/











































































































































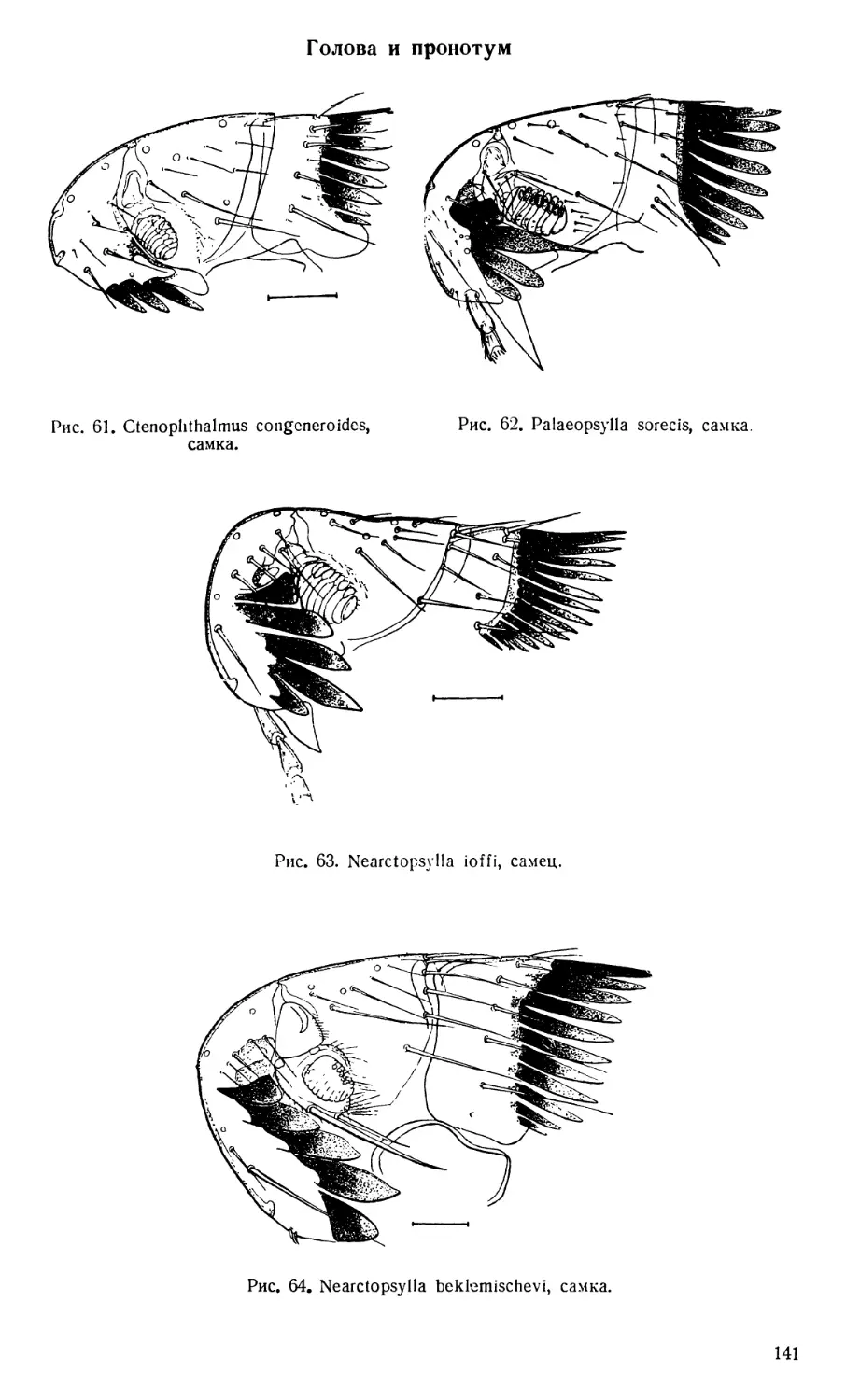





























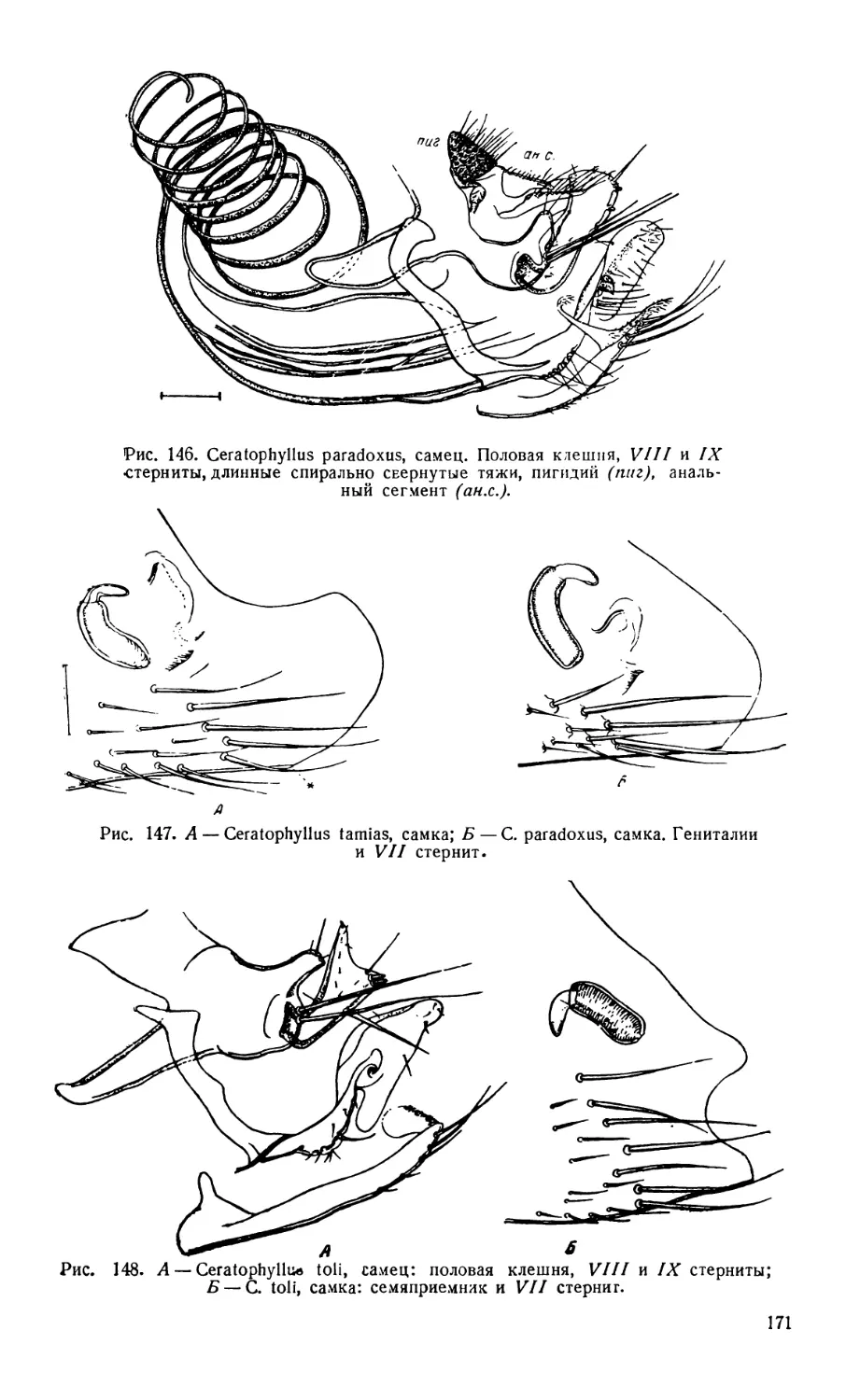





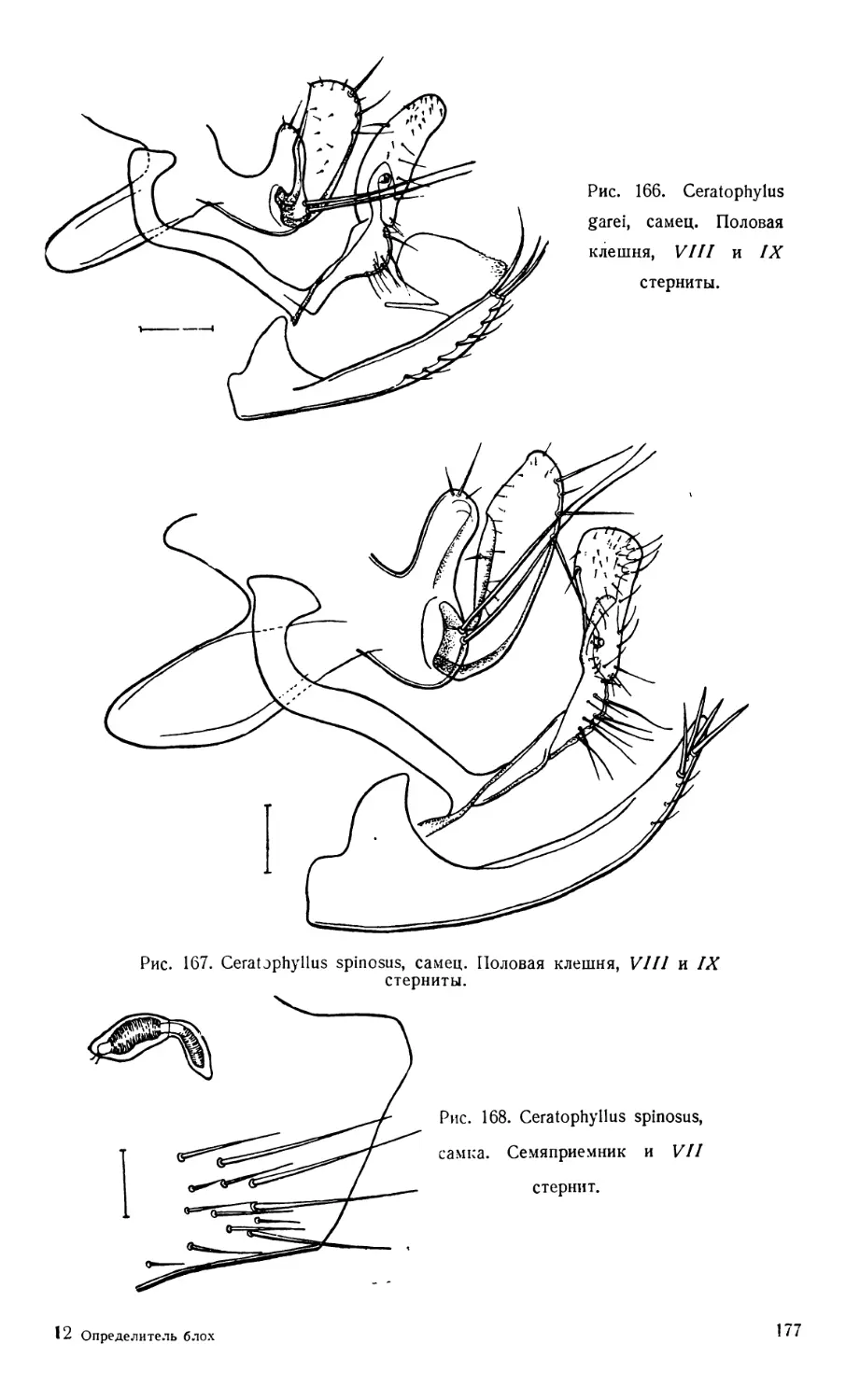




































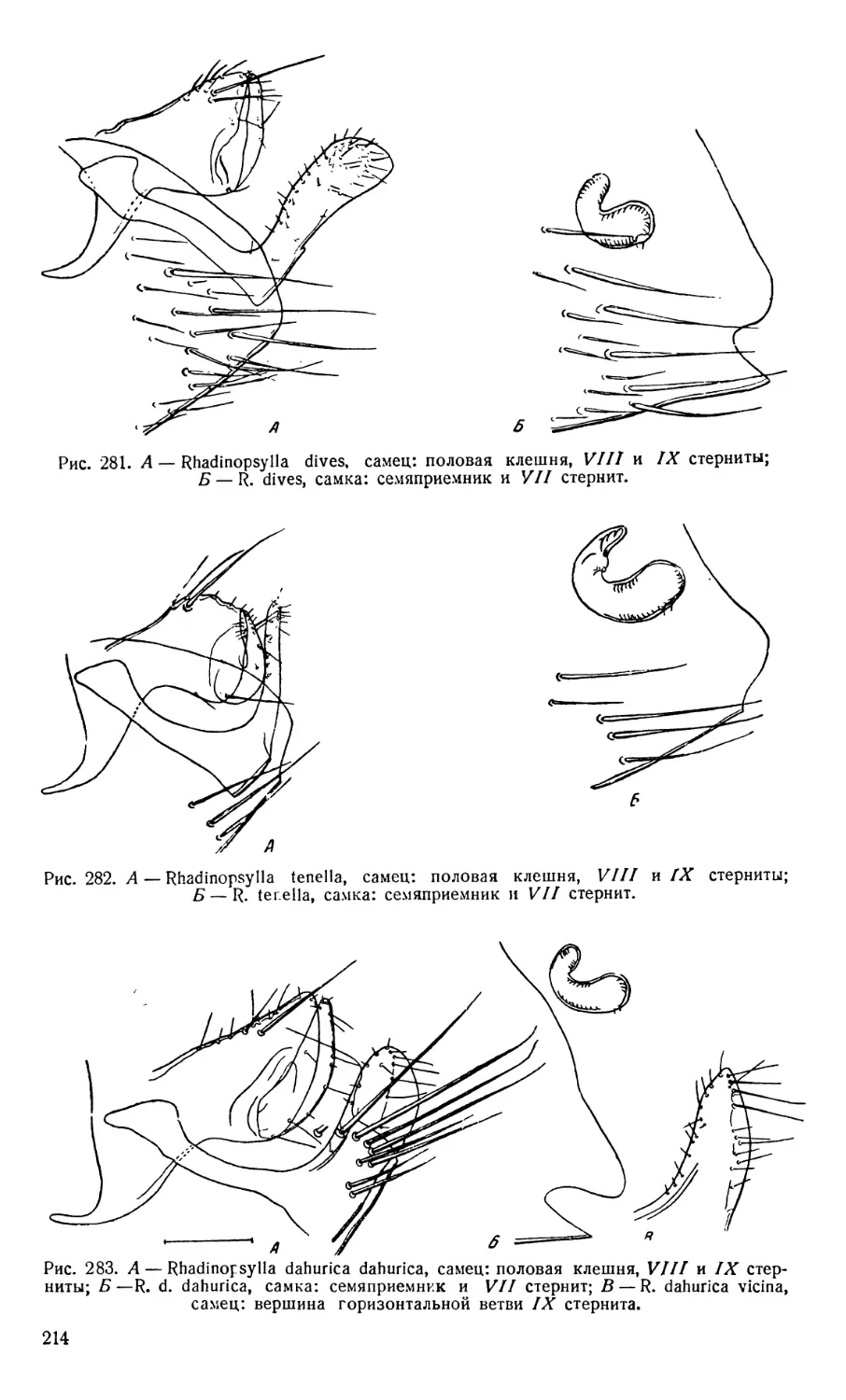
































































Текст
ОПРЕДЕЛИТЕЛЬ
ВОСТОЧНОЙ СИБИРИ,
.ДАЛЬНЕГО ВОСТОКА
И ПРИЛЕГАЮЩИХ
РАЙОНОВ
МЕДГИЗ • 1954
АКАДЕМИЯ МЕДИЦИНСКИХ НАУК СССР
И. Г. ИОФФ и О. И. СКАЛОЙ
ОПРЕДЕЛИТЕЛЬ БЛОХ
ВОСТОЧНОЙ СИБИРИ,
ДАЛЬНЕГО ВОСТОКА
И ПРИЛЕЖАЩИХ РАЙОНОВ
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО
МЕДИЦИНСКОЙ ЛИТЕРАТУРЫ
МЕД Г ИЗ-1954 —МОСКВА
ВВЕДЕНИЕ
В настоящее время исследование блох на зараженность их микро-
бами — возбудителями некоторых инфекций вошло в практику работы
противоэпидемических учреждений. Определение вида исследуемого насе-
комого является обязательным для того, чтобы сделать полнее выводы
из производимого исследования, ибо каждый вид имеет свои физиоло-
гические, экологические и зоогеографические особенности, которые глу-
боко влияют на его эпидемиологическое значение. Достаточно напом-
нить о том, насколько различна активность отдельных видов блох как
переносчиков инфекции, их долговечность, специфичность в выборе хо-
зяина, способность кусать человека и т. д. Если все же в некоторых
случаях массовая обследовательская работа, направленная на выясне-
ние только основного факта — наличия или отсутствия инфекции в той
или иной местности, может проводиться без определения видов иссле-
дуемых объектов, то углубление обследовательских задач, а тем более
научно-исследовательская работа по экологии или эпидемиологическому
значению блох, уже совершенно обязательно требует точного определе-
ния исследуемых насекомых.
Одной из трудных задач, возникших перед нами, было составление
списка видов, подлежащих включению в это пособие. Мы не могли
ограничиться только теми видами, которые фактически были обнаружены
на избранной нами территории Восточной Сибири и Дальнего Востока,
так как всегда приходится считаться с возможностью встретить еще
и другие виды, распространенные в более или менее близких местностях;
кроме того, и границы такой территории нельзя было очертить с полной
точностью, так как интересы соответствующих учреждений нередко про-
стираются за пределы обычно обслуживаемых районов. Поэтому было
решено включить в этот список также и многие виды, которые известны
из других ближайших областей Азии, т. е. из Западной Сибири, с Алтая,
из северо-западных районов Китая, со всей территории Монголии и Ти-
бета. К сожалению, из перечисленных местностей только по Западной
Сибири, Алтаю и ряду областей Монголии имеются в нашем распоря-
жении сколько-нибудь достаточные сведения (по большей части в наших
неопубликованных коллекциях). Особенно недостаточны сведения, ка-
сающиеся стран, лежащих к югу и юго-востоку от избранной нами тер-
ритории, — из пустынь Центральной Азии и нагорий Тибета и Китая.
Несомненно, что их изучение в очень значительной степени увеличит спи-
сок видов интересующей нас фауны, но пока эти виды не известны и по-
этому мы должны предупредить всех, кто будет пользоваться этой
книгой, что, в особенности в отношении представителей пустынно-степной
фауны Центральной Азии, наш определитель не может претендовать на
полноту даваемых сведений.
В некоторых случаях мы пополнили список упоминаемых видов
представителями фауны среднеазиатских пустынь и гор (Тянь-Шаня
Р 3
и др.), учитывая возможность их более широкого распространения на
восток; такие виды приведены в части случаев без рисунков. Мы воздер-
жались от более широкого включения и освещения представителей сред-
неазиатской фауны, так как это чрезмерно увеличило бы объем книги.
Для работы по изучению фауны Aphaniptera пустынь и гор Центральной
Азии будет, вероятно, полезным определитель блох Средней Азии, со-
ставление которого стоит на очереди. Можно упомянуть, что средне-
азиатские представители многих групп (Gerbillophilus, Frontopsylla, Cte-
nophyllus, Pectinoctenus и др.) освещены в монографии «Aphaniptera
Киргизии» (И. Г. Иофф, 1949). Вообще дополнительная литература
о каждой группе или каждом отдельном виде указывается нами во всех
случаях.
Из распространенных в Китае видов включены те, которые были об-
наружены в Маньчжурии и провинциях Жехе, Шеиьси, Шаньси и Ганьсу;
о фауне блох других северных районов Китая в доступной нам лите
ратуре сведений не найдено, а виды, описанные из более южных про-
винций (Сычуань, Гуйчжоу, Фудзянь, Юньнань), не включены, так как
обнаружение их на интересующей нас территории Восточной Сибири
и Дальнего Востока менее вероятно.
В определитель родов включены все роды, известные из СССР и
упомянутых выше соседних районов.
Для того чтобы иметь возможность в дальнейшем изложении крат-
ко называть территорию, для которой рассчитан наш определитель, мы
совершенно условно назовем ее Восточной Азией, не придавая этому
термину точного географического смысла.
Приводимые ниже определительные таблицы построены в основном
по дихотомической системе; в скобках после номера каждой тезы стоит
номер соответствующей антитезы (где даны признаки видов, обладаю-
щих противоположными или отличающимися особенностями). Таблица
читается подряд, пока признаки подходят к определяемому экземпляру,
до того пункта, в котором дано название вида. Если признаки не подхо-
дят, следует перейти к соответствующей антитезе. В некоторых случаях
мы пользуемся в таблицах и буквенной нумерацией, чаще для того, что-
бы выделить пункты, относящиеся к подвидовым различиям, или в тех
случаях, когда представляется более удобным дать параллельно при-
знаки трех или более видов, без противопоставления тез и антитез.
В настоящее время изучающему систематику блох уже нельзя обой-
тись без учета деления родов блох на подроды. Знание признаков
подродов в ряде случаев (в родах Ceratophyllus, Ophthalmopsylla, Rhadi-
nopsylla и др.) не только облегчает определение видов, но иногда совер-
шенно для этого необходимо (это часто относится к более трудно опре-
делимым самкам). Кроме того, надо учитывать, что в литературе назва-
ния некоторых групп блох употребляются то как родовые, то как подро-
довые названия, и потому знание последних необходимо каждому спе-
циалисту. Мы считаем правильным относить к числу родов достаточно
крупные и естественные группы с более доступными для неспециалистов
и достаточно надежными распознавательными признаками и более из-
вестными названиями, а многие недавно предложенные дробные подраз-
деления относим к числу подродов или еще более подчиненных катего-
рий (секций). Для начинающих изучать систематику следует указать,
что, как правило, в практической работе следует употреблять лишь ро-
довые и видовые названия, а названия подродов (и секций) употребля-
ются обычно лишь в специальной работе по систематике. Например,
блоха, относящаяся к роду Ceratophyllus, подроду Nosopsyllus, секции
Gerbillophilus, виду laeviceps, коротко может быть обозначены: Cerato-
phyllus laeviceps. В более специальных работах, где желательно уточнить
положение этого вида, можно его назвать Ceratophyllus (Gerbillophilus)
4
laeviceps. Такой способ обозначений мы и применяем в ниже приведен-
ных таблицах, т. е. в необходимых случаях в скобках ставим название
более узкой группы, к которой относится данный вид, чаще же приводим
лишь родовое название (или его инициал) и видовое.
За последние годы в систематике блох начался довольно интенсив-
ный процесс выделения низших таксономических катего-
рий, лежащих внутри вида — подвидов, рас, племен, морф, аберраций.
Этот процесс вполне законен и дал уже ряд интересных зоогеографиче-
ских и экологических фактов. Однако нужно пожелать, чтобы он остался
на строго фактической основе, т. е. каждая выделяемая форма должна
иметь надежные морфологические особенности, чтобы ее можно было
узнать именно по этим последним, а не по этикетке, указывающей на
происхождение экземпляра. Понятно, что отличия низших категорий мо-
гут быть менее четкими, чем отличия видов. Так, очень часто расцени-
вают как подвиды такие викарирующие (заменяющие друг друга) формы
блох, которые могут быть распознаны лишь по одному полу L Чаще
всего такие формы различаются по половому аппарату самцов, а самки
оказываются неразличимыми. Реже наблюдаются обратные случаи,
когда хорошо удается различить самок (например, мышиную и сурковую
формы блохи Rhadinopsylla ventricosa), а самцы не обнаруживают за-
метных различий. Ясно, что в случаях, когда насекомое может быть
определено до вида по экземплярам обоих полов, а более дробное деле-
ние возможно лишь для одного пола, соображения простого удобства
номенклатуры заставляют считать эти дробные деления лежащими ниже
видовой категории.
Иногда приходится сталкиваться с наличием переходных форм меж-
ду выделяемыми подвидами. Если это происходит в районе соприкосно-
вения или наложения их ареалов, — это вполне законное явление. Одна-
ко вариабильность бывает подчас велика на большом протяжении ареала,
и тогда распознавание подвидов требует больших серий и статистической
обработки, что делает подобные случаи изменчивости мало удобными
для практического использования. Следует пожелать, чтобы систематики
не спешили с выделением низших категорий в подобных случаях и обо-
сновывали свои выводы на хороших сериях и достаточно полных в гео-
графическом отношении материалах.
В определителе приводится ряд примеров деления видов блох на
подвиды (Ceratophyllus tesquorum, Ceratophyllus penicilliger, Amphipsylla
sibirica и многие другие). В отдельных случаях географические, эколо-
гические и морфологические отличия подвидов так четки, что подобные
насекомые могли бы расцениваться как географически или экологически
викарирующие виды. Однако наличие у некоторых подобных форм пере-
ходов на границе ареалов или соображения простого удобства (поскольку
различимы не оба пола) заставляют сохранить их подвидовое обозначе-
ние. Некоторое неудобство подвидового деления заключается в необходи-
мости пользования тройным названием; понятно, что было бы проще пи-
сать два слова Ceratophyllus sungaris вместо трех Ceratophyllus tesquo-
rum sungaris. Поэтому в некоторых случаях при частом повторении
какого-либо подобного названия мы считаем допустимым опускать ви-
довое название и вместо него употреблять подвидовое.
Как известно, после названий животных принято ставить фамилию
автора и год, когда было дано данное название. Это в некоторых слу-
чаях совершенно необходимо, когда читатель должен обратить внимание
1 К сожалению, в распоряжении систематика далеко не всегда имеются более
объективные критерии для решения вопроса о видовом или подвидовом значении
выделяемых форм. Для этого нужны эксперименты по выяснению результатов скре-
щивания ил^ детальные географические материалы по изучению изменчивости, в осо-
бенности материал с границ ареалов, который показал бы наличие или отсутствие
гибридных форм.
а
на приоритет того или другого названия или на запутанность номен-
клатуры из-за синонимики. Во всех других случаях указание автора
усложняет текст без необходимости и мы предпочли такие полные на-
звания с их авторами вынести в особый список в конце книги.
Также в особом списке приведены для справок и некоторые сино-
нимы упоминаемых нами видов; полную синонимику читатель может
найти в Каталоге Ю. Н. Вагнера (с добавлениями к нем^).
Нами включены в определительные таблицы некоторые виды, опи-
сания которых еще не опубликованы; в алфавитном списке видов
(стр. 249) они приведены с указанием на их авторов (in litt.); права этих
авторов, конечно, должны быть сохранены, если определитель будет
напечатан ранее, чем их работы.
В ряде случаев, согласно правилам номенклатуры, допускающим из-
менение одной буквы в названии, мы изменили окончание видового назва-
ния для того, чтобы согласовать его с родовым; например, Ceratophyllus
elatus в настоящее время должен называться Frontopsylla elata. Однако
мы воздержались от такого исправления названий в тех случаях, когда
с самого начала автором было дано название с несогласованным окон-
чанием (Chaetopsylla lasius, Doratopsylla dasycnemus).
После каждого пункта определительной таблицы, дающего опреде-
ление вида (или группы), приводятся сведения о географическом
распространении (местах обнаружения) данного вида, его хозяе-
вах и других важнейших экологических особенностях (сезон, стация,
привязанность к гнезду и т. п.), если они заслуживают упоминания.
Все эти сведения приводятся на основании нашей картотеки, содержа-
щей как все опубликованные, так и многочисленные ранее неопубли-
кованные данные. Мы обращаем особое внимание читателя на необ-
ходимость в каждом случае определения убедиться
в том, что данный вид встречается в том районе, от-
куда поступил определяемый материал. Географические
указания во многих случаях сильно облегчают определение вида, так как
направляют внимание на те виды, которые могут встретиться в данном
районе, или, наоборот, на малую вероятность встретить данный вид, по-
скольку он распространен в удаленных местностях. В ряде случаев гео-
графический критерий введен нами даже в текст определительных таб-
лиц. В тех случаях, когда встречается вид, еще не известный из данного
района, определение обязательно должно основываться на совершенно
бесспорных морфологических признаках (модифицированные сегменты
самца и др.), а при недостаточном материале (например, когда имеются
только самки с недостаточно надежными отличиями) следует воздержи-
ваться от окончательного определения, оставляя решение вопроса до
получения более надежного материала. Гораздо полезнее оставить
экземпляры без видового определения, чем фиксировать ненадежные
фаунистические факты. Экземпляры, которые не могут быть определены
с полной уверенностью до вида, следует обозначать, например, так:
Ceratophyllus sp. или Ceratophyllus (Nosopsyllus) sp., или Ceratophyllus
aff. garei (aff. означает affinis — близкий; иногда употребляется также
выражение: conf. — confinis — близкий; ргоре — близко).
Географические указания даются нами для краткости в возможно
обобщенном и сжатом виде. Только в некоторых случаях, когда геогра-
фия вида требует особого внимания, мы приводим более детальные ука-
зания о местах его обнаружения. Приводя в списке районов обнаруже-
ния обозначение «ДВК», мы имеем в виду те районы Советского Даль-
него Востока, которые лучше обследованы паразитологически и лежат
к востоку от 125 меридиана, примерно между 54 и 42 параллелями.
«Восточное Забайкалье» — обследовались преимущественно степные
районы — Борзинский и соседние и лишь частично лесостепные —
Александрозаводский, Оловяннинский. «Западное Забайкалье» — имеют-
ся в виду главным образом лесо-степные и горно-лесные районы — Се-
ленгинский, Джидинский и Кубанский. «Предбайкалье» — территория,
лежащая к западу от Байкала (часто обозначаемая также как «При-
байкалье»— лесные и лесо-степные местности в районе г. Иркутска,
с. Монды, Болыпеглубоковского). Для Западной Сибири, Монгольской
Народной Республики и многих других мест приводятся указания на
крупные районы обследования: области, аймаки или горные хребты.
Более детальные сведения следует искать в специальных работах.
.Приводя сведения о хозяевах блохи, мы в большинстве случаев
ограничиваемся указанием главных — специфических или наиболее обыч-
ных хозяев, или даем их обобщенную характеристику (например, блохи
мелких лесных грызунов, блохи наземных птичьих гнезд и т. п.). Пере-
числение всех видов животных, на которых встречался тот или другой
вид блох, заняло бы слишком много места, так как обмен блохами и пе-
реход блох с основных хозяев на другие виды животных — очень обыч-
ное явление. Поэтому, указывая, например, что данный вид паразити-
рует на сусликах и мелких степных грызунах, мы не оговариваем, но
подразумеваем, что он может встретиться и на хищниках, охотящихся
за этими грызунами, на насекомоядных, живущих в той же стации, и т. п.
Наоборот, случаи более специфичного паразитизма оговорены спе-
циально.
Заканчивая определение вида, необходимо использовать не только
те признаки, которые перечислены в таблице, но нужно тщательно срав-
нить свой экземпляр со всеми рисунками данного вида, которые приве-
дены в книге (или с препаратами образцовой коллекции, если она
имеется). Такое сравнение предотвратит возможные ошибки. При этом
надо помнить, что многие признаки блох чрезвычайно характерны (мо-
дифицированные сегменты самца, очертания VII стернита самки и мно-
гие другие), но в то же время никогда нельзя забывать о наличии весь-
ма близких видов, подчас надежно различимых только по особям одного
пола (чаще по самцам, иногда по самкам). Нельзя также забывать
о вариабильности признаков. Лишь опыт и внимательная работа позво-
лят каждому накопить необходимые навыки, чтобы во всех случаях на-
ходить правильное решение. Само собой разумеется, что изучение систе-
матики и определение блох должно быть начато с тщательного изучения
(по препаратам) признаков этих насекомых, на основе приводимого
ниже очерка их морфологии.
Для лиц, желающих найти более детальные морфологические све-
дения о блохах того или другого вида или рода, мы приводим необходи-
мые литературные источники — цифры в прямых скобках указывают на
номера по списку литературы. Когда литература для вида не указана
или недостаточна, следует использовать литературу, указанную для рода.
Более исчерпывающие справки о литературе можно найти в каталогах
[1, 2, 3 и другие].
Добавим еще, что в этой книге даны лишь немногие сведения о при-
знаках более крупных таксономических подразделений блох — над-
семейств, семейств и подсемейств. Несмотря на всю важность таких
крупных категорий и их признаков для понимания естественных группи-
ровок блох по их родству, в деле определения видов эти признаки далеко
не всегда бывают удобными, и потому при построении определительных
таблиц мы в ряде случаев игнорировали эти естественные группировки,
подбирая наиболее простые и надежные дифференциальные признаки.
Например, отсутствие гребней на голове (при наличии гребня на передне-
груди) является признаком у представителей из целого ряда подсемейств
(Spilopsyllinae, Ceratophyllinae, Paradoxopsyllinae, Leptopsyllinae, Neopsyl-
liaae, Hystrichopsyllinae и др.). Однако из-за этого не имело смысла от-
7
называться от использования такого четкого признака, и потому таблица
для определения родов не учитывает деления на подсемейства. Вообще,
повторяем, мы в этой книге не ставили себе задачу дать характеристики
всех высших таксономических категорий блох, а поместили лишь неко-
торые, чтобы познакомить читателя с новыми взглядами на систематику
этих насекомых, как она сложилась в последние годы в работах спе-
циалистов (К. Иордан, 1948; И. Г. Иофф, 1946, 1949).
На таблицах под большинством рисунков (или рядом с ними) при-
ведены масштабные линейки в 100 микронов (0,1 мм), что позволяет
судить о степени увеличения каждого данного рисунка. В некоторых
случаях дан общий масштаб для целой серии однотипных рисунков. Без
масштабов приведены некоторые рисунки, заимствованные из лите-
ратуры.
В заключение необходимо указать, что настоящая книга является
результатом труда очень большого числа лиц, принявших участие в из-
учении фауны Aphaniptera Восточной Азии и прежде всего — в соби-
рании материалов по этой фауне. Мы не имеем возможности перечис-
лить всех участников этой большой работы, проводившейся главным
образом на протяжении последних 20 лет силами многих зоологов, па-
разитологов, лаборантов и врачей, участвовавших в противоэпидемиче-
ских обследовательских отрядах, зоологических экспедициях и т. п. Со-
бранные ими материалы лишь частично послужили для составления
специальных -статей о фауне Aphaniptera Азии, приведенных в списке
литературы. Имена авторов этих статей: А. И. Аргиропуло, Ю. Н. Ваг-
нера, 3. М. Вовчинской, Н. Л. Гершкович, Н. Ф. Дарской, В. Б. Дуби-
нина, И. С. Дудченко, Н. Д. Емельяновой, К. Иордана, И. Г. Иоффа,
В. В. Кучерука, Ч. Лю, П. И. Мариковского, М. А. Микулина, Е. И. Пав-
лова, Н. Ротшильда, О. Н. Сазоновой, В. Н. Скалона, О. И. Скалой,
В. В. Сукнева, П. Т. Сычевского, В. Е. Тифлова, О. А. Фединой,
Л. В. Федоровой, П. И. Ширановича, Г. Г. Этмара и многих других,
являются именами лиц, вложивших особенно много труда в нашу об-
ласть исследования. Однако не только опубликованные и оформленные
в виде статей материалы, но также и сведения, учтенные в картотеках
и рукописных фондах учреждений, руководивших работой, представляют
важный вклад в наши знания и нашли полностью свое отражение
в предлагаемом руководстве. Пользующийся им вместе с авторами дол-
жен с благодарностью вспомнить о всех тех, кто помог накоплению кол-
лективных знаний, здесь изложенных.
Эта книга выходит из двух учреждений: Ставропольского противо-
эпидемического института и Иркутского противоэпидемического инсти-
тута. Руководители этих учреждений и многие сотрудники их, а также
другие лица оказали авторам большое содействие при оформлении ра-
боты и подготовке ее к печати. Авторы особенно признательны
Н. Д. Алтаревой, Н. Т. Быкову, В. Н. Тер-Вартанову, 3. С. Брагиной,
О. И. Желудковой, В. М. Жданову, М. А. Ивановой, Л. В. Крекниной,
Н. Ф. Лабунец, И. И. Рогозину, Б. Н. Пастухову, М. П. Покровской,
А. А. Свистельниковой, Н. Т. Соловьевой, М. Н. Споткай, В. Е. Тифлову
и многим другим товарищам, перечислить которых нет возможности.
О всех затруднениях, возникающих при использовании этой книги^
недостатках ее и ошибках просьба сообщать по адресу: Ставрополь-
Кавказский, Советская, 13. Весьма желательна также присылка образцов
блох, вызвавших затруднения при определении, сводных цифровых таб-
лиц для картотеки по фауне и экологии блох, оттисков и рукопи-
сей работ.
СОБИРАНИЕ БЛОХ И ПРИГОТОВЛЕНИЕ ПРЕПАРАТОВ
Сбор блох производят или с хозяев, на которых паразитируют эти
насекомые (преимущественно из шерсти млекопитающих), или в поме-
щениях, логовищах, гнездах и норах, где живут хозяева блох. Сборы из
гнезд дают обычно наибольшее количество насекомых.
Если сбор блох производится в эпизоотическом районе, то вся рабо-
та должна проводиться лишь опытным персоналом с необходимыми ме-
рами предосторожности, как при работе со всяким инфекционным или
подозрительным материалом.
Сбор блох с млекопитающих
Как правило, такие сборы надо производить возможно скорее после
поимки животного, так как многие виды блох быстро уходят даже с жи-
вых хозяев, не говоря уже о трупах. Если возможно, производят сбор на
месте, где животное поймано, быстро перенеся зверька на белую ткань;
не следует забывать осмотреть и самое место поимки — на земле около
капкана, в мышеловке и т. п.
В случае если осмотр животных откладывают до возвращения в ла-
бораторию, то необходимо сделать так, чтобы блохи не могли разбе-
жаться или перейти с одного хозяина на другого. Для этого трупы жи-
вотных, каждый в отдельности, помещают в мешочки из плотной тка-
ни, из которых блохи убежать не могут, а живых зверьков лучше всего
переносить в ловушках, в которых они пойманы, также уложив их в ме-
шочки. Если это невозможно, то для перевозки живых зверьков употреб-
ляют металлические банки, ведра или клеенчатые мешки, которые долж-
ны в лаборатории тщательно осматриваться. Мешочки, употребляемые
для сборов, не должны быть слишком узкими, так как тогда их трудно
выворачивать. Обычно с внутренней поверхности мешка удается собрать
значительную часть эктопаразитов, бывших на трупе животного.
Употребление индивидуальных мешочков для каждого отдельного
зверька важно для точного количественного учета встречаемости
паразитов (т. е. процента зараженных ими зверьков) и учета максималь-
ного числа блох на одном зверьке. Если в один мешок кладутся несколь-
ко зверьков одного и того же вида, то такие сборы могут быть исполь-
зованы лишь для учета индекса обилия (т. е. среднего числа блох
на одного хозяина). При смешивании в одном мешке разных зверьков
сборы эктопаразитов совсем не могут быть использованы для экологиче-
ских выводов (но годны для фаунистических, бактериологических и дру-
гих исследований). Кроме небольших индивидуальных мешочков, иногда
удобно пользоваться большими мешками (наволочками). Собирая в та-
кой мешок несколько животных, их следует отделять одно от другого,
перевязывая мешок веревкой. Мешки рекомендуется использовать рубцом
наружу. Завязывать мешки следует плотно и всегда снаружи (удобнее
всего пришитым шнурком), так как внутренние стягивающие завязки
9
не обеспечивают достаточной плотности. Для дезинфекции мешков
рекомендуется подвергать их кипячению; допустимо также вымачивание
в сулемовом растворе с последующим тщательным выполаскиванием
в проточной воде. Не следует при паразитологической работе (для дезин-
фекции мешков и посуды) пользоваться лизолом и другими пахучими
дезинфицирующими средствами, которые могут повредить блохам. Упо-
требление для сборов клеенчатых мешков также бывает вредным для
эктопаразитов, вероятно, тоже в связи с наличием в них паров дезинфек-
ционных средств.
1.
только
Рис. 1.
Аспиратор. Резиновая
груша снабжена двумя клапа-
нами, позволяющими через ре-
зиновую трубку со стеклянным
наконечником всасывать воздух
в пробирку вместе с мелкими
насекомыми.
Для успешного проведения сборов необходимо помнить еще следующие правила.
Сборы блох со зверьков, пойманных живыми, выгнанных из нор водой или
что застреленных, бывают обычно более обильными, чем с трупов (и даже
живых) зверьков, долго пробывших в капкане С
2. Перенос зверька в мешок должен произво-
диться так, чтобы блохи не успели спрыгнуть; для
этого необходимо положить раскрытый мешок рядом
с ловушкой или, еще лучше, захватить зверька вме-
сте с ловушкой мешком и лишь в нем освободить
его из ловушки. Надо иметь в виду, что в момент
изъятия зверька из ловушки и при переносе его бло-
хи спрыгивают не только с трупов, но и с живых
зверьков; давно замечено, что многие блохи выходят
из шерсти хозяина, когда последний волнуется
и бьется.
3. Сравнительно больше блох удается собрать
со зверьков, пойманных в холодные ночи и в тени-
стых прохладных местах, например, близ речки; при
более высокой температуре блохи двигаются более
активно и скорее покидают труп хозяина. После ноч-
ного лова собирание зверьков из ловушек необходи-
мо производить возможно раньше на рассвете, пока
солнце не пригрело. При дневном лове капканы дол-
жны осматриваться возможно более часто. При
расстановке ловушек и капканов следует по возмож-
ности выбирать места, укрытые от дождя.
4. При сборе зверьков следует также осматри-
вать место, где лежала ловушка; иногда там можно
обнаружить блох; полезно слегка похлопать рукой
по этому месту или подуть на него, тогда блохи вы-
лезают из мусора. Ловить их можно с помощью
аспиратора (рис. 1) или захватывая вместе со ще-
поткой земли. С поверхности земли можно также
собрать блох, накладывая кусок белой ткани, к ко-
торой они прицепляются. Можно также собирать
поверхностный слой почвы и просматривать его на
месте в тазике или на материи, или уносить с со-
бой, но не следует класть землю, мусор вместе
со зверьками, так как это затрудняет сбор эктопаразитов, находящихся на зверьке.
В зимнее время, когда блохи менее подвижны и далеко не уходят, удавалось, напри-
мер, собрать около капканов с песчанками до 30% того количества блох, которые были
сняты со зверьков (В. Е. Тифлов).
Сбор блох с животных производится над белой поверхностью (мате-
рией, бумагой, кюветом и т. п.) путем вычесывания (и стряхивания)
эктопаразитов гребнем, зубной щеткой, пинцетом и т. п. или (при раз-
двигании или раздувании меха) непосредственным вылавливанием пара-
зитов с помощью мягкого пинцета или ватки, смоченной в хлороформе,
1 Ввиду того что для изучения численности блох на зверьках важно обеспечить
проведение возможно более полных сборов, нередко при капканном методе добы-
вания зверьков рекомендуют производить частые осмотры капканов (например,
каждый час) или устанавливать дежурство около них. Само собой разумеется, что
это может быть обеспечено лишь в немногих случаях, и мы полагаем, что получен-
ные таким методом показатели следует использовать главным образом лишь для
проверки и оценки основных цифр, получаемых при обычных массовых сборах, но
при тщательном их проведении. Можно использовать специальные ловушки [183 а],
убивающие сразу и зверьков и их эктопаразитов (с помощью цианистых препаратов)
и не допускающие расползания последних.
10
спирте или бензине. Применение наркотических веществ необходимо при
сборе далеко прыгающих блох, например, блох с хищников, на которых
встречаются Pulex irritans, а также при работе на эпизоотических тер-
риториях. Следует по возможности собрать всех блох, имеющихся на
животном.
Пойманных блох переносят в пробирку с помощью мягкого пин-
цета, сделанного из тонкой эластичной стальной пружины и имеющего
гладкие тупые концы (рис. 2). Такой пинцет не раздавливает блох. Ино-
гда с этой же целью употребляется кисточка, смоченная водой.
Кюветы или тазы, употребляемые при сборах, должны быть со стен-
ками высотой около 10—20 см. Такая высота стенок оказывается в боль-
шинстве случаев достаточной для того, чтобы блохи не могли выбраться.
Рис. 2. Мягкий пинцет.
Удобны эмалированные тазы или тазы из белой жести, по стенкам
которых блохи не могут подниматься вверх.
Если животное не убито, то указанная методика может применяться
лишь к сравнительно спокойным или вялым зверькам (например, удается
вычесать суслика сразу после того, как его выгоняют из норы водой) или
к самым мелким — мышам и т. п. В других случаях животное и блох
приходится наркотизировать. С этой целью мешочек с животным (или
только животное) помещают в плотно закрывающуюся коробку или банку
и бросают туда вату, смоченную хлороформом или лучше эфиром. Если
животное кладется без мешка, то блох собирают не только из шерсти
хозяина, но и со дна банки. Если наркотизирование производить осто-
рожно (небольшой дозой, в течение непродолжительного времени — не-
сколько секунд), то можно сохранить как животное, так и блох жи-
выми — через некоторое время после наркоза они просыпаются.
При отсутствии анестезирующих веществ хозяев убивают уколом
в мозг, удушением и т. п. В этом случае в условиях лаборатории можно
собрать блох и другим способом: труп подвесить над большим тазом
с водой; блохи бегут с трупа, падают в воду и плавают на поверхности,
где их легко собрать. Можно также подвешивать животных над большой
воронкой, под которой ставится стакан.
С собак, ежей и других «спокойных» животных можно собрать блох
следующим образом: напудрить шерсть мелко растертым нафталином
и завернуть животное на несколько минут в материю: блохи оглу-
шаются запахом нафталина и выпадают на материю, где их легко можно
собрать.
Сбор блох в гнездах млекопитающих и птиц
Как указано, сборы, производимые в жилище тех или других хозяев,
бывают особенно обильны. В жилье человека (в одежде, под кошмой
и т. д.), в логовище зверя, в гнезде грызуна или птицы часто удается
собрать десятки, сотни, а иногда и тысячи экземпляров этих паразитов.
Чаще всего приходится производить сборы в подземных гнездах раз-
личных мелких млекопитающих: грызунов, насекомоядных, хищников.
Добывание таких гнезд из норы не представляет в большинстве случаев
особых затруднений, так как многие из этих мелких зверьков делают
неглубокие норы — до 0,5 м, а чаще всего в несколько сантиметров глу-
биной. Отыскивание таких норок тоже не представляет трудностей; так,
11
например, легко найти в поле, в особенности в тех местах, где лежал
собранный хлеб, норы полевок; легко обнаружить норы сусликов и т. д.
Раскопка норы должна производиться лопатой и совком очень осторож-
но; за ходом все время надо тщательно следить, вставляя в него палочку
или проволоку для того, чтобы, во-первых, не терять этого хода, а во-
вторых, не засыпать самого гнезда. Хорошим инструментом при этой
работе является длинный (гинекологический) пинцет, который употреб-
ляется для нащупывания хода и гнезда и для схватывания хозяина, если
он окажется в норке.
Рис. 3.
А — ящик из жести для просеивания гнезд с блохами, раз-
деленный на две разъемные части. Верхняя половина имеет
съемную крышку и дно, затянутое сеткой (изображенной
внизу рисунка); ячейка сетки диаметром в 1 см; Б — термо-
эклектор, боковой разрез. Легкий шкафчик бездна, с полоч-
ками шириной в 8 см. Заднюю стенку образует резервуар
для горячей воды с трубкой для спуска ее (или электриче-
ский согреватель). Через дверку в передней стенке на по-
лочки засыпается мусор с блохами, которые, спрыгивая, по-
падают в жестяной тазик. Поперечные размеры: шкафчи-
ка — 27 см, тазика — 35 см. Чтобы блохи не задерживались
на стенках шкафчика, желательно обить его внутри белой
жестью.
Рис. 4. Термофотоэклектор
Тульгрена (видоизмененный
Сазоновой).
Добравшись до гнездовой камеры и расширив ход в нее, тщательно
собирают оттуда гнездо и весь мусор в мешок и доставляют для раз-
борки в лабораторию. Разборка производится в тазах, кюветах или
больших кристаллизаторах с помощью твердой кисточки или пинцета.
Блохи вылавливаются с помощью мягкого пинцета или мокрой мяг-
кой кисточки.
При наличии большого количества блох в гнезде можно для облегчения работы
воспользоваться особым прибором — термоэклектором; полезно также сито для отде-
ления мелких частиц гнезда вместе с насекомыми от крупных стеблей травы и т. д.
(рис. 3); [более полное описание см. 136].
Для добывания единичных блох из больших количеств сыпучего материала, на-
пример, при поисках блох в зерне или мусоре складов, а также для сбора личинок
блох (как и мелких клещей) можно пользоваться прибором, изображенным на рис. 4,
представляющим большую воронку, освещаемую сверху электрической лампой (или
солнцем); материал, содержащий насекомых, накладывается на сетку, вложенную
в эту воронку. Уходя от яркого света и жара, насекомые проваливаются в подстав-
ленную банку. Другой прибор, применяемый при разборке зерна [167], устроен так,
что блохи выпрыгивают из низкой сковородки на окружающую ее водную поверхность.
Так как в гнездах почти всегда остается много личинок и куколок
блох, то весь мусор следует поместить в большие банки и, завязав их
бумагой, хранить во влажном помещении (подвал, ящик с мокрыми
тряпками). Время от времени оттуда можно опять выбирать блох.
12
В случае, если разборку гнезда нельзя произвести сразу, то, чтобы
сохранить блох живыми (мертвых блох выбирать гораздо труднее), гнез-
да следует беречь от высыхания и согревания — держать в тени, в яме
или подвале, опрыскивать мешки водой и т. д. При пересылке и перевоз-
ке гнезд необходимо для поддержания влажности обильно обложить
мешки снаружи свежей травой.
Сбор мертвых блох из мусора, а также их коконов может быть облегчен при-
менением метода флотации. Для этого мусор смешивается небольшими порциями
с водой. Все смачивающиеся и тяжелые частицы тонут, а легкие частицы, в том числе
блохи и коконы, всплывают на поверхность, откуда их можно легко собрать. При
разборке мусора, содержащего большое число хитиновых остатков насекомых (помет
летучих мышей и т. п.), полезно бывает смешивание мусора со слабым (10—20,%)
раствором спирта.
Сбор блох во входах в норы
В последние годы большое значение в практике обследовательской
работы противоэпидемических учреждений приобрел сбор блох во вход-
ной части нор, где часто имеются блохи, мигрирующие из гнезда к выхо-
Рис. 5. Инструменты для добывания блох из входной части нор.
А — скребок для выгребания земли из косых нор; Б — резиновый шланг, обшитый фланелью;
В — экскаватор.
ду из норы, а также, повидимому, и те блохи, которые мигрируют с по-
верхности земли в норы. У входов в нору нередко удается собрать блох
без помощи каких-либо ловушек или помещая у входов различные ло-
вушки для блох: трубки из липкой бумаги, ватные тампоны или спе-
циальные ящики с водой [В. Е. Тифлов и В. О. Потапов— 159]. Однако
более значительные сборы производятся из глубины первого колена нор
путем выгребания оттуда мусора в тазик, а также путем опускания туда
намотанного на палку ватного тампона или узких длинных лент из фла-
нели (для нор сусликов берутся ленты шириной 4—5 см и длиной до
100—150 см). Наиболее удобны ленты, сшитые вдвое в виде узкого меш-
ка, в который вставляется резиновый шланг (рис. 5, Б), что позволяет
вводить ленту как в вертикальные, так и косые норы (В. Н. Тер-Варта-
нов). Шланг вводят в нору на несколько секунд, слегка пошевеливают
им, затем извлекают и осматривают (см. нижнее фото на стр. 14);
в каждую нору его опускают два-три раза. Блохи прыгают на фланель
и прицепляются к ней в момент движения шланга, так как одним
13
из главных стимулов для прыжка блохи является движение воздуха.
С фланели блохи снимаются мягким пинцетом.
Выгребание мусора и песка из наклонных нор производится с по-
мощью ложки или совочка на длинной ручке или специального скребка
Блоха суслика Ceratophyllus tesquorum.
а самец; б — самка.
(рис. 5, Л — по П. И. Ширановичу), а для извлечения мусора из верти-
кальных нор Б. Е. Лебедевым предложен простой прибор «экскаватор»
(рис. 5, В). Мусор и песок помещаются в таз, откуда и выбираются бло-
хи. В песчаных местностях выгребание часто дает возможность собрать
гораздо больше блох, чем извлечение с помощью шланга.
Сбор блох из норы суслика при помощи резинового ш >анга, обшитого
фланелью.
За один день можно обследовать до 150—200 нор. В среднем из
100 нор сусликов удается собрать от 20 до 100 блох; более высокие ин-
дексы обилия наблюдались весной и в начале июня; к осени блох в но-
14
pax сусликов становится меньше, а в холодное время года сборы из нор
бывают обычно совсем безуспешными L Большое количество блох удается
собрать в норах песчанок, а также иногда и других зверьков Особенно
много блох бывает в норах, оставшихся без хозяина после вылова
зверьков или их гибели. Несомненно, что исследование блох, пойманных
из входов в норы (наряду со сбором трупов грызунов) путем радиальных
и маршрутных обследований может служить одним из самых быстрых
и легких методов обследовательской работы.
Сбор блох в жилищах человека
Для сбора блох в жилищах предложены следующие методы.
1. Осмотр постелей, одежды, ног и т. п.
2. Протаскивание по полу куска белой материи.
3. Выпускание морских свинок и других экспериментальных живот-
ных с последующим их осмотром.
4. Раскладка на ночь липких листов (плотная бумага, смазанная
гусеничным или мушиным клеем, который приготовляют из канифоли —
25 частей, касторового масла— 17 частей и глицерина — 8 частей). Бло-
хи для фаунистических целей могут сниматься с таких листов с помощью
растворителей (скипидар, бензин), а для бактериологического исследо-
вания — без растворителей.
5. Расстановка на полу неглубоких тарелок с жидкостью (водой,
мыльной водой, маслом).
Во время эпидемий в Африке используют этот метод следующим об-
разом: комната полностью затемняется и остается закрытой на 4—5 дней,
после чего дверь быстро открывается и в освещенную полосу ставятся
тарелки с водой. Блохи прыгают на белую поверхность и попадают в во-
ду, откуда их легко собрать.
6. Расстановка тарелок с зерном, на которое положен труп мыши.
Хранение, консервирование, упаковка и этикетировка сборов
Если блохи собираются для бактериологического исследования, био-
логических наблюдений или экспериментальной работы, то необходимо
обеспечить их сохранение в живом виде. Без подкармливания блох мож-
но сохранить живыми в течение нескольких дней и дольше, если пред-
отвратить их высыхание. Для этого следует поместить их в чистые стек-
лянные сосуды — пробирки, банки и т. п. с плотными (не ватными!)
пробками, например, стеклянными, чистыми корковыми или сделанными
из лучших сортов резиновыми, не имеющими запаха. В сосуд для увлаж-
нения воздуха следует положить несколько кусочков зеленой травы. Тра-
ву также следует выбирать без сильного запаха. Для связывания капель
росы на дно пробирки следует положить тонкий слой (не более 1 см)
песка или кусочки сухой фильтровальной бумаги. Надо помнить, что при-
чинами гибели блох бывают: 1) высыхание, 2) избыток влаги (роса),
3) вредные запахи, 4) развитие плесени, 5) выделение аммиака и других
газов из гниющего субстрата. Поэтому надо сосуды через 2—3 дня про-
сматривать, открывать для проветривания и регулировать в них влаж-
ность, добавляя или вынимая траву. Веществ, способных загнивать,
должно быть там возможно меньше, чтобы обеспечить чистоту воздуха;
при этих условиях, несмотря на плотную пробку, блохи не страдают от
недостатка последнего. Наоборот, при ватных пробках они погибают го-
раздо скорее от высыхания.
1 В последнее время рекомендовано (П. С. Козлов) осенью собирать блох из
второго колена нор; для этого в первое колено вставляется палка, и земля вокруг
раскапывается, после чего из второго колена шлангом собирают блох.
15
Высокая температура ускоряет гибель блох. Поэтому желательно
держать их в прохладном месте — в леднике, подвале или хотя бы в яме,
вырытой в тени и хорошо укрытой. Обязательно беречь от прямых сол-
нечных лучей.
Если сборы производятся для фаунистического изучения и т. п., то
для консервирования собранных паразитов следует пользоваться
50—70° спиртом1. Формалин (1:20) ухудшает впоследствии заделку
блох в препараты (блохи из формалиновых сборов в препаратах тем-
неют вследствие плохого проникновения в них бальзама; кроме того,
в них иногда появляются черные зернистые включения). В крайнем же
случае блохи могут быть собраны и в сухом виде — в бумажные
пакетики.
Каждый сбор эктопаразитов, снятых с одного экземпляра хозяина,
должен укладываться в отдельную посуду: пробирку, пузырек и т. п.
Наиболее экономна упаковка в мелкие пробирочки, употребляемые для
оспенного детрита, или просто в обрезки стеклянной трубки, концы кото-
рых плотно закрываются ватой (следует избегать рыхлой ваты, из кото-
рой паразитов потом трудно извлекать). Такие трубочки складываются
в банку или бутылку с 50—70° спиртом.
Каждый сбор обязательно сопровождается паспортом (этикеткой),
написанным простым карандашом или тушью; этикетка вкладывается
в пробирку вместе с блохами. В паспорте должны даваться следующие
сведения: где, когда, с кого (или в чьем гнезде, норе) и кем произведен
сбор. Следует также указать обстановку, где пойманы зверьки и блохи,—
в поле, в лесу, в доме и т. п., а также отметить пол и возраст этих
зверьков. Если сбор был произведен с нескольких зверьков (например,
находившихся в одном мешке), обязательно должно быть отмечено,
сколько было зверьков.
ОБРАЗЕЦ ЭТИКЕТКИ
1/Х 1946 г.
Окрестности ст. Борзя
Тушканчик самец взрослый
№ 27, в степи
П. И. Иванов
Если хозяин не может быть точно определен на месте, то его необ-
ходимо взять для определения (в формалине 1 : 10 или в виде шкурки
с черепом), поместив его номер на этикетке.
В полевом журнале, который ведется при проведении сборов эктопа-
разитов, детально описываются места сборов, обследованные стации,
отмечаются численность животных, метеорологические данные и т. п.
Следует подчеркнуть, что для экологических выводов сбор можно
считать полноценным только при количественном учете собираемого ма-
териала и тщательной его этикетировке. Наиболее частой ошибкой яв-
1 Заметим, что при сборах членистоногих рекомендуют убивать их смесью, со-
стоящей из 97 частей 20° спирта и 3 частей эфира. Это обеспечивает фиксацию ножек
в расправленном состоянии. Однако хранить материал и в этом случае необходимо
б спирте крепостью не ниже 50—70°.
Приведем наиболее простой способ расчета потребного количества спирта и воды
для тех или иных разведений спирта (по Е. Н. Павловскому). Разведение необходимо
производить в измерительном цилиндре. Берется столько объемных единиц спирта,
сколько градусов должно быть в готовом спирте, и доливается дестиллированной во-
дой до объема, содержащего столько единиц, сколько было градусов в исходном
спирте. Например, для приготовления 50° спирта из 95° надо взять 50 мл спирта
и долить водой так, чтобы общий объем получился 95 мл.
15
ляется отсутствие учета тех животных, на которых паразитов не оказа-
лось; не имея данных о всем количестве обследованных животных, нель-
зя вычислить ни процента зараженных эктопаразитами животных (т. е.
встречаемости или показателя экстенсивности инвазии), ни среднего чис-
ла паразитов на одно животное (индекса обилия).
Снаряжение для производства сборов
Лэвушки, капканы и т. п.
Лопата
Сэвок садовый
Частый гребень или зубная щетка
Кисточка твердая, употребляемая для
клея
Тонкие кисточки для акварели № 1
или № 2
Пинцет энтомологический с узким кон-
цом
Пинцет мягкий (из тонкой стальной
ленты)
Пинцет длинный (гинекологический)
Посуда для сбора
Мешочки из плотной материи или боль-
шие мешки (наволочки) с завязками
Веревка
Белая ткань для расстилания
Банки, коробки, таз, кюветы
Шланг, обшитый фланелью
Выгребалка
Спирт 50—70°
Эфир или хлороформ
Формалин
Бумага для этикеток
Карандаш мягкий (не чернильный)
Ножницы
Хранить спиртовые сборы следует в пробирках, заткнутых ватой
и уложенных в банки с 60—70° спиртом, закрытые плотными, лучше все-
го притертыми, пробками. Эти банки должны храниться в темноте, так
как на свету кутикула блох со временем выцветает. Во избежание высы-
хания блох рекомендуется к спирту добавлять немного глицерина.
Микроскопирование блох и приготовление препаратов
Определение видов блох возможно как по живым экземплярам, так
и по заспиртованному материалу. Но легче разбираться в морфологии их
на просветленных препаратах. Тем не менее нет надобности весь сравни-
тельно однообразный материал просветлять и заделывать (монтировать)
в препараты; достаточно изготовить препараты только из некоторой части
материала (выбирая наиболее сохранные и чистые экземпляры), тща-
тельно их изучить, а затем вести определение уже знакомых видов без
изготовления препаратов.
Определение видов блох производится под микроскопом, причем при-
ходится пользоваться как слабым (в 70—100 раз), так и более сильным
(в 140—200 раз) увеличением, при котором рассматриваются отдельные
детали. Для этого нужно иметь объективы X 10 и X 20 и окуляры X 7
и хю.
Микроскопирование блох в живом виде имеет
большое значение при изучении вопросов физиологии и экологии блох
и при определении блох, предназначенных для бактериологического
исследования. Для быстрого просмотра рекомендуется закрепить живую
блоху под микроскопом, положив ее на предметное стекло и покрыв ку-
сочком другого тонкого предметного стекла, удерживающего блоху своей
тяжестью. Однако больше деталей, в особенности во внутренних органах
блохи, можно увидеть при исследовании их в воде под покровным сте-
клом. Пользоваться следует простой, не дестиллированной водой. После
просмотра блоха переносится мягким пинцетом (позволяющим брать
блоху без риска повредить ее) или кисточкой в пробирку, в которую
сверху вложен кусок фильтровальной бумаги для обсушивания блохи.
При длительном пребывании в воде блохи временно теряют подвиж-
ность, но это быстро проходит после их обсыхания. Для ускорения ра-
боты определение живых блох следует производить с помощником, на-
кладывающим блох на стекла. Сухих блох переносят со стекла в про-
бирку, стряхивая их через воронку.
2
Определитель блох
17
Исследуя живых блох следует убирать со стола все лишнее, чтобы
можно было легко найти блоху, если она спрыгнет с кисточки или со
стекла. Изучая блох, обладающих большой длиной прыжка, можно на
время усыпить их эфиром или хлороформом. Для ускорения работы по
определению большого количества живых блох существует специальный
прибор, механизирующий перенос блох под микроскоп и обратно [166].
Спиртовые сборы блох микроскопируются на пред-
метных стеклах, причем блоха должна быть полностью покрыта каплей
Рис. 6. Про-
бирка с ме-
таллической
крышечкой.
жидкости, а иногда для лучшего рассмотрения необходимо
бывает наложить и покровное стекло. Просмотр можно про-
изводить в 50° спирте, но лучше пользоваться глицерином,
смешанным пополам с 50° спиртом; при этом способе рас-
кладку блох на стекла может производить помощник. Про-
питанные глицерином блохи могут оставаться на стекле не-
сколько дней и даже недель, становясь при этом даже
несколько прозрачнее, что облегчает их определение. Эко-
номия времени специалиста, которое он истратил бы для
раскладки сборов на стекле, бывает очень значительной.
Если при лежании в глицерине некоторые поверхностные
тонкие структуры (край сегментов и т. п.) делаются труд-
но различимыми, то добавление капли 50—70° спирта вос-
станавливает хорошую видимость их.
Иногда возникает необходимость улучшить видимость
внутренних органов блохи; это достигается путем надавли-
вания на покровное стекло (например, при использовании
для определения строения внутренних гениталий). При ча-
стом повторении таких манипуляций можно облегчить ра-
боту, используя пружинки из стальной проволоки, надевае-
мой на объектив и автоматически придавливающей покровное стекло.
Наконец, самый простой способ сделать спиртовые сборы блох
более прозрачными — это обработать их щелочью, как указано ниже при
описании техники изготовления препаратов.
Иногда для изучения морфологии отдельных сегментов применяют
отпрепаровку их от остальных сегментов блохи. Но для целей опреде-
ления видов это не требуется.
Можно рекомендовать следующую стандартную технику при-
готовления просветленных тотальных препаратов
блох. В маленькой пробирке с металлической крышкой (рис. 6) бло-
ха, оставаясь в этой же пробирке, последовательно проводится через
ряд сменяемых жидкостей.
1. Раствор едкой щелочи — NaOH или КОН 7% —одни сутки (кипя-
тить не следует).
2. Тщательно отмыть щелочь водой, сменяя ее несколько раз.
3. Спирт 50° — около 1 часа, но можно оставить на сутки и больше.
4. Спирт 70° — » 1 » » » » » » » »
5. Спирт 95° — » 1 » » » » » » » »
6. Спирт абсолютный — от 1 часа до суток.
7. Еще раз промыть свежим абсолютным спиртом.
8. Ксилол — от 1 часа до суток.
9. Гвоздичное масло — от 1 часа до суток.
10. Заделка в канадский или пихтовый бальзам, не особенно густой
(с добавлением ксилола).
Достаточное обезвоживание объектов устанавливается по быстрому
исчезновению мути в ксилоле после его наливания при смене спирта.
Если все же после заделки в бальзам в блохе обнаруживается некоторое
помутнение (мельчайшие капельки воды), то это объясняется чаще все-
го недостаточной отмывкой щелочи. Тогда препарат осторожно раскры-
18
вается при подогревании на пламени и покровное стекло снимается
вместе с блохой. Растворив канадский бальзам в ксилоле, отклеившуюся
блоху вымачивают в последнем, переносят в абсолютный спирт, и снова
проводят через все жидкости (начиная с воды и слабого спирта) и за-
делывают.
При описанном методе сравнительно редко наблюдается последую-
щее потемнение препарата из-за образования пустот, вследствие
пепроникновения канадского бальзама через покровы внутрь блохи. Если
все же такое потемнение произойдет, то для исправления приходится
препарат раскрывать и переделывать, как описано выше.
Предохранению препарата от потемнения способствуют следующие
моменты: 1) достаточное размягчение хитина в щелочи; 2) прокол хити-
на иглой, который делается с правой стороны, вблизи основания брюш-
ка, чтобы не повредить важных деталей; 3) пропитывание блохи перед
заделкой гвоздичным маслом или жидкой смесью канадского бальзама
с ксилолом; 4) употребление для заделки более жидкого бальзама, раз-
веденного ксилолом.
Накрывать препараты следует покровными стеклами малого размера
(примерно 6X9 или 9X9 мм), для чего обычные покровные стекла
режутся алмазным пером на четыре-шесть частей. Употребление ма-
леньких стекол значительно экономит расходование бальзама и облегчает
последующее раскрывание препарата, если в этом возникает необходи-
мость. При отсутствии алмазного пера можно ломать покровные стекла,
положив их на книгу и сильно надавливая толстым предметным стеклом,
вдоль края которого обычно и проходит линия разлома. При отсутствии
покровных стекол их заменяют кусочками прозрачной отмытой от жела-
тины фотопленки.
При обработке сильно загрязненных экземпляров блох можно попытаться перед
выниманием их из щелочи отмыть загрязнения путем взбалтывания окружающей
жидкости или осторожно дотрагиваясь кисточкой до насекомых. В это же время,
пользуясь размягчением тканей в щелочи, можно выправить положение ног, которые
иногда при фиксации блох спиртом загибаются кверху. Однако надо помнить, что
всякое неосторожное прикосновение может повести к поломке ног и щетинок, поэтому
ценные экземпляры лучше не подвергать этому риску.
Если блохи погибли от высыхания, то сегменты их находят один на другой, вся
блоха оказывается сильно сократившейся, что мешает изучению ее морфологии. Рас-
правление достигается одним из следующих способов.
1. После обработки щелочью и промывания в воде в течение 1 часа рекомен-
дуется положить блох на 30 минут в 1—2% раствор уксусной кислоты, которая долж-
на растянуть и уплотнить их, затем промыть водой и, как обычно, обезводить
спиртами.
2. В качестве расправляющего средства используется иногда 2% раствор пере-
киси водорода.
3. После размачивания блохи целесообразно положить ее на 1 час или больше
в 70° спирт и накрыть тяжелым стеклом, чтобы зафиксировать сегменты в растяну-
том положении. Этот же прием применяется иногда для расплющивания слишком
толстых экземпляров блох (самок с яйцами и т. п.).
При сильном наполнении темной кровью кишечника блохи можно сделать пре-
парат более прозрачным путем вымачивания насекомого в растворе нашатыр-
ного спирта, а затем в 2,% растворе перекиси водорода. Для просветления сильно
потемневших старых сборов вшей рекомендуется вымачивать их в 2Q% едком кали
в течение минимум 12 часов, после чего они тщательно отмываются в воде; возмож-
но, что этот метод иногда применим и для блох.
При переносе блох на стекло и нанесении канадского бальзама сле-
дует быть осторожным, чтобы не поломать щетинок, ножек и т. п. Надо
учитывать, что в абсолютном спирте и ксилоле они делаются весьма
хрупкими, особенно при слишком продолжительном пребывании в этих
жидкостях. Переносить блоху следует осторожно маленьким шпателем,
мягкой кисточкой или тонким пинцетом, не зажимая концов его. Повора-
чивать блоху в густом канадском бальзаме, а также надавливать на
покровное стекло не следует.
2*
19
можжевеловым,
C*> H3а моба
№200
3 W/-5O
3 Забайкалье
ОЯрес/п. с 66/дЛа
с C/teffus
Otaar/cfjj
7
4
Для ускорения и упрощения процесса изготовления препарата следует знать
еще следующие приемы.
1. Все стадии обезвоживания и просветления (от 50° спирта до ксилола или
гвоздничного масла включительно) можно заменить вымачиванием блохи в течение
одних суток в креозоте или феноле (acidum carbolicum liquefactum). При этом иногда
обходятся и без обработки щелочью, но темнопигментированные блохи получаются
при этом недостаточно прозрачными. Вообще описанная выше стандартная обработка
дает более прозрачные и отчетливые препараты.
2. Ксилол может быть заменен бензолом или толуолом, а гвоздичное масло —
кайепутовым, или, в крайнем случае, скипидаром.
3. Для заделки блох и в особенности их личинок может
быть применена также среда Фора (Faure), употребляемая для
заделки мелких клещей. Преимуществом этого метода является
его простота, однако многие важные для систематики блох мор-
фологические элементы в этих препаратах бывают хуже разли-
чимы, чем в бальзаме. Кроме того, препараты в среде Фора
недолговечны — в них через несколько месяцев появляются пу-
зыри воздуха. Для переделки таких препаратов их размачивают
в воде, а затем отмывают в 50° спирте. Удлинить срок суще-
ствования этих препаратов можно путем обмазки вокруг по-
кровного стекла (после затвердения среды Фора, на что тре-
буется 2 недели и более) канадским бальзамом. Заделка в сре-
ду Фора производится без всякой предварительной обработки
объектов — живых или зафиксированных в 50—70° спирте. По-
ложив объект в каплю среды на предметное стекло и накрыв
покровным, препарат следует слегка подогреть над пламенем,
не доводя, однако, до кипения; при подогревании объект не-
сколько просветляется, насекомые с мягкими покровами (напри-
мер, личинки блох) расправляются. Рецепт среды Фора: сме-
шать 25 мл дестиллированной воды, 10 мл глицерина, 100 г
хлоралгидрата до полного растворения, затем понемногу доба-
вить 15 г порошка гуммиарабика. Среда бывает готова через
2—3 дня. Хранить ее надо в темном сосуде или в темном месте.
При заделке блоха кладется всегда
на правый бок — для удобства сравне-
н и я. Монтировать следует по одному экземпляру на
каждое стекло. Рекомендуется для препаратов брать
возможно тонкие предметные стекла, позволяющие
> препарат с обеих сторон при достаточно сильном уве-
личении. На стекле тонким чертежным пером с помощью особых чернил
(20% водный раствор таннина, насыщенный спиртовый раствор фуксина
поровну) делается подробная надпись (рис. 7). Для стирания этих чер-
нил со стекла применяется раствор щелочи или соскабливание лезвием
безопасной бритвы. Можно делать надписи также тушью, но это менее
практично. Надписи можно закрепить, покрыв лаком из органического
стекла, растворенного в хлороформе (4—5%).
у frontopsytta
биса £е ata J ef#
det. Иванов
Рис. 7. Образен
надписи на препа
рате.
ОЧЕРК НАРУЖНОЙ АНАТОМИИ (ЭЙДОНОМИИ) БЛОХ
Настоящий очерк приводится для ознакомления с важнейшими мор-
фологическими признаками блох, необходимыми для их определения.
Тело блохи, сплющенное с боков, что, несомненно, является приспо-
соблением, облегчающим движение среди волосяного покрова млекопи-
тающих, удобнее всего рассматривать в боковом положении (в профиль)
как обычно и располагают блох в препаратах, для удобства сравнения
укладывая, как правило, на правый бок1.
Счет члеников (сегментов) ведется от места прикрепления органа.
Первый, или передний, членик называется основной или базальный. Наи-
более удаленный от места прикрепления, т. е. последний членик (или
вообще какой-либо орган или край органа) носит название апикальный
(вершинный), или терминальны/! (концевой); расположенный отступя от
вершины, но ближайший к ней — субапикальный. Поясним еще некото-
рые термины: спинной — дорзальный, брюшной—вентральный, боковой—
латеральный, краевой — маргинальный. Отсюда понятны и термины: суб-
маргинальный, субдорзальный, субвентральный, т. е. расположенный
немного отступя от соответствующего спинного или брюшного края; бли-
жайший к основанию органа или к передней части блохи — проксималь-
ный; обратное этому понятие — дистальный. Приставки: супра, супер —
над, суб — под, анте — перед чем-либо. Самка Q , самец с?.
Рассмотрим отдельно голову, грудь с тремя парами ног и брюшко.
Голова блохи (рис. 8) разделяется ямкой усика на переднюю
и заднюю часть. На лбу, т. е. переднем крае головы, бывает более или
менее развитый лобный зубчик или бугорок1 2. Различают лоб крутой
(рис. 25), пологий (рис. 26), конусообразно выпуклый (рис. 51, 55),
усеченный (рис. 23, 24) и т. д. Необходимо учитывать, что у многих
блох имеется половой диморфизм в отношении формы лба и вообще го-
ловы; обычно более крутой лоб бывает у самцов (рис. 9, 40).
По нижнему краю передней части головы различают ротовой край
(угол его с лобным краем называют предротовым, или преоральным,
углом) и щечный, или генальный, край (ближе к ямке усика). Угол
1 При просмотре непросветленного материала приходится иногда, наоборот,
класть блох на левый бок, если требующий рассмотрения орган лежит ближе к пра-
вой стороне и лучше просвечивает в таком положении; это относится особенно к семя-
приемнику у самок; копулятивная сумка, наоборот, лучше видна слева.
2 Зубчиком (protectum) следует называть особое хитиновое острие, выступающее
из лобного края и отделенное от него (рис. 54, Ж), а бугорком — простое возвышение
этого края (рис. 54, Е). При более сильном развитии зубчику дается иногда назва-
ние mucro, или listron. Резко выраженный излом лобного края можно называть лоб-
ным углом.
Заметим, что по Снодграссу [190] передний край головы у блох правильнее
было бы называть не лбом, а клипеусом, а собственно лбом, по его мнению, правиль-
нее считать отрезок края головы, лежащий позади антенны, который принято называть
теменем (vertex). Однако мы не видим необходимости отказываться от принятой
простой и общепонятной терминологии из-за маловажных соображений, основанных
на гомологизации тех или иных органов.
21
ап. зуб.
жя Й A t\\ m ‘ /*W k \ w' mp 1 Ллг 1 В J 1 Шт. IV m. Vm. Шт^Шт.^Шт^ к^УзУУк' v^//wj/i cm' И) IH \\ H '7/^/^yvn / II \| \ \\ />%z ^cm / jdyt^ i\у/ / / YT—HL £-- *<>‘*^^— J 2yy^_23 Vcm. L^jT\7 т\ Шет. 1 X ЦгадХ jb I— VIIIm— тергиты по порядку;/— V—чле- /Й Wk ники задней лапки,/—VIIIст,—стерниты; yflJxV4 зуб. — лобный зубчик; ус. — усик (антен- чМСЕЧ^ на)’’ мк' п'—максиллярные пальпы; мк. — ШП^\ максиллы; хоб. — хоботок; пр. — спинка 1 JBnk х передней груди (пронотум); кт. — гре- /LukП ^ень (ктенидий); мез. — спинка средпе- гЙ\ груди (мезонотум); мет. — спинка задне- ilrvS^ груди (метанотум); ап. зуб. — апикаль- L5jk ><2/7 ные зубчики тергитов; пиг. — пигидий; IiWk впереди от пигидия изображена стигма 'лЖ тергита (на других сегментах стигмы /йп /Р" изображены в виде маленьких овалов); кА ц. — церки; ан. с. — анальный сегмент; сем. — семяприемник. Членики задней ОК У ноги: косса—тазик или ляжка; тр.— чя£Ду\ вертлуг (трохаитер); бед. — бедро; гол. — голень.
Рис. 8. Самка блохи Ceratophylhts tesquorum.
между щечным краем и ямкой усика образуется щечной лопастью. В зад-
ней части головы различают темя и задний край головы. В ямке усика
с каждой стороны помещается усик, состоящий из трех члеников, или
антенна, причем на третьем членике, носящем название булавы,
в свою очередь более или менее ясно намечается деление на ряд члени-
ков (обычно до 10, включая узкий членик — petiolus — соединяющий бу-
лаву со вторым сегментом усика). Деление булавы на членики может
быть намечено вокруг всей булавы (рис. 9, 40) или только с одной сто-
Рис. 9. Голова и пронотум самца Oropsylla silantiewi.
А — с усиком, лежащим в усиковой ямке, и Б — с поднятым усиком; т, ц. — центральная
трабекула.
роны ее (рис. 21). На этих члениках можно заметить небольшие ямки
органов обоняния. Форма булавы также бывает у разных групп блох
весьма различной — от круглой (рис. 21) до более вытянутой (рис. 56).
Различают еще так называемую асимметричную форму булавы, при ко-
торой базальные членики с одной стороны булавы листообразно расши-
рены (рис. 27, 28). При наличии усиков удлиненной формы ямка усиков
распространяется и на передний сегмент груди. Это наблюдается у сам-
цов, у которых булава чаще имеет вытянутую форму, напоминая еловую
шишку (рис. 9). У самцов нередко усики не лежат в ямке, а подняты
кверху (рис. 9), с чем связана наблюдающаяся у них изогнутая форма
первого членика усика. Ямки усиков у самцов обычно распространяются
выше по направлению к темени, хотя и у самок многих родов блох эти
23
ямки или бороздки, тянущиеся от них (межусиковая бороздка), соеди-
няются на темени.
Таких блох, у которых есть развитая межусиковая бороздка (рис. 51, 55, 70),
разделяющая голову (caput fractum) на обособленные переднюю и заднюю части,
называют «раздельноголовыми» (fracticipita) в отличие от имеющих «цельную» го-
лову (caput integrum — integricipita — рис. 27, 43). Еще недавно «Fracticipita» и «1п-
tegricipita» рассматривались как основные подразделения в систематике блох, но В на-
стоящее время стало ясным, что это деление не имеет систематического значения,
так как среди близко родственных видов блох встречаются оба указанных типа
строения головы, а также и тип промежуточный между ними (caput semifractum),
когда межусиковая бороздка не развита, но вместо нее имеется шсв (рис. 21, 61).
Следует еще отметить, что передний край усиковой ямки образует у некоторых
блох тонкую пластинку, покрывающую переднюю часть этой ямки и часть усика
(рис. 21—28).
Установлено, что у самцов усики несут функцию не только органа чувств, но
служат и для поддержания самок при коитусе; поднимаясь над головой, они захваты-
вают основание брюшка самки. В связи с этим у самцов многих видов блох имеется
на темени продольный желобок — теменная ямка, куда попадает киль брюшка удер-
живаемой самки. Эта ямка видна при рассматривании самцов в обычных препаратах
(сбоку) в виде полосы вдоль теменного края; чем глубже теменная ямка, тем шире
эта полоса (рис. 29,Л—Б). Глубина теменной ямки используется иногда в качестве
видового признака блох. У самцов некоторых блох этот желобок продолжается вдоль
спинки грудных и брюшных сегментов (например, у Chaetopsylla mirabilis).
Перед ямкой усика помещается глаз, который бывает развит весь-
ма различно у разных родов и видов блох. Различают большие темно-
пигментированные глаза (рис. 21, 22), глаза уменьшенного размера
(рис. 40, 44), глаза с развитой верхней пузыреобразной частью и темным
нижним придатком (рис. 45—49, 53; иногда такие глаза кажутся двой-
ными— рис. 48, 49), глаза полурудиментарные (рис. 54, А—Б—В—Г;
60) и рудиментарные (рис. 61, 69), до едва заметных или отсутствующих
(рис. 50; 54, Д; 56).
Кроме того, на поверхности передней и задней части головы можно
заметить несколько небольших круглых «органов Удеманса», являющихся
окончаниями каких-то чувствительных нервов (рис. 21, 22 и др.).
Различные кутикулярные выросты, имеющиеся на поверхности голо-
вы и других частей тела блох, носят разное название в зависимости от
величины и формы. Тонкие иглообразные выросты называют волоска-
ми и щетинками, толстые — шипами, шпорами, плоские вы-
росты — зубцами, зубчиками (иногда они неудачно называются
также иглами). Ряды зубцов образуют гребни (ктенидии). Все эти
элементы «кутикулярного вооружения» блох обычно направлены кзади,
что облегчает движение блох среди шерсти. Особенности развития и рас-
положения щетинок (хетотаксия), а также гребней и других выростов
и их число во многих случаях представляют весьма важные системати-
ческие признаки, однако надо всегда учитывать также значительную
изменчивость (вариабильность) в числе тех или других щетин или зуб-
цов у каждого вида блох.
На голове блох различают ряды щетинок, идущие косо (снизу и спе-
реди, вверх и назад) и в разной степени развитые у разных видов и ро-
дов блох (рис. 45, 47, 69, 70 — примеры хорошо развитых рядов; рис. 21,
43, наоборот,— неразвитых). В передней части головы наиболее постоян-
ным является глазной ряд, идущий непосредственно перед глазом
и состоящий обычно из трех (реже из четырех или одной-двух) щетинок.
Верхняя щетинка этого ряда называется глазной и расположена то на
уровне верхнего края глаза (рис. 42, 44), то выше (рис. 45) или ниже
его (рис. 21); расположение этой щетинки часто является важным при-
знаком. Нижняя щетинка глазного ряда называется ротовой или максил-
лярной. Перед глазным рядом бывает лобный, или фронтальный,
ряд, который иногда совсем отсутствует (рис. 43), или в нем имеются
лишь единичные верхние (рис. 42) или нижние (рис. 41) щетинки; тогда
21
Рис. 10. Наружные ротовые
органы блохи Vermipsylla alacurt
(вид сзади).
эф. — непарный колющий орган; л. —
парный колющий орган; гб. — нижняя
губа; г. п. — губные пальпы; мк. —
максиллы; мк. п. — максиллярные
пальпы.
его называют неполным, или рудиментарным, в отличие от полного ряда,
когда ряд щетинок идет от верхней части ямки усиков до нижнего края
головы (рис. 45, 47, 69, 70). Иногда между глазным и фронтальным ря-
дами сидят единичные вставочные щетинки (рис. 47; 54, А—Б—В—Г);
если при этом фронтальный ряд придвинут к переднему краю лба
(рис. 46), то в этих случаях его называют иногда префронтальным рядом
или говорят о переднем и заднем фронтальных рядах. Иногда отмечается
наличие единичных щетинок, сидящих на щечной лопасти ниже (и сзади)
глаза (рис. 33, 34, 56).
В задней части головы различают два
(иногда три) теменных и задне-
крайний ряд щетинок. Нижняя сильная
щетинка последнего носит название угловой;
иногда она сопровождается добавочной ще-
тинкой. Разные степени развития этих рядов
см. на рис. 40, 44, 45, 56, 69.
Число щетинок в теменных рядах часто учиты-
вается в систематике; при описании иногда приво-
дится сокращенное обозначение этих данных; напри-
мер, фраза: «теменные ряды щетинок: 0(1) -{-2»
обозначает, что в первом ряду щетинок обычно нет
(или редко бывает одна щетинка), а во вторОхМ ряду
имеются две щетинки.
На голове блох (обычно на лбу) могут
быть, кроме щетинок, еще толстые темные
шипы (рис. 46, 51, 52). Большое значение
в систематике блох имеет наличие на голове
плоских выростов — зубцов, образующих бо-
лее или менее развитые ктенидии. В за-
висимости от места расположения эти кте-
нидии носят различные названия: вдоль
всего нижнего края головы — ротовой
(рис. 25, 60); у предротового угла — пред-
ротовой (рис. 75, 76); по щечному краю—
щечный (рис. 51, 65—68); перед ямкой уси-
ка по ее переднему краю — усиковый
(рис. 55) и т. д.
Наружные ротовые органы
блохи (рис. 10, 23, 24) состоят из сле-
дующих частей, названия которым разные
авторы дают не всегда одинаковые (мы при-
водим как более обоснованные, так и неко-
торые другие распространенные названия).
1. Непарный колющий орган — надглоточник, или эпифаринкс (дру-
гие его названия: верхняя губа, иногда неправильно — гипофаринкс),
представляет узкий длинный стилет.
2. Парный колющий орган — лацинии (верхние челюсти, манди-
булы). Они имеют также форму узких длинных стилетов. На большем
или меньшем протяжении от своего конца они покрыты зубчиками и слу-
жат блохам для прокалывания и пропиливания кожи хозяина, а у не-
которых видов — и для длительного прикрепления к коже; тогда они
бывают развиты особенно мощно (рис. 23, 24). Все вместе три колющих
органа образуют трубку, по которой из ранки насасывается кровь. Для
насасывания крови служат пищевой и глоточный насосы, располагаю-
щиеся по ходу пищеварительной трубки в передней части головы.
3. Непарная нижняя губа (labium), имеющая два членистых отростка
(так называемые губные щупики или пальпы), которые, складываясь,
служат как бы футляром для колющих органов — надглоточника и ла-
25
циний, и образуют хоботок (rostrum). У блох с мощными лациниями,
прикрепляющихся надолго к коже хозяина, губные пальпы могут быть
недоразвиты.
4. Две плоские нижние челюсти, или максиллы, имеют обычно тре-
угольную форму и несут членистые максиллярные щупики, или пальпы,
выступающие вперед и вниз из-под предротового угла головы и наиболее
заметные из всех ротовых органов.
5. Непарный короткий подглоточник (гипофаринкс), трудно разли-
чимый на тотальных препаратах. Через него проходит выводной проток
слюнных желез, из которого слюна изливается к основанию колющих
органов.
Длина хоботка и колющих органов, количество члеников, из которых
состоят губные пальпы, форма максилл и длина члеников максиллярных
пальп используются как систематические признаки блох.
Рис. 11. Схема эндоскелета головы (вид сверху).
А — Amphipsylla, т. ц.—центральная трабекула; в. т. — передние ветви; ям. ус. — дно ямки
усика; гл.— глаза; Б—Xenopsylla, ямки усиков соприкасающиеся: В — Coptopsylla, ямки
усиков неглубокие, центральная трабекула отсутствует.
Во многих случаях более длинный хоботок бывает у блох, паразити-
рующих на более крупных животных (на копытных — Vermipsylla,
рис. 35, 36; на медведях — Arctopsylla; на сурках и крупных сусликах —
Oropsylla, рис. 40; Rostropsylla и т. д.), но, конечно, это правило не яв-
ляется абсолютным.
Внутри головы на просветленных препаратах просвечивают внутрен-
ние части головного хитинового скелета (эндоскелета), из которых для
целей систематики необходимо знать так называемые передние ветви
тентория, которые имеются у многих родов блох. Они находятся в перед-
ней части головы по одной с каждой стороны и заметны в виде тонкого
изогнутого тяжа, начинающегося от боковой стенки головы около средней
глазной щетинки и уходящего кзади, скрываясь за глазом (рис. И, А, 45,
52 и др.). У некоторых блох также внутри передней части головы бывает
заметен темнопигментированный рычаг (базальный склерит или аподе-
ма) колющих ротовых органов (рис. 56).
Из других элементов внутреннего скелета головы следует отметить
так называемую центральную перекладину (trabecula centralis), проходя-
щую поперек головы от одной усиковой ямки к другой (рис. 11, А).
В обычных препаратах она бывает видна в области ямки усика, выше
глаза в виде небольшого темнохитинизированного круглого пятна (рис. 9).
У некоторых блох усиковые ямки очень глубоки и соприкасаются одна
с другой своим дном (рис. 11, Б), тогда на месте трабекулы видно боль-
шое светлое пятно. Иногда трабекула совсем отсутствует (рис. И, В).
26
Грудь состоит из трех сегментов (рис. 8, 21, 27, 28, 57): передне-
средне- и заднегруди (pro-, meso- и metathorax). В каждом из грудных
сегментов различают спинное полукольцо (пронотум, мезонотум и мета-
нотум) и грудно-боковые пластинки, или стерноплевры. Каждая стерно-
плевра теоретически состоит из соединенных или слившихся между собой
хитиновых пластинок; ввиду разной степени развития и слияния этих
пластинок они то более, то менее различимы на разных сегментах груди.
При рассматривании блохи сбоку на переднегруди мы видим обычно
только одну нерасчлененную пластинку — плевру. На среднегруди бывает
заметна стернальная часть и нерасчлененная плевра; вертикальный вну-
тренний тяж (рис. 27, 28, 57), пересекающий эту плевру у многих родов
блох, спаян с плеврой только своими концами и не делит пластинку на
две части. На заднегруди, кроме названных частей, различают еще эпи-
стерн и эпимер Г
Эпимер заднегруди (метэпимер, рис. 8, 21, 57), налегающий на пер-
вый сегмент брюшка, называют иногда крыловидной пластинкой; однако
эту пластинку не следует рассматривать как рудимент крыльев, которые
у блох совершенно отсутствуют. У куколок блох такие рудименты отме-
чаются в виде особых бугорков, находящихся по бокам среднегруди.
Вооружение грудных сегментов рядами и группами щетинок также
используется для систематики, но особенно важным признаком являются
гребни, имеющиеся у многих блох на спинке по заднему краю сегмен-
тов — чаще всего на пронотуме (рис. 25, 40 и др.) и иногда на метано-
туме. При описании таких гребней число зубцов подсчитывается обяза-
тельно с обеих сторон тела; для этого следует посмотреть блоху с обеих
сторон, так как гребень не всегда лежит строго симметрично. Кроме
того, отмечается длина зубцов, которые бывают «нормальной» длины
(рис. 51) и «укороченные» (рис. 50). Часто длину зубцов (дорзальных)
сопоставляют с длиной сегмента, на котором они сидят, измеряя его
вдоль дорзального края; при этом измерении границей между зубцами
и сегментами следует считать середину темной полосы, лежащей у осно-
вания зубцов.
Следует обратить внимание на то, что гребень переднегруди (проно-
тума) состоит из большего числа зубцов у тех видов блох, которые пара-
зитируют на птицах. Например, птичьи Ceratophyllus и Frontopsylla
имеют в гребне всегда более 24 зубцов, часто до 30 и больше, тогда как
у представителей тех же родов, живущих на млекопитающих, бывает, как
правило, лишь до 20—22 зубцов.
1 К сожалению, различные авторы по-разному трактуют гомологию частей груд-
ных склеритов и по-разному используют эти наименования [188 (5), 181, 178, 161,
171, 23, 190]. В особенности это касается той пластинки, которая располагается
между метанотумом и стерноплеврой заднегруди и бывает иногда полностью слита
то с метанотумом (например, у Micropsylloides и некоторых отсутствующих у нас
родов блох), то со стерноплеврой (у Synosternus — рис. 28, Eopsylla рис. 57, и др.).
Однако чаще всего она бывает отделена от метанотума внутренним хитиновым тя-
жом, а от стерноплевры — настоящим швом (рис. 21, 27). Эта пластинка имеет боль-
шое значение в систематике. В литературе она упоминается под разными названиями:
«эпистернум», «супраэпистернум», «боковая (вентролатеральная) доля метанотума»;
последнее название, повидимому. наиболее обосновано, но мы сохраняем более при-
нятое название «эпистернум» или «эпистерн» (метэпистерн), используя его только
как условный термин (не затрагивая вопросов гомологии склеритов груди).
На русском языке нет еще установившейся терминологии грудных склеритов;
изображая русскими буквами латинские названия, оканчивающиеся на «ит», их пи-
шут иногда без изменений (мерум, стернум, нотум и т. д.), иногда же придают им
русские окончания — то мужского, то женского рода (парамер, или парамера, и т. п.).
Слово pleura изображается то как плевра, то как плейрит и т. п. Мы полагаем, что
пора придти к единой русской номенклатуре и предлагаем пользоваться преимуще-
ственно мужскими окончаниями слов (парамер, эпистерн и т. п.), которые более при-
вычны. Слово «плевра» удобнее, чем плейрит; слово «нотум», повидимому, придется
сохранить, но при этом допустить его склонение по правилам русской грамматики —
«от нотума», «на нотуме», «с вотумом» и т. д.
27
У многих блох на мезонотуме, а иногда и на пронотуме при отсутствии гребней
отмечается наличие их зачатков в виде бледных тонких острых «щетинковидных вы-
ростов» (псевдосеты) близ заднего свободного края сегмента, на внутренней его по-
верхности. Эти выросты отличаются от настоящих щетинок тем, что у их основания
нет характерного для щетинок кольца (сочленения). На метанотуме у многих блох
вместо гребня имеются единичные короткие апикальные зубчики, такие же, как
и на тергитах брюшка.
К членикам груди прикрепляются три пары ног (рис. 8), из которых
задняя сильнее развита и играет главную роль при прыжке. Нога состо-
ит из нескольких сегментов; считая от груди, они носят названия: кокса
(тазик, ляжка — соха), вертлуг (trochanter), бедро (femur), го-
лень (tibia), пять члеников лапки (tarsus); последний из члеников
лапки — пятый — несет коготки. Элементы кутикулярного вооружения
ног — щетинки, шипики, шпоры — часто используются в качестве отли-
чительных признаков блох; принято обращать особое внимание на во-
оружение задней ноги.
На задней ляжке имеет большое значение группа или косой ряд коротких ши-
пиков, сидящих у некоторых блох на внутренней поверхности этого членика (у Хепо-
psylla и др.— рис. 346, Л—Б). У многих ви-
дов они замещены тонкими щетинками
или волосками (рис. 346, В—Г). Высказы-
валось (не всеми разделяемое) предполо-
жение, что эта группа шипиков может слу-
жить в качестве органа, производящего зву-
ки при трении о боковую поверхность осно-
вания брюшка; соответственно этому, на
поверхности метэпимера у некоторых блох
можно отметить наличие небольшой пло-
щадки с особой шероховатостью или во-
лосатостью покрова (striarium); однако во-
прос о функции указанных образований по-
ка совершенно не изучен.
Fla бедре различают: по дорзальному
краю 1 ряд щегинок и здесь же с наруж-
ной стороны бедра близ вершины имеются
более или менее толстые дорзальные («ко-
ленные») шпоры. Щетинки, расположен-
ные у основания бедер (близ вентраль-
ного края), носят название основных, или
базальных, а близ вершины — субапикаль-
пых вентральных. На боковой поверхности бедер как с внутренней, так и с наружной
стороны почти параллельно вентральному краю бывает расположен боковой, или
субвентральный, ряд щетинок.
По’ заднему краю голени находятся крепкие щетины — так называемые дорзаль-
ные шпоры, сидящие обыкновенно группами пли пучками, по две-три в группе
(рис. 347, А—В; 348, Д, Б); иногда они образуют правильный гребневидный ряд
(рис. 347, Б). На наружной и внутренней поверхностях голени щетинки часто рас-
полагаются продольными рядами.
В старых работах по систематике блох большое значение придава-
лось сравнительной длине члеников лапок (tarsus) по отношению друг
к другу. В дальнейшем выяснилось, что этот признак может быть целе-
сообразно использован для отличия видов лишь в немногих случаях.
Зато большое значение имеет вооружение члеников лапки, в частности,
длина апикальных (т. е. расположенных на вершине) и субапикальных
щетинок II членика задних лапок (рис. 348, Г; 349, В, Г). Эти щетинки
бывают особенно длинными у тех видов блох, которые живут в песчаных
районах (псаммофилы), у многих паразитов песчанок и т. д. Повиди-
мому, такие щетинки облегчают передвижение по песку.
Особенно большое систематическое значение имеет вооружение
пятого членика лапок (рис. 12), которое рассматривается как
один из главных родовых признаков блох. На этом членике надо разли-
чать, кроме коготков, несколько (3—6) пар боковых щетинок, боковые
Рис. 12. Пятый членик лапок.
А — Ccratcphyllus. Пять пар боковых щетпгок
и пара прнк< гтевых шипиков; Б — Dasypsyllus.
Третья пара боковых щетине к передвинута на
подошву. В — Myoxopsylla. Первая (основная)
пара боковых щетинок передвинута на подошву.
1 Дорзальный край ноги — это край, обращенный (при вытянутой вниз ноге)
кзади, а вентральный — кпереди.
волоски, подошвенные щетинки, подошвенные волоски, прикогтевые ши-
пики. Последние в количестве от одного до нескольких расположены на
подошве1 вблизи коготков (рис. 351, А, Б). Что касается подошвенных
щетинок, расположенных обычно парами, то их рассматривают как сбли-
женные между собой и передвинутые на подошву пары боковых щети-
нок; чаще бывает передвинута на подошву первая, основная пара
(рис. 12, В), реже — третья (рис. 12, Б).
Хитиновый покров брюшка блохи состоит из ряда черепицеобразно
расположенных сегментов (рис. 8). Спинное полукольцо каждого сегмен-
та носит название т е р г и т а, а вентральное полукольцо — стернита.
Хитиновые пластинки (склериты) всех сегментов соединяются интерсег-
ментальной перепонкой, или мембраной. Та часть склерита (задний сво-
бодный край), которая черепицеобразно налегает на следующий сегмент,
носит название колларе (collare— воротничок).
Первый сегмент не имеет стернита (его место занято метэпимером),
вследствие чего стернит второго сегмента носит название основного, или
базального, стернита. Сегменты с третьего по шестой называют средни-
ми, а начиная с седьмого сегменты являются модифицированными, т. е.
сильно измененными по форме и вооружению по сравнению с прочими.
Как правило, каждое тергальнос или стернальное полукольцо представляет цель-
ную пластинку, т. е. не разделено на правую и левую половины. Однако в некото-
рых случаях сегмент бывает разрезан полностью или частично вдоль дорзального или
вентрального края па две части. Полное разделение сегментов на правую и левую
пластинки («двойные склериты») наблюдается у тех видов, у которых брюшко при
созревании яиц очень сильно растягивается (рис. 37, А, Б). Полное или чаще непол-
ное разделение (т. е. образование глубокой узкой дорзальной или вентральной вы-
резки апикального края) наблюдается также на модифицированных сегментах.
В некоторых случаях учитываются особенности формы средних
брюшных тергитов, поскольку они могут быть разной ширины, иметь
разные очертания заднего края (рис. 353, А—А4). Следует отметить, что
нижний край III—VI тергитов налегает на верхний край соответствую-
щего стернита; верхний край II стернита, а у самок также и VII стерни-
та, наоборот, налегает на тергит.
На тергитах и стернитах учитываются вертикальные ряды щетинок,
а иногда перед этими рядами — и неправильно разбросанные, так назы-
ваемые «передние добавочные», обычно более мелкие, щетинки. Когда
учитывается число щетинок на том или ином склерите, то для удобства
их подсчитывают обычно лишь с одной стороны блохи, обращенной к гла-
зу наблюдателя, хотя иногда дается цифра, подсчитанная для обеих
сторон, что должно быть в этих случаях оговорено.
Число рядов на каждом из тергитов различно, от одного до несколь-
ких. При этом один из рядов (задний или субмаргинальный, т. е. бли-
жайший к заднему краю) состоит из более крупных щетин и называется
главным; он в большинстве случаев хорошо развит, тогда как передние
ряды бывают неправильные, неполные и т. п.
На средних стернитах брюшка имеется обычно один ряд из сильных
щетинок и иногда перед ними—более мелкие добавочные, которые обычно
не образуют правильных рядов. На основном (втором) стерните имеются
лишь немногочисленные вентральные щетинки и иногда несколько щети-
нок на боковой поверхности. В некоторых случаях характерным призна-
ком является особая склеротизация1 2 некоторых стернитов, например,
1 Подошвенной поверхностью этого членика условно считают ту поверхность,
в сторону которой загнуты боковые щетинки и коготки.
2 Термин «склеротизация» (того же корня, что «склерит») употребляется для
обозначения уплотненных, утолщенных и потому более темных участков кутикулы
и эндоскелета блохи; часто употребляемый термин «хитииизация» признается не со-
вЬем подходящим, так как основные свойства кутикулы — ее твердость, упругость,
непроницаемость — зависят от присутствия кутикулина и других веществ, а не хи-
тина [160].
29
вертикальный, широкий более темный тяж с расплывчатыми краями
у некоторых видов Amphipsylla, более заметный на VI стерпите
(рис. 244).
У многих блох на тергитах имеются по их апикальному, т. е. свобод-
ному заднему краю, короткие апикальные зубчики (рис. 8),
а иногда целые гребни, подобные тем, с которыми мы встречались при
рассмотрении тергитов груди. Если зубья этих гребней сидят не по задне-
му краю членика, а отодвинуты от края к середине членика и представ-
ляют собой лишь расширенные и уплощенные щетинки главного верти-
кального ряда этих члеников, то в таком случае говорят о ложном гребне
(например, Myodopsylla — рис. 76). Зубчики и гребни на тергитах бы-
вают у некоторых родов блох возле спины, у других — на боках брюшка.
Рис. 13. Схема последних сегментов брюшка самцов.
А — Neopsylla; Б — Ctencphthalmus; В — Ceratophyllus.
Часто используются для целей систематики так называемые
стигмы, расположенные на боковой поверхности тергитов и представ-
ляющие собой ямки, в которых открываются дыхальца, т. е. отверстия
трахеальных трубок. С каждой стороны тела имеется две грудных стигмы
и восемь брюшных. Стигмы различаются по форме, размеру и положе-
нию (рис. 319, Б; 320, Б; 353, А—Л4); часто учитывается также количе-
ство щетинок, сидящих на тергитах ниже стигмы.
Важным признаком для распознавания семейств блох является вы-
сота положения первой брюшной стигмы (которая находится у верхнего
края метэпимера). У блох семейства Pulicidae передний край эпимера
заднегруди идет более или менее вертикально кверху, и стигма располо-
жена высоко над уровнем верхнего края метэпистерна (рис. 21, 27).
В семействе Ceratophyllidae, как и в некоторых других, передний край
метэпимера идет от плевры косо кзади и кверху и стигма лежит не вы-
ше или немного выше верхнего края метэпистерна (рис. 8, 57).
Особенно характерные отличительные признаки обнаруживают м о-
дифицированные сегменты (рис. 13, 14, 15).
Верхне-задний угол VII брюшноготергита иногда имеет в той
или иной степени развитый выступ (рис. 257, А; 258, А), на котором
у большинства видов находятся сильные предпигидиальные ще-
тинки, которые защищают так называемый пигидий (pygidium, sensi-
lium — рис. 14; 15; 256, Б; 264) —округлый щиток с чувствительными
органами и волосками (трихоботриями), расположенный на заднем кон-
це брюшка в дорзальной вырезке VIII тергита. Число и степень развития
предпигидиальных щетинок относятся к довольно ценным отличительным
признакам блох; у самцов зачастую их меньше, чем у самок. Особен-
ности формы и размера пигидия, который бывает выпуклым (рис. 264)
или невыпуклым (рис. 15; 111, А; 162), также являются важными при-
знаками блох. Есть данные, указывающие на то, что пигидий является
30
органом, улавливающим движение воздуха; как известно, движения воз-
духа служат основными сигналами, по которым блоха узнает о при-
ближении хозяина или опасности и делает прыжок (анемотаксис).
Форма VIII брюшного тергита и расположение щетинок на
нем представляют довольно характерный систематический признак.
В дорзальной вырезке его мы видим пигидиальную пластинку. В этой же
вырезке у края VIII тергита расположена последняя стигма. Эта стигма
особенно сильно развита и бывает различной формы — узкая, воронко-
Рис. 14. Схема последних сегментов брюшка у блохи-самца.
VII и VIII т.—тергиты, VII—IX ст.—стерниты. В. в.—вертикальная ветвь IX стер-
нита: г. л, — горизонтальная ветвь IX стернита; бр.— брюшко IX стернита; яз.—
язычок IX стернита; ан. т. — анальный тергит; ан. ст. — анальный стерпит;
пи?. — пигидий; стиг. — стигма VII! тергита; к. — тело пологой клешни; д. — дор-
зальная пластинка половой клешни; м.— рукоятка пологой клешни; Р. — непо-
движный палец половой клешни; F—подвижный палец половой клешни или
дигитоид.
видная, напоминающая букву Y или Т (рис. 268, Б), или широкая, «по
душкообразная», покрытая мелкими волосками (рис. 157, Б; 264).
У самок VIII тергит покрывает большую часть заднего конца брюш-
ка, простираясь от самой спинки и до вентрального края. На нем
(рис. 111, А) находятся группы щетинок, сидящих над и под стигмой;
ниже их — боковые или промежуточные щетинки; затем иногда бывает
ряд щетинок вдоль задне-нижнего края — субмаргинальный или субвент-
ральный (рис. 264); несколько генитальных щетинок сидит на внутренней
поверхности свободного края тергита близ его заднего края (на
рис. 267, Б; 271, Б они изображены пунктиром). У некоторых видов
отмечается наличие резко выступающего угла по заднему краю этого
тергита ниже пигидиальной вырезки (рис. 115, Б).
31
У самцов развитие VIII тергита резко вариирует—у некоторых видов
он образует большие боковые лопасти, покрывающее следующие моди-
фицированные сегменты (рис. 13, В), тогда как у других он слаборазвит
(рис. 13, А и Б). Вооружение его также представляет иногда характер-
ные особенности, например, он может быть снабжен пучком длинных ще-
тин, напоминающих гриву (рис. 192). Кзади от стигмы на внутренней
поверхности VIII тергита у самцов некоторых видов (птичьи Ceratophyl-
Рис. 15. Схема последних сегментов брюшка и гениталий блохи-самки.
ц. — церк; сем. — семяприемник; с. п. — семенной проток; сл. п. — слепой проток;
к. с. — копулятивная сумка; к. п.—проток копулятивной сумки; в. — влагалище. Осталь-
ные обозначения см. на рис. 11.
lus и др.) наблюдается на определенном участке большое количество
мелких шипиков (поле шипиков — area spiculosa — рис. 126, Л).
Что касается модифицированных стернитов, то у самок из них хоро-
шо развит и имеет большое систематическое значение лишь VII стер-
н и т, очертания заднего (апикального) края которого бывают часто весь-
ма характерны для данного вида блохи (рис. 147, А; 199; 212, В; 249;
283, В). Различается этот край не всегда легко. Часто приходится про-
слеживать его очень внимательно, с большим увеличением, но без этого
определение самок многих видов невозможно. Нужно иметь в виду так-
же и вариабильность этого признака, благодаря чему приходится давать
изображения нескольких форм края VII стернита, характерных для дан-
ного вида (рис. 123; 147, В; 222 и др.). Различают следующие особенно-
32
ста этого края: по средней вентральной линии он бывает часто рассечен
вентральной вырезкой (рис. 16; 147, А; 160; конец вырезки отмечен звез-
дочкой) ; боковой край его иногда без всяких выступов и вырезок, верти-
кальный— косо срезанный (рис. 114; 240, В) или равномерно округлен-
ный (рис. 219, В). Часто на нем имеются выступы: боковой (рис. 157, В;
212, В; 249; 303), боковой и вентральный, или чаще субвентральный,
т. е. с небольшой вентральной вырезкой под ним (рис. 140, 160); иногда
имеются два боковых выступа: верхний и нижний (рис. 105, 264). Меж-
ду выступами образуются вырезки — боковая (рис. 199; 212, В), или
верхняя и нижняя боковые (рис. 105; 333, Д).
Боковой край VII стернита самок бывает иногда в каком-нибудь ме-
сте слегка утолщен, темнее пигментирован; это тоже приходится исполь-
зовать как признак (рис. 266, В; 268, В), но еще
Рис. 16. Ceratophyllus tur-
bidus. Самка. VII стер-
нит в развернутом виде
с вентральной (а) и бо-
ковой (б) вырезками.
важнее те склеротизованные тяжи и складки, ко-
торые иногда просвечивают сквозь край VII стер-
нита самки1 и бывают весьма характерны
(рис. 262, В; 264; 267, Б, и др.).
Следующие стерниты у самок блох развиты
слабо и особого внимания не привлекают:
VIII стернит обычно узкий и бледный, слегка
выступает под VIII тергитом (рис. 111, Л), или
просвечивает у его основания (рис. 262, В; 264
Рис. 17. Ceratophyllus tesquorum transvolgensis.
Самец. VIII стернит в развернутом виде.
и др.); IX стернит в большинстве случаев совсем слабо склеротизован
или замещен мембраной. Между VIII и IX стернитами располагается
влагалище (vagina — рис. 15).
У самцов модифицированные стерниты лучше развиты и заслужи-
вают большого внимания, кроме VII стернита, который обычно мало
отличается от средних стернитов.
VIII стернит самца бывает развит в различной степени — у одних
родов блох он широкий, у других—узкий (см. рис. 13). В последнем слу-
чае (рис. 13, В) в нем различают треугольную основную пластинку и уз-
кую, направленную кзади горизонтальную часть, на вершине которой
часто расположены концевые щетинки (рис. 108), а с дорзальной сторо- 1 * 3
1 Они относятся обычно к VIII сегменту. Так, например, бывает заметна тем-
ная склеротизация в основании VIII стернита, или склеротизация отдельных участков
VIII тергита, или стенок вагины.
3 Определитель блох 33
ны — тонкий прозрачный бахромчатый вырост разнообразной формы
(рис. 125, 145, 158, 166). В развернутом виде такой узкий стернит имеет
форму, изображенную, например, на рис. 17. У некоторых видов
VIII стернит вследствие недоразвития представлен только основной пла-
стинкой, которая может быть очень бледной, едва различимой — руди-
ментарный VIII стернит (рис. 113).
Между VIII и следующими стернитами соединительная мембрана у многих сам-
цов (Ceratophyllus и др.) образует так называемые интерсегментальные бахромчатые
лопасти (рис. 175) — нежные прозрачные разрастания и складки мембраны причуд-
ливой формы, покрытые мелкими волоскообразными выростами. Ю. Н. Вагнер свя-
зывает функцию этих лопастей с особым «Х-органом», лежащим у самцов некоторых
видов блох возле боковых углов основания VIII стернита (рис. 175). Х-орган пред-
ставляет овальную или шаровидную капсулу, густо покрытую изнутри волосками.
Ю. Н. Вагнер полагает, что в этом органе собирается секрет особых клеток, который,
быть может, распределяется затем по поверхности указанных лопастей и, испаряясь,
производит определенное действие на самку.
Начиная с IX сегмента, последние настолько модифицированы, что
вопрос о их количестве и расположении решается различно отдельными
авторами. До недавнего времени принималось, а большинство специали-
стов и теперь придерживается взгляда, что у блох имеется только 10
брюшных сегментов. Однако Ю. Н. Вагнером [195] разработана теория
о наличии 12 брюшных сегментов у блох, аналогично некоторым другим
насекомым. При этом часть последних сегментов, по его теории, руди-
ментарны, слиты с другими или совсем отсутствуют. По старой номен-
клатуре пигидий относился к IX тергиту; по вагнеровской теории он
должен быть отнесен к X; последний анальный сегмент считался X, по
Вагнеру — XII.
Вопрос о порядковом номере и счете последних сегментов усложняется нали-
чием у многих блох различных так называемых интерсегментальных склеритов
и лопастей. Так, позади VIII стернита самца у многих Ceratophyllinae наблюдаются
бахромчатые лопасти, у Amphipsylla — интерсегментальные вилочки (рис. 242, В;
243,А), у некоторых Neopsylla— настоящий, хотя и узкий склерит (рис. 315) и т. п.
Сложное строение IX стернита самца (у Ceratophyllinae, Wagnerinae и многих дру-
гих) тоже дает основание для сомнений о действительном числе отдельных склери-
тов брюшка блохи. Около пигиднального и анального склеритов у некоторых видов
также имеются особые бахромчатые выросты, тяжи или совсем обособленные так
называемые субпигидиальные и субанальные склериты, или тергоплевриты, при-
чем все подобные образования произвольно относятся разными авторами то к одному,
то к другому сегменту, в зависимости от чего может меняться и счет и нумера-
ция склеритов.
Мы полагаем, что в номенклатуру последних сегментов можно было
бы внести упрощение и единообразие, отказавшись от пользования по-
рядковой нумерацией этих органов и заменив ее словесной номенклату-
рой. Ниже мы приводим все необходимые для этого термины, но, учи-
тывая, что номерные обозначения (по 10-сегментной схеме) прочно во-
шли в литературу и практику, мы параллельно приводим и их Ч
П оловая клешня самца (хватательный или прикрепительный
аппарат, clasper—рис. 14), которая служит для удержания самки при
совокуплении, дает особенно ценные признаки для систематики блох. Ее
принято относить к IX брюшному сегменту.
В клешне различают: тело клешни (corpus), подвижный палец, или
дигитоид, неподвижный палец, рукоятку (manubrium), дорзальную пла-
стинку (называемую еще иногда рукояткой IX тергита), суставный бугор
(сочленовная ямка, acetabulum) — место сочленения тела клешни с по-
движным пальцем. На теле клешни, чаще всего на неподвижном пальце,
бывают щетинки; когда они сидят на суставном бугре, их называют
ацетабулярными. Иногда ацетабулярные щетинки находятся на специ-
альном выступе (рис. 183; 219, Л; 224, Л). Задний край тела клешни не-
редко образует особые выросты в области суставного бугра (рис. 87, Л;
1 Более подробные сведения о 12-сегментной схеме Вагнера интересующиеся
могут найти в «Пособии» И. Г. Поффа и В. Е. Тифлова [14], стр. 22—23.
34
262, A; 268, Л) или ниже его (рис. 89). На подвижном пальце, кроме
щетинок, бывают часто различные шипы, шпоры и т. д. (рис. 121, 139,
238, М—Н, и др.), а иногда особые отростки (рис. 106, 107). Форма
и вооружение клешни представляют собой один из самых характерных
видовых признаков блох.
У некоторых групп блох половая клешня построена по иному типу;
так, иногда имеется два подвижных пальца (рис. 80—85, Pi и Р2) и, кро-
ме того, пластинчатый вырост (Р3).
Ю. Н. Вагнер считает, что половая клешня является гомологом брюшных ко-
нечностей низших насекомых — гоноподой, и предлагает верхний из подвижных паль-
цев считать экзоподитом гоноподы, а нижний — эндоподитом. Тело клешни образует
базиподит. У большинства блох, как говорилось выше, имеется только один подвиж-
ный палец. Ю. Н. Вагнер считает его эндоподитом, основываясь на том, что он
укреплен на внутренней стороне тела клешни. В этом случае экзоподит считается
неразвитым или сросшимся с телом клешни; неподвижный палец тогда может быть
развит за счет экзоподита. Учитывая, что другие авторы до недавнего времени на-
зывали экзоподитом подвижный палец, мы полагаем, что удобнее пользоваться при-
водимыми выше условными терминами (дигитоид, отростки Рь Р2 и Рз, неподвиж-
ный палец), не затрагивающими спорных вопросов гомологизации.
Рукоятка и дорзальная пластинка, по Вагнеру, представляют части эндоскелета
и возникли в результате хитинизации тех тяжей, которыми клешня удерживалась
в своем положении. Степень их развития у разных видов блох бывает весьма
различной.
Так называемый IX стернит самцов, который лучше обозначить
термином субгенитальный стернит, представляет у многих
блох весьма сложное образование; нередко он как бы составлен из не-
скольких частей и всегда дает ценные для систематики признаки. Мы
называем его субгенитальным по той причине, что он всегда примыкает
снизу к половым органам самца, а иногда бывает тесно слит с ними. Он,
несомненно, входит в комплекс органов, участвующих в удержании самки
при совокуплении. Форма этого стернита в общем напоминает изогнутую
под углом ленту (рис. 14), вследствие чего в нем различают две основ-
ные части: 1) вертикальную ветвь, верхний конец которой налегает на
рукоятку половой клешни и сочленяется с последней у ее основания;
нижний конец этой ветви переходит в 2) вентральную, или горизонталь-
ную, ветвь, направленную косо кзади и кверху. Соединение вертикальной
и горизонтальной ветвей бывает у некоторых блох довольно сложным
(рис. 101, 113 и др.); это наблюдается обычно в тех случаях, когда
к IX стерниту прикрепляется один из тяжей полового аппарата. Гори-
зонтальная ветвь нередко в свою очередь делится вентральным синусом
(вырезкой) на дистальную и проксимальную доли (рис. 113; 144, Л).
Проксимальная доля (т. е. так называемое «брюшко» стернита) бывает
сильно вооружена щетинками (рис. 153, 179). Иногда характерный пучок
щетинок бывает и на вытянутом передне-нижнем углу («язычке») ди-
стальной доли непосредственно позади от вентрального синуса (рис. 128,
137)!. На всем многообразии форм субгенитального стернита останав-
ливаться нет возможности, и мы ограничимся только упоминанием
1 У многих блох (Ceratophyllinae и др.) дистальная, т. е. апикальная, доля
этого стернита имеет приспособление (в виде дугообразно загнутого продольного
ребра, или «рельсы» — рис. 155, А, или отверстия — рис. 157, А и т. п.), служащие, пови-
димому, для сцепления с одним из подвижных склеритов эдеагуса, именно с так на-
зываемым «крючком эдеагуса», близ основания которого в таких случаях имеется
темный склеротизированный выступ или «кнопка» («мыщелок»), входящий в соот-
ветствующее углубление IX стернита. Когда этот мыщелок сдвинут вниз, крючок
эдеагуса лежит вдоль апикальной доли IX стернита и скрывается за ней. Когда
мыщелок сдвинут вверх, вершина (конец) крючка выступает позади этого стернита.
Надо полагать, что дугообразная «рельса» IX стернита и подобные приспособления
служат ограничителями передвижений мыщелка, а следовательно, и всего крюч-
ка эдеагуса. *' - ’
На многих наших изображениях IX стернита нарисован этот мыщелок (хотя он
относится к эдеагусу); при этом следует помнить, что расположение его непостоянно.
3* 35
о тех случаях, когда этот стернит бывает построен проще (рис. 263;
286, А; 328) и отсутствует даже деление его на две ветви (рис. 80—85),
или когда он полурудиментарен и почти слит с гениталиями
(рис. 92, Л; 95).
Последний сегмент — анальный (telson, proctiger) обычно густо
покрыт щетинками, развитие которых иногда используется для диферен-
циации видов; форма анального стернита самца приобретает значение
важного группового признака — различают случаи, когда анальный стер-
нит раздвоен и намного превышает длину тергита (рис. 146, 162), и когда
он не заходит за тергит (рис. 109).
Рис. 18. Схема строения полового проводящего аппарата (фаллосома)
самца.
Эд. — эдеагус; энд.—эндофаллус; т. п.—тяжи пениса; в. т.—внутристенные тяжи; а. т.—
аподемальный тяж; а. э. — аподема эдеагуса; ш. а. — шейка аподемы; с. а. — суппорт
аподемы; т. э. — внутренняя трубка эдеагуса; д. п. — дорзальный парамер; в. п. — вен-
тральный парамер; к. э. — крючок эдеагуса. Вверху (Б) — поперечный разрез аподемы
эдеагуса.
У самок с боков анального тергита прикрепляются так называемые
«це р к и» (или «анальные стилеты» — cercus, или stylet — рис. 15; 264,
а также рис. 339,5, В, Г), цилиндрической или конусообразной формы
выросты, по одному с каждой стороны тела; на вершине каждого церка
сидят обычно сильные щетинки. У некоторых групп блох церки отсут-
ствуют (Vermipsyllidae).
Внутри брюшка у блох, в особенности на просветленных препаратах,
видны некоторые склеротизованные части полового аппарата
как у самцов, так и у самок. У самцов хорошо заметны длинные тяжи
полового аппарата, у многих спирально изогнутые (рис. 146; 169, Л, В),
входящие в состав большого и сложно построенного органа — фаллосо-
ма (рис. 18); особенности последнего из-за сложного строения еще не-
достаточно используются в систематике; только в последнее время на
них стали обращать большое внимание.
Фаллосом (проводящий, интромиттирующий орган — рис. 18) состоит из
апикальной наружной части — эдеагуса (aedeagus) и удлиненного внутреннего эндо-
фаллуса (endophallus), представляющего топкостенный мешок, не различимый в про-
светленных препаратах и заключающий в себе пенис и два выдвижных склеротизо-
ванных тяжа пениса; кроме того, в стенках этого мешка бывают заметны два обычно
более коротких внутристенных тяжа — вентральный и дорзальный; под эндофаллусом
идет еще один аподемальный тяж, задний конец которого бывает прикреплен или
к эдеагусу (рис. 18), или к IX стер ниту (рис. 14 и 146). От основания эдеагуса внутрь
брюшка над эндофаллусом идет склеротизованная широкая аподема эдеагуса (ман-
36
тия или футляр пениса); на рис. 18, Б, изображающем ее поперечный разрез,
видно, что аподема представляет жолоб с продольной перегородкой. Суженное место
аподемы называют ее шейкой. Место соединения аподемы с эдеагусом (суппорт или
мыщелок ее) представляет довольно сложное сильно склеротизованное образова-
ние, позади которого находится полость, называемая капсулой эдеагуса. От этой
полости кзади идет склеротизованная внутренняя трубка эдеагуса (трубка пениса)
нередко с воронкообразной концевой частью; через нее при коитусе выдвигаются
семяпроводящие тяжи пениса, проникающие в гениталии самки (в копулятивную
сумку). Апикальные лопасти эдеагуса называют парамерами или фаллитами (рис. 80;
88, А; 331); среди них различают дорзальную лопасть (дорзальный «колпак», «клюв»,
«палец» «коготь»), боковые или наружные лопасти («крылья»), вентральную лопасть
(вентральный «палец», «коготь»). Кроме того, часто имеется апикальный подвижный
склерит или крючок эдеагуса.
Следует учитывать, что все модифицированные сегменты и элементы
копулятивного аппарата самца являются подвижными органами и их
взаиморасположение не является постоянным. Отдельные детали этих
органов (дигитоид, парамеры, крючки) также могут менять свое распо-
ложение. Поэтому на рисунках важно не расположение, а форма отдель-
ных органов. Иногда при рисовании даже допускается некоторое пере-
мещение отдельных элементов, например, VIII и IX стернитов, для того
чтобы один орган не закрывался другим.
У самок особенно сильно склеротизован семяприемник (спер-
матека, receptaculum seminis — рис. 15), просвечивающий примерно
в области VII и VIII брюшных сегментов в виде небольшого колбообраз-
ного мешка. В нем различают более широкую часть — резервуар (тело
или головку) и более узкую — придаток (аппендикс, или хвост). У мно-
гих блох граница между резервуаром и хвостом семяприемника хорошо
заметна (рис. 111, А; 156, Б; 215, А), у других же — один постепенно пере-
ходит в другой (рис. 151, 201). Есть роды блох, имеющие два семяприем-
ника (Hystrichopsylla — рис. 325, Б; Coptopsylla и др.). Форма семя-
приемника часто является ценным родовым признаком, а иногда и видо-
вым. Например, различают резервуары вальковатой (рис. 144, Б; 147, Б;
171), лимонообразной (рис. 165, 168), бочкообразной (рис. 99, Б; 160,
190), почковидной (рис. 239, Б), грушевидной (рис. 111, А; 143, Б),
округлой (рис. 114; 119, Б; 215, А) формы и т. п. Тот конец резервуара,
где в него впадает семенной проток, называют привратниковым (вести-
булярным) отделом; он иногда бывает несколько обособлен и образует
тогда вестибулярную камеру (рис. 333, А). Отверстие семенного протока
бывает расположено симметрично в центре вестибулярного конца резер-
вуара (терминальное отверстие — рис. 197), или сдвинуто к вентральной
стенке резервуара (субвентральное или вентральное отверстие)
(рис. 239, Б; 249; 283, Б), или к дорзальной стенке (дорзальное отвер-
стие). Отмечают также соотношение длины резервуара и придатка, нали-
чие на конце придатка склеротизованной кнопки (рис. 111, А; 140) и дру-
гие особенности. Не следует только забывать о том, что семяприемник
подвижен, и потому очертания его меняются в зависимости от положе-
ния; кроме того, он может несколько сгибаться, а также и деформиро-
ваться при изготовлении препарата.
На удачных препаратах просвечивают у самки и другие менее скле-
ротизованные части полового аппарата (см. рис. 15): влагалище (vagina),
копулятивная сумка (bursa copulatrix), слепой проток (ductus obiuratus)1
и семенной проток (ductus seminalis), ведущий в семяприемник; часть
этого протока, начинающаяся от копулятивной сумки, бывает иногда рас-
ширена (pars dilatata — рис. 157, Б). Проток, соединяющий вагину и ко-
1 Возможно, что слепой проток представляет рудимент второго семенного про-
тока, оставшийся от исчезнувшего у большинства блох второго семяприемника. У не-
которых блох слепой проток совершенно отсутствует (рис. 154,А), у других он бы-
вает склеротизован и представляет ценный признак для распознавания этих видов
(рис. 147,5; 160; 171; 180,5).
37.
пулятивную сумку — копулятивный проток (ductus bursae copulatricis),
в большинстве случаев сливающийся с последней без ясной границы, ча-
сто бывает довольно сильно склеротизован и может представлять ценный
видовой или групповой признак (рис. 241, А—17), но опять-таки нельзя
забывать о его подвижности, некоторой гибкости и возможности де-
формации.
Из отдельных характерных форм копулятивной сумки можно отметить: спираль-
но изогнутую сумку (у Nosopsyllus— рис. 114), мягкую мешкообразную (у Citello-
philus— рис. 123; 126, Б) или подобную ей, но имеющую грубые склеротизованные
складки (у Callopsylla — рис. 129, А и Б\ 133), окруженную железистыми тканями
(у некоторых Megabothris— рис. 154,А). Копулятивный проток бывает простой скле-
ротизованный (у Neopsylla — рис. 312, А), разнообразно изогнутый (рис. 215, А,
241, А—77) и т. д. Иногда используется сравнение длины склеротизованной части
протока у разных видов (например, у Amphipsylla — ср. рис. 241,Ж и М или Б и И)\
или отмечается разная степень склеротизации его (у Chaetopsylla — ср. рис. 88, Б
и 92, Б; у Ceratophyllus — рис. 171 и 181).
Следует еще остановиться на явлении, которое резко отражается на морфологии
гениталий и модифицированных сегментов блох и носит название паразитарной
кастрации; оно было установлено нами у блох впервые в 1939—1940 гг.
Рис. 19. Ctenophthalmus breviatus. Самец. Вид на половую
клешню сверху и сзади; видны также пигидий и восьмые стер-
ниты. На дигигоидах видны крючкообразные бугорки (•*), на-
правленные один к другому.
(см. И. Г. Иофф и В. Е. Тифлов [150], И. Г. Иофф [18]) и позволило объяснить до
того непонятные случаи повторяющихся аномалий, которые приводили иногда даже
к описанию «новых» видов блох. Оказалось, что эти аномалии вызываются заражением
блох еще в их личиночной стадии мелкими нематодами (круглые черви, вероятно, из
отряда Spirurida), которые у взрослых блох обнаруживаются иногда в большом количе-
стве в полости тела. Эти нематоды бывают видны в затемненном поле зрения не толь-
ко при вскрытии блох, но и при рассматривании последних в живом виде, а также и
в монтированных препаратах, но при условии, что препарат изготовлялся без обработки
блох щелочью. Степень кастрации бывает весьма различной — от небольших аномалий
внутренних гениталий до деформации или полного недоразвития всех внутренних
половых органов, в том числе и склеротизованных их частей (семяприемника, фалло-
сома, рукоятки клешни и т. д.) и резкого уродства внешних органов (модифици-
рованных сегментов самца и клешни). До настоящего времени паразитарная кастра-
ция установлена у многих видов следующих родов и подродов блох: Coptopsylla,
Xenopsylla, Gerbillophilus, Citellophilus, Nosopsyllus, Ophthalmopsylla, Frontopsylla,
Amphipsylla, Neopsylla. Кроме того, у Vermipsylla alacurt также обнаружены [142]
повторяющиеся аномалии гениталий, происхождение которых пока не выяснено. Что
касается других индивидуальных уродств и аномалий (деформация головы, аномалии
хетотаксии, уродства гребней и ног, тератомы и т. д.), то и они встречаются неред-
ко, и о них тоже следует помнить, чтобы они не привели к ошибкам при опре-
делении блох.
Изучение столь важных для систематики модифицированных сегментов и поло-
вого аппарата блохи в обычных препаратах, где насекомое находится в боковом
положении, в некоторых случаях бывает полезно дополнить исследованием блохи
38
Рис. 20. Ctenophyllus rigidus.
Лоб молодой (а) и старой (б) самки.
до 15 мм. Как правило, самцы блох
в «стоячем положении», когда мы рассматриваем последние сегменты сверху и сзади
или снизу и сзади. Для этого блоху помешают в особую склеенную из кусочков
стекла небольшую камеру и укрепляют в желаемом положении. На рис. 19 пред-
ставлено, например, то, что видно в таких препаратах при рассматривании самца
Ctenophthalmus breviatus. Хорошо заметны крючкообразные загибы или бугорки на
дигитоидах, которые в обычных «профильных» препаратах едва заметны в виде тем-
ных хитиновых пятен. Подробнее о таких препаратах см. в работе И. Г. Иоффа
и В. Е. Тифлова {146}
Наконец, при описаниях блох отмечается общий тон окраски их
покровов — от соломенножелтого или светлорыжего до ржаво-коричне-
вого, бурого и смоляно-черного цвета; но надо учитывать, что этот цвет
зависит и от возраста — только что вышедшие из кокона блохи обычно
бледнее окрашены. Экологические условия, в которых протекает метамор-
фоз блох, также влияют на
цвет их покровов; например,
паразитирующая на челове-
ке блоха Pulex irritans, име-
ющая в нормальных услови-
ях темную, почти черную
окраску, при выплоде в но-
рах лисиц приобретает более
светлый коричневый цвет, а
в норах хорьков даже жел-
тый или рыжеватый. У не-
которых видов отмечается
неравномерность окраски
склеритов (птичьи Cerato-
phyllus maculatus и др.).
Отмечается также и
размер блох,, которые
встречаются длиной от 0,75
до 5 мм, а у некоторых
видов, у которых брюшко
самок при созревании яиц
сильно растягивается (Ver-
iriipsylla), длина может дохе
меньше самок.
Самки от самцов легко отличаются не только особенностями строе-
ния модифицированных сегментов и гениталий, но нередко также по фор-
ме головы и всего тела (ср. рис. 9 и 40, а также фото на стр. 14).
В последнее время обращают внимание [19] и на изучение возраст-
ного состава популяции блох. «Старые» блохи, давно вышедшие из коко-
на, узнаются по сравнительно более темной пигментации покровов и по
наличию повреждения кутикулярных элементов — поломанных зубцов
ктенидия, поломанных или притупленных щетинок (рис. 20), поломанного
края VII стернита у самок 1 и т. п. «Молодые» блохи распознаются по
светлой кутикуле, все зубчики и щетинки у них целые и острые, отсут-
ствуют скопления частичек пыли под нависающими черепицами сегмен-
тов. У некоторых видов (Xenopsylla) только что вышедшие из коконов
самки хорошо распознаются по светлому, не пигментированному резер-
вуару семяприемника.
1 Частые повреждения этого края у некоторых видов [19], невидимому, свя-
заны с недостаточной «прилаженностью» его формы к форме органов прикрепитель-
ного аппарата самца, которые вдвигаются при коитусе под край VII стернита. Не-
сомненно, что происходящее в процессе эволюции взаимоприспособление соответ-
ствующих органов самца и самки и приводит к развитию различных вырезок и вы-
ступов этого края.
ОПРЕДЕЛИТЕЛЬ РОДОВ БЛОХ
В эту таблицу включены все роды блох, обнаруженные на террито-
рии СССР, и некоторые другие из смежных стран.
1(14). Блохи без гребней на тергитах груди и брюшка.
2(3). Все три членика груди вместе короче спинки первого сег-
мента брюшка. Лоб сильно склеротизован и угловат
(рис. 23, 24)............................Echidnophaga (стр. 52).
' 3(2). Грудь не короче первого тергита брюшка.
4(11). Передний край метэпимера идет от плевры более или менее вер-
тикально вверх; стигма первого тергита брюшка расположена
высоко над уровнем эпистерна заднегруди (рис. 21, 27). На
II—VII тергитах брюшка нет щетинок впереди от главного ряда.
На внутренней поверхности задней коксы имеется ряд шипиков
(рис. 346, Д). Губные пальпы состоят не более чем из четырех
члеников.
5(6). Плевра среднегруди без внутренней хитиновой перекладины,
идущей от места прикрепления коксы к верхнему краю плевры
(рис. 21). Глазная щетинка сидит ниже глаза . . Pulex (стр. 52).
6(5). Упомянутая перекладина плевры среднегруди имеется (рис. 27, 28),
а глазная щетинка сидит против глаза или отсутствует.
7(8). Эпистерн заднегруди слившийся со стерноплеврой (рис. 28)
...........................................Synosternus (стр 57).
8(7). Эпистерн заднегруди отделен от стерноплевры ясным кутику-
лярным швом (рис 27).
9(10). Щечный край головы с большим кинжалообразным острием
(рис. 30). Спинка переднегруди длиннее, чем спинка среднегоу-
ди. Паразиты дикобразов.....................Pariodontis (стр.. 57).
10(9). Выступ щечного края головы слабо развит (рис. 27). Спинка пе-
реднегруди короче, чем спинка срендегруди . . Xenopsylla (стр. 54).
11(4). Передний край метэпимера идет от плевры косо кзади и кверху;
стигма I тергита брюшка лежит не выше или немного выше
уровня эпистерна заднегруди (как на рис. 8 и 57). На внутрен-
ней поверхности задней коксы нет группы шипиков. Губные паль-
пы состоят более чем из четырех члеников.
12(13). На II—VII тергитах брюшка щетинок впереди от главного ряда
нет. Предпигидиальные щетинки имеются. На пятом членике ла-
пок обычно шесть пар боковых щетинок. Самки имеют церки
и два семяприемника. На голове (рис. 32) характерно наличие
бороздки, отделяющей нижнюю часть лба от вышележащей
части.........................................Coptopsylla (стр. 58).
13(12). На II—VII тергитах брюшка впереди от главного ряда имеются
щетинки. Предпигидиальные щетинки отсутствуют. На пятом
членике лапок четыре пары боковых щетинок. У самок церки
отсутствуют и имеется лишь один семяприемник.
40
а) Губные пальпы (хоботок) состоят не более чем из десяти члеников.
Голова — см. рис. 33, 34. Паразиты хищных........................
ч......................................Chaetopsylla (стр. 59).
б) Хоботок длинный, пальпы состоят не менее чем из десяти члеников.
Голова — см. рис. 35, 36. Паразиты копытных......................
.......................................Vermipsylla (стр. 58).
j 1x14(1). На спинке переднегруди (пронотум) имеется гребень (ктенидий),
у-'15 (61). Ктенидий на спинке переднегруди состоит из довольно длинных
зубцов. На голове зубцы или ктенидии имеются или отсутствуют.
16(23). Кроме гребня переднегруди, на тергитах груди и брюшка апи-
кальных зубчиков или гребней не имеется. На II—VII тергитах
брюшка имеется не более чем по одному ряду щетинок. Перед-
ний край метэпимера заднегруди идет от плевры более или ме-
нее вертикально вверх; его стигма расположена высоко над уров-
нем эпистерна заднегруди (как на рис. 21, 27). Глаза хорошо
развиты. Имеется лишь одна развитая предпигидиальная щетин-
ка с каждой стороны.
17(18). Лоб с выдающимся углом выше уровня глаза. На голове вдоль
щечного края имеется почти вертикальный ктенидий из пяти
зубцов (рис. 31)...............................Spilopsyllus (стр. 57).
18(17). Лоб равномерно закруглен, без выдающегося бугорка выше
уровня глаза.
19(20). Нижний край головы без ктенидия или отдельных зуб-
цов ...........................................Hoplopsyllus (стр. 57).
20(19). На голове по щечному краю имеются зубцы или ктенидий.
21(22). Имеются два (реже один-три) щечных зубца. Гребень передне-
груди не более чем из трех-четырех зубцов с каждой стороны
(рис. 22).....................................Archaeopsylla (стр. 53).
22(21). Имеется гребень из семи-восьми зубцов по ротовому краю го-
ловы. Гребень переднегруди приблизительно из восьми зубцов
с каждой стороны (рис. 25, 26) . . . Ctenocephalides (стр. 53).
23(16). Кроме переднегруди, гребни или отдельные апикальные зубчики
имеются еше на некоторых тергитах Г Передний край метэпи-.
мера идет от плевры косо кзади и кверху; его стигма лежит не
выше или немного выше уровня эпистерна заднегруди
(рис. 8, 57).
v 24(64). На голове не имеется ктенидия.
25(26). На пятом членике задней лапки имеются только четыре пары
боковых щетинок, а основная подошвенная пара (имеющаяся на
передней и средней лапках) отсутствует. На внутренней поверх-
ности задней коксы имеется группа шипиков (рис. 346, Б). На
голове хорошо развитые фронтальные и теменные ряды щетинок.
Глаз рудиментарный, но заметный. У самок межусиковая темен-
ная бороздка хорошо развита (рис. 70) . . Catallagia (стр. 119).
26(25). На V членике задней лапки имеется пять пар боковых щетинок
или четыре пары боковых и основная подошвенная пара.
27(28). На V членике лапок пять пар боковых щетинок. Имеются три
теменных ряда щетинок (кроме заднекрайнего). Глаз полуруди-
ментарный. В передней части головы у основания хоботка про-
свечивает темиопигментированный рычаг колющих органов.
Максиллярные пальпы длинные, достигают вершины передней
коксы. На тергитах брюшка имеются зачаточные гребни из ред-
ких зубчиков. У самки два семяприемника и развитая межуси-
ковая бороздка................................Saphiopsylla (стр. 121).
1 Апикальных зубчиков на тергитах брюшка нет у самок некоторых Mesopsylla,
но на спинке заднегруди (метапотум) у них есть зубчики. У некоторых Myodopsylla
нет зубчиков, но имеются расширенные щетины, сидящие по краю тергитов.
41
28(27). Такой комбинации признаков, как указано в п. 27, не имеется.
29(30). Лоб низкий, покатый (рис. 57), без лобного зубчика. Глаз полу-
рудиментарный. Эпистерн заднегруди слабо развит и сливается
с плеврой (рис. 57). На пятом членике лапок четыре пары бо-
ковых и основная подошвенная пара щетинок. На IV—УЯтерги-
тах брюшка лишь по одному ряду щетинок . . Eopsylla (стр. 105).
30(29). Лоб и грудь не такие.
31(44). Верхняя щетинка глазного ряда («глазная щетинка») сидит близ
края усиковой ямки выше уровня глаза (а при отсутствии раз-
витого глаза выше обычного места его расположения). Внутри
головы впереди и выше глаза имеется хитиновый тяж (передняя
ветвь тентория), который начинается вблизи середины глазного
ряда и, образуя дугу, идет кзади, уходя за глаз (рис. 45—49,56).
32(35). V членик лапок с четырьмя парами боковых щетинок, причем
между щетинками первой пары вдвинута подошвенная пара
щетинок.
33(34). Глаз отсутствует или рудиментарный. На щечной лопасти позади
и ниже обычного места расположения глаза имеется сильная ще-
тинка. В передней части головы у основания хоботка просвечи-
вает темнопигментироваяный рычаг колющих органов (рис. 56).
Метанотум без апикальных зубчиков . . Wagnerina (стр. 103).
34(33). Глаз уменьшенный или рудиментарный. Между глазным ря-
дом щетинок и фронтальным рядом имеются вставочные
крупные щетины. Лоб в большинстве случаев заметно вы-
ступает кпереди (рис. 54, А—Д). Метанотум с апикальными
зубчиками ....................................Amphipsylla (стр. 94).
35(32). V членик лапок с пятью парами боковых щетинок, причем пер-
вая (основная) пара может быть несколько сдвинута на подош-
венную поверхность, но не вдвинута между щетинками следую-
щей пары.
36(41). Теменные ряды щетинок недоразвиты — в каждом от нуля до
двух (во втором ряду иногда до трех-четырех) нижних щетинок.
37(40). Глаз развитый, с более бледной пызуреобразной верхней и тем-
ной несколько обособленной нижней частью.
38(39). Развитых предпигидиальных щетинок у самца две, у самки —
две-три. VIII стернит самца большой, широкий. Дигитоид неболь-
шой, без шипов и шпор (рис. 213, 214, 216). Придаток семя-
приемника самки широкий мешкообразный, резко отделенный от
резервуара (рис. 215, А—Б; 217, А).................................
...................................Paradoxopsyllus (стр. 89).
39(38). Развитых предпигидиальных щетинок у обоих полов по три
(у самца иногда две). VIII стернит самца слабо развит. Диги-
тоид довольно широкий, с характерными шипами у заднего края
(рис. 219, А; 221, А; 223, А). Придаток семяприемника постепен-
но переходит в резервуар (рис. 219. Б; 221. Б). Глаз большой
(рис. 48, 49) . ..........................Ophthalmopsylla (стр. 91).
40(37). Глаз не развит. Предпигидиальных щетинок у самца одна-две,
у самки — две-три. VIII стернит самца широкий. Семяприемник
самки, как у Ophthalmopsylla .... Phaenopsylla (стр. 92).
41(36). Имеются два развитых теменных ряда щетинок (рис. 45, 46).
42(43). По заднему краю задней голени многочисленные пучки (не ме-
нее 10 групп) очень длинных щетин (рис. 348, А). У некоторых
видов щетинки, сидящие вдоль лобного края, превращены в ши-
пы (ср. рис. 46 и 47)........................Ctenophyllus (стр. 88).
43(42). По заднему краю голени обычные шпоры, которые не выделя-
ются своей длиной и образуют менее 10 групп (рис. 348, Б). Го-
лова изображена на рис. 45...................Frontopsylla (стр. 84).
42
44(31). Верхняя щетинка глазного ряда сидит перед глазом (рис. 40—44);
она находится не выше его верхнего края (кроме случаев, когда
глаз рудиментарен). Передний тяж тентория (тяж перед глазом,
описанный в п. 31), как правило, не развит1.
45(46). I членик задних лапок длиннее, чем II, III и IV вместе взятые
(рис. 348, В). Лобный зубчик не развит. Глаз хорошо развит.
Паразиты белок................................Tarsopsylla (стр. 63).
46(45). I членик задних лапок не длиннее, чем II, III и IV вместе
взятые.
47(52). На наружной поверхности переднего бедра нет боковых щети-
нок или имеется лишь одна (не считая сидящих у вентраль-
ного края).
48(49). V членик лапок с четырьмя парами боковых щетинок и по-
дошвенной парой, вдвинутой между щетинками первой пары
(рис. 12, В). Паразит сонь (Myoxidae) . . Myoxopsylla (стр. 63).
49(48). V членик лапок с пятью парами боковых щетинок и без подош-
венной пары.
50(51). Наружная поверхность передней коксы довольно равномерно
усеяна щетинками. На II членике антенны у самца имеется вен-
тральная группа сильных щетин, концы которых далеко заходят
за вершину булавы (рис. 345,4), а у самки щетинки этого чле-
ника короткие (рис. 38). Толстых предпигидиальных щетинок
у самки и самца три (иногда четыре). У видов, известных
в СССР, хоботок длинный; за вершину передней коксы обычно
заходит весь последний членик губных пальп или более. У этих
же видов сигмы средних тергитов брюшка круглые и большие;
их вертикальный диаметр равен примерно 1 2/3 поперечного (ко-
роткого) диаметра глаза 2. Характерна половая клешня с боль-
шим широким подвижным пальцем (рис. 101, 102) и семяприем-
ник с относительно узким резервуаром (рис. 103, 104). Пара-
зиты барсуков..................................Рагасегаз (стр. 63).
51(50). Развита передняя ветвь тентория в виде тонкого изогнутого
тяжа внутри головы перед глазом (рис. 39). Значительная часть
наружной поверхности передней коксы свободна от щетинок. Ко-
нец хоботка заходит за вершину коксы (иногда около половины
членика и больше). Стигмы средних тергитов брюшка неболь-
шие. Толстых предпигидиальных щетинок у самки три, а у сам-
ца — одна. Половая клешня и семяприемник изображены на
рис. 100,4—Б................................Aenigmopsylla (стр. 64).
52(47). На наружной поверхности переднего бедра имеется несколько
боковых щетинок.
53(54). Подвижный палец половой клешни (дигитоид) с длинным вен-
тральным отростком, конец которого расширен (рис. 106, 107).
Церки цилиндрические с многочисленными щетинкам на округ-
ленной вершине (рис. 105). В гребне на пронотуме не менее
24 зубцов. Паразиты пищух......................Amphalius (стр. 64).
54(53). Дигитоид без длинного вентрального отростка. Церки кониче-
ские с единичными щетинками на вершине (рис. 111,4).
55(56). Хоботок очень длинный, более чем целый членик, а часто почти
два членика его заходят за коксу передней ноги. У самца стер-
нит анального сегмента намного длиннее тергита. Дигитоид тре-
угольной формы. У самки резервуар семяприемника бочко-
образный. Паразит тонкопалого суслика, обитающего в Сред-
ней Азии.....................................Rostropsylla (стр. 66).
1 Тонкий тяж есть лишь у Aenigmopsylla (рис. 39).
2 В Китае встречаются виды с более коротким хоботком и небольшими стиг-
мами, детальнее не изученные.
43
56(55). Хоботок менее длинный, он не заходит за вершину передней
коксы или заходит не более чем на полтора членика.
57(58). Хоботок длинный, и в большинстве случаев не менее целого
членика его заходит за вершину передней коксы (рис. 40), иногда
около половины членика (рис. 41) С Стернит анального сегмента
самца не длиннее тергита (рис. 109). Дигитоид удлиненно-
овальной формы (рис. 108 и 109). У самки, как правило, три
предпигидиальных щетинки (редко две-пять), а у самца
обычно только одна хорошо развитая. Семяприемник сам-
ки с коротким грушевидным или шаровидным резервуаром
(рис. 111, А—Б).................................Oropsylla (стр. 65).
58(57). Хоботок в большинстве случаев короткий, не достигает вершины
передней коксы (рис. 42), и лишь у некоторых видов несколько
заходит за нее1 (рис. 43). Половая клешня самца и семяприем-
ник самки не такие, как у Oropsylla.
59(60). Третья пара боковых щетинок пятого членика лапок сильно
сдвинута на подошву, а ее место занято тонкими волосками
(рис. 12, Б). В гребне не менее 24 зубцов. Имеются два более
или менее развитых теменных ряда щетинок и два (иногда один)
фронтальных ряда. Глаз очень большой . . Dasypsyllus (стр. 83).
60(59). Третья пара боковых щетинок V членика лапок расположена
обычным образом (рис. 12, А) . . . Ceratophyllus1 2 (стр. 66).
61(15). Ктенидий на переднегруди состоит из коротких зубчиков
(рис. 50). На голове ктенидия нет.
62(63). Хоботок не заходит за вершину передней коксы. Глаз
имеется....................................Mioctenopsylla (стр. 84).
63(62). Хоботок далеко заходит за вершину передней коксы. Глаз от-
сутствует (рис. 50)......................Brachyctenonotus (стр. 92).
v 64(24). На голове имеется ктенидий.
65(74). На голове имеется своеобразный предротовой ктенидий из двух
зубцов (рис. 75, 76). Блохи летучих мышей.
66(73). Предпигидиальные щетинки нормально развиты. VII тергит без
гребня.
67(72). На брюшке нет настоящих ктенидиев.
68(69). На некоторых тергитах брюшка имеются ложные ктенидии,
образуемые несколькими утолщенными щетинками тергитов
(рис. 76)....................................Myodopsylla (стр. 126).
69(68). Брюшко без ложных ктенидиев.
70(71). Основания тергитов//—VII имеют вид темных лент. На заднем
крае головы непосредственно позади ямки усика имеется ряд из че-
тырех или больше толстых острых шипов..............................
......................................Araeopsylla (стр. 126).
1 Пользуясь признаком длины хоботка как основным при диференцировании
Oropsylla от Ceratophyllus, необходимо учитывать, что этот признак в большинстве
случаев очень удобен и прост, но иногда может ввести в заблуждение вследствие
того, что среди Oropsylla и среди Ceratophyllus имеются виды, у которых конец
хоботка заходит за конец коксы примерно на половину последнего членика
(О. asiatica — рис. 41, Cer. lebedewi, С. menzbieri, С. tersus, С. turkmenicus). В не-
которых случаях при неудачном положении хоботка или коксы ошибки могут про-
изойти и при определении таких видов Ceratophyllus, у которых хоботок сравни-
тельно немного (на четверть-треть членика) заходит за конец коксы (Cer. tesquorum
и др. — рис. 43). Поэтому во всех неясных случаях, когда хоботок заходит менее
чем на целый членик за конец коксы, необходимо использовать другие признаки,
главным образом форму половой клешни самца и семяприемника самки (см. также
примечание на стр. 72).
2 Этот обширный род включает много подразделений, принимаемых разными
авторами то за отдельные роды, то за низшие подразделения. Таблица для их опре-
деления дана отдельно (стр. 66—68).
44
71(70). Основания тергитов без особой пигментации. Позади ямки уси-
ка сидят по краю головы у ее задне-нижнего угла обычно лишь
одна-три шиповидных щетинки . . . Rhinolophopsylla (стр. 125).
72(67). На первом и третьем члениках груди и на нескольких тергитах
брюшка нормально развитые гребни (всего шесть или восемь).
Голова изображена на рис. 75 ... . Ischnopsyllus (стр. 123).
73(66). Вместо нормальных предпигидиальных щетинок VII тергит
брюшка с рядом шипов, образующих гребневидный ряд ....
.........................................Nycteridopsylla (стр. 123).
>^74 (65). Предротового ктенидия из двух зубцов не имеется. Ктенидий
находится под глазом на щечной лопасти головы или идет вдоль
всего ротового края.
v 75(94). Гребни на тергитах брюшка отсутствуют, а апикальные зубчики
расположены близко к спине.
v 76(77). Глаз развитый. Непосредственно ниже и сзади него имеется
щечный ктенидий из двух зубцов, направленных кзади и тесно
прилегающих друг к другу (как на рис 53). На пятом членике
лапок имеются четыре пары боковых щетинок и одна пара по-
дошвенных .......................................Mesopsylla (стр. 93).
77(76). Глаз плохо развитый, рудиментарный или вообще отсутствует.
78(81). Пигидий не выпуклый (рис. 111,4). На задней голени вдоль
заднего края у большинства видов имеется правильный гребне-
видный ряд ровных щетин (рис. 347, В). Метанотум с апикаль-
ными зубчиками. Лоб конусообразно выпуклый (рис. 51, 55).
У вершины его часто имеются толстые шиповидные щетины.
79(80). Щечный ктенидий состоит из небольшого числа зубцов — от
двух до шести (рис. 51, 52)..................Leptopsylla (стр. 101).
80(79). Ктенидий из многочисленных зубцов (7—20) идет параллельно
всему переднему краю ямки усика (рис. 55)...........................
.....................................Pectinoctenus (стр. 102).
81(78). Пигидий более или менее выпуклый, во всяком случае у самки
(рис. 264). Щетинки не образуют гребневидного ряда по заднему
краю голени (рис. 349,4—Б). Метанотум без апикальных
зубчиков.
82(85). Щечный ктенидий состоит из двух зубцов, расположенных под
рудиментарным глазом. Последний членик лапок без основной
подошвенной пары щетинок, с четырьмя или пятью парами
боковых.
83(84). Из двух зубцов щечного ктенидия один более короткий направ-
лен вниз, другой более длинный перекрещивает его, направляясь
кзади (рис. 69). Конец максиллярных пальп намного не достигает
до вершины передней коксы. Теменные ряды щетинок развиты
хорошо. Три предпигидиальные щетинки с каждой стороны . .
................................................Neopsylla (стр. 114).
84(83). Оба зубца щечного ктенидия удлиненной формы и направляются
кзади. Максиллярные пальпы длинные, конец их достигает вер-
шины передней коксы. Теменные ряды щетинок развиты слабо.
Две (реже три) предпигидиальные щетинки с каждой стороны
..........................................Paraneopsylla (стр. 118).
85(82). Ктенидий головы состоит более чем из двух зубцов.
86(87). Щечный ктенидий состоит из трех зубцов1 приблизительно оди-
наковой формы, направленных косо кзади и книзу (рис. 61).
V членик лапок с тремя или четырьмя парами боковых щетинок
и с парой основных подошвенных . . Clenophthalmus (стр. 105).
87(86). Ктенидий головы состоит более чем из трех зубцов.
1 В виде редкой аномалии бывает два зубца; такие аномалии наблюдались
с одной стороны головы.
45
88(91). Вертикально расположенный перед усиком ктенидий состоит
из четырех или пяти зубцов различной формы, направленных
кзади.
89(90). Из четырех зубцов головного ктенидия второй сверху длиннее
других, с вытянутым в острие концом (рис. 62). Имеется лобный
зубчик. На V членике лапок четыре пары боковых и основная
подошвенная пара щетинок..................Palaeopsylla (стр. 108).
90(89). Из пяти зубцов головного ктенидия самый длинный второй сни-
зу (рис. 63, 64). Лобного зубчика нет. На V членике лапок пять
пар боковых щетинок (на задней лапке иногда только четыре
пары).....................................Nearctopsylla (стр. 109).
91(88). Головной ктенидий имеет другую форму.
92(93). На V членике лапок четыре пары боковых щетинок и одна пара
основных подошвенных. Губные пальпы состоят из четырех чле-
ников. Щечный ктенидий—из четырех тупых зубцов (рис. 58—60).
..........................................Doratopsylla (стр. 107).
93(92). На V членике лапок четыре или пять пар боковых щетинок, по-
дошвенная пара отсутствует. Губные пальпы состоят не менее
чем из пяти члеников. Щечный ктенидий—обычно из пяти-шести
(реже четыре-восемь) зубцов (рис. 65—68)...........................
...................................Rhadinopsylla (стр. 109).
94(75). На некоторых тергитах брюшка имеются настоящие или хотя бы
зачаточные гребни в виде ряда редко расположенных апикаль-
ных зубчиков на боках, отступя от спины. Щечный гребень со-
стоит из многочисленных (не менее четырех) узких зубцов. Блохи
очень крупные.
95(96). На V членике лапок четыре пары боковых и одна пара подош-
венных щетинок, расположенных между боковыми первой пары.
На I тергите брюшка имеется гребень из многочисленных зуб-
цов. Самки с одним семяприемником. Голова изображена
на рис. 73, 74 ...............................Stenoponia (стр. 122).
96(95). На V членике лапок пять пар боковых щетинок; подошвенная
пара отсутствует. Самки с двумя семяприемниками. Голова —
на рис. 71, 72............................Hystrichopsylla (стр. 120).
УПРОЩЕННЫЙ ОПРЕДЕЛИТЕЛЬ
НАИБОЛЕЕ ЧАСТО ВСТРЕЧАЮЩИХСЯ В ВОСТОЧНОЙ АЗИИ
РОДОВ БЛОХ
Учитывая, что в некоторых случаях определение блох может произ-
водиться лицами, недостаточно хорошо владеющими полными опреде-
лительными таблицами (причем в таких случаях определение чаще все-
го доводится только до рода), мы даем краткую определительную таб-
лицу для наиболее часто встречающихся 24 родов.
1(6). Блохи без гребней на тергитах груди и брюшка.
2(5). На II—VII тергитах брюшка нет щетинок впереди от главного
ряда. Предпигидиальные щетинки имеются у обоих полов. В глаз-
ном ряду не более двух щетинок.
3(4). Плевра среднегруди без внутренней хитиновой перекладины,
идущей от места прикрепления ляжки к верхнему краю плевры
(рис. 21). Глазная щетинка сидит ниже глаза . . Pulex (стр. 52).
4(3). Упомянутая перекладина плевры среднегруди имеется (рис. 27),
а глазная щетинка сидит против глаза или отсутствует
...........................................Xenopsylla (стр. 54).
5(2). На II—VII тергитах брюшка имеются щетинки впереди от глав-
ного ряда. Предпигидиальные щетинки отсутствуют. В глазном
ряду обычно четыре щетинки (рис. 34) . . Chaetopsylla (стр. 59).
6(1). На спинке переднегруди (пронотум) имеется гребень.
7(10). Кроме гребня переднегруди, на тергитах груди и брюшка апи-
кальных зубчиков или гребней не имеется. На тергитах брюшка
с II по VII имеется не более чем по одному ряду щетинок. Пе-
редний край метэпимера заднегруди идет от плевры более или
менее вертикально вверх; его стигма расположена высоко
над уровнем эпистерна заднегруди (как на рис. 21 и 27). Глаза
большие темнопигментированные. Имеется лишь одна развитая
предпигидиальная щетинка с каждой стороны.
8(9). По ротовому краю головы имеется гребень из семи-восьми зуб-
цов (рис. 25, 26)........................Ctenocephalides (стр. 53).
9(8). Имеется лишь два (реже один-три) щечных зубца. Гребень пе-
реднегруди состоит не более чем из трех-четырех зубцов с каж-
дой стороны (рис. 22)......................Archaeopsylla (стр. 53).
10(7). Кроме переднегруди, гребни или отдельные апикальные зубчики
имеются еще на некоторых тергитах груди или брюшка. Передний
край метэпимера идет от плевры косо кзади и кверху; его стиг-
ма лежит не выше или немного выше уровня эпистерна задне-
груди (рис. 8, 57).
11(32). На голове не имеется ктенидия.
47
12(13). Глаз рудиментарный, но заметный (рис. 70). Фронтальный и
теменные ряды щетинок хорошо развиты. На пятом членике зад-
ней лапки имеются только четыре пары боковых щетинок; основ-
ная подошвенная пара на этом членике отсутствует ....
............................................Catallagia (стр. 119).
13(12). На пятом членике задней лапки имеется пять пар боковых
щетинок, или четыре пары боковых и основная подошвенная
пара.
14(23). Верхняя щетинка глазного ряда («глазная щетинка») сидит близ
края усиковой ямки выше уровня глаза, а при отсутствии разви-
того глаза — выше обычного места его расположения (рис. 45—
49, 56).
15(16). V членик лапок с четырьмя парами боковых щетинок, причем
между щетинками первой пары вдвинута подошвенная пара.
Глаз уменьшенный или рудиментарный (рис. 54, А—Д) . . .
............................................Amphipsylla (стр. 94).
16(15). V членик лапок с пятью парами боковых щетинок, причем пер-
вая (основная) пара может быть несколько сдвинута на подош-
венную поверхность, но не вдвинута между щетинками следую-
щей пары.
17(20). Теменные ряды щетинок недоразвиты — в каждом от нуля до
двух (во втором ряду иногда три-четыре) нижних щетинок. Глаз
развитый, с более бледной пузыреобразной верхней частью и тем-
ной несколько обособленной нижней (рис. 48).
18(19). Развитых предпигидиальных щетинок у самца две, у самки—две-
три. VIII стернит самца большой, широкий. Дигитоид небольшой,
без шипов и шпор (рис. 213, 214). Придаток семяприемника сам-
ки широкий, мешкообразный, резко отделенный от резервуара
(рис. 215, А, Б)........................Paradoxopsyllus (стр. 89).
19(18). Развитых предпигидиальных щетинок у обоих полов по три
(у самца иногда две). VIII стернит самца слабо развит. Диги-
тоид довольно широкий, с характерными шипами у заднею края
(рис. 219, А; 221, А). Придаток семяприемника постепенно пере-
ходит в резервуар (рис. 219, Б; 221, Б). Глаз большой (рис. 48).
........................................Ophthalmopsylla (стр. 91).
20(17). Имеется два развитых теменных ряда щетинок (рис. 45—47).
21(22). По заднему краю голени многочисленные пучки (не менее
10 групп) очень длинных щетин (рис. 348, А). У некоторых видов
вдоль лобного края находится ряд толстых шиповидных щетин
(рис. 46)....................................Ctenophyllus (стр. 88).
22(21). По заднему краю задней голени обычные шпоры, которые не
выделяются своей длиной и образуют менее 10 групп
(рис. 348, Б). Голова — см. рис. 45. . . Frontopsylla (стр. 84).
23(14). Верхняя щетинка глазного ряда сидит перед глазом (рис. 40—44);
она находится не выше его верхнего края (кроме случаев,
когда глаз рудиментарен или уменьшен).
24(25). I членик задних лапок длиннее, чем II, III и IV вместе взятые
(рис. 348, Б). Лобный зубчик не развит. Глаз хорошо развит.
Паразиты белок...............................Tarsopsylla (стр. 63).
25(24). I членик задних лапок не длиннее, чем II, III и IV вмест?
взятые.
26(27). На наружной поверхности переднего бедра нет боковых щети-
нок или имеется лишь одна (не считая сидящих у вентрального
края). У видов, известных в СССР, хоботок длинный (рис. 38);
за вершину передней коксы обычно заходит последний членик
губных пальп или более. У этих же видов стигмы средних терги-
тов брюшка круглые и большие; их вертикальный диаметр при-
48
мерно равен 2/3 поперечного (короткого) диаметра глаза. Имеет-
ся три-четыре толстые предпигидиальные щетинки. Характерна
половая клешня с большим широким дигитоидом (рис. 101, 102)
и семяприемник с относительно узким резервуаром (рис. 103,
104). Паразиты барсуков..................Paraceras (стр. 63).
27(26). На наружной поверхности переднего бедра имеется несколько
боковых щетинок.
28(29). Дигитоид с длинным вентральным отростком, конец которого
расширен (рис. 106). Церки цилиндрические, с многочисленными
щетинками на округленной вершине (рис. 105). В гребне на про-
нотуме не менее 24 зубцов. Паразиты сеноставок.......................
.........................................Amphalius (стр. 64).
29(28). Дигитоид без длинного вентрального отростка. Церки конические
с единичными щетинками на вершине (рис. 111, Д).
30(31). Хоботок длинный и в большинстве случаев не менее целого чле-
ника его заходит за вершину передней коксы (иногда около поло-
вины членика !)—рис. 40, 41. Дигитоид удлиненно-овальной фор-
мы (рис. 108, 109). У самки,, как правило, три предпигидиаль-
ных щетинки (редко две-пять), а у самца обычно только одна
хорошо развита. Семяприемник самки с коротким грушевидным
резервуаром (рис. 111,Д и Б)....................Oropsylla (стр. 65).
31(30). Хоботок в большинстве случаев короткий, не достигает вершины
передней коксы и лишь у некоторых видов несколько заходит
за нее1 (рис. 42, 43). Половая клешня самца и семяприемник
самки не такие, как у Oropsylla .... Ceratophyllus (стр. 66).
32(11). На голове имеется ктенидий.
33(34). На голове имеется своеобразный предротовой ктенидий из двух
зубцов (рис. 75, 76). Блохи летучих мышей............................
............................семейство Ischnopsyllidae (стр. 123).
34(33). Предротового ктенидия, состоящего из двух зубцов, не имеется.
Ктенидий находится под глазом на щечной лопасти или идет
вдоль всего ротового края.
35(44). Гребни на тергитах брюшка отсутствуют, а апикальные зубчики
расположены близко к спине.
36(39). На задней голени вдоль заднего края имеется правильный греб-
невидный ряд ровных щетин (рис. 347, В). Лоб конусообразно
выпуклый (рис. 51, 55). У вершины его часто имеются толстые
шиповидные щетины. Метанотум с апикальными зубчиками.
37 (38). Щечный ктенидий состоит из небольшого числа зубцов — от
двух до шести (рис. 51, 52)..................Leptopsylla (стр. 101).
38(37). Ктенидий из многочисленных зубцов (7—20) идет параллельно
всему переднему краю ямки усика (рис. 55)............................
....................................Pectinoctenus (стр. 102).
39(36). Щетинки не образуют гребневидного ряда по заднему краю го-
лени (рис. 349, А, Б). Метанотум без апикальных зубчиков.
40(41). Щечный ктенидий состоит из двух зубцов, расположенных возле
ямки усика под рудиментарным глазом; из этих зубцов один,
более короткий, направлен вниз, а другой, более длинный, пере-
крещивает его, направляясь кзади (рис. 69). Теменные ряды
щетинок развиты хорошо. Три предпигидиальных щетинки с каж-
дой стороны.....................................Neopsylla (стр. 114).
41(40). Ктенидий головы состоит более чем из двух зубцов.
1 См. примечание на стр. 44.
4 Определитель блох 49
42(43). Щечный ктенидий состоит из трех зубцов1 приблизительно оди-
наковой формы, направленных косо кзади и книзу (рис. 61).
V членик задних лапок с тремя или четырьмя парами боковых
щетинок и с парой основных подошвенных.............................
....................................Ctenophthalmus (стр. 105).
43(42). Щечный ктенидий состоит обычно из пяти-шести (реже четырех-
восьми) зубцов (рис. 65—68). На V членике лапок четыре или
пять пар боковых щетинок; подошвенная пара отсутствует
...........................................Rhadinopsylla (стр. 109).
44(35). На некоторых тергитах брюшка имеются настоящие гребни или
хотя бы зачаточные гребни в виде ряда редко расположенных
апикальных зубчиков на боках, отступя от спины. Щечный гре-
бень состоит из многочисленных (не менее четырех) узких зуб-
цов. Блохи очень крупные.
45(46). На V членике лапок четыре пары боковых и пара подошвенных
щетинок, расположенных между боковыми щетинками первой
пары. На 1 тергите брюшка имеется гребень из многочисленных
зубцов. Самки с одним семяприемником. Голова см. рис. 73, 74.
..............................................Stenoponia (стр. 122).
46(45). На V членике лапок пять пар боковых щетинок; подошвенная
пара отсутствует. Самки с двумя семяприемниками. Голова см.
рис. 71, 72 ... .........................Hystrichopsylla (стр. 120).
1 В виде редкой аномалии бывает два зубца; такие аномалии наблюдались
с одной стороны головы.
ПЕРЕЧЕНЬ НАДСЕМЕЙСТВ, СЕМЕЙСТВ, ПОДСЕМЕЙСТВ
И РОДОВ БЛОХ С УКАЗАНИЕМ ИХ РАСПРОСТРАНЕНИЯ
И ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ, ВСТРЕЧАЮЩИХСЯ
В ВОСТОЧНОЙ АЗИИ И СОПРЕДЕЛЬНЫХ РАЙОНАХ
ОТРЯД SUCTORIA RETZIUS 1783= APHANIPTERA
KIRBY \§\7=SIPHONAPTERA LATREILLE 1825
В литературе употребляются все три перечисленные названия отряда
блох. Для ликвидации разногласия по этому вопросу и считаясь с пра-
вилами приоритета, очевидно, лучше будет придерживаться старого на-
звания: Suctoria.
НАДСЕМЕЙСТВО PULICOIDEA [16]
К этому надсемейству принадлежат многие виды блох, привлекаю-
щие к себе особое внимание в связи с тем, что они являются парази-
тами человека, домашних животных (собак, кошек), крыс, песчанок
и многих других млекопитающих и птиц.
Общая характеристика этих -блох следующая. Брюшко сравнительно
короткое и высокое. Метанотум и тергиты брюшка без апикальных
зубчиков. Стигмы круглые. На тергитах II—VII сегментов имеется
не более чем по одному ряду щетинок. Над стигмой VIII тергита
нет щетинок. Метэпимер расширен кверху, его стигма расположена
немного выше, чем метэпистерн. Мезонотум без псевдосет под апикаль-
ным краем. Наружная стенка средней коксы без продольного склероти-
зованного саблеобразного внутреннего тяжа, идущего от сочленения
с грудью вниз.
Для большинства типичных представителей этой группы характерны
еще следующие признаки (не наблюдающиеся, правда, у некоторых ви-
дов семейства Tungidae и у некоторых других не встречающихся у нас
видов). Как правило, глазной ряд состоит из двух щетинок. На V чле-
нике лапок четыре пары боковых щетинок; базальная подошвенная пара
отсутствует. У обоих полов имеется лишь по одной развитой предпиги-
диальной щетипке с каждой стороны тела. Для гениталий самца харак-
терно наличие двух подвижных пальцев половой клешни.
51
СЕМЕЙСТВО PULICIDAE
ПОДСЕМЕЙСТВО PULICINAE
Род Pulex [124, 135, 19]
Этот род представлен в СССР лишь одним видом: Pulex irritans,
распространенным во многих странах земного шара. Это сравнительно
крупная блоха, как правило, темнобурого цвета, обладающая исключи-
тельно большой силой прыжка — до 32 см в длину. Pulex irritans
паразитирует на человеке, обитает в его жилище и нередко обнаружи-
вается на домашних животных (собаках, свиньях и т. д.). Весьма часто
встречается также на хищных животных — волках, лисах, хорьках и др.
При развитии в глубоких темных норах (например, у хорьков) блохи
этого вида несколько изменяются морфологически; они отличаются
сравнительно меньшим размером и более бледно окрашены — желтого
или светло*коричн.евого цвета (Р. irritans var. fulvus). На грызунах (за
исключением сурков) этот вид блох встречается редко. На сурках же
в некоторых местностях (повидимому, более близких к жилью человека)
они встречаются очень часто.
Род Echidnophaga [187, 145, 126, 157]
Из блох этого рода в интересующем нас районе пока обнаружен
лишь один вид, но, учитывая широкое их распространение в Средней
Азии, мы все же приведем таблицу и некоторые рисунки для определе-
ния всех четырех известных в СССР видов. Ряд других видов Echidno-
phaga живут в южном полушарии. Блохи этого рода очень малого раз-
мера (часто менее 1 мм в длину). С помощью колющих органов они
могут надолго прикрепляться к коже животных.
1 (4). V членик всех лапок с двумя прикогтевыми шипиками
(рис. 351, Л).
2(3). Колющие ротовые органы (лацинии или мандибулы) не особенно
длинные (рис. 23), у самца не заходят, а у самки немного захо-
дят за передний вертлуг. Глаз расположен несколько ближе
к щечному краю головы, чем к темени. Язычковидный выступ
на заднем крае головы самки сравнительно слабо развит
.....................................................Е. gallinacea.
Паразит ушастых ежей (Hemiechinus), домашних птиц, домашних животных,
иногда человека. Встречается во многих теплых странах Евоопы, Азии, Африки, Аме-
рики; у нас — в Средней Азии, в Нижнем Поволжье и на Кавказе. Находка в Китае
(Шанхай [27]) требует подтверждения.
3(2). Колющие органы очень длинные и мощные; у самца далеко за-
ходят за вертлуг, а у самки при сравнении с вытянутой передней
ногой — за бедро. Глаз расположен примерно по середине между
теменным и щечным краями головы или даже ближе к темени.
Стигма VIII тергита, а также язычковидный выступ на заднем
крае головы самки сильнее развиты, чем у предыдущего вида.
Лобный угол выражен в большинстве случаев резче, чем у дру-
гих видов...............................................Е. popovi.
Истинный хозяин еще неизвестен, но до сих пор этот вид обнаруживался
преимущественно в норах хищников (лисиц, барсуков). Встречается в полупустын-
ных местностях Ирана, в СССР известен из Восточного Закавказья и Предкавказья,
а также Заволжья [145].
52
4(1). V членик лапок с одним прикогтевым шипиком (рис. 351, Б).
5(6). Колющие ротовые органы длинные; при сравнении с вытянутой
передней ногой конец их у самца достигает основания бедра,
а у самки — почти его вершины. Глаз расположен примерно по
середине между теменным и щечным краем головы или выше.
Язычковидный выступ на заднем крае головы самки довольно
длинный (рис. 24).........................................Е. oschanini.
Паразит песчанок, главным образом Rhombomys opimus. Распространен в Сред-
ней Азии и обнаружен в Монголии (предгорья Гобийского Алтая) [126, 145].
6(5). Кроме отличия в пятом членике лапок, в остальном довольно
сходен с Е. gallinacea (см. выше). Предротовой (нижний) угол
головы выступает вперед обычно несколько сильнее, чем лоб-
ный .......................................................Е. murina.
Паразит крыс (Rattus) известен из Италии и в СССР — из Батуми [187, 154].
ПОДСЕМЕЙСТВО ARCHAEOPSYLLINAE
Род Archaeopsylla [29, 100, 163]
Паразиты ежей (Erinaceus). Встречаются в Европе, Азии и Север-
ной Африке.
1(2). Большая волосатая пластинка половой клешни самца более
длинная, суживающаяся кзади............................... A. erinacei.
Паразит Erinaceus europaeus. Известен от Англии до Поволжья и Закав-
казья (29, 163].
2(1). Большая волосатая пластинка половой клешни более короткая,
тупая (рис. 77, А).........................................A. sinensis.
Паразит ежей. Распространен на востоке Азии: найден в районах Восточного
Забайкалья, Приморья (Сихотэ-Алинь) и Китая (Шеньси) [100].
Род Ctenocephalides [179 (32, 40)]
К этому роду относятся широко распространенные паразиты домаш-
них собак и кошек, нападающие нередко на человека, а иногда встре-
чающиеся и на диких хищных млекопитающих. Другие виды этого рода
живут на хищных преимущественно в Африке.
1(2). Лоб крутой у обоих полов (рис. 25). На заднем крае задней
голени между средней и апикальной группами шпор сидит обыч-
но не менее двух толстых щетин, на метэпистерне — обычно не
менее трех щетин. Хоботок короткий. Стигмы брюшных тергитов
крупные. У самца рукоятка половой клешни сильно расширяется
к переднему ее концу..........................................С. canis.
Блоха собаки. Имеет всесветное распространение. В сухих степных и полу-
пустынных районах с кочевым укладом быта эта блоха на собаках обычно отсут-
ствует, замещаясь Pulex irritans.
2(1). Лоб менее крутой. На заднем крае задней голени между средней
и апикальной группами шпор имеется обычно лишь одна толстая
щетина, а на метэпистерне, как правило, не более двух.
3(4). Лоб пологий, особенно у самок (рис. 26). Хоботок короткий, до-
стигает не далее, чем 2/з передней коксы. Стигмы брюшных тер-
гитов сравнительно узкие. У самца рукоятка половой клешни
слабо расширяется к переднему ее концу........................С. felis.
Блоха кошки. Имеет всесветное распространение. В Южной Азии (Индия, Бир-
ма, Малайя, Цейлон, Новая Гвинея и др.) встречается подвид С. felis orientis, кото-
рый по форме лба приближается к С. canis, но по другим признакам должен быть
отнесен к С. felis [179 (32, 40)].
53
4(3). Лоб менее пологий (т. е. средний по форме между С. canis
и С. felis). Хоботок заходит за 2/3 передней коксы. Стигмы брюш-
ных тергитов крупнее, чем у С. felis. У самца рукоятка половой
клешни, как у С. canis.................................... . С. саргае.
Блоха козы. Найдена в Узбекистане (предгорья Туркестанского хребта)
и в Азербайджане (близ Гадрута).
ПОДСЕМЕЙСТВО XENOPSYLLINAE
Род Xenopsylla [16, 124, 137, 176, 181]
Представители этого обширного рода паразитируют на песчанках
(Gerbillinae), крысах (Rattus, Nesokia) и некоторых других животных
и птицах. Эти блохи наиболее многочисленны в Африке и Средней Азии,
однако некоторые виды расселились со своими хозяевами и во многие
другие страны мира. В пределах рассматриваемого нами района до си^
пор обнаружены лишь два вида (X. cheopis и X. skrjabini), но, учитывая
недостаточное обследование пустынь Центральной Азии, а также воз-
можность заноса крысами некоторых видов этих блох в портовые горо-
да, мы перечислим в таблице все виды, известные в СССР, и ряд других,
занос которых наиболее вероятен.
1(12). От первого и второго теменного ряда сохранилось хотя бы по
одной щетинке в каждом (рис. 27). У самца (рис. 77, Б; 78)
отросток Р2 половой клешни обычно перекрещивается с отрост-
ком Pi и находится выше него, а отросток В3 без щетинок, недо-
развит или мало заметен. У самки VIII тергит с многочислен-
ными щетинками на боковой поверхности. Семяприемник с сильно
расширенной базальной частью придатка, который не менее чем
до середины пигментирован (рис. 86, А—Д).
Сюда относятся паразиты крыс, из числа которых один вид (X. cheopis)
распространился по многим странам и в том числе встречается в Восточной Азии.
2(11). Предпигидиальная щетинка сидит не на особом выступе, а близ-
ко от края VII тергита. Резервуар семяприемника не шире, чем
придаток (рис. 86, А—Г).
3(4). У самца (рис. 77, Б) дорзальный и вентральный край IX стер-
нита почти одинаково склеротизованы. У самки семяприемник
длинный, основание придатка лишь слегка шире, чем резервуар
(рис. 86, А)..........................................X. cheopis.
Паразит крыс рода Rattus, широко распространенный во многих теплых странах
и завезенный в портовые и другие города всех континентов, в том числе в Хабаровск,
Владивосток и многие места Маньчжурии, Кореи и Китая. Активный переносчик
инфекций [16, 18].
4(3). У самца вентральный край IX стернита гораздо сильнее склеро-
тизован, чем дорзальная его часть (рис. 78). У самки основание
придатка семяприемника значительно шире, чем резервуар
(рис. 86, Б, В).
5(10). У самца парамер эдеагуса с узким апикальным дорзальным вы-
ростом в виде отдельного хвостика (рис. 79, А, Б). Семяприем-
ник короче, чем у X. cheopis, и основание придатка сильно взду-
то (рис. 86, Б, В).
Сюда относятся три близкие вида, которые между собой можно различить
безошибочно лишь по самцам.
31
6(7). Дорзальный теменной желобок глубокий, с волнистым дном
(рис. 29, Л). «Воронка» внутренней трубки эдеагуса с выступаю-
щими дорзальным и вентральным уголками и коротким седло-
образным склеритом над ней (рис. 79, Л). . . . . X. astia.
Паразит крыс (Rattus), распространенный в Иране, Индии и прилежащих к ним
странах. Считается менее активным переносчиком инфекций, чем X. cheopis [16, 18].
7(6). Дорзальный теменной желобок самца не такой глубокий
(рис. 29, Б).
8(9). «Воронка» внутренней трубки эдеагуса почти без выступающих
уголков, с длинным седлообразным склеритом над ней (рис. 79, Б)
:.......................................................X. nubicus.
Африканский вид, живущий на крысах, песчанках и т. д. [16, 18].
9(8). Эдеагус примерно такой, как у X. astia. Глаз уменьшен . . .
........................................................X. nesokiae.
Паразит пластинчатозубой крысы (Nesokia indica), распространен в Средней
Азии (Туркменская ССР, Таджикская ССР, Узбекская ССР) [153].
10(5). У самца парамер эдеагуса конической формы без дорзального
«хвостика». Семяприемник длинный, как у X. cheopis, но резер-
вуар его меньше (рис. 86, Г)..........................X. hawaiiensis
Паразит крыс (Rattus) Гавайских островов, где живет преимущественно вне го-
родов, на полевых крысах [16].
11(2). У самца предпигидиальная щетинка сидит на узком конусооб-
разном выступе VII тергита. На отростке Р{ половой клешни
щетины очень толстые, шпоровидные. У самки резервуар семя-
приемника значительно шире, чем придаток (рис. 86, Д) . . .
.......................................................X. brasiliensis.
Паразит крыс (Rattus) тропической и южной Африки, завезен в Бразилию и из-
вестны случаи завоза этого вида в портовые города других стран [16].
12(1). Имеется всего одна теменная щетинка (не считая угловой ще-
тинки заднекрайнего ряда). У самца (рис. 80—85) отросток Рз
половой клешни с апикальными щетинками. У самки VIII тер-
гит с единичными щетинками на боковой поверхности (не считая
краевых щетинок). Семяприемник со слабо расширенным осно-
ванием придатка (рис. 86, Е, Ж).
Сюда относятся главным образом паразиты песчанок. Самки приводимых ниже
видов различимы между собой не всегда; в частности, неразличимы самки трех под-
видов X. gerbilli и X. skrjabini; весьма между собой сходны самки X. conformis
и X. persica. При их определении следует учитывать сопутствующих самцов и геогра-
фический критерий. В Центральной Азии пока обнаружен лишь один вид —
X. skrjabini.
13(14). Щетинки, сидящие вдоль заднего края первого членика задней
лапки, сильно развиты (особенно у самцов); они намного длин-
нее щетинок переднего края и направлены в сторону от членика.
У самца край VII тергита позади предпигидиальной щетины об-
разует сильно развитый выступ. Отростки клешни Р\ и Р2 рас-
положены довольно тесно друг к другу, а Рз примерно той же
длины, что Р\..............................................X. hirtipes.
Паразит песчанок Средней Азии, живущий в сыпучих песках (с чем и связано
удлинение щетинок на лапке) [114].
14(13). Щетинки заднего края первого членика задней лапки (кроме
апикальных) мало удлинены и направлены почти вдоль членика.
15(18). Хоботок далеко не достигает конца передней коксы. Отростки Pi
и Р2 половой клешни тесно прилегают один к другому, а Рз не
55
заходит или слабо заходит за середину Pi (рис. 80). Резервуар
семяприемника небольшой (рис. 86, Е).
16(17). Глаз большой (52—80 у, длиной). Паразит песчанок ....
............................................................X. conformis.
Широко распространен на песчанках рода Meriones (включая подрод Pallasio-
mys), обитающих в Нижнем Поволжье, Закавказье, Средней Азии, Афганистане.
В Африке живет близкий вид или подвид —X. mycerini1 [137, 99а]. На тушканчиках
в пустынях Средней Азии недавно найдены блохи, очень близкие к X. conformis, от-
личающиеся более длинным хоботком.
17(16). Глаз очень мал (34—40 ^). Глазная щетинка часто отсутствует.
Паразит слепушонок........................ X. magdalinae.
Встречается на слепушонках (Ellobius talpinus) в Нижнем Поволжье и Средней
Азии [138].
18(15). Хоботок достигает или почти достигает вершины передней коксы
(кроме X. persica и самцов X. nuttali, у которых он несколько
короче). Глаз, как правило, не очень велик (36—52 ^).
19(20). Половая клешня самца как у X. conformis, но отросток шире,
а трапециевидная фигура парамера эдеагуса заметно суживается
к вершине. Резервуар семяприемника более крупный, его наи-
больший диаметр значительно больше, чем наибольший диаметр
придатка. Дополнительным отличием от X. conformis может слу-
жить отсутствие толстой щетинки на заднем крае задней голени
между четвертой и пятой группами шпор. См. также географию
этого вида............................................. . . X. nuttali.
Паразит песчанок, обитающих в прикаспийских районах Туркменской ССР
и прилежащей части Усть-Урта и Ирана (Шахруд); чаще на Rhombomys
opimus [137].
20(19). Отросток Рз половой клешни не короче или немного короче,
чем Pi (рис. 81). Резервуар семяприемника не особенно велик
(рис. 86, Ж).
21(22). На VIII стерпите самца близ середины вентрального края с каж-
дой стороны имеется группа из семи-восьми щетин (рис. 81).
У самки. (как и у самца) хоботок достигает конца передней
коксы. Семяприемник с небольшим резервуаром (рис. 86,2К);
самки едва ли отличимы от самок X. gerbilli. Глаз не очень мал
(44—54 ц)....................................................X. skrjabini.
Паразит песчанок (Rhombomys opimus). Распространен на Усть-Урте и в райо-
нах, лежащих севернее и восточнее Аральского моря, найденный также в Голодной
степи (Бетпакдала), в Южном Прибалхашье и в Западной Монголии (предгорья
Гобийского Алтая). Все места его обнаружения лежат около широты в 45° или не-
сколько севернее [137].
22(21). У середины вентрального края VIII стернита самца имеются
лишь единичные щетинки. Половая клешня с широким промежут-
ком между отростками Pi и 7Y Самки — см. ниже об отдель-
ных видах.
1 Иордан [16, 99а] сообщает, что в Туркменской ССР (Имам-Баба) в сборах
В. Кожанчикова (1912) с Rhobomys opimus найден один самец X. regis. Этот вид
был ранее известен только из Южной Аравии (с Meriones гех). По форме клешни
и семяприемника он напоминает X. conformis, но хоботок у него достигает вершины
коксы, а отросток Р3 очень короток, немного заходит за основание Щетинки на
тергитах более многочисленны, чем у других близких видов, например, на III тер-
гите брюшка имеется не менее восьми-девяти щетин с каждой стороны тела (у дру-
гих видов — до семи); задне-крайний ряд щетинок головы непрерывный. Отмечается
также сильное развитие поверхностной скульптуры тергитов, вследствие чего дорзаль-
ные очертания тергитов образуют в проходящем свете как бы мелкую пилу. Вентраль-
ный край головы кзади от передней коксы образует развитый треугольный выступ
(примерно, как у Echidnophaga); однако этот важный признак хорошо заметен лишь
в случаях, когда голова слегка отодвинута от переднегруди [16, 99а, 176, 181, 124L
Других сведений о находках этого вида в СССР у нас нет.
56
23(24). У самца IX стернит не расширяется к вершине (рис. 82—84).
Самки едва ли отличимы от самок X. skrjabini. Распространен
на равнинах Средней Азии........................................X. gerbilli.
Паразиты песчанок, преимущественно Rhombomys opimus, широко распростра-
нены в Средней Азии. Вид включает два четко выраженных подвида — западный
и восточный, и третий промежуточный между ними.
а. Отросток Рз обычно не короче чем Pif а часто даже заходит за его конец. Вы-
ступ VII тергита самца позади предпигидиальной щетинки слабо выражен
(рис. 82). Глаз небольшой (36—48 )...................X. g. gerbilli.
Западный подвид, распространенный в южных районах Туркменской
ССР [124].
б. Отросток Рз короче, чем Р\. Указанный выступ VII тергита резко выражен
(рис. 83). Глаз обычно крупнее (48—52 и)........................X. g. minax.
Восточный подвид распространен в Казахстане в пустынях долины р. Или
от границ СССР до оз. Балхаш, а также по среднему течению р. Сыр-Дарья
(Урсатьевская, Пахта-Арал, Джулек Чиилийского района) и в южной части
пустыни Кызыл-Кум (Узбекская ССР).
в. Отросток Рз в большинстве случаев короче, чем Pi, или равен ему, а выступ
VII тергита слабее, чем у X. g. minax (рис. 84) .... X. g. caspica.
Промежуточная форма, распространенная в северных районах Туркменской
ССР, а также на Усть-Урте и в северной части пустыни Кызыл-Кум (Кара-
Калпакия, Узбекистан, Казахстан) [154].
24(23). У самца IX стернит широкий, особенно к вершине, а на апикаль-
ном крае VIII стернита сидит заметная группа сильных щетинок
(рис. 85). Хоботок у самца и у самки далеко не достигает вер-
шины передней коксы. Глаз довольно большой (48—56 д). Оби-
татель горных районов...........................................X. persica.
Паразит песчанок Meriones persicus, обнаружен на Копет-Даге (Туркмен-
ская ССР) [153]. Отличительные признаки самок этого вида от самок X. conformis
требуют дополнительного изучения.
Род Sy по st emus [124, 176, 181]
Представители этого рода встречаются на грызунах и ежах в Афри-
ке и Средней Азии. Нередко переходят и на хищных. В Средней Азии
известны два вида.
1(2). На задней голени шесть групп шпор. По вентральному краю
заднего бедра близ его основания имеется слабый зубчик. У сам-
ца отростки половой клешни короткие..........................S. pallidus.
Блоха ежей, распространена в АсЬрике, Месопотамии и передней Индии. У нас
известна в Туркменской ССР, Кара-Калпакской автономной области и в Таджик-
ской ССР. Обитатель пустынь и полупустынь.
2(1). На задней голени больше чем шесть групп шпор. Вентральный
край заднего бедра близ его основания закруглен, без зубчика.
Отростки половой клешни длиннее, чем у предыдущего вида . .
.......................................................... S. longispinus.
Блоха ежей, распространена во многих районах Средней Азии.
Род Pariodontis [124, 130, 176, 181]
Паразиты дикобразов (Hystrix), обитающих в Африке и южной
Азии. У нас из Туркменской ССР описан Pariodontis riggenbachi tur-
kestanica.
ПОДСЕМЕЙСТВО SPILOPSYLLINAE
Род Spilopsyllus [29]
S. cuniculi — паразит кроликов в Западной Европе.
Род Hoplopsyllus [183]
Паразиты кроликов и зайцев Северной Америки. Один вид известен
также из Азии: Hoplopsyllus glacialis profugus (рис. 87, А, Б), где он
57
обнаружен на Lepus tolai в Казахской ССР (Илийская долина), в Туве
(Саянский хребет) и Монголии (Северо-западный и Южный Хангай, Тай-
шири). Другие подвиды этого вида обнаружены в Гренландии и Канаде.
НАДСЕМЕЙСТВО CERATOPHYLLOIDEA
Это надсемейство включает (по Иордану) большинство видов блох
земного шара. Возможно, что такое объединение несколько искусственно,
и поэтому общая характеристика надсемейства встречает затруднения.
Большинство относящихся сюда видов блох объединено многими об-
щими признаками, но имеются отдельные представители, в отношении
которых приходится делать различного рода оговорки. Можно сказать,
что к этому надсемейству относятся все блохи, которые по совокупности
признаков не могут быть отнесены к Pulicoidea, а отдельные морфологи-
ческие признаки, перечисленные в общей характеристике Pulicoidea
(стр. 51), оказываются у большинства или у многих Ceratophylloidea пря-
мо противоположными.
СЕМЕЙСТВО COPTOPSYLLIDAE
Род Coptopsylla [148, 150, 153, 154]
Представители этого рода являются зимними паразитами песчанок
(Gerbillinae) Африки и Азии и проникли в прикаспийские районы Евро-
пейской части СССР. В Средней Азии известно уже шесть видов и ряд
подвидов, а в Закавказье два вида. В пределах Восточной Азии пока
не обнаружены.
СЕМЕЙСТВО VERMIPSYLLIDAE
Род Vermipsylla [122 (23), 123, 129, 142, 153]
Блохи этого рода живут на копытных как диких, так и домашних,
нападая на них на зимних горных пастбищах Средней и Центральной
Азии. Местному населению эти паразиты известны под названием «ала-
курт» (пестрый червь). Они приносят значительный ущерб животновод-
ству, в особенности овцеводству, так как портят шерсть, вызывают исто-
щение молодняка, а иногда приводят его к гибели. Это редкий пример,
когда блохи, паразитируют на не имеющих постоянного жилья или ло-
гова животных. Предимагинальное развитие у блох происходит летом
на поверхности земли; этому способствуют своеобразные климатические
условия высокогорных пастбищ. Блохи начинают нападать на животных
поздней осенью, откладывают яйца зимой, а к весне погибают. Харак-
терной особенностью этих блох, за которую они получили свое название
«алакурт» является сильное растяжение брюшка у самок во время созре-
вания яиц; такая самка напоминает полосатого червяка. Известны два
вида этого рода, причем каждый из них рассматривается как представи-
тель особого подрода.
1(2). Хоботок состоит из 10—14 члеников (рис. 35). На задней голени
шесть групп шпор (рис. 347, Л). Самка при созревании яиц до-
стигает 6.мм в длину (рис. 37, Л) ..... V. (V.) alacurt.
Паразитирует на многих видах копытных животных (овцах, лошадях, архарах
и др.), а также встречается и на хищных (собаках, лисицах, волках); нападает и на
людей. Распространен в Центральном Тянь-Шане (Казахская ССР, Киргизская ССР)
' на высокогорных пастбищах, расположенных на высоте от 3 000 до 3 500 м над уров-
нем океана. Известен также из Монголии (близ Улан-Батора).
58
2(1). Хоботок очень длинный, состоит из 16—20 члеников (рис. 36).
На задней голени четыре группы шпор (рис. 347, Б). Самка при
созревании яиц (имеющих темносерый цвет) достигает 16 мм
в длину (рис. 37, Б)......................V. (Dorcadia) dorcadia.
Этот вид блох обнаружен на диких копытных (джейране и др.) в Китае
(Шаньси), Монголии (южные склоны Гобийского Алтая около оз. Сого-Нор) и на
овцах Центрального Тянь-Шаня (Киргизская ССР). На Тянь-Шане живет на высоте
от 1 800 до 3 500 м над уровнем океана.
Род Chaetopsylla (=Trichopsylld) [123, 148]
К этому роду принадлежат паразиты хищных — лисиц, барсуков,
куниц, медведей и т. д. Довольно многочисленные виды этого рода рас-
пространены главным образом в Европе и Азии. Выделенные в послед-
ние годы группы Arctopsylla и Achaetopsylla родственны с остальными
представителями и рассматриваются нами как подроды.
Ввиду большого сходства между собой видов этих блох и непостоян-
ства у некоторых из них отдельных важных признаков не всегда легко
можно определить эти виды. Однако, если тщательно сравнивать пре-
параты с приводимыми нами рисунками и учитывать отмечаемые исклю-
чения, то во всех случаях можно будет точно определить тот или иной
вид. Для определения самцов блох этого рода особенно большое значе-
ние имеет строение эдеагуса, а для самок — форма копулятивной сумки.
При рассмотрении эдеагуса нельзя забывать о подвижности отдельных
частей, влияющей на их взаиморасположение и общую конфигурацию
органа (рис. 88, А). На рисунках, на которых изображены самцы
Chaetopsylla, значком (*) отмечен особенно характерный боковой пара-
мер эдеагуса. Что касается копулятивной сумки, то рассмотреть ее
у блох этого рода в большинстве случаев можно только после просвет-
ления препарата.
Общей особенностью блох рода Chaetopsylla является сильное рас-
тяжение брюшка у самок при созревании яиц (до 4—5 мм). Большинство
видов этого рода имеют довольно крупный размер и темнобурый цвет
покровов, но есть и некоторые желтые или светлокоричневые виды
(Ch. homoeus и др.).
Из 14 видов Chaetopsylla, имеющихся в СССР, мы включаем в табли-
цу И обнаруженных в Сибири или в восточных районах Средней Азии,
а остальные виды только упоминаем в примечаниях, так же как и один
южно-китайский вид.
1(4). Средние тергиты брюшка имеют форму, изображенную на
рис. 353, В, Г, т. е. нижняя часть каждого из этих тергитов об-
разует резкий узкий язык, направленный кзади. Стигмы этих
тергитов очень велики. На щечной лопасти сзади глаза имеется
щетинка, на метэпимере — более чем два ряда щетинок. Стерни-
ты IV—VII разделены на правый и левый склерит.
2(3). Лоб с сильно выступающим бугорком, угловатый (рис. 33). Хо-
боток состоит из 7—10 члеников (часто наблюдается ложная,
т. е. неполная, членистость). Хоботок немного заходит за верши-
ну коксы. Число щетинок в теменных рядах: 2 (0—3) + 2—4
(до 6). Апикальные щетинки II, III и IV члеников задней лапки
толстые и не особенно удлиненные. Например, самая длинная
щетинка II членика лишь немного заходит за вершину IV чле-
ника. У самки (рис. 98, Б) семяприемник с большим резервуа-
ром, а проток копулятивной сумки короткий, с частичной скле-
ротизацией. У самца (рис. 98, А) характерной формы эдеагус и
половая клешня с коротким подвижным пальцем, напоминающим
клюв, изогнутый кпереди . . Ch. (Arctopsylla) tuberculaticeps.
59
Паразит медведей. Обнаружен в Италии, Северной Европе, на Тянь-Шане, по-
бережье Охотского моря, Камчатке и в Северной Америке (где именуется Ch. ursi
Roths. 1902, однако отличия американских представителей твердо не установлены) 1
[123, 179, (38), 19].
3(2). Лоб закругленный с небольшим острым бугорком (который ча-
сто бывает стерт). Хоботок состоит из пяти-восьми члеников1 2.
Число щетинок в теменных рядах 1—3 + 2—3. На вершине II,
III и IV члеников задней лапки имеются тонкие и очень длинные
щетинки, самая длинная щетинка II членика достигает вершины
V членика или заходит за нее. У самки резервуар семяприемника
меньше, чем у предыдущего вида, а копулятивный проток длин-
ный и хорошо склеротизован. Эдеагус самца также совершенно
иной формы, с резко выступающим кверху дорзальным параме-
ром, а подвижный палец половой клешни имеет вид клюва, на-
правленного кзади.............................................Ch. lasius.
Этот вид до недавнего времени был известен по описанию двух самок, обна-
руженных в одном из районов Центральной Азии (Анкуза 3) [185]. В настоящее время
мы получили от Р. П. Зиминой много экземпляров блох, которых надо отнести к дан-
ному виду; они собраны с лисиц в Иссык-Кульской области (Киргизская ССР) на
хребте Терскей Алатау.
Ch. lasius можно пока относить к подроду Arctopsylla (куда относится предыду-
щий вид), однако своеобразный эдеагус и другие особенности потребуют, вероятно,
выделения его в особый подрод.
4(1). Средние тергиты брюшка имеют иную форму (рис. 353, Д—Л4).
Стигмы этих тергитов не особенно велики.
5(10). Как правило, хоботок состоит из пяти члеников (иногда у Ch. glo-
biceps, alia, zibellina, mirabilis, как, вероятно, и у других видов,
наблюдается увеличение числа члеников хоботка). Хоботок не
заходит за вершину передней коксы, а часто даже не дости-
гает ее 4.
6(9). На щечной лопасти сзади глаза имеется сильная щетинка. У са-
мок на IV—VII тергитах ниже уровня стигм бывают щетинки
(рис. 353, Д—3 и 7И) хотя бы на некоторых из названных терги-
тов (исключение — Ch. appropinquans, рис. 353, Д).
7(8). На метэпимере, кроме двух рядов щетинок, имеются еще много-
численные щетинки на передней части этого склерита. Форма
средних тергитов изображена на рис. 353, М. Теменные щетинки
многочисленны (4—5 + 5—9). У самки (рис. 90) VII стернит
, с узкой глубокой вентральной вырезкой. Семяприемник довольно
крупный, а проток копулятивной сумки склеротизованный, длин-
ный. Половая клешня самца (рис. 89) с очень характерным
большим вентральным, покрытым волосками и щетинками, отрост-
ком с гривой по краю, которого нет у других блох. Теменная
1 Другой представитель подрода Arctopsylla: Ch. (A.) hyaenae Koi. паразитирует
на гиенах и медведях, обитающих на Кавказе [123], и отличается от Ch. tuberculaticeps
только резко удлиненными апикальными щетинками II, III и IV члеников зад-
ней лапки.
2 У экземпляров из СССР он состоит из шести-восьми члеников, часто с ложной
членистостыо, и достигает вершины коксы или слегка за нес заходит. Однако в ори-
гинальном описании Ротшильда указано, что хоботок состоит из пяти члеников. Во
всяком случае надо иметь в виду, что колебание числа члеников хоботка у Chaeto-
psylla нередкое явление.
3 Где находится Анкуза, остается невыясненным. Гопкинс (Британский музей)
сообщил нам, что, по его сведениям, это место расположено близ оз. Иссык-Куль.
Однако местные работники не знают такого названия.
4 Ниже отмечены случаи, когда хоботок более короток. В остальных случаях
он достигает или почти достигает вершины коксы в выпрямленном ее состоянии, что,
к сожалению, у этих блох наблюдается далеко не всегда, так как хоботок бывает
отогнут кзади.
60
ямка (желобок) самца переходит и на все тергиты груди и брюш-
ка, причем вдоль спины расположено много щетинок, образую-
щих гриву...........................Ch. (Achaetopsylla) mirabilis.
Представители этого вида обнаружены на Кавказе (оз. Севан, Теберда), на
Копет-Даге (близ Ашхабада); в большом числе с куниц (Martes foina) и отдель-
ные экземпляры с лисиц [145].
Этот вид Аргиропуло [10] предложил выделить в особый род Achaetopsylla. Мы
полагаем, что удобнее считать его подродом.
Упомянем еще, что, судя по описанию, южнокитайский вид Ch. hangchowensis
Liu 1939 [27] имеет три ряда шетинок на метэпимере и этим приближается к Ch. mi-
rabilis, но по другим признакам он ближе к Ch. zibellina, giobiceps, trichosa и др.
По хоботку, имеющему шесть-семь члеников, он сходен с Ch. micado.
8(7). На метэпимере имеются лишь два развитых ряда щетинок (при-
чем передний ряд иногда может быть не совсем правильным и с
единичными передними добавочными щетинками). Форма сред-
них тергитов изображена на рис. 353, Д—3.
а) Число щетинок в теменных рядах: 2(1) + 2—3(4). Самка (рис. 96, А)
имеет ровный округленный край VII стернита и длинный дуго-
образный хорошо склеротизованный проток копулятивной сумки.
Для самца (рис. 93) особенно характерен сильно развитый язы-
кообразный боковой парамер эдеагуса . . . Ch. appropinquans.
Блоха хищных Восточной Сибири; обнаружена на куницах (Mustela flavigula),
колонках (Mustela sibirica), барсуках (Meles meles), лесном коте (Felis euptilura)
•и др. в ряде мест Приморской и Амурской областей и в Забайкалье [126, 148].
б) Теменные ряды щетинок: 3—4(5)+ 4(3—5). Хоботок далеко не дости-
гает вершины коксы. У самки (рис. 88, Б) край VII стернита
с довольно широкой вентральной вырезкой. Проток копулятив-
ной сумки короткий, лишь частично и слабо склеротизованный.
У самца (рис. 88, А) боковой парамер эдеагуса округлый, мало
выступающий, а дорзальный — неправильно трапециевидный
(расположение парамеров изменчиво — рис. 88, А)..........................
.................................................Ch. homoeus.
Блоха хищных, преимущественно семейства Mustelidae (хорьки, горностаи, ласки
•и Др.), но часто паразитирует и на лисицах, корсаках, волках и т. д. Изредка попа-
дается на сурках и других грызунах. Широко распространена, в особенности в гор-
ных странах Европы и Азии — в Альпах, на Кавказе, Тянь-Шане, Алтае, в Саянах,
Забайкалье, на Тибете, в Монголии и Бирме. Встречается преимущественно в степ-
ных местностях, как в горах, так и на равнинах. [185, 148, 179, (38)].
•в) Теменные ряды щетинок: 3—4 + 4—6. Очертания тергитов более
узкие и более изогнутые, чем у близких видов (рис. 353, Ж).
У самки (рис. 91,5) проток копулятивной сумки несколько
длиннее и заметнее, чем у Ch. homoeus1. У самца (рис. 91, А)
сочленовная ямка клешни расположена вблизи заднего края и
несколько ниже его середины. Боковой парамер сильнее склеро-
тизован, чем у Ch. homoeus, широкий, а дорзальный клюво-
образно нависает выше него. На обеих сторонах вершины
IX стернита сидят по две сильных щетинки.........................Ch. alia.
Этот вид в небольшом числе найден на соболях в Западной Сибири (Кондо-
‘Сосьвинский заповедник), но настоящий хозяин его и ареал еще не выяснен [153].
г) Теменные ряды щетинок: 3—5 + 3—5. Хоботок не достигает вершины
передней коксы. Очертания тергитов см. рис. 353,3. У самки
(рис. 92, Б) край VII стернита с небольшой вентральной вырез-
кой. Проток копулятивной сумки склеротизованный, разделен
перехватом на две примерно равные по длине части. У самца
1 VII стернит у имеющихся экземпляров самок сломан или измят (как на
рис. 91, Б); видимо, у него нет вентральной вырезки, ошибочно упомянутой в кратком
•описании этого вида (И. Г. Иофф, 1946), и вообще апикальный край его ровный.
61
(рис. 92, Л) дорзальный парамер занимает всю вершину эдеагу-
са, причем его округленный конец заходит за вентральный па-
рамер .............................................Ch. dogieli.
Вид, обнаруженный в большом количестве на колонках (Mustela sibirica) в Суд-
зухинском заповеднике Приморской области [154].
9(6). На щечной лопасти сзади глаза, как правило, нет щетинки (ис-
ключения встречаются среди Ch. trichosa, имеющих иногда эту
щетинку). На метэпимере два ряда щетинок. У самок на IV—VII
тергитах ниже уровня стигм щетинок нет (рис. 353,//, /С)1.
а) Число щетинок в теменных рядах: 3(4) +3. Хоботок не достигает
вершины передней коксы. У самки (рис. 94, Б) край VII стернита
с узкой боковой щелевидной вырезкой. Проток копулятивной
сумки короткий, слабо склеротизованный. У самца (рис. 94, Л)
сочленовная ямка клешни находится в верхней ее половине. Вер-
шина эдеагуса треугольная с характерным острым выступаю-
щим дорзальным парамером и слабо выдающимся боковым
..........................................................Ch. zibellina.
Паразит соболя (Martes zibellina) обнаружен в Западной Сибири (Кондо-Сось-
винский заповедник) и Приморье (Судзухинский заповедник) [153].
б) Теменные ряды щетинок: 0—3+1—3. У самки (рис. 96, Б) край
VII стернита с вариирующей, часто очень слабой боковой вырезкой.
Проток копулятивной сумки дугообразный, хорошо склеротизо-
ванный. У самца (рис. 95) сочленовная ямка клешни находится
вблизи середины заднего края. Дорзальный парамер округленный,
а боковой слабо различим................................Ch. trichosa.
Паразит барсука (Meles meles). Широко распростанен в Европе, на Кавказе,
Урале, Тянь-Шане, Алтае, на горах и равнинах [148].
в) Теменные ряды щетинок: 2—3(0—1)+2—3(1—4). У самки край
VII стернита с широкой боковой вырезкой, обычно более глубо-
кой, чем у предыдущего вида. Проток копулятивной сумки корот-
кий, слабо склеротизованный. У самца сочленовная ямка еще
ближе придвинута к вершине клешни, чем у Ch. zibellina, и по-
движный палец поэтому очень короткий. Дорзальный парамер
округленный, а боковой образует характерный узкий язычок
.......................................................Ch. giobiceps.
Паразит лисицы (Vulpes vulpes). Широко распространен в Европе, на Кавказе,
Усть-Урте и, невидимому, есть на Тянь-Шане [148].
10(5). Хоботок состоит из шести, редко семи, члеников, достигает вер-
шины коксы, но за нее почти не заходит. Позади глаза щетинок
нет. Число щетинок в теменных рядах: 3(2—4) + 3(4). На
метэпимере два ряда щетинок. У самки на IV—VII тер-
гитах ниже уровня стигм щетинок нет (рис. 353, Л). Край
VII стернита (рис. 97, Б) ровный, округленный. Проток копуля-
тивной сумки длинный, хорошо склеротизованный. У самца
(рис. 97, Л) характерный дорзальный парамер в виде широкого
клюва или шлема, нависающего над всей вершиной эдеагуса. Бо-
ковой парамер в виде небольшого узкого язычка . . Ch. mikado.
Этот вид был описан по экземплярам самок, собранных в Японии; после
обнаружения его в Приморском крае (окрестности Владивостока и Судзухинский за-
поведник) он получил у нас название Ch. nocens. Полученный от К. Иордана рисунок
самца позволил установить, что Ch. nocens I. 1946 = Ch. micado R. 1904. Паразити-
рует на колонке (Mustela sibirica) и других хищных животных (Lutra lutra, Felis
euptilura, Vulpes vulpes), а также встречен на зайце [188 (11), 153, 154].
1 В эту же группу, кроме перечисленных ниже трех видов, попадают: Ch. matina
(Пиренеи, Кавказ [179 (32)]), Ch. korobkovi (Усть-Урт, Копет-Даг [158]) и несколько
близко родственных форм, образующих группу Ch. rothschildi (Западная Европа,.
Кавказ, Копет-Даг [156]).
62
СЕМЕЙСТВО CERATOPHYLLIDAE
Сюда относятся многие блохи, паразитирующие на грызунах и дру-
гих млекопитающих, а также на птицах. Многие из относящихся сюда
видов принадлежат к «блохам шерсти», т. е. предпочитают держаться на
самом хозяине, а не в его гнезде (в отличие от блох следующего семей-
ства). Мы принимаем несколько иной состав этого семейства, чем было
до сих пор принято, и потому даем краткую его характеристику.
Как правило, имеются апикальные зубчики на метанотуме1 и на
нескольких тергитах брюшка; однако гребней на тергитах брюшка нет.
У обоих полов пигидий не выпуклый. Задняя кокса без группы шипиков
на внутренней поверхности. У самца IX стернит с аподемальным тяжем,
идущим кпереди под эндофаллусом. У самки на вершине церка, кроме
сильной апикальной щетинки, имеются одна-две (или больше) довольно
длинные щетинки. Семяприемник лишь один.
ПОДСЕМЕЙСТВО CERATOPHYLLINАЕ
Род Tarsopsylla [192]
В Азии и Европе обитает только один представитель этого рода —
Т. octodecimdentata (часто обозначаемый сокращенно: 18-dentata), пара-
зит белок (Sciurus vulgaris), распространенный почти всюду, где имеются
эти грызуны, — от Германии и Швейцарии на западе до лесов Кавказа
(куда этот вид был завезен с белками), Алтая, Хангая, Хэнтэя, Сихотэ-
Алиня и многих других пунктов Европы и Сибири. Признаки самцов
и самок см. на рис. 99, А, Б.
Другой вид этого рода живет в Северной Америке.
Род Myoxopsylla [192]
Паразиты грызунов из семейства сонь (Myoxidae). Известно всего
два вида — западноевропейский и североафриканский М. laverani и кав-
казский— М. jordani [145].
Род Paraceras [29, 100, 125, 179 (32, 38, 41)]
Паразиты барсуков (Meles meles), обитающих в Европе и Азии.
Кроме приводимых нами видов, в Китае и вообще Южной Азии встре-
чаются виды с более коротким хоботком (менее чем целый членик за-
ходит за коксу передней ноги); мы не включаем их в таблицу, так как
они недостаточно изучены и относятся в основном к Индомалайской
зоогеографической области [см. о них 179 (32, 38, 41)]; одна подобная
самка найдена в провинции Шэньси.
1(2). У самцов на вершине II и III члеников лапок имеется ото-
рочка из нескольких длинных тонких волосковидных щетинок
(рис. 350,5). Задний край подвижного пальца половой клешни
выпуклый (рис. 101). Задний край VIII тергита самки (рис. 103)
вариирует, но обычно ниже пигидия образует более или менее
резкий выступ1 2...................................Р. flabellum.
1 В виде исключения у Ceratophyllus, паразитирующих на ласточках, зубчик на
метанотуме может быть или рудиментарным, или отсутствовать вообще.
2 Следует заметить, что самок Р. moles и Р. flabellum отличить нелегко и, может
быть, было бы правильно объединить эти формы в один вид, сохранив их лишь как
подвиды; это соответствовало бы и современным взглядам на систематику их хозяев
барсуков, обитающих в Европе и Сибири. Однако мы оставим решение этого вопроса
до накопления большего материала.
63
Тянь-Шань, Алтай, Кустанайская область, Забайкалье, Приморская область.
Возможно, что восточные представители слегка отличаются более узким дигитоидом.
В Южном Китае обнаружен и описан весьма близкий вид Р. crispus, имеющий,
однако, сравнительно прямой задний край и резко вогнутый передний край дигитоида;
по самкам он, вероятно, неотличим от Р. flabellum. То же надо сказать о Р. sinensis
из Сычуани, который описан только по самкам [125, 100, 27, 105].
2(1). У самцов на вершине II и III члеников задних лапок отдель-
ные длинные волосовидные щетинки не образуют оторочки
(рис. 350, Л). Задний край подвижного пальца половой клешни
(рис. 102) вогнутый или прямой. Выступ заднего края VII тер-
гита самки обычно более закруглен (рис. 104)1 . . . Р. melis.
Блохи этого вида встречаются в Западной Европе, в различных частях СССР
(в Московской, Рязанской и Ленинградской областях, в Крыму, на Кавказе) и в Се-
верном Иране.
В литературе [29] имеются указания о наличии этого вида также в Северной
Азии и Японии. Однако в Японии были обнаружены только самки [111], которые
трудно отличимы от самок близких видов, а относительно упоминаемых находок в Се-
верной Азии никаких подробностей неизвестно. Поэтому распространение Р. melis
в Азии требует дальнейшего изучения.
Род Aenigmopsylla [154]
Единственный известный представитель этого рода — Ае. grodekovi
обнаружен недавно в лесных районах Приморского края (Гродеково, Си-
хотэ-Алинь) и описан по единичным экземплярам, собранным с белок
и других лесных грызунов. Следует подчеркнуть, что эти блохи, обла-
дающие большим сходством с другими представителями подсемейства
Ceratophyllinae, отличаются от них наличием развитых (хотя и тонких)
передних ветвей тентория, что несколько приближает этот вид к Рага-
doxopsyllinae. У самца (рис. 100, А) характерно отсутствие неподвижного
пальца половой клешни, длинный треугольный дигитоид с шипами на
вершине, своеобразная форма и вооружение IX стернита. Признаки са-
мок (округленный край VII стернита и бочкообразный резервуар семя-
приемника) см. на рис. 100,5.
Род Amphalius [179 (39), 180, 173]
Паразиты пищух (сеноставок — Ochotona), обитающих в Азии и Се-
верной Америке. Пока известно три вида, из которых два живут в Азии.
Эти виды (или подвиды?) хорошо отличаются по самцам, а самки могут
быть распознаны по географическому признаку.
1(2). Дигитоид (рис. 106) сравнительно широк; у его верхне-заднего
края сидит широкая, довольно светлая саблевидная шпора. При-
знаки самки изображены на рис. 105...........................A. runatus.
Паразит даурской пищухи (Ochotona daurica), но нередко встречается и на дру-
гих видах пищух, а также на сусликах, тарбаганах и многих других животных. Рас-
пространен в Забайкалье, в верховьях р. Лены (Усть-Курт), на Алтае, в Саянах
(Канск, Тува), Монголии (Хангай, Тайшири, Монгольский Алтай, Убсанурский, Цен-
тральный и Хэнтейский аймаки). Указания на обнаружение в Маньчжурии [57] осно-
ваны, вероятно, на том, что этот вид блохи был найден в районе станции Маньчжу-
рия, около северо-западной границы Маньчжурии {180]. Возможно, блохи этого вида
проникают и в некоторые районы этой страны, поскольку ареал даурской пищухи
доходит до озера Далай-Нор (а может быть, даже до Шаньси) [155], но пока досто-
верных данных о наличии этих блох в Маньчжурии не имеется. По направлению
с севера на юг A. runatus несколько изменяется морфологически, начиная прибли-
жаться к следующему виду.
1 См. примечание 2 на стр. 63.
64
2(1). Дигитоид (рис. 107) более узок и у его верхне-заднего края
сидит не широкая шпора, а обычная щетина. Неподвижный
отросток клешни несколько расширяется к вершине. Самки мор-
фологически плохо отличимы от предыдущего вида . . . A. clarus.
Паразит пищух подрода Conothoa (macrotis, rutila) и др. Описан по экземпля-
рам, найденным на Тибете. Обнаружен также в районе Центрального Тянь-Шаня.
Род Oropsylla [128, 133, 174, 19]
Представители этого рода паразитируют на сурках и сусликах, оби-
тающих в Евразии и Северной Америке. Ниже приводятся сведения
о трех известных у нас видах подрода Oropsylla и об одном виде под-
рода Diamanus, живущем в Китае.
1 (6). VIII стернит самца узкий со щетинками на вершине (рис. 108—110).
Придаток семяприемника с хитинистой кнопкой на конце
(рис. 111, А—Б) ..........................подрод Oropsylla.
2(5). На наружной поверхности заднего бедра одна или единичные
щетинки. Подвижной палец половой клешни с более или менее
прямым передним краем (рис. 108, 109).
3(4). Хоботок длинный; не менее, чем один членик, а чаще и часть
следующего заходит за вершину передней коксы. Глаз неболь-
шой (рис. 40). Лобный зубчик ясно выступает. Зубцы гребня,
как правило, расположены не тесно друг к другу; в большинстве
случаев они значительно короче, чем спинка переднегрудки (ко-
торая удлинена у этого вида), иногда лишь немного короче
(измерять по дорзальной линии). Резервуар семяприемника гру-
шевидный, малый (рис. Ill, А), форма его несколько варьирует.
Модифицированные сегменты самца — см. рис. 108 . . .О. silantiewi.
Паразит сурков (тарбаганов) (Marmota) распространенный всюду, где они встре-
чаются: от Донских и Приволжских степей на западе до Тихого океана на востоке
и от Приуральских степей и Камчатки на севере до Тянь-Шаня и Гималаев на юге.
Разными авторами делались попытки разделить этот вид и выделить в пределах его
ареала ряд подвидов и даже особые виды (О. crassus [100], О. weisskei [198],
О. segregatus [194, 139]); однако до настоящего времени ареалы их и морфологические
различия недостаточно выяснены, и мы предпочитаем не вводить этих названий
в обращение [122, 133, 19].
4(3). По сравнению с предыдущим видом, хоботок короче, за вершину
передней коксы заходит меньше, чем на целый членик. Глаз не-
сколько крупнее и сильнее пигментирован (рис. 41). Лобный
зубчик слабый, часто в виде следа. Зубцы гребня, как правило,
несколько длиннее, чем спинка переднегруди, и расположены
густо. Резервуар семяприемника несколько крупнее (рис. 111, Б).
Модифицированные сегменты самца отличаются мало (рис. 109).
..................................О. asiatica (или О. alaskensis).
Паразит длиннохвостого суслика Citellus undulatus. Распространен во многих
районах Азии (Якутск, Колыма. Камчатка, Восточный Мыс, Забайкалье, Предбай-
калье, Красноярск, Тува, юго-восток Алтая, Джунгарский Алатау, Монголия, Маньч-
журия) [126, 19]. Учитывая, что тот же вид суслика живет и в Северной Америке
(Аляска, Канада), где он известен под именем Citellus parryi, а также принимая во
внимание недавно опубликованные данные по морфологии паразитов этих сусликов
[174, 172], в настоящее время почти не остается сомнений, что рассматриваемый вид
блох идентичен американскому виду О. alaskensis. Если это так, то по закону приори-
тета должно быть принято именно это название как наиболее старое. К числу сино-
нимов должны быть отнесены также: О. elana — Маньчжурия [92] и О. stejnegeri —
Восточный Мыс [179 (35)].
5(2). На наружной поверхности заднего бедра в подавляющем боль-
шинстве случаев имеется развитый боковой ряд — около восьми-
девяти щетинок. Хоботок длинный, как у О. silantiewi. Гребень —
5 Определитель блох
65
как у О. asiatica. Передне-верхняя часть дигитоида срезана, и по-
этому передний край последнего образует угол, направленный
кпереди (рис. ПО)................................О. ilovaiskii.
Паразит крупных сусликов (Citellus fulvus, С. major и др.) [128, 133]. Обнару-
жен во многих степных местностях от Херсонских степей на западе до Чуйской до-
лины (Киргизская ССР) на востоке (а возможно, что до Алтая и Кузбасса, откуда
имеются только самки) и от Куйбышевских (Самарских) степей на севере до Армян-
ской ССР на юге. Восточная граница распространения мало обследована. Отметим
также, что предполагавшийся ранее [140] разрыв ареала этого вида между Волгой
и Доном не подтвердился, но все же в Сальских степях он менее распространен, чем
в других частях его ареала.
6(1). VIII стернит самца (рис. 112, Л) рудиментарный. Дигитоид
узкий и длинный, далеко заходит за тело клешни. Придаток семя-
приемника без хитинистой кнопки на конце, резко изогнутый
у основания (рис. 112, Б) . . . . О. (Diamanus) mandarinus.
Паразитирует на Citellus mongolicus в Китае (Шэньси). Другой вид этого под-
рода живет в Северной Америке [100, 179 (39), 139, 26].
Род Rostropsylla [179 (18), 128[
К этому роду принадлежит блоха среднеазиатского тонкопалого сус-
лика (Spermophilopsis leptodactylus)—R. daca; обнаружена в Туркмен-
ской ССР, Таджикской ССР (низовья Вахша), Узбекской ССР, Кара-
Калпакской АССР, в южных областях Казахской ССР (Балхашский р-н,
Пахта-Арал). Наличие этого вида блох в Закавказье (долина Аракса)
требует подтверждения.
Род Ceratophyllus
Из обширного рода Ceratophyllus (в состав которого еще недавно
входили почти все представители теперешних подсемейств Ceratophyllinae
и Paradoxopsyllinae) разными авторами постепенно выделялись отдельные
группы в самостоятельные роды [128, 192, 197, 179 (39), 139 и др.], при-
чем этот процесс дробления привел к тому, что в настоящее время в ино-
странной литературе к Ceratophyllus s. str. 1 относится лишь срав-
нительно небольшая группа блох, паразитирующих на птицах. Одна-
ко, учитывая нежелательность перемены родового названия многих
широко известных видов блох, паразитирующих на сусликах, крысах
и т. д., а также крайне близкое родство ряда выделенных групп, свя-
занных между собой переходными формами (Ceratophyllus s. str., Mono-
psyllus, Megabothris, Callopsylla, Citellophilus и т. д.), мы считаем целе-
сообразным рассматривать подобные группы как низшие подразделения
(подроды, секции) одного рода Ceratophyllus s. 1.1 2. Ниже приводятся
признаки для определения этих групп. На практике самцы многих ви-
дов легче определяются путем сравнения модифицированных сегментов
с соответствующими рисунками. Для самок же, во избежание ошибок,
всегда следует просмотреть основные групповые признаки, приводимые
в таблице. ~ 'А:‘
Определитель подразделений рода Ceratophyllus s. I.
1(2). У самца VIII стернит рудиментарен, анальный стернит не длин-
нее тергита, и имеются две развитые предпигидиальные щетинки:
кроме главной длинной щетинки, довольно хорошо развита еще
1 s. str. — sensu strict© — в узком смысле.
2 s. 1. — sensu lato — в широком смысле.
66
одна — верхняя. Модифицированные сегменты общего характера
(изображены на рис. 113; 119, Л). У самки копулятивная сумка
спирально изогнута, а семяприемник типа, изображенного на
рис. 114; 119, Б............подрод Nosopsyllus s. 1. (стр. 68).
а) Апикальные щетинки II членика задних лапок не заходят за вершину
IV членика. У самок в большинстве случаев имеются хотя бы
единичные верхние фронтальные щетинки (рис. 42).....................
................................секция Nosopsyllus s. str. (стр. 68).
б) Указанные апикальные щетинки заходят за вершину IV членика
(рис. 348, Г). У самки нет фронтальных щетинок ....................
.....................................секция Gerbillophilus (стр. 69).
2(1). Признаки иные.
3(12). Стигма VIII тергита обычная, небольшая (рис. 140, 146).
4(7). На внутренней поверхности задней и средней ляжки щетинки
имеются в большинстве случаев не только на апикальной поло-
вине ляжки, но и на базальной, хотя бы единичные (не считая
сидящих по переднему краю) — рис. 346, В. У самца анальный
стернит намного длиннее тергита (как на рис. 146, 162). У самки
копулятивная сумка мешкообразно расширена (рис. 123; 129, Б).
5(6). У самцов имеется покрытая шипиками площадка (поле шипиков)
на внутренней поверхности VIII тергита близ дорзального края
(рис. 126, Л). Дигитоид всегда широкий, примерно треугольной
(или трапециевидной) формы, с задним краем, вооруженным
t сильными щетинами или шипами, не сидящими тесно друг к другу.
Строение VIII и IX стернитов показано на рис. 121; 124—126, А,
например, типична конусообразная апикальная доля IX стер-
нита с выступающим острым передне-нижним углом, характерны
также лопасти VIII стернита. У самок, как правило, фронталь-
ные щетинки отсутствуют и имеются лишь две предпигидиальные
щетинки (три бывают только в качестве аномалии). Семяприем-
ник— типа, изображенного на рис. 123; 126, Б; придаток, как
правило, длиннее резервуара, довольно широкий, а резервуар
небольшой, овальный. Копулятивная сумка без сильно склероти-
зованных участков................подрод Citellophilus (стр. 70).
6(5). Поле шипиков VIII тергита самца отсутствует или рудиментар-
но. Дигитоид с расширенной вершиной, на верхне-заднем выступе
его сидят близко друг к другу два коротких шипа (рис. 128, 137).
Характерны общие черты строения VIII и IX стернитов, напри-
мер, бахромчатая лопасть VIII стернита, пучок щетин на вытя-
нутом передне-нижнем язычке апикальной доли IX стернита
и т. д. У самок, как правило, имеются три предпигидиальных
щетинки. Семяприемник с удлиненным бочкообразным или валь-
коватым резервуаром, причем придаток обычно не длиннее
резервуара (рис. 129, Б; 133). Копулятивная сумка в боль-
шинстве случаев с сильно склеротизованными складками . . .
...................................подрод Callopsylla s. 1. (стр. 72 С
а (е). Глаз нормальный.
б (в). В гребне переднегруди менее 24 зубцов. Из теменных щетинок оста-
лась лишь одна. У самки фронтальных щетинок нет (кроме ано-
мальных случаев). В копулятивной сумке видны склеротизованные
складки (рис. 129, А—Б) . . . секция Callopsylla s. str. (стр. 72).
в (б). В гребне переднегруди не менее 24 зубцов. На лбу и темени часто
больше щетинок, чем одна.
г (д). В гребне 24—25 зубцов. На задней голени 10 групп шпор. Щетин-
ки II членика антенны у самца достигают лишь середины була-
вы. В копулятивной сумке очень грубые склеротизованные склад-
ки (рис. 133)......................секция Paracallopsylla (стр. 73).
5* 67
д (г). В гребне около 30 зубцов. На задней голени восемь групп шпор.
Щетинки II членика антенны почти достигают или заходят за
вершину булавы. Копулятивная сумка без склеротизованных
складок (рис. 136, Б). Паразиты птиц . . . секция Orneacus (стр. 73).
е (а). Глаз полурудиментарный, паразит кротов — рис. 137, 138 ....
.....................................секция Typhlocallopsylla (стр. 73).
7(4). На внутренней поверхности задней и средней ляжки щетинки
имеются в большинстве случаев лишь на апикальной половине
(рис. 346, Г). Строение модифицированных сегментов самца и ге-
ниталий самки не соответствует полностью описанному в п. 4,
5, 6. Как правило, фронтальный ряд щетинок развит, или в нем
имеются хотя бы единичные щетинки.
8(11). В гребне переднегруди менее 24 зубцов. Поле шипиков на
VIII тергите самца отсутствует или рудиментарно.
9(10). Во втором теменном ряду больше двух щетинок. Глаз пигменти-
рован нормально, но слегка уменьшен; его продольный диаметр
меньше, чем расстояние от глаза до вершины щечной лопасти
(рис. 44). На дигитоиде выше пары тупых темных шипов име-
ются шпоры (рис. 139; 141, А—Ж). Семяприемник с хитинистой
кнопкой на конце придатка (рис. 140)................................
...............................подрод Amalaracus (стр. 74).
10(9). Во втором теменнохм ряду две щетинки. Наибольший диаметр
глаза больше, чем расстояние от глаза до вершины щечной ло-
пасти ...............................подрод Monopsyllus s. 1. (стр. 75).
11(8). В гребне переднегруди 24 зубца или более (очень редко — 23).
Поле шипиков на VIII тергите самца развито. Вершина верти-
кальной ветви IX стернита самца дугообразно изогнута кзади.
Дигитоид без коротких тупых шипов (рис. 166; 170, А; 182). Ре-
зервуар семяприемника удлиненный вальковатый или лимоно-
образный (рис. 165; 170, А; 180,5). В большинстве случаев пара-
зиты птиц........................подрод Ceratophyllus s. str. (стр. 78).
12(3). Стигма VIII тергита большая (рис. 157, 5; 162), особенно у сам-
ки. Во фронтальном ряду имеется около четырех-шести щетинок,
а во втором теменном ряду три-четыре щетинки. Дигитоид все-
гда с сильными шпорами (рис. 153; 157, А; 158; 159). У самца
стернит анального сегмента, как правило, не заходит за тергит1.
....................................подрод Megabothris (стр. 76).
А. Подрод Nosopsyllus
а. Секция Nosopsyllus s. str. [179 (39), 102]
Паразиты крыс, мышей, полевок. В Восточной Сибири и на Даль-
нем Востоке пока обнаружен лишь один вид — Ceratophyllus (Nosopsyl-
lus) fasciatus, паразит крыс. Два другие вида приводятся в определи-
тельной таблице потому, что встречаются в соседних областях Азии.
Следует упомянуть еще о двух видах, близких к С. fasciatus и паразитирующих
на крысах в южном Китае — С. (N.) nicanus и С. wualis и о двух видах из северной
Индии и Средней Азии (гималайско-тянь-шаньских) С. punjabensis и С. Simla. Эти
паразиты крыс или родственные им виды могут быть занесены в паши дальневосточ-
Егые города или в Монголию [см. о них: 95, 97, 102, 19].
1(2). У самца (рис. 113) вершина IX стернита асимметричная, угло-
ватая; задний край VIII тергита более или менее округленный,
а не угловатый; верхняя короткая предпигидиальная щетинка не
1 В виде исключения очень длинный анальный стернит имеется у самцов
С. (М.) taiganus (рис. 162); этот вид вообще резко отличается от других Megabothris
(см. в определительной таблице видов).
68
толще главной; дигитоид с угловатым передним краем. У самки
фронтальных щетинок, как правило, не более двух (рис. 42); зад-
ний край VII стернита без выступов; придаток семяприемника
не особенно длинный (рис. 114)1...................С. fasciatus
Паразит крыс, обитающих в Европе, имеет всесветное распространение. Обна-
ружен во многих городах Восточной Азии (Иркутск, Чита, Хабаровск, Николаевск,
Благовещенск, Поярково, Ворошилов-Уссурийский, Гродеково, Владивосток, Александ-
ровск на Сахалине), а также и в сельских местностях Западного Забайкалья (Кабан-
ский район), в Хабаровском и Приморском краях, на Сахалине. Имеются сборы
с Курильских островов, из Кореи, Японии, Маньчжурии, а по литературным данным,—
и из некоторых других районов Китая (Шанхай, Сучжоу), где, однако, надо иметь
в виду и упомянутые выше родственные виды. Найден также в Западной Сибири
(Новосибирская, Томская области, Ханты-Мансийск, Алтай). Из числа городов Сред-
ней Азии завезен в Ташкент. С крыс иногда переходит и на других грызунов, но не
особенно часто [102, 146].
2(1). У самца (рис. 115, A; 116, А) вершина IX стернита симметричная
(как профиль сахарной головы); задний край VIII тергита угло-
ватый. У самки фронтальных щетинок обычно больше двух,
а VII стернит с боковым выступом (рис. 115, Б; 116, Б).
3(4). Передний край дигитоида без резкого угла (рис. 115, Л). У сам-
ки верхний угол заднего края VIII тергита не округлен
(рис. 115, Б)................................... .С. mokrzeckyi.
Паоазит домовых мышей Mus musculus степных районов Европейской части
СССР. Проник в Западную Сибирь (Купинский район Новосибирской области) [125].
4(3). Передний край дигитоида с резким углом, находящимся выше
середины этого края (рис. 116, Л). У самки верхний угол заднего
края VIII тергита округлен (рис. 116, Б).....................С. fidus.
Паразит домовых мышей Средней Азии, обнаружен от Ашхабада до Алтая. Как
и предыдущий вид, живет в степях, на полях и в населенных пунктах ’101].
б. Секция Gerbillophilus [197, 141, 19]
Паразиты песчанок Средней Азии и Африки. Представители одного
из видов этой группы обнаружены в Монголии и Китае. В определитель-
ную таблицу введены нами еще некоторые среднеазиатские виды (без
рисунков), чтобы обратить внимание исследователей на возможность
обнаружения родственных видов в Монголии. Для большинства видов
мы приводим только признаки самцов, так как признаки самок без ри-
сунков описать трудно.
1(2). На V членике лапок прикогтевых шипиков больше двух
............................................................С. tersus.
Обнаружен в пустынях Туркменской ССР, Кара-Калпакскон АССР и Илийской
долине на Rhombomys opimus [101].
2(1). Этих шипиков не более двух.
3(4). Самцы не имеют нормальной пары ацетабулярных щетинок
..........................................С. monstrosus и С. aralis.
Обнаружены на Meriones tamariscinus и других песчанках, обитающих в доли-
нах рек Теджен, Мургаб, Аму-Дарья, Сыр-Дарья и Чу [126, 153, 87, 19].
4(3). Ацетабулярные щетинки имеются.
5(8). Неподвижный палец половой клешни короткий и широкий; его
ширина посредине примерно лишь в полтора-два раза меньше,
чем расстояние от его вершины до места прикрепления ацетабу-
лярных щетинок. Конец дигитоида заходит за неподвижный палец
(рис. 117, А; 118, А; 119, А; 120, А).
1 Перечисленные признаки позволяют отличать этот вид также от группы видов,
распространенных в Китае и Индии.
69
6(7). Вершина IX стернита самца клювообразная ... С. laeviceps.
Паразит песчанок, распространенный от Закавказья и северо-западного При-
каспия до Китая. Представлен несколькими подвидами.
а (г). «Клюв» вершины IX стернита резко выражен (рис. 117, А; 118, А).‘
б (в). Половая клешня изображена на рис. 117, А; дигитоид и неподвижный палец
клешни относительно длинные. Самка изображена на рис. 117,5; апикаль-
ный край VII стернита, как правило, с глубокой вырезкой, однако очерта-
ния его варьируют...................................С. laeviceps laeviceps.
Известен от Азербайджанской ССР до Джаркента (Казахстан), но, возможно,
встречается и восточнее. Паразитирует на различных видах песчанок [141].
в (б). Половая клешня изображена на рис. 118, Д; неподвижный палец клешни
и дигитоид сравнительно широкие и короткие. Самка изображена на
рис. 118,5...........................................С. laeviceps kuzenkovi.
Недавно описан с когтистой песчанки из Китая (провинция Жэхэ) [86].
г (а). «Клюв» IX стернита несколько укорочен и округлен (рис. 119, А); дигитоид
относительно короткий. Самка изображена на рис. 119,5....................
................................................ С. laeviceps ellobii.
Встречается в Монголии (северные и южные предгорья Гобийского Алтая близ
озера Сого-Нор и Орон-Нор), а также в Дзабханском аймаке. Название «ellobii» дано
случайно, так как это паразит песчанок, а не слепушонок [141].
7(6). Вершина IX стернита самца более округленная. Дигитоид очень
узкий. Самец и самка изображены на рис. 120, Л, Б).........................
................................................. С. turkmenicus.
Распространен в Средней Азии от Ашхабада до Джаркента, паразитирует на
песчанках [141].
8(5). Неподвижный палец клешни узкий и длинный, ширина его при-
мерно в четыре раза меньше, чем расстояние от вершины до
прикрепления ацетабулярных щетин. Конец дигитоида не заходит
за конец неподвижного пальца..............................С. abramovi.
Обнаружен пока только в некоторых долинах Тянь-Шаня на Meriones tama-
riscinus [19].
Б. Подрод Citellophilus [197, 140, 19]
Паразиты сусликов Азии и Европы (и некоторых сурков Тянь-Ша-
ня), нередко переходящие и на других грызунов, живущих рядом с их
хозяевами. Из включаемых нами в определитель представителей этой
группы только три — С. tesquorum sungaris, С. t. mongoli-cus, С. t. al-
taicus — фактически найдены на интересующей нас территории, осталь-
ные же встречаются лишь в близлежащих местностях.
1(10). На заднем крае дигитоида (рис. 121; 124—126, А) имеются два
шипа (вместо нижнего может быть щетинка). На наружной бо-
ковой поверхности заднего бедра, как правило, имеется не более
четырех щетинок (включая субапикальную).
2(7). Дорзальные зубцы гребня примерно равны длине спинки пе-
реднегруди или длиннее ее. Хоботок не особенно длинный—дости-
гает вершины коксы или середины вертлуга, редко немного длин-
нее L У самок на некоторых из средних стернитов брюшка
(III—VI), как правило, имеются единичные передние добавоч-
ные щетинки.
Блохи сусликов. Относящиеся сюда виды и подвиды достаточно хорошо отли-
чаются по самцам, а самок часто можно распознать лишь по географическому
признаку.
3(6). Дигитоид треугольный, шипы его заостряются к концу (рис. 121;
124—126, А).
1 Об отличиях самок Citellophilus и Oropsylla — см. примечание на стр. 72.
70
4(5). Задне-верхний угол дигитоида прямой или закругленный, а зад-
ний край выпуклый (рис. 121, 122, 124, 125). У VII стернита сам-
ки вентральная вырезка чаще слабо развита или отсутствует
вообще (рис. 123)...........................................С. tesquorum.
Ряд подвидов этой блохи, различающихся по самцам, встречается на сусликах
в степях от Украины (Мариуполь) до Китая. Мы упомянем только азиатские подвиды
[об остальных см. 140]. По самкам подвиды С. tesquorum не различаются, однако
в Монголии обнаружена значительная изменчивость в очертаниях апикального края
VII стернита у самок, требующая дальнейшего изучения (рис. 123).
а (з). Имеются оба шипа дигитоида.
б (д). Шипы заднего края дигитоида сравнительно длинные и сидят не близко друг
к другу. На VIII стерните самца апикально-дорзальная бахромчатая лопасть
не развита (рис. 121).
в (г). Вершина дигитоида расширена за счет передне-верхнего угла, вытянутого
вперед (рис. 121). Самки изображены на рис. 123 . . С. tesquorum sungaris.
Распространен в Предбайкалье (Тункинская долина, Осинский район, Иркутск),
в Восточном и Западном Забайкалье, Монголии (Хэнтэйский, Центральный, Убурхан-
гайский, Баян-Хонгорский, Дзапханский, Гоби-Алтайский аймаки), в Амурской области
(Благовещенск и соседние районы), в окрестностях Якутска, в Китае (Жэхэ). Пара-
зитирует на даурском и длиннохвостом сусликах (Citellus dauricus, С. undulatus)
и других грызунах и хищниках [92, 55, 127].
г (в). Передне-верхний угол дигитоида менее вытянут вперед (рис. 122) . . . .
....................................................С. tesquorum mongolicus.
Найден только в Китае (Шэньси) [100, 128, 55].
д(б). Шипы дигитоида короче. На VIII стерните самца имеется апикально-дор-
зальная бахромчатая лопасть.
е(ж). Шипы дигитоида сидят близко друг к другу (расстояние между ними в сред-
нем около 30 — рис. 124)..................С. tesquorum transvolgensis.
Обнаружен в Заволжье, Западной Сибири (Омская область, Купино Новосибир-
ской области, в гг. Славгород и Камень Алтайского края и в окрестностях Красно-
ярска) [140].
ж (е). Шипы дигитоида далеко раздвинуты (расстояние между ними около 50 ц —
рис. 125)................................................С. tesquorum altaicus.
Встречается в Зайсанской котловине, Джунгарии, на южных склонах Джунгар-
ского Алатау, в Южном Алтае (долина Бухтармы и верховьев Иртыша), в Туве, Мон-
голии (Убсанурский аймак, Монгольский Алтай). Паразитирует на Citellus interme-
dius, С. undulatus и др. [140].
з(а). Нижний шип дигитоида заменен щетинкой. Апикально-дорзальная бахром-
чатая лопасть VIII стернита самца слабо развита или рудиментарна . . .
....................................................С. tesquorum dzetysuensis.
Форма, близкая к С. t. altaicus и заменяющая его в более северных районах
Джунгарского Алатау на Citellus undulatus [62].
5(4). Задне-верхний угол дигитоида вытянутый, острый, а задний край
вогнутый (рис. 126, X). VII стернит самки, как правило, с разви-
той вентральной вырезкой (рис. 126, Б) . . . .С. jenissejensis.
Встречается на юге Красноярского края (Хакасская автономная область). Пара-
зитирует на длиннохвостом суслике (Citellus undulatus) и других грызунах [128]. Ука-
зание на наличие этого вида в Тянь-Шане [121] неправильно и вызвано ошибкой
на этикетке.
6(3). Дигитоид широкий, с округленным задне-верхним краем, имеет
форму полукруга. Шипы его притуплены. VIII стернит самца без
бахромчатог! лопасти. У самки копулятивная сумка с мелкими
хитинистыми складочками более грубыми, чем у блох группы
С. tesquorum................................................С. relicticola.
Паразит реликтового суслика (С. relictus) Тянь-Шань [84].
7(2). Дорзальные зубцы гребня значительно короче, чем спинка пе-
реднегруди, и заметно раздвинуты один от другого. Хоботок
длинный, за вершину передней коксы часто заходит около поло-
1 У самых восточных представителей (из окрестностей Красноярска) это рас-
стояние чаще равно 40 ц.
71
вины членика (у самок хоботок длиннее, чем у самцов)1. На сред-
них стернитах брюшка передние добавочные щетинки отсут-
ствуют. Шипы дигитоида тупые. Блохи сурков Тянь-Шаня.
8(9). Дигитоид трапециевидный. Семяприемник с кнопкой на конце
придатка. На наружной поверхности заднего бедра нет боковых
щетинок..................................................С. lebedewi.
Паразит сурков Тянь-Шаня (Marmota caudata и М. baibacina). Известен в двух
расах, различимых по самкам [196, 19].
а (б). Апикальный край VII стернита самки с глубокой боковой вырезкой . . .
..................................................С. lebedewi lebedewi.
Центральный Тянь-Шань [19].
б (а). Апикальный край VII стернита самки без глубокой боковой вырезки . . .
.................................................. С. lebedewi princeps.
Западный Тянь-Шань и Таджикская ССР [19].
9(8). Дигитоид очень широкий, треугольный, с острым задне-верхним
углом. У самки апикальный край VII стернита без глубокой бо-
ковой вырезки. Семяприемник без кнопки на конце придатка.
На наружной поверхности заднего бедра часто имеются единич-
ные щетинки (одна-две)...................................С. menzbieri.
Паразит -сурка Мензбира. Западный Тянь-Шань [154].
10(1). На заднем крае дигитоида три сильных щетины близ верхнего
угла. На наружной поверхности заднего бедра имеется, как пра-
вило, не менее пяти щетинок................................С. trispinus.
Распространен на равнинах Средней Азии — от Усть-Урта до Чуйской долины;
возможно, что встречается и восточнее. Паразит Citellus fulvus [128].
В. Подрод Callopsylla
а. Секция Callopsylla s. str. [197, 139, 179 (41), 19]
Паразиты преимущественно видов грызунов (Chionomys, Alticola,
Ochotona), живущих в скалах и каменных осыпях горных стран юга
Евразии от Пиренеев до Китая. На интересующей нас территории пока
найден лишь один представитель — С. (С.) gaiskii. В горах Центральной
Азии весьма вероятно обнаружение и других известных и неизвестных
видов 2.
Признаки самок недостаточно изучены, и мы даем таблицу, рассчи-
танную преимущественно для определения самцов пяти видов, распро-
страненных в местностях, ближайших к Восточной Азии.
1(2). Задне-нижний угол дигитоида хорошо выражен (рис 127, 128)
или хотя бы намечается. Боковой угол апикального края
VII стернита самки (рис. 129, А, Б) в большинстве случаев (кро-
ме С. lagomys) более или менее округлен. Склеротизованные
складки копулятивной сумки довольно сильно развиты, сильнее,
* Уместно привести таблич::у признаков самок Citellophilus и Oropsylla, которые
можно легко спутать:
Oropsylla
У О. asiatica тоже меньше,
чем на целый членик.
У других — на целый членик
или больше
Всегда есть одна или не-
сколько из числа нижних.
Меньше, иногда и светлее.
Обычно три (иногда
две-пять)
С грушевидным или шаро-
видным резервуаром, чаще с
кнопкой на конце придатка
(рис. 111, А, Б)
описана под названием Ceratophyllus sparsilis одна самка.
совместно паразитируют на
Хоботок заходит за
вершину коксы
Фронтальные щетинки
Глаз
Предпигидиальные ще-
тинки
Семяприемник
сурках и сусликах и которых
Citellophilus
Немного, меньше чем
целый членик
на
Нет или очень редко имеется
одна верхняя
Больше и темнее
Обычно две (редко три)
С овальным резервуаром
и широким придатком
(рис. 123; 126, Б)
2 Из Тибета (Tingri)
вероятно, также относящаяся к Callopsylla [180].
72
чем у С. saxatilis (Кавказ [145]), и слабее, чем у С. tiflovi (Копет-
Даг [198]).
а. Задне-нижний угол дигитоида выступает лишь немного меньше, чем
его верхний угол (рис. 127)...................................С. caspius.
Найден на Тянь-Шане и Кавказе, где живет на полевках родов Alticola и Chio-
ncmys [82].
б. Указанный угол выступает слабее, чем у предыдущего вида (рис. 128)
..............................................................С. gaiskii.
Известен из Монголии (Хангай: Б а ян-Хон горский и Дзабханский аймаки, хребет
Тайшири, Монгольский Алтай, Убсанурский аймак) и из Тувинской автономной области,
где паразитирует на высокогорных полевках (Alticola semicanus) и сеноставках
(Ochtona pricei) [154]. Близкая форма (С. tiflovi) обитает в Иране и на Копет-
Даге [198].
в. Выступ заднего края дигитоида на месте задне-нижнего угла (близ
середины заднего края) лишь слегка заметен. Длина дигитоида
примерно вдвое превышает ширину его расширенной верхней
части. У самки боковой угол апикального края VII стернита
не округлен...................................................С. lagomus.
Обнаружен на Гиссарском и Ферганском хребтах Средней Азии; паразитирует
па сеноставках [122].
2(1). Задне-нижний угол дигитоида не выражен.
3(4). Задний край дигитоида почти прямой (рис. 130). Боковой угол
апикального края VII стернита самки не округлен......................
.......................................................С. dolabris.
Известен из Китая (провинция Ганьсу), найден на Marmota sp. [100].
4(3). Задний край дигитоида резко вогнутый (рис. 131). Самка неиз-
вестна ....................................................С. kozlovi.
Один самец найден в Тибете в районе верхнего течения реки Хуанхэ, на Micro-
tus sp. [126].
б. Секция Paracallopsylla [139, 19]
Известен один представитель — С. (Р.) kaznakovi, несколько экзем-
пляров которого было собрано во время тибетской экспедиции Козлова
в провинции Кам (р. И-Чу и р. Джа-Чу) с мелких хищников Putorius,
Mustela. На рис. 132, 133 приведены характерные признаки этого пара-
зита, который может встретиться в Монголии [126].
в. Секция Ortieacus [94, 19]
Паразиты, живущие в гнездах ласточек, известны от Англии и Кав-
каза до Тянь-Шаня и Кашмира. Кроме того, на Тянь-Шане один вид
найден в гнездах голубей и других птиц; иногда он встречается
и на людях.
1(2). Паразит ласточек. Хоботок не заходит за конец передней коксы.
Зубцы гребня не короче, чем спинка пронотума. Признаки самок
и самцов изображены на рис. 134, 135 .....................С. oreinus.
Известен из Кашмира и Центрального Тянь-Шаня. Гнезда Delichon urbica.
2(1). Паразит голубей и других птиц. Не менее половины членика хо-
ботка заходит за конец передней коксы. Зубцы гребня укорочены—
спинка пронотума более чем в полтора раза превышает их длину.
Признаки самок и самцов — см. рис. 136, Л, Б .... С. geminus.
Центральный Тянь-Шань. Гнезда Columba sp. и других птиц.
г. Секция Typhlocallopsylla [139, 141]
Один представитель — С. (Т.) semenovi, паразит сибирского крота
(Tapia altaica). Найден в Западном Забайкалье, Новосибирской обла-
сти, Ямало-Ненецком национальном округе (р. Таз), на Саянах и Алтае
(рис 137, 138).
73
Г. Подрод Atnalaraeus [139, 19, 42, 179 (41)]
Паразиты грызунов, преимущественно полевок, лесной, луговой
и горной фауны Европы, Азии и Северной Америки. Главный представи-
тель С. (A.) penicilliger известен в нескольких подвидах. Мы приводим
здесь сведения о всех известных подвидах С. penicilliger. Что касается
кавказского вида С. (A.) improvisus, то сведения о нем см. в соответ-
ствующей литературе [144].
1 (2). Дигитоид резко расширен близ середины и вооружен дву\1я
шпорами и двумя тупыми шипами (рис. 139). У самки хитино-
вый тяж, идущий вдоль VIII тергита (вентральный край IX стер-
нита), склеротизован умеренно (рис. 140) ... С. penicilliger.
Распространен в Евразии и Северной Америке, паразит лесных полевок и других
грызунов, обитателей леса и лугов. Представлен многими подвидами, изучение кото-
рых не закончено. Пока известны следующие формы, хорошо различимые по самцам
(а иногда и по самкам) 119]:
а (е). Клюв эдеагуса длинный, пальцеобразный (рис. 141, А—В).
б (в). Дигитоид длинный и потому кажется узким (рис. 141,4). VIII стернит
самца более узкий, чем у других подвидов. У самки VII стернит с глубокой
вырезкой (рис. 142, В).................................С. penicilliger dissimilis.
Описан по экземплярам, найденным на Аляске. Недавно обнаружен у нас на
Камчатке. Кроме того, в Сибири в верховьях р. Таз (Мотылька Ямало-Ненецкого
округа) собрано несколько экземпляров, весьма близких к этому подвиду. Паразит
лесных полевок (Cleithrionomys) [179 (41)].
в (б). Дигитоид относительно более широкий. У самки VII стернит с неглубокой
боковой вырезкой или совсем без нее.
г (д). Апикальная часть дигитоида развита (рис. 141, В). VII стернит у самки
см. на рис. 142, В............................................С. penicilliger syrt.
Описан по экземплярам, найденным на высокогорных сыртах Тянь-Шаня, но
распространен, повидимому, гораздо шире в горах Центральной Азии. Во всяком слу-
чае в Монголии (Хэнтэй, Тайшири, Южный Хангай) обнаружены представители этого
вида, ближе всего стоящие к данной форме, проявляющие, однако, вариабильность
в направлении к С. р. dissimilis (более узкий дигитоид). Паразит узкочерепных поле-
вок (Stenocranius); возможно, что иногда живет и на лесных полевках (Clethrionomys),
но в этих случаях следует тщательно проверить, нет ли там другого подвида [19].
д (г). Апикальная часть дигитоида укорочена (рис. 141, В). У самки край VII стер-
нита с небольшой боковой и значительной вентральной вырезкой (рис. 142, Г)
.......................................................С. penicilliger arvicolae.
Паразит водяных крыс (Arvicola). Пока обнаружен в Якутии (на р. Лене близ
Якутска) и на Кавказе. Возможно, что представляет особый хорошо диференцирован-
ный вид, распространение которого еще требует изучения [154, 143].
е (а). Клюв эдеагуса короче (рис. 141, Г—Е).
ж (к). Нижняя шпора дигитоида в большинстве случаев находится ближе к шипам,
чем к верхней шпоре (рис. 141, Г, Д). У самок форма края VII стернита
непостоянна (рис. 142,4).
з (и). Неподвижный палец половой клешни не очень широк. Клюв эдеагуса до-
вольно узкий (рис. 141, Г)..............................С. penicilliger penicilliger.
Эта номинальная форма широко распространена в лесах Сибири (Туруханск,
Новосибирская и Томская области, Барнаул, Алтай, Предбайкалье, Забайкалье, Якут-
ская АССР), паразитируя на лесных полевках Clethrionomys и многих других лесных
грызунах; встречается и па водяных крысах. Более детальное изучение вариабиль-
ности этого подвида — дело будущего [122, 19].
и (з). Неподвижный палец половой клешни широкий. Клюв эдеагуса (рис. 141, Д)
короткий, конусовидный, иногда слегка изогнутый . . С. penicilliger vallis.
Тяньшаньская лесная (долинная) форма. На Clethrionomys frater [19].
к (ж). Нижняя шпора дигитоида отодвинута от шипов и находится почти по се-
редине между ними и верхней шпорой (рис. 141, Е). Край VII стернита
самки с глубокой боковой вырезкой (рис. 142, Д)..................................
..................................................С. penicilliger demotus.
Паразитирует на лесных полевках Clethrionomys и других лесных грызунах. Евро-
пейская форма (Лапландия, Ленинградская, Калининская, Московская, Горьковская
области и Татарская АССР); в Западной Европе самцы имеют дигитоид этого же
типа, но у самок нет глубокой вырезки VII стернита. На Урале и в Западной Сибири
также встречается С. р. demotus с некоторыми переходными признаками к типичным
С. р. penicilliger (дигитоид у них напоминает таковой С. р. demotus, но часто варьирует,
а у самок вырезка VII стернита часто менее глубокая) [144, 19]. Заметим, что
С. р. demotus может оказаться идентичным С. pedias Rqths. 1910, описанному из
Финляндии по единственному аномальному самцу (с неразвитой половой клешней);
ввиду этой аномалии и отсутствия другого материала вопрос об идентичности этих
форм пока остается не разрешенным.
74
2(1). Вершина дигитоида шире, чем его середина; у задне-верхнего
угла сидит тупой шип, а около середины заднего края — удли-
ненная шпора с двумя тупыми шипами под ней (рис. 141, Ж).
У самки (рис. 142, Е) склеротизованный тяж вентрального края
IX стернита сильнее развит, чем у других видов ... С. ioffi.
Вид, недавно обнаруженный в горах Северной Кореи на лесных полевках
(Clethrionomys) [42].
Д. Подрод Monopsyllus [179 (39), 139]
К этому подроду относятся довольно разнохарактерные блохи, па-
разитирующие на различных видах грызунов и мелких хищных Европы,
Азии и Америки и разделяемые на несколько более мелких система-
тических групп, которые могут рассматриваться как секции данного под-
рода. Ниже приводятся сведения о тех секциях и видах, которые должны
быть учтены в соответствии с задачами этой книги.
1(12). Стернит анального сегмента самца далеко заходит за тергит
(рис. 146). Апикальный край VII стернита самки без широкой
и глубокой боковой вырезки (рис. 143, Б; 144, Б; 147, А, Б).
2(9). У самцов (рис. 143, А; 144, А; 145; 146) дигитоид без короткого
тупого шипа в нижней части заднего края, а у большинства ви-
дов — вообще без тупых шипов. Тело половой клешни не вытя-
нуто лентообразно, хотя может быть сильно сужено (у С. (М.) pa-
radoxus). Резервуар семяприемника при переходе в придаток
образует резкое сужение (рис. 143, Б), а у большинства видов
придаток вообще значительно уже, чем резервуар (рис. 144, Б;
147, А, Б).................................секция Monopsyllus s. str.
Паразиты белок, бурундуков, соболей и других млекопитающих, обитают в Ев-
ропе и Азии. Сюда относятся следующие виды (пп. 3—8):
3(4). VIII стернит самца (рис. 143, А) относительно слабо развит,
без длинных щетинок. У самки (рис. 143, Б) резервуар семя-
приемника короткий, грушевидный; он не длиннее придатка; при-
даток широкий. Форма края VII стернита несколько варьи-
рует ...................................................С. sciurorum.
Паразит белок (Sciurus) и сонь (Glis. Dyromys), распространен в Европе, на
Урале, Кавказе и в горах Средней Азии, на восток обнаружен до Кондо-Сосьвинского
заповедника (Зауралье), Томской области (Парбигский район) и Центрального Тянь-
Шаня. В Средней Азии представлен подвидом С. sciurorum asiaticus (рис. 143,4),
имеющим более короткий дигитоид и несколько более развитый VIII стернит самца,
чем европейский подвид [122, 141, 99].
4(3). VIII стернит самца сильнее развит, с длинными апикальными
щетинками. Резервуар семяприемника удлиненный, вальковатый,
значительно длиннее придатка.
5(6). Лобный зубчик развит. Теменные ряды щетинок: 0—1 + 2.
Задне-верхний угол тела половой клешни округленный, без раз-
витого неподвижного пальца (рис. 145). VII стернит самки
(рис. 147, А) с глубоким вентральным вырезом и характерным
выступом, имеющим выраженный апикальный дорзальный угол.
Резервуар семяприемника узкий и длинный. Имеется склероти-
зация части слепого протока около копулятивной сумки . . .
........................................................С. tamias.
Паразит бурундука (Eutamias sibiricus), часто встречающийся также на белках.
Обнаружен в гнездах соболей в питомнике. Распространен в лесах от Поволжья
(Костромская область) до Сахалина, включая Урал, Сибирь, Алтай, Тувинскую авто-
номную область, ДВК, лесные районы Монголии (Хэнтэй, Центральный аймак, Хан-
гай, Убсанурский аймак) [34].
Известные из Японии С. argus и С. indages, повидимому, близки к данному
виду, но описаны недостаточно [111].
75
6(5). Неподвижный палец клешни развит. У самки VII стернит и семя-
приемник несколько иной формы.
7(8). Лобный зубчик развит. Теменные ряды щетинок: 0 + 2, т. е. пер-
вый теменной ряд щетинок, как правило, отсутствует (у самцов
изредка в нем бывает одна щетинка). У самца (рис. 144, Л) не-
подвижный палец клешни узкий, конусовидный; дигитоид без
коротких тупых шипов; апикальная мембранозная лопасть
VIII стернита самца слабо развита. VII стернит и семяприемник
самки изображены на рис. 144, Б. Слепой проток без склероти-
зации.......................................................С. anisus.
Паразит крысы карако (Rattus norvegicus сагасо). Обнаружен в Иркутске, За-
байкалье, Амурской области и Еврейской автономной области, Владивостоке, Вороши-
лсве-Уссурийском, Хасанском районе, в Китае (Маньчжурия, Квантунский полу-
остров, Шэньси, Гуйчжоу), в Корее и Японии. Завезен также в Северную Аме-
рику [188, 92].
8(7). Лобный зубчик рудиментарен. В первом теменном ряду, как
правило, имеется одна небольшая щетинка. У самца (рис. 146)
на дигитоиде имеются два коротких тупых шипа; спиральные
тяжи пениса исключительно сильно развиты. VII стернит самки
и семяприемник — см. на рис. 147, Б. Слепой проток частично
склеротизован ................. ...................... С. paradoxus.
Паразит соболя (Martes zibellina). Известен из Кондо-Сосьвинского заповедника
(Зауралье) и Забайкалья (Александровский Завод) [75].
9(2). Дигитоид с коротким тупым шипом в нижней части заднего
края. Тело половой клешни (рис. 149, 150) лентообразно вытя-
нуто. Резервуар семяприемника (рис. 151, 152) узкий и длин-
ный, постепенно переходит в придаток почти той же ширины
..............................................секция Paramonopsyllus.
Эта группа недавно описана и включает два следующих вида паразитов мон-
гольских сеноставок (Ochotona pricei) и высокогорных полевок (Alticola) — обитателей
каменных осыпей [153, 154].
10(11). Дигитоид узкий, полулунной формы — рис. 149. VII стернит сам-
ки и семяприемник — см. на рис. 151...........................С. scaloni.
Найден в Монголии (Южный Хангай, Тайшири, Центральный и Хэнтэйский
аймаки) и в Тувинской автономной области на полевках (Alticola, Phaiomys, Evoto-
mys), сеноставках (Ochtona pricei, О. alpina) и длиннохвостом суслике [154].
11(10). Дигитоид более широкий, а неподвижный палец короче и шире,
чем у предыдущего вида (рис. 150). VII стернит самки и семя-
приемник— см. на рис. 152...................................С. desertus.
Собран с монгольской сеноставки в Карагандинской области Казахской ССР [153].
12(1). Стернит анального сегмента самца не заходит или мало заходит
за тергит. На заднем крае дигитоида имеются шипы или толстые
шпоры. Вершина вертикальной ветви IX стернита самца слабо
изогнута кзади. У распространенного в Азии вида апикальный
край VII стернита самки с широкой и глубокой боковой вырез-
кой ..............................................секция Amonopsyllus.
Блохи этой группы распространены преимущественно в Северной Америке,
а у нас известен лишь один вид С. (A.) toli — рис. 148, А—Б, встречающийся
в северо-восточной Сибири (правый берег Лены против устья Вилюя, Верхоянск, ниж-
нее течение Анадыря, побережье Охотского моря к югу от Аяна, бухта Одян). Име-
ются единичные сборы с северной сеноставки Ochotona hyperborea и летяги Pteromys
yclans [122, 139, 76].
Е. Подрод Megabothris [179 (39), 139]
Паразиты грызунов, преимущественно семейства Muridae, распро-
страненные в лесных и вообще во влажных местностях Европы, Азии
н Северной Америки.
7(3
1(2). На пронотуме, как правило, впереди от щетинок главного ряда
имеется еще ряд из мелких щетинок Ч Стигма VIII тергита мень-
ше, чем у других представителей этого подрода. Признаки самок
и самцов—см. на рис. 153; 154, А. Весьма характерна форма
дигитоида, а у самки форма VII стернита и широкая складча-
тая копулятивная сумка без склеротизации и слепого про-
тока ...............................................С. calcarifer. /
Распространен на полевках, леммингах и других грызунах в очень влажных,
обычно болотистых местностях, в лесах и долинах рек на севере Сибири от Тихого
океана (Камчатка) до Зауралья, найден также на Кольском полуострове (Лапланд-
ский заповедник) и в Монголии (Хэнтэйский аймак). Особенно обычен в Уссурий-
ской долине, Приамурье, Забайкалье. Ближайшие родственные формы живут в Се-
верной Америке; возможно, что они являются расами одного вида [33, 172 а].
2(1). На пронотуме лишь один ряд щетинок (у С. walkeri перед ним
бывают иногда одна-три щетинки).
3(4). Дигитоид широкий с характерным вооружением (рис. 155, Л).
VII стернит самки с очень большой боковой вырезкой и узким
выступом над ней (рис. 154, Б). Гениталии самки, как у преды-
дущего вида...........................................С. walkeri.
!
Паразит водяной крысы (Arvicola terrestris). Широко распространен в долинах
рек в Западной Европе и в Европейской части СССР (Латвийская ССР, Беловежская
пуща, Московская, Ленинградская, Вологодская, Горьковская, Саратовская области,
г. Уральск), а также в Закавказье (Грузинская ССР, Армянская ССР, Нахичеван-
ская АССР), на Урале (районы Клецка, Кустаная, Тюмени, Печорского заповедника,
в Молотовской области и Башкирской АССР), в Казахской ССР (Бетпакдала) и в За-
падной Сибири (Алтайский край, Кемеровская, Омская, Новосибирская и Томская
области) [29, 139].
4(3). Дигитоид самца и VII стернит самки иной формы.
5(6). Дигитоид несколько напоминает С. calcarifer (см. выше), но бо-
лее широк, а отросток, на котором сидят два коротких темных
шипа близ вершины дигитоида, более узок (рис. 155, Б). Отли-
чие строения копулятивной сумки от С. calcarifer показано на
рис. 156, Л, так же как форма края VII стернита . . С. sokolovi.
Найден недавно на Курильских островах (о-в Шпанберга) на лесных полевках
(В. Соколов, Н. Л. Гершкович). Вид до известной степени промежуточный между
неарктическими С. calcarifer и С. asio (имеющим два ряда щетинок на пронотуме)
и нашими С. rectangulatus и С. advenarius.
6(5). Признаки иные.
7(10). Дигитоид в виде узкого прямоугольника. Гениталии самки без
склеротизации в области копулятивной сумки, которая не рас-
ширена; слепой проток имеется; начальная часть семенного про-
тока расширена.
8(9). По заднему краю дигитоида (рис. 157, Л) сидят два коротких
тупых шипа и внизу сильная почти прямая шпора. У самки
(рис. 157, Б) край VII стернита с округленным боковым углом,
выступающим не меньше, чем вентральный угол; вырезка между
ними выражена в разной степени; вентральная вырезка отсут-
ствует ..............................................С. rectangulatus.
Широко распространен на различных видах полевок и других грызунов в лесах
Евразии — в Англии, Скандинавии, Европейской части СССР (Ленинградская, Воло-
годская, Горьковская, Молотовская области, Мурманск, Лапландский и Печорский
заповедники), в Западной Сибири (Омская, Томская, Новосибирская, Красноярская
области), в Туве, на Тянь-Шане и Алтае, в Иркутской области, в Якутии (р. Вилюй)
и в Западном Забайкалье, а из Восточного Забайкалья известен лишь в Бальзиио
и Александрозаводском районе. Обнаружен также в Монголии (Хэнтэйский, Цен-
тральный, Баян-Хонгорский и Дзабханский аймаки) [132, 184].
1 Встречались единичные самки без этого переднего ряда.
77
9(8). По заднему краю дигитоида (рис. 158) сидят два довольно
длинных шипа и внизу кривая шпора. Край VII стернита самки
(рис. 156, Б) в большинстве случаев имеет более или менее замет-
ную вентральную вырезку; боковой угол выступает обычно мень-
ше, чем вентральный (или субвентральный) ... С. advenarius.
Этот вид замещает С. rectangulatus в лесах Восточной Сибири и на Дальнем
Востоке, причем оба вместе они обнаружены пока только в Александрозаводском
районе Читинской области, а промежуточных форм между ними не найдено. Известно
не менее двух рас этого вида [34, 154]:
а) С. advenarius advenarius — форма с двумя острыми шипами на дигитоиде
(рис. 158,А), обнаружена в районе Якутска и с. Наканна (на р. Н. Тунгуска);
экземпляры из Забайкалья (рис. 158 — Александровский Завод) также можно отнести
к этой форме, хотя у них уже намечаются признаки перехода к следующему подвиду.
б) С. advenarius bifallax — указанные шипы не суживаются к концу, имеют за-
кругленную вершину, на которой у молодых блох имеется тончайшее острие
(рис. 158,5). Эта форма найдена в Приамурье, Еврейской автономной области, на
Камчатке и Сихотэ-Алине. Представители этого вида, вероятно, этой же дальнево-
сточной формы, обнаружены и на Сахалине.
10 ( 7). Такого сочетания признаков, как указано в пп. 7—9, не имеется.
11(12). На голове, как и у большинства Megabothris, имеются лишь
обычные развитые ряды щетин — глазной, лобный, два теменных
и задне-крайний. У самца дигитоид треугольный с характерным
вооружением (рис. 159). У самки (рис. 160) VII стернит с не-
большой боковой и ясной вентральной вырезкой. В области
основания слепого протока имеется характерная для этого вида
короткая склеротизация.....................................С. turbidus.
Паразит лесных мышей, полевок и других грызунов. Широко распространен
в лесах от Западной Европы до Урала; весьма обычен также на Кавказе и известен,
хотя и не очень обилен, на Алтае, в Западной Сибири (Омская, Новосибирская и юг
Красноярской области) и в Западном Забайкалье (Кабанский район) [29, 122, 139].
12(11). Лоб сильно выпуклый; кроме обычных рядов щетин, имеется
префронтальный ряд, а впереди него у самки еще добавочные
щетинки; на темени выше рядов также имеются добавочные ще-
тинки. У самца (рис. 162) анальный стернит очень длинный. Тело
половой клешни вытянутое, лентовидное. Дигитоид узкий, пальце-
образный с характерным вооружением. У самки (рис. 161)
VII стернит с острым боковым углом и вырезкой под ним . . .
............................. .............................С. taiganus.
Известны только одна самка и один самец из Кондо-Сосьвинского заповедника
(Зауралье), найденные на лесных полевках. Истинные хозяева и другие эколо-
гические и географические особенности этого вида, резко отличающегося от
других Megabothris, — требуют выяснения [75].
Ж. Подрод Ceratophyllus s. str. [179 (39), 29, 13, 131, 19]
Большинство видов этой группы — паразиты птиц, но имеется также
один вид, паразитирующий на горностаях. Птичьи блохи мало изучались
в Азии и, несомненно, список представителей этой группы значительно
возрастет в будущем, но и теперь мы имеем уже возможность включить
в определитель довольно много видов, имеющихся в интересующих нас
и соседних районах, откуда занос птичьих блох вероятен. Некоторые
из птичьих блох нередко попадают на грызунов и в их норы, так как
паразитируют на птицах, устраивающих свои гнезда под землей в норах
(чеканы) или на земле (жаворонки и др.). Из птичьих гнезд, устраивае-
мых на деревьях, блохи попадают иногда на белок, бурундуков,
сонь и т. п.
1(18). Задняя часть тергита заднегруди (метанотума) и находящийся
на ней зубчик развиты нормально (рис. 352, А).
2(3). Стигмы брюшных тергитов большие. В гребне не особенно много
зубцов, обычно около 25 (от 23 до 28). На наружной поверхно
78
сти задней голени много щетинок — обычно около 18 или боль-
ше. Признаки самца и самки — см. на рис. 163, 164. У самца
тело клешни узкое. У самки копулятивная сумка без склеротиза-
ции, а слепой проток склеротизован на небольшом протяжении.
Резервуар семяприемника удлиненный, вальковатый, но более ши-
рокий, чем у птичьих блох, имеющих резервуар сходной формы
........................................................С. lunatus.
Паразит горностая (Mustela erminea). Обнаружен в Швейцарии, Западной Си-
бири, Якутской АССР, Забайкалье, Предбайкалье, на Тянь-Шане и Камчатке [180,
132, 19].
Американский. С. tundrensis [172] представляет, вероятно, не более как подвид
этого вида.
3(2). Такого сочетания признаков, как указано в п. 2, не имеется.
4(7). У самца (рис. 166, 167) неподвижный палец клешни хорошо
развит, а тело клешни часто сильно сужено. На конце VIII стер-
нита, как правило, имеются толстые укороченные щетины. Спи-
ральные тяжи пениса образуют не более одного оборота (т. е.
полного круга, рис. 169, Л). У самки (рис. 165, 168) резервуар
семяприемника суживается к концам (лимонообразный); копу-
лятивная сумка и слепой проток без ясной склеротизации (слепой
проток обычно совсем не развит).
5(6). На внутренней поверхности заднего бедра в боковом ряду не
более одной-трех щетинок, а на наружной поверхности этот ряд
отсутствует. Глаз большой. У самца (рис. 166) на конце
VIII стернита среди обычных длинных щетин имеются, как пра-
вило, единичные укороченные и утолщенные; мембранозная ло-
пасть этого стернита большая, округлой формы, с характерным
узким передним отростком. Тело клешни сильно сужено. У самки
(рис. 165) боковой угол заднего края VII стернита обычно округ-
лен и не сильно выдается назад. Придаток семяприемника без
ясной апикальной кнопки......................................С. garei Г
Обитатель птичьих гнезд, устраиваемых преимущественно на поверхности земли
или на кустарниках (жаворонки, некоторые чайки, гаги, камышевки, овсянки и др.).
Случайно попадается и на грызунах (суслики, белки, полевки и др.) и в их гнездах.
Встречается во всех странах северного полушария (деление на подвиды еще недо-
статочно разработано). В СССР найден на островах Мурманского побережья, около
Ленинграда, в Вологодской области, на Северном Урале, Кавказе, Алтае, в Западной
Сибири, Иркутской области (Усть-Кут), Якутской АССР (р. Вилюй), Забайкалье
(Даурия, Бырка, Александрозаводский район, Агинский аймак и др.)1 2, в устье р. Ко-
лымы, на Дальнем Востоке (Еврейская автономная область, районы Лазо, Ленин-
1 Очень возможно, что на северном побережье и в горах Сибири встречается
также С. borealis, живущий на морском побережье и в лесах севера Европы, а так-
же на Кавказе. Самцы этого вида отличаются от С. garei более широким полем
шипиков VIII тергита, длинным узким парамером, формой половой клешни и лопасти
VIII стернита и т. п. У самок VII стернит с более узким и резким боковым высту-
пом; семяприемник с кнопкой на конце придатка [13, 29].
2 Приводимые В. Б. Дубининым [46] данные по биологии С. garei в Забай-
калье (размножение в норах) противоречат известным фактам о биологических осо-
бенностях этого вида. Возможно, что в действительности автор имел дело с видом
С. .avicitelli, который в период проведения наблюдений (1943) не был еще описан,
как и другая норовая птичья блоха Забайкалья Frontopsylla frontalis baikal. Пови-
димому, настоящий С. garei упоминается в указанной работе под названием «Сег.
gallinae», среди блох, собранных в массовом количестве в наземных гнездах жаво-
ронков (/\lauda) и т. п. Что касается действительных С. gallinae, то можно пред-
положить, что они могли быть среди тех темных блох, которые были найдены в во-
робьиных гнездах и отмечены под названием «Pulex irritans». Проверить опреде-
ления блох, собранных В. Б. Дубининым, оказалось возможным только в 1945
и 1946 гг., когда сохранилась лишь небольшая часть сборов; получив их от В. Б. Ду-
бинина, мы могли установить среди блох, взятых из птичьих гнезд, наличие следую-
щих видов: С. garei (из гнезд Alauda и Motacilla), С. riparius (из гнезд Riparia),
единичные самки С. avicitelli и F. frontalis baikal, а также ряд видов блох, пара-
зитировавших на грызунах. К сожалению, при печатании упомянутой работы исправ-
ление было внесено только в название блох береговой ласточки (С. riparius).
79
ский, Хасанский, Супутинский заповедник). Найден также в Монголии (Хэнтэйский
аймак) [13, 29].
6(5). На внутренней поверхности заднего бедра ряд из четырех-восьми
щетинок, а на наружной поверхности также бывает несколько
щетинок. Глаз сравнительно небольшой. Стигмы тергитов круп-
нее, чехМ у других видов Ceratophyllus s. str. (кроме С. lunatus).
У самца (рис. 167) на конце VIII стернита три-четыре шипо-
образных щетины, а дорзальные апикальные лопасти не развиты.
Клешня — см. на рисунке. У самки (рис. 168) VII стернит с рез-
ким усеченным боковым выступом. Семяприемник толстостенный,
особенно на конце придатка...................................С. spinosus.
Этот вид известен из Предкавказья (Дагестанская АССР), Средней Азии (Мур-
габ) и Забайкалья (Кяхта). Встречался пока лишь в гнездах хищных птиц (пустельги
Tinnunculus и др.) и случайно на суслике [13, 122].
7(4). У самца неподвижный палец клешни чаще менее развит
(рис. 173, 176), а на конце VIII стернита сидят обычные щетин-
ки. Спиральные выдвигатели пениса образуют более чем целый
оборот (рис. 169, Б). У самки (рис. 174; 180, А—Б\ 181) семя-
приемник с узким вальковатым резервуаром; базальная часть
слепого протока сильно склеротизована.
8(11). На внутренней поверхности заднего бедра ряд из четырех-шести
(редко семи) щетин. У самца (рис. 170, Л; 173; 175), дорзальный
апикальный отросток VIII стернита узкий и высокий, кинжалооб-
разный, без бахромчатой лопасти или с рудиментом ее. У самки
(рис. 171, 174) край VII стернита округлый без резких боковых
выступов и вырезок. Склеротизованный участок слепого протока
короче булавы усика. Копулятивная сумка и ее проток склеро-
тизованы на небольшом протяжении и частично.
9(10). На наружной поверхности II членика средних лапок в базаль-
ной половине имеются щетинки. У самца на вершине VIII стер-
нита обычно по две щетинки с каждой стороны (редко три или
пять с обеих сторон вместе) рис. 170, Л; 173. Вестибулярная
камера резервуара семяприемника слабо развита (рис. 171,
172, 174)....................................................С. gallinae.
Один из самых распространенных в Европе и Азии видов птичьих блох. Ниже
приводятся сведения о трех формах блох, рассматриваемых нами в качестве подви-
дов С. gallinae, так как на границе их ареалов наблюдались переходные между ними
формы. Для самок этих подвидов хорошо выраженных различий не найдено [131].
а (б). Две или три щетинки, сидящие по заднему краю дигитоида, толстые. Ди-
гитоид к вершине не суживается и примерно на половину выступает над
вершиной неподвижного пальца; задний край дигитоида ниже середины
слегка вогнут (рис. 173). Край VII стернита самки чаще довольно равно-
мерно округлен (рнс. 174)...............................С. gallinae gallinae.
Европейская форма. Обитает в гнездах воробьев, скворцов и многих других
птиц, преимущественно устраивающих гнезда в закрытых местах (в дуплах, под кры-
шами и т. п.). Размножается также в больших количествах в курятниках. Встре-
чается в достаточно влажных (лесных и лесостепных) местностях и в расположенных
в этих районах населенных пунктах. Весьма обычна в Западной Европе и в Евро-
пейской части СССР, а также на Кавказе и на Урале; наиболее восточные место-
нахождения, известные до сих пор, — Кондо-Сосьвинский заповедник и окрестности
Ханты-Мансийска (Западная Сибирь) [131].
б (а). Щетинки заднего края дигитоида тонкие. Дигитоид выдается над неподвиж-
ным пальцем менее, чем на половину; задний край его без вогнутости в ниж-
ней половине или слегка выпуклый (рис. 170,4—Б). Край VII стернита
самки имеет выступы и вырезки, хотя и не резкие (рис. 171, 172).
в (г). Дигитоид без значительного расширения в средней части и к вершине не
суживается (рис. 170,4)................................С. gallinae tribulis.
Форма, встречающаяся на юге Европейской части СССР, но, повидимому, более
распространенная в Азии. Найдена в Киеве, в Нижнем Поволжье, * Предкавказье
и Закавказье (Сухуми), на Тянь-Шане, в Забайкалье (Кабанский район, Баргузин-
ский заповедник) и в Якутске. В Монголии (Улан-Батор), пока обнаружены только
самки, вероятно, относящиеся к этому подвиду. В Киеве, Саратове, Предкавказье
встречается вместе с предыдущим подвидом и дает там нередко экземпляры с пере-
80
ходными признакамиВ Забайкалье встречаются экземпляры, приближающиеся
к следующему подвиду.
Эти блохи чаще обнаруживались в гнездах воробьев под крышами в селах и го-
родах; иногда в гнездах других птиц [179 (33) 131].
г (в). Дигитоид в средней своей части заметно расширен, а к вершине суживается
(рис. 170, Б); форма его, однако, несколько вариирует . . С. gallinae dilatus.
Дальневосточный представитель, близкий к предыдущему подвиду. Описан по
экземплярам, обнаруженным в Гродековском районе Приморской области. Найден
также в Корее и на Ляодунском полуострове [131].
10(9). На наружной поверхности II членика средних лапок в базальной
половине обычно нет щетинок. У самца на вершине VIII стернита
лишь одна щетинка с каждой стороны (редко — три щетинки
с обеих сторон), а кинжаловидный отросток сидит апикально
и под тупым углом к стерниту (рис. 175). Дигитоид по направле-
нию к вершине суживается; щетинки на нем тонкие. Резервуар
семяприемника самки с обособленной вестибулярной камерой
............................................................С. fringillae.
Блоха живет в гнездах воробьев, скворцов и других мелких птиц; известна из
Западной Европы, а у нас найдена в южных степных районах (Предкавказье, Араль-
ское море, Усть-Урт) и в Таджикской ССР. У среднеазиатских экземпляров на вер-
шине VIII стернита самца бывает иногда по две щетинки с каждой стороны [131].
11(8). На внутренней поверхности заднего бедра боковой ряд, как пра-
вило, состоит не менее чем из семи щетин. У самца на вершине
VIII стернита с каждой стороны не менее трех щетин (обычно
четыре и более), а апикальный дорзальный отросток с бахромча-
той лопастью. У самки край VII стернита имеет резкие выступы
и вырезки или усечен; если же край более или менее округлый, то
отличием от группы С. gallinae — fringillae служит более развитая
склеротизации копулятивной сумки и слепого протока (рис. 180,
А, Б; 181); длина склеротизованной части последнего больше
длины булавы усика.
12(13). У самца (рис. 176) вершина дигитоида с косо срезанным перед-
ним и вытянутым задними углами. Поле шипиков VIII тергита
уже, чем дигитоид. Брюшко IX стернита сильно выпуклое и несет
обычные щетинки. У самки (рис. 177) край VII стернита равно-
мерно округлый со слабо развитой вентральной вырезкой; пиги-
дий обычно длиннее, чем расстояние от него до церка; анальный
стернит широкий и угловатый...............................С. vagabundus.
Этот вид широко распространен на берегах и островах северных морей и во
многих горных местностях северного полушария и распадается на ряд подвидов;
например, из Красноярского края (район озера Есей) был описан С. vagabundus
orientalis [126], но, к сожалению, описание, сделанное без рисунков, не позволяет
пока распознавать эту форму, что мешает выяснению подвидового состава всех
обнаруженных в СССР представителей, найденных на Мурманском побережье, на
Кавказе, Тянь-Шане, Алтае, в Забайкалье (Александровский Завод) и Монголии
(Центральный аймак). Блохи этого вида живут в гнездах, устраиваемых разными
видами птиц чаще всего в скалах, а иногда и в норах (трехпалые чайки — Rissa
tridactvla, снежные вьюрки и др.) и случайно обнаруживались в норах грызунов
[13, 19, 179 (33)].
13(12). У самца дигитоид иной формы, а поле шипиков очень широкое;
его ширина равна или больше максимальной ширины дигитоида.
У самки край VII стернита с резкими вырезками; если же тако-
вых нет и имеются лишь нерезкие боковые выступы (у С. avici-
telli — рис. 181), то пигидий короче, чем расстояние от него до
церка, а анальный стернит более узкий и менее угловатый, чем
у предыдущего вида.
14(15). Базальный стернит брюшка, кроме обычной вентральной, имеет
многочисленные (5—15) щетинки, причем часть из них сидит на
1 С. gallinae kievensis [126] представляет, повидимому, такую гибридную форму.
6 Определитель блох 81
боковой поверхности, а не у вентрального края. На наружной
поверхности задней голени много щетин (18—33). У самца
(рис. 178) вершина горизонтальной ветви IX стернита узкая
и резко изогнута под углом; брюшко этого стернита с тонкими
щетинками. Дигитоид резко суживается к вершине; вершина
округлая. Парамер широкий, тупой, не уже вершины IX стерни-
та. У самок (рис. 180, Л) край VII стернита своеобразной формы
с развитыми вырезками и выступами; на вентральной половине
VIII тергита сидит около 25—30 щетин....................С. styx.
Обитатель нор береговой ласточки (Riparia riparia). Известен из Западной
Европы. У нас найден в Московской, Горьковской, Кзылординской и Кустанайской
областях и Ставропольском крае [13].
15(14). На боковой поверхности базального стернита нет щетин или бы-
вают как исключение одна-две; впереди от обычной вентральной
щетинки сидит не более одной-четырех мелких добавочных.
У самца вершина IX стернита не изогнута под углом, широкая.
Парамер узкий, заостренный. У самки VII стернит иной формы,
а на вентральной половине VIII тергита сидит не более 20 щетин.
16(17). Хоботок короткий и достигает примерно лишь половины длины
передней ляжки. У самца (рис. 179) брюшко IX стернита выпук-
лое с шипообразными щетинками. У самки (рис. 180, Б) край
VII стернита с глубокой боковой вырезкой. Пигидий длиннее,
чем расстояние от него до церка..............................С. riparius.
Обитатель нор береговой ласточки (Riparia riparia), распространен в Северной
Америке и Восточной Азии, в последнем случае найден пока лишь в районе Восточ-
ного Забайкалья [13, 180].
17(16). Хоботок несколько не достигает вершины передней ляжки. У сам-
ца (рис. 182) брюшко IX стернита слабо выпуклое, с обычными
щетинками. Ширина дигитоида варьирует. У самки (рис. 181)
край VII стернита без резкой боковой вырезки, с округлыми вы-
ступами (варианты изображены на рисунке) . . С. avicitelli.
Обитатель нор чеканов (Oenanthe), нередко обнаруживаемый также в норах
сусликов и других степных грызунов, а иногда и на самих грызунах. Найден в За-
байкалье, Предбайкалье, Монголии (Хэнтэйский, Баян-Хонгорский, Убсанурский айма-
ки), на Алтае и Тянь-Шане. Этот вид описан недавно [19, 13] и ранее в литературе
упоминался как вид из группы С. styx—riparius.
Следует также отметить, что С. sinicus [93], описанный по одной самке, найден-
ной в Китае (Сычуань), может оказаться идентичным с нашим С. avicitelli и тогда
наименование С. sinicus будет иметь право приоритета.
18(1). Паразиты ласточек. Задняя часть тергита заднегруди (метаноту-
ма) и находящийся на ней апикальный зубчик заметно редуциро-
ваны (рис. 352, Б). Семяприемник с узким вальковатым резер-
вуаром и склеротизованной начальной частью слепого протока.
19(20). Брюшко имеет неравномерную полосатую окраску вследствие
того, что основания тергитов и стернитов пигментированы темнее,
чем остальные участки. Задний край тела клешни образует рез-
кий выступ, на котором сидят ацетабулярные щетины (рис. 183).
Резервуар семяприемника сильно суживается к придатку и пере-
ходит в него без резкого уступа; вестибулярный отдел резервуара
обособлен (рис. 185, Л)....................................С. maculatus.
Найден в гнездах ласточек в Якутской АССР (Якутск и берег р. Амга), в Ир-
кутске и в Баргузинском заповеднике, а также несколько самок, вероятно, этого вида
найдено средн ласточкиных блох из Монголии (Улясутай) [34, 13].
20(19). Окраска брюшка без описанных темных полос. Половая клешня
и семяприемник иной формы.
21(22). У самцов (рис. 184) апикальный дорзальный отросток VIII стер-
нита узкий, без бахромчатой лопасти. Дигитоид резко суживается
82
к вершине. У самок (рис. 185, Б) край VII стернита, как прави-
ло, с глубокой вырезкой Ч Копулятивная сумка склеротизована.
На наружной поверхности заднего бедра, как правило, имеется
боковой ряд из трех-четырех щетин....................С. farreni.
Найден в Северной Африке, Англии, Германии, Монголии (Улясутай), в СССР—;
в Киеве, Московской и Воронежской областях, Предкавказье (Ставрополь), Закав;
казьс (Сухуми), Забайкалье (Баргузинский заповедник) и в Южном Приморье. Гнезда
городской ласточки Delichon urbica ЛЗ, 29].
22(21). У самца VIII стернит с широкой апикально-дорзальной бахром-
чатой лопастью. Край VII стернита самки без боковой вырезки
или с небольшой вырезкой. На наружной боковой поверхности,’
заднего бедра нет щетин, кроме одной субапикальной.
23(24). Дигитоид сравнительно простой удлиненной формы (рис. 186).
У самки пигидий равен или больше расстояния от него до осно-
вания церка. Копулятивная сумка не склеротизована. Край
VII стернита с намечающимся округлым субвентральным высту-
пом (рис. 188, А). Блохи мелкие.............................С. caliotes.
Кашмир (Северная Индия), в СССР — Тянь-Шань, Кавказ (Грузинская CCP)j
Забайкалье (Баргузинский заповедник), Иркутск. В гнездах городской ласточки [94, 13],
24(23). Дигитоид резко расширяется или суживается к вершине. У сам;
ки пигидий короче, чем расстояние от него до основания церка.
25(26). У самца (рис. 187) дигитоид сильно расширяется к вершине*
и имеет грушевидную форму; на дорзальном крае VIII тергита
бросается в глаза пучок сильных щетин. У самки (рис. 188, Б)
край VII стернита с ясной неглубокой вырезкой; копулятивная
сумка склеротизована; предпигидиальных щетин обычно две
(редко три). Блохи довольно крупные...........................С. orites.
Кашмир (Северная Индия) и Тянь-Шань. В гнездах городской ласточки [94, 13].
26(25). Дигитоид резко суживается к вершине; на углу заднего края
сидят две длинные шпорообразные щетины. Край VII стернита
самки без ясной боковой вырезки. Копулятивная сумка не скле-j
ротизована. Пигидий очень маленький .... С. hirundinis.1
Северная Африка, Западная Европа, Северная Индия (Кашмир); в СССР —
Киевская. Московская и Воронежская области, Предкавказье и Закавказье. Гнезда|
городской ласточки [13, 29]. <
Род Dasypsyllus [179 (39), 29, 25]
Блохи этого рода живут в гнездах различных птиц, преимуществен-
но в Южной Америке и Южной Азии. Необходимо отметить два вида.
1(2). На передней части головы три ряда щетинок. Дигитоид четырех-
угольный с оттянутым задне-нижним углом; на этом углу, а так-
же выше по краю сидят несколько шпор (рис. 189). У самки раз-
вита лишь одна предпигидиальная щетинка; семяприемник с боч-
кообразным резервуаром, а край VII стернита с глубокой и ши-
рокой боковой вырезкой, разделяющей примерно одинаковые
верхний и нижний выступы (рис. 190) . . . . D. gallinulae.
Паразит, живущий в птичьих гнездах в Западной Европе и Северной Америке.
В СССР обнаружен на Кавказе и на Карпатах. Найден также на Гималаях [29].
2(1). На передней части головы имеются два ряда щетинок. Дигитоид
конический с двумя шпоровидными щетинками у задне-нижнего
угла и одной около середины заднего края (рис. 191, А). У сам-
ки — три предпигидиальных щетинки; край VII стернита с боко-
вой вырезкой, разделяющей небольшой верхний и широкий ниж-
1 Об экземплярах без такой вырезки см. статью Н. Ф. Дарской [13].
6*
83
ний выступы (рис. 191, Б). Семяприемник с широким округлым
резервуаром.....................................D. stejnegeri.
Этот вид был найден в 1882 г. экспедицией Штейнегера на Командорских остро-
вах (остров Беринга — против Камчатки) и описан в 1929 г. Иорданом как Cerato-
phyllus stejnegeri без указания хозяина [179 (35)] по материалам Вашингтонского
музея. Впоследствии вид найден на островах Аляски (Holland [25]; этот автор и отнес
его к роду Dasypsyllus).
Род Mioctenopsylla [26]
Единственный представитель этого рода — М. arctica, блоха трехпа-
лой чайки (Rissa tridactyla), распространена на Новой Земле, Шпицбер-
гене и Аляске. Вероятно, встречается и в арктической Азии.
ПОДСЕМЕЙСТВО PARADOXOPSYLLINAE
Род Frontopsylla [128, 19]
Представители этого рода распространены преимущественно в Азии,
и лишь немногие обитают в Европе. Паразитируют на различных видах
грызунов, причем большинство видов весьма неразборчиво в выборе
хозяина. Одна из групп этого рода (подрод Orfrontia) является парази-
тами птиц. Сведения о разделении этого рода на группы, а также о био-
логии и географии некоторых относящихся сюда видов см. в соответ-
ствующей литературе [19].
1(16). Гребень переднегруди состоит не более чем из 24 зубцов (как
исключение — до 26).
2(15). Апикальные зубчики метанотума не сильнее развиты, чем зубчики
тергитов брюшка.
3(14). Дигитоид у задне-нижнего угла без щетин и шпор (ниже середи-
ны заднего края). Тело клешни короткое. VIII стернит самца ши-
рокий. Семяприемник с терминальным отверстием, без деления
на резервуар и придаток (рис. 197) . . . подрод Frontopsylla s. str.
Паразиты грызунов (сеноставок, сусликов, полевок, тушканчиков и т. п.), мало
разборчивые в выборе хозяина. Мы приводим ниже все известные палеарктические
виды этого подрода, но рисунки даются только для видов, встречающихся на инте-
ресующей нас территории.
Кроме того, в Индии (Бирма) и Китае (Юннань и Сычуань) встречается относя-
щийся, вероятно, к этой же группе вид F. spadix, образующий два подвида [см. 180,93],
характеризующиеся очень длинным и сильным апикальным шипом дигитоида и ко-
ротким неподвижным пальцем клешни; самки, судя по описанию, близки к F. elata
и F. luculenta.
4(5). В глазном ряду имеется четыре щетины (редко три-пять). У сам-
ца (рис. 193, А) дигитоид почти полукруглой формы, далеко вы-
дается за конец короткого неподвижного пальца. Вообще очер-
тания клешни и IX стернита очень характерны. У самки
(рис. 193, Б) VII стернит с широкой боковой вырезкой . . .
..........................................................F. wagneri.
Паразит тушканчиков, реже встречающийся на других грызунах. Распространен
в степях Забайкалья и в Монголии (Хэнтэйский и Центральный аймаки, хребет
Тайшири, Гоби). Имеются также две самки из Тибета (хребет Бурхан-Будда) [132].
5(4). В глазном ряду, как правило, имеется лишь три щетинки (ред-
ко четыре). Половая клешня самца и VII стернит самки иной
формы.
6(7). Дигитоид полукруглый, напоминает очертания шлема с двумя
темными толстыми шипами у заднего края, не выдается за конец
неподвижного пальца. Край VII стернита самки с глубокой вы-
резкой, благодаря чему вентральный угол менее выступает и не
шире, чем боковой.........................................F. semura.
Западный вид, распространен в степях от Украины (Одесса) и Предкавказья
до Казахской ССР (Тургай); восточная его граница еще не выяснена. Встречается
преимущественно в холодное время года (до мая). Паразит сусликов [128, 14].
84
О предполагаемом реликтовом характере распространения этого вида ом. литера-
туру [18, стр. 45].
7(6). Дигитоид примерно треугольной или трапециевидный с одним
тупым шипом у вершины (рис. 194, 195, 198). Боковая вырезка
VII стернита самки меньше, чем у предыдущего вида, или отсут-
ствует (рис. 197, 199).
8(9). В гребне имеется 21—24 зубца. Лобный бугорок расположен
низко. Хоботок в большинстве случаев достигает вершины перед-
ней коксы. У самца (рис. 195) на вершине VIII стернита пучок
из толстых шпорообразных щетин. У самки (рис. 197) край
VII стернита без боковой вырезки............................F. hetera.
Этот вид, описанный по одной самке, добытой в Монголии (Хангай), долгое
время представлялся несколько загадочным. Мы надеемся, что правильно отнесли
к этому виду блох, обнаруженных в Монголии (хребты Хангай, Тайшири и Монголь-
ский Алтай, Убсанурский и Центральный аймаки), в Предбайкалье (Монды), в Чи-
тинской области (Мангут), в Восточном Забайкалье (Кайластуй— только самки),
в Тувинской автономной области, в Абаканской степи и на юго-востоке Алтая, пара-
зитирующих на мелких мышевидных и других грызунах и обладающих характерными
морфологическими особенностями; в общем последние соответствуют описанию F. he-
tera Ю. Вагнера [196].
9(8. В гребне имеется около 20 зубцов (18—22, очень редко — боль-,
ше). Хоботок в большинстве случаев заметно или даже намного
не достигает вершины передней коксы, особенно у самцов*. У сам-
ца на вершине VIII стернита нет такого пучка, как у F. hetera.
У самки край VII стернита чаще с боковой вырезкой.
10(11). Лобный бугорок в большинстве случаев находится выше, чем
у F. hetera. У самца (рис. 194) нет густой щеточки из волосков
на внутренней поверхности дигитоида. Брюшко IX стернита до-
вольно сильно свисает. На вершине VIII стернита некоторые
щетинки немного утолщены; на боковой его поверхности имеется
густая группа щетин. У самки (рис. 196) форма края VII стер-
нита вариирует. Семяприемник чаще с довольно узким концом
придатка. Под стигмой VIII тергита имеется (состоящая обычно
не менее, чем из пяти-шести) группа щетинок. На внутренней по-
верхности задних бедер семь-восемь боковых щетинок . . . .
............................................................F. elatoides.
Паразит сусликов (Citellus undulatus), но переходит и на других грызунов. Най-
ден на Алтае (Катон-Карагай, Сарыбель, Черновая), в горах Джунгарского Алатау,
в окрестностях Красноярска и Минусинска, в Читинской области (Мангут — один
самец), Тувинской автономной области и Монголии (Баян-Хонгорский, Дзабханский,,
Гоби-Алтайский и Убсанурский аймаки) [126, 19].
11(10). Положение лобного бугорка варьирует; чаще он сидит низко.
У самца на внутренней поверхности дигитоида имеется густая
щеточка из мелких волосков, хорошо видная в просветленных
препаратах (рис. 195, 198, 202). На вершине VIII стернита —
тонкие щетинки, а боковая поверхность/как правило, менее ще-
тиниста, чем у F. elatoides. Самки по совокупности признаков,
приведенных в п. 10, не могут быть определены как F. elatoides J.
1 Самки близких видов Frontopsylla (hetera, elatoides, elata, luculenta) в неко-
торых случаях могут быть точно определены лишь по сопутствующим самцам. Еще
труднее распознавание самок многочисленных форм, которые рассматриваются как
расы видов F. elata и F. luculenta (см. ниже). Высказываюсь предположение, что
этих блох можно было бы объединить в одну группу форм под названием F. elata.
Однако от этого приходится воздержаться, так как в некоторых местностях, где
встречаются по две-три формы вместе, скрещивания между ними не обнаруживается,
например, F. luculenta и F. botis в низовьях Амура и Александрозаводском районе;
F. elata и F. elatoides на Алтае и в Красноярске; F. hetera и F. elatoides на Мон-
гольском Алтае, в Дзабхаиском, Убсанурском и Баян-Хонгорском аймаках МНР и в
Тувинской автономной области; F. taishiri и F. hetera в Баян-Хонгорском и Убсанур-
ском аймаках и на хребте Тайшири; F. taishiri и F. elatoides в Убсанурском и Баян-
Хонгорском аймаках; F. parilis, F. taishiri и F. hetera в Сэргулен-Сомоне Централь-
ного аймака МНР.
85
12(13). У самца (рис. 198) волосатое брюшко IX стернита отделено рез-
кой вырезкой от дистальной части стернита; темный шипик этого
брюшка сидит близ середины его. Дигитоид не заходит или мало
заходит за вершину неподвижного падьца. Нижняя ацетабуляр-
ная щетинка сидит выше верхнего края сочленовной ямки. У сам-
ки (рис. 199) край VII стернита, как правило, с боковой вырез-
i кой, причем угол, расположенный над этой вырезкой, острый
и обычно выступает несколько меньше, чем вентральный (в виде
исключения у некоторых экземпляров вырезка может быть слабой
или совсем невыраженной).......................................F. luculenta.
Эти блохи распространены от Центральной Монголии до Тихого океана; опи-
саны две следующие весьма близкие между собой расы, которые могут быть рас-
познаны по самцам, а самки — по географическому критерию. Встречаются часто и в
большом количестве на многих видах грызунов — даурской сеноставке, сусликах, по-
левках, крысах и т. д., в степных и лесостепных районах [103, 92].
а (б). Дигитоид (рис. 198) слегка (но в разной степени) заходит за конец непо-
движного пальца. Под апикальным шипом дигитоида начинается ряд сильных
г щетин, расположенных дугой поперек дигитоида; верхняя щетина этого ряда,
как правило, мало отличается от следующих, но иногда несколько утол-
щена .......................................... F. luculenta luculenta.
Восточносибирская форма, распространенная в Предбайкалье (окрестности Ир-
кутска), Западном и Восточном Забайкалье. Представители этого вида из Якутии
(окрестности Якутска) и из долины Амура (Благовещенск, Мариинское, Николаевск1)
требуют дальнейшего изучения в отношении подвидовой принадлежности [103].
б (а). Дигитоид (рис. 200) не заходит за неподвижный палец. Верхняя щетина,
сидящая под тупым шипом дигитоида, резко утолщена, шипообразна . . .
........................................................F. luculenta parilis.
Монгольская форма, найденная в Кяхтинском районе Забайкалья и в Восточном,
Хэнтэйском и Центральном аймаках МНР [92].
13(12). У самца (рис. 202) волосатое брюшко IX стернита не отделено
вырезкой от дистальной части стернита, а непрерывно переходит
в нее; темный шипик сидит чаще в дистальной части этого брюш-
ка. У самки край VII стернита с боковой вырезкой, развитой
в различной степени, а иногда отсутствующей; боковой угол этого
края, как правило, выступает не меньше, чем вентральный,
и обычно (но не всегда) более широкий и тупой, чем у предыду-
щего вида.......................................................F. elata.
Несколько подвидов распространено от Закавказья до Тихого океана. Возможно,
что в будущем их придется рассматривать как представителей нескольких отдельных
^идов. Все эти блохи живут на различных видах полевок (Microtinae) и на многих
других грызунах, не обнаруживая ясной специфичности в выборе хозяина и стации!
Предпочитают умеренно влажные и влажные местности (лесостепь, лес и горы). Мы
делим этих блох на четыре группы, сведения о которых даются ниже под лите-
рами А, Б, В, Г.
А. У самца (рис. 202; 203, А) дигитоид мало заходит за вершину
подвижного пальца. Передне-верхний угол дигитоида резко вы-
ступает, так что передний край образует дугу, выпуклую кзади.
Под шипом дигитоида поперечно расположенный ряд щетин.
Нижняя ацетабулярная щетина сидит несколько ниже уровня
верхнего края сочленовной ямки. У самки (рис. 203, Б) VII стер-
нит с боковой вырезкой.
а (б). Дигитоид нс очень широкий (рис. 202)...............F. elata elata.
Западносибирская форма — районы Томска, Минусинска, Красноярска, Пред-
байкалье, Алтай, Восточный Тянь-Шань [101].
б(а). Дигитоид относительно короче и шире; он почти не заходит за неподвиж-
ный палец (рис. 203, А)...................................F. elata taishiri.
Монгольская форма (хребет Тайшири, Убсанурский и Баян-Хонгорский аймаки,
Улясутай, Сергулен-Сомон Центрального аймака). Чаще всего на горных полевках —
Alticola semicanus, обитающих на каменных осыпях и в горной степи [154].
1 Ошибочно приводится также для Ворошилова-Уссурийского [59, 61]; там рас-
пространена F. elata botis.
86
Б. Отличается от предыдущей монгольской формы более прямым
передним краем дигитоида, слабым развитием щеточки на внут-
ренней его поверхности, более высоким расположением нижней
ацетабулярной щетинки, находящейся несколько выше верхнего
края сочленовной ямки. Горизонтальная ветвь IX стернита с бо-
лее удлиненной базальной частью и сильно свисающим брюш-
ком. У самки VII стернит с хорошо развитым боковым выступом
и укороченным вентральным..............................F- elata mutata.
Афганистан [99].
В. У самца (рис. 204) дигитоид значительно заходит за вершину
неподвижного пальца. Передний край дигитоида более или менее
прямой или образует дугу, выпуклую кпереди. Ряд щетин, начи-
нающийся под шипом, идет косо вниз. Нижняя ацетабулярная
щетинка сидит выше уровня верхнего края сочленовной ямки или
примерно на этом уровне. У самки VII стернит с боковой вырез-
кой (рис. 205).
а (б). Волосатое брюшко IX стернита сильно свисает (рис. 20) . . . F. elata botis.
Дальневосточная форма — лесостепные районы Восточного Забайкалья (Алексан-
дровский Завод), Биробиджан, Хабаровск и прилежащие районы, Уссурийский край,
Николаевск-на-Амуре, Сахалин, Северо-Восточная Монголия (Хэнтэйский аймак),
Маньчжурия, Корея [92].
б (а). Волосатое брюшко IX стернита не свисает:
1. Дигитоид несколько шире, чем у F. е. botis.......F. elata caucasica.
Кавказская форма [145].
2. Отличается от предыдущей формы значительно более широким дигитоидом
(рис. 202, а)...........................................F. elata popovi.
Известна из Омской области [145а].
3. Отличается отсутствием мелких волосков («щеточки») на внутренней по-
верхности VIII тергита самца близ апикально-дорзального края. Ширина
дигитоида примерно такая же, как у F. е. caucasica . . . . F. elata koksu.
Джунгарский Алатау [19].
Г. Дигитоид в общем, как в группе В, но передний край его с углом,
направленным кпереди. Нижняя ацетабулярная щетинка сидит
примерно на уровне верхнего края сочленовной ямки. У самки
край VII стернита далеко не всегда имеет хорошо развитую
вырезку, вариирует.
а. Угол переднего края дигитоида резко выражен. Брюшко IX стернита слегка
свисает.................................................F. elata humida.
Заилийский Алатау [82, 19].
б. Отличается от предыдущей формы более густой группой щетин на боковой
поверхности VIII стернита самца, более широким дигитоидом и не свисаю-
щим брюшком IX стернита.................................F. elata pilosa.
Терскей Алатау [19].
в. Дигитоид с менее выраженным утлом переднего края и слабо развитой
щеточкой на внутренней поверхности. Щетина, сидящая под апикальным
шипом, сильно утолщена. Вершина неподвижного пальца узкая. На боковой
поверхности VIII стернита группа щетин густая...........F. elata glabra.
Западный Тянь-Шань [19].
14(3). Дигитоид с щетинами или шпорами у задне-нижнего угла. Тело
клешни вытянуто лентовидно. VIII стернит самца уменьшен вен-
тральной вырезкой. Семяприемник с субвентральным отверстием
и намечающимся обособлением резервуара и придатка. Тянь-
шаньские виды............................................подрод Profrontia.
Паразиты горных грызунов Тянь-Шаня — полевок, лесных мышей и т. п. Из-
вестны три вида: F. protera, F. ornata, F. ambigua. Сведения о них см. в соответ-
ствующей литературе [19].
15(2). Апикальный зубчик метанотума особенно сильный, почти вдвое
шире зубчиков тергитов брюшка. Глаз очень большой
. . . . ..........................подрод Mafrontia.
87
Известен один вид — F. macrophthalma из Джаркента, Ашхабада, Аральска,
Северозападного Прикаспия, Баку; чаще встречается на тушканчиках в холодное
время года. Характерными особенностями этой блохи являются: очень большой глаз,
очень маленький неподвижный палец клешни (дигитоид длиннее его во много раз),
вырезанный снизу VIII стернит самца, семяприемник самки с резервуаром, довольно
резко обособленным от придатка [101, 19].
16(1). Гребень переднегруди состоит более чем из 28 зубчиков
.......................................................подрод Orfrontia.
Паразиты птиц. Одна группа видов живет в норах чеканов (Oenanthe) и других
птиц, устраивающих гнезда в норах. Другая группа — паразиты ласточек. Представи-
тели подрода Orfrontia известны от Центральной Европы до Забайкалья. Мы приво-
дим сведения о формах, известных из Азии [19, 10, 188 (16)].
17(18). Количество апикальных зубчиков на тергитах небольшое — один,
редко — два зубчика с каждой стороны тергита. Апикальный шип
дигитоида недлинный (рис. 206). На заднем крае VII стернита
самки имеется боковая вырезка (рис. 207) . . . F. frontalis.
Под названием F. frontalis объединяется ряд форм, живущих в устраиваемых
в норах гнездах птиц. Эти блохи часто бывают и на грызунах (сусликах, тушкан-
чиках и т. п.). Всего известно пять форм блох этой группы, но мы приводим только
три, четвертая—(F. f. gud) известна лишь с Кавказа [10], пятая (F. f. frontalis),—
вероятно, западноевропейская [188 (16)].
а. Неподвижный палец клешни короткий (рис. 206). У самки VII стернит с не-
глубокой боковой вырезкой (рис. 207)................F. frontalis baikal.
Забайкалье, Монголия (Хэптэйский. Баян-Хонгорский и Центральный аймаки),
Юго-Восточный Алтай, Тянь-Шань. Этот подвид ранее упоминался как Fronto-
psylla sp. [19].
б. Неподвижный палец значительно более длинный. У самки VII стернит с глу-
бокой узкой вырезкой................................F. frontalis alatau.
Тянь-Шань, Копет-Даг, Приаральские и Прикаспийские степи, Азербайджан-
ская ССР [19].
в. Форма средняя между приведенными выше — самцы сходны с F. f. alatau,
а самки — с F. f. baikal............................F. frontalis dubiosa.
Тянь-Шань [19].
18(17). На нескольких тергитах брюшка с каждой стороны имеется по
два-пять зубчиков. Метанотум укорочен (особенность блох, пара-
зитирующих на ласточках). Апикальный шип дигитоида длинный.
VII стернит самки без боковой вырезки.........................F. cornuta.
Паразит ласточек Delichon urbica. Известен пока лишь с Тянь-Шаня. Другой
вид этой группы — F. laeta — найден на Кавказе и в Швейцарии [19, 180].
Род Ctenophyllus [192, 19, 154]
Паразиты пищух (сеноставок — Ochotona); встречаются в Азии
и Америке.
1(8). На лбу (рис. 46) имеется ряд шипов (более толстых у самок).
Эти шипы бывают заостренные или тупые, в зависимости от воз-
раста блохи (рис. 20)..........................подрод Ctenophyllus s. str.
Самцы блох этого подрода имеют характерный VIII стернит с конусообразным,
склеротизованным апикальным выступом, на котором сидит пучок щетин (рис. 208, 210).
Семяприемник самок с широким резервуаром, довольно отчетливо отделенным от при-
датка, а VII стерпит без глубоких вырезок (рис. 211, Б, В).
Разделение рода Ctenophyllus на более мелкие группы не связано с чрезмер-
ным обилием относящихся сюда видов; но поскольку оно было начато выделением
Geusibia Jordan 1932 (Сычуань), Cratinius Jordan 1933 (Ява) и Augustsonius
Hubbard 1941 (Северная Америка), нам приходится идти по тому же пути; однако
в этом случае, как и в ряде других, мы предлагаем мелкие группировки считать
подродами, а не отдельными родами [192, 179 (38), 26, 154].
Виды (или подвиды?), относящиеся к подроду Ctenophyllus, достаточно хорошо
различаются по самцам; что касается самок, то отличительные признаки их пока
для ряда форм не выявлены и различать их приходится по сопутствующим самцам
и географическому критерию.
88
2(3).. Шипы лобного ряда очень толстые, часто короткие и тупые.
Дигитоид сравнительно широкий (рис. 211, Л).............................
....................................................С. subarmatus.
Паразит алтайской пищухи Ochotona alpina; обитатель каменных осыпей и скал.
Достоверно известен лишь с Алтая и Саян [122].
3(2). Шипы лобного ряда менее толстые. Дигитоид более узкий.
4(5). Шипы лобного ряда средней толщины. Дигитоид с узким вытя-
нутым передне-верхним углом и вогнутым верхним краем
(рис. 210)................................................С. tarasovi.
Паразит пищухи Ochotona hyperborea Южного Хангая (МНР) [78].
5(4). Шипы лобного ряда тоньше, чем у предыдущего вида; у самца
они похожи на обычные толстые укороченные щетинки. Дигитоид
с выпуклым верхним краем (рис. 208, 209). Склеротизованный
тяж VIII стернита самца сильнее развит.
6(7). Верхне-задний угол тела половой клешни сильно оттянут. Гори-
зонтальная ветвь IX стернита выпуклая книзу и несколько рас-
ширена посредине (рис. 208)................................С. rigidus.
Паразит пищухи Ochotona hyperborea, найден в горах Северной Кореи [43].
7(6). Верхне-задний угол тела половой клешни менее оттянут. Гори-
зонтальная ветвь IX стернита узкая, в большинстве случаев пря-
мая (рис. 209)............................................С. armatus.
Паразит пищух Ochotona hyperborea и О. alpina; как и все предыдущие виды
Ctenophyllus — обитатель каменных осыпей и скал. Обнаружен во многих местах Во-
сточной Сибири: Иркутская область (верховья р. Лены), Забайкалье (Александров-
ский Завод, Селенга, Джида, Кайластуй >, в Якутской АССР (устье р. Вилюя,
Верхоянск), на побережье Охотского моря (Аян, Одян), в Хабаровском крае (Обор)
и в Монголии (окрестности Улан-Батора) [122, 83].
Еще один вид этой же группы С. terribilis известен в Северной Америке (Ка-
нада, США) [173].
8 (1). Щетинки лобного ряда не превращены в шипики (рис. 47).
9(10). Апикальный край VIII стернита самца с рядом плоских ланцето-
видных щетин. У самки семяприемник с довольно узким резер-
вуаром, не совсем отчетливо отделенным от придатка, а VII стер-
нит с глубокой боковой вырезкой .... подрод Ochotonobius.
Сюда относится С. hirticrus — паразит даурской пищухи (Ochotona daurica).
Найден пока лишь в Забайкалье (Восточном и Западном), Тувинской автономной
области и Монголии (Центральный, Баян-Хонгорский, Дзабханский, Гоби-Алтайский
и Убсанурский аймаки). Указания на обнаружение этого вида в Приамурье и Маньч-
журии [56, 57], повидимому, недостаточно обоснованы (см. выше об Amphalius
runatus). Обитатель равнинных и горных степей. Признаки самца и самки см.
рис. 212, А, Б [103].
Другой вид — С. rufesccns — живет в горах Копст-Дага па рыжеватой пищухе
(О. rufescens). Он отличается деталями формы дигитоида и семяприемника, а также
более широким выступом над боковой вырезкой VII стернита самки [153].
10(9). По апикальному краю VIII стернита самца сидят щетины обыч-
ного типа. У самки семяприемник с небольшим округлым резер-
вуаром, резко отделенным от длинного придатка, а VII стернит
без глубокой боковой вырезки......................подрод Conothobius.
Сюда относится С. conothoae — паразит пищух подрода Conothoa (macrotis,
rutila). Тянь-Шань [19, 154].
Род Paradoxopsyllus [116, 27]
Представители этого рода, довольно многочисленные, известны
только из Азии, от Закавказья до Японии; они паразитируют на
многих видах грызунов. В пределах Восточной и Центральной Азии пока
известны четыре вида, но мы включили в определительную таблицу (без
1 Найден в каменистых сопках.
89
рисунков) еще три вида, паразитирующие на песчанках и других грызу-
нах в восточных районах Средней Азии, допуская возможность их обна-
ружения в пустынях или горах Монголии. Совсем не упомянуты нами
в таблице два более западных вида — Р. hesperius [144] и Р. microphthal-
mus [153], а также китайский (Сычуань) —Р. custodis [93].
1(14). Лобный зубчик имеется.
2(13). Концы наиболее длинных апикальных щетинок II членика задних
лапок по меньшей мере достигают, а чаще далеко заходят за вер-
шину IV членика.
3(12). Глаз довольно значительного размера, нормальный для рода.
4 (5). У самки (рис. 217, Б) проток копулятивной сумки дугообразно
изогнут; резервуар семяприемника относительно мал. Самцы не-
известны ..............................................Р. conveniens.
Описан по экземплярам, обнаруженным в Монголии (берег озера Сого-Нор,
южный склон Гобийского Алтая) на грызуне неизвестного вида [126].
5(4). У самок проток копулятивной сумки с угловатым изгибом
(рис. 215, А, Б; 217, А; 218, Б). О самцах — см. в следующих
пунктах.
6(11). У самцов на VIII тергите единичные щетинки. У самок VII стер-
нит имеет форму, изображенную на рис. 215, А, Б\ 217, А.
7(8). У самца (рис. 213) дигитоид сравнительно длинный; передний
край его с выступающим углом, расположенным ниже середины
этого края; вершина дигитоида обычно округлая. Брюшко IX стер-
нита расширено, на нем до трех щетинок более значительных
и единичные слабые; апикальная долька стернита языкообразной
формы. У самки (рис. 215, А) апикальный край VII стернита
с глубокой прямоугольной вырезкой над широким вентральным
выступом; край выступа со склеротизованной складкой, указы-
вающей на расщепление этого края. Проток копулятивной сумки
длинный...........................................Р. scorodumovi.
Найден в степях Восточного Забайкалья (единичные сборы с сеноставок). До-
вольно обычен в некоторых районах Монголии (Тайшири, Баян-Хонгорский, Дзабхан-
ский и Убсанурский аймаки) и Тувинской автономной области (Монгун-Тайгинский,
Овюрский районы), а также на юго-востоке Алтая на горных полевках (Alticola
semicanus), сеноставках и некоторых других высокогорных грызунах, на которых блохи
этого вида появляются с августа—сентября, наиболее обильны в октябре, а с ноября
уже исчезают [73].
8(7). У самцов угол переднего края дигитоида находится на середине
этого края; вершина дигитоида сужена; брюшко IX стернита
с более или менее густым рядом длинных щетинок; оно лишь
немного шире (рис. 214, 216) лежащей кзади от него части гори-
зонтальной ветви; апикальная долька IX стернита узкая. У самки
апикальный край VII стернита без следов расщепления; проток
копулятивной сумки короткий.
9(10). У самца (рис. 214) дигитоид более узкий, чем у Р. scorodumovi,
со слабо выступающим передним углом; вершина его значитель-
но превышает неподвижный палец; на брюшке IX стернита густой
неправильный ряд из шести-десяти длинных щетинок. У самки
(рис/215, Б) апикальный край VII стернита косо срезан; копуля-
тивный проток склеротизован до нижнего изгиба . . . Р. integer.
Имеются единичные сборы из лесостепных районов Забайкалья (Кабанский,
Ононский) с сеноставок и крыс и в большом количестве найден в Монголии (Цен-
тральный и Баян-Хонгорский аймаки) в осенних сборах с высокогорных грызунов —
на Ochotona pricei и в гнездах Alticola semicanus. Найден также в Тувинской авто-
номной области (Овюрский район) на Ochotona alpina и единичные экземпляры на
других грызунах [153].
90
10(9). У самца (рис. 216) дигитоид значительно короче, едва превы-
шает массивный неподвижный палец; передний край дигитоида
с сильно выступающим углом; на брюшке IX стернита около пяти
длинных щетинок образуют правильный ряд по вентральному
краю. У самки (рис. 217, Л) апикальный край VII стернита с не-
большим округлым боковым выступом и слабой вырезкой под
ним. Копулятивный проток склеротизован только в верхней части;
шейка его относительно короче, чем у Р. integer ....
...................................................... Р. dashidorzhii.
Серия экземпляров найдена в Монголии (Баянхонгорский аймак, Цзаг-Сомон)
в каменных осыпях на Ochotona pricei (сборы в октябре и ноябре) [78].
11(6). У самца (рис. 218, Л) на VIII тергите группа многочисленных
щетинок; подвижный и неподвижный пальцы клешни короткие
и широкие; апикальная долька IX стернита треугольная. У самки
апикальный край VII стернита (рис. 218, Б) широко округлый
.......................'...................................Р. curvispinus.
Известен из Китая (Шэньси — Yen-an Fu), Японии, Кореи, Маньчжурии (Чань-
чунь), с крыс [27, 113].
12(3). Глаз малого размера. Дигитоид небольшой, простой овальной
формы. Апикальная долька IX стернита треугольная. У самки
VII стернит и копулятивный проток примерно такие, как
у Р. conveniens............................................Р. repandus.
Равнины Средней Азии (Илийская долина, Урсатьевская, Джусалы, Мангышлак);
зимний паразит песчанок [116].
13(2). Концы апикальных щетинок II членика задних лапок не дости-
гают вершины IV членика. Глаз довольно большой. У самца
дигитоид не длинный, он лишь слегка заходит за вершину непо-
движного пальца. Апикальная долька IX стернита треугольная.
У самки край VII стернита без расщепления и имеет развитый бо-
ковой выступ. Проток копулятивной сумки узкий, без угловатого
изгиба.......................................................Р. naryni.
Найден на Тянь-Шане и Памире. Паразит высокогорных грызунов (Alticola,
Cricetulus и др.) [126].
14(1). Лобный зубчик отсутствует...........................Р. teretifrons.
Песчаные равнины Средней Азии (Илийская долина, Байрам-Али, Репетек, Кара-
Кум, Кызыл-Кум и др.); зимний паразит песчанок [116].
Род Ophthaltnopsylla [128]
К этому роду принадлежат преимущественно паразиты тушканчиков,
распространенные в Азии и в восточной части Европы. Мы рассмотрим
лишь виды, известные из Восточной Азии. Сведения о других видах и под-
видах (montana, intermedia, arnoldi, kasakiensis и др.) см. в соответ-
ствующей литературе [128, 84, 153, 134, 19].
1(6). Лобный бугорок, нередко имеющий форму зубчика, расположен
довольно высоко (рис. 48) . . подрод Ophthalmopsylla з. str.
2(3). У самца (рис. 219, Л) дигитоид треугольный. У самки фронталь-
ный ряд щетинок слабо развит, а лобный бугорок обычно имеет
форму зубчика; апикальный край VII стернита закругленный
(рис. 219, Б) или с одной глубокой вырезкой (рис. 220, Б)
..........................................................О. praefecta.
Блоха тушканчика прыгуна (Allactaga saltator), найдена в Забайкалье, Мон-
голии, на Тибете (Бурхан Будда) и Тянь-Шане. Представлена двумя географическими
расами, различаемыми лишь по самкам [101, 92].
а (б). Край VII стернита самки закругленный (рис. 219, Б).............
.............................................. О. praefecta praefecta.
91
Распространена в степях юго-восточного Забайкалья и проникает в северо-восточ-
ную Монголию. Эта форма впервые описана по экземплярам, обнаруженным в районе
Джаркента (Тянь-Шань) и ареал ее подлежит дальнейшему изучению [101].
б (а). Край VII стернита самки с более или менее глубокой вырезкой (рис. 220, Б)
........................................................О. praefecta pernix
Распространена на юге Забайкалья (Кяхта) и в Монголии (Центральный и Хэн-
тэйский аймаки, хребет Тайшири, Монгольский Алтай, Гоби) [92].
3(2). У самца (рис. 221, А—Б) дигитоид с закругленной вершиной.
У самки несколько нижних фронтальных щетинок хорошо раз-
виты; лобный бугорок часто не имеет формы зубчика; край
VII стернита — см. рис. 221, Б; 222; 223, Б.
4(5). У самца (рис. 221, А) ацетабулярная щетинка сидит на конусо-
образном выступе, направленном кзади и кверху. Брюшко
IX стернита с довольно значительным числом щетинок, а вершина
горизонтальной ветви образует закругленное расширение. У сам-
ки (рис. 221, Б) край VII стернита с двумя боковыми вырезка-
ми; на рис. 222 изображены варианты этого края.........................
. . . ......................................... О. kukuschkini.
Не особенно часто встречается в Забайкалье, в зоне степи и лесостепи, на раз-
ных грызунах; имеется также в Маньчжурии и Китае (Жэхэ) [132].
5(4). У самца (рис. 223, Л) ацетабулярная щетинка сидит на сустав-
ном бугре, не образующем конусообразного выступа, направ-
ленного кверху. Брюшко IX стернита с единичными щетинками,
а вершина горизонтальной ветви узкая. У самки (рис. 223, Б)
апикальный край VII стернита без ясных боковых вырезок
......................................................О. volgensis.
Блохи этого вида живу г в гнездах тушканчиков и распространены от степей
Украинской ССР и Предкавказья до пустынь Западной Монголии, образуя ряд под-
видов [128, 19]. На рис. 223,4—Б изображены модифицированные сегменты О. vol-
gensis extrema ssp. п. Этот подвид обнаружен в Монгольском Алтае и в Зайсанской
котловине. Он отличается от других подвидов угловатым задним краем дигитоида,
большим расстоянием между шпорами, довольно широким и округлым брюшком
IX стернита.
6(1). Лобного зубчика нет; лобный край образует тупой выступающий
угол, расположенный очень низко (рис. 49) . . подрод Cystipsylla.
7(8). Дигитоид с прямым передним краем (рис. 224, Л). Апикальный
край VII стернита самки в нижней части с глубокой вырезкой,
выше которой — узкий длинный выступ (рис. 224, Б) . . . .
. ..................................................О. kiritschenkoi.
Распространен в Монголии (оз. Орок-Нор и Убсанурский аймак), Тувинской
автономной области, на Алтае (Сарыбель). Пески. Паразитирует на тушканчиках
и хомячках (Phodopus songarus) [126].
8(7). Передний край дигитоида выемчатый (рис. 225). Апикальный
край VII стернита самки без узкого длинного выступа (рис. 226)
...........................................................О. jettmari.
Распространен в Южной Маньчжурии. Паразитирует на грызунах — обитателях
песков [92].
Род Phaenopsylla [99, 153, 154]
Паразиты мышевидного хомячка (Calomyscus bailwardi), распростра-
нены в горных районах Туркменской ССР (Копет-Даг) и в Афганистане.
Представители: Ph. kopetdag, Ph. tiflovi, Ph. mustersi.
Род Brachyctenonotus [126]
Единственный представитель этого рода — В. myospalacis — паразит
алтайского цокора Myospalax myospalax (рис. 50, 227, 228). Обнаружен
пока только на Алтае в зимних сборах (октябрь — ноябрь).
92
ПОДСЕМЕЙСТВО MESOPSYLLINAE
Род Mesopsylla [163, 134, 19]
Паразиты тушканчиков (Dipodidae), обитающие в Азии и Европе.
Мы приводим все известные до сих пор виды (кроме закавказского
М. apscheronica) [10], хотя в пределах Восточной Азии обнаружен пока
только один вид, так как тушканчики здесь еще мало обследованы.
Определительную таблицу удобнее было построить отдельно для самок
и самцов; однако, чтобы не повторяться, сведения о географии отдельных
видов приводятся только по одному разу.
1(14). Лобный зубчик имеется. От фронтального ряда у самки оста-
лись лишь единичные верхние щетинки. Дигитоид без темных ши-
пов. У самки на апикальном крае VIII тергита имеются шпорооб-
разные щетинки. Резервуар семяприемника овальный, короче,
чем придаток.
2(9). Самцы.
3(6). На VIII стерните с обеих сторон имеется по паре толстых палоч-
ковидных щетин, сидящих на выступе (рис. 229, 231).
4(5). Угол на переднем крае дигитоида (рис. 229) расположен при-
мерно посредине; по заднему краю дигитоида имеются три (редко
две-четыре) довольно длинных щетинки (кроме двух апикальных)
и несколько нижних коротких, превращенных в шипики . . .
..........................................................М. hebes.
Паразит большого тушканчика (Allactaga jaculus). Распространен в степях Кры-
ма, Предкавказья, Нижнего Поволжья, во многих местах Казахской ССР (от Запад-
ного Казахстана и Усть-Урта до Илийской долины и Алтая включительно), в Киргиз-
ской ССР, Алтайском крае (Бийск), Северной Джунгарии и Монголии (Монгольский
Алтай, Гобийский Алтай, Западный Хангай). Экземпляры из Монголии несколько от-
личаются от остальных (особенно самки) и требуют дальнейшего изучения [101, 134].
5(4). Угол на переднем крае дигитоида (рис. 231) расположен ниже
середины этого края; по заднему краю имеются (кроме двух апи-
кальных) пять-шесть щетинок, из которых верхние очень длинные,
а две нижние немного укорочены, но не превращены в ши-
пики .....................................................М. lenis.
Паразит малого тушканчика (Allactaga elater). Обнаружен в Нижнем Поволжье,
на Усть-Урте и в Илийской долине [101, 134].
6(3). Признаки иные.
7(8). На VIII стерните (рис. 237) с обеих сторон имеется по одной
толстой щетине, сидящей на выступе. На заднем крае дигитоида
несколько нижних щетинок (а иногда даже почти все, кроме двух
апикальных) шипикообразны.................................М. eucta.
Паразит тушканчиков, обитающих в пустынях Средней Азии. Распространен
в Туркменской ССР, Узбекской ССР, южной и юго-восточной части Казахской ССР,
в Киргизской ССР, Афганистане и Синь-Цзяне (Шихо). Образует географические расы:
М. eucta shikho — с Восточного Тянь-Шаня (отличается более широкой вершиной
дигитоида и более выраженным укорочением щетинок его заднего края); М. eucta
kochkor — из Центрального Тянь-Шаня (отличается более острым задне-верхним углом
волосатой части IX стернита); М. eucta afgana—из Афганистана (отличается узкой
волосатой частью IX стернита) [163, 134, 99, 153, 19].
8(7). На VIII стерните отсутствуют описанные выше толстые щетины.
На заднем крае дигитоида все щетинки (кроме двух апикаль-
ных) более или менее укорочены, шипикообразны (рис. 232)
............................................... . М. tuschkan.
Паразит мелких тушканчиков (Alactagulus acontion, Scirtopoda telum и др.).
Найден в Закавказье, Предкавказье, в Нижнем Поволжье и в ряде мест Казах-
ской ССР, в том числе во многих пунктах от Волги до Эмбы, Усть-Урта, Иргнза,
Тургая, Джезказгана. Южнее, в долине Сыр-Дарьи (станция Сауран), заменяется
подвидом М. tuschkan mesa (имеющим более широкий дигитоид с острой вершиной).
93
Восточнее, в Карагандинской области (Ачатау, Кызыл-тау) встречается подвид
М. tuschkan andruschkoi (с более тупой вершниой дигитоида) [128, 134, 153, 19].
Следует отметить, что самки М. eucta и М. tuschkan, повидимому, неразличимы;
поэтому возможно, что в будущем окажется целесообразным объединить эти два
вида в один, образующий много географических рас.
9(2). Самки.
10(11). Над двумя апикальными шпорами VIII тергита (рис. 230) сидит
щетинка, в большинстве случаев утолщенная по сравнению со все-
ми прочими щетинками этого тергита. Апикальный край VII стер-
нита с выраженным боковым выступом. Придаток семяприемника
присоединен к резервуару несколько отступя от его узкой сто-
роны ........................................................М. hebes.
11(10). Указанная щетинка VIII тергита, как правило, не толще прочих
его щетинок.
12(13). Боковой выступ края VII стернита выражен слабо (рис. 234).
Придаток семяприемника присоединяется к резервуару с его
узкой стороны................................................М. lenis.
13(12). Боковой выступ края VII стернита хорошо развит, над ним
стернит глубоко вырезан (рис. 235). Семяприемник такой же,
как у М. hebes.................................М. eucta и М. tuschkan.
14(1). Лобный зубчик отсутствует. Кроме развитого фронтального ряда
имеются еще вставочные щетинки между ним и глазным рядом.
Дигитоид широкий с темным шипом у вершины (рис. 233). У са-
мок (рис. 236) край VII стернита без больших выступов и выре-
зок, а по краю VIII тергита нет особо утолщенных щетин. Резер-
вуар семяприемника длиннее, чем придаток............................
...............................М. (Desertopsylla) rothschildi.
Паразит тушканчиков, обитающих в песчаных пустынях Средней Азии (Кара-
кум, Кызыл-Кум) [153].
ПОДСЕМЕЙСТВО LEPTOPSYLLINAE
Род Amphipsylla [149, 19, 154, 182]
Паразиты мелких грызунов Азии (полевок, хомячков и др.); немно-
гие виды проникли в Европу и Америку. Мы приводим все известные
виды этого рода, но рисунки и более детальные сведения даем только
для 16 видов. Обзор большинства видов Amphipsylla сделан в дру-
гой работе [149]. Самцы легко определяются по характерной для каждого
вида форме и вооружению дигитоида. Для определения самок этого рода
в большинстве случаев необходимо рассмотреть строение лобного зуб-
чика, форму копулятивной сумки с ее протоком и семяприемника, что
требует приготовления просветленных препаратов или надавливания
покровным стеклом. В ряде случаев самки могут быть определены лишь
по сопутствующим самцам или по географическому критерию.
1(16). Лобный зубчик у большинства видов не развит, имеется лишь
слабый бугорок (рис. 54, Б, Е), иногда совсем рудиментарный.
В первом теменном ряду обычно имеется не менее двух щетинок1.
Все темные шипы или шпоры дигитоида находятся выше его
середины (рис. 238, А—Е) 1 2. Копулятивный проток самки не
очень длинный (рис. 241, А—Е), также и резервуар семя-
1 Из относящихся к этой группе видов A. prima часто имеет лишь одну щетин-
ку, реже — две.
2 Исключение представляет недавно найденный на Копет-Даге паразит полевок
A. parthiana; этот вид по высокому выпуклому лбу, наличию двух (трех) щетинок
в первом теменном ряду, по вентральной вырезке VIII стернита самца и строению
гениталий самки явно родственен блохам I группы (ближе всего он к A. phaiomy-
dis), но нижняя шпора дигитоида расположена у него ниже середины заднего края.
94
приемника, имеющий почкообразную форму (как на рис. 239, Л)
......................................................группа I.
Относящаяся сюда группа видов (I по принятой классификации) распадается
на две подгруппы (А и Б), из которых вторая включает переходные формы
К группе II.
2(5). Лобный бугорок расположен сравнительно высоко. Лоб выпук-
лый, т. е. лобный край ниже бугорка, обычно направляется за-
метно кзади (рис. 54, А—Б). В первом теменном ряду имеется
две-четыре щетинки. Вентральный край VIII стернита самца
(рис. 242, Л) с заметной вырезкой, но без склеротизованного
тяжа с пучком щетинок на вершине; интерсегментальная вилочка
отсутствует.............................................подгруппа А.
3(4). Лобный бугорок расположен выше, чем у других видов
(рис. 54, Л). У самца: дигитоид — рис. 238, Л, VIII стернит —
рис. 242, Л. У самки VII стернит с резким боковым выступом
(рис. 240, Л); семяприемник — см. рис. 239, Л, проток копулятив-
ной сумки — рис. 241, В....................................A. rossica.
Паразит обыкновенной полевки (Microtus arvalis); встречается и на других мел-
ких грызунах и хищниках, а иногда и в жилище человека. Распространен от Боге-
мии на западе до Алтая на востоке и от Ленинградской области и Архангельска
на севере до Закавказья и северных хребтов Тянь-Шаня на юге. Обитатель степей
и луголесья. Повидимому, более обилен зимой.
Выделенные географические расы: А. г. contigua (Западная Европа), А. г. 1о-
cuples (Тянь-Шань)—пока трудно отличимы [149, 101].
4(3). Лобный бугорок расположен немного ниже (рис. 54, Б). Между
глазным и лобным рядом нередко бывает больше двух крупных
вставочных щетинок (из других близких видов эта особенность
иногда наблюдалась еще у A. kuznetzovi). Самец: дигитоид — см.
рис. 238, Б. Самка: VII стернит — см. рис. 240, Б, семяприемник
примерно такой же, как на рис. 239, Л, копулятивная сумка —
см. рис. 241, Л, Б............................................A. sibirica.
Паразит лесных полевок (Clethrionomys) и других мелких лесных зверьков. Рас-
пространен во многих лесных районах северного полушария (Западная Европа, Лап-
ландия, Урал, Тянь-Шань, Сибирь, Канада). Представлен несколькими подви-
дами [149, 19]:
a) A. s. sepifera: Швейцария, Австрия, Франция. Отношение ширины дигитоида
К его длине: Д — 14 : 35 [.180].
б) A. s. hetera: Центральная Европа (горы Высокая Татра). Д = 14 : 36; между
шпорами дигитоида сидят лишь два слабых волоска, а у вершины VIII стернита
самца много мелких шипиков. Каков копулятивный проток у названных двух евро-
пейских форм—неизвестно [179 (38)].
в) A. s. sibirica (thoracicus): Финляндия, Лапландия, Урал, Западная Сибирь
(Омская, Новосибирская. Томская, Кемеровская области), север Сибири (Туруханск,
верховья р. Таз), Алтай, Тянь-Шань. Д = 14:30. Склеротизованная часть поотока
копулятивной сумки короткая и образует дугу, выпуклую кпереди (рис. 241, А) [126, 19].
г) A. s. orientalis: Предбайкалье и Западное Забайкалье. Дигитоид иногда
немного длиннее и уже, чем у предыдущей формы, Д = 14:32. Склеротизованный
копулятивный проток заметно длиннее и выпуклая кпереди дуга его обособляется
от копулятивной сумки «шейкой» (рис. 241, Б). Таким образом, этот подвид надежно
отличается от предыдущего лишь по самкам: зато эти самки иногда трудно отличимы
от самок следующей группы видов (A. marikovskii и др.) [19].
д) A. s. pollionis: Канада. Самцы этой формы, повидимому. отличаются от
A. s. orientalis более многочисленными крупными щетинками на VIII стерните самца,
образующими более одного ряда (на рисунке Ротшильда их девять; v наших
A. s. orientalis три-пять щетинок, расположенных в один ряд). Копулятивный проток
у самки A. s. pollionis, судя по рисунку [25], такой же, как у A. s. orientalis
[188 (12), 25].
В общем создается представление, что, начиная от Финляндии через Сибирь
и до Канады, признаки A. sibirica меняются постепенно и не очень значительно;
бпдее заметно удлинение копулятивного протока (Восточная Сибирь и Канада), ме-
нее отчетливо меняются очертания дигитоида. В ряде случаев определение возможно
только до вида.
5(2). Лобный бугорок расположен примерно как у A. sibirica или
ниже, он развит слабо, иногда почти отсутствует (A. kuznetzovi,
95
A. asiatica, A. montana), у некоторых видов переходит в рудимен-
тарный зубчик (A. phaiomydis, A. parthiana, A. petristshevae).
Лоб менее выпуклый, чем в предыдущей группе видов. В первом
теменном ряду обычно две щетинки, реже — три и очень редко —
одна (у A. priina, наоборот, — часто одна, редко — две). Вдоль
вентрального края VIII стернита самца (рис. 242, В) у большин-
ства видов имеется внутренний склеротизованный тяж с пучком
щетинок на вершине (у A. montana и A. parthiana он рудимента-
рен, а у A. phaiomydis исчез почти бесследно). У многих видов
один или оба главных шипа дигитоида тупые (рис. 238, Г; исклю-
чения— см. рис. 238, Д, Е и A. parthiana). У самок блох этой
группы на анальном стерните проксимальные вентральные ще-
тинки часто утолщены и темные (исключения: A. montana и ино-
гда некоторые другие). Задний край VII стернита образует до-
вольно правильную дугу (рис. 240, Д, £), иногда с боковым
выступом (рис. 240, Г). Копулятивный проток однотипной формы
(рис. 241, Г—Е)..................................подгруппа Б.
Сюда относятся азиатские виды, преимущественно обитающие в горах и обра-
зующие по морфологии переходную подгруппу к видам группы II. Этот переходный
характер проявляется в вариабильности важнейших групповых признаков, упомянутых
выше. Ввиду близкого родства относящихся сюда видов распознавать их приходится
в основном по модифицированным сегментам самца путем сравнения с рисунками.
Некоторые особенности самок отдельных видов упомянуты в п. 5, а также приводятся
ниже, но иногда самки отличимы от близких видов лишь по сопутствующим самцам
или по географическому критерию.
6(15). В первом теменном ряду обычно не менее двух щетинок
(очень редко — одна).
7(10). Оба больших шипа (шпоры) дигитоида заострены (рис. 238, Д, £).
Восточноазиатские виды.
8(9). Дигитоид — см. на рис. 238, Д, VIII стернит самца с усеченным
задним краем и резко выступающим нижним углом; ряд из
пяти-семи сильных щетинок идет косо, отступя от этого края.
Самки: край VII стернита — см. рис. 240, Д, копулятивный про-
ток— рис. 241, Д....................................A. marikovskii.
Паразит лесных мышевидных грызунов востока Сибири — север Иркутской об-
ласти (Наканна на р. Н. Тунгуска), Якутская АССР (Якутск, устье р. Вилюй),
Забайкалье (Александровский Завод), Амурская и Хабаровская области, Приморье,
Монголия (близ Улан-Батора) [149].
Описанная с Аляски A. ewingi представляет, судя по описанию [170], не более
как подвид A. marikovskii, а возможно, что вполне идентична с этим видом. Из США
описана еще A. neotomae I. Fox 1940 [25, 26], однако, судя по описанию, едва ли
этот вид относится к Amphipsylla.
9(8). Нижняя шпора дигитоида (рис. 238, Е) расположена выше, чем
у предыдущего вида. Задний край VIII стернита самца округлый
с многочисленными щетинками, рассеянными вдоль края стернита.
Самка: край VII стернита — см. рис. 240, Е . . . A. petristshevae.
Распространен в Корее на лесных полевках (Clethrionomys), обитающих в гор-
ных лесах [42].
10(7). Один или оба большие шипа дигитоида тупые (кроме A. par-
thiana). Западноазиатские виды.
11(12). Дигитоид топорообразный, довольно широкий (рис. 238, Г).
У самки задний край VII стернита с более или менее выражен-
ным боковым выступом (рис. 240, Г), однако очертание края
стернита варьирует, так же как и степень развития проксималь-
ных щетин на анальном стерните (но чаще они сильные); копу-
лятивный проток — рис. 241, Г...........................A. kuznetzovi.
96
Вид, распространенный от Предбайкалья (Болынеглубоковское) и Красноярска
до Урала, Алтая, Тувинской автономной области, Монголии (Тайшири), Тянь-Шаня
и Кавказа, преимущественно на различных полевках в горах. Образует местные расы
(A. k. deminuta на Заилийском Алатау, A. k. glacialis на Кавказе), которые еще
требуют изучения [149, 10, 19].
12(11). Дигитоид более узкий. Живут в горах Средней Азии.
13(14). Из двух шипов дигитоида верхний — тупой, а нижний — острый.
У самки две проксимальные щетинки анального стернита очень
толстые и темные............................................A. asiatica.
Паразит узкочерепных полевок (Stenocranius), обитающих на горных лугах
Тянь-Шаня [149, 132, 19].
14(13). Шипы дигитоида иные. Проксимальные щетинки анального
стернита самок не выделяются толщиной и цветом в такой мере,
как у предыдущих видов этой подгруппы.
Сюда относятся три близких вида, признаки которых мы приведем без деления
на тезы и антитезы. Самки этих видов должны определяться с учетом признаков
сопутствующих самцов, географического критерия и хозяина.
а) Верхний шип дигитоида заменен щетинкой, а нижний тупой.
Край VII стернита самки со значительной выпуклостью кзади,
т. е. вентрально довольно сильно вырезан .... A. montana.
Паразитирует на высокогорных полевках (Alticola), обитающих в скалах и осы-
пях гор Западного Тянь-Шаня и Памира [19].
б) Оба шипа дигитоида тупые. Край VII стернита самки менее вы-
пуклый кзади, чем у предыдущего вида ... A. phaiomydis.
Паразит центральноазиатских полевок (Phaiomys), обитающих в горах Запад-
ного Тянь-Шаня, Алая и Памира [19].
в) Оба шипа дигитоида острые, и нижний из них расположен ниже
середины заднего края. У самки задний край VII стернита слабо
выпуклый................................................A. parthiana.
Паразит полевок Microtus socialis, обитающих в горах Копет-Дага [154].
15(6). В первом теменном ряду обычно одна щетинка, редко — две.
У самца: дигитоид — см. рис. 238, В; у самки: край VII стерни-
та— рис. 240, В, копулятивная сумка — рис. 241, Е ... A. prima.
Паразит степной пеструшки (Lagurus lagurus) и других полевок, найден в За-
падной Сибири (Омская область, Змеиногорск) и в Нижнем Поволжье (Саратов)
и Западно-Казахстанской области [126, 149].
16(1). На лбу имеется не бугорок, а настоящий зубчик (рис. 54, В и Ж)
чаще довольно хорошо развитый, но иногда и рудиментарный
(рис. 54, В); расположен он низко. Выпуклость лба слабо выра-
жена или не выражена (рис. 54, В—Д). В первом теменном ряду
обычно имеется лишь одна щетинка, изредка в виде аномалии
бывает две. На дигитоиде (рис. 238, Ж—/7), кроме темных шипов
и шпор, находящихся в апикальной половине, имеется еще шпо-
ра, сидящая ниже середины заднего края, редко на середине или
несколько выше. Вентральный склеротизованный тяж VIII стерни-
та самца, характерный для подгруппы Б, отсутствует или руди-
ментарен (от него часто остается вершинная группа щетинок
у края стернита)...........................................группа II.
17(36). У самца первый членик антенны без развитого пучка волосков
на дорзальном крае. Вентральный край VIII стернита самца
без развитой вырезки; у его основания обычно имеется «интер-
сегментальная вилочка» — цилиндрический вырост с одной-дву-
мя-четырьмя щетинками на вершине (рис. 242, 5; 243, 5; эта
вилочка у A. dumalis чаще отсутствует). У самки копулятивный
7 Определитель блох 97
проток примерно такой, как в группе I, с некоторыми уклоне-
ниями в форме и длине (рис. 241, Ж—М; исключение — A. kala-
bukhovi). Резервуар семяприемника часто удлиненный, иногда
почкообразный (рис. 239, В—Е)....................подгруппа В.
18(29). Лобный зубчик слабо выступает над краем лба, часто совсем
рудиментарный или незаметный. Глаз обычно слабее пигментиро-
ван или уменьшен по сравнению с видами группы I (рис. 54, Г)
или рудиментарен. IX стернит самца без сильной шпорообразной
щетины на вершине. У самки копулятивный проток (рис. 241,
Ж—/С), как правило, не отличается большой длиной (расстояние
между концами склеротизованной части около 0,1 мм; исключе-
ния— A. argoi и A. montium, у которых это расстояние равно
0,12—0,15 мм).
19(26). Глаз развит. У самки семяприемник с удлиненным резервуаром
(кроме A. tuta?).
20(21). Имеется лобный зубчик. Глаз, как правило, сравнительно боль-
шой, но бледно пигментирован. Дигитоид (рис. 238, Ж) в общем
четырехугольный, задне-нижний угол его сильно выступает, а зад-
ний край вогнутый; по этому краю у верхнего угла сидят две
шпоры и одна сильная — у нижнего угла. Край VII стернита сам-
ки без боковой вырезки, примерно такой, как на рис. 240, Д. Се-
мяприемник с удлиненным резервуаром (рис. 239, Г). Копулятив-
ный проток с дугой малого радиуса (рис. 241,3).........................
.............................................A. schelkovnikovi.
Паразит хомячков Cricetulus migratorius. Известен от Закавказья и Заволжья
до северных предгорий Тянь-Шаня. Описан подвид: A. s. irana — с более узким диги-
тоидом (обнаружен в Иране, Туркменской ССР и в Самарканде). Что касается
A. s. certa (Тянь-Шань), то морфологические отличия этого подвида пока не ясны
[149, 182, 153, 19, 99].
21(20). Лоб очень низкий (рис. 54, Г). Лобный зубчик рудиментарный
или даже не заметен. Глаз уменьшенный. Дигитоид более узкий,
задний край его не вогнутый (рис. 238, 3—И; 243,5). У самок на
стернитах, в особенности на шестом, имеется вертикальный более
темный тяж (рис. 244); край VII стернита с неглубокой боковой
вырезкой (рис. 240, Ж—И). Копулятивный проток (рис. 241,
И—К) отличается тем, что «шейка», не изгибаясь, переходит
в дугу (кроме A. tuta — рис. 241, Ж)-
22(23). Дигитоид (рис. 238,3) с округленным задне-верхним краем;
вместо двух главных шпор заднего края имеются щетинки, а две
шпоры внутренней боковой поверхности сильно развиты. У самки
VII стернит с небольшой боковой вырезкой (рис. 240,3), семя-
приемник и копулятивный проток изображены на рис. 239, Д\
241, И...................................................A. vinogradovi.
Паразит хомячков Cricetulus barabensis (= furunculus), С. longicaudatus и дру-
гих мелких грызунов степей Восточной Азии [Хакасская автономная область, г. Мину-
синск, Тувинская автономная область, Предбайкалье, Забайкалье, Амурская область,
Приморский край (Турий Рог, Гродеково, Ворошилов-Уссурийский), Маньчжурия,
Монголия (Хэнтэйский, Центральный, Баян-Хонгорский, Дзабханский, Гоби-Алтай-
ский и Убсанурский аймаки) [132, 149].
23(22). Дигитоид (рис. 238, И; 243, Б) с выраженным задне-верхним
и задне-нижним углом; верхняя шпора бледная. У самки
VII стернит — см. на рис. 240, Ж, И.
24(25). Дигитоид изображен на рис. 243, Б; особенно характерна длин-
ная шпора, сидящая на середине заднего края. VII стернит (бо-
ковая вырезка которого шире, чем у предыдущего вида), семя-
приемник и копулятивный проток самки см. на рис. 239,5; 240 И\
241, К; 244 . . . .......................................A. longispina.
Распространен в Монголии (Хэнтэйский и Убсанурский аймаки, хребет Тайшири),
паразитирует на хомячках Phodopus songarus [154].
98
25(24). Дигитоид — см. на рис. 238, Я; верхняя шпора его слабо пигмен-
тирована. VII стернит самки с боковым выступом см. на
рис. 240, Ж, а копулятивный проток — на рис. 241, Ж . • .A. tuta.
Вид, известный только с Тибета (верховья рр. Хуанхэ и Бар-Чу), паразитирует
на Microtus и Ochotona [126, 149].
26(19). Глаз сильно редуцирован, почти до полного отсутствия. Семя-
приемник почкообразный.
27(28). Глаз рудиментарен или отсутствует. Дигитоид четырехугольный;
у верхнего и нижнего заднего угла сидит по одной длинной
шпоре, а между ними очень сильные щетины; верхняя шпора со
вздутием посредине (как бы изломана). Край VII стернита сам-
ки со слабо выраженной неглубокой боковой вырезкой. Семя-
приемник и копулятивный проток примерно такие же, как на
рис. 239, Б и 241, Л...................................... . . A. argoi.
Паразит мышевидного хомячка Calomyscus bailwardi, обитающий на Копет-Даге.
В Закавказье имеется близкая, но отличающаяся форма [153].
28(27). В описании указано, что глаз меньше, чем у A. schelkovnikovl.
Самец неизвестен. Край VII стернита самки с небольшой, но
обычно резкой и довольно глубокой вырезкой. Внутренние гени-
талии того же типа, что у предыдущего вида. Отмечается тонко-
стенность копулятивного протока...........................A. montium.
Известно четыре самки, собранные с Calomyscus bailwardi в Северо-Восточном
Афганистане [99].
29(18). Лобный зубчик развит. Глаз развит хорошо или рудиментарен.
У самца на вершине IX стернита имеется шпора. У самки склеро-
тизованная часть копулятивного протока длиннее, чем у большин-
ства ранее рассмотренных видов (расстояние между ее концами
0,13 мм и более — рис. 241, Л, М 1). Резервуар семяприемника поч-
кообразный, но несколько удлиненный (рис. 239, В).
30(35). Дигитоид довольно узкий (рис. 238, К—Л). Копулятивный про-
ток самки изображен на рис. 241, Л, М.
31(32). Глаз обычно сильно редуцирован. Хоботок варьирует, но обычно
почти достигает вершины ляжки. У самца интерсегментальная
вилочка у основания VIII стернита бывает только в редких слу-
чаях. Дигитоид — см. на рис. 238, Л. У самки край VII стернита
изображен на рис. 240, К (боковой выступ не всегда выражен).
Расстояние между концами склеротизованной части копулятивно-
го протока около 0,13—0,14 мм (рис. 241, Л) .... A. dumalis.
Паразит слепушонок Ellobius talpinus, распространен в горах Центральной
и Средней Азии — Монголия (Орон-Нор, Южный Хангай. Тайшири), на Тянь-Шане,
Алае. Вопрос о достаточной диференцировке подвидов (например, A. d. dilatata —
Монголия) еще требует выяснения [101, 149, 19].
32(31). Глаз развит, как у большинства видов этого рода (иногда в виде
аномалии сильно редуцирован у A. anceps). Хоботок достигает
примерно 3/4 ляжки. Интерсегментальная вилочка имеется.
33(34). Дигитоид, как у A. dumalis, но шпоры длиннее. На заднем бедре
меньше боковых щетинок, чем у A. dumalis, — по одной-две на
наружной и внутренней поверхностях. Кавказский вид . . .
..........................................................A. georgica.
Кавказ (Грузинская ССР). Известны единичные экземпляры, собранные с мел-
ких мышевидных грызунов [156].
1 В эту группу по признакам самцов попадает A. kalabukhovi, однако самки
этого вида имеют иную форму копулятивного протока, характерную для следующей
подгруппы (как на рис. 241, Я).
7* 99
34(33). Нижняя шпора дигитоида (рис. 238, Л) ближе к его нижнему
концу, чем у A. dumalis. У задне-нижнего угла VIII стернита
самца (рис. 242, Б) имеется пучок темных щетинок. У самки край
VII стернита примерно такой, как на рис. 240, Д, К. Расстояние
между концами склеротизованной части копулятивного протока
около 0,14—0,16 мм (рис. 241, М)...............................A. anceps.
Паразит хомячков Cricetulus migratorius, распространен в горах Алтая, Тянь-
Шаня, Алая, Памира, Гиссарского хребта. Подвид: A. a. coangusta (Алтай), для
которого считался характерным более узкий дигитоид, оказался географически недо-
статочно обособленным — подобные экземпляры блох встречаются и в других местно-
стях [19, 149, 194].
35(30). Дигитоид более широкий, четырехугольный, на заднем крае
у верхнего и нижнего углов имеется по одной сильной шпоре.
У самок копулятивный проток примерно такой, как на рис. 241, Н
...................................................... A. kalabukhovi.
Паразит хомячков Cricetulus eversmanni и других грызунов, обитающих в сте-
пях западных районов Казахской ССР (р-ны Тургая, Гурьева, Н. Казанки) и в За-
волжье (Куйбышевская область) [149].
36(17). Лобный зубчик развит. Глаз чаще совсем рудиментарный (кро-
ме A. primaris). У самца на дорзальном крае I членика антенны
имеется пучок из длинных волосков. Вентральный край VIII стер-
нита самца с глубокой вырезкой (рис. 242, Г); интерсегменталь-
ная вилочка отсутствует. На вершине IX стернита имеется шпора.
У самки копулятивный проток напоминает дверную ручку, он
длиннее, чем у других видов, и его дуга направлена своей выпук-
лостью кверху и кзади (рис. 241, Н—П). Резервуар семяприем-
ника почкообразный (рис. 239, Б)...........................подгруппа Г.
37(38). Глаз развит, но сравнительно мал (рис. 54, В). Между глазным
и лобным рядом имеется лишь две вставочных щетинки. Хоботок
не достигает вершины ляжки. На средних тергитах брюшка ниже
стигмы сидит лишь одна щетинка. Дигитоид — см. рис. 238,
М—Н. У самки на V и VI стернитах имеется вертикальный более
темный тяж. Край VII стернита — см. рис. 240, Л, копулятивный
проток — рис. 241, Н.......................................A. primaris.
Распространен на многих видах грызунов (полевки, тушканчики и др.) в степных
и горных районах Азии от Якутской АССР через Предбайкалье, Забайкалье, Тувин-
скую автономную область, юго-восток Алтая, Монголию, Карагандинскую область,
Тянь-Шань до Памира. Политопный и полизойный вид, довольно заметно варьирует
в разных местностях. Пока выделено два подвида:
a) A. primaris primaris: западная форма — Тянь-Шань, Памир (рис. 238, М)
[101, 149].
б) A. primaris mitis: восточная форма (Восточная Сибирь, Монголия). Самцы
этого подвида отличаются несколько более узким дигитоидом по сравнению с номи-
нальной западной формой (рис. 238, Н) [92, 149].
Следует заметить, что во многих районах Предбайкалья, Западной Монголии
и несколько западнее встречаются переходные экземпляры с признаками обоих
подвидов. В таких случаях следует ограничиваться определением до вида.
38(37). Глаз рудиментарен или отсутствует (рис. 54, Д). Между глаз-
ным и лобным рядами обычно не менее трех вставочных щетин.
Хоботок почти достигает вершины коксы. На тергитах брюшка
ниже стигмы сидят две щетинки.
39(40). У самца задний верхний угол дигитоида выше переднего и сидя-
щий на нем шип заострен (рлс. 238, П). У самки щетинки// чле-
ника антенны достигают 3Л булавы. На стернитах с /// по VI
имеется вертикальный более темный тяж. Край VII стернита
(рис. 240, //) со значительной вентральной вырезкой. Копулятив-
ный проток с пологой дугой (рис. 241, П) .... A. aspalacis.
Паразит маньчжурского цокора Myospalax psilurus, распространен в степях и ле-
состепях Маньчжурии, Приморья (Гродеково) и Восточного Забайкалья [92, 149].
100
40(39). Задний верхний шип дигитоида короткий, тупой. Край VII стерни-
та самки иной.
41(42). Дигитоид изображен на рис. 238, О. Край VII стернита самки —
см. рис. 240, М. Копулятивный проток изогнут круче (рис. 241,0)
..................................................................A. daea.
Паразит забайкальского цокора Myospalax aspalax. Распространен в Забайкалье
(Оловянинский и Борзинский районы) и Монголии (Хэнтэйский аймак), обитает на
луголесных участках [163, 149].
42(41). Дигитоид более узкий, а задне-верхний угол у него косо срезан
(рис. 243, Л). Для самок надежных отличительных (от A. daea)
признаков в описании не указано...................................A. casis.
Паразит цокора Myospalax fontanieri. Китай. (Шэньси) [100, 27].
Род Leptopsylla (=Ctenopsyllus}
Блохи этой группы являются паразитами мелких грызунов и широ-
ко распространены в Азии, Африке и Северной Америке. Мы включили
в определительную таблицу только шесть видов, относящихся к инте-
ресующему нас району.
1(6). В щечном гребне имеется более двух зубцов.........................
.................................................подрод Leptopsylla.
2(5). В щечном гребне имеется четыре зубца.
3(4). У переднего угла лба имеются два толстых темных шипа
(рис. 51)......................................................L. segnis.
Паразит домовых мышей, завезенный вместе с ними как во многие населенные
пункты Западной (Сибири и Восточной Азии, так и в другие страны мира. На инте-
ресующей нас территории обнаружен в Иркутске, Якутске, Чите, многих селах Забай-
калья, Ворошилове-Уссурийском, Владивостоке, в Амурской и Уссурийской областях,
Еврейской автономной области, а также в Маньчжурии, Китае и Корее [29].
4(3). У переднего угла лба имеются пять-семь толстых коротких чер-
ных шипов. Конусообразная выпуклость лба более резко выра-
жена, чем у предыдущего вида. Половая клешня самца и VII стер-
нит самки изображены на рис. 245, А—Б; дигитоид иной формы,
чем у L. segnis..................................................L. папа.
Паразит мелких грызунов (Alticola, Cricetulus и др.) горных районов Азии, рас-
пространен на Кавказе, Памире, Тянь-Шане и в Монголии (Хангай). Блохи, обнару-
женные в Монголии, отличаются вариабильностыо формы дигитоида, который иногда
весьма узок, а в других случаях расширяется к основанию (рис. 246). Вариабильность
L. папа требует дальнейшего изучения [153, 19, 121].
5(2). В щечном гребне шесть зубцов. На лбу три-четыре шипа. Греб-
невидный ряд щетин заднего края голеней развит слабо . . .
........................................................L. sexdentata.
Паразит домовых мышей, распространен в некоторых горных районах Закав-
казья, на Копет-Даге и Тянь-Шане [126, 84].
^6(1). В щечном гребне два зубца.................подрод Peromyscopsylla.
7(8). У переднего угла лба имеется два толстых шипа, а остальные
щетинки хотя утолщены, но, как правило, все же отличаются от
этих шипов. У самки VII стернит с узким длинным боковым вы-
ступом и глубокой вырезкой под ним (рис. 248, 249). Половая
клешня, VIII и IX стерниты самца — см. рис. 247, 250 ....
.......................................L. bidentata и L. ostsibirica.
Эти два близких вида паразитируют (чаще осенью) на лесных полевках (рода
Clethrionomys) и на других мелких лесных зверьках Европы и Азии. L. bidentata
встречаются во многих лесных районах: Франции, Швейцарии, Югославии, Германии.
Норвегии, Европейской части СССР (от Белоруссии до Нижнего Поволжья, Урала
и Северного Кавказа), в Зауралье, на севере Казахской ССР, в горах Тянь-Шаня,
Алтая, в Томской области (с. Сурово Туганского района), Ямало-Ненецком нацио-
нальном округе (Мотылька).
101
L. ostsibirica распространен в лесных районах Иркутской области (Наканна на
р. Н. Тунгуска, Янды на р. Ангара, Большеглубоковское хозяйство), в Забайкалье,
Якутской АССР, Хабаровском и Приморском краях, а также в Корее.
Самцы этих видов хорошо отличаются друг от друга по форме дигитоида —
у L. ostsibirica—он шире и верхнезадний его край более угловатый (рис. 250),
у L. bidentata этот край более закругленный; из трех сильных щетинок, сидящих на
нем, одна далеко отодвинута книзу от двух других (рис. 247). Самок следует опре-
делять по сопутствующим самцам или по географическому критерию.
К этой Же группе принадлежит американский вид L. hamifer, у которого диги-
тоид еще шире, чем у L. ostsibirica [75, 169, 25, 26].
8(7). У переднего угла лба имеются три шипа. У самки VII стернит
с небольшим боковым выступом (рис. 251, Б) . Половая клешня,
VIII и IX стерниты самца изображены на рис. 251, А . . . .
.........................................................L. silvatica.
Паразит мелких леоных млекопитающих Европы и Западной Сибири; чаще
встречается на полевках (Clethrionomys), но нигде еще не собран в достаточном
количестве, чтобы установить его истинного хозяина. Имеется как в летних, так
и в осенних сборах. Распространен в лесах Западной Европы, Европейской части
СССР, Зауралья, Ханты-Мансийска, Томской (Парбигский район) и Новосибирской
областей (Купинский район). В Западной Европе образует подвиды: L. s. spectabilis,
L. s. fa Пах [186, 179 (38)].
Род Pectinoctenus [126, 19J
Представители этого рода (близкого к предыдущему) живут в Азии
на мелких грызунах — хомячках, лесных мышах и т. п. Мы включили
в таблицу все известные виды, а рисунки даем лишь для тех, которые
живут в Восточной Азии.
1(8). В гребне на голове не менее 11 зубцов.
2(5). Сибирские и центральноазиатские виды. На пронотуме впереди
от главного ряда имеется развитый ряд щетинок. У самки
VII стернит с очень глубокой и широкой вырезкой (рис. 253,
255). Дигитоид не расширяется книзу (рис. 252, 254).
3(4). В гребне на голове у самки имеется 17—20 (редко 15—16) зуб-
цов, а у самца 15—18. У самки (рис. 253) VII стернит с довольно
многочисленными добавочными щетинками, расположенными впе-
реди основного ряда. У самца (рис. 252) на заднем крае диги-
тоида имеется шипик, а на VIII стерните — единичные (обычно
три) щетинки...........................................Р. pavlovskii.
Паразит джунгарского хомячка (Phodopus songarus), а также многих других
степных грызунов. Найден в Западной Сибири (Омская область, Славгород Алтай-
ского края, Хакасская автономная область), Тувинской автономной области, Казах-
ской ССР (Кокчетавская область, близ Щучинска), в районах Западного и Восточного
Забайкалья, в Монголии (Хангай, Тайшири, Монгольский Алтай, Убсанурский и Хэн-
тэйский аймаки) [132].
4(3). В гребне на голове (рис. 55) у самки имеется 14—17 зубцов,
а у самца 13—17. У самки (рис. 255) VII стернит без добавоч-
ных щетинок впереди от главного ряда (или иногда с единичными
такими щетинками). У самца (рис. 254) на заднем крае дигитои-
да нет шипика, а на VIII стерните расположены многочисленные
щетинки.................................................Р. pectiniceps.
Паразит лесных мышей (Apodemus speciosus) и других грызунов, живущих вме-
сте с ними в Восточной Азии. Распространен в лесных и горных районах Забайкалья
(преимущественно Западного), Приамурья, Сахалина, Сихотэ-Алиня, Приморья и Ко-
реи [122, 132].
5(2). Тяньшаньские виды. На пронотуме впереди от главного ряда нет
щетинок или имеются лишь единичные. В гребне на голове около
14—16 зубцов. У самки VII стернит с вентральной вырезкой. Ди-
гитоид расширяется книзу.
102
6(7). У самки VII стернит с широкой вентральной вырезкой и заострен-
ной боковой лопастью над ней. Дигитоид самца имеет неболь-
шой задне-нижний выступ..............................Р. ctenophorus.
Паразит хомячков 'Cricetulus migratorius и других грызунов, живущих в горных
местностях с преобладанием засушливого пустынно-степного ландшафта. Найден
в Тянь-Шаньской области Киргизской ССР [126, 19].
7(6). У самки VII стернит с довольно узкой глубокой вентральной вы-
резкой. Дигитоид с резким клювообразным задне-нижним вы-
ступом ....................................................Р. lautus.
Вид, близкий к предыдущему и заменяющий его в более восточных районах
Центрального Тянь-Шаня (Джунгарский и Заилийский Алатау, Кетмень, Нарын-
кол) [188 (22), 19].
8 (1). В гребне на голове менее 11 зубцов.
9(10). В гребне на голове имеется около восьми-десяти зубцов. У сам-
ки VII стернит с неширокой субвентральной вырезкой. Дигитоид
имеет форму банана.....................................Р. nemorosus.
Паразит среднеазиатских горных лесных мышей (Silvimus silvaticus). Найден
во многих местах Центрального и Восточного Тянь-Шаня (до Джунгарского Алатау
и Синь-Цзяня включительно) и на Гиссарском хребте [82].
10(9). В гребне на голове семь зубцов. Подвижный и неподвижный
пальцы короче, чем у предыдущего вида. Самка неизвестна
........................................................Р. pamirensis.
Один самец этого вида снят с хомячка на Памире (Хорог) [153].
СЕМЕЙСТВО CTENOPHTHALMIDAE
Блохи этого семейства, как и предыдущего, паразитируют на грызу-
нах, насекомоядных и т. д., причем большинство относящихся сюда видов
принадлежит к числу «гнездовых блох» и имеет недоразвитые или совсем
рудиментарные глаза. Морфологическая характеристика этой группы
следующая. Метанотум без апикальных зубчиков. Пигидий у обоих полов
более или менее выпуклый (иногда этот признак слабо выражен, напри-
мер, у некоторых Nearctopsylla, Palaeopsylla, Wagnerina, особенно у сам-
цов). Внутренняя поверхность задней коксы иногда с группой шипиков.
У самки на вершине церка рядом с длинной апикальной щетинкой сидят
мелкие волоски; иногда имеются две сильные апикальные щетинки. Семя-
приемников один или два. У самца IX стернит без тяжа, идущего кпере-
ди под эндофаллусом.
ПОДСЕМЕЙСТВО WAGNERINAE
Род Wagnerina {145, 19, 78]
Паразиты грызунов, живущих в горах Азии, преимущественно в ска-
лах и осыпях. Встречаются обычно осенью и зимой.
1 (6). Хоботок достигает вершины передней коксы или едва заходит
за нее. У самцов вместо предпигидиальных щетинок имеется бо-
лее или менее длинный выступ VII тергита, покрывающий сверху
пигидий. У самок проток копулятивной сумки без резкой склеро-
тизации.
103
2(5). У самцов (рис. 256, А; 257, А; 259) выступ VII тергита над
пигидием очень длинный; пигидий вертикальный; тело половой
клешни лентовидно суженное. У самок (рис. 256, Б; 257, Б)
вентральный край анального стернита прямой или слегка
выпуклый.
3(4). У самцов (рис. 256, Л) выступ VII тергита несколько короче
и шире, чем у следующего вида; на вершил е его сидит несколько
щетинок разной длины; половую клешню и модифицированные
сегменты см. на рис. 256, А; на VIII стерните, близко к краю си-
дит до четырех щетинок и единичные добавочные. У самок
(рис. 256, Б) на VII стерните пять-семь крупных щетинок и впе-
реди них большое число добавочных, образующих до трех
неправильных рядов; апикальный край VII стернита с широкой
боковой вырезкой. Стигма VIII тергита несколько удлиненная
..........................................................W. tecta.
Известен с Памира и Тянь-Шаня, паразитирует на высокогорных полевках
(Alticola argentata) и хомячках (Cricetulus migratorius). Возможно, что экземпляры
из Киргизии несколько отличаются от типичных [19].
4(3). У самцов (рис. 257, Л) выступ VII тергита длиннее и уже, чем
у W. tecta; на его вершине сидит одна совсем маленькая щетинка
и два-три волоска; на VIII стерните, близ апикального края
обычно три щетинки; строение половой клешни и модифицирован-
ных стернитов см. на рис. 257, Л; неподвижный выступ клешни
и треугольный дигитоид более широкие, чем у предыдущего вида.
У самок (рис. 257, Б) на VII стерните впереди от ряда крупных
щетинок имеется лишь небольшое число добавочных; апикальный
край этого стернита имеет неширокую боковую вырезку, глубина
которой варьирует. Стигма VIII тергита короткая; ниже ее сидит
одна крупная щетинка..................................W. longicauda.
Паразит горных сеноставок. Найден в Монголии (Ба:ян-Хонгорский аймак) на
Ochotona pricei совместно с W. antiqua [78].
В Тувинской автономной области на Ochotona alpina найден один экземпляр —
самец, напоминающий W. longicauda, но имеющий ряд отличий. Мы считаем целе-
сообразным выделить его под названием W. tuvensis sp. и. Он отличается наличием
двух светлых шпор на дигитоиде, из которых апикальная очень длинная (у других
видов имейся только одна шпора), и деталями в очертаниях дигитоида, неподвижного
пальца и IX стернита (рис. 259).
5(2). У самца (рис. 258, Л) длина выступа VII тергита не превышает
трех поперечников этого выступа; на вершине его три длинных
щетинки. Пигидий наклонный. Тело половой клешни широкое, ко-
роткое; дигитоид узкий, серповидно изогнутый, с выростом на
задне-нижнем крае; на VIII стерните обычно две щетинки; строе-
ние модифицированных стернитов см. на рис. 258, Л. У самки
(рис. 258, Б) вентральный край анального стернита образует
резкий выступ. На VII стерните несколько крупных щетинок и до
12 добавочных, расположенных двумя-тремя неправильными
рядами; апикальный край имеет неглубокую вырезку и слабый
выступ над ней. На VIII тергите ниже стигмы три (реже две)
длинных щетинки, расположенные косым рядом. Стигма VIII тер-
гита у обоих полов удлиненная..............................W. antiqua.
Паразит горных грызунов Монголии. Найден в Баян-Хонгорском и Дзабханском
аймаках на Ochotonai pricei и Alticola semicanus [78].
6(1). Хоботок длинный, почти на целый членик заходящий за вершину
передней коксы. У самцов на VII тергите одна обычная длинная
предпигидиальная щетинка. У самок проток копулятивной сумки
заметно склеротизован....................... . W. schelkovnikovi.
104
Вид, распространенный в Закавказье (Армянская ССР, Азербайджанская ССР),
в Иране и Hai Копет-Даге. Паразитирует на хомячка!х (Cricetulus migratorius,
Cailomyscus bailwardi), персидских песчанках (Meriones persicus) «и домовых мышах
(Mus musculus) [145].
ПОДСЕМЕЙСТВО ANOMIOPSYLLINAE
Род Eopsylla 1153]
Ряд родов, относящихся к этому же подсемейству, обитает в Север-
ной Америке. У нас известен лишь один представитель — Е. nuda (рис. 57,
260, 261), который пока был найден только в Сихотэ-Алиньском запо-
веднике, в гнезде летяги (Pteromys volans) и на лесной полевке
(Clethrionomys rufocanus).
ПОДСЕМЕЙСТВО CTENOPHTHALMINАЕ
Род Ctenophthalmus [12, 10, 199]
Блохи этого обширного рода распространены преимущественно
в Европе и Африке, и мы в этой книге ограничимся рассмотрением сравни-
тельно немногих видов, встречающихся в интересующем нас районе. Все
они являются паразитами грызунов. Кроме упоминаемых в таблице видов,
в Китае известны еще: Ct. parcus и Ct. dinormus (Сычуань), Ct. junnanus
(Юннань), а в Индии: Ct. crudelis [179 (38)]. В 1940 г. Ю. Н. Ваг-
нер [199] род Ctenophthalmus разделил на ряд более мелких групп, ко-
торые мы считаем ничем иным, как подродами L
Самцы видов этого рода проще всего определяются по форме по-
ловой клешни, путем сравнения с рисунками 262, А; 263; 265, А и др.
Поэтому приводимая ниже таблица нужна в основном для определе-
ния самок.
1(10). Стигма VIII тергита широкая, выпуклая, как на рис. 264; 267, Б.
Самцы —см. рис. 262, А; 263; 265, А; 266, А; 267, А.
2(9). Проток копулятивной сумки короткий (как на рис. 262, Б\ 264);
он значительно короче, чем зубцы гребня переднегруди. Самцы —
см. рис. 262, А; 263; 265, А; 266, А.
1 Приведем краткую определительную таблицу для известных до настоящего
времени групп рода Ctenophthailmus:
а (б). Стигма VIII тергита широкая, выпуклая (рис. 264; 267, Б)..................
.................................................группа Euctenophthalmus.
Сюда относятся: Ct. uncinatus, Ct. assimilis, Ct. congeneroides, Ct. arvalis и др.
б (а). Стигма VIII тергита узкая, Y-образная (рис. 268, Б).
в (г). Конец последнего членика хоботка без искривленной серповидной щетинки
..................................................... группа Palaeoctenophthalmus.
Сюда относятся несколько кавказских видов.
г (в). Такая щетинка имеется (как на рис. 345, Б).
д (е). На III и следующих тергитах брюшка под стигмой сидят две-три крупных
щетинки. Паразиты слепышей........................... группа SpailhcoctenophthaImus
Сюда относятся европейские виды, паразитирующие на слепышах.
е (д). Этих щетинок не более одной.
ж (к). Наиболее длинные щетинки II членика задней лапки заходят за вершину
IV членика.
з (и). Хоботок не заходит или едва заходит за вершину коксы.......................
.............................................. группа Paractenophthalmus.
Сюда относятся: Ct. dolichus и Ct. dux.
и (з). Хоботок длинный — достигает вершины вертлуга или заходит за нее. Пара-
зиты алтайского цокора................................группа Neoctenophthalmus.
Сюда относится Ct. dilatatus.
к (ж). Наиболее длинные щетинки II членика задней лапки далеко не достигают
IV членика............................................группа Ctenophthalmus s. str.
Сюда относится много европейских видов, а из азиатских Ct. pisticus.
105
3(4). Под краем VII стернита самки (рис. 262, Б) видна темная скле-
ротизация в виде двух резких параллельных полос. Самец с ши-
роким дигитоидом, верхне-задний угол которого выступает крюч-
кообразно (рис. 262, Л)................................Ct. uncinatus.
Паразит мелких лесных грызунов (Clethrionomys, Silvimus и др.), обитающих
в Северной Европе и Западной Сибири. Известен из многих мест — от Польши и Нор-
вегии до Ханты-Мансийска и Юргинского района Кемеровской области [122, 12].
4(3). Склеротизации, имеющаяся под краем VII стернита самки, иная.
Самцы — см. рис. 263; 265, А; 266, А.
5(6). Край VII стернита самки (рис. 264) с двумя боковыми выступа-
ми, разделенными развитой вырезкой. Полоски склеротизации,
просвечивающей под этим краем,— слабые, расходящиеся кзади.
У самца дигитоид топорообразной формы, подвержен значитель-
ной индивидуальной вариабильности (рис. 263). Один из наи-
более часто встречающихся видов Восточной Азии..........................
............................................Ct. congeneroides.
Обитатель гнезд мелких грызунов (Microtus, Apodemus и др.) луговых и луго-
лесных местностей и сельскохозяйственных угодий Восточной Азии. Распространен
в Западном и Восточном Забайкалье, Приамурье, Приморском крае, в Маньчжурии
и в Корее. Близкородственные виды обитают в Европе; такое распространение пред-
ставляет один из характерных примеров так называемого «европейско-дальневосточ-
ного разрыва» фауны [126].
6(5). Край VII стернита самки и дигитоид самца иной формы.
7(8). Верхний боковой выступ края VII стернита самки (рис. 265, Б)
более узкий, чем у предыдущего вида. Дигитоид (рис. 265, А)
с усеченным задне-верхним углом..........................Ct. truncus.
Вид, близкий к предыдущему и замещающий его на Сахалине (Александровск,
Ново-Сахалинск). Найден также на Курильских островах (о-в Шпанберга). Парази-
тирует на лесных полевках и крысах ["154].
8(7). Боковая вырезка VII стернита самки (рис. 266, Б), так же, как
и склеротизации, просвечивающая под этим краем, выражена сла-
бее, но сам край заметно утолщен. У самца дигитоид (рис. 266, Л)
имеет форму перевернутого (каблуком вверх) башмака . . .
....................................................... Ct. assimilis.
Паразит полевок (Microtus arvailis) и многих других мелких грызунов, распро-
страненный в луговых и луголесных местностях от Голландии и Италии до Запад-
ного Забайкалья, Алтая и Тянь-Шаня. Из Сибири имеются сборы, произведенные
в Омской, Томской, Новосибирской, Кемеровской, Иркутской областях, Алтайском
и Красноярском краях, на севере Казахской ССР и в Бурят-Монгольской АССР
[122, 12].
9(2). Проток копулятивной сумки длиннее (рис. 267, Б); он примерно
равен по длине или слегка длиннее, чем зубцы гребня передне-
груди. Край VII стернита самки и форму половой клешни сам-
ца — см. рис. 267, А, Б......................................Ct. arvalis.
Обитатель гнезд степных грызунов — сусликов, полевок, хомячков и др. Распро-
странен в Предбайкалье, Алтайском и Красноярском краях, Новосибирской, Восточно-
Казахстанской, Талды-Курганской областях, Тувинской автономной области, Северной
Джунгарии и Монголии (Тайшири, Монгольский Алтай) [128].
К этой же группе принадлежат более западные степные виды: Ct. breviatus,
Ct. pollex и др.
10(1). Стигма VIII тергита узкая, Y-образная (рис. 268, Б). Самцы —
см. рис. 268, Л; 270, Л; 271, Л; 272, Л.
11(12). Наиболее длинные щетинки II членика задней лапки далеко не
достигают вершины IV членика. Край VII стернита самки изобра-
жен на рис. 268, Б, а форма половой клешни самца — на
рис. 268, Л................................................Ct. pisticus.
106
Паразит бурундука (Eutamias sibiricus). Найден во многих местах Азии —
в Приморье, Приамурье, Забайкалье и Предбайкалье, Красноярской и Томской об-
ластях, на Алтае, в Ханты-Мансийске, а также в Корее [180].
Этот вид описан также под названием Ct. dahuricus [132], которое надо считать
синонимом. Дальневосточные представители этого вида (из района Гродеково и др.)
отличаются от всех остальных более расширенным основанием неподвижного отростка
половой клешни и иногда более узким в верхней части дигитоидом с выпуклым
передне-верхним углом (рис. 269); они выделены в особый подвид и названы
Ct. pisticus pacificus [154].
12(11). Наиболее длинные щетинки II членика задней лапки заходят за
вершину IV членика.
13(16). Хоботок не заходит или едва заходит за вершину коксы.
14(15). Внутренняя субапикальная вентральная щетинка задних бедер
не шипикообразная, довольно длинная. Верхний боковой выступ
апикального края VII стернита самки широкий, короче нижнего
бокового (рис. 270,5). Самец — см. рис. 270, Л ... Ct. dux.
Паразит слепушонок (Ellobius talpinus), обнаружен на Тянь-Шане (восток и юг
Казахской ССР и Киргизская ССР), но, вероятно, встречается и восточнее,— одна
самка была найдена на Алтае [101].
15(14). Внутренняя субапикальная вентральная щетинка задних бедер
шипикообразная. Верхний боковой выступ апикального края
VII стернита самки сравнительно узкий и выдается не меньше
нижнего (рис. 271, Б). Самец — см. рис. 271, Л .... Ct. dolichus.
Паразит песчанок, распространенный от северо-западного берега Каспийского
моря до Гобийского Алтая в Монголии. В Средней Азии образует ряд подвидов
[116, 141, 19].
16(13). Хоботок заходит за вершину переднего вертлуга. Самка и са-
мец— см. рис. 272, А, Б......................................Ct. dilatatus.
Паразит алтайского цокора. Найден на Алтае, в Новосибирской и Томской
областях [126].
Род Doratopsylla [179 (19), 180, 193, 163, 43]
Паразиты землероек (Soricidae), живущие в Европе, Азии и Север-
ной Америке. Мы рассмотрим все пять видов и два подрода, известных
в Европе и Азии.
1 (8). Лоб выпуклый, с лобным зубчиком. Гребень расположен на щеч-
ной лопасти и прикрывает ее (рис. 58). Апикальный край VII тер-
гита без выроста над предпигидиальными щетинками.......................
. ...........................................подрод Doratopsylla.
2(5). Верхний (задний) зубец щечного гребня изогнут и немного ко-
роче соседнего зубца (рис. 58).
3(4). Не более трети дигитоида заходит выше неподвижного отростка
клешни (рис. 273, А). У самки край VII стернита с двумя высту-
пами, разделенными довольно глубокой боковой вырезкой
(рис. 273, Б)........................................D. dasycnemus.
Паразитирует на! землеройках (Sorex, Neomys), обитающих в лесах Европы
и Западной Сибири. Распространен до Архангельска на севере и низовьев р. Иртыш
на востоке [163, 29].
4(3). Дигитоид более узкий и почти наполовину своей длины заходит
за конец неподвижного отростка. У самки край VII стернита
с развитым субвентральным выступом, без боковой вырезки . .
...........................................................D. cuspis.
Паразитирует на землеройках в горах Центральной и Южной Европы» (Швейца1-
рия, Венгрия) [180].
5(2). Верхний зубец щечного гребня прямой и значительно короче
соседнего зубца.
107
6(7). Узкая щечная лопасть видна между третьим и четвертым зубца-
ми гребня (рис. 59). Дигитоид суживается к вершине (рис. 274, Л).
У самки край VII стернита с двумя выступами (из которых ниж-
ний узкий) и глубокой вырезкой между ними (рис. 274, Б) . .
.............................................................D. согеапа.
Описан по экземплярам, обнаруженным в Северной Корее на землеройках [43].
7(6). Щечная лопасть находится под последним зубцом гребня. Диги-
тоид не суживается к вершине. У самки выступ под вырезкой
VII стернита не уже верхнего выступа.........................D. dampfi.
Паразит землероек, обитающих на Кавказе [119, 10].
8(1). Лоб довольно покатый, без ясного зубчика. Гребень начинается
от переднего конца нижнего края головы; последний зубец его
находится впереди от щечной лопасти (рис. 60). Апикальный
край VII тергита с резким выростом над предпигидиальными ще-
тинками ......................................подрод Corrodopsylla.
В Европе и Азии встречается только один вид этого подрода D. birulai; моди-
фицированные сегменты его см. на рис. 275, А, Б.
Паразит кутор (Neomys) и землероек (Sorex). Широкий ареал этого вида прости-
рается от Брянских лесов на западе, Архангельска! на севере, Тянь-Шаня Hai юге до
Забайкалья, Якутской АССР, Приамурья и Приморья включительно [132, 193]. Дру-
гие виды живут в Америке.
Род Palaepsylla [162, 163, 29, 153]
Паразиты землероек (Soricidae), живущие в Европе и Азии. Из
многих известных видов этого рода необходимо упомянуть только три,
обнаруженные в интересующих нас местностях или других странах Во-
сточной Азии. Блохи землероек мало обследовались в Азии. О европей-
ских и кавказских видах см. соответствующую литературу [162, 163,
153, 11].
1(4). Второй снизу зубец щечного гребня тупой, третий — с широкой
серединой, за которой сразу резко суживается (рис. 72).
2(3). Зубцы грудного гребня более или менее прямые. Горизонталь-
ная ветвь IX стернита самца без сужения к вершине (рис. 276, Л).
У самки край VII стернита с длинным субвентральным и более
коротким боковым выступом (рис. 276, Б) . . . . Р. sorecis.
Паразит землероек (Sorex), распространен от Англии до Тянь-Шаня, Алтая
и Западного Забайкалья. В Западной Сибири найден в Зауралье, в низовьях
р. Иртыш, в Омской и Кемеровской областях и Красноярском крае. Образует под-
виды: Р. s. sorecis — в Западной Европе, Р. s. starki — в СССР, кроме Кавказа, где
живет Р. s. gromovi [162, 29, 126, 153].
3(2). Зубцы грудного гребня вогнуты книзу. Самец неизвестен. У сам-
ки боковой выступ VII стернита широкий, округленный, т. е. иной
формы, чем у Р. sorecis...................................Р. incurva.
Блохи этого вида паразитируют на Sorex в Бирме [179 (38)]. Две самки, сход-
ные с этим видом, найдены на Сихотэ-Алине (с. Самарка) и в Северной Корее;
однако они отличаются угловатым боковым выступом края VII стернита и другими
деталями и при накоплении большего материала, вероятно, будут выделены в осо-
бый вид.
4(1). Второй снизу зубец щечного гребня заострен. Горизонтальная
ветвь IX стернита самца с вытянутой узкой вершиной. У самки
субвентральный выступ края VII стернита короткий, а верхний
выступ мало развит........................................Р. remota.
Вид, известный из Китая (Chungking) и Сиккима [179 (35, 38, 41)].
108
Род Nearctopsylla [188 (22), 25, 26, 154]
Паразиты насекомоядных (главным образом землероек) и мелких
хищников, распространенные преимущественно в Северной Америке. Два
вида недавно найдены и в Азии.
1 (2). Нижний конец гребня переднегруди изогнут кпереди. Второй сни-
зу зубец щечного гребня значительно длиннее нижнего зубца
(рис. 63). Пронотум и мезонотум с одним рядом щетин. Модифи-
цированные сегменты самца и самки — см. на рис. 277, 278.
У самки край VII стернита с глубокой и широкой вырезкой . .
............................................N. (Beringiopsylla) iofii.
Пара131ит землероек (Sorex), обнаруженный пока! в единичных экземплярах в При-
морье (Гродеково— самец) и в Корее (самка). Другие представители этого подрода
живут в Северной Америке [154].
2(1). Второй снизу зубец щечного гребня короче, чем у предыдущего
вида. У самки на своеобразно измененном II членике антенны,
имеющем вид чаши, в которой помещается маленькая булава, си-
дят сильные щетинки (рис. 64). Пронотум с одним рядом, а ме-
зонотум с двумя рядами щетинок. Край VII стернита самки
(рис. 279) без глубокой вырезки. Резервуар семяприемника
очень мал.............................N. (Chinopsylla) beklemischevi.
Единственный экземпляр (самка) этой своеобразной блохи описан из Китая
(Sian) с белки [154].
ПОДСЕМЕЙСТВО RHADINOPSYLLINAE
Род Rhadinopsylla [151, 19, 179 (35)]
К этому роду принадлежит довольно много видов блох, распростра-
ненных преимущественно в Азии, а также в Европе и других странах,
и живущих в гнездах грызунов, причем более многочисленны они зимой.
Мы включили в определительную таблицу 28 форм, принадлежащих
к пяти подродам (из них некоторые описывались как отдельные роды,
что едва ли целесообразно); рисунки даем для 23 форм, ближе относя-
щихся к интересующим нас районам. Сведения об остальных известных
видах можно найти в обзоре рода, который был составлен в 1941 г. [151]
и в настоящее время требует некоторых дополнений [19, 43].
1 (38). Эпистерн заднегруди не слившийся с метанотумом (отделен
склеротизованным швом).
2(25). Лоб довольно высокий (рис. 65), с нормальным лобным бугор-
ком («зубчиком»)................................подрод Rectofrontia.
К этой группе относится большинство азиатских и европейских видов.
3(4). Хоботок достигает вершины передней ляжки или заходит за нее
и состоит из шести (семи) члеников. Щечный гребень состоит из
шести (пяти-семи) зубцов; гребень переднегруди — из двадцати
четырех-двадцати пяти зубцов. У самца (рис. 280, А) основная
половина горизонтальной ветви IX стернита шире, чем апикаль-
ная половина. У самки VII стернит с раздвоенным боковым вы-
ступом (рис. 280, Б); семяприемник очень большой, с грушевид-
нььм резервуаром........................................R. insolita.
Вид, распространенный в Уссурийском и Приморском краях (близ городов
Ворошилов, Гродеково и южнее), а также в Маньчжурии и Корее на хомячках, по-
левых мышах и других мелких грызунах, живущих на лугах и в степях; в лесу встре-
чается, повидимому, реже [95, 151].
4(3). Хоботок состоит из пяти члеников и в большинстве случаев дале-
ко не достигает вершины передней ляжки.
109
5(6). Гребень переднегруди состоит примерно из двадцати шести-
двадцати восьми зубцов. На последнем членике лапок пять пар
боковых щетинок. У самца (рис. 281, Л) VIII стернит с многочис-
ленными (14—20) щетинками, а вершина горизонтальной ветви
IX стернита с резко выпуклым вентральным углом. У самки
VII стернит с боковым и вентральным выступом (рис. 281, Б)
.............................................................R. dives.
Известен из Южной Маньчжурии, паразитирует на хомячках, мышах и сусликах,
живущих в песчаных дюнах (близ Тунляо) [92].
6(5). Гребень переднегруди содержит менее 26 зубцов.
7 (8). Гребень переднегруди состоит примерно из шестнадцати-семнадца-
ти зубцов. У самца (рис. 282, Л) горизонтальная ветвь IX стерни-
та очень узкая; тело клешни почти шарообразное с округленным
апикально-дорзальным краем. У самки (рис. 282,5) VII стернит
с боковым выступом без вентрального; семяприемник с вдавлен-
ным концом придатка................................... . R. tenella.
Обнаружен там же, где и предыдущий вид, в гнездах хомячков и на мышах [92].
8(7). Гребень переднегруди содержит не менее 17 зубцов, а обычно —
больше. Горизонтальная ветвь IX стернита самца шире, чем вер-
тикальная (в наиболее узком месте последней). VII стернит сам-
ки с боковым и вентральным выступами на заднем крае.
9(10). На V членике лапок, как правило, имеется пять пар боковых ще-
тинок. Конец наиболее длинной апикальной щетинки II членика
задних лапок заходит за вершину IV членика или хотя бы дости-
гает ее. Голова изображена на рис. 65. У самца (рис. 283, Л)
задне-нижний угол VIII стернита сильно вырезан, так что ряд
щетинок расположен близко к заднему краю. У самки VII стер-
нит (рис. 283, 5) с языкообразным боковым выступом и глубо-
кой вырезкой под ним над острым вентральным углом; семяпри-
емник узкий................................................R. dahurica.
Блохи этого вида известны в Азии от Памира и Тянь-Шаня до Забайкалья;
живут в норах полевок, сеноставок, сусликов и других грызунов. По самцам хорошо
различаются четыре подвида (см. ниже). Кроме того, имеются экземпляры с Алтая
и из некоторых других мест Сибири, но ввиду того, что там пока найдены только
самки, эти находки требуют дальнейшего изучения [103, 151, 82, 126].
а (е). Горизонтальная ветвь IX стернита самца с относительно широкой тупой вер-
шиной (рис. 283,Л).
б (д). VIII стернит самца с хорошо развитым боковым выступом (рис. 283,Л). Ди-
гитоид относительно широкий.
в (г). На внутренней поверхности голени, как правило, нет щетинок.......
.............................................. R. dahurica dahurica.
Предбайкалье (Иркутск, Монды), Забайкалье, Тувинская автономная область,
Монголия (Хэнтэйский, Центральный, Баян-Хонгорский и Убсанурский аймаки, хребет
Тайшири). Лесостепи и степи [103, 151].
г (в). На внутренней поверхности голени имеется, как правило, довольно сильная
щетинка. На горизонтальной ветви IX стернита) самца нет продольной скла-
дочки хитина, имеющейся у всех других известных подвидов R. dahurica вбли-
зи основания ветви, ближе к дорзальному краю . . . R. dahurica diclinica.
Обитатель горных долин Тянь-Шаня (около 2000 м над уровнем океана). Луга,
лесостепь По вооружению голени можно узнавать и самок этого подвида [82, 151].
д (б). Боковой выступ VIII стернита самца слабо развит. Дигитоид относительно
узкий. На внутренней поверхности голени, как правило, нет щетинок . .
...................................................... R. dahurica tjanshan.
Обитает на высокогорных сыртах Тянь-Шаня. Горная полупустыня (3000 м
и выше) [151].
е (а). Горизонтальная ветвь IX стернита самца с вершиной гораздо более за-
остренной (рис. 283, В).................................R. dahurica vicina.
Тибетская форма [126, 151].
10(9). На V членике лапок имеется четыре пары щетинок.
11(16). На всех или некоторых средних (II—VII) тергитах брюшка ниже
стигм имеется две щетинки (у самки — как правило, а у самца--
ПО
иногда1)- VZZZ стернит самца довольно сильно вырезанный,
с многочисленными щетинками (часто более десяти), образую-
щими иногда более одного ряда.
12(15). В щечном гребне пять зубцов (реже четыре-шесть).
13(14). Хоботок несколько длиннее, чем у других видов, и иногда почти
достигает вершины коксы. У самца (рис. 284, Д) VIII стернит
с длинным языкообразным боковым выступом и многочисленны-
ми (11 —13) щетинками, образующими более одного ряда. Гори-
зонтальная ветвь IX стернита с узкой заостренной вершиной. По-
ловая клешня длинная, а дигитоид относительно короткий. У сам-
ки (рис. 284, Б) VII стернит с широкой боковой вырезкой и ко-
ротким округлым выступом над ней; семяприемник — см. ри-
сунок 284, Б.................................................R. ioffi.
Паразит алтайского цокора Myospalax myospalax. Алтай, Томская область
[126, 151].
14(13). У самца (рис. 285, Л) VIII стернит с широким боковым выступом
и дугообразным рядом из восьми щетин; IX стернит и клешню
см. на рис. 285, А. У самки (рис. 285, Б) VII стернит с узкой
вырезкой, а семяприемник довольно большой . . R. aspalacis.
Паразит забайкальского цокора Myospalax aspal ах, найден в Забайкалье (Оло-
вянинский район) и Монголии (Хэнтэйский аймак). Лесостепь [151].
15(12). В щечном гребне, как правило, шесть зубцов (реже пять-семь).
У самца (рис. 286, Д) IX стернит с характерным расширением
апикальной части. У самки (рис. 286, Б) VII стернит с усеченным
боковым выступом, семяприемник — см. рисунок 286, Б . . .
....................................................... R. rotschildi.
Распространен в Забайкалье, Приамурье (Еврейская автономная область), Монголии
(Хэнтэйский, Центральный, Баян-Хонгорский и Дзабханский аймаки, хребет Та’йшири).
Живет в гнездах полевок, а иногда и других мелких грызунов и хищников [141, 151].
16(11). На средних (III—VI) тергитах ниже стигмы имеется лишь одна
щетинка. На VIII стерните самца в большинстве случаев не более
семи щетинок (редко 8—14), расположенных в основном
в один ряд.
17(20). На внутренней поверхности задней голени единичные щетинки.
Половая клешня примерно в три раза длиннее дигитоида. Гори-
зонтальная ветвь IX стернита самца узкая, со щетинками по
вентральному краю (рис. 287).
18(19). У самца VIII стернит со значительной вентральной вырезкой.
У самки VII стернит с языкообразным боковым выступом и не
особенно глубокой вырезкой под ним; семяприемник — примерно
такой, как на рис. 286, Б и 289, Б.......................R. angusta.
Блоха гнезд полевок (Stenocranius, Clethrionomys), обитающая в долинах Тянь-
Шаня. Горные леса! и луга [82, 151].
19(18). У самца (рис. 287) VIII стернит без широкой вентральной вырез-
ки. Самка неизвестна.......................................R. accola.
Тибет (р. И-Чю); известен лишь один самец, добытый на хорьке [126, 151].
20(17). На внутренней поверхности задней голени в большинстве слу-
чаев щетинок нет. Половая клешня относительно короткая, лишь
в 2—272 раза длиннее дигитоида. IX стернит самца без щетинок
по вентральному краю, с тупой и довольно широкой вершиной
(рис. 288, Д; 289, Д; 290, Д, В; 291, Д).
1 При пользовании этим признаком следует учитывать возможность встретиться
с экземплярами R. rotschjildi, у которые он не выражен; в этом случае следует руко-
водствоваться другими признаками, указанными в пп. 11—15.
111
21(22). Щечный гребень расположен косо и слабо изогнут. У самца
(рис. 288, А) VIII стернит сильно усеченный; щетинки (от 7—9 и
до 14) образуют на нем дугообразный ряд и сидят близко от зад-
него края. Вершина горизонтальной ветви IX стернита овальная.
У самки (рис. 288, Б) VII стернит с глубокой вырезкой под язы-
кообразным выступом, а семяприемник с небольшим резервуаром,
довольно отчетливо отграниченным от придатка . . R. altaica.
Найден на Алтае, в Тувинской автономной области, Монголии (Баян-Хонгор-
ский аймак), паразитирует на сеноставках Ochotona ailpina [122, 151].
В Монголии встретились экземпляры самцов с некоторыми особенностями (чаще
с шестью зубцами в щечном гребне, с более длинным дипитоидом и с увеличенным
числом длинных щетин на VIII стерпите, с более яркой коричневой пигментацией),
которые заставляют в дальнейшем обратить внимание на изучение изменчивости блох
этого вида.
22(21). Такой комбинации признаков, как указано в п. 21, не имеется.
23(24). Щечный гребень расположен более вертикально, чем у R. altaica.
У самца (рис. 289, Л) боковой выступ VIII стернита несколько
лучше развит, чем у R. altaica. Апикальная часть IX стернита
расширена, вершина округленная. У самки (рис. 289, Б) VII стер-
нит напоминает R. dahurica, а семяприемник примерно такой, как
у R. integella (вообще самки отличаются от R. integella, пови-
димому, лишь слегка менее вертикальным щечным гребнем) . .
.............................................R. pseudodahurica.
Блохи этого вида паразитируют на разных грызунах; они обнаружены в не-
большом числе экземпляров в нескольких удаленных друг от друга местностях Азии:
Забайкалье (Александровский Завод, Баргузинский заповедник), Иркутская область
(Монды, Усть-Кут, Наканна), Якутская АССР (устье р. Вилюй), Западная Сибирь
(Ханты-Мансийск), Монголия (Центральный аймак). Единичные самки, повидимому,
этого же вида найдены на Алтае и в Дзабханском аймаке Монгольской Народной
Республики [75, 151].
24(23). Щечный гребень, как правило, вертикальный, изогнутый. У сам-
ца (рис. 290, А) задний край VIII стернита мало вырезан, так что
щетины находятся сравнительно далеко от края. Апикальная
часть IX стернита менее расширена, чем у предыдущего вида,
и с более выраженным апикально-дорзальным углом, который
иногда округлен, а иногда и заострен. У самок VII стернит и се-
мяприемник — см. на рис. 290, В..............................R. integella.
Блохи этого вида встречаются в гнездах мелких млекопитающих (как и другие
Rhaidinopsylla, преимущественно в зимнее время), чаще в лесах и распространены
от Западной Европы до Тихого океана. Этот вид изучен еще недостаточно и относи-
мые к нему формы потребуют пересмотра. Некоторые из них будут, вероятно, рас-
сматриваться как особые виды. В Азии обнаружен в Зауралье (р. Сосьва|), Западной
Сибири (Ханты-Мансийск, Бийск), Приморье и в Корее [180, 179 (34, 35), 151, 120].
Из семи описанных подвидов (преимущественно европейских) упомянем три.
a) R. integella concava [151]. Эта блоха обитает в луговых местностях Совет-
ского Южного Приморья и Кореи. Она хорошо отличается удлиненным верхним зуб-
цом щечного гребня, вогнутым дорзальным краем IX стернита самца (рис. 290, Б),
большим резервуаром семяприемника и сравнительно менее широким боковым высту-
пом VII стернита самки (рис. 290, Б). В горнолесных районах Кореи найдена другая
форма, детально еще не изученная; повидимому, она близка к следующему подвиду.
б) R. integella casta [179 (34)] — наиболее распространенная лесная форма, встре-
чающаяся в странах Западной Европы и многих лесных районах Европейской части
СССР; в Азии найдена в Зауралье (р. Сосьва) и Западной Сибири (Ханты-Мансийск,
Бийск). Модифицированные сегменты самца этого подвида изображены» на рис. 291, А.
Для самок характерен VII стернит (рис. 291, Б) с широким боковым выступом апи-
кального края, нередко со слегка волнистыми очертаниями.
в) R. integella caucasica [120, 151]—кавказская форма, близкая к R. i. casta, но
отличающаяся по форме VII стернита самки (боковой выступ у нее более узок) и по
форме дигитоида, который заметнее расширяется к основанию.
25(2). Лоб иной формы.
26(31). Лоб нормальный, довольно высокий, но без явно выраженного
бугорка («зубчика»); передняя часть головы несколько вытяну-
112
тая (рис. 66). Имеется тупой лобный угол, расположенный
около середины лба или лоб округлен. В виде очень редкой ано-
малии может быть зачаток лобного бугорка. Хоботок достигает
вершины передней ляжки или заходит за нее и состоит из пяти
или шести-восьми члеников. У самца (рис. 294 и 295, Л) рукоятка
клешни короткая и узкая, обычно образует с телом клешни до-
вольно резкий угол. У самки VII стернит с извилистым задним
краем (рис. 293, А—Д), а семяприемник узкий, с небольшим
впячиванием на самом конце придатка (рис. 293, Л)...................
.................................................подрод Ralipsylla.
Подрод включает исключительно обитающие на территории Азии виды, пере-
числяемые ниже [19].
27(30). Хоботок состоит из пяти-шести (редко семи) члеников. Лоб не-
сколько угловатый (рис. 66).
28(29). У самца (рис. 294, 295, Л) апикальная часть IX стернита не
сильно расширена. У самки VII стернит — см. рис. 293, Л—Б
......................................................................R. И.
Этот алта1йско-монголо-забайкальсюий вид, экология которого еще недостаточно
ясна, встречается на разных грызунах (цокоры, суслики, хомячки, полевки и др.),
мелких хищника» и насекомоядных (Talpa) и известен в двух подвидах [120, 151].
а (б). Хоботок состоит из шести (иногда семи) члеников. Самец и самка изобра-
жены на рис. 293, А и 294 ..........................................R. П 11.
Алтайская форма [120, 151].
б (а). Хоботок состоит из пяти члеников. Самец и самка — см. рис. 293, Б
и 295, А..............................................R. И transbaikalica.
Распространен в Предбайкалье (Монды), Забайкалье (чаще в Западном), Тувин-
ской автономной области, Монголии (Центральный, Убурхангайский, Дзабханский,
Баян-Хонгорский и Убсанурский аймаки, хребет Тайшири) [151].
29 (28). У самца апикальная часть IX стернита (рис. 295, Б) сильно рас-
ширена, с резко выпуклым вентральным краем («брюхатая»).
У самки VII стернит изображен на рис. 293, В, Г.............................
...................................................R. ventricosa.
Тяныпаньский вид, представленный двумя подвидами, которые хорошо отличаются
по их привязанности к определенным хозяевам и по морфологии самок [151, 19].
а (б). Край VII стернита самки с хорошо развитой вырезкой между двумя, рез-
кими боковыми выступами (рис. 293, В)...............R. ventricosa ventricosa.
Паразит сурков, обитающих на Тянь-Шане [151, 19].
б (а). Край VII стернита самки с узкой щелевидной вырезкой между двумя боко-
выми выступами (рис. 293, Г)..........................R. ventricosa murium.
Паразит мышевидных грызунов Тянь-Шаня [151, 19].
30(27). Хоботок длинный, далеко заходит за передний вертлуг и состоит
из восьми (иногда семи-девяти) члеников. Апикальная часть
, IX стернита самца не расширена (рис. 295, В). Край VII стер-
нита — см. на рис. 293, Д.............................R. semenovi.
Памир и Тянь-Шань (в СССР и в Синь-Цзяне); на разных грызунах в сухих
горно-степных местностях [153, 19, 151].
31(26). Лоб с резким выступающим углом. Хоботок состоит из пяти чле-
ников и не достигает вершины передней коксы.
32(37). Лоб низкий (рис. 68) с выдающимся резким углом на незначи-
тельном расстоянии от основания максиллярных пальп; нижняя
щетинка фронтального ряда сидит примерно на уровне лобного
угла или выше него. У самца (рис. 296, А—Б) горизонтальная
ветвь IX стернита ланцетовидная, чаще с острой клювовидной
вершиной; рукоятка клешни длинная, переходящая в передний
край тела клешни без выраженного угла. У самки (рис. 299, А)
семяприемник с резко сплющенным (вдавленным с одной сторо-
ны) концом придатка ..... подрод Rhadinopsylla s. str.
8 Определитель блох 113
Подрод включает несколько видов, распространенных в Средней Азии и запад-
нее. Чаще всего блохи этих видов встречаются в гнездах песчанок в зимнее время.
Мы упомянем три из них.
33(34). Последний членик лапок с пятью парами боковых щетинок.
У самки (рис. 299, Л) край VII стернита с сильно развитым боко-
вым выступом; субвентральная вырезка сильно развита. Модифи-
цированные сегменты самца — см. на рис. 296, Л .... R. cedestis.
Зимний паразит песчанок. Известен из многих мест от Илийской долины
(Тянь-Шань) до Северо-Западного Прикаспия и Закавказья (115, 14, 151].
34(33). Последний членик лапок с четырьмя парами боковых щетинок.
35(36). На II членике задних лапок сильно развиты и резко удлинены
не только апикальные, но и субапикальные щетинки заднего края
(рис. 349, В*); эти субапикальные щетинки обычно заходят
у самки за вершину III членика, а у самца достигают вершины
V членика или заходят за нее. Модифицированные сегменты сам-
ца— см. рис. 296, В. Край VII стернита самки (рис. 299, В) со
слабой субвентральной вырезкой .... . . R. bivirgis.
Паразитирует на разных грызунах зимой. Распространен от долины р. Или до
Северо-Западного Прикаспия (Яндыки) [115, 14, 151].
36(35). Субапикальные щетинки II членика задних лапок не удлинены
(рис. 349, Г *) .Модифицированные сегменты самца — см. рис. 297;
характерен IX стернит с довольно узкой горизонтальной
ветвью и округленной вершиной. Край VII стернита самки
(рис. 298) с маленьким боковым сильнее склеротизованным вы-
ступом . . ......................................R. socia.
Известен из немногих районов Средней Азии (Илийская долина, Прибалхашье,
Каракалпакская АССР), паразитирует на песчанках [126, 151].
37(32). Выдающийся резкий угол лба (рис. 67) расположен выше, чем
у видов предыдущей группы; нижняя щетинка фронтального ряда
сидит значительно ниже уровня лобного угла. Хоботок пятичлс-
нистый, далеко не достигает вершины передней коксы. Зубчики
на тергитах очень короткие и широкие . . . подрод Rangulopsylla.
Известен лишь один вид этого подрода — R. valenti; модифицированные сегменты
самца — см. на рис. 292, А. У самки (рис. 292,5) семяприемник большой, грушевид-
ный. Описан по экземплярам из Кореи, обнаруженным на Apodemus agrarius и Crice-
tulus triton [43].
38(1). Эпистерн заднегруди не отделен склеротизованным швом от ме-
танотум а .....................................подрод Micropsylloides.
Известен лишь один вид этого подрода [168] — R. jaonis, описанный по двум
самкам, обнаруженным в Китае (Шаньси и Маньчжурия) на Scaptochirus и цокоре
[179, (35), 92]. Хоботок у блох этого вида длинный, заходит за вертлуг и состоит из
семи-восьми члеников. В щечном гребне четыре зубца (всегда ли?). VII стернит
и семяприемник самки — см. на рис. 300. Самец неизвестен. Родственные виды живут
в Северной Америке и образуют группу Micropsylla [25, 26]. Мы рассматриваем эти
группы как подроды рода Rhadinopsylla.
(Кстати заметим, что в Китае (Сычуань) есть еще другая блоха этого подсемей-
ства — Stenischi а miraibilis [179 (38)] —с подобным же строением заднегруди, но
отличающаяся расположением гребня, который не поднимается по краю ямки усиков,
а идет по щечному краю; кроме того, она отличается частичной темной склеротиза-
цией брюшных тергитов и стернитов.
ПОДСЕМЕЙСТВО NEOPSYLLINAE
Род Neopsylla [180, 19]
Многочисленные и весьма распространенные представители этого
рода живут на мышевидных и некоторых других грызунах Азии; немно-
гие из них проникли в Европу, и несколько видов известно из Северной
114
Америки. Чаще и в большем количестве эти блохи встречаются в гнез-
дах, чем на самих зверьках. Самки в некоторых случаях могут быть
точно определены лишь при учете географического распространения или
наличия соответствующих самцов в данной местности. В определитель-
ную таблицу включены все палеарктические виды, но рисунки даны
только для видов, известных из Восточной Азии
1(20). На внутренней поверхности задних ляжек нет группы шипиков,
а имеются лишь тонкие мелкие щетинки.
К группе блох, объединяемых этим признаком, относятся почти все встречаю-
щиеся у нас виды.
2(15). На пронотуме впереди от главного ряда щетинок имеется еще
один ряд (от этого переднего ряда остаются иногда лишь еди-
ничные верхние щетинки; только в виде исключения у некоторых
экземпляров N. pleskei ariana они могут совсем отсутствовать).
3(14). Задняя голень (рис. 349, Б) на наружной поверхности имеет
два-три продольных ряда щетинок (не считая передних марги-
нальных или субмаргинальных).
4(11). IX стернит самца с сильно развитыми толстыми, темноватыми,
тупыми или серповидно изогнутыми шипами; VIII стернит с гри-
вой, состоящей из длинных извилистых щетин (рис. 301, 302,
305). Признаки самок приводятся для каждого отдельного вида.
5(6). На пронотуме и на наружной поверхности задней голени у боль-
шинства особей передние щетинки более многочисленные, чем
у других видов (часто семь-восемь щетинок в переднем ряду
пронотума и три продольных ряда на голени, но бывает и гораздо
меньше). У самца (рис. 301) задний край дигитоида с ясным высту-
пом, а IX стернит с двумя группами тупых темных шипов. У сам-
ки (рис. 303) VII стернит с резким и довольно узким боковым
выступом ...... . N. bidentatiformis.
Один из обычных видов блох, живущих в степях и лесостепях Восточной Азии;
иногда попадается также в городах и селах; найден в Предбайкалье (верховья
р. Нижняя Тунгуска, Усть-Удинский и Осинский районы), Забайкалье, Монголии
(Хэнтейский, Центральный, Дзабханский и Убсанурский аймаки), в районах* 'Благо-
вещенска, в Биробиджане, Хабаровске, Уссурийском крае, в Северной и Южной
Маньчжурии, Корее, Китае (Квантунский полуостров, Жэхэ). Паразитирует на хомяч-
ках, сусликах, крысах, полевках и многих других грызунах, а также и на хищниках
[122, 128, 27].
6(5) Приз ШЕИ илы?.
7(10). Вершина IX стернита самца (рис. 302) с одним рядом из тол-
стых шипов и небольшим вентральным отростком. У самки боко-
вой угол VII стернита округлый, слабо выступающий (рис. 304)'.
8(9). У самца (рис. 302) по краю VIII стернита резко выделяете^
нижняя группа длинных щетин. Пальцевидный выступ заднего
края вертикальной ветви IX стернита с кольчатыми насечками
(видными на просветленных препаратах). У самки (рис. 304) бо-
ковой выступ VII стернита очень слабо выражен . . . N. mana.
Живет в гнездах полевок и других грызунов в достаточно прохладных лесо-
степных и луговых, часто горных местностях: Тянь-Шань, Алтай, Красноярский край
(Ермаковский район), Минусинский округ, Тувинская автономная область, Предбай-
калье (Тункинская долина). Якутск. В Монголии известен на хребте Тайшири, а так-
же в Центральном Баян-Хонгорском. Дзабханском и Убсанурском аймаках [34].
9(8). У самца край VIII стернита густо усажен длинными щетинками,
причем верхние не очень уступают по развитию нижним. Паль-
цевидный выступ заднего края вертикальной ветви IX стернита
1 В маньчжурских работах [109, 57] упоминается без описания Neopsylla abei
Ono; нам неизвестно, опубликованы ли где-либо признаки этого вида; пока мы
считаем это название за nomen nudum.
8*
113
без кольчатых насечек. У самки боковой выступ VII стернита ши-
рокий, округлый, обычно несколько более заметный, чем у пре-
дыдущего вида..........................................N. meridiana.
Замещает предыдущего на! высокогорных* сыртах Тянь-Шаня; известен пока лишь
из Киргизской ССР [158, 19].
10(7). IX стернит самца (рис. 305) с двумя рядами шипов; шипы одно-
го ряда длинные и серпообразно загнутые. Боковой угол апи-
кального края VII стернита самки (рис. 307) довольно ясно вы-
ражен, приострен..................................................N. teratura.
Паразит серых хомячков и многих других грызунов распространен на Тянь-
Шане. в Восточном Памире, Бетпакдала и Западной Монголии (Убсанурский аймак
и Монгольский Алтай) [116].
t 11(4). Шипы на IX стерните самца менее отличаются от обычных уко-
роченных щетинок, светлые, не очень толстые (рис. 306; 309, Л;
310, Л). VIII стернит без гривы из длинных извилистых щетин
(кроме N. democratica).
12(13). У самца (рис. 306) нижняя часть заднего края VIII стернита
с гривой. У самки (рис. 308) VII стернит с развитым широко
округленным боковым выступом и вырезкой под ним . . . .
.......................................................... N. democratica.
Этот вид до сих пор был найден лишь в с. Локоть близ Змеиногорска (Алтай-
ский край) на узкочерепных полевках (14 самок, 11 самцов) [191].
13(12). У самца (рис. 309, Л; 310, Л) задняя часть VIII стернита (поза-
ди щетинок) слабо склеротизована и расщеплена на две лопасти
узкой щелью; впереди от этих бледных лопастей имеются не густо
расположенные и не извитые щетины. Дигитоид и апикальная
треть горизонтальной ветви IX стернита довольно узки. У самки
(рис. 309, Б; 312, А) VII стернит с боковым выступом, несколько
более широким и более коротким, чем у N. bidentatiformis . .
...............................................................N. pleskei.
Блохи этого вида живут в норах многих мышевидных и других» грызунов, при-
«тем разные подвиды распространены от Поволжья и Закавказья до Забайкалья
[132, 145, 19]. Мы приведем здесь сведения о трех восточных подвидах, а о двух
западных — N. р. rossica и N. р. armeniaca — см. соответствующую литературу [145].
а) Задний край дигитоида (рис. 309, А) вогнут в нижней, слабее склеротизован-
ной части; граница склеротизации нерезкая; дигитоид относительно корот-
кий. У самки (рис. 309, Б) стигма VIII тергита сравнительно широкая,
а копулятивный проток довольно длинный...................N. pleskei pleskei.
, Западносибирская форма. Известна от реки Оби до озера Байкала (окрестности
Нарыма, Колпашова, Красноярска, Иркутска и др.); паразитирует на полевках и дру-
гих грызунах — обитателях лесостепи [132, 145].
б) Задний край дигитоида (рис. 310, А) без вогнутости; граница! между верх-
ней склеротизованной частью дигитоида и нижней бледной частью резкая;
дигитонд относительно длинный. У самки (рис. 312, А) стигма VIII тергита
значительно меньше, чем у предыдущего подвида, ai копулятивный проток
короче............................................... N. pleskei orientalis.
Забайкальско-монгольская степная форма; распространена в Восточном Забай-
калье, Тувинской автономной области. Восточном, Хэнтэйском, Центральном, Убур-
хангайском, Баян-Хонгорском и Гоби-Алтайском аймаках Монголии; паразитирует на
полевках Брандта и многих других грызунах [145].
Какой подвид N. pleskei встречается в Западном Забайкалье — не выяснено.
в) Дигитоид с легкой вогнутостью в нижней трети заднего края; граница скле-
ротиза1шии дигитоида менее резкая, чем у предыдущего подвида; отличается
также форма и склеротизация дорзального парамера) (рис. 310, Б). Самка —
как у предыдущего подвида................................N. pleskei ariana.
Форма, распространенная в горах Алтая, Тянь-Шаня, Памира, Копет-Дага
Ч' Ирана (Эльбурс); встречается чаще всего на серых хомячках (Cricetulus migra-
t-rius) [19].
14(3). На наружной' поверхности задней голени (рис. 349, А) имеется
лишь один продольный ряд щетинок, впереди от которого щети-
нок нет или имеются единичные (не считая передних маргиналь-
на
ных или субмаргинальных). У самца (рис. 311,71) на VIII стер-
ните гривы нет, а бледные апикальные лопасти довольно слабо
развиты. IX стернит с многочисленными бледноокрашенными ши-
пиками и резким горбом в апикальной трети. Дигитоид с острой
вершиной, узкий; в нижней части его характерная склеротизации
в виде темного узкого язычка, оставляющего у заднего края
светлую полоску. Самки (рис. 311, Б) похожи на N. pleskei
orientals, но отличаются по вооружению голени, а также несколь-
ко более крупной стигмой VIII тергита . . . . N. acanthina.
Сибирский вид, известный из многих местах от р. Оби (Ханты-Мансийск) до
острова Сахалина, обитает в лесных и лесостепных районах; более многочислен-
ные находки имеются из бассейна рек Амура и Уссури (Хабаровск, Николаевск, Бикин,
Турий-Рог, Супутинский заповедник, район г. Лазо и др.). Кроме упомянутых мест,
найден также в Новосибирской области, Предбайкалье, Восточном и Западном Забай-
калье, Якутске, около устья р. Вилюя и в Монголии (окрестности Улан-Батора).
Встречается на бурундуках, лесных мышах и полевках. Возможно, что основным
биотопом окажутся гнезда бурундука [103].
Самка, обнаруженная в Якутской АССР и описанная под названием N. uniseriata,
повидимому, принадлежит к этому же виду [34, 74].
15(2). На пронотуме впереди от главного ряда нет щетинок (лишь
в виде исключения у некоторых экземпляров бывают одна-две,
очень редко три маленькие верхние щетинки). У самца на VIII
стерните нет гривы или она развита гораздо слабее, чем у видов,
описанных в пп. 4—10. Шипики IX стернита не толстые
и светлые.
16(17). На наружной поверхности задней голени развит лишь один про-
дольный ряд щетинок (как на рис. 349, Л). У самца (рис. 313, Л)
задний край дигитоида с резким закругленным' выступом.
IX стернит с горбом у самой вершины и немногочисленными ту-
поватыми светлыми шипиками. Для самок (рис. 313, Б) характер-
ны: очень маленькая стигма VIII тергита, обычно угловатый бо-
ковой выступ VII стернита, маленький резервуар семяприемника,
довольно широкий копулятивный проток.......................N. abagaitui.
Найден в некоторых пунктах Забайкалья (Агинский район, Абагайтуй), а так-
же в окрестностях Красноярска, в Тувинской автономной области, Монголии
(Хэнтэйский, Центральный, Убурхангайский, Баян-Хонгорский аймаки) и в Китае
(Жэхэ). Повидимому, предпочитает лесостепные местности. Встречается па сусликах»,
полевках и др. [153].
17(16). На наружной поверхности задней голени, как правило, впереди
от продольного ряда имеется еще ряд или разбросанные щетин-
ки. У самцов задний край дигитоида без резкого выступа. У са-
мок резервуар семяприемника не укорочен, край VII стернита,
иной, чем у предыдущего вида.
18(19). У самца (рис. 315) дигитоид узкий, с округленным задним
краем; неподвижный выступ клешни широкий и волосатый. На
VIII стерните многочисленные довольно сильные щетинки, более
густо расположенные вблизи вентрального и нижней части зад-
него края; апикальная, не покрытая щетинками лопасть узкая.
Вершина IX стернита с острым дорзальным выступом и единич-
ными мелкими шипиками. У самок (рис. 312, Б) край VII стер-
нита с резким боковым выступом и вырезкой под ним; стигма
VIII тергита небольшая........................................N. galea.
Вид, известный пока по немногим экземплярам, обнаруженным в Забайкалье
(Агинский район) и Монголии (Баян-Хонгорский аймак, Цзаг-Сомон) на Cricetulus
barabensis и в гнезде Microtus brandti. Самка описывается впервые [153].
19(18). На наружной поверхности задней голени не менее двух про-
дольных рядов щетинок. У самца дигитоид с острой вершиной, но
117
широкий в середине (более широкий, чем, например., у N. pleskei).
Апикальная часть VIII стернита позади от щетин мембранозная,
густо покрытая мельчайшими волосками. Вооружение горизон-
тальной ветви IX стернита в общем напоминает N. pleskei, но
впереди шипов имеется ряд тонких длинных щетинок с завитыми
в клубочек концами. У самки край VII стернита со слабо выра-
женным боковым выступом...............................N. setosa.
Блоха гнезд многих видов сусликов, широко распространенная в степях от
Румынии до Тянь-Шаня, где она) проникает и в некоторые горные долины (Иссык-
Куль) [122, 128].
20(1). На внутренней поверхности задней ляжки есть группа шипиков ’.
Блохи, объединяемые этим признаком, принадлежат к фауне Индо-Малайской
зоогеографической области, но из них три вида обнаружены в северных провинциях
Китая, а один у нас в горах Средней Азии. Признаки этих видов приводятся ниже.
21(26). Пронотум с одним рядом щетинок.
22(23). Хоботок очень короткий, достигающий примерно середины коксы.
У самца близ задне-нижнего угла дигитоида имеется шипик; зад-
ний край дигитоида прямой (не вогнутый). У самки край
VII стернита с резким боковым выступом и глубокой вырезкой
под ним. Резервуар семяприемника удлиненный . . . N. hissarica.
Паразит туркестанской крысы (Rattus turkestanicus), обнаружен на Гиссарском
хребте в Таджикской ССР [154].
Родственные N. hissarica виды паразитируют также на крысах, обитающих
в Гималаях (Симла) — N. secura, в Китае — N. honora (Юннань), N. stevensi
(Сычуань) и ряд других [180, 93].
23(22). Хоботок заходит за середину коксы. Дигитоид без шипика на
заднем крае. Резервуар семяприемника короткий.
24(25). Из теменных рядов щетинок развит лишь один. У самца
(рис. 314, А) дигитоид наиболее широк в своей апикальной части.
IX стернит без ряда шипов, с более или менее обычными ще-
тинками. У самки (рис. 314, Б) край VII стернита с резкой боко-
вой вырезкой и округлым выступом над ней . . . N. aliena.
Описан по экземплярам из Китая (Шэньси) с Myospailax [100].
25(24). Имеются оба ряда теменных щетинок (у самок в переднем ряду
лишь две щетинки). У самца (рис. 316, А) дигитоид более широк
в базальной части; IX стернит без шипов. У самки (рис. 316, Б)
край VII стернита с небольшим выступом над боковой вырезкой
...........................................................N. anoma.
Описан по экземплярам из Китая (Шэньси) с Myospalax [113].
26(21). Пронотум с двумя рядами щетинок. Самец неизвестен. У самки
(рис. 314, В) край VII стернита со слабой боковой вырезкой.
Резервуар семяприемника удлиненный (как у N. bidentatiformis
и др.).....................................................N. compar.
Описан по экземплярам из (Китая (Шэньси) с Dignus.
Род Paraneopsylla [82, 84, 144]
Представители этого рода паразитируют на горных грызунах, пре-
имущественно живущих в скалах и каменных осыпях или горных лесах
Кавказа, Тянь-Шаня и Хангая. Известны три вида.
1 (4). Имеются две предпигидиальные щетинки. Нижняя часть дигитои-
да неплотно прижата к телу клешни (рис. 317, А). У самки
1 Относительно наличия их у N. ailiena в литературе имеются противоречивые
сведения [12], но К. Иордан сообщил нам, что они имеются.
118
(рис. 317, Б) край VII стернита с симметричным, широким, округ-
лым боковым выступом; проток копулятивной сумки длинный.
2(3). У самца (рис. 317, Д) на VIII стерните немного (около четырех
щетинок, на вентральном крае IX стернита не имеется шпоры.
Дигитоид довольно узкий. Самка изображена на рис. 317, Б . .
........................’.................................Р. ioffi.
Паразит высокогорных полевок (Alticola) и пищух (Ochotona), найден в Цен-
тральном Тянь-Шане и в Монголии (Дзабханскнй аймак) [82].
3(2). У самца на VIII стерните около 10—12 щетинок, а на вентраль-
ном крае IX стернита имеется шпора. Дигитоид более широкий.
Самка отличается от предыдущего вида менее расширенной стиг-
мой VIII тергита...........................................Р. dampfi.
Паразит снежных полевок (Chionomys), обитающих на Кавказе [144].
4(1). Как правило, имеются три предпигидиальные щетинки. Нижняя
часть дигитоида (рис. 318, Д) плотно прижата к телу клешни.
Между VIII и IX стернитами самца имеется еще один склерит
в форме копыта. Боковой выступ края VII стернита самки
(рис. 318, Б) несимметричный: снизу он круче вырезан, чем
сверху. Проток копулятивной сумки заметно короче, а стигма
VIII тергита значительно менее расширена, чем у Р. ioffi . . .
. . . . ......................Р. tiflovi.
Паразит тяньшаньской лесной полевки (Clethrionomys frater) и других мелких
грызунов Центрального Тянь-Шаня [84].
Род Catallagia [180, 141, 75, 35]
Представители этого рода до недавнего времени были известны
только в Северной Америке. В настоящее время обнаружено четыре вида
этого рода и в Азии. Паразитируют преимущественно на лесных полев-
ках (рода Clethrionomys), но встречаются и на многих других видах
мелких лесных млекопитающих. Блохи этого рода, повидимому, наиболее
обильны весной, затем осенью и зимой, а летом довольно малочисленны.
Распространены на Урале, в Сибири, на Дальнем Востоке, в Тувинской
автономной области и Монголии.
1(2). Стигмы II—VII брюшных тергитов расширенные, яйцевидной
формы (рис. 319, Б); одна из ряда передних, более мелких, щети-
нок на средних тергитах сидит, как правило, ниже стигмы и лишь
в редких случаях вровень с ней. У самца (рис. 319, Д) дигитоид
небольшой; «лапка» IX стернита (апикальная часть горизонталь-
ной ветви) имеет около восьми темных тупых шипов. На
VII стерните брюшка у самки, впереди ряда крупных щетинок
имеется 9—16 мелких (рис. 319, Б)......................С. dacenkoi.
Паразит лесных полевок (Clethrionomys). Найден в Печорском и Кондо-Сосьвин-
ском заповедниках (Урал), на Алтае, в низовьях р. Иртыша и на р. Таз (Мотылька),
в центральной части Якутской АССР, в Забайкалье и лесных районах Монголии
(Улан-Батор) [141, 75].
Близкая форма (известная пока только по единичным самкам) обнаружена на
Сихотэ-Алине (Приморский край) [52]. Недавно С. dacenkoi (возможно, особая раса)
найдены также в Северной Америке (Аляска).
2(1). Стигмы //—VII брюшных тергитов нерасширенные, соскообраз-
но вытянутые назад (рис. 320, Б); нижняя из передних щетинок
на средних тергитах сидит не ниже стигмы; у самок на VII стер-
ните брюшка впереди ряда крупных щетинок имеются лишь
три-семь (у С. fetisovi — до II) мелких.
3(4). У самца (рис. 320, Д) вершина дигитоида слегка превышает не-
подвижный палец; задне-нижний край дигитоида правильно
119
округлый. На «лапке» IX стернита три крупных темных шипа и
ряд более мелких шипов и щетинок, расположенных по нижнему
краю с внутренней стороны. Апикальный край VIII стернита
с небольшим заостренным выступом. У самки (рис. 320, Б) стигма
VIII тергита расширена лишь немного меньше, чем у С. dacenkoi;
резервуар семяприемника овальной формы, отверстие семен-
ного протока мало смещено на нижнюю сторону......................
......................................................С. ioffi.
Паразит лесных полевок (Clethrionomys). Широко распространен в таежной по-
лосе Сибири, встречается в лесах Восточной Монголии. Найден в Западной Сибири.
(Кондо-Сосьвинский заповедник), в низовьях р. Иртыша в Северной Сибири — бассейн
р. Таза (Мотылька), в центральной части Якутской АССР, в верховьях р. Лены, бас-
сейне р. Нижней Тунгуски и в Предбайкалье, Забайкалье, Восточной Монголии
(Улан-Батор) [75].
4(3). Дигитоид самца или короче неподвижного пальца (рис. 321), или
значительно превышает его (рис. 322). У самок стигма VIII тёр-
гита маленькая (рис. 323,А—Б).
5(6). Дигитоид самца (рис. 321) значительно короче неподвижного
пальца; на нижнем его крае две глубокие выемки. «Лапка»
IX стернита миниатюрная, на конце ее сидят три темных, загну-
тых книзу шипа. Апикальная лопасть VIII стернита с глубокой
выемкой, ниже которой — длинный клювообразный выступ.
У самки (рис. 323, А) тело семяприемника удлиненное, слегка
оттянутое в сторону отверстия семенного протока, полностью сме-
щенного на нижнюю сторону. На VII стерните впереди ряда
крупных щетинок — лишь три-пять мелких .... С. striata.
Паразит Clethrionomys. Известны единичные находки этого вида в Восточном
Забайкалье (Александрозаводский район). Амурской области, Приморье (с. Гроде-
ксво, Хасанский район). Обычен в Корее [75].
6(5). У самца (рис. 322) вершина дигитоида значительно превышает
широкий, короткий неподвижный выступ клешни. Апикальная
лопасть VIII стернита с широким тупым выступом. У самки
(рис. 323, Б) тело семяприемника овальное, как у С. ioffi; отвер-
стие семенного протока мало смещено на нижнюю сторону; на
VII стерните до 11 мелких щетинок, расположенных впереди от
ряда крупных................................................С. fetisovi.
Единичные находки в Забайкалье, Предбайкалье и Тувинской автономной обла-
сти на Clethrionomys rutilus, Apodemus speciosus, Ochotona alpina и Mustela alpina.
Распространен в Монголии (Хэнтэйский, Центральный и Баян-Хонгорский аймаки)
на Alticola semicanus (живущих среди камней) [35].
ПОДСЕМЕЙСТВО HYSTRICHOPSYLLINAE
Рол Hystrichopsylla [122, 154, 179 (21), 75]
Очень крупные блохи (до 5 мм длины), живущие в гнездах грызу-
нов и насекомоядных, обитающих в Европе, Северной Африке и Север-
ной Америке.
В систематике относимых нами к этому роду блох до сих пор не
было ясности, так как ряд видов с переходными признаками между
Hystrichopsylla и Typhloceras относились то к первой, то ко второй из
этих групп. Мы считаем целесообразным выделить эти промежуточные
виды в новую группу Hystroceras и всем трем видам придать подродо-
вое значение.
1(2). Щечный гребень состоит из многочисленных (около 10) узких
и длинных зубцов (рис. 71). На II и двух-трех следующих терги-
120
тах брюшка имеются настоящие гребни. У самок — четыре,
у самца — обычно три предпигидиальных щетинки...................
................................подрод Hystrichopsylla s. str.
К этому подроду относится только один вид Н. talpae (рис. 324, А), размножаю-
щийся в гнездах кротов, полевок и других мелких млекопитающих, живущих в лесах
и на лугах. Обнаружен во многих местах: от Англии и Италии на западе до Ерма-
ковского района Красноярского края на востоке и от Архангельска, Кондо-Сосьвин-
ского заповедника и Ханты-Мансийска на севере до Кавказа, Тянь-Шаня и Алтая
на юге. В Западной Сибири собран, кроме упомянутых мест, еще в Томской, Ново-
сибирской и Кемеровской областях, а также в Наурзуме (Кустанайская область)
и на Джунгарском Алатау [29].
2(1). Щечный гребень состоит из четырех-восьми зубцов. На II и сле-
дующих тергитах, на боках, отступя от спины, имеются лишь за-
чаточные гребни из редких зубчиков. У самки и самца три пред-
пигидиальных щетинки.
3(8). Нормального лобного зубчика нет. Глаз полурудиментарный или
рудиментарный. Щечный гребень из пяти-шести (иногда до вось-
ми) зубцов (рис. 72). Дигитоид прикреплен в задне-нижнем углу
тела клешни (рис. 325,Л). У самца VIII стернит отчетливо де-
лится на вертикальную и горизонтальную части.........................
..........................................подрод Hystroceras.
4(7). Сибирские виды. У самца вершина горизонтальной части
VIII стернита конусовидно суживается (рис. 325, Л). На вершине
передних голеней (у обоих полов) — поперечный гребневидный
ряд шпор и утолщенных щетинок.
5(6). В гребне переднегруди 26—31 зубец. Модифицированные сегмен-
ты самца и самки — см. на рис. 325, Л—Б. У самки апикальный
край VII стернита в нижней части с небольшим выступом . . .
..........................................................Н. microti.
Обитатель гнезд мелких лесных млекопитающих Забайкалья, Приамурья,
Сихотэ-Алиня, Приморья и Кореи [75, 154].
6(5). В гребне переднегруди 36 зубцов. У самки (рис. 324, Б) апи-
кальный край VII стернита ниже бокового угла более или менее
ровный. Самец неизвестен..................................Н. nicolai.
Одна самка этого вида снята с белки в Иркутской области (Тулунский район).
Безусловно необходим дальнейший материал для изучения этого вида [73].
Следует отметить, что в Северной Америке известен ряд видов блох, принад-
лежащих к той же группе, что и Н. microti и Н. nicolai [25, 26].
7(4). Кавказский вид. У самца вершина горизонтальной части
VIII стернита более или менее закруглена. На вершине передних
голеней нет сплошного поперечного ряда шпор . . . Н. satunini.
Паразит мелких лесных грызунов п кротов, обитающих на Кавказе [119, 10].
8(3). Имеется лобный зубчик. Глаз развитый. Щечный гребень из че-
тырех, иногда пяти зубцов; кроме того, один зубчик бывает рас-
положен вплотную под глазом. Дигитоид прикреплен выше задне-
нижнего угла тела клешни. У самца VIII стернит не модифици-
рован .........................................подрод Typhloceras.
Представители этого подрода Т. poppei и Т. favosus живут в Западной Европе
и Северной Африке на мелких лесных грызунах [122, 179 (21)].
Род Saphiopsylla [179 (36), 25]
В Евразии обитает только один вид S. ппрега, найденный в Альпах
(Южная Франция, Австрия) и на Кавказе, где встречается на горных
полевках в зимнее время года. Блохи другого вида — S. bishopi — живут
в Северной Америке.
121
Отметим, что этот род крайне близок к североамериканскому роду Atyphloceras,
имеющему более развитый глаз, низко сидящий лобный зубчик и более пяти члени-
ков в губных пальпах. Едва ли была необходимость разделять эти группы. В случае
их соединения они должны получить название Atyphloceras. В зоогеографическом
отношении вся эта группа представляет интерес, как пример европейско-сонорских
связей, т. е.. повидимому, имеет нижнетретичное (палеоген) происхождение.
Род Stenoponia [100, 147]
Крупные довольно темные блохи (2,5—5,5 мм длины), живущие
преимущественно в гнездах грызунов — полевок и других мышевидных,
цокоров и песчанок, на юге Европы, в Азии и Северной Америке. Все
виды этого рода встречаются чаще в холодное время года и выплажи-
ваются осенью. Мы приводим ниже данные о пяти видах, не касаясь
среднеазиатских паразитов песчанок S. conspecta (= solitaria) и S. vla-
sovi и тяньшаньского паразита мышевидных S. suknevi, о которых, как
и о европейских видах, в случае надобности можно найти сведения
в обзоре, посвященном этому роду [147].
1(2). Расстояние от нижнего угла лба до / зубца щечного гребня
большое и почти равно длине гребня (рис. 73). VII стернит самки
и гениталии самца изображены на рис. 326, 329 ...........................
...................................................... S. singularis.
Паразит цокора Myospalax aspalax, обнаружен в Борзинском и Оловяннинском
районах Восточного Забайкалья и на) северо-востоке Монголии [147].
2(1). Указанное расстояние меньше.
3(4). Указанное расстояние не меньше половины длины гребня.
VII стернит самки с двумя глубокими вырезками (рис. 333, А).
Вершина IX стернита самца широкая (рис. 327) . . . . S. ivanovi.
Блоха гнезд полевок. Найдена в Западной Сибири (Абаканская степь), на
Тянь-Шане, Монгольском Алтае, в Забайкалье и на) Кавказе [147, 19].
4(3). Указанное расстояние очень короткое, не больше половины дли-
ны гребня (рис. 74).
5(6). Губные пальпы (хоботок) одночленистые. VII стернит самки,
семяприемник и признаки самцов изображены на рис. 328, 330
.........................................................S. formozovi Г
Блоха гнезд полевок. Известна из Восточного Забайкалья и Благовещенска на
Амуре (из последнего места — лишь одна самка). География и экология этого вида
требуют дальнейшего изучения [147, 83].
6(5). Губные пальпы двучленистые.
7(8). Вершина IX стернита самца (рис. 331) усеченная, с густым пуч-
ком толстых шпорообразных щетин; парамер шлемовидных
очертаний. У самки (рис. 333, Б) VII стернит чаще без высту-
пающего вентрального угла; резервуар семяприемника груше-
видный ........................................................S. sidimi.
Паразит мышевидных грызунов — Apodemus agrarius, Cricetulus sp. и других. Из-
вестен из района Владивостока, Гродеково, Ворошилова-Уссурийского, Лазо, Благо-
вещенска и из Кореи. Живет в сельскохозяйственной лесо-луговой зоне [60].
8(7). Вершина IX стернита самца (рис. 334) округленная и пучок ще-
тин на ней менее развит; парамер крючковидный. У самки
(рис. 335) VII стернит, как правило, с заметно выступающим
вентральным углом, а резервуар семяприемника более округлый
...............................................................S. montana.
Живет на лесных полевках (Clethrionomys) в горных лесах Кореи [42].
1 Китайская S. coelestis, описанная по одной самке из Дацзянлу (Taj-tsien-lu,
провинция Синкан, между Сычуанем и Тибетом), имеет, судя по описанию, одно-
членистый хоботок, как и S. formozovi, но по форме края VII стернита более напо-
минает S. sidimi; семяприемник изображен на рис. 332 [100].
122
СЕМЕЙСТВО 1SCHNOPSY LLIDAE
Блохи летучих мышей. Группа блох, обособившаяся от Ceratophylli-
dae. Характеризуется своеобразной формой головы (рис. 75, 76) с двумя
(или тремя — у некоторых внепалеарктических видов) предротовыми
зубцами.
Род Ischnopsylliis |164, 165]
Блохи летучих мышей, главным образом семейства Vespertilionidae.
Отдельные виды в ряде случаев имеют предпочитаемого хозяина, но
нередко паразитируют и на. других видах летучих мышей. Почти никогда
не встречаются на представителях других отрядов. Учитывая, что ареалы
ряда видов этого рода оказались весьма широкими (быть может, в связи
со способностью их хозяев к перелетам) и география отдельных видов
еще мало выяснена, мы включаем в определительную таблицу все 11 ви-
дов, распространенных на территории СССР и сопредельных стран, но
рисунки даем лишь для некоторых видов.
Как и во многих других группах, самцы лучше всего определяются
по форме и вооружению половой клешни, а также VIII стернита путем
сравнения с рисунками. Для определения самок приходится использовать
многие признаки, причем для этого в большинстве случаев необходимо
изготовление просветленных препаратов.
1(2). Блохи с шестью спинными гребнями I. hexactenus.
Встречается чаще на ушанах (Plecotus auritus). Известно два подвида:
a) I. hexactenus hexactenus — западноевропейская форма, встречающаяся до
Ленинграда. Отличается тем, что у самцов вершина дигитоида и вершина VIII стер-
нита менее расширены (рис. 336,5), чем у следующего подвида [126, 180].
б) I. hexactenus kolenatii--восточная форма; распространенная от Европейской
части СССР до Тихого океана (найдена в Ленинграде, Саратовской области, на
Урале, в Зауралье, Красноярской, Иркутской областях и Забайкалье). Отличается
более расширенной вершиной дигитоида и VIII стернита) (рис. 336, Л) [126].
Неизвестно, какой подвид обнаружен в Москве, Сальском округе, Западном
Казахстане, Закавказье, Приморье, так как найдены только самки.
2(1). Блохи с восьмью спинными гребнями.
3(6). В гребне / членика брюшка (третий гребень) много зубцов
(примерно от 26 до 46). Длина блох обычно не менее 2,5 мм.
4(5). Третий гребень короче второго. Зубцы второго гребня такие же
острые, как первого, в отличие от остальных гребней. У самки
(рис. 338; 339, Г) церк очень широкий, короткий; характерна так-
же склеротизации заднего края VIII тергита. Проток копулятив-
ной сумки очень длинный, начало семенного протока слабо рас-
ширено, а резервуар семяприемника относительно большой
(рис. 338), У самца (рис. 337) характерна половая клешня
с сильно развитым неподвижным пальцем и торчащим перед ним
узким отростком ............................................I. obscurus.
Встречается, повидимому, чаще на кожанах (Vespertilio murinus). Распространен
во многих местах Европейской части СССР (начиная от Белоруссии), в Румынии, на
Северном Кавказе, в Зауралье, Казахской ССР, Средней Азии, Сибири (Новосибир-
ская, Кемеровская области), Забайкалье (Бырка, Борзя, Кайластуй, Агинский район).
Из Приморья (Ворошилов-Уссурийский) и Китая (Квантунский полуостров)
в сборах имеются пока только единичные самки [122, 164, 165].
5(4). Третий гребень примерно такой же длины, как и второй. У сам-
ки церк очень длинный и узкий (рис. 339, Б), длина его примерно
в четыре раза превышает наибольшую ширину. Проток копулятив-
ной сумки короткий; расширенная часть семенного протока в два-
три раза шире, чем копулятивная сумка, резервуар семяприемни-
ка сравнительно небольшой. Характерно также большое число
123
разбросанных на VII стерните щетинок и вытянутый задне-ниж-
ний угол VIII тергита. У самца (рис. 339, Л) дигитоид треуголь-
ный с острым верхне-задним углом и вогнутым верхним краем.
Пигидиальный тергит самца образует языкообразные боковые
лопасти, направленные косо вниз и кзади..............I. elongatus.
Встречается чаще на вечернице (Nyctailus noctula). Широко распространен
в Европе, на Кавказе, в Средней Азии (Ташкент, Алма-Ата, Джаркент). Одна самка,
повидимому, этого же вида найдена в Китае (Квантунский полуостров) [164, 165].
6(3). В третьем гребне обычно менее 20, иногда до 27 зубцов Ч У сам-
ки церк (рис. 339, В) более короткий, чем у предыдущего вида;
его длина превышает ширину примерно в 2—3—3!/2 раза (у I. co-
mans и I. needhamia — обычно в два раза, у других видов —
часто более чем в три раза). Проток копулятивной сумки более
короткий, чем у I. obscurus. Блохи обычно менее 2,5 мм длины,
светлого цвета.
7(8). Предротовой склеротизованный тяж (tuber preorale), идущий
кзади от предротового ктенидия и изогнутый под углом, сравни-
тельно узкий и длинный. У самки резервуар семяприемника до-
вольно велик. У самца резко удлинены верхние щетинки главно-
го ряда мезонотума. Модифицированные сегменты самки и сам-
ца— см. на рис. 340, 341 ...................................I. comans.
Имеются сборы с кожановидного нетопыря Vespertilio savii и др. Восточноазиат-
ский вид. Найден в Приморье (Ворошилов-Уссурийский, Славянка, Краскино) и Китае
(Пекин, Цзинань) [180, 98].
8(7). Предротовой склеротизованный тяж более толстый и короткий.
9(10). У самки (рис. 343, А) стигма VIII тергита расширена и сильно
нависает над пигидием. Резервуар семяприемника мал. У самца
(рис. 342) особенно характере^ пучок длинных, толстых, изогну-
тых под углом щетин на VIII стерните .... I. needhamia.
Имеются сборы с кожана Vespertilio superans и др. Восточпоазиатский вид. Из-
вестен из Забайкалья (Борзинский и Агинский районы), Приморья и Китая (Сучжоу)
[91, 73, 98].
10(9). Признаки иные.
11(12). В третьем гребне довольно много (19—27) зубцов, причем ниж-
ний из них нередко сидит на уровне или даже ниже уровня ниж-
ней щетинки главного ряда этого тергита. Метэпистерн обычно
с двумя щетинками (редко одна-три). У самки начальная часть
семенного протока (pars dilatata) вдвое шире, чем копулятивная
сумка, а резервуар семяприемника мал; задняя половина ниж-
него края головы образует углообразный выступ, более выра-
женный, чем у других видов. У самца VIII стернит широкий.
Ацетабулярные щетинки сидят выше сочленовной ямки. Дигитоид
треугольный с округленным задне-верхним углом. Основание
верхней ацетабулярной щетинки очень сильно утолщено. Дор-
зальные щетинки тергитов средне- и заднегруди у самца удли-
нены и образуют гриву 1 2.................................I. intermedins.
Встречается чаще на позднем кожане Vespertilio (Eptesicus) serotinus. Распро-
странен в Зашадной Европе и многих местах Европейской части СССР; известен так-
же на Кавказе и на Урате [164, 165].
В Средней Азии (Чуйская долина) найден близкий вид I. plumaitus, отличаю-
щийся несколько меньшим развитием третьего гребня (около 15—22 зубцов), более
округленным задним краем дигитоида и более (многочисленными щетинками на
VIII стерните самца [19].
1 В гребне VI тергита также обычно мало зубцов — от 6 до 11—у I. comans,
1. needhamia, I. dolosus, I. simplex; у других видов — до 18.
2 У тех видов, для которых этот признак не отмечен, названные щетинки
не удлинены.
124
12(11),. В гребне I тергита брюшка 6—17 (редко до 22) зубцов, причем
нижний зубец сидит обычно выше уровня нижней щетинки глав-
ного ряда этого тергита. Метэпистерн обычно с одной щетинкой
(редко две). У самки расширенная часть семенного протока не
шире или немного шире, чем копулятивная сумка, а резервуар
семяприемника крупнее, чем у предыдущего вида. У самца
VIII стернит неширокий.
13(16). Предротовой склеротизованный тяж изогнут под прямым углом.
У самца ацетабулярные щетинки сидят против нижней части
сочленовной ямки.
14 (15) .• В гребне I и VI тергитов брюшка по 14—22 зубца. У самки
« • резервуар семяприемника относительно большой. У самца диги-
тоид треугольный с заостренным задне-верхним углом . . . .
.....................................................I. variabilis.
Встречается чаще на нетопырях (Vespertilio nathusii, V. pipistrellus). Известен
из многих районов Европы — от Швейцарии и Германии до Среднего Поволжья
« Предкавказья [164, 165, 17].
15(14). В гребнях I и VI тергитов по 7—16 зубцов. У самки резервуар
семяприемника несколько меньше, чем у предыдущего вида.
У самца дигитоид полулунной формы с округленным задним
краем. Дорзальные щетинки тергитов средне- и заднегруди у сам-
ца удлинены и образуют гриву..............................I. octactenus.
Встречается чаще на нетопыре-карлике (Vespertilio pipistrellus). Распространен
в Западной и Южной Европе, на юге Европейской части СССР, на Кавказе и в Сред-
ней Азии [164, 165].
16(13). Предротовой склеротизованный тяж изогнут под тупым углом.
В гребне I брюшного тергита 6—17 зубцов, в гребне VI тергита
6—11 зубцов. У самки резервуар семяприемника довольно велик.
У самца ацетабулярные щетинки сидят выше сочленения; диги-
тоид полулунной формы с округленным задним краем . . . .
.............................................................I. simplex.
Встречается чаще на ночницах (Myotis). Распространен в Западной Европе,
где описано два подвида) [164, 165, 177].
На Кавказе также на Myotis живет близкий вид I. dolosus, блохи этого вида
отличаются отсутствием сильной щетины на верхнем крае тела половой клешни, тре-
угольным дигитоидом с округленным задне-верхним углом и другими деталями
[165, 179, (35)].
Род Rhinolophopsylla |180]
Единственный известный у нас представитель этого рода Rhinolophop-
sylla unipectinata паразитирует на летучих мышах семейства и рода под-
ковоносов (Rhinolophus). Распространен в Южной Европе, Северной
Африке, Крыму, Закавказье, Средней Азии, на северо-западе Индии.
Образует географические подвиды.
Род Nycteridopsylla [29, 15, 90, 177, 19]
Паразиты летучих мышей семейства Vespertilionidae. Распростране-
ны преимущественно в Западной Европе, где встречаются несколько ви-
дов; на территории СССР найдены лишь единичные экземпляры следую-
щих видов: Nycteridopsylla dictena (Киргизская ССР), N. eusarca (Став-
рополь-Кавказский). Кроме того, следует упомянуть, что из Китая (Шан-
хай, Сучжоу) описана N. galba. Представители этого рода встречаются
чаще зимой.
125
Род Myodopsylla [921
Паразиты летучих мышей, повидимому, преимущественно рода
Myotis. Большинство видов известно в Америке, а в Евразии распро-
странен лишь один вид Myodopsylla trisellis (рис. 343, Б; 344); он об-
наружен в Маньчжурии, а в СССР — в Предбайкалье (с. Янды на
р. Ангаре), на Урале, в Поволжье, Воронежской, Московской и Ленин-
градской областях.
Род Araeopsylla [188 (13), 19J
Паразиты бульдоговых мышей рода Tadarida. В СССР известен
один вид: A. gestroi, описанный из Италии и обнаруженный в За-
кавказье.
АЛЬБОМ РИСУНКОВ
Рис. 21. Pulex irritans, самка. Голова и грудь.
Характерна мезоплевра (мп.) без пересекающего ее вертикального
склеротизованного тяжа. Стигма первого тергита брюшка (стиг.)
расположена высоко, пр. — спинка передней груди (пронотум);
мез. — спинка среднегруди (мезонотум); мет. — спинка заднегруди
(метанотум)); эст. — эпистерн заднегруди; эм. — эпимер заднегруди
(метэпимер). Внизу слева помещена масштабная линейка в 100
(0,1 мм). Линейки, приведенные под большинством рисунков (или
рядом с ними), также равны 100 (л, кроме случаев, специально
оговоренных.
Рис. 22. Archaeopsylla sinensis, самка. Голова и пронотум.
9 Определитель блох
129
Рис. 23. Echidnophaga gallinacea, самка.
Голова, грудь и I тергит брюшка.
Рис. 24. Echidnophaga oschanini, самка.
Голова, грудь и / тергит брюшка.
Рис. 25. Ctenocephalides canis, самка. Голова
и пронотум.
Рис. 26. Ctenocephalides felis, самка. Голова
и пронотум.
130
Голова и грудь самки
Рис. 27. Xenopsylla cheopis.
Затушеваны некоторые склеротизованные тяжи стерноплевры
средне- и заднегруди; мп.— мезоплевра, которую пересекает
характерный вертикальный тяж; эст.— эпистерн заднегруди;
эм.—эпимер заднегруди (метэпимер); стиг.— стигма.
Рис. 28. Synostemus longispinus. мп.— мезоплевра; эст.— эпистерл заднегруди,
слившийся со стерноплеврой.
9-
131
Рис. 29. A—Xenopsylla astia, самец; Б — Xenopsylla nubi-
cus, самец. Теменная ямка.
Рис. 30. Pariodontis nggenbachi, самка. Голова и пропогум.
Рис. 31. Spilopsyllus cuniculi, самка. Голова
и пронотум.
132
Голова и пронотум самки
133
Голова, пронотум и мезонотум самки
Рис. 35. Vermipsylla alacurt.
Рис. 37. Самки Vermipsylla с созревшими яйцами: А — Ver-
mipsylla alacurt; В — Vermipsylla dorcadia. Масштаб — 2 мм.
134
Голова и пронотум самки
.135
Голова и пронотум самки
Рис. 42. Ceratophyllus fasciatus.
Рис. 45. Frontopsylla luculenta.
Рис. 44. Ceratophyllus penicilljger.
136
Рис. 46. Ctenophyllus armatus.
Рис. 47. Ctenophyllus hirticrus.
Рис. 48. Ophthalmopsylla prat*
fecta.
Рис. 49. Ophthalmopsylla ki-
ritschenkoi.
137
Голова и пронотум самки
Рис. 50. Brachyctenonotus myospalacis.
Рис. 52. Leptopsylla ostsibirica.
Рис. 53. Mesopsylla hebes.
.138
Голова и пронотум самки
Рис. 54. Голова и пронотум самки: А — Amphipsylla rossica; Б — A. sibirica;
В—A. primaris; Г—A. vinogradovi; Д — A. daea; Е — лоб A. sibirica;
Ж — лоб A. primaris.
Рис. 55. Peciinoctenus pectiniceps.
Рис. 56. Wagncrina antiqua.
139
Рис. 57. Eopsylla nuda, самка. Голова и грудь. Значение букв см. на рис. 27.
Голова и пронотум самки
Рис. 58. Doratopsylla dasycnemus.
Рис. 60. Doratopsylla birulai.
140
Голова и пронотум
Рис. 62. Palaeopsylla sorecis, самка.
Рис. 61. Ctenophthalmus congcneroides,
самка.
Рис. 63. Nearctopsylla ioffi, самец.
141
Рис. 65. Rhadinopsylla dahurica, самка.
Передняя часть головы.
Рис. 66. Rhadinopsylla li,r'самка. Передняя
часть головы.
Рис. 67. Rhadinopsylla valenti, самка. Рис. 68. Rhadinopsylla cedestis, самка. Голова
Передняя часть головы. и пронотум.
142
Голова и пронотум самки
Рис. 69. Neopsylla bidentatiformis.
Рис. 70. Catallagia ioffi.
Рис. 71. Hystrichopsylla talpae.
Рис. 72. Hystrichopsylla microti.
143
Рис. 73. Stenoponia singulars,
самка. Передняя часть головы.
Рис. 74. Stenoponia for-
mozovi, самка. Передняя
часть головы.
Рис. 75. Ischnopsyllus obscurus, самка. Голова и
пронотум.
Рис. 76. Myodopsylla triscllis, самка. Голова, грудь и / тергит брюшка
с ложным гребнем — л. г.
144
Модифицированные сегменты и гениталии самца
Рис. 77. A—Archaeopsylla sinensis; Б — Xenopsylla cheopis. Обозначения см. рис. 80.
10 Определитель блох
Рис. 79. Эдеагус. Xenopsyl-
la: А —X. astia; Б — X. nubicus;
* — выступающий дорзальный
уголок воронки внутренней
трубки эдеагуса; с — седлооб-
разный склерит над воронкой.
Рис. 80. Xenopsylla conformis.
Рг и Р2 — подвижные пальцы
половой клешни; — пластин-
чатый вырост; эд — парамер
эдеагуса; VII т. — тергит;
♦ WII—IX ст. — стерниты.
145
Модифицированные сегменты и гениталии самца
Рис. 81. Xenopsylla skrjabini. Обозначения см. рис< 80.
Рис. 82. Xenopsylla gerbilli gerbillL
116
Модифицированные сегменты и гениталии самца
Рис. 83. Xenopsylla gerbilli minax.
Рис. 84. Xenopsylla gerbilli caspica.
147
10*
Рис. 85. Xenopsylla persica, самец. Модифицированные сегменты и
гениталии.
Рис. 86. Семяприемник самок Xenopsylla: 'А—cheopis; Б — X. astia; В — X. nubicus;
Г — X. hawaiiensis; Д — X. brasiliensis; Е — X. conformis; Ж — X. skrjabini.
Рис. 87. А — Hoplopsyllus glacialis profugus, самец: половая клешня, VIII и IX
стерниты; Б — Н. g. profugus, самка: VII стернит, семяприемник и копулятив-
ный проток.
Рис. 88. Слева — Chaetopsylla homoeus, самец: половая клешня, IX стернит и эдеагус
(эд)\ справа вверху — вариант эдеагуса; справа внизу—Ch. homoeus, самка: VII стер-
нит, семяприемник и копулятивный проток.
149
Рис. 89. Chaetopsylla mirabilis, самец. Половая к.«шшя и VIII стернит.
Рис. 90. Chaetopsylla mirabilis, самка. VII и VIII стерниты, семяприемник
копулятивный прото*к.
150
Модифицированные сегменты и гениталии самца й самки
Рис. 91. А — Chaetopsylla alia, самец; Б — Ch. alia, самка.
Рис. 92. А — Chaetopsylla dogieli, самец; Б — Ch. dogieli, самка.
151
Модифицированные сегменты и гениталии самца и самки
Рис. 93. Chaetopsylla appropinquans, самец.
152
Модифицированные сегменты и гениталии самца и самки
Рис. 9э. Chaetopsylia trichosa, самец.
5
Рис. 96. А — Chaetopsylia appropinquans, самка; Б — Ch. trichosa, самка.
153
Модифицированные сегменты и гениталии самца и самки
Рис. 97. А — Chaetopsylla mikado, самец; Б — Ch. mikado, самка.
Рис. 98. А — Chaetopsylla tuberculaticeps, самец; Б — Ch. tuberculaticeps,
самка.
154
Рис. 99. А— Tarsopsylla octodecimdentata, самец: половая клешня, VIII и IX стерниты;
Б — Т. octodecimdentata, самка: семяприемник и VII стернит с вариантом.
Рис. 100. А — Aenigmopsylla grodekovi, самец: половая клешня, VII, VIII и IX стер-
ниты; Б — A. grodekovi, самка: семяприемник и VII стернит.
155
Рис. 101. Paraceras flabellum, самец. Половая клешня, VIII и IX стерниты.
Рис. 102. Paraceras melis, самец. Половая клешня, IX стернит, пигидий и
анальный сегмент.
156
1
Рис. 103. Paraceras flabellum, самка.
Семяприемник, VII стернит и VIII
тергит.
Рис. 104. Paraceras melis, самка.
Семяприемник, VII стернит и
VIII тергит.
Рис. 10Й. Amphalius runatus, самка. Семяприем-
ник, копулятивная сумка (яг. с.), VII стернит
\ и церк (ц.).
7\ i; мн им г.
157
Рис. 106. Amphalius runatus, самец. Половая клешня, VIII и IX стерниты,
эдеагус—ad., трубка эдеагуса — т. э.
Рис. 107. Amphalius clarus, самец. Половая клешня и IX стернит.
158
Рис. 108. Oropsylla silantiewi, самец. Половая клешня, VIII и IX стерниты,
парамер эдеагуса — эд. Слева — вариант клешни и парамера самца с Кам-
чатки.
Рис. 109. Oropsylla asiatica, самец. Половая клешня, анальный сегмент (ан. с.%
VIII и IX стерниты. Слева —часть клешни и VIIT стернит экземпляра
с Камчатки (тип).
15&
Рис. ПО. Oropsylla ilovaiskii,
самец. Половая клешня, VII,
VIII и IX стерниты.
Рис. 111. А—> Oropsylla silantiewi,
самка: семяприемник, копулятивный
проток, VII и VIII стерниты; VIII
тергит со стигмой, пигидий, анальный
тергит — ан. т. и церк — ц; Б — Oro-
psylla
asiatica, самка: семяприемник
и VII стернит.
с та г
. ан.т.
шт.
А
VU ст
Рис. 112. А — Oropsylla (Diamanus) mandarinus, самец. Половая клешня и модифициро-
ванные стерниты; Б — О. (D.) mandarinus, самка. Семяприемник, VII и VIII стерниты;
Vi VIII Т^пгыт
Рис. 113. Ceratohpyllus fasciatus, самец. Половая клеши?»
VII, VIII и IX стерниты; VIII тергит, парамер эдеагуса
(эд.).
Рис. 114. Ceratophyllus
fasciatus, самка. Семя-
приемник, копулятивная
сумка, копулятивный про-
ток, VII стернит.
Рис. 115. А — Ceratophyllus mokrzeckyi, самец: половая клешня и IX стернит;
Б — С. mokrzeckyi, самка: семяприемник, VII стернит и VIII тергит.
Рис. 116. А — Ceratophyllus fidus, самец: половая-клешня, IX стернит и VIII тергит;
Б — С. fidus, самка: семяприемник, VII стернит, VIII тергит.
11 161
11 Определитель блох
Рис. 117. А — Ceratophyllus laeviceps laeviceps, самец: половая клешня
и горизонтальная ветвь IX стернита; Б — С. 1. laeviceps, самка:
копулятивный проток и VII стернит с вариантами края.
Рис. 118. А — Ceratophyllus laeviceps kuzenkovi, самец: половая клешня ч
IX стернит; Б — С. 1. kuzenkovi, самка: гениталии и VII стернит.
Рис. 119. Л — Ceratophyllus laeviceps ellobii, самец: половая клешня
и горизонтальная ьегвь IX стернита; Б — С. 1. ellobii, самка:
гениталии и VII стернит.
162
Рис. 120. А — Ceratophyllus turkmenicus, самец: половая клешня ,У7/>
VIII и IX стерниты, часть VII тергита; Б — С. turkmenicus, самка:
гениталии и VII стернит.
Рис. 121. Ceratophyllus tesquorum sungaris, самец. По-
ловая клешня, VIII и IX стерниты.
Рис. 122. Ceratophyllus tes-
quorum mongolicus, самец.
Часть половой клешни.
Рис. 123. Ceratophyllus tesquorum sungaris, самка: гениталии и VII стернит. Справа —
варианты апикального края VII стернита самок из Монголии.
11*
163
Рис. 124. Ceratophyllus tesquorum trans-
volgensis, самец. Часть половой
клешни и VIII стернит.
Рис. 125. Ceratophyllus tesquorum altaicus
самец. Часть половой клешни и VIII стер-
нит-
с. 16.4 —Ceratophyllus jenissejensis, самец: половая клешня, VIII и IX стерниты,
VIII тергит; Б — С. jenissejensis, самка: гениталии и VII стернит.
164
Рис. 127. Ceratophyllus
caspius, самец. Половая
клешня, VIII и IX стер-
нитьи
Рис. 128. Ceratophyllus gaiskii, самец. Половая клешня, VIII и IX
стерниты.
Рис. 129. А — Ceratophyllus caspius, самка: семяприемник, копулятивная сумка — к.
копулятивный проток — /г. л., VII стернит; Б — С. gaiskii, самка.
165
Рис. 131. Ceratophyllus kozlovi, самец. Поло-
вая I "ггг
Рис. 130. Ceratophyllus dolabris, самец,
Пэлэвая клешня, VIII и IX стерниты.
клешня, VIII и IX стерниты, парамер
эдеагуса.
Рис. 132. Ceratophyllus kaznakovi, самец. По-
ловая клешня, VIII и IX стерниты.
Рис. 133. Ceratophyllus kazna-
kovi, самка. Гениталии и VII
стернит.
Рис. 134. Ceratophyllus oreinus, самец. Половая
клешня, VIII и IX стерниты.
Рис. 135. Ceratophyllus oreinus, сам-
ка. Семяприемник и VII стернит.
163
Рис. 136. А — Ceratophyllus geminus, самец: половая клешня, VIII и IX стерниты,
ларамер эдеагуса (эд.); Б —С. geminus, самка: гениталии и VII стернит.
Рис. 137. Ceratophyllus semenovi, самец. Половая
клешня, VIII и IX стерниты.
167
Рис. 139. Ceratophyllus penicilliger, самец. Половая клешня, VIII и IX ciep-
ниты, парамер эдеагуса (эд.).
Рис. 140. Ceratophyllus penicilliger,
самка. Семяприемник, VII стернит и>
стигма VIII тергита (стиг.).
168
Рис. 141. 'Часть половой клешни и парамер эдеагуса самца: А — Cerato-
phyllus penicilliger dissimilis; Б — С. р. syrt; В — С. р. arvicolae; Г — С. р-
penicilliger; Д — С. р. vallis; Е— С. р. demotus; Ж— Ceratophyllus ioffi.
Рис. 142. VII стернит самки; А — Ceratophyllus penicilliger penicilliger, ва-
рианты VII стернита; Б —С. р. dissimilis; В — С. р. syrt; Г —С. р. arvi-
colae; Д — С. р. demotus, с вариантом; Е — Ceratophyllus ioffi.
169»
Рис. 143. А — Ceratophyllus sciurorum asiaticus, самец: половая клешня, VIII и IX стер
ниты; Б — С. s. asiaticus, самка: семяприемник и VII стернит.
Рис. 144. Л — Ceratophyllus anisus, самец: половая клешня, VIII и IX стерниты;
Б — С. anisus, самка: семяприемник и VII стернит.
Рис. 145. Ceratophyllus tamias, самец. Половая клешня, VIII и IX
стерниты.
170
пиг
Рис. 146. Ceratophyllus paradoxus, самец. Половая клешня, VIII и IX
•стерниты, длинные спирально свернутые тяжи, пигидий (пиг), аналь-
ный сегмент (ан.с.).
Рис. 147. А — Ceratophyllus tamias, самка; Б — С. paradoxus, самка. Гениталии
и VII стернит.
Рис. 148. А — Ceratophyllue toli, самец: половая клешня, VIII и IX стерниты;
Б — С. toli, самка: семяприемник и VII стерниг.
171
Рис. 149. Ceratophyllus scaloni, самец. Половая клешня, VIII и IX стерниты, парамер.
эдеагуса (эд.).
Рис. 150. Ceratophyllus desertus, самец. Половая клешня, VIII и IX стерниты, парамее»
эдеагуса (эд.).
172
Рис. 152. Ceratophyllus desertus, самка.
Семяприемник, копулятивная сумка и
VII стернит.
'Рис. 151. Ceratophyllus scaloni,
самка. Семяприемник, VII
<тернит, стигма VIII тергита.
Рис. 153. Ceratophyllus calcarifer, самец. Половая клешня, VIII и IX стерниты.
.Р,ис. 154. Гениталии и VII стернит. Л — Ceratophyllus calcarifer, самка;
Б — С. walkeri самка.
173
Рис. 155. А — Ceratophyllus walkeri, самец. Половая клешня, VIII и IX стерниты;
Б — С. sokolovi, самец. Половая клешня, IX стернит и часть эдеагуса {эд.).
Рис. 156. Л — Ceratophyllus sokolovi, самка: гениталии и VII стернит; Б — С. advenarius,
самка: семяприемник и VII стернит.
Рис. 157. А—Ceratophyllus rectangulatus, самец: половая клешня, VIII и IX стерниты;
Б— С. rectangulatus, самка: гениталии и VII стернит, справа—стигма VIII тергита и
варианты VII стернита.
174
Рис. 158. Ceratophyllus advenarius, самец. Половая клешня, VIII и IX
стерниты. А — С. advenarius advenarius— часть дигитоида с шипиками;
Б—С. advenarius bifallax— часть дигитоида с шипиками.
Рис. 159. Ceratophyllus turbidus, самец. Половая клешня,
VIII и IX стерниты.
Рис. 160.
Ceratophyllus
turbidus,
самка.
Гениталии
и VII
стернит;
*—отмечен
конец
вентральной
вырезки.
Рис. 161.
Ceratophyllus
taiganus,
самка.
Семяприем-
ник и VII
стернит.
175.
Рис. 162. Ceratophyllus taiganus, самец. Половая клешня, VIII и IX
стерниты, стигма VIII тергита, анальный сегмент.
Рис. 163.
Ceratophyllus lunatus,
самец. Половая клеш-
ня, VIII и IX
стерниты.
> Рис. 164. Ceratophyllus lunatus,
«самка. Семяприемник и VII стернит.
Рис. 165.
Ceratophyllus
garei,
самка.
Гениталии
и VII
стернит.
А—вариант
семяприем-
ника.
.176
Рис. 166. Ceratophylus
garei, самец. Половая
клешня, VIII и IX
стерниты.
12 Определитель блох
177
т.п.
Рис. 1691 А— Сеган
tophyllus garei,
самец: половой
аппарат; Б—С. gal-
linae, самец:
половая клешня,
модифицированные
сегменты и поло-
вой аппарат: эд.—
эдеагус; а. э,—апо-
дема эдеагуса;
т. п.— тяжи пени-
са; пиг.— пигидий;
Р — неподвижный
палец; F—дигито-
ид; М — рукоятка
клешни.
Рис. 170. А—Ceratophyllus
gallinae tribulis, самец:
половая клешня, VIII и
IX стерниты; Б — С. di-
latus, самец: часть ' по-
ловой клешни.
Рис. 171. Ceratophyllus gallinae
tribulis, самка. Гениталии и VII
стернит; сл. п. — слепой проток.
Рис. 172.
Ceratophyllus
gallinae
dilatus,
самка.
Гениталии
и VII
стернит.
Рис. 173.
Ceratophyllus
gallinae gallinae,’
самец.
Половая клеш-
ня, VIII и IX
стерниты.
Рис. 174. Ceratophyllus gallinae
gallinae, самка. Гениталии
(сл. и.—слепой пр эго к) и
VII стернит.
Рис. 175. Ceratophyllus fringillae, самец. VIII
и IX стерниты, интерсегментальные бах-
ромчатые лопасти (и.), Л’-орган.
Рис. 176. Ceratophyllus vagabundus, самец.
Половая клешня, VIII и IX стерниты.
Рис. 177. Ceratophyllus vagabundus,
самка. Семяприемник и VII стер-
нит.
12*
179
Рис. 178. Ceratophyllus styx, самец. Пэлэвтя клешня, VIII и IX стернита.
Рис. 179. Ceratophyllus riparius, самец. Половая клешня, VIII и IX стерниты.
130
Рис. 180. Гениталии и VII стернит самки: А—Ceratophyllus styx; Б—С. riparius.
Рис. 181. Ceratophyllus avicitelli, самка. Гениталии, VII
стернит с вариантами, анальный стернит, церк.
Рис. 182. Ceratophyllus avicitelli, самец. Половая клешня
с вариантом, VIII и IX стерниты.
181
Рис. 183. Cerato -
phyllus maculatus,
самец. Половая
клешня, VIII и IX
стерниты.
Рис. 185. Гениталии и VII стернит самки: А—Ceratophyllus maculatus; Б—С. farreni.
182
Рис. 186. Ceratophyllus
caliotes, самец. Половая
клешня, VIII и IX
стерниты.
Рис. 187.’
Ceratophyllus orites,
самец. Половая
клешня и IX
стерниг.
Рис. 183. I ениталии и VII стернит самки: A— Ceratophyllus caliotes; Б — С. orites.
183
Рис. 191. А — Dasypsyllus stejnegeri, самец:
часть половой клешни;# — Dasypsyllus stej-
negeri, самка: VII стернит.
Б
184
185-
Рис."195. Frontopsylla hetera, самец. Часть половой клешни, IX и VIII
стерниты.
Рис. 197. Frontopsylla hetera,
самка. Семяприемник и VII
стернит.
Рис. 196. Frontopsylla elatoides, сам а.
Семяприемник и VII стернит.
Рис. 198. Frontopsylla luculenta luculenta,
самец. Часть половой клешни и IX стернит.
Рис. 199. Frontopsylla luculenta luculenta
самка. Семяприемник и VII стернит
486
Рис. 200. Frontopsylla luculenta parilis, самец. Часть Рис. 201. Frontopsylla luculenta
половой клешни и IX стернит. parilis, самка. Семяприемник и
VII стернит.
Рис. 202. Frontopsylla ehta elata, самец. Часть половой клешни, IX и VIII стерниты;
а—F. data popovi, самец. Часть половой клешни.
А б
Рис. 203. А — Frontopsylla elata taishiri, самец. Часть половой клешни и IX стернит;
Б—F. е. taishiri, самка: варианты апикального края VII стернита.
187
Рис. 204. Frontopsylla elata botis, самец.
Часть половой клешни и IX стернит.
Рис. 206. Frontopsylla frontalis bai-
kal, самец. Часть половой клешни.
Рис. 205. Frontopsylla elata botis*
самка. Семяприемник и VII
Рис. 207. Frontopsylla frontalis baikal*
самка. Семяприемник и VII стернит.
188
Рис. 209. Ctenophyllus armatus, самец. Половая клешня, VIfl и IX стерниты.
Рис. 210. Ctenophyllus tarasovi, самец. Половая клешня, VIII и IX стерниты,
а—вариант дигитоида.
189
Рис. 211. Л—Ctenophyllus subarmatus, самец: часть половой клешни и IX стернит;
Б — С. armatus, самка, семяприемник и VII стернит; В — С. rigidus, самка. Гешь
талии и VII стернит.
Рис. 212. Л—Ctenophyllus hirticrus, самец: половая клешня, VIII и IX Стерниты;
Б—С. hirticrus самка, семяприемник и VII стерни г.
190
Рис. 216. Paradoxopsyllus dashidorzhii, самец. Половая клэшня, VIII и IX
стеониты.
Рис. 217. А — Paradoxopsyllus dashidorzhii, самка: гениталии и VII стернит (с вариан-
тами апикального края); Б — Р. conveniens, самка: гениталии и VII стернит.
И92
Рис. 218. А — Paradoxopsyllus curvispinus, самец: половая клешня и модифицированные
сегменты; Б— Р. curvispinus, самка: гениталии и VII стернит.
Рис. 219. A—Ophthalmopsylla praefecta praefecta, самец: половая клешня
и IX стернит; Б—О. р. praefecta, самка: семяприемник и VII стернит.
13 Определитель блох
193
Рис. 220. A—Ophthalmopsylla praefecta pernix, самец: часть половой клеш-
ни; Б— О. р. pernix, самка: VII стернит.
Рис. 221. А — Ophthalmopsylla kukuschkini, самец: половая клешня и IX
стернит; Б — О. kukuschkini, самка: семяприемник и VII стернит.
Рис. 222. Ophthalmopsylla kukuschkini, самка. Варианты апикального края VII стернита.
194
Рис. 223. Ophthalmopsylla volgensis extrema. A—самец: половая клешня и'/Х
стернит; Б—самка: гениталии и VII стернит.
Рис. 224. A—Ophthalmopsylla kiritschenkoi, самец: половая клешня и IX стернит;
Б — О. kiritschenkoi, самка: VII стернит.
13*
195
Рис. 225. Ophthalmopsylla jettmari,
самец. Часть половой клешни.
Рис. 226. Ophthalmopsylla
jettmari, самка. VII стер-
нит.
Рис. 227. Brachyctenonotus myospalacis, самец. Половая клешня,
VIII и IX стерниты.
195
Рис. 228. Brachyctenonotus myo-
spalacis, самка. Гениталии и
VII стернит.
Рис. 229. Mesopsylla hebes, самец. Половая
клешня, VIII и IX стерниты, парамер
эдеагуса (эд.).
Рис. 230. Mesopsylla hebes, самка. Семяприемник,
VII стернит и VIII тергит. Справа—варианты края VII
стернита.
197
Рис. 231. Mesopsylla lenis,
самец. Половая клешня,
VIII и IX стерниты, па-
рамер эдеагуса (эд.).
Рис. 232. Mesopsylla tu-
schkan, самец. Половая
клешня, VIII и IX стер-
ниты, парамер эдеагуса.
Справа—вариант вершины
дигитоида.
Рис. 233. Mesopsylla rothschildi, самец. Половая клешня,
VIII и IX стерниты.
198
Рис. 234. Mesopsylla lenis,
самка. Семяприемник, VII
и VIII стерниты и VIII
тергит.
Рис. 235. Mesopsylla tuschkan,
самка. Семяприемник, VII и
VIII стерниты, VIII тергит.
Рис. 236. Mesopsylla rothschildi, самка.
Гениталии и VII стернит.
Рис. 237. Mesopsylla eucta, самец.
Часть половой клешни, VIII стернит
и парамер эдеагуса (ad.).
199
Рис. 238. Часть половой клешни самца: А — Amphipsylla rossica; Б — A. si-
birica; В — A. prima; Г — A. kuznetzovi; Д — A. marikovskii; Е — A. pet-
ristshevae; Ж—A. schelkovnikovi; 3—A. vinogradovi; И—A. tuta; К—А. ап-
ceps; Л—A. dumalis; М — A. primaris primaris; Н— A. primaris mitis;
О — A. daea; П — A. aspalacis.
Рис. 239. Семяприемник самки: А — Amphipsylla rossica; Б — A. primaris; В — A. du-
malis; Г — A. schelkovnikovi; Д — A. vinogradovi; Е— A. longispina.
200
Рис. 240. VII стернит самки: А — Amphipsylla rossica; Б — A. sibirica; В — A. prima;
Г —A. kuznetzovi (три варианта); Д — A. marikovskii; Е —A. petristshevae; Ж —
A.*tuta; 3 —A. vinogradovi; И — A. longispina; К — A. dumalis; Л — А. primaris
mitis; М — A. daea; Н — A. aspalacis.
Рис. 241. Копулятивный проток самки: А — Amphipsylla sibirica sibirica; Б — A. sibi-
rica orientalis; В — A. rossica; Г — A. kuznetzovi; Д— A. marikovskii; Е— A. prima;
Ж— A. tuta; 3 — A. schelkovnikovi; И — A. vinogradovi; К — A. longispina; Л.—
A. dumalis; Л4 — A. anceps; Н — A. primaris mitis; О — A. daea; П — A. aspalacis.
201
Рис. 242. VIII стернит самца; А — Amphipsylla rossica; Б—A. anceps;
В — A. kuznetzovi; Г — A. primaris mitis.
Рис. 243. А — Amphipsylla casis, самец: часть половой клешни; Б — Amphi-
psylla longispina, самец: половая клешня, VIII и IX стерниты (‘—интер-
сегментальная вилочка).
Рис. 244. Amphipsylla longispina, самка. Семяприемник, копулятивный
проток, К, VI и VII стерниты.
202
Рис/ 245. Leptopsylla папа. А — самец: часть
половой клешни, VIII стернит; Б — самка: ге-
ниталии и VII стернит.
Рис. 249. Leptopsylla ostsibirica, самка.
Семяприемник и VII стернит.
Рис. 248. Leptopsylla bidentata, самка.
Семяприемник и VII стернит.
203
Рис. 250. Leptopsylla ostsibirica, самеи. Половая клешня, VIII и IX стерниты.
Рис. 251. А — Leptopsylla silvatica, самец: половая клешня, VIII и IX стерниты;
Б — L. silvatica, самка: семяприемник и VII стернит.
Рис. 252. Pectinoctenus pavlovskii, самец.
Половая клешня, VIII и IX стерниты.
Рис. 253. Pectinoctenus pav-
lovskii, самка. VII стернит.
204
Рис. 254. Pectinoctenus pectiniceps, самец. Половая клешня,
VIII и IX стерниты.
Рис. 255. Pectinoctenus pe-
ctiniceps, самка. VII стер-
нит.
Рис. 256. Л — Wagnerina tecta, самец: половая клешня, модифицированные сегменты,
стиг. — стигма VIII тергита; Б — W. tecta, самка: гениталии, модифицированные сег-
менты, стиг. — стигма VIII тергита.
205
стиг.
Рис. 257. А — Wagnerina longicauda, самец: половая клешня, модифицированные
сегменты, стиг.— стигма VIII тергита; Б — W. longicauda, самка: гениталии, моди-
фицированные сегменты, стиг. — стигма VIII тергита.
Рис. 258. А — Wagnerina antiqua, самец: половая клешня, модифицированные сегменты,
стиг. — стигма VIII тергита; Б — W. antiqua, самка: гениталии, модифицированные
сегменты, стиг. — стигма VIII тергита.
206
Рис. 259. Wagnerina tuvensis — самец.
Половая клешня и модифицированные
сегменты.
Рис. 260. Eopsylla nuda, самка. Се-
мяприемник и VII стернит.
Рис. 261. Eopsylla nuda, самец. Половая клешня, VIII и
IX стерниты.
207
Рис. 262. А — Ctenophthalmus uncinatus, самец: половая клешня и. IX стернит;
Б — С.* uncinatus, самка: гениталии и модифицированные сегменты.
Рис. 263. Ctenophthalmus congeneroides, самец. Половая клешня и IX стернит. Справа—
варианты дигитоида.
Рис. 264. Ctenophthalmus congeneroides, самка.
Гениталии, модифицированные сегменты, стиг. —
стигма VIII тергита.
208
Рис. 265. А — Ctenophthalmus truncus, самец: половая клешня и IX
стернит; Б — С. truncus, самка: варианты края VII стернита.
Рис. 266. А — Ctenophthalmus assimilis, самец: половая клешня и IX стернит;
Б — С. assimilis, самка: семяприемник и модифицированные сегменты.
Рис. 267. А — Ctenophthalmus arvalis, самец: половая клешня и IX стернит;
Б — С. arvalis, самка: гениталии и модифицированные сегменты.
14 Определитель блох
209
Рис. 268. А — Ctenophthalmus pisticus pisticus, самец: половая клешня
и IX стернит; Б — С. р. pisticus, самка: семяприемник, модифицированные
сегменты, стигма VIII тергита (с/ипг.).
Рис. 269. Ctenophthalmus pisticus pacificus, самец.
Половая клешня и IX стернит.
Рис. 270. А — Ctenophthalmus dux, самец: половая клешня и IX стернит;
Б — С. dux, самка: семяприемник и модифицированные сегметы.
210
Рис. 271. А — Ctenophthalmus dolichus, самец: половая клешня, VIII и IX
стерниты; Б — С. dolichus, самка: модифицированные сегменты.
Рис. 272. А — Ctenophthalmus dilatatus, самец: половая клешня и IX стернит;
Б — С. dilatatus, самка: семяприемник и модифицированные сегменты.
Рис. 273. А — Doratopsylla dasycnemus, самец: половая клешня, VIII и IX стер-
ниты; Б — Г), dasycnemus, самка: гениталии и VII стернит»
14*
211
л б
Рис. 274. А — Doratopsylla coreana, самец: половая клешня, VIII и IX стерниты,
внизу — вариант части клешни; Б — D. согеапа, самка: гениталии и VII стернит
с вариантом.
Рис. 276. А — Palaeopsylla sorecis starki, самец: половая клешня и IX стер-
пит; Б— Р. s. starki, самка: семяприемник и VII стернит.
212
Рис. 277. Nearctopsylla
ioffi, самец. Половая
клешня, VIII и IX стер-
ниты.
Рис. 280. А — Rhadinopsylla insolita, самец: половая клешня, VIII и IX стерниты;
Б — R. insolita, самка: семяприемник и VII стернит.
213
Рис. 281. А — Rhadinopsylla dives, самец: половая клешня, VIII и IX стерниты;
Б — R. dives, самка: семяприемник и VII стернит.
Рис. 282. А — Rhadinopsylla tenella, самец: половая клешня, VIII и IX стерниты;
Б — R. ter.ella, самка: семяприемник и VII стернит.
Рис. 283. А — Rhadinopsylla dahurica dahurica, самец: половая клешня, VIII и IX стер-
ниты; Б— R. d. dahurica, самка: семяприемник и VII стернит; В — R. dahurica vicina,
самец: вершина горизонтальной ветви IX стернита.
214
Рис. 284. А — Rhadinopsylla ioffi, самец: половая клешня, VIII и IX стерниты;
Б— R. ioffi, самка: семяприемник и VII стернит.
Рис. 285. Л — Rhadinopsylla aspalacis, самец: половая клешня, VIII и IX стерниты;
Б — R. aspalacis, самка: семяприемник и край VII стернита.
215
Рис. 286. А — Rhadinopsylla rothschildi, самец: половая клешня, VIII и IX стерниты;
Б — R. rotschildi, самка: семяприемник и VII стернит.
Рис. 287. Rhadinopsylla accola, самец: половая клешня, VIII и IX
стерниты.
Рис. 288. А — Rhadinopsylla altaica, самец: половая клешня, VIII и IX стерниты;
Б — R. altaica, самка: семяприемник и два варианта края VII стернита.
216
Рис. 289. А — Rhadinopsylla pseudodahurica, самец: половая клешня, VIII и IX стер-
ниты; Б — R. pseudodahurica, самка: семяприемник и VII стернит.
Рис. 290. Rhadinopsylla integella. А — R. i. caucasica, самец: половая клешня, VIII
и IX стерниты; Б — R. i. concava, самец: вершина вертикальной ветви /X стернита;
В — R. i. caucasica, самка: семяприемник и VII стернит; Г — R. i. concava, самка:
семяприемник и VII стернит.
Рис. 291. А — Rhadinopsylla integella casta, самец: половая
клешня, VIII и IX стерниты; Б — R. i. casta, самка: край VII
стернита.
217
Рис. 292. А — Rhadinopsylla valenti, самец: половая клешня, VIII и IX стерниты;
Б — R. valenti, самка: семяприемник и VII стернит.
Рис. 293. Rhadinopsylla. VII стернит самки: А — R. li li; Б — R. li trans-
baikalica; В— R. ventricosa ventricosa; Г—R. ventricosa murium; Д—R. semenovi.
Рис. 294. Rhadinopsylla li li, самец; половая клешня, VIII и IX стерниты.
Справа — вариант горизонтальной ветви IX стернита.
218
Рис. 295. А — Rhadinopsylla li transbaikalica, самец: половая клешня, VIII и IX стер-
ниты; Б — R. ventricosa, самец: горизонтальная ветвь IX стернита; В — R. semenovi,
самец: горизонтальная ветвь IX стернита.
Рис. 296. А — Rhadinopsylla cedestis, самец: Б — R. bivirgis, самец; половая
клешня и IX стернит.
Рис. 297. Rhadinopsylla socia, самец: половая
клешня и IX стернит.
Рис. 298. Rhadinopsylla
socia, самка: VII стер-
нит.
219
Рис. 299. А — Rhadinopsylla cedestis, самка;
В— R. bivirgis, самка: семяприемник и VII
стернит.
Рис. 300. Rhadinopsylla
jaonis, самка. Семя-
приемник и VII стер-
нит.
Рис. 301. Neopsylla bidentatiformis, самец: половая клешня, VIII и IX
стерниты, пигидий.
220
Рис. 302. Neopsylla mana, самец: половая клешня, пигидий, VIII и IX стерниты.
Рис. 303. Neopsylla bidentatiformis,
самка. Семяприемник, копулятивный
проток, VII стернит и стигма VIII
тергита.
Рис. 304. Neopsylla mana, самка. Семя-
приемник, копулятивный проток, VII
стернит и стигма VIII тергита.
221
Рис. 305. Neopsylla teratura, самец. Половая клешня, VIII ji
IX стерниты.
Рис. 30G. Neopsylla democratica, самец. Половая клешня, VIII и IX стерниты.
222
Рис. 308. Neopsylla democratica, самка. Се-
мяприемник, копулятивный проток и VII
стернит.
Рис. 307. Neopsylla teratura, самка.
Семяприемник, копулятивный про-
ток, VII стернит и стигма VIII тер-
гита.
Ри.\ 309. А — Neopsylla pleskei pleskei, самец: половая клешня, пигидий, VIII и IX стер-
ниты; Б — N. р. pleskei, самка. .Семяприемник, копулятивный проток, VII стернит и
стигма VIII тергита.
223
Рис. 310. А — Neopsylla pleskei orientalis, самец: половая клешня, VIII и IX стерниты,
эдеагус; Б — N. р. ariana, самец: часть половой клешни, эдеагус.
Рис. 311. А — Neopsylla acanthina, самец: половая клешня, VIII и IX стерниты;
Б — N. acanthina, самка: гениталии, VII стернит и стигма VIII тергита.
224
Рис. 312. А — Neopsylla pleskei orientalis, самка; Б — N. galea, самка: семяприемник,
копулятивный прото к. VII стернит стигма VIII тергита.
Рис. 313. A — Neopsylla abagaitui, самец: половая клешня, VIII и IX стерниты; Б—Neo-
psylla abagaitui, самка: гениталии, VII стернит, стигма VIII тергита.
Рис. 314. A — Neopsylla aliena, самец: половая клешня, VIII и IX стерниты; Б — Neo-
psylla aliena, самка: семяприемник и модифицированные сегменты; В — Neopsylla com-
par, самка: семяприемник и VII стернит.
15 Определитель блох
225
Рис. 315. Neopsylla galea, самец половая клешня, VIII и IX стерниты.
Рис. 316. А — Neopsylla anoma, самец: половая клешня, IX стернит; Б — N. anoma,
самка: семяприемник и модифицированные сегменты.
А 5
Рис. 317. А — Paraneopsylla ioffi, самец: половая клешня, VIII и IX стерниты; Б — Р.
ioffi, самка: семяприемниК и VII стернит.
226
Рис. 318. А — Paraneopsylla tiflovi, самец: половая клешня, VIII и IX стерниты; Б — Р.
tiflovi, самка: гениталии и VII стернит.
Рис. 319. А — Catallagia dacenkoi, самец: половая клешня, VIII и IX стерниты; Б — С.
dacenkoi, самка: семяприемник, VII стернит, стигма VIII тергита (а), средних
тергитов (о).
15*
227
Рис. 320. А —Catallagia ioffi, самец: половая клешня, VIII и IX стерниты; Б—С. ioffi,
самка: семяприемник, VII стернит, стигма VIII тергита (а), средних тергитов (5).
228
Рис. 323. A — Catallagia striata, самка; семяприемник, VII стернлт с вариан-
том, стигма VIII тергита; Б — Catallagia fetisovi, самка: семяприемник,
VII стернит, стигма VIII тергита.
Рис. 32k А — Hystrichopsylla talpae, самец: пэлозая клешня, VIII и IX стерниты;
Б — Н. nikolai, самка: семяприемники и VII стернит.
229
Рис. 325. А — Hystrichopsylla microti, самец: половая клешня, VIII и IX стерниты;
Б — Н. microti, самка: семяприемники и VII стернит.
Рис. 326. Stenoponia singularis, самец: половая клешня, IX стернит,
вершина эдеагуса (зд.).
230
Рис. 327. Stenoponia ivanovi, самец: половая клешня, IX с терпит,
вершина эдеагуса (эд.).
Рис. 328. Stenoponia formozovi, самец: половая клешня и
IX стернит.
Рис. 329. Stenoponia singularis, самка: Рис. 330. Stenoponia formozovi, самка: се-
семяприемник, VII стернит и VIII мяприемник, VII стернит и VIII тергит.
тергит.
231
Рис. 331. Stenoponia sidimi, самец: половая клешня,
IX стернит и вершина эдеагуса.
Рис. 332. Stenoponia сое-
lestis, самка: семяприем-
ник и VII стернит.
Рис. 333. А — Stenoponia ivanovi, самка: семяприемник и VII стернит; Б — Stenoponia
sidimi, самка: семяприемник и VII стернит.
232
Рис. 334. Stenoponia montana, самец: половая клешня, IX стернит
и вершина эдеагуса (зд.).
Рис. 335. Stenoponia montana, самка: семяприемник и VII стернит
с вариантами.
Рис. 336. А — Ischnopsyllus hexactcnus kolenatii самец: половая клешня и VIII стернит;
Б — I. hexactenus hexactenus, самец: дигитоид и вершина VIII стернита.
233
Рис. 337. Ischnopsyllus obscurus, самец: половая клешня и вершина VIII стернита.
Рис. 338. Ischnopsyllus obscurus, самка: гениталии, модифициро-
ванные сегменты, стигма VIII тергита.
234
Риг. 339. А — Ischnopsyllus elongatus, самец: часть полевой клешни; Б — Г—церки
самок: Б — Ischnopsyllus elongatus; В — intermedins; Г — obscurus.
Рис. 341. Ischnopsyllus comans, самка: семя-
приемник и модифицированные сегменты.
schnorsyllus comans,
самец: часть половой клешни,
VIII и IX стерниты.
235
Рис. 342. Ischnopsyllus
needhamia, самец: поло-
вая клешня, пигидий,
VIII стернит.
Рис. 343. А — Ischnopsyllus needhamia, самка: гениталии и модифицированные
сегменты; Б — Myodopsylla trisellis, самка: семяприемник и VII стернит.
Рис. 344. Myodopsylla trisellis, самец: половая клешня и VIII стернит.
236
Антенна
Конец хоботка
Рис. 345. А — Paraceras crispus, самец; Б — Ctenophthalmus
congeneroides, самец.
Внутренняя поверхность
задней ляжки
Рис. 346. А — Xenopsylla cheopis, самка; Б — Саtallagia ioffi, самка;
В—Ceratophyllus tesquorum sungaris, самка; Г — Ceratophyllus pa-
radoxus, самка.
237
Задняя голень
Рис. 347.
dorcadia,
A -Vermipsylla alacurt, самка; Б- Vermipsylla
о — Leptopsylla ostsibirica, самка.
самка;
Задняя голень
Задняя лапка
р“'
238
Задняя голень
Задняя лапка
Рис. 349. А—Neopsylla acanthina, самка; Б — Neopsylla bidentatiformis, самка;
В—Rhadinopsylla bivirgis, самец; Г—Rhadinopsylla socia, самец; * — субапикаль-
ные щетинки II членика.
//, /// членики задней лапки самца
Рис. 350. А — Paraceras melis; Б — Рага-
ceras flabcllum.
V членик задней
лапки самки
Рис. 351. A —Echidnophaga galli-
пасеа; Б — Echidnophaga oschani n.
Рис. 352. A — Ceratophyllus garei, самка; Б — Ceratophyllus farreni, самка.
239
Четвертый тергит брюшка самки
Рис. 353. А — Vermipsylla alacurt; Б — V. dorcadia; В — Chaetopsylla lasius; Г — Ch. tu-
berculaticeps; Д — Ch. appropinquans; E— Ch. bomoeus; Ж— Ch. alia; 3 — Ch. dogieli;
И — Ch. zibellina; К — Ch. trichosa: JJ — Ch. inicado; M — Ch. mirabilis.
240
СПИСОК РИСУНКОВ С УКАЗАНИЕМ,
ИЗ КАКОЙ МЕСТНОСТИ ВЗЯТЫ ЭКЗЕМПЛЯРЫ БЛОХ
ДЛЯ РИСОВАНИЯ ИЛИ ОТКУДА ЗАИМСТВОВАНЫ РИСУНКИ
Рис. 1—20. Рисунки в тексте.
Рис. 21—76. Голова и грудь.
Рис. 77—344. Модифицированные сегменты и гениталии самца и самки.
Рис. 345—353. Антенна, хоботок, ноги, тергиты груди и брюшка.
На всех рисунках блоха рассматривается в одинаковом положении, лежащей на
правом боку, т. е. передний край каждого органа слева, а задний справа.
Ниже приводятся указания, из какой местности взяты экземпляры блох для ри-
сования, а для рисунков, взятых из литерат
1—7. Рисунки к очерку методики сбора бл
8. Ceratophyllus tesquorum.................
9 . Oropsylla silantiewi...............
10 — 13. По ГО. Н. Вагнеру.
14 — 15. По И. Г. Иоффу и В. Е. Тифлову.
16. Ceratophyllus turbidus................
17. По И. Г. Иоффу.
18. По Снодграссу.
19. По И. Г. Иоффу и В. Е. Тифлову.
20. Ctenophyllus rigidus..................
21. Pulex irritans........................
22. Archaeopsylla sinensis................
23. Echidnophaga gallinacea...............
24. „ oschanini................
25. Ctenocephalides canis.................
26. „ felis7..............................
27. Xenopsylla cheopis....................
28. Synosternus longispinus...............
29, A.—Xenopsylla astia; Б — Xenopsylla
nubicus...............................
30. Pariodontis riggenbachi...............
31. Spilopsyllus cuniculi.................
32. Coptopsylla lamellifer . . . • ....
33; Chaetopsylla tuberculaticeps...........
34. Chaetopsylla homoeus..................
35. Vermipsylla alacurt...................
36. „ dorcadia............................
37, A—Vermipsylla alacurt; Б — V. dorcadia
38. Paraceras flabellum.................... 39 40 41 42 43 44 45 46 47 48
39. Aenigmopsylla grodekovi................
40. Oropsylla silantiewi...................
41. „ asiatica........................
42. Ceratophyllus fasciatus................
43. „ tesquorum..................,.
44. „ penicilliger..................
45. Frontopsylla luculenta..................
46. Ctenophyllus armatus...................
47. „ hirticrus......................
48. Ophthalmopsylla praefecta...............
ры, указаны авторы этих рисунков:
к.
по И. Г. Иоффу п В. Е. Тифлову.
Юго-восточное Забайкалье.
Западное Забайкалье, Кабанский р-н.
Корея.
Саратовская область.
Юго-восточное Забайкалье, Борзинский р-н.
Сальский округ, с. Ремонтное.
Туркменская ССР, Кара-Кала,
юго-восток Европейской части СССР.
по И. Г. Иоффу и В. Е. Тифлову.
по Иордану.
Туркменская ССР, Копет-Даг.
Германия.
по И. Г. Иоффу и В. Е. Тифлову.
Побережье Охотского моря, бухта Одян.
юго-восточная часть Забайкалья, ст. Ма-
циевская
Киргизская ССР, Атбашинский р-н.
Киргизская ССР, Иссык-Кульская область,
по И. Г. Иоффу.
юго-восточная часть Забайкалья, ст. Ма-
циевская.
Приморский край, Гродеково.
юго-восточная часть Забайкалья,
окрестности г. Якутска.
Хабаровский край.
юго-восточная часть Забайкалья, с. Кай-
ластуй.
Восточное Забайкалье, с. Александров-
ский Завод.
юго-восточная часть Забайкалья, Борзин-
ский р-н.
юго-восточная часть Забайкалья, с. Кай-
ластуй.
юго-восточная часть Забайкалья, ст. Ага.
юго-восточная часть Забайкалья, с. Кай-
ластуй.
16 Определитель блох
241
49. Ophthalmopsylla kiritschenkol.........
50. Brachyctenonotus myospalacis..........
51. Leptopsylla segnis....................
52. B ostsibirica.........................
53. Mesopsylla hebes......................
54, A—Amphipsylla rossica; Б — A. sibirica;
B —A. primaris; Г — A. vinogradovi;
Д — A. daea; E—A. sibirica; Ж— A.
primaris................................
55. Pectinoctenus pectiniceps............
56. Wagneria antiqua......................
57. Eopsylla nuda.........................
58. Doratopsylla dasycnemus...............
59. . coreana .................
60. B birulai.............................
61. Ctenophthalmus congeneroides . . . .
62. Palaeopsylla sorecis..................
63. Nearctopsylla ioffi...................
64. „ beklemischevi.......................
65. Rhadinopsylla dahurica................
66. B li.......................
67. , valenti..................
68. „ cedestis.................
69. Neopsylla bidentatiformis.............
70. Catallagi ioffi.......................
71. Hystrichopsylla talpae................
72. 9 microti.............................
73. Stenoponia singularis.................
74. w formozovi...........................
75. Ischnopsyllus obscurus................
76. Myodopsylla trisellis.................
77, A—Archaeopsylla sinensis..............
Б — Xenopsylla cheopis..................
78. Xenopsylla astia......................
79, A—Xenopsylla as'ia; Б—X. nubicus . .
80. B conformis...........................
81. B skrjabini...................
82. B gerbilli gerbilli...................
83. B B minax ............................
84. B B caspica...........................
85. „ persica.............................
Юго-западный Алтай, с. Сарыбель Самар-
ского р-на.
Алтай, Катон-Карагайский р-н.
г. Якутск.
Восточное Забайкалье, с. Александровский
Завод (тип) 1.
по И. Г. Иоффу и В. Е. Тифлову.
по И. Г. Иоффу и В. Е. Тифлову.
юго-восточная часть Забайкалья, Борзин-
ский р-н.
Западная Монголия, Баян-Хонгорский
аймак, Цзаг-Сомон.
Сихотэ-Алиньский заповедник.
Московская область, Михневский р-н.
Северная Корея.
Зауралье, Кондо-Сосьвинский заповедник.
Восточное Забайкалье, с. Александровский
Завод.
по И. Г. Иоффу и В. Е. Тифлову.
Приморский край, Гродеково (тип).
Китай (тип).
но И. Г. Иоффу и В. Е. Тифлову.
по Н. Ф. Дарской (тип).
по И. Г. Иоффу и В. Е. Тифлову.
юго-восточная часть Забайкалья.
Якутия, против устья р. Вилюя.
Зауралье, Кондо-Сосьвинский заповедник.
Восточное Забайкалье, с. Александровский
Завод.
по И. Г. Иоффу и В. Е. Тифлову.
Зауралье, Кондо-Сосьвинский заповедник,
по Иордану и Ротшильду;
Китай.
Индия, Мадрас.
по Иордану.
по И. Г. Иоффу.
по И. Г. Иоффу (тип).
Туркменская ССР, Имам-Баба.
Казахская ССР, долина р. Или.
Казахская ССР, Усть-Урт (тип).
Туркменская ССР, Фирюза (тип).
86, А—Xenopsylla cheopis — по Иордану; Б — X. astia, Иран, Мешхед; В —X. nu-
bicus — по Иордану; Г — X. hawaiiensis — по Иордану; Д — X. brasiliensis — по
Иордану; Е — X. conformis — по И. Г. Иоффу; Ж — X. skrjabini — по И. Г. Иоффу.
87, А и Б—Hoplopsyllus glacialis profugus
88, А и Б—Chaetopsylla homoeus.........
89. Chaetopsylla mirabilis.............
90. „ „ ...............
91, А и Б—Chaetopsylla alia............
92, А и Б— dogieli
93. - appropinquans . .
94, А и Б— 9 zibellina . . . .
95. trichosa
96, А — 9 appropinquans . .
Б — 9 trichosa
97, А и Б— 9 mikado . . . .
98, А и Б— 9 tuberculaticeps . .
Казахская ССР, близ г. Джаркента.
Киргизская ССР, Иссык-Кульская область,
по И. Г. Иоффу и А. И. Аргиропуло (тип).
Тебердинский заповедник.
Зауралье, Кондо-Сосьвинзкий заповедник
(тип).
Приморский край, Судзухинский заповед-
ник (тип).
Приморский край.
Зауралье, Кондо-Сосьвинский заповедник
(тип).
Алтай, Катон-Карагайский р-н.
окрестности г. Благовещенска;
Предкавказье, район г. Кизляра.
Приморский край, Судзухинский заповед-
ник.
Охотское побережье.
1 Типо—(мтип) обозначают экземпляр, послуживший для описания данного вида-
242
99, Л и Б—Tarsopsylla octodecimdentata .
100, А и Б—Aenigmopsylla grodekovi . .
101. Paraceras flabellum..................
102. я metis........................
103. Paraceras flabellum..................
104. „ melis........................
105. Amphalius runatus....................
103. я , .....................
107. „ clarus......................
108. Oropsylla silantiewi..............
109. „ asiatica.....................
110. „ ilovaiskii....................
Ill, A—Oropsylla silantiewi............
Б— „ asiatica..........................
112, А и £>—Oropsylla mandarinus . . .
113. Ceratophyllus fasciatus...........
114. w „ . •.............
115, А и Ceratophyllus mokrzeckyi . .
116, А и Б— fidus...........
117, А и Б— , laeviceps laevi-
ceps
118, А и £—Ceratophyllus laeviceps kuzen-
kovi .......................................
119, A—Ceratophyllus laeviceps ellobii— no
Монголия, Гобийский Алтай, оз. Coro-;
120, А и Б—Ceratophylla turkmenicus . . .
121. „ tesquorum sunga-
ris......................................
122. Ceratophyllus tesquorum mongolicus . .
123. „ „ sungaris . . .
124. „ „ transvolgensis. .
125. „ „ altaicus . . . .
126, A— „ jenissejensis................... Красноярский край, с. Монок; Б —
С. jenissejensis—Красноярский край, д. Светлолобово.
127. Ceratophyllus caspius................ по В. Е. Тифлову (тип).
128. „ gaiskii............................ Западная Монголия, Баян-Хонгорский
аймак.
129, A—Ceratophyllus caspius..............
Б — С. gaiskii........................
130. Ceratophyllus dolabris...............
131. „ kozlovi............................
132. , kaznakovi .........................
133. „ „ .......
134. „ oreinus............................
135. „ , ................
136, А и Б. Ceratophyllus geminus . . . .
137. Ceratophyllus semenovi...............
138. , w ...................
139. , penicilliger.......................
140.
Иркутская область, Тулунский р-н.
Приморский край, Гродеково.
Алма-Атинская область.
по Дампфу.
Алма-Атинская область.
Крым.
Восточное Забайкалье, с. Александровский
Завод.
Юго-восточное Забайкалье, Агинский р-н
Центральный Тянь-Шань (рис. В. Е. Тиф-
лова).
Юго-восточное Забайкалье, Агинский р-н.
Камчатка, Кроноцкий заповедник.
окрестности г. Якутска.
Камчатка (тип).
по И. Г. Иоффу (тип).
юго-восточная часть Забайкалья, Агин-
ский р-н;
окрестности г. Якутска.
по Иордану и Ротшильду (тип).
г. Хабаровск.
по И. Г. Иоффу и В. Е. Тифлову.
по Иордану и Ротшильду (тип).
по И. Г. Иоффу.
Китай, Жэхэ (тип).
И. Г. Иоффу (тип); Б — С. 1. ellobii —
[op (тип).
по И. Г. Иоффу (тип).
Юго-восточное Забайкалье, с. Кайластуй.
по Иордану и Ротшильду.
юго-восточная часть Забайкалья, с. Кай-
ластуй и Западная Монголия, Баян-
Хонгорский аймак.
Чкаловская область, Соль-Илецкий р-н.
Северо-западная Монголия, Убсанурский
аймак.
юг Киргизской ССР, Алайская долина;
Западная Монголия, хребет Тайшири.
по Иордану и Ротшильду.
по Ю. Н. Вагнеру (тип).
Тибет.
но Иордану (тип).
по И. Г. Иоффу.
, (тип).
Кемеровская область, Юргинский р-н.
Юго-восточный Алтай, Катон-Карагай-
ский р-н (тип).
Восточное Забайкалье, с. Александров-
ский Завод.
Восточное Забайкалье, с. Александров-
ский Завод.
141, А — Ceratophyllus penicilliger dissimilis — Камчатка; Б — С. penicilliger syrt — по
И. Г. Иоффу (тип); В — С. penicilliger arvicolae — Ставропольский край, Усть-
Джегутинский р-н (тип); Г — С. penicilliger penicilliger — по И. Г. Иоффу (тип);
Д — С. penicilliger vallis — по И. Г. Иоффу (тип); Е — С. penicilliger demotus — по
И. Г. Иоффу (тип); Ж -— С. ioffi — Корея (гип).
142, A—Ceratophyllus penicilliger penicilliger—по И. Г. Иоффу; Б—С. penicilliger dissimilis-
Камчатка; В —С. penicilliger syrt — по И. Г. Иоффу (тип); Г — С. penicilliger
arvicolae — Ставропольский край, Усть-Джегутинский р-н (тип); Д — С. penicilliger
demotus — по И. Г. Иоффу (тип); Е — С. ioffi — Корея (тип).
143, А и £>—Ceratophyllus sciurorum asiaticus .... Казахская ССР, Кетменские горы.
17 Определитель блох
243
144, А и Б—Ceratophyllus anisus.......... юго-восточная часть Забайкалья, Борзин-
ский р-н.
145. „ tamias. Восточное Забайкалье, с. Александровский
Завод.
146. „ paradoxus ... по О. И. Скалой (тип).
147, А— „ tamias. Восточное Забайкалье, с. Александровский
Завод; Б — С. paradoxus по О. И. Скалой (тип).
148, А и Б—Ceratophyllus toli............ Якутская АССР, против устья р. Вилюя.
149. Ceratophyllus scaloni............... Северо-восточная Монголия, Хэнтэйский
аймак.
150. - desertus
151. scaloni .
152. • desertus
153. calcarifer
154, А—
Казахская ССР, хребет Кызыл-Тас, близ
г. Караганды (топотип).
Северо-восточная Монголия, Хэнтэйский
аймак.
Казахская ССР, хребет Кызыл-Тас, близ
г. Караганды (топотип).
Зауралье, Кондо-Сосьвинский заповедник.
Б— п walkeri....................... Новосибирская область
155, A—Ceratophyllus walkeri—Новосибирская область; Б — С. sokolovi — Курильские
161.
162.
163.
164.
165.
острова.
156, А — Ceratophyllus sokolovi — Курильские острова; Б — С. advenarius — Восточное
Забайкалье, с. Александровский Завод.
157, А и Б.—Ceratophyllus rectangulatus. . Восточное Забайкалье, с. Александровский
Завод.
158. Ceratophyllus advenarius. Восточное Забайкалье, с. Александровский
Завод; А—С. advenarius advenarius—Якутск (тип); Б—С. a. bifallax—Хабаровск (тип).
159. Ceratophyllus turbidus.... Западное Забайкалье, Кабанский р-н.
taiganus................. по О. И. Скалой (тип),
lunatus................. рисунок Ю. Н. Вагнера, Камчатка.
garei ................... Северо-восточная Монголия, Хэнтэйский
аймак; А — С. garei — Юго-восточное Забайкалье, ст. Даурия.
166. Ceratophyllus garei................... Восточное Забайкалье, с. Александров
ский Завод.
167. „ spinosus . . .
168. „ ..............
169, А— я garei . . . .
Б—С. gallinae—Ставропольский край.
170, А— я я tribulis —г. Якутск;
Гродеково (тип).
171. Ceratophyllus gallinae tribulis. . . .
172. „ „ dilatus . . . .
173. я „ gallinae . . .
174. „ , . . .
175. „ fringillae......................
176. „ vagabundus .....................
177. „ „ ..............
178. „ styx...........................
179. „ riparius
180,
А— „ styx..........................
Наурзумский заповедник; Б — С.
ст. Даурия
Ceratophyllus avicitelli...........
Дагестанская АССР, Махачкала.
юго-восточная часть Забайкалья, с. Бырка;
‘—С. gallinae dilatus — Приморский край,
г. Якутск.
Приморский край, Гродеково (тип).
Зауралье, Кондо-Сосьвинский запо-
ведник.
Горьковская область, с. Киселево.
по Ю. Н. Вагнеру.
по Дампфу.
Казахская ССР, Кустанайская область,
Наурзумский заповедник.
юго-восточная часть Забайкалья, ст. Дау-
181.
182.
183.
184.
185,
186.
187.
188,
рия.
Казахская ССР, Кустанайская область,
riparius — юго-восточная часть Забайкалья,
по И. Г. Иоффу (тип).
я maculatus................ по Ю. Н. Вагнеру.
„ farreni.................. окрестности г. Москвы.
А— „ maculatus - по Ю. Н. Вагнеру; Б — С. farreni — по Ротшильду.
, caliotes................. по Иордану (тип).
„ orites...................
А— я caliotes—по Иордану (тип); Б — С. orites —то же.
189. Dasypsyllus gallinulae................... окрестности г. Гамбурга.
190. я я ..................
191, Ап Б—„ я stejnegeri . по Йордану (тип).
192. Frontopsylla wagneri....................... по И. Г. Иоффу.
244
193, А и Б— Frontopsylla wagneri . . . .
194. elatoides
195. hetera
196. •> elatoides
197. hetera
198. •> luculenta luculenta . .
199. w n • •
.00. n „ parilis . . .
201. w W J? * *
202. » elata elata
203, A— elata taishiri
Б — F. elata taishiri Баян-Хонгорский
204. elata botis
205. yy » •>
206. 4t frontalis baikal ....
207. ъ n ъ • • • •
208. Ctenophyllus rigidus...................
209. „ armatus........................
Юго-восточное Забайкалье, Борзин-
ский р-н.
Южный Алтай, Катон-Карагай (тип).
Предбайкалье, с. Монды.
Южный Алтай, Катон-Карагай (тип).
Предбайкалье, с. Монды.
юго-восточная часть Забайкалья, Борзин-
ский р-н.
юго-восточная часть Забайкалья, Борзин-
ский р-н.
Северо-восточная Монголия, Ундурхан.
Алтай, Телецкое озеро; а — F. elata
popovi — Омская область.
Западная Монголия, хребет Тайшири;
аймак.
окрестности г. Хабаровска,
п п п
по И. Г. Иоффу (тип).
юго-восточная часть Забайкалья, ст.
Бырка.
по Н. Ф. Дарской (тип).
Восточное Забайкалье, с. Александров-
ский Завод.
210.
211, А- „
Б — С.
по И.
tarasovi................ по О. И. Скалой (тип).
subarm itus................. Южный Алтай, Катон-Карагай;
armatus — Юго-восточное Забайкалье, Борзинский р-н; В — С. rigidus —
Ф. Дарской (тип).
212, A—Ctenophyllus hirticrus.............
213. Paradoxopsyllus scorodumovi..........
214. „ integer............
215, A— „ scorodumovi ....
Западное Забайкалье, Кабанский р-н.
216. Paradoxopsyllus dashidorzhii ........
217, A- „ „ .........
217, Б— „ conveniens.......................
218, А и Б— , curvispinus.........
219, А и Ophthalmopsylla praefecta prae-
fecta .....................................
220, А и Ophthalmopsylla praefecta
pernix.....................................
юго-восточная часть Забайкалья, Борзин-
ский р-н; Б — ст. Ага.
по О. И. Скалон (тип).
Западное Забайкалье, Кабанский р-н.
по О. И. Скалон (тип.); Б — Р. integer —
по О. И. Скалон (тип).
Юго-западная Монголия, Гобийский Ал-
тай, оз. Сого-Нор (тип).
Китай, Шэньси.
Юго-восточная часть Забайкалья, Борзин-
ский р-н.
по Иордану (тип).
221, А — Ophthalmopsylla kukuschkini — Юго-восточное Забайкалье, Оловяннилский р-н;
Б — О. kukuschkini—Борзинский р-н.
222.
223,
224,
225.
226.
Ophthalmopsylla kukuschkini
А и Б —
А и Б —
volgensis extrema,
kiritschenkoi . . .
jettmari...........
227. Brachyctenonotus myospalacis
228.
Юго-восточное Забайкалье.
.Монгольский Алтай (тип).
Алтай, Сарыбель Самарского р-на.
по Иордану (тип).
Алтай, Катон-Кара гайский р-н.
229.
230.
231.
232.
233.
234.
235.
236.
237.
Mesopsylla hebes
п 1»
по И. Г. Иоффу.
lenis ..............
tuschkan............
rothschildi . . . .
lenis...............
tuschkan............
rothschildi . . . .
eucta...............
рисунок А. И. Аргиропуло (ти
по Й. Г. Иоффу.
Туркменская ССР, Репе тек.
рисунок Иордана (паратип).
238, A—Amphipsylla rossica — по И. Г. Иоффу и В. Е. Тифлову; Б—A. sibirica— Ямалс-
Ненецкий нац. округ, р. Мотылька; В — A. prima — по И. 1'. Иоффу и В. Е. Тифлову;
Г — A. kuznetzovi — тоже; Д — A. marikovskii — то же (тип); Е — A. petri*
stshevie — Корея; Ж— A. sshelkov.iikovi— пэ И. Г. Иоффу и В. Е. Тифлову;
245
3 — A. vinogradovi — то же (тип); И — A. tuta — то же (тип); К — A. anceps —
то же; Л — A. dumalis—то же; М — A. primaris primaris — Центральный Тянь-
Шань; Н — A. primaris mitis — по И. Г. Иоффу и В. Е. Тифлову; О — A. daea—
то же; П — A. aspalacis — то же. .
239, А—Amphipsylla rossica — по И. Г. Иоффу и В. Е. Тифлову; Б — A. primaris mitis —
то же; В—A. dumalis — то же; Г — A. schelkovnikovi — то же; Д — A. vinogra-
dovi— то же; Е — A. longispina — Северо-восточная Монголия, Хэнтэйский аймак.
240, A—Amphipsylla rossica — по И. Г. Иоффу и В. Е. Тифлову; Б— A. sibirica — то же;
В — A—prima—то же; Г — A. kuznetzovi — то же; Д — A. marikovskii—Восточное
Забайкалье, с. Александровский Завод; Е— A. petristshevae—Корея; Ж — A. tuta —
по И. Г. Иоффу и В. Е. Тифлову (тип); 3 — A. vinogradovi — то же; И — A. lon-
gispina— Северо-восточная Монголия, Хэнтэйский аймак; К—A. dumalis—по
И. Г. Иоффу и В. Е. Тифлову; Л — A. primaris mitis — то же; М — A. daea —
то же; Н — A. aspalacis — то же.
241, A—Amphipsylla sibirica sibirica — по И. Г. Иоффу и В. Е. Тифлову; Б — A. sibirica
orientalis — Западное Забайкалье, Кабанский р-н (тип); В — A. rossica — по
И. Г. Иоффу и В. Е. Тифлову; Г — A. kuznetzovi — то же; Д — A. marikovskii —
Восточное Забайкалье, с. Александровский Завод; Е — A. prima—по И. Г. Иоффу
и В. Е. Тифлову; Ж — A. tuta — то же; 3 — A. schelkovnikovi — то же; И — A- vi-
nogradovi — то же; К — A. longispina — Северо-восточная Монголия, Хэнтэйский
аймак; Л — A. dumalis — то же; М — A. anceps — то же; Н — A. primaris mitis —
то же; О — A. daea — то же; П — A. aspalacis—по И. Г. Иоффу и В. Е. Тифлову.
242, A—Amphipsylla rossica — по И. Г. Иоффу и В. Е. Тифлову; Б — A. anceps — то же;
В — A. kuznetzovi — то же; Г — A. primaris mitis — по И. Г. Иоффу и В. Е. Тиф-
лову.
243, A—Amphipsylla casis — по Иордану и Ротшильду (тип); £> —Amphipsylla longi-
spina — Северо-восточная Монголия, Хэнтэйский аймак (тип);
244. Amphipsylla longispina — северо-восточная Монголия, Хэнтэйский аймак (тип).
245, А и Б—Leptopsylla папа............... по И. Г. Иоффу.
246. „ „ ........ Западная Монголия, Баян-Хопгорский
аймак.
247. „ bidentata.............. Зауралье, Кондо-Сосьвинский заповедник.
248. „ п ............ „ w п
249. „ ostsibirica .... по О. И. Скалой (тип).
250. n п .... я „ »
251, А и Б— .. silvatica. Зауралье, Кондо-Сосьвинский заповедник.
ский р-н.
253. ч „ юго-восточная часть Забайкалья, Борзин-
ский р-н.
254. w pectiniceps.......... Юго-восточное Забайкалье, с. Кайластуй
Борзинского р-на.
255. я „ Юго-восточное Забайкалье, с. Кайластуй,
Борзинского р-на.
256, А и Б—Wagnerina tecta........ Памир, Вахан (тип).
257, А и Б— „ longicauda .... по О. И. Скалой (тип).
258, А и Б— „ antiqua...........
259. „ tuvensis.......... Тувинская автономная область.
'260. Eopsylla nuda..................... Сихотэ-Алинь.
261. п - ..................... »» ”
262, А и Б —Ctenophthalmus uncinatus . . Зауралье, Кондо-Сосьвинский заповедник.
263. „ congeneroides Восточное Забайкалье, с. Александров-
ский Завод.
264. п „ Амурская область, с. Поярково.
265, A—Ctenophthalmus truncus.............
остров Сахалин, г. Александровск.
266, А и Б—Ctenophthalmus assimilis . . . .
267, А и Б— , arvalis.............
268, А и Б— „ pisticus pisticus . .
269. „ pisticus pacificus . .
270, А и Б— „ dux.................
271, А и Б— „ dolichus............
272, А и Б— „ dilatatus...........
273, А и Б—Doratopsylla dasycnemus . . .
274, А и Б— „ coreana...........
275, А и Б— „ birulai...........
276, А и Б—Palaeopsylla sorecis starki . .
277. Nearctopsylla ioffi..................
278. , » ....................
279. „ beklemischevi......................
Южно-Сахалинск (тип); Б — Ct. truncus—
Иркутская область, Усть-Удинский р-н.
рисунок Ю. Н. Вагнера (тип).
Новосибирская область, с. Колпашево.
Приморский край, Гродеково (тип),
по Иордану и Ротшильду (тип),
по Ротшильду (тип).
Алтай, р. Катунь, Аскат.
Ленинградская область,
по Н. Ф. Дарской (тип).
Зауралье, Кондо-Сосьвинский заповедник.
Приморский край, Гродеково.
Северная Корея.
Китай, Сиан (тип).
246
280, А и Б—Rhadinopsylla insolita.............
dives.............
tenella...........
dahurica dahurica
dahurica vicina . .
ioffi..............
aspalacis . . . • .
rothschildi ....
accola.............
altaica...........
pseudodahurica . .
integella caucasica
Тифлову.
В. Е.
И. Г. Иоффу и
Иордану (тип).
по
по
281, An
282, А и
283, А и
283, В—
284, A
285, A
286, A
287.
288, A
289, A
290, A—
integella
gella concava — Приморский край, застава Мраморная.
291, А и Б —Rhadinopsylla integella casta . . Зауралье, Кондо-Сосьвинский заповедник.
292, Ап Б — „ valenti........... по Н. Ф. Дарской (тип).
293, А— „ li li — по И. Г. Иоффу и В. Е. Тифлову; Б — R. li transbaika-
lica — то же (тип); В — R. ventricosa ventricosa—то же (тип); Г — R. ventricosa mu-
rium — то же (тип); Д — R. semenovi —
Rhadinopsylla li li.....................
A—Rhadinopsylla li transbaikalica . . .
Б— . ventricosa............
В— - semenovi...............
A— „ cedestis..............
Б— , bivirgis..............
, socia......................
294.
295,
295,
295,
296,
296,
297.
298.
299,
300.
301.
302.
303.
и
и
и
и
и
Б—
Б—
Б—
Б—
Б—
Б—
Б—
Б—
по
И. Г.
Иоффу
и
В. Е.
Тифлову.
(тип).
(тип).
по И. Г.
concava — то же (тип); В — R. integella
по
Тифлову; Б— R.
Е.
В.
Иоффу
caucasica — то же; Г — R. inte-
и
И. Г. Иоффу и
В. Е.
южное
Тифлову.
(тип).
Прибалхашье, не-
Казахская ССР,
ски Тау-Кум.
.......................рисунок Ю. Н. Вагнера (тип).
А— „ cedestis и Б- R. bivirgis—по И. Г. Иоффу и В. Е. Тифлову.
Rhadinopsylla jaonis
Neopsylla bidentatiformis
mana ....
bidenta tiformis
304.
305.
306.
307.
308.
309, А
и
mana.........................
„ teratura ......................
„ democratica....................
„ teratura ......................
„ democratica....................
Б—Neopsylla pleskei pleskei . .
_ „ orientals . . .
по Иордану (тип).
юго-восточная часть Забайкалья, Борзин-
ский р-н.
Алтай, Катон-Карагай (тип).
юго-восточная часть Забайкалья, Борзин-
ский р-н.
г. Якутск.
Монгольский Алтай.
рисунок Ю. Н. Вагнера (тип).
Монгольский Алтай.
рисунок Ю. Н. Вагнера (тип).
Красноярский край, близ г. Минусинска
(тип).
Юго-восточное Забайкалье, ст. Бырка (тип);
310, A— . . . .
Б — N. p. ariana — Памир, Вахан (тип).
Z>—Neopsylla acanthina......... Сахалин.
„ " pleskei orientalis . . юго-восточная часть Забайкалья, ст. Быр-
ка (тип); £—N. galea—Западная Монго-
лия, Баян-Хонгорский аймак, Цзаг-Со-
мон (тип).
” i............ окрестности г. Красноярска (тип).
aliena; В—N. compar—по Иордану и Ротшильду (типы),
galea
311, А и
312, А —
313, А и
314, А и
315.
Б—Neopsylla abagaitui
Б- „ Г'
316, А—
anoma
317, A—Paraneopsylla ioffi
318, А и Б—Paraneopsylla tiflovi
319, А и Б—Catallagia dacenkoi
320, А и Б— „ ioffi . .
321. , striata .
322. , fetisovi .
Читинская область, Агинский нац. округ
(тип).
по Ротшильду (с поправкой Иордана);
Б — N. anoma — по Ротшильду.
Заилийский Алатау, близ г. Алма-Аты
(тип); Б — Р. ioffi — по В. Е. Тифлову
(тип).
по О. А. Фединой (тип).
Якутская АССР, против устья р. Вилюя,
по О. И. Скалон (тип).
Восточное Забайкалье, с. Александровский
Завод.
по О. И. Скалон (тип); Б — С. fetisovi —
Монголия, Центральный аймак.
324, А — Hystrichopsylla talpae — окрестности г. Пржевальска; Б — Н. nicolai —
по О. И. Скалон (тип).
325, А и Б—Hystrichopsylla microti . .
323, А-
striata
по О. И. Скалон (тип).
247
326. Stenoponia singularis
327.
328.
по И. Г. Иоффу и В. E. Тифлову (тип).
юго-восточная часть Забайкалья, Борзин-
ский р-н.
по И. Г. Иоффу и В. Е. Тифлову (тип),
юго-восточная часть Забайкалья, Борзин-
ский р-н.
Амурская область, Благовещенский р-н.
рисунок К. Иордана (тип).
оим, /-1— „ .......................... Ставропольский край, Левокумский р-н;
Б—S. sidimi—Амурская область, Благовещенский р-н.
331. Stenoponia montana................... по Н. Ф. Дарской (тип).
335. „ „ .................. „ „ „
ivanovi .
formozovi
329.
330.
331.
332.
333, A—
singularis
formozovi
sidimi .
coelestis
ivanovi
336, A—Ischnopsyllus hexactenus kolenati— рисунок Ю. Н. Вагнера (тип); Б—I. hexactenus
hexactenus — рисунок Ю. Н. Вагнера.
337. Ischnopsyllus obscurus............. Зауралье, Кондо-Сосьвинский заповедник.
338. „ „ .............. юго-восточная часть Забайкалья, с. Кай-
339, А и Б—Ischnopsyllus elongatus; В -!. in-
termedius; Г — I. obscurus..................
340. Ischnopsvllus comans...................
341. „ „ .................
342. „ needhamia............................
343, A— „ „ ...............
ластуй.
по И. Г. Иоффу и В. E. Тифлову.
по Иордану и Ротшильду (тип).
по О. И. Скалой
юго-восточная часть Забайкалья, с. Кай-
ластуй; Б — Myodopsylla tr’.sellis--Зауралье, Кондо-Сосьвинский заповедник.
344. Myodopsylla trisellis.................. Зауралье, Кондо-Сосьвинский заповедник.
345, A— Paraceras crispus ................... по Иордану и Ротшильду; Б — Ctenoph-
thalmus congeneroides—Восточное Забай-
калье, с. Александровский Завод.
346, A—Xenopsylla cheopis—г. Владивосток; Б — Catallagia ioffi—Якутская АССР, про-
тив устья р. Вилюя; В — Ceratophyllus tesquorum sungaris—Юго-восточное Забай-
калье, Борзинский р-н; Г — Ceratophyllus paradoxus — Зауралье, Кондо-Сосьвинский
заповедник.
347, А—Vermipsylla alacurt — Киргизская ССР, Атбашинский р-н; Б — Vermipsylla dor-
cadia — Киргизская ССР, Иссык-Кульская область; В — Leptopsylla ostsibirica —
Восточное Забайкалье, с. Александровский Завод.
348, A— Ctenophyllus hirticrus — Читинская область, Агинский нац. округ; Б — Fron-
topsylla wagneri — Юго-восточное Забайкалье, Борзинский р-н; В— Tarsopsylla
octodecimentata — Иркутская область, Тулунский р-н; Г — Ceratophyllus laeviceps
ellobii — Монголия, Гобийский Алтай, оз. Сого-Нор.
349, А—Neopsylla acanthina — остров Сахалин; Б — N. bidentatiformis—Юго-восточное
Забайкалье, Борзинский р-н; В — Rhadinopsylla bivirgis—рисунок Ю. Н. Вагнера;
Г — R. socia—то же.
350, А—Paraceras melis. — Закавказье, Курдистан; Б—Р. flabellum — Алма-Атинская
область.
351, А—Echidnophaga gallinacea и Б — Е. oschanini по И. Г. Иоффу и В. Е. Тифлову.
352, A—Ceratophyllus garei.............. юго-восточная часть Забайкалья, ст. Дау-
рия; Б — С. farreni — Западная Монголия, Улясутай.
353, А — Vermipsylla alacurt — Центральный Тянь-Шань; Б — V. dorcadia — то же;
В — Chaetopsylla lasius — по Ротшильду (тип); Г — Ch. tuberculaticeps — Кольский
полуостров, Лапландский заповедник; Д—Ch. appropinquans—окрестности г. Благо-
вещенска; Е— Ch. homoeus — Казахская ССР, Заилийский Алатау; Ж—Ch. alia —
Зауралье, Кондо-Сосьвинский заповедник; 3—Ch. dogieli — Приморский край, Суд-
зухинский заповедник; И—Ch. zibellina — Зауралье, Кондо-Сосьвинский заповедник;
К — Ch. trichosa — Предкавказье, Кизлярский р-н; Л — Ch. mikado — окрестности
г. Владивостока, Седанка; М — Ch. mirabilis — Тебердинский заповедник.
АЛФАВИТНЫЙ СПИСОК
ИЗОБРАЖЕННЫХ НА РИСУНКАХ И ПРИВЕДЕННЫХ
В ОПРЕДЕЛИТЕЛЬНЫХ ТАБЛИЦАХ ВИДОВ БЛОХ
Знаком минус (—) отмечены виды, приведенные для сравнения и пока не обна-
руженные в местностях, для которых рассчитан этот определитель (см. введение).
Сокращения: Arg. = Argyropulo
Jord. = Jordan
Koi. = Kolenati
Roths. = Rothschild
Tasch. ^=-. Taschenberg
Tifl. = Tiflov
Wagn. = Wagner
J. et R. = Jordan et Rothschild
Aenigmopsylla grodekovi Sychevskij 1950 ...................
Amphalius clarus J. et R. 1922 ............................
„ runatus J. et R. 1923 ..............................
Amphipsylla anceps Wagn. 1930 .............................
„ argoi loff 1946 ....................................
„ asiatica loff 1927 .................................
, aspalacis Jord. 1929 ...............................
, casis J. et R. 1911.................................
„ daea Dampf 1910.....................................
, dumalis J. et R. 1915...............................
georgica Savenko 1948 ..........................
kalabukhovi loff et Tifl. 1939..................
„ kuznetzovi Wagn. 1912...........................
„ longispina Scalon 1950 .........................
„ marikovskii loff et Tifl. 1939 .....................
, montana Arg. 1946 ..................................
„ montium Jord. 1944 .................................
parthiana loff 1950 ............................
„ petristshevae Darskaja 1949 ........................
phaiomydis loff 1946 ...........................
prima Wagn. 1929 ...............................
primaris primaris J. et R. 1915.................
„ „ mitis Jord. 1929 .........................
Рисунки №
39; 100, А,Б
107
105, 106
238, K\ 241, M; 242, Б
238 , 77; 240, /7; 241, 77
243, А
54, Д\ 238, О; 240, М;
241, О
238, Л; 239, В, 240, В;
241, Л
238, В; 240, В; 241, В;
242, В
239, Е\ 240 , 77; 241, К\
243, Б\ 244
238, Д; 240, Д; 241, Д
„ rossica Wagn. 1912 . ’................................
„ schelkovnikovi Wagn. 1908 ............................
„ „ irana loff 1946 ..........................
„ sibirica sibirica Wagn. 1893 .........................
— л „ hetera Jord. 1932 ..........................................
„ „ orientalis loff 1946 .........................
— „ „ pollionis Roths. 1905 ........................
— „ „ sepifera J. et R. 1920 .......................
„ tuta Wagn. 1929 ......................................
„ vinogradovi loff 1927 ................................
— Araeopsylla gestroi Roths. 1906 ................................
— Archaeopsylla erinacei Curtis 1832 .............................
„ sinensis J. et R. 1911................................
238, Е\ 240, Е
238, В; 240, В; 241, Е
238, М
54, В, Ж\ 238, /7;
239, В; 240, Л; 241, /7;
242, Г
54, А; 238, А; 239, А;
240, А; 241, В; 242, А,
238, Ж\ 239, В; 241, 3
54, В, В; 238, В; 240, В;
241, А
241, В
238, 77; 240, Ж\ 241, Ж
54, В; 238 , 3; 239, Д;
210, 3; 241, 77
22, 77, А
249
Brachyctenonotus myospalacis Wagn. 1929 ....................... 50, 227, 228
Catallagia dacenkoi loff 1940 ................................. 319, А, Б
„ fetisovi Vovchinskaja 1944...................... 322; 323, Б
„ ioffi Scalon 1950 .................................. 70; 320, А, Б; 346, Б
„ striata Scalon 1950 ................................ 321; 323, A
— Ceratophyllus abramovi loff 1946 .............................
„ advenarius advenarius Wagn. 1927 •..................156, Б; 158; 158, A
„ „ bifallax loff et Scalon 1950 .... 158, Б
„ anisus Roths. 1907 ................................. 144, А, Б
— , aralis Arg. 1946 ...........................................
avicitelli loff 1946 .............................. 181, 182
— „ borealis Roths. 1907 ...................................
calcarifer Wagn. 1913..............................153; 154, A
„ caliotes Jord. 1937 .................................... 186; 188, A
— „ caspius Tifl. 1937 . . ?............................127; 129, A
— , desertus Arg. 1946 ................................. 150; 152
„ dolabris J. et R. 1911 ...........................130
„ farreni Roths. 1905 .............................. 184; 185, Б; 352, Б
„ fasciatus Bose 1801 .............................. 42, 113, 114
„ fidus J. et R. 1915...............................116, А, Б
— „ fringillae Walker 1856 ............................. 175
„ gaiskii Vovchinskaja 1950 .......................... 128; 129, Б
— „ gallinae gallinae Schrank 1803 ................... 169, Б; 173; 174
„ „ dilatus Dudolkina 1946 .................. 170, Б; 172
„ „ tribulis Jord. 1926 ..................... 170, A; 171
„ garei Roths. 1902 ...................................... 165; 166; 169, A; 352, A
— „ geminus loff 1946 .................................. 136, А, Б
— „ hirundinis Samouelle 1819...........................
ioffi Darskaja 1949 ............................... 141, TA*; 1.42, E
„ jenissejensis Wagn. 1902 ........................... 126, А, Б
„ kaznakovi Wagn. 1929 ............................... 132, 133
„ kozlovi Wagn. 1929 ................................. 131
— „ laeviceps laeviceps Wagn. 1908 ......................... 117, A, 5
„ „ ellobii Wagn. 1933 ...................... 119, A, £; 348, Г
, „ kuzenkovi Jagubiants 1953 ............... 118, А, Б
lebedewi lebedewi Wagn. 1932 .....................
„ princeps loff 1946 . . . ....................
lunatus J. et R. 1920 .............• •...........163, 164
maculatus Wagn. 1927 ............................. 183; 185, A
menzbieri loff 1948 ..............................
mokrzeckyi Wagn. 1916..............................115, А, Б
monstrosus Wacn. 1929 ............................
oreinus Jord. 1937 ............................... 134, 135
orites Jord. 1937 ................................ 187; 188, Б
paradoxus Scalon 1950 ............................ 146; 147, Б; 346, Г
penicilliger penicilliger Grube 1852................44; 139; 140; 141, Г;
142, A
„ arvicolae loff 1948 ..................... 141, B\ 142, Г
„ demotus loff 1946 ....................... 141, E; 142, Д
„ dissimilis Jord. 1938 ................... 141, A; 142, Б
svrt loff 1946 ........................ 141, S; 142, В
„ vallis loff 1946 ..................... 141, Д
rectangulatus Wahlgren 1903 ...................... 157, А, Б
relicticola Fedina 1946 ..........................
riparius J. et R. 1920 ........................... 180, A, £
scaloni Vovchinskaja 1950 ........................ 149, 151
sciurorum sciurorum Schrank 1781..................
asiaticus loff 1936 ................... 143, A, £
semenovi loff 1936 ............................... 137, 138
sinicus Jord. 1932 ...............................
sokolovi Gershkovich 1953 ........................ 155, £; 156, A
spinosus Wagn. 1903 .............................. 167, 168
styx Roths. 1900 ................................. 178; 180, A
taiganus Scalon 1950 ............................. 161, 162
tamias Wagn. 1927 ................................ 145; 147, A
tersus J. et R. 1915 .............................
tesquorum Wagn. 1898 .............................8
„ altaicus loff 1936 ......................... 125
n dzetysuensis Mikulin 1951....................
n mongolicus J. et R. 1911......................122
250
Ceratophyllus tesquorum sungaris Jord. 1929 .................. 43; 121; 123; 346, В
transvolgensis loff 1935 .......... 124
— „ tiflovi Wagn. 1936 ..............................
toli Wagn. 1900 ............................... 148, А, Б
— . trispinus Wagn. et loff 1926 ................
, turbidus Roths. 1909 ................................ 16, 159, 160
— n turkmenicus Vlasov et loff 1937 .............. 120, А, Б
— , vagabundus vagabundus Boheman 1865 . . . • 176, 177
, „ orientals Wagn. 1929 .....................
, walkeri Roths. 1902 ........................ 154, Б; 155, A
Chaetopsylla alia loff 1946 ............................. 91, А, Б; 353, Ж
„ appropinquans Wagn. 1929 ................... 93; 96, A; 353, Д
dogieli loff 1950..........................92, A, 5; 353, 3
— „ giobiceps Tasch. 1880 ......................
n homoeus Roths. 1905.........................34; 88, А, Б; 353, E
— n lasius Roths. 1906..........................353, В
„ mikado Roths. 1904..........................97, А, Б\ 353, Л
— „ mirabilis loff et Arg. 1934 ................ 89; 90; 353, M
trichosa Kohaut 1903 ...................... 95; 96, Б\ 353, К
„ tuberculaticeps Bezzi 1890..................33; 98, А, Б-, 353, Г
zibellina loff 1946........................94, A, Б; 353, И
— Coptopsylla lamellifer Wagn. 1895.......................32
Ctenocephalides canis Curtis 1826............................25
„ caprae loff 1953 ...........................
w felis Bouche 1835 .......................... 26
Ctenophthalmus arvalis Wagn. et loff 1926.................267, А, Б
„ assimilis Tasch. 1880 ................... 266, А, Б
„ congeneroides Wagn. 1929.................61; 263; 264; 345, Б
„ dilatatus Wagn. 1929........................272, А, Б
„ dolichus Roths. 1913........................271, А, Б
dux J. et R. 1915..........................270, А, Б
, pisticus pisticus J. et R. 1921............. 268, А, Б
, „ pacificus loff et Scalon 1950 . . . 269
, truncus loff et Scalon 1950 ................ 265, А, Б
„ uncinatus Wagn. 1898 ................... 262, А, Б
Ctenophyllus armatus Wagn. 1900 ............................... 46; 209; 211, Б
„ conothoae loff 1946 ..................................
hirticrus J. et R. 1923 .......................... 47; 212, А, Б; 248, /I
„ rigidus Darskaja 1949 ................................ 20; 208; 311, В
— „ rufescens loff 1946 ..................................
„ subarmatus Wagn. 1900 ........................ 211, A
„ tarasovi Scalon 1953 ................................. 210
—Dasypsyllus gallinulae Dale 1878 ............................... 189, 190
„ stejnegeri Jord. 1929 ................................. 191, А, Б
Doratopsylla birulai loff 1927 ................................ 60; 275, А, Б
„ coreana Darskaja 1949 ................................. 59; 274, А, Б
— „ cuspis Roths. 1915...............................
— w dampfi Arg. 1935 ................................
— „ dasycnemus Roths. 1897 ....................... 58; 273, А, Б
— Echidnophaga gallinacea Westwood 1875 .................. 23; 351, A
— „ murina Tiraboschi 1903 ...............................
„ oschanini Wagn. 1929 ................................. 24; 351, Б
— „ popovi loff et Arg. 1931..............................
Eopsylla nuda Arg. 1946 ....................................... 57, 260, 261
— Frontopsylla ambigua Fedina 1946 .......................
— „ cornuta loff 1946 ....................................
„ elata elata J. et R. 1915........................ 202
n „ botis Jord. 1929 .................................... 204, 205
— „ „ caucasica loff et Arg. 1934 ........................
„ humida Tifl. 1937
„ koksu loff 1946 ..........................
w mutata Jord. 1944 ........................
„ popovi Lebedev 1953 .................... 202a.
„ taishiri Emeljanova 1959 ............... 203, А, Б
elatoides Wagn. 1929 ........................ 194, 196
251
— Frontopsylla frontalis frontalis Roths. 1909 ................
— „ „ alatau Fedina 1946 ........................
, , baikal loff 1946 .......................... 206, 207
— „ « dubiosa loff 1946 .........................
w hetera Wagn. 1932 ................................ 195, 197
„ luculenta luculenta J. et R. 1923 ................ 45, 198, 199
„ w parilis Jord. 1929 ........................ 200, 201
— „ macrophthalma J. et R. 1915...............................
— „ ornata Tifl. 1937 ........................................
— w protera Wagn. 1932 .......................................
— „ semura Wagn. et loff 1923 ................................
— „ ipadix J. et R. 1921 .....................................
„ wagneri loff 1927 ..................................... 192; 193, А, Б\ 348, Б
— Hoplopsyllus glacialis glacialis Tasch. 1889 ...............
„ „ profugus Jord. 1925 ....................... 87, А, Б
Hystrichopsylla microti Scalon 1950 .......................... 72; 325, А, Б
„ nicolai Scalon 1935 .................................. 324, Б
— „ poppei Wagn. 1902 .........................................
— « satunini Wagn. 1916........................................
„ talpae Curtis 1826 ................................... 71; 324, A
Ischnopsyllus comans J. et R. 1921 ........................... 340, 341
— „ dolosus Dampf 1912........................................
„ elongatus Curtis 1832 ................................ 339, А, Б
— „ hexactenus hexactenus Koi. 1857 .............. 336, Б
« „ kolenatii Wagn. 1929 ...................... 336, A
— „ intermedius Roths. 1898 ........................ 339, В
„ needhamia Hsu 1935 ............................... 312; 343, A
„ obscurus Wagn. 1898 .............................. 75; 337; 338; 339, Г
— „ octactenus Koi. 1857 ...........................
— „ plumatus loff 1946 ........................................
— „ simplex Roths. 1906 .......................................
— n variabilis Wagn. 1898 .....................................
Leptopsylla bidentata Koi. 1860 .............................. 247, 248
„ nana Arg. 1946 ................................... 245, А, Б\ 246
„ ostsibirica Scalon 1936 .......................... 52; 249; 250; 347, В
„ segnis Schoncher 1811.......................... 51
— „ sexdentata Wagn. 1929 .....................................
« silvatica Meinert 1896 .......................... 251, А, Б
—Mesopsylla eucta eucta Dampf 1910............................... 237
— „ afgana Jord. 1944 ........................................
— « „ kochkor loff 1950 ..............................
— „ shikho loff 1950 .............................•...........
„ hebes J. et R. 1915 ................................... 53, 229, 230
— „ lenis J. et R. 1915 ................................. 231, 234
— , rothschildi Arg. 1938 ............................... 233, 236
— « tuschkan tuschkan Wagn. et loff 1926 .......... 232, 235
— „ „ andruschkoi Arg. 1946 .....................
— w „ mesa Arg. 1946 ............................
- Mioctenopsylla arctica Roths. 1922 ......................
Myodopsylla trisellis Jord. 1929 ............................. 76; 343, Б\ 344
— Myoxopsylla jordani loff et Arg. 1934 ...................
— „ laverani Roths. 1910...................................
Nearctopsylla beklemischevi loff 1950 ........................ 64, 279
„ ioffi Sychevskij 1950 ............................ 63, 277, 278
Neopsylla abagaitui loff 1946 ................................ 313, А, Б
„ acanthina J. et R. 1923 ............................. 311, А, Б\ 349, A
r, aliena J. et R. 1911................................. 314, А, Б
9 anoma Roths. 1912.................................... 316, А, Б
„ bidentatiformis Wagn. 1893 .......................... 69; 301; 303; 349, Б
, compar J. et R. 1911................................. 314, В
9 democratica Wagn. 1926 .............................. 306, 308
galea loff 1946 .................................... 312, Б\ 315
— , hissarica loff et Sosnina 1950 .........................
mana Wagn. 1927 .................................... 302, 304
meridiana Tifl. et Kolpakowa 1936 ..................
, pleskei pleskei loff 1927 ......................... 309, А, Б
« n ariana loff 1946 ............................ 310, Б
» „ orientalis loff et Arg. 1934 ................ 310, A; 312, A
„ teratura Roths. 1913 ............................... 305, 307
— „ setosa Wagn. 1898 ......................................
252
— „ galba Dampf 1910........................................
Ophthalmopsylla jettmari Jord. 1929 ........................... 225, 226
„ kiritschenkoi Wagn. 1929 ....................... 49; 224, Л, Б
„ kukuschkini loff 1927 ..................... 221, А, Б\ 222
„ praefecta praefecta J. et R. 1915............ 48; 219, А, Б
„ „ pernix Jord. 1929 ..................... 220, А, Б
— „ volgensis volgensis Wagn. et loff 1926 . . .
, „ extrema loff et Scalon ssp. n. . . . 223, А, Б
Oropsylla alaskensis Baker 1904 ...............................
„ asiatica Wagn. 1929 ..................................... 41; 109; 111, Б
— „ ilovaiskii Wagn. et loff 1926 .......................... 110
„ mandarinus J. et R. 1911.............................. 112, А, Б
„ silantiewi Wagn. 1898 ................................ 9; 40; 108; 111, A
— Palaeopsylla incurva Jord. 1932 .............................
— „ remota Jord. 1929 .........................................
— „ sorecis sorecis Dale 1878 .................................
„ „ starki Wagn. 1929 . . . . •................... 62; 276, А, Б
Paraceras flabellum Wagn. 1916................................. 38; 101; 103; 350, Б
— „ melis Curtis 1832 .........................•............. 102; 104; 350, A
— „ crispus J. et R. 1911 ................................ 345, A
— „ sinensis Liu 1935 ....................................
Paradoxopsyllus conveniens Wagn. 1929 ...................... 217, Б
„ curvispinus Miyajima et Koidsuma 1909 . . . 218, А, Б
„ dashidorzhii Scalon 1953 ...................... 216; 217, A
„ integer loff 1946 ...................... 214; 215, Б
— „ naryni Wagn. 1929 ......................................
— „ repandus Roths. 1913....................................
„ scorodumovi Scalon 1935 .............................. 213; 215, A
— „ teretifrons Roths. 1913.................................
w ioffi Tifl. 1937 ....................................... 317, А, Б
— „ tiflovi Fedina 1946 ...................................... 318, А, Б
—Pariodontis riggenbachi riggenbachi Rjths. 1904 ...................
„ „ turkestanica Dubinin 1947 ••.... 30
— Pectinoctenus ctenophorus Wagn. 1929 ..........................
— „ lautus Roths. 1915...........................................
— „ nemorosus Tifl. 1937 ........................................
— w pamirensis loff 1946 ........................................
„ pavlovskii loff 1927 .......................... 252, 253
„ pectiniceps Wagn. 1893 ............................. 55, 254, 255
— Phaenopsylla kopetdag loff 1946 ...............................
— „ mustersi Jord. 1944 .........................................
— „ tiflovi loff 1950 ...........................................
Pulex irritans L. 1758 .......................................... 21
„ „ v. fulvus loff 1929 ..................................
Rhadinopsylla accola Wagn. 1929 ................................. 287
„ altaica Wagn. 1900 ................................. 288, А, Б
— „ angusta Tifl. 1937 .................................
„ aspalacis loff et Tifl. 1946 ....................... 285, А, Б
— „ bivirgis Roths. 1913 ............................... 296, Б; 299, Б\ 349, В
— „ cedestis Roths. 1913................................ 68; 296, A; 299, A
„ dahurica dahurica J. et R. 1923 .......................... 65; 283, А, Б
— r, n diclinica Tifl. 1937.........................
— „ n tjanschan loff et Tifl. 1946 ................
„ „ vicina Wagn. 1929 ........................... 283, Б
„ dives Jord. 1929 ................................... 281, А, Б
„ insolita Jord. 1929 ................................ 280, А, Б
— „ integella integella J. et R. 1921...................
„ „ casta Jord. 1928 ......................... 291, А, Б
— я „ caucasica Arg. 1941 ................. 290, A, В
„ „ concava loff et Tifl. 1946 ............... 290, Б, Г
„ ioffi Wagn. 1929 ..................................... 284, А, Б
„ jaonis Jord. 1929 .................................. 300
. li li Arg. 1941 ................................. 66; 293, A; 294
„ „ transbaikalica loff et Tifl. 1946. ...... 293, Б\ 295, A
„ pseudodahurica Scalon 1950 ......................• ... 289, А, Б
„ rothschildi loff 1935 .............................. 286, А, Б
— „ semenovi Arg. 1941 ................................. 293, Д; 295, В
— „ socia Wagn. 1929 ................................... 297; 298; 349, Г
» tenella Jord. 1929 ....................................... 282, А, Б
253
Rhadinopsylla valenti Darskaja 1949 ......................... 67; 292, Л, Б
— „ ventricosa ventricosa Joff et Tifl. 1946 .... 293, B; 295, Б
— „ „ murium loff et Tifl. 1946 ........... 293, Г
- Rhinolophopsylla unipectinata Tasch. 1880 .................
— Saphiopsylla nupera Jord. 1931.............................
— Spilopsyllus cuniculi Dale 1878 ............................ 31
Stenoponia coelestis J. et R. 1911............................ 332
, formozovi loff et Tifl. 1933 ...................... 74, 328, 330
„ ivanovi loff et Tifl. 1933 ........................ 327; 333, A
„ montana Darskaja 1949 ............................. 334, 335
„ sidimi Marikowsky 1935 ............................ 331; 333, Б
singulars loff et Tifl. 1933 ..................... 73; 326, 329
— Synosternus longispinus Wagn. 1893 ......................... 28,
— „ pallidus Tasch. 1880 .....................................
Tarsopsylla octodecimdentata Koi. 1863 ....................... 99, А, Б\ 348, В
Vermipsylla alacurt Schimkewitsch 1885 ....................... 35; 37, A; 347, A; 353, A
„ dorcadia Roths. 1912.................................. 36; 37, 5; 347, 5; 353, Б
Wagnerina antiqua Scalon 1953 ................................ 56; 258, А, Б
„ longicauda Scalon 1953 ................................ 257, А, Б
„ tuvensis loff et Scalon sp. n...................... 259
— w schelkovnikovi loff et Arg. 1931...................
— „ tecta Joff 1946 ................................... 256, А, Б
— Xenopsylla astia Roths. 1911............................... 29, A; 78,79, A; 86, Б
— „ brasiliensis Baker 1904 ................... 86, Д
„ cheopis Roths. 1903 .................................. 27; 77, Б\ 86, A; 346, A
— „ conformis Wagn. 1903 ................... 80, 86, A'
— , gerbilli gerbilli Wagn. 1903 ................... 82
— „ caspica loff 1950 ........................................ 84
— „ w minax Jord. 1926 ....................................... 83
hawaiiensis Jord. 1932 ........................... 86, Г
— 4 hirtipes Roths. 1913...............................
— 4 magdalinae loff 1935 ..............................
— „ nesokiae loff 1946 ................................
— „ nubicus Roths. 1903 ............................... 29, Б\ 79, Б\ 86, В
— , nuttali loff 1930 .................................
persica loff 1946 ...........................• . . 85
— regis Roths. 1903 ..........................................
» scrjabini loff 1928 .................................. 81; 86, Ж
СПИСОК НЕКОТОРЫХ синонимов
В этом списке до знака равенства поставлены замененные названия (синонимы)
а после него — принятые в настоящее время. Знак вопроса означает, что полная
идентичность названных форм требует еще подтверждения.
Amphipsylla
certa J. et R. = schelkovnikovi Wagn.
contigua J. et R. = rossica Wagn.
ewingi I. Fox = ? marikovskii I. et T.
locuples J. et R. = rossica Wagn.
primaris botis Jord. = primaris mitis Jord.
thoracicus Roths. = sibirica Wagn.
transbajkalica Wagn. = primaris mitis Jord.
Ceratophyllus
anadyrus Jord. = toli Wagn.
baikalensis loff = rectangulatus Wahlgren
caracoi Scalon = anisus Roths.
consor Roths. = laeviceps Wagn.
crassus J. et R. — ? Oropsylla silantiewi Wagn.
dacus J. et R. = Rostropsylla daca J. et R.
elatus J. et R. = Frontopsylla elata J. et R.
famulus J. et R. = ? tesquorum Wagn.
laeviceps mongolicus Wagn. = laeviceps ellobii Wagn.
microti Nordb. = ? rectangulatus Wahlgren
mustelae Schill. = turbidus Roths.
nanodes Arg. = caliotes Jord.
pteromydis Nordb. = tamias Wagn.
sciurorum cophinus Jord. = sciurorum asiaticus loff
silantiewi Wagn. = Oropsylla silantiewi Wagn.
sucinus J. et R. = mokrzeckyi Wagn.
uralensis Wagn. = Tarsopsylla octodecimdentata Koi.
Ctenocephalus = Ctenocephalides
serraticeps Gerv. = canis Curt.
serraticeps Gerv. = felis Bouche
Ctenonotus = Tarsopsylla
Ctenophthalmus
dahuricus loff = pisticus J. et R.
saratovi I. et T. = golovi I. et T.
typhlus Motsch. = ? spalacis J. et R.
Ctenopsyllus = Leptopsylla
monoctena Wagn. = bidentata Koi.
musculi Dug. = segnis Schanch.
sobrinus Roths. = bidentata Koi.
Dermatophilus = Sarcopsylla = Tunga
Echidnophaga
rhynchopsylla Tir. = murina Tir.
Frontopsylla
subula Wagner — Ctenophyllus hirticrus J. et R.
Ischnopsyllus
teres Jord. = dolosus Dampf
wassilii Scalon = needhamia Hsii
Loemopsylla = Xenopsylla
mycerini1 = conformis Wagn.
mycerini1 = magdalinae loff
1 Это относится только к указаниям об обнаружении этого вида на территории
СССР. Африканские X. mycerini Roths, не идентичны нашим.
255
Neopsylla
spinea Roths. = setosa Wagn.
uniseriata Wagn. == acanthina J. et R.
Nycteridopsylla
wui Hsii = galba Dampf
Oropsylla
asiatica Wagn. = ? alaskensis Baker
elana Jord. = ? alaskensis Baker
sinensis Liu = Paraceras sinensis
stejnegeri Jord. = ? alaskensis Baker
weisskei Wagn. — ? silantiewi Wagn.
Paraceras
segregatus Wagn. = ? Oropsylla silantiewi Wagn.
Paradoxopsyllus
subcaecatus Roths. = curvispinus M. et K.
transbaikalicus Tifl. et Pavl. = scorodumovi Scalon
Pectinoctenus
adalis Jord. = pectiniceps Wagn.
Stenoponia
solitaria I. et T. = conspecta Wagn.
Trichopsylla = Chaetopsylla
nocens loff = mikado Roths.
УКАЗАТЕЛЬ ОСНОВНЫХ ЭКОЛОГИЧЕСКИХ
ГРУППИРОВОК БЛОХ
Как известно, многие виды блох относятся к числу паразитов, сравнительно
легко меняющих своих хозяев, т. е. переходящих с одного вида животных на другой.
Тем не менее для большинства видов блох могут быть указаны определенные виды
или группы животных, на которых эти паразиты встречаются значительно чаще, чем
на других животных, не говоря о том, что известны также и весьма специфичные,
связанные с узкими группами хозяев виды блох.
Кроме связи с хозяином, экология каждого вида блох может быть охарактери-
зована привязанностью к определенным местам обитания (стациям, биотопам), большей
или меньшей привязанностью к телу хозяина или его гнезду („блохи шерсти“,
„блохи гнезда*), сезонным характером распространения и т. п.
Ниже приводятся списки видов блох, включенных в наш определитель, сгруппи-
рованные по их основным экологическим особенностям, прежде всего по стациям
и хозяевам. Важнейшие виды, обнаруженные на основной территории, для которой
рассчитан наш определитель, отмечены в этих списках звездочкой (*). Виды, экологи-
ческая характеристика которых недостаточно выяснена, не включены. Некоторые виды
упоминаются в ряде списков, так как они встречаются в различных стациях и на раз-
ных хозяевах, но мы старались свести число таких указаний к минимуму, помещая
большинство видов в наиболее характерные для них группы. Более детальные све-
дения об отдельных видах имеются в тексте книги.
В первых девяти группах дается распределение видов по стациям и хозяевам,
затем по сезонности и, наконец, по особенностям паразитирования.
1. Блохи человека и домашних животных
- Pulex irritans — блоха человека * Vermipsylla alacurt — блоха овец и
* Ctenocephalides canis — блоха собаки других копытных
felis — „ кошки * Vermipsylla dorcadia — блоха овец
„ саргае — „ коз и других копытных
2. Блохи домовых грызунов
а) Блохи крыс (Rattus norvegicus,
Rattus rattus)
* Xenopsylla cheopis
„ brasiliensis
„ hawaiiensis
Echidnophaga murina
- Ceratophyllus (Nosopsyllus) fasciatus
„ „ punjabensis
Ceratophyllus (Nosopsyllus) Simla
„ „ wualis
Ceratophyllus (Nosopsyllus) nicanus
(Monopsyllus) anisus
Paradoxopsyllus curvispinus
Neopsyl a hissarica
б) Блохи домовых мышей (Mus
musculus)
Ceratophyllus (Nosopsyllus) fidus
„ „ mokrzeckyi
* Leptopsylla segnis
„ sexdentata
3. Блохи степных (в том числе горностепных и пустынных) грызунов
а) Блохи сурков — тарбаганов
(Marmota sibirica, Marmota baibacina
и др.)
’ Oropsylla silantiewi
Ceratophyllus (Citellophilus) lebedewi
„ „ menzbieri
Rhadinopsylla (Ralipsylla) ventricosa
б) Блохи сусликов
* Oropsylla asiatica (= O. alaskensis)
, ilovaiskii
Rostropsylla daea
” Ceratophyllus (Citellophilus) tesquorum
„ „ jenisse-
jensis
Ceratophyllus (Citellophilus) trispinus
„ „ relicticola
* Frontopsylla elatoides
„ semura
Neopsylla setosa
257
в) Виды, мало разборчивые в от-
ношении выбора хозяина,
встречающиеся преимущественно в
гнездах и норах разных степных
грызунов (сусликов, полевок, хомяков,
пищух и др.), а также в горах, в ле-
состепи.
* Frontopsylla luculenta
„ ~ hetera
Rhadinopsylla (Ralipsylla) li
Neopsylla bidentatiformis
pleskei
, teratura
„ mana
„ meridiana
г) Блохи с т e ii н ы x (и горн o-c т e n-
ii ы x) полевок (Microtus, Stenocra-
nius, Phaiomys) и пеструшек (Lagurus).
Amphipsylla primaris
w prima
„ rossica
4 asiatica
„ montana
parthiana
phaiomydis
Ctenophthalmus arvalis
Rhadinopsylla (Rectofrontia) dahurica
rothschildi
„ (Ralipsylla) ventricosa
murium
Stenoponia ivanovi
д) Блохи хомяков (Cricetulus, Meso-
cricetus, Phodopus)
: Amphipsylla vinogradovi
longispina
schelkovnikovi
„ kalabukhovi
anceps
Pectinoctenus pavlovskii
e) Б л о x и даурской пищухи
(Ochotona daurica)
Amphalius runatus
“ Ctenophyllus (Ochotonobius) hirticrus
ж) Блохи тушканчиков (Allactaga,
Alactagulus, Scirtopoda, Dipus)
x Ophthalmopsylla praefecta
„ volgensis
„ (Cystipsylla) jettmari
« » kiri-
t sc hen koi
: Frontopsylla wagneri
„ (Mafrontia) macro-
phthalma
" Mesopsylla hebes
„ lenis
„ eucta
„ (Desertopsylla) rotschildi
з) Блохи слепушонок (Ellobius tal-
pinus)
Xenopsylla magdalinae
Amphipsylla dumalis
Ctenophthalmus dux
и) Блохи цокоров (Myospalax)
Brachyctenonotus myospalacis
* Amphipsylla aspalacis
- „ daea
„ casis
Ctenophthalmus dilatatus
Rhadinopsylla (Rectofrontia) aspalacis
. n ioffi
* Stenoponia singulars
к) Блохи песчанок (Rhombomys,
Meriones, Pallasiomys)
* Echidnophaga oschanini
- Xenopsylla scrjabini
„ gerbilli
„ hirtipes
„ • persica
, regis
« conformis
„ nuttali
Coptopsylla lameliifer и др.
* Ceratophyllus (Gerbillophilus) laevi-
ceps
Ceratophyllus (Gerbillophilus) turkmeni-
cus
Ceratophyllus (Gerbillophilus) tersus
„ „ abramovi
„ „ monstrosus
„ « aralis
Paradoxopsyllus conveniens (?)
„ repandus
„ teretifrons
- Ctenophthalmus dolichus
Rhadinopsylla bivirgis
„ cedestis
„ socia
л) Блохи зайцев (Lepus tolai)
* Hoplopsyllus glacialis profugus
м) Блохи кроликов (Oryctolagus
cuniculi)
Spilopsyllus cuniculi
н) Блохи дикобразов
Pariodontis riggenbachi
4. Блохи грызунов, живущих
в скалах и каменных осыпях
а) Блохи пищух (Ochotona pricei,
О. alpina, О. hyperborea, О. rutila,
О. macrotis)
Amphalius clarus
runatus
Ceratophyllus (Callopsylla) lagomys
„ « tiflovi
• Ctenophyllus armatus
„ subarmatus
„ rigidus
„ tarasovi
w (Conothobius) conothoae
„ (Ochotonobius) hirticrus
rufescens
* Rhadinopsylla (Rectofrontia) altaica
б) Блохи высокогорных камен-
ных полевок (Alticola, Chionomys)
:v Ceratophyllus (Callopsylla) gaiskii
„ „ caspius
* Amphipsylla primaris
Paraneopsylla ioffi
„ dampfi
в) Виды, менее разборчивые
в выборе хозяина, встречаю-
щиеся часто как на каменных полев-
ках, так и на пищухах, хомячках
и других грызунах, живущих в ска-
лах, среди камней и в горной степи.
* Ceratophyllus (Paramonopsyllus) sca-
loni
258
Ceratophyllus (Paramonopsyllus) deser-
tus
* Frontopsylla elata
* Paradoxopsyllus scorodumovi
* „ integer
* „ dashidorzii
w naryni
* Leptopsylla nana
Pectinoctenus ctenophorus
„ lautus
* • Wagnerina antiqua
longicauda
„ tuvensis
Wagnerina tecta
w schelkovnikovi
Rhadinopsylla (Ralipsylla) semenov i
Saphiopsylla nupera
г) Блохи мышевидных хомяч-
ков, обитающих в горах Средней
Азии (Calomyscus bailwardi).
Phaenopsylla mustersi
„ tiflovi
„ kopetdag
Paradoxopsyllus microphthalmus
Amphipsylla montium
« argoi
5. Блохи лесных и луголесных грызунов
а) Блохи грызунов, живущих
на деревьях, — белки, бурундука,
летяги, полчка, сонь (Sciurus vulgaris,
Eutamias sibiricus, Pteromys volans, Glis
glis, Dyromys nitedula)
* Tarsopsylla octodecimdentata
Myoxopsylla jordani
Aenigmopsylla grodekovi
Ceratophyllus (Monopsyllus) sciurorum
* Ceratophyllus (Monopsyllus) tamias
Eopsylla nuda
* Ctenophthalmus pisticus
* Neopsylla acanthina
б) Блохи наземныхлесныхгры-
з у н о в, преимущественно лесных
полевок рода Clethrionomys, а также
живущих в лесах полевок из рода
Microtus, лесных мышей (Apodemus
silvaticus, A. speciosus), частью мало
разборчивые в выборе хозяина и встре-
чающиеся на многих лесных млеко-
питающих
* Ceratophyllus (Amalaraeus) penicilliger
„ . ioffi
* „ (Megabothris) rectangu-
latus
* Ceratophyllus (Megabothris) advenarius
„ „ turbidus
„ „ sokolovi
* Frontopsylla elata
Frontopsylla (Profrontia) protera
« „ ornata
„ „ ambigua
* Amphipsylla sibirica
„ kuznetzovi
* „ marikovskii
„ petristshevae
* Leptopsylla ostsibirica
„ bidentata
silvatica
* Pectinottenus pectiniceps
« nemorosus
Ctenophthalmus uncinatus
* Rhadinopsylla (Rectofrontia) integella
pseudo-
daurica
Rhadinopsylla (Rectofrontia) angusta
„ „ dahurica
diclinica
'* Catallagia dacenkoi
* „ ioffi
* „ striata
* Neopsylla mana
Paraneopsylla tiflovi
* Stenoponia sidimi
„ montana
* „ formozovi
Hystrichopsylla talpae
* „ microti
в) Блохи мелких грызунов(ML
_£rQtus, Arvicola, Apodemus), п p e д n o-
читающие наиболее вл а ж-
ные биотопы — луга, болота, пой-
мы pelc
Xenopsylla nesokiae
* Ceratophyllus (Megabothris) calcarifer
„ „ walkeri
„ (Amalaraeus) penicilliger
arvicolae
Ctenophthalmus assimilis
* „ congeneroides
„ truncus
* Rhadinopsylla (Rectofrontia) insolita
6. Блохи насекомоядных (Insectivora)
а) Блохи ежей, живущих в су-
хих местностях (Hemiechinus,
Paraechinus)
Echidnophaga gallincea
Synosternus longispinus
„ pallidus
б) Блохи ежей, живущих в бо-
лее влажных местностях
(Erinaceus)
* Archaeopsylla sinensis
„ erinacei
в) Блохи кротов (Talpa)
Ceratophyllus (Tiflocallopsylla) seme-
novi
Ctenophthalmus bisoctodentatus
Palaeopsylla similis
„ kohauti
г) Блохи землероек(Sзгех,NeDmys)
- Doratopsylla birulai
„ coreana
„ dasycnemus
„ dampfi
„ cuspis
Palaeopsylla sorecis
incurva
„ remota
* Nearctopsylla ioffi
259
7. Блохи хищных (Carnivora)
Помимо перечисляемых ниже видов блох, на хищных часто встречаются блохи че-
ловека и домашних животных: Pulex irritans, Ctenocephalides canis, Ctenocephalides felis,
а также многие паразиты грызунов, которыми питается тот или иной хищник.
а) Блохи мелких хищных
(Mustelidae)
" Chaetopsylla homoeus
„ dogieli
„ zibellina
„ mirabilis
Ceratophyllus lunatus
* „ (Monopsyllus) parado-
xus
Ceratophyllus (Paracallopsylla) kazna-
kovi
б) Блохи барсуков (Meles)
Chaetopsylla trichosa
* Paraceras flabellum
„ crispus
„ melis
в) Блохи лисиц (Vulpes)
* Chaetopsylla homoeus
„ giobiceps
г) Блохи медведей (Ursus)
* Chaetopsylla tuberculaticeps
„ hyaenae
д) Блохи, встречающиеся на
разных видах хищных (ис-
тинный хозяин не выяснен)
Echidnophaga popovi
* Chaetopsylla homoeus
* „ appropinquans
• alia
* „ mikado
„ lasius
8. Блохи летучих мышей (Chiroptera)
* Все виды Ischnopsyllus * Все виды Rhinolophopsylla
„ „ Nycteridopsylla „ „ Araeopsylla
„ „ Myodopsylla
9. Блохи птиц (Aves)
- Ceratophyllus я garei borealis - Ceratophyllus maculatus * „ caliotes
я spinosus „ orites
я gallinae * „ farreni
У) fringillae „ (Orneacus) oreinus
я vagabundus « „ geminus
я avicitelli Myoctenopsylla arctica
я sinicus Dasypsyllus gallinulae
* riparius „ stejnegeri
У) Styx * Frontopsylla (Orfrontia) frontalis
я hirundinis „ „ cornuta
10. Сезонность распространения некоторых блох
Для всех видов блох могут быть установлены более или менее точно сроки вы-
хода их из коконов и большее или меньшее обилие их в те или иные сезоны года.
Однако у многих видов эта сезонность выражена сравнительно слабо, и они встре-
чаются в фазе имаго в течение круглого года, хотя и в разном количестве. Ниже
перечисляются те виды и роды блох, у которых сезонность отчетливо выражена. При
этом необходимо отметить, что приводимые данные относятся к основным или наибо-
лее важным для нас местообитаниям каждого вида, так как при проникновении в иные
местности (например, в горы или в широты с иным климатом) у блох могут сдвигаться
сроки их сезонной приуроченности. Затем следует отметить, что почти все блохи
в местностях с жарким и сухим климатом обнаруживают некоторую депрессию числен-
ности и активности в жаркие летние месяцы.
У видов, не включенных в приводимые списки, сезонность менее выражена или
менее известна.
а) Блох и, встречающиеся в хо-
лодное время года — с осени
до весны, а летом исчезающие (или
почти исчезающие), условно назы-
ваются „зимними видамиа, хотя мно-
гие из них имеют осенний или весен-
ний пик численности.
Vermipsylla alacurt
„ dorcadia
Coptopsylla — все виды
Oropsylla ilovaiskii 1
Frontopsylla semura
Paradoxopsyllus — почти все виды
Brachyctenonotus myospalacis
Wagnerina — все виды
Rhadinopsylla — почти все виды
Paraneopsylla — все виды
1 Исчезает ли этот вид летом в основных районах своего распространения (в пре-
делах ареала крупных сусликов), еще необходимо выяснить.
260
Catallagia — все виды
Saphiopsylla nupera
Stenoponia — все виды
Nycteridopsylla — все известные в
СССР виды
б) Блохи, встречающиеся пре-
имущественно в холодное
время года, имеющие резкий осен-
ний (а часто и весенний) подъем чис-
ленности, но сохраняющиеся и в дру-
гие сезоны.
Tarsopsylla octodecimdentata
Oropsylla silantiewi
Amphipsylla — все виды
Rostropsylla daca
Leptopsylla bidentata
Ctenophthalmus — многие виды
Neopsylla setosa и, вероятно, другие
виды этого рода
в) Блохи, имеющие осенние
или весенние подъемы чис-
ленности, исчезающие зи-
мой.
Echidnophaga — вероятно, все виды
г) Блохи, имеющие подъем чис-
ленности в теплое время г о-
д а („летние виды*).
Pulex irritans
Ctenocephalides — вероятно, все виды
Xenopsylla — „
Ceratophyllus tesquorum и, вероятно,
многие другие виды этого рода
Doratopsylla dasycnemus и, вероятно,
многие другие виды этого рода
Palaeopsylla sorecis и, вероятно, многие
другие виды этого рода
11. Распределение блох по группам: „блохи гнезда* и „блохи шерсти*
В приводимые списки включены некоторые наиболее характерные случаи распре-
деления блох по указанным группам. Для прочих видов это разграничение не может
быть сделано в отдельных случаях из-за недостаточной их изученности. Надо учиты-
вать, что многие виды обнаруживают переходные свойства в отношении большей или
меньшей привязанности к телу хозяина и потому отнесение к той или иной группе
в известной мере условно, тем более что частота и продолжительность пребывания их
на хозяине резко зависят от температурных и других условий среды.
а) Блохи гнезда или жилья хо-
зяина — нападающие на хозяина
лишь на короткие сроки.
Pulex irritans
Ceratophyllus s. str. (птичьи блохи)
Ophthalmopsylla — вероятно, все виды
Ctenophthalmus— большинство видов
Rhadinopsylla — вероятно, все виды
Neopsylla — „
Hystrichopsylla — „ „ „
Stenoponia— „ - •
б) „Блохи шерсти* — находящиеся
на теле хозяина более продолжитель-
ное время, а некоторые виды — значи-
тельную часть жизни (в фазе имаго).
Ctenocephalides — вероятно, все виды
Xenopsylla — „
Synosternus — „
Pariodontis — „
Chaetopsylla — „ „
Tarsopsylla — „
Myoxopsylla — вероятно, все виды
Paraceras — „ , „
Oropsylla — „
Ceratophyllus s. 1. — многие виды из
числа паразитов грызунов и других
млекопитающих
Mesopsylla — вероятно, все виды
Amphipsylla — „ .
Leptopsylla — „
Pectinoctenus — „ „ „
Ischnopsyllus — „ „
Rhinolophopsylla — вероятно, все виды
Nycteridopsylla — > „ „
Myodopsylla — ,
Araeopsylla — „
в) Полустационарные парази-
ты — надолго прикрепляющиеся к ко-
же хозяина
Echidnophaga gallinacea
Vermipsylla alacurt
» dorcadia
18 Определитель блох
УКАЗАТЕЛЬ ЛИТЕРАТУРЫ
А. Каталоги
1. Вагнер Ю. Н. (Wegner J.), b'atalcg der palaearktischen Aphanipteren, 1—55, Wien,
1930.
2. В а г н e p Ю. H. (W’agner J.), Добавления к предыдущему каталогу, I—IV, Kono-
wia, 10, 96-100, 1931; 12, 212-216, 1933; 14, 217-224, 1935; 17, 8-18, 1938.
3. Иофф И. Г., Ростигаев Б. А., Пятое добавление к Вагнеровскому каталогу
палеарктических Aphaniptera. Эктопаразиты, 2, 166—187, 1950.
4. Тиф лов В. Е., Библиография по блохам СССР, Вести, микробиол., эпидемиол.
и паразитол., 15, I, 131—141, 1936.
5. Тиф лов В. Е., Библиография по блохам СССР. Первое добавление. Эктопара-
зиты, 2, 188-197, 1950.
6. Dalia Torre С., Aphaniptera erbis terrarum. Berichte des Naturwiss., Med. Verei-
nes in Innsbruck, 39, 1—29, 1924.
7. Jell i son W., Good N., Index to the literature of Siphonaptera of North America,
National institute of health bull., 178, 1 —193, Washington, 1942.
8. Lima A. da Costa, Hathaway C., Pulgas. (библиография, каталог, хозяева),
Monografias do inst. Oswaldo Cruz, 4, 1—522, Rio de Janeiro, 1946
9. Liu C.. Catalogue of chirese Siphonaptera, Lingnan science journal, Canton, China,
15, 3, 379-390; 4, 583—594, 1936.
Б. Определители
10. Apr ир on у ло А. И., Блохи (Aphaniptera) Закавказья (краткие определитель-
ные таблицы), Тр. Азербайд. ин-та микробиол. и эпидемиол., 5, 1, 119—215, 1935
(поправки — см. [152 и 3]).
11. Ар гиропул о А. И., Aphaniptera — блохи. В кн.: „ Определитель насекомых
Европейской части СССР4*, 599—610, Сельхсзгиз, 1948 (поправки — см. [3]).
12. В а г н е р Ю. Н., Аналитические таблицы для определения видов Aphaniptera,
живущих на Muridae, Параз. сборник, изд. АН СССР, 1, 97—192, 1930 (поправ-
ки — см. [3]).
13. Д арская Н. Ф., Определитель птичьих блох рода Ceratophyllus, Эктопаразиты,
2, 85-105, 1950.
14. Иофф И. Г., Тиф лов В. Е., Пособие для определения блох (Aphaniptera) юго-
востока Европейской части СССР, 1—116, Саратов, 1938 (поправки — см. [3]).
15. Тарбинский С., Ион О., Вагнер Ю., Определитель насекомых, 87—113,
1927.
16. Jordan К., Suctoria (Fleas) в кн.: Smart, „А handbook for the identification of
insects of medical importance, second edition", 211—245, 1948.
17. W a g n e r J., Flohe, Die Tierwelt Mitteleuropas, 17, 1—24, Leipzig, 1936.
В. Монографии и руководства
18. Иофф И. Г., Вопросы экологии блох в связи с их эпидемиологическим значе-
нием, 1—116, Пятигорск, Крайгиз, 1941.
19. Иофф И. Г., Aphaniptera Киргизии, Эктопаразиты, 1, 1—212, изд. АМН СССР,
1949 (поправки — см. [3]).
20. Павловский Е. Н., Наставление к собиранию и исследованию блох (Aphani-
ptera), Наставления для собирания зоол. коллекц., изд. Зоол. муз., АН СССР, 15,
1—40, 1927; Вестн. микробиол. и эпидемиол., 6, 2, 191—220, 1927.
21. П а в л о в с к и й Е. Н., Руководство по паразитологии человека, 2, 668—702,
1948.
22. Шир а но вич П. И., Временная инструкция по сбору и исследоганию экто-
паразитов, 1—22, Ростов н, Д, 1949.
23. Ewing Н., Fox I г., The f’.eas of North America, U. S. dep. of agric. miscellaneous
publication, 500, 1 — 1.42, 1943.
262
24. Fox I г., Fleas of eastern United States, 1 — 191, Ames, 1940.
25. Holland G., The Siphonaptera of Canada, 1—306, Kamloops, 1949.
26. Hubbard C., Fleas of Western North America, 1—533, Ames, 1947.
27. Liu C., The fleas of China, The Philippine journ. of science, 70, 1, 1—122, Manila*
1939.
28. Peus F., Die Flohe, 1 — 106, Leipzig, 1938.
29. Rothschild N., A synopsis of British Siphonaptera, The entom. monthly magazine
third series, 3, 49—112, 1915.
30. Wagner J., Aphaniptera, Klassen und Ordnungen des Tierreichs, 13, f, 1—114,
Leipzig, 1939.
Г. Специальные работы по изучению блох Восточной Азии
и некоторых сопредельных стран
31. Бром И. П., В овчин ск а я 3. М., Федорова Л. В., О роли хищных млеко-
питающих в распространении блох грызунов, Зоол. жури., 27, 2, 167—174, 1948.
32. Бычков В. А., О роли блох в хранении и рассеивании чумного вируса, Сборник,
посвященный Е. Н. Павловскому, „Паразиты, переносчики и ядовитые животные
ВИЭМ, 1935.
33. Вагнер Ю. Н., Ceratophyllus calcarifer sp. п., Ежегод. зоол. муз., АН СССР, 18
263-267, 1913.
34. Вагнер Ю. Н., Материалы по фауне Aphaniptera Якутии. Материалы комиссии
по изучению Якутской АССР, 16, 1—12, изд. АН СССР, 1927.
35. Во вч и иска я 3. М., Новый вид блохи Aphaniptera из Забайкалья, Изв.
Иркутск, гос. противэчум. ин-та Сибири и Дальн. Востока, 5, 192—193, 1944.
36— 37. Вовчи некая 3. М., Материалы к фауне Aphaniptera Западного Забайкалья,
Изв. Иркутск, гос. противочум. ин-та Сибири и Дальн. Востока, 5, 176—183, 1944.
38. Вовчинская 3. М., О л овин а М. Д., Материалы по сезонному изменению
видового и количественного состава блох на тарбагане и в его гнезде, Изз.
Иркутск, гос. противочум. ин-та Сибири и Дальн. Востока, 6, 171 — 177, 1946.
39. Гершкович Н. Л., Материалы по фауне блох серой крысы Хабаровского
края, Изв. Иркутск, гос. противочум. ин-та Сибири и Дальн. Востока, 7, 230—236,
1949.
40. Гершкович Н. Л., Материалы к фауне Aphaniptera Дальнего Востока, 1948.
41. Дар с кая Н. Ф., Блохи даурского суслика (Citellus dauricus), Изв. Иркутск, гос.
противочум. ин-та Сибири и Дальн. Востока, 7, 200—213, 1949.
42. Дар с кая Н. Ф., Новые виды блох лесных полевок (Clethrionomys) из горной
тайги Северной Кореи, ДАН СССР, 67, 5, 949—952, 1919.
43. Д арская Н. Ф., Новые виды Aphaniptera из Северной Кореи, Доклады АН
СССР, 68, 2, 429-432, 1949.
44. Дар ска я Н. Ф., Блохи даурской пищухи и ее нор, 1945.
45. Дубинин В. Б. и Дубинина М. Н., Паразитофауна млекопитающих даурской
степи, Материалы по грызунам, 4, изд. Моск, об-ва испыт. природы, 1951.
46. Дубинин В. Б., Птицы даурской степи и их роль в распространении блох,
Изв. Иркутск, гос. противочум. ин-та Сибири и Дальн. Востока, 7, 237—253, 1949.
47. Дудченко И. С., Вши и блохи у тарбаганов и других грызунов в Забайкаль-
ском очаге эпидемической чумы людей, Мед. обозр.. 5—6, 1916.
48. Емельянова Н. Д., Наблюдения над миграцией блох Oropsylla silantiewi из
нор тарбагана (Marmota sibirica R.), Изв. Иркутск, гос. противэчум. ин-та Сибири
и Дальн. Востока, 5, 194—197, 1944.
49. Емельянова Н. Д., О видовом и количественном составе блох, встречаю-
щихся в Забайкалье во входах нор грызунов и на поверхности земли, Изв.
Иркутск, гос. противочум. ин-та Сибири и Дальн. Востока, 6, 178—183, 1946.
50. Заболотный Д. К., Происхождение эндемичной чумы, A’anales de Flnst. Pasteur,
37, 618-626, 1923.
51. И о ф ф И. Г., К изучению фауны блох Алтая, Изв. АН Казахск. ССР, серия па-
разитол., 8, 41—53, 1950.
52— 54. Иофф И. Г., Дубинин В. Б., Желудкова О. И., К изучению блох Уссурий-
ско-Приханкайской равнины и хребта Сихотэ-Алиня, Эктопаразиты, 2, 30—43,
1950.
55. Иофф И. Г., Эрлих 3. С., О блохе забайкальского суслика, Изв. Иркутск, гос.
противочум. ин-та Сибири и Дальн. Востока, 5, 184 — 191, 1914. .
56. Колосов А. М., Распространение, биология и эпидемиологическое значение крыс
на Дальнем Востоке, Тр. дальневост. базы. АН, Серия .зоол., 1, 1—62, 1945 (по-
правки — см. [3]). ’
57. Кучерук В. В., Заметки о блохах Маньчжурии по иностранной литературе,
Вопр. краевой, общей и эксперимент, паразиюл., изд. АМН СССР, 4, 224—230.
1949 (поправки — см. [3]). 4
18:’ 263
58. Куче рук В. В., Блохи Маньчжурии и их эпидемиологическое значение по Дан-
ным японской литературы, Исслед. по краевой, эксперимент, и описательн. пара-
зитол., изд. АМН СССР, 6, 5—15, 1949.
59. Марико вс кий П. И., Материалы по изучению фауны блох (Aphaniptera)
Дальневосточного края, Вести, дальневост. фил. АН, 13, 124—127, 1935 (поправ-
ки — см. [3]).
60. Марико в ск и й П. И., Новый вид Aphaniptera из Дальневосточного края, Изв.
Гос. протпвочум. ин-та Сибири и ДВК, 2, 39—42, 1935; Вести, микробиол., эпиде-
миол. и паразитол., 15, 3—4, 430—433, 1936.
61. Мариковский П. И., К изучению Aphaniptera Дальневосточного края. Сооб-
щение II. Вести, дальневост. фил. АН, 27, 148—156, 1937 (поправки — см. [3]).
62. Мик ул ин М. А., Материалы к познанию афаниптерофауны юго-востока
Казахстана, Тр. Средне-азиатск. научно-исслед. противочум. ин-та, 1, 1951.
63. Павлов Е. И., Степные грызуны и их естественные вредители забайкальского
эндемического очага чумы, их биология и роль в распространении чумы, Сбор-
ник работ противочум. орган. Вост.-Сиб. края за 1932—1933 годы, Иркутск, 101—130,
1934 (1935).
54— 65. Павлов Е. И., Отчет о зоологической работе междуведомственной полевой
противочумной организации в забайкальском очаге чумы за период с 23. VIII по
20.IX. 1935 г., Изв. Гос. противочум. ин-та Сибири и ДВК, 3, 20—31, 1936.
66. Павлов Е. И., Стадная полевка (Stenocranius gregalis), ее жилище и сезонные
изменения видового и количественного состава блох гнезда, Изв. Иркутск. Гос.
противочум. ин-та Сибири и Дальн. Востока, 7, 259—266, 1949.
67. Попов В., Материалы по изучению фауны блох (Aphaniptera)Западной Сибири.
Эпидемиология и профилактика инфекций, сб. Томск, ин-та эпидемиол.
и микробиол., 80—84, 1945.
68. Рябов Н., Материалы к биологии тарбагана в зимний период, Изв. Иркутск.
Гос. противочум. ин-та Сибири и Дальн. Востока, 6, 184—194, 1946.
69. Сазонова О. Н., О блохах с грызунов и насекомоядных низовьев Иртыша. Пара-
зитология и трансмиссивные болезни, Нов. мед., 5,29—30,1947 (поправки—см. [3]).
70—71. Скалой В. Н., О сборах паразитов птиц, Изв. Гос.противочум. ин-та Сибири
и ДВК, 2, 185-186, 1935.
72. Скалой В. Н., Тарасов П. П., К изучению причин и механизма чумной энзоотии
в Монголии и Забайкалье, Ученые записки Монгольск. Гос. ун-та им. маршала
Чойбалсана, 2, 2, 1—25, Улан-Батор, 1946.
73. Скалой О. И., Материалы к фауне блох (Aphaniptera) Сибири и Дальневосточ-
ного края, Изв. Гос. противочум. ин-та Сибири и ДВК, 2, 24—38, 1935.
74. Скалой О. И., Материалы к фауне блох (Aphaniptera) Сибири и Дальневосточ-
ного края, Изв. Гос. противочум. ин-та Сибири и ДВК, 4, 46—56, 1936.
74а. Скалой О. И., Краткая инструкция по сбору эктопаразитов, Научно - методич.
записки, 6, 100—104, 1940.
75. Скалой О. И., Новые виды блох из Сибири, Эктопаразиты, 2, 74—84, 1950.
76. Скалой О. И., К фауне паразитов промысловых животных Якутии, 1939.
77. Скалой О. И., Материалы по фауне паразитов млекопитающих Кондо-Сось-
винского государственного заповедника. I. Aphaniptera, 1942.
78. Скалой О. И., Новые виды блох с пищух из Монголии, 1950.
79. Слепцов М., К биологии уссурийской мыши-малютки (Micromys minutus).
Фауна и экология грызунов, Материалы по грызунам, 2, 69—100, изд. Московск.
об-ва испыт. природы, 1947 (поправки — см. [3]).
80— 81. Сычевский П. Т., Колосов А. М., Блоха грызунов южного приморья (Даль-
ний Восток), Изв. Иркутск. Гос. противочум. ин-та Сибири и Дальн. Востока,
7, 214—229, 1949.
82. Тиф лов В. Е., К изучению фауны блох (Aphaniptera) Восточной части Казах-
ской ССР. Материалы по вредителям животноводства и фауне преимущественно
Южного Казахстана, Тр. Казахск. фил. АН СССР, 2, 189—219, 1937.
S3. Т и ф л о в В. Е., Павлов Е. И., Материалы к изучению фауны блох Забайкалья,
Вести, микробиол., эпидемиол. и паразитол., 15, 1, 79—88, 1936.
84. Федина О. А., Блохи Алма-Атинской области, Изв. АН Казахск. ССР, 43,
Серия паразитол., 5, 76—91, 1948.
85—86. Федорова Л. В., Посезонное изменение видового и количественного состава
блох на полевке Брандта (Phaiom/s brandti) и в ее гнезде, Изв. Иркутск, гос.
противочум. ин-та Сибири и Дальн. Востока, 7, 194—199, 1949.
87. Ши ран о вич П. И., К фауне Apnaniptera Казахстана, Изв. АН Казахск. ССР,
43, Серия паразитол., 5, 92—99, 1948.
88. Шпрингольц-Шмидт А. И., Материалы по фауне эктопаразитов грызунов
Дальневосточного края, Изв. Гос. противочум. ин-та Сибири и ДВК, 4, 36—45,
89. Этмар Г. Г., Доклад о деятельности отряда по наблюдению за тарабаганьим
промыслом и т. д., Мед. журн. Забайкальск, об-ва врачей, 1—3, 1—40, 1923.J
264
90. Dampf A., Eine neue Nycteridopsylla aus Shanghai, Zool. Anzeiger, 36, 11 15,
1910.
91. Hsu Y., Two new species of insect parasites of the bat in Soochow, Peking nat.
hist, bull., 9, 293—298, 1935.
92. Jordan К.» On fleas collected by dr. H. M. Jettmar in Mongolia and Manchuria
in 1927 and 1928; Novit. zool., 35, 155—164, 1929.
93. Jordan K., Siphonaptera collected by H. Stevens... in Yunnan and Szechuan,
Novit. zool., 38, 276—290, 1932.
94. Jordan K., Three new bird-fleas from Kashmir, Novit zool., 40, 299—306, 1937.
95. Jordan K., A new flea from China, Novit. zool., 40, 295—296, 1937.
96. Jordan K., Siphonaptera collected by dr. Gerd Heinrich in Birma, Novit. zoo’.,
41, 1939.
97. Jordan K-, A new rat-flea from China, The entomologist, 74, 85—86, 1941.
98. Jordan K-, On four eight-combed Chinese bat-fleas of the genus Ischnopsyllus in the
collection of the british museum. Parasitology, 33, 4, 363—372, 1941.
99. Jordan K., Siphonaptera collected by Mr. J. L. Cha worth-Musters in north-eastern
Afghanistan, Proceed, zool. soc., Part III, 114, 3, 360—367, 1944.
99a. Jordan K., On some palaearctic species Xenopsylla, The entomologist, 82,
111-115. 1949.
100. Jordan K. a. Rothschild N., New Siphonaptera from China, Proceed, of zool. soc,
London, 365—393, 1911.
101. Jordan K. a. Rothschild N., On some Siphonaptera collected by W. Riickbeil
in East Turkestan, Ectoparasites, 1—24, 1915.
102. Jordan K. a. R о t h sc h i 1 d N., On Ceratophyllus fasciatus and some allied indian
species of fleas, Ectoparasites, 178—216, 1921.
103. Jordan K. a. Rothschild N., On some Siphonaptera from the eastern hemisphere,
Ectoparasites, 293—308, 1923.
104. Liu C., On the unrecorded male of the bat-flea Ischnopsylla tateishii Sugimoto
(Siphonaptera), The China journal, 23, 306—310, 1935.
105. Liu C., A new Chinese flea, Peking nat. hist, bull., 9, 273—275, 1935.
106. Ono, О блохах, паразитирующих на мышевидных грызунах в районе Ченгочи,
Bull, of the inst. of scient. research. Manchoukuo, 6, 4, 487—497, 1942 (на япон-
ском языке).
107. Ono, Блохи провинции Дуннань уезда Боли (Туашань), Bull, of the inst. of scient.
research Manchoukuo, 6, 6, 903—917, 1942 (на японском языке).
108. Ono, Исследования блох мышевидных грызунов во время чумы в Чанчуне
в 1941 г., Bull, of the inst. of scient. research. Manchoukuo, 6, 6, 919—932, 1942
(на японском языке).
109. Ono, Kato, Блохи в Ченгочи, Bull, of the inst. of scient. research. Manchoukuo,
7, 1, 63—67, 1943 (на японском языке).
110. Ono, Kato, Сезонная динамика крысиных блох в Чангочи, Bull, of the inst.
of scient. research. Manchoukuo, 7, 5, 591—596, 1943 (на японском языке).
Ill. Rothschild N., Siphonaptera collected by Mr. Anderson in Japan in 1904, Proceed,
of the zool. soc., London, 627—629, 1908.
112. Rothschild N., Ceratophyllus silantiewi W., Entomol. monthly magazine, Sec. Ser.,
22, 141, 1911.
113. R о t h s c h i 1 d N., Description of three new species of Siphonaptera, In Clark and
Sowerby’s „Through Shen Kan“, Appendix, IV, 194—203, 1912.
114. R о t h s c h i 1 d N., A new palaearctic species of Xenopsylla, The Ent. Record., 25,
241, 1913.
115. Rothschild N., Two new palaearctic species of Rhadinopsylla, The entomologist,
66, 297, 1913.
116. Rothschild N., Five new Siphonaptera from Asiatic Russia collected by
W. Ruckbeil, The annals and magazine of natur. history (8), 12, 538—544, 1913.
117. Sugimoto M., Note on fleas in Formosa, Trans, nat. hist. soc. Formosa, 23,
116 —145, 1933 (на японском языке с английским резюме).
118. Wu С., Rat flea survey of Shanghai (и др. статьи о крысиных блохах Китая с об-
зором литературы), Reports National Quarantine Service, 3, 4, 5, 6, Shanghai, 1932—
1936.
Д. Некоторые другие работы
119. Аргиропуло А. И., Новые и малоизвестные виды блох (Aphaniptera) Кавказа,
Тр. зоол. сектора грузинск. фил. АН СССР, 2, 185—194, 1938.
120. А р г и р о п у л о А. И., До фауни Aphaniptera УРСР, 36ipHHK праць зоол. муз.
АН УРСР, 24, 15-21, 1941.
121. Аргиропуло А. И., Дубинин В., Новые и малоизвестные Aphaniptera (Insecta)
Средней Азии и Казахстана, Тр. Зоол. ин-та АН СССР, 7, 162—170, 1948
(поправки—см. [3]).
265
122. Вагнер Ю. Н., Афаниптерологические исследования, 1—5, Тр. Русск. энтомол.
об-ва, т.т. 23, 27, 31, 35, 36, 1889-1903.
123. Вагнер Ю. Н., Систематический обзор видов Aphaniptera, описанных до 1904г.,
Сем. Vermipsyllidae, Тр. Русск. энтомол. об-ва, 37, 439—471, 1906.
124. Вагнер Ю. Н.. То же. Сем. Pulicidae, Тр. Русск. энтомол. об-ва, 39, 508—569,
1910.
125. Вагнер Ю. Н., Заметка об Aphaniptera Крымского полуострова, Тр. муз. Тав-
рич. губ. зем. за 1914 г., 4, 1—6 (оттиск), 1916.
126. Вагнер Ю. Н., О новых палеарктических видах блох (Aphaniptera). I и II,
Ежегод. зоол. муз. АН, 30, 21 — 33 и 521—547, 1929.
127. Вагнер Ю. Н., К вопросу о соотношении между фауной паразитов и их хозяев
Тр. IV съезда акад, организаций, 2, 109—119, 1929.
128. Вагнер Ю. Н., Иофф И. Г., О блохах сусликов (и тушканчиков) в связи
с их ролью в распространении чумы в приволжских степях, Вестн. микробиэл.
и эпидемиол., 5, 1—2, 57—116, 1926.
129. Гребенюк Р. В., Вермипсиллез (алакурт)—кожно-эптомозная инвазия овец
высокогорных районов Киргизии, изд. Киргизск. фил. АН СССР, 1—16, 1948.
130. Дубинин В. Б., Географическое распространение и вероятные пути расселения
блох рода Pariodontis в связи с историей их хозяев (дикобразов), Доклады АН
СССР, 58, 7, 1557 — 1560, 1947.
131. Дудолкина Л. А., О паразите кур Ceratophyllus gallinae и близких видах блох,
Эктопаразиты, 2, 106—111, 1950.
132. Иофф И. Г., О новых видах блох в коллекции Зоологического музея АН
СССР, Ежегод. зоол. муз. АН СССР, 28, 407-439, 1927.
133. Иофф И. Г., Блохи сурка и желтого суслика, Вестн. микробиол. и эпидемиол.,
6, 3, 319 -328, 1927.
134. Иофф И. Г., Блохи тушканчиков (Dipodidae), Изв. Гос. микробиол. ин-та,
Ростов-на-Дону, 8, 6—28, 1929.
135. Иофф И. Г., Блохи степных хорьков, Изв. Гос. микробиол. ин-та, Ростов-на-
Дону, 8, 44-52, 1929.
136. Иофф И. Г., О выбирании блох из гнезд грызунов, Лабор. практика, 5, 5—7,
1929.
137. Иофф И. Г., О Xenopsylla conformis и некоторых близких видах блох, Zool.
Anzeiger, 92, 191—206, 1930.
138. Иофф И. Г., Блохи слепушонки, Вестн. микробиол., эпидемиол. и паразитол.,
14, 1, 79-86, 1935.
139. Иофф И. Г., К систематике блох из подсемейства Ceratophyllinae, Zeitschr. f.
Parasitenkunde, 9, 73—124, 1936.
140. Иофф И. Г., О географическом распространении сусликовых блох в связи
с историей расселения сусликов, Параз. сборник, изд. АН, 6, 313—361, 1935.
141. Иофф И. Г., О некоторых новых или малоизвестных видах блох (Aphaniptera),
Параз. сборник, изд. АН, 7, 210—229, 1939.
142. Иофф И. Г., Алакурт, Киргизск. фил. АН СССР, Тр. биол. ин-та, I,
111—135, 1947; Эктопаразиты, 2, 4—29, 1950.
143. Иофф И. Г., Новые афаниптерологические находки, Природа, 10, 80—81,
1948.
144. Иофф И. Г., К изучению фауны и экологии грызунов Кавказа и их эктопарази-
тов, Тр. Зоол. ин-та АН Груз. ССР, 8, 289—315, 1948 (1949).
144а. Иофф И. Г., Новые случаи видообразования у блох при перемене хозяина, Док-
лады АН СССР, 89, 1, 189—192, 1953.
145. Иофф И. Г., АргиропулоА. И., Блохи Армении, Zeitschr. f. Parasitenkunde,
7, 2, 138-166, 1934.
145а. Иофф И., Гершкович Н., ЗагнибородоваЕ., Лабунец Н., Лебе-
де в А., М и к у л и н М.» Скалон О., Т и ф л о в В., Шварц Е., Юркина В.,
Я г у б я н ц И., Новые виды блох (Suctoria-Aphaniptera). Сообщение 3. Мед. пара-
зит. и паразитар. болезни, 5, 460—465, 1953.
146. Иофф И. Г., Тиф лов В. Е., К фауне и экологии блох лесостепи, Параз.
сборник, 1, 193-227, 1930.
147. Иофф И. Г., Тиф лов В. Е., Род Stenoponia J. et R., Вестн. микробиол.,
эпидемиол. и паразитол., 12, 3, 199—210, 1933 (1934).
148. Иофф И. Г., Тиф лов В. Е., Роды Coptopsylla J. et R. и Chaetopsylla Koh.,
Вестн. микробиол., эпидемиол. и паразитол., 12, 4, 303—321, 1933 (1934).
149. Иофф И. Г., Тиф лов В. Е., Род Amphipsylla W., Вестн. микробиол., эпидемиол.
и паразитол., 16, 3—4, 401—437, 1937 (1939) (поправки — см. [150 и 3]).
150. Иофф И. Г., Тиф лов В. Е., Дополнительные замечания о роде Coptopsylla
Явления паразитарной кастрации у блох, Вестн. микробиол., эпидемиол. и пара-
зитол., 19, 1, 98—102, 1940.
151. И о ф ф И. Г., Т и ф л о в В. Е., Род Rhadinopsylla J, et R., Эктопаразиты, 2,
44-73, 1950.
266
152. Иофф И. Г., Тиф лов В. Е., О двух недавно изданных пособиях для опреде-
ления блох, Вестн. микробиол., эпидемиол. и паразитол., 15, 3—4, 485—489,
1936 (1937).
153. Иофф И. Г., Тиф лов В. Е., Аргиропуло А. И., Федина О. А.,
Дудолкина Л. А., Ширанович П. И., Новые вид л блох (Aphaniptera),
Мед. паразитол. и паразитар. бол., 15, 4, 85—94, 1945 (исправление опечаток —
см. в. 6, стр. 94, 1945).
154. Иофф И. Г., Скалой О. И., В о в ч и н с к а я 3. М., Д а р с к а я Н. Ф., Емель-
янова Н. Д., И с а е в а-Г у р в и ч Э., Рост ига ев Б. А., Савенко Р. В.,
Соснина Е. Ф., Сыче вс кий П. Т., Новые виды блох (Aphaniptera). Сообще-
ние П, Мед. паразитол. и паразит, бол., 19, 3, 268—273, 1950 (исправление
опечаток — см. в. 6, 1950).
155. Огнев С. И., Звери СССР и прилежащих стран, 1—7, 1923—1950.
156. Савенко Р. Ф., Два новых вида Aphaniptera из Грузии, Сообщ. АН Груз. ССР, 9,
501-505, 1948.
157. Скрябин К. И., Блохи человека и домашних животных и их санитарно-пато-
логическое значение, Арх. вет. наук, 12, 1178—1191, 1913.
158. Тифлов В. Е., Колпакова С. А., Блохи лисы и т. д., Вестн. микробиол.
эпидемиол. и паразитол., 15, 413—423, 1935 (1937). ’
159. Тифлов В. Е., Потапов В. О., О миграции блох суслика, Вестн. микро-
биол., эпидемиол. и паразитол., 16, 3—4, 438—466, 1937 (1939).
160. Уигглсуорс В., Физиология насекомых, 1—148, 1937.
161. Crampton G., A phylogenetic study of the posterior metathoracic and basal abdo-
minal structures of insects, Journ. New York ent. soc., 39, 323—357, 1931.
162. Dampf A., Eine neue Palaeopsylla-Art aus Ostpreussen, Schrifften der physikalisch-
okonomischcn Gesellschaft zu Konigsberg, 51, 324—330, 1910.
163. Dampf A., Mesopsylla eucta n. g. n. so. ein neuer Floh von der Springmaus (Ala-
ctaga jaculus) nebst Beitragen zur Kenntnis der Gattung Palaeopsylla, Zoolog. Jahrbii-
cher, Suppl 12, 609—664, 1910.
164. Dampf A., Zur Kenntnis der Aphanipteren-fauna Westdeutschlands, mit besonderer
Beriicksichtigung der acht kammigen Ischnopsyllus-Arten. Berichten uber d. Versam-
mlungen d. Botanisch. u. Zoologisch. Vereins f. Rheinland—Westfalen, 73—113, Bonn
(1911), 1912.
165. Dampf A., Eine neue Aphanipteren-Art aus dem Kaukasus (обзор самцов
Ischnopsyllus), Русск. энтомол. обозрение, 12, 41—59, 1912.
166. Edney E., An apparatus for handling small living insects, Bull. ent. research, 37,
83-87, 1946.
167. Es trade F., Technique et appareil pour la capture des puces dans les poussieres
et debris de cereales, Bull. soc. path, exot., 27, 458—451, 1934.
168. Ewing H., Two new genera and one new soecies of Siphonaptera, Proc. ent. soc.
Wash., 40, 93-95, 1938.
169. Fox Ir., New species and a new genus of nearctic Siphonaptera, Proc. ent. soc.
Wash., 41, 2, 45-50, 1939.
170. Fox Ir., Notes on north american Dolichopsyllid Siphonaptera, Proc. ent. soc. Wash.,
42, 3, 64-69, 1940.
171. Fox Ir., Th£ siphonapteran thorax, Proc. ent. soc. Wash., 43, 1, 6—10, 1941.
172. Holland G., Notes on some northern Canadian Siphonaptera, with the description of
a new species, The Canadian entomologist, 76, 242—246, 1944.
172a. Holland G., Notes on Megabothris asio and M. calcarifer, with the description of
a new subspecies, The Canadian entomologist, 82, 126—133, 1950.
173. Jellison W., Siphonaptera: The genera Amphalius and Ctenophyllus in North Ame-
rica, Publ. health reports, 56, 49, 2341—2349, 1941.
174. Jellison W., Siphonaptera: The genus Oropsylla in North America, Journ. of para-
sitol, 31, 2, 83-97, 1945.
175. Jellison W., Kohls G., Siphonaptera: A list of alaskan fleas, Publ. health rep.,
54, 2020-2023, 1939.
176. Jordan K., On Xenopsylla and allied genera of Siphonaptera. Ill Internation. Ento-
mol. Kongress (Zurich, Juli 1925), 2, 593—627, 1926.
177. Jordan K., On four new palaearctic bat-fleas etc., Eos, 18, 243—250, 1942.
178. Jordan K., On some phylogenetic problems within the order of Siphonaptera (Suc-
toria), Tijdschrift voor entomol., 88, 79—93 (1945), 1947.
179. Jordan К., а также R о t h s c h i 1 d N. и их совместные работы в журнале
Novitates Zoologicae, в томах 5—41 за 1898—1939 гг. (при ссылке на этот номер
в скобках указывается том журнала).
180. Jordan К., а также Rothschild N. и их совместные работы в Ectoparasites, 1,
1915-1924.
181. Jordan К. a. R о t h s с h i 1 d N., Revision of the non-combed eyed Siphonaptera,
Parasitology, 1, 1—100, 1908.
182. Jordan K. a. Rothschild N., On the genus Amphipsylla, The Zoologist, 869,
401—408, 1913.
257
183. Kohls G., Siphonaptera: A study of the species infesting wild hares and ratbits
of N. America, National inst. of health bull., 175, 1—34, Washington, 1940.
183a. Nicholson H. a. Vetter M., A lethal trap for capturing small mammals with
their ectoparasites, Joum. of parasitol., 36, 235—237, 1950.
184. Rothschild M., Megabothris rectangulatus a flea new to Great Britain, Novit.
zool., 39, 270—274, 1936.
185. Rothschild N., Some new exotic fleas, Ent. mon. magaz., ser. 2, 17,
221—224, 1906.
186. Rothschild N., On Ctenopsyllus spectabilis and some closely aled species, Ent.
mon. magaz., ser. 2, 20, 184—186, 1909.
187. Rothschild N., A synopsis of the Fleas found on Mus norwegicus etc., Bull,
ent. research, 1, 89—98, 1910.
188. Rothschild N, 1898-1915—[cm. 179].
189. Snodgrass R., The feeding apparatus of biting and sucking insects affecting man
and animals, Smithsonian miscellaneous collection, 104, 7, 83—94, 1944.
190. Snodgrass R., The skeletal anatomy of fleas (Siphonaptera), Smithsonian miscel-
laneous collections, 104, 18, 1 89, 1946.
191. Wagner J., Zwei neue palaearktische Floh-Arten (Stenoponia conspecta und
Neopsylla democratica), Konowia, 5, 79—83, 1926.
192. Wagner J., Uber die Einteilung der Gattung Ceratophyllus Curtis, Konowia, 6,
101-113, 1927.
193. Wagner J., Notiz liber Doratopsylla curvata Roths, und D. birulai loff (Corro-
dopsylla subg. n.), Konowia, 8, 316—318, 1929.
194. Wagner J., Neue Floharten vom Pamir, Konowia, 9, 279—281, 1930.
195. Wagner J., Zur Morphologie der letzten Abdominalsegmente der Flohe. Zool.
Jahrbiich. Abt. f. Anat. u. Ontog. der Tiere, 56, 54—120, 1932; 57, 365—374, 1933.
196. Wagner J., Fiinf neue palaearctische Flohe, Konowia, 11, 273—280, 1932.
197. Wagner J., Weitere Einteilung der Cattung Ceratophyllus, Konowia, 13,
260-263, 1934.
198. Wagner J., Einige neue Oder wenig bekannte Floharten, Zeitschr. f. Parasiten-
kunde, 8, 332—350, 1936.
199. Wagner J., Gattung Ctenophthalmus und ihre Einteilung (Aphaniptera), Zeitschr.
f. Parasitenkunde, 11, 4, 593—606, 1940.
УКАЗАТЕЛЬ НАЗВАНИЙ БЛОХ
Цифры указывают страницы текста. Ссылки на названия, упомянутые в подписях под
рисунками и в списках рисунков, в указатель не внесены.
Atyphloceras...............................122
Augustsonius............................. 88
avicitelli..............................79, 82
П7,
117
115
70
255
111
. 59, 61
. . 256
. . 78
. 43, 64
. . 93
. 38, 58
. . 88
65, 256
. 60, 61
. . 118
. . 112
71
. 70,
. 68, 74
87
76
abagaitui . . .
abei..........
abramovi . .
acanthina . .
accola . . . .
Achae topsy Ila
adalis ....
advenarius . .
Aenigmopsylla
afgana ....
alacurt ....
alatau ....
alaskensis . .
alia..........
aliena ....
altaica ....
altaicus . . .
Amalaraeus
ambigua . . .
Amononsyllus
Amphalius........................ 43, 49, 64
Amohipsylla . . 5, 26,30, 34, 38, 42, 48, 94, 255
anadyrus ....
anceps .........
andruschkoi . .
angusta ....
anisus..........
anoma...........
Anomiopsyllinae
antiqua ....
appropinquans .
apscheronica . .
Araeopsylla . .
aralis .........
Archaeopsylla .
Archaeopsyllinae
arctica.........
Arctopsylla . . .
argoi...........
argus...........
ariana..........
armatus ....
armeniaca . . .
arnoldi ....
arvalis..............................105, 106
arvicolae.....................• . . . . 74
asiatica (Amphipsvlla)................96, 97
„ (Oropsylla) .... 44, 65, 66, 72, 256
asiaticus..............
aspalacis (Amphipsylla)
„ (Rhadinopsylla)
assimilis..............
astia..................
255
100
94
111
255
118
105
104
. . . 60, 61
. . . . 93
. . 44, 126
. . . . 69
.41, 47, 53
. . . . 53
. . . . 84
. 26, 59, 60
. . .98, 99
. . . . 75
. . 115, 116
. . . . 89
. ... 116
. . . . 91
76,
105,
75
100
111
106
55
baikal....................................79, 88
baikalensis.................................255
beklemischevi................................109
Beringiopsylla...............................109
bidentata........................ 101, 102, 225
bidentatiformis....................115, 116, 118
bifallax.................................... 78
birulai......................................108
bishopi.....................................121
bivirgis....................................114
borealis.................................... 79
botis (Amphipsylla).........................255
„ (Frontopsylla)................ 85, 86, 87
Brachyctenonotus.........................44, 92
brasiliensis................................ 55
breviatus.......................... 38, 39, 106
calcarifer................................. 77
caliotes............................... 83, 255
Callopsylla................... 38, 66, 67, 72
canis............................. 53, 54, 255
caprae...................................... 54
caracoi ..........................255
casis......................................101
caspica..................................... 57
caspius.................................... 73
casta.......................................112
Catallagia..........................41, 48, 119
caucasica (Frontopsylla).................... 87
„ (Rhadinopsylla)...................112
cedestis...................................114
Ceratophyllidae.................... 30, 63, 123
Ceratophyllinae........... 7, 34, 63, 64, 66
Ceratophylloidea............................ 58
Ceratoohyllus 4, 5, 6, 27, 32, 34, 38, 39, 44,
49, 66, 68, 78, 80, 255
certa....................................... 98
Chaetopsylla ... 6, 24, 38, 41, 47, 59, 256
cheopis..................................... 54
Chinopsvlla.................................109
Citellophilus............. 38, 66, 67, 70, 72
clarus..................................... 65
coangusta...................................100
coelestis..................................122
co mans....................................124
compar......................................118
concava.....................................112
conformis.................... 55, 56, 57, 255
congeneroides.........................105, 105
conothoae.................................. 89
269
Conothobius............................ 89
consor.....................................255
conspecta........................... 122, 256
contigua............................. 95, 255
conveniens............................ 90 91
cophinus.................................*255
Coptopsylla................. 23, 37, 38, 40, 58
Cop topsy llidae.......................... 58
coreana....................................108
cornuta..................................... 88
Corrodopsylla..............................108
Cratinius................................... 88
crassus.................................... 65
crisp us................................... 64
crudelis...................................105
Ctenocephalides............. 41, 47, 53, 255
Ctenocephalus............................’ 255
Ctenonotus.................................255
ctenophorus.............................103
Ctenophthalmidae........................103
Ctenophthalminae........................105
Ctenophthalmus............ 45, 50, 105, 255
Ctenophyllus................ 4, 42, 48, 88, 89
Ctenopsyllus........................101, 255
cuniculi................................. 57
curvispinus...........................91, 256
cuspis....................................’ Ю7
custodis..................................... 90
Cystipsylla.................................. 92
daca................................. 66, 255
dacenkoi ............................119, 120
dacus......................................255
daea.......................................101
dahurica..............................ПО, 112
dahuricus............................ 107, 255
dampfi (Doratopsylla)......................108
(Paraneopsylla)......................119
dashidorzhii............................... 91
dasycnemus............................6, 107
Dasypsyllus...........................44, 83
deminuta...................................*97
democratica................................116
demotus.................................... 74
Dcrmatophilus
Desertopsylla .
desertus . .
Diamanus . .
dicl mica . .
dictena . .
dilatata . . .
dilatatus . . .
dilatus . . . .
dinormus . . .
dissimilis . . .
dives . . . .
dogieli . . .
dolabris . . .
dolichus . . .
dolosus . . .
Doratopsylla .
Dorcadia . . .
dorcadia . . .
dubiosa . . .
dumalis . . .
dux...........
dzetysuensis .
. . 255
. . 94
. . 76
. 65, 66
. . 101
. . 125
99
105, 107
.... 74
.... 110
.... 62
.... 73
. .105, 107
124, 125, 255
. . 46, 107
.... 59
.... 59
.... 88
... 97, 99
. .105, 107
.... 71
Echidnophaga................ 40, 52, 56, 255
elana................................. 65, 256
elata................ 6, 84, 85, 86, 87, 255
elatoides.................................. 85
elatus.................................6, 255
ellobii................................ 70, 255
elongatus..................................124
Eopsylla.......................... 27, 42, 105
erinacei............................... 53
eucta...................................93, 94
Euctenophthalmus...........................105
eusarca....................................125
ewingi................................. 96, 255
extrema.................................... 92
f al lax......................................102
famulus.......................................255
farreni....................................... 83
fasciatus..................................68, 69
favosus.......................................121
felis................................ 53, 54, 255
fetisovi..................................119, 120
fidus......................................... 69
flabellum .................................63, 64
formoso vi....................................122
fringillae.................................... 81
frontalis..................................... 88
Frontopsylla . . 4, 6, 27, 38, 42, 48, 84, 255
fulvus........................................ 52
gaiskii.................................72, 73
galba................................. 125, 256
galea......................................117
gallinacea..............................52, 53
gallinae . . . ................... 79, 80, 81
gallinulae................................. 83
garei ...................................6, 79
geminus.................................... 73
georgica................................... 99
gerbilli................................55, 57
Gerbillophilus................. 4, 38, 67, 69
gestroi....................................126
Geusibia................................... 88
glabra..................................... 87
glacialis (Amphipsylla).................... 97
„ (Hoplopsyllus)....................... 57
giobiceps..........................60, 61, 62
golovi.....................................255
grodekovi.................................. 64
gromovi....................................108
hamifer..................................102
hangchowensis........................... 61
hawaiiensis............................. 55
hebes..................................93, 94
hetera (Amphipsylla)...................... 95
„ (Frontopsylla)....................... 85
hesperius................................. 90
hexactenus ................................123
hirticrus .
hirtipes .
hirundinis
.......................... 89, 255
............................... 55
............................... 83
hissarica.......................................118
homoeus...............................59, 61
Hoplopsyllus..........................41, 57
honora....................................118
humida................................... 87
hyaenae.................................. 60
Hystrichopsylla .... 37, 46, 50, 120, 121
Hystrichopsyllinae.....................8, 120
Hystroceras...............................121
ilovaiskii .............................. 66
improv isus.............................. 74
incurva...................................108
indages.................................. 75
integella.................................112
integer ..............................90, 91
270
intermedia.................................... 91
intermedins.................................124
insolita.....................................109
ioffi (Catallagia) .........................120
„ (Ceratophyllus)...................... 75
„ (Nearctopsylla)..........................109
„ (Paraneopsylla)..........................119
„ (Rhadinopsylla)..........................Ill
irana...................................... 98
11, 39, 52, 79
... 49, 123
. 45, 123, 255
..........122
irritans . . . .
Ischnopsyllidae
Ischnopsyllus .
ivanovi . . .
jaonis......................................114
jenissejensis............................... 71
jettmari.................................... 92
jordani..................................... 63
junnanus.....................................105
kalabukhovi..................... 98, 99, 100
kasakiensis............................. 91
kaznakovi................................ 73
kievensis................................ 81
kiritschenkoi........................... 92
kochkor.................................. 93
koksu.................................... 87
kolenatii...............................123
kopetdag................................. 92
korobkovi................................ 62
kozlovi.................................. 73
kukuschkini.............................. 92
kuzenkovi................................ 70
kuznetzovi............................95, 96
laeta
88
laeviceps....................... 4, 5, 70, 255
lagomys...................................... 73
lasius....................................6, 60
lautus.......................................103
laverani..................................... 63
lebedewi..................................44, 72
lenis.....................................93, 94
Leptopsylla................ 45, 49, 101, 255
Leptopsyllinae..........................7, 94
li........................................113
locuples............................ 95, 255
Loemopsylla..............................255
longicauda...............................104
longispina.................................. 98
longispinus................................. 57
luculenta........................... 84, 85, 86
lunatus..................................79, 80
macrophthalma.......................... 88
maculatus............................39, 82
Mafrontia.............................. 87
magdalinae.......................... 56, 255
mana ..................................115
mandarinus............................... 66
marikovskii......................... 96, 255
matina................................... 62
Megabothris............. 38, 66, 68, 76, 78
melis..................................63. 64
menzbieri..............................44, 72
meridiana................................116
mesa.................................... 93
Mesopsylla...........................45, 93
Mesopsyllinae........................... 93
microphthalmus.......................... 90
Micropsylla..............................114
Micropsylloides......................27, 114
microti (Ceratophyllus)..................255
„ (Hystrichopsylla)..................121
mikado.......................... 61, 62, 256
minax.................................... 57
Mioctenopsylla........................44, 84
mirabilis (Chaetopsylla)........ 24, 60, 61
„ (Stenischia).......................114
mitis............................... 100, 255
mokrzeckyi.......................... 69, 255
mongolicus (Ceratophyllus laeviceps) . . 255
„ (Ceratophyllus tesquorum) .70, 71
monoctena................................255
Monopsyllus..................... 66, 68, 75
montana (Amphipsylla).................96, 97
„ (Ophthalmopsylla).................. 91
„ (Stenoponia)..................• . 122
montium...............................98, 99
monstrosus............................... 69
murina .............................. 53, 255
murium...................................113
musculi..................................255
mustelae.................................255
mustersi................................. 92
mutata................................... 87
mycerini............................. 56, 255
Myodopsylla..................... 30, 44, 126
myospalacis.............................. 92
Myoxopsylla...........................43, 63
nana.....................................101
nanodes..................................255
naryni................................... 91
Nearctopsylla................. 46, 103, 109
needhamia........................... 124, 255
nemorosus................................103
Neoctenophthalmus........................105
Neopsylla............ 34, 38, 45, 49, 114, 256
Neopsyllinae..................• ... 7, 114
neotomae................................. 96
nesokiae................................. 55
nicanus.................................. 68
nicolai..................................121
nocens............................... 62, 256
Nosopsyllus................... 4, 6, 38, 67, 68
nubicus.................................. 55
nuda ....................................105
nupera....................................121
nuttali.................................. 56
Nycteridopsylla................. 45, 125, 256
Ochotonobius............................. 89
octactenus................................125
octodecimdentata..................... 63, 255
obscurus............................123, 124
Ophthalmopsylla............ 4, 38, 42, 48, 91
oreinus.................................. 73
Orfrontia.............................84, 88
orientalis (Amphipsylla)................. 95
„ (Ceratophyllus)................ 81
„ (Neopsylla)...................116, 117
orientis................................. 53
orites................................... 83
ornata................................... 87
Orneacus..............................68, 73
Oropsylla .... 26, 44, 49, 65, 70, 72, 256
oschanini................................ 53
ostsibirica..........................101, 102
pacificus................................107
Palaeoctenophthalmus.....................105
Palaeopsylla.................... 46, 103, 108
pallidus.................................. 57
271
pamirensis................................103
Paracallopsylla........................67, 73
Paraceras.................... 43, 49, 63, 256
Paractenophthalmus........................105
Paradoxopsyllinae ............. 7, 64, 66, 84
Paradoxopsyllus............... 42, 48, 89 256
paradoxus..............................75, 76
Paramonopsyllus........................... 76
Paraneopsylla........................45, 118
parcus...................................105
parilis................................85, 86
Pariodontis............................40, 57
parthiana.......................... 94, 96, 97
pavlovskii.................................102
pectiniceps........................... 102, 256
Pectinoctenus......... 4, 45, 49, 102, 256
pedias............• •.................... 74
penicilliger.............................5, 74
pernix..................................... 92
Peromyscopsylla............................101
persica............................ 55, 56, 57
petristshevae.............................. 96
Phaenopsyll ............................42, 92
phaiomydis......................... 94, 96, 97
pilosa..................................... 87
pisticus...................105, 106, 107, 255
pleskei................................116, 118
pollex.....................................106
pollionis.................................. 95
popovi (Echidnophaga)...................... 52
n (Frontopsylla)....................... 87
poppei.....................................121
praefecta...............................91, 92
prima...................................94, 97
primaris...................................100
princeps................................... 72
Profrontia................................. 87
profugus................................... 57
protera.................................... 87
pseudodahurica.............................112
pteromydis.................................255
Pulex..................... 11, 39, 40, 47, 52
Pulicidae...............................30, 52
Pulicinae.................................. 52
Pulicoidea..............................51, 58
punjabensis............................... 68
Ralipsylla.................................113
Rangulopsylla..............................114
rectangulatus...........................77, 78
Rectofrontia...............................109
regis...................................... 56
relicticola................................ 71
remota.....................................108
reoandus................................... 91
Rhadinopsylla . 4, 5, 46, 50, 109, 113, 114
Rhadinopsyllinae...........................109
Rhinolophopsylla...........................125
rhynchopsylla..............................255
riggenbachi................................ 57
rigidus.................................... 89
.riparius...............................79, 82
rossica (Amphipsylla).................. 95, 255
я (Neopsylla)..........................116
rothschildi (Chaetopsylla).............. 62
„ (Mesopsylla) 94
я (Rhadinopsylla)..................Ill
Rostropsylia....................... 26, 43, 66
rufescens.................................. 89
runatus.................................64, 89
Saphiopsylla . .........................41, 121
saratovi...................................255
Sarcopsylla...................................255
satunini..................................... 121
saxatilis..................................... 73
scaloni....................................... 76
schelkovnikovi (Amphipsylla) . . 98, 99, 255
„ (Wagnerina)..................104
sciurorum..................................... 75
scorodumovi............................. 90 , 256
secura........................................118
segnis.................................101, 255
segregatus............................. 65, 256
semenovi (Ceratophyllus)...................... 73
я (Rhadinopsylla) ......... 113
semura........................................ 84
sepifera................<..................... 95
serraticeps...................................255
setosa.................................118, 256
sexdentata....................................101
shikho ....................................... 93
sibirica ........................... 5, 95, 255
sidimi........................................122
silantiewi......................... 65, 255, 256
silvatica.....................................102
Simla '.........................l \. . . . 68
simplex................................124, 125
sinensis (Archaeopsylla).................. 53
w (Oropsylla).........................256
n (Paraceras) . . . ............... 64, 256
singularis . ..............,. •. \. '. . 122
sinicus....................................... 82
skrjabini...................... 54, 55, 56, 57
sobrinus.....................................255
socia........................................ П4
sokolovi..................................... 77
solitaria.............................. 122, 256
sorecis......................................108
spadix....................................... 84
spalacis.....................................255
Spalacoctenophthalmus........................105
sparsilis.................................... 72
spectabilis..................................102
Spilopsyllinae.............................7, 57
Spilopsyllus.............................41, 57
spinea.......................................256
spi nosus.................................... 80
starki.......................................108
steinegeri (Ceratophyllus)................... 84
я (Dasypsyllus)......................... 84
(Oropsylla).................. 65, 256
Stenischia...................................114
Stenoponia................. 46, 50, 122, 256
stevensi..................................118
striata.....................................120
styx........................................ 82
subarmatus.................................. 89
subcaecatus..............................256
subula...................................255
sucinus...................................255
suknevi...................................122
sungaris.............................5, 70, 71
Synosternus........................ 27, 40, 57
syrt........................................ 74
taiganus...................................... 78
taishiri..................................85, 86
talpae........................................121
tamias................................... 75, 255
tarasovi...................................... 89
Tarsopsylla.......................... 43, 48, 63
tecta ......................................Ю4
tenella..........................................HO
teratura........................................116
272
teres .............................................255
teretifrons........................................ 91
terribilis......................................... 89
tersus....................................44, 69
tesquorum.................. 5, 44, 70, 71, 255
thoracicus...................................255
tiflovi (Ceratophyllus)...................... 73
„ (Paraneopsylla).........................119
„ (Phaenopsylla).......................... 92
tjanshan....................................110
toli......................................... 76
transbaikalica (Rhadinopsylla).............113
transbaikalica (Amphipsylla).................255
transbaikalicus..............................256
transvolgensis............................... 71
tribulis..................................... 80
Trichopsylla............................ 59, 256
trichosa................................61, 62
trisellis...................................126
trispinus................................... 72
truncus.....................................106
tuberculaticeps..........................59, 60
tundrensis.................................. 79
Tunga.......................................255
turbidus.................................... 78
turkestanica................................ 57
turkmenicus..............................44, 70
tuschkan.................................93, 94
tuta.....................................98, 99
tuvensis....................................104
Typhlocallopsylla........................68, 73
Typhloceras.............................120, 121
typhlus....................................255
uncinatus.............................105, 106
unipectinata................................125
uniseriata............................117, 256
uralensis..................................255
ursi....................................... 60
vagabundus.................................. 81
valenti.....................................114
vallis..................................... 74
variabilis..................................125
ventricosa..............................5, 113
Vermipsylla............... 26, 38, 39, 41, 58
Vermipsyllidae..........................36, 58
vicina.....................................110
vinogradovi................................. 98
vlasovi ....................................122
volgensis................................... 92
wagneri..................................... 84
Wagnerina..............................42, 103
Wagneri пае............................34, 103
walkeri.................................... 77
wassilii...................................255
weisskei.................................... 65
wualis...................................... 68
wui........................................256
Xenopsylla .... 26, 28, 38, 39, 40, 47, 54
Xenopsyllinae .............................. 54
zibellina...........................60, 61, 62
СОДЕРЖАНИЕ
Введение .................................................................... 3
Собирание блох и приготовление препаратов.................................. 9
Сбор блох с млекопитающих............................................. 9
Сбор блох в гнездах млекопитающих и птиц............................. 11
Сбор блох во входах в норы ..........................................• 13
Сбор блох в жилищах человека..........................................15
Хранение, консервирование, упаковка и этикетировка сборов ........... 15
Микроскопирование блох и приготовление препаратов ................... 17
Эчерк наружной анатомии (эйдономии) блох..................................... 21
Эпределитель родов блох...................................................... 40
Упрощенный определитель наиболее часто встречающихся в Восточной Азии
родов блох............................•................................... 47
Перечень надсемейств, семейств, подсемейств и родов блох с указанием их рас-
пространения и определительные таблицы видов, встречающихся в Восточной
Азии и сопредельных районах............................................... 51
Надсемейство Pulicoidea................................................. 51
Семейство Pulicidae, 52
52
52
53
53
54
57
57
57
57
Подсемейство Pulicinae . . . Род Pulex..................................
Род Echidnophaga................• . . .
Подсемейство Archaeopsyllinae. .Род Archaeopsylla.......................
Род Ctenocephalides.....................
Подсемейство Xenopsyllinae . . Род Xenopsylla.........................•
Род Synosternus.........................
Род Pariodontis.........................
Подсемейство Spilopsyllinae . . Род Spilopsyllus........................
Род Hoplopsyllus........................
Надсемейство Ceratcphylloidea.................................................
Семейство Coptopsyllidae .... Род Coptopsylla.............................
Семейство Vcrmipsyllidae .... Род Vermipsylla.............................
Род Chaetopsylla.......................
Семейство Ceratophyllidae
Подсемейство- Ceratophyllinae. .Род Tarsopsylla.........................
Род Myoxopsylla .......................
Род Paraceras..........................
Род Aenigmopsylla......................
Род Amphalius..........................
Род Oropsylla..........................
Род Rostropsylla.......................
Род Ceratophyllus......................
Определитель подразделений рода Ceratophyllus....................
А. Подрод Nosopsyllus ... а. Секция Nosopsyllus....................
б. Секция Gerbillophilus...............
Б. Подрод Citellophilus............................................
В. Подрод Callopsylla ... а. Секция Callopsylla....................
б. Секция Paracallopsylla.............
в. Секция Orneacus....................
г. Секция TyphlocallopsyНа............
Г. Подрод Amalaraeus...............................................
Д. Подрод Monopsyllus..............................................
Е. Подрод Megabothris..............................................
Ж. Подрод Ceratophyllus............................................
Род Dasypsyllus........................
Род Mioctenopsylla.....................
58
58
58
59
63
63
63
63
64
64
65
66
66
66
68
69
70
72
73
73
73
74
75
76
78
83
84
"4
Подсемейство Paradoxopsyllinae.. .Род Frontopsylla....................... 84
Род Ctenophyllus....................... 88
Род Paradoxopsyllus.................... 89
Род Ophthalmopsylla.................... 91
Род Phaenopsylla....................... 92
Род Brachyctenonotus................... 92
Подсемейство Mesopsyllinae . . Род Mesopsylla....................... 93
Подсемейство Leptopsyllinae . . Род Amphipsylla...................... 94
Род Leptopsylla............• . . . . 101
Род Pectinoctenus......................102
Семейство Ctenophthalmidae, Подсемейство Wagnerinae.. .Род Wagnerina..................103
Подсемейство Anomiopsyllinae . . Род Eopsylla.......................105
Подсемейство Ctenophthalminae.. .Род Ctenophthalmus ..................105
Род Doratopsylla.....................107
Род Palaeopsylla.....................108
Род Nearctopsylla....................109
Подсемейство Rhadinopsyllinae ...Род Rhadinopsylla.......................109
Подсемейство Neopsyllinae . . . Род Neopsylla............................114
Род Paraneopsylla......................118
Род Catallagia.........................119
Подсемейство Hystrichopsyllinae. .Род Hystrichopsylla....................120
Род Saphiopsylla.......................121
Род Stenoponia.........................122
Семейство Ischnopsyllidae.........................................................123
Род lscanobsyilus ...................123
Род Rhinolophopsylla.................125
Род Nycteridopsylla..................Г25
Род Myodopsylla......................Г26
Род Araeopsylla......................Г26
Альбом рисунков ................................................................. 127
Список рисунков с указанием, из какой местности взяты экземпляры блох для ри-
сования или откуда заимствованы рисунки...............................241
Алфавитный список изображенных на рисунках и приведенных в определитель-
ных таблицах видов блох......................................................249
Список некоторых синонимов........................................................255
Указатель основных экологических группировок блох ................................257
Указатель литературы..............................................................262
Указатель названий блох ......................................................... 269
Редактор Н, Г. ОЛСУФЬЕВ
Техн, редактор Р. П. Ротермель
Т 03839. Подписано к печати 20/V 1954 г.
Формат бумаги 70 X Юв1/^. Бум. л. 8,63.
Печ. л. 23,63. Уч.-изд. л. 24,60. Тираж 3000 экз.
МН-51. Цена 12 р. 30 к. Переплет 1 р. Заказ 423.
Медгиз, Москва, Петровка, 12
Набрано и отпечатано в 13-й журнальной тип.
Союзполиграфпрома Главиздата
при Министерстве культуры СССР,
Москва, Гарднеровский пер., 1а.
О И Е Ч А Т к и
Стр. Строка Напечатано | । Следует читать
22 Рисунок I ст. эм. (эпимер’заднегруди)
22 2 строка подписи к рисунку I—VIII ст. II—VIII ст.
40 16 сн. срендегруди среднегруди
43 26 св. сигмы стигмы
85 3 „ треугольной треугольный
85 21 . 9 (8. 9 (8).
94 2 . вершнйой вершиной
108 22 , Palaepsylla Palaeopsylla
115 7 сн. Центральном Баян — Хонгорском Центральном, Баян-Хон горском
117 10 св. местах мест
119 3 , (около четырех (около четырех)
164 2 сн. с. 16. А Рис. 126. А
235 3 , j schnopsyllus comans Рис. 340. Ischnopsyllus comans
239 2 строка под- писи под рис. 351 oschan in. cschanini.
240 Подпись под рис. 53 сн. E — Ch. bomoeus Е — Ch. homoeus
242 11 св. Wagneria Wagnerina
242 25 , Catallagi Catallagia
242 1 сн. Типо — (мтип) Типом (тип)
243 31 св. Ceratophylla Ceratophyllus
244 2 си. 191, А и Б— » » stejnegeri 191, А и Б — „stejnegeri
245 17 св. 204. > elata botis i 204. Frontopsylla elata botis
245 9 сн. (ти . (тип).
248 14 св. kolenati kolenatii
248 18 сн. octodecimentata octodecimdentata
250 Ю „ 180, А, Б 179; 180, Б
251 30 , 248, A 348, A
251 29 , 311, В 211, В
253 16 „ 283, Б 283, В