/
















































































































Автор: Нарская Е.В.
Теги: растения биология животный мир ботаника флора животные фауна растениеводство
Год: 1957




Текст
БЮЛЛЕТЕНЬ
МОСКОВСКОГО ОБЩЕСТВА
ОТДЕЛ
БИОЛОГИЧЕСКИЙ
5
19 5 7
БЮЛЛЕТЕНЬ
МОСКОВСКОГО ОБЩЕСТВА
ИСПЫТАТЕЛЕЙ ПРИРОДЫ
НОВАЯ СЕРИЯ
128-й год издания
ОТДЕЛ БИОЛОГИЧЕСКИЙ
Том LXII, вып. 5
СЕНТЯБРЬ — ОКТЯБРЬ
Выходит 6 раз в год
ИЗДАТЕЛЬСТВО МОСКОВСКОГО УНИВЕРСИТЕТА
19 5 7
СОДЕРЖАНИЕ т. LXII (5)
Дубинин Н. П. О некоторых философских вопросах современной генетики ... II
Алпатов В. В. Левизна-правизна в строении растительных и животных ор-
ганизмов ................................................................ . 10
Аверин Ю. В. Зоогеографический очерк Камчатки.........................• . . 29
Арсеньев В. А. Наблюдения за тюленями Антарктики......................... : . 39
ШтегманБ. К. Некоторые особенности строения плечевого пояса у голубей
и рябков и вопрос о функциональном значении ключицы у птиц............. 45-
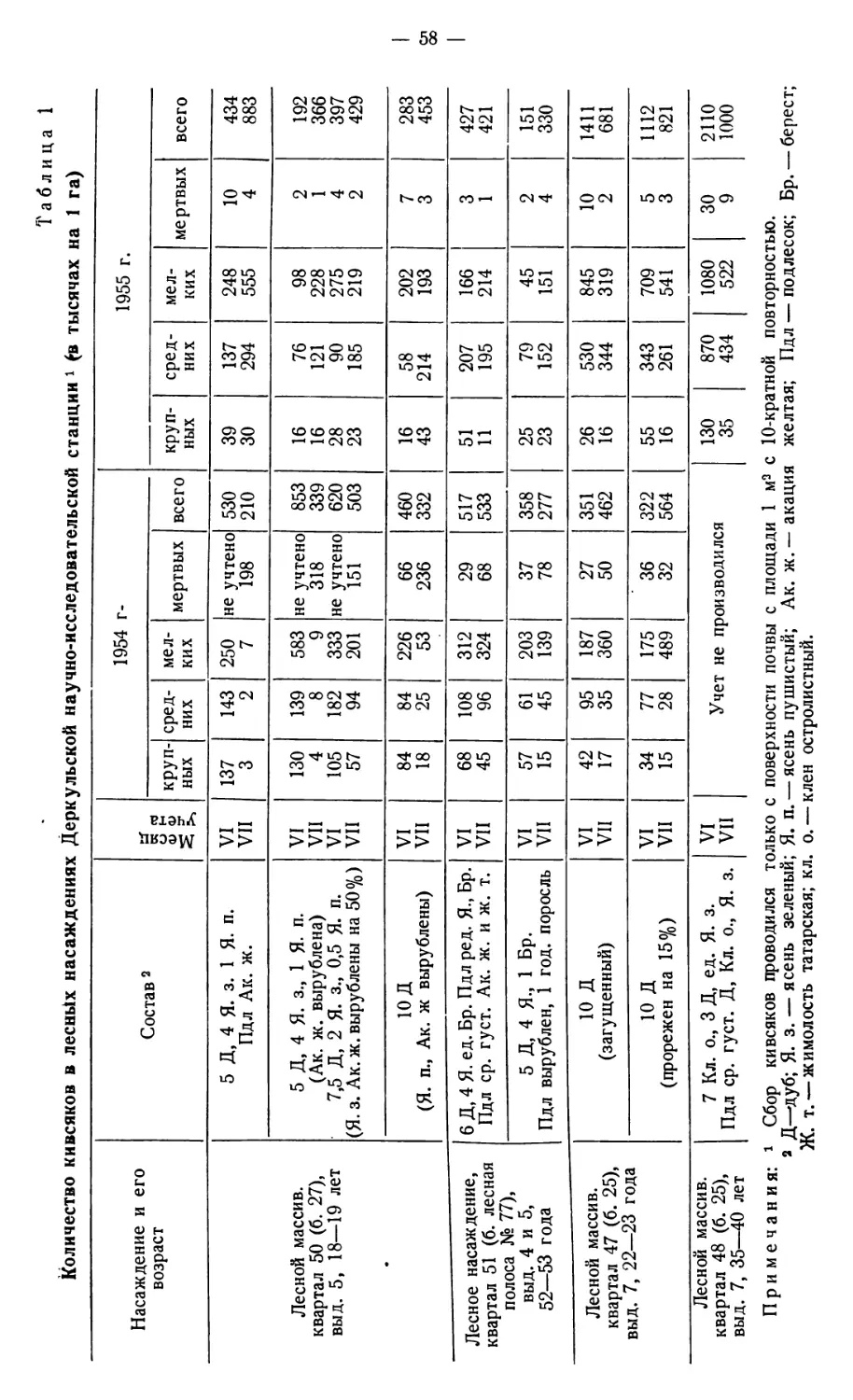
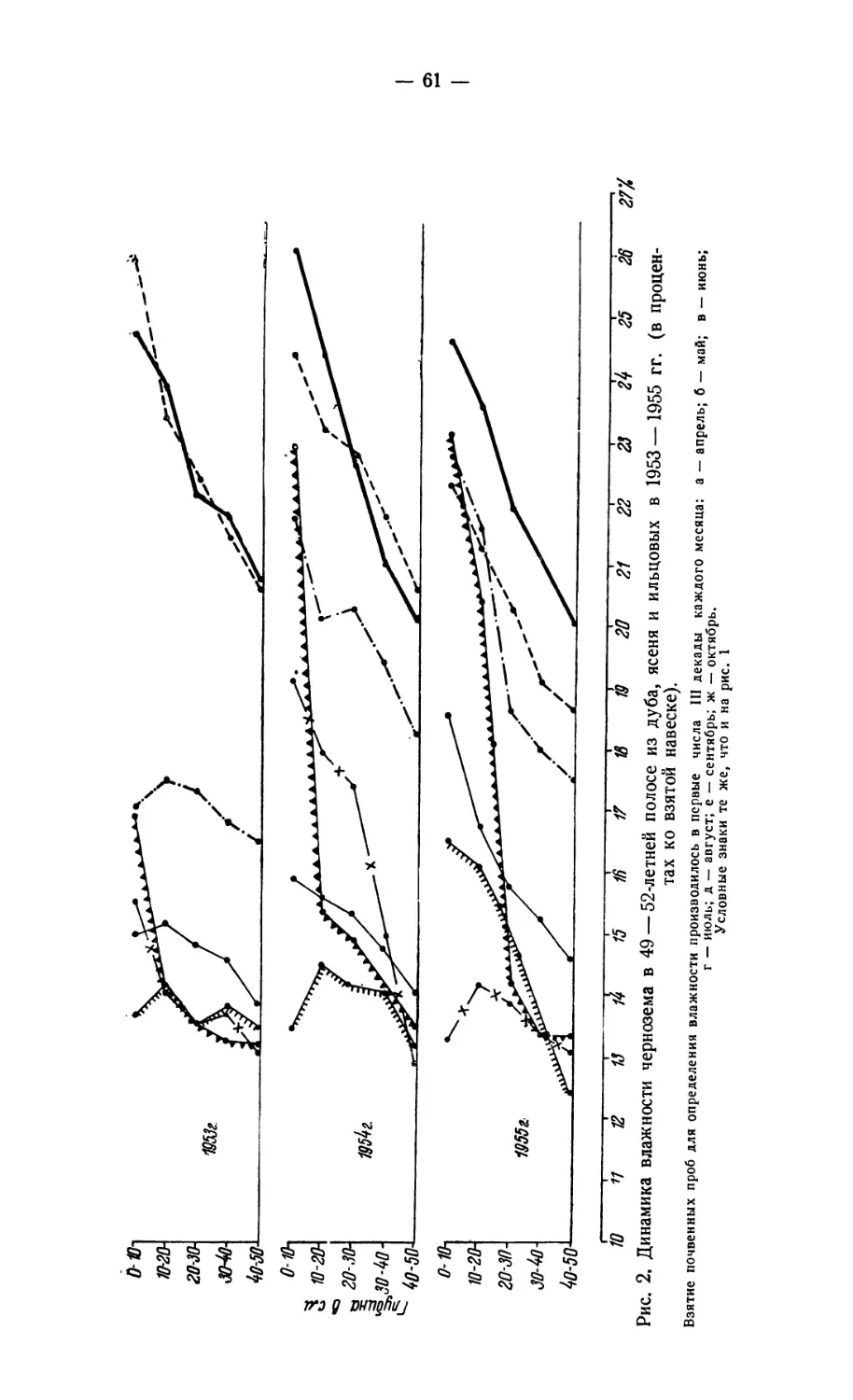
Соколов Д. Ф. О значении кивсяков и муравьев в трансформации органического
вещества под лесными насаждениями в условиях сухой степи................ 57
Г у п а л о П. И. Проблема онтогенетического старения и омоложения растений
и ее значение в растениеводстве........................................ 77
Работнов Т. А. Основные виды изменчивости луговой растительности .... 93
ВороновА. Г. иТагуноваЛ. Н. О стадиях формирования фитоценозов . 105
Хроника.....................................................................113
CONTENTS OF VOLUME LXII (5)
Dubinin N. P. Some philosophical problems of modern genetics......................... 11
Alpatov W. W. Left and right handedness in the structure of plants and animals . 19^
Averin Y. V. Zoogeographic survey of Kamchatka........................................ 29
Arsenyev V. A. Observations on the seals of the Antarctic............................ 39'
Stegmann В. K. Ober einige Eigenheiten im Bau des Schultergiirtels bei den
Tauben und Flughiihnern und die funktionelle Bedeutung der Clavicula bei den
Vogeln............................................................................ 45
Sokolov D. F. The importance of Jvlidae and ants in the transformation of organic
substance under forest timber in conditions of the dry steppe zone................ 57
Gupalo P. J. The problem of senescence and rejuvenescence of plants and its
importance In plant breeding...................................................... 77
Rabotnov T. A. The main forms of changes in meadow vegetation......................... 93
Voronov A. G. and Tagunova L. N. The stages of phytocenoses formation . . 105
Chronicle.............................................................................ИЗ-
Посвящается
XL годовщине
Великой Октябрьской
с оциалистической
революции
БЮЛЛЕТЕНЬ М. О-ВА И СП. ПРИРОДЫ, ОТД. БИОЛОГИИ, Т. LXII (5), 1957
Великая Октябрьская социалистическая революция, сороколетие ко-
торой отмечает в ноябре текущего года все культурное человечество,
открыв новый путь в развитии человеческого общества, создала в то же
время совершенно невиданные ранее возможности для развития науки.
Если при капитализме достижения науки использовались лишь не-
многочисленным господствующим классом общества, подавляющая же
часть народа была далека от всего того, что дает человечеству наука, то
после Великой Октябрьской социалистической революции в нашем оте-
честве завоевания науки стали доступны всему народу. Мало того, само
создание нового социалистического общества и построение величествен-
ного здания коммунизма осуществляются на базе науки с учетом всех ее
достижений. Эта возросшая роль науки, ее, если можно так выразиться,
всенародность — явились не только сильнейшим стимулом к мощному
усилению творческой работы ученых, но и чрезвычайно содействовали
вовлечению в исследовательские работы новых народных сил. Широко
раскрывшиеся двери высших учебных заведений, создание большой сети
разнообразных научно-исследовательских институтов и все увеличиваю-
щиеся ассигнования на научно-исследовательские работы, достигшие не-
виданных масштабов, определили высокие темпы развития науки в Со-
ветском Союзе.
Скрытые ранее творческие силы всех народов, населяющих СССР,
получили возможность проявить себя в полной мере. Теперь в многочис-
ленных научно-исследовательских учреждениях наших республик успешно
трудятся ученые самых различных национальностей.
Особенное же значение имело то, что, если раньше истинные ученые,
открывая объективные законы природы, часто были лишь стихийными
материалистами-диалектиками, то после Великой Октябрьской социали-
стической революции большинство советских ученых основательно озна-
комились с марксизмом-ленинизмом и начали сознательно пользоваться
в своей научной работе единственно правильной методологией — филосо-
фией диалектического материализма.
Четыре особенности характеризуют развитие наук в СССР за по-
следние 40 лет.
Во-первых, создание специальных научно-исследовательских инсти-
тутов. До Октябрьской революции научная работа проводилась по пре-
имуществу на кафедрах высших учебных заведений. В Российской
Академии наук наука была сосредоточена лишь в некоторых музеях
(Ботаническом, Зоологическом, Этнографическом и др.) или в небольших
лабораториях (например, физиологии растений, особой зоологической и
некоторых других). Ныне же центр научной работы переместился из
ВУЗ’ов в специальные институты. Одно только Биологическое отделение
Академии наук СССР имеет 17 больших институтов и 4 самостоятельные
— 6 —
лаборатории, не считая особых научных станций. Многие отраслевые био-
логические институты созданы ведомствами, а также входят во Всесоюз-
ную Академию сельскохозяйственных наук имени В. И. Ленина. Если ка-
федры Высших учебных заведений в дореволюционное время имели
в своем составе лишь по нескольку сотрудников, ведущих научную рабо-
ту, то теперь в штатах научно-исследовательских учреждений насчиты-
ваются сотни научных работников.
Во-вторых, в организацию научной работы введена плановость. Если
в дореволюционное время тематика научных работ как на кафедрах, так
и в музеях и лабораториях определялась личными интересами их сотруд-
ников, то ныне научная работа всех исследовательских учреждений регу-
лируется планами, ежегодно или на известный период составляемыми
данными учреждениями, с учетом важности теоретических проблем и на-
роднохозяйственных запросов практики и вообще жизни человека.
Тесная связь работы научно-исследовательских учреждений, не толь-
ко отраслевых, но и академических, с жизнью, с практикой составляет
третью характерную черту состояния науки в СССР в послеоктябрьский
период.
Наконец, четвертой специфической особенностью развития науки
в СССР особенно в последнее время является все расширяющееся при-
менение комплексного метода решения научных проблем.
Если ранее при кафедрах ВУЗ’ов чаще всего научная работа велась
одним ее заведующим или им с помощью немногих помощников той же
специальности, то ныне крупные проблемы разрешаются целыми коллек-
тивами ученых, в которые входят представители многих различных наук.
Внедрение возможно более широко комплексного метода в научную рабо-
ту обеспечивает более быстрое, а главное, более глубокое разрешение
научных проблем. Однако надо все же отметить, что применение этого
метода находится еще только на первоначальных этапах. Еще предстоит
много сделать в направлении организационных форм коллективной рабо-
ты и путей согласования стремлений и труда различных специалистов.
У ученых еще сильно чувствуется привычный индивидуалистический
подход к научной работе. Необходима известная перестройка психологии
ученого для участия в коллективной работе. Но плодотворность этого ме-
тода не подлежит никакому сомнению. Она доказана работами многих
исследовательских учреждений последнего времени.
В связи с этим стоит явление, на которое обратил внимание Прези-
дент Академии наук СССР, академик А. Н. Несмеянов, в недавнее время
на одном из общих собраний Академии наук СССР. Именно он отметил,
что характерной чертой современности является возникновение и разви-
тие новых промежуточных областей знания среди ранее обособленно суще-
ствовавших наук, например, возникновение таких наук, как физическая
химия, биофизика, биохимия, биогеохимия и др. Появление этих комплек-
сов знания оказалось очень плодотворным для развития науки в целом.
В Советском Союзе в этих контактных, пограничных, науках в послеок-
тябрьский период сделаны большие успехи. Для разработки их созданы
в системе Академии наук СССР и некоторых других академий специаль-
ные институты.
Говоря об условиях развития наук в СССР, нельзя не коснуться зна-
чения научных обществ. Как известно научные общества во всех странах
играли и играют большую роль в развитии науки и в воспитании новых
кадров. Это же имело место и в дореволюционной России. Однако по-
сле Великой Октябрьской социалистической революции у нас был пе-
риод неодооценки научных обществ. Некоторые считали, что при наличии
крупных объединений ученых по исследовательским учреждениям науч-
— 7 —
ные добровольные общества отжили свой век. Но жизнь решительно от-
вергла такое мнение. Ныне мы видим, что научные общества сильно ожи-
вили свою деятельность. Это и понятно. Добровольные научные общества,
являясь вневедомственными организациями, не стеснены в своей деятель-
ности рамками и традициями отдельных исследовательских учреждений.
Они служат прекрасным местом для свободного, объективного, принци-
пиального обсуждения научных вопросов, для организации всякого рода
научных дискуссий. Не отрицая большого значения обществ, объединяю-
щих ученых какой-либо одной научной специальности, например ботани-
ческого, энтомологического, гидробиологического, физиологического и
тому подобных обществ, все же нельзя не отметить особенное значение
обществ более широкого научного профиля, например Московского обще-
ства испытателей природы. Именно подобные общества должны сыграть
большую роль в развитии комплексного метода решения крупных науч-
ных и народнохозяйственных проблем и в развитии «пограничных» наук.
Все, что сказано о развитии науки вообще в послеоктябрьский период
в Советском Союзе, целиком относится и к биологии. За этот период био-
логические науки у нас не только успешно развивались, но установились
и более активные связи их с другими науками. Так, ботаника в целом
чрезвычайно выиграла оттого, что в физиологии растений широко начали
применять физические, химические и биохимические методы. Эти же ме-
тоды много содействовали решению крупнейших проблем филогении, эко-
логии растений, геоботаники и ботанической географии. В этом отношении
особое значение имеет все расширяющееся у нас применение радиоактив-
ных и стабильных изотопов, хроматографического анализа, электронного
микроскопа и других методов микроскопии. Геоботаника и фитоценоло-
гия, особенно укрепив в течение последних десятилетий свои связи с поч-
воведением, климатологией, геоморфологией, а также и с биогеохимией
и используя не только описательный, но и экспериментальный методы,
сильно развились в последнее время и заняли ведущее место в соответ-
ствующих разделах мировой науки. В частности, их развитие привело
к выделению особой, также в некотором роде пограничной области зна-
ния — биогеоценологии.
Успешное развитие работ по общей систематике растений и по во-
просам эволюционной морфологии и филогении растений не только при-
вело у нас к созданию новых, вполне оригинальных систем растительного
мира (А. А. Гроссгейм, Б. М. Козо-Полянский, А. Л. Тахтаджян, А. Я. Ва-
га и др.), но и повлекло за собой составление фундаментальной «Флоры
СССР». Это грандиозное многотомное издание, равного которому по его
масштабам нет в мировой литературе и которое заканчивается в ближай-
шее время, подводит итоги огромной флористической работы, проведен-
ной в значительной степени уже в советский период.
Не меньшее научное значение имеют работы по картографии расти-
тельного покрова, которые весьма интенсивно развернулись у нас в после-
революционный период. До революции, в сущности, у нас не было бота-
нических карт России, если не считать весьма схематических мелкомас-
штабных карт С. И. Коржинского и Г. И. Танфильева. После же Октября
Ботанический институт Академии наук СССР провел большую работу по
составлению разнообразных в значительно больших масштабах карт
растительности сначала Европейской части СССР, а затем и всего СССР.
Эта работа завершилась составлением и изданием в 1954—1956 гг. гео-
ботанической карты СССР в масштабе 1 : 4 000 000, научное и народно-
хозяйственное значение которой трудно переоценить.
К наиболее развитым разделам ботаники в Советском Союзе при-
надлежит и физиология растений, особенно в отношении проблем фото-
— 8 —
синтеза, корневого питания, вопросов роста и развития, водного режима
растений й проч.
Близкая связь науки с практикой в Советском Союзе определила воз-
никновение и развитие таких особых научных дисциплин, как лесоведе-
ние, луговедение, болотоведение, тундроведение и проч.
Не менее успешно развивались за этот период исследования фауны
СССР, охватившие все районы обширной территории страны. Этому зна-
чительно способствовало создание широкой сети научных учреждений
союзной и республиканских академий и отдельных ведомств, организация
системы заповедников и зональных станций.
Развитие работ по систематике животных позволило создать капи-
тальные монографии и сводки, посвященные отдельным группам фауны
СССР. Таковы, например, «Звери СССР и прилежащих стран» С. И. Ог-
нева, «Птицы Советского Союза» и ряд других. Ведется работа по созда-
нию многотомной «Фауны СССР», ряд томов которой в настоящее время
уже опубликован. Все эти труды, подводящие итоги многолетним иссле-
дованиям фауны СССР, вошли в основной арсенал мировой научной ли-
тературы и имеют важнейшее значение для изучения фауны Палеарктики
в целом. Издана целая серия региональных монографий, освещающих
фауны отдельных республик (Азербайджанской, Казахской, Армянской,
Туркменской, Узбекской, Таджикской и др.), которые еще несколько де-
сятилетий тому назад были известны очень мало.
Необходимость удовлетворения запросов, которые ставят перед со-
ветской зоологией промышленность, сельское хозяйство и здравоохране-
ние СССР, вызвала быстрый рост экологических исследований. Большие
успехи достигнуты советскими зоологами в области изучения экологии
промысловых млекопитающих, птиц, рыб и некоторых беспозвоночных,
вредителей сельского хозяйства, переносчиков заболеваний человека и до-
машних животных. Многочисленны и широко известны также работы по
различным вопросам паразитологии, особенно гельминтологии. Прогресс
научных знаний во всех этих областях неразрывно связан с развитием
науки в нашей стране в период после Великой Октябрьской социалисти-
ческой революции.
Однако, говоря о достижениях биологических наук в СССР, которы-
ми советская наука может с полным основанием гордиться, нельзя не от-
метить, что в некоторых ее разделах мы все же отстаем от уровня их
в мировой науке. Это прежде всего относится к генетике и цитологии,
которые связаны между собой.
Также имеется отставание в разработке и другой крупной биологи-
ческой проблемы, проблемы вида и видообразования, имеющей столь
большое теоретическое и практическое значение. Правда, по проблеме
вида и видообразования в самые последние годы в некоторых журналах,
в том числе и в «Бюллетене Московского общества испытателей приро-
ды» (отдел биологический), успешно развернулась дискуссия, в извест-
ной степени исправившая создавшееся положение в этой области, но в то
же время дискуссия выявила недостаточность фактических знаний по
этой проблеме и показала необходимость значительного расширения ис-
следовательских, особенно экспериментальных, работ в этом направлении.
Решения XX Съезда КПСС и последующих пленумов ЦК КПСС
по вопросам развития народного хозяйства СССР и построения комму-
низма в нашей стране — поставили новые и еще более ответственные за-
дачи перед советскими биологами, в частности и перед биологами Мо-
сковского общества испытателей природы и их журналом «Бюллетень
МОИП» (отдел биологический). Общество и журнал должны усилить
— 9 —
свое содействие разработке крупных биологических проблем большого
теоретического и народнохозяйственного значения, повышению научного
уровня биологических исследований и внедрению комплексного метода
в научную работу. Критикуя и разоблачая различные идеалистические и
метафизические биологические теории, все еще имеющие хождение среди
значительной части ученых капиталистических стран, Общество в то же
время должно всемерно содействовать использованию у нас научных до-
стижений всех зарубежных стран. В этих целях необходимы еще более
тесная связь Московского общества испытателей природы с зарубежны-
ми научными обществами, регулярный обмен между ними изданиями и
активное и широкое участие членов Общества в международных научных
конгрессах и конференциях.
БЮЛЛЕТЕНЬ М. О-ВА ИСП. ПРИРОДЫ, ОТД. БИОЛОГИИ, Т. LXII (5), 1957
О некоторых философских вопросах
современной генетики
Н. П. Дубинин
Some philosophical problems of modern genetics
2V. P. Dubinin
Для разработки философских вопросов современной генетики определяющее
значение имеет вопрос о сущности материальных основ наследственности и процессов
их изменений, происходящих на основе единства внутреннего и внешнего. Ф. Энгельс
указывал в «Диалектике природы», что специфика той особо организованной формы
движения материи, которою является жизнь, по сравнению с остальными формами со-
стоит в наличии особого рода взаимодействия простых форм движения, благодаря че-
му возникает новое качество — явление жизни.
Наследственность, как и все другие явления жизни, имеет материальную приро-
ду. В явлениях жизни нет ничего, кроме взаимодействующих молекул и атомов с их
ядрами и электронами. Это показывает, насколько велика роль физических, химиче-
ских и математических методов исследования для биологии в целом и для проблемы
наследственности в частности. Только путем применения этих методов можно исследо-
вать все те простые формы движения, в том числе и квантово-механические, которые
лежат в основе той особой формы движения материи, которая именуется жизнью.
У многих организмов связь между поколениями осуществляется через одну клет-
ку — оплодотворенное яйцо, образуемое слиянием яйцеклетки и спермия. Через него
передаются материальные структуры, физические и химические формы движения кото-
рых своим специфическим взаимодействием определяют свойство наследственности.
Объем этих структур, главной из которых является ядро клетки, вернее — составляю-
щие его хромосомы, в ряде случаев равен всего лишь одному кубическому микрону,
т. е. группе атомов, число которых равно 1012. Вполне понятно, какое громадное зна-
чение для анализа природы таких структур должны иметь методы физики и химии.
Детальными цитологическими исследованиями удалось дифференцировать структуру
хромосомы вдоль по ее длине. Каждая хромосома представляет собою длинную нить,
в основе которой лежат цепи, Составленные из белка и нуклеиновой кислоты, т. е. це-
пи нуклеотидов. Использование дифракции рентгеновских лучей позволило опре-
делить, что диаметр отдельных цепей составляет около 20 ангстремов.
Точно доказано, что отдельные участки, следующие друг за другом вдоль по
длине каждой хромосомы, имеют специфическое влияние на развитие организма. Та-
кие элементарные локусы хромосом получили наименование генов. Путем сопоставле-
ния данных, полученных в экспериментах при использовании предельной разрешаю-
щей способности микроскопа и ультрафиолетовой микроскопии. Меллер и Прокофьева
(1934) определили максимальный размер гена, равный 20X 125 миллимикрон. В на-
стоящий момент имеются серьезные доказательства в пользу того, что в генетическом
веществе хромосом выдающаяся роль принадлежит дезоксирибонуклеиновой кислоте
(ДНК). Химический состав ДНК изучен. Показано, что генетическая специфичность
может быть связана с различием молекул по взаимоположению в них азотистых ос-
нований: цитозина, содержащего 12 атомов, гуанина (15 атомов), аденина (14 атомов)
и тимина (14 атомов). Размер азотистых оснований равен 5—7 ангстремам, вдоль по
цепи молекулы ДНК эти основания отстоят один от другого на расстоянии
-3,4 ангстрема.
— 12 —
Современные методы физического и химического анализов
материальных основ наследственности
Генетическими методами скрещивания и цитологическим исследованием путем
применения светового микроскопа, разнообразных дифференциальных окрасок разных
частей клетки и путем изучения живых структур установлено, что наследственность
в первую очередь связана с ядром клетки, точнее, с хромосомами. Химическими и фи-
зическими исследованиями установлено, что хромосомы и гены являются нуклеопро-
теидами. Это доказано в четырех направлениях работ:
1. Само название хромосом, т. е. красящихся тел, происходит от их способности
избирательно окрашиваться многими основными красками. Специфическое окраши-
вание хромосом по Фельгену показывает, что они содержат большое количество нук-
леиновой кислоты. Установлено также наличие в них некоторого количества рибонук-
леиновой кислоты. Окрашивание хромосом метил-грюнпиронином позволяет отличать
дезоксирибонуклеиновую кислоту (окрашивается в зеленый цвет) от рибонуклеиновой
кислоты /окрашивается в пурпуровый).
2. Применение методов ультрафиолетовой микроскопии показало, что хромо-
сомы абсорбируют волны длиной в 2600 ангстремов, что доказывает наличие
в них нуклеиновых кислот, и волны в 2380 ангстремов, что указывает на нали-
чие белков.
3. Путем центрифугирования и фильтрации было осуществлено изолирование
ядер из спермиев и эритроцитов рыб и из ряда тканей млекопитающих. Подробно
изучались изолированные хромосомы из клеток тимуса теленка. Прямой химический
анализ показал, что хромосомы состоят из нуклеопротеида, в котором 45°/о падает на
дезоксирибонуклеиновую кислоту, а остальное на низкомолекулярные протеины типа
гистона. Небольшая часть падает на белки, содержащие триптофан.
4. Обработка хромосом на цитологических препаратах нуклеазами и пепсином
показала наличие в них нуклеиновых кислот и белков.
На основании всех этих и других данных установлено, что хромосомы клеток
животных, растений и микроорганизмов состоят из нуклеопротеидов.
Вирусы по своим размерам приближаются к макромолекулам. С помощью элект-
ронного микроскопа показано, что диаметр вируса полиомиелита равен всего лишь
22 миллимикронам. Для вируса табачной мозаики электронный микроскоп в сочетании
с химическим анализом, дифракцией рентгеновских лучей и изотопным методом по-
зволили установить структуру частиц этого вируса. Они имеют форму цилиндров диа-
метром в 150 и длиной в 3000 ангстремов. Стенки цилиндров состоят из двойных
белковых молекул, как бы блоков в форме сектора, толщиной в 23 ангстрема. Блоки
связаны в спиральную структуру с полостью внутри. Эта полость занята нуклеино-
вой кислотой.
В результате исследований последних лет вскрылась картина единства физико-
химической организации наследственных структур во всем органическом мире. На-
следственные структуры вирусов, микроорганизмов, растений, животных и человека
представляют собою нуклеопротеиды, т. е. образуются из нуклеиновых кислот и бел-
ков. Среди нуклеиновых кислот главной для этих структур является дезоксирибонук-
леиновая кислота. В нуклеопротеидной организации наследственных структур уловле-
но коренное всеобщее свойство жизни.
Доказательство того, что ДНК играет выдающуюся роль в наследственности, так-
же получены путем использования комплексных методов химии, новых методов изо-
топного анализа, генетики и электронной микроскопии. Было показано, что наслед-
ственность пневмококков может быть направлено изменена путем внесения в среду,
где развиваются пневмококки данного типа, ДНК от пневмококков другого типа.
Активность трансформирующей ДНК настолько велика, что концентрация 6Х10-8 до-
статочна для наследственного преобразования клеток другого типа.
Исследованиями последних месяцев обнаружено, что качественные изменения
в составе ДНК являются Неиссякаемым источником появления новых наследственных
уклонений (мутаций). Как указано выше, в состав ДНК входит азотистое основание —
тимин. Его аналогом служит урацил, входящий в состав рибонуклеиновой кислоты.
Чтобы нарушить синтез тимина, вирусы подвергались воздействию сульфаниламина,
а затем они культивировались на клетках, содержащих соединения урацила, что при-
водило к появлению громадного числа мутаций. Частота мутаций оказалась в 1000 раз
больше естественной. В этом случае установлена такая частота мутирования, которая
до сих пор не была достигнута ни при каких других воздействиях, в том числе и под
влиянием атомной и сходной с ней радиации, химических веществ и т. д. Аналогич-
ные результаты получены также в опытах с бактерией кишечной палочки.
Эта работа представляет собою первый пример изменения наследственных свойств
организмов путем химического вмешательства во внутреннюю структуру ДНК. Сооб*-
щение об этих открытиях было напечатано под названием «Гены создаются челове-
ком» (1957).
Эти и другие факты не оставляют сомнений, что ДНК играет выдающуюся роль
в наследственности организмов.
— 13 -
Физическое и химические исследования молекулярной
и атомной природы ДНК
Открытие громадной роли ДНК в наследственности потребовало знания состава
и строения молекул этого органического соединения.
Дезоксирибонуклеиновая кислота представляет собою уникальное химическое со-
единение в том смысле, что во всем мире оно встречается лишь в наследственных
структурах растений, животных, микроорганизмов и вирусов. Поэтому, когда речь идет
об изучении физики и химии ДНК, практически мы имеем дело с анализом важнейше-
го компонента хромосом. Цитологическое, цитохимическое изучение структуры хромо-
сом, применение методов ультрафиолетовой и электронной микроскопии дали очень
много как для характеристики общей структуры, так и для познания тонких деталей
хромосом. Однако до сих пор неясны многие кардинальные вопросы, такие, как внут-
ренняя картина взаимоотношения в хромосоме белка и нуклеиновой кислоты, вопросы
о том, каким образом молекулярная структура ДНК может приобрести генетическую
специфичность; как осуществляются процессы развития новых хромосом этих гигант-
ских молекул, при которых новые хромосомы воспроизводят физические и химические
особенности исходных молекул (ауторепродукция), без чего не могло бы иметь место
само явление наследственности, и в чем состоит сущность физических и химических
преобразований генетического характера хромосом при мутациях; какова физическая
природа гомологического притяжения, благодаря которому парные хромосомы и даже
их отдельные локусы находят в клетке друг друга и конъюгируют, без чего не было
бы организованного редукционного деления, а стало быть, размножения и менделев-
ского расщепления и т. д.
Громадное значение для всей проблемы наследственности имеет тот факт, что при
наличии глубоких генетических и цитологических знаний о хромосомах как о мате-
риальном субстрате наследственности наряду с этим оказался возможным анализ моле-
кулярной и атомной природы важнейших веществ, входящих в состав генетического
материала. В результате в настоящее время нам многое известно, с одной стороны,
о морфологии и химическом составе хромосом, с другой,— об их молекулярной при-
роде. Одна из главных задач современности состоит в необходимости заполнить брешь
между этими знаниями и показать, как особенности молекулярного строения претво-
ряются в клеточные биологические структуры.
Дезоксирибонуклеиновая кислота, важнейший компонент хромосом, представляет
собою полимерное соединение, состоящее из громадного числа отдельных нуклеотид,
каждый из которых является соединением азотистого основания, и остатков молекул
сахара и фосфорной кислоты. В основе молекулы ДНК лежит длинная цепь из обя-
зательно чередующихся молекул сахара и фосфора. Азотистые основания присоеди-
няются с боку молекулы к сахарам через один из азотов основания. Ширина молекулы
20 ангстремов, а длина равна тысячам и десяткам тысяч ангстремов. Химическим ана-
лизом установлено наличие попарной зависимости между одним пуриновым основанием
(аденин) и одним пиримидиновым (тимин), а также соответственной зависимости меж-
ду другим пурином и пиримидином (гуанин — цитозин).
Решающее значение для установления стереохимической формулы молекул ДНК
имело применение рентгеноструктурного анализа. Рентгенограммы ДНК показали, что
наибольшее скопление атомов отмечается на периферии структур. Это оказались две
извитые цепи из сахаров и фосфатов. Основания расположены внутри в виде пере-
кладин, причем каждая перекладина состоит из попарно соединенных оснований —
аденина с тимином и гуанина с цитозином. Основания соединены водородными связя-
ми, которые объединяют две цепи ДНК в единую структуру. Различия молекул воз-
никают за счет разных взаиморасположений четырех указанных выше оснований. Тео-
ретически можно говорить о любой возможной последовательности оснований вдоль
по цепи ДНК.
Тот факт, что молекулы ДНК обладают генетической специфичностью, требует
истолкования. В настоящее время, исходя из физических и химических особенностей
ДНК, приходится признать, что наследственная информация может быть записана
в структуре молекулы ДНК только на основе различий в специфическом взаимополо-
жении оснований. Однако возникает вопрос, могут ли всего лишь четыре разные ка-
тегорий* оснований обусловить бесчисленное наследственное многообразие таксономи-
ческих и индивидуальных отличий организмов. Математический расчет показывает, что
возможное число специфических взаимоположемий этих четырех оснований исключи-
тельно велико. В отрезке цепи ДНК, содержащем 100 нуклеотидов, мыслимое коли-
чество разных взаимоположений четырех оснований составляет 4100. Это число превы-
шает количество атомов в солнечной системе.
Признание того, что генетическая специфичность записана в молекулярной струк-
туре ДНК столь неспецифическнм образом, ставит кардинальную проблему: каким же
образом столь сравнительно простые молекулы обеспечивают организацию сложней-
ших внутренних процессов развития особи, выступая в качестве материальных носи-
телей наследственных свойств. В первую очередь встает вопрос: каким образом осу-
— 14 —
ществляется регулярное появление в клетке видоспецифических и других наследствен-
но обусловленных белков?
Объяснение этих процессов требует привлечения современной математической
теории информации. Высказана идея, что сущность передачи информации, записанной
в молекулах ДНК, может заключаться в том, что определенная последовательность
оснований приводит к организации определенной последовательности аминокислот, свя-
зывающихся в цепь полипептида. Для проведения соответствующих расчетов были
привлечены электронные быстродействующие счетные машины. Будущее покажет пра-
вомочность этих представлений, однако теперь уже ясно, что математический анализ
этой проблемы так же, как и самого вопроса о формах записи генетической инфор-
мации в молекуле ДНК, является исключительно перспективным.
Современные открытия в области физики и химии наследственных структур имеют
принципиальное значение. Прежде всего это относится к данным, показавшим, что
ДНК является материальным носителем наследственности. Из всех современных дан-
ных следует, что роль ДНК в наследственности исключительно велика, так что в на-
стоящее время мы имеем все основания сказать, что в результате исследований гене-
тиков, цитологов, физиков и химиков, проведенных в самые последние годы, стали
известны молекулярные и атомные основы наследственных структур. Эти открытия,
позволившие материализовать одно из главных свойств жизни — наследственность,
являются великой победой не только для современного естествознания, но и для
марксистского философского материализма.
В настоящее время в качестве первоочередной встает задача выяснить, как осу-
ществляется высший синтез физических и химических форм движения, наличие кото-
рого и обусловливает наследственность. Явления жизни нельзя свести к химии и фи-
зике, ибо жизнь представляет собою высший синтез всех более низких, простых форм
движения материи. Одно ясно, что это новое качество — жизнь — не может быть по-
нято без знания сущности входящих в него простых форм движения материи. По-
этому проблема физических и химических основ наследственности является ныне од-
ной из центральных в генетике. Разработка этого вопроса должна заложить основы
для решения проблемы наследственности во всей сложности ее биологического содер-
жания. Совершенно ясно, что основные вопросы философского материализма связаны
с разработкой этого вопроса. Материалистическая постановка решающих вопросов
проблемы наследственности немыслима без признания того, что явление наследствен-
ности материально обусловлено, что в клетке, которая связует поколения, должны
быть определенные материальные структуры, физические и химические формы дви-
жения которых, благодаря их специфическому взаимодействию, создают явление на
следственности.
Физические и химические методы вызывании наследственной
изменчивости организмов (мутаций) 9-
Источником появления новых наследственных уклонений служит мутационный
процесс, основанный на качественных преобразованиях, идущих в материальных осно-
вах наследственности. В начале XX в. вопрос о появлении новых наследственных
уклонений Вейсманом, Де-Фризом, Лотси и другими решался неправильно. Были раз-
виты метафизические концепции, с одной стороны, о неизменяемости генов, с другой —
о чисто автогенетических факторах их изменений. Аналогично тому как в химии тео-
рия Дальтона привела на некоторое время к учению о неизменности атомов, так и
в генетике в течение некоторого времени широкое распространение получила идея
о неизменности генов.
Серьезный удар по этим воззрениям нанесли открытия Надсона и Филиппова,
Меллера, Стадлера, сделанные 30 лет тому назад. Они заключались в том, что иони-
зирующие излучения, такие, как рентгеновские лучи, атомный распад и другие, вызы-
вают громадное количество мутаций. Эти открытия привели вначале к глубокой раз-
работке биофизики мутационного процесса, а затем к созданию целого направления,
называемого радиационной генетикой. В исследованиях по первичным радиационным
генетическим эффектам наряду с генетикой и цитологией были применены многие до-
стижения физики. Наряду с ионизирующей радиацией широко изучалась природа воз-
действия на хромосомы ультрафиолетового света, началась разработка вопроса о роли
фотодинамических процессов. В самые последние годы принципиальное значение в ра-
диогенетике обещает приобрести принцип миграции и консервации энергии в макро-
молекулах и микроструктурах.
Сейчас, в эпоху наступления атомной эры, проблема вызывания мутаций ионизи-
рующими излучениями приобрела громадное значение. С одной стороны, радиацион-
ная селекция микроорганизмов и растений имеет исключительно важное производ-
ственное значение; с другой — встает задача защитить наследственность человека от
повреждающего воздействия радиации. В свете последней задачи такие, казалось бы,
чисто теоретические принципы биофизики мутационного процесса, как, например, от-
сутствие пороговой дозы радиации при вызывании мутаций, прямопропорциональная
- 15 -
зависимость частоты мутаций от дозы радиации, повышенная генетическая радиочув-
ствительность млекопитающих и другие, приобрели важнейшее практическое значение.
Однако следует отметить, что разработка методов вызывания мутаций действием
радиации хотя и разрушала идею о неизменности гена, но не обусловила отказ от
автогенетических воззрений, в свете которых неправильно решался вопрос о связи
между наследственностью и средой.
После установления мутагенного эффекта радиации историческое значение имело
открытие того, что многие химические вещества, проникая в клетку, вызывают наслед-
ственную изменчивость. Однако и после этих открытий все еще сохранилось, а в ряде
случаев сохраняется и до сих пор, ошибочное представление, что гены являются
особыми высокостабильными образованиями, выключенными из общего метаболизма
клетки и организма как целого, что только прямое воздействие относительно высокой
энергией может вызвать их изменчивость.
В утверждении этой неверной концепции большую роль сыграла попытка свести
весь мутационный процесс к квантовой механике, что было сделано М. Дельбрюком
(Delbruck М., 1935), Э. Шредингером (1947), X. Блумом (Blum Н., 1951) и другими.
Согласно атомно-физической модели гена М. Дельбрюка, поддержанной и развитой
Э. Шредингером, изменчивость генов представляет собой процесс чисто «спонтанный»,
не стоящий ни в какой связи с качественными условиями среды. С этой точки зрения
мутации генов представляют собой квантовые скачки в результате поглощения моле-
кулой энергии случайного теплового движения. Эта энергия, позволяя преодолеть
определенный энергетический уровень, вызывает изменения в конфигурации атомов,
т. е. появление изомерных молекул. Эта концепция возводила принцип случайности
в естественном мутационном процессе в абсолют, полагая, что причины мутаций коре-
нятся в процессах, основанных на принципах теории физического индетерминизма,
разработанной в приложении к микромиру. Согласно этим воззрениям, воздействия,
проникающие в клетку и вызывающие мутации, не оказывают никакого влияния на
качественную сторону происходящих изменений; они только лишь ускоряют спонтан-
ный, принципиально недетерминированный процесс появления мутаций. В последнем
издании самого распространенного американского руководства по генетике Э. Синнот,
Л. Денн и Ф. Добжанский (Sinnot Е., Dunn L. a. Dobzhansky Th., 1950) пишут, что
«имеется достаточно оснований считать, что Х-лучи и другие мутагенные воздействия,
насколько это изучено, просто ускоряют спонтанный процесс мутаций, а не вызывают
новые изменения». Г. Г. Меллер (Muller Н., 1951) в итоговой статье за 50 лет разви-
тия генетики писал, что «общее повышение мутаций при высокой температуре и ра-
диации всех родов как раз таково, какое и следовало ожидать, исходя из концепции
«молекулярного хаоса».
Ныне на микроорганизмах, растениях, на дрозофиле показана специфичность
действия ряда химических веществ. В частности, О. Фахми и М. Фахми (О. Fahmy
а. М. Fahmy) в 1956 г. получили в половой хромосоме дрозофилы при действии алки-
лирующих соединений 200 новых мутаций, не возникавших до сих пор под действием
радиации и других мутагенных факторов. Показано, что мутагенные воздействия в
основном вызывают мутации через изменения метаболических процессов в клетке.
Выяснено, что ионизирующие излучения наряду с прямой ионизацией и возбуж-
дением атомов в наследственных молекулах обусловливают в клетке радиолиз воды,
вследствие чего появляются активные радикалы Н2О2, ОН, НО2 и другие, которые
вызывают окислительные процессы, ведущие к появлению мутаций. О том, что мута-
ции в живой клетке в естественных условиях возникают под влиянием продуктов ее
обмена, говорят, например, опыты с воздействием на каталазу. При аэробном дыха-
нии в клетке образуется перекись водорода (Н2О2). В живых клетках перекись не мо-
жет накапливаться, так как она разлагается каталазой. На нейроспоре показано, что
при добавлении ядов, отравляюпжх каталазу (цианистый калий, азид натрия), частота
естественных мутаций возрастает, так как в этих условиях, вследствие подавления
каталазы, перекись водорода, синтезируемая организмом, накапливается в клетках.
На растениях установлено мутагенное действие путресцина (тетраметилдиамина),
который возникает как продукт декарбоксилизации аминокислоты орнитина. В нор-
мальных условиях растения обладают ферментом, катализирующим окисление путрес-
цина до соответствующего альдегида, чем предотвращается его мутационный эффект.
Появление мутаций в естественных условиях имеет много причин. Часть из них
обусловлена влиянием естественной радиации, слагающейся из космической и земной
радиации, и зависит от радиоактивных элементов, входящих в состав организма. Не-
которое количество мутаций вызывается действием температуры. Однако ныне стало
ясно, что основным фактором появления естественных мутаций служат биохимические
процессы обмена веществ в клетке. Крупной вехой в развитии этого вопроса было
долго недооценивавшееся открытие М. С. Навашина (1933), показавшего, что в старых
семенах растений появляется огромное количество мутаций. Принципиальное значение
имеют факты качественной зависимости характера мутаций от качества воздействую-
щих условий.
Все эти факты отчетливо показывают, что концепция Дельбрюка, Шредингера
и других о принципиальной случайности процессов появления мутаций должна быть
— 16 —
заменена признанием принципиальной зависимости качества мутаций от качества воз-
действующих условий.
Исключительно важно, однако, то обстоятельство, что в обычных условиях ха-
рактер мутаций в первую очередь определяется природой того локуса в хромосоме,
который претерпевает изменение и в результате влияния одного и того же фактора
вызывает массу разнообразных уклонений, так как действует на многие качественно
разные участки хромосом. Однако, каков тонкий механизм влияния данного опреде-
ленного метаболита на ген, мы не знаем. Данные по специфическому мутированию
показывают, что оно может иметь вполне определенный характер. Раскрытие природы
этого влияния необходимо для разработки методов получения направленных
изменений.
Таким образом, новые данные ясно показывают наличие связи материальных
основ наследственности с организмом в целом и через него со всеми материальными
условиями среды. Явление наследственности не обусловлено монопольно генами и хро-
мосомами, так как они представляют собой лишь элементы более сложной целостной
системы — клетки. Благодаря связи со средой и организмом в целом явление наслед-
ственности не может быть автономным, оно базируется на широком взаимодействии
как внутренних факторов жизнедеятельности организма, так и этих последних с мате-
риальными условиями внешней среды.
Применение физических, математических и химических методов не ограничивает-
ся двумя разобранными выше кардинальными областями генетики. На современном
этапе применение этих методов пронизывает многие разделы этой науки: укажем на
математическую» генетику, разработка которой исключительно важна для проблем
селекции и эволюции; на биохимическую генетику, которая раскрыла сущность ряда
важнейших сторон наследственно обусловленного обмена веществ; на радиационную
селекцию, успехи которой во многом базируются на достижениях ядерной физики; на
иммуногенетику и т. д.
Философский анализ основных проблем современной генетики требует учета
громадного роста фактического материала науки, который во многом заставляет сти-
хийно встать на правильные позиции и отвергнуть ошибочные попытки сведения яв-
лений наследственности к закономерностям квантовой механики, отказаться от авто-
генетических теорий наследственности, от неверных представлений о природе наслед-
ственной изменчивости и мн. др. Увеличение фактического материала в генетике идет
стремительными темпами. В 1951 г. в Нью-йо«ке вышел сборник «Генетика в XX сто-
летии», в котором ведущие генетики (Меллер, Стертевант, Гольдшмидт, Бидл, Кас-
персон, Добжанский и другие) подвели итоги за 50 лет, прошедшие после вторичного
открытия законов Менделя. Однако за последние семь лет накоплен такой фактиче-
ский материал, что целый ряд основных вопросов и в первую очередь те, что разо-
браны в настоящей статье, были подняты на новый уровень, чем была создана проч-
ная основа для их разработки с позиций марксистско-ленинского философского мате-
риализма. Это произошло за счет беспрецедентного внедрения в генетику, методов
физики, химии и математики. Новые факты ярко вскрыли методологическую ограни-
ченность в анализе коренных вопросов генетики, на котором многие ведущие генетики
настаивали всего лишь 5—10 лет тому назад, а некоторые из них настаивают
до сих пор.
Вполне понятно, что без знания роста и содержания современной науки фило-
софский анализ становится беспредметным. Это особенно важно в отношении таких
бурно развивающихся наук, как генетика^ которая, как это мы видели выше на при-
мере изучения природы материальных основ наследственности и причин появления
наследственных изменений, вступает в эпоху решающих открытий. На наших глазах
теоретическая генетика из науки, объясняющей явления наследственности, становится
наукой, перестраивающей наследственность органических форм. Это доказано внедре-
нием в практику межлинейных гибридов кукурузы, триплоидной свеклы, гетерозисных
кур, радиационных мутантов у производителей антибиотиков и т. д.
Исследование проблем и задач генетики, разработка вопроса о соотношении
простых (физических и химических) форм движения и особо организованного дви-
жения материи, которым является жизнь и одно из ее основных свойств — наследст-
венность, требуют внимательного методологического анализа в свете марксистско-ленин-
ского философского материализма. От правильного решения этих важных вопросов
зависит будущее развития генетики и ее влияния на практическую деятельность.
Настоящая статья касается лишь двух вопросов среди основных проблем совре-
менной генетики. Необходимо начать систематическую работу по методологическому
анализу всех главных разделов этой науки. Задача эта связана с разоблачением
враждебной идеологии в естествознании, с активной борьбой за утверждение все-
побеждающих принципов философского марксистско-ленинского материализма.
- 17 —
SUMMARY
The article gives and exposition of physical, chemical and mathematical methods,
applied to the problem of heredity as grounds for philosophical dialectical materialistic
concepts in genetics.
Two problems are discussed by the author: the physical and chemical nature of the
foundations of heredity and the interrelation of external and internal factors in the natural
process of mutation. Thanks to contemporary data on the role of desoxyribonucleic acid
in hereditary formations and the established fact that the process of mutation is connected
with general metabolism in the cell, it has been possible to materialize a major feature of
life — heredity. This marks a great victory in modern natural science as well as in marxist
philosophical materialism.
The philosophical analysis of genetic problems must necessarily be supported by
a great array of facts available in modern science.
ЛИТЕРАТУРА
Меллер Г. Г. и Прокофьева А. А. 1934. Непрерывность и дискретивность
наследственного материала. ДАН СССР, т. 4.
Шредингер Э. 1947. Что такое жизнь. Иногиз.
Blum Н. F. 1951. Times arrov and evolution. Prinecton University Press, Prinec-
ton, N. Y.
Delbruck M. 1935. Uber die Natur der Genmutationen und der Genstruktur.
Dritter Teil: Atomphysikalisches Modell der Gennutation. Nachr. Ges. Wiss. Gottingen
(Math. — phys. kl., Biol).
Fahmy O. G. and Fahmy M. Y. 1956. Cytogenetic analysis of the action of car-
cinogens and tumor inhibitors in Drosophila Melanogaster. Your, of Genetics, vol. 54, № 1.
Genetics in the 20-th Century, essays on the progress of Genetics during its first
50 years. New York, 1951.
Muller H. Y. 1951. The development of the Gene Theory, Genetics in the 20-th
Century. New York.
Mane made Genes, Scientefical American, Februar, 1957.
Nawachin M. S. 1933. Altern der Samen als Ursache von Chromosomenmutationen.
Planta, 20.
Sinnot E., Dunn L., Dobzhausky Th. 1950. The principles of Gene-
tics. N. Y.
БЮЛЛЕТЕНЬ М. О-ВА ИСП. ПРИРОДЫ, ОТД. БИОЛОГИИ, Т. LXII (5), 1957
Левизна-правизна в строении растительных
и животных организмов
В. В. Алпатов
Left and right handedness in the structure
of plants and animals
W. W. Alpatov
«Телами должно называть вещества,
ограниченные поверхностями, или
имеющие формы... кристал г, расте-
ние, дом — суть тела».
Д. И. Мен !глеев. Осно-
вы химии. *3 изд., 1906 г.
стр. 406.
Развитие проблемы левизны-правизны
В области биологии можно привести ряд примеров таких открытий, которые в
дальнейшем оказывались открытиями более широкого значения: явления, сходные с
биологическими, обнаруживались в области мертвой природы. Так, например, суще-
ствование левых и правых форм у живых организмов было замечено человеком го-
раздо раньше, чем были найдены левые и правые формы кристаллов.
Первыми объектами, на которых была обнаружена правизна-левизна, оказались
брюхоногие моллюски (Gastropoda). Эти животные издавна привлекали человека
правильностью своей формы и красотой окраски. Особенно богаты и разнообразны
моллюски южных морей и океанов. Неудивительно, что индусы, создавшие в тропи-
ческой Азии одну из древнейших культур, заметили, что среди моллюсков, раковина
которых, как правило, завита по правой спирали, как редкость встречаются раковины
левоспиральные. Эта особенность левых раковин нашла себе отражение в скульптуре.
Статуя индусского бога Вишну держит в левой руке левую крупную раковину мор-
ского моллюска, напоминающего моллюсков рода Turritella.
От индусов знакомство с левыми моллюсками проникло в Европу, куда стали
привозить из Индии всякие диковинные предметы, особенно после путешествий Васко
да Гама (1469—1524). В Европе левизна и правизна моллюсков нашли также отра-
жение в произведениях искусства раньше, чем стали достоянием науки. На картине
Рубенса (1577—1640) «Святой Августин», находящейся ныне в Праге, изображен по-
чтенный старец, стоящий на берегу моря У его ног играет младенец. Правой ногой
он попирает крупного левого моллюска, а на песке рядом лежит правый моллюск.
В XVIII в. замечательным мыслителем и писателем Бернарденом де Сен-Пье-
ром, служившим в 70-х годах в России в качестве строителя дорог, сделана была
первая попытка научного объяснения того факта, что огромное большинство брюхо-
ногих моллюсков имеет тело и раковину, завитые по правой спирали. В своих «Этю-
дах о природе» он писал: «Очень замечательно, что все моря наполненные одно-
створчатыми раковинками бесчисленного множества видов, у которых все завитки
направлены в ту же сторону, т. е. слева направо, подобно движению Земли, если
поставить их отверстиями к северу и их острым концом к земле, лишь очень малое
число видов составляет исключение» (цитировано по В. И. Вернадскому, 1940 6).
- 20 —
Мысль Б. де Сен-Пьера объяснить точно установленный факт преобладания на
земном шаре правых моллюсков должна считаться первой попыткой связать диссимет-
рии моллюсков с явлениями планетарного характера — с вращением Земли вокруг оси
против часовой стрелки.
Лишь в начале XIX столетия X. С. Вейсс (1780—1851) нашел, что среди кри-
сталлов кварца можно наблюдать как правые, так и левые формы. Джон Гершель
(1792—1871) открыл в 1821 г. «зависимость между направлением вращения плоскости
поляризации в кристаллах и их наружной формой» (по Вернадскому, 1903). Следую-
щим этапом в разработке вопроса о правизне и левизне кристаллов были классиче-
ские труды Луи Пастера (1861 и др.), выполненные примерно в 1848—1860 гг. и при-
ведшие к созданию в 1874 г. Лебелем и Вант-Гофом новой науки о расположении
атомов в пространстве, или стереохимии. Здесь надо отметить, что А. М. Бутлерову
принадлежит честь за 11 лет до опубликования работ Лебеля и Вант-Гофа предска-
зать появление стереохимии: «Если же атомы действительно существуют, я не вижу
почему, как думает Кольбе, должны быть тщетными все попытки определить про-
странственное расположение последних, почему будущее не должно научить нас
производить подобное определение» (Бутлеров А. М., 1863).
Определение основных понятий
отражения в плоскости.
Рис. 1. Левый и правый кристаллы
кварца
Учение о симметрии и соразмерности природных тел лучше всего разработано
в применении к кристаллам. Для уточнения понятий правизны и левизны нам надо
поэтому обратиться к кристаллографии.
«Бывают многогранники, один из которых является отражением другого. Ника-
ким движением их совместить нельзя. Совмещение достигается только при участии
совмещающиеся друг с другом только при
помощи зеркально-поворотных преобра-
зований, называются энантиоморфными
(обратноформенными в переводе с гре-
ческого на русский язык.— В. А.). В ви-
де энантиоморфных пар могут суще-
ствовать лишь такие многогранники, ко-
торые лишены зеркально-поворотных
осей, что вытекает из определения этих
фигур». (Болдырев, 1932, стр. 21).
Среди 32 видов симметрии, к кото-
рым относятся все кристаллические фор-
мы, лишь 11 видов имеют только простые
оси симметрии и только среди этих ви-
дов можно встретить правые и левые
формы. На рис. 1 представлено изобра-
жение левого и правого кристаллов
кварца.
Так как энантиоморфные тела не
имеют плоскости симметрии, Л. Пастер
назвал их диссимметричными. Он дает
очень образную картину того, что все реальные тела можно разбить на два класса.
Вот что он пишет (1861):
«Если взять любые материальные тела и рассматривать их формы и проявление
в них сходных частей, можно тотчас установить что оми разделяются «а два боль-
ших класса, отличающиеся следующими признаками: одни, помещенные перед зерка-
лом, дают изображения, совместимые с оригиналом; изображение же других не мо-
жет совместиться с оригиналом, несмотря на то, что первые с точностью воспроиз-
водят все детали оригиналов. Прямая лестница, ветка с супротивным расположением
листьев, куб, человеческое тело... вот тела первого класса. Винтовая лестница, ветка
•со спиральным расположением листьев, винт, рука, неправильный тетраэдр — таковы
формы второго класса. Эти последние не имеют плоскость симметрии».
Какие же группы диссимметрических явлений в области биологии известны в
-настоящее время? В сводке, посвященной явлениям левизны и правизны у животных,
Людвиг (W. Ludwig, 1932) различает 3 основных группы диссиметричных тел или
их проявлений:
1. Двусторонне симметрические тела с вторично неодинаково развитыми, пер-
вично строго симметричными признаками (например, тело человека с преобладающим
развитием правой руки, с перемещением сердца влево, крабы с одной клешней, раз-
витой сильнее другой, и т. д.).
2. Винтообразные и улиткообразные образования: моллюски, спиральные сосу-
ды растений.
— 21 —
3. Спиральные, круговые или циклоидные образования: спиральные колонии бак-
терий Bacillus mycoides, круговые пути животных на плоскости L
Необходимо также остановиться на самом понятии и обозначении правизны и
левизны. Эти термины чисто условны и обязаны своим возникновением тому, что одна
рука носит название правой, а другая левой, что связано в основном с двусторонней
симметрией тела человека. В. И. Вернадский (1903) вспоминает, что на Руси в XV в.
и до последнего времени в языке обитателей русского севера, поморов, сохранились
слова «посолонь» — движение по солнцу и «противусолонь» — движение против
солнца (рис. 2).
Точно так же условно обозначение винтов и винтообразных образований. Вин-
тами правыми называют винты, которые ввинчиваются движением отвертки по часо-
вой стрелке, а левыми — против часовой стрелки. Распространенные в технике и в
противусолонь против чаа стр.
посолонь
по часовой
Стрелка
Рис, 2. Обозначение различных направлений движения в горизонталь-
ной плоскости
быту винты, штопоры и т. п. в подавляющем числе случаев являются правыми вин-
тами. Для получения наглядной картины того, что представляют собой правая и ле-
вая винтообразные линии, полезно взять мягкую проволоку и обматывать ею палочку
снизу по направлению кверху. При вращении свободного конца проволоки по часо-
вой стрелке получается левая винтообразная спираль, а при вращении против — пра-
вая. Важно отметить, что левые и правые спирали и винты при любых положениях
по отношению к наблюдателю сохраняют свое специфическое направление завитков.
У левых спиралей завитки идут снизу справа наверх налево, у правых наоборот.
О встречаемости левых и правых тел в природе
Познакомившись с диссимметричными правыми и левыми организмами, естествен-
но задать себе вопрос о их встречаемости в природе. Мы остановимся лишь на
2-й и 3-й группах диссимметрических образований. Нам при этом придется перейти
от рассмотрения диссимметрии отдельных тел или предметов к рассмотрению их сово-
купностей, скоплений. Мы перейдем, таким образом, от диссиметрии индивидов к дис-
симметрии коллектива. Можно представить себе, что скопления тел могут состоять
лишь из левых тел, либо из правых, либо из смеси правых и левых.
Прежде чем познакомиться с встречаемостью различных совокупностей — скопле-
ний левых и правых тел в мертвой и живой природе, необходимо несколько остано-
виться на уточнении понятия диссимметрии как свойства совокупности тел. Г. Ф. Гау-
зе (1940) считает, что Л. Пастер термин «диссимметрия» применял лишь к отдельным
телам — молекулам, и лишь англичанин Джепп (Japp, 1898) употребил впервые тер-
мин «асимметрия» для выражения свойств совокупностей молекул. Это мнение
Г. Ф. Гаузе, по-моему, ошибочно. Достаточно привести следующие слова Л. Па-
стера (1860): «Таким образом я считаю, что в теорию и практику физиологии введена
важная идея молекулярной диссимметрии естественных органических соедине-
ний» и т. д. (по Дюкло, 1929), чтобы понять, что Пастер и вещества — соединения,
состоящие из совокупностей молекул, называл диссимметричными. Термин «асиммет-
рия» появился в научном обиходе лишь вследствие того, что при переводах на
английский, немецкий и русский языки (1894) сочинений Пастера пастеровский термин
«диссимметрия» переводился словом «асимметрия». По-моему, нет никаких оснований
диссимметрию — свойство отдельных особей или частичек при переходе к совокупно-
1 Людвиг в качестве четвертой группы выделяет еще один тип, свойственный
многокамерным корненожкам; я думаю, что он может подойти к одному из перечис-
ленных выше трех типов.
— 22 -
Рис. 3. Кривые встречаемости правизны скопле-
ний кристаллов кварца (1), колоний Bacillus
mycoides (2) и популяций наземных брюхо-
ногих моллюсков Achatinella mustelina и
Partula suturalis (3, 4); n — число коллек-
ций кристаллов и популяций организмов, по
которым построена кривая (Алпатов, 1953)
стям этих частичек— обозначать новым термином. В самом деле, ведь говорим мы о
весе отдельных людей и в то же время о среднем весе той или иной группы — кол-
лектива людей. В. И. Вернадский (1940а) высказывался также против применения
термина «асимметрия протоплазмы».
Для того чтобы характеризовать степень диссимметрии совокупности тел, я пред-
лагаю выражать число правых тел в процентах общего числа тел, взятых из того или
иного скопления их. Предположим, что в данном месторождении кварца найдено
35 правых и 40 левых кристаллов.
Тогда свойство данного месторожде-
или его декстральность (dexter — по
латыни правый), будет равна
35
утр 100 = 46,7%. При отсутствии пра-
вых кристаллов декстральность бу-
дет равна 0; при наличии только
правых кристаллов она будет равна
100%. С ер ед ин у — 50%, когда левые
и правые тела или особи встречают-
ся поровну, можно назвать рацеми-
ческим состоянием, взяв этот термин
из органической химии, где со вре-
мен Пастера вещества, в состав ко-
торых входят в равном числе пра-
вые и левые диссиметрические моле-
кулы, называют рацемическими. При
этом диссиметрия совокупностей мо-
жет принимать различные количе-
ственные выражения. Правизна, или
декстральность, может выражаться
любым числом от 0 до 100 (это край-
ние пределы диссиметрии).
В работах Г. Г. Леммлейна
(1939, 1944) приведены числа левых
и правых кристаллов, встреченных
в коллекциях кристаллов из 34 ме-
сторождений. Для каждого из них
мною был вычислен показатель дек-
стральности, или показатель правиз-
ны. Эти показатели были разнесены
по классам: 0— 10%, 10—20% и т. д.
до 90—100%. Получены частоты,
т. е. числа местонахождений в пре-
делах каждого класса. Эти частоты
отдельных классов выражены в про-
центах общего числа, т. е. 34, и нане-
сены против середин классов на диа-
грамме (рис. 3). Кривая через эмпи-
рические точки проведена от руки.
Кривая кристаллов кварца
имеет вид так называемой нормаль-
ной, или биноминальной, кривой рас-
пределения. На основании вида этой
кривой можно сделать вывод, что
чаще всего встречаются месторожде-
ния кристаллов, в которых, судя пс
взятым пробам, левые и и правые
тела представлены поровну. Лишь
для одного местонахождения в Гре-
ции А. Б. Вистелиус (1950) показал, что в нем преобладают левые кристаллы над
правыми, и это заключение, судя по достаточному числу взятых на изучение кри-
сталлов, должно считаться статистически обоснованным. Если откинуть этот случай,
для всех остальных местонахождений можно считать нормальным рацемическое со-
стояние, т. е. принять одинаковую встречаемость левых и правых тел косной природы.
Обратимся теперь к встречаемости правизны и левизны в живой природе. Одной
из весьма распространенных споровых бактерий почвы является Bacillus mycoides.
При культивировании этой бактерии на твердой среде с агаром колония имеет вид
спиральной туманности. Обычно у нас в СССР встречаются колонии с завитками, на-
правленными против движения часовой стрелки. Такие колонии называются левоза-
витыми колониями (рис. 4).
— 23 —
Совместно с О. К. Настюковой мною были изучены образчики почв из 127 пунк-
тов Европы, Америки, Азии и Австралии. Была обследована встречаемость правых
колоний у 108 популяций. Из каждой популяции для посева брали кусочки почвы, ко-
торые высевались в чашки Петри. Для обследованных популяций бактерий вычислял-
ся процент встречаемости правых колоний. Полученные характеристики декстраль-
ности 108 популяций были разнесены по классам декстральности, подобно тому как
это делалось для коллекций кристаллов. Полученные ряды изменчивости декстраль-
ности популяций изображены в виде кривой 2 на рис. 3.
Как показывает распределение правизны, наиболее часто встречаются популяции
с небольшой примесью правых форм. Это дает резко выраженную асимметрическую
кривую распределения.
Если обратиться к встречаемости правизны у брюхоногих моллюсков, то для
вида Partula suturalis на о-ве Моорея в Тихом океане (Н. Crampton, 1932) статисти-
Рис. 4. Вид левых и правых колоний Bacillus mycoides
веская обработка дает четко выраженную «чашеобразную» кривую распреде-
ления (рис. 3).
Сходную картину дает распределение правизны популяций моллюска Achatinella
mustelina на о-ве Оаху (Гавайские о-ва) (данные взяты из работы D. Welch
a. A. Alte, 1938).
У нас в Средней Азии широко распространены правые популяции моллюска
Fruticicula lantzi Lnd. Лишь в некоторых районах около Алма-Аты имеются почти
сплошь левые популяции этого вида и крайне редко попадаются смешанные популя-
ции, в которых живут и правые и левые особи.
Таким образом, для популяций моллюсков рацемичность, т. е. существование
популяций с почти равным соотношением левых и правых особей,— явление весьма
редкое.
В итоге изучения кривых встречаемости правизны можно сделать вывод о том,
что тела мертвой природы в отношении правизны скоплений характеризуются коло-
колообразным (биноминальным) распределением, тогда как популяции организмов, у
которых имеются как правые, так и левые особи, характеризуются «чашеобразным»
(для моллюсков) или «получашеобразным» (бактерии) распределением.
В строении высших растений имеется одна очень четко выраженная структура,
которая может быть как левой, так и правой. Это винтообразные утолщения стенок
сосудов. Открыты они были без малого 300 лет назад знаменитым итальянским био-
логом Марчелло Мальпиги. Вот как образно описывает Н. А. Холодковский (1923)
открытие трахей растений — сосудов со спиралями в стенках: «Как-то раз вечером
(в 1662 году) Мальпиги прогуливался в дачном саду недалеко от города. Вдруг
ветка каштана, простиравшаяся поперек дороги на высоте человеческого роста, боль-
но задела его по лицу. С досадой он схватил ее, сломал и на разломе увидел идущие
вдоль нее нити, которые при более внимательном рассмотрении оказались каналами,
наполненными воздухом, трахеями растений». С тех пор как анатомия растений во-
шла в число предметов преподавания в высшей школе, десятки тысяч студентов-био-
логов всего мира изучают на занятиях эти спиральные, точнее, винтообразные утол-
щения сосудов растений, служащих не для проведения воздуха, как думал М. Маль-
пиги, а для проведения соков.
— 24 —
В ботанической литературе до последнего времени не было ни одной специаль-
ной работы, посвященной направлению винта, т. е. правизне и левизне хода винто-
вого утолщения стенок сосудов. Рисунки, даже приводимые в учебниках, в ряде слу-
чаев неправильно указывают направление спиралей (Алпатов, 1951). Мною было об-
следовано 204 вида сосудистых растений (Алпатов, 1952). Огромное большинство
(93.2%) видов относятся к типу растений с левым ходом винтообразных утолщений;
5,4% видов — к типу рацемическому, т. е. к типу, у которого левые и правые сосуды
представлены в одном и том же растении, и 1,5% — к типу с правым ходом утолще-
ний. В отношении приуроченности направления винтообразных утолщений в сосудах
и отдельных семействах можно сказать, что стоящими особняком оказались: сем.
Balsaminaeceae с преобладанием правых видов, сем. Begoniaceae и Orchidaceae напо-
ловину с рацемическими видами. Все эти три семейства характеризуются тем, что в
основном виды, относящиеся к ним, распространены в тропиках. В целом декстраль-
3-100
ность сосудистых растений очень невелика: —jgg- = 1,55%.
Географические закономерности в распространении диссимметричных
и рацемических популяций организмов
В распределении диссимметричных и рацемических популяций в пределах
небольших районов трудно бывает подметить какие-либо закономерности. Как уже
было сказано выше, наземный моллюск Fruticicola lantzi в Средней Азии, в частности
в Казахстане, дает пример любопытного распространения левизны и правизны (Цвет-
ков, 1938). Под г. Алма-Атой и к западу от него встречаются только правые формы.
В 20 км к востоку от Алма-Аты в Талгарском ущелье к правым примешано 6,2% ле-
вых форм, а в Иссыкском ущелье (еще 20 км к востоку) левые формы представлены
уже в количестве 95,5%. Далее к востоку опять идут правые популяции и лишь в
окрестностях с. Чилик опять появляются левые популяции.
Если же обратиться к пространствам большого масштаба, то в распространении
правизны и левизны намечаются более определенные закономерности.
Бактерия Bacillus mycoides представляет собой исключительно благодарный
объект для решения поставленной задачи. Она распространена в почвах всего земно-
го шара и в стадии спор выживает в сухом виде до 90 лет. Можно поэтому для вы-
ращивания колоний брать землю с корней растений, пролежавших много лет в гер-
бариях, и таким образом получить картину изменчивости этой бактерии из самых раз-
нообразных мест по гербарным сборам растений.
О. К. Настюкова (1948) изучила популяции этой бактерии из 58 пунктов разных
районов Союза. В табл. 1 приведены результаты этой работы.
• Таблица 1*)
Группа пунктов Район сбора почв Число пунктов сбора Число пунктов, почвы из которых дали рост бактерий % пунктов с рацемическими популяциями
1 Европейская часть Союза, Северный Кавказ, Запад- ная и Восточная Сибирь, Якутия 25 18 4,6
2 Закавказье 17 16 87,4
3 Приморский край 8 8 50,0
4 Тянь-Шань и Фергана . . . 8 6 83,5
*) Таблица составлена по данным О. К. Настюковой (1948).
В итоге этого обследования О. К. Настюкова делает вывод, что за исключением
случайных находок правых колоний (Игарка по Мишустину и окрестности Москвы
по Настюковой) вся территория Советского Союза занята популяциями левозавитных
и неопределенных колоний В. mycoides. Все Закавказье, часть Средней Азии и Уссу-
рийский край выделяются присутствием там в большем числе правых колоний.
Г. Ф. Гаузе (1949) подтвердил вывод О. К. Настюковой в отношении Европейской
части Союза и Закавказья, добавив к ним данные по тропикам, а именно, по Бра-
зилии, где из 15 проб земли в 8 пробах были обнаружены правые формы. О. К. На-
стюкова объясняет столь своеобразное распределение правых вариантов колоний тем.
- 25 —
что районы с правыми колониями представляют собой места, которые биогеографы
считают убежищами, где сохранилась в почти нетронутом виде третичная флора и
фауна Европы и Сибири, уничтоженная в более северных районах Евразии великим
оледенением. Нахождение правых форм в тропиках ЮЯсной Америки делает толко-
вание О. К. Настюковой еще более вероятным. Добавлю, что на Балканском полу-
острове, который также считается биогеографическим убежищем (средиземноморское
убежище), по моим данным, также встречаются правые колонии.
Обращаясь к высшим растениям, надо указать на явление спирального листо-
расположения. У очень большого числа растений при соединении линией оснований
листьев на вертикальных стеблях получается винтообразная линия. Она может быть
левой или правой. Л. А. Смирнов (1950), обследовав эти спирали у большого числа
тропических растений, нашел «факт широкого и, по-видимому, одинаково широкого
проявления и правых и левых изомеров в листорасположении (многих) тропических
растений» (стр. 397). К сожалению, Л. А. Смирнов не приводит точных цифровых
показателей морфологической рацемичности тропической флоры по сравнению с фло-
рой умеренных широт. С этим выводом Л. А. Смирнова стоит сопоставить отмеченную
мною выше рацемичность и декстральность трех тропических семейств сосудистых
растений по сравнению с другими семействами цветковых растений нашей флоры.
В итоге намечается вывод о том, что древние флоры нашей планеты, приурочен-
ные к тропическому поясу, характеризуются меньшей диссимметрией — рацемичностью
по сравнению с флорой Палеарктики, флорой более молодой.
Корпускулярная основа явлений левизны — правизны
в органическом мире
До сих пор мы придерживались очень строго фактического материала, допуская
лишь время от времени эмпирические обобщения наблюдаемых явлений. В этом же
разделе мы сделаем попытку перебросить воздушный мостик гипотезы между двумя
группами фактов. Здесь стоит вспомнить то, что писал о роли гипотез в науке
Ф. Энгельс: «Если бы мы захотели ждать, пока материал будет готов в чистом виде
для закона, то это значило бы приостановить до тех пор мыслящее исследование и
уже по одному этому мы никогда не получили бы закона», и несколько раньше:
«Формой развития естествознания, поскольку оно мыслит, является гипотеза» ’.
Приведем пример превращения гипотезы в теорию, взяв его из области учения
о кристаллах. Гениальная гипотеза кристаллического сложения М. В. Ломоно-
сова (1749) геометрически закономерного расположения «корпускул» в пространстве
в итоге применения рентгеновских лучей, нащупавших расположение в твердых те-
лах мельчайших элементарных частиц, стала в наше время (с 1912 г.) теорией.
В применении к телам мертвой природы нет сомнений в том, что «само собой
разумеющейся задачей является установление связи между макро- и микроморфоло-
гией» [Ниггли (Р. Niggli, 1950), стр. 131]. Можно подумать, что эта задача в своеоб-
разном виде может стоять и в учении о форме растений и животных.
Остановимся сперва на описанных выше отличиях в характере встречаемости
левизны-правизны кристаллов и живых организмов. Можно подумать, что эти отли-
чия весьма вероятно стоят в связи, во-первых, с весьма существенным отличием в мо-
лекулярном сложении материи косной и материи живой и, во-вторых, с отличиями в
возникновении тел мертвых и тел живых.
Рассмотрим сперва первое отличие. Правизна и левизна кристаллов кварца за-
висят от диссимметрии расположения атомов в кристаллической решетке. Как известно
по В. Браггу, в кристалле кварца атомы кремния и кислорода расположены по правой
и левой спиралям, что определяет правизну и левизну внешней формы кристалла. Опти-
ческая активность кристаллов неорганических веществ не связана с правизной и ле-
визной слагающих их молекул, так как в растворе или расплаве оптическая актив-
ность исчезает. Одинаковая вероятность расположения атомов кремния и кислорода’
либо по левой, либо по правой спиралям, определяет примерно одинаковую встречае-
мость левых и правых кристаллов кварца в месторождениях и колоколообразную,
(биноминальную) форму распределения правизны месторождений. Эта колоколооб-
разная форма распределения возникает следующим образом. В подавляющем боль-
шинстве случаев отклонения месторождений кристаллов кварца от 50-процентной
встречаемости правых и левых кристаллов объясняется тем, что в руки исследова-
телей в порядке случайной выборки попадают правые и левые кристаллы в не абсо-
лютно одинаковом числе. Лишь для одного месторождения в Плакосе (Греция)
А. Б. Вистелиус (1950) статистически доказал «совершенно несомненное преобладание
левых кварцев». Господствуют же на Земле рецемические месторождения кварца.
Иная картина наблюдается при изучении веществ органических. Здесь диссиммет-
рия молекул зависит от присутствия диссимметричного углеродного атома, связанного
с четырьмя различными атомами или группами других атомов.
1 Ф. Энгельс. Диалектика природы. Госполитиздат, 1938, стр. 193.
- 26 -
Встречаемость левых и правых молекул в органических веществах разного про-
исхождения неодинакова:
1. В органических веществах, синтезированных в лаборатории из неорганических
веществ, левые и правые диссимметрические молекулы встречаются в равном числе.
Растворы таких веществ оптически не активны. Кристаллы, выпадающие из рацемат-
ных растворов, представлены, примерно, равным числом левых и правых кристаллов,
а потому, если составить кривую встречаемости правых кристаллов в пробах из ра-
цематов, то кривая получается колоколообразной.
2. Органические же вещества, выделенные из живых организмов (аминокислоты,
жиры, углеводы и т. п.), за редкими исключениями оптически активны (Пастер), что
указывает на то, что они состоят либо лишь из левых, либо лишь из правых молекул.
В частности, известно, что выделяемые из белков аминокислоты за редчайшими
исключениями принадлежат к левому стерическому (пространственному) ряду.
Существование левой и правой форм бактериальных колоний левых и правых
вариантов в пределах отдельных видов моллюсков, а также левых и правых видов
моллюсков в пределах отдельных родов, преобладание видов растений с левыми вин-
товыми утолщениями стенок сосудов позволяет высказать предположение о суще-
ствовании левой (обычной) и правой (редкой) протоплазмы, состоящей из соответ-
ственно инверсных химических веществ.
Перейдем теперь к рассмотрению второго существенного отличия косного и жи-
вого вещества, отличия также имеющего значение для понимания найденной нами
разницы в распределении встречаемости левизны, и правизны. Если бы вещества, из
которых состоят живые организмы, создавались каждый раз вне зависимости от дру-
гих уже существующих организмов, подобно тому как в колбах химиков-синтетиков
возникают органические препараты, то эти вещества были бы, вероятно, рацемически-
ми, и левых и правых организмов не существовало бы вообще.
Тот факт, что «кирпичики», слагающие протоплазму, представлены либо левыми,
либо правыми диссимметричными молекулами, в итоге чего протоплазма оптически
активна, указывает на то, что оптическая активность обладает какими-то жизненными
преимуществами по сравнению с рацемическим состоянием. Г. Ф. Гаузе (1940), обсу-
див теоретические исследования химиков по кинетике экзиматических реакций в дис-
симметрическом и рецематном состояниях, приходит к выводу о том, что «переход от
рацематов к оптически активным веществам должен значительно повысить скорость
всех стереоспецифичных реакций, протекающих в живых системах».
Весьма возможно, что именно этим объясняется показанная мною выше ред-
кость рацемических популяций по сравнению с популяциями, состоящими либо из
левых, либо из правых особей с небольшой примесью противоположных форм. Чаше-
образное и получашеобразное распределение встречаемости энантиоморфных живых
тел выражает особое качество живой материи, в которой господствует преемственная
связь поколений с поколением и закон выживания особей, наиболее приспособленных
к условиям существования. Колоколообразное же распределение характеризует встре-
чаемость левых и правых тел косной природы, возникающих из маточного раствора
вне преемственной связи с телами своего вида.
Высказанные выше соображения могут быть толчком к постановке эксперимен-
тов, в которых должны принять участие как биологи, так и химики, кристаллогра-
фы и физики.
SUMMARY
After giving a description of dissymmetric phenomena in biology the author introdu-
„ , * i л p i number of dextral bodies
ces an index of dissymetry calculated as follows: i. d. = ‘ 100•
An attempt is made to investigate the occurence of populations characterized by different
degree of dextrality. Fig. 1 represents curves of dextrality distribution of different bodies:
curve I belongs to the distribution of the dextrality of quarz crystal samples from diffe-
rent localities; curve 2 represents the dexstrality distribution of populations of colonies of
Bacillus mycoides and the curves 3 and 4 represents the dextrality distribution of popula-
tion of Achatinella mustelina and Partula suturalis, terrestrial molluscs from pacific
islands. While the samples of crystals give a binomial type of the frequency curve, the
curves of living organisms have either a U-type ot L-type of frequency curves of the
dextrality of populations. The direction of coiling of filaments in the walls of vessels of
204 species vascular plants have been investigated. The majority of them (93,2%) has left
spiral filaments, 1,5% have right spirals, while the remaining species (5,3%) have left and
right spirals simultanously or in other word can be consideretate as racemic. The dextrality
of vascular plants is therefore very low and equal to 1,55%.
Populations of organisms with different degrees of dextrality are not proportionately
distributed over the globe. Table I taken from a paper of Nastjukova (1948) shows that the
— 27 —
Harger part of the territory of USSR is inhabited by populations of B. mycoides with a very
low (4,6%) index of dextrality. Transcancasia, a part of Middle Asia and the Ussuri district
of the Far East have populations with a high percentage of dextral forms of B. mycoides
and are therefore characterized by a rather high index of dextrality (50,0—87,4). According
to Nastjukova the high dextrality of population is observed in refugii — regions of survi-
vorship of plants and animals during the glacial period.
Returning to the dextrality of vascular plants in regard to the coiling of spiral vess-
els it must be pointed out that a high dextrality in other words the presence of „left" as
well of „right" species characterizes families of plants of tropical origin (i. e. the Begonia-
ceae, Balsaminaceae and Orchidaceae). In conclusion to his paper the author discusses two
features of living organisms distinquishing them from the inorganic world as a possible
explanation of the dextrality distribution curves of populations of living organisms:
1. In living organisms the optical activity of their protoplasm and the prevalence of
left or right molecules gives evidentely certain advantages in comparison to the race-
’mic state.
2. The living bodies or organisms originate from thoset of the preceeding generation,
while the inorganic bodies — the crystals are formed from the uncristallised matter without
^connection to each other.
ЛИТЕРАТУРА
Алпатов В. В. 1951. О левизне и правизне спиралей в сосудах растений. Приро-
да, № 11.
Алпатов В. В. 1952. О встречаемости левых и правых винтообразных утолще-
-ний в сосудах растений. ДАН, т. 84, № 5.
Алпатов В. В. 1953. О встречаемости левых и правых тел в неживой и живой
лтрироде. Бюлл. МОИП, отд. биол., т. 58, вып. 5.
Болдырев А. К. 1932. Кристаллография. М.
Бутлеров А. М. 1951. Избранные работы по органической химии. М.
Вернадский В. И. 1903. Основы кристаллографии. Вып. 1, М.
Вернадский В. И. 1940а. Биохимические очерки. М. — Л.
Вернадский В. И. 19406. Проблемы биогеохимии. IV. О правизне и ле-
визне. М. — Л.
Вистелиус А. Б. 1950. О распространении энантиоморфных типов кварца. Зап.
Минер, о-ва, т. 79, вып. 3.
Дюкло Н. С. 1929. Химия живого вещества. М. — Л.
Гаузе Г. Ф. 1940. Асимметрия протоплазмы. М. — Л.
Гаузе Г. Ф. 1949. О правизне — левизне у Bacillus mycoides. 1. Географическое
распространение и некоторые физиологические свойства правозавитых и левозавитых
<форм. Микробиология, т. 18, № 2.
Лейммлейн Г. Г. 1939. О числе левых и правых кристаллов в каком-либо
месторождении. Тр. биогеохим. лаборатории, т. 5.
Лейммлейн Г. Г. 1944. Относительное число правых и левых кристаллов. Зап.
Минер, о-ва, т. 73, вып. 2—3.
НастюковаО. К. 1948. О закономерностях географического распространения
лево- и правозавитых колоний Bacillus mycoides Fltigge на территории СССР. ДАН,
т. 59, вып. 9.
Пастер Л. 1891. Винная кислота и ее значение для учения о строении материи
(об асимметрии органических соединений). СПб.
Смирнов Л. А. 1950. Спираль листорасположения и проблема диссиметрии.
Бот. журн., № 4.
Холодковский Н. А. 1923. Марчелло Мальпиги. Берлин.
Цветков Б. Н. 1938. Правые и левые формы моллюска Fruticicala lantzi Lndh.
ti их географическое распространение. Бюлл. МОИП, отд. биол., т. 47, вып. 5—6.
Цветков Б. Н. 1941. Изменчивость Fruticicola lantzi Lndh. (Mollusca, Pulmo-
nata). Сб. тр. Гос. зоомузея МГУ, вып. 6.
Crampton Н. Е. 1932. Studies on the variation, distribution and evolution of the
g. Partula. Garnegie Inst. Publ. May.
Gause G. F. 1941. Optical activity and living matter. Normandy.
Ludwig W. 1932. Rechts — Links Problem im Tierreich und beim Menschen.
Berlin.
Niggli P. 1950. Probleme der Naturwissenschaft. Basel.
Pasteur L. 1861. Recherches sur la dissymetrie moleculaire des produits organi-
ques naturelles. Paris.
Welch D. a. A 11 e A. 1938. Distribution and variation of Achatinella mustelina
Michals in the Waianae mountains Oahu. Bernice P. Bishop Museum, № 152.
БЮЛЛЕТЕНЬ М. О-ВА ИСП. ПРИРОДЫ, ОТД. БИОЛОГИИ, Т. LXII (5), 1957
Зоогеографический очерк Камчатки
Ю. В. Аверин
Zoogeographic survey of Kamchatka
У. V. Averin
I
Настоящая статья представляет собой главный итог зоогеографического исследова-
ния Камчатки на основе изучения автором ее орнитофауны. Для выполнения этой работы
были использованы богатые коллекции птиц с полуострова, хранящиеся в Зоологиче-
ском институте АН СССР и в Зоологическом музее Московского университета, а также
небольшие сборы, находящиеся в Камчатском областном музее краеведения, литератур-
ные источники, ценнейшие рукописи И. Г. Вознесенского и Г. Стеллера, хранящиеся
в Архиве АН СССР, и, наконец, результаты личных шестилетних (1940—1946 гг.) иссле-
дований восточной части полуострова. Помимо орнитофауны Камчатки, которая находи-
лась в центре нашего внимания, попутно рассмотрена фауна птиц северных и средних
Курильских и Командорских островов, а также Корякской Земли и западных островов
Алеутской цепи.
Из общего количества до сих пор приводимых для Камчатки видов исключены
не подкрепленные находками: коршун (Milvus korschun lineatus Gray), желна (Dryoco-
pus martins L.), белоспинный дятел (Dryobates leucotos wosnessenskii But) и крапивник
(Troglodytes troglodytes pallescens Ridgw.). В состав фауны полуострова внесены добы-
тые белая цапля (Egretta alba modesta Gray) и дербник (Aesalon columbarius pad fi-
cus Steg); восстановлены по литературным первоисточникам, сравнительно недавно опуб-
ликованным, пропускаемые в сводках по орнитофауне СССР: кулик черныш (Tringa
ochropus L.), американская свиязь (Anas americana Gm.), гага-гребенушка (Somateria
spectabilis L.) и сибирская завирушка (Prunella montanella badia Port.).
В настоящее время для орнитофауны Камчатки известно 197 видов птиц. Из них
гнездящихся 127 (в том числе 32 оседлых), или 64%, гнездование 9 весьма вероятно,
но еще не доказано, зимующих — 11, пролетных — 32 и залетных — 27. Все они принад-
лежат к 14 отрядам: гагарообразных — 6 видов, трубконосых — 7, веслоногих — 2, аисто-
образных— 1, пластинчатоклювых — 32, дневных хищных птиц — 14, куриных — 3,
журавлеобразных — 2, ржанкообразных — 63 (в том числе куликов — 34, чайковых — 15,
чистиковых — 14), кукушкообразных — 2, сов — 4, дятлов — 3, стрижеобразных — 1 и во-
робьиных — 57 видов.
Явное несоответствие между чрезвычайно разнообразной экологической обстановкой
на полуострове и количеством населяющих его видов видно из того, что авиафауна Кам-
чатки богато представлена лишь видами, привязанными в своем распространении к воде и
относящимися к отрядам пластинчатоклювых, ржанкообразных и некоторым другим. Почти
все остальные группы птиц бедны видами, если сравнивать их соответственно с такими же
группами в близко расположенной Восточной Сибири. Особенно мало видов из семейств
Aquilidae, Falconidae, Tetraonidae, Strigidae, Picidae, Corvidae, Paridae, Turdidae и Frin-
gillidae.
В общем из всего количества живущих на Камчатке форм тяготеют к водоемам
приблизительно 56% (из них 24%— к морским побережьям, 32%—к внутренним во-
доемам), тесно связано с древесно-кустарниковой растительностью — 30%, прочих назем-
ных видов птиц — 14%.
Бедность видового состава (главным образом, лесных птиц) орнитофауны Камчат-
ки, отмеченная еще прежними исследователями, не препятствует, однако, причислению
ее к таежной фауне, наиболее характерной для Восточной Сибири и тяготеющей к по-
бережьям Охотского моря. Так, в таежной фауне птиц Камчатки нет ни одного вида,
— 30 —
который бы не встречался на материке в зоне темнохвойных лесов. Вместе с таежным
комплексом орнитофауны, являющимся для полуострова основным, имеется небольшое
число нетаежных видов, обитающих в -пойменных лесах, морских и других птиц.
Птицы, гнездящиеся на Камчатке, принадлежат к четырем типам фауны (в пони-
мании Б. К. Штегмана, 1938, с небольшими нашими изменениями): сибирскому, тибет-
скому, китайскому и арктической области, причем преобладают виды типов сибирского-
и арктической области.
Сибирский тип орнитофауны, состоящий преимущественно из форм, живущих в
тайге, на Камчатке представлен следующими видами: Anas falcata, Bucephala clangula
Oidemia deglandi Aegolius funereus, Sarnia alula, Picoides tridactylus, Nucifraga caryo-
catactes, Sitta europaea albifrons, Calliope calliope, Tarsiger cyanurus, Pseudaedon sibi-
lans, Tardus naumanni, T.pallidas, Phylloscopus, borealis, Locustella ochotensis, L. lan-
ceolaia, Muscicapa sibirica, Siphia parva, Bombidylla garrulus, Carduelis flammea,
Fringilla montifringilla, Pyrrhula pyrrhula, Pinicola enucleator, Loxia curvirostra,
Emberiza rustica и др. Их количество составляет примерно 35% гнездящихся на полу-
острове форм. Большинство перечисленных видов очень близки к дендрофильным оби-
тателям тайги Восточной Сибири. Птицы пресных .водоемов, находящихся среди хвой-
ного или смешанного лесов, составляют примерно четвертую часть данного списка.
В тех районах Камчатки, где преобладают лиственные леса из каменной березы (Betula
Ermani), обитают те же таежные виды птиц, что и в восточно-сибирской тайге; вместе
с другими таежными элементами ландшафта они придают большое своеобразие этим
участкам местности.
К тибетскому типу орнитофауны принадлежат Charadrias mongolus,, Calpella
solitaria, Anthus spinoletta и представитель рода Leucosticte — L. arctoa, близкий к
тибетскому виду L. brandti. Все эти птицы гнездятся в альпийском поясе, а сибирский
вьюрок селится также на скалистом побережье восточной Камчатки.
Китайский тип фауны экологически связан с широколиственными и отчасти сме-
шанными лесами Восточной Азии. На Камчатке живут следующие виды: Apus pacifi-
cus, Anthus hodgsoni, Lanius cristatus cristatus, Chloris sinica. К этой группе следует
присоединить и японскую овсянку (Emberiza variabilis), которая, впрочем, в Китае не
распространена. Из этих пяти форм четыре встречаются на полуострове в области
сплошного распространения лесов из каменной березы, а белопоясный стриж гнездится*
вероятно, по всей Камчатке. Где-то недалеко от полуострова находится северо-восточ-
ный предел его распространения. Между прочим, этот вид гнездится на скалах побе-
режья Гижигинской губы.
Орнитофауна арктической области полуострова представлена следующими видами:
Somateria mollissima, Falco gyrfalco, Lagopus lagopus, L. mutus, Phalaropus loba-
tus, Stercorarius parasiticus, S. longicaudus, Uria lomvia, U. aalge, Fratercula cor-,
miculata, Anthus cervina, Calcarius lapponicus, Plectrophenax nivalis и др. Птицы
арктической области составляют около 20% всех гнездящихся видов орнитофауны по-
луострова. Они встречаются преимущественно в открытых ландшафтах. Видов, широко,
распространенных в тайге Северной Америки и общих только с Камчаткой (но не с
угими областями Восточной Сибири), нет.
Отсутствие на полуострове многих типичных таежных видов фауны Восточной
Сибири придает его животному миру облик, свойственный островным фаунам. Некоторые
характерные черты лесной орнитофауны Камчатки дали основание Стейнегеру (L. Stej-
neger, 1885) и позже В. Л. Бианки (1909) считать, что она имеет черты островной
фауны. И действительно, из лесных птиц на Камчатке не встречаются желна (Dryoco-
pus martins), кукша (Craetes infaustus), рябчик (Tetrastes bonasia) и другие; из.
млекопитающих — лось (Alces alces), бурундук (Eutamias asiaticus), летяга (Pteromys
volans); рептилий на полуострове вовсе нет, а из амфибий живет только один вид —
сибирский тритон (Hynobius keyserlingii), в то время как на материке Восточной Си-
бири на широте Камчатки обитает несколько видов тех и других. Из настоящих прес-
новодных рыб на полуострове встречается только один хариус (Thymallus arcticus).
Об островных чертах в таежном комплексе орнитофауны Камчатки свидетельст-
вует также присутствие довольно большого числа эндемичных форм из различных
классов животных. Среди птиц известны следующие подвиды: Accipiter nisus pall ens,
Tetrao parvirostris kamtschaticus,Dryobates major kamtschaticus, Picoides tridactylus
albidior, Paras montanus kamtschatkensis и Sitta europaea albifrons. Характерно,
что орнитофауне полуострова свойственны только эндемичные подвиды, эндемичные же
виды остутствуют.
Названные эндемичные подвиды обладают характернейшими признаками — очень
светлой окраской, обычно самой светлой у живущих в Палеарктике подвидов, и часто
крупными размерами.
Имеются также такие формы, ареалы которых в Северо-Восточной Азии весьма
узки, но в них обязательно входит Камчатский п-ов. О камчатском происхождении этих
расселившихся форм говорят отличительные признаки камчатских эндемиков: крупные
размеры и бледная окраска. Так, на Камчатке, в Корякской Земле и в части Анадыр-
- 31 —
ского края обитают сорока (Pica pica kamtschatica) и малый пестрый дятел (Dryobates
minor immaculatus)\ на северных и средних островах Курильской гряды, в Корякской
Земле и в Анадырском крае — кедровка (Nucifraga caryocatactes kamtschatkensi)\ на
северных и средних островах Курильской гряды, на Камчатке, в Корякской Земле,
Анадырском крае и на западном побережье Охотского моря — щур Pinicola enucleator
kamts chat kens is), на Камчатке, в Корякской Земле, Анадырском крае и от него на
запад до среднего течения Колымы — мохноногий сыч (Aegolius funereus magnus); на
северных островах Курильской гряды и на Камчатке — пеночка-таловка (Phylloscopus
borealis xanthodryas), соловей-красношейка (Calliope calliope kamtschatkensis), сне-
гирь (Pyrrhula pirrhula cassini).
ТакЯИГЪбразом, своеобразие комплекса таежной орнитофауны Камчатки выражает-
ся: а) в обедненности видового состава по сравнению с таежной фауной соседних тер-
риторий материка Восточной Сибири; б) в наличии значительного числа эндемичных
подвидов; в) в очень светлой окраске и крупных размерах, присущих камчатским
эндемикам, а также многим видам, распространенным не только на Камчатке, но и бо-
лее широко — в Северо-Восточной Азии; г) в приспособленности эндемиков и некоторых
других таежных видов к жизни исключительно в условиях березовой лиственной „тай-
ги" (имеются в виду формы вроде глухаря, трехпалого дятла, для которых за преде-
лами полуострова обитание в темнохвойной тайге характерно)1; д) в распространении
этого комплекса на территории, имеющей четкие географические границы: весь Кам-
чатский полуостров на север до южной части Парапольского дола и северные средние
острова Курильской гряды.
Поэтому мы считаем, что главному комплексу орнитофауны полуострова — таеж-
ному, имеющему черты, свойственные островным фаунам, можеть быть дано название
камчатского. Г. П. Дементьев (1940), анализируя орнитофауну Корякской Земли и
Камчатки, пришел к выводу, что островные черты своей лесной фауны полуостров имеет
не из-за присутствуя к северу от него преграды в виде тундры Парапольского дола, а
благодаря особенностям ландшафтов рассматриваемых территорий. Он пишет: „В общем
различие между фауной птиц Корякской Земли и Камчатки довольно точно совпадает
с различиями и сменой соответствующих ландшафтов".
Соглашаясь в основном с указанным мнением, мы считаем необходимым уточнить
вопрос о значении Парапольского дола как физической преграды. Этот дол представ-
ляет собой обширную депрессию, простирающуюся от основания полуострова в глубь
материка в северо-северо-восточном направлении примерно на 350 км, и приближается
там к Анадырской низменности. Северная часть Парапольской низменности (около
300 км) проходит через Корякскую Землю между Пенжинским и Корякским хребтами,
а южная вклинивается приблизительно на 50 км в глубь Камчатского перешейка, рас-
полагаясь здесь между хребтами Маметчикинским и северо-западными отрогами Сре-
динного хребта. В северной части камчатского отрезка Парапольского дола одно его
ответвление отходит к берегу Пенжинского залива Охотского моря вблизи бухты Ре-
киники, второе — к заливу Анапка Берингова моря.
Парапольский дол в своей южной части имеет вид плоской, почти идеально ров-
ной сильно заболоченной низменности, прорезанной широкими долинами рек. На этой
поверхности разбросано множество озер. Средние высоты равны 25—30 м. Древесная
растительность отсутствует даже в защищенных от ветра долинах, плоские и низкие
междуречья покрыты обширными сфагновыми бугристыми болотами и кустарниково-
лишайниковой тундрой. Повсеместно на водоразделах развита вечная мерзлота,
д Таким образом, южный отрезок Парапольского дола, так называемый Рекиникский
ол, вместе со Срединным Камчатским хребтом, ограничивающим его с востока и ухо-
дящим своей осью к южной Камчатке, а своими отрогами заполняющим всю восточ-
ную часть Камчатского перешейка вплоть до Карагинского залива Берингова моря, как
°ы закрывает доступ лесным и равнинным формам на Камчатку через Корякскую Зем-
лю. Поэтому Рекиникский дол и северую часть Срединного хребта с отрогами нужно
признать реальной физической преградой.
Заселение Камчатского полуострова с севера через переешеек с преодолением
указанной преграды, в виде исключения, возможно для немногих форм. Пока известна
только одна белка (Sciurus vulgaris jacutensis), которая в 1920 г. проникла на Кам-
чатку и к настоящему времени уже расселилась по всему полуострову вплоть до край-
него юга.
Образование и обособление местных форм, ранга не более высокого чем подвил,
свойственно не только птицам, но и другим группам животных Камчатки. Из млеко-
питающих соболю (Martes zibellina kamtschadalica), снежному барану (Ovis nivicola
nivicola), длинохвостому суслику (Citellus undulatus stejnegeri), черношапочному cyp-
1 На Камчатке елово-лиственичная тайга сохранилась только в Камчатской депрессии
в виде маленького по площади „хвойного острова", населенного некоторыми реликтовы-
ми видами фауны и флоры.
— 32 —
ку (Marmota camtschatica camtschatica) и красно-серой полевке (Cletryonomys rufc*
canus vosnessenskii).
Камчатская флора тоже отличается присутствием в ее составе высокого процента
эндемичных форм, что свидетельствует об интенсивном формообразовании. Е. Л. Люби-
мова (личное сообщение) подсчитала, что из 1032 форм высших растений, найденных на
полуострове, 286, или 28%, Для него эндемичны. Особенно велик эндемиз среди злако-
вых— 50%, ивовых — 48%, бобовых — 35%, сложноцветных — 40% и т. д. Даже среди
немногочисленных древесных пород полуострова имеется ряд эндемичных форм: авачин-
ская береза, камчатская ель, камчатская пихта, камчатский ольховый стланец и некото-
рые другие.
Анализ состава фауны птиц Камчатки и окружающих ее территорий (Корякской
Земли, Курильских и Командорских о-вов и островов западных групп Алеутской гряды)
показал, что некоторые подвиды, свойственные камчатской таежной орнитофауне, входят
как составная часть в наземную фауну птиц северных и, в меньшей степени, средних
островов Курильской гряды; с ними она имеет наиболее близкие систематические и
фаунистические связи, а также связана частично и общей историей формирования.
Можно сказать, что часть комплекса фауны наземных птиц Курильских о-вов
в большей степени дериват наземной орнитофауны Камчатки, как последняя в свою
очередь дериват таежной фауны Восточной Сибири.
Орнитогеографическое районирование Камчатки, Курильских и Командорских о-вов
позволяет судить не только о взаимосвязи и субординации между их фаунами, но и о
месте их среди зоогеографических категорий Палеарктики. В основу нашей работы бы-
ло положено только современное распространение птиц. Что касается применения и
названий крупных подразделений Палеарктической области (подобластей, провинций и
большинства подпровинций), то в этом мы следуем работам М. А. Мензбира (1914 и др.)
с некоторыми изменениями, внесенными П. П. Сушкиным (1925 и др.). Выделение же
дробных орнитогеографических категорий — округов и участков, а также Камчатской
подпровинции с подразделениями — результат нашего исследования. Для Камчатки де-
тальное зоогеографическое районирование дается впервые.
О зоогеографии Курильских о-вов общие соображения высказал Ямасина (Jamasina
1931). По фауне птиц Командорских о-вов нами использована хорошая сводка
Г. X. Иогансена (1934). Общая картина орнитогеографии Северо-Восточной Азии дана
в работе Л. А. Портенко (1937).
Мы предлагаем следующее орнитогеографическое районирование указанных выше
территорий (см. таблицу).
Т а б и а
Палеарктическая область
Подобласти Провинции Подпровин- ции Округа Участки
Европейско- Сибирская Арктиче- ская, или Т ундры и Лесотундры Берингийская Камчатско-Коман- дорский прибрежный Камчатский восточно- бережный Командорский
Восточно- Сибирская (Парапольский)1 Камчатско-Командор- ский равнинный (Чукотско-Камчат- ский альпийский)1 Рекиникский Камчатский западно- приморский Долинно-Камчатский Берингийский Камчатский альпийский
Северная Камчатская Камчатский лесной Северо-Курильский Таежный Центральный
Маньчжур- ская Китайско- Японская Японская (Южно-Курильский и др)1
9 Большие территории округа находятся за пределами Камчатки.
- 33 —
В этой схеме главное место принадлежит выделенной нами Камчатской подпровин-
ции, своеобразные черты орнитофауны которой отмечены выше.
Камчатская под провинция
Камчатский лесной округ
Самый большой из округов, равный примерно половине площади полуострова,
занимает всю его лесную территорию, располагаясь по средним и нижним склонам гор
с юга на север от основания мыса Лопагки почти до соединения перешейка с матери-
ком. С запада на восток этот округ занимает территорию от предгорных лесов, лежащих
в бассейне камчатских рек, впадающих в Охотское море, почти до берега Тихого океана.
Он имеет сложную конфигурацию и граничит со всеми остальными орнитогеографиче-
скими округами и участками полуострова.
Округ населен характерным для Камчатки комплексом видов таежной орнито-
фауны и обитателями широколиственных лесов. Преобладают дендрофильные виды.
Обыкновенны эндемики, только случайно выходящие за границы округа: Accipiter nisus
pal lens, Tetrao parvirostris kamtshsticus, Dry abates major kamtschaticas, Picoides
tridactylus albidior, Paras montanus kamtschatkensis и Sitta europaea albifrons.
Характерны следующие виды, редко встречающиеся за пределами леса: Caculus
optatas, Aegolius fanereus, Sarnia al ala, Dryobates minor, Tarsiger cyanurus, Tardus
naumanni, T. pallid is obsciras, Phylloscopus borealis, Anthus hodgsoni, Muscicapa
sibirica, M. griseisticta, Siphia parva, Cardaelis flammea, Pyrrhula pyrrhula,
Pinicola enucleator, Fringilla montifringilia, Loxia curvirostra, Emberiza vaiiabilis.
Главная ландшафтная древесная порода — каменная береза, которая почти полностью
отсутствует в северной части Камчатского перешейка, замещаясь здесь зарослями кедро-
вого стланика (Pinas pumila), а в южной части Камчатки (южнее 52° с. ш.) ольхового
стланика (Ainas fruticosa).
В средней части округа на дне нижних склонов Камчатской депрессии находится
знаменитый „хвойный остров". Это насаждение — реликт третичной и доледниковой
флоры полуострова, образованный аянской елью (Picea ajanensis) и даурской листвен-
ницей (Larix daurica).
В камчатском лесном округе естественно выделяются два участка: Таежный,
занимающий не менее 95% площади всех лесных угодий полуострова, для которого
вполне применима орнитологическая характеристика, данная всему округу, и Цент-
ральный, на который приходятся остальные 5% лесной территории Камчатки (см. карту).
Центральный участок находится в долинах рек Камчатки и ее левого притока Еловки,
от селения Мильково до деревни Еловки. Он занимает „хвойный остров* вместе с высо-
коствольными пойменными насаждениями, произрастающими вдоль многочисленных рек,
пересекающих участок. Его длина равна примерно 300 км, а ширина — 30—90 км.
Только здесь наряду со многими перечисленными для Таежного участка видами встре-
чаются Aegithalos caadatus, Periparas ater, Bombycilla garrulas и Coccothraastes coc-
cothraustes.
Северо-Курильский округ
В этот округ входят острова северной и средней части Курильской гряды,
от о-ва Шумшу на севере до о-ва Симушир на юге.
Здесь расположены десятки гнездовых колоний трубконосых, веслоногих, чаек
и чистиковых. Большинство из составляющих их видов обычны на всех скалистых бере-
гах восточной Камчатки и Берингова моря. Однако здесь, южнее его, на рубеже Охот-
ского моря и Тихого океана, особой „южный" колорит этим колониям придает гнездова-
ние Laras crassirostris и Cepphas snowi.
Главная особенность фауны птиц Курильского округа заключается в присутствии,
главным образом в северной его части, значительного числа видов таежной камчатской
орнитофауны. Поэтому он принадлежит Камчатской подпровинции. Здесь гнездятся формы
чаще всего тождественные с живущими на Камчатке: Nucifraga caryocatactes kamts-
chatkensis, Calliope calliope kamtschatkensis, Tardus pallidas orii, Phylloscopus
borealis xanthodrias, Locustella ochotensis, L. lanceolata, Muscicapa sibirica, Siphia
parva, Cardaelis flammea flammea, Pyrrhula pyrrhula cassini, Pinicola enucleator
kamtschatkensis и др. Из птиц лиственных камчатских лесов гнездятся Chloris sinica
и Emberiza variabilis.
Кроме отмеченных выше особенностей в видовом составе наземных и морских
птиц северных и средних островов Курильской гряды, необходимо подчеркнуть, что на
ней живут оседло Erolia maritima kurilensis и Troglodytes troglodytes kurilensis.
Полуостров Камчатка. Масштаб 1 : 5 000 000
Схема орнитологических участков:
1—Таежный; 2 — Центральный; 3 — Камчатский альпийский; 4 —Долинно-
Камчатский; 5 — Камчатский западноприморский; 6~— Рекиникский; 7 — Кам-
чатский восточнобережный
- 35 -
Восточно-Сибирская подпровпнци т
Рекинский участок. Этот участок Парапольского округа занимает самую
северную часть Камчатского перешейка, подходя к Пенжинскому заливу Охотского
моря. Северная граница участка находится на прямой, проведенной от бухты Рекиники
до залива Анапка, южная и другие — неясны.
Орнитофауна Рекиникского участка известна плохо, поэтому мы заселяем его веро-
ятно живущими здесь видами, учитывая, что орнитофауны южных частей Камчатки,
а также Корякской Земли и Анадырского края, расположенных к югу и к северу
от него, изучены лучше и могут служить поэтому исходными. Мы основывались на дан-
ных Л. А. Портенко (1939) о фауне Анадырского края, Г. П. Дементьева (1940) —
о Корякской Земле и наших — о Камчатке [Тачановский (L. Taczanowski, 1893), Бергманн
(В. Bergman, 1935), Аверин, 1948 и др.].
Ландшафт Рекиникского участка известен как равнинный, поэтому нельзя предполо-
жить, что он заселен птицами отвесных морских берегов, камчатского леса или альпий-
скими. Мы полагаем, что на этом участке живет комплекс видов, свойственный заболо-
ченной тундре и лесотундре, и видов, обитающих в мелколесье и кустарниках таежной
зоны: Colymbus stellaius, Anser fabatis serrirostris, Nyroca marila, Clangula hyema-
lis, Buteo lagopus, Lagopus lagopus, L. mat as, Erolia alpina, Phalaropus lobatus,
Stercorarius parasiticaus, S. longicaudus, Nucifraga caryocatactes, Phylloscopus
borealis xanthodryas, Motacilla alba lugens, Anthus cervina, Carduelis flammea exili-
pes, Pinicola enucleator, Calcarius lapponpcus и др.
Однако виды птиц широколиственных лесоз, обитающие на Камчатке (Anthus
hodgsoni, Chloris sinica, Emberiza variabilis и т. п.), следует полагать, отсутствуют.
Камчатско-Командорский равнинный округ
Преобладает тундрово-луговой ландшафт. Орнитофауна этого округа). слагается
из чайковых, куликов, пластинчатоклювных, немногих видов воробьиных- и. представите-
лей других отрядов.
Округ состоит из трех участков: два из них находятся на полуострове — Камчатский,
западноприморский и Долинно-Камчатский, а третий — Бсрингийский — на о-ве Беринга
Камчатский западноприморский участок расположен полосой в голь
побережья Охотского моря от р. Голыгиной на юге до р. Пананы на:севере. На востоке
граница участка проходит у лесистых предгорий Срединного хребта, а на западе — по-
берегу Охотского моря. Длина участка равна примерно 900 км, ширина — до 100 км.
Орнитофауна состоит из следующих видов: Podiceps griseigena, Anser fabalis.
serirostris, Anas platyrrhyncha, A. acuta, Nyroca marila, Limosa limosa, Numenius mada~
gas cariensis, Larus conus, L. ridibundus, Sterna hirundo и многих других. Некоторые чз-
перечисленных видов встречаются одиночно или небольшими группками в других мес-
тах Камчатки, но нигде не образуют столь полного комплекса.
Долинно-Камчатский участок находится в средней и нижней частях
долины р. Камчатки, располагаясь узкой лентой в ее пойме от низовьев притока Щапи-
ной до устья р. Камчатки и от него через основание Озерновского п-ва по системе озер
и стариц, находящихся на месте древней поймы р. Камчатки. Площадь этого участка
равна примерно 6000—1200 км3. ‘
Фауна птиц участка в основном такая же, как и в Камчатском западноприморским
участке. Кроме того, только в долине р. Камчатки известно гнездование Podiceps аиsi-
tus, Anas strepera, Nyroca ferina, Mergellus albellus и Cyanosylvia svecica.
Беринг и некий участок находится на о-ве Беринга, занимая его северную,
относительно равнинную часть. Орнитофауна этого участка имеет в своем составе виды,
часть которых распространена в средних и южных районах Камчатки, а часть харак-
терна для более северных территорий — Рекиникского участка, Корякской Земли и более
высоких широт (Colymbus arcticus viridigularis, Branta cacadensis, Anas plaiyrrhyncha,
Nyroca fuligula, N. marila, Clangula hyemalis, Charadrius mongolus, Erolia
sabminata, Stercorarius pomarinus, S. parasiticus, Nyctea scandiaca, Motacilla flava
simillima, Anthus gustavi, Calcarius lapponicus и др.).
На верхних склонах гор, лишь кое-где занятых цветковой растительностью,
на фоне кустистых и накипных лишайников живут оседлые эндемики Lagopus mutus
ridgwayi, Erolia maritima quarto и Plectrophenax nivalis townsendi.
Камчатский альпийский участок. Альпийский пояс расположен в верх-
них частях высоких горных хребтов, на горных плато („долах") и конусах вулкановл
находясь над поясом древесно-кустарниковой растительности. Высотные пределы альпий-
ского пояса лежат в среднем между 900 и 1590 м н. у. м. Этот пояс занимает вершины
всех основных горных систем полуострова: Срединного и Восточного хребтов й Вулка-
нической области, которая начинается на юге полуострова у мыса Лопатки и здкавчип
вается на его севере вулканом Шивелучем. В этой области находятся все,действующие
и большинство потухших вулканов Камчатки. ' * *......* *’
- 36 -
Видовой состав орнитофауны альпийского участка Чукотско-Камчатского аль-
пийского округа небогат, но очень характерен. Здесь гнездятся Lagopus mutus,
Charadrius mongolus, Capella solitaria, Anthus spinoletta japoncus, Leucosticte1
arctoa, past ul ata и Plectrophenax nivalis nivalis. Из них тундряная куропатка и сибирский
вьюрок живут кое-где и близ моря, на скалистых участках побережья восточной поло-
вины полуострова.
К числу обитателей сурового камчатского высокогорья относится и подорожник
(Calcarius lapponicus), не являющийся альпийским видом в других частях своего аре-
ала, а на Камчатке гнездящийся на многих приморских низменностях.
Наибольшая высота распространения горных жиеотных на Камчатке — 2300 м н. у. м.
Выше этой, границы на вершинах некоторых вулканических конусов и многих высоких
гор и хребтов находится безжизненный нивальный пояс с голыми скалами, снежными
полями и ледниками.
Бери игл йс кая подпровинция
Камчатско-Командорский прибрежный округ
Этот округ занимает восточный берег Камчатки и берега обоих командорских
о-вов. Преобладает ландшафт скалистого и обрывистого морских берегов. Населен видами
чистиковых, чаек, бакланов, трубконосых, гнездящихся преимущественно колониально
и некоторыми другими. Мы выделяем в округе два участка:
Камчатский восточнобережный участок захватывает узкую полосу
восточного берега полуострова от мыса Лопатки на юге до залива Анапки на севере,
простираясь в длину примерно на 1200 км. К этому же участку относится небольшой
отрезок берега на крайнем юго-западе Камчатки (мысы Лопатка — Камбальный)
и берег о-ва Карагинского.
Участок населен большим специфическим комплексом видов морских птиц, гнездя-
щихся на обрывистых скалистых берегах: Uria lomvia, Uria aagle, Cepphus columba
columba, Brachyramphus marmoratus, Synthlyboramphus antiquus, Fratercula cornicu-
lata, Lunda cirrhata, Aethia cristatella. A. pygmaea, большинство из которых образует
огромные гнездовые колонии в различных количественных соотношениях друг с дру-
гом и с Larus schistisagus, Rissa tridactyla и Phalacrocorax pelagicus. Также много-
численны обитатели пологих песчаных берегов и отдельные наземные формы, устраиваю-
щие свои гнезда на береговых обрывах и вершинах скал.
Гнездование на обрывистых скалистых берегах в северной половине участка, начи-
ная с мыса Кроноцкого, таких видов, как Fulmarus glacialis, Larus glaucescens, Sterna
camtschatica (гнездится на песчаной косе), Somateria mollissima и S. spectabilis,
свидетельствует о большой неоднородности орнитофауны во всем участке. Это же
подтверждает и неравномернее размещение некоторых видов чистиковых.
Из птиц не морских, но связанных с морскими побережьями, гнездятся на бере-
говых скалах Falco peregrinus и Leucosticte arctoa pustulata, а на песчаных бере-
гах — Haematopus ostralegus.
Командорский участок включает берега о-вов Беринга и Медного вместе
с небольшими надводными скалами у их берегов — о-ва Топоркова и Арьего камня.
Комплекс видов птиц, населяющих Командорский участок, в общем напоминает по своему
составу фауну птиц Камчатского восточнобережного участка, имея, однако, некоторое
количество оригинальных видов.
На скалистых берегах Командорских о-вов гнездятся живущие и на Камчатке
Fulmarus glacialis, Phalacrocorax pelagicus, Somateria mollissima, Larus glauces-
cens, Rissa tridactyla, Uria lomvia, U. aalge, Synthlyboramphus antiquus, Fratercula
corniculata, Lunda cirrhata, Aethia pygmaea. Обитают только на островах, лишь зале-
тая на восточное побережье Камчатки, Oceanodroma furcuta, О. leucorrhoa, Phalacro-
corax urile, Rissa brevirostris, Cepphus columba kaiurca и Cyclorrhynchus psittacula.
К этой же группе принадлежит недавно вымерший Палласов баклан Phalacrocorax
perspicillatus), жившие, вероятно, оседло на о-ве Беринга. Все они, за исключением
Палласова баклана, гнездятся и на Алеутских о-вах.
К сухопутным видам, по-разному использующим береговые скалы и пляжи, отно-
сятся два весьма, характерные оседлые эндемика островов—Troglodytes troglodytes
pallescens и Leucosticte arctoa maxima, и, вероятно, в некотором числе улетающие
на зиму Falco gyrfalco и F. peregrinus pealei.
Выводы
1. Орнитофауна Камчатки гетерогенна, ее составляют виды четырех типов фауны:
сибирского, тибетского, китайского и арктической области, причем первый из них
является основным по числу составляющих его видов; только он имеет черты, свой-
ственные островным фаунам.
— 37 —
2. Комплекс видов орнитофауны сибирского типа представляет собой на Камчатке
обедненный дериват таежной фауны материка Восточной Сибири. На полуострове этот
комплекс появился, вероятно, отступая перед оледенением Северо-Восточной Азии.
3. Черты островной фауны, имеющиеся у таежной орнитофауны Камчатки, заклю-
чаются главным образом в бедности ее видового состава по сравнению с таежной
фауной материковых частей Восточной Сибири и в присутствии ряда эндемичных подви-
дов, характеризующихся очень светлой окраской и обычно крупными размерами. Приз-
наки эти возникли как результат воздействия своеобразных факторов среды в ус-
ловиях длительной изоляции.
4. Таежную орнитофауну полуострова, имеющую островные черты, свойственные
островным фаунам, и рассматриваемую в качестве обедненного деривата таежной фауны
материковой области Восточной Сибири, мы называем камчатской. Некоторое коли-
чество форм камчатской орнитофауны встречается на северных и средних островах Ку-
рильской гряды. Занятая лесом площадь полуостроза вместе с частью территорий
названных островов объединяется в Камчатскую подпровинцию.
SUMMARY
The ornithological fauna of the peninsula consists of representatives of 4 types of
fauna: Siberian, Tibetian, Chinese and Arctic. The first of these has the greatest number
of species. The Siberian type of ornithological fauna, which in the Kamchatka has certain
feat res inherent to insular fauna, is a poorer derivative of the fauna of the continental
part of East Siberia. The main features of insular fauna peculiar to the ornithological fauna
of Kamchatka forests are: the scarcity of species composing it, as compared to the forest
fauna of the continental part of East Siberia, and the presence of a number of endemic
subspecies, with a very light colouring and large dimensions.
We give the name of К a m c h a t k a to the ornithological fauna of the peninsula.
Some forest subspecies peculiar to the Kamchatka are also found on the northern and cent-
ral islands of the Kuril group. The afforested part of the peninsula, together with part of
the territory of these islands, form the Kamchatka subprovince.
ЛИТЕРАТУРА
Аверин Ю. В. 1948. Наземные позвоночные восточной Камчатки. Тр. Кроноцкого
заповедника, вып. 1. Изд. Гл. упр. по заповедникам.
Бианки В. Л. 1909. Отчет о командировке в Камчатку в 1908 г. Изв. Акад. наук.
Дементьев Г. П. 1940. Материалы к авифауне Коряцкой Земли. Изд-во Моск,
о-ва испыт. природы.
Иогансен Г. X. 1939. Птицы Командорских островов. Изд. Томского гос. ун-та.
Мензбир М. А. 1914. Зоологические участки Туркестанского края и вероятное
происхождение фауны последнего. М.
Портенко Л. А. 1937. Орнитогеографические соотношения на крайнем северо-
востоке Палеарктики в связи с особенностями ландшафта. Сб. Памяти М. А. Мензбира.
Изд-во АН СССР.
Портенко Л. А. 1939. Фауна Анадырского края. Птицы, части 1—2. Изд.
Главсевморпути.
Сушкин П. П. 1925. Зоологические области средней Сибири и ближайших
частей нагорной Азии и опыт истории современной фауны Палеарктической Азии. Бюлл.
МОИП, т. 35.
Штегман Б. К. 1938. Основы орнитогеографического деления Палеарктики.
Фауна СССР. Птицы, т. I, вып. 2. Изд-во АН СССР.
Bergman S. 1935. Zur Kenntnis Nordostasiatischer Vogel. Ein Beitrag zur Syste-
matic Biologie und VerbreiUng der V3gel Kamtschatcas und die Kurilen.
Stejneger L. 1885. Results of Ornithological Explorationes in the Commander
slands and in Kamtschatka. Bull. № 29 of the Unit. St. Nat. Mus.
Taczanowski L. 1893. Faune Ornithologie de la Sibirie Orientate. Mem.de
Г Acad. Imp. de SC. de St. Petersbourg (VI Serie), LXXIX.
Jamasina J. 1931. Die Vogel der Kurilen. Journ. f. Ornithol, LXXIX.
БЮЛЛЕТЕНЬ М. О-ВА ИСП. ПРИРОДЫ, ОТД. БИОЛОГИИ, Т. LX1I (5), 1957
Наблюдения за тюленями Антарктики
В. А. Арсеньев
Observations on the seals of the Antarctic
' V. A. Arsenyev
В настоящем сообщении приводятся материалы наблюдений за тюленями, про-
веденные автором во время первого рейса морской Антарктической экспедиции на
судне «Обь» в 1955—1956 гг. Жизнь тюленей тесно связана со льдами — плавучими
и неподвижными — и поэтому в данном случае нас будет интересовать только район
ледового плавания «Оби». Плавание до берегов Антарктиды автор совершил на ди-
зель-электроходе «Лена».
Первые плавучие льды были встречены нами на подходе к морю Дейвиса, омы-
вающему берега Антарктиды в границах от шельфового ледника Шеклтона на во-
стоке до Западного ледника на западе. Пройдя полосу дрейфующего льда, корабль
подошел к неподвижному ледяному припаю, располагавшемуся в районе о-вов Хасуэл.
Здесь наблюдения велись в течение почти полутора месяцев — с 20 января по 28 фев-
раля 1956 г., во время разгрузки кораблей и постройки южнополярной обсерва-
тории «Мирный».
28 февраля «Обь» вышла в море и, обогнув с севера ледник Шеклтона, напра-
вилась к востоку вдоль берегов Антарктиды. 28 марта достигли о-вов Баллени, откуда
повернули к северу, покинув районы льдов. За время плавания у берегов материка
наш корабль неоднократно пересекал пояс паковых льдов, то выходя на прибрежную
полынью, то снова удаляясь в открытое море, причем плавание во льдах занимало
чуть ли не большую часть нашего времени.
Все это время с борта судна велись наблюдения за морскими млекопитающими,
в том числе и за тюленями. В случае обнаружения зверя нами отмечались числен-
ность замеченных животных, видовая принадлежность, условия их залегания и пове-
дение. Эти материалы и послужили основанием для настоящей статьи.
Как известно, семейство настоящих тюленей представлено в Антарктике пятью
видами: тюлень Уэддела (Leptonychotes weddelli), тюлень-крабоед (Lobodon carcinop-
hagus), тюлень Росса (Ommatophoca rossii), морской леопард (Hydrurga leptonyx) и
морской слон (Mirounga leonina), из которых последний обитает на островах субан-
тарктики и в собственно Антарктику не заходит.
Самый малочисленный из антарктических тюленей — тюлень Росса. Судя по ли-
тературным данным, этот тюлень большую часть жизни проводит на льдах открытого
моря, причем распространен он в самых южных, не всегда доступных районах Антарк-
тики. Это определяет чрезвычайно слабую изученность биологии тюленя Росса. Внеш-
ний вид тюленя Росса несколько необычен, так как он имеет относительно короткое
и толстое тело, вследствие чего его нельзя спутать ни с одним из других видов.
За весь рейс «Оби» не удалось наблюдать ни одного тюленя Росса. Однако
начальник австралийской Антарктической экспедиции Ф. Лоу сообщил, что при пересе-
чении пакового пояса на меридиане Западного шельфового ледника на судне «Киста-
Дан» в январе 1956 г. им были замечены два тюленя Росса, лежавшие на пла-
вучем льду.
Почти не наблюдался нами и другой вид антарктических тюленей — морской
леопард. Это наиболее крупный из тюленей Антарктики и единственный хищник среди
•отряда ластоногих. Считается, что большую часть жизни леопард проводит в одиноч-
ку (исключая период размножения).
Мне удалось наблюдать морского леопарда дважды и оба раза на плавучем
льду. Бросаются в глаза более крупные размеры этого тюленя по сравнению с дру-
40
гими видами. Тело его покрыто крупными темными пятнами, ярко выделяющимися на
общем более светлом фоне. В одном случае зверь лежал на большой, длиной до 50 м,
льдине, оторвавшейся от припая. Льдина проходила в нескольких стах метрах от
корабля, и леопард явно беспокоился — он часто поднимал голову и оглядывался.
Два другие вида тюленей Антарктики — тюлень Уэддела и тюлень-крабоед —
наблюдались нами довольно часто, причем крабоедов мы встречали в гораздо боль-
шем количестве, чем тюленей Уэддела.
Тюлени Уэддела (рис. 1) отмечались нами только в районе о-вов Хасуэл в море
Дейвиса во время стоянки для разгрузки кораблей. Чаще их можно было видеть
Рис. 1. Тюлень Уэддела’
на неподвижном припае, но когда в начале февраля в результате двухдневного
шторма припай начал быстро разрушаться и появилось много мелко битого льда, на
нем сейчас же появились тюлени, среди которых было замечено несколько тюленей
Уэддела. Все звери лежали на льдинах как на крупных, так и на мелких. Эти наблю-
дения оставили впечатление, что тюлени охотнее ложатся на плавучий лед, так как
перед штормом на неподвижном припае животных было гораздо меньше.
По литературным данным, тюлень Уэддела считается наиболее многочисленным
видом, не образующим, однако, крупных скоплений. Весной, в период деторождения,
эти тюлени залегают на льдах небольшими группами, но в летнее время выходят
из воды лишь время от времени.
Наши наблюдения производились в середине антарктического лета (январь —
февраль). Никаких, даже мелких, скоплений мы не наблюдали; звери в большинстве
случаев лежали по одному и только один раз; 27 февраля, были замечены три тю-
леня Уэддела, лежавшие на припае метрах в 100 друг от друга. Среди* этих тюленей
было две самки, а пол третьего животного определить не удалось. Как в этом, так
и в других случаях тюлени лежали на крепком гладком льду припая довольно дале-
ко от воды. Так, например, в радиусе более 500 м от замеченных нами трех тюленей
никаких лазок и проталин обнаружено не было. День был тихий, солнечный, и все тю-
лени крепко спали. Следует отметить, что вблизи трех тюленей Уэддела лежал-
один крабоед.
В литературе есть указания, что тюлень Уэддела может достигать длины в 3 м.
Нами было измерено две самки тюленя этого вида, из которых одна оказалась дли-
ной в 214 см, а другая 284 см. У последней обхват за передними ластами оказался
равным 148 см, длина переднего ласта 40 см, длина заднего ласта 52 см и длина,
хвоста 13 см.
Окраска тюленя Уэддела довольно характерна. Спина темно-бурая, почти одно-
тонная, на боках окраска зверя постепенно светлеет и на брюхе принимает желто-
бурый оттенок. Тело покрыто крупными продолговатыми пятнами бурого цвета, вытя-
нутыми продольно, вдоль оси тела. Ярче всего пятна на боках, но на брюхе они так-
же очень хорошо заметны, особенно тогда, когда шерсть сухая. На темной спине-
пятен почти незаметно. Голова, передние и задние ласты темные и совершенно ли-
шены пятен. Вибрисы не очень длинные. У самки длиной в 284 см я насчитал па
26 вибрис с каждой стороны морды. Самые большие вибрисы имели длину в 11 см.
Глаза у тюленя очень крупные, темно-карие.
Тюлень Уэддела — совершенно безобидное животное и в то же время он не ис-
пытывает перед человеком никакого страха. Зверь подпускает к себе вплотную, его
— 41 —
можно трогать руками. Когда тюлень спит, то лежит чаще на боку или даже на спи-
не и не всегда слышит, что к нему подошел человек. Если тюленя спугнуть, толкнув
чем-нибудь, то первое его движение — это поворот на спину. При этом голова и часть
туловища до передних ластов поднимаются кверху и зверь внимательно смотрит на
подошедшего человека своими большими глазами. Поза эта как бы оборонительная,
но никаких агрессивных действий тюлень не предпринимает: пасть не открывает, ни-
каких угрожающих звуков не издает. Через несколько секунд голова снова опускает-
ся на лед, и тюлень крепко зажмуривает глаза. Немного полежит, затем снова по-
смотрит на человека, поморгав глазами, и опять крепко засыпает.
В литературе есть указания на то, что тюлень Росса обладает замечательной
способностью издавать громкие и довольно мелодичные звуки. Относительно других
видов тюленей такого указания «нет. Однако, наблюдая тюленей Уэддела, я обнару-
жил у них такую же способность. В то время, когда зверь принимает оборонительную
позу, неизменно раздается тихий свист. Звук очень тихий, короткий и отрывистый, рот
в этом никакого участия не принимает. Создается впечатление, что звук идет отку-
да-то изнутри. В этот момент по горлу проходят как бы обратные глотательные дви-
жения, кажется, что снизу вверх идет какой-то комок, как это наблюдается, напри-
мер, у коровы при пережевывании жвачки. Мне пришлось так близко обследовать
Рис. 2. Тюлень-крабоед
только одного тюленя Уэддела. Обладают ли все животные этого вида такой способ-
ностью, подлежит тщательной проверке.
Движения у тюленя Уэддела очень неуклюжие. Чтобы заставить зверя сдви-
нуться с места, приходится несколько раз его подтолкнуть. Тогда он переворачивается
на брюхо и ползет. Двигается тюлень очень медленно как бы всем телом, чем-то
отдаленно напоминая движение гусеницы. На ласты совершенно не опирается. Ско-
рость движения равна скорости медленно идущего человека.
Все тюлени, виденные мною относительно близко, были совершенно вылинявши-
ми, волос пальцами не выщипывался. Линяющих тюленей не наблюдалось, не были
отмечены также и молодые тюлени.
Четвертый вид антарктических тюленей — крабоед — также относится к много-
численным видам (рис. 2). Этого тюленя мы наблюдали в гораздо большем количе-
стве, чем всех остальных. Наибольшее число крабоедов наблюдалось восточнее лед-
ника Шеклтона, но и в районе «Мирного», на припае и плавучем льду крабоед, пожа-
луй, по численности преобладал над другими тюленями.
На припае крабоеды лежат почти всегда поодиночке. Однажды наблюдалась па-
ра, которую удалось осмотреть вблизи. Это были самец и самка, лежавшие рядом
метрах в 700—800 от края припая. Оба зверя были, видимо, взрослыми, примерно
одинаковой длины, около двух метров. Самец был несколько светлее самки. Спина
и брюхо обоих зверей имели почти одинаковый серебристый оттенок, никаких пятен
у тюленей не было.
22 февраля вблизи судна на припай вышел крабоед и залег метрах в 200 от
края. Это была самка примерно такого же размера, как и предыдущие особи. Волос
ее был почти серебристого цвета, особенно с брюшной стороны. Были заметны отдель-
ные темные пятна, главным образом у основания задних ластов.
Крабоед не подпускал к себе так близко, как тюлень Уэддела. Уже метров за
30—50 зверь поднимал голову, оглядывался и начинал отползать. На подходящего
— 42 —
совсем близко человека шипел, широко открывая пасть. Крабоед издает негромкий
звук, что-то среднее между шипением и рычанием. Если зверю преградить путь, то
он, раскрыв пасть, делает несколько устрашающих движений навстречу человеку, но
не бросается. Когда двинувшейся таким образом самке крабоеда подставили ногу,
она не схватила зубами, а сразу же начала отползать, продолжая шипеть и
оглядываться.
Интересно реагировал этот тюлень на появление вертолета. Зверь явно забеспо-
коился, все время поднимал голову и шипел. С поднятой головой и раскрытой пастью
зверь начал вертеться на месте, вокруг своей оси, все время следя за пролетающей
машиной. Затем со всей возможной быстротой стал отползать от севшего побли-
зости вертолета.
По льду крабоед передвигается довольно медленно, хотя и быстрее, чем тюлень
Уэддела. При этом зверь как-то подпрыгивает, отталкиваясь передними ластами. Но
долго так бежать не может. Уже через несколько метров он останавливается и отды-
хает. Задняя часть тела при движении зверя остается обычно неподвижной, но ино-
гда тюлень делает какие-то горизонтальные движения, что, однако, ходу нисколько
не помогает. Передвигаясь по суше, крабоед сильно напоминает нашу северную коль-
чатую нерпу (акибу).
Измерить удалось только одну самку крабоеда. Ее длина была 196 см, обхват
за передними ластами 108 см, длина переднего ласта 31 см, длина заднего ласта
34 см, длина хвоста 12 см. Вибрис по 20 штук с каждой стороны морды, их длина
2—3 см, две крайние задние — 6—7 см. Общий тон окраски серый, спина более тем-
ная с бурым оттенком, брюхо серебристо-серое, почти белое. Граница между темным
и светлым проходит по прямой линии на уровне передних ластов. Передние и задние
ласты сверху темные, снизу светлые.
Все обследованные нами тюлени имели крепкий волос и никаких следов линьки
у них обнаружено не было. Из литературы известно, что окраска крабоеда меняется
в течение года. В первое время после линьки мех зверя имеет бурый оттенок, который
постепенно светлеет и к началу следующей линьки тюлень становится серебристо-бе-
лым. Поскольку мех всех виденных нами тюленей имел явно серебристый оттенок,
надо полагать, что линька у них к этому времени уже закончилась.
При плавании в дрейфующем льду мы наблюдали уже настоящие скопления тю-
леней (рис. 3). Интересно, что во всех случаях это были исключительно крабоеды.
Никаких других тюленей вместе с ними замечено не было.
Первое такое скопление было обнаружено 8 марта между 109 и 110° в. д. и, при-
мерно, на 65°30' ю. ш. В течение двух часов при плавании в крупно-мелкобитом льду
наблюдались лежавшие на нем тюлени. Звери располагались как на мелких, так и
на крупных льдинах. Многие лежали поодиночке, очень часто по 3—5 штук вместе,
но отмечались залежки и по 15—20 голов. За это время по курсу корабля было отме-
чено ориентировочно не менее 500 животных.
Цвет преобладающего числа животных был стальной с более темной спиной,
несколько штук были серебристые. При осмотре на расстоянии создалось впечатле-
ние, что зверь уже не линяет, так как все животные были гладкими, а на лежках на
льду выпавшего волоса также не наблюдалось.
Корабля тюлени совершенно не боялись. Даже с тех льдин, которые разворачи-
вались от удара «Оби» зверь в воду не сходил и лишь ползал по льдине из стороны
в сторону. Если же судно проходило метрах в 50 от льдины, тюлени даже не шевели-
лись. За все время было замечено только два нырнувших тюленя.
Глубина в этом районе более 1000 м. На станции, взятой за 3—4 часа до подхо-
да к залежке, планктонные сети принесли много калянусов и бокоплавов, рыб пой-
мано не было. Возможно эти животные и служат пищей крабоедам. Погода стояла
ясная, солнечная, при почти полном штиле.
На другой день, милях в 60 к востоку, было замечено несколько крабоедов, ле-
жавших на плавучем льду. В этом районе была предпринята высадка на небольшой
•скалисты/’ островок, на берегу которого было обнаружено до 15 тюленей — крабоеды
и тюлени Уэддела. С острова удочкой была поймана рыба из трематомусов.
Второе скопление несколько меньшей численности обнаружено 14 марта в районе
сомнительной земли Дисковери. Координаты этого района таковы: широта 65°30'—
65°40' южная долгота 124°30'—125° восточная. Этот район изобиловал громадными
айсбергами, видимо стоявшими на месте, так как у подножья большинства из них
располагались большие поля довольно толстого ровного льда типа припая. На этом
льду и залегали тюлени. Снова наблюдались исключительно крабоеды, представителей
других видов отмечено не было. Тюлени лежали группами голов по 20—25, а иногда
и больше. Реже отмечались мелкие группы и еше реже одиночки. Зверь встречался в
течение нескольких часов хода корабля. Можно считать, что здесь располагалась за-
лежка промыслового типа.
Большая часть тюленей имела темную, буроватую окраску, но встречались
экземпляры и с мехом серебристого тона. Следует напомнить, что в первом встре-
ченном нами скоплении этих тюленей преобладающее большинство особей было
— 43 —
светлым, серебристым. Чем может быть объяснена такая разница в окраске, ска-
зать трудно.
16 марта был сильный шторм с пургой при ветре до 30 и даже более метров в
секунду. Наше судно отстаивалось у припая земли Дисковери. В течение дня группы
из нескольких тюленей много раз подплывали к кромке льда, но на лед не выходили.
Вид этих тюленей определить не удавалось. Однажды два тюленя вылезли на лед
вблизи судна, однако сейчас же сошли в воду. Оба они были крабоедами, что при-
водит к мысли, что и остальные животные принадлежали к этому виду. Тюленей, ле-
жащих на льду, в этот день не наблюдалось, возможно, что сильная пурга препят-
ствовала выходу зверя на лед.
Рис. 3. Карта района наблюдений
При приближении к северному из островов Баллени с запада в течение более по-
ловины дня судно проходило через молодой блинчатый лед, имевший уже порядочную
толщину. Среди молодого льда были вкраплены отдельные старые льдины разного раз-
мера. На многих из этих льдин лежали тюлени-крабоеды. За день удалось заметить
более двух десятков тюленей. Все они без исключения лежали поодиночке, на прохо-
дящее вблизи судно никакого внимания не обращали, даже не шевелились и не под-
нимали голову. Погода была тихая, ясная — полный штиль; в полыньях между раз-
водьями не было даже ряби.
Вот те немногие наблюдения за антарктическими тюленями, которые йам уда-
лось провести во время первого рейса морской Антарктической экспедиции. Эти на-
блюдения показывают, что тюлени Антарктики, не имеющие врагов, кроме леопарда и
косатки, совершенно не боятся ни человека, ни громадных кораблей, ни даже идуще-
го с сильным шумом трактора. Из всех видов тюленей в летнее время толью одни
крабоеды образуют более или менее значительные скопления. Все эти скопления рас-
полагаются на плавучих льдах пакового антарктического пояса. В районах беретового
припая тюлени чаще всего лежат одиночками или, в крайнем случае, по 2—3 особи.
Однако, судя по наблюдениям на острове, который мы посетили, в случае наличия
открытого от льда берега тюлени могут образовывать небольшие залежки. Обнару-
женные нами залежки тюленей-крабоедов могут быть оценены как имеющие промыс-
ловое значение для судов типа зверобойных шхун.
Наиболее доверчивы и флегматичны из трех наблюдавшихся нами видов антарк-
тических тюленей безусловно тюлени Уэддела. Крабоед всегда беспокоится при при-
ближении человека, однако никаких агрессивных действий также не предпринимает.
— 44 —
Леопард, видимо, наиболее осторожный и недоверчивый, а так как это настоящий
хищник, то он. конечно, опасен и для человека.
Надо надеяться, что в следующем рейсе в Антарктику удастся собрать новь^е и
интересные данные об этих еще в сущности очень мало изученных ластоногих.
SUMMARY
Five species of the family of seals proper inhabit the Antarctic: Weddel’s seal (Lepto-
nychotes weddelli), the crab eater (Lobodjn carcinophagus), Rosse’s seal (Onmatophoca,
rossii), the sea leopard (Hydrurga leptonix) and sea elephant (Mir о и ng a leonina). The
latter inhabits subantarctic islands and does not wander into the Antarctic itself. As no
Rosse’s seals were observed during the voyage and leopard seals were seen twice only,
the author of this article confines himself to observations on the two species of seals
most numerous in the Antarctic: the weddel seal and crab eater. They were observed from
on board the electric ship „Ob“, in 1956, in the middle of the antarctic summer (weddel seals
were observed in January and February and crab eaters in January, February and March).
Weddel seals were observed only on solid ice attached to ihe mainland, or on drif-
ting ice — floes that had broken off from it, but never on drifting ice in the open sea.
The seals did not conglomerate in large groups, but were seen either singly, or, more
rarely, in pairs and groups of three. All the animals examined had solid fur (did not mo-
ult). No yoing seals were observed. Crab—eaters Were seen in much greater numbers
than weddel seals. On solid ice near the continent they lay for the most part singly, but
on drifting ice to the East of Shackleton glacier the author observed large groups of seals
(several hundreds in a group). In this case also no young animals were observed.
Antarctic seals, especially of the weddel species, hardly ever ‘how any fear of man,,
whom they do not recognize as an enemy (seal hunting is not practiced in the Antarctic).
The killer and, to a lesser extent, the sea leopard are the only animals to be regarded as-
enemies of the seal.
БЮЛЛЕТЕНЬ М. О-ВА ИСП. ПРИРОДЫ, ОТД. БИОЛОГИИ, Т. LXII (5), 1957
Некоторые особенности строения плечевого пояса
у голубей и рябков и вопрос о функциональном
значении ключицы у птиц
Б. К. Штегман
Шег einige Eigenlieiten im Bau des Schultergiirtels bei
den Tauben und Flughiihnern und die funktionelle
Bedeutung der Clavicula bei den Vogeln
В. K- Stegmann
Изучение морфологии голубей и рябков дало возможность выявить ряд харак-
терных особенностей, отличающих этих птиц от всех родственных групп, в частности
я от куликов. Среди большого количества признаков выделяются естественным комп-
лексом особенности строения плечевого пояса, тесным образом связанные с летными
качествами изучаемых птиц. Исследование особенностей летательного аппарата голу-
бей и рябков в связи с особенностями летных способностей этих птиц дало возмож-
ность определить направление эволюции полета тех и других (Штегман, 1957). Для
лучшего понимания последующих рассуждений мы им предпошлем краткие выводы
из данной работы.
Еще Фюрбрингер (М. Furbringer, 1888) установил родственные связи между
куликами, голубями и рябками, и его выводы не оспаривались, а, наоборот, лишь под-
тверждались последующими исследователями. Так, и в отношении развития летатель-
ного аппарата можно сказать, что кулики представляют собой исходный тип для раз-
вития более узко специализованного полета голубей и рябков. У куликов, приспособ-
ленных к скоростному полету среди открытого ландшафта, крылья узкие, острые,
с довольно длинным скелетом. У голубей, в связи с переходом к жизни среди пересе-
ченной местности или древесной чащи, произошло приспособление к большей манев-
ренности полета, выразившееся в приобретении способности к трепещущему типу его.
Крылья у них с более коротким скелетом, более широкие, с очень жесткими маховыми,
основная летательная мускулатура более мощная, чем у куликов, с особенно силь-
ным развитием подъемных мускулов крыла. Рябки, подобно куликам приспособленные
.к скоростному полету среди открытого ландшафта, имеют летательный аппарат, резко
отличный от летательного аппарата куликов. У них крылья, хотя и очень своеобразной
•формы, широкие у основания, как у голубей, с очень жесткими маховыми и коротким
скелетом. Летательная мускулатура голубиного типа. Характерные особенности лета-
тельного аппарата у рябков ясно указывают на то, что эти птицы вторично приспосо-
бились к жизни среди открытого ландшафта, пройдя стадию приспособления к манев-
ренному полету, следовательно, развились не непосредственно из группы куликов, а из
предков современных голубей, образ жизни которых был уже «голубиным».
Таким образом, и в анатомических признаках можно ожидать большего сход-
ства между голубями и рябками, чем между каждой из этих групп и куликами.
Действительно, скелет крыла у голубей и рябков не только более короткий, чем
у куликов, но и значительно более мощный, так как все кости относительно более тол-
стые. Можно не сомневаться в том. что эта особенность голубей и рябков связана
с приспособлением данных птиц к трепещущему полету, требующему большой затраты
энергии и высокой нагрузки на крылья.
Менее понятной кажется своеобразная форма плечевой кости у голубей. У этих
птиц crista lateralis плеча, место прикрепления грудного мускула, образует острый
'угол, причем проксимальная его часть имеет несколько вогнутую линию (рис. 1,а).
— 46 —
У других птиц, в частности у куликов и чаек, crista lateralis никогда не образует
острого угла; край этого гребня имеет мягкие изогнутые очертания и сам гребень не
бывает таким высоким, как у голубей, но обычно более длинный и подходит ближе
к сочленовной поверхности плечевого сустава (рис. 2, а, б). Между тем у разных ви-
дов голубей вышеописанная форма crista lateralis очень постоянна и повторяется
у рябков (рис. 1,6). Можно даже сказать, что по форме плечевой кости и ее лате-
рального гребня голуби и рябки практически не различимы.
Для понимания функционального значения своеобразного строения плеча у го-
лубей и рябков необходимо изучить и другие особенности морфологии этих птиц, свя-
занные с плечевым суставом. И тут бросается в глаза необычная форма ключицьь
у этих птиц. Тогда как у большинства птиц, в частности и у куликов, ключица, кроме
изогнутости в виде дужки, т. е. в латеральной плоскости, имеет еще и сильный выгиб
Pre. 1. а—плечевая кость сизого голубя
(Columba livia); б— плечевая кость чернобрю-
хого рябка (/ terocles orientalis)
а б
Рис. 2. а — плечевая кость чибиса
(Vanellus vanellus); б — плечевая
кость веретенника (Limosa limosay
вперед, у голубей она в этом направлении не имеет никакого выгиба. Разница эта
очень значительная и не может быть пропущена даже при самом беглом осмотре со-
ответствующих скелетов (рис. 3, 4, 5, 6).
Относительно функционального значения ключицы у птиц долгое время не воз-
никало разногласий. Считалось, что ключица, соединенная своими вершинами с вер-
шинами коракоидов, фиксирует расстояние между плечевыми суставами и создает пру-
жинящую амортизацию при боковом давлении на плечевые суставы, которое возни-
кает при работе мускулов во время полета. Такое объяснение казалось очень вероят-
ным и подкупало своей кажущейся логичностью. В связи с этим и в учебниках было
принято писать, что это действие ключицы — необходимое условие для полета птиц,
и у нелетающих видов ключица, лишенная функции, редуцируется. Однако при этом
упускалось из виду, что имеются и вполне нормально летающие птицы, у которых
ключица настолько редуцирована, что не может выполнять приписываемой ей функции.
Так, например, у многих попугаев сохранились лишь вершины ключицы в местах их
соединения с коракоидами и лопатками, между тем как вся свободная часть, обра-
зующая «дужку», исчезла.
Позднее Си (М. Sy, 1936) было доказало, что расстояние между коракоидами
фиксируется не только ключицей, но. видимо, в большей степени, толстой и эластичной
соединительнотканной перепонкой (membrana sterno-coraco clavicularis), натянутой
между коракоидами, ключицей и грудиной.
Рис. 4. Грудина веретенника (Limosa limosa)
Рис. 3. Грудина чибиса (yanellus vanellus)
— 49 —
В случае редукции ключицы эта перепонка одна выполняет функцию сохранения рас-
стояния между коракоидами. Таким образом, мнимая функция ключицы у птиц на са-
мом деле оказалась побочной, второстепенной и, во всяком случае, не определяющей
ни размеров, ни формы ее.
Какова же основная функция ключицы у птиц? Уже ранее было указано, что
ключица, если она хорошо развита, имеет более или менее ясно заметный выгиб впе-
ред (рис. 3, 4, 5). При этом, как правило, она сплюснута с боков, т. е. имеет большее
сечение в сагиттальной, меньшее — в латеральной плоскости. Из этого следует, что
наибольшую нагрузку ключица в состоянии вынести по продольной линии тела и, если
принять во внимание выгиб ключицы вперед, то именно в каудальном направлении.
Далее, рассматривая мускулатуру птицы, мы видим, что большой грудной мус-
кул, опускающий крыло, своей передней частью прикреплен к ключице и спереди
ограничивается ею. Так как место прикрепления этого мускула очень обширное, то
разные его части имеют неодинаковое действие; значительная часть его при сокраще-
нии не только опускает крыло, но и тянет его назад, другая часть мускула тянет
крыло прямо вниз, а передняя часть, прикрепленная к ключице,— вперед. Следователь-
но, ключицу мускул тянет в обратном, т. е. в каудальном направлении. Таким обра-
зом, становится вполне понятным, почему ключица имеет большее сечение именно
в этой плоскости.
Но какое значение имеет для птиц вышеописанная особенность расположения
большого грудного мускула? Неоднократно высказывалось мнение, что этот большой
мускул иннервируется не весь одновременно, а частями. Если это так, то птица мо-
жет, напрягая ту или иную часть грудного мускула, при опускании крыла то оттяги-
вать его назад, то подтягивать его вперед. Наблюдения над механикой полета птиц
вполне подтверждают такую точку зрения. Во время активного полета птица, опуская
крыло, сильно пропеллирует им. При этом сила тяги прилагается к вершинной части
крыла, которая стремится ускользнуть вперед. Понятно, что в это время компонент
'грудного мускула, тянущий крыло вперед, не имеет значения. Так как при опускании
крыла напрягается весь мускул, то вследствие незначительности его части, тянущей
крыло вперед, преобладает тяга крыла назад.
Иначе обстоит дело при подъеме крыла во время горизонтального полета. Из-
вестно, что при таких обстоятельствах подъем крыла осуществляется пассивно, за
счет летной нагрузки. Конечно, он происходит при известном сопротивлении грудного
мускула для замедления движения крыла и сохранения подъемной силы его. Но так
как во время подъема крыла птица летит за счет инерции, то источником энергии
в данный момент является ее корпус, обладающий главной массой, между тем как
крылья представляют основное сопротивление. Следовательно, крыльям в это время
приходится преодолевать лобовое сопротивление, стремящееся оттолкнуть их назад.
Итак, во время подъема крыла должна действовать сила, препятствующая слиш-
ком быстрому его подъему, и еще вторая сила, противодействующая лобовому сопро-
тивлению, т. е. тянущая крыло вперед. Между тем передняя часть грудного мускула
и опускает крыло и тянет его вперед. Следовательно, вполне вероятно, что во время
«пассивного» подъема крыла в основном работает передняя часть грудного мускула.
Такое же распределение сил и нагрузок несомненно имеет место во время планирую-
щего полета, а также во время парения, представляющего собой то
же планирование с использованием восходящих течений воздуха.
У парителей, летающих в основном без взмахов крыльев, можно
предполагать особенно сильное развитие передней части грудных мускулов. Действи-
тельно, у птиц, много парящих, как, например, альбатросы, фрегаты, хищные птицы,
грудина короткая и часть грудного мускула, тянущая крыло назад, развита слабо.
Ключица же у этих птиц очень крепкая и толстая и имеет особенно большое сечение
в сагиттальной плоскости. Кроме того, она у этих птиц сильно выгнута вперед, в свя-
зи с чем передняя порция грудных мускулов должна быть особенно сильно развита.
В качестве примера такого распределения грудных мускулов можно привести исследо-
ванного мной гималайского сипа. У этой птицы задний край грудины, несмотря на
ее незначительную длину, не был покрыт грудными мускулами. Зато передняя часть
грудных мускулов, прикрепленная к ключице, была очень мощная, составляя больше
половины всего их веса.
Таким образом, большая толщина ключицы и сильный ее выгиб вперед у паря-
щих птиц имеют совершенно определенное функциональное значение. В свете выше-
сказанного становится понятным и значение наличия у некоторых парящих птиц плот-
ного соединения нижней части ключицы (hypocleidium) с вершиной киля грудины.
Этот упор ключицы представляет собой несомненно дальнейшее ее укрепление против
тяги грудного мускула в каудальном направлении.
Однако, как уже было указано, и при активном полете во время подъема крыла
передняя часть грудного мускула, тянущая крыло вперед, получает специальную на-
грузку. Конечно, эта нагрузка не столь велика и длительна, как у парящих птиц, но
с наличием ее все-таки приходится считаться. Очень вероятно, что она у медленно
летающих птиц невелика, но значительно повышается у скоростных летунов с увели-
чением лобового сопротивления крыла. Действительно мы пилим v бигтпп пртяш.
— 50 —
щих куликов довольно толстую, крепкую и сильно выгнутую вперед ключицу. Чайки
и крачки, хотя и летают спокойнее, но в случае необходимости могут сильно уско-
рить полет и, кроме того, много планируют; поморники летают очень быстро.
Вполне понятно, что у всех этих птиц ключица сильно выгнута вперед. Наконец,
у чистиков мы видим еще более сильно выгнутую ключицу. Например, у кайры (Uria)
она от места соединения с акрокоракоидом направляется сначала прямо вперед
(в краниальном направлении) и затем уже поворачивает вниз и назад (в вентраль-
ном и каудальном направлениях) (рис. 5). Такая форма ключицы обусловливает осо-
бенно выгодное расположение передней части грудных мускулов для подтягивания
крыльев вперед, что на первый взгляд представляется мало оправданным, так как
чистики не парят. Если же иметь в виду, что чистики плавают под водой при помощи
крыльев или как бы летают в воде, то такое приспособление у них становится впол-
не понятным. Вода значительно более плотная среда, чем воздух, и, чтобы в ней пре-
одолеть лобовое сопротивление, необходимо большое усилие.
Итак, нам удалось определить основное функциональное значение ключицы
у птиц, а также значение выгиба ее вперед у многих видов. Однако, как уже было
указано, ключица у голубей не имеет никакого выгиба вперед. Это означает, что у го-
лубей большой грудной мускул не имеет отдела, тянущего крыло вперед. Изучение
расположения этого мускула у голубей вполне подтверждает предположение, сде-
ланное на основании изучения скелета: действительно, грудной мускул, ограничивае-
мый спереди ключицей, может тянуть крыло вниз и назад, но не вперед, что, в част-
ности, видно и по направлению волокон этого мускула, идущих от места прикрепле-
ния его к плечу. Между тем голуби относятся к скоростным летунам и несомненно
нуждаются в какой-то силе, подтягивающей крыло вперед во время его подъема.
Так как грудной мускул для этой цели не приспособлен, то приходится полагать, что
у голубей имеется другое эквивалентное приспособление.
У всех птиц имеется мускул, специальная функция которого заключается в отве-
дении плеча от тела, иначе говоря, в подтягивании крыла вперед — это т. сагасо-'
brachialis anterior. Он прикреплен к акрокоракоиду и к плечу и у большинства птиц
очень мал, будучи лишь слегка крупнее малого дельтовидного мускула. Места при-
крепления этого мускула к плечу различны. У одних видов он прикрепляется к прок-
симальной части ребра crista lateralis и тогда ничтожно мал. В других случаях
(в частности, у куликов) он прикрепляется у основания указанного гребня, с вент-
ральной его стороны и прикрыт широким плоским сухожилием, тянущимся от
m. biceps к акрокоракоиду. При таком расположении он бывает несколько больше,
главным образом длиннее.
У голубей проксимальная часть края crista lateralis, как уже было указано, во-
гнута, как бы вырезана. На границе с дистальной частью гребня образуется острый
угол, к которому и прикреплен m. coraco-brachialis anterior. Он заполняет всю шири-
ну вырезки crista lateralis и в несколько раз более мощный, чем у других птиц. Так
как он прикреплен к самой широкой части гребня, то его усилие передается на рычаг
с довольно длинным плечом, что увеличивает эффект его тяги. Это приспособление
несомненно компенсирует у голубей отсутствие тяги крыла вперед при помощи груд-
ного мускула и представляет собой не менее яркий, чем форма ключицы, признак
этих птиц. И, как мы видели, одно с другим связано, представляя собой единый комп-
лекс особенностей плечевого пояса у голубей.
Но каково значение этих приспособлений? Из вышесказанного можно сделать
заключение, что первым звеном в данном комплексе является изменение формы клю-
чицы, обусловившее собой прочие морфологические изменения. Что же касается изме-
нений формы ключицы, то об этом можно сделать следующие предположения. Как из-
вестно, все представители группы Laro-Limicolae, т. е. кулики, чайки и чистики, пи-
таются анимальной пищей и не имеют зоба. У голубей, перешедших на растительную»
пищу, развился большой зоб, который в наполненном виде имеет значительный вес и
должен перемещать центр тяжести этих птиц вперед. Такое перемещение центра тя-
жести затрудняет полет, в особенности взлет. В недавней нашей работе (Штегман.
1950) было подробно описано, применительно к куриным птицам, какое значение для
полета имеет перегрузка передней части тела при наличии большого зоба. Там же бы-
ли описаны изменения в форме грудины у куриных, связанные с необходимостью пе-
ремещения зоба в сторону центра тяжести, для того чтобы он не слишком выдавался
вперед. У голубей мы не видим столь кардинальных изменений плечевого пояса, как
у куриных, вследствие чего наполненный зоб у них не распространяется так далеко
назад, как у последних. Однако в связи с тем что у голубей ключица не имеет выги-
ба в латеральной плоскости, зоб у этих птиц, налегая на нее, в наполненном состоя-
нии выдается не так далеко вперед, как, например, у хищных. Для последних, при-
способленных к парению, сильный выгиб ключицы особенно необходим, а так как они
не приспособлены для быстрого взлета и, кроме того, имеют длинные крылья, то для
них временное перемещение центра тяжести вперед благодаря торчащему зобу не
столь неудобно, как для голубей и куриных.
Что у голубей по сравнению с куриными проблема перемещения зоба назад раз-
решена лишь наполовину, имеет следующие причины. У куриных изменение формы
— 51 —
грудины привело к перемещению грудных мускулов далеко назад, в связи с чем
уменьшен коэффициент их полезного действия. Для голубей, приспособленных к дли-
тельному скоростному полету, необходимо возможно более полное использование мощ-
ности мускулатуры. Кроме того, у голубей масса летательной мускулатуры еще боль-
ше, чем у куриных, и громадный грудной мускул требует много места для своего раз^
мещения. Наконец, для голубей далекое перемещение зоба назад не играет такой
важной роли, как для куриных, так как для них стремительный взлет не столь необхо-
дим. как для последних.
Таким образом, у только что перечисленных групп птиц, имеющих большой зоб,
мы замечаем различное реагирование на наличие этого неудобного для полета приспо-
собления. У хищных птиц видно полное игнорирование неудобств, связанных с нали-
чием зоба, у куриных фундаментальное приспособление для ликвидации указанных
неудобств, между тем как у голубей имеет место известный компромисс при решении
данного вопроса. Все-таки и у голубей приспособления для перемещения зоба оказа-
лись достаточно сложными, охватившими плечевой пояс в целом.
Интересно отметить еще одну особенность строения ключицы у голубей: эта кость
у них очень тонкая и слабая, что бросается в глаза при сравнении с многими другими
птицами, в частности с куликами, чайками и чистиками. Слабость ключицы является
прямым следствием отсутствия у нее выгиба. Как уже было указано, в связи с этим
у голубей грудной мускул не выдается вперед от плечевого сустава, волокна перед-
ней его части, прикрепленные к ключице, располагаются почти параллельно к ней и
при сокращении не тянут ее назад. Ключица не несет механической нагрузки, утратила
свое основное функциональное значение и находится в стадии редукции. Понимание
этого явления имеет большое значение для дальнейших выводов.
В пределах семейства голубей форма ключицы очень постоянна. У разных видов
наших голубей, у изученных экзотических видов, в частности и у плодоядных голубей
(Treroninae), ключица в сагиттальной плоскости лишена изгиба. У венценосных голу-
бей (Goura) мы видим некоторый изгиб* ключицы, но в обратном направлении, т. е.
не вперед, а назад, не выгиб, а прогиб, вмещающий в себя заднюю стенку зоба, оче-
видно. особенно большого у этих птиц.
Что касается рябков, то у них ключица тоже совершенно лишена изгиба в сагит-
тальной плоскости. Причина этого та же, что и у голубей: рябки имеют большой зоб,
перемещение которого в сторону центра тяжести очень важно для облегчения полета,
в особенности взлета. Однако в отличие от голубей ключица у рябков не только тон-
кая, но и сильно укорочена, по крайней мере на одну треть длины, так что вершина
ее достигает лишь половины расстояния от плечевых суставов до киля грудины
(рис. 7, 8). Таким образом, можно сказать, что у рябков ключица еще более редуци-
рована, чем у голубей.
Необходимость приближения зоба к центру тяжести у рябков вызвана еще боль-
шим количеством причин, чем у голубей. Известно, что рябки носят своим птенцам
воду в зобу, зачастую издалека. У чернобрюхих рябков в зобу бывает до одного ста-
кана воды, что составляет почти половину веса этих птиц. Ясно, что такая большая
перегрузка передней части тела.должна сильно затруднять полет, и перемещение зоба
в сторону центра тяжести в данном случае является особенно важным.
Изменение формы ключицы и расположения грудных мускулов привело у ряб-
ков к сильному развитию m. coraco-brachialis anterior, который прикреплен тем же
способом, как и у голубей. Таким образом, можно сказать, что у этих птиц весь-
комплекс приспособлений, связанный с наличием зоба и перемещением его в сторону
центра тяжести, принципиально сходен с таким же комплексом приспособлений у го-
лубей. Более того, развитие m. coraco-brachialis anterior, способ его прикрепления
к плечу и форма плечевой кости у тех и других птиц одинаковы. Имея в виду ори-
гинальность данного комплекса приспособлений и его сложность, приходится считать,
что он вряд ли возник самостоятельно у голубей и рябков. Более вероятным кажется,
что эти признаки являются общими для обоих семейств, подтверждая их филогенети-
ческую близость.
В пределах других отрядов птиц вышеуказанные особенности строения плечевого
пояса, характерные для голубей и рябков, не встречаются даже в приближенной
форме, за одним исключением. У попугаев, по общему облику и по своему образу жиз-
ни ничем не напоминающих голубей и рябков, мы видим почти точное повторение
вышеуказанных морфологических особенностей. Ключица у попугаев совершенно не
имеет выгиба в сагиттальной плоскости, у многих видов даже вогнута, как у венце-
носных голубей. Гребень плеча (crista lateralis) имеет такую же острую вершину и
вырезку в проксимальной части, как у голубей и рябков, и эта особенность морфоло-
гии плеча связана с таким же сильным развитием и таким же прикреплением т. сагасо-
brachialis anterior. Если к этому добавить, что у попугаев имеется большой зоб, то
становится ясным, что вся система морфологических особенностей у попугаев и
у вышеупомянутых семейств связана с аналогичными приспособлениями.
Действительно, главная масса попугаев питается древесными семенами, в осо-
бенности орехами, в поисках которых им приходится совершать значительные переле-
ты. Летят попугаи высоко над лесом, со значительной скоростью, имея приспособлен-
— 53 —
ный для этого летательный аппарат — относительно узкие и острые крылья. Во время
же кормления они перемещаются среди древесной чащи на небольшие расстояния тре-
пещущим полетом. Так как места кормления и отдыха у попугаев не совпадают, то
эти птицы должны иметь большой зоб для хранения запаса пищи.
Из вышесказанного явствует, что у попугаев некоторые особенности образа жиз-
ни совпадают с таковыми у голубей, несмотря на полное несходство в общем облике
этих птиц. И эти особенности образа жизни привели к развитию комплекса приспо-
соблений, во всех подробностях повторяющего аналогичный комплекс приспособлений
у голубей. Конечно, из этого еще не следует, что на основании сходства только дан-
ных признаков можно говорить о филогенетической близости столь различных по об-
лику птиц, как голуби и рябки, с одной стороны, и попугаи — с другой. Но если иметь*
в виду, что в данном случае наблюдается сходство не в одном признаке, а в целом
комплексе их, взаимосвязанных в приспособлении к одинаковым биологическим осо-
бенностям. то совсем независимое развитие их у вышеуказанных групп представляется
тоже мало вероятным.
Впрочем, изучение попугаев, именно в сравнении с голубями, дает материал еще
для некоторых соображений. Как уже было ранее указано, у голубей ключица не вы-
полняет своей основной функции и не имеет той сильной механической нагрузки, кото-
рую она испытывает у большинства других птиц, вследствие чего она заметно реду-
цировалась, став слабой и тонкой. У рябков эта редукция пошла еще дальше, так как
ключица значительно уменьшилась в длине. Что касается попугаев, то мы видим дей-
ствие тех же закономерностей, что и у предыдущих групп. В связи с утратой основ-
ного функционального значения ключица у попугаев стала такой же тонкой и слабой,
как и у голубей. Более того, у ряда видов (Agapornis, Nasiterna, Melopsittacus, Platy-
cercus, Psittacula) она деградировала до такой степени, что нижняя часть ее, образую-
щая собственно дужку, полностью исчезла и сохранились лишь вершинные части в ви-
де рудиментов. Таким образом, вполне естественно объясняется явление редукции
или «отсутствия» ключицы у ряда попугаев и у некоторых других летающих птиц,
ранее казавшееся загадочным.
Впрочем, полная редукция ключицы может произойти и у голубей. В атласе
Мейера (А. В. Меуег, 1889—1897) изображен скелет редкого голубя из Новой Гвинеи
(Otidiphaps nobilis Gould), у которого не осталось и следа ключицы. В тексте
(Мейер, стр. 6) указано, что у этого вида от ключицы сохранилось только по не-
большой костной пластинке с каждой стороны тела, прикрепленной к вершине кора-
коида в области плечевого сустава. По изображению у Гульда (J. Gould, 1850—1883)
видно, что этот голубь приспособлен к наземной жизни, имея короткие крылья и длин-
ные ноги. В особенностях же скелета не видно ничего, что бы могло этот вид резко
противопоставить другим голубям. Что же касается редукции ключицы, то эта осо-
бенность является только дальнейшим развитием той тенденции, которая характерна
для всей группы голубей.
Говоря о птицах с сильно редуцированной ключицей, нельзя не упомянуть о не-
которых отечественных видах. У сов ключица не имеет значительного выгиба вперед,
относительно слабая и тонкая, особенно в нижней части. У некоторых видов, напри-
мер у сычика-воробья (Glaucidium passerinum), у домового сыча (Athene noctua) и
у ястребиной совы (Surnia ulula), нижняя часть ключицы, составляющая «дужку»,
полностью исчезла и сохранились лишь вершинные части, соединенные с коракоидами
и лопатками. На рис. 9 изображена грудина ястребиной совы с коракоидами. Пунк-
тиром обозначена исчезнувшая часть ключицы.
Не подлежит сомнению, что редукция нижней части ключицы у сов смогла про-
изойти лишь по тем же причинам, что у попугаев, т. е. в результате отсутствия рабо-
чей нагрузки. Однако первопричина изменения формы ключицы у них, видимо, иная,
в связи с чем и конечный результат оказался не вполне одинаковым с таковым у по-
пугаев. Как известно, совы не имеют зоба и связь развития этого органа с формой
ключицы у них отпадает. Далее, при рассматривании ключицы сравниваемых птиц
сбоку, мы видим существенную разницу в их форме. У попугаев, голубей и рябков
ключица у вершин тонкая и на всем своем протяжении не имеет изгиба вперед. У сов
она от мест соединения с коракоидами направляется вперед и потом уже образует
резкий изгиб, поворачиваясь в вентральном направлении (рис. 10). Вершинные части
толстые, имея значительную ширину в сагиттальной плоскости, и лишь ниже изгиба
ключица становится более тонкой, в особенности в области «дужки», где она может
совершенно редуцироваться.
Своеобразная форма ключицы у сов связана с особым развитием грудных мус-
кулов у этих птиц. Передняя часть грудных мускулов, прикрепленная к вершинным
отделам ключицы, имеет направление волокон вперед и, несомненно, при сокращении
осуществляет тягу крыльев в этом направлении. Вполне понятно, что вершинные
части ключицы в связи с этим должны быть хорошо развитыми и достаточно крепки-
ми. Что касается нижней, вентральной части ключицы, лишенной выгиба, то части
грудных мышц, прикрепленных к ней, имеют направление волокон, почти параллель-
ное ей. Поэтому вентральная часть ключицы лишена рабочей нагрузки и смогла реду-
цироваться.
— 54 —
Таким образом, мы видим у cqb совершенно оригинальное, «компромиссное» строе-
ние ключицы, имеющей один отдел, несущий определенную функцию и поэтому хоро-
шо развитый, и другой, лишенный функции и подверженный редукции. Эта особен-
ность, связанная также с особенностью развития и расположения грудных мускулов,
резко противопоставляет сов дневным хищным птицам, имеющим сильно выгнутую и
очень крепкую ключицу, и явно противоречит мысли о непосредственном филогенети-
ческом родстве между этими отрядами.
Что касается функционального значения вышеуказанных особенностей строения
ключицы у сов, то определить его не так просто. В отличие от голубей, рябков и по-
Рис. 10. Грудина ястребиной совы (Sarnia
alula) сбоку
Рис/ 9. Грудина ястребиной совы
(Sarnia alula) спереди
пугаев, у сов грудные мышцы имеют некоторый компонент, тянущий крылья вперед,
хотя и более слабый, чем у птиц с сильно выгнутой ключицей. Так как совы летают
медленно, то они, вероятно, не нуждаются в особенно сильных мускулах, подтяги-
вающих крылья вперед. Все-таки у них для осуществления этого движения имеются
еще и другие возможности, кроме вышеописанных. М. coraco-brachialis anterior у сов,
так же как у куликов и многих других птиц, расположен у основания crista lateralis
humeri с вентральной стороны его. Этот мускул у сов небольшой, но поверх него рас-
положено широкое сухожилие, тянущееся от m. biceps brachii к акрокоракоиду. Из
этого следует, что при сокращении бицепса, кроме сгибания локтевого сустава, долж-
но происходить движение крыла в плечевом суставе в том же направлении, что и при
сокращении m. coraco-brachialis anterior, т. е. вперед. Если же локтевой сустав фикси-
рован напряжением других мускулов, то сокращение бицепса должно всецело приво-
дить к подтягиванию крыла вперед.
Следует отметить, что у всех птиц m. biceps brachii имеет двойное прикрепле-
ние; в проксимальной своей части одно сухожилие связано с плечевой костью, другое —
с акрокоракоидом. Таким образом, во всех случаях сокращение этого мускула долж-
но содействовать движению крыла вперед. У птиц с сильно выгнутой ключицей ука-
занное действие m. biceps brachii, вероятно, имеет второстепенное значение так же,
как у голубей и других птиц с сильно развитым m. coraco-brachialis anterior. У сов
это действие двуглавого мускула, несомненно, имеет более важное значение, так как
у них подтягивание крыльев вперед при помощи других мускулов происходит лишь
с незначительной силой.
Итак, мы видим, что ключица у сов лишь частично, в области «дужки», утрати-
ла свое функциональное значение. У тех видов, у которых «дужка» не редуцирована
полностью, она тонкая, подвижная и широко расходится. Имея в виду ее способ-
ность раздвигаться, можно сделать предположение о том, что в данном случае имеется
приспособление, связанное с характерным способом питания сов. Известно, что совы
очень неохотно и лишь в крайних случаях прибегают к расчленению добычи и пред-
— 55 —
почитают ее заглатывать целиком. Так, взрослый филин заглатывает целиком крысу,
-филиненок весом в 300 г — взрослую домовую мышь. Продвижение таких крупных
кусков по пищеводу проследить нетрудно, причем ясно видно, что «дужка» лимитирует
их проходимость. Слабое развитие или полное отсутствие дужки, несомненно, дает воз-
можность совам заглатывать необыкновенно крупные куски, и возможно, что вся вы-
шеописанная цепь адаптаций началась именно с приспособления к такому своеобраз-
ному способу питания этих птиц.
Описывая способы, которыми осуществляется движение крыльев вперед у раз-
ных птиц, необходимо упомянуть и о куриных. У этих птиц передний край киля гру-
дины сильно вырезан с целью максимального приближения зоба к центру тяжести,
в связи с чем передний край грудного мускула отодвинут далеко назад. Соответ-
ственно изменилось и положение ключицы, которая своей нижней частью, т. е. «дуж-
кой», отодвинута далеко в каудальном направлении. Она не имеет явного выгиба впе-
ред и, не неся рабочей нагрузки, стала тонкой и слабой.
Ясно, что при вышеописанном положении грудные мускулы у куриных никак
не могут осуществить тягу крыльев вперед. Между тем форма плеча у этих птиц ни-
сколько не напоминает голубиную, crista lateralis обычного типа. М. coraco-
brachialis anterior у куриных расположен на вентральной стороне плечевого гребня,
как у куликов и сов, при этом чрезвычайно мал, имея, например, у фазана вес 0,1 г
при весе самой птицы в 1200 г. Видимо, наличие этого мускула у куриных практиче-
ского значения не имеет.
Однако у этих птиц сильно развита передняя, пропатагиальная часть дельтовид-
ного мускула. Она пересекает переднюю перепонку крыла (propatagium) и прикрепле-
на широкой сухожильной лентой к разгибателю кисти (m. extensor metacarpi radialis).
При сокращении она подтягивает этот мускул, содействуя, таким образом, разгиба-
нию кисти. Но, будучи прикреплена к предплечью на некотором расстоянии от локтя,
пропатагиальная часть дельтовидного мускула должна содействовать сгибанию локтя,
а в случае фиксирования его другими мускулами — отводить крыло в плечевом суста-
ве от тела вперед.
Описанное движение имеет довольно сложную механику, но оно поддерживается
только что описанным действием m. biceps brachii. Оба мускула совместно, несомнен-
но, развивают тягу крыльев вперед, достаточную для куриных. Эти птицы не относят-
ся к скоростным летунам, обычно не летают на большие расстояния, а во время ин-
тенсивного взлета, характерного для них, крылья пропеллируют при всех фазах дви-
жения, так что в это время их приходится подтягивать лишь назад. Во всяком слу-
чае, других возможностей для движения крыла вперед у куриных не имеется.
Из вышесказанного явствует, что основная функция ключицы у птиц состоит
в том, что эта кость служит местом прикрепления передней части грудного мускула,
тянущего крыло вперед. Чем сильнее выгиб ключицы, тем с большей силой совершает-
ся тяга крыла вперед. В случае отсутствия выгиба вперед ключица теряет свою ра-
бочую нагрузку и редуцируется, вплоть до полного исчезновения вентральной части,
т. е. «дужки». Это никак не связано с редукцией способности к полету, но тяга крыла
вперед осуществляется в таких случаях иным способом, либо за счет сильного раз-
вития m. coraco-brachialis anterior, либо за счет работы m. biceps brachii, прикреплен-
ного одним сухожилием к акрокоракоиду, либо за счет этого же мускула плюс дей-
ствие пропатагиальной части дельтовидного мускула.
Что касается голубей и рябков, то их по вышеописанному комплексу признаков
можно объединить между собой и резко противопоставить ближайшим родственным
группам, в частности куликам. Основная причина развития этого комплекса призна-
ков — переход данных птиц от животной пищи к растительной, что повлекло за собой
развитие большого зоба. Так, изменение способа питания привело к глубоким и свое-
образным изменениям в строении плечевого пояса, единственная функция которого
у птиц заключается в передвижении их в воздухе, т. е. в полете. Такая связь на пер-
вый взгляд кажется парадоксальной, но более глубокое изучение этого вопроса делает
понятной взаимосвязанную цепь изменений от приспособления к растительному пита-
нию до приобретения своеобразной формы crista lateralis плечевой кости и сильного
развития m. coraco-brachialis anterior. Для понимания филогенетических связей голу-
бей и рябков выявление вышеописанных морфологических особенностей их имеет опре-
деленное значение. Несмотря на совершенно различные условия обитания, голуби и
рябки сохранили удивительное сходство в морфологии плечевого пояса, объединяясь
естественным образом в одну группу, резко противопоставляемую куликам, чайкам и
чистикам, объединенным еще Гадовым (Н, Gadow u. F. Selenka, 1891—1893) в группу
Laro-Limicolae. При этом следует отметить, что и по данному комплексу признаков
голуби и рябки являются измененными и резко аберрантными по сравнению с кули-
ками, сохранившими исходный тип, менее специализированный и широко распростра-
ненный среди других отрядов птиц. Впрочем, при близком сходстве между рябками
и голубями намечается и некоторая разница в данном комплексе признаков: у первых
ключица в большей степени редуцирована, чем у последних. Из этого следует, что
и по данному комплексу признаков рябки являются более измененными, чем голуби,
представляя собой дальнейшую специализацию голубиного типа.
— 56 —
Что касается попугаев, то они при большом сходстве с голубями и рябками
в вышеуказанном комплексе признаков в прочих чертах строения Резко отличаются
от них, не исключая также ряда особенностей плечевого пояса. Так, например, форма
коракоидов у Попугаев существенно иная, чем у всех ржанкообразных в широком
смысле. В пределах последних опять голуби и рябки объединяются, противопостав-
ляясь достаточно резко куликам. По форме грудины, в частности по развитию и рас-
положению вырезок ее заднего края, голуби и рябки тоже объединяются, представляя
собой резко аберрантный тип, развившийся путем своеобразной специализации из бо-
лее генерализованного типа куликов. Между тем у попугаев грудина совсем иной
формы, одинаково резко отличающаяся от грудины как голубей и рябков, так
и куликов.
ZUSAMMENFASSUNG
Das Skelett des Schultergiirtels weist bei den Tauben einige Eigenheiten auf, die bei den
Flughiihnern ohne jegliche Abanderung wiederkehren. Die Crista lateralis des Humerus ist
bei diesen Vogeln kurz und in einen spitzen Haken ausgezogen (Fig. 1) und das Schliissel-
bein besitzt keine bogenformige Biegungin sagittaler Ebene nach vorne, wie sie den meisten
anderen Vogeln eigentiimlich ist. Hochstwahrscheinlich ist die eigenartige Form des
Schliisselbeines bei den Tauben und. Flughiihnern durch das Vorhandensein bei dieseri
Vogeln eines grossen Kropfes bedingt, welcher anderenfalls in gefiillten Zustande den
Schwerpunkt weit nach vorne verlagern wiirde und beim Auffliegen vom Boden hinder-
lichsein miisste.
Bei alien Vogeln dient das Schliisselbein als Ansatz fur die vordere Partie des Bru-
stmuskels. Die bogenformige Bieg ng des Schliisselbeines nach vorne bedingt eine Verlagerung
der obengenannten Partie des Brustmuskels vor das Schultergelenk, welche auf diese Weise
den Flugel nach vorne zieht und gleichzeitig auf das Schliisselbein einen Zug nach hinten
ausiibt. Gegen diesen Zug nach hinten ist das Schliisselbein durch die bogenformige Aus-
buchtung und den grossen Durchmesser in sagittaler Richtung gefestigt.
Da bei den Tauben und Flughiihnern das Schliisselbein keine Biegungnach vorne besitzt^
so ist ein Zug des Flugels durch den Brustmuskel nach vorn ausgeschlossen. Die Belastung
des Schliisselbeines durch den Zug nach hinten fallt auch weg, infolgedessen das Schliissel-
bein bei diesen Vogeln schwach und diinn wird und iiberhaupt in Riickbildung beg-
riffen ist.
Das Fehlen einer vorwartszienden Komponente des Brustmuskels wird bei den Tauben
und Flughiihnern durch eine ausserst starke Ausbildung des m. coraco-brachialis anterior
ersetzt. Dieser Muskel zieht den Fliigel direct nach vorne ist aber bei den meisten Vogeln
so klein, dass er praktisch ohne Bedeutung ist. Seine starke Ausbildung bei den Tauben
und Flughiihnern bedingt die eigentiimliche Form der Crista lateralis bei diesen Vogeln^
welche dem obengenannten Muskel als Ansatzflache dient.
ЛИТЕРАТУРА
Штегман Б. К. 1950. Функциональное значение особенностей строения грудины
у куриных птиц. Сб. Памяти акад. П. П. Сушкина, изд-во АН СССР.
Штегман Б. К. 1957. О летных качествах голубей. Зоол. журн., т. XXXVI, вып. 2.
Штегман Б. К. 1957. О летных качествах рябков. Зоол. журн., т. XXXVI, вып. 10.
Fiihrbringer М. 1888. Untersuchungen zur Morphologic und Systematik der
Vogel. Bd. I—II, Amsterdam.
G a d о w H. und S e 1 e n k a F. Bronn’s Klassen und Ordnungen des Tier — Relcu’s,
Vogel, Bd. 6, Anatomie.
Gould J. 1850—1883. Birds of Asia, Bd. VI, pl. 53, London.
Meyer A. B. 1889—1897. Abbildungen von Vogel-Skeletten. Bd. I, Dresden.
S у M. 1936. Funktionell-anatomische Untersuchungen am Vogelfliigel.—Journ. f.
Ornithol., Bd. 84, Heft 2.
БЮЛЛЕТЕНЬ М. О-ВА ИСП. ПРИРОДЫ, ОТД. БИОЛОГИИ, Т. LXII (5), 1957
О значении кивсяков и муравьев в трансформации
органического вещества под лесными насаждениями
в условиях сухой степи1
Д. Ф. Соколов
The importance of Julidae and ants in the
transformation of organic substance under forest
timber in conditions of the dry steppe zone
D. F. Sokolov
Роль почвенных беспозвоночных животных в переработке органического вещества
под пологом леса общеизвестна. Наиболее изучены в этом отношении дождевые черви,
деятельности которых посвящена обширная литература. Из работ по этому вопросу,
заслуживающих особого внимания, следует указать на исследования Ч. Дарвина
(1882), М. П. Архангельского (1929), Н. А. Димо (1938), С. И. Пономаревой, (1948,
1949), М. С. Гилярова (1951), А. И. Зражевского (1954) и др.
В работах по изучению состава органического вещества почв под лесными на-
саждениями Деркульской научно-исследовательской станции Института леса
АН СССР, расположенной в условиях сухой степи, было обращено внимание на роль
кивсяков и некоторых видов муравьев в переработке растительных остатков.
Кивсяки и их значение в превращении растительных остатков
Кивсяки, или многоножки (Julidae), значительно распространены под лесными
насаждениями и в естественных лесах степной и лесостепной зон. Жизнь и развитие
многоножек зависят от наличия мертвого органического субстрата, являющегося источ-
ником питания (опад, подстилка, корни леоной растительности), и от его влажности.
Двухлетние наблюдения за численностью кивсяков под разновозрастными и разнопо-
родными лесными насаждениями показали, что количество этих животных, находя-
щихся в активном состоянии, не остается постоянным (табл. 1). Численность кивсяков
при более детальных раскопках под пологом посадок лесной полосы № 77 превышала
500 экземпляров на 1 м2 (Гиляров, 1956).
Собранные (за оба периода времени) с учетных площадок (10 м2) кивсяки бы-
ли подразделены по их размеру на 3 группы. Вес 1 экземпляра для первой группы
варьировал от 328 до 448 мг, для второй — от 223 до 316 мг, для третьей —
от, 25 до 61 мг.
Масса живых кивсяков, как установлено лабораторными исследованиями, состоит
из зольных элементов (26,1%), воды (29,3%), летучих веществ (44,6%). В составе по-
следних на долю С приходится 30,8%, а на долю N — 7,43%.
В зольном составе кивсяков, согласно данным А. К. Алешиной (Зонн, 1954)
преобладают соединения кальция (в процентах к золе):
SiO2...........6,92; Fe2O3............3,47;
CaO............ 66,40; А13О3..........0,97;
MgO.............2,05; SO3............2,96.
P2O5...........3,86;
1 Работа выполнена в лаборатории лесного почвоведения Института леса
АН СССР, руководимой проф. С. В. Зонном. Значительная часть анализов проведена
старшими лаборантами С. С. Францкевич и А. С. Дубковой, которым приношу глубо-
кую благодарность.
Таблица 1
количество кивсяков в леснь!х насаждениях Дернульской научно-исследовательской станции1 (в тысячах на 1 га)
Насаждение и его возраст Состав3 Месяц учета 1954 г- 1955 г.
круп- ных сред- них мел- ких мертвых всего круп- ных сред- них мел- ких мертвых всего
5 Д, 4 Я. з. 1 Я. п. VI 137 143 250 не учтено 530 39 137 248 10 434
Пдл Ак. ж. VII 3 2 7 198 210 30 294 555 4 883
Лесной массив. 5 Д, 4 Я. з., 1 Я. п. (Ак. ж. вырублена) VI 130 139 583 не учтено 853 16 76 98 2 192
квартал 50 (б. 27), VII 4 8 9 318 339 16 121 228 1 366
выд. 5, 18—19 лет 7,5 Д, 2 Я. з„ 0,5 Я. п. (Я. з. Ак. ж. вырублены на 50%) VI 105 182 333 не учтено 620 28 90 275 4 397
VII 57 94 201 151 503 23 185 219 2 429
10 Д VI 84 84 226 66 460 16 58 202 7 283 |
(Я. п., Ак. ж вырублены) VII 18 25 53 236 332 43 214 193 3 453 Сл QO
Лесное насаждение, 6 Д, 4 Я. ед. Бр. Пдл ред. Я-, Бр. VI 68 108 312 29 517 51 207 166 3 427 1
квартал 51 (б. лесная полоса № 77), Пдл ср. густ. Ак. ж. и ж. т. VII 45 96 324 68 533 11 195 214 1 421
5 Д, 4 Я., 1 Бр. VI 57 61 203 37 358 25 79 45 2 151
выд. 4 и 5,
52—53 года Пдл вырублен, 1 год. поросль VII 15 45 139 78 277 23 152 151 4 330
Лесной массив. 10 д VI 42 95 187 27 351 26 530 845 10 1411
квартал 47 (б. 25), выд. 7, 22—23 года (загущенный) VII 17 35 360 50 462 16 344 319 2 681
10 Д VI 34 77 175 36 322 55 343 709 5 1112
(прорежен на 15%) VII 15 28 489 32 564 16 261 541 3 821
Лесной массив. 7 Кл. о., 3 Д, ед. Я. з. VI 130 870 1080 30 2110
квартал 48 (б. 25), выд. 7, 35—40 лет Пдл ср. густ. Д, Кл. о., Я. з. VII «учет не производился 35 434 522 9 1000
Примечания: 1 Сбор кивсяков проводился только с поверхности почвы с площади 1 м3 с 10-кратной повторностью.
з Д-^дуб; Я. з. — ясень зеленый; Я. п. — ясень пушистый; Ак. ж.—акация желтая; Пдл — подлесок; Бр. — берест;
Ж. т. — жимолость татарская; к л. о. — клен остролистный.
— 59 —
Следует указать, что наибольшее накопление неорганических компонентов обна-
ружено в хитинизированном покрове животных (до 46%), который периодически сбра-
сывается по мере их роста.
С целью выяснения характера питания кивсяков и их отношения к качественно-
му составу энергетического материала и состоянию увлажненности последнего были
проведены лабораторные опыты, сущность которых заключается в следующем:
Опыт первый. В сосуды была помещена воздушно-сухая почва (чернозем),
на поверхность которой внесена такого же состояния влажности дубово-ясенево-клено-
вая подстилка в количестве 100 г.
Сосуды содержались в лаборатории при температуре 20—25°. Из помещенных
в сосуды при закладке опыта относительно однородных по развитию 25 кивсяков че-
рез 9 суток погибло 3 особи, или 12%. Через 13 суток остальные углубились в почву и
свернулись в клубочки. Через 18 суток погибло еще 20 кивсяков, что составляет 80%
от всех помещенных в сосуды. Гибель последних двух особей была обнаружена через
21 сутки. Из сказанного вытекает, что недостаток влаги в субстрате привел перво-
начально к прекращению активности (в том числе и питания), а в дальнейшем к ги-
бели кивсяков.
Необходимо отметить, что в условиях сухой степи в течение вегетационного пе-
риода очень часто наблюдается резкое снижение влаги в почве (А. Н. Ступникова,
личное сообщение) (рис. 1 и 2), благодаря чему значительно варьирует количество
кивсяков, находящихся на поверхности почвы.
Опыт второй был поставлен с целью выявления потребности кивсяков в пи-
тательных веществах. В сосуды с почвой было внесено по 100 г листьев древесно-
кустарниковых пород и по 40 особей однородных кивсяков: влажность почвы и ра-
стительного материала в течение опыта поддерживалась на уровне 60% от их полной
ълагоемкости.
Через месяц листовая пластинка была объедена: у клена — на 95—97%, у ясе-
ня — приблизительно на 85% и у дуба — на 60%. У акации желтой объедания листьев
не наблюдалось, и в сосудах с этими растительными остатками кивсяки в скором
времени начали погибать.
По истечении указанного времени осталось живых кивсяков:
в сосуде с кленовыми листьями ... 38 особей, или 95%,
в сосуде с ясеневыми листьями ... 35 особей, или 87,5%,
в сосуде с дубовыми листьями .... 32 особи, или 80%.
в сосуде с листьями акации желтой .. 3 особи, или 7,5%.
В процессе своей жизнедеятельности кивсяки перерабатывают растительные
остатки, заглатывая при этом и почвенные частицы. При нормальном развитии они
ежедневно выделяют массу экскрементов, равную около 7з1 части их живого веса,
с содержанием влаги около 47%.
Относительная подвижность воднорастворимых и легко гидролизуемых форм
углерода и азота как в отмерших кивсяках, так и в их копролитах значитель-
на (табл. 2).
Таблица 2
Содержание зольных и биогенных элементов в кивсяках и копролитах
и подвижность в них форм углерода и азота (Соколов, 1955)
Объект иссле- дования pH (водная суспензия) В % на сухое вещество В % на сухое вещество к общему содержанию С и N
гигроскопи- ческая Н2О зольность С N
водная вытяжка кислотная вытяжка
С N С N
Кивсяки . . . 7,8 6,08 26,1 30,78 7,433 4,52 14,7 1,008 13,6 5,54 18,0 2,166 29,1
Копролиты . . 6,7 6,11 38,9 37,77 2,655 2,68 7,9 0,404 15,2 6,60 19,5 0,762 25,3
Из массы кивсяков в водную вытяжку переходит от 13,6 до 14,7% растворимых
«форм углерода и азота, а в кислотном гидролизате, соединения которого усваиваются
Глубина в см
8
io т/ 7г и « 7з 7о Т' 7s 7о lb 7з Tk г§ То Th
А •------------•----------------•б --------------9г ............iii»Z? •------X-----9 в 9*-*
Рис. 1
Взятие
. Динамика влажности чернозема в 17 — 19-летнем дубово-ясеневом насаждении в 1953 —1955 гг. (в процентах ко
взятой навеске).
почвенных проб для определения влажности производилось в первые числа П1 декады каждого месяца: а — апрель; б — май; в — июнь;
г — июль; д — август; е — сентябрь; ж — октябрь
/о v 2 7з 7r & 7a To Tt Тг а г4 To T Tx
Рис. 2. Динамика влажности чернозема в 49 — 52-летней полосе из дуба, ясеня и ильцовых в 1953 —1955 гг. (в процен-
тах ко взятой навеске).
Взятие почвенных проб для определения влажности производилось в первые числа III декады каждого месяца: а — апрель; б — май; в — июнь;
г — июль; д — август; е — сентябрь; ж — октябрь.
Условные знаки те же, что и на рис. 1
Таблица 3
Влияние кивсяков на ход разложения и минерализации лесного опада (листьев) в лабораторных условиях
Внесено в 1 сосуд при закладке опыта (4/IX 1953 г.) Определено при снятии опыта (5/Х 1953 г.) Уменьшение (—) или увеличение Потеря от прока-
Варианты опыта CQ CQ в том числе: кивсяков С-, CQ CS CQ в том числе: кивсяков ( + ) за время опыта в % от начального коли- ливания в % к сухому веществу при:
О О О О) CD особей CQ О чества содержания
сухого в С особей общий вес в г сухого в< С N живых мертвых общий В( С N закладке опыта снятии опыта
Почва 500,0 22,135 2,3350 — — 500,0 21,158 2,2350 — — — — 4,5 — 4,3 12,8 10,8
Почва + кивсяки 500,0 22,135 2,3350 100 32,33 500,0 21,345 2,1400 74 26 31,90 — 3,6 — 8,4 12,8 11,3
Почва + кивсяки) + листья дуба 525,0 32,923 2,5625 100 37,56 520,16 30,307 2,3353 92 8 36,82 —19,4 —16,6 12,8 93,1 11,0 88,3
Листья дуба + + кивсяки . . . 25,0 10,788 0,2275 100 36,57 20,17 9,655 0,2474 87 13 36,24 —10,5 + 8,5 93,1 91,6
Листья луба . . . 25,0 10,788 0,2275 — — 21,07 9,920 0,2299 — — — — 8,2 + М 93,1 91,3
Примечание: В графе „Потеря от прокаливания“
потери от прокаливания листьев дуба.
в числителе даны величины потери
от прокаливания почвы, в знаменателе —
- 63 —
как высшими растениями, так и микроорганизмами, количество их возрастает
в 1,5—2 раза.
Исходя из зольности экскрементов, а также из потери от прокаливания почвы
и подстилки, можно предполагать, что кивсяки заглатывают около Чз почвенных ча-
стиц и 2/з растительных остатков. Относительная подвижность воднорастворимых и
легко гидролизуемых углеродистых и азотистых веществ копролитов примерно та же,
что и у мертвых кивсяков. Исключение составляет растворимость в воде соединений
углерода, выход которых определяется половиной величины в сравнении с раствори-
мостью этих же веществ в сухой массе кивсяков.
Лабораторные опыты, поставленные с целью выявления участия кивсяков в раз-
ложении разнородного лесного опада, показали, что для жизнедеятельности этих жи-
вотных необходимы и почва и остатки лесной растительности. Так, наибольшее коли-
чество сохранившихся кивсяков отмечено в сосуде с почвой и листьями дуба, и, наобо-
рот, наибольшая гибель их установлена в сосуде с одной почвой без листьев (табл. 3).
Если дождевые черви довольствуются энергетическим веществом органоминеральной
части почвы, то кивсяки нуждаются еще и в органическом веществе опада древесных
насаждений.
Выживаемость кивсяков в лабораторных условиях не превышала 6 месяцев.
К концу первого месяца лабораторного опыта в сосудах погибло от 8 до 26% жи-
вотных \ причем наименьшая гибель их выявлена в сосудах с почвой и листьями
дуба и, наоборот, наибольшая — в сосудах с одной почвой без внесения каких-либо
растительных остатков.
Потеря углерода и азота в растительных остатках при их разложении и мине-
рализации тесно связана с выживаемостью и сохранностью подопытных животных.
Потеряно указанных веществ больше там, где созданы наиболее благоприятные усло-
вия для существования кивсяков. Но, наряду с этим, в двух вариантах опыта (в со-
судах с листьями дуба и кивсяками и в сосудах с листьями дуба) отмечена незначи-
тельная прибавка азотистых веществ. В первом случае, очевидно, это связано с нали-
чием копролитов, являющихся прекрасным субстратом для оптимального развития
микрофлоры, и в том числе азотфиксирующих микроорганизмов. Во втором варианте,
прибавка этого компонента незначительна и находится в интервале отклонений точно-
сти опыта. С увеличением разложения растительных остатков естественно уменьшается
как абсолютное, так и относительное содержание углерода и увеличивается процентное
содержание азотистых и зольных элементов, в особенности это заметно в сосудах
с кивсяками (табл. 4).
Из приведенных данных следует, что в результате превращения органических
веществ происходит подщелачивание последних, с более ярко выраженным показателем
в сосудах без почвенного фона (pH с 5,5 до 7,7). В связи с чем происходит подщела-
чивание? Это, по всей вероятности, связано как с освобождением (при увлажнении
опытного материала) органических кислот, распадом и окислением их до СО2 и Н2О,
так и с одновременным образованием углекислых солей кальция, в основном в форме
бикарбоната кальция. С уменьшением углеродистых соединений в процессе разложе-
ния органического вещества отмечается и изменение подвижности воднорастворимых
и легко гидролизуемых биогенных элементов (табл. 5).
В сосудах с кивсяками наблюдается незначительное увеличение легко гидроли-
зуемых азотистых соединений, в то время как воднорастворимые формы этого компо-
нента уменьшены наполовину и больше.
По углероду получена несколько иная картина. Во всех случаях в присутствии
кивсяков падает выход этих соединений, растворимых в воде и гидролизуемых в кис-
лоте. Однако следует иметь в виду, что легко гидролизуемые соединения углерода
определяются в несколько раз большими величинами, чем воднорастворимые.
Изменение группового состава растительных остатков в процессе
* их разложения и минерализации
Данные по изменению группового состава органических остатков представлены
в табл. 6.
С разложением растительных остатков в естественных или в лабораторных усло-
виях происходит относительное нарастание количества зольных элементов, азотистых
веществ и негидролизуемого остатка.
Гемицеллюлозы (пентозы, гексозы, соединения сахаров с уроновыми кислотами)
исследованного материала относительно мало изменяются, зато количество клетчатки
как в сосудах с кивсяками, так и в сосудах с листьями дуба без почвенного фона
резко снижается (почти наполовину, в среднем на 49%). За счет уменьшения рас-
сматриваемой группы веществ, а также и за счет группы воскосмол наблюдается
1 В сосуды при закладке опыта было помещено по 100 особей кивсяков, относи-
тельно Однородных по размерам и весу.
Таблица 4
Изменение pH, относительного содержания биогенных и зольных элементов при разложении и минерализации органического
вещества в лабораторных опытах
Варианты опыта Количество вещества pH (водная суспензия) Содержится в % на сухое вещество
углерода азота зольных элементов и ми- нерализованного остатка
а б в а б в а б в а б в
Листья дуба + почва 100 г сухого вещества 1000 г . 5,9 7,3 6,2 7,17 6,8 7,6 51,03 4,43 50,16 4,46 42,68 4,19 0,918 0,476 1,241 0,429 1,353 0,420 6,83 87,47 10,02 83,26 16,1 87,21 | 2
Листья дуба + 100 кивсяков + почва 100 г сухого вещества 1000г . 5,9 7,3 7,1 7,5 6,9 7,5 51,03 4,43 40,62 4,45 40,37 4,26 0,918 0,467 1,807 0,505 2,201 0,429 6,83 87,47 18,59 83,88 1 19,44 87,15
Почва 1000 г сухого вещества 7,3 7,8 7,6 4,43 Не опре- делено 4,15 0,467 Не опре- делено 0,417 87,47 Не опре- делено 87,01
Листья дуба 100 г сухого вещества 5,9 Не опре- делено 7,7 51,03 Не опре- делено 41,51 0,918 Не опре- делено 1,418 6,83 Не опре- делено 13,08
Примечание: а — показатели величин в исходном, взятом для опыта материале;
б — „ «в момент снятия опыта через 100 дней;
в— > > „ „ 305 дней.
— 65 —
Таблица 5
Подвижность биогенных элементов в листьях дуба за время опыта
Время и виды определений По почвенному фону Без фона
листья дуба листья дуба + кивсяки листья дуба
При закладке опыта определено:
воднорастворимого С (г на сосуд) (а) 10,278 10,278 9,118
легко гидролизуемого С (г на сосуд) (б) 18,602 18,602 12,592
воднорастворимого N (г на сосуд) (в) 0,258 0,258 0,118
легко гидролизуемого N (г на сосуд) (г) . . . . 0,720 0,720 0,250
По снятии опыта через 100 дней определено:
г на сосуд 2,299 1,314 Не опре-
воднорастворимого Св 22,4 12,8 делено
г на сосуд 7,577 8,314
легко гидролизуемого СЗ в 6u •••••• 40,7 44,7 » •
т г на сосуд 0,048 0,132
воднорастворимого N в 0/ „ 70 ОТ „ В 18,6 51,2 п п
г на сосуд 0,511 0,810
легко гидролизуемого N в о/ Q-p р« 70,0 112,9 п »
По снятии опыта через 305 дней определено:
г на сосуд 0,985 0,818 0,770
воднорастворимого С в— 9,6 8,0 8,5
г на сосуд 7,500 8,494 1,945
40,3 45,6 15,5
легко гидролизуемого СЗ в q/ qt 6u ••••••
wT г на сосуд 0,083 0,034 0,040
воднорастворимого N в 0/ Л и /0 от „ в 32,1 13,2 33,9
жт г на сосуд 0,559 0,813 0,144
легко гидролизуемого N в qt——— 77,6 112,9 57,6 •
увеличение белковых (протеиновых) соединений, в особенности в сосудах с кивсяка-
ми (в 2,2 раза).
Таким образом, в течение 305-дневного разложения и минерализации листьев
дуба в лабораторной обстановке произошли следующие изменения (табл. 7).
Максимальные потери органического вещества, в том числе и биогенных эле-
ментов. установлены в сосуде с листьями дуба без почвенного фона, и, наоборот, ми-
нимальные — в сосудах с одной почвой. Но отсюда еще не следует, что в сосудах с
кивсяками разложение и минерализация растительных остатков шли менее интенсивно,
чем в сосудах без этих животных и почвенного фона. Кивсяки произвели большую
работу: листья значительно ими измельчены, а нижние слои органической массы в
какой-то мере перемешаны с почвенными частицами, благодаря чему и происходит
более интенсивное поглощение и закрепление растворимых соединений органомине-
ральных веществ. Одновременно необходимо отметить, что в сосудах с кивсяками
потеряно ничтожное количество азотистых соединений (2,7% от общего содержа-
ния N), тогда как в сосудах без животных эти потери возрастают до 10,2%.
Одним из наиболее распространенных методов определения интенсивности разло-
жения органических остатков под пологом леса является метод учета углекислоты с
определенной площади и за определенный промежуток времени. Исходя из этой пред-
посылки, был заложен в вакуум-эксикаторах лабораторный опыт по схеме: 1) сосуд
с почвой; 2) сосуд с почвой и кивсяками; 3) сосуд с почвой, листьями дуба и кив-
сяками; 4) сосуд с листьями дуба и кивсяками; 5) сосуд с почвой, листьями дуба и
акации желтой и 6) сосуд с листьями дуба.
— 66 —
Таблица 6
Изменение в составе листьев дуба за время опыта с 12/VI 1953 г. по 17/IV 1954 г.
(в % на сухое вещество)
Перечень определений При за- кладке опыта В конце опыта (17/IV 1954 г.)
по почвенному фону листья дуба без почвен- ного фона.
листья дуба листья дуба листья дуба + кив- сяки
Зольные элементы Сухой остаток в том числе ( зодчих веществ Вещества, экстрагируемые спиртобензолом . . Гемицеллюлозы Клетчатка Негидролизуемый остаток (лигнин, кутин и пр.) Протеин * 6,83 28,35 1,01 2,93 6,15 11,40 16,92 16,34 4,72 16,10 14,13 0,09 3,20 4,00 9,43 15,64 31,62 7,88 19,44 11,00 0,4 2,90 3,76 8,48 8,04 32,90 10,48 13,08 15,88- 0,09 3,88 4,23 11,59 9,17 34,09 8,76
Сумма 87,78 95,60 91,20 92,92
В течение 10 суток велись наблюдения за разложением органического вещества
по количеству выделившейся углекислоты, учет которой производился специально в
утренние часы и один раз круглосуточно, через каждые 4 часа.
Наиболее интенсивное выделение СО2 наблюдалось, как и следовало ожидать,,
в варианте «почва 4- листья дуба + кивсяки», где величины двуокиси углерода ино-
гда достигали свыше 45 мг за время 15-минутного наблюдения в течение 10 суток
(рис. 3). Интенсивность «дыхания» при разложении и минерализации органического-
вещества зависит не только от породы, состава и количественного соотношения его,
но и от увлажненности субстрата и колебания температуры, при которой протекают
биологические процессы и биохимические реакции. Отсюда понятны и те резкие коле-
бания в выделении углекислоты: ее максимумы и минимумы не совпадают в отдель-
ных вариантах опыта.
По продуцированию углекислоты в среднем за весь период наблюдения (правый
график, рис. 3) варианты опыта располагаются в • следующем убывающем порядке:
почва с листьями дуба и кивсяками > почва с листьями дуба и акации желтой >
листья дуба с кивсяками > листья дуба > почва с кивсяками > почва.
Из этого ряда распределения отдельных вариантов опыта по выделению угле-
кислоты следует, что в сосудах «листья дуба + кивсяки» биохимические процессы
несколько замедлены в сопоставлении с сосудами «листья дуба и акации желтой».
При последнем сочетании смешения опада древесно-кустарниковых пород происходит
более энергичное разложение и минерализация органических остатков (Рунов и Со-
колов, 1956).
Суточная динамика выделения СО2 показала, что наибольшее продуцирование
углекислоты падает на утренние часы и, наоборот, наименьшее — на полуденное и
ночное время (рис. 4).
Таким образом, роль кивсяков в переработке растительных остатков под поло-
гом естественных лесов и лесных насаждений довольно значительна. В результате их
жизнедеятельности происходит не только перемешивание и перетирание подстилки и
рыхление поверхностных слоев почвы, чем создаются благоприятные условия для
микронаселения и воздушного режима лесных почв и почв под лесными насажде-
ниями, но и обогащение последних копролитами, которые выделяются в течение суток,
в зависимости от численности и возраста животных, в следующем количестве (по уче-
ту 4/VII 1955 г.), приводимом в табл. 8.
Исходя из этих данных, можно утверждать, что кивсяками при указанных усло-
виях будет выделено в течение вегетационного периода от ПО до 686 кг/га экскремен-
тов в пересчете на сухую массу. Конечно, это количество выделенич может быть
Таблица 7
Изменения в составе органического вещества в процессе его разложения
и минерализации
Варианты опыта и перечень определений Внесено при закладке опыта в г (100%) Определено по снятии опыта, г/% Потеряно за время опыта, г/%
93,2 45,68 47,52
органического вещества 49,0 51,0
Листья дуба С 51,03 21,28 41,7 29,75 58,3
в том числе 0,92 0,75 0,17
N 81,5 18,5
218,5 179,8 38,70
органического вещества 82,2 17,8
Листья дуба 95,33 67,39 . 29,94
+ почва С 70,7 29,3
в том числе • 5,59 5,02 0,57
N 89,8 10,2
218,5 171,0 45,50
Листья дуба органического вещества 78,2 21,8
+ кивсяки + почва в том числе С 95,33 63; 97 67,1 5,44 31,36- 32,9 0,15
N . . . . 5,59 97/3 2,7
125,3 119,6 5,70
органического вещества 95,4 " 4,6
Почва 44,30 41,04 3,26
С 92,6 7,4
в том числе 4,12 0,55
N 4,67 88,2 11,8
Таблица 8
Суточное выделенйе копролнтов кивсяками
Насаждение Копролитов в кг/га
свежих сухих
Массивное (квартал 50) { уясень у акация у ; ; Лесная полоса № 77 j Дуб--ясень - ильмовые по- Массивное (квартал 48) 1 ™е« 4 1 | дуб — ясень зеленый 3,75 2,28 1,55 J9,74 (4/VI) 1 4,39 1,77 1,07 0,73 4,59 2,07.
В среднем за 15 минут
Рис 3. Динамика выделения С02 в лабораторных опытах с почвой, листьями и кивсяками (в мг за 15 минут наблюдения).
rnrvnbi* 1 - с почвой; 2 — с почвой и кивсяками; 3 - с почвой, листьями дуба и кивсяками; 4 - с листьями дуба и кивсяками; 5 - с листьями дуба и
сосуды» акации желтой; 6 — с листьями дуба
Рис. 4. Суточные изменения СО2 в лабораторных опытах с почвой, листьями дуба и кивсяками (в мг за 15 минут наблю-
дений).
Сосуды: 1 — с почвой; 2 — с почвой и кивсяками; 3 — с почвой, листьями дуба и кивсяками; 4 — с листьями дуба и кивсяками; 5 — с листьями
дуба и акации желтой; 6 — с листьями дуба
— 70 —
значительно увеличено за счет не только численности, но и нормального развития
кивсяков, зависящих исключительно, как уже отмечалось, от погодных условий (тепла
и увлажненности субстрата). Экскременты, благодаря подвижности в них воднораство-
римых и легко гидролизуемых форм углерода и азота, являются прекрасным энерге-
тическим материалом для микроорганизмов и источником питательных веществ для
высших растений.
Роль муравьев в переработке растительных остатков
«Польза муравьев заключается в том, что они уничтожают много вредных насе-
комых и рыхлят почву, способствуя ее аэрации, снижают, благодаря выделениям
слюнных желез, кислотность почвы» (Гринфельд, 1939).
Рис. 5. Вертикальный разрез гнезда Formi-
ca pratensis
Рис. 6. Вертикальный разрез гнезда Formi-
ca pressilabris
Мы не ставили перед собой задачи изучить влияние муравьев вообще на почво-
образовательный процесс под пологом лесной растительности, а ограничились только
небольшими исследованиями по выяснению роли двух видов муравьев: Formica pra-
tensis Retz. и F. pressilabris Nyl.,— в переработке растительных остатко-в. Названные
виды муравьев широко распространены в лесостепной и степной зонах. Из литера-
туры известно (Леббок, 1884), что пищей для муравьев служат насекомые, мед, мед-
вяная роса тлей и плоды, в связи с чем нет никаких оснований предполагать воз-
можность использования этими животными опада в качестве энергетического мате-
риала, как это имеет место у дождевых червей или кивсяков.
- 71 —
Для выяснения значения муравьев в изменении найденных в их гнездах расти-
тельных остатков и их воздействия на органо-минеральный комплекс почвы были
сделаны вертикальные разрезы двух муравейников, откуда послойно взяты для ана-
лиза образцы в количестве: 5 проб из первого гнезда (рис. 5) и 3 проб с этой же
площади вне гнезда (№ 9—11); 4 проб из второго гнезда (рис. 6) и 2 проб вне
гнезда (№18—19).
Следует оговориться, что муравьи различных видов по своим размерам резко
различаются между со’бой, благодаря чему используемый для строительства мура-
вейника растительный материал также различен. Более крупному виду муравьев
(F. pratensis), естественно соответствуют более крупные растительные остатки, меха-
нически перемешанные с почвенными частицами.
Соотношения между минеральной частью и растительными остатками в пробах
были неодинаковы (табл. 9). Особенно эти различия заметны в количестве органиче-
ской массы отдельных фракций. Так. наибольшие количества указанных веществ в
пробах, взятых из муравейника F. pratensis, падают на крупную фракцию (диамет-
ром больше 3 мм), в то время как на долю такой же фракции муравейника F. pressi-
labris приходится лишь от 7,3 до 15,4°/о. Зато в пробах из последнего муравейника
значительно преобладает более мелкая фракция полуразложившегося органического
вещества (диаметром менее 2 мм), достигающая для верхнего слоя 61,6%>, для ниж-
него — 79,5°/о от общего его содержания.
Таблица 9
Механический состав почвенных проб, взятых для анализа из муравейников
№ мура- вейника № пробы Глубина в см № проб иЭ мура- вейников Составная часть пробы Соотно- шение состав- ных частей в % Удельный вес расти- тельных остатков в %
диаметром больше диаметром меньше 1 мм
3 мм 2 мм 1 мм
№ 1 1 0—10 1 почва 67,93
{Formica 2 растительные остатки 32,07 52,8 13,4 14,3 19,5
praten- 3 10—20 2 почва 79,75 — — — —
sis) 4 растительные остатки 20,25 51,3 13,9 17,8 17,0
5 20—30 3 почва 82,48 — — — —
6 растительные остатки 17,52 58,0 12,2 11,8 18,0
7 30—40 4 почва 100,0 — — — —
8 40—50 5 почва 100,0 — — — —
Вне мура- 9 0—10 почва 100,0
вейника 10 10—20 — почва 100,0 —
№ 1 11 25—35 — почва 100,0 — — — —
№ 2 12 0—10 6 почва 70,76
(Formica 13 растительные остатки 29,24 14,4 24,0 37,6 24,0
pressi- 14 10—20 7 почва 82,58 — — — —
labris) 15 растительные остатки 17,42 7,3 13,2 32,7 46,8
16 20—30 8 почва 100,0 — — — —
17 30—40 9 почва % 100,0 — — — —
Вне мура- 18 0—10 почва 100,0
вейника 19 10—20 — почва 100,0 —
№ 2
Данные физико-химического анализа исследованных почв представлены в
табл. 10, из которой видно, что органические остатки в качественном отношении да-
Таблица 10
Краткая физико-химическая характеристика растительных остатков и почв, взятых из муравейников
№ муравейника № пробы Глубина в см № пробы из муравейника Анализируемая часть пробы В % на сухое вещество Легко гидролизуемые соединения
потеря от про- калива- ния со3 общий С общий N С от N от
сухого вещества общего содержа- ния С сухого вещества общего содержа- ния N
1 1 0—10 1 почва 19,72 0,81 5,51 0,525 0,30 5,44 0,04 7,62
(Formica 2 растительные остатки 70,09 — 37,23 1,241 2,01 5,40 0,41 33,03
pratensis) 3 10—20 2 почва 21,20 0,91 5,74 0,535 0,29 5,05 0,06 11,21
4 растительные остатки 70,08 — 30,99 1,302 2,36 7,62 0,33 25,34
5 20—30 почва 16,32 0,68 5,42 0,562 0,08 1,47 0,06 10,67
6 растительные остатки 67,40 — 37,12 1,513 2,56 6,90 0,19 12,55
7 30—40 4 почва 9,41 0,63 3,05 0,338 0,11 3,61 0,01 2,95
8 40—50 5 почва 15,00 0,55 2,62 0,249 0,07 2,27 0,01 4,01
Вне муравей- 9 0—10 — почва 16,36 0,28 3,22 0,283 0,06 1,86 0,02 7,07
ника № 1 10 10—20 — почва 13,48 0,94 3,35 0,304 0,06 1,79 0,01 3,28
11 25—35 — почва 13,84 0,50 2,29 0,260 0,07 3,05 не определялось
*
№ 2 12 почва 24,05 0,22 6,28 0,581 0,44 7,01 0,05 8,60
( Formica 13 0—10 6 растительные остатки 65,11 33,29 1,327 3,03 9,10 0,23 12,59
pressilabris) 14 почва 16,43 0,33 5,46 0,559 0,62 11,35 0,05 8,94
15 10—20 7 растительные остатки 51,11 — 26,40 1,468 2,49 9,43 0,27 18,39
16 20—30 8 почва 15,89 0,58 3,45 0,354 0,13 3,77 0,01 1,73
17 30—40 9 почва 15,48 0,53 3,65 0,284 0,09 2,47 0,01 1,89
J3nc муравей- 18 0—10 — почва 13,41 0,38 3,74 0,372 0,11 2,94 0,01 2,63
ника № 2 19 10—20 — почва 15,24 0,66 3,04 0,276 0,03 2,68 0,01 1,57
— п —
леко не однородны. Например, соотношения между зольными элементами и азотисты-
ми веществами варьируют в следующих пределах:
Толщина слоя в см Разрезы
муравейников
№ 1 № 2
0—10 .............. 24,1 26,2
10—20 ............. 22,9 .33,3
20—30 ............. 14,9 нет
Это указывает на различный состав органических остатков, использующихся жи-
вотными в качестве строительного материала для муравейников. В разрезе муравей-
ника № 1 по мере углубления наблюдается уменьшение показателей соотношений, что
говорит о неравномерном нарастании неорганических веществ в сравнении с азоти-
С7ЫМИ соединениями. Зольность в данном случае увеличивается от поверхности в
глубину, достигая 8% прироста, тогда как азот в этой же толще почвенного профиля
возрастает до 21%. Совершенно иначе получается с растительными остатками мура-
вейника № 2. где зольные компоненты в них увеличиваются на 40%, а азот только
на 10%, в связи с чем и возросло отношение между ними на 7 единиц.
Почва, находящаяся в сфере деятельности муравьев, значительно обогащена
углеродом и азотом, в сопоставлении с поверхностным слоем, взятом вне муравейников:
Муравейник Гнездо (а) Вне гнезда (б)
№ 1 С, % N, % В % б от а
С N
0—10 см 5,51 0,525 58,4 53,9
о-зо „ 5,56 0,541 59,7* 52,3*
№ 2 0—10 см 6,28 0,581 59,6 64,0
0-20 „ 5,87 0,570 63,7* 65,2*
Почвенные слои, непосредственно примыкающие к муравейнику, по содержанию
рассматриваемых компонентов почти не отличаются от таких же почвенных профи-
лей рядом расположенных участков.
Легко гидролизуемых форм углерода и азота в почве и растительных остатках
муравейников имеется в несколько раз больше, чем в почве контрольных участков.
Относительно высокий выход азотистых веществ выявлен в органических остатках
гнезда F. pratensis. Большей подвижности легко гидролизуемого азота органических
остатков муравейника № 1 соответствует пониженная растворимость углерода и, на-
оборот, с увеличением растворимости последнего, наблюдается некоторое снижение
этих же форм азотистых соединений (муравейник № 2).
Групповой состав органических веществ исследованных остатков и их количе-
ственный выход далеко не всегда зависят от степени разложения и минерализации
последних (табл. 11). На долю органических соединений рассмотренных групп при-
ходится от 77 до 93% от суммарного их содержания при пересчете на обеззоленное
вещество. Меньшие количества вещества группового состава отмечены в растительных
остатках на глубине 10—20 см муравейника № 1. Чем же вызваны указанные пони-
жения по сравнению с выше или ниже лежащими слоями органических остатков
в профиле муравейника? Ведь зольных элементов как в первом 10-сантиметровом, так
и в следующем за ним слое содержится совершенно одинаковое количество. Дело
заключается в различной подвижности указанных компонентов. Так. в остатках, при
определении лигнина, кутина и других негидролизуемых веществ, было обнаружено
золы (в процентах от начального количества, муравейник № 1):
на глубине 0—10 см ... .69,4%,
. 10—20 .......... 88,2%,
. 20-30 ............ 65,2%.
За зольными элементами по удельному значению следует группа лигнина и ку-
тина, составляющая около 7з, а в некоторых случаях и большую часть органических
веществ (35,52%). Характерно, что с увеличением разложения и минерализации ра-
стительных остатков уменьшается относительное содержание гемицеллюлоз и белко-
♦ Приведенные цифры показывают процентное содержание углерода и азота в
слое 0—10 см почвы вне гнезда, отнесенное к этим же компонентам 20- и 30-санти-
метровой толщи муравейников.
Таблица 11
Групповой состав органических остатков исследованных муравейников (в процентах на сухое или обеззоленное вещество)
№ мура- вейника и слой разреза в см Произведен расчет на вещество Зола Воднорастворимые соединения Воско- смолы Гемицел- люлозы Клет- чатка Негидро- лизуемый остаток (лигнин, кутин и пр.) Белковые вещества (NX6.25) Сумма
сухой остаток в том числе
зольных элемен- тов сахаров С N
№ 1 сухое 29,91 7,14 0,54 0,55 2,55 0,49 2,29 12,37 8,94 21,21 8,39 89,72
0—10 обеззоленное — 9,89 — 0,81 3,82 0,73 3,43 18,53 13,39 31,77 12,57 89,58
10—20 сухое 29,92 5,70 0,60 0,69 1,88 0,11 1,80 10,12 10,64 19,06 7,49 84,13
— обеззоленное — 7,27 — 0,98 2,68 0,16 2,57 14,43 15,17 27,18 10,68 77,30
20—30 сухое 32,60 5,08 0,42 0,64 1,69 0,15 1,84 10,02 11,37 23,95 7,32 91,76
— обеззоленное — 6,91 — 0,95 2,51 0,22 2,73 14,86 16,86 35,62 10,86 87,74
№ 2 сухое 34,89 5,82 0,72 1,23 1,58 0,18 2,28 10,66 9,78 18,60 11,16 90,19
0—10 обеззоленное — 7,83 — 1,89 2,43 0,28 3,50 16,36 15,01 28,55 17,13 88,38
10—20 сухое 48,89 8,43 1,05 0,97 1,69 0,24 2,98 5,79 7,65 14,79 8,90 96,34
— обеззоленное — 14,44 — 1,90 3,31 0,47 5,83 11,33 14,96 29,32 17,41 93,29
— 75 —
вых веществ. Последние, как правило, при гумусообразовании количественно увеличи-
ваются, что в данном случае (в связи с своеобразным процессом разложения орга-
нического вещества в муравейниках) не наблюдается.
Исходя из рассмотренных данных, можно утверждать, что и муравьи указанных
видов оказывают положительное влияние на обогащение почвы самого гнезда угле-
родистыми и азотистыми веществами.
Заключение
В статье рассмотрено значение кивсяков и двух видов муравьев в переработке
растительных остатков под пологом леса сухой степи. Большое влияние на трансфор-
мацию органического вещества оказывают кивсяки, значительно меньшее — муравьи.
В результате жизнедеятельности кивсяков растительные остатки частично из-
мельчаются и перемешиваются с верхними слоями почвы, частично заглатываются
вместе с почвенными частицами самими животными и, пройдя через их кишечный
тракт, выбрасываются в форме копролитов, значительно обогащенных углеродистыми
и азотистыми соединениями.
Роль муравьев в этом процессе незначительна, но она положительна не только
в смысле обогащения почвы гнезда биогенными элементами, но также и в смысле
некоторого механического измельчения органических остатков, благодаря чему проис-
ходит относительно равномерное разложение и минерализация последних с освобож-
дением из них питательных органоминеральных веществ.
SUMMARY
Julidae аге the most wide — spread representatives of invertebrate fauna In forest
black soil of the steppe zone. It has been established, as a result of research conducted
both in the laboratory and under conditions of the natural environment, that in the course
of their vital activity julidae perform a great deal of work connected with the transforma-
tion of vegetable remains. Unlike rain worms, these insects use as energetic material»
besides the organic matter of the soil, also the dead litter of forest vegetation, from which
they choose parts of the most suitable quality. They digest maple leaves first, then ash
tree leaves and lastly oak leaves. The dead leaves of yellow laburnum, that contain the
largest quantity of nitric substances, are not digested by these insects.
Besides the composition of the vegetable litter covering the forest soil, the number
of julidae also depends on weather conditions (heat and moisture). It varies between
21 and 211 and, according to the observations of M. S. Guilyarov (1956) even 500 speci-
mens per square metre of forest. It has been established by observations conducted over
a period of two years that a live julidae weighs from 25 to 448 milligrammes.
Together with vegetable remains, julidae swallow particles of soil, which, according
to coprolite ash composition, from about one third of the energetic material consumed by
them. Normally developing julidea cast out a mass of excrement (coprolite) equal to a
thirihy first part of their live weight.
And so, the forest litter is mixed and ground as a result of the julidae’s vital acti-
vity and the upper layers of soil loosened, thus providing favourable conditions for the
micropopulation and for the stpply of air in the soil of natural forests and planted woods.
The soil is, moreover, enriched with coprolite, which is excreted daily to the amount of
from 0.7 to 4.6 kilogrammes of dry mass per hectare, depending on the number and age
of the insects (abo t 110 to 686 kg. per hectare during the vegetation season). Due to the
mobility of water soluble and easily hydrolized forms of carbon and nitrogen, contained
in coprolite, the latter is an excellent energetic material for microorganisms and a source
of nutritive substances for the higher plants.
The role played by 2 species of ants (Formica pratensis and Formica pressilobris)
in the transformation of vegetable remains was established at the same time as that of
julidae. Their role in the process of decomposition and mineralization of organic matter
under cover of the forest was found to be negligible. The good results of their activity
consist in enriching the soil of and nests with biogeneous elements and in the mechanical
grinding of vegetable remains. This helps the more regular decomposition of the litter,
from which organic and mineral substances are released.
ЛИТЕРАТУРА
Архангельский M. П. 1929. К вопросу о влиянии дождевых червей на уро-
жай озса и ячменя в связи с внесением в почву некоторых удобрительных веществ.
Научно-агрономический журн., № 12.
Гиляров М. С. 1951. Роль почвенных животных в формировании слоя печвы
Усп. совр. биол., т. XXXI, вып. 2.
— 76 —
Гиляров М. С. 1956. Почвенная фауна лесных насаждений и открытых степ-
ных пространств бассейна р. Деркул. Тр. Ин-та леса АН СССР, т. XXX.
Гринфельд Э. К. 1939. Экология муравьев заповедника „Лес на Ворскле"
и его окрестностей. Уч. зап. ЛГУ, № 28, вып. 7.
Дарвин Ч. 1882. Образование растительного слоя деятельностью дождевых
червей. Перевод с англ.
Ди мо Н. А. 1938. Земляные черви в почвах Средней Азии. Почвоведение, № 4.
3 о н н С. В. 1954. Влияние леса на почвы. Изд. АН СССР.
Зражевский А. И. 1954. О роли фауны беспозвоночных в образовании плодо-
родия лесных почв. Тр. Ин-та леса АН СССР, т. XXIII.
Леббок Дж. 1884. Муравьи, пчелы и осы. Перевод с англ.
Пономарева С. И. 1948. Скорость образования в почвах кальцита дождевыми
червями. Докл. АН СССР, т. 61, № 3.
Пономарева С. И. 1949. Влияние деятельности дождевых червей на создание
устойчивой в эрозионном отношении структуры почв. Тр. Юбилейной сессии, посвящ.
столетию со дня рождения В. В. Докучаева.
Рунов Е. В. и Соколов Д. Ф. 1956. Исследование влияния опада на био-
химические и микробиологические процессы в почвах под лесными насаждениями.
Тр. Ин-та леса АН СССР, т. XXX.
Соколов Д. Ф. 1955. О роли кивсяков в разложении органического вещества
в почвах под лесными насаждениями в степи. Докл. АН СССР, т. 100, № 3.
БЮЛЛЕТЕНЬ М. О-ВА ИСП. ПРИРОДЫ, ОТД. БИОЛОГИИ, Т. LXII (5), 1957
Проблема онтогенетического старения и омоложения
растений и ее значение в растениеводстве
П. И. Гупало
The problem of senescence and rejuvenescence of plants
and its importance in plant breeding
P. I. Gupalo
Обсуждение проблем онтогенеза растений, начатое в «Бюллетене МОИП (отдел
биологический)» статьей В. В. Скрипчинскогоимеет большое теоретическое и прак-
тическое значение.
Жизнь давно уже настоятельно требует этого обсуждения. Оно было начато в
журнале «Селекция и семеноводство» еще в 1952 г.1 2, а также в журнале «Успехи
современной биологии»э, однако ни в первом, ни во втором не получило надлежа-
щего развития и не было завершено. Причиной этого, по-видимому, явилось догмати-
ческое отношение редакций этих журналов к теории стадийного развития: дискуссия
о научном наследстве Н. П. Кренке, начатая в указанных журналах, затронула тео-
рию стадийного развития, и это побудило редакции прекратить начатую дискуссию.
Ряд рукописей, поступивших в редакцию «Успехи современной биологии», не бы-
ли опубликованы и итоги дискуссии не подведены. Будем надеяться, что обсуждение
проблем онтогенеза, начатое «Бюллетенем МОИП». окажется более плодотворным.
Нельзя не согласиться с высказыванием, изложенным в программной редакцион-
ной статье журнала «Физиология растений», т. I, № 1 (1954): «Именно вследствие
догматизма мало развивалась за последнее время теория стадийного развития расте-
ний. Многие физиологи ограничивали свою задачу лишь характеристикой уже откры-
тых стадий (яровизации и световой), хотя эти стадии еще не охватывают всех эта-
пов онтогенеза растений, в том числе и очень важных как с теоретической, так и с
производственной стороны» (стр. 11).
Одним из таких этапов онтогенеза растений, не охватываемых теорией стадий-
ного развития Т. Д. Лысенко, является этап их старости, общего одряхления. Несмот-
ря на очевидное общеб!?ологическое значение онтогенетического старения организмов
и их омоложения, эта сторона жизни растений еще не получила должного призна-.
ния. В частности, работы крупного советского ученого Н. П. Кренке по этой проблеме
в последние годы игнорировались и даже были объявлены «идеалистическими» и «ме-
тафизическими» 4.
Отрицание самостоятельного значения возрастных качественных изменений ра-
стений, трактовка их, как изменений чисто количественных, в противоположность ка-
чественным стадийным изменениям (Казарян, 1952; Володарский, 1954), а также
оценка старения как простого следствия эколого-физиологических стадийных измене-
ний (Казарян, 1952; Кружилин и Шведская, 1950) — все это приводило по существу
к игнорированию проблемы старения и омоложения растений и ее приложения в прак-
тике растениеводства.
Совершенно иначе выглядит эта проблема в свете учения Ф. Энгельса
и И. В. Мичурина о жизненных фазах организмов, если положить его в основу пони-
1 Бюллетень МОИП (отдел биологический), т. 61, вып. 4 и 5, за 1956 г.
2 «Селекция и семеноводство» за 1952 г. № 2, 8, 9 и за 1953 г. № 2, 4.
3 «Успехи современной биологии». 1951 г., т. 32, вып. 1 (4); 1953 г., т. 36,
вып. 1 (4) ; 1954 г., т. 37, вып. 3; 1954 г., т. 38, вып. 1 (4); 1955 г., т. 39. вып. 1.
4 БСЭ, т. 23, статья «Кренке».
— 78 —
мания закономерностей онтогенеза растительных индивидов, как предлагает
В. В. Скрипчинский.
Общеизвестно огромное практическое значение проблемы возрастных качествен-
ных изменений человеческого организма. Большое значение имеют возрастные изме-
нения организмов также в животноводстве, что ярко было показано специальной науч-
ной конференцией, проведенной Московской сельскохозяйственной академией им. Ти-
мирязева в 1953 г. Не меньшее значение имеет эта проблема и в растениеводстве.
В последней статье, которой было закрыто обсуждение вопросов о научном на-
следстве Н. П. Кренке в журнале «Успехи современной биологии» основным аргу-
ментом Ф. А. Дворянкина в защиту монополии теории стадийного развития было то,
что эта теория много дала для растениеводческой практики. Поэтому в целях обосно-
вания необходимости расширения теоретических представлений о закономерностях он-
тогенеза растений мы и поставили своей задачей рассмотреть в данной статье, глав-
ным образом, вопрос о практическом значении проблемы старения и омоложения
растений в растениеводстве.
Краткий исторический обзор теоретической
разработки данной проблемы
Еще в 30-х годах XIX столетия русский агроном В. В. Беликов, рассматривая
в своих статьях вопросы индивидуального развития сельскохозяйственных растений,
обсуждал вопросы долголетия растений и вырождения сортов плодовых культур при
длительном вегетативном размножении. В. В. Беликов считал прививку на молодой
дичок, средством временного омоложения и подчеркивал, что полное омоложение ра-
стений происходит только при семенном размноженини (по Максимову и Кал-
мыкову, 1954).
В зарубежной литературе мысли о старении и омоложении растений, как суще-
ственных сторонах их жизни, были в свое время четко сформулированы ботаником
Брауном (А. Braun, 1851, 1872). Работы Брауна привлекли к себе пристальное вни-
мание А. Н. Бекетова (1858, 1862).
В отличие от животных, растительные организмы имеют своеобразную особен-
ность, выражающуюся в том, что вегетативный рост их в определенных точках
не прекращается до самой смерти и поэтому существование растительного индивида
может быть продлено на неопределенно долгий срок путем вегетативного размноже-
ния. Это вызвало у ряда ботаников неправильное представление о потенциальном бес-
смертии растительных индивидов (Декандолль, Клебс и др.). Обзор и критика этих
неверных представлений даны Н. П. Кренке (1928, 1950).
Коршельт (Е. Korschelt, 1924), а затем Молиш (Н. Molisch, 1929) подчеркивают
ограниченность индивидуальной жизни растений, особенно высших, и практическое
значение процессов их старения. Молиш провел также опыты по продлению жизни
растений, их омоложению.
И.В. Мичурин считал возрастные качественные изменения основной чертой онто-
генеза растений. Он широко разработал эту проблему, особенно в отношении измене-
ния наследственности, и придавал большое значение онтогенетическому старению ра-
стений, признавая также и старение клонов при длительном искусственном вегета-
тивном размножении.
Оригинальную общебиологическую разработку проблемы старения и омоложения
организмов, в том числе и растений, дал Чайльд (С. Child, 1915).
Он рассматривает старение и омоложение как диалектически связанные физио-
логические процессы, свойственные организмам и являющиеся следствием жизнен-
ного процесса как такового. При этом термином «старение» (senescence) Чайльд
обозначает процесс снижения уровня метаболизма, который, по его экспериментальным
данным, начинается уже на самых ранних этапах онтогенеза и вызывается измене-
ниями субстрата организма. Омоложением (rejuvenescence) Чайльд называет
противопбложный процесс — повышение метаболизма, связанное также с качествен-
ным изменением субстрата организма. По Чайльду, омоложение ярко проявляется
при всяком образовании молодых организмов в процессе размножения, но оно имеет
место также и в ходе онтогенеза каждого организма в зависимости от условий жизни.
Четкое понимание значения внешних условий для жизни организмов, а также
острая критика вейсмановской концепции организма, пронизывающая всю книгу, де-
лают указанную выше работу Чайльда весьма интересной для советских биологов, ра-
ботающих над развитием науки на основе мичуринского направления. Приходится
пожалеть, что труд Чайльда «Senescence and rejuvenescence» остается мало извест-
ным для ’ многих советских биологов, так как он не переводился на русский язык и
даже его содержание до сих пор не было достаточно освещено в нашей биологиче-
ской литературе.
1 «Успехи современной биологии», т. 39, выл. 1, 1955.
— 79 —
Н. П. Кренке (1928, 1940, 1950) критически рассмотрел и обобщил все накоп-
ленные в науке данные о проблеме возрастности, выполнил ряд оригинальных экспе-
риментальных работ и наблюдений и вновь привлек внимание широкого круга бота-
ников и растениеводов к проблеме старения и омоложения растений. Сознательно при-
менив философию диалектического материализма, Н. П. Кренке сделал набросок тео-
рии возрастной цикличности растений. Несмотря на недостаточную разработанность
этой теории автором вследствие его преждевременной смерти, она вызвала большой
интерес как v советских ученых, так и за рубежом, в частности в странах народной
демократии (Шикула, 1955).
О значении теоретической работы Н. П. Кренке свидетельствует тот факт, что
его концепции приняты, как одна из теоретических основ садоводства, коллективом
авторов — работников Мичуринского плодоовощного института и Научно-исследова-
тельского института плодоводства в недавно вышедшем учебнике по плодоводству
(Курындин, Малинковский, Беньяминов, Белохонов, 1954).
Не останавливаясь в данной статье на полном критическом разборе теоретиче-
ских концепций Н. П. Кренке, считаем необходимым подчеркнуть, что много неяс-
ностей и даже противоречий в его теории возрастной цикличности растений, отмечен-
ных, в частности, А. А. Сапегиным (1944), обусловлено недостаточно четкой термино-
логией, что является отражением недостаточной разработанности ряда понятий, отно-
сящихся к проблеме старения и омоложения.
В разговорной речи в русском языке принято называть «старением» одряхление
организма или его частей, ослабление их. С таким содержанием этот термин вошел в
широкую биологическую и растениеводческую литературу. Сохраняя значение термина
«старение» как синонима комплекса одряхления, наблюдаемого на нисходящей ветви
онтогенеза, Н. П. Кренке нередко употребляет его также в смысле общего онтогене-
тического (возрастного) продвижения организма вообще, аналогично термину «aging»
у Чайльда.
Поэтому у Н. П. Кренке встречаются такие формулировки: «Здесь большее ее
старение на восходящей ветви онтогенеза ведет не к старению, а к большей вегета-
тивной мощности индивидуумов в их целом» (1940, тезис 41).
Употребляемый им термин «омоложение» Н. П. Кренке определяет так: «Омоло-
жение есть новообразование и развитие молодых веществ и структур, а также за-
держка старения существующих элементов, но не возврат индивидуума или его ча-
стей к прошлому» (1940, стр. 7).
Этот термин также требует уточнения. Н. И. Дубровицкая и А. Н. Кренке (1954)
дали критический обзор употребления термина «омоложение» у разных авторов и на
основе анализа фактических данных по этому вопросу предлагают различать два типа
омоложения: а) глубокое омоложение частей организма, протекающее со снятием ста-
дийных, т. е. качественных онтогенетических изменений, которые для целого организма
являются необратимыми; б) омоложение целого организма или его частей в том смыс-
ле, как это указано в определении Н. П. Кренке.
Глубокое омоложение, наблюдающееся при половом процессе, естественном веге-
тативном размножении и при регенерации, Н. И. Дубровицкая и А. Н. Кренке пред-
лагают называть обновлением, а термин «омоложение» сохранить для всех остальных
случаев (при образовании молодых клеток и тканей в процессе роста, при искусствен-
ном вегетативном размножении, при некоторых воздействиях на растения, таких, как
поливы, подкормки и т. д.).
Мы полагаем, что следует согласиться с этим предложением, хотя резкой грани-
цы между этими двумя типами омоложения нет.
Глубокую теоретическую и практическую разработку проблемы старения и омо-
ложения плодоягодных растений сделал П. Г. Шитт. В его книге «Биологические
основы агротехники плодоводства» (1952), вышедшей после смерти автора под редак-
цией В. И. Эдельштейна, термин «старение» в отдельных случаях также применяется
как синоним возрастного продвижения организма вообще (стр. 88. 91 и др.), но в
основном он употребляется для обозначения одряхления клеток, органов и организма
в целом. При этом часто преимущество отдается термину «отмирание» (стр. 126 и др.).
Противоположный процесс — зарождение нового, повышение жизнеспособности — в
книге Шитта называется «обновлением» (стр. 97. 126, 129 и др.), хотя употребляется
также и термин «омоложение» («омолаживающая подрезка»).
Весьма показательным является тот факт. что. подводя итоги своей полувековой
работы в области плодоводства, П. Г. Шитт пришел к концепции индивидуального
развития растений, как диалектического единства и противоположности старения и
омоложения (отмирания и обновления), т. е. к той же концепции, которую развивал
Н. П. Кренке, исходя из другого ряда наблюдений и экспериментальных данных.
В ряде мест своей книги П. Г. Шитт подчеркивает свое понимание индивидуаль-
ного развития растений как единства противоположностей — старения и обновления
(1952, стр. 49, 93, 97, 99, 129, 159, 172, 181, 201, 204). Эта концепция явилась резуль-
татом длительного и кропотливого труда по изучению онтогенетического развития
растений в связи с задачей управления жизнью плодового дерева и его урожаем.
— 80 —
П. Г. Шитт, как и Н. П. Кренке, рассматривает растение как сложный организм,
каждый метамерный орган которого имеет свой собственный онтогенез, участвуя так-
же и в онтогенезе организма в целом, что подчеркивал в свое время еще К. А. Ти-
мирязев (1942).
Диалектическое сочетание противоположных процессов — старения (отмирания)
и омоложения (обновления) — П. Г. Шитт понимает в их конкретном морфологиче-
ском выражении в сложном растительном организме, как системе клеток, тканей и
органов (1952, стр. 144—145). В процессах омоложения он учитывает корреляционное
влияние корневой системы, на что не обратил достаточного внимания Н. П. Кренке.
Следует признать, что П. Г. Шитту по существу принадлежит приоритет в экс-
периментальном доказательстве старения корневой системы, качественные изменения
которой не учитываются теорией стадийного развития. Мы имеем в виду опыты
П. Г. Шитта с прививкой концевых веточек яблони 15-летнего возраста на концевых
корнях той же яблони. В отличие от обычных прививок черенков на молодых дичках
такая прививка дала карликовые растения, которые по морфолого-анатомической
структуре, этапу возрастных изменений и физиологическим функциям соответствовали
возрастному состоянию маточного растения.
Существенный вклад в разработку проблемы возрастных изменений растений
внесен Т. А. Работновым (1946, 1949, 1950), который изучал жизненный цикл много-
летних травянистых растений в луговых ценозах в целях более глубокого познания
этих ценозов и управления ими.
Непосредственно исходя из мичуринских представлений о жизненных фазах,
Т. А. Работнов конкретизировал эти представления применительно к жизни травяни-
стых многолетников и однолетних растений. Он указывает, что явления онтогенетиче-
ского старения у травянистых растений распространены достаточно широко, но мало
известны благодаря отсутствию соответствующих наблюдений. Т. А. Работнов подчер-
кивает, что длительность жизни луговых растений, а следовательно, и наступление
старческого периода, существенно и притом очень резко, меняются в зависимости от
условий произрастания. Поэтому многие растения, которые в естественных луговых
ценозах имеют продолжительность жизни 5—10—15 лет, в питомниках ведут себя как
типичные двухлетники (например, бодяк болотный, порезник горный и др.).
Большое внимание проблеме возрастных качественных изменений растений и их
старения уделил в своем оригинальном труде И. Г. Серебряков (1952), а также в
ряде своих опытов и в докторской диссертации А. К. Ефейкин (1956).
Убедительно доказывая, что меристема по своей природе всегда является носи-
телем молодого состояния организма и развитие из нее тех или иных (вегетативных
или репродуктивных) органов зависит не от узко локальных изменений меристемы, а
от состояния растительного организма в целом, А. К. Ефейкин, на наш взгляд, до-
пускает существенную ошибку, отрицая старение клонов при искусственном вегета-
тивном размножении. Эта ошибка обусловлена тем, что, сосредоточив свое внимание
на меристеме, как носительнице молодого состояния организма и непрерывности
жизни вида, А. К. Ефейкин не учитывает, что при вегетативном размножении отде-
ляется не одна меристема, а довольно значительная часть растения, несущая в себе
признаки и свойства соответствующего онтогенетического уровня, достигнутого этой
частью на материнском организме.
Проблема жизненности организмов, выдвинутая в последние годы Т. Д. Лысенко
(1949), по существу совпадает с проблемой старения и омоложения, поскольку
Т. Д. Лысенко подчеркивает, что жизненность организма в целом по мере старения
отдельных участков тела и всего организма затухает, но вновь возобновляется при
оплодотворении и размножении.
Мы показали выше, какими разнообразными путями подходила научная мысль к
обсуждению проблемы старения и омоложения растений. Значимость теоретических
представлений определяется в конечном счете их применением в производственной
деятельности человека. Поэтому перейдем к рассмотрению практического значения
данной проблемы.
Старение и омоложение плодоягодных культур
Из практических наблюдений давно было известно, что с возрастом плодовых
деревьев и ягодных кустарников их продуктивность сначала возрастает, а потом
снижается. Садоводы давно уже выработали приемы поддержания плодоношения
у стареющих деревьев путем обрезки старых ветвей и формирования плодоносящих
частей кроны из более молодых побегов. Приемы эти были созданы на основе чисто
эмпирических наблюдений за естественным ходом отмирания и обновления кроны
деревьев и только в последние годы им начали уделять серьезное теоретическое
внимание (Кренке, 1928, 1940; Шитт, 1936, 1952; Гусева, 1946; Иванов, 1949).
Особенно подробно разработаны вопросы агротехники плодоводства на основе
учета возрастных изменений плодового дерева П. Г. Шиттом.
- 81 —
Развивай положение И. В. Мичурина о возрастных этапах онтогенеза плодового
дерева, П. Г. Шитт разделил весь жизненный цикл плодового дерева на девять воз-
растных периодов, которые различаются по соотношению роста вегетативных частей,
плодоношения и усыхания кроны.
По ходу этих последовательных периодов изменяются все биологические и хо-
зяйственные свойства плодового дерева, а также и задачи агротехники ухода за
растениями.
Омолаживающая подрезка, по П. Г. Шитту, должна начинаться с третьего воз-
растного периода, причем в пятом периоде дополнительно вводится омолаживание
корневой системы путем полосной плантажной вспашки, сочетаемой с внесением
органических и минеральных удобрений, а если возможно, то и с поливом.
Являются ли все эти приемы ухода за плодовыми деревьями действительным
омоложением их? Да, являются, если понимать омоложение не в смысле полного
обновления организма в целом, а как новообразование более молодых частей.
Существенные практические результаты получили П. Г. Шитт и его сотрудники,
применив также концепцию о старении и омоложении растений к вопросам агротех-
ники земляники. Наблюдениями в производственных условиях установлено, что уже
на четвертом году жизни на плантациях земляники появляется тенденция к сниже-
нию количества и качества урожая с единицы площади: общий урожай снижается,
ягоды становятся мельче, кусты больше страдают от зимнего обмерзания. Поэтому
сохранение плантации земляники свыше 4—5, реже 6 лет, уже является производ-
ственно невыгодным. Применив омолаживающую обрезку земляники, П. Г. Шитт
получил эффективные производственные результаты (1952, стр. 350—353). Эффектив-
ные результаты дал также и метод черенкового размножения земляники для полу-
чения более раннего цветения и созревания ягод, поскольку растения, полученные от
взрослых стеблевых частей, являются более онтогенетически продвинутыми в своем
развитии.
Большое практическое значение имеет омолаживающая подрезка в культуре
винограда (Барский, 1952; Машинская, 1953 и др.). Л. П. Машинская сообщает, что
в опытах, проведенных Всероссийским научно-исследовательским институтом вино-
градарства и виноделия с 1944 по 1952 г., правильно проведенное омолаживание при
веерной форме куста дало повышение урожая до 33%.
Наблюдения над процессами старения у малины (Герасимова, 1953) выявил»
разнокачественность кустоз малины в зависимости от возраста корневой системы.
Разновозрастность корней в пределах сорта сказывается на времени начала плодо-
ношения, величины и продолжительности его, качестве ягод, а также на морозо-
стойкости. Оптимальным для плодоношения является средний возраст корневой си-
стемы, причем скороспелые сорта имеют менее долговечные корни, чем позднеспелые.
На основе изучения возрастных изменений малины 3. А. Герасимова рекомендует ряд
приемов для увеличения урожая, улучшения качества ягод и получения более друж-
ного плодоношения.
На возрастное снижение урожайности кустов смородины и необходимость свое-
временного омоложения их указывают П. И. Житнева (1948) и М. А. Лисавенко (1950).
Таким образом, возрастные качественные изменения имеют весьма существенное
значение в онтогенезе плодоягодных культур. Хотя теория стадийного развития
Т. Д. Лысенко, разработанная на данных по однолетним семенным растениям, может
быть частично применена к многолетним древесным и кустарниковым растениям, одна-
ко совершенно очевидно, что процессы возрастных изменений она не охватывает.
Старение травянистых многолетников
У травянистых многолетников процессы онтогенетического старения выражены
менее ясно, чем у древесных растений и кустарников. Причина этого заключается
в том, что у многих травянистых многолетников ежегодное нарастание индивида при-
нимает характер вегетативного обновления и размножения.
Так, С. П. Смелов (1947) отмечает, что у луговых злаков при вегетативном
возобновлении каждое поколение побегов при возникновении из почек образует не
только новые надземные, но и подземные органы, т. е. происходит полное обновление
организма, вследствие чего признаки старения могут сниматься почти в такой же
мере, как это происходит при семенном возобновлении в условиях самоопыления.
Для таких растений проблема онтогенетического старения становится в основном
проблемой старения клона, т. е. проблемой старения при длительном вегетативном
размножении.
В отличие от луговых злаков, люцерна и клевер при ежегодном вегетативном
возобновлении, как правило, самостоятельных молодых корней у новых побегов не
образуют, т. е. у них корневая система при обычных условиях культуры обновляется
недостаточно. Поэтому можно ожидать, что у этих растений онтогенетическое старение
будет резко выражено.
Таблица 1
Основные Данные по физиологической характеристике клевера второго и третьего годов жизни
Дата взятия пробы Фаза развития и части растений, взятые для анализа Максимальная водоемкость ткани листьев (в % воды к сухому веществу) % золы к сухому веществу % кальция в золе % калия в сухом веществе Активность каталазы
2-й г. 3-й г. 2-й г. 3-й г. 2-й г. 3-й г. 2-й г. 3-й г. 2-й г. 3-й г.
20/VI Начало стеблевания; один верх- ний лист, сидящий ниже мо- лодого, но уже раскрытого листа; взяты пластинки листьев 359,0 343,4 10,72 9,20 28,1 30,5 0,844 0,775 22,4 21,5
17/VII Фаза хорошо развитых бутонов (начало цветения):
а) первый лист сверху, не считая листьев, отверты- вающих головку .... 330,4 309,6 10,25 8,59 40,7 39,8 не о пр еделяли — —
б) второй лист не опр< гделяли 16,21 9,6 33,6 40,8 0,681 0,549 — —
в) третий лист 16,65 9,73 34,8 41,2 не опр еделяли — —
5/VIII Разгар цветения; листочки, об- вертывающие головку .... — — — — — — 21,4 20,3
- 83 —
М. Я. Трегубенко (1940, 1947, 1950) установлено резкое снижение морозостой-
кости люцерны в связи с ее старением и предложены агротехнические мероприятия,,
направленные к тому, чтобы предотвратить преждевременное старение люцерны и
снижение ее зимостойкости. Старение тканей люцерны доказано Трегубенко путем
определения гидрофильности коллоидов (содержания связанной воды) в однолетних,
двухлетних и трехлетних растениях.
Е. Н. Синская (1949) показала, что устойчивость люцерны к заболеваниям
тесно связана с конституцией растений и процессами их старения и омоложения.
Ф. Филатов (1950) установил, что высокие урожаи семян люцерны получаются толь-
ко на возрастно молодых посевах: по данным обследования 687 колхозов, урожай
семян на молодых посевах был на 47,9°/о выше, чем на старых. А. Пустовойт (1950)
предложил производить перепашку старовозрастных участков люцерны с дискованием
для их омоложения и повышения урожая сена и семян. Прием этот получил приме-
нение в практике растениеводства.
Онтогенетическое старение красного клевера впервые экспериментально устано-
вил М. А. Лохин (1948), который на основе своих опытов, проводимых в течение
8 лет, показал, что старение является одной из основных причин изреживания посе-
вов клевера. М. А. Лохин указывает, что к постановке этих опытов его побудили
работы Н. П. Кренке.
Многолетними наблюдениями Красноуфимской государственной селекционной
станции было установлено, что участки клевера на третьем году от посева (т. е. на
втором году пользования), как правило, дают гораздо меньше семян, чем участки
первого года пользования, даже при одинаковой густоте травостоя. Это явление было
мало понятным, так как считалось, что естественное долголетие позднеспелого перм-
ского клевера равно 5—8 годам. Кроме этого, сельскохозяйственные органы рекомен-
довали колхозам оставлять на семена участки позднеспелого (одноукосного) клевера
только второго года пользования, чтобы обеспечить отбор более долговечных растений.
Это побудило нас заняться изучением процессов старения у красного клевера (Гупало,
Савченюк, Мохнаткин, 1950; Гупало, 1951, 1953). Результаты изучения подтвердили
наличие отчетливой картины онтогенетического старения у растений клевера на
третьем году жизни.
Изучение проводилось на полевых опытных участках, которые выбирались так,
чтобы они различались только годом пользования, а все почвенные и агротехнические
условия были одинаковы.
В 1948 и 1949 гг. определяли оводненность и процент белка в пробах листьев
клевера второго и третьего годов жизни, а также нектарность цветов методом центри-
фугирования по А. Ф. Губину. Были обнаружены отчетливые различия: у клевера
третьего года жизни показатели оводненности листьев и содержание белка в них,
а также нектарность цветов оказались ниже, чем у клевера второго года жизни.
В 1950 г. изучение было проведено по более широкому комплексу показателей:
оводненности листьев, проценту золы, содержанию кальция и калия, активности ката-
лазы и содержанию лигнина в стеблях. Полученные данные приведены в табл. 1 и 2.
Содержание лигнина определяли в стеблях, взятых к концу цветения клевера
1/IX, а именно в таких, у которых верхушечные головки находились в одинаковой
фазе развития: самые верхние цветы головки еще не расцвели. Стебли при этом еще
имели зеленую окраску. В анализ шла верхняя часть стебля длиной 15 см. В резуль-
тате анализа в стеблях клевера второго года жизни найдено 19,4% лигнина от веса
сухого вещества, а у растений третьего года — 22,6%.
Таблица 2
Содержание нектара (в мг на головку) в цветах
клевера второго и третьего годов жизни
Дата определения Второй год жизни Третий год жизни
11/VIII 17,3 13,5
23/VIII 42,4 20,6
Цветение клевера третьего года жизни началось на 5 дней раньше, и цветонос-
ные стебли в этом травостое начали отмирать с 25/VIII, в то время как клевер вто-
рого года жизни цвел и зеленел до октября, когда он был скошен. Все эти данные
свидетельствуют о наличии у клевера на третьем году жизни отчетливо выраженных
признаков онтогенетического старения.
Наши данные были подтверждены наблюдениями, проведенными во Всесоюзном
институте растениеводства. Так, Т. А. Коломиец (1955) в условиях Ленинградской
— 84 —
обл. нашла, что растения одноукосного красного клевера на третий год жизни по
своей мощности значительно уступали растениям второго года жизни.
Таким образом, прежние взгляды относительно естественной продолжительности
жизни растений позднеспелого культурного красного клевера не подтвердились, и по-
этому сельскохозяйственное производство должно считаться с неизбежно наступаю-
щим старением клевера на третьем году жизни (втором году пользования) и сделать
отсюда соответствующие практические выводы.
Поэтому в зоне деятельности Красноуфимской селекционной станции в агро-
правила по семеноводству клевера были внесены изменения, а именно было указано,
что при отводе семенных участков надо принимать во внимание, главным образом,
состояние растений, собирая семена как с участков второго, так и первого года
пользования.
В последнее время против этого выступил Д. Федюнькин (1955, 1956), который
требует прекратить сбор семян с участков клевера первого года пользования, моти-
вируя это тем, что такая практика приведет к отбору более рано зацветающих скоро-
спелых и менее зимостойких морфобиотипов, а следовательно, и к ухудшению при-
роды пермского клевера.
Известно, что популяции красного клевера состоят из морфобиотипов разной
скороспелости, долговечности и зимостойкости. Однако Д. Федюнькин и другие за-
бывают, что определенное и закономерное различие в сроках зацветания этих морфо-
биотипов наблюдается только при обособленном размещении отдельных растений.
В густом же хозяйственном посеве клевера цветение растений по годам пользования
будет обусловлено не столько наследственными особенностями их, сколько случайны-
ми условиями загущенности или разреженности стояния растений: при густом стоянии
наступление цветения клевера задерживается, а при редком — ускоряется (Сукачев,
1941; Работнов, 1950). Именно этим и объясняется тот факт, что при сборе семян
клевера с участков первого года пользования в течение периода свыше 10 лет, про-
изведенном в специальном опыте, никакого ухудшения наследственной природы этих
семян не было обнаружено: урожай сена от них был получен даже более высокий
(Мохнаткин, Гупало и Савченюк, 1952). Конечно, при решении практического вопроса
о том, на каком году пользования следует собирать семена с красного клевера, сле-
дует учитывать все стороны этого вопроса. Однако при этом ни в коем случае
нельзя упускать из виду такой существенный фактор, как старение растений, которое
совсем не учитывается Д. Федюнькиным.
В последнее время в сельскохозяйственном производстве складывается опреде-
ленная тенденция к одногодичному использованию клевера в полевых севооборотах
(мы считаем эту тенденцию вполне обоснованной), но для кормовых севооборотов
проблема старения клевера сохраняет свое значение.
Описанные нами возрастные изменения травянистых многолетников не могут
быть сведены к прохождению ими эколого-физиологических стадийных изменений
(стадии яровизации и световой), а являются качественно своеобразным общебиоло-
гическим процессом, как и у древесных растений.
Старение п омоложение картофеля и овощных культур
Т. Д. Лысенко (1935) показал, что причиной вырождения посадочного материала
картофеля на юге в основном является онтогенетическое старение (одряхление), вы-
званное влиянием высокой температуры на молодые формирующиеся клубни, особенно
на их почки. На этой основе был предложен метод летних посадок картофеля, ока-
завший огромную помощь народному хозяйству нашей страны в деле повышения
урожаев картофеля на юге.
В ботанической литературе и учебниках обычно пишут, что теоретической осно-
вой метода летних посадок является теория стадийного развития. Это утверждение
неправильно. Теоретической основой этого приема является положение о старении,
одряхлении растений и их частей, протекающем в связи с определенными условия- >
ми жизни.
С теорией стадийного развития здесь общим является только принцип экологи-
ческой обусловленности жизненных процессов, протекающих в растительном организ-
ме. Старение — это особый физиологический процесс качественных изменений, кото-
рый нельзя отожествлять со стадийными изменениями, как эколого-физиологическими:
приспособлениями в ходе развития, т. е. конкретно с прохождением стадий яровиза-
ции и световой.
Правильное теоретическое представление о причинах вырождения картофеля дает
возможность найти и другие меры борьбы с ним.
Н. С. Бацанов (1952, 1953) предложил проводить, наряду с методом летних
посадок, также посадку семенных участков картофеля на окультуренных торфяниках^
т. е. на низких и достаточно влажных местах с большим количеством органических
веществ. Доказано, что такие посадки также предупреждают старение клубней и>
улучшают качество посадочного материала.
-85 —
Н. С. Бацанов отмечает, что длительное хранение картофеля при температуре
ъыше 3° также вызывает старение почек глазков клубня, равносильное южному
вырождению.
Г. X. Молотковский (1945, 1948) предложил бороться с вырождением картофеля
путем срезания ботвы, чем вызывается омолаживание растений и задержка клубне-
образования, которое отодвигается на более поздний период лета. Позже Г. X. Мо-
лотковский и В. Т. Бубырева (1954) провели физиологическое исследование картофеля
с обрезанной ботвой и установили, что «глубокая обрезка ботвы повышает жизне-
деятельность организма, омолаживая его». Так, по данным анализов, у вновь появив-
шихся после обрезки стеблей повышались содержание хлорофилла в листьях, актив-
ность каталазы и содержание аскорбиновой кислоты.
Сопоставляя анатомическое строение клубней картофеля, выросшего в условиях
севера и на юге, О. А. Муравьева (1949) обнаружила характерные признаки старе-
ния, которые заключаются в изменении соотношений тканей мягких паренхимных
и одревесневших с преобладанием последних. В клубнях от летних посадок на юге
признаки старения не были обнаружены.
Г. Н. Линник (1955), изучавший вопрос о вырождении картофеля в течение
многих лет, пришел к выводу, что это явление необходимо рассматривать в основном
как внешнее проявление процесса старения. Конечно, нельзя отрицать роль вирусов
в комплексе явлений вырождения картофеля, но совокупность фактических данных
свидетельствует о том, что влияние вирусов накладывается на процессы онтогенетиче-
ского старения, одряхления растений.
Овощеводы давно установили (Смерткин, 1940), что корнеплоды моркови на юге
СССР, например в условиях Кубани, оставляемые для хранения с целью получения
в будущем году семян, плохо хранятся в зимний период именно вследствие их пере-
зревания, т. е. старения. Поэтому было предложено для получения маточных корне-
плодов моркови производить летние посевы, чтобы к осени корнеплоды не успели
постареть. Это предложение дало вполне положительные результаты (Сагалович,
1949; Гойко, 1954).
В этом случае особенно наглядно видно, что процессы старения нельзя свести
к стадийным изменениям: ведь корнеплоды к осени еще не прошли ни стадии ярови-
зации, ни световой. Это не исключает того, что в других случаях процессы старения
резко усиливаются генеративным развитием. Например, старение огуречных растений
тесно связано со сроками снятия плодов. Они стареют и отмирают очень рано, если
плоды оставляются на семена, т. е. до полного созревания. Если же не допускать
этого и снимать «зеленец», т. е. 6—10-дневные завязи, то огуречные растения будут
расти «неопределенно долгое время» (Эдельштейн, 1953, стр. 26).
Клинские огородники применяют особый прием омолаживания огуречных расте-
ний, а именно: когда плеть внизу, вследствие отмирания листьев, оголится, ее укла-
дывают на землю и слегка присыпают землей. В результате образуются добавочные
корни и ^сизнь растений удлиняется. Такой прием позволяет колхозникам Клинского
р-на, Московской обл., получать урожай огурцов в течение 13 месяцев, считая от
посева. Резкое влияние оказывает удаление плодов в молодом возрасте и у овощ-
ного гороха: при обычном созревании плодов отмирание растений происходит через
2,5 месяца, т. е. после вызревания плодов, а при удалении молодых завязей или
цветов растения продолжают вегетировать до самых заморозков. При этом контроль-
ные растения образовали в среднем 51 лист на один экземпляр, а растения, у которых
обрывались цветки — по 264 листа, т. е. в пять раз больше (Эдельштейн, 1953,
стр. 344).
Противоположное влияние оказывает на растение удаление молодых растущих
побегов или их верхушек. В практике садоводства давно применяется прищипка
концов побегов при буйном росте молодого деревца, чем обеспечивается прекращение
роста и вызревание древесины. Пасынкование помидоров вызывает более раннее
созревание плодов, но в то же время, по нашим наблюдениям, оно приводит и к ран-
нему отмиранию растений.
Старение и омоложение у других сельскохозяйственных растений
Возможность задержки старения растений путем удаления органов плодоношения
давно привлекала к себе внимание растениеводов (Егоров, 1923; Тарановская, 1927).
М. А. Егоров установил, что «кастрированные растения», кроме увеличения пе-
риода вегетации, обнаруживают более сильный рост, накапливают большую массу
сухого вещества, причем эта масса более богата белками, чем у контрольных расте-
ний. Это побудило ряд агрономов применить данный прием для увеличения урожая
кормовой массы у силосных растений, например у подсолнечника. Результаты полу-
чены вполне положительные (Савута, 1948).
Старение однолетних хлебных злаков, наступающее к концу их онтогенеза, долго
оставалось вне поля зрения исследователей и растениеводов-производственников, так
как после плодоношения эти растения уже не представляют интереса для произвол-
— 86 -
ства. Однако работы Н. В. Цицина и А. И. Державина по выведению многолетних
форм хлебных злаков вызвали теоретический и практический интерес также и к ста.-
рению этой группы растений.
В. В. Скрипчинский (1952, 1954) получил ряд существенных экспериментальных
данных по этому вопросу. Так, например, им было установлено, что в условиях
Ставропольского края устранение плодоношения у однолетних хлебных злаков (ка-
стрирование) почти не задерживает их старения и отмирания. Больше того, в тех же
условиях при весеннем посеве озимой пшеницы неяровизированными семенами наблю-
далось, что полученные растения, давшие сильное кущение и не прошедшие стадийное
развитие, к осени все же полностью отмирали, в то время как в северных, более
влажных и прохладных районах такие посевы обычно переносят лето и могут разви-
ваться в следующем году. Многолетняя рожь в тех же условиях сохранялась живой.
Эти факты показывают, что процесс старения и отмирания растений может про-
текать в определенных условиях и без перехода к плодоношению, без прохождения
растением генеративного (стадийного) развития, хотя обычно генеративное развитие
и плодоношение является важнейшим фактором старения и естественной смерти
растений.
Старение и отмирание без стадийного развития наблюдается при длительном
хранении семян. При этом происходят закономерные качественные изменения семян:
опыты разных авторов показали, что растения из старых семян обнаруживают более
слабый рост и быстрее переходят к цветению (Barton a. Garman, 1946; Чирков-
ский, 1953).
Конечно, процессы старения семян, старения листьев, старения растений при
действии засухи и других неблагоприятных условий и старения после цветения и
плодоношения имеют разное конкретное физиолого-биохимическое содержание. Но
вместе с тем все эти процессы имеют также и некоторые общие черты, общую осно-
ву, установленную рядом физиолого-биохимических исследований, о которых кратко-
будет сказано дальше.
Много примеров практического значения учета возрастных качественных изме-
нений растений и управления ими приведено Н. П. Кренке (1940), который указывал
также на перспективы значительного расширения применения этих закономерностей,
в практике растениеводства. Указания Н. П. Кренке полностью подтвердились. Рас-
крыто решающее значение возрастных качественных изменений растений и их органов
в стойкости их к грибным и бактериальным поражениям (Дунин, 1946; Трофимович^
1948 и др.), при укоренении черенков (Дубровицкая, 1953), при скрещивании систе-
матически отдаленных растений (Долгушин, 1946; Кузьмин, 1948) и т. д.
Физиолого-биохимические исследования процессов
старения и омоложения растений и их органов
В последние два десятилетия в физиологии и биохимии растений получено много-
данных, характеризующих возрастные качественные изменения растений и значение
этих изменений для отдельных физиологических процессов.
Мы не имеем возможности в рамках данной статьи дать критический обзор
этих данных, хотя в этом имеется настоятельная потребность. Укажем лишь неко-
торые основные моменты.
Н. П. Кренке, уделявший большое внимание вопросам физиолого-биохимической
сущности процессов старения и омоложения, хотя сам лично и не успел организовать
соответствующие исследования, считал наиболее глубокой основой процессов старения
качественные изменения протоплазмы, сводящиеся в основном к явлениям синерезиса
и гистерезиса, присущим всем коллоидным системам, на что указывали Ружичка и
Бакхайзен (Кренке, 1950, стр. 490—492).
Многочисленные последующие исследования, как и данные, полученные раньше,
подтвердили, что снижение оводненности тканей является одним из самых характер-
ных показателей их старения (Максимов, 1948: Алексеев, 1948; Гупало, 1949 и др.).
Н. А. Максимов (1941) показал, что засуха и вообще всякое обезвоживание резко
ускоряют процесс старения растений и их органов. Установлено также закономерное
снижение интенсивности дыхания растений и их органов в связи с возрастом (Саби-
нин, 1955), изменение вязкости и проницаемости протоплазмы (Максимов и Можаева,
1944), смещение ИЭТ протоплазмы (Конарев, 1948), изменение содержания хлоро-
филла в листьях (Гюббенет, 1951), изменение активности ферментов и направленности
их действия (Курсанов и Брюшкова, 1940; Сисакян, 1954), уменьшение содержания
свободных нуклеиновых кислот (Lindner, Kirkpatrik, Weeks, 1956) и т. д.
Показано значение возраста листьев и растений в целом в отношении восприим-
чивости их к фотопериодическим воздействиям (Жданова, 1947; Мошков, 1950 и др.).
В. И. Разумов (1954) получил цветение красного клевера в условиях короткого
дня при выдерживании его в этих условиях в течение 2—3 лет, в то время как
в первом году жизни клевер в условиях короткого дня не цветет.
— 87 -
Работами А. А. Табенцкого (1947, 1953) показан закономерный процесс старе-
ния пластид в листьях, а также установлено, что старение хлоропластов замедляется
при подкормке растений азотом.
А. В. Благовещенский (1934, 1950) отмечает, что при старении растений про-
исходит снижение качества ферментов и усиливаются экзотермические процессы цик-
лизации и полимеризации, что ведет к накоплению стойких продуктов специализиро-
ванного обмена, таких, как лигнин, каучук и т. д.
Ф. В. Турчин, М. А. Гуминская и Е. Г. Плышевская (1955), применив изотоп
азота N 15, показали, что в растениях происходит непрерывное обновление азотного
состава белка и пиррольных групп хлорофилла, но по мере старения растений и их
органов интенсивность обновления конституционных белков и хлорофилла падает.
Эти данные совпадают с результатами исследований А. В. Нагорного (1953) и его
сотрудников, которые считают одним из основных показателей ’старения снижение
самообновления белка.
Физиолого-биохимические процессы, протекающие в тканях растений при про-
хождении ими стадий яровизации и световой, еще мало изучены. Однако те данные,
которые уже имеются (Коновалов, 1942; Сисакян, 1954; Конарев, 1954), свидетельству-
ют о том, что это процессы иного порядка, чем процессы старения. Достаточно ука-
зать на то, что при яровизации содержание нуклеиновых кислот увеличивается и ин-
тенсивность дыхания возрастает, в то время как при старении содержание нуклеино-
вых кислот уменьшается и интенсивность дыхания снижается. Следовательно, и с этой
точки зрения нет основания к тому, чтобы сводить возрастные изменения к стадийным
изменениям по Т. Д. Лысенко.
Заключение
Индивидуальное развитие (онтогенез) растений — это сложный и многогранный-
процесс, в котором наряду с другими сторонами отчетливо выступают явления старе-
ния и омоложения отдельных клеток, тканей, органов и организма в целом. Старе-
ние— это качественное изменение организма или его частей, проявляющееся в сни-
жении интенсивности ростовых процессов и физиологической активности. При репро-
дукции происходит омоложение (обновление) вновь образующегося организма по
сравнению с материнским, возврат к исходной точке онтогенеза, хотя и не полный,
так как идет филогенетическое развитие по спирали. Возможно частичное омоложение
и для организма в целом, которое следует понймать как неполное и временное по-
вышение уровня обмена веществ, новообразование более молодых структур и задерж-
ку старения.
Старение и омоложение обусловлено в основном внутренними закономерностями
роста и развития организмов, однако интенсивность этих процессов в значительной
мере зависит от внешних факторов.
Все процессы, наблюдающиеся в ходе онтогенеза растений, взаимосвязаны
между собой. Также взаимосвязаны возрастные и эколого-физиологические стадийные
(по Т. Д. Лысенко) изменения. Однако нельзя сводить возрастные изменения к эко-
лого-физиологической стадийности или отрицать качественный характер возрастных
изменений.
В философии диалектического материализма категорией «качество» обозначаются
коренные свойства предметов, явлений, процессов материального мира. Качественные
изменения — это изменения коренных свойств предметов. Переход растения от обра-
зования вегетативных органов к образованию органов генеративных и прохождение
эколого-физиологических стадий в процессе этого перехода — это, конечно, каче-
ственные изменения. Но переход клетки, органа или целого организма от состояния
активной жизнедеятельности (явной или потенциальной) к состоянию одряхления
и отмирания, или восстановление жизнедеятельности дочернего организма при раз-
множении — это тоже качественные переходы. Старение и омоложение клеток, тканей,
органов и организма в целом, как диалектически противоположное проявление воз-
растных качественных изменений, составляет коренную черту онтогенеза, вытекаю-
щую из жизненного процесса как такового.
1Чы показали многообразное практическое значение возрастных качественных из-
менений растений в растениеводстве. Несомненно, что дальнейшее углубленное изу-
чение этих процессов даст науке не меньше, чем дала теория стадийного развития.
Но для успешности этого изучения необходимо изжить догматическое отношение к
теории стадийного развития, полностью использовав ее фактическую часть и обще-
биологические положения о связи организма с условиями его жизни.
«Чтобы действительно знать предмет, надо охватить, изучить все его стороны, все
связи и «опосредствования». Мы никогда не достигнем этого полностью, но требова-
ние всесторонности предостережет нас от ошибок и от омертвения» 1.
Попытки задержать учение об онтогенезе растений в рамках теории стадийного
развития Т. Д. Лысенко (Дворянкин, Володарский, Казарян и многие другие) должны
быть отметены бурно развивающейся советской биологией.
1 В. И. Ленин. Соч., изд. 4-е, т. 32, стр. 72.
— 88 —,
SUMMARY
The article gives a short historical survey of the theoretical development of the
problem of senescence and rejuvenescence of plants. The practical importance of these proces-
ses, as applied to fruit and berry yielding plants, perennial grass, potatoes, vegetables and
other agricultural crops, is also discussed by the author.
Particular attention is given in the historical survey to the works of C. Child, I. V.
Michurin, N. P. Krenke, P. G. Schitt, T. A. Rabotnov abd other scientists.
The author provides a more exact definition of the terms „senescence* and „rejuven-
escence*. He points out the various meanings of the term „aging* according to Krenke and
agrees with the suggestion made by N. I. Dubrovitskaya and N. P. Krenke regarding the
delimitation of the terms „reneval* and „rejuvenescence*.
That section of the article which deals with the process of senescence in perennial grass
contains a detailed statement of the work done by the author at the Krasnoufimsk plant
breeding station in 1948—1952, in connection with the study of that process in clover.
The factual data quoted in the article shows that senescence is a special physiological
process involving changes of quality. It must neither be identified with the changes due
to different stages of development in the sense proposed by T. D. Lysenko, nor regarded
as a simple consequence of these changes.
The dogmatic attitude adopted by some Soviet scientists in regard to the theory of
development by stages prevents their getting a correct conception of the substance and
importance of the processes of senescence and rejuvenescence of plants, since the theory of
development by stages does not reveal that side of ontogenesis. Recognition of the principles
advanced by F. Engels and I. V. Michurin concerning the life phases of organisms, as a
basis for understanding the laws of ontogenesis, is the right premise to a profound and
fruitful study of the whole many — sidea process of individual plant development and to
the control of that process in the interests of man.
ЛИТЕРАТУРА
Алексеев A. M. 1948. Водный режим растения и влияние на него засухи,
Казань.
Алексеев А. М. 1953. Влияние минеральных удобрений на водный режим
растений. Тр. ИФР им. К. А. Тимирязева, т. 8, вып. 1.
Барский Я. С. 1952. Омолаживание винограда. Сад и огород, № 12.
Бацанов Н. С. 1952. Возделывание картофеля для семенных целей в нечерно-
земной полосе. Изв. ТСХА, вып. 1.
Бацанов Н. С. 1953. Возделывание картофеля в районах нечерноземной по-
лосы. Изд. «Знание», М.
Бекетов А. Н. 1858. Обновления и превращения в мире растений. Русск.
вестник, 15, Май, кн. 1.
Бекетов А. Н. 1862. Курс ботаники. СПб.
Благовещенский А. В. 1934. Биохимия растений. Госхимтехиздат, М.
Благовещенский А. В. 1950. Биохимические основы эволюционного про-
цесса у растений. Изд. АН СССР, М.
Бурмистрова Н. Д. 1949. Омолаживание персика. Сад и огород, № 12.
Васильев И. М. 1953. Зимостойкость растений. Изд. АН СССР, М.
Вобликова Т. В. 1941. Интенсивность фотосинтеза и дыхания листа в зави-
симости от возраста. ДАН СССР, т. 33, № 1.
Володарский Н. И. 1954. Теория стадийного развития и возрастность
растений. Усп. совр. биол., т. 37, вып. 3.
Володарский Н. И. 1955. Влияние листьев разных ярусов на сроки буто-
низации табака. ДАН СССР, т. 100, № 4.
Герасимова 3. А. 1953. Некоторые биологические особенности малины.
Тр. Всесоюзн. н-иссл. ин-та консервн. промышленности, т. 2.
Гой ко В. А. 1954. Влияние возраста семенников на урожай семян двухлетних
овощных культур. Укр. н.-иссл. ин-т овощеводства. Научн. труды, т. 3.
Голицынский Д. А. 1939. Влияние возраста и сроков посева на зимостой-
кость озимой пшеницы в различных условиях зимовки. Тр. Белорусск. СХИ, т. VIII (30).
Головко Д. М. 1955. Влияние калийного питания на рост и продолжитель-
ность жизни листьев подсолнечника. Физиол. растений, т. 2, вып. 2.
Гупало П. И. 1949. Оводненность листьев в связи с динамикой онтогенетиче-
ского развития растений. ДАН СССР, т. 67, № 2.
Гупало П. И. 1950. О значении сроков посева озимой пшеницы для ее зимо-
стойкости. ДАН СССР, т. 74, № 2.
Гупало П. И. 1951. Новые данные о старении клевера в хозяйственных посе-
вах. ДАН СССР, т. 79, № 3.
— 89 -
Гупало П. И. 1952. Об использовании и развитии научного наследства'
Н. П. Кренке. Селекция и семеноводство, № 9.
Гупало П. И. 1953.. К вопросу о физиологической характеристике морфобио-
типов красного клевера. ДАН СССР, т. 91, № 4.
Гупало П. И. 1954. Пути дальнейшего развития теории онтогенеза растений.
Усп. совр. биологии, т. 38, вып. 1 (4).
Гупало П., Савченюк Д., Мохнаткин И. 1950. Возрастные измене-
ния красного клевера в хозяйственных посевах. ДАН СССР, т. 72, № 1.
Гусева Е. И. 1946. Биологическое обоснование обрезки мандарина. Бюлл.
Всес. ин-та чайн. промышленности и субтропич. культур, № 1.
Гюббенет Е. Р. 1951. Растение и хлорофилл. Изд. АН СССР, М.— Л.
Долгушин Д. А. 1946. О некоторых особенностях оплодотворения растений.
Агробиология, № 3.
Дубровицкая Н. И. 1953. Возраст и регенерация растений. Автореферат
дисс. на соиск. уч. степ. докт. биол. наук. Главн. бот. сад АН СССР.
Дубровицкая Н. И. и Кренке А. Н. 1954. О критике теории возрастной
цикличности. Усп. совр. биол., т. 36, № 1 (4).
Дунин М. С. 1946. Иммуногенез и его практическое использование. Тр. Моск.
СХ академии им. К. А. Тимирязева, вып. 40, Рига.
Е фей кин А. К- 1956. Онтогенез и меристема у покрытосеменных растений.
Автореф. дисс. на соиск. уч. степ. докт. биол. наук.
Егоров М. А. 1923. Вопросы зольного питания растений. Харьков.
Жданова Л. П. 1947. Фотопериодическая реакция коротко дневных и длинно-
дневных растений в связи с их возрастом. ДАН СССР, т. 58, № 3.
Житнева П. И. 1948. Биологические основы обрезки смородины. Докл. Моск.
СХ акад. им. Тимирязева, вып. 7.
Иванов П. П. 1949. История и теория обрезки плодоносящей яблони. Дисс. М.
Казарян В. О. 1952. Стадийность развития и старения однолетних растений.
Изд. АН АрмССР, Ереван.
Коломиец Т. А. 1955. Вегетативное возобновление красного клевера. Автореф.
дисс. (ВИР), Л.
Конарев В. Г. 1948. Возрастные изменения в клетках растения и изоэлектри-
ческая точка протоплазмы. ДАН СССР, т. 59, № 4.
Конарев В. Г. 1954. Влияние яровизации на поведение нуклеопротеидов и
нуклеиновых кислот в зародышах злаков. Биохимия, 19, вып. 2.
Коновалов И. Н. 1942. Физиологические особенности развития яровизиро-
ванных растений. Дисс., М.
Кренке Н. П. 1928. Хирургия растений. Изд-во «Новая деревня», М.
Кренке Н. П. 1940. Теория циклического старения и омоложения растений
и практическое ее применение. СХ ГИЗ, М.
Кренке Н. П. 1950. Регенерация растений. Изд. АН СССР, М.— Л.
Кружилин А. С., Шведская 3. М. 1950. Об индивидуальной роли листьев
при прохождении стадий. Агробиология, № 5.
Кузьмин А. Я. 1948. Влияние возраста цветка на скрещиваемость отдален-
ных растений. ДАН СССР, т. 59, № 2.
Курсанов А. Л. и Брюшкова Н. 1940. Действие ферментов в листьях
различных ярусов в связи с их индивидуальным развитием и общим развитием рас-
тения. Биохимия, 5, № 2, 187.
Курындин Н. И., Малинковский В. В., Беньяминов А. Н.,
Белохонов И. В. 1954. Плодоводство. М.
Линник Г. Н. 1955. О причинах вырождения картофеля. Бот. журн., т. 40,№4
Лисавенко М. А. 1950. За дальнейший расцвет сибирского садоводства.
Сад и огород, № 1.
Л ох ин М. А. 1948. Биологическое старение — одна из основных причин изре-
живания красного клевера. Агробиология, № 4.
Лысенко Т. Д. 1935. Теория развития растений и борьба с вырождением
картофеля на юге. Яровизация, № 22, 3—23.
Лысенко Т. Д. 1949. И. В. Сталин и мичуринская агробиология. Агробио-
логия, № 6.
Максимов Н. А. 1941. Влияние засухи на физиологические процессы в рас-
тениях. Сб. работ по физиологии растений памяти К. А. Тимирязева, изд. АН СССР.
Максимов Н. А. 1948. Краткий курс физиологии растений. СХ ГИЗ, М.
Максимов Н. А., Можаева Л. В. 1944. Возрастные изменения коллоидно-
химических свойств протоплазмы растительных клеток. ДАН СССР, т. 42, -№ 5 и 6.
Машинская Л. П. 1953. Значение омолаживания многолетних рукавов куста.
Виноградарство и виноделие СССР, № 10.
Максимов Н. А. и Калмыков К. Ф. 1954. Краткий очерк истории фи-
зиологии растений (Ботаника. Книга для учителя, изд. Ак. пед. наук РСФСР, М.)<
Мичурин И. В. 1939. Сочинения. Изд. 1-е. Сельхозгиз, М.
Молиш Г. 1933. Физиология растений как теория садоводства. Сельхозгиз, М.
— 90 —
Молотковский Г. X. 1945. К вопросу о борьбе с вырождением картофеля
на юге. ДАН СССР. т. 46, № 9.
Молотковский Г. X. 1948. Обрезка ботвы картофеля как способ борьбы
с его вырождением. Уч. зап. Черновицкого гос. ун-та, т. 1, серия биол. наук, вып. 1.
Молотковский Г. X. 1950. Об удлинении жизни некоторых однолетних
растений путем трансплантации. ДАН СССР, т. 71, № 6.
Молотковский Г. X., Бубырева В. Т. 1954. К вопросу о физиологии
растений картофеля с обрезанной ботвой. Бот. журн. АН УССР, т. 11, № 2.
Мошков В. С. 1950. Физиологическая природа фотопериодической реакции
листа. Пробл. ботаники, т. 1, М.— Л.
Мохнаткин И. П., Гупало П. И. и Савченюк Д. М. 1952. Урожай
семян одноукосного клевера на участках второго года пользования. Сов. агроно-
мия, № 12.
Муравьева О. А. 1949. Анатомическое строение столона картофеля в онто-
генезе и при явлениях израстания. Труды Ленинградск. о-ва естествоиспытателей,
т. 19, вып. 3.
Нагорный А. В. 1953. Некоторые закономерности возрастной эволюции жи-
вотного организма. (В кн.: Научн. конференция по вопросам значения возраста при
разведении с.-х. животных Моск. СХ акад. им. Тимирязева).
Р а б о т н о в Г. А. 1946. Длительность виргинильного периода жизни травяни-
стых растений в естественных ценозах. Бюлл. МОИП, отд. биол., т. 51, вып. 2.
Работнов Т. А. 1949. Луговые сорняки и меры борьбы с ними. Сельхозгиз.
Работнов Т. А. 1950. Жизненный цикл многолетних травянистых растений
в луговых ценозах. Тр. БИН им. В. Л. Комарова, серия III (геоботаники), вып. 6.
Разумов В. И. 1954. Некоторые вопросы стадийного развития растений. Вопр.
ботаники, т. II.
Сабинин Д. А. 1955. Физиологические основы питания растений. М.
Сапегин А. А. 1944. Критика теории возрастной цикличности. Юбилейный
сб. АН УССР, г. Уфа.
Саву та А. С. 1948. Декапитация подсолнечника на силос. Сов. агрономия, № 3.
Сагалович Е. Н. 1949. Использовать для отбора на семена морковь летних
посевов. Сад и огород, № 9.
Серебряков И. Г. 1952. Морфология вегетативных органов высших растений.
Изд-во «Сов. наука», М.
Синская Е. Н. 1949. Устойчивость люцерны к заболеваниям в зависимости от
конституции растений, условий существования и процессов старения и омоложения.
Тр. по прикл. бот., ген. и сел., т. 28, вып. 3—18.
Сисакян Н. М. 1954. Биохимия обмена веществ. Изд-во АН СССР, М.
Скрипчинский В. В. 1952. Отмирание многолетних озимых злаковых расте-
ний при различной продолжительности яровизации семян. ДАН, 87, № 5.
Скрипчинский В. В. 1954. Рост, развитие и отмирание однолетних и много-
летних злаков при разных сроках посева. Тр. Ставропольского СХИ, вып. 6.
Смелов С. П. 1947. Биологические основы луговодства. М.
Смелов С. П. 1954. О научных проблемах луговодства в нечерноземной поло-
се. Бот. журн., т. 39, № 1.
Смерткин В. Д. 1940. Борьба со старением корнеплодов моркови на юге
Союза. Овощеводство, № 11—12.
Сукачев В. Н. 1941. О влиянии интенсивности борьбы за существование
между растениями на их развитие. ДАН СССР, т. 33, № 8.
Табенцкий А. А. 1946. Структура хлорофиллового зерна как показатель
жизнедеятельности листа. Изв. АН СССР, серия биол., № 5.
Табенцкий А. А. 1953. К вопросу об управлении процессами образования
зеленых пластид. Изв. АН СССР, серия биол., № 1.
Тарановская.В. Г. 1927. Об изменениях, вызываемых приемом кастрирова-
ния в растениях. Журн. оп. агрономии, т. 23, кн. 2.
Тимирязев К. А. 1942. Исторический метод в биологии. Изд. АН СССР, М.
Трегубенко М. Я. 1940. Снижение морозостойкости люцерны в связи с воз-
растом. Докл. ВАСХНИЛ, вып. 23—24.
Трегубенко М. Я- 1947. Снижение морозостойкости люцерны в связи со
старением. Докл. ВАСХНИЛ, вып. 4.
Трегубенко М. Я. 1950. О приемах повышения зимостойкости люцерны.
Сб. «Травосеяние и семеноводство многолетних трав», МСХ СССР, Сельхозгиз, М.
Трофимович А. Я. 1948. Возрастная приуроченность в паразитизме пероно-
споровых грибов. Докл. Моск. СХ академии им. Тимирязева, вып. 8.
Турчин Ф. В., Гуминская М. А. и Плышевская Е. Г. 1955. Иссле-
дование азотного обмена растений с применением изотопного азота N15, Физиология
растений, т. 2, № 1.
Чирковский В. И. 1953. Влияние старения семян на развитие растений
у табака. ДАН СССР, т. 92, № 2.
— 91 —
Шику л а Я. 1955. Жизнь и труды морфолога растений Н. П. Кренке. «За
социалистическое земледелие» № 5, Прага (на чешском языке).
Шитт П. Г. 1936. Введение в агротехнику плодоводства. Сельхозгиз, М.
Шитт П. Г. 1940. (В соавторстве с 3. А. Метлицким). Плодоводство. Сельхоз-
гиз, М.
Шитт П. Г. 1952. Биологические оснозы агротехники плодоводства. Сельхоз-
гиз, М.
Федюнькин Д. 1955. Пермские клевера. Газета „Сельское хозяйство*
от 6/V 1955 г., № 106.
Федюнькин Д. 1956. Восстановить семеноводство пермского клевера. Газета
„Сельское хозяйство* от 13/VI 1956 г.
Филатов Ф. 1950. Колхозная практика выращивания семян многолетних трав
на Юго-Востоке. Газета „Соцземледелие* № 83 от 17/IV 1950 г.
Эдельштейн В. И. 1953. Овощеводство, М.
Barton L. V. a. Garman Н. R. 1946. Effect of age and storage condition of
seeds on the yields of certain plants. Contrib. from B. Thomps. Jnstitute, vol. 14, N 4.
Braun A. 1851. Betrachtungen uber die Erscheinungen der Verjungung in der
Naturgeschichte. Leipzig.
Braun A. 1872. Veber der Bedeutung der Entwickelung in der Naturgeschichte.
Berlin.
Child С. M. 1915. Senescence and rejuvenescence. Chicago. Jllinois.
Korschelt E. 1924. Lebensdauer, Altern und Tod. 3 te Auflage, Jena, G. Fischer.
Lindner R. S., К i r k p a t r i k H. S. a. Weeks T. E. 1956. Free ribonucleic
acid particles in Homogenates of plant tissue (Plant Physiology, vol. 31, N 1).
Molisch H. 1929. Die Lebensdauer der Pflanze Jena.
Paech K. 1940. Ursache und Verlauf des Alterns bei Pflanzen. I (Zeitschrift fur
-Altersforschung, Band II, Heft III).
БЮЛЛЕТЕНЬ М. О-ВА ИСП. ПРИРОДЫ, ОТД. БИОЛОГИИ, Т. LX1I (5), 1957
Основные виды изменчивости луговой растительности
Т. А. Работное
The main forms of changes in meadow vegetation
T. A. Robotnov
Луг — сложное природное образование, подверженное непрерывным изменениям.
Изучение изменчивости лугов и луговой растительности имеет очень большое значе-
ние для разрешения многих теоретических вопросов, для практики геоботанического
исследования лугов [выделение ассоциаций (типов), картирование] и, в особенности,
для разработки наиболее эффективных приемов рационального использования и улуч-
шения лугов. Ниже рассматриваются некоторые вопросы изменчивости луговой расти-
тельности и намечаются пути управления ею *.
Формы и особенности изменчивости луговой растительности
Луговые ценозы образованы многими видами травянистых, преимущественно
многолетних, мезофильных растений. Каждый вид в ценозе представлен, как правило,
многочисленными особями. В связи с возникновением и отмиранием особей и с изме-
нением их возраста и жизненного состояния меняются численность и состав популя-
ций компонентов ценозов и как следствие этого происходит изменение самих ценозов
.Это всегда сопровождается: 1) увеличением обилия (числового, весового, проектив-
ного) одних видов; 2) уменьшением обилия других видов вплоть до полного исчез-
новения их из ценоза. Кроме того, может происходить внедрение в ценоз новых
видов. При этом всегда меняются соотношение в обилии видов, а иногда и состав
компонентов.
Основная причина изменчивости луговых ценозов — изменение среды. Оно может
быть связано: 1) с воздействием внешних по отношению к ценозу условий (метеоро-
логические условия, заливание полыми водами, отложение наилка, деятельность чело-
века и пр.); 2) с жизнедеятельностью растений и других организмов, образующих
ценозы. .
Среда может меняться: Повременно, на непродолжительный срок (1—2 года),
нередко в различных направлениях (например, при чередовании сухих и влажных
лет), обусловливая лишь временные, обычно не очень крупные изменения, главным
образом, в соотношении обилия видов; 2)\ непрерывно, в определенном направлении
и приводить через тот или иной срок к созданию иных, по сравнению с исходными,
условий произрастания и как следствие этого — к смене одного ценоза другимЛЗ) ка-
тастрофически (распашка, погребение луга овражным выносом и пр.), в результате
чего происходит уничтожение ценоза, с последующим его восстановлением через
известный срок или с последующим образованием нового ценоза.
Нередко между этими тремя группами изменений нет резких границ.
Формы изменчивости одни и те же у различных типов растительности. В то же
время в изменчивости каждого типа растительности имеется своеобразие; на лугах
оно выражается в следующем.
1. Луга на значительных площадях — вторичные образования, возникшие в ре-
зультате деятельности человека на месте сведенных лесов и кустарников, осушенных
болот, спущенных озер, в результате орошения степей и пр. Они, как правило,
1 Краткие обзоры огромной работы, проведенной в нашей стране по изучению
лугов за советский период, даны Т. А. Работновым (1950) и А. П. Шенниковым
(1954).
— 94 —
используются под сенокосы и пастбища. Большинство типов лугов при прекращении
использования их человеком постепенно возвращается к исходным типам раститель-
ности (превращаются в леса и пр.). Следовательно, их происхождение, либо их
современное состояние, или то и другое тесно связаны с деятельностью человека.
Нельзя поэтому изучать изменчивость лугов вне связи с деятельностью человека.
Сенокошение и выпас оказывают очень большое и в то же время различное влияние
на луговую растительность. На месте однородной растительности при различных
формах использования (число стравливаний или скашиваний, высота скашивания или
стравливания, нагрузка, вид скота и пр.) может возникать большое количество суще-
ственно отличающихся друг от друга растительных группировок. При этом в зависи-
мости от форм и интенсивности использования меняется изменчивость растительности,
обусловленная другими причинами (жизнедеятельностью растений, метеорологическими
условиями и пр.). • .
2. Луговая растительность на значительных площадях — не только вторичное, н(>
и молодое образование. Нередко возраст лугов измеряется всего двумястами-
тремястами, а иногда даже несколькими десятками лет. Естественно, что в связи
с относительной непродолжительностью периода отбора растения, образующие луго-
вые ценозы, менее приспособлены к местным условиям, нежели растения коренных
типов растительности, на месте которых возникли луга. Кроме того, луговые травыг
образуя травостои, создают свой фитоклимат, в значительно большей степени завися-
щий от изменения метеорологических условий отдельных лет, нежели фитоклимат
лесов и кустарников. Поэтому на лугах разногодичная изменчивость проявляется
особенно резко.
3. Значительные площади лугов расположены в пониженных элементах рельефа
(в поймах, низинах, западинах), водный режим которых более резко колеблется
по годам (и в течение вегетационного сезона), нежели на положительных элементах
рельефа. Это определяет возможность резких изменений условий произрастания от
года к году.
4. Длительность жизни луговых растений большей частью невелика, причем, как
правило, многие из них не достигают предельно возможного в данных условиях воз-
раста, отмирая в значительных количествах в годы метеорологических аномалий или
в годы массового размножения мышевидных грызунов или вредных насекомых.
Многие виды луговых растений размножаются исключительно или преимущественно
семенами. Семенное размножение на лугах затруднено и условия, благоприятные для
него, реализуются периодически. Поэтому на лугах нередко наблюдается «циклич-
ность» в обилии ряда компонентов, связанная с периодическим массовым отмиранием
(либо в связи с достижением предела жизни или в связи с иными причинами) и перио-
дически реализующимися условиями, благопрятными для семенного размножения.
Для лугов наиболее универсальны следующие формы изменчивости:
/ 1) сезонная изменчивость;
2) разногодичная изменчивость1;
3) смены и изменения ценозов, связанные с деятельностью человека;
1 4) смены ценозов в связи с изменением среды в результате жизнедеятельности
луговых растений;
5) изменчивость, связанная с жизненным циклом растений.
Экзодинамические смены, помимо смен, вызванных деятельностью человека,
на лугах происходят редко. Наиболее широко распространены экзо-эндодинамические
смены, обусловленные изменением среды в связи с жизнедеятельностью луговых рас-
тений и под влиянием внешних по отношению к ценозам причин.
Все причины изменчивости луговой растительности действуют одновременно.
При этом происходят то более медленные, то более быстрые смены одних ценозов
другими, а в пределах периода существования ценозов — более или менее значитель-
ные изменения в соотношении компонентов ценозов от года к году и в течение веге-
тационных периодов. Процесс смены ценозов редко бывает плавным, более часто он.
нарушается временными изменениями, связанными с своеобразием метеорологических
условий отдельных лет, массовым размножением насекомых, мышевидных грызунов,
сменой формы использования человеком и пр.
^-Для каждого типа луга существует свой ход развития, обусловленный направле-
нием' изменения физико-географической среды (например, с постепенным увеличением
высоты пойменной террасы в связи с ежегодным отложением наилка), формой и ин-
тенсивностью использования луга и особенностями воздействия компонентов ценозов
на среду. Скорость смен зависит от совместного воздействия всех трех групп факто-
ров. Воздействие одной группы факторов может ослабить или, наоборот, усилить влия-
ние другой группы факторов. Например, отложение наилка ослабляет или полностью-
предотвращает накопление в почве неразложившихся остатков растений. Наоборот,
ежегодное скашивание и отчуждение с урожаем значительных количеств питательных
веществ приводит к обеднению почвы, ухудшает условия разложения отмерших остат-
1 Вопросы разногодичной изменчивости рассмотрены нами в другой работе
(Работнов, 1955).
— 95 —
ков растений и тем самым ускоряет смену ценозов. Из всех групп факторов особенно
велико значение деятельности человека. Применяя рациональные формы использования,
внося удобрения, регулируя водный режим, можно предотвратить изменение условий
произрастания и удержать луг в желательном состоянии неопределенно долгое время.
Однако для предотвращения смен ценозов и поддержания луговой растительности в
определенном состоянии недостаточно создания условий, устойчивых и благоприятных
для ценных кормовых растений. Необходимо, помимо того, обеспечить возможность
размножения растений, чтобы вместо отмерших возникали новые особи. Разрешение
этой задачи на постоянных лугах (или лугах длительного использования), по-видимэ-
му, представляет большие трудности, чем создание соответствующих условий увлаж-
нения, обеспечения питательными веществами и аэрации.
Обеспечение непрерывного размножения растений в луговых ценозах наиболее
легко осуществимо для видов, возобновляющихся вегетативно. Правда, по-видимому,
при непрерывном вегетативном размножении жизненное состояние растений и способ-
ность создавать урожай постепенно снижаются (Вильямс, 1922; Кренке, 1940 и др.).
Однако снижение жизненного состояния при непрерывном вегетативном размножении
у разных видов проявляется в различной степени и, несомненно, зависит от условий
произрастания. Некоторые виды луговых растений, во всяком случае, в определенных
условиях могут размножаться вегетативно, без утраты способности давать высокие
урожаи, в течение десятков лет, а возможно, даже свыше ста лет. Известны интен-
сивно-используемые высокопродуктивные пастбища, где возможность семенного раз-
множения исключена или весьма ограничена, существующие в течение десятилетий.
Значительно труднее на интенсивно используемых высокопродуктивных лугах (сено-
косах и пастбищах) поддержать в течение длительного периода на должном уровне
обилие растений, размножающихся исключительно семенами, в особенности таких
недолговечных растений, как клевер красный и розовый. Чем выше урожайность
луга, тем, обычно, менее благоприятны условия для семенного размножения растений.
Сезонная изменчивость луговой растительности
Ценное обобщение данных о сезонной изменчивости луговой растительности дано
А. П. Щенниковым (1941); им указано на необходимость разностороннего и более
глубокого изучения этого явления. Однако до сих пор изучались, главным образом,
два вопроса: смена аспектов и динамика изменения урожайности (массы надземных
органов) в течение вегетационного сезона. Между тем для геоботаники и луговодства
большой интерес представляет изучение сезонного изменения обилия компонентов
ценозов и структуры травостоев, а также условий семенного и вегетативного размно-
жения и пр. По этим вопросам имеется мало данных.
Луговые ценозы образованы видами, отличающимися друг от друга по динамике
изменения массы надземных органов в течение вегетационного периода (по динамике
абсолютного обилия). В связи с этим сезонные изменения в соотношении обилия
компонентов являются характерными для луговых ценозов. У отдельных ценозов они
выражены в различной степени, завися от следующих условий:
1) Состав компонентов; численность и состав группы доминантов (сезонные
изменения состава травостоев наиболее значительны в полидоминантных груп-
пировках).
2. Динамика надземной массы отдельных доминантов. Наиболее велики изме-
нения в составе травостоев при наличии доминантов, резко отличающихся друг от
друга по сезонной динамике, в особенности при наличии доминантов, вегетирующих
не весь вегетационный сезон [весенние эфемероиды, растения весенне-летней вегета-
ции (Veratrum lobelianum, Polygonum bistorta)].
3) Доля участия доминантов в сложении травостоев. Наиболее устойчив состав
монодоминантных группировок, в которых преобладающее растение образует (по мас-
се) 80—90% и более травостоя (например, некоторые костровые луга).
4) Воздействие человека. Динамика изменения обилия по сезону существенно
меняется в зависимости от воздействия человека. При двуукосном использовании
от первого ко второму укосу обыкновенно происходят значительные изменения в от-
носительном обилии луговых растений. Большое значение также имеет весенний вы-
пас на сенокосах и пр.
При резко выраженной разногодичной изменчивости сезонные изменения оби-
лия компонентов ценозов в отдельные годы могут проявляться по-разному. При про-
ведении геоботанических исследований лугов, в особенности при установлении ассо-
циаций, необходимо учитывать сезонную изменчивость луговых травостоев. В одних
случаях для установления ассоциаций достаточно однократного описания раститель-
ности, в других — однократное описание может дать ошибочное представление об уча-
стии отдельных видов в ценозе. Нами изучался субальпийский луг (Северный Кавказ),
который на основании описаний, проведенных в середине июня, следовало отнести
к чемеричникам, поскольку в это время в травостое преобладала чемерица (Veratrum
lobelianum), а по описаниям, проведенным в середине августа, когда чемерица за-
— 96 -
сохла и ее проективное и весовое обилие резко снизилось, а в травостое преобладал
аконит восточный (Aconitum orientate), образовавший почти сплошной покров,—
к аконитникам. Крупные ошибки в выделении ассоциаций можно допустить, если
в одних случаях луга характеризуются на основании изучения травостоев перед
первым укосом, а в других случаях — перед вторым укосом.
Изучение сезонной изменчивости луговых травостоев имеет большое значение
для определения рациональных сроков использования. Такие наблюдения должны
быть длительными в связи с необходимостью учета влияния использования в от-
дельные сроки на урожайность и состав травостоев в последующие годы. Необходимо
также изучение сезонных изменений эффективности приемов улучшения лугов. Луго-
воды часто ставят опыты для выяснения рациональных сроков внесения удобрений,
подсева и пр. Однако, как правило, такие опыты краткосрочны и проводятся без учета
состояния травостоев.
Омены луговой растительности под влиянием изменения условий произрастания
в результате жизнедеятельности растений
Наиболее полно этот вопрос рассмотрен в работах В. Р. Вильямса (1922, 1926).
Си показал значение жизнедеятельности растений как фактора смен луговой расти-
тельности и предложил теорию, объясняющую эти смены (теорию дернового процес-
са). По В. Р. Вильямсу, в результате жизнедеятельности луговых многолетних травя-
нистых растений в почве происходит постепенное накопление мертвого органического
вещества и, как следствие этого, снижение обеспеченности растений доступными для
них элементами минеральной пищи, увеличение влажности, ухудшение условий
аэрации. Все это приводит, по В. Р. Вильямсу, к последовательным сменам одних
ценозов другими. Учение В. Р. Вильямса о дерновом процессе представляет большую
ценность для почвоведения, геоботаники и луговодства. Впервые в. работах
В. Р. Вильямса был осуществлен комплексный биоценотический подход к изучению
луга как развивающегося явления. К сожалению, теория дернового процесса, после
ее опубликования, не разрабатывалась ни самим В. Р. Вильямсом, ни его последо-
вателями. В большинстве случаев эта теория излагалась догматически. Лишь в немно-
гих работах отдельные положения теории дернового процесса рассмотрены критически
(Сукачев, 1916; Шенников, 1941 и др.) Геоботаники мало использовали работы
В. Р. Вильямса при изучении смен луговой растительности. Очень мало было прове-
дено углубленных наблюдений и экспериментов для изучения изменения среды в ре-
зультате деятельности растений в условиях луга. Между тем изучение смен луговой
растительности в результате жизнедеятельности луговых растений и теоретическое
обоснование приемов управления сменами естественных и сеяных ценозов имеет
огромное значение для практики луговодства. Помимо проведения специальных иссле-
дований, необходимо обобщить большей частью отрывочный фактический материал,
имеющийся в работах геоботаников, почвоведов, луговодов, микробиологов и др.
Впредь до проведетия такой работы представляется целесообразным рассмотреть основ-
ные положения теории дернового процесса В. Р. Вильямса и наметить вопросы для
дальнейшего изучения.
Прежде всего следует отметить, что воздействие луговых растений на почву
весьма разносторонне и не может быть сведено только к накоплению мертвого органи-
ческого вещества и к связанным с этим изменением пищевого, водного и воздушного
режимов. В изменении среды принимают участие и живые органы растений, погло-
щающие из почвы влагу и питательные вещества и выделяющие в почву различные
соединения. Большое значение в изменении среды имеют также продукты разложения
отмерших остатков растений. Растения могут изменять почву не только в сторону
ухудшения обеспеченности пищей, влагой и воздухом, но и в сторону улучшения
условий произрастания; обогащая почву азотом, улучшая структуру почвы (и тем са-
мым улучшая водно-воздушные свойства почвы), перенося зольные элементы из ниж-
них слоев почвы в верхние, растворяя труднорастворимые соединения, и освобождая
их после минерализации отмерших остатков в форме соединений, легко доступных
для других видов растений, содействуя размножению полезных микроорганизмов и
дождевых червей и пр. Таким образом, воздействие луговых растений на почву мо-
жет быть очень разносторонним. Необходимы углубленные исследования для выясне-
ния разнообразных воздействий луговых растений на почву. Особенно важно выявле-
ние и изучение растений, непосредственно или косвенно содействующих повышению
плодородия почвы: бобовые растения, растения, благоприятствующие жизнедеятель-
ности свободно живущих азотфиксирующих организмов, виды, благоприятно влияю-
щие на жизнедеятельность дождевых червей, и пр. Луговодам нужно иметь данные
не только по урожайности и кормовой ценности отдельных видов луговых растений,
но и по влиянию их на почву. Только тогда представится возможным более созна-
тельно подходить к регулированию видового состава луговых сообществ.
Несомненно, однако, что прогрессивное накопление в почве неразложившихся
и полуразложившихся остатков растений имеет большое значение в смене луговых
— 97 —
ценозов. Поэтому все, что содействует этому процессу, ускоряет смены, и, .наоборот,
все, что препятствует проявлению дернового процесса, замедляет или даже пред-
отвращает смены.
Накопление мертвого органического вещества в почвах под луговой раститель-
ностью зависит от многих причин, а в частности от: 1) деятельности человека; 2) ха-
рактера экотопа; 3) видового состава растительности; 4) почвенной фауны.
В. Р. Вильямс рассматривал дерновый процесс и смены луговой растительности
вне зависимости от деятельности человека. Между тем луга, в преобладающей своей
части, возникли в результате деятельности человека и в настоящее время используют-
ся как сенокосы и пастбища. Форма и интенсивность использования лугов, а также
характер ухода за ними оказывают огромное влияние на условия произрастания
растений. Поэтому нельзя рассматривать смены луговой растительности вне связи
с деятельностью человека. Накопление в почве мертвого органического вещества и
обеднение ее элементами минерального питания растений наиболее ярко выражено на
сенокосах, где обеднение почвы усиливается в связи с ежегодным отчуждением с уро-
жаем значительных количеств элементов пищи растений, в особенности азота. Коли-
чество ежегодно отчуждаемых с урожаем питательных веществ существенно варьирует
от одного типа луга к другому в зависимости от урожайности и химического состава
сена. Так, по данным Г. Я. Бронзовой (1936), в Сафоновском р-не, Смоленской обл.,
при рациональных сроках использования на отдельных типах лугов ежегодно отчуж-
дается от 11,0 до 58,0 кг/га азота.
Исходя из данных среднего содержания Са, Р и К в сене (Томмэ, Ксанфопуло
и Сементовская, 1948), можно считать, что с суходольных лугов при средней урожай-
ности 10 ц/га с урожаем ежегодно уносится из почвы (в кг/га): Са — 70, Р—1,6',
К— 13,5. Ежегодно повторяемое отчуждение питательных веществ в местах, где нет
обогащения ими за счет отложения наилка, приноса грунтовыми водами, внесения
удобрений и пр., приводит к прогрессирующему обеднению лугов доступными для
луговых растений минеральными элементами пищи растений. Это не только усиливает
ухудшение обеспеченности луговых трав минеральной пищей, но, ослабляя жизнедея-
тельность микроорганизмов, затрудняет разложение отмерших органов растений, со-
действует накоплению мертвого органического вещества в почве L
Обеднение почвы элементами минеральной пищи растений можно, однако, легка
приостановить путем регулярного внесения соответствующих минеральных удобрений.
К сожалению, до сих пор, несмотря на большое число опытов с удобрением лугов,
нет достаточно точных данных о том, как влияет регулярное внесение удобрений на
изменение среды в результате жизнедеятельности луговых растений и, в частности,
на накопление в почве мертвого органического вещества. В качестве косвенного noj
казателя условий произрастания, правда, можно использовать урожайность. В лите-
ратуре имеются многочисленные и довольно противоречивые данные об изменении
урожайности лугов при регулярном внесении удобрений. Имеющаяся противоречи-
вость, несомненно, связана с тем, что отдельные исследователи изучали различные
луга (естественные, сеяные), вносили различные удобрения и проводили опыты в те-
чение различного срока. Опытов, длившихся долгое время, проведено, к сожалению,
мало. Особый интерес представляет опыт, заложенный в 1856 г. Лоозом и Гильбер-
том на Ротамстедской опытной станции и продолжающийся по настоящее время.
В этом опыте урожайность делянок, ежегодно удобряющихся азотом (в форме селит-
ры— 96,5 кг/га), фосфором и калием за 70 лет не снизилась, в то время как уро-
жайность неудобрявшихся делянок снизилась вдвое (W. Е. ВгепсЫеу, 1926). Таким
образом, результаты этого опыта показывают, что при регулярном внесении соответ-
ствующих удобрений можно не только добиться высокой урожайности, но и поддер-
живать ее в течение длительного времени на высоком уровне (68,9 ц/га).
В литературе, однако, имеются многочисленные указания на быстрое снижение
урожайности сеяных лугов даже при регулярном внесении удобрений (Смелов,
1936, и др.).
Очевидно, что снижение урожайности сеяных лугов связано в этом случае не
только и не столько с обеднением почвы и накоплением в ней мертвого органического
вещества, сколько с несовершенством созданного сообщества, с малой приспособлен-
ностью высеянных видов или форм растений к местным условиям произрастания,
с неспособностью их к размножению при данных формах использования.
При пастбищном использовании лугов при круглосуточном содержании скота на
пастбищах 75—96% азота и 90—96% зольных элементов, содержащихся в съеденной
1 Косвенным доказательством этого могут служить результаты исследований
Кмоха (Н. G. Kmoch, 1952). Изучая подземные органы растений на лугах, он уста-
новил, что количество их возрастает по мере обеднения почвы Р2О5; так, в среднем
при изучении большого числа образцов он получил следующее содержание подземных
органов в граммах на 1000 см3 почвы: на почвах, богатых Р2О5,— 58,5, умеренно
богатых — 84,0; бедных — 145,1. Несомненно, что Кмох определял массу не только
живых, но и отмерших корней.
- 98 -
животными траве, возвращаются на луг с экскрементами скота (Р. D. Sears, 1950; Ра-
ботнов, 1956). Таким образом, на пастбищах с урожаем отчуждается несравненно
меньшее количество питательных веществ, чем на сенокосах. Кроме того, в связи
с отложением на поверхности почвы значительных количеств легко минерализуемого
органического вещества создаются более благоприятные условия для деятельности
микроорганизмов и дождевых червей, способствующих минерализации отмерших
остатков растений в почве. Все это препятствует накоплению в почвах пастбищ мерт-
вого органического вещества; эти почвы, как правило, содержат меньше мертвого
органического вещества, нежели почвы сенокосов (М. L. Hart, 1950). При правиль-
ном использовании пастбищ и уходе за ними со временем на них не происходит ухуд-
шения условий произрастания для ценных кормовых растений, требовательных к фак-
торам среды; об этом можно судить по тому, что с возрастом повышается продук-
тивность пастбищ, увеличивается содержание ценных кормовых растений на них,
улучшаются условия аэрации (увеличение объема пор) (Е. Klapp, 1954). Все это
дает основание утверждать, что изменение среды под влиянием жизнедеятельности
луговых растений на пастбищах идет иначе, чем на лугах, используемых как сено-
косы. Следовательно, правильно используя луга, в особенности используя их как
пастбища, и внося удобрения, можно контролировать дерновый процесс и управлять
сменами луговой растительности в желательном направлении. Необходимо дальней-
шее углубленное изучение отдельных форм воздействия человека на луговую расти-
тельность.
Исходное состояние, в частности механический состав и химические свойства
почвы, и условия увлажнения имеют большое значение для хода дернового процес-
са. Все, что благоприятствует деятельности микроорганизмов, разлагающих отмершие
части растений (нейтральная или близкая к ней реакция почвы, обеспеченность каль-
цием и фосфором, благоприятные условия аэрации й увлажнения) замедляет темпы
дернового процесса или даже вовсе устраняет накопление мертвого органического ве-
щества в почве. Наоборот, наличие условий, затрудняющих разложение микроорга-
низмами отмерших частей растений (повышенная кислотность, недостаток фосфора,
повышенное увлажнение, плохая аэрация), ускоряет ход дернового процесса.
Общеизвестно, что в поймах, в местах ежегодного отложения обильного наилка,
накопления в почве неразложившихся остатков растений не происходит. Таким обра-
зом, проводя известкование кислых почв, осушение заболачивающихся пространств
и прочее, можно также воздействовать на ход дернового процесса.
Несомненно, что дерновый процесс зависит и от видового состава растительных
сообществ. Отдельные растения, при произрастании в одних и тех же условиях, мо-
гут существенно различаться друг от друга по количеству подземных органов, от-
мирающих ежегодно, по их химическому составу и по скорости разложения. Послед-
нее находится в связи с различием в содержании веществ (в частности, азота), не-
обходимых для нормальной жизнедеятельности микроорганизмов, а также веществ,
трудно поддающихся разложению или затрудняющих жизнедеятельность микроорга-
низмов (лигнин, дубильные вещества и пр.).
Несомненное значение также имеет глубина размещения подземных органов
луговых растений. По наблюдениям ряда исследователей (Bates, 1948; Куркин, 1952),
разрастание низкорослых злаков с поверхностной сильно разветвленной корневой си-
стемой (полевицы, мятлики и пр.) создает неблагоприятные условия для произраста-
ния более урожайных высокорослых злаков. В связи с этим для поддержания высо-
кой урожайности переменных лугов стремятся не допускать разрастания мелкоукоре-
няющихся малоурожайных злаков (Prince, 1956). Выше было отмечено, что изучение
средоизменяющих свойств отдельных видов представляет большой интерес для прак-
тики. При проведении таких исследований необходимо обратить должное внимание
на свойства растений, определяющие накопление в почве мертвого органического ве-
щества.
Почвенная фауна, в особенности дождевые черви, может оказать большое влия-
ние на ход дернового процесса. Поедание отмерших частей растений дождевыми
червями и другими животными, обитающими в почве, содействует быстрой минерали-
зации органического вещества, препятствует его накоплению. Дождевые черви (а так-
же и другие животные), пронизывая почву своими ходами, улучшают ее водно-воз-
душные свойства и тем самым ускоряют разложение отмерших остатков растений
микроорганизмами. Дождевые черви содействуют образованию прочной комковатой
структуры. Чем больше дождевых червей, чем выше их активность, тем большие ко-
личества отмерших частей растений они переработают, тем в большей мере они пре-
пятствуют накоплению неразложившихся остатков растений. Создавая благоприят-
ные условия для дождевых червей на лугах можно существенно изменить ход дерно-
вого процесса. Дождевые черви являются постоянным компонентом луговых биоцено-
зов. К сожалению, в нашей стране роль их в жизни луговых ценозов почти не изуче-
на. Очень мало известно о взаимоотношениях дождевых червей с отдельными видами
луговых растений, в частности поедаемость отмерших подземных органов различных
видов растений дождевыми червями. Исследования в этом направлении представляют
несомненный интерес.
— 99 —
Смены луговых ценозов в результате жизнедеятельности луговых растений
(а также в связи с другими причинами) в существенной степени зависят от внедре-
ния в ценозы новых видов растений. Как только в связи с изменением среды в це-
ноз внедряется новый вид (или новые виды), более приспособленный к создавшимся
условиям, он (при наличии соответствующих биологических свойств) может быстро
размножиться и тем самым существенно воздействовать на среду. Внедрение новых
видов, таким образом, ускоряет смену ценозов или даже служит причиной смены це-
нозов. Поэтому все, что затрудняет или предотвращает внедрение новых растений,
задерживает, а иногда даже исключает смену ценозов. Примером могут быть неко-
торые лесные суходольные поляны, которые не превратились в белоусники лишь по-
тому, что не было заноса сюда семян белоуса. Следовательно, путем контроля режи-
ма зачатков также можно воздействовать на смены ценозов и в комплексе с другими
приемами управлять сменами. Представляет несомненный интерес изучение факто-
ров переноса зачатков, процесса внедрения новых растений в ценозы и влияния их
внедрения на другие виды растений.
Изменчивость луговой растительности, связанная с жизненным
цивлом компонентов
В. Н. Сукачев (1942) выделил как особую форму динамики растительности «из-
менение фитоценозов в связи с онтогенезом 1 эдификаторов». Этот вид изменчивости
часто свойствен определенным типам лугов. Однако он часто связан с прохождением
жизненного цикла растений, которые вряд ли можно отнести к эдификаторам1 2.
К числу их относится ряд моно- и олигокарпичных растений, характеризующихся не-
большой длительностью жизни, в частности клевера красный и розовый и некоторые
монокарпичные зонтичные (порезник промежуточный, дягиль лекарственный и др.).
У этих растений нередко хорошо выражена цикличность в их участии в создании це-
нозов. Их обилие и характер его изменения по годам определяются возможностью
обсеменения, приживанием всходов, а в дальнейшем — условиями произрастания при-
жившихся растений.
л Особенно резко цикличность выражена у клеверов красного и розового на
почвах, благоприятных для их роста и развития (нейтральная или слабокислая реак-
ция, обеспеченность Р и К). На таких лугах возникающие из семян особи клевера
развиваются быстро и обыкновенно на третий год (а иногда даже на второй год) при
благоприятных метеорологических условиях превращаются в пышно развитые расте-
ния с многочисленными генеративными побегами. На таких лугах клевера красный и
розовый являются либо моно- либо дикарпичными растениями. Поэтому пышно раз-
витые особи клеверов обычно отмирают к Весне следующего года или цветут в тече-
ние двух лет и затем отмирают. Массовый переход клеверов из вегетативного состоя-
ния в генеративное на естественных лугах происходит лишь при благоприятных соче-
таниях метеорологических условий (в предшествующий и в данные годы). Эти усло-
вия ближе не изучены. По-видимому, массовый переход клевера от вегетативного
состояния в генеративное может происходить при различных сочетаниях гидротерми-
ческих режимов в предшествующий и в данный годы. В некоторые годы пышное
развитие клеверов наблюдается лишь на лугах определенных типов, в то время как
в другие годы клевера обильны на лугах всех типов, где они входят в состав цено-
зов. Вероятно влияние метеорологических факторов сказывается* на клеверах не столь-
ко непосредственно, сколько косвенно, обусловливая то увеличение, то уменьшение
обилия других растений в ценозе, в частности злаков (Работнов, 1950). Некоторое
подавление групп растений, сдерживающих рост клеверов в определенный год, может
привести в последующий год к резкому увеличению клеверов. Цикл участия клеверов
в сложении ценозов включает следующие периоды (Работнов, 1950):
1. Наличие большого числа особей клевера, находящихся в вегетативном, не-
редко в угнетенном состоянии; роль клеверов в сложении ценоза незначительна.
2. Превращение большей части особей в пышно развитые растения с многочис-
ленными генеративными побегами. Численность особей обычно не меняется, но весо-
вое и проективное обилие и участие в сложении ценоза резко (в десятки и даже сот-
ни раз) возрастает.
3. Резкое снижение численности популяций в результате массового отмирания
пышно развитых растений.
4. Увеличение численности популяций за счет возникновения новых особей из
семян.
1 Эмбриональный этап онтогенеза не имеет значения для изменчивости расти-
тельности, поэтому лучше говорить о жизненном цикле растений, понимая под ним
часть онтогенетического развития растения, от возникновения особи до отмирания
(после завершения жизненного цикла).
2 Изучение эдификаторов имеет огромное значение (Лавренко, 1947). Однако
выделение их на лугах часто сопряжено с большими трудностями. Нельзя относить
к эдификаторам любой доминирующий вид растения в ценозе.
— 100 —
Период преобладания клеверов обычно равен одному-двум годам, но в некото-
рых условиях (что случается редко) может быть и более длительным (в наших опы-
тах на субальпийских лугах Северного Кавказа до трех лет). При этом, вероятно,
имеет большое значение характер использования луга. Раннее (до образования семян)
скашивание, по-видимому, увеличивает длительность жизни клеверов. Массовое отми-
рание клеверов может происходить не только в связи с завершением жизненного
цикла, но также в результате деятельности мышевидных грызунов и других причин
(Работнов, 1950). Однако, вероятно, оно большей частью связано с их моно- и ди-
карпичностью. Не ясен вопрос о значении «клевероутомления» как причины циклич-
ности клеверов на лугах.
Интервалы между «клеверными» годами сильно варьируют в зависимости от
возможностей обсеменения и наличия всхожих семян клеверов в почве и от условий
произрастания. Особенно важно подчеркнуть значение обсеменения клеверов или при-
носа их семян извне (в том числе и в виде подсева).
Внесение удобрений (Са, Р, К, Мо) там, где имеется в том потребность, со-
действует приживанию всходов и превращению молодых (а также угнетенных взрос-
лых) растений во взрослые пышно развитые особи. Однако увеличение устойчивости
клеверов (красного, розового) при этом может быть достигнуто лишь при обеспече-
нии их обсеменения (что зависит от характера использования) или при регулярном
подсеве их семян.
Несомненно, что периодическое преобладание клеверов в травостоях сказы-
вается на обилии других растений. Следовательно, даже при относительно устойчи-
вых условиях произрастания, исключающих возможность смены одного ценоза дру-
гим, растительность луга в связи с «цикличностью» участия клеверов, может претер-
певать существенные изменения. Это, несомненно, осложняет разработку классифика-
ций луговой растительности. Возникает вопрос—происходит ли на лугу периодиче-
ская смена «клеверных» и «неклеверных» ассоциаций или происходит смена состояний
одного и того же ценоза в связи с особенностями жизненного цикла клеверов. Пра-
вильнее не выделять «клеверные луга» в самостоятельные ассоциации1 или типы,
а рассматривать их как временное состояние, характерное для многих лугов (пой-
менных, низинных, суходольных, субальпийских), отмечая периодичность преоблада-
ния клеверов при наличии их обсеменения или приноса их семян извне. В названии
таких типов лугов (ассоциаций) должно быть отражено чередование состояний с пре-
обладанием и без преобладания клевера. Это может быть выражено следующим об-
разом: А + С + D А 4- В + Trifolium.
Если есть возможность указать длительность периодов, то это следует ввести
в название, например: А + С + D (2—4 rojia)^Trifolium + А + В (1—2 года). Воз-
можен также несколько иной способ обозначения таких лугов; А+ (Trifolium + В) +
(С + Д). Для обоснованного выделения и для точной характеристики таких типов
необходимы многолетние наблюдения.
Иной характер имеет изменчивость растительности, связанная с прохождением
жизненного цикла монокарпичных зонтичных, в частности дягиля лекарственного
(Archangelica officinalis).
Наблюдениями и опытами, проведенными в пойме р. Оки близ Дединова (Мос-
ковской обл.), установлено, что отсутствие или малое обилие дягиля на больших пло-
щадях центральной поймы, где он широко распространен, вызвано не условиями про-
израстания, а затруднением обсеменения и отсутствием заноса его семян. Периоди-
чески, во всяком случае на отдельных участках, в связи с запозданием со скашива-
нием травы, дегиль обсеменяется. Обсеменение обычно происходит во время сушки
травы и сгребания сена. При этом на небольших участках на поверхность почвы
нередко осыпается значительное количество семян дегиля. На следующий год по-
являются массовые всходы. Молодые растения развиваются медленно, постепенно,
с каждым годом увеличивая массу надземных органов, и лишь в возрасте 5—10 лет
достигают взрослого состояния и могут стать преобладающими растениями в ценозе.
Обычно ежегодно зацветает лишь часть особей. Благодаря этому преобладание
дягиля может длиться ряд лет, пока большая часть особей не зацветет и не отомрет
(если при этом не происходит нового обсеменения дягиля). Таким образом, в отли-
чие от клеверов, у монокарпических зонтичных наблюдается постепенное увеличение
их обилия, а после достижения определенного максимума — более или менее посте-
пенное снижение его. При этом одно и то же обилие на восходящей и нисходящей
части кривой имеет различное значение в жизни ценоза. Несомненно, что в связи
с изменением обилия дягйля меняются условия произрастания для других компонен-
тов ценозов и, следовательно, меняется их обилие. Такого рода изменчивость лугов
в связи с прохождением жизненного цикла монокарпичных зонтичных особенно резко
1 При выделении луговой растительности с преобладанием клевера красного или
розового в самостоятельные ассоциации необходимо указывать, что такие «ассоциа-
ции» периодически возникают, существуют один-два года, а затем сменяются други-
ми группировками. Их нельзя признать равнозначными длительно существующим
группировкам.
— 101 —
выражена на отдельных небольших участках, площадью в несколько десятков или
сотен квадратных метров, на которых особенно обильно осыпались семена при убор-
ке сена
В менее резких формах она широко распространена. Как особую форму сле-
дует рассматривать изменчивость луговых травостоев, обусловленную колебанием по
годам численности полупаразитных однолетников (погремков и др.).
О длительности существования и степени изменчивости луговых ценозов
При стационарном изучении луговой растительности можно установить су-
щественные признаки ценозов: длительность существования, возраст ц степень выра-
женности сезонной и разиогодичной изменчивости. Эти признаки обычно ие учитыва-
ются в практике геоботанических исследований и при разработке классификаций.
Каждый ценоз возникает, существует определенное время, а затем сменяется
другим ценозом. Отдельные ценозы несомненно, характеризуются различной дли-
тельностью существования. К сожалению, наши знания по этому вопросу очень скуд-
ны. Длительность существования ценозов определяется: 1) устойчивостью условий
произрастания, что зависит а) от устойчивости режимов экотопа, б) от средоизменяю-
щей способности компонентов ценоза, в) от деятельности человека; 2) длительностью
жизни компонентов ценоза и способностью их возобновляться при данных условиях
использования; 3) возможностью заноса зачатков видов растений, более приспособлен-
ных к данным условиям по сравнению с видами, образующими ценозы.
Основными моментами управления изменчивостью луговой растительности
должны быть ускорение смен менее ценных ценозов более ценными и удержание на
возможно более длительный срок ценозов, наиболее отвечающих запросам производ-
ства, что может быть достигнуто путем:
1) создания благоприятных и в то же время устойчивых условий произрастания
для ценных кормовых растений (внесение удобрений, регулирование водного режима);
2) обеспечение желательного состава ценозов (уничтожения вредных или мало-
ценных в кормовом отношении растений, а также видов, неблагоприятно влияющих
на условия произрастания ценных кормовых растений, подсев); очень важно создать
биологически сбалансированные ценозы, чтобы изменения среды, вызываемые одними
компонентами, устранялись в результате жизнедеятельности других компонентов;
3) обеспечения возможности возобновления (семенного, вегетативного) для ви-
дов, признаваемых желательными для данных условий (рациональные формы ис-
пользования и ухода, обеспечивающее возможность обсеменения и приживания всхо-
дов, подсев).
В пределах периода существования ценозов они изменяются с возрастом. Опре-
ределение возраста и возрастных изменений ценозов представляет значительный инте-
рес. Несомненно, что возрастные изменения у отдельных ценозов проявляются различ-
но. В связи с различиями в длительности существования отдельных ценозов и в ходе
их возрастных изменений при одном и том же возрасте одни ценозы могут быть «мо-
лодыми», а другие «старыми».
Отдельные ценозы отличаются друг от друга и по степени выраженности сезон-
ной и разногодичной изменчивости. Существуют ценозы, весьма устойчивые в тече-
ние вегетационного периода и от года к году, и ценозы с резко выраженной сезон-
ной и разногодичной изменчивостью. Целесообразно различать сезонные и разного-
дичные состояния ценозов, которые закономерно сменяются в течение вегетационного
периода, от года к году или от одного периода лет к другому. При этом меняется не
состав компонентов, а их обилие, что обыкновенно сопровождается изменением уро-
жайности, а иногда и структуры травостоев. Меняется лик ценозов (Раменский,
1938). Мо-кно поэтому говорить об одноликих и многоликих ценозах (сезонноодноли-
ких и сезонномноголиких ценозах, а также разнйгодично одноликих и многоликих
ценозах).
Для выявления признаков ценозов необходима не только определенная пло-
щадь— площадь выявления (Раменский, 1925), но и определенное время — время
(или период) выявления. Для ценозов, у которых не выражена сезонная и разного-
дичная изменчивость (например, у Pinetum cladinosum), это «время выявления» мало,
для других оно равно вегетационному сезону или нескольким годам.
Длительность существования, возраст и степень выраженности сезонной и раз-
ногодичной изменчивости, а иногда также изменчивость, связанную с прохождением
жизненного цикла компонентов ценозов, необходимо учитывать при установлении
ассоциаций и при разработке классификаций луговой растительности. Для обосно-
ванного выделения ассоциаций (типов) и для разработки классификаций недостаточно
материалов, получаемых при маршрутных геоботанических исследованиях, когда • ис-
следователь может судить об изучаемой растительности лишь на основании одно-
1 Неравномерность обсеменения ряда растений при сеноуборке нередко обуслов-
ливает существенную неоднородность травостоев, увеличивает их пестроту, создает
ложную комплексность.
— 102 -
кратных описаний отдельных участков. При сопоставлении и анализе отдельных опи-
саний не исключена возможность неправильной трактовки изучаемых объектов,,
в частности отнесение к различным ассоциациям разногодичных, а в некоторых слу-
чаях даже сезонных состояний одной и той же ассоциации. Настала пора бэлее углуб-
ленного изучения луговой растительности и разработки ее классификаций с учетом
длительности существования ценозов и степени выраженности их сезонной разно-
годичной и прочей изменчивости. Следует выделять и классифицировать типы лугов
(типы биогеоценозов — В. Н. Сукачев, 1942, 1945, 1954), понимая под ним.и стадии
развития растительности в пределах определенных экотопов (Работнов, 1955). В пре-
делах типа следует различать возрастные, сезонные и разногодичные и прочие со-
стояния.
SUMMARY
The most common forms of changes in meadow vegetation are: alteration of co-
mposition during vegetation period and from year to year; successions due to human
activity; successions due to changes in the environment as a result of the activity of
meadow plants; changes connected with the life cycle of the components of cenoses.
Cenoses differ from each other as regards the length of their existence, their age, the
degree of seasonal and differentyear alteration and the changes connected with the course
of their components’ life cycle. All this must be taken in consideration in establishing
associations and classifying meadow vegetation. Not only definite area but definite time
(definite observation period) are required to determine the attributes of a cenosis. The
different types of meadow must be classified as stages in the development of vegetation..
The different age, seasonol etc. states must be distinguished within the limits of every *
particular type.
ЛИТЕРАТУРА
Бронзова Г. Я. 1936. К вопросу о методах уточнения геоботанических показа-
телей. Сб. „О методике комплексных обследований “.
Вильямс В. Р. 1922. Общее землеведение. Часть вторая. Естественно-истори-
ческие основы луговодства. М. ч
Вильямс В. Р. 1926. Почвоведение. М.
Кренке Н. П. 1940. Теория циклического старения и омоложения растений. М.
Куркин К. А. 1952. Луга Барабы и их омоложение. Автореф. дисс. на соиск.
уч. степ. канд. биол. наук.
Лавренко Е. М. 1947. Об изучении эдификатороз растительного покрова.
Сов. ботаника, т. XV, вып. 4.
Работнов Т. А. 1950а. Советское луговедение. Бюлл. МОИП, отд. биол.,
т. 55, вып. 1.
Работнов Т. А. 19506. Жизненный цикл многолетних травянистых растений
в луговых ценозах. Тр. БИН АН СССР, геоботаника, 6.
Работнов Т. А. 1955. Разногодичная изменчивость лугов. Бюлл-. МОИП,
отд. биол., т. 60, вып. 3.
Работнов Т. А. 1956. Улучшение пастбищ в районах влажного умеренного
климата. Сб. иностр, с.-х. информации, № 8.
Раменский Л. Г. 1925. Основные закономерности растительного покрова.
Воронеж.
Раменский Л. Г. 1938. Введение в комплексное почвенно-геоботаническое
исследование земель. М.
Смелов С. П. 1936. Теоретические основы лугопастбищного хозяйства. Про5л-,
животноводства, № 11.
Сукачев В. Н. 1916. О „теории дернового процесса* В. Р. Вильямса. Почво-
ведение, № 2.
Сукачев В. Н. 1942. Идея развития в фитоценологии. Соз. ботаника, № 1—3.
Сукачев В. Н. 1945. Биоценология и фитоценология. Докл. АН СССР, т. 47,
вып. 6. •
Сукачев В. Н. 1954. Некоторые общие теоретические вопросы фитоценологии.
Вопр. ботаники, № 1.
Томмэ М. Ф., КсанфопулоО. И. и Сементовская Н. М. 1948. Мине-
ральный состав кормов СССР. М.
— 103 —
Шен ни ков А. П. 1941. Луговедение. Л.
Щенников А. П. 1954. Изучение лугов в СССР. Вопр. ботаники, № 1.
Bates G. Н. 1948. An investigation into cause and prevention of deterioration of
leys. J. british. grassl. soc. 3.3.
Brenchley W. E. 1926. Die Rothamste der Wiesendiingungsversuche von 1856
bis 1919.
Hart M. L. 1950. Organishe stofen Grasland. Landbouwk. Tijdschr. 62. Ref. Herbage
Abstracts 21, I. 1951.
Klapp E. 1954. Wiesen und Weiden. 2. Auflage, Berlin.
К moc h H. J. 1952. Uber den Umfang und einige Gesetzmassigkeiten d. Wurzel-
massenbildung unter Grasnarben. Zeitschr. f. Acker u. Pflanzenbau, 95,4.
Prince F. S. 1956. Grassland farming in the humid Northeast. New Vork—London.
Sears P. D. 1950. Soil fertility and pasture growth. J. British, grasse. soc., 5,4.
БЮЛЛЕТЕНЬ М. О-ВА И СП. ПРИРОДЫ, ОТД. БИОЛОГИИ, Т. LXII (5), 1957
О стадиях формирования фитоценозов
Д. Г. Воронов и Л. И. Тагунова
The stages of phytocenoses formation
A. G. Voronov and L. N. Taganova
Растительный покров изменяется под влиянием изменений условий среды й
внутренней перестройки самих фитоценозов. Анализ смен растительного покрова по-
казывает, что в этом процессе эндо- и экзогенез тесно переплетаются между со-
бой. Гак, при отступании моря изменяется базис эрозии, понижается уровень грун-
товых вод, на обсохших участках побережья поселяются однолетние (солерос) и мно-
голетние (сарсазан) солянки, которые, накопляя соли, способствуют уменьшению засо-
ления грунтов, увеличению их опресненности и, следовательно, поселению менее га-
лофильных видов — полыней, пырея, бескильницы. Если же в прибрежной полосе раз-
виты заросли тростника, то при обсыхании такого участка процесс почвообразования'
идет по иному пути, так как тростник создает специфические условия и для форми-
рования почвенного покрова и для развития растительности. Условия, создающиеся;
под влиянием изменения среды, оказываются, таким образом, благоприятными для1
определенных растительных сообществ; под влиянием жизнедеятельности образую-'
щих эти сообщества растений условия существования продолжают изменяться и ста-
новятся уже более благоприятными не для живущих здесь, а для других видов, что1
и приводит к новой смене растительных ассоциаций. При этом растения, обитающие
в сообществе, приспособлены не только к физико-химическим особенностям почв и‘
к другим абиотическим факторам, но и к определенной фитоценотической среде. По-
этому имеют место не только смены некоторых видов, входящих в состав сообщества,
другими, но и смены целых сообществ другими.
Особый интерес представляет вопрос о формировании фитоценоза, т. е. о посте-
пенной выработке его основных особенностей. Этот процесс можно наблюдать на
освобождающихся от воды побережьях морей й~днйщах озер. При всех конкрет-
ных особенностях хода этого процесса, наблюдающихся в зависимости от характера
грунтов, в частности степени увлажнения и степени и характера засоления, можно
выделить определенные стадии развития растительного покрова. Этому вопросу были
посвящены специальные исследования (Гроссгейм, 1929; Воронов, 1940; Шахов, 1946);
Мы также попытаемся охарактеризовать эти стадии.
При этом мы будем придерживаться определения фитоценоза, данного В. Н. Су-
качевым (1938): «Под фитоценозом (растительным сообществом) надлежит понимать
всякую совокупность растений на данном однородном участке территории, находя1
щуюся в состоянии взаимозависимости и характеризующуюся как определенным со^
ставом и строением, так и определенным взаимоотношением со средой... Там, где нет
указанной взаимозависимости растений друг от друга, нет фитоценоза. Такого рода
растительные покровы мы встречаем на самых первых стадиях заселения не занятой
еще растениями территории» (стр. 19).
Распределение растений на той или иной территории и их сочетания в опре1-
деленные фитоценозы являются, таким образом, результатом: а) действия комплекса
факторов среды, б) особенностей различных видов растений и в) взаимовлияния рас-
тений при совместном обитании. При этом, как указывал Л. Г. Раменский (1938),
чем более суровыми являются условия среды, тем меньшее значение приобретает кон-
куренция и тем более заметно выступает на передний план прямое подавляющее
действие внешних условий. Однако и здесь в подборе сочленов биоценоза будет играть
огромную роль приспособленность их к данным условиям, их экологические осо-
бенности.
Уже на самых ранних стадиях формирования фитоценоза на лишенных расти-
— 106 —
тельного покрова участках поверхности литосферы при поселении единичных экземп-
ляров растений, довольно заметно удаленных один от другого, условия произрастания
несколько меняются: изменяется влажность приземного слоя воздуха, падает ско-
рость ветра, извлекаются из грунта солевые частицы и т. д., т. е. начинается посте-
пенное формирование фитосреды. Вопреки мнению И. К. Пачоского (1925), который
полагал, что только на почвах развиваются настоящие растительные сообщества,
а остальные субстраты допускают развитие лишь зарослей, мы полагаем, что при
длительном существовании одновидовой заросли (например, в течение хотя бы года,
а тем более — нескольких лет), начинается почвообразовательный процесс и форми-
рование пусть вначале слабо выраженного почвенного покрова со свойственными ему
особенностями (так существуют заросли тростника на плавневых почвах, сарсазан-
ники на мокрых солончаках). В определенных условиях чистая заросль может пред-
ставлять собой одну из начальных стадий формирования сообщества (фитоценоза).
Таковы заросли тростника или солероса на северном побережье Каспия, чистые пио
нерные группировки шведки на высохших днищах озер Наурзума и т. д.
Рассматривая вопрос о смене фитоценозов во времени при заселении ранее не
занятой территории, В. Н. Сукачев (1938) указывал, что «первая стадия этого про-
цесса характеризуется случайным составом растений, не образующих сомкнутого
ковра, растущих отдельно друг от друга. Вначале нет фитоценоза (сообщества), так
как нет еще между растениями сколько-нибудь ясно выраженного взаимного влия-
ния» (стр. 24). Этот процесс первоначального формирования растительного покрова
в связи с вселением растений на данную территорию В. Н. Сукачев (1938) называет
сингенезом. В процессе приспособления к существующим условиям среды происходит
Отбор определенных видов (эцезис). В дальнейшем, под влиянием изменения фито-
ценозом среды, изменяется и сам фитоценоз, т. е. происходит экогенез.
Таким образом, в процессе формирования фитоценоза (ценозогенеза) очень
важна степень замкнутости этого фитоценоза, характеризующая выраженность его
среды, т. е. взаимовлияние слагающих его видов (Шахов, 1941, 1946).
В. Н. Сукачев характеризует три стадии экогенеза, четко прослеживающиеся
при формировании фитоценоза. Для первой — открытый фитоценоз — характерно
раздельно-зарослевое сложение растительного покрова. Для . следующей — закрытый
невыработавшийся фитоценоз — обычно смешанно-пятновое сложение, й, наконец, для
третьей стадии — выработавшийся фитоценоз — типично равномерное смешение ви-
дов, диффузное сложение, и прочное сохранение за собой территории в течение до-
вольно продолжительного времени. Очевидно, совершенно правильная схема, данная
В. Н. Сукачевым, является общей, недетализированной.
В более новых руководствах по фитоценологии стадиям формирования фито-
ценозов почти не уделяется внимания. П. Д. Ярошенко (1953) ограничивается опре-
делениями сингенетических, эндоэкогенетических и экзогенных смен (стр. 125).
Б. А. Быков (1953), уделяющий этим сменам около 1,5 страницы (276—278), рисует
следующую общую схему, в дальнейшем им несколько детализируемую: «...во всех
случаях прохождения первичных сукцессий, как, впрочем, и в случае вторичных
сукцессий, весь процесс смены может быть сведен к следующим явлениям: занос за-
чатков воздушными или водными течениями или животными (продуцирование их
в самом ценозе в случае вторичных сукцессий); прорастание и развитие тех из них,
для которых условия оказываются достаточно благоприятными; образование зачи-
нающегося ценоза; развитие установившегося ценоза; превращение его в угасающий
ценоз» (стр. 277). Эта схема еще более обща. Нам представляется, что не следует
говорить об угасающем ценозе. В природной обстановке ухудшение условий существо-
вания для одного ценоза есть улучшение их для другого ценоза, поэтому мы не на-
блюдаем в чистом виде угасания растительного ковра, а видим цепь сукцессий. Все
первичные сукцессии автор почему-то относит в раздел эндодинамических смен.
Дальнейшую детализацию стадий формирования фитоценоза Б. А. Быков про-
водит преимущественно на основе классификации А. А. Гросгейма (1929). Этот автор,
давший одну из первых (по времени опубликования) схем начальных стадий форми-
рования фитоценоза, называл чистые заросли одного вида на освободившейся терри-
тории аггрегациями, группировки из нескольких однородных в экологическом отно-
шении видов, представляющие следующую стадию,— аггломерациями; следующей фа-
зой он считал семиассоциацию, представляющую собой сочетание не связанных аггло-
мераций, в которые могут вселяться новые виды, и, наконец, последней стадией, по
А. А. Гроссгейму, является ассоциация, представляющая собой связанные аггломера
ции, иными словами — выработавшийся фитоценоз.
Уточняя и перерабатывая классификацию А. А. Гроссгейма, А. Г. Воронов
(1940) предложил прежде всего отказаться от терминов «семиассоциация» и «ассо-
циация», так как ассоциация является таксономической единицей в фитоценологии
и поэтому применение такого и сходных терминов к стадиям формирования сообще-
ства (фитоценоза) недопустимо. Автор также указал, что ход процесса формирова-
ния фитоценоза (по наблюдениям на высыхающих днищах озер Наурзума) весьма
резко различается в зависимости от того, каков характер заселяемого грунта: является
ли он засоленным или пресным.
— 107 —
Так при зарастании соленых озер последовательно сменяют друг друга: 1) чистые
пионерные группировки (Salicornia herbacea или Suaeda maritima s. 1.); 2) чистые
группировки тех же солянок; 3) смешанные группировки (образованы теми же видами);
4) сложные группировки с участием многолетников (Artemisia solina, Agropyrum repens)
и однолетников. Все эти стадии могут быть объединены под общим названием группи-
ровок; следующие стадии уже являются стадиями развития фитоценозов. Это: 5) заросли
многолетников (например, Н alocnemum strobilaceum, Phragmites communis), которые
в свою очередь подразделяются на чистые и смешанные; 6) сложный фитоценоз, обра-
зованный многими видами.
Границы пионерных группировок передвигаются за год на сотни метров, чистых
и смешанных группировок однолетников — на десятки метров, сложных группировок —
на один-два метра, редко' на 10—20 м. Фитоценозы устойчивы, они медленно изменяют
свои границы. Несколько по-иному проходят начальные стадии формирования фитоценоза
на обсыхающих днищах пресных озер.
Так- как условия существования на таком днище благоприятны для ра-
стений с самыми различными требованиями к среде, резко возрастает роль случая в
формировании пионерной группировки; если на засоленных днищах из попавших сюда
семян могут прорасти и развиться лишь семена немногих видов солянок, то на пресных
и слабо засоленных грунтах могут практически развиться семена и плоды всех видов,
которые произрастают поблизости. Поэтому-то возможность развития здесь того или
иного вида определяется почти исключительно возможностью заноса сюда его семен-
ных зачатков. Прежде всего сюда попадают семена и плоды растений, обитающих по-
близости и особенно те, которые характеризуются высокой летучестью. В качестве приме-
ра приведем список растений, развившихся на высыхающем днище о. Байназар-копа. Здесь
А. Г. Вороновым и А. А. Шаховым 23 июня 1937 г. на 92 площадках по 1 м2 были об-
наружены проростки семенного происхождения, изредка (у Atropis convoluta) наряду
с проростками цветущие экземпляры, принадлежащие 18 видам. Далее мы указываем
после названия вида общее число экземпляров на 92 м2, а в скобках — число метровых
площадок, на которых этот вид встречен. Виды расположены в порядке убывания числа
экземпляров: Phragmites communis—132(56), Panicum miliaceum— 26(13), Polygonum
scabrum — 25 (4), Scirpus maritimus—\1 (13). Chenopodium chenopodioides—14 (10),
Aster tripolium—13 (8), Atropis convoluta—9 (9), Polygonum patulum—9 (4), Alope-
curus ventricosus — 1 (7), Agropyrym re pens — 3 (1), J uncus Gerardi — 3 (2), Crypsis
spec. — 1 (1), Nasturtium palustre—1 (1), Scolochloa festucacea — 1 (1), Salicornia her-
bacea — 1 (1), Rumex maritimus—1 (1), Oenanthe aquatica—1 (1), Glycyrrhiza ura-
lensis—1 (1). Кроме того, были обнаружены не попавшие на эти площадки проростки:
Salsola Kali s. Matricaria inodora, Statice caspica, Sonchus arvensis. Senecio pa-
luster, Atropis distans, Ranunculus sceleratus.
Различия экологических особенностей растений, входящих в приведенный список,
бросаются в глаза. Та же картина наблюдалась одним из авторов на оз. Южная Сарпа,
в северо-западном Прикаспии, 4 сентября 1954 г. Здесь на обсохшем днище озера, сре-
ди сухого войлока, образованного нитчатками, с остатками водных растений Lemna tri-
sulca, Potamogeton pectinatus, Ceratophyllum demersum и семенами Ceratophyllum
demersum, были обнаружены Chenopodium rubrum, Alhagi camelorum, Atriplex patens,
Agropyrum repens, Crypsis aculeata, Abutilon avicennae, Tournefortia sibirica, Convol-
vulus arvensis и другие, многие в цвету.
Позже количество видов резко падает, так как возрастающая сухость грунта спо-
собствует отбору определенных, немногих растений. Таким образом, здесь сначала воз-
никает смешанная пионерная группировка, затем чистая пионерная группировка (в усло-
виях Наурзума обычно образованная Chenopodium chenopodioides или Aster tripolium).
Дальнейшие этапы сходны со стадиями смен на днищах соленых водоемов (не столько
по составу, сколько по характеру проходимых стадий). Отчасти это связано с высыха-
нием днищ пресных озер и капиллярным подтягиванием солей к поверхности их грунта,
отчасти — с выносом солей ветрами с днищ соленых озер; поэтому условия существова-
ния сближаются и чистые или смешанные группировки Petrosimonia triandra и Bassia
hyssopifolia могут развиваться в обоих случаях.
В более суровых, чем в Наурзуме (Кустанайская обл.), условиях северо-восточного
Прикаспия роль случая в распределении растений на зарастающих участках грунта умень-
шается: проростки многих видов засыхают на ранних стадиях развития, и группировки
образуют растения, наиболее выносливые по отношению к сухости и засолению, поэтому
здесь даже на самых ранних стадиях заселения мы наблюдаем развивающиеся про-
ростки всего нескольких видов, а не десятков их.
А. А. Шахов (1946) дает свою классификацию стадий формирования фитоценоза.
Он принимает следующие понятия: 1) колония (одновидовая или разновидовая); 2) коло-
ния-группировка (ничтожное, но большее влияние растений друг на друга); 3) развив-
шаяся группировка (одновидовая или разновидовая); косвенное взаимное влияние — за-
держание песка и пр.; 4) несомкнутый фитоценоз (создается фитосреда, синузии);
5) сомкнутый фитоценоз; вселение других видов затруднено; наиболее^ соответствует
современным почвенно-климатическим условиям, но развивается.
— 108 —
Следует прежде всего отметить крайне неудачное применение термина „колония"
(и, следовательно, „колония-группировка"). Как известно, в биологии колонией называет-
ся сложное объединение ©предельных организмов (губки, кораллы, мшанки, сальпы), а
также разводки бактерий на питательной среде. Зачем же вводить этот термин для
обозначения совершенно иного понятия? Нам представляется, что для ранних стадий
развития фитоценоза наиболее приемлем термин, данный в свое время В. В. Алехиным
(1938)—„группировка". Под группировкой мы вслед за В. В. Алехиным понимаем бес-
порядочную, еще не сложившуюся комбинацию растений, в той или иной степени слу-
чайную. Группировки могут состоять из многих, немногих или одного вида растений,
по большей части однолетних. Заросли многолетников не обязательно являются груп-
пировками. В длительно существующих зарослях (фитоценозах) многолетников (А. Г. Во-
ронов, 1940) какие-то условия существования (например, весьма сильное засоление почвы
или значительное увлажнение) подавили развитие большинства видов и способствовали
мощному развитию немногих или даже одного вида. При этом чистые заросли (моно-
ценозы), как и одновидовые группировки, мы называем так, конечно, условно; ибо, как
показал В. Н. Сукачев (1950), в состав фитоценозов следует включать все раститель-
ные организмы, обитающие на данном участке, в том числе и низшие растения и мик-*
роорганизмы в воздухе и в почве, поэтому моноценозов в строгом смысле слова не
существует.
Затем, как правильно указывает Б. А. Быков (1953), А. А. Шахов совершенно не-
прав, считая, что ни в „колонии", ни в „колонии-группировке", ни даже, в сущности,
в группировке (если не считать косвенного взаимовлияния растений) нет фитоценоти-
ческой среды. Это, конечно, не так: даже в пионерных группировках начинается воз-
действие растений на среду, т. е., создание фитосреды.
Замкнутость (выработанность) фитоценоза не обязательно должна соответствовать
его сомкнутости, поэтому совершенно необязательно в природе проходят стадии раз-
вития фитоценоза в том порядке, как на это указывается А. А. Шаховььм (1946, схема
на стр. 133; см. также примечание к нашей табл. 1), — сначала сомкнутый ценоз, затем
замкнутый. Если, как полагает А. А. Шахов (1946), „фитоценотическая среда создается
в период смыкания подземных частей растений", то будет непонятным, как сарсазан
(Halocnemum strobilaceum) может образовывать замкнутый ценоз при отсутствии смы-
кания его подземных частей. Здесь замкнутость в первую очередь определяется харак-
тером местообитания, которое .насыщается" определенным количеством нес*\ ы сающихся
экземпляров растений, а не смыканием надземных или подземных органов сарсазана.
В то же время существование в моноценозе этого вида фитоценотической среды не
подлежит сомнению. Могут в то же время иметь место и случаи, соответствующие
схеме А. А. Шахова.
/ Сопоставление различных классификаций начальных стадий развития сообщества
'(табл. 1) показывает, что и В. Н. Сукачев, и А. А. Гроссгейм, и А. Г. Воронов прежде
всего выделяют самую первую стадию зарастания свободных территорий как стадию с
пятнисто-зарослевым сложением растительного покрова. Пионерная растительность об-
разует обычно несомкнутые чистые или иногда смешанные заросли более или менее
случайно попавших сюда видов, часто присутствуют растения, заведомо не выносящие
конкуренции с другими видами, иногда развиваются вегетативно-подвижные растения
из многолетников (с ползучими корневищами, отпрысками и т. п.); последнее бывает
обычно при разрастании зарослей многолетников (ползучий пырей, тростник), развитых
почокраине свободных от растительного покрова участков. Здесь еще сначала нет- вза-
имного влияния, борьбы за существование, нет фитоценоза, позже таксе влияние начи-
нает слабо проявляться при сохранении случайного состава зарослей. Очевидно, стадия
отсутствия фитоценоза В. Н. Сукачева соответствует частй стадии чистой и смешанной
пионерных группировок А. Г. Воронова. Открытый фитоценоз В. Н. Сукачева охватыва-
более поздние периоды развития пионерных (чистых и смешанных), а также его чистые
и смешанные группировки, что соответствует аггрегациям и аггломерациям А. А. Гросс-
гейма. Закрытый невыработавшийся фитоценоз В. Н. Сукачева охватывает сложную груп-
пировку (конгломерацию) А. Г. Воронова и соответственно „семиассоциацию" А. А. Гросс-
гейма. Наконец, выработавшемуся фитоценозу В. Н. Сукачева соответствует стадия „ас-
социации" А. А. Гроссгейма и фитоценоза А. Г. Воронова.
Таким образом, если принять терминологию А. Г. Воронова, то возникновение от-
крытого фитоценоза происходит на стадии пионерных группировок, переход к закрытому
невыработавшемуся фитоценозу соответствует началу стадии сложной группировки (конг-
ломерации) и переход к выработавшемуся фитоценозу соответствует переходу от груп-
пировок к фитоценозам.
Эта выработка-фитоценозов включает: приспособление первых растений-поселенцев
к условиям среды; преимущество,получаемое одними видами над другими в процессе
конкуренции; отбор и приобретение господства определенными видами (В. Н. Сукачев, 1938).
Формирование сложных фитоценозов, лишь медленно уступающих свое место дру-
гим, продолжается длительное время. На обсыхающем побережье моря этот срок равен
10—20 годам (Ревердатто, 1935). Видовая насыщенность возрастает от чистых пионерных
группировок и чистых группировок к смешанным и сложным группировкам, достигая
Классификация начальных, стадий развития сообщества
Таблица 1
По В. Н. Сукачеву (1938) По А. А. Гроссгейму (1929) По А. Г. Воронову (1940) По А. А. Шахову (1946)
1. Отсутствие фитоценоза (слу- чайный состав растений, нет взаим- ных влияний, нет борьбы) 2. Открытый фитоценоз (случай- ный состав растений, слабое взаим- ное влияние, нет сомкнутого ковра) 1. Аггрегация (чистые заросли одного вида на грунтах) (многолетняя аггрегация = псевдо- аггрегации) I. Группировки 1. Пионерные группировки а) чистые ( = аггрегации) б) смешанные ( = аггломера- ции) 1. Колония (одновидовая или разновидовая)
2. Аггломерация (группировка из нескольких однородных в экологи- ческом отношении видов) 2. Группировки однолетников а) чистые ( — аггрегации) б) смешанные ( = аггломера- ции) 2. Колония-группировка (одно- видовая и разновидовая)
3. Закрытый невыработавшийся фитоценоз 3. Семиассоциация (состоит из несвязанных аггломераций) 3. Сложные группировки ^кон- гломерации) 3. Группировка (одновидовая и и разновидовая)
4. Выработавшийся фитоценоз, прочно удерживающий завоеван- ную территорию 4. Ассоциация (состоит из свя- занных аггломераций) II. Фитоценозы 1. Заросли многолетников (=псевдоаггрегации) а) чистые б) смешанные 2. Сложные фотоценозы (из мно- гих видов) 4. Несомкнутый фитоценоз (од- новидовой и синузийный) 5. Сомкнутый фитоценоз (одно- видовой и синузийный) 6. Незамкнутый фитоценоз (од- новидовой и синузийный) 7. Замкнутый фитоценоз (одно- видовой и синузийный)1 8. Фитоценоз-климакс
1 Автор указывает, что несомкнутый и сомкнутый ценозы, с одной стороны, незамкнутый и замкнутый — с другой, самостоятельные и неза-
висимые стадии ценрзогенезиса, но..в с^еме (стр. 133) они даны последовательно.
- по -
невысокого максимума при переходе от сложных группировок к смешанным зарослям.
На опресненных участках наблюдается еще один максимум насыщенности — в смешанных
пионерных группировках.
Число экземпляров (или стеблей многолетников) на единицу площади достигает
максимума в чистых, реже в смешанных группировках однолетников, очевидно, в силу
меньшей „конкрурентной мощности" однолетников по сравнению с многолетниками, а так-
же лучших условий увлажнения. Количество стеблей на единицу площади может в за-
рослях одного и того же вида резко меняться в различные годы (см. рисунок). На
Количество стеблей солероса (Salicornia herbacea)
на I м3 в зарослях солероса у оз. Кутун-тал (На-
урзум, Кустанайская обл. Казахстана): 1 — первый
участок; 2 — второй участок
рисунке показано, что в густо населенном участке число стеблей на втором году наблю-
дений было крайне незначительным, в то время как на участке с меньшим обилием соле-
роса на второй год число стеблей даже несколько повысилось. Очевидно, в первом слу-
чае имело место использование густыми зарослями питательных веществ и превращение
благоприятных для вида условий в неблагоприятные.
В чистых группировках однолетников стебли распределяются по кл^сам высоты,
аналогично стволам деревьев в лесных ценозах (табл. 2).
Можно различать „угнетенные" и „господствующие" экземпляры.
Одиночные, выросшие вне густых участков группировок стебли однолетников Bas-
sia hyssopifolia, Petrosimonia triandra, Salicornia herbacea, Suaeda maritima s. 1,
Aster tripolium и других видов ветвятся от основания, у многих видов образуя пере-
кати-поле; в густых участках группировок они или слабо ветвятся (Salicornia, Suaedai
Aster) или их ветви вытянуты вверх почти вертикально (Bassia hyssopifolia).
В ходе смен, соответствующих намеченным нами стадиям, наблюдалось выпадение
поясов и их инверсця. Выпадение поясов, неоднократно наблюдавшееся на обсыхающих
водоемах Наурзума, находит себе объяснение в различной экологической амплитуде раз-
ных видов. Выпадают обычно пояса, образованные видами, имеющими малую экологиче-
скую амплитуду. Так, например, если в первый год по высыхании водоема по его ок-
раине были развиты пояса Salicornia herbacea, Suaeda maritima s. Bassia hyssopi-
- ш -
folia, то на второй год пояс Suae da часто выпадает и пояс Bassia следует непосред-
ственно за поясом Salicornia.
Таблица 2
Распределение солероса Salicornia herbacea (Наурзум, Кустанайской обл.)
по классам высоты у о. Кутантал 22 августа 1936 г.
Классы высоты (в см) 1—5 6—10 11—15 . 16—20 1 21-25 26-30 31—35 36—40 41—45 46—50 51—55 56—60
Число стеб- лей 0 0 1 6 12 14 12 13 3 3 1 1
Инверсия поясов происходит благодаря отложению по окраинам озерной котловины
золовых наносов, засыпающих стебли Salicornia и Suaeda, препятствующих развитию
проростков этих видов и создающих условия значительно большей сухости. Поэтому на
участках, заваленных наносами, развиваются виды, не стойственные существовавшим
здесь поясам Salicornia и Suaeda. (Ofaiston monandrum, Petrosimonia triandra, Ar-
temisia salina). Следовательно, пояса с господством Artemisia salina или Petrosimonia
triandra, ранее расположенные к периферии от поясов Salicornia и Suaeda, в следую-
щем году могут предшествовать им.
В зарослях тростника, камышей и рогоза, располагающихся по окраинам обсыхаю-
щих днищ водоемов, также происходит ряд изменений. В первое время, пока грунт еще
влажный, заросли медленно надвигаются по направлению к центру озера. В дальнейшем
это поступательное движение останавливается. Одни из видов, образующих заросли,
сравнительно быстро перестают развивать свои надземные части (Scirpus lacustris, S. та-
ritimas, Typha angustifolia). Другие,, как Phragmites communis, еще долгое время
дают вегетирующие побеги. Под пологом сухих стеблей тростника среди все более из-
реживающихся зарослей его угнетенных побегов как бы проходит ряд самостоятельных
смешанных группировок. У уреза воды здесь развиваются Rumex Marschallianus, Che-
nopodium chenopodioides, Scutellaria gal eric al at а, далее от воды — Saussurea amara,
на еще более сухой почве — Artemisia salina. Наконец, сухие стебли тростника изрежи-
ваются очень сильно и на месте его зарослей возникает новый фитоценоз, например ас-
социация Artemisia salina, существующая до нового наполнения озера водой. Укажем,
что важное биоценотическое значение зарослей тростника сохраняется им на весь период
существования сухих стеблей, только вместо водоплавающих птиц, усатых синиц, водя-
ной крысы здесь поселяются дрофы, зайцы и другие представители степной фауны, ис-
пользующие защитные свойства этих зарослей.
Что касается изменения границ прибрежных фитоценозоз, то одни из них, образо-
ванные видами с небольшой экологической амплитудой, при высыхании озер довольно
быстро уменьшают занятую ими площадь (например, мезофильная ассоциация Agropyrum
repens, заросль Scirpus maritimus), другие, образованные видами с более широкой эко-
логической амплитудой, изменяют свои границы, наступая в сторону озерного днища и,
следовательно, расширяя свою площадь, причем это расширение площади идет бок о
бок с изменением строения фитоценоза: вселение новых видов, изменение обилия отдель-
ных компонентов и т. д.; это может привести к превращению одной ассоциации в дру-
гую (например, ассоциации Artemisia salina в ассоциацию A. salina 4- Saussurea amara).
ЗАКЛЮЧЕНИЕ
Пои завоевании растительностью освобожденных из-под воды территорий (озерные
днища, морские побережья) наблюдаезся постепенное развитие фитоценозов, при кото-
ром растительный покров проходит стадии: чистых пионерных группировок, смешанных
пионерных группировок, чистых и смешанных группировок однолетников, сложных груп-
пировок (конгломераций), чистых и смешанных зарослей многолетников и сложных фито-
ценозов. В процессе становления фитоценозов некоторые стадии могут отсутствовать;
процесс развития может в некоторых условиях долго задерживаться на той или иной
стадии, .например на стадий чистой заросли многолетников, смешанной заросли много-
летников. В условиях опресненных грунтов днищ пресноводных озер перечисленным
стадиям предшествует стадия смешанной пионерной группировки.
В проникновении растений на освободившуюся территорию значительную роль иг-
рает случай — наличие поблизости тех или иных видов растений, а также способы пере-
носа семенных зачатков различных видов. Затем начинается процесс приспособления
растений к определенным условиям среды—процесс отбора видов как физико-географи-
- 112 —
ческими факторами, так и в результате конкуренции, причем роль последней возрастает
по мере выработки фэрмирующимся ценозом своей внутренней фитоценотической среды.
Фитоценотическая среда начинает формироваться еще на стадии пионерных груп-
пировок. В процессе формирования фитоценоза уменьшается роль однолетних растений
и возрастает роль многолетников.
При смене во времени различных поясов, наступающих на освободившуюся терри-
торию, наблюдается выпадение поясов, образованных видами с узкой экологической амп-
литудой, а также инверсия поясов, связанная с образованием на этих участках, напри-
мер, эбловых наносов.
В чистых группировках однолетников мы можем наблюдать, как в лесу, диффе-
ренциацию растений по классам высоты и выработку характерных для густых группи-
ровок форм роста—почти без ветвей или с обращенными вверх прижатыми к стеблю ветвями.
Стадии формирования фитоценоза заслуживают дальнейшего изучения.
SUMMARY
The penetration of vegetation into territories drained of water (lake bottoms, sea costs>
is accompanied by the gradual development of phytocenoses, in the course of which the
vegetable cover passes through the following stages: purely pioneering groups, mixed pio-
neering groups (agglomerations), pure and mixed thickets of perennial plants and compli-
cated phytocenoses. Some stages may be absent in the process of phytocenosis formation
and the process of development may, under certain conditions, be very much delayed in.
some stage, for example in the stage of pure thicket of perennials, or mixed thicket of pe-.
rennials. In the none-saline ground of fresh water lakes these stages are preceded by the
stage of mixed pioneering groups.
Chance, that is — the proximity of certain plant species and methods of transportation
of the seed germs of various species, plays a considerable role in the penetration of plants
into drained territory. This is followed by the process of the plants’ adaptation to definite-
environmental conditions, and the process of species selection, both according to physico-
geographical factors and as a result of competition. As the cenoses gradually develop their
own internal phytocenotic meditm the role of competition rises accordingly.
The phytocenotic medium begins to form already in the pioneering group stage. In
the process of phytocenosis formation the roje of annual plants gradually diminishes an±
that of perennials gradually rises.
In connection with changes in the time of penetration of different belts into drained!
territory, it can be observed that belts formed of species with a narrow oecological ampeli-
tude drop .out and an Inversion of belts can also be observed, due to the formation on these
territories of eolic deposits, for example.
The differentiation of plants according to height classes and the development of forms
growing in a way characteristic for thickly grouped plants: with hardly any branches or bran-
ches directed upwards and pressed close to the stem, can be observed in pure annual gro-
ups, as they are observed in forests.
The stages of phytocenosis formation deserve to be studied more closely.
ЛИТЕРАТУРА
Алехин В. В. 1938. География растений. Учпедгиз.
Быков Б. А. 1953. Геоботаника. Алма-Ата.
Воронов А. Г. 1940. Водная и прибрежная растительность Наурзумского за-
поведника. Дисс. на соиск. уч. степ. канд. биол. наук, М.
Гроссгейм А. А. 1929. Введение в геоботаническое обследование зимних
пастбищ ССРАзербайджана, Баку.
Пачоский И. К. 1925. Социальный принцип в растительном царстве. Бот.
журн., т. 10, № 2.
, Раменский Л. Г. 1938. Введение в комплексное почвенно-ботаническое
исследование земель. М.
Ревердатто В. В. 1935. Введение в фитоценологию. Часть I. Томск.
Сукачев В. Н. 1938. Главнейшие понятия из учения о растительном покрове
Растительность СССР, т. I, М.— Л.
Сукачев В. Н. 1950. О некоторых основных вопросах фитоценологии. Сб.
«Проблемы ботаники», М.— Л.
Шахов А. А. 1941. О зарастании безводных озерных котловин Наурзумского
заповедника. Сб. научных студенческих работ, вып. 24, Ботаника, М.
Шахов А. А. 1946. Формирование фитоценоза (ценозогенез). Бюлл. МОИП,
отд. биол., т. 51, вып. 4—5, М.
Ярошенко П. Д. 1953. Основы учения о растительном покрове. Изд. 2-е, пере-
работанное и дополненное, М.
БЮЛЛЕТЕНЬ М. О-ВА И СП. ПРИРОДЫ, ОТД. БИОЛОГИИ, Т. LXII (5), 1957
ХРОНИКА
CHRONICLE
Деятельность зоологической секции Московского общества испытателей природы
с октября 1956 г. по март 1957 г.
(председатель А. Н. Формозов, член Бюро секции А. А. Насимович)
25 октября 1956 г. Доклад В. А. Арсеньева «Зоологические работы в Ан-
тарктической экспедиции» (с демонстрацией диапозитивов).
15 ноября 1956 г. Доклады И. Л. Кулик «Структура популяций больших
песчанок в северо-западной части их ареала» (см. автореферат, стр. 118) и М. В. Ше-
х а н ов а «Географические и стадиальные особенности размножения больших пес-
чанок» (см. автореферат, стр. 118).
В обсуждении докладов приняли участие Н. В. Башенина, В. Г. Кривошеев,
В. В. Кучерук, В. Лобачев, Н. П. Наумов и В. Я. Шубин.
29 ноября 1956 г. (совместно с секцией географии МОИП и Московским
филиалом Географического общества СССР). Доклад А. Н. Формозова «Впечат-
ления о поездке в Бразилию и о работе секций биогеографии и медицинской географии
XVIII Международного географического конгресса в Рио-де-Жанейро» (с демонстра-
цией коллекций тропииеских птиц, зарисовок автора и местных изданий).
6 декабря 1956 г. Доклады Н. В. Б а ш е н и н о й «К вопросу о сущности
критической температуры у мелких полевок» и Н. Н. Воронцова «Значение изу-
чения хромосомных наборов для систематики млекопитающих» (будет опубликован
в Бюллетене МОИП, отдел биологический).
Выступивший по первому докладу А. А. Насимович отметил необходимость
тщательного изучения на большом материале физиологических показателей для от-
дельных половых и возрастных групп в пределах вида, без чего вопрос о межвидо-
вых различиях не может быть решен убедительно. Работу необходимо сочетать
с накоплением данных для разных сезонов, различных кормовых рационов и разных
состояний зверьков.
Н. Н. Воронцов в докладе, построенном на большом литературном материале,
почерпнутом, главным образом, из зарубежных источников, рассказал о значительных
успехах, достигнутых в изучении хромосомных наборов и применения этих знаний
в систематике млекопитающих для выяснения родствейных связей в пределах низших
и высших таксономических подразделений, а также для установления центров про-
исхождения и последующего расселения некоторых групп животных, в том числе
и одомашненных видов (доклад сопровождался демонстрацией большого числа
диапозитивов).
Выступившие по докладу В. Г. Гептнер, А. А. Прокофьева-Бельговская,
В. И. Цалкин и А. Н. Формозов отметили значительный интерес сообщения, обстоя-
тельность данного обзора и желательность его опубликования в бюллетенях МОИП,
что было поддержано всем собранием. Вместе с тем А. А. Прокофьева-Бельговская
указала, что данные Н. Н. Воронцова о полиплодии хомячков неверны: докладчик
фактически отрицает влияние внешних условий на хромосомный аппарат, что, однако,
неверно. В. И. Цалкин считает, что данные о монофилитическом происхождении со-
бак, без участия шакала, спорны; остеологический и другой материалы свидетель-
ствуют об обратном. В. Г. Гептнер подробно остановился на вопросе о значении
различных признаков для систематики позвоночных.
- 114 -
20 декабря 1956 г. (совместно с Московским филиалом Географического*
общества СССР). Доклад С. М. Успенского «Остров Беннет, его фауна и история
исследования».
На основании поезАки летом 1956 г. С. М. Успенский рассказал о природе
о. Беннет и дал обзор истории изучения этого острова. На острове были сделаны
новые находки следов пребывания здесь в 1902 г. экспедиции Э. Толя.
27 декабря 1956 г. Доклад А. Н. Формозова «Впечатления о посещении на-
учных учреждений Бразилии» и отчет и перевыборы Бюро секции.
В обсуждении деятельности Бюро зоологической секции за 1954—1956 гг. приня-
ли участие В. Я. Шубин, В. В. Кучерук, В. И. Цалкин и другие, сделавшие ряд кри-
тических замечаний по повестке докладов (преобладание самотека, редкость проблем-
ных докладов, недостаточное внимание обсуждению выходящих книг и т. д.). В це-
лом деятельность Бюро секции за отчетный период признана вполне удов-
летворительной.
24 января 1957 г. Доклады Л. В. Заблоцкой «Распределение и динами-'
ка численности мышевидных грызунов в борах левобережья Оки и Московской обла-
сти» (опубликован в Бюллетене МОИП, отд. биологический, т. 62, вып. 4, за 1957 г.)
и А. Д. Бернштейн «К изучению биологии черной крысы в Абхазии».
В обсуждении докладов приняли участие В. И. Цалкин, М. А. Рубина,
Л. П. Никифоров, В. Я. Шубин, В. В. Груздев, И. Л. Кулик и Л. Н. Елисеев.
7 февраля 1957 г (совместно с секциями биологических основ животновод-
ства и истории естествознания) К столетию со дня рождения проф. А. А. Браунера.
Доклады И. И. Пузанова «Памяти А. А. Браунера», С. Н. Боголюбского
«Воспоминания о проф. А. А. Браунере» и М. А. Заблоцкого «Последний период
работы А. А. Браунера (1936—1938 гг.) в Аскания-Нова».
Отмечена желательность издания избранных трудов А. А. Браунера и ценная
инициатива, проявленная в подготовке к этому И. И. Пузановым.
11 февраля 1957 г. (совместно с Московским филиалом Географического
общества СССР). Доклад А. П. Кузякина «Схема ландшафтного районирования
для биогеографических целей».
Выступившие по докладу А. Г. Банников и другие отметили, что схема райони-
рования, предложенная А. П. Кузякиным, страдает формализмом, неудачна в целом
и не может быть использована для биогеографических целей.
14 февраля 1957 г. Доклады С. А. Ш ил о в о й, В. Б. Троицкого,
Г. Б. Малькова и В. М. Бельковича «Значение подвижности лесных мыше-
видных грызунов в распределении клещей в очагах клещевого энцефалита» (см. авто-
реферат, стр 119) и П. П. Тарасова «Внутривидовые отношения у соболя
и горностая».
Доклады вызвали оживленный обмен мнениями, в котором приняли участие
В. В. Кучерук, Л. П. Никифоров, М. Я. Лаврова, Е. В. Карасева, В. Лобачев,
Э. И. Коренберг, Л. Г. Динесман, К- М. Эфрон, С. Е. Клейненберг и А. В. Рюмин.
Оба доклада, как имеющие значительный теоретический и практический интерес, ре-
комендованы к опубликованию в бюллетенях МОИП.
20 февраля 1957 г. (совместно с Комиссией по охране природы МОИП).
Доклад В. В. Криницкого «О некоторых вопросах научной работы в запо-
ведниках».
Докладчик отметил, что научные исследования в заповедниках должны прово-
диться, исходя из задач заповедников по сохранению исторически обусловленных
взаимоотношений между отдельными элементами ландшафта; они должны быть на-
правлены на комплексное изучение климата, водного режима, почв, растительности
и животного мира. Основные методы исследовательской работы в заповедниках —
многолетние стационарные наблюдения, проводящиеся во все сезоны года. Научные
исследования в части заповедников не увязаны с их профилем, некоторые экспери-
менты (например в Астраханском заповеднике) причиняют серьезный ущерб природе
заповедников. Биотехнические мероприятия (например, неумеренное истребление не-
которых хищников) проводятся без серьезного изучения природных процессов, вслед-
ствие чего эти мероприятия не только не достигают цели, но и приносят в отдельных
случаях вред природному комплексу заповедника.
Выступивший по докладу А. Г. Банников подробно остановился на специфике
работы небольших заповедников, являющихся островками нетронутой природы среди
вторичного ландшафта. В подобных заповедниках сильно размножившиеся дикие
копытные нередко не находят в смежных районах подходящих условий для расселе-
ния, вследствие чего в заповеднике (Воронежский и др.) создается опасность исто-
— 115 —
щения кормовой базы. А. Г. Банников и С. Е. Клейненберг отметили неоправданное
увлечение заповедников узко утилитарными исследованиями, не увязанными с зада-
чами заповедников и охраной природы в целом; подобные исследования усиленно
насаждались в свое время А. В. Малиновским, однако они более подстать отраслей
вым институтам. С. Е. Клейненберг и В. Б. Дубинин подчеркнули ошибочность аккли^
матизации некоторых новых видов животных в заповедниках (по мнению первого,,
акклиматизационные работы вообще несовместимы с задачами заповедников).
В. Б. Дубинин подробно рассмотрел, что следует понимать под комплексным подхо-
дом к исследованиям в заповедниках, и высказал ряд предположений о причинах
деградации в Астраханском заповеднике птичьих колоний {частые нарушения режи-
ма заповедности, старение ивовых зарослей и т. д.) и массовой гибели в «зимоваль-
ных ямах» сазана (отравление продуктами распада промышленных вод и других
нечистот). В обсуждении доклада приняли участие также А. А. Насимович
и Г. Е. Бурдин.
21 февраля 1957 г. (совместно с секцией ботаники МОИП и Московским
филиалом Географического общества СССР). Доклады Е. А. Дороганевской
«Проблемы географической изменчивости химизма растений» см. автореферат, стр. 120
и Ю. Н. Куражсковского «Некоторые результаты и задачи изучения географи-
ческих закономерностей изменчивости состава кормов в СССР».
7 марта 1957 г. Доклады Е. В. Карасевой, Е. В. Нарской и
А. Д. Бернштейн «Полевка-экономка, обитающая в окрестностях озера Неро
Ярославской области» (опубликован в Бюллетене МОИП, отд. биологический, т. 62,
вып. 3, за 1957 г.) и М. Я. Л а в р о в о й, Е. В. П р о х о р о в а и А. П. В и ш н я к о-
в а «Итоги изучения природного очага лептоспироза в Шаховском районе Москов-
ской области» (см. автореферат, стр. 120).
На заседании были оглашены результаты перевыборов Бюро зоологической сек-
ции; в состав нового бюро вошли А. Н. Формозов, С. Е. Клейненберг, А. А. Насимович,
А. Г. Воронов и С. А. Шилова, дополнительно кооптирован В. В. Кучерук.
По докладам выступили А. Н. Формозов, Е. В. Карасева, В. В. Кучерук,
В. Я. Шубин и другие. Большинство выступавших отметило значительный научный
интерес обоих сообщений и желательность продолжения начатых исследований; отпу-
щенные МОИП средства для работы М. Я. Лавровой были использованы Главным
образом на привлечение коллектива студентов и юннатов, т. е. на расширение
объема исследований. Вместе с тем Е. В. Карасевой и В. В. Кучеруком отмечено,
что число анализов мочи, по которым судили о зараженности зверьков лептоспирозом,
было недостаточно, методика работ, применявшаяся М. Я. Лавровой, требует улучше-
ния в целом; необходимо уделить большее внимание параллельному сбору материалов
по фауне млекопитающих Московской обл.
14 марта 1957 г. Доклады Т. В. Кошкиной «Динамика численности мыше-
видных грызунов на Кольском полуострове» и А. А. Кищинского «Некоторые
черты.биологии хищных птиц в’тундре севера Кольского полуострова».
Доклад Т. В. Кошкиной вызвал большое число вопросов и оживленный обмен
мнениями. А. Н. Формозов, как и большинство выступавших, отметил значительный
интерес доклада, построенного на обширном фактическом материале; в будущем не-
обходимо учитывать не только семеношение хвойных, но и березы, семена которой
также служат важным кормом для мышевидных. В. В. Кучерук подчеркнул, что из
данных Т. В. Кошкиной наглядно видно, что даже при известной монотонности ланд-
шафта, как это имеет место в некоторых районах Кольского полуострова, динамика
численности полевок в разных районах не однотипна. По мнению В. П. Теплова, до-
кладчик недооценивает значение метеорологических факторов, в то время как именно
они обусловливают четырехлетний цикл колебаний численности полевок на Кольском
п-ве. А. А. Насимович считает, что вопрос значительно сложнее, чем его представляет
В. П. Теплое, колебания численности грызунов зависят от взаимодействия многих
факторов: численности и состояния популяции самих мышевидных и их врагов, оби-
лия корма, метеорологических условий и ,т. д.
Секция вынесла решение о желательности дальнейшей разработки данного во-
проса в эколого-географическом аспекте. В интересах непрерывности и накопления
многолетних данных по динамике мышевидных целесообразно, чтобы и в будущем
поддерживались и сохранялись ранее заложенные стационары по учету мышевидных,
особенно стационар в Лапландском заповеднике, заложенный в 1936 г.
По второму докладу выступили В. Я. Шубин и Т. В. Кошкина, отметившие
интерес доложенных данных.
21 марта 1957 г. Доклады Е. В. Карасевой, Л П. Семеновой и
Н. М. Окуловой «Ландшафтная приуроченность мелких млекопитающих на Алтае
и в Северном Казахстане и их значение в природной очаговости лептоспироза»
— 116 —
и Н. А. Никитиной «Особенности использования территории полевыми мышами,
изученные методом мечения».
В обсуждении докладов приняли участие М. Я. Лаврова, А. Н. Формозов,
В. В. Кучерук, С. Д. Перелешин и Е. В. Карасева. Отмечен значительный интерес
обоих сообщений. Данные Е. В. Карасевой и ее соавторов показали, что, вопреки
распространенному мнению об исключительном значении в засушливых районах ско-
та в рассеивании лептоспироза, в Акмолинской обл. большое эпидемиологическое
значение имеют и мелкие млекопитающие. В связи со вторым докладом обсуждался
вопрос о том, что следует понимать под «инстинктом дома», насколько изучен этот
вопрос и удачен ли сам термин.
ПРИЛОЖЕНИЯ
Структура популяций больших песчанок в северо-западной части их ареала
И. Л, Кулик
(Автореферат доклада, прочитанного 15 ноября 1956 г.)
Помимо экологических популяций, в понимании их Н. П. Наумовым (1955),
можно различать еще микропопуляции — низшие группировки населения одного вида,
из которых складываются группировки более высокого порядка — экологические попу-
ляции. Микропопуляция — эта группа зверьков, населяющая непрерывный участок
одной стации, обеспечивающий существование микропопуляции в течение нескольких
сезонов. Зверьки одной микропопуляции имеют сходное питание, единые сроки и ин-
тенсивность размножения, в результате чего в микропопуляции наблюдается опреде-
ленный ход сезонной динамики численности. Несколько нетождественных микропопу-
ляций составляют экологич-ескую популяцию, которая населяет разнородную террито-
рию (комплекс, мозаику стаций), имеет единый ритм многолетней динамики числен-
ности, образует поселение того или иного типа и обеспечивает существование вида
на данной территории.
В юго-восточном Приаралье для мозаичного ландшафта характерны поселения
песчанок, состоящие из микропопуляций, населяющих массивы песков и прилежащие
участки такыровидной равнины. Эти микропопуляции имеют различия: песчанки на
такыровидных участках питаются более однообразным кормом, так как видовой со-
став растений здесь беднее, чем в песках. В связи с более поздним началом вегета-
ции растений на такыровидных участках зверьки начинают здесь питаться свежей
зеленью позднее, чем в песках. Соответственно и начало размножения здесь песча-
нок по сравнению с песками запаздывает (в 1953 г. на 2 недели). На такыровидных
участках песчанки более привязаны к своим норам (Кулик, 1955). В период кочевок
выводки на такыровидных участках чаще перемещаются из одной части норы в дру-
гую ее часть, в песках же они перемещаются, главным образом, из одной норы
в другую. На такыровидных участках больше, чем в песках нор, в которых песчанки
живут постоянно или продолжительное время; норы, в которых выловлены все зверь-
ки, интенсивнее, чем в песках, заселяются песчанками. Все это свидетельствует о том,
что на такыровидных участках, в отличие от песков, значительно меньше мест, удоб-
ных для поселения песчанок, и те места, где зверьки могут жить постоянно, редко
пустуют. В результате песчанки «а такыровидных участках больше заражены бло-
хами. Все перечисленные особенности показывают, что такыровидные участки и мас-
сивы песков заселены различными микропопуляциями, в комплексе составляющими
одну популяцию с поселением мозаичного типа. В таком поселении в микропопуля-
циях такыровидных участков, где много постоянно обитаемых и интенсивно заселяе-
мых зверьками нор и где наблюдается высокая численность блох, создаются необхо-
димые условия для возникновения элементарных очагов инфекций (Наумов, 1955).
Географические и стадиальные особенности
размножения больших песчанок
М. В. Шеханов
(Автореферат доклада, прочитанного 15 ноября 1956 г.)
В результате многолетних наблюдений различными исследователями, в том числе
и автором (северное Приаралье, 1946—1950 гг.; Туркмения, 1952—1953 гг.), был со-
бран обширный материал по размножению больших песчанок, показавший, что оно
косвенно во многом зависит от количества выпадающих осадков, влияющих на раз-
витие растительности; не менее важное значение могут иметь и биотические факторы.
— 117 —
При анализе автор пользуется двумя показателями: интенсивностью размножения
(средний размер выводка 100 половозрелых самок) и плодовитостью (средний размер
приплода 100 половозрелых песчанок обоего пола).
На интенсивность размножения и плодовитость песчанок влияет качественный
состав корма. Наибольшие величины вышеуказанных показателей повсеместно и во
все годы наблюдаются в апреле и мае, т. е. в период, когда основной пищей песча-
нок служат эфемеры и эфемероиды. В остальные месяцы периода размножения даже
максимальные показатели интенсивности размножения значительно ниже, чем такие
же показатели в апреле и мае: в июне — в 2 раза, в марте и июле — в 2,5, в августе
и сентябре — в 4, в октябре — в 10 раз (Прибалхашье, 1939—1948 гг.). В северной
пустыне в один и тот же год (1947) интенсивность размножения с марта по май во
все декады была в 1,3—3 раза выше в стациях с относительным преобладанием эфе-
меров (глинистая пустыня), чем в стациях с меньшим количеством их (песчаная
пустыня). В песчаной пустыне во всех районах более низкая интенсивность размно-
жения компенсируется более поздними сроками окончания его. Общее увеличение
количества эфемеров в растительном покрове при продвижении с севера на юг опре-
деляет значительное возрастание плодовитости песчанок весной, что обусловлено
в основном увеличением числа самок, носящих 2—3 выводка (в северном Приаралье
до 40%, в северных Кызыл-Кумах до 60%, в южных пустынях до 100%). Меньшая
весенняя плодовитость песчанок в северных пустынях компенсируется в некоторые
годы летне-осенним размножением.
Колебание урожайности кормов по годам в зависимости от метеорологических
условий (особенно от количества зимних и весенних осадков и хода температур вес-
ной и летом) вызывает значительные изменения в интенсивности размножения. Одна-
ко урожайность кормов зависит не только от условий текущего, но и предыдущего
года, обеспечивающего запас семян, в связи с чем находится и более высокая интен-
сивность размножения весной в относительно менее благоприятные годы (1942, 1947,
1950, 1953), следующие за более благоприятными. Соответственно урожайности кор-
мов имеют место стадиальные и географические отличия в размножении.
Необходимо помнить, что при высоком уровне численности грызунов интенсив-
ность размножения ниже, чем в годы (и в стациях) с низким уровнем ее (при про-
чих равных условиях). Указанные особенности размножения следует учитывать при
анализе и прогнозировании изменений численности больших песчанок.
Значение подвижности лесных мышевидных грызунов в распределении
клещей в очагах весенне-летнего энцефалита
С. А. Шилова, В. Б. Троицкий, Г. Б. Мальков, В. М. Белъкович
(Автореферат доклада, прочитанного 14 февраля 1957 г.)
Авторы ставили своей целью выяснить, как отражается характер использования
территории мелкими лесными грызунами на формировании клещевых очагов. Работа
проводилась в 1954—1956 гг. в Пермской обл. Для изучения подвижности зверь-
ков применялась методика мечения. За период работы было помечено 706 зверьков,
которые ловились 4226 раз. Помимо мечения в разных биотопах добыто и исследо-
вано на заклещевение около 6 тыс. зверьков (набрано 90 тыс. ловушко-суток).
В год, последовавший за периодом глубокой депрессии численности (1955), по-
пуляция рыжих полевок характеризовалась исключительно высокой подвижностью.
В 1956 г., когда численность зверьков возросла, их подвижность заметно уменьши-
лась. В 1956 г. зверьки равномерно заселили всю территорию, при этом оседло живу-
щие особи преобладали в популяции в течение всего года. В стациях переживания
в год депрессии (1954) зверьки также в основном жили оседло.
Во всех стациях, где проводились наблюдения, клещи Ixodes persulcatus — основ-
ные переносчики клещевого энцефалита — встречались постоянно и в больших коли-
чествах. На площадке мечения их численность доходила до 20 особей на 1 человеко-
час. Личинки и нимфы кормились преимущественно на рыжих полевках, лесных лем-
мингах, полевках-экономках и землеройках-бурозубках. На всех меченых зверьках
в течение обоих сезонов проводили учет присосавшихся личинок и нимф. Таким обра-
зом, авторы имели возможность учесть, сколько личинок и нимф прокормил каждый
зверек в течение сезона, где он заразился ими и где их сбросил. При этом удалось
подметить, что существуют участки, где постоянно наблюдается высокое заклещевение
зверьков, в том числе мало подвижных (самки в период беременности). Таким обра-
зом было установлено несколько зон заражения в пределах площади, где происходит
нападение голодных личинок на зверьков. В местах концентрации личинок жили пре-
имущественно оседлые зверьки, которые разносили их в пределах своих индивидуаль-
ных участков. Нимфы встречались на зверьках в различных местах площадок, но
основная масса их регистрировалась именно там, где в предыдущем год} были сбро-
шены личинки.
— 118 —
Ограниченные участки территории, которые наиболее благоприятны для суще-
ствования грызунов (для рыжих полевок это захламленные понижения с зарослями
малины), заселены в основном оседлыми зверьками, которые живут здесь группами.
В том случае, если на этом участке самка /. persulcatus составляет кладку, личинки
прокармливаются в основном на оседлых зверьках и распределяются они в пределах
той же территории. Часть выплодившихся на следующий год нимф также остается
на этом участке и прокармливается на зверьках. В этом ограниченном участке на-
блюдается значительная численность половозрелых клещей. Такие микроучастки среди
заклещевленных территорий, на которых наблюдается очень высокая численность
клещей, мы называем локальными клещевыми очагами. Большей частью эти участки,
располагаются в местах с повышенной численностью грызунов.
Таким путем при участии грызунов возникают локальные клещевые очаги.
Проблемы географической изменчивости химизма растений
Е. А. Дороганевская
(Автореферат доклада, прочитанного 21 февраля 1957 г.)
Географическая изменчивость биохимических процессов — общебиологическая*
закономерность, свойственная животным и растениям. Географическая изменчивость
химического состава растений имеет большое значение для животных, питающихся
растениями. Количество ценных питательных веществ в растениях непостоянно, оно
меняется в процессе развития растений, а сезонная динамика их содержания меняет-
ся в географическом разрезе. Проблема географической изменчивости химизма расте-
ний возникла в связи с изучением вопросов качества растительной продукции, но
из-за недостаточного развития методов массового анализа химизма растений развитие
этой области науки, находящейся на стыке интересов нескольких дисциплин, сильно
задержалось. В настоящее время с развитием в биологии взглядов на роль химических
превращений в процессах жизнедеятельности все больше выявляются связи рассмат-
риваемой проблемы с вопросами продуктивности, растений и географии. В основе
продуктивности растений лежит их способность вырабатывать различные органические
вещества, составляющие растительную массу вегетативных частей растений, или от-
лагаемые в семенах, клубнях и т. п. Масштабы продуктивности как одних и тех же,
так и разных видов растений варьируют по различным зонам и районам в зависи-
мости от природных условий. С биогеографией эту проблему связывает: 1) простран-
ственная изменчивость биохимических реакций, лежащих в основе жизни, 2) причин-
ная связь этих изменений с различиями природных условий и 3) их роль в форми-
ровании ландшафтов.
Развитие исследований географической изменчивости химизма культурных и
дикорастущих растений обусловлено заинтересованностью в этом разнообразных от-
раслей сельского хозяйства: животноводства, звероводства и т. д.; оно облегчается
с разработкой новых методов массового анализа (хроматографического, спектрогра-
фического и др.). Необходимо обобщение фактических материалов, накопленных в-
практической работе отраслевых опытных учреждений и организация дальнейших
исследований, подчиненных общей программе. Комплексный характер проблемы тре-
бует участия в ее развитии различных специалистов — химиков, физиологов растений,,
почвоведов, климатологов и других. Разработка этой проблемы может сочетаться с
комплексными исследованиями природных ресурсов. Необходима инициатива научных
и хозяйственных организаций, заинтересованных в прогрессе данной области знания.
Автор изучал: 1) количественные связи между продуктивностью культурного
растения и климатическими условиями, 2) ландшафтные особенности химизма кор-
мовых растений.
Итоги изучения природного очага лептоспироза в Шаховском районе
Московской области
М Я. Лаврова, Е. В, Прохорова и А. П. Вишняков
(Автореферат доклада, прочитанного 7 марта 1957 г.)
В 1953—1956 гг. в Шаховском р-не Московской обл. в природном очаге лепто-
спироза сотрудники Института им. Мечникова при содействии МОИП проводили изу-
чение лептоспирозной эпизоотии среди мелких млекопитающих. Смена аспектов массо-
вых видов зверьков — носителей инфекции — влечет за собой преобладание одного
из двух, свойственных данному очагу серологических типов лептоспир. В 1953 и
1956 гг. в Шаховском р-не наблюдалась высокая численность полевок и других видов-
мышевидных грызунов, в связи с этим преобладала Leptospira grippotyphosa. В 1954—
1955 гг. при депрессии численности полевок и высокой численности насекомоядные
— 119 -
сем. Soricidae преобладала L. sorex. Для Шаховского р-на типичен диффузный ха-
рактер поселений полевок-экономок — главного источника L. grippotyphosa — возбуди-
теля водной лихорадки людей. Болотистые угодья служат основным местообитанием
полевок-экономок, а элементарные очажки L. grippotyphosa бывают диффузно разбро-
саны по территории района, составляя около 5% его площади. Диффузная структура
очагов L. grippotyphosa, по-видимому, широко распространена в условиях сельско-
хозяйственного ландшафта с преобладанием пахотных земель. Такой тип поселений
полевок-экономок, а также способность местных почв долго удерживать влагу обу-
словливают сравнительно высокий показатель зараженности лептоспирозом обыкно-
венных полевок Microtus arvalis на пахотных землях, который в сырую осень 1953 г.
достигал 5О°/о. Обыкновенные полевки — главный источник заражения людей водной
лихорадкой в Шаховском р-не. Эпидемиологический прогноз в очаге данного типа
нужно строить с учетом эпизоотологической ситуации среди популяции этих зверьков.
Третий многочисленный вид мышевидных грызунов — рыжие полевки — в циркуляции
лептоспирозной инфекции в шаховском очаге играют второстепенную роль.
Среди насекомоядных — носителей L. sorex — на первом месте по зараженности
лептоспирозом стоит обыкновенная бурозубка (6—15% зараженных особей). У зверь-
ков этого вида лептоспирами заражены преимущественно перезимовавшие, интенсивно
размножающиеся особи (50—70%); инфицированные прибылые составляют всего
1—2°/о. Общие показатели интенсивности лептоспирозной эпизоотии среди бурозубок
определяются численностью в популяции группы перезимовавших особей и зависят от
времени установления и высоты снежного покрова предшествующей зимы. Изучение
сезонных изменений интенсивности лептоспирозной эпизоотии в 1954—1956 гг. пока-
зало, что пики кривых эпизоотии типа L. sorex приходятся на наиболее жаркие ме-
сяцы лета и не всегда связаны с периодами обильного выпадения дождей, как это
имеет место в случае эпизоотии типа L. grippotyphosa среди грызунов. Не отмечено
и сколько-нибудь отчетливой стадиальной приуроченности эпизоотии типа L. sorex.
Факт наличия в природе очень интенсивной и широко разлитой лептоспирозной эпи-
зоотии типа L. sorex в условиях засушливого лета и высокой плотности популяции
бурозубок говорит о независимости развития этой инфекции от воды внутри популяции
основного хозяина и о несомненном преобладании контактного пути передачи L. sorex
от зверька к зверьку. Вода сохраняет свое значение в передаче L. sorex молодым
бурозубкам и неспецифическим хозяевам — другим видам землероек и грызунам.
В засушливом 1955 г. при наличии интенсивной эпизоотии типа L. sorex в Ша-
ховском р-не заболевания людей лептоспирозом этого типа отсутствовали, так как
был сильно ограничен единственно возможный — водный — путь передачи инфекции.
Наблюдения за мечеными зверьками в 1955—1956 гг. (с целью выяснения механизма
распространения лептоспирозной инфекции в природе очага) показали, что высокие
плотности популяции и степень индивидуального контакта между отдельными полев-
ками-экономками, а также между ними и зверьками других видов не достаточны
для распространения лептоспирозной эпизоотии типа L. grippotyphosa вне болот. Даже
при обильном выпадении дождей эпизоотия распространяется среди зверьков только
при наличии полевок-экономок старших возрастных групп — главных носителей
инфекции. В связи с тем, что наиболее старые полевки-экономки в Шаховском р-не
обитают преимущественно на болотах, только там в годы среднего и малого увлаж-
нения происходит вовлечение в лептоспирозную эпизоотию молодых особей этого ви-
да, а также и других зверьков. Последнее обстоятельство в значительной степени и
обеспечивает диффузную .структуру природного очага L. grippotyphosa в Ша-
ховском р-не.
Редакционная коллегия:
Р. И. Белкин, А. Г. Воронов, Л. А. Зенкевич, Б. А. Кудряшов, К- И. Мейер,
Т. А. Работнов, И. Г. Серебряков, акад. В. Н. Сукачев (главный редактор), А. Н. Фор-
мозов, В. И. Цалкин (зам. главного редактора)
* *
*
Адрес редакции: Москва, К-9, ул. Герцена, 6
Московское общество испытателей природы
Тел.: Б 9-99-14, доб. 2-16
Редактор Е, В. Нарекая
Технический редактор Г. И. Георгиева
Сдано в набор 19/VIII 1957 г.
Подписано в печать 1/XI 1957 г.
Л-37642. Изд. № 879.
Печ. л. 10,27.
Бум. л. 3,375.
Тираж 2500.
Цена 10 р.
Формат 70X108r/i6.
Физич. л. 7,5.
Уч.-изд. л. 11,68.
Заказ 1705.
Издательство Московского
университета
Москва, Ленинские горы
13 типография
Московского городского Совнархоза.
Москва, ул. Баумана,
Гарднеровский пер., д. 1а.