],
у Cyclopidae состоят из 1 ветви, в огромном большинстве случаев
4-членистой (у р. Halicyclops — 3-членистая вследствие слияния 2 дис-
тальных члеников в 1). 1-й членик обычно несколько толще прочих
и, помимо косорасположенных рядами или рассеянных мелких шипиков,
в своей дистальной части на внутренней поверхности несет всегда 1
длинную оперенную щетинку; на внешнем крае 2-го членика имеется
1 щетинка, а на внутреннем крае этого и следующих за ним 2 члени-
ков обычно имеются ряды коротких шипиков. 3-й членик вдоль внешнего
края вооружен несколькими щетинками и, кроме того, снабжен дисталь-
ной группой щетинок, более длинных, чем расположенные на внешнем
1 Обозначения: А — сензорные колбы, С — сензорные цилиндры, F — сензорные
волоски.
МОРФОЛОГО-AH АТОМИЧЕСКИЙ ОЧЕРК
25
крае; у р. Halicyclops длинная щетинка прикрепляется почти на середине
этого края. Дистальный членик несет группу из 7, обычно изогнутых,
щетинок различной длины. О функциональном назначении задних антенн
Cyclopidae говорится ниже (см. стр. 38—39).
Ротовое отверстие сверху прикрывается маленькой, на заднем
крае расщепленной на несколько мелких зубцов, хитиновой пластинкой —
верхней губой (фиг. 5з, 4). Зубцы занимают серединное положение
и, кроме них, справа и слева губа образует еще по 1 крупному грубому
J
Фиг. 5. Задние антенны самки. (По Gurney);
7 — Macrocyclops allidus (Jur.); 2—Halicyclops aequoreus (Fisch.).
Верхняя губа. 3—Eucyclops serrulatus var. speratus (Lili.); 4 — Microcyclops
gracilis (Lili.).
зубцу. На внешней поверхности губа несет 1 или несколько рядов^очень
тонких волосков. Снизу ротовое отверстие ограничено нижней ' губой
с зазубренным свободным краем.
1-я пара околоротовых конечностей представлена верхними
челюстями (мандибулы). Они состоят из довольно крупной пластинки
и подвижно соединенного с ней очень короткого щупальца, которое
у Speocyclops demetiensis совершенно отсутствует, а у всех прочих видов
семейства Cyclopidae сравнительно с семейством Cyclopinidae и Oitho-
nidae1 очень сильно редуцировано, будучи представлено коротким при-
датком, вооруженным 3 щетинками, из коих 1 (у р. Euryte\ а обычно
2, — длинные, сильно оперенные. Hansen (1925) считает основную пла-
1 У этих 2 семейств мандибулярное щупальце хорошо развитое, многоиленистое.
26
ВВЕДЕНИЕ
стинку мандибул прекоксальным члеником, а щупальце—:коксальным,
тогда как Gurney (1931) полагает, что основная часто соответствует
коксе, при полном отсутствии прекоксы. Спереди основная пластинчатая
часть мандибул образует ряд довольно крупных заостренных зубцов,
служащих для размельчения захватываемой пищи; передний из них обычно
заметно крупнее остальных. На противоположном (заднем) конце сво-
бодного края пластинки, близ заднего зубца, имеется 1 короткая опе-
ренная щетинка» возможно, сензорного характера.
2-я пара околоротовых конечностей (фиг. б2) представлена
1-й парой нижних челюстей (передняя, верхняя, внутренняя максилла,
максиллула). Эти челюсти состоят из основной широкой пластинчатой
части (гнатобаза), на переднем (внутреннем) крае снабженной несколь-
кими крупными коническими хитиновыми зубцами, и из одно-членистой
придаточной части (щупальца), подвижно сочлененной с основной. Прида-
Фиг. 6. (По Gurney).
1 — мандибула Maarocyclops distinctus (Rich.); 2—максиллула М. fuscus (Jur.);
3—максиллярная нога М. fuscus (Jur.); 4 — максилла' М. fuscus (Jur.).
точная часть вооружена несколькими щетинками и у некоторых видов
(С. strenuus) образует 2 коротких выроста (лопасти), несущих 4 щетинки.
У некоторых видов (Mesocyclops leuckarti) имеется 1 такой вырост;
рассматриваемые выросты представляют рудименты экзо- и эндоподита.
Свободный конец придаточной части у рода Euryte зазубрен и несет
щетинки только в проксимальном отделе. У некоторых видов (Paracyclops
fimbriatus) щупальце явственно 2-членистое, у большинства видов оно
1-членистое.
3-я пара околоротовых конечностей (фиг. 64) представлена
2-й парой нижних челюстей (= задняя, нижняя, наружная максилла).
Каждая из этих челюстей состоит из 4 явственно дифференцированных
члеников; 5-й членик редуцирован до степени маленького выроста. Первые
2 членика (протоподит) крупнее прочих, из них передний (коксоподит,
кокса) имеет внутренний отросток с 2 оперенными щетинками, а задний
(базиподит, базис) на внутреннем крае несет 1 щетинку и близ конца,
МОРФОЛОГО-АН АТОМИЧЕСКИЙ ОЧЕРК
27
на особом удлиненном отростке, несет другую щетинку, в основании
которой отросток продолжается в длинный шиповидный придаток,
у некоторых форм (род Euryte) неотчлененный от самого отростка,
у большинства же вполне от него отчлененный и образующий утолщенную
шипообразную щетинку. У некоторых видов (Ectocyclops phaleratus) оба
членика протоподита очень короткие и широкие, причем базиподит
снабжен поперечным рядом сильных шипов. Следующие за протоподи-
том 2 членика представляют собой эндоподит. Из них наиболее развит
1-й членик, внутри образующий клювообразно изогнутый вырост, близ
основания которого прикрепляется толстая оперенная щетинка, а в самом
основании имеется, повидимому постоянно, еще 1 очень короткая
и тонкая щетинка. Конечный членик вндоподита вооружен 4—5 щетин-
ками, из которых 2—3 особенно крупные.
4-я пара околоротовых конечностей (фиг. 63) представлена
челюстными (максиллярными) ногами. Как правило, эти ноги состоят
из 4 члеников. У Cyclops первые 2 членика (коксоподит и базиподит),
особенно коксоподит, наиболее объемистые. Коксоподит с внутренней
стороны вооружен 3 оперенными щетинками (у Acanthocyclops sensitivus
и у видов рода Euryte — 2), а базиподит —2 оперенными щетинками (у рода
Euryte лишь 1). За протоподитом следует 2 (или 1 —Ectocyclops phaleratus,
Paracyclops affinis) членика эндоподита. Из них 1-й снабжен внутри
1 щетинкой (близ конца членика), а 2-й несет 3 щетинки, из которых
внешняя маленькая и тонкая, а 2 прочие длинные. Единственный членик
эндоподита Ect. phaleratus и Par. affinis снабжен сильным, заостренным,
изогнутым выростом н 3 щетинками. У Graeteriella unisetigera, ведущей
подземный образ жизни, вся максиллярная нога несет лишь 4 щетинки.
У Speocyclops demetiensis максиллярная нога 3-членистая, снабженная
5 щетинками (дистальный членик несет 3 щетинки). У рода Halicyclops
максиллярная нога состоит лишь из 2 члеников, вероятно представляю-
щих эндоподит; из них 1-й членик особенно удлиненный. Максиллярная
нога рода Euryte 4-членистая, с изогнутым внутрь дистальным отделом
(эндоподит), причем конечный членик несет пару когтевидных придатков,
и вся конечность имеет хватательный тип.
4 пары следующих за околоротовыми грудных конечностей
носят название плавательных ног. Они прикрепляются к брюшной поверх-
ности I, II, III и IV торакального сегмента и построены по типу двувет-
вистой конечности, одинаково (или почти одинаково) у обоих полов.
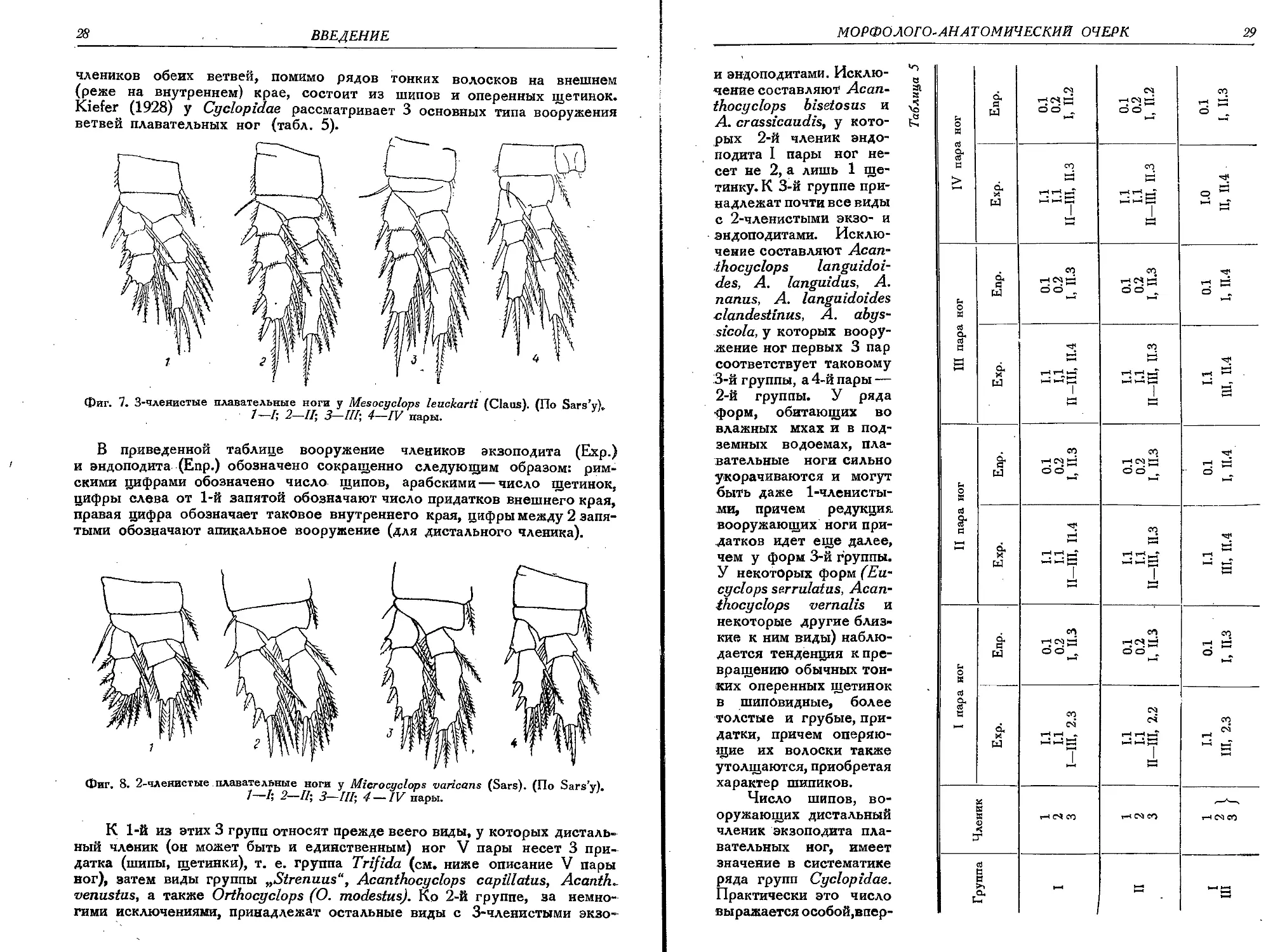
Каждая плавательная нога (фиг. 7—9) состоит из 2 широких основ-
ных члеников (коксоподит и базиподит) и из 2 ветвей — внешней (экзо-
подит) и внутренней (эндоподит). Прекоксальный членик имеется лишь
в очень слабо выраженном состоянии (Cyclops).
Коксоподит на внутренне-заднем углу обычно несет крупную опе-
ренную щетинку, направленную кзади. Базиподит, как правило, несет
тонкую короткую щетинку на внешнем крае, а у ног первой пары близ
внутренне-заднего угла несет или оперенный шип, или оперенную щетинку.
Коксоподиты ног правой и левой стороны каждой пары внутри соеди-
няются особой тонкой соединительной пластинкой, причем для некото-
рых Cyclopidae (например род Mesocyclops) строение и вооружение
свободного края этой пластинки у ног IV пары имеют существенное
систематическое значение. Каждая из 2 ветвей плавательных ног состоит
не более как из 3 члеников (фиг. 7), но может быть 2-х (фиг. 8) и даже
1-членистой. Иногда сокращенное число члеников наблюдается только
у эндоподита, тогда как экзоподит остается 3-членистым. Вооружение
28
ВВЕДЕНИЕ
члеников обеих ветвей, помимо рядов тонких волосков на внешнем
(реже на внутреннем) крае, состоит из шипов и оперенных щетинок.
Kiefer (1928) у Cyclopidae рассматривает 3 основных типа вооружения
ветвей плавательных ног (табл. 5).
Фиг. 7. 3-членистые плавательные ноги у Mesocyclops leuckarti (Claus). (По Sars’v).
7-7; 2-ZZ; 3-III; 4—IV пары.
В приведенной таблице вооружение члеников экзоподита (Ехр.)
и эндоподита (Епр.) обозначено сокращенно следующим образом: рим-
скими цифрами обозначено число щипов, арабскими — число щетинок,
цифры слева от 1-й запятой обозначают число придатков внешнего края,
правая цифра обозначает таковое внутреннего края, цифры между 2 запя-
тыми обозначают апикальное вооружение (для дистального членика).
Фиг. 8. 2-членистые плавательные ноги у Microcuclops varicans (Sars). (По Sars’v).
1—h 2—II; 3—HI; 4 — IV пары.
К 1-й из этих 3 групп относят прежде всего виды, у которых дисталь-
ный членик (он может быть и единственным) ног V пары несет 3 при-
датка (шипы, щетинки), т. е. группа Trifida (см. ниже описание V пары
ног), затем виды группы „Strenuus", Acanthocyclops capillatus, Acanthi
venustus, а также Orthocyclops (O. modestus). Ко 2-й группе, за немно-
гими исключениями, принадлежат остальные виды с 3-членистыми экзо-
МОРФОЛОГО-АНАТОМИЧЕСКИЙ ОЧЕРК
29
и эндоподитами. Исклю-
чение составляют Асап- ®
thocyclops bisetosus и
A. crassicaudis, у кото- й
рых 2-й членик эндо-
подита I пары ног не-
сет не 2, а лишь 1 ще-
тинку. К 3-й группе при-
надлежат почти все виды
с 2-членистыми экзо- и
эндоподитами. Исклю-
чение составляют Acan-
thocyclops languidoi-
des, A. languidus, А.
nanus, A. languidoides
clandestinus, A. abys-
sicola, у которых воору-
жение ног первых 3 пар
соответствует таковому
3-й группы, а 4-й пары —
2-й группы. У ряда
форм, обитающих во
влажных мхах и в под-
земных водоемах, пла-
вательные ноги сильно
укорачиваются и могут
быть даже 1-членисты-
ми, причем редукция,
вооружающих ноги при-
датков идет еще далее,
чем у форм 3-й группы.
У некоторых форм (Еи-
cyclops serrulatus, Acan-
thocyclops vernalis и
некоторые другие близ-
кие к ним виды) наблю-
дается тенденция к пре-
вращению обычных тон-
ких оперенных щетинок
в шиповидные, более
толстые и грубые, при-
датки, причем оперяю-
щие их волоски также
утолщаются, приобретая
характер шипиков.
Число шипов, во-
оружающих дистальный
членик экзоподита пла-
вательных ног, имеет
значение в систематике
ряда групп Cyclopidae.
Практически это число
выражается особой,впер-
IV пара ног Епр. 0.1 0.2 I, II.2 0.1 0.2 I, П.2 S4I ‘I го
Ехр. и 1.1 II—III, П.З 1.1 1.1 П-Ш, П.3 1.0 II, П.4
JOH Епр. 0.1 0.2 I, П.З 0.1 0.2 I, П.3 0.1 I, П.4
III пара Ехр. 1.1 1,1 II-III, П.4 1.1 1.1 П-Ш, П.З 1.1 III, П.4
НОГ Епр. 0.1 0.2 I, И.З 0.1 0.2 I, П.З 0.1 I, П.4 1
II пара Ехр. 1.1 1.1 П-Ш, 11.4 1.1 1.1 П-Ш, П.3 1.1 III, П.4
НОГ Епр. 0.1 0.2 I, П.З 0.1 0.2 I, П.З СО О *”1 нм
I пара Ехр. 1.1 1.1 I—III, 2.3 1.1 1.1 П-Ш, 2.2 1.1 III, 2.3
Членик г-4 еч со t-M CSJ со г-4 (М СО
1руппа нм IM
30
ВВЕДЕНИЕ
вые предложенной Vosseler’oM (1836), „формулой шипов" (Dornformel).
Так формула 2’3"3-3 показывает, что дистальный членик экзоподита ног
I пары ног вооружен 2, а остальных 3 ног — 3 шипами. Формула З^^’З-
(например Paracyclops fimbriatus) показывает, что дистальные членики
экзоподита I и IV пар ног снабжены 3, а II и IV пар ног—4 шипами.
Число щетинок в формуле шипов не обозначается, оно выражается
отдельной „формулой щетинок" (Borstenformel). Так формула щетинок
4.4.4.4 (например у некоторых видов рода Acanthocyclops) показывает,
что дистальные членики экзоподита всех 4 пар плавательных ног несут
по 4 щетинки.
По числу шипов дистального членика экзоподита 2 первых пар
плавательных ног различают 2 основных типа вооружения: тип Bini
(формула шипов 2-3-3-3) и тип Terni (формула шнпов 3-4-3-3). Особое
Фиг. 9. Нога IV пары. (Но Gurney).
1 — Eucyclops macrarus (Sars); 2 — Acanthocyclops vemalis (Fisch.); 3 — Microcyclops
bicolor (Sars); 4 — M. minutus (Claus).
значение эти 2 типа вооружения имеют для систематики группы Stre-
nuus, где, однако, нередко встречается и смешанный тип вооружения
(подробнее см. ниже, в описании рода Cyclops).
Для систематики некоторых групп Cyclopidae далее имеет значение
строение и апикальное вооружение (а равно и относительные размеры)
дистального члени.ка эидоподита IV пары ног (фиг. 9). Оно состоит из
2 шипов, обычно разной длины, редко 1 из них заменяется щетинкой
или вообще имеется лишь один апикальный шнп {Microcyclops minutus).
Для некоторых групп (род Mesocyclops) принимается во внимание
относительная длина обоих апикальных шипов, внешнего и внутреннего.
Плавательные ноги служат главным органом передвижения животного,
функционируя наподобие весел. При ударе ног кзади щетинки внутрен-
него края экзоподита располагаются перед эндоподитом той же ноги.
Образующимся противодействующим давлением воды эти щетинки тесно
прижимаются к эндоподиту, таким образом образуя поверхность, функ-
ционирующую как весло. Обе ноги каждой отдельной пары, вследствие
их прочной связи соединительной пластинкой, двигаются одновременно
и однотипно. При быстром ударе плавательных ног рачок делает быст-
МОРФОЛОГО-АН АТОМИЧЕСКИЙ ОЧЕРК
31
рое скачкообразное движение, постоянно меняя положение тела, — брюш-
ной поверхностью то вверх, то вниз, то занимая косое или вертикаль-
ное положение. Эти порывистые скачкообразные движения крайне харак-
терны для Cyclopidae. Плавание циклопов весьма образно описывается
Исаевым (1934): „передвигаясь ритмическими скачками, циклоп может
хорошо держаться на одном уровне, подниматься вверх и опускаться
вниз под углами различной крутизны. Циклоп может плавать с одинако-
вой легкостью, перевернувшись на спину. Циклоп хорошо описывает
дуги, делает мертвые петли, ординарные и множественные, прямые
и обратные. Циклоп может делать поворот под углом 90°, вращаться
вокруг оси не только со снижением, напоминающим витки «штопора»
самолета, но и с поднятием вверх. Циклоп может скользить на антенну,
делать через нее переворот, пикировать вниз головой под углом в 90°
и скользить на хвост. Характер «фигур», выполняемых циклопом, очень
сходен с фигурами высшего пилотажа. Обладание фигурами высшего
пилотажа, необходимыми для самолетов-истребителей, несомненно, облег-
чает циклопу — активному хищнику—возможность обеспечить себе суще-
ствование охотой за водными обитателями, служащими ему пищей".
При остановке работы плавательных ног рачок начинает погружаться,,
и тело из первоначального, приблизительно горизонтального, положения
постепенно меняет последнее на косое, смещаясь цефалотораксом все
ниже и ниже, в конце концов принимая почти вертикальное положение,
головой вниз. Затем следуют новые удары плавательных ног, и живот-
ное поднимается опять кверху. В фазе погружения передние антенны
простираются приблизительно перпендикулярно к длинной оси рачка.
Следующая за плавательными V пара ног прикрепляется к брюш-
ной поверхности заднего отдела 5-го торакального сегмента. Рассматри-
ваемая пара ног рудиментарна и имеет совершенно иное, чем плаватель-
ные ноги, строение, одинаковое у обоих полов. Нога V пары Cyclopoida
(фиг. 10) состоит обычно из 1 или 2 члеников, очень редко она 3-члени-
стая {Orthocy clops, морской род Euryte). У некоторых видов говорить
хотя бы об 1 членике этой ноги, в сущности, возможно лишь условно,,
так как последняя представляет собой очень узкую маленькую пластинку,
не отчлененную от последнего торакального сегмента. Основной членик
(базиподит) 2-членистой ноги V пары на внешнем угле несет всего 1,
большей частью оперенную щетинку. У рода Euryte, где V пара 3-чле-
нистая, этому членику соответствует 2-й членик, тогда как 1-й является
коксоподитом. 2-й членик 2-членистой ноги, гомология которого окон-
чательно не выяснена (эндо- или экзоподит?), у пресноводных Cyclopidae
снабжен 2 или 3 придатками, иногда только 1 придатком. У морского
подсемейства Halicyclopinae (рода Euryte, Halicyclops) гомолог этого
членика имеет 4 придатка (дистальный членик у Euryte, единственный
членик V ноги у Halicyclops). Комбинация придатков, вооружающих
дистальный членик V ноги, у разных родов и подродов Cyclopidae
может быть очень разнообразной. У видов с 3 придатками (подсемейство
Eucyclopinae) или оба крайних шиповидны, а срединный щетинковидный
(Macrocyclops), или только внутренний является шипом, а остальные
2 — щетинками {Eucyclops)'. одна щетинка на конце членика (апикальная),
другая—на его внешнем крае (латеральная). A. Graeter (1903) всех
Cyclopidae с 3 придатками на дистальном членике V ноги выделяет
в группу Trifida ( — подсемейства Eucyclopinae в смысле Kiefer),
а с 2 придатками — в группу Bifida. У последних апикальный придаток
представлен щетинкой, а другой придаток, прикрепленный внутри рядом:
32
ВВЕДЕНИЕ
с этой щетинкой, является шипом, иногда очень похожим на щетинку
(щетинко видный шип; например, у Mesocyclops). A. Graeter считает
этот 2-н придаток настоящей щетинкой, выделяя особую подгруппу Bifida-
Chaetophora (= Mesocyclops s. lat.), характеризующуюся присутствием
2 „щетинок" на конце дистального членика ног V группы, тогда как
подгруппа Bifida Acanthophora объединяет формы, у которых этот чле-
ник вооружен шипом и щетинкой. Едва ли однако можно оспаривать
соображения Kiefer‘а (1928), по которым оба придатка у Chaetophora
Фиг. 10. (По Gurney).
7— ноги V и VI самца Paracyclops fimbriatus (Fisch.); 2—то же Eucy clops serrulatus
(Fisch.); 3—то же Cyclops abyssorum (Sars); 4—то же Mesocyclops leuckarti (Claus);
5—то же Microcyclops minutus (Claus); 6 — генитальные придатки (нога VI) самки —
Cyclops abyssorum (Sars); 7—то же Acanthocyclops viridis (Jur.); 8—то же самца
M icrocyclops varicans (Sars); 9 — ноги V и VI самца Acanthocyclops viridis (Jur.).
(например у Mesocyclops leuckarti) достаточно дифференцированы, при-
чем внутренний из них не имеет структуры нормальной щетинки и соот-
ветствует шипу прочих представителей группы Bifida. Очень часто рас-
сматриваемый придаток заметно короче апикальной щетинки или весьма
сильно редуцирован до степени мелкого шипика (например у Acanthocy-
clops viridis). В редких случаях он совершенно редуцируется, и можно
рассмотреть лишь соответствующий ему пункт прикрепления (например
у Microcyclops varicans var. rubellus), так что дистальный членик ока-
зывается вооруженным лишь 1 апикальной щетинкой. Взаимное располо-
жение шипа и щетинки может быть очень различно. В некоторых случаях
МОРФОЛОГО-AH АТОМИЧЕСКИМ ОЧЕРК 33
шип прикреплен весьма близко от щетинки, занимая почти апикальное
положение (например у Acanthocyclops bicuspidatus, Microcyclops anceps),
в других же случаях он прикреплен почти на середине внутреннего края
членика, занимая, таким образом, латеральное положение (например
у Cyclops strenuus, Acanthocyclops viridis). Вполне точное апикальное
положение шипа однако представляет редкий случай (у некоторых видов
подрода 7h.erm.ocyclops).
У ног с 4 придатками дистального членика (роды Euryte, Halicy-
clops) 3 из них (2 внешних и внутренний) являются шипами, а 1 (2-й,
считая с внутренней стороны) — щетинкой. Если базиподит 2-членистой
ноги V пары совершенно сливается с торакальным сегментом, так что от
этого членика остаются лишь следы или он вполне исчезает, то гомоло-
гичная ему внешняя щетинка принадлежит уже. торакальному сегменту
(например у Microcyclops varicans var. rubellus). По бокам заднего
отдела генитального сегмента обеих полов, у самцов несколько ближе
к брюшной поверхности, имеется рудимент ног VI пары, построенных
по типу сильно редуцированных ног предыдущей пары (особенно значи-
тельна редукция VI пары ног самок). У взрослых самок каждая нога
VI пары (фиг. 106,7) состоит из очень короткого и маленького выроста
кутикулы, а у самцов она в виде маленькой неотчлененной от сегмента
пластинки (фиг. 101_5,8_9). Вооружение рассматриваемой ноги у самок
представлено большей частью 2 шипами и 1 щетинкой, у самцов — 1
шипом и 2 щетинками, причем эти придатки особенно сильно развиты
у самцов. В редких случаях (например у самок Graeteriella unisetigera,
у самцов Microcyclops minutus) имеется только 2 придатка. Основной
отдел ноги у самок прикрывает выводные отверстия половых ор-
ганов.
Тело всех пресноводных Cyclopidae покрыто хитинозной кути-
кулой, которая в области переднего и заднего отдела пищеваритель-
ного аппарата (рот, глотка, передний и задний отдел кишки) заворачи-
вается внутрь, выстилая внутренний просвет этого отдела тонким слоем.
В местах сочленения сегментов тела кутикула значительно тоньше
в виде тонкой эластичной мембраны. Покровы особенно утолщены не
брюшной стороне тела (исключая задний торакальный сегмент), где
образуют прочную опору для прикрепления конечностей и мускулатуры.
Кутикула утолщена также в пластинке мандибул и частью в других
конечностях. Под кутикулой лежит гиподермальный слой, состоящий
из полигональных, главным образом гексагональных, клеток с овальными
ядрами. Среди клеток гиподермы особенного внимания заслуживают
так называемые Ришаровские железы, представляющие собою изменен-
ные овальные, сильно вакуолизированные гиподермальные клетки, снаб-
женные округлыми ядрами и открывающиеся наружу особыми порами
в кутикуле. Железы расположены главным образом группами, по 2—-3
в каждой, но имеются и одиночные железы. Они округлой или груше-
видной формы, снабжены эксцентрично расположенным крупным ядром,
1 или несколькими вакуолями и зернистым содержимым. Кожные железы
многочисленны в покровах торакса, где Они имеют главным образом
форму округлых пузырьков. В области покровов абдомена и в конеч-
ностях, включая и рудиментарную пару ног, они овальной, более или
менее вытянутой вдоль члеников формы. Фуркальные ветви снабжены
обычно 2 парами кожчых желез. Кожные железы выделяют клейкий
секрет. Секрет кожных желез играет крупную роль в образовании
покоящихся состояний циклопов.
3 Фаувл СССР
34
ВВЕДЕНИЕ
Мускулатура Cyclopidae до настоящего времени изучена очень
неполно; наиболее подробо она рассматривается лишь в старой работе
Hartog’a (1888). Мускулы Cyclopidae имеют резко выраженную попереч-
ную полосатость и к внутренней поверхности кутикулы, в месте при-
крепления обычно несколько утолщенной, прикрепляются с помощью
тонких эластичных сухожилий. Мускулатура тела представлена, главным
образом, продольными, парно расположенными мускулами, обслуживаю-
щими движение отдельных сегментов относительно друг друга, сгиба-
ние и разгибание циклопа. Сокращение этих мускулов проходит весьма
быстро; в связи с этим стоят необычайно быстрые и энергичные дви-
жения тела рачка в различных направлениях.
В тораксе, справа и слева от срединной линии, в косом направле-
нии и расходясь кзади, симметрично расположены большие парные
экстензоры. Они берут начало от боковых участков передней части
стенок цефалоторакса и идут к переднему отделу 3-го и 4-го торакаль-
ных сегментов. Пара больших флексоров начинается от пост-максилляр-
ной области и до 4-го торакального сегмента, продолжаясь и в абдомен.
Ближе к серединной линии имеются парные группы мелких V-образных
экстензоров, вершиной направленные кзади, в области 2—4 торакальных
сегментов. Большие продольные мускулы имеют точки прикрепления
в пределах каждого сегмента. В абдомене имеется 4 группы мускулов:
2 супра-латеральных (экстензоры) и 2 инферо-латеральных; у последней
группы нижние мускулы являются флексорами. По Hartog’y эти мускулы
в фуркальные ветви не заходят. Однако Claus (1858) упоминает о муску-
лах, из абдомена вступающих в фуркальные ветви и дающих тонкие
мускульные нити к фуркальным щетинкам. По Hartog’y в торакальных
сегментах, кроме упомянутых выше, имеются короткие плевральные
мускулы, соединяющие стенки „плевры", т. е. складок покровов по
бокам торакальных сегментов, на местах их границ относительно друг
Друга.
Конечности снабжены, в основном, 2 типами мускулов — внешними
и внутренними. Первые мускулы обслуживают конечность снаружи, одним
концом прикрепляясь к покровам тела, 2-ые лежат в пределах самой конеч-
ности. Верхняя губа снабжена 6 констрикторами, располагающимися
в 2 группы, сверху и снизу, по 3 мускула в каждой. Спереди губы,
кроме того, имеются леваторы. В антеннулах самки различают группу
из 3 коротких экстензоров, длинный, прикрепленный к основанию
17-го членика, экстензор и тянущийся от 1-го до начала 17-го членика
длинный флексор. В антеннулах самцов основной отдел обслуживает так
называемая базальная группа мускулов, состоящая из короткого и
длинного экстензора и короткого флексора. Далее у самцов различают
группу мускулов, обслуживающих 9-й — 14-й членики антенну л, затем
крупный толстый флексор среднего подвижного отдела антеннул,
берущий начало в 9-м и заканчивающийся в 15-м членике, и флексор
дистального отдела, начинающийся в основании 16-го и заканчива-
ющийся в 17-м членике. 2-й паре антенн свойственны: флексор
2-го членика, экстензор 3-го членика, мускул, берущий начало
в середине базального членика и заканчивающийся в дистальной
части 3-го членика, толстый мощный мускул, берущий начало рядом
с основанием предыдущего и заканчивающийся в конце дистального
членика антенны. Рассматриваемые антенны снабжены кроме того 5 внеш-
ними экстензорами и флексорами, прикрепляющимися к внутренней
поверхности кутикулярных покровов цефалоторакса. Наружную муску-
Морфолого-анатомический очерк
35
латуру имеют также мандибулы, снабженные парой аддукторов и 3 абдук-
торами. Внутри максиллул находятся мощный флексор и экстензор
дистального членика и пара маленьких мускулов в основании подвижной
придаточной ветви; снаружи максиллулы обслуживаются 7 мускулами,
в том числе 3 абдукторами и парой аддукторов. Внутри задних максилл
находятся 3 пары экстензоров и 3 пары флексоров, а снаружи эти
максиллы снабжены длинным аддуктором, парой коротких аддукторов
и 1 экстензором-абдуктором. Внутренняя мускулатура максиллярных
ног состоит из мускула-леватора и выроста 1-го членика и из 3 экстен-
зоров и 3 флексоров, попарно расположенных в следующих 3 члениках,
а снаружи эти конечности обслуживаются парой аддукторов и 1 абдук-
тором-экстензором.
Плавательные ноги имеют весьма сложную мускулатуру, распреде-
ляющуюся следующим образом.
Базиподит. 3 экстензора — внутренний (аддуктор), внешний
(абдуктор) и средний (вероятно аддуктор); они начинаются в основании
коксоподита и прикрепляются к последнему отделу базиподита; 3 силь-
ных, косорасположенных флексора-абдуктора. От внутреннего края
базиподита к внутреннему углу основной части 1-го членика экзоподита
идет аддуктор экзоподита, а от проксимального угла внутреннего края
базиподита идет косорасположенный абдуктор экзоподита, прикрепляю-
щийся другим концом к внешнему углу основания экзоподита.
Эндоподит. Пара аддукторов, идущих от основания базиподита
до основания 2-го и 3-го члеников эндоподита, к которым и прикреп-
ляется близ внешних углов. От основания базиподита до внешнего угла
основания 2-го членика эндоподита идет абдуктор этого членика.
Экзоподит. 2 аддуктора, соединяющих внешние углы основной
части 1-го членика и внутренние углы этой части 2-го и 3-го члеников.
У ног V пары, у Cyclopidae сильно редуцированных, имеются лишь
внешние мускулы — экстензор и флексор, прикрепляющиеся во внутрен-
ней поверхности покровов заднего торакального сегмента, справа
и слева.
Рот спереди снабжен констрикторами, сзади — аперторами. Глотка
обслуживается передними и задними мускулами-диварикаторами и, кроме
того, так называемыми угловыми мускулами, расположенными между
основанием рострального отдела головы и основанием антеннул.
Кишечник снабжен следующими внешними мускулами.1 Спереди —
2 передними леваторами (протракторами), идущими от верхней стенки
желудочного отдела до срединной линии спинных покровов. Кзади сле-
дует пара задних леваторов, начинающихся в месте перехода желудоч-
ного отдела в собственно кишку, до срединной линии передней части
3-го торакального сегмента. От нижней (брюшной) поверхности кишки
отходит 2 пары расходящихся депрессоров, направленных косо — 2 кпе-
реди и 2 кзади. Задний отдел кишечника, прикрепленный к внутренней
поверхности покровов 2 латеральными парами мезентериев, обслужи-
вается внешней мускулатурой только сзади, где расположены парные,
очень короткие медианные и латеральные мускулы (ретракторы и про-
тракторы). Передняя медианная пара свободными концами прикрепляется
к местам соединения 2-го и 3-го абдоминального сегментов. Задняя
медианная пара идет от передней стенки 4-го абдоминального сегмента
к верхней стенке ректума, прикрепляясь перед анальным отверстием
1 О его мускулатуре см. также нике, стр. 37 и след.
3*
36
ВВЕДЕНИЕ
(ретракторы и дилятаторы этого отверстия). Латеральные мускулы
имеют поперечное расположение и прикрепляются к вентральной поверх-
ности кишки и к стенке 4-го абдоминального сегмента (передние ретрак-
торы и протракторы); кроме того в области анального отверстия имеется
2 мускула апертора, латеральный и вентральный. Имеется маленькая
связка мускулов, обслуживающая специально анальную пластинку (Vos-
seler, 1886).
Пищеварительный аппарат (фиг. 11 наиболее тщательно
у Cyclopidae изучен Farkas’oM (1923). Кишечник Cyclopidae лишен при-
даточных желез. В области верхней губы имеются крупные парные
„слюнные" железы, функция которых не выяснена. Gicklhorn (1930)
показал, что они гомологичны кожным железам. По Richard’y (1891)
у Acanthocyclops viridis они одноклеточны, расположены по бокам губы
и с каждой стороны представлены 3 группами желез (всего их 8), впа-
Фиг. 11. Продольный раврез самки Acanthocyclops viridis (Jur.). (По Farkas’у).
а — ротовое отверстие; Ъ — ротовая полость (атриум); с—верхняя губа; d— глотка;
е —пищевод; /—зитинозный отдел желудка; g—клеточный отдел желудка; Л — тонкая
кишка: i — толстая кишка; к — сфинктер толстой кишки; I — прямая кишка; т — фурка
дающих в общую ампулу, которая особым коротким каналом открывается
наружу. По Farkas'y железы эпистома пронизаны особой капиллярной
сетью канальцев и секрет выделяется через них.
По исследованиям Farkas’a и Исаева (1923, 1934) кишечник Cy-
clopidae состоит из следующих отделов: ротовой полости (atrium, b),
глотки (pharynx, о?), пищевода (oesophagus, е), желудка (ventriculus, f, g),
тонкой кишки (intestinum tenue, Л), толстой кишки (intestinum crassum,
z) и прямой кишки (rectum, 1). Первые 3 отдела и последний имеют экто-
дермальное происхождение и внутри выстланы довольно толстым хити-
новым слоем, представляющим прямое продолжение наружных покровов
(см. выше). Средняя часть кишечника эндодермального происхождения.
Ротовая полость объемиста. В месте перехода полости в глотку на вен-
тральной стороне имеется особое, ближе еще не изученное, повидимому
вращающееся, образование в виде плоского округлого хитинового
диска, снабженного острыми шипиками; диск снабжен особой мускула-
турой и служит для размельчения пищи. Спереди ротовой полости имеется
верхняя губа (labrum, с) с рядом острых зубцов, сзади и снизу находится
особый вырост — нижняя губа (labium), вдоль наружного края вооружен-
ная мелкими зубчиками. Мандибулы, расположенные с боков ротового
отверстия, ориентированы таким образом, что их свободный зазубрен-
ный край вдается в ротовую полость, куда вдаются также и острые
МОРФОЛОГО- АН АТОМИЧЕСКИЙ очерк
37
концы расположенных ниже (кзади) максиллул. Ротовая полость про-
ходит в глотку, трудно различимую у живых циклопов. Глотка может
очень сильно расширяться, и выстилающий ее довольно толстый хити-
новый слой образует складки. Глотка снабжена сильными мускулами-
дилататорами. В месте перехода глотки в пищевод хитиновая выстилка
утолщается и состоит из 2 слоев — очень тонкого внешнего, сильно
окрашивающегося железным гематоксилином, и неокрашивающегося
им, в 2—4 раза более толстого внутреннего, к которому прилегают
рассеянные, образующие хитин, клетки с большим круглым ядром
и плотной протоплазмой. Кутикулярная выстилка очень длинного пище-
вода также образует складки; он снабжен сильными мускулами, главным
образом констрикторами, расположенными спирально. Напротив диля-
таторы пищевода развиты слабо и лишь в месте перехода пищевода
в желудок имеются сильные мускулы-дилататоры. Хитиновая внутренняя
выстилка пищевода, вместе с упомянутыми Образующими хитин клетками,
отчасти продолжается и в желудок, занимая его передний отдел, около
г/з—г/4 всей длины, за которым следует эпителиальный слой желудка,
почти совершенно лишенный хитинового покрова. Желудок очень объ-
емистый. В его передней части, выстланной эпителием, находится слой
так называемых „начальных клеток", являющихся очень низкими эпи-
телиальными клетками. Высота клеток увеличивается по направлению
к заднему отделу желудка.
Стенка желудка, таким образом, разделяется на 2 отдела — перед-
ний, хитинозный, и задний, клеточный. В передней части клеточного
отдела клетки слабо дифференцированы и имеют фибриллярную струк-
туру; среди них лишь редко расположены клетки железистого характера.
Последние свойственны задней части желудочного отдела. Они сильно
вакуолизированы, снабжены довольно крупным ядром и выделяют сероз-
ный секрет. Между фибриллярными и вакуолизированными клетками
имеются клетки переходного характера, так называемые альвеолярные
клетки. Поверхность эпителиальных клеток желудка снабжена „палочко-
вым" слоем. Среди эпителиальных начальных клеток желудка имеются
еще особые клетки, также фибриллярной структуры, у которых между
фибриллами, однако, находится система канальцев; эти клетки образуют
выросты, очень похожие на псевдоподии корненожек. Проходя через
наружную оболочку (membrana propria) и между сетью мускульных
волокон желудочной стенки в полость тела, эти псевдоподии захватывают
находящиеся близ желудка яйцевые клетки. При этом псевдоподии
внедряются внутрь в молодые яйцевые клетки. Farkas считает, что
образующие псевдоподии клетки желудка играют известную роль в пи-
вании яйцевых клеток. Membrana propria желудка не покрывает его
сплошным слоем, но образует местами просветы, что, очевидно, облег-
чает прохождение псевдоподий в полость тела.
Функция отдельных типов эпителиальных клеток желудка, по Farkas’ у,
распределяется следующим образом: „начальные" клетки — трофические
клетки, фибриллярные — клетки, резорбирующие воду и растворенные
в ней вещества, альвеолярные—клетки, резорбирующие жиры, вакуо-
лизированные— клетки, содержащие секреты и экскреты.
Поперечный разрез просвета тонкой кишки имеет звездчатое
очертание. В тонкой кишке эпителий образован цилиндрическими,
одинакового типа клетками, снабженными палочковым слоем и довольно
большим овальным ядром. Тонкая кишка образует перед началом абдо-
мена сильный загиб и переходит в толстую кишку, причем цилиндри-
38
ВВЕДЕНИЕ
ческий эпителий становится более низким, так что в пределах толстой
кишки он состоит уже из очень плоских клеток, ширина которых в б—
8 раз более высоты. Толстая кишка простирается до конца 3-го абдо-
минального сегмента, где она сильно суживается; в этом месте кишка
снабжена сильно развитым сфинктером (£). Прямая кишка (rectum)
находится в пределах 4-го и 5-го абдоминального сегментов; она несколько
шире толстой кишки. Внутренняя поверхность прямой кишки выстлана
довольно толстым складчатым слоем кутикулы, а снаружи стенка кишки
снабжена сильно развитым мускульным слоем, причем мускулы имеют
не спиральное, а продольное расположение. Помимо выталкивания непе-
реваренных пищевых частиц, прямая кишка совершает пульсирующие
сокращения, у Calanus finmarchicus установленные Dakin’oM (1908).
Благодаря этим сокращениям происходит постоянное вхождение и выхож-
дение воды через анальное отверстие (см. дыхание). Сзади прямая кишка
открывается дорзально в заднем абдоминальном сегменте в супрааналь-
ную полость, прикрытую анальной пластинкой. Dakin (1908) отрицал
присутствие в кишечнике Copepoda мерцательных клеток, найденных
однако у Cyclopidae Farkas’oM (1923). Число этих клеток непостоянно,
причем они совсем не наблюдаются у циклопов с набитым пищей кишеч-
ником, появляясь, однако, при недостаточном питании рачков в клеточ-
ном отделе желудка, в тонкой кишке и в начальном и конечном отделе
толстой кишки. Очертание мерцательных клеток, в общем, удлиненно-
кубическое, их обращенная в просвет кишки поверхность снабжена
очень тонкими щетинками (стероцилии), совершающими неправильные
бьющие движения (не ритмические). Длина стероцилий колеблется
от 1—2 до 10—15 р.; они длиннее всего (до 25 р.) в тонкой и толстой
кишке. Некоторые из стероцилий вовсе лишены способности к движе-
нию. Число их у 1 мерцательной клетки равно 30—40. В базальном
конце такой клетки лежит ядро довольно разнообразной, иногда округ-
лой формы. С помощью активных движений стероцилий происходит
возобновление питательной жидкости, омывающей клетки, что имеет
особенно существенное значение при недостаточном питании циклопов.
Питание пресноводных Cyclopidae изучено еще далеко не полно.
Способ захвата пищи достаточно изучен только для науплиальной фазы,
с 3 парами функционирующих конечностей — 2 парами антенн и парой
мандибул. Тщательный анализ функции этих конечностей и их значение
для восприятия пищи у Cyclops strenuus дает Storch (1928). Антеннулы
у науплиусов являются исключительно локомоторным оргавом. Передняя
губа прикрывает околоротовую полость, а на место отсутствующей
задней губы расположены 1—2 ряда длинных оперенных щетинок,
составляющих несколько групп, по 5—8 щетинок в каждой. 1-я из этих
групп достигает до края верхней губы и своими оперенными щетинками
ограничивает околоротовую полость справа и слева, причем образуется
окруженное сеткой из волосков пространство, напоминающее вершу.
На коксальном членике II пары антенн с каждой стороны находится
по 2 сильных крючка (крючковидных выроста), снабженных волосками.
Длинные волоски вентрального крючка образуют вертикально располо-
женные, близко стоящие друг к другу поверхности. При ударе этих
антенн их крючки всовываются в околоротовое пространство, своими
концами достигая ротового отверстия. Проксимальный членик эндопо-
дита мандибул на брюшной стороне снабжен подвижной треугольной
пластинкой с 3—4 сильными щетинками, из которых 3 особенно длин-
ные, направленные кпереди1.; и достигающие до губы. Эти 3 щетинки
МОРФОЛОГО-АНАТОМИЧЕСКИЙ ОЧЕРК 39
ограничивают в разрезе треугольное пространство, образуя род черпа-
ка. В противоположность Diaptomus, 2-я пара антенн и мандибулы
науплиусов Cyclops не совершают быстрых вибраций, а производят силь-
ные толчкообразные движения, обычно одновременно с такими же дви-
жениями антеннул. Результатом является быстрый скачок науплиуса.
Во время такого скачка мандибулы функционируют наподобие черпака,
причем вода проходит через вышеуказанные их щетинки, которые задер-
живают внутри ограниченного ими пространства взвешенные в воде
пищевые частицы. При дальнейших толчкообразных ударах мандибул
задержанные пищевые частицы попадают в упомянутую выше ограни-
чивающую околоротовую полость вершу. Одновременно работают зад-
ние антенны, которые с помощью своих крючков проталкивают достав-
ленные мандибулами пищевые частицы глубже в полость верши и, наконец,
в ротовое отверстие; затем сокращением пишевода (акт заглатывания)
пищевые частицы проводятся в среднюю кишку. Таким образом у нау-
плиусов Cyclops пища захватывается активно с помощью черпающих
движений мандибул, причем приносящих сестон токов воды (вызываемых
околоротовыми конечностями у Diaptomus) вовсе не образуется. Такой
способ захвата пищи имеет место на всех науплиальных фазах, до 1-й
копеподитной фазы, когда антеннулы теряют свою весельную функцию
и начинают служить в качестве органов балансирования, а задние антенны
утрачивают свои крючковидные придатки и совершенно исключаются
из процесса захвата пищевых частиц. Вместе с тем у мандибул появля-
ются жевательные зубцы, треугольные придатки их эндоподита реду-
цируются, а равно сильно редуцируется и сам эндоподит. Окончательно
формируются обе пары максилл и максиллярные ноги, которые сразу
же начинают функционировать как аппарат для активного захвата пищи.
Эта функция приписывалась рассматриваемым конечностям уже давно,
как и хватательная функция задней пары антенн. Подробности акта
хватания пока не изучены. Размалывание пищи происходит в ротовой
полости с помощью зазубренных концов мандибул, а также указанного
выше дисковидного образования, находящегося в этой полости; в процес-
се размельчения пищи принимают участие также снабженные зубчиками
верхняя и нижняя губы. Функция отдельных пар околоротовых конеч-
ностей у взрослых Cyclopidae выяснена пока лишь в общих чертах.
Циклопы, по способу захвата пищи, являются типичными „хватателями"
и относятся к группе „преследователей" (Spiirer) по терминологии
Lohmann’а; по Dietrich’y (1915) вообще все Podoplea принадлежат к актив-
ным хищникам, схватывающим добычу в различных пунктах водной
массы, с помощью характерных скачкообразных движений, быстро меняю-
щих в ней их положение. Захват пищи производится, по Naumann’y
(1923), с выбором, а не чисто механически. Содержимое кишечников
циклопов исследовалось целым рядом авторов. Существует множество
указаний на хищный образ их жизни, поедание ими коловраток, инфу-
зорий, в частности Paramaecium, на захват циклопами турбеллярий.
олигохет, молоди Cyclopidae, науплиусов и кладоцер. Ряд авторов
наблюдал захватывание других циклопов у Macrocyclops fuscus, М. al-
bidas, Acanthocyclops viridis, у которых мы наблюдали случаи канниба-
лизма. Wolf (1905) описывает случай, когда гидра, пытавшаяся захва-
тить Macrocyclops fuscus, сама стала жертвой этого рачка. Все эти
факты, бесспорно, доказывают хищный способ питания ряда видов
Cyclopidae. Вместе с тем ряд авторов отмечает для них растительную
пищу в виде различных диатомовых (в том числе Melosira и Cyclotella),
40
ВВЕДЕНИЕ
десмидиевых, перидиниевых и синезеленых водорослей (у науплиусов).
Вопреки предположениям Colditz’a (1914) взрослые Cyclopidae, по на-
блюдениям Naumann’a (1923), лишь в очень малых количествах загла-
тывают наннопланктические водоросли, но могут заглатывать более круп-
ные водоросли (величины микро-и мезотипа). По нашим исследованиям
(Рылов, 1930в) Mesocyclops leuckarti быстро набивает кишечник круп-
ными пандоринамн (Pandorina morum, диаметр колоний 50:—75 у.), тогда
как мелкие Chlamydomonas даже при их количестве около 400 000
в 1 см3 захватываются лишь в единичных экземплярах. Из приведенных
данных можно заключить, что пелагические Cyclopidae, помимо хищного
образа питания, могут захватывать и растительную пищу в виде сравни-
тельно крупных планктических водорослей, очевидно хватаемых активно,
т. е. являются всеядными, причем, вероятно, у них преобладающее
значение имеет животная пища. На всеядность вообще всех Cyclopidae
указывал ряд авторов (см. сводку у Steuer, Planktonkunde, 1910). Неко-
торые авторы отмечают также пищевое значенине для Cyclopidae орга-
нического детрита, нередко находимого в их кишечниках. Возможно,
что в данном случае дело касается сравнительно крупных частиц
и скоплений детрита, доступных для активного захвата. Вероятно такой
детрит, а также обрастания водорослей на макрофитах, представляет
важный элемент питания у видов, обитающих на обрастаниях и на верх-
нем слое богатого растительными остатками ила (например у Ectocy-
clops, Paracyclops affinis, Acanthocyclops abyssicola, у некоторых Mic-
rocyclops). Захват при этом происходит на самом субстрате; ряд авто-
ров отмечает, что циклопы ищут пищу не только плавая в воде, но
и лазая по субстрату, отрывая от него куски. Совершенно недостаточно
изучено значение в питании Cyclopidae бактерий. Интересные наблю-
дения были сделаны Horhammer’oM (Arch. f. Hygiene, 73, 1910, см.
также: Reitz, Mikrokosmos, V, 1912) над питанием бактериями тифа
С. strenuus. Применяя метод окраски метиленовой синью, этот автор
наблюдал как после двухдневного голодания циклопы набивали кишеч-
ник массой окрашенных бацилл, активно нападая на скопления послед-
них; захватываются также и неокрашенные бациллы. Однако равномерно
рассеянные в воде бациллы циклопами не захватывались. Эти экспери-
менты, произведенные в явно искусственной постановке, однако отнюдь
не проливают света на питание циклопов бактериями в естественных
условиях, когда налицо имеется обеспеченность животной пищей.
Специальных органов кровообращения (в частности сердца) у Cy-
clopidae не имеется. К р о в ь (целомическая жидкость, циркулирующая
в полости тела, у Crustacea являющейся псевдоцелем) весьма текуча
и бесцветна. По Hartog’y (1888) она содержит богато вакуолизирован-
ные и гранулированные амебовидные элементы (лейкоциты) с тонкими
псевдоподиями. Циркуляция целомической жидкости, по предположению
Zenker’a (1854) и Hartog’a (1888), обусловливается движениями кишеч-
ного тракта. На дорзальной стороне жидкость перемещается кпереди
(к голове), на вентральной — в обратном направлении. По старым наблю-
дениям Zenker’a движение целомической жидкости у Cyclops quadricor-
nis (= Macrocyclops) происходит очень медленно, и она омывает весь
псевдоцель лишь 3 раза в минуту. Наблюдения эти нуждаются в про-
верке.
Дыхание Cyclopidae совершается поверхностью тела, диффузно,
как и у прочих Copepoda. Особенно интенсивно дыхание происходит
в области 1-го торакального сегмента. Частично имеет мёсто и аналь-
МОРФОЛОГО-АНАТОМИЧЕСКИЙ ОЧЕРК
41
ное дыхание, совершающееся задним отделом задней кишки, насчет
ее ритмических сокращений.
Органы выделения (фиг. 12м) у науплиусов представлены
антеннальными железами, которые в конце науплиальной фазы начинают
редуцироваться, и у взрослых Cyclopidae, как и у прочих Copepoda,
исчезают (остается лишь их рудимент) и заменяются хорошо видимыми,
Фиг. 12.
/ — Мозг Mesocyclops oithonoides (Sars): ck — центральное тело, pbr— протоцеребраль-
ный мост, sm — зрительная масса, ма — медианный глаз, gz— слой ганглиозных клеток,
Нур. — гиподермис, А1. — нерв к антенну лам, Fr. — фронтальный орган; 2, 3 — нефридий
Cyclops strenuus, схематично: с — целомический мешочек; 4— передний отдел головы
С. strenuusi 5—to же, схема иннервации корнеальных линз: а — корнеальная линза,
п1> п2 — нервы, N—пигмент глаза, К—хрусталики глаза, R — рострум, Ь — щетинка,
возможно, относящаяся к фронтальному органу. (7 — по Hanstrom’y, 2—4—по Gickl-
horn’y).
начиная с 1-й копеподитной стадии, максиллярными железами. Их 2—
справа и слева по 1, в области максилл и максиллярных ног. В по-
следнее время эти железы подробно изучены Gicklhorn’oM (1930) мето-
дом прижизненной элективной окраски. У Cyclops они были описаны
под названием раковинных желез (Schalendriise) Zenker’oM и Leydig’oM
(1859), а особенно точно и полно Claus’oM (1877), Richard’oM (1891)
и Plenk’oM (1911).1 Каждый максиллярный нефридий состоит из конеч-
1 Сводку см. у Giesbrecht’a: Lang, Handbuch d. Morphol. etc., 1921.
42
ВВЕДЕНИЕ
ного мешочка, от которого отходит изогнутый выводной проток, откры-
вающийся наружу особым отверстием на задней поверхности 2-й (зад-
ней) пары максилл. Конечный мешочек имеет форму тетраэдра и содержит
внутри выстланную несколькими (не более 5) плоскими клетками,
довольно объемистую полость. Возле этого мешочка, по Gicklhorn’y,
имеется лишенный выводного канала рудимент целомического мешочка
прежней антеннальной железы, который крупнее у молоди, чем у взрослых
рачков.1
Выводной проток имеет вид извитой трубки и состоит из неболь-
шого числа клеток; просвет канала образован насчет прободения клеток
(интрацеллюлярный просвет), которые снабжены особенно мощным
в проксимальном отделе канала палочковидным кутикулярным слоем.
Функция максиллярных желез была изучена рядом авторов (см. сводку:
Winterstein, Hdb. vergl. Physiol., 2, 1924). По исследованиям Gicklhorn’a,
выводной канал служит не только для пассивного выведения продуктов
экскреции, но его клетки имеют самостоятельную функцию, точно
не выясненную (экскреция или резорбция).
Центральная нервная система Cyclopidae изучена неполно.
Спереди головного отдела лежит крупная ганглиозная масса, так называе-
мый „мозг". От мозга по бокам пищевода отходят 2 параэзофагеальных
цепочки (Hartogf, 1888), no 1 с каждой стороны, а на брюшной стороне
проходит брюшная нервная цепочка, доходящая до начала заднего
торакального сегмента, кзади от которого она разветвляется надвое.
По Richard’у, брюшная цепочка заходит и в 1-й абдоминальный сегмент,
далее разветвляясь. Спереди мозг дает очень тонкие оптические нервы,
нервы к фронтальному органу (Fr.), нервы к голове, нервы к антен-
нулам и к верхней губе. Довольно подробно мозг был изучен Hanstrom’oM
(1924) у Mesocyclops oithonoides (фиг. 12). Непосредственно на мозге
лежит глаз, причем ганглиозный слой мозга (gz) окружает глаз с боков.
Под глазом находится непарная, так называемая „зрительная" масса
нервных клеток (sm), которая сзади разделена на 3 отростка. В сообще-
нии с этой массой находятся нервы фронтального органа, а также
нервы самого глаза (см. ниже). Зрительная масса окружает протоцере-
бральный мост (pbr), под которым находится центральное тело мозга
(ск). Брюшная нервняя цепочка лежит под кишечником, и ее задние
ветви заканчиваются в фуркальных ветвях. Она имеет ряд ганглиозных
утолщений с отходящими от них нервами. Отходящие от ганглиев пар-
ные нервы очень тонки и, как показал Richard (1891), у Cyclopidae
снабжены неврилеммой. Richard подробно исследовал центральную
нервную систему у Macrocyclops fuscus, Acanthocyclops viridis и Cyclops
strenuus. По этому автору, в их тораксе имеется 5 ганглиев (торакаль-
ные ганглии), которые соответственно иннервируют все 5 пар конеч-
ностей; задней (рудиментарной) паре ног соответствует 5-й торакальный
ганглий, который иннервирует также мускулы, причем от него с каждой
стороны отходит пара нервов — один к рудиментарной ноге, другой
к мускулам. В генитальном сегменте имеется абдоминальный ганглий,
от которого отходят нервы к receptaculum seminis и к генитальным
отверстиям. Кзади от этого ганглия брюшная нервная цепочка делится
на 2 ветви, заходящие в фурку. Всю нервную цепочку автор делит
1 Ранее этот рудимент был обнаружен у взрослых Cladocera. Наличие у взрослых
циклопов рудиментов пары нёфридиев (максиллулы), о которых упоминает Gicklhorn,
не доказано.
МОРФОЛОГО-АН АТОМИЧЕСКИЙ ОЧЕРК
43
на 3 отдела — околопищеводный, торакальный и абдоминальный. Ман-
дибулы и максиллы иннервируются 1-м нз этих отделов (по 2 пары
нервов), 5 пар ног—2-м из них. Кроме центральной нервной системы,
у Cyclopidae описаны еще мало изученные элементы симпатической
нервной системы, рассеянные в гиподерме верхней губы и в области
ротового отверстия.
Помимо описанных выше сензорных придатков (сензорные колбы
и цилиндры), особенно многочисленных на антеннулах самцов Cyclopidae,
важнейшим органом чувств у Copepoda вообще является непарный
медианный глаз. Его центральная масса состоит из 3 крупных,
наполненных темнокоричневым пигментом, клеток, разделенных 2 взаимно
перпендикуляоными мембранами, в виде перегородок. В центральную
пигментную массу погружены 3 глазка (ocelli) — 2 боковых и 1 вен-
тральный. Каждый глазок состоит из удлиненных цилиндрических клеток,
собранных в группы. Каждая из этих клеток имеет сильно преломляю-
щее свет прозрачное светлое содержимое, и лишь в основании клеток
имеются зерна пигмента. Каждая цилиндрическая клетка снабжена
лежащим в ее базальном отделе ядром. Оптические нервы подходят
не к пигментной центральной массе, а к глазкам. По Zenker’y (1854)
светопреломляющая сила глазков взрослых Cyclopidae = 1.50, неполово-
зрелых—1.36. По исследованиям Erhard’a (1922) глаз Cyclopidae
не менее чувствителен к изменениям силы света, нежели человеческий
глаз, реагируя очень тонко на малейшие ее изменения; циклопы
реагируют на ультрафиолетовые лучи, но не различают цветов. У пещер-
ных Cyclopidae глазной пигмент иногда (далеко не у всех видов)
совершенно исчезает, и глаз, очевидно, теряет функцию органа зрения,
хотя светопреломляющее тело сохраняется. У живущих в пещерных
водах популяций широко распространенного в наземных водах Рага
cyclops fimbriatus глазной пигмент не исчезает бесследно, но очень
сильно редуцируется. Chappuis (1920) показал, что лишенные глазного
пигмента пещерные Eucyclops teras и Е. graeteri при культивированих
на свету восстанавливают этот пигмент.
Над медианным глазом лежит пара так называемых „корнеаль-
ных линз" (фиг. 124,5), впервые открытых у Cyclopidae Claus’ом
(1863) и недавно подробно изученных Gicklhorn’oM (1930а) у Cyclops
strenuus и Macrocyclops fuscus. Эти линзы с трудом заметны на живых
циклопах, но хорошо различаются при окраске животных конгоротом;
у Cyclopidae корнеальные линзы являются не плотным телом, а округ-
лыми участками очень тонкой мембрановидной кутикулы а, находящи-
мися справа и слева впереди медианного глаза; эти мембраны гораздо
тоньше остальной кутикулы цефалоторакса. К каждой линзе подходит
тонкий нерв, отходящий от вентральной части переднего отдела мозга.
Нервы заканчиваются униполярными нервными клетками, тесно при-
легающими к внутренней поверхности мембраны. Таких клеток наблю-
дается от 1 до 3. Существуют ли клетки самих корнеальных линз, — пока
не установлено. По Gicklhorn’y (193Эа) корнеальные линзы, вопреки
предположению Claus’a, не имеют никакого отношения к зрительной
функции и, вероятно, являются конечными пластинками органов, вос-
принимающих химические раздражения.
В области цефалоторакса на уровне антенны II пары лежит так
называемый фронтальный орган. Фронтальный орган является
парным образованием и представлен латерально и симметрично отно-
сительно продольной оси расположенными группами клеток, иннервирую
44
ВВЕДЕНИЕ
щихся непосредственно отходящими от мозга нервами. Каждый орган
состоит только из пары кеглеобразных клеток, лежащих непосредственно
под кожными покровами. Ядро лежит в тонко оттянутом конце клетки,
оно маленькое; этот нитевидный конец клетки представляет нервное
волокно, отходящее от мозга (см. выше). Таким образом фронтальный
орган представлен униполярными чувствительными нервными клетками,
прямо сообщающимися с мозгом (см. также Hanstrom, 1924, у Meso-
cyclops oithonoid.es}. На роструме Cyclops strenuus имеется всегда 2 пары
очень тонких коротких волокон (Richard, 1891; Gicklhom, 1930b), отно-
шение которых к фронтальному органу не выяснено. Функция этого
органа неизвестна.
ОК
Фиг. 13.
7— Половые органы самки Cyclops sp.: ov— оварий, od— яйцевод, rs—reeeptaculum
seminis; 2 — половые органы самц-: а — а2 vasa deferentia (парный), а2 и — 1-й секре-
торный отдел vas deferens (до места, обозначенного буквой е, он имеет железистую
структуру), &2 — 2-й секторный отдел, с — изогнутый к спиниой стороне отдел vas defe-
rens, d—его конечный отдел, переходящий в тонкий канал ведущий в генитальный
сегмент (отрезки с и d представляют 3-й секреторный отдел), g—гонады (семенник).
Отрезок а — а2 представляет vas def. recurrens superior (см. текст), 6, и b-> — vas def.
procurrens superior, с и d — vas def. descendens и recurrens inferior.
Bee Cyclopidae раздельнополы. Половые органы самки (фиг. 13!)
состоят из непарного яичника (оо), лежащего в переднем отделе торакса,
от которого с каждой стороны (справа и слева) отходит по 1 яйцеводу
(od), направленному кзади и открывающемуся сбоку (несколько ближе
к спинной поверхности) 1-го абдоминального сегмента, иногда на гра-
нице со 2-м сегментом. По исследованиям Matschek’a (1910) у всех
видов родов Macrocyclops и Ectocyclops, у Eucyclops serrulatus, Tropocyc-
lops prasinus и Paracyclops affinis яичник двураздельный (по терминоло-
гии этого автора „парный"), что, вероятно, свойственно вообще под-
семейству Eucyclopinae, тогда как у видов подсемейства Cyclopinae
(исследованы Cyclops strenuus, С. insignis, Acanthocyclops b 'cuspidatus,
A. vemalis, Microcyclops gracilis, Mesocyclops leucKarti, M. dybowskit)
МОРФОЛОГО-АН АТОМИЧЕСКИЙ ОЧЕРК 45
он имеет вид простого, неразделенного мешка. Яичник занимает средин-
ное положение между кишечником и спинным покровом тела, причем
в зависимости от физиологического состояния (наполнение половыми
клетками) оно может несколько изменяться. У Ectocyclops яйцеводы
заходят далеко в абдомен. Лопастевидный широкий отдел яйцеводов
нередко называют маткой (см. Hartog1, 1888), тогда как задний их отдел
всегда лишен выростов и имеет вид простой изогнутой трубки. Конеч-
ный отдел яйцеводов снабжен железистыми клетками, выделяющими
особый секрет для оболочек яйцевых мешков (Gruber, 1878, 1879; Mat-
schek, 1910); по Walter (1922) этот секрет выделяется кожными железами
в области генитального отверстия, т. е. непосредственно у выходов
яйцеводов, а железы конечного отдела яйцеводов дают секрет лишь
ддя образования оболочек яиц. От заднего конца каждого яйцевода
отходит по очень короткому тонкому протоку; протоки впадают справа
и слева в особый, чрезвычайно характерный для Cyclopidae, объемистый
орган — семенной мешок, reeeptaculum seminis (фиг. 13, rs; фиг. 14).
Он нередко занимает значительную часть генитального сегмента
и имеет очень различное строение.
Весьма подробное исследование строения и функции reeeptaculum
seminis было произведено Walter (1922) у Acanthocyclops viridis. Орган
служит для сохранения изливаемой из сперматофоров спермы. Он занимает
в генитальном сегменте медианное положение, располагаясь близко
от брюшной поверхности генитального сегмента, на которой открывается
особым непарным отверстием, лежащим на срединной продольной оси
сегмента. С боков это отверстие выстлано хитином, образующим маленькую
папиллу, заворачивающуюся в полость семенного мешка. Упомянутые
выше короткие протоки яйцеводов впадают в мешок вблизи его наруж-
ного отверстия. Каждый проток укрепляется с помощью особого хити-
нозного тяжа, идущего от покровов сегмента. Конечный отдел яйцево-
дов снабжен сильной кольцевой мускулатурой, причем специальный
кольцевой мускул имеется, кроме того, у самого отверстия яйцевода.
Эти мускулы способствуют выталкиванию из яйцеводов зрелых яиц.
Стенки reeeptaculum seminis лишены железистых клеток и выстланы
тонкой кутикулой — орган служит исключительно для сохранения спермы;
Claus неправильно приписывал ему также секреторную функцию.
Проходя через конечный отдел яйцеводов и при прохождении сокращая
их стенки, яйца засасывают находящуюся в семенном мешке сперму,
поступающую через короткий проток и попадающую на яйцо, которое
и оплодотворяется; наружу яйцо выходит уже оплодотворенным, причем
несколько вышедших оплодотворенных яиц склеивается в яйцевые
мешки, по одному справа и слева. У Acanthocyclops viridis образова-
ние мешков происходит очень быстро, в течение около 1 минуты. Число
яиц в каждом мешке может быть очень различным. Иногда мешок
содержит лишь по 1—2 яйца, иногда число их достигает до 100.
У одного и того же вида число яиц в мешках непостоянно, колеблясь
в зависимости от ряда условий. Одна и та же самка образует яйцевые
мешки несколько раз, причем первые мешки содержат меньшее коли-
чество яиц, чем образующиеся позже. Самка A. viridis, по Walter,
образует яйцевые мешки до 12—13 раз (вид полицикличен, размножается
весь год), откладывая всего до 800—1000 яиц, из них лишь около х/з
дает половозрелых рачков. По Manfredi (1923) одна самка Tropocyclops
prasinus дала 13 пар яйцевых мешков в течение 2г/2 месяцев, самка
Eucyclops serrulatus за 3 месяца дала 12 пар мешков, а самка
46
ВВЕДЕНИЕ
Acanthocyclops bicuspidatus за 1г/2 месяца образовала 7 пар яйцевых
мешков.
Фиг. 14. Receptaculum seminis.
7 — Cyclops abyssorum Sars; 2— Acanthocyclops gigas (Claus); 3 — A. bise-
tosus (Rehb.), 4—A. abyssicola (Lili.); 5—Mesocyclops crassus (Fisch.); 6—
Microcyclops gracilis (Lili.); 7—Macrocyclops jascus (Jur.); 8— Eucyclops
macruroides (Lili.); 9—Acanthocyclops sensitivus (Graet. et Chapp.);
10 — A. bicuspidatus (Claus); 11 — Macrocyclops albidus (Jur.); 12— Tropo-
cyclops prasinus (Fisch.). (7—8—no Sars’y, 9 и 12—no Kiefer'y, 10—11 —
no Gurney).
Из сперматофоров сперма изливается непосредственно в recepta-
culum seminis через отверстие последнего. Порция сперматозоидов,
МОРФОЛОГО-АН АТОМИЧЕСКИЙ ОЧЕРК
47
попавших в receptaculum seminis из пары сперматофоров, служит для
оплодотворения нескольких серий яиц (Wolf, 1905). По исследованиям
Claus’a (1899) и Walter (1922) к восприятию спермы способны только
половозрелые самки, причем их оплодотворение происходит сразу после
последнего линяния, но образование яйцевых мешков наступает только
через 2—3 недели (у A. vzrzdzs).
Половые органы самцов Cyclopidae (фиг. 132) исследованы
рядом авторов (Claus, 1863; Gruber, 1878; Hartog, 1888; Heberer, 1926).
Особенно тщательно изучены они НеЬегег'ом у Cyclops strenuus
и Acanthocyclops viridis. В области 1-го торакального сегмента лежит
непарный, нередко двулопастный семенник (§), сильно суживающийся
кзади; форма задней части гонады очень вариирует, резко выраженное
разделение ее на 2 лопасти особенно характерно для видов с большим
числом хромозом. Задняя часть гонады содержит сперматогонии. Перед-
ний ее отдел снабжен парой семепроводов (vasa deferentia, а — d), рас-
положенных симметрично. От гонады каждой семепровод отходит
в виде узкого тонкостенного канала, который у взрослых самцов в основа-
нии ампулообразно вздут. Семепровод направляется кзади приблизи-
тельно параллельно дорзальной поверхности тела и образует резко
выраженный загиб, в пределах которого внутренний эпителиальный
слой становится толще, а его клетки содержат крупные светлые вакуоли;
этот отдел семепровода имеет секреторную функцию (1-й секреторный
отдел). В конце 3-го или в 4-м торакальном сегменте семепровод
загибается кпереди (к головному отделу); эпителий его стенок здесь
также железистого характера, но кпереди он становится тоньше и снабжен
более мелкими вакуолями (2-й секреторный отдел). Дальнейшее направ-
ление семепроводов стоит в связи с возрастом и половым состоянием
самца. У деятельных в половом отношении самцов каждый семепровод
образует новый загиб, направленный почти отвесно книзу (к брюшной
стороне) и, изгибаясь, в виде тонкого канала продолжается кзади,
в область генитального сегмента, где под вентрально расположенной
генитальной складкой кутикулы (Genitalklappe) открывается гениталь-
ным отверстием. Отрезок семепровода от последнего загиба до гени-
тального сегмента представляет 3-й секреторный отдел. Конечная часть
каждого семепровода расширена в особую сперматофорную сумку
(Kittdruse старых авторов), причем ее вентральная часть, лежащая
в генитальном сегменте, образует 4-й секреторный отдел. Спермато-
форные сумки, лежащие симметрично, справа и слева сегмента, очень
хорошо видны у половозрелых самцов. В этих сумках происходит окон-
чательное формирование эллиптических крупных сперматофоров, в общем
имеющих бобовидную форму. Как видно из вышеизложенного, различ-
ные отделы семепровода имеют секреторную функцию. Heberer (1926)
предлагает для них следующую терминологию: передний отдел — vas
deferens recurrens superior (a—a2), в задней части железистого харак-
тера; 2-й отдел — vas. def. procurrens superior (6X — 62), его передняя
часть функционально соответствует задней части предыдущего отдела;
3-й отдел — vas def. descendens и recurrens inferior (с—d), которые
у молодых циклопов снабжены высоким эпителием, а у взрослых —
плоским (вследствие излияния секрета в просвет семепровода). В заднем
(4-м) отделе, в пределах генитального сегмента, только часть стенок
vas deferens имеет секреторную функцию. По исследованиям цитируе-
мого автора в образовании сперматофора участвуют 3 различных сектора,
соответственно 3 передним отделам семепроводов. Сперматофор обра-
48
ВВЕДЕНИЙ
зуется, главным образом, насчет зернистого секрета, выделяемого 2-м
отделом семепровода (в задней части vas. def. procurrens superior).
В области перехода этого отдела в 3-й (vas. def. descendens и recurrens
inferior) выделяется секрет пенистой структуры, образующий наружную
оболочку сперматофора. Особый секрет, состоящий из шаровидных
скоплений, выделяется передней частью vas. def. procurens superior и зад-
ней частью vas. def. recurrens superior (передний отдел семепровода);
в начале последнего секрет отсутствует, и проникшие сюда из гонады
сперматозоиды плавают свободно в тканевой жидкости. В самом заднем,
Фиг. 15. Расположение сперматофоров на генитальном
сегменте самки. (По Sars’y).
7 — у Cgclbps strenuus Fisch.; 2—у С. scutifer Sars;
3—у С. insignis Claus; 4 — у Acanthocgclopscras-
sicaudis (Sars),
4-м отделе семепровода (в области сумки) выделяется лишь очень
небольшое количество секрета, в построении самого сперматофора
не участвующего.
Заключающие сперматозоиды, вполне сформированные спермато-
форы, в процессе копуляции всегда парой прикрепляются самцом на
брюшную поверхность генитального сегмента самки (фиг. 15). У самцов
окончательное сформирование обоих сперматофоров занимает довольно
продолжительное время — по Walter (1922) у Acanthocyclops viridis оно
продолжается в течение 10—15 дней существования самца после 5-й
копеподитной стадии. Акт копуляции у свободноживущих Copepoda
неоднократно наблюдался уже старыми исследователями (Jurine, 1820;
Siebold, 1839; Zenker, 1854; Leydig, 1859; Claus, 1857, 1893; Gruber,
1879; Schmeil, 1892). Особенно подробно копуляция у Cyclopidae
описана .Wolf’oM (1905) у Macrocyclops fuscus, Eucyclops serrulaius,
МЕТАМОРФОЗ
49
Cyclops strenuus, Acanthocyclops viridis, A. vernalis; Holmes’oM (1909)
у Paracyclops fimbriatus и Walter (1922) у Acanthocyclops viridis.
При копуляции самец обхватывает своими передними антеннами
(изгибая их в подвижном сочленении среднего и дистального отделов)
IV пару плавательных ног самки; обхват именно этой пары ног
наблюдался уже Jurine’oM (1820). Обхватываются обе ветви ног.
Пара циклопов in copula плавает, причем самец производит быст-
рые удары ногами этой же пары в области отверстия receptacuhim
seminis самки. Генитальные сегменты самца и самки в этот момент
находятся на одном уровне, прикладываясь друг к другу. При этом оба
сперматофора, без помощи каких-либо других конечностей, быстро
прикрепляются (приклеиваются) симметрично по бокам отверстия гесер-
taculum seminis, тесно прилегая к брюшной поверхности генитального
сегмента самки. Schmeil (1892) наблюдал, как при копуляции самец
Paracyclops fimbriatus своими передними антеннами охватывал также
верхнюю часть абдомена самки, что Wolf (1905) наблюдал также у
Р. affinis и Ectocyclops phaleratus. Holmes (1909) упоминает, что во время
копуляции Paracyclops fimbriaius самка более пассивна, чем самец,
который оживленно плавает, увлекая с собой удерживаемую самку,
причем иногда самец копулирует с самкой, уже вынашивающей яйцевые
мешки. Прикрепленные сперматофоры быстро опоражниваются, и их пустые
оболочки отпадают. Один и тот же самец может оплодотворить несколько
самок. Так Manfredi (1923) наблюдала, как 1 самец Tropocyclops prasinus
оплодотворил 7 самок. Walter (1922) упоминает, что 1 самец Acanthocy-
clops viridis оплодотворял самок несколько раз, каждые 1—2 дня.
Сперматозоиды у Cyclopidae имеют различную форму, изученную
пока лишь у немногих видов. В общем они веретенообразны, но при
набухании в воде становятся округлыми. По Walter (1922), попав в recepta-
culum seminis самки, они становятся ромбовидными и квадратными;
в receptaculum seminis сперматозоиды сохраняют свою жизнеспособность
очень долго, у Acanthocyclops viridis не менее 1 месяца.
Эмбриональное развитие пресноводных Cyclopidae, главным образом
родов Cyclops и Acanthocyclops {viridis), изучалось многими авторами
(Urbanowitsch, 1884, 1886; Haecker, 1892—1901; Ruckert, 1894,
1895; Matschek, 1909, 1910; Amnia, 1911; Fuchs, 1913, 1914;
Jacobs, 1925; Heberer, 1925, 1927, 1931). Как у прочих Copepoda,
развитие яйца Cyclopidae имеет резко выраженный детерминативный
характер. Дающие начало зародышевым пластам клетки обособляются
уже на 1-х фазах дробления. Дробление яйца тотальное, большей
частью адеквальное. 9-я фаза дробления приводит к гаструляции. В резуль-
тате 1-го дробления, которое меридионально, образуется 2 бластомеры,
из которых одна является 1-й зародышевой клеткой (I Keimbahnzelle,
Stammzelle), содержащей зачатки всех зародышевых пластов, а другая
бластомера заключает лишь соматический материал. Деление начинается
всегда уже на вышедших из яйцеводов наружу яйцах.
МЕТАМОРФОЗ
Развитие Cyclopidae, как и прочих свободноживущих Copepoda,
после выхода из яйца личинки протекает путем сложного метаморфоза.
У Cyclopidae имеется 5 науплиальных и 5 копеподитных стадий; взрос-
слому индивиду соответствует б-я копеподитная стадия. Некоторые
4 Фауна СССР
50
ВВЕДЕНИЕ
авторы считают возможным выделить еще 6-ю1 науплиальную стадию.
На всех стадиях развития Cyclopidae находятся в свободно плавающем
состоянии. Появлению каждой новой стадии предшествует линяние.
Науплиусы Cyclopidae, как и вообще Cyclopoida, несегментиро-
ваны; намечающиеся на последних науплиальных стадиях границы между
тораксом и абдоменом еще не приводят к сегментации, которая имеет
место только в копеподитной фазе развития.
Фиг. 16. Науплиальные стадии Acanthocyclops viridis (Jur.). (По Walter).
7—1-я (ортонауплиус); 2—2-я (метанауплиус I); 3—3-я (метанауплиус II);
4—4-я (метанауплиус III).
Тело науплиуса (фиг. 16) яйцевидно-округлой формы, более или
менее сплющенное в спинно-брюшном направлении. Сзади тело несет
сперва 1 пару симметрично расположенных фуркальных щетинок; затем
появляются еще 2 пары щетинок.
Науплиус снабжен передней (верхней) губой, в виде почти круглой
крупной дисковидной пластинки; задняя (нижняя) губа отсутствует.
1-я науплиальная стадия (ортонауплиус, фиг. 161). 3 пары конечно-
стей: пара одноветвистых 3-членистых антенну л, пара 2-ветвистых
антенн (задние антенны), пара 2-ветвистых верхних челюстей (мандибул).
Имеется верхняя губа в виде округлой плоской крупной пластинки.
1 Zigelmayer (1925), работа которого имеет ряд крупных недостатков, ошибочно
устанавливает даже 7-ю стадию.
МЕТАМОРФОЗ 51
2-я науплиальная стадия (метанауплиус I, фиг. 162). Кроме указан-
ных конечностей и губы, имеется пара зачаточных нижних челюстей
(максиллул) в виде мелких бугорков.
3-я науплиальная стадия (метанауплиус П, фиг. 1б3). Отличается
от предыдущей появлением 2-й пары фуркальных щетинок.
4-я науплиальная стадия (метанауплиус III, фиг. 1б4). Отличается
от 3-й появлением 3-й пары фуркальных щетинок. Максиллулы получают
уже значительное развитие.
5-я науплиальная стадия (метанауплиус IV, фиг. 17). Появляется
4 пары новых конечностей: 2-я пара нижних челюстей, пара челюстных
(максилярных) ног, 2 пары (1-я и 2-я) плавательных ног (еще в зачаточ-
ном состоянии).
Копеподитные стадии развития (фиг. 172,3) отличаются от нау-
плиальных прежде всего явственной сегментацией тела. Отдельные стадии
имеют следующие признаки. 1-я копеподитная стадия (фиг. 172). Тело
состоит из 5 сегментов, число члеников антеннул обычно более 3,
у ряда видов оно равно 6; иногда имеется только 3 членика (Halicyclops)
или 5 члеников (Paracyclops, fimbriatus). Базиподит антенн II пары еще
имеет рудимент экзоподита. Околоротовые конечности уже достаточно
развиты и, в общих чертах, близки к таковым взрослого животного;
однако мандибулы еще имеют (у всех видов?) следы экзоподита. 3 пары
плавательных ног, из них 2 передние 2-ветвистые, хорошо развитые,
а 3-я пара в зачаточном состоянии. Экзо- и эндоподиты обеих передних
пар плавательных ног 1-членистые. Хорошо развиты фуркальные ветви,
у которых имеются латеральные и дорзальные и все 4 апикальные
щетинки; последние еще не вполне развиты и оперены не полностью.
2-я копеподитная стадия. Тело состоит из б сегментов. Антеннулы
состоят из б (например Eucyclops serrulatus) или 7 (например Cyclops
strenuus), редко из 5 (Paracyclops fimbriatus) члеников. Антенны II пары
и околоротовые конечности, в общем, как у предыдущей стадии. Губа
уже явственно зазубрена. 4 пары плавательных ног, из них передние
3 пары хорошо развиты, задняя — зачаточна. Экзо-и эндоподиты первых
2 пар плавательных ног 2-членистые. Все щетинки хорошо развитых
фуркальных ветвей получают полное оперение.
3-я копеподитная стадия (фиг. 17s). Тело состоит из 7 сегментов.
4 пары хорошо развитых плавательных ног, у ног I, II и III пар экзо-
и эндоподиты 2-членистые, у ног IV пары ветви 1-членистые, ноги
V пары в зачаточном состоянии. Антеннулы уже с большим числом
члеников (например у Acanthocyclops b 'cuspidatus и у Cyclops strenuus 9,
у Tropocyclops prasinus 8, у Eucyclops serrulatus 7, у Paracyclops
fimbriatus б, у Halicyclops только 4 членика). У антенны II пары
вооружение соответствует таковому взрослых индивидов. Окончательно
формируются околоротовые конечности. Имеется различие в величине
самок и самцов, причем самки более крупны.
4-я копеподитная стадия. Тело состоит из 8 сегментов. Различие
в величине обоих полов выражено еще значительнее. Дальнейшее уве-
личение числа члеников передних антенн (у Cyclops strenuus, Microcyclops
minutus и Acanthocyclops bicuspidatus 10, у самцов C. strenuus 9 или 10,
у Tropocyclops prasinus 9, у Eucyclops serrulatus 8, у Paracyclops б,
у Halicyclops 5). Все плавательные ноги с 2-членистыми экзо- и эндо-
подитами. У ряда видов ноги V пары состоят из 2 члеников, 1-й членик
уже несет наружную щетинку, 2-й вооружен, как и у взрослых', шипом
и щетинкой; у других видов ноги V пары 1-членистые.
4*
ВВЕДЕНИЕ
5-я копеподитная стадия. Тело состоит из 9 сегментов. Наблюдается
явственное различие у обоих полов в толщине некоторых члеников
передних антенн, а также и в их числе. У самок Cyclops strenuus,
Acanthocyclops bicuspidatus, A. vernalis a Tropocyclops 11 члеников,
у самцов 10. У некоторых видов на 5-й копеподитной стадии
у обоих полов передние антенны состоят из одинакового числа
МЕТАМОРФОЗ
53
члеников; у Eucyclops serrulatus 10, у Halicyclops 5 члеников, у Масго-
cyclops albidus 11 члеников. Резко выраженное различие в величине
самцов и самок. Плавательные ноги или с 2-членистыми экзо- и эндо-
подитами (например у Acanthocyclops bicuspidatus), или обе ветви
становятся уже 3-членистыми (например у Cyclops strenuus, у Eucyclops
serrulatus). Окончательно сформировываются ноги V пары.
При переходе к половозрелости копеподитная личинка 5-й стадии
линяет, у ней окончательно формируется генитальный сегмент, вполне
утрачивающий следы расчленения на 2 сегмента, и образуется гесер-
taculum seminis, отсутствующий у 5-й копеподитной стадии. Вместе
с тем заканчивается окончательная дифференцировка обоих полов.
Продолжительность метаморфоза Cyclopidae зависит
от условий водной среды, из которых исследовано влияние температуры.
Развитие яйца Acanthocyclops viridis при 22—23° занимает всего 2—3
дня, но зимсй, при низких температурах, оно продолжается 10—15 дней.
При оптимальной температуре весь метаморфоз этого вида длится
нередко только 3—4 недели, а при низкой температуре — несколько
месяцев. 1-я линька науплиусов наступает уже через 2—3 дня, причем
время между 2 линьками на последующих фазах, в общем, удлиняется.
Созревание самцов после 5-й копеподитной стадии у A. viridis происхо-
дит быстрее, чем самок. По Walter поколения этого вида, относящиеся
к разному времени года, имеют неодинаковую продолжительность жизни.
С ноября по февраль самка дает поколения, живущие около 9 месяцев,
но в марте появляется поколение с коротким (4—6 месяцев) жизненным
периодом. Отношение числа долгоживущих циклопов к числу живущих
короткое время в ноябре до февраля = 4:1, в марте = 2:1, в апреле
= 1:1 и 1:2, в мае = 1:4, причем самцы живут меньше, чем самки.
В общем мелкие виды Cyclopidae. живут, вероятно, 4—б месяцев, круп-
ные— не свыше 10—14 месяцев, что совпадает с предположением
Burckhardt’a (1900), по которому пелагические циклопы живут 8—14
месяцев. Eucyclops serrulatus развивается гораздо быстрее, чем A. viridis-,
зимой его метафорфоз занимает около 1.5 месяцев, при температуре
10° — всего 2 недели. Дицикличные виды (Tropocyclops prasinus, Para-
cyclops, некоторые виды Cyclops) живут, вероятно, 10—12 месяцев.
Сколь-либо точных исследований над влиянием других факторов
на метаморфоз пока не имеется.
Исследования над метаморфозом пресноводных Cyclopidae показы-
вают, что науплиальные стадии различных видов различаются по ряду
признаков, причем, по крайней мере в некоторых случаях, вид циклопа
можно определить уже по его науплиусу. Нужно, однако, заметить, что
для подавляющего большинства видов науплиус в различных стадиях
его развития совершенно не изучен. Даже для большинства родов
мы пока не можем дать вполне надежной морфологической характери-
стики всех стадий науплиальной фазы метаморфоза.
Для родов Eucyclops, Tropocyclops Ectocyclops (Е. phaleratus) общим
признаком является длина дистального членика экзоподита II пары
антенн, которая равна или более длины 3 предыдущих члеников этой
ветви, взятых вместе. Признак этот, однако, не характеризует всего
подсемейства Eucyclopinae, так как у Macrocyclops albidus длина ука-
занного членика менее таковой 3 предыдущих члеников. У рода Tropo-
cyclops из щетинок среднего членика антеннул наиболее длинная дисталь-
ная, а у Eucyclops vl Ectocyclops все щетинки этого членика прибли-
зительно одинаковой длины. Ectocyclops характеризуется очень сильно
54
ВВЕДЕНИЕ
развитым жевательным крючком коксоподита задних антенн, тогда как
у рода Eucyclops этот крючок гораздо меньше. У следующих видов
дистальный членик экзоподита II пары антенн короче общей длины его
3 предыдущих члеников: Mesocyclops leuckarti, М. dybowskii, М. oithono-
ides, Acanthocyclops bicuspidatus, A. vernalis, A. viridis, Macrocyclops
albidus. У видов рода Mesocyclops и у Acanthocyclops bicuspidatus
длина внешних фуркальных щетинок на науплиальных стадиях 1-й — 3-й
почти равна или более половины длины тела науплиуса, в противном
случае внешняя щетинка максиллул (=передние максиллы) в 2 (и более)
раза превышает длину их прочих щетинок, выдаваясь за нижний край
тела личинки. У Mesocyclops leuckarti внешняя апикальная щетинка
дистального членика эндоподита мандибул вдвое длиннее соседней
внутренней щетинки (у 1-го науплиуса) или равна, или более длины
2 соседних щетинок этого членика, взятых вместе (у 2-го — 5-го нау-
плиусов), тогда как у Mesocyclops oithonoides и М. dybowskii, а также
у Acanthocyclops bicuspidatus внешняя щетинка не длиннее прочих
щетинок дистального членика (на всех науплиальных стадиях). У М. oitho-
noides наиболее длинная из апикальных щетинок этого членика эндо-
подита мандибул не имеет оперения, а у М. dybowskii и A. bicuspidatus
она оперенная. Науплиусы этих 2 видов отличаются длиной внешних
фуркальных щетинок на первых 3 стадиях (у A. bicuspidatus они менее,
а у М. dybowskii равны или более длины тела) и числом фуркальных
щетинок с каждой стороны на 3-й и 4-й стадиях (у A. bicuspidatus
их 3, у М. dybowskii только 2, на месте 3-й щетинки имеется короткий
внутренний шип). В противоположность приведенным 4 видам, у Acan-
thocyclops viridis, A. vernalis и Macrocyclops albidus длина внешних
фуркальных щетинок на 3 первых науплиальных стадиях заметно менее
половины длины тела науплиуса, в противном случае внешняя щетинка
максиллул не длиннее их остальных щетинок и за край тела личинки
не выдается. Науплиусы A. viridis отличаются присутствием на поверх-
ности верхней губы поперечного ряда длинных волосков, которых нет
у М. albidus и A. vernalis. Эти два циклопа отличаются расположением
волосков на брюшной стороне тела науплиуса: у М. albidus эти волоски
собраны в 2 боковых пучка, между которыми находятся поперечные
ряды более мелких волосков, а у A vernalis волоски расположены
петлеобразно, ве образуя пучков. Ортонауплиусы (науплиусы 1-й стадии)
Eucyclops serrulatus и Е. macrurus отличаются вооружением дистального
членика антеннул и эндоподита мандибул: у 1-го вида этот членик
мандибул вооружен 4 щетинками (дистальный членик антеннул только
3), у 2-го — 3-щетинками (дистальный членик антеннул 2). Эти виды
во 2-й науплиальной стадии отличаются числом щетинок (4 щетинки
у 1-го и 3 у 2-го) дистального членика антеннул и числом щетинок
(у 1-го 6, у 2-го 5) экзоподита мандибул. На 3-й науплиальной стадии
(метанауплиус 2) эти виды отличаются вооружением нижнего края тела:
у Е. serrulatus здесь с каждой стороны присутствует 1 тонкая неопе-
ренная и 1 толстая оперенная щетинка, а у Е. macrurus 2 оперенных
щетинки, т. е. оперена и внешняя из них. На 4-й и 5-й стадии (равняю-
щимся 3-му и 4-му метанауплиусам) науплиусы Е. serrulatus имеют почти
одинаковой длины фуркальные щетинки, а у Е. macrurus внутренние
из них заметно длиннее внешних.
Исследования Cyclopidae пресных вод в последнее время все более
и более уделяют внимание постэмбриональному развитию. Работы в этом
направлении чрезвычайно ценны для разрешения ряда проблем. Только
ЦИКЛ РАЗВИТИЯ
55
при детальном изучении морфологии отдельных стадий метаморфоза
будет возможно построение достаточно убедительной схемы генеалоги-
ческих отношений между родами и видами пресноводных Cyclopidae.
Предложенные до сих пор филогенетические схемы носят спекулятивный
характер; их крупным недостатком является отсутствие опоры на данных
онтогенеза. Изучение влияния различных факторов водной среды
на длительность отдельных стадий развития и на процесс метаморфоза
в его целом, с другой стороны, совершенно необходимо для выяснения
циклов развития (цикличности) в их зависимости от экологических
условий. В частности, разрешение этого вопроса является непременным
условием для выяснения изменений цикличности пресноводных Cyclopidae
в связи с географическим фактором, — проблема, по которой мы пока
имеем крайне скудный фактический материал.
ЦИКЛ РАЗВИТИЯ
Различают виды моноцикличные (1 генерация в год), дицикличные
(2 генерации в год) и полицикличные (более 2 генераций в год). Так
как у Cyclopidae нет смены полового и партеногенетического размно-
жения,1 то термин цикличность здесь понимается в ином смысле,
нежели, например, для Cladocera, и основывается на числе генераций
в год, причем каждой новой генерации предшествует фаза интенсивного
размножения. Темп развития Cyclopidae и число генераций, падающее
на год, находится в очень тесной связи с экологическими условиями
водоема. Один и тот же вид циклопа может иметь неодинаковое число
генераций, в зависимости от того, обитает ли этот вид в постоянном
или во временном водоеме. Виды, полицикличные в постоянном водоеме,
могут быть ди- и моноцикличными в очень мелком, существующем лишь
короткое время, водоеме. Так, например, в мелких, сравнительно дли-
тельно существующих водоемах Средней Европы, A. languidus полицик-
личен, а в быстро усыхающих он дицикличен, так как 3-я генерация
не имеет времени, необходимого для прохождения всех фаз метаморфоза.
Cyclops strenuus во временных водоемах имеет лишь 1 генерацию,
а в постоянных мелких водоемах и прудах встречается весь год и дает
2, а вероятно и более генераций. Cyclops scutifer в мелких промерзающих
лужах горной области Северной Швеции моноцикличен, а в озерах
он моно- и дицикличен.
Возможность изменения числа генераций даже в соседних водоемах,
в зависимости от экологических условий, точно установить которые
весьма часто пока мы не можем, свойственна особенно видам непела-
гическим, обитающим в мелких водоемах, где мы находим большое
разнообразие в сочетании факторов водной среды. Виды пелагические,
в общем, характеризуются гораздо большим постоянством цикла раз-
вития. Так, например, Cyclops abyssorum и С. bohater в озерах средней
Европы строго дицикличны. У видов моноцикличных в некоторых озерах
наблюдается дициклия, иногда лишь тенденция к образованию 2-й гене-
рации {Cyclops scutifer var. wigrensis, отчасти C. scutifer typ.), тогда
как полициклия у этих видов пока не установлена и, вероятно, отсут-
ствует. Исключением является нередко Mesocyclops leuckarti, в некоторых
озерах дицикличный, в других — полицикличный; вид этот, однако,
встречается и в литорали озер и в планктоне более мелких водоемов.
1 Некоторые авторы (Olofsson, 1918) применительно к Copepoda воздерживаются
от термина цикличность, говоря лишь о числе генераций.
56
ВВЕДЕНИЕ
В общем характеристика цикличности родов пресноводных Cyclopidae,
поскольку она изучена у палеарктических видов, сводится к следую-
щему:
Род Cyclops. Преимущественно моно- и дицикличные виды, поли-
циклия редка (с достоверностью установлена лишь для С. strenuus
s. str.).
Род Acanthocyclops. Главным образом ди- и полициклия, в кратко-
временных усыхающих водоемах весьма часто моноциклия — дициклия.
Род Microcyclops. Палеарктические, достаточно изученные виды
дицикличны, М. minuius, вероятно, главным образом моноцикличен.
Род Mesocyclops. Главным образом моно- и дициклия.
Род Ochridacyclops. Цикл развития не изучен.
Род Graeteriella. Цикл развития не изучен. Замедленный темп мета-
морфоза и очень ограниченная продукция яиц (троглобионты!).
Род Speocyclops. Тоже.
Род Macrocyclops. Дициклия и полициклия, моноциклия с точностью
не установлена.
Род Paracyclops. Дициклия и полициклия. Р. fimbriatus, возможно,
также моноцикличен.
Род Ectocyclops. Дициклия, возможно, также полициклия.
Род Eucyclops. Главным образом полициклия.
Род Tropocyclops. Дициклия; в некоторых водоемах, возможно, три
генерации в год.
Явление изменения цикличности в связи с географическим положе-
нием водоема, столь резко и определенно выраженное у Cladocera,
у которых имеет место смена полового и партеногенетического поколе-
ний, у Cyclopidae, судя по имеющимся данным, выражена далеко не столь
отчетливо. Напротив местные экологические факторы для цикличности
Cyclopidae приобретают решающее значение, так что в двух рядом лежащих
мелких водоемах число генераций одного и того же вида может быть,
в зависимости от экологических условий, далеко не одинаковым. Такая
разница в картине цикла развития этих групп ракообразных стоит в тес-
ной связи с характером их размножения. У Cyclopidae отсутствует пар-
теногенез, и широко обеспечена возможность переживания неблагопри-
ятных условий среды в состоянии стадий покоя. Весьма существенно,
что таковые относятся к копеподитной фазе метаморфоза к взрослому
животному и, наконец, к яйцевым мешкам рачка: для Cyclops strenuus
установлены особые покоящиеся яйца (см. главу по экологии). Едва ли
подлежит сомнению, что латентные стадии свойствены всем видам
Cyclopidae. Образование фаз покоя у Cyclopidae, таким образом,
по существу имеет совершенно иной характер, чем у Cladocera,
где оно реализуется исключительно в образовании эфиппия и специаль-
ных покоящихся яиц, которому непременно предшествует относительно
длительная фаза только партеногенетического размножения (при полной
невозможности в этой фазе образовывать покоящиеся яйца) и появление
самцов. Cyclopidae обладают гораздо большими возможностями в обра-
зовании покоящейся фазы, вполне обеспечивающей переживание небла-
гоприятных колебаний экологических условий, причем число генераций
сразу сокращается, и характер цикличности изменяется.
Весьма важным фактором, определяющим темп метаморфоза Cyclo-
pidae, как сказано выше, является температура. Если взять специально
этот фактор, то, в общем, надо принять, что в широтах, где лето
коротко (крайний север Палеарктики, арктические острова) и господ-
КЛАССИФИКАЦИЯ
57
ство низких температур охватывает очень значительную часть года,
будет иметь место тенденция к сокращению числа генераций. Виды,
в таких же водоемах умеренных широт обнаруживающие ди- и полицик-
лию, в условиях арктики становятся моноцикличными. Так на Шпиц-
бергене оба найденных там вида циклопов (A. crassicaudis, Cyclops
strenuus s. lat., вероятно = C. kolensis Lili.) во всех водоемах, не исклю-
чая и постоянных, моноцикличны, причем фаза покоя занимает около
10 месяцев, и только 2 месяца падает на активный период, тогда как
в западной части УССР С. kolensis дицикличен, a A. crassicaudis
в средней Европе ди- и полицикличен (в быстро усыхающих мелких
водоемах может быть и моноцикличен). Литературные указания на раз-
личие в цикличности в высокогорных и арктических водоемах и в водое-
мах равнин умеренных широт касаются, к сожалению, почти исключительно
„Cyclops strenuus", вида сборного, в настоящее время разделенного
на несколько видов с различной биологией; все эти указания нуждаются
в систематической проверке.
КЛАССИФИКАЦИЯ
Семейство Cyclopidae Sars относится к подотряду Cyclopoida, при-
надлежащему отряду Copepoda — веслоногих раков. Здесь мы следуем,
таким образом, системе, приблизительно соответствующей системе
G. О. Sars (1901—1921) и принятой большинством авторов. Классификация
V. Brehm’a (1927) является слишком искусственной и спорной и справед-
ливо была подвергнута основательной критике со стороны Gurney (1933);
эта классификация не нашла последователей вплоть до настоящего
времени. Gurney (1931—1933) склоняется к разделению веслоногих
раков на 7 отрядов (а не подотрядов), причем Cyclopoida рассматрива-
ются в качестве одного из этих отрядов. Вместе с тем отряд Copepoda
получает уже высшее таксономическое значение. Система Gurney, однако,
принципиально не противоречит системе Sars’a и исходит из последней,
внося в нее лишь некоторые изменения.
Подотряд Cyclopoida подразделяется на 3 секции: Siphonostoma
(задняя пара антенн с 1-членистой придаточной ветвью), Gnathostoma
(эти антенны без придаточной ветви, мандибулы имеются, ротовые
конечности не сосательного типа) и Poecilostoma (нет ни мандибул, ни
придаточной ветви задних антенн, сосущие ротовые органы).
Свободноживущие Cyclopoida пресных вод относятся к секции
Gnathostoma. Последняя заключает 3 семейства — Cyclopidae Sars,
Oithonidae Sars и Cyclopinidae Sars.
К семейству Oithonidae принадлежат морские формы (подсемейство
Oithoninae Kief.). Только 1 род — Limnoithona Burckh., содержащий вид
L. sinensis и выделенный Kiefer’oM в подсемейство Limnoithonidae,
представлен в пресных водах Китая (оз. Таху, р. Ян-цзы-цзян). Семей-
ство Cyclopinidae является исключительно морским. Семейство Cyclopi-
dae заключает как морские, так и пресноводные и солоноватоводные
формы. Kiefer (1929) подразделяет семейство Cyclopidae Sars на 3 под-
семейства — Halicyclopinae, Eucyclopinae, Cyclopinae. Это подразделение,
однако, не принимается Gurney’eM, указывающим, что положение родов
Euryte и Halicyclops, относимых Kiefer’oM к первому подсемейству,
неясно, причем, кроме вооружения ног V пары (4 придатка у дисталь-
ного членика), эти 2 рода ничем не сближаются и между собою морфо-
логически они менее близки, нежели род Halicyclops и Cyclops (s. lat.).
58
ВВЕДЕНИЕ
Неопределенность положения Euryte, однако, едва ли вообще может
быть принципиальным возражением против установления подсемейства
и может указывать лишь на спорность положения рода в пределах по-
следнего; в дальнейшем его придется выделить из Halicyclopinae.
Что касается положения рода Halicyclops, резкая обособленность кото-
рого совершенно ясна и никем не оспаривалась со времени его описа-
ния, то соображение Gurney (1933:7) отнюдь не может служить аргу-
ментом против выделения Halicyclops в подсемейство того же семейства,
к которому принадлежит и род Cyclops s. lat., ибо указываемые автором
генетические отношения остаются в силе и для подсемейств. Вместе
с тем нужно заметить, что группа Trifida характеризуется присутствием
3, а не 4 придатков на дистальном членике ног V пары и что наличие
4 придатков является признаком, абсолютно отсутствующим у пресновод-
ных Cyclopidae.1 Род Halicyclops, кроме его морфологических
особенностей, характеризуется совершенно особой биологией — род
является чисто морским и его присутствие в континентальных водах
представляет бесспорно вторичное явление; как раз обратные соотноше-
ния мы имеем для пресноводных Cyclopidae. Эволюция рода Halicyclops
и пресноводных Cyclopidae (=род Cyclops s. lat.), в частности группы
Trifida, очевидно, происходила независимо друг от друга и в
различных направлениях. Поэтому, думается, выделение рода Halicyclops
(и, во всяком случае, близкого к нему рода Neocyclops) в отдельное
подсемейство имеет полное основание.
Gurney (1933:17) отрицает также возможность деления сем. Cyclo-
pidae на подсемейства Eucyclopinae и Cyclopinae. Род Cyclops s. lat.
подразделяется на 2 секции, установленные в 1903 г. Graeter’oM — Tri-
fida и Bifida, отличающиеся присутствием 3 или 1—2 придатков у дисталь-
ного членика ног V пары.
Секция Trifida целиком соответствует подсемейству Eucyclopinae,
секция Bifida целиком укладывается в подсемействе Cyclopinae. О том,
почему секции Trifida и Bifida нельзя рассматривать как подсемейства,
Gurney, в сущности, ничего определенного не говорит, но, очевидно,
невозможность такого подразделения им основывается на том обстоя-
тельстве, что генетические отношения между этими двумя подсемействами
и их родами (подродами) в данное время недостаточно ясны. Однако
Gurney принимает подразделение старого рода Cyclops на ряд подро-
дов, хотя генетические отношения между последними также еще далеко
не выяснены, что, конечно, всецело относится и к принимаемым им
родам Halicyclops, Neocyclops и Cyclops s. lat. При таком положении
систематики Cyclopidae нам кажется, вопреки Gurney, принципиально
вполне возможным, не предрешая в данное время истинных генетических
отношений между подсемействами и родами (подродами) этого семейства,
сохранить систему, предложенную Kiefer'oM и уже принятую целым
рядом авторов. Практическое подразделение на 3 подсемейства гораздо
более удобно, чем подразделение рода Cyclops s. lat на 2 секции с 2
подсекциями, при котором роды Halicyclops и Neocyclops занимают
изолированное положение и, вместе с тем, объем рода получает в системе
всего семейства крайне различное и неравномерное содержание. Разде-
ление прежнего рода Cyclops на указанные секции и подсекции отпа-
дает при разделении этого рода на ряд отдельных родов, независимо
1 Если не считать уродства у Par. fimbriatus (f. quadrispinusa Kief.J и некоторых
других тератологических случаев.
КЛАССИФИКАЦИЯ
59
от их количества. При этом разделении, принимаемом нами в настоящей
работе, каждая из секций будет заключать ряд родов, группируемых
в секцию по общему для них признаку (признакам), а, вместе с тем,
в систематическом отношении объем секции должен приравниваться
таковому подсемейства и этот последний термин должен заменить собою
термин „секция”.
Попытки разделить род Cyclops s. lat. на естественные группы,
не придавая им, однако, значения родов и подродов, делались неодно-
кратно начиная с Rehberg-’a, который в 1880 г. разделил Cyclops на 3
группы, опираясь на метаморфоз, — 1-я группа этого автора заключала
только С. affinis, 2-я — С. fimbriatus, С. poppei и’ С. phaleratus, 3-я —
остальные виды Cyclops s. lat. Fric (1882) рассматривает 2 группы —
Brachypoda (науплиусы, сдавленные с боков, мандибулы крючковидные —
С. serrulatus, С. fimbriatus, С. phaleratus) и Dolichopoda (науплиусы
овальные, мандибулы с длинной плавательной ветвью — С. viridis,
С. albidus, С. fuscus и др.). Vosseler (1886) предложил разделение Cyclops
на группы, положив в основу строение ног V пары и вооружение перед-
них антенн самцов. Schmeil (1892), исходя из подразделений Vosseler’а,
установил 8 следующих групп пресноводных Cyclopidae на основании
членистости и вооружения ног V пары: 1-я группа strenuus-insignis, 2-я
группа leuckarti-oithonoides, 3-я группа bicuspidatus, 4-я группа gracilis-
diaphanus, 5-я группа varicans-bicolor, 6-я группа fuscus-albidus, 7-я
группа serrulatus-prasinus, 8-я группа affinis-fimbriatus. Schmeil разде-
лил эти группы на 2 больших отдела на основании следующих признаков:
1) дистальный членик ног V пары с 3 придатками, задний торакальный
сегмент по бокам всегда с группами щетинок, передние антенны
самцов вооружены сензорными цилиндрами (группы б, 7 и 8);
2) дистальный членик ног V пары с 1—2 придатками, указанный
сегмент по бокам без групп щетинок, передние антенны самцов
вооружены сензорными колбами. Позднее A. Graeter (1903) дал
свое подразделение по тем же признакам на группы Trifida и Bifida,
упомянутое выше, причем группа Trifida соответствует 1-му отделу
Schmeil’H а группа Bifida — его 2-му отделу. Система Schmeil’я
и Graeter’a, однако, не рассматривает указанные группировки как родо-
вые или подродовые. Впервые разделение Cyclops s. lat. на роды
и подроды было предложено Claus’oM (1893), который, учитывая историю
развития антенн и в связи с этим и характер их членистости, устано-
вил 4 рода: 1) Cyclops с подродами Cyclops, s. str. и Macrocyclops’,
2) Microcyclops-, 3) Eucyclops’, 4) Paracyclops. В другой работе Claus
(1893a) изменил эту классификацию в том смысле, что все свои роды
низвел до подродов одного рода Cyclops, причем выделил еще подрод
Heterocyclops (Н. affinis). Система цитируемого автора, однако, имела
тот существенный недостаток, что, исходя из неверного принципа,
в корне разошлась с вполне естественным подразделением A. Graeter’a
на Trifida к Bifida-, она подверглась весьма обстоятельной критике со сто-
роны Mrazek’a (1893) и была принята лишь Forbes’oM(1897), который уста-
новил еще 2 подрода — Orthocyclops (О. modestus) и Homocyclops (Н. ater).
В 1904 г. Brady устанавливает род Ectocyclops (Е. rubescens). В упо-
мянутой обширной монографии Cyclopoida Sars’a автор разделяет род
Cyclops s. lat. на следующие 5 родов: Cyclops s. str., Mesocyclops, Pa-
chycyclops, Leptocyclops и Platycyclops. Непонятным образом Sars совер-
шенно игнорировал работы Claus’a и Graeter’a, ничего не упоминая
О подродах 1-го автора и вообще крайне неполно учитывая литературу
60
ВВЕДЕНИЕ
по Cyclopidae до 1913 г. Вследствие этого, исключая род Mesocyclops,
остальные 3 рода Sars’a отходят в синонимы: Pachycyclops Sars = Macro-
cyclops Claus, Leptocyclops Sars = Eucyclops Claus, Platycyclops Sars =
= Paracyclops Claus. Род Cyclops в понимании Sars’a остается крайне
гетерогенным. Позже Sars (1927) выделяет еще 2 рода — Cryptocyclops
и Afrocyclops, из них 1-й соответствует подроду Microcyclops Claus,
а 2-й — Eucyclops Claus.
Из приведенного краткого исторического обзора можно видеть,
что, в сущности, уже с 1893 г. систематика пресноводных Cyclopidae
встала на путь более дробного деления обширного старого рода Cyclops,
причем основные группировки разных авторов достаточно полно соот-
ветствуют друг другу. Принцип естественного разделения Cyclops
на основании филогенетически весьма важного признака — строения
и вооружения V рудиментарной пары ног, с учетом и других морфоло-
гических признаков был впервые особенно тщательно разработан A. Gra-
eter’oM, и неудивительно, что затем работа этого автора легла в основу
системы Kiefer’a и Сигпеу’я, несмотря на ряд расхождений систем этих
двух авторов. Равным образом и система Sars’a вполне укладывается
в систему A. Graeter’a.
В 1927 г. выходит статья Kiefer’a, в которой автор, опираясь
в основном на систему Graeter’a, намечает ряд родов и подродов,
частью уже установленных предыдущими авторами. Подробное развитие
своей системы Kiefer дает в ряде последующих работ (см. особенно
1928, 1929 и 1937), причем вносит в нее некоторые изменения и допол-
нения. Предложенная цитируемым автором система пресноводных
Cyclopidae разработана им весьма тщательно, и здесь едва ли стоит
повторять приводимые им основания необходимости подразделения ста-
рого рода Cyclops на ряд родов, что, в сущности, является естествен-
ным завершением изложенной выше истории системы этого рода и едва ли
может встретить серьезные принципиальные возражения. Поэтому едва
ли можно согласиться с Gurney’eM (1933), отрицающим возможность
разделения Cyclops на ряд родов, но принимающим разделение на под-
роды, исходя из признаков, положенных в основание групп Trifida
и Bifida. Подобное положение всей системы у этого автора создалось
вследствие отрицания возможности подразделения семейства Cyclopidae
(в смысле Sars’a) на подсемейства, на что мы указывали выше. Вместе
с тем нельзя не согласиться с Gurney’eM, что Kiefer в ряде случаев
допускает слишком узкое понимание рода (подрода) и особенно вида,
явно переоценивая значение некоторых морфологических признаков
и не учитывая их изменчивости, в особенности индивидуального харак-
тера, значительность амплитуды которой вполне доказана работами
ряда авторов.
Подразделение группы видов на ряд родов практически сводится
к установлению определенной для каждого рода комбинации признаков,
которая должна быть общей для всех видов данного рода. Особенную
ценность здесь представляют наиболее постоянные в пределах данного
семейства (подсемейства) признаки. У Cyclopidae к таким принадлежит
рудиментарная, не имеющая определенной функции, V пара ног — обра-
зование, которому всеми систематиками этой группы придается
исключительно важное филогенетическое значение.
Едва ли прав, однако, Kiefer, придавая консервативности вооруже-
ния ног V пары слишком широкое значение вообще в пределах всего
семейства Cyclopidae. В пределах подсемейства Eucyclopinae мы, дей-
КЛАССИФИКАЦИЯ
61
ствительно, в праве говорить о чрезвычайной консервативности строе-
ния и вооружения этой пары ног, но в пределах подсемейства Cyclopi-
пае у целого ряда видов, как наземных, так и свойственных пещерам,
мы наблюдаем значительную вариантность ног V пары, идущую
в направлении редукции одного из придатков дистального членика,
именно внутреннего шипа, который может совершенно редуцироваться,
а также в направлении изменения положения этого шипа. Весьма зна-
чительной изменчивостью, далее, в пределах некоторых групп видов
этого подсемейства отличается членистость рассматриваемой пары ног,
до полного слияния базального членика 2-членистой ноги с задним
торакальным сегментом, причем щетинка, вооружающая этот членик,
переносится на указанный сегмент. В пределах подсемейства Eucyclo-
pinae такое слияние мы наблюдаем лишь у Ectocyclops, причем здесь
одновременной редукции вооружения V пары ног не происходит. В пре-
делах этого подсемейства членистость всех 4 пар плавательных ног
является абсолютно константным признаком (3-членистые экзо- и эндопо-
диты), тогда как в пределах подсемейства Cyclopinae она весьма сильно
вариирует; у некоторых видов эти вариации (редукция числа члеников
с 3 до 2) носят индивидуальный характер {Acanthocyclops languidus,
A. languidoides, некоторые пещерные виды). Эго же мы наблюдаем и отно-
сительно формулы шипов, которая в подсемействе Eucyclopinae отличается
весьма большим постоянством, в противоположность другому подсемей-
ству. Приведенные факты, число которых можно было бы увеличить
и за счет других морфологических признаков, приводят к заключению,
что рассматриваемые подсемейства в данное время находятся в неоди-
наковой фазе вариантности (в широком смысле) и видообразования,
причем особенной константностью характеризуется подсемейство Eucy-
clopinae. Это вполне согласуется с филогенетическими соображениями
Kiefer’a, согласно которому группы Trifida и Bifida, имея общий корень
происхождения, развиваются независимо друг от друга, причем развитие
обеих групп шло различно, отчасти параллельно и конвергентно. К этому
надо прибавить, что, конечно, и темп развития в обоих случаях был
неодинаков. Из этих положений, однако, естественно вытекает неравно-
ценность таксономического значения различных признаков в пределах
этих 2 подсемейств. Там, где в данное время наблюдается широкая
амплитуда изменчивости какого-либо признака, последний может
не иметь того таксономического значения, которое он имеет там, где.
получает характер константного, филогенетически законченного образо-
вания. Эти соображения, конечно, в полной мере относятся и к видам,
в установлении которых, как вытекает из самого понятия „вид", необ-
ходимо руководствоваться наиболее константными признаками (для дан-
ного момента филогенетического развития), всегда отличающими все
особи данного вида от всех особей другого, хотя бы и близкого к первому,
вида.
Принимая во внимание приведенные соображения, вытекающие
из самой динамики эволюции морфологических признаков рассматривае-
мого семейства, объединение групп видов в роды едва ли может встре-
тить возражение в тех случаях, когда каждая из таких групп морфоло-
гически является хорошо обособленной, т. е. между устанавливаемыми
родами имеется определенно выраженный морфологический hiatus. Так
обстоит дело с подсемейством Eucyclopinae. В тех случаях, однако,
когда этот hiatus не может быть установлен достаточно отчетливо,
а вместе с тем в пределах данной группы намечается возможность
62
ВВЕДЕНИЕ
комбинирования видов в отдельные подгруппы, hiatus между которыми,
вследствие неполной законченности (в данный момент исторического
развития всей группы) морфологической дифференцировки признака
(признаков), не может быть достаточно резким и абсолютным, — едва ли
правильно говорить о родах, так как в обоих случаях мы имеем груп-
пировки различного таксономического значения. Во втором случае можно
было бы говорить не о родах, а о подродах, т. е. о категории низшего
таксономического значения, вместе с тем, конечно, расширяя объем
рода, как суммы подродовых группировок. Дальнейшая эволюция такого
рода широкого содержания может привести к дальнейшему обособлению
входящих в него группировок (подродов) и к установлению между
ними достаточно резкого морфологического hiatus’a, допускающего
разделение на морфологически хорошо очерченные роды. Подсемейство
Cyclopinae в современной фазе эволюции его некоторых групп видов
представляет нам пример таких группировок.
Подсемейство Eucyclopinae Kiefer первоначально (1927, 1928, 1929)
подразделил на 4 рода — Macrocyclops Claus, Eucyclops Claus (с подродом
Tropocyclops Kief.), Paracyclops Claus и Ectocyclops Brady; род
Afrocyclops Sars (1927) считался автором синонимом подрода Eucyclops
s. str., подрод Homocyclops Forbes (1897) — синонимом рода Macrocyclops.
Затем Kiefer (1930) выделяет род Thaumasiocyclops, восстанавливает
роды Afrocyclops и Homocyclops, придает родовое значение подроду
Tropocyclops и описывает новый род Ochridacyclops (1937d). Таким
образом, в данное время подсемейство, по Kiefer’y, заключает 9 родов,
тогда как Gurney (1933:35) считает возможным всех Trifida объединить
в один род Cyclops с 5 подродами — Macrocyclops, Eucyclops, Tropocyclops,
Paracyclops и Ectocyclops", подрод Homocyclops. таким образом, сли-
вается с подродом Macrocyclops, подрод Afrocyclops — с подродом
Eucyclops, род Thaumasiocyclops сближается с подродом Paracyclops,
хотя определенно об этом Gurney не высказывается. Устанавливаемые
этим автором 5 подродов настолько резко обособлены, что я, в сущно-
сти, не вижу никаких оснований против придания им значения родов,
тем более, что их разделение здесь может быть строго проведено
уже на основе генеалогически столь важного признака, как характер
и структура вооружения ног V пары обоих полов, очень константного
в пределах видов каждого из этих родов. Особенно гомогенной является
группа видов, объединенных в род Macrocyclops, где даже такой вари-
антный признак, как членистость передних антенн самки, сохраняется
с абсолютным постоянством. Неудивительно поэтому, что Gurney выде-
лил один этот род в самостоятельную подсекцию Polyarthra.
Вместе с Kiefer’oM мы придаем родовое значение и подроду Homocy-
clops Forbes, у которого имеются 1-членистые ноги V пары, при полном
исчезновении базального членика, тогда как у рода Macrocyclops эти
ноги 2-членистые, причем тенденция к сокращению числа члеников
абсолютно отсутствует. Кроме того Homocyclops ater, как отмечают
Forbes, (1897) и Marsh (1909), имеет необычайно широкий передний
отдел тела, тогда как у всех Macrocyclops он значительно уже и строй-
нее. Вследствие выпадения базального членика ног V пары у рода
Homocyclops, эта пара получает сходство с этой же парой ног у Eucy-
clops, но совокупность прочих признаков, в частности строение recepta-
culum, ставит вне сомнений близость первого рода к роду Macrocyclops,
причем сходство в строении и вооружении указанной пары ног у родов
Homocyclops и Eucyclops бесспорно представляет случаи конвергенции
'КЛАССИФИКАЦИЯ
63
и отнюдь не может служить указанием на непосредственные генетиче-
ские отношения этих двух родов. Группе циклопов, объединяемых в подрод
Tropocyclops, вместе с Kiefer’oM вполне можно придать родовое значе-
ние. Группа заключает 8 видов, характеризующихся вполне определен-
ной комбинацией признаков, резко отличающей всю группу от рода
Eucyclops в узком смысле слова. Особенно следует отметить 2 таких
признака, как важнейшие. Во-первых, у Tropocyclops мы постоянно
наблюдаем иное, чем у Eucyclops, вооружение ног V пары — у послед-
него рода внутренний придаток представлен грубым, толстым, иногда
ножевидным крупным шипом, а у Tropocyclops этот шип стройный, тон-
кий, напоминающий щетинку. Во-вторых, у Tropocyclops имеется совер-
шенно своеобразный reeeptaculum seminis, крайне резко отличающийся
от такового всех Eucyclops и представляющий отдельный самостоятель-
ный тип этого органа вообще среди всех представителей подсемейства
Eucyclopinae, отсутствующий в пределах подсемейства Cyclopinae. Уже
эти 2 признака, всегда комбинированные друг с другом, без всяких
затруднений позволяют отличить любой вид Tropocyclops от видов
Eucyclops s. str. Однако имеется еще ряд признаков, свойственных той
или иной из этих 2 групп Eucyclopinae — длина фуркалькых ветвей
у Tropocyclops чаще всего в 1.5—2.5 раза (и не более чем в 4 раза)
превышает их ширину, ряд шипиков на их внешних краях всегда отсут-
ствует, внутренний апикальный шип дистального членика эндоподита ног
IV пары у Tropocyclops обычно очень длинный и пр. Наконец сущест-
венно отметить и то обстоятельство, что уже в пределах рода Tropocy-
clops намечается дифференциация на 2 группы форм Quaterni (формула
шипов 3-4-4-3) и Terni (формула шипов 3-4-3-3), причем, как показал
Kiefer, обе группы имеют совершенно различное географическое рас-
пространение.
Выделение рода Afrocyclops едва ли правильно. Sars выделил
в этот род африканского Eucyclops gibsoni (Brady), основываясь на воору-
жении плавательных ног. У Afrocyclops, по Sars’y, эти ноги сильно разви-
тые и у ног I—III пары дистальный членик экзоподита на внешнем крае
несет не 3, а 2 шипа, так что у Е. gibsoni общее число придатков
этого членика на 1 менее, нежели у остальных Eucyclops. Что касается
числа придатков дистального членика экзоподита ног IV пары (именно 8),
то оно одинаково у Е. gibsoni с таковым Е. serrulatus и Е. macrurus,
тогда как у Е. macruroides этот членик несет 9 придатков. Выделение
рода только на основании числа шипов у экзоподитов плавательных
ног неприемлемо; признак этот не фигурирует в качестве родового
у всех прочих Eucyclopinae, а также и в подсемействе Cyclopinae. Ука-
зываемые Sars’ом различия в вооружении ног V пары у Afrocyclops
и Eucyclops s. str. (направление изгиба внутреннего шипа) явно недо-
статочны для выделения рода;1 ссылка цитируемого автора на морфоло-
гические взаимоотношения самок и самцов у Eucyclops и Afrocyclops
отнюдь не является аргументом в пользу признания последнего само-
стоятельным родом. Как упоминалось выше, Kiefer (1935), на основании
нахождения трех восточно-африканских видов с таким же, как у Е. gibsoni,
вооружением плавательных ног, сохраняет за всей этой группой цикло-
пов значение подрода {Afrocyclops}, с чем можно согласиться вопреки
недавно предложенному тем же автором (1937) толкованию рассматри-
1 К тому же признак этот едва ли реален (см. особенно описание Е. gibsoni var.
abbreviates у Kiefer’a, 1933),
64
ВВЕДЕНИЕ
ваемой группы в качестве самостоятельного рода. Следует категориче-
ски возражать против попытки Lindberg-’a (Rec. Ind. Mus., 37, 1935 :408)
выделить виды Eucyclops без ряда шипиков на внешних краях фуркаль-
ных ветвей (Е. multicolor Lindb. 1935) в отдельный подрод, проме-
жуточный между родами Eucyclops и Tropocyclops. Строение receptaculum
seminis, вооружение V пары ног и плавательных ног таких видов Eucy-
clops явно указывает на их принадлежность к этому роду (подроду
Eucyclops s. str.). Ряд шипиков на внешних краях фуркальных ветвей
в пределах Eucyclops s. str. является признаком, находящимся в фазе
интенсивных вариаций в направлении редукции. Достаточно указать
на эти вариации у Е. serrulatus (см. ниже описание этого вида); вообще
у видов Eucyclops, у которых этот ряд шипиков присутствует, он выра-
жен в весьма различной степени, причем последняя не может быть
положена даже в основу различения видов этого рода.
Подобно роду Macrocyclops, морфологически весьма резко очерчен-
ным является род Ectocyclops, куда относятся типичные бентонические,
лазающие по субстрату, очень близкие друг другу виды, характери-
зующиеся весьма сплющенным в спинно-брюшном направлении передним
отделом тела, укороченным толстым абдоменом, своеобразным воору-
жением внешних краев экзоподитов плавательных ног, особым строением
ног V пары и рядом других признаков. Равным образом и группа видов,
входящих в род Paracyclops, является настолько дифференцированной
от прочих Eucyclopinae, что родовое значение всей этой группы едва ли
может возбуждать сомнения. Вся группа резко и притом вполне опре-
деленными признаками, и в первую очередь строением и вооружением
ног V пары, не слитой с задним торакальным сегментом, отличается
от рода Ectocyclops. В общем род Paracyclops представлен видами,
морфологически более уклоняющимися друг от друга, чем, например,
роды Macrocyclops и Ectocyclops, причем Р. fimbriatus является сильно
вариирующим видом, а Р. eucyclopoid.es приближается к видам рода
Eucyclops', принадлежность этого вида к роду Paracyclops, однако, судя
по структуре и вооруженно ног V пары, может считаться бесспорной
У недавно установленного Kiefer’oM (1930, 1933) рода Thaumasio-
cyclops (1 вид— Th. insulanus. о. Бали) нога V пары напоминает тако-
вую рода Eucyclops, но вооружена лишь двумя придатками. Принадлежность
рассматриваемого циклопа к подсемейству Eucyclopinae бесспорна.
Несмотря на неполноту его описания, совокупность характерных для
него признаков не позволяет отнести этот вид ни к одному из указан-
ных выше родов подсемейства; ближе всего он стоит к родам Eucyclops
и Paracyclops. Kiefer отмечает его близость к Paracyclops eucyclopoides,
но нам кажется, что оба вида очень резко отличаются по ногам V пары
причем ноги Th. insulanus скорее напоминают редуцированные ноги
рода Eucyclops. Однако у всех наземных Eucyclops число придатков (3)
у ног V пары, несмотря на изменчивость их относительной длины),
сохраняется с абсолютным постоянством и тенденции к слиянию един-
ственного членика этих ног с задним торакальным сегментом не наблю-
дается. Таким образом рассматриваемый признак у Th. insulanus нахо-
дится в состоянии, которое едва ли можно сопоставить с изменчивостью
этого признака у всех видов Eucyclops из наземных вод.
Наконец род Ochridacyclops Kief, бесспорно является хорошо диф-
ференцированным родом, на что указывает строение и вооружение ног
V пары, фуркальных ветвей и плавательных ног, а равно и переднего
отдела тела.
КЛАССИФИКАЦИЯ
65
В основу подразделения подсемейства Cyclopinae на роды мы кла-
дем, как и Kiefer, строение ног V пары. При этом ряд группировок
представителей этого подсемейства получает объем, соответствующий
родам подсемейства Eucyclopinae, разделенного выше на роды в основ-
ном по этому же признаку. Такие группы видов подсемейства Cyclopl~
пае вполне могут считаться родами, так как морфологический hiatus
между отдельными группами выражен настолько резко, что всякая воз-
можность их смешивания безусловно отпадает. Таковы роды Cyclops
s. str., Orthocyclops, Mesocyclops, Speocyclops, Muscocyclops, Bryocy-
clops, Graeteriella. Иначе обстоит дело с группами, в системе Kiefer’a
фигурирующими в качестве родов Acanthocyclops, Diacyclops, Megacy-
clops, Microcyclops, Metacyclops, Neutrocyclops, Allocyclops и Thermocy-
clops. Критический обзор признаков, положенных в их основание указан-
ным автором, приводит нас к заключению, что содержание этих родов
является далеко не одинаковым, причем лишь при условии слияния
некоторых из них в один род мы получаем группировки видов, соответ-
ствующие по содержанию родам семейства, вполне дифференцированным
и резко очерченным морфологически.
Вместе с Kiefer’ом всей группе „strenuus", бесспорно, надо придать
-родовое значение — род Cyclops. Он характеризуется прежде всего
характерным строением и вооружением ног V пары. Их дистальный
членик никогда не бывает цилиндрическим и имеет угловатые очертания.
Внутренний шип этого членика всегда прикреплен строго на средине
края, отличаясь своим положением даже от Mesocyclops s. str. и Mega-
cyclops (sensu Kiefer), где этот шип может переноситься на значитель-
ное расстояние кпереди от конца членика, не занимая, однако, столь
определенного положения, как у рода Cyclops. В основании апикальной
щетинки у видов этого рода всегда имеется снаружи группа мелких
шипиков, отсутствующих у прочих Cyclopinae. На спинной поверхности
•фуркальных ветвей постоянно присутствует продольная складка. Перед-
ние антенны с полным числом члеников (17), receptaculum seminis,
исключая лишь С. iisignis, характерной округлой формы, без придатков.
Планктические, частью пелагические виды.
Установленный Forbes’oM род OrthocydopS должен быть сохранен
в его родовом значении, на что прежде всего указывают состоящие из
3 члеников ноги V пары, тогда как у всех прочих пресноводных Cyclo-
pidae число члеников этих ног не превышает 2. По фуркальным ветвям
Orthocyclops напоминает Mesocyclops, дистальный членик ног V пары
с парой очень длинных придатков. Рассматриваемый род был восстано-
влен Kiefer’oM (1927). Orthocyclops ни с каким родом практически
смешиваться не может.
К вполне дифференцированным, хорошим родам надо отнести также
род Mesocyclops Claus (s. lat.). Резкая морфологическая обособленность
Mesocyclops была принята уже Claus’ом, Schmeil’eM, и Graeter’oM, причем
последний объединил все виды этого рода в филогенетически обособлен-
ную подсекцию Chaetophora, противопоставляя ей всех прочих Cyclopi-
nae. Выделение Kiefer’oM подрода Thermocyclops в самостоятельный
род без каких-либо убедительных доводов надо признать безусловно
неверным, так как оно основано на признаке, в высокой степени вари-
антном (место прикрепления внутреннего придатка дистального членика
ног V пары). Непостоянство этого признака Kiefer (1928) отмечает как
раз применительно к „oz'iAonozcfes-Gruppe s. str.“. Kiefer делает ошибку,
кладя в основу рода признак, находящийся в фазе широкой изменяй-
5 Фауна СССР
66
ВВЕДЕНИЕ
вости в пределах группы Thermocyclops, которая таксономически не может
быть эквивалентной 3 предыдущим родам, причем морфологический
hiatus, например, между родами Cyclops и Mesocyclops совершенно иного
характера и иной широты, нежели между Mesocyclops и Thermocyclops.
Вполне установившаяся комбинация других признаков определенно-
указывает на то, что Mesocyclops и Thermocyclops составляют в родовом
отношении единую группу. Согласно изложенным выше признакам родо-
вого подразделения семейства, в данном случае мы имеем пример, когда
можно говорить лишь о 2 подродах, и только в таком содержании весь-
род Mesocyclops получает положение, таксономически равноценное поло-
жению в системе Cyclopinae остальных родов.
Обращаясь к группе циклопов, разбиваемой Kiefer’oM на роды.
Acanthocyclops, Megacyclops и Diacyclops, прежде всего необходимо
отметить, что именно в пределах этой группы мы нередко находим:
чрезвычайно широкую изменчивость целого ряда признаков в пределах
одного и того же вида. Достаточно указать, что именно сюда относятся
столь вариантные виды, как A. vernalis (robustus), A. bicuspidatus,.
A. crassicaudis (ряд вариететов), A. languidus (atavus, disjunctus),.
A. languidoides (ряд вариететов), A. nanus (incertas), A. viridis и A. gigas..
Совершенно не изучены вариации вне-палеарктических видов, но несо-
мненно, что, по крайней мере у многих из них, они имеют место
в широкой степени. Вместе с тем виды, входящие в рассматриваемую
обширную группу циклопов, представляют в ряде признаков очень боль-
шое разнообразие.. При таком морфологическом разнообразии и вариант-
ности признаков разделение на группы, характеризующиеся вполне
определенными в каждом случае комбинациями признаков, является
весьма затруднительным.
Приведем сравнительный анализ признаков родов Megacyclops,
Acanthocyclops и Diacyclops, выдвигаемых Kiefer’oM (1929) в его диагно-
зах.
Членистость ног V пары. У Megacyclops и Acanthocyclops 2 членика,,
у Diacyclops 1—2.
Базальный членик ног V пары. У Megacyclops очень широкий,,
у Acanthocyclops большей частью более суженный, но у некоторых,
видов столь же широкий, у Diacyclops —то же самое.
Дистальный членик ног V пары. У Megacyclops вооружен мелким
шипом близ середины внутреннего края, но шип может прикрепляться:
и близ дистального конца края (субапикально), причем положение шипа
очень вариирует уже в пределах вида (Д. viridis, A. gigas). У Acantho-
cyclops этот шип обычно (но не всегда) крупнее, прикреплен близ самой
вершины членика, иногда ближе к середине, причем шип может занимать
положение, одинаковое с таковым у Megacyclops. Длина рассматривае-
мого шипа у Diacyclops очень вариирует, его положение — близко от
конца внутреннего края членика (субапикальное), что, однако, свой-
ственно и целому ряду видов Acanthocyclops.
Анализ строения ног V пары, таким образом, приводит к заключению,
что разделение трех рассматриваемых групп как родов решительно непри-
емлемо. Разделение невозможно уже по членистости ног, тем более, что
1-членистая нога в пределах группы Diacyclops наблюдается лишь
у вариетета D. nanus (var. incertas), у которого базальный членик цели-
ком погружается в кутикулу торакального сегмента и исчезает. Суще-
ственно, однако, отметить, что Gurney (1933) у D. nanus находил все
переходы от 2-членистой (т. е. с базальным члеником) до 1-членисгой
КЛАССИФИКАЦИЯ
67
ноги, что имеет место также и у D. languidoides var. hypnicola, у которого,
однако, следы базального членика все же остаются, так что о полном
его выпадении говорить нельзя. Равным образом и вооружение ног
V пары не дает оснований к разделению на 3, вполне определенно
характеризующихся по этому признаку, группы; мне, в сущности, непо-
нятно, как мог сам Kiefer, указывающий в ряде работ на крайнюю
вариантность этого признака у циклопов рассматриваемой группы,
положить его едва ли не в основу разделения на 3 рода (первоначально —
подрода).
Членистость плавательных ног. У Megacyclops и Acanthocyclops всегда
по формуле З-З/З-З/З-З/З-З, у Diacyclops З-З/З-З/З-З/З-З или у первых
двух пар число члеников редуцируется до 2. Так формула ног D. langui-
dus 2-2/3-2/3-3/3-3, у var. atavus З-З/З-З/З-З/З-З, у D. languidoi-
des 2-2/3-2/3-3/3-3, тоже у D. abyssicola и D. tantalus. У большинства
Diacyclops формула плавательных ног З-З/З-З/З-З/З-З. Несмотря на частич-
ную редукцию, у Diacyclops обе задние пары плавательных ног все же
сохраняют 13-членистость экзо- и эндоподитов.
Вооружение дистального членика экзоподита плавательных ног.
У I—IV пар ног Megacyclops, кроме шипов, имеется по 4 щетинки,
у Acanthocyclops большей частью 4, иногда 5 щетинок, у Diacyclops
чаще всего по 4 щетинки, у I пары ног иногда имеется 5-я щетинка.
Вооружение эндоподитов, в частности дистального членика ног IV пары,
у всех 3 групп весьма однотипно.
Передние антенны у Megacyclops 17-. у Acanthocyclops 11—17(18)-,
у Diacyclops 10—17-членистые, у всех 3 без гиалиновых пластинок
на дистальных члениках. Этот признак, как и предыдущие 3, совершенно
очевидно, в основу разделения 3 родов положен быть не может.
Вооружение внутренних краев фуркальных ветвей. Этот признак
Kiefer’oM кладется как один из главнейших в основу отделения Mega-
cyclops от 2 остальных групп. У Megacyclops края несут ряд волосков,
которых, как правило, нет у 2 других групп. Однако сам Kiefer описал
A. troglophilus, A. stammeri и A. pilosus, у которых ряды волосков
имеются. Едва ли в данном случае этот признак можно рассматривать
как родовой. Для рода Cyclops s. str. он, действительно, весьма характе-
рен, хотя у разных видов выражен в неодинаковой степени. Однако
в основу рода Cyclops s. str. этот признак не кладется, и весь род
характеризуется по ногам V пары, т. е. по филогенетически весьма
убедительному признаку, который, однако, не может быть положен
в основу разделения на роды группы Megacyclops — Diacyclops. Весьма
существенно, что в других случаях волоски на внутренних краях
фуркальных ветвей являются отнюдь не родовым признаком. Таков,
например, род Mesocyclops, где их отсутствие или присутствие наблю-
дается даже у одного и того же вида. В пределах рода Macrocyclops
их нет у М. albidus, но они имеются у остальных 3 видов. Существенно
отметить, что рассматриваемый ряд волосков у A. viridis, по нашим
наблюдениям, появляется лишь на последней (5-й) копеподитной стадии;
это же нашел Gurney (1933:44, рис. 1261) у С. strenuus.
Строение receptacnlum seminis. В пределах Acanthocyclops и Dia-
cyclops строение кэайне разнообразно. Пользоваться рассматриваемым
признаком для разделения группы Megacyclops — Diacyclops тем более
рисковано, что в пределах всей группы строение receptaculum seminis
может быть совершенно различным даже у близко стоящих видов,
причем, например, в пределах рода Diacyclops (sensu Kiefer) мы находим
5*
68
ВВЕДЕНИЕ
явно различные типы этого органа (сравни, например, D. bicuspidatus,
D. crassicaudoides, D. langaidus), а в роде Acanthocyclops (sensu Kiefer)
встречаем столь различные типы reeeptaculum seminis, как, например,
у A. vernalis и A. sensitivus. В то время как, например, род Mesocy-
clops действительно характеризуется общим для всех видов типом
reeeptaculum seminis, группа Megacyclops—Diacyclops характеризуется
у различных видов столь различным строением этого органа, что считать
последний родовым признаком в пределах каждого из 3 разделов этой
группы решительно невозможно. >
Приведенный разбор морфологических признаков группы Megacy-
clops— Diacyclops приводит, таким образом, к заключению, что разбить
ее на 3 установленных Kiefer’oM рода не представляется возможным.
Мы считаем поэтому единственно правильным рассматривать всю группу
целиком, как отдельный род Acanthocyclops. Несмотря на разнообразие
входящих в него компонентов, весь род Acanthocyclops в таком толкова-
нии не может быть смешан ни с одним из других родов подсемейства
Cyclopinae, причем получает содержание, гораздо более соответствую-
щее и равноценное другим родам этого подсемейства, нежели каждый
из 3 вышеприведенных родов Kiefer’a. Мы в то же время не считаем
возможным, согласно приведенному анализу признаков, разбивать этот
род на подроды, так как в данном случае не находим сколь-либо
определенного признака (или их комбинации), который мог бы точно
охарактеризовать каждый из таких подродов в целом. Мы рассматриваем
весь род Acanthocyclops как род, в данный момент находящийся в фазе
интенсивного видообразования, причем сколь-либо четкое разделение на
морфологически уже достаточно обособленные группировки, которые
таксономически можно было бы оценивать как подродовые, в пределах
этого рода еще не имеет места.
Принимаемые Kiefer’oM роды Microcyclops, Metacyclops и Neutro-
cyclops составляют группу циклопов, охватывающую около 50 видов.
Вся группа в целом хорошо характеризуется всегда 2-членистыми ногами
первых 4 пар и всегда 1-членистыми ногами V пары, причем принадлежа-
щая выпавшему базальному членику этих ног щетинка сохраняется
на заднем торакальном сегменте (торакальная щетинка). Вооружение
дистального членика V пары ног представлено апикальной щетинкой
и внутренним, как правило, очень коротким шипом, у некоторых видов
совершенно редуцированным; шип занимает субачикальное положение,
иногда переносясь значительно кпереди, к середине внутреннего края
членика. Для всей группы Sars предложил (1927:129) родовое название
Cryptocyclops, причем обратил внимание на отличие этого рода от про-
чих Cyclopidae1 по членистости плавательных ног.
Принимая за основу строение ног V пары и 2-членистость всех Пар
плавательных ног, мы, соответственно тому, что сказано о предыдущем
роде, всю эту группу циклопов (1933) рассматриваем как один род
Microcyclops. В таком объеме род Microcyclops резко отличается от близ-
кого к нему рода Acanthocyclops и вместе с тем, принимая во внимание
строение фуркальных ветвей, присутствие торакальной щетинки и вооруже-
ние ног V пары, от других родов Cyclopinae с редуцированным числом
члеников плавательных ног. Наиболее близким к Microcyclops в роде
Acanthocyclops является A. nanus var. incertus Wolf, у которого базаль-
1 Здесь Sars впадает в некоторую ошибку, так как 2-членистые ноги I—IV пар
свойственны и некоторым другим родам.
КЛАССИФИКАЦИЯ
69
ный членик ног V пары совершенно слит с торакальным сегментом.
Однако, как указывалось выше, у A. nanus встречаются все переходы
от вполне дифференцированного базального членика до полного его
выпадения, в то время как формула ног (2-2/3-2/3-3/3-3) сохраняет свое
постоянство и обе задние пары всегда 3-членистые.
Вместе с Gurney’eM роду Microcyclops мы придаем расширенное
толкование, включая в него весь род Metacyclops Kiefer. Сравнение
двух диагнозов, даваемых Kiefer’oM для родов Microcyclops и Metacyclops,
убеждает нас в том, что речь в них идет об одном и том же роде.
Замечание Kiefer’a в первом диагнозе о reeeptaculum seminis у Metacyclops
лишено значения, так как сильно развитой задний отдел этого органа
мы находим и у некоторых видов, Kiefer’oM относимых к роду
Microcyclops" отметим, что строение reeeptaculum seminis у целого
ряда видов Microcyclops неизвестно. Несомненно лишь, что оно в преде-
лах этого рода очень разнообразно, так же, как в пределах рода Acantho-
cyclops й даже Cyclops s. str., куда Kiefer сам относит С. insignis
Cis., несмотря на совершенно иной, чем у прочих Cyclops, reeeptaculum
seminis. Строение этого органа также разнообразно и у видов рода
Metacyclops (sensu Kiefer). Достаточно упомянуть М. gracilis Lili. (rec.
seminis по типу Mesocyclops) и М. dengizicus Lep. (rec. seminis по типу
A. bicuspidatus). Разделение Microcyclops и Metacyclops по этому при-
знаку безусловно невозможно.
В другой работе (1929) Kiefer пытается установить для рода
Metacyclops еще 1 по мнению этого автора характерный признак —
значительная длина внутреннего апикального шипа дистального членика
эндоподита ног IV пары. Очевидно, однако, что сам Kiefer не настаи-
вает на этом признаке, так как в одновременно вышедшем обзоре
Cyclopidae в „Tierreich" в диагнозах Microcyclops и Metacyclops о нем
уже не упоминает. Отметим, что у М. arnaudi Sars, относимого
Kiefer’oM к Metacyclops, указанный шип короче дистального членика,
тогда как у М. cunnigtoni Sars, относимого к М crocyclops, он заметно
длиннее и почти равен длине дистального членика эндогодита. Длинный
внутренний апикальный шип имеется также у М. inopinatus Sars
(= Microcyclops по Kiefer’y), где он заметно длиннее дистального членика,
причем у Meiacyclops necessarius Kief, он и абсолютно и относительно
членика нисколько не длиннее, чем у М. inopinatus. В последнее время
Kiefer (1936а, 1937) склоняется к разделению рода Microcyclops на 2 под-
рода— Microcyclops s. str. и Cryptocyclops Sars, — невидимому, главным
образом, на основании строения reeeptaculum seminis и вооружения ног
IV пары. О невозможности такого подразделения на основании строения
reeeptaculum seminis достаточно сказано выше, и сам Kiefer, как мы
думаем, с полным на то основанием, ранее не считал возможным по
этому признаку разделять этот род на 2 подрода. Разделение на основа-
нии вооружения плавательных ног, при одинаковой формуле шипов
экзоподитов (3-4-4-3) и одинаковом числе шипов и щетинок (2 ч-4)
у дистального членика эндоподита ног IV пары у М. bicolor и М. vari-
cans и близких к ним видов, крайне рискованно, тем более, что устано-
вить корреляцию со строением reeeptaculum seminis в пределах всего
рода невозможно. Отметим, кроме того, что нам крайне мало известно
о вариациях длины обоих шипов дистального членика эндоподита ног
IV пары у внеевропейских видов; возможность разделения по этому
признаку М. bicolor и М. varicans сама по себе еще недостаточна
для подразделения всего рода в целом на 2 подрода.
70
ВВЕДЕНИЕ
Описанный Lowndes’oM (1934) из Бразилии Cryptocyclops brevifurca.
Kiefer’oM выделяется в отдельный род Neutrocyclops. Признаки этого
рода, по Kiefer’y (1936а), следующие: 1) 11-членистые антенны; 2) из
апикальных шипов дистального членика эндоподита ног IV пары внутрен-
ний длинный, внешний короткий; 3) соединительная пластинка и базопо-
диты ног этой пары, по мнению Kiefer’a, особого строения; 4) ноги V
пары имеют очень маленький членик с 1 апикальной щетинкой, торакаль-
ная щетинка присутствует; 5) receptaculum seminis близок к молото-
образному; 6) фуркальные ветви,очень короткие, их вооружение близко
к таковому у Thermocyclops.
11-членистые передние антенны свойственны и Microcyclops (вклю-
чая Metacyclops). В строении и вооружении эндоподита ног IV пары
я не вижу абсолютно никакой разницы с М. gracilis Lili., где имеется
столь же удлиненный дистальный членик и совершенно такое же отноше-
ние длины его апикальных шипов, из которых внутренний очень длин-
ный. Равным образом у рассматриваемых видов нет никакой, сколь-либо
существенной разницы в структуре соединительной пластинки базипо-
дитов и в их вооружении. Строение и вооружение ног I пары также
•одинаковы у обоих видов. Ноги V пары у С. brevifurca построены,
безусловно, по их типу у Microcyclops и отличаются лишь величиной
членика; очень близкое строение этих ног мы имеем, например,
у М. varicans var. rubellus, где членик лишь относительно длиннее.
Однако длина этого членика не может служить ни в качестве родового,
ни в качестве подродового признака, а редукция его внутреннего шипа
свойственна и ряду видов Microcyclops (sensu Kiefer). Receptaculum
seminis у C. brevifurca построен по типу М. gracilis; такой же receptacu-
lum seminis имеется и у некоторых других видов Microcyclops {Metacy-
clops sensu Kiefer). Повидимому Kiefer придает в роде Neutrocyclops
особое значение укорачиванию и вооружению фуркальных ветвей —
он пишет даже, что подобного соотношения длины их апикальных
щетинок, исключая Thermocyclops, мы вообще не имеем у всех других
Cyclopinae. Однако вооружение фуркальных ветвей у С. brevifurca
крайне близко к таковому у М. gracilis. Вместе с тем необходимо иметь
в виду, что длина фуркальных ветвей и соотношение длины их апикаль-
ных щетинок у близких видов одного и того же рода могут вообще
быть различными. Признаки эти во всей системе Cyclopidae лишены
родового и подродового значения. Анализируя признаки С. brevifurca,
мы, таким образом, не находим достаточных оснований выделять этот
вид из рода Metacyclops (в смысле Kiefer’a), который нами относится
выше к роду Microcyclops Cis.
До последнего времени в крайне запутанном положении оставалась
систематика некоторых групп Cyclopinae, обитающих в условиях специфи-
ческих биотопов, — влажные мхи, скопление вод в листьях Bromeliaceae,
некоторые троглобионты. В 1926 г. Brehm предлагал выделить в особый
род Graeteriella Cyclops operculatus Chapp. и 3 троглобионтов —
Cyclops racovitzai Chapp., C. troglodytes Chapp. и C. unisetiger Graeter.
Kiefer в 1927 г. установил род Bryocyclops, охватывающий специально
обитателей влажных мхов и отчасти скоплений вод в листьях наземных
высших растений. Остальные виды рассматриваемой группы циклопов
относились, главным образом, к подроду Diacyclops. Недавно Kiefer
(1937) дал весьма обстоятельную полную ревизию всех перечисленных
форм, внеся значительную ясность в их систематику, тщательно им
Разработанную. В основу группировок этих форм Kiefer, с полным
КЛАССИФИКАЦИЯ
77
на то основанием, положил строение и вооружение ног V пары, по
возможности в комбинации с некоторыми другими признаками. В оконча-
тельной форме система Kiefer’a сводится к установлению 4 родов —
Graeteriella, Bryocyclops, Speocyclops Kief, и Muscocyclops Kief.
Перечисленные роды резко дифференцированы по структуре ног
V пары, так что их разделение без труда проводится уже по этому,
для систематики Cyclopidae столь важному, признаку. Родовое значение
этих групп едва ли может встретить принципиальные возражения; оно
подкрепляется и иными морфологическими признаками, а также дан-
ными географического распространения. Изолированность и специфич-
ность биотопов, населяемых этими группами циклопов, дает основание
предполагать образование новых родов и видов в таких специальных
условиях.
Принимая систему Kiefer’a, при которой содержание установленных
им 4 родов не противоречит объему других родов Cyclopinae, в частно-
сти родов Acanthocyclops и Microcyclops, мы должны, однако, сделать
следующие к ней замечания.
Ввиду совершенно одинакового с С. unisetigera строения ног V пары
(1 членик с 2 придатками, отсутствие торакальной щетинки) и притом
однотипного строения receptaculum seminis, принадлежность к роду
Graeteriella описанного Kiefer’oM из пещер Верхнего Рейна Diacyclops
laisi можно считать достоверной, — только у этих 2 видов мы находим
-такие ноги V пары. Однако по другим признакам оба вида значительно
отличаются, на что обращает внимание и Kiefer. Оба вида имеют раз-
личное строение генитального сегмента и вообще абдомена, различное
вооружение фуркальных ветвей, различное развитие анальной пластинки,
неодинаковую членистость плавательных ног. Мы полагаем, поэтому,
-что имеются достаточные основания к выделению G. laisi в отдельный
подрод, называемый нами Paragraeteriella; выделение рода, ввиду пол-
ной идентичности структуры и вооружения ног V пары, здесь, конечно,
невозможно. Признаки, отличающие подроды Graeteriella s. str. и Para-
. graeteriella, следующие (no J 5).
. Г енитальный
сегмент
Graeteriella s. str.
Очень широкий, его длина
значительно менее ширины
Paragraeteriella mihi
Нормального строения, сильно
суженный кзади, его длина при-
близительно равна его ширине
в переднем отделе и значитель-
но более таковой в заднем
•Фуркальные
ветви
Анальная
пластинка
С 3 апикальными придатками,
из них средний очень развит,
другая срединная щетинка отсут-
ствует, крайние придатки в виде
толстых шипов, из них внутрен-
ний гораздо короче внешнего.
Треугольная, очень сильно раз-
витая, выдающаяся далеко кзади
между фуркальными ветвями,
иногда с зазубренным краем
отделе.
С нормальным вооружением из
4 придатков, обе средние щетин-
ки нормально развитые, из край-
них щетинок внутренняя почти
вдвое длиннее внешней.
Умеренно развитая, полукруглая,
кзади не доходящая до основа-
ния ветвей, гладкая
Членистость
плаватель’
ных ног
У О 2-2/2-2/2-2/2-2,
•у 5 2-2/2-2/2-2/3-2
У J 2-2/3-2/3-3/3-3
Gr. unisetigera является видом гораздо более измененным (специа-
-лизованным), нежели Gr. laisi. По членистости плавательных ног подрод
72
ВВЕДЕНИЕ
Paragraeieriella близко напоминает род Acanthocyclops, с которым обнару-
живает известное сходство и в строении абдомена и в вооружении,
фуркальных ветвей. К сожалению, Kiefer не дает рисунка общего видя
Gr. laisi, который, вероятно (судя по абдомену, в частности по гениталь-
ному сегменту), не имеет столь резко выраженного гарпактикоидного
типа, как у Gr. unisetigsr.
Описанной Kiefer’oM (1933) Allocyclops chappuisf, как указывает
сам автор, близок к Bryocyclops и Metacyclops, особенно к 1-му роду..
У Allocyclops и Bryocyclops совершенно одинаковые ноги V пары: отсут-
ствие обоих члеников, прикрепление 3 придатков к заднему торакаль-
ному сегменту, причем только у этих родов мы находим столь свое-
образные рудиментарные ноги; у обоих сильно развитая анальная пла-
стинка, очень близкое по типу строение receptaculum seminis, очень-
короткие фуркальные ветви. Указывая на сходство обоих родов в этих
признаках, Kiefer, однако, считает, что по величине циклопа (самка:
Allocyclops 0.72 — 0.74 мм), по менее редуцированным плавательным
ногам и по вооружению фуркальных ветвей род Allocyclops не может
считаться идентичным с родом Bryocyclops. Таким образом эти признаки,
по Kiefer’y, будут в данном случае родовыми. Длина самок Bryocyclops:
очень различна, чаще всего от 0.32 до 0.5 мм; однако длина самок
Br. difficilis Kief. 0.5—0.68 мм, а самки Br. phyllopas Kief. 0.70 мм
(везде без фуркальных щетинок), что уже очень близко к длине самок.
Allocyclops- chappuisi. Ссылка Kiefer’a на этот признак поэтому лишена
основания. Членистость плавательных ног у Allocyclops по формуле
2-2/2-2/2-2/2>2; 6 видов Bryocyclops имеют такую же формулу
ног, у меньшинства наблюдается редукция числа члеников ног IV пары,
до 1. Признак этот, следовательно, не может служить аргументом про-
тив объединения Allocyclops и Bryocyclops в 1 род. Если обратиться
к вооружению ног IV пары, то у Allocyclops дистальный членик эндопо-
дита вооружен 6 придатками (2 апикальные шипа -+- 4 щетинки), а у Bryo-
cyclops 4 или 5 (как правило, 1 шип -+- 3 или 4 щетинки). Однако у очень
обстоятельно описанного Kiefer’oM Br. africanus этот членик (IV ноги)
несет 2 крупных апикальных шипа и 3 щетинки, т. е. лишь на 1 щетинку
менее, чем у Allocyclops. Вместе с тем вооружение экзоподита IV пары
ног у Br. africanus также отличается от прочих Bryocyclops — дисталь-
ный членик вооружен 8 (а не 7) придатками. Как указывалось выше,.
Kiefer отмечает различие обоих родов по фуркальным ветвям. Несо-
мненно, однако, что различия в этом признаке у Allocyclops и Bryocy-
clops гораздо менее выражены, чем у 2 упомянутых выше видов рода
Graeteriella, что не мешает Kiefer’y без оговорок объединять их в 1 род..
У видов одного и того же рода фуркальные ветви могут быть очень
различными. У Allocyclops строение и вооружение фуркальных ветвей,,
в общем, конечно, менее специализовано, нежели у Bryocyclops, но
различать здесь 2 рода по этому признаку едва ли возможно. У раз-
личных видов Bryocyclops анальная пластинка развита далеко не одинаково
сильно. В пределах рода этот признак, однако, может вариировать в весьма
значительной степени. Достаточно упомянуть род Speocyclops и род
Acanthocyclops. Приведенные соображения приводят к тому, что на пере-
численных признаках едва ли можно настаивать как. на родовых, когда
имеется полное сходство в строении HorVпары, прямо указывающее на поло-
жение в пределах одного и того же рода. Принимая во внимание, что решаю-
щее значение здесь должен иметь именно этот, столь важный в генеалоги-
ческом отношении, признак, в данном случае к тому же коррелятивный:;
КЛАССИФИКАЦИЯ
73'
со строением receptaculum seminis и с членистостью всех 4-х пар
плавательных ног, мы относим Allocyclops chappuisi к роду Bryocyclops.
Среди видов рода этот вид является наименее специализованным,
наиболее сохраняющим морфологические черты „нормальных" циклопов,,
напоминающие Microcyclops (sensu mihi, Metacyclops sensu Kiefer), —
строение и вооружение фуркальных ветвей, относительно слабое разви-
тие анальной пластинки, сохранение щетинки на внутреннем углу 1-го
базального членика ног IV пары, отсутствие выростов на соединитель-
ной' пластинке этой пары ног. Вместе с тем необходимо принять, что-
в пределах рода Bryocyclops последние 2 признака не имеют абсолют-
ного значения, что, в сущности, относительно 2-го из них признает
и Kiefer, описав Br. africanus. Несомненно род Bryocyclops охватывает
еще мало изученную группу циклопов, и дальнейшие исследования
населенных ими весьма специфических биотопов дадут ряд новых видов;,
очень вероятно, что присутствие щетинки на базальном членике IV пары
окажется признаком, помимо „Allocyclops", свойственным и некоторым
другим видам Bryocyclops. Зоогеографически отнесение All. chappuisi
к Bryocyclops препятствий не встречает. Этот вид найден в Западной
Африке, где найден и Br. africanus (в Восточной Африке найдено-
несколько видов), в источнике близ Daloa. Очень вероятно, что
All. chappuisi обитает также в формации мхов, так как, подобно другим:
Bryocyclops, это, очевидно, типично бентический циклоп.
Вследствие слияния Allocyclops с Bryocyclops, видовое название
chappuisi может быть оставлено лишь для 1 вида последнего рода.
Оставляя таковое за Br. chappuisi Kiefer (Zool. Anz., бб), описанным
в 1928 г. с о-ва Явы, мы изменяем видовое название американского
вида на Bryocyclops kieferi nom. nov (= Allocyclops chappuisi Kiefer,
Arch. f. Hydr. 26., 1933).
Kiefer (1927—1937)
Подсем. Eucyclopinae
Род Macrocyclops Claus
„ Homocyclops Forbes
„ Eucyclops Claus
„ Afrocyclops Sars
„ Tropocyclops Kief.
„ Paracyclops Claus
„ Ectocyclops Brady
„ Thaumasiocyclops Kief.
. Ochridacyclops Kief.
Подсем. Cyclopinae
Род Cyclops O. F. Mull.
„ Megacyclops Kief.
„ Acanthocyclops Kief.
„ Diacyclops Kief.
„ Microcyclops Claus
„ Metacyclops Kief.
„ Nc.utrocyclops Kief.
„ Bryocyclops Kief.
„ Allocyclops Kief.
„ Muscocyclops Kief.
„ Speocyclops Kief.
„ Graeteriella Brehm
„ Orthocyclops Forbes
„ Mesocyclops Claus
„ Thermocyclops Kief.
Наша система
Подсем. Eucyclopinae
Род Macrocyclops Claus
„ Homocyclops Forbes
„ Eucyclops Claus
„ Tropocyclops Kief.
„ Paracyclops Claus
„ Ectocyclops Brady
„ Thaumasiocyclops Kief..
„ Ochridacyclops Kiefer
Подсем. Cyclopinae
Род Cyclops O. F. Mull.
„ Acanthocyclops Kief.
„ Microcyclops Claus
„ Bryocyclops Kief.
Muscocyclops Kief.
„ Speocyclops Kief.
„ Graeteriella Brehm
„ Orthocyclops Forbes
„ Mesocyclops Claus
.74
ВВЕДЕНИЕ
В вышеприведенной таблице показано соотношение системы обоих под-
семейств пресноводных Cyclopidae Kiefer’a и предлагаемой намина основа-
нии соображений, высказанных выше.
РАСПРОСТРАНЕНИЕ И ИСТОРИЯ ФАУНЫ
В противоположность пресноводным Calanoida, где отдельные роды
и виды имеют резко выраженный, нередко весьма суженный ареал оби-
тания, пресноводным Cyclopoida до последнего времени не придавалось
сколь-либо существенного зоогеографического значения, причем указы-
валось на космополитичность и убиквизм многих видов этой группы
ракообразных. Нужно заметить, что уже данные литературы конца
XIX — начала XX века едва ли оправдывали подобное мнение. Наряду
с широко распространенными видами, уже в указанное время были изве-
стны виды, свойственные какой-либо одной зоогеографической области.
Исследования последних двадцати лет показали, что в целом ряде
случаев мнение о космополитизме Cyclopidae основывалось на неверных
определениях,—-более или менее близкие морфологически виды прини-
мались ошибочно за один, известный ранее йз Палеарктики. За последнее
время из различных областей, в частности южного полушария, описано
большое количество новых видов, а частью и новых родов. Часть этих
видов, возможно, представлят таксономические единицы низшего порядка,
часть должна быть отнесена в синонимы. Несомненно, однако, что для
целого ряда вновь описанных форм видовое их значение можно считать
бесспорным; вместе с тем наши современные данные по фауне Cyclopoida
отдельных областей уже таковы, что, если не в деталях, то по крайней
мере в общей форме позволяют отрицательно ответить на вопрос
о космополитизме видов, а отчасти и родов этой группы ракообразных.
В нескольких работах Kiefer дает попытку зоогеографически характери-
зовать роды и отчасти виды пресноводных Cyclopoida (1931, 1932, 1933).
Нижеследующая табл. 6 представляет собой несколько измененную
Таблица 6
Космо- политы Северное полушарие Южное полушарие
Macrocyclops .... Homocyclops .... Eacyclops s. str. . . . Подр. Afrocyclops. . Tropocyclops .... Paracyclops .... Ectocyclops Thaumasiocyclops . . Ochridacyclops . . Cyclops ...... Acanthocyclops . . . Microcyclops .... Bryocyclops Muscocyclops .... Speocyclops . ... . Graeteriella . ... Orthocyclops .... Mesocyclops s. str. . Подр. Thermocyclops • —
РАСПРОСТРАНЕНИЕ И ИСТОРИЯ ФАУНЫ
75
и дополненную таблицу этого автора, предложенную в одной из его работ.
Толстая черта показывает, какому полушарию исключительно или глав-
ным образом свойствен данный род, тонкая короткая черта обозначает
лишь присутствие данного рода в том или ином полушарии; в левой
графе отмечен космополитизм.
В следующей таблице 7 дается зоогеографический анализ видового
состава отдельных родов.
Таблица 7
Общее число видов Число ви- дов с кос- мополит- ным рас- простране- нием Процентное соотношение космополитов к общему числу видов Примечания
Macrocyclops ...... 4 2 50 Космополиты М. albi-
dus и М. disiinctus.
Homocyclops ...... 1 — — Известен из Неаркт. и Неотроп, областей
Eucyclops s. str. .... (Бразилия).
ок. 40 1 2.5 Космополит Е. serru-
5 0 0 latus.
.Подр. Aphrocyclops . . .
Tropocyclops . ... . 8 1 12.5 Космополит Т. prasi- nus, который однако
отсутствует на севере
Голарктики.
Paracyclops ...... Ectocyclops ...... 6 3 50 Космополит Е. phale-
6 1 16.7
ratus.
Thaumasiocyclops . . . 1 0 0 Европа (оз. Охрида).
Ochridacyclops . ... . 1 0 0
Cyclops ........ Acanthocyclops .... ок. 20 ок. 40 0 4 0 10
.Microcyclops ...... ок. 50 2 4 Космополиты М. va-
10 0 0 ricans и М. bicolor.
Bryocyclops ......
Muscocyclops . ... 2 0 0 Европа, троглобионты.
Speocyclops ...... 12 0 0
Graeteriella 2 0 0
Orthocyclops ...... 1 0 0 Неарктика.
Mesocyclops s. str. . . . 13 1 8.3 Космополит М. leu-
ckarti.
Подр. Thermocyclops . . ок. 30 2 6.7
Приведенная таблица показывает, что можно говорить о космополи-
тизме лишь 8 родов пресноводных Cyclopidae-. из подсемейста Eucyclo-
pinae— 5, из подсемейства Cyclopinae — 3. Основываясь на видовом
составе (число видов в данном роде), можно установить различную сте-
пень космополитизма отдельных родов. Только 2 рода (Macrocyclops
и Paracyclops) отличаются высокой степенью (50% видового состава)
космополитизма. В остальных 6 родах число космополитных видов
очень невелико, не свыше 17%.
Анализ всего семейства и его обоих подсемейств приводит к сле-
дующему результату (см. табл. 8).
76
ВВЕДЕНИЕ
Таблица 8'
Число родов Число космопо- литных родов % родов с космопо- литизмом В 50% Число видов (около) Число космополитных видов (в % к общему их числу)
Подсемейство Eucyclo- pinae 8 5 25 72 5.5
Подсемейство Cyclopinae 9 3 0 180 5
Все семейство Cyclopidae 17 8 11.8 252 5.2
Приведенные цифры, конечно, приблизительны — нахождение новых
видов в дальнейшем будет их несколько изменять. Едва ли однако может
вызывать сомнения основной результат этой таблицы, показывающий^
что говорить о значительном космополитизме даже отдельных родов
семейства едва ли возможно — космополитных родов в нем, приблизи-
тельно, 50% от всего числа родов. Что касается до видового состава
семейства, то лишь около 5°/0 всего числа входящих в него видов могут
быть отнесены к космополитам.
Ниже мы даем краткую зоогеографическую характеристику фауны
пресноводных Cyclopidae основных зоогеографических областей земного
шара. Несмотря на известную неполноту данных (особенно для Австра-
лийской области), такая характеристика может наметить основные черты
состава фауны Cyclopidae каждой отдельной области.
Палеарктическая, область. Богато представлены видами
космополитические роды Macrocyclops (3 вида из 4) и Paracyclops
(3 вида из б). По преимуществу голарктический род Acanthocyclops
особенно богато представлен именно в Палеарктике: из 40 видов рода
в ней обитает 23 вида, из которых 14 только палеарктических (из них
10 троглобионтов). Другой чрезвычайно характерной чертой Палеарк-
тики является присутствие рода Cyclops s. str., насчитывающего в ней
около 18 форм, из которых только 2—3 формы (1 вид с достовер-
ностью) указаны для Северной Америки; 2 вида этого рода известны
из Ориентальной области; в других областях род отсутствует. Только
в Палеарктике пока найдены все виды троглобионтных родов Speocy-
clops и Graeteriella (оба лишь в Европе) и род Ochridacyclops. Отрица-
тельные черты фауны Cyclopidae Палеарктики: гораздо беднее, чем
в других областях (Южное полушарие), представлены роды Eucyclops
(только 5 видов из 40, отсутствует подрод Afrocyclops), Tropocyclops
(1 вид из 8), Ectocyclops (1 вид из 6), Microcyclops (б видов из 50),
Mesocyclops с подродом Thermocyclops (7 видов из 42), отсутствуют
роды Homocyclops, Thaumasiocyclops, Bryocyclops, Muscocyclops,.
Orthocy clops.
Неарктическая область. Все 3палеарктических видаMacrocy-
clops известны и из Северной Америки. В ней широко распространен
монотипный род Homocyclops. Фауна рода Eucyclops еще мало изучена.
В Северной Америке показан космополит Ей. serrulatus и Ей. priono-
phorus, известный также из Неотропической области (Уругвай). Eucyclops
elegans Herrick, по нашему мнению, является членом вариационного
РАСПРОСТРАНЕНИЕ И ИСТОРИЯ ФАУНЫ
77
ряда serrulatas — proximus. Род Tropocyclops представлен 2 видами —
•общим с Палеарктикой космополитическим, Т. prasinus и Т. extensus,
•который пока больше нигде не найден. Роды Paracyclops, и Ectocyclops
представлены космополитами Р. fimbriatus, Р. poppei, Р. affinis и Е. pha-
leratus. Из рода Cyclops с достоверностью показан лишь С. scutifer
(возможно также С. kolensis и С. strenuus; указания требуют проверки);
судя по имеющимся данным, этот род в Неарктике далеко не имеет
разнообразия, свойственного Палеарктике, и представлен только в север-
ной ее половине (Аляска, Канада, примерно до широты Нью-Йорка).
Сравнительно богато представлен в Неарктике род Acanthocyclops —
около 12 видов, из них 7 общие с Палеарктикой, а 5 (и 1 подвид —
A. bicuspidatus var. thomasi) найдены только в ее пределах: A. nearcti-
cus, A. haueri, A. venustoides, A. jeanneli, A. donnaldsoni (из них 2
последние троглобионты). Характерный для пелагиали озер Северной
Америки A. americanus известен из Англии и очень близок к широко
распространенному в Палеарктике A. vernalis; эндемики Неарктики
также очень близки, главным образом, к палеарктическим видам рода.
Род Microcyclops в Неарктике представлен, судя по имеющимся данным,
необычайно бедно; в ее пределах найдено всего 3 вида — М. varicans
(включая var. rabellus), М. dengizicus и М. bicolor, все 3 космополиты.
Эндемиком Неарктики является род Orthocyclops с 1 видом О. modestus,
•близким к роду Mesocyclops; последний в Неарктике представлен 2 видами:
-М. leuckarti (с эндемическим подвидом edax) и М. tenuis, последний
известен также из Южной Америки (Парагвай). Подрод Thermocyclops
в Неарктике отсутствует (если к нему не относить М. tenuis).
Ориентальная область. Изучена менее полно, чем Пале-
арктическая. Наиболее полные данные имеются для Малайского
архипелага, однако и их нельзя считать исчерпывающими. Характерной
особенностью Малайский подобласти является, прежде всего, присут-
ствие рода Bryocyclops (4—5 видов), который вне Ориентальной области
представлен лишь в Экваториальной Африке. Род Macrocyclops пред-
ставлен всеми 4 видами, из которых М. neuter известен вообще только
с Явы. Из рода Eucyclops в Ориентальной области известно 8 видов,
из них Е. thienemanni, ruttneri и gibsoni var. abreviatus — только
в Малайской подобласти, тогда как Е. serrulatus и Е. permixtus найдены
и в Индии, Е. euacanthus— также на Формозе; исключительно для Индии
показаны Ё. indicus и Е. productus. Из приведенных видов Е. eyacanthus
и Е. gibsoni (typ.) обитают также в Эфиопской области. Род Tropocy-
clops, помимо Т. prasinus, в Ориентальной области представлен еще 2
видами — Т. confinis (с var, frequens, Ява) и Т. nigroviridis (Формоза).
Tropocyclops prasinus широко распространен во всей области. Из рода
Paracyclops в Ориентальной области широко распространены космопо-
литы Р. fimbriatus и Р. affinis; только с Формозы известен Р. sitiseien,
sis и только из Малайской подобласти Р. eucyclopoides. Ectocyclops pha-
leratus известен из Британской Индии, Е. polyacanthus—только с Фор-
мозы, других видов этого рода для рассматриваемой области не указано.
Эндемиком является монотипный род Thaumasiocyclops, известный
с о-ва Бали, т. е. с северной границы Меланезийской подобласти Австра-
лийской области. Из рода Cyclops на севере Индийской подобласти,
на границе с Палеарктикой (Ладак), найдены еще недостаточно полно
описанные С. ladakanus и С. hutchinsoni; оба, однако, известны только
в горных водоемах (1582—4000 м над ур. м.). Южнее род отсутствует.
Род Acanthocyclops в Ориентальной области представлен очень бедно —
78
ВВЕДЕНИЕ
известно всего 6 видов, из них 5 широко распространены в Палеарк-
тике. Эндемиком является A. alticola, найденный только в высокогор-
ных водоемах (4252—4977 м над ур. м.) северной Индии. С Формозы
описан проблематичный A. formosanus, вероятно идентичный с A. viridis.
Таким образом по видовому составу фауна этого рода в Ориентальной
области является сильно обедненной по сравнению с палеарктической. Род.
Microcyclops в Ориентальной области насчитывает около 15 видов. Эндеми-
ками являются 10 видов, общими с Палеарктикой будут М. varicans, М. bico-
lor, М. dengizicus и М. minutus, общим с Эфиопской областью — М. Нп-
janticus и, вероятно, М. davidi. В Ориентальной области отсутствуют
роды Muscocyclops, Speocyclops, Graeteriella, но довольно богато пред-
ставлен преимущественно тропический род Mesocyclops с подродом
Thermocyclops. В ней, повидимому, широко распространены космополиты
Mesocyclops leuckarti и М. crassus. Из подрода Mesocyclops s. str.
М. meridianus известен из западной Индии (также из Неотропической
области), М. tobae — с Явы н Суматры, М. thermocyclopoides — с Фор-
мозы. Помимо М. crassus, из подрода Thermocyclops в Индии найдены
М. rylovi и М. mongolicus (по всем вероятиям идентичный с М. asiatucus),
т. е. 2 свойственных югу Палеарктики вида. Исключительно в Малайской
подобласти1 пока найдены М. operculifer, М. trichophorus, М. philippi-
nensis (Филиппины); М. taihokuensis, М. brevifurcatus и М. ianthinus
пока известны только с Формозы.
Приведенные данные показывают некоторое сходство Ориентальной
фауны Cyclopidae с палеарктической (присутствие рода Acanthocyclops
в сильно обедненном видовом составе, присутствие на севере рода Cy-
clops), причем это сходство фауны касается северной части Ориенталь-
ной области, главным образом горной (т. е. климатически обособленной),
а в остальном сводится к присутствию в обеих областях космополитич-
ных видов. Отношение к австралийской фауне пока неясно. Зато опре- ,
деленно устанавливаются черты сходства ориентальной и эфиопской
фауны; только из этих областей известны Eucyclops euacanthus, Ей*
gibsoni, Tropocyclops confinis, Macrocyclops albidus var. oligolasius, Mi-
crocyclops linjanticus, M. davidi, Mesocyclops leuckarti aequaiorialis, род
Bryocyclops, тогда как только в Ориентальной и Неотропической обла-
стях найден лишь 1 Mesocyclops meridianus. Сходство Ориентальной
и Эфиопской областей замечается также в фауне Diaptomidae (группе
orientalis) и Harpacticoida (род Elaphoidelldy, Chappuis (1930, 1931)
пытается объяснить это сходство бывшим в палеозое сообщением обеих
областей материком Гондвана и устанавливает „гондванскую" группу
гарпактицид (р. Elaphoidella, Aitheyella, Anipodiella), с которой Kiefer
(1933, 1933а) сопоставляет и указанные выше виды родов Eucyclops,
Tropocyclops и Microcyclops.
Австралийская область. Для нее мы имеем крайне скудные
данные. Все подсемейство Eucyclopinae представлено 7 космополитич-
ными видами, известными также из Палеарктической и Ориентальной
областей. Род Cyclops отсутствует. Из рода Acanthocyclops указаны
A. bicuspidatus, A. bisetosus (там же), A. crassicaudoides (близок к A. cras-
sicaudis) и A australis. Эндемиками являются только 2 последних вида.
Из рода Microcyclops в области известны М. varicans, М. bicolor, М. mona-
canthus и М. amaudi, из них 2 последних вида больше нигде не най-
дены. Из рода Mesocyclops найдены только М. leuckarti и М. crassus
М. analogus Kief., вероятно, идентичен с М. rylovi.
РАСПРОСТРАНЕНИЕ И ИСТОРИЯ ФАУНЫ
79
Судя по этим, далеко не полным, данным, фауна пресноводных Cyclopi-
dae Австралийской области, в противоположность таковой Calanoida,
не имеет сколь-либо резко выраженного эндемизма и близко примыкает
к фауне Палеарктической области и только в комплексе палеарктиче-
ских форм — к Ориентальной.
Никакого сходства в фауне пресноводных Cyclopidae Австралийской,
Эфиопской и Неотропической областей не наблюдается.
Эфиопская область. В Южном полушарии это, пожалуй, наи-
более исследованная по фауне Cyclopidae область. Из 4 видов Macro-
cyclops в ней с достоверностью показан лишь М. albidus (с var. oligola-
sius, который известен также с о. Суматры). Для рассматриваемой обла-
сти в высшей степени характерно необычайное богатство видов рода.
Eucyclops — больше 20 видов (из них 5 эндемичного подрода Afrocy-
clops'), из которых 2 известны также из Ориентальной области, а все
остальные свойственны только Эфиопской области. Из них 4 вида оби-
тают лишь в оз. Танганьика и 2 вида известны только с о. Мадагаскара.
Космополитом является Е. serrulatus ( — Е. agiloides Sars). Богато пред-
ставлен также род Tropocyclops — 5 видов, из которых 2 (Т. varicoides
и Т. tenellus) свойственны только Африке. Т. parvus известен также
из Южной Америки и 1 (Т. confinis) — из Ориентальной области; кроме
того показан Т. prasinus. Из рода Paracyclops в Эфиопской области
указано 4 вида, из них 3 космополита и 1 (Р. oligarfhrus) эндемик
оз. Танганьика. Очень богато представлен также род Ectocyclops-. 4 вида,
из вих 3 эндемика (Е. coperes, Е. compactus и Е. hirsutus); Е. phaleratus
представлен вариететом rubescens, известным также из Неотропической
области. Род Cyclops в Эфиопской области отсутствует (известен из
палеарктической части Африки), род Acanihcyclops в ней представлен
только 1 видом A. viridis (4 вида известно из палеарктической части
Африки). Род Microcyclops, напротив, в Эфиопской области насчитывает,
помимо космополитов М. varicans и М. bicolor, около 20 видов, из кото-
рых 18 свойственно только этой области, а 1—2 Вида общи с Ориен-
тальной областью {М. davidi для о. Явы указан под вопросом). Помимо
2 указанных космополитов, остальные палеарктические виды Microcy-
clops заходят лишь в палеарктическую часть Африки. Своеобразной
чертой Эфиопской области (экваториальной ее части) является присут-
ствие рода Bpyocyclops, с 5 эндемическими видами; род этот свойствен
еще только Ориентальной области. Подрод Mesocyclops s. str. в Эфиоп-
ской области насчитывает только 2 вида: М. leuckarti и М. аппае.
Напротив, подрод Thermocyclops в Эфиопской области представлен,,
подобно роду Eucyclops, необычайно богато; из 15 известных в ней
видов 12 (или 13) свойственны только ей, а 2 вида (М. crassus и М. dybow
skii) являются широко распространенными.
Н е о т р о п и ч е ск ая облает!^. Из рода Macrocyclops найден
только М. albidus, что, вероятно, не может быть объяснено простою слу-
чайностью, так как, например, в Ориентальной области, которая едва ли
изучена более полно, чем Неотропическая, известно 4 вида этого рода, а из
совсем слабо изученной Австралийской области известно 2 вида. Заме-
чательно присутствие в Неотропической области Homocyclops ater
(Парагвай), который, кроме того, известен лишь из Неарктики. Из Нео-
тропической области известно 6 видов Eucyclops (подрод Afrocyclops
отсутствует), из них Е. prionophorus показан также для Неарктики,
а остальные 5 видов больше нигде не найдены; палеарктических видов
Eucyclops в рассматриваемой области вовсе нет. Фауна этого рода в ней
80
ВВЕДЕНИЕ
несравненно беднее эфиопской и ничего общего с последней не имеет.
Род Tropocyclops представлен, кроме Т. prasinus, 2 видами, из кото-
рых Т. schubarti больше нигде не найден, а Т. parvus известен также
из Эфиопской области. Из рода Paracyclops для Неотропической обла-
сти показаны лишь широко распространенные Р. fimbriatus и Р. poppei,
из рода Ectocyclops, помимо Е. phaleratus (с var. rubescens), показан
лишь Е. bromelicola, больше нигде не найденный и обитающий в микро-
водоемах листьев Bromeliaceae. Род Cyclops в Неотропической области
отсутствует, род Acanthocyclops представлен в ней очень бедно: помимо
космополитов A. viridis и A. vernalis, известен A. uruguayensis (Уруг-
вай), A. lobulosus (только в Антарктике) и A. michaelseni (там же).
Напротив, род Microcyclops в Неотропической области, помимо космопо-
литов М. varicans и М. dengizicus и палеарктического М. gracilis (най-
ден в Аргентине), представлен еще 13 видами, отличными от эфиопских
и больше нигде не найденными. Род Bryocyclops отсутствует, вместо
него присутствует близкий биологически (обитатели влажных мхов) род
Muscocyclops (2 вида — М. operculatus и М. staheli, оба в экваториаль-
ной части Южной Америки). Значение обоих подродов рода Mesocyclops
в Неотропической области обратное тому, что имеет место в Эфиопской.
В первой подрод Mesocyclops представлен 7 видами, из них только
М. leuckarti космополит, М. tenuis — вид, известный из Неарктики,
М. meridianus известен также из западной Индии, а 4 вида являются
эндемиками. Напротив, из подрода Thermocyclops для Неотропической
области описано, помимо космополита М. crassus, всего 2 вида (М. breh-
mi и М. minutus). Неотропическая фауна Cyclopidae имеет очень мало
общего с ориентальной и, несомненно, обнаруживает черты сходства
с эфиопской, однако при резко выраженном эндемизме видовом, а отча-
сти н родовом {Muscocyclops). Общими с эфиопской фауной являются
Tropocyclops parvus, Ectocyclops phaleratus var. rubescens, затем весьма
большое разнообразие видов Microcyclops, однако эндемичных. Соедине-
ние Африки с Южной Америкой имело место не только в палеозое
(Гондвана), но и в мезозое (Бразильско-Эфиопской континент). Если
•считать, что в верхнем мезозое фауна Cyclopidae обоих материков была
более или менее однообразной, то можно допустить, что род Microcy-
clops обитал уже в верхнем мезозое на обоих континентах и затем, уже
в третичное время, развились параллельно неотропическая и эфиопская
фауны этого рода, совершенно различные по своему видовому составу;
повидимому так обстояло дело и с родом Eucyclops, представленным
в Южной Америке 5 эндемичными видами (из 6 в ней вообще извест-
ных).
Остановимся подробнее на характере и происхождении палеаркти-
ческой фауны пресноводных Cyclopidae.
Ввиду полного отсутствия палеонтологических данных, разрешение
проблемы истории фауны пресноводных Cyclopoida наталкивается на
значительные затруднения и является делом весьма сложным. Данные
.зоогеографического порядка позволяют считать несомненным, что в тре-
тичное время существовала богатая пресноводная фауна Cyclopidae,
и можно думать, что она имелась уже в мезозойскую эпоху. Все сооб-
ражения по истории фауны этих ракообразных мы можем базировать
лишь иа данных зоогеографического, отчасти морфологического и эко-
логического характера; и здесь необходимо также учитывать еще далеко не
лолную и, главное, неравномерную обследованность фаун пресноводных
Cyclopoida отдельных областей. Необходимо также иметь в виду, что
РАСПРОСТРАНЕНИЕ И ИСТОРИЯ ФАУНЫ
81
генеалогические взаимоотношения родов и видов этой группы пресно-
водных ракообразных ни в коем случае не могут считаться выяснен-
ными даже в главных своих чертах, а существующие схемы этих отно-
шений далеко не лишены искусственности и почти совершенно не бази-
руются на данных онтогенетического развития.
Как отмечено выше, только 2 рода {Macrocyclops и Paracyclops)
могут считаться космополитическими. Все остальные роды явственно
группируются в 2 комплекса — комплекс родов Северного полушария
и комплекс родов Южного полушария (тропический). К первому принад-
лежат роды Ochridacyclops, Cyclops, Acanthocyclops, Speocyclops, Grae-
ieriella, Orthocyclops, t. e. 5 родов подсемейства Cyclopinae и только
1 род подсемейства Eucyclopinae. Тропический комплекс насчитывает
4 рода 2-го подсемейства {Eucyclops, Tropocyclops, Ectocyclops, Thau-
masiocyclops) и 4 рода 1-го {Microcyclops, Bryocyclops, Muscocyclops,
.Mesocyclops).
Роды Cyclops и Acanthocyclops, как указывалось выше, чрезвычайно
разнообразно представлены в Голарктике и их бесспорно можно считать
ее автохтонами. Существенно отметить, что процесс видообразования
в пределах этих обоих родов весьма интенсивно происходит и в совре-
менную нам эпоху, на что мы уже указывали в главе, посвященной система-
тике, причем толчком к этому, очевидно, явились условия ледникового
периода. Оба эти рода тесно связаны генетически и стоят особняком в общей
системе подсемейства Cyclopinae. Установить центр образования этих
.родов едва ли возможно. Во всяком случае едва ли можно наметить
два центра, как это имеет место, например, для пресноводных Harpacti-
coida (западный и восточный центры, по Боруцкому; европейско-
сонорское и ангаро-американское царство для Trichoptera, по Мартынову,
1924). Современное географическое распространение пресноводных Cy-
clopidae в Голарктике не дает никаких сколько-нибудь убедительных
доказательств в пользу установления двух центров (западный и восточный)
многообразия видов. Правда фауна Cyclopidae восточной Азии изучена
значительно менее, чем Европы; однако накопившиеся за последнее время
данные показывают, что на востоке мы находим общепалеарктический
комплекс видов, наблюдающиеся же различия видового состава касаются
южного комплекса (род Mesocyclops). Присутствие только на западе
родов Ochridacyclops, Speocyclops и Graeteriella объясняется специаль-
ными условиями их образования (см. ниже), равно как и присутствие
в Западной Европе ряда троглобионтных видов рода Acanthocyclops.
Для целого ряда групп палеарктической фауны зоогеографы склонны
считать очагом образования древний Ангарский материк, вовсе не покры-
вавшийся морем с палеозойской эры (сводку см. у Н. Я. Кузнецова,
1938), причем Reinig (1937) считает даже, что Ангарский континент
являлся центром образования всей голарктической фауны. Если этот
взгляд справедлив для родов Cyclops и Acanthocyclops, то можно при-
нять, что, как и для многих других групп голарктической фауны, в тре-
тичный период эти два рода или их родоначальники были более или менее
однообразно распространены до Арктики включительно. Современное
географическое распространение этих родов, насколько оно нам теперь
известно, а равно генеалогические отношения их ныне живущих видов,
правда, не дают никаких указаний специально на восточное происхожде-
ние. Однако доледниковая умеренная фауна Cyclopidae Ангарского
континента, свойственная третичному периоду, в четвертичное время,
•под влиянием оледенения, претерпела весьма существенные изменения
6 Фауна СССР
82
ВВЕДЕНИЕ
первоначального распределения, в общем вытесняясь к югу и юго-во-
стоку. Можно допустить, что в это время, благодаря ледникам, западный
и восточный комплекс фауны Cyclopidae был более или менее изолиро-
ван, и развитие обоих комплексов шло независимо. Но затем, когда
ледник отступил к северу, произошло настолько полное слияние обоих
комплексов, что в настоящее время уже нет никакой фактической воз-
можности установить хотя бы следы двух центров формообразования.
Такому смешению благоприятствовала способность пресноводных Cyclo-
poida в Голарктике к распространению в широтном направлении, у этой
группы ракообразных выраженная в гораздо большей степени, нежели у мно-
гих других групп палеарктической фауны. Однообразие фауны Cyclopoida
в широтном направлении особенно выражено в зоне тундры; примеры
этому дают также Calanoida (например Heterocope borealis). Однако
это замечается и для родов Cydops и Acanthocyclops. Исключая Acantho-
cyclops languidoides var. gracilicaudatus, известного только с Даль-
него Востока, нам неизвестны специально восточные подвиды обоих
родов. Указанному вариетету, без сомнения, надо приписать лишь моло-
дой возраст, во всяком случае послеледниковый. Нет, однако, уверен-
ности, что этот вариетет не будет найден и на западе. A. crassicaudis
var. cretensis известен с запада (о. Крит), из Японии и с о. Формозы.
Арктический комплекс видов в Голарктике представлен исключи-
тельно двумя рассматриваемыми родами. К нему мы относим Cyclops scuti-
fer, С. rolensis, Acanthocyclops capillatus, возможно — Cyclops lacustris^
Едва ли всех их можно рассматривать в качестве эварктов, в смысле
Н. Я. Кузнецова (1938). Первый из этих видов на юг доходит до широты
Сувалок, а в Северной Америке — до Нью-Йорка. Не исключена воз-
можность принадлежности этого вида к борео-альпийской фауне, хотя
в западноевропейских Альпах пока с достоверностью он не найден.
Отнесение этого вида в средней Европе к ледниковым реликтам едва
ли может быть, доказано. Он распространен во всей Скандинавии и на
всем Кольском полуострове. Нахождение его в озерах (олиготрофных —
слабо эвтрофированных) района западной части СССР между Ладож-
ским озером и оз. Вигры (Сувалки) не представляется невероятным.
В таком случае последнее местонахождение явится лишь наиболее южным
пунктом его сплошного ареала распространения на северо-западе и утра-
тит свой изолированный характер. Принимая во внимание наличие
в южной части этого ареала достаточно глубоких озер с низкими тем-
пературами гиполимниона, становится возможным расселение этого вида
в пределах рассматриваемого ареала уже в послеледниковое время.1
Если бы подтвердилось нахождение С. scutifer в субальпийских озерах
Альп; то там этот вид можно было бы рассматривать как ледникового
реликта (подобно Heterocope borealis). Распространение Cyclops kolensis
и Acanthocyclops capillatus, арктический характер которых можно счи-
тать бесспорным, еще настолько мало изучено, что толковать нахожде-
ние их, например, в районе около 55—60° с. ш. (южная Швеция, Карело-
Финская ССР) в духе ледниково-реликтовой теории едва ли
возможно. Так как распространение A, abyssicola также еще очень мало
изучено, то пока мы воздерживаемся от причисления этого вида, неви-
димому резко стенотопного, даже к комплексу арктических форм.
Попытки некоторых авторов считать A. crassicaudis ледниковым реликтом
1 Существенно отметить, что в Вигерских озерах обитает не только var. wigrensis,
но и типичная форма С. scutifer.
РАСПРОСТРАНЕНИЕ И ИСТОРИЯ -ФАУНЫ
8з
в Западной Европе бесспорно ошибочны и опровергаются всеми дан-
ными его распространения. Из перечисленных видов в сущности только
2, Cyclops kolensis и Acanthocyclops capillatus, могли бы быть отнесены
к эварктам. Они, действительно, весьма характерны для водоемов тунд-
рового ладшафта. Однако оба они встречаются и в зоне тайги, а первый из
них известен и из зоны смешанных лесов.
Переходим к южному комплексу форм Палеарктики. Едва ли под-
лежит сомнению, что троглобионтный род SpeocyclopS является релик-
том третичного времени (Kiefer, 1937), и его история, в общем, анало-
гична таковой гарпактицид Elaphoidella, Parasternocaris и некоторых
других (Chappuis, 1927, 1931). Генетически род Speocyclops связан
с эфиопским и ориентальным тропическим родом Bryocyclops. Проник-
новение предков рода Speocyclops в Палеарктику имело место в мио-
цене, при климатических условиях этого времени, когда Африка была
связана с Европой материком (район нынешнего Эгейского моря). В это
время попавшие в Европу тропические иммигранты жили в наземных
водоемах, как это теперь свойственно роду Bryocyclops. Охлаждение
в плиоцене и особенно в ледниковый период вызвало переселение этих
форм из наземных в подземные водоемы, где колонии и сохранились
до настоящего времени, причем образовался род Speocyclops, давший
ряд видов. Судя по нахождению иа Кавказе и в Англии, где обитают
самостоятельные виды Speocyclops (Sp. colchidanus, Sp. demetiensis),
можно думать, что род был сформирован очень давно, вероятно не
позднее плиоцена, а затем произошло изолированно образование этих
2 видов. Третичное время сопровождалось мощными горообразователь-
ными процессами, и именно в это время возникли Карпаты, Альпы,
Кавказ. Представляется вполне вероятным, что переход тропических
иммигрантов в подземные воды мог произойти уже в верхне-третичное
время (в конце неогена), так как в это время подземные воды, несо-
мненно, уже существовали. Существенно отметить, что охлаждение
имело место уже в верхнем миоцене (меотическое время), в плиоцене
оно прогрессировало и в это время реки европейской части СССР уже
покрывались льдом (см.: Берг. Климат и жизнь, 1922:26). Едва ли
есть необходимость приурочивать отщепление рода Speocyclops от общего
предка с Bryocyclops специально к четвертичному времени. Более слож-
ным представляется происхождение рода Graeteriella. Он встречается
исключительно в подземных водах Западной Европы. По Kiefer’y (1937)
из 13 его местонахождений 3 лежат в южной Европе (Румыния, Юго-
славия, северная Испания) и 10 к северу от Альп. На юге ареалы оби-
тания этого и предыдущего родов совпадают. Генеалогические отноше-
ния рода Graeteriella крайне неясны. Отнесение его к подроду „Diacy-
clops" (т. е. к голарктическому роду Acanthocyclops) Kiefer’oM (1928,
1929) оказалось столь же ошибочным, как и отнесение к нему тем же
автором видов рода Speocyclops. Никаких оснований к утверждению
близости Graeteriella к обоим голарктическим видам не имеется. Напро-
тив, гораздо более вероятным нам кажется предположение Chappuis
(1927), по которому предков Graeteriella („Cyclops unisetiger"), как
и предков Speocyclops („Cyclopidae troglodytes"), надо искать в тропи-
ческой фауне Cyclopidae. Если этот взгляд подтвердится дальнейшими
исследованиями, то Graeteriella, подобно другому роду, может быть
также отнесена к третичным реликтам палеарктической фауны Cyclopi-
dae. Факт более северного главного ареала распространения Graeteriella
(если он действительно имеет место), сравнительно с основным компле-
6*
84
ВВЕДЕНИЕ
ксом видов рода Speocyclops, едва ли противоречит такому положению,
тем более, что в последнем роде мы имеем 1 вид (Sp. dementiensis)
с еще более северным ареалом (Англия). Род Ochridacyclops занимает
совершенно изолированное положение, по вооружению фуркальных вет-
вей напоминая Mesocyclops. Как отмечает Kiefer (1937d), Ochridacyclops
однако несомненно примыкает генетически к группе Eacyclops — Afro-
cyclops— Tropocyclops, т. e. к тропическому комплексу. Чрезвычайно
существенно при этом, что Ochridacyclops найден в оз. Охрида, в кото-
ром мы находим ряд третичных реликтов. Вероятно к таковым же надо
отнести и рассматриваемый род, южное происхождение которого едва
ли может подлежать сомнениям.
Известно, что подземная фауна Европы (например десятиногий рак
Troglocaris schmidti, из Syncarida — Bathynella и Parabathynella) обна-
руживает родство с наземной фауной Ориентальной (Индо-малайской)
и Австралийской областей. Происхождение Speocyclops однако можно
связать специально с Эфиопской областью, где ныне обитает половина
видов рода Bryocyclops (см. выше). Исключительный зоогеографический
интерес представляет фауна Cyclopoida подземных вод азиатской части
СССР, особенно Дальнего Востока, о которой, к сожалению, нам пока
ничего не известно.
Приведенный зоогеографический анализ шести родов Cyclopidae,
свойственных Северному полушарию, показывает, что „северный" ком-
плекс состоит из 2 групп совершенно различного происхождения — из
автохтонной голарктической группы, центром образования которой пред-
положительно являлся Ангарский континент (роды Cyclops, Acanthocy-
clops), и из третично-реликтовой группы, генеалогически связанной
с современной фауной тропических областей (роды Speocyclops, Ochri-
dacyclops, вероятно Graeteriella). Дальнейший состав палеарктической
фауны относится уже к родам, исключительно богатым видами специально
в Южном полушарии, напротив в Палеарктике (и вообще в Голарктике)
представленным чрезвычайно бедно. Таковы роды Eucyclops, Тгоросу-
cldps, Ectocyclops, Microcyclops, Mesocyclops (с подродом Thermocyclops).
Распространение этих родов в Палеарктике имеет некоторые своеобраз-
ные черты. Род Tropocyclops имеет резко выраженное южное распро-
странение, он отсутствует в зоне тундры и тайги и очень редок в зоне
смешанных лесов (встречается в ее южной части); основной ареал его рас-
пространения в Палеарктике лежит южнее 50° с. ш. Из рода Microcy-
clops исключительно в Средиземноморской подобласти обитает М. planus,
главным образом в южной части Палеарктики распространены М. minu-
tus (на север не заходит далее широты Костромы, отсутствует в Скан-
динавии и во всей Северной Америке), М. gracilis (к северу не заходит
далее широты Кострома — Молотов, нет в Северной Америке, но присут-
ствует в Южной Америке) и М. dengizicus (степная и пустынная зона юга
СССР, в Северной Америке найден лишь в аналогичных условиях ланд-
шафта около 40° с. ш., известен также с о. Гаити и из Индии). Даже
столь космополитические виды этого рода, как М. bicolor и М. varicans,
в зоне тундры отсутствуют или там встречаются лишь крайне редко,
отсутствуют они и на арктических островах (оба тенловодны). Ectocy-
clops phaleratus севернее Пудожа (Карело-Финская ССР) у нас вообще
не найден, на арктических островах он также отсутствует. В зоне тундры
отсутствуют Mesocyclops dybowskii, М. crassus, вероятно М. oithonoides,
тогда как остальные известные в Палеарктике виды подрода Thermo-
cyclops (asiaticus, rylovi) встречаются исключительно на юге и юго-
РАСПРОСТРАНЕНИЕ И ИСТОРИЯ ФАУНЫ
85
востоке (проблематичный М. mongolicus Kief, найден только в восточ-
ной Монголии). Наиболее распространены в северной части Палеарктики
Mesocyclops leuckarti (на севере, однако, он, в общем, встречается реже,
чем в умеренных широтах) и виды рода Eucyclops, особенно Е. serrula-
tus и Ё. macruroides.
О происхождении перечисленных родов в Палеарктике, насколько
нам известно, пока никто не высказывался. Принимая во внимание дан-
ные их современного распространения, изложенные выше, нет основа-
ний считать эти роды автохтонами Северного полушария. Напротив,
нужно предполагать центр образования этих родов где-то в Южном
полушарии. Если считать, что в мезозое существовала уже фауна прес-
новодных Cyclopidae, то, может быть, образование этих родов связано
с фауной Cyclopidae бывшего в мезозое бразильско-эфиопского мате-
рика, объединявшего в верхнюю эпоху юрского периода Южную Аме-
рику и Африку и частью нынешнюю Ориентальную область (Индию)
и охватившего частью тот обширный район, который в палеозое был
занят Гондваной, простиравшейся до Австралии и Индо-Малайского
архипелага включительно. В третичное время произошло проникновение
этих родов из Эфиопской и Ориентальной областей, возможно в мио-
цене, в Палеарктику, где при теплом климате этого времени они имели
обширное распространение. Существенно отметить, что исключительное
обилие видов родов Eucyclops, Tropocyclops, Ectocyclops и Mesocyclops
(подрод Thermocyclops) мы находим именно в Эфиопской области, в кото-
рой очень богато представлен и род Microcyclops. Затем уже в плио-
цене могло произойти смещение ареала обитания к югу (роды имеют
явно выраженный тепловодный характер), а в ледниковое время ареал
обитания этих родов был совершенно изменен — часть видов тепловод-
ной фауны Cyclopidae вообще вымерла, часть же оттеснилась на юг
Палеарктики. В четвертичное время, после отступания ледника, произо-
шло новое заселение оставшимися видами прежней территории, и сфор-
мировалась картина их современного распространения. При этом, разу-
меется, не исключалось образование уже в конце третичного времени
в умеренных широтах новых видов, которые ныне мы находим только
в Северном полушарии. Таковы Eucyclops macruroides, Е. macrurus,
пещерные Eucyclops graeteri и Е. teras, Microcyclops planus, Mesocy-
clops oithonoides (все известны только из Палеарктики).
Приведенные соображения о происхождении рассматриваемых родов
(тропиче’ский комплекс) в Голарктике являются, конечно, лишь гипоте-
зой. Думается, однако, что они приемлемы, тогда как обратное предпо-
ложение об автохтоном происхождении этих родов в Северном полуша-
рии и затем их позднейшее внедрение в фауну тропических областей
настолько расходится с данными современного географического распро-
странения этих родов, что вряд ли вообще допустимо.
Фауна пресноводных Cyclopoida Неарктической области весьма
тесно примыкает к фауне Палеарктики. Если взять эндемические формы
Северной Америки, которые среди Cyclopoida в ней гораздо менее
обильны, нежели, например, среди пресноводных Calanoida, то можно
видеть крайнюю близость этих эндемиков к палеарктическим видам
(троглобионты отмечены звездочкой):
Acanthocyclops americanas
„ donnaldsoni *
„ jeanneli*
,, nearcticus
близок к A. vernalis
„ „ A. viridis (latipes)
„ „ A. crassicaudis и A. bisetosas
„ „ A. langaidas
86
ВВЕДЕНИЕ
Acanth ocylops haueri
„ bicuspidatus thomasi
„ venustoides
Eucyclops prionophrus
Tropocyclops extesus
близок к A. bicuspidatus
„ „ A. venustus
„ „ E. serrulatus
„ „ T. prasinus
Из этих видов A. americanus недавно указан и для Англии, но глав-
ный ареал его распространения лежит в Северной Америке. Фауну
пресноводных Cyclopoida последней, насколько она нам пока известна,
вообще можно рассматривать как обедненную и частью измененную
фауну Палеарктики. В обеих областях преобладание остается за родом
Acanthocyclops (15 видов в Неарктике), в обеих бедно представлены
роды Microcyclops, Eucyclops и Mesocyclops. Несомненно в прошлом
должен был происходить обмен фаунами обеих областей, всего вероят-
нее посредством Берингии, соединявшей на востоке оба материка в тре-
тичное время, причем, возможно, этот обмен шел особенно деятельно
в направлении с запада на восток. Обмен мог продолжаться, возможно,
и позже. На западе обмен фаун Cyclopoida восточных частей Северной
Америки (амфиатлантические элементы; Eidmann, 1934) мог происходить
по обширному материку Северной Атлантиды, существовавшему вплоть
до плиоцена. В фауне пресноводных Calanoida (Epischur а, некоторые
Diaptomidae) наблюдается сходство северо-восточной Азии и западных
берегов Северной Америки (амфипацифическое распространение), тогда
как оно вовсе не имеет места на западе Европы. Это говорит за мигра-
ционный путь Calanoida именно на востоке. Однако для пресноводных
Cyclopidae мы имеем интересный случай нахождения A. americanus
именно на западе (Англия), а северо-американский A. venustoides близок
к западноевропейскому A. venustus, известному только из Швейцарии,
Германии и Англии. К сожалению, огромное большинство данных по
фауне Cyclopidae Северной Америки относится к атлантическому побе-
режью, тогда как по Тихоокеанскому побережью пока имеются лишь
очень скудные данные. Поэтому вопрос об амфиатлантических и амфи-
пацифических компонентах этой фауны пока остается открытым; для
фауны Calanoida такие компоненты устанавливаются для немногих, правда,
видов (например род Epischura) с большей достоверностью. После окон-
чательной изоляции Неарктической и Палеарктической областей пресно-
водная фауна стала развиваться независимо, что привело к известной
дифференциации фауны Cyclopidae обеих областей, и выработались,
в частности, характерные черты специально североамериканской фауны
пелагических Cyclopoida (также Diaptomidae и Cladocera—’Daphnia
группы pulex—-parapulex), отличной от палеарктической (пелагический
A. americanus, несомненно примыкающий к A. vernalis, A. bicuspidatus
thomasi, присутствие в пелагиали Tropocyclops prasinus). Едва ли, однако,
можно отрицать, что выработка специфических черт североамериканской
фауны пресноводных Copepoda происходила уже в третичное время;
состав фауны Diaptomidae достаточно определенно говорит именно за
относительную древность этой дифференциации. Весьма характерной
чертой неарктической фауны пресноводных Cyclopidae является присут-
ствие в ней совершенно чуждых Палеарктике Homocyclops ater (род
монотипичен) и Orthocyclops modestus (род также монотипичен). Первый из
них, однако, найден недавно также в Бразилии и, возможно, принадлежит
в североамериканской фауне к элементам тропической группы. К этой
же группе мы склонны отнести Orthocyclops, несомненно родственный
тропическому роду Mesocyclops, затем Mesocyclops tenuis Marsh, извест-
ный также из Парагвая и Панамы и в Северной Америке найденный
БИОЛОГИЯ
87
только на юге (Аризона). Эти формы, надо думать, проникли в Север-
ную Америку с юга, а не являются дериватом неарктической фауны.
БИОЛОГИЯ
Семейство Cyclopidae представлено в пресных и солоноватых
водоемах очень разнообразного характера, как в стоячих проточных
и непроточных, так и в текучих. Виды, свойственные только последним
(реобионты), отсутствуют. В состав планктона рек входят виды, свой-
ственные стоячим водоемам, откуда они и проникают в реки; специально
потамопланктических родов и видов, а равно и вариететов, не имеется.
Для потамопланктона неоднократно указывалось в качестве характерной
черты особенно крупная роль личиночных стадий Cyclopidae. Причины
этого явления пока нельзя считать окончательно выясненными. Едва ли,
однако, можно сомневаться в том, что весьма крупную роль в этом
явлении играет, во-первых, значительная длительность метаморфоза
Cyclopidae и, во-вторых, влияние на него механического фактора тече-
ния и, вероятно, иных факторов, с этим связанных. Фауна Cyclopidae
прибрежных участков рек й ручьев складывается из бентических
и факультативно-планктических (Рылов, 1922) видов, свойственных
стоячим водоемам. Для дна фарватера рек в качестве обычного элемента
фауны нередко указывается Paracyclops fimbriatas, некоторыми авто-
рами причисляемый даже к реофилам; отсюда рачок нередко попадает
и в планктон реки. Пресноводные Cyclopidae, таким образом, по существу
свойственны ленитической области.
Фауна Cyclopidae озер, т. е. водоемов достаточно глубоких, с резко
дифференцированной пелагической областью, характеризуется прежде
всего присутствием специфических элементов пелагического планктона.
Для пелагиали озер Палеарктики характерны представители исключи-
тельно подсемейства Cyclopinae, притом только родов Cyclops, Meso-
cyclops и Acanthocyclops. Среди видов первого рода мы насчитываем
9 пелагических. Планктическимй являются также все виды рода Mesocy-
clops. Из них для пелагиали озер Палеарктики очень характерны
Mesocyclops leuckarti, встречающийся также в планктоне более мелких
водоемов, М. oithonoides, М. crassus— последний также в планктоне
прудов. Иногда, повидимому, специально в эвтрофных озерах, в пела-
гиали встречается М. dybowskii, количественно никогда не играющий
столь крупной роли, как первые 3 вида, и свойственный, главным
образом, планктону мелких водоемов. В пелагиали озер Дальнего
Востока, вероятно, крупную роль играет М. asiaticus. Из рода Acantho-
cyclops, представленного, главным образом, непланктическими видами,
A. americanus характерен для пелагиали озер, он широко распространен
в озерах Северной Америки, но найден также в Англии, где, однако,
обитает и в мелких водоемах. В пелагиали озер Неарктической области
крупную роль играет, кроме того, A. bicuspidatus thomasi. Известен
лишь один случай присутствия A. viridis в качестве постоянного компо-
нента в пелагиали озер (оз. Иссык-куль). Фауна литорали и сублиторали
озер населена видами, встречающимися и в мелких водоемах. Она
резко приурочена к зоне зарослей прибрежных макрофитов, где мы
и находим особенное разнообразие форм Cyclopidae. Некоторые видь!
(например Paracyclops affinis, Ectocyclops phaleratus, Microcyclops bico-
lor, M. var icons) встречаются также в различных обрастаниях —
в налетах синезеленых и диатомовых водорослей, в клубках различных
88
ВВЕДЕНИЕ
нитчатых и проч. Только 2 вида до сих пор были найдены исключи-
тельно в озерах—Acanthocyclops abyssicola (озера севера Европы)
и Ochridacyclops arndti. Последний найден только в оз. Охрида
и является, вероятно, коменсалистом губки Ochridaspongia. Только
этот вид показан исключительно для профундали (глубина около 30 м),
но неизвестно, является ли он батибионтом. Нельзя также отнести
к батибионтам A. abyssicola. Все остальные виды Cyclopidae, найденные
в профундали озер, надо отнести к эврибатным, частично проникающим
в профундаль (до эвпрофундали включительно) из зоны прибрежных
макрофитов. Таковы, однако, очень немногие виды — Paracyclops fim-
briatus (в Thunersee и Brienzersee показан до глубины 200 м), A. gigas
(найден до глубины 200 м), A. bisetosus (в Brienzersee был найден
на глубинах до 100 м), A. vernalis (известен с глубин до 40 м). Наибо-
лее постоянными компонентами профундальной области являются 2 пер-
вых вида, присутствие же в ней двух последних носит лишь случайный
характер. В противоположность некоторым Harpacticoida и Cladocera,
среди Cyclopidae мы вовсе не находим специфических обитателей
иловых отложений профундали; указанный выше A. abyssicola, по иссле-
дованиям Ekman’a (1916), приурочен не к илистым, а к песчаным и глини-
сто-песчаным грунтам, что отмечалось и для Paracyclops fimbriatus^
Остальные найденные в профундали виды нормально обитают в лито-
рали среди макрофитов и к илистым грунтам не приурочены; некоторые
виды (Paracyclops, Ectocyclops) факультативно обитают на прибрежных
отложениях, богатых растительными остатками.
В противоположность Calanoida, Cladocera и Rotatoria, среди Cyclo-
poida специально прудовых форм мы не находим. Некоторые виды
(Cyclops insignis, Mesocyclops leuckarti, M. Crassus, M. dybowskii) очень
обычны в прудовом планктоне, но едва ли их можно считать гелеопланк-
тическими, так как они частью встречаются и в пелагиали озер, частью,
и в мелких водоемах. Характеристика Cyclopidae специально гелеопланк-
тона сводится более к его отрицательным чертам — к отсутствию
эвпелагических, свойственных только пелагиали озер, видов (см. выше),
а также видов, свойственных планктону специально временных, весен-
них мелких водоемов.
Население Cyclopidae более мелких водоемов — болота, лужи —
отличается большим разнообразием, что стоит в связи с чрезвычайным
разнообразием таких водоемов. Наиболее богато представлены Cyclo-
pidae в мелких, богатых водной растительностью, водоемах, исключая
некоторые водоемы со специализованными условиями существования.
Две группы мелких наземных водоемов отличаются характерным соста-
вом фауны Cyclopidae. Во-первых, богатые гуминовыми веществами мелкие
сфагновые и гипновые болота-лужи резко выраженного дистрофного
типа, постоянные или пересыхающие. Об их фауне упоминается ниже.
Во-вторых, временные, наполняющиеся снеговой и дождевой водой,
лужи, пересыхающие летом, резко астатического характера. Фауна
таких временных водоемов вообще отличается присутствием ряда специ-
фических элементов. Из Cyclopidae к ним принадлежат Cyclops furcifer,
С. strenuus (typ.), Acanthocyclops vernalis, A. bicuspidatus, A. bisetosus,
A. crassicaudis, которые, однако, встречаются и в постоянных мелких
водоемах, а также Microcyclops minutus, свойственный исключительно
временным усыхающим водоемам, характер которых может быть очень
разнообразен (снеговые весенние лужи, дождевые эфемерные водоемы
степного, полупустынного и пустынного ландшафта). Наиболее приуро-
БИОЛОГИЯ
89
чены специально к астатическим весенним лужам, наполняемым снего-
вой водой, С. strenuus (typ.) и С. furcifer. Для временных, усыхающих
мелких водоемов Средиземноморской подобласти очень типичен Micro-
cyclops planus. Cyclopidae способны переносить полное высыхание
водоема в особых покоящихся стадиях, что было установлено уже
старыми авторами (Sars, 1889; Schmeil, 1891; Brauer, 1891; Claus, 1894).
Claus нашел, что Microcyclops minutus переносит высыхание на различ-
ных копеподитных стадиях метаморфоза, до 5-й копеподитной включи-
тельно, но не в фазе науплиальной. Kammerer (1907) выводил Acan-
thocyclops vernalis из ила, в сухом виде пересланного из Швеции в Вену
и сохранявшегося около 3 лет, причем копеподитные стадии появлялись
очень быстро. Н. Nordquist (1921) упоминает, что в спускных рыбовод-
ных прудах южной Швеции половозрелые Cyclops strenuus появляются
очень быстро после наполнения этих прудов свежей водой. Olofsson (1918)
также отмечает, что в мелких водоемах Шпицбергена у A. crassicaudis,
по наполнении весной этих водоемов талой водой, сразу же появляются
взрослые самки и самцы, быстро приступающие к копуляции, что нами
наблюдалось для этого вида в мелких весенних лужах Старого Петер-
гофа. Интересно отметить, что, по Olofsson’y, A. crassicaudis переносит
полное промерзание луж. Вопрос о покоящихся стадиях Cyclopidae
особенно полно освещен работами Wolf’a (1905), Lowndes’a (1930)
и Roy (1932). Первый автор установил способность переносить усыха-
ние водоема у Cyclops strenuus, A. bicuspidatus, A. bisetosus и A. dia-
phanus (вероятно = А. nanus). У этих видов, по Wolf’y, не образуется
специальных покоящихся яиц, снабженных плотными толстыми оболоч-
ками, как, например, у некоторых Diaptomidae, но яйцевые мешки
обволакиваются особым секретом, выделяемым кожными железами;
секрет этот может обволакивать и всего рачка. Такие рачки становятся
совершенно непрозрачными и иначе относятся к окраске гематоксилин-
эозином, нежели нормальные, не покрытые секретом. При обволакива-
нии секретом как весь рачок целиком, так и его яйцевые мешки могут
пе°реносить не только пересыхание водоема, но также и вмерзание
в лед. Wolf отмечает, что уже приступившие к размножению, вполне
половозрелые циклопы высыхания переносить не могут, но это для них
возможно до наступления половозрел ости и на последней копеподитной
стадии. Однако по исследованиям Roy (1932), Cyclops furcifer переносит
при обволакивании секретом высыхание в состоянии яиц и в IV—V
копеподитной стадии, но также и в половозрелом состоянии (самки).
По наблюдениям этого автора A. bicuspidatus и A. bisetosus
переносят усыхание на V копеподитной стадии (самки и самцы), причем
тело обволакивается секретом; яйца этих 2 видов переносят высыхание
только в прикрепленных к телу самки мешках, но, будучи оторванными
от самки, погибают. Образование особого „кокона", описанного Birge
и Juday (1908), у Acanthocyclops bicuspidatus в озерах Северной Америки
(без сомнения, данные этих авторов относятся к A. bicuspidaius var.
thomasi) Roy не наблюдал.
Lowndes (1930) для Cyclops strenuus ( = C. pictus) описал покоя-
щиеся яйца. Они чрезвычайно крупные, образуются в незначительном
числе (не более 6) и снабжены более толстой, нежели у обычных яиц,
наружной оболочкой. Такие яйца имеют длительную фазу развития, что
вообще характерно для покоящихся яиц Entomostraca и Rotatoria. Не
доказано, способны ли эти яйца переносить высыхание или служат
для переживания неблагоприятных условий только в постоянных водо-
DO
ВВЕДЕНИЕ
пммх. Не исключена возможность образования подобных покоящихся
лиц и у некоторых других Cyclopidae.
Совершенно специфичным биотопом Cyclopidae являются скопления
атмосферной воды в листьях наземных высших растений. Такие „мйкро-
лужи“ образуются па листьях Bromeliaceae в тропиках. В таких водо-
емах 1 обитает 1 вид (Br. bogoriensis) свойственного южным широтам
рода Bryocyclops, живущий в скоплениях воды в листьях Bilbergia
pyramidalis на Яве.
Среди палеарктических пресноводных Cyclopidae есть виды, обитаю-
щие в мелких заросших моховых лужах не только в свободной толще
воды, но и в погруженных в воду подушках густо расположенных мхов;
при выжимании таких подушек, образованных мхами Sphagnum и Hypnum,
вместе с водой выжимаются некоторые мелкие виды рода Acanthocy-
clops—A. muscicola, A. vernalis, A. languidus, A. nanus, некоторые
вариететы A. languidoides, затем A. venustus, отчасти A. crassicaudis.
Только в подушках погруженных мхов обитает A. languidoides var.
hypnicola, вероятно A. languidus var. disjunctus-, A. languidoides var.
eriophori обитает среди густых зарослей Eriophorum. Специально влаж-
ным наземным подушкам мхов свойственны почти все виды тропического
рода Bryocyclops и виды рода Muscocyclops (М. staheli, М. operculatus).
В пределах Палеарктики (Англия) во влажных растительных наземных
налетах скал был найден Speocyclops demefiensis, который, как все про-
чие виды этого рода, вероятно, является подземным циклопом.
Горизонтальное распределение пресноводных Cyclopidae в пределах
одного и того же водоема в настоящее время изучено чрезвычайно
мало. Как мы уже упоминали выше, в озерах огромное большинство
видов Cyclopidae свойственно литорали и отчасти сублиторали, и только
очень немногие виды проникают в профундаль. В пределах литорали
и сублиторали горизонтальное распределение состоит в тесной связи
с распределением водной растительности. Лишенные макрофитов участки
литорали, с каменистым и песчаным дном, черезвычайно бедны цикло-
пами; в таких условиях нередко встречается Paracyclops fimbriatus,
очевидно приуроченный'здесь к налетам микрофитов и к участкам дна
с иловыми отложениями наносного характера, а также некоторые дру-
гие, очень немногие, виды, очевидно проникающие сюда из зарослей
макрофитов. Вопрос о приуроченности тех или иных видов Cyclopidae
к определенным формациям прибрежных зарослей макрофитов остается
почти не изученным. Большинство видов, обитающих в этих условиях,
очевидно, весьма эвритопно и встречается в различных формациях
зарослей. Резко очерчен в литорали биотоп прибрежных участков сфаг-
новых и гипновых мхов, где мы находим характерный для таких усло-
вий комплекс сфагнофильных форм, при отсутствии ряда видов, в мохо-
вой формации вообще не обитающих. По нашим наблюдениям в ряде
озер европейской части СССР фауна Cyclopidae прибрежных зарослей
Carex и Phragmites качественно отличается значительной бедностью,
а виды, свойственные только зарослям этим макрофитов, совершенно
отсутствуют. Напротив, качественно очень разнообразную фауну Cyclo-
pidae мы находим в зарослях Myriophyllum, Ceratophyllum, Elodea, Utri-
cularia, Potamogeton и некоторых других макрофитов, особенно при
значительном развитии на них обрастаний различного рода (синезеле-
ные, нитчатки, налеты диатомовых).
1 В них же обитает Ectocyclops bromelicola Kief. (Бразилия),
БИОЛОГИЯ
91
Для озер СССР вопрос о распределении Cyclopidae в связи с рас-
пределением прибрежной водной растительности наиболее полно освещен
в работах Ласточкина (1926а, 1927), данные которого, однако, отнюдь
не могут распространяться вообще на все озера. Для дистрофированного
Валдайского озера (Ивановской области) этот автор намечает следую-
щие биоценетические группировки литоральной области: 1) сфагновые
зыбуны — Acanthocyclops nanus; 2) покрытый водой сфагновый мох —
A. nanus, A. muscicola, A. viridis, Mesocyclops leuckarti, сопровождаю-
щие виды—Macrocyclops albidus, Ectocyclops phaleratus, Paracyclops
affinis; 3) заросли Hypnum и Calliergen, примыкающие к сфагновым
зарослям, — A. nanus, Eucyclops macruroides var. denficulatus { — Euc.
lilljeborgi), сопровождающие виды Macr. albidus, Par. affinis, A. viridis,
.Mes. leuckarti; 4) прибрежные придонные мхи в пределах распростра-
нений Equisetum, Polygonum и Potamogeton—A. nanus, Macr. albidus,
сопровождающие виды—Par. affinis, A. viridis, Euc. macruroides var.
denficulatus; 5) толща воды в пределах зарослей Carex и Equisetum —
Euc. macruroides var. denticulatus, Mes. leuckarti, Euc. macrurus,
Macr. albidus; в зарослях Polygonum и Sparganium эти же виды,
но здесь чаще встречается Euc. macrurus и значительно реже первый
и последний вид; 6) открытый (без зарослей) грунт типа дьи, с примесью
глины или песка, — Par. fimbriatus, сопровождающие виды — A. nanus,
Mes. leuckarti. В эвтрофном оз. Переславском (той же области), дно
которого сплошь покрыто Chara, Zannichellia и Heleocharis, водная
толща в зарослях Equisetum -+- Polygonum и Potamogeton perfoliatus харак-
теризуется присутствием Mier, bicolor, которого нет в зарослях Chara -+-
Heleocharis и Potamogeton -ь Zannichellia, а также нет в сублиторальной
зоне. На самом дне этих видов нет, но здесь широко распространен
Par. fimbriatus, обитающий и на местах с песчаным грунтом. В зарос-
лях нитчаток этого вида иет, но здесь обитает Mier, bicolor. В водной
толще среди всех упомянутых зарослей широко распространены Еис.
serrulatus и A. viridis. Характер горизонтального распределения расти-
тельных зарослей играет очень важную роль и в горизонтальном рас-
пределении свойственного этим зарослям комплекса видов в прудах
мелких водоемах. В прудах, где дно сплошь покрыто макрофитами,
и ил богат их остатками, господствующий комплекс видов Cyclopidae
может занимать всю территорию водоема. Такое обширное распростра-
нение качественно весьма разнообразного комплекса Cyclopidae наблю-
дается, например, в некоторых исследованных нами прудах г. Пушкина
(окрестности Ленинграда), дно которых, сплошь покрыто зарослями
Chara. Комплекс видов, обитающий в озерах только в пределах лито-
рали и сублиторали, в таких случаях занимает обширное распростране-
ние, иногда охватывающее дно всего водоема.
Вопрос о горизонтальном распределении пелагических видов Cyclo-
pidae изучен также очень неполно. Горизонтальное распределение вообще
пелагического планктона до сих пор служит предметом дискуссии,
и едва ли можно сказать, что оно нашло себе в современной планкто-
логической литературе, касающейся эвлимнопланктона, достаточно ясное
освещение. Несомненно в водоемах с изрезанной береговой линией, а также
в очень крупных озерах, различные области которых лежат в различных
физико-географических условиях (например оз. Байкал, Ладожское
озеро), отдельные участки пелагиали могут представлять биотопы,
характеризующиеся далеко не одинаковыми физико-химическими усло-
виями. Установлено, что в таких случаях и планктон различных участ-
92
ВВЕДЕНИЕ
ков пелагиали озера и качественно и количественно не одинаков. Это
различие относится и к фауне пелагических Cyclopidae. Приведем
несколько примеров. По Верещагину (1924) в олиготрофном Сегозере
(Карело-Финская ССР) пелагиаль северо-восточной и западной его
части отличается достаточно резко по химизму, в частности по окисляе-
мости и содержанию CaO; Mesocyclops leuckarti обитает только
в северо-восточной части озера. По нашим исследованиям (Рылов,
1933а) в пелагиали оз. Балхаша, где химизм, в частности содержание
хлоридов и карбонатная жесткость, сильно изменяется, увеличиваясь
в направлении с запада на восток, — Cyclops vicinus, встречаясь в участ-
ках пелагиали с CI 90—880 мг на литр и при жесткости 8.3—32.2°,
совершенно отсутствует в пелагиали самого восточного, наиболее мине-
рализированного района, при С1 свыше 880 мг на литр и жесткости
более 34.6°, тогда как Mesocyclops crassus в значительных количествах
встречается и в этом последнем районе, где одновременно мы находим
типичного галобионта, коловратку Pedalion охуига. В оз. Иссык-куль
(Рылов, 1930а) Cyclops vicinus совершенно отсутствует в пелагиали
глубин свыше 25—50 м, встречаясь, однако, в пелагическом планктоне
изобаты 10—25 м. Весьма вероятно, что такое распределение рассмат-
риваемого вида, особенно характерного для эвтрофных озер, стоит
в связи с эвтрофированием пелагиали прибрежных областей этого озера.
Вопрос о возможности образования „роев", т. е. местных скоплений,
в пределах физико-химически однородных участков пелагиали, т. е.
в пределах однотипного пелагического биотопа, до известной степени
освещенный для некоторых Cladocera, для Cyclopidae совершенно
не выяснен, и факт бесспорного образования роев в таких условиях
у этих рачков не установлен.
Явление „избегания берегов" (Uferflucht) у Cyclopidae изучено
еще очень мало. По исследованиям Burckhardt’a (1910) в Женевском озере
и в оз. Лугано оно очень сильно выражено у самцов Cyclops sp. группы
strenuus (С tatricus ?), в меньшей степени у самцов Mesocyclops leu-
ckarti, в минимальной степени у самок этих видов. В Нижнем Лунцском
озере (юго-восточная Германия) Ruttner (1929/1930) у самок и молоди
С. tatricus не нашел сколь-либо в заметной степени выраженного избегания
прибрежной зоны. Данные этих авторов едва’ли, однако, могут иметь
общее значение, так как рассматриваемое явление настолько тесно
связано с горизонтальным распределением экологических факторов,
что один и тот же вид циклопов в разных озерах, при различных
условиях, может дать совершенно различную картину этого яв-
ления.
Вертикальное распределение планктических пресноводных Cyclopidae
изучено для очень многих водоемов, особенно для озер и прудов.
Сколь-либо подробный обзор обширной литературы по этому вопросу
невозможен в рамках нашей главы, так как данные по Cyclopidae
мы находим почти во всех, очень многочисленных, статьях, касающихся
вообще вертикального распределения зоопланктона. Ограничимся здесь
лишь общей характеристикой этого распределения.
Все планктические виды рода Mesocyclops в озерах обитают в зоне
эпилимниона, в пределах которого происходят и вертикальные суточ-
ные миграции. Нижним пределом распределения является зона металим-
ниона, в которой Mesocyclops встречается лишь в единичных экзем-
плярах. В гиполимнион проникают, очевидно, лишь случайные индивиды.
Приуроченность видов этого рода к верхним слоям воды характерна
БИОЛОГИЯ
93
также и для прудов. Днем рачки держатся обычно не у самой поверх-
ности, а на некоторой глубине, которая, как и для других планктических
рачков, в различных озерах и прудах стоит в связи с физико-химиче-
скими факторами, в первую очередь в связи со световыми условиями,
в разных водоемах часто далеко не одинаковыми.
Ввиду смешивания систематиками различных близких видов рода
Cyclops s. str., многочисленные данные по их распределению нуждаются
в проверке. Это особенно относится к С. Strenuus, заведомо ошибочно
очень часто указываемого для пелагиали озер. В противоположность
роду Mesocyclops, среди рассматриваемого рода имеются виды с явно
выраженной тенденцией к образованию днем максимума в глубоких
слоях воды, до гиполимниона включительно. Это наблюдалось у С. Scutifer
в олиготрофных озерах Северной Норвегии, у С. abyssorum. в ряде озер
севера Европы, у С. bohater в слабо эвтрофированных озерах окр.
Сувалок, у С. tatricus в Нижнем Лунцском озере (зимой) в юго-восточной
Германии, у С. lacustris в южной части Ладожского озера. Образование
максимума в глубоких слоях, повидимому, гораздо менее выражено
у С. vicinus.
Отношение Cyclopidae к различным физико-химическим факторам
водной среды изучено пока лишь в общей форме, и наши представления
по экологии того или иного вида циклопов основываются почти исклю-
чительно на данных прямого наблюдения в природе и лишь в крайне
редких случаях на экспериментальном материале (влияние температуры,
активной реакции, отчасти солености); при этом экспериментальные
данные еще очень неполны даже в отношении таких факторов водной
среды, как, например, температура и соленость. Без риска впасть
в ошибку можно утверждать, что разработка проблемы экологии Cyclo-
pidae на физиологической основе в настоящее время находится лишь
в самой начальной стадии и является всецело делом будущих исследо-
ваний. При таком положении проблемы крайне рискованным представ-
ляется попытка дать экологические спектры для различных родов
и видов Cyclopidae. Для некоторых факторов среды мы можем в отдель-
ных случаях указать приблизительные пределы амплитуды колебания,
в которых данный вид встречается в естественных условиях. Однако
каковы оптимальные пределы фактора, при которых данный вид находит
себе реальные возможности наиболее интенсивного размножения и нор-
мального существования, каким образом изменяются эти пределы в зави-
симости от весьма сложного комплекса других факторов водной среды,
постоянно изменяющегося во времени, — пока сказать мы не можем.
Между тем разрешение именно этих вопросов, которые можно было бы
кратко формулировать как „проблему оптимума", является необходимой
предпосылкой для освещения вопроса о продукции того или иного вида
Cyclopidae в данных конкретных физико-химических и биологических
условиях биотопа и водоема в целом.
В настоящей главе на основании фактического материала, изло-
женного более подробно в систематической части, мы даем сводную
картину отношения Cyclopidae континентальных вод Палеарктики, глав-
ным образом видов, встречающихся в пределах СССР, к различным
факторам водной среды, как она рисуется в естественных условиях.
Такая картина, разумеется, не может претендовать на исчерпывающую
полноту и точность; однако она достаточно ясно намечает основные
элементарные экологические черты рассматриваемой группы низших
ракообразных.
94
ВВЕДЕНИЕ
Световые условия. Огромное большинство Cyclopidae континен-
тальных вод живет в наземных водоемах и подвергается суточным
и сезонным изменениям фактора света. В таких условиях живут все
виды родов Macrocyclops, Ectocyclops, Cyclops, Mesocyclops и Microcy-
clops. 1 Род Ochridacyclops найден на глубине 30 м в оз. Охрида, кото-
рое, однако, отличается значительной прозрачностью (олиготрофный
водоем). Нахождение О. arndii только на этой глубине, однако, едва
ли связано со световыми условиями (вероятно коменсалист придонных
губок). Нахождение на значительных глубинах некоторых прибрежных
Cyclopidae (см. выше) указывает лишь на возможность жизни этих
видов и в условиях олиготипа света, т. е. на широкую амплитуду при-
способленности к этому фактору. Среди рода Acanthocyclops целый ряд.
наземных видов способен к продолжительной жизни в пещерных усло-
виях. Таковы A. bicuspidatus, A. viridis, A. languidoides, A. languidus
(в пещерах редко), A. crassicaudis (типичная форма в пещерах отсут-
ствует), A. bisetosus. При этом некоторые из них в условиях полного
отсутствия света образуют особые вариететы (частично, быть может,
морфического характера) — A. languidoides var. clandestinus, var. nagy-
salloensis, var. zschokkei, var. putealis, var. italianus, var. goticus, var.
tantalus. Acanthocyclops crassicaudis в пещерной фауне представлен
особым вариететом — var. cretensis. Широкой возможностью длительно
жить в условиях пещерных водоемов, при отсутствии света, обладает
род Paracyclops, особенно Р. fimbriatus, многими авторами относимый
к троглофилам. У этого вида при переходе к подземному образу жизни
также наблюдается тенденция к образованию вариететов (var. imminutus,
var. finitimus).
Род Eucyclops представлен, главным образом, видами, обитающими,
в наземных водоемах, но Е. serrulatus обладает весьма широкой при-
способленностью к изменению световых условий и часто встречается
в пещерных водах, а 2 вида и 1 вариетет являются троглобионтами.
Tropocyclops prasinus—вид наземных вод, частично обитающий и в под-
земных водах.
Следующие виды являются троглобионтами. Род Acanthocy-
clops: kieferi, venustus (var. crinitus), stammeri (включая и его var. west-
falicus), rhenanus, phraeticus, karamani, scophensis, stygius, hypogeus.
Кроме того ряд вариететов, указанных выше, найден только в подзем-
ных водах.1 2
Род Eucyclops: teras, graeteri (= inarmatu), serrulatus var. nagasakai.
Род Speocyclops: 12 видов, только подземные воды (Sp. demetiensis
найден во влажных налетах на скалах, но, вероятно, является также
троглобионтом).
Род Graeteriella: 2 вида, оба троглобионты. Последние 2 рода,
таким образом, свойственны исключительно подземным водам.
В связи с подземным образом жизни у циклопов-троглобионтов
наблюдается ряд характерных особенностей — тенденция к редукции
некоторых морфологических признаков (членистость плавательных ног,
вооружение фуркальных ветвей и др.), потеря пигмента (окраски) тела,
1 М. planus и М. minutus иногда были находимы в колодцах, но, бесспорно, при-
надлежат к наземной, а не к пещерной фауне. Нахождение (очень редко) Mesocyclops
leuckarti и М. dybowskii в пещерах объясняется чисто случайным заносом. Из рода
Macrocyclops таков же М. albidus.
2 В последнее время описано (Kiefer, 1938) еще несколько специально пещерных
видов Acanthocyclops и Speocyclops, которые в этой работе не учтены.
БИОЛОГИЯ
95
иногда полная редукция глазного пигмента (но не самого глаза).
Последнее, очевидно, стоит в связи с отсутствием в пещерных водах
света, так как на свету пигмент восстанавливается. Однако у Cyclopi-
dae этих вод отсутствие пигмента отнюдь не указывает на длительность
пребывания данного вида в условиях отсутствия света. Так, например,
циклопы рода Graeteriella, несомненно, издавна обитающие в подземных
водах, снабжены глазным пигментом, тогда как гораздо более молодые
по происхождению Eucyclops teras и Е. serrulatus var. nagasakai, близкие
к наземному Е. serrulatus, глазной пигмент утратили, что наблюдается
также у Е. graeteri, близкому к Е. macrurus. У рода Speocyclops,
в пещерных водах несомненно древнего происхождения, глазной пигмент
у некоторых видов сохранился, у других редуцировался до полного
исчезновения.
Виды Cyclopidae, живущие в наземных водоемах, в высокой сте-
пени фототаксичны. В связи с неподвижностью глаза (в противополож-
ность Cladocera) на изменения в направлении и интенсивности освеще-
ния циклопы реагируют изменением положения всего тела относительно
источника света. Затем происходит активное перемещение рачка в напра-
влении к источнику света или в 'обратном направлении. Знак фото-
таксиса может меняться в зависимости от предварительной световой
адаптации и от некоторых химических факторов водной среды. Так,
присутствие свободной СО2 изменяет положительный фототаксис
на отрицательный.
В естественных условиях реакция Cyclopidae, как и других свободно-
живущих Copepoda, на световые раздражения реализуется в явлении
вертикальных суточных миграций. Особенно резко оно выражено у пела-
гических видов Cyclopidae (роды Cyclops, Mesocyclops, Acanthocyclops
americanus, A. bicuspidatus var. thomasi). Из палеарктических видов эти
миграции особенно хорошо изучены у Cyclops tatricus, С. scutifer,
С. vicinus (упоминаем точно определенные виды группы strenuus),
Mesocyclops leuckarti, М. oithonoides и М. crassus. Для всех этих видов,
в общем, характерно появление в сумерки и ночью в самых верхних
слоях воды, тогда как при дневном освещении они держатся главной
массой на некоторой глубине с пониженной интенсивностью освещения.
Время появления вечером у поверхности и положение слоя дневного
максимума зависит от ряда факторов, последнее — главным образом от
условий прозрачности воды. В оченб прозрачных олиготрофных озерах
циклопы, как и прочие пелагические рачки, днем держатся глубже
и имеют гораздо большую амплитуду вертикальной миграции, нежели
в мало прозрачных эвтрофных водоемах. Приведем некоторые цифры,
показывающие амплитуду этих миграций. Burckhardt (1900) в олиготроф-
ном Фирвальдштедском озере для Mesocyclops leuckarti наблюдал
амплитуду в 30—40 м; в эвтрофных озерах, с сильными цветениями
воды, амплитуда миграций этого вида, по нашим наблюдениям, может
быть всего в 2—4 м, в Люцернском озере, по Worthinglon’y (1931), она
колеблется в разное время года от 3 до 15 м, а у молоди Cyclops
strenuus (auct.)—от 12 до 46 м. По нашим наблюдениям амплитуда
суточных миграций взрослых самок scutifer в олиготрофных озерах
Мурмана не превышает 10 м. В прозрачном Нижнем Лунцском озере взрос-
лые самки С. tatricus имеют амплитуду около 10 м, копеподитные стадии
мигрируют также интенсивно, но науплиусы вовсе не мигрируют. Послед-
нее наблюдается и для науплиальных стадий других пелагических
Cyclopidae. По нашим наблюдениям (Рылов, 1927), в прудах окрестно-
96
ВВЕДЕНИЕ
стей Старого Петергофа (ныне Петродворца) самцы Mesocyclops oitho-
noides мигрируют не менее интенсивно, чем самки, копеподитные стадии
(главным образом 3-я и 4-я) — слабее, чем последние, а науплиусы
вовсе лишены способности к вертикальным суточным миграциям.
По Burckhardt’y (1900), в Фирвальдштедском озере самцы Cyclops (stre-
nuus auct.), однако, мигрируют слабее самок. Один и тот же вид
циклопа в разных водоемах и в одном и том же водоеме в разные фазы
своей жизни может дать различную картину миграции.
О вертикальных суточных миграциях планктических Cyclopidae,
свойственных мелким водоемам (например Cyclops furcifer, Tropocyclops
prasinus) ничего не известно, но таковые, вероятно, происходят,
при очень узкой амплитуде. Не выяснено также, имеют ли место эти
миграции у прибрежных, факультативно-планктических и бентических
форм (например роды Eucyclops, Macrocyclops и др.). Доказано, однако,
что явление фототаксиса свойственно и этой группе форм (например
Macrocyclops aldidus; эксперименты Esterly, 1907).
Температурные условия. Влияние температуры на метамор-
фоз Cyclopidae рассмотрено в главе .о метаморфозе. Приведенные в ней
данные относятся к эвритермичным или к тепловодным видам, у кото-
рых понижение температуры ведет к замедлению, а повышение — к уско-
рению развития. Влияние температуры на метаморфоз видов Cyclopidae,
вообще приуроченных к низким температурам воды (холодноводно-сте-
нотермичные циклопы), экспериментально не изучено. Весьма вероятно,
что в этом случае повышение температуры при прочих равных условиях
может приводить и к замедлению темпа метаморфоза. За это говорят
наблюдения над циклом развития, например, Acanthocyclops gigas, у кото-
рого метаморфоз занимает весьма длительный период, падающий на
время господства высоких температур (лето — осень), когда взрослые
самки и самцы этого строго мотоцикличного вида отсутствуют.
Среди Cyclopidae континентальных вод Палеарктики достаточно
резко выделяются 2 основные группы видов — группа эвритермичных,
широко приспособляющихся к колебаниям температуры видов, и группа
стенотермичных, приуроченных к определенным температурным условиям
видов. В последней группе различают холодноводно-стенотермичные
и тепловодно-стенотермичные виды. Ряд видов, вследствие спорадичности
нахождения (некоторые найдены только 1 раз), пока не может быть
отнесен ни к одной из этих групп.
Эвритермичными являются: Cyclops strenuus (s. str.), C. vicinus,
Acanthocyclops viridis, A. vernalis, A. bicuspidatus, A. biseiosus, A. cras-
sicaudis, A. languidus, A. languidoides typ. (?), A. nanus, Macrocyclops
fuscus, M. distinctus, M. albidus, Paracyclops fimbriatus, P. affinis, Eucy-
clops serrulatus, M. minutus.
Тепловодно-стенотермичны: Mesocyclops leuckarti, M. oithonoides,
M. crassus, M. dybowskii, M. rylovi, вероятно M. asiaticus, затем Eucy-
clops macrurus, Tropocyclops prasinus, Microcyclops gracilis, M. bicolor,
M. varicans; резко выраженной склонностью к теплым водам, далее,
отличаются Euc. macruroides (включая var. denticulatus), экология
которого еще мало изучена, и Ectocyclops phaleratus. Холодноводно-сте-
нотермичны: Cyclops insignis, С. abyssorum, С. tatricus, С. strenuus
var. landel, по всем вероятиям С. scutifer и С. kolensis, затем Acantho-
cyclops gigas и A. capillatus. Отношение к температуре у различных
родов характеризуется следующим образом:
БИОЛОГИЯ
97
Род Cyclops. Тепловодно-стенотермичные виды отсутствуют, весьма
значительный процент видов холодноводных. Только 2 вида с достовер-
ностью можно считать эвритермичными. Род отсутствует в южных
областях, распространен только в Палеарктике и частично на севере
Ориентальной и Неарктической области.
Род Acanthocyclops. Если исключить троглобионтов, по крайней
мере часть которых, вероятно, холодноводно-стенотермична (низкие
температуры подземных вод, при слабой амплитуде годового колеба-
ния), то большинство палеарктических видов эвритермично. Тепловодно-
стенотермичных видов пока не указано (возможно таковы некоторые
вне-палеарктические виды). 2 вида холодноводно-стенотермичны. Род свой-
ствен, главным образом, Голарктике, вообще космополитичен.
Род Microcyclops. Почти все виды тепловодно-стенотермичны.
Холодноводных нет, М. minutus эвритермичен. Основной ареал распро-
странения рода лежит в Южном полушарии, в арктических областях род
почти отсутствует.
Род Mesocyclops (включая подрод Thermocyclops). Исключительно
тепловодно-стенотермичные виды; род отсутствует в высокоарктических
областях, основной ареал — в Южном полушарии.
Род Macrocyclops. Все виды, встречающиеся в Палеарктике, эври-
термичны. Род космополитичен.
Род Paracuclops. Как предыдущий род; 2 тропических вида.
Род Ectocyclops. Эвритермичные и тепловодные виды. Род космо-
политичен.
Род Eucyclops. Эвритермичные и тепловодные виды. Основной
ареал распространения — Эфиопская область.
Род Tropocyclops. Исключительно тепловодные виды. Один вид
-свойствен югу Палеарктики и Неарктике, 1 вид — Неарктике, остальные
виды — тропикам.
Отношение к температурам трогдобионтных родов Speocy-
dops и Graeteriella не вполне ясно. Вероятно большинство их видов
приурочено к постоянно низким температурам, что характерно для насе-
ляемых ими биотопов.
Установить границы температурного оптимума для отдельных
видов Cyclopidae с достаточной убедительностью пока невозможно даже
относительно наиболее распространенных и часто встречающихся видов.
Наблюдения непосредственно в природе позволяют предполагать, что,
например, для холодноводного Acanthocyclops capillatus оптимум лежит
при температуре не выше 14—15°, а у A gigas верхняя граница опти-
мума, вероятно, выражается еще более низкими температурами. Ruttner
(1929/1930) оптимальные условия существования взрослых Cyclops tat-
ricas определяет в 12—13°. Очевидно весьма низкий температурный
оптимум имеет С. insignis и, особенно, С. strenuus var. landei, встре-
чающийся только зимой. Оптимальные температуры для Mesocyclops
leuckarti можно принять в 15—30°. Весьма высокие границы оптимума
наблюдаются для Tropocyclops prasinus и Microcyclops varicans —
последний вид прекрасно размножается даже при температуре 34—37°.
Судя по имеющимся данным, верхняя граница амплитуды колебаний
температуры, в пределах которой вообще возможно существование
палеарктических Cyclopidae, лежит около 40°; нижняя граница немного
выше 0°. Весьма существенным для понимания экологии отдельных
видов Cyclopidae является вопрос о значении температуры для разных
стадий метаморфоза. Недавно Боруцкий (1937) показал для Bryocamptus
7 Фауна СССР
98
ВВЕДЕНИЕ
arcticus, что оптимальные температурные условия для отдельных стадий
его метаморфоза совершенно различны — оптимум для ортон ауплиуса.
лежит в пределах 8—10°, при более высокой температуре ортонау-
плиусы погибают, но последующие личиночные стадии переносят тем-
пературу около 20°. Насколько нам известно, аналогичных данных
по пресноводным Cyclopidae не имеется. В литературе имеются указа-
ния, что величина взрослых самок и самцов Cyclops strenuus в различ-
ные сезоны года не одинакова. Одни авторы констатируют увеличение
длины этого вида зимой, другие, напротив, — в летние месяцы. Едва
ли подлежит сомнению, однако, что данные эти относятся к разным
видам группы „strenuus". Высказывалось предположение, что одним из
факторов, от которого зависит окончательная величина циклопов,
является температура. Специально этому вопросу посвящено обширное
экспериментальное исследование Coker’a (1933) над Acanthocyclops
vernalis, A. viridis и Eucyclops serrulatus (все 3 вида эвритермичны).
Самые крупные самки A. vernalis, воспитанные при 28—30°, оказались
меньше, чем самые мелкие из воспитанных при 18—19°, а воспитанные
при 18—19° оказались мельче, чем воспитанные при 7—10°. Обратная
корреляция температуры и величины рачка установлена Сокег’ом также
для самцов этого вида, в более узких температурных границах'—для
обоих полов Е. serrulatus, а в менее ясной форме и для A. viridis.
Наиболее крупные самки (2.20—2.27 мм) и самцы (1.42—1.43 мм)'
последнего вида получены при 7—15°. В другой работе Coker (1934)
показал, что температура оказывает влияние и на формирование морфо-
логических признаков циклопов. Так, у A. vernalis при воспитании при
низких температурах (7—10°) самцы и самки получат гораздо более
длинные фуркальные ветви, нежели при высоких температурах (28—30°).
Температура оказывает влияние также на число и строение шипов,
вооружающих дистальные членики обеих ветвей IV пары плавательных
ног. Исследования цитируемого автора, касающиеся .только одного этого
фактора из целого ряда других, в широких размерах изменяющихся
локально и во времени, представляют особое значение для систематиков,
показывая, насколько осторожным надо быть с таксономической оценкой
признаков, иногда ошибочно понимаемых даже в качестве видовых,
тогда как эксперимент доказывает явно морфический их характер.
Термический фактор играет крупную роль в вертикальном распределении
пелагических Cyclopidae. В озерах с резко выраженной летней стратифи-
кацией температуры (озера умеренного типа) и сильно прогреваемым
эпилимнионом, холодноводно-стенотермичные виды (например Cyclops
abyssorum) держатся в глубоких холодных слоях воды, в слое гиполим-
ниона. В холодных озерах, где летние температуры верхних слоев эпи-
лимниона не высоки (например озеро Телецкое на Алтае), упомянутый
вид держится гораздо ближе к поверхности, и здесь положение слоя
его максимума на некоторой глубине определяется уже не термическими,
а иными условиями, в частности световыми. Тепловодно-стенотермичные
виды, обитающие в пелагиали озер (род Mesocyclops), имеют резко
выраженную приуроченность к верхним слоям эпилимниона, где господ-
ствует наиболее высокая летняя температура.
Соленость. До сих пор мы не имеем полной сводки по отноше-
нию к общей солености видов Cyclopidae континентальных вод Пале-
арктики. На основании данных, которые могут более или менее полно
характеризовать отношение Cyclopidae к степени общей солености,
может быть предложена следующая группировка.
БИОЛОГИЯ
99
Эвригалинная группа. Cyclops strenuus (typ.), С. vicinus,
Acanthocyclops viridis, A. bicuspidatus, A. bisefosus, Mesocyclops leuc-
karti, M. crassus, Macrocyclops albidus, Paracyclops fimbriatus, Eucyclops
serrulatus. Из этой группы A. bicuspidatus и особенно A. bisefosus неко-
торыми авторами относятся к галофилам и даже к галобионтам (A bise-
tosus), что едва ли правильно, так как оба вида в очень больших коли-
чествах встречаются и в мягких, относительно бедных электролитами,
водах (например в весенних лужах зоны смешанных лесов). Массовое
развитие этих 2 видов в соленых континентальных водоемах отнюдь
не указывает, что именно в этих водоемах оба вида находят наиболее
благоприятные, оптимальные условия существования. Эвригалинность
Mesocyclops leuckarti подчеркивается наблюдениями Таусон (1934)
и нашими (Рылов, 1935), а равно и неоднократным нахождением
этого вида в солоноватых морских водах (Финский и Ботнический
заливы). Отношение к общей солености перечисленных видов, конечно,
не одинаково, так что говорить об их эвригалинности можно только
в общей форме. Мы не видим, однако, оснований выделять специальную
группу галофильных видов. В качестве таковых в литературе фигури-
рует лишь A. bicuspidatus и A. bisefosus, о которых упомянуто выше.
Галофильность в данном случае есть просто неудачное выражение
широких границ эвригалинности, которые у прочих видов рассматривае-
мой группы, в общем, более сужены.
Стеногалинная группа. Мы подразделяем ее на 2 подгруппы —
агалобионтов и галобионтов. Первые обитают в нормальных пресных
водах (агалинные — слабо олигогалинные воды) и в гидробиологии отно-
сятся к галоксенам. Вторые обитают исключительно в соленых конти-
нентальных водоемах, совершенно отсутствуя в агалинных (олигогалин-
ных) водах.
К 1-й подгруппе относятся Cyclops kolensis, С. furcifer, С. scutifer
(включая var. wigrensis), С. tatricus, С. lacustris, С. abyssorum, С. boha-
ter, C. insignis, C. ochridanus, Acanthocyclops gigas (при повышенном
содержании NaCl очень редок), A. vernalis (нахождение в слабо соле-
ных водах, вероятно, объясняется случайным заносом), A. abyssicola,
A. languidus, A. languidoides (во всяком случае вся группа eriophori-hyp-
nicola), A. nanus, A. muscicola, A. capillatus, A. crassicaudis, Microcyclops
gracilis, M. minutus (данные Gauthier, 1928), Mesocyclops oithonoides,
M. rylovi, Macrocyclops fuscus, M. distinctus, Paracyclops affinis, Ecto-
cyclops phaleratus, Ochridacyclops arndti. По всем вероятиям к агало-
бионтам надо отнести Mesocyclops dybowskii, Microcyclops bicolor,
Eucyclops macrurus, Tropocyclops prasinus. Ряд видов рассматриваемой
подгруппы относится к „сфагнофилам" (A languidus, A. languidoides —
группы eriophori— hypnicola, A. nanus, A. muscicola).
К группе галобионтов относится лишь 1 палеарктический вид —
Microcyclops dengizicus. Вид свойствен соленым водоемам пустынного
и полупустынного, отчасти степного ландшафта, встречаясь в них сов-
местно с другими галобионтами (Brachionus miilleri, Lecane grandis
и др.). Вместе с некоторыми другими галобионтами-планктерами является
важным элементом специально салинопланктона (Rylov, 1935b). В соле-
ных водоемах Муганской степи (Вейсиг, in litt.) М. dengizicus обитает
в массах в водах с сухим остатком 25.924 г на литр и при содержании
С1 10.426 г на литр; Kiefer находил его при S-41°/00. Рачок характерен
для лимно-мезогалинных и димно-полигалинных вод.
т
1.00
ВВЕДЕНИЕ
Чрезвычайно крупное экологическое значение имеет, как известно,
не только степень общей минерализации воды (S pro mille), но также
и ее солевой состав и количественное взаимоотношение растворенных
солей (антагонистическое действие солей).
Наши сведения о физиологическом действии ионов отдельных эле-
ментов на пресноводных Cyclopidae, однако, настолько еще незначи-
тельны (см. Беклемишев, 1923, 1924, Acanthocyclops viridis-, Таусон,
1924, Cyclops insignis; Скадовский, 1928, Acanthocyclops languidus), что
едва ли могут объяснить с физиологической точки зрения присутствие
или отсутствие упомянутых видов (не говоря уже о других, по кото-
рым аналогичных данных не имеется) в водах с различным качественным
и количественным солевым составом в естественных условиях, когда
экологическая картина еще более усложняется одновременным влиянием
других факторов водной среды. Известную картину отношения видов
Cyclopidae к солевому составу воды нам дают, однако, наблюдения
в самой природе, в особенности в водоемах со специфическим химиз-
мом (воды аберрантного типа, идиотрофные водоемы). Нужно заметить,
что и здесь наши сведения далеко не полны и нередко опираются
лишь на очень элементарные химические данные.
Несомненно ряд видов пресноводных Cyclopidae широко приспосо-
бляется к содержанию СаО. Так, например, Acanthocyclops vernalis очень
обычен и в больших количествах развивается в бедных Са (и вообще
электролитами) дистрофирйванных водоемах, встречаясь также и в водах
с высоким содержанием Са (высшие ступени мезотрофности, примерно
до 100 мг СаО на литр), A. viridis встречается в Са-политрофных
(свыше 100 мг СаО на литр) водоемах и широко распространен в мягких
водах. По Козминскому (Kozminski, 1936) Cyclops kolensts живет
и в очень мягких водах и при жестокости в 19 немецких градусов, что,
вероятно, справедливо также для типичного С. strenuus и С. vicinus и,
несомненно, для вообще весьма эвритопного вида Eucyclops serrulatus.
Напротив, имеющиеся данные указывают на приуроченность С. scu-
tifer к водам с олиго-мезотипом СаО, а С. strenuus var. landei резко
приурочен к олиготипу СаО (не более 2 мг на литр; Kozminski, 1936).
Судя по факту массового размножения так называемых „сфагнофильных"
циклопов в бедных электролитами мягких дистрофных водах, можно
думать, что оптимальные условия таких форм лежат в области олиго-
типа Са. Фауна Cyclopidae алкалитрофных водоемов (СаО свыше 100 мг
на литр) изучена еще очень недостаточно. Судя по имеющимся данным,
качественно она бедна, что вообще характерно для зоопланктона Са-
политрОфных вод. В них обитают некоторые широко эвритопные Cyclo-
pidae, упомянутые выше {Eucyclops serrulatus, Acanthocyclops viridis
и некоторые другие). Циклопов, приуроченных специально к водам
с богатым содержанием Са, не имеется.
Данные для содовых (резкое преобладание Na над К, при очень
высокой цифре НСО3-ьСО3) озер центральной Якутии, с резко щелоч-
ной реакцией (Семенович, 1935; Рылов, 1935), относящихся к 1-му
классу озер по классификации Palmer’a, (1911), показывают чрезвычайно
широкую амплитуду приспособленности к химизму таких вод некоторых
видов CyclopidaeAecK. Mesocyclops crassus в них развивается в значитель-
ных количествах (в планктоне), преобладая над всеми прочими Entomo-
straca при общей минерализации 0.39 —1.99 г на литр, М. leuckarti—
при 0.39 —1.77 г, Cyclops vicinus и С. strenuus typ. — при 0.62 —1.77 г;
однако 3 последних вида уже не встречаются при 1.99 г на литр
БИОЛОГИЯ
ЮГ
(оз. Атахан). В водах хлоридного типа, с преобладающим значением
NaCl, мы наблюдаем особенно значительное качественное обеднение
фауны Cyclopidae. При высоких содержаниях NaCl совершенно отсут-
ствуют агалобионты, а из эвригалинных форм выживают только очень
немногие виды, например Acanthocyclops bicuspidatus, A. bisetosus,
A. viridis. Особенно характерны для таких вод два первые вида. По дан-
ным Tauson (1933) в соленом оз. Чебаркуль A. viridis выпадает при повы-
шении содержания С1 с 4 до 8.98 г на литр, тогда как A. bisetosus
при последней концентрации еще присутствует; однако в 1931 г., когда
озеро значит ельно осолонилось (содержание возросло с 3.55 до 7.82 г,
а С1 — с 8.98 до 18.40 г на литр), выпал и этот вид, и планктон принял
характер типичного салинопланктона (появление и развитие галобионтов
Artemia salina, Moina microphthalma и др.). По Schafer’y (1933) в хло-
ридных водах A. bisetosus встречается еще при содержании 5.61 г
на литр, тогда как в хлоридных водах окрестностей Oldesloe
(Голыптиния), по Klie (1925), типичный A. bicuspidatus живет при
содержании NaCl до 10.24 г в литре; по Schafer’y он обитает
в хлоридных водах при S = 0.562 —1.589%, но в Каспийском море,
где мы имеем специфический химизм воды, с иным соотношением
ионов Са и Mg1, этот вид отсутствует уже при С1 свыше 1.72 г
на литр. В южной части Каспийского моря он обитает лишь в опреснен-
ных участках (Мурд-аб; см. Рылов, 1928а). Значительные содержания
NaCl выносят Mesocyclops leuckarti и М. crassus. Оба, особенно послед-
ний, очень характерны для пелагиали открытой части Аральского моря,
нормально размножаясь при 0 = 3.3— 3.6 г на литр (S — до 1О°/оо). Оба,
однако, отсутствуют в пелагическом планктоне Каспийского моря (leu-
ckarti встречается лишь в сильно опресненном северном его участке,
М. crassus совсем отсутствует), что может найти свое объяснение
в соотношении Mg и Са в воде Аральского моря. Равным образом
в последнем Acanthocyclops viridis обитает при содержании С1 в коли-
честве 3.58 — 3.62 г на литр (Бенинг, in litt.), тогда как в Каспийском
море не встречается уже при содержании С1 = 1.172 г.
Влияние свободного С1 на Cyclopidae почти не изучено. Соответ-
ственные наблюдения имеются лишь для Mesocyclops leuckarti. По экспе-
риментальным исследованиям Таусон (1934) этот вид, сравнительно
с кладоцерами Daphnia, Ceriodaphnia и некоторыми другими пресно-
водными организмами, относительно свободного хлора довольно устой-
чив. При содержании С1 в количестве 1.0 —1.25 мг в литре рачок
живет 2 суток, при 1.5 мг — 3. 5 часа (остаточный С1 около 0.4 мг),
при 2 мг в литре (остаточный С1 составлает 0.6, мг) — 2 часа 40 мин.,
а при содержании С1 4.00 мг в литре (остаточный С1 составляет 1 мг)
умирает через 1 час 10 мин.
Отношение Cyclopidae к солям железа не выяснено. Богатые желе-
зом сидеротрофные водоемы (свыше 1 мг Fe2O3 на литр), несомненно,
имеют обедненную фауну Cyclopidae. Thienemann (1907) для таких вод
Рюгена (Голштиния) отмечает присутствие вообще весьма эвритопного
Paracyclops fimbriatus; Klie (1925) отмечает Microcyclops varicans var.
rubellus для железистого источника в Oldesloe. В богатых железом
мелких водоемах неоднократно был находим Eucyclops serrulatus.
Сидеротрофные воды весьма часто имеют дистрофный характер, при
резко кислой реакции, причем специфичность экологических условий,
обусловливающих состав населения таких водоемов, выражается одно-
временно не только в высоком содержании железа, но и в комбинации
102
ВВЕДЕНИЕ
других факторов водной среды, имеющих важное экологическое значение
(присутствие значительных количеств гуминовых веществ, pH и проч.).
Отношение пресноводных Cyclopidae к активной реакции (pH) рас-
сматривается в работе Lowndes’а (1928), который на основании своих
исследований дает лимиты pH для ряда видов. Экспериментально это
отношение изучено особенно Таусон (1924) и Скадовским (1926, 1928).
Влияние pH на организмы может быть прямым (например воздействие
Н-ионов) и косвенным (например воздействие свободной СО2) и в есте-
ственных условиях, при сложной комбинации влияния целого ряда других
факторов водной среды, является весьма сложным. Весьма крупную роль
здесь играет солевой состав воды — фактор, вариирующий в очень широ-
ких пределах. Приведенный ниже общий обзор отношения к pH различных
видов Cyclopidae показывает лишь те границы pH, в которых данный вид
встречается в естественных условиях и не предрешает вопроса о прямом
или косвенном влиянии активной реакции среды. Фактический материал
(наблюдения в природе) пока еще невелик. Он позволяет, однако,
наметить основные группировки циклопов, характерные для вод кислых,
нейтральных и щелочных. В табл. 9 горизонтальная черта показывает
границы pH, в которых данный вид был вообще находим, двойная
черта отмечает приблизительные границы оптимума, которые пока
могут быть с достаточной степенью вероятности установлены лишь для
очень немногих видов. Приведенные данные позволяют наметить 3 группы
видов по их отношению к pH.
1. Эврионная группа; виды, широко приспособляющиеся
к условиям активной реакции. Таков, прежде всего, эвритопный вид
Paracyclops fimbriatas, который встречается даже при pH менее 4.
К этой же группе надо отнести Eucyclops serrulatus,1 Е. macruroides,
Tropocyclops prasinus, Macrocyclops albidus (вероятно также M. fuscus),
Paracyclops affinis, Acanthocyclops viridis (вероятно и A. gigas), A. bicu-
spidatus, A. bisetosus, A. vernalis, Microcyclops varicans. Все эти виды,
встречаясь и, повидимому, нормально размножаясь в кислой, нейтральной
и щелочной среде, однако отсутствуют при pH около 4, а некоторые
уже при pH около 5 {Macrocyclops fuscus, Paracyclops affinis, Eucyclops
macruroides, Tropocyclops, Microcyclops varicans).
2. Группа, характерная для кислых в о до е м о в. Среди
подсемейства Eucyclopinae вовсе нет видов, которых можно было бы
отнести к этой группе. Из подсемейства Cyclopinae к ней принадлежат
Acanthocyclops languidus, A. languidoides (группа eriophori—hypnicola),
A. nanus и, судя по еще очень скудным данным, Cyclops strenuus var.
landei. Особенно характерны для рассматриваемой группы 3 указанных
вида Acanthocyclops. Они могут встречаться в слабо кислых — нейтраль-
ных водах, но, судя по всем данным, оптимальные условия существо-
вания находят себе в резко выраженной кислой среде (кислые полионные
водоемы, при pH примерно 3.5 — 5, A. languidus и A- nanus встреча-
ются даже при pH = 3), при условиях pH, которые, например, для Meso-
cyclops leuckarti являются уже летальными (pH = 3 — 4). Эти виды
Acanthocyclops чрезвычайно характерны для дистрофных, богатых гуми-
новыми веществами, мелких водоемов с зарослями Sphagnum и Hypnum
(так называемые сфагнофильные виды).
3. Группа, характерная для нейтральных (слабо кислых)
и нейтрально-щелочных вод (нейтральные мезоионные и близкие
1 Пещерный Euc. serrulatus v. nagasakai Ueno встречается даже при pH = 3.4.
БИОЛОГИЯ
103
к таковым водоемы). Типичными представителями этой группы являются
Mesocyclops leuckarti, М. oithonoides, М. crassus, М. rylovi и, по всем
вероятиям, все прочие виды рода Mesocyclops. Оптимальные условия
для этих видов лежат в пределах pH примерно 6.5 — 8 (главным обра-
зом, вероятно, около 7). Для Mesocyclops leuckarti критическим интер-
валом является pH — 4 — 5. На основании пока еще немногочисленных
данных, к рассматриваемой группе относятся также Macrocyclops distin-
ctus (?), Microcyclops gracilis, M. minutus, M. bicolor и, вероятно, Cy-
clops strenuus typ., C. vicinus и C. scutifer, которые, однако, оптимальные
условия находят и при несколько более кислой реакции, но, во всяком
случае, не ниже pH = 6. Повидимому к этим последним видам примы-
кают Ectocyclops phaleratus и Eucyclops macrurus, которые наиболее
часто (и в значительных количествах) встречаются в нейтральной
и щелочной среде.
Видов, приуроченных исключительно к высокощелочным водам
(pH —8), среди Cyclopidae не имеется. В этих водах (см. выше, са-
политрофные водемы) представлена 1-я и 3-я группа видов, однако
в обедненном составе; 2-я группа в них вовсе отсутствует.
Таблица 9
pH
з
10
А.
Macrocyclops fuscus ....
М. distinctas ........
М. albidus .........
Paracyclops fimbriatus . . .
P; affinis .........
Ectocyclops phaleratus . . .
Eucyclops serrulatus ....
Euc. macrurus .......
Euc. macruroides ......
Tropocyclops prasinus : . .
Cyclops strenuus (typ.) . . .
C. v. landei ........
C. furcifer\ . ... . . .
C. vicinus...............
C. scutifer .............
Acanthocyclops languidus . .
languidoides (rp. eriophori-
hypnicola) .......
nanus .........
vernali ..............
viridis................
gigas .................
bisetosus ........
bicuspidatus (typ.) ....
crassicaudis..........
Microcyclops gracilis ....
M. minutus...............
M. bicolor .........
M. varicans (typ)........
Mesocyclops leuckarti ....
M. oithonoides .......
M. crassus ...... ...
M. rylovi .........
А.
А.
А.
А.
А.
А.
А.
104
ВВЕДЕНИЕ
Наши данные по экологическому значению для Cyclopidae растворен-
ных в воде органических веществ базируются на цифре окисляемости воды,
которая, однако, не вскрывает их химической природы и дает известное
представление (далеко не всегда полное) лишь о суммарном количестве
растворенных веществ, физиологическое значение отдельных раство-
ренных органических веществ, которое может быть далеко не оди-
наковым, при таком положении дела не выясняется. Для водоемов
специально дистрофного характера, где крупную роль играют гуминовые
вещества, в общем, характерно обеднение качественного состава фауны
Cyclopidae при переходе от мезотрофии этих веществ (окисляемость.
в КМпО4 = 25—100 мг на литр) к политрофии (окисляемость свыше
100 мг на литр). В очень богатых этими веществами болотистых водо-
емах, с бурой непрозрачной водой, это обеднение выражено весьма резко,
и здесь мы находим почти исключительно ацидофильные виды циклопов.
Такие водоемы характеризуются очень кислой реакцией, и говорить
здесь с уверенностью о прямом влиянии растворенных органических
веществ едва ли возможно. О положении Cyclopidae в системе сапробов.
говорится ниже, в главе о значении этой группы ракообразных в хозяйстве,
человека.
Отношение Cyclopidae к растворенным в воде газам изучено мало.
В естественных условиях недостаток растворенного в воде кислорода
обычно сопровождается накоплением свободной углекислоты, так что
на организм одновременно воздействуют оба эти фактора. Эксперимен-
тальные исследования Никитинского (1928) показывают, что в насыщен-
ном растворе СО2 Cyclopidae погибают очень быстро, Cyclops strenuus —
через 10—15 сек., С. insignis — через 30—40 сек., молодь Cyclops sp.—
через 15—20 сек., причем гибель вызывается ядовитым воздействием сво-
бодной СО2, а не недостатком кислорода и не водородными ионами.
Исследования того же автора над влиянием недостатка кислорода,
при отсутствии свободной СО2, показали, что при почти полном
отсутствии С2 С. strenuus и молодь Cyclopidae живут 1—3 дня, ,
а С. insignis живет 18 дней. Наблюдения в природе над типичной формой
С. strenuus указывают на широкую эвриоксибионтность этого вида,
одновременно способного некоторое время жить в присутствии серо-
водорода. Так, по нашим наблюдениям (Rylov, 1924; Рылов, 1927а),:
в прудах окрестностей Старого Петергофа (ныне Петродворец)
этот вид живет несколько недель в период заражения H2S (до 11 мг
на литр) даже верхнего слоя воды (вторая половина зимы), когда
анализ в этом слое не дает указаний на следы свободного О2; раз-
множения в таких условиях, однако, не происходит, и рачки встре-
чаются лишь в единичных экземплярах. Huss (1913) экпериментально
показал, что циклопы (С. strenuus?) в полном отсутствии О2 в течение
33 мин. выживали при содержании H2S 70 мг на литр и около 10 мин.
при H2S около 140 мг на литр, а в придонном слое водоемов могут долго
жить при H2S до 4.4 мг на литр. По исследованиям Bigre и Juday
(1908) в озере Mendota, облеченный в „кокон" Acanthocyclops bicuspi-
datus (var. thomasi?), выживает в покоящемся состоянии несколько-
месяцев в придонном слое воды при полном отсутствии кислорода,
причем в таких условиях некоторое время живут и молодые, вышедшие
из кокона, циклопы; повидимому этот вид переносит без вреда содер-
жание О2 в 0.1 см3 на литр. По данным Зверевой (1921), A. gigas в оз. Чаны
(Барабинская степь) зимой живет при содержании О2 в 0.4—1.72 см8
на литр. Сильное обеднение кислородом может выносить также Cyclops.,
БИОЛОГИЯ
105
kolensis (Kozminski, 1933, 1936). По Ziegelmayer (1923) в каменноуголь-
ных шахтах Саарбрюкена несколько1 видов Cyclopidae, в том числе
Eucyclops serrulatus и Acanthocyclops viridis, обитают в условиях полного
отсутствия света, при содержании О2 у поверхности не более 1.49 см®
на литр и в присутствии свободной СО2, метана и сероводорода.
Для огромного большинства пресноводных Cyclopidae вопрос об отно-
шении к растворенным газам остается совершенно не освещенным. Судя
по некоторым данным местонахождений, в высокой степени эвриокси-
бионтны Paracyclops fimbriatus, Eucyclops serrulatus, Acanthocyclops
viridis, A. bicuspidatus. По всем вероятиям сюда же относится ряд
троглобионтов (может быть, даже все троглобионты), обитающих в усло-
виях, при которых имеет место сильное понижение содержания растворен-
ного кислорода и накопление свободной углекислоты, иногда также амми-
ака, метана и сероводорода. Отношение к газовому режиму, в частности?
к растворенному кислороду пелагических видов, почти не выяснено.
В эвтрофных озерах, со значительным дефицитом кислорода в гиполимни-
оне в периоды стагнаций, обитающие в пределах гиполимниона пелагические
виды (например Cyclops abyssorum, С. bohater), несомненно, должны пере-
носить содержание кислорода менее 5О°/0 нормального насыщения, при
некотором накоплении свободной углекислоты. Длительное заражение
сероводородом гиполимниона, однако, ведет к полному выпадению в этом
слое всех Cyclopidae. Так, по наблюдениям Borner’а (1921), в Ritomsee
в Швейцарии мощная зона с глубины 12.5 м до дна постоянно заражена серо-
водородом и совершенно азойна. Существование полиоксибионтных видов.
Cyclopidae не доказано. Если таковые имеются, то в состоянии покоящихся
стадий полиоксибионтные виды (и вообще виды, не переносящие значитель-
ного дефицита кислорода) свободно могут переносить длительное время
крайне неблагоприятные условия газового режима — полное отсутствие
кислорода, накопление свободной углекислоты, аммиака, сероводорода.
Это фактически имеет место в сильно заиленных мелких водоемах,
когда зимой вся толща воды, отрезанная от атмосферы ледяным покро-
вом, нацело теряет кислород и обогащается вредными газами, а также
в эвтрофных, богатых иловыми агильными отложениями, озерах, где
на зимнее время ряд пелагических Cyclopidae (например род Mesocy-
clops) из планктона выпадает, тогда как покоящиеся стадии опускаются
на дно, где и переносят зимний период несмотря на исключительно
неблагоприятные газовые условия придонного слоя зимой. Весной,
с наступлением полной весенней циркуляции, рачки переходят в активное
состояние и появляются в зоне эпилимниона (Рылов, 1927а). Наличие
у Cyclopidae покоящихся стадий, таким образом, имеет огромное значе-
ние в формировании в пределах данного водоема определенных биоце-
нозов Cyclopidae, так как таким путем из года в год сохраняется гене-
тическая преемственность популяций, в частности планктических видов
Cyclopidae, причем комплекс последних, Как и вообще зоопланктон
в целом, получает тот' стойкий, определенный в основных чертах, харак-
тер, который мы находим не только в пелагиали озер, но также в прудах
и, в большей или меньшей степени, даже во временных мелких вод о-
емах.
В заключение настоящей главы остановимся на вопросе о характере
фауны Cyclopidae в водоемах различного трофического типа- Для зоо-
1 Кроме указанных, автор приводит Eucyclops macrurus и Macrocyclops albidus;
что требует проверки.
J06
ВВЕДЕНИЕ
планктона этот вопрос пока разработан вообще крайне недостаточно. Наблю-
дения над зоопланктоном, главным образом озер, показывают, что каждая
из общепринятых в лимниологии 3-х основных групп—олигротрофные,
эвтрофные и дистрофные озера — имеет некоторые специфические черты
состава зоопланктона. Здесь мы будем иметь в виду 3 этих группы
в их крайнем, наиболее типическом выражении, оставляя в стороне
водоемы переходного характера.
Несомненно некоторые виды Cyclopidae широко приспособляются
к условиям водоемов эвтрофной и олиготрофной группы. Так, например,
Cyclops abyssorum. в средней Европе обитает в эвтрофных озерах, но он
в больших количествах развивается и в пелагиали ультраолиготррфного
Телецкого оз. Eucyclops serrulatus л Acanthocyclops viridis встречаются
в литорали как эвтрофных (до крайних ступеней эвтрофности), так
и ультраолиготрофных (стеноолиготрофных) озер. Некоторые виды,
однако, обнаруживают приуроченность только к одной из указанных
групп.
Для пелагического планктона высоко олиготрофных озер средней
Европы весьма характерно нередко полное отсутствие или лишь слабое
количественное развитие Mesocyclops leuckarti и, вероятно, М. crassus.
В таких озерах севера Европы и Азии характерно сильное развитие
Cyclops scutifer и С. lacustris, которые еще встречаются в слабо эвтро-
фированных озерах, но отсутствуют в нормальных эвтрофных, где заме-
няются в пелагиали другими видами этого рода. В эвтрофной группе
озер и более мелких водоемов, напротив, род Mesocyclops, в частности
М. leuckarti, играет очень крупную роль, нередко до преобладания
над прочими Entomostraca в летнем планктоне. Далее, главным образом
для эвтрофных озер, характерны Cyclops kolensis, С. scutifer var. wigren-
sis, C. vicinus (Kozmifiski, 1933, 1936), причем последний вид, невиди-
мому, выпадает из планктона на высоких ступенях эвтрофирования.
Дистрофный тип озер и прудов, при резко выраженной степени
дистрофности (сильная гумификация, до полигумозноети включительно,
значительная кислотность воды, очень малая ее прозрачность, бедность
электролитами), отличается вообще специфическим и весьма обедненным
составом зоопланктона. Качественный состав планктических Cyclopidae
таких водоемов характеризуется полным отсутствием (или лишь совер-
шенно ничтожным значением) Mesocyclops leuckarti и вообще рода
Mesocyclops, а нередко и полным отсутствием эвпланктических Cyclopi-
dae. Так, например, в дистрофном Неклюдовском оз. (Московской области),
при pH = 5, по Скадовскому (Skadowski, 1926), из Copepoda обитает
только Diaptomus graciloides. Выше мы отмечали, что pH = 5 является
критическим пунктом для Mesocyclops leuckarti, так что уже один этот
фактор объясняет отсутствие этого вида в высокодистрофных водоемах.
По данным Н. Nordquist’a (1921) при эвтрофировании (искусственное
удобрение) дистрофных прудов, когда, несомненно, происходит сдвиг
спектра активной реакции в сторону щелочности, этот вид начинает
сильно развиваться. О видах Cyclopidae, характерных для прибрежной
зоны дистрофных озер и для мелких водоемов этого типа, мы упоминали
выше („сфагнофильные" циклопы). Из приведенной выше табл. 9 (см.
стр. 1G3) можно видеть, насколько крупное экологическое значение
имеет pH (прямое и косвенное влияние) для Cyclopidae в тех случаях,
когда происходит резкий сдвиг от нейтрального (нейтрально-щелочного)
интервала в сторону кислотности. Несомненно в сильно дистрофирован-
дгых водоемах, имеющих резко выраженный специализированный характер,
хозяйственное значение
107
специфический состав фауны Cyclopidae (присутствие и высокое разви-
тие „сфагнофилов", резкое общее обеднение качественного состава)
находит себе объяснение уже в кислотности среды, что подтверждается
и экспериментально. Иначе обстоит дело, однако, в водоемах двух других
групп, где пока едва ли возможно точно установить, какие .именно усло-
вия определяют особенности состава фауны Cyclopidae олиготрофной
и эвтрофной группы.
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ .
Циклопы являются промежуточными хозяевами целого ряда пара-
зитических червей (лентецов Cestodes и круглых червей Nematodes),
паразитирующих в различных позвоночных животных, в частности
в домашних, а также в человеке, и вызывающих весьма серьезные забо-
левания (злокачественное глистное малокровие, спарганоз, лигулез,
дракункулез и др.).
В качестве промежуточных хозяев известны следующие роды и виды
Cyclopidae (наиболее полная сводка у Neveu-Lemaire, Traite d’Helmin-
thologie, 1936; нами внесены необходимые исправления согласно новей-
шей синонимике): Macrocyclops fuscus, М. albidus, Eucyclops serrulatus,
Tropocyclops prasinus, Paracyclops affinis, Ectocyclops phaleratus,
Mesocyclops leuckarti, M. oithonoides, Cyclops strenuus, C. vicinus,
C. insignis, Acanthocyclops viridis, A. bicuspidatus, A. vernalis (включая
var. robustus,), Microcyclops gracilis. Ряд указаний на присутствие личинок
паразитических червей относится к ближе не определенным Cyclopidae
.(„Cyclops sp."/ Таким образом промежуточным хозяином являются
не менее 15 видов, относящихся к 9 родам; едва ли подлежит сомнению,
что и некоторые другие виды обоих подсемейств Cyclopidae также могут
быть промежуточными хозяевами, особенно для паразитических цестод.
Заслуживает внимания факт почти полного отсутствия (одно указание —
Microcyclops gracilis) данных на присутствие личинок паразитических
червей у видов рода Microcyclops, к которому принадлежат, главным
образом, мелкие циклопы. Большинство перечисленных выше видов рас-
пространено чрезвычайно широко, а указанные роды, исключая Cyclops
^Палеарктика, отчасти Ориентальная область) обладают также весьма
широким распространением. Южное распространение имеет Tropocyclops
prasinus.
Для различных видов паразитических червей в качестве промежуточ-
ных хозяев были указаны1 следующие Cyclopidae (личинки червей живут
всегда в полости их тела).
Cestodes. Ленточные черви
Diphyllobothrium latum Liihe. Лентец широкий. Окончательный хозяин—
человек (кишечный канал), также в тонких кишках собаки, кошки и неко-
торых других млекопитающих. Может вызвать злокачественное глистное
малокровие вследствие действия глистного токсина. Первый, проме-
жуточный хозяин: Eucyclops serrulatus, Tropocyclops prasinus Cyclops
1 Кроме того Cyclopidae в качестве промежуточных хозяев были еще указаны
и для некоторых других видов: Philometra (Ichtyonema) sanguineum, Eubotrium, Pro-
tocephalas (Cyclops strenuus), Ancylostoma, Strongyloides и др. К вышеприведенному
списку Cyclopidae — промежуточных хозяев — эти указания новых видов не добавляют.
108
ВВЕДЕНИЕ
strenuus, Acanthocyclops viridis, A. vernalis (включая var. robustus),
A. bicuspidatus, также Diaptomidae. Вторым промежуточным хозяином
являются многие рыбы (щука, окунь, ерш, сиговые и др.), употребляемые
в пищу человеком.
Diphyllobothrium (Sparganum) mansoni Cobbold. Окончательный
хозяин — собака, волк, кошка и некоторые другие млекопитающие.
Взрослый лентец живет в тонких кишках. Первый промежуточный хозяин:
Macrocyclops albidus, М. fuscus, Eucyclops serrulatus, Paracyclops
affinis, Ectocyclops phaleratus, Mesocyclops leuckarti, M. oithonoides,
Cyclops vicinus, Acanthocyclops viridis, A. bicuspidatus. Вторым проме-
жуточным хозяином является человек (также крысы, кошки, лягушки,
домашняя птица, змеи, песцы на Командорских островах), в подкожной
клетчатке которого, а также в брюшной и плевральной полости, в шейных
мускулах и в глазу живет плероцеркоид этого лентеца, вызывая болезнь
спарганоз — появление воспалительных опуханий в области залегания
плероцеркоидов. В СССР известен из Дальнего Востока.
Hymenolepis fasciculata (Rud.). Окончательный хозяин — домашний
и дикий гусь и некоторые другие водяные птицы. Живет в тонкой кишке.
Промежуточный хозяин цистицеркоид — в полости тела Eucyclops serrula-
tus, Acanthocyclops viridis и Diaptomus. Вся Европа.
Hymenolepis setigera (Frol.) Взрослый червь—в кишках лебедя,
дикого гуся и др. водоплавающих птиц. Цистицеркоид — в полости тела
Cyclops strenuus и Diaptomus. Вся Европа.
Hymenolepis tenuirostris (Rud.). Взрослый червь — в кишках гуся
и домашней утки. Цистицеркоид — в полости тела Eucyclops serrulatus
и Acanthocyclops bicuspidatus, также Diaptomus, Gammarus, Astacus.
Европа.
Hymenolepis anatina (Krabbe). Взрослый червь — в кишках домаш-
ней утки. Цистицеркоид — в полости тела Eucyclops serrulatus, Acantho-
cyclops viridis, A. vernalis, Diaptomus, Gammarus.
Hymenolepis (Weinlandia) gracilis (Zeder). Окончательный хозяин —
как у предыдущего вида. Цистицеркоид—в полости -тела. Acanthocyclops
viridis, Diaptomus, Ostracoda.
Schistocephalus solidus (O. F. Mull.). Взрослый червь — в кишках раз-
личных диких водоплавающих птиц. Первый промежуточный хозяин —
Acanthocyclops viridis, вероятно A. bicuspidatus и Eucyclops serrulatus;
процеркоид —: в полости тела. Вторым промежуточным хозяином служат
рыбы (колюшки, Coitus, Salmo), тюлень, в брюшной полости которых
живет плероцеркоид (2-я личиночная стадия). Распространен, главным
образом, на севере Европы.
Triaenophorus nodulosus (Pall.). Взрослый червь — в кишечнике рыб,
особенно щуки. Первым промежуточным хозяином является Paracyclops
fimbriatus, Cyclops strenuus и, вероятно, другие Cyclopidae; в полости
их тела живет цистицеркоид. Второй промежуточный хозяин — рыбы
(окунь, форель, ерш, колюшка, налим, щука), в которых плероцеркоид
живет обычно в печени, также в других органах. Наносит сильный вред
рыбному населению, особенно в форелевом прудовом хозяйстве.
Ligula intestinalis (L). Ремнец. Взрослый червь живет в кишечном
канале водных птиц (Podiceps, Larus и др.). Процеркоид — в полости
тела Cyclops strenuus и Diaptomus gracilis. Вторым промежуточным
хозяином служат очень многие пресноводные рыбы (щука, многие кар-
повые, сиговые, окуневые, миноги), в брюшной полости которых живет
плероцеркоид, вызывая болезнь рыб — лигулез. Иногда лнгулез вызывает
ХОЗЯЙСТВЕННОЕ значение
109
целые эпизоотии н наносит существенный вред рыбному хозяйству.
У зараженных рыб чрезвычайно сильно нарушается функциональная
S б
Фнг. 18. 1 — процеркоиды Scfiistocephalus solidas в Cyclops
sp.; 2—процеркоид Sckistocephalus solidus; 3—Cyclops stre-
nuus auct. e процеркоидом Diphyllobothrium latum в полости
тела; 4 — процеркоид Diphyllobothrium latum; 5 — процеркоиды
Diphyllobothrium latum в полости тела Paracyclops affinis (Sars);
6—личинка Dracunculus medinensisB полости тела Mesocyclops
(Thermocyclops); 7—свободноживущие личинки Dracunculus
medinensis. (/—2— no Calio t et Desports, 3—4—no Janicki
et<Rosen, 5—по B. Li, 6 — по E. H. Павловскому, 7—-no
Neveu-Lemaire).
деятельность, брюхо вздувается, и рыба не может уйти на глубину
плавая у самой поверхности; рост рыбы очень сильно замедляется.
Drepanotaenia lanceolata (Bloch.). Взрослый червь — в кишках гуся,
утки, лебедя, очень редко человека. Вызывает кишечный гельминтиазис.
110 ВВЕДЕНИЕ
Цистицеркоидв полости тела Macrocyclops sp., Eucyclops serrulatus,.
Cyclops strenuus, C. insignis, Acanthocyclops bicuspidatus.
Eimeriaria fascicularis (Pall.) Взрослый червь — в тонких кишках
домашней и дикой утки и гуся. Цистицеркоид — в полости тела Cyclopi-
dae (n. det.).
Nematodes. Круглые черви
Dracunculus medinensis (L.). Ришта. Взрослый червь живет в под-
кожной клетчатке (главным образом ног) человека, вызывая чрезвычайно
тяжелую болезнь дракункулез, распространен исключительно в жарких:
странах — тропическая Африка, Иран, Индия. В СССР очаг ее был
в Старой Бухаре. При болезни покровы изъязвляются, иногда нарушается
функция суставов ног. Обычно у человека бывает только 1 червь. Кроме
человека, окончательным хозяином могут быть лошади, собаки, быки,,
бараны, шакалы, дикие кошки, леопарды и некоторые другие млекопи-
тающие. Промежуточным хозяином является Macrocyclops fuscus,.
Eucyclops serrulatus, Tropocyclops prasinus, Mesocyclops leuckarti»
M. oithonoides, Cyclops strenuus, C. vicinus, Acanthocyclops bicuspidatus,
A. viridis, в полости тела которых живут личинки — микрофилярии.
Ghathostoma spinigerum (Owen). Взрослый червь живет в узлах желудка
кошки, собаки, диких Felidae, в качестве гостепаразитов у человека,
у последнего — в коже и под ней, вызывая отеки и дерматоз. Паразит
свойствен Азии и западной и средней Америке. Личинка живет в полости
тела ближе не определенных Cyclopidae.
Camallanus lacusiris (Zoega). Взрослый червь в кишечнике рыб
(окунь, ерш, налим и др.). Первый промежуточный хозяин — Macrocyclops
и другие Cyclopidae, также Asellus и личинки Agrion.
Процесс проникновения личинок паразитических червей в полость
тела циклопов и дальнейшая судьба личинок особенно тщательно изучены
у лентеца широкого и у ришты. Яйца широкого лентеца вместе с экскре-
ментами окончательного хозяина попадают в воду — в различные водо-
емы, в которых обитают циклопы. Из яиц выходит личинка-корацидий,
снабженная б-ю крючками и ресничным покровом, с помощью которого
они свободно плавают в воде. Циклопы, питаясь, заглатывают корацидии,
которые в кишке рачка, сбрасывая реснички, превращаются в личинку-
онкосферу; эта личинка проникает через стенку кишки и попадает
в полость циклопа. В ней онкосфера живет 2—3 недели, превращаясь
в удлинённую личинку — процеркоид, у которой на заднем конце тела
имеется шаровидный придаток с 6-ю крючками; длина такой личинки дости-
гает 50—60 р.. Циклоп редко содержит более 1 —2 процеркоидов. Циклопы
заглатываются рыбами; освобождающиеся из рачков процеркоиды про-
никают через стенки желудка в различные органы; превращаясь в пле-
роцеркоид, они оседают в мышцах, печени, яичниках и в других органах
рыбы, служащей пищей человека. При съедании рыбы в сыром, полу-
сыром или вяленом состоянии плероцеркоиды, сохранившие жизнеспо-
собность, попадают в тонкие кишки человека, прикрепляются к их стенке
и превращаются во взрослого лентеца.
У ришты внутри тела самки находится весьма объемистая матка,.
заполненная личинками-микрофиляриями, длиной 0.5—0.7 мм и шириной
0.015—0.025 мм; взрослая самка имеет длину от 32 до 120 см и толщину
всего 1—1.7 мм. Микрофилярии живут несколько дней в сырой почве
и до 3 дней в воде. Попадая в воду, они плавают в ней и заглатываются
хозяйственное, значение
1JT
циклопами. Процесс захвата личинок последними недавно был весьма
подробно изучен у нас Л. М. Исаевым (1934). Обычно циклоп хватает
личинку с хвостового конца или за середину тела, реже с головного
конца. Захваченная ротовыми конечностями (максиллярными ногами)
личинка обычно свернута в клубок. Затем циклоп заглатывает личинку,
и она проходит в среднюю кишку. Часть личинок при захвате подвер-
гается травматическим повреждениям. Циклопы хватают, по наблюдениям
Исаева, только плавающих личинок; осевших на дно сосуда личинок
они не трогают. Необходимо иметь, однако, в виду, что Исаев ставил
свои опыты с чисто планктическйми видами циклопов — Mesocyclops
oithonoides1 и Cyclops vicinus; возможно, что виды непланктические
хватают и осевших личинок (например Macrocyclops, Eucyclops) Загло-
ченные личинки передвигаются в полости кишечника, причем их движе-
ния постепенно усиливаются; иногда личинкам удается добраться
до задней кишки и выйти через заднепроходное отверстие наружу, при-
чем, по Chatton’y, они остаются живыми. Переход личинок из кишеч-
ника в полость тела, по наблюдениям Исаева, происходит только
в области переднего, желудочного отдела средней кишки. При этом
личинка принимает форму вытянутого кольца, так что ее концы сходятся.
Затем, опираясь на стенку кишки, личинка выпрямляет головной конец
и надавливает им на противоположную стенку желудка, причем послед-
няя несколько выпячивается наружу (в полость тела), и в конце концов,
после движений личинки в этом положении, стенка его пробуравливается,
и она, головным концом вперед, проникает в полость тела, при прохож-
дении сильно изгибаясь. После прохождения личинки раздвинутые ею
клетки стенки желудка быстро смыкаются. Если циклоп проглотил
б—8 и более личинок, то последние своими движениями могут совер-
шенно разорвать желудок, что ведет к гибели циклопа. Обычно переход
личинки в полость тела циклопа происходит через 5—б часов после
ее заглатывания, иногда же уже через несколько минут. Предположение
Федченко, что личинки ришты способны пробуравливать наружные
покровы циклопов, решительно опровергается новейшими исследова-
ниями— внедрение микрофиллярий в тело циклопов происходит исклю-
чительно путем заглатывания (Исаев, 1934).
Ветвистоусые рачки (Cladocera) — активные фильтраторы, как, напри-
мер, Daphnia, Simocephalus, Ceriodaphnia, Moina и др„ не могут захва-
тывать микрофилярий в силу устройства их фильтрационного аппарата.
Исаев показал, что среди Copepoda Phyllodiaptomus bland, очень рас-
пространенный в водоемах Бухары и относящийся также к активным
фильтраторам, может лишь в виде исключения заглатывать микрофиля-
рий, но в таких случаях последние всегда выходят наружу через анальное
отверстие. Таким образом промежуточными хозяевами ришты являются
только Cyclopidae.
В полости тела циклопов микрофилярии развиваются в продолжение
1—1 у месяцев, линяют и достигают длины около 1мм, теряя тонкий
хвостовой отдел. Употребляя для питья воду вместе с планктоном,
в котором имеются зараженные риштой циклопы, человек проглатывает
и этих циклопов. В пищеварительном канале человека микрофилярии,
после переваривания циклопов, освобождаются. Затем они мигрируют
по телу и проникают в подкожную клетчатку, главным образом ног.
1 Возможно, что автор имел не этот вид, а М. crassus (Fisch.) или М. rylovi Smirn»
.112
ВВЕДЕНИЕ
•Способ и путь проникновения личинок в последнюю остается невыяс-
ненным.
Меры борьбы против заражения лентецом широким сводятся, прежде
всего, к профилактическим и направленным, главным образом, к обезвре-
живанию второго промежуточного хозяина — рыбы. Ее необходимо хорошо
проваривать или прожаривать, так как при высокой температуре плеро-
церкоиды погибают. Это достигается также выдерживанием рыбы в холо-
дильниках; наконец плероцеркоиды быстро убиваются солением и мари-
нованием рыбы. Очень важны также меры санитарной охраны водоемов,
при которых они оберегаются от попадания в воду экскрементов. Унич-
тожение первого промежуточного хозяина — циклопов — здесь невозможно,
так как в данном случае приходится иметь дело, главным образом,
с крупными водоемами рыбохозяйственного значения. Существенно также
отметить, что первым промежуточным хозяином лентеца является не один
какой-либо вид циклопа, специально на уничтожение которого могли бы
быть направлены меры борьбы, а целый ряд видов Cyclopidae, как
пелагических, так и литоральных, т. е. свойственных весьма различным
биотопам и имеющих различные циклы развития. К тому же хозяевами
процеркоидов могут быть и Diapiomidae. Если для небольших мелких
водоемов и можно ставить себе задачу уничтожения Cyclopidae, в тех
случаях, конечно, когда это не наносит вреда рыбному хозяйству,
то в крупных водоемах (озера, рыбоводные пруды) такой метод борьбы
не приложим. Необходимо также иметь в виду весьма крупное кормовое
значение Entomostraca и в частности Cyclopidae для мальков рыб и для
целого ряда взрослых рыб. В таком же положении находится вопрос
о мерах борьбы с лигулезом, при котором первым промежуточным хозяи-
ном является Cyclops strenuus и Eudiaptomus gracilis, очень обычные
компоненты озерного (также пруды) планктона, развивающиеся в очень
больших количествах (Е. gracillis обычно держатся в планктоне круглый
год). Уничтожение переносчиков процеркоидоз здесь также технически
невыполнимо. Так как при лигулезе пораженные им рыбы для человека
безвредны, то борьба с лигулезом, очевидно, должна быть направлена
специально к охрана рыбного населения, в тех случаях, когда лигулез
принимает эпизоотический характер и грозит гибелью значительной
части рыбных запасов. Единственной мерой является отстрел водопла-
вающих птиц (чайки, крачки и т. д.), которые служат окончательным
хозяином паразита. При заражении Triaenophorascnn борьба с циклопами
также отпадает и направляется на уничтожение окончательного хозяина
паразита.— рыб, в первую очередь щук. Иначе обстоит дело с мерами борьбы
против дракункулеза (ришты). Профилактической мерой здесь является,
прежде всего, употребление для питья исключительно профильтрованной
или прокипяченой воды. При фильтровании необходимо иметь в виду
фильтры, заведомо не пропускающие циклопов, не только взрослых,
но и находящихся и в предыдущих стадиях копеподитной фазы, когда
рачки меньше взрослых, так как активный захват пищи у Cyclopidae
начинается с 1-й копеподитной стадии, когда происходит быстрое пере-
формирование ротовых конечностей науплиальной фазы и прекращается
науплиальный способ захвата пищи. Кипячение воды убивает всех нахо-
дящихся в ней рачков. Одновременно необходимо запрещать купанье
и мытье ног в водоемах (в Бухаре — хаузы), чтобы предотвратить попа-
дание труда личинок ришты. Необходимо также вести учет всех лиц,
зараженных риштой, а также обратить внимание на собак и других
домашних животных, так как для них доказана возможность заражения
ХОЗЯЙСТВЕННОЕ значение
113
риштой. Вместе с тем при борьбе с риштой весьма успешно применяются
методы, направленные к уничтожению циклопов. В данном случае при-
ходится иметь дело с мелкими водоемам (в Бухаре — система хаузов),
лишенными сколь-либо большого рыбохозяйственного значения, а обычно
и вовсе не имеющими такового. В Бухаре большую роль в уничтожении
дракункулеза сыграла регулярная очистка и осушение, а главным обра-
зом полное уничтожение водоемов, в планктоне которых присутствуют
Cyclopidae. Необходимо, однако, иметь в виду, что для ряда Cyclopidae
известны стадии покоя, в которых рачок легко переносит полное усыха-
ние водоема. Так как эти состояния покоя относятся к последним ста-
диям копеподитной фазы, то при новом наполнении водоема циклопы
появляются весьма быстро и притом сразу в стадии, допускающей воз-
можность активного захвата пищи. Leiper (1907) считает надежным
подвергать осушаемые (в периоды засухи естественным порядком) водо-
емы действию пара, под давлением вводимого в водоем, при температуре
не ниже 35°, кроме того в воду вводится марганцево-кислый калий
в пропорции 0.07 на 100. Меры этого порядка должны предусматривать
не только свободно плавающих рачков, но также и стадии покоя, тем
более, что их образование крайне вероятно в условиях усыхающего
мелкого водоема. Из указанных в качестве промежуточного хозяина
ришты циклопов стадии покоя установлены у Acanthocyclops bicuspidatus
и Cyclops strenuus, но возможность их образования и у прочих видов
едва ли подлежит сомнениям. Так как для развития микрофилярий в полости
тела циклопа необходимо довольно продолжительное время (1—
1 g месяца), то вопрос о длительности жизни циклопов, как взрослых,
так и на копеподитной фазе, является весьма существенным. В литера-
туре по этому вопросу имеется ряд данных. Burckhardt (1900) полагал,
что Cyclopidae живут 8—14 месяцев. У Acanthocyclops viridis, по Walter
(1922), весь метаморфоз занимает, в зависимости от условий темпера-
туры и питания, от 3—4 недель до нескольких месяцев, а взрослые
циклопы живут до 10—14 месяцев: этот автор полагает, что более мелкие
виды Cyclopidae живут 4—6 месяцев, дицикличные виды, например
Tropocyclops prasinus, Paracyclops fimbriatus, некоторые виды группы
strenuus, живут 10—12 месяцев, Eucyclops serrulatus проделывает свой
метаморфоз, в общем, гораздо быстрее, чем Acanthocyclops viridis. Весь
цикл развития у Acanthocyclops bicuspidatus, по Manfredi (1923), при 13—
15°, занимает около 1—месяцев, а при 20—27° около 3 недель.
В водоемах юга СССР и южных широт, где имеет место весьма силь-
ное прогревание воды, метаморфоз, в частности копеподитная фаза,
очевидно, должен происходить в ускоренном темпе; однако индивидуаль-
ная жизнь циклопа в половозрелом состоянии продолжается значительно
дольше, чем время, необходимое для развития микрофилярии в полости
рачка, причем это относится ко всем видам—промежуточным хозяевам
ришты. Рассматривая биологию циклопов в связи с вопросом о дракунку-
лезе, необходимо обратить внимание на очень важный факт существования
в пределах одного и того же водоема целого ряда видов Cyclopidae, ыспу-
щих служить в качестве промежуточного хозяина ришты. Некоторые из этих
видов (например Eucyclops serrulatus, Macrocyclops) в высокой степени
эвритермичны, другие (Tropocyclops, Mesocyclops) характерны для теплого
времени года; Cyclops vicinus встречается нередко круглый год, С. stre-
nuus —форма весенняя. Цикличность этих видов различна, но вследствие
8 Фауна СССР
114
ВВЕДЕНИЕ
присутствия биологически и экологически разнородного комплекса видов
в одном и том же водоеме, в любое время имеется наличие циклопа-
(или их комбинации), могущего стать промежуточным хозяином ришты,
и в этом смысле водоем в любое время года может представлять опас-
ность как очаг заражения. Исключением являются, помимо периода
полного высыхания, лишь те моменты в жизни водоема, когда в нем,
в силу тех или иных обстоятельств, возникают условия, неблагоприятные
для развития циклопов, например сильное понижение активной реакции
(рН = 3—4), недостаток кислорода, слишком сильное органическое загряз-
нение. Однако, например, в условиях сильного дефицита расстворенного-
кислорода и сапробности воды, близкой к а-мезосапробности, некоторые
Cyclopidae (например С. strenuus), способны выживать более или менее
продолжительное время. Безусловно летальным для всех Cyclopidae
является понижение активной реакции в кислую сторону (рН = 3, для"
Mesocyclops, вероятно, уже pH около 5.) Неизвестно, однако, в какой
мере кислую реакцию могут переносить циклопы на стадиях покоя.
Весьма возможно, что регулирование спектра активной реакции, которое-
в мелком водоеме практически достижимо наряду с прочими мерами
борьбы, направленной специально к уничтожению циклопов-переносчи-
ков ришты, окажется весьма существенной радикальной мерой.
Говоря о вредном значении пресноводных Cyclopidae, следует еще
упомянуть о вреде, наносимом ими вместе с остальным планктоном
различного рода фильтрационным сооружениям водопроводов, в частности-
снабжающих население городов питьевой водой. Попадая на фильтры,,
планктон закупоривает их своей массой и может сильно понизить про-
пускную способность. При очень большом преобладании Cyclopidae
в пелагическом планктоне водоема (озера, водохранилища), что, например,
может иметь место зимой, когда фитопланктон обычно развит очень
слабо, участие специально циклопов в засорении фильтров может оказа-
ться весьма значительным. Особенную опасность в этом отношении
могут иметь пелагические Cyclopidae, например Cyclops scutifer (в озе-
рах севера), С. vicinus, С. abyssorum и другие формы группы „strenuus",.
из летних форм — виды рола Mesocyclops (leuckarti, oithonoides, crassus).
Пресноводные Cyclopidae, несомненно, играют крупную роль в пита-
нии рыб. Как показывают анализы, Copepoda вообще содержат довольно-
значительное количество жиров и, особенно, белков. У морских Copepoda
Brandt (1898) нашел 59% белков и 7% жиров, а у пресноводных —
57.25°/0 белков и 6.40% жиров; около 9—100/0 приходится на зольные
вещества. По Knauthe (1907) веслоногие рачки из хорошо удобренного
пруда (в Германии) содержат 52% белков и 12—14% жиров. Специально
для Cyclopidae (ближе не определенных) данные химического анализа
приводятся Birg-e и Juday (1922, планктон озер Северной Америки).-
белков 59.81%, жиров 19.80%, золы около 6%, содержание азота дости-
гает 9.57—10.15%. Эти цифры указывают на высокую пищевую ценность
Cyclopidae, не уступающую ценности других пресноводных и морских
ракообразных.
Весьма многочисленные работы по питанию различных пресновод-
ных рыб, имеющих хозяйственное значение, нередко содержат указания-
на питание циклопами; обычно в этих работах не дается определения»
вида, а родовое название дается в духе старой систематики—„Cyclops".1
Несмотря на несовершенство определений, литературные указания
1 Точное определение затрудняется тем, что его часто приходится производить,
по сильно переваренным остаткам.
хозяйственное значение
115
на питание рыб циклопами в ряде случаев свидетельствуют о поедании
(и переваривании) рыбами циклопов в весьма значительном количестве,
причем некоторые рыбы питаются ими во взрослом состоянии, поедая
циклопов вместе с другими планктическими ракообразными, другие же
рыбы питаются ими в состоянии малька или сеголетка. К рыбам-планкто-
ноядам относятся, например, сиги (в значительной степени), ряпу-
шка, снеток, уклейка и некоторые другие. Из пелагических Cyclopidae
особое значение в питании таких рыб имеют виды, в пелагиали разви-
вающиеся в больших количествах, особенно же крупные циклопы —
различные виды рода Cyclops (группа „strenuus" — С. abyssorum, С. lacu-
stris, С. vicinus, С. scutifer), из мелких циклопов — виды рода Mesocyclops
{leuckarti, oithonoides, crassus). Необходимо иметь в виду, что планктоно-
ядные рыбы, например сиги, могут питаться и донной фауной, и здесь
крупное питательное значение могут иметь специально придонные циклопы.
Так, например, по нашим наблюдениям, пересаженный в оз. Севан (Армян-
ская ССР) ладожский сиг-лудога во взрослом состоянии пожирает в огром-
ных количествах придонного Acanthocyclops gigas, так что кишечники
сигов нередко сплошь набиты остатками этого крупного циклопа.
В пелагиали оз. Севан сиг-лудога, помимо дафний и диаптомусов, хва-
тает Cyclops (гр. strenuus).
Едва ли можно указать на специальное значение отдельных видов
Cyclopidae для питания рыб придонной и прибрежной зоны, в котором
главную роль играют бентические и факультативно-планктические орга-
низмы. Едва ли можно сомневаться, что в таких случаях объектом
питания может стать любой вид циклопа, причем встречающиеся в боль-
ших количествах и притом крупные циклопы должны представлять осо-
бенно значительную пищевую ценность. Таковы, например, виды рода
Macrocyclops (особенно albidas как вид, встречающийся особенно часто,
нередко в значительных количествах), в прудах — Cyclops strenuus,
С. insignis, затем более мелкие циклопы родов Eucyclops, Acanthocyclops
(в озерах Северной Америки также и пелагические формы этого рода,
например A. americanus) и некоторых других родов.
Говоря о полезном значении пресноводных Cyclopidae, отметим
их роль в качестве показательных организмов при санитарно-биологиче-
ской оценке воды. В качестве индиктаторов в известной системе са-
пробов Kolkwitz’a и Marsson’a (1908,1909; см. также: Вислоух, 1916; Kolkwitz,
1922; Долгов и Никитинский, 1927) приводятся как факультативные:
а-мезосапробы Cyclops strenuus и Acanthocyclops viridis, как р-мезоса-
пробы — эти же два вида и Eucyclops serrulatus, Ectocyclops phaleratus,
Paracyclops fimbriatus и Mesocyclops leuckarti, как олигосапробы —
Acanthocyclops viridis, A. bicuspidatus, Eucyclops serrulatus, Macrocyclops
albidus M. fuscus и Mesocyclops oithonoides.
4 основных сапробных зоны (полисапробная, а- и Р-мезосапробная,.
олигосапробная) характеризуются целым рядом признаков. Основными
являются химический состав воды (богатство белковыми веществами'
и продуктами их распада, до полной их минерализации включительно)1
и, в связи с ним, способность воды к загниванию. В связи с этими
признаками далее стоят кислородные и вообще газовые (H2S, свободная
СО2, аммиак) условия и общее направление биохимических процессов
(восстановление — окисление). Таким образом каждая зона (ступень)
сапробности выражается в сложном комплексе экологических факторов,
которым и определяется возможность существования того или иного
вида Cyclopidae в условиях той или иной степени сапробности.
8*
176
ВВЕДЕНИЕ
Среди Cyclopidae мы не знаем видов, встречающихся в условиях
полисапробной зоны. Обилие способных к распаду органических (бел-
ковых) веществ, интенсивные процессы их разложения, с образованием
и аккумуляцией продуктов разложения (свободная СО2, H2S), анаэроб-
ность,— все эти характерные для полисапробной зоны условия в сово-
купности не могут переноситься даже самыми стойкими, широко эври-
топными видами Cyclopidae.
В |3-мезосапробной зоне, где господствуют уже частью окислитель-
ные процессы и полуанаэробные условия, могут выживать лишь очень
немногие виды. В упомянутой шкале сапробов к таковым отнесены
Cyclops strenuus и Acanthocyclops viridis; оба в высокой степени эври-
топны и широко приспособляются к неблагоприятным условиям газового
режима, в частности к пониженному содержанию кислорода и присут-
ствию сероводорода и аммиака. К этим 2-м видам надо прибавить Cyclops
insignis, несомненно встречающегося в условиях, соответствующих
а-мезосапробным. Все эти виды, однако, в таких условиях в массах
не развиваются, а характерны для ^-мезосапробной зоны, в а-мезосапроб-
ной зоне встречаясь лишь факультативно как виды с широкой амплиту-
дой приспособленности к изменениям газового режима в неблагоприят-
ную сторону. Присутствие этих трех видов в а-мезосапробных водах, таким
образом, отнюдь не может служить само по себе индикатором специально
на эти воды.
В [i-мезосапробной зоне, где господствуют окислительные процессы
и вполне нормальные аэробные условия, a H2S, благодаря интенсивным
процессам окисления, отсутствует или имеется лишь в малых количест-
вах, мы находим уже вообще качественно богатую флору и фауну. Состав
фауны Cyclopidae резко обогащается, и ряд видов в условиях рассмат-
риваемой зоны развивается в больших количествах. Как указано выше,
в качестве p-мезосапробов в системе сапробов упоминается 6 видов.
Бесспорно, их число значительно больше. Не говоря уже о Cyclops
insignis (факультативно также а-мезосапроб), экологические данные
позволяют отнести к видам, встречающимся в ^-мезосапробных условиях,
еще следующих циклопов: С. abyssorum (в гиполимнионе эвтрофных озер),
Acanthocyclops gigas (тоже), A. vernalis, A. bicuspidatus, A. bisefosus,
Mesocyclops crassus, М. dybowskii, Macrocyclops albidus, Paracyclops
affinis’, вероятно также, что факультативно в условиях низших ступеней
tB-мезосапробности могут жить Cyclops furcifer, Acanthocyclops crassicau-
dis, A. abyssicola, Mesocyclops oithonoideS, Macrocyclops fuscus, Eucyclops
macruroides, E. macrurus, Tropocyclops prasinus, Microcyclops bicolor,
M. varicans, M. planus.
Только 3 вышеупомянутых вида из этого перечня видов могут встре-
чаться также в условиях а-мезосапробной зоны, тогда как все прочие
виды в ней совершенно отсутствуют.
Целый ряд видов, встречающихся в условиях [В-мезосапробной зоны,
прекрасно развивается в условиях олигосапробной зоны, в практически
вполне чистых водах. К таковым относятся, в сущности, все перечис-
ленные виды, исключая, вероятно, Cyclops strenuus (typ.) и С. insignis.
Mesocyclops leuckarti также может считаться видом, характерным, глав-
ным образом, для JB-мезосапробных вод, но частично он встречается
и в олигосапробных водах, например в эпилимнионе олиготрофных озер,
где, однако, не развивается в сколь-либо значительных количествах,
а нередко и вовсе отсутствует. Для p-мезосапробных вод очень харак-
терны также Paracyclops fimbriatus и Ectocyclops phaleratus, но едва ли
ЛИТЕРАТУРА
117
можно утверждать (Долгов и Никитинский, 1927), что эти 2 вида отсут-
ствуют в олигосапробНых водах. Судя по имеющимся данным, облигат-
ными компонентами олигосапробных вод являются лишь некоторые
формы рода Cyclops {Cyclops strenuus var. vranae, C. tatricus). Далее
для олигосапробных вод весьма характерно сильное развитие Mesocy-
clops oithonoides и Cyclops scutifer. Отнесение Macrocyclops albidus
и M. fuscus исключительно в группу олигосапробов (Долгов и Ники-
тинский, 1927) едва ли правильно.
Вообще в методе санитарио-биологической оценки хозяйственных
вод, по существу методе комплексном, Cyclopidae едва ли имеют в каче-
стве индикаторов абсолютное значение; эти рачки, однако, должны
учитываться наравне с рядом других животных организмов для выясне-
ния общей картины состава населения, что в санитарно-биологическом
анализе имеет уже весьма большое показательное значение.
ЛИТЕРАТУРА
Али-Заде, А. 1934. Гидрофауна Апшеронского полуострова. Тр. Азербайдж. отд.
Закавк. фил. Акад. Наук СССР, сектор зоол., 7:3—13.
Амелина, Л. 1927. Личинки пресноводных Cyclopidae (Copepoda). Тр. Косинской
биол. ст., 5:31—39.
Амелина, Л. 1929. Copepoda Севанского озера. Тр. Севанск. озерн. ст., 2:31—37.
Бенинг, А. 1928. Некоторые данные по фауне колодцев г. Саратова. Раб. Волжск,
биол. ст., X (1):3—2.
Бенинг, А. 1928а. О планктоне оз. Чалкар. Русек, гидроб. журн., 7:219—228.
Бен ни г, А. 1934. Гидрологические н гидробиологические материалы к составлению
промысловой карты Аральского моря. Тр. Аральек. отд. Всес. Инет, морск. рыбн.
хоз., 3:183-200.
Бенинг, А. и Медведев, Н. 1926. О микрофауне водоемов окрести. Эльтона
и Баскунчака. Изв. краев, инет. изуч. южно-волжек. обл. при Саратов, унив.,
1:1—39.
Бенинг, А. и Никольский, Г. 1933. Материалы по рыбному хозяйству озер
низовьев Сары-су. Тр. Аральск, отд. Всес. научн.-иссл. ннст. Морск. рыбн. хоз.,
2:105—148.
Боруцкий, Е. 1937. Географическое распространение Bryocamptus (Arcticocamptus)
arcticas Lili, на основе его экологии. Сб. тр. Гос. Зоол. муз. при МГУ, 4:135—189.
Бродский, А. 1929. Фауна водоемов пустыни Кара-кум. Тр. Среднеазиатск. Гос. унив.
(сер. ХПа), 5:1-42.
Верещагин, Г. 1923. Заметка о пресноводной фауне Памира. Изв. Рос. гидр, инет.,
6:1—20.
Гальцов, П. 1913 (1914). Исследование Косинских озер. Ч. I. Дневн. зоол. отд. Общ.
люб. ест., ант. и этн., 3:1—142; ч. II — стр. 1—107.
Грезе. Б. 1933. Лимнологический очерк Валдайских озер н их предварительная рыбо-
хозяйственная оценка. Изв. Всесоюзн. Научн.-иссл. ин-та рыбн. хоз., 16:66—128.
Дементьева, Т. 1927. Материалы к точному познанию изменчивости низших рако-
образных. Тр. Косинской биол. ст., 5:3—43.
Долгов, Г. и Никитинский, Я. 1927. Гидробиологические методы исследования.
Станд. методы исследов. питьевых и сточных вод. Изд. Пост, бюро Всесоюзн.
водопров. н санит.-технич. съездов. Москва 1—76.
Зернов, С. 1903. О животном планктоне Аральского моря по материалам, собранным:
Л. С. Бергом в 1900 г. Изв. Туркест. отд. Русек, геогр. общ., 3:1—38.
Зиверт, М. 1927. Обзор фауны пресноводных Eucopepoda Украины. Тр. Харьков
общ. исп. прир., 50 (2):3—22.
Зиверт, М. 1929. Опыт изучения фауны пресноводных Eucopopeda бассейна
р. Донца. Тр. Харк. тов. доел, прир., 52:169—241.
Зиновьев, А. 1931. Водоемы Троицкого лесостепного заповедника и их фауна
(Copepoda и Phyllopoda). Тр. Пермск. биол. и научн.-исслед. инет., 3:281—-364.
Зиновьев, А. 1933. Периодические водоемы Троицкого лесостепного заповедника
и его окрестностей. Фауна Copepoda и Р11у11оро8а.Изв. Пермск. биол. научн.-
нссл. ннст., 8:319—360.
Зиновьев, А. 1934. Соленые и солоноватые водоемы Троицкого района Уралобласти.
Изв. Пермск. биол. научн.-иссл. инет., 9:271—295.
118
ВВЕДЕНИЕ
Зиновьев, А. 1935. Постоянны г пресные водоемы Троицкого лесостепного заповед-
ника и его окрестностей. Фауна Copepoda и Phyllopoda. Тр. Пермск. биол.
научн.-иссл. инет., 6:1—118.
Зверева, О. 1930. Опыт рекогносцировочного обследования озер по Омскому
и Славгородскому округам Сибкрая. Тр. Сиб. научи, рыбн. ст., 5:1—90.
Исаев, Л. 1934. О способе проникновения личинок ришты (Dracunculus medinensis)
в циклопов. Мед. даразитол. и паравит. болевни, 3:212—230.
К е й в е р, Н. 1923. К фауне Copepoda и Cladocera р. Сыр-дарьи и ее прибрежных
водоемов. Бюлл. 1 Среднеаз. Гос. Уиив., 2; также Русек, гидроб. жури.,
4 (1925):7б—83.
Ласточкин, Д. 1924. Новые и редкие Copepoda и Oligochaeta в фауне Ив.-Вознес.
губ. Изв. Гос. Гидрол. инет , 9:1—22.
Ласточкин, Д. 1926. Веслоногие Плещеева озера. Докл. Переясл.-Залесск. научн.-
проев. общ., 14:32—33.
Ласточкин, Д. (в сотр. с Кордэ Н. и др.), 1926а. Прибрежные сообщества Валдай-
ского озера. Зап. Гос. Гидрол. инет., 1:1—70.
Ласточкин, Д. 1928. Ракообразные. Crustacea. Мат. по фауне Плещеева овера. VIII.
Тр. Переясл.-3алесск. истор, худож. краев, музеи, 8:32—35.
Ласточкин, Д. (в сотр. с Кордэ, Н., Рубинович, А. и Уломским, С.). 1934.
Типологический анализ иепойменных озер Вязииковского, Южского и Семенов-
ского районов. Рыби. хоз. Ивановск. пром. обл. и его перспективы, 2:7—55.
Ласточкин, Д. и Кордэ, Н. 1921. Copepoda района исследования. Исследовании
по фауне Ив.-Вознес. губ., произведенные Сельскохозяйственным факультетом
Ив.-Возиес. биол. инет. Изв. Ив.-Вознес. политехи, инет., 4:3—5.
Лепешкин, В. 1900. К фауне Copepoda Акмолинской области. Изв. Общ. люб. ест.,
антр. и этногр., 98 (2), 1—29.
Марковский, Ю. 1935. Овера Конча та його зооплаиктон. 2. Тр. Гидроб. ст. ВУАН
8/9:86—215
Мир ошниче нк о, О. 1930. Зам1тка про Copepoda водоймищ нетеч долины р. П1вд.
Бог. 36ipH. прац. Дншр. бюл, ст., 5:327—330.
Морозов, А. 1915. Река Цивнль и ее обитатели. Тр. Общ. ест. Каваиск. унив.,
47(3^:1—194.
Неизвестнова-Жадина, Е. 1929. К изучению микрофауны р. Оби и ее бас-
сейна. Изв. Гос. гидр, инет., 25:59—68.
Па ж и т н о в а, 3. 1935. Материалы к изучению мнкрофауны рисовых полей. II. Микро-
фауна типового рисового поля Самаркандского оазиса. Тр. Среднеазиатск. Гос.
унив., VIII, Зоология, 183:62.
Рузский, М. 1889. О пелагической фауне оз. Кабана. Тр. Общ. ест. Казанск. унив.,
19:1—31.
Рувскнй, М. 1926. Заметка о ракообразных озер Госкурорта Карачи. Курорт
оз. Карачи. Изд. Бальн.-физ.-терап. инет., Томск: 37—38.
Ру век ий, М. 1929. Курорти. оз. Горькое. Сб. кур. Урала, I. (Цит. по Таусон, 1933).
Рылов, В. 1915. К планктону некоторых озер Витебской губернии. Фауна Copepoda
н Cladocera. Тр. Петр. общ. естеств., 44:111—162.
Рылов, В. 1916. Материалы к познанию фауны Copepoda и Cladocera Тверской губ;
Тр. Бородинской биол. ст., 4:1—92.
Рылов, В. 1917. Материалы по фауне свобэдноживущих пресноводных Copepoda Сев.
России, I. Ежег. Зоол. муз. Росс. Акад. Наук, 22:247—310.
Рылов, В. 1918. Материалы по фауне свобэдноживущих пресноводных Copepoda Сев.
России. II. Ежег. Зоол. муз. Росс. Акад. Наук, 23:43—96.
Рылов, В. 1920. К сведениям о фауне водоемов г. Петрограда и его окрестностей.
Журн. Петрогр. агрон. инет., 2:82—104.
Рылов, В. 1921. Пресноводные свободноживущие Copepoda Петроградской губернии.
Фауна Петрогр. губ., (ивд. Петр, агрон. инет.), 2:2—11.
Рылов, В. 1922. Свободноживущие веслоногие ракообразные (Eucopepoda). Преснов,
фауна Европ. России, изд. Природы и Научрыббюро Главрыбы, 1:1—126.
Рылов, В. 1923. К фауне Crustacea-Eucopepoda Манчжурии. Ежег. Зоол. мув. Росс.
Акад. Наук, 24:52—95.
Рылов, В. 1927. К познанию фауны Eucopepoda некоторых водоемов Олонецкого
края. Тр. Олонецкой эксп. Гос. гидр, инет., Зоология, 6:1—44.
Рылов, В. 1927а. Исследования над планктоном прудов окрестностей Петергофского
• еетеств.-научн. инет. Изв. Петергофск. естеств.-научн. инет., 4:1—253.
Рылов, В. 1928. Материалы к фауне пресноводных свободноживущих веслоногих
ракообразных северной Сибири. Тр. Комисе, по изуч. Якутск. АССР, 11:1—33.
Рылов, В. 1923а. К фауне Eucopepoda Кавказа, Закавказья и северной Персии. Раб.
Сев.-Кавказской гидроб. ст., 2:1—17.
ЛИТЕРАТУРА
119
Крылов, В. 1930а. Некоторые данные о зоопланктоне озера Исеык-куль. Мат. Комиее.
эксп. исел. Акад. Наук, СССР, II, Иссык-кульская эксп. 1928, 1:111—135.
Рылов, В. 1930b. Некоторые наблюдения над захватом сестона у Diaptomus eoeruleup
Fisch. Тр. Ленингр. общ. естеств., 60:149—176.
Рылов, В. 1933. Пресноводные Eucopepoda Уссурийского края. Тр. Зоол. инет. Акад.
Наук СССР, 1:243—280.
Рылов, В. 1933а. К сведениям о планктоне озера Балхаш. Иссл. озер СССР, 4:57—69.
Рылов, В. 1933b. К сведениям о зоопланктоне горных озер окр. Телецкого озера
(Алтай). Исел. озер СССР, 3:121—130.
Рылов, В. 1935. Состав и вертикальное распространение зоопланктона некоторых
озер центр. Якутии. Иссл. озер СССР, 8:85—124.
Рылов, В. 1935а. Некоторые количественные и качественные данные о зоо-
планктоне озера Иссык-куль. Тр. Киргизской компл. экспед. Акад. Наук СССР,
3:87—96.
Рылов, В. 1936. К фауне Eucopepoda озер поймы низовьев р. Амура. Вести. Дальне-
вост. фил. Акад. Наук СССР, 21:143—160.
Рылов, В. 1936а. К лимнологии Кавказа. III. Некоторые данные по химизму и зоо-
планктону водоемов окр. Хосты и Красной Поляны. Тр. Зоол. инет. Акад. Наук
СССР, 3:79—88.
Рыло в, В. 1937. Зоопланктон некоторых горных водоемов Байкальского хребта. Тр.
Байк, лимнол. ст., 7:75—86.
С к а д о в с к и й, С. Н. 1928. Влияние изменений pH в растворах различных электро-
литов на выживание личинок Cyclops languidas Sars. Тр. Звенигор. гидрофизиол.
ст., 1:82—95.
'Скадовский, С., Савич, В. и Брюхатова, А. 1928. Биологические и физико-
химические исследования оз. Бисерова и Неклюдова, Московской губ. Тр. Звени-
гор. гидрофизиол. ст., 1:167—214.
Скадовский, С., Кортиков, А., Амалинскин, И. и Брюхатова, А. 1933.
Биология планктона и физико-химич. режим Петровских озер Оршанского тор-
фяника Московской обл. Зоол. журн., 12,3:3—107.
"Смирнов, С. 1926. К фауне веслоногих ракообразных окрестностей г. Костромы. Тр.
Костр. научн. общ., 37:7—18.
Смирнов, С. 1928. Заметка о фауне Copepoda Кавказа. Тр. Ленингр. общ. ест.,
58, 1:11-28.
Смирнов, С. К фауне Eucopepoda Средней России. Тр. Костромск. научн. общ. по
изуч. мести, края, 43:1—12.
'Смирнов, С. 1930. Заметки по фауне Copepoda СССР. II. Русск. гидроб. журн.,
9:8—22.
Смирнов, С. 1930а. О двух новых формах Copepoda из Амурского края. Докл.
Акад. Наук СССР, V:281—286.
Смирнов, С. 1933. Материалы к познанию воопланктона озер Карелии. I. Зоопланк-
тон Кончезерской группы озер. Тр. Бородинской биол. ст., 7, 1:27—55.
Смирнов, С. 1935. Зоопланктон озер Катунских альп. Иссл. озер СССР, изд. ГГИ.,
8:254—262.
Совинский. В. 1888. Очерк фауны пресноводных ракообразных окрестностей Киева
и северной части Киевской губернии, Зап. Киевск. общ. ест., 9:225—298.
Совинский, В. 1891. Материалы по фауне пресноводных ракообразных Юго-западного
края. Зап. Киевской общ. ест., 11:107—177.
Соколова, М. 1927. Зоопланктон восточной части Финского залива. Иссл. р. Невы
и ее бассейна. Гос. Гидр, инет., 2 (5):3—45.
Соколова, М. 1930. К зоопланктону. Ладожского озера, Исслед. р. Невы и ее басе.
Гос. Гидр, инет., 6:53—67.
Соколова, М. 1935. Планктон Ладожского озера в районе Осиновецкого маяка. Тр.
Петергофск. биол. инет., 13—14:124—149.
Соловьев, М. 1923. К познанию известковых сапропелитов. Изв. Сапроп. ком.,
4:61-124.
Тау'с о н, А. 1934. Изучение влияние хлора на организм при хлорировании воды. Изв.
Пермск. биол. научн.-иссл. инет., 9:251—269.
Таусон, А. 1935. Озеро Дикое. Уч. зап. Пермск. Гос. унив., 1:3—-55.
Ульянин, В. 1875. Ракообразные. Путешествие Федченко в Туркестан, 2, 6:23—41.
Уломский, С. 1928. Материалы к познанию фауны Eucopepoda Ярославской губ.
Тр. Яросл. ест.-истор. и краевед., общ., 4, 2:41—46.
Фадеев, Н. 1925. К сведениям о фауне озер Закавказья. Раб, Сев.-Кавказской гидроб.
ст., 1:19—26.
Харин, Н. 1925. К изучению фауны Eucopepoda Воронежской губ. Русск. гидроб.
журн., 4:162—163.
120
ВВЕДЕНИЕ
Шмаикевич, В. 1875. Некоторые ракообразные соляно-озерных и пресных вод.
и отношение их к среде. Зап. Новоросс. общ. ест., 3,2:1—391.
Яшнов, В. А. 1925. Crustacea Новой: Земли. Тр. Пловуч. морск. научн. инет.,
12:51 7 б.
Яшнов, В. А. 1927. Зоопланктон Карского моря. Тр. Пловуч. морск. научн. инет.,.
2:1—59.
Атта, К. 1911. Ueber die Differenzierung der Keimbahnzellen bei den Copepoden.
Arch. Zellforsch., 6:497—576.
Birge, E. 1897. Plankton-studies on Lake Mendota. II. Trans. Wiseons. Acad. Sei.
a. Let., 11:274—448.
Birge, E. a. Ju day, Ch. 1908. A summer resting stage in the development of Cyclops
bicuspidatus Claus. Trans. Wiseons. Acad. Sei., 16:1—9.
Borutzky, E. 1927. Cyclops muscicola Meuzel and Cyclops mnscicolus Last. Zook
Anz, 71:63.
Borutzky, E. 1930. Zur Kenntnis der uuterirdischen F auna der Kutais-Hohlen am Rion
(Transkaukasus, Georgien). Copepoda-Cyclopoida. Zool. Anz., 89:331—335.
Bowkiewicz, J. 1928. Cyclops scutifer Sars in dem Krzyzaki-See bei Wilno. Arch.
Hydrob. i Rybact., 3:39.
Braun, H. 19C9. Die spezifischen Chromosomenzahlcn der einheimischen Arten der
Gattung Cyclops. Arch. Zellf., 3:449—482.
Brehm, V. 1918. Ergebnisse einiger im Marienbader Moor unternommener Exkursionen..
Arch. f. Hydrob., 12:422—453.
Brehm, V. 1919. Aus dem Kaiserwald. Uuser Egerland. 23. (Цитир. no Kiefer'y, 1923).
Brehm, V. 1926. Zoologische Ergebnisse der von Prof. Dr. F. Klute nach Nordpatago-
nien unternommener Forschungsreise. I. Die Entomostraken. Arch. f. Hydrob.,.
16:302—329.
Brehm, V. 1927. 3. Ordnung der Crustacea Entomostraca. Copepoda. Handb. d. Zool..
v. W. Kiickenthal u. O. Krumbach, 3:435—496.
Burckhardt, G. 1913. Wiss. Ergebnisse einer Reise pm die Erde von M. Pernod und.
C. Schroter. 111. Zooplankton aus ost- und siid-asiatischen Binnengewassern. Zool.'
Jahrb, Syst, 34:341—472.
Chambers, R. 1912. A discussion of Cyclops viridis Jur. Bull. Biol., Woods Hole Mass.,
22:291—296.
Chambers, R. 1912a. Egg maturation, chromosomes and spermatogenesis in Cyclops.
Toronto Stud. Univ. Biol., 14:1—37.
Chappuis, P. 1920. Die Fauna der unterirdischen Gewasser der Umgebungen Basel.
Arch. Hydrob., 14:1—88.
Chappuis, P. 1925. Sur les Copepodes et les Syncarides des eaux souterraines de Cluj;
et des Monts Bihar. Bull. Soc. Stiinte Cluj, 2:157—182.
Chappuis, P. 1931. Crustaces Copepodes. Campagne speologique de C. Bolivar et
R. Jeannel dans 1’Amerique du Nord (1928). Arch. Zool. Exper. et Gener, 71:345—-360.
Chappuis, P. 1933. Copepodes, avec Г enumeration de tons les Copepodes cavernicoles
connus en 1930. Arch. Zool. Exper. et Gener., 76:1—57.
Chat ton, E. 1918. Prenne experimentale de 1’infestation des Cyclopes par vue digestive..
Bull. Soc. path. exot. Paris, 11:338—348.
Claus, C. 1857a. Das Genus Cyclops und seine einheimische Arten. Arch. Natnrgesch.,
23:1—40.
Claus, C. 1357. Weitere Mitteilungen fiber die einheimischen Cyclopiden. Ibid., 23:205—211..
Claus, C. 1858. Zur Anatomie und Entwicklungsgeschichte der Copepoden. Arch. Natur-
gesch., 24:1—76.
Claus, C. 1863. Die freilebenden Copepoden mit besonderer Berucksichtigung der Fauna
Deutschlands, der Nordsee und des Mittelmeeres, Leipzig:!—230.
Claus, C. 1877. Die Schalendrfise der Copepoden. Sitzungsber. Akad. Wien, Math. Nat.
KI., 74:717—721.
Claus, C. 1893. Ueber die Antennen der Cyclopiden und die Auflosung der Gattung
Cyclops in Gattungen und Untergattungen. Akad. Anz. Wien, 9:79—83; 13:116—120.
Claus, C. 1893a. Neue Beobachtungen fiber die Organisation nnd Entwicklung von
Cyclops. Arb. Zool. Inst. Wien, 10:283—356.
Claus, C. 1894. Ueber die Wiederbelegung im Schlamm eingetrockneter Copepoden und
Ostracoden-Eier. Arb. Zool. Inst. Wien, 17:1—12.
Claus, C. 1899. Ueber die Maxillarffisse der Copepoden und die morphologische Deutung-
der Cirripedien:Gliedmassen. Arb. Zool. Inst. Wien., 11:49—65.
Coker, R. 1933. Influence of temperature on size of freshwater Copepods (Cyclops).
Int. Rev. ges. Hydrob. u. Hydrogr, 29:406—435.
Coker, R. 1934. Influence of temperature on form of the freshwater Copepod Cyclops:
vernalis Fischer. Int. Rev. ges. Hydrob. u. Hydrogr., 30:411—427.
ЛИТЕРАТУРА
121
D a d а у, Е. 1908. Entomostraea et Hydrachnidae. е Tibet. Rec. Ind. Mus., Calcutta,
2:323—341.
Dietrich, W. 1915. Die Metamorphose der freilebenden Copepoden. I. Zeitschr. wiss.
Zool., 113:252—324.
Ek man, S. 1904. Die Phyllopoden, Cladoceren und freilebenden Copepoden der nord-
schwedischen Hochgebirge. Zool. Jahrb., Syst., 21:1—170.
Ekman, S. 1907. Ueber das Crustaceenplancton des Ekoln (Malaren) und fiber verschie-
dene Kategorien von marinen Relicten in schwedischen Binnenseen. Zool. Studieh
till Prof. T. Tullberg:42—65.
Ewers, L. 1930. The larval development of fresh-water Copepoda. Diss. Ohio St.
Univ., 1:60—70. (Цитировано no Gurney, 1933).
Farkas, B. 1923. Beitrage zur Kenntnis der Anatomie und Histologie des Darmkanals
der Copepoden. Acta lit. Acad, scient. Reg. Univ. Francisco-Josephinae, Sect. Sc.
Nat., 1:47—76.
Far wick, B. 1917. Neuere Untersuchungen fiber die Verbreitung der freilebenden Cope-
poden am Niederrhein und ihre Biologie. Arch. Naturg.:7—27.
Fischer, S. 1851. Beitrage zur Kenntnis der in der Umgegend von St. Petersburg sich
findenden Cyclopiden. Bull. Soc. Imp. Nat. Moscou, 24:409—-438.
Fischer, S. 1853. Beitrage zur Kenntnis der in der Umgegend von St. Petersburg sich
findenden Cyclopiden. Bull. Soc. Imp. Nat., Moscou, 2o:74—100.
Fischer, S. I860. Beitrage zur Kenntnis des Entomostraceen. Abh. Bayer. Akad. Wiss.,
8:645—682.
F c r b e s, E. 1897. A contribution to a knowledge of N. A. freshwater Cyclopidae. Bull.
Illinois State Labor. Nat. Hist., 5:27—82. ,
Fric, J. 1882. Note preliminaire sur 1’ontogenie de nos Copepodes d’eau douce. ZooL
Anz„ 5:498—503.
Fuchs, K. 1914. Die Keimblatterentwickluug von Cyclops viridis Jurine. Zool. Jahrb.,
Anat., 38:103—156.
G a g e r n, E. 1924. Beitrage zur Kopepodenkunde Deutschlands. 2. Zool. Anz., 60:334—336.
Gauthier, H. 1928. Recherches sur la faune des eaux continentales de I’Algerie et de la
Tunisie. Alger, Minerva: 1—419.
Gelmini, G. 1928, Contribute alia couoscenza dello sviluppo larvale di Cyclops leuckarti
Claus. Natura, Milano, 19:89—96.
Gicklhorn, J. 1930. Bau und Funktion der Excretionsorgane von Cyclops strenuus
Fischer. Zeitschr. wiss. Zool., 127:120—149.
Gicklhorn, J. 1930a. Notiz fiber die sogen. Cornealinsen von Cyclops strenuus. Zool.
Anz., 90:250—258.
Gicklhorn, J. 1930b. Zur Kenntnis der Frontalorgane von Cyclops strenuus. Zool. Anz.,
90:209—216.
Graeter, A. 1903. Die Copepoden der Umgebung Basels. Rev. Suisse Zool., 11:419—541.
Graeter, E. 1911. Die Copepoden der unterirdischen Gewasser. Arch. Hydrob., 6:1—48,
111—152.
Grander i, R. 1926. Noune specie di Copepodi della Laguua Veneta. Boll. 1st. Zool.
Roma, 3:38—70.
Gurney, R. 1916. On some fresh-water Entomostraea from Ceylon. Proc. Zool. Soc..
London : 333—343.
Gurney, R. 1927, Some forms of Cyclops allied to C. languidus Sars, with notes on
C. minutus Claus. Ann. Mag. Nat. Hist., (9) 19:497—509.
Gurney, R. 1929. Dimorphism and rate of growth in Copepoda. Int. Rev. ges. Hydrob.
u. Hydrogr., 21:189-207.
Gurney, R. 1931. British fresh-water Copepoda. London, Ray Soc., 1:1—238.
Gurney, R. 1933. British fresh-water Copepoda, London, Ray Soc., 3:1—384.
Haberbosch, P. 1920. Die Sfisswasser-Entomostraceu Gronlands. Zeitschr. Hydrol.,
1:130—184, 245—349.
Haecker, V. 1897. Die Keimbahn von Cyclops. Arch. mikr. Anat., 49:35—91.
Handlirsch, A. 1914. Eine interessante Crustaceenform aus der Trias der Vogesen.
Verb, zool.-bot. Ges. Wien, 64:1—8.
Hansen, H. 1825. Studies on Arthropoda. II. Ou the comparative morphology of the
appendages in the Arthropoda Crustacea. Copenhagen:!—176.
H ans t г о m, B. 1924. Beitrag zur Kenntnis des zentralen Nerveusystems der Ostracoden
und Copepoden. Zool. Anz., 61:31—38.
Harada, I. 1931. Studien fiber die Sfisswasserfauna Formosas. Annot. Zool. Japon.,
13:149—168. * .
Hartmann, O. 1917. Ueber die tcmnorale Variation bei Copepoden [Cyclops, Diaptomus]
und ihre Beziehunv zu der bei Cladoceren. Zeitschr. f. indukt. Abst. u. Vererbungs-
iehre, 18:22-43.
122
ВВЕДЕНИЕ
Н а г t о g, М. 1888. The morphology of Cyclops and the relations of the Copepoda. Trans.
Linn. Soc. London, 5:1—46.
H e b e r e r, G. 1926. Zur Kenntnis der mannlichen Generationsorgane der Cyclopiden.
Verb. Deutsch. Zool. Ges.: 141—148.
Herrick, C. a. Turner, C. 1895. Synopsis of the Entomostraca of Minnesota. Rep.
Surv. Minnesota, Zool. Ser., 2:1—525.
.Holmes, S. 1909. Sex recognition in Cyclops. Biol. Bull., 16:313—315.
Huss, H. 1913. Konnen die Cyclopiden intramolekular atmen., lut. Rev. ges. Hydrob.
u. Hvdrogr., 6:38—43.
Hutchinson, G., Pickford, G. a. Seh uur m a n, M. 1932. A contribution to the
hydrobiology of pans and other inland waters of South-Afrioa. Arch. f. Hydrob.,
24:1—154.
Jacobs, M, 1925. Entwicklungsphysiologische Untersuchungen am Copepodenei (Cyclops
viridis Jurine). Zeitschr. wiss. Zool., 124:487—541.
Janicki, C. A. et Rosen, L. 1918. Le cycle evolutif du Dibotriocephalus latus L. Bull.
Soc. Sc. Nat. Neuchatel, 42:19—53.
Jurine, L. 1820. Histoire des Monocles, qui se trouvent aux environs de Geneve. Paris:
1—260.
Kai m us, H. 1928. Ueber die Bodenfauna der Moldau im Gebiete von Prag. Ein Jahres-
zyklus. II. Int. Rev. ges. Hydrob. u. Hydrogr., 19:349—429.
Keiser, N. 1231. Ueber die Cladoeeren und Copepoden der Wiiste Kara-Kum. Int. Rev.
ges. Hydrob. u. Hydrogr., 25:355—372.
Kiefer, F. 1923. Beitrag zur Kenntnis von Cyclops crassicaudis Sars. Zool. Anz.,
56:283—289.
Kiefer, F. 1927. Versuch eines Systems der Cyclopiden. Zool. Anz., 73:302—308.
Kiefer, F. 1928. Ueber Morphologie und Systematik der Siisswasser-Cyclopiden. Zool.
Jahrb., Syst., 54:495—556.
Kiefer, F. 1929. Crustacea Copepoda. IL Cyclopoida Gnathostoma. Das Tierreich,
53:1—102.
Kiefer, F. 1929a, Zur Kenntnis einiger Artengruppen der Siisswasser-Cyclopiden.
Zeitschr. wiss. Zool., 133:1—56.
Kiefer, F. 1930. Mitteilung uber die Cyclopiden der Deutschen Limnolog schen Sunda-
Expedition. Zool. Anz., 89:319—322. *
Kiefer, F. 1931. Die Untergattung Tropocyclops der Gattung Eucyclops (Copepoda,
Cyclopoida). Zeitschr. wiss. Zool., 138:487—514.
Kiefer. F. 1931a. Neue Siisswassercopepoden aus Jugoslawien. Zool. Anz., 101:49—60.
Kiefer, F. 1931b. Zur Kenutnis der freilebenden Siisswassercopepoden, insbesondere der
Cyclopiden Nordamerikas. Zool. Jahrb., Syst., 61:579—620.
Kiefer, F. 1932. Zur Kenntnis der Copepodenfauna der Sunda-Insel. Arch. Naturgesch.,
N. F„ 1:225-274.
Kiefer, F. 1932a. Neue Diaptomiden und Cyclopiden aus Franzosisch-Westafrika. Bull.
Soc. Stiinte Cluj, 6:523—528.
Kiefer, F. 1932b. Ein neuer Mesocyclops (Copepoda Cyclopoida) aus der Mandschurei.
Zool. Anz., 100:234—237.
Kiefer, F. 1933. Freilebende Binnengewasser-Copepoden. Voyage de Ch. Alluaud et
P. A. Chappuis en Afrique Occidentale Fran^aise. Arch. f. Hydrob., 26:121—142.
Kiefer, F. 1933a. Die freilebenden Copepoden der Biunengewasser von Insuliude. Arch,
f. Hydrob., Suppl. 12:519—621.
Kiefer, F. 1935. Neue Susswasser-cyclopiden aus Ostafrika. Bull. Soc. Stiinte Cluj,
8:237—242.
Kiefer, F. 1936. Ueber die Krebstiere, insbesondere die Ruderfusskrebse des Eichener
Sees- Beitr. naturkundl. Forseh. Siidwestdeutsehland, 1:157—162.
Kiefer, F. 1336a. Brasilianische Ruderfusskrebse, gesammelt von Herrn Dr. Otto Schu-
bart. П. Zool. Anz., 114:129—133.
Kiefer, F. 1937. Ueber Systematik und geographische Verbreitung eiuiger Gruppen stark
verkiimmerten Cyclopiden. Zool. Jahrb., Syst., 70:419—442.
Kiefer, F. 1937a. Freilebende Ruderfusskrebse (Crustacea Copepoda) aus Angola.
I. Diaptomiden u. Cyclopiden. Arch. f. Hydrob., 32:470—485.
Kiefer, r. 1937b. Cyclopiden (Crustacea Copepoda) aus nordspanischen Hohlen. Zool.
Anz., 119:321—327.
Kiefer, F. 1937c. Eine kleine Copepoden-Ausbeute aus der ostlichen Mongolei, Zool.
Anz., 119:293—298.
Kiefer, F. 1937d. Ein- neuer Cyclopide (Crustacea Copepoda) aus dem Ochridasee. Zool.
Anz., 120:137—143.
Kleiber, O. 1911. Die Tierwelt des Moorgebietes von Junpholz im siidlichen Schwarz-
wald. Areh. Naturg., Suppl. 3:1—115.
ЛИТЕРАТУРА
123
KoimiHski, Z. 1927. Ueber die Variabilitat der Cyclopiden aus der sfrenuns-Gruppe
auf Grund von quantitativen Untersuchungen. Bull, Int. Acad. Pol. Sc. Lettr., Suppl.:
1—114.
Kozminski, Z. 1932. Ueber die systematische Steliung von Cyclops strenuus aus den
Gebirgsseen. Arch. Hydrob. i Rybactwa, 6:140—151.
Kozminski, Z. 1933. Badania morfometryczne i ekologiczne nad oczlikami {Cyclopidae)
z grupy strenuus. Arch. Hydrob. i Rybactwa, 7:59—140.
Ko^mihski, Z. 1934. Ueber die morphologiche Gruppierung der Arten des Subgenus
Cyclops. Mem. Acad. Polon. Sc. Lettr., B, 3:105—121.
Ko^mihski, Z. 1934a. Ueber die okologische Verteilung einiger limnetischer Cyclopiden
in den Wigryseen. Verb. Int. Ver. theor. u. angew. Limnol., 6:299—307.
Kozminski, Z. 1935. Ueber die Eigentiimlichkeiten des Zooplanktons des Ochridasees.
Verb. Int. Ver. theor. u. angew. Limnol., 7:245—254.
Kozminski, Z. 1936. Morphometrische und okologische Untersuchungen an Cyclopiden
der strenuus-Gruppe. Int. Rev. ges. Hydrob. u. Hydrogr., 33:161—240.
Lande, A. 1890. Materyjaiy do fauny skorupiakow nidlonogicb (Copepoda) Krolestwa
Polskiego. Widlonogi swobodnie Zyjace, I. Rodzina Cyklopy. Pamiet. Fizyjogr,
10:307—398.
Lastockin, D. 1927. Biosooziologische Studien in der Litoralregion eiuiger russischen
Seen. Verb. Int. Ver. theor. u. augew. Limnol., 3:262—276.
Lilljeborg, W. 1901. Synopsis specierum hue usque in Suecia observatorum generis
Cyclops. Kngl. Svenska Vet. Acad. Handl., 35:1—118.
Lindberg, K. 1936. Note sur les Cyclopides de 1’Iran. Bull. Mus. Hist. Nat. Belg.,
12 (17):1—26.
Lint, G. M. 1922. Untersuchungen uber Plankton-Copepoden, in niederlandischen Gewas-
sern. Int. Rev. ges. Hydrob. u. Hydrogr., 10:76—90.
Lowndes, A. 1926. Cyclops americanus March. Ann. Mag. Nat. Hist., (9), 17:616—619.
Lowndes, A. 1928. Freshwater Copepoda and hydrogen ion concentration. Ann. Mag.
Nat. Hist., (10) 1:457-460.
Lowndes, A. 1928a. Cyclops americanus Marsh, a discussion and description of its
specific characteristics and its occurence in Europe. Int. Rev. Ges. Hydrob. u. Hyd-
rogr., 19:11—20.
Lowndes, A. 1928b. The result of breeding experiments and other observations on
Cyclops vernalis aud Cyclops robusius G. O. Sars. Int. Rev. ges. Hydrob. u. Hyd-
rogr., 21:171-188.
Lowndes, A. 1930. Some Entomostraca from Skye, with notes on the resting eggs
of Cyclops pictus Koch. Ann. Mag. Nat. Hist., (10), 5:209—217.
Lowndes, A. 1932. The results of further breeding experiments on four species of
Cyclops. Ann. Mag. Nat. Hist., (10) 9:265—297.
Lowndes, A. 1932a. The results of breeding experiments on the geuus Leptocyclops
G. O. Sars, with some general notes on the results of culture experiments. Ann.
Mag. Nat. Hist., (10), 10:45—80.
Lowndes, A. 1934. Report of an expedition to Brazil and Paraguay in 1926—7. Cope-
poda. Journ. Linn. Soc., Zool., 39:83—131.
Lucks, R. 1927. Zur Entwicklungsgeschichte des Cyclops viridis Jur. und seine Stellung
zum Cyclops clausii Heller. Sehr. nat. Ges. Danzig, 17:128—169.
Manfredi, P. 1923. Etude Sur le developpement larvaire de quelques especes du genre
Cyclops. I. Ann. Biol. Lac., 12:272—303.
Manfredi, P. 1925. Etude sur le developpment larvaire de quelques especes du genre
Cyclops. II. Ann. Biol. Lac., 14:111—129.
Marsh, C. 1909. A revision of the North American species of Cyclops. Trans. Wisconsin
Acad. Sc. Arts a. Let., 16:1067—1134.
Marsh, C. 1912. Notes on fresh-water Copepoda in the United States National Museum.
Proc. Un. St. Nat. Mus., 42:245—255.
Marsh, C. 1913. Report on fresh-water Copepoda from Panama, with descriptions of
new species. Smithson. Misc. Coll., 61:1—30.
Marsh, C. 1920. The fresh-water Copepoda of the Canadian Arctic Expedition. Rep.
Canad. Arct. Exped., 7, G :1—25.
Marsh, C. 1926. Crustaces Copepodes recoltes par M. Henri Cadeau de Kerville pendent son
voyage zoologique enSyrie. Voyage zool. d’Henri Cadeau de Kerville enSyrie, 1:171~185.
M a t i 1 e, P. 1897. Contribution a la faune des Copepodes des environs de Moscou. Bull.
Soc. Imp. Nat. Moscou, N. S., 11:113—139.
M a t s c h e к, H. 1909. Zur Kenntnis der Eireifung und Eiablage bei Copepoden. Zool.
Anz., 34:42—54.
Matschek, H. 1910. Ueber Eireifung und Eiablage bei Copepoden. Arch. Zellforsch.,
5:36—119.
124
ВВЕДЕНИЕ
Menzel, R. 1925. CyclopideS muscicoles et bromelicoles de Java. Ann. Biol. Lac.,
14:209—216.
Mrazek, A. 1893. Prispevky к pozndni sladkovodnich Copepoda. Vestn. krdl. ceske
spol. nSuk, 8:1—74.
Mr&zek, A. 1913. Androgyne Erseheinungen bei Cyclops gigas Cis. Zool. Anz,
43:245—250.
N e u b a u r, R. 1912. Ueber Beziehungen zwischen Cyclops fuscus (Jur.), Cyclops albidus
(Jur.) and dem angenommenen Bastard Cyclops distinctus (Richard.). Zool. Jahrb.,
Syst., 34:1—-69.
Okul, A. 1937. Copepoden- and Phyllopoden-Fauna der Umgegend von Odessa. Int.
Rev. ges. Hydrob. u. Hydrogr, 34:87—101.
Olofsson, O. 1917. Siisswasser-Entomostraken and Rotatorien von der Murmankuste
and aus dem nordlichsten Norwegen. Zool. Bidr. Uppsala, 5:259—294.
Olofsson, O. 1918. Studien fiber die Siisswasserfauna Spitzbergens. Zool. Bidrag
Uppsala, 6:183—648.
P est a, O. 1914. Euthycarcinus kessleri Handlirsch und die reeenlen Copepoden. ZooL
Anz., 45:44—47.
P e s t a, O. 1928. Krebstiere-Crustacea. I. Ruderfiissler oder Copepoda. 1. Calanoida,
2. Cyclopoida. Die Tierwelt Deutsehlands, 9:1—-136.
P e s t a, O. 1937. Beitrage zur Kenntnis der Tierwelt (Entomostrakenfauna) des Zicklacken-
gebiets. Zool. Anz., 118:177—192.
Pine, R. 1934. Metamorphosis of Cyclops viridis. Trans. Amer. Mier. Soc. Menasha,
53:286-292.
Plenk, J. 1911. Zur Kenntnis der Anatomie und Histologie der Maxillardrfise bei Cope-
poden. Arb. Zool. Inst. Wien, 19:29—56.
R e h b e r g, H. 1880. Beitrag zur Kenntnis der freilebenden Sfisswasser-Copepoden. Abh.
Natur. Ver. Bremen, 6:533—554.
Rehberg, H. 1884. Beitrage zur Naturgeschichte niederer Crustaceen (Cyclopiden und
Cypriden). Abh. Nat. Ver. Bremen, 9:1—18.
Richard, J. 1891. Recherches sur le systeme glandulaire et sur le systeme nerveux des-
Copepodes libres d’eau douce. Ann. Sc. Nat., Zool., 12:113—270.
Richard, J. 1895. Cladoeeres et Copepodes recueillis par M. Kavraisky pres de Tiflis
et dans le lac Goktseha. Bull. Soc. Zool. France, 20:91—92.
Richard, J. 1896. Sur la faune de quelques lacs eleves du Caucase d’apres les recoltes
de M. Kavraisky. Bull. Soc. Zool. France, 21:183—185.
Roy, J. 1932. Copepodes et Cladoeeres de 1’ouest de la France :1—224.
Rut n er, F. 1929/1930. Das Plankton des Lunzer Untersees. Int. Rev. ges. Hydrob..
u. Hydrogr.. 23:1—287.
Rylov, W. 1930. Cladocera und Copepoda. Памирск. экспед. Акад. Наук СССР,
Зоология, 2:105—130.
Rylov, W. 1932. Zur Kenntnis der Copepoden- und Cladocerenfauna der Insel Sachalin.
Zool. Anz., 99:101—108.
Rylov, W. 1932a. Ein neuer Bryocamptus aus dem Kaukasus. Zool. Anz, 99:171—174.
Rylov, W. 1935b. Das Zooplankton der Binnengewasser. Die Binnengewasser, 15:1—
272.
Rylov, W. u. Jarnefelt, H. 1936. Ueber das Vorkommen von Acanihodiaptomus
tibetanus (Dad.) in Nordfinnland. Ann. Zool. Soc. Vanamo, 2:3—9.
R z 6 s k a, J. 1925. Contribution a I’etude des Copepodes de la Grande Pologne. Bull..
Soc. des amie des sciences de Poznan, B, 1:34—43.
Rzoska, J. 1927. Einige Beobachtungen fiber temporale Grossenvariation bei Copepoden
und einige andere Fragen ihrer Biologie. Int. Rev. ges. Hydrob. u. Hydrogr.,.
17:99—114.
Rzoska, J. 1930/1931. Biometrische Studien fiber die Variabilitat einer Cyelopidengruppe
(C. strenuus s. 1.). Arch. Hydrob. i Rybactwa, 5:193—220.
Rzoska, J. 1932. Some general remarks on the faunistics and variability of some Cyclo-
pida. Int. Rev. ges. Hydrob. u. Hydrogr., 26:424—430.
Sars, G. O. 1898. The Cladocera, Copepoda and Ostracoda of the Jana-Expedition.
Ежег. Зоол. муз. Акад. Наук СССР, 3:224—359.
Sars, G. О. 1903. On the Crustacean Fauna of Central Asia. Ежег. Зоол. муз. Акад.
Наук СССР, 8:195—264.
Sars, G. О. 1913—1918. An account of the Crustacea of Norway, Cyclopoida, 6:1—
225.
Sars, G. O. 1927. The fresh-water Entomostraca of the Cape province. 3. Copepoda.
Ann. S. AMc. Mus, 25:85—149.
Sars, G. O. 1927a. Notes on the Crustacean Fauna of the Caspian Sea. Сб. в Честь
проф. H. М. Книповича, Москва: 315—329.
ЛИТЕРАТУРА
/25
Schafer, Н. 1933. Unsere Kenntnis der Copepodenfaima des Braekwassers, nebst Notizen
iiber die Brackwasser-fauna von Hiddensee. Int. Rev. ges. Hydrob. u. Hydrogr.,
29:264—273.
Schafer, H. 1934. Entomostrakenfunde aus dem Riesengebirge. Arch. f. Hydrob.,
27:271—276.
Schiklejev, S. 1931. Einige neue Arten und Varietaten der Eucopepoda aus den Siiss-
wassern des Kaukasischen Naturschutzgebietes und seiner nachsten Umgebung.
Zool. Anz., 94:185—194.
Schmeil, O. 1892. Deutsehlands freilebende Siisswasser-Copepoden. I. Cyclopidae. Bibl.
Zool., 11:1-191.
Schmeil, O. 1893. Copepoden des Rhatikon-Gebirges. Abh. Naturf. Ges. Halle, 19:1—-40.
Schmeil, O. 1898. Nachtrag. Bibl. Zool., 8, 21:145—188.
Sewell, R. B. 1934. Studies on the bionomics of fresh-waters in India. II. Int. Rev. ges.
Hydrob. u. Hydrogr., 31:203—238.
S i e w e r t h, M. 1928. Nachtrag zur Uebersieht der Siisswasserfauna Eucopepoda aus der
Ukraine. Zool. Anz., 77:145—156.
Skadowsky, S. 1926. Ueber die aktuelle Reaktion der Siisswasserbecken und ihre
biologische Bedeutung. Verh. Int. Ver. theor. u. angew. Limn., 3:109—144.
Smirnov, S. 1929a. Mesocyclops rylovi n. sp., ein neuer Siisswasser-Cyclopide aus dem
Kaukasus. Zool. Anz., 80:37—42.
Smirnov, S. 1929b. Beitrage zur Copepodenfauna Ostasiens. Zool. Anz., 81:317—329.
Smirnov, S. 1931. Ein Beitrag zur Copepoden-Fauna des Amur-Gebietes. Arch. f. Hyd-
rob., 23:618—638.
Smirnov, S. 1932. Notizen fiber einige Siisswasser-Copepoden Japans. Annot. Zool.
Japon., 13:283—296.
Spandl, H. 1922. Zur Artberechtigung von Cyclops clausii Heller. Zool. Anz., 54:273—
275.
Spandl, H. 1926. Copepoda. Ruderfusskrebse. Biol, der Tiere Deutsch!., 19:1-—82.
Stella, E. 1931. La citologia delle cellule sess. di alcuni Ciclopidi in relazione con la
loro sistematica. Int. Rev. ges. Hydrob. u. Hydrogr., 36:112—142.
-Steue r, A. 1898. Die Entomostraken der Plitvicer-Seen und des Blata-Sees (Croatien),
gesamm. von Dr. R. Sturany (1895). Ann. naturhist. Mus. Wien, 13: 159—-188.
Storch, O. 1928. Der Nshrungserwerb zweier Copepoden-Nauplien. Zool. Jahrb, Alig.
Zool., 45:385—436.
Tauson, A. 1933. Plankton des Sees „Gorjkoe" und die Biologie desselben. Arch,
f. Hydrob., 25:351—364.
Thallwitz, J. 1926. Ueber Varietatbildung bei Cyclops ’vernalis uud C. robustus Sars.
Arch. Hydrob., 17:366—380.
Thiebaud, M. 1915. Copepodes. Catal. des Invertebres de la Suisse, 8:1—125.
Ueno, M. 1934. A new subterranean Copepod from Japan. Proc. Imp. Acad. Jap.,
10:229—232.
Ueno, M. 1934a. Subterranean Crustacea from Kwantung. Annot. Zool. Japon., 14:445— 450.
•Ueno, M. 1936. Zooplankton of Lake Taraika and its neighbouring waters, Southern
Sakhalin. Trans. Sapporo Nat. Hist. Soc., 14:173—178.
Van-Breemen, P. 1907. Vrijlevende zoetwater Copepoden van Nederland. Tijdschr.
Ned. Dierk. Ver. (2), 10:303—368.
Van-Douwe, C. 1905. Copepoden von Transkaukasien, Transkaspien und Turkestan.
Zool. Jahrb., Syst., 22:673—700.
Voss el er, J. 1886. Die freilebenden Copepoden Wiirtembergs und angrenzender Gegen-
den. Jahresh. Ver. Vaterl. Naturk. Wiirtemberg, 22:167—-204.
Wahlberg, A. 1913. Bidrag till kannedomen om Littois Trask med sarskild Hansyn till
dess plankton. Acta Soc. pro Fauna et Fl. Fennica, 38:1—201.
Walter, A. 1888. Tr’anskaspische Binnenkrustaceen. Zool. Jahrb., Syst., 3:987—1014.
Walter, E. 1922. Ueber die Lebensdauer der freilebenden Siisswasser-Cyclopiden und
andere Fragen ihrer Biologie. Zool. Jahrb., Syst., 44:375—420.
Wesenberg-Lund, C. a. Ostenfeld, C. 1905. A regular fortnightly exploration
of the plankton of the Icelandic lakes, Thingvallavathn and Myvatn. Proc. R. Soc.
Edinburgh., 25:1092—1167.
Wierzbicka, M. 1934. Les resultats du croisement de certaines formes du groupe
de Cyclops strenuus (sensu lato) Mem. Acad. Polon. Sc., 13:189—206.
Wierzejski, A. 1883. Zarys fauny stawow tatrzaiiskich. Pamietn. Tow. Tatrz., Krakow,
8:95—123.
Willey, A. 1923. Notes on the distribution of free-living Copepoda in Canadian waters.
Contr. Canad. Biol., Ottawa, N. S. 1:303—334.
Willey, A. 1925. Northern Cyclopidae and Cantocamptidae> Proc. Trans. R. Soc. Canada,
Ottawa, 19 (5):137—159.
126 ВВЕДЕНИЕ
Willey, А. 1934. Some Laurentian Copepoda and their variation. Trans. Canad. Inst..
Toronto, 20:77—98.
Wilson, Ch. 1932. The Copepods of the Woods Hole region Massachusetts. Bull. Uri.
St. Nat. Mus. Washington, 158:1'—635.
Wolf, E. 1905. Die Fortpflanzungsverhaltnisse unserer einheimischen Copepoden. Zool.
Jahrb., Syst., 22:101—280.
Zenker, W. 1854. Ueber die Cyclopiden des sussen Wassers. Arch. Naturgeseh.,
20:88—102.
Ziegelmayer, W. 1925. Metamorphose und Waehstum der Cyclopidae. Zeitschr. wiss.
Zool., 126:493—570.
Zykoff, W. 1904. Zur Crustaceenfauna der Insel Kolgujev. Zool. Anz.. 28:337—345.
Z у к о f f, W. 1906. Ueber das Plankton des Saison-Sees. Zool. Anz., 29:477—482.
Zykoff, W. 1908. Das Plankton des Flusses Irtiseh unds einer Nebenfliisse Bukon und
Tobol. Zool. Anz., 33:103—112.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Сем. CYCLOPIDAE
Голова сращена с передним сегментом торакса в один общий отдел —
цефалоторакс. Очень сильно редуцированные мандибулярные пальпьи
Задние антенны большей частью 4-членистые, редко 3-членистые,
антеннулы 6—21-членистые, ноги V пары, как правило, сильно редуци-
рованные. — 3 подсемейства.
Подсем. Halicyclopinae заключает 3 рода, из которых Euryte Phil,
и Neocyclops Gurn. исключительно морские; ряд видов рода Halicyclops
Norm, обитает в солоноватых континентальных водах, некоторые виды
обитают только в морях. 2 другие подсемейства являются исключительно
пресноводными.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ
1(2). Дистальный1 членик ног V пары (g) с 4 придатками (3 шипа
и 1 щетинка) (Подсем. Halicyelopinae).
2(1). Этот членик с 1—3 придатками.
3(4). Этот членик с 3 придатками, задний торакальный сегмент
по бокам почти всегда вооружён щетинками........................
........................................ 1. Подсем. Eucyclopinae^
4(3). Этот членик с 1—2 придатками, задний торакальный сегмент
обычно без групп щетинок по бокам ... 2. Подсем. Cyclopinae
1. Поде. EUCYCLOPINAE
Eucyclopinae Kiefer, 1927:303; Kiefer, 1929:26. — Cyclops section Trifida subsec-
tion Polyarthra et Oligarthra Gurney, 1933 : 35.
Задний торакальный сегмент по бокам с группой щетинок, лишь
в редких случаях отсутствующих. Фуркальные ветви очень различной
длины, часто с рядом мелких шипиков (в виде „пилы") вдоль внешних
краев или с косыми рядами их на спинной поверхности. Обе ветви
плавательных ног всегда 3-членистые. Ноги V пары 1-членистые или
2-членистые, обычно с 3 придатками (шипы, щетинки) на конце. Передние
антенны б—17-членистые.
К подсемейству принадлежат 8 родов. Род Homocyclops Forbes
заключает лишь 1 вид — Н. ater (Herrick), известный из Северной
Америки и Бразилии. Род Ochridacyclops Kief. (оз. Охрида в Югосла-
вии) заключает также лишь 1 вид—Ochr. arndtiKiet. Роды Ectocyclops„
Paracyclops, Eucyclops, Tropocyclops и Macrocyclops имеют обширное
1 У ряда видов этот членик является единственным.
128
EUCYCLOPINAE
распространение (см. ниже). Род Thaumasiocyclops, недайно установлен-
ный Kiefer’oM (1933), заключает 1 вид—Th. insulanus Kief. (Малай-
ский архипелаг — Бали). В подсемействе известно около 100 видов.
Пресноводные рачки, обитающие в литорали озер, особенно в зарос-
лях макрофитов, а также в разнообразных мелких водоемах (пруды,
болота, лужи и проч.). Ряд видов известен и из текучих вод (реки,
ручьи, особенно род Paracyclops). В пелагиали озер отсутствуют. При-
уроченность специально к бентическому образу жизни (лазание по
субстрату) особенно выражена у родов Ectocyclops и Paracyclops,
у которых наблюдается, в связи с этим, значительная сплющенность
тела в спиннобрюшном направлении и сильное укорачивание передних
антенн. В родах Paracyclops и Eucyclops имеются некоторые виды,
помимо нормальных пресных вод обитающие и в водах со значительной
степенью минерализации (гораздо менее характерно это для рода
Macrocyclops). Некоторые виды рода Eucyclops обитают исключительно
в подземных водах южной Европы.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (4). Передние антенны 17-членистые.
2(3). Ноги V пары 1-членистые................(Homocyclops Forbes)
3(2). Ноги V пары 2-членистые................1. Macrocyclops Claus
4(1). Передние антенны б—12-членистые, ноги V пары всегда 1-члени-
стые.
5(6). Ноги V пары в виде очень широкой в основании и очень корот-
кой пластинки, неотчлененной от заднего торакального сегмента;
передние антенны 6—10-членистые, редко 11-членистые1 . . . .
.............................................6. Ectocyclops Brady
6(5). Ноги V пары явственно отчленены1 2 от торакального сегмента,
угловатых очертаний, неширокие, передние антенны б—12-члени-
стые.
7 (8). Передний отдел тела заметно сплющенный в спинно-брюшном
направлений, передние антенны б—11-членистые3....................
.......................................5. Paracyclops Claus
В (7). Передний отдел тела нормальной толщины, лишь слабо сплющен-
ный, передние антенны 11—12-членистые.4
9(10). Ноги V пары с 2 придатками.........(Thaumasiocyclops Kief.)
10(9). Ноги V пары с 3 придатками.5
11 (14). Receptaculum seminis без трубчатых выростов.
1 В виде исключения 11-членистые (нормально 10-членистые) передние антенны
иногда бывают у Ed. phaleratus Koch. Они всегда (?) 11-членистые у мадагаскарского
Ed. hirstttus Kief.
2 Лишь у Thaumasiocyclops insulanus Kief, пластинка этих ног слита с задним
торакальным сегментом.
3 Лишь у Par. eucyclopoid.es Kief, (о-ва Индо-малайского архипелага) передние
антенны 12-членистые (всегда?); у встречающихся в СССР Paracyclops они имеют не
более 11 члеников.
4 У Euc. echinatus Kief. (Мадагаскар) и Euc. stuhlmanni (Mraz.) (восточная Африка)
передние антенны 11-членистые (см. также Euc. inarmatus Kief., пещеры Югославии),
то же у р. Ochridacyclops.
5 У пещерного Euc. teras (Graeter) ноги V пары с 2 придатками; вид очень бли-
зок к Euc. serrulatus (Fisch.) и, несомненно, с ним связан генетически, обладая явными
признаками редукционного характера.
1. MACROCYCLOPS
129
12(13). Членик ног V пары удлиненный, все 3 его придатка расположены
апикально, фуркальные ветви как у Mesocyclops, без шипов на
внешних краях................................ 4. Ochridacyclops Kief.
13 (12). Членик ног V пары короткий, внутренний и внешний его при-
датки расположены не апикально, лишь средний придаток
(щетинка) занимает открытое положение. Фуркальные ветви
обычно с рядом шипиков на внешних краях.............................
...............................................2. Eucyclops Claus
14(11). Receptaculum seminis с трубчатыми S-образно изогнутыми
выростами.....................................3. Tropocyclops Kief.
1. Род MACROCYCLOPS CLAUS
Monoculus (part.) Jurine, 1820 :44. — Cyclops (part.) Koch, 1838, Deutschl. Crust.,
21:3; Lande, 1890:10; Schmeil, 1892:15, — Cyclops (Macrocyclops) Claus, 1893:82.—
Cyclops (part.) Lilljeborg, 1901:3; Marsh, 1909:1070. — Pachycyclops Sars, 1914:64.—
Macrocyclops (part.) Kiefer, 1927:303; Kiefer, 1929:26.— Macrocyclops Kiefer, 1931b:
609, 617 (выделен p. Homocyclops Forbes); Kiefer, 1933, Zool. Anz., 105: 42. — Macrocy-
clops (part.) subgenus, Gurney, 1933 s 67.
Передний и задний отделы тела резко дифференцированы, передний
отдел значительно расширенный. Генитальный сегмент постепенно
суживающийся кзади, объемистый. Receptaculum seminis у разных видов
неодинаковой формы, его задний отдел состоит из 2 мешковидных
частей, у М. albidus относительно коротких, у прочих видов удлинен-
ных. Анальная пластинка не развитая. Фуркальные ветви относительно
толстые, очень короткие, их длина самое большее в 2.3—2.6 раза более
их ширины1 (у М. neuter Kief, только в 1.5—1.75 раза), крайние апи-
кальные щетинки относительно длинные, внутренние края ветвей глад-
кие или с рядом более или менее густо расположенных волосков.
Передние антенны всегда 17-членистые, 3 дистальных членика вдоль
внутреннего края снабжены узкой, хорошо различимой гиалиновой
пластинкой, иногда зазубренной. Плавательные ноги сильные, хорошо
развитые. Ноги V пары очень крупные, дистальный членик вооружен
3 придатками — на конце длинной оперенной щетинкой, по бокам,
немного кзади от конца членика, парой шипов, из которых внешний
короче внутреннего. У самцов передние антенны сильные, богато воору-
женные длинными сензорными цилиндрами. Науплиусы не сплющенные,
с поперечными рядами коротких волосков на вентральной поверхности
(особенно у III и IV стадии). Главным образом крупные циклопы, длина
самки 1.7—4 мм, лишь яванский М. neuter Kief, принадлежит к мелким
видам (длина £ не более 1.3 мм).
Род имеет космополитическое распространение и заключает 4 вида
и 1 вариетет, в СССР известно 3 вида. Все виды рода не принадлежат
к планктическим, обитают в зарослях макрофитов, в литорали озер
и в мелких, богатых водной растительностью, водоемах. Эвритермичны,
пресноводны; специально пещерных видов нет.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Внутренние края фуркальных ветвей длинные, без волосков . . .
. ................................... . . *3. М. albidus (Jur.)
1 Kiefer (Zool. Jahrb. Syst., 65, 1934: 55) отмечает, что у M. albidus из Палеарк-
тики отношение длины фуркальных ветвей к их ширине = 2.3—2.6:1, а у популяций
этого вида из тропиков (субтропиков) лишь 2—2.1:1. Положение это, однако, требует
проверки иа достаточно обширном материале.
9 фауна СССР
13&
EUCYCLOPINAE
2(1). Внутренние края фуркальных ветвей с рядом волосков.
3(4). Эти волоски густо расположенные, гиалиновая пластинка дисталь-
ного членика передних антенн в проксимальной половине края
грубо зазубрена....................................*1. М. fuscus (Jur.)
4(3). Эти волоски расположены редко, густого ряда не составляют,
указанная пластинка или очень тонко зазубрена или цельнокрайняя
...............................................*2. М. distinctus (Rich).
1. Macrocyclops fuscus (Jurine) (фиг. 19, 205).
Monoculus quadricornis fuscus Jurine, 1820:47, t. 2, f. 2. — Cyclops signatus Koch’
1838, Deutsehl. Crust., 21, t. 8.— C. coronatus Claus, 1857 a: 29, t. I, f. 5; t. II, f. 1—11;
Lubbock, 1863, Trans. Linn. Soc., 24: 199. — C. signatus Brady, 1880, Monogr. Brit. Cop.,
1:100, t. 17, f. 4—12. — C. palusiris (juv.), Совинский, 1887, Зап. Киевск. общ. ест., 8:52
и- 8, ф. 11.— С. fuscus Schmeil, 1892:123, t. I, f. 1—7 6, t. IV, f. 16. — Macrocyclops
coronatus Claus, 1893 a: 347, t. I, f. 5; t. II, f. 2; t. IV, f. 7—9; t. VII, f. 1—3, 12.—
Cyclops signatus var. coronatus Herrick, 1895 :105, t. 15, f. 1—4. — C. fuscus Lilljebarg,
1901:44, t. 3, f. 12—15. — Macrocyclops fuscus Kiefer, 1929:27—23, f. 11. — Cyclops
(Macrocyclops) fuscus Gurney, 1933:67, f. 1292—1313.
Самка. Передний отдел тела овальный, кзади несколько более
суживающийся, чем кпереди, длина цефалоторакса и торакальных сег-
ментов приблизительно в 1% раза превышает наибольшую ширину,
задний сегмент приблизительно такой же ширины, как основание гени-
тального сегмента. Генитальный сегмент объемистый, умеренно расши-
ренный впереди, его длина в 1.12—1.2 раза более его ширины. Recepta-
culum seminis с полуовальным, иногда несколько вдавленным впереди
передним отделом и с задним отделом в виде 2 отвисающих мешков.
Фуркальные ветви короткие и толстые, расходящиеся под значительным
углом, их длина приблизительно вдвое более ширины, внутренние края
со сплошным рядом густо расположенных волосков. Латеральная щетинка
прикреплена близко от заднего конца ветвей, крайние апикальные
щетинки относительно длинные, из них внешняя на 3/з—г/г короче вну-
тренней, средние апикальные щетинки хорошо развитые, из них внутрен-
няя лишь немного более чем вдвое превышает длину крайней внутренней
щетинки; все апикальные щетинки густо оперены. Передние антенны
почти достигают заднего конца торакса, 12-й членик с очень малень-
кой сензорной колбой, близ дистального конца 8-го — 11-го и 12-го —
14-го члеников имеются поперечные ряды (по одному на каждом членике)
очень мелких шипиков, не образующих вокруг членика сплошного
кольца. Внутренние края 15-го—17-го члеников снабжены тонкой
гиалиновой пластинкой; пластинка 17-го членика своим концом выдается
немного кпереди от конца членика, очень тонко зазубрена в дистальной
половине и глубоко зазубрена в проксимальной половине свободного
края; края пластинок 15-го и 1б-го члеников без зазубренности. Задние
антенны очень крупные, их оба конечные членика удлиненные, почти
одинаковой длины, 3-й от конца членик короткий, утолщенный. Щупальце
мандибулы с 3 щетинками. У максилл относительно длинный 2-й членик,
его длина почти вдвое более ширины. Максиллярные ноги стройные,
удлиненные, их дистальный членик заметно длиннее проксимального.
Обе ветви всех 4 пар плавательных ног 3-членистые, формула шипов
3 • 4 • 4 • 3. Соединительная пластинка ног IV пары с выпуклым килем,
вооруженным рядом густо расположенных тонких шипиков; на задней
поверхности этой пластинки имеется еще 2 поперечных ряда шипиков,
которые в дистальном ряду относительно длинные и редко расположен-
ные, а в проксимальном — очень мелкие и густо расположенные. Ноги
1. MACROCYCLOPS
131
V пары мощные, длинные, их проксимальный членик внутри с мелкими
волосками, сзади с поперечным рядом мелких шипиков, щетинка внеш-
него угла этого членика длинная, хорошо развитая. Дистальный членик
короче : предыдущего, с длинной передней апикальной щетинкой
Фиг. 19. Macrocyclops fascus (Jur.).
7—II антенна самки; 2—нога V самки; 3—фуркальная ветвь самки;
4 — reeeptaculum seminis; 5'— дистальный членик антенны I самки. (5 — по
Schmeil’io).
и с 2 тонкими шипами по бокам ее, из которых внешний в Р/о—2 раза
короче внутреннего. Длина 1.8—4.0 мм. Окраска значительно вариирует,
обычно очень яркая* нередко темнозеленая с голубыми включениями,
reeeptaculum seminis обычно яркокрасный или красно-оранжевый. Иногда
самки очень темного цвета, почти черные, редко они прозрачны и бес-
цветны. Яйцевые мешки овальные, крупные, обычно темнокоричневые или
д*
/52
eucyclopinae
черные, состоящие из значительного количества яиц, тесно прилегающие
к абдомену, нередко отчасти прикрывающие его спинную поверхность.
Самец. Передние антенны богато вооружены длинными, притуплен-
ными на конце, сензорными цилиндрами, у 1-го членика 2 цилиндра,
у 2-го — б-го, у 8-го — 9-го члеников—по 1. Ноги VI пары с 3 придат-
ками: внешней оперенной щетинкой и парой голых шипов, из которых
внешний короткий, менее ’/а длины внутреннего. Длина 1.2—2.5 мм.
Сперматофоры крупные, бобовидные. Сперматозоиды очень мелкие,
в виде укороченных веретенец.
Распространение. Вся Европа, северная Азия, Япония, Суматра,
Северная Африка, Северная Америка.1 Широко распространен
в европейской части СССР, в которой найден во многих местах, от
крайнего севера (Кольский полуостров, Соловецкие острова) до окре-
стностей Одессы включительно, на восток до Челябинской области
включительно, в бассейне р. Волги известен до Саратова, но, без
сомнения, распространен и южнее, вероятно, до северной части Каспия.
На Кавказе известен из окр. Дзауджикау, из Теберды (оз. Каракель,
высота около 1300 м над ур. м.), из оз. Абрау близ Новороссийска
и Махач-кала. В азиатской части СССР найден в р. Сыр-дарье, до низовьев
р. Сарысу, в р. Нуре, близ Акмолинска, в окр. Мары и Пржевальска.
В горах известен до высоты около 2000 м.
Биология. Обитает в очень разнообразных водоемах, от глубоких
крупных озер до мелких луж и болот включительно. Как правило,
отсутствует во временных, летом пересыхающих мелких лужах, однако
найден в крупном, но мелководном периодическом Blata-See в Кроации
(Steuer, 1898). Неоднократно указывался для текучих вод, где обитает
в прибрежных зарослях макрофитов, при слабом течении; в быстро текучих
ручьях отсутствует. Известен из холодных ключевых водоемов типа
гело- и лимнокренов (Klie, 1925). В озерах обитает в зоне макрофитов
литорали, лишь случайно попадая в пелагиаль1 2 и лишь в виде исклю-
чения и в единичных экземплярах проникая на значительные глубины;
наибольшая глубина, где он найден, равна 45 м. Троглоксен. В подзем-
ных водах лишь в виде редкого случайного компонента. Эвритермичен,
в мелких водоемах был находим при температуре до 31°. Указания
на холодноводность (Thiebaud, 1908) надо считать ошибочными. В соло-
новодных водоемах отсутствует. Обычен в водах с pH = 5.0—8.4, по
Harnisch (1925), отсутствует в сильно дистрофировацных моховых боло-
тах с pH менее 5.
Встречается обычно в течение всего года, полицикличен, вероятно
до 3 максимумов в год, в некоторых водоемах, возможно, и моноцикли-
чен (Borner, 1917, St. Moritzer-See), с максимумом летом — осенью.
Указание Kolkwitz’a (1909) и Долгова и Никитинского (1927) на
облигатную олигосапробность М. fuscus ошибочно, рачок, несомненно,
нормально встречается и в ₽-мезосапробных условиях.
*2. Macrocyclops distinctus (Richard) (фиг. 20i_3).
Cyclops bistriatus Koch, 1832, Deutschl. Crnst., 21, t. 7. — C. tenuicornis var. distin-
eta Richard, 1887, Bull. Soc. Zool. France, 12:162. — C. gracilicornis Lande, 1890:345,
t. 15, f. 13—20; t. 16, f. 21. — C. annalicornis (non Koch, 1832) Richard, 1891, Ann. Sc.
1 Нахождение в Южной Америке (Daday, 1905) сомнительно.
2 В старой литературе есть указания аа пелагический образ жизни М. fascas,
которые, безусловно, ошибочны.
1. MACROCYCLOPS
133
Nat., 12 : 225. — C. distinct us Lilljeborg', 1901 : 47, t. 3, f. 16—20. — C. bistriatus Scour-
field, 1903, Journ. Quek. Mier. Club, 3:535.— C. distinctus Van Breemen, 1907:321,
t. 6, f. 6s — C. distinctus Neubaur, 1912:117, t. 6, f. 25, textf. C, F, K, N, O, R, T, B1,
D1, F1, J1. — P achy cy clops bistriatus Sars, 1914:67, t. 41—Macrocyclops distinctus Kiefer,
1929:28. — C. (Macrocyclops) distinctus Gurney, 1933 : 79, f. 1332—1348.
Самка. Передний отдел тела правильной овальной формы, отношение
длины цефалоторакса -+- торакса к ширине около 1.7:1. Генитальный сегмент
широкий, отношение его длины к ширине приблизительно 1:1 или 1:1.2.
Receptaculum seminis напоминает таковой М. fuscus, но в заднем отделе
с более объемистыми боковыми выростами. Фуркальные ветви короткие,
Фиг. 20. (По Neubaur’у).
Macrocyclops distinctus (Rich.): 7 — фурка самки (ще-
тинки не изображены), 2 — генитальный сегмент с re-
ceptaculum seminis, 3—сензорный придаток на 12-м
членике антенны I самки; 4 — М. albidus (Jur.), сензор-
ный придаток на 12-м членике антенны 1 самки; 5'— М. fus-
cus (Jur.), сеизориый придаток на 12-м членике ан-
тенны 1 самки.
их длина в 2г/2—3 раза превышает их ширину, внутренние края
с рядом из немногих, редко расположенных, волосков. Апикальное во-
оружение фуркальных ветвей как у М. f ascus, наиболее длинная из апи-
кальных щетинок превышает половину длины цефалоторакса и иногда
в заднем отделе несколько изогнута ковнутри (лирообразно). Передние
антенны как у М. albidus и т. fascas, но сензорная колба на 12-м
членике длиннее, чем у М. fascas, и короче, чем на 13-м членике, чле-
ники 8-й — 10-й и 12-й — 14-й без поперечных рядов шипиков, гиалино-
вая пластинка 17-го членика не имеет глубокой зазубренности, характер-
ной для М. fascas. Задние антенны как у М. albidus, их предпоследний
членик заметно короче дистального. Щупальце мандибул без короткого
выроста, свойственного М. fuscus. Строение и вооружение ног I—IV
пар и соединительной пластинки ног IV пары как у М, fuscus, но апи-
134
EUCYCLOPINAE
кальные шипы дистального членика эндоподита несколько более корот-
кие. Ноги V пары как у М. fuscus, но из апикальных придатков дисталь-
ного членика боковые более удлиненные, чем у последнего вида (осо-
бенно внешний придаток). Длина 1.5—2.25 мм. Окраска обычно голубо-
ватая или темносиняя, нередко с зелеными включениями, абдомен
и передние антенны нередко красноватые, яйцевые мешки темнокорич-
невые, крупные, мало расходящиеся, менее тесно прилегающие к абдо-
мену, чем у М. fuscus.
Сам-ец, как у М. fuscus, но более удлиненный, стройный; длина
1.2—1.4 мм.
Schmeil (1892) предполагал, что М. diSfinctus является помесью М. fus-
cus и М. albidus, но позже (1898) высказался за его видовую самосто-
ятельность. Ряд авторов, однако, считает его помесью этих двух видов.
Экспериментальные исследования Braun’а (1909), Neubaur’a (1912)
и Lowndes’a (1928) над скрещиванием М. fuscus и М. albidus дали,
однако, отрицательные результаты.
Распространение. В виду смешивания с М. fuscus, оно выяс-
нено далеко не полно. Широко распространен в Европе, известен
с о-вов Суматры, Явы, Цейлона,1 из Лагора, с Новой Зеландии
и из Северной Америки. В пределах СССР пока показан с достовер-
ностью лишь для Челябинской области (оз. Кукай близ г. Троицка;
Зиновьев, 1935), без сомнения, должен иметь широкое распространение
в СССР.
Биология изучена крайне недостаточно. Обитает в зарослях
макрофитов в литорали озер и в мелких стоячих и проточных водоемах.
Известен из холодных ключей. Нередко встречается вместе с М. fuscus
и М. albidus. Эвритермичен. По Lowndes (1928) обитает в водоемах
с рН=7.б—8.4. Встречается, вероятно, круглый год. По исследованиям
Neubaur’a период максимального развития не совпадает с таковым
М. fuscus и М. albidus (в одном и том же водоеме), вероятно ди-(поли-)
цикличен.
*3. Macrocyclops albidus (Jurine).
Monoculus quadricornis albidus Jurine, 1820:44, t. 2, f. 10—11. — Cyclops annuli-
cornis Koch, 1838, Deutschl. Crust., 21:6. — C. ienuicornis Claus, 1857a:31, t. 3, f. 1—11.—
— C. pennatus Claus, 1857a:35, t. 3, f. 12—17. — C. clausi. Поггенполь, 1874 (non C. cla-
usi Lubbock, 1863, non C. clausi Heller, 1871), Изв. Общ. люб. ест., антр. и эти., 10:70,
т. 15, рис. 4—14. — С. latissimus Поггенполь, 1874, ibid., 10:71, т. 15, рис. 16—17, т. 16,
рис. 1, 10.— С. signatus Ульянин, 1875 (non С. signatus Koch, 1838), ibid., 11:29, т. 9,
рис. 6—11, т. 11, рис. 8. — С. korostyschevi (juv.) Совинский, 1887, Зап. Киевск. общ.
ест., 8:51, т. 8, рис. 7. — С. gyrinus Forbes, 1887, Rep. U. S. Fish. Comm., 707, t. 2,
f. 9—12; t. 3. f. 14. — C. albidus Schmeil, 1892:128, t. I, f. 8—14,b; t. 4, f. 15, — Cyclops
(Macrocyclops) ienuicornis Claus, 1893a:347, t. 2, f. 1. — C. signatus Marsh, 1893 (non
C. signatus Koch, 1838), Tr. Wiscons. Acad., 9:21, t. 5, f. 7—9. — C. viridosignatusByrn.es,
1909, Cold. Spr. Harb. Mon.:23, t. 9, f. 1—8. — C. signatus var. annulicornis Byrnes,
1909, ibid.: 10, t. 4, f. 1—6. — Pachycyclops annulicornis Sars, 1914:68, t. 42. — Macrocy-
clops albidus Kiefer, 1929:28. — C. (Macrocyclops) albidus Gurney, 1933:74, f. 1314—1331.
Самка. Передний отдел тела более суженный и стройный, чем
у М. fuscus и М. distinctus, его наибольшая ширина лежит приблизи-
тельно на середине его длины, отношение длины цефалоторакса -ь тора-
кса к их ширине =1.8:1. Генитальный сегмент удлиненный, заметно
1 Kiefer (Rec. Ind. Mus., 1928 г 333) ставит нахождение на Цейлоне под сомнение,
но Gurney (1933) приводит его вторично. Нахождение на о-вах Яве, Суматре и в Лагоре
(Индия), можно думать, окончательно решает вопрос в положительном смысле.
1. MACROCYCLOPS
135
расширенный спереди, кзади равномерно суживающийся. Receptaculum
seminis состоит из поперечно расположенного овального переднего
отдела и из короткого заднего, в виде 2 широких и коротких полукруг*
лых мешков. Фуркальные ветви слабо расходящиеся, их длина прибли-
зительно в 2.5—'3 раза более их ширины, их внутренние края без волосков. Из
апикальных крайних щетинок внешняя не более половины длины внутрен-
ней, не превышающей половину длины внешней средней щетинки,
а нередко бывает и еще короче; латеральная щетинка прикрепляется
очень близко от внешнезаднего угла ветвей. Передние антенны дости-
гают заднего конца торакса, членики 8-й—10-й и 12-й—14-й с рядами
шипиков, более мелких, чем у М.. fuscus, и расположенных только
на брюшной поверхности члеников. Сензорная колба 12-го членика
сильно развитая, своим концом достигающая основания 14-го членика,
такая же колба на 1б-м членике очень тонкая и короткая. Вооружение
15-го—17-го члеников как у М. fuscus, но гиалиновая пластинка 17-го
членика лишь с едва различимой тонкой зазубренностью, или цельно-
крайняя. Задние антенны относительно короче, чем у М. fuscuS,
их предпоследний членик заметно короче дистального и лишь немного
короче 3-го от конца членика. 2 щетинки мандибулярных щупальцев
очень длинные, 3-я их щетинка очень короткая. Максиллы и максилляр-
ные ноги, как у М. fuscus, но отличаются лишь относительной длиной
вооружающих их щетинок. Плавательные ноги такие же, как у этого
вида, но у соединительной пластинки ног IV пары шипики свободного
края более нежные, а проксимальные шипики тонкие, в виде волосков.
Апикальное вооружение дистального членика эндоподита ног IV пары
очень похоже на таковое М. distinctus. Строение и вооружение ног
V пары как у М. fuscus1. Длина 1.5—2.5 мм. Обычно светлосерая, ино-
гда коричневая (до темнокоричневой включительно) Окраска; иногда
самки почти бесцветны, Фуркальные ветви, задний отдел абдомена
и отчасти передние антенны обычно темносиней (черно-синей, черной)
окраски. Яйцевые мешки значительно расходящиеся, не прилегающие
к абдомену, содержащие значительное (нередко свыше 50 в каждом)
количество яиц.
Самец. Очень похож на самца М. fuscus, длина 1—1.3 мм. Спер-
матофоры более удлиненные, а сперматозоиды гораздо более крупные
(в виде длинных веретенец), чем у указанного вида.
Распространение. Космополит в широком смысле слова.
В СССР, исключая арктические острова (показан лишь для Соловецких
островов), распространен, буквально, везде и принадлежит к наиболее
обычным циклопам. Неоднократно указывался для высокогорных водоемов
Европы и Северной Америки, причем в горы, повидимому, заходит
выше, чем М. fuscus (до 2600 mJ.
Биология. Характерен для зарослей макрофитов литорали (частью
и сублиторали) озер и самых разнообразных мелких водоемов, до луж
и маленьких болот включительно. Встречается также в периодически
высыхающих мелких fастатических,) водоемах. Неоднократно указывался
для текучих вод, где держится, главным образом, в зарослях прибреж-
ных макрофитов. В озерах лишь случайно проникает в глубинную зону,
где показан до глубины 60 м. Очень часто встречается совместно
с М. fuscus. В пелагиаль озер попадает лишь случайно, выносясь ветром
1 О постоянстве видовых признаков М. fuscus и М, albidus см. экспериментальные
исследования Byrnes (Biol. Bull., 1919).
136
EUCYCLOPINAE
и волнением из литорали.1 Указывался1 2 3 для подземных водоемов,
и колодцев, но едва ли является троглофилом. Эвритермичен, обитает
как в сильно прогреваемых водоемах (в южной области распространения),
Фиг. 21. Macrocyclops albidus (Jur.), самка.
7 — нога V; 2 — reeeptaculum seminis; 3 — фуркальная ветвь со спинной
стороны; 4— антенна II, 5—сперматофоры; 6—дистальный членик ан-
тенны I. {6— по Schmeil’io).
так и в холодных ключевых и родниковых водах. По Lowndes’y (1928)
обычен в водах с pH = 4.4—9.8. В нейтральных — слабощелочных водах
очень обычен, но оптимальные условия находит и в высокощелочных
водах. В противоположность М. fuscus (см. выше), рассматриваемый
вид в значительной степени эвригалинен. В большинстве постоянных
1 Старые литературные данные о нахождении этого вида в пелагиали в качестве
постоянного компонента пелагической фауны безусловно ошибочны.
2 Под вопросом указан Лебединским (1900, 1904: С. ten.uicorn.is) для пещер Крыма
и Боруцким (Borutzky, 1930) для пещер Кавказа (близ Кутаиси).
2. EUCYCLOPS
737
водоемов M. albidus встречается круглый год, полицикличен, с тремя
половыми периодами — зимой (январь — февраль), летом и осенью.
Отнесение М. albidus к облигатным олигосапробам, бесспорно,
ошибочно, — он нормально обитает и в [3-мезсапробных условиях.
2. Род EUCYCLOPS CLAUS
Cyclops (part.) Fischer, 1851:423; Claus, 1863:96; Schmeil, 1892:15. — Cyclops (subgen.
Eucyclops) Claus, 1893:82. — Cyclops (part.) Lilljeborg, 1901:3; Marsh, 1909:1070. — Lepto-
cyclops Sars, 1914:70.—Eucyclops (s. str.) Kiefer, 1927:308; Kiefer, 1928:541; Kiefer,
1929:29. — Eucyclops (part.) (subgen.) Gurney, 1933:97.
Передний отдел тела лишь немного сплюснутый в спиннобрюшном
направлении, явственно дифференцированный от заднего. Задний тора-
кальный сегмент по бокам с группой сильных щетинок (у пещерного
Е. teras они редуцированы). Генитальный сегмент спереди широкий,
кзади сильно суженный, причем его задний отдел цилиндрический,
стройный. Reeeptaculum seminis из двух поперечно расположенных
один над другим широких отделов, обычно с заметным вдавлением
посредине, спереди и сзади. Анальная пластинка не развита. Длина
фуркальных ветвей очень различна, отношение их длины к ширине
от 2.5:1 до 11:1, большей частью превышает 4:1. У большинства
видов внешние края фуркальных ветвей снабжены продольным рядом
шипиков, крайние апикальные щетинки относительно короткие, средние
апикальные щетинки нередко с гетерономным вооружением. Передние
антенны большей частью 12-, редко 11-членистые,1 3 дистальных
членика с очень узкой гиалиновой пластинкой вдоль внутреннего
края. На конце дистального членика эндоподита ног IV пары вну-
тренний шип длиннее внешнего. У ног V пары, явственно отделенных
от заднего торакального сегмента и 1-членистых, не в виде широкой
пластинки, вооружение состоит из 3 придатков — из хорошо развитого
толстого, иногда ножевидного внутреннего шипа и из 2 оперенных щети-
нок — апикальной и внешней (у пещерного Е. teras имеется лишь 2 при-
датка). Относительно мелкие (g 0.5—1.7 мм, обычно около 1 мм),
стройные циклопы.
Род подразделяется на 2 подрода — Eucyclops s. str. и Afrocyclops
Sars. В СССР последний подрод (5 видов) отсутствует. Род заключает
пресноводные виды (некоторые эвригалинны), литораль (заросли макро-
фитов) озер, всевозможные мелкие водоемы. Некоторые виды подрода
Eucyclops свойственны только подземным водам.
К роду Eucyclops в настоящее время относится около 40 видов.
Подрод Космополитен, некоторые виды (например Е. serrulatus) распро-
странены чрезвычайно широко. В Европе обитают1 2 Е. serrulatus (Fisch.)
Е. macrurus (Sars), Е. macruroides (Lili.), E. graeteri (Chapp.), E. teras
(Graet.), из них три первых вида обитают также в Азии и палеаркти-
ческой части Африки. Только Эфиопской области свойственно около
16 видов (кроме того 4 вида Afrocyclops), т. е. около половины всего
числа видов подрода; 2 вида (Е. madagascariensis Kief, и Е. echinatus
Kief.) известны только с Мадагаскара, 1 (Е. euacanthus Saxs — E. cognatus
1 У всех встречающихся в СССР видов они 12'членястые.
2 Е. inarmatus Kief. (Zool. Anz., 1932, Югославия) вид крайне сомнительный и,
как полагает и сам Kiefer, вероятно, идентичен с Е. graeteri (Chapp.). Описание Lepto-
cyclops brevifurca Grandori (Boll. 1st. Zool., Roma, 3, 1936) осталось мне недоступным,
E. porrectus Kief, для меня также очень сомнителен.
EUCYCLOPINAE
Kief.) — из Африки, с о. Формозы и с Малайского архипелага, только
островам которого (в пределах Ориентальной области) свойственны
3 вида. Только в пределах Индийской и Цейлонской подобласти най-
дено 2 вида. В общем, специально Ориентальной области свойственно
5 видов (и 1 подвид подрода Afrocyclops). Только в Неотропиче-
ской1 области найдено 5 видов. Из Северной Америки1 2 известен
Е. prionophorus Kief., найденный в штате Коннектикут и обнаруженный
также в Уругвае. Несмотря на неодинаковую степень исследованности
фауны Copepoda различных областей земного шара, все же устана-
вливается факт чрезвычайного богатства видами подрода Eucyclops
специально Эфиопской области, тогда как в палеарктической части
Африки подрод представлен уже гораздо беднее.
Виды всего рода Eucyclops является непланктическими циклопами,
обитающими в зоне макрофитов литорали озер и в разнообразных мелких
водоемах (пруды, лужи, болота и проч.). Большинство видов тепловодно-
стенотермичны, частью эвритермичны, холодноводных нет. К троглоби-
онтам принадлежат европейские Е. teras (Graet.), Е. graeteri (Chapp.)
и японский Е. serrulatus var. nagasakai (Ueno). В СССР известно 3 вида.
Sars (1914) и Kiefer (1929) считают „С. varius var. speratus
Lili." самостоятельным видом; на сомнительность этого я уже указывал
ранее (Рылов, 1918, 1927). Lowndes (1932а) полагает, что самки
Е. speratus ввиду наличия переходных форм не всегда отличимы от
таковых Е. serrulatus s. str., но самцы отличаются длиной внутренней
средней апикальной щетинки фуркальных ветвей: у Е. serrulatus s. str.
эта щетинка длиннее фуркальных ветвей (или равна им), у Е.
speratus — короче. Однако после разъяснений Сигпеу’я (1933:105),
на абсолютности этого признака самцов едва ли можно настаивать.
Мы полагаем единственно возможным рассматривать Е. speratus
в качестве вариетета Е. serrulatus (Fisch.). Можно считать бесспор-
ным, что под названием „С. serrulatus var. denticulata“ A. Graeter
(1903) описал форму, идентичную с „Leptocyclops lilljeborgi" Sars (1914).
С этим согласен и Kiefer (1929:35), и непонятно, почему этот автор
не признает в названии приоритета Graeter’a, давшего очень подробное
описание var. denticulata. Вместе с тем изучение материалов из СССР
приводит к заключению, что, кроме удлиненных и менее расходящихся
фуркальных ветвей, Е. macruroides (Lili.) ничем не отличается от Lepto-
cyclops lilljeborgi Sars, причем между обоими существуют переходные
формы. Совершенно то же мы находим у E.serrulatus (Fisch.), где име-
ются все переходы между короткими (var. brachyurus) и удлиненными
и более параллельными (var. proximus) фуркальными ветвями. В данном
случае у 2 видов мы наблюдаем совершенно одинаковые по направле-
нию вариации, касающиеся двух бесспорно коррелятивных признаков.
Попытки различать Е. macruroides и Е. lilljeborgi по самцам (Thallwitz,
1926; Lowndes, 1932), как показал Gurney (1933:114), лишены основа-
ния. Вследствие объединения этих двух- форм в один вид, вместе с Gur-
ney’eM (1933:109), необходимо оставить за видом данное Lilljeborg-’ом
(1901) название macruroides, тогда как за вариететом оставить, как более
1 Описанный из Аргентины Е. silvestris (Brian) остается сомнительным видом
(см. Kiefer, 1929:31).
2 Описанный Herrick’oM (1884) С. serrulatus var. elegans (Миннезота) по Kiefer’y
(Zool. Anz., 80,1929:305) является самостоятельным видом, что этим автором доказывается
крайне неубедительно. Мы полагаем, что эта форма является просто одним из членов
вариационного рода serrulatus — proximus.
2. EUCYCLOPS
139
раннее, название denticulata (A. Graeter, 1903), отнеся Leptocyclops
lilljeborgi Sars’a (1914) в синонимы последнего.1
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (6) Глазной пигмент отсутствует (пещерные виды).
2 (3) Ноги V пары лишь с 2 придатками (шип и щетинка)..............
............................................2. Е. teras (Graet.)
3 (2) Ноги V пары с 3 придатками (1 шип и 2 щетинки).
4 (5) Дистальный конец шипа ног V пары щетинкообразно оттянут . .
.........................................5. Е. graeteri (Chapp.)
5 (4) Этот шип ножевидный; без щетинкообразного окончания ....
........................* 1с. Е. serrulatus var. nagasakai (Ueno)
б (1). Глазной пигмент имеется. F
7 (10). Гиалиновые пластинки вдоль внутреннего края 10-го — 12-го чле-
ников передних антенн пилообразно зазубрены.
8 (9). Фуркальные ветви длинные, не менее длины 3 задних абдоми-
нальных сегментов, взятых вместе, параллельные или почти парал-
лельные • • -............................* 3. Е. macruroides (Lili.) typ.
9 (8). Фуркальные ветви короче, их длина менее длины 3 задних абдо-
минальных сегментов (взятых вместе), сильно расходящиеся. . . •
...............*3а. Е. macruroides var. denticulatus (A. Graet.)
10 (7). Гиалиновые пластинки вдоль внутреннего края 10-го — 12-го
члеников передних антенн не зазубрены.
11 (12). Фуркальные ветви очень длинные, параллельные, их длина
не менее, чем в 10 раз более их ширины, на внешних их краях
имеется очень короткий ряд шипиков близ латеральной щетинки,
10-й — 12-й членики передних антенн укороченные.....................
..........................................* 4. Е. macrurus (Sars)
12 (11). Длина фуркальных ветвей обычно не более, чем в б—6.5 раз,
редко в 8 раз, превышает их ширину, обычно они расходящиеся,
на внешних краях с почти сплошным рядом шипиков, иногда более
или менее редуцированным, 10-й — 12-й членики передних антенн
удлиненные.
13 (16). Шипики занимают почти весь внешний край (исключая его
передний отдел) фуркальных ветвей, хорошо различимые, или лишь
мало редуцированные.
14 (15). Фуркальные ветви короткие, их длина не более длины 2 задних
абдоминальных сегментов (взятых вместе)...............................
. . . . •.........................* 1. Е. serrulatus (Fisch.) typ.
15 (14). Длина фуркальных ветвей несколько более длины 2 задних
абдоминальных сегментов (взятых вместе).............................
. . . .............*la. Е. serrulatus (Fisch.) var. proximus Lili.
16 (13). Ряд шипиков на внешних краях удлиненных и почти параллель-
ных (или параллельных) фуркальных ветвей очень сильно редуци-
рован, причем шипики остаются лишь в задней ]/3 длины края.
.................* lb. Е. serrulatus (Fisch.) var. speratus (Lili.)
*1. Eucyclops (s. str.) serrulatus (Fisch.) (фиг. 22, 23).
Cyclops agilis Koch, 1838, Deutschl. Crust., 21:3. — C. serrulatus Fischer, 1851:423,
t. 10, f. 22—23, 26—31. — C. serrulatus var. montana Brady, 1878, Mon. Brit. Cop.,
1 К сожалению, мне остались недоступными работы по вариациям европейской
группы serrulatus, опубликованные Thallwitz’eM (Sitzsber. Isis, Dresden, 1927) и SrAmek-
Husek (Mem. Soc. Roy. Sc. Boherne, 7, 1934, по-чешски).
140
EUCYCLOPINAE
1:109, t. 22, f. 1—14. — C. novae-zealandiae Thomson, 1879, Tr. New Zeal. Inst., 11:258,
t. 11, f. 1. •—C. agilis (non C. agilis Koch) Rehberg, 1880, Abh. Nat. Ver. Bremen; 6:545.—
C. pectinifer Cragin, 1883, Tr. Kansas Acad., 8:71, t. 4, f. 1—7. — C. serrulatus f. typica,
+- f. brevicaudata Совинский, 1888, Зап. Киевск. Общ. ест., 9:233. — С. serrulatus
(part.) Schmeil, 1892:141, t. 5. — Eucyclops serrulatus Claus, 1893:82. — C. varius var.
brachyurus Lilljeborg, 1901:89, t. 6, f. 5 — 9. — C. agiloides Sars, 1909, Proc. Zool. Soc.
London:59, t. 20, f. 183—188. — Leptocyclops agilis Sars, 1914:71, t. 43. — C. serrulatoides
Labbe, 1927, Arch. Zool. Exp., 66:200, f. 184—186. — Eucyclops (s. str.) serrulatus Kiefer,
1929:31, f. 12 a — b. — Leptocyclops agilis Lowndes, 1932a:45. — Eucyclops (s. str.) serru-
latus Рылов, 1933:256, т. 2, рис. 12; т. 3, рис. 1. — Cyclops (Eucyclops) agilis Gurney,
1933:99, f. 1374—1394.
Самка. Тело стройное, туловище правильной овальной формы,
его наибольшая ширина лежит приблизительно на середине его длины
(считая задний тор.акальный сегмент). Задний торакальный сегмент
по бокам с группами мелких волосков. Абдомен тонкий, генитальный
сегмент в основном отделе сильно расширенный, его задний отдел почти
цилиндрический, лишь незначительно суживающийся кзади; длина гени-
тального сегмента приблизительно равна его наибольшей ширине. Recep-
taculum seminis состоит из 2 широких отделов, из которых передний1
вдавлен посредине спереди, а задний — сзади, оба отдела в средней
части сообщаются узким протоком. Длина фуркальных ветвей и степень
их расхождения весьма вариирует. Обычно ветви расходятся значительно
(у var. speratus почти параллельны), их длина в 4—5 (у var. speratus
в б—‘8) раз более ширины. Внешние края ветвей с частым рядом мел-
ких, хорошо различимых шипиков (наподобие пилы), иногда сильно
редуцированных (var. speratus); обычно этот ряд лишь немного не
доходит до основания ветвей. Их латеральные щетинки 'Прикреплены
очень близко от заднего угла. Из апикальных крайних щетинок внешняя
шипообразна, обычно лишь немного короче (или равна) внутренней
крайней, тонкой щетинке. Средние апикальные щетинки хорошо разви-
тые, из них внешняя более :,/2 длины внутренней. Передние антенны
12-членистые, едва доходящие до середины или до заднего конца
2-го торакального сегмента, 3 дистальных членика очень длинные
и тонкие, с очень узкой цельнокрайней гиалиновой пластинкой вдоль
края. На 10-м членике сензорный придаток в виде тонкой щетиики.
У задних антенн дистальный членик заметно короче предыдущего.
Максиллярные ноги короткие, тонкие, с не вполне разделенными
2 дистальными члениками эндоподита. Формула шипов плавательных
ног 3-4-4-3- Дистальный членик эндоподита ног IV пары узкий и длин-
ный, его длина приблизительно втрое более его ширины, внутренний
из апикальных шипов этого членика в 1.2—1.5 раза длиннее внешнего,
нередко сильно изогнутый. Ноги V пары в виде угловатой пластинки,
несущей внутри мощный ножевидный шип, а снаружи и посредине зад-
него края по тонкой относительно длинной щетинке. Длина1 2 0.8—1.5 мм.
Окраска сильно вариирует, обычно однообразная соломенножелтая,
иногда коричневая или розоватая (красноватая). Яйцевые мешки сильно
расходящиеся, удлиненно-овальные, обычно содержащие значительное
количество яиц.
Самец. Внешние края фуркальных ветвей без ряда шипиков,
гладкие, ветви короче, чем у самки, обычно почти параллельные. У ног
1 По Gurney (1933:103) передний отдел крупнее заднего, что, однако, наблюдается
далеко не всегда.
2 В? Coker’y (1933) при высоких температурах (28—30°) получается поколение
22 и оо меньшей величины, нежели при низких температурах.
2. EUCYCLOPS
HI
Фиг. 22. Eucyclops serrulatus (Fisch.).
/—задний отдел торакса и генитальный сегмент с reeeptaculum seminis;
2—нога V самки; 3—3 дистальных членика антенны I самки; 4—фурка
самки var. brachyurus Lili, с брюшной стороны; 5— фурка самки var.
proximus Lili, с брюшной стороны; б — фурка самки (промежуточная между
brachyurus и var. proximus форма) с брюшной стороны.
142
EUCYCLOPfNAE
VI пары внутренний шип длиннее 2 абдоминального сегмента и обеих
щетинок. Длина 0.65—0.8 мм.
Распространение (включая var. proximus, встречающегося
почти всегда совместно с типичной формой).1 Космополит. Вся Европа
и Азия (включая и Малайский архипелаг), Северная Америка, Новая
Зеландия, вся Африка. В СССР обширно распространен, встречается
на арктических островах и в пределах всех ландшафтных зон, на восток
до Дальнего Востока включительно (известен с севера о. Сахалина;
Rylov, 1932). В горных водоемах встречается до высоты 4000 м над
ур. м. (Памир; Rylov, 1930). Принадлежат к самым обыкновенным
циклопам.
Биология. Вид бентический. Обитает в самых разнообразных
стоячих и текучих водоемах, в зарослях водных микро- и макрофитов.
Обычен в литорали озер, где, как правило, глубже сублиторали почти
не опускается; в глубоких озерах единичные экземпляры были найдены
на глубине до 95 м. Широко распространен в различных мелких водо-
емах, в том числе и в астатических. В реках и ручьях обитает в при-
брежных зарослях, редко встречаясь на фарватере и избегая открытых
мест с сильным течением. Нередок в наземных ключевых водах. Целый
ряд нахождений относится к подземным водоемам — пещеры, глубокие
колодцы. В СССР указан под вопросом для окрестностей Кутаиси (Во-
rutzky, 1930); известен из колодцев Саратова, Баку и окрестностей Троицка.
Эвритермичен, амплитуда приспособляемости к изменениям температуры
очень широка. В качестве постоянного компонента отмечался неоднократно
для фауны родников и ручьев с постоянно низкой температурой. Вместе
с тем очень обычен в сильно прогреваемых летом мелких водоемах
(до 30° и выше). Прямых наблюдений над отношением Е. serrulatus
к содержанию растворенного кислорода не имеется. Есть, однако, осно-
вания предполагать его высокую приспособленность к колебаниям
и этого фактора. Так, как упоминалось выше, рачок обычен в подземных
водах. A. Graeter (1903) упоминает о нахождении его Richard’oM в серных
источниках Энгиена (Франция), а Дексбах (1931) находил его зимой
в придонном слое зараженного сероводородом пруда в окрестностях
Москвы (Косино). Во всех этих случаях едва ли можно говорить о бла-
гоприятных кислородных условиих. По Lowndes’y (1928) Euc. serrulatus
обычен в водах с pH = 4.6—9.8, указания для СССР относятся к водам
с pH = 5.6—9.2. В дистрофных водах с pH менее 4.5 рачок не встре-
чается. Что касается окисляемости воды, то он встречается как в гумус-
олиготрофных чистых водах (например оз. Кордывач на Кавказе), так
и в водах с окисляемостью 70—80 мг 02 %0 (вероятно и при более
высокой окисляемости). В высокой степени эвригалинен. Цикл размно-
жения изучен рядом авторов. По Wolf у (1905), Зиверту (1929) и некото-
рым другим авторам полицикличен, встречается круглый год; в окрест-
ностях Вюртемберга Wolf наблюдал в постоянных водоемах до 6 половых
периодов в течение года (также зимой, подо льдом). Самки с яйцевыми
мешками зимой были находимы неоднократно. Точных указаний надици-
клию, сколько нам известно, не имеется.
Рассматриваемый вид является, главным образом, /?-мезосапробой;
несомненно он встречается, однако, и в олигосапробной и, повидимому,
отчасти (редко) в а-мезосапробной зоне. Можно согласиться с Зивертом,
1 Из очень большого числа литературных указаний часть сомнительна (смешивэ
нив с Euc. macruroides).
2. EUCYCLOPS
143
по которому Е. serrulatus, являясь убиквистом в широкой степени, едва ли
может иметь значение индикатора в санитарно-биологическом анализе.
Fischer (Bull. Soc, Nat. Moscou, 1851) под C. serrulatus описал
форму с относительно короткими фуркальными ветвями, сильно воору-
женными на внешних краях шипиками. С ней идентионой надо признать
описанную Lilljeborg-’ом, 1901 (Svenska Vet. Akad. Handl., 35:90,
t. 6, f. 5—9) C. varius var. brachyurus.
Типичная форма отличается сравнительно короткими фуркальными
ветвями, длина которых приблизительно равна длине 2 задних абдоми-
нальных сегментов, взятых вместе, или несколько менее последней;
ветви относительно толстые.
* la. Eucyclops (s. str.) serrulatus (Fisch.) var. proximus Lili.
Cyclops varius var. proximus Lilljeborg- 1901:89, t. 6, f. 1—4. — Eucyclops (Eucy-
clops) serrulatus var. proximus Рылов 1933:257, т. 3, p. 2—3.
От типичной формы самка отличается более длинными фуркальными
ветвями, превосходящими длину 2 задних абдоминальных сегментов,
взятых вместе; нередко ветви почти параллельны, шипики на их внешних
краях хорошо развитые, иногда редукцированные, но в незначительной
степени, их ряд занимает почти весь внешний край, исключая лишь его
передний отдел.
Очень часто встречается совместно и одновременно с типичной
формой, причем между обеими встречаются все переходы.
* lb. Eucyclops (s. str.) serrulatus (Fisch.) var. speratus (Lili.) (фиг. 23).
Cyclops varius var. speratus Lilljeborg, 1901:88, t. 5, f. 12—15. — Leptocyclops
speratus Sars. 1914:72, t. 44. — Cyclops speratus Рылов, 1918:70, рис. 27—28. — Eucyclops
(s. str.) speratus Kiefer, 1929:33. — Leptocyclops speratus Lowndes, 1932a : 50—53, 56.—
Cyclops (Eucyclops) agilis speratus Gurney, 1933 :104, f 1395—1405.
Самка. Ряд шипиков на внешних краях фуркальных ветвей очень
сильно редуцирован, причем шипики (притом очень мелкие) остаются
лишь в задней 1/3 длины края. Фуркальные ветви почти параллельные
(иногда вполне параллельные), стройные, длинные, их длина в б—8 раз
превышает их ширину, внутренняя крайняя апикальная щетинка обычно
достигает 60% длины ветви. Длина 1—1.7 мм.
Самец. Длина фуркальных ветвей в 4—б раз более их ширины,
внутренняя из средних апикальных щетинок обычно короче самих ветвей
(иногда равна им, иногда длиннее их). Длина 0.7—0.9 мм.
Как указывалось выше, считать speratus самостоятельным видом
нет оснований. Таксономическое положение вариетета неясно; едва ли
оно аналогично таковому var. proximus. У var. speratus изменчивость
идет в направлении не только длины и расхождения фуркальных ветвей,
но и в направлении редукции вооружения их внешнего края, что почти
не имеет места (во всяком случае не характерно) для ряда typicus-
proximus.
Распространение. С достоверностью вариетет найдеи
в Европе, Азии и северной Африке. Распространение совпадает с тако-
вым типичной формы и var. proximus. Вероятно встречается и высоко
в горах.
Биология изучена неполно. Обитает, главным образом, в мелких
стоячих и слабопроточвых водоемах, встречается также в заро-
слях макрофитов литорали озер и в реках, при слабом течении, также
144
EUCYCLOPINAE
в ключах (Klie, 1925). Эвритермичен. Указан для вод с pH = 4.8—9.
Пресноводная форма, в солоноватых водах редка (Schafer, 1933, при
S=0.603 — 0.725%, Пирожников, 1929 — в оз. Сартлан). Цикл размно-
жения не изучен. По Зиверту (1929) в окр. Харькова встречается круглый
год. Вероятно, как и типичная форма, полицикличен.
Фиг. 23. Eucyclops serrulatus var. speratus
(Lili.), самка.
7—фурка; 2—дистальный членик антенны I;
3— нога V самкн. (7—2—по Рылову, .2—по
Sars’y).
*lc. Eucyclops (s. str.) serrulatus var. nagasakai (Ueno).
Eucyclops nagasakai Ueno, 1934:229, f. 1, A—J; ib., 1934a: 449, f. 3, A — C.
Самка. Строение тела как у типичной формы. Глазной пигмент
отсутствует. Фуркальные ветви заметно расходящиеся, короткие, их длина
приблизительно в 4 раза более их ширины, внешние края несут ряд
мелких шипиков лишь в задней половине, из крайних апикальных щети-
нок внутренняя приблизительно на % длиннее внешней, шипообразной,
средние апикальные щетинки хорошо развитые, как у типичной формы.
Передние антенны 12-члени-
стые, доходящие приблизитель-
но до середины 2 торакаль-
ного сегмента, гиалиновые пла-
стинки 3 дистальных члеников
редуцированы. Плавательные
ноги и ноги V пары вооружены,
как у типичной формы. Дли-
на =0.9—1.2 мм. Бесцветны.
Яйцевые мешки содержат 8 яиц.
Самец. У VI пары 1 из 3
придатков сильно редуцирован,
в виде мелкого шипика. Внеш-
ние края фуркальных ветвей
без ряда шипиков. Длина 0.8—
0.85 мм.
Вопреки мнению Ueno, я
не могу признать за рассма-
триваемой формой видового
значения. Сам Ueno указывает
на ее чрезвычайную близость
к Е. serrulatus (Fisch.), приводя в качестве признаков отличия от
последнего „отсутствие глаза", длину и вооружение краев фуркальных
ветвей и отсутствие шипиков на этих краях у самцов. Что здесь имеет
место не отсутствие глаза, а глазного пигмента, — не подлежит сомнениям.
Однако экспериментально доказано, что у пещерных Cyclopidae с отсут-
ствующим пигментом, в частности у Е. graeteri, гораздо более дифферен-
цированного морфологически, при культивировании на свету пигмент
восстанавливается (Chappuis, 1920). Этот признак, следовательно,
не является видовым. Длина и вооружение фуркальных ветвей самки
Е. serrulatus очень сильно вариирует вообще, в частности вариирует
вооружение их внешних краев, которые у Е. serrulatus из наземных
водоемов может быть даже еще более редуцированными, чем это описано
Ueno для Е. nagasakai. Отличие самцов, отмечаемое этим автором,
фактически неверно, — шипики на внешних краях фуркальных ветвей
отсутствуют и у самцов типичных Е. serrulatus (см., например Gurney,
1933, f. 1393). В виду чрезвычайной изменчивости последнего вида,
с систематической оценкой перечисленных признаков надо быть особенно
2. EUCYCLOPS
из
осторожным при определении его популяций из пещерных водоемов,
в условиях которых, как мы знаем, ряд видов (например р. Acanthocy-
clops) склонен к образованию форм, которым систематика с полным на
то основанием не придает видового значения.
Распространение. Центральная Япония (Shinano), подземный
водоем в пещере у Hirogawara, Манчьжурия (провинция Квантунь),
водоем близ города Пи-тен-во.
Биология. Троглобионт, найден при pH от 3.4 до 7.6 (особенно
многочислен при щелочной реакции) и содержании кислорода в ЗО°/0
нормального насыщения; годовые колебания температуры воды 6—10°.
2. Eucyclops (s. str.) teras (E. Graeter) (фиг. 24ь 2).
Cyclops teras E. Graeter, 1907, Zool. Anz., 31:349, f. 1—2; E. Graeter, 1911:31,
f. 13—14. — Eucyclops teras Kiefer, 1929:89.
Глазной пигмент отсутствует. Задний торакальный сегмент по бокам
без щетинок. Фуркальные ветви несколько искривленные, очень короткие
Фиг. 24. (По Е. Graeter’у).
Eucyclops teras (Е. Graet.): 7 — фурка самки, 2—генитальный сегмент, V пара ног
и receptaculum seminis; 3—Е. graeteri (Chapp.), нога V самки.
(их длина приблизительно в 3 раза более их ширины), на внешних краях
без ряда шипиков, внутренний из крайних апикальных придатков (щетин-
ка) приблизительно вдвое длиннее внешнего (шипа). Передние антенны
12-членистые, достигающие середины 1-го торакального сегмента.
Ноги V пары только с 2 придатками — с внутренним шипом и внешней
щетинкой. Длина 1.0 мм. Бесцветны и очень прозрачны. В каждом
яйцевом мешке не более 8 яиц. Самцы не описаны.
Распространение. Швейцария.
Биология. Троглобионт, исключительно в пещерных водах. Исход-
ная форма — Е. serrulatus (Fisch.).
* 3. Eucyclops (s. str.) macruroides (Lili.) (фиг. 254—6).
Cyclops macruroides Lilljeborgi, 1901:85, t. 5, f. 7—11.— C. serrulatus (part.)
Graeter, 1903:494. — Leptocyclops macruroides Sars, 1914:74, t. 46. — C. macruroides
Рылов 1918:73, рис. 29—30. — Eucyclops (». str.) macruroides Kiefer, 1929:35. — Lepto-
10 Фауаа СССР
146
EUCYCLOPINAE
cyclops macruroides Lowndes, 1932a: 53. — C. (Eucyclops) macruroides Gurney, 1933:109.
f. 1406—1419.
Самка. Габитус тела такой же, как у Е. serrulatus. Фуркальные
ветви очень длинные, стройные, почти параллельные (у var. denticulata
расходящиеся), их длина не менее (у этого вариетета несколько менее)
длины 3 задних абдоминальных сегментов, взятых вместе, приблизительно
в 8 —9 раз (иногда в 9 раз слишком) более их ширины (у var. denticu-
lata в б—8 раз); внешние края ветвей с рядом сильных шипиков, крайняя
внутренняя апикальная щетинка приблизительно в 1 1/2 раза длиннее
внешнего крайнего апикального шипа, средние апикальные щетинки
вооружены тетерономно. Генитальный сегмент и reeeptaculum seminis
как у Е. serrulatus. Передние антенны достигают середины 1-го
торакального сегмента, 10-й—12-й членики с тонко зазубренной узкой
гиалиновой пластинкой вдоль внутреннего края; нередко на месте этой
зазубренности образуется ряд чрезвычайно тонких шипиков, которые
крупнее и толще в проксимальной половине пластинки. У плавательных
ног IV пары длина дистального членика эндоподита почти в 3 раза
превышает его ширину, внутренний апикальный шип приблизительно
в l1^ раза длиннее внешнего. Ноги V пары со сравнительно тонким
и коротким внешним шипом. Длина 1.2—1.5 мм. Яйцевые мешки как
у Е. serrulatus.
Самец. Длина фуркальных ветвей в 5—7 раз более их ширины,
их внешние края без ряда шипиков, у ног VI пары внутренний шип
очень короткий, почти одинаковой длины с вооружающими членик щетин-
ками. Длина 0.7—0.9 мм.
Распространение. Европа, Северная Африка, Азия. В СССР
известен с Соловецких островов, из Болыпеземельской тундры, Карело-
Финской ССР, с Кольского полуострова, из ряда центральных областей.
На Кавказе указан для Краснодарского края и Армении, в азиатской
части СССР найден в среднем течении р. Енисея, близ Барабинска,
в центральной части Якутской АССР, в оз. Кара-куль на Памире,
в р. Мургаб, в литорали Телецкого озера. В Швейцарских Альпах
известен до высоты 1000 м над уровнем моря, на Памире—на высоте
4000 м. Распространение Е. macruroides пока нельзя считать выясненным
в достаточной степени; очень вероятно, что он встречается и на Даль-
нем Востоке.
Биология. Подобно Е. serrulatus, вид не планктический; обитает
в разнообразных мелких водоемах, в зоне макрофитов литорали озер,
в прудах. В мелких временных водоемах гораздо реже. Большинство
имеющихся данных определенно указывает на обитание, главным обра-
зом, в более или менее крупных водоемах (озера) и в связанных с ними
текучих и стоячих водоемах (заливные водоемы); в реках обитает в заро-
слях прибрежных макрофитов. Троглоксен, в подземных водах лишь
случайно. Вероятно пОлиоксибионт, эвритермичен, невидимому, с склон-
ностью к более или менее высоким температурам воды. Встречается
в водах с рН=5.2—9.8, невидимому, особенно в нейтральных (слабо-
кислых) слабощелочных водах. Вид пресноводный, но может жить
в сильно минерализированных водах. По данным Зиверта (1929) является
олигосапробом, /?-мезосапробом.
Цикл размножения изучен неполно. В старицах Кубани (Калита,
1930) на зиму выпадает, по Зиверту (1927, 1929) яйценосные $>$> только
летом, но рачок встречается круглый год (небольшие озера поймы
р. Донца). Вероятно полицикличен.
2. EUCYCLOPS
147
Фиг. 25.
Eucyclops macruroides var. denticulatus (Graet.): 7 — фурка самки, 2 — нога V самки,
3 — дистальные членики антенны I самки.
Е. macruroides (Lili.) typ.: 4— фурка самки, 5 — йога V самки, 6—дистальный членик
антенны I самки; Е. macrurus Sars: 7—фурка самки, 8—нога V самки,
9—3 дистальных членика антенны I самки.
10'
148
EUCYCLOPINAE
Типичная форма характеризуется очень длинными, почти параллель-
ными, узкими фуркальными ветвями, длина которых не менее длины
3 задних сегментов абдомена, взятых вместе, и в 8 — 9 (или несколько
более) раз превышает их ширину.
* За. Eucyclops (s. str.) macruroides var. denticulatus (A. Graeter)
(фиг. 25j,2).
Cyclops sorrulatus (non C. serrulatus Fischer) Lilljeborg, 1901:81, t. 5, f. 1—6,—
C. serrulatus var. denticulatus A. Graeter, 1903:491. — Leptocyclops lilljeborgi (part.)
Sars, 1914:73, t. 45. — C. lilljeborgi Рылов, 1918:65, рис. 72—73; Рылов, 1923:79,
т. 2, рис. 24—25. — С. denticulatus Thallwitz, 1927, Sitzungeber. Ges. Isi-,.: 4. — Eucy-
clops (s. str.) lilljeborgi Kiefer, 1929:35. — ? Eucyclops fimbriatus var. anyschghizara1
Schiklejev, 1931, Zool. Anz., 94:188, f. 4, a — c. — E. (E.) lilljeborgi Рылов, 1933 : 257,
т. 3, рис. 4—6.— Cyclops (Eucyclops) macruroides denticulatus Gurney, 1933: 111,
f. 1410—1423.
Самка. Фуркальные ветви сильно расходящиеся, их длина в б — 8
раз превышает их ширину и несколько менее длины 3 задних абдоми-
нальных сегментов, взятых вместе; внешние края ветвей нередко несколько
вогнутые, их вооружение состоит всегда из сильных шипиков, внутрен-
няя крайняя апикальная щетинка обычно вдвое длиннее внешнего апи-
кального шипа, иногда почти равняется длине самой ветви. Передние
антенны и вооружение плавательных ног IV пары как у типичной формы.
У ног V пары внутренний шип относительно короче и тоньше, чем
у последней. Длина 0.9 —1.2 мм.
Самец — как у типичной формы.
Распространение. С достоверностью var. denticulatus указан
для Европы, Азии и северной Африки. В СССР найден на Мурманском
побережье, в Карело-Финской ССР; на Кавказе известен близ
Новороссийска и, вероятно, в окрестностях Пицунды (= С. fimbriatus
var. anyschghtzara Schiklejev, 1931), в азиатской части СССР — из окр.
Барабинска, из оз. Марка-куль на Алтае, с Дальнего Востока. В общем
распространение вариетета и типичного Е. (s. str.) macruroides совпадает,
ограничиваясь Палеарктикой; обе формы встречаются нередко совместно
и одновременно.
Биология изучена недостаточно. Как и типичная форма, обитает
в зарослях макрофитов, главным образом, литорали озер и в связанных
с ними стоячих и текучих водах, также в прудах. Повидимому, эвритер-
мичен, со склонностью к тепловодности. По Lowndes’у (1928) обычен
в водах с pH = 4.6 — 7.6, но Зиновьевым (1935) в водоемах Челябинской
области найден при pH = 7.8 — 8.2; обыкновенен в нейтральных (слабо-
кислых) и слабощелочных водах. Вместе с типичной формой найден
в солоноватом озере Сартлан. Судя по имеющимся данным, биология
типичной формы и рассматриваемого вариетета одинакова.
* 4. Eucyclops (s. str.) macrurus (Sars) (фиг. 257)9).
Cyclops macrurus Sars, 1863, Forh. Selsk. Christ., (1862): 254. — C. maarensis
Vosseler, 1839, Arch. Nature-., 55:110, t. 6, f. 1—7.— C. macrurus Schmeil, 1892: 146,
t. 5, f. 15—17. — Leptocyclops macrurus Sars, 1914:75, t. 47. — Eucyclops (s. str.) macru-
rus Kiefer, 1929: 36. — C. macrurus var. caucasicus Schiklejev, 1931, Zoo). Anz., 94:190,
f. 51. — C. (Eucyclops) macrurus Gurney, 1933:115, f. 1424—1437.
1 Относительно этой формы Gurney (1933) впал в непонятную для меня ошибку,
отнеся ее в синонимы Par. fimbriatus. Описание Шиклеева основано на грубейшей
ошибке в определении. Как видно из рис. 4 а—с этого автора, он имел, бесспорно,
неполовозрелый экземпляр циклопа из рода Eucyclops s. str. и, судя по рисунку перед-
них антенн, именно Е. macruroides var. denticulatus.
2. EUCYCLOPS
149
Самка. Тело более стройное, нежели у Е. serrulatus, задние углы
цефалоторакса и 2-го — 4-го торакальных сегментов закругленные, у 4-го
сегмента более заметно оттянутые кзади, задний сегмент по бокам
с группами тонких волосков. Генитальный сегмен и receptaculum seminis
как у Е. serrulatus. Фуркальные ветви параллельные, очень длинные,
их длина приблизительно в 10 раз (иногда немного более) превышает
их ширину, внешние края в задней 1fi длины вооружены группой из 4—6
мелких шипиков (близ латеральной щетинки). Из крайних апикальных
придатков внутренняя щетинка почти вдвое длиннее внешнего придатка,
который является шипом, средние апикальные щетинки хорошо развиты,
с гетерономным вооружением. Передние антенны едва достигают зад-
него конца цефалоторакса, их дистальный членик относительно короче,
чем у Е. serrulatus, с очень узкой, с трудом различимой, гиалиновой
цельнокрайней пластинкой вдоль внутреннего края. У максиллярных
ног оба дистальных членика резко дифференцированы. Плавательные
ноги как у Е. macruroides, соединительная пластинка ног IV пары
с выпуклым краем. У ног V пары внутренний шип короткий и относи-
тельно тонкий. Длина 1.1 —1.4 мм. Окраска обычно желтоватая, до корич-
невой. Яйцевые мешки несколько меньше, чем у Е. serrulatus, тесно
прилегающие к абдомену.
Самец. Длина фуркальных ветвей приблизительно в 8 раз более
их ширины. У ног VI пары внутренний шип почти одинаковой длины
со 2-м абдоминальным сегментом, несколько более длинный, чем
соседняя с ним (средняя) щетинка. Длина 0.7—0.9 мм.
Распространение. Вся Европа и часть Азии (запад азиатской
части СССР, Палестина), северная Африка. В европейской части СССР
распространен очень широко. В азиатской части СССР показан для
окрестностей Красноярска, Акмолинска, Барабинска, из озер курорта
Карачи, в низовьях р. Сыр-дарьи. На Кавказе известен лишь из окрест-
ностей Краснодара и Адлера (Schiklejev, 1931, С. т. var. caacasicus).
В горной области Кавказа и азиатской части СССР отсутствует. В Запад-
ной Европе также не заходит высоко в горы (показан до высоты около
1000 м над уровнем моря).
Биология. Обитает, главным образом, в зарослях макрофитов
литорали озер, также в прудах. Гораздо реже был находим в мелких
водоемах (болотца, лужи), притом, вероятно, главным образом, в свя-
занных с крупными водоемами. В реках исключительно в зарослях при-
брежных макрофитов, при слабом течении. Обычен в стоячих водоемах
речной поймы. Сколь-либо глубоко в озерах не встречается, повидимому,
отсутствуя уже в верхних горизонтах сублиторали. Троглоксен, в под-
земных водоемах1 указывался как редкое исключение (колодцы). Боль-
шинство авторов считает, что Е. macrurus является хорошо выраженным
тепловодным рачком, на что указывает и цикл его размножения. Судя
по населяемым биотопам, вид обитает исключительно в богатых кисло-
родом водах. Встречается в водах с pH = 5.4—9.2, причем в водах с pH
более б он особенно распространен. Типично пресноводный вид. В соло-
новатых водах был находим лишь очень редко. Е. macrurus следует
отнести к олигосапробам — [3-мезосапробам; высоких степеней /9-мезоса-
пробности, повидимому, избегает.
1 Е. macrurus var, subterranea есть синоним троглобиота Е. graeteri (Chapp.)
(см. ниже).
150
EUCYCLOPINAE
Цикл размножения изучен недостаточно полно. Как считает Зиверт
(1929), вероятно, полицикличен. На зимние месяцы выпадает; указания
на нахождение в середине зимы сомнительны.
5. Eucyclops (s. str.) graeteri (Chappuis) (фиг. 243).
Cyclops macrurus var. subterranea E. Graeter, 1907, Zool. Anz., 31:850; E. Graeter,
1911:32, f. 15, t. 1, f. 2. — Cyclops graeteri (nom. nov.) Chappuis, 1927, Die Binnengew.,
3:42.— Eucyclops graeteri Kiefer, 1929:36; Kiefer, 1932, Zool. Anz., 101:53, f. 9.—
? Eucyclops inarmatus Kiefer, 1932, ibid.: 51, f. 3—8.
Глазной пигмент отсутствует.1 Длина фуркальных ветвей приблизи-
тельно в б раз более их ширины, их внешние края без шипиков, из край-
них апикальных щетинок внутренняя вдвое длиннее внешней. Передние
антенны 12-членистые, достигающие заднего края цефалоторакса.
У дистального членика эндоподита ног IV пары внутренний апикальный
шип почти вдвое длиннее внешнего. У ног V пары внутренний шип
в основании широкий, в дистальном отделе тонкий, щетинкооэразный.
Длина 1.7 мм (с фуркальными щетинками). В каждом яйцевом мешке
не более 2 яиц. Самцы не известны.1 2
Распространение. Швейцария и Франция.
Биология. Троглобионт, исключительно в пещерных водах. Родо-
начальной формой вида Chappuis предполагает Е. macrurus (Sars).
Е. inarmatus Kief., описанный из пещер Герцеговины, по всем
вероятиям, идентичен с рассматриваемым видом, от которого отличается
лишь более короткими фуркальными ветвями, 11-членистыми (всегда ?)
передними антеннами с очень длинной щетинкой на их 4-м членике,
а отчасти апикальным вооружением фуркальных ветвей; самец не изве-
стен.
3. Род TROPOCYCLOPS KIEF.
Tropocyclops (subgenus) Kiefer, 1927:303; Kiefer, 1928:541, Kiefer, 1929:39; Kiefer, 1930,
Zoo). Anz., 87:118; Kiefer, 1931:437, 505; Gurney, 1933:86. — Tropocyclops (genus) Kiefer,
1937:421.
Фуркальные ветви короткие, их длина самое большее в 3—4, обычно
лишь в 1.5—2.5 раза превышает ширину, внешние края без ряда шипиков.
У ног V пары внутренний придаток в виде длинного тонкого шипа. На
конце дистального членика эндоподита ног IV пары внутренний шип
очень длинный, обычно почти вдвое длиннее внешнего шипа. Передние
антенны всегда 12-членистые, относительно длинные. Receptaculum seminis
по бокам переднего отдела с трубчатыми придатками приблизительно
cv-образно изогнутыми. Мелкие циклопы, длина самок 0.4—0.9 мм;
обычно яркозеленой окраски. Науплиусы не сильно сплющенные.
По вооружению плавательных ног Kiefer (1931:506) различает
2 группы: 1) Формула шипов 3-4-4*3, группа Quaterni; сюда относятся
Т. prasinus (Fisch.) с его вариететами (см. ниже), Т. extensus Kief,
(с f. longispina Kief.), T. nigroviridis Harada и T. schubarti Kief.
1 По исследованиям Chappuis (Arch. f. Hydrob., 1920:24) при воспитании E. grae-
teri на свету получается поколение (уже в 1-й генерации) с красным глазным пигментом.
Вообще у всех подземных Cyclopidae глаз сохраняется, редуцируется только его
пигмент.
2 Kiefer (1929: 36) в диагнозе упоминает, что 33 Е. graeteri не известны, и затем,
по какому-то недоразумению, приводит размеры о (длина 1.5 мм.), В другой работе
(1932 ; 53) автор указывает, что лично этот вид он не находил.
3. TROPOCYCLOPS
151
2) Формулы шипов 3-4-3-3, группа Terni; к ней принадлежат Т. conf inis
Kief, (с f. frequens Kief.), T. parvus Kief., T. tenellus (Sars). Формула
шипов T. varicoid.es (Brady) не известна.1
Всего известно 8 видов.1 2 Наиболее богато род представлен в тро-
пической зоне, особенно в Эфиопской области (см. выше. Eucyclops).
На севере Палеарктики и Неарктики род отсутствует. Прибрежные виды,
литораль озер, планктон мелких водоемов разнообразного типа. Тепло-
водно-стенотермичиы. Троглобионты отсутствуют.
* 1. Tropocyclops prasinus (Fisch.) (фиг. 26).
Cyclops prasinus Fischer, 1860, Abh. Bayer. Akad., 8:652, t. 20, f. 19—26, a; Schmeil,
1892:150, t. 5, f. 1—5. — C. (Eucyclops) prasinus Claus, 1893a:348.— C. magnoctavus
Kokubo, 1912, Ann. Zool. Japon., 8:93, t. 2, f. 8 —12. — Leptocyclops viridis Henry, 1919,
Joum. Roy. Soc. N. S. Wales, 53:40, t. 2, f, 8, 9. — Leptocyclops prasinus Sars, 1927:119,
t. 12, f. 11—20.— Eucyclops (Tropocyclops) prasinus Kiefer, 1929:39, f. 13; Kiefer, 1931:506,
f. 1—2. — E. prasinus candidiusi Harada, 1931:150, f. 1—3. — E. (Tr.) prasinus Kiefer,
1932:244, f. 43—-50; Рылов 1933:257, т. 3, рис. 7—9. — Cyclops (Tropocyclops) prasinus
Gurney, 1933:37, f. 1349-1373.
Самка. Тело более широкое и укороченное, нежели у Е. serrulatus,
задний сегмент торакса по бокам с группами волосков. Генитальный
сегмент мало расширенный спереди. Receptaculum seminis 3 показанной
на фиг. 26 характерной формы (см. также определительную таблицу
подродов). Фуркальные ветви очень мало расходящиеся, их длина
не более как в 2.2—2.5 раза превышает их ширину, латеральная щетинка
прикрепляется немного кзади от середины их внешнего края, из крайних
апикальных щетинок внутренняя лишь немного короче внешней, имеющей
шипообразный вид. Средние апикальные щетинки относительно короткие,
утолщенные. Передние антенны 12-членистые, достигающие заднего
края 2-го торакального сегмента, щетинки 1-го и 4-го члеников очень
длинные, 3 дистальных членика с очень узкими, с трудом различимыми,
вдоль края иногда очень тонко зазубренными гиалиновыми пластинками,
9-й членик с мелким сензорным цилиндром. Задние антенны суженные,
их 4-й членик немного длиннее 3-го. Мандибулы с парой очень длинных
щетинок. У максиллярных ног оба дистальных членика явственно расчле-
нены. У плавательных ног I—III пар свободный край соединительной
пластинки вогнутый, у ног IV пары — выпуклый. Дистальный членик
эндоподита ног IV пары длинный, его длина в 2.3—2.6 раза более
ширины, внутренний апикальный шип длиннее самого членика и прибли-
зительно в 2—2.3 раза длиннее его внешнего апикального шипа. Ноги
V пары похожи иа таковые у Е. (s. str.) serrulatus, но внутренний шип
гораздо тоньше, стройнее, а внутренний край членика несет группу
мелких шипиков. Длина 0.7—0.9 мм. Обычно яркозеленая, реже серая
окраска; иногда рачок бесцветен. Яйцевые мешки содержат (каждый)
по 10—20 яиц; они тесно прилегают к абдомену, округлые.
Самец, Передние антенны 16—17-членистые, богато вооруженные
щетинками. Ноги VI пары с тонким внутренним шипом, несколько более
коротким, чем 2 соседние с ним оперенные щетинки, которые почти
равной длины. Длина 0.5—0.7 мм.
1 С ним идентичен „С. virescens“ Brady (Ann. Trop., Med., I, 1907; см. об этом у
Kiefer, Zool. Anz., 1930).
2 Систематика рода недостаточно выяснена. По Lowndes’y (1932) и Gurney (1933)
некоторые виды сомнительны (быть может, являются подвидами?).
3 О вариациях формы этого органа у Tr, prasinus см. Kiefer (1932).
152
EUCYGL0P1NAE
Kiefer различает1 четыре „формы" и „подвида", которые, по нашему
мнению, представляют, по веем вероятиям, чисто индивидуальные вариа-
ции: Tr. prasinus forma guwana Kiefer, 1931, найдена на Яве (пещеры);
7r. prasinus subsp. jerseyensis1 2 Kiefer, 1931, в Северной Америке; Tr.
prasinus subsp. meridionalis Kiefer, 1931, в Западной Африке и Южной
Америке (Ла-Плата, Уругвай); 7r. prasinus forma pelagica Kiefer, 1933,
Суматра.
Фиг. 26. Tropocyclops prasinus (Fisch.), самка.
7 — фурка co спинной стороны; 2—3 дистальных членика антенны I; 3— йога V;
4 — абдомен с reeeptaculum seminis; 5—дистальный членик эндоподита ноги IV.
Распространение.3 Европа (исключая север!), восточная, юго-
восточная и южная Азия, Новая Зеландия, Северная и Южная Америка,
Африка, Новые Гебриды. В СССР известен из окрестностей Москвы,4
из Одесской области, из р. Сыр-дарьи, из мелких водоемов Дальнего
Востока. На Кавказе найден в мелких водоемах близ Хосты. Отсутствует
в зонах тундры и тайги и, очевидно, очень редок в зоне смешанных
лесов, где найден только в южной части. Главный ареал его распро-
странения у нас, повидимому, охватывает степную зону, южнее 50° с. ш.;
1 Только по самкам. Недавно Kiefer (Zool. Anz. 123, 1938) из Мексики описал еще
подвид mexicanus (только по <?!), крайне проблематичный.
s С этим вариететом надо признать идентичным Tr. prasinus candidiusi Harada
(1931, рис. 1—3), описанного с о. Формозы.
3 Включая и указанные проблематичные „формы" и „подвиды" Kiefer’a.
4 Указания на нахождение в Калининской обл. (окрестности оз. Селигера) требуют
Проверки-
3. TROPOCYCLOPS, 4. OCHRIDACYCLOPS
153
присутствие в зоне субтропиков (Колхида, Талыш) едва ли подлежит
сомнениям. В горах встречается до высоты около 903—1200 м над
уровнем моря.
Биология. Обитает в хорошо прогреваемых мелких водоемах
типа луж, гораздо реже в литорали (заросли макрофитов) озер. Неодно-
кратно указывался для зарослей макрофитов медленно текучих рек.
Планктичность этого вида подтверждается наблюдениями Kiefer’a (1933).
Имеется единственное указание на присутствие его в пелагиали озер
(Green Lake в Северной Америке. Marsh, 1897). Неоднократно был
находим в подземных водах, возможно, является троглофилом. Экспери-
менты, произведенные Roy (1932), показывают, что при высоких темпе-
ратурах воды развитие Tr. prasinus идет ускоренным темпом. Особняком
стоят данные Kleiber’a (1911), по которым в мелких водоемах южного
Щварцвальда (на высоте около 740 м) размножение происходит и зимой,
подо льдом, тогда как другими авторами максимум наблюдался в теп-
лое время года, хотя в южных областях рачок встречается и в
зимние месяцы (указания такого же рода есть и для средней
Европы, где половой цикл имеет место, однако, в теплое время года).
По Lowndes'y (1928) обычен в водоемах с рН-4.б—9.8. Скудные дан-
ные для СССР относятся к нейтральным и слабощелочным водам.
Главным образом в пресных водах, лишь очень редко указывался
в солоноватых.
Дицикличен (возможен 3-й половой период), половые циклы в теплое
время года, на которое падает и максимум развития.
Вероятно олигосапроб — [3-мезосапроб.
4. Род OCHRIDACYCLOPS KIEF.
Ochridacyclops Kiefer, 1937, Zool. Anz., 120:137.
Самка. Цефалоторакс очень объемистый, удлиненный. Задние углы
2-го — 4-го торакальных сегментов сильно выступают кзади, по бокам
предпоследнего торакального сегмента имеются группы тонких щети-
нок, которые хорошо развиты и по бокам заднего торакального
сегмента. Генитальный сегмент толстый, лишь слабо суживающийся
кзади. Reeeptaculum seminis построен по типу такового у рода Eucyclops.
Фуркальные ветви толстые и короткие, мало расходящиеся, их длина
приблизительно в 2г/2 раза превышает их ширину, внутренние края без
волосков, внешние края без шипиков, латеральная щетинка прикреплена
в начале задней трети их длины, средние апикальные щетинки относи-
тельно короткие, изогнутые в дистальном отделе, гомономно вооружен-
ные, из крайних апикальных щетинок внутренняя заметно длиннее
нешипообразной внешней, дорзальная щетинка очень короткая. Передние
антенны 11-члениетые, очень укороченные, значительно не достигающие
заднего края цефалоторакса, их 3 дистальных членика короткие.
Членистость плавательных ног по формуле 3-3/3-3/3-3/3-3, формула
шипов экзоподита 3-4-3-3, щетинок 5-5-5-5. У ног I пары на внутренней
стороне 2-го базального членика нет ни шипа, ни щетинки. Дистальный
членик эндоподита ног IV пары короткий, внутренний из апикальных
шипов почти вдвое длиннее внешнего (1.75 : 1). Ноги V пары состоят
из удлиненного прямоугольного членика, вооруженного 3 апикальными
придатками—внутренним, очень коротким и тонким шипом и 2 оперен-
ными длинными щетинками. Длина 0.75 мм.
/54
EUCYCLOPINAE
Самец. Фуркальные ветви, плавательные ноги, V пары ног как
у самки. У ног VI пары 3 придатка — мелкий шип и 2 хорошо развитые
щетинки. Длина около 0.79 мм.
Единственный вид с признаками рода.
1. Ochridacyclops arndti (Kief.) (фиг. 27).
Ochridacyclops arndti Kiefer, 1937, Zool. Anz., 120:137, fig. 1—9.
Местонахождение. Оз. Охрида в Югославии.
Биология не изучена; повидимому обитает на губке Ochridaspongia
rotunda Arndt в качестве комменсалиста.
5. Род PARACYCLOPS CLAUS
Cyclops (part.) Fischer, 1853:94; Schmeil, 1892:15.— Cyclops (subgen. Paracyclops,
part.) Claus, 1893:33.— Cyclops (part.) Lilljeborg, 1901:3; Marsh, 1909:1070. — Platycyclops
(part.) Sars, 1914:76.— Paracyclops Kiefer, 1927:704; Kiefer 1923:542; Kiefer, 1929 : 40;
Kiefer, 1929a:47.— Cyclops (subg. Paracyclops) Gumey, 1933:97.
Передний отдел тела заметно сплющен в спинно-брюшном направ-
лении, абдомен толстый, причем
дифференцировка этих 2 отделов
выражена менее, чем у рода Ей-
cyclops. Задний торакальный сег-
мент по бокам на задних углах
с группой мелких щетинок. Ге-
нитальный сегмент сравнительно
широкий, утолщенный.1 Recepta-
culum seminis очень похож на тако-
вой рода Eucyclops. Фуркальные
ветви различной длины, отноше-
ние которой к ширине от 2:1
до 6:1, внешние края без пило-
образного продольного ряда ши-
пиков, но обычно с поперечным
или косым рядом на спинной
поверхности ветвей, кпереди от
основания латеральной щетинки.
Передние антенны 6 — 11-члени-
стые, у Р. eucyclopoides 12-чле-
нистые. Внешние края экзопо-
дитов плавательных ног с силь-
ным вооружением из мелких ши-
пиков. Ноги V пары 1-членистые,
единственный членик явно отчле-
нен от заднего торакального сег-
мента и имеет угловатые очер-
тания (не в виде широкой пла-
стинки), его вооружение состоит
Мелкие циклопы, длина самки не
Фиг. 27. Ochridacyclops arndti Kief., самка.
(По Kiefer’y).
/ — со спинной стороны; 2 — нога V; 3 —
эндоподит ноги IV.
из внутреннего шипа и 2 щетинок,
более 1.5 мм, обычно около 1 мм.
Известно б видов: Р. fimbriatus (Fisch.), Р. poppei (Rehb.), Р. affinis
(Sars), P. sitiseiensis Harada, P. eucyclopoides Kief., P. oligarthrus Sars.
1 Исключением является P. eucyclopoides, имеющий более стройное тело (как
у рода Eucyclops) и занимающий как бы промежуточное положение между последним
родом и родом Paracyclops,
5. PARACYCLOPS
155
После исследований Kiefer’a видовая самостоятельность Р. poppei
не подлежит сомнениям, хотя ранее последний считался вариететом
fimbriatus^ Систематика Р. fimbriatus является довольно неясной. Kiefer
выделил из него 2 очень близкие формы — abnobensis Kief, и finitimus
Kief, в качестве самостоятельных видов, но позже сам высказался об
этом с большим сомнением. Рассматриваемые 2 формы отличаются
от Р. fimbriatus (Fisch.) длиной фуркальных ветвей, относительной
длиной их средних апикальных щетинок и придатков ног V пары
и незначительным различием в длине внутреннего апикального шипа
эндоподита ног IV пары. Едва ли, однако, возможно на основании таких
признаков говорить об отдельных видах. Длина фуркальных ветвей
вообще у Eucyclopinae является признаком очень вариирующим,
в частности у Р. poppei; это относится также к длине придатков ветвей
и ног V пары, значительная амплитуда вариаций которых установлена
у целого ряда видов. Невозможность пользоваться признаком длины
апикальных придатков эндоподита ног IV пары в качестве видового
вполне выяснена экспериментально (Lowndes, 1928). Поэтому нам
кажутся достаточно убедительными высказанные Gurney’eM сомнения
в видовом значении формы abnobensis и finitimus, и ниже мы рассмат-
риваем эти формы в качестве вариететов — таксономических единиц
низшего порядка (возможно это аберрации).1 2
Род имеет космополитическое распространение. Р. oligarthrus
известен только из средней Африки (оз. Танганайка), Р. eucyclopoides —
с Малайского архипелага, Р. sitiseiensis — со. Формозы. Остальные 3 вида
имеют весьма обширное распространение и известны из СССР.
Все виды рассматриваемого рода принадлежат к бентическим цикло-
пам. Обитают в литорали (Р. fimbriatus также на дне глубинной области
озер) и в мелких стоячих и проточных водоемах (в реках — особенно
указанный вид).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Передние антенны 11-членистые .... • . . 3. Р, affinis (Sars)
2(1). Передние антенны 8-членистые.
3 (10). Ряд шипиков расположен лишь в заднем отделе спинной
поверхности фуркальных ветвей, занимая приблизительно косое
положение.
4(5). Фуркальные ветви длинные, их длина в 5—б раз превышает их
ширину.................................*1. Р. fimbriatus (Fisch.) typ.
5 (4). Фуркальные ветви относительно короткие, их длина, самое
большее, в 4 раза превосходит их ширину.
6(9). Внешняя из средних апикальных щетинок фуркальных ветвей
приблизительно на х/3 длиннее внутренней.
7 (8). У ног V пары боковая щетинка чрезвычайно длинная..........
......................lb. Р. fimbriatus var. abnobensis (Kief.)
8(7). Эта щетинка гораздо короче, чем у типичной формы ......
......................* la. Р. fimbriatus var imminutus Kief.
1 Такого мнения держится Gurney (1933:129), с которым едва ли можно
согласиться.
2 Р. fimbriatus f. quadrispinosa Kiefer (Zool. Anz., 79, 1923;24б), без сомнения,
является уродливой самкой этого вида (4 придатка у ног V пары); о f. imminuta
см. ниже.
156
EUCYCLOPINAE
9(6). Внешняя из средних апикальных щетинок фуркальных ветвей
в 2 с лишком раза длиннее внутренней..............................
.................................1с. Р. fimbriatus var. flnltlraus Kief.
10(3). Ряд шипиков на спинной поверхности фуркальных ветвей начи-
нается от внешнего края их заднего отдела и затем загибается
кпереди, охватывая и их передний отдел.................................
...............••...........................* 2. Р. poppei (Rehb.)
* 1. Paracyclops fimbriatus (Fisch.) (фиг. 28, 29).
Cyclops fimbriatus Fischer, 1353:94, t. 3, 1. 19—23, 30.— C. crassicornis Sars, 1353,
Forh. Selsk. Christ., (1352):255; Brady, 1373, Monogr. Brit. Cop., 1:113.— C. margoi Daday,
1832, Orv. Term. Ertes., 7:226.— C. fimbriatus Schmeil, 1392:161, t. 7, £.8 —13.— C.
(Paracyclops) fimbriatus Claus, 1893a:348, t. 5, f. 12—13.
Самка. Тело сплющенное в спинно-брюшном направлении, передний
отдел правильной овальной формы, его наибольшая ширина лежит,
приблизительно, на середине его длины, 5-й торакальный сегмент
по бокам с группами мелких щетинок. Абдомен широкий, кзади мало
суживающийся. Ширина генитального сегмента более его длины; аналь-
ная вырезка с шипиками по бокам. Reeeptaculum seminis широкий,
с поперечно-овальным, посредине спереди не вдавленным передним
и с узким, в виде сегмента, задним отделом. Фуркальные ветви у типич-
ной формы широко расставленные, слабо расходящиеся, их длина
в 5—б раз (у var. imminutas в 2х/2—4, у var. abnobensis менее чем в 4,
у var. jimtimus в 3—Зг/2 раза) превышает их ширину. На спинной
поверхности фуркальных ветвей имеется поперечно расположенный ряд
из нескольких мелких шипиков, частью заходящий и на внешние края
ветвей. Обе крайние апикальные щетинки почти равной длины (у var.
abnobensis внутренняя из них немного короче внешней), из них внешняя
шипообразная толще внутренней, средние апикальные щетинки обычно
несколько изогнутые, гетерономно вооруженные, из них внутренняя
приблизительно вдвое длиннее внешней. Передние антенны 8-член истые,
очень короткие, достигающие, приблизительно, середины цефалото-
ракса. Максиллярные ноги 4-членистые, с короткими шипами на 3-м и 4-м
члениках. Формула шипов плавательных ног 3-4-4-3. Щетинка внутрен-
него угла 2-го базального членика ног I пары очень длинная. У ног IV пары
длина дистального членика эндоподита немного менее, чем вдвое пре-
вышает его ширину, из его апикальных шипов внутренний немного более,
чем вдвое (у var. abnobensis Kief, этот шип особенно длинный), длиннее
внешнего шипа и несколько длиннее самого членика. Ноги V пары
в виде расширенной сзади пластинки с хорошо развитым внутренним
шипом; внешняя щетинка несколько длиннее (у var. abnobensis почти
вдвое длиннее) внутренней.1 Длина 0.8—1.5мм.1 2 3 Обычно прозрачны
и бесцветны, иногда светлорозовой окраски. Яйцевые мешки содержат
по 5—8 яиц (в каждом) и прилегают к абдомену.
Самец. Фуркальные ветви относительно короче, чем у самки,
передние антенны сильно развитые, 1-й их членик с парой крупных
сензорных цилиндров, одна из его щетинок сильно утолщена в основа-
нии, прочие членики несут 8 сензорных придатков (по Gurney, 1933),
12-й членик снабжен хитиновым крючковидным шипом с мелкими
1 По Gurney (1933:124) у британских экземпляров эти щетинки обычно одинаковой
длины; этого мы не нашли у экземпляров из СССР.
2 Длина у 1.5 мм указана Daday (= С. margoi, 1. cit., 1882). Исследованные нами
экземпляры (самки) не превышали 1.2 мм.
5. PARACYCLOPS
/57
зубцами. Ноги VI пары со стройным прямым внутренним шипом
и с 2 щетинками, из которых внешняя длиннее внутренней. Длина
0.7—1 мм.
Науплиусы очень сильно сплющенные, широкие.
Распространение. Космополит в широком смысле слова. В СССР
широко распространен всюду, исключая арктические острова. Неодно-
Фиг. 28. Paracyclops fimbriatus (Fisch.) typ., самка.
7— генитальный сегмент с reeeptaculum seminis; 2—фурка со спин-
ной стороны; 3 — нога V; 4 — антенна II.
кратно указывался для высокогорных водоемов, в западноевропейских
Альпах до высоты около 2700 м над уровнем моря.
Биология. Является типично бентическим рачком, хорошо лазает
по субстрату, обитает главным образом в зарослях макрофитов, предпо-
читая песчаный грунт, в литорали и сублиторали озер, но встречается,
однако, и в глубинной области, известен до глубины 200 м. Судя
по имеющимся данным, на значительных глубинах находит себе вполне
158
EUCYCLOPINAE
нормальные условия существования и едва ли там является только
случайным элементом. Вместе с тем Р. fimbriatus очень обычен в самых
разнообразных мелких стоячих и проточных водоемах и В текучих
водах, причем обитает и на дне фарватера, а не только у берегов.
Встречается также в моховых подушках1 быстро текучих горных ручьев.
Из всех Cyclopidae рассматриваемый вид наиболее реофилен. A. Graeter
(1903:505) указывает на его способность цепко лазать по субстрату
и крепко удерживаться на последнем даже при сильном течении.
Встречается также в ключах и подземных водах, некоторыми авторами
относится к троглофилам. Указан для Адельбергской пещеры, в подзем-
ных водоемах (в частности в пещерах) Франции, Бельгии, Швейцарии,
Испании, Германии, Италии, Румынии, Югославии, Англии; в СССР
найден в колодцах Саратова, в Бакинском водопроводе, в гроте близ
Карадага в Крыму. Амплитуду приспособления к световым условиям
для рассматриваемого рачка надо признать чрезвычайно широкой. Это же
относится и к температурам. Указания некоторых авторов (Thiebaud,
1908) на склонность Р. fimbriatus к теплым водам нельзя считать
верными, так как максимум развития и половой период для него
неоднократно отмечались при низких температурах воды. Он известен
из ключей с постоянной температурой не более 12° и из сильно прогре-
ваемых водоемов зоны полупустыни и пустыни; близ Байкала мы нашли
его в мелкой луже при 31.1°, образуемой горячим ключом (71°). Судя
по обширному распространению в пещерных водоемах, рачок, очевидно,
в значительной степени эвриоксибионтная форма, способная жить при
небольших содержаниях растворенного кислорода; распространение
рачка в реках, таким образом, едва ли объяснимо потребностью в высо-
ком содержании кислорода. По Lowndes’y (1928) Р. fimbriatus обычен
в водоемах при pH = 4.1—8.6. Данные по отношению к активной
реакции пока еще неполны, но, по всем вероятиям, нижняя граница pH
для рачка лежит около 4; для var. finitimus отмечено нахождение даже
при pH =3.7. Обитает в пресной воде, но встречается и в солоноватых
водах,— эврнгалинен в значительной степени. Судя по данным Thiene-
mann’a (1907), может обитать в водоемах с высоким содержанием
железа.
Главным образом дицикличен и полицикличен, в некоторых водоемах,
возможно, моноцикличен. Максимумы н половые периоды нередко
наблюдаются в теплое время года, но также и зимой.
Кажется все авторы согласны с тем, что рассматриваемый вид
является Р-мезосапробом.
* la. Paracyclops fimbriatus (Fisch.) var. immmutus Kief. (фиг.Л9г_3).
Cyclops fimbriatus var. Kiefer, 1926, Schr. Ver. Gesch. u. Naturo-esch. Baar, 16:274,
f- 1—4.— Paracyclops fimbriatus f. imminuta Kiefer, 1929a:49, f. 54—55; Kiefer, 1929:41;
Borutzky, 1930:332, f. 1—2.
От типичного P. fimbriatus отличается (самки) следующими при-
знаками:
1. Более короткими фуркальными ветвями, длина которых не более
чем в 3—4 раза превышает ширину; иногда (пещеры Риона) их длина
всего в 21/2 раза более ширины.
2. Более сближенными фуркальными ветвями.
1 Schneider (1886) предполагал способность Р. fimbriatus к амфибиальной жизни,
что надо признать ошибочным (см. об этом у A. Graeter, 1903:507).
5. PARACYCLOPS
159
Распространение. Северная Германия (водопровод в Oefing-er)
и на о. Рюген мелкие водоемы Силезии (Schafer, 1936). В СССР
найден в Рионской пещере близ Кутаиси.
Биология не изучена. Судя по имеющимся пока данным, обитает
главным образом в связанных с подземными водами водоемах (гелокрены,
пещеры). Весьма возможно, является морфой основного вида.
Фиг. 29.
Paracyclops fimbriatus (Fisch.) var. imminvtus Kief.: 7 — фурка самки, 2— дистальный
членик эндоподита ноги IV самки, 3—нога V самки; Р. /. var. abnobensis (Kief.):
4__ноГа V самки, 5—фурка самки, 6 — дистальный членик эндоподита ноги IV самки;
Р. poppci (Rehb.): 7 — удлиненная фурка самки, 8—укороченная фурка самки.
(1—6—по Kiefer’y, 7—по Marsh’y, 8—по Gurney).
lb. Paracyclops fimdriatus (Fisch.) var. abnobensis Kief. (фиг. 294_6).
Paracyclops abnobensis Kiefer, 1929a:50, f. 57; Kiefer, 1929:11.
От типичного P. fimbriatus отличается следующими признаками:
1. Фуркальные ветви короткие, их длина менее чем в 4 раза пре-
вышает их ширину.
2. Внутренние края фуркальных ветвей несколько выпуклые.
3. Их внутренняя крайняя апикальная щетинка немного короче
внешней крайней апикальной щетинки.
4. Дистальный членик эндоподита ног IV пары с более длинным,
нежели у типичной формы, внутренним апикальным шипом.
5. У ног V пары внешняя щетинка очень длинная, почти вдвое
превышающая длину внутренней.
160
EUCYCLOPINAE
Распространение. Германия (Северный Шварцвальд, Муммель-
ское озеро около 1100 м над уровнем моря). Нахождение в СССР
вероятно.
Биология не изучена.
1с. Paracyclops fimbriatus (Fisch.) var. finitimus Kief.
Paracyclops finitimus Kiefer, 1928. Bull. Soc. Nat. Maroceo, 8:99, f. 24—26; Kiefer,
1929a:51, f. 60—61; Kiefer, 1929:42; Kiefer, 1934, Zool. jahrb, Syst, 65:160.
От типичного P. fimbriatus отличается1 следующими признаками:
1. Фуркальные ветви еще короче, чем у var. abnobensis, их длина
лишь в 3— З1/, раза более их ширины.
2. Внутренняя из средних апикальных щетинок фуркальных ветвей
немного длиннее, чем у типичных экземпляров, вдвое длиннее внешней
средней щетинки.
Распространение. Северная Америка (Мамонтова пещера,
Chappuis, 1931), Южная Америка (Kiefer, 1936а, по Brehm in lift.),
северная и южная Африка, Новая Зеландия, Ява (Kiefer, 1933а).
Нахождение в СССР возможно. Биология не изучена.
2. Paracyclops poppei (Rehb.) (фиг. 297_8).
Cyclops poppei Rehberg', 1880, Abh. Ver. Bremen, 6:550, t. 6, f. 9—11.— C. fimbriatus
Herrick, 1884, Rep. Geol. Suiv. Minnesota, 12:162, t. Q, 3, f. 9—14, t. R, f. 11.— C. fimbriatus
var. poppei Schmeil, 1892:168, t. 7, f. 14—16. — Paracyclops poppei Kiefer, 1929. — Cyclops
(Paracyclops) fimbriatus poppei Gurney, 1933:129, f. 1459.
Самка. От P. fimbriatus отличается строением и вооружением
фуркальных ветвей. Они расположены гораздо ближе друг к другу, чем
у последнего, их длина обычно1 2 лишь в 3—Зт/2 раза более их ширины,
ряд шипиков на их спинной поверхности расположен не поперечно,
а направляется наискось кпереди и затем изгибается, располагаясь
почти параллельно краям ветвей, лишь немного не достигая их осно-
вания.3 У йог V пары внешняя щетинка лишь немного длиннее внутрен-
него шипа. Длина самки 0.75—0.90 мм. Яйцевые мешки содержат
незначительное количество яиц. Длина самца 0.70 мм.
Распространение. Вид, несомненно, неоднократно смешивался
с предыдущим. С достоверностью он указан для Европы (Германия,
Голландия, Англия, Швейцария^ Франция, СССР), Северной Америки,
Южной Америки (Бразилия, Парагвай), южной Африки, Гавайских
островов (Sars, 1901), южного Сахалина (Ueno, 1936). В СССР известен
из окр. Костромы (Смирнов, 1926, 1930); под вопросом указан Совинским
(1888) для окрестностей Киева. Несомненно, у нас должен иметь
широкое распространение, охватывающее и азиатскую часть СССР.
Для высокогорных водоемов не указывался.
Биология изучена крайне неполно. Обитает в литорали озер
и в мелких водоемах. Вид бентический, характерный для зарослей
1 Kiefer (1939) указывает некоторые другие отличительные признаки, которые мы,
ввиду их явной неприемлемое!и, опускаем. Эти признаки свойственны в типичным
Р. fimbriatus; их упоминание цитируемым автором является просто каким-то недора-
зумением.
2 Повидимому столь короткие ветви у Р. poppei ие представляют постоянного
признака; Marsh (1909, t. 81, f. 6) для североамериканских экземпляров изображает
более удлиенные фуркальные ветви.
3 Такое расположение шипиков у Р. poppei является вполне постоянным признаком
позволяющим легко отличить этот вид от Р. fimbriatus.
5. PARACYCLOPS
161
макрофитов. Известен также из текучих вод. Неоднократно был находим
совместно с Р. fimbriatus, но встречается гораздо реже последнего.
Вероятно эвритермичен. По Ueno (1936) на южном Сахалине обитает
в мелких тундровых водоемах, при pH = 4.5; известен также из нейтраль-
ных слабощелочных вод.
Цикл развития не изучен. По Farwick (1917) встречается весь год,
причем самки с яйцевыми мешками найдены в январе.
* 3. Paracyclops affinis (Sars) (фиг. 30).
С. pygmaeus Rehberg, 1380, Abh. Ver. Bremen, 6; 546, t;6,'f. 3 — 6. — C. affinis
Schmeil, 1892:157, t. VII. f.l.— Paracyclops cffinis Claus, 1893:83. — C. affinis Lilljeborg,
1901:98, t. 6, f. 15—16.— Platycyclops affinis Sars. 1914/15:80, t. 49. — Paracyclops affin s
Kiefer, 1929:42. — Platycyclops affinis Lowndes, 1932, Ann. Mag. Nat. Hist., X:395, f. 1—9,
t. 16, f. 1—3.— Cyclops (Paracyclops) affinis Gurney, 1933:130—136, f. 1460—1478.
Самка. Передний отдел тела заметно сплющенный в спинно-брюш-
ном направлении, его наибольшая ширина лежит несколько кзади от его
середины, задний торакальный сегмент по бокам с группами мелких
волосков. Генитальный сегмент очень широкий, его ширина несколько
более его длины. Абдомен укороченный, толстый, очень мало суживаю-
щийся кзади, анальная вырезка с группами мелких волосков. Receptaculum
seminis широкий, показанной на рисунке формы, его передний отдел
немного уже заднего. Фуркальные ветви почти параллельные, короткие,
их длина в 2—2г/2 раза более их ширины, на спинной поверхности их
задней половины имеется несколько расположенных косым рядом мелких
шипиков, латеральная щетинка очень мелкая, прикрепленная не на крае, .
а вблизи его, на спинной поверхности ветвей, внутренняя крайняя
апикальная щетинка немного (примерно на 1/3) короче внешней крайней,
средние апикальные щетинки с гетерономным вооружением, из них
внутренняя приблизительно вдвое длиннее внешней. Передние антенны
очень короткие, не достигающие заднего края цефалоторакса,
11-членистые. У задних антенн оба дистальных членика почти равной
длины. Формула шипов плавательных ногЗ-4-З-З, илиЗ-З-З-З. У плаватель-
ных ног I пары очень длинные щетинки на задних углах обоих базальных
члеников. Соединительная пластинка IV пары ног сзади выпуклая, с рядом
тонких волосков вдоль свободного края, длина дистального членика
эндоподита почти вдвое более его ширины, его внутренний шип почти
вдвое длиннее внешнего. Ноги V пары 1-членистые, в виде расширяю-
щейся кзади пластинки, вооруженной 3 придатками, из которых внутрен-
ний является длинным шипом, а средний — короткой и тонкой щетинкой.
Длина 0.6—0.9 мм. Преобладает светлая красно-коричневая окраска,
фуркальные ветви и конечности обычно с голубоватым оттенком. Яйцевые
мешки тесно прилегают к абдомену и содержат 4—8 очень крупных яиц.
Самец. Передние антенны короткие и толстые, загнутые
ко внутренней стороне, в проксимальном отделе с 6 длинными сензор-
ными цилиндрами. Ноги VI пары с длинным внутренним шипом и с 2 щетин-
ками, из которых внешняя короткая, а внутренняя лишь немного короче
шипа. Длина около 0.6—0.7 мм.
Распространение. Вся Европа, средняя и южная Азия, Канада,
западная, восточная и северная Африка, Мадагаскар, Новый Южный
Уэльс. В СССР широко распространен, от крайнего севера (Соловецкие
острова) до Одессы и Ташкента включительно; нахождение на Кавказе
и в Крыму вероятно; известен с Дальнего Востока. В горных областях
11 Фауна СССР
162
EUCYCLOPINAE
отмечается вообще редко, наивысшая точка нахождения —1600 м ^над
уровнем моря.
Биология. Типичная бентическая форма. Обитает в зоне макро-
фитов в литорали озер и мелких стоячих водоемах разнообразного
типа. В озерах, повидимому, не спускается глубже литорали (сублито-
рали?). В медленно текучих водах указывался редко, при этом отме-
чался в условиях прибрежной зоны (заросли макрофитов). Троглоксен,
Фиг. 30. Paracyclops affinis (Sars).
1— фурка самки; 2—генитальный сегмент и reeeptaculum
seminis; 3—нога V самки; 4 — антенна I самки; 5—антенна!
самца. (5—по Schmeil’ro).
имеется указание на нахождение (1 5) в глубоком (20 м) колодце близ
Красноводска (van Dauwe, 1905). Эвритермичен, вероятно приурочен,
главным образом, к повышенным температурам. Встречается в водах
с pH = 4.7— 9.2. Отсутствует в солоноватых водах; указание Daday
(1893) на нахождение в натронных водоемах Alfoldes (Венгрия) сомни-
тельно. Вероятно, главным образом, Р-мезосапроб, отчасти олигосапроб.
Цикл размножения изучен недостаточно. Вероятно в некоторых
водоемах дицикличен. Е. Graeter, Wolf и некоторые другие авторы
отмечают внезапное появление этого рачка, после чего следует очень
краткий половой период и выпадение на несколько месяцев.
6. ECTOCYCLOPS
163
6. Род ECTOCYCLOPS BRADV
Cyclops (part.) Koch, 1838, Deutschl. Crust., 21:9; Fischer, 1851:426; Claus, 1863:96;
Schmeil, 1892:170. — Paracyclops (part., subgen) Claus, 1893:83. — Cyclops (part.).
Lilljeborg,. 1901:3.— Ectocyclops Brady, 1904, Proc. Zool. Soc. London, 2:124. — Cyclops
(part.) Marsh, 1909:1070. — Platycyclops (part.) Sars, 1914:76.—Ectocyclops Kiefer,
1927:304; Kiefer, 1928:543; Kiefer, 1929:43. — Ectocyclops (subgenus) Gurney,
1933.137•
Тело, сильно сплющенное в спинно-брюшном направлении, перед-
ний и задний отделы дифференцированы еще менее, чем у рода Рата-
cyclops, так что рачки имеют близкий к гарпактикоидному габитус.
Абдомен очень широкий, грубый. Reeeptaculum seminis широкий, весьма
похожий на таковой рода Paracyclops. Фуркальные ветви очень корот-
кие, их длина самое большое в 2т/2—23/4 раза превосходит их ширину,1
обычно с несколькими рядами шипиков, косо заходящих на спинную
и внутреннюю поверхность ветвей. Передние антенны очень короткие,
6—11-членистые.1 2 3 Внешние края экзопрдитов плавательных ног с рядами
сильных шипиков. Ноги V пары в виде короткой и широкой пластинки,
слитой с задним торакальным сегментом и вооруженной 3 придатками.
К роду принадлежат 5 точно установленных видов: Е. phaleratus
(Koch), Е. compactus (Sars), Е. hirsutus (Kief.), E. coperes (Gurney),
E. bromeltcola (Kief), (j не известна). Описанный Brady (1904:124)
E. rubescens Kiefer’oM сначала считался синонимом E. phaleratus (Koch),
но позже им выделен в отдельный вид. Мы, однако, разделяем мнение
Gurney, по которому Е. rubescens является лишь подвидом Е. phale-
ratus, к которому морфологически весьма близок./;, medias3 Kief, отли-
чается 6т Е. phaleratus лишь немного более длинной внешней щетинкой
ног V пары и 11-членистыми передними антеннами, что, однако, нередко
наблюдается у Е. phaleratus; вместе с Gurney мы относим Е. medias
Kief, в синонимы этого вида. Видовая самостоятельность Е. polyacanthus
Harada сомнительна.
Род Космополитен. Е. compactus (Sars) известен лишь из Централь-
ной и Западной Африки, Е. coperes (Gurney) — только из Центральной
Африки (из Танганайки), Е. hirsutus Kief. — с о. Мадагаскара, Е. brome-
licola Kief. — из Бразилии. Е. phaleratus (Koch) широко распространен;
его подвид rubescens пока известен из Парагвая и Африки (Южная
Африка, Абиссиния). Kiefer (1935) высказывает предположение, нуждаю-
щееся, однако, в проверке, что в южных широтах обитает не типичный
Е. phaleratus, а только этот подвид, отсутствующий в Европе.
К роду относятся типично бентические, хорошо приспособленные
к лазанию по субстрату циклопы. Обитают в литорали озер и в мелких
водоемах различного типа (пруды, болота, лужи и пр.). Е. bromelicola
найден исключительно в скоплениях воды в листьях Bromeliaceae. Глав-
ным образом тепловодные формы (Е. phaleratus эвритермичен, но со склон-
ностью к теплым водам), троглоксены. В СССР обитает 1 вид.
* 1. Ectocyclops phaleratus (Koch) (фиг. 31).
Cyclops phaleratus Koch, 1838, Deutschl. Crust., Myr., Arachn., 21, № 9; Schmeil.
1892: 170, t. 8, f. 1—11,—C. (Paracyclops) phaleratus Claus, 1893a, 10:348, t. 5, f. 14.—
C. phaleratus japonicus (o juv.) Kokubo, 1912, Ann. Zool. Japon., 8:103, t. 2, f. 27—30.—
Platycyclops phaleratus Sars, 1914:73, t. 43. — Ectocyclops phaleratus Kiefer, 1929:43,
f. 15. — C. (Ectocyclops) phaleratus Gurney, 1933 :137, f. 1479—1501.
1 У E. bromelicola Kiefer (Zool. Anz., 112, |1935 :23) их длина не более ширины.
2 У Е. coperes Gurney они 6-членистые.
3 Западная и южная Африка, Малайский архипелаг.
11
164
EUCYCLOPINAE
Самка. Тело толстое, передний отдел значительно сплющенный
в дорзо-вентральном направлении, наибольшая ширина лежит на средине
его длины, задний торакальный сегмент сзади на брюшной стороне
вооружен поперечным рядом (прерванным посредине) шипиков. Гени-
тальный сегмент очень широкий, его ширина несколько более его длины.
Receptaculum seminis сильно расширенный. Относительно переднего
отдела тела абдомен очень широк, слабо суживающийся кзади, анальная
вырезка с группой мелких щетинок. Фуркальные ветви толстые и корот-
кие, заметно суживающиеся кзади, их наибольшая ширина приблизительно
равна половине их длины, на спинной поверхности ветви вооружены
несколькими (иногда лишь 3) поперечными рядами шипиков, заходящих
отчасти и на брюшную поверхность ветвей. Латеральная щетинка
маленькая, близ нее прикрепляется несколько мелких шипиков, на брюш-
ной стороне ветвей, близ их заднего конца, также имеется группа
из 3—5 мелких шипиков. Крайние апикальные щетинки фуркальных
ветвей короткие, из них внутренняя (более тонкая) обычно лишь немного
длиннее шипообразной внешней, но может быть и короче последней,
или обе щетинки равной длины. Средние апикальные щетинки с гете-
рономным вооружением, из них внутренняя заметно толще внешней
и приблизительно вдвое длиннее ее. Передние антенны короткие,
не достигающие заднего конца цефалоторакса, 10-членистые, иногда,;
вследствие слияния 7-го и 8-го члеников, 9-членистые, а иногда, вслед-
ствие разделения б-го членика, — 11-членистые. Задние антенны очень
короткие, утолщенные. Максиллы очень укороченные, максиллярные
ноги с 1-членистым сильно редуцированным эндоподитом. Плавательные
ноги с очень широкими базиподитами. Формула шипов 3-4-4-3. Соеди-
нительная пластинка IV пары ног без шипиков, гладкая, на свободном
крае вооруженная лишь 2 изогнутыми щетинками (по 1 с каждой стороны).
У ног IV пары дистальный членик эндоподита с внутренним шипом,
который приблизительно вдвое длиннее внешнего. Ноги V пары состоят
из широкой пластинки, в основании совершенно слитой с торакальным
сегментом и на свободном крае вооруженной 3 придатками — парой шипов
и щетинкой. Длина 0.9—1.2 мм. Обычно самки желтого или красно-корич-
невого цвета, концы передних антенн, плавательных ног и конец
торакса и абдомена часто голубоватой (синей) окраски. Яйцевые мешки
тесно прилегают к абдомену и содержат небольшое (5—20) количе-
ство яиц.
Самец. 17-членистые передние антенны, сильно загнутые ковнутри
(клещневидные) и богато вооруженные цилиндрическими сензорными
придатками. Ноги VI пары сильно развитые, напоминающие ноги V пары
обоих полов, вооруженные очень длинным мощным внутренним шипом
и 2 внешними короткими и тонкими щетинками почти одинаковой длины.
Длина равна 0.7—0.1 мм.
Распространение. Космополит.1 В СССР широко распро-
странен, но не найден на крайнем севере и на арктических островах.
Известен с Дальнего Востока (бассейн Амура). Известен из горных
водоемов до высоты 2400 м над уровнем моря.
Биология. Типично бентический вид, живет на субстрате,
по которому ползает, напоминая этим Harpacticoida, с которыми имеет
сходство по ряду признаков (сплюснутое тело, очень толстый абдомен,
короткие, своеобразно вооруженные, фуркальные ветви, далеко захо-
1 Принимая во внимание и Е. phaleratus rubescens (Brady).
6. ECTOCYCLOPS
165
дящие в абдомен яичники и некоторые другие признаки). Не относится,
однако, к обитателям специально иловых отложений и некоторыми авто-
рами даже считается обитателем песчаных грунтов. Обитает в литорали
(сублиторали) озер, а главным образом в разнообразных стоячих мелких
водоемах, повидимому, преимущественно в постоянных; в состав фауны
астатических водоёмов нормально не входит. Редко в ключах. В озерах
показан до глубины 40 м.1 Обычен в зарослях макрофитов, повидимому
Фиг. 31. Ectocyclops phaleratus (Koch), самка.
1 — генитальный сегмент с receptaculum seminis; 2—фурка с брюш-
ной стороны; 3—антенна I; 4—-нога V; 5 — фуркальная ветвь
со спинной стороны и несколько сбоку.
предпочитая дно, покрытое свежими растительными остатками. Обитает
при pH = 5.8—8.6, нередко он указывался для болотистых водоемов
(в частности с сфагновыми мхами). В сильно дистрофированных водоемах
(при pH = 4.5 и ниже), однако, отсутствует. По Kolkwitz’y является мезо-
сапробом, но, вероятно, может обитать и в олигосапробных условиях.
Троглоксен. Эвритермичен, но с определенной склонностью к повышен-
ным температурам. Дицикличен, в некоторых водоемах, возможно, имеет
3 половых периода в год; периоды максимального развитии падают
на теплое время года.
1 Thiebaud (1908) склонен считать Е. phaleratus даже глубоководным видом, что
безусловно ошибочно.
lf>6
CYCLOPINAE
2. Подсем. CYCLOPINAE
Cyclopinae (part,) Dana, 1852, Un. Rt. Esplor. Exped., 13 : 1177. — Cyclopinae Kiefer,
1927 : 304; Kiefer, 1929 : 45. — Cyclops section Bifida Gurney, 1933 : 36.
Задний торакальный сегмент, за редкими исключениями, по бокам
без группы щетинок. Фуркальные ветви очень различного строения
и различной длины, иногда сильно измененные, укороченные, их внеш-
ние края всегда без пилообразного ряда шипиков, плавательные ноги
3-членистые или 2-членистые, редко одночленистые. Ноги V пары глав-
ным образом 1—2-членистые, у одного рода совсем лишенные членика,
совершенно слитые с задним торакальным сегментом, причем свойствен-
ные ноге придатки прикрепляются непосредственно к этому сегменту.
Вооружение дистального членика состоит из 2 придатков, из коих
внутренний шип иногда сильно редуцируется, до полного его исчезно-
вения. При полном слиянии базального членика V пары ног с торакаль-
ным сегментом его щетинка прикрепляется на этом сегменте, очень
редко исчезает бесследно.
К подсемейству принадлежат 9 родов. Род Orthocyclops Forbes
заключает один вид — О. modestus (Herrick), известный только из Север-
ной Америки. Роды Acanthocyclops, Microcyclops и Mesocyclops имеют
обширное распростанение в Голарктике (особенно в Палеарктике).
В подсемействе известно около 180 видов. Пресноводные рачки обитают,
главным образом, в литорали озер и в разнообразных мелких водоемах,
ряд видов планктических, частью характерных для пелагиали озер
(роды Cyclops, Mesocyclops). Некоторые роды свойственны специальным
биотопам: влажные, частью наземные мхи, скопления воды в листьях
наземной растительности (Bromeliaceae); таковы роды Mascocyclops
и Bryocyclops. Только пещерным водам свойственны роды Graeteriella
и Speocyclops (исключая 1 вид), к троглобионтам принадлежит также
ряд видов рода Acanthocyclops.
В пределах СССР известны виды родов Cyclops, Acanthocyclops,
Microcyclops, Mesocyclops и Speocyclops.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Ноги V пары 3-членистые...................(Orthocyclops Forbes)
2 (1). Ноги V пары состоят менее чем из 3 члеников.
3 (10). Ноги V пары 2-членистые.1
4 (5). Членики этих ног расположены не один на другом, а рядом,1 2 3
у всех 4 пар плавательных ног обе ветви 2-членистые..................
...........................................10. Speocyclops Kief.
5(4). Членики этих ног расположены один на другом. Обе ветви всех
4 пар ног 3-членистые, редко (Acanthocyclops) число члеников
ног I—II пар редуцируется до 2, тогда как ноги III—IV пар
остаются 3-членистыми.
6 (7). На спинной поверхности фуркальных ветвей имеется продольная
складка; внутренний придаток дистального членика ног V пары
в виде сильного шипа, прикрепленного всегда строго на середине
1 Только у A. nanus (var. incertas) иногда замечается полное слияние базального
членика с торакальным сегментом.
2 У 5. demetiensis (Scourf.) оба членика сливаются в 1 пластинку, по которой,
однако, можно заключить о слиянии ее из 2 члеников; у самок других видов рода оба
членика различимы, иногда весьма резко дифференцированы.
7. CYCLOPINAE
167
края членика, в основании (снаружи) его апикальной щетинки
всегда имеется группа мелких шипиков . . 1. Cyclops О. F. Mull.
7 (6). На спинной поверхности фуркальных ветвей нет продольной
складки; внутренний придаток дистального членика ног V пары
расположен апикально или субапикально, редко несколько при-
ближаясь к середине внутреннего края;1 близ основания (снаружи)
апикальной щетинки этого членика нет группы шипиков.
8(9). 2 дистальных членика всегда 17-членистых передних антенн вдоль
внутреннего края с узкой гиалиновой пластинкой; внутренний при-
даток дистального членика ног V пары представлен длинным
щетинкообразным шипом, длина которого обычно лишь немного
менее таковой апикальной щетинки; receptaculum seminis всегда
молотообразный...............................12. Mesocyclops Claus
.9(8). 2 дистальных членика 10—18-членистых передних антенн без гиа-
линовой пластинки; внутренний придаток дистального членика ног
V пары в виде настоящего короткого шипа, длина которого,
за очень редкими исключениями, менее половины длины апикаль-
ной щетинки; receptaculum seminis не молотообразный ...............
.......................................8. Acanthocyclops Kief.
.10(3). Ноги V пары 1-членистые или вовсе лишены членика, причем
тогда свойственное им вооружение прикрепляется к заднему тора-
кальному сегменту.
11(12). Ноги V пары без членика, все 3 их придатка прикрепляются
непосредственно к заднему торакальному сегменту. У ног IV пары
1-й базальный членик, за одним исключением,1 2 внутри без щетинки
..................................................(Bryocyclops Kief.)
12(11). Ноги V пары состоят из 1 членика, вооруженного 2 при-
датками. 1-й базальный членик ног IV пары внутри снабжен
щетинкой.
13(14). Щетинка, соответствующая базальному членику ног V пары,
отсутствует, так что задний торакальный сегмент ее не имеет
.............................................11. Graeteriella Brehm.
14(13). Задний торакальный сегмент снабжен щетинкой, соответствующей
исчезнувшему базальному членику ног V пары.
15(16). Генитальный сегмент очень широкий, его ширина приблизительно
в 11/2 раза более его длины, членик ног V пары несет 2 почти
одинаковой (или одинаковой) длины придатка, фуркальные ветви
сильно измененные, очень широкие и короткие .......................
....................................... (Muscocyclops Kief.)
16(15). Генитальный сегмент нормальный, удлиненный, его ширина
не превышает длины, оба придатка, за крайне редкими исключе-
ниями, резко различной длины, причем внутренний из них гораздо
менее внешнего и иногда совсем редуцирован. Фуркальные ветви
удлиненные, нормального строения..............9. Microcyclops Claus
7. Род CYCLOPS О. F. MULL (s. str.)
Cyclops (part.) Muller, 1776, Zool. Dan. Prodr.: 201. — Cyclops strenuus — insignis
' Gruppe Schmeil, 1892 : 39.— Cyclops s. (str. Kiefer, 1927:305; Kiefer, 1928:545; Kiefer,
1929 : 49. — Cyclops s. str. Gurney, 1933:147. — Cyclops Kiefer, 1936, Zool. Anz., 113:5;
Kiefer, 1937 : 442, 443.
1 У Mesocyclops s. str., иногда у Acanthocyclops viridis, A. gigas.
2 У Br. kieferi mihi (= Allocyclops chappuisi Kiefer, 1933).
168
CYCLOPINAE
На спинной поверхности фуркальных ветвей имеется продольная
кутикулярная складка, их внутренние края с рядом волосков. Передние
антенны 14—17- (редко 18-)членистые, 3 дистальных членика вдоль вну-
треннего края снабжены в более или менее резкой степени выраженным
рядом очень тонких, тесно расположенных шипиков. Формула члени-
стости плавательных ног З-З/З-З/З-З/З-З. Дистальный членик экзоподи-
тов всех 4 пар ног вооружен 5 щетинками. Вооружение шипами
по формуле Terni, Bini или по смешанному типу. Ноги V пары 2-чле-
нистые, базальный членик всегда явно отчленен от заднего торакального
сегмента, шире дистального членика; с хорошо развитой щетинкой
на внешне-заднем углу, дистальный членик удлиненный, не прямоуголь-
ный, угловатых очертаний, его внутренний, обычно крупный, заострен-
ный шип прикреплен строго на средине внутреннего края (а не субапи-
кально); на конце членик вооружен довольно длинной оперенной щетин-
кой, близ основания которой, в конечном отделе внешнего края, имеется
группа из нескольких хорошо различимых коротких шипиков. Recepta-
culum seminis, за исключением 1 вида (С. insignis Claus), очень простого
строения, округлый, без выростов. Крупные циклопы, длина самок
до 3.5 мм., главным образом более 1.5 мм.
К роду Cyclops s. str. относится около 20 форм, из которых хорошо
очерченными видами являются С. tatricus Койт., С. strenuus Fisch.,
С. kolensis Lili., С. furcifer Claus, C. scutifer Sars, C. vicinus Uljan.,
C. insignis Claus. Вследствие чрезвычайно широкой изменчивости ряд
форм рассматриваемого рода, описанных в качестве самостоятельных
видов (например С. lacustrts Sars, С. abyssorum Sars), до последнего
времени занимал спорное таксономическое положение. За последние
10 лет группой „strenuus" особенно много занимались Rzoska (1925,
1927, 1930, 1931, 1932) и Kozminski (1927, 1932, 1933, 1934, 1936), при-
менившие биометрический метод исследования на обширном материале..
Работами этих авторов систематика и таксономическое положение спор-
ных „видов“ группы „strenuus" в значительной степени разъясняется.
К сожалению, до сих пор не произведено подобных исследований над
материалом из всей Палеарктики, что, конечно, является задачей чрез-
вычайно обширной, потребующей труда целого ряда специалистов.
Нужно заметить, что ряд форм группы, описанных как самостоятельные
виды, описан лишь по одному — нескольким экземплярам, без приме-
нения вариационно-статистического метода, в лучшем случае лишь
с несколькими измерениями некоторых немногих признаков. Таксономи-
ческое положение таких форм не может считаться окончательно выяснен-
ным до детального их исследования на достаточно обширном материале,
и ниже мы лишь провизорно сохраняем за ними видовое значение.
Род является, главным образом, палеарктическим. Только в преде-
лах Палеарктики обитают С. abyssorum Sars, С. bohater Kozm., С. lacu-
stris Sars, C. scutifer var. wigrensis Kozm., C. furcifer Claus (известен
и из палеарктической части Африки), С. tatricus Kozm., С. ochridanus
Kief., С. insignis Claus; пока только для Палеарктики указаны С. smir-
novi sp. n., C. vicinus var. klkuchii Smirnov, C. strenuus var. landei
Kozm., C. s renuus var. vranae Kozm. и спорный C. strenuus var. diver-
gens Lindberg- (1936). Кроме того в пределах Палеарктики обширно
распространен С. strenuus Fisch, (typ.), присутствие которого в Север-
ной Америке пока с достоверностью не установлено. С. kolensis Lili.,
помимо севера Палеарктики, возможно, обитает и на севере Неарктиче-
ской области (С. vicinus? Marsh 1920, Аляска), С. vicinus Uljan с досто-
7. CYCLOPS
169
верностью показан также только для Палеарктической области, но, воз-
можно, будет найден в Ориентальной (Индийская подобласть). Присут-
ствие в Северной Америке широко распространенного на севере
Палеарктике С. scutifer Sars можно считать достоверным. Пока только
в пределах Ориентальной области (в Индийской подобласти — Ладак,
Кашмир, на высоте 1600—4000 м над уровнем моря) найдены С. lada-
kanus Kief, и С. hutchinsoni Kief., описанные очень неполно. В прочих
зоогеографических областях род Cyclops s. str. отсутствует.
Фиг. 32. (По Kozminsk’oMy).
1 — основные индексы измерений Cyclops', 2, 3— то же,
для заднего отдела торакса и генитального сегмента.
Объяснение см. в тексте.
Род заключает планктические виды, частью характерные для пела-
гиали озер. Троглоксены. Некоторые виды (С. vicinus), помимо пресных
вод, обитают в водах повышенной минерализации, в частности богатых
NaCl. В сильно дистрофированных водоемах (pH = 3.2— 4) род отсут-.
ствует. Вероятно, главным образом, эвритермические формы, С. insignis
холодновоцно-стенотермичен. Предлагаемая определительная таблица
составлена с небольшими изменениям ии дополнениями, по Козминскому.
Упомянутый выше С. strenuus var. divergens, описанный Lindberg-’ом:
(1936) из Ирана, ввиду неполноты его диагноза, нами в эту таблицу
не вводится.
Для точного определения форм р. Cyclops необходимо изложение
целого ряда признаков.
.170
CYCLOPINAE
Согласно Козминскому (Ko:zminski, 1936), разработавшему метод
измерений для группы strenuus, применимый и для прочих циклопов,
измерения отдельных элементов тела Cyclopidae, производятся
следующим образом и имеют следующие обозначения (фиг. 32, 33):.
1. Общая длина цефалоторакса (= длина головы -+- длина слитого
с ней 1-го торакального сегмента) и 4 остальных (2-ьЗ-ь4-ь5) торакаль-
ных сегментов. Long-, cphth. -+- thor, 2—5. Сумма ab -+- cdef -+- gh
(фиг. 33).
Фиг. 33. (По Kozmiask’oMy).
7 — основые индексы измерений абдомена Cyclops-,
2 — то же, V ноги. Объяснения см. в тексте.
12. Максимальная ширина цефалоторакса: Lat. max. cphth. (фиг. 32, if).
.За очень редкими исключениями эта величина не совпадает с макси-
мальной шириной переднего отдела тела, которая в таких случаях
.лежит несколько позади цефалоторакса.
3. Ширина предпоследнего сегмента торакса: lat. IV. s. thor.
(фиг. 32, kt).
4. Ширина последнего (заднего) сегмента торакса: Lat. V. s. thor.
(фиг. 32, kl).
; -j 5. Максимальная ширина переднего абдоминального (генитального)
•сегмента: Lat. max. gensgt. (фиг. 32, op).
6. Общая длина абдомена, не считая фурки: Long. abd. (фиг. 33, ab -+-
~ь cd -+- ef-i- gh).
7. Длина генитального сегмента: Long, gensgt. (фиг. 33, ab).
7. CYCLOPS
171
8. Общая длина 3 задних абдоминальных сегментов: Long. tr. ult.
ss. abd. (фиг. 33, cd-i- ef gh).
. 9. Длина тела: Long. corp. Представляет сумму Long, cphth. -+- thor.
2—5~bLong. abd.
10. Длина фуркальных ветвей: Long. furc. (фиг. 33, z'Z).
11. Ширина фуркальных ветвей: Lat. furc. (фиг. 32, iu).
12. Расстояние между местом прикрепления мелких щетинок в перед-
нем отделе внешнего края фуркальных ветвей и основанием последних:
.sp. furc. I (фиг. 33, ij).
13. Расстояние между местом прикрепления латеральной щетинки
фуркальных ветвей и основанием последних: sp. furc. II (фиг. 33, ik).
14. Длина фуркальных щетинок (фиг. 33), а именно:
Длина внешней крайней апикальной щетинки (Long. set. apic. ext.).
Длина внутренней крайней апикальной щетинки (Long. set. apic. int.).
Длина внешней из средних апикальных щетинок (Long. set. apic.
med. ext.).
Длина внутренней из средних апикальных щетинок (Long. set. apic.
med. int.).
Длина дорзальной щетинки (Long. set. dors.).
15. Степень расхождения фуркальных ветвёй (sp. interfurc., фиг. 32, rs),
т. е. расстояние между окончаниями их внутренних краев. Передние
антенны измеряются по кривой (т. е. их длина — сумме длины каждого
отдельного членика и выражена в °/00 длины тела). Обозначение длины
I антенны: long. ant. I par.
У дистального членика Епр. ног IV пары измеряется длина апикаль-
ных шипов: Long. sp. ext. end. IV par. и Long. sp. int. end. IV par.
У ног V пары измеряется длина шипа дистального членика (Long, spi-
nae ped. V par., фиг. 33, b) и длина щетинки этого членика и преды-
.дущего. Щетинки измеряются по прямой линии, не считаясь с их кривиз-
ной, которая у придатков V пары ног лишь незначительна. Обозначения
длины щетинок: Long. , set. maj. ped. V par. и long. set. min.
_ped. V par.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1(2). Reeeptaculum seminis не округлый, очень широкий, с очень узким
задним отделом, передние антенны 14-членистые...................
........................................... 12. С. insignis Claus
2 (1). Reeeptaculum seminis округлый, неширокий, передние антенны 17-
(редко 16- или 18-) членистые.
-3(6). Предпоследний торакальный сегмент сзади гораздо шире, по бокам
с крупными, направленными кзади заостренными лопастями; задний
торакальный сегмент с боков вытянут в длинные заостренные
лопастевидные выросты.
-4 (5). Задний торакальный сегмент лишь немного уже, чем предыдущий.
Генитальный сегмент равномерно и значительно суживающийся
кзади. Вооружение плавательных ног по типу Terni. Sp. furc. в °/0
long. furc. 62 (59—65)1 ......................*10. С. scutifer Sars
1 Цифра перед скобкой есть среднее арифметическое величины данного признака.
Цифры в скобках показывают границы вариаций данного признака, в которых нахо«
.дится теоретически 95.5% измеренных экземпляров.
172
CYCLOPINAE
5(4). Задний торкальный сегмент значительно уже предыдущего. Гени-
тальный сегмент кзади мало суживающийся. Вооружение плаватель-
ных ног по типу Bini. Sp. furc. в °/0 long. furc. 76 (73—79) . . .
.........................................*11. С. vicinus Uljan.
6 (3). Предпоследний торакальный сегмент сзади уже или лишь немного
шире, чем спереди, его задние углы в виде коротких отростков
не лопастные, иногда не отличающиеся от таковых предыдущего
сегмента: задний сегмент с боков без лопастных выростов, лишь
с короткими тупыми или заостренными отростками.
7 (12). 2-й торакальный сегмент с очень сильно развитыми задними углами,
значительно выступающими кнаружи, иногда загибающимися
на брюшную сторону сегмента. Вооружение плавательных ног
по типу Terni.
8 (9). Цефалоторакс удлиненный, не расширенный в среднем отделе,
равномерно расширяющийся кзади..................5. С. tatricus Kozm.
9 (8). Цефалоторакс очень короткий, его длина лишь немного превышает
его наибольшую ширину, лежащую, приблизительно, на средине
его длины или лишь немного кзади от середины, так что сзади,
сегмент несколько суживается.
10 (11). Фуркальные ветви заметно расходящиеся, снабженные волосками
вдоль всего их внутреннего края, а не только в задней его поло-
вине. Long. set. min. ped. V par. в % long. set. maj. ped. V par-
81 (72—90)..................................... 3. C. bohater Kofm.
11(10). Фуркальные ветви параллельные, волоски имеются только
в задней х/3 длины их внутреннего края. Long. set. min. ped. V par.
в °/o I011?, set. maj. ped. V par. 65.........................
..................................*6. C. smirnovi, sp. nov.
12 (7). 2-й торакальный сегмент co слабо развитыми задними углами,
не выдающимися или едва выдающимися кзади, похожими на углы
3-го торакального сегмента. Вооружение плавательных ног по типу
Terni или Bini или по смешанному типу.„
13 (18). Задние углы 4-го (предпоследнего) торакального сегмента,,
слабо выступающие кзади, одинакового строения с задними углами,
предыдущего сегмента. Вооружение плавательных ног по типу
Terni.
14(17). Тело стройное, удлиненное, задний торакальный сегмент лишь
немного шире основного отдела значительного удлиненного гени-
тального сегмента, внутренний шип дистального членика ног V пары
короткий, слабый.
15(16). Фуркальные ветви короткие, их длина не более чем в 5—6.2 раза
превышает их ширину............................*4. С. iacustris Sars
16(15). Фуркальные ветви очень длинные, их длина приблизительно
в 9 раз более их ширины.......................7. С. ochridanus Kief.
17 (14). Тело укороченное, довольно толстое, задний торкальный сегмент
значительно шире основного отдела сильно укороченного гениталь-
ного сегмента, внутренний шип дистального членика ног V пары
крупный, сильный................*1в. С. strenuus var. vranae Kozm.
18(13). Задние углы 4-го торакального сегмента с затупленными, заметна
выдающимися кзади и иногда загибающимися кпереди оттянутыми
углами, сильно отличающимися от задних углов предыдущего»
сегмента.
19 (20). Из крайних апикальных щетинок фуркальных ветвей внутренняя
очень длинная; фуркальные ветви стройные, удлиненные. Long. set.
7. С7 CLOPS
173
apic. int. в °/00 Ions', corp. 195 (154—214), long. abd. в % long,
cphth-i-thor (2—5) 36.5 (34—42), редко более 33 . . . . ~ .
................................................*2. C. abyssorum Sars
20(19). Внутренняя крайная апикальная щетинка фуркальных ветвей
короче. Long. set. apic. int в %0 long. corp, в среднем не более 138
(максимум около 160), long. abd. в °/0 long, cphth. ч- thor. (2—5)
в среднем не менее 42 (38—50).
21 (22). Фуркальные ветви очень узкие и длинные [lat. furc. в % long,
furc. в среднем около 11 (9.3—13)], генитальный сегмент не рав-
номерно, а сразу суживается кзади, так что его задний отдел
цилиндрический, суженый. Вооружение плавательных ног по типу
Bini, Terni или по смешанному типу. ... *9. С. furcifer Claus
22(21). Фуркальные ветви короче и шире [lat. furc. в °/0 long. furc.
не менее 15 (12.6—25)], генитальный сегмент равномерно суживаю-
щийся кзади.
23(24). Вооружение плавательных ног по типу Bini.......................
............................................*8. С. kolensis Lili.
24(23). Вооружение плавательных ног по типу Terni.
25 (26). Передний отдел тела сильно суживающийся кзади, задний тора-
кальный сегмент лишь незначительно шире переднего отдела гени-
тального сегмента Lat. thor4 в °/оо I011? corp. 199 (177—221), lat.
thor3 в %» I011?- copr. 166 (152—181), long. tot. 1.514 мм (1.346—
1.681)................................la. C. strenuus var. iandei Kozm,
26 (25). Передний отдел тела слабо суживиющийся кзади, задний тора-
кальный сегмент значительно шире переднего отдела генитального
сегмента. Lat. thor. в °/00 long. corp, около 238 (213—264, иногда
до 282), lat. thor. в °/00 long. corp. 187 (158—216), иногда немного
более, long. tot. 1.890 мм (1.458—2.322). ............................
.....................................*1. С. strenuus Fisch, (typ)
*1. Cyclops strenuus Fisch, (фиг. 34).
Cyclops strenuus Fischer, 1851 :419, t. 9, f. 12—21. — C. strenuus (part.) Schmeil,
1892:39; Lilljeborg’, 1901:28.— C. strenuus Sars, 1913:32, t. 16; I Rzoska, 1925:35, t. 2,
f. 3—4. — C. strenuus f. strenuus Kozminski, 1927 : 70, 11—13. — C. strenuus var. abyssorum.
(part.) Рылов, 1928:11. — C., (s. str.) strenuus Kiefer, 1929 : 49; Gurney, 1933 : 153, f. 1502—
1525. — C. strenuus subsp. strenuus Kozminski, 1933:106; Kozmiilski, 1936:219.
Самка. Тело относительно толстое, мало суживающееся кзади,
наибольшая ширина переднего отдела тела лежит приблизительно на
середине его длины (считая и задний торакальный сегмент). Цефалото-
ракс лишь немного длиннее следующих 4 сегментов или приблизительно
равен общей длине последних. Задние углы 2-гй и 3-го торакальных
сегментов у типичной формы одинакового строения, лишь слабо выдаю-
щиеся кзади, реже у 2-го сегмента они значительно развиты. У 4-го
сегмента эти углы образуют вариирущей величины и очертаний
выросты, иногда несколько загнутые кпереди. Задний торакальный
сегмент по бокам с хорошо выраженными, обычно затупленными,
иногда заметно укороченными выростами. Абдомен относительно длин-
ный, его длина в % длины тела, в среднем, =42 (36—47), обычно
более 40. Генитальный сегмент расширенный спереди, кзади равномерно
суживающийся. Receptaculum seminis широко овальный, иногда почти
круглый, у некоторых экземпляров несколько расширенный в переднем
отделе. Фуркальные ветви обычно заметно расходящиеся, очень вари-
ирующей длины, в среднем их ширина составляет не менее 16.0 (12.6—
т
CYCLOPINAE
19.4) % их длины. Из крайних апикальных щетинок внутренняя отно-
сительно короткая, ее длина в °/Оо длины тела не более 158 (73—158),
средние апикальные щетинки сравнительно короткие, из них внутренняя
Фиг. 34. Cyclops strenuus Fisch, (typ.), самка.
7— задний отдел торакса; 2—задний сегмент торакса, генитальный сегмент
и receptaculum seminis; 3—йога V; 4— фурка со спинной стороны; 5—эк-
зоподит ноги (первый членик не изображен).
лишь немного длиннее внешней. Дорзальная щетинка короткая, лате-
ральная прикрепляется близко от заднего конца ветвей, внутренние
края последних с почти сплошным рядом волосков. Передние антенны
17-членистые, достигающие приблизительно средины 2-го торакаль-
ного сегмента. Вооружение плавательных ног по типу Terni. У ног
7. CYCLOPS
175-
V пары щетинка базального членика вариирующей длины, иногда дости-
гающая середины щетинки дистального членика, шип на внутреннем
крае последнего крупный, сильный, выступающий кзади от конца членика;
близ основании дистальной щетинки имеется группа из нескольких
мелких шипиков. Длина 1.45—2.32 мм. Окраска сильно вариирует, нередко-
желтоватая, до яркооранжевой и красной включительно; рачки мало
прозрачны. Яйцевые мешки удлиненно-овальные, реже округлые, при-
легающие к абдомену, обычно заключающие значительное количество
(иногда свыше 100 в каждом) яиц.
Самец. Гораздо стройнее и тоньше самки, фуркальные ветви
короче, чем у последней, без продольной складки на спинной поверх-
ности. Ноги VI пары с длинной внешней щетинкой с 2 шипами, из которых
один лишь немного длиннее другого. Плавательные ноги с вооружением
по типу Terni или Bini. Сперматофоры небольшие, иа генитальном
сегменте самки они располагаются, не заходя своими концами за края
сегмента. Длина 1.1—1.6 мм.
Основные индексы приводятся ниже.
Под названием „С. strenuus" в литературе фигурирует как типичная
форма, так и ряд близких к ней форм. За типичную форму мы прини-
маем, вместе с Kozminsk’nM, впервые описанную в 1851 г. Fischer’oM.
Вопреки Козминскому (1936), мы считаем бесспорной идентичность
последней с описанным Багз’ом (1913) С. strenuus, который во всех
основных признаках вполне укладывается в диагноз Fischer’a. Отличия
от С. abyssorum и других близких форм приводятся ниже.
Распространение. Типичная форма. Европа, Малая Азия,
Северная Африка и, возможно, Северная Америка. Большинство указа-
ний для СССР требуют проверки. Известна из следующих районов;
Карело-Финская ССР, Коми АССР, окр. Ленинграда; области: Горьковская,
Ярославская, Ивановская, Московская, Харьковская, Киевская, Днепро-
петровская; бассейн Южного Буга; Кавказ (окрестности Махач-Кала);
Крым (Карадаг); Новосибирские острова; окр. Атбасара, Памир; цен-
тральная часть Якутской АССР; Дальний Восток.
Биология. Очень характерен для мелких, в частности для времен-
ных астатических водоемов (лужи, болотца и тому подобное), но встре-
чается также в прудах и в более или менее крупных постоянных-
водоемах; повидимому гораздо реже в литорали озер. Принадлежа
к настоящим планктическим циклопам, он, однако, повидимому вовсе
не встречается нормально в пелагиали озер. В мелких лужах развивается
в массах. Относить С. strenuus (typ.) к холодноводным космополитам
едва ли возможно. Его выпадение на лето из мелких временных луж,,
очевидно, связано отнюдь не с термическими, а с иными условиями,
вообще резко изменяющимися при усыхании таких водоемов. Рачок
неоднократно был находим и при высоких температурах воды в постоян-
ных водоемах и летом (см. ниже). Равным образом нет оснований счи-
тать рассматриваемый вид ледниковым реликтом. С. strenuus s. str.
может довольно продолжительное время существовать при отсутствии
растворенного кислорода и при наличии сероводорода; несомненно —
это в широкой степени форма эвриоксибионтная. Широко распро-
странен в водах с слабокислой, нейтральной и слабощелочной реак-
цией, при pH =6—7.5 (8.0). Ласточкин (1934) отмечает его для дистроф-
ных водоемов, однако при pH = 5.75 — 5.95. Повидимому в сильно
дистрофированных водоемах, при pH около 4 и ниже, типичный
С. strenuus отсутствует (см. ниже var. landei). Как отмечает
176
CYCLOPINAE
Kozminski (1936). C. strenuus s. str. широко приспособляется к коле-
баниям содержания хлоридов. Повидимому он переносит и весьма
широкие колебания содержания Са. Так, в мелких водоемах г. Пушкина
он обитает при содержаниях СаО близких к политипу, но вместе
с тем неоднократно был найден и в очень мягких водах, при олиго-
типе СаО.
Цикл размножения изучен далеко не полно. Максимум весной, при-
чем в высыхающих летом водоемах рачок моноцикличен и исчезает
нередко уже в мае. В постоянных водоемах встречается, однако, весь
год, летом менее многочислен; самки меньших размеров, нежели
весной.
Главным образом [3-мезосапроб, однако отчасти и а -мезосапроб
(см. выше).
Типичная форма Cyclops strenuus Fisch, характеризуется следую-
щими основными индексами (самка): общая дл. тела = 1890 (1458—2322);
шир. тор. сегм.4 в °/00 к дл. тела — 238 (313—282)); шир. тор. сегм5
в °/00 к дл. тела =187 (148—239); шир. тор. сегм.4 в °/0 к макс. шир.
цефалот. = 66 (58—79); дл. антенн. 1 пары в °/(0 к дл. тела = 488 (430—
547); дл. внутр, ап. щет. в °/оо к дл. тела = 116 (87—150); дл. внутр,
ап. щет. в % к дл. внеш. ап. щет. = 140 (122—219).
На табл. 10 даются подробные измерения типичной формы по мате-
риалам из СССР.
la. Cyclops strenuus var. landei Kozm. (фиг. 35).1
Cyclops strenuus landei Kozminski, 1933:108, t. 4, f. 7—8; Kozminski, 19’4:10, t. 4,
f. 6; Kozminski, 1938:221.
Самка. Цефалоторакс очень широкий и укороченный, заметно
суживающийся кзади, следующие 2 сегмента с одинаково развитыми
задними углами, которые у 4-го сегмента оттянуты кзади и несколько
в стороны, задний торакальный сегмент лишь немного шире гениталь-
ного сегмента, с мало выступающими боковыми выростами. Гениталь-
ный сегмент укороченный, мало и равномерно суживающийся кзади.
Reeeptaculum seminis округлый, как у типичной формы. Фуркальные
ветви относительно короткие, слабо расходящиеся, с почти сплошным
рядом волосков на внутренних краях. Из крайних апикальных щетинок
внутренняя менее чем вдвое длиннее внешней, дорзальная щетинка
короче внешней крайней. Передние антенны 17-членистые, относительно
короткие, едва достигающие середины 2-го торакального сегмента.
Плавательные ноги вооружены по типу Terni. У ног V пары щетинка
базального членика относительно короткая, внутренний шип дистального
членика сильный, длинный, заходящий за задний край членика. Яйцевые
мешки тесно прилегают к абдомену и содержат обычно не более
30 яиц (в каждом).
Основные индексы (по Козминскому, 1933,1936): общ. дл. тела = 1514
(1345—1681); шир. тор. сегм.4 в °/0 к макс. шир. цефалот. = 56 (49—63);
шир. тор. сегм.Б в °/оо к Дл- тела = 166 (152—181); шир. тор. сегм.4
в %о к дл- тела = 199 (177—221); дл. антенн 1 пары в °/00 к дл.
(135 =168)^ (511—ДЛ’ BHyT₽’ ап- Щет- в °/о к дЛ" внешн- ап- щет. = 151
1 Средние величины по Kozminsk’oMy (1933, 1936), крайние предельные величины
нами несколько изменены согласно нашим измерениям (на средние это существенно
не влияет).
12 Фауна СССР
Фиг. 35. (По Ко zminsk’oMy).
Cyclops strenuus var. landei Kozm., самка; 2—C. bohater Kozm., самка; 3—C. tatricus Kozm., самка.
Таблица JO
Измерения C. strenuus Fisch, (typ.).
Экз. № Крым (Карадаг) Новосибирские острова
1 2 3 4 5 1 2 3 4 5
Дл. цефалоторакса 776.4 739 758 696 741 696 660 668 650 708
176.4 186 187 180 180 152 132 144 156 136
180 138 137 174 208 132 126 144 148 138
„ тор. сегм. 4+5 „ цефалоторакса -+- тор. сегм. 2—5 240 282 240 202 226 199 200 128 185 203
1373 1345 1322 1352 1355 1179 1118 1154 1139 1185
Макс. шир. цефалоторакса 720 732 715 688 679 662 630 634 бЗо 678
.. ,, „ В °/пп к дл. тела . . . 368 375 375 359 357 398 386 385 385 395
Шир. тор. сегм. 4 518.4 523 517 501 538 419 • 427 370 410 451
• „ „ „ в °/оо к Дл. тела 265 268 271 262 282 251 263 225 251 263
„ „ „ в ®Iq к макс. шир. цефалот. . . 72 71 72 73 79 63 67 58 65 66
м тор. сегм. к 434.4 428 438 430 430 344 347 330 343 367
„ „ „ в °/ап к дл. тела 222 219 229 225 226 206 213 201 210 214
„ ,, ,, в 0/л к шир. тор. сегм. 4 . . . * 84 78 85 86 80 82 81 89 84 81
314.4 320 300 271 274 252 260 2ч0 245 254
Макс. шир. ген. сегм 312 311 312 313 301 249 259 236 238 238
„ „ „ „ в к дл. ген. сегм. . . . 99 97 104 116 113 99 100 98 97 94
270 286 283.2 290 274 236 252 250 252 279
.. » » в °М к дл. фурк. ветви • • • 88 100 102 103 100 102 105 106 105 113
584 606 583 561 548 488 512 490 497 533
„ „ в 0/0 к дл. цефалот. -+- тор. сегм. 2-в • Дл. тела Дл. фурк. ветви „ ,, в °/пл к дл. тела • • 43 1957 45 1951 44 1905 42 1913 40 1903 41 1567 46 1630 42 1644 44 1636 45 1718
306 285 277 282 273 231 241 235 240 247
156 146 145 147 143 139 148 143 147 144
44.7 50 50 46 47 40 39 37 37 40
,/ „ в °/о к дл. фурк. ветв. . . . . • • 15 18 18 16 17 17 16 16 15 16
*si
Продолжение таблицы 10
Экз. № Крым (Карадаг) Новосибирские острова
1 2 3 4 5 1 2 3 4 5
Расст. А (фиг. 33. ik) ............... 253 231 234 240 230 183 185 191 189 200
„ „ в °/о к дл. фурки 83 81 85 85 84 79 77 81 79 81
Дл. внутр, кр. ап. щет 277 293 250 269 259 170 162 162 178 191
„ „ в О/оо к ДЛ. тела 142 150 131 141 136 102 99 99 109 111
Дл. внеш. ср. ап. щет 460 440 430 480 453 322? 367 340 350 407
„ „ „ В с/оо к ДЛ. тела 235 226 226 251 238 193? 224 207 214 231
Дл. внутр, ср. ап. щет 502 518 552 547 534 425 410 390 398 443
„ „ „ » В °/оо К дл. тела 256 266 289 286 281 255 245 237 243 252
Дл. внеш. ап. щет 138 148 114 153 125 108 120 109 110 108
„ „ » » в °/00 к дл. тела 71 76 59 80 66 65 73 66 67 63
Дл. латер. щет. 59 48 48 49 47 39 40 38 40 36
„ „ » в °/оо к ДЛ. тела 30 25 25 26 25 24 25 23 24 21
Дл. внутр, ап. щет. в °/о к дл. внеш. ап. щет. . . . 201 200 219 176 206 157 135 149 163 178
Дл. внеш. ср. ап. щет. в °/о к дл. внутр, ср. ап. щет. 92 85 78 88 85 76? 89 90 88 92
Дл. антенн. I пары 959 850 906 960 884 780 852 780 817 810
„ „ » „ в о/оч к дл. тела 490 435 475 501 464 469 522 474 499 471
Дл. эндоп. з IV пары ног 112 127 119 126 121.5 115.5 126 115 110 121
Шир. эндоп. з IV пары ног 54.6 52 48.3 47.6 46 38 41 38 38' 42
Дл. внутр, ап. щет. эндоп. 3 IV пары ног 84.7 85 100 111 105 94 99 96 91 95
Дл. внеш. ап. щет. эндоп. 3 IV пары ног 28? 56 51.1 54.6 52 57 58 61 57 58
Дл. экзоп. з IV пары ног 110.6 1032 114 115.5 115 119 125 117 126 127
Шир. экзоп. з IV пары ног 52 46.2 58 49 55 49 50 45 54 53,
Дл. мал. щет. V пары ног 70 78.4 66 83 70 54 62 52 52 56,
„ „ в °/0 к дл. болып. щет.
V пары ног 65 65 60 66 62 77? 76 58 62 60
Дл. болып. щет. V пары ног 108.5 120 110 125 ИЗ 70? 82 89 84 93
Общая длина тела 2263 2236 2182 2195 2176 1898 1871 1879 1876 1965
Колич. яид 46-50 34 40-42 52-55 42-45 12-16 8 19 12-12 9—10;
180
CYCLOPINAE
От C. strenuus s. str. отличается более сильно суженным кзади
цефалотораксом и менее широким (относительно генитального сегмента)
задним торакальным сегментом.
Рас простр анение. Пока известен лишь в районе оз. Вигры
(Сувалки).
Биология. По Козминскому обитает в мелких дистрофных водо-
емах, бедных электролитами, в частности Са; в водах с содержа-
нием СаО более 2 мг 0/00 не найден. Повидимому отсутствует в сильно
дистрофированных (pH — 4 и менее) водоемах. Моноцикличен; молодь
появляется осенью, половозрелость наступает в ноябре—декабре,
максимум обычно в феврале, в апреле исчезает. Является, таким
образом, специально зимним циклопом.
* 1в. Cyclops strenuus vranae Kozm. (фиг. 36).
Cyclops strenuus var. vranae Kozminski, 1927:71, f. 6. — C. strenuus vranae Kozminski,
1933:107; Kozmiiiski, 1934:110, t. 4, f. 3; Kozminski, 1936:221.
Самка. 2-й торакальный сегмент с тупо заканчивающимися зад-
ними углами, лишенными выроста (как у С. strenuus s. str./ задние
углы 3-го и 4-го торакальных сегментов одинаковые, оттянутые кзади,
хорошо развитые. Задний торакальный сегмент, каку С. tatricus (см. ниже),
значительно шире генитального сегмента. Генитальный сегмент короткий,
слабо и равномерно суживающийся кзади. Фуркальные ветви довольно
длинные, сильно расходящиеся, латеральная щетинка прикрепляется
близко от заднего конца внешнего края, дорзальная щетинка обычно
относительно длиннее, чем у С. strenuus s. str., и относительно длиннее,
чем у последнего, щетинки ног V пары. Передние антенны 17-члени-
стые, вооружение плавательных ног по типу Terni
Основные индексы (самки, по Козминскому 1933, 1936): общ. дл.
тела = 1895 (1664—2125); макс. шир. цефалот. в °/00 к дл. тела = 340
(323—358); шир. тор. сегм.4 в °/00 к дл.тела = 214—278; шир. тор. сегм.5
в °/оо к дл. тела = 175—219; шир. тор. сегм.4 в °/о к макс. шир. цефа-
лот. =64—81; шир. тор. сегм.5 в °/0 к шир. тор. сегм.4 = 72—88; дл. задн.
абд. сег. в % к дл. фурк. ветв. = 83—118; расст. А (фиг. 33. ik)
в °/0 к дл. фурк. ветв. = 75.0—83.4; дл. фурк. ветв. в °/Оо к дл. те-
ла =148 (135—161); шир. фурк. ветв. в °/0 к дл. фурк. ветв. = 13,8—19.2;
дл. внутр, ап. щет. в °/00 к дл. тела 133 (120—146).
Яйцевые мешки содержат небольшое количество яиц. Окраска
(оз. Севан) желтоватая, до оранжевой (красно-оранжевой) включительно.
Наиболее характерными признаками рассматриваемого вариетета
являются: довольно грубое, толстое тело, ширина заднего торакального
сегмента, заметно превышающая длину генитального сегмента, который
относительно очень толст и короток, довольно длинные, заметно расхо-
дящиеся фуркальные ветви, относительно крупный шин на дистальном
членике ног V пары. Этими признаками var. vranae хорошо отличается
от С. lacustris Sars, который Также обитает в пелагиали озер. К С. stre-
nuus var. vranae мы относим форму strenuus из оз. Севан, которая
в общем укладывается в таблицу измерений, опубликованную
Козминским (1933) и соответствует краткому диагнозу этого автора
для vranae. Ни с какой другой формой группы strenuus севанская
форма идентифицироваться не может (в частности с С. abyssorum,
который для оз. Севан ошибочно приводится Амелиной, 1929). Измере-
ния севанских С. strenuus var. vranae приводятся в табл. 11.
7. CYCLOPS
/SX ;
Фиг. 36. Cyclops strenuus var. vranae Kozm., самка.
1—передний отдел тела со спинной стороны; 2—фурка со спинной сто-
роны; э— абдомен и задний сегмент торакса; 4—фурка со спинной сто-
роны; 5— задний отдел торакса и генитальный сегмент; б и 7—нога V;
8 — задний отдел торакса и генитальный сегмент, форма из оз. Врана. .
(7—7—по экземплярам из оз. Севана, 8—по Kozminsk’oMy).
182
CYCLOPINAE
Таблица 11
Измерения С. strenuus var. vranae Kozm.
Оз. Севан. Экземпляры № 1—5 собраны в VII 1931 (е глубины 15—30 м), экзем-
пляр № 6—19 XI 1931 (с глубины 17—47 м).
Экз. № ad.)
1 2 3 4 5 6
Дл. цефалоторакса .........................
„ тор. сегм. 2....................... . .
„ тор. сегм. з..................•.........
„ тор. сегм. 4+5..........................
„ цефалот. -+- тор. сегм. 2—в.............
Макс. шир. цефалоторакса...................
» „ „в °/00 к дл. тела . .
Шир. тор. сегм. 4 ..............
„ „ „ в °/00 к ДЛ. тела.............
Шир. тор. сегм. 4 в °/о к макс. шир. цефалот .
Шир. тор. сегм. 5.............•.............
» „ „ В °/оо К ДЛ. тела...........
„ „ „ в °|о к шир. тор. сегм. 4 . .
Дл. ген. сегм...............................
Макс. шир. ген. сегм. . . ..................
» „ „ „ в % к дл ген. сегм. . . .
Дл. задн. абд. сегм...........•............
„ „ „ „в e/оо к дл. фурк. ветви . .
Дл. абд. . . ..............................
„ „ в к дл. цефалот. ч- тор. сегм. о-в .
Дл. тела...................................
Дл. фурк. ветв.............................
„ „ „ в 0/0 к дл. тела.............
Шир. фурк. ветв............................
„ „„ в °/0 к дл. фурк. ветв. . . .
Расст. А (рис. 33, ik) ............
п ,, в °/0 к дл. фурки...................
Дл. внутр, кр. ап. щет.....................
Дл. внутр, кр. ап. щет. в °/оо к дл. тела . . .
Дл. внеш. ср. ап. щет...............• ...
» » „ „в °/оо к дл. тела . . .
Дл. внутр, ср. ап. щет......................
» « ,, ,, „ в °/оо К ДЛ. тела ....
Дл. внеш. ап. щет. .... ....................
» » » » в ^/оо к дл. тела.........
Дл. латер. щет..............................
» » В к дл. тела..................
Дл. внутр, ап. щет. в к дл. внеш. ап. щет.
Дл. внеш. ср. ап. щет. в °/о к дл. внутр, ап.
ср. щет. .... ............................
Дл. антенн I пары...................• • . .
» » ,, в °(оо к дл. тела.........
Дл. эндоп. з IV пары ног....................
Шир. эндоп. з IV пары ног..................
Дл. внутр, ап. щет. эндоп. 3 IV пары ног . . .
Дл. внеш. ап. щет. эидоп. 3 IV пары ног . . .
Дл. экзоп. з IV пары ног................ .
Шир. экзоп. з IV пары ног...................
619 588 596 638 650 680.4
120 128 124 162 153 150
138 130 136 180 178 178
185 168 165 176 174 188
962 914 921 1156 1155 1196
540 520 500 600 600 632
379 384 369 357 352 368
343 338 330 392 390 411
241 250 243 233 229 240
64 65 66 65 65 65
284 291 280 333 320 349
200 215 206 198 188 203
83 86 85 85 82 85
231 210 201 259 251 253
218 210 210 259 245 262
94 100 105 100 98 104
230 229 234.5 267 298 266
119 112 111 108 110 111
461 439 436 526 549 519
48 48 47 46 48 42
1424 1353 1357 1682 1704 1716
192 204 210 248.5 270 240
135 151 155 147 159 140
38 35 35 41 39 41
20 17 17 17 15 17
158 164.5 169 197 217 190
82 80 80 80 80 79
204 203 185 246 252 226
143 157 136 146 148 132
450 436 422 516 513 410
316 322 311 307 301 239
523 558 516 638 620 550
368 412 381 379 364 321
112 109 113 144 130 114
78 81 83 86 76 66
41 39 41 50 51 46
29 29 30 29 29 27
182 186 164 171 194 198
86 78 82 82 83 75
761 730 725 912 810 896
535 539 535 542 475 522
116 108 108 125 133 109
39 38 36 41 43 40
123 119 116 124 135 112
53 46 48 67 66 58
112 103 102 131 132 129
35 40 38 50 52 40
7. CYCLOPS
183
Продолжение таблицы 11
Экз. № ad.)
Дл. эндоп. з III пары ног....................
Шир. экзоп. з III пары ног...................
,Дл. мал. щет. V пары ног ...................
„• „ „ „ „ „ в 0/0 к дл. болып.
щет. V пары ног....................... . .
Дл. болып. щет. V пары ног...................
Общая длина тела........................ . .
Колич. яиц............................ , . .
1 2 3 4 5 6
102 123 109
— — 40 51 — 38
— 70 70 80 70 64
55 62 56 62 64
—. 127 113 144 130 100
1615 1557 1567 1931 1974 1956
9-10 — 8-8 — — —
Распространение. Оз. Врана (о. Херсо в Адриатическом
море); оз. Севан в Армянской ССР.
Биология. Пелагическая форма глубоких крупных озер. Цикл
размножения не изучен. В оз. Севан половозрелые самки и молодь
найдены нами в июле — сентябре и ноябре.
*2. Cyclops abyssorym Sars (фиг. 37).
Cyclops abyssorum Sars, 1863, Forh. Selsk. Christ. (1862): 238. — C. strenuus var.
gracilipes Sars, 1903 : 217. — C. abyssorum Sars, 1913, t. 17; Kozmiiiski, 1933:103, f. 3, 4,
t. 2; Kozminski, 1934:110; Kozminski 1936:214.
Самка. Тело относительно утолщенное и укороченное, передний
отдел равномерно суживающийся кзади, 4-й торакальный сегмент уже
преыдущего, с хорошо развитыми, нередко заметно оттянутыми кзади
и в стороны задними углами. Задние углы 2-го торакального сегмента
развиты слабо, не загибаются на брюшную сторону. 5-й торакальный
сегмент с хорошо развитыми, нередко заостренными выростами. Гениталь-
ный сегмент заметно расширенный спереди, относительно короткий,
кзади равномерно и лишь незначительно суживающийся. Reeeptaculum
seminis как у С. strenuus. Абдомен относительно короток, его длина
составляет в среднем 36.50/0 (обычно менее 39 %) длины переднего
отдела тела (включая задний торакальный сегмент). Фуркальные ветви
заметно расходящиеся, обычно очень длинные и стройные, с почти
сплошным рядом волосков на внутренних краях. Длина внутренней
из крайних апикальных щетинок в °/00 длины тела в среднем не ме-
нее 200 (154—214), наичаще более 160—170. Латеральная щетинка
прикреплена близко от заднего конца ветвей, длина дорзальной
щетинки обычно несколько менее таковой внешней крайней апикальной
в °/00 длины тела = 79—108; иногда эти обе щетинки почти одинаковой
длины. Передние' антенны 17-членистые, достигающие конца 2-го
торакального сегмента, иногда несколько длиннее. Вооружение плава-
тельных ног по типу Terni. У ног V пары щетинка базального чле-
<ника = 35—55% длины щетинки дистального членика, обычно 1-я щетинка
менее 1/2 длины 2-й. Шип внутреннего края дистального членика
сильный, выступающий заметно кзади от конца членика. Длина 1.49—
2.24 мм, обычно около 2 мм. Окраска серовато-белая, иногда рачки
бесцветны. Яйцевые мешки овальные, реже округлые, прилегающие
184
CYCLOPINAE
к абдомену, у типичной формы небольшие, заключающие небольшое-
количество крупных яиц.
Самцы — как у С. strenuus. Сперматофоры крупные, не выступаю-
щие за края генитального сегмента самки.
Фнг. 37. Cyclops abyssorum Sars.
1 — передний отдел тела самки со спинной стороны; 2—2 задних,
сегмента торакса и абдомен самки; 3—экзоподит ноги I; 4—
5— фурка самки со спинной стороны.
Результаты измерений экземпляров С. abyssorum из водоемов СССР'
приведены в табл. 12. Основные индексы (самки): общ. дл. тела =1486—2243,
(обычно около 2000); дл. абд. в °/0 к дл. цефалот. тор. сегм.2_5=3б.5
(34—42.8); дл. фурк. ветв. в %о к дл. тела = 155—190, обычно около 175;;
шир. фурк. ветв. в °/0 к дл. фурк. ветв. = 10.6—16, обычно около 12—14;,
7. CYCLOPS
785:
Таблица T2
Измерения С. abyssorum Sars.
Экз. № (2 ad.) Оз. Телецкое, 21 VIII 1930 Оз. Бол. Миа- СОВО, . 13 VIII 1928
‘ 1 2 3 4 2 3
Дл. дефалоторакса 588 600 544 608 800 817 785
Дл. тор. сегм. 120 121 117 132 153 150 156
127 138 111 132 139 164 163
Дл. тор. сегм. 4+5 150 156 146 130 202 207 213
Дл. цефалот. ч- тор. сегм. 2_g 985 1015 918 1052 1294 1338 1317
480 497 450 528 685 701 685-
„ „ „ В %0 к ДА- 350 344 355 370 389 382 375
Шир. тор. сегм. 4 ............... 295 300 288 332 403 390 374
„ „ „ в °1по к дл. тела .... • . 215 208 227 233 229 212 204.
„ п „ в о/о к макс. шир. цефалот. . 61 60 64 63 59 56 55
Шир. тор. сегм. 240 249 230 280 357 350 340'
176 172 181 196 203 191 186
„ „ „ в °|о К шир. тор. сегм 4 . . 81 83 80 84 89 90 91
Дл. ген. сегм 196 204 186 207 215 243 244
Макс. шир. ген. сегм 1'96 196 184 204 274 272 270
„ „ „ „ в % к дл. ген. сегм. . . 100 96 99 ок.100 127 112 111
Дл. задн. абд. сегм 189 225 165 168 248 254 269
„ „ „ „ в °|0 к дл. фурк. ветв. . . 89 97 76 76 83 83 99
385 429 351 375 463 497 513
„ „ в % к ДА- дефалот. ч- тор. сегм. 2_g . 39 42 38 36 37 37 39
Дл. тела 1370 1444 1269 1427 1757 1835 1830
Дл. фурк. ветви 212 233 217 222 299 307 271
„ „ „ В 0/00 к дл. тела 155 161 171 156 170 167 148
Шир. фурк. ветв. 32 33 26 35 43 43 43
» » » В °!о К дл. фурк. ветв. . . . 15 14 12 16 14 14 16-
Расст. А (фиг. 33, ik) . 173 182 164 172 235 237 223
„ „ в °/о к ДА. фурки 82 79 76 78 79 79 82
Дл. внутр, кр ап. щет 239 222 211 233 341 322 321
„ » „ » п В о/оо К дл. тела . . . 174 154 166 163 194 175 175
Дл. внеш. ер. ап. щет 509 504 453 480 504 — 504
„ и „ в °|оо к дл. тела .... 372 349 357 336 287 — 275
Дл. внутр, ср. ап. щет ••.... 594 617 592 643 646 575 570
„ „ я » » В °/оо К дл. тела.... 434 427 467 493 368 313 311
Дл. внеш. ап. щет 96 108 96 120 160 — 140
„ „ „„ в °/оо к дл. тела 70 75 76 84 91 — 76
Дл. латер. щет. ............... 43 52 54 52 — — 60
„ „ „ В 0/оо к дл. тела 31 36 43 37 — •— 32
Дл. внутр, ап. щет. в о/0 к дл. внеш. ап. щет. 249 206 219 194 213 — 229
Дл. внеш. ср. ап. щет. в °/0 к дл. внутр, ср. ап. щет. 86 82 76 75 78 — 88
Дл. антенн I пары 828 798 783 852 991 1031 1002
„ I пары в °/оо к дл. тела 604 553 617 597 564 562 548
Дл. эндоп. з IV пары ног 104 122 112 120 132 140 135
Шир. эндоп. з IV пары ног 35 32 28 36 48 46 42
Дл. внутр, ап. щет. эндоп. 3 IV пары ног . . . 111 131 120 127 157.5 155 147
Дл. внеш. ап. щет. эндоп. 3 IV пары ног . . . 39 52 35 38.5 77 70 86
Дл. экзоп. з IV пары ног 116 116 109 121 136 140 125
Шир. экзоп. з IV пары ног . 35 34 33 37 52.5 50 52.5
Дл. эндоп. з III пары ног 98 90 100 105 — — —
Шир. эндоп. з III пары ног 39-8 42 36 45 — — —
Дл. мал. щет. V пары ног 78 80 70 74 85 85 77
„ ,, » » „ в О/о к дл. больш. щет. V пары ног 52 53 49 56 50 49 46
Дл. больш. щет. V пары ног 149 151 143 150 170 176 166
Общая длина тела 1582 1677 1486 1649 2056 2142 210L
.186
CYCLOPINAE
дл. задн. абд. сег. в °/0 к дл. фурк. ветв. — около 80 (70—99); дл. внутр,
ап. щет. в °/оп к дл. тела = 154—214 (преимущественно 160—200);
дл. мал. щет. V пары ног в °/0 к дл. болып. щет. V пары ног обычно
не более 53.
Рассматриваемый вид очень близок к С. strenuus Fisch, и рядом
авторов считается вариететом последнего. Однако он хорошо отличается
от указанного вида определенной комбинацией признаков — иным соотно-
шением длины переднего отдела тела и абдомена, иной длиной крайней
внутренней апикальной щетинки (относительно длины тела), иной длиной
фуркальных ветвей (относительно длины тела); биология обеих форм
также различна (см. ниже).
Распространение. С. abyssorum, несомненно, нередко смеши-
вается с другими видами группы strenuus, вследствие чего ряд литера-
турных данных требует проверки. Следующие указания можно считать
достоверными: Норвегия, Швеция, Германия, СССР; вероятно, также
Финляндия. В СССР известен из Карело-Финской ССР, из Свердлов-
ской области, из оз. Телецкого, Ниже-Кочурлинского и Кара-коль (все
озера лежат на Алтае, на высоте до 1785 м.). Вероятно нахождение
в Ладожском озере.
Биология изучена неполно. Обитает в пелагиали озер,
до очень глубоких (оз. Телецкое) включительно: вопреки Козмин-
скому (1936), встречается не только в умеренно эвтрофных и слабо
дистрофированных, но также и в олиготрофных озерах (Телецкое, Кара-
коль). В озерах днем держится, вероятно, в придонных слоях воды.
По Ekman’y в оз. Malaren днем скопляется на глубине 21—36 м, близ
дна (лето, конец зимы), в оз. Кара-коль (глуб. 29.5) — на глубине 15—22 м.
По нашим данным в Телецком оз. в июле — сентябре С. abyssorum
днем держится на глубине 20—40 м (молодь и взрослые самки и самцы),
причем в единичных экземплярах встречается до глубины 180—200 м
(молодь и на глубине в 300 м), так что в этом озере рачок отнюдь
не является глубоководным. До сих пор указывался для слабокислых,
нейтральных и слабощелочных озер. Возможно, что приуроченность
этого вида в некоторых относительно неглубоких озерах специально
к гиполимниону стоит в связи с его холодноводной стенотермичностью;
в Телецком озере рачок также избегает верхней, наиболее прогреваемой
зоны эпилимниона. Судя по наблюдениям в некоторых озерах С. abys-
sorum дицикличен, максимум зимой (февраль — март) и летом (июль —
август); дициклия, по нашим данным, наблюдается и для формы
из Телецкого озера, где однако 2-й максимум запаздывает приблизи-
тельно на месяц.
3. Cyclops bohater Kozm. (фиг. 352).
Cyclops bohater Kozminski, 1933:105, t. 3, f. 5, 6; Kozminski, 1934:110, t. 4, f. 5;
Kozminski, 1936:218.
Самка. Цефалоторакс очень широкий, кзади несколько суженный,
2-й торакальный сегмент с очень сильно развитыми задними углами,
несколько загнутыми на брюшную сторону. Следующие 2 сегмента
заметно уже 2-го, с почти одинаковыми задними углами. Задний тора-
кальный сегмент с сильно развитыми заостренными боковыми выростами.
Генитальный сегмент как у типичного С. strenuus. Фуркальные ветви
довольно длинные, расходящиеся, внутренняя из крайних апикальных
щетинок приблизительно вдвое длиннее внешней крайней, дорзальная
7. CYCLOPS
187
щетинка почти равна по длине последней. Передние антенны 17-, редко
18-членистые. У ног V пары щетинки базального членика длинные,
в среднем 81 (72—90) °/0 длины щетинки дистального членика, шип
внутреннего края последнего как у типичного С. strenuus. Дли-
на 2.288 — 2.583 мм. Яйцевые мешки удлиненно-овальные, тесно при-
легающие к абдомену, содержащие значительное количество сравнительно
мелких яиц. Основные индексы (самки) (по Козминскому, 1933, 1836):
общ. дл. тела в среднем 2435, дл. абд. в °/0 к дл. цефалот. -+- тор.
• сегм., + 5 = 41 (37—45); дл. внутр, ап. щет. в °/о к Дл- внеш. ап. щет. = 209
(192—226), дл. мал. щет. V пары ног в % к дл. болып. щет V пары
ног = 81 (72 -90).
Рассматриваемый вид чрезвычайно близок к С. abyssorum, но хорошо
отличим от него, главным образом, по углам второго торакального
сегмента.
Распространение. Известен лишь из Вигерских озер близ
«Сувалок (Польша). Нахождение в пределах СССР вероятно.
Биология. По Козминскому обитает в пелагиали умеренно
:эвтрофных озер, встречается и в олиготрофных озерах (последнее, как
мы указывали выше, свойственно и С. abyssorum). Вертикальное распре-
деление С. bohater и С. abyssorum, вероятно, однотипно, равно как
л цикл размножения одинаков (2 максимума в год—зимой и летом).
* 4. Cyclops lacustris Sars (фиг. 38).
Cyclops lacustris Sars, 1863, Forh. Selsk. Christ. (1862): 239.— C. strenuus f. lacustris
Lilljeborg;, 1901:31, t. 2, f. 23. — C. lacustris Sars, 1913 : 35, t. 18; Kiefer, 1929: 51;
Kozminski, 1936:219.
Самка. Tело стройное, относительно длинное, наибольшая ширина
переднего отдела лежит приблизительно на середине его длины (считая
задний торакальный сегмент). Цефалоторакс— удлиненный, в виде
полуовала. Задние углы торакальных сегментов (включая и задний)
лишь незначительно выдаются кзади, почти одинакового строения.
Генитальный сегмент относительно длинный, заметно суженный в заднем
отделе. Reeeptaculum seminis нормального для С. strenuus строения, его
передний отдел обычно несколько меньше заднего. Фуркальные ветви
короткие и относительно толстые, значительно расходящиеся, с рядом
волосков почти на всем внутреннем крае. Из крайних апикальных
щетинок внутренняя приблизительно вдвое длиннее внешней, внутренняя
из средних щетинок лишь немного длиннее внешней, дорзальная щетинка
заметно длиннее внешней крайней. Передние антенны 17-членистые,
достигающие середины или заднего края 2-го торакального
сегмента. Вооружение плавательных ног по типу Terni. У ног V пары
дистальный членик удлиненный, шип его внутреннего края очень малень-
кий, значительно не доходящий до конца членика. Длина 1.5—2.3 мм,
длина самцов 1.3—1.5 мм. Рачок очень прозрачен, голубоватой или
зеленоватой окраски. Яйцевые мешки округлые, тесно прилегающие
к абдомену, содержащие обычно небольшое количество яиц (по 4—7,
у экземпляров из Онежского озера по 7—14, из Ладожского — по 9—18
в каждом).
Измерения самок из Онежского и Ладожского озер приведены
в табл. 13 (см. ниже С. smirnovi, средняя и правая графы).
В группе strenuus рассматриваемый циклоп/ пожалуй, занимал
наиболее неопределенное положение. Лишь в 1913 г. Sars дал очень
188
CYCLOPINAE
точные изображения С. lacustris. За отсутствием материала Rzdska
и Козминский не ввели С. lacustris в свои биометрические исследования
Фиг. 38. Cyclops lacustris Sars, самка.
7 — передний отдел тела, со спинной стороны; 2—фурка со спинной
стороны; 3—абдомен, нога V и генитальный сегмент; 4—экзоподит
ноги II.
группы strenuus. В литературе нередко он фигурирует в качестве
вариетета С. strenuus, но Козминский вместе с Sars’oM склонен считать
его самостоятельным видом. Исследования материала из Онежского
7. CYCLOPS
189
и Ладожского озер произведенные нами, заставляют присоединиться
к этому мнению; идентичность формы из этих двух озер с С. lacustris
Sars не вызывает сомнений. Рассматриваемый вид наиболее близок
к С. strenuus, но от него, как и от его вариететов landei и vranae,
отличаетея следующими признаками: 1) у С. lacustris гораздо более
стройное, удлиненное тело, в частности цефалоторакс; 2) генитальный
сегмент удлиненный, сильно суженный в заднем отделе; 3) шип дисталь-
ного членика V пары ног у С. lacustris чрезвычайно тонок и короток;
4) процентное соотношение длины фуркальных крайних щетинок у этого
вида больше (174—235), чем у типичного С. strenuus (ИЗ—166) и у var.
landei (126—176) и близко к таковому у var vranae (153—205); однако
от последнего С. lacustris весьма резко отличается 3 вышеотмеченными
признаками, а также меньшей шириной 2 задних торакальных сегментов
(у С. lacustris lat. thor.4 в °/0 lat. max. cphth -+- thor.2_5 = 51—61, в сред-
нем 55.4, редко более 56, а у var. vranae — 64—81).
Распространение. С. lacustris, без сомнения, нередко смеши-
вался с другими формами группы strenuus; можно предполагать, что ряд
указаний на „С. strenuus“ для пелагиали озер северной Европы относится
именно к С. lacustris. С достоверностью последний известен из озер
Швеции, Норвегии и СССР, где он найден в Онежском (Кондопожская
губа, ЗИН) и Ладожском оз. (ЗИН), в 3 озерах Карело-Финской ССР
и в оз. Нижне-мультинском на Алтае.
Биолог ия. Изучена очень неполно. Судя по немногочисленным
вполне достоверным данным С. lacustris обитает в пелагиали олиготроф-
ных (слабо эвтрофированных) озер. По Sars’y (1913), населяет верхние
слои эпилимниона, но, по нашим наблюдениям, в пелагиали открытого
района южной половины Ладожского озера днем главной массой держится
глубже 20 м. Смирнов в Нижне-мультинском озере нашел максимум
в слое 0.5 м. Цикл размножения не изучен. Возможно моницикличен,
с максимумом летом — осенью.
5. Cyclops tatricus Ко£га. (фиг. З53).
Cyclops strenuus f. tatricus Kozmiiiski, 1927:71, f. 1—5.—C. strenuus f. Rzoska,
1930:208—214, t. 1, 2. — C. strenuus Baldi, 1931, Mem. Mus. Stor. Nat. Ven. Trid., 1:9,
f. 1—3.— C. tatricus Kozmiiiski, 1932:140, t. V; Kozmiiiski, 1933:104. — C. strenuus
tatricus Gurney, 1933:167, f. 1540. — C. tatricus Kozmiiiski, 1934:110. — C. strenuus lagora-
ensis Baldi,1 1935, Boll. Pesca di Pise, e di Idrob., 13:40, f. 10, a — e. C. tatricus
Kozmincki, 1936:216.
Самка. Тело стройное, цефалоторакс относительно удлиненный,
его наибольшая ширина лежит сзади, задние углы 2-го торакального
сегмента чрезвычайно сильно развитые, отвисающие кзади, не заострен-
ные, нередко загибающиеся на брюшную сторону. Задние углы обоих
следующих торакальных сегментов развиты менее, но также оттянуты
кзади, задний торакальный сегмент уже предыдущего сегмента, по бокам
со слабо развитыми заостренными' выростами. Генитальный сегмент
широкий и укороченный, мало, и равномерно суживающийся кзади.
Receptaculum seminis округлый, как у С. strenuus. Фуркальные ветви
заметно расходящиеся, довольно длинные, на внутренних краях со сплош-
ным рядом волосков. Дорзальная щетинка хорошо развитая, несколько
длиннее внешней из крайних щетинок. Антенны 17- (иногда 18-)
1 Рисунки Baldi относятся явно к неполовозрелым самкам; идентичность С. strenuus
lagoraensis с С. tatricus, однако, кажется мне очень вероятной.
1У0
CYCLOPINAE
членистые, доходящие приблизительно до середины 2-го торакаль-
ного сегмента. Вооружение плавательных ног по типу Terni. У ног
V пары щетинки обоих члеников относительно длинны, шип внутрен-
него края дистального членика также длиный, значительно выступающий
над задним краем этого членика. Длина самки 1.35—2.05 мм, самца
1.30—1.40 мм. Окраска, по Minkiewicz’y (1917, Татры, около 2000 м),
красная. Яйцевые мешки обычно содержат лишь незначительное коли-
чество яиц, округлые, тесно прилегающие к абдомену.
Основные индексы самки (по Kozminsk’oMy 1933, 1936): общ. дл.
тела 1702 (1353—2052); дл. абд. в °/о к дл. цефалот. -+-тор. сегм. 2_5 =
34.8 (32.0—37.6):, дл. фурк. ветв. в %о к Дл. тела 150 (134—166); дл.
внутр, ап. щет. в %о к Дл- тела = 164 (141—188); шир. фурк. ветв. в °/о
к дл. фурк. ветв. = 15.3 (13.7—16.9); дл. мал. щет. V пары ног в %
к дл. больш. щет. V пары ног = 54 (46—63).
С. tatricus является морфологически хорошо очерченным видом.
От С. strenuus и С. abyssorum он резко отличается очень сильным
развитием задних углов 2-го торакального сегмента, что наблюдается
также у С. bohater и С. smirnovi. От 1-го однако, С. tatricus
хорошо отличается слабо развитыми боковыми выростами заднего
торакального сегмента и иным процентным соотношением длины,
щетинок ног V пары (у С. bohater оно, в среднем, 81 (72—-90), у С. tatri-
cus— лишь 54 (46—63). От С. smirnovi рассматриваемый вид без труда
отличается строением и вооружением фуркальных ветвей, а также вели-
чиной шипа дистального членика ног V пары.
Распространение. Выяснено крайне неполно (часто смешивался
с близкими видами). Указан с достоверностью для горных озер Татр,
для 2 горных озер Швейцарии, для озер южного склона Итальянских
Альп, для горных озер восточных Альп, Нижнего Лунцского озера в юго-
восточной Германии. Нахождение в СССР вероятно. Повидимому широко-
распространен вообще в горных (показан на высоте от 607 до 2144 м
над уровнем моря) озерах юга Западной Европы.
Биология. Обитает в пелагиали озер, встречается также и в лито-
рали; биология изучена особенно Minkiewiez’eM (1917) в Татрах и Ruttner’oM
(1929/30) в Нижнем Лунцском озере (высота 607 м). Встречается весь год,
зимой и осенью размножается особенно интенсивно, летние самки,,
по Rzoska (1927), крупнее зимних. Температурный оптимум для взрослых
рачков, по Ruttner’y, лежит, вероятно, между 12—13°. Летом в Нижнем
Лунцском озере максимум взрослых самок наблюдался в слое 1—10 м,.
особенно много в слое 3—5 м, зимой вертикальное распределение гораздо
равномернее, но иногда максимум наблюдается зимой у самого дна (около .
30 м глубины). Науплиусы в этом озере летом образуют максимум:
в слое 10—20 м, обитая во всей толще воды, начиная с глубины 1—3 м;
зимой они распределяются равномерно. Суточные вертикальные миграции
взрослых самок интенсивны.
* 6. Cyclops smirnovi, sp. п. (фиг. 39).
Самка. Цефалоторакс очень крупный, широкий, несколько суживаю-
щийся кзади; его наибольшая ширина лежит, приблизительно, на середине
его длины. 2-й торакальный сегмент с весьма значительно развитыми
задними углами, выдающимися кнаружи и отчасти на вентральную
сторону. У следующих 2 торакальных сегментов задние углы развиты
гораздо менее, у 4-го сегмента они несколько вытянуты в стороны,.
7. CYCLOPS
191
наподобие отростков. Боковые выросты заднего торакального сегмента
хорошо развитые. Генитальный сегмент широкий, равномерно и мало сужи-
Фиг. 39. Cyclops smirnovi, sp. nov., самка.
/ — передний отдел тела со спинной стороны; 2 — задний отдел торакса н генитальный-
сегмент; 3 — фурка со спинной стороны; 4 — нога V; 5—экзоподит ноги I;
6 — receptaculum seminis.
вакицийся кзади, боковые выросты его переднего отдела слабо развитые._
Receptaculum seminis округлый, относительно небольшой. Фуркальные
ветви стройные, длинные, параллельные. Латеральная щетинка хорошо-
.192
CYCLOPINAE
развитая, прикрепляющаяся значительно кпереди от заднего конца ветвей.
Внутренние края последних снабжены рядом волосков лишь в задней
—73 своей длины. Из крайних апикальных щетинок внутренняя
приблизительно вдвое длиннее внешней. Средние апикальные щетинки
длинные, хорошо развитые, из них внутренняя лишь немного длиннее
внешней. Дорзальная щетинка очень длинная, превышающая длину
внешней крайней апикальной щетинки. Передние антенны 17-членистые, до-
ходящие до конца 2-го или середины 3-го торакального сегмента, т. е. отно-
сительнодлинные. Вооружение плавательных ног по типу Terni. У ног.У па-
ры щетинка базального членика относительно короткая. Дистальный членик
очень удлиненный, его апикальная щетинка очень длинная, шип его
внутреннего края чрезвычайно короткий, не доходящий до конца
членика. Длина 1.81—1.93 мм. Самец не известен. Измерения приведены
в табл. 13.
По строению переднего отдела тела, в частности его 2-го сегмента,
С. smirnovi напоминает С. bohater Kozm.; на близость именно к послед-
ней форме указывают и индексы отдельных признаков (табл. 13).
Однако по следующим признакам рассматриваемый вид отличается
от С. bohater весьма резко:
1. У С. bohater длина внутренней из средних апикальных щетинок
в °/00 длины тела=232—321, у С. smirnovi— 392—444 (в среднем 418).
2. У С. bohater длина меньшей щетинки ног V пары в % длины
большей щетинки = 72—90 (в среднем 81), а у С. smirnovi—53—63
(в среднем лишь 58).
3. Шип внутреннего края дистального членика ног V пары у С. bohater
гораздо длиннее и толще, чем у С. smirnovi, у которого он очень мал
и тонок, как у С. lacustris. Дистальный членик у С. smirnovi гораздо
длиннее, чем у С. bohater.
4. Фуркальные ветви у С. bohater значительно расходящиеся, снаб-
женные волосками вдоль всего внутреннего края, а у С. smirnovi
параллельные, снабженные волосками лишь в заднем отделе края.
Отличия от С. tatricus указаны при описании последнего. С. smirnovi
близок к С. ochridanus Kiefer (1931)—пелагической форме оз. Охрида.
Сходство обоих видов выражается в длине и расположении фуркальных
ветвей, в вооружении ног V пары, отчасти в строении 3 задних торакаль-
ных сегментов и генитального сегмента; у обоих видов плавательные
ноги вооружены по типу Terni. Однако Kiefer (1931:53) определенно
говорит в диагнозе, что „das 2 Thoraxsegment ist nicht besonders ver-
starkt und umfasst nicht das 3 Segment auf der Bauchseite"; рис. 10
(стр. 54) Kiefer’a соответствует этому диагнозу и определенно указывает,
что говорить об идентичности сравниваемых двух форм не приходится.
К сожалению, Kiefer не дает измерений С. ochridanus, а результаты
биометрического исследования этого вида, о котором упоминает Kozmifiski
(1935:250), насколько нам известно, не опубликованы. Отметим также,
что у сравниваемых видов различно вооружение внутреннего края фур-
кальных ветвей и строение дистального членика ног V пары.
Распространение. С. smirnovi в большом количестве найден
нами в пелагических сборах Седельникова (18—19 VI 1916) из оз. Таль-
меньего на Алтае, лежащего на высоте 1460 м над ур. м. близ Бийска.
Биология не изучена. В 1-й половине июня найдены, главным
образом, неполовозрелые самки, единично взрослые. Сборы относятся
к глубине 0—30 и 0—60 м (максимальная глубина озера 73 м, средняя
глубина 38 м). Температура поверхности 18 VI была равна 8°.
<3
Sft
ft
$
Измерения С. smirnovi, йр. п. и С. lacustris Sars
С. lacustris Sars. (Ладожское озеро) ио сч ио оо о оо со г-н сч СО о со ио ио U0 <Р 1П 04 СЧ О СО СО гЧ г* г* о СО хР О' СЧ СЧ 40 со О'* ио оо 4© оо сч со о оо г-н о СЧ З3 хр г-н О> СО Г* <ч г-«г-1 СЧ 40 СО СО ГН СЧ г-1 СЧ СЧ г-1 СЧ г-н U0 Г” СЧ г-4 т—1 т—1 г-Ч
О 40 г-н О> 40 СХ 04 Г-СО U0 00 со СО 40 04 40 О 40 40 04 СЧ ио Г-«-и Г—со о г.Ч . • 40 Г* СО со 40 хР О и*>00.4>00СЧСПО1НОГ’« СО т—« СО СО хР t—«ООО© г- г-1 сч 40 СО СО СЧ СЧ г-1 СЧ СЧ 1-4 СЧ г-1 Хр г-.ЧгЧ r-i Г-Н t-H
со Ч* Г- 4© 00 U0 О <Р -С? Г* г-Ч 04 © 04 СО хР 00 Q t-ч СО хР СО U0 Г—4© О СО СЧ иОсои^чОг-нОГ^чОСЧ чОСОсОГ”СЧСЧО'ч5г-<сО хР О СО <!-хР СЧ 04 оО чО Г-4 г-н 1—1 гЧ 40 СО СО СЧ СЧ г-н СЧ СЧ СЧг-с^ 40 СЧ t-ч i г-н г-Ч
сч СО U0 00 U0 U0 ’чр о \ПСОГ’«СОчОчОчОООгЧ’^ и004’чРОГ-0404С4 СО Г* Г* 40 U0 ч0 40 40 О 1П О 40 00 1П -хР 04 О СЧ ЧО • хр >-н UO хР^ч ©4 г— (О4Ч0СО со СЧ СО 1-4 СЧ СЧ СОгЧиО оО СЧ 1-4 гч
Т-—< чПчОчПОО'П г-’гр Г—о Г— сч ЧО хР О 00 U0 О 04 СО хР 04 СО СЧ хР СО 04 гЧ<Пч©1П0401ПчС>гЧ 40Г-40Г^СОСО0>ООСОг-ч хр ( т-ч СЧ хР гЧ рч i — г-1-ч гЧ ,-4 r-ч 4D СО СО СЧ СЧгЧ СЧ СЧ СЧ гЧ U4 Г-СЧ т-ч t-ч гЧ г-Ч
С. lacustris Sars. (Онежское озеро) U0 ио ио СО 00 ОхР ОхРгЧГ^ .хР гЧСЧСЧгЧГ-1ПСЭчд1ПС0 Г'-хРхРгчСЧгНхРО Г—40хРГ-40 40 40ООоО иОиО40СОСЧОО>Г—СОО хР40ОСОхРСЧ40ОО иОт-ЧгЧгЧфчПСОСЧ 1-4 СЧгЧ СЧ СЧ СЧ т-Ч <0 иО СЧ i—< т-ч чЧ t—|
хр 4040 ТГ СО хР СО СО хР СЧ г—«041-4 СЧСОгЧО4ОхРсОхР40хР xP0Qc0O44©O4O4CO t-н хР СХ U0 СО СЧ U0 40 оО 1П СО чО СО 1-4 04 04 СЧ 1П хр 00 04 СЧ СО t-Н U0 СО 40 т-Ч t-H i—i О U0 СО СЧ t-H СЧг-Ч СЧ гЧ СЧ г-Ч хР Хр r-Ч ^ч г-Ч
со Г— хр Т хР 0 40 СО 00 Г- о> Г* СЧСО- СЧ об СЧ СО Г-СО 04 04 Г-4 Ч© 00 СО S г-4 сб СЧ Г-Ч сч 40 U0 СЧ 40 О СО 00 Г— 1П хР 1П 00 СЧ О 00 04 1П сч хр СО 04 СЧ’Ч1 СЧ 4© оО 40 t-Ч т-ч г-Ч t-Ч U0 СО СЧ t-ч СЧ Г-Ч СЧСЧ СЧг-HUO 40 г-Ч г-4 г-Ч
сч хР хр Ь- U0 тРСОСОГ-СЧобг-ОСЧ ’З'чПчО'-Чг-ЧхПГ-СЧСЧхР гЧ 40 00 мА оо 00 о г— СЧГ*СЧ40(0Чг-чсООООО U01xP00 04C004UOC4xP хРСООСОСОг-Ч40Г— КО 1—• ₽ч о мп СО СЧ 1-4 СЧгЧ г-Ч t-ч U0 СЧ гЧ гЧ
т^ 4© 40 40Г-хРсОО'’ЧсО4©Г- гЧ4©хриОхРгЧоОООхРСО счсоооио^сооо Q ио СО 40 40 СО U0 СЧ Г-4 ч© хр 40 Г* О О О' СО —'Хр хРОг-Ч^хРСЧ40г- 40 т-ч гЧ т-ч О Щ СО СО СЧ С. 11—4 СЧ . Ч СЧ *4 хр ио СЧ Т-ч т-ч —Ч Т-Ч
С. smirnovi sp. n. Оз. Тальменье, Алтай. U0 хр 04 хР 04 г* 04 о Г-О 40 40 ч© со ио ио Т-ч 04 Г-ч хР СЧ »-ч 1—(, оо Г- 4© ио ГЧ 40 00 М* UO U0 СО СЧ in т-ч . 40 04 СО 00 • 4 СЧ 04 ^Р О Г— хр СЧ СО хР СО «*“« СО Г- 40 Т-ч t-ч Г-Ч г-Ч ио СО СО СЧ СЧг-ч СЧСЧ СЧ т-ч ХР 40.4 1-4 г-Ч »-Ч г-Ч
ХР Хр О 40 О О Хр UO U0 хР О СО Г- СЧ СО т-ч 04 ХР UO U0 »-Ч U0 О U0 Г-Ч© 00 40 ио Г—ЧП 00 40 40 Тр СО О 40 Г* 40 со СЧ СЧ 04 U0 ГЧ г- Тр со ' Ч со со т-ч ч© Г*. Ч© t-Ч г-Ч Т-ч Т-ч ио со СО . Ч СЧг-ч СЧСЧ СЧ >-ч тр 4© СЧ чч гч т-Ч т-Ч
со 'Р 4© 40 СО'Р О U0 О ио Т-ч г* 00 г* Т-ч т-ч о чч 40 СЧ СО 4© U0 СО Г* 4© U0 Чр W0 ^Р тр U0 О'О U0 хР t-Н ч© 04 оо 00 СЧ СЧ О ио О Г- ’ЧР 1> СО хр со г-Ч 1> г* 40 т-4 t--« t—<—<U0C0C0.N СЧгЧ СЧ .4 г-< •. ч т-ч ^р U0 СЧ т—< ,-ц
сч -оо 00 40 40 00 00 СЧ *«т Хр -Чр СЧ Т-ч Т-ч 40 О Г- МО Чр т-ч СЧ 04 гЧ хР 00 40 00 00 о UO ио U0 г-оо U0 СЧ 4© Q 00 00 1-4 СО-О 1> г-Ч 04 хРч©-чр’5рСО,-чСОГ'- г-Ч ^ч ^-4 ^ч ио СО СО СЧ СО т-Ч СЧ СЧ т-ч СЧ т-Ч хР ч© СЧ т-ч т-ч т-Ч
г-Ч Хр 4© U0 00 Г* 40 00 04 СЧ т-ч О сч ио 40 00 со 00 Т-ч СЧ 4© 4© U0 со хр Т-ч 40 ч© тр СО 00 O4UOxp\04©r-xPI>C4 40ОООООСЧСООГ*ОО xPI>40UOcOt-lOl> 40 t-Ч т-ч Т-ч т-ч ио СО со СЧ СО т-Н СЧ СЧ г-Ч СЧ г-Ч U0 4© СЧ *-Ч СЧ г-Ч т-ч
d о+ со 1 СП л Дл. цефалоторакса Дл. тор. сегм. 2 Дл. тор. сегм. з Дл. тор. сегм. 4-|_5 Дл. цефалот. -+- тор. сегм. 2—5 • • Макс. шир. цефалот Я я . В °/00к ДЛ. тела . Шир. тор. сегм. 4 „ В °/оо к ДЛ. тела . . „ „ „ в о/о к макс. шир. цефалот Шир. тор. сегм. 5 » я » в О/оо к дл. тела . . я „ я в °/0 к шир. тор. сегм. 4 Дл. ген. сегм. Макс. шир. ген. сегм я „ я я в % к ДЛ. ген. сегм. Дл. задн. абд. сегм „ „ я » в % к дл. фурк. ветв. Дл. абд , „ в % к дл. цефалот-ь -+- тор. сегм: g—5 • Дл. тела „ фурк. ветв Дл. фурк. ветв. в °/оо к дл. тела . . Шир. фурк. ветви „ я я в % к дл. фурк. ветви Расст. А (рис. 33, ik) ....... . „ „ в °/0 к дл. фурки
13 Фауна СССР
Продолжение таблицы 13
С. lacustris Sars. (Ладожское озеро) чЛ СО- r-«d«04»—(0404чНОГ'-СО СО r*O44Hd*O4r*04OO404d*4© Г-04 Г-г-Н О г* чП к© 04 О СО 00 чП СО 04 00 чП чН 04 СО 04 СО 04 чП 04 d< 04 Г* 40 Г* | СП Г-< d* 04 чН СО «—< 04 own l-Н J—* 1—4 »—< !—< г-4 04 1 г-4 ч© гЧ
d1 О;; к© О СО U0 00 СО СО г-4 04 СО Г* СП О d СО Г- СО чО Г* d< 00 4©i—«Г-J-H ч© чП чН к© d* r-t d* 00 чП со 00 00 d* чН СО СО СО чП 04 d* i-^ чН СО Г* 00 d* | 04 i—1 d1 04 ЧН СО г—1 г-4 ОЧЧПг-4 г-4 t—( г-4 г—( г-4 04 । г-4 00 г—4
со чН dj 04 i—( ЧЛ чП чН Г-04 04 о чН О СО О 04 04 СО 40 СО чП ч© d* 04 40 ОО г-4 СТчСООч^ноО'-Оч/ООЧЦОСО О'. OO4n0xcOd*d*d1O4d*O4d1cn О- СО СО | 04 г-н со 1/0 СО г-4 г-4 044Hi—1 i i—1 »—4 i—IQO 1 i—i чН т—<
сч U000^0»’^fi/0’^00404 ч0 т—lOOQ^DO^r^r-i 04 СО Q 40 О Г*. чП О 4© Оч U0 04 00 ©Ч т—< <© СП СО 04 ч© 04 ЧП СО d СП Г-00 1 СО •—i U0 04 ЧН СП >—1 г-4 Olflr-1 i-4 ,—i t—i ,—< нООО «-» 04 г-ч
г-4 d1 О СО 40 d* 04 00 г-н 04 О 04 04 г— СО СО 04 Оч 2 00 1 1 1 1 I | 1 1 1 1 2 r'£g,J:
С. lacustris Sars. (Онежское озеро) _ . ' ЧП d in • о ^ООООГтЧ^чОч© 04 Г-040ч\©О00^1ПС0ГГ>сП0ч <О О ОО,г? СО d 04 4© 00 О О \О чП СО СО СО d СО г-i d i—ICO»—i d О ЧЛ 04 оО чП ч© 1 ’ 04 г-i d 04 d СО г-4 04 OOlHi—i i—i i—i г-i i—i «—‘Г^оО
d " d d d «4*Г-040ч©г-ОчсОГ-04 О чП О d 04 СО 04 ч© d* 04 04 СО СО d 04 i—1 ! 4© Г- ч© »—i СО 04 СО 04 Г- чН 0ч 04 г-< d 04 СО О СО О d О d О ч© чН СО 04 d "d СО d* СО i—i • i-ч 00 чЛ 1—i i-и 1—i г-< 1—14©Г— 1-4
со чП d 1—1СООО4О4Г-ОС")ОЧ40 .-и С0ОС0С0ОС0С0ОчО04О40 ООО ddd4©OO©4C*u0C0 О 00 СО О О d* i—1 о «t О in М • г* чО со 04 i—i d 04 чП СО г-1 04 ОО чп 1—i t—l ,—i r-1 i—i i—i CO 04 1-И i-4
04 ЧП d O4©dO'O©4 014©04O4 m d4nOc6cOOG4COCOdCO40 40 uo d . d чО> i—<CO0>i—i©4©<fco CO СО-^чПОСО'ОСНг-!^O^r-< co CO d 04 i—l d 04 d СП i—( 04 004Hi—i 1—1 r-4 r—< i—i 1 1-4
Т—I 1П1П \Л чЛ чП i—i 1—i СЭ © Оч ЧП CO 40 4© г— d 04 40 04 СО СО 40 СО чН ч© ч© 04 04 о 1—1 co’T* r-COCOt—(ООчПОчЛСО О Г*чЛч©0<Л04СО»—idOdO’ 00 40 Г-н 1 04 1—« d со 4© d 1—11—( 1—1 оо чП 1—1 1—1 1—< 1—1 1—1 Г-,Г','С)
С. smirnovi sp. n. Оз. Талъменъе, Алтай 5" 04 04 сч СО О 4© 04 04 04 чН 04 О 4© d d Г- СО Г- d Г- 00 40 04 ч© ч© СО ч© 04 d i—i Г—ЧЛ СО 04 00 О ЧН СО СО d О» 04 d | 1 04 чН 4© чП I CM i—i U0 СО Г-d d 04 О4чПт—< .-и г-н II t-400 1 1-4
d* 1—1 Г-04 '<4 чП 1—1 о Г—00 04 04 40 О 1—1 СО Г* чП чП ч© СО 00 г* 04 чП d d СТ' о ЧП о 1—1 ч© d 04 1—1 Г* чП 00 ЧП со чП 00 СО 1 104 чП Г- чП | 041—1 d* СП 4© -d* т-Ч 04 04 ЧЛ 1—Е г-4 Т-Ч II 1—* 00 1 J
со чЛ Ч0ч©ООчЛОО4О4©С01—1 чЛ О О 1—1 О 00 чП О чЛ 04 . , О СО i—> i—i , ’d* чЛ 04 CO 4© i—i 04 Г* М* СО О 00 О Г* «ч* СО d1 04 СО‘«J1 04 чЛГ-н сч t-н чЛ СО 4© «d* »—1 04 ©4 ЧЛ !—1 ,—1 ,—1 11 1—(СО 1 1-4
04 04 U0 чЛ М* 04 04 i—i 04 i—i СО О СО О 0-СО ЧЛ СО О чЛ 0- СО 4© О •d* d1 СЧ г-l чЛ 04 1-4 0- ЧЛ СП О 00 1—11—1 СО со чЛ со d* d1 | |н 4© СО i—i 1 04 1—1 чЛ со 4© t.o 1—1 СЧ О 4© 1-4 г-н | | т—1 1—1 04 |
г—4 чЛ ч© 00 d1 Оd* 4© d* О'N 40 СО чЛ 1-1 d* 04© о о о о со . . ₽+* 00 О \Л . чЛ ЧЛ d* 4© 04 СО 04 Г* чЛ со i—i 4© О \© СО d1 d* Г* d* чЛ О чЛ СО СО 04 rH d* 04 Г* d1 гН -.4 г—1 4© 1—1 ,—i ,—i 1 1 1—1 г-Н 04 1 1“^ 1“Н
( рв б) Дл. внутр, кр. ап. щет „ ,, „ „ » в °/оо к Дл.тела . Дл. внеш. ер. апик. щет ,, » ,, „ » в%окдл-тела Дл. внутр, кр. ап. щет „ „ „ „ „ в °/оо к дл. тела Дл. внеш. ап. щет » „ ,, ,, в %о к дл. тела . Дл. латер. щет „ „ „ в °/оо к ДЛ. тела . . . Дл. внутр, ап. щет. в % к дл. внеш. Дл. внеш. ср. ап. щет. в °/0 к дл. внутр, ср. ап. щет Дл. антенн I пары „ „ ,, ,, в °/оо к дл. тела . . Дл. эндоп. з IV пары ног Шир. эндоп. з IV пары ног Дл. внутр, ап. щет. эндоп.д IV пары ног п внеш. » » » » » „ „ экзоп. з IV пары ног Шир. ft п it „•••••• Дл. энд. з III пары ног Шир. „ » » » •••••• Дл. мал. щет. V пары ног ..... » » it » >» » в °/о к ДЛ. болып. щет. V пары ног Дл. болып. щет. V пары ног .... Общая длина тела Количество яиц ..........
7. CYCLOPS
195
7. Cyclops ocbridanus Kief.
Cyclops ochridanus Kiefer, 1931a:53, f. 10—13.
Самка. Тело стройное, задние углы 2-го торакального сегмента
слабо развитые, не загибающиеся на брюшную сторону. Задние углы
3 задних торакальных сегментов слабо выступающие кзади. Генитальный
сегмент объемистый, мало и равномерно суживающийся кзади. Recep-
taculum seminis несколько вытянутый вдоль генитального сегмента.
Фуркальные ветви очень длинные, их длина, в среднем, в 9 раз превы-
шает ширину, внутренние края снабжены редко расположенными
волосками, не доходящими до основания ветвей, дорзальная щетинка
прикреплена очень близко от их конца, длина внутренней из крайних
апикальных щетинок немного менее чем вдвое превосходит длину внеш-
ней щетинки. Передние антенны достигают, приблизительно, до заднего
края 2-го—или до середины 3-го торакального сегмента. Формула
шипов 3-4-3-3 (группа Terni). Длина дистального членика эндоподита
ног IV пары приблизительно в 3.5 раза более ширины, внутренний
апикальный шип этого членика несколько более, чем вдвое, длиннее
внешнего. Ноги V пары как у С. strenuus, но внутренний шип дисталь-
ного членика короткий, слабо развитый. Длина 1.6—1.64 мм. Самцы
не известны.
Распространение. Известен только из оз. Охрида в Югославии.
Биология. Изучена очень мало. Вид пелагический, в указанном
озере встречается круглый год.
8. Cyclops kolensis Lili. (фиг. 40).
Cyclops kolensis Lilljeborg-, 1901:21, t. 2, f. 11—12. — C. strenuus (part.) Рылов,
1928:10, рис. 5—6. — C. kolensis Rzoska, 1930:199, 206, f. 2; Kozminski, 1933:110, f. 2—-10,
t. 5.— C. kieferi Schafer, 1933, Zool. Anz., 104:102, f. 1—6. — C. silesicus Schafer, 1934,
ibid., 105:224, — C. kolensis Kozminski, 1934:110, 115, 117, t. 4, f. 8; Kozmiiiski, 1936:223.
Самка. Туловище относительно толстое, укороченное, цефалоторакс
очень объемистый, задние углы 2 передних торакальных сегментов
одинаковые, незначительно выдающиеся кзади, у предпоследнего сегмента
значительно оттянутые кнаружи, иногда немного загнутые кпереди.
Задний торакальный сегмент с хорошо развитыми боковыми выростами.
Генитальный сегмент очень узкий, стройный, receptacnlum seminis круп-
ный, округлый, очень близкий к таковому С. strenuus. Фуркальные
ветви весьма укороченные, толстые, мало расходящиеся, на внутренних
краях вооруженные немного не доходящим до основания ветвей рядом
волосков. Внутренняя из крайних апикальных щетинок относительно
короткая, менее двойной длины внешней, внутренняя из средних апикаль-
ных щетинок лишь немного длиннее внешней, дорзальная щетинка при-
близительно равна внешней крайней. Передние антенны 17-(иногда 18-)
членистые, обычно доходящие до заднего края цефалоторакса, иногда
достигающие конца 2-го или середины 3-го торакального сегмента.
Вооружение плавательных ног всегда по типу Bini. У ног V пары
щетинка базального членика относительно очень короткая, значительно
не достигающая середины апикальной щетинки дистального членика,
хорошо развитой боковой шип которого своим концом выдается кзади
от конца самого членика. Длина 1.03—1.77 мм. Окраска светложелтая,
более темная у цефалоторакса. Яйцевые мешки небольшие, тесно при-
13*
196
CYCLOPINAE
легающие к генитальному сегменту, содержащие небольшое количество
яиц (по 7—18 в каждом).
Самец. Фуркальные ветви относительно короче, чем у самки,
их латеральная щетинка прикреплена несколько далее от их конца,
а внутренняя крайняя апикальная щетинка длиннее, чем у самки. Ноги
Фиг. 40. Cyclops kolensis Lili., самка.
1 — передний отдел тела со спинной стороны; 2—задний отдел тора-
кса и генитальный сегмент со спинной стороны; 3—receptaculum semi-
nis; 4 — фурка с брюшной стороны.
VI пары вооружены внутри сильным шипом и парой оперенных щетинок,
которые длиннее шипа; внешняя из них более чем вдвое превышает
длину шипа. Сперматофоры на генитальном сегменте самки располагаются,
расходясь под значительным углом. Длина около 1 мм.
На табл. 14 даны измерения (в микронах) 5 самок с яйцевыми
мешками с о. Новая Сибирь.
Рассматриваемый вид долгое время смешивался с другими видами
группы С. strenuus, в частности с С. strenuus Fisch., пока Rzoska
и Козминский не выяснили его видовой самостоятельности. Он, бесспорно,
является „хорошим" видом. По строению и вооружению фуркальных
7. CYCLOPS
197
Таблица 14
Измерения С. kolensis u. С. furcifer
Экз, № (g ad.) С. kolensis (О. Новая Сибирь) С. furcifer (Днепропетров- ская область)
1 2 3 4 5 1 2 3
Дл. цефалоторакса 441 528 458 448 499 699 764 655
Дл, тор, сегм. 83.3 120 108 101 110.6 194 209 181
Дл. тор. сегм. 87.5 120 120 94 112.7 219 223 194
Дл. тор. сегм. 88.2 169 142 132 149 240 250 222
Дл. цефалоторакса тор. сегм, 2_g . 700 937 828 775 871 1352 1446 1252
Макс. шир. цефалоторакса 425 512 426 437 443 704 758 698
» „в °100 к дл. тела 402 381 334 379 345 349 359 374
Шир. тор. сегм. 327 388 350 345 350 486 511 460
» » в °/оо К дл. тела . . 309 288 283 299 272 240 242 247
» „ » в и/0 к макс. шир.
цефалотор 77 76 82 79 79 69 67 66
Шир. тор. сегм. 244.3 274.4 259 307 251 364 365 371
» » в °/оа к дл. тела . . 231 203 209 266 196 179 173 199
» » » в »/о к пгир. тор. сегм. 4 75 71 74 89 72 75 71 81
Дл. ген. сегм 191 210 210 196 208 337 340 299
Макс. шир. геи. сегм . . 176 211 196 179 186 274 286 266
» » » » в °/q к дл. ген.
сегм. 92 100 93 92 90 82 84 89
Дл. задн. абд. сегм 164.5 198 199.7 183 206 335 325 314
» >> » „ в °/о к дл. фурк. ветв. 118 103 129 120 147 103 103 105
Дл. абд 355.5 408 410 379 414 672 665 613
» » в °/а к дл. цефалот. -+-
н- тор. сегм. 50 44 49 49 48 50 46 49
Дл. тела . . 1056 1345 1238 1154 1285 2024 2111 1865
Дл. фурк. ветв 140 192 154 152 140 326 317 298
» » » в °/оа к дл. тела . . 133 143 124 132 110 161 150 159
Шир. фурк. ветв 31 35 29 32 35 36.4 40 39
_ » » >• в % к дл. фурк. ветв. 24 18 19 22 25 11 13 13
Расст. А (фиг. 33, ik) 106.4 147 111 112 107 255 274 228
» „ в к дл. фурки 76 77 72 74 76 78 86 77
Дл. внутр, кр. ап. щет 99.4 127 104 113 120 226 202 200
» » » » » в к дл. тела . 94 94 84 98 93 111 96 107
Дл. внеш. ср. ап. щет. 294 350 300 350 305 520 485 532
’> » » » » в °/оок ДЛ. тела . 279 260 242 217 237 257 230 285
Дл. внутр, ср. ап. щет 400 430 400 400 375 632 546 654
» ” »» » » в Уоо к дл. тела 379? 320 323 347 300 314 259 351
Дл, внеш. ап. щет 70 85.4 74 78 86 182 150 156
» » ” » в °/оо к Дл. тела . 66 63 60 67 67 90 71 84
Дл. латер. щет. 38 39.2 36 39.2 34.3 55 53 46
" " » в °1оп к дл. тела . . Дл. внутр, ап. щет. в % к дл. внеш. 36 29 29 34 26 22 25 25
ап. щет 141 147 141 145 140 124 136 128
Дл. внеш. ап. ср. щет. к о/о к дл.
внутр, ср. ап. щет 74 82 75 87 82 82 88 81
Дл. антенн I пары 380 539 547 399 — 867 996 810
» » » » в °/аа к дл. тела . . 366 401 442 346 — 428 472 434
Дл. дорэ. щет, — — — — — 104 116 80
» » >, в °/оо к дл. тела . . . — — — — 51 55 43
Дл. эндоп. з IV пары йог 60 — 75.3 70 — 123 140 100
Шир. „ „ „ „ 22.5 27 26.7 — 44 50 43
198
CYCLOPINAE
Продолжение таблицы 14
Экз. № (g ad.) С. kolensis (О. Новая Сибирь) С. furcifer (Днепропетров- ская область)
1 2 3 4 5 1 2 3
Дл. внутр, ап. щет. эндоп. я IV пары ног 60 70.5 83 115.5 115 108
24.6 — 25.8 30 — 56.7 73 51
Дл. экзоп. з IV пары ног 71.7 — 83.7 91 — 120 148 100
Шир. „ „ „ „ 30 — 28 33 — 54 54 51
Дл. мал. щет. V пары ног 40 49 40 38 39 75 66 62
» » » » » В °/о К ДА. больш. гцет. V пары ног .... 54 64 58 54 70 70 58 62
Дл. больш. щет. V пары ног .... 74 77? 69 71 56 107 113 100
Общая длина тела 1196 1537 1392 1306 1425 2350 2428 2163
Колин, яиц 11—12 15-18 7—7 13—13 —
ветвей н плавательных ног он хорошо отличим от С. strenuus, С. abys~
sorum, С. tatricus и близких к ним форм. Отличия от С. furcifer указаны
при описании последнего. От С. vicinus, у которого плавательные ноги
также вооружены по типу Bini, С. kolensis без труда отличается
по габитусу туловища, особенно по строению 2 задних торакальных
сегментов, а также по строению генитального сегмента и фуркальных
ветвей.
Распространение выяснено очень неполно. С достоверностью
найден в средней и южной Швеции, северовосточной Германии и на севере
СССР. Возможно, что этот вид был найден ClofssonoM (1918; С. strenuus,
part.) на Шпицбергене и Marsh’eM (1920; С. vicinus) на Аляске. В пре-
делах СССР известен с Кольского полуострова (Lilljeborg, 1901, оз. Пере-
сяр близ Мурманска) и с о. Новая Сибирь (из группы Новосибирских
островов; Рылов, 1928, С. strenuus part.).
Биология. Обитает в пелагиали и литорали преимущественно
эвтрофнрованных озер, в олиготрофных озерах спорадически и в незна-
чительных количествах. Обитает также в мелких водоемах, исключая,
повидимому, сильно дистрофироваяные. Переносит сильное обеднение
растворенным кислородом и высокое содержание Са, обильное развитие
наблюдалось при 19° (немецких) жесткости. Цикл развития изучен
довольно полно. Дицикличен, главным образом зимой. В Вигерских
озерах (северовосточная Германия) половой период наблюдался с декабря
до конца марта, максимум обычно в феврале — марте. В апреле — мае
встречается лишь молодь (новой генерации), причем старые самки и самцы
отмирают. Новая генерация достигает половозрелости в июне, причем
наступает новый максимум (начало лета), менее высокий, однако, чем
зимой. К августу рачок исчезает и снова появляется в сентябре — октябре.
* 9. Cyclops furcifer Claus (фиг. 41).
Cyclops furcifer Claus, 1857:208, t. II, f. 14—16. — C. brevicaudatus (part.) Claus,
1863:100. — C. miniatus Lilljeborg, 1901:24, t. II, I. 13—15. — C. lacunae Lowndes, 1926,
Ann. Mag. Nat. Hist., (9), 18—142, t. 7, i. 1—13. — C. strenuus f. furcifer Kozminski,
1927:72, f. 14. — C. miniatus Kiefer, 1927, Arch. Balat., 1:412, f. 33—35. — C. (s. str.)
furcifer Kiefer, 1929:51; Rzoska, 1930, t. 1—2; Rzoska, 1932:428; Roy, 1932, Bull. Soc.
Agric. Sarthe, 53:9; Roy, 1932, Cop. Clad. Ouest France: 163, f. 77; Kozminski,1933:109;
Qurney, 1933:170, f. 1542-1548; Kozminski, 1934:110, t. 4, f. 7.
7. CYCLOPS
199
Самка. Туловище толстое, сильно расширенное в переднем отделе,
.цефалоторакс относительно короткий, внешне-задние углы передних
Фиг. 41. Cyclops furcifer Claus, самка.
1 — передний отдел тела со спинной стороны; 2—задний отдел и гениталь-
ный сегмент;1 3 — абдомен и receptaculum seminis; 4 — нога V самки;
5 — фурка со спинной стороны.
2 торакальных сегментов лишь слабо выдаются кзадн, у 4-го сегмента
они очень сильно вытянуты кнаружи, в виде отростков, нередко загнутых
кпереди, боковые выросты заднего торакального сегмента короткие,
тупо заостренные. Генитальный сегмент довольно широкий спереди,
сильно суженный и почти цилиндрический в задней половине. Recepta-
200
CYCLOPINAE
culum seminis обычно удлиненный, похожий на таковой у С. strenuus..
Фуркальные ветви относительно тонкие, очень длинные, обычно умеренно
расходящиеся. Из крайних апикальных щетинок внутренняя лишь немного
длиннее внешней, иногда почти равна ей. Средние апикальные щетинки
хорошо развитые, нз них внутренняя немного (примерно на */в—Vs)*
длиннее внешней. Дорзальная щетинка почти вдвое короче внешней
из крайних апикальных щетинок (ее длина равна 43—71 % длины
туловища). Внутренние края фуркальных ветвей с немного не доходящим
до их основания рядом волосков. Передние антенны 17-членистые, очень
короткие, доходящие лишь до заднего конца цефалоторакса или.
лишь немного длиннее. Вооружение плавательных ног непостоянно1 —
по типу Bini, или Terni, или по смешанному типу. У ног V пары щетинка
базального членика относительно короткая, не доходящая до середины
апикальной щетинки дистального членика, боковой шип последнего
сильно развитой, значительно выдающийся кзади от конца этого членика;
группа шипиков в основании апикальной щетинки хорошо развитая.
Длина 1.5—2.4 мм. Окраска обычно желтоватая, иногда оранжевая.
Яйцевые мешки небольшие, содержащие обычно значительное количе-
ство яиц.
Измерения экземпляров из СССР (Днепропетровская обл., воору-
жение ног по типу Terni) приведены в табл. 17 (см. С. kolensis, в таблице
справа).
Самец. Фуркальные ветви немного короче, чем у самки. У ног
VI пары внутренний шип короткий, из 2 щетинок внешняя немного
длиннее соседней. Сперматофоры прикрепляются на генитальном сегменте
самки косо и расходятся под значительным углом. Длина 1.1—1.5 мм.
Долгое время С. furcifer смешивался с другими формами группы
strenuus и, без сомнения, нередко указывался как С. strenuus Fisch.
От С. kolensis он без труда отличим по строению туловища (особенно
по форме цефалоторакса), по форме 2 задних торакальных сегментов,
а особенно по длинным фуркальным ветвям и их апикальному вооружению
и по очень стройному абдомену.
Распространение выяснено очень недостаточно. Известен
из Великобритании, Германии, Венгрии, Франции, Швеции, СССР
и Северной Африки (Алжир). В СССР найден близ Полтавы, в Днепро-
петровской области и в низовьях р. Колымы. Вероятно у нас распро-
странен очень широко.
Биология. Очень характерен специально для мелких водоемов
(лужн, болота, мелкие пруды), особенно для астатических, пересыхающих
летом; нередко совместно с С. strenuus s. str. По Lowndes’y (1928)
обитает в водах с pH = 6.6—8.2. Встречается в средней Европе зимой,
весной и осенью, максимум ранней весной. По экспериментам Roy (1932)
пересыхание водоема может переносить и в состоянии яйца (фаза раз-
вития не выяснена), и во взрослом состоянии (также на 4-ой копеподит-
ной стадии). Специального „кокона" при этом не образуется, а рачок
приобретает опаковый оттенок, покрываясь обильно слизью (секретом
кожных желез).
*10. Cyclops scutifer Sars (фиг. 42).
Cyclops scutifer Sars, 1863, Forh. Selsk. Christ. (1862):237; Sars, 1828:349, t. 9, f. 1—5?
Lilljeborg’, 1901: 33, t. 2, f. 26—27; 36, t. 19; Рылов, 1918:234, фиг. 12—13; Bowkiewiez,
1 О формуле шипов С. furcifer см. особенно у Lowndes’a (1927:170) и Kiefera,
(1936:160).
7, CYCLOPS
201
1928, Arch. Hygrob. iRybact., 3:40,1.1; Kiefer, 1929:51; Gurney, 1933:169, f. 1541; Kozminski,.
1933:101, t. 1, f. 1—2; Kozminski, 1934:110, 114—117, t. 4, f. 1; Kozminski, 1936:208,—
Cyclops strenuus Marsh, 1920:10, t. 4, f. 2, 3, 7. — Cyclops scutifer f. asc Kozminski,
1927:68, f. 15.
Фиг. 42. Cyclops scutifer Sars, самка.
7 —фурка с брюшиой стороны; 2—задний отдел торакса и абдомен; 3—дистальный;
членик эндоподита ноги IV; 4—генитальный сегмент с receptaculum seminis; 5—геиг-
тальный сегмент с прикрепленными сперматофорами; б—экзоподит ноги I;
7—3 дистальных членика аитени I; 8—нога V.
202
CYCLOPINAE
Самка. Тело стройное, удлиненное, задние углы цефалоторакса
и следующих за ним 2 торакальных сегментов не выдаются кзади.
Предпоследний торакальный сегмент сильно расширен сзади, его наиболь-
шая ширина лежит на линии, соединяющей заостренные на конце,
весьма сильно оттянутые кзади и в стороны задние углы, причем сзади
справа и слева сегмент образует мощные крыловидные лопасти. Задний
торакальный сегмент с сильно развитыми заостренными боковыми выро-
стами, лишь немного уже предыдущего сегмента. Генитальный сегмент
сильно расширенный спереди, значительно и равномерно суживающийся
кзади. Receptaculum seminis овальный, вариирующих очертаний, иногда
заметно удлиненный. Фуркальные ветви относительно короткие, очень
мало расходящиеся, со сплошным рядом волосков на внутренних краях.
Латеральная щетинка прикреплена значительно кпереди от конца ветвей.
Внутренняя из крайних апикальных щетинок в 2—2'/2 раза длиннее
внешней, внутренняя из средних апикальных щетинок заметно (самое,
меньшее на */4) длиннее внешней. Длина дорзальной щетинки, в % длины
тела, в среднем 93 (78—108). Передние антенны 17-членистые, доходя-
щие до середины или до заднего конца 2-го торакального сегмента,
иногда немного длиннее. Вооружение плавательных ног по типу Terni.
У ног V пары щетинка базального членика относительно короткая,
значительно не доходящая до середины апикальной щетинки дисталь-
ного членика, шип на внутреннем крае последнего длинный, хорошо
развитый, выступающий за конец членика. Длина 1.135—1.40 мм (var.
wigrensis— до 1.86 мм). Самки очень прозрачны, обычно бесцветны,
иногда голубоватой или зеленоватой окраски; последняя иногда смешан-
ная, например в олиготрофных озерах Мурманского побережья у самок
и самцов туловище, абдомен и фуркальные ветви светлоголубые, с оранже-
выми включениями жира в тораксе; яйцевые мешки темнофиолетовые,
округлые, тесно прилегающие к генитальному сегменту, содержащие
небольшое количество яиц (по 4—10, редко более 10 в каждом; по Ekman’y,
1904, в водоемах крайнего севера Скандинавии — до 40).
Самец. Фуркальные ветви лишь немного превышают длину 2
задних абдоминальных сегментов. Длина 1—1.3 мм. Сперматофоры
очень крупные, бобовидные, на генитальном сегменте самки располагаются
расходясь под значительным углом, задними концами выступая за края
сегмента.
С. scutifer близок к С. vicinus, но хорошо от него отличается
по вооружению плавательных ног (Terni, вместо Bini), по относительной
ширине 2 задних торакальных сегментов, по форме генитального сегмента,
по строению и вооружению фуркальных ветвей, в частности по положе-
нию места прикрепления латеральной щетинки относительно начала
внешнего края — у С. scutifer она прикреплена ближе к началу, нежели
у С. vicinus.
Известен 1 вариетет — var. wigrensis Kozm., описание которого дано
ниже.
Распространение.1 Европа (Скандинавия, север Финляндии,
Германия, север СССР), север азиатской части СССР, Северная Америка
[Канада (Marsh, 1920 — С. strenuus, non vicinus; Willey, 1923 — C. scutifer)
близ Нью-Йорка (Marsh, 1912 — С. strenuus Gurney, 1933 — C. scutifer)).
1 Нахождение в Татрах (Wierzejski, 1833, brevicaudatus) и в Альпах (Zsehokke,
1900, и др., С. strenuus part.; Schmeil, 1893, С. scutifer) требует проверки (см. Kozminski,
1936:210). Указание Ekman’a (1904) на нахождение в Гренландии и Исландии ошибочно
(см. Wesenberg-Lund, 1905; Haberbosch, 1920; Рылов, 1928).
7. CYCLOPS
203
В СССР известен с достоверностью с о. Вайгач, Соловецких остро-
вов, с Новой Земли, с Мурманского побережья, из ряда озер Кольского
полуострова, из Выпочозера и окрестностей Умбы (ЗИН) на северном
берегу Белого моря, из ряда озер Карело-Финской ССР, с севера
Якутской АССР, из озер бассейна Хатанги. Наиболее южное нахождение —
область озер Сувалок и Вильнюса. В СССР распространение резко
ограничено зоной тундры и тайги (до 60° с. ш.), однако на западе
(Вильнюс) встречается и в зоне смешанных лесов. В горных областях
севера Европы показан до высоты 1200—1500 м. По Ekman’y (1904)
в горах Шведской Лапландии встречается до высоты 1350 м (наивысшее
положение исследованных этим автором водоемов).
Биология. Пелагический озерный вид, в водоемах крайнего севера
живет также в мелких болотах и лужах. Обитает, главным образом,
в олиготрофных (и слабо эвтрофированных) озерах (типичная форма).
Отсутствует в высокоэвтрофных и в полигумозных, очень кислых
дистрофных озерах, встречаясь, однако, при олиго-мезогумозности.
Вертикальное распределение в пелагиали озер изучено пока мало. В олиго-
трофных озерах Мурманского побережья в полдень обитает в больших
количествах в слое 2—7 м, лишь единично встречаясь в слое 0—2 м,
куда мигрирует с наступлением темноты. В карельском олиготрофном
Сайозере, по нашим исследованиям, в августе днем рачок держится
на глубине 5—9.5 м (глубина озера до 11 м), отсутствуя в слое 0—5 м,
а в оз. Утренняя Ламба (там же, максимальная глубина 15.3 м) в слое
О—4 м днем мы нашли только самцов, на глубине 4—8 м — самцов
и самок. По Strom (1931) в олиготрофном оз. Feforvain (Северная
Норвегия, максимальная глубина 53 м) днем держится в слое 0—20 м
и в нижних слоях гиполимониона (30—50 м). С. scutifer, во всяком слу-
чае, в озерах имеет явственную вертикальную стратификацию и днем
избегает верхней зоны эпилимниона; суточные вертикальные миграции,
по всем вероятиям, интенсивны, со значительной амплитудой. Отношение
к факторам водной среды выяснено мало. Некоторые авторы
считают С. scutifer холодноводным рачком, что, однако, оспаривает
Козминский (1936), отмечающий развитие его в летние месяцы; однако
вопрос может быть решен лишь при условии изучения вертикального
распределения в связи с температурой. Повидимому приурочен к слабо-
кислым— нейтральным — слабощелочным водам. По Таусон (1933) в ста-
рице найден при pH = 6.5—7.7. Вероятно в высокой степени полиокси-
бионт (точных данных по отношению к кислороду не имеется). Характерен
для вод с олиго- и мезотипом СаО.
Типичная форма, по Козминскому (1936), моноциклична: молодь
появляется в марте — апреле, половозрелость наступает в июне, максимум
обычно в августе, в сентябре резкая убыль количества и в октябре
выпадение из планктона. По Ekman’y (1904) в горной области северной
Швеции в озерах дицикличен, в мелких водоемах моноцикличен. Вероятно
дициклия имеет место и в оз. Ессей (в бассейне р. Хатанги; Рылов,
1928). Судя по имеющимся данным, вероятно, олигосапроб (лишь в слабой
степени Р-мезосапроб).
Типичная форма Cyclops scutifer Sars, помимо обычно небольшой
величины, отличается следующими индексами (по Kozminski, 1933, 1936):
расст. А (фиг. 33, ik) в % к Дл- ФУРК- ветв. = 62 (59—65); дл. внешн.
ср. ап. щет. в % к дл. внутр, ср. ап. щет. = 68 (62—73); дл. фурк. ветв.
в °/00 к дл. тела=120 (111—129); шир. фурк. ветв. в °/0 к дл. фурк.
ветв. = 19,5 (17—22); шир. тор. сегм. 5 в %0 к дл. тела = 232 (209 —
204
GYCLOPINAE
255); дл. дорз. щет. в °/00 к дл. тела=93 (78—108); дл. больш. щет. V
пары ног в %о к дл. тела = 74 (61—88).
10а. Cyclops scutifer var. wigrensis Kozm.
Cyclops scutifer forma wigrensis Kozminski, 1927:68, f. 10, 16; Kozminski, 1933:102,
t. I, f. 2; Kozminski, 1934:110; Kozmihski, 1936:210.
От типичной формы отличается (самка) следующими признаками:
тело более утолщенное, предпоследний торакальный сегмент спереди
более расширенный, фуркальные ветви относительно длиннее, длина
дорзальной щетинки ветвей относительно несколько менее длины тела,
щетинка дистального членика ног V пары короче.
Основные индексы (по Козминскому, 1933,1936): дл. внешн. ср. ап. щет.
в °/о к ДЛ- внутр, ср. ап. щет. =71 (66—77); дл. фурк. ветв. в °/оо к Дл.
тела = 122 (114—130); шир. тор. сегм. 5 в °/00 к дл. тела = 236 (220—252);
дл. дорз. щет. в °/оо к ДА. тела = 77(65—89); дл. больш. щет. V пары
ног в %0 к дл. тела=61 (50—73); общ. дл. тела 1612 (1360—1865).
Самки var. wigrensis менее прозрачны, яйцевые мешки крупнее, обычно
содержат значительное количество яиц (по 20 в каждом).
Распространение. Известен из оз. Вигры и из нескольких
с ним соседних озер (северовосточная Германия). Нахождение в СССР
вероятно.
Биология. Характерен для эвтрофных озер (нет в высокоэвтро-
фированных). Половозрелость наступает иногда ранее, чем у типичной
формы, уже в мае, но главным образом в июне (как и у типичной
формы), моноцикличен, возможно с наклонностью к дипиклии.
* 11. Cyclops vicinus Uljan. (фиг. 43).
Cyclops vicinus Ульянин, 1875:30, т. 10, рис. 1—7; Lande, 1890:47, t. 16, f. 33—
41, t. 21, f. 164; Lilljeborg, 1901:26, t. 11, f. 16—19; Sars, 1903:216, t. 15, f. 2, a-k; Sars,
1913:37, t. 20; de Lint, 1922:76, 78—80, t. 4, f. 2-5; Kozminski, 1927:44, 74, 79, 91, f. 8,
7; Kiefer, 1929:52; Rzoska, 1930:200, 204, 210—214; Rzoska, 1932:428; Gurney, 1933:175,
f. 1549—1562; Kozminski, 1933:102; Kozminski, 1934:115, t. 4, f. 2; Koz'mmski, 1936:211.—
Cyclops strenuus Schmeil, 1892:39, t. 2, f. 12 (part.). — Cyclops strenuus var. vicinus Frio
u. V&vra, 1894, Arch. Land. Bohmens, 9:57. — Cyclops vicinus var. glacialis Lilljeborg,
1901, Bih. Svenska Vet. Ak. Handl., 26(4):12.
Самка. Тело стройное, задние углы первых 3 сегментов одинакового
строения, без выростов, предпоследний торакальный сегмент очень
крупный, его наибольшая ширина лежит на линии, соединяющей весьма
сильно развитые, оттянутые в заостренные лопасти задние углы. Зад-
ний торакальный сегмент значительно уже предыдущего, с очень сильно
развитыми, заостренными боковыми выростами. Генитальный сегмент
умеренно расширенный спереди, равномерно и относительно мало
суживающийся кзади. Receptaculum seminis округлый, обычно немного
удлиненный. Фуркальные ветви длинные (у var. kiku.ch.ii очень длинные),
узкие, обычно мало расходящиеся (у var. kiku.ch.ii параллельные), со
сплошным рядом волосков на внутренних краях. Дорзальная щетинка
длинная, приблизительно равная длине внешней крайней апикальной
щетинки, латеральная щетинка прикрепляется недалеко от конца внеш-
него края ветвей. Из крайних апикальных щетинок внутренняя прибли-
зительно вдвое (или немного более чем вдвое) длиннее внешней, средние
апикальные щетинки хорошо развитые, из них внутренняя лишь немного
длиннее внешней. Передние антенны 17-членистые, редко 16-членнстые
(неполное разделение 8-го членика), очень редко 18-членистые, дохо-
7. CYCLOPS
205
Фиг. 43. Cyclops vicinus Uljan., самка.
1 — фурка co епинной стороны; 2 — задний отдел торакса и генитальный сегмент;
.3—4 — нога V; 5—дистальный членик эндоподита ноги IV; 6—экзоподит ноги !;
7 — задний сегмент торакса и генитальный сегмент с receptaculum seminis;
8— три дистальных членика антенны I.
206
CYCLOPINAE
дящие приблизительно до середины 2-го- торакального сегмента, иногда
немного более короткие, ряды тонких шипиков вдоль краев 3 дисталь-
ных члеников, за редкими исключениями, выражены явственно. Вооруже-
ние плавательных ног по типу Bini. У ног IV пары внутренний апикаль-
ный шип дистального членика эндоподита в 2—3 раза длиннее внешнего..
Ноги V пары как у С. strenuus, щетинка базального членика довольно
длинная, шип внутреннего края дистального членика хорошо развит,
своей вершиной заходящий за конец членика (у var. klkuchii он короче).
Длина 1.25—2.18 мм. Окраска вариирует, обычно однообразная бело-
ватая, иногда с коричневыми или красными включениями, или светло-
голубая. Яйцевые мешки овальные, обычно заключающие небольшое:
количество яиц, более или менее тесно прилегающие к абдомену.
Самец. Тело стройнее, чем у самки, 2 задних торакальных сегмента
без выростов сзади и сбоков, ноги VI пары вооружены очень длинной
внешней щетинкой, более чем вдвое превышающей длину соседней
с ней внутренней щетинки, и шипом, который короче последней щетинки..
Длина 1.15—1.5 мм. Сперматофоры на генитальном сегменте самки
располагаются расходясь под значительным углом, своими концами,
однако, не заходя наружу за края сегмента.
Известен один вариетет — var.klkuchii Smirnov, описание которого
следует ниже. В 1901 г. Lilljeborg' описал с Медвежьих островов варие-
ч'ет glacialls, отличающийся (самки) сравнительно незначительной длиной
(около 1.4 мм), более слабым развитием боков обоих задних торакаль-
ных сегментов, 16-членистыми передними антеннами и редукцией рядов
шипиков на краях и 3 дистальных члеников; самцы ничем не отличаются
от типичных. Описываемую форму, однако, нет оснований выделять
в отдельный вариетет, так как она представляет просто весеннюю,
не вполне развитую (антенны) генерацию; сам автор высказывает пред-
положение, что самки позже, летом, достигают нормального развития
и не отличаются от типичных.
Распространение.1 Европа, северный Иран, М есопотамия,
Япония (окрестности Токио, var. klkuchii), Китай, Манчжурия, Монголь-
ская Народная республика. Известен с Медвежьих островов (Lilljeborg,
1901). Указания для Аляски (Marsh, 1920) и Шпицбергена (Olofsson,
1918) требуют проверки (см. С. kolensis). В СССР С. vicinus обитает
в пределах всех ландшафтных зон, исключая лишь зону субтропиков,
доходя на восток до бассейна р. Амура. Характерно, что в зоне пустынь.
(Кизыл-кумы) он обнаружен в ключе (определение Ульянина не подле-
жит сомнениям) и в пределах этой зоны, повидимому, вообще не пред-
ставляет редкости (также и в зоне полупустыни). В пределах зоны
тундры он, по всем вероятиям, встречается гораздо реже, нежели
С. scutifer.
Биология. Характерен для пелагиали озер, главным образом,
эвтрофных; обитает также в мелких стоячих водоемах, указывался для
планктона медленно текучих рек. В литорали озер, повидимому,
в меньших, чем в пелагиали, количествах. Данные о вертикальном
распределении очень скудны. По данным Скорикова (1904) в оз. Пестове
летом днем максимум рачка наблюдается в слое 8—12 м:, в декабре
он распределен равномерно по всей толще воды. В озерах централь-
ной части Якутской АССР (Рылов, 1935) днем максимум лежит в слое
0—3 и 0—7 м, но в одном озере резко выраженный максимум находился,.
1 Часть указаний, возможно, относится к var. klkuchii (см. ниже).
7. CYCLOPS
207
в слое 4—8 м (в слое 0—4 м отсутствует; оз. Мюрю, максимальная
глубина 14 м). В прибрежных районах оз. Иссык-куль (Рылов, 1930а)
днем держится особенно в слое 7.5—16 м, встречаются лишь единичные
экземпляры. Во всяком случае в пелагиали озер С. vicinus обитает,
главным образом, в пределах эпилимниона.
Эвритермичен, отношения к кислородным условиям не выяснены,
возможно полиоксибионт. Обитает в водах в pH = 5.9—9.13. Рассматри-
ваемый рачок, бесспорно, избегает сколь-либо значительно дистрофи-
рованных водоемов. Помимо нормальных пресных олиго- и мезоагалинных
водоемов, С. vicinus неоднократно указывался для вод повышенной
минерализации, в частности и для солоноватых вод. Амплитуда колеба-
ний содержания СаО и MgO для него может быть значительной. Так
по Дексбаху (1931) он обитает в водоеме (пруд в Косине), где содер-
жание СаО колеблется от 2.6 до 25.4 мг-°/00 (олиго-мезотип), а в озере
Балхаше (pH = 8.0—9.1) — при общей жесткости 13.4—48.7° немецких
и карбонатной жесткости 8.3—32.2° (преобладает магний). Равным
образом широко приспособляется к колебаниям в содержании хлоридов.
В эвтрофных водоемах средней Европы половозрелые индивиды
встречаются весь год, максимум падает на зиму и весну (декабрь,
апрель), причем зимние самки несколько мельче летних и содержат
меньшее количество яиц в мешках. Ряд авторов отмечает зимнее нахо-
ждение рачка и в водоемах СССР. Рачок, однако, обычен и летом, при
высоких температурах воды.
Судя по местонахождениям, С. vicinus следует отнести преимуще-
ственно к |3-мезосапробам; отметим, что в Англии, по Gurney (1933),
в мелких водоемах он обычно встречается совместно с Daphnia pulex.
Типичная форма Cyclops vicinus Uljan. характеризуется (самки) сле-
дующими индексами (по Козминскому, 1933, 1936): общ. дл. тела = 1717
(1409—2024); шир. тор. сегм.4 в °/00 к макс. шир. цефалот.=86 (80—92);
дл. внутр, ап. щет. в °/00 к дл. тела = 190 (164—216); дл. внутр, ап. щет.
в °/0 к дл. внешн. ап. щет. = 227 (196—257); дл. фурк. ветв. в °/00 к дл..
тела = 160 (141—178); дл. внутр, ср. ап. щет. в °/00 к дл. тела 313
(271—354).
* Ila. Cyclops vicinus var. kikuchii Smirnov.
Cyclops kikuchii Smirnov, 1932:233, f. 1—5. — C. vicinus var. brachismerinthos.
Wierzbieka, 1933, C. R. Mem. Aead. Polon. Se., 8; Kozminski, 1933:103, 111. — C. vicinus .
var. kikuchii Wierzbieka, 1934:t. 7, f. 1, 5, t. 8, t. 1—9; Kozminski, 1936:200, 204, 212.
От типичной формы самки этого варнетета отличаются иным соот-
ношением длины крайних апикальных щетинок фуркальных ветвей
и иным отношением длины внутренней крайней апикальной щетинки
к длине тела. Фуркальные ветви очень длинные, тонкие, параллельные,
шип на внутреннем крае дистального членика ног V пары короче
(всегда), чем у типичного С. vicinus, едва доходящий до конца этого
членика.
Индексы (в среднем): общ. дл. тела = 1744; дл. внутр, ап. щет..
в °/00 к дл. тела = 140; дл. внутр, ап. щет. в °/00 к дл. внешн. ап. щет.=
171; длина фурк. ветв. в °/оо к дл. тела = 171; дл. внутр, ср. ап. щет. в °/00,
к дл. тела = 393.
Распространение. Найден в северовосточной Г ермании
и в Японии (окрестности Токио). Возможно, что литературные указания,
на „С. vicinus" частично относятся к var, kikuchii.
208
CYCLOPINAE
Биология изучена очень мало. Возможно встречается исключи-
тельно в пресной воде. Мелкие водоемы (до луж включительно), лито-
раль озер. Встречается отдельно от типичной формы, но также и совместно
с последней, в одном и том же водоеме одновременно. Легко скрещи-
вается с типичным С. vicinus, потомство достигает половозрелости. Был
находим зимой и осенью.
* 12. Cyclops insigriis Claus (фиг. 44).
Cgclops insignis Claus, 1857:209, t. II, f. 8—13; Schmeil, 1892:52, f. 2, f. 16—19,
t. 8, f. 15; Sars, 1913:39, t. 21; Kiefer, 1929:52.
Самка. Туловище стройное, кзади равномерно суживающееся,
наибольшая ширина переднего отдела тела лежит приблизительно на
Фиг. 44. Cgclops insignis Claus, самка.
/ — фурка с брюшной стороны; 2 — задний сегмент торакса и гени-
тальный сегмент с receptaculum seminis; 3 — нога V; 4 — дисталь-
ный членик антенны I.
середине его длины (считая задний торакальный сегмент). Задние углы
II—IV торакальных сегментов немного оттянуты кзади, задний торакаль-
ный сегмент уже предыдущего, с заостренными и сильно выступающими
кнаружи и кзади задними углами. Генитальный сегмент очень сильно
расширенный спереди, сильно и сразу суживающийся в заднем отделе;
боковые выросты его переднего отдела затупленные, хорошо развитые.
Receptaculum seminis характерной формы (фиг. 44), расположенный
поперек переднего отдела сегмента (в самом его начале), в виде
неправильно очерченного, кзади суживающегося короткого овала;
7. CYCLOPS
209
отвисающий кзади мешковидный отдел, свойственный С. strenuus
и близким к нему видам, отсутствует. Фуркальные ветви параллельные
(или почти параллельные), тонкие, чрезвычайно длинные, их длина
в 7.5—8 раз более их ширины, продольная хитиновая складка на их
дорзальной поверхности явственно выражена, внутренние края несут
лишь немного не доходящий до основания ветвей ряд тонких волосков.
Латеральная щетинка прикрепляется относительно далеко от конца
внешнего края ветвей. Из крайних апикальных щетинок внутренняя
лишь немного длиннее внешней, очень утолщенной, шипообразной.
Средние апикальные щетинки хорошо развитые, из них внутренняя лишь
немного длиннее внешней. Латеральная щетинка не длиннее внешней
крайней апикальной щетинки, часто гораздо короче ее. Передние антенны
14-членистые, приблизительно доходящие до середины 2-го сегмента
торакса, 8-й членик особенно длинный, 9-й членик вооружен дистальным
коротким копьевидным сензорным придатком, ряды мелких шипиков,
свойственные краю 3 дистальных члеников антенны С. strenuus и близ-
ких к нему видов, у С. insignis различимы лишь с трудом и, повидимому,
могут отсутствовать.1 Вооружение плавательных ног по типу Bini. Ноги
V пары как вообще в группе strenuus: внутренний шип дистального
членика мощный, выступающий за конец членика. Длина 2.5=5.0 мм.
Окраска: самки большей частью бесцветны, иногда зеленоватой, желто-
ватой— до красной (включительно) окраски. Яйцевые мешки очень
крупные, удлиненно-овальные, содержащие весьма значительное количе-
ство яиц, тесно прилегающие к абдомену.
Самец. Стройнее самки. Ноги VI пары с коротким шипом и парой
длинных щетинок, из которых латеральная особенно длинная. Длина
2.2—2.5 мм. Сперматофоры относительно узкие, вытянутые, прикрепляясь
поперек генитального сегмента самки справа и слева на одной прямой
линии (а не под углом), они своими внешними концами выдаются
за края сегмента.
От остальных форм группы strenuus рассматриваемый вид резко
отличается, главным образом, своими 14-членистыми передними антеннами,
чрезвычайно длинными параллельными фуркальными ветвями, строением
генитального сегмента и receptaculum seminis. По длине фуркальных
ветвей напоминает С. furcifer, от которого легко отличим по форме
туловища, членистости передних антенн, строению 2 задних торакальных
сегментов и генитального сегмента, а также по строению receptaculum
seminis.
Распространение. Европа (Скандинавия, Дания, Германия,
СССР, Австрия, Швейцария, Франция, Болгария). В СССР достоверно
известен из областей: Ленинградской, Горьковской, Московской, Сверд-
ловской, Воронежской, Харьковской, Киевской.
Биология. Обитает, главным образом, в мелких водоемах (лужи,
болотца, пруды, мелкие заливные водоемы), гораздо реже указывался
для литорали (заросли макрофитов) озер. Все указания на нахождение
в пелагиали озер, в качестве постоянного компонента пелагического
планктона, крайне еомнительны (случайный занос из литорали, конечно,
возможен). Холодноводно-стенотермичен. Повидимому встречается
в нейтральных (слабо кислых) и слабо щелочных водах, избегает сильно
дистрофированных водоемов. По исследованиям Никитинского (1928),
в широкой степени приспособляется к колебаниям содержания раство-
1 Не обнаруживаются при очень сильных увеличениях.
14 .Фауна СССР
210
CYCLOPINAE
ренного кислорода и может до 18 дней жить при отсутствии последнего
и, повидимому, в присутствии H2S.
Цикл развития изучен неполно. Многочисленные данные указывают,
что С. insignis встречается1 в холодное время года (летом лишь в виде
редкого исключения. Можно считать вполне установленным, что максимум:
количественного развития всегда падает на холодное время года (в част-
ности на период ледостава). Возможно, что в постоянных водоемах
дицикличен.
Положение в системе сапробов неясно. Вероятно, главным образом,
р-мезосапроб.
8. Род ACANTHOCYCLOPS KIEF.
Subgen. Acanthocyclops, Megacyclops, Diacyclops (part.) Kiefer, 1927:305; Kiefer,.
1928:545; Kiefer, 1929:55. — Subgen. Acanthocyclops Gurney, 1933:182. — Acanthocyclops
Megacyclops, Diacyclops Kiefer, 1937:421.
Фуркальные ветви без продольной складки на спинной поверхности^
за редкими исключениями, без ряда волосков на внутренних краях.
Анальная пластинка у большинства видов слабо развита, у некоторых
пещерных видов (Л. stygius Chapp., A. phraeticus Chapp.) значительно-
развитая. Передние антенны 10—17- (редко 18-)членистые, обычно не
менее чем 12-членистые, их 2 дистальных членика без гиалиновых пла-
стинок и без ряда шипиков вдоль внутреннего края. Членистость
плавательных ног по формуле З-З/З-З/З-З/З-З, редко 2-2/3-2/3-3/3-3
(A. languidus typ., A. languidoides Lili., A. abysslcola Lili., A. hypogeus
Kief., A. tantalus Kief., A. nanus Sars) или 3-2/3-2/33/3-3 (A. stygius
Chapp.), у 2 задних пар экзо- и эндоподиты всегда 3-членистые.1 2 Основ-
ная формула щетинок экзоподитов 4-4-4-4 или 5-5-5-5. Ноги V пары
2-членистые, иногда наблюдается частичное слияние базального членика
с задним торакальным сегментом, полное исчезновение этого членика
иногда наблюдается лишь у A. nanus (var. Incert a Wolf). Дистальный
членик этих ног внутри с шипом, прикрепленным субапикально или
апикально, редко несколько ближе к середине внутреннего края, апи-
кальная щетинка членика хорошо развитая, близ ее основания (снаружи)
нет группы шипиков. Длина внутреннего шипа, как правило, не более
3/2 длины апикальной щетинки. Receptaculum seminis очень разнообразной
формы, никогда не бывает молотообразным. Главным образом мелкие
виды, длина самок редко более 2 мм.
Как указывалось выше (см. введение), к роду Acanthocyclops нами
относятся роды Megacyclops Kief, и Diacyclops Kief, (в современном
объеме последнего). В таком положении род в данное время насчиты-
вает около 40 видов. На обособленность положения Cyclops australis
King (Proc. R. Soc. Van-Diemen’s Land, 3, 1855) уже обратил внимание
Sars (Arch, naturv. Krist., 29, 1908:16). Ноги V пары этого, известного
лишь из Австралии (окр. Сиднея), вида соответствуют таковым рода
Microcyclops, к которому и отнес этот вид Kiefer (1929). Однако прочие
признаки, в частности членистость плавательных ног, определенно
указывают на близость к роду Acanthocyclops. Отметим, что С. australis
Является крупным видом (длина 2—2.2 мм), тогда как к роду Microcy-
1 По Seligo (1890, 1900) в озерах Германии встречается весь год, но данные автора,,
по всем вероятиям, относятся не к этому виду (пелагиаль озер!).
2 Лишь у A. stugius Chapp. описан случай 2-членистости эндоподита ног III пары
(var. deminut a Chapp.. 1925), требующий, впрочем, проверки.
& ACANTHOCYCLOPS
2H
clops относятся очень мелкие (преимущественно не более 1 мм) видьи
Внутренние края фуркальных ветвей у С. australis снабжены сплошным
рядом волосков, что наблюдается у некоторых Acanthocyclops (Megacy-
clops sensu Kiefer). Если положение рассматриваемого вида можно
считать несколько спорным в роде Acanthocyclops, то, несомненно, оно-
гораздо более сомнительно в роде Microcyclops.
Род в целом обладает космополитным распространением, но лишь
очень немногие виды могут считаться космополитами (A. viridis неиз-
вестен лишь из Австралии, A. bicuspidatus—Палеарктика и Новая
Зеландия, A. bisefosus, A. vernalis). Только Палеарктике свойственно
14 видов, из них не менее 10 являются пещерными.1 Кроме того из
Палеарктики известно 9 видов, найденных и в других областях. Только
Неарктической области свойственны A. venustoides Coker, A. bicuspidatus
var. thomasi Forbes, A. nearcticus Kief., A. haueri Kief, и троглобионт
A. jeanelli Chapp., Неотропическая область насчитывает 3 специфических
вида — A. uruguaensis Kief., A. lobulosus Ekm. (Огненная Земля, Фаль-
клэндские острова) -и A. michaelseni Mraz. (Фальклэндские острова),
кроме того, в ее пределах найдены широко вообще распространенные
A. viridis и A. vernalis.
В последнее время мы имеем целый ряд работ по Copepoda Эфиоп-
ской и Ориентальной областей, которые доказывают значительную
бедность видами Acanthocyclops обеих этих областей; это подтверждается
и более старыми работами. В Эфиопской области показан лишь
A. viridis, в палеарктической части Африки известны, кроме этого вида,
A. donnaldsoni var. algericus Kief.,1 2 A. bicuspidatus m. odessana Schm.,
A. crassicaudis Sars. A. vernalis Fisch.; специально эфиопских Acantho-
clops пока неизвестно. Только Ориентальной области свойственны
A. viridis var. acutulus Kief. (Бали) и A. alticola Kief. (Индия). Кроме
них, в этой области показаны A. bisetosus, A. vernalis var. robustus,
A. crassicaudis (о. Формоза),3 A. languidus (о. Цейлон). Исключительно
Австралийской области свойственны A. crassicaudoides Kief. (Новая
Зеландия) и А. (?) australis King (окрестности Сиднея), кроме того для
нее указаны A. bicuspidatus и A. bisetosus. Таким образом род Acaniho-
cyclops является преимущественно голарктическим и, главным образом,
свойствен Палеарктике.
Род заключает, главным образом, наземные виды, обитающие
в мелких водоемах различного типа, некоторые виды (например
A. languidoides) характерны для дистрофных, богатых мхами (Hypnum,
Sphagnum) болотистых водоемов и отличается чрезвычайной изменчивостью.
A. bicuspidatus, A. bicuspidatus m. odessana, A. bisetosus широко распро-
странены в солоноватых континентальных водоемах (особенно 2 последние
формы). В роде обширно представлены троглобионты—10—11 видов
и ряд вариететов A. languidoides', дальнейшие новые виды и вариететы
в пещерных водах очень вероятны. Главным образом непланктические
1 Из описанных в последнее время Kiefer’oM некоторых пещерных видов мы не
упоминаем здесь виды явно сомнительные (о них см. ниже, в описании пещерных
видов).
2 Эта форма крайне поверхностно описана Kiefer’oM, ее отношение к американ-
скому виду очень сомнительно. Быть может, он является одной из форм A. viridis.
Описываемый Lindberg- Megac. viridis var. deserticola (Иран), по нашему мнению, выде-
ления в особый вариетет не заслуживает.
3 Описанный с о. Формозы С. (Megac.) formosanus Harada (1931:157), вероятно,
идентичен с A. viridis (возможно отдельный вариетет).
14*
212
CYCLOPINAE
циклопы, редко в пелигиали озер (A. bicuspidatus v. thomasi Forb.—
в Северной Америке, A. vernalis var. pelagica Lint1 —в Голландии,
A. americanus Marsh — в Северной Америке и Англии; очень редко
A. viridis — см. ниже описание этого вида). В условиях, повидимому
'близких к таковым пелагиали озер, иногда были находимы (в солоно-
ватых водах) в массовом количестве A. bicuspidatus и A. bisetosus.
Вероятно все виды Acanthocyclops, исключая A. gigas (холодноводный),8
эвритермичны в широкой степени.
Всего в пределах Палеарктической области констатировано 23 вида,1 2 3
из них 12 известны в СССР. Составить определительную таблицу для
всех указанных для Палеарктики видов рассматриваемого рода весьма
затруднительно, ввиду большой изменчивости некоторых из них; особен-
ные затруднения представляет A. languidoides, заключающий 12 варие-
тетов. Их определительная таблица дается ниже (см. описание этого
вида). Однако в определительную таблицу видов необходимо было
внести отдельно A. languidoides var. eriophori, резко противопоставля-
ющийся (положение литеральной щетинки фуркальных ветвей) всей
группе остальных форм этого вида. Это не пришлось сделать для
A. bicuspidatus m. odessana, A. stammeri var. westfalicus и A. languidus
var. belgicus (у последнего число члеников передних антенн может быть
16 и 13—14). Внесение всех 12 вариететов A. languidoides, а также
вариететов A. crassicaudis, A. vernalis, A. languidus и A. venustus,
чрезвычайно загромоздило и усложнило бы таблицу, в ущерб ее прак-
тичности. Имеющиеся в литературе определительные таблицы видов
этого рода (Kiefer, Gurney и др.) нужно признать неудовлетворяющими
своему назначению.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (14). Передние антенны 17-членистые.4
2(3). Генитальный сегмент очень широкий, его ширина превышает
длину, receptaculum seminis характерной бабочковидной формы .
...................................12. A. sensitivus (Graet. et Chapp.)
3(2). Ширина генительного сегмента не превышает его длины, recep-
taculum seminis не бабочковидной формы.
4(7). Внутренние края фуркальных ветвей с рядом волосков.
5(6). Длина самки обычно менее 2.5 мм, отношение длины фуркаль-
ных ветвей к их ширине обычно менее 4:1, отношение длины
дистального членика эндоподита ног IV пары к его ширине
обычно 2(2.2):1....................................*1. A. viridis (Jur.)
6(5). Длина самки 2.7—4.5 мм, обычно более 3.5 мм, отношение
длины фуркальных ветвей к их ширине всегда более 4:1 (обычно
6:1), отношение длины дистального членика5 эндоподита к его
ширине обычно более 2.4:1 (до 2.7:1).................................
...................................... . *2. A. gigas (Claus) typ.
1 Мы разделяем мнение Gurney (1933:191), по которому С. viridis var. pelagica
Lint (1922) относится к виду A. vernalis. Для нас остается сомнительным, идентична
ли описанная Lint пелагическая форма A. vernalis, как это думает Gurney, е A. ameri-
canus Marsh.
2 Возможно, что холодноводяыми являются и некоторые пещерные виды.
3 Некоторые из указанных для Палеарктики видов, вследствие их сомнительности,
нами не учитываются.
4 У A. vernalis иногда 18-членистые.
5 У A. gigas var. talipes (Lownd.) этот членик укороченный, длина самки менее 2.5 мм.
8. ACANTHOCYCLOPS
213
7(4). Внутренние края фуркальных ветвей без волосков.
8(9). Из крайних апикальных щетинок фуркальных ветвей внутренняя
короче шипообразной внешней.................*15. A. bisetosus (Rehb.)
9(8). Из крайних апикальных щетинок внутренняя длиннее (редко
равна) внешней.
10(11). Задний отдел receptaculum seminis в виде очень объемистого,
отвисающего кзади мешка1............................................
............................ *13. A. bicuspidatus (Claus) typ.
11 (10). Задний отдел receptaculum seminis маленький, узкий.
12(13). Расширенный передний отдел генитального сегмента имеет
более или менее резко выраженные угловатые очертания боков.
Передний отдел receptaculum seminis без широкой прозрачной
каймы. Вид не пелагический.....................*3. A. vernalis (Fisch.)
13(12). Этот отдел генитального сегмента с боков равномерно закруг-
лен. Передний отдел receptaculum seminis с широкой прозрачной
каймой. Пелагический вид....................5. A. americanus (Marsh)
14(1). Передние антенны состоят менее чем из 17 члеников.
15(16). Передние антенны 16-членистые...............................
.......................*18. A. languidus (Sars) (включая var.
disjunctus Thall. и var. belgicus Kief, с 16-членистыми I антеннами).
16 (15). Передние антенны состоят менее чем из 15 члеников.
17 (40). Передние антенны 12—14-членистые.
18(19). Фуркальные ветви очень длинные, их длина приблизительно
в 9 раз превышает их ширину....................14. A. karamani (Kief.)
19(18). Фуркальные ветви короче, их длина лишь в 2.5—6 раз более
их ширины.
20(21). У эндоподитов ног IV пары шипы'по бокам с тонкой прозрачной
перепонкой.....................’...............*4. A. muscicola (Last.)
21 (20). Эта перепонка на шипах отсутствует.
22 (23). Поверхность покровов тела (на абдомене всегда) покрыта распо-
ложенными в отдельные ряды и группы очень мелкими шипооб-
разными складками. Передние антенны 12-членистые....................
................................8. A. venustus (Norm, et Scott)
23(22). Поверхность покровов тела гладкая или однообразно и равно-
мерно покрытая1 2 мелкими шипиками.
24(27). Передние антенны 13—14 членистые.3
25 (26). Длина дистального членика эндоподита ног IV пары самое
большее в 1.25 раза превышает его ширину. Receptaculum semi-
nis с маленьким задним отделом .....................................
. ............... 18в. A. languidus (Sars) var. belgicus Kief.
26(25). Длина дистального членика эндоподита ног IV пары в 2—3 раза
превышает его ширину, задний отдел receptaculum seminis
в виде далеко заходящего кзади объемистого мешка, передние
антенны 14-членистые................................................
.........*13а. A. bicuspidatus (Claus) m. odessana (Schmank.)
27(24). Передние антенны 12-членистые.
1 Этот признак свойствен и A. bicuspidatus m. odessana, у которой, однако,
передние антенны 14-членистые (см. ниже).
2 У A. bicuspidatus m. odessana, у которого нередко покровы совершенно гладкие,
без шипиков.
3 В виде очень редкого исключения 12-членистые передние антенны бывают
и у Л. languidoides var. zschokkei.
214 CYCLOPINAE
28(35). Длина дистального членика эндоподита ног IV пары равна его
ширине или превосходит последнюю менее чем вдвое.
29(30). Генитальный сегмент стройный, удлиненный, его длина заметно
более его ширины ...........................17. A. scopliensis (Kief.)
30(29). Генитальный сегмент короткий и толстый, его длина приблизи-
тельно равна его ширине или лишь очень немного более последней.
31(32). Из крайних апикальных придатков фуркальных ветвей внутрен-
ний заметно короче внешнего (редко) или почти одинаковой
длины......................................*16. A. crassicaudis (Sars)
32(31). Из этих придатков внутренний вдвое (или почти вдвое) длиннее
внешнего.
33(34). Формула шипов экзоподитов плавательных ног 3-4-4-4, щети-
нок 5-5-5-5 .... 9а. A. stammer! Kief. var. westfalicus Kief.
34(33). Формула шипов экзоподитов плавательных ног 2-3-3-3, щети-
нок 4-4-4-4................................. 10. A. rhenanus Kief.
35(28). Длина дистального членика эндоподита ног IV пары вдвое
и более чем вдвое превышает его ширину.
36(39). Из крайних апикальных придатков фуркальных ветвей внутрен-
ний более чем в 2 раза длиннее внешнего. Латеральная
щетинка прикрепляется в начале задней 73 длины внешнего
края ветвей или .еще далее кзади.
37 (38). Длина фуркальных ветвей в 4 и более раза превышает их
ширину..................................... 9. A. stammer! Kief. typ.
38(37). Длина фуркальных ветвей в 3.5 раза более их ширины ....
.....................................7. A. phraeticus (Chapp.)
39(36). Из крайних апикальных придатков фуркальных ветвей внутрен-
ний лишь немного длиннее внешнего. Литеральная щетинка
прикрепляется почти на середине внешнего края ветвей . . .
......................................*11. A. capillatus (Sars)
40(17). Передние антенны 10—11-членистые.
41 (42). У ног V пары апикальные придатки дистального членика почти
одинаковой длины.............................21. A. stygius (Chapp.)
42 (41). Эти придатки резко различаются по длине; внутренний шип вдвое
короче внешней щетинки или еще более короткий.
43(46). Латеральная щетинка фуркальных ветвей прикрепляется на сере-
дине (или лишь немного кзади от нее) их внешнего края.
44(45). Фуркальные ветви длинные, их длина в 4—4.5 раза более их
ширины, длина дистального членика эндоподита ног IV пары
вдвое или более чем вдвое превышает его ширину ......
........................................*20. A. nanus (Sars)
45(44). Фуркальные ветви более короткие, их длина не более как
в 3.5 раза превышает их ширину, длина дистального членика
эндоподита ног IV пары приблизительно равна его ширине . .
...............19а. A. languidoides (Lili.) var. eriophori Gurney
46(43). Латеральная щетинка фуркальных ветвей прикрепляется в начале
задней 7з длины их внешнего края или еще далее кзади.
47(48). Внешние края эндоподитов всех пар плавательных ног вместо
щетинок вооружены шипами.....................*23. A. abyssicola (Lili.)
48(47). Внешние края эндоподитов плавательных ног вооружены
не шипами, а щетинками.
49(52). Внутренний придаток (шип) дистального членика ног V пары
чрезвычайно короткий, значительно менее половины ширины этого
членика.
8. ACANTHOCYCLOPS
21
50(51). Апикальные шипы дистального членика эндоподита ног IV пары
равной или почти равной длины и короче этого членика; из
средних апикальных щетинок фуркальных ветвей внутренняя
приблизительно вдвое длиннее абдомена (включая фуркальные
ветви) .......................................22. A. hypogeus (Kief.)
51 (50). Эти шипы резко различной длины, из них внутренний более
чем в 1.5 раза длиннее внешнего и длиннее самого членика;
из средних апикальных щетинок фуркальных ветвей внутренняя
лишь немного длиннее абдомена .(включая фуркальные ветви) .
. ....................................6. A. kieferi (Chapp.)
52 (49). Внутренний придаток (шип) дистального членика ног V пары
сильный, длинный, гораздо длиннее ширины этого членика, более
половины его длины......................... *18. A. languidoides (Lili.)
1. Acanthocyclops viridis (Jur.) (фиг. 45).
Monoculus quadricornis viridis Jurine, 1820:45, t. 3, f. 1.— Cyclops viridis Fischer, 1851:
412, t. 9, f. 1—11. — C. brevicornis Glaus, 1857a : 32, t. 3, f. 12—17. — C. clausi 1 (non
C. clausi Lubbock, non C. clausi Pog-g-enpohl), Heller, 1871, Ber. Nat. Med. Ver. Innsbruck,
7:73, f. 1—2. — C. viridis Schmeil, 1892:97, t. 8, f. 12—14. — C. viridis europaeus Herrick,
1895:90. — C. viridis Lilljeborg-, 1901:8, t. 1, f. 6—11. — C. vulgaris (non C. vulgaris Koch)
Sars, 1913:40, t. 22. — C. viridis Рылов, 1917:305. — C. teres Wilson, 1925. Pr. U. St. Nat.
Mus., 64:17. — C. viridis Рылов, 1928:14. — C. (Megacyclops) viridis Kiefer, 1929.: 53,
J. 17, a. — b. — C. (Acanthocyclops) viridis Gurney. 1933:185, f. 1563—1582.
Самка. Тело относительно толстое, наибольшая ширина переднего
отдела лежит приблизительно на середине его длины (считая и задний
торакальный сегмент). Задние углы обоих задних торакальных сегмен-
тов не выступают кнаружи. Генитальный сегмент короткий, умеренно
расширенный спереди. Reeeptaculum seminis очень вариирующих очер-
таний, передний отдел объемистый, обычно вдавленный впереди, задний
отдел шире переднего, часто с глубоким вдавлением сзади. Фуркаль-
ные ветви слабо расходящиеся, иногда почти параллельные, их длина
весьма вариирует, обычно в 2.5—4 раза более ширины, внутренние
края с рядом довольно крупных волосков. Из крайних апикальных
щетинок внутренняя обычно приблизительно в 2 с лишком раза длин-
нее внешней, но иногда более короткая; средние апикальные щетинки
хорошо развитые, довольно толстые, из них внутренняя обычно немного
длиннее внешней; относительная длина этих щетинок вариирует в зна-
чительной степени. Латеральная щетинка прикреплена к задней %—1/3
длины внешнего края ветвей. Передние антенны 17-членистые, дохо-
дящие приблизительно до заднего края цефалоторакса. Обе ветви
всех 4 пар плавательных ног 3-членистые, формула шипов 2-3-3-3.
Длина дистального членика эндоподита ног IV пары обычно лишь не-
много более чем вдвое превышает его ширину,, апикальные шипы этого
членика довольно толстые, из них внутренний немного длиннее внеш-
него и обычно короче самого членика; длина этих шипов относительно
длины последнего очень вариирует. У ног V пары базальный членик
очень широкий, дистальный членик удлиненный, его внутренний шип
вариирующей длины, обычно очень маленький; он прикрепляется иногда
-близ самого конца1 2 внутреннего края членика (но не апикально), иногда
же лишь немного кзади от его середины. Длина 1.5—2.5 мм, обычно
1 Является молодым A. viridis (см. Spandl, 1922; Lucks, 1927; Kalmus, 1928).
2 Такая форма была описана Lmderg-’ом (1936) под названием „Megac. viridis var.
tdeseriicola" (Иран).
216
GYCLOPINAE
Фиг. 45. Acanthocyclops •viridis (jur.).
7— фурка самки с брюшной стороны; 2—генитальный сегмент
с receptaculum seminis; 3—дистальный отдел фуркальной ветви
самки; 4—часть генитального сегмента самца с сперматофором
и ногой VI (вид сбоку); 5 — дистальный членик эндоподита ноги,
самки; 6—7—нога V самки.
& ACANTHOCYCLOPS
217
менее 2.5 мм, редко больше. Окраска очень вариирует, иногда зеленая,
нередко беловато-серая или желтоватая. Яйцевые мешки крупные, удли-
ненно-овальные, содержащие значительное количество яиц (по Wolf,
1905, 20—50 в каждом), обычно сильно расходящиеся.
Самец. Длина фуркальных ветвей немного менее длины 3 задних
абдоминальных сегментов, взятых вместе. Сензорные придатки передних
антенн очень длинные, у ног VI пары внутренний (медиальный) шип
немного длиннее средней щетинки, длина внешней щетинки вариирует,,
иногда лишь немного длиннее средней щетинки. Длина 1.4—1.6 мм.
В таблице 15 даются измерения (в микронах) фуркальных ветвей, их
крайних апикальных щетинок, дистального членика эндоподита ног IV
пары и его апикальных шипов.
Acanthocyclops viridis является видом весьма вариирующим в целом
ряде признаков (см. Рылов, 1917, 1928; Lowndes, 1932), что нередко
затрудняет его определение; он легко может быть смешан с очень
близкими к нему A. gigas Claus. На взаимоотношениях этих двух видов
мы останавливаемся ниже (см. A. gigas). Наиболее характерными при-
знаками рассматриваемого вида являются следующие:
1. Незначительная величина, обычно менее 2.5 мм. Однако иногда,
встречаются и более крупные самки. Coker (1933) показал, что при
высоких температурах получается поколение A. viridis меньшей вели-
чины, нежели при низких температурах.
2. Отношение длины фуркальных ветвей к их ширине у типичного
A. viridis менее 4:1, иногда лишь 2.5:1 (см. табл. 15).
3. Внутренняя крайняя апикальная щетинка фуркальных ветвей
обычно более чем вдвое длиннее внешней крайней апикальной щетинки.
Исключения редки.
4. Как правило, у A. viridis имеется относительно короткий дисталь-
ный членик эндоподита ног IV пары, причем отношение длины этого
членика к его ширине редко превышает 2.0(2.2);1.
5. Некоторые особенности биологии, в особенности полициклич-
ность A. viridis, его эвритермичность, встречаемость в течение круглого
года, темп метаморфоза.
Распространение.1 Космополит. Вся Европа и Азия, Северная
Америка, Южная Америка, Африка, Азорские острова. В СССР распро-
странен всюду, населяя все ландшафтные зоны. В горных водоемах
Альп показан до высоты около 2000 м над уровнем моря, в СССР —
до высоты около 1900 м. Указание Daday (1908) для Тибета, на высоте
4480 м, требует проверки (см. ниже A. gigas).
Биология. Обитает в самых различных, постоянных и временных,
водоемах. Обычен в литорали (заросли макрофитов) и сублиторали
озер, где отчасти заходит и на значительные глубины, что, однако, для.
этого вида далеко не столь характерно, как для A. gigas. Очень обы-
чен в лужах, болотцах, прудах и т. п., водоемах, до искусственных
вклкчительно, известен из ключей, из периодически высыхающего
Blata-See в Кроации, из ряда рек, где обитает на дне береговой области
и фарватера. Держится преимущественно в зарослях различных водных
растений (Sphagnum, Scirpus, Nuphar, Polygonum, Chara и др.), но также
1 Несомненно рассматриваемый вид нередко смешивался с A. gigas. Мы приводим:
данные, которые можно считать вполне достоверными. Указание для Новой Зеландии.
(Thomson, 1892; см. Gurney, 1933) сомнительно. Указание для Гренландии, Медвежьих:
островов и Исландии (Richard, 1898; Gurney et Richard, 1892) требует проверки (см._
Haberbosch, 1920).
8. ACANTHOCYCLOPS
219
Измерения (в микронах) Acanthocyclops viridis Jur. из различных местностей СССР.
Длина апикальных шипов этого членика внутренних внешних 1.2: 1 1.2:1 1.1 : 1 1.2:1 1.2: 1 1.1: 1 14:1 1.1:1 1.3:1 1.1:1 1.1:1 1.1:1
хиншонд 'СОГ-‘СОО>ОСОСОГ-С01>СО
хиннэблЛна £2 1—< GO О --- т—< .—< 'хГ1 О СО СО
Дистальные членики Enp. IV пары ног длина ширина с00909»-;09 09\С'^^0909г-< 04 04 09 04 09 09 09 09
внибипг ООО1ЛООСЭО1ЛО1Л\П
вниу? 140 135 100 122 : 120 130 130 150 110 120 100 120
Крайние апикальные щетинки внутренние внешние 09 г-н 09 г-Чг-;09»-’*^1ПС00909 09 04 09 09 09 04 09 09 09 09 04
эиншена ООО1Л\Л1ПСЭООООО 1П’^^^О9\ОГ'С>г-'С0т—1ТГ
эиннэблЛна СП СП 09 СИ 09 СИ СИ 09 СП 04 СП
Фуркальные ветви длина ширина СИ СП СП СО СИ СП СП 09 СО
внидшп 50 60 50 60 55 70 60 60 50 55 50 50
гниуГ ОООООООООООО 09 СМ г-< 09 »—< 04 С9 04 г—< •—<—<09
(ии а) вниу'р' СО СО 09 04 СО 04 09 >—< 09 09 09 09 0-. г-< 09 <04 09 V V V V
Местонахождение Новосибирская область . » » * Бассейн Хатанги .... Коми АССР „ ,, • а » » „ „ ...... Окрестности Харбина . . » » • •
и на илистом (илисто-песчаном) дне. В пелагиали озер является вообще
случайным компонентом; Gurney (1933) сомневается в возможности нор-
мального существования этого вида в условиях пелагической области.
Однако в солоноватом оз. Иссык-куль (Рылов, 1930а, 1935т) мы нашли
морфологически не отличающегося от A. viridis циклопа в качестве
постоянного компонента планктона пелагиали. В широкой степени эври-
термичен. Очень обычен в подземных водах. Повидимому выносит зна-
чительные колебания в содержании растворенного кислорода, Встре-
чается в водах с pH =4.4—9.8. В Лузинском болоте под Москвой
Скадовский (1923) находил этого рачка в зоне рН=4.38—4.50, в зоне
-3.7—3.8 он, однако, отсутствовал. В бедных электролитами водах ниж-
няя граница pH лежит, очевидно, около 4.5, т. е. она выше, чем для
A. nanus и A. languidus (см. ниже). В значительной степени, эвригали-
нен. Являясь видом, характерным для нормальных пресных олиготроф-
ных, эвтрофных и дистрофных (исключая высокой степени дистрофии)
вод, рачок неоднократно был находим в водах с высокой минерализа-
цией, в частности в водах с повышенным содержанием NaCL. Вместе
с тем он обилен всюду в оз. Иссык-куль, где сульфаты (повышенное
содержание Mg) преобладают над хлоридами.
Цикл размножения изучен подробно. В мелких постоянных водое-
мах и в литорали озер полицикличен, встречается весь год; для литорали
озер установлена и дициклия. Половые периоды падают на различное
время года, в том числе на лето и зиму.
Kolkwitz’oM и Marsson’oM (1909) ошибочно отнесен к олигосапробам.
Зиверт (1929) отмечает чрезвычайно широкую его приспособленность
к изменениям сапробности — от а-мезосапробности до олигосапробности.
Особенно характерен для ₽-мезосапробных условий.
*2. Acanthocyclops gigas (Claus) (фиг. 46).
Cyclops gigas Claus, 1857:207, t. П, f. 1—5. — C. ingens Herrick, 1832, Rep. Geol.
Surv. Minnesota, 10:228, t. 4, f. 1—8. — C. viridis var. gigas Sostaric, 1888, Rad. Jugosl.
.Ak. Znan. i Umjetn., 92:64, t. I, f. 7. — C. viridis (part.) Schmeil, 1892. — C. gigas Lilljeborg',
1900, Bih. Svenska Vet. Ak. Handl., 26:4; Lilljeborg, 1901:5, t. I, f. 1—5; Sars, 1913:41,
t. 23; Рылов, 1918:307. — C. viridis var. gigas Рылов, 1928:14. —: C. (Megacyclops) gigas
(part.) Kiefer, 1929:53. — C. (Acanthocyclops) gigas (part.) Gurney, 1933:191, 194,
f. 1583-1585.
Самка. Габитус тела как у предыдущего вида. Фуркальные ветви,
-за редкими исключениями, почти параллельные, длинные: их длина
до 6 раз превосходит их ширину. Внешняя из крайних апикальных
щетинок относительно длинная, отношение ее длины к длине внутрен-
ней крайней щетинкв 1:1.6—1:1.9, редко 1:2 (1:2.1). Receptaculum semi-
nis как у A. viridis. Дистальный членик эндоподита ног IV пары удли-
ненный, отношение его длины к ширине от 2.3:1 до 2.7:1, обычно
более 2.4:1, редко менее; апикальные шипы этого членика нередко
относительно длиннее, чем у A. viridis. Ноги V пары—-как у послед-
него. Длина 2.7 — 4.5 мм, обычно более 3.5 мм. Окраска обычно жел-
товатая или желто-коричневая, иногда с зелеными включениями. Яйце-
вые мешки очень крупные, содержащие значительное количество яиц,
обычно прилегающие к абдомену.
Самец. Длина фуркальных ветвей приблизительно равна (или
немного более) длине 3 задних сегментов абдомена, взятых вместе.
.Длина 2—'2.5 мм.
Систематическое положение A. gigas долгое время оста-
валось спорным. Ряд авторов не считал возможным отделять его от
220
CYCLOPINAE
Фиг. 46. Acanthocyclops gigas (Claus), самка.
7 —фурка с брюшной стороны; 2—дистальный членик эндоподита ноги IV-
J дистальный отдел фуркальной ветви; 4 — нога V. ’
8. ACANTHOCYCLOPS
221
предыдущего вида, считая вариететом последнего. Обратного мнения
держались Sars и Lilljeborg. В последнее время, однако, выясняется
с достаточной определенностью видовая самостоятельность A. gigas (см.
особенно Kiefer, 1929; Roy, 1932; Lowndes, 1932; Gurney, 1933). Послед-
ний автор (см. также Рылов, 1928, 1933) установил, что ряд признаков
этого вида не может служит критерием для его отделения от A. viridis.
Однако некоторые признаки являются для ’него достаточно характер-
ными. Таковы длина фуркальных ветвей, соотношение длины их край-
них апикальных щетинок, длина и ширина дистального членика эндопо-
дита ног IV пары. Нужно заметить, что, в виду чрезвычайной изменчи-
вости A. viridis, его вариации могут отчасти налагаться на таковые
A. gigas и в этих признаках. Повидимому это особенно имеет место
в мелких водоемах севера, где оба вида встречаются и совместно.
В таких случаях определение обеих форм встречает затруднения и тре-
бует изучения всего цикла их развития. Однако обычно популяции
A. gigas указанными признаками без труда отличимы от таковых
A. viridis (табл. 15 и 16). Исследования последнего времени вместе
с тем доказывают, что биология обоих видов совершенно различна.
Распространение. Европа (Скандинавия, Финляндия, СССР,
Голландия, Германия, Венгрия, Франция, Великобритания); известен
с Исландии и Медвежьего острова, из азиатской части СССР, Северной
Америки, вероятно Северной Африки (Roy и G uthier, 1927). Указание
для Шпицбергена требует проверки.
В СССР, как и предыдущий вид, встречается во всех ландшафтных
зонах, исключая лишь зону субтропиков. Наивысшая точка местонахож-
дения— 4000 м (Памир), в Альпах — 2000 м (несомненно смешивался
с A. viridis).
Биология, Обитает как в мелких водоемах, так и в литорали,
сублнторали и профундали озер.1 В целом ряде озер отмечен как очень
характерный компонент придонной фауны значительных глубин (до 180 м),
Указание Zschokke (1907) на нахождение A. viridis в Фирвальдштет-
тском озере на глубине 200 м относится, вероятно, к A. gigas. В СССР
характерен для глубинной области ряда озер, в оз. Телецком, Ладож-
ском, Онежском, Севан, в ряде озер Карело-финской ССР и Кольского
полуострова. На холодноводную стенотермию указывает ряд авторов.
Половозрелые самки были находимы, за редкими исключениями, в холод-
ное время года (зима, ранняя весна, поздняя осень; см. ниже, цикл
развития), но в глубокой области озер, где летом господствуют низкие
температуры, половэзрелость наступает ранее, даже в июле. Повиди-
мому, как и предыдущий вид, широко приспособляется к колебаниям
в содержании растворенного кислорода. На глубинах олиготрофных
озер он выдерживает понижение содержания О2 до 50% нормального
насыщения; для оз. Чаны (Зверева, 1927) он показан в период ледо-
става (март) при О2 всего 0.41—1.72 см3 на литр. Lowndes (1928) ука-
зывает A. gigas исключительно, для щелочных вод, с pH =7.4—8.2.
Однако он был находим и в кислых водах, при рН=5.2—6.1. Вероятно
амплитуда pH для рассматриваемого вида достаточно широка, не отли-
чаясь существенно от таковой для A. viridis- в очень кислых, дистроф-
ных водах (pH менее 4.5—5), однако, A. gigas отсутствует. Форма
пресноводная, однако изредка была находима и в солоноватых
1 В пещерных водоемах, насколько известно из литературы, отсутствует; имею-
щиеся указания относятся лишь к A. viridis (Jur.).
8. ACANTHOCYCLOPS
221
змерения (в микронах) Acanthocyclops gigas (половозрелые £ £>) и в различных местностей СССР
Апикальные шипы этого1 членика (V S № О & И И внешние о ” '" р н
9ИНШ9НЯ
ОИК -модлХня
Дистальные членики Епр. IV пары ног длина ширина г-, -г vr; г- -г co u-i к, ,r> 45г- чо см co CM CO cm cm cm cm cm cm CM cm cm CM cm cd cm <n’ cm cm cm cm cm od см сч см
внидиш
вниуГ гЧ H r-i r-( H CN H H i-l
Крайние апикальные щетинки Внутренние внешние 1.7:1 1.7:1 1.8: 1 1.7:1 1.7: 1 1.8: 1 1.8: 1 1.8:1 1.7:1 1.9:1 1.8:1 1.78:1 1.8: 1 1.9:1 1.76: 1 1.7:1 2:1 1.6:1 1.7 : 1 1.8:1 1.7: 1 1.7: 1 1.8:1 1.6:1 1.6:1
эиншоня 512 M OOOOOOQQOOOOOGOOae*1 n n
9ИН -нэблЛна S2S88°S888S8S28RS8gg§§gg^ co co мг co co co co co N co N co co см ом cm cm cm °
Фуркальные ветви длина ширина >r> u-> 45 CO Г- OM CO O' CO — 1 1 1 1 1 1 1 | 1 1 45 co 45 CO —
внибиш 1 1 1 1 1 1 1 1 1 1Sgggg
внигГ © tn о О О О О tn о О , , , . tn гП 40 £. о Г' ч© I I | I I [SS222 cncncocn-'tfcnoocococn1 1 1 । 1 1 1 1 1 । S Я Й 5
(ии a) bhhv^/ <^ОО^Ми^Оч00ООО1>|>ч0’^\П['-.\ПС<|СЧ\ПО1ПОа-)0\
Местонахождение Окр. Ленинграда (г. Пушкин) . То же » •• » » * • п » • • » Оз. Севан То же . . , ,, • >1 'и •••••••••.,, » Коми АССР То же Окр. Архангельска бассейн Хатанги
водах. Повидимому эвригалинность у A. gigas гораздо менее выражена,
чем у A. viridis, который для солоноватых вод указывался гораздо чаще.
Рачок широко приспособлен к колебаниям в содержании Са. Так весной обы--
чен в богатых кальцием водах окр. Ленинграда (воды Силурийского плато),
но неоднократно был находим И в водоемах очень мягких, бедных Са.,
Цикл размножения. По Lilljeborg’y (1900), в мелких маленьких
водоемах и в литорали озер Швеции моноцикличен, самцы только
осенью и в начале зимы (сентябрь — декабрь), в озерах, на глубине
30—100 м, половозрелые самки и самцы появляются уже в июле—-
августе. По Sars’y, в мелких прудах окр. Осло половозрелые самки,
и самцы встречаются еще подо льдом, ранней весной, а к концу лета,
рачок, вероятно, совсем выпадает; с этим согласуются наблюдения:
в г. Пушкине, где в мелких водоемах рачок в массе встречается весной,
(апрель, март, в мае уже реже) и исчезает на лето, снова появляясь
(молодь) в ноябре; однако осенью полового периода не наблюдалось.
Судя по приведенным данным, единственный половой период у A. gigas
в мелких водоемах может иметь место или весной или поздней осенью
(начало зимы), т. е. в обоих случаях при низких температурах воды,,
причем темп метаморфоза является весьма замедленным, и продолжи-
тельность индивидуальной жизни весьма значительна, около года
(Roy, 1932).
Вероятно A. gigas является, главным образом, ^-мезосапробом.
2а. Acanthocyclops gigas var. iatipes (Lowndes).
Cyclops Iatipes Lowndes, 1927, Ann. Mag. Nat. Hist. (9), 19:266, £. 1—4. — C. (Megacy-,
clops) Iatipes Kiefer, 1929:54.— C. (Acanthocyclops) gigas Iatipes Gurney, 1933:195,
f. 1589—1579.
Самка. Близок к A. gigas. Длина почти параллельных фуркаль»
ных ветвей в 5—б раз более их ширины. Из крайних апикальных щети-
нок внутренняя менее чем вдвое длиннее внешней. Дистальный членик
эндоподита ног IV пары очень короткий, его длина не более чем вдвое
(обычно менее) превышает его ширину, апикальные шипы этого чле-
ника относительно короткие, из них внутренний не превышает длины
самого членика. Длина 1.85—2.25 мм. Окраска серовато-белая. Яйцевые
мешки крупные, расходящиеся.
Рассматриваемая форма чрезвычайно близка к A. gigas и отли-
чается от последнего, главным образом, величиной и размерами дисталь-
ного членика эндоподита ног IV пары, чем она сближается с A. viridis
Jur; однако по Lowndes (1932) A. viridis и A. gigas. var Iatipes не скрещивают-
ся. Таксономическое положение A. Iatipes отнюдь нельзя считать выяснен-
ным. Мы следуем здесь Gurney (1933), который не находит возможным
выделять A. Iatipes в отдельный вид и сближает его с A. gigas Claus,
Распространение. Великобритания (Lowndes, Gurney) и Фран-
ция (Roy). Нахождение в пределах СССР возможно.
Биология. Обитает в мелких водоемах (лужи, болотца и т. п.),
особенно в усыхающих летом, при pH =6.5—8.2; цикл размножения
не изучен; вероятно эвритермичен, хотя Gurney находил его только
весной и зимой.
*3. Acanthocyclops vernalis (Fisch.) (фиг. 47х_4).
Cyclops vernalis Fischer, 1853:9. t. 3, f. 1—5.— C. lucidulus (non C. lacidalas Koch,
1838) Sars, 1363, Forh. Selsk. Christ. (1862):245.— C. (parcus Herrik, 1832, Rep. Geol.
Surv. Minnesota, 10:229, t. 4, f. 1—8.— C. vernalis (part.) Schmeil, 1892:88, t. 2, f. 4—7.—.
.22Z
CYCLOPINAE
C. vernalis var. acaleata Mrizek, 1893:30, t. 6, f. 2.— C. vernalis Lilljeborg, 1901:17, t. 2,
f. 5—7.— C. lu.cidu.lus Sars, 1913:44, t. 25.— C. vernalis var. ornatus Jungmayer, 1914,
Math. Term. Kozl., 33:68, f. 17, a — c.— C. vernalis-t-C. vernalis var. ambigua -+- C. ver-
nalis var. infesta-t-C. vernalis v. acaleata, Thallwitz, 1926, Arch. f. Hybrob., 17:366, f. 4,
6—8, 15—19.— C. vernalis (part.) Lowndes, 1928b:171. — C. (Acanthocyclops) vernalis
Kiefer, 1929:54, f. 18, a — b.— C. (Acanthocyclops) vernalis (part.) Gurney, 1933:198.
f. 1598—1615. — Acanthocyclops vernalis Kiefer, 1936, Bull. Mus. R. Hist. Nat. Belg.,
12:12.
Самка. Тело относительно толстое, длина переднего отдела
(считая и задний торакальный сегмент) менее его двойной ширины,
которая лежит, приблизительно, на середине этой длины. Задние углы
заднего торакального сегмента значительно вытянутые кнаружи. Гени-
тальный сегмент короткий, сильно расширенный спереди, где его бока
имеют угловатые очертания, его наибольшая ширина приблизительно
равна его длине. Receptaculum seminis с поперечно-овальным передним
и с очень узким маленьким задним1 отделом, иногда вдавленным сзади.
Фуркальные ветви очень мало расходящиеся или параллельные, их
длина1 2 в 4—5 раз (редко в 5.5—6 раз) более их ширины, латеральные
.щетинки прикреплены приблизительно в начале задней 1/i длины внеш-
него края. Из апикальных крайних щетинок внутренняя иногда заметно
длиннее внешней или же обе эти щетинки одинаковой длины. Средние
апикальные щетинки хорошо развитые, очень длинные, из них внут-
ренняя на 74—г/3 длиннее внешней. Передние антенны 17-членистые
(иногда, вследствие разделения надвое 7-го членика, 18-членистые,
доходящие до заднего конца цефалоторакса или немного короче. У ног
I—IV пар обе ветви 3-членистые, формула шипов 2-3-3-3 или 3-4-4.-4
(последний особенно у var. robust ns), на внешнем крае дистального
членика эндоподита ног IV пары имеется нормальная щетинка или
(у var. robusius) шип, длина этого членика обычно немного более чем
вдвое (реже менее чем вдвое) превышает его ширину, апикал! ные шипы
этого членика относительно толстые, почти равной длины или внешний
из них немного длиннее внутреннего. У ног V пары базальный членик
широкий, с относительно короткой щетинкой, дистальный членик
маленький, обычно .с довольно толстым шипом и с относительно корот-
кой щетинкой; длина этого членика приблизительно вдвое превышает
его ширину. Длина3 1.0—1.8 мм. Окраска желтоватая или красноватая,
до яркокрасной (особенно в высокогорных водоемах). Яйцевые мешки
удлиненные, овальные, крупные, очень слабо расходящиеся, иногда
прилегающие к абдомену, обычно содержащие большое (до 100 в каж-
. дом, но иногда не более 10) количество яиц.
Самец. У ног VI пары внутренний шип хорошо развитый, несколько
короче обеих щетинок, которые почти одинаковой длины. Сперматофоры
овальные, на генитальном сегменте самки они приклеиваются продольно.
..Длина 1.0—-1.2 мм.
Описанный в 1863 г. Sars’oM С. robusius (см. ниже) некоторыми авто-
рами считается самостоятельным видом (Lilljeborg, 1901; Sars, 1913;
Kiefer, 1924, 1929; Thallwitz, 1926). Экспериментальные исследования
1 Иногда задний отдел вовсе неразличим.
2 По Coker’у (1934) у этого вида длина фуркальных ветвей экспериментально
может быть изменена; при низкой температуре (7—10°) получается поколение с удли-
ненными ветвями.
3 По экспериментальным исследованиям Coker’а (1933), выведенные при низких
температурах экземпляры отличаются большей величиной (почти в 1.5 раза длиннее),
нежели при высоких (28—30°) температурах.
5. Acanthocyclops
225
Lowndes’a (1928в), однако, убедительно показали возможность скрещи-
вания A. robustus л A. vernalis и, вместе с тем, полную невозможность
их разделения на основании формулы шипов; этот автор из культур
A. robustus получил поколение A. vernalis, так что говорить здесь
о 2 самостоятельных видах, конечно, нельзя. Gurney (1933) подтверж-
дает исследования Lowndes’a. Несмотря на чрезвычайную изменчивость
целого ряда признаков A. vernabs, выделение A. robustus в особый
вариетет все же возможно по строению придатков, вооружающих плава-
тельные ноги (см. ниже), — признак, на основании которого Gurney
(1933) и считает возможным выделить var. robustus. Описанного в 1893 г.
С. americanus Marsh (Trans. Wise. Acad., 9) Kiefer (1929:55) ставит
в синонимы C. robustus, a Gurney (1939:25) считает подвидом A. vernalis.
Однако Lowndes (1926, 2028а) считает A. americanus самостоятельным
видом, близким к A. vernalis var.robustus, что надо признать правиль-
ным, принимая во внимание особенно строение receptaculum seminis,
генитального сегмента самки и вооружение ног VI пары самца. A. ame-
ricanus — вид пелагический (широко распространен в Северной Аме-
рике, пелагиаль озер), в противоположность типичному A. vernalis и var
robustus. Есть предположение, что описанный из Голландии Lint
(1922:80) пелагический A. viridis var. pelagica идентичен с A. america-
nus Marsh (Gurney, 1933). В старой литературе последний совершенно
ошибочно относился к вариететам A. viridis.
Распространение (включая и var. robustus). Вся Европа, Азия,
Северная Америка, Канарские острова, Кергуелен. Известен из Грен-
ландии и Исландия. В СССР известен из очень многих местностей, от
крайнего севера (включая арктические острова) до крайнего юга (окре-
стности Одессы, Крым, Кавказ, включая и его горную область); широко
распространен в азиатской части СССР, включая весь север Сибири
и Дальний Восток (бассейн Амура, Камчатка). Повидимому отсутствует
в зоне пустыни и полупустыни; широко распространен во всех осталь-
ных ландшафтных зонах. Очень распространен в высокогорных водое-
мах, где является обычным компонентом фауны мелких луж и показан
до высоты около 3000 м над урознем моря.
Биология. Обитает в очень разнообразных водоемах — в лужах,
болотах, прудах, в прибрежной и придонной области медленно текучих
рек и ручьев. Обычен в мелких временных водоемах и в литорали
(сублиторали) озер, где спускается до довольно значительных глубин.
Особенно характерен для мелких болотистых, в частности сфагновых и
гипновых луж, а также для весенних снеговых и дождевых водоемов. Живет,
очевидно, не только в свободной толще воды, но и в моховых подуш-
ках (выжимки мхов). Указания на пелагический образ жизни A. vernalis
(и var. robustus) надо считать ошибочными. Троглоксен. Эвритермичен,
найден (половозрелые индивиды) при температурах от 1° до 28—30°.
Указание некоторых авторов (Graeter, 1903; Thiebaud, 1908) на холодно-
водность ошибочно и основано на выпадении его на лето в усыхающих
весенних водоемах, тогда как в постоянных водоемах он размножается и
летом, при высокой температуре воды. Встречается в водах с pH =4.4.—8.2.
По Скадовскому (1928), ядовитое воздействие на A. vernalis Н-ионов
парализуется ионами Са (но не Na). Очевидно он переносит самые
различные содержания СаО — он обычен в бедных электролитами
дистрофных водах, но известен также в водах с содержанием СаО
100 мг на литр. Очевидно также, что рачок переносит высокую оки-
сляемость воды [например, по данным Кузнецова и Щербакова (1925)
15 (Cavua ГГСО
226
CYCLOPINAE
он найден при окисляемости 115 мг О2 на литр в гипновом болоте
близ Москвы]. Вид пресноводный, в сколь-либо осолоненных водах
встречается лишь случайно.
Цикл размножения. В средней Европе встречается в постоян-
ных водоемах круглый год, что отмечено для водоемов Гренландии.
Полицикличен, в некоторых мелких водоемах 2 половых периода в год;
в постоянных водоемах 1 из половых периодов наблюдается нередко
зимой. Возможно, однако, что в мелких, быстро усыхающих весенних
лужах имеет не более 1 (2) половых периодов. Несомненно, что A. ver-
nalis имеет стадии покоя, в которых переносит высыхание водоема.
Kammerer (1907) выводил его из ила, взятого в мелком водоеме
в Швеции; ил был взят 31VIII 1901 и помещен в воду лишь 10 V 1904,
причем уже 18VI 1904 в аквариуме появилась молодь. Вероятно рачок
переносит высыхание на последних стадиях метаморфоза (см. A. bicus-
pidatus), а не в состоянии покоящихся яиц.
Вероятно, главным образом, Р-мезосапроб, отчасти олигосапроб.
Типичная форма Acanthocyclops vernalis Fisch, отличается от var.
robustus наличием щетинок (а не шипа) на внешних краях дистального
членика эндоподита ног II—IV пары. Апикальные щетинки фуркальных
ветвей обычно не утолщенные.
*3а. Acanthocyclops vernalis var. robustus (Sars) (фиг. 475-,).
C. robustus Sars, 1863, Forh. Selsk. Christ. (1862):245.— C. brevispinosus Herrick,
1884, Rep. Geol. Survey Minnesota, 12:148, t. S, f. 7—12. C. vernalis (part.) Schmeil,
1892.— C. robustus Lilljeborg', 1901:19, t. 2, f. 8—10; Sars, 1913, t. 26.— C. viridis var.
dives Juno-mayer, 1914, Math. Term. Kozl., 33:61, f. 14.— C. vernalis var. robustus, Рылов
1918:50.— C. vernalis var. tetracaniha ч- tetracantha f. robusta var. Kiefer, 1926, Arch,
f. Hydrob., 16: 504, f. 16—19.— C. robustus var. saxonica ч- var. aphanes -+ var. armata 4-
ч-var. setiger Thallwitz, 1926, ibid., 17:366, f. 1—3, 5,9—14. — C. robustus Lowndes,
1928b:184, f. 1—2,7—12, 15—20.— C. vernalis var. robustus Gurney, 1933:201, 203, f. 1612.
От типичных A. vernalis самка отличается присутствием шипа
(а не щетинки) на внешнем крае дистального членика эндоподита ног
IV пары (нередко и ног II и III пары). Апикальные щетинки фуркаль-
ных ветвей обычно несколько толще, чем у типичного A. vernalis.
Формула шипов, как правило, 3-4-4-4, но может быть и 2>3-3-3.
Распространение. Как у типичной формы (см. выше). В СССР
известна с о. Кильдина, с Новой Земли, с севера Кольского полуост-
рова, из Коми АССР, из областей Московской, Калининской, Одесской,
из окрестностей Махач-кала, с Дальнего Востока, Камчатки. Нередко
встречается с типичной формой совместно и обитает в тех же усло-
виях, что и последняя.
*4. Acanthocyclops muscicola (Lastotschkin).
Cyclops muscicolus Ласточкин, 1924:4, рис. 1—3.— C. muscicola Borutzky,
1927:63.— Cyclops (Acanthocyclops) muscicola Kiefer, 1929:56.
Самка. Фуркальные ветви немного (в 1.1—1.3 раза) длиннее 2
задних абдоминальных сегментов, взятых вместе; внутренняя из край-
них апикальных щетинок в 2—2.1 раза длиннее внешней, латеральная
щетинка прикреплена на расстоянии 0.59—0.65 длины внешнего края,
считая от основания ветви. Генитальный сегмент спереди значительно
расширенный, receptaculum seminis с очень объемистым поперечно-оваль-
ным передним и с очень узким задним отделом, очень близок по форме
& ACANTHOCYCLOPS
227
Фиг. 47.
Acanthocyclops vernalis (Fisch, typ.): 7 — фурка самки с брюшной стороны,
2—задний сегмент торакса и абдомен с receptaculum seminis, 3—нога
V самки, 4 — эндоподит ноги IV самки; A. vernalis var. robustus (Sars):
j — фурка самки с брюшной стороны, б—2 задних членика эндоподита ноги
IV самки, 7 — дистальный членик экзоподита иоги IV самки.
228
CYCLOPINAE
к таковому A. vernalis. Передние антенны 12—13-членистые (последнее
вследствие расщепления надвое 9-го членика), немного не достигаю-
щие до заднего конца цефалоторакса. Членистность плавательных ног
как у A. vernalis. Дистальный членик экзоподита несет 3 или 4 шипа,
апикальные шипы дистального членика эндоподита ног IV пары одина-
ковой длины (или почти одинаковой). У ног II—IV пар шипы обеих
ветвей в дистальной ]/2—3/t Длины окружены каемкой из тонкой про-
зрачной перепонки с зазубренными краями. Ноги V пары очень похожи на
таковые A. vernalis, шип их дистального членика толстый, хорошо разви-
той. Длина 0.9—1.0 мм. Окраска однообразная, сероватая иди светлого-
лубая, с синими включениями, иногда самки бесцветны. Самец не описан.
Распространение. Ивановская область (оз. Валдайское; Ласточ-
кин, 1924, 1925, 1926, 1927). Больше пока нигде не найден.
Биология. Вид очень характерен для покрытых водой сфагновых
и гипповых мхов мелководной прибрежной области указанного озера;
также и в мелких дистрофных лужах. Цикл развития неизвестен.
5. Acanthocyclops americanus (Marsh).
Cyclops amerikanus Marsh, 1893. Wiss. Trans. Wise. Acad., 9:202, t. 4, f. 8—10.— C. viri-
dis amerikanus Herrick and Turner, 1895:92, t. 14.— C. viridis var. insectus Forbes, 1897:41,
t. 11, f. 3—6.— C. viridis var. amerikanus Marsh, 1909:1076, t. 79, £. 6.— C. amerikanus
Marsh, 1920:9j; Lowndes, 1926:616; Lowndes, 1926, Rep. Marlbor. Coll.. 74:86, t. 4., Lown-
des, 1928a:12, f. 1—8.— Acanthocyclops robustus (part.) Kiefer, 1929:55.— C. vernalis-
amerikanus Gurney, 1933:205, f. 1616—1625.
Самка. Тело более стройное и узкое, чем у A. vernalis, задние
углы 2 задних сегментов торакса выступают кнаружи. Длина фуркаль-
ных слабо расходящихся ветвей приблизительно в б раз превышает
ширину, из их крайних апикальных щетинок внутренняя почти вдвое
длиннее внешней; внутренние края ветвей без волосков. Плавательные
ноги как у A. vernalis, формула шипов 2-3-3-3 или 3-4-4-4. Дисталь-
ный чЛеник эндоподита ног IV пары очень удлиненный, его длина
приблизительно в 2.75 раза более ширины, его апикальные шипы почти
равной длины, придаток внешнего края представлен шипом. Ноги V
пары как у A. vernalis. Генитальный сегмент сильно расширенный
в переднем отделе, где его бока закруглены, цилиндрически суженный
в заднем. Receptaculum seminis отличается от такового A. vernalis
отсутствием вдавления спереди, присутствием (всегда?) сосковидных
выростов по бокам переднего отдела, который окружен бесцветной
каймой, и очень узким задним отделом. Длина 1.2—1.5 мм. Бесцветны,
с голубыми включениями в области ротового отверстия и абдомена
(иногда по всему телу). Яйцевые мешки очень крупные, довольно зна-
чительно расходящиеся, содержащие значительное количество (по 60)
яиц.
Самец. У ног VI пары 2 почти равной длины шипа и щетинка,
которая очень длинна (более чем вдвое длиннее шипов).
Распространение.1 Озера Северной Америки и Англии.
Биология. Вид пелагический, в озерах Северной Америки
весьма распространенный также и в мелких водоемах. Половое размно-
жение, повидимому, в холодное время года (осень — зима — начало
весны).
1 В Германии и Венгрии (С. viridis var. dives Jung-mayer, Math. Term. KozL, 33, 1917)
пока с точностью не установлен.
8. ACANTHOCYCLOPS
223
6. Acanthocyclops kieferi (Chapp.) (фиг. 484_8).
Cyclops kieferi Chappuis, 1925, Bull. Soe. Stiint. Cluj, 2:159, f. 1—3; Kiefer, 1925
Mierosk. f. Naturfreunde, 3, f. 5; Kiefer, 1926, Zool. Anz., 67:103, f. 1—5. — Cyclops
(Acanthocyclops) kieferi Kiefer, 1929 : 56. — Acanthocyclops hispanicus Kiefer, 1937b : 323,
f. 1—3
Самка. Глаз крупный, с пигментом. Тело немного сплющенное
в спинно-брюшном направлении. Генитальный сегмент спереди сильно
расширенный. Receptaculum seminis поперечно-овальный. Анальная
пластинка умеренно развитая, трапециевидная, с немного вогнутым задним
краем, лишенным вооружения. Длина почти параллельных фуркальных
ветвей не более чем в 2—2.5 раза превышает их ширину, из крайних
апикальных щетинок внутренняя короче шипообразной внешней, из сред-
fl
Фиг. 48. (По Kiefer’y).
Acanthocyclops hypogeus Kief.: 1 — абдомен с фуркой самки ео спинной стороны,
2 — дистальный членик эндоподита ноги IV самки, 3 — нога V самки; A. kieferi (Chapp.)
3—абдомен с фуркой самки, 5—генитальный сегмент и receptaculum seminis, 6—ди-
стальный членик эндоподита ноги IV самки, 7 — нога V самки, 8 — анальная пластинка,
230
CYCLOPINAE
них апикальных щетинок внешняя почти равна длине всего абдомена,
а внутренняя вдвое длиннее внешней. Передние антенны 11-членистые,
немного не доходящие до заднего конца цефалоторакса. Обе ветви
всех плавательных ног 3-членистые, с очень короткими члениками,
у дистального членика эндоподита ног IV пары внутренний апикальный
шип почти вдвое длиннее внешнего. Ноги V пары 2-членистые, внутрен-
ний шип дистального членика очень короткий, апикальная щетинка
хорошо развитая. Длина 0.6—0.84 мм.
Самец. Генитальный сегмент очень похож на таковой A. crassicau-
dis, ноги VI пары с 1 шипом и 2 щетинками. Строение и вооружение
фуркальных ветвей и плавательных ног как у самки. Длина около
0.68 мм.
Распространение. Подземные воды Венгрии и Г ермании.
Троглобионт.
Вероятно с A. kieferi идентичен A. reductus (Chapp., 1905), у кото-
рого анальная пластинка несколько более развитая, членистость ног
по формуле 3-2-/3-2/3-2/3-3, апикальные шипы дистального членика
эндоподита ног IV пары немного более длинные, и базальный членик
ног V пары немного более узкий. Описан неполно.1 Acanthocyclops
hispanicus Kief., по всем вероятиям, есть также синоним A. kieferi.
Из описания Kiefer’a никаких существенных отличий от последнего
вида усмотреть нельзя
7. Acanthocyclops phraeticus (Chapp.).
Cyclops phraeticus Chappuis, 1928, Bull. Soc. Stiint. Cluj, 4: 20, f. 1—4. — Cyclops (Acan-
thocyclops) phraeticus Kiefer, 1929: 56.
Самка. Анальная пластинка умеренно развитая. Длина фуркаль-
ных ветвей в 3.5 раза более их ширины, из крайних апикальных щети-
нок внутренняя в 2.5 раза длиннее внешней, латеральная щетинка при-
креплена в задней 1/3 длины внешнего края. Receptaculum seminis как
у A. capillatus. Передние антенны 12-членистые, не заходящие за задний
конец цефалоторакса. Обе ветви всех пар плавательных ног 8-членистые,
длина дистального членика эндоподита ног IV пары вдвое более его
ширины, апикальные шипы почти равной длины, около 3/4 длины чле-
ника. У ног V пары широкий базальный членик, вооружение дистального
членика как у A. vernalis. Длина 0.85 мм. Самцы не известны.
Распространение. Колодцы Румынии.
8. Acanthocyclops venustus (Norm, et Scott) (фиг. 493 4).
Cyclops venastas Normann and Scott, 1906, Crust. Devon Cornwall, 189, t. 16, f. 11;
t. 18, f. 9; t. 20, f. 1—3; Gumey, 1923, Joum. Linn. Soc. London, 35 :437; Klie, 1928,
Zool. Anz., 78; 62, f. 1—2. — Cyclops (Acanthocyclops) venastas Gurney, 1933: 210,
f. 1626—1639.
Самка. Передний и задний отделы тела утолщенные, задние края
абдоминальных сегментов зазубрены. Глазной пигмент имеется. Поверх-
ность кутикулы тела (включая отчасти и фуркальные ветви) покрыта
рядами очень мелких складок, которые, повидимому, постоянно отсутствуют
на тораксе у var. crinitus (см. ниже). Генитальный сегмент крупный,
его ширина почти равна его длине. Receptaculum seminis объемистый,
1 Об A. reductus см, также Kiefer (1928:525, f. 37 и 1929:57).
8. ACANTHOCYCLOPS
231
Фиг. 49.
Acanthocyclops rhenanus Kief.: 1 — абдомен и фурка еамки с брюшной стороны, 2— эндо-
подит ноги IV самки; A. venustus Norm, et Scott: 3— генитальный сегмент самки, ноги V
и receptaculum seminis, 4 — фурка самки с брюшной стороны; A. languidus var. disjun-
ctus Thallw.: 5—фурка самки с брюшной стороны, 6—соединительная пластинка ног IV,
7—2 членика эндоподита ног IV; A. languidus var. belgicus Kief.: 8—фурка самки
со спинной стороны, 9—2 членика эндоподита ноги IV.
(/—2 и 5—9—по Kiefer’y, 3—4 — по Gurney).
232
CYCLOPINAE
с крупным передним и с более мелким задним поперечно расположен-
ными отделами. Анальная пластинка умеренно развитая, без придатков.
Фуркальные ветви заметно расходящиеся, их длина в 3—4 раза более
их ширины, внутренние края с неправильно расположенными группами
мелких волосков, из крайних апикальных щетинок внутренняя почти
вдвое длиннее внешней, обе апикальные срединные щетинки очень
длинные, из них внутренняя равна длине переднего отдела тела, а у var.
crinitus даже всего тела (без фуркальных ветвей). Передние антенны
12-членистые, немного не доходящие до заднего конца цефалоторакса.
Членистость плавательных ног по формуле З-З/З-З/З-З/З-З. Длина дисталь-
ного членика эндоподита ног IV пары вдвое более его ширины, его
апикальные шипы почти равной длины; формула шипов 3-4-4-4. Ноги
V пары очень похожи на таковые A. viridis, внутренний шип дисталь-
ного членика короткий. Длина типичной формы 1.0—1.3 мм. Яйцевые
мешки содержат небольшое количество яиц и тесно прилегают к абдо-
мену. Самец не известен.
Вид наиболее близок к A. phraeticus (Chapp.) и A. stammeri (Kief.).
Типичная форма A. venustus (Norm, et Scott) известна из Англии
и Германии.
Биология почти не изучена. Обитает в мелких болотистых водоемах
(сфагновые лужи), известен из Wollingster-See (Германия, озеро с Lobe-
lia и Isoetus).
8а. Acanthocyclops venustus var. crinitus (E. Graeter) (фиг. 50e_8).
Cyclops crinitus E. Graeter, 1903, Zool. Anz., 33:45, f. 1—13; E. Graeter, 1911,
t. 1, f. 3, 4, t. 2, f. 5, textf. 16, 17; Chappuis, 1920 (1922), ibid, 16 :24.
Самка. От типичной формы отличается отсутствием (всегда?)
шипиков на поверхности кутикулы торакса,1 очень длинной (почти
равна длине всего тела, исключая фуркальные ветви) внутренней сред-
ней апикальной щетинкой фуркальных ветвей и- гораздо большей вели-
чиной— длина 2.0—2.3 мм.
Все авторы считают рассматриваемый вариетет вполне идентичным
с типичной формой, лишь Gurney (1933:212) в этом сомневается. При-
нимая во внимание весьма обстоятельное описание Е. Graeter, нет
оснований смешивать эти 2 формы, так как их различия касаются
нескольких признаков и довольно значительны.
Распространение. Швейцария.
Биология. Троглобионт (типичная форма, вероятно, в подземных
водах отсутствует), обитает в пещерных водах и колодцах.
9. Acanthocyclops stammeri Kief. (фиг. 51t_4).
Cyclops (Acanthocyclops) stammeri Kiefer, 1930, Zool. Anz., 87:223, f. 1—4.—
Acanthocyclops troglophilus Kiefer, 1932, ibid, 101 : 55, f. 14—18.
Самка. Задние края абдоминальных сегментов зазубрены, аналь-
ная пластинка умеренно развитая, с гладким, закругленным задним
краем, не заходящая за основание фуркальных ветвей. Длина последних,
параллельно поставленных, в 4 (или несколько более) раза превышает
1 По данным Graeter и Gurney у A. crinitus эти шипики имеются только на
абдомене (включая фуркальные ветви).
8. ACANTHOCYCLOPS
233
их ширину (у var. westfalicus ветви более короткие), внутренние края
или гладкие, или лишь с немногими очень короткими волосками, из
Фиг. 50.
Acanthocyclops sensitivus (Graet. et Chapp.): 1 — абдомен самки и receptaculum seminis,
2—дистальный членик эндоподита ноги IV; A. karamani Kief.: 3—генитальный сегмент
с receptaculum seminis и ноги V, 4 — фурка самки с брюшной стороны, 5 — дистальный
членик эндоподита ноги IV; A. venustus var. crinitus (Graet.): 6—нога V самки,
7—фурка самки со спинной стороны, 8—генитальный сегмент самки с receptaculum
seminis; Graeteriella (Paragraeteriella) laisi Kief.: 9 — генитальный сегмент самки
c receptaculum seminis, 10— фурка самки со спинной стороны.
(7—5 и 9—10 — по Kiefer’у, 6—8—по Graeter’y).
крайних апикальных щетинок внутренняя более чем вдвое длиннее
внешней, средние щетинки очень длинные, из них внутренняя почти
равна длине всего рачка, латеральная щетинка прикреплена в начале
234
CYCLOPINAE
задней г/3 длины внешнего края. Reeeptaculum seminis с объемистым
поперечно-овальным передним отделом. Передние антенны 12-членистые.
Членистость плавательных ног по формуле З-З/З-З/З-З/З-З, формула
шипов экзоподитов 3-4-4-4, щетинок 5-5-5-5. Длина дистального чле-
ника эндоподита ног IV пары более чем вдвое превышает его ширину,
у var. westfalicus членик несколько короче; апикальные шипы почти
одинаковой длины, немного короче самого членика, на внешнем крае
которого имеется шипообразная щетинка.1 Дистальный членик ног V
пары крупный, его внутренний шип короткий, щетинка длинная, хорошо
развитая. Длина около 1.2 мм.
Вид близок к A.venustus var. crinitus (Е. Graet.).
Распространение. Карстовые пещеры северной Италии (St.
Ganzian), пещеры в Герцеговине (A. troglophilus Kief.). Биология неизу-
чена. Троглобионт.
9а. Acanthocyclops stammeri var. westfalicus Kief. (фиг. 515_6).
Cyclops (Acanthocyclops) stammeri westfalicus Kiefer, 1931, Mitt. Hohl-u. Karst,
forsch., 3, f. 1—2. — Acanthocyclops stammeri westfalicus Kiefer, 1936, Bull. Mus. R. Hist-
Nat. Belgique, 12 :4, f.l—3.
От типичной формы самка отличается более короткими фуркаль-
ными ветвями, длина которых лишь в 3.1 раза более ширины, более
короткими фуркальными щетинками и более коротким дистальным чле-
ником эндоподита ног IV пары. Дорзальная щетинка фуркальных ветвей
рчень длинная, превышающая их длину.
Распространение. Пещеры Вестфалии; пещеры Бельгии. Тро-
глобионт.
-10. Acanthocyclpos rhenanus Kief. (фиг. 491_2).
Acanthocyclops rhenanus Kiefer, 1936, Zool. Anz., 115:168, f. 1—5.
Самка. Генитальный сегмент и reeeptaculum seminis как у A. stam-
meri Kief. Задние края абдоминальных сегментов слабо зазубрены,
анальная пластинка слабо развитая. Фуркальные ветви параллельные,
их длина лишь в 2.5 — 3 раза более их ширины, внутренние края без
волосков, из крайних апикальных щетинок внутренняя вдвое длиннее
внешней, средние апикальные щетинки хорошо развитые, их длина —
внутренней менее половины длины тела, а внешней равна половине
длины внутренней. Дорзальная щетинка очень длинная (как у A. stam-
meri var westfalicus), почти втрое длиннее внешней крайней щетинки
и значительно длиннее ветвей. Передние антенны 12-членистые, на 9-м
членике очень длинный сензорный придаток. Членистость плавательных
ног по формуле З-З/З-З/З-З/З-З, формула шипов экзоподитов 2-3-3’3,
щетинок 4-4-4-4. Длина дистального членика эндоподита ног IV пары
менее чем в 1.5 раза превосходит его ширину, внутренний апикальный
шип приблизительно в 1.5 раза длиннее внешнего, внешний край членика
снабжен нормальной щетинкой. Ноги V пары, как у A. vernalis. Длина
0.75 мм. Самец не известен.
Вид очень близок к A. stammeri var. westfalicus Kief., от которого
отличается более короткими средними апикальными щетинками фуркаль-
ных ветвей, иной формулой шипов и щетинок экзоподитов, иным соотно-
1 На рис. 3 (Bull. Mus. R. Hist. Nat. Belgique, 12,1936) Kiefer рисует у var. west-
falicus здесь не шип, а нормальную щетинку, что и отмечает в тексте.
8. ACANTHOCYCLOPS
235
Фиг. 51.
Acanthocyclops stammeri Kief.: 1 — фурка самки co спинной стороны, 2—генитальный
сегмент с reeeptaculum seminis, 3—дистальный членик эндоподита ноги IV, 4 — нога V
самки; A. stammeri var. westfalicus Kief.: 5— дистальный членик эндоподита ноги IV,
6 — фурка самки со спинной стороны; A. scopliensis Kief.: 7 — фурка самки с брюшной
стороны, 8 — генитальный сегмент с reeeptaculum semiuis, 9 — дистальный членик эндо-
подита ноги IV; A. stygius (Chapp.): 10— нога V самки, 11— генитальный сегмент
с reeeptaculum seminis.
(7—9 — по Kiefer’у, 10—11 — по Chappuis).
236
CYCLO R INAE
шением длины апикальных шипов дистального членика эндоподита ног
IV пары.
Распространение. Германия (область Верхнего Рейна, также
близ Карлсруе); подземные воды, колодцы.
*11. Acanthocyclops capillatus (Sars) (фиг. 52).
Cyclops capillatus Sars, 1863, Forh. Selsk. Christ., 1862: 248; Schmeil, 1898:152, t. 13,
f. 1; Lilljeborg', 1901 :54, t. IV, f. 1—4; Sars, 1913:43, t. 24; Рылов, 1918:43, фиг. 14—16;
Marsh, 1920, I: II, t. IV, f. 4, 6—8—11. — Cyclops (Acanthocyclops) capillatus Kiefer,
1929:57.
Самка. Общий габитус тела очень близок к таковому A. viridis,
но цефалоторакс более удлиненный. Наибольшая ширина цефалоторакса
2-го — 5-го сегментов торакса лежит на середине его длины. Гениталь-
ный сегмент укороченный, мало расширенный спереди, его наибольшая
ширина приблизительно равна его длине. Receptaculum seminis характер-
ной формы, с объемистым передним поперечно-овальным отделом
и с полуовальным менее объемистым задним отделом. Фуркальные вет-
ви параллельные или почти параллельные, их длина приблизительно
в 4—4.5 раза превышает ширину, латеральная щетинка прикреплена
почти на середине внешнего края, хорошо развитая. Внутренняя из край-
них апикальных щетинок лишь очень немного длиннее внешней крайней,
средние апикальные щетинки длинные, из них внутренняя лишь немного
длиннее внешней. Передние антенны 12-членистые, обычно немного
не доходящие до заднего конца цефалоторакса, 8-й и 9-й членики
(особенно 8-й) относительно длинные. У задних антенн дистальный
членик заметно длиннее предыдущего членика (3-го от начала антенны).
Обе ветви плавательных ног всех 4 пар 3-членистые, формула шипов
3-4*4-4, дистальные членики обеих ветвей удлиненные, стройные,
у дистального членика эндоподита ног IV пары апикальные шипы отно-
сительно длинные, из них внутренний немного длиннее внешнего, длина
этого членика не менее чем вдвое превышает его ширину. Ноги V пары
очень похожи на таковые A. viridis, но базальный членик менее широ-
кий. Длина 1.7—1.8 мм. Окраска желтоватая, иногда с коричневым или
зеленоватым оттенком. Яйцевые мешки весьма удлиненные, сильно рас-
ходящиеся, обычно заключающие значительное (12—20 и более) коли-
чество яиц.
Самец. Ноги VI пары с 1 коротким шипом и с 2 щетинками,
из которых средняя вдвое длиннее шипа и почти вдвое короче внешней
щетинки. Длина 1.1—1.2 мм.
Распространение. Европа (Скандинавия, СССР), Аляска.
В СССР показан для Мурманского побережья, Большеземельской тундры,
севера Кольского полуострова, Карело-Финской ССР, для северного
острова Новой Земли.1 Таким образом A. capillatus имеет хорошо
выраженный северный ареал распространения, охватывающий зоны
тундры и тайги; едва ли подлежит сомнениям, что в пределах этих зон
(особенно первой) широко распространен и в азиатской части СССР
(на Аляске найден Marsch, 1920).
Биология изучена очень неполно. Обитает преимущественно
в мелких маленьких водоемах (лужи, болота), а также в литорали
1 Следующие указания требуют подтверждения: лесостепная часть Омской области
(Зверева, 1930), Калининская обл. (Степанова и Белоголовая, 1934). Указание Совин-
ского (1888) на нахождение в окрестностях Киева безусловно ошибочно.
8. ACANTHOCYCLOPS
237
и в донной области озер, в крупных озерах Норвегии живет на илистом
грунте, на глубинах 3—10 м. Холодноводно-стенотермический вид,
оптимум, вероятно, не выше 14—15°. Судя по имеющимся пока данным,
обитает в относительно бедных электролитами олиготрофных (быть
Фиг. 52. Acanthocyclops capillatus (Sars).
/—2— фурка самки со спинной стороны; 3—два дистальных членика экзопо-
дита ноги IV; 4— генитальный сегмент с receptaculum seminis; 5—нога V
самки; 6—дистальный членик эндоподита ноги IV.
может отчасти в слабо эвтрофированных) водоемах; по данным для
севера Новой Земли найден при pH = 7.5 и в кислых водах отсутствует,
во всяком случае не входя в состав населения сильно дистрофирован"
ных водоемов, с pH = 4 и менее.
Цикл развития не исследован. В постоянных водоемах, по всем
вероятиям, встречается круглый год и полицикличен (быть может дицик-
"238
CYCLOPINAE
личен). Olofsson (1917) на Мурмане в мелком водоеме находил самок
с яйцами и самцов в июне, нами самки с прикрепленными сперматофо-
рами в низовьях р. Колы (заросли макрофитов) были находимы в конце
июля.
12. Acanthocyclops sensitivus (A. Graether et Chapp.) (фиг. ЗО^г).
Cyclops sensitivus Graether und Chappuis, 1914, Zool. Anz., 43:507, f. 1—5. —
Cyclops (Acanthocyclops) sensitivus Kiefer, 1929:58; Gurney, 1933:215, f. 1643—1654;
Kiefer, 1935, Verb. Nat. Ver. Karlsruhe, 31:133, f. 1—5.
Самка. Тело стройное, ширина генитального сегмента несколько
более его длины, receptaculum seminis характерной „бабочковидной“
формы, задний его отдел в виде 2 крупных мешков. Задние края абдо-
минальных сегментов не зазубрены. Фуркальные ветви короткие, их длина
приблизительно в 2.5 раза превышает их ширину, латеральная щетинка
прикрепляется лишь немного кзади от середины их внешнего края,
из крайних апикальных щетинок внутренняя в 2 с лишком раза длин-
нее внешней, из хорошо развитых средних апикальных щетинок внутрен-
няя приблизительно на 1/3 длиннее внешней. Передние антенны 17-чле-
нистые, немного не доходящие до заднего края 2-го торакального сег-
мента, иногда несколько более короткие, у 12-го членика очень длинная
сензорная щетинка. У задних антенн дистальный членик немного тоньше
и короче предыдущего членика. Членистость плавательных ног по фор-
муле З-З/З-З/З-З/З-З, формула шипов 2-3-3-3. Длина дистального чле-
ника эндоподита ног IV пары вдвое более ширины, внутренний апи-
кальный шип короче самого членика. Ноги V пары 2-членистые, базальный
членик очень широкий, дистальный членик с коротким внутренним
шипом, длина которого приблизительно равна длине членика, и с хорошо
развитой, на конце оперенной внешней щетинкой. Глазной пигмент
имеется. Длина 0.72 мм. Бесцветный, чрезвычайно прозрачный. В яйцевых
мешках, расходящихся и удлиненных, заключается по 4—8 яиц. Самец
не известен.
Распространение. Франция, Бельгия, Германия, Швейцария,
Англия.
Биология. Троглобионт, подземные воды (колодцы).
*13. Acanthocyclops bicuspidatus (Claus) (фиг. 53).
Cyclops bicuspidatus Claus, 1857b: 209, t. II, f. 6—7. — C. pulchellus Sars, 1863,
Forh. Selsk. Christ., (1862): 246.— C. enizi’-t-C. roseus Daday, 1882, Orv. Term. Erdl.
Mus., 7 : 220. — C. gigas (non C. gigas Claus) Thomson, 1833, Tr. New-Zeal. Inst.,
15:96, t. 9, f. 8—10. — C. bicuspidatus Schmeil, 1892:75, t. 2, f. 1—3.— C. pulchellus
Sars, 1913:47, t. 27. — C. pulchellus var. synarthrus Lowndes, 1926, Rep. Marlbor. Soc.,
74:88, t. 25, f. 1—7.— C. (Diacyclops) bicuspidatus Kiefer, 1929:58, f. 19, a — b.—
C. (Acanthocyclops) bicuspidatus Gurney, 1933:219, f. 1655—1657, 1659—1671.
Самка. Тело стройное, наибольшая ширина цефалоторакса —
торакса лежит на середине их длины, задние углы заднего торакаль-
ного сегмента слабо вытянутые кнаружи. Генитальный сегмент объемистый,
постепенно суживающийся кзади, у receptaculum seminis задний отдел
очень обширный, в виде отвисающего кзади широкого мешка, передний
отдел меньше заднего, широкий. Фуркальные ветви почти (или вполне)
параллельные, их длина в 6—7 раз (иногда менее чем в 6 раз),—
modessana) превышает их ширину, латеральная щетинка прикреплена
обычно близ середины внешнего края, иногда несколько кзади от сере-
8. Acanthocyclops
239
дины, в конце передней ]/4—У3 длины этого края прикрепляется 1
(иногда 2—3) мелкий шипик. Внутренняя крайняя апикальная щетинка
лишь немного длиннее внешней крайней, шипообразной; иногда обе
крайние щетинки одинаковой длины. Средние апикальные щетинки
хорошо развитые, из них внутренняя приблизительно на 2/3 длиннее
внешней. Кутикула всего тела, включая фуркальные ветви и передние
Фиг. 53. Acanthocyclops bicuspidatus (Claus), самка.
/ — генитальный сегменте receptaculum seminis; 2—фурка с брюшной
стороны; 3—дистальный членик эндоподита ноги IV; 4—5— нога V.
антенны, нередко покрыта очень мелкими шйпиками. Передние антенны
14-(m. odessana) или 17-членистые (типичная форма), обычно доходя-
щие лишь до заднего конца цефалоторакса, на 12-м членике имеется
длинный тонкий сензорный придаток. Дистальный членик задних антенн
лишь немного длиннее предшествующего. Обе ветви ног I—IV пары
3-членистые.1 Формула шипов 2-3'3-3. Длина дистального членика
эндоподита ног IV пары в 2—3 раза более его ширины, апикальные
шипы этого членика хорошо развитые, из них внутренний обычно заметно
1 У var. synathrus Lown. 2-членистые; эта форма от типичного A. bicuspidatus
больше ничем не отличается.
240 CYCLOPINAE
короче внешнего.1 У ног V пары дистальный членик удлиненный,
узкий, его длина приблизительно втрое более его ширины, внутренний
шип прикрепляется близко от конца внутреннего края, мощный, заострен-
ный, равный длине самого членика, иногда длиннее последнего; базаль-
ный членик крупный, его длина почти равна его ширине. Длина 0.95—
1.7 мм. Окраска очень вариирует, обычно желтоватая, иногда серо-белая,
нередко с розовым или красным оттенком. Яйцевые мешки удлиненные,
обычно сильно расходящиеся и содержащие значительное число яиц.
Самец. Строение фуркальных ветвей и плавательных ног .как
у самки, 1-й членик передних антенн с длинным сензорным придатком.
Ноги VI пары с хороио развитым внутренним шипом и 2 щетинками,
из которых внутренняя (средняя) почти равна длине шипа, а внешняя
почти вдвое длиннее внутренней. Длина около 1.0 мм.
Распространение. Вся Европа, Азия, Новая Зеландия. Неод-
нократно типичная форма ошибочно указывалась для Северной Америки,
где в пелагиали озер обитает var. thomasi Forbes (1882); указание
последней (Brady, 1892) для Шотландии ошибочно (Gurney, 1933), ука-
зание типичного A. bicuspidatus для Канады требует подтверждения.
В пределах СССР распространен весьма широко. Указан для многих
местностей, начиная с Соловецких островов до Крыма и Кавказа
включительно; ряд местонахождений в азиатской части СССР (бассейн
Хатанги, р. Енисея, окрестности Томска, Славгорода, Казахстан).
Распространение типичного A. bicuspidatus, исключая зону субтро-
пиков и, возможно, тундры, охватывает все прочие зоны. В Альпах
указан до высоты 2300 м, в СССР до высоты 2000 м (Армени).
Биология. Обитает как во временных пересыхающих (например
в весенних снеговых), так и в постоянных разнообразных водоемах
(лужи, пруды, литораль озер и проч.). Очень характерен для весенних
луж. В озерах типичный A. bicuspidatus не спускается глубже 10 м.
Известен из медленно текучих рек и ручьев и из ключей типа лимно-
кренов. Не входит в состав пелагического планктона пресных озер
(в Северной Америке пелагический var. thomasi Forbes), но в солоно-
ватых озерах, подобно A. bisetosus, в больших количествах обитает
и в условиях пелагиали (см. ниже). Известен из пещерных вод и колод-
цев. Тенденция к троглофилии, в общем, выражена достаточно резко,
причем в пещерах обитает очень близкий вид рода Acanthocyclops,
генетически несомненно связанный с рассматриваемым видом (Л. karamani
Kief.). Указания на холодноводную стенотермичность A. bicuspidatus
едва ли правильны и основаны, очевидно, на факте высокого развития
этого вида в весенних водоемах; однако в постоянных водоемах он
встречается и летом, при высоких температурах воды (например при 20°).
Распространение в подземных водах указывает на способность A. bicus-
pidatus жить в условиях низкого содержания растворенного кислорода;
по исследованиям, Birge и Juday (1908, данные, по всем вероятиям,
относятся к var. thomasi), в североамериканских озерах он выживает
в придонных слоях в инцистированном состоянии (см. ниже) даже при
отсутствии кислорода. Широко приспособляются к колебаниям pH,
обычен в водах с pH — 4.1—8.6; в очень кислых водах (pH = 3.5)
отсутствует. Типичный A. bicuspidatus чрезвычайно эвригалинен. Помимо
1 Длина этих шипов очень вариирует даже у одной и той же популяции; шипы
могут быть одинаковой длины, но иногда внешний шип более чем вдвое длиннее
внутреннего.
8. ACANTHOCYCLOPS
241
нормальных пресных вод, часто был находим в водах высоко минера-
лизованных, в частности при значительном содержании NaCI. К сожале-
нию в большинстве случаев такие нахождения не сопровождаются точ-
ными химическими данными. Эксперименты Шманкевича (1875) над отно-
шением A. bicuspidatus к содержанию NaCI требуют повторения,
но, думается, 3 факта устанавливаются с достаточной убедительностью.
Во-первых, из этих экспериментов следует, что A. bicuspidatus с 17-
членистыми передними антеннами (возможно — в 2 формах) обитает
и в пресных водах и в водах е повышенным содержанием NaCI. Во-вторых,
форма с 14-членистыми антеннами (m. odessana) при известных условиях
эксперимента может быть превращена в таковую с 17-членистыми
антеннами. Кроме того эти эксперименты показывают, что A. bicuspi-
datus с 17-членистыми передними антеннами может обитать и при
гораздо более высоких содержаниях NaCI, нежели m. odessana.
По исследованиям Wolf (1905) A. bicuspidatus встречается круглый
год, полицикличен, периоды особенно интенсивного размножения в постоян-
ных водоемах наблюдаются весной и осенью, причем половозрелые
самки и самцы встречаются и зимой, подо льдом. По Murray (1911),
в мелких, усыхающих летом, прудообразных водоемах в окрестностях
Глазго он встречается зимой и также полицикличен. Однако в очень
кратковременных весенних лужах рачок, по всем вероятиям, имеет
не более 2 (быть может и 1) половых периодов. В таких лужах он
появляется обычно вскоре после заполнения водоема водой. Пересыха-
ние последнего переносит, как и A. bisetosus, в состоянии последних
стадий метаморфоза и во взрослом состоянии, облекаясь секретом,
выделяемым кожными железами (Roy, 1932).1 Отнесение A. bicuspidatus
к олигосапробам (Kolkwitz и Marsson, 1909) безусловно неверно (см.
Зиверт, 1929). Свойственные ему биотопы определенно указывают
на fi-мезосапробный характер, так что можно говорить лишь о факуль-
тативной олигосапробности этого вида.
Известно 2 вариетета1 2—var. thomasi Forbes (Amer. Nat., 16, 1882),
считаемый Kiefer’oM (1929), без достаточных оснований, самостоятель-
ным видом, и var. odessana Schmank. 1-й обитает лишь в озерах Север-
ной Америки (пелагиаль). Вышеуказанные эксперименты Шманкевича
(1875), несмотря на их неполноту, показали, что при изменении условий
температуры и солевости типичный A. bicuspidatus в ряде поколений
может быть превращен в описанного этим автором „С. odessanus",
почему имеются основания последнего считать морфой.
*13а. Acanthocyclops bicuspidatus m. odessana (Schmank.).
C. lubbocki Brady, 1869, Tr. Nat. Hist. Soc. Northumb., 3:127, t. 4, f. 1—8.—
C. odessanus Шман,<'евич, 1875.1:3—6. — C. helgolandicus Rehberg, 1880, Zool. Anz.,
3: 301. — C. lubbocki Canu, 1892, Rr. Labor. Wimeraux, 6:182, t. 3, f, 11—18; f. 4,
£.1—5. — C. bicuspidatus var. odessanus Schmeil, 1892 : 80. — C. bicuspidatus var. lubbocki
Gurney, 1904, Tr. N rfolk Soc., 7 :645.— C. pulchellus var. Sars, 1913:48, t. 27, f. A—
F. — C. bicuspidatus odessanus Kiefer, 1928, Bull. Soc. Nat. Marocco, 8:99. — C. (Diacy-
clops) bicuspidatus odessanus Kiefer, 1929 :59. — C. (Acanthocyclops) bicuspidatus lub-
iocki Gurney, 1933:222, f. 1658.
От типичной формы самка отличается 14-членистыми антеннами
и обычно несколько более короткими фуркальными ветвями.
1 Специального „кокона" не образуется, но тело циклопа просто обволакивается
секретом.
2 Указанный выше „synarthrus“ Lowndes (1926), кроме 2-членистости плаватель-
ных ног, от типичной формы ничем, в сущности, не отличается.
16 фауна СССР
242
CYCLOPINAE
Распространение. Европа (Великобритания, Скандинавия,
СССР, Германия, Южная Италия, Франция), Азия (Ирак, Иран, Памир),
Северная Африка (Алжир, Марокко). В СССР известен1 из окрестностей
Одессы, из Крыма (близ Карадага; Уломский, in lift), из соленого
озера Шор-куль на Памире (высота около 3700 м). Характерен для
солоноватых вод пустынной и полупустынной зон.
Биология выяснена очень недостаточно. Обитает, главным обра-
зом, в континентальных солоноватых водоемах (Одесские лиманы).
Известен также из пресных вод. Так, он обитает в мелких пресных
водоемах окрестностей Одессы, в мелких приморских пресных водоемах
во Франции; известен также из пресных вод Германии (Wolf, 1905} и
Англии (Gurney, 1933). В общем в пресных водах редок, и указанные
нахождения в них, по крайней мере в большинстве случаев, повидимому,
объясняются случайными заносами из ближайших соленых вод, частью
же относятся к водам с повышенной минерализацией. По Lundbeck
(1932) в солоноватых водоемах Голштинии обитает при 3 — 8%о (водоемы
осолоняются водами Балтийского моря), что соответствует мезогалинной
степени солености по классификации Redeke (1922). Помимо солености,
на членистость передних антенн A. bicuspidatus влияют и другие факторы,
в частности, вероятно, температура (Шманкевич). В окр. Одессы
m. odessana, по этому автору, обитает при солености менее 1° Бомэ
и температуре воды 14° R; по Okul (1937), он обитает в холодных
(постоянная температура 7—8°) глубоких колодцах (пресные воды),
также в дождевых и снеговых быстро высыхающих мелких водоемах.
Однако, по Richard’y (1895), он встречается как зимой, так и в теплое
время года.
14. Acanthocyclops karamani Kief. (фиг. 503_s).
Cyclops {Diacyclops) karamani Kiefer, 1932, Zool. Anz., 101: 56, 19—23.
Самка. Габитус тела как у A. bicuspidatus. Задние края абдоми-
нальных сегментов зазубрены. Генитальный сегмент значительно рас-
ширенный спереди и сильно суженный в задней половине. Receptaculum
seminis очень похож на таковой A. bisetosus. Анальная пластинка как
у A. crassicaudis, сзади закругленная. Фуркальные ветви стройные,
параллельные, их длина почти в 9 раз превышает их ширину, латеральная
щетинка прикреплена близко от их конца, крайние апикальные придатки
приблизительно одинаковой длины. Передние антенны 12-членистые,
доходящие до заднего конца цефалоторакса. Плавательные ноги как
у A. bicuspidatus, длина дистального членика эндоподита ног IV пары
вдвое более его ширины, внутренний апикальный шип на х/3 длиннее
внешнего и приблизительно равен длине членика. Ноги V пары как
у A. bicuspidatus. Длина около 1 мм.
Самец. Фуркальные ветви и вооружение эндоподита ног IV пары
как у самки. Ноги VI пары с 3 придатками. Длина 0.82 мм.
Распространение. Пещеры в Югославии.
1 Возможно, что A. bicuspidaias, указанный Рузским (1929) для оз. Горького
(Свердловской обл.), есть m. odessana (см. об этом у Таусон, 1933). Это предположе-
ние относится также к указанию того же автора (1926) для курорта Карачи (близ
Томска) и к указанию Зверевой (1930) для Западной Сибири.
8. ACANTHOCYCLOPS
243
15. Acanthocyclops bisetosus (Rehb.) (фиг. 54).
Cyclops bisetosus Rehber, 1880, Abb. Natur. Bremen, VI: 543; Schmeil, 1892 : 94, t. II,-
f. 8—11; Lilljeborg-, 1901: 14, t. II, f. 2—4; Sars, 1913 : 48, t. 28. — Cyclops bicuspidatus (non
Claus) Sars, 1863, Forh. Vid. Selsk. Christ., 1862:247. — Cyclops {Diacyclops} bisetosus
Kiefer, 1929:60, f. 20. — Cyclops {Acanthocyclops) bisetosus Gurney, 1933 : 227, f. 1675—1685.
Самка. Тело более удлиненное и стройное, чем у A. bicuspidatus,
немного сплющенное в спинно-брюшном направлении. Задние углы зад-
него торакального сегмента мало выступающие кнаружи. Длина гениталь-
ного сегмента приблизительно равна его ширине.Задний отдел recepta-
culum seminis обширнее переднего, в виде отвисающего кзади мешка,-
относительно более короткого, чем у A. bicuspidatus, по периферий
rec. seminis окружен довольно широко, сильно преломляющей свет;
бесцветной или желтоватой каемкой, передний отдел rec. seminis спереди
образует 2 бугорка вариирующей формы, иногда в виде пары рожек.
Фуркальные ветви параллельные, их длина в 5—7 раз более их ширины,
латеральная щетинка прикреплена значительно кзади от середины внеш-
него края, близко от его заднего конца. Из крайних апикальных щети-
нок внутренняя заметно короче внешней шипообразной, средние апикаль-
ные щетинки хорошо развиты, из них внутренняя лишь немного (при-
близительно на 1/4) длиннее внешней. Как у A. bicuspidaias, кутикула
тела, включая и фуркальные ветви, нередко мелко пунктирована. Перед-
ние антенны 17-членистые, достигающие до основания следующего
за цефалотораксом сегмента. Обе ветви I—IV пары плавательных ног
3-членистые, формула шипов 2-3-3-3, длина дистального членика
эндоподита ног IV пары приблизительно в 1т/2 раза превышает его
ширину, из апикальных шипов этого членика внутренний заметно длин-
нее внешнего. Ноги V пары очень похожи на таковые A. bicuspidatus,
базальный членик довольно широкий, его длина несколько менее
ширины. Длина 1.0—1.5 мм. Окраска серо-белая, иногда с более или
менее значительным красным, розовым или коричневым оттенком, иногда
яркокрасная. Яйцевые мешки крупные, овальные, содержащие обыкно-
венно значительное количество яиц (до 30), мало расходящиеся.
Самец. Внутренний шип ног VI пары очень маленький, внешняя
щетинка почти в 2х/2—3 раза длиннее этого шипа, средняя щетинка
(внутренняя) заметно короче внешней, но длиннее шипа. Длина около
1.0 мм.
Распространение. Вся Европа, Северная Америка (Канада)
вероятно вся Азия, Северная Африка (Алжир, Египет), Новая Зеландия.
Известен с Новой Земли и Шпицбергена. В СССР указан для неболь-
шого числа мест, но, без сомнения, у нас имеет очень широкое распро-
странёние. Пока известен из Карело-Финской ССР, из областей Яро-
славской, Горьковской, Ленинградской, Кировской, Московской, Саратов-
ской, Челябинской, Киевской, Харьковской, из окрестностей оз. Эльтон,
из окрестностей Карадага в Крыму. На Кавказе указан для окрестно-
стей Тбилиси и окрестностей Новороссийска. В азиатской части СССР
известен с Новосибирских островов, из бассейна р. Енисея (близ
Енисейска и Красноярска) и из Актюбинской области. В горах высоко
не встречается.
Биология. Характерен для мелких, особенно для временных,
весенних, к лету высыхающих водоемов, обитает также в прудах, реже
в озерах, где встречается до глубины 100 м. В текучих водоемах был
находим редко. Неоднократно указывался для пещерных вод и колодцев.
16*
244
CYCLOPINAE
В состав планктона прудов и пелагиали пресных озер не входит, но
в мелких водоемах и в солоноватых озерах ведет планктический образ
жизни. Температурная амплитуда для рассматриваемого вида, очевидно,
очень широка. Он был, например, находим в ключах с постоянной
температурой?0 и в очень мелких, прогреваемых свыше 20—25°, водоемах.
Фиг. 54. Acanthocyclops bisetosus (Rehb.), самка.
Z — фурка с брюшной стороны; 2—3—генитальный сегмент с recepta-
culum seminis;^ — нога V; 5—дистальный членик эндоподита ноги IV.
Отношение к кислороду не выяснено; вероятно, способен выносить
содержания раствора О2 менее 50% нормального насыщения. Обычен
в водах с pH = 5.4—8.4; в очень кислых, сильно дистрофированных
водоемах, однако, отсутствует, и, вероятно, pH около 4.5 является
нижней границей колебаний активной реакции. Широко распространен
в нормальных пресных водах, но обитает также в солоноватых водах
и рядом авторов относится к галофилам (Klie, 1925, неправильно относит
его к галобионтам). Очень характерный компонент планктона многих
сильно минерализованных, в частности богатых NaCl водоемов. A. bise-
8. ACANTHOCYCLOPS
245
tosus может переносить полное высыхание обитаемых им водоемов,
появляясь затем при новом заполнении их дождевой и снеговой водой.
По исследованиям Roy (1932) покоящихся яиц не образует, перенося
высыхание в состоянии последних стадий метаморфоза, а равно и во
взрослом состоянии, причем тело рачка обволакивается довольно плот-
ной оболочкой секрета, выделяемого кожцыми железами. Цикл размно-
жения изучен неполно. По Wolf’y (1905) в ряде водоемов встречается^
весь год, полицикличен; в мелких временных водоемах (лужи) едва ли
имеет более 2 (вероятно, иногда лишь 1) половых периодов.
*16. Acanthocyclops crassicaudis (Sars) (фиг. 55).
Cyclops crassicaudis Sars, 1863, Forh. Selsk. Christian. (1862): 249; Lilljeborg’,
1901:57, t. 4, f. 5—7; Van-Douwe, 1903, Zool. Anz., 26:463, f. 1—3; Sars, 1913:49, t. 29;
Рылов, 1918:54, рис. 20—23; Kiefer, 1923:283. — C. brucei Scott T’n., 1889, Joum. Linn.
Soc. London, 27: 93, t. 6, f. 1—6. — C. bissextilis Willey, 1929, Tr. Roy. Soc. Canada,
19:140. — Cyclops (Diacyclops) crassicaudis Kiefer, 1929:60. — Cyclops (Acanthocyclops)
crassicaudis Gurney, 1933:233, f. 1868—-1696.
Самка. Передний отдел тела довольно стройный, длина цефало-
торакса -ь2-го — 5-го сегментов торакса почти в 13/4—2 раза превышает
ширину, наибольшая ширина лежит немного кпереди от середины длины.
Задние углы торакального сегмента значительно выступают кнаружи.
Генитальный сегмент чрезвычайно широкий и толстый, его ширина
приблизительно равняется длине, по бокам переднего отдела сегмент
образует вдавление, кзади он суживается незначительно. Receptaculum
seminis очень широкий, окруженный сильно преломляющей свет прозрач-
ной (иногда желтоватой) каемкой, которая обычно особенно широка
в переднем отделе. Фуркальные ветви очень мало расходятся или
параллельные, их длина в 4.2—5 раз (у var. brachycercus Kief, лишь
в 3.2—3.4 раза, у var. cretensis Kief, в 4—4.5 раза) превышает их ширину,
латеральная щетинка прикрепляется приблизительно в задней 1/3 длины
края, из крайних апикальных щетинок внутренняя заметно короче внеш-
ней, редко почти равна ей, средние апикальные щетинки хорошо раз-
витые, из них внешняя значительно длиннее половины длины внутрен-
ней. Передние антенны 12-членистые, достигающие приблизительно
заднего конца цефалоторакса. Обе ветви ног I—IV пар 3-членистые.
У ног II—IV пар внутренний край базиподита вооружен группой мелких
шипиков, длина дистального членика эндоподита ног IV пары обычно
приблизительно равна его ширине (у типичной формы и var. brachycercus),
иногда более последней (у var. cretensis), из 2 апикальных шипов этого
членика внутренний заметно длиннее внешнего. У ног V пары базаль-
ный членик широкий, дистальный членик стройный, удлиненный, его
апикальный шип хорошо развитый, равный длине самого членика.
Длина 0.72—1.12 мм. Окраска однообразная, серовато-белая, иногда
молочно-белая, ноги розоватые. Яйцевые мешки длинные, мало расходя-
щиеся, обычно содержащие значительное количество яиц (по Kleiber,
1911, в каждом по 15—20, по Смирнову, 1930, по 23—32).
Самец. У ног III пары дистальный членик эндоподита с направлен-
ным ковнутри и перекрещивающимся с соседней апикальной щетинкой
апикальным шипом. Генитальный сегмент очень широкий, сперматофоры
очень крупные, на сегменте самки они прикрепляются вдоль срединной
линии, а не поперек. У ног VI пары 1 короткий шип и 2 щетинки, из
них врутренняя немного длиннее шипа, а внешняя приблизительно вдвое
длиннее внутренней. Длина 0.75—0.90 мм.
246
CYCLOPINAE
Kiefer различает 2 вариетета — var. brachycercus Kief, и var. cretensis
Kief. — основываясь на длине фуркальных ветвей и дистального членика
°эндоподита плавательных ног IV пары.1 Таксономическое значение
обеих неясно, — вероятно, в данном случае вариации длины фуркальных
ветвей параллельны таковым рода Eucyclops.
Фиг. 55. Acanthocyclops crassicaudis (Sars).
7 — фурка самки со спинной стороны; 2— абдомен самки с
receptaculum seminis; 3— эндоподит ноги IV; 4—5 — нога V
самки; б — сперматофоры.
Распространение A. crassicaudis Sars, в виду того, что
в литературе под таким названием, по всем вероятиям фигурируют
отчасти и оба вариетета (особенно var. brachycercus), рассматривается
нами для всех 3 форм совместно, с указанием на вариетет в тех случаях,
когда оно имеется. Европа (Скандинавия, Финляндия, СССР, Франция,
Далмация, Германия, Великобритания, о. Крит (здесь показан var. cre-
tensis), арктические острова (Шпицберген, Земля Франца-Иосифа,
Новая Земля, Медвежий остров, Соловецкие острова, о. Кильдин
1 Kiefer упоминает еще н о других различиях (вооружение ног Ш пары самца,
строение дистальной щетинки внутреннего края дистального членика эндоподита
у самки); как показал Gurney, признаки эти свойственны н типичной форме.
8. ACANTHOCYCLOPS
247
на Мурмане, Новосибирские острова), Северная Америка (в окрестностях
Нью-Йорка, var. brachycercus)- Канада, Северная Африка, Япония, о. Фор-
моза (var. cretensis). В СССР, помимо указанных арктических островов,
известен с Мурманского побережья, окрестностей Костромы и Ленин-
града, Московской и Харьковской1 областей. Считать A. crassicaudis
исключительно „северным" видом, как видно из приведенных данных,
нет оснований. Основной ареал обитания его лежит, очевидно, на севере,
охватывая зону тундры и тайги, но отдельные нахождения лежат далеко
к югу, в СССР, например, до широты Харькова, на западе — до широты
северной Африки и Крита, на востоке — до широты о. Формозы. Brehm
(1918, 1919) и Gagern (1924) высказывали мнение, что в средней Европе
A. crassicaudis является ледниковым реликтом. Ошибочность этого
была убедительно показана Kiefer’oM (1923, 1925; см. также Schaffer,
1934) и вряд ли нуждается в дальнейших комментариах.
В горы A. crassicaudis высоко не заходит, наивысшее местонахо-
ждение— около 750 м над уровнем моря.
Биология. Близка к таковой A. bisetosus Rehb. Крайне характерен
для очень мелких маленьких водоемов типа луж, очень обычен во вре-
менных (дождевых и снеговых), летом пересыхающих водоемах, но,
например, на Новой Земле обитает также в литорали небольших
мелких озер, с нейтральной — слабощелочной реакцией воды. В мелких
лужах—нередко в массах. В литературе неоднократно указывалось
на холодноводную стенотермию A. crassicaudis, что отчасти и послужило
основанием к неправильному толкованию его распространения в духе
ледниково-реликтовой теории. Он, действительно, неоднократно конста-
тировался в весенних снеговых лужах, при низких температурах воды
(см., например, Смирнов, 1929, 1930, в лужах при 1,3—9°), на которые
на севере и падает период размножения. Однако рачок неоднократно
был находим и при гораздо более высоких температурах воды (16°
и выше) в мелких лужах в 1-й половине лета. С другой стороны,
Kiefer (1923, 1925) показал, что рачок прекрасно живет и размножается
в аквариумах летом, при комнатной температуре. Этот автор справед-
ливо отрицает связь распространения A. crassicaudis специально с тем-
пературным фактором. Очевидно здесь играют решающую роль иные
условия среды, которые надо искать в таковых мелких луж и, прежде
всего, астатических водоемов (см. выше, A. bisetosus). В пещерах рассматри-
ваемый вид был найден лишь однажды (о. Крит, var. cretensis). Обитает
только в нормальных пресных водах. По Lowndes’y (1928), встречается
при pH = 6.2—8.7. Повидимому в очень кислых (pH = 4 и менее) дистроф-
ных водоемах всегда отсутствует. Цикл размножения изучен достаточно
полно (все данные относятся к лужам). На Шпицбергене (Olofsson,
1918) моноцикличен, половой цикл вскоре после образования водоема
(моховые лужи) весной, причем к размножению приступали половозре-
лые самки и самцы, которые перед этим перезимовывали; затем они
отмирали. Первая генерация, достигнув половозрелости, к копуляции
не приступает, а перезимовывает в фазе покоя в промерзшей до дна
луже. Рачок, таким образом, несомненно, имеет фазу покоя не в виде
покоящихся яиц, а на последних копеподитных стадиях и во взрослом
состоянии. В мелких водоемах южного Шварцвальда (высота около 750м
1 Зиверт (1929) отмечает некоторые уклонения найденных им экземпляров от
описанных Lilljeborg-’oM (1901) и мною (1918). Повидимому они, однако, не выходят
ва пределы индивидуальных вариаций. Указания для окрестностей Казани (Рузский,
1916) и Красноярска (Тюшняков и Воробьев, 1918) требуют проверки.
248
CYCLOPINAE
над уровнем моря), по Kleiber’y (1911), рачок дицикличен. В мелких
водоемах верхнего Дуная (Villingen), по Kiefer’y (1923), рачок встречается
в июне, июле, августе, январе и феврале, причем цикл размножения
весьма зависит от -характера водоема, от продолжительности существо-
вания последнего; числа половых периодов автор не установил. Оче-
видно в тех водоемах средней Европы, которые не высыхают продолжи-
тельное время, а тем более в постоянных, рачок имеет во всяком случае
более одного полового периода (см. также A. bisetosus, A. bicuspidatus
и A. vernalis). Вероятно олигосапроб — [3-мезосапроб.
У типичной формы A. crassicaudis фуркальные ветви самки от-
носительно длинные, их длина в 4.2—5 раз превышает их ширину. Длина
дистального членика эндоподита ног IV пары самки приблизительно
равна его ширине, его апикальный (внутренний) шип почти равен длине
самого членика.
1ба. Acanthocyclops crassicaudis var. brachycercus Kief.
Cyclops crassicaudis Kiefer, 1927, Zool. Anz., 72: 263, f. 1—2. — Cyclops [Diacyclops)
crassicaudis brachycercus Kiefer, 1929 : 61.
Фуркальные ветви самки укороченные, их длина лишь в 3.2—3.4 раза
более их ширины. Длина дистального членика эндоподита ног IV пары
самки не более его ширины (у типичной формы может быть и немного
более его ширины).
16b. Acanthocyclops crassicaudis var. cretensis Kief.
Cyclops crassicaudis cretensis Kiefer, 1928, Zool. Anz., 79 :245, f. 1—3.— Cyclops
(Diacyclops) crassicaudis cretensis Kiefer, 1929 : 61; — Cyclops crassicaudis taipehensis
Harada, 1931:158, f. 17—18.
Фуркальные ветви самки очень близки к таковым у типичной
формы, их длина в 4—4.5 раза превышает их ширину. Дистальный
членик эндоподита IV пары ног самки, однако, заметно более удлинен-
ный, его длина в 1.56—1.65 раза более его ширины, внутренний апикаль-
ный шип этого членика очень длинный, на 1/3 длиннее последнего и
почти вдвое длиннее внешнего апикального шипа.
17. Acanthocyclops scopljensis Kief. (фиг. 517__9).
Cyclops (Diacyclops) scopljensis Kiefer, 1932, Zool. Anz., 101 :58, f. 24—27.
Самка. Генитальный сегмент удлиненный, его длина почти вдвое
более его ширины, задний отдел лишь немного уже переднего. Recep-
taculum seminis очень похож на таковой A. crassicaudis. Задние края
абдоминальных сегментов не зазубрены, анальная пластинка мелкая.
Фуркальные ветви параллельные, их длина в 4 раза превышает их
ширину, из апикальных щетинок внутренняя крайняя несколько более
чем вдвое длиннее внешней крайней. Передние антенны 12-членистые,
приблизительно достигающие заднего края цефалоторакса. Длина
дистального членика эндоподита ног IV пары в 1т/3 раза более его
ширины, из апикальных шипов этого членика внутренний на х/з—7г
длиннее внешнего. Ноги V пары как у A crassicaudis. Длина 0.88—
0.92 мм.
8. ACANTHOCYCLOPS
249
Самец. Фуркальные ветви короче, чем у самки, дистальный
членик эндоподита ног IV пары почти как у самки, длина 0.74 мм.
Распространение. Подземные воды Югославии (Скоплие).
Троглобионт. Очень близок к A. crassicaudis var. cretensis Kief., но от-
личается более стройным удлиненным генитальным сегментом, удли-
ненной внутренней апикальной крайней щетинкой фуркальных ветвей и
несколько иным вооружением указанного членика эндоподита ног IV пары,
*18. Acanthocyclops languidus (Sars) (фиг. 495_9) 56).
Cyclops languidus Sars, 1863, Forh. Selsk. Christ. (1862): 249; Schmeil, 1892:84,
f. 3, f. 9—17; Lilljeborg-, 1901:59, t. 4, f. 8—9; Sars, 1913:50, t. 30. — Cyclops languidus
f. atava Thallwitz, 1922, Zool. Anz., 54: 266. — Cyclops {Diacyclops) languidus Kiefer,
1929:61, f. 22. — Cyclops {Acanthocyclops) languidus, Gurney 1933:236, f. 1697—1712.
Самка. Тело удлиненное, стройное, наибольшая длина его перед-
него отдела лежит посредине его длины (считая и задний торакальный
сегмент), по бокам сзади с умеренно выступающими углами. Гениталь-
ный сегмент значительно расширенный спереди, с неглубокой перетяжкой
несколько кпереди от середины его длины. Receptaculum seminis очень
вариирующей формы, с поперечно-овальным широким передним отделом;
задний отдел гораздо меньше переднего, иногда совсем маленький.
Фуркальные ветви стройные, почти параллельные, их длина в 4—5 раз
(у var. disjunctus хишъ 2.5—2.7 раза) превышает ширину, латеральная
щетинка прикреплена к задней 1/3 длины их внешнего края, средние
апикальные щетинки хорошо развитые, очень длинные; из них внешняя:
лишь немного короче внутренней, из крайних апикальных щетинок
внутренняя очень маленькая, иногда равная длине внешней крайней,
обычно несколько короче последней. Передние антенны 16-членистые
(у var. belgicus иногда лишь 13—14-членистые), приблизительно дости-
гающие заднего конца цефалоторакса, иногда более короткие. Экзо-
подиты ног I пары и эндоподиты ног I и II пары 2-членистые, остальные
ветви плавательных ног 3-членистые (членистость 2-2/3-2/3-3/3-3),
реже обе ветви всех пар плавательных ног 3-членистые (f. atava
Thallw.). Формула шипов З-З-З-З. Дистальный членик эндоподита ног
IV пары с толстыми апикальными шипами, из которых внутренний лишь
немного длиннее внешнего или оба шипа равной длины; длина этого
членика 1.5—2 раза (у var. disjunctus и var. belgicus лишь в 1.2—
1.25 раза) более его ширины. Ноги у V пары 2-членистые, базальный
членик широкий, дистальный членик длинный, стройный, с длинным
внутренним шипом (его длина иногда равна длине членика) и с хорошо
развитой внешнёй щетинкой. Длина 0.7—1.0 мм. Окраска однообразная,
серовато-белая. Яйцевые мешки сильно расходящиеся, очень крупные,
содержащие значительное количество яиц.
Самец. Передние антенны с длинными цилиндрическими сензор-
ными придатками. У ног VI пары внешняя щетинка очень длинная,
шипы очень короткие, из них средний немного крупнее внутреннего.
Длина 0.5—0.8 мм.
Распространение. Вся Европа, Азия, о. Цейлон, Китай, вероятно
Малая Азия, Япония, возможно, Канада (Willey, 1925, С. languigulus).
В СССР пока не указан для азиатской части, где его присутствие, однако,
очень вероятно. Известен из областей: Кировской, Ярославской, Иванов-
250
CYCLOPINAE
ской, Московской, Ленинградской, Киевской, Воронежской, Харьковской.1
В горах показан до высоты 1200—1300 м над уровнем моря.
Биология. Вид характерен для мелких сфагновых (гипновых)
дистрофных водоемов (лужи, болотца); встречается также в заболоченных
(сфагновое, гипновое заболачивание) участках литорали озер. Обычен
во временных, летом пересыхающих, лужах указанного типа. Подобно
A. nanus, является сфагнофильной формой, но не сфагнобионтом,
Фиг. 56. Acanthocyclops languidus (Sars).
.1—фурка самки с брюшной стороны; 2—дистальный членик эндоподита ноги IV
самки; 3—нога V самки; 4—б—генитальный сегмент самки с reeeptaculum seminis;
7 — нога V самки.
(4—по Lilljeborg-y, 5—по Kiefer'y, б—7—по Sars'у).
так как частично обитает и в иных биотопах. Изредка встре-
чается и в подземных водоемах, являясь троглоксеном; показан
также для ключей. Отнесение A. languidus к тепловодно-стено-
термичным циклопам ошибочно. Он встречается круглый год,
Kleiber (1911) находил его как подо льдом, так и при температуре воды
в 30°. Согласно экспериментальным исследованиям Скадовского (1928),
однако, температура в 25° отрицательно влияет на развитие яиц этого
1 Указание для Татарской АССР (Морозов, 1915) требует проверки. Возможно, что
рассматриваемый вид у нас ошибочно отмечался частью как „С. diaphanus".
8. ACANTHOCYCLOPS
251
вида, причем 20° является верхней границей оптимума, а при 10° яйца
^развиваются очень быстро. По Lowndes’y (1928) A. languidus встречается
в водах pH от 3.0 до 7.2. По Скадовскому (Skadowski, 1926) в Лузин-
ском болоте (Московской обл.) он приурочен, главным образом, к зоне
с pH = 3.8, единично встречаясь при pH = 4.2—6.4 и отсутствуя
в водах с pH = 7.5—8.4. Эксперименты этого автора (1928) показывают,
что слабые концентрации Н-ионов в отсутствии или при, очень малом
содержании Са- и Na-ионов отрицательно влияют на личиночные стадии
рассматриваемого вида, причем pH = ± 6.0 для них является уже леталь-
ной концентрацией; в растворах NaHCO3-ь NaOH они отмирают при
pH = 9—9.2. В отсутствии слабых кислот и оснований (СН3 СООН
и NH4OH) яйца нормально развиваются при pH = 3.0—9.2. В противо-
положность A. vernalis, вредное влияние на A. languidus Н-ионов пара-
лизуется не только Са, но и Na. Цитируемый автор предполагает, что
в естественных дистрофных очень кислых (pH, например, 3,5—4) водах,
бедных электролитами, в частности Са и Na, парализующее влияние
на ядовитое воздействие Н-ионов оказывают ионы и молекулы раство-
ренных органических веществ, а возможно и коллоидов, которыми богаты
дистрофные воды.
Рассматриваемый вид известен только из пресных вод. Несомненно
он нормально живет при высоких содержаниях растворенных органиче-
ских веществ (богатые гуминовыми веществами естественные дистроф-
ные водоемы).
Цикл размножения изучен неполно. Wolf (1905, окрестности Вюртем-
берга) считает вероятной полициклию, Kleiber (1911, южный Шварцвальд,
высота около 750 м) устанавливает полициклию в мелких водоемах
(яйценосные самки весь год). Однако в быстро высыхающих лужах
рачок, по всей вероятности, имеет не более 1—2 половых периодов.
Бесспорно, он переносит усыхание водоема, вероятно, на последних
стадиях метаморфоза и во взрослом состоянии. Известно 2 вариетета,1
присутствие их в фауне СССР очень вероятно.
18а. Acanthocyclops languidus var. disjunctus Thallw.
Cyclops languidus var. disjuncta Thallwitz, 1927, Zool. Anz., 71:59, £. 1—2. —
C. (Diacyclops) languidus disjunctus Kiefer, 1929. — Diacyclops disjunctus Kiefer,
1937b: 324, f. 4—7.
От типичной формы самка отличается очень укороченными фуркаль-
ными ветвями, длина которых лишь в 2.5—2.7 раза более их ширины,
и очень коротким дистальным члеником эндоподита ног IV пары, длина
которого лишь в 1.2 раза превышает ширину; латеральная щетинка
фуркальных ветвей прикрепляется значительно кзади от начала задней
Vg длины внешнего края. Длина самки 0.7 мм, самца около 0.6 мм.
Силезия, мелкие водоемы.
18b. Acanthocyclops languidus Sars var. beigicus Kiefer (фиг. 498_9).
Diacyclops languidus beigicus Kiefer, 1936, Bull. Mus. Hist. Nat. Belg1., 12:8,
f. 4—7.
Длина фуркальных ветвей в 4—5 раз более их ширины, внутренняя
крайняя апикальная щетинка не длиннее внешней крайней, иногда короче
1 Кроме того Thallwitz (1. cit., 1922) выделяет в особую форму (f. atava) самок,
у которых обе ветви всех пар плавательных ног 3-членистые. Выделение Kiefer ом
.(1937/38) var. disjunctus Thallw. в самостоятельный вид сомнительно.
252
CYCLOPINAE
последней. Передние антенны 16- или 13—14-членистые. Плавательные
ноги как у типичной формы, но дистальный членик эндоподита ног .
IV пары очень короткий, его длина не более как на 2/4 превышает его
ширину, внутренний из апикальных шипов этого членика немного короче
последнего и на 1/5—1/4 короче внешнего шипа. Длина самки 0.720—
0.780 мм, самца 0.50. Колодцы Бельгии.
*19. Acanthocyclops languidoides (Lili.) (фиг. 57x_s).
Cyclops languidoides Lilljeborg-, 1901:61, t. 4, f. 10—12; Kiefer, 1926, Int. Rev. ges.
Hydrob. u. Hydrogr., 14 : 344, f. 2, a — c. — Cyclops (Diacyclops) languidoides Kiefer,
1929:62; Kiefer, 1931, Zool. Jahrb., Syst., 61: 697. — Cyclops (Acanthocyclops)
languidoides Gurney, 1933. — Diacyclops languidoides Kiefer, 1936, Bull. Mus. R. Hist.'
Nat. Belg., 12:11.
Самка. Габитус тела очень близок к таковому A. languidus.
Фуркальные ветви слабо расходящиеся или почти параллельные, их
длина приблизительно в 5 раз (у типичной формы), а у отдельных
вариететов в 2т/2—8 раз более их ширины. Латеральная щетинка при-
креплена приблизительно в начале задней х/з (У типичной формы) длины
их внешнего края, внутренняя из крайних апикальных щетинок обычно
заметно короче внешней, но у вариететов может быть и длиннее послед-
ней. Передние антенны 11-членистые.1 Формула членистости плаватель-
ных ног 2"2/3-2/3-3/3-3. Дистальный членик эндоподита ног IV пары
у типичной формы короткий, его длина лишь в 1.2—1.4 раза более
ширины. Ноги V пары, генитальный сегмент и receptaculum seminis
очень похожи на таковые A. languidus, причем очертания receptaculum
seminis, как у последнего, весьма вариируют в зависимости от степени
наполнения спермой. Длина 0.72—0.75 мм. Яйцевые мешки содержат
по 10—12 яиц.
Самец. Апикальные щетинки фуркальных ветвей короче, чем
у самки, длина фуркальных ветвей приблизительно равна длине 3 задних
абдоминальных сегментов. Длина 0.65 мм. Рассматриваемый вид чрез-
вычайно изменчив в целом ряде признаков. Описано 12 его вариететов,,
таксономическое значение которых совершенно не выяснено; возможно,
частью они являются аберрациями. Все эти вариететы экологически
группируются в 2 характерных биотипа (см. ниже — экология). Нахожде-
ние еще новых вариететов крайне вероятно. Kiefer и Gurney склонны
придавать этим варйететам значение подвидов (очевидно в смысле эко-
типов, infraspecies, по Бергу), что, однако, едва ли может быть доказан-
ным; очень возможно, что частью они являются и морфами. Пока из них
лишь 1 известен в пределах СССР (var. gracilicaudaius Smirn.); нахо-
ждение у нас описанных из Западной Европы вариететов, однако,
весьма вероятно, почему ниже мы даем описание и последних.
Распространение (включая и вариететы, распространение
которых дается отдельно ниже при их описании). Европа (Швеция,
СССР, Германия, Северная Италия, Швейцария, Бельгия, Англия, Ирлан-
дия, Венгрия, Франция, Югославия, Румыния), Малая Азия (Aleppo),
Дальний Восток. В СССР известен с Мурманского побережья (типичная
форма; Olofsson, 1917) и с Дальнего Востока (бассейн Амура, Смирнов,
1930а, 1931). Распространение нельзя считать выясненным достаточно.
Вероятно в Палеарктике вид распространен очень широко, и в СССР
1 У var. zschokkei также 12-члеинстые.
8. ACANTHOCYCLOPS
253
<он будет еще найден во многих местностях, в частности в подземных
водах.
F ||Биология. Места его обитания сводятся к двум следующим
основным биотопам.
1. Мелкие наземные болотистые моховые водоемы с зарослями
Eriophorum, Sphagnum, Hypnum и др. Главным образом бедные электро-
Фиг. 57.
Acanthocyclops languidoides (Lili.) f. typica: 7 — фурка самки, 2—дистальный членик
эндоподита ногн IV самки, 3—фуркальная ветвь самки, 4 — дистальный членнк эндо-
подита IV самки, 5— базальный членнк ногн IV самки; A. languidoides var. clandesti-
nus Kief.: 6— фурка самкн, 7 — дистальный членик эндоподита ноги IV самки; A. lan-
guidoides var. zschokkei (Е. Graet.): 8—фурка самки, 9—дистальный | членик
эндоподита ноги IV самки.
(7—2—по Olofsson’y, 3—9— по Kiefer’y).
литами кислые воды, богатые гуминовыми веществами, в частности
коллоидами. Подвергаются значительным суточным колебаниям темпе-
ратуры, характеризуются, главным образом, мезо- и политипом света;
очень: часто такие водоемы являются астатическими, высыхающими
летом. В рассматриваемых водах обитает ряд вариететов (var. eriophori,
badeniae, hypnicola, gracilicaudaius), а также типичная форма, которая
в водоемах следующей группы очень редка (показана, сколько нам
254
CYCLOPINAE
известно, лишь для 2 каптажных источников в Швейцарии, Chappuis,
1933).
2. Подземные воды, повидимому, с весьма различным солевым
составом, до значительно минерализованных включительно. Олиготип
света, в общем низкие и постоянные температуры. В таких условиях:
обитают вариететы: clandestinus, nagysalloensis, putealis, zschokksi, На-
lianas, goiicus, возможно hiberniae, tantalus.
Цикл размножения не выяснен. Вероятно от моно- до полициклии
включительно. Для определения вариететов приводится нижеследующая
таблица.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВАРИЕТЕТОВ
1 (4). Длина фуркальных ветвей в 2.5 раза более их ширины.
2(3). Генитальный сегмент удлиненный, его длина немного более его
ширины..........................••• 19b. A. I. var. hypnicola Gurney
3(2). Генитальный сегмент очень короткий, его длина менее его ширины
..................................... 19с. А. 1. var. badeniae Kief..
4 (1). Длина фуркальных ветвей в 3 раза и более превосходит их
ширину.
5(6). Латеральная щетинка фуркальных ветвей прикреплена лишь немного
кзади от середины их внешнего края ...............................
...............................19а. А. 1. var. eriophori Gurney.
6(5). Эта щетинка прикрепляется в начале задней 1/3 длины внешнего
края или еще далее кзади.
7 (8). Апикальные придатки дистального членика эндоподита ног
IV пары представлены шипом и щетинкой.............................
.................................19m. А 1. var. tantalus (Kief.)
8(7). Эти придатки в виде шипов.
9(10). Фуркальные ветви очень длинные, их длина в 8 раз превышает
их ширину.................19d. А. 1. var. gracilicaudatus Smirnov
10(9). Фуркальные ветви короче, их длина не более чем в 7 раз пре-
вышает их ширину.
11 (22). Их длина в 4—7 раз более их ширины.
12 (13). Длина дистального членика эндоподита ног IV пары в 2.4—
2.5 раза более его ширины, длина фуркальных ветвей в 5 раз
более их ширины .............19g-. А. 1. var. nagysalloensis Kief.
13(12). Длина дистального членика эндоподита ног IV пары менее чем
в 2 раза превышает его ширину или равна последней.
14 (15). Длина этого членика равна его ширине, членик очень короткий
.....................................19е. A. I. var. hiberniae Gurney
15(14). Длина этого членика в 1.2—1.7 раза более его ширины.
16 (17). Длина фуркальных ветвей в 6 раз более их ширины.........
.................................19i. A. I. var. putealis Chapp-
17 (16). Их длина не более чем в 4—5 раз превышает их ширину.
18(19). Их латеральная щетинка прикреплена в начале 1/3 Длины внеш-
него края, длина фуркальных ветвей в 5 раз более их ширины,
длина дистального членика эндоподита ног IV пары в 1.2—1.4 раза
превышает его ширину..................19. A. languidoides (Lili.) typ.
19(18). Латеральная щетинка прикрепляется в начале задней т/4 Длины
внешнего края фукальных ветвей.
20(21). Длина дистального членика эндоподита ног IV пары лишь
несколько более его ширины, из его апикальных шипов внутренний
заметно длиннее внешнего...............19k. А. 1. var. italianus Kiefi
8. ACANTHOCYCLOPS
255
21(20). Длина этого членика в 1.5 раза более его ширины, его апикаль-
ные шипы почти равной длины .... 191. А. 1. var. goticus Kief.
22 (11). Длина фуркальных ветвей в 3—3.5 раза более их ширины.
23(24). Длина дистального членика эндоподита ног IV пары лишь
в 1.3 раза превышает его ширину, крайние апикальные щетинки
фуркальных ветвей почти равной длины . . ........................
................• •..........19f. А. 1. var. clandestinus Kief,.
19а. Acanthocyclops languidoides (Lili.) var. eriophori Gurney (фиг.
58б—э).
Cyclops languidoides eriophori Gurney, 1927, Ann. Mag. Nat. Hist. (9) 19:503,
f. 3, a—k.— Cyclops (Diacyclops} languidoides eriophori Kiefer, 1929 — Cyclops (Acan-
thocyclops} languidoides eriophori Gurney, 1933:246, f. 1726—1735.
Самка. Длина иногда сильно расходящихся фуркальных ветвей,
в 3—З1^ раза более их ширины. Латеральная их щетинка прикрепляется
лишь немного кзади от середины внешнего края. Генитальный сегмент
значительно короче, чем у типичной формы. Длина дистального членика
эндоподита ног IV пары не более его ширины, длина внутреннего апи-
кального шипа этого членика приблизительно равна длине последнего,,
или несколько превосходит ее.
Распространение. Англия (Норфольк), мелкие болотца, зарос-
шие Eriophorum.
19b. Acanthocyclops languidoides Lili. var. hypnicola Gurney (фиг.
58з-5).
Cyclops languidoides hypnicola Gurney, 1927, Ann. Mag. Nat. Hist., ( 9), 19:501, .
f. 2, a—d. — Cyclops (Diacyclops} languidoides hypnicola Kiefer, 1929:62. — Cyclops
(Acanthocyclops} languidoides hypnicola Gurney, 1933:244, f. 1719—1725.
Самка. Фуркальные ветви очень короткие, их длина лишь
в 2т/2 раза более их ширины, внутренняя крайняя апикальная щетинка,
немного короче внешней крайней. Длина дистального членика эндоподита
ног IV пары лишь очень немного превышает его ширину, апикальные
шипы почти одинаковой длины, заметно короче самого членика. Базальный
членик ног V пары более или менее тесно слит с задним торакальным,
сегментом. Длина самки 0.42 мм.
Распространение. Англия; гипновые болотные мхи.
19с. Acanthocyclops languidoides Lili. var. badeniae Kief.
Cyclops (Diacyclops) languidoides badeniae Kiefer, 1933, Beitr. Nat. Erforsch. Badens, .
11:184, t. I, f. 4-6.
Самка1. Наибольшая ширина генитального сегмента немного более
его длины, фуркальные ветви почти параллельные, их длина приблизи-
тельно в 2.5 раза превышает их ширину, латеральная щетинка прикреп-
лена приблизительно в начале задней 1/s длины внешнего края, крайние
апикальные щетинки почти одинаковой длины, тонкие и короткие. Длина
дистального членика эндоподита ног IV пары равна его ширине, из апи-
кальных шипов внутренний немного длиннее самого членика, внешний
заметно короче внутреннего. Длина 0.5 мм.
Распространение. Шварцвальд, сфагновые болота (найдена.
1 самка).
256
CYCLOPINAE
* 19d. Acanthocyclops languidoides Lili. var. gracilicaudatus Smirnov
{фиг. 59).
Cyclops 1 languidoides var. gracilicaudatus Смирнов, 193Oat225, рис. Id; Smirnov,
1931:624, f. 8—11.
Фиг. 58.
Acanthocyclops languidoides var. nagysalloensis Kief.: 7 — фурка самки co
спинной стороны, 2 —• дистальный членик эндоподита ноги IV самки,
A. languidoides var. hypnicola Gum. 3—фурка самки со спинной стороны,
4— дистальный членик эндоподита ноги IV самки, 5— ноги V и reeeptaculum
seminis; A. languidoides var. eriophori Gum.: 6— фурка самки co спинной
стороны, 7 — эндоподит ноги IV самки, 8 — нога V самки, 9 — генитальный
сегмент и reeeptaculum seminis; A. languidoides var. hiberniae Gum.: 10 —
фурка самки co спинной стороны, 77 — нога IV самки.
(7—2—по Kiefer’y, 3—77 -—по Gurney).
Самка. Фуркальные ветви стройные, параллельные, очень удли-
ненные, их длина почти в 8 раз превышает их ширину, латеральная
щетинка прикрепляется лишь немного кзади от начала задней J/3 длины
8. ACANTHOCYCLOPS
257
их внешнего края, внутренняя крайняя апикальная щетинка очень корот-
кая, приблизительно вдвое короче внешней крайней. Членистость плава-
тельных ног 2-2/3-2/3-3/3-3. Длина дистального членика эндоподита
ног IV пары приблизительно вдвое более его ширины, апикальные щетинки
почти одинаковой длины, равняются приблизительно 3/4, длины самого
членика. Длина 1.01 мм.
Близок к var. putealis Chapp., но отличается еще более длинными
фуркальными ветвями и иными размерами и вооружением дистального
членика эндоподита задней пары плавательных ног.
Распространение. Дальний Восток, бассейн р. Амура, мелкий
водоем на берегу р. Зеи.
Фиг. 59. Acanthocyclops languidoides var. gracilicaudatus Smirn.
(По Смирнову).
1 — абдомен и фурка самки с брюшной стороны; 2—нога
IV са1мки; 3—нога V самки; 4—нога VI самки.
19е. Acanthocyclops languidoides Lili. var. hiberniae Gurney
(фиг. 58io-ii).
Cyclops languidoides hiberniae Gurney, 1927, Ann. Mag. Nat. Hist., (9), 19:493,
f.?l, a — e.— Cyclops (Diacyclops) languidoides hiberniae Kiefer, 1929. — Cyclops (Acan-
thocyclops) languidoides hiberniae Gurney 1933:243, f. 1713—1713.
Самка. Фуркальные ветви удлиненные, их длина в 53/2 — 7 раз
более их ширины, латеральная щетинка прикрепляется близко от заднего
конца их внешнего края. Длина дистального членика эндоподита ног IV
пары приблизительно равна его ширине, из апикальных шипов внутрен-
ний гораздо длиннее самого членика и заметно длиннее внешнего шипа.
Оба членика ног V пары резко разделены. Длина самки 0.84—0.94 мм,
самца 0.72 мм. 4
Распространение. Ирландия, мелкие водоемы (лужи, цистерна,
ручей).
17 фауна СССР
258
CYCLOPINAE
19f. Acanthocyclops languidoides Lili. var. clandestinus Kief,
(фиг. 57б-?).
Cyclops languidoides clandestinus Kiefer, 1926, Schr, Ver. Gesch. u. Naturgesch.
Donaueschingen, 16:276, f. 5—7; Kiefer, 1926, Int. Rev. Ges. Hydrob. u. Hydrogr., 14:345,
f. 3, a — b. — C. nanus (part.) Chappuis, 1922, Arch. f. Hydrob. 14:32. — Cgclops {Diacy-
clops) languidoides clandestinus Kiefer, 1929:62. — Diacyclops languidoides clandestinus
Kiefer, 1936, Bull. Mas. R. Hist. Nat. Belg., 12:11.
Самка. Длина фуркальных ветвей приблизительно в 3 раза более
их ширины, крайние апикальные щетинки почти одинаковой длины.
Дистальный членик эндоподита ног 1V пары короткий, его длина при-
близительно в 1.3 раза превышает ширину, его апикальные шипы очень,
короткие, короче, чем у типичной формы. Длина 0.8—0.9 мм.
Распространение. Найден в водопроводах в Offingen (Баден),
в Бельгии, в гротах Семигорья, Венгрии, Италии (близ Сото),
в колодцах Aleppo (Малая Азия), Румынии и Югославии. Троглобионт.
19g. Acanthocyclops languidoides Lili. var. nagysalloensis Kief,
(фиг. 58 i-г).
Cyclops languidoides nagysalloensis Kiefer, 1927,- Zool. Anz., 71:6, f. 1—Kiefer,,
1927, Arch. Balat., 1:418, f. 40—42. — Cyclops {Diacyclops) languidoides nagysalloensis
Kiefer, 1929:63.
Самка. Длина фуркальных ветвей в 5 раз более их ширины,,
их крайние апикальные щетинки почти одинаковой длины, или внутрен-
няя немного длиннее внешней. Дистальный членик эндоподита ног IV
пары удлиненный, его длина в 2.4—2.5 раза превышает ширину, апикальные
шипы этого членика короче последнего из них внутренний немного длин-
нее внешнего.
Самец. Фуркальные ветви короче, чем у самки, их длина в 4т/2 раза
более их ширины, длина дистального членика эндоподита ног IV пары,
вдвое более ширины.
Длина самки и самца не известна, вероятно самец около 0.8 мм.
Распространение. Колодцы в Венгрии (в Nagysallo), совместно1
с подземными Niphargus dudichi и Ostracoda.
19h. Acanthocyclops lagnidoides Lili. var. zschokkei (E. Graeter):
(фиг. 57g-g).
Cyclops zschokkei E. Graeter, 1911:42, t. 2, f. 6, Textif. 18—19.— C. nanus (part.}
Chappuis, 1922, Arch. f. Hydrob., 14:36. — Cyclops languidoides zschokkei Kiefer, 1926,
Int. Rev. ges. Hydrob. u. Hydrogr., 14:347, f. 4, a — b. — Cyclops {Diacyclops) languidoi-
des zschokkei Kiefer, 1929:63; Kiefer, 1935, Verh. Nat. Ver. Karlsruhe, 31:134, f. 6—8.
Самка. Длина фуркальных ветвей в 3—3.5 раза более их ширины,
внутренняя из крайних апикальных щетинок приблизительно вдвое длин-
нее внешней. Длина дистального членика эндоподита ног IV пары
в 1.5—1.55 раза превышает ширину, апикальные шипы короткие, внут-
ренние короче самого членика и немного длиннее внешнего. Передние
антенны 11-членистые, редко 12-членистые. Длина около 0.5 мм; экзем-
пляры очень прозрачны, глазной пигмент отсутствует.
Распространение. Пещеры Швейцарии и подземные воды
в верховьях Рейна. Троглобионт.
8. ACANTHOCYCLOPS
259
19i. Acanthocyclops languidoides Lili. ver. putealis (Chapp.).
Cyclops languidoides putealis Chappuis, 1928, Bull. Soc. Stiint. Cluj, 4:21. — Cyclops-
(Diacyclops) languidoides putealis Kiefer, 1929:63.
Самка. Длина фуркальных ветвей в 6 раз более их ширины,.,
из крайних апикальных щетинок внутренняя немного длиннее внешней,,
латеральная щетинка прикреплена в начале задней */4 длины внешнего
края. Длина дистального членика эндоподита ног IV пары в 1.7 раза
превышает ширину, апикальные шипы короче самого членика, из них
внутренний немного длиннее внешнего. Длина 0.72 мм.
Распространение. Колодец во Франции (департамент Эн).
19k. Acanthocyclops languidoides Lili. var. italianus Kiefer.
Cyclops (Diacyclops) languidoi les f. italiana Kiefer, 1931, Zool. Jahrb., Syst.
61:697, f. 1—3.
Самка. Длина генитального сегмента равна его ширине или не-
много менее последней. Фуркальные ветви слабо расходящиеся, их длина
в 4—4.3 раза более их ширины, латеральная щетинка прикрепляется
приблизительно в начале задней % длины их внешнего края, внутренняя
крайняя апикальная щетинка немного короче внешней крайней. Длина
дистального членика эндоподита ног IV пары немного более ширины,
апикальные шипы хорошо развиты, из них внутренний заметно длиннее
внешнего и обычно немного длиннее самого членика. Длина 0.60—0.64 мм.
Морфологически занимает среднее положение между типичной формой
и var. clandestinus.
Распространение. Северная Италия, мелкий водоем в рас-
щелине скалы, в затененном месте (совместно с Nlphargus).
191. Acanthocyclops languidoides Lili. var. goticus Kiefer.
Cyclops (Diacyclops) languidoides f. gotica Kiefer, 1931, Zool. Jahrb., Syst.,
61:699.
Самка. Длина фуркальных ветвей в 4.4—4.7 раза более их ширины.
Длина дистального членика эндоподита ног IV пары в l1^ раза превы-
шает его ширину и более внутреннего апикального шипа, почти равного
внешнему апикальному шипу. Длина не известна. По длине фуркальных
ветвей близок к типичной форме, по дистальному членику эндоподита
ног IV пары сближается с var. zschokkei.
Распространение. Северная Италия, мелкий водоем в Адельс-
бергской пещере.
19m. Acanthocyclops languidoides var. tantalus (Kief.).
Diacyclops tantalus Kiefer, 1937, Zool. Anz., 119:16, f. 1—4.
Фуркальные ветви длиннее, чем у zschokkei, их длина приблизи-
тельно в 5.7 раз превышает ширину; вооружение ветвей как у указанного
вариетета, в частности их дорзальная щетинка очень длинная. Передние
антенны 11-членистые, членистость плавательных ног по формуле
2-2/3-2/3-3/3-3. Дистальный членик эндоподита ног IV пары удлинен-
ный, его длина вдвое больше его ширины, из пары апикальных придат-
17*
260
CYGLOPINAE
ков внешний представлен шипом, а внутренний — щетинкой (а не шипом,
как у прочих форм languidoides). Ноги V пары как у типичной формы.
Длина самки 0.62—0.72 мм. Найден в пещерах Югославии.
Kiefer считает tantalus самостоятельным видом, с ним едва ли можно
согласиться, так как кроме присутствия апикальной щетинки (вместо
внутреннего шипа) на дистальном членике эндоподита ног IV пары
он ничем не отличается от A. languidoides, вида крайне изменчивого.
Удлиненный членик эндоподита этой пары ног мы находим, например,
у var. nagyssaloensis Kief, (здесь членик даже еще длиннее). Принимая
во внимание, что вообще у одного и того же вида шип этого членика
может заменяться щетинкой (см. A. vernalis var. robustus),—с этим
признаком, когда он остается единственным для различия двух форм, надо
быть особенно осторожным, и мы считаем правильным отнесение tantalus
к виду A. languidoides.
*20. Acanthocyclops nanus (Sars) (фиг. 60i—7).
Cyclops nanus Sars, 1863, Forh. Selsk. Christ. (1862):251.— C. languidus var. nanus
Schmeil, 1898:151. — C. diaphanus Lilljeborg, 1901:63, t. IV, f. 13—15. — C. nanus Scour-
field, 1903, Queq. Mier. Club, (2), IX:63. — C. diaphanus var. diaphanoides A. Graeter,
1903:535, t. 15, f. 11, 12, 24, 28 —C. nanus Kleiber, 1911:36, f. 13—16.—C. diaphanus
Sars, 1913:52, t. 31. — C. languidoides Klie, 1913, Int. Rev. Hydrob. u. Hydrogr., Biol.
Suppl., VI:6. — C. diaphanus Рылов, 1918:58, рис. 24—26. — C. languidus var. intermedia
Kiefer, 1923, Zool. Anz., 58:278. — C. nanus Kiefer, 1925, Mikrokosmos, 18:243, fig.—
Kiefer, 1926, Int. Rev. Hydrob. u. Hydrog-r., 14:348, f. 5, a — e. — Gurney, 1927, Ann.
Mag. Nat. Hist., (9), 19:504, f — 4, a — c. — C. (Diacyclops) nanus Kiefer, 1929:63. —
C. (Acanthocyclops) nanus Gurney, 1933:248, f. 1736—1746.
Самка. Тело несколько более суженное, чем у A. languidus, наи-
большая ширина переднего отдела лежит приблизительно на середине
его длины. Генитальный сегмент стройный, значительно расширенный
спереди, его длина немного более его ширины.1 Reeeptaculum seminis
поперечно-овальный, очень похожий на таковой A. languidus. Фуркаль-
ные ветви почти параллельные, их длина приблизительно в 4—4’/2 раза
более их ширины,8 латеральная щетинка прикреплена на середине внеш-
него края, из крайних, апикальных щетинок внутренняя немного короче
шипообразной внешней, средние апикальные щетинки хорошо развитые,
из них внутренняя почти вдвое длиннее внешней. Передние антенны
11-членистые, не доходящие до заднего конца цефалоторакса, на 8-м
членике имеется очень длинная сензорная щетинка. Членистость плава-
т?льных ног как у A. languidus, но вооружающие их шипы более длин-
ные. У ног III и IV пар 2-й членик эндоподита только с 1 внутренней
щетинкой, причем у ног IV пары спереди ее на внутреннем крае иногда
присутствует очень короткий волосок.1 2 3 * У ног IV пары ширина дистального
членика эндоподита более чем вдвое превышает его длину, внутренний
апикальный шип немного длиннее внешнего. У ног V пары типичных
A. nanus базальный членик отделен от заднего торакального сегмента,
дистальный членик удлиненный, стройный, обычно резко дифференциро-
1 По Gurney (1933:248) длина почти равна ширине, что противоречит рис. 1737
этого автора.
2 По Gurney (1933) в 5 раз (но по его рис. 1738 в 4 раза), по Sars’y (1913) —
в 4% раза.
3 Он изображается на рисунках Sars’a и Gurney; у экземпляров с Мурмана
мы не всегда обнаруживали его, у некоторых самок он отсутствовал.
8. ACANTHOCYCLOPS
261
Acanthocyclops nanus (Sars): 7 — генитальный сегмент самкн с
reeeptaculum seminis, 2—экзоподит ноги IV самки, 3—эндоподит
ноги IV самки, 4—- нога I самки, 5—фуркальная ветвь самки со
спннной стороны, 6—7 — нога V самки; A. nanus var. incertus
(Wolf): 8— нога V самки, 9— задний сегмент торакса и гениталь-
ный сегмент самки с reeeptaculum seminis, 10— антенна I ?самки.
(8—10— по Wolf’y).
262
CYCLOPINAE
ванный, но иногда без явственного отчленения от базального членика;
вооружение дистального членика состоит из длинной внешней щетинки
и из довольно длинного внутреннего шипа, не доходящего до середины
щетинки и прикрепленного очень близко от конца членика. У var. incertus
базальный членик совершенно сливается с задним торакальным сегмен-
том и ноги V пары 1-членистые. Длина 0.66 — 0.9 мм. Рачок очень
прозрачен, бесцветен, иногда окрашен в нежный желтоватый или крас-
новатый (розовый) цвет. Яйцевые мешки довольно крупные, овальные,
мало расходящиеся.
Самец.1 Цефалоторакс несколько более удлиненный, чем у самки,
2-й и 3-й абдоминальные сегменты почти равной длины, фуркальные
ветви немного короче 3 задних абдоминальных сегментов, передние
антенны 14-членистые, длина около 0.6 мм.
Рассматриваемый вид, бесспорно, неоднократно приводился в лите-
ратуре как „С. diaphanus Fisch". После разъяснения Kiefer’a (1926, 1929)
полная неопределенность этого синонима едва ли подлежит сомнениям.
В подавляющем большинстве случаев невозможно решить, какой именно
вид приводит автор под названием С. diaphanus. Сравнительно в очень
немногих случаях можно установить, что указание относится именно
к A. nanus Sars (см. выше синонимический список). Только такие, вполне
надежны?, указания приняты нами во внимание в нижеследующем обзоре
географического распространения.
Распространение. Европа (Великобритания, Скандинавия,
Голландия, Германия, Швейцария, Румыния), Азия (южный Сахалин).
В СССР известен с Мурмана (Рылов, 1918, С. diaphanus), из Карело-
Финской ССР, из окрестностей Костромы, из областей Московской (?),1 2
Ивановской, Ленинградской. Присутствие на севере Сибири очень вероятно.
Распространение A. nanus в СССР, таким образом, ограничено север-
ной его половиной, охватывая зоны тундры, тайги и северную часть
зоны смешанных лесов. В горах показан до высоты 750 м над уровнем
моря.
Биология. Вид чрезвычайно характерен для мелких дистрофных
водоемов (сфагновые и гипновые лужи, болота), в том числе и для
высыхающих; реже в литорали заболачивающихся озер, в зоне при-
брежных сфагновых и гипновых мхов. Ласточкин (1924) в Валдайском
озере отмечает до 2000 экз. на 1м2 моховых зарослей. Был находим
и в колодцах, так что его правильнее относить к сфагнофилам, а не
к сфагнобионтам. В пещерах, при полном отсутствии света, отсутствует.
Эвритермичен. По Lowndes (1928) обитает в водах с pH — 4.4—7.2, по
Gessner’y (1931) характерен специально для вод с pH = 3 — 4, при
высокой окисляемости (282—243 мг. КМпО^ на литр) и ничтожной
жесткости (общая жесткость лишь 0.336° немецких), — условия, кото-
рые, в общем, надо признать оптимальными. Цикл размножения изучен
очень неполно. В болотах Jungholz в южном Шварцвальде (Kleiber,
1911), на высоте около 750 м над уровнем моря, полицикличен, встре-
чается круглый год; Wolf (1905) также предполагает полицикличность,
по Gessner’y (1931) в болотистых водоемах (высота около 750 м) Шварц-
вальда встречается с конца марта до ноября включительно и, во всяком
случае, имеет не менее 2 половых периодов. Что касается отношения
1 По Lilljeborg,,y (1901).
2 Указан Россингким (1892), данные требуют проверки.
8. ACANTHOCYCLOPS
263
к планктону, то, по наблюдениям Kleiber’a, A. nanus способен к свобод-
ному плаванию по субстрату (т. е. факультативно-планктичен).
*20а. Acanthocyclops nanus var. incertus (Wolf) (фиг. бОв-ю).
Cyclops incertus Wolf, 1905:161, t. 8, f. 18, a — c. — Cyclops (Diacyclops) nanus
incertus Kiefer, 1929:64.
От типичной формы отличается полным слиянием базального чле-
ника ног V пары с задним торакальным сегментом, так что эта пара
ног оказывается 1-членистой, причем единственный членик не имеет
явственного отчленения от сегмента. A. Graeter (1903) описал форму
A. nanus, у которой у ног V пары базальный членик еще имеется, но
он погружен в кутикулу заднего торакального сегмента (= С. diaphanus
var. diaphanoides Graet., no Kiefer’y, 1929, — C. (D.) nanus f. diaphanoides).
Gurney (1933) отмечает, что у A. nanus встречаются все переходы от
вполне дифференцированного до вполне слитого с торакальным сегмен-
том базального членика ног V пары, причем нога теряет всякие следы
2-членистости. Цитируемый автор полагает, что в данном случае имеются
вариации чисто индивидуального характера. Говорить здесь о подвиде,
как это делает для incertus Kiefer, едва ли возможно. Вместе с тем
едва ли правильно incertus систематически не отделять от типичных
A. nanus, так как полное изменение членистости ног V пары, что харак-
терно для крайней степени вариационной амплитуды рассматриваемого
признака, без труда позволяет морфологически отличить incertus от
типичной формы, еще сохраняющей базальный членик ног V пары.
Поэтому вместе с Kiefer’oM мы выделяем описанную Wolf’oM форму
в отдельный вариетет, не предрешая его таксономического значения
и, вопреки Kiefer’y, воздерживаясь от придания ему значения подвида.
Распространение. Европа (Шварцвальд, СССР). В СССР
найден близ гг. Мурома и Иванова. Указание Морозова (1915) на нахож-
дение в окрестностях Казани (плактон р. Цивиль) сомнительно.
Биология. Как у типичной формы; встречается совместно с по-
следней, но также и изолированно.
21. Acanthocyclops stygius (Chapp.) (фиг. 51ю-ц).
Cyclops stygius Chappuis, 1924, Bull. Soc. Stiint. Cluj, 2:27, f. 1—2; Kiefer, 1928:
527, f. 40—45. — Cyclops stygius var. deminutus Chappuis, 1925, Bull. Soc. Stiint. Cluj,
3:162. — Cyclops (Diacyclops) stygius Kiefer, 1929:54.
Самка. Глазной пигмент отсутствует. Тело несколько сплющенное
в спинно-брюшном направлении, с мало дифференцированными передним
и задним отделом, задние края абдоминальных сегментов не зазубрены.
Анальная пластинка хорошо развитая, но не заходящая кзади за основа-
ние фуркальных ветвей, с закругленным гладким задним краем. Фур-
кальные ветви слабо расходящиеся, их длина немногим более чем
в 2 раза превышает ширину. Из крайних апикальных щетинок внутрен-
няя очень маленькая, вдвое меньше шипообразной внешней. Генитальный
сегмент очень широкий. Передние антенны 11-членистые, с сензорным
цилиндром на середине 8-членика. Членистость плавательных ног по
формуле 3 • 2/3 • 2/3 • 3/3 • 3, у var. deminutus Chapp. 3 • 2/3 • 2/3 • 2/3 • 3.
У ног V пары дистальный членик с внутренним шипом лишь немного
бодее коротким, чем соседняя с ним щетинка. Длина 0.55 мм. Самцы
не известны.
Распространение. Пещеры Югославии, подземные воды Венгрии.
Троглобионт.
264
CYCLOPINAE
22. Acanthocyclops hypogeus Kief. (фиг. 48i_3).
Cyclops (Diacyclops) hypogeus Kiefer, 1930, Zool. Anz., 78:224, f. 5—7.
Самка. Тело несколько сплющенное в спинно-брюшном направле-
нии, анальная пластинка кзади за основание фуркальных ветвей не
выступает, с гладким закругленным краем. Длина фуркальных ветвей
приблизительно вдвое превышает их ширину, из крайних апикальных
щетинок внутренняя лишь немного длиннее внешней. Reeeptaculum semi-
nis не описан. Передние антенны 11-членистые. Членистость плавательных
ног по формуле 2 • 2/3 • 2/3 • 3/3 • 3. Длина дистального членика эндопо-
дита ног IV пары почти равна его ширине, апикальные шипы этого
членика почти одинаковой длины, короче самого членика. Ноги V пары
2-членистые, внутренний шип дистального членика очень маленький,
гораздо короче соседней с ним щетинки. Длина около 0.5 мм. Самец
не известен.
Распространение. Пещеры в Италии (St. Ganzian). Троглобионт.
Очень близок к A. stygius (Chapp.), от которого отличается, помимо
членистости плавательных ног, чрезвычайно коротким внутренним шипом
дистального членика ног V пары.
*23. Acanthocyclops abyssicola (Lili.) (фиг. 61).
Cyclops abyssicola Lilljeborg-, 1901:66, t. 4, f. 16—19; Sars, 1913:53, t. 32; Kiefer,
1926, Int. Rev. Hydrob. u. Hydrogr., XIV.-354, f. 6, a — d. — Cyclops (Diacyclops) abyssicola
Kiefer, 1929:64.
Самка. Передний отдел тела широкий и толстый, длина цефалото-
ракса не более как в 1х/2 раза превосходит ширину, наибольшая ширина
лежит посредине длины, внешние задние углы задних торакальных
сегментов закругленные. Абдомен очень толстый, укороченный, гени-
тальный сегмент чрезвычайно широкий, в переднем отделе заметно рас-
ширенный, более толстый, нежели задний торакальный сегмент; длина
генитального сегмента приблизительно равна его ширине. Прочие абдо-
минальные сегменты толстые, весьма укороченные. Reeeptaculum seminis
состоит из поперечно расположенного овального крупного переднего
отдела и из маленького мешковидного заднего, похож на этот орган
у A. languidus. Фуркальные ветви толстые и короткие, параллельные,
их длина приблизительно в 2.5—3 раза более их ширины, латеральная
щетинка прикреплена в задней 2/з длины их внешнего края, крайняя
внутренняя апикальная щетинка короче крайней внешней, средние апи-
кальные щетинки хорошо развитые, относительно короткие, внешняя
из них несколько более г/2 длины внутренней. Передние антенны обычно
10-членистые, но у старых самок (по Lilljeborg’y, 1901) могут быть
и 11-членистые; они очень короткие, едва достигающие т/2 длины
цефалоторакса, богато вооруженные щетинками. Эндо- и экзоподиты
I пары плавательных ног и эндоподиты II пары 2-членистые, остальные
ветви всех плавательных ног 3-членистые. Дистальный членик эндопо-
дитов вместо щетинки снабжен внешним апикальным шипом, внутри
рядом с ним у ног I—II пар имеется щетинка, а у ног JII—IV пар шип.
Формула шипов З-З-З-З. Дистальный членик эндоподита IV пары пла-
вательных ног короткий, его длина приблизительно в l1^ раза более
ширины, оба апикальные шипа превышают длину членика, из них внут-
ренний заметно длиннее внешнего. Ноги V пары 1-членистые, но иногда
не явственно 2-членистые, причем базальный членик более или менее
полно слитый с торакальным сегментом, дистальный членик удлинен-
9. MICROCYCLOPS
265-
Фиг. 61. Acanthocyclops abyssicola (Lili.),,
самка.
7— фурка самки с брюшной стороны; 2 —
генитальный сегмент и reeeptaculum seminis;
3 — дистальный членик эядоНоднта ноги IV;
4 — нога V.
до 28 м, а в оз. Сейтес-явр — на:
ный, стройный, вооруженный довольно крупным шипом (субапикально)
и щетинкой, длина которой в 2—3 раза более шипа. Длина 0.75—0.9 мм.
Яйцевые мешки маленькие. Окраска беловатая, с лиловым и красноватым
оттенком.
Самец. Отличается также значительной толщиной тела, что менее
выражено, однако, чем у самки. Длина 0.7 мм.
По членистости плавательных ног (2-2/3-2/3-3/3-3) A. abyssicola
близок к A. languidus typ., по фуркальным ветвям—к A. languidus var.
disjunctus Thalw. Однако от этого
вида и вообще от видов группы
languidus по плавательным ногам
A. abyssicola отличается весьма
резко (присутствие длинных ши-
пов), равно как по чрезвычайно
утолщенному телу и по очень ко-
ротким передним антеннам.
Р а с п ро странение.
Пока известен лишь из Скан-
динавии и с севера Кольского
полуострова, где он найден в
Умбозере и оз. Сейтес-явр.
Биология. Почти совер-
шенно не изучена. Был находим
только в озерах (вероятно толь-
ко в олиготрофных), где ведет
бентический образ жизни, обитая,
главным образом, на глубинах.
В шведском оз. Веттерн обитает
на глубине 25—113 м на песча-
ном и глинисто-песчаном грунте
(до 14 экз. на 5 дм2). В оз. Ifosjon
(в Швеции) обитает на глубине
около 45 м, причем не находили
его ни в литорали, ни в мелких
водоемах. Однако (1913) в нор-
вежском озере Mjosen находили
его на глубине 7—11 м, в Умб-
озере он присутствует на или-
стых грунтах, на глубинах от 3
мелководьи у берега. Отнесение A. abyssicola специально к сублито-
рально-профундальной фауне (Ekman, 1916), таким образом, едва ли пра-
вильно. Цикл размножения не изучен, половозрелые самки были
находимы летом и осенью, в Умбозере—во 2-й половине сентября.
Возможно, моноцикличен.
9. Род MICROCYCLOPS CLAYS
Microcyclops Claus, 1893:82; Claus, 1893:34 7. — Cryptocyclops Sars, 1927:129.—
Microcyclops и Metacyclops Kiefer. 1929a: 41; Kiefer, 1929:65—75. — Microcyclops (part.,
subg.) Gurney, 1933:251. — Neutrocyclops Kiefer, 1936a: 130.
Фуркальные ветви на спинной поверхности без продольной складки,
их длина обычно в 3—5 раз (редко в 6—7 раз) более их ширины.
Анальная пластинка слабо развитая. Передние антенны 9—12-, большей
частью 11—12-членистые, дистальные членики без гиалиновой пластинки,.
.266
CYCLOPINAE
и без продольного ряда шипиков. Членистость плавательных ног по
формуле 2-2Z2-2/2-2/2-2. Ноги V пары одночленистые, базальный чле-
ник слит с задним торакальным сегментом, до полного его исчезновения,
свойственная ему щетинка прикрепляется на торакальном сегменте.
Единственный членик почти у всех удлиненный, приблизительно прямо-
угольный, на конце вооруженный щетинкой и ковнутри от нее мелким
шипиком, прикрепленным субапикально, в задней 1/3 длины внутреннего
края членика, близко от его конца, редко — апикально. У некоторых
видов шипик редуцирован. Receptaculum seminis различной формы, округ-
' лый, редко близкий к молотообразному (например у М. gracilis).
В принятом нами объеме рода к нему в настоящее время принад-
лежит около 50 видов.
Род в целом имеет космополитическое распространение, но космо-
политами в тесном смысле слова являются лишь М. varicans (все обла-
сти) и М. bicolor (не указан лишь в Неотропической области). Видов,
свойственных только Палеарктике, исключая М. planus (северная Африка,
Сирия, Кроация, Крым), не имеется. Нет также специально неарктиче-
ских видов. В пределах Палеарктики известны М. varicans Sars, М. bico-
lor Sars, М. gracilis Lili., M. dengizicus Lep., M. minutus Claus, M. pla-
nus Gurn., т. e. всего 6 видов.1 Специально Неотропической области
свойственно 13 видов, кроме того в пределах этой области найдены
М. varicans, М. gracilis, М. dengizicus, всего, следовательно, 16 видов.
Только в Эфиопской области известно 18 видов, кроме того М. varicans,
М. bicolor, М. linjanticus Kief (также на Малайском архипелаге), т. е.
всего 21 вид. Только в Ориентальной области пока показано 9 видов,
кроме того М. varlacans, М. bicolor, М. dengizicus, М. minutus, М. linjan-
ticus и М. davidi (?). В Австралийской области, помимо космополитов
-М. varicans и М. bicolor, известны лишь М. monacanthus Kief. (Новая
Зеландия) и М. arnaudi Sars (Виктория). Несмотря на неравномерность
исследованности Cyclopidae различных областей, из которых, конечно,
наиболее изучена Палеарктика (особенно Европа), все же можно заклю-
чить, что род Microcyclops является свойственным, главным образом,
Неотропической и Эфиопской областям, на которые приходится свыше
30 эндемичных видов, а всего 34 вида, т. е. более 3/5 всего видового
состава рода. Род заключает пресноводные виды, но 2 вида — М. dengi-
zicus Lep. и М. panamensis Marsh. — являются галобионтами (солоноватые
и соленые континентальные водоемы). Обитатели, главным образом,
мелких водоемов, до пересыхающих летом луж включительно (особенно
.М. minutus Claus), а также литорали (заросли макро- и микрофитов)
озер. Пелагических видов нет. Большинство видов тепловодны, некоторые
эвритермичны, холодновато-стенотермических видов нет. Некоторые ука-
заны и для подземных вод (колодцы — М. planus, М. minutus), но тро-
глофилов и троглобионтов в роде нет.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1(8). Латеральная щетинка прикреплена на середине или лишь немного
кзади от середины внешнего края фуркальных ветвей.
2(5). Длина фуркальных ветвей приблизительно в 3—3.5 раза превышает
их ширину, receptaculum seminis молотообразной или близкой
к таковой формы.
1 Описанный Lindberg’oM (1936) Metacyclops unacanthus, по нашему мнению, бес-
спорно идентичен с М. minutus Claus.
9. MICROCYCLOPS
267
3(4). Передние антенны 11—12-членистые, генитальный сегмент строй-
ный, значительно удлиненный.......................*3. М. gracilis (Lili.)
4(3). Передние антенны 9-(редко 10-) членистые, генитальный сегмент
толстый, его длина лишь незначительно превосходит ширину . . .
............................................... * 6. М. planus( Gurney)
5(2). Длина фуркальных ветвей в 4—6 раз более их ширины, recepta-
culum seminis не молотообразный.
6(7). Членик ног V пары очень широкий, его длина почти вдвое менее
его ширины, генитальный сегмент сильно расширенный спереди
и значительно суженный в заднем отделе . . * 4. М. dengizicus (Lep.)
7(6). Членик ног V пары удлиненный, его длина заметно более его
ширины. Генитальный сегмент лишь мало суживающийся кзади .
................................................ * 5. М. minutus (Claus)
8(1). Латеральная щетинка прикрепляется в начале задней х/з внешнего
края фуркальных ветвей или еще далее кзади, близко от их зад-
него конца.
'9(10). Внутренний апикальный шип дистального членика эндоподита ног
IV пары гораздо (в 3. 5—6 раз) длиннее внешнего апикального
шипа................................... ..........* 2. М. bicolor (Sars)
.10(9). Этот шип лишь 1.2—2 раза длиннее внешнего апикального шипа
............................................* 1. М. varicans (Sars)
*1. Microcyclops varicans (Sars) (фиг. 62).
Cyclops varicans Sars, 1863, Forh. Selsk. Christ., (1862) : 252.— C. orientalis Ульянин,
1875:33, t. 7, f. 10—11; t. 10, f. 8—13; t. 11, f. 1—2. —<7. varicans Schmeil. 1392:116,
t. 6, f. 1—5. — C. (Microcyclops) varicans Claus, 1893a:347, t. 3, f. 7; t. 5, f. 7—9. —
C. africanus Bourne, 1893, Pr. Zool. Soc. London, 165, t. 6, i. 1—8. — C. varicans Lillje-
borg, 1901:72, t. 4, f. 23, 24; Sars, 1913:54, t. 33. — C. caeibaensis Marsh, 1919, Proc. U.
St. Nat. Mus., 55:546, t. 49, f. 1—5. — C. subaequalis Kiefer, 1927, Faune Col. Franc.,
1:558, f. 39—40. — C. varicans Kiefer, I929a:29, f. 24—26. — C. subaequalis Kiefer, 1929a:
31, f. 27—28. — C. ’(Microcyclops) varicans С. (M.) subaequalis Kiefer, 1929:66, 67, f. 24,
a—c. — С. (M.) varicans subaequalis Kiefer, 1932:249. — C. (Microcyclops) varicans Gur-
ney, 1933:255, f. 1747—1764; Рылов, 1933:261, т. 4, ф. 4—6; т. 6, ф. 1.
Самка. Передний отдел тела овальный, относительно широкий,
более удлиненный, чем у М. bicolor, задние углы предпоследнего тора-
кального сегмента закругленные, задний торакальный сегмент по бокам
с длинной щетинкой, позади которой этот сегмент не образует округ-
лого выроста, направленного кзади. Генитальный сегмент удлиненный,
спереди мало расширенный, receptaculum seminis очень вариирующих
очертаний, в его типичной форме с приблизительно равными по вели-
чине передним и задним отделами, в виде овальных мешков; нередко
receptaculum seminis образует объемистые округлые боковые выросты,
принимая крестообразные очертания. Фуркальные ветви почти парал-
лельные, их длина в 3—4 раза (у var. rubellus в 23/2—3 раза) более
их ширины, латеральная щетинка прикрепляется приблизительно в начале
.задней 1/3 длины внешнего края, внутренняя крайняя апикальная щетинка
заметно длиннее внешней крайней, средние апикальные щетинки хорошо
.развитые, нормальной толщины, из них внутренняя в 4х/2—53/2 раз длин-
нее самой ветви и заметно длиннее внешней средней щетинки; оперение
средних щетинок менее густое, нежели у М. bicolor. Передние антенны
12-членистые, иногда, вследствие неполного расчленения З’-го (от начала)
членика, — 11-членистые (Рылов, 1923, 1933; Gurney, 1933), 2-й от конца
членик заметно длиннее предыдущего и лишь немного короче дисталь-
ного. Обе ветви ног I—IV пары 2-членистые, формула шипов 3-4-4-3.
268
CYCLOPINAE
Длина дистального членика эндоподита ног IV пары в 2—21/2 раза
(у var. rubelliis в 2 и менее раза) более его ширины, внутренний апикаль-
ный шип этого членика хорошо развитой, очень вариирующей длины,
иногда он почти равен длине членика, иногда менее х/2 этой длины,»
внешний апикальный шип короче внутреннего (обычно около 3/4 длины
последнего), но всегда не менее т/2 его длины. Ноги IV пары немного*
Фиг. 62. Microcyclops varicans (Sars), самка.
/—2— фурка с брюшной стороны; 3—нога IV; 4-—генитальный сег-
мент с receptaculum seminis; 5—дистальная часть антенны I; 6—7—ьнога V.
менее ног III пары. Ноги V пары 1-членистые (базальный членик совер-
шенно слит с торакальным сегментом), единственный членик стройный,
цилиндрический, длинный, на конце вооруженный длинной щетинкой
и сбоку, несколько кзади от основания щетинки, очень мелким, нередко
трудно различимым шипиком (у var. rubellus он совершенно редуцирован).
Длина 0.6—1.0 мм. Окраска серо-белая, однообразная, иногда с желтым
9. MICROCYCLOPS
269
шли красноватым оттенком, иногда соломенно-желтая. Яйцевые мешки
крупные, очень удлиненные, обычно содержащие значительное количе-
ство яиц, расходящиеся под значительным углом.
Самец. Фуркальные ветви короче, чем у самки, внутренняя край-
няя апикальная щетинка длиннее, чем у последней, передние антенны
как у самца М. bicolor. У ног VI пары внешняя щетинка длинная, более
чем вдвое длиннее внутреннего шипа, срединные щетинки заметно длин-
нее последнего и короче внешней щетинки. Длина 0.5—0.6 мм.
Распространение. Вся Европа, Азия, Африка, Северная и
Южная Америка, Новая Зеландия, Новый Южный Уэльс. В европей-
ской части СССР известен (типичная форма) из Карело-Финской ССР,
из областей центральной, юго-восточной и южной, также из Челябинской
области. В азиатской части СССР указан из ряда местностей, до Даль-
него Востока включительно. В горах показан до высоты 1000 м. На
арктических островах и на крайнем севере СССР (зона тундры), подобно
М. bicolor, отсутствует.
Биология. Характерен для мелких заросших макрофитами водо-
емов, нередко во временных, летом высыхающих, лужах, встречается
также в зарослях макрофитов литорали озер. Указан для ключевых вод,
где, очевидно, является редким видом. Тепловодно-стенотермичен (см.
ниже о размножении), рядом авторов отмечается как специально летний
циклоп. По исследованиям Пажитновой (1935), близ Самарканда М. vari-
cans очень характерен для рисовых полей, где летом в больших коли-
чествах встречается при температуре воды 34—37°. Обитает преимуще-
ственно в водах с рН==7.0—9.0. Однако по Себенцову и Адовой (1928)
обитает также в сфагновых торфяных лужах (карьеры) Московской
области при pH не менее 5.06 и при высокой окисляемости (100—152.4
мг О2 на литр), т. е. в значительно дистрофированных водах; возможно,
эти данные относятся к vai. rubellus, который, по Gurney (1933), совме-
стно с Acanthocyclops nanus встречается в больших количествах и
в дистрофных водоемах (моховые болота). В пещерных водоемах отсут-
ствует. Повидимому широко приспособляется к разнообразному химизму
воды, так как широко распространен в мягких, бедных электролитами
водах, но Sars’oM (1909) найден в богатом магнием оз. Танганайке (цен-
тральная Африка), а Зверева (1930) находила этот вид в рч. Китерме
(Омской обл.) — стоке оз. Ик (сухой остаток 598.4 мг на литр, слабо
соленая вода, СаО до 80 мг на литр).
Цикл размножения изучен неполно. По Wolf'y (1905) в мелких
водоемах окрестностей Вюртемберга дицикличен. Зимой отсутствует,
появляется весной, максимум в летние месяцы, осенью выпадает.
*la. Microcyclops varicans Sars var. rubellus (Lili.) (фиг. 633_10).
Cyclops rubellus Lilljeborg, 1901:75, t. 4, f. 25—26. — C. varicans var. rubens Wolf,
1905:173, t. 8, f. 9—11. — C. davidi Chappuis, 1922, Rev. Suisse Zool., 29:172, f. 2; Kiefer,
1927, Faune Colon. Franc., 1:560, f. 41—43. — C. rubellus Lowndes, 1928, Nat. Hist.
Wicken Fen, 4:338, t. 8, f. 1—6. — C. (Microcyclops) rubellus Kiefer, 1929:67, f. 25.—
C. rubellus Смирнов, 1930:10, рис. 1—3. — C. (Microcyclops) varicans rubellus Gurney,
1933:260, f. 1765—1770.
От типичного M. varicans самки отличаются следующими призна-
ками:
1. Фуркальные ветви короче, их длина в 2х/2—3 раза превышает
их ширину.
270
Cyclopinae
2. У ног IV пары дистальный членик эндоподита короче, чем
у типичной формы, его длина не превышает его двойной ширины.
3. У членика ног V пары всегда отсутствует мелкий шипик, так что-
членик вооружен лишь щетинкой.
Длина самки 0.6—0.9 мм. В яйцевых мешках по 4—12 яиц.
Фнг. 63.
Microcyclops planus (Gurney), самка: 7 — генитальный сегмент и reeeptaculum seminis:
2—нога V; Microcyclops varicans var. rubellus (Lil!.), самка: 3—фурка, 4 — recepta-
culum seminis, 5 — нога IV, 6 — нога V, 7 — фурка с брюшной стороны, 8— reeeptaculum
seminis, 9—10—дистальный членик эндоподита ноги IV.
(7—2—по Chappuis, 3—6 н 10— по Lowndes’y, 7—9—по Kiefer’y).
Sars (1913) уже оспаривал видовую самостоятельность описанного
Lilljeborg-’oM С. rubellus. Lowndes и Kiefer последнего считают за само-
стоятельный вид. Исследования Gurney, однако, говорят убедительно
против этого. Придавать значение форме reeeptaculum seminis здесь
едва ли возможно, так как этот орган чрезвычайно вариирует у типич-
ного М. varicans. Равным образом отсутствие или присутствие мелкого
шипика на членике V пары ног едва ли можно считать видовым призна-
ком; у М. varicans этот шипик находится явно в фазе сильной редукции.
9, MICROCYCLOPS
277’
Наконец размеры дистального членика эндоподита IV пары ног у типич-
ных М. varicans очень вариируют, что, очевидно, носит индивидуальный
характер и не дает оснований к строгому различению обеих форм.
Поэтому мы считаем rubellus лишь вариететом, таксономическое поло-
жение которого относительно типичного varicans пока остается неясным.
В приведенном кратком описании rubellus мы опускаем некоторые ука-
зываемые Kiefer’oM и Gurney’eM признаки как явно непригодные для
различения обеих форм (очертания reeeptaculum seminis, длина внутрен-
ней крайней апикальной щетинки фуркальных ветвей, положение их;
латеральной щетинки, наличие ряда шипиков в их основании, длина
апикальных шипов дистального членика эндоподита ног IV пары).
Распространение. Известен из Германии, СССР, Швеции,.
Голландии, Англии, Северной Америки. В СССР найден в окрестностях
Костромы.
Биология. Характерен, главным образом, для заросших мелких,
водоемов, до значительно дистрофированных включительно (Gurney, 1933)..
Смирнов (1930) находил этот вариетет в зарослях нитчаток в р. Кост-
роме, Klie (1925) — в богатом железом источнике в Голштинии. По Lown-
des’у (1928), обитает в водах с рН=6.3—7.2; несомненно встречается
и в более кислых водах. В пещерных водоемах не найден. Летняя,,
дицикличная форма, цикл размножения изучен неполно; Wolf и Gurney
предполагают наличие, как и у типичной формы, стадии покоя (она не
описана), в состоянии которой рачок должен переносить высыхание:
водоема и, вероятно, условия зимнего режима.
*2. Microcyclops bicoior Sars (фиг. 64).
Cyclops bicolor Sars, 1863, Forh. Selsk. Christ, (1862): 253. — C, longicaudatus Пог-
генполь, 1874, Изв. Моск. общ. люб. ест., антр. и эти., 10, 2: 72, t. 15, f. 19, 20; t. 16,
f. 5, 6. — C. diaphanus (non Fischer) Rehberg, 1880, Abh. Ver. Bremen, 6 : 547.— C.breui-
seiosus Daday, 1884, Math. term. Kozl., 19:225, t. 3, f. 3—5, 10. — C. tenuicauaus Daday,.
1884, ibid., 6:258, t. 3, f. 18—19. — C. diaphanus (non Fischer) Richard, 1891, Ann. Sc.
Natur., ser. 7, 12:236, t. 6, f. 26.— C. bicolor Schmeil, 1892:118, t. 6, f. 6—13. — Cy--
clops (Microcyclops) bicolor Claus, 1893a: 347, —C. bicoior Lilljeborg, 1901:78, t. IV, f. 27,
28; Sars, 1913: 56, t. 34. — Cyclops (Microcyclops) bicoior Kiefer, 1929 : 70, f. 26, a. —
Crypiocyclops bicoior Lowndes, 1930, Proc. Zool. Soc. London, 174, t. II, f. 8—9. — Cy-
clops (Microcyclops) bicoior Gurney, 1933:362, f. 1771—1784.
Самка. Передний отдел тела широко овальный, несколько сплющен-
ный в спинно-брюшном направлении, относительно короткий и толстый,,
длина цефалоторакса ч- длина 2-го—5-го сегментов торакса приблизи-
тельно в l’/г раза более ширины. Задние углы предпоследнего тора-
кального сегмента равномерно закругленные, задний сегмент по бокам
с довольно крупной щетинкой, позади места прикрепления которой'
сегмент образует (справа и слева) небольшое округлое выпячивание,,
направленное кзади. Генитальный сегмент довольно длинный, сравни-
тельно мало расширенный спереди. Reeeptaculum seminis объемистый,
очень широкий, его передний отдел крупнее заднего, с небольшим
вдавлением спереди, задний отдел в виде короткого мешка, иногда вда-
вленного сзади. Фуркальные ветви мало расходящиеся, иногда параллель-
ные, их длина в 4—5 раз превышает ширину, из крайних апикальных
щетинок внутренние в 2 (иногда и более) раза длиннее внешней край-
ней, средние апикальные щетинки очень толстые и короткие, густо
оперенные, из них внутренняя лишь немного длиннее внешней и лишь
незначительно превышает длину ветвей. Латеральная щетинка прикре-
пляется близ конца внешнего края последних. Передние антенны 11-чле-
-272
CYCLOPINAE
нистые, очень укороченные, не доходящие до середины цефалото-
ракса, 7-й и 8-й членики удлиненные 8-й членик снабжен тонким щетин-
кообразным сензорным придатком, 11-й членик немного длиннее 2 пре-
дыдущих, которые очень коротки и почти равной длины. Обе ветви
ног I—IV пары 2-членистые, формула шипов 3-4-4-3. Ноги IV пары
несколько длиннее ног предыдущих пар, длина дистального членика
эндоподита ног IV пары в 2—21/2 раза превышает его ширину, внутрен-
ний апикальный шип этого членика хорошо развит, довольно толстый,
.заметно короче самого членика, внешний апикальный шип очень малень-
Фиг. 64. Microcyclops bicolor (Sars), самка.
/ — фурка со спинной стороны; 2—генитальный сегмент с reeepta-
culum seminis; 3—нога V; 4 — дистальный членик эндоподита ноги IV;
5—дистальные членикн антенны I.
жий, не более 1/3—т/5 длины внутреннего. Ноги V пары цилиндрические,
удлиненные, 1-членистые (базальный членик совершенно слит с тора-
кальным сегментом), вооруженные длинной щетинкой и, несколько кзади
•от ее основания, на внутреннем крае членика — очень мелким, часто
с трудом различимым шипиком. Длина 0.6—0.8 мм. Передний отдел
тела иногда бесцветен, иногда с голубым оттенком, абдомен и перед-
ние антенны яркой золотисто-желтой окраски; иногда весь рачок бес-
цветен. Яйцевые мешки овальные, прилегающие к абдомену, содержащие
обычно небольшое количество яиц (от 4 до 20).
Самец. Фуркальные ветви относительно короче, чем у самки.
Внутренняя из крайних апикальных Щетинок относительно длиннее, чем
у последней, сензорные придатки на 1-м и 4-м члениках передних антенн
9. MICROCYCLOPS
27'i
очень крупные, цилиндрические. У ног VI пйры внутренний шип круп-
ный, средняя щетинка очень короткая, в виде мелкого шипика, внешняя
щетинка немного длиннее внутреннего шипа. Длина 0.5—0.7 мм.
Распространение. Вся Европа и Азия, Северная Америка,
Африка, Новые Гебриды. В СССР широко распространен, до Дальне-
восточного края включительно; не встречается, однако, в зонах тундры,
полупустыни и пустыни. В горах показан до высоты около 2000 м над
уровнем моря.
Биология. Обитает, главным образом, в мелких заросших водо-
емах, встречается также в зоне макрофитов литорали озер, но в субли-
торали отсутствует. Неоднократно был находим в усыхающих летом
водоемах. В реках обитает исключительно в местах со слабым течением,
в прибрежных зарослях макрофитов. Очень обычен в обрастаниях нит-
чаток и др. водорослей. По данным всех авторов, тепловодно-стенотер-
мичен, встречается в теплое время года, размножается и при очень
высоких температурах (рисовые поля близ Самарканда, при температуре
воды 34—37°). Троглоксен. Встречается при рН=6.4—9.2, особенно в ней-
тральных и слабощелочных водах; в дистрофных водоемах, при pH менее
6, отсутствует. /9-мезосапроб, вероятно также олигосапроб. Вид чисто
пресноводный, единственное мне известное нахождение в солоноватой
воде (мелкая лужа в окрестностях Троицка, Зиновьев, 1934), по всем
вероятиям объясняется случайным заносом из пресного водоема.
Цикл размножения изучен детально; вид дицикличен.
*3. Microcyclops gracilis (Lili.) (фиг. 65).
Cyclops gracilis Lilljeborg, 1853, Crust, ord. trib. : 208. — C. diaphanus (non Fischer)
Schmeil, 1891. Zeitschr. Naturw., 64:33. — C. gracilis Schmeil, 1892:110, t. 6, f. 14—16;
Lilljeborg, 1901:69, t. IV, f. 20—22; Graeter A., 1903: 514, t. 15, f. 13, 29; V. Breemen,
1907 : 345, t. VI, f. 56—57. — Mesocyclops gracilis Sars, 1914 : 63, t. 39. — C. (Metacyclops)
gracilis Kiefer, 1927 :303; Kiefer, 1929 : 72. — C. (Microcyclops) gracilis Gurney, 1933:272—
276, f. 1801—1817.
Самка. Тело стройное, его передний отдел значительно суживаю-
щийся кзади, его наибольшая ширина лежит несколько кпереди от его
середины, ширина заднего торакального сегмента не более (или лишь
незначительно более) ширины переднего отдела генитального сегмента;
его задние углы закругленные. Абдомен относительно длинный, гениталь-
ный сегмент мало расширенный в переднем отделе, очень удлиненный,
его длина почти вдвое более его ширины. Receptaculum seminis молото-
образный, фуркальные ветви более или менее расходящиеся, иногда
почти параллельные, относительно очень подвижные, их длина прибли-
зительно втрое более их ширины, латеральная щетинка хорошо развитая,
прикрепляется приблизительно на середине внешних краев, внутренняя
из крайних апикальных щетинок лишь немного менее чем вдвое превы-
шает длину внешней крайней, обе средние апикальные щетинки корот-
кие, сильно утолщенные в передней длины, из них внешняя прибли-
зительно на г/4—1/5 короче внутренней. Передние антенны 11-членистые,
редко 12-членистые, почти доходящие до заднего края 3-го торакаль-
ного сегмента, иногда несколько более короткие. Обе ветви I—IV пар
плавательных ног 2-членистые, формула шипов 3-4-4-3. У IV пары ног
1-й членик экдоподита внутри без щетинки (имеется лишь 1 внешний
шип), 2-й членик эндоподита очень длинный, его длина приблизительно
в 3 раза превышает ширину, из апикальных шипов внутренний очень
длинный, иногда длиннее самого членика, внешний очень короткий.
18 Фауна СССР
274
CYCLOPINAE
Ноги V пары одночленистые, базальный членик совершенно слит с зад-
ним торакальным сегментом, остающийся единственный членик удлинен-
ный, на конце вооруженный длинной щетинкой и коротким шипиком.
Длина 0.76—0.82 мм. Окраска буроватая, иногда розово-красная. Яйце-
вые мешки небольшие, обычно тесно прилегающие к абдомену, содер-
жащие небольшое (4—5) число яиц.
Фиг. 65. Microcyclops gracilis (Lil!.).
/—фурка самки со спинной стороны; 2—генитальный сегмент и receptaculum seminis;
у—4—нога V самки; 5—дистальный членик эндоподита ноги IV; 6—Tettataxusaxi
сегмент и recptaculum seminis; 7—антенна I самки.
(7—3—по Sars’y 4—7 — по Gurney).
Самец. У ног VI пары лишь 2 придатка — довольно длинный тон-
кий внутренний шип и более короткая внешняя щетинка. Длина 0.63—
0.7 мм.
Науплиус последней стадии очень удлиненный, с сильно развитыми
выступающими кзади фуркальными выростами, перед которыми тело
науплиуса заметно сужено.
Распространение. Европа, Азия (Монголия), Южная Америка
(Аргентина). В СССР известен из областей: Горьковской, Ярославской,
Калининской, Свердловской, Харьковской. На крайнем севере Европы
и на арктических островах отсутствует, высоко в горы не заходит.1
1 Данные R. Monti (1906, Итальянские Альпы, до высоты 2500 м над уровнем
моря) сомнительны.
9. MICROCYCLOPS
275'
Биология. Обитает в более или менее заросших макрофитами
мелких водоемах — болотцах, старицах, лужах, прудах и проч., также
в зоне макрофитов литорали озер. Тепловодно-стенотермичен. По Таусон
(1935) в мезотрофном оз. Диком (близ г. Молотова) живет при рН =
6.8—7.5; отсутствует в сильно дистрофированных кислых водах, опти-
мум, вероятно, в нейтральных и слабощелочных водах. Для подземных
вод не указан. Чисто пресноводный вид. Судя по данным Wolf’a (1905),
для окрестностей Вюртемберга и Зиверта (1929) для окрестностей Харь-
кова,— дицикличен. Все авторы отмечают М. gracilis как летнюю форму,
появляющуюся весной — в начале лета — и на зиму выпадающую. Guiney
(1933) полагает, что зиму рачок проводит в состоянии покоящихся яиц
(они не описаны), так как весной появляются науплиусы и 1-е копепо-
дитные стадии.
*4. Microcyclops dengizicus (Lepeschkin) (фиг. 66).
Cyclops diaphanus var. dengizicus Лепешкин, 1900:26, t. 3, f. I, 17, 19,21,25,26.—
C. buxtoni Gurney, 1921, Joum. Bombay Soe., 27 : 840, t. I, f. A — E. — C. dengizicus
Kiefer, 1926, Zool. Anz., 67:106, f. 6—8.— C. (Metacyclops) dengizicus Kiefer, 1929:73,
f. 27; 1936, Arch. f. Hydrob., 30 : 296, f. 71—77.— C. (Metacyclops) dimorphus Kiefer,
1934, Zool. Anz., 107:271, f. 5—6.
Сам к а. Передний отдел тела равномерно суживающийся кпереди
и кзади, наибольшая ширина лежит приблизительно на середине его
длины, предпоследний торакальный сегмент с несколько выступающими
кнаружи задними углами, вооруженными группой очень мелких щетинок.
Генитальный сегмент очень широкий спереди и значительно суженный
в задней половине, его длина приблизительно равна его наибольшей
ширине. Receptaculum seminis похож на таковой A. bicuspidatus,передний
отдел узкий, иногда вдавленный спереди, задний отдел мешковидный,
очень объемистый, передний отдел окружен сильно преломляющей свет
каймой, которая имеется и вдоль переднего края мешковидного отдела.
Фуркальные ветви удлиненные, параллельные, их длина приблизительно
в 41/2 раза более их ширины, латеральная щетинка прикреплена обычно
на средине внешнего края или немного кзади от нее, ближе к заднему
концу ветвей, крайние апикальные щетинки короткие, почти одинаковой
длины, средние апикальные щетинки хорошо развитые, нормальной тол-
щины, из них внутренняя лишь немного длиннее внешней. Передние
антенны 11-членистые, едва доходящие до заднего конца цефалото-
ракса. Обе ветви всех пар плавательных ног 2-членистые, формула
шипов 3-4-4-3, длина дистального членика эндоподита ног IV пары
приблизительно в 1% раза более ширины, внутренний край вооружен 3
длинными оперенными щетинками (они длиннее самого членика), внеш-
ний— 1 более короткой оперенной щетинкой; апикальное вооружение
этого членика состоит из длинного (более 1/2 длины членика) внешнего
шипа и внутренней щетинки (а не шипа!), длина которой приблизительно
на 1/3 более таковой шипа и почти равняется длине самого членика.
Экзоподит немного короче эндоподита, его 1-й членик снабжен 1 внеш-
ним шипом, а внутри 1 щетинкой, 2-й членик — соответственно 3 шипами
и 5 щетинками. Ноги V пары весьма характерного строения, 1-членистые,
базальный членик совершенно слит с задним торакальным сегментом,
принадлежащая этому членику щетинка относительно короткая. Единст-
венный членик чрезвычайно широк, его ширина приблизительно вдвое
более его длины, так что членик имеет вид широкой и короткой пла-
стинки; его вооружение состоит из сильного шипа на внутренне-заднем
18*
2/6
cyclopinae
углу и из длинной (в несколько раз длиннее шипа) оперенной щетинки
на внешне-заднем углу. Длина 0.9—1.2 мм.
Самец. У ног V пары внутренний шип вдвое длиннее, чем у самки,
ноги VI пары с 3 придатками — длинным внутренним шипом и 2 щетин-
Фиг. 66. Microcyclops dengizicus (Lepeschk.), самка.
7— фурка со спинной стороны; 2 — абдомен с reeeptaculum seminis; 3—нога V;
4—дистальный членик эндоподита ноги IV; 5—нога IV.
ками, из которых внешняя немного длиннее средней; обе щетинки слабо
оперены. Длина 0.9—0.94 мм.
Рассматриваемый вид был описан Лепешкиным как новая разновид-
ность, под именем Cyclops diaphanus var. dengizica. Автор дал хорошие
рисунки, но не вполне удовлетворительный диагноз; в частности им не
была описана IV пара ног. Позже Sars (1903) высказался за видовую
самостоятельность С. dengizicus, не дав, однако, его описания. Kiefer
9. MICROCYCLOPS
277
(1936), основываясь на рисунках Лепешкина (русское описание этому
автору осталось недоступным), подтвердил мнение Sars’a и доказал иден-
тичность С. dengizicus с С. buxtoni Gurney (1921). Позднее Kiefer (1936),
по материалу с о. Гаити, дал весьма обстоятельное описание М. dengi-
zicus, сопровождаемое рядом рисунков, которое вполне подтверждается
нашими исследованиями материала, определенного Sars’oM (оз. Денгиз;
Sars, 1903). От прочих видов рода Microcyclops рассматриваемый вид,
помимо строения ног V пары, хорошо отличим по вооружению эндопо-
дита ног IV пары. По строению reeeptaculum seminis М. dengizicus при-
ближается к М. minutus, от которого, однако, легко отличается по во-
оружению дистального членика эндоподита ног IV пары, по строению
ног V пары и по вооружению фуркальных ветвей. Как показал сам
Kiefer (1936), описанный им М. dimorphus (Калифорния) идентичен с рас-
сматриваемым видом. Наиболее близок к М. dengizicus описанный Marsh’eM
(1913) из солоноватых водоемов Панамы М. panamensis.1
Распространение. Ирак (р. Тигр; Gurney, 1921—С. bux-
toni), Индия (оз. Чилка близ Калькутты; Sewell, 1924), о. Гаити (Kiefer,
1936), Египет (Кабрет, Gurney, 1927), Калифорния (Соленое оз.; Kiefer,
1934 — С. dimorphus). В СССР найден в солоноватом оз. Денгиз (Кара-
гандинской обл.) и в солоноватом болотце близ этого же озера совме-
стно с Moina microphthalma, Diaptomus salinas и Cletocamptus retro-
gressus. Таким образом M. dengizicus характерен для зоны пустынь.
Биология. Очень специально для солоноватых вод и дол-
жен быть отнесен к галобионтам. Обитает как в крупных солоно-
ватых озерах, так и в солоноватых мелких лужах. Единственное место-
нахождение, которое можно было бы отнести к пресной воде, предста-
вляет мелкий водоем (pH —8.3) на о. Гаити, но Kiefer (1936) полагает,
что и этот водоем получает соленую воду из рядом лежащего обширного
соленого пруда. Отношение к температурам не выяснено; во всяком
случае М. dengizicus способен переносить высокие температуры (25—30°;
Kiefer, 1936:264).
Цикл размножения не известен.
*5. Microcyclops minutus Claus (фиг. 67).
Cyclops minutus Claus, 1863:102, t. 10, f. 6—8; — C. longicaudatus Brady, 1892,
Trans. Nat. Hist. Northumb., XI, 1:88, t. 10, f. 5—1.— C. diaphanus (non Fischer) Schmeil,
1892:112, t. 5, f. 18. — C. diaphanus (non Fischer) Richard, 1893, Rev. Biol. Nord. Er.,
V : 404, f. 5—6. — Microcyclops diaphanus (non Fischer) Claus, 1894:12, t. I, f. 1—8.—
C. minutus A. Graeter, 1903 : 424. — C. diaphanus (non Fischer) Gurney, 1921, Journ.
Bomb. Nat. Hist. Soc., 27 : 90. — C. minutus Thallwitz, 1922, Zool. Anz., 54: 263; Kiefer,
1926, Int. Rev. Hydrob. u. Hydrogr., 14 357, f. 7, a—-f.— C. diaphanus (non Fischer)
Marsh, 1926, Voyage zool. H. Gadeau de Kerville, 1:179, t. 26, f. 3—6. — C. minutus
Gurney, 1927, Ann. Mag. Nat. Hist., (9), 19:506, f. 5 a—f.; Смирнов, 1929:2,
рис. 1, a — d. — C. (Microcyclops) minutus Kiefer, 1929 : 74, f. 28; Gurney, 1933 : 266—271,
f. 1785—1800. — Metacyclops unacanthus Lindberg, 1936:9, f. 13—16.
Самка. Длина цефалоторакса плюс длина 2-го — 5-го сегментов
торакса приблизительно в 1 3/4 раза превосходит ширину, торакс довольно
толстый, задние углы предпоследнего торакального сегмента закруглены,
закругленные задние углы последнего торакального сегмента по бокам
с длинной, обычно загнутой кзади щетинкой. Генитальный сегмент
очень объемистый, почти не суживающийся кзади, его длина лишь
1 Кроме Панамы, этот вид известен из солоноватых водоемов о. Гаити, о. Аруба
И северного побережья Южной Америки (отсюда описан как <5 С, vidnus Kief.)
278
CYCLOPINAE
немного более ширины; несколько кпереди от середины этот сегмент
по бокам неглубоко вдавлен. Receptaculum seminis очень крупный, его
передний отдел маленький, очень узкий, задний отдел в виде широкого,
далеко заходящего кзади обширного мешка. Фуркальные ветви парал-
лельные или почти параллельные, их длина в 4—5 раз более их ширины,
Фиг. 67. Microcyclops minutus (Claus).
I—Фурка самки с брюшной стороны; 2—фурка самки со спинной стороны; 3—4 — гени-
тальный сегмент и receptaculum seminis; 5 — дистальный членик Эндоподита ноги IV;
б—нога IV самки; 7—нога V самки; 8—антенна! самки; 9—нога I.
(1—3, 5 и 7—по Kiefer’y; 4, б, 8 и 9 — по Gurney).
латеральная щетинка хорошо развита и прикреплена почти на середине
внешнего края, внутренняя из крайних апикальных щетинок несколько
короче внешней крайней, средние апикальные щетинки относительно
очень короткие, утолщенные, густо оперенные, из них внутренняя лишь
немного длиннее внешней и не более чем в 3 раза длиннее фуркальной
ветви. Торакс, абдомен и фуркальные ветви обычно пунктированы
(очень мелкие ямкообразные вдавления кутикулы). Передние антенны
11-членистые, не доходящие до заднего края цефалоторакса. Обе ветви
9. MICROCYCLOPS
279
I—IV пар ног 2-членистые, формула шипов 3 • 4 4 • 3. Ноги IV пары
несколько короче ног III пары. Эндоподит ног IV пары короче экзопо-
дита, длина дистального членика эндоподита в 2—2 1/2 раза (иногда
несколько более) превышает его ширину, на конце этот членик воору-
жен лишь 1 апикальным шипом, длина которого более длины членика.
Ноги V пары 1-членистые, базальный членик совершенно слит с задним
торакальным сегментом, остающийся членик очень маленький, его длина
приблизительно в 1 х/2 раза более ширины, на конце он вооружен длин-
ной щетинкой, близ которой в конце внутреннего края членика при-
крепляется довольно крупный шип, длина которого вариирует, — иногда
он несколько длиннее членика. Длина 0.7—0.98 мм. Окраска сероватая.
Яйцевые мешки крупные, не прилегающие к абдомену.
Самец. Фуркальные ветви и их апикальные щетинки длиннее, чем
у самки, передние антенны с очень короткими сензорными цилиндри-
ческими придатками, 1-й членик снабжен лишь 1 таким придатком. Ноги
VI пары снабжены лишь 2 придатками — шипиком и щетинкой, прибли-
зительно одинаковой длины. Длина 0.7 — 0.92 мм.
Едва ли можно сомневаться, что под названием „ Cyclops diaphanus"
неоднократно указывался рассматриваемый вид; однако под этим же
названием в литературе приводились и другие виды {Acanthocyclops
nanus Sars, Microcyclops bicolor Sars, M. gracilis Sars, M. dengizicus Lep.),
так что в целом ряде случаев нельзя сказать, какой именно из этих
видов приводится автором в действительности. Некоторые указания
на нахождение „С. minutus Claus", имеющиеся в старой литературе,
безусловно ошибочны. Таковы указания РогеГя, 1884,1885 (определение
Н. Vernet, указан как пелагический вид субальпийских швейцарских
озер), Pavesi, 1879, 1883 (озера Италии), Fric’a, 1871 (Богемия), Heller’a
1870 (Тироль).
Распространение. Бесспорные данные имеются для Европы
(Англия, Германия, Португалия, Болгария, СССР), Азии (Палестина,
Сирия, Ирак, Синай, Индия, азиатская часть СССР), северной
и северозападной Африки. Под вопросом недавно приведен Ueno (1936)
для южного Сахалина. В пределах СССР указан с достоверностью
для окрестностей Костромы и Одессы, Апшеронского полуострова,
долины Маныча, для мелких водоемов пустыни Кара-кумы. Известен
из Крыма (Карадаг).
Биология. Чрезвычайно характерен для временных, мелких, летом
высыхающих луж, причем встречается и в очень мутных лужах. Особенно
характерен для таких водоемов (также в колодцах) области степей,
полупустынь и пустынь. Так, в Кара-кумах он является обычным весло-
ногим рачком (обычно единственным из Cyclopidae) временных луж
(их местное название „куймы" и ,,каки“) и колодцев, причем здесь встре-
чается с большим постоянством; в водоемах такого же типа М. minutus
очень распространен в Малой Азии и Северной Африке. Однако на севере
(окрестности Костромы) рачок обитает во временных лужах совершенно
другого типа; Gurney в Англии находил его в болотистых усыхающих
водоемах, a Gauthier (1928) в Северной Африке неоднократно констати-
ровал его в пресных, проточных и стоячих, временных, заросших макро-
фитами, водоемах. По Okul (1937) в окрестностях Одессы М. minutus
обитает в пересыхающих водоемах с Lemna и с зарослями Typha, Scir-
pus, Phragmites. Его приуроченность специально к эфемерным временным
водоемам резко выраженного астатического типа является в высшей
степени характерной.
280
CYCLOPINAE
«>
Судя по ряду данных, М. minutus безусловно эвритермичен. По дан-
ным Gauthier (1928) в Алжире М. minutus встречается только в пресных
водах, при pH = 6.6—7.6, отсутствуя в солоноватых водоемах. Цикл
размножения не изучен. Очень вероятно, что в ряде случаев рачок
моноцикличен. Рачок переносит высыхание водоема не в виде покоя-
щихся яиц или науплиуса, а в состоянии последних фаз метаморфоза
(последние копеподитные стадии), причем при новом заполнении бывшего
водоема снеговыми и дождевыми водами очень быстро достигает поло-
возрелости (появляются самки и самцы) и сразу же приступает к раз-
множению.
*6 Microcyclops planus (Gurney) (фиг. 631-2).
Cyclops planus Gurney, 1909, jouru. R. Mier. Soc.: 293, t. 12, f. 24—27. — C. hale-
pensis Chappuis, 1922, Zool. Anz., 55:28, f. 1—4. C. planus Kiefer, 1924, Zool. Anz.,
61:220. — C croaticus Krmpotic, 1924, Arch. f. Hydrob., 15:42. t. 5, f. 29—31. — C. (Mic-
rocyclops) planus Kiefer, 1929 : 175. — Microcyclops planus (o) Stephauides, 1937, Arch,
f. Hydrob., 31: 329, f. a — e.
Самка. Тело заметно сплющенное в спинно-брюшном направлении.
Генитальный сегмент короткий и широкий, его длина лишь немного
более ширины, кзади сегмент почти не суживается. Receptaculum seminis
объемистый, с очень крупным мешковидным, далеко продолжающимся
кзади, задним отделом и 2-раздельным передним отделом. Длина очень
мало расходящихся фуркальных ветвей равна (или несколько более)
длине 2 задних абдоминальных сегментов, взятых вместе, из крайних
апикальных щетинок внутренняя заметно короче шипообразной внешней,
средние апикальные щетинки, хорошо развитые, из них внутренняя более
чем вдвое превышает длину внешней, латеральная щетинка крупная,
прикрепленная почти на средине внешнего края ветвей. Передние антенны
лишь 9-(редко 10-)членистые, очень короткие, д'оходящае лишь до
середины цефалоторакса. Дистальный членик эндоподита ног IV пары
снабжен лишь одним апикальным шипом, другой апикальный придаток
представлен щетинкой. Ноги V пары с очень маленьким члеником,
вооруженным очень мелким внутренним шипом и довольно длинной
внешней щетинкой; торакальная щетинка сильно развитая. Длина 0.65 —
0.80 мм. Яйцевые мешки округлые, очень тесно прилегающие к абдо-
мену.
Самец. Фуркальные ветви и передние антенны относительно длин-
нее, чем у самки, последние достигают заднего конца цефалоторакса,
14-членистые. Ноги VI пары сильно развитые, с 2 довольно длинными
оперенными щетинками. Длина 0.7—0.76 мм. Сперматофоры удлиненные.
Распространение. Северная Африка (Тунис, Алжир, Египет),
Сирия, Кроация, Греция (Корфу). В СССР найден в Крыму (Карадаг).
Биология. Обитает в мелких водоемах типа луж, в частности
пересыхающих; известен из колодцев (Aleppo). В Алжире и Тунисе,
по Gauthier (1928), обитает в мелких водоемах (pH 7.2—7.6) степной
и полустепной зоны.
10. Род SPEOCYCLOPS KIEF.
Graeteriella (part.) Brehm, 1926:134. — Cyclops (subg. Diacyclops, part.) Kiefer>
1929:65, 66. — Cyclops (subg. Bryocyclops, part.) Scourfield, 1932, Aun. Mag. Nat. Hist.,
10:559. — Cyclops (subg. Microcyclops, part.) Gurney, 1933 : 253, 281. — Speocyclops Kiefer,
1937b: 325; Kiefer, 1937 : 431, ' " ppp
10. SPEOCYCLOPS
28/
Габитус тела более или менее гарпактикоидный, абдомен короткий
и очень толстый, генитальный сегмент очень широкий, объемистый,
у некоторых видов на спинной поверхности с поперечным швом, так
что при рассматривании рачка со спины его абдомен оказывается 5-чле-
нистым. Фуркальные ветви очень короткие, средние апикальные щетин-
ки хорошо развитые, длинные, крайние апикальные щетинки очень
короткие. Анальная пластинка полукруглой или треугольной формы,
иногда зазубренная, у большинства видов довольно сильно развитая.
Членистость плавательных ног по формуле 2-2/2-2/2-2/2-2, дистальный
членик экзоподита ног IV пары с 3 шипами и 3 щетинками, эндоподита —
с 4 придатками (1 апикальным шипом, 1 апикальной щетинкой и 2 суба-
пикальными щетинками). Щетинка на внутреннем крае 1-го базального
членика этой пары ног имеется, выросты на их соединительной пластин-
ке отсутствуют. Ноги V пары состоят из 2, лежащих рядом (а не один
над другим), маленьких члеников, из них внешний (гомологичный базаль-
ному 2-членистой нормальной ноги) несет 1 щетинку, а внутренний
(гомологичный дистальному) — 2 придатка, обычно различной длины.
Иногда оба членика (или 1 из них) настолько тесно сливаются с задним
торакальным сегментом и друг с другом, что от них остаются лишь
следы, по которым однако можно всегда заключить о 2-членистости
ног (Sp. plutonis, Sp. demetiensis). Передние антенны 11-членистые,
очень короткие. Мелкие циклопы, длина самок не более 0.4—0.6 мм,
частью без глазного пигмента.
Троглобионты. В наземных условиях (влажные налеты на скалах)
найден лишь Sp. demetiensis (Scourf.), но и этот вид, по всем вероятиям,
относится к троглобионтам.
Известно 12 видов. Род распространен в пещерных водоемах
Венгрии, Югославии, Испании, Франции, Италии, Германии, 1 вид
(Sp. demetiensis) в Англии, 1 вид (Sp. colchidanus) на Кавказе (близ
Кутаиси). Исключительно палеарктический род.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (10). Генитальный сегмент на спинной поверхности со сплошным
поперечным швом.
2 (3). Анальная пластинка очень маленькая, значительно не доходящая
до основания фуркальных ветвей.
......................................1. Sp. racovitzai (Chapp.)
3 (2). Анальная пластинка крупная, заходящая кзади за основание фур-
кальных ветвей.
4 (5). Крайние апикальные щетинки фуркальных ветвей почти одинако-
вой длины.............................. 2. Sp. iafernus Kief.
5 (4). Эти щетинки заметно разной длины.
6 (7). Анальная пластинка сзади узкая, удлиненная, гладкая или тонко
зазубренная лишь в дистальном отделе.. . 3. Sp. sisypbus Kief.
7 (6). Анальная пластинка сзади широкая, грубо зазубренная.
8 (9). Оба членика ног V пары крупные, на внутреннем членике
внутренний придаток короче внешнего................................
.................................. 4. Sp. troglodytes (Chapp.)
9 (8). Внешний членик ног V пары гораздо короче внутреннегоа у кото-
рого внутренний придаток длиннее внешнего .........................
5, Sj». sebastiamis Kief,
282
CYCLOPINAE
10 (1). Генитальный сегмент на спинной поверхности без поперечного
шва или лишь с боков со следами такового.
11 (14). Оба членика ног V пары почти вполне слиты с задним тора-
кальным сегментом, так что от них остаются лишь следы в виде
плоских бугорков.
12 (13). Длина дистального членика эндоподита ног IV пары не-
сколько более его ширины......................б. Sp. plutonis Kief.
13 (12). Длина этого членика не превышает его ширины..............
.........................................7. Sp. demetiensis (Scourf.)
14 (11). Оба членика ног V пары резко дифференцированы.
15 (16). Длина фуркальных ветвей в 2.5 раза более их ширины . .
.........................•.........• ... 8. Sp. cerberus (Chapp.)
16 (15). Их длина в 1.7—2 раза более их ширины.
17 (18). Членики ног V пары крупные, удлиненные...................
. ........................................9. Sp. spaeleus Kief.
18 (17). Эти членики маленькие, укороченные ......................
. . ...................................10. Sp. proserpinae Kief.
1. Speocyclops racovitzai (Chapp.) (фиг. 68 1_2, 6_7).
Cyclops racovitzai Chappuis, 1923, Bull. Soc. Sciint. Cluj, 1 :587, f. 1, 4, 8; Kiefer,
1928 : 527, f. 54—57. — Cyclops (Diacyclops) racovitzai Kiefer, 1929 : 66. — Speocyclops
racovitzai Kiefer 1937:425, t. 7, f. 30—32.
Самка. Генитальный сегмент на спинной поверхности с поперечным
швом, receptaculum seminis очень крупный, с поперечно-овальным перед-
ним отделом, окруженным прозрачной каймой и с редуцированным
задним отделом. Анальная пластинка маленькая, треугольная, с зазуб-
ренным задним краем. Длина фуркальных ветвей немного менее чем
вдвое превышает их ширину; из крайних апикальных щетинок внутрен-
няя немного короче внешней; средние апикальные щетинки длинные.
Длина дистального членика эндоподита ног IV пары немного менее чем
в 1.5 раза превышает ширину, апикальный шип хорошо развит, в 1.5 раза
длиннее членика, щетинки короткие. Оба членика ног V пары сильно
сближены, хорошо дифференцированы, внешний снабжен длинной щетин-
кой, внутренний — парой неодинаковой длины придатков. Длина 0.40—
0.45 мм. Самец не известен.
Распространение. Югозападная Франция, пещеры. Трогло-
бионт.
2. Speocyclops infernus Kief. (фиг. 68 3_s).
Cyclops (Diacyclops) infernus Kiefer, 1930, Zool. Anz., 87: 225, f. 8—10; Kiefer,
1931, Zool. Jahrb., Syst., 61 : 705, f. 9—11. — Speocyclops infernus Kiefer, 1937: 425,t. 7,
f. 33—36.
Самки. Генитальный сегмент на спинной поверхности с поперечным
швом. Receptaculum seminis как у Sp. troglodytes. Анальная пластинка
крупная, треугольная, грубо зазубренная по краям, значительно захо-
дящая кзади за основание фуркальных ветвей. Длина последних прибли-
зительно в 1.5 раза превышает их ширину, крайние апикальные щетинки
почти одинаковой длины, средние щетинки длинные, из них внешняя
приблизительно вдвое короче внутренней. Дистальный членик эндоподита
ног IV пары короткий, его длина лишь немного более его ширины,
апикальный шип приблизительно в 1.5 раза длиннее этого членика, его
щетинки относительно короткие. Оба членика ног V пары удлиненные,
10. SPEOCYCLOPS
283
хорошо дифференцированные, очень тесно сближенные, из них внешний
очень узкий, снабженный длинной щетинкой, внутренний более широкий,
вооруженный парой почти равных коротких придатков. Длина 0.40 мм.
Самец не известен.
Распространение. Северная Италия, пещеры St. Ganzian.
Троглобионт.
Фиг. 68.
Speocyclops racovitzai (Chapp.): 1 — генитальный сегмент
с receptaculum seminis, 2—нога V самки; S. infernus Kief.:
3—дистальный членик эндоподита ноги IV самки, 4—фурка
самки со спинной стороны, 5— нога VI самца; S. racovitzai
(Chapp.): 6— анальная пластинка самки, 7 — нога VI самца.
(7—2— по Chappuis’y, 3—7—по Kiefer’y).
3. Speocyclops sisyphus Kief. (фиг. 69г _2).
Speocyclops sisyphus Kiefer, 1937:435, t. 9, f. 67—73.
Самка. Генитальный сегмент на спинной поверхности с попереч-
ным швом, анальная пластинка очень удлиненная, треугольная, нево-
оруженная или лишь на конце очень тонко зазубренная, задним своим
углом заходящая за основание фуркальных ветвей. Длина последних
в 1.75 раза более их ширины, внутренняя из крайних апикальных щетинок
заметно короче внешней, средние апикальные щетинки длинные. Дисталь-
ный членик эндоподита ног IV пары несколько удлиненный, его апикаль-
ный шип длинный, почти в 1.5 раза длиннее самого членика. Оба
284
CYCLOPINAE
Фиг. 69.
Speocyclops sisyphus Kief.: 1— абдомен самки co спинной стороны, 2 — нога V самки;
S. plulonis Kief.: 3— нога IV самки, 4 — нога V самки; 5—S. cerberus Kief, абдомен
самки с брюшной стороны; 6-—S. spelaeus Kief., нога V самки; 7—S. sebastianus Kief.,
нога V самки; 8— S. proserpinae Kief., нога V самки; S. demetiensis Scourf.: 9—фурка
самки со спинной стороны, 10—reeeptaculum seminis, 77 —нога V самки.
(7—8—ПО Kjefer’y, 9—11 — пр Scourfield’y),
10. SPEOCYCLOPS 285
членика ног V пары явственно дифференцированы, из них внутренний
значительно крупнее внешнего и снабжен 2 придатками — длинным
внутренним и более коротким внешним; придаток внешнего членика
приблизительно равен внутреннему придатку другого членика. Длина
0. 42 мм. Самец не известен.
Распространение. Франция, грот в департаменте Арьеж.
Совместно с Sp. racovitzal (Chapp.).
4. Speocyclops troglodytes (Chapp.) (фиг. 70g_10).
Cyclops troglodytes Chappuis, 1923, Bull. Soc. Sci. Cluj, 1:585, f. 3, 5, 7, 9, 10;
Kiefer, 1928:527, f. 51—53. — Cyclops {Diacyclops) troglodytes Kiefer, 1929:65, f. 23.—
Speocyclops troglodytes Kiefer, 1937:425, t. 7, f. 26—29.
Самка. Генитальный сегмент на спинной поверхности с попереч-
ным швом, задние края абдоминальных сегментов тонко и неравномерно
зазубрены. Reeeptaculum seminis с объемистым передним отделом.
Анальная пластинка треугольная, по краям грубо зазубренная, лишь
немного выдающаяся кзади за основание фуркальных ветвей. Длина
последних приблизительно в 1.5 раза превышает их ширину, из крайних
апикальных придатков внутренний приблизительно вдвое короче внешнего,
средние апикальные щетинки хорошо развитые. У ног IV пары дисталь-
ный членик эндоподита очень короткий, апикальный шип приблизительно
в 1.5 раза длиннее этого членика, щетинки относительно короткие.
У ног V пары оба членика резко дифференцированы, из них внутренний
вооружен 2 придатками, из которых внешний длиннее внутреннего,
а внешний членик несет 1 еще более длинную щетинку. Длина 0.5—
0.55 мм. Самец такой же длины.
Распространение. Пещеры Венгрии.1 Троглобионт.
5. Speocyclops sebastianus Kief. (фиг. 697).
Speocyclops sebastianus Kiefer, 1937b: 327, f. 11—13, Kiefer, 1937:436, t. 9,
f. 73—82.
Самка. Генитальный сегмент на спинной поверхности с попереч-
ным швом, reeeptaculum seminis близок к таковому Sp. spaeleus, с очень
объемистым поперечно-овальным передним отделом. Длина фуркальных
ветвей в 1.75 раз более их ширины, внутренняя из крайних апикальных
щетинок приблизительно вдвое короче внешней, средние апикальные
щетинки длинные, густо оперенные, из них внутренняя почти вдвое
длиннее внешней. Анальная пластинка треугольная, немного заходящая
кзади за основание фуркальных ветвей, зазубренная. Дистальный чле-
ник эндоподита ног IV пары очень короткий, его длина не превышает
ширины, апикальный шип заметно длиннее членика, щетинки короткие.
У ног V пары внутренний членик явственно дифференцированный,
на конце с парой неравной длины придатков, внешний членик очень
короткий, значительно слитый с торакальным сегментом, его щетинка
очень длинная. Длина около 0.40 мм. Самец не известен.
Распространение. Северная Испания, пещерный водоем близ
Сан-Себастьяна. Найдена 1 самка.
1 Chappuis (1933) приводит вид также для пещеры в Югославии, но Kiefer (1937)
в материале из этой пещеры нашел другой вид — Sp. proserpinae Kiefer.
286
CYCLOPINAE
6. Speocyclops plutonis Kief. (фиг. 693_4).
Speocyclops plutonis Kiefer, 1937: 434, t. 8, f. 49—60, t. 9, f. 61—63.
Самка. Генитальный сегмент широкий, без поперечного шва
на спинной поверхности, анальная пластинка хорошо развитая, заходящая
кзади за основание фуркальных ветвей, сзади грубо зазубренная. Длина
фиг. 70. Speocylops colchidanus (Вог.), самец. (По Боруцкому).
/—общий вид со спинной стороны; 2—абдомен со спинной стороны; 3 — абдомен
с брюшной стороны; 4 — нога I; 5—нога III; б—нога IV.
фуркальных ветвей несколько менее чем вдвое превышает их ширину,
крайние апикальные щетинки короткие, почти одинаковой длины, средние
щетинки хорошо развитые, длинные. У ног IV пары длина дистального
членика эндоподита немного более его ширины, апикальный шип заметно
длиннее этого членика. У ног V пары оба рядом расположенных членика
слиты с задним торакальным сегментом; однако задний отдел обоих
члеников, вроде короткого выроста, остается свободным, так что при-
сутствие обоих члеников устанавливается без труда. Внешний членик
вооружен 1 довольно длинной щетинкой, внутренний — парой более
коротких щетинок. Длина 0.50 мм.
10. SPEOCYCLOPS
287
Самец. Ноги VI пары с 3 придатками — с очень толстым внутрен-
ним шипом и с парой коротких щетинок.
Распространение. Югославия, пещера близ Злот.
7. Speocyclops demetiensis (Scourf.) (фиг. 699_u).
Cyclops (Bryocyclops) demetiensis Scourfield, 1932, Ann. Mag-. Nat. Hist., 10:559. —
Cyclops (Microcyclops) demetiensis Gurney, 1933 : 221, f. 1836—1852. — Speocyclops deme-
tiensis Kiefer, 1937:425, t. 7, f. 37—41.
Самка. Генитальный сегмент на спинной поверхности без попе-
речного шва, receptaculum seminis с очень объемистым поперечно-
овальным передним и с очень маленьким задним отделом; по бокам
последний снабжен парой коротких выростов. Анальная пластинка
полукруглая, грубо зазубренная, доходящая до основания фуркальных
ветвей или лишь немного заходящая кзади от их основания. Длина
фуркальных ветвей приблизительно вдвое более их ширины, из крайних
апикальных щетинок внутренняя почти вдвое короче внешней, средние
апикальные щетинки длинные. Дистальный членик эндоподита ног IV пары
очень короткий, его длина не превышает его ширины, апикальный шип
крупный, в 1.5—2 раза длиннее членика. Оба членика ног V пары
почти совершенно слиты с торакальным сегментом, так что от них
остается лишь пара низких плоских бугорков, из них более узкий
внешний вооружен длинной щетинкой, а более широкий и укороченный
внутренний несет 2 придатка, из которых внутренний, шиповидный,
немного длиннее внешнего. Длина 0.50 мм. Окраска беловатая. Яйца,
вероятно, откладываются прямо в воду.
Самец. У ног VI пары 3 придатка, из них внешний в виде щетинки,
длина которой почти вдвое более длины внутренних шиповидных при-
датков. Длина 0.48 мм.
Распространение. Найден в Англии (близ Тенби), во влажных
налетах на скалах. Вероятно, троглобионт.
8. Speocyclops cerberus (Chapp.) (фиг. 69s).
Cyclops (Diacyclops) cerberus Chappuis, 1934, Bull. Soc. Sd. Cluj, 8.—Speocyclops
cerberus Kiefer, 1937: 426, t. 8, f. 42—44.
Самка. Генитальный сегмент на спинной поверхности без попереч-
ного шва, leceptaculum seminis как у Sp. spaeleus, анальная пластинка
маленькая, треугольная, с гладкими краями, не доходящая до основа-
ния фуркальных ветвей. Длина последних приблизительно в 2.5 раза
более их ширины, крайние апикальные щетинки хорошо развитые, из них
внутренняя в 1.5 раза длиннее внешней. Дистальный членик эндоподита
ног IV пары очень короткий, его длина лишь немного более его шири-
ны, апикальный шип длинный, почти вдвое длиннее этого членика,
апикальная и внутренняя щетинки относительно очень длинные (также
и у дистального членика экзоподита). Оба членика ног V пары (особенно
внутренний) резко дифференцированы, из них внутренний несет 2 нерав-
ной длины придатка, внешний — 1 довольно длинную щетинку. Длина
около 0.5 мм.
Распространение. Пещера близ Лунца, юго-восточная Гер-
мания.
286
CYCLOPINAE
9. Speocyclops spaeieus Kief. (фиг. 69e).
Speocyclops spaeieus Kiefer, 1937b: 326, f. 8—10; Kiefer, 1937:435, t. 9, f. 74—77.
Самка. Генитальный сегмент на спинной поверхности без попереч-
ного шва (его следы лишь с боков), receptaculum seminis с очень
объемистым передним поперечно-овальным и с очень маленьким мешко-
видным задним отделом; передний отдел окружен широкой прозрачной
каймой. Анальная пластинка треугольная, сзади тонко зазубренная,
не выдающаяся за основание фуркальных ветвей. Длина последних
вдвое более их ширины, крайние апикальные щетинки почти равной
длины, средние хорошо развитые, длинные, густо оперенные. У ног
IV пары длина дистального членика эндоподита в 1.3 раза более шири-
ны, апикальный шип лишь немного длиннее этого членика, обе внутрен-
ние щетинки (щетинки внутреннего края и апикальная) очень длинные.
Оба членика ног V пары длинные, явственно дифференцированные,
придаток внешнего членика гораздо длиннее 2 придатков внутреннего
членика, из которых внутренний немного длиннее внешнего. Длина
0.52—0.56 мм.
Самец. Длина 0.420 мм. Строение плавательных ног и ног V пары —•
как у самки.
Распространение. Северная Испания, пещерные воды.
10. Speocyclops proserpmae Kief. (фиг. 69g).
Speocyclops proserpinae Kiefer, 1937 :434, t. 9, f. 64—66.
Самка. Генитальный сегмент широкий, без поперечного шва
на спинной поверхности, анальная пластинка немного заходит кзади
за основание фуркальных ветвей, на заднем конце тонко зазубренная.
Длина фуркальных ветвей приблизительно в 1.75 раза превышает
их ширину, крайние апикальные щетинки как у Sp. plutonis, средние
хорошо развиты. Дистальный членик эндоподита ног IV пары как
у Sp. plutonis. Оба членика ног V пары явственно различимы, короткие,
но длиннее, чем у названного вида, вооружение состоит из 3, почти
равной длины, придатков. Длина 0.54—0.56 мм.
Самец. Длина 0.44 мм.
Распространение. Югославия, пещерные воды.
11. Speocyclops orcinus Kief.
Speocyclops orcinus Kiefer, 1937: 436, t. 9, f. 83—85.
Самка не известна.
Самец. Анальная пластинка маленькая, с полукруглым, тонко
зазубренным задним краем, едва заходящая за основание фуркальных
ветвей. Последние очень короткие, их длина менее чем в 1.5 раза пре-
восходит их ширину, из крайних апикальных щетинок внутренняя более
чем вдвое короче внешней, средние апикальные щетинки относительно
очень короткие, из них внешняя лишь немного короче внутренней. Обе
ветви всех пар плавательных ног 2-членистые.1 Внутренний членик ног
V пары явственно дифференцированный, с 2 неравной длины придатками,
1 В описание вооружения дистального членика эндоподита ног IV пары у Kiefer’a
вкралось явное противоречие. Автор, ссылаясь на рис. 84 табл. 9, в диагнозе пишет,
что членик вооружен лишь 1 апикальным шипом, а на рис. 84 изображает 2 апикаль-
ных шипа, из которых 1 почти вдвое длиннее другого.
11. GRAETERIELLA
289
внешний членик очень короткий, в значительной степени слитый с тора-
кальным сегментом, вооруженный длинной щетинкой. Длина 0.40 мм.
Распространение. Франция, пещеры в департаменте Basses
Pyrenees, совместно с Sp. racovitzai (Chapp.).
*12. Speocyclops colchidanus (Borutzky) (фиг. 70).
Cyclops colchidanus Borutzky, 1930:333, f. 3—8. — Speocyclops colchidanus Kiefer,
1937:427, 431.
Самка не известна.
Самец. Генитальный сегмент очень широкий, задние края абдоми-
нальных сегментов тонко зазубрены, анальная пластинка неправильной
треугольной формы, короткая, асимметрично зазубренная, не доходя-
щая до основания фуркальных ветвей. Длина последних менее чем вдвое
превышает их ширину, из крайних апикальных щетинок внутренняя лишь
немного короче внешней, средние апикальные щетинки хорошо развитые,
из них внешняя приблизительно вдвое короче внутренней. Дистальный
членик эндоподита ног IV пары очень короткий, его длина почти равна
ширине, апикальный шип очень мощный, почти вдвое длиннее членика,
апикальная щетинка немного длиннее шипа, 2 прочие (боковые) щетинки
короткие. Ноги V пары описаны недостаточно полно. По Боруцкому,
они одночленисты и снабжены парой придатков. По всем вероятиям,
однако, имеются и следы 2-го членика, а равно и 3-я щетинка (Kiefer,
1937:431). Ноги VI пары, как у прочих видов Speocyclops, с 3 придат-
ками, из которых средний заметно короче обоих крайних. Длина 0.324 мм.
Несмотря на неполноту описания (найден лишь 1 экз.), принадлеж-
ность рассматриваемого вида к роду Speocyclops бесспорна (см. особенно
вооружение ног IV пары).
Распространение. Кавказ, пещера на берегу р. Ухаль-цители
(на Рионе, близ г. Кутаиси).
11. Род GRAETERIELLA BREHM
Graeteriella Brehm, 1926:313 (part.); Kiefer, 1937:429.
Передние антенны 11-членистые, очень короткие, достигающие лишь
в середине цефалоторакса. Членистость плавательных ног самок по фор-
муле 2-2/2-2/2'22-2 или 2-2/3-2/3-3/3-3. Ноги V пары 1-членистые,
с 2 апикальными придатками — внутренним коротким шипом и длинной
(в 2—2 1/2 раза длиннее шипа) оперенной щетинкой: щетинки, соответ-
ствующей совершенно отсутствующему базальному членику, не имеется,
так что задний торакальный сегмент ее вовсе лишен. Receptaculum
seminis очень широкий, поперечно расположенный, в переднем отделе
с прозрачной каемкой, задний отдел не развитой. Мелкие циклопы,
длина самок менее 1 мм, троглобионты. Род представлен исключительно
в Европе.
2 подрода: Graeteriella s. str. и Paragraeteriella.
1. Подрод Graeteriella s. str.
Graeteriella (part.) Brehm, 1926:313; Kiefer, 1937:429.
Резко выраженная гарпактикоидная форма тела. Генитальный сег-
мент очень широкий, его длина значительно менее ширины. Фуркальные
ветви с 3 апикальными придатками, причем из 2 средних щетинок имеется
19 Фауна СССР
290
CYCLOPINAE
только 1. Анальная пластинка очень сильно развитая. Членистость
плавательных ног у самок 2 • 2/2 - 2/2 - 2/2-2, у самцов 2-2/2-2/2-2/3'2.1
1 вид.
Фиг. 71.
Graeteriella (s. str.) unisetigera (E. Graet.): 1 — фурка самки, 2 — фурка
самца, 3—общий вид самки со спинной стороны, 4—5—нога V самки,
б—нога V самца, 7 — абдомен, reeeptaculum seminis и фурка самки с брюш-
ной стороны; Speocyclops troglodytes (Chapp.); 8—reeeptaculum seminis,
9— фурка самки co спинной стороны, 10— нога V самки. (/ — по Е.
Graeter’y, 2—4—по Gurney, 5—7—по Kiefer’y, 8—10 — по Chappuis).
1. Graeteriella (s. str.) unisetigera (E. Graet.) (фиг. 71!_7).
Cyclops unisetiger E. Grae'er, 1908, Zool. Anz., 33 : 49; E. Graeter, 1911:44, textfig,
20, t. 2, f. 7, 8; t. 3, f. 9; Chappuis, 1923, Bull. Soc. Stiint. Cluj, 1:585, t. 2, 6, 11; Kiefer,
1928:526, t. 58—62. — Cyclops unisetiger f. biarticulata Kiefer, 1926, Verb. Geseh. Nat.
Baar, 16:277, f. 8. — C. (Diacyclops) unisetiger Kiefer, 1929:65. — C. (Diacyclops) unisetiger-
f. biarticulata Kiefer 1929:65. — C. (Microcyclops) unisetiger Gurney, 1933:278, f. 1818—
1835. — Graeteriella unisetiger Kiefer, 1937:424, t. 6 f. 1—16.
1 О членистости ног (исправление прежних данных автора) см. Kiefer, 1937:424
(примечание 1).
11. GRAETERIELLA.
291
Самка. Глаз и пигмент его имеются. Тело утолщенное, несколько
сплюснутое в спинно-брюшном направлении, абдомен очень толстый,
границы между передним и задним отделами тела мало выражены. Аналь-
ная пластинка очень крупная, задним краем заходящая за основание
фуркальных ветвей, сзади иногда вооруженная 2—3 короткими зубцами.
Фуркальные ветви сильно расходящиеся, их длина приблизительно
в 1.5—2 раза более ширины, единственная средняя апикальная щетинка
сильно развитая, в переднем отделе утолщенная, латеральная щетинка
короткая, она прикрепляется несколько отступя от внешнего края,
приблизительно против его середины. Из апикальных крайних шипооб-
разных придатков внутренний значительно (в 2—5 раз) короче внешнего.
Дорзальная щетинка хорошо развитая, длиннее внешнего крайнего
апикального придатка. На 8-членике передних антенн имеется длинный
сензорный цилиндр, своим концом доходящий до конца 9-го членика.
Дистальный членик эндоподита ног IV пары с 3 придатками — 1 апи-
кальным шипом и, по бокам его, 2 субапикальными оперенными щетинками.
Дистальный членик экзоподита этой пары ног с 8 придатками —4 шипами
на внешнем крае, 3 оперенными щетинками на конце и 1 оперенной
щетинкой на внутреннем крае. Членик ног V пары очень маленький,
прямоугольный, резко отчлененный от торакального сегмента, оба при-
датка хорошо развитые. Ноги VI пары с парой крайне мелких шипиков,
без оперенных щетинок. Длина 0.4—0.7 мм; бесцветны, очень прозрачны,
яйцевые мешки содержат лишь по 1 яйцу.
Самец. Ширина генитального сегмента почти вдвое превышает его
длину. Ноги VI пары с 2 очень короткими шипиками, без оперенных
щетинок. Передние антенны на 1-м, 4-м, 9-м и 13-м члениках с крупными
сензорными цилиндрами. Длина 0.7—0.8 мм.
Распространение. Европа (Германия, Швейцария, Югославия,
Венгрия, Франция, Бельгия, Северная Испания, Англия).
Биология почти не изучена. Троглобионт, пещерные водоемы,
водопровод и колодцы.
2. Подрод Paragraeteriella, subg. nov.
Graeteriella Kiefer, 1937:424 (part.).
Генитальный сегмент нормального строения, его ширина в переднем
отделе приблизительно равна его длине, а в заднем значительно менее
последней. Фуркальные ветви короткие, их длина немного более чем
вдвое превосходит их ширину, их апикальное вооружение нормальное —
4 придатка. Анальная пластинка умеренно развитая, кзади не заходящая
за основание фуркальных ветвей, не вооруженная. Членистость плава-
тельных ног самки 2-2/3-2/3-3/3-3. 1 вид.
2. Graeteriella (Paragraeteriella) laisi (Kief.).
Diacyclcps laisi Kiefer, 1936, Zool. Anz., 113:85, f. 1—8. —Graeteriella laisi Kiefer,
1937:424, t. 6, f. 17—21; t. 7, f. 22—24.
Самка. Генитальный сегмент укороченный, спереди сильно рас-
ширенный, задние края абдоминальных сегментов не зазубрены, аналь-
ная пластинка с закругленным гладким задним краем, не доходящая
до основания фуркальных ветвей, последние короткие, параллельные,
из крайних апикальных придатков внутренний почти вдвое длиннее внеш-
него, латеральная щетинка прикреплена несколько отступя от внешнего
19*
292
CYCLOPINAE
края (как у Gr. unisetigera), немного кзади от его середины, дорзальная
щетинка немного длиннее внешней из крайних апикальных, из средних
апикальных щетинок внутренняя очень длинная, внешняя немного менее
чем вдвое короче внутренней. Передние антенны 11-членистые. У ног
IV пары дистальный членик эндоподита с 5 придатками —2 апикальными
шипами (из них внутренний приблизительно на 1/3 длиннее внешнего)
и 3 щетинками. Формула шипов 3-3-3»3. Членик ног V пары очень
короткий, резко отчлененный от торакального сегмента, его внутренний
апикальный шип тонкий, приблизительно вдвое короче соседней апи-
кальной щетинки. Длина 0.5 мм. Яйцевые мешки не описаны.
Самец не известен.
Распространение. Пока найден лишь близ Фрейбурга (Баден)
и Страсбурга (Эльзас).
Биология не изучена. Найден только в подземных водах (источ-
ники, колодцы).
12. Род MESOCYCLOPS SARS
Cyclops (part.) Claus, 1863:96; Schmeil, 1892:15; Lilljeborg, 1901:3. — Mesocyclops
Sars, 1914:57; Kiefer, 1927:307; Kiefer, 1928:552; Kiefer, 1929:77; Kiefer, 1929a:3. — Cyclops
(Mesocyclops) Gurney, 1933:286.
Тело большей частью стройное, удлиненное, с резко дифференци-
рованными передним и задним отделами. У некоторых видов задний
торакальный сегмент по бокам со слабым вооружением из группы корот-
ких щетинок. Генитальный сегмент удлиненный, равномерно суживаю-
щийся кзади. Receptaculum seminis молотообразный, передний отдел
из 2 поперечно расположенных удлиненных частей, задний в виде
отвисающего кзади более или менее удлиненного мешка. Фуркальные
ветви относительно короткие, их длина обычно в 2.5—3.5 раза (редко
в 4—5 раз) более их ширины, лишь у немногих видов с рядом волосков
на внутренних краях. Из крепких апикальных щетинок внутренняя обычно
сильно развита. Передние антенны всегда 17-членистые, 2 дистальных
членика вдоль края с узкой гиалиновой пластинкой, иногда зазубренной.
Обе ветви всех 4 пар плавательных ног 3-членистые. Оба членика ног
V пары всегда резко расчленены, дистальный членик вооружен длинной
внешней апикальной щетинкой и длинным щетинкообразным шипом, длина
которого не менее половины длины, а обычно лишь немного менее всей
длины внешней щетинки; этот шип прикреплен или апикально, или
медиально близ конца внутреннего края членика. Мелкие циклопы, длина
самок преимущественно около 1 мм, редко больше (до 1.8 мм). Известно
около 40 видов, несколько вариететов. Планктические виды, ряд видов
в пелагиали озер. Род Космополитен, но отсутствует на арктических
островах. Специально пещерных видов нет. Главным образом в южных
широтах, виды тепловодно-стенотермические.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ
1 (2). Внутренний щетинкообразный шип1 дистального членика ног
V пары прикреплен не апикально, а на внутреннем крае членика .
................................1. Подрод Mesocyclops s. str.
1 Kiefer называет внутренний придаток шипом (Dorn, Staehel), Gurney (1933:36
и 286) — щетинкой или шипом (seta or spine). Наиболее подходящим нам кажется
термин „щетинкообразный шип“; несомненно в данном случае мы имеем измененный
шип, как Можно видеть из положения этого придатка.
12. MESOCYCZOPS
293
2(1). Этот шип прикреплен апикально, около внешней апикальной
щетинки................................2. Подрод Thermocyclops Kief.
1. Подрод Mesocyclops s. str.
Kiefer, 1927:307; Kiefer, 1928:552; Kiefer, 1929a:3; Kiefer, 1929:79; Gurney; 1933:286
(part.).
Характерный признак подрода — положение внутреннего придатка
дистального членика ног V пары. Наиболее крупные виды рода Mesocy-
clops принадлежат к рассматриваемому подроду. К нему относится около
13 видов (и ряд вариететов вида М. leuckarti). Отнесение Kiefer’om
(Zool. Anz., 114, 1936:133) М. tenuis Marsh к подроду Thermocyclops,
судя по рисункам Marsh’a (1909), нам кажется неправильным. Отнесение
Kiefer’oM (1929) М. tenuisacus Sars (1927) в синонимы М. (s. str.) annula-
tus (Woerz.), судя по описанию Sars’a, не внушает сомнений.
Подрод в целом Космополитен, но из видов к космополитам можно
отнести только М. (s. str.) leuckarti, известного из всех зоогеографических
областей. В Палеарктике встречаются только 2 нижеописываемых вида.
В Неарктической области, кроме него, обитает М. (s. str.) tenuis Marsh,
известный и из Южной Африки, в Эфиопской — М. (s. str.) апае Kief.,
в Ориентальной1 — М. (s. str.) tobae Kief. (Ява, Суматра), М. (s. str.)
thermocyclopides Hasada (Формоза) и M. (s. str.) meridianus Kief., который
известен также из Южной Америки. Только Неотропической области
свойственно 4 вида, и эта область наиболее богата видами (всего 7 видов).
Из Австралийской области пока известен лишь М. (s. str.) leuckarti.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Длина фуркальных ветвей в 3—3.5 раза более ширины, щетинко-
образный шип дистального членика ног V пары лишь немного
короче апикальной щетинки . . . . . *1. М. (s. str.) leuckarti Claus
2 (1). Длина фуркальных ветвей почти в 4 раза превышает ширину,
щетинкообразный шип дистального членика ног V пары короткий,
приблизительно вдвое короче апикальной щетинки .....................
.................................2. М. (s. str.) bodanicola Kief.
*1. Mesocyclops (s. str.) leuckarti Claus (фиг. 72, 73^,)
Cyclops leuckarti Claus, 1857:35, t. I, f. 4; t. 2, f. 13—14. — C. simplex Поггенполь,
1874, Изв. Общ. люб. ест., антр. и этн., 10:70, т. 15, рис. 1—3. — С. tenuicornis (non
С. tenuicornis Claus) Ульянин, 1875, ibid., 11:30, т. 9, рис. 12—13. — С. leeuwenhockki
Hoeck, 1878, Tijdskr. Nederl. Dierk. Ver., 3:19, t. 3, f. 1—12. — C. pectinatus Daday, 1884,
Math. Term. Kozl., 19:223, t. 1, f. 713. — C. lucidus Рузский, 1889, Тр. Общ. ест. Казанск.
ун-та, 19 : 27, рис. 7—12. — С. leuckarti Schmeil, 1892:57,- t. 3, f. 1—8. — C. leuckarti
var. australiensis Sars, 1908, Ark. Nat. Christ., 29 : 16, t. 3, f. 19—24. — C. pulchellus
(non C. pulchellus Koch) Byrnes, 1909, Cold Spr. Harb. Mon., 7 : 24, t. 10, f. 1—3. —
Mesocyclops obsoletus (non C. obsoletus Koch) Sars, 1914:58, t. 35. — Mesocyclops major
Sars, 1927:116, t. II, f. 16-—20. — Mesocyclops (s. str.) leuckarti Kiefer, 1929a:4, f. 1—6;
Kiefer, 1929:79, f. 31, a—• b. — C. (Mesocyclops) leuckarti Gurney, 1933:287, f. 1853—1877.
Самка. Туловище относительно широкое, его наибольшая ширина
лежит приблизительно на середине его длины (считая задний торакаль-
ный сегмент). Генитальный сегмент стройный, удлиненный, равномерно
и мало суживающийся кзади, спереди слабо расширенный. Receptaculum
seminis объемистый, с мешковидным задним отделом. Задние края абдо-
1 М. iranicus Lindberg- (1936), возможно, идентичен с М. leuckarti var. pilosa Kief.
294
CYCLOPINAE
минальных сегментов не зазубрены. Фуркальные ветви очень мало рас-
ходящиеся, иногда почти параллельные, их длина в 3—3.5 раза (у var.
aequatorialis Kief, лишь в 2.5—2.75 раза) превышает их ширину, внутрен-
Фиг. 72. Mesocyclops (s. str.) leuckarti (Claus), самка.
7—2—фурка co спинной стороны; 3—генитальный сегмент с recep-
taculum seminis; 4 — нога IV; 5—дистальный членик I.
ние края без волосков (они имеются у var. pilosa Kief.), латеральная
щетинка длинная, прикрепляющаяся приблизительно на середине внеш-
него края ветви, дорзальная щетинка короткая, короче внешней из край-
них апикальных. Внутренняя крайняя апикальная щетинка обычно немного
12. MESOCYCLOPS
295
более чем вдвое превышает длину внешней крайней, средние апикальные
щетинки хорошо развитые, длинные. Передние антенны обычно доходя-
щие до заднего конца 2-го торакального сегмента или немного длиннее.
Оба дистальных членика с хорошо развитой гиалиновой пластинкой,
Фиг. 73.
Mesocyclops (s. str.) leuckarti (Claus): 1 — дистальный членик антенн I самки,
2 — нога самки; М. leuckarti var. pilosus Kief.: 3—дистальный членик
антенны I самки, 4 — дистальный членик эндоподита ногн IV самки, 5 — фурка
самки с брюшной стороны.
(3—5— рисунки С. С. Смирнова по экземплярам из Бухары).
пластинка 17-го членика с глубокой вырезкой и нередко с тонко зазуб-
ренным краем1 (у var. edax Forb, пластинка грубо зазубрена), иногда
пластинка снабжена еще 1—3 глубокими вырезами. Задние антенны
стройные, короткие. Плавательные ноги всех 4 пар с 3-членистыми
обеими ветвями. У ног IV пары длина дистального членика эндоподита
в 3—3.75 раза (обычно в 3—3.2, у var. aequatoria/is Kief, лишь
1 Такая зазубренность иногда имеется и у пластинки 1б-го членика.
296
CYCLOPINAE
в 2.5—2.75 раза) более ширины, дистальные шипы этого членика хорошо
развитые, из них внешний немного длиннее или короче внутреннего
(у v. aequatorialis Kief, разница в длине шипов более значительна).
Соединительная пластинка ног IV пары сзади по бокам с более или
менее крупным шипиком. Ноги V пары крупные; внутренний щетинкооб-
разный шип дистального членика длинный, лишь немного короче апи-
кальной щетинки; щетинка базального членика относительно короткая,
доходящая приблизительно до середины апикальной щетинки. Длина
0.9—1.3 мм. Обычно сероватая окраска, иногда самки бесцветны и про-
зрачны. Яйцевые мешки обычно крупные, расходящиеся, с очень различ-
ным количеством яиц (от 4 до 30 в каждом).
Самец. Передние антенны с длинными сензорными цилиндрами
на 1-м и 4-м члениках и с мелким сензорным придатком на 9-м членике.
Ноги VI пары с коротким внутренним шипом и с 2 длинными щетинками,
из которых внешняя несколько длиннее средней. Длина 0.8—1.0 мм.
Mesocyclops leuckarti—вид значительно вариирующий. Kiefer (1929а)
подразделил его на подвиды и описал несколько „форм". Их таксоно-
мическое положение не вполне ясно; с большой степенью вероятности
действительно подвидами (subspecies) можно считать М. (s. str.) leuckarti
aequatorialis Kief. (1929a), Экваториальная Африка, Индия, Малайский
Архипелаг, М. (s. str.) leuckarti edax Forbes, Rep. H. St. Fish. Comm.,
1891, пелагиаль озер Северной Америки.
М. (s. str.) leuckarti deccanensis Lindberg (Rec. Ind. Mus., 1935),
описанный из Индии (Декан), возможно, является самостоятельным
видом, М. (s. str.) leuckarti f. pilosa Kief., возможно, является подвидом;
ниже мы рассматриваем эту форму как вариетет, таксономическое зна-
чение которого пока неясно.
Распространение. Космополит в широком смысле слова.
В СССР широко распространен. Встречается от крайнего севера
(Мурманское побережье, Соловецкие острова) до широты южного Кавказа
включительно. В азиатской части СССР известен от севера Сибири
(бассейн р. Хатанги, устье р. Оби, Якутская АССР) до широты Самар-
канда, на востоке — до Сахалина и бассейна р. Амура включительно.
На арктических островах отсутствует, в зоне тундры гораздо реже, чем
южнее. Неоднократно указывался для планктона горных озер, где, однако,
не найден выше 1240 м над уровнем моря.
Биология. Эвпланктический виде обитает в самых различных
водоемах, от мелких луж и болот1 до крупнейших озер включительно.
В озерах является характерным компонентом пелагического планктона,
но обитает и в условиях литорали. Гораздо реже в мелких водоемах;
особенно редко в астатических; обычен в планктоне рек, лишь в виде
редкого исключения в ключах (Klie, 1925). Обычный компонент прудо-
вого планктона. В пелагиали озер обитает в зоне эпилимниона, суточные
вертикальные миграции интенсивны. В некоторых глубоких озерах их
амплитуда доходит до 40 м. Судя по ряду данных, в некоторых водое-
мах достигает значительного количественного развития и в прибрежной
зоне. Троглоксен в подземных водах встречается очень редко и слу-
чайно. Тепловодный рачок. По исследованиям Roy (1932), при 15—20°
размножается особенно интенсивно, температура в 4° оказывает явно
неблагоприятное влияние и вызывает гибель науплиусов. Вместе с тем
1 Так Thiebaud (1906) в окр. Невшателя. находил этот вид в маленьком болотце,
глубиной всего около 0.5 м.
12. MESOCYCLOPS
297
прекрасно развивается и при температуре выше 25°. Так, на рисовых
полях и в их оросителях близ Самарканда (Пажитнова, 1935) является
обычнейшим и постоянным компонентом планктона при 34—37°. Кисло-
родный оптимум, вероятно, лежит в пределах, близких к нормальному
насыщению. Обычен в водах pH =7.2—9.8, отсутствует в озерах
с pH =4.2—5.6.1 Скадовский (1926) показал, что при pH =3 яйца М. leuc-
karti могут развиваться нормально, но науплиусы быстро погибают при
концентрациях водородных ионов <4.5. На основании имеющихся
данных можно заключить, что М. leuckarti оптимальные условия находит
при pH = 6—8.5 (быть может и в несколько более щелочной среде)
и что критическим интервалом для него является pH = 4 — 5, а летальный
исход наступает при pH около 4. В сильно дистрофированных водах
не встречается, характерен для гумус-олиготрофных и гумус-мезотрофных
водоемов. Обитает в Са-олиготрофных, до Са-мезотрофных вод вклю-
чительно, в Са-политрофных водах, повидимому, встречается гораздо
реже и значительного количественного развития не достигает. В значи-
тельной степени эвригалинен. Найден в солоноватых водах восточной:
части Финского залива, в Ботническом заливе. В континентальных соло-
новатых водоемах СССР отмечался неоднократно.1 2 Согласно экспери-
ментальным исследованиям Таусон (1934), вообще очень вынослив отно-
сительно содержания остаточного С1. По нашим наблюдениям (Рылов,
1935), в богатых натрием щелочных озерах центральной части Якутской
АССР М. leuckarti обитает в больших количествах в 7 озерах при общей
минерализации от 0.39 до 1.77 г на литр и содержании НСО3-ьСО3
от 266.16 до 1094 мг на литр; для натронных озер Венгрии (Alfoldes)
давно уже указан Daday’eM (1893). Цикл размножения изучен с доста-
точной полнотой. По данным почти всех авторов, М. leuckarti встречается
в теплое время года, выпадая на зиму; это отмечалось и для мелких
постоянных водоемов, где рачок полицикличен. В озерах установлена,
дициклия, но, по всем вероятиям, в некоторых озерах и более 2 половых
периодов в год. Периоды размножения падают на весну, лето и осень..
Лишь редко М. leuckarti был находим в Европе в озерах и зимой как
форма постоянная, встречающаяся круглый год.
В шкале Kolkwitz’a и Marsson’a (1909) М. leuckarti показан как
р- мезосапроб, что подтверждается данными Зиверта (1929). Однако, хотя?
и сравнительно редко, он указывался и для олигосапровных условий
(например пелагиаль Боденского озера).
*la. Mesocyclops (s. str.) leuckarti Claus var. pilosus Kief. (фиг. 733_s).
Mesocyclops leuckarti f. pilosus Kiefer, 1930, Zool. Anz., 87:45.
От типичной формы отличается присутствием более или менее
густо расположенных волосков на внутренних краях фуркальных ветвей.
Распространение. Мадагаскар (Тананариво). В СССР найден
Смирновым в Бухаре (мелкие водоемы, май, 1929).
2. Mesocyclops (s. str.) bodanicola Kief. (фиг. 74).
Mesocyclops leuckarti bodanicola Kiefer, 1929a:17, f. 7—10; Kiefer, 1929:30.— M. boda‘
nicola Kiefer, 1938, Zool. Anz. 124:150, f. lb—5b.
1 Для Ленинградской области (Сяберские озера) есть указание на нахождение
при pH =4.6—5.2 (Са-олиготрофные и гумус-олиготрофные озера!) и на отсутствие его-
при pH —4 (оз. Ершовичи). Данные эти, возможно, нуждаются в проверке.
2 В Аральском море обширно распространен, представляя один из важнейших.,
компонентов планктона пелагиали.
298
CYCLOPINAE
От M. leuckarti самка отличается следующими признаками. Тело
прозрачное и бесцветное, более стройное и удлиненное. Длина фуркаль-
ных ветвей почти в 4 раза более их ширины, дорзальная щетинка
длинная, достигающая или почти достигающая конца внешней апи-
кальной щетинки. Вырезка на гиалиновой пластинке 17-го членика перед-
них антенн неглубокая, очень маленькая, иногда ее вовсе нет. Длина
дистального членика эндоподита ног IV пары приблизительно в 3.75 раза
превышает его ширину, на конце этот членик вместо внутреннего шипа
иногда несет оперенную щетинку. Внутренний щетинкообразный шип
.дистального членика ног V пары вдвое короче апикальной щетинки
этого членика, щетинка базального членика заметно длиннее, чем
Фиг. 74. Mesocyclops (s. str.) bodanicola Kief.,
самка. (По Kiefer’y).
/ — фурка co спинной стороны; 2—дистальный
членик эндоподита ноги IV; 3—йога V; 4—два
дистальных членика антенны I.
у М. leuckarti. В яйцевых мешках небольшое количество яиц (по Wolf’y,
1905, лишь по 4—5 яиц в каждом).
Распространение. Боденское оз., Neuenburger и Bielersee
в Швейцарии в пелагиали и литорали; нахождение в озерах СССР вероятно.
2. Подрод Thermocyclops Kief.
Mesocyclops (Thermocyclops) Kiefer, 1927:308; Kiefer, 1938:552, Kiefer, 1929:82.
— Cyclops (Mesocyclops) Gurney, 1933:286 (part.). — Thermocyclops Kiefer, 1937a:479.
Подрод характеризуется апикальным положением щетинкообразного
шипа дистального членика ног V пары. Мелкие формы, длина самок
не более 1—1.1 мм, обычно менее 1 мм (самка южноамериканского
М. retroversus Kief, лишь 0,6 мм). У большинства видов длина фуркаль-
ных ветвей лишь в 2.5 —3 раза превышает ширину, самое большее
в 4 раза (южноамериканский М. brehemi Kief.) Известно около 30 видов.
„Cyclops philippinensis“ Marsh (Tr. Wise. Acad., 1932) должен быть
«отнесен к этому подроду; положение в роде Mesocyclops описанного
12. MESOCYCLOPS
299
из Бразилии Th. inversus Kiefer (Zool. Anz., 114, 1936) остается невыяс-
ненным (нет описания ног V пары), М. (Th.) analogus Kiefer (Zool.
Anz., 113, 1936, Индия) очень близок к М. (Th.) rylovi Smirn.,
М. (Th.) decipiens Kief. (Zool. Anz., 80, 1929) едва ли отличим от
М. (Th.) crassus (Fisch.) и нами относится к видам сомнительным,
к которым принадлежит и М. (Th.) potamius Burckh. (Китай). В последнее
время Kiefer’oM описан целый ряд видов подрода, очень близких
к М. (Th.) crassus. Полная ревизия всего подрода весьма желательна,
так как отношения между рядом описанных форм не ясны и возможно,
что, вопреки мнению Kiefer’a (1937а:479), некоторые из описанных им
видов придется выделить в единицы низшей таксономической категории.
Географическое распространение целого ряда видов поэтому нельзя
считать выясненным. Подрод в целом Космополитен. Наиболее обшир-
ное распространение имеет М. (Th.) crassus, известный из всех обла-
стей, исключая Нёарктическую, где пока подрод вообще не констати-
рован. В пределах Палеарктики, помимо этого вида, обитает М. (Th.)
oithonoides (Sars), М. (Th.) dybowskii (Lande), M. (Th.) asiaticus Kief.,
M. (Th.) rylovi Smirn. и M. (Th.) mongolicus Kief, (восточная Монголия).1
Из Неотропической области, помимо М. (Th.) crassus, известны 2 вида,
из Ориентальной—7 видов. Чрезвычайно обильно подрод представ-
лен в Эфиопской области, только из ее пределов известно 12 видов
(всего 14 или 15), частью очень близких к М. (Th.) crassus. Из Авст-
ралийской области (Новые Гебриды) пока известен только М. (Th.) crassus
(Fisch.).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Из апикальных щетинок фуркальных ветвей средняя внешняя почти
втрое длиннее внутренней крайней. Внутренний апикальный шип
дистального членика эндоподита ног IV пары короче внешнего .
... •..........................*7. М. (Th.) dybowskii (Lande)
2(1). Из апикальных щетинок фуркальных ветвей средняя внешняя
не более чем вдвое превышает длину внутренней крайней. Внут-
ренний апикальный шип дистального членика эндоподита ног
IV пары длиннее внешнего.
3(6). Внутренний апикальный шип дистального членика эндоподита ног
IV пары в 4—5 раз длиннее внешнего.
4 (5). Латеральная щетинка фуркальных ветвей прикреплена приблизи-
тельно на середине внешнего края. Боковые выросты переднего
отдела receptaculum seminis очень мало загнутые кзади или прямые,
расположенные почти перпендикулярно к продольной оси гени-
тального сегмента......................*3. М. (Th.) oithonoides Sars
5(4). Латеральная щетинка фуркальных ветвей прикреплена значительно
отступя кзади от середины их внешнего края. Боковые выросты
переднего отдела receptaculum seminis очень сильно загнутые
кзади..................................*4. М. (Th.) asiaticus Kief.
6(3). Внутренний апикальный шип дистального членика эндоподита ног
IV пары не более чем в 2—3 раза длиннее внешнего.
7(9). Из крайних апикальных щетинок фуркальных ветвей внутренняя
не более чем в 2—2.5 раза превышает длину внешней и лишь
1 Самостоятельность этого вида для меня очень сомнительна. По всем вероятиям
он идентичен с М. (Th.) asiaticus Kief. М. (Th.) tinctus Lindberg (1936; Иран), no нашему
мнению, требует повторного описания.
300
CYCLOPINAE
немного более половины длины внешней из средних апикальных
щетинок. Задний край выростов соединительной пластинки ног
IV пары вооружен мелкими тонкими шипиками...................
....................................*6. М. (Th.) rylovi Smirn.
8 (7). Из крайних апикальных щетинок фуркальных ветвей внутренняя
в 3—Зт/3 раза длиннее внешней и лишь немного короче внешней
из средних апикальных щетинок. Задний край выростов соедини*-
тельной пластинки ног IV пары вооружен грубыми толстыми шипи-
ками..................................*5. М. (Th.) crassus (Fisch.)
*3. Mesocyclops (Thermocyclops) oithonoides Sars (фиг. 75).
Cyclops oithonoides Sars, 1863, Forh. Selsk. Christ., (1862):21; Schmeil, 1892:64, t. 4,
f. 6—11.—-Mesocyclops oithonoides Sars 1914:59, t. 36. — Mesocyclops {Thermocyclops}
oithonoides Kiefer, 1929:82, f. 35, — Cyclops (Mesocyclops) oithonoides Gurney, 1933:300.,
f. 1897—1900.
Самка. Тело очень стройное, удлиненное, длинацефалоторакса плюс
длина торакса несколько более чем вдвое превышает их наибольшую ши-
рину, которая лежит немного кпереди от середины длины. Абдомен строй-
ный, относительно тонкий. Генитальный сегмент весьма удлиненный, посте-
пенно суживающийся кзади, его длина приблизительно в 1г/2 раза
(или несколько более) превышает его наибольшую ширину. Receptaculum.
seminis с боковыми выростами переднего отдела, лишь незначительно
загнутыми кзади, иногда почти прямыми, задний отдел объемистый, очень
удлиненный. Фуркальные ветви обычно значительно расходящиеся, строй-
ные, удлиненные, их длина приблизительно в 31/2 раза более их ширины,
латеральная щетинка прикреплена на середине внешнего края, хорошо
развитая, из крайних апикальных щетинок внутренняя в Зх/2 раза длиннее
внешней и почти равна 3/5 длины внутренней из средних апикальных
щетинок; внешняя средняя апикальная щетинка лишь немного длиннее
внутренней крайней, дорзальная щетинка приблизительно вдвое длиннее
внешней крайней апикальной щетинки. Передние антенны достигают
конца 3-го или середины 4-го торакального сегмента, оба дисталь-
ных членика с очень узкой цельнокрайней гиалиновой пластинкой вдоль
внутреннего края. Соединительная пластинка ног IV пары сзади по бокам
с крупными закругленными выростами, на свободном (заднем) крае воору-
женными очень мелкими волосообразными шипиками, число которых
равно 5—6 или немного более. Дистальный членик эндоподита ног IV пары
удлиненный, его длина приблизительно в 3J/4 —31/2 раза более его ширины,
внутренний апикальный шип этого членика очень длинный, стройный,,
немного изогнутый, в 4—5 раз длиннее внешнего апикального шипа.
Ноги V пары с удлиненным дистальным члеником, его шипообразная
внутренняя щетинка прикреплена очень близко от конца внутреннего
края, почти одинаковой длины с внешней апикальной щетинкой этого
членика; обе щетинки хорошо развитые, длинные. Длина 0.9—1.0 мм.
Рачок очень прозрачен, нередко бесцветен или окрашен в нежный
желтоватый или голубоватый цвет. Яйцевые мешки крупные, прилегают
щие к генитальному сегменту, содержащие очень небольшое (нередко
только по 4—5 в каждом) количество яиц.
Самец. Тело более удлиненное, чем у самки, апикальные щетинки
фуркальных ветвей немного короче, чем у последней, длина 0.6—0.7 мм.
Распространение. Вследствие частого смешивания этого вида
с близким к нему М. (Th.) crassus (Fisch.) (— С. hyallnus auct.), ряд
указаний в литературе на нахождение М. (Th.) oithonoides сомнителен.
12. MESOCYCLOPS
301
С достоверностью этот вид показан для Европы и Азии. В пределах
СССР1 он известен во многих местностях, от северного побережья
Фиг. 75. Mesocyclops (Th.)' oithonoides (Sars), самка.
И— фурка eo спинной стороны; 2—абдомен и receptaculum seminis; 3—нога V;
4— соединительные пластинки ног IV; 5—дистальный членик эндоподита ноги IV.
Белого моря до окрестностей Одессы включительно; повидимому широко
распространен в азиатской части СССР (окрестности Новосибирска,
Акмолинска, Сыр-дарья и Аму-дарья, Дальний Восток).
1 Целый ряд данных в русской литературе должен быть отброшен. Здесь мы
приводим местонахождения, не возбуждающие сомнений, тщательно проверенные или
на основании личных исследований, или по литературным данным. Указания тех авторов,
"которые в своих работах различают М. (Th.) oithonoides и М. (Th.) crassus (— hyalinus)
мы считаем за достоверные.
302
CYCLOPINAE
Отсутствует в зоне тундры и обширно распространен в зоне тайги,,
смешанных лесов и лесостепи, встречаясь, однако, также в бассейнах рек
степной, полупустынной1 и пустынной зоны. На арктических островах
отсутствует. В Западной Европе в Альпах указан до высоты не свыше
500 м над уровнем моря, в СССР до высоты около 1000 м (Байкаль-
ский хребет); в состав планктона высокогорных озер Кавказа* 2 и азиат-
ской части СССР не входит.
Биология. Чрезвычайно характерный компонент озерного планк-
тона, главным образом пелагиали; в условиях литорали также встре-
чается, но, невидимому, не достигает столь высокого, как в пелагиали,
количественного развития. Обитает также в планктоне чистых (в част-
ности проточных) водоемов прудового типа. Гораздо реже встречается
в более мелких (главным образом в пойменных) водоемах. Обычен
в планктоне рек, куда проникает из связанных с ними озер и поймен-
ных водоемов озерного типа. В пелагиали озер обитает в пределах
эпилимниона, в границах которого имеют место вертикальные суточные-
миграции с незначительной амплитудой. В подземных водоемах не встре-
чается. Как показывает цикл размножения и в частности образование
максимумов в теплое время года, рассматриваемый вид является в доста-
точной степени выраженным тепловодным рачком. Voigt (1905) в Б. Плён-
ском озере наблюдал,3 что в ноябре и декабре, после периода полной
циркуляции, рачок скопляется в глубоких, более теплых слоях воды,,
тогда как летом он держится исключительно в верхних слоях эпилим-
ниона. Повидимому оптимальные условия рачок находит при высоких
содержаниях растворенного кислорода. Судя по литературным данным,,
он обитает при pH = 6.4—7.6, главным образом при pH = 6.8—7.3.
Нами исследованы материалы по М. (Th.) oithonoides из 40 озер,
откуда имеются данные по pH (озера Ленобласти, Свердловской обл.,.
побережья Белого моря, с Кольского полуострова, озера низовьев
Амура и др.), причем границы pH определились в 6.9—7.6. В сколь-
либо значительно дистрофированных водоемах отсутствует, обитая, глав-
ным образом, в эвтрофных (мезотрофных) и в близких к олиготрофным
озерах. Повидимому обитает лишь при олиготипе и мезотипе кальция.
Типично пресноводный вид.4 5 Цикл развития еще не изучен достаточно
полно. Судя по ряду данных, в некоторых озерах встречается в планк-
тоне весь год. Вместе с тем почти все авторы отмечают, что зимой
рачок встречается лишь в единичных экземплярах и что максимум его
количественного развития падает на теплое время года, причем рачок
дицикличен — максимумы летом и в конце осени; однако в некоторых
озерах он моноцикличен. В мелких водоемах (пруды, старицы и проч.)
держится почти везде только в теплое время года, выпадая на зиму;
повидимому это имеет место и в некоторых неглубоких озерах. Voigt
(1905) упоминает, что в Б. Плёнском озере М. (Th.) oithonoides в сентябре
образует темноокрашенные покоящиеся яйца;® это наблюдение пока
не нашло подтверждения.
• 1 Указания для Аральского моря (Виркетис, 1927; Бенинг, 1933, 1934) ошибочны;-
в материалах из различных частей этого моря (сборы А. Л. Бенинга) я нашел исклю-
чительно М. (Th.) crassus (Fisch.).
2 На Кавказе и в Крыму вообще не найден.
3 Эти наблюдения, к сожалению, не проверены.
4 Редкие данные о нахождении в солоноватых водах надо отнести не к этому
виду, а к М. (Th.) crassus (см. ниже), с которым он часто смешивался.
5 Они не описаны.
12. MESOCYCLOP S
303
В шкале сапробов Kolliwitz’a и Marsson’a рассматриваемый вид.
помечен олигосапробом. Едва ли, однако, подлежит сомнениям, что
он обитает и в р-мезосапробных условиях, исключая высшие степени
последних.
*4. Mesocyclops (Thermocyclops) asiaticus Kief. (фиг. 76).
Mesocyclops oithonoides Smirnov, 1929 в: 318. — Mesocyclops (Thermocyclops\
asiaticus Kiefer, 1932в: 234, f. 1—4; Рылов, 1936:154, рис. V, 1—4, рис VI.— Th. топ-
golicus Kiefer, 1937 c : 297, f. 11—-15.
Самка. Тело относительно утолщенное, похожее на таковое
М. (Th.) crassus. Генитальный сегмент равномерно суживающийся кзади,,
его длина приблизительно на х/5 более ширины его переднего отдела..
Reeeptaculum seminis с очень сильно загнутыми кзади боковыми выро-
стами переднего отдела. Задний абдоминальный сегмент приблизительно-
вдвое короче предыдущего сегмента. Фуркальные ветви стройные*,
расходящиеся под значительным углом, их длина приблизительно
в 3 раза превышает ширину, из крайних апикальных щетинок внутрен-
няя Приблизительно в 21/2 раза длиннее внешней, средние апикальные
щетинки хорошо развитые, из них внешняя приблизительно на '/4 длин-
нее крайней внутренней. Все апикальные щетинки очень густо оперены и,
исключая крайнюю внешнюю, в заднем отделе обычно сильно изогнуты —
средние внутрь, крайняя внутренняя — в противоположном направлении.
Дорзальные щетинки необычайно длинные, густо оперенные, их длина
равняется половине длины внешней из средних щетинок; иногда дор-
зальная щетинка еще более длинная. Латеральная щетинка фуркальных
ветвей прикрепляется значительно кзади от середины их внешнего
края. Передние антенны почти достигают до заднего края 3-го тора-
кального сегмента, на дистальном конце 12-гО членика имеется очень,
крупный копьевидный сензорный придаток. Длина дистального членика
эндоподита ног IV пары приблизительно в 3 -Зг/3 раза более его ширины,
внутренний из его апикальных шипов очень длинный, в 4—4.5 раза
длиннее внешнего. Соединительная пластинка ног IV пары по бокам,
с довольно крупными, сильно выступающими кзади выростами, на сво-
бодном крае вооруженными очень тонкими шипиками. У ног V пары
придатки стройного и удлиненного дистального членика почти одина-
ковой длины. Длина 0.84—1.0 мм. Яйцевые мешки округлые, с очень-
различными (от 4 до 20 в каждом) количеством яиц; обычно их 14—20.
При значительном содержании яиц мешки очень значительно удлиняются.
Окраска не известна.
Самец. Фуркальные ветви несколько короче, чем у самки,
ноги VI пары с 3 придатками, из них внешний щетинкообразный,
в 2х/2—3 раза превышает длину каждого из 2 других придатков, которые
почти одинаковой длины. Длина 0.68—0.70 мм.
Рассматриваемый вид близок к М. (Th.) oithonoides, но хорошо
отличим от него, главным образом, по строению reeeptaculum seminis.
(у oithonoides боковые выросты его переднего отдела никогда не заги-
баются столь сильно кзади), по вооружению фуркальных ветвей, по более
грубому утолщенному телу, а также и по апикальному вооружению
дистального членика эндоподита IV пары ног.
Распространение. Известен из окрестностей Мукдена (Kie-
fer, 1932 Ь), из притока р. Суйфуна в Уссурийском крае (Smir-
nov, 1929 b = 717. oithonoides). По нашим исследованиям (Рылов, 1936)
304
CYCLOPINAE
очень распространен в озерах поймы нижнего течения р. Амура, где
встречается совместно с М. {Th.) crassus, но, повидимому, викарен
относительно М. {Th.) oithonoides.
Фиг. 76. Mesocyclops {Th.) asiaticus Kief., самка.
-7 — фурка co спинной стороны; 2—йога V; 3—дистальный членик эндоподита ноги IV;
'4 — генитальный сегмент с receptaculum seminis; 5—соединительная пластинка ног IV;
б—11-й — 13-й членики антенны I.
Биология не изучена. Вид планктический, в некоторых озерах
поймы низовьев Амура летом развивающийся в очень большом коли-
честве. Kiefer (1932) нашел этот вид в пробе из мелкого водоема
‘(ноябрь).
12. MESOCYCLOPS
305
* 5. Mesocyclops (Thermocyclops) crassus (Fisch.) (фиг. 77).
Cyclops crassus Fischer, 1353:92, t. 3, f. 14—16 (non f. 29). — C. hyalinus Reh"
berg', 1880, Abh. Nat. Ver. Bremen, 6:542, t. 6, f. 1—2. — C. cabanensis Рузский, 1889,
Тр. Общ. ест. Казанск. унив., 19 : 29, рис. 1—6.- С. oithonoides var. hyalina Schmeil, 1892 : 68,
t. 4, f. 12—14. — C. hyalinus Lilljeborg, 1901 : 40, t. 3, f. 4—-7. — Mesocyclops crassus
Sars, 1914:61, t. 37. — C. crassus Рылов, 1923:75, т. 2, рис. 21—22.—Mesocyclops
{Thermocyclops} hyalinus Kiefer, 1929:83, i. 36, a — c. — M. (Th.} crassus Рылов, 1933:263,
т. 5, рис. 7—8; т. 6, рис. 2. — C. (Mesocyclops) hyalinus Gurney, 1933 : 295, f. 1880—1896. —
M. (Th.) hyalinus Rylov, 1935b: 212, t. 30, f. 324b. — M. (Th.) crassus Рылов, 1936:156,
рис. V, 2—3.
Самка. Тело более утолщенное и короткое, чем у М. (Th.) oitho-
noides', наибольшая ширина торакса лежит приблизительно на середине
длины переднего отдела тела, длина которого почти вдвое более его
ширины. Генитальный сегмент более короткий и широкий, чем у М. (Th.)
oithonoides, его длина несколько менее чем в 2г/2 раза превышает его
ширину. Receptaculum seminis с почти прямыми или лишь мало загну-
тыми кзади боковыми выростами переднего отдела. Фуркальные ветви
заметно расходящиеся, укороченные, их длина в 21/2 раза более
их ширины. Внутренняя из крайних апикальных щетинок приблизительно
в 3—Зг/2 раза длиннее внешней крайней и приблизительно на короче
внешней из средних апикальных щетинок. Латеральная щетинка при-
креплена немного кзади от середины внешнего края фуркальных ветвей,
их дорзальная щетинка приблизительно вдвое длиннее внешней крайней
апикальной щетинки. Передние антенны достигают не более как до зад-
него конца 2-го торакального сегмента, т. е. значительно короче, чем
у М. (Th.), oithonoides', оба дистальные членики с узкой цельнокрайней
гиалиновой пластинкой вдоль края. Соединительная пластинка IV пары
сзади по бокам с крупными выростами, на свободном закругленном
крае несущими по 3—5 сильных, довольно толстых и крупных шипиков.
Длина дистального членика эндоподита ног IV пары приблизительно
в 2г/2—3 раза превышает его ширину, внутренний из апикальных шипов
этого членика гораздо короче, чем у М. (Th.) oithonoides, и обычно
лишь в 2У2—3 раза (иногда только в 2 раза) превышает длину внешнего
шипа. Ноги V пары, как у М. (Th.) oithonoides. Длина 0.8—1.0 мм.
Окраска желтоватая, до желто-коричневой включительно, иногда с голу-
быми включениями (особенно в фуркальных ветвях); обычно самки очень
прозрачны. Яйцевые мешки крупнее, чем у М. (Th.) oithonoides; обычно
со значительным (самое меньшее по 6 в каждом) количеством яиц,
в основании прилегающие к абдомену.
Самец. У передних антенн нет сензорных придатков на 1-м
и 13-м члениках, у 4 го и 9-го члеников эти придатки очень маленькие.
У ног VI пары внешняя щетинка очень длинная, более чем вдвое длин-
нее внутреннего шипа, средняя щетинка короче последнего. Длина
около 0.6. мм.
Sars (1914) определенно высказался за идентичность1 „С. crassus"
Fischer’a с С. oithonoides var. hyalina Schmeil и других авторов. Ранее
я уже указывал (Рылов, 1923, 1933) на правильность этого мнения,
вопреки Kiefer’y и Gurney, которые не считают возможным вообще
отождествлять С. crassus, описанного Фишером, с каким-либо из видов
рода Mesocyclops. На путаницу в синонимике вида, приведенной Kie-
1 Sars, вместе с тем, полагал, что описанный Rehberg’om (1880) С. hyalinus иден-
тичен е С. (Th.) oithonoides, но не идентичен с С. oith. var. hyalinus, описанным
в монографии Schmeil’a. Ошибочность этого мнения показана Kiefer’oM и Gurney’eM.
306
CYCLOPINAE
fer’oM (1929), я уже указывал ранее (1933). Gurney (1933:298) относит
С. crassus Фишера в группу species incertae sedis, в сущности, на том
только основании, что разными авторами эта форма отождествлялась
с различными видами, — Schmeil (1882) отождествлял ее с С. {Acanth.)
viridis Jur., Lande (1890) — c Mes. (s. str.) leuckarti, Lilljeborg (1901)—
4
Фиг. 77. Mesocyclops (Th.) crassus (Fisch.), самка.
1—фурка с брюшной стороны; 2—абдомен с receptaculum
seminis; 3—еоедявптехьъая пластинка ног IV; 4 — дистальный
членик эндоподита ноги IV.
с М. (77г.) dybowskii. Г рубая ошибка в сближении С. crassus с первыми
двумя видами становится очевидной, если внимательно прочесть диагноз
Фишера (1853) и тщательно изучить его рисунки. Вообще С. crassus
этого автора можно было бы отождествлять с 2 видами — М. {Th.) hya-
linus (Rehb.) и M. {Th.) dybowskii (Lande), a все прочие сближения
здесь безусловно отпадают. Вместе с тем рис. 14, 15, 16 Фишера
(рачок in toto, фуркальные ветви) совершенно определенно говорят
за то, что Фишер описал именно М. {Th.) hyalinus; особенно следует
отметить, что соотношение длины апикальных щетинок фуркальных
12. MESOCYCLOPS
307
ветвей этим автором вполне точно и совершенно одинаково изобра-
жается на 2 рисунках (рис. 14 и 15). Единственное сомнение мог бы
вызывать рис. 29 (ноги IV пары), где апикальные шипы эндоподита
очень близки к таковым М. (Th.) dybowskii. Весьма возможно, что
на этом рисунке Фишер ошибочно нарисовал ногу какого-либо другого
вида; во всяком случае диагноз этого автора и его рис. 14—16 могут
относиться исключительно к М. (Th.) hyalinus Rehberg’a. Cyclops caba-
nensis Рузского (1889) в литературе под знаком вопроса ставится
в синонимы С. hyalinus Rehb. (см., например, Kiefer, 1929). Русский
диагноз цитируемого автора, очевидно недоступный иностранным спе-
циалистам, однако едва ли может оставить сомнение в том, что С. саЬа-
nensis идентичен с М. (Th.) crassus (— С. hyalinus Rehb.). Особенно
существенно, что Рузский сам отмечает отличия описываемого им циклопа
от М. (Th.) oithonoides, выдвигая как раз те признаки, которые харак-
терны для С. hyalinus Rehb. С другой стороны, прекрасно исполненный
в красках рис. 7 Рузского не может относиться ни к oithonoides,
ни к dybowskii (а тем более к leuckartii), вполне соответствуя С. (Th.) cras-
sus (см. особенно соотношение длины и ширины переднего отдела
тела). На идентичность С. cabanensis с С. hyalinus Rehb. уже ранее
указывал Matiie (1897), что, к сожалению, не было принято во внимание
всеми прочими авторами.
Распространение.1 Европа, Азия (включая Японию и остро-
ва Малайского архипелага), Африка, Центральная Америка, Филиппины,
Новые Гебриды. Отсутствует на крайнем севере Европы и Азии.
В СССР с достоверностью известен из целого ряда местностей,
от центральной части Карело-Финской ССР (окрестности Пудожа)
и средней части Кольского полуострова до крайнего юга и юго-востока
включительно. На Кавказе с достоверностью не указан (? Richard, 1895,
Дагестан), в азиатской части СССР, повидимому. широко распространен
(низовья Иртыша, центральная часть Якутской АССР, Казахстан, оз. Бал-
хаш, Аральское море, р. Сыр дарья, р. Сары-су).
Биология. Очень характерен для пелагического планктона эвтроф-
ных и мезотрофных озер; отсутствует в водоемах высокой дистрофности
(полигумозных). Обитает также в прудовом планктоне, гораздо реже
встречается в очень мелких водоемах, куда, вероятно, проникает из более
крупных. Неоднократно указывался для планктона рек, проникая в них
из стоячих водоемов. Обитает также в планктоне прибрежной области
озер. В пелагиали характерен для верхней зоны эпилимниона. Ампли-
туда вертикальных суточных миграций очень невелика. Тепловодно-
стенотермичен, в планктоне озер и прудов появляется обычно в конце
апреля—в мае и выпадает поздней осенью — в начале зимы. В подзем-
ных водах отсутствует. Обитает в слабокислых, нейтральных и щелоч-
ных водах, по Lowndes’y (1928), при pH = 7.6—8.4. Однако по Ласточ-
кину (1934), помимо нейтральных озер, он встречается также при
pH = 5.9—6.4 (оз. Б. Гарава близ Иванова олиго-дистрофное, по тер-
минологии этого автора). В общем границы pH для рассматриваемого
вида определяются как 5.9—8.4, причем, вероятно, оптимум лежит
при pH = 7—8. Гораздо более эвригалинен, нежели М. (Th.) oithonoides.
С достоверностью показан для слабосоленых вод восточной части
1 См. выше замечание о достоверности данных М. (Th.) oithonoides Sars.
М. (Th.) hyalinus /ar. persicus Lindberg’ (1936, Иран) едва ли Заслуживает выделения
в особый вариетет,
20’
308
CYCLOPINAE
Финского залива, в северной части Каспия, в Аральском море, в ряде
солоноватых озер низовьев р. Сары-су, в озерах низовьев р. Сыр-дарьи.
Исследования в озерах центральной части Якутской АССР (Рылов, 1935)
показали чрезвычайно высокую приспособленность М. (Th.) crassus
к колебаниям общей минерализации; здесь рачок найден в значительных
количествах в 9 озерах, при общей минерализации от 0.39 до 1.99 г
на литр. *
Повидимому, главным образом, дицикличен; в некоторых водоемах,
возможно, полицикличен; максимальное развитие падает на теплое
время года.
Положение М. (Th.) crassus в системе сапробов пока не выяснено.
Повидимому, главным образом, [S-мезосапроб, частично олигосапроб.
Есть основание полагать, что рачок может переносить более высокую
степень [3-мезосапробности, нежели М. (Th.) oithonoides.
*6. Mesocyclops (Thermocyclops) rylovi Smirnov (фиг. 78).
Mesocyclops rylovi Смирнов, 1928 :17, рис. 5—8; Smirnov, 1929a : 38, f. 1—5.
— Mesocyclops (Thermocyclops) rylovi Kiefer, 1929:85; Lindberg, 1935, Rec. Ind.
Mus., 37 : 412, t. VIII, f. 6.
Самка. Тело широкое, напоминающее таковое М. (Th.) crassus,
наибольшая ширина переднего отдела лежит на границе цефалоторакса
и торакса, отношение длины этого отдела к его ширине от 1.73:1
до 1.86:1. Генитальный сегмент слабо расширенный спереди, мало
суживающийся кзади, его длина лишь немного более его ширины.
Reeeptaculum seminis очень похож на таковой М. (Th.) dybowskii, боко-
вые выросты его переднего отдела мало загнуты кзади. Фуркальные
ветви расходятся под значительным углом, их длина приблизительно
в 3 раза превышает их ширину, латеральная щетинка прикреплена зна-
чительно кзади от середины внешнего края, нередко близко от его
заднего конца. Внутренняя апикальная щетинка лишь очень немногим
более чем в два раза превосходит длину внешней крайней щетинки
и немного более половины длины внешней из средних апикальных
щетинок. Дорзальная щетинка фуркальных ветвей относительно очень
короткая, едва достигающая половины длины внутренней из крайних
апикальных щетинок. Передние антенны обычно достигают до начала
или до середины 2-го торакального сегмента, 2 дистальных членика
вдоль края с узкой цельнокрайней гиалиновой пластинкой. Длина
дистального членика эндоподита ног IV пары приблизительно вдвое
превосходит его ширину, из апикальных шипов этого членика внутрен-
ний приблизительно вдвое длиннее внешнего. Соединительная пластинка
этой пары ног по бокам с полукруглыми выростами, вдоль свободного
края вооруженными 5—9 очень мелкими тонкими шипиками. Ноги V пары
с относительно очень короткой щетинкой на базальном членике, внутрен-
ний апикальный шип дистального членика хорошо развит, с мелкими
шипиками по бокам, очень непохожий на нормальную щетинку.
Длина 0.90—1.07 мм. По наблюдениям Seymour Sewell’a (1934:229),
зимой самки и самцы несколько крупнее (самка 1.022 мм.), нежели
летом — осенью (самка 0.796— 0.864 мм.). Яйцевые мешки удлиненно-
овальные, очень длинные, заходящие иногда даже за концы фуркальных
ветвей, содержащие значительное количество (до 27—30 в каждом) яиц.
Самец.1 Ноги VI пары с 3 придатками, из которых внутренний
1 Описан Landberg’ом (1935). По этому автору длина самца лишь 0.209 мм (!),
что, очевидно, ошибочно,
12. MESOCYCLOPS
309
(шип) немного длиннее среднего (щетинка), внешний придаток (щетинка)
почти в Зг/2 раза длиннее внутреннего.
Окраска обоих полов, по Lindberg’y, серая или желтоватая.
Lindberg (1935:415, t. 8, f. 7—9) описывает, кроме типичной формы
этого вида, его вариетет — var. vermifer, который отличается от типич-
ной формы более длинными передними антеннами, достигающими
заднего края 4-го торакального сегмента, иным соотношением длины
апикальных щетинок фуркальных ветвей и иным вооружением боковых
выростов соединительной пластинки IV пары ног — у var. vermifer эти
выросты снабжены 3—5 сильными, довольно крупными шипиками.
©
Фиг. 78. Mesocyclops (Th.) rylovi Smirn., самка. (По Смирнову).
1 — общий вид; 2— фурка со спинной стороны; 3 — генитальный сегмент
и reeeptaculum seminis; 4—нога V; 5—соединительная пластинка ног IV.
К сожалению, рисунки цитируемого автора, относящиеся к var. vermifer,
далеко не безупречны и очень неполны (дан лишь рисунок ноги V пары,
генитального сегмента с reeeptaculum seminis и соединительной пла-
стинки задней пары плавательных ног). Прилагаемые автором много-
численные измерения отдельных признаков, однако, не дают достаточ-
ного основания для безусловного отнесения var. vermifer к рассматри-
ваемому виду; очень возможно, что этот „вариетет“ окажется
самостоятельным видом.
Распространение. Найден в окрестностях Тбилиси и Махач-
калы (Смирнов, 1928), в Индии (Декан — Lindberg, 1935, окрестности
Калькутты; Gurney, 1907, С. oithonoides*, Seymour Sewell, 1934), в Иране
(Lindberg, 1936).
Биология. Планктический вид, до сих пор был найден лишь
в мелких водоемах (пруды, лужи), при pH =7.65—8.8. По наблюдениям
Seymour Sewell в пруде в Калькутте держится в планктоне в ноябре
ЗЮ
CYCLOPINAE
и декабре (самки с яйцевыми мешками), отсутствует в феврале, затем
появляется в марте и держится весь апрель, май, июнь (много), август
(много, самки с яйцевыми мешками), по январь включительно; зимой
редко. Вероятно дицикличен и тепловодно-стенотермичен.
*7. Mesocyclops (Thermocyclops) dybowskii Lande (фиг. 79).
Cyclops dybowskii Lande, 1890 : 363, t. 17, f. 60—68; Schmeil, 1892 : 72, t. IV, f.l—5.—
Cyclops crassus Lilljeborg', 1901 (non Fischer) : 38, t. 3, f. 1—3. — Mesocyclops dybowskii
Sars, 1914:62, t. 38.—Mesocyclops (Thermocyclops) dybowskii Kiefer, 1929:84.—
Cyclops (Mesocyclops) dybowskii Gurney, 1933: 302, f. 1901—915.
Самка. Тело менее стройное, чем у М. (Th.) oithonoides, передний
отдел более широкий, чем у последнего. Габитус рачка близок к тако-
вому М. (Th.) crassus Ficsh. Генитальный сегмент относительно корот-
кий и широкий, кзади равномерно суживающийся; длина этого сегмента
лишь немного превышает его ширину. Receptaculum seminis обычно
значительно вдавленный спереди, боковые выросты' переднего отдела
заметно загнуты кзади. Фуркальные ветви сравнительно мало расходя-
щиеся, их длина в 21/2—3 раза более их ширины, латеральная щетинка
прикрепляется несколько кзади от середины внешнего края. Внутрен-
няя из крайних апикальных щетинок менее чем вдвое превышает длину
внешней крайней и заметно менее половины длины внешней из средних
апикальных щетинок. Передние антенны достигают до заднего края
2-го торакального сегмента, оба дистальные членика очень короткие
(особенно 17-й), с очень узкой, цельнокрайней гиалиновой пластинкой
вдоль края. Соединительные пластинки ног I—IV пар с каждой стороны
с закругленным выростом, на краю несущим ряд очень мелких шипиков.
У ног IV пары длина дистального членика эндоподита приблизительно
втрое более его ширины, внутренний апикальный шип этого членика
немного короче внешнего. Ноги V пары очень похожи на таковые
М. (Th.) oithonoides, шипообразная внутренняя апикальная щетинка
немного длиннее внешней апикальной щетинки. Длина 0.85—1.1 мм.
Окраска обычно коричневая, иногда частично или сплошь с фиолетовым
оттенком. Яйцевые мешки крупные, расходящиеся.
Самец. Передние антенны с длинными сензорными Цилиндрами
на 1-м, 4-м и 8-м члениках. У ног VI пары средняя щетинка гораздо
короче обоих боковых придатков и почти вдвое короче внутреннего
из них, который является шипом; внешний придаток в виде относительно
очень длинной щетинки. Длина около 0.7 мм.
Распространение. Широко распространен в Европе, Азии
(исключая север); северная и восточная Африка. Для СССР пока
имеется небольшое число вполне достоверных местонахождений.1 Пока-
зан для ряда западных, восточных и центральных областей европейской
части СССР, на юге — для бассейна Южного Буга, для Кавказа, Крыма;
в азиатской части СССР указан из ряда мест, исключая северную
Сибирь, известен с Дальнего Востока.
Биология. Обитает, главным образом, в зарослях макрофитов,
литорали озер и в различных мелких богатых растительностью водоемах.
Известен из астатических водоемов (весенние лужи), а также из при-
1 Указание для Северной Двины у Архангельска (Величковский, 1927) мало
вероятно. Gurney (1933:306) ошибочно приводит М. dybowskii, ссылаясь на Van-
Douwe (1905), для Туркестана; последним автором этот вид указан для Ленкорани
Эту же ошибку делает и Kiefer (1929),
12. MESOCYCLOPS
брежных зарослей медленно текущих рек. В некоторых озерах входит
в состав пелагического планктона, в котором, однако, обычно встре-
чается в небольших количествах, не достигая такого развития, как
М. (Th.) oithonoides и М. (Th.) crassus. Равным образом иногда при-
сутствует и в прудовом планктоне. Повидимому встречается в пелагиали
исключительно эвтрофных (и близких к таковым) озер. Троглоксен,
Фиг. 79. Мesocy clops (Th.) dybowskii (Lande), сайка.
/ — фурка с брюшной стороны; 2—генитальный сегмент с receptaculum seminis
3 — нога V; 4 — эндоподит ноги IV; 5—3 дистальных членика антенны I.
в подземных водах показан с достоверностью лишь в Адельсбергской
пещере в Италии. Вид стенотермический, тепловодный. Насколько нам
известно) нет ни одного указания на нахождение М. (Th.) dybowskii
в зимнее время. Можно думать, что температурный оптимум для
М. (Th.) dybowskii лежит, примерно, между 15—25°. Судя по место-
нахождениям, рачок нормально обитает лишь при высоких содержаниях
растворенного кислорода. Данные по отношению к концентрации водо-
родных ионов очень незначительны. В водоемах СССР он показан при
pH от 5.2—5.8 до 8.4. Очень вероятно, что границы pH для рассматри-
ваемого вида, в общем, соответствуют таковым для М. (s. str.) leuckarti’,
312
CYCLOPINAE
во всяком случае M. (Th.) dybowskii характерен для вод нейтральных
и щелочных. Нахождение последнего в солоноватых водоемах представ-
ляет нехарактерное явление для этого вида, ведущего вообще чисто
пресноводный образ жизни. Так, Зиновьев (1933) указывает его для
болотца в Челябинской области, а Пирожников (1929) — для солонова-
того оз. Сартлан. Указания эти, равно как и указание Зверевой (1927)
для оз. Чаны, впрочем, нуждаются в проверке.
В мелких водоемах рачок дицикличен, с максимумами весной
(в 1-й половине лета) и во 2-й половине лета — осенью; вероятно,
дицикличен и в пелагиали озер.
Вероятно, по преимуществу [3-мезосапроб.
ДОПОЛНЕНИЕ
После представления рукописи в редакцию появились описания
следующих новых палеарктических форм Cyclopidae.
1. Microcyclops stammeri (Kief.). 1938 {Metacyclops stammeri Kiefer,
Zool. Anz., 123:7, f. 11—17), близок к M. minutus Claus, но на конце
эндоподита IV ноги самки имеется 2 придатка — длинная щетинка и шип,
длина которого приблизительно равна половине эндоподита; последний
значительно удлинен. Receptaculum seminis гораздо короче, чем у М. minu-
tus. Длина самки 0.62—0.64 мм. Пещеры в южной Италии (Otranto),
самцы и самки в массе.
2. Microcyclops subdolus (Kiefer), 1938 {Metacyclops subdolus Kiefer,
Zool. Anz., 123:9, f. 18—24), очень близок к M. minutus Claus, с кото-
рым имеет одинаковое строение и вооружение дистального членика
эндоподита IV ноги самки. Receptaculum seminis более короткий, чем
у М. minutus, из средних апикальных щетинок фуркальных ветвей
внутренняя почти вдвое длиннее внешней. Длина самки около 0.62 мм.
Найден вместе с предыдущим видом в пещерах южной Италии. Очень
возможно, является вариететом М. minutus Claus.
3. Speocyclops itallcuS Kief., 1938 (Zool. Anz., 123:11, f. 25—28),
близок к S. sebastianus Kief., но анальная пластинка закругленная,
фуркальные ветви более длинные, щетинка совершенно редуцированного
базального членика V ноги очень короткая. Длина самки 0.40 мм.
Южная Италия, грот в Салерно.
4. Mesocyclops {Thermocyclops) siephanidesi Kief., 1938 (Zool.
Anz., 123:96, f. 1—5), близок к M. {T.) oithonoides и М. (Г.) crassus,
но у V ноги самки субапикальный шип значительно длиннее апикаль-
ного придатка, щетинка базального членика короткая, не достигающая
до середины апикального придатка, боковые выросты receptaculum semi-
nis сильно загнуты кзади, на конце с маленьким бугорком. Длина самки
1.0—1.1 мм. Греция, о. Корфу.
Описанный Kiefer’oM (Zool. Anz., 1938, 123:5, f. 6—10) из грота
близ Салерно Acanthocyclops agamus (найден 1 самец!) этим автором
относится к названному роду с сомнением. Мы полагаем, что этот
самец действительно принадлежит к Acanthocyclops. но является ли
он новым видом, очень проблематично; возможно, что, как допускает
и Kiefer, он относится к какому-либо из уже известных видов рода
(например A. hypogeus. A. rhenanus), у которых самцы не описаны.
Алфавитный указатель латинских названий животных
abnobensis, Paracyclops 159
abyssicola, Acanthocyclops 214, 264
abyssicola, Cyclops 264
abyssicola, Cyclops (Diacyclops) 264
abyssorum, Cyclops 173, 183
Acanthocyclops 167, 210
affinis, Cyclops 161
affinis, Cyclops (Paracyclops) 161
affinis, Paracyclops 55, 161
affinis, Platycyclops 161
africanus, Cyclops 267
agilis, Cyclops 139, 140
agilis, Cyclops (Eucyclops) 140
agilis, Leptocyclops 140
agilis speratus, Cyclops (Eucyclops) 143
agiloides, Cyclops 140
albidus, Cyclops 134
albidus, Cyclops (Macrocyclops) 134
albidus, Macrocyclops 129, 134
americanus, Acanthocyclops 213, 228
americanus, Cyclops 228
annulicomis, Cyclops 132, 134
annulicomis, Pachycyclops 134
arndti, Ochridacyclops 127, 154
asiaticus, Mesocyclops (Thermocyclops) 299,
301, 303 ater. Homocyclops 127
bicolor, Cripiocyclops 2?1
bicolor, Cyclops 271
bicolor, Cyclops (Microcyclops) 271
bicolor, Microcyclops 267, 271
bicuspidatus, Acanthocyclops 213, 238
bicuspidatus, Cyclops 238, 243
bicuspidatus, Cyclops (Acanthocyclops) 238
bicuspidatus, Cyclops (Diacyclops) 238
bicuspidatus var. lubbocki, Cyclops 241
bicuspidatus lubbocki, Cyclops (Acantho-
cyclops) 241
bicuspidatus m. odessana, Acanthocyclops
213, 241
bicuspidatus odessanus, Cyclops 241
bicuspidatus odessanus, Cyclops (Diacy-
clops) 241
bisetosus, Acanthocyclops 213, 243
bisetosus, Cyclops 243
bisetosus, Cyclops (Acanthocyclops) 243
bisetosus, Cyclops (Diacyclops) 243
bissextilis, Cyclops 245
bistriatus, Cyclops 132, 133
bistriatus, Pachycyclops 133
bodanicola, Mesocyclops (s. str.) 293
bohater, Cyclops 172, 186
hrevicaudatus (part.), Cyclops 198
brevicornis, Cyclops 215
brevisetosus, Cyclops 271
brevispinosus, Cyclops 226
brucei, Cyclops 245
Bryocyclops Kief. 167
buxtoni, Cyclops 275
cadanensis, Cyclops 305
caeibaensis, Cyclops 267
capillatus, Acanthocyclops 214, 236
capillatus, Cyclops, 236
capillatus, Cyclops (Acanthocyclops) 236
cerberus, Cyclops (Diacyclops) 287
cerberus,, Speocyclops 282, 287
clausi, Cyclops 134, 215
colchidanus, Cyclops 289
colchidanus, Speocyclops 289
coronatus, Cyclops 130
coronatus, Macrocyclops 130
Cpalustris (juv.) 130
Craeteriella (part.) 280
crassicaudis, Acanthocyclops 214, 245
crassicaudis var. brachycercus, Acanthocy-
clops 248
crassicaudis brachycercus, Cyclops (Diacy-
clops) 248
crassicaudis var. cretensis, Acanthocyclops
248
crassicaudis cretensis, Cyclops 248
314
Алфавитный указатель латикских названий
crassicaudis cretensis, Cyclops (Diacyclops)
248
crassicaudis, Cyclops 245, 248
crassicaudis, Cyclhops (Acanthocyclops) 245
crassicaudis, Cyclops (Diacyclops) 245
crassicaudis iaipehensis, Cyclops 248
crassicornis, Cyclops Sars 156
crassus, Cyclops 305, 310
crassus, Mesocyclops 305
crassus, Mesocyclops (Thermocyclops) 300,305
crinitus, Cyclops 232
croaticus, Cyclops 280
Cryptocyclops 265
Cyclopidae 127
Cyclopinae 127, 166
Cyclops 127, 167
Cyclops (part.) 129, 137, 154, 163, 167, 292
Cyclops 167
Cyclops (subg. Bryocyclops) 280
Cyclops (subg. Diacyclops) 280 f
Cyclops (subgen. Eucyclops) 137
Cyclops (Macrocyclops) 129
Cyclops (Mesocyclops) 292, 298
Cyclops (subg. Microcyclops, part.) 280
Cyclops (Paracyclops) 154
davidi, Cyclops 269
demetiensis, Cyclops (Bryocyclops) 287
demetiensis, Cyclops (Microcyclops) 287
demetiensis, Speocyclops 282 287
dengizicus, Cyclops 275
dengizicus, Cyclops (Metacyclops) 2Tb
dengizicus, Microcyclops 275, 267
deniiculatus, Cyclops Thallwitz Sitzungeb 148
Diacyclops 210
diaphanus, Cyclops 260, 271, 273, 277
diaphanus var. dengizic., Cyclops 21b
diaphanus var. diaphanoides, Cyclops 260
diaphanus, Microcyclops 277
dimorphus, Cyclops (Metacyclops) 275
disjunctus, Diacyclops 251
distinctus, Cyclops 133
distinctus, Cyclops (Macrocyclops) 133
distinctus, Macrocyclops 130, 132, 133
dybowskii, Cyclops 310
dybowskii, Cyclops (Mesocyclops) 310
dybowskii, Mesocyclops 310
dybowskii, Mesocyclops (Thermocyclops) 299,
310
Ectocyclops 128
Ectocyclops 163
entzi, Cyclops-t-roseus, Cyclops 238
Eucyclopinae 127
Eucyclops 127, 128, 129, 137
fimbriatus, Cyclops 156, 160
fimbriatus var. poppei, Cyclops 160
fimbriatus var., Cyclops 158
fimbriatus, Cyclops (Paracyclops) 156
fimbriatus var. anyschohtzara Eucyclops 148
fimbriatus poppei Cyclops (Paracyclops) 160
fimbriatus, Paracyclops, 155, 156
fimbriatus var. abnobensis, Paracyclops 155,
159
fimbriatus var. finitimus, Paracyclops 156
fimbriatus var. imminutus Kief, Paracyclops
155, 158
finitimus, Paracyclops 160
furcifer, Cyclops 173, 198
fuscus, Cyclops 130
fuscus, Cyclops (Macrocyclops) 130
fuscus, Macrocyclops 130
gigas, Acanthocyclops 212, 215
gigas, Cyclops 219, 238
gigas, Cyclops (Acanthocyclops) 219
gigas, Cyclops (Megacyclops) 219
gigas var. Iatipes, Acanthocyclops 223 •
gigas Iatipes, Cyclops (Acanthocyclops) 223
gracilicornis, Cyclops 132
gracilicornis var. distincta, Cyslops 132
gracilis, Cyclops 273 Lilljeb.
gracilis, Cyclops (Metacyclops) 273
gracilis, Cyclops (Microcyclops) 273
gracilis, Mesocyclops 273
gracilis, Microcyclops 267, 273
graeteri, Cyclops 150
graeteri, Eucyclops 139, 150
Graeteriella 167, 289
Graeteriella 289, 291
gyrinus, Cyclops 134
halepensis, Cyclops 280
helgolandicus, Cyclops 241
hispanicus, Acanthocyslops 229
Homocyclops 127, 128, 129
hyalinus, Cyclops 305
hyalinus, Cyclops (Mesocyclops) 305
hyalinus, Mesocyclops (Thermocyclops) 305
hypogeus, Acanthocyclops 215, 264
hypogeus, Cyclops (Diacyclops) 264
inarmatus, Eucyclops 150
incertus, Cyclops 263
infernus, Cyclops (Diacyclops) 282
infernus, Speocyclops 281, 282
Алфавитный указатель латинских названий
315
ingens, Cyclops 219
insignis, Cyclops 171, 208
insulanus, Thaumasiacyclops 128
karamani, Acanthocyclops 213, 242
karamani, Cyclops (Diacyclops) 242
kieferi, Acanthocyclops 215, 229
kieferi, Cyclops 195, 229
kieferi, Cyclops (Acanthocyclops) 229
kikuchii, Cyclops 207
kolensis, Cyclops 173, 195
korostyschevi, Cyclops 134
lacunae, Cyclops 198
lacustris, Cyclops 172, 187
laisi, Diacyclops 291
laisi, Graeteriella 291
laisi, Graeteriella (Paragrieteriella) 291
languidus Acanthocyclops 213, 249
languidus, Cyclops 249
languidus, Cyclops (Acanthocyclops) 249
languidus, Cyclops (Diacyclops/249
languidus f. atava, Cyclops 249
languidus var. beigicus Acanthocyclops 213,
251
languidus beigicus, Diacyclops 251
languidus var. disjunctus, Acanthocyclops 251
languidus var. disjuncta, Cyclops 251
languidus disjunctus, Cyclops (Diacyclops)
251
languidus var. intermedia, Cyclops 260
languidus var. nanus, Cyclops 260
languidoides Acanthocyclops 215, 252, 254
languidoides, Cyclops 252, 260
languidoides, Cyclops (Acanthocyclops) 252
languidoides, Cyclops (Diacyclops) 252
languidoides, Diacyclops 252
languidoides var. badeniae, Acanthocyclops
' 254, 255
languidoides badeniae, Cyclops (Diacyclops)
255
languidoides var. clandestinus Acanthocy-
clops 255, 258
languidoides clandestinus, Cyclops 258
languidoides clandestinus, Cyclops (Diacy-
clops) 258
languidoides clandestinus, Diacyclops 258
languidoides var. eriophori Acanthocyclops
214, 254, 255
languidoides eriophori Cyclops 255
languidoides eriophori, Cyclops (Acantho-
cyclops) 255
languidoides eriophori, Cyclops (Diacy-
clops) 255
languidoides var. goticus, Acanthocyclops
255, 259
languidoides f. gotica, Cyclops (Diacy-
clops 259
languidoides var. gracilicaudatus, Acantho-
cyclops 254
languidoides var. gracilicaudatus, Cyclops
257
languidoides var. hiberniae Acanthocyclops
254, 257
languidoides hiberniae, Cyclops 257
languidoides hiberniae, Cyclops (Acantho-
cyclyps) 257
languidoides hiberniae, Cyclops (Diacy-
clops) 257
languidoides var. hypnicola, Acanthocyclops,
254, 255
languidoides hypnicola, Cyclops 255
languidoides hypnicola, Cyclops (Acantho-
cyclops) 255
languidoides hypnicola, Cyclops (Diacy-
clops) 255
languidoides var. italianus, Acanthocyclops
254, 259
languidoides f. italiana, Cyclops (Diacy-
clops) 259
languidoides var. nagusalloensis, Acantho-
cyclops 254, 258
langyidoides nagysalloensis, Cyclops 258
languidoides nagysalloensis Cyclops (Dia-
cyclops) 258
languidoides, var. putealis, Acanthocyclops
254, 259
languidoides putealis, Cyclops 259
languidoides putealis, Cyclops (Diacyclops)
259
languidoides var. tuntalus Acanthocyclops
254, 259
languidoides var. zschokkei, Acanthocyclops
258
languidoides zschokkei, Cyclops 258
languidoides zschokkei, Cyclops (Diacy-
clops) 258
Iatipes, Cyclops 223
Iatipes, Cyclops (Megacyclops) 223
latissimus Cyclops 134
leeutoenhockki, Cyclops 293
Leptocyclops 137
leuckarti, Cyclops 293
leuckarti, Cyclops (Mesocyclops) 293
leuckarti, Mesocyclops (s., str.) 293
316
Алфавитный указатель латинских названий
leuckarti, var. australiensis 293
leuckarti bodanicola, Mesocyclops, 297
leuckarti var. pilosus, Mesocyclops, 297
lilljeborgi, Cyclops 148
lilljeborgi, Eucyclops 148
lilljeborgi, Leptocyclops Sars 148
longicaudatus, Cyclops 271, 277
lubbocki, Cyclops 241
lucidulus, Cyclops 223
lucidus, Cyclops 293
maarensi, Cyclops 148
Macrocyclops 127, 128, 129
Macrocyclops 129
macruroides, Cyclops 145
macruroides, Cyclops (Eucyclops) 146
macruroides, Eucyclops 139
macruroides, Eucyclops 145
macruroides denticulatus, Cyclops (Eucy-
clops 148
macruroides var. denticulatus, Eucyclops 139,
148
macruroides, Leptocyclops 145, 146
macrurus var. caucasicus, Cyclops 148
macrurus, Cyclops (Eucyclops) 148
macrurus, Eucyclops 139
macrurus, Eucyclops (s. str.) 148
macrurus, Leptocyclops 148
macrurus var. subterranea, Cyclops 150
magnoctavus, Cyclops 151
major, Mesocyclops 293
margoi, Cyclops 156
Megacyclops 210
Mesocyclops 167, 292
Mesocyclops s. str. 292, 293
Mesocyclops (Thermocyclops) 298
Microcyclops 167, 265
Microcyclops (part, subg.) 265
Microcyclops u Metacyclops 265
miniatus, Cyclops 198
minutus, Cyclops 277
minutus, Gyclops (Microcyclops) 277
minutus, Microcyclops 267, 277
mongolicus Thermocyclops 303
Monoculus 129
muscicola, Acanthocyclops 213, 226
muscicola, Cyclops 226
muscicolus, Cyclops 226
muscicola, Cyclops (Acanthocyclops) 226
Muscocyclops 167
J
nanus, Acanthocyclops 214, 260
nanus. Cyclops 258
nanus, Cyclops 260
nanus, Cyclops (Acanthocyclops) 260
nanus, Cyclops (Diacyclops) 260
nanus var. incertus, Acanthocyclops 263
nanus incertus, Cyclops (Diacyclops) 263
nagasakai, Eucyclops 144
Neutrocyclops 265
novae-zealandiae, Cyclops 140
obsoletus, Mesocyclops 293
Ochridacyclops 129, 153
ochridanus, Cyclops 172, 195
oihonoides, Cyclops 300
oithonoides, Cyclops (Mesocyclops) 300
oithonoides, Mesocyclops 300, 303
oitonoides, Mesocyclops (Thermocyclops) 299,
300
oithonoides var. hyalina, Cyclops 305
odessanus, Cyclops 241
Oligarthra 127
orcinus, Speocyclops 283
orientalis, Cyclops 267
Orthocyclops 166
Pachycyclops 129
Paracyclops 128, 154
Paracyclops (part, subgen) 163
Paragraeteriella subg. nov. 291
parcus, Cyclops 223
pectinifer, Cyclops 140
pennatus, Cyclops 134
phaleratus, Cyclops 163
phaleratus, Cyclops (Ectocyclops) 163
phaleratus, Cyclops (Paracyclops) 163
phaleratus, Ectocyclops 163
phaleratus japonicUs Cyclops 163
phaleratus Platycyclops 163
phraeticus, Acanthocyclops 214, 230
phraeticus, Cyclops, 230
phraeticus, Cyclops (Acanthocyclops) 230
planus, Cyclops 280
planus, Cyclops (Microcyclops) 280
planus, Microcyclops 267, 280
Platycyclops (part.) 154, 163
plutonis, Speocyclops 281, 286
Polyarthra 127
poppei, Cyclops 160
poppei, Paracyclops 156, 160
prasinus, Cyclops 151
prasinus Cyclops (Eucyclops) 151
prasinus, Cyclops (Tropocyclops) 151
prasinus, Eucyclops (Tropocyclops) 151
prasinus, Leptocyclops 151
prasinus, Tropocyclops 151.
Алфавитный указатель латинских названий
317
prasinite candidiusi, Eucyclops 151
proserpinae, Speocyclops 282, 288
pulchellus, Cyclops 238, 293
pulchellus var., Cyclops 241
pulchellus var. synarthrus, Cyclops 233
pygmaeus, Cyclops 161
quadricornis albidus, Monoculus 134
quadricornis fuscus, Monoculus 130
quadricornis viridis, Monoculus 215
racovitzai, Cyclops 282
racovitzai, Cyclops (Diacyclops) 282
racovitzai, Speocyclops 281, 282
rhenanus, Acanthocyclops 214, 234
robustus Acanthosyclops 228
robustus, Cyclops 226
robustus var. saxonica -+- var. aphanes -+- var.
armata-t-var. setiger, Cyclops 226
rubellus, Cyclops 269
rubellus, Cyclops (Microcyclops) 269
rylovi, Mesocyclops 308
rylovi, Mesocyclops (Thermocyclops) 300, 308
scopliensis, Acanthocyclops 214, 248
scopliensis, Cyclops (Diacyclops) 248
scutifer, Cyclops 171, 200
scutifer f. asc, Cyclops 201
scutifer var. wigrensis, Cyclops 204
sebastiinus, Speocyclops 281, 285
Sensitivus, Acanthocyclops 212, 238
sensitivus, Cyclops 238
sensitivus, Cyclops (Acantocyclops) 238
serrulatoides, Cyclops 139, 140, 148
serrulatus (part.) Cyclops 140, 145
serrulatus, Eucyclops typ. 139, 140
serrulatus, Eucyclops (s. str.) 139
serrulatus (s. str.) Eucyclops 140
serrulatus var. denticulatus, Cyclops 148
serrulatus var. montana, Cyclops 139
serrulatus var. nagasakai Eucyclops 139, 144
serrulatus var. proximus, Eucyclops 139, 143
serrulatus var. proximus, Eucyclops (Eucy-
clops) 143
serrulatus var. speratus, Eucyclops, 139, 143
serrulatus f. iypica-t- f. brevicaudata, Cy-
clops 140
signatus, Cyclops 130, 134
signatus var. annulicornis, Cyclops 134
signatus var. coronatus, Cyclops 130
silesicus, Cyclops 195
simplex, Cyclops 293
Sisyphus, Speocyclops 281, 283
smimovi sp. nov., Cyclops 172, 190
spaeieus, Speocyclops 282, 288
spaeieus, Speocyclops 288
Speocyclops 166, 280
speratus, Cyclops 143
speratus, Eucyclops (s. str.) 143
speratus, Leptocyclops 143
stammeri, Acanthocyclops 214, 232
stammeri, Cyclops (Acanthocyclops) 232
stammeri var. westfalicus, Acanthocyclops
214, 234
stammeri westfalicus, Cyclops (Acanthocy-
clops) 234
strennus, Cyclops 173, 189, 201, 204
strenuus (part.), Cyclops 173, 195
strenuus, Cyclops (s. str.) 173
strenuus (typ.), Cyclops 173
strenuus var. abyssorum, Cyclops 173
strenuus f. furcifer, Cyclops 198
strenuus var. gracilipes, Cyclops 183
strenuus-insignis, Cyclops 167
strenuus f. ,,L“, Cyclops 189
strenuus f. lacustris, Cyclops 187
strenuus lagoraensis, Cyclops 189
strenuus var. landei, Cyclops 173, 176
strenuus subsp. strenuus, Cyclops 173
strenuus tatricus, Cyclops 139
strenuus var. vicinus, Cyclops 204
strenjius vranae, Cyclops 172, 180
stygius, Acanthocyclops 214, 263
stygius, Cyclops 263
stygius Cyclops (Diacyclops) 263
stygius var. deminutus, Cyclops 263
subaequalis,. Cyclops 267
tantalus, Diacyclops 259
tatricus, Cyclops 172, 189
tenuicaudus, Cyclops 271
ienuicornis, Cyclops 134, 293
ienuicornis var. distincta, Cyclops 132
ienuicornis, Cyclops (Macrocyclops) 134
teras, Cyclops 145, 215
teras, Eucyclops 139, 145
Thaumasiocyclops 128
Thermocyclops 293, 298
Trifida 127
troglodytes, Cyclops 285
troglodytes, Cyclops (Diacyclops) 285
troglodytes, Speocyclops 281, 285
troglophilus, Acanthocyclops 232
Tropocyclops 129, 150
Tropocyclops 150
Tropocyclops 150
318
Алфавитный указатель латинских названий
unacanthus, Metacgclops 277
unisetiger, Graeteriella 290
unisetiger, Cgclops 290
unisetiger, Cgclops (Microcgclops) 290
unisetiger f. biatriculata, Cgclops 290
unisetiger, Diacgclops 290
unisetiger f. biatriculata (Diacgclops) 290
unisetigera, Graeteriella 290
varicans, Cgclops 267
varicans, Cgclops (Microcgclops) 267
varicans, Microcyclops 267
varicans rubellus, Cgclops (Microcgclops) 269
varicans rubellus, Microcyclops 269
varicans var. rubens, Cgclops 269
varicans subaequalis, Cgclops (Microcg-
clops) 267
varius var. brachgurus Cgclops 140
varius var. proximus, Cgclops 143
varius var. speratus, Cgclops 143
venustus, Acanthocyclops 213, 230
venustus var. crinitus, Acanthocyclops 232
venustus, Cgclops 230
venustus, Cgclops (Acanthocgclops) 230
vernalis, Acanthocyclops 213, 223, 224
vernalis, Cgclops 223, 226
vernalis Cgclops 224
vernalis, Cgclops (Acanthocgclops) 224
vernalis (part.), Cgclops (Acanthocgclops)
vernalis var. robustus, Acanthocyclops 226
vernalis var. aculeata, Cgclops 224
vernalis-americanus, Cgclops 228
vernalis var. ocnatus, Cgclops 224
vernalis var. robustus, Cgclops 226
vernalis var. tetracantha, Cgclops -+- tetra-
cantha f. robusta var. 226
vicinus, Cyclops 172, 204
vicinus var. brachismerinthoa, Cgclops 207
vicinus var. glacialis, Cgclops 204
vicinus var. kikuchii, Cyclops 207
viridis, Acanthocyclops 212, 215
viridis, Cgclops 215
viridis (part.) Cyclops 219
viridis, Cyclops (Acanthocyclops) Gurney 215
viridis, Cyclops (Megacyclops) Kiefer 215
viridis, Leptocyclops Henry 151
viridis var. americanus, Cyclops 228
viridis europaeus, Cyclops 215
viridis var. dives, Cgclops 226
viridis var. gigas, Cgclops 219
viridis var. insectus, Cgclops 228
viridosignatus, Cyclops 134
vulgaris, Cyclops 215
schokkei, Cgclops 258
Содержание
Стр.
Вячеслав Михайлович Рылов (1889—1942)..................................... 3
Предисловие............................................................... 11
Систематический указатель видов.......................................... 13
Введение.................................................................. 17
Морфолого-анатомический очерк....................................... 17
Метаморфоз.......................................................... 49
Цикл развития..................................................... 55
Классификация....................................................... 57
Распространение и история фауны . .................................. 74
Биология............................................................ 87
Хозяйствеиное значение............................................. 107
Литература........................................................ 117
Систематическая часть.................................................... 127
Дополнение............................................................... 312
Указатель латинских названий............................................. 313
ГТ вчитается по постановлению редакционно-издательского совета
РИСО АН СССР № 3032. Подписано к печати 7)VI 1948 г. М-00926. Печ. л. 20.
Уч.-изд. л. 28,5. Тираж 2000 экз. Зак. № 1170.
1-н Тип. Изд. АН СССР. Ленинград, В. О., 9 линия, д. 12.
ОПЕЧАТКИ
Страницу Строка Напечатано Следует читать
3 14 св. науки наук н
8 21 „ Infusors Infusoriums
10 1 он.' 1—000,000 фиг. 1—319,79 фиг.
104 23 сн. С, о2
122 22 св. 1231 1931
122 14 сн. 1336а 1936а
123 20 св. Biosooziologische Biosoziologische
126 3 сн. Saison-Sees Saisan-Sees
Фауна СССР> Ракообразные, т, Ш, вып. 3.