/

































































































































Текст
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
АКАДЕМИЯ НАУК СССР
НАУЧНО-ПОПУЛЯРНАЯ СЕРИЯ
К. Е. ОВЧАРОВ
ВИТАМИНЫ
В ЖИЗНИ
РАСТЕНИЙ
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
Москва —1955
О' ............
Ответственный редактор
В. Н. БУКИН
ВВЕДЕНИЕ
Осенью 1741 г. русские моряки во главе со своим ка-
питаном — командором Берингом, борясь с непогодой и
преодолевая огромные лишения, отстаивали славу рус-
ского флота и честь недавно созданной Российской Ака-
демии наук.
Длинные ночи с дождем и снегом, изнуритель-
ный труд сильно подрывали здоровье моряков экспе-
диции. Но самым ужасным и губительным бедствием
была цынга.
В ноябре, когда корабль пристал к берегу, несколько
офицеров и матросов были уже мертвыми, а восьмого де-
кабря умер и капитан корабля «Святой Петр» Беринг.
Такие же бедствия потерпела и команда корабля «Святой
Павел», входившая в состав экспедиции Беринга.
Моряки знали, что избавиться от цынги можно, упо-
требляя травы, но они не могли найти их на Крайнем
Севере, на земле, занесенной снегом. Участник экспедиции
Беринга ботаник и зоолог Стеллер научился предупреж-
дать появление цынги приготовлением отваров из травы,
собранной на Шумагинских островах, но зимой этой тра-
вы не было.
Цынга была постоянным спутником экспедиций и в
течение столетий поражала человечество, особенно наро-
ды северных областей, где недостаток овощей и фруктов
был обычным явлением. Только в одном 1849 г. в России
болело цынгой 260444 человека, из них около 70 тыс.
человек умерло.
В Японии, Китае, Индии, Африке и Южной Америке,
т. е. в тех странах, где рис являлся основным продуктом
питания, население страдало от другого заболевания,
3
известного под названием «бери-бери». В 1878—1889 гг.
бери-бери болело около половины всего личного состава
японского флота. Хотя причина этого заболевания и была
найдена около 50 лет назад, тем не менее оно и в
наше время приносит значительный ущерб. Так, например,
в английской колонии Гонконге в 1941 г. из 350 тыс. че-
ловек населения болели бери-бери 10 тыс., из них более
1000 человек умерло. В Японии за период 1920—1929 гг.
от бери-бери умерло около 200 тыс. человек.
Населению Италии, Франции, Испании, южных штатов
США хорошо известно заболевание под названием пеллаг-
ра, или шершавая кожа. Тяжелые страдания людей, ча-
стые смертные случаи заставляли ученых искать пути
борьбы с этой болезнью. Чтобы представить, как велико
число заболеваний пеллагрой, стоит, например, сказать,
что в 1917 г. только в США зарегистрировано 165 тыс.
случаев заболевания, а в 1930 г. эта цифра превысила чет-
верть миллиона.
В течение тысячелетий дети страдали от рахита.
В наше время рахит является широко распространенным
заболеванием в капиталистических странах.
Гениальное открытие Луи Пастером инфекционной
природы многих болезней, казалось, должно было объяс-
нить и причины указанных выше заболеваний. Однако
многочисленные попытки ученых выделить бактерии, вы-
зывающие эти болезни, не имели успеха. Безуспешны
были и исследования, основанные на замечательном от-
крытии Д. И. Ивановским вирусов. Оказалось, что эти
заболевания вызываются не вирусами. Неосновательны-
ми были и объяснения появления цынги, бери-бери и дру-
гих болезней наличием в пище ядовитых веществ.
Некоторое время считалось вероятным, что одной из
причин заболевания цынгой является недостаток пищи.
Но объяснение подобного рода опровергалось практикой.
Хорошо известно, что в России цынга считалась «благо-
родной болезнью», так как ею обычно болели дворяне и
купцы, а не крестьяне, употреблявшие много лука, Ка-
пусты, репы. Известно также, что заключенные в тюрьмах
на голландских островах, питавшиеся полированным ри-
сом, заболевали бери-бери чаще, и болезнь у них проте-
кала более тяжело, чем у тех, которые получали полу-
очищенный рис.
4
Неосновательными и ошибочными были также пред-
ставления, согласно которым для нормального функцио-
нирования организма необходим рацион, состоящий лишь
из белка, жира, углеводов и минеральных солей. Эти
представления в конце прошлого века опроверг наш со-
отечественник Н. И. Лунин, создавший теоретические
основы нового учения, которое мы теперь называем вита-
минологией. В 1880 г. Н. И. Лунин убедительными опыта-
ми показал, что для жизни организма совершенно недо-
статочно употребление белков, углеводов и минеральных
солей и что, кроме них, необходимо наличие в пище каких-
то еще невыявленных, но жизненно важных веществ.
Н. И. Лунин проводил свои опыты на мышах. Одну
группу мышей он кормил обычным молоком, другую —
искусственным, т. е. изготовленным из очищенных белков,
жиров, сахаров и солей, входящих в состав молока и
взятых в тех же самых количествах. В результате мыши
второй группы погибли, а первой — остались вполне здо-
ровыми. На основании этого Лунин заключил: «Если...
невозможно обеспечить жизнь белками, жирами, саха-
ром, солями и водой, то из этого следует, что в молоке,
помимо казеина, жира и молочного сахара и солей, со-
держатся еще и другие вещества, незаменимые для пи-
тания».
Установив столь важный факт, Лунин решил подробно
изучить природу этих незаменимых для питания веществ.
Талантливый исследователь в своей диссертации, защи-
щенной им 18 сентября 1880 г. на Совете медицинского
факультета Юрьевского университета, указывал: «Пред-
ставляет большой интерес исследовать эти вещества и
изучить их значение для питания». Но осуществить наме-
ченный план исследований ему не удалось. Через полго-
да после защиты, когда после 1 марта 1881 г. в универ-
ситетах был введен полицейский режим, Лунин вынуж-
ден был покинуть Юрьевский университет. Оставив на-
чатые исследования, Лунин перешел на практическую вра-
чебную работу и больше не имел возможности возвра-
титься к своему замечательному открытию. Через 10 лет
открытие Н. И. Лунина было подтверждено русским уче-
ным С. Сосиным. Многочисленные опыты, проведенные
на собаках и мышах, привели и его к аналогичному выво-
ду.- В 1891 г. он писая: «Эти недостающие-вещества,- как
5
совершенно ясно, содержатся в молоке и яичном желтке:
их обнаружение является первой задачей, которую необ-
ходимо поставить прежде чем проводить новые опыты по
кормлению».
В. В. Пашутин в 1901 г. указывал, что вещества, из-
лечивающие цынгу, в изобилии находятся в растениях и
имеют органическую природу. «Итак,— писал В. В. Па-
шутин,— скорбут есть проявление одной из форм голо-
дания и именно органического голодания». В. В. Пашу-
тин тогда же высказал мысль, что эти вещества имеются
не только в высших растениях, но и в бактериях, и что при
помощи последних происходит обогащение пищевых про-
дуктов необходимыми факторами питания.
За границей после замечательного открытия Н. И. Лу-
нина с каждым годом появлялось все больше и больше
исследований, подтверждающих его классические опыты.
Через 30 лет после опытов Лунина английский ученый
Гопкинс пришел к тем же выводам. Он писал: «Ни одно
животное не может жить на смеси чистых белков, жиров
и углеводов, даже если ему будет доставлен весь необхо-
димый неорганический материал».
Мечта Лунина изучить природу и физиологическое
значение открытых им новых веществ была успешно осу-
ществлена польским биохимиком Казимиром Функом.
В 1911 г. Функ впервые выделил из дрожжей и отру-
бей кристаллическое вещество, которое в дозе 0,004 г
исцеляло голубя от заболевания (полиневрита), напоми-
нающего бери-бери. Определяя химический состав этого
вещества, Функ нашел, что оно содержит азот в виде
аминной группы. Желая подчеркнуть жизненно важное
значение открытых Луниным веществ и предполагая, что
все они содержат аминную группу, он предложил их на-
зывать витаминами (жизненный аминами), а заболева-
ния, возникающие на почве недостатка в пище витами-
нов — авитаминозами. Заслуга Функа состоит не только
в том, что он впервые получил кристаллический препа-
рат, излечивающий полиневрит у птиц, но и в том, что, обоб-
щив имеющиеся факты, далее развил открытие Н. И. Лу-
нина. Функ писал: «Витамины находятся в нашей пище в
весьма малых количествах и, тем не менее, они самым
поразительным образом доминируют над всей картиной
обмена...». «Таким образом,— делает вывод Функ, к из-
6
вестному перечню элементов пищи — белкам, жирам,
углеводам, пуринам, липоидам и минеральным солям —
нужно прибавить новую группу, а именно — «вита-
мины».
Представления Лунина и Функа вначале не нашли
широкого признания. Многие исследователи продолжали
объяснять развитие цынги, бери-бери, рахита не отсут-
ствием витаминов, а изменением качества белка, появле-
нием в процессе пищеварения ядовитых веществ и т. п.
Однако дальнейшими иссследованиями была установле-
на необходимость наличия в пище витаминов для нор-
мальной жизнедеятельности животных и человека, но мне-
ние Функа о том, что все витамины имеют в своем составе
азот, не подтвердилось.
Накопленный материал позволяет оха-
рактеризовать «витамины» как группу ор-
ганических веществ разнообразной хими-
ческой природы, необходимых в ничтож-
ных количествах для нормальной жизне-
деятельности организма.
Чем больше мы узнаем о природе и биологическом
действии витаминов, тем яснее становится огромное
значение открытия Н. И. Лунина, сделанного 75 лет на-
зад. Только после Великой Октябрьской социалистиче-
ской революции, когда русская наука вышла на широкий
простор, открытие Н. И. Лунина нашло не только все-
общее признание, но и получило необычайно быстрое раз-
витие. Наряду с крупными исследованиями по физиоло-
гии и биохимии витаминов, советскими учеными сделан
большой вклад и в разработку способов искусственного
получения витаминов путем химического синтеза. Успехи
советской витаминологии быстро и широко внедряются в
народное хозяйство. Витамины стали неотъемлемой
частью высококачественной пищи советских людей.
Учение о витаминах содействовало и более быстрому
развитию многих разделов биологии и особенно медици-
ны. Академик А. Н. Бах писал: «Трудно найти такой раз-
дел физиологии и биохимии, который не соприкасался бы
с учением о витаминах. Обмен веществ организма, дея-
тельность органов чувств, функции нервной системы, фер-
ментативные процессы, явления роста и размножения
и т. д.? все эти разнообразные и коренные по своей важ-
7
ности области биологических дисциплин теснейшим обра-
зом связаны с витаминами».
Одновременно с изучением роли витаминов в жизне-
деятельности человека и животных, в последнее время
значительное внимание исследователей было обращено на
условия образования витаминов в растениях и на выяс-
нение значения этих веществ для микроорганизмов и
высших растений.
Накопленный за последние годы значительный факти-
ческий материал указывает на важную роль витаминов в
жизнедеятельности не только человека и животных, но и
растений. Вместе с тем этот материал позволяет расши-
рить область применения витаминов и еще более деталь-
но познать многие стороны жизни высших и низших ра-
стений.
В настоящей брошюре мы и поставили перед собой
задачу — дать краткий обзор данных о биосинтезе и зна-
чении витаминов в жизнй* растений — основных постав-
щиков витаминов для человека и животных.
Гл а в а 1
ОБЩИЕ СВОЙСТВА ВИТАМИНОВ
1. Характеристика свойств витаминов
Успешное использование ряда растительных продук-
тов для борьбы с авитаминозами побудило химиков,
биохимиков и физиологов развернуть широкие исследо-
вания по выделению из растений веществ, обладающих
витаминными свойствами, и всестороннему их изуче-
нию. Эти исследования требовали огромных усилий ввиду
ничтожно малого содержания витаминов в растительных
материалах, но они принесли человечеству замечательные
успехи — были выделены и подробно изучены многие ви-
тамины. Получение витаминов в чистом виде и подробное
изучение их физико-химических свойств позволило при-
ступить к синтезу витаминов и принять радикальные ме-
ры борьбы с различными авитаминозами. Вместе с тем
эти успехи обогатили наши знания в области обмена ве-
ществ как у животных, так и у растений.
Витамины обозначаются большими буквами латин-
ского алфавита: витамин А, витамин С, витамин D и т. д.
Кроме того, после установления химической структуры
некоторые витамины получали соответствующие хими-
ческие названия. Например, тиамин (витамин Bi), рибо-
флавин (витамин В2), никотиновая кислота (вита-
мин РР) и т. д. Витамины принято классифицировать
по их растворимости в воде и в жирах. К воднораствори-
мым относятся, например, витамины Bi, В2, В6, РР, Н,
Р, С, фолиевая кислота, пантотеновая кислота, инозит,
пара-аминобензойная кислота и др.; к жирораствори-
мьш — вйтаминьгА,'D, Е и К.
9
В настоящее время обнаружено более пятнадцати
витаминов. Здесь мы рассмотрим лишь те из них, био-
синтез и значение которых в жизнедеятельности расте-
ний наиболее изучены.
Воднорастворимые витамины
К воднорастворимым витаминам относятся многочис-
ленные витамины, известные как «комплекс витаминов В».
Наиболее изучены витамины Bi, В2, Вб, В12, РР, И, панто-
теновая кислота, инозит, пара-аминобензойная кислота,
фолиевая кислота.
Витамин Bi, или тиамин
Около 60 лет назад было установлено, что в рисовых
отрубях содержится вещество, легко растворимое в воде,
предохраняющее птиц от заболевания полиневритом.
В 1911 г. Функ выделил из отрубей риса это вещество в
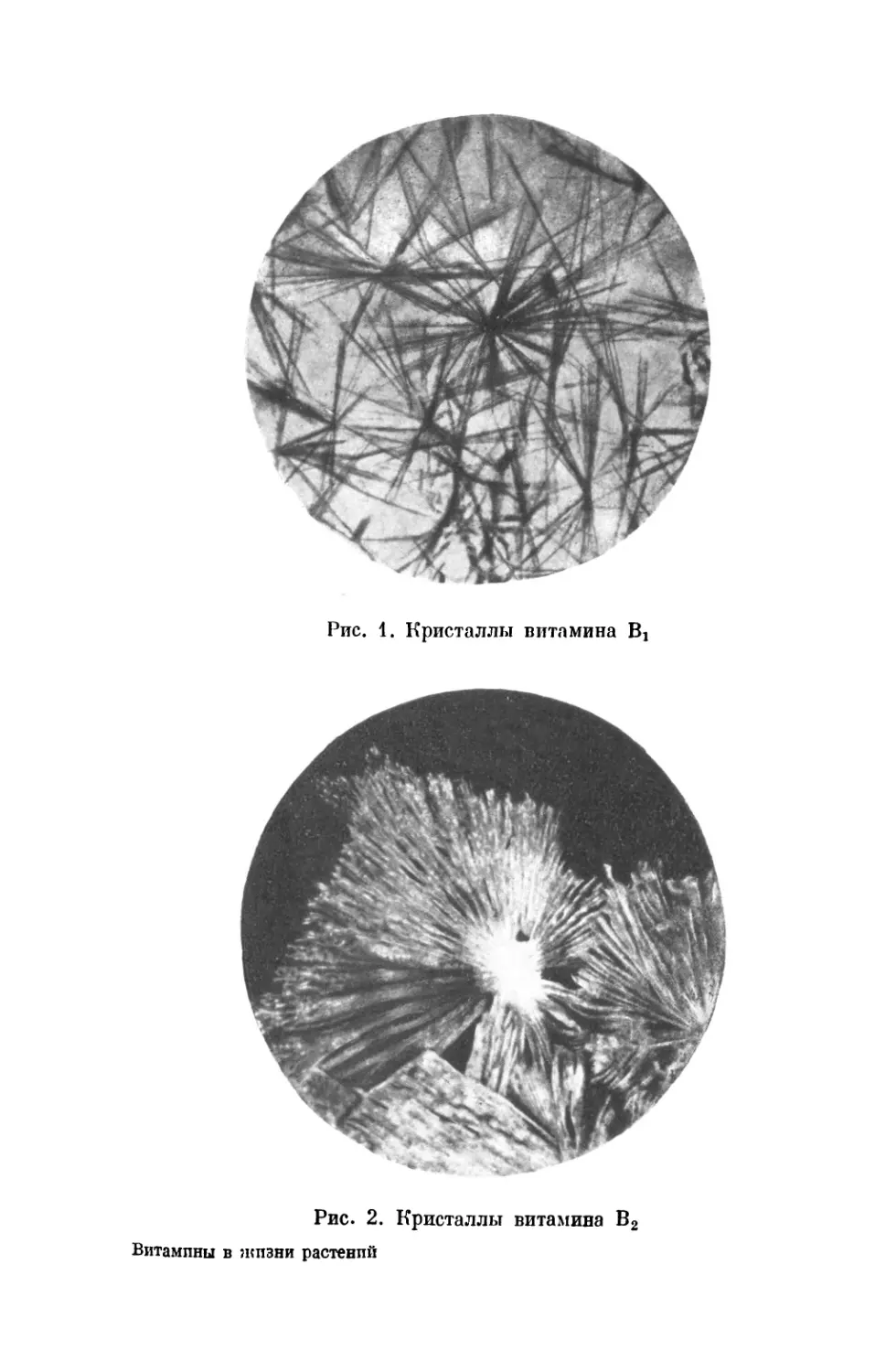
кристаллическом виде (рис. 1) и показал, что его добав-
ление в диету излечивает голубей от полиневрита. Через
год активные препараты витамина Bi были получены
Функом из дрожжей, молока, лимона, мозга быка.
Витамин Bi имеет следующее строение:
Витамин Bi
Как видно из структурной формулы, витамин Bi со-
стоит из пиримидиновой (слева) и тиазоловой части
(справа).
Витамин Bi хорошо растворим в воде. В кислой сре-
де он устойчив к действию высокой температуры, тогда
как в щелочной — быстро разрушается при нагревании.
Синтез этого витамина осуществлен в лаборатории
более 15 лет назад. В настоящее время предложено не-
сколько промышленных способов оинтеза витамина В|.
10
Среди этих способов оригинальным и простым является
синтез, осуществленный советскими химиками.
Витамин В2, или рибофлавин
Давно отмечалось, что у мышей при содержании их
на очищенных диетах в некоторых случаях наблюдается
облысение мордочки, помутнение роговицы глаз и при-
остановка роста. У человека появлялись заболевания, со-
провождающиеся понижением температуры тела, ослабле-
нием пульса, частоты дыхания и, наконец, потерей созна-
ния.
Вначале считалось, что это заболевание у человека и
животных вызывается отсутствием в пище витамина Вь
Однако вскоре выяснилось, что введение в пищу витами-
на В1 не устраняло заболевания, тогда как добавление к
диете автоклавированных дрожжей и мяса предохраняло
от него. Эти факты позволили прийти к заключению, что
в дрожжах и мясе, кроме витамина Bi, имеется еще
какой-то другой витамин. Выделенное из сыворотки мо-
лока, мяса, дрожжей, печени и растительных тканей ве-
щество, введенное в рацион, полностью излечивало жи-
вотных и человека. Это вещество было названо витами-
ном В2. Его формула Си H2oN406.
Рибофлавин представляет собою игольчатые кристал-
лы (рис. 2) желто-оранжевого цвета, растворимые в во-
де и не растворимые в обычных жирорастворителях.
Витамин В2 имеет следующее строение:
Витамин В8
И
Пути биологического синтеза рибофлавина до сих пор
недостаточно выяснены, хотя в лаборатории он был синте-
зирован около 20 лет назад.
Витамин Вб, или пиридоксин
20 лет назад этот витамин впервые нашли в дрожжах,
а в 1939 г. была установлена его элементарная и струк-
турная формула:
Витамин Вб
Витамин В6 представляет собою бесцветные мелкие
кристаллы, хорошо растворимые в воде и в этиловом
спирте. Этот витамин устойчив к действию высокой тем-
пературы, но разрушается на свету. Витамин Вб синтези-
рован в лаборатории. Выяснена его биологическая роль.
При недостатке в диете пиридоксина у животных
отмечаются припадки, напоминающие эпилептические.
При Вб-авитаминозе у крыс, кроме того, наблюдалось по-
ражение кожи.
Витамин В12
Хотя около 30 лет назад было установлено наличие в
печени вещества, оказывающего поразительное действие
при пернициозной анемии (злокачественном малокро-
вии) , прошло около 20 лет, прежде чем его удалось выде-
лить в кристаллическом виде.
В отличие от всех других известных витаминов, этот
витамин содержит в своем составе микроэлемент кобальт.
Витамин В12 кристаллизуется в иглах или призмах орто-
ромбической системы. Он весьма устойчив к воздействию
12
высокой температуры. Кристаллический витамин Bi2 не
разлагается при нагревании до 100° в течение продолжи-
тельного времени. При действии на этот витамин веществ,
обладающих сильными окислительными и восстанови-
тельными свойствами, а также тяжелых металлов он бы-
стро теряет активность.
Витамин В12 в настоящее время получают из бакте-
рий и грибов, где он накапливается иногда в значитель-
ных количествах.
Витамин В12 содержится также в продуктах живот-
ного происхождения и почти отсутствует в тканях выс-
ших растений.
Витамин Вс, или фолиевая кислота
Этот витамин получил свое название от латинского
слова фолиум — лист. Давно было замечено, что для ро-
ста молочнокислых бактерий необходимо вещество, на-
ходящееся в значительных количествах в листьях. Дей-
ствительно, это вещество было выделено из листьев и на-
звано фолиевой кислотой.
В настоящее время витамин Вс получен в кристалли-
ческом виде и синтезирован. Фолиевая кислота представ-
ляет собой желтоокрашенные игольчатые кристаллы.
Витамин Вс и несколько его производных успешно
синтезированы А. В. Труфановым и В. А. Кирсановой.
Кристаллический витамин Вс слабо растворим в воде.
Витамин РР, или никотиновая
кислота
Этот витамин широко распространен в растительном и
животном мире. Изучение природы витамина РР пока-
зало, что он представляет собою никотиновую кислоту
(C6H5O2N). Витаминной активностью обладает также ве-
щество, легко образующееся из никотиновой кислоты —
амид никотиновой кислоты.
Витамин РР получают путем окисления никотина,
а амид никотиновой кислоты образуется из никотиновой
кислоты при взаимодействии ее с аммиаком. Витамин
13
Pf> слабо растворим в воде. Он имеет форму белых
кристаллических игл (рис. 3).
Никотиновая кислота
Н
Амид никотиновой кислоты
Синтез никотиновой кислоты был осуществлен более
60 лет назад путем окисления никотина перманганатом,
хромовой или азотной кислотой, но лишь в 1937 г. было
установлено, что никотиновая кислота и ее амид облада-
ют витаминной активностью и могут излечивать челове-
ка и животных от пеллагры.
Пара-аминобензойная кислота
Открытие этого витамина связано с выяснением дей-
ствия сульфамидных препаратов (белого стрептоцида,
сульфидина и других) на микроорганизмы. Было установ-
лено, что введение этих веществ в питательную среду
сильно угнетает рост бактерии — гемолитического стреп-
тококка. Однако при введении в культуру дрожжей это
угнетающее действие белого стрептоцида устраняется, бак-
терии вновь начинают нормально расти. Анализ благо-
приятного действия дрожжей на рост гемолитических
бактерий показал, что дрожжи выделяют в среду пара-
аминобензойную кислоту, которая и нейтрализует вред-
ное действие сульфамидных препаратов. Прямые опыты
14
Подтвердили это предположение, введение в питательную
среду синтетической пара-аминобензойной кислоты дей-
ствительно снимало губительное действие белого стреп-
тоцида и других сульфамидных препаратов на бакте-
риальную клетку. Аналогичное явление было обнаружено
и у высших растений. Так, например, в 1%-ном раство-
ре сульфаниламида семена риса совершенно не прораста-
ют, тогда как при одновременном введении в семена
этого препарата и пара-аминобензойной кислоты семена
прорастают так же энергично, как и в воде. Этот вита-
мин, принимающий участие в синтезе аминокислот, не-
обходим для человека, животных и растений. Природа
пара-аминобензойной кислоты установлена. Витамин
имеет следующее строение:
соон
I
НС сн
нс сн
V
NH,
Пара-аминобензойная кислота
Бесцветные кристаллы пара-аминобензойной кислоты
хорошо растворяются в спирте и эфире и плохо раствори-
мы в воде.
Пантотеновая кислота, или пантотен
Этот витамин был выделен из рисовых отрубей более
15 лет назад. Вскоре выяснилось, что он встречается у
всех растений и животных. Широкое распространение и
необходимость этого витамина для живых существ отра-
жается и в самом его названии «пантос», что по-грече-
ски. означает «всюду», или «вездесущий». Пантотеновая
кислота является необходимой для нормального роста
многих микроорганизмов и изолированных органов выс-
ших растений. Этот витамин имеет формулу C9H17O5N и
следующее строение:
15
Н3сч
Н3С-С - СНОН -CONH - сн2- СН2СООН
HOH2CZ
Пантотеновая кислота
Пантотеновая кислота — светлое вязкое маслообраз-
ное вещество. Она и ее кристаллические соли хорошо ра-
створимы в воде и не растворяются в жировых раствори-
телях.
ВитаминН, или биотин
При изучении азотфиксирующей бактерии было уста-
новлено, что эта бактерия требует для своего роста какое-
то вещество, выделяемое другой бактерией — азотобакте-
ром. Однако оказалось, что подобное вещество присутству-
ет не только в культурах азотобактера, но и во многих
других микроорганизмах и в тканях высших растений.
Интересно отметить, что еще до установления этих
фактов было известно, что сырой белок куриного яйца,
будучи введен в рацион в значительных количествах, при-
водит к характерному заболеванию кожи у мышей — облы-
сению вокруг глаз. В том случае, если в пищу вводились
дрожжи с яичным белком, токсическое действие яичного
белка не проявлялось. Вещество, нейтрализующее токси-
ческое действие этого белка, было названо витамином Н.
Получение биотина в кристаллическом виде позволило
изучить потребность разных животных и микроорганизмов
в витамине и установить характерные симптомы этого
авитаминоза. Синтез витамина Н осуществлен в лабора-
тории и установлена его химическая природа.
О
KN NH
НС — сн
I I
н2с сн-сн2-сн2-сн2-сн2соон
Витамин Н
16
Рис. 2. Кристаллы витамина В2
Витамины в жизни растений
Рис. 3. Кристаллы никотиновой кислоты
Рис. 4. Кристаллы витамина Н (биотина)
Рис. 5. Кристаллы витамина С
Рис. 6. Кристаллы каротина (провитамина А)
Витамины в жизни растений
Рис. 7. Кристаллы витамина Д2 (кальциферола)
Витамин Н представляет собой тонкие иглообразные
кристаллы (рис. 4). Он хорошо растворяется в воде и ал-
коголе и весьма устойчив к действию высокой темпера-
туры.
Инозит
Этот витамин широко распространен у растений и жи-
вотных. В 1935 г. была доказана необходимость инозита
для нормального роста некоторых микроорганизмов, а
через пять лет — для животных.
/CJHOH
НОНС СНОН
нонс^ СНОН
cflOH
Инозит
Инозит — шестиатомный спирт. Трудно растворим в
воде, а в спирте и эфире совсем не растворяется. В ра-
стениях этот витамин находится в связанном с фосфорной
кислотой соединении, известном под названием фитин.
Инозит синтезирован в лаборатории.
Витамин Р
Витамин Р в настоящее время выделен из многих ра-
стений. При недостатке витамина Р у животных и чело-
века наблюдается повышенная проницаемость и хруп-
кость кровеносных сосудов. Р-витаминной активностью
обладает целый ряд находящихся в растениях полифено-
лов. Наиболее активными из них являются катехины и, в
частности, эпикатехин.
Эпикатехин
2 Витамины в жизни растений
17
А. Л. Курсановым и М. Н. Запрометовым подробно
изучены катехины чайных листьев, что позволило более
полно представить биосинтез этих чрезвычайно важных
для медицинских целей соединений и разработать способы
их промышленного получения.
Витамин С, или аскорбиновая
кислота
Витамин С выделен в кристаллическом виде из ра-
стений ранее того, как стало известно, что именно это
вещество может избавить человека от заболевания цын-
гой. В 1927 г. Сцент-Гиорги, выделив из капусты, апель-
синов и надпочечников животных вещество, названное
им гексуроновой кислотой, лишь четыре года спустя уста-
новил, что эта кислота и витамин С — одно и то же. Ви-
тамин С представляет собою бесцветные кристаллы
(рис. 5), очень хорошо растворимые в воде:
I
С —ОН
S-OH '
н-{------
но—сн
СН2ОН
Витамин С
Изучение химической природы и физико-химических
свойств витамина С показало, что этот витамин пред-
ставляет собой соединение простого химического соста-
ва. (СбНвОб). Благодаря наличию в его структуре группы
А гчи витамин С обладает сильно выраженными
—с—он
восстановительными свойствами. В настоящее время пред-
ложен ряд способов промышленного синтеза аскорбино-
вой кислоты.
18
Жирорастворимые витамины
Витамин А
Еще в прошлом столетии из растений был выделен
пигмент каротин (рис. 6), а в начале нашего века под-
робно изучалась его физиологическая роль в растении.
Особенно оригинальные работы по изучению роли каро-
тина в растении выполнены нашими соотечественниками
Д. И. Ивановским и М. С. Цветом.
H3Qj
с
Н2С
СНз СНз
с-сн=сн-с=сн—сн=сн—с=сн-сн=ск С.н=
н2с
с-снч
/ *
СНз
СНз
=с-сн=сн -сн=с-сн=сн-с сн2
HSC-C сн2
\н2
р-каротин
Однако лишь в начале 30-х годов было эксперимен-
тально доказано, что широко распространенный в расте-
ниях пигмент желтого цвета — каротин, будучи введен-
ным в организм животного, превращается в витамин А.
Вещества, являющиеся, подобно каротину, предшествен-
никами витаминов, обычно называют провитаминами.
Н3С СНз
С СНз СНз
К,С/ХС-СН=СН-С=СН-СН = СН-С=СН-СНзОН
I II
Н2С С-СНз
хсн2
Витамин А
2*
19
Витамин А представляет собою светложелтые кри-
сталлы, хорошо растворимые в жирах и органических ра-
створителях.
Механизм превращения каротина в витамин А в орга-
низме животных до сих пор окончательно не расшифро-
ван. Значительное внимание уделяется изучению биосин-
теза каротина, однако до сих пор не удалось установить-
те звенья реакции, которые приводят к его образованию.
Витамин D, или антирахитический
витамин
В настоящее время установлено, что способностью
излечивать рахит обладает ряд веществ. Они обознача-
ются как витамины группы D: D2, D3, D4, D5. Эти вита-
мины в растениях не обнаружены. В растениях образу-
ются провитамины, из которых при соответствующих
условиях происходит синтез витаминов группы D. Так,
например, при облучении ультрафиолетовым светом эрго-
стерина — вещества, широко распространенного в расте-
ниях, грибах и дрожжах, образуется витамин D2. Другие
витамины группы D (D3, D4, D5) также образуются при
облучении соответствующих провитаминов ультрафиоле-
товыми лучами.
Особенно много эргостерина накапливается в дрож-
жах и низших грибках, откуда он и добывается для про-
мышленных целей.
Витамин D2 представляет собою бесцветные кристал-
лы (рис. 7).
Эргостерин
20
СН3 СНз СНз
Cf-к I । I
3 сн —сн=сн—сн—сн—сн3
н
Витамин D2
Витамин Е, или токоферол
Витамин Е широко распространен в растениях. Осо-
бенно много его находится в пыльце, в зародышах пше-
ницы и других злаков. При недостатке витамина Е у крыс
нарушаются процессы формирования половых клеток,
оплодотворение и развитие зародыша. Поэтому витамин Е
часто называют антистерильным. В 1937 г. была уста-
новлена структурная формула витамина Е, который полу-
чил название токоферола, а через год был осуществлен
синтез этого витамина. Свое название витамин Е, или то-
коферол (tocos — роды; phero — производить) получил
в связи с той важной ролью, которую он выполняет в про-
цессах размножения. Установлено, что биологическим
действием обладает не только а-токоферол, но и два дру-
гих токоферола: ₽- и у-токоферол.
“СН—(CHJ3—СН—(СН2)3—СН—СН3
СНз СНз СНз
а-токоферол
Все три вещества, обладающие биологическими свой-
ствами витамина Е, представляют собой маслообразные
вещества, совершенно не растворимые в воде и хорошо
21
растворимые в ацетоне, в серном и петролейном эфирах
и в других органических растворителях. Витамин Е устой-
чив к действию высокой температуры, но весьма чувстви-
телен к воздействию ультрафиолетовых лучей.
Витамин К
Известны три вещества, обладающие активностью
витамина К: Ki, Кг и Кз.
О
Н3
Н2СН=С(СН2)з-СН(,СН2)з-СНГСН2)з-СН-СН3
СН3 СПз СНз СН3
Витамин Ki
Витамин Ki — светложелтое вещество маслянистой
консистенции, не растворимое в воде, но хорошо раство-
римое в эфире, бензоле и в других жирорастворителях.
Биосинтез этого витамина в растениях осуществляется
при участии фитола. Некоторые бактерии синтезируют
витамин Кг, имеющий вместо фитола несколько более
удлиненную боковую цепь. Активной оказалась и одна
циклическая часть молекулы витаминов Ki и Кг, назван-
ная витамином Кз-
Советские ученые внесли большой вклад в выясне-
ние природы витаминов группы К* А. В. Палладии и
А. А. Шмук синтезировали витамин Кз, а М. М. Шемя-
киным был расшифрован механизм действия этого вита-
мина.
Из весьма краткого обзора следует, что за последние
два десятилетия выяснена химическая природа значи-
тельного числа известных в наше время витаминов. Боль-
шие успехи достигнуты и в осуществлении их синтеза.
Эти успехи позволили не только найти эффективные ме-
тоды лечения авитаминозных заболеваний, но также
22
содействовали решению другой важнейшей стороны уче-
ния о витаминах — выяснению роли витаминов в процес-
сах обмена веществ в организме.
2. Роль витаминов в обмене веществ
организма
«Короли и капуста одинаково нуждаются в аскорбино-
вой кислоте»,— говорил крупнейший венгерский биохимик
Сцент-Гиорги, установивший химическую природу вита-
мина С. Этот афоризм справедлив не только по отноше-
нию к витамину С, но и к другим витаминам. Хорошо
известно, что все организмы нуждаются в витаминах, хо-
тя они по-разному обеспечивают себя этими незаменимы-
ми элементами питания. Одни организмы синтезируют
витамины в достаточном количестве и даже выделяют
в окружающую среду; другие — совсем не обладают спо-
собностью синтезировать витамины самостоятельно или
образуют их в недостаточном количестве и требуют вве-
дения витаминов извне.
Так почему же витамины нужны организму? Как
известно, формирование свойств и особенностей растений
и животных связано с совершающимися в них процесса-
ми обмена веществ. Ф. Энгельс указывал, что из обмена
веществ вытекают все проявления жизни. Ведущую роль
обмена вещества в жизни организма подчеркивал и
К. А. Тимирязев. Он писал: «Основное свойство, характе-
ризующее организмы, отличающее их от неорганизмов,
заключается в постоянном обмене между их веществом и
веществом окружающей среды. Организм постоянно вос-
принимает вещество, превращает его в себе подобное
(усвояет, ассимилирует), вновь изменяет и выделяет.
Жизнь простейшей клеточки, комка протоплазмы, суще-
ствование организма слагается из этих двух превраще-
ний: принятия и накопления — выделения и траты веще-
ства»1. '
Хотя витамины содержатся в организме в незначи-
тельных количествах, они тем не менее выполняют в нем
многообразные, важные функции. Отсутствие или не-
достаток того или иного витамина приводит к нарушению
1 К. А, Тимирязев. Соч., т. V, 1940, стр. 146.
23
течения физиологических процессов, к появлению харак-
терных признаков витаминной недостаточности — авита-
миноза и гиповитаминоза. Последние обычно обозначают
буквами, соответствующими названию того витамина, от-
сутствие которого приводит к появлению специфического
заболевания (например, Bi-авитаминоз, О2-авитаминоз
и т. д.).
Остановимся на тех звеньях процесса обмена веществ,
в которых витамины принимает непосредственное уча-
стие. В живых организмах широко распространены биоло-
гические катализаторы — ферменты, ничтожно малые ко-
личества которых способны в сотни и тысячи раз ускорять
те или иные биохимические реакции. При непосредствен-
ном участии ферментов происходит превращение веществ:
образование и разрушение углеводов, жиров, белков, ор-
ганических кислот и аминокислот, осуществление взаимо-
связи между организмом и окружающей их средой. В от-
сутствии ферментов многие жизненно необходимые реак-
ции в организме или вообще не могут протекать, или про-
текают так медленно, что не в состоянии обеспечить нор-
мальную жизнедеятельность организма. Так, А. Н. Бах
указывал, что без содействия пищеварительных фермен-
тов на превращение крахмала в сахар, белка в пептон
под влиянием воды потребовались бы не десятки минут,
а десятки месяцев. Можно сказать, что без ферментов не
было бы жизни, так как без них организм не в состоя-
нии усваивать и перерабатывать питательные вещества.
В образовании многих ферментов огромную роль игра-
ют витамины.
Академик Н. Д. Зелинский в своем предисловии к
русскому изданию монографии К. Функа «Витамины»
писал в 1922 г.: «Химическая природа витаминов, как по-
казали исследования Функа и других авторов, начинает
понемногу выясняться, и, может быть, мы скоро будем в
состоянии определенно ответить на вопрос,— что пред-
ставляет та группа органических соединений, присутствие
которой в комплексной форме с другими телами столь
важно и необходимо для всего обмена и роста организ-
ма...
Связь между ферментами и витаминами, возможно, и
выражается в том, что последние необходимы как строи-
тельный материал для первых».
24
Предсказание нашего выдающегося химика было эк-
спериментально подтверждено последующими исследова-
ниями.
Как известно, ферменты в большинстве случаев пред-
ставляют собою сочетание активной группы (так называе-
мого кофермента, вступающего в химическое взаимодей-
ствие с субстратом) и коллоидального белкового «носи-
теля». Оказалось, что активной группой многих фермен-
тов являются витамины.
В зависимости от того, с каким белком соединяется
кофермент, образуется тот или иной фермент. Так, на-
пример, витамин Вг образует кофермент, который, соеди-
няясь более чем с 10 белками, дает начало образованию
соответствующего числа ферментов с совершенно различ-
ными физиологическими функциями.
Данные о том, что витамины являются составной ча-
стью ферментов, впервые были получены около 20 лет
назад. Удалось, например, экспериментально установить,
что при соединении витамина Bi с двумя молекулами фос-
форной кислоты образуется кофермент карбоксилазы —
кокарбоксилаза.
Карбоксилаза — фермент, широко распространенный
в растениях и животных,— принимает непосредственное
участие в превращении углеводов. Как известно, посту-
пившие или образовавшиеся в организме углеводы пре-
терпевают глубокие изменения, прежде чем они могут
быть использованы для построения белков и других
жизненно важных соединений. Из углеводов при помо-
щи ферментов образуется пировиноградная кислота —
СНз • СО • СООН, служащая источником образования
аминокислот, альдегидов, органических, в том числе жир-
ных кислот и других соединений, необходимых для нор-
мальной жизнедеятельности организма. Иначе говоря, пи-
ровиноградная кислота является узловым звеном обмена
веществ в живой клетке, обусловливающим взаимное
превращение углеводов, белков и жиров. В ее превраще-
ниях существенную роль играет фермент карбоксилаза,
в состав активной группы которого входит витамин Вь
При отсутствии или недостатке витамина Bi задер-
живается образование кофермента, что приводит к рез-
кому снижению активности карбоксилазы. В результате
25
этого углеводный обмен в организме останавливается на
стадии пировиноградной кислоты, и дальнейшие процес-
сы образования жизненно необходимых соединений также
задерживаются. Кроме того, накопление в тканях значи-
тельных количеств этой кислоты оказывает вредное дей-
ствие на организм. Такое нарушение обмена веществ
приводит к задержке ростовых процессов как у выс-.
ших растений, так и у микроорганизмов. Многочис-
ленными опытами было доказано, что при недостатке
витамина Bi клетки бактерий, дрожжей и грибов
уменьшаются в размере и размножение их задержи-
вается.
При участии витамина Bi происходит не только от-
щепление углекислоты от пировиноградной кислоты, но и
противоположная реакция, т. е. реакция присоединения
углекислого газа к пировиноградной кислоте- Установле-
ние этого факта имеет огромное научное значение. Как
известно, А. Ф. Лебедев 30 лет назад установил возмож-
ность фиксации СО2 микроорганизмами. Это выдающееся
открытие вызвало вначале большие сомнения, так как
считалось общепризнанным, что усвоение углекислого га-
за возможно только на свету при участии хлорофилла зе-
леного растения. Однако последующие работы полностью
подтвердили справедливость выводов нашего соотече-
ственника и внесли значительный вклад в выяснение при-
роды этого процесса. Оказалось, что «темновая фиксация»
углекислоты свойственна не только микроорганизмам, но
также и зеленым растениям и что роль витаминов в этой
реакции весьма велика. А. Л. Курсанов (1953) показал,
что усвоение углекислого газа корнями высших растений
происходит без участия света и играет существенную роль
в общем балансе усвоения СО2 зелеными растениями. Это
далеко не полный перечень функций, выполняемых в
обмене веществ витамином Вь
Еще более многообразны функции других витаминов,
особенно тех, которые принимают участие в образовании
и превращении белков.
Белки являются основой жизни. Еще на заре развития
химии и биохимии белка Ф. Энгельс писал: «Повсюду,
где мы встречаем жизнь, мы находим, что она связана
с каким-либо белковым телом, и повсюду, где мы встре-
чаем какое-либо белковое тело? которое не находится в
26
процессе разложения, мы без исключения встречаем *и
явления жизни»
Витамины, сами являющиеся продуктами обмена бел-
ков, принимают непосредственное участие в образовании
и превращении белков в организме.
Классическими работами академика Д. Н. Прянишни-
кова давно была выяснена общая картина образования и
превращения белков в растениях. Он показал, что так
называемые дикарбоновые аминокислоты (аспарагиновая
и глютаминовая) и их амиды (аспарагин и глютамин) яв-
ляются первыми продуктами синтеза азотистых веществ.
При образовании этих веществ одновременно происходит
обезвреживание аммиака в организме, что имеет важное
значение для нормальной жизнедеятельности растений-
Дальнейшие исследования, посвященные выяснению
механизма образования аминокислот, показали, что в
этом процессе непосредственное участие принимают ви-
тамины. Советские ученые А. Е. Браунштейни М. Г. Криц-
ман, открыв реакцию переаминирования, т. е. переноса
аминогруппы (NH2) с одного соединения на другое, пока-
зали, что витамин Вб принимает участие в этой реакции.
При недостатке витамина Вб снижается синтез амино-
кислот из пировиноградной кислоты или других кетоки-
слот и аммиака. При необеспеченности организма вита-
мином Вб задерживается и обратная реакция, т. е. обра-
зование кетокислот из аминокислот.
Витамин Вб принимает также непосредственное уча-
стие в образовании аминокислот из органических кислот
и аммиака путем прямого присоединения к ним аммиака.
Так, в результате присоединения аммиака к щавелевоук-
сусной кислоте образуется широко распространенная в
растениях аминокислота — аспарагиновая.
СООН-СН2СО-СООН+ NH3 + 2Н = COOH-CH2.CH-NH2-COOH+H2O
Шавелевоуксусная
кислота
Ам- Во-
ми- до-
ак род
Аспарагиновая
кислота
Вода
Когда же происходит присоединение аммиака к дру-
гим органическим кислотам — пировиноградной и кето-
1 Ф. Энгельс. Анти-Дюринг, 1953, стр. 77.
27
глютаровой — образуются соответственно новые амино-
кислоты — аланин и глютаминовая.
Важное значение витамин В6 имеет также и в обра-
зовании аминокислот путем так называемого переамини-
рования, т. е. переноса аминогруппы (NH2) из амино-
кислот на органические кислоты, в результате чего исход-
ная аминокислота превращается в органическую кислоту,'
а исходная органическая кислота в соответствующую ами-
нокислоту.
Примером переаминирования является следующая ре-
акция:
СООН
СН2 СН3
HG — NH24-CO
/ I
СООН СООН
Глютаминовая Пировиноград-
кислота ная кислота
СООН
сн2
I
СНа СНз
со + нс — nh2
I /
СООН СООН
Кетоглютаро- Аланин
вая кислота
В синтезе аминокислот и белков также принимают
участие витамины В2, В!2, РР, Н и др.
В свете последних представлений о биологической роли
корневой системы у растений, развиваемых А. Л. Кир-
сановым, витаминам принадлежит весьма важная роль.
Как известно, согласно этим представлениям, в кор-
нях растений имеет место значительное образование ща-
велевоуксусной и кетоглютаровой кислот. Последние,
присоединяя к себе азот аммонийных удобрений при по-
мощи ферментов, содержащих витамины, образуют со-
ответствующие аминокислоты. В настоящее время извест-
но до 14 важнейших аминокислот, образующихся в кор-
нях. Из корней эти кислоты поднимаются вверх по расте-
нию, поступают в растущие побеги и плоды, где и исполь-
зуются для образования белков.
Таким образом, синтез и превращение аминокислот —
составных частей белка, происходят при непосредствен-
ном участии витаминов. Без витаминов не может обра-
зоваться белок и не могут проявиться его жизненные
свойства.
28
Распад сахаров в тканях растений и животных приво-
дит также к образованию жиров. В синтезе жиров из са-
харов принимают участие витамины Вь В2, РР и пан-
тотеновая кислота, а при превращении в жиры белков
необходим еще пятый витамин — Вб. Установление
факта непосредственного участия пантотеновой кислоты
в образовании жиров из более простых соединений было
использовано для синтеза жиров вне организма.
Огромная роль принадлежит витаминам и в процессе
дыхания. Как известно, дыхание является важнейшим
источником энергии у растений и животных, используемой
для Осуществления синтеза жизненно необходимых со-
единений, а также для процессов роста и движения. Вме-
сте с тем дыхание является источником образования со-
единений, которые служат исходным материалом для мно-
гих реакций синтеза. Процесс дыхания осуществляется в
организме при помощи ферментов. По образному выра-
жению А. Н. Баха, без дыхательных ферментов организм
задохнулся бы в атмосфере чистого кислорода.
20 лет назад был впервые выделен фермент, прини-
мающий непосредственное участие в дыхании. Изучение
химической природы фермента показало, что он состоит из
белка и кофермента. Последний представляет собой со-
единение витамина В2 с фосфорной кислотой. Вслед за
этими работами появился ряд других исследований
по выделению новых ферментных систем, в которых ви-
тамин В2 выполняет окислительно-восстановительные
функции.
В настоящее время доказано, что ни одна живая клет-
ка микроорганизма, высших растений или животных не
может существовать, если в ней отсутствуют биохимиче-
ские системы, в которые входит витамин В2. Это и понят-
но, ибо там, где есть жизнь, должно быть дыхание, а там,
где есть дыхание, должны быть ферментные системы, осу-
ществляющие окислительные реакции.
Большую роль в дыхании растений выполняет и фоли-
евая кислота. Не случайно, что при прорастании семян,
когда особенно интенсивен обмен веществ, идет быстрое
накопление в проростках фолиевой кислоты, витамина В2
и витамина С. Одновременно с возрастанием содержания
этих витаминов повышается и интенсивность дыхания про-
ростков. В том случае, если прорастание семян произво-
29
дится на растворе фолиевой кислоты, то рост проростков
ускоряется, их дыхание возрастает более чем на 40% по
сравнению с проростками, не получившими этого витами-
на. При введении фолиевой кислоты в листочки капусты
в последних также значительно увеличивается энергия
дыхания (табл. 1).
Таблица 1
Дыхание} листочков* капусты при введении
различных количеств фолиевой кислоты
(мм3 О2 на 1г сырого веса за 10 мин.)
(По Андреевой, 1953)
Вода (контроль) Концентрация фолиевой кислоты, уг
2 5 10 20
3,3 3,5 3,8 4,62 5,46
Витамины не только принимают непосредственное уча-
стие в превращении веществ, находящихся в организме, но
они участвуют также в усвоении растением веществ, по-
ступающих из окружающей среды. Раньше мы указывали
на важную роль некоторых витаминов в усвоении угле-
кислого газа. Имеющиеся материалы показывают, что ви-
тамины принимают участие и в процессах усвоения расте-
нием минеральных веществ. Установлено, например, что
витамины Bi, В2, Вб и другие участвуют в поглоще-
нии растениями из почвы фосфорной кислоты. Кроме
утилизации фосфорной кислоты, при участии витаминов
осуществляется улавливание и обезвреживание аммиака
в растении. Некоторые дрожжи при отсутствии вита-
мина Н не в состоянии использовать аммиак из питатель-
ной среды. При введении же в питательную среду этого
витамина потребление дрожжами аммиака резко воз-
растает.
В процессах усвоения азота растением существенную
роль играет и витамин С. По мнению Д. М. Михлина
(1936), этот витамин принимает участие в восстановлении
нитратов в растительных тканях.
30
Таким образом, витамины в растениях играют исклю-
чительно важную роль в превращениях веществ
и в усвоении растениями минеральных и органических
соединений. Отсутствие или недостаток того или иного
витамина приводит к существенным нарушениям обмена
веществ, снижению интенсивности физиологических про-
цессов, к анатомо-морфологическим изменениям, а иногда
и к приостановке роста растений. В. А. Энгельгардт счи-
тает, что витамины являются биохимическими факторами,
которые связывают между собой и растительный мир, и
мир животных, и мир микроорганизмов.
Изучение природы действия витаминов шло и идет
параллельно с выяснением механизма действия фермен-
тов. Такое изучение взаимно оплодотворило оба эти на-
правления, способствовало быстрому и успешному их раз-
витию.
Как указывалось выше, физиологическая активность
витаминов тесно связана с белками. Поэтому при наличии
даже значительных количеств витаминов, но при недо-
статке специфического белка, с которым связывается тот
или иной витамин, активность последнего не проявляется,
и организм страдает соответствующим авитаминозом.
С другой стороны, установлена зависимость между нали-
чием в организме витаминов В2, В6 и РР и рациональ-
ным использованием животными пищевых белков, между
содержанием в тканях витаминов А и К и использовани-
ем жиров.
Эти факты указывают на пути более эффективного
использования витаминов в практике. К сожалению, по-
добных исследований еще не проведено в отношении выс-
ших растений. Однако хорошо известно, что необеспе-
ченность растений минеральным питанием, неблагоприят-
ные условия водного режима оказывают существенное
влияние на содержание белка в растении.
Витамины в организме не изолированы. Образуясь или
поступая в организм, они вступают в соединение с дру-
гими группами веществ. Одни из таких веществ являются
необходимыми для проявления функциональной активно-
сти витаминов, другие, наоборот, инактивируют вита-
мины.
В настоящее время известен ряд антивитаминов, при-
чем для одного и того же витамина таких антагонистов
31
может быть несколько. Антивитаминами пара-аминобен-
зойной кислоты являются лечебные препараты — сульфа-
миды (стрептоцид, сульфидин). Сульфамиды активно
соединяются с белком, с которым должна соединяться
пара-аминобензойная кислота, и препятствуют проявле-
нию ее витаминной активности.
Пиридин-З-сульфокислота
Антивитамином никотиновой кислоты является ее ана-
лог — пиридин-3-сульфокислота, которая отличается от
витамина тем, что карбоксильная группа витамина заме-
нена сульфогруппой.
При введении в организм пиридин-3-сульфокислоты
или других антивитаминов последние блокируют фер-
ментный белок, вытесняя витамин РР из жизненно важ-
ных реакций. В растениях обнаружен естественный анта-
гонист витамина РР. Так, например, в некоторых злаках
содержится ацетил-пиридин — вещество, инактивирующее
витамин РР. В сладком клевере обнаружен дикумарин,
инактивирующий витамин К, а в папоротнике — вещество,
являющееся антагонистом витамина Вь Имеется ряд дру-
гих антивитаминов. Так, например, близкое по строению
к фолиевой кислоте вещество аминоптерин тормозит ее
действие, и это объясняется более прочным присоединени-
ем антивитамина к белковой основе фермента и вытесне-
нием из белковой молекулы фолиевой кислоты.
Уже отмечалось, что некоторые витамины, соединяясь
с белками, образуют активные ферментативные системы,
обеспечивающие течение сложных процессов в организме.
К таким витаминам относится и биотин. Соединяясь с бел-
ками, витамин Н образует ферменТы, принимающие уча-
стие в углеводном и белковом обмене. Но биотин жадно
связывается и с другим белком — авидином.
32
Авидин получен из белка сырых яиц, и его присутствие
в тканях приводит к Н-авитаминозу. Выяснение механиз-
ма инактивации витамина Н авидином позволило прийти
к заключению, что соединяясь с авидином в отношении
1:1, витамин Н выводится из дальнейших реакций. Та-
кой связанный витамин не в состоянии соединиться с дру-
гим белком, а потому не может образовать активные фер-
ментативные системы. Блокирование авидином витамина
Н также приводит к авитаминозу, несмотря на то, что жи-
вотные получили, казалось бы, достаточное количество
биотина.
Антивитамины не только инактивируют поступившие
в ткани витамины, но они могут и задерживать их образо-
вание. Так, например, в присутствии органического веще-
ства пиритиамина — антивитамина Bi — синтез этого ви-
тамина из пиримидина и тиазола резко задерживается.
Анализ этого явления показал, что антивитамин тормозит
активность фермента, соединяющего пиримидин и тиазол
в молекулу витамина, и тем самым снижает биосинтез ви-
тамина Вь
Как указывалось выше, источником образования вита-
мина РР является аминокислота триптофан. Оказа-
лось, что превращение триптофана в витамин задержи-
вается 3-метоксиантраниловой кислотой. Таким образом,
усиленный синтез в организме антивитаминов либо вве-
дение их извне могут явиться причинами необеспеченности
растений витаминами.
Интересно отметить, что иногда один витамин может
инактивировать действие другого. Так, например, витамин
К является антивитамином витамина РР. Было доказано,
что введение в питательную среду дополнительного ко-
личества витамина РР снимает токсическое действие
витамина К.
На судьбу витаминов в организме оказывают сущест-
венное влияние и ферменты. Так, содержание витамина
В] в некоторых растениях обусловливается Не только его
синтезом и потреблением, но и зависит от активности фер-
мента, переводящего витамин в неактивную форму (Тита-
ев, 1950). В отношении витамина С также установлено
наличие в организме специфического фермента — аскор-
биноксидазы, разрушающего этот витамин (Энгельгардт
и Букин, 1937). Следовательно, наряду с ферментами,
3 Витамины в жизни растений 33
способствующими синтезу витаминов, в организме встре-
чаются и такие ферменты, которые разрушают эти веще-
ства.
Участие того или иного витамина в обмене веществ
строго специфично и определяется его химической приро-
дой. Недостаток витамина не только препятствует осу-
ществлению биохимических реакций, в которых витамин
принимает непосредственное участие, но и течению ряда
смежных реакций, что может привести к расстройству
жизнедеятельности организма. Значительное разнообра-
зие витаминов в организме позволяет им принимать уча-
стие в самых различных процессах обмена веществ. По-
этому витамины и являются обязательными и незаменимы-
ми участниками обмена веществ, без них организм не
может существовать.
Глава II
БИОСИНТЕЗ ВИТАМИНОВ В РАСТЕНИЯХ
1. Образование витаминов растениями
Много столетий назад было известно, что цынга может
быть излечена употреблением в пищу отваров из расте-
ний. Не случайно поэтому поискам таких растений уде-
лялось исключительное внимание. Русский академик
П. С. Паллас в «Описании растений Государства Россий-
ского» в 1785 г. сообщал, что «Собираемые по концам ве-
ток молодые сосновые и кедровые вершинки похваляются
от всех наших в Сибири промышленников и мореходов
как лучшее противоцынготное и бальзамическое средство
и составляют в лечебной науке преизрядное от цынготных
болезней лекарство. Таковых сосновых вершинок выво-
зится из Государства Российского в иностранные аптеки
великое количество».
Из вывозимых за границу сосновых шишек приготов-
ляли эссенции и продавали по дорогой цене русским как
антицынготный препарат. Такой эссенцией, в частности,
пользовалась русская экспедиция Крузенштерна, совер-
шившая свое замечательное кругосветное плавание.
Начальник экспедиции снабжал своих людей не только
этой эссенцией, но два раза в день матросы «Надежды» и
«Невы» получали напиток с лимонным соком. Когда же
корабли приставали к берегу, закупалось такое количество
овощей, которого хватило бы до следующих остановок.
Успешно закончив экспедицию, ее начальник с радостью
сообщал: «За весь длинный переход от Канарских остро-
вов до Южной Америки в лазаретах не было ни одного
больного».
35
РаСтенйя йздавйа использовались для лечёнйя не
только цынги, но и других авитаминозов — заболеваний,
вызванных недостатком витаминов в пище. Однако, не
зная причин такого исцеляющего действия растительной
пищи, человек, естественно, не мог ставить ни вопроса
с количестве витаминов, необходимом для излечивания
заболеваний, ни о содержании их в тканях растений и
животных.
Лишь после получения в 1911 г. Казимцром Функом
активного препарата, излечивающего бери-бери, были
предприняты большие исследования во всех лабораториях
мира по выделению витаминов, установлению химиче-
ской природы и выяснению возможности их синтетиче-
’ с кого изготовления.
О путях синтеза некоторых витаминов
в растениях
Разработка биологических и химических методов опре-
деления витаминов способствовала поискам витаминно-
го сырья, изучению судьбы поступивших в организм
витаминов, установлению количества витаминов, необхо-
димого для предупреждения и лечения авитаминозов.
Одновременно с этим были предприняты крупные ис-
следования по выяснению образования витаминов в ра-
стительных и животных организмах. Было установлено,
что почти все витамины образуются в растениях. Лишь
витамины А и D в микроорганизмах и в высших расте-
ниях не синтезируются, но в них образуются так называ-
емые провитамины — предшественники витаминов. По-
падая в организм животного, провитамины служат ис-
точником образования^ соответствующих витаминов. Так,
например, обстоит дело с каротином — провитамином А,
который превращается в витамин А уже в теле животных.
Большинство витаминов животные не в состоянии син-
тезировать даже из провитаминов и потому требуют вве-
дения витаминов в готовом виде. Растения в этом отно-
шении отличаются от животных — они способны образо-
вывать витамины из простых соединений.
Синтез витаминов в лабораторных условиях осущест-
вляется совершенно иными путями, чем в растительных и
животных клетках. В лабораториях прибегают к воздей-
36
ствию на исходные составные части витаминов высокими
температурами, большими концентрациями щелочей или
кислот, применением веществ, совершенно не встречаю-
щихся в тканях живых существ. Не случайно поэтому,
что путь лабораторного синтеза многих витаминов резко
отличается от образования этих веществ в живой лабора-
тории — клетке.
Биологи в своих исследованиях широко используют
успехи химиков-органиков. Так, установление того фак-
та, что витамин Bi состоит из тиазола и пиримидина, а со-
единение этих компонентов в молекулу витамина Bi осу-
ществляется в лаборатории сравнительно легко, было
использовано биологами для выяснения синтетических
возможностей растений. Оказалось, что, если в питатель-
ную среду вместо витамина Bi дать его составные части —
тиазол и пиримидин, то некоторые микроорганизмы не
растут совсем, другие же растут так же успешно, как и в
случае получения целой молекулы витамина. Корни поми-
доров и гороха нормально растут, если они получают ви-
тамин Bi не в виде целой молекулы, а в виде его ком-
понентов.
Выяснение этого факта привело к открытию нового
фермента, осуществляющего соединение тиазола и пири-
мидина в молекулу витамина Вь Это в свою очередь по-
могло объяснить разную' реакцию организмов на введе-
ние в питательную среду составных частей витамина Вь
В последние годы удалось установить, что широко
распространенная в растениях и животных аминокислота
триптофан является исходным веществом для образования
витамина РР.
О важной роли этой аминокислоты в биосинтезе ви-
тамина РР свидетельствует, например, тот факт, что
при недостатке в пище животных триптофана возникает
заболевание, сходное с РР-авитаминозом. Витамин
РР образуется не непосредственно из триптофана, а
через ряд промежуточных соединений. Образование его
из триптофана доказано как для низших и высших ра-
стений, так и для животных. Иные аминокислоты явля-
ются предшественниками витамина, называемого пантоте-
новой кислотой. В биосинтезе этого витамина значитель-
ную роль играет аминокислота (3-аланин, входящая в
состав его молекулы. Таким образом, изучение путей об-
37
разования этих витаминов в организме помогло одновре-
менно выяснению важной физиологической роли широко
распространенных в растениях аминокислот.
Более сложное строение, чем пантотеновая кислота,
имеет витамин Н, поэтому и пути его биосинтеза также
сложнее. Биосинтез витамина Н протекает через образо-
вание близкого к витамину вещества — дестиобиотина.
Позднее было установлено, что биосинтез этого предшест-
венника витамина Н значительно усиливается, когда в
питательную среду вводится пимелиновая кислота. При
одновременном введении в питательную среду дестиобио-
тина и аминокислоты цистеина, образование витамина Н
идет особенно успешно.
В низших и высших растениях в значительных коли-
чествах образуется эргостерин — провитамин D, однако
дальнейшее превращение этого вещества в витамин в тка-
нях растений не происходит. Детальное изучение биосин-
теза витамина D позволило прийти к заключению, что
превращение предшественников витамина D в витамин
происходит лишь под влиянием ультрафиолетовых лучей.
Этот факт был установлен 30 лет назад. Освещая светом
кварцевой лампы льняное и хлопковое масла, Хесс и
Стинбок получили масла, содержание витамин D. Даль-
нейшие исследования биосинтеза витамина D позволили
разработать более рациональные способы его промышлен-
ного получения.
Другие исходные вещества служат источником образо-
вания витамина С. Наличие прямой зависимости между
содержанием сахаров в растении и накоплением витами-
на С, а также сходство в их строении, указывало на воз-
можность образования этого витамина из углеводов. Од-
нако нужны были прямые доказательства — опытным пу-
тем. Такие исследования были успешно осуществлены
в нашей стране. К. Л. Поволоцкая (1937), вводя в пита-
тельную среду глюкозу, фруктозу и другие сахара, пока-
зала, что из них проростки гороха обоазуют витамин С.
При непосредственном введении сахаров в листья образо-
вание в них витамина С также значительно возрастает
(Рубин, 1940). В результате исследования В. А. Девятки-
на (1950) было установлено, что материалом для обра-
зования витамина С в растениях являются такие сахара,
которые содержат шесть углеродных атомов и близки по
38
расположению отдельных группировок к аскорбиновой
кислоте (витамин С). Установив исходные для синтеза
витамина С продукты, биохимики напряженно работают
над выяснением ферментных систем, принимающих
участие в биосинтезе витамина С в живой растительной
клетке. Выяснение этого вопроса позволит правильно
представить сложный путь образования аскорбиновой
кислоты и объяснить, почему животные не в состоянии
синтезировать витамин С из тех же продуктов, что и рас-
тения.
Инозит также образуется из сахаров, но пути образо-
вания этого витамина резко отличны от биосинтеза вита-
мина С. О важном значении сахаров для инозита могут
свидетельствовать такие опыты. Вводя в листья чая са-
хара, А. Л. Курсанов (1949) получил прирост до 40%
инозита за полтора часа, причем из сахарозы биосинтез
витамина происходил быстрее, чем из глюкозы. Еще более
интенсивно шел синтез в том случае, когда в листья вво-
дились более сложные вещества — арбутин и салицин.
Все эти материалы показывают, что пути образования
разных витаминов весьма различны. Неодинаковы и ис-
ходные вещества, и ферментные системы, необходимые
для образования того или иного витамина.
Содержание витаминов в растениях
Некоторые растения издавна известны как богатые ис-
точники витамина С.
Не случайно, например, шиповник в течение несколь-
ких веков считается наилучшим противоцынготным сред-
ством. Для сбора плодов шиповника еще в XVI в. посы-
лали специальные экспедиции в степи нынешней Чкалов-
ской области. С течением времени человек перестал
довольствоваться дикими видами шиповника. Из них он
отбирал виды, наиболее эффективно излечивающие цын-
гу.. Благодаря естественному отбору и селекции, витамин-
ность шиповника значительно повысилась. • Одновремен-
но с улучшением витаминных качеств шиповника изыски-
вались новые источники витамина С. С этой целью
проведены многочисленные исследования, показавшие со-
держание аскорбиновой кислоты (витамина С) во многих
низших и высших растениях. Результаты исследований
39
свидетельствуют о том, что у бактерий и грибов этот ви-
тамин либо совсем отсутствует, либо находится в очень
малых количествах. У мхов содержание витамина С до-
стигает 1 мг на 1 кг их веса; водорослей и хвощей —
2 мг, у папоротников и голосемянных — 4 мг, у по-
крытосемянных— 10 мг на 1 кг веса (Девятнин, 1946),
у покрытосемянных растений содержание витамина С
по разным семействам также неодинаково. Так, напри-
мер, у растений семейства Пастуховых содержание вита-
мина С в среднем — 850 мг на 1 кг веса, у крестоцвет-
ных— 4 тыс. мг, у первоцветных — 9410 мг, у ирисо-
вых— 14 тыс. мг (Девятнин, 1946). Внутри каждого
семейства есть виды, отличающиеся один от другого по
содержанию этого витамина. Этот анализ помогает иссле-
дователям объяснить своеобразие обмена веществ в ра-
стениях и указывает на те классы и семейства растений,
представители которых могут быть использованы как ис-
точники витамина С.
В шиповнике в значительных количествах содержит-
ся также провитамин А. По содержанию этого провита-
мина среди плодов и ягод шиповник уступает только
абрикосам, а среди овощей — близок к лучшим сортам
моркови. В плодах шиповника найдены в заметных коли-
чествах и витамины В2, К, Р, Bi и др.
Ценным витаминным продуктом является также чер-
ная смородина. Высокие витаминные качества черной
смородины давно ’привлекали внимание исследователей.
При исследовании более 60 сортов черной смородины ока-
залось, что содержание витамина С в них колеблется от
498 до 3890 мг на 1 кг плодов. Ряд высоковитаминных
сортов черной смородины широко внедряется в практику
сельского хозяйства (Степанова, 1950). Однако при вы-
соком содержании витамина С черная смородина обычно
содержит лишь незначительное количество провитамина
А — каротина (до 7 мг на 1 кг). Недавно получен такой
сорт смородины, в котором содержание каротина дости-
гает более 50 мг на 1 кг плодов. Чтобы представить, на-
сколько богат этот сорт смородины каротином, вспомним,
что красная морковь — корнеплод, наиболее богатый ка-
ротином — содержит до 170 мг этого провитамина на 1 кг
корнеплодов. Следовательно, этот сорт черной смородины
может быть использован не только для получения каро-
40
тина, но и явиться исходной формой для создания новых,
высоковитаминных сортов.
Концентраты из плодов черной смородины и шипов-
ника оказались не только эффективным средством борь-
бы с цынгой, но и играют большую роль как профилак-
тическое средство и против других заболеваний. Богаты
витамином С незрелые плоды грецкого ореха и актини-
дии. Значительно меньше этого витамина в перце и осо-
бенно мало в тыкве, арбузе, сливе и чернике.
В годы Великой Отечественной войны Н. А. Максимов,
Ю. В. Ракитин и Р. X. Турецкая (1948) детально иссле-
довали еще один источник витамина С — гладиолус.
В некоторых сортах гладиолусов в 1 кг листьев содержит-
ся более 17 тыс. мг витамина С, что позволяет с каждого
гектара плантации при среднем урожае, равном 6 т, по-
лучить свыше 100 кг чистого витамина.
Содержание каротина — провитамина А — также раз-
лично у разных растений. По среднему содержанию каро-
тина А. Д. Егоров (1954) разбивает изученные им семей-
ства (высших и низших растений) на четыре большие
группы. К первой группе он относит очень богатые про-
витамином А растения, куда входят семейства березовых,
крестоцветных, пасленовых; вторую группу составляют
растения, относительно богатые каротином — гречишные,
губоцветные, осоковые, злаки, зонтичные, маревые, розо-
цветные, бобовые, сложноцветные; к третьей группе при-
надлежат средние каротиноносители: хвощевые, ивовые,
лютиковые, мареновые, камнеломковые; наконец, к
четвертой — растения, бедные каротином: лилейные, со-
сновые.
‘ Схематически содержание каротина у отдельных се-
мейств приведено на рис. 8.
Содержание других витаминов в различных растениях
также неодинаково. В одних они представлены богато, и
потому такие растения являются надежными источника-
ми витаминов; в других — их содержание крайне низко.
Важнейшим источником витаминов являются также
фрукты, ягоды и овощи. Как следует из табл. 2, некото-
рые из них (смородина, томаты, капуста, шпинат, карто-
фель и другие) содержат много витамина С; другие,—
например, морковь — богаты каротином, но витамина С в
них мало. Некоторые фрукты, ягоды и овощи богаты так-
41
же и другими витаминами. Особенно много в них инози-
та и в значительных количествах содержатся витамины
РР, Bi, В2, пантотеновая и фолиевая кислоты. В табл. 2
1 — березовые; 2 — крестоцветные: 3 — пасленовые- 4 — гречишные;
5 — губоцветные; 6 — осоковые; 7 — злаки; 8 — зонтичные; 9 — маре-
вые; 10 — розоцветные; 11— бобовые; 72—сложноцветные; .3—иво-
вые; 14— хвощевые; 15—лютиковые; 16 — мареновые; 17 — камне-
ломковые; 18 — лилейные; 19 — сосновые
приведены средние данные. В зависимости от сорта куль-
туры, места ее произрастания и физиологического состо-
яния растений содержание витаминов в них может быть
большим или меньшим.
В семенах некоторые витамины также представлены в
значительных количествах. Особенно много в них вита-
минов РР и пантотеновой кислоты, в ничтожно малых
42
Таблица 2
Содержание витаминов в фруктах, ягодах и овощах
(мг на 1 кг)
Фрукты, ягоды И ОВОЩИ Bi в9 РР с Каротин Пантотеновая кислота вв Витамин Н Инозит Фолиевая ки- слота
Апельсин .... 0,84 0,38 1,36 530 3,4 0,8 0,019 2100 0,83
Вишня 0,50 — 1,36 100 5,4 — — — — —
Виноград .... 0,56 0,56 8,40 63 1,25 — — .— — —
Груша 0,35 0,63 1,16 60 — — — — — —
Земляника .... 0,28 2,60 2,42 53 — 2,6 0,44 0,04 600 0,23
Лимон 0,2 0,035 1,72 430 .— — — — — —
Малина 0,27 — — 21 — — — — — —.
Персик 0,18 0,57 6,4 14 — 1,7 0,16 0,017 960 0,17
Смородина черная 0,30 — — 2500 20,0 •— — — — —
Томат 1,10 0,53 4,30 300 16,0 3,7 0,60 0,04 460 0,75
Морковь .... 1,15 0,60 8,35 40 80 2,5 1,2 0,025 480 0,97
Картофель .... 1,05 0,57 5,70 150 Следы 3,2 2,2 0,006 290 0,83
Капуста белокоч. 1,50 0,56 2,50 300 Следы 1,8 1,2 0,024 950 0,65
Шпинат 1,58 0,24 6,46 570 150,0 1,7 0,83 0,064 270 0,17
количествах содержится витамин Н и совершенно нет
аскорбиновой кислоты (табл. 3).
Недавно (Поволоцкая, 1953) было установлено, что
витамин В2 в тканях растений и животных находится не
только в свободном состоянии, но и в состоянии прочной
связи с белковыми молекулами. Обычно применявшими-
ся методами определения учитывался лишь свободный
или слабо связанный с белком витамин В2, другая же
форма этого витамина не обнаруживалась. Новый метод
определения витамина В2 позволяет учитывать и эту фор-
му рибофлавина (витаминВ2).Так, например, в 1 кг семян
пшеницы обычным способом найдено витамина В2 —
1.82 мг. Применив новый метод, удалось в этих семенах
дополнительно обнаружить 1,68 мг витамина В2. Таким
образом, общее содержание рибофлавина в 1 кг семян
пшеницы достигает 3,50 мг, а в кукурузе 5,8 мг.
43
Таблица 3
Содержание витаминов в семенах
(мг на 1 кг)
Растение Каротин (провита- мин А) рр Bi в, в. Пантоте- новая ки- слота Витамин Н
Ячмень 0,42 56,9 4,8 0,13 6,25 0—02
Пшеница 2,53 49,8 5,1 1,32 6,06 12,36 0—06
Гречиха — 44,0 4,5 — — — —
Чечевица — 31,0 4,9 2,6 — — —
Соя — 31,6 14,5 4,9 4,5 1,20 0,5-0,6
Рожь 0,09 15,1 4,5 2,74 — 10,38 —
Овес 0,11 13,5 6,5 0,9 — 9,90 0,02
Кукуруза 4,89 14,1 4,53 1,32 6,68 7,39 —
Как следует из табл. 3, содержание витаминов (Вь
Вб, пантотеновой кислоты, РР) в указанных семенах
заметно выше, чем витамина В2 и каротина. Овес, соя, яч-
мень, рожь и пшеница богаты витамином Вь В пшенице,
ржи и овсе в значительных количествах представлен и ви-
тамин В6. Пшеница и ячмень, по сравнению с другими
культурами, содержат много и витамина РР. Кукуруза,
которая по постановлению Январского Пленума ЦК КПСС
должна получить широкое распространение во всех рай-
онах страны, выгодно отличается от других зерновых
культур высоким содержанием каротина (в случае желто-
мясых сортов) и, как указано выше, высоким содержани-
ем рибофлавина. По содержанию витамина РР кукуруза
уступает другим культурам и потому ее рекомендуется
применять в сочетании с другими продуктами, богатыми
этим витамином. Этот факт, однако, нисколько не умаля-
ет огромной ценности кукурузы как неисчерпаемого ис-
точника белков и углеводов и как достаточно богатого
источника витаминов (Вь В2, В6 и др.).
Таким образом, в семенах содержатся не только зна-
чительные количества белков, углеводов и жиров, но и
много витаминов. Витамины в семенах распределяются
неравномерно: в одних частях семени их больше, в дру-
гих— меньше. Так, например, в рисе витамин Bt сосре-
44
доточен главным образом в оболочке. Поэтому отходы
от очистки риса (отруби) являются богатым источником
этого витамина. В отходах мукомольной промышленности
и в жмыхах в больших количествах находятся и другие
витамины. Широко используемая в животноводстве мука
хлопковых семян не только богата белком, но и является
важным источником витаминов Bi, В2, РР и пантоте-
новой кислоты (табл. 4).
В пшеничных отрубях в значительных количествах
представлены витамины Bi, В2, РР и пантотеновая ки-
слота. Сено люцерны является важнейшим источником
как каротина, так и других витаминов.
Советские ученые приготовили специальные витамин-
но-белковые препараты, которые находят все большее и
большее применение в сельском хозяйстве. Передовики
Таблица 4
Содержание витамина в некоторых кормах
(мг на 1 кг корма)
Корм Каротин (про- витамин А) Витамин Bi Витамин В3 Витамин РР Пантотеновая кислота Витамин В,
Мука хлопковых семян (бел-
ка 38—43%) Мука из льняного семени 0,20 13,48 8,98 44,88 13,97 —
(белка 33—38%) .... 0,26 12,85 6,05 48,95 7,04 —
Мука из орехов арахис 5,15
(белка 38—43%) .... — 7,23 170,50 53,02 —
Соевая мука (белка 38—
43%) 0,22 5,77 4,11 38,72 13,79 —
Рисовые отруби — 22,70 3,04 284,02 22,73 32,03
Рисовые отходы при по-
лировке — 19,45 1,92 715,00 — 23,73
Пшеничные отруби .... 2,60 7,13 2,95 307,93 24,42 —
Высевки Сено люцерны молотое (об- 3,06 15,40 1,53 116,11 15,12 —
лиственное, солнечной
сушки) . . ’ 42,68 2,47 11,79 38,94 26,51 —
Люцерна зеленая 63,26 — 4,88 17,47 11,18 —
животноводства, рационально применяя в своей практике
успехи витаминологии, значительно повысили продуктив-
ность скота.
Изучая образование витаминов и значение отдельных
звеньев реакций и условий внешней среды в этом важном
процессе, биохимики и физиологи успешно решают во-
просы биосинтеза витаминов и указывают практике пути'
повышения витаминное™ культурных растений.
Советские селекционеры получили ряд совершенно но-
вых сортов растений, отличающихся богатым содержани-
ем витаминов. Так, например, удовлетворяя запросы жи-
вотноводов, селекционеры вывели новый сорт тыквы —
«Витаминная», содержащий до 40 мг каротина на 1 кг
веса, тогда, как обычно тыква содержала в 1 кг не выше
5 мг каротина. На Воронцовской станции Всесоюзного ви-
таминного института путем скрещивания географически
отдаленных форм шиповника получены сорта с содержани-
ем в них витамина С от 150 до 270 тыс. мг на 1 кг плодов.
В настоящее время обращено внимание на получение та-
ких сортов, которые бы сочетали в себе высокое содер-
жание не одного, а нескольких витаминов.
Источником витаминов для человека и животных яв-
ляются не только продукты питания. Для витаминного ба-
ланса имеет большое значение деятельность кишечных
бактерий. Доказано, что в желудке коров, овец и других
животных микроорганизмы синтезируют витамины Bb Н,
РР, В2, Вб, К и пантотеновую кислоту. Последними ра-
ботами установлено, что в кишечнике человека при по-
мощи бактерий может осуществляться синтез витаминов
Bi, В2, Н, РР, К и фолиевой кислоты. Этим обстоя-
тельством можно объяснить и различную потребность
организма в добавочном введении витаминов извне и бла-
гоприятное косвенное действие некоторых пищевых ве-
ществ на излечение различных заболеваний. Так, напри-
мер, увеличение в диете молочного сахара способствует
развитию полезной микрофлоры в кишечнике и тем самым
содействует дополнительному обеспечению организма ви-
таминами. Становится понятной и увеличенная потреб-
ность в витаминах при лечении человека сульфамидными
препаратами, которые наряду с уничтожением вредных
бактерий, оказывают губительное действие и на полезную
бактериальную флору.
46
Однако не все микроорганизмы в одинаковой степени
образуют витамины. Установлено, что наряду с микро-
организмами, которые совершенно не синтезируют
некоторые витамины, имеются виды и расы, обладающие
высокой синтетической способностью. Так, например, сре-
ди молочнокислых бактерий найдены виды, которые не в
состоянии синтезировать витамин Н, РР и пантотеновую
кислоту, но в достаточном количестве образуют витамины
Bi, В2, В6 и фолиевую кислоту. Некоторые виды молоч-
нокислых бактерий синтезируют либо витамин Н, либо
витамин РР или пантотеновую кислоту, но не способны
образовать другие витамины.
Дрожжи также обладают неодинаковыми синтетиче-
скими возможностями. Из исследованных 163 штаммов
дрожжей, принадлежащих к ПО видам, не обладали спо-
собностью синтезировать витамин Н 76 видов, витамин
Bi—33 вида, пантотеновую кислоту—30 видов, ино-
зит— 15 видов, витамин РР — 13 видов и витамин Ве —
13 видов.
Наряду с видами дрожжей, не синтезирующими неко-
торые витамины, есть и такие дрожжи, которые образуют
значительные количества витаминов. Последнее свойство
микроорганизмов нашло широкое использование для по-
лучения витаминов. Дрожжи давно используются как на-
дежный источник получения витамина Вь Однако ученые
не ограничились установлением и использованием того ко-
личества витамина Вь которое синтезируют дрожжи в
обычных условиях. Изучив биологию этих организмов,
М. Н. Мейсель и Е. Одинцова в 1943 г. предложили метод
обогащения хлебопекарных дрожжей витаминами. При
добавлении к бродящей взвеси специальных веществ или
экстрактов из пшеничных зародышей, люцерны, карто-
фельных очистков и других естественных обогатителей
количество витамина Bi в дрожжах увеличивалось в 8—
10 раз. Из дрожжей в значительных количествах также
получаются витамин В2 и эргостерин — провитамин D.
Как указывалось выше, в зеленых растениях витамин D
не синтезируется, но они являются поставщиками матери-
ала, необходимого для образования витаминов D в теле
животных.
Витамины иногда находятся в дрожжах и в других ор-
ганизмах в очень малых количествах. Так, например, для
47
получения 1 г кристаллического витамина Н из обыкно-
венных пекарских дрожжей понадобилось бы перерабо-
тать 360 т дрожжей, а для получения 10 мг этого вита-
мина из сухого яичного желтка пришлось бы переработать
свыше 250 кг, что соответствует 30 тыс. шт. яиц. Нахо-
дясь в ничтожно малом количестве в организме, витамин
Н выполняет очень важную роль. Его действие на размно-
жение дрожжей проявляется даже при разведении
1 : 400 000 000 000.
При помощи селекции удалось получить микроорганиз-
мы, обладающие новыми возможностями образования
витаминов. Так, например, при скрещивании дрожжей,
синтезирующих пантотеновую кислоту, но не синтезиру-
ющих витамин Bi, с дрожжами, обладающими противо-
положными свойствами, получены дрожжевые культуры,
способные синтезировать и тот и другой витамины.
Изучение распределения витаминов в растениях и в
продуктах их переработки позволило разработать рацио-
нальные приемы использования растительного сырья.
Вместе с тем выяснение биосинтеза витаминов дало но-
вые материалы для более полного познания процессов,
лежащих в основе жизнедеятельности растений.
2. Изменение содержания витаминов
в процессе развития растений
Значительное влияние на содержание витаминов в ра-
стениях оказывают не только природные свойства орга-
низма, но и его физиологическое состояние. Вместе с из-
менением обмена веществ, обусловливающего процессы
роста и развития растений, изменяются биосинтез и на-
копление витаминов. Так, например, в покоящихся се-
менах капусты не найден витамин С, но при прорастании
их, когда в семенах возрастает обмен веществ, образова-
ние этого витамина резко повышается (табл. 5).
Биосинтез витамина С при прорастании семян отме-
чен для всех исследованных культур. Содержание вита-
мина В2 по мере прорастания семян также возрастает.
Так, например, на 3-й день прорастания ячменя количе-
ство витамина В2 по сравнению с непроросшими семена-
ми возрастает на 40%, на 7-й день — на 240% и на 12-й
день — на 320%. При прс^растании семян фасоли и пше-
48
Таблица 5
Изменение содержания витамина С
. при прорастании семян
капусты различных сортов
(по Рубину, 1940)
Дни проращи- вания Содержание витамина С, мг на 1 кг
Амагер М 1
0 0 0
3 255 122
7 290 273
10 590 315
13 590 320
ницы содержание витамина РР увеличивается еще боль-
ше, чем витамина В2.
По мере прорастания семян начинается и новообразо-
вание каротина. Так, к моменту, когда корешок грейп-
фрута достигнет длины 2,5—5,0 см, содержание каротина
в зародыше увеличивается более чем в 13 раз. Еще энер-
гичнее происходит биосинтез фолиевой кислоты. За ко-
роткий срок ее содержание в прорастающих семенах ов-
са возрастает почти в 40 раз.
При прорастании семян происходит заметное перерас-
пределение витаминов внутри семени. Витамины из за-
пасных частей прорастающего семени устремляются к ра-
стущим частям. В опытах с пшеницей, культивируемой
в темноте, было показано, что в то время, как общее со-
держание витамина Bi в семени осталось одним и тем
же, количество этого витамина в зародыше за 18 дней
уменьшилось в три раза. Из эндосперма в зародыш, т. е.
туда, где при прорастании семян происходит особенно
энергичный обмен веществ, передвигаются также вита-
мины В2, Вб, РР и другие. Существенным образом
в этот момент изменяется и качество витаминов. В то вре-
мя, как в покоящемся семени витамин Bi находится в сво-
бодном состоянии, при прорастании семян он присоеди-
няет две молекулы фосфорной кислоты и переходит в ак-
тивную форму — кокарбоксилазу.
4 Витамины в жизни растений 49
Увеличение количества витаминов при прорастании
семян имеет важное биологическое значение. Этот период
в жизни растений, как известно, связал с усиленными
процессами перестройки белков, углеводов, жиров, с ново-
образованием клеток и тканей проростков. Принимая
непосредственное участие в синтезе белков, аминокислот,
сахаров и других жизненно необходимых веществ, вита-
мины способствуют нормальному росту молодых расте-
ний.
Увеличение содержания витаминов при прорастании
семян пшеницы, ячменя и овса нашло и практическое
применение. Передовики совхозного и колхозного живот-
новодства начинают использовать проросшие семена для
обогащения кормов витаминами.
При подготовке растений к цветению содержание ви-
таминов достигает максимума, что связано с существенной
перестройкой питательных веществ в организме и транс-
портировкой их в органы плодоношения. Так, например,
содержание витамина С в 1 кг свежих листьев гладиолу-
са в момент появления цветоноса было равно 9940 мг, в
период массового цветения— 16 050 мг и в период нача-
ла пожелтения листьев—12 710 мг. Аналогичная картина
наблюдается и в накоплении провитамина А-
Образование каротина также начинается сразу после
прорастания семян и уже в проростках он содержится в
больших количествах. Установлено, что в период макси-
мальной скорости роста растений кукурузы концентрация
каротина также является максимальной, а затем, по ме-
ре старения растения, она заметно падает. Об этом мож-
но наглядно судить по данным, приведенным в графике
(рис. 9).
Как следует из кривых графика, наибольшая концен-
трация каротина (на единицу веса) наблюдается в рас-
тениях 45-дневного возраста; в 80-дневных растениях кон-
центрация провитамина А снижается в 2 раза, а в 120-
дневном возрасте — более чем в 10 раз. В этот период и
общее содержание каротина (в пересчете на одно расте-
ние) падает. Максимальное же количестве каротина со-
держат растения 80-дневного возраста.
По мере роста красного клевера, тимофеевки, люцерны
и других культур содержание каротина в листьях увели-
чивается, достигая максимума до начала бутонизации, и
S0
затем резко снижается. Было найдено, что в красном кле-
вере содержалось до бутонизации 43 мг каротина, перед
Рис. 9. Изменение концентрации каротина и об-
щего содержания каротина в развивающемся
растении кукурузы
1— концентрация каротина; 2—общее содержание каротина
цветением — 29 мг, во время цветения — только 19 мг в
1 кг зеленой массы. В зеленой массе тимофеевки содер-
жание каротина в период цветения и особенно при созре-
вании семян снижается еще больше. Снижение содержа-
ния каротина в листьях в период массового цветения и
образования завязей обусловливается, с одной стороны,
усиленным его расходом на синтетические процессы, свя-
занные с оплодотворением, а с другой,— снижением интен-
сивности образования этого витамина, вызванного старе-
нием листьев. В. Р. Вильямс указывал, что «на акт цвете-
ния многолетние травы затрачивают половину — 50 про-
центов всего количества белков, которое они накопили со
времени их цветения в прошлом году. Разрушаются при
4* 51
этом наиболее ценные — подвижные белки, в том числе
огромный процент витаминов».
В кормовых травах, отрастающих после первого уко-
са, концентрация каротина обычно восстанавливается
сравнительно быстро. И задержка со вторым укосом, как
и с первым, приводит к значительным потерям вита-
минов.
Так, если при скашивании пырея якутского до цветения
в 1 кг сухого сена содержалось 339 мг каротина, а в пе-
риод цветения — 220 мг, то в период вегетации, когда
началось пожелтение листьев,— всего 137 мг (Егоров,
1954). Таким образом, изучение изменения содержания
витаминов в различные периоды развития растений имеет
большое практическое значение, так как указывает, в ка-
кой период необходимо убирать растения, чтобы получить
корм, богатый витаминами. В настоящее время твердо
установлено, что необеспеченность животных витамином
А резко снижает их сопротивляемость различным заболе-
ваниям. Недостаток каротина и витамина Е в корме при-
водит к яловости или резкому уменьшению количества
приплода, к снижению его жизнеспособности. Поэтому
передовики животноводства уделяют большое внимание
своевременной уборке и высококачественному приготов-
лению кормов.
Содержание каротина в листьях изменяется по фазам
развития неодинаково в растениях разного пола. В листь-
ях мужских особей конопли в период бутонизации каро-
тина больше, чем в женских, тогда как во время цветения
это соотношение изменяется: в женских особях содержа-
ние каротина заметно возрастает, а в мужских, наоборот,
падает.
Содержание витаминов в разных органах вегетирую-
щего растения зависит как от места синтеза витаминов
И интенсивности оттока их в другие части растения, так
и от напряженности обмена веществ и от физиологического
состояния отдельных органов. В листьях, как правило, со-
держится больше витаминов, чем, например, в корнях.
В качестве примера приведем содержание витаминов в
листьях и корнях свеклы (рис. 10).
Как следует из приведенной диаграммы, содержание
витаминов Bi, В2, В6, пантотеновой кислоты, фолиевой
кислоты и особенно витамина Н в корнях во много раз
52
меньше, чем в листьях. Меньшая разница наблюдается
в содержании витаминов РР и инозита.
У шиповника, вики и фацелии максимальное количе-
ство витамина РР содержится в листьях, меньше в
Рис. 10. Содержание витаминов в листьях и корнях свеклы
(мг на 1 кг сухого веса)
1 — листья; 2 — корни
цветах, значительно меньше в стеблях и совсем мало —
в корнях.
Исследования А. Д. Егорова (1954), проведенные
на большом количестве различных растений, свидетель-
ствуют, что, чем больше витамина» С содержится в
листьях, тем больше его и во всех других частях ра-
стений.
Содержание аскорбиновой кислоты и других витами-
нов в растении зависит также от физиологического состоя-
ния его отдельных органов.
В молодых, растущих частях растения витаминов обра-
зуется значительно больше, чем в частях старых, закон-
чивших рост. В старых листьях томатного растения со-
держание витаминов Bi, РР, В2 и пантотеновой кислоты
значительно снижено, по сравнению с листьями молоды-
ми, и наибольшее количество этих витаминов находится
в листьях средних ярусов (6—8). В молодых, энергично
растущих частях стебля, содержание витаминов повыше-
но, по сравнению со старыми, менее жизнедеятельными
53
частями стебля. В корнях содержатся большие количества
пантотеновой кислоты, тогда как другими витаминами
корни томатов бедны. Аналогичное распределение вита-
мина Bi было обнаружено в растениях кукурузы, гороха и
хлопчатника. Значительное увеличение содержания вита-
минов в наиболее жизнедеятельных частях растения
объясняется как повышенным биосинтезом, так и пере-
движением синтезированных витаминов в эти части расте-
ния.
Давно были высказаны предположения о передвиже-
нии витаминов в растении. Однако лишь недавно прямыми
опытами было доказано, что витамины, как и другие орга-
нические вещества, передвигаются по коре. На это ука-
зывает, например, такой факт. Если в нижней части стеб-
ля томатного растения сделать кольцеобразный вырез
коры (произвести кольцевание), то обнаруживается бы-
строе накопление витамина Bi непосредственно над коль-
цом, тогда как в стебле, ниже кольца, и в корнях количе-
ство витамина резко падает. У древесных растений вита-
мин Bi из надземных частей растения также передвигается
в корни по коре- Передвижение витамина Bi в вегетирую-
щих растениях происходит как по направлению к корням,
так и к растущей верхушке.
Величина оттока зависит от возраста листьев. Обра-
зование и передвижение витамина Bi идет чрезвычайно
интенсивно в зрелых, сформировавшихся листьях, тогда
как в молодых, энергично развивающихся листьях пере-
движение витамина не наблюдается или происходит
в очень незначительной степени. В этих листьях биосинтез
витамина Bi значительно меньше, чем в листьях средних
ярусов. В связи с усиленными процессами обмена веществ
в молодых листьях, потребность их в витаминах повышена.
Таким образом, сниженный биосинтез и повышенная по-
требность в витамине Bi энергично растущих листьев
являются одной из причин, задерживающих его отток
в другие части растения.
Совершенно иная картина наблюдается во взрослых,
сформировавшихся листьях. В последних процессы ново-
образования тканей закончены, расход витамина в них
понижен, а биосинтез этих веществ резко увеличен. Сле-
довательно, синтезированный в этих листьях витамин Bi
не потребляется на месте и оттекает в другие части расте-
54
ния. Подобно витамину Bi, витамины С, В6 и РР
образуются главным образом в листьях и оттекают из них
по направлению к корням. Так, например, было показа-
но, что при кольцевании стеблей шиповника витамин С,
как и витамин Bi у томатов, накапливается в значитель-
ных количествах в листьях и коре, лежащих выше кольца.
Из листьев витамин С передвигается в корни. Установле-
но, что клубни картофеля, лишенные связи с листьями,
не накапливали, а даже значительно снижали содержание
этого витамина.
Витамины Н и пара-аминобензойная кислота, анало-
гично витаминам В2 и патнотеновой кислоте, передвигают-
ся из листьев в корни в незначительных количествах.
В листовой пластинке происходит не только синтез са-
харов, но в ней энергично идут процессы образования
и других веществ, в том числе и витаминов. Черешок ли-
ста служит тем каналом, через который эти вещества пе-
ретекают в другие части растения. Не случайно поэтому
содержание многих витаминов в листовой пластинке боль-
ше, чем в черешке. Так, содержание провитамина А в
листовой пластинке иногда в 50 раз больше, чем в сред-
ней жилке или черешке. Причем, если пересчитать содер-
жание каротина на все растение, можно обнаружить, что
у многих растений в листьях содержится до 90% всего
провитамина А.
Установление факта передвижения витаминов помогло
объяснить причины большего или меньшего их накопления
в отдельных органах растений. Перераспределение пи-
тательных веществ, в том числе и витаминов, происходит
в растении во все периоды его жизнедеятельности.
Витамины, подобно белкам, сахарам и другим физиологи-
чески важным веществам, оттекают в молодые, растущие
части, а в период плодообразования — главным образом
в плоды. В качестве примера можно привести данные по
изменению содержания витамина С в листьях и плодах
томата (табл. 6).
Обращает на себя внимание тот факт, что в период
после отцветания наибольшее содержание витамина С
обнаруживается в листьях и незначительное — в завязях.
По мере роста плодов картина меняется. В зрелых пло-
дах содержится максимальное количество витамина С.
Подобные изменения в содержании этого витамина в
55
Таблица 6
Сезонные колебания содержания витамина С
в листьях и плодах томатов
(по Поповской, 1950)
Фазы развития Количество витамина С (мг на 1 кг)
в листьях в плодах
До цветения 274
Цветение 413 —
Завязи 407 180
Зеленые плоды .... 456 400
Начало созревания . . — 430
Зрелые плоды 224 450
В лежке через 20 дней — 444
плодах и листьях были установлены для шиповника,
кормовых трав и других растений. Об интенсивности нако-
пления витамина С по мере созревания плодов шиповни-
ка можно судить по данным, приведенным в табл. 7.
Таблица 7
Накопление витамина С в процессе созревания
плодов шиповника
(по Валовой, Бейдеру и Янишевской, 1941)
Состояние плодов
мг на 1 кг
сухой мякоти
плодов
Плоды зеленые (26.VII)...............
„ бледножелтые (4.VIII)...........
„ желтые (15.VIII)................
„ оранжево-желтые (26.VIII) . . .
„ оранжево-красные (15.IX) . . .
„ красные (15.Х)..................
2500
4040
4890
5150
6480
6780
56
Как наглядно показывают данные таблицы, по мере
созревания плодов шиповника содержание в них витами-
на С возрастает, достигая максимума в период их полной
зрелости- Следует отметить, однако, что накопление вита-
минов в плодах при созревании обусловлено не только их
поступлением из других частей растения, но и биосинте-
зом в самих плодах.
Содержание каротина в
органах растения подверже-
но значительным изменениям
в различные периоды его
жизнедеятельности. В период
цветения — плодообразова-
ния в листьях резко сни-
жается содержание этого
провитамина, тогда как в
цветах и завязях количество
его значительно возрастает.
В некоторых частях цветка,
например в пыльце, каротина
и витамина Е накапливается
в сотни раз больше, чем в
листьях. Значительное накоп-
ление витаминов в пыльце и
в завязи обусловливается
не только биосинтезом этих
Рис. 11. Передвижение вита-
мина Вх из стеблей и листьев
в семена пшеницы
веществ, но и притоком их
из других частей растения.
Много витаминов в завязь
/—витамин Bj вТлистьях и стебле;
2— витамин Вх в семенах
притекает из листьев и стеблей. Наглядным примером
оттока витаминов из стеблей и листьев в семена являют-
ся данные по витамину Bi (рис. 11).
По мере увеличения возраста и размера зерен пшени-
цы содержание витамина Bi в листьях ц стеблях резко
снижается и на 28-й день после цветения количество вита-
мина Bi в них падает до 1 мг в кг (пунктирная кривая).
Одновременно в семенах идет нарастание количества
витамина Вь достигая максимума в возрасте 21 дня
(сплошная кривая).
Формирование семян связано со значительной пере-
стройкой поступивших в них питательных веществ. В пре-
вращении этих веществ принимают непосредственное
57
участие и витамины. И не случайно, что в период наиболее
интенсивного обмена веществ в семенах содержание в них
витаминов заметно возрастает. При дальнейшем созрева-
нии семян, когда интенсивность обмена снижается, содер-
жание витаминов в них снова падает. Об интенсивности
снижения содержания витаминов Bi и фолиевой кисло-
ты по мере созревания семян пшеницы можно наглядно
судить из следующей диаграммы (рис. 12) (по Андре-
евой, 1953).
Рис. 12. Изменение содержания фолиевой
кислоты и тиамина при созревании семян
пшеницы (мг на 1 кг сухого веса)
I — тиамин; 2 — фолиевая кислота
При созревании семян овса, ржи и гороха также отме-
чается падение в них количества фолиевой кислоты.
В этот период снижается и количество каротина, витами-
нов С и В2. Это уменьшение вызвано усиленным расхо-
дом витаминов ра процессы, связанные с формированием
и созреванием семян, с уменьшением притока витаминов
из листьев и переходом их в другие соединения.
Существовавшее ранее мнение, что в семенах содер-
жится больше витаминов, чем в плодах, оказалось несо-
стоятельным. Полученные факты свидетельствуют о том,
что в некоторых плодах содержание витаминов во много
раз превышает их содержание в семенах. Особенно много
каротина, витаминов Bg, С и РР и очень небольшое коли-
5? л
чество витамина Bi содержится в плодах. Характер на-
копления того или иного витамина в плодах зависит от
природы растения и от физиологического состояния пло-
дов. Так, в зрелых плодах красного перца каротина в
30 раз больше, чем в незрелых зеленых плодах. Хорошо
выяснен биосинтез каротина в плодах томатов. Былс
установлено, что в зеленых плодах томатов каротина
меньше всего, в белесоватых количество каротина воз-
растает в два раза и в спелых — в три раза, по сравне-
нию с зелеными. В переспелых плодах содержание каро-
тина вновь падает. Большое влияние на содержание ка-
ротина в плодах оказывает продолжительность их созре-
вания. Плоды томатов, созревшие в течение 1—8 дней,
содержали 3 мг, а созревшие в течение 13—19 дней —
лишь 1,9 мг каротина в 1 кг свежих плодов. В связи с
этим следует отметить, что ускорение созревания плодов
при помощи газа этилена не только повышает вкусовые
свойства плодов, но и улучшает их витаминные качества
(Ракитин, 1938).
Накопление витамина С и фолиевой кислоты по мере
созревания плодов несколько отличается от изменения со-
держания каротина. В зрелых плодах содержание этих
витаминов не уменьшается, а возрастает. Так, например,
если зеленые плоды томатов содержали 66,0 мг витами-
на С, то зрелые — 219,0 мг в 1 кг. Подобное изменение
в содержании витамина С найдено и для ягод (смороди-
ны, малины, земляники и Др.).
Ю. В. Ракитин и А. В. Крылов (1950), разработав ме-
тод резкого повышения продуктивности томатов при по-
мощи стимуляторов роста, который в настоящее время на-
шел широкое практическое применение, установили, что
полученные при обработке цветов бессемянные или мало-
семянные плоды содержат больше сухого вещества, са-
харов и витамина С. У томатов сорта Спаркс Эрлиана,
например, содержание этого витамина при обработке сти-
муляторами роста увеличивается почти в два раза.
Неодинаково содержание витаминов в разных частях
одного и того же органа. В кожице крыжовника количе-
ство витамина С в два раза больше, чем в мякоти, в ко-
жице лимона в три раза и в кожице сливы в десять
раз больше, по сравнению с мякотью этих плодов. В ко-
жице редиски, тыквы, моркови, томатов,- репы также
59
содержится больше витамина С, чем в мякоти тех же
овощей.
Витамины в значительных количествах представлены
не только в надземных органах, но и в корнях, клубнях и
корнеплодах. Некоторые из них являются богатыми
источниками витаминов. Корнеплоды моркови особенно
выделяются высоким содержанием каротина. В самом
названии «каротин» отражен тот факт, что он был впер-
вые получен из моркови, имеющей латинское назва-
ние Daucus carota, С тех пор прошло более 120 лет. За это
время были выявлены сорта моркови с высоким содержа-
нием этого провитамина, установлены наилучшие агро-
технические условия культуры, сроки уборки корней мор-
кови, распределение каротина в корне. Было установлено,
что не во все периоды развития корней содержание в них
каротина велико: в молодых корнях его мало, а по мере
старения количество провитамина заметно возрастает
(табл. 8). Эта закономерность широко учитывается прак-
Таблица 8
Динамика накопления каротина в сортах
моркови во время их роста
(по Мурри, 1948)
тиками при заготовке сырья для получения провитамина
А из моркови.
Интересно отметить, что в разных частях корнеплода
содержание каротина неодинаково: в коре концентрация
каротина на 30% выше, чем в его сердцевине. Корни
моркови богаты и фолиевой кислотой (6,9 мг в 1 кг су-
хого вещества), но значительно больше ее в ботве морко-
ви (34,8 мг в 1 кг сухого вещества). Несколько меньше,
60
чем в корнях моркови, содержится каротина в клубнях
батата. Однако среди других корне- и клубнеплодов ба-
таты выделяются необычайно высоким содержанием это-
го провитамина. В клубнях картофеля, например, содер-
жится в 50 раз меньше каротина, чем в клубнях батата.
Однако клубни картофеля могут служить примером рас-
тительного продукта, в котором сочетается высокое со-
держание нескольких витаминов. По данным М. С. Про-
кошева, пищевой картофель в нашей стране доставляет
ежегодно примерно 1500 тонн витамина С. Клубни кар-
тофеля содержат также значительные количества вита-
минов Bi, В2, РР.
Изучение изменения содержания витамина С по мере
роста клубней картофеля показало, что с момента обра-
зования клубней начинается быстрое накопление в них
этого витамина. В период цветения растений в клубнях
содержится наибольшее количество витамина, а затем
происходит резкое снижение его содержания (табл. 9).
Таблица 9
Содержание аскорбиновой кислоты в клубнях картофеля
(по А. Д. Егорову, 1954)
Фаза развития
Содержание аскор-
биновой кислоты,
мг на 1 кг сырого
веса клубней
Клубнеобразование (13.VII)
Цветение (27.VII)........
Конец цветения (13.VIII) .
Созревание (29.VIII) . . .
Полная зрелость (10.IX) .
282
425
350
201
142
При прорастании картофеля в хранилищах имеет ме-
сто значительная потеря как углеводов, так и витаминов.
Не случайно поэтому практикой давно была поставлена
задача изыскания эффективного метода задержки про-
растания картофеля при его длительном хранении. Такой
метод был разработан Ю. В. Ракитиным и А. В. Крыло-
61
вым. Он состоит в том, что клубни картофеля при заклад-
ке на хранение опыливаются специальными химическими
препаратами (М-1 и др ). Такой обработкой удается за-
держать прорастание клубней на значительный срок. При
обработке картофеля химическими препаратами сокра-
щаются потери сухого веса клубней, в последних же луч-
ше сохраняются витамины и крахмал, уменьшается на-
копление вредного глюкозида — соланина.
В настоящее время этот метод широко внедряется в
практику народного хозяйства.
Много накапливается витамина С также в корнях хре-
на — до 9000 мг в 1 кг сухих корней.
Приведенные материалы показывают, что большее
или меньшее содержание витаминов в растениях зависит
как от природы организма, так и от физиологического
состояния его органов. В периоды жизни растения, свя-
занные с усиленным превращением веществ, обусловли-
вающим новообразование тканей, усиливаются как био-
синтез, так и потребление витаминов.
3. Влияние факторов внешней среды
на содержание витаминов в растениях
Многочисленными исследованиями было установлено,
что условия произрастания оказывают существенное влия-
ние на содержание витаминов в растении. У растений,
выросших в северных районах и в горах, витамина С, как
правило, больше, чем у растений южных или выросших
в низменных местах. В качестве примера приведем содер-
жание этого витамина в плодах розы иглистой (табл. 10).
Противоположная закономерность наблюдается в со-
держании витамина Вь Так, мягкая озимая пшеница
«Украинка», произрастающая на Крайнем Севере (Коми
АССР), содержала 2,6 мг витамина Bi, в Кировской об-
ласти — 3,7 мг, в Ростовской области — 4,8 мг и в Дне-
пропетровской области — 5,8 мг в 1 кг сухого зерна
(Мурри, 1948).
Эти примеры показывают, что одни и те же климати-
ческие условия оказывают различное действие на биосин-
тез разных витаминов. Различные синтетические возмож-
ности растений в этих условиях зависят как от обеспечен-
ности растений исходными веществами для биосинтеза
62
Таблица 10
Содержание витамина С в плодах розы иглистой
из различных районов СССР
(по Розановой,1946)
Место сбора мг на 1 кг сырого веса мякоти плода мг на 1 кг сухого веса мякоти плода
Северные районы 17 290 50830
Алтайский край (горные районы) 17 280 46 650 '
Сибирь 13 730 33 060
Дальневосточный край . . 8 690 23 460
витаминов и наличия ферментных систем, осуществляю-
щих этот процесс, так и от той роли, какую выполняет
тот или иной витамин в жизнедеятельности растений.
Минеральное питание также является мощным факто-
ром, обусловливающим синтез и накопление витаминов
в растении. Высокая агротехника и своевременное внесе-
ние минеральных удобрений увеличивают валовой выход
витаминов с единицы посевной площади не только за счет
увеличения массы урожая, но и за счет повышения содер-
жания витаминов в растениях. Органо-минеральное пита-
ние является важным фактором воздействия на растение.
Своевременное и полное удовлетворение растений мине-
ральным питанием создает наилучшие условия для обме-
на веществ в организме. Усиление физиологических
процессов в растении, в свою очередь, способствует более
энергичному росту и развитию растений. Не удивительно
поэтому, что, как правило, в условиях низкой агротехники,
при несвоевременном и неполном обеспечении растений
органическим и минеральным удобрением, содержание
витаминов в растениях падает. Так, например, рост овса
при недостатке нитратов в питательной среде резко за-
держивается и более чем в два с половиной^аза снижает-
ся содержание в нем пантотеновой кислотьг й витамина Н.
Недостаточная обеспеченность растений азотом и фос-
фором отрицательно сказывается на содержании витами-
63
на С. Так, при внесении полного минерального удобрения
урожай черной смородины составил 5185 кг с 1 га, при
содержании в ней 1827 мг витамина С в 1 кг ягод, тогда
как без удобрения получен урожай, равный 3199 кг с
1 га, с содержанием в ней витамина 1599 мг в 1 кг. В пер-
вом случае валовой выход витамина С с 1 га составлял
9,52 кг, во втором — только 5,1 кг. Одностороннее вне-
сение азотно-фосфорных удобрений также снижает со-
держание этого витамина в черной смородине (Степа-
нова, 1950). Усиление азотного питания на тяжелом опод-
золенном суглинке увеличивает содержание каротина в
злаках и других растениях. По данным Е. Дьяковой
(1945), внесение одинарной дозы азота повысило содер-
жание каротина в стадии выхода овса в трубку с 10 до
28 мг, а тройная доза азота — с 10 до 53 мг каротина в
1 кг сырой массы. Увеличение содержания каротина при
усилении азотного питания было показано также для
костра безостого и тимофеевки. У последней три этом
отмечалось значительное уменьшение количества усыха-
ющих листьев. Внесение навоза повышает содержание
каротина в растениях, особенно в ранние периоды их жиз-
ни. Так, в люцерне, произрастающей на почве, удобрен-
ной навозом, содержалось перед бутонизацией 81 мг ка-
ротина, а в растениях, не получивших навоз,— всего 27 мг
в 1 кг. При дальнейшем развитии эти различия сглажи-
ваются, но содержание каротина во всем урожае в опы-
тах с навозом значительно больше, чем в контроле без
удобрений. При известковании почв также увеличивается
содержание каротина в растениях.
Заметное влияние на биосинтез витаминов оказывает
и удобрение растений микроэлементами. Есть указания,
что внесение в почву ограниченного количества железа
повышает содержание витамина С в ананасе и в других
растениях. Увеличение содержания витамина С в шпина-
те, овсе и суданке наблюдается и при внесении в почву
марганца. Опыты также указывают на стимулирующее
действие марганца на синтез витамина С. Введение
в листья сернокислого марганца значительно усиливает
биосинтез этого витамина. Внесение в почву меди повы-
шает содержание витамина С, тогда как магний и молиб-
ден уменьшают его содержание в томатах и турнепсе.
Цинк усиливает биосинтез витаминов С, Bi, и Р.
64
Благоприятное влияние на биосинтез витамина Р оказы-
вают микроэлементы: медь, железо, молибден и особенно
никель. Один и тот же микроэлемент оказывает неодина-
ковое влияние на синтез разных витаминов. Так, при не-
достатке железа синтез витаминов Вь В2, Вб и РР микро-
организмами повышается, а образование витамина Н,
инозита и пара-аминобензойной кислоты понижается.
Присутствие в среде марганца тормозит образование ви-
тамина В2.
Подробное изучение действия микроэлементов на
биосинтез витаминов помогло выяснить физиологическую
роль некоторых микроэлементов в жизни растений. Так,
например, установление того факта, что кобальт входит
в состав витамина BJ2, позволило более точно предста-
вить значение этого микроэлемента в жизни растений.
Дальнейшее изучение взаимосвязи между витаминами и
микроэлементами поможет подробнее вскрыть причины
необходимости микроэлементов для растений и животных
и объяснить высокую эффективность некоторых микро-
элементов в повышении урожайности сельскохозяйствен-
ных культур.
Большое влияние на биосинтез витаминов оказывают
кислотность среды и концентрация солей в питательном
растворе. Выше мы указывали, что понижение кислотно-
сти почвы путем известкования способствует увеличению
содержания каротина. В отношении витамина Bi доказа-
но, что у некоторых грибов теряется способность синтези-
ровать этот витамин в кислой среде. Установлено также,
что при увеличении концентрации минеральных солей в
питательной среде с 1,64 до 16,4 на 1 л плесневый гриб
прекращает синтез одной из частей витамина Bi—пири-
мидина.
Значительное внимание исследователями уделено вы-
яснению роли света в биосинтезе витаминов. Накоплен-
ный материал показывает, чтр свет является мощным
фактором в образовании витаминов. Давно было замече-
но, что в годы с небольшим количеством ясных дней
содержание витамина С в плодах уменьшается. Это под-
тверждают и опыты. Плоды томатов, развивавшиеся
в темноте, содержат значительно меньше витамина С, чем
выросшие на свету. Резкое снижение образования этого
витамина наблюдается и в проростках при выращивании
5 Витамины в жизни растений 65
их в темноте. Установлено, что в укропе, помйдорах,
петрушке, капусте, салате, редисе в условиях закрытого
грунта содержание витамина С в листьях, стеблях, пло-
дах и корнеплодах значительно снижается, по сравнению
с растениями открытого грунта- Изучение изменения со-
держания витамина С в течение суток показало, что
в ночные часы накопление витамина С в листьях, как пра-
вило, резко снижается. В качестве одного из примеров
приведем данные по изменению содержания этого вита-
мина в листьях томатов.
Таблица 11
Суточная динамика витамина С в листьях томатов
(по Поповской, 1950)
Фаза развития Часы суток
5 9 13 17 21 1 1 а
До цветения . . . 21,5 27,4 34,5 24,6 22,0 20,1
Цветение .... 37,5 40,7 45,6 42,8 45,6 36,6 32,2
Зрелые плоды . . 20,8 22,4 27,3 26,1 27,3 20,2 —
Аналогичная закономерность получена для листьев
шиповника и картофеля. В отличие от этих культур,
в листьях гороха содержание витамина С падает днем
и возрастает ночью. Установлено также, что в период мглы,
когда фотосинтез подавлен, содержание этого витамина
в листьях хлопчатника и других растений возрастает в де-
сятки раз (Брегетова, 1951).
Изучение действия отдельных лучей солнечного спек-
тра на образование витамина С в проростках пшеницы
показало, что наиболее интенсивно биосинтез этого ви-
тамина происходит при действии на проростки белого
света, хуже — на красном свете и еще хуже — в зеленой
части спектра.
В проростках, культивируемых в темноте, биосинтез
витаминов Р и Вь как и биосинтез витамина С, значи-
тельно уменьшается. В отсутствии света и в корнях, и в
надземных частях гороха витамин Bi почти не образуется,
66
тогда как на свету за то же время его содержание увели-
чивается более чем в три раза. Подобные результаты по-
лучены для проростков помидоров и пшеницы.
Если зеленые листья поместить в темноту, в них не
только уменьшается содержание крахмала и сахаров, но
быстро падает количество витаминов. Об этом говорит
такой опыт. Из листьев брюквы вырезались диски. Часть
из них помещалась в темноту, а другая — под лампы
дневного света. По истечении некоторого времени в них
определялось содержание витаминов С, В2 и фолиевой
кислоты (табл. 12).
Таблица 12
Изменения содержания витаминов в листьях
брюквы, выдерживаемых в темноте и при лам-
пах дневного света
(по Андреевой, 1953)
Изменения, % от исход-
ного
Витамин
в темноте на свету
Фолиевая кислота . .
Витамин С............
Витамин В2...........
91,6 129,1
41,6 116,2
95,0 125,0
Приведенные данные показывают, что биосинтез фо-
лиевой кислоты, витамина С и витамина В2 происходит
на свету. В темноте же витамины потребляются, но не
синтезируются. Эти опыты вместе с тем свидетельствуют
о том, что синтез указанных витаминов происходит в ли-
стьях, ибо в изолированные диски витамины не могли по-
пасть из других частей растения.
В плодах накопление витаминов происходит не только
за счет притока их из листьев, но и за счет их новообра-
зования, причем и в этом случае свет оказывает благо-
приятное действие. Так, например, при искусственном до-
зревании плодов шиповника в темноте в них содержалось
20 мг каротина на 1 кг сухого веса, тогда как в плодах,
&♦ 67
находившихся на свету,— 77 мг. Так как прочие условия
опыта были одинаковыми, то можно сделать вывод, чго
увеличение содержания каротина в плодах вызвано дей-
ствием света.
Повышение содержания витаминов на свету позволи-
ло некоторым авторам прийти к выводу, что свет прини-
мает прямое участие в биосинтезе витаминов. Однако та-
кое утверждение является ошибочным. Многочисленными
опытами давно было доказано, что микроорганизмы, куль-
тивируемые в темноте, образуют витамин Bi в значитель-
ных количествах. Известно также, что изолированные
корни гороха, хлопчатника и многих других культур син-
тезируют витамины Bi, В2, РР и другие витамины в тем-
ноте. Подробное изучение изменения содержания витами-
нов при прорастании бобов сои в темноте показали, что
в этих условиях биосинтез витаминов Вь В2, Вб, пантоте-
новой кислоты и особенно витамина РР, инозита и био-
тина весьма значителен. Лишь содержание фолиевой кис-
лоты заметно снижается.
Таким образом, нельзя говорить о прямом, непосред-
ственном действии света на образование витаминов. Снш
жение биосинтеза некоторых витаминов в темноте пра-
вильнее поэтому объяснять глубоким нарушением обмена
веществ, уменьшением синтетических процессов в рас-
тениях. Такое направление обмена, вызванное не свойст-
венным для зеленого растения фактором — продолжи-
тельной темнотой,— задерживает биосинтез не только
витаминов, но и многих других жизненно необходимых
веществ, играющих значительную роль в образовании ви-
таминов. Этими же причинами объясняется изменение
содержания витаминов в низших и высших растениях,
культивируемых при неблагоприятных температурных
условиях. Так, у некоторых грибов, при температуре 25°
резко снижается образование витамина В2. Накопление
витамина С значительно повышается в проростках расте-
ний, выросших при низкой температуре. Большое влияние
на содержание каротина оказывает температура. При
температуре 25—35° биосинтез провитамина А в надзем<
ных частях растения снижается, по сравнению с растения-
ми, культивируемыми при 10—15°.
Образование каротина в корнях моркови и свек-
лы лучше всего происходит при 15—20°. Морковь,
68
выросшая при более низких температурах, окрашена ме-
нее интенсивно вследствие низкого содержания провита-
мина А.
Исследованиями К. Л. Поволоцкой (1937) и
С. М. Прокошева (1944) было установлено значительное
повышение биосинтеза витамина С при раневой реакции.
Позже К- Л. Поволоцкая (1953) показала, что на пора-
нение растения отвечают усиленным биосинтезом не толь-
ко витамина С, но и витамина Вг, а также увеличением
энергии дыхания.
Приведенные примеры убедительно показывают, что
условия внешней среды оказывают огромное влияние на
•образование витаминов в растении. Следовательно, куль-
тивируя растения в условиях, благоприятных для биосин-
теза витаминов, можно значительно повысить содержание
этих веществ в растениях. Дальнейшее выяснение усло-
вий образования витаминов является необходимым как
для познания роли витаминов в жизнедеятельности рас-
тений, так и для рационального использования человеком
витаминного сырья.
Содержание витаминов в организме обусловливается
интенсивностью их образования и степенью их потребле-
ния, которое в свою очередь зависит от состояния орга-
низма и от условий внешней среды. Таким образом, изу-
чение действия факторов внешней среды на биосинтез ви-
таминов в растении не только дает ключ к управлению
образованием витаминов, но и помогает выяснению на-
правленности обмена веществ в растениях в тех или иных
условиях их произрастания. Вместе с тем изучение зави-
симости образования витаминов от условий внешней сре-
ды способствует более детальному установлению путей
биосинтеза витаминов и выяснению причин необеспечен-
ности растений этими жизненно необходимыми вещест-
вами.
Глава III
ПОТРЕБНОСТЬ РАСТЕНИЙ В ВИТАМИНАХ
1. Потребность микроорганизмов в витаминах
В 1860 г. французский ученый Луи Пастер сообщил,
что при посеве небольших количеств дрожжей (объемом
с «булавочную головку») на искусственную среду из
золы, аммонийной соли и сахара наблюдается хороший
рост и размножение этих организмов. Сообщение Пасте'-
ра привлекло внимание многих исследователей. В их
числе был и немецкий ученый Либих. Как известно, ра-
боты Луи Пастера, поставленные с исключительной тща-
тельностью и аккуратностью, изложенные ясным и об-
разным языком, были легко повторимы в любой лабора-
тории. Исследователи и практики, проверявшие работы
Пастера, всегда получали тот же результат, который был
получен автором.
Либих, проверявший работу Пастера, не получил ре-
зультатов, о которых сообщал выдающийся французский
биолог. Либих не мог добиться развития культуры
дрожжей на питательной среде, предложенной Пастером.
Когда же к среде Пастера он добавлял мясной экстракт,
рост дрожжей проходил нормально. Сообщение Либиха
о том, что ему не удается получить по предложенному
Пастером рецепту ни роста, ни брожения дрожжей уди-
вило гениального ученого, и он предложил повторить
опыты в присутствии Либиха на любом количестве пив-
ных дрожжей, но при этом поставил условие — Либих
должен оплатить расходы, связанные с постановкой это-
го опыта. Вскоре Либих умер и потому не мог убедиться
в достоверности опытов Луи Пастера.
70
Через 30 лет их спор был разрешен русским ученым
Я. Никитинским и бельгийским ученым Вильде. Я. Ни-
китинский точными опытами показал, что грибы выде-
ляют в питательную среду вещества, активирующие раз-
множение. Аналогичные результаты были получены
и Вильде. Засевая достаточное количество дрожжей, т. е.
«с булавочную головку», как рекомендовал Луи Пастер,
’он получил нормальное размножение их и брожение на
искусственной среде. В том же случае, если посев произ-
водился меньшим количеством дрожжей, как это делал
Либих, развитие культуры не происходило. Прибавляя
к среде органические экстракты из тканей, Вильде, как
и Я. Никитинский, получал нормальное развитие дрожжей
и в том случае, когда при посеве было внесено неболь-
шое число клеток. Вещество, находящееся в экстрактах
и в дрожжах и усиливающее рост микроорганизмов, было
названо биосом (от греческого слова — жизнь).
Исследования Никитинского и Вильде показали, что
и Луи Пастер, и Либих были правы. Пастер, засевая
множеством клеток питательную среду, вносил вместе
с культурой дрожжей небольшое количество биоса, доста-
точное для нормального размножения микроорганизмов.
Либих же при высеве небольшого количества дрожжей
вносил вместе с ними слишком малое количество биоса
в питательную среду.
Позднее было установлено, что не все микроорга-
низмы одинаково реагируют на внесение биоса. Одни из
них совершенно не растут без биоса и потому нуждают-’
ся в его введении извне. Другие сами обеспечивают себя
биосом. Биос был найден не только в микроорганизмах
и высших растениях, но и во всех животных тканях. По-
дробное изучение природы этого фактора позволило
прийти к заключению, что биос представляет собою не
одно вещество, а состоит из ряда витаминов. В его состав
входят инозит, витамин. Вг, витамин Н, пантотеновая
кислота, витамин Вб, витамин РР, пара-аминобензой-
ная кислота.
Выяснение химической природы биоса, получение в
чистом виде отдельных его компонентов позволило изу-
чить значение каждого2 из-них для жизнедеятельности
организмов. Было установлено; что для роста некоторых-
бактерий и грибов на синтетической среде не требова--
П
лось добавления веществ группы биоса, другие организ-
мы требовали введения всех компонентов биоса, и, нако-
нец, некоторые организмы нуждались лишь в одном или
нескольких компонентах биоса. Так, например, было по-
казано, что нематоспора — паразит, поражающий хлоп-
чатник, не способен к синтезу составных частей биоса
и потому для нормального роста гриба необходимо поступ-
Рис. 13. Прирост сухого вещества мице-
лия грибка нематоспоры на разных источ-
никах витаминов (%) (по Кеглю и Фризу,
1937)
1—инозит; А— витамин В,; В — витамин Н; 14-А—
инозитН-витамин Вх; А •+ В—витамины Вж 4-Н; l-f-B—
инозит-рвитамин Н; 1 +-А 4-В— инозит+витамин Вх-|-
витамин Н
ление указанных веществ извне. При этом следует отме-
тить, что введение одной из составных частей биоса не
оказывает почти никакого действия; при одновременном
введении инозита и витамина Bi рост гриба несколько
улучшается. Еще лучше развитие нематоспоры идет при
одновременном добавлении витаминов Bi и Н, и особенно
хорошо — при внесении в среду инозита и витамина Н.
Когда же в питательную среду добавлялись одновремен-
но все три компонента биоса, т. е. витамин Bi, инозит и
витамин Н, рост гриба был наилучшим (рис. 13).
72
Гриб фитофтора является представителем такой
группы организмов, которая способна синтезировать ви-
тамины Н и инозит и потому не отзывается увеличени-
ем роста на добавление в среду этих веществ, тогда как
внесение в питательную среду витамина Bi—вещества,
не синтезируемого самим грибом, значительно увеличи-
вает его развитие. В эту группу входит большое количе-
ство микроорганизмов. В настоящее время известно бо-
лее 100 видов бактерий, дрожжей и грибов, нуждающих-
ся в снабжении витамином Bi извне. Большинство из них
паразитирует в живых тканях других организмов и лишь
некоторые поселяются на отмерших остатках растений и
животных. I
По отношению к витамину Н микроорганизмы могут
быть разбиты на три группы: одни — неспособны синте-
зировать пимелиновую кислоту—источник образования
этого витамина — и потому нуждаются во введении ее
или витамина Н, другие — синтезируют пимелиновую
кислоту, но не могут образовать из нее предшественник
биотина — дестиобиотин, наконец, третьи — синтезируют
дестиобиотин, но не могут его превращать в витамин Н.
Известны также организмы, которые способны синте-
зировать не целую молекулу витамина, а только ее часть.
Так, например, некоторые грибы и дрожжи способны
строить одну половину молекулы витамина Bi — пири-
мидин, но не в состоянии синтезировать другую часть —
тиазол. Поэтому для нормального развития этих орга-
низмов достаточно ввести в питательную среду только
тиазол. Представителей такой группы известно немного.
Значительно многочисленнее группа растений, способных
синтезировать тиазол и нуждающихся в снабжении лишь
одним пиримидином. Есть и такие организмы, которые
не способны синтезировать ни тиазол, ни пиримидин, но
могут из этих составных частей строить молекулу вита-
мина Вь Введение этих частей тиамина или целой моле-
кулы витамина в одинаковой степени (рис. 14) способ-
ствует росту таких организмов.
Наконец, известны и такие организмы, как, например,
приб фитофтора, которые не могут синтезировать моле-
кулу витамина Bi из введенных пиримидина и тиазола и
поэтому, подобно высшим животным, требуют целой мо-
лекулы витамина Вь
73
Даже морфологически близкие организмы обладают
различной способностью синтезировать витамины и пото-
му неодинаково реагируют на внесение не только целой
молекулы витамина, но и составных ее частей. Устано-
вив, что некоторые дрожжи успешно синтезируют одну
часть витамина Bi—пиримидин — и плохо образуют
другую часть витамина — тиазол,— удалось повысить со-
держание витамина Bi на
Рис. 14. Гриб (Phycomyces nitens)
Слева без витамина В,; справа — с вита-
мином Bi
40—100% путем введения
в питательную среду тиа-
зола или вытяжки из пше-
ничных зерен, экстрактов
хвои, содержащих тиазо-
ловое кольцо, т. е. той ча-
сти витамина, которую
дрожжи синтезируют в
меньшей степени.
Различная реакция
близких между собою ор-
ганизмов на добавление
отдельных витаминов или
их составных частей по-
зволяет не только выяс-
нить роль того или иного
витамина в обмене ве-
ществ организма, но может
быть также использована для распознавания морфологи-
чески близких форм, что часто невозможно сделать дру-
гими методами. Кроме того, установленные факты позво-
лили выяснить некоторые'вопросы взаимоотношения одних
организмов с другими. После того как было выяснено, что,
например, один вид дрожжей синтезирует витамин Вь но
не способен строить витамин Н и потому нуждается в
снабжении- последним извне, другой же — наоборот син-
тезирует только витамин Н, но не в состоянии строить мо-
лекулу витамина Вь задались целью путем совместного
культивирования заставить их нормально развиваться без
добавления извне витаминов. Попытка увенчалась успе-
хом. Совместное культивирование таких дрожжей дало им
возможность обмениваться недостающими веществами,
что создало условия для успешного развития каждого из
симбионтов. Такое же взаимное положительное влияние
74
двух организмов наблюдается в случае совместного
культивирования некоторых дрожжей с грибами. При
культивировании раздельно в отсутствии витамина Bi
эти организмы не развиваются. Однако при совместном
их посеве развитие дрожжей и грибов идет нормально,
так как дрожжи снабжают гриб одной частью витамина
Bi — тиазолом, а гриб обеспечивает дрожжи другой со-
ставной частью этого витамина — пиримидином.
Микроорганизмы не только синтезируют, но и разру-
шают витамины. На это, например, указывают такие
факты. При введении витаминов в почву, богатую микро-
организмами, содержание этих веществ быстро умень-
шалось, тогда как в стерильной почве, т. е. без микро-
организмов, содержание витаминов сохранялось продол-
жительное время. Позднее были установлены и пути раз-
ложения некоторых витаминов микроорганизмами. Так
было показано, что особый микроорганизм расщепляет
активную часть витамина В2 до углекислоты и воды и
тем самым инактивирует этот витамин. Другая бактерия,
выделенная из почвы, окисляет пара-аминобензойную
кислоту до углекислоты, аммиака и воды. Выделены бак-
терии, разлагающие витамины Вь РР, С и др.
Подробное изучение физиологических и морфологиче-
ских изменений в микроорганизмах при недостатке или
отсутствии витаминов привело к заключению, что 6 этих
условиях у них также наступают явления авитаминоза
(рис. 15).
Впервые обстоятельные исследования морфолого-фи-
зиологических изменений в бактериальной клетке при не-
достатке витаминов были выполнены советскими учены-
ми. М. Н. Мейсель (1950) показал, что у микроорганиз-
мов при авитаминозе резко нарушается обмен веществ
и происходят глубокие изменения в строении клеток. В без-
витаминных культурах клетки мелкие, имеют нетипич-
ную форму, размножение задерживается, урожай куль-
туры резко снижается (рис. 16).
Выяснение потребностей микроорганизмов в том или
ином витамине не только позволяет установить синтети-
ческие возможности организма и создать необходимые
питательные среды для его культуры, но и дало легкие и
надежные методы определения витаминов. В настоящее
75
Рис. 15. Реакция Endotnyces tnagnusii на витамин В! (фиксирован-
ные и окрашенные препараты)
1—клетки, выросшие на среде Ридер—аспаоагин—агар без витамина Bj; 2—то
же с витамином Вх; 3 - клетки, выросшие на среде Ридер с сернокислым аммо-
нием без витамина ; 4 — то же с витамином Bj (по Мейселю, 1950)
Рис. 16. Морфологические проявления активирующего действия
никотиновой кислоты на клетки дрожжей Zygosaccharotnyces таг-
x'anus
1 — на среде без никотиновой кислэты; 2 — над улетучивающейся никотиновой
кислотой (по Мейселю, 1950)
время микроорганизмы широко используются для опреде-
ления многих витаминов.
Таким образом, витамины, принимая непосредствен-
ное участие в обмене веществ бактерий и грибов, необ-
ходимы им так же, как и животным. В отличие от живот-
ных, некоторые микроорганизмы обеспечивают себя ви-
таминами за счет синтеза, однако известно весьма много
бактерий и грибов, нуждающихся в снабжении вита-
минами.
Так установлено, что лишь немногие почвенные ми-
кроорганизмы не нуждаются в дополнительном их обес-
печении витаминами, а большинство для своей нормаль-
ной жизнедеятельности получают витамины из почвы.
Обстоятельные исследования по витаминному пита-
нию микробов помогли также выяснению потребности
высших растений в витаминах.
2. Потребность высших растений в витаминах
Долгое время считалось, что лишь человек и живот-
ные требуют введения витаминов извне. Однако, как мы
видели выше, многие бактерии, дрожжи и грибы или не
в состоянии синтезировать некоторые витамины совер-
шенно, или образуют их в недостаточном количестве
и поэтому нуждаются в введении их. В последние годы
исследования показали, что и у высших растений имеется
потребность в витаминах, что может представлять не
только научный, но и практический интерес.
Влияние витаминов на рост корней
Исследования последних лет все больше и больше
вскрывают причины давно установленной зависимости,
что растения, выращенные на почвах с обильной микро-
флорой и там, где даны зеленые и органические удобре-
ния, развиваются значительно лучше, чем выращенные на
почвах, бедных органическим веществом. Оказалось, что
микроорганизмы не только способствуют созданию
структуры почвы, но в процессе своей жизнедеятельности
выделяют в окружающую среду физиологически актив-
ные вещества, благоприятно действующие на рост и раз-
витие растений. В числе этих веществ витамины
78
занимают значительное место. Так, например, в почвах
Вахшской долины- в Таджикской ССР, на которых куль-
тивируется хлопчатник, по теоретическим подсчетам
М. Н. Мейселя (1950) на 1 га площади синтезируется ми-
кроорганизмами в течение года около 400 г витамина
eBb около 300 г витамина В6 и около 1 кг витамина РР.
Особенно много накапливается витамина Bi в поч-
вах, в которых культивируется азотобактер. Этот микроб
синтезирует и выделяет в почву значительные количества
витамина Вь
Выделение многочисленными почвенными микроорга-
низмами витаминов в почву является одним из звеньев
сложных взаимоотношений между низшими и высшими
растениями. Такое выделение обеспечивает возможность
получать недостающие организму витамины и тем самым
способствует лучшему развитию.
Опыты показывают, что в наиболее жизнедеятельном
(пахотном) слое почвы содержится больше витаминов,
чем в нижележащих горизонтах. В качестве примера
приведем результаты одного из многочисленных анали-
зов (табл. 13).
Таблица 13
Количественное распределение витаминов
Bi и Н в почве на различных глубинах
(по Шопферу, 1950)
Глубина, см Количество витамина Bt мг на 1 кг почвы Количество витамина Н, мг на 1 кг почвы
10 19,3 0,0062
20 8,6 0,0032
30 6,2 0,0027
50 2,9 0,0023
Микроорганизмы не являются единственным источник
ком витаминов в почве. Витамины в почве образуются
также при разложении остатков животных и растений.
79
Большое количество витаминов вносится в почву с наво-
зом и навозной жижей.
Сами растения отдают в почву значительное количе-
ство витаминов. Часть из них попадает в почву из над-
земных частей растений, но особенно много витаминов
выделяется в почву корнями. Так, например, в почве из
зоны распространения корней табака найдено 0,01—
0,15 мг витамина Bi в 1 кг почвы, тогда как в почве
вне зоны корней содержалось только 0,001—0,004 мг.
Выделение корнями высших растений витаминов в почву
имеет огромное биологическое значение как для расте-
ния, так и для микроорганизмов (Красильников, 1952).
Возникает вопрос, усваиваются ли витамины, находя-
щиеся в почве, корнями растений и усиливают ли они
рост последних?
На этот вопрос ученые могли ответить не сразу. Каза-
лось бы, что внесение в почву витаминов и учет роста
и развития растений должны разрешить этот вопрос.
Такие опыты были поставлены многими исследователями
и лишь некоторые из них получили положительные ре-
зультаты. Проверка этих работ другими учеными, как пра-
вило, не имела успеха. Исследователи, вносившие растворы
витамина Bi в черноземные почвы, не получили усиления
роста корневой системы, тогда как те же концентрации
этого витамина, будучи добавлены в подзолистые или
песчаные почвы, увеличивали рост растения космоса.
Расхождение полученных результатов, как выяснилось
позднее, объяснялось тем, что раньше не учитывалось
содержание витаминов в почве. Оказалось, что в черно-
земах содержатся значительные количества витамина Вь
образовавшиеся в результате жизнедеятельности микро-
организмов. Поэтому дополнительное внесение витамина
не давало никакого эффекта. В песчаных же почвах ви-
тамин Bi почти отсутствует, и его введение способствует
усилению роста корневой системы.
Но даже в том случае, если опыт ставился на почвах,
лишенных витаминов, исследователь не мог ответить на
вопрос — поглощаются ли корнями внесенные в почву
витамины? В почве, кроме корней, находятся в огромном
количестве бактерии и грибы, большинство которых, как
мы видели выше, потребляют витамины. Следовательно,
витамины, внесенные в такую почву, являются достоя-
80
нием не столько корней, сколько микроорганизмов. Кроме
того, сами микробы и грибы выделяют в среду некото-
рые количества того или иного витамина.
Опыты, поставленные в стерильной почве, в которой
микроорганизмы отсутствовали, также не ответили
с определенностью на этот вопрос, так как витамины, син-
тезируясь в листьях, оттекают в корни. В разных усло-
виях в листьях образуются и из них передвигаются
в корни неодинаковые количества витаминов. Поэтому
в каждом конкретном случае трудно было учесть, сколько
же витаминов поступило в корни.
Использование методов меченых атомов и стерильных
культур помогло решить эту задачу. Выращивая гречи-
ху, горох и кукурузу на питательной среде, содержащей
витамин Bi, меченый радиоактивным изотопом серы S35,
удалось отчетливо показать, что этот витамин погло-
щается корнями.
Поглощенный корнями витамин затем в значитель-
ных количествах накапливается в других частях расте-
ния (в листьях и стеблях). Даже в том случае, когда
растение само синтезирует этот витамин, оно продолжает
поглощать витамин Bi из питательной среды (Шавлов-
ский, 1954). При помощи метода меченых атомов была
также выяснена передача высшему растению витаминов,
синтезированных микроорганизмами. Опыт ставился так.
Меченый витамин Bi вводился в микробные клетки,
после чего последние наносились на простерилизованные
семена гречихи. Через 11 дней в проростках гречихи было
определено содержание меченого витамина В]. Опреде-
ление показало, что те микробные клетки, которые выде-
ляют витамин Bi в окружающую среду в больших коли-
чествах, способствуют и большему накоплению этого
витамина в проростках гречихи (Шавловский, 1954).
Поглощение корнями витаминов было доказано и
другим способом. Семена исследуемых растений прира-
щивались в стерильных условиях. Когда корни пророст-
ков достигли длины 1—2 см, от них отрезались вер-
хушки, которые переносились в стерильную питательную
среду, содержавшую минеральные соли, аминокислоты и
сахар. Оказалось, что на этой среде корни не могут
расти продолжительное время. Корни некоторых расте-
ний прекращают рост через 1—2 педели и лишь корни
6 Витамины в жизни растений 81
немногих растений нормально «росли продолжительное
нремя.
Так, например, рост корней томата заметно увеличи-
вался, когда в среду добавлялся витамин Вь Корни, куль-
тивировавшиеся в питательном растворе, содержащем ви-
тамин Bi, аминокислоты и другие вещества, хотя и росли
с достаточной интенсивностью, однако были тонкими,
скрюченными и обладали пониженной жизнеспособ-
ностью. Это указывало на то, что для нормального роста
корней не хватало еще каких-то важных питательных ве-
ществ. Оказалось, что введение в питательную среду, кро-
ме витамина Bi, витамина В6 значительно улучшало рост
корней томата (рис. 17).
Еще лучше рост корней томата в том случае, если в
среду добавить, кроме витаминов Bi и В6, также вита-
мин РР. Из этого можно заключить, что корни томата
мало образуют витаминов РР и В6 и особенно плохо син-
тезируют витамин Вь Вместе с тем эти опыты показы-
вают, что внесенные в среду витамины потребляются ра-
стущими корнями. В дальнейшем было установлено, что
корни разных растений для своего нормального роста на
питательной среде требуют различных витаминов. Так,
корни люцерны, получавшие только витамин Bi или
только витамин РР, переставали расти после двух ежене-
дельных пересадок на свежую питательную среду.
Те же корни, которые получали оба эти витамина, про-
должали расти с первоначальной для них скоростью,
сохраняя прирост по 98 мм за неделю в течение 25 недель.
Прибавление к среде витамина Be не увеличивало ско-
рость роста.
Корни белого клевера хорошо росли на среде, содер-
жащей витамины Bi и РР, и их рост даже несколько
увеличивался при непрерывном переносе в свежую пита-
тельную среду. Так, в течение первых пяти недель после
переноса в среду, содержавшую эти витамины, корни
увеличили свой размер на 84 мм, за вторые пять недель
давали прирост в среднем по 109 мм, за третьи пять не-
дель— по 129 мм, за четвертые пять недель — по 136 мм
и за пятые пять недель — в среднем по 151 мм в неделю.
Прибавление витамина В6 не увеличивает больше ско-
рость роста корней белого клевера. Корни, которым
давался витамин РР, продолжали расти с приблизи-
82
тельно постоянной скоростью, давая прирост по 22 мм
в неделю.
Корни хлопчатника росли с одинаковой скоростью
в продолжение 15 недель в среде, содержащей витамины
Bi и РР. К концу этого времени корни хлопчатника
были того же диаметра, какими они были в первую не-
делю культуры. Прибавление витамина Be не увеличи-
вало больше скорость роста корней хлопчатника. Корни
дурмана, не получившие витаминов, очень бы-
стро прекращали рост. Корни же, обеспеченные
витаминами Bi, В6 или РР, росли нормально, f6j
но при перенесении в безвитаминную среду их I
рост замедлялся, а затем и прекращался.
Корни дурмана, получавшие все три вита-
мина, росли хорошо и сохраняли среднюю ско-
рость роста в размере 38 мм за неделю в те-
чение 15 недель. Корни моркови, не получав-
шие витаминов или получавшие только вита-
мин Вб, значительно уменьшали скорость сво-
его роста. Когда же эти корни получали вита-
мин Bi, их рост улучшался, но особенно хоро-
ший рост наблюдался
в том случае, если в
питательную среду
одновременно вводи-
лись витамины Bi и В6.
Корни подсолнеч-
Рис. 17. Влияние витаминов Вх и В6
на рост отчлененных корней томата
(вес корня в мг)
ника, не получавшие
никаких витаминов или
получавшие витамин
РР или витамин В(5,
прекращали рост после семи последовательных ежене-
дельных переносов. Корни, получавшие либо витамин
Bi, либо витамины Bi и РР, либо витамины РР и В6, да-
вали прирост в неделю 7 мм или меньше. Корни, полу-
чавшие витамины Вь РР и В6, энергично росли в тече-
ние пяти еженедельных переносов.
Так, скорость роста корней подсолнечника в течение
первых пяти недель была 74 мм, в течение вторых пяти
недель — 96 мм, в течение третьих пяти недель — 120 мм
и в течение последующих двух недель—150 мм в не-
делю.
6* 83
Потребность в витаминах изолированных корней деся-
ти видов растений приводится в табл. 14.
Таблица 14
Потребность в витаминах изолированных корней растений
Потребность в витаминах Вид
Витамин Bj Витамины Вх и РР ... Лен Горох, редис, люцерна, кле- вер, хлопчатник
Витамины Bi и В6 .... Витамины Вь РР и Вб . . Морковь Томат, дурман, подсолнечник
Все исследованные виды требуют введения в питатель-
ную среду витамина Вь Хотя изолированные корни льна
и клевера и растут без добавления в среду витамина Bi,
но скорость их роста увеличивается на 400—500% при
введении этого витамина. Восемь из этих десяти видов
требуют добавления витамина РР. Однако степень потреб-
ности в этом витамине у разных видов неодинакова. Так,
корни томата и подсолнечника растут и без витамина РР,
нс их рост увеличивается на 35—140% в присутствии это-
го витамина.
Следовательно, корни десяти изученных видов расте-
ний по их отношению к витаминам могут быть разбиты на
три группы: одни не реагируют на добавление в среду
того или иного витамина, другие — усиливают свой
рост и, наконец, для третьих — витамины являются
абсолютно необходимыми, без них рост корней невоз-
можен.
Для объяснения причин разного действия витаминов
на рост различных корней было высказано предположе-
ние, что корни, которые сами не способны синтезировать
тст или иной витамин, требуют для нормального роста
в искусственной среде введения этих витаминов. Те же
корни, которые синтезируют малые количества витамина,
84
не обеспечивающие нормальный обмен веществ и макси-
мальный рост, нуждаются в дополнительном введении
витаминов.
Это предположение нашло в дальнейшем и экспери-
ментальное подтверждение.
Известно, что корни льна хорошо растут, если им да-
вать витамин Bi. С этими корнями и был поставлен спе-
циальный опыт. Корни льна культивировались в течение
тридцати шести еженедельных переносов. В присутствии
витамина Bi скорость роста при последних десяти пере-
носах была равна 185 мм в неделю, а в отсутствии вита-
мина она достигала всего лишь 31 мм в неделю. Медлен-
ный рост корней в отсутствии витамина Bi указывал на
то, что корни льна способны к ограниченному синтезу это-
го витамина. Проверка высказанного предположения про-
изводилась таким образом: один корень льна, который
подвергался 36 еженедельным переносам в среду, свобод-
ную от витамина Bi, вновь был перенесен в такую же
•питательную культуру. В течение пяти недель этот отре-
зок корешка синтезировал примерно в сто раз больше
витамина Bi, чем его было первоначально в верхушке
корня. Однако синтезированного витамина Bi было не-
достаточно для нормального роста корней. Если рассчи-
тать синтез витамина Bi по неделям, то получим, что в
среднем синтез этого витамина корнем был приблизи-
тельно равен 0,02 микрограмма в неделю, потребность же
равнялась 2 микрограммам. Таким образом, синтетиче-
ские возможности корня льна отставали от его потребно-
сти в витамине Bi почти в сто раз. Поэтому, когда в пи-
тательную среду дополнительно вносили этот витамин,
корни льна значительно усиливали свой рост. Корни кле-
вера обладают значительно более слабой способностью
синтезировать витамин Вь если в корешках льна за пять
недель его содержание увеличилось в 100 раз, то в ко-
решках клевера — только в 6 раз. Поэтому дополнитель-
ное введение витамина В] в питательную среду вызывает
более значительную прибавку скорости роста корней кле-
вера, чем корней льна.
Изучение причины медленного роста корней в отсут-
ствии витамина Bi показало, что в этом случае резко сни-
жается и даже прекращается деление клеток. Хотя корни
страдали прежде всего от недостатка витамина Bi, но
85
наблюдавшиеся симптомы могли быть приписаны отчасти
и недостатку витамина РР.
Для проверки этого был поставлен такой опыт. В од-
ном варианте корни гороха получали только витамин Вь
в другом — только витамин РР, в третьем — контроль-
ном — одновременно оба витамина. Результаты опыта до-
казали, что недостаток
Рис. 18. Влияние витаминов Bi и
РР на рост отчлененных корней
гороха
1 — Вх 4- РР; 2 — Вь 3 — РР
того или другого вита-
мина приводит к умень-
шению скорости роста
(рис. 18).
Это снижение темпа
роста корней сопровож-
дается как задержкой
деления клеток в расту-
щей зоне корня, так и
уменьшением размера
клеток. При недостатке
витамина РР умень-
шаются диаметр клеток
корня и число рядов
клеток в корне, вслед-
ствие чего эти корни
были тоньше, чем кор-
ни, получившие вита-
мин РР.
Систематические на-
блюдения за ветвле-
нием корней в искусст-
венной культуре пока-
зали, что характер его оставался типичным для данного
вида растений. Так, томат, имеющий волокнистую корне-
вую систему, обнаружил обильное ветвление и быстрый
рост боковых корней, тогда как у растений с главным кор-
нем, таких, как горох, люцерна и лен, в этих условиях
корни ветвились меньше, и рост боковых корней у них
был значительно задержан.
Корни растений одного и того же вида неодинаково
отвечают на введение витаминов. Так, например, было
установлено, что рост изолированных корней томатов
одного сорта значительно ускорялся прибавлением в сре-
ду витамина Во и не изменялся при добавлении витамина
86
РР, тогда как корни другого сорта томатов нуждались
не в витамине Вб, а в витамине РР. При скрещивании
этих сортов получался гибрид, корни которого не нужда-
лись ни в том, ни в другом витамине.
В естественных условиях, при наличии листьев, свои
потребности в витаминах корни покрывают не только пу-
тем биосинтеза, но и за счет их притока из надземных
частей растения. Большой интерес представлял вопрос,
как будет идти образование корней, если у растений уда-
лить листья, но корням дать витамины. Для решения этой
задачи нами был поставлен такой опыт. 14 июня, т. е. в
период бутонизации, у растений хлопчатника обрывались
все листья, их корни переносились в питательную среду,
содержащую витамин РР.
Наблюдения за образованием корней показали, что в
варианте с витамином РР, взятом в концентрации
(0,001%), черёз шесть дней после начала опыта отмеча-
лось массовое образование корневых бугорков. 6 августа
был произведен учет числа образовавшихся корней и их
рост. Результаты учета приводятся в табл. 15.
Таблица 15
Интенсивность образования корней у хлопчатника
под влиянием обработки растении витамином РР
Вариант, концентрация (%) Число корней на 1 растение Длина корней, см Число вторичных корней
Контроль 1 2 о
Витамин РР (0,1) . . . растения погибли
» (0,01) . . . 6 3 4
» (0,001) . . . 3 4 3
Из данных таблицы следует, что подкормка растений
витамином РР усиливает образование и рост корней
у хлопчатника. Однако это усиление неодинаково при вве-
дении в питательную среду разных доз витамина. Замет-
ное образование корней наблюдается в варианте с 0,01%
концентрацией витамина. При снижении дозы витамина
87
(0,001%) образование корней уменьшается по сравнению
с предыдущей концентрацией витамина, но их число и раз-
мер больше, чем у растений, не получивших витамин
(контроль). При избытке витамина РР (0,1%), т. е. при
гипервитаминозе, растения погибли. Отметим, кстати, что
в физиологии животных также известны случаи, когда
значительное повышение в тканях этого витамина вызы-
вает вредное действие. Таким образом, не только недо-
статок, но и избыток витаминов может привести к глу-
боким расстройствам физиологических процессов в орга-
низме.
Благоприятное действие некоторых витаминов на рост
корневой системы находит практическое применение
в растениеводстве. Как известно, для повышения образо-
вания и усиления роста корней у трудно укореняющихся
черенков нашли широкое применение стимуляторы
роста — гетероауксин, альфанафтилуксусная кислота,
2—4-дихлорфеноксиуксусная кислота и др.
Прием улучшения укоренения черенков при помощи
стимуляторов роста, разработанный в Институте физио-
логии растений им. К. А. Тимирязева Академии наук
СССР академиком Н. А. Максимовым и Р. X. Турецкой,
нашел в настоящее время широкое применение при вы-
ращивании различных культур. Несколько лет назад в
этом же институте В. Ф. Верзилов использовал стимуля-
торы роста для усиления образования корневой системы
у саженцев и взрослых деревьев.
Представляло большой интерес изучение совместного
действия стимуляторов роста и витаминов на образование
корней. Такие исследования были проведены во многих ла-
бораториях. В результате было, например, установлено
значительное усиление образования корней у гороха при
обработке черенков витамином Bi и индолилуксусной кис-
лотой (гетероауксином). Опыт ставился таким образом.
Этиолированные стебли гороха помещали в растворы ин-
долилуксусной кислоты (20—200 мг на 1 л) на 20 часов
и затем переносили в склянки, содержавшие 5 мл 2%-ного
раствора сахарозы. Через различные сроки после этой об-
работки к растворам сахарозы прибавлялся витамин Bi
в различных концентрациях. Добавление витамина Bi в
эти растворы вызывало значительное увеличение как
числа, так и размера корней. Без применения стимул я-
88
торов роста витамин Bi не оказал положительного дей-
ствия. Такие же результаты были получены и для черен-
ков лимона. Обработанные 0,02 %-ной индолилуксусной
кислотой черенки на неделю помещали в парники, затем
вынимали и на сутки нижней частью погружали либо в
воду (контроль), либо в 0,0001 %-ный раствор витамина
Вь Через 13 дней в варианте с витамином число корней,
по сравнению с контролем, увеличивалось вдвое, при этом
длина корней также значительно возросла. Черенки, не
обработанные стимулятором роста, имели в среднем 0,3
корня на черенок, тогда как при обработке индолилуксус-
ной кислотой их было 8,1; а после воздействия на них
стимулятором роста и витамином Bi — 16,3.
Еще более эффективным оказалось совместное дейст-
вие этих веществ на образование корней у облиственных
черенков камелии.-Через неделю после обработки черен-
ков камелии индолилуксусной кислотой у них образо-
валось в среднем три коротких и слабых корня, в то вре-.
мя как при совместном воздействии кислоты и витамина
Bi на каждом черенке образовалось по 67 корней. Паль-
мы также весьма отзывчивы на этот витамин. Манговые
чувствительны к обработке витаминами Bi и РР лишь
в молодом возрасте. Путем совместной обработки стиму-
лятором роста и витамином Bi удалось также значитель-
но повысить процент укоренения черенков красной ка-
занлыкской розы, кокаинового куста, березы, чая и других
трудно укореняемых растений. Есть данные о благопри-
ятном действии смеси витамина Bi и индолилуксусной
кислоты на укоренение, и рост бегонии, сливы и других
растений.
Чем же объяснить благоприятное действие на укоре-
нение черенков совместного применения витамина Bi и
стимуляторов роста? Специальные исследования показали,
что закладка корней происходит под влиянием индолилук-
сусной кислоты, но некоторые корни дальше не могут
развиваться в отсутствии витамина Вь Добавляя этот ви-
тамин, удается значительно усилить рост зачаточных кор-
ней. Обработка же черенков одним витамином Bi у неко-
торых растений не приводит к заметному образованию
корней. Эти исследования, кроме значительного теорети-
ческого интереса, имею? и большое практическое значе-
ние, так как они указывают новые пути ускорения обра-
89
зования и роста корневой системы у трудно укреняющих-
ся растений.
Дополнительное обогащение растений витаминами
усиливает во многих случаях обмен веществ и ускоряет
их рост. В связи с этим часто говорят о том, что витами-
ны оказывают на растение стимулирующее действие. Сти-
муляция — это усиление обмена веществ, вызываемое
действием факторов, не свойственных норме требований
организма. Наблюдаемое при стимуляции усиление обме-
на Ю. В. Ракитин (1953) рассматривает как защитную
реакцию организма, направленную на обезвреживание
вредных для организма химических агентов, на преодоле-
ние нарушенного обмена веществ. При усиленном обмене
вредные начала скорее обезвреживаются, и организм ско-
рее приходит к норме. Совсем по-другому действуют на
растение витамины, являющиеся необходимыми для ор-
ганизма факторами, т. е. условиями существования. По-
падая в растение, эти соединения включаются в обмен
веществ как необходимые его компоненты. При недостатке
витаминов наблюдаются нарушения обмена, а при по-
полнении содержания этих веществ до нормы нарушения
в обмене веществ ликвидируются, и организм приходит
к нормальному состоянию. Усиление обмена веществ, на-
блюдаемое при обогащении растений витаминами, не яв-
ляется результатом стимуляции и должно рассматривать-
ся как следствие более полного удовлетворения потреб-
ности организма.
Витамины и рост надземных частей
растений
Тот факт, что витамины в семенах находятся в зна-
чительных количествах, позволяет предположить, что они
играют существенную роль в развитии зародыша, в про-
растании семян и в ранний период роста растений. Полу-
ченные экспериментальные данные действительно указы-
вают на важную роль витаминов в этих процессах.
В опытах с проростками гороха, из которых были удале-
ны семядоли, было показано, что на среде, содержащей
только минеральные соли и сахар, рост этих проростков
был весьма слабым. При добавлении же витамина Н ско-
рость роста проростков значительно увеличивалась, а вве-
90
дение витамина Bi заметно ускоряло рост корней. Панто-
теновая кислота, подобно витамину Н, увеличивает рост
надземной части проростка гороха, а витамин РР способ-
ствует росту как корней, так и надземной части пророст-
ка. Однако даже при применении оптимальных комбина-
ций витаминов Bi, Н, РР и С, пантотеновой кислоты рост
таких проростков менее интенсивен, чем рост проростков
с сохраненными в них семядолями. Этот факт свидетель-
ствует о том, что для нормального роста зародышей горо-
ха нужны другие, пока еще не известные вещества. Зре-
лые зародыши дурмана легко культивируются на искус-
ственной среде без витаминов. Однако незрелые зароды-
ши, составляющие 73 от конечного размера, не разви-
ваются в обычной среде. Их развитие начинается только
при добавлении к обычной среде витаминов РР, Вь Вб,
С и других веществ. Еще более сложной среды требуют
более молодые зародыши.
Добавление к питательной среде витаминов РР, Bi и
Во позволило культивировать незрелые зародыши в искус-
ственных условиях. Благоприятное действие оказывает
пантотеновая кислота на рост люцерны в стерильных пес-
чаных культурах. Этот витамин, а также витамины Bi,
РР и Н заметно усиливали рост мятлика.
В последнее время началось подробное изучение дей-
ствия витаминов на интенсивность прорастания семян.
Хотя и допускалось, что семена могут довольствоваться
собственными витаминами, запасы которых значительны,
все же не исключена возможность повышения продуктив-
ности растений при помощи витаминных подкормок. Уже
первые работы в этом направлении дали положительные
результаты. Прибавляя в питательную среду витамин С,
удалось заметно увеличить интеноивность роста пророст-
ков гороха в стерильных условиях. Однако не во всех
случаях этот витамин оказывал положительное действие.
Так, оказалось, что сорта гороха, синтезирующие при про-
растании семян значительные количества витамина С,
при их обработке витамином увеличивали скорость роста
всего на 4%, тогда как сорта со слабым биосинтезом ви-
тамина С в этих условиях увеличивали рост на 163—
213% по сравнению с контролем.
Благоприятное влияние на прорастание семян оказы-
вают и другие витамины. Обработка семян винограда
91
0,01%'-ным раствором витамина РР ускоряет прораста-
ние семян. У обработанных семян корешки и надземные
части развивались также значительно лучше, чем у семян,
не получивших витамин (Флеров и Коваленко, 1947).
Есть указания на благоприятное действие предпосевной
обработки семян моркови витаминами. Вводя в семена
моркови витамины РР, Bi и другие, удалось вызвать усй-
ление ростовых процессов и повышение урожая моркови
на 30%.
Предпосевная обработка семян хлопчатника витами-
нами усиливает энергию прорастания семян и улучшает
рост проростков. При обработке семян хлопчатника ви-
таминами Bi и РР значительно увеличивается содержа-
ние в семенах сахаров и небелковых форм азота, что яв-
ляется следствием усиления активности ферментов. Все
это способствует более быстрому прорастанию семян. Так,
например, если на пятый день после обработки у кон-
троля взошло 45% семян и средняя длина проростков
составляла 1,35 см, то при обработке семян витамином
РР всхожесть составляла соответственно 75% и пророст-
ки достигали 1,65 см, а при обработке витамином Bi
всхожесть — 64% и длина — 1,74 см. Значителтное по-
вышение всхожести семян хлопчатника сорта С-460 при
предпосевной обработке семян витаминами получили и
другие исследователи (Захарьянц, Горбачева и Зглин-
ская, 1950). Обработка семян фолиевой кислотой увели-
чивает образование и рост корешков у проростков пше-
ницы.
Значительный практический интерес представляют ре-
зультаты опытов по предпосевной обработке семян фасо-
ли витаминами Bi и РР. При дополнительном обеспе-
чении этих семян витамином Bi заметно усиливаются ро-
стовые процессы растений и повышается урожай (Дагис,
1954). Так, например, рост фасоли в высоту увеличился
на 18%, прирост вегетативной массы — на 39%, урожай
семян — на 20,8%. Заметная прибавка урожая фасоли
наблюдается и при предпосевной обработке семян вита-
мином РР (табл. 16).
Как видно из данных таблицы, особенно значитель-
ное увеличение урожая фасоли наблюдается при воздей-
ствии на семена витамином РР, взятым в концентрации
5 мг на 1 л воды. При обработке же семян меньшими до-
92
Таблица 16
Влияние предпосевной обработки семян витамином РР
на урожай фасоли
(по Дагису, 1954)
в ариант Общий вес одного растения Урожай семян с одного растения Число бобов с одного растения Число семян с одного растения
г ' /о г 7о г /о г 7о
Контроль 39,1 100,0 20,5 100,0 14,0 100,0 46,5 100,0
Витамин РР (5 МГ/Л) 45,9 117,5 24,2 118,0 17,8 127,1 61,4 129,8
» (1 мгул) 38,4 98,2 23,5 114,6 15,0 107,1 49,2 105,7
(0,2 мг/л) 39,0 99,7 23,2 113,1 15,6 111,1 52,1 112,0
зами витамина РР общий вес растения остается почти
таким же, как у растений контрольного варианта, и имеет
место незначительное увеличение числа бобов и семян.
Приведенный материал свидетельствует о благоприят-
ном действии некоторых витаминов на энергию прораста-
ния семян и рост молодых растеньиц. К сожалению, об-
стоятельных исследований в этом отношении до настоя-
щего времени не проводилось, что затрудняет дать ответ
на вопрос о практическом использовании предпосевной
обработки семян витаминами. Несомненно, результаты,
полученные в лабораторных условиях, нельзя перенести
в поле, так как в последнем случае на эффективность
предпосевной обработки могут оказывать влияние другие
факторы внешней среды и содержание в почве вита-
минов.
Как отмечалось выше, биосинтез витаминов происхо-
дит главным образом в надземных частях. Это дало ос-
нование считать, что листья, стебли и побеги не нуждают-
ся в дополнительном обеспечении их витаминами. Однако
при этом упускали один важный момент. Не при всех
условиях внешней среды происходит нормальный процесс
биосинтеза витаминов. При одних,— как мы видели выше,
93
биосинтез витаминов усиливается, при других — наоборот,
он тормозится. Усиленный обмен веществ, в котором при-
нимает участие тот или иной витамин и связанный с
этим повышенный его расход, также может явиться одной
из причин необеспеченности растения витаминами. Таким
образом, хотя биосинтез витаминов и осуществляется
главным образом в листьях и в плодоэлементах, но тем
не менее может быть такое их состояние, что надземные
части не полностью обеспечены этими жизненно необхо-
димыми соединениями.
Некоторые исследователи потребность растений в ви-
таминах ставят в прямую связь с наличием этих витами-
нов в том или ином растении. Так, американские иссле-
дователи Боннар и Грин, определив содержание витами-
на Bi в листьях различных растений и установив реак-
цию этих растений на обработку их витаминами, пришли
к категорическому выводу, что чем меньше содержится
витамина В в листьях, тем сильнее растения реагируют
на такую обработку. Так, например, горох и томат, со-
держащие 13—18 мг витамина Bi в 1 кг сухих листьев,,
не ускоряют рост при добавлении витамина Вь тогда
как капуста, космос, камелия японская и другие, имею-
щие низкое содержание этого витамина, положительно
реагируют на такую обработку.
Однако другие данные показывают, что низкое содер-
жание витамина Bi в том или ином растении еще не мо-
жет указывать на потребность его в этом витамине. Об-
мен веществ у растения может быть таким, что для его
нормального осуществления совсем не нужно повышенное
содержание витамина. Вегетирующие растения могут
осуществлять нормальный обмен и рост при невысоком
содержании в них того или иного витамина, если это ко-
личество витамина является свойственным природе ра-
стения, произрастающего в определенных условиях. Со-
вершенно естественно поэтому, что положение указанных
авторов, сделанное ими на основании реакции капусты
и космоса на введение витамина Вь не отражает общей
закономерности и не нашло подтверждения в дальнейших
работах. Так, например, мятлик, содержащий 4,2 мг ви-
тамина Bi в 1 кг сухого веса листьев, отзывается менее
значительно на витаминную подкормку, чем полевица,
содержащая 6,0 мг Bi в 1 кг листьев. С другой стороны,
94
Рис. 19. Влияние витамина Bi на
накопление сухого веса космоса
(г на 1 побег)
содержание витамина Bi в растениях при подкормках мо-
жет значительно увеличиваться, а усиление роста может
быть незначительным. Например, в предыдущем опыте
содержание витамина Bi в тканях мятлика увеличилось
всего на 2,8 мг (менее чем в два раза), а рост этих рас-
тений за два месяца увеличился в 6 раз, по сравнению
с растениями, не получавшими витамин. У белой же гор-
чицы количество витамина возросло в 9,4 раза, а накоп-
ление сухого вещества
увеличилось всего в
4 раза.
Дополнительная под-
кормка витамином Bi
оказала благоприятное
действие и на другие
растения. На обработку
витамином Bi хорошо
реагирует космос и тун-
говое дерево. Растения,
получавшие по 0,5 мг
витамина через каждые
два дня, за 70 дней
дали почти в два раза
больший прирост, чем
контрольные, * не полу-
чавшие этого витамина
(рис. 19).
Еженедельные под-
кормки мака снотворного витамином Bi от момента про-
растания и до сбора урожая увеличили вес коробочек
и повысили урожай этой культуры. Систематическое вне-
сение раствора витамина в почву оказало благоприятное
действие и на рост шпината. Растения, получавшие за
время вегетации ряд поливов водой, содержащей витамин
Bi, имели большее количество листьев, причем средний
вес листьев также был выше, чем в контроле (табл. 17).
Подкормки растений витамином Bi ускоряют рост
стебля у разных растений неодинаково. Так, введение ви-
тамина Bi в почву увеличило длину стебля у сливы и
японской камелии на 131%, у ливанского кедра на 247%
по сравнению с контролем.
Есть указания о благоприятном действии витамина
95
Таблица 17
Сухой вес листьев шпината через 63 дня опыта, после 13
орошении раствором витамина В1
(по Шопферу, 1950)
Вариант Положение листьев
1 2 3 4 5 6 7 8 9
Контроль, мг 32,4 58,1 56,2 35,8 9,6 — — — —
Витамин Bi . . 28,5 103,9 255,6 390 504,1 534,9 375,9 205,4 45,7
Во на рост баклажан. Денисон, вводя в питательный рас-
твор вита'мин В2 в количестве 2,5 мг на 1 л, получил зна-
чительное увеличение роста корней, стеблей и ботвы у
баклажан. Так, например, если у 77-дневного контроль-
ного растения длина стебля составляла 7,1 см, вес бот-
вы— 48,5 г и вес корней— 12,5 г, то у растений, полу-
чавших витамин В2, эти показатели составляли соответ-
ственно: 12,2 см, 121,0 г и 22,9 г. Эти опыты, правда, не
дают окончательного ответа о действии витаминов на рас-
тение, так как введенные в почву витамины могли дей-
ствовать не только непосредственно на растение, но и ак-
тивировать размножение почвенных микроорганизмов,
благоприятно действующих на рост шпината и снотвор-
ного мака. Поэтому наиболее точный ответ на вопрос
о влиянии витаминов на ростовые процессы растения мо-
жет быть получен при введении витаминов через стебель
или лист. Обработка черешков винограда раствором ви-
тамина РР заметно усиливает рост надземных частей
и особенно листьев (Флеров и Коваленко, 1947). Отзыв-
чивы к действию этого витамина соя и лимонное деревцо.
Под влиянием витамина РР уже через 12 дней после
укоренения сои сырой вес обработанных проростков уве-
личился на 63% по сравнению с контролем. Сеянцы
лимона, смазанные ланолином, в котором содержался ви-
тамин РР, заметно увеличили прирост побегов (табл.
18).
Усиление ростовых процессов под влиянием обработки
растений витамином РР наблюдается и у бухарского
96
Таблица 18
Увеличение скорости роста сеянцев лимона под влиянием
витамина РР
(по Кочерженко и Снегиреву, 1946)
Вариант Средняя высота растений» см
В день обработки— 15. VIII 15.IX 15.x • 15. XI
Контроль (чистый ланолин) 27,2 29,2 35,5 38,3
Витамин* РР (10,5%) . . 27,0 33,2 40,5 42,9
. (1,0%). . . 24,6 34,3 39,0 44,0
(1,5%). . . 23,8 34,7 44,5 47,7
(2,0%). . .• 24,1 33,0 38,2 44,2
миндаля (Матвеев и Овчаров, 1949). 13 июля у бухар-
ского миндаля, произрастающего в необеспеченных вла-
гой условиях, удаляли листья, и оголенные побеги опры-
скивали водными растворами либо витамина РР, либо
аденина. Контрольные деревья опрыскивались водой. На-
блюдения за контрольными и обработанными деревьями
показали, что при введении витамина, раньше вскрывают-
ся почки. Так, у контрольных деревьев 25 июля вскры-
лись немногие верхушечные почки, тогда как в варианте
с витамином РР и аденином в это время появилось зна-
чительное количество зеленых листочков. В дальнейшем
эта разница стала еще большей (табл. 19).
Из данных таблицы следует, что число листьев, по-
явившихся под влиянием витамина РР, в четыре раза
превышает контроль. По другим показателям — число и
вес листьев в розетке — контрольные деревья сильно от-
ставали от деревьев, обработанных витамином РР
и аденином. В начале октября листья, появившиеся на
деревьях, получивших витамин, оставались яркозелеными
и свежими, тогда как листья контрольных деревьев по-
краснели и несколько подсохли.
7 Витамины в жизни растение 97
Таблица 19
Влияние витамина РР и аденина на образование и рост
листьев и побегов у бухарского миндаля
(по состоянию на 8 сентября)
Вариант Число листьев на 1 растение Число листьев в розетке 6 8 « См SSx Побега
число общая длина, см число листьев на побегах
Контроль 631 3-5 0,100 Побегов не образовалось
Витамин РР 2464 7—11 0,190 9 108,3 147
Аденин 575 8—12 0,580 7 44,0 132
Под влиянием витамина РР и аденина в конце ав-
густа началось образование новых побегов. Число побе-
гов, их общая длина и число листьев на побегах у де-
ревьев, обработанных витамином РР и аденином, в сен-
тябре достигли значительных размеров. В следующем
году деревья, обработанные этими веществами, нормаль-
но вегетировали, тогда как контрольные деревья погибли.
Гибель деревьев в контрольном варианте мы склонны
объяснить тем, что эти деревья образовали новую ассими-
ляционную поверхность в столь малых количествах, что
она не могла заменить собой потерянную. Это привело
к резкому спаду синтеза в растении и к истощению запа-
сов, необходимых для обеспечения вегетации следующего
года. Наоборот, деревья, обработанные витамином РР
и аденином, взамен удаленных листьев образовали боль-
шое количество молодой листвы (рис. 20). Это позволило
обработанным деревьям накопить запасы ассимилятов,
достаточных для вегетации следующего года.
Как известно, кольцевание саженцев плодовых древес-
ных и кустарниковых пород способствует ускорению цве-
тения и усилению плодоношения окольцованных ветвей.
В целях ускорения цветения и плодоношения сеянцев
цитрусовых М. X. Чайлахян и Т. В. Некрасова (1954)
предприняли исследования по изучению влияния кольце-
вания на рост и развитие сеянцев лимона. При этом было
98
установлено, что в том случае, если место кольцевания
не срастается, рост ветвей прекращается, листья опадают
и, наконец, отмирают ветви. Естественным желанием
исследователей было найти такие условия, при которых
происходили бы быстрое срастание кольца и более энер-
гичный рост окольцованных побегов. Нанося на места
кольцевания ланолин в смеси с витамином РР,
М. X. Чайлахян и Т. В. Некрасова получили наиболее
Рис. 20. Влияние витамина РР и аденина
на рост листьев бухарского миндаля
1 — контроль; 2 — никотиновая кислота; 3 — аденин
(по Матвееву и Овчарову, 1949)
быстрое и хорошее срастание кольца и более энергичный
рост окольцованных ветвей. Так, например, если в кон-
троле без обработки прирост побегов составлял 439 мм
и число новых листьев на них 21, то при нанесении на
кольцо витамина РР прирост побегов увеличился до
716 мм, а число листьев — до 34.
Большую роль витамины играют в жизни орхидей.
Известно, что в искусственных условиях семена орхидей
могут нормально прорастать только в том случае, если
на них поселяется особый грибок. Впоследствии выясни-
лось, что в семенах орхидей содержится очень мало вита-
мина РР, которого не хватает для нормального прора-
7* 99
стания семян. Грибок же, поселяясь на орхидеях и полу-
чая из ее тканей питательные вещества, в свою очередь,
снабжает растеньице орхидеи недостающим ей витами-
ном РР. При добавлении витамина РР семена легко про-
растают и в отсутствии грибка. Установление этого
факта позволило создать высокоэффективные питатель-
ные среды для орхидей, в состав которых наряду с дру-
гими питательными веществами входят и витамины. Ор-
хидеи полностью обеспечивают себя витаминами С и В5
и потому их рост не усиливается при введении этих вита-
минов в питательную среду. Образование же витамина Bi
и особенно витамина РР не удовлетворяет потребность
растения и поэтому добавление этих веществ, как показал
Геприксон, значительно увеличивает рост корней и над-
земных частей растения (табл. 20).
Таблица 20
Влияние витаминов на рост орхидей
Вариант
Контроль ...............
Bi......................
Вв......................
С •.....................
Р ......................
30,8 5 48,0 42,0 5,5 60,5
59,1 6 83,5 65,0 8,5 92,5
18,1 6 48,0 25,0 4,5 58,5
30,1 6 55,5 42,0 5,0 55,5
86,6 8 101,0 73,5 9,0 176,5
я
я
В варианте с витамином РР почти в три раза уве-
личились сухой вес растения, высота надземных частей
и длина корней и почти в два раза — размер листьев, по
сравнению с контролем. Этот факт все больше и больше
используется для ускоренного получения цветов орхидей.
Таким образом, здесь мы имеем один из ярких при-
меров так называемого симбиоза—полезного сожитель-
ства совершенно различных организмов, снабжающих
друг друга недостающими питательными веществами.
Симбиоз имеет широкое распространение в живой при-
100
роде, и тот факт, что витамины являются одной из причин
такого сожительства, наглядно демонстрирует огромное
значение их в жизни растений, во взаимоотношениях ор-
ганизмов.
Значительный интерес представляло изучение дейст-
вия витаминных подкормок на культуре хлопчатника.
Как известно, в начальные периоды хлопчатник растет
медленно, особенно в случае низких весенних температур.
Это обстоятельство побудило пас (Ракитин и Овчаров,
1948) испытать действие некоторых витаминов и аденина
на рост и плодоношение хлопчатника. Систематическая
обработка хлопчатника витамином РР или аденином
усиливает образование и рост листьев. По мере*роста ра-
стений число листьев в варианте с витамином РР и
аденином все больше и больше возрастает по сравнению
с растениями контрольного варианта. Размер листьев при
внекорневой подкормке растений витамином РР и адени-
ном так же был несколько большим.
Значительное увеличение листьев у хлопчатника в пе-
риод бутонизации и начала цветения имеет весьма суще-
ственное значение для дальнейшего роста и развития все-
го растения. Практикой хлопководства давно было уста-
новлено, что внешние условия, задерживающие рост ли-
стьев в начальный период, приводят к резкому отстава-
нию новообразования и роста симподиев и к понижению
урожая хлопка-сырца. Наоборот, агротехнические усло-
вия, способствующие более дружному и раннему образо-
ванию листьев, ускоряют закладку плодовых побегов,
улучшают рост симподиев и приводят к более раннему
созреванию коробочек.
Добавочное введение в растения витамина РР и
аденина не только увеличивает интенсивность образования
листьев, по и способствует усилению роста других частей
растения. Высота главного стебля и его диаметр, число
симподиев и их рост также заметно возрастают. Одновре-
менно с усилением роста симподиев происходит и энергич-
ное образование коробочек. Вследствие этого наблюда-
лось ускорение созревания коробочек и повышение уро-
жая хлопка-сырца. Аналогичные результаты получены
и другими авторами (Захарьянц, Горбачева и Зглинская,
1950), которые, опрыскивая хлопчатник витамином РР
и тиамином, получили увеличение числа и размера ли-
101
стьев. Особенно благоприятное действие оказала обработ-
ка растений витамином РР. В случае обработки хлоп-
чатника витамином РР число листьев на растении уве-
личилось почти на 50%, заметно увеличился их размер.
Так, например, если у контроля размер листа был равен
11,06 см (длина) и 14,78 см (ширина), то при обработке
витамином РР размеры листа составляли соответствен-
но 11,90 и 16,17 см. По данным этих авторов, обработка
хлопчатника витамином РР увеличила урожай хлопка-
сырца на 34,1 % по сравнению с контролем.
Таким образом, своевременное обеспечение растений
витаминами может представлять значительный практиче-
ский инте{Яес.
Витаминные подкормки растений могут иметь боль-
шое значение для повышения продуктивности ценных
сельскохозяйственных культур. Особое внимание при
этом заслуживают витамины группы В, т. е. витамины Bi,
В2, Вб, РР, Н, а также аденин.
В противоположность витаминам группы В, так назы-
ваемые жирорастворимые витамины (А, Е, К), оказы-
вают задерживающее действие на рост растений.
М. X. Чайлахян (1950), нанося на место срезов верхушки
хризантемы ланолиновую пасту в смеси с растворенным
в рафинированном масле кристаллическим каротином
или сухой измельченный порошок рылец шафрана, весьма
богатых каротиноидами, установил, что под влиянием этих
веществ рост побегов задерживался, пластинки листьев
становились меньше и уже, черенки укорачивались. Вита-
мин К также задерживает рост растений. Он подавляет
рост некоторых грибов, бактерий и корней высших расте-
ний. Введение в питательную среду витамина РР сни-
мает тормозящее действие витамина К. Дополнительное
обеспечение растений витамином Е не усиливает, а, на-
оборот, сильно тормозит ростовые процессы (табл. 21).
Как следует из данных таблицы, введение в щелкунец
витамина Е резко подавляет рост растения и особенно —
образование цветов.
Приведенные примеры показывают, что хотя зеленые
растения сами способны синтезировать витамины, но
в некоторые периоды своей жизни они находятся в со-
стоянии необеспеченности этими веществами. К сожале-
нию, подробных исследований по изучению потребностей
102
Таблица 21
Влияние витамина Е на рост щелкунца
(по Шопферу, 1950)
Вариант Высота растения, см Число междоузлий Число цветов
Витамин Е 6,14 3—7 10,0
Контроль без витамина . . 35,72 7—14 80,40
в витаминах у различных групп растений, произрастаю-
щих в разных агротехнических и климатических условиях,
в настоящее время нет. Дальнейшее изучение роли вита-
минов в жизнедеятельности высших растений еще больше
углубит наши сведения в этой интересной и важной обла-
сти знания и поможет выявить новые возможности управ-
ления ростом и развитием растений.
3. Участие витаминов в процессе
оплодотворения у растений
Оплодотворение является одним из сложнейших про-
цессов в растительном организме. Не случайно поэтому
проблема оплодотворения многие годы была наименее
разработанным разделом биологии. Разработка И. В. Ми-
чуриным метода опыления смесью пыльцы, метода мен-
тора и вегетативной гибридизации открыла новые пер-
спективы для понимания сущности явления оплодотворе-
ния. Представление о ведущей роли обмена веществ
в процессах оплодотворения открывает широкие возмож-
ности для направленного изменения свойств организма
и для управления этими процессами.
Витамины как важные звенья обмена веществ играют
существенную роль и в процессе оплодотворения.
Давно известно целебное действие пчелиного меда.
Химический анализ состава меда показал, что в нем, кро-
ме сахаров и других веществ, содержится значительное
количество разнообразных витаминов. Так, например,
в 1 кг меда было найдено: витамина Bi —до 0,1 мг, вита-
мина В2 — до 1,5 мг, витамина В6 — до 5 мг, пантотено-
103
вой кислоты—до 2 мг, витамина РР — до 1 мг, а ви-
тамина С — до 54 мг.
Откуда же появились витамины в меду?
Исследования о происхождении витаминов в этом про-
дукте показали, что они появляются из растений. Имею-
щиеся материалы позволяют сделать вывод, что витами-
ны Bi, Вг, РР, В6, витамин С и каротин содержатся
в большом количестве в цветах. Пчелы, посещая цветы,
вместе с сахарами и другими физиологически важными
соединениями доставляют в ульи и значительные количе-
ства витаминов. Содержание витаминов в цветах разных
растений неодинаково. Так, в цветах капусты содержание
витамина Bi невелико и еще меньше содержится витами-
на В2. В табл. 22 приводятся данные Кравнато и других
о содержании некоторых витаминов в 1 кг цветов ряда
растений.
Таблица 22
Содержание витаминов в цветах
Растение Витамин В|, мг Витамин В2, мг Витамин РР, мг
Тыква .... 20,0 25,0 122
Капуста . . . 4,6 1,5 73
Агава .... 12,2 5,6 23,2
Вика .... 14,0 9,0 58,0
В цветах агавы содержится небольшое количество ви-
тамина В2, больше витамина Bi н особенно много вита-
мина РР. Цветы тыквы отличаются высоким содержа-
нием всех трех' витаминов. Витамины в цветах распреде-
лены неравномерно: в некоторых частях цветка их содер-
жится много, другие весьма бедны этими веществами.
Так, например, у гороха значительное количество витами-
на Bi найдено в пестиках, чашечке и меньше — в тычин-
ках и венчике. В цветах тюльпана, как было установлено
Ритцем, особенно много этого витамина в тычинках
(табл. 23).
Изучение динамики содержания витаминов в цветах
хлопчатника разного возраста позволило К. Т. Сухоруко-
104
Таблица 23
Содержание витамина Вх в разных частях цветка
Растение Содержание витамина, мг на 1 кг сухого веса
незрелые бутоны чашечка венчик тычинки пестики
Горох . . 23 21 14 13 20
Тюльпан . . — 4 4 10 5
ву и Филиппову (1945) прийти к заключению, что по
мере старения венчиков содержание в них витаминов рез-
ко падает, а в завязях их количество возрастает.
Особенно богата витаминами пыльца некоторых рас-
тений. Так, например, в пересчете на 1 кг в пыльце жел-
той акации содержится 2300 мг каротина, в пыльце под-
солнечника— 1460, а в кукурузе всего 10 мг. Различно
содержание и других витаминов. Если в 1 кг пыльцы
осота найдено 1700 мг витамина Е, то у гречихи — всего
210 мг. В пыльце дудника и сныти витамина В2 содер-
жится лишь в 2 раза меньше, чем в сухих пивных дрож-
жах, весьма богатых этим витамином. В пыльце других
растений, как видно из табл. 24, содержание витамина
В2 меньше, чем у дудника и сныти.
Наличие витаминов в пыльце, как и во всем расти-
тельном организме, не является случайным и бесполезным
для растения. Прямые факты показывают, что находя-
щиеся в пыльце и пестике витамины играют важную роль
в жизнедеятельности этих частей цветка и в процессах
оплодотворения.
С. И. Лебедев (1953) установил, что пыльца с желтой
иди оранжевой окраской, т. е. содержащая значительное
количество каротина, подвергнутая воздействию света,
быстро обесцвечивается и теряет жизнеспособность. Так,
например, пыльца аморфы и пальмы, обесцвеченная на
рассеянном свету, в четыре-пять раз снижала способность
к прорастанию, а пыльца коровяка совсем потеряла спо-
собность к прорастанию. Аналогичное явление наблюдает-
ся и в том случае, если обесцвечивание пыльцы произво-
8 Витамины в жизни растений 105
Таблица 24
Содержание витаминов в пыльце
(по Грунт, Девятнину и др., 1948)
Растение Содержание, мг на 1 кг пыльцы
витамин В витамин С витамин Bi витамин в2
Акация желтая 1184 14 15 15
Борщевик обыкновенный 600 18 5 13
Гречиха посевная • . . . . 210 35 13 16
Дудник аптечный 600 20 12 21
Ежа сборная 423 — 7,5 13
Кипрей (Иван-чай) .... 255 14 5 12
Клевер белый 545 — 14 12
Липа 750 — 7,5 12,5
Лютик едкий — — 7,4 15
Одуванчик лекарственный — — 7,5 16
Осот желтый 1700 41 И 14
Пеон желтый — — 2,5 15
Полынь 455 57 7 10
Синюшник 900 27 10 12
Сныть съедобная . . . . • — — 9 23
Таволга вязолистная . . . 445 36 6 И
Яблоня 900 36 10 18
Рябина обыкновенная . . . 327 — — —
Фацелия 400 — •— —
дилось извлечением каротина арахисным маслом. Напри-
мер, у пыльцы жасмина наблюдалось 62,21% прораста-
ния необеспеченной пыльцы при длине пыльцевых тру-
бок 6,73 ьтогда как у обесцвеченной — проросло 52,54%
при длине пыльцевых трубок 4,96 р. Непосредственное
изучение действия каротина на прорастание пыльцы и
рост пыльцевых трубок показало, что если на контроль-
ной среде, т. е. без каротина, проросло 39,0% пыльцы ко-
нопли и длина пыльцевых трубок в среднем равнялась
100 то при добавлении каротина в концентрации
106
0,005%' эти показатели соответственно увеличивались до
53,5% и 312 р. (рис. 21).
Рис. 21. Прорастание пыльцы конопли
1 — среда без каротина; 2 — среда с добавлением О,ОО57о каротина
(по Лебедеву, 1953)
Аналогичные результаты получаются и при смешива-
нии пыльцы, богатой этим витамином, с пыльцой, в кото-
107
рой его чрезвычайно мало. Когда на питательную среду
одновременно высевается пыльца бегонии или бузины,
содержащая очень мало каротина и обычно плохо прора-
стающая, и пыльца акации, или коровяка с высоким со-
держанием каротина, то прорастание пыльцы бегонии и
бузины заметно улучшается (Лебедев, 1953). Нами по-
добное явление было установлено на разных сортах хлоп-
чатника. При совместном культивировании на питатель-
ной среде пыльцы сортов хлопчатника, бедных каро-
тином, с пыльцой сортов, богатых провитамином А,
отмечалось лучшее прорастание пыльцы и более энер-
гичный рост пыльцевых трубок у пыльцы, бедной ка-
ротином.
Интересно отметить, что в естественной обстановке,
как в пыльце, так и в пестиках к моменту оплодотворения
содержание каротина заметно возрастает. Однако в пыль-
це во все периоды ее жизни значительно больше этого
провитамина, чем в пестиках. Возрастание содержания
каротина в пестиках во время оплодотворения происхо-
дит в значительной степени за счет попадания его из
пыльцы. Как показали наши исследования, при нанесении
пыльцы на рыльца содержание каротина в последних
увеличивается более чем в три раза, а в столбиках —
в два раза.
Увеличение содержания каротипа в рыльцах и столби-
ках происходит, однако, не только за счет попавшей на
рыльце пыльцы, но и за счет притока каротина из других
частей цветка. В лепестках и прицветниках в момент от-
крытия бутонов содержится максимальное количество ка-
ротина. На второй день после оплодотворения содержа-
ние этого провитамина в указанных частях цветка резко
падает, в то же время количество каротина в рыльце и
пестике заметно возрастает (табл. 25).
Накопленный материал не оставляет сомнения в том,
что для нормального прорастания пыльцы и роста пыль-
цевых трубок, являющихся важным звеном сложного
процесса оплодотворения высших растений, необходимо
наличие определенного количества каротина. Этот прови-
тамин оказывает существенное влияние и на половой
процесс у низших растений. Имеющиеся данные показы-
вают, что при недостатке каротина и каротиноидов за-
держивается половой процесс у водорослей и грибов.
108
Таблица 25
Содержание каротина в разных частях цветка хлопчатника
(мг в 1 кг)
Состояние цветка Пыльца Пести- ки Лепестки Прицвет- ники
Перед открытием бутонов 400 130 105 160
В момент открытия цветков .... 951 284 210 257
На 2-й день после оплодотворения — 336 148 132
Р. Кун (1941), получив интересный материал о влиянии
некоторых каротиноидов на образование и копуляцию
мужских и женских половых клеток у водоросли хламидо-
монад пришел к выводу, что исследованные им каротино-
иды являются специфическими гормонами: сафронол —
мужским, а пикрокроцин — женским. Задавшись целью
объяснить, почему в разных условиях происходит синтез
того или другого вещества, Кун приходит, однако, к не-
правильному заключению, что этот процесс регулируется
особым набором генов.
На важную роль каротина и каротиноидов в образова-
нии половых форм, и половой функции у микроорганиз-
мов указывают многочисленные данные других исследо-
вателей. Таким образом, провитамин А и каротиноиды
являются важными факторами в процессах оплодотворе-
ния как у животных, так и высших и низших растений.
Значительное влияние на половой процесс оказывают
и другие витамины. В отношении бактерий, грибов и
дрожжей было установлено благоприятное действие на
этот процесс витаминов Вь В2, РР и др. Имеются обстоя-
тельные исследования и о действии витаминов на форми-
рование и взаимодействие мужских и женских половых
элементов у высших растений.
Установлено, что на прорастание пыльцы многих рас-
тений благоприятно влияет дополнительное введение ви-
тамина Вь При этом было показано, что для пыльцы
разных сортов табака потребность в этом витамине ко-
леблется от 0,0002 до 0,005 мг на 1 л раствора (Поляков,.
1949). Пыльца разных сортов дынного дерева также со-
вершенно по-разному реагирует на добавление витами-
10Э
на Вь Так, для одних сортов введение в питательную
среду этого витамина увеличивает прорастание пыльцы
на 10—12%, для других — на 35% и для третьих — на
52%. Неодинаковая потребность разной пыльцы в вита-
мине Bi может обусловливаться не только тем, что для
осуществления нормального обмена веществ необходимо
больше или меньше этого витамина, но она зависит так-
же от того, что содержание и активность фермента, раз-
рушающего витамин Вь в пыльце разных растений не-
одинаковы.
Положительное влияние на прорастание пыльцы неко-
торых растений оказывают также витамины В2, С, Be и
РР. Так, например, если на среде без витамина РР
проросло 31,0% пыльцы сосны и длина пыльцевых трубок
была равна 0,608 мм, то при введении этого витамина
проросло 54,0% и длина пыльцевых трубок была равна
0,721 мм.
Приведенные примеры показывают, что хотя содержа-
ние провитамина А, аскорбиновой кислоты (витамина С)
и других витаминов в пыльце и значительно, но их иног-
да недостаточно для нормального прорастания пыльцы
в искусственных условиях. В природной обстановке при
попадании пыльцы на рыльце может и не наблюдаться
этого явления, так как пыльца может получить недостаю-
щее количество витаминов из пестиков, в которых вита-
мины также представлены в значительных количествах.
Установление потребности прорастающей пыльцы в ви-
таминах помогло создать эффективные питательные сре-
ды для культивирования пыльцы, а также содействовало
выяснению жизнедеятельности пыльцы и пестика и их
взаимодействия.
Давно было установлено благоприятное действие ры-
лец на прорастание пыльцы. Пыльца некоторых растений
на питательной среде совершенно не прорастает, если нет
в этой среде рыльца; пыльца других растений хотя ц про-
растает без рылец, но очень плохо, тогда как в присутст-
вии рылец этот процесс идет значительно быстрее и луч-
ше. Изучение этого явления позволило прийти к заключе-
нию, что в благоприятном действии рылец на прораста-
ние пыльцы, наряду с другими веществами, значительная
роль принадлежит витаминам. Рыльца выделяют в пита-
тельную среду витамины и другие жизненно необходимые
НО
вещества и тем самым обеспечивают пыльцу недостаю-
щими для ее прорастания веществами. Так, нами было
установлено выделение рыльцем хлопчатника витами-
нов Bi, РР, Вб, Н и пантотеновой кислоты и показано,
что дополнительное введение этих витаминов в питатель-
ную среду, на которой культивировалась пыльца хлопчат-
ника, усиливает как прорастание, так и рост пыльцевых
трубок.
Высокое содержание в рыльцах витаминов и большая
отзывчивость пыльцы на введение их извне позволяет
прийти к заключению, что в благоприятном действии
рылец на прорастание пыльцы витаминам принадлежит
значительная роль. Пыльца, в свою очередь, дополни-
тельно снабжает пестик теми веществами, которые необ-
ходимы ему для нормальной жизнедеятельности. Такой
взаимный обмен растущей пыльцы с тканями пестика со-
здает благоприятные условия для развития пыльцы и пе-
стика, для формирования и роста семяпочки. Как извест-
но, после оплодотворения семяпочек к ним резко увели-
чивается приток сахаров, аминокислот и органических
кислот, азота, фосфора и др. В это время в оплодотворен-
ных семяпочках значительно повышается также и содер-
жание витаминов. Так, например, в наших опытах через
сутки после оплодотворения содержание витамина С в
семяпочках возрастало более чем на 40%. Увеличение
количества витаминов в семяпочках способствует усиле-
нию превращения поступающих в зародыш веществ, что
создает необходимые условия для более энергичного роста
и формирования семени.
Дальнейшее выяснение роли витаминов — одного из
важных звеньев обмена веществ — в оплодотворении ра-
стений не только позволит более полно выяснить взаимо-
действие репродуктивных и вегетативных органов расте-
ний, влияние внешних условий на процесс оплодотворе-
ния, взаимосвязь между мужскими и женскими частями
цветка, но и откроет новые возможности управления
этим сложнейшим процессом жезнедеятельности расте-
ний.
* *
*
Таким образом, изучение роли витаминов в жизни
растений привело к выяснению новых сторон участия этих
111
веществ в сложных физиологических процессах, происхо-
дящих в организме низших и высших растений. Вместе
с тем вскрыты многие тайны жизнедеятельности растений,
которые были недоступны исследователю в те времена,
когда он не имел в своих лабораториях витаминов. Выяс-
няя действие того или иного витамина на физиологические
процессы, исследователь все больше и больше открывает
взаимосвязь организма с внешней средой, взаимоотноше-
ния организмов между собою и взаимосвязь отдельных
частей высшего растения. Прошло немного лет с тех пор,
как физиологи растений обратили внимание на важную
роль витаминов в жизни зеленых растений. Но и за этот
короткий срок были достигнуты значительные резуль-
таты. Дальнейшее, более подробное изучение роли вита-
минов в жизни зеленых растений может указать на новые
пути повышения продуктивности культурных растений
при помощи широкого применения витаминов в растение-
водстве.
ЛИТЕРАТУРА
Андреева Н. А. О характере биосинтеза фолиевой кислоты в рас-
тениях. «Биохимия», т. 18, вып. 6, 1953.
Брегетова Л. Г. О содержании аскорбиновой кислоты в листь-
. ях хлопчатника. «Ботан. журн.», т. 36, № 1, 1951.
Браунштейн А. Е. Биохимия аминокислотного обмена. М., 1949.
Будницкая Е. В. Современные представления о биосинтезе и
физиологической роли каротиноидов. «Изв. АН СССР» серия
биол.», № 4, 1952.
Букин В. Н. Витамины. Пищепромиздат, М., 1940.
Букин В. Н. и Водолазская Н. А. Метод определения и не-
которые данные о качестве белка пищевых продуктов в связи с
витаминизацией. «Биохимия», т. 15, вып. 1, 1950.
Букин В. Н., Поволоцкая К. Л. и Онохова Н. Т. Содержа-
ние антицынготного витамина в сортах плодов, ягод, овощей
Севера. Проблема витаминов. Сб. эксперим. работ витаминной
лаборатории ВИР’а, под ред. проф. Н. Н. Иванова. Л., 1934.
Верзилов В. Ф. Применение стимуляторов роста при пересадке
деревьев. АН СССР и Всес. хим. об-®о им. Д. И. Менделеева.
М., 1949.
Грунт Е. В., Девятнин В. А., 3 а х а р о в а М. П., Зворы-
кина В. В., Кравчина Л. Н. и Рыбаков М. Н. Витамины
в пыльце. «Пчеловодство», № 10, 1948.
Д а г и с И. К. Опыты по применению стимуляторов роста и микро-
элементов для повышения урожайности культурных растений
«Изв. АН Латв. ССР», № 7(84), 1954.
Девятнин В. А. Распределение аскорбиновой кислоты в листьях
растений. Сб. «Новое в пауке и технике витаминов», вып. 1.
Пищепромиздат, М., 1946.
113
Девятнин В. А. Витамины. Пищепромиздат, М., 1948.
Д е в я т н и и В. А. Об исходных веществах для биосинтеза аскор-
биновой кислоты в растениях. «Биохимия», т. 15, вып. 4, 1950.
Дьякова Е. Влияние условий питания на содержание каротина
в кормовых травах. «Докл. ВАСХНИЛ», вып. 6, 1945.
Егоров А. ,Д. Витамин С и каротин в растительности Якутии.
Изд. АН СССР, 1954.
Жуковский П. М. и Медведев. Ж. А. Значение световой
энергии и каротиноидов для развития бесполого и полового
поколений в растительном мире. «Усп. соврем, биол.», т. 26,
вып. 1 (4), 1948.
ЗахарьянцИ. Л., ГорбачеваЗ. И. и ЗглинскаяН. А.
Влияние ростактивирующих веществ на хлопчатник. «Тр. Ин-та
ботаники и зоологии АН Узб. ССР», вып. 3, 1950.
Иерусалимский Н. Д. Азотное и витаминное питание микро-
бов. Изд. АН СССР, 1949.
Кочерженко Е. И. и Снегирев Д. П. Влияние задерживаю-
щих рост веществ на морозоустойчивость лимона. «Бюлл. по
культурам влажных субтропиков», № 14—15, 1946.
Красильников Н. А. Роль микроорганизмов в дополнительном
питании растений. «Усп. соврем, биол.», т. 32, вып. 3, 1952.
Кудряшев В. А. Витамины, их физиологическое и биохимическое
значение. Моск, об-во испыт. природы, 1953.
Кун Р. Вещества, стимулирующие оплодотворение и определяющие
пол у растений и животных. «Усп. соврем, биол.», т. 14, 1941.
Курсанов А. Л. Значение изотопов и других новейших методов
исследования в биологии для решения вопросов сельского хо-
зяйства. «Вести. АН СССР», № 12, 1953.
Курсанов А. Л., Выскребенцева Э. И., Воробьева М. В.
Участие m-инозита в углеводно-фенольном обмене чайного листа.
«ДАН СССР», т. 68, № 5, 1949.
Курсанов А. Л., Крюкова Н. Н. и Пушкарева М. И. Тем-
новая фиксация и освобождение углекислоты, поступающей в
растения через корни. «ДАН СССР», т. 88. № 5, 1953.
Лебедев А. Ф. Об ассимиляции углерода сапрофитами. «Изв.
Донск. ун-та», 1921.
Лебедев С. И. Физиологическая роль каротина в растении. Изд.
АН Укр. ССР, Киев, 1953.
Леутский К. М. Витамины (комплекс Вг). Изд. Львовск. гос.
ун-та, 1949.
114
Леутский К. М. и Стульникова Р. И. Содержание никоти-
новой кислоты и триптофана в семенах фасоли при прорастании
и созревании. «ДАН СССР», т. 80, № 6, 1951.
ЛунинА. И. Ueber die Bedeutung der anorganischen Salze fur die
Ernahrung des Tieres. «Ztschr. physiol. Chem.», m. 5. 1881.
Максимов H. А., Ракитин Ю. В., Турецкая P. X. Гладио-
лус как новый вид сырья для получения витамина С. Химиздат
МОВХО им. Д. И. Менделеева, 1948.
Матвеев М. И. и Овчаров К. Е. Влияние аденина и никотино-
вой кислоты на вторичную вегетацию бухарского миндаля
(Amygdalus bucharica К о г s h.), «ДАН СССР», т. 65, № 3, 1949.
Мейсель М. Н. Функциональная морфология дрожжевых орга-
низмов. Изд. АН СССР. М.—Л., 1950.
Мейсель М. Н., Помощникова Н. А. и Трофимова Н. П.
Биологический синтез пантотеновой кислоты из Р -аланина и
некоторых его производных. «Биохимия», т. 14, вып. 4, 1949.
Михлин Д. М. О роли аскорбиновой кислоты при восстановле-
нии нитратов в растительных тканях. «Биохимия», т. 1, вып. 6,
1936.
М у р р и И. К. Проблема витаминов в растениеводстве. «Биохимия»,
т. VIII, 1948.
Никитинский Я. Ueber die Beeinflussung der Entwicklung einiger
Schimmelpilze durch ihre Stoffwechselprodukte. Inaugural Disser-
tation, Leipzig, 1904.
Прокофьев At, А. Дополнительные факторы роста и их роль
в жизни растений. «Усп. соврем, биол.», т. 26, 1948.
Прокошев С. М. Раневое образование витамина С в картофеле.
«Биохимия», т. 9, вып. 1, 1944.
Прокошев С. М. Биохимия картофеля. Изд. АН СССР. М.— Л.,
1947.
Поволоцкая К. Л. Витамин С в прорастающих семенах. Про
блемы витаминов, сб. 2, 1937.
Поволоцкая К. Л. О новой, связанной с белком форме рибо-
флавина. «Биохимия», т. 18, вып. 5, 1953.
Папандопуполо П. X. Витамин А и его роль в животновод-
стве. Сельхозгиз, 1952.
Поповская Е. М. К вопросу об образовании и передвижении
аскорбиновой кислоты в растении. «Биохимия», т. 15, вып. 3,
1950.
115
Ракитин Ю. В. Практическое руководство по ускоренному созре-
ванию плодов. Изд. АН СССР, 1938.
Ракитин Ю. В. Проблема стимуляции растений в связи с зада-
чами сельского хозяйства. «Усп. соврем, биол.», т. 36, вып. 3 (6)^
1953.
Ракитин Ю. В. и Крылов А. В. Применение стимуляторов,
роста для повышения продуктивности культуры томатов. Изд.
АН СССР. М,—Л., 1950.
Ракитин Ю. В. и Овчаров К- Е. Влияние аденина и никотино-
вой кислоты на рост и плодоношение хлопчатника. «ДАН
СССР», т. 61, № 5, 1948.
Розанова М. А. Влияние климатических и почвенных условий на
содержание аскорбиновой кислоты в растениях. Сб. «Новое в
науке и технике витаминов». Пищепромиздат, М., 1946.
Рубин Б. А. Условия образования витамина С и каротина и их
роль в растении. «Тр. Моск. Дома ученых и Ин-та биохимии
АН СССР», вып. 4, 1940.
Сисакян Н. М. и Васильева Н. А. Механизм стабилизации
аскорбиновой кислоты сернистой кислотой. «Биохимия», т. 10,
вып. 2, 1945.
Сисакян Н. М., Глущенко И. Е., Васильева Н. А. и
Кобякова А. М. Изменения биохимических признаков в се-
менных потомствах томатов от прививок. «Биохимия», т. 11,
вып. 2, 1946.
Степанова Е. М. Черная смородина — витаминная культура.
Пищепромиздат. М., 1950.
Сухоруков К. и Филиппов В. Гормоны клеточного деления
в цветках хлопчатника. «ДАН СССР», т. 67, № 5, 1945.
Титаев А. А. Тиаминдегидраза, ее свойства, распространение и
функции. «Биохимия», т. 15, вып. Э, 1950.
Труфанов А. В. Витамины и антивитамины. Пищепромиздат, М.,.
1950.
Турецкая Р. X. Приемы ускорения размножения растений путем
черенкования, 1949.
Уайт Ф. Р. Культура растительных тканей. Изд. иностр, лит-ры,
М.» 1949.
Флеров А. Ф. и Коваленко Е. И. Влияние ростовых веществ
и алкалоидов на развитие черенков и прорастание семян вино-
града. «ДАН СССР», т. 58, № 4, 1947.
116
Функ К. Витамины, их значение для физиологии и патологии. Гос-
издат, М., 1922.
Чайлахян М. X. Влияние каротина на рост и формирование рас-
тений. «ДАН СССР», т. 74, № 2, 1950.
Чайлахян М. X. и Некрасова Т. В. Влияние кольцевания на
рост и развитие сеянцев лимона. «ДАН СССР», т. 96, № 2, 1954.
Шавловский Г. М. Участие микроорганизмов ризосферы в
снабжении растений витаминами. «ДАН СССР», т. 95, №5, 1954.
III о п ф е р В. X. Витамины почвы. Биохимия и физиология витами-
нов. I. Биосинтез и функции витаминов, 1950.
Энгельгардт В. А. и Букин В. Н. Устойчивость аскорбиновой
кислоты и ее дегидроформы. «Биохимия», т. 2. 1937.
ОГЛАВЛЕНИЕ
Введение ................................................ 3
Глава I. Общие свойства витаминов ....................... 9
1. Характеристика свойств витаминов.................. 9
2. Роль витаминов в обмене веществ организма .... 23
Глава II. Биосинтез витаминов в растениях............... 35
1. Образование витаминов растениями................. 35
2. Изменение содержания витаминов в процессе развития
растений............................................ 48
3. Влияние факторов внешней среды на содержание вита-
минов в растениях................................... 62
Г лава III. Потребность растений в витаминах............ 70
1. Потребность микроорганизмов в витаминах.......... 70
2. Потребность высших растений в витаминах.......... 78
3. Участие витаминов в процессе оплодотворения у растений 103
Литература.............................................. ИЗ
Печатается по постановлению
Редакционно-из )ателъского совета
Академии Наук СССР
*
Редактор издательства Е. А. Критиков
Технический редактор Г. А. Астафьева
Корректор В, Раскопова
*
РИСО АН СССР № 15—108 В. Сдано в набор
31/III 1955 г. Подп. к печ. 4/VI 1955 г. Формат
бум. 84хЮ81/за. Печ. л. 3,75-6,15. 4-2 вкл.
Уч.-издат. л. 5,64-2 вкл. (0,2 уч-изд. л.) Тираж 8000
Т-04488 Издат. № 1037. Тип. заказ № 1200
Цена 1 р. 90 к.
Издательство Академии Наук СССР
Москва, Подсосенский пер., д. 21
2-я тип. Издательства Академии Наук СССР
Москва, Шубинский пер., д. 10
СПИСОК ОПЕЧАТОК К КНИГЕ ОВЧАРОВА
.ВИТАМИНЫ В ЖИЗНИ РАСТЕНИЙ'
Стра- ница Строка Напечатано Следует читать
б 10 строка снизу жизненный аминами жизненными аминами
40 6 строка сверху (Девятнин, 1946), у (Девятнин 1946). У
53 рис. 10 Пантогенная Пантотеновая
Цена 1 р. 90 к.