/


































































































































































































































































































































































































































































































































































































































































Автор: Дементьев Г.П. Спангенберг Е.П. Гладков Н.А. Птушенко Е.С. Судиловская А.М.
Теги: птицы фауна ссср птицы советского союза
Год: 1952




Текст
Г. П. ДЕМЕНТЬЕВ, Н. А. ГЛАДКОВ,
Ю. А. ИСАКОВ, Н. Н. КАРТАШЕВ, С. В. КИРИКОВ,
А. В. МИХЕЕВ, Е. С. ПТУШЕНКО
ПТИЦЫ
СОВЕТСКОГО
СОЮЗА
Том IV
Под редакцией
Г. П. Дементьева и Н. А. Гладкова
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО
«СОВЕТСКАЯ НАУКА»
Москва— 19 52
ОТРЯД КУРИНЫЕ
RASORES или GALLIFORMES
Характеристика отряда
Морфология. Внешние признаки. Птицы главным образом
средней, реже крупной или мелкой величины. Самый крупный вид в фауне
СССР—глухарь—достигает веса 5000 г; самый мелкий—перепел—100 г. Тело-
сложение плотное, шея короткая, крылья короткие и широкие, хвост разно-
образной формы, ноги умеренной длины, четырехпалые. У многих видов
самцы имеют на цевке шпору. Для некоторых характерно наличие окаймля-
ющих пальцы роговых бахромок, способствующих удержанию на ветвях и
передвижению по заснеженной земле. Цевка у одних голая, у других оперен-
ная; у некоторых видов оперены и пальцы. Клюв короткий, выпуклый по
хребту надклювья, с тупой, слегка загнутой вершиной.
Оперение плотное и жесткое. Перья с хорошо развитым побочным ство-
лом (кроме гоацина). Десять первостепенных маховых. Пух только по апте-
риям. Число рулевых варьирует от 12 (и даже 10 у гоацина) до 24. Окраска
разнообразная. У многих видов резко выражен половой диморфизм.
Анатомические признаки. Череп схизогнатический, голо-
ринальный, сошник развит очень слабо. Гемиптеригоидной кости нет. Неб-
ные кости без lamina interna. Шейных позвонков 16 (исключение гоацин).
Вилочка с хорошо развитым гипоклейдием. В позвоночнике хорошо выра-
жена os dorsale. На processus lateralis posterior грудины имеются processus
obliquus. Кольца трахеи окостеневают. Желчный пузырь имеется. Слепые
кишки хорошо развиты и функционирующие. Зоб большой, мускульный желу-
док хорошо развит (исключение гоацин). Крыло обычно эвтаксического типа.
Образ жизни. Разнообразие форм и условий жизни куриных позволяет
дать характеристику их биологии лишь в самых общих чертах. Гокко и гоа-
цины—древесные виды. Остальные хотя и присаживаются на деревья, но
гнездятся на земле. Многие виды всю жизнь проводят на земле Громадное
большинство видов оседлые или совершают только небольшие перекочевки,
но немногим, как, например, перепелу, свойственны настоящие перелеты.
Пища преимущественно растительная и разыскивается на земле, причем пти-
ОТРЯД КУРИНЫЕ
цы роются в земле сильными ногами. В выкармливании молодых, однако,
значительное место занимают мелкие беспозвоночные.
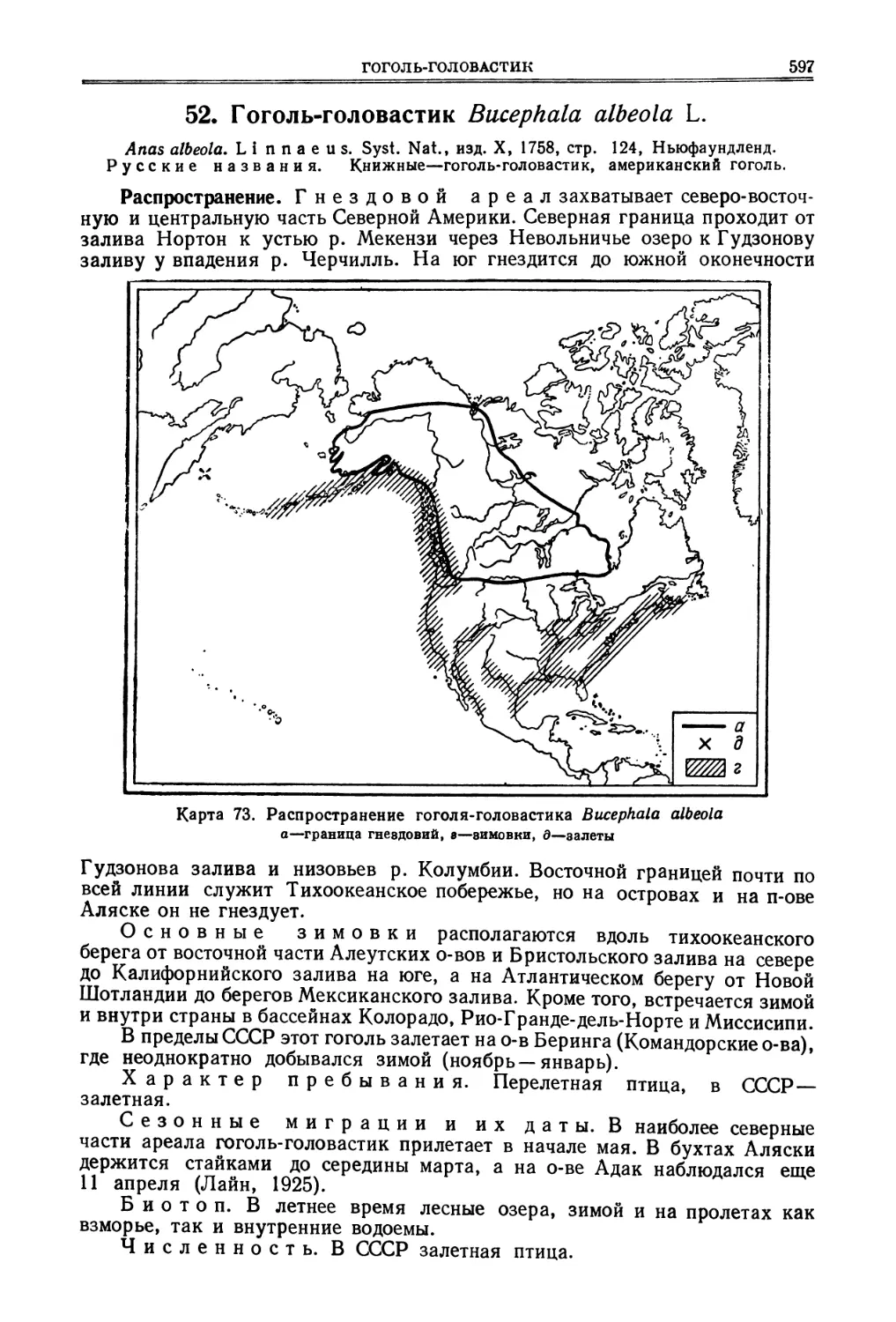
Половая зрелость у видов мелкой и средней величины наступает в годо-
валом возрасте, у крупных видов—позднее. Брачный период у многих харак-
теризуется токами, в которых принимает участие несколько самцов. Неко-
торые виды разбиваются на пары, другие пар не образуют. В последнем слу-
чае самцы в заботах о выводке участия не принимают. У видов, не образую-
щих пар, самцы, как правило, не только больше по размерам, но и ярче окра-
шены, чем самки. У видов, образующих пары, самцы и самки, за некоторыми
исключениями, окрашены одинаково.
Вне брачного времени многие виды ведут стайный образ жизни.
Яйца (за исключением гокко и большеногих кур) относительно мелкие,
но количество их в кладке большое. Скорлупа белая или пестрая. Исключение
составляют гокко и гоацины, кладущие по 2—3 яйца. Как правило, один
репродуктивный цикл в году. Эмбриональное развитие протекает быстро —
от 12 до 28 дней. Развитие птенцов—по выводковому типу.
Линька—раз в году, полная, в конце брачного периода. Исключение—
белые куропатки, у которых самец имеет четыре, а самка три наряда
в году. У некоторых видов имеется, повидимому, частичная предбрачная
линька весной, захватывающая оперение шеи. Возрастные изменения окрас-
ки незначительны, но у крупных видов прослеживаются до двухлетнего
возраста. Кроме того, довольно длительное время сохраняются и остеологи-
ческие (краниологические) особенности. Весьма характерно для куриных
(кроме гокко) и развитие маховых у молодых птиц. Передние два первостепен-
ных маховых развиваются только тогда, когда птенец меняет пуховой наряд
на первый контурный и сохраняются в течение первого года жизни до пер-
вой полной годовой линьки—по окраске этих перьев можно, таким образом,
всегда определить возраст этих птиц. Остальные первостепенные маховые
развиваются]у пуховиков и сменяются уже в первую осень жизни при ювеналь-
ном линянии. У взрослых птиц первостепенные маховые сменяются от
заднего (внутреннего) края партии к переднему (наружному). Рулевые ли-
няют то центробежно, то центростремительно. У некоторых тетеревиных
при линянии сменяются не только оперение, но и когти, и бахромки на лапах,
и клюв (белые куропатки, глухари и др.).
Куриные хорошо передвигаются по земле—ходят и бегают. Полет их
в высшей степени специализирован. Как правило, птицы взлетают в послед-
ний момент, стремительно и с шумом. Фазаны и кеклики поднимаются при
этом вверх «свечкой», потом полого планируют. Прибегают к полету обычно
на очень короткое время, но перепел способен перелетать Черное море от
Крыма к берегам Малой Азии; в соответствии с этим он имеет более острое
крыло.
Систематика. Куриные распадаются на два подотряда: гоацинов Opis-
thocomi и собственно куриных Galti.
К первому принадлежит одно семейство Opisthocomidae с единственным
родом и видом Opisthocomus hoatzin, распространенным в Южной Америке
от Суринама до Колумбии и Боливии. Морфологически и экологически гоа-
цины представляют собою резко уклоняющуюся от собственно куриных
группу; некоторые исследователи предлагали рассматривать их в качестве
самостоятельного отряда. Гоацин — древесная птица, которая редко летает
и почти не спускается на землю. Морфологически гоацины характеризуются
сильным развитием подвижности верхней челюсти (в силу особенностей со-
членения лобных и носовых костей), срастанием грудины с вилочкой и во-
роньими костями и слабым развитием киля грудины (он выражен, в сущности,
только в каудальной части); последнее связано с сильным развитием и круп-
ными размерами зоба, служащего для перетирания основной пищи гоацина—
ХАРАКТЕРИСТИКА ОТРЯДА
жестких листьев. В связи с последним мускульный желудок мал. Слепые
кишки, в отличие от собственно куриных, развиты слабо и укорочены.
Пух, в отличие от всех остальных куриных, расположен не только по апте-
риям, но и по птерилиям. Цевка спереди покрыта мелкими чешуйками
(у остальных куриных двумя рядами поперечных щитков). Шейных позвон-
ков 19. На груди имеется кожное утолщение (мозоль), которому соответст-
вует утолщение грудины—на это место гоацин опирается при отдыхе. Птенцы
развиваются, в сущности, по типу, промежуточному между выводковым и птен-
цовым: рождаются они слабо развитыми, с редким пуховым нарядом, который
впоследствии сменяется вторым пуховым нарядом. Первый и второй пальцы
передних конечностей у них весьма развиты и подвижны, несут когти—пте-
нец пользуется ими при лазании по ветвям деревьев (у взрослых гоацинов
строение пальцев крыла обычное). Свои гнезда гоацины устраивают над во-
дою на деревьях из небрежно сложенных веточек. При опасности птенцы
гоацина или вылезают из гнезд и лазают при помощи крыльев и ног по вет-
вям, или бросаются в воду, ныряют и плавают. Характерно медленное раз-
витие маховых у птенцов гоацина. Выкармливают их родители отрыжкой
из зоба, как это делают голуби.
Подотряд собственно куриных разделяется на четыре семейства: боль-
шеногов, или сорных кур Megapodiidae, гокко Cracidaey павлиньих, или фаза-
новых Pavonidae (Phasianidae), и тетеревиных Tetraonidae.
У первых двух семейств, в отличие от двух последних, первый (задний)
палец прикреплен на одном уровне с передними, а основная его фаланга
примерно равна по длине основной фаланге среднего (третьего) пальца.
Они поэтому выделяются в группу голубеногих, или равнопалых Peristero-
podes. Павлиньи и тетеревиные—с первым пальцем, прикрепленным выше
остальных; основная его фаланга заметно короче основной фаланги сред-
него пальца. Эта группа носит название куроногих, или неравнопалых Ale-
ctoropodes.
К семейству большеногов, или сорных кур, относится 10 видов. Это—
крупные и средней величины неярко окрашенные птицы, ведущие наземный
образ жизни. Вырезки грудины у них развиты слабо; кости слабо пневма-
тичны; трахея удлинена, как у гокко, но в отличие от последних не имеет изви-
вов; побочный ствол развит слабо, копчиковая железа голая или почти голая;
имеется только правая сонная артерия; пальцы длинные, хорошо приспособ-
ленные к разрыванию земли, куч, опавших листьев и т. п., когти прямые;
у двух родов—Megapodius и Megacephalon—крыло аквинтокубитального
типа. Распространены в Юго-Восточной Азии от Филиппин и Индо-Австра-
лийского архипелага до Австралии. Семь родрв—Catheturus, Leipoa, Tale-
gallus, Aepypodius, Megacephalon, Eulipoa, Megapodius. В биологическом
отношении интересно, что большеноги откладывают свои очень крупные
яйца в большие сгребаемые ими кучи гумуса и растительных остатков,
развитие яиц происходит без насиживания, за счет тепла субстрата, а отча-
сти—развиваемой эмбрионами теплоты. Некоторые виды зарывают яйца
в песок. Птенцы выходят из яиц покрытые перьями и могут перепар-
хивать.
Гокко—38 видов, относимых к родам Crax, Mitua, Penelope, Ortalis и др.
Распространены в Центральной и Южной Америке. У них ряд морфологи-
ческих черт, сближающих их сбольшеногами: строение ноги, слабое развитие
вырезок грудины, слабое развитие побочного ствола перьев, голая копчико-
вая железа. Трахея длинная и у самцов имеет петлеобразные извивы; имеется
половой член; крылья эутаксического типа. В отличие от других куриных,
у гокко при ювенальной линьке сменяются все, включая два передних,
первостепенные маховые. Древесные птицы, гнездящиеся также на деревьях.
Кладки небольшие, из 2—3 яиц, насиживаемых в течение 22—29 дней.
ОТРЯД КУРИНЫЕ
Фазановых — Pavonidae, или Phasianidae,—174 вида, широко распростра-
ненных по всему свету, кроме холодной зоны южного и северного полушария.
Кроме общих куроногим особенностей строения задней конечности, для
фазановых характерна неоперенная цевка, неприкрытые перьями ноздри,
отсутствие бахромок или оторочек по краям пальцев. Грудина с глубокими
вырезками и длинными боковыми отростками. Сонные артерии парные.
Побочные стволы хорошо развиты. Гнездятся на земле, откладывают срав-
нительно большое количество некрупных яиц. Внешность весьма разно-
образная, как и величина. Семейство делится обычно на три подсемейства:
американских индюков Meleagrinae с двумя монотипическими родами Meleag-
ris и Agriocharis\ африканских цесарок Numidinae, всего 7 видов, относимых
к родам Numiduy Guttera, Acryllium и др.; наконец, фазановых в узком смыс-
ле—Phasianinae всего 165 видов, относимых к родам Odontophorus, Lophor-
tyxy Colinus, Phasianus, Gennaeus, Crossoptilon, Ithaginis, Lophophorus, Chryso-
lophuSy Polyplectron, Tragopan, Diardigallus, Reinchardtia, Syrmaticus, Pavo,
Afropavo, Gallus, Perdix, Alectoris, Ammoperdix, Francolinus, Coturnix, Tet-
raogallusy Arborophila, Lerwa и др.
Тетеревиные—относительно небольшое семейство, всего 18 видов, отно-
симых к родам глухарей Tetrao, тетеревов Lyrurus, рябчиков Tetrastes, дикуш
Falcipennis, белых куропаток Lagopys, а также американских Dendragapusy
Tympanuchus, Centrocercus, Bonasa, Canachites, Pedioecetes. В морфологиче-
ском отношении они, в сущности, очень близки к фазановым, но отличаются
от последних прикрытыми перьями ноздрями, оперенной цевкой и нали-
чием бахромок, оторачивающих пальцы.
В систематическом отношении отряд куриных представляет собою
резко очерченную группу. Он в известной мере приближается к тинаму—
с одной стороны, а к трехперсткам—с другой. Тем не менее никаких
палеонтологических данных в пользу общего происхождения этих групп не
имеется.
В мезозое ископаемые куриные неизвестны, но уже в среднем эоцене
Северной Америки появляются представители двух семейств этого отряда.
Существенно, что здесь представлено семейство Gallinuloididae со смешанны-
ми признаками, промежуточными, до известной степени, между большеногами
и гокко. В Европе куриные найдены с позднего эоцена или раннего олигоцена.
Заслуживает внимания, что позднеэоценовый европейский вид Filkolornis
также обнаруживает известные черты сходства с гокко и даже в известном
отношении напоминает гоацина.
Несомненные гокко, повидимому, идентичные с современным родом
Ortalis, известны из миоцена—плиоцена Северной Америки. В Южной
Америке ископаемые остатки гокко известны только с плейстоцена.
Описанный из эоцена штата Уайоминг Palaeophasianus meleagroides пред-
ставлен, в сущности, столь фрагментарными остатками, что говорить о его
месте в системе куриных трудно.
Семейство фазановых в третичных отложениях Европы и Америки пред-
ставлено довольно богато, но и для этих остатков трудно определить с до-
стоверностью их место в системе. Можно только сказать, что близкие
к современным фазанам птицы жили и в Америке в олигоцене и миоцене, но не
позднее.
Индюки, в сущности, известны только с позднего плиоцена и плейстоце-
на Северной Америки. Описанный из миоцена вид Meleagris antiqua едва ли
представляет собою индюка.
Современные роды появляются в плиоцене—Alectoris, Francolinus в Ев-
ропе, Ortalis, Colinus—в Америке. В плейстоцене современные роды куриных
уже многочисленны, встречаются и современные виды, как глухарь, тетерев,
рябчик, кеклик, перепел, серая куропатка, обыкновенный фазан и др.
ХАРАКТЕРИСТИКА ОТРЯДА
Географическое распространение. Как указывалось в очерках семейств,
куриные птицы распространены по всему свету, за исключением только Ант-
арктики и некоторых островов. В биотопическом отношении распространение
куриных, как отряда, также чрезвычайно широкое. Они встречаются и в откры-
тых ландшафтах, например тундрах и степях, и в лесах; хорошо уживаются
и в культурном ландшафте. Обитают как на равнинах, так и в горах, дости-
гая значительных высот в альпийском поясе.
Вместе с тем для куриных, как и для других групп оседлых птиц, харак-
терно наличие эндемизмов высокого ранга. Так, в тундрах Старого и Нового
Света представлен только один род—белые куропатки.
Гоацин и гокко характерны для Неотропики, индюки—для Неарктики.
Тетеревиные населяют только Палеарктику и Неарктику, причем и тут и там
представлены эндемичными родами. Большеноги характерны для Австралии
и Папуасии.
В фауне СССР имеется 20 видов куриных, или 8,3 % от общего числа ныне
живущих видов отряда. В том числе 8 видов семейства тетеревиных и 12 видов
семейства фазановых.
Тетеревиные в основном распространены в лесной зоне, выходя из нее
в пределы лесостепи. Наибольшее число видов характерно для тайги:
обыкновенный и каменный глухарь, рябчик, дикуша (последняя распро-
странена только на востоке Сибири и имеет своих ближайших родственников
среди лесных тетеревиных Северной Америки).
Тундреная куропатка имеет типичное аркто-альпийское распростране-
ние, встречаясь, с одной стороны, в каменистых тундрах и на островах север-
ных океанов, а с другой—в альпийской зоне гор. Белая куропатка, не заходя
в столь высокие широты, встречается в субарктических тундрах, а также
по болотам таежной полосы, выходя в лесостепь в Северном Казахстане
и на юго-западе Сибири; встречается она и в горных местностях.
Среди фазановых все улары—птицы азиатского высокогорья, прони-
кающие в пределы Европы только на Кавказе. Характерно, что они предста-
влены рядом близких экологически и сходных морфологически викарирующих
видов. Горная, широко распространенная в Палеарктике птица, но менее
связанная с большими высотами,—кеклик.
С западной Палеарктикой, в основном, связана пустынная куропатка,
также держащаяся на расчлененном вертикально рельефе, но не выходя-
щая, как правило, за пределы нижнего пояса гор и предгорий.
Перепел, серая и бородатая куропатки—птицы степей и культурных
ландшафтов Палеарктики.
Фазаны в целом тяготеют к восточной области. Обыкновенный фазан—
эндемик Палеарктики, встречающийся у нас преимущественно в речных доли-
нах Кавказа, Средней Азии и Дальнего Востока. Местами в Азии проникает
по долинам довольно высоко в горы.
Наконец, турач принадлежит к группе африканско-переднеазиатского
происхождения. В СССР он находится у крайнего северного предела видо-
вого ареала в Азии и встречается только на незначительной части территории
Закавказья и Юго-Западной Туркмении.
Практическое значение. Практическое значение куриных очень велико.
Как известно, это лучший объект и промысловой и спортивной охоты. В этом
отношении из тетеревиных на первом месте стоит рябчик, за ним следует
тетерев. Для населения севера большое значение имеют куропатки—тунд-
реная и в особенности белая. Глухари уступают перечисленным видам по вку-
совым качествам, и охота на них имеет спортивное, а в хозяйственном отно-
шении лишь местное значение.
Из фазановых главное значение как предмет охоты имеют фазаны, пере-
пела, серая и бородатая куропатки. Турач имеет на нашей территории столь
8
ОТРЯД КУРИНЫЕ
ограниченное распространение, что в советское время стал полностью
охраняться как на Кавказе, так и в Средней Азии. Местами в Средней Азии
и на Кавказе довольно значительна роль в охотничьем хозяйстве кеклика,
чего нельзя сказать о пустынной куропатке и уларах: первая немногочис-
ленна, вторые—малочисленны и труднодоступны.
Надо еще отметить, что куриные хорошо подвергаются акклиматиза-
ции. Во многих охотничьих хозяйствах нашей страны успешно разводятся
в парковом состоянии фазаны. В Крым несколько раз завозились кеклики,
в последний раз после Великой Отечественной войны. В средних частях
Туркмении, у Ашхабада, делались опыты вольного разведения турача.
Нет сомнения, что при достаточном контроле за результатами акклимати-
зации имеются достаточно широкие возможности обогащения ценными в охот-
ничьем отношении видами куриных. Этот вопрос возникает, в частности, в свя-
зи с осуществлением плана преобразования природы нашей страны.
Надо еще иметь в виду, что многие куриные, в особенности серые куро-
патки, приносят большую пользу сельскому хозяйству истреблением вред-
ных насекомых.
Еще более велико значение куриных в качестве домашних сельскохо-
зяйственных птиц.
Таблица для определения видов куриных птиц СССР
1B). Цевка оперена полностью или, по крайней мере, на три четверти длины; пальцы
ног окаймлены роговой бахромой; ноздри прикрыты перьями 3
2A). Цевка не оперена; пальцы ног без роговой бахромы; ноздри не прикрыты
25
перьями
11A2
1201
13A4
ИОЗ
3D). Нижняя четверть цевки не
оперена . . . рябчик Tetrastes bonasia
4C). Цевка оперена полностью . 5
5F). Пальцы оперены; маховые бе-
лые 7
6E). Пальцы не оперены; маховые
темной окраски 9
7(8). Крыло самцов обычно длин-
нее 210 мм, самок—длиннее 190 мм;
клюв сильнее, выпуклее и выше (вы-
сота его у основания обычно превышает
8 мм); черной полосы у глаза в зимнем
наряде нет
. . белая куропатка Lagopus lagopus
8G). Крыло самцов обычно менее
210 мм, самок —менее 190 мм; клюв
слабее, площе и ниже (высота его у
основания обычно менее 7,5 мм); перед
глазом в зимнем наряде обычно имеется
черная полоса т у н д-
реная куропатка Lagopus mutus
9A0). Рулевых 8 пар; вершины
передних маховых резко сужены . .
. .дикуша Falcipennis falcipennis
10(9). Рулевых 9 пар; вершины
передних маховых без резких суже-
ний И
Боковые рулевые короче средних; длина крыла превышает 250 мм .... 13
Рис. 1. Характер оперения цевки куриных
птиц. Наверху—фазан, внизу—тетерев
Боковые рулевые длиннее средних; длина крыла короче 250 мм 19
Окраска спины бурая, хвост без поперечных полос 15
Окраска спины пестрая, хвост поперечнополосатый 17
15A6). Клюв черный, рулевые черноватые без белых пестрин, плечевые с белыми
пятнами каменный глухарь Tetrao parvirostris (самцы)
16A5). Клюв беловатый, рулевые с белыми пестринами, плечевые без белых пятен
обыкновенный глухарь Tetrao urogallus (самцы)
17A8). Зоб рыжевато-бурый без поперечного рисунка
обыкновенныйглухарь Tetrao urogallus (самки)
ХАРАКТЕРИСТИКА ОТРЯДА
Рис. 2. Голова белой куропат-
ки (наверху) и тундреной ку-
ропатки (внизу)
18A7). Зоб иной окраски, поперечнополосатый
каменный глухарь Tetrao parvirostris (самки>
19B0). Общая окраска однообразная с преобладанием черноватого цвета .... 21
20A9). Общая окраска пестрая с преобладанием
рыжеватых или сероватых тонов 23
21B2). Боковые рулевые изогнуты лирообразно
(снаружи); нижние кроющие хвоста, задние (внутрен-
ние) первостепенные маховые и основные части второ-
степенных маховых белые
тетерев Lyrurus tetrix (самцы)
22B1). Боковые рулевые изогнуты внизу; нижние
кроющие хвоста и все маховые черноваты ....кав-
казский тетерев Lyrurus mlokosiewiczi (самцы)
23B4). Длина хвоста менее 2/з длины крыла и
обычно менее 130 мм. Темный рисунок оперения груди
и боков широкий . . тетерев Lyrurus tetrix (самки)
24B3). Длина хвоста более 2/3 длины крыла и обыч-
но превышает 130 мм; темный рисунок оперения груди
и боков узкий и неправильный кавказ-
ский тетерев. Lyrurus mlokosiewiczi (самки)
25B6). Величина мелкая, длина крыла менее 120 мм;
рулевые перья скрыты кроющими хвоста
перепел Coturnix coturnix
26B5). Величина иная, длина крыла превышает
120 мм; рулевые перья не скрыты кроющими хвоста 27
27B8). Хвост заметно длиннее крыла; оперение или
яркое с сильно развитыми металлическими отливами
(самцы) или пестрое охристо-серовато-буроватое (сам-
ки) фазан Phasianus colchicus
28B7). Хвост короче крыла, оперение без ярких ме-
таллических отливов 29
29C0). Размеры средние: крыло короче 200 мм,
хвост короче 130 мм; рулевых не более 9 пар . . .31
30B9). Размеры крупные: крыло длиннее 200 мм>
хвост длиннее 130 мм; рулевых более 10 пар 39
31C2). Окраска рулевых, кроме средней их пары, черная с узкими беловатыми по-
перечными полосками турач Francolinus francolinus-
32C1). Окраска рулевых иная, кроме средней пары, они рыжеватые, без резкого
поперечного рисунка 33
33C4). Первостепенные маховые,
кроме первого, окрашены на наруж-
ных опахалах в поперечнополосатый
рисунок, на внутренних опахалах
одноцветны; рулевых 6 пар; крыло-
не длиннее 140 мм
пустынная ку-
рочка Ammoperdix griseigularis
34C3). Первостепенные маховые,
кроме первого, или все одноцветные,
или окрашены и на наружных и на
внутренних опахалах в поперечнопо-
лосатый рисунок; рулевых более
6 пар; крыло длиннее 145 мм . . 35
35C6). Первостепенные маховые,
кроме первого, одноцветные; ноги и
клюв красные; шея с черной полосой
на передней стороне
к е к л и к Alectoris graeca
36C5). Первостепенные маховые,
кроме первого, поперечнополосатые;
ноги и клюв буроватые, шея без чер-
^ , ной полосы на передней стороне . . 37
Рис. 3. Хвост обыкновенного тетерева (слева) 37C8). Зоб одноцветный, серый;
и хвост кавказского тетерева (справа) пятно на передней части брюха каш-
таново-бурое в разных оттенках; чер-
ной полосы под глазом нет серая куропатка Perdix perdix
38C7). На зобе желтое пятно; пятно на передней части брюха черное или черновато-
бурое; под глазом черная узкая полоса . ..бородатая куропатка Perdix daurica
39D0). Первостепенные маховые белые с темными вершинами 41
10 ОТРЯД КУРИНЫЕ
40C9). Первостепенные маховые бурые; если белый цвет на них имеется, то только
«а вершинах или же у оснований внутренних опахал 45
41D2). От угла рта вокруг горла идет каштановая, рыжая или серая полоса ... 43
42D1). Окраска иная, без каштановой, рыжей или серой полосы у горла
каспийскийулар Tetraogallus caspius
43D4). Полосы у горла рыжие или серые; зоб с узким и правильным поперечным ри-
сунком кавказский улар Tetraogallus caucasicus
44D3). Полосы у горла каштановые; зоб иной окраски, серий с черными предвершин-
ными пестринами гималайский улар Tetraogallushimalayensis
45D6). Первостепенные маховые бурые со светлыми вершинами; перья груди с чер-
ными каемками тибетский улар Tetraogallus tibetanus
46D5). Первостепенные маховые бурые с беловатыми полями у оснований внутрен-
«их опахал; перья груди беловатые . . . алтайский улар Tetraogallus altaicus
СЕМЕЙСТВО ТЕТЕРЕВИНЫЕ TETRAONIDAE
РОД БЕЛЫЕ КУРОПАТКИ LAGOPUS BRISSOH A760)
Тип Tetrao lagopus
1. Белая куропатка Lagopus lagopus Linn.
Tetrao lagopus. Linnaeus. Syst. Nat., изд., X,I, 1758, стр. 159. Шведская Лапландия.
Распространение. Ареал. Кругополярная птица. В Старом Свете
встречается от Скандинавии и тундр Евразии к югу до Южного Казахстана
и Северной Монголии, а в Северной Америке от Гренландии, Ньюфаундленда,
Арктического архипелага и Аляски к югу через Канаду, залетая иногда зи-
мою в Соединенные Штаты. В Северной Америке (Бент, 1932) гнездовая
область простирается: к северу—до побережий Аляски (мыс Лисберн,
Уэнрайт, мыс Барроу и мыс Демаркейшн), Франклина (порт Чайлю) и Лаб-
радора (Оккак); на восток—до Лабрадора (бухта Гамильтон) и Ньюфаундлен-
да (Ролей и Сент-Джонс); на юг—до Ньюфаундленда, Центрального Квебека,
Северного Онтарио D0 миль к югу от мыса Генриетта-Мария), Северного
Манитоба (форт Черчилль), Южный Мекензи (форт Резолюшен), Централь-
ной Британской Колумбии (Оленье ущелье, горы Ича и гора Девяти миль)
и Юго-Восточной Аляски (о-в Сант-Хуан); на запад—до Аляски (Ледниковый
залив, о-в Нельсон, Аскинукский хребет, Ном, р. Минг., мыс Лисберн). В За-
падной Европе населяет субальпийскую зону и морской берег от района
Ставангер до Нордкапа в Норвегии, обитает от Вермланда до Лапландии
в Швеции, встречается в Финляндии. Прежде водилась и была нередка на
болотах северо-восточной части б. Восточной Пруссии, особенно у Мемеля
и Тильзита. Исчезла там примерно в 30—40-х годах прошлого столетия
(Хартерт, 1921—1922). В небелеющей на зиму расе (L. /. scoticus L a t h.)
встречается на Британских о-вах: в Англии и Уэльсе — в Глеморгане,
Монмаузе, Гересфорде, Лэлоне, Дерби, Йоркшайре, кочующие отмечены
в Серрее, Девоне и Сомерсете; в Шотландии—всюду, включая Гебридские
и Оркнейские о-ва, но не Шетландские; встречается в Ирландии (Уайзерби,
1941). Наибольшая же часть ареала приходится на СССР (см. ниже).
Характер пребывания. В лесной и лесостепной зонах—
оседла, слабые кочевки изредка бывают в неурожайные для кормов годы.
В тундрах Крайнего Севера и в альпийском поясе гор совершает регуляр-
ные сезонные перекочевки в лесотундру или в нижние пояса гор и долины.
Биотоп. В гнездовое время: открытые тундры с наличием зарослей
ивняка, карликовой березы и ягодников; альпийский и субальпийский поя-
са гор; моховые болота лесной полосы; березовые и осиновые колки и забо-
лоченные ольшаники в лесостепной зоне. Зимою—заросли ивняков, березы
БЕЛАЯ КУРОПАТКА
11
и ольхи, преимущественно в поймах рек, где больше всего концентрируются
эти кустарники.
Подвиды. На территории СССР семь подвидов, различающихся размерами и окрас-
кой: L. /. lagopus Li п п а е u s, 1758—север Европы; L. L rossicus Sserebrowsky,
1926—центральные районы европейской части СССР; L. /. koreni Thayeret Bangs,
1914—север Сибири до Томска, Красноярска, Якутска; L. /. major L о г е n z, 1904—
Западная Сибирь к югу от Ишима и Тары, Северный Казахстан, Башкирия на север до
Свердловска; L. I. brevirostris Hesse, 1931—Алтай, Саяны, Танну-Ола, Хангай,
Кентэй; L. /. sserebrowsky Domaniewsky, 1933—горы Восточной Сибири южнее
Якутска и Верхоянского хребта; L. /. okadai Moraiyama, 1928—Сахалин, Камчатка.
Карта 1. Распространение белой куропатки Lagopus lagopus
1 — L. I. scoticus, 2 — L. /. variegatus, 3—L. I. lagopus, 4—L. I. rossicus, 5—L. I. koreni, 6—L. I.
major, 7—L. I. brevirostris, 8—L. I. sserebrowsky, 9—L. L okadai, 10—L. I. alexandrae, 11—L. I.
•alasceneis, 12—L. L albus, 13—L. /. ungamis, 14—L. I. alleni; a—граница гнездовий, б—не вполне
выясненная граница гнездовий
Выделяемого некоторыми авторами в самостоятельный 'вид грауса с Британских
островов правильнее считать лишь хорошо обособленной небелеющей на зиму расой белой
куропатки L. /. scoticus Lath. Это подтверждается географическим викариатом этих форм,
полным сходством биологии, а также существованием в Норвегии как бы переходной
к граусу формы L. I. variegatus Salomonsen, 1936, оперение которой белеет на зи-
му не полностью.
В Северной Америке различают пять географических рас: L. I, albus G m e I i n,
1789— L. I. ungavus R i 1 e y, 1911—Северный Квебек—Гудзонов залив; L. /. alleni
Stei neger, 1884—Ньюфаундленд; L. /. alascensis Swarth, 1909—Аляска севернее
бассейна Юкона; L. I. alexandrae Gri nnel 1, 1926—о. Баранов, о. Шумагин,Аляска
до Британской Колумбии.
Лапландская белая куропатка Lagopus lagopus lagopus L.
Синонимы. Tetrao lapponicus. Gm e 1 i n. Syst. Nat., 1, 2, 1789, стр. 751. Лапландия.—
Lagopus albus auctorum nee G meli n, 1789.—Lagopus brachydadylus. Gould. Birds of
Europe. IV, 1836, стр 256. Северная Россия.—Lagopus lagopus kapustini. Sserebrow-
ski. Journ. fur Ornith., 1926, стр. 512. Кольский п-ов.— Lagopus lagopus septentrional is.
Бутурлин. Сборник трудов Зоологического музея Московского университета, 1,
1934, стр. 83, низовья Таза.
Распространение. Ареал. Скандинавия, Финляндия, Прибалтика,
север европейской части СССР.
В СССР на север проникает до побережья Кольского п-ва, Канинской,
Тиманской (Михеев, 1948) и Большеземельской тундр, за исключением узкой
прибрежной полосы Югорского Шара, на о-в Колгуев (Портенко, 1937); к югу
12
ОТРЯД КУРИНЫЕ
по тундре, лесотундре и северной части лесной полосы через Прибалтику (Эсто-
ния, Латвия, Литва; Лоудон, 1909) до Белоруссии (без ее южных частей),
Ленинграда, вероятно Вологодской обл. и Камского бассейна, может быть
только до Вычегды; экземпляр с Вычегды по рыжей окраске больше сходен
со среднерусской куропаткой, хотя имеет выраженные черные центры, сбли-
жающие ее несколько с лапландской (Михеев, 1948); к востоку—до Урала.
Характер пребывания. В лесной полосе в основном оседла,
делает местные кочевки в зимнее время, особенно при неурожае кормов.
В тундрах и на островах северных морей совершает регулярные перелеты
в лесотундру и прилегающие
районы лесной полосы. Степень
выраженности перелетов зави-
сит от характера зимы, особен-
ностей местности и урожайно-
сти кормов. В годы, когда ку-
старники вовсе не заносятся сне-
гом, почти вся масса птиц оста-
ется зимовать в тундре. Из
районов с высокими зарослями
ивняков улетает птиц меньше и
отлет происходит позднее, чем
из арктической зоны тундры и
с островов, где ивняков вовсе
нет или они низкорослы; оттуда
куропатки улетают раньше и
почти полностью. Изредка от-
мечаются массовые переселения
из тундр, во время которых пти-
цы движутся по кустарникам в
определенном направлении в те-
чение всей зимы или летят гро-
мадными стаями почти без пере-
рыва (Соловьев, 1927). Эти пе-
реселения связаны, видимо, с
особой неурожайностью кормов
в тундре и являются крайним
выражением обычных сезонных
миграций.
Места массовых зимовок птиц тундры—лесотундра и верховья тундро-
вых рек с обширными и высокими ивняками. При наличии последних на тунд-
ровых реках ежегодно зимует значительное количество куропаток. В Канин-
ской тундре, например, к таким рекам относятся Губистая, Двойник, Косты-
лиха, Жемчужная; в Тиманской—Вельть, Икча, Черная, Великая, Волонга
(Михеев, 1948, см. карту 2). Часть птиц заходит на зимовке в северные районы
таежной зоны. Глубина проникновения в лесную зону зависит от характера
зимы и в разные годы колеблется. Куропатки из Большеземельской тундры
чаще всего достигают среднего и нижнего течения р. Усы, иногда только ее
верховьев, а в наиболее суровые и многоснежные зимы проникают по Печоре
даже в хвойные таежные лесные массивы на 200—250 км—до рек Абрамской
и Шуроге (Соловьев, 1927). Из Канинской тундры обычно доходят до г. Мезе-
ни, а по долине р. Мезени на 50 км южнее (до д. Кильцы, Михеев, 1948).
В Северном Крае наблюдаются три очага массового скопления зимующей
куропатки: район р. Усы и низовьев Печоры с их притоками, где зимуют пти-
цы из Большеземельской и отчасти Малоземельской тундры; район р. Сулы—
из Малоземельской, Тиманской и отчасти Канинской тундр; район рек Несь,
Карта 2. Схема зимних перелетов белой куропатки
БЕЛАЯ КУРОПАТКА 13
Вижас, Ома, Пеша и прилегающих к ним березников—из Канинской тундры,
а иногда и с Кольского п-ва. Наиболее мощный очаг первый; значительные
массы птиц скопляются и в последнем.
Время отлета из тундры и появления в лесах зависит от характера зимы
и сильно колеблется в разных районах и в разные годы. В окрестностях
с. Несь (Канинская лесотундра) в 1914 г. куропатки появились в октябре,
в 1922 г.—в ноябре, в 1931 г. в феврале, в 1937 г.—в конце февраля—начале
марта, в 1939 г. первые стайки в конце октября, основная масса в начале де-
кабря. В окрестностях с. Вижас (Канинская лесотундра) в 1935, 1936,
1937 гг.—в конце февраля—начале марта. В низовьях реки Пеши отмечаются
обычно две волны прилета: в сентябре—октябре и массовый—в ноябре—
декабре, а иногда в январе. В окрестностях с. Паново (верховья р. Индиги,
Тиманская тундра)—обычно в январе (Михеев, 1948). В притундровую полосу
на Печоре—в среднем между 1 ноября и 1 декабря (Соловьев, 1927); в зиму
1931/32 г. первая волна здесь в начале декабря, вторая в конце января
(Брюханов, 1935).
Решающим фактором, влияющим на сроки отлета, являются кормовые
условия в тундре и на пути перелетов; имеют значение направление и сила
ветра: куропатка плохой летун, не способна летать против ветра и пользуется
попутными ветрами.
В зависимости от состояния погоды и времени зимы куропатки движутся
или прямым полетом, изредка опускаясь в кустарниках на отдых и кормежку,
или кочуют по зарослям; так они, кормясь, перебегая и делая короткие пере-
леты, постепенно передвигаются на юг. Так, на Печоре с первозимья 1931/32 г.
слет в притундровые леса проходил медленно; табунки вылетали из тундры
и останавливались у первых высоких зарослей. Позднее наблюдался резко
выраженный пролет: в продолжение 2—5 дней в одном направлении летел
табун за табуном по 100—300 птиц на высоте от 50 до 200 м. Птицы следо-
вали вдоль реки и редко удалялись в сторону. Изредка они опускались на кор-
межку и рассыпались по кустарникам, бегали и перелетали вперед, создавая
картину «перекатывания» табуна (по материалам Архангельской зональной
охотничье-промысловой станции).
Из тундры в леса птицы летят по наиболее удобным в отношении корма
местам, в первую очередь по заросшим ивняками долинам тундровых рек.
Но там, где отсутствуют идущие с севера на юг реки, на направление
перелетных путей, а в конечном счете и на выбор зимовок, большое влияние
оказывает направление и сила ветра. Зимой в тундре почти всегда дуют сильные
ветры, и куропатки пользуются тем попутным ветром, который застанет
их в момент исчезновения корма. Так, в район с. Нижняя Пеша они прилетают
с Канинского п-ва только тогда, когда дуют северо-западные ветры. Если же
дует северный или северо-восточный ветер, то основная масса летит вдоль
западного берега полуострова к лесам в район с. Несь и только при дальней-
шем попутном юго-западном ветре иногда продвигается в район с. Нижней
Пеши. Точно так же бывают годы, когда с попутными ветрами куропатка
прилетает с Кольского п-ва в окрестности с. Несь, а изредка с Канинского
п-ва на Терский берег Белого моря (Михеев, 1948). Наиболее краткий путь
с о-ва Колгуева на материк проходит через устье р. Вельть в Тиманской
тундре, куда чаще всего в начале зимы и прилетают колгуевские птицы. Но
нередко они прибывают в район устья р. Индиги и даже на северную оконеч-
ность Канинского п-ва (Соловьев, 1927).
Тем не менее имеются более или менее постоянные направления, по ко-
торым чаще всего происходят сезонные миграции из тундр в леса и обратно.
Так, из Тиманской- тундры в Канинскую лесотундру куропатки никогда не
мигрируют. С Канина же они ежегодно летят в Тиманскую тундру и далее,
в пограничные с нею лесные районы. При этом летят сюда несколькими путя-
14 ОТРЯД КУРИНЫЕ
ми. Часть птиц, повидимому из северных районов полуострова, перелетает
Чешскую губу в наиболее узкой ее части (Микулин мыс—мыс Бармина) и,,
следуя далее в юго-восточном направлении, уходит в леса и кустарники по
р. Сула, а по некоторым сведениям даже в район Печоры. Часть пересекает
губу в южной ее оконечности и попадает в леса окрестностей с. Нижняя Пеша.
Большинство же куропаток движется сюда по восточному берегу Канина
через села Вижас и Ома. Значительный пролет идет вдоль западных берегов
полуострова в район с. Несь, откуда часть спускается к окрестностям г. Ме-
зени, а часть повертывает на восток в сторону с. Нижняя Пеша. Как отме-
чалось, с о-ва Колгуева в большинстве случаев перелет идет в Тиманскую
тундру через устье р. Вельть.
Наличие определенных направлений пролета, проходящих иногда через
довольно широкие морские проливы,—показатель того, что сезонные мигра-
ции куропаток тундры не являются местными кочевками птиц, постепенно
двигающихся в поисках корма от одного кормного места к другому, а пред-
ставляют собой довольно хорошо выраженное явление перелета, связанное
с определенными, выработавшимися исторически инстинктами.
Возвращаются куропатки в тундру ранней весной, когда начинают вы-
таивать из-под снега кустарники и появляются первые проталины. В зави-
симости от характера весны птицы или летят на значительной высоте (вне
досягаемости выстрела) почти без задержки, или же, как и осенью, пере-
двигаются постепенно с длительными остановками. Обратный отлет происхо-
дит, в общем, теми же путями, что и осенью, но не всегда. Так, птицы, про-
шедшие зимой через окрестности с. Несь, как правило, весной сюда не воз-
вращаются, а продвинувшись на восток, летят в тундру через район сел Сно-
па—Нижняя Пеша, где наблюдается регулярный осенний и весенний проле-
ты. Через с. Нижняя Пеша возвращается куропатка из Тиманской тундры
на Канин. В Тиманскую и Канинскую тундры прилет происходит в конце
марта—начале апреля, в Большеземельскую (по данным 1932 г.)—в 20-х чис-
лах апреля (материалы Архангельской зональной охотничьё-промысловой
станции).
Сезонные миграции белой куропатки вызывают миграции кречета и белой
совы, для которых куропатка служит основным зимним кормом.
Явлениями миграций нередко объясняют колебание численности про-
мысловых птиц, причем об урожайности последних судят по размерам промы-
сла. Учитывая особенности миграций белой куропатки и зависимость от них.
численности прилетающих на зимовку в лесотундру птиц, где происходит
основной промысел, можно определенно высказаться о неприменимости
подобного метода в отношении описываемого вида, по крайней мере для севера
Биотоп. Гнездовый—открытая тундра и моховые болота в северной
части лесной зоны, преимущественно по долинам рек. Внегнездовый—заро-
сли ивняков по тундровым рекам, ивняки, низкорослые березники и ольша-
ники в лесотундре, чистые высокоствольные березовые, а также хвойные
насаждения с примесью кустарников и березников в лесной полосе.
Наиболее характерная гнездовая стация—среднеувлажненная кочко-
ватая тундра с ягодниками, карликовой березой и куртинками ивняка. На
участках, где кустарники часто перемежаются кочковатой тундрой с ягод-
никами, отмечается наибольшая плотность птиц в гнездовый период. Густых
и обширных зарослей ивняков во время гнездования избегает и предпочитает
держаться по их опушкам или на соседних участках тундры. Гнездится и в со-
вершенно открытой местности в значительном удалении от ивняков, на
при этом обязательно на кочковатой тундре, поросшей ягодниками и хотя бы
редкими кустиками карликовой березы. Обширных голых участков тундры
с лишайниками или низкорослой редкой травой на песчаных почвах со-
вершенно избегает. Примиряясь с чрезмерной влажностью заболоченных
БЕЛАЯ КУРОПАТКА 15
мест с сухими кочками, она вовсе избегает их в тех случаях, когда кочки по-
крыты густым, плотным покровом мха, сохраняющим влагу и вытесняющим
ягодники и всякую травяную растительность. В незначительном количестве
гнездится и на лайдах, но обязательно при наличии ивняков.
Численность. На гнездовье в тундрах—высокая. Средняя плот-
ность в Тиманской тундре 20—25 гнезд на 1 км2, максимальная 30 и более.
В лесотундре и лесной полосе—значительно ниже. На зимовках в тундре ко-
леблется по годам, но в общем незначительная. В лесотундре численность
зимующих птиц высокая, о чем свидетельствуют и цифры промысла. На Мур-
мане один охотник добывает за зиму до 600—800 птиц, в Канинско-Тиманском
районе—1000—1500 и даже до 3000—5000, в Печорском крае до 4000—
6000 птиц (Ивлев, 1930), а нередко и до 10 000 (Соловьев, 1927).
Во время сезонных миграций нередки случаи массовой гибели куропа-
ток. Особенно много гибнет их от штормов при перелете с островов на мате-
рик и обратно. По сообщению местных жителей в 1926 или 1927 г. в устье
р. Индиги было обнаружено много куропаток, погибших при перелете с о-ва
Колгуева на материк. Соловьев A927) приводит случай особенно грандиоз-
ной гибели птиц в 1915—1916 гг., когда выброшенные на берег Большеземель-
ской тундры птицы местами образовали вал высотой до 2 м. Гибель птиц, по
его словам, имела место также в 1921, 1922 и в 1925 гг. Немало куропаток
при перелетах разбивается о телеграфные провода. Так, зимой 1937 г. жите-
ли с. Несь ежедневно собирали под телеграфной линией по 70—80 птиц на
протяжении 10—15 км> а в течение месяца отдельные лица подобрали до
300 шт. По 15, 20, а иногда по 100 шт. в день находили под проводами
в 1925 г. в Большеземельской тундре (Соловьев, 1927). Есть основания пола-
гать, что разбиваются в первую очередь слабые птицы, не способные доста-
точно хорошо управлять своим полетом при сильном ветре. В литературе
есть указания, что белая куропатка по каким-то непонятным причинам в не-
которые годы осенью устремляется в Северный Ледовитый океан где и поги-
бает массами (Ивлев, 1930). Наши данные не подтверждают это. Здесь или
смешивают весенние миграции с осенними, или же неправильно истолковы-
вают могущие быть случаи уноса стай куропаток сильными штормами в от-
крытое море в направлении к Северному полюсу во время их осенних пере-
летов на материк.
Экология. Размножение. Куропатки моногамные птицы, дости-
гающие половой зрелости в первую же весну. По нашим наблюдениям в Канин-
ской и Тиманской тундрах признаки наступления брачного периода (весен-
ние крики самцов, увеличение бровей, первые следы весенней линьки) замет-
ны еще на местах зимовок. По прилете в тундру птицы сразу рассыпаются
по проталинам. При похолодании и выпадении снега вновь сбиваются в стаи
и переселяются в ивняки, причем самцы образуют обособленные от самок
стаи численностью до 100—150 шт. С наступлением вновь теплой погоды и
появлением проталин снова быстро распределяются по тундре. Начало брач-
ного периода тесно связано с ходом весны и состоянием снежного покрова
и в разных районах наступает в разные сроки. Так, весной 1938 г. в западной
части Тиманской тундры (р. Великая) куропатки рассыпались по проталинам
и оглашали тундру громкими брачными криками в начале апреля, а в восточ-
ной (р. Черная—р. Вельть), где снег еще лежал сплошным покровом, более
чем на декаду позднее. Рассыпавшись по оголенным от снега участкам, самцы
сразу приступают к ухаживанию за самками. В течение всего дня слышны
громкие крики и видны токовые полеты самцов, во время которых они взле-
тают на 15—20м и опускаются с характерными для брачного периода кри-
ками. Первое время пар не заметно, птицы держатся поодиночке, лишь изред-
ка сходятся по две-три вместе и тут же расходятся. Самки ведут себя спо-
койно, в течение всего дня кормятся на проталинах и по кустикам карлико-
16 ОТРЯД КУРИНЫЕ
вой березы с частыми перерывами на 15—25 мин. Самцы кормятся мало,
чаще преследуют самок, при этом вытягивают и опускают вниз шею, подни-
мают хвост, напоминая в такой позе курицу. Приближаясь к самке, изредка
наклоняют голову, а подойдя вплотную, распускают веером хвост, оттопы-
ривают одно крыло и, как петухи, забегают боком кругом. Самка в этот период
всегда убегает от самца. Иногда самец бросается то к одной, то к другой, то
к третьей самке, но везде его постигает неудача. Нередко одну самку пресле-
дуют два самца, но драк и столкновений между самцами в это время не наблю-
дается. Крики самцов и преследования самок в течение дня то разго-
раются, то прекращаются. После непродолжительного затишья, во время
которого птицы кормятся и отдыхают, достаточно раздаться только одному
крику самца, как в ответ ему с разных сторон тундры доносятся ответные
крики других самцов, начинаются перелеты с места на место, возобновляется
преследование самок, и снова наступает шумное оживление. Будучи молча-
ливой в другие сезоны года, в брачный период белая куропатка довольно
Рис. 4. Схема токового полета самца белой куропатки
криклива. От криков самцов в тундре временами стоит сплошной гул. Крики,
в общем, однотонны, но имеют немало вариаций. Наиболее типичен для
•самца громкий крик, издаваемый при опускании на землю во время токового
полета, который можно изобразить так: очень высокого тона «кою», ниже
«крр», выше «ква-ква», очень низко «кува-кува-кува-кува». Первые два слога
издаются в воздухе при опускании на землю, остальные на земле. Самки зна-
чительно молчаливее самцов, но и они в этот период подают голос, чаще всего
по утрам и вечерам. Наиболее обычен негромкий крик, напоминающий голос
домашней курицы: «та-та-та-та», иногда довольно продолжительный.
С продвижением весны, с потеплением погоды и увеличением проталин
брачная деятельность проходит интенсивнее. Самцы упорно придержи-
ваются определенной самки, все чаще и чаще поблизости от самца можно
обнаружить и самку. Образуются пары. В течение 3—4 дней разбивка на
пары почти полностью заканчивается. Самцы с тихими гортанными, вор-
кующими голосами, оттопыренными крыльями и распущенными веером
хвостами расшаркиваются, кланяются в такт шагам перед самками, бегают
за ними с поднятыми вверх хвостами, преследуют в воздухе. С этого момента
начинают замечаться враждебные отношения между самцами. Самец, уха-
живающий за самкой, завидев поблизости другого самца, оставляет подругу
и с вытянутой вниз шеей и опущенным хвостом бежит к сопернику. Тот при-
нимает такую же позу, и обе птицы, слегка покачивая шеями и покланиваясь,
молча наступают друг на друга, перебегая с места на место. При этом до драк
дело еще не доходит и один из самцов скоро улетает.
Одновременно с образованием пар начинается размещение птиц по гнез-
довым участкам. Каждая пара придерживается определенного места, на ко-
тором кормится и отдыхает. Вспугнутый с этого места самец вскоре возвра-
щается обратно на него. После того как большинство птиц разобьется на па-
БЕЛАЯ КУРОПАТКА \J
ры и значительная часть удобной территории окажется занятой, начинаются
драки самцов. Это дает основание предполагать, что дерутся самцы не из-за
самок, а из-за гнездовых территорий. И чем меньше остается незанятой терри-
тории, тем чаще и ожесточеннее драки. Драки возникают тогда, когда на за-
нятый одним самцом участок прилетает другой. Самец—хозяин участка—
немедленно вступает в драку с конкурентом и прогоняет его. Сходясь для
драки, самцы издают свой боевой громкий клич: «ваак, ваак», после которого
набрасываются друг на друга. Позы самцов во время схватки напоминают
петушиные, птицы также часто клюют землю перед броском на врага, под-
скакивают в воздух в момент драки. Но бросаются друг на друга не спере-
ди, как домашние петухи, а сбоку, стараясь наносить удары в голову и шею.
Птицы придерживаются избранных ими мест чрезвычайно упорно, и ни
частое спугивание, ни выстрелы, ни близость жилья с бегающими собаками
не в состоянии вынудить их покинуть облюбованные места. Объясняется это
необходимостью иметь хоть где-нибудь гнездовую территорию и жестокой
борьбой из-за нее между птицами в брачный период. На гнездовом участке
самец держится по высоким кочками и буграм, с которых легче совершать
обзор своих владений. Если гнездовый участок расположен на низинной
лайде с кустарниками, то в качестве сторожевого поста избирается высокий
куст.
Пока не все птицы разбились на пары и не вся гнездовая территория
занята ими, отдельные самцы или пары свободно перемещаются по незаня-
тым участкам. Но это становится с каждым днем труднее и труднее, и вскоре
вовсе прекращается. Закрепившиеся на гнездовых участках самцы тотчас
изгоняют всякого прилетевшего на их участок другого самца.
Каждый из них вынужден держаться на том участке, на котором он
оказался к этому времени.* В отличие от самцов, самки свободно перелетают
с одного гнездового участка на другой и никакой вражды к ним со стороны
самцов и самок других пар не замечается. Отсюда следует, что в выборе
и закреплении гнездового участка ведущую роль играет самец, а не самка.
Самец, последовавший за перелетевшей на соседний гнездовый участок сам-
кой, никогда не вступает в драку на чужом участке, а всегда при первом же
враждебном действии со стороны самца—хозяина участка—возвращается
на свой участок. Драки происходят главным образом с самцами, ищущими
гнездовую территорию, а также на границах участков.
При образовании пар самки первое время не реагируют на ухаживание
самцов, убегают и улетают, не выказывая никакой привязанности к участку.
Но с момента, когда вся территория уже занята самцами, они начинают упор-
но держаться близ самца и, отлетев на другой участок, вновь возвращаются.
Это изменение в поведении самок совпадает по времени с интенсивным раз-
витием яичников и созреванием половых желез. Вскоре перемещение по тунд-
ре самок также прекращается и они оседают по гнездовым участкам в парах
с самцами. Вся гнездовая территория оказывается поделенной на гнездовые
участки, зорко охраняемые самцами. Запоздавшие с началом брачного перио-
да птицы вынуждены довольствоваться худшими местами или оставаться во-
все без них.
Неоднократные опыты с манщиком (чучело самца в брачном наряде),
проделанные автором в Тиманской тундре, подтверждают ожесточенность,
с которой самцы изгоняют соперников со своих гнездовых участков и полное
безразличие к чужим самкам. Однажды легко раненный самец был заклеван
в драке насмерть. В другом случае спугнутый со своего гнездового участка
самец в течение значительного времени летал над тундрой и не мог найти
места, где бы мог опуститься и отдохнуть: всюду его встречали самцы и мо-
ментально изгоняли со своих участков. Он вынужден был вернуться на
свой гнездовый участок и сесть поблизости от спугнувшего его человека.
2 Птицы СССР, 1. IV
18 отряд куриные
Величина гнездового участка зависит от характера стаций и плотности
населения куропаток. В Тиманской тундре в стациях со средней плотностью
сторожевые посты самцов располагались на расстоянии 150—250 м один
от другого и размеры гнездовых участков колебались от 30 до 70 тыс. м2.
Средняя плотность составляла 20—25 пар на 1 км2, максимальная —
30 пар и более. В начале брачного периода плотность выше, чем позднее. Так^
на обследованном участке Тиманской тундры площадью 0,5 км2 14 мая оби-
тали 15 пар, а 1 июня—только 12. Отход происходит за счет гибели от хищ-
ников. Гнезда устраиваются не обязательно в центре участка, а нередко на
его краю, и расстояния между двумя соседними гнездами бывают значительно
короче, чем между сторожевыми постами самцов. Охрана гнездовых террито-
рий происходит не только в брачный период, но и в течение всего периода
насиживания и заканчивается только за несколько дней до появления птен-
цов. Это совпадает с интенсивной линькой самцов и появлением темного
оперения на верхней стороне тела. В это время самцы оставляют сторожевые
посты и затаиваются между кочками, плотно прижавшись к земле. В период
выкармливания птенцов никакого обособления гнездовых территорий и враж-
ды между самцами не существует, а наоборот, наблюдается приуроченность
нескольких выводков к одним местам и нередко даже соединение их в общее
стадо. Образование гнездовых участков и борьба из-за них в брачный период
есть приспособление вида к равномерному распределению популяции на
гнездовой территории и наиболее успешному воспроизведению потомства.
Начало весеннего возбуждения самцов совпадает по времени с увеличе-
нием половых желез и с началом линьки. По мере роста семенников увели-
чивается половая активность птиц, интенсивнее проходит линька. У самок
наблюдается отставание в развитии яичников, чему соответствует отста-
вание в появлении половой активности и более позднее начало линьки.
Весь брачный период, т. е. время от рассеивания зимних стай и начала
проявления брачной активности до момента кладки и насиживания яиц,
можно разделить на три стадии. Первая—от начала весеннего возбуждения
до разбивки на пары, вторая—от момента разбивки на пары до спаривания,
третья—от спаривания и кладки яиц до начала насиживания. Первая стадия
относится к ранней весне, когда наряду с проталинами в тундре еще много-
снега, ясные теплые дни сменяются холодными, часто выпадает снег, бывают
дни с пургой и метелями. Птицы имеют увеличенные гонады и в теплые дни
проявляют активную брачную деятельность. При возврате холодов и снего-
падах иногда наблюдается временное возвращение птиц к стайному образу
жизни. В низовьях р. Вельть (Тиманская тундра) в 1938 г. эта стадия имела
место с 18 апреля по 4 мая. Вторая стадия совпадает с освобождением значи-
тельной площади тундры от снега, потеплением, прекращением сильных
холодов и снегопадов, с массовым прилетом куликов и водоплавающих птиц
с юга. В это время у куропаток происходит интенсивное развитие гонад,
заканчивающееся к концу периода, образование пар и закрепление гнездовых
участков за ними. Эта стадия наблюдалась с 4 по 26—27 мая. Третья стадия
начинается тогда, когда гнездовые места полностью освободятся от снега,
кочки и почва пообсохнут, установится теплая летняя погода. Этот период
совпадает с появлением в тундре мелких насекомоядных птиц. К этому вре-
мени заканчивается созревание половых желез, самки приобретают почти
полный летний наряд, хорошо маскирующий их на темном фоне почвы.
В этот период характерна еще более жестокая борьба за гнездовые участки,
спаривание, устройство гнезд и откладывание яиц. Этот период имел место-
меж ду 26—27 мая и 8—10 июня.
Таким образом, продолжительность брачного периода в Тиманской тун-
дре в 1938 г. равнялась 50—55 дням, т. е. почти двум месяцам. Это составляло
около 40% всего гнездового периода против 25% у степной расы из Северного
БЕЛАЯ КУРОПАТКА \д
Казахстана (Ульянин, 1939). Столь большая длительность брачного периода
на севере с его и без того коротким летом объясняется тем, что на севере весна
развивается значительно медленнее, в связи с чем и развитие половых желез
у куропаток происходит не так быстро, как у южных. Зато затяжка брач-
ного периода на севере компенсируется более быстрым темпом роста молодых,
и весь гнездовый период как южные, так и северные птицы заканчивают
примерно в 4,5 месяца.
При выборе места для гнезда существенную роль играют характер ра-
стительности, характер микрорельефа, влажность почвы. Для гнезда куро-
патка выбирает место с такой растительностью, которая, с одной стороны,
надежно маскирует гнездо (для чего при хорошей покровительственной окра-
ске птицы достаточно малейшего прикрытия одиночных веток), а с другой—
не скрывает от сидящей на гнезде птицы окружающую местность и позволяет
во-время заметить опасность. Свойство куропатки при надвигающейся опас-
ности крепко затаиваться на гнезде делает последнее обстоятельство крайне
важным для выбора момента взлета. В силу этого куропатки по возможности
избегают гнездиться в густых зарослях ивняка и березы и явное предпоч-
тение отдают участкам открытой тундры. Из 14 найденных в Тиманской тун-
дре гнезд среди зарослей ивы помещалось одно, под прикрытием одиночных,
но сравнительно густых кустиков карликовой березы и ивы—3, на открытой
тундре с нависшими над гнездом мелкими одиночными ветками карликовой
березы—9 и на голом участке тундры без всякого прикрытия—1. Типичным
местоположением гнезд в отношении микрорельефа является кочковатая
тундра; на ней располагается большая часть гнезд. Устраиваются они чаще
всего между кочками или на кочках, но всегда так, что гнездо с одной или двух
сторон защищено не слишком высокими выступами кочек, не мешающими
птице иметь под надзором окружающую местность. Встречаются гнезда
и на участках ровной песчаной или лишайниковой тундры в низинах между
кочками и на ровных лугах лайд, но, как правило, под значительным при-
крытием той или иной растительности. Из отмеченных выше гнезд, помещав-
шихся в открытой тундре, 7 располагались среди кочек, 3 на участках с ров-
ным микрорельефом и 3 в понижениях тундры. Почти всегда гнезда распо-
лагаются на сухом месте и этим в значительной мере объясняется тяготение
к кочковатой тундре: кочки весной обсыхают раньше, а нередко предста-
вляют единственные сухие места на довольно обширных площадях. Однако
гнезда бывают и в сыроватых низинках и даже на мокрых лугах лайд.
В этих случаях замечается более толстая подстилка из сухой травы под
яйцами.
Устройство гнезда весьма простое. Это небольшое углубление в земле,
бедно выстланное окружающей растительностью—сухими стеблями, тон-
кими ветками и листьями трав и кустарников и иногда несколькими соб-
ственными перышками. Ямки, как правило, овальной формы 4—8 см глуби-
ной (средняя 6 см), 9—19 см в диаметре (средний 17 см). На увлажненной
почве подстилка гуще, размер нередко больше. Так, гнездо, найденное авто-
ром на мокром лугу лайды в Тиманской тундре, представляло не углубление
в почве, а сооружение, дно и стенки которого были свиты из длинной сухой
травы. Глубина его 12 см, диаметр лотка 18 сму толщина дна свыше 1 см, тол-
щина стенок до 2 см. Благодаря искусной маскировке гнезд, покровитель-
ственной окраске яиц и самих птиц и привычке самки крепко затаиваться на
гнезде (не только в период насиживания, но и в момент кладки) обнаружить
гнездо белой куропатки в тундре чрезвычайно трудно. На время отсутствия
на гнезде птица прикрывает яйца кусочками окружающей растительности.
Яйца откладываются ежедневно. Число яиц в полной кладке Толмачев
A927) указывает для о-ва Колгуева 6—10, Семенов Тян-Шанский A937) для
Лапландии 7—10. В Тиманской тундре в 1938 г. кладки колебались от 8
2*
ОТРЯД КУРИНЫЕ
до 15 яиц, а в среднем (по 14 гнездам)—11. Поданным жителей тундры, число
яиц в гнезде достигает иногда 20 и более, а в некоторые годы в большинстве
гнезд встречается всего по 5—7 яиц. Размеры яиц: длина 44—52 мм, средняя
47 мм (по обмеру 20 шт.); вес ненасиженных 19—22 г, насиженных 17—19 г.
Окрашены в бледножелтоватый основной тон, испещренный коричневатыми
и буроватыми крапинками и пятнами разной величины. Пигментация зна-
Рис. 5. Схема расположения гнезд белой куропатки
чительно большая, чем у тетерева. Мелкие крапинки, сливаясь, образуют
немало крупных пятен неправильной формы, в результате чего темный цвет
занимает больше половины плещади скорлупы. Окраска только что снесенного
яйца свежая и сочная, основной тон, крапинки и пятна красного оттенка.
Полежавшее яйцо имеет более бледную окраску основного тона и более темную,
темнобурую и черно-коричневую, а также больше крапинок и пятен. Кладка
яиц обычно в первой половине июня (Семенов Тян-Шанский, 1937—Лаплан-
дия; Михеев, 1948—Тиманская тундра). В Тиманской тундре начало кладки
с 27 мая по 7 июня, конец с 7 по 17 июня, общая продолжительность времени
БЕЛАЯ КУРОПАТКА 21
кладки 21 день. Большое число яиц в полной кладке указывает на малую про-
должительность откладывания яйцу птиц одной популяции. К насиживанию
приступают после окончания всей кладки. На незаконченную кладку не са-
дятся и во время ночных заморозков. Морозы до —3,5° С никакого влияния
на результаты вывода птенцов в Тиманской тундре в 1938 г. не оказали.
Насиживает только самка, самец находится неотлучно близ гнезда, охраняя
гнездовый участок и защищая гнездо от нападения чаек и поморников. Начало
массового насиживания в Тиманской тундре в 1938 г.—около середины июня.
К этому времени зазеленела трава на лугах и сырых низинах, начали распу-
скаться листья березки на расположенных у земли ветках, появились цветы
и много насекомых. С этого времени самцы становятся молчаливее, больше
затаиваются, тундра не оживляется их голосами. Самка сидит на гнезде
очень крепко, вплотную подпускает человека, иногда до нее можно дотро-
нуться даже рукой. Спугнутая, быстро возвращается на гнездо. Сидит на гнез-
де круглые сутки, слетая на короткое время для кормежки. При поспешном
взлете с гнезда птица нередко выталкивает часть яиц на 10—20 см и более.
При возвращении самка вкатывает их обратно. В одном гнезде мы несколько
раз откатывали от 4 до 7 яиц на 5—20 см от гнезда, некоторые слегка прикры-
вали травой, но каждый раз птица водворяла все яйца на место. Даже
яйцо чирка, подложенное в кладку, ею аккуратно вкатывалось обратно. Как
правило, яйца располагаются в гнезде в один ряд, но в крупных кладках—
в два ряда, причем всегда острым концом вниз, тупым кверху. Для равномер-
ности согревания яйца регулярно перекладываются птицей с краев в середи-
ну и наоборот, в чем мы неоднократно убеждались, делая пометки на яйцах. Это
способствует одновременному вылуплению птенцов, наблюдающемуся в гнез-
дах куропатки. В период насиживания птица худеет. Весной вес самок (в Ти-
манской тундре в 1938 г.) от 600 до 700 г, к концу насиживания не более 550г.
В этот период наблюдается наибольшая зараженность птиц пухоедами и гли-
стами; зараженность последними иногда достигала 50% всех бегло осмотрен-
ных птиц (начало июля 1938 г.). В кишечнике одной особи—до десятка круп-
ных глистов. Наибольшая зараженность падает на конец июня—июль, т. е.
на период, когда в кормовом рационе птиц отсутствуют ягоды.
Продолжительность насиживания по Греббельсу A937) 22, 24 и 26 дней,
по нашим данным для Тиманской тундры —19 дней, возможно в некоторых
гнездах 20. В зависимости от характера почвы, на которой расположено
гнездо, температуры воздуха в период насиживания, влажности воздуха
и почвы, а также интенсивности насиживания сроки насиживания в разных
гнездах могут колебаться в пределах 1—2 дней и равны 18—20 дням. Более
же длительные сроки, указываемые Греббельсом, нуждаются в проверке.
Появление птенцов отмечается: в Лапландии—в конце июня—начале
июля (Семенов Тян-Шанский, 1938), на Канине (Банников, 1934)—в первой
половине июня, на Колгуеве (Толмачев, 1937)—в середине июля. В 1938 г.
в Тиманской тундре первые птенцы—в 20-х числах июня, массовый выход—
в последних числах июня, а в запоздалых кладках до середины июля.
Массовый выход птенцов совпал с началом отцветания морошки, концом
распускания листьев ивы и карликовой березы, массовым цветением голу-
бики, массовым появлением комаров. Продолжительность выхода птенцов
и$ яиц в одном гнезде 20—28 часов, в среднем одни сутки. Число одновре-
менно вылуплявшихся птенцов в наших гнездах было 1, 3, 4, 5, 9. Процент
выхода птенцов у белой куропатки высок: из 107 яиц, находившихся в 10 гнез-
дах, вывелись 97 птенцов, т. е. 90,7 %. Результаты вывода по отдельным гнез-
дам: в 7 гнездах—100%, в 1—80%, в 1—72%, в 1—63,6 %. Последние три гнез-
да часто нами посещались, что, очевидно, сказалось на результатах вывода.
Из 10 погибших яиц 1 похищено, 1—болтун, 5—с недоразвитыми зародышами
и 3 погибли в момент выклевывания из яйца вследствие нашего прямого вме-
22 ОТРЯД КУРИНЫЕ
шательства. Помимо отхода отдельных яиц, нередка гибель полных кладок.
Из 16 найденных нами гнезд погибли 3 A8,8%), два из которых разорены мох-
ноногим канюком (погибла самка) и длиннохвостым поморником (похитил
яйца), одно, повидимому, человеком. Нормально самка сидит до тех пор,
пока не вылупится последний птенец; но если самка согнана с гнезда в то время,
когда еще остается несколько яиц даже с проклюнувшимися зародышами,
а вместе с самкой выскочили и разбежались полуобсохшие птенцы, то обратно
птица в гнездо не возвращается, и зародыши в яйцах погибают.
Обычно самка уводит птенцов с гнезда сразу после того, как они обсох-
нут, т. е. через несколько часов после вылупления. К выводку сразу присое-
диняется самец, у которого к тому времени летний наряд полностью покры-
вает верхнюю сторону тела. Размещение куропаток в период выкармливания
птенцов резко меняется. Выводки переселяются к зарослям карликовой бере-
зыи ивнякам, на опушки травянистых лайд. Наибольшая плотность выводков
наблюдается в местах, где среди ивняков располагаются участки более или
менее сухой кочковатой тундры, поросшей карликовой березой и ягодниками.
Родители очень привязаны к птенцам и смело защищают их, пренебре-
гая смертельной опасностью. Как взрослые, так и птенцы при опасности
стараются прежде всего затаиться, а если это невозможно, пытаются убежать.
Только позднее, когда птенцы достигнут величины примерно половины взрос-
лых, родители при тревоге поднимаются и улетают молча. Самец и самка
в одинаковой мере заботятся о птенцах. При гибели самки все функции ухода
за птенцами принимает самец, а при гибели обоих родителей—птенцы присое-
диняются к соседнему выводку.
Смертность птенцов белой куропатки, по сравнению с другими вывод-
ковыми птицами, невелика, и по наблюдению в Тиманской тундре, до двух-
месячного возраста, когда молодые по величине уже не отличимы от взрос-
лых, доживают 80% птенцов (у глухарей смертность 60%, несколько меньше
у тетеревов, Северцев, 1932). Незначительный процент гибели молодняка
у белой куропатки объясняется рядом биологических особенностей: моно-
гамность и большая привязанность родителей к птенцам, обыкновение под-
бирать осиротевшие выводки, группирование нескольких выводков в общее
стадо и коллективная защита от врагов, способность затаиваться при опас-
ности и вести скрытый наземный образ жизни, чему способствует хорошо
выраженная покровительственная окраска. Кроме того, разнообразие и оби-
лие кормов в тундре исключает более или менее массовую гибель птенцов от
заболеваний. Враги куропаток в тундре: песец, поедающий яйца, птенцов
и взрослых; серебристая чайка и поморники (короткохвостый и длиннохво-
стый), особенно опасные для кладок и птенцов; мохноногий канюк, охотя-
щийся за птенцами и взрослыми, а также сапсан, кречет, белая сова, ястреб-
тетеревятник. Для белой совы и кречета белая куропатка служит зимой
основным кормом; вместе с нею они откочевывают на зиму в лесотундру.
Преследуют куропаток орлы, филины, длиннохвостая и каменная неясыть
и даже ястребиная сова. Таскают их яйца вороны и сороки. При таком обилии
врагов только плодовитость белой куропатки да отмеченные выше биологи-
ческие особенности позволяют этому виду успешно воспроизводить потомство
и поддерживать свое существование.
Анализы темпа роста птенцов (по весу) показывают, что первые 15—
20 дней увеличение веса идет сравнительно медленно: с 15—16 г в возрасте одно-
го дня до 60—80 г в возрасте 20 дней. Затем вес быстро возрастает, к 40 дням
достигая 300—350 г; после этого рост снова замедляется. К 60 дням вес птен-
цов равен 500—600 г, т. е. почти полному весу взрослых. Периоды замедлен-
ного увеличения веса совпадают с интенсивным формированием оперения
и линькой. В течение первых 15—20 дней происходит закладка и бурное
развитие перьев, заменяющих пух почти на всех частях тела. После 40 дней
БЕЛАЯ КУРОПАТКА
23
начинается линька маховых, идет интенсивный рост рулевых, происходит
формирование осеннего, а почти одновременно и зимнего нарядов. Размеры
птенцов в одном выводке колеблются: из пяти птенцов, добытых нами 14 июля
1937 г., самый маленький весил 22,4 г, самый крупный 43,1 г.
В Тиманской тундре птенцы достигают величины взрослых в конце авгу-
ста—в начале сентября, и весь период развития птенцов равен в среднем
2 месяцам, в то время как в Северном Казахстане у степной расы—3 меся-
цам. В краткости сроков развития птенцов на севере можно видеть при-
способление к короткому полярному лету, причем это связано с обилием
в тундре пищи и длительностью полярного дня, позволяющего птенцам
кормиться в течение круглых
суток.
Суточная жизнь куропаток
в период выкармливания птен-
цов в общем сходна с таковой
в брачный период: кормежка пе-
ремежается частым коротким
отдыхом и лишь в течение не-
скольких ночных часов насту-
пает период покоя. В конце
июля—начале августа ночи на-
рушаются частыми и громкими
криками самцов и перелетами,
напоминающими токовые в брач-
ный период, что связано, пови-
димому, с пробуждением поло-
вой активности и сходно с осен-
ним токованием тетеревов.
Выводки в течение лета ве-
дут довольно оседлый образ жиз-
ни и суточная деятельность од-
ной даже взрослой семьи обыч-
но ограничена территорией, не
превышающей 1—2 гектаров.
Родители находятся при птен-
цах до тех пор, пока те не до-
стигнут величины взрослых и
не наденут зимнего оперения.
Взрослые за это время претерпе-
вают все последующие линьки.
Рис. 6. Белая куропатка (самец в зимнем опе-
рении)
Линька. В течение года самцы носят четыре, самки—три хорошо
выраженных и резко отличных один от другого сезонных наряда (Михеев,
1948). В соответствии с этим куропатка линяет три или четыре раза в году.
Две линьки, послебрачная и предзимняя,—полные, в результате их появ-
ляется летний и зимний наряды; две другие—предбрачная и осенняя, даю-
щие весенний и осенний наряды,—частичные. У самок отсутствует пред-
брачная линька. В линьке куропаток характерно то, что одна линька по
времени налегает на другую: не успевает сформироваться один наряд, как
начинает развиваться другой, а осенний и зимний наряды начинают заклады-
ваться почти одновременно. По существу, беспрерывная линька идет с ранней
весны до самой зимы, т. е. 7—8 месяцев в году. Возможно с этим связано то, что
у белой куропатки не накопляется подкожного слоя жира, так как все за-
пасы питательных веществ идут на воспроизведение новых перьев при линьке.
Все сезонные наряды, за исключением зимнего, никогда не бывают чи-
стыми, они всегда перемешаны. От весны до зимы в оперении каждой птицы
24
ОТРЯД КУРИНЫЕ
имеются одновременно перья 2—3 нарядов. Ранней весной на горле у самцов
среди белых зимних перьев появляются каштаново-рыжие перья весеннего на-
ряда; вначале они скрыты белыми, а во второй половине апреля пробиваются и
образуют на белом фоне зимнего наряда первые яркие пятна весеннего наряда.
Вслед за горлом весенним нарядом охватываются бока шеи, головы и зоб,
а затем верх шеи и головы, а вскоре грудь и спина. Эта стадия наступает
примерно в конце мая. На этом линька не приостанавливайся, как полагает
Бутурлин A935), а продол-
жает развиваться, и уже в
начале июня весенние перья
появляются последователь-
но в удлиненных плечевых
и кроющих верх крыла, на
пояснице, надхвостье, верх-
ней части боков туловища,
на спине и нередко брюхе;
иногда последовательность
несколько иная. На всех
этих частях тела весенний
наряд распространен не
сплошь, а в виде отдельных
перьев или партий, переме-
шанных сначала с зимними,
а потом с летними перьями.
Весенним нарядом охваты-
ваются не все части тела,
а лишь голова, шея, зоб,
передние части груди и спи-
ны, часть удлиненных плече-
вых и кроющих верх крыла,
часть верха боков тулови-
ща, отдельные перья пояс-
ницы, надхвостья, спины и
изредка подхвостья и брю-
ха. Перья летнего наряда на-
чинают показываться вскоре
после появления весенних
перьев на верхней стороне
тела—обычно в первой по-
ловине июня. На передних
частях груди и спины они
бывают видны до появле-
ния весенних перьев на спи-
не, пояснице и надхвостье.
Обычная последовательность появления перьев летнего наряда: передние
части груди и спины, удлиненные плечевые и кроющие верх крыла, поясни-
ца, надхвостье, бока туловища и спина, верхняя сторона шеи, бока шеи,
зоб, горло, голова и позднее всего нижняя часть груди и брюха. Брюхо
в нижней части нередко остается белым в течение всего лета. Белые зимние
перья ноздрей и подбородка заменяются летними, окрашенными после пол-
ного формирования летнего наряда остальных частей оперения. Развитие
летнего наряда проходит быстро и полностью заканчивается в середине июля.
Отдельные перья весеннего наряда (чаще на горле и зобу) держатся иногда
до полного осеннего наряда. По окончании развития летнего наряда начи-
нается линька маховых и рулевых, затягивающаяся до поздней осени. В это
Рис. 7. Белая куропатка; сверху—самец в весеннем
наряде, снизу—самка в летнем наряде
БЕЛАЯ КУРОПАТКА 2S
же время начинают линять клюв и когти. Оперение цевки и пальцев начи-
нает изменяться еще в весеннем наряде, к полному летнему наряду почти пол-
ностью сбрасывается и вскоре вновь появляется в виде новых белых зимних
перьев. Вскоре после формирования полного летнего наряда появляются
первые перья осеннего наряда, окрашенные в более интенсивный и темный
рыжий тон. Пеньки осенних перьев закладываются в середине июля вначале
на зобу и горле, почти одновременно на боках нижней части шеи, а вскоре
на верхней стороне шеи и головы. Затем последовательно на груди, брюхе,
боках туловища, в удлиненных плечевых, кроющих крыла, спине, пояснице,
надхвостье. Осенним нарядом охватываются не все указанные части оперения
и больше всего он развит на зобу и груди, выражен на голове, шее,.брюхе
и боках туловища, но всюду перемешан с летним. Хорошо развитый осенний
наряд наблюдается с первой половины августа. Зимний наряд начинает
развиваться почти одновременно с осенним. В конце июля среди полного»
летнего наряда на брюхе, зобу, боках и задней части верхней стороны шеи
появляются распускающиеся из пеньков белые перья. В середине—во второй
половине августа под летними перьями в большом количестве встречаются
зимние на всех частях тела. Первые белые перья пробиваются наружу из-под
окрашенных в середине августа. Последовательность развития зимнего наря-
да: брюхо, подхвостье и нижняя часть боков туловища, грудь, зоб, подборо-
док, ноздри и одновременно поясница, верх крыла, надхвостье; позднее спина,
верхняя часть боков туловища, удлиненные плечевые, горло, шея, голова.
Полный зимний наряд—во второй половине октября.
У самок весной начинает развиваться сразу летний наряд и лишь у отдель-
ных особей на голове, зобу, горле, шее и верхней стороне иногда встречаются
одиночные рыжеватые перья, впоследствии заменяющиеся типичными лет-
ними. Первые распускающиеся из пеньков летние перья охватывают одно-
временно все части головы и шеи, за исключением подбородка, затем зоб и позд-
нее подбородок, вершина которого остается белой до полного летнего наряда.
К концу мая голова, зоб, шея полностью в летнем наряде, а спина, верх крыла,
поясница, передняя часть боков, надхвостье—пестрые, где летние перья уже
преобладают над зимними. К этому времени пеньки закладываются на груди,
на остальных частях боков туловища, брюха и на подхвостье. В первой поло-
вине июня наряд на этих частях тела пестрый, а полный летний наряд поя-
вляется в середине—конце июня. В отличие от самцов, брюхо у самок в летнем
наряде обычно полностью покрыто окрашенными перьями, за исключением
иногда одиночных. Осенний наряд начинает формироваться с конца июля.
Порядок его развития в общем сходен с таковым у самцов и охватывает те же
части тела, но выражен слабее. Зимний наряд, как и у самцов, появляется
почти одновременно с осенним, а первые зимние перья на наседных пятнах
даже раньше. Порядок развития как и у самцов. Начиная появляться в сере-
дине августа, полностью формируется во второй половине октября. Таким
образом, в течение года оперение белой куропатки 4—5 месяцев состоит из
одного наряда (зимний), iy2—2 месяца—из двух (зимний, весенний), 5—б ме-
сяцев из трех нарядов (зимний, весенний, летний и летний, осенний, зимний).
Рулевые перья сменяются один раз в году со второй половины июля до се-
редины сентября, выпадая постепенно. Надрулевые (Михеев, 1948)—два раза
в году; в июне и во второй половине сентября—в октябре. Маховые претер-
певают одну линьку. Начинается она в середине июля и полностью закан-
чивается в конце сентября—первой половине октября. Линька маховых
начинается с внутренних первостепенных и наружных второстепенных
и идет в двух направлениях: к наружным первостепенным и внутренним
второстепенным. Последними из первостепенных линяют второе и первое
маховые. Также один раз линяет оперение ног. Вследствие обнашивания
и обламывания бородок к июлю пальцы и цевка становятся полуголыми,
2tt ОТРЯД КУРИНЫЕ
с редкими упругими щетинками, представляющими собой сохранившиеся
стержни перьев, которые позднее исчезают совсем. В середине августа закла-
дываются пеньки новых зимних перьев, полностью одевающие плюсну и паль-
цы в конце сентября—первой половине октября. Когти линяют в июле. Отра-
стание новых когтей начинается вместе с появлением осеннего наряда и закан-
чивается к моменту появления полного зимнего наряда. По Ионсену A929),
линька клюва происходит в июле и начале августа; на нашем материале
сроки не прослежены, но отмечалось линяние в конце мая—начале июня.
Летний, осенний и зимний наряды связаны с сезонными изменениями
ландшафта и являются покровительственными. Весенний наряд—охрани-
тельный. Яркая окраска сидящего на сторожевом посту самца в брачном
наряде делает его заметным издали и отпугивает от занятого гнездового
участка другие пары, а также отвлекает внимание врагов от гнезда. Последо-
вательность формирования каждого сезонного наряда подчинена сохранению
наилучшей маскировки птицы. Помимо защитной роли, сезонные наряды
имеют значение в приспособлении к температурным условиям. Зимние перья
длиннее, их пуховая часть шире, побочный ствол развит лучше, чем у летних.
Длина и развитие пуховой части осенних перьев в общем сходные таковыми
зимних. Зимние перья дольше всего остаются весною и раньше всего появля-
ются осенью на брюхе, в чем можно видеть приспособление к защите от низ-
кой температуры снега и земли (при заморозках).
Белая куропатка носит два возрастных наряда: первый—пуховой, вто-
рой—ювенальный перьевой или птенцовый. Окончательный наряд—первый
зимний. Порядок формирования птенцового наряда в общем такой же, как
летнего наряда взрослых. Первыми начинают расти маховые первого порядка,
затем махи второго порядка и перья на хвосте, плечах, верхней части спины
и боках туловища. Все эти перья быстро отрастают, особенно маховые,
в результате чего полупуховые птенцы хорошо перепархивают на большое
расстояние. Дальше перья появляются на боках головы сзади глаз, на гру-
ди, верхней стороне головы и шеи, брюха, ноздрях, по средней линии спины,
затем на остальных частях спины и позднее всего—на горле, подбородке
и боках шеи. В начале—середине августа птенцы в Тиманской тундре бывают
полностью покрыты перьями. Вес их в это время достигает х/з—lU веса взро-
слых и представляют они собой маленьких птичек с хорошо развитым опере-
рением, но короткохвостых ввиду отсутствия рулевых. К этому моменту чер-
ные с белыми вершинами рулевые начинают только появляться. Сначала
показываются крайние, а затем постепенно средние. Развитие их затягивается
и они долго остаются короче надрулевых и кроющих хвоста, удлиняющихся
но мере роста птенцов. Средняя пара рулей начинает развертываться,
когда крайние почти достигают нормальной длины. Маховые перья первого
контурного наряда темные. Позднее они заменяются белыми, входящими
в состав окончательного зимнего наряда. Замена окрашенных маховых белы-
ми начинается еще у полупуховых птенцов. Порядок линьки маховых отли-
чен от взрослых. Первыми заменяются два наружных первостепенных махо-
вых (сначала первое, вскоре второе) и одновременно самые крайние внутрен-
ние первостепенные. Движение линьки от наружных к внутренним на первых
двух махах заканчивается, а от внутренних к наружным продолжается.
Первое время замена внутренних первостепенных махов идет быстро и к мо-
менту полного развития перьевого наряда большинство маховых оказывается
белыми, а темными остаются только 3—4 пера (третье—шестое первостепен-
ные маховые). Линька их затягивается и последнее, третье, первостепенное
маховое перо заменяется белым поздней осенью. Поздним летом и осенью
уже взматеривших молодых легко отличить от взрослых по наличию одного-
двух окрашенных в темный цвет первостепенных махов (не перелинявших),
я j первозимья—по третьему первостепенному маху, более короткому, чем вто»
БЕЛАЯ КУРОПАТКА 27
рое и четвертое маховые (у взрослых третье маховое всегда длиннее второго).
Вместе с развитием птенцового наряда происходит замена ювенального
пуха на цевке и пальцах. Зеленовато-желтоватый пух заменяется бело-
ватыми пуховыми перьями сначала на цевке, а затем на пальцах. К моменту
появления полного перьевого наряда ноги полуголые с наличием белых пень-
ков; несколько позднее: и они покрываются пухом, но редким и не повсемест-
но. Это соответствует летнему наряду ног взрослых. К тому периоду, когда
в оперении молодых появятся зимние перья, пуховое оперение цевки, а затем
и пальцев, вновь заменяется белыми перьями, которые и остаются на зиму.
Первый перьевой наряд никогда не бывает чистым, к нему всегда примеши-
ваются перья осеннего и зимнего нарядов. Еще у полупуховых птенцов, по-
мимо белых маховых, появляются белые зимние перья среди мелких крою-
щих крыла. Вскоре на груди и зобу закладываются пеньки с развивающимися
рыжими перьями осеннего наряда. К середине августа у тиманских птиц
осенний наряд бывает выражен хорошо. Развит он обычно на груди и зобу;
на боках туловища и. верхней стороне тела проявляется в виде отдельных
перьев. У самцов осенний наряд выражен значительно лучше, чем у самок,
покрывая почти сплошь грудь и зоб (у самок в виде одиночных перьев). Это
позволяет легко отличить самца от самки в довольно раннем возрасте. Одно-
временно с осенними закладываются пеньки с зимними перьями на груди
и брюхе, а позднее на боках туловища. Развитие пеньков идет медленно и
лишь вместе с появлением хорошо развитого осеннего наряда одиночные
белые перья пробиваются наружу, создавая известную пестроту в окраске
оперения. Дальнейшее развитие зимнего наряда у молодых, повидимому,
сходно с-таковым у взрослых. Когти и клюв у птенцов также подвергаются
линьке.
Питание. Типично растительноядная птица. Насекомые в питании
взрослых фигурируют редко. По Михееву A948) в Тиманской тундре зареги-
стрированы 37 кормовых видов растений, из которых 16 видов занимают
значительный удельный вес или в отдельные сезоны, или в течение всего года:
ивы F видов), подбел, толокнянка альпийская, водяника, клюква мелкоплод-
ная, голубика, береза карликовая, гречишник живородящий, кукушкин лен,
осока водяная и морошка. Наибольшее значение имеют ивы, средний про-
цент встреч которых за шесть месяцев (март—август) в зобах равен 52,8%.
Их удельный вес в кормовом рационе по месяцам: март 100%, апрель 56,8%,
май 58,8%, июнь 50%, июль 40,6%, август 44,4%. Поедаются почки, ветки
и зеленые листья. На втором месте стоит подбел (средний процент встреч
за шесть месяцев 32,4%), у которого поедаются преимущественно почки и вет-
ки, а также зеленые листья, прекрасно сохраняющиеся под снегом; реже упо-
требляются цветы и коробочки. Удельный вес по месяцам: март—во всех
зобах в значительном количестве, апрель—42, май—52,2, июнь—16,6%;
в июле и августе редок. На третьем месте—толокнянка (средний процент встреч
26,4%). Обнаружен в зобах только в июне, июле и августе с удельным весом
соответственно 16,6, 30,6, 22,2%. Употребляются в пищу листья и ягоды,
реже цветы. Далее следует поставить водянику, встреченную в 22,8% всех
осмотренных зобов. Значительный удельный вес имеет лишь в апреле и мае,
достигая 28,4 и 23,2%, а отчасти в июле. Поедаются ягоды, сохраняющиеся
под снегом до весны, а отчасти и листья. Являясь массовым кормом в период
конца зимы и весны, водяника тем самым представляет особенно ценный
объект питания. Не менее важным кормом служит голубика, приобретающая
особенное значение в летнем питании (средний процент встреч 22,8). Удель-
ный вес становится заметным в июне A6,6%), увеличиваясь в последующих
двух месяцах до 23,8 и 22,2 %. В пищу идут главным образом листья и ягоды,
отчасти цветы. Значительное место в питании занимает карликовая береза
(средний процент встреч 20,4), но удельный вес ее невелик: в июле 10,2,
28
ОТРЯД КУРИНЫЕ
в августе 11,1%. В пищу употребляются ветки, почки, листья и сережки. То-
локнянка альпийская играет существенную роль в весенний и раннелетний
период. Ее удельный вес: апрель 21,3, май 11,6, июнь 16,6%. В пищу идут
почти исключительно ягоды и только отчасти зеленые листья. Все остальные
корма употребляются на протяжении весьма ограниченного периода времени.
Количество растений, употребляемых в пищу в разные сезоны года, далеко
не одинаково. У птиц, добытых в марте, обнаружено 5 видов, в апреле—15,
в мае—14, в июне—18, в июле—28 и в августе—18. Наибольшее значение
в питании имеют три группы кормов: 1) сережки, почки и ветки растений,
являющиеся основным, а часто и единственным кормом зимой и ранней
весной, 2) зеленые листья, преобладающие в кормовом рационе летом, 3) яго-
ды, занимающие весьма большое место в питании ранней весной, а также
в конце лета и осенью. Количество поедаемого.корма колеблется по месяцам,
достигая максимума в период питания грубыми кормами (почками, ветками,
сережками), т. е. зимой, и снижаясь летом, когда употребляется более нежная
и питательная пища. Так, средний вес содержимого зоба в апреле равен 13,6 г,
в мае 6,6 г, в июне 3 г, в июле 1,9 г, в августе 2,5 г. В период питания грубы-
ми кормами в желудке больше содержится мелких камешков и непереварен-
ных костянок, способствующих механическому перетиранию кормов.
Видовой состав кормов птенцов всех возрастных групп несколько раз-
нообразнее, чему взрослых, и насчитывает 42 вида, из которых 31 относится
к растительным и 11— к животным кормам, состоящим исключительно из
насекомых. Последние обнаружены только .у птенцов ранних возрастов;
позднее птенцы, как и взрослые, становятся почти исключительно раститель-
ноядными. Общий состав кормов птенцов виден из следующих данных про-
цента встречаемости в зобах: насекомые 35,5, голубика 37,5, береза карли-
ковая 27,2, осоки 20,9, толокнянка 13,8, ивы 18,8, кукушкин лен 14,6, гречиш-
кин живородящий 12,5. В первые же дни жизни птенцы потребляют все виды
кормов, фигурирующие у взрослых, вплоть до грубых. Это свидетельствует
о ранней физиологической приспособленности к перевариванию и усвоению
самых разнообразных кормов, что увеличивает выживаемость птенцов при
неблагоприятных кормовых условиях. Но наибольший удельный вес имеют
нежные корма—насекомые и цветы. Это характерно для птенцов до 20-днев-
ного возраста. В последующих возрастах характер питания приобретает
сходство со взрослыми.
Удельный вес групп кормов в разных возрастных группах птенцов (в %)
Возрастные
группы птенцов
I
II
III
IV
Во всех воз-
растных груп-
пах в целом
Сережки,
почки и
веточки
18,8
37,5
28,6
16,7
Листья
и другие
зеленые
части
растений
18,8
47,2
75,0
86,0
47,9
Цветы
50
17,7
16,7
Семена
и плоды
травяных
растений
31,5
35,4
37,5
43
33,3
Ягоды
12,5
41,8
62,5
71,4
37,5
Насеко-
мые
(встречае
мость)
63
41,8
35,5
Состав кормов этого подвида в разных районах может иметь некоторые
различия, основанные на различной растительности этих районов. Так,
в Норвегии (Нордхаген 1928) куропатки больше, чем в Тиманской тундре,
БЕЛАЯ КУРОПАТКА 29
употребляют в пищу ветки, почки и листья черники, почки и сережки березы,
и используют вереск, вовсе не употребляющийся тиманскими птицами.
В то же время последние поедают ряд кормов, не отмеченных для Норвегии.
Небольшие различия имеются в кормовом рационе лапландских птиц, опи-
санном Семеновым Тян-Шанским A937). Однако общий характер питания
и смены кормов по сезонам в разных географических районах в общем один
и тот же. В зимние месяцы поедаются почти исключительно почки, ветки
и сережки древесных растений. Весною заметную роль играют перезимовав-
шие под снегом ягоды и зеленые листья. В течение всего вегетационного перио-
да имеют крайне важное значение листья и другие зеленые части растений.
В период цветения кормовой рацион разнообразится цветами, а позднее пло-
дами и семенами травянистых растений. В конце лета и осенью в массе по-
требляются ягоды нового урожая, составляющие одну из основных групп
кормов белой куропатки.
Полевые признаки. От других куриных, как и тундреная куропатка,
хорошо отличается в зимний период белоснежным нарядом и густым опере-
нием цевки до пальцев включительно. Среди белого оперения контрастно
выделяются лишь черные глаза, клюв, а при полете—темные рулевые и стерж-
ни маховых перьев. Весною самцы легко отличимы от других видов наличием
каштановой-рыжей окраски передних частей тела, а осенью самцы и самки—
наличием интенсивно рыжих пятен, вкрапленных в более бледную окраску
летнего наряда, преимущественно на передних частях тела. Летом отличается
рыжеватыми и желтоватыми тонами оперения с характерным чередованием
светлых и темных полос и пятен на пере. От тундреной куропатки отличается
более крупными размерами, более крупным клювом, а также отсутствием
черной уздечки в зимнем наряде и отсутствием ярких каштаново-рыжих,
рыжеватых и желтоватых тонов в оперении других сезонных нарядов; общий
окрас тундреной куропатки летом более темный и сероватый
Промысел. Обширная область распространения, богатейшие естественные
запасы и хорошие товарные качества ставят белую куропатку в число важней-
ших промысловых птиц страны. Из всех подвидов более других хозяйственно
используется популяция описываемой формы, населяющая более или менее об-
житые районы, а поэтому и более доступная для массовой добычи, особенно при
скоплении на зимовках в лесотундре. Промысел на куропатку в этих районах
имеет большое экономическое значение. Во многих районах лесотундры про-
мыслом занимается зимою поголовно все население. До Октябрьской револю-
ции, когда на север завозилось мало хлеба, мясо куропаток заменяло насе-
лению хлеб (Соловьев, 1927). Лет 20—30 тому назад ежегодный вылов белой
куропатки только в одном Усть-Усинском районе (на Печоре) колебался от
400 до 1000 тыс. шт. (Соловьев). За последний период вследствие ослабления
внимания к дичезаготовкам размер промысла этого вида снизился. Но и
при былом промысле естественные запасы белой куропатки использо-
вались далеко не полностью. Недопромысел связан с трудностями транспор-
тировки добытой птицы из глубинных районов в промышленные центры
и в особенности со слабой организацией промысла.
Описание. Размеры и строение. Тело, как и у других куриных, плотного
сложения, с небольшой головой, средней длины шеей, слегка выпуклым клювом; ноги
средней длины с крепкими пальцами, с короткими, слегка изогнутыми тупыми когтями;
по краям пальцев бахромка из роговых пластинок; цевка и пальцы оперены. Рулевые
сверху прикрыты парой надрулевых перьев и кроющими хвоста. Крылья округленные
и недлинные. Формула крыла: 3>4>2^>5>6> 1. Длина крыла самцов (86) 198—231,
самок (87) 194—215, в среднем 215,5 и 205 мм; длина клюва самцов (86) 9,5—13, самок (87)
8—11,6, в среднем 11,3 и 10,4 мм; высота клюва самцов (86) 9—12,2, самок (87) 9—12,
в среднем 10,9 и 10,4 мм. Вес самцов C2) 550—715, самок B9) 400—700, в среднем 658
и 616 г.
Окраска. Птенец в первом пуховом наряде от беловатого, иногда с коричневым от-
тенком, до желтого, иногда с зеленоватым оттенком, основного цвета; на этом общем фоне
30
ОТРЯД КУРИНЫЕ
с боков и верхней стороны тела разбросаны темные полосы и пятна от черного до рыжего-
и охристо-рыжего цвета. На голове коричневая или темнокоричневая шапочка, окай-
мленная черным; на боках головы сзади глаз черная изогнутая полоска или пятна; вдоль
спины тянутся две продольные черные (иногда слабо выраженные) или черно-бурые
полосы, на крыльях темные пятна с коричневым, а иногда охристым цветом вокруг них.
Надклювье желтоватое; когти беловато-желтоватые, иногда с темными краями.
Птенцовый наряд сходен с летним нарядом взрослых самок (см. ниже), имеются лишь
некоторые отличия: светлые полосы на перьях верхней стороны тела, следуя за вытянутым
черным центром, часто образуют концентрической формы рисунок. Темные поперечные
полосы на перьях брюха или слабо, или почти совсем не выражены. Больше проявляется
желтоватый или рыжеватый тон. Вершины перьев верхней стороны тела имеют белые тре-
угольники, а не каемки во всю ширину пера, как у взрослых. Маховые не белые, а серо-
вато-бурые с узкими беловатыми белыми полосками. Надклювье черное, подклювье с тем-
ными (или черными) краями и вершиной, с желтоватым или беловатым основанием.
Зимний наряд у самцов и самок белый, лишь хвост и стержни маховых черные.
Рулевые сверху прикрыты белыми надрулевыми и кроющими хвоста.
Молодые в первом зимнем пере не отличимы от взрослых. Распознать их можно лишь
по беловатому или желтоватому основанию подклювья, каковое у взрослых темное.
Весенний наряд самца: голова, шея, зоб и верхняя часть груди каштаново-рыжие;
перья верха головы и шеи с черными центрами, иногда на шее светлые вершинные каемки;
на зобу и груди нередко узкие темные
поперечные полоски, особенно выражен-
ные на передней части спины. Весенние
перья остальных темных частей тела
несколько светлее зоба и груди и варьи-
руют от темнорыжего с многочисленными
темными крапинками и полосками до*
темнобурого однотонного цвета. Бровь
красная. Оперение всех прочих частей
тела белое. У самок весенний наряд вы-
ражен слабо и не у всех особей, он пред-
ставлен в виде отдельных рыжеватых
или темнокоричневатых перьев, главным
образом на голове, зобу и шее.
Летний наряд самца: голова и шея
от охристо-бурого до бледнорыжего и
даже рыжевато-желтоватого цвета, не-
сколько более темного на голове, чем на
шее; зоб и грудь немного темнее, интен-
сивно рыжий цвет испещрен черными или
черно-бурыми поперечными полосами и
пятнами. Перья спины, удлиненные пле-
чевые, кроющие верх крыла, поясницы
и надхвостья—черные или черно-бурые с бледными желтоватыми или рыжеватыми попереч-
ными полосками и беловатыми вершинными каемками. Бока туловища несколько свет-
лее. Перья брюха от беловатого и бледнорыжеватого с темными буроватыми полосками
и пятнами до темнобурого и даже черного однотонного цвета. Маховые, большие кроющие
и верхние ряды малых кроющих верх крыла—белые, как и все кроющие низ крыла. Окра-
шенные кроющие верх крыла длиннее белых и полностью скрывают их на сложенном кры-
ле. Рулевые черные с яркобелыми вершинными каемками у новых после линьки, исчезаю-
щими по мере обнашивания. Цевка и пальцы голые. Когти темного или желтоватого цвета
с более темным основанием; клюв темный. Для самцов описываемого подвида характерен
хорошо развитый черный цвет на верхней стороне тела. Интенсивно черные перья изрезаны
сравнительно редкими узкими светлыми поперечными полосами, чем отличаются от севе-
росибирской и среднерусской форм. Окраска светлых полос верха интенсивно желтая,
рыжеватая и коричневатая в отличие от более бледных желтоватых тонов северосибир-
ской и более светлых интенсивно рыжих тонов у среднерусской. Белые вершинные каемки
развиты слабо, довольно узкие, создающие нередко на темном фоне спины характерный
тонкий струйчатый рисунок. Интенсивно черный цвет, узкие светлые полоски и сравни-
тельно темный окрас их, а также слабая развитость белых вершинных каемок, создают
характерный для этого подвида темный окрас верха. Окраска низа также более темного
рыжеватого тона с более узкими черными поперечными полосами на боках туловища, чем
у северосибирской. Окраска оперения самок в летнем наряде отличается от окраски сам-
цов следующим: 1) рыжие и рыжеватые тона выражены значительно слабее или вовсе
отсутствуют и заменены желтоватыми, охристо-желтоватыми или бледными тонами, осо-
бенно на горле, зобу, груди и брюхе, на которых у самцов рыжий цвет обычно развит
интенсивно; в результате общая окраска самок более бледная, чем у самцов; 2) поперечно-
полосатость на верхней стороне тела выражена слабее, а зачастую и вовсе отсутствует»
так как черные центры перьев крупнее, светлые полосы не разрезают их поперек, а не-
Рис. 8. Пуховой птенец белой куропатки
БЕЛАЯ КУРОПАТКА 31
редко, следуя за формой центра, образуют концентрический рисунок, характерный и для
молодых; 3) брюхо, как правило, полностью покрыто окрашенными перьями; 4) ниж-
няя сторона тела испещрена черными широкими поперечными полосами, чем самки до-
вольно резко отличаются от самцов. Самки данного подвида характеризуются, подобно
самцам, интенсивно черным основным цветом перьев верха, значительно более узкими
(чем у северосибирских) светлыми полосами, в окраске которых также преобладают
коричневатые и рыжеватые тона и более интенсивные (не бледные) желтоватые. Черные
центры перьев верха менее выражены, белые вершинные каемки развиты слабее.
В общем верхняя сторона тела более темная и значительно менее пестрая, чем у северо-
сибирских.
Осенний наряд самца: подбородок, горло, зоб, бока головы и шеи рыжевато-коричне-
ватые, иногда с узкой черной каймой на перьях подбородка и шеи или с темными крапин-
ками и пятнами на перьях зоба. Грудь такого же или более темного цвета, иногда с более
развитыми крапинками и пятнами и светлыми узкими вершинными каемками. Брюхо
и бока окрашены как и грудь, но всегда осенние перья перемешаны с летними, а на брюхе
и с белыми зимними, благодаря чему эти части обычно имеют пестрый окрас. На осенних
перьях боков туловища разбросаны темные крапинки и пятна. Подхвостье такого же цве-
та, как и остальной низ, с черными крапинками, а иногда полосками. Верхняя сторона
головы и шеи темнорыжевато-коричневатая с широкими черными полосами и пятнами.
Разбросанные на остальных частях верха тела осенние перья такого же цвета, с много-
численными черными крапинками, нередко сливающимися в центре пера в сплошное перо.
Окраска осеннего наряда самок сходна с самцами. Среди более бледных, не имеющих
почти совершенно рыжеватого оттенка летних перьев самок осенние перья выделяются
более резко, чем у самцов.
Среднерусская белая куропатка Lagopus lagopus rossicus Sserebrowsky
Lagopus lagopus rossicus. Sserebrosky. Journ. fur Ornith., 1926, стр. 511.
Егорьевск, б. Рязанская губ.
Распространение. Ареал. Центральные районы европейской части
СССР от Ленинграда и Вологодского края до долины рек Оки и Цны, к за-
паду включая Белоруссию, а к востоку, вероятно, Камский бассейн. На
юге граница проходит по южным районам Белоруссии, несколько севернее
Гомеля, но южнее Слуцка и Рогачева, т. е. приблизительно по 52° 30' с. ш.
(Мензбир, 1902). В Смоленской обл. доходит до широты Рославля, или 54°
с. ш. (Станчинский, 1915). Была добыта у Ивниц Житомирской обл., есть
указания на присутствие ее в Черниговской обл. (устное сообщение В. И. Зу-
баревой), изредка встречается в северных частях Засурских лесов (Бутур-
лин- 1935). В Тульской обл. прежде доходила до г. Одоева и района Епифани
(Сушкин, 1892), позднее держалась лишь в северо-западном .углу области
в районе Козельска, где очень редка (Филатов, 1915). В Московской обл.
гнездится в северной половине, включая и Ногинский район (Поляков, 1924),
заходит до района Спасска в Рязанской обл. (Хомяков, 1900), указывается
для района Темникова Мордовской АССР. В Горьковской обл. гнездится
только по левому берегу Волги (Серебровский, 1918), в Кировской—до райо-
на Халтуринска (Лобачев, 1930), т. е. приблизительно до 58° с. ш. До той же
примерно широты доходит и в Молотовской обл., где гнездится в районе устья
р. Обвы (Ущков, 1927). Северная и восточная границы достаточно не про-
слежены. Экземпляры из района станции Тундра (в 40 км к югу от Архан-
гельска) относятся к лапландской, с Вычегды—к среднерусской формам;
район Свердловска и Башкирию населяет степная форма.
X а р а.к тер пребывания. В основном оседла, но местами, осо-
бенно при неурожае кормов, совершает небольшие осенние и зимние переко-
чевки. Александров A903), описывая распространение белой куропатки
в б. Романово-Борисоглебском уезде Ярославской губ., пишет, что она по-
падается по окраинам болот Золотого и Варегова исключительно осенью.
Рузский A899) указывает, что в б. Казанской губ. летом куропатки нет, а зи-
мой изредка встречается в Козмодемьянском, Царевококшайском, Свияж-
ском и Казанском уездах. Хомяков A901) отмечает, что в б. Спасском уезде
32 ОТРЯД КУРИНЫЕ
Рязанской губ. куропатка водится только в западной его части, зимой же
встречается по всему уезду. Переходя зимою к питанию почками древесных
растений, куропатки вынуждены выбираться из своих летних местообита-
ний—моховых болот и переселяться в березовые леса, заросли ивняков по бе-
регам рек, отстоящие иногда на значительном расстоянии от районов гнез-
дования.
Биотоп. Гнездовый—преимущественно моховые болота, негнездо-
вый—открытые, поросшие кустарником места. По Битриху A926) на севере
европейской части СССР куропатка избегает сплошных массивов хвойного
леса, обитая по обширным моховым болотам и лесным площадям, испещрен-
ным сфагновыми формациями различного рода. Наиболее характерны места,
где чередуются чистые моховики-торфяники с торфяными болотами, по-
крытыми низкорослой сосной с типичным для них моховым покровом и под-
леском из карликовой березы, багульника, подбела, голубики и болотного
мирта. Часто встречается по краям моховых болот, по соседству с остров-
ными борами, веретьями, среди подлеска и полукустарников и низкорослых
ив. Во время осенней линьки забивается в крепь заболоченных березняков,
в кочковатые согры, поросшие ивняками, и избегает появляться в белом
наряде на открытых пространствах до выпадения снега. Зимою живет табу-
нами по порубам, пожарищам и тальникам.
В Ленинградской обл. летом куропатка держится чаще в больших боло-
тах, поросших молодым кустарником; реже наблюдается на песчаных, порос-
ших вереском горках, граничащих с болотами. В другие времена года изби-
рает вблизи лежащие открытые места, поросшие кустарником (Бихнер, 1884).
В Смоленской обл. куропатка гнездится в березовом мелколесье по торфя-
ному болоту и в сосновых насаждениях по моховому болоту (Станчинский,
1927). В Калининской обл. обитает по моховым болотам, поросшим редкой
низкорослой сосной. По окраинам таких болот, а местами на возвышенных
сухих участках среди них растут береза, осина, ива и другие кустарники.
По самым болотам встречается вереск, багульник, голубика, брусника,
клюква, а также морошка и черника. Болота зачастую вплотную подходят
к возделываемым полям, которые белая куропатка нередко посещает. Летом
держится на самом болоте, зимой концентрируется по их опушкам среди
кустарников.
Численность. Не высокая, с продвижением от севера к югу замет-
но падает и у южных границ ареала—низкая, что связано с меньшим распро-
странением болот в южных районах. На сокращение численности известное
влияние оказывает изменение стаций под воздействием хозяйственной дея-
тельности и непосредственное истребление самих птиц. Нами A948) в Кали-
нинской обл. определена плотность не выше одной пары на 1—2 км2. Низкую
плотность в значительной мере определяет ограниченность зимних кормов.
Характер кустарниковых зарослей в окрестностях обследованного нами
болота позволяет заключить, что для прокорма одной птицы в течение зимы
требуется от 4 до 9 га сплошных густых зарослей. Площадь последних была
такова, что позволяла держаться не более чем одной паре на 1 км2.
Экология. Размножение. Общие закономерности сходны с номи-
нальной расой. Весною табунки развиваются в марте (север европейской
части СССР, Битрих, 1926), токованье с апреля до конца мая—начала июня
(Костромская обл., Кирпичников, 1915; Ленинградская обл., Бихнер, 1884).
Полная кладка в середине—конце мая (Бихнер, 1884). Появление выводков
Битрих A926) отмечает около 15—25 июня, Бихнер A884)—в середине—
конце июня. К середине августа птенцы почти достигают размера взрослых
{Битрих, 1926).
Линька. Как у номинальной расы, но в несколько другие сроки,
детально не прослеженные.
БЕЛЛЯ КУРОПАТКА 33
Питание. Характер тот же, что и у номинальной расы, но набор
кормов несколько иной. Зимой—почти исключительно почки и верхушки
побегов ивы, березы, изредка осины, летом нежные листья, цветы, побеги
и ягоды голубики, черники, морошки, брусники, клюквы, иногда и насе-
комые (Михеев, 1948, Бихнер, 1884; Петрункевич, 1929). Мошнин A905) ука-
зывает на питание травой и что куропатка выедает в день в среднем не менее
2 кв. аршин луга. Это может быть и преувеличено, но факт вылета куропаток
осенью на поля и кормежки на них отмечает также Петрункевич A929).
Промысел* Этот подвид представляет интерес главным образом как
объект охоты. Добывается ружьем.
Описание. Размеры. Чуть помельче номинальной расы. Длина крыла самца D)
205—208, самки D) 192—218, в среднем 206 и 203 мм\ длина клюва самца D) 11—12,
самки D) 9,6—12, в среднем 11,4 и 11,1 мм; высота клюва самца D) 10,5—11, самки D)
10—12, в среднем 10,8 и 10,3 мм.
Окраска. В летнем наряде характеризуется более светлой окраской и сильно
выраженным чистым интенсивно рыжим тоном, особенно на верхней стороне тела, чем
резко отличается от других рас. Черный цвет на верхней стороне развит слабо. Черновато-
бурые (а не интенсивно черные, как у лапландской) перья верха изрезаны частыми и широ-
кими интенсивно рыжими полосами, пересекающими и центр пера. В результате черные
центры на перьях обычно отсутствуют, сравнительно хорошо выражена поперечнополоса-
тость, а рыжий цвет в общем покрывает даже большую часть пера, чем черный. На
боках туловища рыжая окраска особенно преобладает над темной, и рыжие полосы
значительно шире, чем разделяющие их темные, что не всегда наблюдается у других рас.
Бросается в глаза также отсутствие или слабая выраженность темных полос и пятен на
перьях груди и зоба. В других сезонных нарядах не отличима от лапландского подвида.
Северосибирская белая куропатка Lagopus lagopus koreni Thayer
et Bangs
Lagopus lagopus koreni Thayer et Bangs. Proceed. New-England. Zool. Club., V, 1914,
стр. 4, Нижне-Колымск.
Синонимы. Lagopus lagopus birulai. Sserebrowsky. Journ. fur Ornith., 1926,
стр. 513. Новая Сибирь.— Lagopus lagopus septentrionalis. Бутурлин. Сборник трудов
Зоологического музея Московского государственного университета, 1, 1934, стр. 83,
низовья Таза.
Распространение. Ареал. Север Сибири—от УРала до Анадыря. К югу
до Томска, Красноярска, Якутска и верховьев бассейна Колымы. К северу
на Ямале и Гыданском п-ве приблизительно до 71° 30' с. ш. (Житков, 1912;
Наумов, 1931), по другую сторону Енисейского залива до о-ва Диксон и севе-
ро-восточных побережий, т. е. почти до 74° с. ш. (Кирпичников, 1936); до той
же широты на Восточном Таймыре (Тугаринов и Толмачев, 1934); распростра-
нена в бассейнах Лены (Романов, 1934), Индигирки, Колымы, и на Чукот-
ском п-ве (Прртенко, 1939) до морских побережий; встречается также на Но-
восибирском архипелаге (Бируля, 1907). На восток до побережий Берингова
и Охотского морей. Довольно обычна по прибрежной полосе Анадырского
залива (Портенко, 1939) и в южных частях Анадырского края вообще (Бело-
польский, 1934), в Коряцкой Земле (Дементьев, 1941), в районе Охотска
(Харитонов, 1915). На запад распространена до Ямала включительно,
по Уралу граница не прослежена.
Характер пребывания. Как и у номинальной расы, из тунд-
ры совершают регулярные перелеты на зиму в прилегающие леса; в лесной
полосе—сравнительно небольшие перекочевки преимущественно в неуро-
жайные на корма годы. Подобно лапландской, нередки случаи зимования
некоторого числа птиц в Арктической тундре, что отмечено для Ленско-
Хатангского района (Романов, 1934,), о-ва Диксон (Кирпичников, 1936)
и даже Новосибирских о-вов (Бируля, 1907)
Время отлета из тундры: с побережья и островов моря Лаптевых
в 1926 г. в конце сентября—октябре и до второй половины ноября, момента
3 Птицы СССР, т. IV
34 ОТРЯД КУРИНЫЕ
наступления полярной ночи (Романов, 1934); с Новосибирских о-вов—
в середине ноября (Бируля, 1907); с о-ва Диксон—во второй половине октя-
бря—начале ноября (Кирпичников, 1936). По Енисею стаи продвигаются
к югу в лесную зону до 66—67° с. ш., а в Колымском крае резко выра-
женный весенний пролет наблюдался под 68° 30' с. ш. (Тугаринов и Бутур-
лин, 1911); по р. Индигирке куропатки добираются до г. Аллаихи A00—
120 км от устья) и вероятно еще выше (Михель, 1935). Большие скопления
зимующих стай отмечены в низовьях Оби (реки Сосьва, Казым, Полуй),
бассейна р. Таза, бассейна Нижней Тунгузки, на водоразделе между реками
Вилюем и Оленеком, в местах вокруг среднего течения Лены и Олекмы,
верховьях Яны, бассейна Индигирки, Колымы (Насимович, 1934), а также
в бассейне Анадыря (Портенко, 1938). Время весеннего прилета в тундру:
в районе о-ва Диксон в 1930 г. 23 апреля, в 1932 г. 18 апреля; в районе
побережья моря Лаптевых в 1927 г. появились: на возвышенной тундре
среднего течения р. Попигая 1 апреля, в дельте Лены 10—15 апреля, в рав-
нинной тундре около 1 мая, в окрестностях с. Быково первые птицы 11 апреля,
массовый прилет во второй половине апреля (Романов, 1934).
Биотоп. Сходен с биотопом номинального подвида.
Численность. В зоне тундры высокая, не ниже чем у номиналь-
ного подвида. По Дементьеву A935), расстояние между сторожевыми по-
стами двух соседних гнездовых участков определяется в 200 м. В лесной
полосе средняя. В связи с значительно более широким распространением
болот в лесной зоне Сибири по сравнению с европейской частью СССР чи-
сленность и общие запасы северосибирской расы в лесной зоне выше, чем
описанных двух рас в лесной полосе европейской части Союза.
Экология. Размножение. Начало брачного периода несколько
более позднее, чем у лапландской: в Гыданской тундре—первая половина
апреля (Наумов, 1931), в Ленско-Хатангском районе—середина апреля
(Романов, 1934), в дельте Индигирки—в апреле (Михель, 1935), на Ново-
сибирских о-вах—во второй половине апреля (Бируля, 1907).
Разбивка на пары в Гыданской тундре—в середине мая, в Ленско-Ха-
тангском районе в первой половине мая, на Новосибирских о-вах в сере-
дине мая. Спаривание и кладка в Ленско-Хатангском районе в первой по-
ловине июня, в дельте Индигирки—в середине июня, на Новосибирских
о-вах—в первой половине июня. К насиживанию приступают сразу же по
окончании полной кладки.
Места для гнездовий—те же ранние проталины, на которых птицы оби-
тают с прилета. Они раньше обсыхают, давая возможность делать гнездо
и класть яйца в то время, когда вокруг, по низинам, еще лежит снег. Как
и у номинальной расы, гнездо—небольшое углубление в почве, выстлан-
ное прошлогодней травой с примесью собственных перышек. Количество
яиц в кладке отмечается: для Ленско-Хатангского района 10—14 (Романов,
1934), для Новосибирских о-вов 8—10 и до 12 (Бируля, 1907). Сроки появле-
ния птенцов: на Ямале, в Гыданской тундре—в середине июля (Житков,
1913; Наумов, 1931), на северо-восточном побережье Енисейского залива—
во второй половине июня (Кирпичников, 1936), в Ленско-Хатангском райо-
не—в конце июня—начале июля (Романов, 1934). Молодые достигают вели-
чины взрослых в конце августа.
Линька. Ход и развитие, как у номинальной расы. Сроки также
близки.
Питание. Сходно с питанием номинального подвида. На почки ивы
и березы, а также на сережки последней, как нз основной корм индигир-
ских птиц, указывает Михель A935). По Наумову A931), основную летнюю
пищу куропаток Гыданской тундры составляют луковички гречишника
живородящего {Polygonum viviparum) и отчасти листья ив; в сентябре почти
БЕЛАЯ КУРОПАТКА 35
исключительно листья и почки ив и луковички камнеломки (Saxifraga
cernna); в ноябре, по выпадении снега,—почки ив.
Промысел. На обширной территории ареала естественные запасы опи-
сываемого подвида велики и превышают запасы всех остальных географи-
ческих рас. Однако экономически используются они весьма незначительно
и опромышляются лишь местами. По подсчетам Куклина A927), в тундровых
и лесотундровых районах Ямальского и Остяко-Вогульского округов еже-
годно добывается до 300 тыс. белых куропаток; на южном Ямале один охотник
добывает ежегодно в среднем до 1000 птиц. В большинстве же районов Сибир-
ской лесотундры ввиду слабой заселенности и отсутствия заготовительных
пунктов белая куропатка вовсе не опромышляется. Добыча производится
почти исключительно самоловами.
Описание. Размеры. Несколько мельче номинальной расы.
Длина крыла самцов A03) 196—226, самок (82) 186—216, в среднем 210 и 201 мм:
длина клюва самцов A03) 9—13, самок (82) 9—12,5, в среднем 10,8 и 10,6 мм; высота клюва
самцов A03) 9—12, самок (82) 9—11,7, в среднем 10,8 и 10,1 мм.
Окраска. Самцы в летнем наряде выделяются более светлой окраской спины,
чем у номинальной расы, но более темной, чем у среднерусской, а также значительно мень-
шим присутствием рыжих тонов, чем у обеих рас. Черный цвет перьев верхней стороны тела
менее интенсивен, чем у лапландской и приближается к черновато-бурому, как и у средне-
русских. Светлые полосы такой же ширины, но несколько реже расположены и окраска
их характеризуется как более бледножелтоватая и рыжеватая, чем у предыдущих рас.
Черные центры перьев верхней стороны выражены хорошо, более обширны, с широкими
белыми вершинными каемками, чем ясно отличаются от других рас. Белые вершинные
каемки, широкие светлые поперечные полосы и обширные темные центры создают ту Не-
строту рисунка оперения, которая так^характерна для птиц этих районов. Окраска груди
и зоба несколько темнее (но менее рыжая), чем у среднерусской, но чуть светлее лапланд-
ской. Темные поперечные полосы на боках туловища более широки и яснее выражены,
чем у обеих указанных рас. Самки в летнем наряде по окраске верха более свет-
лые, чем номинальная раса, но несколько темнее среднерусских. Характеризуются хороша
выраженными широкими светлыми полосами на верхней стороне тела, обширными черными
центрами, что создает характерную пестроту оисунка. Чаще всего у птиц именно этой расы
появляется концентрический рисунок пера. Светлые полосы на спине более бледного цве-
та, чем у типичной и среднерусской форм, с значительно меньшим количеством рыжего*
тона и с преобладанием бледножелтоватых тонов. Основной цвет перьев верха не интен-
сивно-черный, а черновато-бурый, как у самцов. Белые вершинные каемки выражены сла-
бее, чем у самцов. В других сезонных нарядах неотличимы.
Большая белая куропатка Lagopus I ago pus major Lorenz
Lagopus albus major. Lorenz. Orn. Monatsber 1904, стр. 177, степи у Акмолинска.
Распространение. Ареал. Населяет Западную Сибирь примерно
до района Ишима и Тары на север до последних островков леса в Северном
Казахстане на юг и до подножий Алтая на восток; в европейской части Со-
ветского Союза населяет Башкирию и заходит на север приблизительно до
Свердловска. Южная граница проходит в низовьях Илека, Губерлинских
горах, районе Орска, у истоков Тобола (близ озера Айке), в Наурзумском
заповеднике E1° с. ш.) и даже еще южнее—в урочище Каинды-Сай (Тур-
гайская степь) около 50° с. ш. (Сушкин, 1908); в Кокчетавском и Атбасар-
ском районах Северо-Казахстанской обл. проходит на юг до Акмолинска,
Каркаралинска (несколько южнее 50° с. ш.) и Семипалатинска (Штегман, 1934;
Сушкин, 1938); западная, северная и восточная границы точнее не прослежены.
Характер пребывания. Оседла, лишь местами совершает нерегу-
лярные сезонные перекочевки, появляясь в зимний период далеко за предела-
ми ареала. Сушкин A908) отмечает, что зимою куропатки держатся в камышах
Чалкар-Игыз-Кара, отдельные стайки их наблюдались поздней осенью под
Карабутаком, а зимби 1896/97 г. одна куропатка была добыта под Иргизом.
Аксаков A856) писал, что в западных степных уездах б. Уфимской губ*
белая куропатка бывала только зимой, летом ее там нет.
3*
36 ОТРЯД КУРИНЫЕ
Биотоп. По Сушкину, в Зауралье белая куропатка обыкновенно
держится в заболоченных березовых колках или ольшаниках. В подобных
же условиях, если только они имеются, можно встретить ее и в Северо-
Восточном Казахстане, но чаще всего она держится там в таких же березо-
вых колках, разбросанных по степи, или березовых участках боров, как
и тетерев. Иногда такие березовые участки растут среди боров и на пес-
чаной почве. Этот же автор указывает, что, подобно тетереву, куропатка
часто выбегает на полянки купаться в песке и довольно далеко уходит кор-
миться в ковыльную степь. Кириков A935) отмечает куропатку в качестве
весьма обыкновенной птицы в Предуральской и Зауральской лесостепях
лля следующих стаций: мелколесья колковых березников, березово-осо-
ковых болот, осоко-тальниковых мочажин и степных березников с кустар-
никовым подлеском. По нашим наблюдениям A948), характерным место-
обитанием в Северном Казахстане являются березовые, а местами осиновые
колки и перелески с густыми кустарниковыми зарослями и высокой густой
травой, перемежающиеся многочисленными большими и мелкими степными
полянами с низкой разреженной травянистой растительностью. Встречается
и в березовых участках на опушках сосновых боров. Кормящиеся птицы
нередко наблюдаются в открытой степи и на просяных полях близ мелких
кустиков таволги на расстоянии до 7—8 км от ближайшего леса и кустарни-
ковых зарослей. В жаркую летнюю пору, когда все мочажины в борах пере-
сыхают, в Наурзумском заповеднике куропатки регулярно вылетают на
водопой к озерам и карасу на расстояние до 10 км от постоянного места
обитания. В сухих березовых колках с кустарниковым подлеском и много-
численными полянами держатся куропатки и в Барабинской степи. В отличие
от лесной полосы, белая куропатка и тетерев в лесостепи обитают в сходных
условиях. Однако и здесь проявляется тяготение белой куропатки к откры-
тым местам, что особенно сказывается в приуроченности гнезд к полянам,
опушкам колков и даже открытой степи.- По данным Ульянина A939) для
Северного Казахстана, из 19 отмеченных им гнезд только 6 находились в лесу,
остальные вне его. Из последних 6 гнезд располагались на расстоянии до
5 м от леса и отдельных деревьев, 7 гнезд—эт 50 до 10Э лс и 1 гнездо-
до 20 км. Почти все гнезда располагались на пегчанэа почве и лишь
одно на глинистой, что свидетельствует о тяготении птиц к сухим почвам.
Чаще всего и сами птицы держатся в открытых местах, не забираясь в отличие
от тетерева со своими выводками в густые кустарниковые заросли, окружен-
ные густой травой. Больше всего шансов встретить куропатку в таких
местах, где мелкие куртинки березника и кустарников перемежаются обшир-
ными полянами с разреженной низкорослой травяной растительностью.
Численность. Довольно высокая, но ниже, чем у лапландской
и северосибирской форм в тундрах. На небольших площадях изолирован-
ных березовых колков гнездится значительное количество пар, располагая
гнезда иногда на 50—60 м одно от другого. Но небольшой размер территории
лесостепи, заселенной куропаткой, ставит эту область в отношении запасов
на одно из последних мест.
Экология. Размножение. Брачный период в Северном Казах-
стане в 1936 г. с 10 апреля по середину мая, т. е. 35 дней, весь период размно-
жения с 10 апреля по конец августа, т. е. 140 дней (Ульянин, 1939, 1949).
Таким образом, брачный период у степного подвида составляет лишь 25%
от гнездового периода. По Ульянину A949), семенники начинают увеличи-
ваться в длине с 1 апреля, наиболее резкое увеличение с 10 апреля, а к 8 мая
достигают наибольших размеров A6 мм)\ с 8 мая до 10 июня постепенно
уменьшаются до размеров, характерных для периода покоя G—8 мм). Автор
выделяет три стадии в первом этапе периода размножения: 1) птицы концен-
трируются в гнездовых стациях, самцы надевают брачный наряд, семенники
БЕЛАЯ КУРОПАТКА 37
начинают увеличиваться; стаи разбиваются на пары; самки сохраняют
зимний наряд; птицы летают группами по 3—4 и парами, между самцами
происходят драки; 2) куропатки окончательно устанавливают места гнездо-
вья, самки надевают неполный летний наряд (на голове, шее, спине, верх-
них кроющих крыла и хвоста) и начинают откладку яиц; становятся менее
подвижны, поднимаются лишь в исключительных случаях; у самцов—
полный брачный наряд, семенники достигают максимальной величины;
по утрам самцы обычно сидят на березах недалеко от гнезда, кричат реже,
перелетов группами на наблюдается, пары целыми сутками держатся в пре-
делах своего гнездового участка, 3) самки целыми сутками сидят на гнездах,
самцы находятся поблизости в ивняке или таволге. По данным того
же автора, начало кладки в 1936 г. в Северном Казахстане наблюдалось
с 2 по 8 мая, конец с 14 по 20 мая, весь период кладки равнялся 18 дням.
К насиживанию птица приступает после того, как снесет последнее яйцо.
Гнезда в углублениях на земле, размеры несколько мельче, чем у лапланд-
ской в Тиманской тундре: глубина от 2 до 5, диаметр от 16 до 17 см. Подстил-
ка—сухая трава, тонкие ветки таволги и изредка перья самой куропатки.
В части гнезд подстилки вовсе не было, и яйца лежали на сухом песке. Самка
откладывает ежедневно по одному яйцу. Полная кладка из 9—13, большей
частью 11—12 яиц. Из 19 гнезд 9 яиц было в двух, 10—в трех, 11—в семи,
12—в шести и 13—в одном. Окраска яиц сходна с таковыми номинальной
формы. Вес яиц сильно варьирует как в одной кладке, так и в разных: све-
жих от 16 до 21 г, насиженных от 15 до 20,5 г. Сроки насиживания с 14—
20 мая и по 9—10 июня. Продолжительность насиживания 18, в двух гнез-
дах—20 дней. Процент выхода птенцов: из 211 яиц, находившихся в 19 гнез-
дах, вывелось 173 птенца, или 82,5%. Уничтожено хищниками 18 яиц
(8,5%), болтунов 7 C,2%), птенцов, задохнувшихся в скорлупе, 13 E,8%),
из них 11 в одном гнезде, т. е. кладка полностью погибла. В сравнении со
степной расой тетерева процент гибели яиц у белой куропатки значительно
ниже: у тетерева 41,5, у куропатки 17,5. Темп роста птенцов быстрый; дан-
ные взвешивания птенцов из разных гнезд характеризуют прирост в весе
следующим образом: 5 июня (день вылупления из яйца)—14 г, 21 июля—
118 г, 22 августа—416 г, т. е. 63% от среднего веса взрослых самцов. В пер-
вые дни выводки держатся в березнике с разреженным травостоем, через
3—5 дней переходят в лощинки, заросшие зеленой травой. По мере подра-
стания птенцов чаще выходят на открытые места, а к концу августа или же
в начале сентября, когда молодые достигают 70% среднего веса старых сам-
цов и хорошо летают, родители уводят выводки в открытую степь за 2—3 км
и лишь к вечеру возвращаются в лес. В сентябре выводки начинают сбиваться
в стаи, численность которых в Наурзумском заповеднике достигает 30 и реже
70 штук. Как выводки, так и стаи держатся в районе летнего гнездования.
Линька. Общий ход линьки, как у номинальной расы. Сроки линьки
у самцов: появление первых перьев весеннего наряда конец марта—начало
апреля; передние части тела (голова, шея, зоб) полностью в весеннем наряде—
начало мая; появление первых весенних перьев на спинной стороне тела—
первая половина мая; появление первых летних перьев—первая половина
мая; спинная сторона тела полностью в летнем наряде—конец мая—начало
июня; полный летний наряд—конец июня; появление первых пеньков перьев
осеннего наряда—конец июня; появление первых осенних перьев—начало
июля, хорошо развитый осенний наряд—вторая половина июля, появление
пеньков первых зимних перьев—первая половина июля; появление первых
зимних перьев—первая половина августа; брюхо, подхвостье, грудь
полностью белые — вторая половина сентября; полный зимний наряд —
первая половина ноября; начало линьки махов—начало июля, выпадение
последних махов—вторая половина августа, конец линьки рулевых—начало
38 ОТРЯД КУРИНЫЕ
июля, выпадение последних рулей—конец июля, конец линьки рулей—
первая половина сентября, конец первой линьки надрулей—начало июля,
конец второй линьки надрулей—вторая половина сентября—октябрь.
У самок: спинная сторона полностью в летнем наряде—первая половина
мая; полный летний наряд—конец мая—начало июня; первые пеньки осен-
них перьев—первая половина июля; хорошо развитый осенний наряд—со
второй половины июля—начала августа; пеньки первых зимних перьев—
начало июля, первые зимние перья — первая половина августа; брюхо,
подхвостье, грудь полностью белые — вторая половина сентября — начало
октября; полный зимний наряд — первая половина ноября; линька крыла
и хвоста в те же сроки, как и у самца. Таким образом, у степного под-
вида весенний, а также летний и осенний наряды формируются несколько
раньше, чем у северных форм из тундр Евразии. Первые признаки зимнего
наряда южных птиц также появляются чуть раньше, но развивается он
медленнее, и полный зимний наряд появляется позднее, чем у северных.
Длительность формирования каждого сезонного наряда у северных и южных
форм примерно одинакова, за исключением летнего и особенно зимнего,
которые у южных птиц развиваются медленнее.
Питание. Общий характер питания и смена по сезонам сходны
с таковыми номинальной расы. Но состав кормов резко отличен, что связано
с различиями растительности северных тундровых и южных лесостепных
районов. Так, в Наурзумском заповеднике (Ульянин, 1939) из 40 зарегистри-
рованных видов кормовых растений 20 свойственны только для лесостепи
и не отмечены вовсе для северных птиц; из всех основных кормовых видов
75% относятся к специфичным только для этой полосы видам. По данным
анализа 56 зобов и 68 желудков главными растительными кормами явля-
ются: шиповник B9,4% встреч), употребляются плоды и листья с января
по сентябрь включительно; просо D5,5% встреч), поедается в течение
8 месяцев; лапчатка B9,4 % встреч)—в течение 5 месяцев; в отдельных случаях
составляет 90% съеденного корма (по объему); ива B5%)—в течение 7 ме-
сяцев, зимой почки и побеги—главнейший корм, летом—листья; гречиха
A9,1 %)—в течение 3 месяцев, в отдельных случаях составляет 50% съеден-
ного корма; осина G,3%)—в течение 4 месяцев; почки, сережки в осенне-
зимний и ранневесенний период составляют в зобах до 95 % содержимого;
таволга A7,6%); семена и листья поедаются в весенне-летние месяцы (май—
60%, июль—30% встреч); береза F%); почки и сережки в марте в отдельных
зобах занимали до 100% содержимого. К второстепенным кормам относятся,
полынь морская (Artemisia maritima), хвощ (Equisetum sp.), василек (Centau-
via sp.), осока (Carex supina), солодка (Glyсуrrhiza glabra), спаржа {Asparagus
sp.), щавель кислый (Rumexhaplorhizus), хондрилка (C/io/zdr/Z/a juncea), лютик
(Ranunculus sp.). Кроме того, зарегистрирован ряд видов случайных кормов,
как солонечник, дикий миндаль, гвоздика, ковыль и др. Животные корма
не играют существенной роли; всего зарегистрировано 9 видов, в том числе
чернотелка (сем. Tenebrionidae), итальянский прус (Calliptamus italicus),
муравьи (Formicidae), долгоносики (Curculionidae). Только итальянский прус
в двух случаях занимал 90% содержимого зоба (по объему) да муравьи—
иногда до 80%. В зимнее время (с декабря по март включительно) фигури-
рует исключительно растительная пища и главным образом шиповник,
береза, ива, лапчатка, просо, осина, а в глубокую зиму—исключительно
ветки и почки ивы, ягоды шиповника, почки осины и изредка березы.
Сравнительно с тетеревом, обитающим в тех же местах, кормовой рацион
белой куропатки беднее: у тетерева отмечено 54 вида растений и 14 видов
животных.
Промысел. Имеет значение не только для спортивной, но и промысло-
вой охоты. По данным Куклина A927), только в одной Челябинской обл.
БЕЛАЯ КУРОПАТКА 39
ежегодно добывалось приблизительно 200—250 тыс. белой куропатки. Спо-
соб добывания—стрельба ружьем и ловля шатром.
Описание. Размеры. Несколько крупней номинальной формы. Длина крыла самцов
C6) 208—235, самок A6) 208—222, в среднем 224 и 215 мм; длина клюва самцов C6) И —
13, самок A6) 11,3—12,2, в среднем 12,4 и 11,8 мм; высота клюва самцов C6) 10,7—12,5,
самок A6) 11—12,7, в среднем 11,8 и 11,6 мм; вес самцов A7) 450—872, средний 593 г; са-
мок (добытых в сентябре) 640—666 г.
Окраска. Самцы в летнем наряде имеют черный цвет перьев спинной стороны
более густой и интенсивный, чем у среднерусских и северосибирских, сближаясь этим с ла-
пландской расой. Черные центры выражены хорошо, чем отличаются от среднерусских и
сходны с северосибирскими. Светлые полосы бледнее, чем у предыдущих, с меньшим коли-
чеством рыжего; окраска их бледнорыжего, желтоватого и даже беловатого цвета. У боль-
шинства птиц они немного уже, чем у северосибирских, и расположены несколько реже.
Черный цвет, интенсивнее, крупнее черные центры и более редкое расположение более
узких световых полос создают окраску оперения верхней стороны тела темнее, чем у пре-
дыдущих двух форм. Белые вершинные каемки выражены не хуже, чем у северосибирской
формы, что обусловливает пестроту оперения, сходную с таковой у последней. Горло, зоб,
грудь, бока туловища более светлые, чем у всех предыдущих рас, бледнорыжеватого и да-
же желтоватого цвета. Самки в летнем наряде характеризуются теми же признаками.
Кроме того, от лапландской расы отличаются меньшим развитием поперечной полосатости
и более широкими светлыми полосами. Черный цвет на брюхе выражен слабее, чем у севе-
росибирских и лапландских птиц. В осеннем наряде перья бледного, желтовато-рыжевато-
го окраса, напоминающий окрас перьев на зобу и груди у самцов в летнем наряде. В дру-
гих сезонных нарядах неотличимы.
Малоклювая белая куропатка Lagopus lagopus brevirostris Hesse
Lagopus tagopus brevirostris. Hesse. Ornith. Monatsber, 1912, стр. 101, Алтай.
Синоним: Lagopus lagopus kozlowae. Portenko. Mitteil. aus dem Zoolog. Mus.
Berlin. 1931, стр. 416, Хангай.
Распространение. Ареал. Алтай, Саяны, Танну-Ола, Хангай, Кен-
тэй. Южная граница проходит по хребту Сарым-Сакты и Тау-Текель
в Южном Советском Алтае, на Укёке, у Коб до в Монгольском Алтае, на Чу-
лышманском плоскогорье и хребте Танну-Ола на перевале Улуч-арт к северу
от озер Убса-Нор (Сушкин, 1938), через Хангай (приблизительно 48° с. ш.)
и Кентэй (Козлова, 1930, 1932, 1933). Северная, западная и восточная гра-
ницы детально не прослежены.
Характер пребывания. Совершают регулярные вертикаль-
ные сезонные перекочевки. Обитая летом среди низкорослой горной расти-
тельности, скрывающейся зимой под снежным покровом, птицы, лишенные
корма и укрытий, вынуждены спускаться в нижние пояса гор и долины,
где держатся в течение всего зимнего периода. О регулярных вертикальных
кочевках и иногда на большие расстояния говорит Мензбир A902); то же
для Алтая отмечают Фолитарек и Дементьев A938).
Биотоп. По Сушкину A938), куропатка на Алтае связана с заросля-
ми альпийской березки. Особенно много этой птицы на Чулышманском
плоскогорье, где приземистая березка покрывает сплошь целые квадратные
километры, причем держатся они преимущественно там, где ручьи образуют
промоины, почва заболочена и местами в ней обнажены каменные обломки.
Характерной стадией куропаток Алтайского заповедника Фолитарек A938)
считает формацию мохово-кустарниковой тундры. Здесь куропатка держится
в типичных для этой формации обширных зарослях низкорослой и корявой
круглолистной березки, обычно достигающей высоты не более 1 м. Автор
отмечает особенное обилие куропаток в верхнем течении р. Чулышмана,
на р. Еш-Кечи, где на заболоченной долине реки заросли ивняков резко
преобладали над березкой и травянистый покров был более разнообразен.
Любимыми летними стациями в других местах являлись комбинации склонов
с россыпями и щебенкой и пониженных мест в тундре, занятых зарослями
ивняков, прерывающихся травянистыми буграми, лишенными кустарниковой
40 ОТРЯД КУРИНЫЕ
растительности. Их становится мало в чистых и высоких зарослях березки
и даже ивняков.
Численность. В разных районах различна, местами—высокая.
В Алтайском заповеднике Фолитарек A938) поднимал с площади в 20 га до
10 выводков. Учитывая сравнительную обширность площадей с подходя-
щими стациями, к одной из ландшафтных зон, хранящих значительные
запасы белой куропатки, можно отнести горные районы.
Экология. Размножение. Откладывание яиц на Алтае в середине
июня (Фолитарек и Дементьев, 1938), в высокогорном Хангае (Козлова,
1932)—в первой половине июня. Появление выводков на Алтае (Фолитарек
и Дементьев, 1938)—в середине июля A8 июля птенцы весом 39,5 г,
8—12 июля 242—291 г).
Линька. Характер линьки, как и у номинальной расы, но детально
сроки не прослежены. По замечанию Сушкина A938), линька взрослых, а так-
же последующая линька молодых носит иной, несколько сокращенный
характер по сравнению с среднерусской расой: у взрослых не развивается
полностью летний наряд, у молодых—осенний наряд. Автор предполагает,
что эти особенности вызываются коротким и поздним летом альпийской
зоны Алтая.
Питание. Данные о питании отсутствуют.
FОписание. Размеры. Мельче других рас, особенно в размере клюва. Длина крыла
самцов B1) 187—216, самок A0) 182—208, в среднем 205 и 193 мм; длина клюва самцов
B1) 8,5—11, 4, самок A0) 7,5—10,5, в среднем 10,3 и 9,4 мм; высота клюва самцов B1) 9—
10,4, самок A0) 8,5—9,4, в среднем 9,3 и 8,9 мм.
Окраска. Темнее остальных рас, за исключением номинальной. Темный тон
верха интенсивно черный. Светлые полосы более узкие и частые, лучше выражена погте-
речнополосатость на верхней стороне тела. Окраска светлых полос более бледная, чем
северосибирских, с меньшим количеством или полным отсутствием рыжеватых тонов,
но чуть темнее, с меньшей выраженностью белого тона, чем у степной. Черный цвет на
нижней стороне выражен лучше, чем у всех предыдущих, особенно на зобу, груди и пле-
чах; лучше, чем у других рас, выражена поперечнополосатость на боках туловища
и брюха. Самки в осеннем наряде сходны с северосибирскими, но окраска чуть потусклее,
с меньшим оттенком красноватого и большим оттенком бурого. В других нарядах неотли-
чимы.
Восточносибирская белая куропатка Lagopus I ago pus sserebrowsky
Domaniewski
Lagopus lagopus sserebrowsky. Domaniewski. Acta Orn. Mus. Zool. Polon.
1, 1933, № 6, Забайкалье.
Синоним. Lagopus lagopus dybowskii.S t e g m a n n. Orn. Mon., 1934, стр. 150—152
Распространение. Ареал. Горы Восточной Сибири южнее Якутска и
Верхоянского хребта: хребты Становой и Тукурингра, северная часть
Малого Хингана и Сихотэ-Алиня (Штегман, 1934), низовья Амура до притока
Горин (Шульпин, 1936), все горы Забайкалья и Байкальский край на запад
до западного берега Байкала, на юг до Хамар-Дабана и южной оконечности
Байкала.
Характер пребывания. По аналогии с предыдущей расой
можно полагать, что совершают вертикальные сезонные перекочевки, хотя
фактическим материалом мы не располагаем.
Биотоп. На Яблоновом хребте найдена на гольцах, поросших кедро-
вым стланцем, березкой Миддендорфа, даурской лиственницей, рябиной, водя-
никой, брусникой, малиной, шиповником, пахучей смородиной, багульни-
ком. Птицы держались на самой вершине гольцов и несколько ниже, избегая
густых зарослей стланца и придерживаясь мест, густо поросших водяникой
(Павлов, 1948).
БЕЛАЯ КУРОПАТКА 41
Численность. В разных местах различна, но, повидимому, не-
высокая. На сравнительно небольшой площади гольца (менее 1 км2) Яблоно-
вого хребта Павловым были обнаружены 4 выводка.
Экология. Размножение. На Яблоновом хребте 30 июня Павлов
наблюдал выводки слуховыми: птенцами 12 июля птенцы были величиной
с перепела и при тревоге дружно поднялись и далеко улетели. В последних
числах августа и в начале сентября в выводках насчитывалось по 8—10 птиц.
Линька. Сроки линьки не прослежены.
Питание. Просмотренные Павловым зобы и желудки куропаток,
добытых 30 июня, а также в конце августа и начале сентября, содержали:
ягоды водяники, брусники, малины, шиповника, пахучей смородины (еди-
нично), а также почки березки Миддендорфа, цветы и орехи кедрового стлан-
ца; орехи обнаружены лишь у одного самца в сентябре.
Описание. Размеры. Значительно меньше других рас и равна примерно мало*
клювой. Длина крыла самцов F) 195—210, самок A6) 180—197, в среднем 200 и 189 мм;
длина клюва самцов F) 10,1—11,2, самок A6) 9,5—11,8, в среднем 10,7 и 10,7 мм; высота
клюва самцов F) 9—11, самок A6) 8,3—9,5, в среднем 10,5 и 8,9 мм.
Окраска. В летнем наряде отличается от северосибирской куропатки несколь-
ко более узкими и частыми светлыми полосками на перьях верха, более мелкими черными
центрами и лучше выраженными рыжеватыми и коричневатыми тонами в светлых полосах.
У самцов эти же тона хорошо проявляются и в оперении низа. От малоклювой куропатки
отлична также большим количеством рыжих тонов (у самок на верхней, у самцов на ниж-
ней стороне тела), чуть более широкими и частыми светлыми полосками и отсутствием или
слабой выраженностью поперечнополосатости. Перья осеннего наряда самок светлее, чем
у упомянутых рас, но темнее у большой куропатки, светлокоричневатого или рыжевато»-
оранжевого цвета. Птиц в весеннем наряде мы не имели, в зимнем наряде от других рас
не отличима.
Сахалинская белая куропатка Lagopus lagopus okadai Mom i у am a
Lagopus lagopus okadai. M о m i у a m a. Annotationes Orn. Orient 1928, 21. XII,
I, № 3, стр. 236 Сахалин.
Синоним. Lagopus lagopus kamtschatkensis. Momiyama. Annot. Orn. Orient.
1928. 28. XI, № 3, стр. 238, западная часть Камчатки, около 51°50' с. ш.
Распространение. Ареал. Камчатка, Сахалин. На Камчатке распро-
странена ст побережья до верхних частей кустарникового пояса, т. е. до
высоты 800—900 м (Аверин, 1948).
Характер пребывания. По наблюдениям Аверина A948)
сахалинская куропатка на Камчатке совершает регулярные сезонные вер-
тикальные перекочевки из высокогорья в средние и нижние кустарни-
ковые пояса, где ольховый стланец и березовый лес дают корм и защиту.
Зимой в высокогорье совсем не встречается из-за отсутствия корма. Пере-
селение в нижние пояса гор происходит постепенно, начиная с конца октября
и в зависимости от снежности зимы. В местах зимовок концентрируется по
речным долинам, где ивняки и ольховый стланец не заносятся снегом. Обратное
передвижение в верхние пояса происходит в половине мая. Перекочевки
в верхние пояса наблюдаются также осенью, после окончания гнездового пе-
риода. Зимой крупных стай не образует, держится группками до 20 шт.;
только однажды в марте отмечена стая численностью 150 шт.
Биотоп. В гнездовое время держится по окраинам тундрочек вблизи
кустарников и в речных долинах, особенно в их верховьях, где чередуются
открытые высокотравные поляны и ягодниковые тундрочки с зарослями ивня-
ков и ольховника. Кустарники летом представляют только защиту. Придер-
живаясь последних, проникают на открытые окраины высокогорья. С высо-
коствольным лесом летом связана мало. Гнезда встречены на тундровых
кочках, на опушках приречных травянистых полей, на окраинах тундр под
нависшими кустами кедрового стланца. Зимой, помимо речных долин, охот-
но живет по опушкам березового леса, где много молодняка и кустарника.
42 ОТРЯД КУРИНЫЕ
Численность. На Камчатке и Сахалине куропатка обычна, но не
многочисленна.
Экология. Размножение. Начало токования на Камчатке наблю-
дается в половине апреля; распадение стаек на побережье—в 20-х числах
апреля, вдали от моря—в конце мая. Откладка яиц начинается в первых
числах июня: 3 июня найдены два гнезда с двумя яйцами в каждом. Кладка
чаще состоит из 7—9 яиц, но встречались по 17 и даже по 23 яйца. Массовый
вылет птенцов происходит 5—10 июля. В половине августа уже наблюдается
много хорошо летающих выводков, хотя 10 сентября отмечен выводок из
пяти птенцов, которые едва летали.
Линька. Не прослежена.
Питание. В зимний период, с половины ноября по апрель включи-
тельно, пищу составляют почки и тонкие веточки главным образом ив, мень-
ше—почки ольхового стланца и ольхи и изредка почки каменной березы.
В апреле в пищу идут ягоды шикши и брусники, в мае присоединяется моло-
дая зелень, а в конце этого месяца веточки вовсе выпадают из кормового ра-
циона. Со второй половины июля до половины сентября поедаются исклю-
чительно зелень и ягоды голубики, шикши, жимолости, брусники, шипов-
ника; в этот период ягоды составляют основную пищу. С конца сентября по
октябрь включительно—период перехода от летних кормов к зимним: наряду
с ягодами поедается и главный веточный корм—ивы. Кварцевые зерна в же-
лудках летом редки, зимой же обычны. Наибольшее количество гравия—
5,84 г—зарегистрировано у самца, добытого 8 апреля в березовом лесу.
Описание. Размеры. Крупнее восточносибирской расы. Длина крыла самцов A4)
196—220, самок A6) 193—215, в среднем 208 мм\ длина клюва самцов A4) 10,5—12,2,
самок A6) 10 12,4, в среднем 11,4 и 10,4 мм; высота клюва самцов A4) 10—12, самок A6)
9,5—11,3, в среднем 11 и 10,2 мм.
Окраска. Сходна с окраской восточносибирского подвида.
2. Тундреная куропатка Lagopus mutus Montin
Tetrao mutus, Montin. Phys. Saelsk. Handl. 1, 1776, стр. 155, Швеция.
Распространение. Ареал. Кругополярная птица. Населяет полосу
тундр и альпийскую зону горных хребтов от Исландии, Шпицбергена,
Шотландии до Северо-Восточной Азии и Японии в восточном полушарии,
северные части Северной Америки (Ньюфаундленд, Алеутские о-ва) и Грен-
ландию—в западном. В Западной Европе идет к югу до Альп и Пиренеев,
а в Скандинавии до 581/2° с. ш; в Азии—до Северной Монголии.
В СССР—полоса тундр (сухих и каменистых) от Лапландии до Чукот-
ского п-ва, Коряцкой земли и Камчатки, к югу—до Северного Урала
F1°с. ш.), Тарбагатая, Алтая иСаяна, Станового и Яблонового хребта, Коман-
дорских о-вов. Нет ее на Колгуеве, Новой Земле, Вайгаче и Канине, не
найдена на Новосибирских о-вах.
Характер пребывания. Оседла, местами зимой совершает
незначительные вертикальные перемещения.
Биотоп. Каменистая и горная тундра с бедной кустарниковой и травя-
нистой растительностью.
Подвиды. Систематика тундреной куропатки в значительной мере запутана, большое
число подвидов было описано по явно недиагностическим признакам, на очень небольшом
материале. Просмотренная коллекция Зоологического музея Московского государственного
университета и отчасти Зоологического института Академии наук СССР в количестве
112 экземпляров позволила выделить на территории СССР только 4 подвида: L. m. mutus
Montin, 1776 —Скандинавия, Кольский п-ов; L. т. kelloggae G г i n n е 1 1,
1910—Урал, Сибирь, Аляска, Северная Америка до залива Коронации, Арктических о-вов
и Северо-Западной Гренландии; L. т. nadezdae Sserebrowsky, 1926—Тарбага-
тай, Алтай, Саяны и Хангай; L. т. ridgwayi Stejneger, 1884—Командорские о-ва.
ТУНДРЕНАЯ КУРОПАТКА
43
Кроме того, за пределами СССР описаны подвиды: L. m. rupestris G meli n, 1789—
север Британской Колумбии, Южный Юкон, бассейн Мекензи, западное побережье
Гудзонова залива, юг Баффиновой Земли; L. т. reinhardi; В rehra, 1824—Юго-Западная
Гренландия до 66° с. ш.; L, т. welchi Brewster, 1885—Ньюфаундленд; L. т. nelsoni
S t e j n e g е г, 1884—Алеутские о-ва: Уналашка, Акутан и Унимак; L. т. atkhensis
Turner, 1882—о-в Атка (из группы Алеутских о-вов); L. /я. chamberlaini Clark, 1907—
о-в Адах (из группы Алеутских о-вов); L. т. sanfordl Bent, 1912—о-в Танага (из груп-
пы Алеутских о-вов); L. /п. townsendi Elliot, 1896—о-в Кыска (из группы Алеутских
Карта 3. Распространение тундреной куропатки Lagopus mutus
l—L. m. islandorum, 2—L. m. millaisi, 3—L. m. pyrenaicus, 4—L. m. helveticxie, 5—L. m. mutus,
6—L. m. helloggae, 7—L. m. nadezdae, 8—L. m. lidgwayi, 9—L. m. japonicus, JO—L. m. evermanni,
11—L. m. townsendi, 12—L. m. sanfordi, 13—L. m. chamberlaini, 14—L. m. athhensis, 15—L. m.
nelsoni, 16—L. ?n. dixoni, 17—L. m. rupestris, 18 — L. m. welchi, 1U — L. m. reichardi; а—граница
гнездовий
о-вов), L. т. evermanni Elliot, 1896—о-в Атту (из группы Алеутских о-вов); L. т. dixoni
-Q г i n n e 1 1, 1909—о-в Баранов, Чичагов, Адмиралтейство и прилежащие части мате-
рика; L. т. helveticus Thienemann, 1837—Альпы от Савойи до Южной Баварии,
Зальцбург, Каринтия и Штирия; L. т. rnillaisi Hartert, 1923—Шотландия; L. т. pyrenaicus
Н a rert, 1917—Пиренеи; L. т. islandorum F a b е г, 1822—Исландия; L. т. japonicus
•Clark, 1907—о-в Хондо.
Лапландская тундреная куропатка Lagopus mutus mutus Mont.
Распространение. Ареал. Населяет Скандинавию и Кольский п-ов.
На Кольском п-ве поднимается до 1100 м над уровнем моря в Лапландском
заповеднике, а в Хибинах—выше (Семенов Тян-Шанский, 1939).
Характер пребывания. В основном оседла, лишь в зимний
период совершает незначительные вертикальные перекочевки. В Лаплан-
дском заповеднике еще с осени стайки тундреной куропатки спускаются с вер-
шин гор пониже, в районы, богатые ягодами и побегами, где они и остаются
на зиму. Зимой обычно держатся между 300 и 600 м над уровнем моря (Семе-
нов Тян-Шанский, 1939).
Биотоп. Любимое местообитание—каменистые россыпи с растущими
на них тощими мхами и лишаями, вообще же горные или по крайней мере
каменистые тундры. На Кольском п-ве встречается по скалистым участкам
у берегов Баренцова моря и заселяет все горные тундры в центральных частях
полуострова. В разные сезоны года тундреная куропатка живет в тундрах
разного типа, поднимаясь летом в область гольцов. В лес залетает редко.
В конце зимы, когда снег покрывается крепким настом и куропатки не имеют
44 ОТРЯД КУРИНЫЕ
возможности докопаться до пищи, стайки иногда спускаются в зону березо-
вого криволесья, садятся на деревья, как это делают белые куропатки, и клюют
березовые почки. Выводки встречаются в самых разнообразных стациях,
но там, где поблизости есть вода или снег. Их можно встретить в сыроватых
лощинах с низкой травкой лугового характера, недалеко от тундровых ручьев
в долинах с зарослями карликовой березки, в ущельях со снегом и даже
среди пустынных скал, если только здесь есть лужи воды; на сухих ягель-
никах они редки, но на тундровых гарях кормятся охотно. Зимой попадаются
повсеместно, но чаще на ягельниках.
Численность. Не высокая. По данным Семенова Тян-Шанского
A939), в Лапландском заповеднике на 1 км маршрута приходится встреч
от 0,2 до 1,9, а в среднем за 5 лет 0,8 тундреной куропатки. В Архангельской
обл. и Коми АССР ежегодно добывается в пределах 500 шт., а общее пого-
ловье исчисляется 12—15 тыс. шт. (Паровщиков, 1937).
Экология. Размножение. По наблюдениям Семенова Тян-Шан-
ского A939) в Лапландском заповеднике, токовать куропатки начинают
в конце апреля. В это время они живут у верхней границы леса, где птицы
раньше всего и без затруднений находят корм. К закату солнца петухи ожив-
ляются, начинают перелетать и токуют всю короткую и светлую полярную
ночь. Стремительно взлетает петух и, опустив немного крылья и распущен-
ный хвост, сразу задерживает движения и садится на снег, пробегает не-
сколько шагов с приподнятым и полураспущенным хвостом и останавливается.
Обычно сейчас же является и садится поблизости другой петух, и оба они
перебегают друг перед другом, временами приостанавливаясь и исполняя
токовую песню. Поза петуха в этот момент весьма характерна: он немного
опускает хвост и шею так, что вместе с линией спины они образуют пологую
дугу, а оканчивая песню, одновременно поднимает шею и принимает обычную
позу. Брачный крик буквами можно изобразить примерно так: «кррраааэ-крэ-
ке-крээ-э-э-э». Правда, вспугнутый петух кричит также и в другие времена
года. Крик этот несколько варьирует, часто первое колено выпадает и в об-
щем соответствует первой части крика белой куропатки; но ничего анало-
гичного второй части брачного крика последней у самца тундреной куропатки
нет. Курочки ведут себя в это время тихо, малозаметны и издают такие же
негромкие крики, как и белая куропатка: «ме...», или «мя...», или «мяу...».
Лучше всего токуют куропатки в тихие и сравнительно теплые ночи.
Около 1 мая куропатки разбиваются на пары. Среди дня, когда птицы
находятся в покое, самец обыкновенно сидит или, точнее, лежит грудью на
выступающем камне, головой на ветер, а курочка прячется у подножья.
Петух с большого расстояния кажется белоснежной точкой на темном фоне
скалы. При приближении человека он взлетает с обычным брачным криком,
но садится тут же поблизости на камень или кустик. Самка подпускает к себе
ближе, а когда она взлетит, петух присоединяется к ней, и оба улетают.
Живут ли парами тундреные куропатки в течение всего периода размно-
жения—остается неясным. Некоторые наблюдения указанного выше автора,
как преследование самцом другого самца, подсевшего к паре, двукратная
встреча выводков вместе с самцом, усиленно отводившим от птенцов, как
будто подтверждают это. Но вместе с тем автор указывает, что как только
самки сядут на яйца, самцы встречаются поодиночке в тех же местах, где
проводили весну, а в июле поднимаются к вершинам гор в область гольцов,
где заканчивают смену пера. Кроме отмеченных двух случаев, автор не встре-
чал при выводках самцов. Однако отсутствие групповых токов и факт суще-
ствования определенных пар в брачный период дает возможность полагать,
что тундреная куропатка—скорее птица моногамная. О жизни парами,
подобно белой куропатке, указывает и Мензбир A902). В Лапландии кладка
яиц начинается не ранее конца первой декады июня, иногда в середине
ТУНДРЕНАЯ КУРОПАТКА 45
июня, а как исключение даже в августе (Мензбир, 1902). В Лапландском
заповеднике кладка и насиживание происходят в июне (Семенов Тян-Шан-
ский). Гнездо представляет небольшое углубление в почве, устланное
стебельками трав, мохом и иногда небольшим количеством перьев. Кладка
содержит от 3 до 10,а иногда и 12 яиц. Основной цвет их от грязно-
белого до буровато-охристого, с крупными и мелкими яркошоколадно-
бурыми пестринами. По наблюдениям Семенова Тян-Шанского A939), на
Кольском п-ве птенцы появляются в первых числах июля E июля в
1930 г., 3 июля 1933 г.), а при ранней весне даже в конце июня
B4 июня 1934 г.); В 34 зарегистрированных в Лапландском заповеднике (Семе-
нов Тян-Шанский, 1939) выводках число птенцов колебалось от 1 до 10,
чаще 3—6, а в среднем за 5 лет—4,6. По отдельным годам средняя величина
выводков изменялась от 1,6 A932 г.) до 6,2 A934 г.), что, как справедливо
предполагает автор, связано со снижением плодовитости куропаток в небла-
гоприятные в отношении зимних кормов годы и с значительной гибелью кла-
док и птенцов при летних волнах холода. Выводки встречались на разных
высотах от 400 до 830 м над уровнем моря. Выводок, имевший 5 июля пухо-
вых птенцов, 14 июля недурно перепархивал. «Поршки» обычно стараются
улетать как можно дальше под склон или по ветру, а самка остается на месте
и отвлекает на себя внимание врага. Птенцы постарше не взлетают, а вместе
с маткой убегакгГ и прячутся в камняк; как только все успокоится, подают
голос. Призывный крик птенцов—довольно громкое «тиу» или «пиу». Самка
сзывает цыплят негромким «куо», или «куа», или «квэ», причем, смотря по
обстоятельствам, придает ему различный оттенок. К середине августа птенцы
достигают 70% веса взрослой самки A2 августа—от 290 до 335 г), а в сентябре
они по размерам от матки не отличимы.
Линька. Число и порядок смены возрастных и сезонных нарядов,
как у белой куропатки. По Семенову Тян-Шанскому A939), схема линьки
лишь в деталях отличается от таковой у белой куропатки.
У самцов тундреной куропатки первые темные перья весеннего наряда
появляются на затылке, и, когда последний весь почернеет, линька преры-
вается до окончания периода токов. По сравнению с белой куропаткой пауза
наступает на более ранней фазе линьки и поэтому брачный наряд у тундре-
ной куропатки мало отличается от зимнего. У самца в брачном наряде на
теле под белым оперением скрываются пеньки и подрастающие летние перья.
Брачный наряд он носит приблизительно с 10 мая по 20 июня. В июне линька
переходит на шею и голову, а также на кроющие крыла и верхнюю часть спи-
ны. Этими частями оперения и ограничивается распространение весеннего на-
ряда, перья которого (более темного окраса) указанный автор ошибочно при-
нимает за особый тип летних перьев. По крайней мере в просмотренной нами
коллекции Зоологического музея Московского государственного университета
ни у одного из летних или осенних самцов не было обнаружено тех темнокаш-
тановых перьев, которые характерны для весеннего наряда. Таким образом,
полный весенний наряд охватывает голову и шею сплошь, верхнюю часть
спины, плечи и локти в виде отдельных перьев. Почти одновременно с фор-
мированием второй стадии весеннего наряда бурно развивается и летний
наряд, который вскоре становится полным. В летнем наряде остаются белыми
маховые, нижние и большая часть верхних кроющих крыла, кроющие ног,
брюха и задней части груди, центральная пара рулевых. Летний наряд
самец носит все лето, приблизительно с 5 июля по 15 сентября. За это время
он сменяет все перья, опадающие один раз в году, т. е. маховые, рулевые
и все те кроющие, которые на лето оставались белыми. В сентябре среди
летнего оперения появляются белые зимние перья, всюду почти в одно время,
и около 15 октября петух надевает полный зимний наряд. Дольше всех задер-
живаются отдельные летние перышки на шее и затылке. Весенняя линька
46 отряд куриные
у самок начинается почти одновременно с почернением затылка у самцов.
Летние перья пробиваются одновременно на голове, шее и плечах и к на-
чалу июня—к моменту насиживания—она получает полный летний наряд.
Ход осенней линьки у самца и самок одинаков. Осенняя линька, как и у сам-
цов, происходит в среднем с 15 сентября по 15 октября.
Зимняя жизнь. По Семенову Тян-Шанскому A939), зимою тун-
дреные куропатки живут большей частью стайками по 3—6 шт., реже попа-
даются большими стайками или одиночками. В первую, наиболее темную
половину зимы они—почти единственные обитатели снежных гор Лаплан-
Рис. 9. Тундреная куропатка (самец в зимнем наряде)
дии; кроме них, здесь можно встретить только редкие следы зайца, лисы
и горностая. Пищу добывают, раскапывая снег в тех местах, где он неглу-
бок. Ночуют в снежных ямках не далее нескольких метров одна от другой.
Чем толще, плотнее и ледянистее становится снежный покров в тундре, тем
труднее добывать пищу из-под снега. Поэтому во второй половине зимы,
когда северный олень из лесов переселится в тундру, отдельные стайки ку-
ропаток охотно держатся на местах кормежки оленей, где снег взрыт копы-
тами и куропаткам легко добраться до корма. В крайнем случае они спу-
скаются в пояс березников, где кормятся березовыми почками. Автор пред-
полагает, что многоснежные зимы губительны для тундренои куропатки и что
снеговой режим зимы является главным регулятором численности популя-
ции. В качестве примера указывает, что в начале 1932 и 1935 гг. наблюдался
самый глубокий (80—100 см) и плотный снежный покров за ряд лет. Именно
в эти зимы отмечено резкое падение численности куропаток @,3 и 0,2 шт.
на 1 км учетного маршрута вместо 1,7 и 1,9 шт. в 1931 и 1934 гг., отличавшихся
малоснежностью), а в последующее лето резкое уменьшение приплода.
Питание. В Лапландии (Семенов Тян-Шанский, 1939) зимою куро-
патки питаются ягодами вороники, толокнянки альпийской, толокнянки обык-
новенной, добываемыми из-под снега, а также, вероятно, почками и зелеными
частями тех же и других растений горной тундры. Нередко едят почки и
ТУНДРЕИАЯ КУРОПАТКА
47
концы побегов карликовой и простой березы и ивняков. Те же корма состав-
ляют основу питания в конце апреля—начале мая, когда появятся первые
проталины. В июне кормовой рацион значительно разнообразней и состоит
из почек, листьев, цветов и ягод различных видов растений. Однако прош-
логодние ягоды брусники и толокнянки альпийской, а также почки неко-
торых видов ив попрежнему стоят на первом месте. В июле зобы наполнены
по большей части цветами и соцветиями черники, ястребинки альпийской,
гречишника живородящего, нежными листьями ив, осоки, черники, новыми
плодами ивы травянистой, осоки, а прошлогодние ягоды попадаются редко;
у птиц, добытых на тундровых гарях, зобы наполнены преимущественно
коробочками кукушкина льна. По-
падаются насекомые—жуки, мухи,
перепончатокрылые. В августе то
же разнообразие кормов, но добав-
ляются ягоды нового урожая; в од-
ном зобу отмечены слизни A0 шт.).
В сентябре на первом месте ягоды
черники, голубики, вороники, за-
тем сухие плоды осоки, отчасти зе-
леные листья черники и вороники,
изредка насекомые, а с октября на-
чинают попадаться грубые почки ив
и карликовой березки. Всего заре-
гистрировано 27 видов кормовых
растений и несколько видов живот-
ных кормов.
Промысел. Ввиду малой чис-
ленности промысловое значение
небольшое.
Полевые признаки. Похожа на
белую куропатку, но мельче. Отли-
чается от последней в зимнем пере широкой черной уздечкой у самцов (иногда
и у старых самок), в летнем—меньшим развитием рыжих тонов.
Описание. Строение и размеры. Строение, как и белой куропатки, но
чуть длиннее крылья и более мелкий—низкий и короткий—клюв. Длина крыла самцов A1)
195—220, самок (9) 190—206, в среднем 196 и 209 мм. Вес (по Семенову Тян-Шанскому,
1939) самцов A3) 472—610, самок D) 435—480, в среднем 536 и 458 г.
Окраска. Птенцы и молодые похожи на белых куропаток, но светлее. Зимний
наряд взрослых птиц белый, обычно с черной уздечкой, с черными рулевыми. Весенний,
летний и осенний—пестрые. Перья весеннего наряда темнокаштановые, иногда чернобурые.
Летний наряд в общих чертах сходен с таковым белой куропатки, но отличается отсут-
ствием или очень слабым развитием рыжих и охристых тонов и присутствием сероватых.
Верхняя сторона тела самки темная, перья с крупными черными центрами, светлые каем-
ки и полосы узкие, беловатые, сероватые или светлоохристые. Нижняя сторона тела с хо-
рошо выраженными черными поперечными полосами и слаборыжими тонами. Верхняя
сторона тела самцов темная, черные центры перьев разрезаны узкими беловатыми и свет-
лоохристыми полосками, благодаря чему создается общий сероватый окрас со струйчатым
рисунком. Поперечные полосы на нижней стороне тела уже, более рыжеватые. В целом
отличается от других подвидов более темной окраской, меньшим развитием рыжих и боль-
шим серых тонов.
Рис. 10. Пуховой птенец тундреной куро-
патки
Сибирская тундреная куропатка Lagopus mutus kellogae G г i n n e 11
Lagopus rupestris kelloggae. Gri n n e 11. Univ. Calif. Publ. Zool. 1910, V, стр. 383. Аляска.
Синонимы. Lagopus mutus komensis. Sserebrowsky. Journ. fur Ornith.,1929,
стр. 522, Малая Харута, Северный Урал,—Lagopus mutustransbaicalicus. Sserebrows-
ky. Journ. fur Ornith., 1926, стр. 694, Тукурингра.—Lagopus mutus bargusinensis, Бу-
турлин. Птицы СССР, т. II, стр.175, Баргузин.— Lagopus mutus pleskeL Sserebro-
wsky. Journ. fur Ornith., 1926, стр. 696, Таймыр.—Lagopus mutus Kurilensis Kuroda.
Bull. Brit.Ornith. Club, 1934, стр, 15, Парамушир.
48 ОТРЯД КУРИНЫЕ
Распространение. От Урала через Сибирь, Аляску, Северный Юкон,
западное арктическое побережье Америки до залива Коронации и арк-
тических островов (кроме Баффиновой Земли) и Северо-Западной Грен-
ландии. В СССР—Урал, вся Сибирь (за исключением Алтая, Саян и Тар-
багатая), Дальний Восток, Камчатка, Северные Курилы. На север идет до
границ ареала вида, на юг—до 60° с. ш. по Уралу (Портенко, 1937), до Алтая
и Саян в Сибири; населяет Яблоновый, Становой и хребет Тукурингра на
Дальнем Востоке.
Характер пребывания. В зимнее время в поисках корма
местами совершает как горизонтальные, так и вертикальные перекочевки.
На Северном Урале в летнее время держится только к северу от 71-й парал-
лели, зимой же откочевывает к югу приблизительно до Полярного круга
(Куклин, 1937). На Камчатке летом распространена по высокогорью, не ниже
2300 м, зимой спускается в нижние части долов и в пояса леса вплоть до
морского побережья. Спускаться вниз начинают около половины сентября,
т. е. когда высоты закрываются снегом; заканчивается зимнее размещение в се-
редине ноября. Массовый подъем в горы происходит в апреле (Аверин, 1948).
Биотоп. На Северном Урале каменистые россыпи, чередующиеся
с пятнами лишайниковой тундры, сухие бугры и площадки тундры, порос-
шие карликовой березой и ягодниками (Портенко, 1937). На Ямале держится
всегда в высокой тундре: на склонах и вершинах обнаженных от кустар-
никовых зарослей и вообще бедных растительностью холмов, на перевалах
между речными бассейнами и т. п. (Житков, 1912). На Восточном Таймыре
встречается большей частью в сухих участках тундры, нередко слегка порос-
ших кустиками ивы, реже—на более высоких холмах и совсем случайна
в болотистых низинах (Тугаринов и Толмачев, 1934). В Коряцкой Земле
гнездится в каменистой горной тундре, зимой держится в речных поймах
(Дементьев, 1940). Типичное местообитание на Камчатке (Аверин, 1948)—
каменистая горная тундра на склонах хребтов, старые задернованные лаво-
вые потоки; по соседству всегда каменистые осыпи, нагромождение туфовых
обломков или отдельные скалы на склонах хребтов, которые дают возмож-
ность куропаткам спрятаться. Эти стации находятся выше пояса сомкнутых
кустарниковых зарослей и лишь иногда по разреженной их окраине. Зимой
откочевывают вниз. Держатся там, где по условиям рельефа мало снега
и есть бесснежные выдувы, на которых они кормятся.
Численность. В большей части ареала обычна, но немногочислен-
на; в численности далеко уступает белой куропатке*
Экология. Размножение. Токование на Восточном Таймыре
(Тугаринов и Толмачев 1934) отмечено 21 апреля. Образование пар 4—5 июня,
насиживание—во второй половине июня—первой половине июля. Гнезда
с 11 и 13 яйцами находили 23 и 25 июня, 4 июля, в последнем гнезде яйца
были еще 7 и 10 июля. Гнездо представляет неглубокую ямку, скудно выло-
женную перьями; расположено на сухой тундре под защитой ветвей ивы.
Благодаря защитной окраске самку разглядеть на гнезде трудно. Сидит
на гнезде так крепко, что ее можно снять руками с гнезда. Первый выводок
с пуховыми птенцами встречен 18 июля; выводок, вспугнутый 28 июля, под-
нялся на крыло, но полет неуверенный; к 3—4 августа птенцы заметно подросли
и неплохо летали; 11 августа птенцы летали хорошо и величиной были почти
со взрослых. На Среднем Урале гнездо с 5 яйцами найдено около 10 июня.
На Камчатке (Аверин, 1948) птенцы выводятся в середине июля. Встречен-
ные в период с 18 по 30 июля птенцы весили 22,5—40 г; в конце августа
молодые хорошо летали (вес 150—200 г), в сентябре не отличались от взрос-
лых (вес 310—400 г). За лето происходит большой отход молодняка.
Линька. Характер линьки, как у номинального подвида. На Восточ-
ном Таймыре самки начинают линять в первых числах июня; первые перья
ТУНДРЕНАЯ КУРОПАТКА 49
появляются в беспорядке на спине, шее и голове. Линька идет интенсивно
и к середине июля почти полностью заканчивается, остаются белые перья
лишь на брюхе. К концу июня летний наряд становится полным. Первые
линяющие самцы с немногими темными перьями на задней стороне шеи встре-
чены 11 июня,- а вообще без следов линьки попадались до 18 июня. В конце
июня все самцы линяли; у многих из них темные перья были лишь на голове
и шее, а у некоторых и на горле и у основания крыльев. К концу периода
насиживания A0 августа) самцы успели заметно перелинять, но далеко не
полностью; 18 июля сверху они почти вылиняли и на груди проглядывали
темные перья. В первой половине августа они были в полном летнем наряде.
В августе же начинается и осенняя линька. У добытого 25 августа самца на
нижней стороне тела имелось немало белых перьев. Во встреченной 24 сентября
стайке все птицы как будто перелиняли полностью (Тугаринов и Толмачев,
1934). На Северном Урале нередко куропатки белеют до выпадения снега
и становятся легкой дсйбычей охотников.
Зимняя жизнь. На Восточном Таймыре в конце августа сбиваются
в стайки по 15—20 шт., которые держатся весь сентябрь. Начиная с октяб-
ря встречаются только одиночками и по 2—Зшт. вместе (Тугаринов и Толмачев,
1934). На Камчатке с сентября и всю зиму куропатки чаще держатся пооди-
ночке, выводки объединяются редко, на зимовках в лесу встречаются только
небольшими стайками. Ночуют в снегу, иногда в нишах под скалами или кам-
нями. Иногда несколько куропаток обитает на бараньей или оленьей зимовке,
пользуясь растительностью, которую эти животные выбрасывают, раскапывая
копытами снег. Кормежка на оленьих копках отмечена и для Таймыра. В вер-
ховьях Колымы в апреле отмечены большие стаи (Дементьев и Шохин, 1939).
Питание. Состав пищи и смена ее по сезонам сходны с белой куро-
паткой- На Среднем Урале летом и весною тундреная куропатка питается
исключительно листьями и цветами альпийских растений. Ко времени
созревания ягод переходит на питание последними, в частности голуби-
кой. На Камчатке (Аверин, 1948) наиболее постоянным кормом являются
почки и тонкие веточки ив. В различные сезоны года к ним присоединяются
другие корма. Зимой—ягоды шикши и брусники, сухая травянистая рас-
тительность в сильно измельченном виде, встречающаяся во всех зимних
зобах и желудках; в нижнем поясе леса—главным образом почки и семенные
сережки каменной березы, почки и ветки ив, почки ольхового стланца.
Летом к ивам присоединяются листья и цветы остролодки, горной голубики,
куропаточьей травы, мятлика и других растений. По мере созревания ягоды
и семена растений приобретают все большее значение. В августе и сентябре
они встречаются во всех желудках.
Промысел. Незначителен. Добывают ружьем, силками, загоняют зимой
в сети, расстанавливаемые в кустах тальника.
Описание. Размеры. Длина крыла самцов C1) 190—213, самок D8) 182—210,
в среднем 205 и 194 мм; вес самцов A1) 360—750, самок A0) 310—660, в среднем 530 и 470 г.
Окраска. Отличается от типичного подвида большим развитием рыжих и охри-
стых тонов в оперении, меньшим—черного цвета. Черные центры перьев верха мельче, свет-
лые каемки шире, черные поперечные полосы низа—уже. Светлые полосы и каемки на
перьях не сероватые, а светлоохристые или рыжеватые. В целом окраска более светлая,
чем у типичного подвида. Наличие охристых и рыжеватых тонов сближает описываемый
подвид с белой куропаткой.
Алтайская тундреная куропатка Lagopus rnutus nadezdae Sserebr.
Lagopus mutus nadezdae. Sserebrowsky. Journ. fur Ornith., 1926, стр. 692,
Курайские горы, Алтай.
Синоним: Lagopus mutus macrorhynchus. Sserebrowsky. Journ. fur Ornith.,
1926, стр. 693, Тарбагатай.
Распространение. Тарбагатай, Алтай, Саяны и Хангай. К востоку идет
до озера Косогол и Отхон-Тенгри, к северу—до верховьев Маны, к югу—до
4 Птицы СССР, т. IV
50 ОТРЯД КУРИНЫЕ
Урюгнора и Тау-Теке-Лу в Монгольском Алтае, Западного Саяна, Хангая
и Хамар-Дабана. Вертикальное распространение на Алтае от 2200 и по край-
ней мере до 3000 м.
Характер пребывания. Данных нет. По аналогии с преды-
дущими подвидами можно полагать, что совершает сезонные вертикальные
перекочевки.
Биотоп. Характерная стация на Алтае—каменные россыпи, крупные
с камнями до метра в поперечнике, или площадки мелкого щебня, разбросан-
ные среди альпийского луга или крупных валунов. На кормежке иногда
далеко выходит на кочковатый альпийский луг, но никогда не забирается
в березовые заросли (Сушкин, 1938). В Алтайском заповеднике встречается
выше пояса мохово-кустарниковой тундры, в щебенистой и каменистой тундре
(Фолитарек и Дементьев, 1938). В Хангае придерживается исключительно
области каменистой тундры вблизи нижней границы вечных снегов (Козлова,
1932).
Численность. Невысокая, в Хангае редка.
Экология. Размножение. Сведения об экологии крайне
скудны. По Сушкину A938), вывод птенцов на Алтае происходит разно-
временно и на больших высотах вообще запаздывает. 26 июня на Алтын-Ту
встречены птенцы немного крупнее перепелки с пухом, сохранившимся на
горле, щеках и по средней линии зоба. На Маргалинских белках в те же числа
добыты пуховички. В Сайлюгеме у верховьев Хара-Джаматы пуховички
ростом меньше перепелки найдены 18 июля, а неподалеку отсюда, у верхо-
вья Чаган-Бургазы, 5 и 6 августа добыты птенцы уже крупные, размером
в две трети самки, почти перелинявшие в осеннее перо. По наблюдениям тога
же автора, в охране выводка самец не принимает участия, птенцов водит
только самка; самца можно иногда найти неподалеку, но лишь просто пото-
му, что он держится в тех же стациях.
Л и н ь к а. По Сушкину A938), весенняя у самцов на Алтае начинается
в начале июня: у добытого 9 июня только начинали пробиваться темные
перья на голове. В конце июня голова и шея темные, но на уздечке и лбу еще
имеются старые перья зимнего наряда; верхняя сторона тела в это время
у одних сплошь темная, у других только начинает линять; на зобу и боках,
а иногда и на брюхе одиночные летние перья. В середине июля самцы почти
полностью в летнем пере, кроме груди и брюха, которые, за исключением
отдельных перьев, остаются белыми. К концу июля у некоторых уже появ-
ляются на брюхе новые зимние перья; к 10 августа их может быть и довольно
много. Самки в конце июня уже одеты в летний наряд, в котором отдельные
зимние перья остаются только на нижней стороне, среди кроющих крыла.
К концу июля появляются перья осеннего наряда (принимаемые автором за
перья второго летнего наряда), но их бывает не более 1/б общей массы опе-
рения; иногда этот наряд представлен лишь немногими отдельными перьями.
В это же время у некоторых на нижней стороне появляются отдельные перья
нового зимнего наряда, происходит линька махов и рулей и зарастают
белыми перьями наседные пятна. На Хангае (Козлова, 1932) весенняя линька
отмечена во второй половине июня. Указанный автор справедливо пола-
гает что куропатка не успевает целиком сменить зимний наряд на летний.
Зимняя жизнь. Данные отсутствуют.
Питание. Не изучено.
Описание. Размеры. Длина крыла самцов F) 191—216, самок D) 193—198,
в среднем 207 и 194 мм.
Окраска. Занимает промежуточное положение между лапландской и сибир-
ской: темнее, чем сибирская, но светлее, чем лапландская; рыжих и охристых тонов в опе-
рении больше, чем у лапландской, но меньше, чем у сибирской.
ТУНДРЕНАЯ КУРОПАТКА
51
Командорская тундреная куропатка Lagopus mutus ridgwayi Stejneger
Lagopus ridgwayi. Stejneger, Proceed Biol.lSoc Wash., 1884, 1, стр. 98, о-в Беринга.
Распространение. Населяет только группу Командорских о-вов.
Характер пребывания. Оседла. Совершает незначительные
вертикальные перемещения.
Биотоп. Излюбленная стация зимой—малоснежные обветренные
края высокой каменистой и кочковатой тундры. Весною поднимается выше
в горы.
Численность. Обыкновенна и очень многочисленна как на о-ве
Беринга, так и на о-ве Медном.
Карта 4. Распространение тетерева Lyrurus tetrix
l—L. t. britannicus, 2—L. t. tetrix, 3—L. t. viridanus, 4—L. t. tschusii, б—L. t. mongolicua, 6—L. t.
baicalensi3, 7—L. t. ussuriensis; a—граница гнездовий, б—не вполне выясненная граница гнездовий
Экология. Размножение. По Иогансену A934), в конце апреля
разбивается на пары. Токование самцов—с апреля до половины июня. Откла-
дывание яиц в июне: 15 июня 1927 г. в южной части о-ва Медного на высоте
около 500 м найдена неполная кладка в 4 яйца. По данным местных жителей,
полная кладка до 15 шт. Полные кладки в конце июля (Бианки, 1909).
Птенцы выводятся в июле, а в середине августа попадаются уже летные
выводки.
Линька. Предбрачная линька самцов начинается в конце апреля—
начале мая. Полный летний наряд птица надевает в середине августа; в это
время наступает и смена махов. Первые белые перья зимнего наряда проби-
ваются почти одновременно с появлением последних перьев летнего наряда;
к концу первой декады октября распределение белых и окрашенных участков
оперения почти такое же, как в конце мая. Полный зимний наряд появляется
около середины ноября (Бианки, 1909). По данным Иогансена A934),
у самца, добытого 5 июня, почти весь верх, большая часть груди и боков тем-
ные, брюхо, подбородок, маховые и ноги еще белые.
Зимняя жизнь. Зимой встречаются стаями, иногда до 20 шт.
и более, в которые сбиваются в октябре. Благодаря большой доверчивости
птиц к стае можно подойти почти вплотную; по рассказам местных жителей,
иногда птиц бьют палками.
4*
52 ОТРЯД КУРИНЫЕ
Питание. Зимой питаются в основном листьями и ягодами вороники,
которые достают из-под снега.
Промысел. Незначителен. На о-ве Беринга ежегодно добывается около
400 шт. и столько же приблизительно на о-ве Медном.
Описание. Размеры. Длина крыла самцов 187—205, самок 179—197 мм (по
Бутурлину, 1935).
Окраска. Отличается очень темным, почти совсем однообразным черно-бурым
летним оперением самца, причем в отличие от других подвидов даже брюхо и оперение
голени не остаются белыми, а бывают дымчато-черными.
3. Тетерев Lyrurus tetrix L.
Tetrao tetrix. Linnaeus. Syst. Nat., изд. X, I, 1758, стр. 159 Швеция.
Русские названия. Тетерев-косач, березовик, черныш.
Распространение. Ареал. Лесная и лесостепная полоса Евразии от
Шотландии и восточной части Пиренеев до Уссурийского края; но на Саха-
лине, Камчатке, в Крыму и на Кавказе отсутствует. На север распространен
до 70° с. ш. в Скандинавии, до 67—69° с. ш. в европейской части СССР,
до 66—68° в Сибири; на юг до Альп, Черногории, Карпат, Волын-
ского полесья, Черниговской, Курской, Воронежской, северной части
Саратовской обл., в районе рек Урала и Илека до 51° с. ш., Тургая до49°с.ш.,
до Тышкана (Семиречье), Тянь-Шаня, Чжунгарского Ала-Тау, южной Дау-
рии и Южно-Уссурийского края.
Характер пребывания. Оседлая птица, местами совершает
незначительные сезонные перекочевки, более регулярно выраженные в гор-
ных районах.
Биотоп. Лиственные и смешанные леса, особенно осветленные поля-
нами и вырубками; темной тайги избегает.
Подвиды. Существование значительного числа географических рас, выделяемых ря-
дом авторов (Бутурлин, 1935; Штегман, 1932), требует проверки, так как индивидуальные
колебания в окраске оперения и размерах у тетерева значительны и в ряде случаев пере-
крывают приводимые указанными авторами диагностические признаки. Однако недоста-
точность материала по Средней Сибири и Дальнему Востоку не позволила нам это сделать
и ниже приводятся подвиды, выделяемые Штегманом A932), как более близкие к действи-
тельности. L. t. britannicus Witherbyet Lonnberg, 1913—Великобританские
о-ва; L. t. tetrix Linnaeus, 1758—Западная Европа, северная полоса Евразии*.
L. /. viridanus L о г е n z, 1891—южные части Восточной России и Западной Сибири;
L. t. tschusii I о h a n s e n, 1898—Южная Сибирь от Иртыша до Иркутска, к северу до
Томска и Красноярска, к югу до западных и северных частей Алтая и бассейна верховьев
Енисея; L. t. mongolicus Lonnberg, 1904—от Тянь-Шаня до северных склонов
Тарбагатая и южных частей Алтая; L. t. baicalensis L о г е nz, 1911—Забайкалье и Прибай-
калье, северная часть Приамурья, Северная Монголия; L. t. ussuriensis L о г е n z, 1911 —
Уссурийский край, низовья Амура, восточная часть Маньчжурии.
Североевропейский тетерев Lyrurus tetrix tetrix L.
Распространение. Ареал. Скандинавия, Финляндия, Германия, Поль-
ша, Западная Литва, Белоруссия, северная часть СССР к югу по крайней
мере до Оки, Камы, Подкаменной Тунгуски и среднего течения Лены вклю-
чительно. Северная граница проходит в Скандинавии у 70° с. ш., на Коль-
ском п-ве у 68° с. ш., захватывая северные берега озера Имандры (Семенов
Тян-Шанский, 1938), у низовьев рек Мезени, Пёши (около 67е с. ш., Михеев,
1949), у 68° на Печоре (Портенко, 1937), у 67° на Урале, у 66° на Оби, у68 72о
на Енисее, у 67° на Лене, у 68° на Колыме. Имеются указания, что северная
граница распространения тетерева на Кольском п-ве за последние 80 лет
несколько продвинулась на север (Семенов Тян-Шанский, 1938).
ТЕТЕРЕВ 53
Характер пребывания. Оседлая птица, местами зимой в по-
исках корма совершает перекочевки, принимающие в горных районах харак-
тер регулярных сезонных миграций протяженностью в сотни километров
(Мензбир, 1902); изредка бывают массовые переселения тетеревов, связан-
ные, по всей видимости, с большим неурожаем кормов. Такое переселение
с севера на юг, напоминавшее отлет летних птиц, имело место в 1895 г. в* Коми
АССР; в это время наблюдались стаи в 300—400 шт. (Резцов, 1904). Необы-
чайное обилие тетеревов в окрестностях Усть-Сысольска (Вычегда) отмеча-
лось осенью 1882 г.: птицы вылетели из сосновых и еловых лесов и концен-
трировались стаями по 200—300 шт. в березниках (Арсеньев, 1883). Мас-
совое появление наблюдалось в 1881 г. в Новоград-Волынском районе Жито-
мирской обл. (Завилейский, 1882), в 1928 г. в Боровичском районе Псковской
обл.(Нечаев, 1928). В обоих последних случаях в стаях отмечалось явное
преобладание самок над самцами A : 3 и 1 : 7).
О налете тетеревов в Печоро-Илычский заповедник в 1941 г. сообщает
Теплов A941); в результате налета поголовье косачей увеличилось там
в 14 раз.
Биотоп. Гнездовой—березовые леса, чередующиеся с хлебными
полями, осинники и липняки по соседству с обширными вырубками и гарями,
лесные опушки и редкое мелколесье с обязательным присутствием ягодни-
ков и сухих мест, необходимых для устройства гнезда; глухих высокостволь-
ных лесов тетерев избегает. Во время линьки косачи выбирают крепкие
места на окраинах болот с березовым мшарником, в густом мелком березнике,
а нередко и в густом ельнике (Мензбир, 1902). По нашим наблюдениям, ха-
рактерным летним местообитанием тетерева в Приволжско-Дубнинском за-
поведнике (Московская обл.) являются мелкие березняки с полянами, опушки
лиственных и смешанных насаждений с кустарниками, заросли ольхи по
опушкам болот, кустарники поймы р. Дубны, разреженные хвойные и сме-
шанные участки леса с мелким лиственным подлеском или куртинками мел-
ких сосенок. В Лапландии тетерев встречается в ельниках и смешанных за-
болоченных лесах; наиболее многочислен вдоль линии железной дороги,
в полосе, сравнительно густо населенной, с выгоревшими и вырубленными
лесами, в близком соседстве с северными форпостами земледелия—совхозом
«Индустрия» и Полярным отделением Всесоюзного института растениевод-
ства в Хибинах. Таким образом, и на северном пределе распространения
тетерев остается по преимуществу жителем полей или открытых пространств
с перелесками (Семенов Тян-Шанский, 1938). В Печоро-Илычском заповед-
нике летом населяет в борах беломошники и зеленомошники, прибрежные
ельники с примыкающими лугами, березники с лесными полянами, сухие,
сильно захламленные гари. В период линьки косачи и холостые самки дер-
жатся здесь главным образом на гарях, поросших кипреем и малинником,
а в период питания брусникой чаще встречаются в брусничниках, в борах
беломошниках или на открытых гарях. Зимнее (ноябрь—апрель) размещение
тетерева зависит в первую очередь от наличия его основных кормов—березо-
вой сережки и «озими» сосны. В большинстве тетерева сосредоточиваются
в чистых березниках; темнохвойных насаждений, даже с значительной
примесью березы, избегают и встречаются в них только в том случае, если
в березниках они не находят сережек, а тут они есть. В сосновых лесах
тетерев чаще встречается в разреженных насаждениях, в беломошниках и на
сосновых болотах, но перебирается сюда только при урожае сосновой «озими».
Сомкнутых сосновых насаждений избегает, повидимому, из-за их сильной
заснеженности. С уплотнением снега обычно концентрируется в участках
леса, богатых зарослями можжевельника, ягодами которого охотно пи-
тается (Теплов, 1947). На Северном Урале населяет леса из березы и ели,
сосновые леса, чистые березники (Портенко, 1937). На севере Кольского
54 ОТРЯД КУРИНЫЕ
Приуралья придерживается речных долин; в елово-липовой подзоне—всюду,
где есть вырубки и гари, поросшие вторичным лесным кустарником (Ворон-
цов. 1940). По наблюдениям Раевского, в Кондо-Сосьвенском заповеднике
(Тюменская обл.) в гнездовый период тетерев держится в березниках или
в смешанном лесу с преобладанием березы, на кромках обширных болот и по
рекам. Основная масса приурочена к большим площадям сгоревших елово-
кедрово-лиственничных ассоциаций, сменяемых березовым возобновлением
в возрасте 20—40 лет; таковы гари на реках Тугр и Ем-агань, а также на
реке Нюрух севернее Выреи.
На юге ареала вследствие распашки степей и уменьшения лесов рас-
пространение тетерева сокращается, на севере же благодаря вырубке
лесов занятая им территория постепенно все увеличивается.
Численность. На большой части ареала значительная, в густо
населенных районах местами низкая. На основании количественного учета
разными авторами приводятся следующие показатели плотности на 1000 га:
Лапландский заповедник (Мурманская обл.) 100 шт., Центрально-лесной
заповедник (Великолукская обл.) 30 шт., район Верхней Вычегды 73 шт.,
Лисинское учебно-опытное охотничье хозяйство (Ленинградская обл.)
146 шт., Печоро-Ылычский заповедник 26 шт. (Теплов, 1947). В Приволжско-
Дубнинском заповеднике (Московская обл.) на площади около 3 тыс. га в 1948 г.
учтено около 30 выводков. Для ряда районов отмечено колебание числен-
ности тетерева. Так, по данным количественного учета в Печоро-Ылычском
заповеднике, установлены следующие показатели (в процентах от показа-
теля за год максимальной численности): 1937 г.—100, 1938 г.—70, 1939 г.—
22, 1940 г.—4, 1941 г.—61, 1942 г.—74, 1943 г.—72. Колебания численности
отмечены для Лапландского заповедника, Кондо-Сосьвенского заповедника
(Тюменская обл.) и ряда других районов. Для севера эпизоотии у тетеревов
не отмечено и причина колебаний численности лежит, видимо, в колебании
урожаев зимних кормов (Теплов, 1947).
Экология. Размножение. Срок наступления половой зрелости,
повидимому, около года (Северцев, 1935). Пар не образуют, полигамы.
В конце зимы стаи приближаются к местам будущих токов, что в Печоро-
Ылычском заповеднике наблюдается во второй половине марта (Теплов,
1947), в Кондо-Сосьвенском заповеднике (Тюменская обл.) 4 апреля (наблю-
дения Раевского).
Вскоре зимние стаи разбиваются и с первыми признаками весны птицы
приступают к токованию. Как правило, каждая зимняя стая имеет свое то-
ковище, и там, где стаи зимою малочисленны, как, например, у населенных
центров, малочисленны бывают и тока, и наоборот. В малонаселенных местах
на тока вылетают до 150—200 птиц (Теплов, 1947, Коми АССР; Мензбир,
1902, Урал).
В густо населенных районах, где вследствие постоянной охоты тетерева
не собираются в стаи, а также в районах с низкой плотностью птиц, как,
например, на севере, токование происходит в одиночку. Места токовищ
весьма постоянны, и только чрезмерная охота или коренные изменения обста-
новки на них заставляют птиц покидать старые и перемещаться на новые
места. Обычно местом для тока в средней полосе служит поросшая редкими
кустами и деревьями опушка леса, лесная поляна с отдельными кустиками
и деревьями, а иногда и расположенный поблизости от леса участок поля
или луга. В долинах лесных рек, заливаемых водой, как, например, в Яро-
славской обл., ток происходит на поросших редкими лесами песчаных гри-
вах, а нередко и на деревьях. В крупных лесных массивах местом для тока
выбирается моховое болото, поросшее редкими чахлыми сосенками, берез-
ками и кустами, или же редкое мелколесье. Известно токование на чистых,
удаленных от леса болотах, поросших камышом, иногда вовсе без кустар-
ТЕТЕРЕВ 55
ников, (Мензбир, 1902). В Печоро-Ылычском заповеднике (Коми АССР) по
месторасположению тока бывают двух типов: на открытых сфагновых боло-
тах и на невозобновившихся лесом гарях. Тока на болотах характерны своей
мощностью: в некоторых случаях на них регистрировалось до 200 птиц,
в то время как на гарях не более 20—30, а очень часто и одиночные, на рас-
стоянии нескольких десятков и даже сотен метров одна от другой. В районе
темнохвойных лесов заповедника тетерева токуют чаще всего на лесных
полянах в березниках и на мелкоразнотравных и пустотных лугах, в гор-
ном районе—на полянах в субальпийской зоне (Теплов, 1947).
В Лапландском заповеднике токование одиночных косачей отмечалось
исключительно на обширных сфагновых болотах или по окраинам болотных
массивов, иногда на льду озер (Семенов Тян-Шанский, 1938). По наблю-
дениям Раевского, в Кондо-Сосьвенском заповеднике ток происходит обык-
новенно на льду озер или же на открытом болоте и лишь изредка на лесных
полянах. Одиночные тетерева токуют на берегах, травянистых лайдах, на
гарях (поведение птиц во время тока см. в описании следующего подвида).
Сроки токования в сильной степени колеблются в зависимости от гео-
графического положения местности и общего хода весны. Так, по данным
Хомченко A930), начало токования в 1930 г. было: в Ярославской обл.
7 марта, в Псковской 12 марта, в Вологодской 16 марта, в Мозырской 21 мар-
та, в Калужской 22 марта По наблюдениям Раевского, в Кондо-Сосьвен-
ском заповеднике в 1943 г. тетерева начали токовать 27 марта, а в 1939 г.—
20 апреля. По наблюдениям Семенова Тян-Шанского A938), в районе Хибин
и Апатит токование начинается на неделю раньше, чем в расположенном не
столь далеко Лапландском заповеднике, и растягивается на более продол-
жительный срок- Вообще же в средней полосе России первое бормотанье тете-
ревов обычно наблюдается около середины марта, в южной—в начале, на
севере—в конце этого месяца и даже гораздо позднее, в апреле (Мензбир,
1902) и мае.
В Лапландском заповеднике средняя дата начала токования за 6 лет—
6 мая (крайние сроки 1—16 мая), конец токования 23 мая B0—27 мая).
8 Архангельской обл. в 1913 г. тетерева начали бормотать с 17 марта, тока
в разгаре были с 30 марта (Яблонский, 1914). В Кондо-Сосьвенском заповед-
нике начало тока в среднем 10 апреля B7 марта—20 апреля), разгар 20 апре-
ля—15 мая, конец 28 мая (самая поздняя дата 5 апреля наблюдения Раев-
ского).
Насимович A936) приводит начало тока в 1935 г.: в Максатихинском
охотхозяйстве (Калининская обл.) 30 марта, в Долголуговском охотхозяй-
стве 22 марта; в 1934 г. разгар тока здесь был 22 апреля, конец 8 июня.
В Приволжско-Дубненском заповеднике (Московская обл.) первое бормо-
тание в 1947 г. 8 марта, массовое—конец марта; в 1948 г. начало тока 6 марта,
массовое—с начала апреля; отдельные токующие петухи отмечались до
середины апреля (наблюдения автора). В Минской обл. самая ранняя дата
начала тока отмечена 15 марта (Шнитников, 1913). Серебровский A918)
отмечает конец токования в Горьковской обл. 18 мая.
В среднем весеннее токование длится в одной местности 6—7 недель
(Мензбир, 1902), но нередко растягивается до 21/»—3 месяцев. Последними
токуют молодые тетерева, которых можно слышать еще в июне и как исклю-
чение даже в июле.
В первое время бормотанье тетеревов в средней полосе начинается часу
в шестом, незадолго до восхода солнца; позднее, в начале апреля—около
четырех часов утра, в конце апреля—в третьем часу, в мае—во втором часу
ночи. В Лапландии, где в мае солнце почти перестает скрываться за горизонт,
тетерева начинают токовать вечером и токуют всю ночь. В разгар тока тете-
рева токуют не только утром, но и вечером, часов с 7 и до заката солнца.
56 ОТРЯД КУРИНЫЕ
Ночуют птицы недалеко от токовища. Ежедневная продолжительность токо-
вания с разгаром тока увеличивается. Сначала петухи перестают токовать
через 2—3 часа после восхода сонца, но в самый разгар тока при хорошей
погоде токуют до 10 час. утра, после чего улетают на кормежку в чащу леса.
В начале токования самки прилетают к окраинам токовища ненадолго;
позднее, когда начинается настоящий ток, они прилетают до солнечного вос-
хода или на самом восходе и держатся на окраине токовища, издавая квох-
танье. Спаривание происходит как на току, так и за его пределами. В период
токования самки ведут более однообразную и спокойную жизнь, чем самцы,
больше кормятся и отдыхают. Самцы же за это время сильно изнуряются
и худеют (Мензбир, 1902). Данные о половом составе популяций почти отсут-
ствуют, лишь по Печоро-Ылычскому заповеднику указывается соотношение:
57% самцов и 43 Уо самок.
В средней полосе к гнездованию тетерева приступают в конце апреля—
начале мая, когда молодая трава успеет подрасти и на деревьях появится
молодая листва. Большинство самок садится на яйца в середине мая, в юж-
ной полосе—в начале мая, в северной—в конце мая—начале июня. Старые
самки начинают гнездиться раньше молодых.
В Новгородской обл. полные кладки с почти ненасиженными яйцами
в 1879 г. были найдены 27 мая и 2 июня (Бианки, 1910), в Минской обл.
полные, ненасиженные кладки с 20 апреля до 13 мая (Шнитников, 1913),
в Витебской обл. насиженная кладка 5 июня (Федюшин), в б. Келецкойгуб.»
самое раннее нахождение полных кладок в 1908 г.—1 мая (Катин, 1912),
в Горьковской обл. гнезда с яйцами отмечались не позднее 14 мая (Серебров-
ский, 1918), в Кондо-Сосьвенском заповеднике первое гнездо с кладкой в 1 яй-
цо в 1931 г. было найдено 22 мая (наблюдения Раевского).
Место гнездовья обыкновенно находится недалеко от токовища и всегда
по соседству с ягодниками земляники и костяники, на которых впоследствии
кормятся выводки, следовательно преимущественно по березовым лесам.
Гнездятся также в осинниках и липняках, но недалеко от ягодников. Для
устройства гнезда выбираются сухие места, так как сырость для яиц губи-
тельна. Гнездо обычно располагается под кустом или деревцем—березкой,
липой, можжевельником. Оно представляет собой небольшую ямку в земле,
слегка выстланную сухой травой, небольшим количеством моха и перьев.
По Мензбиру A902), количество яиц в кладке обычно 4—8, иногда
10 и даже 12 и 14; вторая кладка из 2—4 яиц; по Серебровскому A918), для
Горьковской обл. от 7 до 9, по Катину A912), для б. Келецкой губ. 9—12.
Теплов A947) среднее число яиц в полной кладке для Печоро-Ылычского
заповедника определяет в 7,3 шт. (по 16 кладкам). Размер яиц: 4,7—52 х
X 34—36 мм (вес и окраску см. в описании следующего подвида). Малочислен-
нее кладки в годы неурожая кормов, а также у молодых птиц. К насиживанию
самка приступает после откладки последнего яйца. При гибели первых
кладок самка откладывает вторую, в результате чего яйца можно встретить
до конца июня (Мензбир, 1902).
По Мензбиру, насиживание продолжается три недели, но с некоторыми
колебаниями в разных широтах: в средней полосе 19—22 дня, на севере 23
и даже 25 дней. Несомненно, что небольшие колебания в продолжительности
насиживания в отдельных гнездах тетеревов имеются, как это точно установ-
лено и для белой куропатки. Но увеличение срока насиживания по мере
продвижения на север сомнительно и требует проверки. По крайней мере
достоверные наблюдения Ульянина A949) определяют срок насиживания
тетерева в Северном Казахстане в 23 дня, т. е. несколько даже больший,
чем указывается для средней полосы.
Первые тетеревята появляются приблизительно через месяц от самого
разгара тока. В средней России массовое появление выводков наблюдается
ТЕТЕРЕВ 57
в середине июня, запоздалые—до середины июля (Мензбир, 1902). В При-
волжско-Дубненском заповеднике (Московская обл.) первые выводки
в 1947 г. начали встречаться в первой половине июня, в массовом количе-
стве—во второй половине июля. Встреченный 10 июня выводок состоял из
пуховых птенцов с распускающимися перьями на крыльях, а 1 июля они
хорошо летали. В 1948 г. первый выводок встречен 14 июня, птенцы были
величиной с галку. Во встреченном 5 июля 1946 г. выводке птенцы хорошо
летали и достигали по величине примерно 2/з величины взрослых (наблю-
дения автора). В Долголуговском охотхозяйстве первые выводки в 1934 г.
появились 12 июня, в 1935 г. 3 июня (Насимович, 1936). В 1928 г. первые
выводки в окрестностях Ленинграда 18 июня, близ Вологды 20 июня, близ
г. Омутнинска (Кировская обл.) 4 июня (Смирнов, 1920); в середине августа
в Ленинградской обл. молодые петушки уже начинают «мешаться пером».
В Новгородской обл. 15 июня отмечены молодые величиной с коростеля,
10 июля—с серую куропатку (Бианки, 1910). В б. Ковенской губ. Баранов-
ский A907) в конце июля нашел много выводков, частью в черном пере,
частью поршки, очевидно второй кладки. В Архангельской обл., по Але-
ксандрову A906), к середине сентября молодые взматерели, и выводки раз-
бились. В Горьковской обл., по Серебровскому A918), летающие молодые
наблюдаются в конце мая. В Печоро-Ылычском заповеднике первые выводки
в среднем появляются 3 июля. В конце августа вес молодых самцов состав-
ляет 70%, а самок 80% от веса взрослых (Теплов, 1947). В Кондо-Сосьвен-
ском заповеднике в конце июля 1931 г. птенцы были ростом с куропатку,
а добытый 10 августа молодой самец носил уже пестрый наряд; в первых
числах сентября тетеревята находились в своем первом взрослом оперении
и только небольшое рыжее пятно имелось на шее и голове, немного на спине
и крыльях и отсутствовали косицы на хвосте. К тому времени все выводки
уже разбились (наблюдения Раевского).
О сроках вывода птенцов дают также некоторое представление следую-
щие материалы из коллекций Зоологического музея Московского готудар-
ственного университета.
Кандалакшский залив—5 августа молодой самец величиной со взрослого,
оперение почти полностью черное, за исключением головы и отдельных пятен
на туловище, имеются косицы на хвосте. Лениградская обл. (Лужский
район)—28 августа молодой самец такого же размера и с таким же характе-
ром оперения, как и предыдущий. Латвия—27 июля полный ювеналь-
ный наряд. Калининская обл.—16 июня пуховой птенец, начинает
оперяться; 26 июня полностью оперенный, размером в половину вели-
чины взрослого; 2 августа молодой самец величиной в три четверти взрослого,
оперение «мешается»—черные перья пробились на нижней части тела и пояс-
нице; два молодых самца, добытых 4 и 14 августа, примерно такой же вели-
чины и с таким же характером оперения. Вологодская обл. —два птенца,
добытых 21 июля, величиной в половину взрослого и полностью оперены.
Печоро-Ылычский заповедник—20 сентября самец размерами со взрослого,
полностью черный, за исключением отдельных серых пестрин. Заповедник
«Денежкин Камень» (Свердловская обл.)—18 сентября самец в таком же сос-
тоянии, как и предыдущий. Ярославская обл.—11 июня пуховой птенец.
Горьковская обл.—13 июля полуоперенный птенец. Московская обл.—
25 июля оперенный, но меньше, чем в половину взрослого; 10 августа моло-
дой самец имеет уже более половины оперения черной. Рязанская обл.—
2 августа самец размером более половины взрослого, оперение «мешается».
Тульские засеки—10 августа полностью оперенный, величиною почти в поло-
вину взрослого.
По Мензбиру A902), количество тетеревят в выводке в средней полосе
чаще всего 7—8, изредка 10, в виде исключения 12—14. По наблюдениям
58 ОТРЯД КУРИНЫЕ
Теплова A947), средний размер выводков, определенный путем подсчета
птенцов в период до середины сентября, колеблется по годам от 4 до 5,4 шт.,
а в среднем за 7 лет A937—1943 гг.) равен 4,5 шт. Отход молодых с момента
вылупления до середины сентября тот же автор устанавливает равным 26—
45%. Прирост поголовья у тетерева, как и других куриных, сильно сни-
жается наличием в некоторые годы значительного количества холостых
самок. Так, по данным Теплова A947), в Печоро-Ылычском заповеднике
в 1939 г. холостые самки тетерева составляли 54%, по данным Белова и
Полубояринова A934), в Ленинском учебно-опытном охотхозяйстве Ленин-
градской обл. летом 1932 г. их было зарегистрировано 20%, в Ильменском
заповеднике в 1940 г. (по наблюдениям Ушкова)—50%, в Башкирском за-
поведнике (по данным Северцова, 1932) летом 1930 г. до 30%. Теплов
наличие холостых самок связывает с гибелью насиженных кладок от июнь-
ских заморозков.
Первые дни тетеревята растут очень медленно, через неделю темп ренета
сильно увеличивается; на 3—4-й день пробиваются маховые, потом рулевые
перья; в недельном возрасте птенцы могут припархивать и подлетывать,
в двухнедельном—перелетать значительное расстояние, а в месячном не
отстают от матки. Примерно к этому времени у них полностью формируется
перовый птенцовый наряд. Тетеревята взлетают на деревья уже в недельном
возрасте, но подолгу оставаться на них и тем более ночевать начинают, только
достигнув 1,5-месячного возраста. Суточная жизнь выводков проста: ранним
утром птенцы выходят кормиться в лесные покосы, на сечи, гари и т. п.,
где держатся до 8—9 час. утра; затем отдыхают на лесной опушке, в кустар-
никах и в других защищенных местах. Во второй половине дня, в 4—5 час,
вновь кормятся на лесных покосах, а к закату возвращаются в защищенные
места на ночевку. Первые 1—1,5 месяца семья ночует на земле, причем
в течение 1—1,5 недели матка прячет птенцов под крылья. С возрастом вы-
водки начинают кочевать на большей площади, позднее переселяются в ягод-
ники, отстоящие нередко за много сот метров от первоначальных мест кор-
межек. С исчезновением ягод и созреванием яровых хлебов выводки по утрам
и вечерам совершают регулярные переходы на хлебные поля, возвращаясь
в полдень и на ночь на лесные опушки. В конце августа—начале сентября
в средней полосе молодые косачи отбиваются от матки, хотя и придержи-
ваются друг друга, а полмесяца спустя они окончательно отбиваются от
•семьи. Молодые тетерки продолжают держаться с маткой до образования
зимних стай (Мензбир, 1902).
В конце лета и осенью у тетеревов наблюдается осеннее токование.
По Мензбиру, осеннее токование происходит лишь местами, но, судя по лите-
ратуре и личным наблюдениям, распространено это явление весьма широко.
Отмечено оно для средней полосы, западных и северных областей европей-
ской части СССР, для севера Сибири и других районов.
По Мензбиру при ранней весне осеннее токование начинается уже
в конце августа, но лучшее время—с конца сентября по конец октября;
бывает оно также в ноябре, декабре и январе. Начинается токование обык-
новенно незадолго до восхода солнца и длится в течение всего дня, наиболее
же дружно идет при восходе солнца, а в глубокую осень часу в десятом.
Охотнее всего тетерев токует на земле, иногда на голом дереве. Токуют почти
исключительно старики. Чуфыканье бывает слышно редко, остальная часть
песни короче и слабее весенней. Позы во время токования те же, что и весной,
но не так сильно оттопыриваются крылья и не в такой степени распускается
хвост. Самки относятся к токованию безразлично. В Приволжско-Дубнен-
ском заповеднике (Московская обл.) осеннее токование в 1946 г. отмечено
27 сентября, в 1947 г. 12 и 26 октября, в 1948 г. 12 сентября (наши наблюде-
ния. В Максатихинском охотхозяйстве (Калининская обл.) осенний ток
ТЕТЕРЕВ 59
«ачался в 1934 г., 7 августа, разгар в октябре. В Долголуговском охотхозяй-
стве тетеревиные тока в 1934 г. продолжались в ноябре, а бормотанье одиноч-
ного косача отмечено 25 декабря (Насимович, 1936). В Кондо-Сосьвенском
заповеднике (Тюменская обл.) начало осеннего бормотания 1 августа—10 сен-
тября, оживленный осенний ток наблюдался в последних числах сентября;
ток происходил по вечерам на деревьях (наблюдения Раевского). Токование
изредка наблюдается и в зимние месяцы. Так, Рыков A930) сообщает о бор-
мотании тетерева в окрестностях Свирьстроя 28 января; Хомченко A930)
в Костромской обл. 24 ноября и в Ярославской обл. 21 февраля.
Линька. Смена нарядов: пуховый—первый птенцовый—второй
птенцовый, он же первый взрослый—второй взрослый наряд; окончатель-
ный наряд—третий.
Вскоре после того как закончится формирование первого птенцового
наряда, тетеревята начинают линять во второй птенцовый наряд, являю-
щийся в то же время первым взрослым нарядом. В средней полосе начало
линыки наблюдается в среднем в конце июля. Начинается линька с брюха
и оканчивается головой. Вместе с линькой появляются первые отличия
молодых самцов от самок: пробивающиеся на крыльях самцов покровные
перья темнобурые, почти черные, на задней части спины сине-стальные,
по бокам груди черные; брови у самцов заметно крупнее и шире, чему самок.
Позднее (в сентябре) у молодых самцов отрастают косицы, в середине сен-
тября тетеревенок уже почти весь покрыт перьями второго птенцового на-
ряда и только голова и часть шеи еще покрыты гнездовыми перьями; но окон-
чательно первый взрослый наряд надевается лишь в конце сентября—октябре
и носится до июня следующего года.
Взрослые косачи начинают линять вскоре после окончания тока, для
чего обычно переселяются в крепи, где и перелинивают. В средней полосе
линька начинается обычно с конца июня и продолжается довольно долго—
около 2,5 месяца и больше, заканчиваясь в сентябре и даже в октябре.
Линька мелкого пера у косачей начинается одновременно на плечах,
зобе и надхвостье и оканчивается шеей и головой; последними сменяются
маховые и рулевые. Холостые самки приступают к линьке примерно на месяц
раньше маток одновременно со старыми косачами, т. е. в средней полосе
с конца июня, и оканчивают линьку к середине августа. Порядок линьки тот
же, что и у тетеревят, т. е. начинается с брюха и оканчивается головой.
Тетерки, имеющие птенцов, начинают линять вместе с последними, в средней
полосе—в конце июля. Прежде всего пеньки перьев появляются на насед-
ных пятнах, затем линька распространяется на грудь и надхвостье и закан-
чивается на голове, как у молодых; последними сменяются маховые и руле-
вые перья: в сентябре в средней полосе линька полностью заканчивается
'(Мензбир, 1902). Следовательно, у взрослых бывает только одна полная годо-
вая линька.
Зимняя жизнь. Зимой тетерева живут смешанными стаями, кото-
рые образуются еще осенью. Каждая стая упорно придерживается извест-
ного участка, выбирая такие места, где поля и перелески расположены неда-
леко от болотистой чащи—место их ночлега. Участок этот обычно занимает
несколько C—6) квадратных километров. С выпадением глубокого снега
-стаи перестают держаться исключительно одной местности и начинают вести
более бродячий образ жизни. В годы неурожая березовой почки тетерева еще
с осени переселяются в другие, иногда весьма отдаленные местности, концен-
трируясь там в крупные стаи до 200 и даже 500 птиц. Осенью тетерева ночуют
в болотистой чаще в густых уремах по берегам озер и рек, либо в густой
осоке, либо под хворостом и кустами; на севере местом ночлега нередко изби-
рают моховые болота. Зимою, особенно в большие холода, они всю ночь и даже
значительную часть дня проводят, зарывшись в снег. Для этого тетерев или
60
ОТРЯД КУРИНЫЕ
с дерева, или прямо с полета бросается в снег и пробивает его верхний за-
твердевший слой. Затем подсовывает голову под этот пробитый слой и, двигая
крыльями, образует углубление, вроде пещеры, куда и забивается. Таким
образом тетерев уходит иногда под снегом на 10 м и более. Вылетает из-под
снега тетерев очень быстро и с шумом. В сильные метели стая может оставаться
под снегом по нескольку дней (Мензбир, 1902). Бывают случаи, когда после
оттепели наступают сильные морозы, и снег покрывается такой твердой
коркой, из-под которой тетерева не могут выбраться и в массе погибают»
По сообщению местных охотников, подобный случай массовой гибели тете-
ревов наблюдался в Талдомском районе Московской обл. в 1944 г., когда
после выпавшего мокрого снега ночью ударил сильный мороз и образовался
крепкий наст, державшийся две недели. До этого в лесах наблюдались стаи
по 20—40 птиц, после их уже не стало. Бродячую жизнь тетеревиные стаи
ведут до конца зимы, а в конце февраля—начале марта (средняя полоса)
снова перемещаются в район токовища.
Питание. В основном растительноядная птица; животные корма
имеют небольшое значение. Исследованием содержимого зобов и желудков,
добытых в нескольких областях европейской части СССР в различные сезоны
года, установлено поедание 79 растительных и 26 животных видов кормов
(Наумов и Лавров, 1948). Характер питания по сезонам года не одинаков:
наиболее разнообразен кормовой рацион в летний период, наиболее одно-
образен зимой. Значение наиболее часто поедаемых кормов по сезонам года
видно из нижеприведеной таблицы (по Лобачеву и Щербакову, 1933).
Наименование корма
% встреч
шма
100
55
33
0
0
0
0
0
0
0
весна и лето
57
14
0
28
57
57
28
0
28
71
осень
Можжевельник (ягоды)
Береза (сережки, почки, побеги)
Ольха (сережки)
Ива (почки и побеги)
Брусника (ягоды и листья)
Черника и голубика (ягоды и листья) . . .
Ястребника, луговик (листья)
Лютик (семена)
Семена полевых культур
Насекомые (главным образом муравьи и гусе
ницы)
71
50
29
21
29
21
29
21
14
36
По данным исследования 14 зобов (Семенов Тян-Шанский, 1938), главными кормами
тетерева в Лапландском заповеднике являются: хвоя и побеги сосны (зарегистрированы
в ноябре и марте), цветочные сережки и побеги с почками березы (октябрь, ноябрь, де-
кабрь, март), плоды, побеги и листья черники (май, сентябрь, октябрь, ноябрь), плоды
и листья брусники (март, сентябрь, октябрь, ноябрь), плоды и листья голубики (сен-
тябрь), листья подбела (май, сентябрь, октябрь, ноябрь), побеги и плоды вороники (март,
сентябрь, октябрь), плоды и листья марьяника (сентябрь), плоды осоки (август,
сентябрь), побеги с почками и листья карликовой березы (май, сентябрь, октябрь).
В качестве второстепенных и случайных кормов отмечены листья луговика, плоды клюквы,
плоды дерена. Из животных кормов встречены муравьи, гусеницы, кобылки, жуки.
В результате анализа содержимого 47 зобов тетеревов, добытых в Печоро-Ылыч-
ском заповеднике в период с ноября по апрель включительно, Теплов A948) дает следую-
щую картину зимнего питания: «озимь» хвойных составляет 20% встреч, сережки березы—
45%, почки и побеги деревьев и кустарников—25%, побеги черники—5%, ягоды—5%
встреч; сережки березы и «озимь» хвойных (сосны) составляют по встречаемости 65%,
а по объему около 80%. Соотношение между березовой сережкой и «озимью» сосны в зобах
тетерева довольно сильно изменяется в отдельные годы. Создается впечатление, что они
заменяют друг друга. Почки и побеги (исключительно березовые) встречаются, как пра-
вило, в виде примеси к березовой сережке и, очевидно, компенсировать недостаток послед-
ТЕТЕРЕВ
61
ней не могут. Автор связывает урожайность сосновой озими и березовых почек с колеба-
ниями численности тетерева и его периодическими миграциями, имеющими место в районе
заповедника.
В зобах тетеревов, добытых в Кондо-Сосьвенском заповеднике, в сентябре преоблада-
ли ягоды брусники и черники, листья этих растений, кусочки стеблей с почками черники,
коробочки мха. В октябре, с перекочевкой птиц в березники, в зобах появились почки
и сережки березы, но преобладающим кормом оставались те же ягоды, к которым стали
примешиваться ягоды и листочки клюквы.Зимой корм состоит преимущественно из сережек
и веточек с почками березы, молодых сосновых шишечек (урожай будущего года) и не-
большого количества хвои. С середины ноября количество сосновых шишечек в зобах воз-
растает и в среднем в зобе равно 6, а
2 декабря в одном зобу достигло 31 шт.
Окончательно на зимний корм птицы
переходят в середине декабря, с появ-
лением глубокого снега (наблюдения
Раевского).
В зобах и желудках 5 тетеревов,
добытых в апреле, сентябре и ноябре
1948 г. в Московской обл., содержа-
лось 22 вида растительного корма: яго-
ды можжевельника, шиповника, кру-
шины, малины, черемухи, рябины, ко-
стяники, брусники, семена осоки паль-
чатой, воробейника полевого, горошка
заборного, мятлика лугового, майника
двулистого, фиалки, лапчатки серебри-
стой, иван-да-марьи, овса, листья кле-
вера лугового, вероники дубравной,
сережки и ветки березы бородавчатой,
ивы, цветочные головки кульбабы
осенней, коробочки иван-да-марьи.
Кроме того, в четырех желудках
содержались камешки. Наиболее разно-
образны корма в сентябре: 18 ви-
дов преимущественно ягод и семян
трав. В ноябре и апреле пища состояла
из сережек и кончиков ветвей березы
бородавчатой, ивы и ягод можжевель-
ника.
Промысел. Благодаря обшир-
ному ареалу и высокой числен-
ности описываемый подвид имеет
большое промысловое значение и
составляет основу промысла все-
го вида. Немаловажное место за-
нимает он и в спортивной охоте.
Добывают тетерева весьма разнообразными способами. Весной охотятся
с подхода и на току, в конце лета — за выводками и на манку. Осенью
на чучела, зимой с подъезда и из ямок. Кроме этих ружейных способов,
существует много промысловых способов добычи с помощью ловушек: вен-
телей, ковшей, коробов, давушек, сильев, кружков и т. д. Все эти способы
довольно полно описаны в охотничьей литературе.
Полевые признаки. Птица средней величины, с короткими изогнутыми
крыльями: поднимается с земли с большим шумом, полет быстрый, часто
машет крыльями. Самец черный, с белыми подмышечными, подбоем крыла
и подхвостьем и с более или менее заметным «зеркалом»—белой поперечной
полосой на крыле: крайние рулевые сильно загнуты лирообразно наружу.
Самка рыжевато-серая, немного мельче самца; подмышечные перья белые,
подхвостье тоже, но с крупными темными полосами и пятнами; хвост выре-
зан. Молодые похожи на самку.
Описание. Размеры и строение. Птицы с совершенно оперенной цев-
кой, но голыми, покрытыми сверху поперечными пластинками, пальцами, имею-
Рис. 11. Тетерев (самец и самка)
62
ОТРЯД КУРИНЫЕ
щими боковые роговые выросты или гребневидную бахрому. Хвост вырезан у сам-
ки меньше, у самца больше, нижние кроющие хвоста обыкновенно длиннее средних
рулевых, крайние же рули у взрослых самцов сильно изогнуты вбок. Над глазом имеется*
голое пространство красной, сильно шершавой кожи. Длина крыла самцов C0) 250—286,
самок C0) 227—255, в среднем 273 и 239 мм; вес птиц, добытых в период сентябрь—апрель,
колеблется: в Печорском заповеднике самцов B6) 1000—1400; самок C5) 765—1050,
в среднем 1255 и 910 г; в Лапландском заповеднике самцов E) 1120—1333, самок F>
934—1056, в среднем 1236 и 975 г.
Окраска. Пуховой птенец желтовато-охристый, верх головы, плечевые и широ-
кая срединная полоса на спине охристо-коричневые с черными пятнами. Птенцовый наряд
у самца и самки одинаков и сходен со взрослой самкой, только с более светлой поперечной
полосатостью. Первый годовой наряд самца в общем сходен со взрослым, но имеет ряд отли-
чий. По бокам головы и на шее остаются рыжевато-охристые перья с поперечным рисунком.
У заднего угла глаза небольшое охристо-беловатое пятно с мелкими более темными пестри-
нами. Окраска верхней и нижней стороны тела черная. Металлический блеск развит слабо.
Кое-где вокруг шеи, на спине и зобе
встречаются рыжевато-охристые перья
птенцового наряда. Второстепенные ма-
ховые, верхние кроющие крыла и верх-
ние кроющие хвоста испещрены мелкой
рыжеватой струйчатостью. Окончатель-
ный наряд надевается только на третьем
году. Во втором годовом наряде самца
поперечный рыжеватый рисунок стано-
вится менее заметен, в особенности на
верхних кроющих хвоста. Голова чер-
ная. Металлический блеск развит силь-
нее. В третьем годовом наряде самца по-
перечнорыжеватого рисунка на верхних
кроющих совсем нет, а на второстепен-
ных маховых и верхних кроющих кры-
ла едва заметен. Металлический блеск
развит, как у взрослых. Взрослый са-
мец имеет общую окраску оперения чер-
ную с металлическим синим блеском на
голове, шее, зобе и нижней части спины.
Маховые темнобурые. Основание второ-
степенных маховых, больших и средних
кроющих крыла белое. Нижние кроющие
хвоста, подмышечные и нижние крою-
щие крыла белые. Над глазом красная
Рис. 12. Пуховой птенец тетерева
бровь. Летом голова черно-бурая в мел-
ких пестринах. Клюв черный, пальцы темнобурые. Крыловое зеркало сравнительно узкое
и на сложенном крыле мало заметно. На больших верхних и кроющих кисти нет белого»
пятна. Рулевые перья обычно вовсе не имеют корневой белизны. У взрослой самки верх-
няя сторона тела с черно-бурой и охристо-рыжеватой поперечной полосатостью. Верхние
кроющие крыла с белыми вершинами, исчерченными черными продольными штрихами.
Второстепенные маховые с белыми основаниями и широкими бурыми вершинами, испещрен-
ными охристыми или беловатыми черточками. Нижние кроющие крыла и подмышечные
белые. Верхние кроющие первостепенных маховых с белым основанием. Шея спереди
охристая с темной поперечной полосатостью. Зоб рыжевато-охристый с редкой и узкой
темной поперечной полосатостью. Грудь, верхняя часть брюха и бока тела с охристо-рыжи-
ми и черно-бурыми поперечными полосами и широкими беловатыми вершинами, испещ-
ренными продольными черными штрихами. Середина брюха черноватая с узким попереч-
ным охристым и белым рисунком. Кроющие хвоста белые с рыжевато-коричневыми ред-
кими поперечными полосами. Отличается от других подвидов более сильно выраженным
рыжим тоном и узким зеркалом, обычно скрывающимся под кроющими перьями при сло-
женном крыле.
Степной тетерев Lyrurus tetrix viridanus Lorenz
Tetrao tetrix viridanus. Lorenz. Journ. fur Ornith.,; 1891, стр. 306, Саратов.
Распространение. Ареал. Населяет юг восточных районов европей-
ской части СССР и лесостепь Западной Сибири, от бассейнов Дона и Суры
на западе до Иртыша на востоке и к северу до низовьев Камы, верховьев
Урала и до района Тюмени. Южная граница проходит в районах Курска,
ТЕТЕРЕВ 63»
Воронежа, по северной части Саратовской области, в области рек Урала
и Илека до 51° с. ш., в районе Тургая до 49° с. ш. Прежде, в конце XVIII
и начале XIX века, ареал степного тетерева был гораздо обширнее. Пови-
димому, именно эта форма занимала почти весь юг европейской части СССР
до Бессарабии, Южноукраинских, Донских степей и Северного Кавказа
включительно. В южных частях Воронежской облв и в Донских степях он
окончательно исчез в конце XIX в., в Харьковской, Киевской, Полтавской
обл.—в середине XIX в., в Бессарабии и Новороссийских степях—в конце
XIX в. На Северном Кавказе тетерев прежде был распространен как в Став-
ропольских и Кумских лесах, так и в Кубанских и Терских степях.
На юг он проникал до предгорий Скалистого хребта. Особенно его много
было в окрестностях Майкопа, Каладжинской, Упорной, Отрадной и Псе-
бая. В 60-х годах численность начала быстро сокращаться, и последние
тетерева исчезли в 80-х годах прошлого столетия (Мензбир, 1902; Аверин
и Насимович, 1938). Основной причиной исчезновения тетеревов на юге
европейской части СССР являются, повидимому, как отмечают Лоренц и
Мензбир, не охота, а усиленная распашка степей, вырубание лесов и
быстрое общее хозяйственное освоение края.
Характер пребывания. Как и номинальная раса, совершает
местные сезонные перекочевки, особенно резко заметные в годы неурожая
кормов. По сообщению Кротова A928), в б. Тюменском округе летом была
немного тетеревов, но за вторую половину ноября и начало декабря они
заметно начали прибывать и в каждом лесном колке можно было видеть
табунки по 15—25 шт.; по предположению местных охотников,* тетерева
налетели из Тобольского округа. О появлении в б. Тарском уезде в некоторые
годы больших стай залетных тетеревов пишет Ушков A913). В нормальные
годы на Южном Урале осенью и в начале зимы тетерева перемещаются иа
вишенников, бобовников и других кустарниковых и травянистых местооби-
таний в уремные и колковые леса; на южной же окраине широколиственной
лесостепи тетерева, наоборот, покидают на зиму уремные и дубово-липовые
леса и перебираются на вершину горного массива, где березы значительна
больше, чем в нижней части склонов (Кириков, 1948).
Биотоп. По Кирикову A948), в лесных ландшафтах Южного Урала
наиболее густо населяет березовые леса, перемежающиеся с полями и поля-
нами, менее густо—разреженные сосново-березовые леса с участками мел-
колесья, с полянами и безлесными склонами и еще реже встречается в гнез-
довое время в старых густых широколиственных лесах и старых борах. В ле-
состепи по восточным предгорьям Урала основными местообитаниями тетере-
вов служат березовые колки, кустарниковая степь и заболоченное березовое
мелколесье по западинам. На южной окраине широколиственной лесостепи
чаще всего встречаются в колках широколиственного мелколесья и в зарос-
лях степной вишни, бобовника и чилиги по безлесным склонам. Колочки
березового мелколесья, вязово-черемуховая урема по дну долов и кустар-
никовая степь—основное местообитание в нагорно-степных ландшафтах
южной оконечности Урала. В равнинных степях Южного Урала, где нет
березового мелколесья и крупных участков кустарниковых степей, тетерева
очень редки. Примерно тот же характер биотопа отмечает для б. Уфимской
губ. Сушкин A897), в больших лесах гнездится редко. За пределы лесной
растительности, несмотря на обилие ягод, не идет.
В б. Бугурусланском уезде наиболее характерен для молодняков и ку-
старников, избегает старого леса (Исполатов, 1910), в южных районах Кам-
ского Приуралья характерен для смешанных насаждений, представленных
колками (Воронцов, 1949), в Мордовском заповеднике—для сильно разре-
женных рубкой сосновых боров с примесью ели, березы и осины в подлеске,
придерживаясь близости вырубок, опушек и полян (Птушенко, 1938). В сте-
64 ОТРЯД КУРИНЫЕ
пях европейской части СССР тетерев распространен по долинам рек и там,
где большие или меньшие участки степи покрыты ягодными кустарниками,
дикой вишней и диким персиком (Мензбир, 1902). В Наурзумском заповед-
нике (Северный Казахстан) наиболее многочислен в березовых колках
и перелесках с густыми кустарниковыми зарослями и высокой густой травой,
перемежающихся с многочисленными степными большими и мелкими поля-
нами; меньше их в сосновых борах Терсеке и Наурзум-Карагае, где они
держатся преимущественно в участках с наличием подлеска из березы и ку-
старников (Михеев, 1936). Березовые колки характерны для тетерева в лесо-
степи Западной Сибири (наблюдение автора).
Численность. В большинстве районов ареала высокая. В Наур-
зумском заповеднике на площади 300 км2 (леса Наурзум-Карагай и Сыпсын-
Агач) в марте 1936 г. подсчитано в стаях 1140 тетеревов, т. е. в среднем около
0,4 шт. на 1000 га, весной того же года на токах учтено 400 шт. (Ульянин,
1949). Лет 20 тому назад в районе того же заповедника один охотник в течение
зимы добывал в среднем около 100 шт., а за день 4—6 шт. (Михеев, 1936).
На Южном Урале за одну утреннюю или вечернюю охоту охотник добывает
от 5 до 20 тетеревов. Там же в березовых лесах величина зимних стай дости-
гает 500—1000 шт. (Кириков, 1948).
Экология. Размножение. Токование начинается с первыми при-
знаками весны, когда, несмотря на утренние 30-градусные морозы, днем ка-
пает с крыш, начинают чернеть большие дороги и на южных склонах пока-
зываются проталины. Сроки начала и конца токового периода сильно изме-
няются в зависимости от характера весны, района и даже ландшафта мест-
ности. По Кирикову, в Башкирском заповеднике (Южный Урал) первое весен-
нее бормотание в 1938 г. отмечено 3 апреля, в 1939 г.—22 марта, в 1940г.—
21марта, в 1941г.—25марта, в 1942 г.—4 апреля, в 1943 г.—20марта,в 1944 г.—
4 марта, в 1945 г.—30 марта; средняя дата за 8 лет—24 марта; разница
между крайними датами—30 дней. Окончание весеннего токования на южной
окраине широколиственной лесостепи в конце мая B6 мая в 1932 г.), а в ланд-
шафте нагорных сосново-березово-лиственничных лесов заповедника—в пер-
вой половине июня- В Мордовском заповеднике (Птушенко, 1938) в 1936 г.
первое бормотание 12 марта; разгар токов—во второй половине апреля
и начале мая, конец—19 мая: продолжительность периода токования 68 дней.
По данным Ульянина A949), в Наурзумском заповеднике (Северный Казах-
стан) массовое токование с 10 апреля по 10 июня, всего 61 день, одиночные же
особи с 30 марта до 15 июня, т. е. 78 дней.
В самом начале токового периода тетерева бормочут на случайных
местах, будучи еще в стае. В солнечную погоду по утрам во время кормежки
или после нее тетерева начинают чуфыкать, одиночки бормотать, а другие
гоняться друг за другом и драться. Токование в неразбившейся стае длится
несколько дней, затем стайки разбиваются, тетерки отделяются, и токование
происходит на постоянных токовищах. Первыми вылетают на ток и бормо-
чут на нем старые черныши, вскоре к ним присоединяются сильные молодые
тетерева прошлогоднего выводка, слабые же молодые даже в разгаре токо-
вого периода хотя и вылетают на ток, но держатся только на его окраинах;
они или молчат или чуфыкают, но не бормочут (Кириков, 1948). Выделяют
три стадии в развитии токов (Ульянин, 1949). Первая стадия—ранневесенние
предбрачные игры, в течение которой происходит рост семенников и бро-
вей и устанавливаются места токования. Стаи полностью слетаются на места
тока, но брачные крики издают далеко не все петухи. Первое время преоб-
ладает бормотанье, чуфыканье слышится редко, драки очень редки, самки
держатся особой группой, несколько в стороне от тока. В 1936 г. эта стадия
продолжалась с 10 по 28 апреля, всего 18 дней, если считать ее с момента
увеличения семенников. Если же за начало ее принять первое бормотанье,
ТЕТЕРЕВ 65
то она занимала 30 дней (с 29 марта по 28 апреля). Вторая стадия—разгар
тока. В течение этого времени семенники и брови достигают предельных
размеров, происходит спаривание большинства самок. Вся стая слетается
на токовища, токует большинство прилетевших на тока петухов, продолжи-
тельность тока наибольшая, часты драки; самки держатся одиночками во-
круг тока, в 200—300 м от петухов. В 1936 г. эта стадия продолжалась
17 дней—с 28 апреля по 15 мая. Третья стадия—угасание тока. В это время
уменьшаются размеры семенников, понижается интенсивность брачных игр.
Число прилетающих на ток петухов уменьшается, самки на ток вовсе не
прилетают, так как сидят на яйцах. Продолжительность тока снижается
до двух часов в сутки, начинаясь и кончаясь в более ранние часы; реже драки,
голоса поющих самцов более глухи и слабы. Наблюдения того же автора за
числом одновременно поющих на току самцов показывают, что в первую
стадию токовало не сыше 12% из числа присутствующих петухов, во вторую
стадию—до 75%, в третью стадию—до 60%. Количество собирающихся на
тока чернышей на Южном Урале от нескольких штук до нескольких десят-
ков (Кириков, 1948), в Северном Казахстане до 3—4 десятков (Ульянин,1949).
В Мордовском заповеднике места токов располагаются на полянах
среди кустарников и на буграх среди заливного луга, покрытого кустарни-
ками (Птушенко, 1938). На Южном Урале в нагорно-лесостепных ландшаф-
тах большинство токов наблюдается на степных вершинах водораздельных
сыртов и отходящих от них отрогов, на участках, поросших типчаком, степ-
ным мятликом, горной гвоздикой и т. д.; в нагорно-лесных ландшафтах—на
безлесных вершинах хребтов и увалов, реже—на полянах и совсем редко—
в редколесье, перемежающемся с участками березово-сосново-лиственничных
молодняков (Кириков, 1948). В Наурзумском заповеднике (Ульянин, 1949)
тока располагаются в довольно разнообразных местах: в типично степных
стациях далеко от леса E—6 км), на степных полянах среди Наурзумского
бора, в зарослях таволги на гриве между озерами, окруженной тростником,
на опушке зарослей ивовых кустов и березника; большинство токов A7 из
21) располагается на песчаной степи с ковыльным покровом, вдали от леса.
По наблюдениям того же автора, ранней весной тетерева слетаются на
ток в 8—9 час. утра, когда уже пригревает солнце; позднее, в разгар тока,
появляются при полной темноте C час.—3 час. 30 мин.), а к концу мая токуют
с 2 час. утра и до восхода солнца. Во второй половине периода токования
слетаются и вечером. Прилетевший на ток петух поднимает голову и ставит
тело вертикально. Взмахивая крыльями и подпрыгнув 1—2 раза, издает
громкое чуфыканье. Это—вызов другим тетеревам и сигнал начала драки:
начинают чуфыкать другие и вскоре следуют драки. После первой схватки
или после чуфыканья тетерев наклоняется, вытягивает горизонтально шею,
поднимает на ней перья и, развернув веером хвост, начинает топтаться на
месте или, медленно передвигаясь, делает небольшие круги. Принимая такие
позы, он непрерывно бормочет. Обычно какой-нибудь наиболее крупный
самец, раскрыв веером хвост и бормоча, проходит по всему току, отгоняя
всех встретившихся ему на пути тетеревов или вступая с ними в драку.
Схватка между самцами в большинстве случаев продолжается 2—7 мин., после
чего петух делает минутную передышку и снова взъерошивает перья на
шее, распускает веером хвост и непрерывно бормочет минут 10. Затем он
сидит спокойно 3—5 мин., а потом начинает свою весеннюю игру сначала.
До отлета с тока каждый самец многократно успевает повторить чуфыканье,
драку и бормотанье. Во время тока некоторые самцы улетают в сторону от
токовища отыскивать держащихся по соседству самок и потом снова воз-
вращаются. Поведение самок в ходе токования меняется. В начале апреля,
т. е. в период предбрачных игр, самки держатся группами метрах в 200—
300 от токующих самцов. В разгар токования самки держатся поодиночке,
5 Птицы СССР, т. IV
66 ОТРЯД КУРИНЫЕ
каждая в районе своего гнезда. Только утром и вечером они случайно соби-
раются иногда по 2—3 вместе недалеко от токовища. Как только в гнезде
появится полная кладка яиц, тетерки приступают к насиживанию, и коли-
чество их вблизи токов быстро сходит на-нет. Косачи некоторое время про-
должают токовать и без самок, но игры становятся все более вялыми и вскоре
прекращаются совсем. Спаривание, как утверждает вышеуказанный автор,
в условиях Наурзумского заповедника происходит всегда в стороне от тока.
Встреча самцов с самками обусловливается не только тем, что тетерки при-
ближаются на 200—300 м к току, но и активными поисками самок самцами,
отлетающими с тока. Роль самок при этом ограничивается только тем, что
они показываются на виду у самца. Подманивания самками самцов голосом
автор никогда не слышал. Наиболее крупные (вероятно, старые) самцы про-
водят на току больше времени, улетая последними; более мелкие петухи
разлетаются с тока в поисках самок раньше и имеют больше шансов оста-
вить потомство (Ульянин, 1949).
Насколько сильно стремление тетеревов к токованию, можно судить
по тому факту, что однажды Кириков A948) встретил чуфыкавшего на току
петуха, который накануне был тяжело ранен (перебито крыло).
По Ульянину A949), период гнездования рисуется в следующем виде.
В начале мая самки выбирают места гнездования неподалеку от тока. С мо-
мента оплодотворения они останавливаются на избранном месте, устраивают
гнезда и откладывают яйца. Гнезда располагаются в кустарниках, ковыль-
ной степи, зарослях осоки и в небольших лесных колках. Гнездо представ-
ляет собою небольшую ямку 16—20 см в диаметре и 4—6 см глубиной, вы-
стланную сухой травой, иногда одними сухими ветками таволги или почти
вовсе без подстилки. Кладка яиц в Наурзумском заповеднике в 1936 г.
происходила с 2 по 26 мая, т. е. 24 дня; наиболее ранние кладки в Башкир-
ском заповеднике (Кириков, 1948)—в середине мая, начало кладки в б. Сим-
бирской губ. (Богданов)—конец апреля—начало мая, полная кладка—
10 мая; чем моложе курочка, тем позже она несется и меньше яиц; свежие
яйца встречаются до конца мая, изредка до половины июня. Сроки размно-
жения в степях на восток от Волги на две недели позже, чем в лесной полосе
б. Симбирской и Казанской губ., что связывается с более ранним появ-
лением и большим количеством пищи в лесах. Кладка яиц в окрестно-
стях Уфы начинается в конце апреля, в окрестностях Златоуста на 2 недели
позже (Сушкин, 1896). Как правило, самка кладет одно яйцо в сутки. Вес
яиц в одной кладке варьирует от 28 до 35 г, а в среднем A0) 30,17 г.Окраска
яиц бледноохристая с темнокоричневыми пятнами различной величины
и формы (от макового зернышка до зерна чечевицы). Размер кладки в Наур-
зугАском заповеднике от 4 до 10 шт., в среднем (по 9 кладкам) 7,3 шт., в Баш-
кирском заповеднике (Кириков^ 1948)—от 5 до 11 шт., но чаще всего 6—8 шт.
Насиживание начинается с момента откладки последнего яйца и продол-
жается 23 дня, а по Богданову, 19—22 дня, что, видимо, преуменьшено.
В период кладки одни самки при уходе с гнезда закрывают его сухой травой,
другие оставляют яйца совершенно открытыми.
Суточная жизнь тетеревов в период кладки и насиживания: утром
самка откладывает яйцо и улетает или уходит к току; на заре, до встречи
с самцами, некоторое время кормится, а найдя самца, улетает с ним в сторону
от токовища, и происходит спаривание; днем она находится вблизи гнезда
и на этом же участке остается на ночь. В период насиживания целые сутки
сидит на гнезде и лишь утром (иногда также перед закатом солнца) сходит
на несколько минут покормиться поблизости. Процент гибели яиц до-
вольно велик и главным образом от хищников. Из 9 гнезд, находившихся
под наблюдением Ульянина в Наурзумском заповеднике в 1936 г.,
3 гнезда были полностью разорены хищниками; из общего числа 65 яиц
ТЕТЕРЕВ 67
в этих гнездах вывелись 38 птенцов E8,5%), похищено хищниками 25 яиц
C5,5%), оказались болтунами 2 яйца C%). Период гнездования (от откладки
первого яйца до нахождения последнего насиживаемого яйца) продолжался
с 3 мая по 16 июня, т. е. 44 суток. Первые выводки в Башкирском заповед-
нике (Кириков, 1948) отмечены: в 1937 г.—10 июня, в 1940 г.—1 июня,
в 1941 г.—20 июня, в 1944 г.—17 июня, в 1946 г.—28 июня; в среднем за 5 лет
15 июня в Мордовском заповеднике (Птушенко, 1938); в 1936 г. 12 июня;
в б. Симбирской губ. ранние в конце мая, в среднем—в первой половине
июня, поздние—до конца июня.
В Наурзумском заповеднике выводки держатся в различных местах:
на степных полянах и в понижениях, поросших березником, таволгой,
шиповником, боярышником среди соснового бора; в ковыльной степи и в ку-
стах таволги в 2—3 км от границы леса; среди березовых колков в кустах
таволги, шиповника и миндаля; на склонах соров, поросших высокой густой
травянистой растительностью; среди ковыльной растительности по залежам,
поросшим густым бурьяном, и т. д. Выводки не уходят далеко от места гнездо-
вания и держатся здесь до конца лета даже в засушливые годы, когда в степ-
ных районах пересыхают водоемы. В Мордовском заповеднике с 15 июня
(т. е. вскоре после вывода) выводки держатся на ягодниках, сначала земля-
ники^ 10 июня—на чернике, а в августе иногда на костянике. В первые
дни птенцы держатся при матери, не отходя от нее дальше чем на 1 м\ при-
мерно к 1 июля уже удаляются от самки на 15—20 м, а в первой половине
августа—до 100—200 м.
Суточный режим в первые дни жизни выводка разнообразен. В теплую
солнечную погоду птенцы выходят кормиться рано утром, в пасмурные дни
или после выпадения обильной росы—позднее, когда потеплеет. Днем они
скрываются в тени—в кустах таволги или березнике; перед заходом солнца
выходят кормиться на открытые места песчано-ковыльной степи; на ночлег
скрываются в кустах таволги и березнике. Когда птенцы подрастут (до 200 г)
и смогут перелетать на 40—50 м, их суточный режим несколько меняется.
С восходом солнца они бродят по березовому колку или по склонам бархана
с разреженной травянистой растительностью; днем больше в тени, под кусти-
ками березы или таволги, а после полудня семья расходится по степи и кор-
мится здесь до сумерек, после чего выводок приближается к кустам или
березам, где и остается ночевать. Ночью тетеревята размещаются на земле
неподалеку друг от друга; на деревьях ночуют только в исключительных
случаях. Повидимому, определенных мест ночлега нет, птенцы ночуют близ
тех кустов, где их застала ночь.
Тетеревята растут очень быстро, но темп роста у отдельных птенцов
различен. Отстрелянные в разные сроки из разных выводков птенцы весили:
8 июня—23 г; 11 июня—24 г и 26 г; 1 июля—54 и 94г; 2 июля—146 г; 18 июля—
526 г, 23 июля—509 г, 24 июля—625 г, в августе—221 и 758 г; в сентябре—
840 г. Птенцы, только что вылупившиеся из яйца, весят 24—26 г, через день вес
их уменьшается до 23 г (за счет поглощения желточного пузыря), а потом
быстро увеличивается: к середине августа отдельные молодые достигают 70%
среднего веса взрослых самцов. По данным Птушенко A938) для Мордов-
ского заповедника, вес молодых изменялся так: самцы 8 августа—748,772 г,
19 августа—873 г, 20 августа—974, 962 и 955 г; 22 августа—962 г, 23 авгу-
ста—970 г; самки 8 августа—623 г, 20 августа—815 г; 23 августа 750 г, 26 сен-
тября—917 г; взрослые самки 13 августа—870 г, 20 августа—816 г. Но попада-
лись отдельные выводки, которые даже 4 сентября достигали всего лишь
3Д величины взрослых. По нашим наблюдениям, в Наурзумском заповед-
нике в 1934 г., в конце июля птенцы почти достигали величины взрослых,
а в середине августа начинали собираться в стаи и вылетать на кормежку
на хлебные поля.
5*
68 ОТРЯД КУРИНЫЕ
После окончания токов самцы, не принимающие участия в воспитании
птенцов, линяют. Во второй половине лета тетерева иногда токуют, но не
каждый год. В Башкирском заповеднике (Кириков, 1948) летнее токование
в 1940 г. наблюдалось в первой половине августа на нескольких весенних
токовищах. В Мордовском заповеднике (Птушенко, 1937) токование отме-
чено во второй половине сентября. 26 сентября автор наблюдал, как два
петуха дрались, наскакивая друг на друга, принимая те же позы, что и при
весеннем токовании; близ них на токовище находились три самки, следив-
шие за дракой самцов.
Линька. Последовательность смены наряда, как у номинального
подвида. По Богданову, птенцовый наряд начинает формироваться сразу,
как только подрастут маховые перья. Сначала появляются перья на крыльях,
потом на плечах и по бокам зоба, затем по спине, надхвостью, груди
и бокам туловища, а к концу четвертой недели оперяются голова и брю-
хо, и тетеревенок достигает величины домашнего голубя. В это время
самца уже можно отличить от самочки: он рослее, брови шире, пестрины на
перьях чаще и мельче, рыжий цвет значительно бурее, чем у последней.
Птенцовый наряд носится недолго. Уже в конце июля у молодых самцов
он начинает заменяться первым взрослым нарядом. Черные перья появ-
ляются сначала по бокам зоба и на плечах, затем распространяются посте-
пенно к середине зоба, по груди, шее и брюху, а от плеч на крыло,
спину, надхвостье и заднюю сторону шеи к голове, которая линяет послед-
няя. Во время смены гнездового наряда темнеют ноги и клюв. Смена махов
происходит одновременно с линькой головы, когда птенцы достигают почти
полного роста. С окончательной сменой махов начинается смена рулей,
наконец появляются косицы, на чем линька и заканчивается. Самые ранние
птенцы с косицами отмечены 18 августа, самые поздние с недоросшими ко-
сицами—27 сентября. Линька старых самок начинается и заканчивается
одновременно с молодыми, т. е. происходит с конца июля по конец августа.
Последовательность линьки: наседные пятна, грудь, надхвостье и далее
достепенно передние части тела. Старые косачи начинают линять около сере-
дины июня и заканчивают в середине или второй половине июля. Чем моложе
косач, тем позже начинается линька; линяющие годовалые самцы раньше
конца июня не встречаются, чаще же в июле; оканчивают линьку они к концу
августа. Линька начинается одновременно на плечах, зобе и надхвостье.
От зоба линянье распространяется на грудь, брюхо, бока, шею, горло, ниж-
ние кроющие крыльев и хвоста; от плеч—на верхние кроющие крыла и от
надхвостья к переду по спине, задней стороне шеи и кончается головой. По-
следними линяют голова и горло, одновременно с ними махи, а затем и рули.
Холостые курочки начинают линьку одновременно с молодыми (годовалыми)
косачами с конца июня, т. е. на месяц ранее, чем самки с выводками.
Зимняя жизнь. В общем сходна с таковой предыдущего подвида.
Особенно велики—до 500—1000 шт.—бывают стаи в березовых лесах по
восточному склону Южного Урала. В сосново-березово-лиственничных
и в широколиственных лесах таких больших стай не бывает. В начале зимы
продолжаются перемещения и перекочевки тетеревов, начало которых наблю-
дается еще осенью. Как только снег станет настолько глубоким, что они
могут зарыться в него, птицы ночуют под снегом; в очень большие морозы
сидят под снегом также значительную часть дня.
Питание. Ульянин A941), исследовавший 62 зоба и 92 желудка тете-
ревов, добытых в Наурзумском заповеднике, зарегистрировал 54 вида расти-
тельных и 14 видов животных кормов. Из растительных кормов к главным
относятся: 1) береза—почки, сережки, ветви; употребляются в течение
8 месяцев—с января по май, в октябре—декабре; средняя частота встреч за
год 32,6%, в отдельных зобах зимой занимает до 100% объема; 2) шиповник,
ТЕТЕРЕВ 69
плоды и отчасти семена; встречен в январе—феврале, в апреле—октябре,
а практически входит в рацион, видимо, круглый год; в отдельных зобах
и желудках до 90% объема; 3) ястребинка—соцветия; встречена в мае, июле,
августе, октябре в 16 случаях A7,3%); 4) просо—зерно, листья; отмечено
в феврале, апреле, мае и с июля по октябрь в 17,3% случаев, в отдельных
зобах до 100% по объему; 5) козлобородник—цветы, корзинки с плодами;
встречен в 16,3% случаев в июле—сентябре, в июле из 24 данных—6 встреч,
по объему в отдельных зобах до 60%; 6) лапчатка—листья и стебли; зареги-
стрирована в марте, апреле, июле, августе и составляет 6,4% всех встреч,
но в марте и апреле в зобах и желудках занимает 100% объема пищи, почему
ее можно считать основным ранневесенним кормом; 7) подорожник—цветы
и листья; 8,1% встреч главным образом в июле и реже в апреле, августе,
сентябре; 8) молочай—соцветия; 8,1% встреч преимущественно в июле и еди-
ничные случаи в августе и сентябре; 9) таволга—цветы, листья, плоды; 8,6%
встреч в мае и особенно в июле; в отдельных случаях достигала 50% объема;
10) мятлик—листья; 5,4% встреч в мае, июне, сентябре, октябре, иногда
до 50% объема; 11) осина—листья, почки, сережки—5,4% встреч в марте,
апреле, июле; 12 ) солнечник—листья; 9,7% встреч, главным образом в июле
и редко в сентябре и ноябре; 13) спаржа—ягоды; 9,7% встреч в июле—сен-
тябре, иногда до 40% объема пищи; 14) латук—листья, цветочные корзинки;
6,4% встреч в мае, июле, августе и октябре; 15) вишня—плоды; 4,3% встреч
в июле и августе, в отдельных зобах до 90% объема; 16) степной миндаль—
плоды; в июле 8,3% встреч, в августе 25% встреч, в сентябре 22,6% встреч,
в отдельных случаях до 90% объема; 17) полынь морская—листья; встречена
в апреле C7,5%) и в ноябре, в апреле до 80% объема. К второстепенным
кормам следует причислить паслен черный, ковылок Лессинга, василек,
ивы, хвощ зимний, костер прямой, воробейник и др., всего 12 видов. Слу-
чайных кормов зарегистрировано 23 вида (осот полевой, щавель, гвоздика,
горошек, кровохлебка, вероника настоящая, анис дикий, боярышник и др.)-
Из животных кормов к главным относятся: 1) саранча, 10,8% встреч:
в июле 29,1%, в августе37,5%; 2) полосатая кобылка,6,4% встреч: в июне
25%, в июле 20,8%; 3) итальянский прус, 5,4% встреч: в июле 16,4%, в ав-
густе 12,5%; 4) гусеницы (ближе не определены): в июле 20,8% встреч; 5) чер-
нотелка в июле и сентябре; в отдельных случаях до 70% объема; 6) муравьи,
9% встреч: в июле 20,8%, в августе 25%, в сентябре 22,2%. В качестве
второстепенных кормов отмечены 4 вида (мягкотелка, бронзовка, кобылка
зеленая и божья коровка) и случайных 4 вида (златка, щитоноска, долго-
носик, муха). В 1935 г., обильном мелкими саранчевыми, тетерева охотно
ими питались. В некоторых зобах и желудках эти насекомые составляли
до 100% объема съеденного корма. По сезонам кормовой рацион сильно ме-
няется. В январе и феврале в него входят: береза (сережки, почки, ветки),
шиповник (плоды), ивы (ветви и почки), изредка веточки сосны. Первое
место занимает береза, второе в январе ива, в феврале шиповник. В марте
к старым кормам прибавляется в значительном количестве лапчатка и почки
осины. В апреле насчитывается до 10 видов, из них наибольшее значение
имеют береза, лапчатка (листья) и осина (почки), несколько меньший—
листья полыни и плоды шиповника. В мае и июне кормовой рацион еще более
разнообразен. В мае основным кормом продолжают оставаться береза и ши-
повник, но к ним добавляются в значительном количестве соцветия василька
и таволги, а в июне мятлик, полосатая кобылка, златки. Наибольшего разно-
образия достигают корма в июле: 39 видов растительных и 9 животных. По
частоте встреч занимают первое место: зеленые ягоды и семена шиповника
F6,6%), козлобородник D1,7%), ястребинка C5,4%), саранча B9,2%)>
спаржа и молочай B5%). По объему в отдельных зобах преобладали следую-
щие виды: итальянский прус (до 100%), другие насекомые (до 90%), шипов-
70 ОТРЯД КУРИНЫЕ
ник (до 95%), козлобородник (до 100%). В августе состав кормов менее раз-
нообразен: 16 видов растительных и 10 животных. Важнейшие корма—ши-
повник, ястребинка, козлобородник, спаржа, саранча. В сентябре число кор-
мовых видов резко падает: 12 растительных, 2 животных. Преобладают просо
(87,5% встреч) и ягоды шиповника F2,5%). В октябре встречены только
растительные корма; из них важнейшие—березовые почки и сережки, лю-
церна, ковелец. В ноябре (по одному данному)—5 растительных видов, из
которых основными являются верхушки побегов полыни и листья вероники.
Таким образом, в зимние месяцы в рацион входят 2—3 вида корма (береза,
шиповник, ивы), летом же, как, например, в июле, число кормовых видов
доходит до 39 растительных и 9 животных.
В Башкирском заповеднике (Кириков, 1948) тетерева, держащиеся зимой
в чистых березниках, кормятся почти исключительно сережками и почками
берез; зимующие же в сосново-березовых лесах питаются также и молодыми
(первогодними) сосновыми шишечками. В летних кормах южноуральских
тетеревов наибольшее значение имеют ягоды, зеленые части растений и бес-
позвоночные (преимущественно кузнечики и жуки). В зависимости от место-
обитания состав ягод, употребляемых в пищу, сильно изменяется: в сосно-
во-березовых лесах чаще всего кормятся ягодами клубники, костяники,
в ольхово-черемуховой уреме—черемухой, а в кустарниковой степи—степ-
ной вишней. Осенний корм сильно различается в зависимости от того, где
держатся тетерева. В южноуральских сосново-березово-лиственных лесах
главнейшими кормами являются сережки, почки и побеги березы, хвоя
лиственницы, первогодние шишечки сосны, листья клевера лесного, соцве-
тия ястребинки зонтичной, ягоды костяники, цветочные почки сон-травы,
плоды можжевельника, почки и побеги осины, побеги черники, листья ман-
жетки. В смешанных дубово-сосново-березовых лесах большое значение име-
ют жолуди. В нагорно-степном ландшафте, где основными кормовыми угодь-
ями в это время года служат кустарниковая степь, поля и вязово-ольховая
урема, главнейший корм—зерна пшеницы и овса, плоды черемухи, почки,
и побеги вяза, плоды степной вишни, сережки черной ольхи, плоды (засох-
шие) клубники, плоды шиповника, листья полыни, сережки березы, соцве-
тия ястребинки зонтичной.
В Мордовском заповеднике (Птушенко, 1938) в июле и августе отмечено
поедание в значительном количестве проса, костяники и разнообразной
травяной растительности.
Промысел. В начале зимы, когда снег еще не глубок и тетерева довольно
долго придерживаются одних и тех же участков, на Южном Урале на них охо-
тятся с подъезда. Прекращается эта охота с установлением глубокого снежно-
го покрова. В Северном Казахстане зимой добывают петлями или капканами.
В лесу на полянке, в обычных местах кормежки тетеревов, втыкают наклон-
но в снег пучок свежесломанных веток березы с сережками так, чтобы сереж-
ки свешивались и птицы могли их достать. Перед пучком с той стороны, на
которую свисают сережки, протягивают веревочку с висящими на ней пет-
лями. Тетерев, подходя к пучку и пытаясь клевать сережки, попадает
головой в петлю и затягивается. Вместо петель иногда расставляют у пучка
капканы. Прежде (лет 20—30 тому назад) этими способами охотник за зиму
добывал в среднем около 100 шт., а за день обычно 4—6 шт.
Осенью на Южном Урале и в Западной Сибири применяют охоту на те-
теревов с чучелами. Заметив места, куда прилетают кормиться стаи тете-
ревов, охотники устраивают под одиночными березами, разбросанными среди
поля, несколько шалашей на расстоянии 200—300 м один от другого.
На вершине дерева, под которым поставлен шалаш, на легком шестике уста-
навливается чучело тетерева. Пролетающие тетерева, заметив чучело, при-
саживаютс! на это же дерево или на соседнее. Иногда в такой охоте уча-
ТЕТЕРЕВ 71
ствует верховой загонщик, который объезжает ближайшие к полю участки
леса и нагоняет тетеревов на поле. Более удачна охота бывает группой
в 5—6 человек, когда тетерева, отлетевшие от одного чучела, замечают сосед-
ние и присаживаются к ним. Кириков A948) приводит случай, когда на од-
ной охоте, где было 5 охотников и 1 загонщик, за одно утро и два вечера
•было убито 152 тетерева. С установлением прочного снежного покрова охота
с чучелами прекращается
Прежде в указанных выше районах в зимнее время тетеревов крыли щат-
рами, одновременно накрывая иногда по нескольку десятков птиц.
Описание. Размеры. По размерам равен примерно номинальному подвиду.
Длина крыла самцов F8) 250—282, самок E9) 225—250, в среднем 268 и 239 мм\ вес сам-
цов (И) 810—1363, самок A1) 615—1100, в среднем 1247 и 754 г.
Окраска. Отличается от номинального, как и от других подвидов, более широ-
ким зеркалом крыла, которое при сложенном крыле хорошо выдается (около 2 см) из-под
верхних кроющих перьев и развито даже на внутренних из первостепенных маховых.
У самца оперение имеет слегка зеленоватый отлив металлически-стально-голубого блеска,
а не слегка вишнево-синий, как у типичного подвида. Третьестепенные маховые и большие
кроющие крыла даже у старых самцов не сплошь черны, а с неясными светлыми крапин-
ками. Самки и молодые значительно светлее, охристее и сероватее типичных, почти без
ржавчатого оттенка; на спине и надхвостье беловатые вершины каймы и белая поперечная
предвершинная пестрина; зоб белесоват; на основании верхних больших кроющих кисти
•белое пятно, прикрытое крылышком. Плечевые верхние кроющие крыла, перья груди
и боков с большими белыми вершинными каемками.
У молодых самцов в первом зимнем наряде плечи и темные части второстепенных
и третьестепенных маховых сильнее испещрены, пестрины светлее—охристо-бурые, а не
темнобурые, как у типичной расы; третьестепенные маховые с беловатыми вершинными
краями; средние части перьев поясницы также сильнее испещрены светлыми крапинками.
Енисейский тетерев Lyrurus tetrix tsckusii Johansen
Tetrao tetrix tschusii. Johansen. Ornith. Jahrb., 1898, Томск.
Синоним. Lyrurus tetrix jenisseensis. С у ш к и н. Спис. и распр. пт. Русск. Алт.,
1925, стр. 62, Минусинский район.
Распространение. Ареал. Распространен по Южной Сибири от
Иртыша до Иркутска, к северу до районов Тары, Томска, Красноярска
и Киренска, к югу до западных и северных частей Алтая и бассейна верхо-
вьев Енисея (Минусинск, Тувинская авт. обл.): Птицы из верховьев Нижней
Тунгуски занимают промежуточное положение между енисейской и но-
минальной формами (Штегман, 1932).
Характер пребывания. Оседлая птица, при неурожае
кормов в послегнездовый период совершает явно выраженные перекочевки.
О появлении в некоторые годы больших стай залетных птиц в б. Тарском
уезде сообщает Ушков A913). Небывало массовый налет тетеревов в окрест-
ностях Томска наблюдался в зиму 1927/28 г. Косачи появились даже в горо-
де и, по свидетельству очевидцев, «небольшими стайками сидели на тополях,
а часть из них расположилась на крыше кинотеатра Арос. Птицы были
весьма смирны. Территория, подвергшаяся налету, повидимому, была зна-
чительной, так как масса тетеревов наблюдалась и на расстоянии 180 км
к северо-западу от г. Томска» (Кареев, 1928; «Уральский охотник», № 11,1928)
Биотоп. В районе Красноярска особенно охотно населяет места,
где сухие березники или колки смешанного леса чередуются с пашнями,
заливными лугами или болотами. Пашни и ягодники на лесных полянах
дают тетереву обильный корм, в соседнем ельнике он находит ночной приют,
я по окраинам болот или в приречных чащах проводит наиболее жаркое
время дня (Тугаринов и Бутурлин, 1911). В Минусинском крае встречается
в лесостепной зоне и березово-боровом окрайке тайги; в тайгу не заходит
(Сушкин, 1914).
Численность. В большей части ареала обыкновенен.
72 ОТРЯД КУРИНЫЕ
Экология. Размножение. Начало весеннего токования даже
в одном и том же районе неодинаково: в лесостепных местностях раньше,
чем в подтаежных, а тем более лежащих внутри тайги, что связано с разными
сроками общего хода весны в этих районах. В районе Красноярска начало
тока в среднем приходится на конец марта—начало апреля, разгар—не
ранее 20-х чисел апреля. Токовища располагаются или на сухих гривах среди
болот с немногими редкими деревьями, или на обширных полянах среди
леса, или же просто на участке мелкого редкого березника. В последнем
случае птицы не концентрируются на каком-либо определенном месте и
токуют разбросанно. Оканчивается токование в середине мая, но не
редкость слышать одиночных токовиков до первых чисел июня. Утром ток
начинается с первыми проблесками зари и кончается часа два спустя после
восхода солнца, отдельные же особи поют до 10—11 час. утра. Вечерние тока
обычны, начинаются с заходом солнца и оканчиваются с наступлением тем-
ноты. По вечерам слетаются реже и токуют вразброд (Тугаринов и Бутур-
лин, 1911). В окрестностях Минусинска Сушкин A914) наблюдал разгар
тока 15 мая. В Красноярском крае кладка яиц заканчивается иногда к 10 мая,
но обычно позднее. Выход молодых начинается с первой декады июня и про-
должается до первых чисел июля. После 10 августа молодые начинают наде-
вать первый взрослый наряд (Тугаринов и Бутурлин, 1911). В Минусин-
ском крае в начале августа птенцы достигают 1/г—2/з величины взрослых,
а в конце августа почти полностью надевают взрослое перо (Сушкин, 1914).
Линька. В сентябре, к моменту образования стай, линька взрослых,
как и птенцов, заканчивается. Сроки линьки не прослежены.
Зимняя жизнь. В общих чертах сходна с таковой номинального
подвида: детально не прослежена.
Питание. Детально не изучено. ПоТугаринову и Бутурлину A911),
в первую половину лета пищу выводков составляют главным образом насе-
комые, гусеницы, мягкие листья растений, семена. С появлением ягод
клубники, черной смородины, кислицы и др. птицы начинают поедать их.
Осенью, начиная с сентября, ежедневно кормятся на хлебных полях.
Описание. Р а з м е р ы. Длина крыла самцов B3) 261—279, самок A6) 233—249
в среднем 269 и 240 мм (по Штегману, 1932).
Окраска. Весьма сходна как со степным, так и с тяньшанским подвидами.
Косач от тяныыанского даже вовсе не отличим, от степного же отличается несколько менее
заметным зеленоватым блеском (скорее чистым голубовато-синим без зеленого, но и без
вишнево-синего, заметного у северных птиц) и более темной окраской темных частей кры-
ла и темным цветом верхней стороны лапы. Самка едва заметно менее серовата, чем степ-
ная, и пестрее: все каемки перьев верха узкие, с темным и мелким крапом, нижняя сто-
рона с более узкими светлыми вершинными каемками, подхвостье с широкими темными,,
неприкрытыми поперечными полосами вместо некрупных пятен, нередко и вовсе отсутст-
вующих у степной формы; по сравнению с тяньшанскими, самка светлее, с резче выделяю-
щимися темными пестринами и более редким крапом на светлых каймах перьев; подхво-
стье более пестро.
Тяньшаньский тетерев Lyrurus tetrix mongolicus Lonnberg
Tetrao tetrix mongolicus. Lonnberg. Orn. Monatsber, 1904, стр. 108, Баимгол.
Распространение. Ареал» Центральный Тяныыань к югу от Заи-
лийского Ала-Тау и Иссык-Куля, Джунгарский Ала-Тау, Тарбагатай до топо-
левых рощ по р. Аягузу, Западный Алтай, Южный и Юго-Восточный
Советский Алтай, западная часть Монгольского Алтая (Черный Иртыш),
Северо-западная Монголия на север до хребта Танну-Ола, Западный Хангай.
В указанной области тетерев распространен неравномерно, местами
спорадично. В Семиречье в наибольшем количестве встречается к югу
и юго-западу от Иссык-Куля, где в подходящих местах он очень обыкнове-
нен. Местами многочисленен в Заилийском Ала-Тау и в Джунгарском Ала-
ТВТЕРВВ 73
Тау; наиболее редок в Киргизском хребте (Шнитников, 1949). В Джун-
гарском Ала-Тау указанный автор в большом количестве нашел тетеревов
в ущелье Юн-Ки (к югу от с. Маловодного), в районе верховьев Карталы,
в верховьях речек Алмалы, Коктал, Аще-Булак, на р. Балыкты, а также
в верховьях рек Саркан и Баскан> Пункты нахождения туркестанского тете-
рева на Алтае. Южный Алтай: Курчумские горы—урочище Сая-Терек на
р. Бала-Кольджир; по Курчуму; южный берег озера Map ка-Куль; поселок
Александровский, поселок Алексеевский; верхнее течение Бухтармы у пе-
ревала Коко-даба; устье Берели, окрестности дер. Каменки на Бухтарме;
окрестности Катон-Карагая на Бухтарме; дер. Яры; по Иртышу. Западный
Алтай и окраина: окрестности Усть-Каменогорска. Центральный Алтай:
по пути из Верхнего Уймона через Кату некий хребет к Фыкалке (Сушкин,
1938)» По утверждению Долгушина A938), к этому же подвиду, а не к степ-
ному, относятся тетерева, встреченные им в следующих пунктах Центрального
Казахстана: баян-аульские и каркаралинские сосновые массивы; горы Кы-
зыл*рай; горы Нуртай, долина р. Токрау. По предположению Сушкина
A938), граница распространения туркестанского и енисейского подвидов
в Алтае проходит в районе Котанды, Уймона и по пути отсюда к Фыкалке.
На Алтае тетерев распространен в сравнительно низких местах до 1000 м
на уровнем моря и только местами заходит значительно выше, к верхней
границе леса (Сушкин, 1938). В Семиречье никогда не спускается ниже
1000 и даже 1200 му чаще всего обитает на высоте 2000—2400 м, а нередко
поднимается на высоту до 3000 м (Шнитников, 1949).
Характер пребывания. Видимо, ведет оседлый образ жизни,
по крайней мере литературными данными о кочевках мы не располагаем.
Биотоп. На Алтае тетерев характерен для лесостепной местности
и лесных кол ков.. Многочислен по перелескам с обилием ягод. Там, где
лес хотя и прерывается полянами, но носит характер тайги, тетерев или
редок, или не встречается вовсе (Сушкин, 1938). В Семиречье в общем при-
урочен к лесной зоне, и частности к зоне елового леса, но не исключи-
тельно, и встречается в самых различных условиях. Любимые места—сла-
боволнистые предгорья на высоте 2000—2400 м, поросшие травой и изредка
группами ели. Однако охотно живет и в настоящем еловом лесу и в арчев-
никах, встречается и на совершенно открытых местах лугового характера,
вдали от всяких деревьев. Селясь на открытых луговых местах, тетерева
выбирают такие, где трава достаточно высока и густа или где есть кустики.
Поэтому их можно встретить в арчевниках, но на альпийском лугу нельзя.
По р. Аягузу тетерев встречен в необычной обстановке—среди типично пу-
стынных кустарников буквально рядом с Джеком. Раньше по Аягузу росла
тополевая урема, но теперь она вырублена, и тетерев вынужден был приспо-
собиться к жизни среди низкорослых зарослей и ивняков (Шнитников, 1949).
В Алма-Атинском заповеднике (Заилийский Ала-Тау) держится почти исклю-
чительно в еловой зоне. Иногда встречается и токует в предгорьях, обычно
на разнотравных верхних склонах или хребтах, будучи приурочен к участ-
кам, где имеются небольшие перелески осины и березы. В яблоневых лесах,
в кустарниках речных долин тетерева нет, в чистых ельниках довольно
обычен. Выводки держатся в ельниках, смешанных с осиной и березой, даже
предпочтительно в осинниках. Следовательно, и здесь, на юге, тетерев при-
держивается главным образом осин, берез и елей подобно обитателям севера
(Шульпин, 1939). Осенью тетерева массами появляются на полях далеко от
ближайших ельников, десятками покрывая скирды хлеба (Шнитников, 1949).
Численность. Колеблется по районам, в целом относительно
невелика.
Экология. Размножение. Сведения о размножении, как и вооб-
ще по экологии, скудны. По данным Шнитникова A949), в Семиречье токо-
74 ОТРЯД КУРИНЫЕ
вание начинается еще при обильном снеге и продолжается очень долго:
им встречен ток 4 мая, когда даже на довольно значительной высоте весна
была в полном разгаре и ни о каком снеге помину не было. В Алтайском запо-
веднике в 1936 г. начало токования отмечено 20 марта, последняя песня 1 июля
(Фолитарек и Дементьев, 1938). Для токовища птицы выбирают открытое место
на горке, обращенное к солнцу, и пользуются им из года в год. Количество
собирающихся на тока птиц в разных районах различно. Шнитников A949)
приводит случай, когда на одном из токов в Джунгарском Ала-Тау было
убито 11 петухов, прежде чем птицы покинули избранное ими токовище.
По Шульпину A939), в Заилийском Ала-Тау крупных токов не бывает,
косачи токуют по голым хребтам, на подходящих по площади возвышенных
пунктах, зачастую токуют разбросанно и самое большее собираются по 12—
15 самцов. По данным Шнитникова A949), в урочище Карамула в начале
мая 1944 г. найдено гнездо с 6 свежими яйцами в зарослях шиповника. 1 сен-
тября 1913 г. встреченные молодые еще только начали «мешаться», а в пер-
вых числах сентября 1912 г. в Каракольском ущелье встречались моло-
дые, почти совершенно не отличимые от стариков.
Линька. Сроки не прослежены.
Зимняя жизнь. Детально не прослежена.
Питание. По опросным данным, собранным Шульпиным A939),
в Алма-Атинском заповеднике, тетерев не ест березовую сережку, а питается
главным образом арчевой ягодой и осиновой почкой; при этом арчевой яго-
дой питается и всю зиму, так как она долго висит на кустарнике и снегу
в арчевнике бывает обычно немного. Ягоды арчи придают смолистый и вя-
жущий привкус мясу птиц. Осенью тетерев кормится здесь в основном яго-
дами можжевельника. Шнитников A949) указывает, что зоб тетерева, добы-
того в начале сентября около Алма-Атинского озера, был набит главным обра-
зом плодовыми головками смолевки с примесью листочков какого-то мел-
кого астрагала и ягод можжевельника. В Алтайском заповеднике отмечено
поедание тетеревами кедровых орешков (Фолитарек и Дементьев, 1938).
Описание. Размеры. Длина крыла самцов B4) 273—296, самок B0) 242—263,
в среднем 277 и 250 мм (по Штегману, 1932).
Окраска. Петух от енисейского окраской не отличается, от степного отличен
более темным оттенком темных частей крыла и верхних частей лапы. Самка потем-
нее енисейской, сверху более тусклокоричневатого оттенка с менее резко выступающими
черными отметинами, середина брюха темнее, светлые охристо-серые края перьев верхней
стороны гуще и мельче испещрены; поясница менее пестра. Беловатые вершины перьев
верхней стороны развиты очень слабо. От типичной формы отличается менее заметным
ржаво-рыжим тоном, а от степной—менее сероватым оттенком, так как в общем более ох-
ристого тона.
Байкальский тетерев Ьугигиь tetrix baikalensis Lorenz
Tetrao tetrix baikalensis. Lorenz. Birkhuhner Russl., 1911, стр. З, Забайкалье.
Распространение. Ареал. Населяет Прибайкалье к востоку от Ир-
кутска, Забайкалье, северную часть Приамурья к северу до хребта Туку-
рингра, Северо-Восточную Монголию (горы Кентэй, район Урги) и Западную
Маньчжурию. В районе Иркутска имеются переходы к енисейской форме
(Сушкин, 1938).
Характер пребывания. Ввиду отсутствия сведений—
неясен. По аналогии с другими формами можно предполагать, что имеются
сезонные перекочевки, хотя бы вертикальные.
Биотоп. В Юго-Западном Забайкалье и Кентэе держится в смешан-
ных и по речным уремам. В Юго-Западном Кентэе многочислен в горах
Ноин-ула, где склоны речных долин густо заросли сосновыми лесами
с примесью березы. В верховьях р. Сунгур, ближе к гольцам, где, кроме сое-
ТЕТЕРЕВ 75
новых и лиственничных лесов, встречаются густые кедровники, тетеревов
значительно меньше. В горах Ноин-ула почти в каждой обособленной реч-
ной долине держится своя стая, численностью иногда до 30—50 особей
(Козлова, 1930).
Численность. В Юго-Западном Забайкалье и Кентэе обычен.
По другим районам сведения отсутствуют.
Экология. Размножение. Данные по экологии весьма скуд-
ны. По наблюдениям Козловой A930) в Юго-Западном Забайкалье и Север-
ной Монголии весеннее токование начинается в середине марта; с 20-х чисел
апреля оно в полном разгаре и кончается к середине июня. Токовища
в Кентэе наблюдались в долине речки, где птицы проделывают узкие тропы
среди многочисленных болотистых кочек, поросших лозой и черной березой;
в долине р. Сунтур ток происходил по каменисто-луговым вершинам и
южным склонам падей, рано освобождающимся от снега и совершенно
лишенным лесной и кустарниковой растительности. На токовищах самок
наблюдалось мало; в середине мая на утренних зорях часто встречались стаи
самок, кормившихся в лесах вблизи гребней увалов. При токовании целой
стаи на высоких деревьях сидят 2—3 нетокующих косача, как бы сторожа.
Токование 2—3 особей изредка наблюдалось на хребтах, в высокоствольном
лесу и в зарослях молодняка; на этих мелких токах присутствовало иногда
до 8 самок. Гнезда с кладками в 6—11 яиц встречаются начиная с первых чисел
июня. 12 июня найдено гнездо с 11 яйцами, располагавшееся на гребне
хребта неподалеку от каменистых россыпей среди редких сосен; свито оно
было из сухой травы и березовых листьев, а лоток выстлан пухом. 27 июня
обнаружен выводок, состоявший из пуховых птенцов. С первых чисел ок-
тября на утренних и вечерних зорях начинается осеннее токование, достига-
ющее к середине месяца полного разгара. Интересны данные о наличии зим-
него тока, происходящего в самые холодные месяцы. С первыми лучами
солнца тетерева оставляют ночлег и перелетают к месту весеннего тока, где
сначала немного бормочут на деревьях, а затем перемещаются на занесенное
снегом токовище; здесь они ходят среди кочек, растопорщив крылья и рас-
пустив хвосты. Через 2—3 дня на токовище образуются узкие протоптанные
тропинки, по которым они могут бегать и подпрыгивать с весенним оживле-
нием. Чуфыканье и бормотанье в это время слышатся непрерывно. Токо-
ванье продолжается до 9 час. утра, а после кормежки на пашнях и в лесу
перед заходом солнца самцы вновь бормочут на деревьях вблизи токовища
до сумерек, после чего удаляются на ночлег в глубину уремы. Зимний ток
продолжается с удивительной интенсивностью в течение самых холодных
месяцев до последних чисел января. Все самцы, добытые во время зимнего
тока, были старые, весом 1,5—1,8 кг.
Линька. Не прослежена.
Зимняя жизнь. Детально не прослежена.
Питание. Данные отсутствуют. Имеется лишь указание Козловой
A930) о том, что весной она находила в зобах целиком проглоченные бутоны
Pulsatilla vulgar is.
Описание. Размеры. Длина крыла самцов B5) 276—291,-самок B1) 245—257,
в среднем 282 и 252 мм (по Штегману, 1932).
Окраска. Самец имеет более зеленоватый металлический блеск, чем у енисейских,
туркестанских и даже, пожалуй, чем у степных. Плечевые и кроющие перья крыла посиль-
нее испещрены светлым, чем у степной формы; следы таких пестрин заметны даже на верх-
них кроющих хвоста. Молодой самец в первом зимнем наряде сильнее испещрен, чем степ-
ная и енисейская расы, так что на плечах и кроющих крылах охристо-бурый цвет места-
ми преобладает; перья горла с широкими белыми концами; поясница и верхние кроющие
хвоста сильно испещрены и тонко поперечно исполосованы охристо-серым. Самка окраше-
на, как у енисейской расы, но темная, бурая форма, которая редко встречается среди ени-
сейских самок, здесь составляет почти треть всего населения.
76 ОТРЯД КУРИНЫЕ
Уссурийский тетерев Lyrurus tetrix ussuriensis Kohts
Tetraotetrixussuriensis. Kohts in Lorenz. Birkhuhner Russl. 1911, стр.3, Уссурийский
край.
Распространение. Ареал. Населяет северные и средние части Уссу-
рийского края к югу до окрестностей Ворошилова, Южное Приамурье
к северу до 51° с. ш. (Воробьев, 1938) и к западу до Хингана, Восточную
Маньчжурию. В указанной области распространен спорадично, приурочен
в основном к сравнительно узкой полосе по Уссури и Амуру; встречается
вниз по Амуру по меньшей мере до Ирри (устье р. Челасо); близ устья
р. Горин Воробьев добыл 2 августа молодую самку из выводка. По Зее
доходит к северу до Домбиков. По Уссури летом на гнездовье довольна
малочислен. На Нижней Уссури гнездование отмечено у ст. Дьяченковой,
под Дормидонтовской, в долине Даубиха под Яковлевкой, в небольшом
количестве в верховьях Сунгачи, в окрестностях Новорусановки, в среднем
течении Сиянхе, на Среднем Мо, под Ворошиловом. В низовьях Имана
обычен, в среднем течении не гнездится; граница распространения вверх
по реке—с. Картун. Вообще же к востоку от Сихотэ-Алиня на гнездовье
не встречается. Спангенберг A941) отмечает, что в районе Картун и Пещер-
ный тетерева появились после вырубки леса. Довольно обычен в лесах Боль-
шого Хингана, а за последние годы начинает шире распространяться и в
лесах восточной части Маньчжурии.
Характер пребывания. Совершает сезонные перекочевки.
Маак A861) отмечает, что на Уссури тетерева ежегодно появляются
с первым снегом, потом попадаются все чаще и чаще, и в ноябре, когда
реки покрываются льдом, делаются наиболее многочисленными; автор пола-
гает, что прилетают сюда тетерева из долины Сунгари. На увеличение тете-
ревов в Приморье в зимний период указывает и Шульпин A936). По его дан-
ным, зимой они доходят до д. Анучино (верховья Даубихе), бывают много-
численны под Раздольным и даже в нижней части р. Сучан; Пржевальский
зимой встречал тетеревов около р. Ванцин (почти под Псухуном). Это указы-
вает на то, что тетерева откочевывают от границы гнездовой области по край-
ней мере на 250—300 км, притом через хребты и сплошные леса. В зимнее
время тетерева также наблюдались под Владивостоком, в устье Суйфуна,
близ ст. Иман, у ст. Шереметьевской, у ст. Розенгартовка, у Хабаровска,
в устьях рек Хор и Мурени. На Сунгари летом редки, осенью и зимой весьма
обильны; многочисленны зимой в Южной Маньчжурии и в лесах Боль-
шого Хингана.
Подобно типичной форме, у уссурийского тетерева изредка наблюдаются
необычно массовые зимние налеты. Об одном из таких налетов в районе
р. Хор сообщает Воробьев A938). Он указывает, что здесь, в районе
р. Бичевой, тетерев редок; осенью же 1929 г. появился вдруг в большом коли-
честве; птицы держались большими стаями, встречаясь даже в глухой тайге,
например по р. Кетынь (притокр. Хор), в 30—35 км от устья. Автор полагает,
что птицы летали, видимо, с юго-запада из бассейна р. Сунгари.
По Радде A863), в зиму 1857/58 г. тетерева в исключительно большом
количестве встречались в долине Амура и особенно его островов к востоку
от устья Уссури, а также на островах между устьем Уссури и Сунгари;
на луговых пространствах в районе Нижней Бурей в октябре и ноябре мест-
ные охотники добыли около 2000 тетеревов.
Обилие тетеревов зимой в Приморье на Среднем Амуре и Сунгари объяс-
няется, с одной стороны, откочевкой к югу птиц, гнездящихся в северной
половине Амурской обл. и с другой—концентрацией всего немногочисленного
местного населения на узких зимних стациях, главным образом в области
ТЕТЕРЕВ 77
заливных долин рек. Наиболее массовые скопления зимующих птиц, в наших
пределах расположены в области правых нижних притоков Бурей и в районе
Ханкайской низменности, в китайских пределах—в области широкой долины
Сунгари и левых притоков Уссури (Шульпин, 1936).
Биотоп. Пржевальский весенние тока по Уссури чаще наблюдал
на луговой стороне реки, реже по травянистым островам реки; Шульпин
A936)—обычно на пашнях, по заимкам, большей частью вблизи кустовых
опушек лесочков, иногда даже на обсохших участках болот, расположенных
среди невысоких, далеко одна от другой отстоящих сопок, покрытых лист-
венным лесом; иногда на сфагновых болотах с угнетенной древесной расти-
тельностью. Выводки и гнезда отмечались в березниках; в сырых густотрав-
ных, частью кочковатых лугах; на пологом склоне горы, поросшей мелким
кустарником; в степной полосе (Ханкайский бассейн). В низовьях Имана
в период тока излюбленными местами являются сухие участки низменности,
занятые прошлогодними посевами; гнездятся по мелким дубовым перелескам
и березникам (Спангенберг, 1940).
В зимний период основная стация—острова и берега крупных рек,
поросшие зарослями кустарниковых ив. В некоторых местах (например под
Дормидонтовкой) тетерева зимуют в ином ландшафте—среди полей, у опу-
шек тайги, по горам (Шульпин, 1936).
Численность. В большинстве районов ареала низкая, выше—
«а Хингане. Большая концентрация наблюдается в скоплениях на зимов-
ках: на Уссури Пржевальский встречал стаи до 100 шт и более, на Суйфуне
один крестьянин в зиму 1866 г. поймал около 200 шт.
Экология. Размножение. Первое токование на р. Сунгаче
в 1868 г. отмечено 23 марта, когда днем самцы и самки держались еще вместе
в одних стаях, 27 марта токование уже слышалось во многих местах, но тете-
рева все-таки еще продолжали держаться стаями в тальниках у берегов рек.
Токование замолкло в середине мая. В холодную весну 1869 г. первые звуки
токующего тетерева замечены только 9 апреля; здесь ток окончился также
в середине мая, но в бассейне р. Сиянхе по утрам тетерева токовали до конца
мая. В 1859 г. в окрестностях Буссеевой ток начался 27 марта (Шульпин,
1936). По сообщению Воробьева, первое токование в Уссурийском крае
в последних числах марта, последнее токование—во второй половине мая,
а иногда до начала июня. В низовьях р. Имана последнее бормотание под
Вербовкой отмечено 5 июня (Спангенберг, 1940). Шульпину A936) не при-
ходилось видеть больших токов тетерева в Уссурийском крае. По непрове-
ренным им расспросным данным в окрестностях Дормидонтовки на тока
иногда собираются до 150 косачей, но в окрестностях Новорусановки птицы
токуют разбросанно, самое большее по два вместе. То же самое для Сунгача
и Уссури отмечает и Пржевальский. Он указывает, что правильных токов
здесь нет, каждый тетерев токует поодиночке, или по два без определенного
места, и все на деревьях; само токование происходит как-то неправильно,
неподолгу, отрывисто. По Спангенбергу A940), в окрестностях Вербовки
(низовья Имана) на тока собирается до 12—15 петухов, чаще 5—7.
О сроках гнездования имеются скудные сведения: на Сунгаче самка
от гнезда была вспугнута 1 мая, на р. Сиянхе найдено 9 довольно сильно
насиженных яиц 2 июня (Шульпин, 1936), в низовьях Имана обнаружено
гнездо с 11 насиженными яйцами; выводок маленьких, но уже летающих
тетеревят встречен там же 14 июня (Спангенберг, 1940); на Суйфуне разби-
вающийся выводок из 5 почти взрослых молодых, но еще при матке,
поднят 15 сентября, причем молодые самцы примерно наполовину перели-
няли в черные перья (Шульпин, 1936); молодой величиной в половину взрос-
лого добыт в Амурской обл. 2 августа (коллекция Зоологического музея
Московского государственного университета).
78 ОТРЯД КУРИНЫЕ
Линька. Сроки не прослежены. Имеются указания Спангенберга
о том, что 15 июня в низовьях Имана им встречено два линяющих самца
с отрастающими перьями на голове; держались они в густом кустарнике
дубняка среди сырого луга. На Средней Мо 10 июля добыт сильно линяющий
самец (Шульпин, 1936).
Зимняя жизнь. В общих чертах сходна с таковой номинального
подвида.
Питание. Изучено слабо. По наблюдениям Шульпина A936) под
Дормидонтовкой, зимний помет тетеревов «березовый, светлозеленой окрас-
ки; когда поспеет голубика, он становится черный благодаря ягодам, а по
вызревании хлебов становится светлый зерновой». Тот же автор отмечает,
что забайкальский и уссурийский тетерева, повидимому, имеют различный
пищевой режим в период кочевок. Уссурийский связан в основном с таль-
никами, почками которых он питается (но ест местами и березовую почку),
забайкальский питается главным образом яблочками сибирской яблони
и почками душистого тополя. Это различие в поведении обеих птиц было
подмечено еще Радде.
Промысел. В 70-х годах прошлого века русское население по Уссури
ловило тетеревов петлями на снегу, а для приманки клались ветки
талины. Иногда же их промышляли и ружьем. Этот же промысел был раз-
вит и на Суйфуне, где один крестьянин из д. Дубнинской поймал
пленками в зиму 1866 г. около 200 тетеревов. Под Новорусановкой русское
население часто специально охотится на тетеревов с ружьем зимой и поздней
осенью, когда эти птицы становятся здесь многочисленными вследствие при-
кочевки стай с севера. Осенью и зимой обильны тетерева на Сунгари, куда
приезжают добывать их охотники из Харбина. Но количество тетеревов,
добываемых на востоке, ничтожно по сравнению с убиваемыми на Хингане.
Здесь был сильно распространен промысел тетерева петлями на токах, совер-
шенно опустошавший ближайшие к железным дорогам районы (Шуль-
пин, 1936).
Описание. Размеры. Длина крыла самцов (8) 266—283, самок E) 232—256,
в среднем 274 и 243 мм (по Штегману, 1932).
Окраска. В общем похож на степного, белое зеркало крыла развито чуть силь-
нее; оперение цевки чуть беловатее. Самец позеленее, почти без примеси синеватого
в металлическом блеске. Общая окраска самки чуть потемнее, чем у степного и даже
темных особей енисейского и байкальского подвидов, ноне так рыжевата, как у типичной
формы; середина горла обыкновенно белая.
4. Кавказский тетерев. Lyrurus mlokosiewiczi Taczan,
Tetrao mlokosiewiczi. Tacza nowcki. Proc. Zool. Soc. Lond., 1875, стр. 267, Лагодехи.
Распространение. Ареал. Альпийский пояс Главного Кавказского
хребта и Малого Кавказа на высоте от 1500 до 3000 м над уровнем моря.
На Главном хребте встречается всюду от бассейна Пшехи (Западный Кав-
каз) до бассейна Самура (Дагестан); распространение к югу от последнего
еще не выяснено. На боковых хребтах южного склона Большого Кавказа
найден от Гагринского (Западный Кавказ) до Кахетинского (водораздел
Иоры и Алазани) хребта. На Малом Кавказе тетерев распространен от
Гурийских гор до Карабахского хребта. Встречается на всем протяжении
Месхийского хребта, соединяющего Большой Кавказ с Малым. Ни в север-
ном Иране, ни в Турции не встречается (Аверин, 1938).
Характер пребывания. В полосе верхнего предела лесов
и на субальпийских лугах тетерева более или менее оседлы; населяющие аль-
пийские луга совершают вертикальные перекочевки, спускаясь зимой к верх-
нему пределу леса или заходят в него. В очень снежные зимы тетерева спу-
КАВКАЗСКИЙ ТЕТЕРЕВ
79
скаются до 700—800 м над уровнем моря. Вертикальное распространение
тетеревов по сезонам видно из таблицы, составленной по данным встреч
по Аверину, 1938):
""^^.^ Пояса
^^^•^^^ растительности
Сезоны ^^^-^^
Зима (декабрь—март)
Весна (апрель—май)
Лето (июнь—август)
Осень (сентябрь—ноябрь)
Пихтово-буко-
вый лес (в %)
28,1
7,9
Нет
4,3
Верхний предел
лесов и субаль-
пийские луга
(в %)
56,3
60,5
48,0
65,2
Альпийские
луга (в о/о)
15,6
31,6
52,0
30,5
Биотоп. Летом альпийские луга, покрытые богатой растительностью,
заросли рододендрона и низкорослой березы; зимой—субальпийское вер-
холесье, выгревы нижней части альпийской зоны, а также высокоствольные
пихтовые леса (Мензбир, 1902;
Аверин, 1938). 3
Численность. В целом
невысокая. Плотность заселения
тетеревом различных частей ареа-
ла далеко не одинакова и нахо-
дится в большой зависимости от
выпаса скота. Там, где скот па-
сется в большом количестве, те-
терев сократился в числе вслед-
ствие вытаптывания кладок ско-
том и уничтожения молодых цып-
лят пастухами и многочисленны-
ми собаками. Видимо, по этим
Карта 5. Распространение кавказского тете-
рева Lyrurus mlokosiewiczi
а—граница гнездовп*
причинам тетерев исчез на хребте
Гернегем и сильно уменьшился
в количестве на Лагодехском пла-
то. Редок он на Скалистом хреб-
те, в верховьях Пшехи, где производится массовый выпас скота. Районы,
закрытые для выпаса скота, например Главный хребет с отрогами в пределах
Кавказского заповедника, населены наиболее плотно. Здесь тетерева обычны,
и за несколько часов ходьбы иногда можно поднять до двух десятков птиц.
Там, где выпас скота прекращается, численность тетеревов заметно увели-
чивается. Так, на хребте Пшекиш раньше тетерев был редок, но с прекра-
щением выпаса после организации Кавказского заповедника Пшекиш стал
одним из богатых тетеревом районов заповедника (Аверин, 1938).
Экология. Размножение. Половой зрелости достигают, видимо,
на втором году жизни. Полигамы. Весной, вместе с быстрым таянием снегов
и появлением больших площадей проталин, тетерева из высокоствольного
леса поднимаются в полосу верхнего предела лесов, в заросли кавказского
рододендрона и на проталины среди альпийских лугов—на свои гнездовые
местообитания. В начале второй половины апреля самцы собираются на тока
и токуют до начала июня. По наблюдениям Аверина A938), различаются
два типа токовищ: 1) небольшие по размеру, где петухи имеют постоянные
места, расположенные одно вблизи от другого; токовище связано с лесом,
и птицы чаще приходят из леса на ток «пешком», чем прилетают; 2) занимаю-
щие огромную площадь, на которой петухи разбиваются на отдельные групп-
80 ОТРЯД КУРИНЫЕ
ки, перелетают из одной в другую и не имеют постоянных мест; токовище
не связано с лесом, поэтому птицы на него прилетают.
Первый тип токовища наблюдался на склоне, поросшем субальпийским
разнотравьем. С одной стороны токовища расположена полукругом полоса
верхнего предела леса в виде субальпийского высокотравья, с другой—узкий
язык букового леса, оканчивающийся двумя-тремя угнетенными соснами,
несколькими ивовыми деревьями и мелким березником; вверх по склону
продолжается субальпийская растительность. Размер токовища прибли-
зительно 60X100 м. Петухи на токовище имели более или менее постоянные
места, которые покидали лишь в моменты наибольшего возбуждения—для
встреч друг с другом или в погоне за самками.
Второй тип токовища встречен на альпийском лугу на крутом склоне
с большим числом обнаженных скал в полукилометре выше леса. Птицы
прилетали поодиночке и небольшими группками и располагались в разных ча-
стях склона партиями от 1 до 4 шт. Между крайними группками было не
менее 500 м. Группки были все время в движении: то из одной в другую
перелетали самцы, то 1—3 петуха из разных групп собирались в погоне за
самкой.
Токование происходит утром и вечером. Вечером самцы появляются
на токовище обычно около 5 час, но токовать начинают не ранее чем через
час; до этого же сидят неподвижно в разных концах токовища в траве, ощипы-
ваются, иногда догоняют насекомых. Самки приходят на ток в сумерках,
и с их приходом напряженность тока сразу возрастает и вновь падает спустя
5—10 мин. после их ухода. Оканчивается вечерний ток около 8 час. вечера.
Утренний ток начинается вскоре после прилета на токовище самцов около
2 час. и продолжается до 3 час. 30 мин—4 час 30 мин, а иногда и до 8 час.
Самки появляются на рассвете и очень редко после восхода солнца. Пове-
дение птиц на утренних и вечерних токах одинаково. За появившейся на току
самкой пускается в погоню обычно один или два ближайших петуха осталь-
ные только быстрее подпрыгивают, сидя на прежних местах. Самка уводит
петуха с тока в ближайший кустарник, откуда он возвращается через 5—
7 мин. обратно на свое место; самка больше не показывается. На тока тетерева
собираются в количестве до 20 шт. Драки между самцами возникают, видимо,
редко. Однажды Аверин A938) видел, как два самца, бросившиеся за самкой,
вступили в драку . Сначала столкнулись грудь с грудью, разошлись и снова
начали сходиться. Подойдя друг к другу, долго стояли почти неподвижно,
надувшись и подняв хвосты. Потом, вытянув шею и щелкая клювом, как
это делают совы, норовили клюнуть друг друга в голову и неожиданно
разошлись на свои постоянные места. Вся «драка» продолжалась около 5 мин.
Однако по наблюдениям Лоренца, кавказские тетерева очень драчливы и
дерутся даже с большим ожесточением, нежели обыкновенные косачи.
Наиболее оживленно токование проходит в утренние и вечерние сумерки,
когда на току бывают тетерки.
Поведение петухов на току отлично от поведения обыкновенного тетерева:
они или спокойно сидят, или подпрыгивают вверх на высоту около метра,
причем прыжок сопровождается характерным хлопаньем крыльев. Частота
прыжков выражает степень возбужденности птицы. С появлением на току
каждого нового петуха или самки частота прыжков увеличивается, с уходом
с тока самки—уменьшается. Если подпрыгивает один петух, то поочередно
редко вместе, прыгают и все остальные. Частота прыжков изменяется также
от метеорологических условий, времени дня и месяца. Так, 10 мая 1935 г.
за весь ток в среднем на одного самца приходилось 135 прыжков, а 2 июня
1935 г.—только 1.
Поза токующего кавказского тетерева постоянна и своеобразна: он отки-
дывает далеко назад голову, втягивает шею, выпячивает зоб, опускает крылья
КАВКАЗСКИЙ ТЕТЕРЕВ 81
и почти вертикально поднимает хвост, не распуская, однако, его веером.
В этом или ином положении тетерев делает прыжки вверх. Несколько при-
седая, птица отталкивается от земли, и быстро взмахивая крыльями с неко-
торой паузой, поднимается на высоту 75—100 см, опускаясь потом на землю
почти на прежнее место. В воздухе птица круто поворачивается, отчего,
опустившись на землю, оказывается повернутой относительно исходного
положения на 180°. Во время прыжка никаких звуков горлом не издает.
Мягкое «цир-цир», описанное Лоренцом A887) как слабый писк, не горловой
звук, а звук, издаваемый крыльями. Обычно ток проходит в молчании, если
не считать хлопанья крыльев при прыжках. Изредка петухи щелкают клю-
вами или издают короткое хрипение, напоминающее приглушенный и мягкий
крик коростеля. В токе участвуют как старые самцы, так и молодые, еще
в пестром сером, невзрослом, наряде. Разгар токов в Кавказском заповеднике
падает в среднем на период с последней пятидневки апреля по четвертую-
пятую пятидневку мая. Наиболее интенсивно токование проходит в безве-
треную погоду, но при небольшом дожде и снеге—вяло. В ветреную по-
году тока даже в период разгара не бывает.
К гнездованию самки приступают в мае; в конце этого месяца они са-
дятся на яйца. Гнездо представляет собой неглубокую, едва выстланную
сухой травой и несколькими перышками ямку, располагается в зарослях
рододендрона, можжевельника или в субальпийском высокотравье, иногда
в нише под камнями. Гнезда искусно замаскированы и найти их—большая
редкость. Размер гнезда: диаметр 21 см, диаметр лотка 17 см, глубина лотка
5 см. Число яиц в кладке от 2 до 10, в среднем (по 10 кладкам) 6. Средний
размер яиц ( по 7 шт.) 52,8x35,6 мм, вес 30,9 г. Продолжительность
насиживания не установлена, по косвенным данным она равна приблизи-
тельно 20—25 дням.
Первые выводки появляются в половине июня, массовый выход птен-
цов в конце июня—начале июля. Вылупившиеся тетеревята, повидимому,
сейчас же покидают гнездо и начинают кочевать вместе с самкой, уводящей
их на открыты епросторы альпийских лугов. В возрасте 2—3 дней они весят
от 21,8 до 23,5 г. При опасности птенцы крепко затаиваются в густой траве,
самка пытается отвести врага прихрамывая, волоча крылья и т. д., как это
делает и обыкновенный тетерев. Гибель птенцов значительна, особенно в пер-
вые дни жизни. С 20 по 30 июня число птенцов в выводке в среднем равня-
лось только 3,8 (по 6 встречам), в июле—3,7 (по 31 встрече), в августе—2,8
{по 6 встречам). Важнейшие причины смертности в раннем возрасте—суро-
вые климатические условия высокогорья: обильные дожди, холодные туманы,
заморозки, град, а также, вероятно, хищники. С половины августа—момента
созревания ягод и семян альпийских растений—выводки кочуют по ягодни-
кам черники и брусники. К концу августа молодые тетеревята достигают
величины взрослых и отделяются от маток.
Линька. Смена нарядов: пуховой, птенцовый, первый годовой,
взрослый. Линька не прослежена. Самцы начинают линять вскоре после
окончания токов. Они забираются в густые заросли рододендрона или берез-
ника и сидят там очень крепко. Самки в сильной линьке находятся, повиди-
мому, в 20-х числах августа.
Зимняя жизнь. В сентябре, с наступлением заморозков и выпа-
дением быстро стаивающего затем снега, тетерева начинают сбиваться в стаи
и концентрироваться вблизи верхней границы лесов. В октябре и ноябре
все чаще выпадает снег и на более долгий срок закрывает сплошным покро-
вом землю. В поисках корма тетерева спускаются ниже в лес, заходя уже
и в его высокоствольную часть. Важнейшими факторами в жизни тетерева
зимой являются распределение и мощность снегового покрова и размещение
растительности по склонам гор. От них зависит доступность корма, они
6 Птицы СССР, т. IV
82
ОТРЯД КУРИНЫЕ
определяют вертикальное распределение тетерева по поясам растительности.
Основная масса птиц придерживается склонов северной экспозиции, так
как здесь обильнее зимний корм и глубже, чем на южных склонах, снеговой
покров, в который они зарываются от ночных морозов. Часть птиц все же
остается на южных склонах, ночуя в кустарниках или под защитой скал.
Держатся зимой тетерева как стаями, так и одиночками. По одним данным,
величина стай достигает 15—20 шт. (Беме, 1925), по другим даже 50 шт.
(Серебровский, 1918). Аверин A938) наблюдал стаи по 13 шт., но чаще в
2—5 шт. и одиночки.
It
Рис. 13. Кавказский тетерев (самец и самка)
Суточный цикл жизни зимой чрезвычайно однообразен. С началом рас-
света стайки покидают места ночлега и в поисках корма разбредаются по
склону . Корм клюют с земли, редко забираются в крону дерева. Днем тете-
рева сидят на солнцепеке или в кустах можжевельника, а во время снего-
пада или больших ветров—в лунках. На вечернюю кормежку вылетают
вскоре после полудня и с наступлением сумерек слетаются на ночлег, ближе
к лесу или в лес. Перед тем как зарыться в снег, обычно сидят на деревьях
или скалах. В лунки, как правило, садятся слета. Глубина лунок 15—25 еле,
ширина 15—20, длина 30 см. В наст, когда птицы не могут пробить ледяной
корки, ночуют под навесами скал, под снеговыми надувами и кустарниками.
Места ночлега более или менее постоянны.
Питание. Данные анализа зобов и желудков тетерева позволили
зарегистрировать 47 видов растительных и 6 видов животных кормов. Расти-
КАВКАЗСКИЙ ТЕТЕРЕВ 83
тельные корма в питании взрослых занимают первое место во все сезоны года.
Животные корма (насекомые и пауки) встречаются очень редко. Зимой
в качестве главных кормов служат почки и сережки березы, хвоя и ягоды
можжевельника, ягоды шиповника, побеги и почки ивы. Первое место среди
них принадлежит березовым сережкам и хвое можжевельника. Среди вто-
ростепенных кормов наиболее обычны листья рододендрона, хвоя пихты
и листья черники и брусники. Весной береза, шиповник и можжевельник
остаются еще главными кормами. Зеленые части травянистых растений,
побеги и почки деревьев и кустарников по мере их развития приобретают
все большее значение. С конца мая главным кормом становятся стебли*
цветы, незрелые семена и коробочки альпийских растений. С начала августа
пища состоит из зрелых семян альпийских растений, а с половины месяца
из ягод волчьего лыка, черники и брусники. Насекомые встречаются доволь-
но редко. Основная пища осенью ягоды брусники и черники, а также семена
альпийских растений. К пище тетеревов, спустившихся в лес, примешиваются
хвоя пихты, листья кислицы, плоды бука и др.
Пища тетеревят отличается от пищи взрослых только в июне и июле,
когда они растут. Уже в августе выводки переходят на ягодники, где питаются
теми же кормами, что и взрослые. Всего у птенцов обнаружено 17 видов
животных и 4 вида растительных кормов. В первые 10—15 дней тетере-
вята питаются почти исключительно насекомыми: из 17 желудков только
6 содержали растительный корм. Для всех выводков характерно преобла-
дание в пище жуков—долгоносиков и листогрызов. В питании тетеревят
одного выводка заметно большое сходство в качественном и количественном
составе насекомых, что свидетельствует об отсутствии избирательной спо-
собности у выводков в отношении тех или иных видов насекомых. Помимо
отмеченных жуков, птенцы поедают пилильщиков, мух, пядениц, нимфалид,
улиток, пауков и др.
Промысел. Ввиду малочисленности промыслового значения не имеет>
служит предметом спортивной охоты. Добывают тетерева ружьем с лега-
вой собакой, на току и случайно при охоте на крупного зверя.
Полевые признаки. Похож на обыкновенного тетерева, но чуть помельче;
самец отличается матово- или бархатисточерным, почти без блеска опере-
нием, отсутствием зеркала, загнутыми больше вниз, чем в бока, крайними
рулевыми; самка имеет более мелкие и однообразные пестрины, образующие
струйчатый рисунок
Описание. Размеры и строение. По строению, кроме формы рулевых
(см. выше) сходен с обыкновенным тетеревом. Длина крыла самцов A2) 209—232, самок
A0I95—220, в среднем 217 и 204 мм.
Окраска. Птенец покрыт блестящим соломенно-желтым пухом. По бокам голо-
вы черные неправильной формы пятна. Верхняя сторона тела со смешанными рыжими
и черными вершинами перьев. Самец в первом годовом наряде сходен оперением с самкой.
Верхняя сторона тела охристая стойким темным струйчатым рисунком. Шея спереди в мел-
ких пестринах. Нижняя сторона тела, плечи, задняя часть верха и бок охристые с серо-
ватым налетом и узкими темными пестринами. Середина брюха темнобурая. Рулевые рыже-
ватобурые с светлыми пестринами. Самец на второй год жизни надевает взрослый наряд.
Взрослый самец черный с легким зеленовато-голубоватым металлическим блеском на го-
лове, шее, груди и спине. Маховые темнобурые, подмышечные белые. Рулевые черные,
изогнуты кнаружи и вниз. Оперение плюсны темнобурое. Клюв черный. Над глазом голое
красное пятно. У самки верх головы и тела с узкой темной и охристой поперечной поло*
сатостью, иногда переходящей в струйчатость. Шея спереди в темных пестринах. Зоб
с охристыми и темными поперечными полосами. Грудь, верхняя часть брюха и бока имеют
струйчатый рисунок. Середина брюха темнобурая. Рулевые темнобурые с охристыми по-
перечными полосами. Оперение плюсны сероватое.
84 ОТРЯД КУРИНЫЕ
РОД ГЛУХАРИ TETRAO LINNAEUS. 1758
5. Глухарь Tetrao urogallus L.
Тип. Tetrao urogallus
Tetrao Urogallus. Linnaeus. Syst. Nat. изд. X, 1758, стр. 159. Швеция.
Русские названия. Самец—глухарь (большая часть страны), глушак (Смолен-
ская обл.), глушень (Осташковский район), глухой тетерев, моховик (Северный Край);
самка—глухарка (большая часть страны), глушица (Смоленская обл.), копалуха (евро-
пейский север), курица (Южный Урал).
Белорусское, украинское и польское названия: глушец.
Названия глухарь, глушак, глушец даны птице, вероятно, потому, что он не слышит во
время песни. Есть и другое объяснение: Аксаков A852) полагал, что имя глухарь
дано ему потому, что он водится в глухих и крепких местах.
Распространение. Ареал. От Скандинавии, Шотландии (где глухарь
был истреблен и затем восстановлен), Кантабрийских гор и Пиренеев глу*
харь распространен к востоку до южного побережья Байкала, низовья р.Муи,
Патомского нагорья, Олекминска, Вилюйска. Восточная граница распро-
странения глухаря изучена слабо, так же как и северная. Дальше всего
на север глухарь заходит по еловым долинным лесам (до 67° с. ш. в районе
р. Шапкиной и до 70° с. ш. в Скандинавии), но в Ямальской лесотундре
(Сдобников, 1937) с ее разреженными лиственничными лесами отсутствует.
Южная граница проходит по Кантабрийским горам, Пиренеям, южной окра-
ине Альп, Балканам и Южным Карпатам, Волыни (Дубно, Овруч), север-
ным районам Киевской и Черниговской обл., через Севск, Серпухов, Рязань,
по южной окраине лесов по рекам Цне и Суре, Самарской Луке, Бузулук-
скому бору, южной окраине Уральской лесостепи с широколиственными
колками E1°40' с. ш.), Санарскому бору, прииртышскцм и приобским лен-
точным борам, южной окраине Алтая, хребту Танну-Ола, Тункинским горам,
Южному Забайкалью.
Область распространения глухаря в прошлом—лесная зона и лесной
горный пояс Европы, Западной и Средней Сибири и крупные острова и вы-
ступы леса в европейской и западносибирской лесостепи. К настоящему
времени область распространения глухаря сильно сократилась, и он стал
редкой птицей в юго-западной и центральной части Европы, сохранившись
там преимущественно в лесах Пиренеев, Юры, Альп, горных массивов Юж-
ной и Средней Германии, Балкан и Карпат; в равнинных частях Западной
Европы уцелел лишь кое-где, в крупных лесах Польши и Германии. В связи
с вырубанием лесов исчез в Шотландии в середине XVIII в., но затем был
завезен в Шотландию из Скандинавии в 1837 г. и успешно там расселяется
(Ричи, 1920). Сильно сократилась область распространения глухаря в лесо-
степной части нашей страны и на южной окраине лесной зоны. Во второй
половине XVIII века глухарь еще жил в лесостепи между Днепром и Доном
у Багалея A890) в списке птиц, живущих в конце XVIII столетия в тог-
дашнем Харьковском комиссарстве, помещены «тетеревы большие и малые»1,
а в Хотомлянском—«тетеревы глухие». Гмелин A771) писал, что в Воро-
нежской губ. глухарь почитается за редкость, и высказывал предположе-
ние, что глухари лишь залетают в нее, «заблудившись». Но правильнее
считать, что в то время глухари жили постоянно в Воронежской губ.,
хотя й были редки, так как и Северцов A855) включил глухаря в список
воронежских птиц, хотя и по расспросным сведениям. В 70-х годах XIX
1 Несомненно что «тетеревами большими» названы глухари, а «малыми»—тетерева.
Любопытно, что на сербском языке глухарь называется «тетреб велики», а тетерев
«тетреб мали» (Д. Стоичевич, 1938).
ГЛУХАРИ
85
века в Харьковской и Воронежской губ. глухарей уже не было. (Саба-
неев, 1876). В эти годы глухарь еще встречался кое-где по отдельным борам
в долине р. Хопра до с. Сестренцев, а на перевале между р. Медведицей и
р. Терешкой самым южным местонахождением глухаря был бор у с. Черкас-
ского (Богданов, 1871).
В лесостепном Зауралье южная граница глухаря также значительно
отступила к северу за последние 80—100 лет. Есть указания, что в конце
прошлого столетия глухари водились в Ара-Карагае (И. П-ий, 1880) и дру-
гих кустанайских борах (Полферов, 1896). Но Сушкин писал A908), что
в начале XX в. глухарей в кустанайских борах уже не было, и ближай-
шее местонахождение глухаря отмечено в Санарском бору (недалеко от
Карта 6. Распространение глухаря Tetrao urogallus
1—Т. и. uralensis, 2—Т. и. urogallus, 3—Т. и. major; a—граница гнездовий
г. Троицка). По данным Штегмана A934), в Кокчетавских горах глухарь во-
дился еще в начале XX столетия, а в борах у озера Щучьего последний глу-
харь был добыт приблизительно в 1918 году1.
В полосе европейских широколиственных лесов, а также в полосе
смешанных темнохвойных широколиственных лесов, глухарь стал редкой
и малочисленной птицей (в Киевской, Черниговской, Гомельской, Смолен-
ской, Калужской, Московской, Рязанской, Тамбовской, Пензенской, Ульянов-
ской, Куйбышевской и некоторых других областях, а также в Татарской,
Марийской, Чувашской АССР). В Тульских засеках глухарь изредка встре-
чался еще в конце 70-х годов прошлого столетия (Мензбир, 1879), но Сушкин,
исследовавший фауну птиц б. Тульской губ. через 12 лет после Мензбира,
уже не застал там глухаря.
Характер пребывания. Оседлая птица. В некоторых мест-
ностях, где расположены не только хвойные и смешанные хвойно-листвен-
ные леса, но и крупные лиственные леса без примеси хвойных пород, отме-
чены ежегодные сезонные перекочевки глухарей (подробнее об этом далее).
Биотоп. Глухарь населяет как хвойные и смешанные хвойно-лист-
венные леса, так и чистые лиственные леса, предпочитая из хвойных лесов
1 «Лет пятнадцать тому назад»,—писал Штегман в сборнике, который подписан
к печати в 1933 г.
86
ОТРЯД КУРИНЫЕ
боры, а из лиственных—дубравы. Живет в лесах различных типов—от болот-
ных сосняков до сухих песчаных и каменистых боров, от корявых низкоро-
слых южноуральских дубняков до корабельных дубрав Волыни и Полесья
и т. д. При выборе местообитания большое значение имеют возраст, полнота
леса, так как глухарь избегает лесных молодняков и редколесья. В лесных
молодняках (до 20—25 лет), хотя бы они занимали и крупные по площади
Рис. 14. Глухарь (самец и самка)
участки, глухари встречаются лишь в том случае, если молодняки примы-
кают к старому лесу, да и то встречаются лишь летом. Имеет значение также
и величина лесного участка.
Подвиды и варьирующие признаки. Окраска оперения глухарей сильно изменяется
в различных частях его области распространения; в меньшей степени изменяются размеры
(вес) этих птиц. Основываясь на этом было описано несколько подвидов глухарей. По
мнению автора, большинство описанных подвидов в действительности представляет собой
не хорошо охарактеризованные подвиды, а узко местные изменения, отличающиеся неко-
торыми, очень незначительными особенностями окраски или даже только различиями
в количественном соотношении тех или иных типов окраски (таких узко местных измене-
ний у глухаря может быть выделено гораздо больше, чем их описано до настоящего вре-
мени].
ГЛУХАРИ 87
В пределах СССР, по мнению, автора, можно выделить три подвида; Т. и. urogallus
(Li n n a e u s, 1758—таежная полоса СССР; Г. ы. uralensis S e v. et M e n z b i e г,
1887—лесостепь, Южный Урал и южные лесные районы Западной и Средней Сибири;
Т. и. major В г е h m, 1831—Западная Европа и юго-западные части СССР (о местных
изменениях см. замечания по систематике).
Белобрюхий глухарь Tetrao urogallus uralensis Sev. et Menzb.
Tetrao urogallus v. uralensis Sev. et. Menzb. В статье Назарова, Бюллетень Моск.
о-ва испыт. природы, т. 62, № 4, 1887, стр. 365.
Синоним. Tetrao urogallus grisescens. К i г i k о v. Alauda, 1932, стр. 239, Южная
Башкирия.
Местные изменения. Т. и. uralensis taczanowskil. Stejneger. Bull. U. S. N,
Mus., 1885, стр. 193, Иркутск.—Г. и. uralensis volgensis. B u t ur 1 i n. Orn. Monatsb. 1907.
стр. 81. Боры между низовьем Барыша и Сурой (см. замечания по систематике).
Распространение. Ареал. Белобрюхий глухарь распространен от
Севского и Касимовского районов на западе до Алтая и Минусинской лесо-
степи на востоке1; южная граница его совпадает с южной границей распро-
странения глухаря как вида. Северная же плохо изучена.
Характер пребывания. Глухари, населяющие хвойные
(за исключением чистых лиственничников) или смешанные хвойно-листвен-
ные леса, живут обычно в пределах небольшого района, где и совершают те
или иные перемещения. Птицы, живущие в чистых лиственных лесах, в не-
которых местностях (широколиственный лесостепной массив Шайтан-Тау,
Кириков, 1937; Чувашия, Першаков, 1932) совершают регулярные сезонные
перекочевки, отлетая из лиственных лесов на зиму в боры или в те участки
леса, где есть хотя бы небольшая примесь сосны, а весною возвращаются
в родные места.
Из южноуральских дубрав на зиму откочевывают не все глухари; часть
их остается и зимуют в дубравах же (Кириков, 1947). По данным Разевига
<1880), глухари, водившиеся в 80-х годах прошлого столетия в б. Сапож-
ковском уезде Рязанской губ., держались оседло в широколиственных лесах.
Сходное явление отмечено Житковым и Бутурлиным A906) в б. Симбирской
губ.
Биотоп. Хвойные и лиственные леса (характер леса см. биотоп вида).
Численность. В настоящее время мы располагаем крайне скуд-
ными сведениями о численности глухарей в тех или иных угодьях, населяе-
мых юго-восточным белобрюхим глухарем.
Некоторые данные по учету глухарей-самцов на токах указываются
в таблице 1.
Приведенные в таблице 1 данные указывают на низкую плотность засе-
ления глухарей не только лесного широколиственного массива Шайтан-Тау,
но и нагорных сосново-березово-лиственничных лесов Башкирского запо-
ведника; невелика плотность населения глухарей и в Бузулукском бору.
По отношению к Башкирскому заповеднику низкая плотность населения глу-
харей объясняется несколькими причинами. Во-первых, еще в 1940 г. (год
учета) сказывался мор, имевший место в 1934 г. и погубивший большинство
глухарей. Во вторых, в той части заповедника, где производился учет,
около половины всех лесов представляет собой мелколесье и редины (обра-
зовавшиеся после опустошительных рубок и пожаров, имевших место
до организации заповедника), где глухари почти не живут.
1 Систематическое положение глухарей, населяющих заенисейскую часть Сибири,
«е ясно, так как в коллекциях крупнейших зоологических музеев нашей страны (Зооло-
гическом институте Академии наук СССР и Зоологическом музее Московского государ-
ственного университета) имеется еще очень мало заенисейских глухарей.
88
ОТРЯД КУРИНЫЕ
Таблица 1
Количество глухарей (самцов), учтенных на токах
Природная зона (или
переходная полоса),
ландшафт и район
Лесная зона; ландшафт
южноуральских сосно-
во-березово-листвен-
ничных лесов; Башкир-
ский заповедник . . .
Лесостепь; ландшафт
южноуральской нагор-
ной лесостепи с широ-
колиственными лесами;
Шайтан-Тау
Лесостепь; ландшафт
заволжской лесостепи
с широколиственными
и сосновыми лесами;
Бузулукский заповед-
ник
Площадь
участка (в
гектарах),
где велся
учет
общая
10 000
4 000
5 241
покры-
тая
лесом
9 000
3 400
5 200
Коли-
чество
токов
6
5
Общее
количе-
ство
учтенных
глухарей
56
18
47
Количество
глухарей
на 1000 гек-
таров по-
крытой ле-
сом площади
6,2
5,3
9,0
Год
учета
1940
1936
1939
По чьим
данным
Кирикоа
A952)
То же
Даркшевич
A940)
В массиве Шайтан-Тау, расположенном у южного предела лесостепи,
состоящем из одних лиственных пород условия существования глухарей
искони мало благоприятны, и низкая численность их не связана с рубкой
леса.
Повсюду значительная часть глухарей, особенно молодняка и глуха-
рок, гибнет от хищных птиц ( и меньше от хищных зверей). Главными при-
чинами небольшой плотности населения южнолесных глухарей следует счи-
тать следующие: гибель их от болезней (мало изученных), от хищных птиц
и зверей, вырубание лесов и превращение их в редколесье и малолесье, лес-
ные пожары (особенно в Сибири) и чрезмерно большой промысел птиц (осо-
бенно в густо населенных людьми районах).
Низкая плотность населения глухарей в малонаселенных районах и
заповедниках особенно удивительна, если принять во внимание, что эти
птицы не только летом, но и зимой в избытке обеспечены естественным кормом.
Экология. Образ жизни значительно различается в зависимости от
того какую географическую область южной полосы лесной зоны или лесо-
степи он населяет, а также от того, в каком лесу (хвойном или лиственном)
он живет. Особенно существенны различия в характере пребывания и в пи-
тании.
Размножение. Полигамная птица. Для токованья собираются
весной из года в год на одних и тех же местах (токовищах). Рано утром, еще
до восхода солнца, один из старых глухарей начинает петь, сидя на дереве.
Песня начинается с глухого щелканья, похожего на звуки «ка-ду», если их
произносить с открытым ртом, приложив язык к небу и затем опустив его.
Сначала глухарь щелкает редко «ка-ду», «ка-ду», и останавливается,
настороженно прислушиваясь. Если его ничто не испугало, щелканье ста-
новится продолжительнее, а перерывы между ними все сокращаются. И вдруг
ГЛУХАРИ 89
вслед за учащенным «ка-ду», «ка-ду» слышится вторая часть песни, похожая
на звуки «скжиги-скжиги-скжиги...» Эту часть песни называют «точеньем»,
«стрекотаньем», «скжигиканьем»; она длится несколько секунд, и именно
в этих мгновенья, когда глухарь «скжигикает», он ничего не слышит. В это
время шея глухаря вытянута вверх, «как свечка», крылья несколько отста-
влены от туловища и опущены, перья на шее и подбородке оттопырены,
а хвост распущен веером и приподнят кверху.
Глухари поют не только на деревьях, но и на снегу или на земле, куда
они слетают в разгар токованья. Распустив веером хвост, оттопырив крылья
и чертя вершинами их по снегу или по земле, глухари ходят и бегают на
прогале, время от времени подскакивая и взлетая, как тетерева. Во время
«точенья» («скжигиканья») они приостанавливаются вытягивая шею вверх
и тряся бородой. Там же, на земле, происходят и ожесточенные драки между
самцами, начинающиеся после того как на тока станут прилетать глухарки.
Сроки некоторых периодических явлений в жизни глухаря, в том числе
и сроки явлений, непосредственно связанных с размножением, значительно
изменяются в связи с географическими различиями жизненной среды (см.
таблицу 2).
Весеннее токование в южноуральских нагорных борах и лесостепных
дубравах имеет следующие основные различия (Кириков, 1947).
Глухари возвращаются с зимовки в нагорную лесостепь с широколист-
венными колками значительно позднее, чем начинается ток в нагорных борах.
Глухари и глухарки появляются весною в нагорной лесостепи в одно
время, и тотчас же после их возвращения начинается ток. Возвращение глу-
харок в широколиственную лесостепь и начало вылета их на тока в нагор-
ных сосново-березовых лесах происходят приблизительно в одно и то же
время. Тока в нагорной лесостепи с широколиственными колками бывают
из года в год на одних и тех же местах. Это служит доказательством, что
глухари возвращаются с зимовки в свои гнездовые места, подобно тому как
возвращаются на них и многие из настоящих перелетных птиц.
Наблюдения над токованьем глухарей в нагорных приуральских борах
велись автором в течение нескольких лет. Из них следует, что первыми на
токах начинают петь глухари старше двух лет; постепенно к первым токо-
викам присоединяются и начинают петь остальные. На тех токах, где были
выбиты все певшие там глухари, на следующую весну первыми начинали
петь двухлетние глухари (точнее—глухари в возрасте 22 месяцев). На таких
токах начало токованья отмечено значительно позднее, чем на токах, где
были и старые глухари. В разгар токов на токовищах наблюдались две группы
глухарей, отличающихся по поведению. В центре тока держались хорошие
токовики—старше двух лет и двухлетки. На окраинах тока держались
глухари-годовики; часть их пела, другие же только «кряхтели« («крэкатни»).
Иногда наблюдалось, что годовалые глухари токовали в одиночку, на отдель-
ных токовищах.
Если на токах были выбиты все глухари старше двух лет, то в центр
тока прилетали годовалые глухари. Повидимому, не все глухари-самцы
становятся половозрелыми и вылетают на весенний ток в возрасте одного
года; часть глухарей становится половозрелой и вылетает на весенний ток
в возрасте двух лет. Драки между глухарями на токах бывают ожесточен-
ными и прекращаются после того как глухарки садятся на гнезда и перестают
вылетать на тока. В конце весеннего токованья (в конце мая—начале июня)
на токах наблюдались как глухари старше двух лет, так и двухлетние и го-
довики.
Глухарки начинают прилетать на весенние тока значительно позже,
а покидают их значительно раньше, чем самцы. По наблюдениям в нагор-
ных сосново-березовых лесах Южного Урала (Башкирский заповедник),
Таблица 2
Сроки весеннего токованья глухарей (самцов) и вылета глухарок на тока
Природная зона (или
переходная полоса);
ландшафт; район
Лесная зона; ландшафт
южноуральских нагор-
ных сосново-березово-
лиственничных лесов;
Башкирский заповед-
ник; 33° 15' с. ш.;
500—600 м над уров-
нем моря
Лесостепь; ландшафт
южноуральской нагор-
ной лесостепи с широ-
колиственными леса-
ми; Шайтан-Тау;5Р40'
с. ш.; 400—500 м над
уровнем моря ....
Лесостепь; ландшафт
заволжской лесостепи
с широколиственными
лесами; Бузулукский
бор: 53Э с. ш. 50 м
над уровнем моря . .
SS»
о * Д
В S ч 5
Л) 33 flj *>
о н з 2
0
3
5
Начало токованья
глухарей на посто-
янных токах
амое
аннее
7/1II
16/IV
2/Ш
0J
амое
оздне
о в
4/IV
19/IV
27/Ш
редне
о
27/Ш
17/IV
17/111
Окончание токованья
глухарей
амое
аннее
О Q«
27/V
си
амое
оздне
о с
8/VI
Наблюдений
На
5людений
CD
редне
о
1/VI
нет
нет
5 о я
ч S к
о ? ч 5
ij ж (у S
S3 rtj ^ j*Zr
§ д 2 2
7
3
Начало вылета
глухарок на тока
амое
аннее
о сх
11/1V
16/IV
амое
оздне
о с
18/IV
19/IV
редне
о
14/IV
17/IV
Окончание вылета глухарок
на тока (за исключением
единичных случаев)
амое
аннее
О Q.
11/V
/
О)
si
2 со
СО О
о в
22/V
CL)
К
О
15/V
Наблюдении нет
Наб;
1юдений н
ет
ГЛУХАРИ 91
сроки первых вылетов глухарок на тока колеблются в пределах от 11 до 18 ап-
реля (в среднем 14 апреля), а последние вылеты отмечены между 11 и 22 мая.
(в среднем 15 мая). Позже этих дат глухарки наблюдались очень редко, по-
одиночке и на очень немногих токах. Следовательно, продолжительность
участия глухарок в весенних токах примерно в два раза короче продолжи-
тельности токованья самцов. Окончание вылетов глухарок на тока совпа-
дает с началом насиживания, которое отмечено в Башкирском заповеднике
между 8 и 21 мая (в среднем 19 мая).
Наблюдений над количеством яиц в кладках южнолесных глухарок
немного. По наблюдениям Ушкова A887), на Среднем Урале кладка чаще
всего состоит из 6—8 яиц; «в хорошую весну у некоторых глухарок бывает
до 12 яиц, а в дурную—не больше». На Южном Урале (Башкирский заповед-
ник) количество яиц в кладке колеблется от 5 до 16, равняясь в среднем (из
22 кладок) 7,6.
Различия в величине кладки зависят от разных причин. Наблюдения
в Башкирском заповеднике (Кириков, 1947) дают основание предполагать
существование связи между зимней и весенней погодой и количеством яиц
в глухариных кладках. Если малоснежная, суровая зима сопровождается
затяжной, холодной и ненастной весной, то такое сочетание будет, повиди-
мому, наиболее неблагоприятным для кладки глухарей1. Наоборот, если
снег ложится рано и достаточно мощным слоем и если такая зима сопро-
вождается достаточно теплой и дружной весной, то следует ожидать, что
количество яиц будет повышенным.
Яйца глухарей по размерам сходны с куриными. Окраска их желто-
вато-белая с бурыми пестринами.
Гнездо—небольшое углубление в земле. Глухарка устраивает его в лесу,
обычно не дальше чем в 1/2—1 км от токовища. Глухарка сидит на гнезде
неподвижно, и оперение ее так сливается с ближайшим окружением гнезда,
что даже подойдя к знакомому гнезду на 5—8 шагов приходится всматривать-
ся несколько минут, чтобы заметить глухарку.
Наблюдений над тем, через какие промежутки откладываются яйца,
очень немного. По наблюдениям Ушкова A887), глухарки на Среднем Урале
откладывали яйца через день. Сходные наблюдения были произведены и на
Южном Урале (Башкирский заповедник), где 6 мая 1941 г. было найдено
гнездо с одним яйцом; 11 мая в этом гнезде было 4 яйца, 14 мая—5 яиц и
17 мая, когда глухарки уже насиживали,—6 яиц. Литературные указания
о продолжительности насиживания несколько расходятся. По Сабанееву
A876), глухарка сидит на яйцах около 4 недель, даже несколько более (до
30 дней), но Ушков A887) указывал на меньшую продолжительность наси-
живания, равную 20—24 дням. В одном из гнезд, за которым велись наблю-
дения в Башкирском заповеднике, последнее яйцо было отложено между
15 и 16 мая, глухарята же в этом гнезде вывелись 9 июня, т. е. на 24-й или
на 25-й день.
Глухари-самцы (как и у других видов полигамных птиц) не принимают
никакого участия ни в насиживании, ни в уходе за птенцами. В середине авгу-
ста молодые глухари отбиваются от матки; глухарки держатся с нею несколь-
ко дольше.
Летнее токованье. Весеннее токованье глухарей неразрывно
связано с периодом размножения этих птиц. Но, кроме весеннего токованья,
у юго-восточных светлосерых глухарей бывает и летнее токованье, наблю-
давшееся на Южном Урале с начала августа или конца июля и заканчивав-
1 Малоснежные суровые зимы вероятно ослабляют организм глухарок, так как
в такие зимы они не могут укрываться под снегом; ослабление организма, несмотря на оби-
лие корма, может, повидимому, привести к ослаблению функций размножения.
92 ОТРЯД КУРИНЫЕ
шееся в конце августа; следовательно, летнее токованье значительно короче
весеннего. Помимо этого, имеются другие особенности летнего токованья,
Отличающие его от весеннего. Все глухари A7 особей), добытые на летних то-
ках, оказались сверстниками: возраст их равен 14 месяцам; глухарей старше
этого возраста на летних токах добыто не было.
Глухарки на летних токах наблюдались крайне редко (за 7 лет наблюде-
ний в Башкирском заповеднике глухарки наблюдались всего лишь три раза).
Драк между глухарями на летних токах не наблюдалось, как не наблюдалось
и того, чтобы они токовали на земле, что бывает обычно весною. Семенники
глухарей, токовавших летом, были увеличены в такой же степени, как и
весной (при сравнении глухарей одной и той же возрастной группы)»
Гистологическое исследование семенников, произведенное Третьяковым,
дало возможность установить, что у глухарей, добытых на летних токах,
семенники находились в стадии активного образования сперматозоидов, но
сперматогенез выражен несколько слабее, чем у глухарей двух лет и старше,
добытых на весенних токах. Все это позволяет рассматривать летнее токова-
нье глухарей, как период половой активности, проявляющейся вне периода
размножения.
Осеннее и зимнее пение. Отличается от весеннего и летнего
токованья прежде всего тем, что обычно происходит не на постоянных токови-
щах и не в определенное время суток, а на местах кормежки и одинаково
часто как утром и днем. Кроме этого, осенью глухари держатся обычно сме-
шанными стайками из глухарей и глухарок, и среди этой стайки поет один:
или несколько глухарей. Зимою юго-восточные глухари поют гораздо реже,
чем осенью, нам известно всего лишь несколько наблюдений над зимним
пением в Башкирском заповеднике, где наблюдения велись в течение 10 лет.
Линька. Имеющийся материал по линьке юго-восточных глухарей
позволяет описать лишь летнюю линьку глухарей (по сборам с южной око-
нечности Урала). У глухаря, добытого 18 июля 1948 г., голова и шея пол-
ностью в летнем оперении (светлосерые перышки покрывают верх головы
и шею, на подбородке и нижней части горла светлосерые перья с белой рябью).
Половина рулевых выпала и сменяется новыми.
Летнее токованье южноуральских глухарей совпадает со сменой летнего-
оперения на зимнее. У глухаря, добытого на току 4 августа 1948 г. (на южной
окраине лесостепи), голова и шея полностью в летнем оперении, появилось
всего лишь 5—6 новых перьев. То же самое заметно и у глухаря, добытого на
току в Башкирском заповеднике? августа 1941 г. У глухаря, добытого 10авгу-
ста 1941 г., новых перьев значительно больше C0—50); на подбородке и ниж-
ней части шеи они еще в пеньках. У глухарей, токовавших 16 августа 1940 г.
и 20 августа 1942 г., новых перьев еще больше, но большинство их еще
в пеньках. Глухарь, токовавший 17 августа 1942 г., полностью сменил лет-
нее оперенье на зимнее.
Питание южно-лесных глухарей, судя по наблюдениям на южной
оконечности Урала (Кириков, 1937 и 1947), резко различается в зависимости
от того, в каких лесах—сосновых или чистых широколиственных—живут
эти птицы. Основной весенний корм глухарей, живущих в южноуральских
сосново-березовых лиственничных лесах,—сосновая хвоя. До конца мая
у глухарей, добытых на токах, зобы продолжали оставаться наполненными
сосновой хвоей, хотя в это время имелся в достаточном количестве и другой
корм—молодые листья, бутоны и цветы различных растений. Из других дре-
весных пород лиственница занимает следующее после сосны место (глухари
весной едят ее почки и молодую хвою); сережки березы и хвою можжевель-
ника глухари едят реже. Из травянистых растений наибольшее пищевое
значение имеют следующие: цветы сон-травы (Pulsatilla patens), листья лупи-
нолистного клевера {Trifolium lupinaster), черники (Vaccinium myrtillus)>
ГЛУХАРИ 93
горного очитка (Sedum hybridum), володушки (Bupleurum aureum), клуб-
ники (Fragaria collina), полыни {Artemisia sp.), лапчатки (Potentilla sp.), чины
(Lathyrus sp.), перезимовавшие плоды костяники и майника.
Пища глухарей, живущих в южноуральских широколиственных лесо-
степных колках, сильно отличается от пищи боровых глухарей. В начале
весны, когда травяной зеленой растительности еще мало, лесостепные дубра-
вные глухари едят главным образом почки побеги дуба, козьей ивы (Satix
capraea) и березы (Betula verrucosa).
В связи с быстрым ходом весны этот корм вскоре заменяется соцветиями
дуба, ивы и осины, цветами сон-травы и хохлатки, листьями и молодыми
стеблями башкирской кислицы (Polygonum alpinum), шелковистой полыни
(Artemisia sericea), горного очитка и некоторыми другими.
Летом как боровые, так и дубравные глухари кормятся преимуществен-
но на земле. Для молодых глухарят, живущих в некоторых борах Южного
Урала, очень большое пищевое значение имеют беспозвоночные животные—
гусеницы и куколки некоторых бабочек (бражников, пестрянок, ночниц
и др.), нимфы и личинки цикад, некоторые виды жуков, клопов и пауков;
из травянистых растений довольно большое значение в пище молодых глуха-
рей имеют семена осок. По мере поспевания ягод (клубники, земляники, чер-
ники, малины, костяники) они становятся самой важной пищей как молодых,
так и взрослых глухарей. Кроме того, боровые южноуральские глухари едят
летом листья лупинолистного клевера и гороховидной чины, соцветия ястре-
бинки, семена перловника (Melica nutans) и речного гравилата (Geum rivale).
Во второй половине лета глухари нередко поднимаются на деревья и едят
хвою лиственницы и листья осины. В годы полного неурожая всех ягод глуха-
ри переходят на питание лиственничной хвоей раньше, чем в «ягодные»
годы.
Летняя пища лесостепных дубравных глухарей более сходна с пищей
боровых глухарей, чем весенняя.
Ягоды малины, степной вишни, клубники и костяники—важнейший
корм дубравных глухарей во второй половине лета; из беспозвоночных
чаще всего попадают в пищу глухарям кузнечики и жуки, реже—клопы и
науки. В очень засушливые годы, когда все ягоды пропадают уже в июле,
глухари необычно рано (в начале августа) начинают кормиться листьями осины
и дуба и семенами березы. Осенью различия в пище боровых и дубравных глу-
харей вновь нарастают. В пище глухарей, живущих в южноуральских лесах
«с примесью лиственницы, очень большое значение имеет хвоя лиственницы.
Глухари нередко едят ее и летом, но регулярные ежедневные вылеты на
лиственницы начинаются позднее и зависят от начала сильных осенних утрен-
ников: чем раньше они начинаются, тем раньше начинают вылетать на лист-
венницы глухари. Хвоей лиственницы глухари кормятся до тех пор, пока
остаются деревья с зеленой хвоей (пожелтевшую они не едят). В первой поло-
вине осени, кроме хвои лиственницы и листьев осины, глухари кормятся так-
же и на земле, собирая ягоды костяники, обрывая листочки лупинолистного
и лесного красного клевера, коробочки колокольчиков, лилиелистных и т. д.
Животный корм осенью сходит на нет. В тех местах Южного Урала, где
среди леса на полянах есть посевы, глухари выходят кормиться пшеницей
и овсом.
Лесостепные дубравные глухари осенью кормятся листьями осины, желудя-
ми, ягодами рябины и калины. При урожае желудей они становятся важней-
шей пищей глухарей.
С наступлением зимы глухари, живущие в сосновых и сосново-листвен-
ных лесах, питаются преимущественно осиновой хвоей; более или менее су-
щественную примесь к ней составляют молодые, первогодние шишечки
сосны и березовые почки; там, где в лесах, кроме сосны и березы, есть листвен-
94
ОТРЯД КУРИНЫЕ
ница, почки последней также иногда поедаются глухарем. Ряд исследова-
телей (Сабанеев, 1876; Кириков, 1937) обратил внимание на то, что глухари
по каким-то, еще невыясненным, причинам едят хвою не на всякой сосне,
а выбирают определенные деревья, стоящие более или менее открыто, а не
в густом лесу. Иногда они даже вылетают кормиться на одиночные сосныУ
стоящие среди поля.
В Западной и Средней Сибири, там, где есть кедр, его хвоя поедается
глухарями зимою, весною и осенью не менее охотно, чем хвоя сосны, а может
быть даже предпочитается. В «Охотнике Сибири» за 1935 г. (№ 5—6) Тузовский
сообщал, что в Прокопьевском районе зимою, весной и осенью глухари едят
преимущественно кедровую хвою; там же указывалось на наблюдения Гольц-
майера в Горно-Шорском районе,
где глухари тоже кормятся глав-
ным образом кедровой хвоей.
Желудок глухарей не может
сам по себе справиться с перетира-
нием грубой древесной пищи, и:
они заглатывают много мелких ка-
мешков, помогающих перетиранию
хвои и побегов. Поздней осенью вес
камешков в желудках южно-ураль-
ских глухарей-самцов колеблется
от 48,5 до 58,9 г, а у глухарок от
24,1 до 31,8 г. Летом количество ка-
мешков в желудках сильно умень-
шается, доходя до 0,35 г.
Полевые признаки. Самая
крупная из куриных птиц на-
шей страны; ростом почти с ин-
дюка. С земли поднимается тяже-
ло, громко хлопая крыльями. Ле-
тит обычно над самым лесом или
в полдерева; лишь осенью совер-
шая более значительные перемеще-
ния, летит высоко над лесом. Молодые глухарята, отъединившись от матки
и друг от друга, протяжно свищут. Мать созывает детей особым голосом,
похожим на протяжное унылое гуденье; на току издает звуки, похожие на
«бак-бак-бак». «Песня» глухарей описана в разделе о токованье.
Описание. Размеры и строение. Длина крыла самцов (85) 373—422,
самок E) 307—315, среднее 392 и 311 мм. Вес самцов двух лет и старше G5) 3600—5050,
самок A0) 1700—1920, среднее 4100 и 1800 г.
Изучение строения черепа показало, что возрастные изменения черепа протекают так
длительно и отдельные стадии этих изменений отличаются одна от другой так ясно, что
по изменениям черепа можно точно определить возраст годовалых, полуторагодовалых,
двухлетних и более старых глухарей (Кириков, 1939, 1944 и 1947).
Окраска. Пуховик. Снизу светложелтый со слабым охристым оттенком; подбо-
родок и шея более белесы, чем остальной низ. Верх желтовато-бурый сдсумя темнобуры-
ми узкими полосками на спине и надхвостье. Голова светложелтая с темными пятнышками.
Первый наряд после пухового полностью не описан. У глухаренка, добытого 1 июля
1946 г. в возрасте около двух недель на Южном Урале, на крыльях уже не осталось пуши-
нок. Первостепенные маховые темнобурые со светлобурыми каемками и такими же пятны-
шками, разбросанными по перу; второстепенные маховые светлобуроватые с широкими бе-
лесыми каемками. Голова светложелтая с темными пятнышками. На нижней стороне тела
преобладает еще пух, но на боках уже обозначилась узкая полоска из светлобурых перьев.
У взрослого самца голова и шея темносерые со струйчатыми или зигзагообразными
черными поперечными черточками, передняя часть спины и поясница пепельно-серые
вследствие наличия сильно развитых беловатых поперечных пестрин, бока тела окрашены
сходно со спиной, нижняя часть груди и брюхо белые с довольно редкими черно-бурыми
пятнами. Соотношение белой окраски и темных пестрин очень изменчиво в пределах не
Рис. 15 Пуховой птенец глухаря
ГЛУХАРИ
95
только подвида в целом, но даже в пределах отдельных стад глухарей, населяющих какой-
либо естественный район. Наиболее характерным для этого подвида является его белобрю-
хость. Верхние кроющие крыла серые с белыми пятнами основных половин и широкими
вершинными краями; плечевые и верхние кроющие крыла светлорыже бурые и пепельные.
Первостепенные маховые темнобурые со светлыми краями передней части наружных
опахал; концы второстепенных маховых с узкими белыми каемками; подбой крыла белый.
Рулевые черные с большим или меньшим развитием неясных беловатых мраморновидных
как бы растушеванных пятен; концы длинных верхних кроющих хвоста с белыми ободка-
ми. Радужина бурая, клюв желтовато-рогового цвета, ноги красновато-бурые.
Окраска глухарок (описываются по экземплярам с Южного Урала). Сверху темно-
бурые с желтыми поперечными полосками; на подхвостье и нижней части спины попереч-
ные полоски не желтые, а белые. Снизу грудь и брюхо белесые от широких поперечных
о д
Рис. 16. Изменчивость окраски глухарей: а—западноевропейский черно-
брюхий глухарь, б—глухарь переходный между юго-восточным и таеж-
ным, в—юго-восточный белобрюхий глухарь
вершинных каемок, перемежающихся с узкими черными полосками и более широкими
желтыми, но последние прикрыты белыми вершинами. Подхвостье чисто белое. Зоб желто-
вато-оранжевый с узкими черно-зелеными поперечными полосками.
Систематические замечания. Описание и латинское название юго-восточного бело-
брюхого (или светлосерого) глухаря впервые было опубликовано в статье Назарова П887),
указавшего, что как описание, так и название Т. и. uralensis принадлежат не ему, а Север-
цову и Мензбиру (tTetrao urogallus v. uralensis S e v. et M e n z b.»). В том же году A887)
и под тем же латинским названием белобрюхий глухарь был описан Мензбиром, но это
описание было опубликовано позднее, чем вышла из печати статья Назарова. Поэтому
должно сохраниться первое название Т. и. uralensis Sev. et M e n z b.
Первое краткое описание Северцова и Мензбира и второе более подробное Менэ-
бира, а также тип описания1 показывают, что Северцев и Мензбир выбрали для описания
глухаря с наибольшим развитием белой окраски на брюхе и груди. Такой тип'окраски
свойствен сравнительно небольшой части южноуральских глухарей; несколько чаще
на Южном Урале встречаются глухари, у которых темная окраска на брюхе и груди пре-
обладает под белой; чаще всего попадаются глухари, у которых белый фон и темные пест-
рины занимают приблизительно одинаковую площадь2.
1 Глухарь, послуживший Мензбиру типом описания, находится в Зоологическом
институте Академии наук СССР. На оригинальной этикетке написано: «Coll. Severtzow.
on ¦ ¦*
Е Baschkiria adlatus, urbe Orenburge emptus. 18 ^т\ 74. subad., 1619».
1 Такие глухари были описаны (Кириков, 1932) под названием Т. и. grisescens.
96
ОТРЯД КУРИНЫЕ
Поволжские глухари, описанные Бутурлиным как особый подвид (Г. и. volgensis),
должны рассматриваться, как узко местная раса юго-восточного светлосерого глухаря.
Судя по небольшой коллекции глухарей (9 экземпляров) из б. Симбирской губ., в этой
местности не было добыто совсем таких глухарей, у которых на брюхе преобладала бы
белая окраска; были добыты только глухари второго D4%) и третьего E6%) типа окраски.
Южносибирские глухари, как уже было замечено и подчеркнуто Сушкиным A914),
чрезвычайно сходны с поволжскими. На этикетках минусинских глухарей Сушкин писал
Т. и. uralensis natio taczanowskii; следовательно, он считал минусинских глухарей мест-
ным изменением белобрюхого глухаря.
У глухарей из прииртышских и приобских ленточных боров, судя по осмотренным
9 экземплярам, преобладает второй тип окраски.
Среди пяти глухарей, добытых в Алтайском заповеднике, у Телецкого озера, лишь
у одного1 белая и темная окраска занимает примерно одинаковую площадь на брюхе ниж-
ней части груди; у остальных же четырех темная окраска преобладает над белой, при-
чем один более темнобрюх, чем самые темнобрюхие уральские и поволжские.
В каком количественном соотношении встречаются глухари того или иного типа
окраски на южной оконечности Урала, в присурских борах, прииртышских и приобских
борах и на Алтае, показано в таблице 3.
Таблица 3
Количественное соотношение глухарей различных типов окраски на южной оконечности
Урала, в присурских и прииртышских борах и в Алтайском заповеднике
Место добычи глухарей
Южная оконечность Урала;
Башкирский заповедник,
окрестности Крепостного
Зилаира и Каноникольска
Поволжье, присурские леса
Прииртышские и приобские
боры
Алтайский заповедник . . .
Количество
осмотрен-
ных тушек
58
9
9
5
Тип окраски
на брюхе и
ьижней части
груди белая
окраска преоб-
ладает над'
черной
11A9%)
белая и темная
окраска зани-
мает примерно
одинаковую
площадь
28D8%)
4D4о/о)
7G7о/о)
1 B0%)
темная окраска
преобладает
над белой
19C3%)
5E6%)
2B3о/о)
4(80о/0)
Таежный темный глухарь Tetrao urogallus urogallus L.
Местные изменения. Г. а. и. karelicus. Lonnberg. Fauna och Flora, стр. 71,
Каяна в Финляндии.—Г. м. и. obsoletus. Snigirewsky. В книге Л. А. Портенко.
Фауна птиц Северного Урала, 1937, стр. 50. Архангельск.—Г. и. и. kureikensis.
Бутурлин Uragus, III, № 2, 1927, стр. 1, Курейка в Туруханском крае.
Синоним. Т. и. pleskei. Stegmann. Доклады Академии наук СССР, серия А,
1926, стр. 65, Луга, Гдов.
Распространение. Ареал. Вся северная часть ареала вида от Сканди-
навии до бассейна р. Курейки, к югу до соприкосновения с северной границей
светлосерого глухаря. Переходная темнохвойно-лиственная полоса европей-
ской части нашей страны населена глухарями, признаки которых переходны
между таежным и светлосерым глухарем или же (в западных частях ареала)
между таежным и западноевропейским глухарем. Из восточной, заенисейской
части ареала имеется очень мало материала, поэтому остается не вполне
ясным, следует ли относить птиц оттуда к этому подвиду или, быть может,
к светлосерому белобрюхому глухарю.
1 О нем упоминал Дементьев A938).
ГЛУХАРИ
97
Характер пребывания. Таежный глухарь обычно дер-
жится оседло в пределах небольшого района, внутри которого и перемещает-
ся. Таких регулярных сезонных перекочевок, которые наблюдались у юго-
восточных глухарей, населяющих чистые лиственные леса, у таежных глу-
харей не отмечено. Но непериодические отлеты глухарей из своих родных
мест и появление их даже на окраинах поселков отмечались неоднократно.
Совершенно не известен характер пребывания глухарей в светлой лист-
венничной тайге Средней и Восточной Сибири, а также в крупных листвен-
ных массивах без примеси хвойных пород.
Биотоп. Таежный глухарь населяет как светлохвойные (сосновые
и лиственничные), так и темнохвойные (еловые, пихтовые и кедровые)
леса, а также смешанные хвойно-лиственные. Таежный глухарь (так же, как
и юго-восточный) избегает молодняков и не живет в мелколесье, если оно рас-
положено вдали от крупного леса.
Численность. О плотности заселения глухарями различных таеж-
ных лесов известно крайне мало. Наиболее точный и сравнительно простой
способ учета глухарей на токах производился лишь в полосе переходных
темнохвойно-дубравных лесов в бывшей Гатчинской Охоте. Данные по учету
глухарей на участке, занятом этой охотой, указаны в таблице 4.
Таблица 4
Учет глухарей (самцов) на токах в Гатчинской Охоте в 1908 г.
Местность
Ландшафт прибал-
тийских хвойно-
лиственных ле-
сов, бывшая Гат-
чинская Охота
Площадь участка
(в гектарах), где
велся учет
общая
98 370
покрытая
лесом
Почти
равна
общей
площади
Количе-
ство
токов
45
Общее
количество
учтенных
глухарей
536
Количество
глухарей, при-
ходящееся на
1000 гектаров
покрытой лесом
площади
5,5
По чьим
данным
Диц
A911)
Незначительная плотность заселения глухарями Гатчинской Охоты веро-
ятно объясняется тем, что на территории этой охоты, кроме еловых и сосно-
вых лесов, было довольно много крупных моховых болот и березово-осино-
вого мелколесья.
На основании многолетних данных (Диц, 1911) выясняется, что в Гатчин-
ской Охоте (где ежегодно производился отстрел глухарей в количестве
9—18% от Btex токовиков) nocjie небольшого уменьшения численности токо-
вавших глухарей, происшедшего в 1896 г., наблюдался постепенный прирост
в течение пяти лет—с 1897 по 1901 г.; затем произошел значительный спад
в 1902 г., продолжавшийся и в 1903 г., и вновь постепенное увеличение в те-
чение трех лет (с 1903 по 1906г.). В течение следующих четырех лет было
двукратное падение численности—в 1907 и 1909 гг.
Экология. Образ жизни таежного глухаря значительно отличается от
образа жизни юго-восточного глухаря, особенно в сроках и продолжитель-
ности некоторых жизненных явлений.
Несомненно также и то, что существуют различия в образе жизни
таежных глухарей, населяющих различные части таежной полосы. Особенно
существенны должны быть различия между птицами, живущими в темно-
7 Птицы СССР, т. IV
ОТРЯД КУРИНЫЕ
хвойной и лиственничной тайге. К сожалению, о жизни глухарей, обитающих
в восточно-сибирской лиственничной тайге, почти ничего неизвестно.
Размножение. О сроках начала и конца весеннего токованья
глухарей, начала и конца вылета глухарей на тока в различных частях таеж-
ной полосы известно очень мало. Наблюдения над токованьем глухарей
в Лапландском и Печорском заповедниках указываются в таблице 5.
Наблюдений над сроками кладки и насиживания очень немного. Семе-
нов Тян-Шанский считает, что в Лапландском заповеднике глухарки на-
чинают насиживание около 1 июня и выводят 20—25 июня (следовательно,
примерно, на три недели позже, чем в южноуральских борах). Количества
яиц в кладках таежных глухарок меньше, чем у юго-восточных птиц. В Лап-
ландском заповеднике оно колеблется в пределах от 4 до 8 (среднее из 10—
6,6), а в Печорском—от 4 до 9 (среднее из 27—6,2). Часть яиц, как и у юго-
восточных глухарок,—болтуны.
О летнем токе таежных глухарей в литературе нет указаний.
Осеннее пение таежных глухарей, по данным Семенова Тян-Шанскога
A939) и Теплова A947), в Лапландском и Печорском заповедниках наблю-
далось очень редко.
Линька таежных глухарей наиболее полно описана Леннбергом
A927) (на скандинавских птицах).
У глухарей (как самцов, так и самок) линяет не только оперение, но и
клюв, когти и бахромки, оторачивающие пальцы.
Бахромки сбрасываются весною после тока и отрастают к осени. Зимок>
глухарь сидит на деревьях гораздо больше времени, чем летом, и поэтому
когти изнашиваются зимою сильнее, чем летом. За лето к осени отрастают
свежие, неизношенные вершины когтей, и когти становятся длиннее на 2 мм
(в среднем) по сравнению с весною.
Линька клюва протекает или частично, или же вся внешняя покрышка
сбрасывается целиком. Во время линьки клюва, проходящей летом, глухари
не едят грубой хвои и кормятся главным образом на земле.
В начале лета происходит частичная линька в летний наряд. Грант,
на которого ссылается Леннберг, описал его следующим образом. Бока головы
и шеи покрыты короткими черноватыми перьями со светлыми песочно-корич-
неватыми черточками. На подбородке и шее расположены короткие, закру-
гленные перья, слегка закрапленные серым. Летние перья, в отличие от зим-
них, не имеют добавочного стволика и менее пушисты.
В августе—сентябре проходит полная (зимняя) линька.
Питание таежного глухаря существенно отличается в различных
ландшафтах и ландшафтных районах тайги. В Лапландском заповеднике
(Семенов Тян-Шанский, 1939) глухарь питается зимою приблизительно на
85% сосновой хвоей и на 15% можжевеловыми побегами. В боровом районе
Печорского заповедника сосновая хвоя—основной, почти единственный вид
пищи глухаря с декабря по апрель (Теплов, 1947). В другом районе заповед-
ника—в районе темнохвойных печорских лесов—кедровая хвоя имеет для
глухаря такое же пищевое значение, как хвоя сосны в печорских борах.
В удмуртских елово-пихтовых лесах, между Камой и Вяткой, глухари зимой
и весной обходятся без сосны, питаясь преимущественно еловой хвоей (Пер-
шаков, 1937).
Хвоя составляет основу питания таежного глухаря и весною. По наб-
людениям Цедлица A929), шведские глухари до конца мая кормятся преиму-
щественно сосновой хвоей Она же—основная пища лапландских глухарей
(Семенов-Тян-Шанский, 1939); кроме того, лапландские глухари едят весной
побеги черники, побеги и листья голубики, вороники и подбела, почки ипобе-
ги березы и осины, листья ожики (Luzula pilosa) и соцветия пушицы (Erio-
phorum vaginatum). На Северном Урале, в Печорском заповеднике, глухари:
Таблица 5
Сроки начала и окончания весеннего токованья глухарей и вылета глухарок иа тока
Природная зона, ландшафт
и район
Лесная зона; таежная поло-
са; ландшафт лапландской
нагорной тайги; Лапланд-
ский заповедник ....
Лесная зона; таежная поло-
са; ландшафт Северно-
уральской нагорной тайги
Начало токования
на постоянных
самое
раннее
16/IV
?
самое
позднее
14/V
?
самцов
токах
среднее
27/IV*
?¦¦
Последнее токованье самцов
самое
раннее
20/V
15/V
самое
позднее
9/VI
«В конце
первой дека-
ды июня»
среднее
3/VI
сОколо
20 мая»
Начало вылета глухарок
на тока
самое
раннее
самое
позднее
среднее
Данных нет
28/IV
11/V
По чьим данным
Семенов-Тян-Шан
ский A939)
Теплое A947)
¦ Наблюдения, как указывал сам Тян-Шанский A939), велись нерегулярно, и потому точность их сомнительна.
¦¦ По словам Теплова A947), в марте и в две первые декады апреля ток еще не носит регулярного характера.
100 ОТРЯД КУРИНЫЕ
питаются весною преимущественно хвоей сосны и кедра и, кроме того, побе-
гами черники и подбела, хвоей можжевельника, бутонами пушицы, перези-
мовавшими ягодами брусники и толокнянки (Теплов, 1947).
Описание. Размеры и строение. Длина крыла самцов B6) 378—422,
самок A2) 300—320, среднее 396 и 310 мм. Вес самцов A70) 3470—4900, среднее 3980 г.
Вес скандинавских, лапландских и северноуральских глухарей (самцов) сходен
между собой, равняясь, в среднем 4 кг.
Окраска. Пуховик (по Хартерту, 1921) сверху ржаво-желтый, на передней
части головы черное пятно. Спина и крылья ржаво-бурые с черными неправильной формы
пестринами. Нижняя сторона сернисто-желтая с охристо-желтоватым налетом; на зобу
светлоржавая полоса.
Взрослые таежные глухари (самцы) хорошо отличаются от юго-восточных глухарей
темной окраской оперения. Брюхо и нижняя часть груди у северноевропейских, ураль-
ских и западносибирских глухарей очень темные с небольшим количеством белых пест-
рин. Окраска низа, по сравнению с юго-восточным глухарем, очень однообразна.
Сверху все таежные глухари (от Скандинавии до Восточной Сибири) гораздо темнее
юго-восточных светлосерых глухарей. Вместе с тем легко заметить, что у таежных глуха-
рей соотношение серого и черного цвета увеличивается по мере движения на восток и не-
сколько изменяется интенсивность бурой окраски верха.
Таежные глухарки также более темны, чем юго-восточные. Нижняя часть груди
и брюха не белеса, как у юго-восточных, а яркотрехцветна, так как белый и оранжевый
цвета почти «равноправны», а черные поперечины значительно шире, чем у юго-восточных.
Зоб яркооранжевый.
Систематические замечания. Среди таежных глухарей можно выделить несколько
местных изменений. Некоторые из них были описаны, как особые подвиды (см. ниже).
Переходная темнохвойно-лиственная полоса европейской части нашей страны и Запад-
ной Сибири населена глухарями, признаки которых переходны между таежным и юго-
восточным светлосерым глухарем, а на крайнем западе нашей страны—между таежным
и западноевропейским чернобрюхим глухарем. Среди этих глухарей также можно выделить
местные изменения (одно из них было описано Штегманом, как особый подвид 7\ и.
plesKet).
Чернобрюхий западноевропейский глухарь Tetrao urogallus
major Brehm
Tetrao urogallus major. Brehm. Handb. Nat. Vog. Deutschl. 1831, стр. 503, Средняя
Германия.
Местные изменения. Т. и. rudolfi. Domaniewski (карпатский глухарь).
Распространение. Ареал. Юго-западная окраина нашей страны—Во-
сточные Карпаты, Волынь, северная Украина, западная окраина Белоруссии
(к востоку по крайней мере до Беловежской пущи), Литва (на восток до Виль-
нюса); вне СССР область распространения его охватывает горйо-лесные
ландшафты Центральной Европы (Германии, Чехословакии, Польши, Фран-
ции) и некоторые лесные массивы на равнинах Польши и Германии.
Характер пребывания. В настоящее время чернобрюхий
западноевропейский глухарь ведет оседлый образ жизни (в прошлом, когда
Западная Европа была покрыта лесами, среди которых было много чистых
широколиственных массивов, глухари, может быть, совершали периодиче-
ские сезонные перекочевки, откочевывая на зиму из дубрав).
Биотоп. Населяет хвойные, хвойно-лиственные и кое-где чистые ши-
роколиственные леса.
Численность этого глухаря и плотность заселения им лесных
угодий очень малы (см. таблицу стр. 35); населяет еловые и пихтовые леса,
поднимаясь до верхней границы тайги и проникая отчасти в пояс криво-
лесья; высотное распространение его здесь от 800 до 1400—1500 м.
Малая плотность заселения глухарем Беловежской пущи объяснялась
главным образом разреживанием лесов вследствие их вырубания, уничто-
жением кладок кабанами и присутствием значительного количества людей
в пуще (особенно летом, во время сбора ягод) (Доманевский, 1933).
ГЛУХАРИ
101
Таблица 6
Природная зона (или
переходная полоса),
ландшафт и район
Лесная зона; полоса ев-
ропейских широколи-
ственных лесов; ланд-
шафт полесских дуб-
равно-хвойных лесов
и болот; Беловежская
пуща
Лесной вертикальный
пояс; ландшафт кар-
патских нагорных
хвойно-широколиствен-
ных лесов; Ясенское
лесничество
Площадь (в гек-
тарах) участка,
где велся учет
общая
125 820
покрытая
лесом
105 726*
15 000
Коли-
чество
токов
?
10
Общее
количе-
ство
учтенных
глухарей
100
20
Количество
глухарей на
1000 гекта-
ров покры-
той лесом
площади
0,95
1,3
Год
учета
1928
По чьим
данным
Дилев-
ский**
Доманев-
ский
A933)
В Германии плотность населения глухарей также очень мала, и перед
началом второй мировой войны отстрел глухарей во всей стране равнялся
в среднем 1410 особям в год (Нитхаммер, 1942).
Данные по учету глухарей на токах показывают, что плотность населе-
ния глухарей в густо населенных людьми странах Западной и Центральной
Европы как в горных, так и в равнинных лесах значительно уступает плот-
ности населения и таежного и юго-восточного светлосерого глухаря (плотность
населения последнего даже в лесостепных дубравах южной оконечности
Урала в несколько раз выше плотности глухарей в Беловежской пуще и на
Карпатах).
Экология. Размножение. Сроки начала и конца токования разли-
чаются довольно сильно в нагорных и равнинных районах, но длительных,
многолетних наблюдений над сроками токования западного глухаря в нашей
стране, повидимому, не производилось (или они не были опубликованы).
Однажды в предгорьях Альп, в окрестностях Вены, глухарь был убит на
току 19 февраля, но все же тока в феврале начинаются лишь в исключительные
годы (Вурм, 1885). Во Французской и Швейцарской Юре, по предположению
Энара и Мейлана A935), тока заканчиваются между 20 и 25 мая. На терри-
тории Советских Карпат ток, по сообщению Страутмана, начинается в начале
апреля и длится до середины мая, но в некоторые годы, при очень ранней
весне, тока бывают уже в середине марта; так, в 1950 г. первые токующие глу-
хари отмечены 13—14 марта. Глухарки начинают готовить гнездо задолго до
окончания тока и приступают к откладке яиц. В Советских Карпатах (Рахов-
ское лесничество) неполные кладки находили в первой половине мая; птенцов
встречают иногда в конце мая, а чаще в начале июня: 14 июня 1950 г. добыты
глухарята величиной с перепелку (Страутман).
В Германии (Нитхаммер, 1942) полные кладки находили между середи
ной апреля и началом июня (в зависимости от погоды и высоты местности)
* По съемке 1890 г. (Энциклопедический словарь Брокгауза и Ефрона, 1891).
** Ссылка на работу Дилевского по Доманевскому A933).
102 отряд куриные
О летних токах имеется лишь упоминание у Вурма A885) о том, что
в Альпах один наблюдатель слыхал летом, начиная с 4 августа, токовавших
глухарей, и что в 1879 г., за время с 4 по 22 августа на токах в Тюрингии
были убиты 4 глухаря. В противоположность таежным и юго-восточным
глухарям, чернобрюхие центральноевропейские глухари цередко поют в теп-
лые зимы, которые довольно часто бывают в Центральной Европе (Вурм,
1885).
Количество яиц в кладках чернобрюхого глухаря колеблется в следую-
щих пределах. В Галиции и Буковине (Завадский, 1840) в гнездах глухарок
находили от 6 до 16 яиц, такие же данные указывает для Карпат Водзицкий
A853); по данным Альтума A873), в Германии глухарки несут от 5 до 12 яиц,
иногда даже 16; в Австрии (Вурм, 1885) глухарки чаще всего кладут от 8 до
12 яиц, иногда даже 161.
Сопоставляя эти данные с наблюдениями над кладками южноуральских
глухарок, можно сделать вывод, что предельные наибольшие и наименьшие
количества яиц в кладках южноуральских и среднеевропейских глухарок
равны. О том, что количество яиц в кладке зависит от возраста, сообщал Вурм
A885), заметивший, что в Австрии молодые глухарки кладут 6—8 яиц, в то
время как взрослые несут чаще всего 8—12 яиц.
Линька. Неполная летняя линька взрослых проходит в мае или
июне, а окончательная полная линька длится (в условиях Германии) с июля
до сентября—октября. Как и у таежных и юго-восточных глухарей, роговые
пластинки, оторачивающие пальцы, сбрасываются после тока и отрастают
вновь осенью. Летом линяют когти и клюв. При линьке глухарят из первого
наряда в окончательный первыми (в возрасте 4 недель) появляются вну-
тренние маховые, а в возрасте 4—5 недель рулевые; в возрасте 6—7 недель
появляются зеленые перья на груди. При смене первого наряда на оконча-
тельный два крайних внешних маховых остаются, не сменяясь (Нитхам-
мер, 1942).
Питание большого глухаря изучено недостаточно, сведения о его
питании даются в самых общих чертах. Указывалось (Вурм, 1885), что зимой
и во время весеннего токования чернобрюхий глухарь питается преимущест-
венно хвоей (чаще всего сосновой). Кроме того, зимою глухари (особенно
глухарки) едят почки и сережки бука, ивы, березы, ольхи, орешника (Вурм,
там же).
В желудках двух глухарей, добытых Щербиной A915) на току в Волын-
ском Полесье, оказались главным образом остатки сосновой хвои; кроме
того, были обнаружены остатки перезимовавших ягод черемухи и клюквы
и ольховых почек. Летом питаются различными ягодами (земляникой, черни-
кой, брусникой, клюквой, малиной, рябиной), листьями клевера, одуванчика
и некоторых других растений и беспозвоночными животными. Осенью боль-
шое пищевое значение имеют жолуди и буковые орешки. У птиц, добытых
в конце апреля на току Страутманом (Советские Карпаты), в зобу обнаруже-
ны лишь кучки еловой хвои, оторванные вместе с кончиками веток, почки
и просто хвоя, а в желудках—хвоя ели на разных стадиях переваривания
и много хорошо окатанных белых кварцевых гастролитов.
Описание. Размеры и строение. Длина крыла самцов C) 405—415 сред-
нее 410 мм. Вес самцов (Померания, Цедлиц, 1924) наибольший 6500, средний 5500 г.
Окраска. Первый наряд после пухового напоминает наряд самки (как и у дру-
гих подвидов). J *J
Для взрослых чернобрюхих западноевропейских глухарей характерно резкое пре-
обладание чисто черного цвета на брюхе и нижней части груди. Сверху западноевропей-
ские глухари также очень темны. Западноевропейским глухарям свойствен более крупный
1 Хартерт A921—1922) и вслед за ним Нитхаммер A942) предполагали, что 16 яиц
несет не одна глухарка, а две, откладывающие яйца в одно гнездо* но нет никаких осно-
вааний согласиться с этим предположением.
КАМЕННЫЙ ГЛУХАРЬ
103
вес, чем юго-восточным и таежным глухарям. Впрочем, о последнем признаке следует
сделать оговорку, так как неясно, свойствен ли крупный вес лишь отдельным местным
изменениям или характерен для этого подвида в целом. Карпатское изменение чернобрю-
хого западноевропейского глухаря было описано как особый подвид G\ и. rudolfi D о т.).
6. Каменный глухарь Tetrao urogalloides Middend.
Tetrao urogalloides. Mi dde ndorf. Sibirische Reise, 1851. II, 2, стр. 195, табл. XVIII,
Становой хребет
Синонимы. Tetrao parvirostris. Bonaparte. Compt. Rend. Acad. Paris, 1856,
т. XII, стр. 880, Становой хребет.
Русское название. Каменный глухарь; было известно еще Мессершмидту,
на которого ссылается Паллас A811)—cKussis Kamenoi gluchar vocatur».
Распространение. А р е а л. В пределах нашей страны населаяет лесную
зону и лесотундру Восточной Сибири, Дальнего Востока, Камчатки и Саха-
лина; в Южном Забайкалье встречается в нагорной лесостепи. За пределами
Карта 7. Распространение каменного глухаря Tetrao
urogalloides
1—Т. и. urogalloides, 2—Т. и. taczanowskii: а—граница гневдовий
нашей страны населяет нагорно-лесные районы Северной Монголии и Маньч-
журии. Тачановский A893) ссылается на сообщение Давида A877) о том, что
в горно-лесных районах Северного Китая (во время пребывания там Давида)
изредка встречались каменные глухари и что одну глухарку ему прислали
оттуда в декабре 1864 г.
Наиболее западные местонахождения каменного глухаря известны из
следующих мест: хребет Хангай—верховье р. Орхон (Козлова, 1933), сред-
нее течение р. Оки (Радде, 1863), северо-западное побережье Байкала, у мыса
Котельникова (Штегман, 1936) среднее течение Нижней Тунгуски—от
д. Ербогачена до устья р. Туры (Ткаченко, 1924), р. Кочечумо—приток р. Ту-
ры (сборы Наумова, 1929, Зоологический музей Московского государствен-
ного университета), р. Курейка—приток Енисея (Штегман, 1936).
На значительном пространстве заенисейской Сибири живут оба глухаря—
каменный и обыкновенный. Отношения этих видов глухарей друг к другу
особенности их образа жизни там, где они встречаются вместе не изучены.
104 ОТРЯД КУРИНЫЕ
Северная граница распространения каменного глухаря изучена слабо.
Известны следующие наиболее северные местонахождения: верховье р. По-
пигай (Тугаринов, Смирнов, Иванов, 1934), с. Говорово на Лене (упомяну-
тые выше авторы, 1934), Крест-Майор на Индигирке (Михель, 1935), с. Ниж-
неколымск (Тугаринов, Смирнов, Иванов, 1934), верховья рек Анадыря и Пен-
жины (Портенко, 1939).
Наиболее южные местонахождения расположены за пределами нашей
страны—хребты Хангай и Кентэй в Монголии, Большой Хинган в Маньчжу-
рии.
Характер пребывания. Оседлая птица, перемещающаяся
в пределах небольшого района.
Биотоп. Каменный глухарь, повидимому, чаще всего встречается
в лиственничных лесах, но не избегает сосновых и кедровых, а на Восточной
Камчатке (Аверин, 1948) населяет преимущественно светлые парковые леса
из каменной березы и (реже) лиственничники.
Подвиды и варьирующие признаки. У каменного глухаря довольно резки различия
в окраске оперения, свойственные взрослым особям из различных частей области распро-
странения этого вида. Направление изменчивости окраски оперения каменного глухаря
в основном сходно с направлением изменчивости обыкновенного глухаря (Tetrao urogal-
lus), а именно: очень темная окраска оперения, свойственная одному подвиду каменного
глухаря, переходит в светлосерую окраску со значительным развитием белых пестрин,
свойственную другому подвиду.
Основываясь на различиях в окраске оперения и отчасти на различиях в длине хво-
ста, было описано несколько подвидов каменного глухаря, но большинство из них, по на-
шему мнению, не подвиды, а местные изменения (часть была описана необоснованно).
У каменного глухаря только два подвида—населяющий Камчатку—Т. и. kamtschaticus
Kittlitz, 1858, и занимающий весь остальной ареал вида—Т. и. urogalloides Mi d d e n-
dorff, 1851.
Камчатский каменный глухарь Tetrao urogalloides kamtschaticus К i 111 i t ъ
Tetrao kamtschaticus. К i t t 1 i t z. Denkw. Reis. Russ. Amer, II, 1858, стр. 353, Камчатка.
Распространение. Ареал. Камчатка.
Характер пребывания. Летом встречается как в лесах, так
и в кустарниковом поясе, но на зиму в ноябре покидает последний и спускает-
ся в березовые леса (Аверин, 1948).
Биотоп. В восточной части Камчатки (в Кроноцком заповеднике)
населяет преимущественно сухие и светлые леса из каменной березы и реже—
лиственничники. Предпочитаются леса на спокойном рельефе широких
водоразделов, крутые и короткие склоны долин малых речек избегаются
(Аверин).
Численность. Немногочисленная птица.
Экология. Размножение. О размножении (так же как о питании
и остальных чертах образа жизни камчатского глухаря) известно очень ма-
ло. Токованье начинается, повидимому, в начале апреля и заканчивается
в начале июня (Аверин). Глухарь токует от рассвета до 9—12 час. дня,
начиная токованье на деревьях, а затем обычно слетает на землю, как и
обыкновенные глухари (Т. urogallus). Песня состоит только из щелканья
сначала редкого, затем все учащающегося и переходящего в своеобразную
трель. Ею заканчивается одна песня и после короткого перерыва начина-
ется новая.
Есть ли летнее, осеннее и зимнее токованье,—неизвестно. Почти ничего
неизвестно и о гнездованье. Аверин встретил в Кроноцком заповеднике за
несколько лет лишь двух глухарок с глухарятами—27 июля и 28 августа.
Линька. Не изучена.
Питание камчатского глухаря изучено слабо. Зимою, по наблю-
дениям Аверина, питается преимущественно сережками, почками и кон-
КАМЕННЫЙ ГЛУХАРЬ
105
чиками ветвей каменной березы. Почки и побеги ивы и почки ольхового
стланца играют очень малую роль в питании камчатского глухаря. У глуха-
рей, убитых на весенних токах в апреле и мае, были найдены почки каменной
березы, ягоды можжевельника, перезимовавшие ягоды брусники и вороники,
стебли хвоща, листья голубики и травянистая растительность (виды растений
не определены). В зобах и желудках глухарки и двух глухарят, добытых
летом в Кроноцком заповеднике были обнаружены ягоды и семена вороники
Рис. 17. Каменный глухарь
(Empetrum nigrum), перезимовавшие ягоды брусники, листья ивы, голубики,
жимолости, цветы брусники и некоторых других растений, гусеницы, мошки.
Полевые признаки. Крупная птица, лишь немного уступающая обыкно-
венному глухарю. «Песня» иная, чем у обыкновенного глухаря, состоящая
из одного щелканья.
Описание. Размеры и строение. Длина крыла самцов D) 382—402; сред-
нее 396 мм. Вес самцов D) 3445—3800, среднее 3640 г.
Окраска. Маленькие, едва перепархивающие каменные глухарята (весом 40 г)
хорошо отличаются от обыкновенных глухарят, в том числе и от светлосерых юго-восточ-
ных. Низ каменных глухарят желтый с сернистым оттенком. Спина буровато-желтая, бо-
лее темная, чем у юго-восточных глухарят. Первостепенные маховые светлобуроватые
с частыми более темными пестринками. На вершинах второстепенных маховых широкие
белые каймы, образующие две светлые перевязки. Голова оранжево-бурая.
Взрослый самец. По степени развития светлосерой и белой окраски камчатский глу-
харь напоминает юго-восточного светлосерого глухаря (Т. и. uralensis), но в отличие от
последнего у камчатского глухаря светлосерая окраска и белые пестрины расположены
преимущественно на верхней части тела, а не на нижней. Спина камчатского глухаря (сам-
106
ОТРЯД КУРИНЫЕ
ца) светлосерая с небольшой приме-
сью желтовато-бурого налета между
лопатками и на верхних кроющих.
Второстепенные маховые С белыми
широкими D—6 мм) вершинами; на
вершинах верхних кроющих крыла
широкие белые треугольники, обра-
зующие широкую белую перевязь;
вторая белая перевязка—из белых
вершин второстепенных маховых. Го-
лова и шея черно-серые. Низ пепель-
но-серый с редкими мелкими белыми
пестринками. У самок также белая
окраска сильнее всего развита на
крыльях: широкие D—5 мм) белые
вершины верхних кроющих крыла
еще шире —до 1 см; на подхвостье
также много белого. Поперечная рябь
нижней части тела (брюха и груди)
трехцветна: черные и желтые попе-
речные полоски почти равны, а бе-
лые немного уже.
Темный каменный глухарь Tetrao
urogalloides urogalloides
Mi ddendorff
Рис. 18. Пуховой птенец каменного глухаря Синонимы. Tetrao parvirostris
macrurus. Stegmann. Доклады
Академии наук СССР, серия А, стр. 231, 1926. — Tetrao urogalloides kolymensis. В u-
t u г 1 i n. Alauda, № 3, 1932, стр. 269, район Верхне-Колымска.
Местные изм енения. Т. и. v. sachalinensis. Bogdanov. Consp. Av. Imp.
Ross.; I, 1884, стр. 122, Сахалин.—Г. и. j an en sis. Tugari nov. Ornith. Monatsb., 1922,
район Верхоянска.—Г. и. turensis. Buturlin. Alauda, № 3, 1932, стр. 269, низовья
рек Туры и Кочечумо.
Распространение. Ареал. Совпадает с ареалом вида, за исключением
Камчатки.
Характер пребывания. Оседлая птица.
Биотоп. Темный каменный глухарь, по наблюдениям Годлевского
(Тачановский, 1898) и Павлова A948), живет в Южном Забайкалье преимуще-
ственно в лиственных лесах, но не избегает сосновых и кедровых, а на Кентее
(Козлова, 1933) встречается и в смешанных лесах, и в борах, и в кедровни-
ках. В бассейне р. Анадыря по сведениям, собранным Портенко A939), зимой
встречается в лиственничниках.
Численность. Плотность населения темного глухаря очень мала.
Некоторые из исследователей (Шульпин, 1936; Павлов, 1948) объясняют это
лесными пожарами, уничтожающими хвойные леса. По наблюдениям Павло-
ва в Читинской области, 1940 г., когда было много весенних лесных пожаров,
отличался особенно малым количеством глухариных выводков.
Экология. Почти совсем не изучена.
Размножение. Песня каменного глухаря резко отличается от пе-
сни обыкновенного глухаря {Tetrao urogallus) и состоит из щелканья (см.
описание песни камчатского глухаря). На Кентэе, по наблюдениям Козловой,
ток начинается в апреле и продолжается до 10—12 июня. В каком возрасте
начинают токовать каменные глухари, неизвестно. Осенью забайкальские
глухари поют утром и вечером (Тачановский). Токуют ли каменные глухари
летом* неизвестно. В Забайкалье глухарки начинают нестись в конце мая,
кладка состоит из 5—8 яиц (Тачановский, 1873 и 1893). Гнезда любят
устраивать под вершиной срубленного дерева.
ДИКУША 107
Л и н ь к а. Не изучена. Частичная линька происходит летом, как у обы-
кновенного глухаря.
Питание. По наблюдениям Годлевского (Тачановский, 1893),
главнейший зимний корм каменного глухаря в Юго-Восточном Забайкалье—
почки лиственницы. В зобу глухарки, добытой зимою в нижнем течении Аму-
ра, обнаружены почки березы и побеги лиственницы (Воробьев, 1951). Осенью
и в начале зимы глухарь любит кормиться ягодами брусники и достает их
из под снега (Радде, 1863). В связи с этим он осенью чаще всего встречается
в тех лесах, где есть брусника. В зобах двух глухарок, добытых в Читинском
округе 16 и 18 июля 1938 г., были обнаружены ягоды голубики, муравьи
и их яйца.
Описание. Размеры и строение. Длина крыла самцов A5) 300—310,
среднее 348,2 мм. Вес самцов C)—2695, 3230 и 4000 г.
Окраска. Первый пуховой наряд не описан. Молодые глухари, сменившие пухо-
вой наряд, в общем похожи на взрослых. Весь низ их в поперечных полосах—черных,
белесых и светловатожелтых.
Взрослый самец хорошо отличается от камчатского каменного глухаря более темной
окраской. Спинная сторона самцов этого подвида темносерая с узкими A—2 мм) белыми
вершинами второстепенных маховых и редкими каплевидными белыми пятнами на верх-
них кроющих крыла; брюшная сторона почти сплошь черная. Самки имеют на брюшной
стороне тела белые узкие поперечные полосы; шире их—черные полосы и самые широкие—
желтые.
Замечания по систематике. Среди темных каменных глухарей можно выделить
местные изменения. Некоторые из них были описаны как особые подвиды (см. выше).
7. Дикуша Falcipennis falcipennis Hartlaub
Tetrao falcipennis. Hartlaub. Journ. fur Orn. (III). 1855, стр. 39, Становой хребет.
Синонимы. Falcipennis falcipennis muratai. Momiyama. Annot. Orn. Orientalis, I,
1928, стр. 234. Южный Сахалин.
Русские названия. Дикуша, черный рябчик, иногда в Приморье—«смиренный
рябчик». Все названия местные, охотничьи; дикуша—от крайней доверчивости и неосто-
рожности птицы.
Распространение. Ареал. Восточные части Забайкалья, южные райо-
ны Якутии, Охотское побережье, бассейн Амура, Уссурийский край, Саха-
лин. Границы ареала приблизительно такие: на западе по южному склону
Станового хребта почти до среднего течения р. Шилки (Шульпин, 1936; Во-
робьев, 1938); на севере—до верховьев Олекмы, Алдана, Уяны и, вероятно,
Май и Юдомы (Бутурлин, 1935); на востоке до берегов Охотского моря, но не
севернее Аянского района (Розов, 1938), низовьев Амура; на юге до Южного
Сахалина, верховьев рек Имана, Тетюхе и вероятно даже Малазы в бассейне
Сучана (Шульпин, 1936). К западу южная граница не прослежена.
Будучи связана с определенными типами леса, дикуша внутри ареала
распространена неравномерно. Детали распространения таковы. По тракту
от Якутска на Удск Миддендорф впервые встретил ее в северных отрогах Ста-
нового хребта на р. Уян; часто встречалась ему на восточных склонах этих
гор и в окрестностях Удска. На север отсюда дикуша добыта у Аяна; по
сведениям, полученным от населения, встречается по тропе между Якутском
и Охотском, а также по наиболее глухим частям Станового и Янканского
хребтов и отчасти хребта Тукурингра. Добыта на Становом хребте в верховьях
Зеи, встречена в верховьях р. Безымянной (левый приток Ольдоя); по южно-
му склону Станового хребта доходит почти до Шилки, встречается по Тикси,
по истокам Киле и по среднему течению Ура, но на реках Лима и Ольдоя не
наблюдается. По расспросным же данным встречается и в истоках Ольдоя
(Маак, 1859) и даже в северо-восточных истоках р. Горбицы (приток Ама-
зара). К востоку от Станового хребта дикуша добывалась в долине р. Бама
(системы р. Норы), в хребте Джагды, в верхнем течении р. Амгуни, в окре-
108
ОТРЯД КУРИНЫЕ
стностях Николаевска, в верховьях р. Кура в южной части Буреинского
хребта. По расспросам встречается также в бассейне Горина и Хунгари.
Обычна в Тунгуро-Чумиканском районе, по р. Горин и в бассейне озера
Эворон; многочисленна по рекам Немилан и Керби, а также в районе озера
Кизи и на побережье залива де-Кастри. На Сихотэ-Алине по Ботчи и Копи
встречается почти от устья этих рек (Емельянов, 1929); в Сихотэ-Алиньском
заповеднике нередка в высокогорном поясе среднего и верхнего течения рек
Та-Кемы и Колумбэ (западные склоны Сихотэ-Алиня) на высоте от 700—
800 до 1500 м над уровнем моря. Найдена в истоках р. Бегаму (бассейн Бики-
на) и в самых верховьях Имана; по расспросам встречается в верховьях
р. Тетюхе и даже в верховьях р. Малазы
в бассейне Сучана.
Вне пределов СССР не встречается,
за исключением, может быть, погранич-
ных с Амуром районов Китая.
Характер пребывания.
Оседлая птица. В горных районах со-
вершает незначительные вертикальные
сезонные кочевки, детально по датам
не прослеженные. В верховьях Тетюхе
и Иодзыхе осенью спускается несколь-
ко ниже по хребту и концентрируется
в местах, богатых ягодами, появляясь
на Брусничной сопкевШубинской пади,
где нормально не живет летом (Шуль-
пин, 1936). В высокогорной части Сихотэ-
Алиня, где летом обычна, зимой не на-
блюдается (Капланов, 1938).
Биотоп. Ограничен зоной охот-
ской растительности, с общей террито-
рией которой совпадает распростране-
ние дикуши. Гнездовой—елово-пихто-
вые леса, ельники, лиственничники, перемежающиеся мелкими еловыми
и лиственными чащами и полянками. Всюду для стаций дикуши характер-
но сочетание тенистых и сырых участков леса с прогалинами и полянками,
густой моховой покров почвы, обилие ягод: голубики, вороницы, морошки,
дикой смородины и др. (Шульпин, 1936). В Сихотэ-Алиньском заповеднике
в гнездовое время дикуши обитают по гольцам, занимаемым участками высо-
когорной тундры у верхней границы лесной растительности, по высокогон-
ным и горным мшистым ельникам, по гарям, покрытым осиново-березовыми
лесами (старые гари), а также по скалистым хребтам, поросшим дубовым ле-
сом с даурской березой. При этом для стаций дикуши характерны обильные
брусничники, криволесье из каменной березы, березовый стланец, большие
заросли (Pinus pumila, Rododendron chrysanthum, бадана и других высокогор-
ных растений (Капланов, 1938).
Численность. В большинстве районов ареала невысокая, хотя
в ряде мест (например, в Охотско-Аянской тайге) может считаться обычной
птицей.
Экология Размножение. Изучено слабо. Брачные игры в местах
гнездовья—в мае. Токуют одиночками. Капланов A938) наблюдал ток дику-
ши 17 мая 1937 г. в Сихотэ-Алиньском заповеднике: «Самец сидел на дереве
распушившись и принимал позу токующего глухаря, приподняв вверх рас-
крытый веером хвост и голову (голова была запрокинута менее круто, чем
у глухаря во время тока), на дереве сидел беззвучно, лишь хвостом издавал
шелест при поднимании и раскрывании его; затем слетал на землю, распушал-
Карта 8. Распространение дикуши Fal-
cipennis falcipennis
а—граница гнездовий, б—не вполне выяснен-
ная граница гнездовий
ДИКУША 109
ся, и, издав протяжный вибрирующий звук вроде «у-у-у-р-р-р», подлетал сан-
тиметров на 20—30, издавая при этом двойное щелканье, вроде глухариного.
Это повторялось несколько раз подряд через равные промежутки времени—
минуты через две-три. Сперва протяжный звук, затем подскакивание на
месте и одновременно двойное щелканье. Невдалеке на дереве сидела самка».
Ток происходил на горелом гольце, поросшем Ledum decumlens, Betula
middendorfiy баданом и обильно брусникой, на высоте 1500 м над уровнем моря.
Сходны наблюдения Миддендорфа, который видел эту птицу токующей
на земле среди небольшой лесной прогалины. Не более как в 15 шагах от него,
на открытой поляне, самец важно выступал, как индюк, распускал и волочил
по земле хвост, моргал побагровевшими глазами, щелкал и по временам даже
фыркал. Затем наступала пауза, в продолжение которой самцы других ку-
риных пород обыкновенно бывают очень осторожны, но самец дикуши под-
пустил наблюдателя на несколько шагов, прежде чем вспорхнул. По на-
блюдениям на Южном Сахалине, самцы токуют на пнях, распустив крылья
и издавая звук вроде «коо» или «куу» (Така-Тсуказа, 1935).
Срок наступления половой зрелости не выяснен, но, видимо, не отли-
чается от других видов семейства тетеревиных.
Весной птицы, повидимому, разбиваются на пары, но самцы не прини-
мают участия в вождении птенцов и все лето держатся обособленно.
Период размножения—между маем и июлем. Гнездо делается на земле,
у подножья деревьев. В гнезде, найденном на Южном Сахалине 8 июня 1934 г.
в густом еловом лесу, были насиженные яйца. Само гнездо помещалось среди
мха, кустиков Vaccinium chamissois и Vaccinium Smallii. Гнездо крайне
простого устройства—ямка в земле на небольшой кочке, выстланная веточ-
ками хвойных деревьев и Vaccinium chamissois. В гнезде было 8 яиц; окра-
ска бледнобурая с оливковым оттенком, по всей поверхности скорлупы мел-
кие бурые пестрины; размеры 48x32; 48,5x32; 46x32; 45x32; 46x32; 48х
Х31,5; 46x32; 43x31 мм (Така-Тсуказа, 1935).
Маленький птенец при самке встречен в верховьях Та-Кунжи на Сихотэ-
Алине 25 июня (Капланов, 1938). Выводок из 8 молодых величиною с серую
куропатку и хорошо летавших, наблюдался в бухте Таба 4 августа. Вывод-
ки предпочитают держаться опушек травянистых полян (Шульпин, 1936).
Подобно тетеревам, у дикуши бывает осеннее токованье. 31 августа
в верховьях Колумбэ (Сихотэ-Алиньский заповедник) Капланов A938) на-
блюдал под пологом хвойного леса на земле осенний ток дикуши, который
очень напоминает весенний ток.
Линька происходит, повидимому, в период июнь—сентябрь, причем
у самцов раньше, чем у самок. Добытый 29 июня самец нес следы интенсив-
ной линьки всего мелкого пера (с кроющими крыла), за исключением перьев
и верхней части шеи (Шульпин, 1936). У добытых 24 и 25 августа в районе
озера Кизи (Амурский край) двух самцов оперение вылиняло, за исключе-
нием верха головы и горла: перья на этих частях тела наполовину вышли
из пеньков. Третий самец, добытый там же в августе (точно дата неизвестна),
следов линьки оперения не имел; наблюдалась лишь линька клюва. Две
самки из тех же мест, добытые 24 августа, несли следы линьки мелкого опе-
рения спины; кроме того, у одной самки линяли рулевые перья, достигшие
половины длины нормальных, а у другой перья горла и клюв (коллекции
Зоологического музея Московского государственного университета). По
Шренку A861), у экземпляров, добытых 1 октября и даже 17 сентября
осенняя линька была закончена.
Питание. Как и другие тетеревиные, кормится преимущественно расти-
тельной пищей. В 11 желудках и зобах, добытых в конце августа, найдены
были почти исключительно растительные остатки и только в двух отмечены
насекомые: одна кобылка (Acrididae) и один муравей (Camponotus). Из ра-
по
ОТРЯД КУРИНЫЕ
стительных остатков главную массу составляла хвоя лиственницы (Larix
dahurica), затем ягоды черники, малины, семена костяники (Rubus saxatilis)
и колоски осоки (Сагех lasicarpa). Во всех желудках имелись камешки,
общим весом от 2,5 до 13 г в одном желудке, в большинстве же 8—12 г. Вес
самых крупных камней 250 мг. У некоторых птиц обнаружены камни и в зобу
(Воробьев, 1938). В зобу самца, добытого 29 июня в верховьях Имана, 75%
по массе—листья брусники и 25% —крупная и жесткая хвоя пихты {Abies
nephrolepis); в желудке 30%—белые блестящие кварцевые камешки, 65%—
Рис. 19. Дикуша
хвоя лиственницы и 5%—остатки жуков, 1 гусеница и 1 крупный черный
муравей. У самца от 3 августа из бухты Таба в полном зобу 90 %—хвоя листвен-
ницы, часть целыми пучками, и 10%—незрелые ягоды морошки; в желудке:
60%—хвоя лиственницы, 15%—незрелые ягоды клюквы и 25%—масса мелких
камней (Шульпин, 1936).
В желудках трех добытых в летнее время в Сихотэ-Алиньском заповед-
нике птиц содержалась главным образом брусника и меньше—хвоя белокорой
пихты (Капланов, 1938). По устному сообщению Воробьева, зимние экзем-
пляры, доставленные ему из бассейна Нижнего Амура, имели в желудках
исключительно хвою пихты. Зоб одной из птиц был туго набит хвоей пихты,
общий вес которой достигал 56 г. Молодые кормятся преимущественно насе
ДИКУША
111
комыми, подросшие переходят на ягодные корма (Така-Тсуказа, 1935). У трех
птенцов от 5 августа из бухты Таба, величиной с маленькую серую куропатку,
в желудках только розово-красная масса с остатками оболочек от ягод воро-
ницы (Empetrum nigrum), а в зобу у двух экземпляров немного хвои пихты
(Шульпин, 1936). Обилие хвои, притом в летнее время и даже у молодых
птиц, сближает дикушу до характеру питания с глухарем.
Полевые признаки. По сложению и общему поведению представляет
нечто среднее между тетеревом и рябчиком, но ближе к последнему. Как
и рябчик, искусно скрывается в ветвях хвойных деревьев, неподвижно сидя
на одном месте. От рябчика отличается более крупными размерами и темной
окраской. Малоподвижная и скрытная птица, держащаяся в чаще и в диких
таежных уголках на земле или
на нижних ветвях деревьев до
2—3 м высоты.
Вследствие малой подвиж-
ности и темной окраски мало
заметна. Потревоженная не под-
нимает шума, не стремится
скрыться, а взлетает на ниж-
нюю ветку дерева и наблюдает
виновника тревоги, продолжая
сидеть даже в 3—5 шагах от че-
ловека. Дикуша известна уди-
вительной доверчивостью, отсут-
ствием осторожности и боязни
человека, за что у населения
Приморья получила широко рас-
пространенное название «сми-
ренного рябчика» (Шульпин,
1936). Снимание птицы петлей
с веток—обычный способ охоты у местного промыслового населения. Свиста
и звуков, подобных рябчику, дикуша не издает. Самка дикуши при тревоге
тихо и довольно грубо квохчет подобно самке тетерева-косача, но менее
громко (Капланов, 1938).
Промысловое значение невелико вследствие малочисленности, спорадич-
ности распространения и обитания в отдалении от населенных мест, в трудно
доступных лесных массивах. Кроме снимания петлями, добывают ружьем
В летнее время (июнь—август) мясо дикуши бело и приятно на вкус, но более
жестко и горьковато, чем мясо рябчика (Шульпин, 1936).
Описание. Размеры и строение. Крыло относительно короткое; наруж-
ные первостепенные маховые короткие, суженные на концах, серповидной формы. Фор-
мула крыла: первое первостепенное маховое короткое, немного длиннее восьмого и короче
седьмого; второе, третье, четвертое, пятое и шестое примерно равной длины и образуют
вершину крыла. Хвост закругленный, из 16 рулевых. Цевка длинная, оперена до пальцев;
особенно развиты перья в нижней части цевки: они длинные и прикрывают пальцы. Пальцы
голые с характерной для тетеревиных бахромой по сторонам. Средний палец, измеренный
без когтя, примерно равен длине плюсны. Величиной крупнее рябчика, но меньше тетере-
ва; вес около 600 г. Крыло 175—200 мм, хвост 100—135 мм, плюсна 32—38 мм, длина
клюва (от ноздрей) 9,5—12 мм, высота клюва 6—7 мм.
Окраска. Летний наряд. Самец. Общая окраска оперения значительно темнее,
чем у обыкновенного рябчика. Основной фон каштаново-черный, на котором раз-
бросаны серые, белые, бледноохристые и рыжеватые пестрины и пятна, обусловливающие
различия в окраске разных частей оперения. Темя и затылок черные, с едва заметными
сероватыми поперечными пестринами, более светлыми у основания клюва. Верхняя часть
спины такого же цвета, испещрена редкими светлыми вершинными поперечными каемками.
Остальная часть верхней стороны тела несколько светлее за счет более частых охристых
поперечных и белых стержневых пестрин. Последние более развиты на концах плечевых
на пояснице. Бока головы и шея с белыми и светлоохристыми пятнами. Зоб и грудь от
Рис. 20. Пуховой птенец дикуши
112
ОТРЯД КУРИНЫЕ
темнокаштанового до черного цвета, без пестрин. Остальная часть низа (бока и брюхо)
окрашены наиболее светло благодаря наличию на перьях широких белых поперечных пред-
вершинных полос. Махи черно-бурые, рули—тоже, но кроме средней пары с широкими
белыми вершинными полосами; нижние кроющие хвоста с белыми вершинами. Клюв
черный. Радужина темная желтовато-бурая.
Самка. Окрашена значительно светлее самца благодаря более развитым светлым
пестринам и пятнам как на нижней, так и на верхней сторонах тела. В светлых пестринах
и пятнах хорошо развиты рыжеватые тона, в том числе на груди и горле. Верх головы
и передняя часть спины с широкими охристо-рыжими поперечными полосами. На осталь-
ной части верхней стороны тела рыжий цвет развит сильнее, чем у самцов. Шея спереди
в серых пестринах. Зоб и грудь с охристыми и темными поперечными полосами. Бока тела—
рыжеватого цвета.
Зимний наряд. Различий в окраске от летнего наряда незаметно. Цевка оперена
более густо; волосовидные перья длинные, достигают концов пальцев.
Молодой самец. Спинная сторона тела с рыжевато-бурыми пятнами и светло
охристыми наствольями. На брюшной стороне тела белые пятна.
Систематические замечания. Выделение двух географических рас дикуши: материко-
вой (F. /. falcipennis Н а г t.) и сахалинской (F. /. muratai Momiyaraa), основанное
якобы на небольших различиях в размерах и окраске (Бутурлин, 1935), едва ли можно
признать достоверным. Более поздние данные о размерах хвоста и цевки сахалинских
птиц (Така-Тсуказа, 1935) свидетельствуют о том, что они ничем не отличаются от разме-
ров материковых птиц. Приводимые различия в окраске настолько незначительны, что их
следует отнести к индивидуальным отклонениям, тем более, что число исследованных птиц
было крайне незначительно.
8. Рябчик Tetrastes bonasia L.
Tetrao Bonasia. Li n n a e u s. Syst. Nat. изд. X, I, 1758, стр. 160, Швеция.
Русские названия. Рябчик, рябец, рябок—народное, от пестрой окраски.
Распространение. Ареал. Населяет леса от Пиренеев на западе до
Колымы, Охотского побережья и Сахалина на востоке, к югу—до Се-
Карта 9. Распространение рябчика Tetrastes bonasia
l—T. b. rupestris, 2—Т. b. bonasia, 3—T. b. sibiricus, 4—T. b. amurensie; a—граница гнездовий
верной Италии, Югославии, Болгарии, Волыни, лесов Черниговских, Брян-
ских, Моршанских, Самарской луки, среднего течения Урала, Алтая, Север-
ной Монголии, Южно-уссурийского края и Северной Японии (о-в Хоккайдо);
к северу идет до 69° с. ш. в Скандинавии и Лапландии, 67° на Урале, 69°
на Енисее, 68%°на Колыме. В лесах Кавказа, Тарбагатая, Танну-Ола и Кам-
чатки отсутствует.
РЯБЧИК ИЗ
Характер пребывания. Оседлая птица. В осенне-зимний
период делает местные перекочевки в поисках корма.
Биотоп. Еловые или елово-лиственные леса, местами лиственные
насаждения, преимущественно сырые, с густым подлеском. Сухих сосновых
боров избегает.
Подвиды. Выделение большого числа подвидов рябчика, описанных рядом авторов,
основано на явно недиагностических признаках; слабые отличия в окраске у большинства
описанных форм перекрываются индивидуальными колебаниями. Просмотр свыше
300 шкурок коллекции Зоологического музея Московского университета позволил выде-
лить на территории СССР только три подвида: 7\ b. bonasia L., 1758—Скандинавия,
европейская СССР к востоку до Нижней Камы; 7\ b. sibiricus Buturlin, 1916—от
бассейна Печоры, Урала и Приуралья на западе, до Охотского побережья и бассейна
Зеи на востоке, до Южного Алтая Северной Монголии и Б. Хингана на юге; Т. Ь. ати-
rensis R П еу,—1916—Восточное Приамурье, Уссурийский край, Сахалин, Маньчжурия
и Северная Корея. Помимо того, для Западной Европы описаны: Т. b. rupestris Brehm,
1831—центральная часть Западной Европы к северу до Померании, к востоку до Карпат,
Трансильвании, Буковины и Болгарии.
Восточноевропейский рябчик Tetrastes bonasia bonasia L,
Синонимы. Tetrastes bonasia volgensis. Бутурлин. Орн. Вестн., 1916, № 4,
стр. 227. Сурские боры у устья Барыша.—Tetrastes bonasia grassmanni. Z e d 1 i t z.
Journ. fur Orn, 1920, стр. 234, Шара, бассейн Припяти.
Распространение. Населяет европейскую часть СССР от западных гра-
ниц, к востоку до бассейна Печоры и Приуралья исключительно. Вне СССР—
северные районы Скандинавии. К северу идет до 69/4° с. ш. в Норвегии, север-
ных частей Финляндии, до 68&° с. ш. на Кольском полуострове (до г. Колы),
северной границы крупного леса, т. е. примерно 6972° с- ш- на остальной
части европейского севера. На юге доходит до лесов Волынской, севера Чер-
ниговской, западных частей Орловской обл., Тульских засек, северо-восточ-
ных районов Рязанской обл., до лесов Суры и Жигулевских гор.
Характер пребывания. Оседлая птица, в гнездовое время
строго привязанная к определенным участкам леса. В послегнездовой период
совершает короткие, но отчетливо выраженные перекочевки, связанные
с изменением кормовых условий.
В Архангельской и Вологодской обл. осенью и зимой держится близ
воды, в березовых и ольховых логах, где сначала кормится ягодами рябины,
а потом почками; к весне же оставляет эти места и на весь гнездовой период
перебирается в хвойные леса. Перелеты в другие стации совершает по зорям
и не сразу, а в течение нескольких дней. Только изредка, при каких-то еще
не выясненных обстоятельствах, рябчик предпринимает более дальние стран-
ствования (Мензбир, 1902).
Биотоп. Коренное местожительство рябчика—старые еловые или
елово-лиственные леса; в юго-западных частях ареала он круглый год дер-
жится в старом лиственном лесу, в других местах в лиственные леса рябчик
перебирается только на весну и лето. Будучи равнинной птицей он заселяет
леса низменностей или речных долин всегда с сырым низом, поросшим гус-
той порослью. Сухих сосновых боров избегает и лишь в виде исключения
встречается в таких сосновых борах, низ которых покрыт папоротником.
Не встречается и на чистых моховых болотах, поросших редким сосняком.
На Кольском полуострове самой излюбленной стацией рябчика, особенно
зимою, являются долины ручьев или лощины с зарослями серой ольхи,
сережки и почки которой составляют его обычную зимнюю пищу. Помимо
ольшаников, нередко встречается в ельниках, предпочтительно в более
густых и захламленных, и даже в борах (чернично-ибруснично-ягельных),
в последнем случае невдалеке от лощин, дающих птице убежище. На чис-
тых ягельниках встречается только поздней осенью или рано весной: пови-
8 Птицы СССР, т. IV
114 отряд куриные
димому, его привлекают сюда ягоды брусники. Выводки можно встретить
также в березниках по берегам рек, среди травы. На болотах не живет, а в
горную тундру не поднимается ни при каких обстоятельствах; не попадается
и на гарях (Семенов Тян-Шанский, 1938). В Подмосковье рябчик придержи-
вается преимущественно хвойных и смешанных лесов с подлеском из моло-
дых елок или из густой поросли осинника, березника и др. Еловые насаж-
дения предпочитает другим. Встречается и в лиственных насаждениях,
главным образом осиновых и ольховых. Увлажненность, легкая захламлен-
ность, хорошо выраженный травяной покров, а также наличие ягодников
положительно сказываются на численности рябчика (наблюдения автора).
По наблюдениям Формозова A934) в Горьковской области, сочетание двух
факторов определяет пригодность какой-либо стации для рябчика: наличие
ели как защитной породы и наличие корма. Количество рябчика в угодьях,
если откинуть значение кормов, прямо пропорционально степени сомкну-
тости крон ели. В этом отношении еловые чащи, возникающие на месте
старых гарей, под пологом временных березников в результате смены по-
род, а также «шохры»—заболоченные еловые или елово-пихтовые леса—наи-
более благоприятные для рябчика типы угодий. Его совершенно нет в со-
сняках-ягельниках, в сосняках-долгомошниках, в сосняках по большим бо-
лотам и на молодых гарях; малочислен в сосняках-зеленомошниках, а
на гари 15—20 лет с частой порослью осинового и березового молодняка
изредка залетает осенью из прилегающих раменей. По Воронцову A949),
в Молотовской обл. рябчик наиболее многочислен в елово-пихтовой и елово-
липовой надзонах.
Численность. В северной половине европейской части СССР до-
вольно высокая, у северных и южных границ ареала, а также вокруг круп-
ных селений и в густонаселенных районах—вообще низкая. В Приволжско-
Дубненском заповеднике, имеющем площадь 3800 га, в 1948 г. было учтено-
34 выводка рябчика (данные автора). В Лапландском заповеднике числен-
ность рябчика составляет всего лишь 10% по отношению к численности дру-
гих куриных птиц.
Экология. Размножение. Половая зрелость наступает, повиди-
мому, в годовалом возрасте. В брачный период рябчики образуют пары, кото-
рые существуют и в период насиживания. Роль самцов в выкармливании
птенцов не ясна и разными авторами трактуется по-разному (см. описание
следующего подвида). Брачный период в Средней России начинается с кон-
ца марта или начала апреля и продолжается до половины мая. В Прибал-
тике гнезда с полными кладками обычны уже в конце апреля (Мензбир„
1902). На Кольском п-ве 22 мая была убита самка с почти готовым к сносу
яйцом; насиживание начинается приблизительно в последних числах мая,.
появление птенцов во второй половине июня (Семенов Тян-Шанский, 1938).
В Минской обл. ненасиженные полные кладки в конце апреля (Шнитников,
1913). В Московской обл. гнездо с девятью насиженными яйцами найдено
12 мая; помещалось оно на земле под основанием ствола толстой березы у
края болотца и по соседству с куртинкой густого ельничка (наблюдение
автора). Устройство гнезда рябчика очень просто; представляет оно собою
небольшую ямочку или углубление под кустом, деревом, валежником или
даже просто во мху или гнилом дереве, выстланное травинками, листьями,
иногда мелкими прутиками. Гнездо обычно хорошо скрыто и найти его мож-
но с большим трудом. Самец с самого начала брачного периода держится
вблизи самки; позднее же, когда самка начнет нестись, почти не отходит
от нее и в это время обращает гораздо меньше внимания на других самцов.
Полная кладка из 6—10 яиц, у более старых самок даже из 14—15 яиц.
Скорлупа яиц гладкая, блестящая, буровато-желтого цвета с редкими красно-
бурыми пятнышками и точками, а иногда и без них. Размер яиц 41—42x28—
РЯБЧИК
29 мм. Самка сидит на гнезде настолько крепко, что иногда позволяет взять
себя руками; при опасности старается незаметно сойти с гнезда и отвести
от него. Насиживание продолжается около трех недель; птенцы могут бегать
сейчас же после того, как матка обсушит их под крыльями. На другой день
обыкновенно уводит их на светлые травянистые лужайки и опушки, где
птенцы находят корм в изобилии. Очень травянистых полян выводки избе-
гают, напротив, лесные дороги с их скудной травянистой растительностью и
колеями, где долго держится дождевая вода, охотно ими посещаются (Менз-
бир, 1902). В Московской обл. первые выводки с пуховыми птенцами встре-
чены в 1947 г. в середине июня, в 1948 г.—9 июня; в середине июля 1947 г.
птенцы достигали двух третей размера взрослых, а в 1948 г. одни в это время
были размером с перепелку, а другие со взрослых. В конце июля и в авгу-
сте молодые, по нашим наблюдениям, уже не отличимы от взрослых. В Дол-
голуговском охотхозяйстве (Московская обл.) первые выводки отмечены
в 1934 и 1935 гг. 10 июня (Насимович, 1936 ). Птенцы превосходно пря-
чутся, затаиваясь в ямках, под сучьями и т. д. Первое время они растут
медленно и начинают перепархивать в возрасте нескольких дней; трех-
недельные уже обыкновенно ночуют на деревьях. Месячные величиной с пе-
репелку, но после этого растут быстро и в двухмесячном возрасте достигают
величины взрослых, отличаясь от последних лишь перьями птенцового на-
ряда на голове. Полностью линька у молодых заканчивается к середине сен-
тября, после чего выводки разбиваются и молодые начинают вести само-
стоятельный образ жизни (Мензбир, 1902).
Осеннее токование рябчика и в Долголуговском охотничьем хозяйстве
начинается в конце августа и продолжается до октября включительно
(Насимович, 1936). В это время происходит образование пар.
Линька. Смена нарядов: пуховой, птенцовый, первый годовой,
взрослый. Линька взрослых ежегодная полная (общий ход линьки см. в опи-
сании амурского рябчика). Старые самцы в средней полосе начинают линять
в конце июля, заканчивают линьку к концу сентября; к этому времени только
кое-где еще остаются не совсем развившиеся перья. У самок линька начи-
нается несколько позднее, но оканчивается одновременно с самцами, т. е.
в конце сентября К этому времени молодые успевают надеть взрослый
наряд; лишь перья плюсны еще коротки, но к середине октября и они от-
растают (Мензбир, 1902).
Зимняя жизнь. Осенью рябчики живут парами или в одиночку
в районе своего летнего местообитания. С началом сильных морозов и выпа-
дением глубокого снега они переселяются к ручьям и речкам, где растет
ольха и береза; на севере это наблюдается в конце октября, в средней по-
лосе—не ранее конца ноября. В местах обилия корма рябчики концентри-
руются и образуют иногда временные стайки по 5—10 шт., которые позднее
рассыпаются (Мензбир, 1902).
Питание. По данным Формозова A934), исследовавшего питание ряб-
чика в Горьковской обл. (всего 124 вскрытия, относящиеся к 9 месяцам года;
не охвачены вскрытиями январь, март, декабрь), наряду с растительными кор-
мами рябчики питаются и животными. Из последних преобладают насеко-
мые, составляя за 8 месяцев 20% встреч. Насекомые встречены в апреле в
1 зобу (из 1 осмотренного), в мае в 1 (из 3), в июне в 1 (из 3), в июле в 3
(из 11), в августе в 5 (из 6), в сентябре в 11 (из 42); совсем не найдены в ок-
тябре и ноябре. По частоте встреч и количеству съеденных экземпляров на
первом месте стоят жуки (жужелицы, иванов-червячок, навозники, листогры-
зы); относительно часто встречаются перепончатокрылые (мелкие наездники,
крупный древесный муравей, личинки наездника), нередко бабочки (гусе-
ницы пяденицы, перламутровицы). За насекомыми на втором месте стоят
пауки, исключительно мелкие наземные формы; из моллюсков имеется толь-
8*
116
ОТРЯД КУРИНЫЕ
ко одна встреча. В общем животные корма можно встретить в желудках
рябчика с весны B2 апреля) до поздней осени B5 ноября), но обычно еди-
ничными экземплярами. Только в одном зобу насекомые (листогрызы) со-
ставляли по объему около 25% всего съеденного. Количественно раститель-
ные корма резко преобладают над животными. Исключение составляют пу-
ховые птенцы в первые дни жизни. Птенчик 41 г весом имел значительное
количество насекомых в зобу и желудке, но немало и мелких семян, и неж-
ные зеленые части растений. Двумя неделями позднее у птиц, выросших
вдвое (весом 71 и 87 г), в зобу и желудке были исключительно ягоды чер-
ники и мелкие семена.
Из растительных кормов охотно поедаются зеленые части растений, как
пуховыми птенцами, так и взрослыми, с весны до выпадения снега. Из травя-
нистых растений отмечены листья
душистой таволги, земляники, мали-
ны, кислицы, лютика и селезеночни-
ка, из полукустарников—листья чер-
ники и брусники. Преобладают над
травянистыми листья древесных по-
род, отмеченные главным образом
осенью. Чаще всего поедаются листья
осины, затем ольхи, березы, ивы и
реже липы и рябины; хвоя—случай-
ный корм. Семена лесных травяни-
стых растений бывают в желудке в
небольшом количестве летом и осенью;
более заметную роль играют семена
осоки, душистой таволги и марьяника
лугового и меньшую—фиалки и щаве-
лей. Важную роль в летнем и осен-
нем питании играют ягоды. Наиболь-
шее значение из них имеют ягоды
малины (в августе 83,3%), черники
(июнь 33%, июль 69%, август 33%,
сентябрь 9,7%), брусники (сентябрь
75,6%, октябрь 19,9%, ноябрь 83%);
по объему брусника составляет в сред-
нем 77,5% содержимого зобов в пер-
вой половине сентября, 31 %—во вто-
рой, соответственно 9% и 3,4% в
октябре и 3,7% в начале ноября. От-
мечены также ягоды костяники, зем-
ляники, майника, клюквы, рябины, калины, жимолости, можжевельника.
К важной группе кормов относятся сережки и почки древесных и кустар-
никовых пород. На первом месте стоят сережки ольхи: в сентябре 24,4%
встреч, в октябре 74,3%, в ноябре 66,6%; по объему от 35 до 65%, в одном
зобу до 164 шт. (вес 20 г). Часто поедаются почки ольхи. На втором месте
сережки почки и части ветвей березы; встречаемость сережек 24% в сен-
тябре, в октябре 35,8%, в ноябре 50%, почек с ветвей соответственно 2,4;
12,8 и 33,3%; по объему от 1,6 до 23,5%. Наибольшее количество сережек
в одном зобу 72, почек 140 шт. Значительное место занимают сережки
и почки ивы, почки осины и черники; реже встречаются почки рябины,
брусники и липы.
Питание по сезонам года изменяется следующим образом. В апреле
и мае основу составляют распускающиеся почки, сережки березы, ивы, лис-
тья осины и ивы; с земли собираются некоторые семена, оставшиеся ягоды,
Рис. 21. Рябчик (самец и самка)
РЯБЧИК
117
листья травянистых растений и, видимо, насекомые. Летом поедаются пре-
имущественно зеленые части растений, семена и насекомые, а позднее—ягоды.
Уже в июле рябчик склевывает зеленую чернику и поспевающую землянику,
а в августе и малину. Некоторые семена и зелень—дополнение к основному
ягодному рациону. В сентябре питается в значительной мере брусникой,
а также ягодами майника, рябины, почками и ветками черники, семенами
таволги и марьяника, листьями кислицы и осины, сережками и почками
ольхи. По характеру питания можно назвать август чернично-брусничным
периодом, первую половину сентября—брусничным, вторую—бруснично-
сережковым. Октябрь и последующие месяцы характеризуются сережковым
питанием: сережки, почки, веточки ольхи и березы вместе заполняют в ок-
тябре более 77% содержимого
зоба. В зобах рябчиков, добы-
тых в августе и октябре в Мос-
ковской обл., нами обнаружены
следующие корма: ягоды брус-
ники, малины, костяники, чере-
мухи, земляники, семена май-
ника, осоки польчатой, семена
и коробочки иван-да-марьи,
сережки и концы веток березы.
У добытых в октябре пища в зна-
чительной мере состояла из се-
режек и кончиков веток березы;
общая длина последних в одном
желудке достигала 70 см, коли-
чество сережек в зобу—9, ягод
брусники—14.
Промысел Промысловое зна-
чение рябчика было велико: еже-
годно добывалось в России до
5—6 МЛН. ШТ. (Мензбир, 1902); Рис- 22« Пуховой птенец рябчика
значительно оно и в настоящее
время. Добывается весьма разнообразными способами: ружьем с легавой
и лайкой, с загонщиками, с пищиком, сильями, давилками и многими дру-
гими самоловами, хорошо описанными в литературе.
Полевые признаки. Птица величиной с галку или чуть крупнее и более
плотного (характерного для куриных) сложения, серого, с поперечными тем-
ными пестринами цвета и с слабо заметным хохолком на голове. У самца
горло черное, окаймленное белой полоской, проходящей по щекам ниже глаза
и затем через лоб. Поднимается с земли и деревьев с большим шумом, при
полете часто машет крыльями, издающими слышимый издали шум.
Описание. Размеры и строение. Средней величины тетеревиные птицы
с густым, но мало заметным лежачим теменным хохолком. Цевка оперена приблизительно
на 2/з длины, хвост слабо округлый на вершине из 8 пар рулей. Крыло тупое. Длина крыла
самцов E3) 162—180, самок C0) 161—178, среднее 172—170 мм.
Окраска. Пуховой птенец. Спинная сторона тела рыжевато-каштаново-корич-
невая, более темная на темени и задней части спины. Уздечка черная. Брюшная сторона
светложелтая, несколько более темная на зобе. Птенцовый наряд—как у взрослой самки,
лишь на спине узкие белые настволья и брюхо посветлее, почти без темных пятен.
Молодая птица в первую осень: горло беловатое в темных пестринах. Перья спинной
стороны тела с белыми наствольями. Зоб бледноохристый. Черные пятна на брюшной сто-
роне тела тусклые, белый цвет не чистый, а беловатосерый. Самки сходны с самцом, но
у самцов черное горловое пятно только намечено, темные пестрины низа яркие, верх охри-
стый. Взрослый самец: верх головы, шеи и передняя часть спины серые с черными и рыжи-
ми поперечными полосами. Спина и кроющие хвоста серые с узким поперечным черными
рыжеватым рисунком и темными наствольями перьев. Маховые бурые с беловато-охристым
118 ОТРЯД КУРИНЫЕ
наружными опахалами перьев. Кроющие крыла пестрые с серыми, охристыми и белыми
пятнами. Рулевые, кроме средней пары, с темной широкой предвершинной полосой и бе-
лой вершиной. Шея спереди черная, окаймленная белой полосой, идущей под глазом до
угла рта и затем на лоб. Зоб в поперечных темных и беловато-охристых полосах, переходя-
щих постепенно на груди и брюхе в пятна. Подхвостье беловато-охристое, нижние кроющие
хвоста охристые с темным рисунком и белыми вершинами. Пальцы буровато-серые,
клюв серовато-черный. Взрослая самка отличается более сильно развитым охристо-рыжим
тоном. Горло белое. Окраска описываемого подвида в целом несколько темнее, чем у дру-
гих, с более выраженным рыжим тоном на верхней стороне тела.
Сибирский рябчик Tetrastes bonasia sibiricus But.
Tetrastes bonasia sibiricus. Бутурлин. Орн. Вестн., 1916, № 4, стр. 224. Енисей.
Синоним. Tetrastes bonasia kolymensis. Бутурлин. Орн. Вестн. 1916, № 4, стр. 226,
Колыма.
Распространение. Ареал. Населяет бассейн Печоры, Урал и При-
уралье, всю Сибирь до Охотского побережья; на север—до границы вида,
на юг до Южного Алтая, Северной Монголии, Б. Хингана; от Б. Хингана
южная граница идет к бассейну Зеи и далее выходит к северному устью
Амура. На Анадыре и Камчатке отсутствует.
Характер пребывания. Оседлая птица, строго привязанная к опре-
деленным участкам леса. Совершает лишь короткие миграции, вызываемые се-
зонной сменой кормов (см. биотоп). Обычно эти перемещения бывают осенью.
В Печоро-Ылычском заповеднике из прибрежных ельников рябчики вре-
менно откочевывают на сезонные корма в другие стации (Донауров, 1947).
Большая часть южноуральских рябчиков (Кириков, 1948) с наступлением
осени перебирается в черемухово-ольховую урему, где зимой встречается
гораздо реже, а в иные годы всю зиму вовсе отсутствуют. На Среднем Урале
с первым снегом рябчики перебираются в березник и ольшаник по речкам,
где и остаются до весны (Мензбир, 1902). В Башкирии (Сушкин, 1897)
поздней осенью одиночные птицы или парочки делают перекочевки и могут
быть встречены в таких лесах, где не гнездятся.
Биотоп. В Печоро-Ылычском заповеднике основной район обитания
рябчика—полоса темнохвойных лесов, причем биотопом постоянного их
обитания служат наиболее широко распространенные типы леса—ельники
заболоченные, зеленомошные и прибрежные. Густые и сомкнутые кроны
деревьев и сильная захламленность ельников создают прекрасные защитные
условия. В хорошо развитом лесном покрове много осоки, кислицы и чер-
ники, составляющих основной корм этих птиц летом и осенью. Береза, ива
и рябина, на которых рябчик кормится поздней осенью и зимой, встречаются
в темнохвойных лесах в большом количестве. В горах густо заселены ряб-
чиком только долины рек, ручьев и нижние склоны. В этих местах характер
древесной растительности и степень обилия кормовых растений такие же,
как и в районе темнохвойных лесов. Но чем выше к вершинам гор, тем лес
становится реже, примесь березы увеличивается и численность рябчика
снижается. В полосе низкорослого редколесья и выше, на склонах горных
тундр рябчик не встречается. В районе боров в Припечорской низменности
места обитания рябчиков связаны с наличием ели; наибольшее число их дер-
жится в еловых лесах по долинам рек: в густых, сильно захламленных при-
брежных ельниках рябчик находит хорошие кормовые и защитные условия.
В сосновых лесах постоянно обитает лишь в том случае, если в них, кроме
ягодников, есть и примесь ели. Но в чистых борах встречается в ничтожном
количестве, главным образом на участках, граничащих с ельниками в
речных долинах. Сезонное размещение рябчика по стациям, установленное
на основании регистрации встреч рябчиков в различных типах леса Печоро-
Ылычского заповедника в течение трех лет (общее число встреч 4100),
характеризует значение отдельных стаций. В темнохвойных лесах в марте
РЯБЧИК 119
и апреле основная масса птиц D9%) сосредоточена в прибрежных ельниках;
значительное их количество C0%) обитает и в заболоченных еловых лесах
на водоразделах. Обе эти стации обеспечивают рябчику наилучшие условия
существования как в отношении корма, так и в отношении защиты.
В зеленомошных ельниках встречается не часто (8%), чаще в березовых ле-
сах с сильным возобновлением ели, покрывающих гари B3%). В мае боль-
шинство птиц остается в прибрежных ельниках, что обусловливается нали-
чием удобных для гнездования мест. Увеличивается число встреч в заболочен-
ных ельниках, за счет откочевки из ельников зеленомошников. Появляется
на гарях, снижается процент встреч в березовых лесах. В летний период
(июнь—август) самцы и одиночные «холостые» самки продолжают держаться
преимущественно в прибрежных и заболоченных ельниках, а со второй по-
ловины июля перекочевывают на ягодники в ельники зеленомошники и бере-
зово-еловые леса. Выводки с момента появления птенцов до середины июля
держатся главным образом на участках леса с хорошо развитым травяным
покровом: на лугах и по краям травяных болот, в березовых лесах и забо-
лоченных ельниках; в густых прибрежных ельниках выводки держатся
редко. С середины июля начинается массовая перекочевка на ягодники, глав-
ным образом в наиболее богатые черникой зеленомошные еловые леса. В ок-
тябре продолжают держаться на ягодниках: в зеленомошных ельниках-чер-
ничниках, в прибрежных ельниках, в заболоченных еловых лесах. В октябре
в темнохвойных лесах исчезает основной корм—черника и основная масса
птиц концентрируется по берегам рек, куда их привлекает обилие почек
и сережек березы и ольхи, почки ив, а также неопавшие плоды шиповника
и рябины. С установлением постоянного снежного покрова рябчики пере-
ходят на питание древесными кормами, с размещением которых связано и
зимнее (ноябрь—февраль) размещение рябчика. В темнохвойных лесах ряб-
яики концентрируются главным образом в прибрежных и заболоченных ель-
никах; в березовых лесах встречаются чаще, чем осенью, в зеленомошных
ельниках редки ввиду малочисленности кормов. Таким образом, в темнохвой-
ных лесах наибольшее значение для рябчиков имеют прибрежные ельники,
где концентрируется большое количество птиц в течение почти всего года,
и обширные заболоченные ельники, где достаточно корма во все сезоны года.
Ельники-зеленомошники—место концентрации в ягодный сезон. В борах
«большинство рябчиков обитает в прибрежных ельниках, лишь временно отко-
чевывая отсюда на сезонные корма в другие стации; только небольшая часть
рябчиков постоянно живет в борах-зеленомошниках, где среди высокостволь-
ных сосен есть густой подлесок из елей (Донауров, 1947 ). На Южном Урале
летом рябчики встречаются в не слишком густых участках сосново-березо-
вых лесов, особенно в местах, где бывает хороший урожай костяники, чер-
ники, малины и брусники; вылетают кормиться в вишенники на безлесные
склоны гор, но очень далеко от леса не отлетают. С наступлением осени боль-
шая часть птиц перебирается в черемухово-ольховую урему, а где ее по-
близости нет, встречаются в сосново-березовых лесах. В последних преиму-
щественно держится и в зимний период (Кириков, 1948). В Алтайском за-
поведнике размещение рябчика тесно связано с наличием в лесных насаж-
дениях березы и ольхи, дающих осенний и зимний корм; в заметных коли-
чествах встречается он здесь лишь в нижнем поясе леса, близ березовых
и ольховых насаждений. Особенно обильны рябчиком леса Прителецкой части
заповедника. Здесь на месте вырубок хвойных лесов обычны временные на-
саждения из березы и осины, чередующиеся с участками редколесья, из ли-
ственницы, осины, березы и других пород. Покрытые лесом участки сменя-
ются яркоцветными злаково-разнотравными полянами, выходами скал и рос-
сыпями, а в пониженных элементах рельефа развиваются поляны разнотра-
вья. В подлеске и подросте леса обычна ольха. Прителецкие леса богаты
120 отряд куриные
ягодными кустарниками, из которых многие виды служат для рябчика кор-
мом. По долинам многочисленных в этом районе речек спускается вниз
темнохвойная пихтово-кедровая тайга. По долинам рек рябчик доходит
до верхней границы леса, но по мере подъема в горы количество его резко
падает, и на верхней границе леса он крайне редок (Фолитарек и Дементьев,
1938). Вертикальное распространение местами за 2 000 м (Сушкин, 1938). В
югозападном Кентэе встречается вуремах горных речек, в густых сиверах по
гребням хребтов и в редколесье—везде, где к хвойному лесу примешивается
береза или где встречаются брусничные кочки (Козлова, 1930). В верховьях
Колымы рябчик встречается в поймах рек, там, где на илистых островах,
берегах и окрайках проток встречаются густые заросли ивняка и особенно
излюбленной рябчиком кустарниковой ольхи. Тут, в непролазной чаще оль-
шаников, рябчик держится стайками всю зиму и большую часть весны, выби-
раясь для гнездования в смежный лиственный лес (Дементьев и Шохин,
1939).
Численность. Набольшей части территории ареала сравнительно
высокая. По данным весеннего и осеннего учетов рябчика на пищик в при-
брежных ельниках Печоро-Ылычского заповедника на площади 10 га прихо-
дится: в боровом районе 3,7 шт., в районе темнохвойных лесов 1,9 шт.,в гор-
ном районе 1,7 шт. (Донауров, 1947). В Алтайском заповеднике при учете
в конце марта и начале апреля в Прителецких лесах на 10 км маршрута
отмечено около 25—30 птиц; осенью за границами заповедника в сходных
угодиях охотник нередко добывает в день 20—25 птиц (Фолитарек и Де-
ментьев, 1938).
Экология. Размножение. Наиболее полные данные из Печоро-
Ылычского заповедника (Донауров, 1947). Весеннее токование начинается
там в первой декаде марта; его начало связано с общим потеплением и уста-
новлением ясных, теплых и уже длинных дней. В это время на обогреваемых
солнцем склонах берегов рек утрами слышны не частые еще брачные песенки
самцов, а на снегу видны следы волочащихся крыльев. Разгар токования
приходится на вторую половину апреля и первую половину мая, а конец—
на вторую декаду июня. Рябчик—моногамная птица, и весь период тока са-
мец постоянно держится около своей, еще с осени выбранной самки. Такая
пара рябчиков в то время, как и в остальную часть года, постоянно держится
на занятых ею строго ограниченных участках леса. Токование самца выра-
жается частым пением и преследованием самки. Бегая за самкой, он рас-
пускает хвост веером и волочит по земле крылья. На интенсивность весен-
него токования влияет плотность населения рябчиков: чем их больше и тес-
нее соприкасаются участки, занятые отдельными парами, тем азартнее поют
самцы. Активнее проходит ток в тихие и ясные утра и вечера; в пасмурные
дни рябчики токуют слабо, а в холодные дни с дождем и снегом токование пре-
кращается вовсе. В 1938 г. период спаривания начался со второй декады мая.
В это время семенники самцов достигли максимального размера, а у всех
добытых самок в яйцеводе отмечались готовые к кладке яйца. Самок, несущих
яйца, добывали только в течение второй декады; с третьей декады мая все сам-
ки сели на яйца. Таким образом, продолжительность периода спаривания
и кладки яиц в 1938 г. равнялась 10 дням. Учитывая, что среднее число яиц
в 16 учтенных гнездах равно 7,3, можно считать, что самки откладывают яйца
каждый или почти каждый день. В 1939 г. общий ход весны запоздал по срав-
нению с предыдущим годом на 10—15 дней, в связи с чем и начало периода
спаривания отодвинулось на декаду. Интересно, что задержка весны отра-
зилась на более позднем развитии половых органов только у самок, семенни-
ки же у самцов достигли максимального размера в те же сроки, что и в 1938 г.,
т. е. во вторую декаду мая. Можно полагать, что на половой цикл у самок
рябчика, как это отмечалось другими исследователями и для глухаря, влияет
РЯБЧИК 12 i
состояние гнездовых стаций и в первую очередь появление обширных про-
талин.
На Южном Урале весенний свист рябчиков начинается с конца марта;
весь апрель и половину мая они держатся парами, самцы сильно возбуждены,
быстро подлетают на манок и при встрече друг с другом легко вступают в дра-
ки. Кладка яиц начинается в мае. В Башкирском заповеднике в раннюю весну
1940 г. полные кладки найдены 16, 23 и 28 мая, в позднюю весну 1942 г.
16 мая обнаружено гнездо только с одним яйцом (Кириков, 1948). В Алтай-
ском заповеднике свист самцов в 1935 г. отмечен 23 марта, разгар тока и спа-
ривание—во второй половине апреля, носка яиц—в конце апреля и в пер-
вой декаде мая (Фолитарек и Дементьев, 1938). В Юго-Западном Кентэе
первый свист в десятых числах апреля, особенно оживленный—в мае (Коз-
лова, 1930). В верховьях Колымы первые яйца рябчиков найдена 4 июня
(Дементьев и Шохин, 1939). В Красноярском крае начало брачного периода
около середины апреля (Тугаринов и Бутурлин, 1911).
Гнездо устроено очень просто и представляет собою углубление в слое
отмершего лесного покрова почти без всякой выстилки. Диаметр лотка ос-
мотренных Донауровым в Печоро-Ылычском заповеднике гнезд равнялся
в среднем 20 см, глубина лотка 4—5 см. Как правило, все гнезда располо-
жены на возвышенных, наиболее сухих и ранее всего освобождающихся из-
под снега местах. От дождя гнездо обычно защищено молодыми пихтами,
елями и валежником. Гнездо, найденное Фолитареком A938) в Алтайском
заповеднике, помещалось на редине с ветровалом, под камнем, и было при-
крыто сверху плетями папоротника. По Донаурову, число яиц в кладке на
Печоре колеблется от 4 до 9, в среднем (по 16 гнездам) 7,3; Кириков A948)
находил гнезда на Южном Урале с 7 (в одном гнезде), 9 (в двух гнездах)
и 11 (в одном гнезде) яйцами; Фолитарек A938) на Алтае —с 11 яйцами.
Размер яиц (8) 37—40,5x25—27,6 мм (Донауров, 1947). Весной самцы ряб-
чиков плохо питаются и к концу брачного периода сильно теряют в весе.
Падение веса происходит с первых чисел апреля до середины мая, причем
за этот период вес птиц снижается в среднем на 42 г, или на 10—11 % ис-
ходного веса. В последней декаде мая брачный период заканчивается и на-
чинается повышение веса самцов. Весеннее падение веса у самцов сопровож-
дается ростом семенников, а увеличение веса в последней декаде мая сов-
падает с периодом их увядания. Весеннее падение веса у самцов связано
с недостаточным питанием их в это время, о чем можно судить по легким
или пустым зобам птиц, добытых в глубоких сумерках. Напротив, самки
весной ведут себя иначе—они усиленно кормятся, предпочитая в это время
насекомых, и вес их в брачный период заметно повышается: с первой декады
апреля до второй декады мая на 56 г, или на 14% исходного веса. Нараста-
ние веса идет параллельно с развитием яичника и яйцеводов. Достигнутый
весной максимальный вес самки снижается в летний период, видимо, глав-
ным образом во время насиживания (Фолитарек и Дементьев, 1938). Период
насиживания в Печоро-Ылычском заповеднике (Донауров, 1938) в 1938 г.
начался с третьей декады мая, в 1939 г. —с первой декады июня, первые вы-
водки были встречены соответственно в начале второй декады и в третью
декаду июня. Таким образом, период насиживания равен примерно 20 дням.
Относительно роли самца в воспитании потомства существуют различные
мнения. Многие наблюдатели отрицают роль самца в выкармливании моло-
дых и уходе за ними, но Богданов A868) и Мензбир A902) указывают, что
самец держится при выводке и разделяет с самкой труд выкармливания моло-
дых. Гейм де Бальсак A935) наблюдал, что при выводке всегда держится
пара родителей. По наблюдениям Кирикова A948) на Южном Урале, самец
не принимал участия в насиживании яиц, при выводке чаще всего бывает
одна самка, но нередко встречаются и самка и самец; при гибели матки на
122 ОТРЯД КУРИНЫЕ
следующий день с молодыми рябчиками часто оказывался самец. Поданным
Донаурова для Печоро-Ылычского заповедника, самцы не покидают самок
в период насиживания и нередко держатся около них почти до вылупления
птенцов. Указанному автору неоднократно приходилось, вспугивая с гнезда
самку, обнаруживать поблизости и самца, который обычно сидел до этого
на нижних ветках дерева. Донауров допускает возможным кормление в пе-
риод насиживания самцом самки, но утверждает, что после вылупления птен-
цов самцы отделяются от самок.
15 Среднее количество рождающихся птенцов по сравнению с числом яиц
в кладке сильно разнится по годам. На основании 181 встречи выводков
в первое время массового появления птенцов в Печоро-Ылычском заповеднике
среднее их количество в выводке в 1938 г. было равно 6, в 1939 г.—4,9,
в 1940 г.—6 шт. Учитывая, что число яиц в кладках существенно по годам
не изменяется, можно полагать, что разница в числе рождающихся птенцов
по годам зависит от гибели яиц. В 1938 г. погибло 12% яиц, в 1939 г.—
33%, в 1940 г.—12%. Подсчет среднего количества птенцов в выводках по
месяцам показывает, что отход молодняка у рябчика к сентябрю составлял:
в 1938 г. не менее 24%, в 1939 г.—37%, в 1940 г.—32%. В течение 4 лет—
с 1937 по 1940 г.—среднее количество птенцов, приходящееся на одну самку
в конце периода размножения, колебалось в пределах от 4,7 до 5,3. Таким
образом, можно считать, что население рябчиков в результате размножения
может ежегодно увеличиваться в 2,5—3 раза, если допустить, что у рябчика
не наблюдается массовой «яловости». Молодые рябчики растут довольно
быстро и через 6 недель достигают размера взрослых. В первую декаду птен-
цы слабо прибывают в весе, увеличиваясь с 8,5 до 10,3 г; мало увеличивается
и длина тела, крыла, хвоста и плюсны. В первые десять дней жизни пуховые
птенцы довольно быстро бегают, скрываясь под корнями и валежинами дере-
вьев, но весьма редко и невысоко поднимаются на деревья при вспугивании.
Со второй декады жизни начинается интенсивный рост птенцов. К концу
второй декады вес увеличивается до 39 г, т. е. более чем в 3 раза, а длина тела
и крыла—в 2 раза; пуховой наряд довольно быстро заменяется ювенальным.
В третью и четвертую декады птенцы растут также интенсивно, вес в каждую
декаду увеличивается в2—2,5раза и впервой декаде августа достигает 191 г;
увеличение размеров тела в это время идет медленнее: длина тела—в 1,5 раза,
а длина крыла—на одну треть. В последние две декады августа молодые
рябчики увеличиваются главным образом в весе: во вторую декаду достигают
255,6 г, в третью—323,5 г; размеры тела и крыла увеличиваются незначи-
тельно. В эти две декады меняется ювенальное перо на наряд взрослых.
В первую декаду сентября молодые рябчики по размерам не отличимы от
старых, а в конце этого месяца они наравне с остальными участвуют в осен-
нем токовании.
В Башкирском заповеднике 13 июня 1937 г. было найдено гнездо с вы-
ходящими из яиц птенцами (Кириков, 1948). В Алтайском заповеднике пер-
вые, уже хорошо летающие выводки в 1935 г. были отмечены 28 июня. Добы-
тые 10 июля 1934 г. два молодых птенца весили 112 и 112,5 г, а шесть птен-
цов в том же самом месте 10 и 11 августа весили от 293 до 305 г. В послед-
ней декаде августа в Прителецкой части молодые по размерам и весу тела
не отличались от взрослых. По мере подъема в горы календарные сроки раз-
множения запаздывают. Так в 1935 г. молодые птицы, добытые на устье
р. Шавлы 10 июля, весили 195 г; добытые 7 июля в вершине р.Чел-Коль (на
500 м устья Шавлы) весили в среднем 50 г. Примерно на той же высоте—
около 1 500 м, в среднем течении р. Сайгоныш 13 августа 1935 г. молодые
имели вес около 150 г (Фолитарек и Дементьев, 1938). В северо-восточных
предгорьях Алтая пуховые птенчики найдены 26 июня, оперенные 8 и 10 июля.
В горах у устья Башкауса, птенцы, немного крупнее перепелки, добыты
РЯБЧИК 123
1 августа; видимо, вывод происходит здесь недели на две позднее, чем в ни-
зинах. 1 сентября добыт молодой с остатками птенцового наряда только на
шее (Сушкин, 1938). В Юго-Западном Кентэе первые выводки в 20-х числах,
июня, совсем оперенные птенцы в начале июля (Козлова, 1930). В верховьях
Колымы только что вылупившиеся птенцы 1—2 июля (Дементьев и Шохин,
1939). В Красноярском крае выводмолодых в середине июля (Тугаринов и Бу-
турлин, 1911). В Якутии пуховые птенцы—в конце июня, летние, достигшие
величины половины взрослых,—в середине июля (Иванов, 1929).
Явление осеннего токования у рябчиков выражено в не менее яркой форме,
чем у тетерева, глухаря и ряда других куриных птиц. По наблюдениям Дона-
урова A947), в Печоро-Ылычском заповеднике во второй половине сентября —
первой половине октября можно часто слышать песни токующих самцов
и отклики самок. В этот период почти так же азартно, как и весной, самцы
отзываются и подлетают на манок. Каких-либо брачных игр самца и самки
не отмечено. Незаметно и увеличения семенников у самцов. Автор подтвер-
ждает высказанную Мензбиром мысль, что осеннее токование есть не что иное
как разбивка рябчиков на пары после прошедшего размножения. В Юго-За-
падном Кентэе разбивка на пары с начала сентября, причем взрослый самец
иногда выбирает молодую самку этого года. С 10-х чисел сентября свист
возобновляется, и в ноябре, несмотря на сильные морозы, слышится так же
часто и с такой же интенсивностью, как весной. Это зимнее оживление про-
должается до половины января, когда рябчики умолкают до весны (Козлова,
1930).
Линька. Последовательность смены нарядов, как и у предыдущего
подвида. В Печоро-Ылычском заповеднике замена пухового наряда птенцо-
вым происходит во второй декаде июля (на второй декаде жизни), формиро-
вание взрослого наряда—во второй половине августа. У самцов линька
начинается в июле, заканчивается к началу сентября. В Якутии линька
у взрослых начинается с конца июня и затягивается до августа (Иванов,
1929).
Зимняя жизнь. По Донаурову A947), рябчик никогда не обра-
зует больших стай. Чаще всего стайки встречаются в сентябре и октябре;
в это время на ягодниках, на шиповнике и ольхе иногда наблюдаются
стаи до 20 птиц. В последующие зимние месяцы величина стай постепенно
уменьшается; птицы собираются по 4, 5, б шт. и более только на время
кормежки на оголенных деревьях — березах, ольхах и ивах, а ночуют
по отдельности. Семь месяцев в году — с ноября по май — рябчик ве-
дет древесный образ жизни. В ноябре, в первый месяц снежного по-
крова, абсолютное большинство птиц (91,6%) кормится уже на де-
ревьях; на поверхности снега зарегистрированы только 4,2% птиц. Ночь
и дневной перерыв между утренней и вечерней кормежкой рябчики в ноябре
проводят в ветвях густых елей или на земле, под их кронами. В декабре,
с наступлением морозов, рябчики ночуют уже в снежных лунках. В январе
и феврале встречаются или спящими в снегу, или кормящимися на деревьях.
В эти неблагоприятные для них месяцы зимы они вылетают лишь на короткое
время на березы и ольхи, чтобы набрать в зоб необходимую порцию корма,
и тотчас же скрываются под снегом. Под снегом на глубине 20 см температура
снега значительно выше температуры воздуха и, кроме того, она здесь более
постоянна. Если температура воздуха изменялась от—16 до—39°С, то в снегу
она колебалась между—10 и—16°С, причем чем ниже температура воздуха,
тем разность ее с температурой снега становится большей. Следовательно, ряб-
чики, проводя зимой большую часть суток (до 20 час.) под снегом, не испыты-
вают влияния сильных морозов, достигающих в условиях Печоро-Ылычского
заповедника—45—50°С. Зимой деятельность рябчика наиболее ограничена.
Продолжительность дня в это время 5—6 час, и рябчики кормятся только
124
ОТРЯД КУРИНЫЕ
один раз в сутки—в наиболее теплое время дня, проводя остальные 18—19
темных часов под снегом.
Питание. Рябчик преимущественно растительноядная птица. По дан-
ным Донаурова A947), основанным на анализе зобов и желудков 307 взрослых
и 54 молодых птиц, употребляемые рябчиком в Печоро-Ылычском заповед-
нике растительные корма довольно разнообразны. Кроме сережек и почек
деревьев и кустарников, а также ягод, в пищу идут стебли и листья трав,
изредка даже мхи и лишайники. К основной группе кормов относятся прежде
всего ягоды, которые имеют существенное значение с июля по октябрь.
Большой удельный вес занимают зеленые части растений, употребляемые
весной и осенью, а также сережки и цветочные почки деревьев и кустарни-
ков—основной корм в снежный период года. Семена трав имеют существен-
ное значение с апреля по ноябрь. Плоды и семена кустарников хотя и упо-
требляются во все сезоны года, но не часто и служат добавочным кормом.
Значение отдельных групп кормов по сезонам года видно из нижепомещеннойг
таблицы.
Группы растительных кормов рябчиков и изменение их значения по сезонам
Месяцы
XII, I,
II, III
IV
VI
VII
VIII
IX
XI
Сережки и почки дере-
вьев и кустарников .
Семена и плоды кустар-
ников
Семена травянистой рас-
тительности
Ягоды
Зеленые части растений
Мхи и лишайники . . .
100
4
75
5
20
35
35
48
19
35
31
73
33
100
17
54
96
46
4
34
73
88
5
3
30
36
59
90
25
4
89
16
16
73
76
70
7
7
63
72
Группу основных кормов составляют следующие виды: береза, ива, ши-
повник, черника, брусника, марьяник, осока. Наибольшее значение имеет
береза, сережками и почками которой рябчики питаются в течение шести
зимних месяцев (ноябрь—апрель). Встречаемость сережек по месяцам: но-
ябрь 71%, декабрь—март 84%, апрель 60%, май 22,9%, сентябрь 16,8%,
октябрь 59,4%; почки поедаются реже и видное место занимают лишь зимой
и весной F0 и 50%), в октябре же встречаемость 8,1 %, в ноябре 28,6%. Ли-
стовые и цветочные почки ивы используются в пищу в течение восьми меся-
цев (октябрь—май), но гораздо реже, чем почки березы; встречаются в зобах
преимущественно осенью (октябрь 10,8%, ноябрь 28,6%) и весной (апрель
15%, май 14,6%); зимой—редко D%). Ягоды и семена шиповника идут в пи-
щу только осенью: август 27,1%, сентябрь 33,8%, октябрь 16,2%; в конце
октября ягоды опадают и в ноябре встречаемость падает до 7,1%. Из ягод
наибольшее значение имеет брусника, употребляемая в летне-осенний и ве-
сенний периоды. Начиная питаться зелеными ягодами в июле и августе
D,2 и 10,2%), рябчики в наибольшем количестве поедают созревшие ягоды
в сентябре—ноябре F4,2%); с выпаданием снега они исчезают из рациона w
вновь появляются в апреле—мае C5 и 27,1 %). Листья и почки брусники упо-
требляются в пищу в апреле C5%) и октябре E4,1 %). Ягодами черники кор-
мятся с июля по ноябрь: июль 54,1 %, август 67,8%, сентябрь 32,5%, октябрь
8,1 %, ноябрь 1,2%. Листья и почки черники употребляются в течение всего
бесснежного периода, но наибольшее значение имеют весной (апрель 55 %„
рябчик 125
май 25%, июнь 33%) и осенью (октябрь 67,6%, ноябрь 100%). Семена осоки
в заметном количестве фигурируют в питании главным образом в июле, ав-
густе и сентябре (встречаемость от 25,9 до 37,5%), семена марьяника в ав-
густе й сентябре D2,4 и 30,1 %). К второстепенным кормам относятся почки
рябины, охотно поедаемые весной (апрель 35%), ягоды можжевельника,
листья кислицы, ягоды костяники, ягоды красной смородины, плоды май-
ника, наземные части хвоща. Из случайных кормов следует упомянуть сережки
и почки ольхи, ягоды рябины, ягоды и почки черемухи, сережки и почки
осины, ягоды крушины, морошки, голубики, клюквы, толокнянки, водяники,
малины, поляники, семена и листья лютика, листья ожики, жимолости и др.
Животные корма встречены у 30 исследованных птиц. Существенное зна-
чение имеют лишь насекомые; пауки, улитки и многоножки поедаются весь-
ма редко C,4, 0,3 и 0,6% встреч). Из насекомых основное место занимают
перепончатокрылые B3,1%) и жуки A1,7%). Бабочки и мухи служат кор-
мом лишь изредка. Первостепенное значение имеют муравьи, которыми ряб-
чики наиболее интенсивно питаются в августе и сентябре E4,2, и 28,6%).
Значение остальных животных кормов, отнесенных к числу основных—листо-
грызы, личинки пилильщиков, гусеницы пяденицы, по сравнению с муравьями
невелико—не более 3,9%. Все случаи поедания этих кормов относятся к ию-
лю—сентябрю, листогрызы были найдены один раз в мае. В группе второстепен-
ных кормов зарегистрировано 9 видов, значение которых в годовом рационе
не превышает 0,9%; случаи их поедания отмечены почти исключительно в сен-
тябре. Таким образом, рябчик питается преимущественно нелетающими вида-
ми насекомых и пауками, причем главным образом в июле, августе и сентябре.
Для перетирания пищи и особенно для измельчения грубых зимних
кормов в желудке рябчика всегда находится мелкий гравий, зимой в большом
количестве (до 3—3,5 г в одном желудке), летом в несколько меньшем A,8—
2,9 г). Питание молодых отличается от взрослых большим потреблением жи-
вотных кормов и большим разнообразием видового состава. В первую декаду
жизни птенцы питаются, видимо, исключительно животной пищей—мелкими
паучками, личинками пилильщиков и цикад, комарами и муравьями.
Во второй декаде в кормовом рационе в небольшом количестве появляются
мелкие семена травянистых растений—кислицы, осоки, марьяника, фиалки
и лютика; основой же корма остаются животные виды пищи; мелкие пауки
A00%), личинки пилильщиков F0%), гусеницы пядениц D0%) и мелкие
жуки D0%). В третьей декаде сразу резко увеличивается удельный вес рас-
тительных кормов, главным образом ягод черники G7,8%) и семян кислицы
B2%); состав животных кормов остается без изменений. В первой декаде
августа значительно расширяется видовой состав поедаемых ягод: черники,
малины, костяники, красной смородины, морошки; важное место занимают
семена травянистых растений: лютика F1,5%), кислицы C8,4%), осоки
C0,7%), седмичника A5,8%), фиалки A5,8%); животная пища встречается
только у 76,9% птиц. Во второй и третьей декадах августа питание молодых
по составу растительных кормов сходно с питанием взрослых птиц; живот-
ная пища остается более разнообразной и встречаемость ее значительно выше
<88,9 и 83,3%), чем у взрослых; в состав ее входят муравьи, пауки,гусеницы
и личинки пилильщиков, мух и комаров. Животная пища, видимо, имеет боль-
шое значение для молодых и в сентябре. Механические примеси в желудках
молодых рябчиков в первую декаду жизни отсутствуют и появляются только
со второй декады. Количество поедаемой рябчиком пищи в разные сезоны
года сильно колеблется: в снежный период (ноябрь—февраль) в сутки съе-
дает 41 г, весной (март—май)—58 г, летом (июнь—август) 28 г, осенью (сен-
тябрь—октябрь) 27 г.
Фолитарек A939), исследовавший 35 желудков и 16 зобов птиц, добытых
в период с 18 августа по 5 сентября 1928 г. в районе р. Кан Красноярского
126 отряд куриные
края, зарегистрировал поедание 30 видов растительных и 26 животных кор-
мов. Можно предполагать, что разнообразие животных кормов относится
к молодым рябчикам. Наибольший удельный вес в кормовом рационе в этот
период у канских рябчиков занимают: ягоды боярышника B6,7% от веса
содержимого зобов), черники B2%), шиповника A2,5%), бузины A,2%),
почки, листья и семена березы E,8%), ягоды костяники C,6%), коробочки
с семенами грушанки зонтичной C,6%); животные корма составляют 9,2%
от веса содержимого зобов. Из животных кормов следует выделить коричне-
вого таежного муравья C1,2% встреч в зобах), пауков E6,2%), личинки и
коконы пилильщиков C1,2%), цикадку A8,7%), тлей A8,7%), листоедов.
A8,7%), кобылку A2,5%), совку A2,5%).
На Южном Урале (Кириков, 1948) главным зимним кормом являются
сережки и почки березы и ольхи. Весной поедаются бутоны и соцветия ивы
и осины, молодые листья черемухи, клубники, селезеночника, семена сос-
ны, костяники и майника, гусеницы и черви. Летом большое значение
имеют гусеницы насекомых, жуки, кузнечики и др., а из растительных
кормов—ягоды костяники, клубники, черники, малины, степной вишни,
брусники, а также семена злаков, осок, фиалок, марьяника. Осенью количест-
во кормовых растений резко снижается и в октябре состоит главным образом
из сережек и почек березы и ольхи и почек черемухи.
По данным Цвеленьева A938), вскрывшего 160 зобов и 220 желудков,
в питании рябчика на Алтае большую роль играют животные корма, употреб-
ляемые в течение 7 месяцев в году A9 апреля—15 ноября). Из общего числа
исследованных автором в этот период птиц (около 190 шт.) у трех насекомые
составляли 100% содержимого зобов, у семи—от 21 до 89%. Общее количество
встреч животных кормов в зобах за апрель—июнь 20,6%,за июль—сентябрь
45,5%; в желудках соответственно 40,6 и 44%. Подавляющее большинство
составляет отряд жуков, главным образом из семейства листогрызов и дол-
гоносиков, а также божьи коровки. Видное место занимают клопы, в част-
ности Acantosoma chemorchoidaiis, ежегодно в сентябре в массе появляющийся
в исследованной местности; в трех случаях было зарегистрировано в зобах
по 54, 55 и 71 клопу. Кроме того, в зобах встречены щелкун, жужелица,
кузнечик, цикадка, наездник, мухи, личинки бабочек, личинки пяденицы,,
пауки, в одном случае улитка. Муравьи, отмеченные Фолитареком в боль-
шом количестве у канских рябчиков, встречены только один раз. Говоря
о животных кормах, Цвеленьев не выделяет возрастные группы рябчиков,
между тем как значительная доля этих кормов несомненно приходится на долю
птенцов. Растительных кормов у алтайского рябчика зарегистрирована
59 видов. Характер питания и смена растительных кормов по сезонам сходны
с таковыми у печорского рябчика. Однако набор кормов и удельный вес
отдельных их видов довольно различен.
Так, в зимнем питании наряду с сережками и почками березы большое
место занимают сережки и почки ольхи (декабрь 50% встреч, январь и фев-
раль—еще больше), входящие у печорских рябчиков в число случайных
кормов. В некоторых зобах подсчитано 95,. 106, 125 сережек, составлявших
75—80% содержимого зобов. Значительно больший удельный вес зимой зани-
мает рябина, составляющая до 50—70% содержимого зобов; она относится
к группе основных кормов. Зимний рацион более разнообразен, чем на
Печоре, и включает 15—16 видов растений. В весенний период поедают-
ся 25 видов, из которых большую роль играют цветы и листья ветреницы
(86,7% встреч, или 52% к общему весу содержимого зобов), вовсе не фигури-
рующей у печорских птиц. В летне-осенний период, в отличие от печорских
птиц, важное место в питании играют ягоды, почки и ветки рябины D2,5%
встреч), таволга A8%), ягоды красной смородины A7%), клевер ползучий
A8%), земляника A2%), малина (9%). Как и в предыдущие сезоны, в этот
РЯБЧИК 127
период корма более разнообразны по сравнению с Печорой: как правило,
все исследованные зобы содержали до 15 видов растений, а всего за летне-
осенний сезон зарегистрированы 44 вида растительных кормов.
Промысел. Из всего количества добываемых в СССР рябчиков большая
часть падает на описываемый подвид, населяющий обширные территории
страны, в том числе и важнейшие промысловые районы. Способы добывания,
как и у предыдущего подвида.
Описание. Размеры. Длина крыла самцов G5) 163—195, самок E4) 161—182,
в среднем 174 и 173 мм. Вес самцов в позднеосенний и зимний периоды C2) 360—500,
самок B4) 340—490, в среднем 435 и 422 г (по Донаурову, 1947).
Окраска. От номинального подвида отличается более светлым оперением как
верхней, так и нижней частей тела за счет более светлого (а не тусклого) окраса и более
широких белых вершинных каемок и стержневых центров, а следовательно, и более узких
и реже расположенных темных полос и пятен. Спина светлопепельно-серая с узкими тем-
ными поперечными полосами, без рыжего (как у номинального подвида) или охристого (как
у амурского) оттенка. Плечевые партии значительно светлее, чем у других подвидов, обра-
зуя почти сплошную белую полосу. Более широка и ярче выражена белая полоса вокруг,
темного горла у самцов.
Амурский рябчик Tetrastes bonasia amurensis Riley
Tetrastes bonasia amurensis. Riley. Proc. Biol. Soc. Wash., 1916, XXIX, январь, стр. 17,
Средняя Маньчжурия (Гирин).
Синонимы. Tetrastes bonasia ussuriensis. Бутурлин. Орн. Вестн., 1916, № 4,
стр. 222, ст. Бочарове р. Бикин и озеро Ханки. — Tetrastes bonasia yamashinai.
М о mi у a m a. Annot. Orn. Orien. 1928, I, ДО> 3, стр. 232, Южный Сахалин.
Распространение. Ареал. Населяет Приамурье к востоку от бассейна
р Зеи, Уссурийский край, Сахалин, Маньчжурию, Северную Корею.
Характер пребывания. Местами совершает значительные
зимние кочевки. Так, Шульпин A936) указывает, что на Федоровской сопке
в устье Сейфуна, поросшей дубовым лесом и отделенной от остальных лесных
массивов широким безлесным пространством, рябчик летом очень редок,
но довольно обыкновенен поздней осенью и зимой. По наблюдениям Шамы-
кина в Сихотэ-Алиньском заповеднике, рябчики совершают правильные вер-
тикальные перекочевки: населяющие восточные склоны Сихотэ-Алиня на
зиму переселяются в верхние пояса гор и на северные склоны сопок; живу-
щие по западным склонам—в нижние пояса гор. По данным указанного автора
сезонные перемещения вызываются не кормовыми условиями, ибо на зиму
птицы нередко переселяются в менее кормные места, а условиями снего-
вого покрова, с которым тесно связано наличие благоприятных мест зимних
кочевок. В предгорьях и средних высотах Западного Сихотэ-Алиня, а также
на южных экспозициях сопок, снеговой покров до февраля или вовсе отсут-
ствует, или настолько незначителен, что не может представить надежного
убежища ночующим в снегу рябчикам; последние вынуждены подниматься
в верхние пояса гор или на северные склоны сопок с более мощным снего-
вым покровом. О большом значении снегового покрова как защитного фак-
тора свидетельствует ряд случаев массовой гибели рябчика в малоснежные
зимы. На западных склонах Сихотэ-Алиня зима, как правило, сопровождает-
ся более ранними снегопадами и относительно равномерным распределением-
снегового покрова. Осенние и зимние перекочевки происходят здесь в об-
ласть предгорий: в поймы больших горно-таежных рек (Колумбэ, Арму,
Татба, Иман) и в низовья ключей, где рябчики находят самые благоприят-
ные условия для зимовки.
Биотоп. По Шренку A861), на Нижнем Амуре рябчик держится
преимущественно по краю леса по берегам рек, там, где к лесным массивам
примешиваются береза, осина, тополь, ольховые и ивовые кустарники, юж-
нее же—в светлом лиственном лесу с обильным подлеском по склонам гор
128 ОТРЯД КУРИНЫЕ
и скалистым берегам рек. Нередко как летом, так и зимой встречается по
поросшим ивами островам и берегам Амура, Горина и Уссури. Шульпин
< 1936) на Нижнем Амуре встречал рябчиков летом главным образом по ли-
ственным участкам леса, по просекам, по опушке хвойной тайги; в моховых
лесах, густых пихтовниках, моховых, хотя и ягодных болотах рябчик дер-
жится сравнительно редко и главным образом по травянистым полянам, по
ручьям среди травянистых берегов. Гнездится и в Охотской тайге по Сихотэ-
Алиню в верховьях Имана; выводки попадаются здесь в довольно глухих
уголках с пухлым моховым ковром и порослями багульника, но все же ряб-
чик предпочитает солнечные места и более лиственный и травянистый ха-
рактер растительности. В южной половине края наиболее плотно населены
смешанные леса Маньчжурской тайги, причем, несмотря на горный харак-
тер местности, рябчики придерживаются не только ручьев, а даже больше
склонов гор, лесов по широким падям и т. п. В чисто лиственных участках
встречается в сравнительно небольшом числе, чаще в дубовых с густым
подлеском. По наблюдениям Шамыкйна, в Сихотэ-Алиньском заповеднике
рябчик населяет все вертикальные ландшафты, начиная от дюнных всхолм-
лений берега моря, поросших крупноплодным шиповником и зарослями
кустарниковой формы монгольского дуба, вплоть до угнетенных подголь-
цёвых ельников. Как в поясе высокогорной тайги, так и в прибрежной зоне
обычен. Часто встречается даже в таких малокормных угодиях, как листвен-
ничные мари, где под пологом редколесья господствуют заросли багуль-;
ника и сфагновые мхи. Придерживается нижних и средних поясов гор и
редко выходит за их пределы во время суточных и сезонных миграций. Коли-
чественный учет показал, что основная масса рябчика сосредоточена в пре-
делах, пояса смешанных маньчжурских лесов, где в годы с нормальным уро*
жаем кормов численность его никогда не падает ниже 80%; в поясе Охотской
тайги обитает, в зависимости от времени года, от 9,5 до 19,9% всего поголо-
вья. Особенно высока плотность птиц в пойменных лесах и уремах нижних
течений горных ключей. Смешанная тайга привлекает рябчика богатством
травянистой растительности, дающей неограниченный выбор разнообразных
зеленых кормов, ранних плодов, ягод и семян, массу насекомых и моллю-
сков. По среднему и нижнему течению Имана населяет лесистые острова из
высоких широколиственных деревьев с густым покровом из папоротника,
часто встречается также в смешанной тайге по сопкам и гораздо реже в темно-
хвойной тайге из кедра, пихты и ели; в последних случаях держится не в глу-
хих участках, а по краям тайги, где наряду с хвойным лесом попадается мел-
колесье по берегам речек и заболоченных низин родникового происхожде-
ния (Спангенберг, 1941). По наблюдениям Воробьева в Уссурийском крае,
начиная от светлых дубовых лесов с подлеском из различных кустарников
широко населяет тайгу маньчжурского типа, а также заходит в горные елово-
пихтовые леса с богатым моховым покровом почвы.
Чисденность. На большей части ареала обыкновенен.
Экология. Размножение. По наблюдениям Шамыкйна в Сихотэ-
Алиньском заповеднике, весенний свист—-с первой декады марта, когда стоят
еще большие морозы. Примерно с этого времени начинается постепенная
разбивка на пары. Вместе с пробуждением половой активности замечается
увеличение гонад. В конце марта длина яичника в среднем 10 мм, к 5 ап-
реля—16—24 мм, диаметр отдельных яиц 6—10 мм\ к середине апреля
яичник достигает почти полного развития—35—37 мм9 диаметр яйца 17—
19 мм\ в конце апреля—полностью сформированные со скорлупой яйца,
диаметр отдельных 19—20 мм, У самцов длина левого семенника в конце
марта 7,8—8,1 мм, в конце апреля 11,2—11,9 мм, во второй декаде мая 9,4—
9,7 мм, в состоянии покоя—только к середине июня (к разгару насиживания).
Ток рябчика распадается на два периода: первый—с начала апреля до се-
РЯБЧИК 129
редины мая, второй—с середины мая до начала насиживания. В первый
период птицы ведут себя более активно. Токующие пары в это время часто
сходятся вплотную, образуя как бы общее токовище; происходит погоня друг
за другом, драки и спаривание. Токуют с восходом солнца до 10—11 час.
утра и вечером с 5—6 час. до вечерней зари. Нормально каждая пара имеет
свой район токования, и самец не терпит присутствия на своем участке дру-
гой пары. На манок токующий самец моментально отзывается посвистом,
а при повторном звуке манка оставляет самку и летит к предполагаемому со-
пернику. Желая обнаружить место расположения вторгшейся пары, он на-
чинает токовать: распустив хвост, раздувшись и бороздя крыльями, он быстро
переходит по отлогому суку дерева, делая резкие повороты и издавая харак-
терный свист. Самка не покидает самца и бежит на его посвист, периодиче-
ски издавая более грубый и отрывистый свист. Самец в период тока большую
часть времени проводит на деревьях, перелетая с одного на другой; часто
залетает на вершины самых высоких деревьев. Спускается на землю только
при спаривании или драках. Самка больше находится на земле, взлетая
на дерево только в крайних случаях.
В первый период тока самцы кормятся мало, главным образом в полуденный
перерыв, и на протяжение всех суток имеют почти пустые зобы; вес содер-
жимого зоба не превышает 5—10 г, в то время как осенью и зимой соста-
вляет 30—35 г. Вес самцов в этот период заметно снижается. Как было от-
мечено и для сибирского рябчика, падение веса самцов совпадает с увели-
чением семенников, снижение веса содержимого зобов идет параллельно
с ростом половой активности. С увяданием семенников вес стабилизируется
и к началу линьки (начало июня) заметно повышается. Самка, наоборот,
в период тока много кормится и вес ее увеличивается до начала насижива-
ния, после чего снижается.
Вторая половина тока протекает менее активно. В это время птицы обо-
собляются парами и держатся в районе гнезда. К концу мая некоторые сам-
ки приступают к насиживанию, но ток еще продолжается. По утрам, вече-
рам и в полдень пары сходятся на жировках. Кладка яиц в нижнем поясе
гор Сихотэ-Алиня начинается с последней трети апреля и растягивается до
конца мая, а в ряде случаев до начала июня; в яйцеводах готовых
к откладке яиц находили в 1935 г. 1 июня, в 1938 г. 2 июня, в 1939 г. 4 июня.
8 верхнем поясе гор сроки начала и конца кладки запаздывают на 10—
12 дней.
По Черскому A915), в окрестностях Нововладимировки (бассейн Имана)
в 1911 г. рябчики начали свистеть 28 марта, когда южные склоны возвышен-
ностей освободились от снега и стали покрываться первыми цветами; ко вто-
рой половине апреля песня смолкла. Несущаяся самка была добыта 25 апреля.
Шульпин A936) в мае 1926 г. уже не слышал трелей рябчика, но в устье Сей-
фуна в холодную и пасмурную весну 1927 г. рябчик впервые начал сви-
стеть только 17 апреля. Спангенберг A940) в низовьях Имана в 1938 г. до-
был 4 и 12 мая двух самцов во время песни, а 27 мая из яйцевода самки
извлек яйцо, готовое к сноске. По наблюдениям Воробьева в Уссурий-
ском крае, первый свист—в конце марта, начало кладки в 20-х числах ап-
реля. Гнезда, по наблюдениям Шамыкина, устраиваются в захламленных
и заросших кустарником участках тайги, под валежинами или у стволов
больших сырорастущих деревьев. Присутствие поблизости человека мало
смущает птицу. Шамыкин приводит пример, когда два гнезда располагались
рядом с тропой в 13 и 21 ^ от лесной избушки. Полная кладка 7—10 (Во-
робьев) и до 14 яиц (Шамыкин). Календарные сроки размножения в условиях
различных высот значительно колеблются, что видно из таблицы, состав-
ленной Шамыкиным по наблюдениям в Сихотэ-Алиньском заповеднике
в период с 1937 по 1945 г.
9 Птицы СССР, т. IV
130
ОТРЯД КУРИНЫЕ
Поелгооья
Средние высоты ....
Высокогорья
Кладка яиц
нача-
ло
21/IV
26/1V
3/V
разгар
15/V
24/V
27/V
конец
2/VI
5/VI
9/VI
Насиживание
нача-
ло
11/V
19/V
26/V
разгар
1/VI
9, VI
18/VI
конец
6/VTI
11/VII
17/VII
Выход птенцов
нача-
ло
2, VI
9/VI
20/VI
разгар
13/VI
16/VI
29/VI
конец
6/VII
11/VII
24/VII
Сроки размножения в одних и тех же вертикальных поясах гор сильно»
растянуты. Воробьев связывает это с гибелью кладок у части птиц; в 1948 г.
им найдено 16 июня гнездо с надклеванными яйцами, в первых числах июня —
еще нелетные выводки, 20 июля—едва летающие птенцы. В Сучане самка
с наседными пятнами была добыта 1 июня 1926 г., 20 июня здесь встречен
выводок пуховичков примерно трехдневного возраста; такого же возраста
птенцы были встречены 26 июня 1928 г. в верховьях Имана; в Пхусуне вы-
водок птенцов с хвостом в 5 см отмечен 3 июля 1927 г.; на озере Кизи
птенцы величиной в полматки найдены 29 августа 1928 г. (Шульпин, 1936).
В низовьях Имана Спангенберг A940, 1941) гнездо с проклеванными яйцами
нашел 3 июня, птенцов двух-трехдневного возраста—8 июня, размером с дят-
ла—17 июня.
Смертность в выводках рябчика довольно значительна и особенно в пер-
вые дни жизни. Шамыкин указывает, что в первые 5—10 дней число птен-
цов в одном выводке падает с И—14 до 6—10 шт.; в нормальные годы отход,
птенцов в первые дни составляет 20—25%, а в неблагоприятные годы 40—
50%. По данным количественного учета в Сихотэ-Алиньском заповеднике
за 7 лет, с 20 до 30 июня отход птенцов составляет 19,4%, к 10 июля 31,2%,
к 1 августа—33,6%, к 1 сентября—36%, к 1 октября—40%. В отдельные годы
гибель птенцов колеблется от 22,8 до 55,8%, а в среднем за 7 лет A937 —
1945) равна 37,4%. Причина большой смертности птенцов состоит в небла-
гоприятных метеорологических условиях первой половины лета, особенна
на восточных склонах Сихотэ-Алиня: преобладающие южные, восточные
и юго-восточные ветры сопровождаются резкими похолоданиями, туманами
и холодными затяжными дождями. Похолодания и затяжные дожди резка
снижают активность насекомых, составляющих основу питания птенцов.
в первые дни их жизни; к тому же птенцы вынуждены большую часть суток
находиться под маткой. Чрезмерная сырость, холод, а главное голодовка вызы-
вают массовую гибель цыплят. На западных склонах Сихотэ-Алиня, изолиро-
ванных от отрицательного воздействия моря, климатический режим более
благоприятен и смертность птенцов ниже, чем на восточных склонах. В бо-
лее благоприятных условиях оказываются и выводки, обитающие в поясе
Охотской тайги, где благодаря более поздним срокам размножения появление
птенцов совпадает с прекращением ненастья. Немалую роль в снижении чис-
ленности молодняка играют мелкие хищники, и в первую очередь колонок.
Темп роста птенцов характеризуется следующими средними весовыми по-
казателями: в первые дни жизни 12—14 июня средний вес птенцов равен 9,1 г,
через 10 дней—22 июня—19,4 г, 30 июня—30,2 г, 10 июля—97,9 г, 20 июля
161,4 г, 31 июля—192,1 г, 15 августа—238,5 г, 31 августа—310,8 г, 15 сен-
тября—342,9 г, 30 сентября—369 г, 5 октября—372,5 г (Шамыкин, Сихотэ-
Алиньский заповедник). Наиболее быстро рост птенцов происходит в пер-
вый месяц жизни. Прирост веса, как и размеров тела, идет неравномерна
и тесно связан с ходом линьки и формированием оперения: бурная линька
снижает темп прироста. Первый сравнительно резкий прирост веса наблю-
дается в первые дни жизни, когда птенцы имеют пуховой наряд. В последую-
РЯБЧИК 131
щие дни в связи с формированием первого перьевого наряда темп прироста
снижается, а по окончании развития первого контурного пера F—10 июля)
вновь увеличивается; в дальнейшем некоторое увеличение прироста заме-
чается в конце августа, когда ход линьки замедляется. Молодые заканчивают
свое развитие и надевают первый годовой наряд в октябре: самцы к середине,
самки—к концу месяца.
Осеннее токование рябчиков, по Шульпину A936), начинается с поло-
вины сентября.
Линька. Последовательность смены наряда, как и у предыдущего
подвида. Птенцовый наряд начинает формироваться с первых же дней жизни
и заканчивается, по наблюдениям Шамыкина, к 17—20 июля; первый годо-
вой наряд формируется с июля до конца октября. В течение свыше четырех
месяцев оперение молодых сменяется почти беспрерывно. По Шамыкину,
линька взрослых начинается еще до окончания тока и длится в течение всего
лета, осени и зачастую захватывает начало зимы. Явные признаки ее обнару-
живаются уже в первой половине мая, и нередки случаи, когда линные особи
добывались в первой декаде декабря. Сроки линьки у самцов и самок различ-
ны. Самцы начинают линять со второй декады мая и заканчивают к середине
октября. У самок линька начинается одновременно с самцами, но на период
насиживания прекращается и вновь возобновляется вскоре по выходе птен-
цов, полностью заканчиваясь к началу ноября. Порядок линьки у самцов
и самок одинаков. В первую очередь линяют крайние верхние и нижние
кроющие хвоста, задняя и средняя часть брюха, подбородок и верх мантии.
Дальнейшее распространение линьки идет за счет расширения указанных
первоначальных очагов. По той же схеме идет формирование первого взрос-
лого наряда у молодых птиц. Рулевые перья начинают выпадать у самцов,
в начале июня, а в единичных случаях в середине мая. Первым выпадает
четвертое или пятое перо, а затем линька идет к внутренней паре и к наруж-
ным перьям, при этом к последним значительно быстрее. Смена маховых
перьев идет от внутренней части крыла к наружной, причем очаги линьки
возникают в каждой партии махов отдельно. Весной, до начала тока, у ча-
сти рябчиков замечается частичная линька боков головы, шеи и туловища; при
этом линьке подвержена только часть перьев. Такая линька наблюдается
в период с середины—конца марта до 10—15 апреля. Шамыкин полагает,
что ранневесенней линьке подвергаются молодые особи, в которых не пол-
ностью вылиняли перья осенью. По Воробьеву A938), линька взрослых
самцов начинается с половины июня и продолжается до октября; самки ли-
няют позднее.
Питание. По наблюдениям Шамыкина в Сихотэ-Алиньском заповед-
нике, корма уссурийского рябчика характеризуются большим разнообра-
зием и насчитывают около 80 видов, из которых свыше 20 относится к живот-
ным. Все корма можно разбить на пять групп, каждая из которых состав-
ляет основу питания в определенный сезон года.
1. Грубые корма—сережки, почки и концевые ветки деревьев и кустар-
ников. Эта группа составляет основу питания с середины октября до конца
апреля, т. е. 6—7 месяцев в году. В октябре указанные корма занимают
53,6% содержимого зобов, зимой—98,5% (почки и концевые ветки 86,7%,
сережки 13,3%), ранней весной 74,5%. Удельный вес за год 50,4%.
2. Зеленые корма (листья, цветы) имеют большое значение в весеннем
питании, но встречаются с конца марта (появление проталин) до глубокой
осени. В весеннем рационе (начало мая—конец июня) зелень занимает 67,2%,
к середине июля с появлением плодов, семян и ягод удельный вес ее падает
до 17,4%, а в осеннем питании до 5,4%. Удельный вес за год 15,4%.
3. Плоды и семена травянистых, древесных и кустарниковых растений —
обычный корм в летнем и осеннем питании. Употребляются с конца июня
9*
132 отряд куриные
вплоть до первых снегопадов, а в бесснежные зимы до конца ноября—начала
декабря. Во второй половине лета (июль—август) удельный вес этой группы
составляет 44,7%, с массовым появлением ягод снижается до 17,4%, средний
годовой 10,6%.
4. Ягоды деревьев и кустарников—важнейший корм во второй половине
лета и осенью. В июле ягоды занимают в кормовом рационе 27,9%, в ав-
густе 31,2%, в сентябре67,3%, в октябре 25,5%, в ноябре 24,7%. Удельный
вес в летне-осеннем питании 30,3%, в годовом 15,8%.
5. Животные корма занимают большое место в летнем питании, особенно
птенцов и линяющих взрослых особей. Насекомые встречаются с конца апре-
ля и обычны еще в сентябре. Наибольшее значение приобретают с конца
июня до середины июля, составляя в это время 28,5% содержимого зобов
и 30—40% по частоте встреч. Удельный вес за год 7,6%.
В состав зимних кормов входит 17—22 вида. В зоне кедрово-широко-
лиственных лесов к ним в первую очередь относятся: актинидия, лимонник,
виноград, реже ребристая береза, маньчжурская лещина, козья ива, чозе-
ния; в полосе приморских стаций маньчжурская лещина заменяется обык-
новенной лещиной, а по мере продвижения в пояс темнохвойной тайги обе
лещины вытесняются березами и ольхами. Грубые зимние корма дают также
аянская ель, белоКорая пихта, клен, липа. Из ягод в зимнее время употре-
бляется шиповник, можжевельник, омела. Количество поедаемого зимой
корма велико: содержимое зоба добытых в январе и феврале рябчиков весит
в среднем 33—40 г, а в отдельных случаях свыше 50 г (против 12—15 г лет-
них птиц). Весной пищу рябчика составляют как грубые, так и зеленые кор-
ма, включающие от 17 до 24 видов. Группа грубых кормов представлена
главным образом ивой, сережки которой дают обильный и легко доступный
корм. В состав зеленых кормов входят 19 видов, в том числе кислица, лесной
мак, ветреница, хохлатка, черемуха обыкновенная. Кроме того, в весеннем
питании фигурируют 5—6 видов насекомых: красный клоп, трубковерт
осиновый, цикадки, слоники. В летнем питании фигурирует около 60 видов.
Из них 19 видов ягод главным образом жимолости, смородины, черемухи,
голубики, малины, терна татарского, актинидии. Большое место занимают
семена травянистых растений, а также насекомые, насчитывающие свыше
20 видов: личинки пилильщиков, гусеницы бабочек (преимущественно пяде-
ниц), листоеды, златки, лубоеды, долгоносики, хрущи, цикадки, лесные
клопы, саранчевые, грибные мухи и др. Часто также поедаются сухопутные
моллюски. В наибольшем числе животный корм поедается в июле, и не толь-
ко птенцами, но и взрослыми самками и линяющими самцами. Осенние
корма также разнообразны и также насчитывают около 60 видов. Главная
масса их падает на ягоды и семена растений. Из грубых кормов поедаются
мужские сережки березы, позднее ольхи и ивы; почки встречаются реже.
Среди большого числа поедаемых рябчиком растений имеется ряд
таких видов, которые фигурируют в течение всего года. К таким в первую
очередь относятся актинидия, лимонник и виноград, почки, зелень и ягоды
которых птицы употребляют во все сезоны года.
Шульпин A936) в зобах рябчиков находил жолуди монгольского дуба,
семена бобовых, кончики веток хвойных, цветы амурского горицвета; у пу-
ховых птенцов—исключительно нежные насекомые, мелкие гусеницы. На-
блюдения Воробьева дают сходную с описанной выше картину питания
амурского рябчика. В апреле большое значение имеют почки деревьев и
кустарников. По мере стаивания снега птицы переходят на зелень весенней
эфемерной растительности, в частности охотно склевывают цветы амурского
горицвета, а несколько позднее—развертывающиеся почки черемухи, амур-
ской сирени и зеленую хвою пихты. В конце июня у молодых в зобу и же-
лудках обнаружены различные гусеницы. В июле и августе приобретают
ОБЫКНОВЕННЫЙ ПЕРЕПЕЛ J33
важное значение ягоды. Особенно охотно поедаются ягоды черемухи, лимон-
ника» винограда, жимолости, бересклета, калины и амурской сирени; ягоды
продолжают играть значительную роль в сентябре и октябре. Осенью в зобах
и желудках часто попадаются различные кобылки и реже гусеницы пядениц.
В ноябре в зобах обнаружены мелкие жолуди. С наступлением зимы пище-
вой режим резко меняется, становится однообразным. Основу его составляют
почки деревьев и кустарников, особенно почки граба, мелколистого клена,
мелкоплодника, лещины маньчжурской и березы серебристой. В долинных
же лесах поедаются главным образом почки черемухи и различных ив.
Кедровые орехи, отмеченные Сушкиным в качестве корма рябчика на Алтае,
у амурского рябчика не встречены. Для амурского рябчика характерно
большое разнообразие кормов, что, несомненно, связано с разнообразием
и богатством растительности края, а также с разнообразием стаций, в кото-
рых обитает рябчик.
Промысел. Промысловое значение амурского рябчика потенциально
значительно, но используется охотниками совершенно недостаточно ввиду
богатства края другими видами дичи, особенно зверем.
Описание. Размеры. Длина крыла самцов B2) 168—185, самок A3) 164—184.
в среднем 175 и 173 мм.
Окраска. Чуть светлее номинального подвида, но темнее сибирского. Кроме того
хорошо отличается охристыми тонами в оперении верха и низа вместо рыжеватых у номи-
нального подвида и более нейтрального светлосерого—у сибирского.
СЕМЕЙСТВО ФАЗАНОВЫЕ PHASIANIDAE
РОД ПЕРЕПЕЛЫ COTU RNIX BONN ATE RRE, 1791
Тип Tetrao coturnix
9. Обыкновенный перепел Coturnix coturnix L.
Tetrao coturnix. Li nnaeus. Syst. Nat., Изд. X, 1, 1758, стр. 161, Швеция.
Синонимы. Coturnix communis. Bonnaterre. Tabl. Enc. et Meth., 1, 1791, стр. 217
новое имя для Tetrao coturnix.—Coturnix dactylisonans. Meyer. Vog. Liv.—und Esth-
lands, 1815, стр. 167 новое имя для Tetrao coturnix.
Русские названия. Перепел, перепелка—почти повсеместно распространенное,
народное название. В некоторых местах (р. Кама) звукоподражательное—подполюшка.
Распространение. Ареал. На гнездовье распространен от о-вов-
Атлантического океана (Азорские, Канарские, Британские, Фарерские)
через всю Евразию до Северного Китая, Японии и Кореи. К северу гнездится
до 65° с. ш. в Скандинавии и Финляндии, до 64° на Урале и до 63°10' на Ени-
сее; к югу область гнездования перепела включает Африку (до мыса Доброй
Надежды), Мадагаскар, Палестину, Месопотамию, северную часть Белуджи-
стана, Северо-Западную Индию, Монголию, Северный Китай (Кукунор,
бассейн Желтой реки).
Характер пребывания. Повсюду перелетная птица, только
в Южной Африке и, видимо, на Мадагаскаре—оседла.
Биотоп. Долины рек и луговые поляны в таежной и лесной зонах,
лесостепь, степь, долины рек и луга в предгорьях и горах, культурный ланд-
шафт (посевы хлебов, клевера, картофеля, сады, огороды и т. п.).
Подвиды и варьирующие признаки. В Европе, Марокко и Северной Африке, в Азии
до Байкала, подножья Кентэя и Северо-Западной Индии распространен д. с. coturnix
Li nnaeus, 1758; в Забайкалье, Приамурье, Корее, Северном Китае и Японии—С. с.
japonica TemmincketSchlegel, 1842; на Канарских о-вах и на Мадере—С. с.
confusa Н а г t е г t, 1917; на Азорских о-вах С. с. conturbans Н а г t e г t, 1917 (очень
плохо отличимый от следующего подвида); в Южной Африке и на Мадагаскаре—С. с. afri-
сапа Temminck et Schlegel, 1849.
134
ОТРЯД КУРИНЫЕ
Отличия подвидов сводятся к незначительным особенностям окраски и величины;
С. с. japonica хорошо отличим от С. с. coturnix по голосу; С. с. africana ведет оседлый
образ жизни. Географические изменения экологии (календарь периодических явлений,
особенности размножения и трофических связей) изучены недостаточно.
Карта 10. Распространение перепела Coturnix coturnix
1—С. с. coturnix, 2—С. с. japonica, 3—С. с. confusa, 4—С. с. conturbans, 5—С. с. africana;
а—граница гнездовий; б—не вполне выясненная граница гнездовий; г—зимовки; цифры
в кружочках—номера зимующих подвидов
Обыкновенный перепел. Coturnix coturnix coturnix L.
Синоним. Coturnix communis orientalis. Bogdanow. Consp. Av. I. Rossici, 1884,
стр. 44, Сибирь.
Распространение. Ареал. Населяет Британские и Фарерские о-ва,
всю Европу, острова Средиземного моря. В Скандинавии и Финляндии про-
никает до 65° с. ш. На юге область распространения охватывает Марокко
и Северную Африку до Египта (где редок), Палестину, Месопотамию, Иран,
северную часть Белуджистана и Северо-Западную Индию навосток до долины
р. Хара в Северо-Восточной Монголии (подножья Кентэя).
В пределах СССР обыкновенный перепел гнездится очень широко. Се-
верная граница области его распространения в Карелии точно не установле-
на; гнездится у Онежской губы Белого моря и у Архангельска; далее навос-
ток гнездится в бассейне Вычегды и Печоры, спускаясь по последней до
Савинобора F3с30' с. ш.; Дмоховский, 1933); Сабанеев A874) на основании
опросных сведений указывает на находки перепела на р. Ижме у 64° с.ш.,
а Портенко A937) слышал однажды бой перепела на восточном склоне Сум-
мях-нёра (Приполярный Урал, 64° с. ш.). Резцов A904) слышал крик пере
пелов в районе Чердыни и по лугам р. Кольвы (Северный Урал). Граница
на Нижней Оби не установлена; гнездится по долине р. Туры (Ларионов
ОБЫКНОВЕННЫЙ ПЕРЕПЕЛ 135
1926), в большом количестве в окрестностях Тобольска (Тарунин, 1928),
обычен в районе Тары (Ушаков, 1913) и довольно многочислен в Нарым-
ском крае (Аникин, 1902). В бассейне Енисея доходит до 63°10' с. ш.; Скалой и
Слудский A941) слышали крик перепелов на р. Елогуе, а 22 июля 1932 г.
добыли самца и самку у ее устья. Далее на восток обычен в верхнем тече-
нии р. Нижней Тунгуски от истоков до с. Ербогачен, т. е. примерно до61°20'
с. ш. (Ткаченко, 1937). В Якутии перепел встречен под Олекминском (Туга-
ринов, 1934), хотя для более юго-западных районов он не указывается;
повидимому, эта самая крайняя северо-восточная точка встречи этого под-
вида. На юге гнездовая область перепела повсюду выходит за государствен-
ные границы Советского Союза. Восточная граница гнездовой области этого
подвида ограничена западным побережьем оз. Байкал и долиной р. Иркут.
В пределах этого большого ареала обыкновенный перепел, даже при
наличии подходящих биотопов, распространен неравномерно. Так, его нет
на Среднем Урале, причем по восточным склонам перепел ближе подходит
к хребту, чем по западным (Сабанеев, 1874); в районе Ильменского заповед-
ника он гнездится повсеместно, кроме Златоустовского участка (Снигирев-
ский, 1929); только на пролете одиночки отмечены в степях Мангышлакского
п-ова и не найдены на гнездовье на Усть-Урте (Долгушин, 1948) и т. п.
В небольшом количестве зимует в Средиземноморье, в Юго-Западной
Европе и в южных частях СССР. Нерегулярно встречается зимой на юге
Украины: добывались в январе 1911 г. под Херсоном (Пачосский, 1911),
в конце ноября уже при значительном снеговом покрове встречены в низовьях
р. Дона (Альфераки, 1910). Регулярно, но в небольшом количестве зимует
на Кавказе: в долине р. Риона и у Батуми (Вильконский, 1897; Домбровский,
1913); по всей низменности Восточного Закавказья (Сатунин, 1907), причем
в Гельской котловине в январе за несколько дней было поднято 12 шт., а до-
бытые экземпляры были жирные и упитанные (Вейцман, 1933); одиночка
добыт у Дзауджикау 6 января 1916 г. (Беме, 1926); зимуют в долине р.Ала-
зани (Чхиквишвили, 1930), крайне редок зимой в Астраханском крае (Хлеб-
ников, 1930), по опросным сведениям зимует в долине р. Сулака (Предгорный
Дагестан; Туров и Красовский, 1933), под Кюрдамиром (Азербайджан;
Спангенберг и Туров, 1950), в Мильской степи (Формозов, 1946), в Талыше
и на прилегающих островах Каспия (Тугаринов и Козлова, 1935, 1938;
Козлова, 1950). Единичные особи зимуют в Волжско-Уральских песках
(Бостанжогло, 1911; Формозов, 1950), иногда пытаются зимовать по неубран-
ным участкам проса в Среднем Поволжье, в "Донских степях и Воронежской
обл. (Формозов, 1946).
Крайне сомнительны и требуют подтверждения указания о том, что пе-
репела нередко в большом количестве зимуют по ельникам Среднего Урала
Щолевская и Нязенетровская лесные дачи; Сабанеев, 1874).
Далее на восток одиночные экземпляры зимуют в Бадхызе и Карабиле
(Туркмения, коллекционные экземпляры); изредка зимует в дельте р. Или
(Штегман, 1949) и в Гиссарской долине (Таджикистан; Иванов, 1945, 1949);
Северцов A873) указывает, что перепела зимуют по саксаульникам и
солонцам на высоте 200—350 м над уровнем моря в III и IV участках Турке-
станского края (Кара-Тау, Западный Тянь-Шань, долина Зеравшана, горы
между ним и Сыр-Дарьей и т. п.) . Эти данные, как давние, требуют проверки.
В ноябре—декабре одиночные птицы встречались в степях под Барнаулом
(Велижанины, 1929), в долине Чулышмана (Алтай; Сушкин, 1938). Прже-
вальским и Козловым встречен зимой в долине Тарима и Хайду-Гол (Северо-
Западный Китай).
Основная масса птиц описываемого подвида зимует в северной части
тропической Африки (Южная Сахара, Гамбия, район озера Чад, Египет,
Абиссиния; отдельные экземпляры в Северном Конго, Нигерии, Кении,
136 ОТРЯД КУРИНЫЕ
Учанде, т. е. до экватора; Гроте, 1930—1937), в Аравии, в Индии (на юг до
Траванкора и Мадраса).
Характер пребывания. Перелетная птица. Особи, зимую-
щие в южных частях1 ареала, в том числе и в пределах СССР, вероятно не
местные, а прикочевавшие с севера птицы (что требует подтверждения путем
кольцевания). Данные кольцевания скудны: молодой самец окольцован
17 августа 1939 г. в пойме р. Дона (с. Троицкое) и добыт там же 30сентября
1939 г.; взрослая птица окольцована 20 мая 1935 г. в долине р. Роны (Фран-
ция) и добыта 30 июня 1935 г. в Ленинградской обл. (с. Колчаново); птица
окольцована в Италии (дата и место неизвестны) и добыта 18 июля 1950 г.
вблизи г. Проскурова.
Даты. На местах зимовки в Африке держатся с конца августа по
конец марта—конец апреля (в Северной Нигерии появляются уже в сентяб-
ре, в Кении в октябре, в тропической Африке в феврале-марте; Гроте, 1937);
на Иерихонской равнине в марте и в первые дни апреля собираются в сотен-
ные стаи, почти полностью улетая в начале мая (Палестина; Давыдов, 1898);
в Индии перелетные птицы появляются в августе и остаются до конца
апреля; особенно многочисленны перепела с октября по начало апреля
(Бекер, 1928).
Сроки пролета колеблются по годам в зависимости от условий погоды.
В Северо-Восточном Иране пролет хорошо заметен уже в конце марта—нача-
ле апреля (Зарудный, 1900), в Кашгарии в конце марта (Судиловская, 1936;
по сборам Дивногорского). В первой половине апреля прилетают в Молда-
вию (Остерман, 1912), Крым (Молчанов, 1906), Одесскую (Куликовский,
1913), Херсонскую обл. (Браунер, 1894), в окрестностях Мариуполя первые
птицы встречены 20 марта 1905 г. и 25 марта 1906 г. (Боровиков, 1907), мас-
совый пролет под Николаевым отмечен 15 апреля 1913 г. (Токарь и Иванов,
1913). С конца первой декады апреля до половины мая идет пролет по Черно-
морскому побережью Кавказа (Аджария; Вильконский, 1897), в середине
апреля птицы появляются на Северном Кавказе (высота до 2750 м над
уровнем моря; Беме, 1926); в начале третьей декады апреля начинается «бой»
по р. Куре (Сатунин, 1914).
В Туркмении весенний пролет продолжается весь апрель и начало мая
(Шестоперов, 1937); на Мангышлаке пролетные птицы наблюдались с 1 по
20 мая (Долгушин, 1948). В Киевской и Черниговской обл. перепела появ-
ляются обычно во второй декаде мая (Чернай, 1853; Данилович, 1932;
Емельяненко, 1913), иногда в конце апреля—начале мая (Шарлемань, 1913,
1930). В Смоленской обл. пролет отмечен в среднем между 10—20 мая (Граве,
1926; однако в 1911 г. появились уже 27 апреля, Гржибовский, 1912, а
в 1912 г.—3 мая, Меландер, 1913), в Московской обл. первый крик (в среднем
за 20 лет) зарегистрирован 16 мая, самый ранний—8 мая, самый поздний—
24 мая, многолетняя амплитуда 16 дней (Долгошов, 1947), в Ярославской
обл. (Плещеево озеро) первые птицы появились между 10 и 15 мая (за пять
Лет; Пгушенко, 1937), в Тульской и Калужской обл. первый «бой» в первой—
второй декаде мая; в Тамбовской обл. пролет заканчивается уже в первой
декаде мая (Предтеченский, 1928). В Среднем Поволжье появляются в конце
апреля—в первые дни мая. В окрестностях Пскова птицы появляются в на-
чале мая (Нестеров и Никандров, 1913); на севере Рыбинского водохрани-
лища первый «бой» отмечен лишь в конце мая—начале июня (Спангенберг
и Олигер, 1949), а в Кайском районе Кировского края лишь в начале
июня (Душин, 1935); в Молотовской обл., однако, появляется значительно
раньше—в первой декаде мая (Воронцов, 1949); в окрестностях Тюмени—
во второй половине мая (Словцов, 1892). В Ленинградской обл. появляется
в третьей декаде мая (Бианки, 1923)—в первой декаде июня (Кайгоро-
дов, 1922).
ОБЫКНОВЕННЫЙ ПЕРЕПЕЛ 137
На северных берегах Каспийского и Аральского морей весенний пролет
начинается во второй половине апреля (Бостанжогло, 1911), однако на
Теджене первые птицы появились уже 1 апреля 1886 г. (Зарудный,1896);
в низовьях Сыр-Дарьи вялый пролет был с 29 апреля до середины мая 1927г.,
одиночки летели и в июне (Спангенберг и Фейгин, 1936); в окрестностях
Чкалова первые птицы появляются во второй половине апреля, валовой про-
лет в мае (Зарудный, 1888, 1897), однако в Северном Казахстане (Наурзум-
ский заповедник) первый «бой» отмечен уже в третьей декаде апреля (Улья-
нин, 1941). Через верховья Аму-Дарьи пролетают в середине мая (Келиф;
Иванов, 1940), хотя в низовьях р. Чу появляются уже в середине апреля
(Долгушин, 1939), а в долине р. Или 10 мая уже было много птиц и «бой»
был в разгаре (Шестоперов, 1929). В Барабинской степи первый «бой» отме-
чен в первой декаде мая (оз. Чаны; Данилович, 1935), в районе Новосибир-
ска—во второй (Гвоздев, 1937;Щепотьев, 1934), в Кузнецкой степи и Салаире
прилет в середине—второй половине мая (Хахлов, 1937), в Минусинском
округе в середине мая (Зверев, 1930).
Таким образом, перепела прилетают на места гнездовий в числе послед-
них перелетных птиц, позже многих мелких насекомоядных (пеночки и т. п.);
в более северных участках равнины появляются раньше, чем в горах
(Памир, отчасти высокогорные районы Кавказа и т. п.). Весной самцы и самки
летят одновременно (Спангенберг и Фейгин, 1936).
Осенний отлет начинается постепенно и мало заметен в северных районах;
перелетных стай на севере не образуется и птицы летят поодиночке. В б. Петер-
бургской губ. последние особи были добыты 3 сентября 1892 г. и 13 сентября
1859 г. (Бианки, 1923); в Молотовской обл. отлет в первой декаде сентября
(Воронцов, 1949), на Урале в конце августа—начале сентября (Куклин,
1938). В районе Плещеева озера (Ярославская обл.) местные птицы исчезают
в первой декаде сентября, а последние пролетные—в конце сентября—начале
октября (Птушенко, 1937). Из пределов Московской обл. улетают во вто-
рой—третьей декаде сентября—начале октября (Смолин, 1948), но отдель-
ные особи задерживаются позже A4 октября одиночный перепел встречен
нами на овсяном поле в Коммунистическом районе Московской обл.)-
Из Минской обл. улетают в течение сентября, одиночки задерживаются до
середины октября (Шнитников, 1913), в Смоленской обл. отлет начинается
в сентябре и заканчивается в начале октября (Граве, 1926); в Тульской обл.
исчезают к концу сентября, часть уже после начала заморозков (Сушкин,
1892). В Мордовии (заповедник) в 1936 г. было много перепелов в первые дни
сентября, затем число их уменьшилось, а 21—22 сентября не встречены
вовсе (Птушенко, 1938). Из пределов Горьковской обл. начинают отлетать
в конце августа и полностью исчезают в первой половине октября, иногда
задерживаются даже до декабря (Пузанов, Кипарисов, Козлов, 1942); в эти
же сроки идет отлет и в Среднем Поволжье (Доброхотов, 1937; Житков и
Бутурлин, 1906). В Полтавской обл. самая поздняя пролетная птица была
добыта 1 ноября 1926 г. (Гавриленко, 1929), под Киевом одиночки также
задерживаются до начала ноября (Кеслер, 1851).
Наиболее отчетливо выражен пролет на юге, где перепела летят уже
стаями. Местные птицы начинают отлетать уже в августе, а интенсивный
пролет на юге Украины (на пространстве Херсон—Мариуполь) занимает
весь сентябрь и начало октября.
На Крымских плоскогорьях перелетные перепела начинают скопляться
в больших количествах, уже в августе—начале сентября (Пузанов, 1931).
Сроки перелетов через Южное побережье Крыма изменяются по годам в за-
висимости от погоды: в 1933 г. пролет наблюдался с 27 сентября по 10 ноября;
в 1934 г.—с 20 августа по 27 октября; в 1935 г. пролет проходил с 31 августа
по 20 октября. Возрастной состав стай меняется: первые пролетные стаи
138 отряд куриные
состоят из 90% взрослых самцов, затем численность самок возрастает и, на-
конец, превышает количество самцов, одновременно начинают попадаться
жирные молодые птицы обоего пола, в конце массового пролета и после него
летит поздняя молодь с низким весом (Спангенберг, 1948).
На Черноморском побережье Кавказа первые одиночки появляются
уже в июле A0 июля 1910 г.; Колхида; Домбровский, 1913); в августе
начинается слабый пролет под Батуми, увеличивающийся к концу месяца
(Нестеров, 1910; Дерюгин, 1900), в конце августа—начале сентября заметно
увеличивается за счет прилетных количеств птиц в горах (Кавказский запо-
ведник—Аверин и Насимович, 1938; Северный Кавказ—Беме 1926; Армения—
Ляйстер и Соснин, 1942). В предгорьях Предкавказья и особенно по Черно-
морскому побережью пролет продолжается в сентябре—октябре (Верещагин,
1947), в окрестностях г. Дзауджикау до 10-х чисел ноября (Беме, 1950).
Интенсивность пролета в Крыму и на Кавказе зависит от погоды. В без-
ветреные ясные ночи или при слабых ветрах с юга и севера птицы летят
через Черное море и перевалы Кавказа не останавливаясь; при сильных
штормах, в туманные и дождливые ночи птицы не летят дальше и число их
резко возрастает за счет прилетевших с севера; при улучшении погоды вновь
возобновляется интенсивный пролет, и все скопившиеся за время ненастья
птицы отлетают на юг.
В Чкаловской обл. валовой пролет заканчивается в первой трети сен-
тября, но одиночки задерживаются значительно позднее (Сушкин, 1908);
в Кызыл-Кумах по опросным сведениям в большом числе бывают в конце
августа—первой половине сентября (Зарудный, 1915), в низовьях Сыр-
Дарьи последние птицы отмечены 15 сентября 1927 г. (Спангенберг и Фейгин,
1936). В Туркмении большие пролетные группы птиц в районе Ашхабада—
Багира и у Теджена были отмечены в первой половине августа 1889 г. (За-
рудный, 1896); в районе Гасан-Кули (Восточный Каспий) первые птицы
появляются в конце сентября, последние—в конце октября (Исаков и Во-
робьев, 1940); в это же время заканчивается пролет по всей Туркмении
(Шестоперов, 1937). В Сей стане (Иран) повидимому первые пролетные птицы
были встречены 15—20 сентября (Зарудный, 1903).
В долине р. Или наиболее интенсивный пролет отмечен в сентябре (Ше-
стоперов, 1929; Корелов, 1948). Повидимому, в конце августа—начале сен-
тября заканчивается пролет птиц по долинам Памира и Тянь-Шаня. Отлет
в Западной Сибири не прослежен; вероятно птицы отлетают в Индию; мертвая
перепелка найдена в начале декабря на р. Зайсан-Сайту между Русским
хребтом и западной оконечностью хребта Алашань (Пржевальский, 1888).
Биотоп. Гнездовой—поля с посевами хлебов, картофеля, клевера,
люцерны и других культур, пары, суходольные и заливные луга, большие
поляны и опушки лесов у полей, поросшие травой лесосеки и гари, лесо-
степи, степи и окраины песков, часто вдали от воды. К концу лета при сено-
косах и уборке хлебов уходит на бахчи, в подсолнухи, кукурузу, заросли
кустарников.
На юге охотно поселяется в полезащитных лесных полосах как моло-
дых, так и сомкнувших кроны (Мельниченко, 1949). По разнотравно-зла-
ковым степям с мозаикой лесных колков, озер и болот более равномерно рас-
пространен и более многочислен, чем в сухих степях (Северный Казахстан;
Формозов, 1950). Заходит в солончаки с зарослями солянок и в глинистые
полынные степи (Зарудный, 1888), проникает в луговые участки пустыни,
но отсутствует в песках (Киргизские степи, Сушкин, 1908; Туркмения, JX&-
ментьев, 1952); гнездится и по каменисто-щебнистой пустынной степи (Ту-
винская авт. обл.; Янушевич, 1948).
Заходит в горы и гнездится по культурным участкам в полосе предго-
рий, по большим полянам в зоне леса и по пятнам лугов, посевов и нагорных
ОБЫКНОВЕННЫЙ ПЕРЕПЕЛ 139
степей субальпийской и, местами, альпийской зоны, по кустарникам речных
долин (до 2810 м над уровнем моря в Даралагезском хребте—Даль, 1949;
до 2575 м в Памбакском хребте—Даль, 1948; до 2100 м в районе Кавказского
заповедника—Аверин и Насимович, 1938; до 2990 м в Зеравшанском иТур-
кестанском хребтах—Даль, 1936).
В горах часто поселяется на сырых и даже заболоченных лугах хотя
на равнине обычно увлажненных мест избегает.
Под Харьковом, вероятно под влиянием энергичного преследования,
еще в конце прошлого века перепела стали чаще поселяться в кустарниковых
зарослях, в балках и оврагах (Сомов, 1897); в меньшей степени это же,пови-
димому, можно видеть и в других частях ареала.
Во время перелетов встречаются преимущественно по открытым участ-
кам; дневки в садах, по огородам, зарослям кустов и тростников, в тугаях,
травянистой степи. Через горы перелетают по речным долинам и перевалам.
За счет увеличения пригодных биотопов (в первую очередь продвижение
земледелия на север) перепела постепенно распространяются на северо-
запад и север.
Численность. Неравномерна в разных частях ареала и в разных
биотопах: на заливных лугах у г. Кузнецка встречена 1 шт. на гектар, в степи
от 2 до 16 шт. на 1 км2, в районе сосновых лесов в предгорьях Салаира 4 шт.
на 1 км2, а на соответствующих стациях в тайге 1 шт. на 1 км2 (Хахлов,
1937); по северному склону Сарайбулагского хребта на высоте до 1920 м над
уровнем моря плотность была 0,4 шт. на 1 га посевов (Кавказ—Даль, 1944);
в июне на 800 км маршрута в Северном Казахстане встречено 112 перепелов,
или 0,12 перепела на 1 км, из них в песчано-ковыльной степи 0,04, в ковыль-
ной степи с кустами таволги 0,3, в понижениях с пырейным покровом 0,5,
по залежам и посевам 0,2 по луговым солонцам 0,08 перепела на 1 км (Наур-
зумский заповедник—Ульянин, 1941).
Установлены значительные изменения численности по годам: в Калуж-
ской обл. в 1907 г. на 3 верстах встречалось до 10 выводков (чаще 4—8),
в 1909 г. птиц почти не было, в 1910 г. стало вновь довольно много (Филатов
1915);врайонеМинскав1899г. на 300 десятин было до 5 гнезд, в 1900—1903 гг.
очень редко удавалось слышать крик, в 1904—1905 гг. ни разу не слышно
крика (Шнитников, 1913). Подобные колебания численности отмечены для
Тульской обл. иСреднеказахстанских степей (Сушкин, 1892, 1908), для Новго-
родской и Псковской обл. (Бианки, 1910), Смоленской обл. (Граве, 1926) и т.п.
Во время засух в южных степных частях ареала (Поволжье, Украина)
массами откочевывают на гнездовья севернее: в сильную засуху 1920 и осо-
бенно 1921 г. резко увеличилось число птиц в Горьковской обл. (Волчанец-
кий, 1924; Формозов, 1935), в северных районах Ивановской обл. (Добро-
хотов, 1937), в окрестностях г. Костромы (Шуммер, 1923) в Московской
{Поляков, 1924) и Витебской обл. (Федюшин, 1926); гнездовой период при
этом затянулся до сентября (Доброхотов, 1937). В засушливое на юге лето
1946 г. вновь отмечено обилие перепелов в Московской обл., а в 1947 г. их
опять стало мало (Смолин, 1948).
Повидимому, на численности перепелов также существенно отражаются
и условия погоды при перелетах (массовая гибель птиц на море во время
штормов и туманов; Формозов, 1935; Исаков и Воробьев, 1940) и на зимовках
(Спангенберг и Туров, 1950; Бостанжогло, 1911).
На гнездовьях много молодых и взрослых птиц гибнет при сенокосах
и уборке хлебов (попадая под косы и режущие части машин). В Наурзумском
заповеднике после 40—50D жары 17—19 июня 1936 г. температура упала
до 8—10°С, давление до 726 мм, шел сильный грозовой дождь с ветром до
17 м/сек; после этого найдены мертвые перепела, повидимому погибшие от
голода и переохлаждения (Рябов, 1949).
140 ОТРЯД КУРИНЫЕ
В ряде районов за последние десятилетия наблюдается сокращение
количества гнездящихся птиц (Закарпатье—Портенко, 1950; Западные обла-
сти—Суслова, 1937; Тульская обл.—Сушкин, 1892; и т. п.), что, повидимому,
объясняется массовым истреблением птиц на перелетах (особенно в Ита-
лии и т. д.).
Экология. Размножение. Повидимому, полигам, не образующий
постоянных пар. Вскоре после прилета (на второй или третий день, Птушен-
ко, 1937; на следующий день после прилета, Герхнер, 1928; в день прилета,
Ульянин, 1941) по утренним зорям самцы начинают «бить» (т. е. издавать то-
ковый крик: сначала хриплое и тихое «ва-вва», «ва-вва», повторяемое обычно-
не более трех раз, а затем чисто и громко начинает собственно «бой», который
можно передать звукоподражательно «подьполоть», «подьполоть», или «фить-
пиль-вить»; этот крик повторяется от четырех до десяти раз). Самка отвечает
на крик тихим и хриплым «тюрюрю» или «брибит». Наиболее интенсивен
«бой» в конце мая—начале июня, когда крик слышен вечером, ночью, утром
и даже днем. В июле интенсивность «боя» уменьшается, а полностью он пре-
кращается даже в средней полосе и на Алтае лишь в середине августа. Во-
время «боя» самцы бегут на крик самки и в это время происходит спаривание.
Ряд авюров утверждает, что у перепелов в году бывают две нормальных
кладки; однако последующие наблюдения опровергают эти утверждения и
показывают, что не только в средней полосе, но и на юге нормально бывает
лишь одна кладка (Ульянин, 1941). Растянутость сезона размножения в по-
пуляции, повидимому, объясняется неодновременной физиологической готов-
ностью к размножению самок различных возрастных групп (Ульянин, 1941)
и тем, что взамен утраченных первых кладок часть самок откладывает допол-
нительные вторые кладки. Так, на склоне Памбакского хребта 15 августа
1940 г. были найдены три гнезда: в одном птенцы недавно вылупились, во-
втором—эмбрионы вполне сформированы, в третьем—эмбрионы в половину
яйца; на следующий день добыты молодые весом в 60 и 67 г, самец с сильно
вздутыми семенниками и самка со сформированным яйцом в яйцеводе, а 5 и
6 сентября 1939 г. там же добывали птенцов весом 37,5 г. (Армения—Даль,
1948); 27 июня встречены птенцы размером с матку, а 14 июля—сильно наси-
женные яйца (Крым—Никольский, 1891); 14 июля найдена кладка из
14 яиц в долине р. Суры (Житков и Бутурлин, 1906); Спангенбергом кладка
из 8 свежих яиц найдена на жнивье 10 сентября 1920 г. у железнодорожной
станции Канкриновка (Запорожская обл.).
Первые яйца откладывает в Чкаловской обл. в конце первой декады
мая, массовая кладка—с середины мая (Зарудный, 1897), полные кладки
во второй половине мая в Туркмении (Дементьев, 1950), по побережью Азов*
ского моря (Боровиков, 1907), в первой половине июня в Северном Казах-
стане (Ульянин, 1941), в июне у Красноярска (Тугаринов и Бутурлин,
1911) и т. д. Самка откладывает по одному яйцу в сутки (Ульянин).
Гнездо устраивается в небольшой ямке на лугах, в степи, часто в хле-
бах, иногда под защитой кустов бурьяна, кочек и т. п. Небольшая ямка
выстилается сухой травой и растительной ветошью, причем в некоторых
гнездах подстилка достигает 1,5 см толщины и хорошо свита (Ульянин),
иногда внутри бывает и скудная выстилка из перьев наседки. Число яиц
в кладке доходит до 9—15 (Наурзумский заповедник—Ульянин), 10—14
(р. Висла—Катин, 1912), до 16 (оз. Чаны—Иогансен, 1911); 10—12, редко 15
(Московская обл.—Лоренц, 1892); много реже встречается в кладках до
20 яиц (Молдавия—Остерман, 1912; Тульская обл.—Сушкин, 1892; Чкалов-
ская обл.—Зарудный, 1888; Астраханская обл.—Хлебников, 1930). Яйца
слабо грушевидные, форма их варьирует даже в одной кладке: некоторые
более вздуты и остроконечны, у других почти не выражен острый и тупой
концы; встречаются и резко грушевидные яйца (куличиного типа). Скорлупа
ОБЫКНОВЕННЫЙ ПЕРЕПЕЛ 141
гладкая и блестящая, со слабо выраженной пористостью. Основной фон из-
меняется от известково-белого до глинисто-буроватого и даже оливково-
охристого цвета; по нему разбросаны пятна: крупные поверхностные бурые
или рыжеватые, иногда с размытыми краями, создающими мраморный рису-
нок; между крупными пятнами разбросаны такого же цвета мелкие пятнышки
и точки; между ними просвечивают более глубокие светлобурые пятнышки.
Размеры яиц A00) 25—32,2x20,6—25, в среднем 29,41 х 22,79 мм (Хартерт,
1921—22). Величина яиц варьирует даже в пределах одной кладки: размеры
15 яиц колебались от 28 до 31 мм по длине и от 22 до 24 мм по ширине (сред-
нее 29x23); вес яиц (за двое суток до выклева) колеблется от 5,6 до 7 г (Наур-
зумский заповедник—Ульянин, 1941).
Продолжительность насиживания (после откладки последнего яйца)
равна 15 суткам (Ульянин, 1941)—17 суткам (Хейнрот, 1932). Насиживает
яйца и водит птенцов, повидимому , одна самка; однако в конце мая 1900 г.
на р. Алатырь на гнезде добыт насиживающий самец с хорошо развитым
наседным пятном, самка поблизости не найдена (Житков и Бутурлин, 1906);
в Красноярском крае добыт самец, отводивший от гнезда (Тугаринов и Бутур-
лин, 1911), вероятно эти исключения объясняются функциональными нару-
шениями гормональной системы. Насиживающая самка сидит на гнезде
очень плотно, почти не сходя.
Процесс вылупления птенцов из яиц в одном гнезде занимает около
двух суток, считая от первых наклевов до выхода последних птенцов. Самка
во время вылупления сидит на гнезде, обогревая птенцов. Обсохшие птенцы
хорошо передвигаются, и самка уводит их от гнезда; через 4 часа после вы-
лупления выводок уходит от гнезда далее 100 м. Тотчас по вылуплении птен-
цы весят 5—6 г (Ульянин, 1941).
Пуховички под Киевом (ранние) встречены уже 21 мая 1924 г. (Шарле-
мань); в Молдавии первые выводки в первой половине июня (Остерман,
1912), под Москвой массовый выклев в конце июня—начале июля, хотя уже
летавшие птенцы встречены 24 мая 1892 г. (Лоренц, 1894), в Тульской обл.
с июня (Мензбир, 1879), в Северном Казахстане во второй половине июня
{Ульянин, 1941), на Алтае в конце июня (Сушкин, 1938). Молодые, почти
достигшие размеров взрослых птиц, встречены под Калугой 19 августа 1907 г
{Филатов, 1915), под Минском 7 июля 1897 г. (Шнитников, 1913), в конце июня
в Чкаловской обл. (Зарудный, 1888, 1897), в начале августа в Северном Ка-
захстане (Наурзумский заповедник—Ульянин, 1941), 24 июля в окрестно-
стях Теджена (Туркмения—Шестоперов, 1936). Растянутость периода раз-
множения в популяции подтверждается тем, что пуховых птенцов находили
еще 2 сентября 1906 г. под Мариуполем (Боровиков, 1907), в конце августа
и даже начале сентября под Киевом (Кеслер, 1851), 21 августа в Подолии
(Герхнер, 1928), 29 августа в Московской обл. (Лоренц, 1894), 17 августа
1907 г. под Калугой (Филатов, 1915), в середине сентября в б. Симбирской
губ. (Житков и Бутурлин, 1906), 28 июля на Алтае (Уймон—Сушкин, 1938),
14 сентября под Минусинском (Зверев, 1930). Чуть перепархивающие птенцы
размером в полматку встречены на Даралагезском хребте 5 августа 1946 г.
{Армения—Даль, 1949), 8—12 июля в Северном Прибалхашье (Долгушин,
1947), 15 августа 1926 г. под Киевом (Шарлемань, 1930). Половозрелыми
молодые птицы становятся на следующий год после рождения (Нитхам-
мер, 1942).
По наблюдениям в неволе рост птенцов идет довольно быстро (Хейнрот,
1924-32):
Выводок водит только самка, повидимому она и улетает на юг вместе
с молодыми (Ульянин, 1941). Вначале выводки держатся в районе гнезд;
подросшие уходят в хлеба (особенно в просо), если поблизости есть посевы;
после уборки хлебов до самого отлета кочуют по убранным полям, кормясь
142
ОТРЯД КУРИНЫЕ
Рост птенцов перепела по наблюдениям в неволе
Возраст
в днях
Вес самца
в г
Вес самки
в г
0
5
5,5
2
6,5
7
4
8,5
10
6
13
15,5
8
17,5
20,5
12
27
28
14
31
34
16
38
41
18
46
51
20
50
57
24
62
67
28
—
77
35
87
97
42
105
129
49
100
120
на жнивье, по лугам и степи. Во время засух выводки уходят к воде, скоп-
ляясь у луж, в болотах и поймах речек (Мордовский заповедник—Птушенко,
1938). Перепелки, гнездящиеся на равнине Ахала и Атэка, с подросшими
выводками откочевывают в предгорья (Туркмения—Зарудный, 1896); эта
указание нуждается в дополнительной проверке.
Линька. Полная линька один раз в году. В соответствии с растяну-
тым сроком размножения линька в популяции также сильно растянута.
У взрослых самцов и самок линька мелкого пера начинается во второй поло-
вине июня—начале июля (Кавказ, Поволжье, Мордовия, озеро Зайсан,
Туркмения). Однако взрослые птицы, только что начавшие линьку, в тех
же местах добывались и в первой половине августа.
Линька начинается со смены мелкого оперения по всему туловищу.
В разгар линьки, когда мелкое перо уже покажется над старым, начинается
смена маховых и рулевых перьев. Первостепенные маховые сменяются от
десятого к первому, т. е. центробежно, второстепенные—центростремительно.
Линька рулевых идет одновременно или лишь с небольшим запозданием со
сменой первостепенных маховых. Взрослые особи, полностью завершившие
линьку, встречаются обычно в конце июля—начале августа (коллекционный
материал Зоологического музея Московского государственного универси-
тета); из ранних встреч перелинявших птиц следует указать самца перепела,
добытого под Красноярском 9 июля 1894 г. (Тугаринов и Бутурлин, 1911).
У птиц, добытых в конце сентября—октябре, следов линьки не обнаружено,
хотя смена мелкого пера у отдельных птиц затягивается до января (Уайзерби,
1946). В феврале—апреле отмечена частичная линька мелкого пера
(Уайзерби, 1946).
Пеньки первого птенцового наряда появляются у пуховиков в первую
неделю жизни. На 11-й день птенцы начинают уже перепархивать. Перво-
степенные маховые вырастают в возрасте трех недель, кроме первого—треть-
его, которые еще в нераскрывшихся пеньках. Через два дня выпадает десятое
первостепенное маховое; на шестой неделе сменяется шестое маховое и пол-
ностью сформировываются пятое—третье маховые (первой, т. е. птенцовой,
генерации), наружные первое—второе меховые у основания еще в пеньках.
На седьмой-восьмой неделе в основном завершается формирование контур-
ных перьев первого годового наряда; первое и второе маховое достигают
окончательной длины, сформировывается четвертое (у самцов) и пятое
(у самок) маховые. В таком состоянии, т. е. с первым—третьим (самцы) или
с первым—четвертым (самки) маховыми первой генерации, молодые птицы
начинают перелет. Дальнейшая линька идет на зимовке, а первое и второе
маховые, повидимому, как и у других куриных, сменяются лишь при полной
линьке на будущий год (по наблюдениям в неволе; Хейнрот, 1924—32).
Молодые птицы в таком состоянии оперения начинают встречаться со второй
половины июля до конца сентября (коллекционные материалы Зоологического
музея Московского государственного университета).
Питание. В основном растительноядная птица. Питается семенами
и побегами растений; в летнее время одним из существенных компонентов
ОБЫКНОВЕННЫЙ ПЕРЕПЕЛ 143»
корма становятся насекомые и, возможно, другие наземные беспозвоночные.
Основными кормовыми растениями для перепелов Памбакского хребта слу-
жили Ballota nigra, Chenopodium album, Polygonum persicara; в зобах и желуд-
ках встречены муравьи (Formicidae), чешуекрылые (Tineidae), двукрылые
(Muscidae) и другие насекомые (Армения—Даль, 1948). В Наурзумском за-
поведнике излюбленный корм перепела—семена Polygonum (в 8 желудках
из 12 и очень много по объему), меньшее место по объему занимают насеко-
мые (9 из 12 желудков); в годы обилия саранчи перепела поедали ее в боль-
шом количестве, концентрируясь у ее скоплений; в августе переходят на
питание культурными растениями, особенно просом (Ульянин, 1941). На
Южном побережье Крыма в зобах у осенних перелетных птиц найдены остат-
ки богомолов и кузнечиков, реже семена злаков; многие желудки содержали
только мелкие камешки (Спангенберг, 1948).
Занесение перепелов в разряд вредителей сельскохозяйственных куль-
тур (пшеница, рожь, овес и просо; Западная Сибирь—Зверев и Климов,
1931) мало оправдано, так как они поедают в основном «падалицу» зерен
культурных злаков. Одновременно следует указать на некоторую пользу
перепела в уничтожении семян сорняков (Воронцов, 1949).
Суточный цикл. Наиболее деятельны ночью и в сумерках;
днем, особенно в жару, отдыхают.
Враги. Лисица, горностай, хорь и хомяки уничтожают кладки, а иногда
и взрослых птиц. Ловят перепелок и хищные птицы: ястреб-перепелятник,
мелкие сокола (дербник, чеглок, меньше пустельга) и др. За перелетными
стаями перепелов следуют перепелятники и сапсаны. Добытые Север-
цовым A873) в 1864 г. C сентября—6 октября) на пролете между Аулье-Ата
и Чимкентом степные орлы A2 шт.) были очень жирны и у всех в зобах най-
дены перепела, иногда заглоченные целиком.
В засухи, смертность перепелов, повидимому, увеличивается не только
от недостатка питания и хищников, но и от сильных инвазий; добытые в
1936 г. в Мордовском заповеднике птицы были сильно истощены и у всех
в желудках и тонких кишках обнаружены ленточные глисты; особенно их
много было у молодых птиц (Птушенко, 1938).
Хозяйственное значение. Местами на Северном Кавказе, в лесостепных
районах европейской части СССР и в Западно-Сибирских степях летом ло-
вят сетью, подманивая самца ручной самкой или манком (вабиком), имити-
рующим голос самки; пойманные птицы откармливаются и идут в продажу
(в том числе и на экспорт); иногда ловят силками, петлями или наволочной
сетью (Силантьев, 1898). В Средней Азии (Туркмения, Узбекистан) перепелов
также ловят сетьми и держат в клетках, как излюбленную комнатную птицу;
местами развит спорт «перепелиных боев».
Повсюду осенью—объект спортивной охоты с ружьем из-под собаки.
В южных районах СССР—Крым, Черноморское побережье, Кавказ, Сред-
няя Азия—спортивная охота вовремя перелетов перерастает в промысловую.
Перепелов добывают ружьем из-под собаки, сбивают сучковатой палкой или
накрывают сачком на взлете, ловят ночью сачком с фонарем (даже на улицах
городов; Сухуми—Верещагин, 1947; Дзауджикау—Беме, 1950). В б. Симбир-
ской губ. (Курмышский и Сергачский уезды) в XIX в. ловили в большом ко-
личестве дрессированными дворняжками и перепелятниками; птиц засали-
вали в бочках впрок (Житков и Бутурлин, 1806). На Кавказе и в Средней
Азии до сих пор местами очень распространена охота с выношенными ястре-
бами-перепелятниками.
В окрестностях Дзауджикау (Северо-Осетинская АССР) за вечер во
время осеннего перелета с фонарем ловят до 25—30 шт.; ружьем в среднем
добывают 30—60 шт. в день, иные охотники добывают до сотни (Беме, 1950).
Общая добыча перепелов за осень на Черноморском побережье Кавказа
144
ОТРЯД КУРИНЫЕ
достигает нескольких десятков тысяч штук (Верещагин, 1947). На Крымском
побережье, несмотря на запрещение охотничьего законодательства, широко
практикуемая охота с фонарем дает за ночь от 40 до 200 птиц на ловца (Спан-
генберг, 1948). В районе Евпатории (Крым) в 1896 г. добыто более 20000 птиц
(Силантьев, 1898).
Для предотвращения уменьшения количества птиц необходимо фактиче-
ское осуществление повсеместного (в южных районах) запрещения ночной
охоты с фонарем и введение предельных норм добычи перепелов за день и за
сезон на ружье.
Полевые признаки. Размером с дрозда; по повадкам и манере держаться
напоминает миниатюрную курочку. Бегает легко и быстро; при добывании
Рис. 23. Перепел (самец и самка)
корма копается в земле, разбрасывая и разгребая ее ногами; охотно купается
в пыли. Летает очень быстро, часто взмахивая крылышками; перед посадкой
некоторое расстояние пролетает планирующим полетом. При опасности за-
таивается или перебегает; вспугнутая —летит низко над землей и невдалеке
опускается на землю, скрываясь в траве. На деревья не садится.
Описание. Размеры и строение. Крыло более острое, чем у других кури-
ных; на первом первостепенном маховом небольшая вырезка на внутреннем опахале,
на втором-т ретьем небольшие вырезки на наружном опахале, на четвертом она чуть замет-
на. Обычно формула крыла 1 > 2 > 3, реже 2 > 1 > 3; разница в длине между первы-
ми тремя маховыми обычно не более 1—2 мм; разница в длине между последующими около
5—8 мм. Шпор нет ни у самцов, ни у самок. Рулевые мягкие, скрыты под кроющими хво-
ста и мало от них отличаются.
Длина крыла у самцов (84) 99—114, у самок E0) 101—112, в среднем соответственно
104, 8 и 106, Б мм. Длина тела у самцов C1) 175—215, у самок D) 157—210, в среднем
соответственно 197,1 и 193 мм. Размах крыльев у самцов B4), 327—390, у самок A0)
ОБЫКНОВЕННЫЙ ПЕРЕПЕЛ
145
352—389, в среднем соответственно 363 и 367,6 мм. Вес самцов (9) 76—106, самок E) 78—
96 г, в среднем соответственно 90,5 и 87,6 г. Во время осеннего перелета вес перепелов
достигает 146 г. (Крым—Спангенберг, 1948); вес местных птиц в августе в среднем 99,4 г,
перелетных в сентябре до 123,3 г (Армения—Даль, 1948). Таким образом, половой димор-
физм размеров отчетливо не выражен.
Окраска. У пуховичка спинная сторона тела рыжевато-охристая с тремя черно-
вато-бурыми полосами по спине и по бокам туловища и двумя темными полосами по верху
шеи и затылку, сливающимися на лбу; более размытые темные полоски есть на крыльях
и бедрах. Брюшная сторона тела, включая горло, шею и брюшко, желтоватая с чуть серо-
ватым оттенком.
Молодые птицы в первом птенцовом наряде сходны по окраске с взрослыми самками,
но оперение молодых более тусклое. Пестрины с зоба дальше, чем у самок распростра-
няются на грудь и бока тела.
'ff4л. {(.iffctoUH
Рис. 24. Пуховые птенцы перепела
Первый годовой наряд сходен по окраске с нарядом взрослых птиц; у молодых сам-
цов полоса на подбородке и горле почти не выражена.
Окраска взрослого самца сверху очень сложного пестрого рисунка. Общая окраска
охристо-бурая с темными охристо-рыжими (а иногда и коричневато-бурыми) поперечны-
ми полосами более или менее выраженными светлоохристыми наствольными полосами.
Верх головы коричневато-бурый, с узкой светлой полосой посредине, образованной светлы-
ми наствольями перьев; над глазом беловатая полоса, проходящая от ноздри за ухо. Под-
бородок, горло и шея спереди охристо-рыжие или темнобурые, отделенные от белых или
красно-бурых щек и от охристых с узкими белыми наствольями перьев зоба узкой белой
полосой; у многих самцов подбородок, горло и шея в осеннем пере, спереди беловатые.
Грудь бледноохристая, брюхо беловатое. По бокам тела более или менее отчетливые про
дольные рыжие или бурые полосы. Первостепенные маховые темнобурые с буровато-охри-
стой поперечной исчерченностью на наружных, опахалах; на наружном опахале первого
первостепенного махового эта исчерченность обычно заменена узкой продольной светлой
полосой.
Оперение взрослой самки сверху очень сходно с оперением самца. Подбородок, гор-
ло и шея спереди беловатые (никогда не бывают охристо-рыжими или бурыми). Зоб и пе-
редняя часть груди с менее выраженным, чем у самцов, охристым налетом, чуть заметными
светлыми настволиями и хорошо выраженными отчетливыми округлыми темнобурыми пят-
нами. По бокам тела, помимо менее резкого продольного рисунка на перьях, более или
менее отчетливо выражены поперечные рыжевато-бурые полосы или пятна.
У отдельных экземпляров как самцов, так и самок встречается более темный, чем
обычно, общий тон окраски; при этом самые светлые участки оперения—грудь, брюшко,
светлые настволия поверху туловища—кажутся грязносерыми (неполный меланизм).
10
Птипы СССР, т. IV
146 ОТРЯД КУРИНЫЕ
Немой, или восточносибирский перепел Coturnix coturnix japonica
Temm. et Schleg.
Coturnix vulgaris japonica. Temminck etSchlegel. Fauna Japon., Aves, 1849V
стр. 103, табл. 61, Япония.
Синонимы. Coturnix ussuriensis. Bogdanow. Consp. Av. J. Rossi cae, 1884, стр. 45.
Уссурийский край.— Coturnix muta. D у b о w s k i. Journ. f. Orn., 1868, стр. 337. Даурия.
P у с с к о.е название. Книжное, данное Палласом, так как токовый крик гораздо
тише, глуше и более хриплый, чем крик обычн го перепела.
Распространение. Ареал. В пределах Советского Союза гнездится
на Витимском плоскогорье и в долине р. Читы (Павлов, 1948), в долине рек
Селенги, Орхона, Чикоя, Иро (Моллесон, 1895), в Даурских степях (р.Онон—
Долгушин, 1941; Александровский завод—Скалой, 1936), в Приаргунье
(Скалой, 1935), по р. Амуру и его притокам (на север до верхнего течения
р. Зеи; Штегман, 1928), в Приморском крае (нижнее течение р.Имана; Спан-
генберг, 1940; повсюду в Южно-Уссурийском крае; хребет Сихотэ-Алиня—
Иогансен, 1927); на пролете встречены на Шантарских о-вах (Дулькейт и
Шулейкин, 1937). Северная граница ареала немого перепела в пределах
нашей страны точно не установлена (самые северные находки: Витимское
плоскогорье, верховья р. Зеи—без указания координат; Шантарские о-ва).
В подходящих биотопах, повидимому, повсюду на о-ве Сахалине.
Вне пределов Советского Союза западная граница проходит через юго-
западную часть Кентэя (долины рек Сунгур, Тола, Хара и т. д.) и даже
до бассейна оз. Убса-Нур (Шульпин, 1936); последнее сомнительно. Далее
он распространен в Маньчжурии, Северном Китае (Куку-нор, Ганьсу, бас-
сейн Желтой реки, т. е. Хуанхэ), Корее и Японии (о-ва Хоккайдо, Хондо,
Кюсю—Ян, 1942).
Отдельные особи или малочисленные группы зимуют в малоснежных.
Забайкальских степях (у рудника Абагайтуй, р. Аргунь; Скалой, 1935;
в окрестностях Кяхты—Бианки, 1915 и т. п.), в Приамурье (район Благо-
вещенска—Шульпин, 1936), в Приморье (в низовьях р. Сучана—Пржеваль-
ский, 1870; в окрестностях бухты Славянка—Медведев, 1914); зимуют и в
Маньчжурии. Основная масса птиц зимует на Японских о-вах—особенна
южных, в Южном Китае, Индо-Китае, Бирме и Ассаме на юг до Брамапутры.
В наших пределах зимуют, в зависимости от метеорологических условий,
не ежегодно: так, совсем не оставались на зиму 1912—1913 г. в окрестностях
бухты Славянка, хотя в другие годы были там обычны (Медведев, 1914).
Характер пребывания. Перелетная птица. Особи, зимую-
щие в пределах СССР, повидимому, принадлежат не местным, а более север-
ным популяциям. Данные по кольцеванию совершенно отсутствуют; приме-
нение кольцевания к особям этого подвида крайне желательно.
Даты. Сроки прилета и отлета очень изменчивы и зависят от метеоро-
логических условий (Шульпин, 1936). Первые одиночки появились в окрест-
ностях бухты Славянка (Приморье) 13 марта 1913 г. (Медведев, 1914),
на Верхней Сунгаче в начале апреля 1869 г. и во второй половине апреля
1868 г. (Пржевальский, 1870), в низовьях р. Имана встречены в середине мая
(Спангенберг, 1940); на Витимском плоскогорье первые птицы отмечены
1 июня 1940 г. (Павлов, 1948). Таким образом, прилет в пределах СССР про-
ходит с конца марта по начало июня.
Даты осеннего отлета выяснены недостаточно точно; отлет, повидимому,
идет в сентябре-октябре: на пролете одиночка добыт на Б.Шантарском о-ве
26 августа 1926 г. (Дулькейт и Шульпин, 1937), со 2 октября птицы парами
и одиночками стали встречаться у Новгородской гавани (Пржевальский,
1870), небольшие стайки встречены на о-ве Аскольд в первой половине,
ноября (Воробьев, 1938).
ОБЫКНОВЕННЫЙ ПЕРЕПЕЛ 147
Биотоп. Гнездится в степях, особенно разнотравных, по пашням,
в кустах по сухим склонам сопок, по сырым и сухим лугам речных долин,
по полянам на лесных болотах, по кустарникам и травянистым зарослям
падей; встречен по полянам в сосновых с примесью осины борах без подлеска
(р. Онон—Долгушин, 1941), а в июне найден в дубовых перелесках по р. Има-
ну (Спангенберг, 1940). Более часто, чем обычный перепел встречается по
сырым участкам лугов, явно предпочитая влажные места (Шульпин,
1936).
Численность. Повидимому, везде невелика. Изменения числен-
ности по годам не зарегистрированы. В Приморье в снежные зимы гибнут
зимующие здесь птицы; во время палов, пускаемых для выжигания
прошлогодней травы, гибнут кладки и, возможно, взрослые птицы (Шуль-
пин, 1936).
Экология. Размножение. Повидимому, полигам, не образующий
постоянных пар; однако Шульпин предполагает, что весной пары почти
неразлучны. Брачный крик, повидимому, вскоре после прилета. Характер-
ного для европейского перепела «боя», следующего вслед за «вавчением»,
у немого перепела нет. Брачный крик самца тихий и глухой, издали напоми-
нающий жужжание; звуками его можно передать, как «чжу-чжир-чжирр-
чжирр-чжиррр» (Пржевальский) или «чу-пит-тррр» (Спангенберг); у неко-
торых, может быть молодых, крик двусложен: «чуть-тррр». Весной крик
слышен с утренних сумерек и до темноты; в июле еще кричат довольно часто;
крик прекращается в конце августа (последний крик 27 августа и даже 10 сен-
тября 1926 г., Приморье—Шульпин, 1936).
Откладка яиц, повидимому, в конце апреля—июне. 26 мая 1928 г. на
озере Ханка добыты две самки, только что окончившие кладку, 23 мая
1927 г. у Сиваковки встречены уже насиженные кладки (Приморье—Шуль-
пин, 1936); самок со сформированными яйцами в яйцеводе добывали до 29 ию-
ня (долина р. Читы—Павлов, 1948). Сезон размножения, повидимому, как
и у европейского перепела, очень растянут: совершенно свежая кладка най-
дена 10 августа в окрестностях г. Спасска (Иогансен, 1928). Утверждения
о наличии, хотя бы в некоторые годы, нормальных двух и даже трех кладок
в году (Шульпин, 1936) крайне сомнительны; повидимому, в году одна нор-
мальная кладка, а взамен утерянных первых кладок могут быть дополни-
тельные вторые кладки.
Гнездо устраивается обычно по сухим местам в траве, и рыхло обклады-
вается небольшим числом сухих травинок (Шульпин, 1936); иногда гнездо
свивается из травинок (Павлов, 1948). Число яиц в полных кладках 9—10
(Шульпин, 1936); кладка от 10 августа содержала 11 совершенно свежих яиц
(Иогансен, 1928). Яйца по форме и характеру рисунка в осмотренной кладке
не отличимы от яиц европейского перепела, но более мелкие по размерам.
Продолжительность насиживания не установлена. Насиживает кладку
и водит выводок, повидимому, только самка; однако в начале июля у гнезда
был задавлен вероятно насиживающий самец (Читинская обл.—Павлов,
1948); подобные случаи, как и у европейского перепела, очевидно крайне
редки.
23 июля в долине р. Читы встречены летные молодые размером в полматки
(Павлов, 1948); с 5 августа 1869 г. в долине Мо молодые хорошо летали и были
в половину маток и больше, однако в конце августа 1867 г. на озере Ханка
молодые были лишь в половину маток (Шульпин, 1936). Эти сведения под-
тверждают растянутость периода размножения в популяции. Птенцы, почти
достигшие величины взрослых птиц, добывались в июле—октябре A4 июля
на р. Янчи-Хе, 20 сентября в Маньчжурии, 12 ноября у с. Сиваковки, 24 октяб-
ря у бухты Кит в Судзухинском заповеднике; коллекция Зоологического
музея Московского государственного университета).
10*
148 ОТРЯД КУРИНЫЕ
Линька. Достаточно не изучена, последовательность ее сходна с ев-
ропейским перепелом. В конце апреля—мае взрослые птицы в обношенном
пере, обычно не линяют; однако у отдельных экземпляров в это время еще
заканчивается частичная линька мелкого пера (на горле; самка от 9 мая из
Хабаровска, самец от 1 апреля 1909 г. с озера Ханка). В июне птицы без
следов линьки, лишь у самца от 26 июня 1939 г., ср. Санчикон появились на
туловище первые тычки мелкого пера; особи, добытые в сентябре (самец от
16 сентября 1909 г. созера Ханка, самец от 9 сентября 1932 г. с о-ва Аскольд),
и, наконец, самец от 26 ноября 1943 г. из Судзухинского заповедника
уже в свежем пере (коллекция Зоологического музея Московского госу-
дарственного университета).
Молодые птицы начинают линять в первый годовой наряд, повидимому,
уже в июле: молодой от 14 июля 1947 г. с р. Янчи-Хе в энергичной линьке по
всему туловищу, первое—третье маховые в чуть раскрывшихся пеньках.
Молодой самец, заканчивающий линьку мелкого пера, с полностью вырос-
шими первым и вторым первостепенными маховыми и со сменяющимися ма-
ховыми птенцовой генерации (восьмое еще у основания в тычке) добыт
24 октября 1943 г. у бухты Кит в Судзухинском заповеднике; молодая птица
от 12 октября 1932 г. из окрестностей с. Сиваковки в энергичной линьке мел-
кого пера, первое—третье первостепенные маховые у основания еще в чех-
ликах, причем первое лишь в половину, второе в три пятых и третье в две
трети окончательной длины.
Питание. Растительноядная птица, летом в больших количествах
поедает насекомых и возможно других беспозвоночных. В июле в долине
р. Читы зобы многих растительноядных птиц были переполнены слониками
(Павлов, 1948); в конце мая на Рябо коне (озеро Ханка) у птиц в желудках най-
дены камешки, множество разнообразных семян, преимущественно злаковых,
много остатков насекомых, одна гусеница и несколько стебельков; у перепе-
ла, добытого Мааком в середине июля, в зобу были небольшие черные гусени-
цы (Шульпин, 1936).
Хозяйственное значение. Несмотря на широкое распространение промыс-
ловое значение невелико. Объект спортивной охоты с ружьем. Кроме этого,
немого перепела ловят сетями и силками, расставленными по канавкам
(Шульпин, 1936).
Полевые признаки. Не отличим по поведению от европейского перепела
(кроме различий в голосе).
Описание. Размеры и строение. Сходен с европейским перепелом, но
несколько меньше по размерам. Длина крыла самцов B2) 90—105, самок E) 90—101,
в среднем соответственно 96 и 96,3 мм; среднее для обоих полов B7) 96 мм.
Окраска. У пухового птенца не описана. Молодые птицы в птенцовом
наряде похожи на самок, но окраска оперения более тускла. Первый годовой наряд сходен
с окраской взрослых птиц; в этом наряде самцы уже отличимы от самок.
Окраска взрослых особей немого перепела по типу очень сходна с окраской евро-
пейского подвида, но более яркая и контрастная. У взрослых самцов бурый
цвет на верхней стороне туловища более темный (почти черный) и яркий, причем он пре-
обладает над охристо-рыжим; светлые настволья несколько шире. У всех самцов в весен-
нем и летнем наряде щеки, горло и зачастую передняя сторона шеи кирпично-рыжие
или буровато-красные; иногда на горле слабо выделяется продольная буроватая полоса
с размытыми краями; продольные полосы по бокам груди и брюха более яркого корич-
невато-рыжего цвета. После осенней линьки горло и щеки (может быть, только у мо-
лодых самцов?) значительно светлеют.
У самок немого перепела, по сравнению с самками европейского подвида, пят-
нистость на груди более темнобурого цвета, а пятна расположены несколько более густо;
охристый налет на груди выражен слабее, продольный и поперечный рисунок боков туло-
вища более яркий.
После осенней линьки у обоих полов на подбородке вырастают удлиненные заострен-
ные перышки (сменяются во время частичной весенней линьки). У отдельных особей как
самцов, так и самок, общий тон оперения, в том числе и брюха, бывает грязноватосерый
(разлитой неполный меланизм, встречающийся и у европейского перепела).
ПУСТЫННАЯ КУРОПАТКА
149
РОД ПУСТЫННАЯ КУРОПАТКА AMMOPERDIX GOULD, 1851
Тип. Perdix griseogularis
10. Пустынная куропатка Ammoperdix griseogularis
Brandt
Perdix griseogularis. Brandt. Bull. Classe Physico-Math. Acad. I. St. Petersbourg,
1, апрель, 1843, стр. 365, Северная Персия.
Синонимы. Caccabis Bonhami. Gray. Ann. and Mag. Nat. Hist., XI, май 1843, стр. 372.
Русское название. Пустынная куропатка—книжное и несовсем удачное, так
как собственно в пустынях эта птица не встречается. Местное население Средней Азии
называет ее чиль—звукоподражатель-
но, по голосу птицы.
Распространение. Ар е а л.
Пустынная куропатка распрост-
ранена от Месопотамии и Ирана
до Инда и Пенджаба; к северу
проникает в Туркмению и Таджи-
кистан.
Характер пребыва-
ния. Повсюду оседлая птица.
Биотоп. Каменистые мест-
ности со скудной и чахлой расти-
тельностью, преимущественно в
предгорьях.
Подвиды и варьирующие признаки.
Почти весь ареал занимает номиналь-
ный подвид A.g. griseogularis Brandt,
1843. Птицы из Месопотамии и Юго-
Западного Ирана были выделены из-за
более рыжеватого тона окраски, как
A. g. ter-meuleni Zarudny et
L о u d о п, 1904; однако реальность этого подвида вызывает сомнение (Хартерт, 1938).
В экологическом отношении все пустынные куропатки сходны между собой.
Карта 11. Распространение пустынной куро-
патки Ammoperdix griseogularis
1—А. д. griseogularis, 2—А. д. ter-meuleni; а—гра-
ницы гнездовий
Туркестанская пустынная куропатка Ammoperdix griseogularis griseogularis
Brandt
Синоним. Ammoperdix bonhami bucharensis. S а г u d n y. Orn. Monatsber., 1911,
стр. 83, Южная Бухара.
Распространение. Ар е а л. Обитает в сухих каменистых местностях
от северо-восточных окраин Месопотамии и северо-западных частей Ирана до
долины р. Инда, восточнее заходя в район Соляного хребта. В пределах Совет-
ского Союза ареал крайне невелик: гнездится на п-ве Мангышлак и на плато
Усть-Урта (в пределах Казахстана; Долгушин, 1948); в горах Куба-Даг и
Кюрен-Даг, на Больших и Малых Балханах; по Копет-Дагу (на запад до хол-
мистых отрогов Сонгу-Дага между Дузлу-Олумом и Чатом), откуда по Атре-
ку доходит до Яглы-Олума (Зарудный, 1898); далее на восток населяет область
Туркменских предгорий Паропамиза (в Бадхызе в частности в хребте Гязь-
Гедык, а также в центре междуречья у озера Ер-Айлан-Дуз—Дементьев,
Спангенберг и Рустамов, 1948; Гептнер, 1949; возможно и Карабиль). На Аму-
Дарье встречена у Келифа, в горах Кулан-Ашар и Кундузляр (Заруд-
ный, 1896). Обычна в хребте Кугитанг. Далее населяет горы между Ка-
фирниганом и Вахшем, начиная от долины Пянджа и кончая холмами Гис-
150 отряд куриные
сарской долины; к северу от г. Сталинабада проникает в долину р. Гюль-
бисты, на р. Варзоб (не выше устья р. Харангон), и в долину р. Люгоб;
населяет горы около Чубека и Богорака и далее по Пянджу до Хирманджоу
и Шуроб-Дара (Иванов, 1940, 1949). Изредка залетает в Присырдарьинские
пустыни (Долгушин, 1948); Зарудный A915) встретил, повидимому залетную,
птицу в горах Аристан-бель-Тау в Кызыл-Кумах.
Характер пребывания. Ведет оседлый образ жизни. Сезонные
вертикальные миграции выражены незначительно (много слабее, чем у кек-
лика и других видов).
Биотоп. Держится обычно в нижних поясах пустынных и полупустын-
ных гор (крайне редко поднимаясь выше 1000 м над уровнем моря; Иванов,
1940), по склонам лессовых, конгломератовых или песчанистых холмов пред-
горий, прорезанных промоинами и оврагами. В общем заселяет открытые
ландшафты с редкой и чахлой растительностью степного или полупустынного
типа; избегает нагромождений скал, явно предпочитая глинистые участки
с выходами камней и осыпями галечника, песчаника и известняка, со сглажен-
ным рельефом. В узкие скалистые ущелья обычно не заходит (Лаптев, 1945).
В арчевники, как правило, не заходит; только один раз Зарудный A896)
встретил пустынных куропаток в июле 1884 г. в зоне арчи между реками
Ширин-Чай и Мергень-Улья (Копет-Даг). Встречается в кустарниках в зоне
горной полупустыни (Салихбаев, 1939). Вблизи гор иногда проникает в буг-
ристые пески (Зарудный, 1903).
Необходимое условие для жизни пустынной куропатки—близость воды:
реки родника и т. п. (редко, но встречается вблизи соленых водоемов; За-
рудный, 1900), так как она не совершает дальних перелетов на водопои.
Численность. В пределах ареала по подходящим биотопам обыч-
ная птица, однако численность повсюду невелика. На маршруте в 428 км
D8 км удобных стаций) в Копет-Даге встречено всего 100 шт., или 5250 шт.
в пересчете на всю обследованную площадь (а кекликов 123 000; Лаптев,
1936). Указание Радде и Вальтера A889) о том, что в Куба-Даге пустынные
куропатки встречаются чаще кекликов, не подтверждается последующими
исследователями. Заметных изменений количества по годам не отмечено.
Возможно, что, не заходя высоко в горы, она меньше, чем родственные
виды (кеклик и др.), подвержена сезонным метеорологическим воздействиям
(обильные снегопады, резкая смена температур и т. д.), а приуроченность
к малонаселенным полупустынным районам избавляет ее от уничтожения
человеком при неблагоприятных условиях, как это имеет место у турача
(весенние пожары, истребление при обильном снеге и т. п.).
Даже при наличии подходящих биотопов распределена по ареалу нерав-
номерно, рассеянными группами (колониями).
Экология. Размножение. Моногамы. Вопрос о постоянстве пар
не решен. Уже 3 апреля у Кизил-Арвата наблюдались в парах, а в Гязь-
Гедыке во второй половине апреля птицы держались парами и устраивали
гнездовые ямки (Дементьев, Спангенберг, Рустамов). Биология гнездового
периода изучена плохо. Период размножения в популяции растянут. Свежие
яйца находили уже с середины апреля (Туркмения—Шестоперов, 1928;
Северная Персия—Зарудный, 1900), в конце апреля встречали неполные
кладки со свежими яйцами и полные свежие и уже насиженные кладки; но
еще 17 мая 1886 г. в отрогах Сончу-Дага у Чата добыта самка со зрелым
яйцом в яйцеводе (Радде и Вальтер, 1889); Иванов добыл подобную самку
около Микоянабапа 2 июля, а Зарудный добыл на Пяндже самку с яйцом
20 июля (Иванов, 1940).
В связи с растянутостью периода размножения высказывались сообра-
жения о возможности нормальных двух кладок в лето (Зарудный, 1896).
Однако большинство наблюдателей полагает, что нормально у пустынных
ПУСТЫННАЯ КУРОПАТКА 151
куропаток в году одна кладка, а поздние кладки и выводки принадлежат пти-
цам, потерявшим первые кладки (Туркмения—Тишкин по Дементьеву,
1952; Таджикистан—Иванов, 1940). Вопрос этот требует выяснения.
Гнездо устраивается на голых склонах холмов, обычно под прикрытием
камня или кустика травы. Почти всегда это плоская ямка, слабо выстланная
-сухими травинками и растительным мусором (Зарудный, 1896; Шестоперов,
1928).
Полные кладки содержат 8—12 яиц (в Хальват-Даге было найдено одно
гнездо с 16 яйцами; Зарудный, 1896). Форма яйца овальная, несколько
заостренная к одному концу (Дементьев, 1952). Скорлупа матово-блестя-
щая, бледного желто-охристого (Зарудный, 1900) или сливочно-желтоватого
(Шестоперов, 1937) цвета без пятен. Размер яиц из Северной Персии 34х
Х24,6 мм (Зарудный, 1900), из Туркмении D) 35,4—38x25—26,5, в сред-
нем 36,3x25,7 мм (Шестоперов, 1928).
Продолжительность насиживания не установлена; повидимому, несколь-
ко более трех недель. Продолжительность постэмбрионального роста птен-
цов неизвестна. Только что вылупившиеся птенцы найдены в Туркмении
во второй половине июня (Дементьев, 1952) и встречаются даже до конца
июля (Зарудный, 1900). Птенцы размером с воробья добыты во второй
половине июля (Радде и Вальтер, 1889); пгенцы с только начинающимися
пробиваться пеньками перьев на крыльях встречены 17 июля у устья
р. Ханака-Дарьи, а к 27 августа они уже достигли величины перепелки (Тад-
жикистан—Иванов, 1940); в горах у Кизил-Арвата 4 сентября 1916 г.
найдены птенцы размером менее перепелки (Шестоперов, 1928). Молодые,
достигшие величины трех четвертей старки и почти закончившие линьку
в первый осенний наряд, встречены 22 сентября у устья р. Ханака-Дарьи
(Иванов, 1940).
Самцы во время насиживания держатся, повидимому, вблизи от гнезда;
встречены самцы и при выводках; однако в период размножения встречались
группы самцов до 20 шт. (Зарудный, 1896). Выводки от погибших самок при-
соединяются к другим выводкам; иногда одна самка водит до 50—60 разно-
возрастных птенцов; встречаются объединенные выводки с несколькими сам-
ками, при них иногда бывают и самцы (Зарудный, 1896). Птенцы довольно
самостоятельны; Зарудный A900) наблюдал как выводок, потерявший самку
через несколько лней после вылупления, самостоятельной стайкой кормился,
ходил на водопой и тесной кучкой ночевал в каменистой расщелине.
Линька. Изучена недостаточно. Ежегодно одна полная линька. Сро-
ки линьки растянуты, что связано с растянутым периодом размножения.
У просмотренных экземпляров взрослых птиц в апреле и в мае следов линьки
нет. Самцы, добытые 11 и 19 июня в Кугитанге, 18 июня в Б. Балханах,
6 июня и 24 июня у Чубека (Бухара), не линяют. Самец от 6 июня
(Б. Балханы) меняет мелкое оперение на туловище; на горле и подбородке
свежее перо, первые пять первостепенных маховых старые, шестые-седьмые
растут, задние маховые новые. Самец от 23 июня (Б. Балханы) меняет мелкое
перо, первые четыре маховых старые, пятое растет, шестые—десятые новые;
самец от 25 июня меняет мелкое перо, растут пятое и шестое маховые, задние
маховые перелиняли. Самец от 9 июля (Ходжа-Дере у Курру-Гоудана, цен-
тральный Копет-Даг) меняет мелкое перо, первые шесть маховых (на правом
крыле пять) старые, седьмое растет, задние восьмое—десятое новые. В такой же
стадии линьки (смена мелкого пера, рост пяти маховых, свежие шестое—деся-
тое маховые) почти на месяц позже, 27 июля добыт самец у Кене-Берме (близ
Ашхабада) и 31 июля у Фирюзы. Самки, добытые 22 и 25 июня (Б. Балханы),
24 июня у Чубека (Бухара) и 9 июля у Ходжа-Дере (Центральный Копет-Даг),
не линяют. У самки от 27 августа из Куркулаба (Центральный Копет Даг)
сменяется мелкое перо, первые пять первостепенных маховых старые, шестое
152
ОТРЯД КУРИНЫЕ
растет, задние седьмое—десятое новые. Таким образом, намечается отстава-
ние линьки у самок (половой диморфизм в линьке; Дементьев, 1952). В сен-
тябре большинство, а в октябре все птицы в новом пере.
Смена пухового наряда на первый птенцовый наряд растягивается в по-
пуляции с июня по август. Молодой самец из Даг-Дага (Б. Балханы) от
25 июня еще сохранил пух на подбородке, горле, шее, брюхе, и темени;
маховые первой генерации (без первого—третьего), длина крыла 56 мм. Пте-
нец, добытый 27 августа у Куркулаба, почти оперился, немного пуха сохра-
нилось только на голове (темя, щеки, подбородок), передние первое—третье-
маховые растут, длина крыла 85,5 мм. Молодой самец из Кене-Берме (Ашха-
бад), добытый месяцем раньше, 27 июля еще крупнее, растут перья на темени
и щеках, передние маховые еще в пеньках, крыло 97 мм.
Рис. 25. Пустынная куропатка (самец и самка)
Смена птенцового наряда на первый годовой наряд, повидимому, проис-
ходит в популяции в августе—октябре. Молодой самец от 6 августа (родник
Кендерли, Б. Балханы) начинает менять перо по туловищу; на горле и зобе
уже светлосерое перо, на лбу и в надглазничной полоске растут черные перья,
маховые старые, длина крыла 114 мм. Найденные 22 сентября у устья Ханака-
Дарьи (Таджикистан) молодые заканчивали линьку в первый годовой наряд
(Иванов, 1940).
Питание. Изучено недостаточно, особенно сезонная смена кормов.
В желудках и зобах добытых птиц обнаружены вегетативные части и семена
травянистых растений и насекомые—муравьи, жуки, мухи (Дементьев, 1952).
Пустынные куропатки регулярно и помногу пьют воду, посещая водопой
2—3 раза в день; обычно утром, при восходе, и вечером, при закате солнца
(Зарудный, 1896; Иванов, 1940). Когда птенцы еще малы и не могут ходить
на водопой, самка поит их, принося воду в клюве (Зарудный, 1896).
ПУСТЫННАЯ КУРОПАТКА 153
Суточный цикл. Не изучен. Даже в негнездовое время не обра-
зуют значительных скоплений: в стайках обычно не бывает более 5—20 птиц,
часто меньше. Враги не изучены.
Хозяйственное значение. Вследствие распространения в малонаселенных
пустынных районах и относительной малочисленности промысловое значение
пустынной куропатки (несмотря на превосходный вкус мяса) крайне невели-
ко. Это же ограничивает ее значение как объекта спортивной охоты.
В прежнее время ее добывали ловчимии птицами (балобан, ястреб),
сейчас добывается преимущественно ружьем (стрельба легкая). Из-за несо-
блюдения сроков охоты и перепромысла численность местами довольно силь-
но сократилась (окрестности г. Сталинабада и другие места Таджикистана—
Иванов, 1948; некоторые районы Узбекистана—Султанов, 1939 и т. д.).
Полевые признаки. По внешности и повадкам очень напоминает миниа-
тюрного кеклика. У самцов даже издали бросается в глаза исчерченность
боков туловища. При приближении опасности хорошо затаиваются как птен-
цы, так и взрослые птицы. Когда враг совсеме близко—взлетает и, пролетев
небольшое расстояние (на другой склон, к обрыву и т. п.), опускается и быстро
бежит (обычно вверх по склону). Бегает пустынная куропатка быстро и ловко
как по равнине, так и по крутым скалистым обрывам. Полет быстрый, порыви-
стый, прямой; всегда непродолжительный. Голос в виде легкого посвиста;
его можно передать словами «силь-силь» или «чиль-чиль» (Дементьев 1952).
Описание. Размеры и строение. В хвосте 12 рулевых. Крыло тупое,
вершину его образуют 2—4 маховых, причем разница в их длине очень невелика; первое
маховое обычно длиннее шестого и седьмого. Плюсна без шпоры даже у самцов. Длина кры-
ла у птиц из Советского Союза: у самцов A5) 125—133, у самок (9) 124—131,5, в среднем
соответственно 128.2 и 126,3 мм. Вес пустынных куропаток достигает примерно 200 г A82—
205 по шести взвешиваниям; Дементьев, 1952). Разница в размере полов крайне незначи-
тельна, и различия сводятся преимущественно к особенностям окраски.
Окраска. Пуховой наряд птенцов неизвестен.
Птенцовый наряд сходен по окраске с нарядом самки. Общий тон окраски песоч-
но-серый, светлее на нижней стороне. Беловатые крапинки образуют вершинные пятна, более
или менее сливающиеся в поперечный рисунок; особенно отчетливо он выражен на второ-
степенных маховых и верхних кроющих крыла. Первостепенные маховые желтовато-охри-
стые, с черновато-бурым рисунком и сероватым краном у вершины. Уши и уздечка темно-
серые, по сторонам темени светлые охристые полоски. Ноги желтоватые, клюв бледно-
роговой.
Первый годовой наряд сходен с нарядом взрослых птиц.
У взрослого самца лоб и полоса над глазом черные, уздечка и крою-
щие уха белые, позади уха небольшое желтоватое пятно. Верх головы серый, щеки и гор-
ло спереди и с боков чистого серо-стального цвета. Верх тела серый с более или менее вы-
раженным охристо-розовым налетом, плечевые и передняя часть спины с едва заметным по-
перечным рисунком, задняя часть спины и верхние кроющие хвоста с мелким темным кра-
пом. Первостепенные маховые светлобурые. Наружные опахала их (кроме первого) с попе-
речной исчерченностью. Перья по бокам туловища в широких продольных коричневых по-
лосах с черным окаймлением. Зоб и грудь розовато-охристые, брюхо беловатое. Рулевые,
кроме средней пары, буровато-рыжие, нижние кроющие хвоста светлоохристые. Раду-
жина от светлобурой до оранжевато-желтоватой, клюв оранжевый, ноги грязножелтые.
Оперение самки более тусклое, серое с желтоватым или розоватым оттенком;
с очень мелким поперечным охристым или темным рисунком. Нижняя сторона тела светлее
верхней. По бокам тела черноватые поперечные пестрины; широких коричневых полос нет
(изредка встречаются самки с полосами по бокам тела как у самцов—частичная «петухо-
перость»; Зарудный, 1915; в коллекциях Зоологического музея Московского государ-
ственного университета таких птиц нет). Голова без черных и белых полос. Клюв оран-
жевый, ноги грязножелтые.
Систематические замечания. По Зарудному A911) и Шестоперову A937) в Восточной
Туркмении (Кугитанг и предгорья западнее его) и в Таджикистане встречается подвид A. g.
bucharensiSy отличающийся будто бы от копет-дагских и североиранских птиц большей
величиной и розовым, а не желтоватым и сероватым тоном окраски. Однако уже Хартерт
A921—22), Бутурлин A935) и Иванов A940) считали этот подвид нереальным и сводили его
в синонимику A.g. griseogularis. Воробьев A941) подтвердил, что в Кугитанге встречаются
и сероватые и розоватые пустынные куропатки: первые встречаются по склонам из плот-
ного серого известняка, а вторые—по легко разрушающимся железистым песчаникам.
154
ОТРЯД КУРИНЫЕ
Эта железистая пыль пропитывает оперение птицы, придавая ему розоватый оттенок
различной интенсивности. Подобный смеханический диморфизм» свойствен пустынным
куропаткам и из других районов (сборы Бобринского из Бухары).
Дементьев A952) говорит о диморфности самок: более темные серые (Б. Балханы,
Ашхабад, Кене-Берме, Кельтечинар) и более светлые желтоватые (Меана, Кугитанг). При-
чина этой диморфности неясна. Вообще этот вопрос требует дальнейшего исследования;
вероятно, здесь имеет место сильно выраженная индивидуальная изменчивость.
РОД КАМЕННАЯ КУРОПАТКА ALECTORIS KAUP, 1829
Тип. Perdix b arbor a.
Синоним. Caccabis Каир, 1829.
11. Кеклик, или каменная куропатка Alectoris graeca
Meisner
Perdix Graeca. Meisner. Syst. Verg. Vog., welche die Schweiz bewohnen etc., 1804,
стр. 41, ex Daubenton PI. Enul., 231, Греция.
Синонимы. Caccabis saxatilis. Meyer. Naturg. Vog. Deutschl., 1, 1805—1809,
стр. 86.—Tetrau kakelik. F a 1 k. Beytz. z. topogr. Kenntn. Russ. Reichs, III, 1786,
стр. 390 (nomen nudum).— Perdix Chukar. Gray. 111. Ind ZooL, 1, 1832, табл. 54.
Русские названия. Каменная куропатка—название книжное. Кеклик—широко
распространенное у народов Средней Азии звукоподражательное название этого вида,
в конце XVIII в. перешедшее в русский язык.
Карта 12. Распространение каменной куропатки Alectoris graeca
1—А. д. saxatilis, 2—А. д. дгаеса, 3—А. д. whitakeri, 4—А. д. Cypriotes, 5—А. д. sinaica,
6—А. д. caucasica, 7—А. д. shestoperovi, 8—А. д. laptevi, 9—А. д. kurdestanica, 10—
А. д. werae, 11—А. д. koroviacovi, 12—А. д. subpallida, 13—А. д. fa'ki, 14—А. д.
pallescens, 15—А. д. pallida, 10—А . д. chukar, 17—А. д. dzungarica, 18—А. д. potanini,
19—А. д. pubescens, 20—А. д. тадпа; а—граница гнездовий
Распространение. Ареал. Южная Европа, Передняя, Средняя и
Центральная Азия: от Пиренеев и Альп, Сицилии, Балканского п-ва и остро-
вов Средиземного моря через Малую Азию, Кавказ, Копет-Даг, Гиндукуш,
Памир, Тянь-Шань и Алтай до Маньчжурии, Тибета и Гималаев.
Характер пребывания. Оседлая птица. При выпадении
снега в горах предпринимает незначительные вертикальные кочевки, переходя
на малоснежные участки
КЕКЛИК, ИЛИ КАМКННАЯ КУРОПАТКА 155
Биотоп. Глубокие ущелья и каменистые склоны гор от верхней гра-
ницы леса до вечных снегов, каньоны рек в горах и предгорьях; по каменистым
россыпям, выходам скал и вертикально расчлененным участкам рельефа
проникает в предгорья и на прилежащие равнины, по скалистым останцам
и глинистым холмам заходит и в песчаные пустыни.
Подвиды и варьирующие признаки. По всем Альпам на восток до Юго-Восточной
Венгрии распространена Л. g. saxatilis Meyer, 1805—1809; от Апеннинских гор (Ита-
лия) до Югославии, Греции, Албании, Западной Болгарии и Ионических о-вов распростра-
нена Л. g. graeca M e i sner, 1804; в Сицилии—A. g. whitaneri S с h i e b e 1. 1934; на
о-вах Эгейского моря, на о-ве Кипр, в европейской части Турции и Юго-ВосточнойБолга-
рии, в западных и южных частях Малой Азии, включая Тавр,—Л. g. Cypriotes Н а г t e г t,
1917; от Синайского п-ва до Сирийского побережья—Л. g. sinaica Bonaparte, 1856,
Среднюю и Восточную Малую Азию, Кавказ и Закавказье населяет Л. g. caucasicaS us с ti-
le i п, 1927; по горам южного берега Каспийского моря и в Копет-Даге живет Л. g. she-
stoperovi S u s с h k i n, 1927; западную часть Туркмении, Большие и Малые Балханы,
Мангышлак и Западный Узбой заселяет Л. g. laptevi Dementiew, 1945; в Южном
Курдистане—Л. g. Kurdestanica Mei nertzhagen, 1923; в Юго-Западном Иране
и прилегающих районах Месопотамии—Л. g. werae Sarudny etLoudon, 1904;
в Восточном Иране, включая Паропамиз и его отроги, в Белуджистане и в Синде—Л. g.
koroviakovi Sarudny, 1914; в Кызыл-Кумах, Кугитанге и Баба-Таге—Л. g. sub-
pallida Sarudny, 1914; горы системы Тянь-Шаня населены Л. g. falki Н а г t e г t,
1917; в Гиндукуше от Гильгита, через Балтистан до Ладака—Л. g. pallescens Hume,
1873; в Кашгарии и возможно в пограничных участках Памира и Семиречья, в Западном
Куэньлуне, Алтын-Таге и Западном Нань-Шане—Л. g. pallida Hume, 1873; в Гималаях
и Непале Л. g. chukar Gray, 1830—1832; Северное Семиречье, Тарбагатай, Алтай, За-
падный Саян, Танну-Ола и прилегающая Монголия населены Л. g. dzungarica S u s с h-
ki n. 1927; от Алашаня до Монгольского Алтая и Хангая распространен Л. g. potanini
S usch ki n, 1927; в Гань-су и прилегающих районах Китая вплоть до Маньчжурии—
Л. g. pubescens Swi n h о е, 1871; в Цайдаме, Куку-норе, Южно-Куку норском хребте,
Бурхан-Будда и в истоках р. Хуанхэ—Л. g. magna Przewalski 1876.
Отличия подвидов сводятся к удовлетворительно или плохо заметным вариациям
особенностей окраски и к небольшим различиям в размерах. Географические изменения
экологии (календарь периодических явлений, трофические связи и т. "•) известны не-
достаточно.
Кавказский кеклик Alectoris graeca caucasica ^ us<\iikin
Alectoris chukar caucasica. S u s h k i n. Bull. B. O. Club., NCCCXVH, 1927, стр. 22.
Ахалцых, Закавказье.
Синонимы. Alectoris graeca armenica. Бутурлин. Полный определитель птиц
СССР, II, 1935, стр. 201, Каре, Арарат.— Лlectoris graeca daghestanica Бутурлин.
Птицы Северного Кавказа, 1929, стр. 12, Карабудахкент, Дагестан.
Распространение. Ареал. По обоим склонам Большого Кавказа и по
всему Закавказью, включая нагорный Талыш. По хребту Большого Кавказа
на западе доходит до района Красной Поляны (Дороватовский, 1913); встре-
чена на горах Аибга, Ачишхо, Чугуш и в хребте Герпегем около Псебая
(Аверин и Насимович, 1938; Пузанов, 1949) на востоке—до Каспийского
моря (Билькевич, 1893). До Крымской войны A855—1856 гг.) птицы, видимо
этого подвида, успешно акклиматизировались в Крыму (район Симферополя —
Никольский, 1891), где встречались до 1898 г. (Бутурлин, 1901) и, возможно,
позднее, но затем были начисто выбиты (Пузанов, 1949). Попытка вновь ак-
климатизировать кекликов в Крыму была предпринята Розановым после
Великой Отечественной войны; результаты ее положительны. Указание на
гнездование кекликов в XVII в. в Днепро-Бугском районе Украины (Шарле-
мань, 1937) кажется в высшей степени сомнительным.
Вне пределов Советского Союза по Армянскому нагорью проникает
в среднюю и восточную часть Малой Азии; граница с другими подвидами
достаточно не выяснена
Характер пребывания. Повсюду оседлая птица. Зимой при
выпадении снегов спускаются в долины в горах и в предгорьях (Беме, 1926),
156 ОТРЯД КУРИНЫЕ
подходят к селениям (Дерюгин, 1900) или перемещаются на более крутые,
малоснежные склоны.
Биотоп. Предпочитает крутые склоны гор или предгорий и ущелья
горных рек с редкой и чахлой ксерофитной растительностью, выходами скал
и каменистыми россыпями; в меньшем числе заселяет можжевеловое редко-
лесье и луговины северных склонов (Даль, 1949), изредка встречается в гор-
ных лесах (хребет Цихис-Джавари выше г. Бакуриани—Сатунин, 1907);
сырых участков с пышной растительностью обычно избегает. На безлесных
скалистых хребтах живет в полосе 1000—2500 м над уровнем моря (Аверин
и Насимович, 1938); в Северной Осетии подымается выше 3500 м над уровнем
моря (Беме, 1926); гнездится по россыпям и вблизи ледников (Сатунин, 1907).
По безлесным каменистым участкам спускается почти до равнины (Сушкин,
1914), где иногда даже встречается на гнездовье в кустарниках вместе с тура-
чом (с. Кадели в долине р. Куры—Сатунин, 1907).
Численность. Неравномерна в пределах ареала: на южных скло-
нах гор птиц больше, чем на северных; в средней полосе гор более обычен
чем на большой высоте, и т. п. В Сарайбулагском хребте, Карабахское на-
горье, плотность колеблется от 0,05 до 1,3шт. на гектар (Даль, 1944).
Изменения численности по годам для этого подвида не отмечены, что,
повидимому, объясняется недостаточной изученностью биологии кавказского
кеклика: условия погоды несомненно отражаются на численности (глубина
снега, гололедица, ранние заморозки и т. п.).
Резкое уменьшение числа птиц в Азербайджане с 20-х годов XX в.
Верещагин A947) объясняет применением отравленного зерна для истребле-
ния полевок (отравление птиц) и широким применением истребительной охоты
с «кобылкой» (см. ниже).
Экология. Размножение. Изучена очень плохо. Повидимому,
моногамы. Разбиваться на пары птицы начинают уже в конце февраля—в на-
чале марта (Мензбир, 1900—1912). Участие самца в устройстве гнезда, наси-
живании и в уходе за выводком точно не установлено. Для балканского кек-
лика указывалось, что самка откладывает две кладки в два гнезда: в одном
насиживает самец, в другом—самка (Портал и др.); однако наблюдения над
кекликами в Советском Союзе этого не подтверждают.
Полные кладки обнаружены в мае; они содержат до 12 яиц. В году не-
сомненно одна кладка, но взамен утерянных возможны вторичные кладки,
что удлиняет период размножения в популяции: 17 мая 1895 г. найдена кладка
уз 14 сильно насиженных яиц (Джеванширский уезд, Сатунин, 1907),
14 июня 1925 г.—6 совсем свежих яиц (Юго-Западный Дагестан, Спанген-
берг), а кладка из 9 сильно насиженных яиц найдена даже 1 августа (Дарала-
гезский хребет, Даль, 1949). Форма яйца несколько вздутая, с хорошо выра-
женным острым и тупым концом. Окраска яйца глинисто-охристая (более
светлая, чем у Л.Я. falki); по основному фону разбросаны слабо выраженные
светлые рыжевато-коричневые пятна.
Гнездо—небольшая ямка, слабо выстланная растительной ветошью,
обычно под защитой нависающего камня, кустика вереска или можжевель-
ника, иногда в трещине скалы. Самка сидит на гнезде очень плотно. Продол-
жительность насиживания и время постэмбрионального роста птенцов не
установлены. В начале августа встречались птенцы размером в полматки
и птенцы, почти достигшие величины взрослых птиц (склоны Арарата,
Сушкин, 1914); птенцы размером в половину матки добыты у Еревана еще
24 сентября 1936 г. (коллекция Зоологического музея Московского госу-
дарственного университета).
Весной птицы держатся парами и одиночками, летом и в начале осени —
стайками-выводками; осенью и зимой стаями в 10—20 шт. и более. Продол-
жительность жизни до 7 лет (по содержанию в неволе; Московский зоопарк).
КЕКЛИК. ПЛИ КАМЕННАЯ КУРОПАТКА 157
Линька. Детально не изучена. В связи с растянутостью периода
размножения в популяции растянут и период линьки. По коллекционным
материалам Зоологического музея Московского государственного университета
в марте—начале июня птицы в обношенном пере без следов линьки. Повидимо-
му, самцы начинают линьку несколько раньше самок. Самцы от 9 июня
1930 г. из Сарай-Булага (Армения) и от 23 июня 1926 г. из Карабудахкента
(Дагестан) в сильно обношенном пере; по телу появились еще нераскрывши-
еся пеньки новых контурных перьев, первое—девятое первостепенные махо-
вые старые, десятое выпало и на его месте еще нераскрывшийся пенек; рули
старые. У самца от 13 августа 1924 г. из того же района энергичная линька
мелкого пера по всему телу, еще не полностью выросшие новые перья уже вы-
ступают над старыми; первостепенные маховые сменяются центробежно: пер-
вое и второе—старые, третье—в три пятых, а четвертое—в шесть седьмых
окончательной длины, пятое—десятое новые, полностью выросшие; второ-
степенные маховые сменяются центростремительно, последние 3—4 пера еще
старые; начинают сменяться рули. В сходном состоянии оперения самцы
из района Тифлиса от 10 августа 1916 г.: по всему телу новое контурное перо,
сохранившее еще чехлики у основания; первое первостепенное маховое ста-
рое, второе—три четверти, а третье—шесть седьмых полной длины, четвер-
тое—десятое полностью выросшие новые; сменяются плечевые; хвост сильно
обношен, выпали средняя и предпоследняя пары рулевых. Взрослые самцы
от октября 1948 г. из района Сарай-Булага и Арпа-Чая (Армения) пол-
ностью перелиняли в свежее перо; у части птиц первое и второе первосте-
пенные маховые еще на 40—10 мм не доросли до полной длины.
Самка от 26 июня 1923 г. из Карабудахкента с большим наседным пятном,
без признаков линьки. Самка от 8 июля 1924 г. из окрестностей Гуниба
(Дагестан) с первыми признаками линьки: отдельные редкие пеньки контур-
ных перьев чуть пробивают кожу на большом наседном пятне и на спине;
десятое первостепенное маховое выпало и на его месте едва, пробил кожу
пенек нового пера. У самки от 25 августа 1908 г. из Мцхета (Грузия) заро-
стает наседное пятно, по всему телу раскрываются пеньки контурных перьев;
первое—шестое первостепенные маховые старые, седьмое в половину, а вось-
мое—в пять восьмых полной длины, девятое—десятое выросли; началась
линька хвоста. Самки от 18 и 26 сентября 1948 г. из района Еревана полно-
стью в новом пере, но первое первостепенное маховое еще в три четверти,
а второе—шесть седьмых окончательной длины; продолжается рост рулевых,
наружная пара лишь чуть больше половины нормальной длины. Самка от
9 октября 1948 г. из Арпа-Чая (Армения) в новом, но еще не полностью вырос-
шем контурном пере (у основания сохраняются чехлики); первое первостепен-
ное маховое в две пятых, а второе—в три четверти окончательной длины.
Ноябрьские и декабрьские экземпляры взрослых птиц в новом пере без
следов прошедшей линьки.
Молодая птица от 16 июля 1907 г. из Армении интенсивно линяет в первый
годовой наряд: по всему телу еще нераскрывшиеся и уже наполовину рас-
крывшиеся пеньки; энергично растут кроющие хвоста и рулевые; первое—
второе первостепенные маховые уже достигли полного роста, третье—четвер-
тое птенцовой генерации, пятое в нераскрывшемся пеньке, шестое в три пятых,
а седьмое в шесть седьмых полной длины, восьмое—десятое уже полностью
выросли. Однако в таком же состоянии оперения молодые птицы добывались
«ще в октябре B, 6, 29 октября; Армения) и в то же время встречены полно-
стью закончившие линьку молодые F и 10 октября 1948 г., р. Арпа-Чай,
Армения). 24 сентября 1936 г. у Еревана добыта молодая птица еще целиком
в птенцовом наряде; по всему телу еще нераскрывшиеся пеньки контурного
пера годового наряда; первое первостепенное маховое еще в нераскрывшемся
пеньке, второе—едва распустилось и достигло лишь трети окончательной
158
ОТРЯД КУРИНЫЕ
длины. Все данные по линьке молодых птиц также свидетельствуют о растя*
нутости периода размножения в популяции.
Питание. Основу питания кеклика составляют семена бобовых и зе-
лень различных трав (особенно Medicago sp.); живущие вблизи посевов птицы
склевывают проросшие зерна злаков (май; Армения; Даль, 1944), в июле-
августе в желудках и зобах—моллюски, гусеницы, муравьи, зерна ячменя,
бобы, семена мятлика и ковыля (Даль, 1949). Осенью кормятся по хлебным
полям, собирая падалицу; по дорогам расклевывают помет (Сатунин, 1907).
Зимой питаются исключительна
растительной пищей, кормясь
на местах пастьбы козлов и ба-
ранов и по выдувам снега на
крутых склонах. Ранней весной
поедают почки и молодые по-
беги кустарников.
Суточный цикл. Кек-
лики просыпаются на рассвете
и начинают кормиться. После
кормежки посещают водопой; в
жаркое время дня отдыхают в
тени скал или кустарников, ча-
сто вблизи от воды. После спада
жары опять кормятся. Охотно
посещают дороги, купаясь в пы-
ли и роясь в помете скота. При
приближении опасности рассы-
паются по склону и затаивают-
ся. После захода солнца устраи-
ваются на ночлег, обычно в тре-
щинах скал, пещерках или вы-
моинах под скалами, реже по
выступам больших камней и
скал.
Враги. В Сарайбулагском
хребте встречены в погадках
филина (Армения—Даль, 1944).
Повидимому, добывают кекли-
ков беркуты, волки, лисы и дру-
гие хищники.
Хозяйственное значение.
Объект спортивной охоты, ме-
стами перерастающей в промы-
словую. Обычно кекликов добы-
вают ружьем, реже с ловчими птицами и ловят сетями и силками на кри-
ковую птицу; в заморозки после оттепелей обмерзших птиц бьют палками
и ловят руками (Силантьев, 1898). Особо губительна запрещенная, но все
еще широко применяющаяся охота с «кобылкой»: охотник подходит к птицам,
прикрываясь рамой, затянутой материей пестрых ярких цветов; любопытные
птицы подпускают близко, и с выстрела охотник берет 5—12 шт. (Цветков).
Кеклики в СССР в благоприятные годы добываются в больших количе-
ствах. Так, только через Ленинградскую экспортную базу прошло кекликов
(всех подвидов) в 1927—1928 гг. 166,7 тыс. шт. A3,6% от всей обработанной
дичи), в 1928—1929 гг.—198,1 тыс. шт. A7,9%), в 1929—1930 гг.—78 тыс.
(8%), в 1930—1931 гг.—233,2 тыс. (9,2%) и в 1931—1932 гг.—78,7 тыс. шт.,
или 4,3% от обработанной дичи (Рудановский и Насимович, 1933).
Рис. 26. Каменная куропатка
КЕКЛИК, ИЛИ КАМЕННАЯ КУРОПАТКА 159
Полевые признаки. По внешнему виду и поведению несколько напоминают
домашних кур; очень живые, подвижные и осторожные птицы. Превосходно
бегают, легко взбираются по крутым скалам. При опасности молодые птицы
затаиваются, взрослые же вначале бегут, а затем с шумом и квохчущим
криком срываются и летят торопливым и порывистым, но не скорым поле-
том и вскоре опускаются на землю, обычно на противоположный склон, убегая
дальше пешком. Кормясь, бегут вверх или вдоль по склону, но никогда
не бегут вниз; при кормежке часто подают квохчущий голос («каккалик»).
Описание. Размеры и строе л и е. В хвосте 14 рулевых. Наружное опахало
первого и внутренние опахала всех первостепенных маховых без заметных вырезок*
крыло тупое. У самцов короткая тупая шпора, отсутствующая у самок.
По величине самки чуть меньше самцов; длина крыла у самцов A5) 160—176, у са-
мок A4) 149—170, в среднем соответственно 167,2 и 159,5 мм. Вес кекликов колеблется
в пределах от 370 до 770 г (Бутурлин, 1935); вес четырех самцов этого подвида был
в среднем 557 г E39—573 г; Сарайбулагский хребет, Армения—Даль, 1944).
Окраска. Пуховой наряд не описан; повидимому, сходен с другими подвидами. У мо-
лодой птицы в первом гнездовом наряде верх буровато-песочный с малозаметным оливко-
вым оттенком; плечевые, малые и средние кроющие крыла и в меньшей степени перья спи-
ны с концевыми тусклобеловатыми пятнами треугольной формы; первостепенные махо-
вые бурые с охристо-песочной поперечной исчерченностью на наружных опахалах; руле-
вые слабо рыжеватого цвета; низ тела более или менее серовато-белый с песочным отли-
вом; на боках тела перья с отчетливо выраженными узкими поперечными полосами на
опахалах.
Первый годовой наряд сходен с нарядом взрослых птиц, отличаясь лишь несколько
более тусклой окраской и более узкими первым—вторым первостепенными маховыми.
Сезонный и половой диморфизм окраски у взрослых птиц отчетливо не выражен;
самки отличаются от самцов лишь отсутствием шпор и несколько меньшей величиной.
У взрослых птиц лоб черный. Верх головы и шеи серый с отчетливым винно-розовым
отливом, отсутствующим у некоторых особей. От лба через глаз к уху идет черная поло-
са, которая спускается на зоб и окружает шею спереди и снизу. У угла рта черное пятно.
Передняя часть спины, плечевые, верхние кроющие крыла и второстепенные маховые
темносерые с густым винно-розовым отливом. Задняя часть спины, поясница и верхние
кроющие хвоста дымчато-серые с оливковым отливом. Первостепенные маховые бурые;
на наружных опахалах продольные охристые пятна, образующие при сложенном крыле
светлую предвершинную полоску. Подбородок и шея спереди охристо-белые, отделенные
от серых с розоватым отливом зоба и груди черной полосой. Брюхо охристо-желтое; ниж-
ние кроющие хвоста также охристо-желтые, но несколько более темного оттенка. Рулевые
каштаново-рыжие; у средних пар в основании более или менее развит дымчато-серый цвет.
Перья на боках туловища с отчетливо выраженными черными поперечными полосами на
опахалах (примерно 4—6 мм шириной) и более или менее выраженными каштановыми
вершинами. Ноги бледнокрасные; клюв и узкое голое кольцо вокруг глаз красные.
От других подвидов, в том числе и от копетдагского, кавказские кеклики отличаются
более темным общим тоном окраски, темя розоватое без буроватого оттенка, на пояснице
отчетливее проступает оливковый цвет, черные поперечные полосы на боках тела
слегка шире.
Систематические замечания. Количество подвидов кекликов, описанных на терри-
тории Советского Союза, очень велико, а различия между ними (в величине и тонких дета-
лях окраски) малозаметны; в пограничных участках ареалов подвидов часто встречаются
особи переходного типа (например, переходные особи между A. g. subpallida и A. g. fa-
Iki встречаются в Нура-Тау, Баба-Дага и т. д.). При изучении этого вида мы были вынуж-
дены часть подвидов свести в синоним. Однако очень вероятно, что дальнейшее накопление
коллекционного материала и сравнительное подвидовое изучение экологии приведет к еще
большему сокращению числа подвидов. Повидимому, кавказские и копетдагские кеклики
должны быть объединены в одну группу, тяньшанский и джунгарский—в другую; перехо-
дом между ними будет равнинный кызылкумский кеклик, объединяющий и западно-турк-
менского равнинного кеклика; вероятно сохранит свое подвидовое значение восточно-
иранский кеклик, объединяющий восточно-иранских и афганских птиц. Сейчас такая
полная ревизия невозможна из-за недостатка коллекционного материала.
Следует отметить, что при частом купании в пыли (что характерно для кекликов)
мелкие частицы почвы остаются на перьях, придавая им специфический оттенок (красно-
ватый на красных глинах, буроватый на лессовых почвах и т.п.); этим достигается точная
«юдгонка» оперения оседлых птиц под цвет почвы (Формозов, 1950). Поэтому при анали-
зе подвидов у кеклика необходимо диференцировать отличия в окраске, обусловленные
физиологическими особенностями отдельных популяций, от цветовых отличий, обуслов-
ленных чисто механическим насыщением перьев мельчайшими частицами почвы, более^
или менее заметно изменяющих общий тон окраски.
160 отряд куриные
Копетдагский кеклик Alectoris graeca shestoperovi Suschkin
Alectoris chukar shestoperovi. S u s h k i n. Bull. B. O. Club, N CCCXVII, 1927, стр. 23. хре-
бет Асыльма (близ Ашхабада), Копет-Даг.
Распространение. Ареал. Встречается в Западном и Центральном
Копет-Даге, на восток, повидимому, до Каакха (восточнее ее замещает A. g.
koroviakovi); гирканские кеклики—из системы Эльбурса—принадлежат
к этому же подвиду, а не к кавказскому (Дементьев, 1948).
Характер пребывания. Оседлая птица, совершающая не-
большие сезонные вертикальные кочевки: весной (в начале марта) поднимает-
ся выше в горы к родникам; с наступлением холодов и выпадением снега спу-
скается вниз вплоть до предгорной равнины; миграции совершаются стаями,
иногда достигающими до 500 шт.; зимой 1934 и 1935 гг. наблюдались массовые
перекочевки в Кара-Кумы (Рустамов, 1945).
Биотоп. Небольшие ущелья#с осыпями камней, крутыми склонами
и родниками во всех участках Копет-Дага, от подгорной равнины до зоны
арчи на высоте от 500 до 2000 м над уровнем моря и выше. В арчевниках
более многочислен, чем в других поясах.
Численность. В пределах Туркменской ССР довольно многочис-
лен. На маршруте в 428 км встречен (за два года учета) 3721 кеклик, что
в переводе на всю обследованную площадь (между Фирюзой и Сулюкли,
около 15 000 га) составляет около 120 000 шт. (Лаптев, 1934). Колебания чис-
ленности в зависимости от условий года (отсутствие воды летом, суровые,
многоснежные зимы и т. п.) несомненны, однако детально они не изучены.
Так, в Туркмении очень «урожайным» был 1934 г.
Экология. Размножение, (преимущественно по Рустамову,
1945). Повидимому, моногамы. Разбиваются на пары в первой половине марта;
при этом самцы иногда вступают в драку друг с другом. Сформировавшиеся
пары в середине марта уже занимают гнездовые участки и приступают к по-
стройке гнезд A2—18 марта 1942 г., горы Хунча). Гнездо—небольшое углуб-
ление, выстланное растительной ветошью или без нее,—устраивается в тре-
щине скалы, под камнями на каменистых откосах, у отдельных кустов арчи;
в постройке гнезда участвуют и самец и самка.
Начало откладки яиц приходится на первую декаду апреля, но свежие
кладки встречаются и в мае и в июне. Полная кладка содержит обычно от 8
до 14 яиц, реже до 18—19. Окраска яиц варьирует (повидимому, сходна с ок-
раской яиц кавказского кеклика); довольна часто встречается сливочно-
(желтовато-) охристый основной тон с мелкими красно-бурыми пятнышками
(Рустамов).
В лето нормально лишь одна кладка; повидимому, возможны вторичные
кладки взамен потерянных, что находит свое выражение в растянутости перио-
да размножения в популяции; возможно сказывается и разновременная
физиологическая готовность отдельных особей к размножению. Насиживание
продолжается около месяца; при этом самец помогает самке (Рустамов,
1945); эти данные требуют дополнительного уточнения.
Первые птенцы встречены в первой декаде мая; однако еще в конце июня
и в первой половине июля попадаются молодые ростом с перепелку, а в сен-
тябре, наряду с молодежью, достигшей размеров взрослых птиц, встречены
птенцы ростом вдвое менее перепелки. Выводок водят оба родителя, бросая
заботу о птенцах лишь тогда, когда они достигнут размеров взрослых
птиц; в августе-сентябре выросшие выводки сбиваются в стаи до 100 шт.
и более.
Линька. Материалы по линьке очень скудны. Добытые между концом
апреля и июнем не линяют. Полная годовая линька начинается во второй
декаде июля: самец, добытый 11 июля 1927 г. у Ашхабада (Багир), интенсивно
КЕКЛИК, ИЛИ КАМЕННАЯ КУРОПАТКА 161
сменял мелкое перо, первое—третье первостепенные маховые старые, четвер-
тое новое в пять шестых окончательной длины, пятое—десятое достигли пол-
ного роста, второстепенные маховые сменились полностью, плечевые еще ста-
рые, предпоследняя пара рулей достигла шести седьмых полной длины, две
средние пары в пеньках, остальные старые; самец от 21 июля 1942 г., добытый
на горе Душак (Центральный Копет-Даг), на сходной стадии линьки. В сен-
тябре встречены уже полностью перелинявшие взрослые птицы. Возможно,
что весной может иметь место частичная линька (?).
Линька пуховиков в первый птенцовый наряд начинается вскоре после
вылупления; она растянута в популяции по времени, что стоит в связи с рас-
тянутостью периода размножения. Линька в первый годовой наряд проходит
во второй половине июня—августе, затягиваясь у отдельных птиц до октября.
Молодые птицы, уже полностью перелинявшие в первый годовой наряд,
встречены в середине августа.
Питание. Пищу копетдагского кеклика составляют различные части
растений (плоды, семена, луковицы, клубни, листья, тонкие веточки) и на-
секомые. Набор кормов довольно обширен и тесно связан с био-фенологиче-
скими стадиями развития основных растительных группировок Копет-Дага
(Федоров, 1949; Коган, 1950). Летом и осенью в зобах и желудках птиц встре-
чены луковицы мятлика Роа bulbosa, плоды и семена шиповника и винограда,
мелкоплодной вишни, кизильника Cotoneaster, жимолости Lonicera persica,
ягод морены Rubia florida, семена ячменя и пшеницы; в меньшем числе
найдены плодики Poterium sanguisorba, семянки Echinops sp., целые бобы
Astragalus pitetocladus9 Onobrychis sp. и т. п., мешочки Carex sp., семена и лу-
ковицы Tulipa sp. и А Шит sp., кусочки клубней Bongardia chrysogonum и
Leontica sp., листочки Astragalus sp., Colutea persica, Thymus Kotschyanus
и т. п.;, в зобах встречены также кусочки камеди трагакантовых астрагалов;
роль жерновков, облегчающих перетирание пищи в желудке, выполняют не
только камешки, но и твердые косточки видов рода Cerasus и, возможно, семян-
ки шиповников (Рустамов, 1945; Федоров, 1949; Коган, 1950). Из беспозво-
ночных в желудках и зобах встречены прямокрылые Orthoptera, жуки Coleop-
tera, клопы Hemiptera, муравьи Formicidae, паукообразные Arachnoidea
(Рустамов, 1945) и др.; однако насекомые в питании играют лишь второсте-
пенную роль и встречаются лишь летом (Коган, 1950). Птицы, особенно в лет-
нее время, нуждаются в регулярном водопое и поэтому обычно держатся
вблизи водных источников.
Зимой, в январе—феврале, основу питания составляют как луковички
горного лука А Шит vavilovi и тюльпана Tulipa Micheliana, которые птицы
выкапывают из земли (Рустамов, 1945), так и зеленые листья, преимуществен-
но мятлика и пустынной осочки; меньшее значение имеют в этот период семе-
на и плоды; количество камешков в желудке птиц зимой увеличивается
(Коган, 1950).
Интересно, что кеклики поедают плоды, семена и цветы ряда таких ядо-
витых растений как: Arum Korolkowi, Hyoscyamus sp., Merendera robusta и
др. (Коган, 1950).
Суточный цикл. Рано утром, еще до восхода солнца, начинается
утренняя жировка и посещение водопоев (ручьи, лужи, ключевые болотца);
к это время птицы много кричат (крик напоминает кудахтанье домашних
кур). К 10—11 час. кормежка прекращается и голоса птиц стихают; в середине
дня, до 4—5 час. вечера, птицы скрываются от солнца в трещинах скал,
в кустарниках и арче, отдыхают и купаются в пыли. После спада жары вновь
кормятся до захода солнца, в сумерках устраиваясь на ночлег (Рустамов,
1945). Ночуют в скалах; также охотно ночуют под «подушками» трагаканто-
вых астрагалов, где ночью температура на несколько градусов выше темпе-
ратуры наружного воздуха (Федоров, 1949).
11 Птицы СССР, т. IV
162 ОТРЯД КУРИНЫЕ
Враги. Старых и молодых птиц на земле и на лету берет беркут;
молодых птиц и птенцов уничтожают коршуны, чеглоки, реже пустельга;
на ночлегах на птиц нападает филин; летящих птиц изредка ловят крупные
соколы. Молодых и взрослых птиц и яйца уничтожают лисицы и куницы.
Хозяйственное значение. Объект любительской спортивной охоты и един-
ственная промысловая птица в горах Туркмении. Особенно добычлива
(и должна ограничиваться, как истребительная) охота на местах водопоя,
когда одним выстрелом из засидки снимают до 10—15 птиц. Широко распро-
странена также охота с ружьем с подхода, под прикрытием щита, затяну-
того яркими кусками материи. Помимо ружейной охоты, кекликов добывают
также петлями и ловчими птицами—сейчас эти виды охоты распространены
мало. В заморозки после оттепелей обмерзших кекликов избивают палками
и ловят руками.
Лучшее время для промысловой охоты в Туркмении—ноябрь, когда
птицы очень жирны и держатся небольшими стаями; наиболее добычлива
охота на утренних и вечерних водопоях (Рустамов, 1945).
В 1929 г. шесть пар кекликов вероятно этого подвида были выпущены на
остров Барса-Кельмес и приступили к размножению, однако вскоре были
полностью уничтожены корсаками (Слудский, 1948); этот опыт показывает,
что при создании благоприятных условий жизни может успешно проводиться
акклиматизация этих птиц.
Описание. Размеры и строение. Очень близки к кавказским кекликам,
отличаясь от них лишь несколько меньшими размерами (отчетливо заметно лишь на
сериях) и небольшими отличиями в окраске.
У самцов копетдагских кекликов F) крыло 160—168 мм, у самок (8) 152—160, в сре-
днем соответственно 162,3 и 155,9 мм. Вес птиц осенью (когда кеклики очень жирны)
достигает 400—500 г (Рустамов, 1945).
Окраска. Копетдагские кеклики очень похожи по окраске на кавказских и от-
личаются от последних лишь тем, что сверху они чуть светлее и серее, темя слегка буро-
ватое, оливковый оттенок на пояснице менее интенсивен, черные поперечные полосы
на боках тела несколько уже C—5 мм шириной).
Западнотуркменский равнинный кеклик Alectoris graeca laptevi
Dementiev
Alectoris graeca laptevi Дементьев. Изв. Туркменского филиала АН СССР, № 5—6, 1945,
стр. 178—179. Бугдайли, Туркмения.
Синоним. Alectoris graeca dementievi. Рустамов. Сборник сОхрана природы»,
№ 5, Москва, 1948, стр. 106—108, колодец Кавешем, Юго-Западный Усть-Урт.
Распространение. Ареал. Равнинные участки Западной Туркмении
и обрывы Атрека, массив Больших Балханов, Западный Узбой (район озер
Ясха и Каратегелек, колодца Ян-Аджи и т. д.), чинк Усть-Урта (в районе
колодцев Кавешем, Аяк-Депме, Баш-Депме); этот же подвид, повидимому,
встречается на Малых Балханах, на Красноводском плато и на Мангышлаке,
на восток проникает до Сарыкамышской впадины. Этот ли или кызылкумский
подвид кеклика населяет останцевые бугры в районе Дарвазы,—остается
невыясненным.
Характер пребывания. Оседлая птица.
Биотоп. Останцы, бугры и небольшие глинистые холмы и обрывы
или отдельные небольшие горные участки в пустынных равнинах; встре-
чается и в бугристых песках (например, в 35—40 км севернее Гасан-Кули),
Необходимое условие—наличие воды, хотя бы и солоноватой (дождевые
лужи, колодцы, родники, озера).
Численность. Менее многочисленны, чем копетдагские кеклики;
в изолированных, но относительно больших горных массивах (например,
Большие Балханы) более обильны, чем по холмам и бугристым пескам.
КБКЛИК, ИЛИ КАМЕННАЯ КУРОПАТКА 163
Экология. Размножение. Биология этого подвида почти совер-
шенно неизвестна. У самцов, добытых 4 мая у колодца Кавешем (ю.-з.Усть-
Урт), семенники были почти максимальных размеров A7,5x10,5 мм; 21,5х
X 10,5 мм), однако в это же время уже встречены молодые размером с пере-
пелку (Рустамов, 1948). На Мангышлаке большинство самок к 20 мая окон-
чило кладку; 1 июня встречены выводки; 13 июня найдено гнездо с 21 сильно
насиженным яйцом; 7 июня добыта самка с готовым яйцом в яйцеводе, птица
сильно упитана, а наседное пятно развито слабо; в это же время добывались
сильно истощенные самки (вес 320 и даже 300 г) с громадными наседными
пятнами (Долгушин, 1948). Все это свидетельствует о растянутости периода
размножения в популяции (что свойственно и другим подвидам). В году,
вероятно, лишь одна кладка; в случае ее гибели возможны повторные, вы-
нужденные, кладки.
В период размножения птицы держатся парами, но самцы, повидимому,
не насиживают (ни у одного из 15 добытых самцов не обнаружено наседных
пятен; Долгушин, 1948). Одно из найденных на Мангышлаке гнезд предста-
вляло ямку глубиной 8—10 см и до 25 см в поперечнике, обвитую травой
и выстланную внутри мягкой, сухой травой и перьями кеклика; оно было рас-
положено в узкой затененной части ущелья у дождевой ямы среди густых
зарослей ежевики, крапивы, мяты и лопухов; второе гнездо найдено на силь-
но опесчаненном месте с редкими кустами терескена, под одним из кустов.
В гнезде с 21 яйцом в 15 яйцах были уже крупные зародыши, а 6 яиц были
болтунами (Долгушин).
Продолжительность насиживания не установлена. Молодые птицы раз-
мером с перепелку встречены у колодца Кавешем 4 мая (Усть-Урт—Рустамов,
1948); в Малых Балханах в выводке из 20 птенцов молодые почти достигли
размеров взрослых птиц (Шестоперов, 1935). До взматерения молодых при
выводке обычно встречаются и самец и самка; число молодых в выводках—
от 8 до 18 шт. (Долгушин, 1948).
Линька. Изучена крайне недостаточно. В январе—феврале птицы
в новом пере без признаков линьки. В апреле—мае птицы уже в заметно об-
ношенном пере, но без следов линьки. Взрослые птицы, вероятно, начинают
полную годовую линьку после того, как подрастут молодые. У самки, добы-
той 10 июня 1948 г. в Секи-Даге (Б. Балханы), на большом наседном пятне
редкие, наполовину раскрывшиеся пеньки; нераскрывшиеся или чуть рас-
крывшиеся пеньки есть по всему телу; первое—четвертое первостепенные
маховые старые, пятое выпало, шестое новое до одной трети полной длины,
седьмое—десятое новые, полностью выросшие; сменяются второстепенные
маховые, плечевые все старые; началась смена рулевых.
Линька молодых птиц из птенцового в первый годовой наряд, повиди-
мому, идет в июле—августе. Молодой самец от 3 августа 1947 г. из окрестно-
стей родника Сакка (Б. Балханы) в стадии энергичной смены мелкого пера
по всему туловищу, новое перо уже показалось над старым; первое—второе
первостепенные маховые уже достигли полного роста, третье—четвертое
птенцовой генерации, следующие уже годового наряда: пятое в половину,
шестое в четыре пятых, а седьмое—десятое полной длины; рулевые все но-
вые, но большинство еще далеко не достигло полного роста.
Питание. Пищу, повидимому, составляют преимущественно раз-
личные части растений и в меньшей степени насекомые; детально пита-
ние не изучено. В связи с обитанием в пустынных районах видимо резко
выражена сезонная смена кормов. В желудках и зобах птиц, добытых
в апреле—мае 1947 г. у колодца Кавешем, обнаружены лишь эфемеры:
коробочки с семенами Gagea reticulata, корзинки Scorzonera pusilla, стручки
Goldbachia laevigato и коробочки Roemeria hybrida (Усть-Урт—Рустамов,
1948).
11*
164
ОТРЯД КУРИНЫЕ
Хозяйственное значение. Ввиду относительной малочисленности и оби-
тания в безлюдных районах имеет крайне малое, по сравнению с другими
подвидами, промысловое значение и
добывается лишь случайно. В связи
со строительством Главного Туркмен-
ского канала и орошением больших,
ныне безводных площадей числен-
ность популяции этого подвида долж-
на возрасти и тем самым, несомнен-
но, увеличится его значение как
охотничьего и промыслового объек-
та. Для этого необходимо выполне-
ние простейших биотехнических ме-
роприятий (запрещение охоты в бли-
жайшие годы, регулирование разме-
ров заготовок и точное соблюдение
сроков охоты и т. п.).
Описание. Размеры и строе-
ние. Очень похожи на копетдагских птиц,
но несколько меньше. Длина крыла сам-
цов 145—163 мм, самок 145—159 мм (Де-
ментьев, 1945); самцов A0) 150—167, у са-
мок (9) 147 — 157 мм, в среднем соответ-
ственно 158,7 и 151,9 мм (коллекция Зооло-
гического музея Московского университе-
та). По сравнению с копетдагскими, рав-
нинные кеклики имеют несколько более
мелкий клюв и более короткую цевку.
По окраске очень сходны с ко-
петдагскими птицами. Некоторые особи от-
личимы чисто серым или серо-сизым окра-
сом поясницы, хотя у других из тех же
мест отчетливо выражен и оливковый налет.
Пуховой птенец: верх головы и за-
шеек каштаново-песочного цвета; от боков
шеи на спину идут узкие охристо-бурова-
тые полоски, сливающиеся на спине в ши-
рокую полосу, идущую до конца туловища,
охристо-буроваты и бока тела; промежу-
ток между спинной полосой и боками свет-
лый серовато-охристый. Подбородок и низ
тела белые; щеки желтовато-белые. По об-
щему тону окраски пуховички западно-
туркменского равнинного кеклика более
Рис. 27. Птенцы каменной куропатки светлопесочные в отличие от тяньшанских,
A. g. laptevi (сверху) и A. g. jalci (сниз>) более темных, рыжевато-бурых.
Восточноиранский кеклик Alectoris graeca koroviakovi Sar.
Caccabis kanelik koroviakovi. Зарудный. Орнит. Вестник, № 1, 1914, стр. 55, горы
Бирджан в Восточном Иране.
р
Н а г t е г t. Vog Palaanct. Fauna, III, 1921—22,
е.—Alectoris graeca koriakovi. Искаженное название
Faun. Br. Ind., V, 1928, стр. 1928.
Бирджан в Восточном Иране.
Синонимы. Alectoris graeca kirthari. Н а г t е г t. Vog Palaan
стр. 1908, хребет Киртар в Синде.—Alectoris graeca koriakovi
в книге Baker. Faun. Br. Ind., V, 1928, стр.
Распространение. Ареал. Населяет Восточный Иран и Белуджистан,
проникая на восток за низовья Инда в Синд. По отрогам Паропамиза про-
никает в пределы Советского Союза, населяя возвышенности Бадхыза (хре-
бет Гязь-Гедык, Акар-Чешме, Фисташковый хребет, окрестности впадины
Ер-Айлан-Дуз, Кызыл-Джар и т. п.) и, повидимому, встречается по гли-
нистым холмам Карабиля, по обрывам верхнего течения Теджена и Мургаба.
КЕКЛИК, ИЛИ КАМЕННАЯ КУРОПАТКА I6S
Характер пребывания. Оседлая птица.
Биотоп. Скалистые участки и выходы камней, пологие глинистые
холмы с обрывами, глинистые береговые обрывы на равнине (долиныр. Тед-
жена и р. Мургаба).
Численность. По подходящим биотопам обыкновенная птица
всего района распространения. Численность неравномерна; так, в Акар-
Гешме они более многочисленны по западным ушельям, чем в других участках
(Лаптев, 1945) и т. д.
Экология. Размножение. В наших пределах не изучено. В Бе-
луджистане и Иране яйца с конца апреля по июнь; размеры яиц E0) 35,2—
43,0 х 26,4—30,2, в среднем 39,0 х 29,3 мм; яйца в среднем меньше
и тусклее , чем у других подвидов (Бекер, 1928).
Линька. Не изучена. Самец от 5 июня 1946 г. из района впадины
Ер-Айлан-Дуз (Бадхыз) в сильно обношенном пере без следов линьки; самка
от 11 мая 1949 г. из Акар-Чешме (Бадхыз) в сильно обношенном пере без
следов линьки, на животе громадное наседное пятно—повидимому, птица
в разгаре насиживания. Самка, добытая в октябре в Акар-Чешме, уже пол-
ностью перелиняла в новый наряд, лишь у основания первого первостепенного
махового сохранились остатки перьевого чех лика.
Молодой самец, добытый 14 октября 1945 г. в хребте Гязь-Гедык, закан-
чивает линьку в первый годовой наряд: завершается рост нового мелкого
пера по всему телу, на подбородке часть перьев еще в пеньках; первое—вто-
рое маховые перья первой генерации достигли полного роста, третье новое
в шесть седьмых, а четвертое—шестое—полной длины; рулевые новые, но
часть их еще в четырех пятых окончательной длины. Одновременно добытый
там же второй молодой самец находится в таком же состоянии линьки, но
две средние пары рулей еще старые, птенцовой генерации.
Скудный материал по линьке все же свидетельствует о значительной
растянутости периода размножения в популяции (самка от 11 мая с большим
наседным пятном, молодые еще в середине октября не закончили первую
годовую линьку).
Питание. Не изучено.
Хозяйственное значение. Ввиду относительной безлюдности районов,
населенных этим подвидом кеклика, имеет лишь ограниченное местное про-
мысловое значение.
Описание. Размеры и строение. Отличаются от других подвидов и в пер-
вую очередь от копетдагских кекликов своей меньшей величиной и более темной
окраской.
Длина крыла у самцов восточноиранских кекликов A9) 144—163,5, у самокFI40,5—
148, в среднем соответственно 152,2 и 143,5 мм (Зарудный, 1914); промеры птиц из коллек-
ции Зоологического музея Московского университета дают большие величины: крыло
у самцов C) 165—169, у самок B) 155—158, в среднем соответственно 166,6 и 156,5 мм;
эти промеры мало показательны из-за недостаточного количества материала.
Окраска. Более темная и яркая, чем у копетдагских птиц. По верху головы
более или менее отчетливо развит буровато-рыжий цвет, зачастую заходящий и на лоб.
Передняя часть спины, плечевые и верхние кроющие крыла ржаво-красные с более или
менее выраженным серым налетом.
Кызылкумский кеклик Alectoris graeca subpallida Sar.
Caccabis kakelik subpallidus. Зарудный. Орнит. Вестн., № 1J1914, стр. 59, горы в пес-
ках Кызыл-Кум и горы Южной Бухары.
Распространение. Ареал. Живет в горах пустыни Кызыл-Кум (Ари-
стан-Бель-Тау, Кюйгень-Тау, Ак-Тау, Тохта-Тау; Зарудный, 1914), в горах
Юго-Восточной Бухары (Ходжа-Баба, Байсун) и в горах между Кафирнига-
ном и Вакшем (южнее Гиссарской долины—Иванов, 1940). На северо-восто-
ке, повидимому, не проникает далее левобережья Сыр-Дарьи, северная гра-
166 ОТРЯД КУРИНЫЕ
ница этого подвида совсем неясна. Среди птиц из Байсуна, Баба-Тага, Хир-
шанджау встречены переходные особи, приближающиеся к A. g. falki\ в Ну-
ра-Тау все особи промежуточного типа, а в Кугитанге птицы очень сходны
с тяньшаньской формой (Мекленбурцев),.
Детали распространения этой формы требуют уточнения: в частности,
следует проверить, не проникает ли она северо-восточнее Сыр-Дарьи, как
далеко идет на восток по Пянджу и т. п.
Характер пребывания. Оседлая птица.
Биотоп. Держится по скалистым щебенистым склонам безлесных
голых гор и в меньшем числе по глинистым и лессовым обрывам; местами
проникает в зону фисташки и в арчевники. Обычно живет на высоте 400—
600 м над уровнем моря, однако в мае в водораздельной части Гардани-
Ушты A00 км на юг от Сталинабада) встречалась не ниже 1500 м над уровнем
моря (Иванов, 1940). Обычно тяготеет к родникам, но от воды встречалась
и в 5—6 км.
Численность. По подходящим биотопам обычен во всех участках
ареала. Многочислен в горах Аристан-Бель-Тау и Ак-Тау (Зарудный,
1915), значительно истреблен в восточных участках Нура-Тау (Мекленбурцев,
1937).
Экология. Размножение. Цикл периодических явлений в жизни
этого подвида совершенно не изучен. В водораздельной части Гардани-Ушты
A00 км южнее Сталинабада) 9—13 мая держался преимущественно парами,
у добытого самца сильно увеличены яичники; гнезд не найдено (Иванов).
Выводки с молодыми в полматки и больше встречены в горах Аристан-бель-
Тау 20 мая, а в горах Тоха-Тау молодые в три четверти роста взрослых птиц
и до размеров дрозда найдены 8 и 9 июня (Зарудный, 1915); молодые в поло-
вину взрослых встречены у Микоянабада еще в конце июня (Иванов, 1940).
Это свидетельствует о растянутости периода размножения в популяции.
Линька. Совершенно не изучена. Птица из Кафирнигана от 1 сен-
тября 1934 г. заканчивает полную линьку в годовой наряд: по всему телу
свежее перо, еще не полностью закончившее рост; все первостепенные махо-
вые новые, но первое лишь в три четверти полной длины; второстепенные
маховые и рули новые, завершившие рост. Птицы от конца апреля и мая без
следов линьки, в слегка обношенном пере.
Питание. Детально не изучено. У птиц, добытых 9—13 мая 1948 г.
в водораздельной части Гардани-Ушты A00 км южнее Сталинабада), в зобах
найдены отцветшие головки Taraxacum, Gagea, плоды Bellivalia (Hyacinthus)
и побеги Polygonum viviparum (Иванов). Регулярно летают на водопой, при-
чем пьют воду и из горько-соленых источников (Зарудный, 1915).
Хозяйственное значение. Объект любительской охоты, имеет местное
промысловое значение. Добывается, повидимому, в меньших количествах,
чем другие подвиды.
Описание. Размеры и строение. Отличаются от тяньшаньских кекликов
преимущественно более бледной окраской и, повидимому, несколько меньшей величиной.
Длина крыла самцов E) 161—165, самок A4) 148—157 мм, в среднем соответственно
163,4 и 151,6 мм (Зарудный, 1914); длина крыла самок C) 150—161) в среднем 155,3 мм
(коллекция Зоологического музея Московского государственного университета).
Окраска. Верхняя сторона с песчаным оттенком, заметно более светлая, чем
у тяньшаньских птиц; розовато-рыжеватый цвет по верху головы и винно-розовый отлив
по бокам передней части груди выражен слабее.
Систематические замечания. Кызылкумские кеклики по размерам и окраске прибли-
жаются к западнотуркменским равнинным кекликам. Однако объединение этих подвидов
сейчас невозможно ввиду скудости коллекционного материала и недостаточно изученной
биологии.
КЕКЛИК, ИЛИ КАМЕННАЯ КУРОПАТКА 167
Тяньшаньский кеклик Alectoris graeca falki Hartert
Atectoris graeca falki. Hartert, Nov. Zool., 1917, стр. 280, Русский Туркестан.
Сти н о н и м ы. Tetrao kakelik. F a i k. Beytz. z. topogr. Kenntn. Russ. Reichs. Ill, 1786,
стр. 390, Бухара; описание птицы крайне неточное и неполное.—Caccabis hyemalts. Sever-
t z о f f. Ibis 1883, стр. 80 (nomen nudum), альпийская зона Туркестана.
Распространение. Ареал. По подходящим биотопам повсюду jb гор-
ной системе Тянь-Шаня, выходя несколько за ее пределы на западе. В пре-
делах Советского Союза населяет Гиссарский, Зеравшанский, Туркестанский,
Чоткальский хребты, Кара-Тау и Таласский Ала-Тау. На восток через
Тер скей-Ала-Тау и Заилийский Ала-Тау доходит до левобережья р. Или,
не проникая, повидимому, через нее далее на северо-восток. Гнездится
в Причуйских песках и Чу-Илийских горах, возможно заходит и в Присыр-
дарьинскую пустыню (Долгушин, 1948). Повидимому, к этому подвиду при-
надлежат кеклики, в небольшом числе встречающиеся в Улу-Тау и в Мугод-
жарах. Птицы из Кугитанга более сходны с тяньшаньской, чем с кызылкумской
формой; в западных участках Туркестанского хребта также встречаются
переходные формы между тяньшаньским и кызылкумским кекликами (Мек-
ленбурцев). Граница подвида на Памире точно не выяснена: он населяет
Дарвазский хребет, водится на р. Беик, в Сардыме и Алмаяне(Иванов, 1940),
но птицы переходного типа между A. g. falki и A. g. pallescens зимой также
встречены по долине р. Пяндж между устьем р. Ванч и Поршневым
(Мекленбурцев).
Характер пребывания. Оседлая птица, предпринимающая
небольшие сезонные кочевки. В Заилийском Ала-Тау зимой перемещается
на южные склоны, где мало снега; иногда поднимается выше мест гнездовий,
так как чем выше, тем колебания температуры меньше (Долгушин, 1949),
в Гиссарской долине перемещается на южные склоны холмов северной окраи-
ны долины (Иванов, 1945). При обильных снегопадах спускается с гор: со
склонов Гиссарского хребта к берегам р. Варзоб и на южные склоны гор,
где между кустами и в тростниках есть участки талой земли (Козлова, 1949;
Иванов, 1949); это явление отмечено также в Юго-Восточном Узбекистане
(Салихбаев, 1939) и других районах.
Биотоп. Каменистые и щебенистые осыпи и выходы скал со скудной
степной и пустынно-степной растительностью, мягкие землистые склоны
с травянистой растительностью и отдельными кустами арчи, скалы над
альпийскими лугами (летом доходят до границы снегов), склоны и дно уще-
лий; щебенистые и резко пересеченные участки предгорий; останцевые и гли-
нистые крутые холмы в предгорных пустынях. Обычно всегда вблизи воды
{лужи, колодцы, родники); однако местами встречены и в 6—7 км от ближай-
ших водных источников (маленькие чинки в пустыне у железнодорожной
станции Дарбаза, в 40 км севернее Ташкента; Мекленбурцев).
В Зеравшанском и Туркестанском хребтах встречены на высоте 2040—
3070 м над уровнем моря (Даль, 1936), в Таджикистане обычно не ниже
1200—1300 и до 3000—3500 м над уровнем моря (Иванов, 1940).
Численность. Различна в разных участках ареала и меняется
по годам. Так, в западных отрогах Кара-Тау он более редок, а далее на юго-
запад, в глубь гор, становится обычным (Спангенберг и Фейгин, 1930); на
южном склоне Гиссарского хребта (верховья р. Ханаки) очень обычен, но
далее к востоку в долине р. Сарда-и-мион и у Гарма редок; редок он и на
Памире (Иванов, 1940); из-за перепромысла стал редок у крупных населен-
ных пунктов.
Численность кеклика заметно меняется по годам, особенно резко умень-
шаясь после многоснежных и холодных зим. В необычайно холодную и мно-
госнежную зиму 1918—1919 гг. кеклики массами появились в окрестностях
168 ОТРЯД КУРИНЫЕ
Алма-Аты, в горах находили мертвых птиц, а весной 1919 г. в Заилийском
Ала-Тау они стали редкостью (Шнитников, 1949). В 1944 г. на маршруте
в 10 км было встречено 25 выводков, а в 1945 г., после зимы с обильными
снегопадами, буранами и падением температуры до —35°С, встречены лишь
три выводка; в ноябре 1944 г. птицы весили 637—650 г, а самец от 15 марта
1945 г.—всего 260 г. (Таласский Ала-Тау; Шевченко, 1948). В суровую зиму
1944—1945 гг. кеклики тысячами спускались с гор к р. Варзоб, где снег был
менее глубок, чем в горах, а наследующий год их численность резко упала:
на р. У ноу от устья до Тангимура встречен всего один выводок; в такую же
суровую зиму 1948—1949 гг. много кекликов появилось в окрестностях Стали-
набада, где добывались массами (один охотник добыл до 300 шт.), осенью
1949 г. выводков почти не было (Иванов).
В «урожайные» годы местами встречаются в громадных количествах:
в Тандаре на 1 км2 встречено 120 шт. (Северный Таджикистан; Даль, 1936).
Экология. Размножение. Повидимому, моногам. Многие исследова-
тели утверждают, что птенцов водит лишь самка (Козлова, 1949, и др.);
однако вблизи гнезда с насиживающей самкой встречали самца, а у выводков
добывали и самца и самку (Шнитников, 1949). По наблюдениям Кузьми-
ной, самец наряду с самкой активно участвует в насиживании, и у многих
самцов бывают наседные пятна; при выводках добывались самцы и самки;
встречены выводки, которых водили только самцы (Заилийский Ала-Тау,
по устному сообщению Корелова).
С конца февраля уже встречаются пары и одиночки (преимущественно
самцы), в середине марта стайки встречаются крайне редко, в мае встречают-
ся почти всегда одиночки, ведущие себя очень тихо и скрытно (Гиссарский
хребет; Козлова, 1949). Самец, «ухаживая» за самкой, бегает вокруг нее, рас-
пустив крылья, подобно домашнему петуху (Шнитников, 1943). Одиночки
и пары встречаются и в середине июля (Иванов).
Период размножения, повидимому, в основном за счет вынужденных
повторных кладок, очень растянут. Самка с готовым яйцом в яйцеводе добыта
на Курдае 20 апреля 1907 г.; в Заилийском Ала-Тау гнездо с 8 свежими
яйцами найдено 18 мая 1907 г., а с 14 яйцами—15 июня; в хребте Борохоро
30 мая найдены два гнезда с одним яйцом в каждом, 13 июня в одном из этих
гнезд уже было 11 яиц (следовательно, в сутки откладывается по одному
яйцу); 2 июня гнездо с 17 яйцами; в ущелье Карабалты 10 июня 1934 г. най-
дено гнездо с 13 слабо насиженными яйцами, 15 июня—с 11 свежими
яйцами, 30 мая 1936 г.—с 15 свежими яйцами, 3 июня—с 15 насиженными
яйцами и 4 июня—24 яйца, из которых начали проклевываться птенцы (Се-
миречье; Шнитников, 1949); в Гиссарском хребте гнездо с 6 свежими яйцами
было найдено 22 мая 1942 г. (Иванов); в верховьях Исфары, на Чичаке и
у Варуха Зарудный 23—27 июня нашел 4 гнезда с 12 яйцами в каждом (в двух
гнездах свежие, в двух—накануне вылупления), Герхнер на озере Искан-
дер-Куль нашел гнездо со свежими яйцами на р. Варзобе 23 июня, в урочище
Щуроб-Дара Зарудный 20 июля 1910 г. добыл самку с готовым яйцом в яйце-
воде и в это же время встречал молодняк ростом со взрослых (Таджикистан;
Иванов, 1940).
Величина кладок сильно варьирует: от 6 до 24 яиц в гнезде. Большинство
яиц сливочно-охристого цвета различной интенсивности, с коричневыми или
серо-коричневыми крапинками; реже встречаются светлые сливочно-охри-
стые яйца без пятен или почти белые (Иванов, 1940); обычно яйца несколько
темнее, чем у кавказского кеклика.
Гнездится в каменистых осыпях, на щебенистых склонах ущелий и овра-
гов со скудной растительностью и отдельными кустиками, часто довольно
далеко от воды (Шнитников, 1949), в лишенных растительности скалах и
кручах, по разреженным местам в лесистых участках (Козлова, 1949).
КЕКЛИК, ИЛИ КАМЕННАЯ КУРОПАТКА 169
Гнездо—вырытая птицей ямка глубиной до 8—9 см и диаметром до 20 см—
обычно расположено под защитой нависшего камня или кустика; ямка
выстлана тонким слоем растительной ветоши, коротких сухих былинок и
перьями птицы; по краям ямки из земли, веточек и травинок устраивается
валик (Иванов, 1940). На яйцах самка сидит очень плотно; спугнутая, отво-
дит от гнезда: раскрыв крылья и припадая к земле, бежит прихрамывая,
иногда почти ползет на брюшке (Козлова, 1949).
Продолжительность насиживания и время постэмбрионального роста
птенцов не установлены. Из-за растянутости периода размножения в одно
время встречаются очень разновозрастные птенцы. У Копала 22 июня 1907 г.
встречен выводок с очень маленькими птенцами, а через несколько дней—
молодые ростом с взрослую птицу (Семиречье; Шнитников, 1949); пуховички
найдены 18 июля (Зеравшанский и Туркестанский хребты; Даль, 1936);
в Киргизском хребте 2—3-дневные пуховички встречены в конце июня —
начале июля (Спангенберг и Дементьев, 1935); на Искандер-Куле 25 августа
были молодые почти со взрослых птиц и лишь в половину перепелки, в конце
июля—молодые в половину матки, а в конце августа—уже кончающие линьку
в первый годовой наряд (Таджикистан; Иванов, 1949); в Гиссарском хребте
первый выводок встречен 5 июня, 16 июня—выводок с птенчиками размером
с воробья и тогда же—размером в полматки, 30 августа встречены молодые
ростом в полматки и молодые, уже заканчивающие линьку в годовой наряд
(Козлова, 1949), в районе Сталинабада молодые в полматки встречены
25—30 июня и 14—17 июля, тогда же—птенцы размером с цыпленка; моло-
дые ростом со взрослых птиц найдены 15—20 августа (Иванов; Салихбаев,
1939), что свидетельствует о значительной смертности пуховиков. Вспугнутая
от выводка самка не отводит, а бросает птенцов и улетает (Козлова, 1949)
или убегает, а молодые затаиваются; если птенцы уже на крыле, то они
вначале бегут, а затем улетают вместе со стариками (Шнитников, 1949).
Л и н ь к а. В соответствии с растянутостью периода гнездованья период
линьки в популяции также растянут. По коллекционным материалам Зооло-
гического музея Московского государственного университета, тяньшаньские
кеклики в ноябре—январе и апреле—мае не линяют. Взрослые самцы начи-
нают полную годовую линьку несколько раньше самок. У самца от 8 июня
из Бурулдая (окрестности Чимкента) на теле отдельные пеньки, выпали две
средние пары рулей, перо сильно обношено; у самца от 9 июня из того же
места редкие, еще нераскрывшиеся пеньки по телу, первое—восьмое первосте-
пенные маховые старые, девятое—десятое в нераскрывшихся пеньках, выпали
две средние пары рулей, началась смена нижних кроющих хвоста; у самца
от 10 июня выпало восьмое первостепенное маховое, девятое в нераскрыв-
шемся пеньке, десятое около трети длины. Самец от 20 июня из Кара-Тау
(Бугунь) и от 22 июня из Киргизского хребта (Сосновка) на сходной
стадии линьки, а от 24 июня (Кара-Тау) в сильно обношенном пере без сле-
дов линьки. Два самца от 14 августа из Кара-Тау (Кара-Мурун) в энергич-
ной линьке: по всему телу свежее контурное перо, у основания кое-где еще
сохранились чехлики; первое первостепенное маховое старое, второе в по-
ловину, третье—десятое новые, полностью выросшие; все второстепенные
маховые новые, плечевые еще старые; часть рулей старых, часть выпала,
часть новых в три четверти полной длины; сменяются верхние и нижние
кроющие хвоста. Самец из Покровского района Иссык-Кульской обл. от
25 сентября завершил полностью линьку контурного пера; первостепенные
маховые новые, но первое еще в одну треть, а второе в пять шестых полной
длины; рули новые, полностью выросшие, но наружная пара еще в половину
полного роста.
Самки от 30 мая из Киргизского хребта, от 5 июня из долины
Сурхан-Дарьи (Гиссарский хребет), от 23 и 27 июня из Кара-Тау в обношен-
170 ОТРЯД КУРИНЫЕ
ном пере, с большими наседными пятнами и без следов линьки. У самки от
2 июля из Киргизского хребта на наседном пятне и на всем теле появи-
лись еще нераскрывшиеся редкие пеньки; первое—восьмое первостепенные
маховые старые, девятое выпало, десятое в нераскрывшемся пеньке. У самки
от 7 августа с берегов Искандер-Дарьи начинается энергичная линька:
тычки появились по всему телу, в том числе и на наседном пятне; первое—
четвертое первостепенные маховые старые, пятое выпало, шестое в нерас-
крывшемся пеньке, седьмое—десятое новые, почти полного роста; в хвосте
четыре пары старых рулевых, остальные новые, в одну треть длины, все
верхние кроющие хвоста в чуть раскрывшихся пеньках. Самка от 7 сентяб-
ря из Гиссарского хребта энергично линяет: по всему телу уже показалось
над старым свежее перо; первое—второе первостепенные маховые старые,
третье в четыре пятых, а четвертое—десятое полного роста; энергичная
смена рулей, причем новые уже до двух третей полной длины. Самка от
25 сентября из Покровского района Иссык-Кульскойобл. завершает линьку:
по всему телу свежее перо, у основания некоторых перьев еще сохраняются
остатки чехликов; рули все новые, последняя и предпоследняя пара еще
в половину длины; первое—второе первостепенные маховые старые, третье
в половину, а четвертое—десятое окончательной длины.
Перья птенцового наряда начинают появляться над пухом, видимо,
уже на 4—6-й день. Продолжительность линьки в птенцовый наряд не уста-
новлена.
Молодые птицы, начавшие линьку в первый годовой наряд, добыты
в Киргизском хребте в середине июля (Спангенберг и Дементьев, 1935).
У молодого самца от 11 августа (окрестности Алма-Аты) энергичная линька
мелкого пера по всему телу; первое первостепенное маховое в половину,
а второе в четыре пятых полной длины, третье—пятое птенцовой генерации,
шестое в половину, седьмое в шесть седьмых, восьмое—десятое в полную
длину первого годового наряда; растут рулевые, еще не достигшие полной
длины. Молодая самка от 7 сентября с берегов Искандер-Куля энергично
линяет: по всему телу свежее перо годового наряда, еще не полностью за-
кончившее рост; плечевые еще птенцовой генерации; первое первостепенное
маховое в одну треть, а второе в полную длину, третье старое (птенцовой
генерации), четвертое в половину, пятое в шесть седьмых, а шестое—десятое
полной длины годового наряда; все рули новые, достигли полного роста.
Закончившие полностью линьку молодые добывались в сентябре—октябре,
но в середине октября еще встречены птицы (р. Алмаатинка) в полном раз-
гаре линьки мелкого пера с сохранившимся еще от птенцовой генерации
третьим (а иногда и четвертым) первостепенным маховым.
Питание. Охотно использует и растительную и животную пищу.
В июле в зобу встречены листья шиповника, в августе зоб битком набит
семенами, остатками саранчевых и раковинами наземных моллюсков (Иванов).
Летом в больших количествах поедает личинок саранчевых; осеннюю пищу
составляют клубни и семена растений и муравьи (Козлова, 1949). В ноябре
в зобу у птиц найдены побеги Роа bulbosa, отдельные зерна ячменя и про-
росшие семена какого-то бобового; в зобах птиц зимой встречены зерна
ячменя и различных бобовых, зеленые свежие побеги злаков и осок; в же-
лудках—камешки, твердые семена и перетертая трава; птицы кормятся
по солнцепекам и у ручьев, где есть талая земля в кустах и тростниках;
много копок найдено у основания кустов Rosa lutea\ в очень снежные дни
садятся на кусты и, повидимому, поедают почки; зимой встречаются как
очень жирные, так и худые птицы (Иванов, 1949). На северных склонах
Заилийского Ала-Тау после выпадения снега косули кормятся опавшими
яблоками, добывая их из-под снега; по копкам косуль яблоками кормятся
кеклики; эти взаимоотношения кеклика и косули продолжаются до января,
КЕКЛИК, ИЛИ КАМЕННАЯ КУРОПАТКА
пока не будут съедены все яблоки (Корелов, 1947); кормятся кеклики зимой
и по копкам других диких копытных.
Суточный цикл. Сходен с другими подвидами. В жаркую пого-
ду отдыхают днем среди лесной растительности (Козлова, 1949), часто вблизи
родников, под навесом камней»
Враги. В Киргизском хребте кекликов добывают беркуты (Спанген-
берг и Дементьев, 1935); в холодные многоснежные зимы процент кекликов
в пище наземных хищников резко увеличивается (Шевченко, 1948).
Хозяйственное значение. Важнейшая промысловая птица в ряде районов
(Таджикистан—Иванов, 1940; Казахстан—Формозов, 1950). Местами добы-
вается в больших количествах: в Таджикистане массами вылавливается
весной силками на манных самцов—за весну на одного самца поймано
150 кекликов (Пенджикент—Иванов, 1940); в многоснежные зимы спускаю-
щихся с гор кекликов бьют палками и добывают ружьем, причем один охотник
добывает до 500—600 шт. (Гузарский район Бухарской обл., Салихбаев,
1939).
Хищническая добыча привела в ряде мест к резкому уменьшению чис-
ленности птиц, чему способствовал и ряд неблагоприятных зим; в других же
районах недопромышляются. Большое промысловое значение кекликов
требует организации планомерных и равномерных заготовок по всей терри-
тории их ареала. Несомненно заслуживает широкого применения предло-
жение Формозова A950) об организации культурных дичных хозяйств
<с основным объектом кекликом) в комплексе с отгонным животноводством.
Полевые признаки. Сходны с другими подвидами. Кормятся стайками
на ходу, быстро двигаясь; по малокормным местам пробегают цепочкой;
вспугнутые снимаются (Козлова, 1949), обычно перелетая на другой склон.
Описание. Размеры и строение. Темнее и несколько крупнее кызылкум-
ского кеклика. Длина крыла самцов B1) 154—176, самок A1) 151—163, в среднем соответ-
ственно 165,2 и 155,4 мм. Длина тела до 370 мм, размах крыльев 500—550 мм. Вес самцов
осенью 535—600, самок 475—530 г (Туркестанский и Зеравшанский хребет; Даль, 1936).
Окраска. Более темный, чем кызылкумский кеклик. Бока шеи и грудь темные
серо-сизые; шапочка на голове, передняя часть спины, плечевые, а часто и верх шеи с ро-
зовато-каштановым отливом; темнооливковый отлив на пояснице частично переходит и на
плечевые перья.
Пуховой птенец по характеру окраски неотличим от пуховиков западнотуркменского
равнинного кеклика, однако общий тон окраски верха головы и туловища более темный,
рыжевато-бурый .
Кара-Корумский кеклик Alectoris graeca pallescens He 11 ma у г
Alectoris graeca pallescens. H e 1 1 m а у г. Field Mus. Nat. Hist. Publ., Zool. Ser., XVII,
1929, стр. 137, Южный Памир.
Распространение. Ареал. Населяет Малый Тибет (Балтистан),
хребет Каракорум, Гильгит, южные и восточные отроги Памира и, вероятно,
проникает с востока в хребет Гиндукуш. В пределах Советского Союза
встречен в Западном Памире в долине р. Пянджа и в среднем течении р. Шах-
Дара; на Восточном Памире встречен лишь в единственном месте—выше
Сарезского озера в обрывах Джаман-Тала (Мекленбурцев, 1946). По долине
р. Пянджа между Поршневом и устьем р. Ванч зимой встречены птицы про-
межуточного типа, приближающиеся по окраске к тяньшаньской форме
(Мекленбурцев, 1946).
Характер пребывания. Оседлые птицы. В снежные зимы
переходят на южные склоны, где солнце и ветер быстрее сгоняют снег.
Биотоп. Склоны гор с каменистыми осыпями, выходами скал и скуд-
ной растительностью. На Шах-Даре обычно на высоте в 3000—3100 м над
уровнем моря, выше 4000 м не встречены (Мекленбурцев).
172 ОТРЯД КУРИНЫЕ
Численность. Меняется по годам. После многоснежных зим чис-
ленность заметно снижается: после бесснежной зимы 1942—1943 г. летом
в ботаническом саду у Хорога кекликов было в 5—6 раз больше, чем после
многоснежной зимы 1941 г. (Гурский, по Иванову). Мекленбурцев A946)
в среднем течении р. Шах-Дара встречал 3—4 пары на 1 км маршрута.
Экология. Размножение. Совершенно не изучены. 9 мая у Хо-
рога найдено гнездо со свежими яйцами, 20 июня встречены молодые ростом
с перепела (Мекленбурцев, 1946); 8 сентября у озера Друм-Куль добыты
молодые ростом со взрослых птиц (Гурский, по Иванову).
Линька. Неизвестна.
Питание. Зимой кормятся на выдувах южных склонов, поедая
плоды облепихи Hippophae rhamnoides, семена разных полыней Artemi-
sia, замерзшие плодики и семена шиповника Rosa (Гурский, по Иванову).
Суточный цикл. Сходен с другими подвидами. В жаркие
полдни летом отдыхают в тени; зимой греются под защитой камней на солнце-
пеках.
Хозяйственное значение. До последнего времени планомерное про-
мысловое использование кеклика не организовано. Местным населением
истребляется во множестве; птенцов ловят руками, взрослых—силками:
в сезон размножения привязывают пойманного кеклика, и идущие на крик
другие птицы запутываются в расставленные вокруг силки (Мекленбур-
цев, 1946).
Описание. Размеры и строение. Крыло самцов и самок 158—172 мм.
Окраска—заметно светлее тяньшаньского и восточноиранского кекликов, общий тип
окраски тусклый, серовато-песочный с нерезко выраженным розоватым отливом. От
кызылкумского кеклика отличается тем, что розоватый оттенок захватывает и бока зоба,
а охристо-розовый налет на кроющих крыла выражен менее отчетливо.
Джунгарский кеклик Alectoris graeca dzungarica Sushkin
Alectoris chukar dzungarica. Sushkin. BulT ВЛ O. Club, XLVIII, 1927, стр. 26, хребет
Монрак в Тарбагатае.
Синонимы. Alectoris chukar obscurata. Sushkin Bull. В. О. Club, XLVIII,
1927, стр. 27, западная часть хребта Танну-Ола,
Распространение. Ареал. Населяет Джунгарский Ала-Тау, Тар-
багатай и склоны Зайсанской котловины, Алтай, Танну-Ола и Северо-
Западную Монголию. Границы ареала и детали распространения в преде-
лах Советского Союза крайне неясны; в Алтае, повидимому, только в запад-
ных его участках; в Центральном Алтае и в Тувинской авт. обл. нет
(Сушкин, 1936).
Характер пребывания. Оседлая птица. Зимой, особенно
в многоснежные зимы, спускается до предгорной степи.
Биотоп. Останцы в предгорьях; склоны гор, где выходы камня
чередуются с зарослями шиповника, барбариса и т. п. и участками степной
растительности. Встречается на высоте 1000—2500 м над уровнем моря.
Численность. В Зайсанской котловине и Тарбагатае много-
числен (Хахлов, 1929), в Джунгарском Ала-Тау обычен (Кузьмина,
1948), в других участках ареала в пределах Советского Союза скорее мало-
числен.
Экология. Размножение. Джунгарский Ала-Тау (по Кузьминой,
1948). Разбиваются на пары во вторую половину марта, в позднюю
весну стайки встречаются до половины апреля. В апреле уже добывались
самки с яйцами. Первый выводок на р. Биже встречен 19 июня. При выводке
встречаются как одна, так и две взрослые птицы; 13 июля добыт самец
с наседным пятном.
НЕКЛИК, ИЛИ КАМЕННАЯ КУРОПАТКА 173
Птенцы на б—7-й день уже подлетывают, к двум месяцам достигают
величины взрослых птиц. В конце августа—начале сентября выводки сби-
ваются в стайки.
Линька. Подробно не изучена. Птицы в ноябре и декабре не ли-
няют. Самец от 4 июня из Сарыбулака (Тарбагатай) в сильно обношенном
пере, начала линьки мелкого пера незаметно; первое—шестое первостепен-
ные маховые старые, седьмое—десятое выпали, вместо них еще неразвер-
нувшиеся пеньки длиной до 4 см\ выпала средняя пара рулей. Самец
от 2 августа с р. Кок-Су (Джунгарский Ала-Тау) энергично линяет: сме-
няется контурное перо по всему телу; первое—третье первостепенные махо-
вые старые, четвертое в половину, пятое в три четверти, седьмое—десятое
новые в полную длину; сменяются второстепенные маховые; идет линька
хвбста, часть рулей в наполовину раскрывшихся пеньках, часть рулей
старые; сменяются верхние кроющие хвоста.
Молодая самка от середины сентября из Зайсанской котловины в све-
жем пере по всему телу, часть перьев у основания еще в чехликах; первое
и второе первостепенные маховые новые, полной длины, третье еще птен-
цовой генерации, четвертое—десятое новые, четвертое в три четверти, а
пятое—десятое полной длины; растут рулевые, сменяются кроющие хвоста.
Питание. В пище преобладают растительные корма: семена,
цветы, листья, луковицы 90 видов растений; летом в пище много и живот-
ных компонентов: насекомые—фитофаги и их личинки, жуки, клопы,
муравьи, саранчевые, реже наземные моллюски (Кузьмина, 1948).
Суточный цикл. Ночуют в расщелинах. С восходом солнца
кормятся и посещают водопой, в это же время купаются в пыли. В середине
дня отдыхают в скалах, почти всегда с криком. В пасмурную погоду и в сне-
гопады не кормятся (Кузьмина, 1948).
Описание. Размеры и строение. Длина крыла самцов C) 157—160,
самок B) 152—160 мм, в среднем, соответственно, 159 и 156 мм.
О к р а с к а. По общему тону окраски еще темнее, чем тяньшаньский кеклик. Серое
пространство на задней стороне шеи развито слабо, красновато-буроватая шапочка зани-
мает весь верх головы, оставляя лишь узкую серую полоску над беловатой бровью;
розовато-винный оттенок с передней части спины распространяется и на бока зоба; на
крыльях слабо заметен темнооливковый налет.
Кашгарский кеклик Alectoris graeca pallida Hume
Caccabis pallidus. Hume. In. Henderson and Hume, Lahore to Jarkand,
1873, стр. 284, Яркенд.
Синонимы. Caccabis kakelik humei. Зарудный. Орнит. Вестн., № 1, 1914, стр.54,
р. Каракаш.— Alectoris chukar fallax. Sushki n Bull. В. О. Club, XLVIII, 1927, стр. 23,
Курук-Таг.
Распространение. Ареал. Встречается в южной части Кашгарии
(р. Каракаш), в западных частях Куэнь-Луня и в Западном Нань-Шане,
включая Алтын-Таг и, может быть, западные отроги хребта Гумбольдта;
к северо-востоку встречается почти до верховьев Таушкан-Дарьи. Из Каш-
гарии, возможно, проникает в юго-восточные участки Киргизии (Шнит-
ников, 1949) и, может быть, на восточные склоны Памира.
Детали распространения этой формы в пределах Советского Союза,
биология и периодические явления в жизни кашгарского кеклика совер-
шенно не изучены. 12 мая у убитой самки в яйцеводе найдено готовое яйцо;
массовая кладка в июне; 6 августа птенцы с воробья ростом (Кашгария;
Судиловская по сборам Дивногорского, 1936).
Описание. Очень бледный подвид с тусклым общим тоном окраски: верх головы и пе-
редней части спины светлой дымчато-серой окраски; черные полосы на боках туловища
заметно уже, чем у других подвидов B—3 мм шириной).
174
ОТРЯД КУРИНЫЕ
РОД ТУРАЧ FRANCOLINUS STEPHENS, 1819
Тип. Tetrao francolinus
12. Турач Francolinus francolinus L.
Tetrao Francolinus. Li nnaeus. Syst. Nat., изд. XII, 1766, стр. 275, Италия, Малая
Азия.
Русские названия. Заимствовано из иранского языка, где эту птицу называют
дуррач; у туркменов—турач, трач; также заимствовано из иранского языка.
Распространение. Ареал. Область распространения—восточная
часть Средиземного моря (о-в Кипр), Малая Азия, Восточное Закавказье,
Юго-Западная Туркмения, Иран, Ирак, Сирия, Индия. В недавнее время
встречался в Юго-Восточной
Испании (Валенсия), в Сицилии
(последние особи добыты в
1869 г.), откуда проникал в Ита-
лию, и по островам Эгейского
моря; сейчас в этих местах пол-
ностью истреблен.
Характер пребы-
вания. Повсюду оседлая
птица.
Биотоп. Заросли ку-
старников по берегам водоемов
(реки, озера, арыки) на равни-
Карта 13. Распространение турача Francolinus не, участки культурного ланд-
francolinus шафта. В горы не проникает.
1—F. /. francolinu8,2 —F. /. asiae, 3—F. /. melanonotus;
а-граница гнездовий Подвиды и варьирующие приз.
наки. На о-ве Кипр, в Малой Азии,
Северном Иране, Туркмении, Закавказье, Палестине и Сирии—/7. /. francolinus L i n-
п а е u s, 1766; в Восточном и Южном Иране, в Южном Афганистане и Белуджистане,
в Индии на восток до Непала и Бехара, а на юг до Диза, Серана и Западной Бенгалии—
F. /. asiae Bonaparte, 1856; в Восточном Непале и Восточной Бенгалии, Манипуре и
Ассаме—/7. /. melcnonotus Hume, 1899.
Подвиды характеризуются незначительными отличиями в величине и характере окра-
ски птиц и в размерах яиц. Географические изменения экологии, касающиеся календаря
периодических явлений, трофических связей и т. п. изучены крайне плохо.
Европейский турач Francolinus francolinus francolinus L.
Синонимы. Francolinus orientalis europaeus. В u t u г 1 i n. Ornithol. Monatsb., 1907,
стр. 81, Греция. —Francolinus orientalis coucasicus. Buturli n, там же, стр. 81, Закав-
казье.—Francolinus orientalis zarudnyi. В u t u г 1 i n, там же, стр. 81, южная часть
Закаспийского края.
Распространение. Ареал. На о-ве Кипре, в Малой Азии, Сирии,
Палестине, Северном Иране. В СССР область распространения невелика.
В Восточном Закавказье сейчас населяет Куро-Аракскую низменность от
Акстафы до Каспия (проникая в пределы Грузии по долине р. Алазани
Верещагин, 1947); обычен в подходящих биотопах в Муганской степи.
По Араксу на запад доходит до Джебраильского района (Худаферское
ущелье, Млокосевич; селение Бегманлу, Сатунин, 1907). По непроверен-
ным данным, возможно, встречается в Мегранском районе Армении (Ляй-
стер и Соснин, 1941). По побережью Каспия регулярно встречается от
Аляты до южной части Кировского (Кызыл-Агачского) залива и далее-—
у Ленкорани и Астары. В 80-х годах XIX в. выпущен на о-ве Сара, но не
ТУРАЧ 175
прижился; в 1939 г. проник туда самостоятельно и необычайно размножился,
в 1941 г. встречался по всему острову (Верещагин, 1947).
В Туркмении населяет узкую полосу вдоль иранской границы по
р. Атреку (длиной до 80 и шириной от 5—6 до 15—20 км): на восток до
Кызыл-Атрека, на запад доходит до аула Аджи-Яб (обычен у озер
Делили и у Челаюка; Дементьев, 1952). В конце XIX в. встречался
в нижнем течении р. Чандырь и в долине р. Сумбара почти до Кара-Кала
(Зарудный, 1896); сейчас турача там нет (Исаков и Воробьев, 1940). К югу
от нашей границы обычен в бассейнах Атрека и Горгана (к западу до Бендер-
Шаха; Дементьев, 1948).
Характер пребывания. Ведет оседлый образ жизни. Сезон-
ные перемещения отсутствуют, а смена биотопов отчетливо не выражена.
Биотоп. Распространен на равнине; в полосу предгорий, как пра-
вило, не заходит. Держится вблизи водоемов (реки, озера, арыки) по густым
кустарниковым зарослям (тамариск, ежевика, держи-дерево, верблюжья
колючка и т. п.) или камышовом плавням, чередующимся с полянами, по-
крытыми высокой травой; обычен в травянистых зарослях полыни (Artemisia
maritime, и др.), солянки (Salsola laricina и др.), селитрянки (Nitraria Scho-
beri) и тому подобных местах. Также обычен и в культурном ландшафте (па
посевам ячменя и пшеницы вблизи зарослей кустарников, на хлопковых
полях, в садах, огородах, виноградниках, по лесным гарям и т. п.). В сухих
степях не встречается.
Численность. В пределах СССР невелика. Плотность популяции
сильно колеблется по годам. Особенно губительны неумеренная охота и мо-
розные зимы с обильными снегопадами.
Зимой 1924—1925 гг. в Северной Myгани (степь) сохранилось лишь 1—5%
поголовья, после морозов в—18°С встречались неспособные двигаться тура-
чи с отмороженными ногами; в районах с густыми кустарниками легче пере-
носят многоснежные зимы; зиму 1924—1925 гг. пережили 60—80% популяции
(Витович, 1926). Это объясняется тем, что в густых кустах снег не проникает
до земли, и турачи могут кормиться под засыпанными снегом кустами (Азер-
байджан; Спангенберг и Туров, 1950). В многоснежные зимы смертность
резко увеличивается, птицы становятся легкой добычей домашних собак
и кошек, камышовых котов и шакалов; даже после 2—3-дневных снегопадов
ослабевших птиц массами бьют палками и ловят руками—все это резко сни-
жает их численность. Поэтому после особо многоснежных и холодных зим
турачи сохраняются в Закавказье и в Иране лишь в незначительных количе-
ствах (Формозов, 1946); однако численность обычно восстанавливается срав-
нительно быстро, в три—четыре года (Дементьев, 1950). Выжигание кустов,
камышей и сухой травы ранней весной (Закавказье, Туркмения) также сильно
сокращает численность птиц. В благоприятные годы в некоторых местах
плотность может достигать 20—25 птиц на 1 га (Азербайджан; Верещагин,
1947). Обычно плотность значительно меньше: около 6 птиц на 1 км2 в За-
кавказье (Бардинский район, Азербайджан; Беме, 1935) и 7—8 птиц на 1 км2
в Туркмении (озеро Б. Делили; Дементьев, 1945).
В связи с созданием полезащитных лесных полос в Закавказье и с осу-
ществлением строительства Главного Туркменского канала следует ожидать
расширения ареала турача в СССР; при вмешательстве человека расселение
турача будет итти значительно быстрее.
Экология. Размножение. Моногамы. Вопрос о постоянстве пар
не выяснен, но, повидимому, пары длительные, так как самцы и самки дер-
жатся вместе и вне периода размножения. Уже в марте держатся парами.
Самцы токуют с конца марта до первой половины июня включительно (у ту-
рачей в Туркмении семенники и яичники начинают увеличиваться в разме-
рах уже в первую декаду марта; Воробьев, 1946).
176 ОТРЯД КУРИНЫЕ
Токующий самец крутится перед самкой, запрокидывает назад голову,
распускает хвост веером и чертит по земле полураспущенными крыльями
(Сатунин, 1907); часто подскакивает вверх, хлопая крыльями, вертит голо-
вой и издает своеобразные крики, далеко разносящиеся по равнине (Заруд-
ный, 1900). Токовый крик из двух протяжных и одной короткой строфы.
Ранней весной самцы кричат только по утрам, перед восходом солнца; позд-
нее, в период спаривания и насиживания, крик начинается задолго до восхо-
да солнца и продолжается весь день, несмотря на жару (Воробьев, 1946).
Наиболее интенсивен крик по утрам, при восходе солнца, и вечером, с 17—
18 час. (Дементьев, 1950); интенсивное токование продолжается и в пасмур-
ные дни (Сатунин, 1907). Самец токует вблизи самки на земле по полянам
среди кустов, на открытых буграх вблизи кустарниковых и травянистых
зарослей; при пении вскакивает на кочку; часто токует, сидя на кусте (Иса-
ков и Воробьев, 1940). При опасности птицы быстро убегают, скрываясь в за-
рослях. Обычно вблизи токующего самца находится лишь одна самка (Са-
тунин, 1907). Крики отдельных самцов можно слышать даже в последней
декаде июля (Зарудный, 1903).
Полные кладки в Закавказье в конце апреля—мае (в My ганской степи
25 апреля кладка из 8 уже слегка насиженных яиц; Спангенберг), в Туркме-
нии с середины апреля, но свежие яйца встречали и в июне. Существует мне-
ние, что в Закавказье (Сатунин, 1907) и в районе Атрека (Тишкин по Дементь-
еву, 1948) бывает нормально две кладки. Однако вопрос этот до сих пор не
выяснен. В июне встречаются свежие яйца и птенцы разного возраста—от
пуховиков до почти достигших размеров взрослых птиц (Зарудный, 1900).
Более вероятно, что растянутость периода размножения в популяции объяс-
няется наличием дополнительных «вынужденных» кладок взамен утраченных
первых весенних кладок, как это характерно и для других куриных и более
поздним началом размножения у молодых птиц; возможно, что в особо бла-
гоприятные годы у отдельных пар бывает нормально две кладки. Для выяс-
нения этого необходимы дополнительные точные наблюдения с использова-
нием кольцевания и маркировки птиц.
Гнездо устраивается обычно в зарослях кустарников, реже на открытых
местах, всегда под прикрытием куста. Чаще всего это небольшая ямка в зем-
ле, ничем не выстланная (Зарудный, 1900) или со скудной выстилкой из су-
хой травы и веточек (Дементьев, 1948). Воробьев A946) в долине Атрека
находил гнезда на глинисто-солончаковой равнине в середине небольшого
кустика солянки; яйца были отложены прямо на землю без всякой под-
стилки, не было заметно и ни малейшего углубления в почве. Гнезда чаще
вблизи водоемов (озера, арыки).
В Закавказье полные кладки содержат около 10 яиц A0—12 и до 14 по
Сатунину; 8—12 по Верещагину), в Туркмении также около 10 (8—15 по
Шестоперову, 7—9 и до 18 по Воробьеву).
Форма яйца яйцевидно вздутая, с нерезко выраженным острым и тупым
концами. Скорлупа, как и у других турачей, блестящая, пористая и необы-
чайно толстая. Окраска яиц равномерного оливково-бурого или молочно-
кофейного тона с редкими, но хорошо заметными светлыми или почти белыми
пятнами, иногда последние бывают бурого цвета. Размеры яиц из Закавказья
40,0—42,1x32,9—33,5 мм (Спангенберг), из Туркмении B2) 38,0—44,2х
У 31,8—34,0 мм, в среднем 41,14x33,12 мм (Воробьев, 1946).
Продолжительность насиживания и постэмбрионального роста птенцов
неизвестна. Самец во время насиживания держится вблизи гнезда, вместе
с самкой сопровождает выводок. Первые молодые в Закавказье встречены
уже во второй половине мая (Витович, 1926); однако однодневные птенцы
добывались и в половине июля A6 июля 1948, коллекция Зоологического
музея Московского государственного университета). Птенцы ростом в полматки
ТУРАЧ 177
добывались в Туркмении 18—19 августа, в это же время встречены птенцы
величиной с перепела (Воробьев, 1946); птенцы ростом в полматки были
встречены и 7 октября (Дементьев, 1945). В Закавказье в сентябре (ст. стиль)
встречались молодые размером с перепела (Сатунин, 1907), а во второй поло-
вине октября добывались молодые размером в полматки (Беме, 1935). Все
это свидетельствует о значительной растянутости периода размножения
в популяции.
Линька. Изучена недостаточно. Указание Исакова и Воробьева
на то A940), что турачи линяют «в течение всего года», не подтверждается
просмотренным коллекционным материалом и, повидимому, обусловлено
тем, что, в связи с растянутостью периода размножения, в популяции (а не
у отдельных особей) соответственно растянут и период линьки. Ошибочным
кажется и указание Зарудного A900) на то, что сильная линька мелкого пера
была замечена у токовавших самцов и спаривавшихся самок. По коллекцион-
ным материалам Зоологического музея Московского государственного универ-
ситета взрослые птицы в конце июня еще не линяют, в сентябре все они в све-
жем пере. Первостепенные маховые сменяются от десятого к первому.
Птенцовый наряд заменяется первым годовым нарядом в октябре-ноябре,
у отдельных особей, возможно, и несколько позже. Февральские особи
с линяющим мелким оперением каштанового ошейника (Талыш, Тугаринов
и Козлова, 1935), повидимому, принадлежат к молодым птицам.
Питание. В основном кормится зелеными побегами, ягодами, семе-
нами, насекомыми. Пищу собирает на земле; ягоды на кустарниках склевы-
вает не только с земли, но и садясь на ветви (Исаков и Воробьев, 1940).
Анализ содержимого 140 зобов и желудков турачей из Туркмении пока-
зал, что летом преобладают животные корма* зимой—растительные (Воробьев,
1946). Из насекомых преобладают жуки: Haltica gtycyrrhizae, Opatroides
punctiilatus, Lophosis punctata, Bothynoderus crispicollus, Dendarus vaga-
bundus, Broscus sp. и др.; кроме жуков, встречены медведки (Gryllotatpa),
кузнечики, клопы, муравьи, зеленые гусеницы. Рано утром в сильные туманы
в большом количестве поедают нелетающих стрекоз Sympetrum meridionale.
Кроме того, летом и осенью большое значение в питании имеют ягоды селит-
рянки NitrariaSchoberi. Зимой набор кормов меняется. Из насекомых в декаб-
ре поедаются почти только одни муравьи. Основным компонентом корма
служит молодая трава, зеленые листочки и зерна ячменя, корневища рогоза
(Typha), земляные черви. Механические примеси (гравий) в желудках встре-
чаются сравнительно редко и преимущественно в зимние месяцы; повиди-
мому, их роль выполняют твердые семена селитрянки (Воробьев, 1946).
В Закавказье к растительным кормам турача принадлежат листья веро-
ники, зеленые побеги разных трав, семена винограда, пшеничные зерна
(Радде), зерна риса, пшеницы, ячменя, проса (Сатунин), кунжутное семя
(Верещагин, 1947). Зерна культурных растений птицы собирают во время
кормежек на убранных полях (падалица), на дорогах, токах и вблизи селе-
ний, куда подходят во время морозов. Осенью в 26 желудках и зобах турачей
из Азербайджана встречены ягоды черного паслена (Solarium nigrum) и чер-
ной бузины (Sambucus nigra), зерна пшеницы и фасоли; насекомые были встре-
чены в меньшем количестве (главным образом жужелицы и муравьи, меньше
саранчовые; Дементьев, 1948). Желудки и зобы турачей, добытых летом
в Сеистане, были набиты преимущественно крупными цикадами (Зарудный,
1900). Изредка в желудках встречаются мелкие ящерицы, моллюски и т. п.
Хорошо упитанные, с жировыми отложениями птицы встречаются осенью
(октябрь—ноябрь).
Суточный цикл. Ночуют обычно в густых зарослях (кустарни-
ки, камыши). С восходом солнца кормятся по полянам и опушкам зарослей.
В жаркое время дня отдыхают в зарослях, обычно вблизи воды; нередко (как
Птицы СССР. т. IV
178
ОТРЯД КУРИНЫЕ
\
и на ночёвке) присаживаются на ветви. После спада жары вновь вылетают
на жировку, кормясь до сумерек. На кормежке, как и другие куриные,
роются в земле (Дементьев, 1948). Весной и летом держатся парами и вывод-
ками; осенью и зимой—небольшими стайками (Сатунин, 1907). После заверше-
ния линьки в теплые солнечные ноябрьские дни иногда слышен токовый крик
турачей—явление повидимому, аналогичное осенним токам тетеревиных
(Закавказье; Спангенберг).
Враги. Турачей и их кладки уничтожают шакалы, лисицы, коты, хорьки,
ласки и некоторые другие хищники; из птиц преимущественно болотные
луни и ястребы.
Хозяйственное значе-
ние. Объект спортивной
охоты; промыслового зна-
чения не имеет из-за огра-
ниченного распростране-
ния в пределах СССР и,
вследствие этого, малой
численности. Нежное бе-
лое мясо выдвигает турача
в категорию наиболее цен-
ной дичи. Добывается
ружьем (с собакой); раньше
в большом количестве про-
мышлялся с охотничьим»
ястребами - тетеревятника-
ми (Станчинский, 1914) и
ловлей западнями-ямами
(Силантьев, 1898). В особо
больших количествах обыч-
но уничтожается в холод-
ные многоснежные зимы.
Общий выход битого тура-
ча в Азербайджане и Гру-
зии в благополучные годы
достигал 50 000 шт. (Вере-
щагин, 1947).
Представляет большой
интерес для разведения в-
охотничьих хозяйствах За-
кавказья (Абхазия и дру-
гие районы). Создание полезащитных лесных полос в Прикаспийской рав-
нине и строительство Главного Туркменского канала, несомненно, позво-
лит расширить ареал турача в пределах СССР путем его интродукции в
районах вновь создаваемых лесопосадок. Об этом свидетельствует выпуск
трех самок и двух самцов в долине р. Агри-Чай (Нухинский район Азер-
байджанской ССР) в 1932 г.; птицы успешно размножились и стали обыч-
ными в долине р. Агри-Чай и р. Алазани на протяжении 100 км (Вере-
щагин, 1947). Успешны были попытки интродукции турача в между-
речье Куры и Аракса (Сатунин, 1914), в Багире (Туркмения; Билькевич,
1902). Для предотвращения резкого уменьшения численности птиц после
суровых многоснежных зим следует использовать опыт суровой зимы
1924—1925 гг., когда в Муганской степи союзом охотников под руководством
Витовича было отловлено и закуплено у населения большое количество
турачей; птицы содержались в теплом помещении, а затем были выпущены*
в охотничьи угодия (Спангенберг и Туров, 1950).
Рис. 28. Турач (самец и самка)
ТУРАЧ
Г/9
Значение турача как ценной охотничье-промысловой птицы и как истре-
бителя наземных насекомых требует более строгой охраны и ограничения
охоты (многолетние запреты охоты не дали должного эффекта лишь потому,
что надлежащий контроль за их выполнением отсутствовал и поэтому про-
цветало браконьерство; не проводилось мер по спасению турачей в много-
снежные холодные зимы—отлов, подкормка).
Полевые признаки. По размерам несколько крупнее куропатки.
Довольно осторожен. При приближении опасности сначала быстро бежит
по земле, затем «свечкой» с шумом взлетает на высоту 2—3 м (иногда
с криком) и переходит на горизонтальный полет. Летит с быстрыми и частыми
взмахами крыльев и скоро опускается на землю; перед спуском планирует.
Опустившись на землю, убегает в густые заросли. От собаки бежит, затем
взлетает.
Описание. Размеры истроение. В хвосте 14 рулевых. Первостепенные махо-
вые без вырезок. Крыло тупое, формула его 4=3>2>5; первое маховое обычно короче
седьмого. У самцов на плюсне всегда шпора.
Рис. 29. Птенцы турача: а—пуховой, б—оперяющийся, в—в первом наряде
Длина крыла птиц из Закавказья: самцов A1) 168—179, самок A2) 164—172 в сред-
нем соответственно 172,5 и 168,5 мм; из Туркмении: самцов B7) 168—181, самок B4) 160—
171, в среднем соответственно 173,7 и 166,2 мм. Вес турачей (измерения в ноябре при наи-
большем весе в году) из Закавказья: самцов 450—500, самок 400—450 г (Верещагин, 1947);
из Туркмении: самцов 425—550; самок 400—450 г (Воробьев, 1946; Дементьев, 1948).
Приведенные размеры свидетельствуют о невозможности разграничения закавказской
и туркменской популяции как подвидов. Таким образом средняя длина крыла у самцов
C8) 173,3 A68—181) и у самок C6) 167 мм A60—172), при длине тела в 360—380 мм и раз-
махе крыльев около 550—600 мм. Как видно, разница в размерах полов незначительна;
однако половой диморфизм окраски выражен отчетливо.
Окраска. Пуховой наряд: общий тон окраски головы охристо-белый, от лба до за-
тылка идет широкая черно-бурая полоса с едва заметным ржавчатым отливом на затылке;
по ее бокам от ноздрей назад идут узкие буроватые полоски; такие же полоски идут назад
от заднего края глаза и от угла рта; подбородок желтовато-белый. По спине проходят три
отчетливых продольных чернобурых полосы, сливающиеся на передней части спины;
такая же полоса проходит по крылу в области предплечья. Нижняя сторона тела желтова-
то-белая.
Общая окраска птенцового наряда охристо-бурая. Перья верха туловища, большие
кроющие крыла, плечевые и маховые бурые с широкими охристыми полосами, более яр-
кими на маховых. Рулевые охристые с поперечной бурой и коричнево-бурой исчерченно-
стью и бурым крапом. Грудь и бока тела бледные охристо-серые с буроватыми продоль-
ными пестринами, которые в области бедер замещаются поперечными полосами; брюхо бе-
ловато-серое с неясно выраженными продольными буроватыми пестринками.
12*
180 ОТРЯД КУРИНЫЕ
Первый годовой наряд сходен с окончательным нарядом взрослых птиц, и лишь
у самцов отличается от последующих белой надбровной полосой и «усами»; иногда сохра-
няются в виде узкой полосы светлые с темной поперечной исчерченностью перья на груди.
У взрослого самца темя и затылок темнобурые с ржаво-охристыми краями перьев,
лоб черноватый; уздечка и полоса над глазом черные, кроющие уха белые с мелким чер-
ным крапом. Подбородок и горло черные. На шее ржаво-коричневый ошейник (ширина его
индивидуально изменчива). Передняя часть спины черная с белыми округлыми пятнами,
постепенно переходящими в продольные беловато-охристые полосы на спине и кроющих
крыла. Задняя часть спины, поясница и надхвостье черные с белой поперечной исчерчен-
ностью. Маховые и задние плечевые темнобурые с широкими охристо-желтыми светлыми
поперечными полосами. Верхние кроющие крыла темнобурые с охристо-беловатыми про-
дольными полосами на опахалах и темными краями перьев. Низ блестящего черного цвета
с белыми округлыми пятнами на брюхе и боках тела; середина брюха каштаново-рыжая.
Подкрылья бледнобурые с белой поперечной исчерченностью; подхвостье каштаново-
рыжее с черными пятнами и белой полосой. Рулевые буровато-черные с узкими белыми
поперечными полосами; все они с широкими темными без поперечной исчерченности верши-
нами. Радужина орехово-коричневая, веки белые, клюв черный, ноги буровато-красные.
Взрослая самка сверху значительно бледнее самца. Голова буроватая, со светлыми
каемками перьев и темными наствольями. Задняя сторона шеи беловатая, за ней узкая
каштаново-рыжая полоса. На спине, крыльях и рулевых более сильно выражен охристо-
желтоватый оттенок, особенно на рулевых. Подбородок и щеки белые, уздечка и бровь
черноватые; грудь и брюхо светлые с черноватым и буроватым темным рисунком, перехо-
дящим на брюхе в поперечные полосы Подхвостье ржавчато-рыжее. Радужина коричне-
во-бурая; клюв черный, у основания желтоватый; ноги буровато-красные.
РОД УЛАРЫ TETRAOGALLUS GRAY, 1834
Тип. Tetraogallus himalayensis
Синоним. Megaloperdix. Brandt, 1841 тип Tetrao caucasicus.
13. Кавказский улар Tetraogallus caucasicus Pall.
Tetrao caucasicus. Pallas. Zoogr. Rosso-Asiat., II, 1811, стр. 76, Кавказ.
С и н о н и м. Chourtka alpina. Moischoulski. Bull. Soc. I. Nat. Moscou, 1839,
стр. 95, табл. VIII, Кавказ.
Русские названия. Улар—тюркское слово, обозначающее этих птиц (и, вероят-
но, другие крупные виды куриных, например глухаря у якутов). Книжное название—
горная индейка (крайне неудачно).
Распространение. Ареал. Населяет альпийские области Главного
Кавказского хребта и его отрогов. На запад не идет далее горы Чугуш и от-
сутствует на крайних западных вершинах Ачишхо, Фиште, Оштена (Аверин
и Насимович, 1938); однако Пузанов A949) полагает, что кавказский улар
по Главному хребту доходит до вершин Фиште и Оштена, но не проникает
на вершины отрогов (Ачишхо, Аибга, Агбеста). На восток доходит до г. Гю-
мюшлю (Азербайджанская ССР; Верещагин, 1947). Детали распространения
в пределах указанного ареала до сих пор еще выяснены недостаточно четко.
Характер пребывания. Во всех участках ареала оседлые.
Предпринимают незначительные вертикальные миграции; во второй половине
лета подымаются вслед за границей вечных снегов (Аверин и Насимович,
1938), а зимой спускаются ниже, в Нагорном Дагестане вылетая даже на степ-
ные склоны хребтов (Формозов, 1946). Зимой избегают мест с глубоким сне-
гом и держатся в районах выпасов туров, разрушающих снежный покров.
Биотоп. Распространен в высокогорьях от верхней границы альпий-
ских лугов до пределов вечного снега, охватывая районы от 1800 до 4000 м
над уровнем моря. Особо предпочитает верховья ущелий с нагромождениями
скал и крутые каменистые россыпи со скудной растительностью, лужай-
ками альпийского низкотравия и пятнами сохраняющегося снега; в зону родо-
дендронов и альпийского высокотравия, как правило, не заходит.
Численность. В пределах ареала численность неравномерна, что,
повидимому, хотя бы отчасти связано с глубиной снежного покрова зимой;
КАВКАЗСКИЙ УЛАР
181
так, в малоснежных горах, окружающих долину р. Б. Лабы, улар более
обычен и многочислен, чем в более снежных горах в верховьях
р. Киши и т. д. Плотность популяции кавказских уларов в отдельных
частях ареала неизвестна. Колебания численности по годам, как и вообще
детали биологии, достаточно не изучены.
Экология. Размножение. Периодические явления в жизни улара
изучены крайне мало. Повидимому, моногамы; но, вероятно, самцы не при-
нимают участия в выводе птенцов, держась небольшими группами, отдельно
от выводков (Беме, 1926). Разбивка на пары проходит в самом начале весны;
первые тока в начале апреля, причем значительны колебания в сроках
по годам; холостые молодые и старые птицы продолжают держаться стайками
(Носка, 1896).
Ток обычно проходит с утра до ночи на свободных от снега полянах,
прекращаясь в середине дня. Самец преследует самку, перебегая в гору и под
гору, иногда перелетая неболь-
шое расстояние; обе птицы часто
свистят. Во время остановок са-
мец крутится вокруг самки, под-
нимая вверх распущенный хвост,
опуская до земли крылья и рас-
пушая все оперение. Наиболее
интенсивен ток в конце апреля
(Носка, 1896).
Самки приступают к отклад-
ке яиц уже в апреле (Мензбир,
1900). Повидимому, взамен ут-
раченных бывают вторые, до-
полнительные кладки, так как
Сатунин A907) на основании
опросных сведений указывает,
что кладки бывают еще в начале
июля. Гнездо устраивается обыч-
но под защитой камней, лоток
лишь слегка выстилается травой. По указанию Радде A884), в полных
кладках бывает до 20 яиц, хотя Носка A896) не находил в гнездах более
8 — 10 яиц. Сатунин A907) на основании опросных данных говорит о нормаль-
ных кладках в 2—4 яйца, что мало вероятно. Основной тон окраски скорлупы
яиц желтовато- или голубовато-серый, иногда с зеленоватым отливом, испещ-
ренный светлобурыми или коричневатыми пятнами. Средние размеры яиц
69x46 мм\ наибольшие 68x56 мм, наименьшие 46x44 мм (Радде, 1884).
Продолжительность насиживания и время постэмбрионального развития
птенцов не установлены. Самец в насиживании участия, вероятно, не при-
нимает. Первые птенцы появляются в конце мая—начале июня; пуховики
встречены еще 21 июня, хотя 13 июня находили уже хорошо летавших птен-
цов (Носка, 1896). Птенцы размером с куропатку встречаются с конца июля
(Беме, 1925). Эти данные также свидетельствуют о растянутости периода
размножения в популяции. Выводки иногда могут объединяться: при десяти
цыплятах величиной с перепела 9 августа были добыты две взрослые самки
с хорошо выраженными наседными пятнами (Цветков, по Мензбиру, 1900).
При опасности самка улетает, а птенцы затаиваются; иногда самка пытается
отводить от выводка; хорошо летающие птенцы взлетают вслед за старой;
разбитый выводок самка сзывает криком, несколько напоминающим квох-
танье, и уводит вверх по склону (Цветков, по Мензбиру, 1900).
Линька. Изучена плохо. Очевидно, полная линька лишь однажды
в году. Весной взрослые птицы не линяют. У взрослых птиц линька начи-
Карта 14. Распространение кавказского улара
Tetraogallus caucasicus
а—граница гнездовий
182 отряд куриные
нается в конце июня ст. стиля (Носка, 1896). У самца, добытого 5 июля
(верховья Кубани), энергичная линька мелкого оперения по всему телу;
первое—четвертое первостепенные маховые старые, пятое в нераскрывшемся
пеньке, шестое достигло половины окончательной длины, седьмое трех чет-
вертей, восьмое—десятое полностью выросли; энергично идет линька руле-
вых: сохраняется часть старых, а новые достигли примерно трех четвертей
окончательной длины. У самца, добытого 27 июля у г. Баталпашинска, раз-
гар линьки мелкого пера ио всему телу; молодое перо частью почти полно-
стью раскрылось, частью в пеньках; первое—третье первостепенные маховые
старые, четвертое в нераскрывшемся пеньке, пятое около одной трети, шестое
около половины, седьмое около четырех пятых окончательной длины, вось-
мое—десятое достигли полного роста; большинство рулевых сменилось новы-
ми перьями, достигшими трех четвертей полной длины, часть рулей еще
старая; сменяются также верхние и нижние кроющие хвоста. Линяющие
самцы держатся группами по нескольку штук, проводя весь день на голых
скалах; линька заканчивается в сентябре (Беме, 1926).
У самок, вероятно, линька, должна протекать несколько позднее чем
у самцов. Однако скудность коллекционного материала не позволяет выяс-
нить этого точно. У самки, добытой 7 июня (ст. стиля ?) на наседном пятне
показались пробившие кожу пеньки; начинается смена мелкого пера по туло-
вищу (наполовину распустившиеся пеньки); первое—пятое первостепенные
маховые старые, шестое в пеньке, седьмое в половину, а восьмое—десятое
около трех четвертей окончательной длины; кроющие хвоста и рули еще
не сменяются. Самка, добытая в сентябре в Дагестане, завершила линьку,
однако еще не все рули полностью закончили рост.
Молодые птицы заканчивают линьку в конце сентября, повидимому,
одновременно со взрослыми птицами (Беме, 1925).
Питание. Достаточно не изучено. Повидимому, в основном кормятся
побегами, ягодами, почками, семенами, клубнями и луковицами, при случае
насекомыми. Зимой кормятся на местах выпаса туров.
Суточный цикл. Ночуют в скалах. При первых проблесках зари
поднимают громкий крик, а затем слетают вниз (Цветков поМензбиру, 1900).
Кормятся на травянистых лужайках; отыскивая корм, постепенно подни-
маются вверх, скрываясь в 10—11 час. в скалах (Беме, 1925). Самка с вывод-
ком маленьких птенцов держится вначале всего на площади в 0,25 га (Носка,
1896). После полудня так же вылетают на кормежку. В жаркие августов-
ские дни уходят на вершины к границам снега и ледников (Аверин и Наси-
мович, 1938).
Обычно встречаются группами в 3—9 шт. (Беме, 1925).
Враги. Помимо человека, уларов добывают беркуты (Цветков по Мен-
збиру, 1900), возможно, каменная куница, волк и другие хищники. Указа-
ния о нападении на уларов бородача мало вероятны.
Хозяйственное значение. Вследствие ограниченности ареала и относи-
тельной малочисленности, а также трудностей охоты в горах, добывается лишь
случайно; основная масса добытых птиц оседает на месте (Верещагин, 1947).
Добывается преимущественно пулевыми ружьями; на дробовой выстрел
подпускает очень редко. Охота ведется путем сочетания скрадывания и под-
карауливания. В неволе (зоопарки) выживает плохо.
Полевые признаки. По внешнему виду и поведению напоминают домаш-
них кур, но заметно превосходят последних по величине. Отличаются
от других видов уларов хорошо заметной резкой черноватой исчер-
ченностью зоба и передней части груди. Кормятся рассыпной стайкой, обычно
двигаясь вверх по склону; бегают очень быстро; необычайно крикливы и из-
дают громкое квохтанье и мелодичный громкий свист, особенно по утрам
и вечерам (Беме, 1925). Мелодичный свист кавказского улара более чист
КАВКАЗСКИЙ УЛАР
18 3
и звонок, чем у других уларов, и слышен почти за километр (Сушкин, 1914),
При опасности всегда бегут вверх к вершине хребта. Достигнув перевала,
взлетают и скрываются в ближайшем ущелье. Летят очень быстро с частыми
взмахами крыльев; в полете громко свистят.
Описание. Размеры и строение. Округлый хвост образован 18—20 руля-
ми. Наружное опахало первого первостепенного махового без вырезки, второго—пятого
с вырезкой. Крыло тупое; обычно формула крыла 2>3>1>4>5, реже 3>2>4>5>1;
разница в длине самых длинных перьев обычно не превышает 2—4 мм. У самцов тупые,
короткие шпоры; у некоторых, повидимому старых, самок, равно как и у молодых самцов,
есть, зачатки шпор. Длина крыла у самцов C) 273—285, у самок (8) 245—260, в среднем
Рис. 30. Кавказский улар
соответственно 279 и 254 мм [среднее для обоих полов A4)— 245 лмс]. Как видно, различия
полов по величине незначительны; половой диморфизм окраски также не выражен.
Окраска. Пуховой наряд не описан, и соответствующие экземпляры отсутствуют
в доступном коллекционном материале. Окраска птиц в первом птенцовом наряде (раз-
мером с куропатку) по общему тону напоминает окраску взрослых птиц, но значительно
бледнее и тусклее. Охристый струйчатый рисунок повсеместно заметно светлее; коричнева-
тый тон на зашейке отсутствует, коричневато-черная исчерченность боков туловища выра-
жена неясно. В этом возрасте начинают разворачиваться рулевые (хвост образован верхни-
ми кроющими); растут первостепенные маховые и их нижние кроющие.
Окраска молодых в первом годовом наряде сходна с окраской взрослых птиц.
У взрослых птиц верх головы, щеки, уздечка и верх шеи серые, с более или менее
выраженным рыжеватым налетом на шее (варьирует индивидуально). Полоса над глазом,
кроющие уха и бока шеи белые. Передняя часть спины, зоб и передняя часть груди в рез-
ко очерченных поперечных черных и светлоохристых полосках. От щек по бокам горла
идут назад темносерого или каштанового цвета полосы; повидимому, каштановый цвет
появляется у старых птиц обоего пола. Остальная часть спины, поясница, верхние крою-
щие крыла и хвоста темносерые с мелким светлоохристым струйчатым рисунком и светло-
184
ОТРЯД КУРИНЫЕ
охристыми пятнами по бокам опахал (пятен нет на кроющих хвоста); на верхних крою-
щих крыла эти боковые пятна рыжие и развиты сильнее. Маховые белые с темнобурыми
вершинами; белый цвет в виде широкой каймы выступает из-под кроющих. Брюхо
аспидно-серое с мелким охристым крапом и бледноохристыми окаймлениями опахал.
Перья боков тела в широких продольных коричневых полосах с черными окаймлениями
опахал. Рулевые коричневато-бурые с каштановыми вершинами; на некоторых перьях
вершина иногда окаймлена узкой белой полосой (варьирует индивидуально). Нижние
кроющие хвоста белые. Оперение ног серое с охристым налетом.
Клюв буровато-роговой, ноги оранжево-желтые.
Систематические замечания. Все виды уларов очень близки между собой. Различия
сводятся преимущественно к небольшим отличиям в размерах, некоторым различиям в го-
лосе и небольшим вариациям деталей окраски, при сохранении сходства в общем ее харак-
тере. Вариации окраски представляют собой последовательное, от вида к виду,замещение
черного цвета белым (кавказский, гималайский, каспийский, алтайский, тибетский улары
Нестеров, 1911); все это крайне напоминает географическое изменение окраски в пределах
вида. По поведению, приуроченности к определенным стадиям и прочим чертам биологии
(правда, еще крайне недостаточно изученной) все виды также очень близки друг к другу.
Каждый из видов приурочен в своем распространении к одной из крупных горных систем
(кавказский улар—к Большому Кавказу; алтайский—к Алтаю и т. п.). Поэтому можно
полагать, что виды этого рода генеалогически очень близки, а их видовая специфичность
обусловлена преимущественно локализацией их ареалов, связанных с древними горными
системами.
14. Каспийский улар Tetraogallus caspius Gm.
Tetrao caspius. Q m e 1 i n. Reise durch Russland, IV, стр. 67, 1784, Гилянь в Северной
Персии.
Распространение. Ареал. Область распространения каспийского
улара охватывает горы Малой Азии (от Тавра до Армянского нагорья,
Малого Кавказа и Ирана; на
востоке проникает в Копет-Даг
(до Ашхабадского меридиана).
Характер пребы-
вания. Повсюду оседлая
птица.
Биотоп. Каменистые
россыпи и луга с выходами скал
в альпийской и субальпийской
зонах гор.
Подвиды и варьирующие приз-
наки. От гор Тавра по всей Малой
Азии и по хребтам Малого Кавказа
обитает Т. с. tauricus Dresser,
1896; в Северном Иране и Копет-
Даге живет Г. с. caspius Gme I i n,
возможно населяющий и Талышские
горы; из Юго-Западного Ирана (хре-
бет Загрош) описан Т. с. semenow-
tianschanskii S а г u d п у, 1908.
Границы между ареалами подвидов
точно не установлены. Подвиды отличаются лишь незначительными различиями в общич
тонах окраски и степенью интенсивности рыжеватых оттенков верхней стороны тела.
Географические различия в биологии до сих пор совершенно не изучены.
Армянский каспийский улар Tetraogallus caspius tauricus Dres.
Tetraogallus tauricus. Dresser. Proc. Lool. Soc. London, 1876, стр. 675, Булгар-Даг
и Ала-Даг в восточной части Тавра.
Синоним. Tetraogallus Challayei. О u s t a 1 е t. Bull. Soc. Philom. Paris, ser., 6, XII,
1877, стр. 54, Эрзерум.
Распространение. Ареал. Населяет высокогорные районы Малой
Азии, от гор Тавра до Понтийских гор и Армянского нагорья. В пре-
улара
Карта 15. Распространение каспийского
Tetraogallus caspius
1—Т. с. tauricus, 2—Т. с. caspius, 3—Т. с. semenow-
tianschanskii; a—граница гнездовий
КАСПИЙСКИЙ УЛАР J 85
делах СССР область распространения армянского улара крайне невелика
и ограничена высокогорьями Армении (от Бзовдальского и Памбакского
хребтов и Алагеза на восток по горам между озером Севан и р. Араке до
Зангезурского хребта; Ляйстер и Соснин, 1942; Даль и Соснин, 1947). Воз-
можно, что по отрогам Карчхальского и Арсианского хребтов проникает
в южные районы Грузинской ССР (Домбровский, 1913). Детали распростране-
ния улара на нашей территории до сих пор достаточно точно не выяснены.
Характер пребывания. Во всех участках ареала оседлая
птица. Армянские улары предпринимают слабо выраженные вертикальные
миграции, связанные с сезонным состоянием снегового покрова в горах;
в августе—сентябре держатся около перевалов, посещая северные склоны
с пятнами снега (Спангенберг, 1948). Небольшие миграции предпринимают
и летом: так, например, в период массового выпаса скота на склонах Пам-
бакского хребта перелетают на соседний Акстафинский хребет (Даль, 1948).
Биотоп. Распространены в альпийской и субальпийской зонах на высоте
1800—3000 м и более над уровнем моря. Держатся преимущественно по кру-
тым склонам, где скудные ксерофитные луга чередуются с каменистыми рос-
сыпями и отвесными скалами (Спангенберг, 1948), заходят и в заросли родо-
дендронов (Нестеров, 1911). Интересно, что на большинстве гор, обитаемых
уларами, вечного снега нет; он лежит лишь с половины октября до половины
июля. Однако в тех участках, где пятна снега сохраняются долго, улары явно
к ним тяготеют (Бобринский, 1915—1916; Даль, 1949). С середины октября
выпавший снег заставляет птиц спускаться ниже, но даже в самые суровые
зимы они не доходят до верхней границы леса (Спангенберг, 1948); однако
в августе армянских уларов иногда удавалось добывать и у верхней границы
леса (Нестеров, 1910).
Численность в пределах ареала неравномерна и повсюду неве-
лика. Вне пределов СССР улар, повидимому, более многочислен. Динамика
изменения популяции совершенно неизвестна.
Экология. Размножение (преимущественно по Спангенбергу,
1948). Экология армянского улара не изучена. Повидимому, моногамы.
В апреле голос-свист уларов слышен более часто; повидимому, в этом месяце
начинается ток, затягивающийся до конца мая. Первые кладки возможны
в апреле B1 апреля добыта самка с готовым яйцом в яйцеводе), хотя в это
время в горах еще много снега (Спангенберг, 1948). 24 апреля нийдено гнездо
с 6 яйцами; редко в кладках бывает до 9 яиц (Хартерт, 1921—1922).
Размеры яиц D0) 63,2—70x43,8—48,6, в среднем 65,4x45,9 мм; окраска
скорлупы светлоглинистая с матовым зеленоватым отливом и отдельными
матовыми красновато-коричневыми пятнами (Хартерт, 1921—22; место сбора
яиц неизвестно, поэтому описание их может относиться как к Т. с. tauricus,
так и к Т. с. caspius).
Продолжительность насиживания не установлена. Самец в насиживании
и выкармливании птенцов участия, вероятно, не принимает. В выводках
встречали до 10 птенцов, сопровождаемых одной взрослой птицей (Нестеров,
1910). 29 июня на горе Кала (близ Зора, Армянское нагорье) встречен выводок
из 7—8 хорошо летавших птенцов, ростом с кеклика (Бобринский, 1915—1916).
В первое время после вылупления птенцов выводки держатся в районе
гнезд (т . е. относительно низко), а затем поднимаются к горным перевалам;
в конце октября молодые достигают размеров взрослых птиц, а в начале
ноября выводки объединяются в крупные зимние стаи (Спангенберг, 1948).
Линька. Совершенно не изучена. Очевидно, в году одна полная
линька. У самца, добытого 21 октября (в 16 км от Кировокана), энергичная
линька по всему телу; часть мелкого пера еще в чуть раскрывшихся пеньках;
первое—третье первостепенные маховые старые, четвертое достигло поло-
вины окончательной длины, пятое почти выросло, шестое—девятое новые>
ОТРЯД КУРИНЫЕ
полной длины; энергично сменяются рули: часть старых еще сохранилась,
а молодые не достигли окончательного роста. У самца, добытого в тот же день,
все туловище в свежем пере, часть их еще не достигла полного роста; первое—
третье первостепенные маховые старые, четвертое достигло трех четвертей
окончательной длины, пятое—десятое новые, полностью выросшие; новые
рули еще не достигли полного роста, сохраняется часть старых рулевых.
Самка, добытая там же 7 ноября, полностью перелиняла в новое перо; пер-
вое первостепенное маховое и некоторые из рулевых перьев еще не достигли
окончательной длины. Повидимому, в ноябре молодые и взрослые птицы
полностью заканчивают линьку.
Питание армянского ула-
ра совершенно неизучено; вероят-
но, сходно с другими уларами.
Суточный цикл однооб-
разен (Спангенберг, 1948). Осенью
и зимой ночуют на недоступных
для четвероногих хищников ска-
лах. Просыпаются с наступлением
рассвета, тихо пересвистываются
(«уйль, уйль»), затем одновременно
срываются с места и тесной стай-
кой планирующим полетом спу-
скаются вниз. Летом обычно при-
держиваются теневых склонов.
Кормящиеся птицы медленно под-
нимаются вверх по склону, бес-
прерывно перекликаются, купают-
ся в пыли и вновь идут вверх. До-
стигнув скал, располагаются на
дневной отдых. Во второй половине
дня вновь спускаются вниз на кор-
межку. Ночуют обычно на одних
и тех же скалах, под которыми
скапливается много помета.
Враги армянского улара не выяснены.
Хозяйственное значение. Вследствие ограниченного ареала (распростра-
нения), малой численности и трудности охоты в высокогорных районах добы-
ваются лишь случайно (Верещагин, 1947). При охоте к птицам нужно под-
ходить сверху, стараясь разбить стаю, и скрадывать укрывшихся одиночек
(Спангенберг, 1948).
Полевые признаки. Общий характерный облик улара. Кормятся по скло-
нам рассыпной стайкой, часто перекликаются. При опасности затаиваются
меж камней или бегут всегда вверх по склону. Бег легкий и быстрый. Спуг-
нутые, взлетают всегда с криком и летят вниз, облетая гору. Летают превос-
ходно; иногда долго планируют, не* махая крыльями. У летящих птиц
бросается в глаза белая перевязь на крыльях; бегущие и сидящие птицы мало
заметны, сливаясь по окраске с окружающим фоном.
Описание. Размеры и строение. Хвост округлый из двадцати—двадцати двух
рулевых. Наружное опахало первого махового без вырезки, второе—пятое с вырезкой.
Крыло тупое, обычно его формула 2>3>4>1>5, иногда 2>3>4 или 4>5>1; разница
между этими перьями не превышает 1 см, У самцов короткая и толстая шпора. Длина
крыла самцов B) 310 и 315, у самок A) 305 мм.
Окраска. Пуховой наряд не описан. Первый годовой наряд, повидимому, схо-
ден с нарядом взрослых птиц.
Взрослые самцы и самки по окраске неотличимы. Верх головы и зашеек, уздечка,
щеки и бока шеи серые; от заушной области на бока шеи спускается широкая белая поло-
Рис. 31. Каспийский улар
КАСПИЙСКИЙ УЛАР 287
са; подбородок и горло белые. Передняя часть спины серая с охристым оттенком. Осталь-
ная спина, плечевые, поясница и верхние кроющие крыла темносерые с мелким струйчатым
охристым рисунком и светлоохристыми продольными краевыми пятнами на опахалах;
на кроющих крыла эти пятна охристо-рыжие. Первостепенные маховые белые с темнобу*
рыми вершинами; второстепенные маховые белые с бурыми вершинами, испещренными гус-
той мелкой струйчатостью светлоохристого цвета. Зоб и передняя часть груди голубовато-
серые со слабым охристым налетом и редкими черными пятнами. Грудь и брюхо серые
с густым и мелким струйчатым охристым рисунком. Перья боков тела с узкими продоль-
ными охристо-рыжими полосами и черными окаймлениями. Рулевые буровато-черные
с мелким рыжевато-охристым крапом и более или менее выраженными рыжеватыми окон-
чаниями; верхние кроющие хвоста темносерые с мелким струйчатым охристым рисунком;
нижние кроющие хвоста белые.
Клюв светлороговой с голубоватым оттенком и темной вершиной. Ноги желтовато-
оранжевые, радужина глаза темнокоричневая.
Систематические замечания. Бобринский A933), а за ним и Бутурлин A935) указы-
вали, что популяция каспийских уларов, населяющих высокогорья Малого Кавказа, не
отличима от птиц из Эрзерума (Т. с. tauricus), а от южнокаспийской популяции (Т. с.
caspius) отличаются более темным и более чистосерым, без рыжевато-охристого оттенка,
тоном верхней стороны тела и кроющих крыльев. Однако уже Даль и Соснин
A947) относили армянских птиц к подвиду Т. с. caspius; добытые Далем A948) два зимних
экземпляра из Акстафинского хребта имели хорошо выраженный рыжевато-охристый от-
тенок на кроющих крыла и на плечевых. Недостаток коллекционного материала не позво-
ляет сделать пока окончательных выводов, но вполне возможно ожидать, что в дальнейшем
придется объединить Т. с. caspius и Т. г. tauricus вместе под общим названием 7\ с. cas-
piuc Gm. Реальность Т. с. semenowtianschanskii пока также остается сомнительной (опи-
сана Зарудным лишь по одному экземпляру).
Южнокаспийский улар Tetraogallus caspius caspius Gm
Синоним. Tetraogallus transcaspius. Reichenow. Journ. f. Orn. 1916, стр. 161,
Копет-Даг.
Распространение. Ареал. Населяет Эльбурский хребет и Туркмено-
Хорасанские горы (Северный Иран); может быть, заходит и в Талышские
горы. В пределах СССР ареал крайне невелик и ограничивается участком
Копет-Дага между Фирюзой и Гоуданом (Дементьев, 1951).
Характер пребывания. Оседлая птица, предпринимающая
незначительные сезонные вертикальные миграции: при обильном снеге
спускается несколько ниже обычных мест обитания и переходит на более
крутые склоны.
Биотоп. Держится обычно на высоте около 2000 м над уровнем моря
по крутым склонам с выходами скал и каменистым россыпям и по мало доступ-
ным глубоким ущельям с отвесными склонами, где летом сохраняется снег.
Численность. В пределах СССР крайне немногочислен; зна-
чительно обильнее в Эльбурском хребте и Иранской части Туркмено-Хора-
санских гор.
Экология. Размножение. Периодический цикл в жизни южно-
каспийского улара совершенно не изучен. Начало кладки, повидимому,
относится к первой половине апреля: в Ашхабадском музее хранится яйцо от
17 апреля 1916 г. Продолжительность насиживания и выкармливания птенцов
не выяснена. 30 июня 1938 г. в Центральном Копет-Даге (Шах-Шах) добыта
молодая птица уже во вполне развитом оперении (Акад. наук Туркм. ССР).
Линька. Не изучена.
Питание. Не выяснено.
Хозяйственное значение. Несмотря на крупные размеры и вкусное
мясо, промыслового значения в Туркмении не имеет: улар слишком малочис-
лен и трудно доступен (Дементьев, 1952).
Полевые признаки. Кормятся по склонам рассыпной стайкой. Двигаются
вверх по склону, часто перекликаясь. Бегают легко и свободно; полет быстрый;
могут планировать, не взмахивая крыльями. У летящих птиц бросается
в глаза белая перевязь на крыле.
188
ОТРЯД КУРИНЫЕ
Описание. Размеры и строение. Сходен с армянским уларом. Длина кры~
ла у самцов 296—312, у самок 280—290 мм (Хартерт, 1921—22).
Окраска. Пуховой наряд не описан. Первый годовой наряд сходен по окраске
с нарядом взрослых птиц.
Взрослые птицы отличаются от армянского улара более сильным рыжевато-охри-
стым оттенком верхней стороны тела и более светлой и рыжевато-охристой исчерченностью
кроющих крыла.
Систематические заметки. Повидимому, при накоплении материала будет возможна
объединить с армянским уларом в один подвид.
15. Гималайский, или темнобрюхий, улар Tetraogallus
himalayensis Gray
Tetraogallus himalayensis. Gray. Proc. Zool. Soc, London, 1842, стр. 105, Гималаи-.
Распространение. Ареал. Область распространения охватывает гор-
ные системы Гиндукуша, Памира, Тянь-Шаня, Куэнь-Луня. На север про-
ходит вплоть до Ала-Тау и Тар-
багатая. На юге проникает в
Гималаи.
Характер пребыва-
ния. Повсюду оседлая птица.
Биотоп. Обычно луга с
выходами скал и каменистые
россыпи в альпийской и субаль-
пийской зоне гор B000—5000 м
над уровнем моря) часто вблизи
границы вечных снегов. Зимой
иногда спускается в верхние
участки арчевой зоны.
Подвиды и варьирующие приз-
наки. В Западных Гималаях, по Гин-
Карта 16. Распространение гималайского улара
Tetraogallus himalayensis
1—Т. h. Himalayeneie, 2—Г. h. дготbczewskii, 3—Т. h.
ko8lowi; a—граница гнездовий
дукушу, Памиру, Тянь-Шаню, Ала-
Тау, Тарбагатаю и Сауру обитает
Т. A. himalayensis Gray; систему
Куэнь-Луня, от Русского хребта на
востоке и на запад примерно до
78° в. д. (включая хребет Кэрийский
и бассейн Каракаша) населяет Т. h. grombczewskii Bianchi, 1898; из северных
окраин Тибета—Алтын-Таг, хребет Гумбольдта (западная часть Нань-Шаня), хребет
Южно-Кукнорский (включая хребет Сарлык-ула) описан Т. h. koslowi Bianchi,
1898; некоторые авторы позднее выделяли его в самостоятельный вид (Бианки, 1907;
Бутурлин, 1935). Различия между подвидами сводятся преимущественно к небольшим
отличиям в общем тоне и некоторых деталях окраски (перья зоба, характер шейных
коричневых полос и т. п.). Географические изменения экологии (трофические связи,
цикл периодических явлений и т. п.) совершенно не выяснены.
Туркестанский темнобрюхий, или гималайский, улар Tetraogallus
himalayensis himalayensis Gray
Синонимы. Tetraogallus himalayensis severzowi. Зарудный. Орнит. Вестн., 1910,
стр. 108, Семиречье.—Tetraogallus himalayensis incognitus. Зарудный. Орнитологи-
ческий вестник, 1911, стр. 311, Куляб и Дарваз в Горной Бухаре.
Распространение. Ареал. Населяет Азию от Восточного Афганистана до
Кашмира, Ладака и Гарвала, Западные Гималаи (на запад до Кумаона), хребет
Гиндукуш, хребет Тянь-Шань (на восток примерно до 94 ° в. д.). В пределах
СССР область распространения туркестанского темнобрюхого улара отно-
сительно невелика. В пределах Таджикистана он широко распространен
по всему высокогорью (исключая низкие горы Юго-Западного Таджикистана
и Кугитанг; Иванов, 1940). Живет в Гиссарском, Зеравшанском, Турке-
станском и Алайском хребтах. В Западном Памире населяет Дарвазский>
ИМАЛАЙ СКИЙ, ИЛИ ТЕМНОБРЮХИЙ, УЛАР 189
Язгулемский, Рушанский, Шуганский, Ваханский хребты и восточную часть
Памирского нагорья до Заалайского хребта включительно (Дементьев, 1935).
Далее к северо-востоку населяет все горные районы Семиречья, включая
Киргизский хребет, Таласский Ала-Тау, Терскей-Ала-Тау, Кунгей-
Ала-Тау, Заилийский Ала-Тау и Джунгарский Ала-Тау (Шнитников, 1949;
Корелов, 1948, Шестоперов и др.). Многочислен в системе Саура и Тар-
багатая и в горах, окружающих Зайсанскую котловину (Сушкин, 1925;
Долгушин, 1948, и др.). Далее на восток расположена область распростра-
нения алтайского улара.
Детали распространения темнобрюхого улара в пределах ареала выясне-
ны еще недостаточно четко и требуют уточнения. Так, Зарудный A915)
предположительно говорит о его отсутствии в Центральном Памире; Меклен-
бурцев A936) указывает, что он редок в рййонах Восточного Памира.
Характер пребывания. Оседлая птица во всех участках
-ареала. Предпринимает незначительные сезонные вертикальные миграции,
спускаясь зимой в зону кустарников (Спангенберг и Дементьев, 1935),
в арчевники (Формозов, 1950) и даже в ельники (Шульпин, 1939); изредка
встречается на высоко лежащих хлебных полях (Шестоперов, 1929); предпри-
нимает незначительные вертикальные миграции и в другие сезоны года,
особо в ненастную погоду (Шульпин, 1939).
Биотоп. Распространен в альпийской и субальпийской зонах, обыч-
но на высотах 2000—5000 м над уровнем моря. Однако в Кульджинских
горах отмечен в гнездовое время ниже обычных мест обитания, на высоте
900—1500 м над уровнем моря (Шестоперов, 1929). Держится преимуще-
ственно на крутых каменистых склонах и осыпях со скудной травянистой
растительностью и мелкими, редкими кустарниками или на лугах, загромо-
жденных каменными глыбами и выходами скал; иногда встречается даже на
почти голых гранитных и мраморных склонах (Шнитников, 1949). Наиболее
часто встречается вблизи нижней границы вечных снегов (Даль, 1936; Спан-
генберг и Дементьев, 1935).
В непогоду и в суровые многоснежные зимы спускается несколько ниже
обычных мест обитания. При этом всегда придерживается более крутых,
скалистых участков, не выходя на равнину.
Численность. Неравномерна в пределах ареала. Он более редок
в районах Восточного Памира (Мекленбурцев, 1936) и, повидимому, в Цен-
тральном Памире (Зарудный, 1916); многочисленнее в Семиречье, особенно
в долине р. Куёлю и на сыртах Сарджаса (Шнитников, 1949). Данные по
примерной плотности уларов в том или ином районе отсутствуют. Вероятно
изменение численности по годам, но все данные по этому вопросу ограничи-
ваются указанием Спангенберга A937) о том, что в июне-июле 1935 г. в рай-
оне озера Сары-Чилек среди уларов наблюдался падеж, причем встреча-
лись трупы хорошо упитанных птиц (эпизоотия). В то же время суровая
зима 1944—1945 гг. с обильными снегопадами и суровыми морозами при силь-
ных ветрах не отразилась на численности уларов в Таласском Ала-Тау;
птицы зимой кормились по копкам горных козлов (Шевченко, 1948).
Экология. Размножение. Экология изучена совершенно недоста-
точно. Повидимому, моногамы: во второй половине апреля—первой половине
мая в долине Пянджа и Шах-Дары птицы держались только парами (Меклен-
бурцев, 1946). Однако уже в последние дни мая (Семиречье; Шнитников, 1949)
в июне (Зеравшанский и Туркестанский хребты; Даль, 1936)—июле (озеро
Сары-ЧиЛек; Кашкаров, 1927) встречены стайки птиц в 6—17 особей. Шесто-
перов A929) полагает, что в стайки собираются самцы и холостые или потеряв-
шие кладки самки; по мнению местных охотников, это исключительно сам-
цы. Можно предполагать, что самцы не принимают участия в высиживании
яиц и выкармливании молодняка (см. биологию алтайского улара). Наблю-
190 ОТРЯД КУРИНЫЕ
дений над уларами в период спаривания нет, возможно, что совокупление
начинается уже в марте. Начало откладки яиц и время появления полных
кладок не установлено.
Гнездо обычно устраивается в скалистых участках с отдельными низко-
рослыми кустиками. Небольшая ямка, выстланная стебельками злаков
и иногда собственными перьями самки, помещается обычно под защитой
куста или нависающего камня. Полные кладки содержат 8—16 яиц (Шестопе-
ров, 1929; Шульпин, 1939). Повидимому, довольно часто в больших кладках
встречаются неоплодотворенные яйца: в кладке из 16 яиц было два болту-
на, а в кладке из 15 яиц—один (Шнитников, 1949). Основной тон окраски
скорлупы яиц варьирует от бледного желтовато-сероватого до довольно
интенсивного красновато-охристого цвета, у некоторых яиц со слабым
сероватым или зеленоватым оттенком; по основному фону разбросаны раз-
личной величины красновато-коричневые пятна. Размеры яиц из районов
Советского Союза (9): 68,0—69,4x43,5—47,0, в среднем 68,5x45,6 мм (раз-
ные авторы). Размеры яиц из Гималаев F8) 62,0—72,8x43,1—48,2, в сред-
нем 65,4x45,4 мм (Бекер, 1928); оттуда же D5) в среднем 68,2—45,3 мм
(Хартерт, 1921—22).
Продолжительность насиживания и постэмбрионального роста птенцов
не установлены. Вылупление птенцов отмечено уже в конце апреля Киргиз-
ский хребет; Спангенберг и Дементьев, 1935) —мае A4 мая, долина р. Или;
Шестоперов, 1929), хотя и 8 июня еще находили яйца (Семиречье; Шнитни-
ков, 1949). Пуховые птенцы наблюдались 10 мая у г. Хорога (Мек-
ленбурцев, 1946). В конце июня на р. Кудара (Памир) добыты птенцы раз-
мером примерно с перепелку, и такие же птенцы добыты месяцем позже
на р. Ягноб (Иванов, 1940), но 2 и 10 июня в Семиречье встречены хороша
летавшие птенцы размером с кеклика (Кореев и Зарудный, 1906; Шнитников,
1949). Молодые размером в полматки встречены на Памире в начале августа
(Иванов, 1940), а птица, уже достигшая размера взрослых, добыта 19 сентября
(Терскей-Ала-Тау; коллекция Зоологического музея Московского государст-
венного университета). Эти факты свидетельствуют о растянутости периода
размножения улара; несомненно, что это связано с микроусловиями обитания
птиц (высота и освещенность склона, ветры, температура, осадки и т. п.); воз-
можно, имеют место дополнительные вторые кладки взамен утерянных.
Самки при опасности пытаются «отводить» от выводка, отбегая в сторону
с опущенными крыльями и поднятым и распущенным хвостом; при этом
совершенно не проявляется свойственная уларам осторожность (Шесто-
перов, 1929).
Линька. Изучена недостаточно. Очевидно, одна полная линька
в году. В апреле—мае птицы одеты в обношенное перо, но не линяют. Самец,
добытый 18 июня (Киргизский хребет) находится в интенсивной линьке:
сменяется мелкое перо на голове, зобе, спине; первое—пятое первостепенные
маховые старые, шестое еще в совершенно нераскрывшемся пеньке, седьмое
достигло половины .длины, восьмое—десятое в три четверти окончательной
длины; выпала часть рулевых перьев. У самца от 18 июля (Тарбагатай) опере-
ние туловища уже новое (но часть перьев еще в пеньках). Первое маховое
старое, второе меньше половины длины, третье и четвертое в три четверти
длины, пятое почти выросло, шестое—десятое достигли окончательных
размеров; крайние рулевые новые, достигли полного роста, средние от одной
трети до трех четвертей длины; средняя пара рулей старая; старые верхние
и нижние кроющие хвоста почти все выпали, новые в пеньках.
У самок линька, повидимому, начинается несколько позднее. У самки,
добытой 8 июля (Киргизский хребет) от выводка, большое наседное пятно
с едва пробивающимися отдельными пеньками новых перьев, начинается
смена мелкого оперения по всему туловищу; первое—пятое маховые старые,
ГИМАЛАЙСКИЙ, ИЛИ ТЕМНОБРЮХИЙ, УЛАР
191
пенек шестого махового чуть пробил кожу, седьмое едва достигло одной чет-
верти, а восьмое—десятое—половины окончательной длины; рулевые, верхние
и нижние кроющие хвоста старые. У самки от 31 июля (Тарбагатай) часть
свежих контурных перьев туловища уже показалась над старым оперением;
часть второстепенных маховых и кроющих крыла новые; первое—четвертое
маховое старые, пятое едва пробило кожу, шестое—седьмое достигли поло-
вины, а восьмое—десятое—двух третей окончательной длины; нижние крою-
щие крыла в пеньках; рули старые, а нижние и верхние кроющие хвоста
начали сменяться. Взрослые птицы, добытые в октябре—ноябре, все в све-
жем, полностью перелиняв-
шем оперении.
У самца из Кулябских
гор от 18 апреля (коллекция
Зоологического музея Мо-
сковского государственного
университета) все первосте-
пенные маховые растут одно-
временно и едва достигли тре-
ти окончательной длины; по-
видимому, эта птица долгое
время жила в неволе и у нее
были вырваны все маховые.
Линька в первый птен-
цовый наряд начинается с до-
стижением пуховиками раз-
меров перепелки (июнь-июль).
В первый годовой наряд мо-
лодые птицы линяют в авгу-
сте— сентябре. У молодой
самки, добытой 10 сентября
(хребет Терскей-Ала-Тау) и
достигшей размера взрослой
птицы, по всему туловищу
идет энергичная смена мел-
кого пера; первое — третье
первостепенные маховые но-
вые окончательной длины,
Рис' 32'
У«Р
четвертое старое, пятое до!
стигло половины, а шестое
трех четвертей окончательной длины, седьмое — десятое маховые новые,
достигли полной длины; рули все новые, но часть их еще только половин-
ной длины; верхние кроющие хвоста также не достигли полной длины.
Таким образом, как и у других куриных, первое—второе маховые в соб-
ственно птенцовом наряде отсутствуют, появляясь позднее, и не сменяются
при линьке в первый годовой наряд.
Питание. Северцов A873) указывает, что этот улар кормится,
смотря по времени года, почками и семенными головками альпийских трав,
летом—еще насекомыми, зимой можжевеловыми ягодами. Весной в пище
преобладают луковицы дикого чеснока; весной же в большом количестве
поедает осыпающиеся семена ели (Шнитников, 1949). В районе Хорога
основа питания зимой—семена Prangos pabularia, плоды и семена шиповника
и т. п. (Иванов по Гурскому).
Суточный цикл. Наиболее интенсивно кормятся утром и вече-
ром, днем отдыхают в нагромождениях скал. Ночуют в скалах, часто в одном
и том же излюбленном месте.
192
ОТРЯД КУРИНЫЕ
Весной держатся парами, летом и ранней осенью выводками. Когда
птенцы окрепнут и подрастут, выводки поднимаются по склонам вверх,
где держатся до октября; после снегопадов и похолодания спускаются не-
сколько ниже (Шестоперов, 1929). Поздней осенью выводки объединяются
в небольшие табунки.
Враги. Данные о врагах отсутствуют.
Хозяйственное значение. Вследствие ограниченного ареала распростра-
нения, очень большой осторожности птиц и трудности охоты в высокогорьях
промысел темнобрюхого улара имеет лишь местное значение. Птицы добы-
ваются преимущественно ружьем. Охота ведется главным образом путем
скрадывания и подкарауливания на местах кормежки и ночевок.
Полевые признаки. По внешнему виду напоминают других уларов;
от тибетского улара, с которым встречается в восточных участках Таджики-
стана, издали отличаются темной ок-
раской нижней стороны тела. Кор-
мятся по склонам рассыпной стай-
кой, часто перекликаясь громким
свистом. При преследовании моло-
дые, еще не вполне выросшие птицы
затаиваются в камнях или у кустов
арчи. Взрослые птицы при опасности
обычно издают громкий предупрежда-
ющий свист и перелетают на другую
сторону ущелья, почти всегда ниже
по склону; при энергичном пресле-
довании улетают через хребет в дру-
гое ущелье (Шнитников, 1949). По са-
мым крутым склонам двигаются очень
.. легко и быстро, при этом на бегу
иногда взмахивают крыльями для со-
хранения равновесия.
Рис.
Пуховой птенец
улара
гималайского
Описание Размеры и стро-
ение. Хвост округлый, образованный
двадцатью — двадцатью двумя рулевыми.
Наружное опахало первого махового без
вырезки, второго—пятого с вырезкой. Крыло тупое обычно, его формула 2>3>4>1>5;
разница между ними не превышает 0,5—1 см. У самцов короткая и толстая шпора; у
некоторых самок, повидимому старых, есть очень низкие, но широкие в основании зачатки
шпор. Длина крыла у самцов F) 320—340, у самок A1) 275—315 в среднем соответственно
328,3 и 297,7 мм (среднее для обоих полов 305,3 мм). Улары, добытые Зиминой в
Нссык-Кульской обл. на северном склоне Терскей-Ала-Тау в сентябре—январе, весили:
самцы GJ200—3100, самки G) 2000—25/0, в среднем соответственно 2529 и 2326 г
(средний вес для обоих полов 2427 г); в это же время молодые птицы (самцы и самки)
достигли веса в 1800—1900 г. Таким образом различия полов по величине незначительны;
половой диморфизм окраски отчетливо не выражен.
Окраска. Верх тела пухового птенца в коричневато-черном и буром крапе; темя
более темное; хорошо выражена черно-коричневая полоса у глаза, вторая темная полоса
идет от основания клюва и раздваивается в ушной области; низ тусклого серовато-белого
цвета (Бекер, 1928).
Первый птенцовый наряд напоминает наряд взрослых птиц, но имеет более выра-
женный тусклосерый оттенок. Голова буровато-серая с мелкой исчерченностью; верхняя
боковая полоса из Коричневато-бурых перьев чуть намечена, нижняя выражена лучше.
Перья нижней стороны тела (в том числе и задней части зоба) буровато-серые со светлыми
охристыми вершинками. Продольные коричневые полосы по бокам тела узки и не окай-
млены черным.
Первый годовой наряд сходен с нарядом взрослых птиц.
У взрослых птиц (самцы по окраске сходны с самками) верх головы, затылок
и зашеек серого цвета; уздечка и щеки светлосерые; лоб и полоса над глазом беловатые;
бока шеи, подбородок и передняя часть зоба белые. От голого пятна за глазом и от осно-
вания нижней челюсти идут назад (окаймляя белые бока шеи) коричневые полосы; они
АЛТАЙСКИЙ УЛАР
193
соединяются друг с другом поперечной полосой, более широкой и отчетливой на задней
части зоба и менее выраженной на зашейке; этот коричневый или темнорыжий ошейник
у разных особей варьирует по ширине. Передняя часть спины светлосерая с редкими
точечными темными крапинками. Остальная часть спины, поясница, плечевые и верхние
кроющие хвоста темносерые с мелким густым струйчатым охристым рисунком и охристыми
каймами перьев; последние более сильно развиты на пояснице, а на верхних кроющих
крыла еще более широкие и рыжие. Первостепенные маховые белые с темными оконча-
ниями; концы рыжеватые с мелким темным крапом; белый цвет в виде широкой белой
перевязи выступает из-под кроющих перьев. Второстепенные маховые буровато-серые
со струйчатым охристым рисунком, более заметным к вершинам. Перья задней части зоба и
передней части груди с крупными предвершинными черными пятнами и широкими белыми
вершинами, образующими черно-белую исчерченность (несколько слабее у самок); послед-
ние ряды перьев белые. Остальная часть груди и брюха темного голубовато-серого цвета
с очень мелким охристым струйчатым рисунком; у некоторых птиц оперение брюха с ясным
черновато-бурым оттенком. Перья боков тела с широкими продольными коричневыми
полосами с черным окаймлением. Оперение ног темносерое с мелким струйчатым охри-
стым рисунком. Внутренние опахала оулевых темносерые, наружные—коричневые, с тем-
ным мелким струйчатым рисунком, более развитым от наружных к внутренним перьям;
вершины рулевых темнорыжие с неясно выраженной предвершинной черноватой полосой.
Клюв буровато-роговой, ноги оранжево-красные, радужина глаза коричневато-бурая*
16* Алтайский улар Tetraogallas altaicas G e b 1 е г
Perdix altaica+Gt bier. Bull. Sc. Acad. I. St. Petersb., P. 1, 1836, стр. 31, Катунские
белки*
Распространение. Ареал. Область распространения алтайского улара
невелика и охватывает преимущественно альпийские и субальпийские зоны
Центрального, Южного и Юго-
Восточного Алтая и Северо-Запад-
ной Монголии (Сушкин, 1938).
На западе встречается в Кал-
бинском хребте (Казахская ССР;
Долгушин, 1948), но не заходит в
Тарбагатай и на склоны Зайсан-
ской котловины (Сушкин, 1925),
где замещается гималайским ула-
ром. Далее на восток в пределах
нашей страны широко распростра-
нен в Алтайской горной стране.
Населяет Теректинский хребет (на
север до р. Урусула и района Он-
гудая; Кащенко, 1899), притоки
верховий р. Кату ни и горы системы
Кату неких белков (Геблер, terra
typica) и далее к югу Курчумские
горы (Сары-Тау) и горы Топай-Ко-
зень (Поляков, 1912), район озера
Map ка-Куль и хребет Сарымсакты
Карта 17. Распространение алтайского ула-
ра Tetraogallus altaicus
\а—граница гнездовий
(Сушкин," 1938)."Встречен в Чуй-
ских горах, Айгулакском и Курайском хребтах и в горах в верховьях
у. Чаган-Бургазы (Сушкин, 1938); от озер Ак-Куль и Джулу-Куль по
склонам Чулышманского хребта до устья р. Башкаус и далее на север
через Прителецкий хребет Корбу до юго-западных отрогов Абаканского
хребта по верховьям р. Чоот и его притоку Харанак (Фолитарек и Де-
ментьев, 1938); встречается и в верховьях р. Абакана в пределах Горной
Шории (Залесский, 1930; Скалой, 1936). На востоке проникает в горную си-
стему Западного Саяна, по р. Кемчик доходя до р. Енисея и по его правому
берегу до устья р. Ус (Нестеров, 1909). Зимой встречен еще восточнее, по
р.Элегестина горе Хаирхан (Сушкин, 1914). Южнее гнездится по хребту
13 Птицы СССР. т. IV 37
194 ОТРЯД КУРИНЫЕ
Танну-Ола и в котловине Успа-Нур; возможно, населяет Восточно-Тувин-
ское нагорье (Янушевич, 1948). Указание Радде, основанное на опросных све-
дениях, о нахождении уларов в Восточном Саяне между верховьями Оки и Ир-
ку та нуждается в проверке.
Вне территории СССР по Монгольскому Алтаю доходит почти до его
восточного конца (г. Гурбун-Сайхан), через котловину оз. Убса-Нор прони-
кает в Хангай (добыт под Улясутаем, сборы Козлова);.далее к востоку добыт
Козловым у Косогола (Сушкин, 1938).
Характер пребывания. Ведет оседлый образ жизни. Местами
отчетливо выражены сезонные вертикальные миграции, вызванные метеоро-
логическими условиями (состояние снегового покрова в горах, господствую-
щие ветры, освещенность склонов и т. п., обычно эти миграции охватывают
небольшие пространства (Сушкин, 1938).
Биотоп. Распространен в альпийской и субальпийской зоне гор
B000 м над уровнем моря и выше), по открытым каменистым склонам
и россыпям в таких местах, где альпийские участки отделены друг от друга
открытыми безлесными пересечениями рельефа (луговые долины, каменистые
склоны, скалистые ущелья); отдельные вершины гор, отделенные друг от друга
обширными сплошными массивами тайги (что характерно для таежного
Северо-Восточного Алтая), как правило, не заселяет. Обычно предпочи-
тает крутые склоны гор, где участки щебня и альпийских лугов чере-
дуются с обрывистыми выходами скал; в больших количествах держится
в местах, где к концу лета совсем не остается снега, но встречается и у гра-
ницы вечных снегов (Сушкин, 1938). При выпадении глубокого снега спут
скается в нижележащие области гор; зимой появляется в степных предгорьях
и у верхней опушки леса.
Численность, повидимому, нигде не бывает очень высокой. Веро-
ятны колебания численности по годам, но достоверные наблюдения по этому
важному вопросу, как и по другим чертам биологии алтайского улара, прак-
тически в литературе отсутствуют. Повидимому, в Западном Саяне (вер-
ховья Енисея) численность его несколько выше, чем на Алтае.
Экология. Размножение (преимущественно по Сушкину, 1938).
Периодические явления жизни улара выяснены совершенно недостаточно.
Повидимому, моногамы; однако на Алтае в начале июня Сушкин встречал вы-
водки (добытые при них взрослые птицы оказались самками) и стайки из
одних взрослых птиц до 30 особей (добытая из стаи птица оказалась самцом);
наседного пятна у самца не образуется—все это, наряду сданными по линьке
самцов и самок, косвенно свидетельствует о том, что самец не принимает
участия в заботе о потомстве.
Спаривание предположительно относят на апрель; возможно, что в пе-
риод спаривания проходят турнирные бои самцов («ток»), на что указывает
весенняя добыча птиц с выщипанным на горле пером (Сушкин, 1938). Пенье
или крик птиц (ряд следующих друг за другом отрывистых звуков), вероятно,
не связано непосредственно с периодом спаривания, так как его слышно
ив конце лета; кричат, повидимому, и самцы и самки. Точные сроки откладки
яиц, равно как и размеры полных кладок, у этого вида не известны. Яйца
находили в мае; их размеры A4) 65,6—73,5x44,0—48,5 мм, в среднем
70,0x47,13 мм.; по окраске они несколько бледнее яиц гималайского улара
(Хартерт, 1921 — 1922).
Продолжительность насиживания, время постэмбрионального роста птен-
цов и поведение самца по отношению к выводку не выяснены. В половине
июня добывались птенцы, почти достигшие размеров куропатки (Кушканур),
а в половине июля—размерами с крупную куропатку (верховья р. Чаган-
Бургазы), здесь же добытый в начале августа птенец был ростом с тетерку;
это свидетельствует о запаздывании сроков вывода птенцов высоко в горах
АЛТАЙСКИЙ УЛАР 195
(Сушкин, 1938) и о растянутости периода размножения у особей даже
одного района.
Линька. Совершенно не изучена. У взрослых птиц в половине июня
оперение сильно обношено. У самца, добытого в половине августа, голова,
шея и бедра еще не линяют, по остальным участкам мелкое перо сменилось
и частично окрепло, рули сменились и почти достигли полной величины,
маховые сменяются; у добытой в это же время негнездящейся самки (без
наседного пятна) линька продвинулась еще больше; у насиживающих самок
линька еще не начинается, если не считать пробивающихся перьев в области
наседного пятна (Сушкин, 1938). У самки (Зоологический музей Московского
государственного университета), добытой 13 июля 1915 (ст. стиль?) на р. Ча-
ган-Бургазы, сменяется оперение по всему телу, линяют кроющие хвоста, рули
старые, первое—пятое маховые старые, пятое—десятое выпали и вместо них
показались тычки с едва раскрывшимися верхушками. Птицы, добытые в
октябре—ноябре (коллекция Зоологического музея Московского государ-
ственного университета) полностью в свежем пере.
Птенцы, добытые в половине июля в верховьях Чаган-Бургазы (ростом
с крупную куропатку), уже полностью одеты в первый птенцовый наряд,
пух остался лишь на голове и в области пахов, маховые уже опять начали
сменяться. Птенцы, добытые там же в половине августа (ростом с тетерку),
полностью одеты в птенцовый наряд, появляются перья первого годового
наряда на спине, надхвостье и боках, все рулевые уже годового наряда, но
еще не полной длины; среди махов (у кистевого сгиба) появляются махи взрос-
лого наряда (Сушкин, 1938).
Питание. Повидимому, в основном кормится побегами, ягодами,
семенами, почками, клубнями и луковицами, насекомыми; при случае хва-
тает мелких грызунов. Поедает побеги и почки карликовой березы ВеШа папа
и ягоды Berberis sibirica (Хартерт, 1921—1922); в зобах найдены побеги и лу-
ковицы дикого полевого лука (Фолитарек и Дементьев, 1938). Зимой держится
по наименее многоснежным склонам, кормясь на местах пастьбы горных
баранов Ovis argali, разбивающих снег (Тугаринов, 1916).
Суточный цикл. Ночуют в скалах. По утрам слетают вниз и,
начиная кормиться, двигаются вверх по склону. Туров по наблюдениям
в районе Алтайского заповедника пишет, что в июле птицы стараются дер-
жаться на тейевых участках склонов, меняя склоны в течение дня (Фоли-
тарек и Дементьев, 1938).
Весной держатся парами. В июле встречаются отдельные пары, группы
поЗ—4 шт. и табунки от 10—12 до 20 особей (Бианки, 1915). Зимой помет
обнаружен под кедрами на границе леса среди альпийских и субальпийских
лужаек A800 м над уровнем моря) (юго-западные отроги Абаканского хребта;
Фолитарек и Дементьев, 1938).
Враги. Кроме человека, уларов преследует беркут (Бианки, 1915). Пови-
димому, кладки могут иногда уничтожать бурые медведи и волки.
Хозяйственное значение. Вследствие ограниченности ареала и труд-
ности охоты в высокогорьях имеет лишь местное промысловое значение.
Однако в некоторые годы заготовляется в довольно значительных коли-
чествах и завозится в крупные центры (Москва, Ленинград и т. п.); все же
основная масса добытых птиц оседает на месте. Добывается преимущественно
пулевыми ружьями. Чаще всего охота ведется путем сочетания скрадыва-
ния и подкарауливания птиц у мест ночлега и кормежки. Птицы, особенно
самцы, держатся осторожно и с трудом подпускают на дробовой выстрел.
Полевые признаки. Издали по манере держаться напоминают очень
крупных домашних кур; при ходьбе часто взмахивают хвостом. Издали хоро-
шо отличимы от уларов других видов по белой нижней стороне туловища.
Кормятся рассыпной стайкой, быстро перебегая или вверх или вдоль по скло-
13*
196
ОТРЯД КУРИНЫЕ
ну, но не вниз; через нагромождения камней перебегают или перелетают;
почти непрерывно перекликаются тихим, далеко слышным посвистом; вре-
менами отдельные птицы взбираются на скалы и издают громкий, свистового
тона крик. Кричат чаще всего утром и вечером в хорошую погоду, в дождь
обычно молчат (Бианки, 1915). Голос алтайского улара сильнее и грубее,
чем у кавказского, но мелодичнее, чему гималайского (Сушкин, 1938). При
Рис. 34. Алтайский улар
одасности птицы стараются убежать вверх по склону; помогая крыльями,
ловко взбираются на почти отвесные скалы. Достигнув гребня, взлетают,
почти всегда с криком. Полет быстрый, с частыми взмахами крыльев и фазами
скольжения на распростертых крыльях. Обычное направление полета—вдоль
или вниз по склону. Спускаясь с большой высоты, летят, планируя, с очень
большой скоростью, со свистом; обычно не пролетают далее 2—Зкм и садятся
(Сушкин, 1938).
Описание. Размеры и строение. Хвост округлый, образован 22 руле-
выми. Наружное опахало первого первостепенного махового без вырезки, второе—пятое
с вырезкой. Крыло тупое; обычно его формула 2>3>4М У самцов тупые короткие
и толстые шпоры; у некоторых старых самок есть зачатки шпор, большие по основанию,
но очень низкие. Длина крыла у самцов G) 290—325, у самок (9) 280—323, в среднем
ТИБЕТСКИЙ УЛАР 197
соответственно 311,4 и 294,2 мм (среднее для обоих полов—301,7 мм). Как видно» различия
полов по величине незначительны; половой диморфизм окраски также не выражен.
Окраска. Пуховой и птенцовый наряд не описаны и отсутствуют в доступном кол-
лекционном материале. Первый годовой наряд, повидимому, сходен с окончательным
нарядом взрослых птиц.
У взрослой птицы (самцы по окраске сходны с самками) верх головы и шеи, уздечка,
щеки и передняя часть спины седые с буроватым налетом и плохо заметным редким и мел-
ким точечным крапом. Кроющие уха буровато-серые с серебристым отливом. Над глазом
белая полоса. Серая окраска передней части спины прерывается в межлопаточной области
поперечной аспидночерной полосой (ширина варьирует индивидуально), передняя гра-
ница которой резкая, а задняя постепенно переходит в темносерую окраску спины. Пояс-
ница, верхние кроющие крыла и хвоста темносерые со струйчатым мелким бледноохри-
стым рисунком; на пояснице и особенно на кроющих крыла по бокам опахал крупные
белые пятна. Первостепенные маховые бурые с черноватыми вершинами; в основной
половине может быть более или менее развит белый цвет (обычно белые участки прикрыты
кроющими крыла). Второстепенные маховые серые, более светлые у основания; наруж-
ные опахала и вершины с мелкой буровато-охристой струйчатостью. Подбородок и горло
грязносерые с темными наствольями. Зоб и передняя часть груди серые со слабым буро-
вато-охристым налетом и более или менее развитыми белыми вершинными и черными
предвершинными пятнами. Грудь и брюхо грязнобелые с узкими слабобуроватыми
кромками опахал, образующими мало заметный крупный чешуйчатый рисунок; посреди
брюха более или менее выражено черное пятно. Подхвостье белое. Бока тела буровато-
черные с неясно выраженным темноохристым струйчатым рисунком. Оперение ног буро-
вато-черное. Рулевые серые с широкими черными вершинными частями и светловатыми
узкими полосками на концах (последние заметны не у всех экземпляров даже из одного
района). Клюв буровато-роговой, ноги оранжево-красные.
Систематические замечания. Сушкин выделил птиц из восточных частей Монголь-
ского (Гобийского) Алтая (от Улясутая до Дунду-Сейхана) как восточный подвид
алтайского улара Tetraogallus altaicus orientalis S u s h k i n (Bull. Br. Ornithol. Cl.,
№ CCCVIII, 1926, стр. 36), противопоставив его, таким образом, западному алтайскому
улару Г. a. altaicus 6 e b 1 е г, населяющему Саяны, Большой Алтай и западные части
Монгольского Алтая (к востоку до озера Косогол); птицы из хребта Баин-Цаган-Ула имеют
промежуточный тип окраски (Сушкин, 1926). Различия указанных подвидов плохо заметны
и сводятся к тому, что Т. a, orientalis имеет более светлый общий тип окраски, менее
густой темный крап и более широкие светлые концы рулей. Однако (по коллекционным
материалам Зоологического музея Московского государственного университета) все эти
признаки подвержены индивидуальной изменчивости и у особей западного подвида; в
частности, светлые концы рулей варьируют по ширине и могут совсем отсутствовать. По-
этому, принимая во внимание изменчивость подвидовых признаков, слабые подвидовые
различия (на что уже указывал Бутурлин, 1935), скудость коллекционного материала
и совершенно неизвестные подвидовые различия в биологии, в настоящий момент выде-
ление подвидов этого вида не может быть оправдано.
17« Тибетский улар Tetraogallus tibetanas Gculd
Tetraogallus libetanus. Go ul d. Proc» Lool. Soc. London, 1853, стр. 47, Ладак.
Распространение. Ареал. Область распространения тибетского улара
простирается от Восточного Памира, Кашгарии и Ладака до Тибета и Запад-
ной Сычуани, на юг доходя до Центральных Гималаев (Кумаон, Сикким,
хребет Гумбольдта).
Характер пребывания. Оседлая птица.
Биотоп. Обычно каменистые россыпи и скалы альпийского пояса
гор от 3000 до 6000 м над уровнем моря (до снеговой линии); зимой спу-
скаются до 3000—4000 м (Бекер, 1928).
Подвиды и варьирующие признаки. В настоящее время описано 6 подвидов: 7\ t. ti-
belanus Gould, 1853—в Памире, Яркенде и Кашгарии, вплоть до Ладака и Северо-
Западного Тибета; Т. t. centralis S ush ki n, 1926—в Южном Тибете, проникает на север
до хребта Тангла; Т. /. aquilonifer Mei nertzhagen, 1926—-в Сиккиме и при-
легающих участках Южного Тибета; 7*. t. tschimenensis S u s h k i n, 1926—населяет си-
стему Куэнь-Луня (хребты Московский, Чиментаг и Колумба); Г. t. przewalskii^ i a n с hi
1907—в восточном Тибете от Камбаянга до хребта Гумбольдта и Нань-Шаня; 7\ t. henrici
О u s t a 1 е t, 1891—по высокогорьям Сычуани (Та-цзин-лу, Сунь-нань и т. п.). Границы
подвидов до сих пор достаточно ясно не установлены. Подвиды отличаются преимущест-
венно малозначительными деталями окраски.
198
ОТРЯД КУРИНЫЕ
Западный тибетский улар Tetraogallus tibetunus ttbetanus Gould
Распространение. Ареал. Населяет Восточный Памир, Яркенд, Каш-
гарию. Ладак и Северо-Западный Тибет. В пределах СССР область распро-
странения крайне невелика и захватывает лишь восточную часть Таджики-
стана; в горах по правой стороне долины р. Башгумбез, по Мургабу (Памир-
скому)—на перевале Харгуш в Уч-Коле, —в горах по р. Кара-Су у Тах-
тамыша в среднем течении р. Ак-Су; Грум-Гржимайло нашел его у перевала
Муз-Курук в Сарыкальском хребте (Мекленбурцев, 1946).
Характер пребывания. Оседлая птица. В середине зимы
снега вынуждают уларов или держаться по пастбищам архаров или спускать-
ся ниже обычных мест обитания (Мекленбурцев, 1946).
Биотоп. Держатся по каменистым россыпям обычно очень высоко
в горах: летом выше 5000 м над уровнем моря, в первой половине зимы (без
снега)—выше 4700 м (Мекленбур-
цев, 1946).
Численность. Достаточ-
но точно не установлена; в горах
в долине Башгумбез многочислен
(Мекленбурцев, 1946).
Экология. Размножение.
Тибетский улар—наименее изучен-
ная птииа Памира. Данные по его
экологии отсутствуют. Сроки раз-
бивки на пары, время откладки
яиц, величина кладок в пределах
СССР не установлены. В Ладаке
и Северо-Западном Тибете период
размножения приходится на конец
мая—начало июля; кладка из семи
яиц была обнаружена 27 мая на
высоте около 4 500 м над уровнем
моря; число яиц в кладке обычно
4—7; размеры яиц 58,4—68.6 X
х41,1— 46,9, в среднем 63,8x44,1 мм
(Бекер, 1928).
Карта 18. Распространение тибетского улара
Tetraogallus tibetanus
l—T. t. tibetanus, 2—Т. t. centralis, 3—T. t. aquilo-
nifer, 4—T. f. tschimenensis, 5—T. t. przewalskii,
6—T. t. henrici; a—граница гнездовий w
Продолжительность насижи-
вания, сроки вылупления и время
роста птенцов не установлены. Период размножения в популяции, растянут:
18 июня в Кашгарии Дивногорский встречал при взрослых пицах по
4—6 летавших птенцов (Судиловская 1936), а еще в августе Пржевальский
встречал птенцов, не превышающих по размерам перепела (Бекер, 1928).
Линька. У тибетского улара не изучена.
Питание. Повидимому, питаются в основном растительной пищей,
но при случае поедают насекомых и, может быть, мелких позвоночных.
В желудках и зобах птиц из Восточного Памира обнаружены ростки, почки
и веточки Stellarча, Роа, Testuca, Smelowska, Nepeta,Saxifraga, Oxytropis,
Potentilla, Alopecurus, Primula sp. (Мекленбурцев, 1946).
Суточный цикл. Подробно не изучен; вероятно, сходен с другими
уларами. Летом встречаются выводками (Дивногорский, по Судиловской,
1938; бывает ли самец при выводке—не выяснено), а в первую половину зимы
стаями по 10—30 шт. (Мекленбурцев, 1946).
Хозяйственное значение. Вследствие -крайне незначительного распро-
странения в пределах СССР и трудностей охоты на высоте 4000—5000 м над
уровнем моря добывается лишь случайно.
ОБЫКНОВЕННЫЙ ФАЗАН
199
Полевые признаки. По внешнему виду и поведению сходен со стальными
уларами. Легко отличим от гималайского улара, с которым, может быть,
встречается в некоторых районах Памира, по белому цвету зоба, груди и брюха.
При преследовании очень быстро и легко бежит вверх по склону; если
охотник спускается к стае сверху, то птицы затаиваются между камней,
а затем шумно и стремительно взлетают и улетают вдоль склона; при взлете
издают резкий и отрывистый свист, более глухой и тихий, чем у гималай-
ского улара (Мекленбурцев, 1946).
Описание. Размеры и стро-
ение. По Хартерту A921—1922) дли-
на крыла у самцов 275—290, у самок—
258—270 мм; по Бекеру A928) общая
длина около 500 мм, длина крыла
255—276 мм.
Окраска. Пуховой наряд не опи-
сан. Птицы в первом годовом наряде
похожи на самок, но черные и охристые
пятна на груди и шее грубее, а перья
спины с бледными стержневыми поло-
сками.
У взрослого самца верх и бока
головы и верхняя часть шеи темносе-
рые, уздечка беловатая, кроющие уха
светлоохристые. Передняя часть спины
светлая, охристо-серая с бурым крапом,
остальная часть спины охристо-серая с
бурым крапом и охристыми окаймле-
ниями перьев; плечевые и верхние кро-
ющие крыла с широкими бледноохристы-
ми окаймлениями. Первостепенные ма-
ховые светлобурые с белыми вершина*
ми (не всегда выраженными на первом—
четвертом маховых); второстепенные ма-
ховые с большими белыми вершинами,
образующими на крыле зеркальце. Под-
бородок, шея спереди и зоб белые; меж-
ду зобом и шеей серая полоса, такая
же полоса проходит поперек передней
части груди; грудь, брюхо и нижние
кроющие хвоста белые с черными боко-
выми окаймлениями опахал (особенно
широки они на нижних кроющих хвоста
и боках груди), создающими продольную исчерченность. Оперение ног светлосерое с
охристым налетом. Рулевые темнобурые с каштановыми вершинами и охристо-розовыми
пятнами у основания. Голые участки кожи вокруг глаз, клюв и ноги красные.
Самка сходна по окраске с самцом, но бока шеи и серые полосы на зобе и груди
испещрены черными и охристыми пятнами. Клюв и голые участки кожи вокруг глаз
красные (Мекленбурцев, 1946).
Систематические замечания. Систематика вида требует дальнейшей разработки
на основе увеличения коллекционных материалов и детального изучения экологии под-
видов.
РОД ФАЗАНЫ PHASIANUS Linnaeus, 1758
Тип Phasianus colchicus
18. Обыкновенный фазан Phasianus colchicus L.
Phasianus colchicus. Li n n a e u s. Syst. Nat., изд. X, 1758, стр. 158, Рион (Колхида).
Русское название. Фазан, на русском, как и на других современных европей-
ских языках, происходит от латинского названия этой птицы phasianus, или phasiana
(avis), так как птица завезена в Европу, по преданию, еще аргонавтами с реки Фазис
(Phasis), ныне Риона в Колхиде.
Распространение. Ареал. Дельта Волги, Кавказ, Болгария (у Ям-
«бола и на реке Тундтс до Елховской и к югу до Бургаса, Патев 1950),
Рис. 35. Тибетский улар
200 ОТРЯД КУРИНЫЕ
Передняя Азия от Южного побережья Черного моря в Анатолии, Ирана
до Средней Азии и Афганистана. Центральная и Восточная Азия от Синц-
зяня и Монголии до Приамурья и Приморья, Японии, Маньчжурии, Кореи
и Китая, на юг до Северной Бирмы и Индокитая (Тонкий); Формоза.
Кроме того, фазаны акклиматизированы в ряде мест Западной Европы и
в Северной Америке.
Еще при Перикле фазан был завезен в Грецию (позднее истреблен там
почти полностью), потом в Италию и затем расселен во многих странах
Карта 19. Распространение фазана Phasianus colchicus
j ph. с. colc/iicus, 2—Ph. с. septentrionatis, 3—Ph. c. talischensis, 4—Ph. c. persicus,
,5 ph. c. principalis, 6—Ph. c. zarudnii, 7—Ph. c. bianchii, 8—Ph. c. chrysnmelas,.
9—ph. c. zerafshanicus, 10—Ph. с turcestanicus, 11—Ph. c. mongolicus, 12—Ph.
с pallasi, 13—Ph. с shawii, 14—Ph. с tarimensis, 15—Ph, с hagenbecki, 16—Ph.
c. vlangalii, 17—Ph. с satscheuensis, 18—Ph. с edzinensis, 1Г—Ph. с strauchi, 20—Ph.
c. 8chokhoten8is, 21—Ph. c. alaschanicus, 22—Ph. c. suehschanensie, 23—Ph. с elegansy
24—Ph. с karpowi, 25—Ph. c. decoLlatus, 26—Ph. c. torquatus, 27—Ph. c. takatsuha*aet
28—Ph. с formosanus, 29—Ph. с ellioti, 30—Ph. с kiusiuensis; а—граница гнез-
довий, б—не вполне выясненная граница гнездовий, в—изолированные гнездования
Европы (особенно в придунайских) к северу до Англии, Южной Швеции
я Осло.
Разводится во многих охотничьих хозяйствах европейской части СССР
к северу до Прибалтийских республик и Ленинградской обл.
Естественная граница распространения фазанов в общих чертах совпа-
дает с полосой малоснежных равнин, предгорий и континентальных сухих
плато с малым количеством зимних осадков и с глубиной снежного покрова
не выше 10—20 см (Формозов, 1946).
Характер пребывания. Оседлый.
Биотоп. Преимущественно кустарниковые и камышевые заросли
речных долин, а также культурный ландшафт. Местами в кустарниках па
речным долинам в горах, где достигает высоты 1500 и даже 2500—2600 м.
Подвиды и варьирующие признаки. Описано много подвидов, различающихся
окраской (главным образом у самцов), в некоторых случаях размерами, а также некото-
ОБЫКНОВЕННЫЙ ФАЗАН 201
рыми деталями биологии (биотопическое распределение, трофические связи, даты перио-
дических явлений)*
В Малой Азии, Закавказье живет Ph. с. colchicus Li п n a e u s, 1758; в дельте
Волги, на Северном Кавказе в бассейнах Кубани и Терека—Ph. с. septentrionalis L о-
г е n z, 1888, в юго-восточной части Закавказья (Талыш) и в южнокаспийских провин-
циях Ирана на восток, повидимому, до Мазандерана,— Ph. с. talischensis L о г е п z,
1888; в Копет-Даге, в бассейнах Гюргеня и Атрека в пределах Ирана—Ph. с. persicus-
Sewertzow, 1875; в бассейнах Теджена и Мургаба, в низовьях Кушки—Ph. с.
principalis S с 1 a t е г, 1885; по среднему течению Аму-Дарьи— Ph. с. zarudnyi В и-
t и г 1 i п, 1904; в дельте Аму-Дарьи—Ph. с. chrysomelas Sewerzow, 1875; в Юго-
Западном Таджикистане—Ph. с. bianchii В u t и г 1 i n, 1904; в долине Зеравшана—
Ph. с. zerafshanicus Tarnovski, 1891; в бассейне Сыр-Дарьи— Ph. с. turcestanicus
L о г е n z, 1896; в Семиречье—Ph. с. mongolicus Brandt, 1844; в Приамурье и При-
морье, в Северной Маньчжурии— Ph. с. pallasi Rothschild, 1903; в Кашгарии—
Ph. с. shawii Elliot, 1870; в Восточном Синцзяне в бассейне Тарима и у Лоб-Нора—
Ph. с. tarimensis Р 1 е s k e, 1888; в долине Кобдо—Ph. с. hagenbecki Rothschild,
1901; в Цайдаме, между Южно-Кукунорским хребтом и горами Толай-Ула и Бурхан-
Будда—Ph. с. vlangalii Przewalski, 1876; в западных частях Ганьсу к северу от
Нань-Шаня, в долинах Булунзира и Дан-Хе, у Халачи-Нор, в оазисе Сачжоу—Ph. с. sat-
scheuensis PI e s k e, 1892; в низовьях реки Эдзингол в Монголии—Ph. с. edzinensiz
S u s с h k i n, 1926; в южных частях Ганьсу на север до р. Тетун-Гол, на восток до сред-
них и южных частей Шеньси (Циньлинские горы)—ЯЛ. с. strauchi Przewalski,
1876; в оазисе Сого-Хото в 100 км северо-восточнее хребта Рихтгофена—РА. с. schokho-
tensis В u t ii г 1 i n, 1908; в оазисе у западных отрогов средней части Алашанских гор—
Ph. с. alaschanicus Alpheraki et Bianchi, 1908; у Сунпана и Тунпея в Сюешане
в северо-восточной части Сычуаня—Ph. с. suehschanensis В i а п с h i, 1906; в горах Юж-
ного Сычуаня (Тацзинлу) и Юннани до Северной Бирмы и Северо-Западного Тонкина—
Ph. с. elegans Elliot, 1870; в Южной Маньчжурии и Северном Китае (Чжили, Шань-
дунь, Шаньси), в Корее, на островах Квельпарт—Ph. с. karpowi В u t и г 1 i n, 1904;
в западных частях Среднего Китая—в Гуй-Чжоу, в западном Хунане, восточных частях
Юннани, на север до равнинных частей Сычуаня (Гуан/—РЛ. с. decollatus Swi nhoe,
1870; в Юго-Восточном Китае от Кантона до восточных частей Хунаня, на север до лево-
бережья Яньцзы, на юг до Северо-Восточного Тонкина—Ph. с. torquatus Gmeli n,
1789; в Восточном Тонкине у Лангсона— Ph. с. takatsukasae D е 1 а с о и г, 1927; на о-ве
Формрза—Ph. с. formosanus Elliot, 1870; в Японии на о-вах Хондо и Сикоку,
на Семи островах—Ph. с. versicolor Vieillot, 1825; на о-вах Кюсю, Танегасима.
и Якусима— Ph. с. kiusiuensis Kuroda, 1919.
Закавказский фазан Phasianus colchicus colchicus L.
Синоним. Phasianus colchicus lorenzi. В u t u r 1 i n. Ibis, 1904, стр. 386. Кура.
Распространение. Ареал. Малая Азия от южного побережья Анатолии
и Закавказья без самого юго-восточного угла его, где обитает уже другой
подвид. Подробности распространения в СССР таковы. Раньше населяя
в сущности всю Грузию, Армению и Рионскую низменность (по побережью
Черного моря не севернее Сухум-Кале); также северо-восточные части Азер-
байджана. В настоящее время есть в Абхазии (Вейцман, 1933), в низменностях
Западной Грузии, в Кахетии и Гареткахетии (Марков, 1950), по Алазан»
не идет на восток далее с. Анвами (согласно Чхиквишвили, 1930,—здесь
водится уже предкавказский подвид, что сомнительно), на восток на севере
Грузии до восточных границ Мелавского района. По Маркову A950), гнез-
дится в Закатало-Измаилинской и Кубино-Хачмасской низменностях Восточ-
ного Закавказья; в Армении встречается, повидимому (и то по расспросным
сведениям), только в Кафанском районе (Ляйстер и Соснин, 1942).
Характер пребывания. Оседлая птица.
Биотоп. Леса с густым подседом, перевитым колючими вьющимися
растениями, заросли лоха; по берегам рек (например, Кура и Араке) держится
в густых Камышевых зарослях, лишенных древесной растительности; может
быть встречен и на хлебном поле; в Абхазии держится также на кукуруз-
ных полях, чайных плантациях и нередко встречается вблизи домов (Вейц-
ман, 1933).
202 ОТРЯД КУРИНЫЕ
Численность. Сильно уменьшившаяся в числе и в ряде мест исчез-
нувшая птица. Однако в местах, где охраняется, увеличивается в числе
и даже появляется там, где раньше его не было (например, в Цхалтубо; Мар-
ков, 1950). Согласно Верещагину, наибольшая плотность населения наблю-
дается в Кубино-Хачмасской и Закатало-Измаилинской низменностях. Чис-
ленность сильно изменяется в зависимости от условий зимовок. Так, фазан
почти совершенно исчез на некоторых реках бассейна Риона после продол-
жительных снегопадов в зиму 1895—1896 гг. (Дандуров, 1896). Неблагоприят-
ное влияние на численность птиц в Западной Грузии оказали в частности
зимы 1910—1911 и 1924—1925 гг. Основные факторы, уменьшающие поголовье
фазанов,—это устойчивый снеговой покров свыше 30—40 см, наст и гололе-
дица, длительные морозы при недостатке корма; в снежные зимы особенно
сказывается также и преследование хищниками; летом уменьшению числен-
ности фазанов способствуют засухи, разливы рек, выжигание кустарников и
тростника (Насимович).
Экология. Размножение. Половая зрелость наступает в возрасте
около года. Вопрос о моногамии остается невыясненным, так как мнения
наблюдателей противоречивы; возможно, в этом отношении существует
различие между подвидами. Привязанность самца к своему гнездовому уча-
стку несомненна, но хорошо известны случаи, что после того как самка при-
ступила к насиживанию, самец привлекает на тот же участок другую самку
и даже третью. Весной между самцами наблюдаются драки, которые, однако,
никогда не бывают ожесточенными. В это время, а также и летом слышны
их громкие крики. Полные кладки из 10—15 яиц. В Арешском районе
кладки находят уже в начале мая, в Джеванширском районе, в предгорьях
позже—яйца с начала июня, а полные кладки в середине этого месяца (Сату-
нин, 1907). Яйца иногда обычной «яйцевидной» формы, иногда укороченно-
овальные. Скорлупа с небольшим блеском, оливково-бурого цвета, часто так-
же бурая, или серовато-зеленая или голубовато-зеленая. Размер яиц B0)
39,0—47,8x34,5—37,0, мм среднее 45,21 х35,5 мм (Хартерт, 1917; данные
относятся к охотничьему фазану—более или менее чистому colchicus).
Длительность насиживания 23—27 дней, насиживает самка. Птенцы
размером в половину взрослой самки отмечены у Иссмаилов 17 июня (Спан-
генберг).
Линька. Общая схема смены оперения у фазанов такова. Пуховой
птенец начинает оперяться очень рано, примерно на третий день жизни,
причем первыми появляются первостепенные маховые, последней опери-
вается голова. Судя по линьке таджикского фазана, возможно появление
элементов промежуточного наряда, перья которого начинают расти еще
до того, как птенец полностью оперился, и вскоре исчезают, заменяясь перь-
ями первого взрослого наряда. Таким образом, смена гнездового наряда
на взрослый происходит осенью; начинается она с перьев головы и шеи,
маховые сменяются едва ли не последними. В дальнейшем линька раз в году
полная, происходит во второй половине лета, у самцов несколько раньше,
чем у самок, что связано с заботой о потомстве у последних. Более точно
сроки линьки не изучены (общий ход линьки в подробностях—см. таджик-
ского фазана).
Питание. Растительная и животная пища. Сколько-нибудь подроб-
ных наблюдений нет. В большинстве мест, где этот фазан акклиматизирован,
самостоятельно прокормиться зимою не может, нуждается в искусственной
подкормке.
Суточный цикл. См. лучше изученные подвиды.
Враги. В низменных районах в основном—камышевые коты и шакалы.
Промысел. Промысловое значение этого подвида (так же, как и других
подвидов) сильно изменчиво в зависимости от численности. Местами эта птица
ОБЫКНОВЕННЫЙ ФАЗАН
203
теряет значение даже как объект любительской охоты, местами может еще
рассматриваться и как объект промысловой охоты. В 1929—1930 г. через
ленинградскую экспортную базу прошло 33 400 особей фазана разных под-
видов (Рудановский и Насимович, 1933). Способы охоты разнообразны. В Азер-
байджане еще до 20-х годов этого столетия сохранилась охота с ловчим соколом
или с ястребом. По тугаям рек Сулака и Алазани их ловят в ямы на травя-
«ых полянах (Верещагин, 1947); охотятся на фазана также с собакой; по ста-
рым данным Силантьева A898), их стреляли на деревьях в лунные ночи с фона-
Рис. 36. Фазан (самец и самка)
рем или даже сдергивали волосяной петлей; местами употребляют щит с яркой
материей, за которым скрывается охотник с ружьем.
Полевые признаки. Довольно крупная, размером примерно с курицу,
птица; общий склад тела, как у курицы, но хвост очень длинный, особенно
у самцов. Самцы отличаются яркостью оперения, самки окрашены скромно.
Взлет шумный, стремительный. От опасности обычно стремится уйти по
земле, в случае необходимости взлетает круто вверх «свечкой», потом очень
полого планирует. Голос самца громкий, несколько напоминает пение
молодого петуха домашней курицы. Этот подвид, как и все кавказские, садит-
ся на деревья, часто на них ночует.
Описание. Размеры и строение. Телосложение плотное. Хвост очень
длинный (даже у курочек превышает длину крыла) из 9 пар рулевых, но бывает из 8 или
10 пар. Рулевые постепенно сильно суживаются к вершине, средняя пара самая длинная,
крайняя пара значительно короче средней. Ноги сильные, цевка покрыта поперечно вытя-
нутыми пластинками и у самцов имеется шпора. У самцов перья ушной области слегка
удлинены, образуя суши», бока головы почти лишены перьев, перья по бокам надхвостья
и края рулевых ближе к их основанию рассучены и волосовидны. Крыло широкое, закруг-
ленное, сильно выпуклое, формула его 4^5>3>6>2>7>1...
204
ОТРЯД КУРИНЫЕ
Половой диморфизм довольно хорошо выражен. Он проявляется в размерах,
в относительной длине хвоста, в различии характера оперения боков головы, в строении
рулевых и перьев поясницы, в отсутствии или наличии шпоры на ноге и очень резко
выражен в расцветке оперения.
Длина крыла самцов C) 250—253, среднее 251 мм. Согласно Далю A947), длина
крыла самцов до 280, самки A) 235 мм (Зоологический музей Московского государствен-
ного университета). Вес самцов в ноябре (по Верещагину, 1947) до 1300, в среднем
1150 г, самок—до 950, в среднем 850 г.
Окраска. (Пуховой птенец—см. таджикский подвид.) Голова и шея взрослого
самца ярко, металлически блестящие, темнозеленого цвета, переходящего в бронзово-
зеленый на темени и в сине-фиолетовый на боках и передней стороне шеи. Зашеек и перед-
няя сторона спины золотисто-оранжевые с черными краешками и черной вершинной стерж-
невой полоской перьев; задняя часть спины с поясницей, плечевыми и кроющими
хвоста меднокрасные с пурпурным отливом;
у плечевых перьев меднокрасные края и бурые
центры, окаймленные бледноохристой и черно-
бурой полосой, и меднокрасные края опахал»
вследствие чего плечевые партии имеют пестрый,
чешуйчатый вид; на пояснице и надхвостье не-
ясные бурые с блеском отметины; рулевые бу-
ровато-оливково-охристые с меднокрасными кра-
ями; на средних рулевых поперечные черные по-
лосы, очень узкие у основания пера и более ши-
рокие ближе к его концу. Зоб и передняя часть
груди меднокрасные с черными краешками
перьев, бока туловища золотисто-оранжевые, с
черными вершинными каймами перьев; нижние
кроющие хвоста каштановые с черными блестя-
щими пятнами; брюхо матовое, спереди полу-
окаймленное полосой блестящих вершин приле-
гающих к нему перьев груди.
Самка в общем песочно-бурая, на голове,
шее, мантии и боках груди с черно-бурыми цен-
трами перьев, с рыжеватым оттенком и очень лег-
ким фиолетовым отливом; на пояснице и крою-
щих хвоста с бурыми отметинами и охристыми
пестринами; горло бледнорыжевато-охристое.
Грудь и брюхо бледнорыжевато-бурые без пяте»
или с неясно проглядывающими бурыми отмети-
нами; средние рулевые рыжевато-бурые с пе-
сочно-оливковыми краями и с неправильными
черно-бурыми и охристыми поперечными полосами. Ноги у обоих полов грязновато-
серые или светловато-бурые, клюв светлобуровато-желтый, радужина светлая красно-
вато-бурая. г~лая кожа головы у самцов красная.
Северокавказский фазан Phasianus colchicus septentrionalis Lor.
Phasianus colchicus septentrionalis. Lorenz. Journ. f. Orniih. 1883, стр. 571, северная
сторона Кавказа.
Распространение. Ареал. Северный Кавказ, к югу по побережью
Каспийского моря, не доходя до Кубинского района, где уже водится закав*
казский подвид, к северу до дельты Волги. Водился раньше в дельте Урала,
но с середины прошлого столетия истреблец. Вследствие резких колебаний
численности и частичного сокращения, а временами расширения ареала
и изменения размещения внутри его указать подробности распространения
этого фазана довольно затруднительно. Раньше он был распространен на се-
веро-западе почти до самого впадения Кубани в море, но теперь по Черно-
морскому побережью истреблен почти повсеместно (Аверин и Насимович,
1938). В окрестностях Дзауджикау к концу прошлого столетия он исчез
совсем, но после первой мировой войны, размножившись в Дагестане, вновь
заселил к 20-м годам и средние части Предкавказья до предгорий, теперь
опять сильно уменьшился там в числе (Беме, 1929), по Тереку он и сейчас
обычен; в дельте Волги почти исчезал и вновь восстанавливался, в 1949 г.
был там обычен (Спангенберг).
Рис. 37. Птенец фазана в возрасте
10 дней
ОБЫКНОВЕННЫЙ ФАЗАН 205
Характер пребывания. Оседлая птица.
Биотоп. Держится по берегам рек и островов в густых зарослях
ивняка, облепихи и лоха, по пойменным рощам, в терновниках, тростниках
и по опушкам лесов. Открытых мест избегает, но во второй половине лета
и поздней осенью его можно увидеть на кукурузных и других полях. В горах
распространен до мест, где долины сильно суживаются и окружены высокими
горами, т. е. примерно до 700 м над уровнем моря.
Численность. Сильно колеблется. Нерациональная охота привела
местами в полному истреблению этой птицы. Большое влияние на численность
имеют неблагоприятные условия зимы (глубокий и длительный снеговой
покров) и наводнения. В низовьях Кубани суровые многоснежные зимы 1875—
1877 гг. привели к почти полному исчезновению этой птицы. В долине Сулака
огромное количество их погибло во время сильных морозов 1927 г., в тех же
местах численность птиц ограничивается также весенними разливами из-за
неправильного пользования оросительными каналами (Туров и Красовский,
1933). Во время высокого половодья большая гибель фазанов наблюдается
также в дельте Волги.
Экология. Размножение. Вероятно, моногам, хотя иногда при
одном петухе бывают две курочки. Приступает к размножению, вероятно,
в возрасте около одного года. Токование (начинается в начале или в середине
апреля) состоит из громкого крика, который птица подает через известные
промежутки времени, главным образом утром и вечером; брачный крик напо-
минает крик петуха, только более хриплый и короче, он может быть передан
звуками «кор-орк», за криком обыкновенно следует хлопание крыльями.
Петух не принимает участия в устройстве гнезда и в насиживании, но дер-
жится в общем недалеко от гнезда и выводка (Мензбир, 1902). Гнездо устра-
ивается на земле и густых зарослях кустарника и представляет собой неболь-
шое углубление в почве, слегка выстланное сухой травой и перьями. Согласно
Беме A926), полные кладки содержат 10—17 яиц, цвет их—как у закавказ-
ского подвида. Полные кладки в Северной Осетии находят обычно в средних
числах мая (Беме, 1926), на Сулаке в конце мая найдено гнездо с яйцами
в последней стадии насиживания (Туров и Красовский, 1933), а у Кизляра
полная кладка (гнездо с 15 свежими яйцами) найдена 4 мая (Беме, 1925).
Длительность насиживания, вероятно, 25 или 26 дней (Мензбир, 1902). В году
одна кладка.
Линька. Общая схема как и у всех фазанов. Точные сроки не
изучены.
Питание. Смешанная растительная и животная пища; более детально
у этого подвида не изучено; осенью и в первую половину зимы поедает ягоды,
в особенности ягоды облепихи, весной—семена и зерна (Россиков, 1884).
Суточный цикл. Как и другие кавказские подвиды, выходит
на кормежку два раза в день—утром и вечером; днем в жаркую погоду отды-
хает и купается в пыли.
Промысел. Как и у других кавказских подвидов. Особенно много
этой птицы добывалось в начале 80-х годов прошлого столетия вскоре после
открытия железнодорожного сообщения Ростов—Владикавказ. В 1882—
1883 гг. было вывезено с Малки, Терека и прилежащих мест (бывш. Нальчик-
ский окр.) свыше 20 000 фазанов; кроме того, большое количество их было
использовано на месте (Россиков, 1884).
Описание. Размеры и строение. Длина крыла самцов A3) 250—267,
самок C) 228—205, среднее 258,5 и 231,3 мм.
Окраска. При большой близости к закавказскому фазану, этот подвид довольно
хорошо отличается от последнего: общая окраска его светлее, больше золотисто-оранже-
вая, а на черных отметинах спины, зоба и боков преобладает зеленый блеск. Самка не-
сколько бледнее самки номинального подвида.
206 ОТРЯД КУРИНЫЕ
Талышский фазан Pftasianus colchicus talischensis Lor.
Phasianus persicus talischensis. L о г е n z. Journ. f. Ornith. 1888, стр. 572, Ленкорань.
Распространение. Ареал. Населяет юго-восточную часть прилегаю-
щей к южной части Каспия низменности, т. е. Талыш, Гилян и, очевидно,
западную часть Мазандерана. Населяет некоторые острова Кизыл-Агач-
ского залива (на о-в Сара завезен в конце XIX в.), к северу доходит до ниж-
него течения Куры.
Характер пребывания. Оседлая птица.
Биотоп. Равнинные леса (главным образом мелколесье), тростниковые
заросли и заросли ежевики; в дубово-грабовых лесах и в лесах предгорий
отсутствует.
Численность. Довольно обыкновенная птица.
Экология. Размножение. Сведений очень мало. Вопрос о моно-
гамности неясен; Тугаринов и Козлова A938) наблюдали в зимнее время
иногда группки птиц, состоящие из одного самца и трех самок. Брачные
крики начинаются с середины марта. Сведений по гнездовому периоду почти
нет. Гнездо, найденное Кузякиным в 1948 г. поблизости от Ленкорани, поме-
щалось в заболоченном лесу на сухой возвышенности около низкорослых
кустов. Поблизости были густые заросли ежевики, а крупные деревья-
(главным образом ольха) нередко были увиты колючей лианой. Гнездовая
ямка была выстлана сухими стеблями трав (главным образом злаков) и сухими
листьями ольхи. 5 мая в гнезде было 4 свежих яйца. Вес трех из них: 24, 24,5
и 27,5 г. По величине, форме и окраске они ничем существенно не отличаются
от яиц других подвидов (во всяком случае, кавказского) фазана. Размер их
C) 43,2—43,5x33,5—34 мм.
Линька. Как и у других подвидов. Смена мелкого пера у старых птиц
наблюдается и в средних числах ноября. В ноябре присходит интенсивная
линька и у молодых (начинается она, вероятно, тотчас после того как молодые
оперятся в гнездовой наряд или, возможно, даже несколько раньше—по
аналогии с таджикским подвидом).
Питание. В зобах и желудках Тугаринов и Козлова A938) находили
зимой ягоды ежевики и небольшое количество других ягод.
Суточный цикл. Как и у других фазанов.
Враги. На о-ве Сара большое значение в ограничении численности фазана
имеют шакалы (Верещагин, 1947).
Полевые признаки. Этот подвид (как и другие кавказские) нередко взле-
тает на деревья. Ночь каждая птица проводит на строго определенном дере-
ве, куда прилетает сразу же после захода солнца. Голос (брачный крик)
более чем у других подвидов напоминает пение молодого петушка и состоит
из трех коротких и одного удлиненного слогов: «ко-ко-ко-коооу» (Спанген-
берг).
Описание. Размеры и строение. В отличие от предыдущих подвидов,
перья зоба в вершинной части сужены и с глубокой выемкой у конца стержня. Длина
крыла самцов B) 237 и 240 (Хартерт, 1917), у самки в коллекции Зоологического
музея Московского государственного университета 22,5 мм.
Окраска. Взрослый самец до известной степени промежуточен между закав-
казским и персидским подвидами, но от последнего хорошо отличается более темной окра-
ской верхней стороны тела и более темными верхними кроющими крыла. От закавказского
отличается более узкими темными каемками перьев зоба. Темный цвет брюха окаймлен
спереди блестящим пурпурно-меднокрасным цветом. Самка несколько темнее самок номи-
нального подвида.
ОБЫКНОВЕННЫЙ ФАЗАН 207
Персидский фазан Phasianus colchicus persicus Sew.
Phasianus persicus. Sewertzow. Bull. Soc. Imp. Nat. Moscou XLVIII, 1874 (в дей-
ствительности 1875), стр. 208, Южное побережье Каспия.
Распространение. Ареал. Западный Копет-Даг и прилежащие райо-
ны Ирана. Раньше доходил к востоку до Ашхабада, окончательно исчез
оттуда около 30 лет назад, теперь встречается еще в Геок-Тепинском районе
(Прохладное). Северная граница распространения более или менее совпадает
с водоразделом Атрека и рек, текущих с Копет-Дага на север в Ахал-Текин-
ские и Атэкские равнины. Западная граница в Туркмении проходит в настоя-
щее время в Сонгу-Даге у нижнего течения Сумбара, на Атреке теперь фазана
нет. Старые сведения (Богданов, 1884), что фазан встречается в низовьях
Атрека, недостоверны (Дементьев, 1952). Довольно обычен фазан у Кара-Кала,
Кейне-Касыра, Терса-кана, Йолдере, вХасар-Даге на Сумбаре и также по
Чандыру. В Иране он населяет прилежащие к Туркмении районы к западу
до Бендер-Шаха и низовий Гюргеня.
Характер пребывания. Более или менее оседлая птица,
но, возможно, совершает на зиму небольшие перекочевки.
Биотоп. Кустарники подгорной полосы и нижнего пояса гор
(Дементьев, 1952), по Зарудному A896)—держится в долинах рек, где эти
долины покрыты Камышевыми плавнями и местами прерваны полянами,
поросшими высокой травой.
Численность. В конце прошлого столетия очень высокая, в настоя-
щее время этот фазан сильно истреблен и местами исчез.
Экология. Размножение. Известно очень мало. Гнездо с 9 сильно
насиженными яйцами найдено было в 1886 г. 19 мая.
Линька. Во второй половине лета. В начален середине сентября можно
видеть как старых, так и молодых птиц, закончивших линьку; однако большая
часть молодых еще продолжает линять. Во время линьки птицы часто встре-
чаются по убранным пашням, среди оставшейся соломы и бурьянов.
Питание. Данных по этому подвиду нет.
Описание. Размеры и строение. Перья зоба—как у талышского фазана,
т. е. с глубокой выемкой. Длина крыла трех самцов 232, 240 и 300, трех самок 220,
225, 230 мм.
Окраска. Занимает как бы промежуточное положение между талышским и мур-
габским подвидами. От первого хорошо отличается верхними кроющими крыла, которые
у персидского фазана белесовато-серые с явственно выраженным буланым оттенком;
на межлопаточной области, зобе, груди и перьях боков преобладает золотистый отлив;
металлически-зеленый цвет и блеск на открытой части перьев передней части надхвостья
почти отсутствует.
Мургабский фазан Phasianus colchicus principalis Sclater
Phasianus principalis. Sclater. Proc. Zool. Soc. London, 1885, стр. 322, Бала-Мургаб
в Северном Афганистане.
Синонимы. Phasianus Komarowii. Bogdanow. Bull. Acad. Sc. Petersb. 1886,
crp. 356, Ашхабад.—Phasianus principalis bogdanowi. В ut urli n, Ibis. 19C4., стр. 390,
новое взамен предыдущих.
Распространение. Ареал. Восточные предгорья Копет-Дага (Каах-
кинский район), долины Теджена и Мургаба. Современные границы распро-
странения на Теджене неясны, во всяком случае фазан сохранился там,
где есть еще тугаи; его много между аулом Ата и Серахсом и далее к югу,
где Теджен течет вдоль нашей государственной границы, к северу доходит
до Карабента, а возможно, и дальше. Западная граница этого подвида
непостоянная. После 1905 г. он был потреблен в Ах ал-Текинском оазисе,
где раньше, судя по данным Варенцова A907), доходил почти до Ашхабада.
208 ОТРЯД КУРИНЫЕ
Однако в последнее время вновь найден у аулов Акча-Теле и Чаача
у иранской границы (Дементьев, 1952). Повидимому, он вновь дохо-
дит теперь на западе до Баба-Дурмаза, а быть может, и до Каахка в Атекском
оазисе. По Мургабу этот подвид населяет тугаи по среднему течению реки
от Тахта-Базара и Таш-Кепри, у Сары-Язы, Имам-Баба, Иолатани вМарый-
ском оазисе (Мары, Байрам-Али), а также низовья Кушки у Кала- и -Мора.
Характер пребывания. Оседлая птица, но в зимнее время
может быть встречена довольно далеко на север в песках.
Биотоп. Травянистые, покрытые кустарниками пространства среди
камышей и невдалеке от хлебных полей; густо заросшие лесом и тамариском
речные долины и прилежащие к ним равнины, покрытые кустарником и вер-
блюжьей колючкой. Зарудный писал об исключительном случае гнездования
фазана около Репетека, т. е. в удаленной от всякого влияния рек пустыне.
Равнинная птица.
Численность. Местами еще значительная, в Марыйском оазисе
очень сильно уменьшилась, в местах, где проводится запрет охоты, вновь воз-
растает. Согласно Шестоперову A936), в 1929 г. численность резко уменьши-
лась, возможно в связи с питанием отравленными приманками против саранчи.
Экология. Размножение. На основании своих наблюдений за этим
подвидом Зарудный A896) считает фазанов полигамными птицами, у которых
самец спаривается с несколькими самками. Судя по другим подвидам, это
не так, хотя случаи, когда самец водит двух, а иногда даже и трех курочек,
достоверно известны. Токование в середине апреля уже в полном разгаре;
Мрачный крик несколько отличается от крика кавказских подвидов; это
двусложное «кох-кох» (Спангенберг); крики можно слышать даже до сере-
дины июля. Токующий самец большей частью располагается вблизи воды,
при этом часто пьет и купается; иногда садится на вершину куста или на невы-
сокое деревцо, иногда на какую-нибудь выдающуюся кочку. Крик фазана
сопровождается шумом крыльев, как при взлете. Заметив появившуюся
курочку, петух преследует ее, и происходит спаривание. В случае появления
другого петуха происходит драка, которую Зарудный A896) характеризует
как остервенелую и рассказывает о случае, когда два петуха были пойманы
во время драки руками.
Гнезда устраиваются на земле, часто под кустиками прошлогодней верб-
люжьей колючки; они представляют собой плоскую ямку, выстланную сухи-
ми стебельками, пухом и перьями. По сообщению Радде и Вальтера A889),
10 августа ими найдено два гнезда с 9 и 5 ненасиженными яйцами. Зарудный
находил гнезда с 9—12 сильно насиженными яйцами с 22 мая по 1 июня,
в то же время он встречал цыплят размером уже с пустынную курочку, в конце
июля в Мервском оазисе встречались цыплята размером с водяную курочку,
а в последнюю треть июля по старому стилю он видел около Серахса молодых,
лишь недавно выклюнувшихся из яиц.
Первые дни после вылупления птенцов выводок держится уединенно,
потом отдельные выводки соединяются вместе и как описывает Зарудный,
бродят под присмотром своего петуха. В таких соединенных стадах может
быть до 50 птенцов (Зарудный, 1896).
Л и н ь к а. С конца июня по начало сентября, общая схема—как у дру-
гих подвидов. В начале августа молодые птицы находятся в состоянии интен-
сивной линьки в наряд взрослых, а старики большей частью кончают смену
мелкого пера, рулевые в это время еще не достигают нормальной длины.
Маховые частично новые, частично отсутствуют и частично старые (Заруд-
ный, 1896). УУ
Питание. По Зарудному A896)—всевозможные семена, хлебные
зерна и различные насекомые, гусеницы и личинки. Часто появляется
на бахчах, где вредит арбузам и дыням.
ОБЫКНОВЕННЫЙ ФАЗАН 209
Описание. Размеры и строение. Длина крыла самцов (9) 238—250,
самок E) 208—225, среднее 244 и 218 мм.
Окраска. Взрослый самец отличается от предыдущего подвида почти чисто
белыми верхними кроющими крыла. Передняя часть спины с преобладанием золотистой
основной окраски, с хорошо развитым чешуйчатым рисунком, благодаря наличию широ-
ких (до 1,5 мм) каемок перьев, которые в межлопаточной области обычно сливаются сере-
динным темным пятном на конце пера. Плечевые также широко окаймлены, на большин-
стве их серединный матовочерный и сливочнобелый рисунок скрыт под другими перьями,
вследствие чего эти участки оперения лишены характерной для кавказских подвидов
пестроты. Белого ошейника нет, но, разбирая перья, иногда можно обнаружить следы
его, совершенно незаметные снаружи. Багряно-красный цвет на зашейке то имеется, то
отсутствует. Багряно-красные каймы перьев середины зоба и груди широки (до 3 мм)
и сравнительно слабо отличаются от прилегающей части пера. На боках груди перья
имеют металлически-фиолетовые или зелено-черные ободки.
Самка бледнее самки номинального подвида.
Аму-Дарьинский фазан Phasianus zarudnyi But.
Phasianus principalis zarudnyi. В u t u r 1 i n. Ibis, 1904, стр. 390, новое имя для Ph.
medius.
Синонимы. Phasianus medius. Зарудный. Орнитолог, фауна Закаспийского
края, 1896, стр. 481, среднее течение Аму-Дарьи. Phasianus tschardjuensis. Бутурлин.
Наша охота, 1908, VII (июль), стр. 10, то же 6 Ibis, 1908, стр. 571, Чарджои.— Phasia-
nus gordius. Алфераки и Бианки, Ежегод. Зоологического музея Академии
наук, XII, 1907 (на самом деле 1908), стр. 440, Карнас.
Распространение. Ареал. Долина Аму-Дарьи от г. Керки на север
до Дарган-Ата. Указания на более северные нахождения (Шейх—Абас-
Вали и Юмалак-тугай) очень сомнительны—в названных местах распростра-
нен уже Ph. chrysomelas (Мекленбурцев, 1941).
Характер пребывания. Оседлая птица.
Биотоп. Заросли кустарников высокой травянистой растительности,
не слишком удаленные от каких-либо водоемов; густая чаща Камышевых
зарослей, иногда совершенно лишенная примеси другой растительности,
иногда с добавлением невысокой осоковой поросли, заросли чингиля, солодки
и др.; тугаи, искусственные насаждения джиды и окраинные участки полей
между культурной полосой и барханами. Довольно близко подходит к киш-
лакам и с окраин заходит внутрь их.
Численность. Довольно высокая, но колеблющаяся. Согласно
подсчетам Мекленбурцева в 1939 г., у Карабекаула, в Дейнау и в Саяте за
2,5—3 часа удавалось отметить по 16—18 фазанов (петухов); непосредственно
у Чарджоу плотность фазаньего населения в 2—2,5 раза меньше. В Кара-
бекаульском районе ежегодно добывается около 25 000 птиц. По данным экспе-
диции Туркменского филиал а Академии наук в августе 1941 г. в пойме Аму-
Дарьи от Чарджоу до Керков было 75 000 птиц (Гизенко, 1946). Временный
запрет охоты вызывает быстрое повышение численности, тогда фазаны встре-
чаются и вблизи крупных городов (Чарджоу) и даже проникают зимой внутрь
их (сады в г. Керки).
Экология. Размножение. Слабо изучено. Массовое начало кладки,
повидимому, в конце апреля и, как указывает Мекленбурцев, происходит,
видимо, в довольно сжатые сроки. Гизенко A946), однако, нашел гнездо
фазана и добыл из него самку с готовым к откладке яйцом в Денаусском районе
в 1941 г. 24 марта. Гнездо было устроено под большим кустом эриантуса
у обочины арыка, его закрывали отогнувшиеся от куста листья, лоток пред-
ставлял собой небольшое углубление в почве, выстланное стеблями и листьями
растений. Яиц в гнезде еще не было. К середине мая кладка, очевидно, уже
заканчивается. 19 мая найдено гнездо с 24 яйцами, 20 мая добыта самка,
имевшая в яйцеводе яйцо в скорлупе и несколько увеличенных желтков
в яичнике, 1 июня самка имела одно вполне сформировавшееся яйцо в яйце-
воде, повидимому, последнее. Запоздалая кладка, видимо вторая, найдена
14 Птицы СССР, т. IV
210 ОТРЯД КУРИНЫЕ
16 июня, состояла из 9 слабо насиженных яиц. Первые птенцы замечены
8 июня, они могли перепархивать не далее как на 2—3 м. 16 июня
встречен выводок хорошо летавших птенцов, величиной с перепела,
18 июня—три выводка, в одном из них птенцы размером с перепела, в других
несколько более крупные.
Начало насиживания приходится на вторую декаду мая.
Линька. Общая схема линьки—как и у других подвидов, детали еще
не изучены.
Питание. В первую половину лета в кормовом режиме самцов пре-
обладает растительная пища, главным образом зерна диких растений, в ча-
стности зерна диких бобовых. Затем следуют остатки зеленых частей расте-
ний и ягоды тута. Попадаются также зерна диких злаков, иногда зернышки
паслена, ячменя, как исключение—фасоль. Животную пищу составляют
муравьи, мелкие жуки (например долгоносики), нимфы цикад и прямокры-
лые, палочники и некоторые другие членистоногие. В желудке у одной самки
преобладала животная пища, также и у птенца, преимущественно насеко-
мые, кроме того паукообразные и некоторое количество растительной пищи
(Мекленбурцев, 1941).
Суточный цикл. Петухи в гнездовое время проводят ночь на земле—
в зарослях солодки, камыша, колючки и др. С рассветом начинают кричать.
Когда становится совсем светло, переходят кормиться на поляны с мелко-
рослой травой, охотно посещают при этом песчаные бугры. В это время
к самцам присоединяются самки, причем у одного петуха можно видеть
до трех самок. В жаркое время куры переходят, видимо, к гнездам, а петухи
к воде. Вечером вновь повторяется выход на кормежку, которая затягивается
почти до сумерек (Мекленбурцев, 1941).
Враги. Шакал, лисица, камышевый кот, домашняя кошка, бродячие
собаки; из птиц—филин, балобан, болотный лунь, коршун. Сколько-нибудь
заметного значения в ограничении численности фазана названные хищники
не имеют (Гизенко, 1946).
Промысел. Диференцированных сведений по подвидам фазана из Турк-
мении нет, но известно, что в 1940—1941 гг. Туркмензаготживсырьем была
заготовлено 9 625 фазанов, что составило, по сведениям Гизенко A946),
7700 кг мясной продукции и почти на 50 000 руб. пера. Помимо сдачи госу-
дарству, охотники используют для себя примерно вдвое большее коли-
чество птиц. Главную роль в заготовках играет аму-дарьинский подвид.
Описание. Размеры и строение. По размерам не отличим от соседних
подвидов. Длина крыла самцов (8) 227—244, в среднем 234 мм, самок 213 и 224 мм.
Окраска. Очень изменчивый подвид. Окраска спины, как правило, ярче и свет-
лее, чем у мургабского фазана, но у отдельных птиц она неотличима, а иногда даже и более
темная. Черные каемки спины иногда совершенно отсутствуют, иногда развиты очень
заметно, но с темным конечным пятном не сливаются; на плечевых эти темные каймы
в большинстве случаев отсутствуют. Белый ошейник развит лучше, чем у мургабского
фазана; он бывает хорошо заметен снаружи в виде двух серпообразных полосок по бокам
шеи, иногда обнаруживается лишь при разбирании перьев и лишь в редких случаях реду-
цирован, как у мургабского. Багряный цвет зашейка у многих экземпляров развит очень
незначительно и в некоторых случаях исчезает совсем. Багряно-красные каймы перьев
зоба и груди в среднем уже, чем у других туркменских подвидов—около 2 мм ширины*
иногда 1,5, иногда до 4 мм. Они резко отличны по цвету от прилегающей части пера,
которая светлая—соломенно-золотистая у летних и желто- или красновато-золотистая у
зимних экземпляров; каймы эти светлее, чем у мургабского подвида. Указанные признаки
сочетаются в различных комбинациях. Некоторые особи имеют широкие багряные каймы
низа, но узкие, почти незаметные темные края перьев спины. Багряный цвет зашейка
встречается у птиц как с хорошо развитой, так и с почти незаметной чешуйчатостью.
Систематические замечания. Описанный Бутурлиным Ph. tschardjuensis есть лишь
резкое индивидуальное уклонение аму-дарьинского фазана, весьма, как сказано выше,
изменчивого. Принять эту форму вслед за Бутурлиным в качестве вида невозможно из-за
большого числа промежуточных особей, в качестве подвида она тоже не может быть при-
ОБЫКНОВЕННЫЙ ФАЗАН 211
знана по причине полного территориального смещения с Ph. с. zarudnyi и отсутствию
какой бы то ни было экологической изоляции.
Дементьев A952) считает возможным признать за Ph. с. zarudnyi гибридное
происхождение.
Таджикистанский фазан Phasianus colchicus bianchii But.
Phasianus chrysomelas bianchii. В u t u г 1 i n. Ibis, 1904, стр. 393, реки Кафирнигая
и Вахш.
Синонимы. Phasianus jabae. Зарудный. «Семья Охотников», 1909, стр. 128, Аму-
Дарья, значительно выше Келифа (вероятно, помесь, синоним условно относится к этому
подвиду).—Phasianus michailowskii. Зарудный. «Семья Охотников», 1909, стр. 129,
Памир.
Распространение. Ареал. Населяет верхнее течение Аму-Дарьи,
будучи распространен в юго-западной части Таджикистана. Подробности тако-
вы: по течению Пянджа он гнездится от Термеза до Чубека, выше Чубека,
возможно, держится в небольших прибрежных тугаях, недоступных ни
с берега, ни со стороны реки. Из долины этой реки он поднимается по ее
правым притокам к северу. По Сурхану распространен до Денау, откуда
область распространения заходит в Гиссарскую долину, достигая Сталина-
бада и Янги-Базара. По Кафирнигану фазан держится и несколько севернее
38-й параллели, по Вахшу несколько севернее Курган-Тюбе до места выхода из
гор этой реки. Обитание на Кизыл-Су и Ях-Су тоже ограничивается долинами
этих рек, причем в их нижнем течении. Как птица низменностей, не прони-
кает сколько-нибудь высоко в горы, и северная граница его ареала очерчи-
вается системой хребтов Гиссара и Каратегина с их отрогами. По Аму-Дарье
вниз от Термеза фазанов, повидимому, нет. За пределами Союза этот фазан
обитает в Северном Афганистане, но границы его распространения здесь
точно не установлены.
Характер пребывания. Строго оседлая птица. Отдельные
особи фазанов, пары, а затем и выводки держатся на некоторых совершенно
определенных участках, не совершая перекочевок.
Биотоп. Тугаи с преобладанием древесной растительности и с раз-
личной густотой насаждений от трудно проходимой чащи до заметно разре-
женных участков с открытыми, занятыми травяной растительностью поля-
нами. Чистые тополевые насаждения, приуроченные к более возвышенным
участкам лолины реки; тугаи с преобладанием злаков; заросли тамириска по
солонцу и тростники (Тугаринов, 1945). Согласно Иванову A940), фазан селит-
ся в тростниковых и древесных тугаях или на заброшенных и заросших бурья-
ном пустырях и залежах. Необходимыми элементами каждого гнездового
участка фазана служат: наличие защитной растительности, причем флорис-
тический состав ее и ярусная ее расчлененность не имеют существенного
значения, кормовые угодья (в данном случае, повидимому, вертикальная
расчлененность и разнообразие видового состава растительности играют
большое значение), наличие водопоя, независимо какого происхождения, и
наличие места, которое может быть названо токовищем; резкое нарушение
защитного или кормового фактора делает биотоп непригодным для заселения
фазанов и заставляет их выселяться оттуда, даже покидая насиженные
кладки (Строганов, 1946).
Численность. В густо населенной долине Сурхана фазан сохра-
нился в небольшом количестве, а в Гиссарской долине представляет большую
редкость, так как истреблен охотниками. По Вахшу, особенно ниже Джили-
куля, а кое-где и выше (Куйбышев) обитает в большом количестве до самого
Пянджа, и местами очень многочислен по Пянджу, но у Чубека уже редок.
Вообще встречаемость фазана находится в прямой зависимости от заселенности
районов (Тугаринов, 1945). У Старой Пристани (в 12 км от Джиликуля)
14*
212 ОТРЯД КУРИНЫЕ
Тугаринов определяет 20 или несколько больше выводков на 1 км2, а число
особей фазана к концу гнездового периода (молодых со старыми)—140—150 шт.
При норме 140 птиц на 1 км2 в долине Вахша должно быть 440 000 птиц,
по правобережью Пянджа и Аму-Дарьи от Пархара до Термеза—450 000 осо-
бей, в долине Кафирнигана несколько более 200 000 шт., по Ях-Су и Кизыл-
Су—300 000 и в долине Сухана 100000 особей. Всего численность фазана
в Таджикистане может быть определена к концу гнездового времени в
1 500 000 голов (Тугаринов, 1945).
Осушка и распашка болот, уничтожение камышей и бесконтрольная
охота приводят к уменьшению численности фазанов (Иванов, 1949); проло-
жение оросительной сети в безводных до того местах способствует увеличе-
нию их численности и приводит к расширению ареала (Строганов, 1946).
Экология. Размножение. По наблюдениям Строганова, а также
Тугаринова и Козловой—моногам, однако в некоторых случаях бывает, что
при одном петухе держатся две курочки и соответственно этому бывает
два выводка. Уже в начале марта самцы перестают держаться обществами
и занимают свои токовые территории, в середине марта весенние тока уже
в полном разгаре. Крик петухов слышен от утренней зари до восхода солнца,
иногда и ночью. Токовый крик всегда двух- или трехсложный, нечто вроде
«ке-ке» или «ке-кере». Тотчас после крика слышно хлопанье крыльев. Токо-
вый крик вместе с хлопаньем повторяется с известными промежутками;
в период особенного оживления, примерно в середине апреля, петушок кри-
чит каждые 5—6 мин., позднее интервалы увеличиваются до 10 минут. Голоса
петухов не вполне одинаковы, и при известном навыке отдельных птиц можно
различать по крику. Кроме того, самцы издают еще своеобразное «гуканье»,
это тихое, очень нежное «гу-гу-гу», повторяемое неопределенное число раз
подряд и прерываемое иногда таким же тихим и протяжным завыванием
«гууу», с повышениями и понижениями тонов. Во время тока фазан непре-
рывно бродит в пределах избранного им участка по более или менее опреде-
ленному маршруту, который в отдельных случаях равен по длине 400—500 м
и совершается с остановками для кормежки в 30—40 мин. В первый период
токов—весь март и до начала апреля—фазаны токуют в одиночестве, самки
в это время держатся отдельно небольшими группками по 3—4 особи. С пер-
вой десятидневки апреля можно видеть уже пары. Самец и самка в такой паре
держатся, по первому взгляду, как бы независимо друг от друга, но как только
удаляются за пределы взаимной видимости, самка начинает откликаться на
каждый токовый крик самца. Копуляция происходит во время токового
маршрута. Козлова в районе своих наблюдений не замечала драк между сам-
цами, хотя токовые маршруты их пересекались, но Строганов A946) пишет,
что между самцами бывают ожесточенные драки, причем дерутся они на манер
петухов, победитель преследует побежденного, потом возвращается на старое
место. Тока продолжаются до начала июня, когда самки уже начинают во-
дить птенцов.
Самка устраивает гнездо на том же участке, гдеона держалась с самцом.
Одним из условий помещения гнезда является наличие укрытого местечка
для его маскировки—густого сплетения стеблей, кучи хвороста, нависших
или упавших стволов дерева и т. п. Иногда гнездо помещается почти открыто.
Строганов описывает гнездо фазана как шарообразную полость, диаметром
около 27 см, с круглым боковым входом, достигавшим в поперечнике 17 см.
Стенки и свод этой полости были весьма плотными и вполне предохраняли
постройку от ветра и дождя. Лоток гнезда имел вид правильной чашеобраз-
ной лунки, глубиной в 6 и диаметром в 20 см. Он был выстлан травянистой
ветошью и перьями, повидимому, самой насиживающей птицы. Размеры лотка
варьировали в пределах— в диаметре 20—23 см, в глубину 5—7 см. Однако
Козлова подобных «закрытых» гнезд не наблюдала. По ее описаниям,
ОБЫКНОВЕННЫЙ ФАЗАН 213-
гнездо сверху обычно хорошо скрывается наклоненными стеблями злаков
или обступающими со всех сторон ветвями кустарника.
Нормально каждая самка имеет одну кладку в лето и лишь в случае
гибели первой может быть вторая—дополнительная. полной кладке от 8
до 14 яиц, по расспросным сведениям бывает до 15 и даже 17. Размеры их
B5L2—46,2x33,7—37 мм (Тугаринов и Козлова, 1945); D2) 42,1—45,3х
Х34—35,1 мм (Строганов, 1946); вес яиц от 25,5 до 29,5 г. Окраска яиц
весьма однотонная. Они очень слабого по интенсивности оливково-бурого
цвета с зеленоватым оттенком, слегка блестящим и без пятен. Форма
их сильно изменчива: одни вытянуты в длину, другие укорочены и вздуты,
некоторые более или менее равномерно эллиптичны на обоих концах, другие
имеют сильно округленное расширение к одному концу и коническое заостре-
ние к другому (Строганов, 1946).
Самка сидит очень крепко, оставляет гнездо для кормежки после восхода
солнца, возвращаясь к гнезду ко времени наступления жары, т. е. в 9—10 час;
между 5 и 8 час. вновь сходит, после захода солнца возвращается и ночь
проводит в гнезде. В первые дни насиживания самка покидает гнездо при
каждом подозрительном шуме, потом сидит все крепче и крепче, в послед-
ние дни подпускает к себе чуть ли не вплотную. Длительность насиживания,
очевидно, 21—23 дня. Во время насиживания птица сильно худеет, теряя
около 40% своего зимнего веса.
Самец в период насиживания держится в пределах того же гнездового
участка и, найдя пищу, нередко призывает к себе голосом самку. Однако он
может в это время привлечь на участок и вторую самку, которая тоже устра-
ивает гнездо. Позднее два разновозрастных выводка могут соединяться
вместе.
Начало откладывания яиц не прослежено; очевидно, в Таджикистане оно
начинается в десятых числах апреля и на откладку яиц самке нужно в сред-
нем 10—12 дней (Тугаринов и Козлова, 1946). Первая полная кладка най-
дена Строгановым 10 мая, последняя (ненасиженная) 27 июня, очевидно,
это уже повторная кладка. Фазанята появляются на свет не ранее 8—10 мая,
а большинство выводится около середины этого месяца, последних пухови-
ков (очевидно, второй кладки) Козлова наблюдала 15 июня, но, судя по дате
нахождения кладок, бывают и более поздние выводки. Выход птенцов из яиц
в одном гнезде происходит примерно в течение суток.
После вывода птенцов семья первое время держится в ближайшем к гне-
зду районе, впоследствии бродит более широко, но все же в пределах одного
и того же гнездового участка. Выводок не разбивается в течение долгого
времени, молодые в полном юношеском наряде с вполне доросшими
крыльями держатся еще с матерью.
Первый день жизни фазанята весят 21,5, 22 и 23 г. На второй день жизни
у них исчезают следы яйцевого зуба, на третий день появляются пеньки
маховых, и уже в это время, по наблюдениям Козловой, птенец может плани-
ровать с полуметровой высоты и взлетать приблизительно на 30 см. В 30-днев-
ном возрасте птенец может взлетать вертикально на высоту до 3 м, а вдоль
пролетает 50—60 шагов. В этом возрасте он весит около 127 г, основания махо-
вых и рулевых у него еще в пеньках. Маховые достигают полного роста
к 50 дням от рождения.
Растянутость гнездового периода у фазана хорошо иллюстрируется раз-
новозрастностью выводков. В сентябре можно встретить птенцов величиной
с перепела и вполне взматеревших птиц, в октябре встречаются еще выводки
с птенцами размером в пол-матки (Строганов, 1946).
Линька. Рост и смена оперения у молодого фазана происходит непре-
рывно, начиная с третьего дня жизни и до пятимесячного возраста. Первыми
у пухового птенца вырастают маховые, позднее всего оперяются шея и голова.
214 ОТРЯД КУРИНЫЕ
На 14-й день жизни появляются перья лопаточных партий и показываются
из пеньков рулевые, в возрасте одного месяца оперяется голова, а в 40 дней
начинается уже новая линька, которая начинается со смены маховых, хотя
полного развития гнездовой наряд достигает к полуторамесячному возрасту,
следовательно, после начала смены маховых на взрослые. Повидимому,
у фазана существует еще какой-то промежуточный, не вполне, однако, выра-
женный наряд, отдельные партии которого появляются у молодой птицы еще
ло того, как вполне развился юношеский гнездовой наряд; перья переход-
ного наряда (отличного от гнездового и от взрослого) достигают наибольшего
развития на нижней стороне тела.
Линька в окончательный наряд начинается, как сказано, со смены пер-
востепенных маховых; смена юношеских рулевых происходит в возрасте
несколько больше двух месяцев. Нижняя и верхняя сторона тела вылинивают
почти одновременно. Позже всего появляются перья взрослого наряда навер-
ху головы, последними развертываются пучки сильно удлиненных зеленых
перьев за глазом, по бокам затылка («уши»).
Дальнейшая ежегодная линька взрослых совершается в иной последо-
вательности: линька взрослых начинается со смены оперения головы и шеи.
У петухов она начинается как только они прекращают токовать—с самого
начала июня в Таджикистане. Смена крупного и мелкого пера происходит
одновременно. Смена маховых начинается одновременно со сменой оперения
головы, несколько позже сменяются перья хвоста. На правом и левом крыле,
также на правой и левой половине хвоста смена отдельных соответствующих
друг другу перьев происходит часто неодновременно. Первостепенные махо-
вые сменяются от внутренних к внешним, второстепенные могут выпадать,
как самые внешние, так и самые внутренние, одновременно. Верхние кроющие
крыла сменяются одновременно с маховыми, среди рулевых последней отра-
стает (но не всегда) средняя пара. Самка начинает линять, когда птенцы дости-
гнут, примерно, величины двух третей взрослой птицы. Линька взрослой
птицы тянется долго—около трех месяцев. Интересно, что во-время линьки
вес тела птицы увеличивается и появляются (у обоих полов) жировые отло-
жения.
Питание. Фазан может быть охарактеризован как всеядная, очень
нетребовательная к составу пищи птица. В пищу входят как различные
животные организмы, так и растения в виде их плодов, семян или зеленых
частей. Избирательной способности фазаны не проявляют, все зависит от
наличия поблизости того или иного рода пищи.
Молодые фазаны, напротив, очень разборчивы в пище; по наблюдениям
в неволе, они предпочитают личинок мелких саранчовых, тлей, муравьев
пауков, от мертвых животных отказываются, насекомых с жесткими покро-
вами начинают есть только с 7-го дня жизни.
Основная масса пищи склевывается птицей с поверхности земли. Зеле-
ные части растений срываются клювом, насекомые с поверхностных слоев
почвы и мокрицы берутся клювом после ее разрыхления клювом и лапами.
Фазаны заглатывают также камешки. По весу их может быть в одном желудке
до 5 г. Ту же роль, что и камешки, играют в желудке фазана косточки джиды.
Кормясь в своем участке, фазан ходит определенными более или менее
постоянными путями, в результате чего в густых зарослях протаптываются
хорошо выраженные тропы.
Суточный цикл. Выход выводка на кормежку после ночного
сна происходит до восхода солнца, когда еще очень прохладно и вся трава
покрыта обильной росой. Около 9 час. утра самка собирает птенцов на днев-
ной отдых в густую траву или под кусты (птенцы не выносят жаркого солнца,
и не могут бегать по раскаленной почве). Вечерняя кормежка выводка начи-
нается около 6 час. вечера и продолжается до 8—8% час.
ОБЫКНОВЕННЫЙ ФАЗАН 215
Примерно таков же и суточный цикл взрослых птиц. Утром и вечером—
жировка, днем—купанье в пыли или в песке, либо отдых где-либо у воды.
Зимняя жизнь. В зимнее время птицы нередко выходят за пре-
делы своих участков, собираются (в январе) группами, предпочитая держать-
ся по более открытым местам—опушкам леса, полям и даже по солончакам
с кустами; в таких группах бывают или одни самцы или самцы с самками
<Иванов, 1945).
Враги. Гибель взрослых птиц от четвероногих хищников явно незначи-
тельна вряд ли какое-нибудь значение имеют и хищные птицы. Кладки
•фазанов уничтожаются шакалами и котом, болотным лунем, вараном. Моло-
дых поедают уже названные четвероногие хищники, кроме того, гюрза и
полоз. Тугаринов и Козлова полагают A945), что в среднем в каждом
выводке погибает около половины птенцов.
Промысел. При правильно поставленном промысле этот фазан, оче-
видно, может иметь большое промысловое значение, так как общее количе-
ство живого веса птицы составляет в Таджикистане, по подсчетам Тугари-
нова, около 1800 т. Однако промысел не регулируется в достаточной сте-
пени, вследствие чего в Таджикистане широко практикуется один из самых
истребительных приемов охоты на фазана—массовая облава во время выпа-
дения снега. Охотники, выстроившись в ряд проезжают по зарослям и ударами
палок о кусты выпугивают птиц. Уже при незначительной толщине рыхлого
снегового покрова фазан бежать не может, а пролететь может около 200 м и
последвух-трех взлетов,пролетая каждый разоколо 150—200 лс, садится потом
в снег, где охотники бьют его палкой. При таком способе охоты в многоснеж-
ные зимы гибнут тысячи фазанов, причем самцов добывается значительно
больше, чем самок. Быть может этим обстоятельством следует объяснить,
что в некоторых местах летом самок значительно больше, чем самцов, и неко-
торые из них остаются холостыми (Строганов, 1946).
Полевые признаки. Внешний облик, как и у других подвидов. Молодые
птицы, будучи вспугнутыми, взлетают на деревья, но старые, как правило,
«а деревья не садятся.
Описание. Размеры и строение. Длина крыла самцов B0) 240—260,
в среднем 249,7 мм, самок D) 219 лле.Вес самцов зимой D) 956—1300, в среднем 1106,5г,
самок B) 710 и 850 г (Иванов, 1945).
Окраска. Пуховой птенец (по описанию Козловой) имеет сверху бледножелто-
ватую окраску. От подклювья до затылка через лоб проходит темнобурая полоска,
расширяющаяся на темени в виде треугольного пятна. По сторонам темени, над глазами—
такие же узкие темные полосы. На щеках под глазом до уха намечается очень узкая темная
черточка, в области уха заканчивающаяся черным пятнышком. От затылка вдоль спины
до поясницы идет буровато-черная полоса; по краям спины—из-за крыла до надхвостья—
идут более узкие темные полосы. На лопатках два рыжевато-бурых пятна. Брюшная
сторона тела однообразно желтовато-белая. На пяточном сочленении сзади—темно-
бурые пятна. Радужина темнобурая, клюв светлороговой, по хребту надклювья темный.
Ноги бледнотелесной окраски.
Гнездовой наряд. Верх головы и зашеек серовато-палевые с черноватым
мелким крапом. Уши грязнопалевые, подбородок и горло беловатые. Бока головы вокруг
глаза в очень мелких беловатых перышках. Под глазом овальное почти белое пятно. Шея
•сзади черноватая с мелкими белыми пятнами. Лопаточные и плечевые буровато-черные
с бледнопалевыми центральными штрихами и широкими охристыми каймами. Спинная
птерилия дымчато-серая, оперение ее рыхлое. Зоб палевый с размытыми округлыми более
темными буроватыми пятнышками. Брюшко посредине беловатое; бока тела палевые,
с буровато-черными пятнами. Крылья—как у взрослой самки. Рулевые палевые с темно-
бурым продолговатым пятном в основной части пера и с крошечным пятнышком перед
его вершиной. Ноги серые, клюв бледнороговой, радужина зеленовато-серая. Перья
зоба и груди у взрослого самца с блестящими черными краями, которые почти
сплошь закрывают темнозолотой цвет на груди и особенно на зобе, придавая этим
частям тела черно-зеленый цвет. Черные концы имеют и перья боков туловища. Спин-
ная сторона тела сходна с аму-дарьинским фазаном, но несколько темнее (примерно, как
более темные аму-дарьинские, описанные Зарудным в качестве особой формы—гордиев
фазан Ph. с. gordius).
216 ОТРЯД КУРИНЫЕ
Хивинский фазан Phasianus colchicus chrysomelas Sew.
Phasianus chrysomelas. Sewertzow. Bull. Soc. I. Nat. Mosc. XLVIII, 1874 в (дей-
ствительности 1875), стр. 207, низовья Аму-Дарьи.
Синоним. Phasianus oxianus и Phasianus Dorandti Sewertzow. Journ. f. Ornth
1875, стр. 225 (автор колебался между несколькими именами и остановился на
Ph. chrysomelas).
Распространение. Ареал. Низовья Аму-Дарьи, вниз примерно от
Дарганата. Область распространения идет сначала довольно узкой полосой
вдоль долины реки, расширяясь потом на весь Хивинский оазис и на дельту
реки. К северо-востоку распространен до границ влияния вод Аму-Дарьи
на пустыню и до юго-восточного угла Аральского моря, где держится как
на островах, так и по берегу моря. Нами в Зоологический музеи Московского
государственного университета доставлен один экземпляр (взрослый самец
от 6 октября 1935 г.) из местности между Акпектами и Бару-Чайнаганом,
Зарудный A915) указывал его для о-вов Аталык, Дамбалы, Силява и др.
На северо-западе граница распространения почти упирается а южный чинк
Усть-Урта (Судочье озеро; Гладков, 1935); на востоке найден Молчановым
A913) в северо-восточных частях Айбугирской впадины; встречается и в
окрестностях Ташауза.
Характер пребывания. Оседлая птица.
Биотоп. Тугаи, тростниковые заросли, заросли гребенчука, проникает
и в сухую степь, поросшую зарослями кустарника (Никольский, 1892).
Численность. Во многих местах обычная птица, но численность „
как и у других подвидов, подвержена значительным колебаниям.
Экология. Размножение. Изучено очень слабо. Во второй поло-
вине июля молодые хорошо летают, самые маленькие из них не больше, чем
дрозд-деряба, самые крупные в это время не менее двух третей роста взрос-
лой самки.
Линька. Схема линьки, очевидно, как и у предыдущего подвида.
Самцы, добытые во второй половине июля, находятся в состоянии интенсив-
ной линьки. Молодые петухи (наиболее выросшие) имеют в это время уже
некоторые перья взрослого наряда.
Питание. Сведений, относящихся специально к этому подвиду, нет.
Описание Размеры и строение. Длина крыла двух самцов 235 и 250,
самки 228 мм. По Хартерту A917), длина крыла самцов 236—247 мм.
Окраска. Взрослый самец очень сходен с таджикским, но хорошо отли-
чается от него тем, что черный или черно-зеленый цвет нижней стороны тела слабее развит,
вследствие чего меднокрасный цвет на груди преобладает над черным, тогда как у таджик-
ского этот цвет только проглядывает среди черного и имеет притом не меднокрасный,
а скорее темнозолотой оттенок. Верхняя сторона тела у хивинского фазана значительно
темнее, чем у таджикского, не желтовато-оранжевая, а бронзово-красная. Кроме того,
у хивинского фазана зеленый цвет на задней стороне спины очень слабо развит,
практически отсутствует.
Зеравшанский фазан Phasianus colchicus zerafschanicus Tarn.
Phasianus principalis zerafschanicus. fTarnovski. The Field, XXVII, 1891, стр. 409,
Катта- Курган.
Синонимы. Phasianus zerafschanicus sive Klossowskii. Tarnovski. The Field,
XXVII, 1891, стр. 409, Катта-Курган. —Phasianus tarnowskii. S e e b о h m. Proc. Zool.
Вое. о! London, 1892, стр. 271, Зеравшан.
Распространение. Ар е а л. Долины рек Зеравшана и Кашка-Дарьи
(есть сведения, что фазаны расселились по Зеравшану совсем недавно—
в 80-х годах прошлого столетия; Мекленбурцев, 1940). Самое юго-западное
нахождение указано Лоудоном A910) в окрестностях ст. Фараб, т. е. уже
в Аму-Дарьинской равнине, где должен быть другой подвид (Ph. с. zarudnyi);
названное местонахождение требует подтверждения, но в окрестностях Ходжа-
ОБЫКНОВЕННЫЙ ФАЗАН 217
Девлета и у озер Сунгур и Каранга безусловно водится уже зеравшанский
фазан (Даль, 1936). К востоку от названных мест распространен вверх по-
лолине Зеравшана до кишлака Педжикент (т. е. до выхода реки из гор).
В стороне от долины реки этот фазан известен у кишлака Дюрт-Куль и у
кишлака Шафрикан (Мекленбурцев, 1940). В долине Кашка-Дарьи он рас-
пространен к западу до Карши, а вверх по реке поднимается до самых гор
и даже несколько заходит в ущелья рек (был отмечен на Джаны-Дарье
и у места выхода из гор Чияль-Дарьи, Мекленбурцев, 1940).
Характер пребывания. Оседлая, в высшей степени привя-
занная к своей территории птица.
Биотоп. Густые камышевые заросли, то более сухие, то сильно увла-
жненные; прибрежные тугаи, образованные главным образом джидой и
в меньшей степени ивами и другими деревьями с густым подседом из солодки
и других высоких трав, в меньшей степени сады и рощи, в которые назван-
ные тугаи почти незаметно переходят; густые заросли трав, перемежающиеся
с тамарисковыми кустарниками на границе речной поймы и пустыни. Зача-
стую поселяется и вдали от древесной растительности, среди полей пшеницы
и хлопка, особенно если вблизи есть участки, заросшие густым камышом.
Наличие воды в виде реки, озера, болота и т. п. обязательно при всех
условиях.
Численность. По мнению Мекленбурцева A940), количество фаза-
нов даже в наиболее густо заселенных ими местах очень далеко от насыщения.
Наиболее бедно населены участки Кашка-Дарии между кишлаками Каучин
и Чиракчи, также мало заселена и область, лежащая на правом протоке
Зеравшана в районе кишлака Кара-Дарья. Значительно выше плотность
населения фазанов у Шахризябса, в садах его окрестностей и на полях у неко-
торых ближних кишлаков. Наиболее богато фазанами место у впадения
Даргома в Зеравшан.
Экология. Размножение. Откладывание яиц и выведение птенцов
происходит, позидимому, в довольно сжатые сроки, но бывают и запоздалые
выводки, очевидно, повторные, так как 6 мая отмечены совсем маленькие птен-
цы—меньше перепела. Обычно же птенцы в конце июля и начале августа
имеют размеры от горной куропатки до серой куропатки.
Число молодых в одном выводке очень непостоянно. Чаще всего встре-
чаются 10—15 фазанят при одной курице, но бывает и больше. Иногда встре-
чается только 3—4 птицы в выводке (возможно, часть выводка погибла).
Каждый выводок держится на сравнительно небольшой строго определен-
ной площади, которая должна включать в себя достаточно густую и высокую
растительность, чтобы птенцы могли скрываться; в ней должны находиться
площадки с невысокой травой, где они могли бы кормиться; также должен быть
участок густого и высокого камыша или кустарника, служащего местом ноч-
лега, и, наконец, вблизи должна быть питьевая вода. Размер такого участка
не превышает 2 га, но с подрастанием птенцов площадь, на которой они дер-
жатся, может увеличиваться.
Л и н ь к а. Не изучена. Молодые начинают перелинивать из гнездового
наряда во взрослый с начала августа или несколько раньше; к десятым чис-
лам этого месяца можно встретить самцов, у которых спина, грудь и зоб по-
крыты золотистыми перьями, а длина хвоста до 15—20 см. Взрослые самцы
линяют тоже с начала августа. Общая схема линьки, очевидно, как и у дру-
гих подвидов.
Питание. Животная и растительная пища. Животная пища пред-
ставлена в подавляющем большинстве случаев насекомыми, главным образом
прямокрылыми (видовой состав поедаемых фазаном саранчовых довольно
разнообразен). Растительная пища состоит почти исключительно из семян
(преобладают Centaurea, затем следуют культурные растения и некоторые
218 ОТРЯД КУРИНЫЕ
другие), гораздо реже попадаются ягоды и в виде исключения—клубни
и листья. На хлопковых полях фазан из-за истребления им саранчи должен
быть признан одной из полезнейших птиц; на зерновых полях, возможно,
приносит некоторый вред, но надо учесть, что он высоких колосьев не трогает
и подбирает семена с земли (возможно, некоторое исключение составляет
низкорослый маш).
Маленькие птенцы едят небольших жуков, более взрослые саранчу и куз-
нечиков, зерна встречаются в зобах и желудках взрослых птиц (Мекленбур-
цев, 1940).
Суточный цикл. Петухи просыпаются по утрам, видимо, раньше,
чем самки с птенцами, так как крик слышится еще до рассвета, но на кормежку
петух выходит, когда становится уже совершенно светло, выводки выходят,
видимо, в это же время. Покормившись около 2—3 час, птицы идут на водо-
пой к ближайшему водоему, выбирая места, где к воде можно подойти неза-
метно. Пьют фазаны много. На местах водопоя не задерживаются и быстро
убегают в места дневного отдыха. Отдых длится приблизительно от 11 до 3 час.
Для отдыха выбираются сухие места у густых кустов или камышей, здесь
птицы любят купаться в пыли, выбивая, подобно курам, небольшие ямки.
На вечернюю кормежку выходят как только спадает дневной жар, вечером
воду не пьют, а прямо отправляются на ночлег. Ночевать предпочитают
в густом камыше, а если его поблизости нет, петухи, видимо, забираются
на деревья, а выводки прячутся в густой кустарник, совершенно выросшие
молодые тоже иногда садятся на ветви. Иногда ночуют в камышах, затоплен-
ных водой, захватывая ногами по нескольку стеблей (Мекленбурцев, 1940).
Описание. Размеры и строение. Сведений в литературе нет.
Окраска. Спинная сторона тела взрослого самца почти так же изменчива,
как и у аму-дарьинского фазана, но попадаются особи более яркие и светлые. Черные
вершинные каемки перьев передней части спины отсутствуют или плохо развиты. Белый
ошейник развит сильнее, чем у предыдущих форм, достигая у некоторых особей ширины
0,5 см. Пурпурные (или багряно-красные) полосы узки—около 11*/2 мм ширины, горло
обыкновенно пурпурно-красное, поясница кирпично-рыжая. За исключением указанных
отличий, сходен с аму-дарьинским.
Сыр-Дарьинский фазан Phasianus colchicus turcestanicus Lorenz
Phasianus mongolicus turcestanicus. Lorenz. Orn. Monatsb., 1896, стр. 189, Сыр-Дарья.
Синонимы. Phasianus mongolicus triznae. Зарудный. Семья Охотников, 1909,
стр. 352. Сев. часть Наманганского у. —Phasianus kvaskovskii. Зарудный. там же
1909, стр. 353, окр. Гульчи.— Phasianus mongolicus bergii. Зарудный. Орнитол.
Вестн. 1914, стр. 277, о-ва Аральского моря.
Распространение. Ареал. Долина Сыр-Дарьи и восточные берега (острова)
Аральского моря. Согласно Эверсману, фазан водился в камышовых полях
к северу от Аральского моря, на самом деле на северном берегу Аральского
моря фазана нет и указание Эверсмана, быть может, следует отнести к Камыш-
лы-Башским озерам. На восточном берегу он, вероятно, встречается еще
между дельтой реки и местностью Кара-Бура, а также и на о-ве Аталык
{Спангенберг, 1936); вероятно, выводит в небольшом количестве по камышам
юго-западного берега озера Камыш л а-Баш; нами гнездо этой птицы найдено
в 1935 г. в дельте реки недалеко от моря. Сведения о нахождении фазана
в устьях Куван-Дарьи ни нашими исследованиями, ни исследованиями Спан-
генберга не подтверждаются. Во время работ Зарудного фазаны были на Узун-
Каире. Южнее они найдены были Бергом на Уялах, но Зарудный их там
уже не нашел. На островах юго-восточной части Аральского моря фазаны
•есть, но относятся уже к хивинской форме (chrysomelas).
По Сыр-Дарье полоса, населенная фазанами, тянется по обоим берегам
реки (в долине р. Арыс—этот же подвид; Шевченко, 1948), не заходя особенно
ОБЫКНОВЕННЫЙ ФАЗАН 219
далеко в прилегающие пустыни; фазан из низовий Сары-Су (очень редок там,
быть может, теперь отсутствует) относится, вероятно, к семиреченской форме,
хотя материалом для решения этого вопроса мы не располагаем. Остается неяс-
ным вопрос о восточной границе этого подвида. В настоящее время он водится
до самых западных границ Ферганской долины (в самой Ферганской долине
он почти истреблен), потом вновь появляется восточнее уже в горах, окай-
мляющих Ферганскую долину с востока. Подвидовая принадлежность этого
горного фазана остается невыясненной. Бутурлин в 1905 г. относил фазанов
с этой части Тянь-Шаня к сыр-дарьинской форме, а не к семиреченской,
позднее Зарудный описал оттуда новый подвид (triznae), реальность суще-
ствования которого, видимо, не подтверждается. Мы принуждены относить
фазана из Тянь-Шаня к югу от Терскей-Ала-Тау условно к сыр-дарьинскому
(хотя есть разница и в биотопе: равнинная форма и горная). Распространение
фазана здесь в настоящее время рисуется следующим образом. В Джалала-
бадской области он распространен спорадически по всему ее протяжению, в
частности юго-восточным склонам Чаткальского хребта, в Арсланбобе, в Джа-
лалабадском районе и т. д.) есть и в бассейне Кара-Дарьи. Водится он и в
Ошской обл., в 1927 г. был известен еще в Гульче, раньше был (этот ли под-
вид?) и по р. Кзыл-Су в Алайской долине, теперь отсутствует, нет его и в Ош-
ском районе (Дементьев, 1943).
Совершенно особняком стоит указание Сушкина A908), что около 1880 г.
один фазан был убит под Иргизом. Слова Бутурлина A905), что фазан про-
никает в пределы Тургайской обл., следует, вероятно, относить к низовьям
Сары-Су, т. е. уже к семиреченскому подвиду.
Характер пребывания. В долинах оседлая птица, в горах
совершает вертикальные миграции, спускаясь на зиму в больших количествах
в долины.
Биотоп. В долине—камышевые заросли, кустарники, тугаи. Охотнее
всего держится в участках, изобилующих озерами и протоками, где характер
местности всего разнообразнее; сплошные тугаи заселяются в меньшей сте-
пени. В горах—прибрежные заросли и кустарниковые заросли орехового
леса. Распространен от самых низких мест до высоты в перевале Гульча
C416 м. н. у. м.) и озера Сыры-Чилек.
Численность. Довольно сильно зависит от преследования человеком.
Большое влияние на численность оказывают условия погоды, сильные
паводки, а также пожары. В ряде мест (некоторые острова Аральского моря,
местами в Фергане и в горах Тянь-Шаня) истреблен, но местами (по Сыр-
Дарье между Джусалами и Тюмень-арыком, в горах у озера Сыры-Чилек;
Спангенберг) и сейчас водится в большом количестве.
Экология. Размножение. Размножение начинается в последней
трети апреля (сведения относятся к равнинной части ареала). Гнездо устра-
ивается или среди мелкого кустарника или среди густой травы и в таких
местах, которые редко затопляются во время летних разливов реки. Лоток
гнезда бывает до 16 см в поперечнике, но в некоторых случаях настолько мал,
что кладка с трудом умещается в гнезде, и яйца располагаются в нем, обра-
зуя как бы перевернутый конус (средние ниже остальных). Гнездо выстлано
чежными стебельками трав, а к концу насиживания в нем имеются и перья
птицы. Количество яиц в кладке в большинстве случаев 8—10, реже встре-
чаются кладки до 16 яиц; Спангенберг A936) указывает случай нахождения
гнезда с 22 яйцами и предполагает, что в одно гнездо неслись две птицы.
По форме и окраске яйца очень сходны с яйцами серой куропатки. Раз-
мер яиц F8) 41,7—48,2x34—37 мм (Спангенберг, 1936).
Сроки кладки яиц разнообразны. Первые яйца найдены (по 1—2 в кладке)
в первых числах мая, но и в середине июня нередки кладки, содержащие
совсем свежие яйца, а в середине июля—яйца, очень сильно насиженные.
220 ОТРЯД КУРИНЫЕ
Первое появление птенцов отмечено уже в конце мая, в середине июня вывод-
ки могут уже свободно летать.
Линька. Общая схема—как у других подвидов. Мелкое оперение взрос-
лых начинает сменяться в начале июля.
П и т а н и е. В общем аналогично питанию других подвидов. Зарудный
A915) находил зимой в зобах птиц из окраины Кызыл-Кумов плоды саксаула
и джузгуна. По наблюдениям Спангенберга, осенью поедает семена дивной
колючки, а в ноябре полностью переключается на ягоды лоха, которые скле-
вывает, сидя на дереве; в теплое время года ест много насекомых и особенно-
саранчи; в горах Ферганы в большом количестве поедает ягоды кашгарской
боярки. Произведенный Кузьминой анализ содержимого осенних и зимних
желудков фазана с Сыр-Дарьи показал, что в сентябре и октябре в пище фаза-
нов играют большую роль насекомые, особенно саранча, позднее (уже в октяб-
ре) растительная пища приобретает все большее значение, причем кроме веге-
тативных частей разных растений поедается рис, служащий в это время пре-
обладающей пищей, в ноябре большое значение приобретают ягоды джиды
(Грачев и Кузьмина, 1947).
Полевые признаки. Крик (брачный) двухсложное «кох-кох». Ночует,
повидимому, только на земле.
Описание. Размеры и строение. Длина крыла самцов B4) 240—263„
самок A0) 220—240,в среднем 253 и 226,8 мм. Вес самцов D) 1170—1477, самок E) 740—
1018, в среднем 1353 и 838 г (сентябрь).
Окраска. Взрослые птицы очень сходны с семиречснским подвидом. От более
южных подвидов хорошо отличается преобладанием меднокрасного тона в передней
части спины и наличием хорошо развитого белого ошейника, который в передней части
может быть прерван на незначительном протяжении.
Семиреченский фазан Phasianus colchicus mongolicus Brandt
Phasianus mongolicus. Brandt. Bull. Phys. Nat. Acad. Sc. Petersb., Ill, 1844,
стр. 61, Область Алтая и Монголия. (Типичное место указано Брандтом непра-
вильно. По мнению Сушкина A938), типами для описания послужили птицы, купленные
на рынке в Семипалатинске, куда фазана нередко привозят с Ала-Куля и впадающей
в него р. Эмиль.)
Синоним. Phasianus brandti. Rothschild. Bull. Orn. Club. XII, 1901, стр. ?0
(новое имя взамен предыдущего).
Русское название. В научной литературе этого фазана называют нередко в со-
ответствии с его латинским именем монгольским, что совершенно неверно, так как в Мон-
голии этот подвид не встречается.
Распространение. Ареал. Юго-восточные части Казахстана к востоку
от Киргизского хребта и, быть может, до Алакуля (Семиречье). Сведения
о нахождении фазана на Зайсане и Кара-Иртыше, а тем более на Алтае
неверны (Сушкин, 1938). Более детально распространение фазана этого под-
вида таково. Он населяет р. Чу по всему ее протяжению вверх до Токмака
и поднимается в горы по ущельям притоков этой реки (Большой и Малый
Кебин, как редкость Шамси и Кегеты; Дементьев, 1943), хотя в настоя-
щее время выше Токмака в долине этой реки встречается как редкость; в на-
чале 30-х годов был обыкновенен около Фрунзе (Дементьев); доходит до Коч-
корского района, а в Иссык-Кульской обл. распространен по берегам озера
Иссык-Куль и поднимается по долинам горных рек (Тюп, Джергалан и др.),
где только есть заросли камыша и кустарниковые (преимущественно обле-
пихи) заросли. Есть фазан и на Джумгале и некоторых его притоках, на
р. Нарын (возможно, завезен туда)—приблизительно до устья р. Атбаш.
К северо-востоку есть по реке Или от Балхаша вплоть до китайской границы
или дальше до Кульджи; по Талкару, Тургени и другим рекам проникает
в горные леса Заилийского Ала-Тау; есть по всему южному побережью Бал-
ОБЫКНОВЕННЫЙ ФАЗАН 221
хаша и по низовьям впадающих в него рек, но не поднимаясь (за исключе-
нием, как указано, Или) по ним до бывшего почтового тракта (Шнитников,
1949). Самое восточное нахождение фазана в Семиречье—р. Лепса (не выше
с. Романовского). По свидетельству Шнитникова A949), в Алакульской рав-
нине фазана нет, хотя Сушкин, ссылаясь на старые сведения от Тихонова,
говорит о гнездовании этой птицы на Алакуле (р. Эмиль).
Характер пребывания. Не вполне оседлая птица. Во вся-
ком случае, в начале зимы спускаются с северного подножья гор в равнину,
уходя от глубокого снега (Зверев); зимние миграции отмечены и в низовьях
р. Или (Штегман, 1946). На ней кочующие стаи начинают появляться при
первых морозах, задолго до появления снега; исчезают стаи (незаметно рас-
сасываясь) с окончательным потеплением, которое наступает уже после
вскрытия рек. Кочующие фазаны держатся на р. Или стаями, но, кроме того,
там можно встретить и одиночных птиц, явно местных (Штегман).
Биотоп. Камыши и тугайные заросли рек; кустарниковые заросли
по склонам хребтов в зоне лиственного леса; близость воды обязательна.
Фазан населяет также густые заросли степной растительности («курайные»
заросли), а также вблизи посевов, не избегая близости человека.
Численность. Подвержена резким колебаниям. Быстрое умень-
шение количества происходит обычно в результате резкого ухудшения усло-
вий существования. Чаще всего падение численности вызывается массовой
гибелью приплода—кладок и маленьких птенцов—вследствие несвоевремен-
ного и чрезвычайно высокого подъема воды в реках низменностей; большое
значение имеют также суровые многоснежные зимы (например, в Северном
Семиречье зимы 1929—30 и 1930—31 гг.; Шнитников, 1949). Местами чис-
ленность фазана очень высокая. При регулировании промысла и охоты чис-
ленность фазана можно держать на высоком уровне. Наблюдается некоторая
тенденция к расселению фазана в горы, причем птица расселяется по реч-
ным долинам там, где по берегам рек имеются кустарниковые заросли, наблю-
даются также и периодические появления фазана в горах, не приводящие
к расширению ареала (Дементьев, 1943).
Экология. Размножение. Моногам (Дементьев, 1943). Брачный
период начинается рано. Хриплый двусложный крик можно слышать на
р. Или с середины февраля, а во второй половине этого же месяца и в на-
чале марта в яйцеводах самок начинают попадаться яйца, готовые к сносу
{Долгушин).
Гнездо фазана (по наблюдениям Дементьева в Киргизии) устраивается
в зарослях облепихи или в зарослях травянистых растений на краю обле-
пиховых зарослей, обычно по краю самих зарослей в густом кусте, иногда
среди кустов экстрагоновой полыни. Оно представляет собой ямку в земле,
слегка выложенную травой с добавлением небольшого числа перьев. Коли-
чество яиц в кладке сильно колеблется, их может быть от 7 до 14, а по рас-
спросным сведениям и больше, даже до 20.
Размер яиц E) 45—47,5x36—36,8 мм, среднее 45,7x36,2 мм (Иоган-
сен, 1906).
В районе озера Иссык-Куль фазанки начинают нестись в первой поло-
вине апреля, в период с 5 по 10 мая большинство птиц заканчивает кладку,
я с 1 по 5 июня происходит вылупление цыплят. Бывают и более ранние клад-
ки (так, в первых числах июня добыты птенцы не менее 6-недельндго воз-
раста) и более поздние (отмечено выведение цыплят 25 июня, а 20 июля най-
дено гнездо с малонасиженными яйцами). Повидимому, поздние кладки
являются вторыми—дополнительными. На р. Или ближе к Джаркенту
20 июня отмечены выводки ростом с перепелку (Шестоперов, 1926), в низовьях
Или в последней декаде июня молодые были в половину величины взрослых
Долгушин).
222 ОТРЯД КУРИНЫЕ
Птенцы выклевываются из яиц очень развитыми; едва обсохнув, начи-
нают быстро бегать. Рост их вначале идет очень быстро. Первые перья (ма-
ховые, кроющие крыла, рулевые и перья на боках груди), по Дементь-
еву, появляются на 8—10-й день жизни (мы думаем, что маховые перья начи-
нают расти несколько раньше).
В недельном возрасте птенцы весят около 20 г, а двухмесячные самцы—
от 320 до 390 г. В одном выводке могут быть очень различные по величине
цыплята, что, вероятно, связано с растянутостью кладки, но чаще в выводке
бывают очень сходные по размерам птенцы.
Первоначально выводки держатся отдельно друг от друга в пределах
своих гнездовых участков, позднее (примерно с первых чисел августа) про-
исходит скопление их.
В возрасте одного месяца цыплята имеют пух только на голове, к 40—
45 дням пуха уже не остается, а еще через две недели появляются половые
различия в окраске.
Линька. Общая схема—как и у других подвидов. Самцы начинают
линять вскоре после того как самки садятся на яйца. Самцы, добытые в пер-
вой десятидневке июля, находятся уже в состоянии интенсивной линьки:
сменяется контурное перо на всех частях тела, причем линька на голове
заканчивается, рулевые отрастают новые, идет смена маховых. Самки
линяют позднее, когда птенцы уже несколько подрастут. У добытых в июне
самок признаков линьки не обнаруживается, только в самом конце месяца
появляются немногочисленные новые перья на голове и на шее; у некоторых
начинается смена маховых и крайних пар рулевых. У большинства самок
линька начинается с середины июля, интенсивная линька в конце июля—
начале августа. Маховые сменяются начиная с десятого; после того как
сменилось пятое, начинают сменяться и внутренние маховые, начиная с
одиннадцатого.
Линька у взрослых птиц заканчивается в начале—середине октября,
у молодых птиц затягивается до середины ноября.
Питание. Пища как растительная, так и животная. Особенно охот-
но поедают ягоды—облепиху, эфедру, барбарис и др., едят как семена, так
и вегетативные части растений, в частности охотно поедают листья солодки.
Животная пища также разнообразна—фазаны ловят различных насекомых,
пауков, очень часто едят моллюсков. По словам Дементьева A943),
растительную пищу едят и маленькие цыплята. Пища собирается большей
частью на земле, но за ягодами птицы (также и небольшие цыплята) взлетают
и на деревья.
Суточный цикл. Регулярно два раза в день выходят на кормеж-
ку: утром около 7—8 час, когда сойдет роса, но в 11—12 час, по наступлении
сильной жары, прячутся в более густую растительность и отдыхают; новая
кормежка начинается около 16—17 часов и продолжается до наступления
темноты. Осенью могут оставаться на ночь и на местах кормежек, довольно
далеко от зарослей облепихи или камыша.
Выводки проводят с матерью большую часть суток в зарослях облепихи,
откуда выходят утром на кормежку на край зарослей, а по мере подрастания
птенцов—все дальше от них.
Зимняя жизнь. Известна очень мало. Известно, что к зиме
птицы сильно жиреют, слой подкожного жира достигает 1, иногда 1,5 см
толщины, имеется и внутренний жир. Сердце хорошо упитанного фазана
весит почти вдвое больше, чем сердце летнего худого. Однако с наступлением
зимних холодов (в низовьях Или около 30°) жировые отложения быстро исче-
зают. Повидимому, фазан очень боится холода,—прячется в самых густых,
защищенных от ветра местах, выходит на кормежку после восхода солнца
и возвращается до его захода; вполне вероятно, что он не успевает собрать
ОКЫКНОВЕННЫЙ ФАЗАН 223
за это время необходимого количества корма (Штегман, 1945). По среднему
течению Или фазаны держатся стадами на рисовых полях.
Враги. Врагов у фазана сравнительно немного. Шнитников на первое
место среди них ставит камышевую кошку, но она имеется не по всему аре-
алу птицы; по Дементьеву, на птенцов, а быть может, и на взрослых птиц
нападает ласка; для гнезд представляет опасность еж; цыплят нередко ловят
бродячие собаки; для молодых фазанов может быть опасен также камышевый
лунь, известную опасность представляют вороны и сороки.
Описание. Размеры и строение. Длина крыла самцов A8) 250—273,
среднее 260 мм.
Вес самцов из окр. Алма-Аты, по Звереву, не превышает 1400 г, среднее 1100 г.
самок—не больше 1000 г, среднее около 800 г; в низовьях Или упитанные самцы весят
от 1200 до 1720 г, доходя до 2 кг, а худые 1000—1400 г; упитанные самки 900—1350 г,
худые—750—975 г (Штегман, 1945).
Окраска. Пуховой птенец (по Дементьеву, 1943) окрашен очень пестро.
Основной цвет пуха желтый, рыжеватый на верхней и беловатый на брюшной стороне
тела. На боках груди пух слегка сероватый от просвечивающих здесь темных основных
частей его. От лба (но не от самой границы оперения) посреди головы тянется темная по-
лоса, постепенно расширяющаяся, — на лбу черного цвета, на темени темнобурого, на
затылке переходящая в рыжевато-коричневое пятно, за которым на задней стороне шеи
расположено черное пятно. По бокам головы, сзади глаз, тянутся две более узкие черные
полосы; такие же, но еще более узкие полосы, на ушах. Посредине спины тянется черная
полоса, доходящая до конца туловища, у заднего конца полосы по сторонам ее по черному
пятнышку. На плечах по большому черному пятну и на локтевом сгибе по пятну шоколад-
ного цвета. На месте хвоста пух яркорыжий. Глаза карие, ноги бледнобурые, клюв светло-
роговой с черным пятном у основания надклювья. Имеются возрастные изменения в
окраске неоперенных частей у птенцов: глаза постепенно светлеют, в возрасте 40—45 дней
они светлокарие, у двухмесячных—желтовато-коричневые, позднее—грязножелтые
с коричневым оттенком; черноватые пятна у основания надклювья постепенно увеличи-
ваются, занимая постепенно все надклювье, кроме светлой полосы по ребру его,—полу-
чается заметная разница в окраске верхней и нижней (светлой) частей клюва. В окраске
ног появляются серые оттенки, которые, постепенно усиливаясь, вытесняют первоначаль-
ный общий бурый тон окраски.
Взрослый самец отличается от ранее рассмотренных подвидов тем, что в передней
части спины преобладает меднокрасная основная окраска; имеется довольно широкий
(около 1 см ширины) белый ошейник, который, однако, может быть и не замкнут спереди.
Спина, плечи, зоб и широкая кайма передней части брюха с преобладанием зеленого
блеска. Перья зоба и середины груди без черных вершинных каемок.
Бутурлин считал необходимым называть этот подвид Ph. mongolicus mongolicusr
полагая, что он описан Палласом в 1811 г.; однако Паллас, как выясняется, употреблял
слово mongolicus по отношению к фазану из Аргуни как прилагательное, а не как соб-
ственное имя; в номенклатурном смысле mongolicus употреблено позднее Брандтом A844)
для фазанов из Семиречья.
Маньчжурский фазан Phasianus colchicus pallasi R о t h s с h.
Phasianus torquatus pallasi. Rothschild. Bull. Brit. Orn. Club, XIII, 1903, стр. 43,
Сидеми.
Синонимы. Phasianus alpherakyi. В u t u г 1 i n. Ibis, 1904, стр. 399, Сунгари, район
Харбина.—Phasianus alpherakyi ussuriensis. В u t u г 1 i п. Там же, стр. 403, Уссури,
Сидеми.
Распространение. Ареал. Бассейн Уссури и Южноуссурийский край.
Расчистка тайги и зарастание лесосек кустарником способствует расселению
фазана на север и северо-восток; за последние 20 лет он распространился
к северу на 300 км от своих старых границ (Рудановский и Насимович,
1933). На Амуре он распространен до Черняево, у Кумары есть и в стороне
от реки и доходит до 52°30' с. ш. по долине р. Селемджи (Шульпин, 1936);
есть на о-ве Аскольд, куда завезен русскими. В Маньчжурии этот фазан рас-
пространен через всю эту страну с востока на запад до Большого Хингана,
а во времена Палласа был известен также на Аргуни и у Абагатуя. В насто-
¦224 ОТРЯД КУРИНЫЕ
•ящее время так далеко на запад он не проходит. Южная граница подвида
проходит вне СССР, так что Южная Маньчжурия, Северный Китай и Корея
находятся уже в области распространения другого подвида (Ph. с. carpowi).
Характер пребывания. Не вполне оседлая птица—в отдель-
ные годы совершает более или менее заметные перекочевки. Пржевальский
предполагал возможность A870) перекочевки фазанов на зиму в горы, где
их будто бы меньше беспокоят морозы и метели. На самом деле, повидимому,
происходит более или менее ясно выраженное передвижение северной и се-
веро-западной популяции за зиму к югу, при этом часть фазанов уходит
за пределы нашей страны. С другой стороны, по наблюдениям Горчаковской
и Слепцова, в конце лета и осенью происходит иногда перекочевка фазанов
к нам из Маньчжурии, что вызвано, повидимому, меньшим количеством корма
в это время в Маньчжурии.
Биотоп. Уремовая и кустарниковая растительность по широким
долинам рек, дубовые леса, обладающие богатым кустарниковым подседом,
высокотравные луга, поля и огороды; заселяет иногда и довольно сырые
места. В осеннее время—поля, в особенности засеянные просом и бобами
(соей), гумна и огороды, вообще держится ближе к человеческому жилью.
Численность. Сильно колеблется в зависимости or условий по-
годы, резко понижается после зим с длительным и глубоким снеговым покро-
вом, а также после продолжительных дождей в период размножения. Веро-
ятно, известное значение имеют и болезни птиц (Шульпин, 1936). Влияние
человека на численность фазанов двоякое. С одной стороны, в результате
неумеренной охоты происходит некоторое уменьшение численности, а с дру-
гой, в результате расчистки лесов под пашни и широким развитием в расчи-
щаемых местах кустарников с высокой травой происходит увеличение чис-
ленности птиц и их некоторое расселение.
Экология. Размножение. Период размножения растянут с ап-
реля по конец июня. В Южном Приморье тока начинаются в начале апреля,
а в более северных с середины этого месяца или в конце его. В холодную
весну 1941 г. в Хабаровском районе ток петухов начался лишь в первых чис-
лах мая. Крик самца напоминает крик молодого петуха, но короче и отры-
вистее; состоит из двух слогов, и местными жителями передается словами
«шаг вперед». В разгар тока самец кричит каждые 5—10 мин. Брачные крики
слышны как только начинает рассветать, достигают наибольшей силы в 10—
11 час. утра и потом в 4—5 час. вечера. Токующий самец нередко вскакивает
на кочку, особенно часто на пень, иногда даже на низкий сук дерева и, крича,
хлопает несколько раз крыльями, как обыкновенный петух. Во время токо-
ванья птица перемещается с места на место, по свидетельству Пржевальского,
иногда даже на полкилометра.
Гнезда помещаются в низком кустарнике, обычно у берега реки под гу-
стым кустом или под деревом, в естественном углублении или в углублении,
выскребленном в почве приблизительно на 4 см вглубь самой птицей. По
одним наблюдениям, в гнезде совсем не бывает подстилки, по словам Гор-
чаковской и Слепцова—несложная подстилка (сухие стебли травы и листья)
все же имеется, хотя и не всегда. Число яиц в кладке колеблется от 4 до 12,
один раз теми же наблюдателями было найдено гнездо с 18 яйцами.
Первая полная кладка (в 6 яиц) найдена Горчаковской и Слепцовым
в 1944 г. 2 мая, последняя кладка A8 яиц) в 1945 г. 15 июня, однако 6 свежих
-яиц были найдены и 19 июня. Длительность насиживания остается неизвест-
ной. Выход птенцов из яиц происходит в конце июня и в июле, но Спанген-
берг встретил 7 июля уже хорошо летавших птенцов, а Годлевский (по Тача-
новскому, 1883) находил очень маленьких цыплят и в конце июля. Молодые
фазаны величиной с вальдшнепа встречены на Средней Мо 20 июля 1869 г.,
там же молодые ростом с перепела 22 июля. В сентябре и середине октября
ОБЫКНОВЕННЫЙ ФАЗАН 225
основная масса молодых фазанов по размерам близка к взрослым, но встре-
чаются и некоторые только в половину веса и размеров взрослой птицы (Гор-
чаковская и Слепцов). Первое время выводки держатся семьями без петухов,
самцы присоединяются к ним только осенью, причем нередко можно видеть
семью фазанов с двумя и даже тремя старыми петухами.
Линька. Схема линьки—как и у других подвидов фазана. В конце
июля у старых птиц замечается интенсивная смена пера на головной птери-
лии, более слабая на шейной, грудной и спинной, начинается смена
пера также на крестцовой .и бедренных птерилиях. В августе линька
на грудной и спинной птерилиях усиливается, захватывает брюшную пте-
рилию, меняются маховые и рулевые перья. В первой половине октября
линька на головной, шейной и плечевых птерилиях постепенно ослабевает
и к середине октября заканчивается, к началу ноября линька заканчивает-
ся полностью. Молодые птицы линяют примерно в те же сроки, с некоторым
ускорением сроков у более ранних выводков и запозданием у поздних
(Горчаковская и Слепцов). У самцов сроки начала линьки связаны с окон-
чанием токования, а самки приступают к линьке после того как птенцы зна-
чительно подрастут.
Питание. По свидетельству Пржевальского A875) и по словам охот-
ников, обычной пищей фазанов осенью служат жёлуди. Кроме того, они
поедают осенью семена культурных растений, забираясь для этого в хлеба,
нередко когда они еще на корню; едят и картофель, который величиной до
грецкого ореха заглатывают целиком. Едят семена бобовых растений, летом
питаются улитками (Тачановский, 1893). По характеру питания видно, что
эти птицы могут приносить местами известный вред на полях.
Зимой фазаны выходят на проезжие дороги и расклевывают лошадиный
навоз, извлекая из него непереваренный овес; в суровые и снежные зимы
фазаны держатся у колхозных ферм и систематически кормятся остатками
пищи, находимыми в навозе. По наблюдениям Горчаковской и Слепцова,
летом и осенью фазаны разоряют иногда гнезда мышей-малюток (часто сделан-
ные из колосьев риса или пшеницы) и при этом поедают маленьних мышат.
По тем же авторам, молодые фазаны, а иногда и старые, охотно поедают
муравьев, мелких улиток, гусениц пядениц, совок, нимф и имаго кобылок,
жуков-навозников, бронзовок, щелкунов и других беспозвоночных.
Враги. Лиса и колонок уничтожают гнезда и истребляют молодых
птиц, зимой они скрадывают взрослых птиц у скирд необмолоченных хле-
бов. Из пернатых хищников фазанов преследуют белые совы, мохноногие
канюки, степные и луговые луни и ястреб-тетеревятник, причем луни напа-
дают на молодых птиц, остальные на взрослых.
Промысел. Фазанов добывают, раскидывая на жнивье сети, под-
манывают зимой на приваду и стреляют; в некоторых местах фазанов при-
кармливали намоченными в корейской водке зернами чумизы, после чего
птицу можно брать руками. По непроверенным сведениям, из пределов
Приморья в 20-х годах вывозили от 10 до 40 тысяч в год (Шульпин, 1936).
Описание. Размеры и строение. Длина тела самцов A5) 760—860, самок
A4) 630—850, среднее соответственно 827 и 820,3 мм; размах самцов A7) 720—820,
самок A5) 660—770, среднее 791,2 и 708,7 мм; длина крыла самцов A6) 245—260, самок
A4) 215—250, среднее 246,2 и 223,2 мм (Горчаковская и Слепцов); самцов A1J47—278,
самок F) 225—243, среднее 259,5 и 233 мм (коллекция Зоологического музея Московского
государственного университета). Вес, по Горчаковской и Слепцову, самцов A4) 1090—1330,
самок (9) 970—1220, в среднем 1174,4 и 1081,5 г.
Окраска. Пуховой птенец серого цвета с буроватыми крапинками и полосками.
От других подвидов фазанов из СССР уссурийский отличается тем, что у него окраска
поясницы и верхних кроющих хвоста матовосерая, голубоватая или зеленоватая, без
примеси меднокрасных, рыжих или охристых оттенков (только по бокам надхвостья
имеется по партии волосовидных матовых рыжих или оранжевых перьев). Мелкие и сред-
15 Птицы СССР, т. IV
226
ОТРЯД КУРИНЫЕ
ние кроющие крыла не белые, но светлые голубовато-серые или с легким оливковым оттен-
ком. На основной половине средней пары рулей темные толстые поперечные полосы, около»
3—4 мм ширины. Передняя часть спины и бока тела золотисто-соломенно-желтые. Белый
ошейник имеется.
РОД КУРОПАТКИ PERDIX BRISSON, 1760
Тип. Tetrao perdix
19. Серая куропатка Perdix perdix L.
Tetrao Perdix. Li nnaeus. Syst. Nat., изд. X, 1758, стр. 160, Швеция.
Русское название. Почти повсеместно распространенное русское название;
повидимому, дано из-за сходства по внешнему виду и поведению с домашними курами.
Распространение. Ареал. Распространена от Британских о-вов и се-
верных районов Португалии до Алтая. На юге область ее распространения
Карта 20. Распространение серой куропатки Perdix perdix
I—P. p. hispaniensis, 2—P. p. armoricana,3—P. p. sphagnetorum, 4—P. p. perdix, 5—P. p.
6—P. p. canescensy 7—P. p. robusta; a—граница гневдовий,
б—не вполне выясненная граница гнездовий
lucida,
охватывает Северную Португалию, Среднюю Испанию, Францию, Италию,
Македонию, Северную Грецию, Малую Азию и Северный Иран, Тянь-Шань,
Тарбагатай и Алтай; на север в Скандинавии проникает до 64° с. ш., в Фин-
ляндии до 66° с. ш., в районе Белого моря до 6,5° с. ш., на Урале до 5872°
и в Зауралье до 61° с. ш.
Характер пребывания. Оседлая птица; однако куропатки
из открытых степных районов с суровыми зимами (между Средней Волгой
и Иртышом) отлетают на зимовку в близлежащие малоснежные районы
с более мягким климатом (в пустыни Средней Азии, степи Предкавказья и
Приазовья и т. п.); в других районах предпринимают местные кочевки,
приближаясь к населенным пунктам и т. д.
СЕРАЯ КУРОПАТКА 227
Биотоп. Хлебные поля, чередующиеся с кустарниками и мелко-
лесьем, сады, сухие луга, степи с колками леса и с поросшими кустарни-
ками балками, большие поляны и лесные вырубки, поймы рек, степные пред-
горья с зарослями кустарников.
Подвиды и варьирующие признаки. Север Португалии, Северную и Среднюю Испа-
нию, Пиренеи и прилегающие участки Французской равнины, а также всю Италию
населяет P. p. hispaniensis Reichenow, 1892; в Арденнах и во всем Арморикан-
ском массиве, включая Нормандию и Бретанию (Франция),—P. p. armoricana Наг-
t e г t, 1917; на Британских о-вах, в Северо-Восточной и Восточной Франции, в Бель-
гии, Голландии и Северо-Западной Германии—P. p. sphagnetorum A I t u m, 1894;
в Швейцарии, Дании, Германии и далее на восток до Балканского п-ова, возможно,
до устья Днепра на юге и до Швеции, Норвегии и, вероятно, Прибалтийских республик
Советского Союза—P. p. perdix Li n na e us, 1758; в Польше (от р. Вислы), в восточных
районах Прибалтийских республик и далее на Восток до Урала—P. p. lucida A I t u m,
1894; на Кавказе и в Закавказье, в Малой Азии и Иране—P. p. canescens В u t и г 1 i п,
1906; от Урала, северо-восточных и восточных берегов Каспия до Тарбагатая и Алтая—
P. p. robusta Homeyer et Tancre, 1883.
Отличия подвидов сводятся к вариациям окраски и крайне небольшим и трудно
уловимым различиям в размерах. Очень значительные индивидуальные вариации
окраски часто перекрывают подвидовые отличия, чем объясняются указания на то, будто
в одном и том же районе встречаются две формы (P. p. robusta и P. p. arenicola в Наурзум-
ских степях; де Ливрон, 1938; Сушкин, 1908; P. p. robusta и P. p. cinerea в б. Харьков-
ской губ.; Аверин, 1910 и т. п.); кроме этого, промежуточные между подвидами особи
занимают значительные территории. Все это в значительной степени усложняет систе-
матику вида. В качестве общей закономерности следует отметить, что в окраске опере-
ния при движении с запада на восток общий рыжий или каштаново-рыжий тон окраски
постепенно все более и более заменяется более чистым и серым тоном, рыжие и каштаново-
рыжие пятна и полосы на верхней стороне тела становятся уже или совсем исчезают.
Среднеевропейская серая куропатка Perdix perdix perdix L.
Распространение. Ареал. Населяет Швейцарию, Германию (за исклю-
чением ее северо-западных районов). Балканский п-ов (Македония, Северная
Греция, Болгария), Румынию, Венгрию. Юго-Западную Польшу и Закар-
патскую Украину, проникая до низовьев Днепра, а местами, вероятно, и
переходя через Днепр (самецот 28 марта 1929 г. из Полтавской обл. неот-
личим от типичной формы); на севере населяет Швецию и Норвегию (до се-
верной границы земледелия, т. е. примерно до 64° с. ш.), Финляндию (до
66° с. ш.). Из Германии, повидимому, проникает в Прибалтийские респуб-
лики Советского Союза (или хотя бы в их западные районы; экземпляры
из Литовской ССР очень близки к номинальной расе).
Характер пребывания. Оседлая птица. Зимой предприни-
мают незначительные кочевки: реже местами и не ежегодно совершает и
перелеты (Клайпеда; Нитхаммер, 1942).
Биотоп. Гнездится по хлебным полям, большим лесосекам, в степях,
прорезанных заросшими кустарниками оврагами, по сухим лугам (и очень
редко по мокрым), по опушкам колков и больших лесных полян, реже в
садах и огородах.
Численность. Неодинакова в разных районах; на севере менее
многочисленна, чем на юге. Численность меняется погодам, особенно резко
снижаясь после суровых многоснежных зим (после зимы 1914—15 г. почти
исчезли в Западном Закарпатье и стали увеличиваться в числе лишь в послед-
ние годы; Портенко, 1950 и т. д.).
Экология. Размножение. Моногамы; самец принимает участие
с насиживании и наравне с самкой водит выводок. Разбивка на пары в конце
марта—начале апреля (Латвия; Тауриньш, 1950). Откладка яиц происходит
в конце апреля—начале мая; полные кладки из свежих яиц встречены во
второй половине мая (Молдавия, Остерман, 1912). Количество яиц в кладке
15*
226
ОТРЯД КУРИНЫЕ
12—20 реже до 22; самка откладывает по одному яйцу в сутки. Размеры
яиц (90) 32—38x24,5—28,6, в среднем 35,2x26,8 мм\ вес свежего яйца
около 14 г (Нитхаммер, 1942).
Гнездо в виде небольшого углубления в земле, выстланного раститель-
ной ветошью с примесью куропаточьих перьев, располагается обычно в гу-
стой траве под защитой куста или кочки. Самка сидит на гнезде очень плотно;
самец находится вблизи гнезда и сменяет самку. Насиживание продолжается
24—25 дней. Птенцы начинают бегать и кормиться в первый день после
вылупления* на 9—11-й день уже перепархивают. Рост птенцов, как и у дру-
гих куринных, относительно медленный (по Хейнроту, 1924—1932):
В озраст
в днях
Вес в г
1
8,5
5
17
10
34
16
55
21
80
26
105
38
160
49
220
56
265
63
300
78
315
91
330
105
350
119
350
При воспитании в неволе четырехмесячный самец весил 332 г, а самка
343 г. Половозрелыми молодые птицы становятся в возрасте одного года,
на следующее лето.
Лин ь к а. У пуховичков пеньки маховых и плечевых перьев начинают
развертываться уже на 5—6-й день после вылупления. В возрасте 24—28
дней птенцы заканчивают линьку в птенцовый наряд.
В первый годовой наряд молодые птицы линяют в августе—октябре.
В возрасте 38—40 дней начинают сменяться маховые перья, затем линька
переходит на туловище; последними сменяются второе и первое первосте-
пенные маховые, рост которых у некоторых особей еще не заканчивается
полностью и в конце октября. В первом годовом наряде отличаются от взрос-
лых птиц более узкими с заостренными (а не округлыми) вершинами первого
и второго первостепенных маховых.
У взрослых птиц полная годовая линька проходит между концом
июля—серединой сентября, затягиваясь у отдельных особей до начала
октября. В марте-апреле отмечена неполная предбрачная линька, охваты-
вающая лишь голову и шею (Нитхаммер. 1942).
Питание. Основу питания составляют семена, клубни, корешки
и свежие побеги многих растений; в летнее время используют и животные
корма: насекомых, их личинок и гусениц, дождевых червей, моллюсков и т. п.
Полевые признаки. Курица небольших размеров; хвост короткий, тон
оперения серый, на груди коричневое подковообразное пятно. Наземная
птица. Взлет сильный, шумный.
Описание. Размеры и строение. Короткий, слегка округленный хвост
образован 16—18 рулевыми. Первостепенное маховое без вырезки. Крыло короткое,
тупое; обычная формула крыла 3>4>5>2>6^1. Шпоры на цевке отсутствуют как
у самцов, так и у самок.
Длина крыла у самцов от 150 до 163 мму обычно 156—160 мм, у самок 148—159 мм\
вес самцов 400— 450 г, самок—около 375 г (Нитхаммер, 1942). Длина крыла самцов C)
154—161, самок C) 149—151,5, в среднем, соответственно, 157,5 и 150,1 мм (коллекция
Зоологического музея Московского государственного университета).
Окраска. Пуховик. Верх головы красновато-коричневатый с черноватыми
пятнами; горло, бока головы и уздечка бледножелтые с буроватыми пятнышками; осталь-
ной верх тела в больших партиях темнобурого, грязновато-сливочного и красновато-
коричневатого пуха, образующих две неясные продольные черноватые полоски; нижняя
сторона шеи и туловища светлые грязновато-желтые (Хартерт, 1921—22).
В птенцовом наряде лоб, верх головы и щеки буроватые со светлыми наствольями;
шея, спина и поясница желтовато-бурые с узкими светлыми наствольными полосками;
плечевые и кроющие крыла бурые со светлыми наствольями и узкими охристыми попе-
речными полосками; подбородок и горло беловатые, грудь светлая желтовато-бурая со
светлыми наствольями, брюхо грязнобелое.
СЕРАЯ КУРОПАТКА
229
Первый годовой наряд молодых сходен с окраской взрослых
птиц; у многих птиц в этом наряде уже отчетливо выражено брюшное пятно, развитое
обычно лишь несколько меньше, чем у взрослых птиц.
У взрослого самца над клювом через лоб проходит узкая черная полоска;
лоб и полоса над глазом, подбородок и прилежащие участки шеи спереди ржавчато-рыжие,
кроющие уха коричневато-бурые. Темя и затылок рыжевато-бурые со светлыми узкими
наствольными полосками и пятнами, отчетливо выраженными не у всех экземпляров.
Передняя часть спины и шеи серые с мелким темнобурым струйчатым рисунком и отчет-
ливо выраженным коричневато-рыжим отливом, на опахалах перьев спины, кроме этого,
Рис. 38. Серая куропатка (самец и самка)
узкие поперечные каштановые и охристые полоски; более отчетливы они на задней части
спины и на верхних кроющих хвоста. Плечевые и крющие крыла с резко очерченными
белыми наствольными полосками (с рыжеватым отливом у большинства особей этого
подвида). Маховые бурые со светлоохристым поперечным рисунком, особенно отчетливо
выраженным на наружных опахалах первостепенных маховых. Рулевые рыжие с более
или менее развитым у основания серовато-бурым цветом. Грудь и отчасти бока тела серые
с густым темнобурым мелким струйчатым рисунком, образующим мало заметную попе-
речную исчерченность; на боках тела широкие поперечные темнорыжие (или каштаново-
рыжие) полосы. На передней части брюха расположено обычно подковообразное пятно
каштанового или темнокаштанового цвета, отделенное от серой груди варьирующей
по ширине узкой белой полоской; задняя часть брюха и подхвостье грязнобелые, на по-
следнем иногда неясно выражен грязноохристый налет.
Взрослая самка по окраске очень сходна с самцом. Лишь на плечевых
и кроющих крыла (лучше всего на средних кроющих) обычно есть по две-три узкие охри-
стые поперечные полоски (у некоторых птиц отсутствуют). Брюшное пятно обычно чуть
меньше по величине брюшного пятна самца, редко отсутствует.
230 ОТРЯД КУРИНЫЕ
Радужина каряя, клюв темнобурый; ноги желтовато-бурые, а у старых самцов голу-
бовато-серые.
Отличается от других подвидов более темным общим тоном окраски и более выражен-
ным рыжевато-каштановым тоном окраски верхней стороны туловища. Грудь темносерая;
каштановый оттенок со спины и кроющих крыла заходит и на верх шеи. На пояснице
и верхних кроющих хвоста отчетливо выделяются более широкие, чем у других подвидов,
поперечные рыжевато-каштановые полосы, чуть окаймленные сверху охристым.
Среднерусская серая куропатка Perdix perdix lucida Altum
Starna cinerea var. lucida. Altum. Journ. f. Orn., 1894, стр. 286, Восточная Пруссия.
Синоним. Perdix perdix caucasica. Rei chenow. Journ. f. Orn., 1903, стр. 543,
Северный Кавказ (сравнивалось лишь с P. p. perdix, но не с P. p. lucida и P. p. robusta).
Распространение. А р е а л. От Восточной Польши (от р. Вислы) до Ураль-
ского хребта. В пределах европейской части Советского Союза по подходя-
щим биотопам встречается почти повсеместно.
В Прибалтийских Советских республиках, повидимому, встречаются
формы, переходные между P. p. perdix и P. p. lucida, это же возможно и для
Правобережья Днепра в пределах Украинской ССР. На севере куропатка
доходит в Карело-Финской ССР до Медвежьегорска включительно (Марвин,
1950); в июне 1950 г. пара серых куропаток встречена на севере Ругозерского
района в окрестностях озера Нюк, а по опросным сведениям она встречается
и на юге Калевальского района в окрестностях северного побережья озера
Нюк и у Юшкозера (Гибет). В Архангельской обл., повидимому, местами
доходит до берегов Онежского залива; далее на восток граница ее ареала
проходит около г. Шенкурска F2° с. ш.; Бобринский, 1934), затем по пра-
вому берегу р. Камы до с. Юрлы E9 1/2° с. ш.; Ушков, 1927) и по западным
склонам Урала до 58 г/2° с. ш. (Резцов, 1904).
На юге населяет Крым, побережье Азовского моря, Северный Кавказ
до поросших кустарниками предгорий, Ставрополье, низовья р. Волги.
Склоны Уральского хребта и районы по р. Уралу вплоть до Каспия и Вол-
жско-Уральское междуречье населены птицами переходного типа (между
P.p. lucida и P. p. robusta).
В связи с продвижением земледелия на север и расширением посевных
площадей в лесных районах расширяется ареал серой куропатки и увели-
чиваются занимаемые ею площади внутри ареала. В 60-х годах XIX в.
стала встречаться в Ярославской обл; в 1913 г. в Мышкинском районе впер-
вые обнаружен один выводок, а в 1936—37 г. за день встречали до 12 вывод-
ков (Кузнецов, 1947). В окрестностях Петрозаводска появилась в 60-х
годах XIX в., сейчас же обычна по берегам Онежского озера (Красовский,
1933) и дошла до Юшкозера. В 30-х годах XX в. проникла на север Киров-
ской обл. (Кайский район; Пузанов и др., 1942); в это же время стала более
многочисленной севернее г. Молотова и у г. Надеждинска (Урал; Куклин,
1948). Существует предание, что во второй половине XVIII в. ввезены в
б. Петербургскую губ., откуда расселились в Финляндию (Мензбир, 1893).
Характер пребывания. В большей части ареала оседлая или
предпринимающая кочевки птица. Зимой приближается к селениям, уходит
в заросли кустов и лозника, в лесные колки, в пересеченные участки мест-
ности (Харьковская обл., Чернай, 1853; Николаевская обл. Воронцов,
1940; юг Горьковской обл., Формозов, 1935). В суровые, многоснежные зимы
откочевки выражены яснее (Московская обл.; Поляков, 1924). Во время
засух из выжженных солнцем степей уходит в поймы рек и лесные массивы
(Бузулукский бор; Доброхотов, 1937). В восточных районах европейской
части СССР перелетна (в условиях многоснежной зимы со средним максиму-
мом снегового покрова 50—60 см и выше); часть птиц пытается зимовать, но
•бычно погибает, особенно в суровые зимы (Формозов, 1946).
СЕРАЯ КУРОПАТКА 231
Осенний пролет куропаток к югу (октябрь-ноябрь) стайками по 40—50
шт. хорошо заметен в лесных районах Южного Приветлужья (Шарьинский
район Горьковской обл.; Формозов, 1935, 1946); отлетают птицы и из Саратов-
ской обл. и б. Самарской губ. (Житков и Бутурлин, 1906). В Саратовской обл.
пролет начинается после первых заморозков; через Р/г—2 недели птицы
появляются у Камышина, еще через несколько дней у Сталинграда; вначале
летят редкие стаи, в конце октября—начале ноября массовый пролет, закан-
чивающийся под Камышином в первой половине декабря; птицы летят от
захода солнца до глубокой темноты и по утрам сотнями табунов от 4 до
20 шт. на юго-восток; в теплые зимы часть птиц оседает у Камышина, а боль-
шинство долетает до юга Астраханской обл. и степей Северного Кавказа.
В Ставрополье первые пролетные птицы появляются в конце августа; вало-
вой пролет у Ростова в конце сентября—начале ноября, по берегу Азовского
моря (мимо Мариуполя, Таганрога и т. п.)—до последней трети ноября;
часть птиц достигает Одесской обл. Весенний пролет назад незаметен (Житков
и Бутурлин). Пролет на побережье Азовского моря в 80-х годах прошлого
века был очень велик и проходил ежегодно, затем количество пролетных
птиц постепенно уменьшалось (истребление во время пролетов и на зимов-
ках) и в первом десятилетии XX в. пролет стал носить случайный характер
{Алфераки, 1910; Вальх, 1911). Раньше перелетные птицы достигали и
Балканского п-ова (Сербия, Хаджи-Ильич, 1910).
Необходимо проведение изучения перелетов в настоящее время с широ-
ким применением метода кольцевания.
Биотоп. Поля и суходольные луга, особенно прилегающие к кустар-
никам, оврагам и балкам, опушкам лесов; большие лесосеки и можжевеловые
поляны на кромках больших лесных массивов; в степи по балкам с зарослями
кустов или бурьянов, островные лески, кустарниковые и травянисто-камы-
шевые заросли в поймах рек и по берегам прудов и озер; заросли ситника
и других растений в бугристых песках Северного Прикаспия, в зарослях
лоха, ивы, гребенчука и т. п.; в меньшей степени огороды и сады. Охотно
поселяется в молодых полезащитных лесных полосах 5—15-летнего воз-
раста; в более старые полосы заходит реже, но обильна на прилегающих
полях (Мельниченко, 1949).
Зимой преимущественно по кустарниковым и тальниковым зарослям
в поймах рек, по опушкам леса, в островных лесах, колках и садах, по овра-
гам и пятнам кустов в степи, в камышах и бурьянах. В глубокие снега под-
ходят к населенным пунктам, кормясь по озимям (особенно по заячьим жи-
ровкам; Формозов, 1946), на токах, у скирдов и по проезжим дорогам.
Численность. Неодинакова в разных районах: например, в Кара-
чевском районе редка, в Трубчевском—обыкновенна, в Поченском—много-
численна (Брянская обл.; Суслова, 1937), в северных районах реже, чем
в южных и западных (Смоленская обл.; Станчинский, 1916) и т. п. Неравно-
мерность распределения связана с наличием подходящих биотопов, глуби-
ной снежного покрова зимой и т. п. В благоприятные годы местами обильна:
у Богдинских полезащитных лесных полос на 2—3 га полей в августе дер-
жалось по выводку (Мельниченко, 1949). В долине р. Иловли пара от пары
в 10—12 км, в степи, изрезанной балками, по 2—3 пары на 1 км2 (Спанген-
<5ерг), в Дубовском районе Сталинградской обл. в мае 1949 г. за день
экскурсии встречалось до 25 птиц (Птушенко, 1949).
Численность сильно снижается после холодных, особенно многоснеж-
ных зим, когда резко увеличивается смертность не столько от переохлажде-
ния, сколько от голода (трудность добывания корма из-под глубокого снега);
увеличивается и смертность от хищников и от охотников.
В б. Калужской губ. после суровой зимы 1907—08 гг., в течение трех лет
не встречены там, где в предыдущие годы выводки летом были обычны (Фила-
232 ОТРЯД КУРИНЫЕ
тов, 1915). Даже в Крыму после суровых зим 1874—75, 1878—79, 1910—11 и
1928—29 гг. численность куропаток резко снижалась (Пузанов, 1932). Анало-
гичные наблюдения есть для окрестностей Мариуполя (Рудневич, 1924),
Подолии (Подольский, 1924), Киевской и Черниговской обл. (Щербина, 1916),
Белоруссии (Федюшин, 1928), Смоленской (Граве, 1926), Ленинградской
(Исполатов, 1907) и других областей.
После крайне холодной зимы 1939—40г., когда морозы доходили до 52°С,
отмечено крайне резкое уменьшение куропаток на Украине, в Калининской,
Ярославской и других областях европейской части Союза (Дементьев и
Шимбирева, 1941); в Молого-Шекснинском междуречье (Рыбинское водохра-
нилище) ранней весной повсюду находили мумифицированные трупики
и остатки растерзанных лисами птиц (Исаков, 1949); гибли от холода и
массами уничтожались хищниками птицы на побережье Каспия A,8%
встреч от найденных мертвых птиц всех видов; Исаков и Сазонова, 1947).
Повидимому, резко сказываются на численности серых куропаток и эпизо-
отии, в частности глистные инвазии (Формозов, 1935).
После ряда мягких и малоснежных зим численность куропаток резко
увеличивается (Белоруссия, Шнитников, 1913).
Количество куропаток в угодьях Гатчинской Охоты с 1886 по 1909 г.
колебалось в широких пределах (ежегодный отстрел обычно не превышал
5—7% летней численности птиц при максимальном отстреле в 17% в год):
в 1886 г. отмечено 363 молодых птицы, в 1887г.—1903, в 1821 г.—2968,
в 1892 г.—950, затем шло постепенное нарастание прироста, и максимум
численности молодых B934 шт.) был достигнут в 1898 г.; в 1900 г. число
молодых птиц снизилось до 1421, в 1903 г. до 1031, затем в течение 3 лет
численность поднялась до 1499 и в 1907 г. вновь упала до 921 молодой птицы
(Север цов, 1941).
Экология. Размножение. Моногамы. Разбивка на пары проис-
ходит в марте-апреле (Сандомирский край, Томкевич, 1914; Среднее Повол-
жье, Доброхотов, 1937), часто еще до появления проталин. Начало кладки
зависит от условий года и приходится обычно на конец апреля—май. Пол-
ные кладки из 16—22 яиц найдены 2 июня 1905 г. (Сандомирский край;
Катин, 1912); гнездо с 25 сильно насиженными яйцами встречено 5 мая 1895 г.
(Тарха-Сунак в Крыму; Сеницкий, 1898), во второй половине мая (Поволжье,
Доброхотов, 1937 и др.; Южный Урал, Куклин, 1938). Сезон размножения
в популяции растянут: 22 апреля найдено гнездо с 5 свежими яйцами, 17—
мая—с 9 свежими яйцами, 4 июня—с 23 насиженными яйцами, 5 июня—
с 19 свежими яйцами, 6 июня—с 18 насиженными яйцами (б. Минская губ.;
Шнитников, 1913); в июне встречены яйца, поршки и хорошо летающие
птенцы (Сандомирский край; Томкевич, 1914).
Количество яиц в кладке до 15—20, реже до 24 (Хлебников, 1930),
25 (Сенецкий, 1898). Яйца правильной яйцевидной формы с хорошо выра-
женным острым и тупым концом. Окраска скорлупы одноцветная серовато-
песчаная, темная охристо-оливковая или глинисто-буроватая; поры мало
заметны; скорлупа лишь с небольшим блеском. Размеры яиц A3) 33—
39x26,5—29 в среднем 35,3x27,2 мм (Сомов, 1897).
Гнездо в небольшой ямке среди хлебов, в густой траве под защитой
куста, дерева или кочки. Лоток скудно выстлан сухой травой и растительной
ветошью. Повидимому, в насиживании принимают участие самец и самка;
однако некоторые авторы утверждают, что самец не насиживает, хотя и
держится около выводка (Пузанов, Кипарисов, Козлов; 1942). Самка сидит
на гнезде плотно.
Длительность насиживания—как у других подвидов. В соответствии
с растянутым периодом откладки яиц растянут и период вылупления:
первые птенцы обычно встречаются в последние дни мая—в первой половине
СЕРАЯ КУРОПАТКА 233
июня (в Карелии в июне; Марвин, 1950). Недельные птенцы найдены
18 мая, выводок с недельными птенцами и гнездо с необсохшими, только что
вылупившимися птенцами—14 июня (б. Минская губ.; Шнитников, 1913),
едва летающие птенцы добыты в половине июня и в начале августа (Куй-
бышевская обл.; Житков и Бутурлин, 1906); 7 июля встречены только что
вылупившиеся пуховики, молодые размером с перепела и молодые, достиг-
шие размеров взрослых птиц; однодневные птенцы найдены и 4 августа
(окрестности Мариуполя; Боровиков, 1907); 1—2-дневные птенцы встреча-
лись в пойме р. Еруслан (Саратовская обл.) с 31 мая до конца июня (Вол-
чанецкийи Яльцев, 1934), молодые ростом в половину взрослых птиц найдены
в половине августа (Рязанская обл.; Хомяков, 1900). Размеров взрослых
птиц молодые в массе достигают в сентябре (Московская обл.; Лоренц,
1892)—в октябре (Карело-Финская ССР; Марвин, 1950).
Выводки охотно держатся по хлебным полям, после уборки переходят
на картофельные поля, в огороды, сады, на опушки лесов и в кустарники.
С половины августа выводки начинают соединяться в стаи по 30—80 шт.
и более и совершают по зорям довольно значительные перелеты на кормежку
(Формозов и Осмоловская, 1950).
Линька. Проходит как у других подвидов. Молодые перели-
нивают в первый годовой наряд в августе-сентябре, отдельные особи закан-
чивают линьку в октябре. Молодые самец и самка от 20 августа 1921 г. (Воро-
нежская обл.) в энергичной линьке мелкого пера по всему туловищу, све-
жее перо проступило под старым; первое и второе первостепенные маховые
в еще нераскрывшихся пеньках, третье птенцовой генерации, четвертое в
нераскрывшемся пеньке, пятое в половину, шестое в три четверти, седьмое—
десятое полной длины. У молодого самца от 12 сентября 1939 г. (Ростовская
обл.) по всему телу свежее перо первого годового наряда, часто еще не пол-
ностью выросшее; первое и второе первостепенные маховые новые, почти
полностью выросшие, третье в чуть раскрывшемся пеньке, четвертое в три
четверти, пятое—десятое новые, полной длины; в сходной стадии линьки
молодые птицы от середины сентября из Ростовской обл. и б. Сувалкинской
губ. Молодой самец от 10 октября 1904 г. (б. Пензенская губ.) закончил
линьку в годовой наряд; перья на груди еще не полностью закончили рост
В октябре заканчивают линьку молодые птицы в Карелии (Марвин, 1950).
Полная линька взрослых птиц проходит в июне—сентябре; сроки
ее в популяции растянуты в соответствии с растянутостью периода раз-
множения. В мае птицы, как правило, не линяют, но одна самка от 18 мая
1922 г. из Боровского района б. Воронежской губ. с большим наседным пят-
ном уже начала линьку: отдельные нераскрывшиеся пеньки появились на
брюхе, спине и голове. В середине сентября встречены как полностью закон-
чившие линьку взрослые птицы, так и в разгаре линьки: с еще не полностью
развернувшимися пеньками контурного пера по туловищу, с первым перво-
степенным маховым в половину, вторым в три четверти и третьим—десятым
полной длины (Восточная Польша, Московская, Воронежская и другие
области).
В марте, а на юге уже в январе (Аскания-Нова), проходит частичная
предбрачная линька, охватывающая, повидимому, только голову и шею
птицы (коллекция Зоологического музея Московского государственного
университета).
Питание. Основу питания составляют семена сорняков, падалица
хлебных злаков, зеленые листья, клубни и корни травянистых растений,
ягоды шиповника и т. п.; в летне-осенний период охотно поедают членисто-
ногих и, вероятно, наземных моллюсков. В малоснежных местах добывают
насекомых из подстилки и в зимнее время (гусениц совок, вредную черепаш-
ку и т. п.).
234 ОТРЯД КУРИНЫЕ
Птенцы вначале питаются нежной животной пищей (зоб нелетного птен-
ца был набит куколками и личинками земляных муравьев; Формозов и
Осмоловская, 1950), лишь постепенно переходя на семена диких растений
и падалицу культурных злаков. В желудках птиц A6), добытых в Тима-
шевских полезащитных лесных полосах в мае—сентябре 1938 г., найдены
10 экземпляров Hemiptera, 13—Curcutionidae, 48—Cassidini, 4—Еlateri-
dae,6—Carabidae>7l—прочих Coleoptera, 35 зерен пшеницы, 102 зерна диких
злаков и других диких растений, 15 зерен проса и 89 листьев злаков (Мель-
ниченко, 1949). В полезащитных лесных полосах Сталинградской обл. серые
куропатки в массе истребляют куколок лесной пяденицы (делая прикопки
на глубину 5—6 см) и несколько меньше ильмового ногохвоста и березового
пилильщика; выводки концентрируются на участках заражения; в меньшем
количестве птицы поедают клопов, жуков, проволочников и цикад; часто
в зобу находили много десятков кобылок (Формозов и Осмоловская, 1950).
В периоды массовых размножений наземных насекомых во множестве их
поедают (вредная черепашка, саранча и т. п.).
Зимой питаются зелеными проростками, семенами и насекомыми, рас-
капывая верхний слой подстилки в малоснежных местах, кормятся на токах
и дорогах; едят березовые сережки (Богачев, 1927) и, возможно, почки различ-
ных кустарников.
Враги. Яйца, птенцов и взрослых птиц истребляют бродячие кошки
и собаки, лисы, хорьки и другие наземные хищники. Сильно уничтожает их
и ястреб-тетеревятник; в течение 10 дней декабря тетеревятник регулярно рано
утром появлялся на одном из берегов озера, где держалась стайка куропа-
ток: сначала их было 8 шт., через 5 дней—3, через неделю исчезли все (Смо-
ленская обл.; Плющенков, 1949); они ловили и сбившихся в суровые морозы
у стогов птиц (Киевская обл., Шарлемань, 1930). Кроме этого, куропаток,
повидимому, берут камышевые луни и другие пернатые хищники.
Хозяйственное значение. Повсеместно объект спортивной охоты, места-
ми перерастающей в промысловую (Поволжье, Северный Кавказ). В зимних
заготовках дичи на территории б. Уральской обл. серая куропатка занимала
отЗ до 15%, в 1931—32г. там было заготовлено свыше70тысяч птиц (Куклин,
1938); местами недопромышляется (южные районы Молотовской обл.;
Воронцов, 1949).
Добываются преимущественно ружьем. Применение промысловых спо-
собов лова (силки в кустах, накрывание шатром, загоны в вентеря и мережи
и т. п.), как истребительные, должны быть запрещены и могут применяться
лишь в организованном порядке под соответствующим контролем. В прош-
лом веке, когда на осенних пролетах была многочисленной, ее ловили под
Камышином и Сталинградом перевесами, устанавливаемыми на оврагах,
идущих из степи в долину Волги (Гринберг, 1934).
Высокая плодовитость и приуроченность к культурному ландшафту
делают ее прекрасным объектом дичеразведения. Несомненная полезность
серой куропатки как истребителя вредных насекомых травянистого яруса,
подстилки и верхнего слоя почвы и куколок вредителей древесных пород
требует привлечения ее в полезащитные лесные полосы. Применение неслож-
ных биотехнических мероприятий (рациональный отстрел, временные за-
преты охоты, охрана в гнездовой период, уничтожение бродячих собак и ко-
шек, подкормка в неблагоприятные по метеорологическим условиям годы,
отлов и содержание в сараях в особо многоснежные зимы с последующим
выпуском птиц весной в угодья, что успешно применялось в Гатчинской
Охоте; Диц, 1917, и т.д.) позволит превратить полезащитные лесные полосы
и прилегающие поля в рассадники этой полезной для сельского хозяйства
и ценной промысловой птицы и резко увеличит размеры промысловых загото-
вок этого вида.
СЕРАЯ КУРОПАТКА 235
Полевые признаки Напоминает курицу небольших размеров и плотного
сложения. Рассыпной стайкой перебегают в поисках корма, роются в земле,
подобно домашним курам; охотно купаются в пыли. На деревья садятся
исключительно редко (Шарлемань, 1930). При опасности стремительно взле-
тают с шумом и громким хлопаньем крыльев; в полете быстро машут крыль-
ями, летят невысоко над землей; перед посадкой планируют, спускаясь на
землю по пологой кривой .
Описание. Размеры и строение. Длина крыла самцов B5) 148—162,5, самок
A7) 144—162, в среднем, соответственно, 155 и 154 мм. Длина тела 313—356 мм, размах
крыльев 500—550 мм. Средний вес самцов (8)—413,7, самок B) 395,5 г (Казань, Руданов-
ский и Насимович, 1933); вес самцов E23) 315—510, самок E14) 310—470, в среднем
соответственно 413 и 400 г (Уральская научно-исследовательская пушнопромысловая стан-
ция; Куклин, 1938). Самки лишь чуть меньше самцов.
Окраска. По общему тону чуть светлее и серее среднеевропейской серой
куропатки. Темя и затылок более серые. Верх тела с рыжеватым отливом, менее интен-
сивным, чем у P. p. perdix, верх передней трети шеи совсем без рыжины, наствольные
полосы белые или с едва заметным охристым отливом; поперечные полосы на кроющих
поясницы и хвоста более узкие. Грудь светлосерая с более редким мелким темнобурым
крапом, образующим плохо заметную поперечную исчерченность. Полосы на боках
тела несколько уже, рыжее и тусклее.
В популяции общий тон окраски (степень развития рыжего оттенка по верху,
тела и т. п.) варьирует довольно сильно: отдельные особи совершенно не имеют рыжины
и почти не отличимы от P.p. robusta).
Закавказская серая куропатка Perdix perdix canescens But.
Perdix perdix canescens. В u t u г 1 i n. Jbis, 1906, стр. 411, Ахалцых.
Синонимы. Perdix perdix furvescens. Бутурлин. «Наша охота», 1908, сентябрь,
стр. 6, Талыш.—Perdix perdix fulvescens. S а г u d n у. Journ. f. Orn. 1911, стр. 204,
Южнокаспийские области Ирана.
Распространение. Ареал. Кавказский хребет, Закавказье и далее
на юг в Малой Азии и северных районах Ирана. Популяция северных пред-
горий Большого Кавказа представлена формами переходного типа (между
P. p. lucida и P. p. canescens).
Характер пребывания. Оседлая птица. В осенне-зимний
период живущие в высокогорьях птицы совершают кочевки (с октября-ноября):
перелетают с северных склонов гор на южные (с Памбакского хребта на
Акстафинский; Даль, 1948), спускаются с гор в долины рек и предгорные
равнины (верхнее течение р. Куры, Цветков), появляются в садах, огородах
и задворках населенных пунктов (Россиков, 1884); в марте вновь возвраща-
ются на места гнездовий в горы (Цветков).
В зимнее время на Кавказе (преимущественно предгорья Дагестана)
встречаются птицы, неотличимые от P. p. lucida, повидимому, залетают во
время осенних кочевок с низовьев Волги и прилежащих районов.
Б и от о п. Окаймленные кустарниками луга и поля в долинах рек, участ-
ки лесостепи и посевов в предгорьях, посевы в лесной зоне, степи и луго-
вины в горах вплоть до субальпийских лугов (местами встречаются в одних
стациях с кекликом; окрестности б. Темир-Хан-Шуры, Дагестан; Бильке-
вич, 1893), реже по травянистым склонам среди можжевельника. В лесу
встречаются лишь во время кочевок, на пролете (Цветков). В период гнездо-
ванья в Даралагезском хребте встречены на высотах от 1660 до 2315 м над
уровнем моря, в Сарайбулагском хребте до 2400 м\ в долине р. Мисханы
встречены в конце ноября на высоте 2580 м над уровнем моря (Даль, 1944,
1948, 1949).
Зимой птицы держатся обычно близ хлебных полей и рисовых планта-
ций в зарослях кустарников и держи-дерева (Сатунин, 1907).
Численность. Различна в разных участках ареала. Более обиль-
на в долине р. Куры, менее в долинах Малки и Баксана (Россиков, 1884).
236 ГОТРЯД КУРИНЫЕ
Плотность колеблется от 0,06 шт. на 1 га в горной степи до 0,9 шт. на 1 га
в кустарниках (Даль, 1944). Колебания численности изучены мало. Причины
их, повидимому, в основном, как и у других подвидов, зависят от степени
морозности и многоснежности зимы. В Кизлярском радоне в 1921 г. была
крайне редка, летом 1922 г. многочисленна (Дагестан; Беме, 1925).
Экология. Размножение. Слабо изучено. Разбиваются на пары,
повидимому, в конце марта—начале апреля. В горах выше древесной расти-
тельности гнездо с 12 совершенно свежими яйцами было найдено 30 мая,
а с 15 слабо насиженными яйцами—2 июня 1937 г. (Спангенберг), однако
первые кладки, вероятно, бывают уже в начале мая. Гнездо—неглубокая
ямка в земле со скудной выстилкой из травы и корешков. Яйца осмотренных
кладок очень сходны с яйцами P. p. lucida, отличаясь лишь несколько более
заостренным концом; окраска светлооливковая или такая же, как и у пре-
дыдущего подвида.
Птенцы ростом чуть больше перепела встречены 27 июня (Дагестан;
Билькевич, 1893), величиной в полматку—10 августа (с. Имырлез, Армения;
Ляйстер и Соснин, 1942), летные птенцы добыты 22 июля (Даралагезский
хребет; Даль, 1949); птенцы, достигшие размеров взрослых птиц, встречены
в июле (Кизляр; Беме, 1925); обычно на плоскогорьях молодые достигают
полного роста в середине августа, а в горах на 2—3 недели позже (Осетия,
Беме, 1926). В конце лета птицы держатся выводками. Осенью сбиваются
в большие табуны (Россиков, 1884).
Линька. Молодые заканчивают линьку в первый годовой наряд
в конце августа на плоскогорье, а в горах на 2—3 недели позже (Северная
Осетия, Беме, 1926).
Взрослые птицы в мае-июне не линяют. У самки от 26 июня 1923 г.
(Карабудахкент, Дагестан) на туловище следов линьки незаметно, выпало
девятое первостепенное маховое, а десятое в нераскрывшемся пеньке.
Самка от 16 июля из Кизляра в энергичной линьке. У самки от 28 августа
1924 г. из района Кизляра все туловище в новом, частью еще не вполне вырос-
шем пере; первое первостепенное маховое в нераскрывшемся пеньке, второе
в две трети, третье в пять шестых, а четвертое—десятое полной длины, закан-
чивается рост рулевых. Взрослые птицы от сентября—октября в новом,
полностью выросшем пере.
Питание. Сходно с другими подвидами. В мае в желудках и зобах
птиц встречены стебли злаков, семена бобовых и почвенные усачи Dorcadion
sp. (Сарайбулагский хребет; Даль, 1944); летом—саранчовые, муравьи,
зерна ячменя, листья дикой люцерны и семена некоторых бобовых; в одном
зобу было 5 г ячменя и 3 г семян бобовых (Даралагезский хребет; Даль,
1949). Повидимому, птицы регулярно посещают водопой; в Сарайбулагском
хребте отмечены полеты на водопой в верховья р. Хагачала (Даль, 1944).
Враги. Остатки куропаток встречаются в погадках филина (Сарай-
булагский хребет; Даль, 1944).
Хозяйственное значение. Объект спортивной охоты. Добывается пре-
имущественно ружьем. Местами на Кавказе практикуется охота с соколами
и ястребами (Каверзнев, 1935). В результате несоблюдения сроков охоты
и отсутствия регулирования отстрела местами сильно уменьшается в чис-
ленности (Северная Осетия, Беме, 1929).
Описание. Размеры и строение. Длина крыла самцов A1) 150—162,5,
самок 149—158,5, в среднем, соответственно, 154,9 и 154,2 мм (коллекция Зоологического»
музея Московского государственного университета); длина крыла 145—170 мм (Армения;
Даль и Соснин, 1941).
Окраска. Очень сходна с P. p. luclda, отличаясь от последней очень неясно.
Еще более тусклосерого общего тона окраски с меньшим развитием рыжего цвета по верху
туловища; рыжеватый оттенок по верху шеи и на боках груди обычно почти не выражен.
СЕРАЯ КУРОПАТКА 237
Рыжие поперечные полосы на пояснице и верхних кроющих крыла выражены несколько
слабее, а у некоторых особей совсем отсутствуют.
Среди зимних экземпляров (верховья Терека и Кубани, Дзауджикау) встречаются
птицы, совершенно неотличимые от Р, p. lucida; повидимому, это особи более северных
популяций, залетевшие сюда на зимовки. В это же время встречаются птицы, видимо
тоже залетные, неотличимые по внешнему виду от западносибирских птиц (самец
от 12 марта 1916 г., с. Скобелевка б. Елизаветпольской губ.).
Сибирская серая куропатка Perdix регйщ rob us ta Homeyer et Tancre
Perdix robusta. Homeyer und Tancre. Mitth. Orn. Verein. Wien, VII, 1883,
стр. 92, Алтай.
Синонимы. Perdix perdix arenicola. В u t u г 1 i n. Orn. Monatsber., 1904, стр. 148,
пески Туссун, 65 км к юго-западу от Тургая.-— Perdix perdix buturlini. S а г u d n у und
L о u d о п. Orn. Monatsber., 1907, стр 47, восточная часть Сыр-Дарьинского бассейна.
Распространение. Ареал. От*Уральского хребта (цепь Приуральских
озер), р. Урала и восточных берегов Каспийского моря до Алтая и Западного
Саяна. Северная граница проходит через Тюменскую обл. по долине р. Туры
(куда впервые проникла в середине прошлого века; Словцов, 1892), далее
встречена в б. Сургутском уезде, т. е. севернее 61° с. ш. (Чугунов, 1915).
Восточнее есть по левобережью р. Тары и, повидимому, доходит до г. Тара
(Шухов, 1928), т. е. до 57° с. ш. Дальше северная граница ареала проходит
несколько севернее г. Томска и круто спускается к юго-востоку, проходя
через Хакасскую и Тувинскую автономные области (г. Минусинск,
с. Уинское, р. Быстрая, Бейгрединская, Туранская и Иджимская степи;
Нестеров, 1909).
Южная граница гнездовой области этой серой куропатки проходит через
низовья р. Урала на северное побережье Аральского моря. Далее на восток
населяет долину Сыр-Дарьи, предгорья Кара-Тау, долины р. Чирчика и
Келеса, долину р. Чу и низовья Или; заходит в пустыню Бет-Пак-Дала
(Селевин, 1933) и Балхаш-Алакульскую пустыню (Долгушин, 1948). В этом
районе проникает и в предгорья (по северным склонам Таласского Ала-Тау
поднимается до 1800—2000 м над уровнем моря; Шевченко, 1948). Встречена
в Джунгарском Ала-Тау, в Тарбагатае и Зайсанской котловине, Алтае (где
кочующие птицы в октябре встречены у Телецкого озера; Фолитарек и
Дементьев, 1938) и далее до Хакассии.
Характер пребывания. Оседлая или кочующая в осенне-
зимний период птица. Зимой придерживается малоснежных участков
с зарослями бурьянов, терна, по озимям, в поймах рек, в осиновых колках,
на бесснежных склонах холмов, на полях у поселков и овечьих тырл.
Из многоснежных районов полностью или частично отлетает в более
южные малоснежные участки. Зимует в Волжского-Уральских песках, тугаях
Сыр-Дарьи, в предгорьях Западного Тянь-Шаня и т. п. (разные авторы).
Проникает на окраины Кара-Кумов, где держится по саксаульникам и кус-
тарниковым зарослям (Зарудный, 1915). Зимой увеличивается в числе (пови-
димому, за счет зимующих) в Зайсанской котловине (Хахлов, 1929). Из
Таласского Ала-Тау откочевывает на равнину и в предгорья (Шевченко,
1948). Осенью сибирские серые куропатки встречены на западном побережье
озера Байкал (Туров, 1940), повидимому, залетевшие из Западного Саяна.
В нижнем течении Сыр-Дарьи число зимовавших птиц начинает заметно
уменьшаться с половины апреля (СпангенбергиФейгин, 1936); осенью прико-
чевавшие птицы появляются в последней трети сентября. (Зарудный, 1916)
Биотоп. Наиболее обычна в разнотравно-злаковых степях с мозаи-
кой колков, кустарников, озер и пашен, в тугаях и на лугах (Казахстан;
Формозов, 1950); встречена на гнездовье в открытой ковыльной степи, на
полянах в сосновом бору и на поросших вершинах песчаных барханов (Се-
верный Казахстан; Ульянин, 1949). Повсеместно встречается в зарослях
238 ОТРЯД КУРИНЫЕ
кустарников по берегам рек и озер, в мелколесье осиновых и березовых кол-
ков, по оврагам в степи и ущельям в предгорьях, по лугам и хлебным полям.
Зимой встречается по гумнам и у стогов, у поселений, в зарослях по
берегам рек и озер. В холода ночует, зарывшись в снег (Щепотьев, 1934).
Численность. Неравномерна в разных участках ареала. Много-
численна в Тюменской обл. (Ларионов, 1926), окрестностях Барнаула
(Велижанины, 1929), Тарбагатае (Хахлов, 1929), Алтае (Формозов, 1950)
и т. д. Спорадично распространена в окрестностях Акмолинска (Штегман,
1928), не найдена у Кокчетава (Залесские, 1931), обычна по Среднему Уралу,
но малочисленна по Илеку (Зарудный, 1888). На численности резко сказы-
вается вырубание кустарников: в 1933 г. в долине р. Кульдемен-Темир
после вырубки кустарников не встречено ни одного выводка, а в полосе лоха
длиной 200 м найден большой выводок (Актюбинская обл.; Формозов, 1950).
Численность птиц резко снижается после многоснежных холодных зим
и гололедицы. После тяжелой зимы 1897—98 г. почти исчезли куропатки по
Иргизу (Сушкин, 1908); в малоснежную с частой гололедицей и снежными
буранами зиму 1936 г. отмечена гибель куропаток в Северном Казахстане;
после двухдневного бурана с температурой до—25° С 18 марта в кустах
ивняка найдены 6 замерзших птиц (Наурзумский заповедник; Ульянин,
194ь^ В Таласском Ала-Тау в 1944 г. за двухчасовую экскурсию встре-
чали 4—6 выводков, а после многоснежной зимы с буранами и моро-
зами до—35°С осенью за две недели поисков найдены всего 2 выводка
(Шевченко, 1948). После повсюду необычайно суровой и морозной зимы
1939—40 г. в окрестностях Тюмени за месяц охоты встречен лишь один выво-
док; под Омском зимой находили замерзших птиц, а летом 1940 г. птиц было
в два раза меньше, чем осенью 1939 г. (Дементьев и Шимбирева, 1941).
Отмечены случаи гибели выводков и взрослых птиц летом. В Наурзум-
ском заповеднике после жарких дней с температурой до 40—50° С сильно
похолодало; 17—19 июня 1936 г. температура упала до 8—10° С, давление
до 726 мм, все время шли грозовые дожди, а ветер достигал 17 м/сек\ после
этого была найдена мертвая самка с выводком, повидимому, птицы погибли
от переохлаждения (Рябов, 1949). В ряде районов численность птиц умень-
шилась в результате перепромысла (Кокчетавские боры; Штегман, 1928).
Экология. Размножение. Моногамы, держатся парами большую
часть года. Только в парах птицы встречены уже в начале апреля (р. Чу,
Долгушин, 1939; окрестности Новосибирска; Щепотьев, 1934). Брачные игры
проходят рано утром еще до восхода солнца: самец вылетает на небольшое
возвышение и начинает издавать своеобразный призывный крик, самка
тихо отвечает на него и приближается к самцу, после чего происходит спа-
ривание; первое спаривание отмечено 10 марта 1936 г., а окончание перио-
да спаривания—10 июня; период спаривания растягивается в популяции на
90 дней, причем максимальная величина семенников и откладка яиц прохо-
дит у разных особей в разные сроки; семенники у самцов увеличиваются
с начала марта, наибольшей величины достигают во второй половине мая
A9 мм 28 мая) и уменьшаются до размеров, характерных для периода
покоя, у некоторых птиц лишь к концу августа (Северный Казахстан;
Ульянин, 1949); повидимому, это связано с разновременным достижением
физиологической готовности к размножению в разных возрастных группах.
Массовая кладка проходит с конца апреля и по конец мая. Число яиц
в полной кладке от 17 до 26; ежедневно откладывается по одному яйцу,
ежедневно повторяется и спаривание. Форма яиц несколько варьирует от
правильной яйцевидной до почти грушевидной; скорлупа бледнооливкового
цвета, варьирующая от бледнозеленоватого до бледного зеленовато-серого.
Размеры яиц чаще 36x26 мм, вес (в период полной кладки)от8,75 до 13,75 г
(Северный Казахстан; Ульянин, 1949).
СЕРАЯ КУРОПАТКА 239
Гнездо обычно под защитой кустика травы или кочки; вырытая в земле
ямка хорошо выстлана сухой травой, к концу насиживания в лотке есть
и перья самки. Длительность насиживания колеблется в разных гнездах
от 21 до 26 дней. Самка сидит на гнезде очень плотно, сходя на кормежку лишь
на несколько минут; самец держится в 10—15 м от гнезда, охраняя самку
(Ульянин, 1949).
Вылупление птенцов в одном гнезде проходит в течение суток. Самка
уводит обсохших птенцов от гнезда; птенцы очень подвижны и за 2 часа ухо-
дят на 200 м и более. Массовое вылупление в Северном Казахстане начинается
с конца первой декады июня (Ульянин, 1949); в Канинском районе Томской
обл. птенцы встречены 16 июня (Иогансен, 1907). Хорошо летающие птенцы
размером со взрослых птиц встречены в середине августа (Северный Казах-
стан—Михеев, 1938; Кузнецкая степь—Зверев, 1927).
Выводки уходят в участка разнотравья, где густой травянистый покров
и обилие насекомых. Кормятся обычно по утрам и вечерам; в полдень отды-
хают в тени, купаются в пыли. При опасности птенцы затаиваются, а самка
отводит от выводка. Летающие птенцы при опасности улетают вместе с роди-
телями. Осенью (август-сентябрь) выводки объединяются в стайки.
Линька. Общее течение линьки сходно с другими подвидами.
Молодые линяют в первый годовой наряд в июле—сентябре, отдельные особи
полностью заканчивают линьку лишь в октябре.
У молодых самца и самки, добытых 23 июля (Северный Казахстан;
вес 195 и 212 г) по телу отдельные полураскрывшиеся пеньки первого
годового наряда; первое первостепенное маховое в чуть раскрывшемся
пеньке, второе в три пятых, третье почти полной длины, у основания в чех-
лике, четвертое—шестое полной длины (птенцовая генерация), седьмое
(годового наряда) в нераскрывшемся пеньке, восьмое в половину, девятое
в три четверти и десятое полной длины, энергично растут новые рулевые.
В сходном состоянии линьки птенцы от 15 августа (р. Сакмара), от середины
сентября (Юго-Восточный Урал), от 9—15 августа (Северный Казахстан),
от 10 августа (Караганда). Полностью завершившие линьку в первый годовой
наряд птенцы встречены 14 августа (Юго-Восточный Урал), 24 сентября
(Северный Казахстан), 30 июля (р. Чу).
Полная линька проходит один раз в году, сроки ее растянуты в связи
с растянутостью периода размножения. В мае—первой половине июня
большинство птиц не линяет; линька начинается обычно в конце июня—
в июле. Самка от 24 мая (Северный Казахстан) без следов линьки с боль-
шим наседным пятном, а у самца от 28 мая (оттуда же) в сильно обно-
шенном пере, по телу пробиваются отдельные, еще не раскрывшиеся пеньки,
выпало десятое первостепенное маховое. Птицы от начала июля (самцы
и самки) начинают энергично линять: наполовину раскрывшиеся опахала
контурных перьев по телу еще не показались над старым пером, девятое
(или восьмое) первостепенное маховое в нераскрывшемся пеньке, десятое
чуть больше половины полной длины; у самок появляются чуть раскрыв-
шиеся пеньки по наседному пятну. Птицы от конца июля энергично линяют:
свежее перо по туловищу уже показалось над старым; первое—четвертое
первостепенные маховые старые, пятое в нераскрышемся пеньке, шестое
в половину, седьмое в три четверти, восьмое в пять шестых, девятое—деся-
тое новые, полного роста; сменяются рулевые и кроющие хвоста. На такой
стадии линьки птицы добывались 12 августа (Юго-Восточный Урал),
21 августа и 2 сентября (Северный Казахстан).
Полностью перелинявшие взрослые птицы с вполне выросшими перьями
встречены в конце сентября—октябре (коллекции Зоологического музея
Московского государственного университета). Повидимому в феврале—в на-
чале марта (по аналогии с другими подвидами) проходит частичная пред-
240 ОТРЯД КУРИНЫЕ
брачная линька (однако соответствующий коллекционный материал (в Зооло-
гическом музее Московского государственного университета отсутствует).
Питание (преимущественно по Ульянину, 1949; Северный Казах-
стан). Основу питания составляет растительная пища, летом рацион попол-
няется животными кормами. В марте (и повидимому всю зиму) в зобах и же-
лудках обнаружены семена и вегетативные части пырея, астрагала, ковыля,
бобовых. В гололедицу, когда другие корма недоступны, птицы поедают
зерновки ковыля и тырсы; у шести найденных мертвыми птиц острые зер-
новки ковыля воткнулись под язык, в стенки пищевода и местами, пройдя
через стенки зоба, достигли кожи или застряли в мышцах. В апреле в желуд-
ках найдены пырей, листья каких-то злаков, полынь, соцветия кермика и зер-
на гречихи. В мае—июле состав корма более разнообразен и включает листья,
соцветия, стебли и корни многих растений (в том числе и тюльпаны); в июле
рацион пополняется животными (муравьи и итальянский прус, редко саран-
ча, чернотелки и другие насекомые). В августе рацион растительных кормов
еще более разнообразен (до 18 видов—гречиха, просо, васильки, солодка,
бобовые, спаржа и т. п.). Зачисление куропатки в разряд вредителей пше-
ницы, ржи, овса и проса (Западная Сибирь; Зверев и Климов, 1931) мало
обосновано, так как птицы, в основном, поедают падалицу культурных злаков.
Питание сочными зелеными листьями и луковицами растений (тюльпаны,
мятлик и т. п.) и насекомыми позволяет выводкам жить в 10—12 км от бли-
жайших водопоев, используя лишь случайные водоемы (лужи на солонцах,
в выбоинах дорог и т. п.), наполняющиеся водой лишь 2—3 раза в лето
(Северный Казахстан; Ульянин, 1949).
Враги. В Северном Казахстане серая куропатка служит добычей ястре-
ба-тетеревятника, орла-могильника, балобана, в меньшей степени луней и
коршунов; вероятно добывают их также волк и лиса (Ульянин, 1949).
Хозяйственное значение. Повсеместно ценный объект спортивной охоты,
местами объект значительного зимнегсУ промысла (лесостепная часть Казах-
стана, Алтай; Формозов, 1950; меньше на севере—Тарский округ, Шухов,
1928 и другие районы Западной Сибири). За зиму 1931—32 г. только на Ленин-
градской экспортной базе было переработано 216 000 серых куропаток (всех,
подвидов), составивших 12% от всей поступившей дичи; в 1930—31 г.—116 400
или 4,6%, в 1929—30 г.—168 700, или 17,2%; в общих заготовках дичи
куропатка на Урале составляла 16%, в Западной Сибири—9,7%, в Казах-
стане—59,8% (Рудановский и Насимович, 1933).
Птицы добываются преимущественно ружьем; в зимнее время часто
устраивают привады из снопов проса (б. Тургайская обл., Полферов, 1896).
На Урале, в Западной Сибири и ряде районов Казахстана ловят шатром
(Гринберг, 1934), на приваде, у скирдов и на гумнах—волосяными петлями
(Западная Сибирь, Зверев, 1930). В Казахстане местами до настоящего вре-
мени добывают ловчими птицами (тетеревятник, реже чеглок).
Описание. Размеры и строение. Длина крыла самцов B5) 147—163,
самок A9) 150—159, в среднем, соответственно, 157 и 154,5 лик. Средний вес (по взвеши-
ваниям на Ленинградской экспортной базе в марте~!1932 г., по Рудаковскому и Наси-
мовичу, 1933) птиц из Кунгура—самцов B6) 420,9, самок B1) 397,5 г, из Омска—самцов
C4) 395,2, самок C1), 377 г, из Татарска—самцов A7) 408, самок A2) 390,7 г, из
Семипалатинска—самцов F6) 402,5, самок A4) 388,8 г, из Новосибирска—самцов A0)
415,2, самок A3) 405,2 г, из Томска—самцов C2) 397,7, самок A0) 392,6 г; средний вес
птиц из всех этих пунктов самцов A85)—404,1 и самок A01) 389,7 г.
Самки по длине крыла и по весу лишь чуть меньше самцов; весовые различия птиц
из разных участков большого ареала подвида также невелики.
Окраска. По общему тону окраски еще серее, чем среднерусская куропатка.
Передняя часть спины и шея без рыжего оттенка; белые наствольные полосы на спине
и кроющих крыла с чуть заметным сероватым отливом. Поперечные полосы на кроющих
поясницы и хвоста более узкие, каштанового или каштаново-бурого цвета. Общий тон
окраски верха чистосерый с мелким темнобурым крапом. Грудь более чистосерого цвета
БОРОДАТАЯ КУРОПАТКА
241
с несколько более грубым темнобурым крапом. Полосы на боках туловища рыжевато-
каштанового или каштаново-бурого цвета.
Вариации в окраске в популяции довольно значительны. Так, среди наурзумских
чистосерых сверху птиц изредка встречаются особи с более или менее выраженным
рыжеватым оттенком на спине; одна птица из Зеренджинского лесничества (Кокчетав-
ский район, Северный Казахстан) неотличима по общему тону окраски от среднерусских
птиц; птицы из Тургая не отличимы от наурзумских, но один экземпляр очень близок
к среднерусскому подвиду.
20. Бородатая куропатка Perdix daurica Pall.
Tetrao perdix var. daurica. Pallas. Zoogr. Rosso-Asiat., II, 1811, стр. 78, Даурия.
Синоним. Perdix bar bat a. Verreaux etDes Murs. Proc. Zool. Soc. London,
1863, стр. 63, табл. IX. Нерчинск.
Русское название. Книжное, данное Палласом: удлиненные, заостренные пе-
рышки по бокам горла несколько напоминают бороду.
Распространение. Ареал. От Ферганы, Семиречья, северо-западных
и северных лесостепных участков Алтая через южные районы Краснояр-
ского края (Минусинские степи), Прибайкалье и Забайкалье вплоть до
Карта 21. Распространение бородатой куропатки
Perdix daurica
1—P. d. daurica, 2—P. d. suschkini, 8—P. d. przewalshii;
a—граница гнездовий, б—не вполне выясненная граница
гнездовий
устья р. Уссури; на юг через Джунгарию, Монголию и Маньчжурию до
Восточного Тянь-Шаня, Северо-Восточного Тибета (верховья Желтой и Го-
лубой рек) и Северо-Западного Китая.
Характер пребывания. Оседлая птица; в осенне-зимний
период совершает небольшие кочевки.
Биотоп. Кустарниковые заросли в степях и на лугах, поймы рек,
степные колки, опушки лесов, лесные поляны, пашни, безлесные склоны
гор, субальпийские луга. Избегает крутых скалистых склонов гор, не захо-
дит в пустыни.
Подвиды и варьирующие признаки. От Ферганы, Семиречья и южных склонов
Восточного Тянь-Шаня через Тарбагатай, Алтай, Монгольский Алтай и Хангай, Запад-
ный и Восточный Саяны (на север до Красноярска) и Тувинскую автономную область
до степей Прибайкалья, Забайкалья и прилегающих районов Северной Монголии распро-
странена P. d. daurica Pallas; Цайдам, район Куку-Нора, прилегающие части
16 Птицы СССР, т. IV
242 ОТРЯД КУРИНЫЕ
Восточного Нань-Шаня и, вероятно, верховья Желтой и Голубой рек населяет P. d. prze-
walskii S u s с h k i n, 1926; в Маньчжурии, Уссурийском крае и Северном Китае живет
P. d. suschkini P о 1 j a k о v, 1915.
Подвиды, как и у серой куропатки, слабо различимы по вариациям окраски и не-
большим, различиям в величине. Пограничные районы, заселенные промежуточным»
формами, занимают большие территории. Очень велика и индивидуальная изменчивость,
иногда перекрывающая подвидовые различия. Особенности биологии подвидов совер-
шенно не выяснены.
Систематические замечания. По образу жизни, строению и окраске бородатая куро-
патка очень близка к серой куропатке. Однако перекрывающиеся на большой площади
ареалы (Семиречье, Алтай) позволяют считать их самостоятельными, но очень близкими
видами.
Сибирская бородатая куропатка Perdix daurica daurica Pall.
Синонимы. Perdix daurica turcomana. S t о 1 z m a n n. FBull. Soc. I. Nat. Moscou
1897, стр. 79, Гульча в Восточной Фергане.—Perdix daurica Occidentalis. B u t u г 1 i n.
Orn. Monatsb., 1908, стр. 46, Семиречье (Кульджа, Иссык-Куль, Нарынкол).
Распространение. Ареал. От Ферганы, Семиречья и южных скло-
нов Восточного Тянь-Шаня через Тарбагатай, Алтай, Монгольский Алтай,
Хангай, Саяны, Танну-Ола и Кентэй до Забайкалья включительно;
может быть доходит до западных склонов Большого Хингана.
Детали распространения в пределах Советского Союза подробно не
выяснены. На западе по предгорьям хребта Кара-Тау доходит до системы озер
р. Аса (Кашкаров, 1928). Далее (на восток и на юг) встречена в Таласском
Ала-Тау (Шевченко, 1948), в западной части Туркестанского хребта
(Мекленбурцев, 1951), в восточной части Ферганской долины, в Киргиз-
ском хребте (Спангенберг и Дементьев, 1935), по всему Семиречью (Шнит-
ников, 1949), в горных участках долины р. Или, в Джунгарском Ала-Тау,
в Тарбагатае, Зайсанской котловине, в лесостепных районах Алтая (Таш-
тып, реки Урусул, Чулышман, Бухтарма, Чуйская степь и т. п.; по Суш-
кину, 1938), в котловинах и лесостепи Западного и Восточного Саяна (на
север до Красноярска; коллекция Зоологического музея Московского Госу-
дарственного университета), в Тувинской автономной области, в степях южной
части Иркутской обл. и в Бурято-Монгольской автономной республике, в Дау-
рии (на север до южных частей Витимского плоскогорья; Штегман, 1928).
Повидимому, залетевшая из Ферганы во время осенних кочевок птица
добыта в Заалайском хребте F ноября; Таджикистан; Иванов, 1940). В ав-
густе 1936 г. куропатка, повидимому также залетная, была добыта в Бара-
бинской степи южнее ст. Чаны (Рузский, 1940). Под Томском одиночные боро-
датые куропатки иногда встречаются осенью в стайках серых куропаток
(Залесский, 1917).
Характер пребывания. Оседлая птица. Зимой стайки пред-
принимают незначительные кочевки: уходят с гор в предгорья и на рав-
нину (Киргизский хребет; Спангенберг и Дементьев, 1935), появляются
в нижней половине еловой зоны гор (Алма-Атинский заповедник; Шульпин,
1939), из узких долин уходят на малоснежные склоны (Алтай; Фолитарек
и Дементьев, 1938), приближаются к поселениями аулам, кормясь на местах
тебеневки скота (Северная Монголия; Формозов, 1946). Указание Северцова
A873) о том, что в горах окрепшие выводки поднимаются до верхнеальпий-
ской зоны, как давнее, нуждается в проверке.
Биотоп. Лесостепи и степи, даже каменисто-щебенистые, на равнине
и в предгорьях, долины рек, луга, посевы, опушки леса и небольшие лес-
ные массивы (Забайкалье). В степи предпочитает участки разнотравья
и кустарниковые заросли. По открытым склонам гор, логам и ущельям,
полянам, зарослям кустарников и опушкам лесов поднимается до субаль-
пийских лугов (в Семиречье до 3000 м над уровнем моря; Шнитников, 1949).
Скалистых и очень крутых склонов гор избегает; в участки пустыни не про-
БОРОДАТАЯ КУРОПАТКА 243
никает. В полосе предгорий распространена значительно шире, чем серая
куропатка.
Численность. В разных участках ареала различна. Обыкновенна
в степях и лесостепях Урянхайского и Минусинского краев (Сушкин, 1914),
в Забайкалье (Штегман, 1929); редка в междуречье Чилика и Чарына (Семи-
речье; Корелов, 1948), в Илийской долине (Шестоперов, 1929) и т. п. На
степных склонах и горных лугах Алтая и Тарбагатая бородатая куропатка
распространена шире серой (Формозов, 1950); в Киргизском хребте менее
многочисленна, чем кеклик и даже улар (Тянь-Шань; Спангенберг и Дементьев,
1935). Изменение численности по годам несомненно существует, но совершен-
но не изучено. Весной держится парами, летом выводками, осенью стайками
в 15—30 шт. и более; вначале зимы в стае отмечали до 200 птиц (Северная
Монголия; Бианки, 1915).
Экология. Размножение. Почти не изучено. Повидимому, моно-
гамы. В середине марта разбиваются на пары (Алтай; Фолитарек и Демен-
тьев, 1938). Начинают откладку яиц вероятно в конце апреля—начале мая.
Самка со зрелыми фолликулами добыта 8 мая (Читинская обл; Павлов, 1948).
с готовым яйцом в яйцеводе 19 мая (Заилийский Ала-Тау; Шнитников,
1949). 20 мая 1907 г. в предгорьях у Алма-Ата найдено гнездо с 20 яйцами;
гнездо помещалось под кустиком верблюжьей колючки (Шнитников; 1949);
16 июня 1936 г. была найдена кладка из 16 насиженных яиц; гнездо поме-
щалось на склоне поросшей шиповником неглубокой щели и представляло
собой небольшую ямку, скудно выстланную сухой травой (Киргизский хре-
бет; Спангенберг).
В осмотренной кладке (Киргизский хребет) яйца по форме очень
сходны с яйцами серой куропатки; они правильной яйцевидной формы с хо-
рошо выраженным острым и тупым концом; в той же кладке были яйца,
несколько приближающиеся по форме к грушевидным. Окраска яиц гли-
нисто-бурая с буроватым или коричневатым отливом; видимо, в некоторых
случаях яйца бородатой куропатки не отличимы от яиц серой куропатки.
Размеры яиц B2) 32,4—35x25,4—27,5, в среднем 33,6x26,6 мм (Хартерт,
1921—22).
Повидимому, период размножения в популяции растянут: 19 июня
1898 г. (ст. стиля?) пуховые птенцы добыты в долине р. Урусула (Онгудай,
Алтай; Кащенко, 1899), 15 июля 1913 г. совсем крошечные птенцы встре-
чены на берегу у р. Текес (долина р. Или; Шестоперов, 1929), а в середине
июля птенцы ростом в половину взрослых добыты в Киргизском хребте
(Тянь-Шань; Спангенберг и Дементьев, 1935).
Линька. Не изучена; ее последовательность, повидимому, сходна
с таковой у серой куропатки. У взрослых птиц годовая линька начинается
в конце июня; весной проходит частичная линька, охватывающая перья шеи.
Птенцы сбрасывают все остатки пуха, достигнув трех четвертей величины
взрослых птиц; линьку в первый годовой наряд заканчивают в сентябре
(Алтай; Сушкин, 1938).
Взрослые птицы от начала июня еще не начали линьку (Тува); птицы
от середины сентября и от октября уже в полностью выросшем свежем пере
(Тянь-Шань); молодые птицы от конца сентября уже в полностью выросшем
первом годовом наряде (Иссык-Куль; коллекции Зоологического музея
Московского государственного университета).
Питание. Не изучено; повидимому, сходно по характеру с пита-
нием серой куропатки. До снега кормятся опавшими хлебными зернами,
семенами дикой конопли, охотно поедают ягоды облепихи и шиповника,
копаются в опавшей листве; в глубокие снега кормятся по токам (Алтай;
Фолитарек и Дементьев, 1938), в местах тебеневки скота, по дорогам и на
жнивье (Сушкин, 1938). В летнее время вероятно питаются, как и серая куро-
16*
244 ОТРЯД КУРИНЫЕ
патка, насекомыми. Более интенсивно кормятся утром и перед вечером;
днюют в кустах по склонам сопок (Некипелов и Скалой, 1936), в оврагах.
Хозяйственное значение. Повсеместно объект спортивной охоты. В ряде
районов имеет важное промысловое значение. В заготовках дичи в Восточной
Сибири составляет до 7,2%, в Западной Сибири до 4,9%, в Казахстане до 1%
(Рудановский и Насимович, 1933). В Тувинской авт. обл. промысел
резко возрос в связи с постройкой железной дороги и за зиму 1899—1900 г.
лишь с Кемчика вывезено не менее 12 000 пар бородатых куропаток
(Сушкин, 1914).
Рис 39. Бородатая куропатка (самец и самка)
Добывается преимущественно ружьем. При промысловой добыче при»
меняют лов капканами и петлями на тропках в кустах и шатрами на привадах
Полевые признаки. Очень похожа на серую куропатку. На свободе
с трудом отличается от последней по охристо-желтоватому налету на зобе
и на передней части груди.
Описание. Размеры и строение. Очень сходны по внешнему виду
истроениюс серыми куропатками. Крыло тупое; его формула обычно 4^3^2>5>1,
реже 3^4>5<6^2>7>1.
Длина крыла у самцов A2) 143—151, у самокA3) 140—152, в среднем соответственно
147,3 и 145,5 мм. Длина тела 300—320 мм, размах крыльев 480—520 мм. Средний вес
птиц, поступивших из различных районов на Ленинградскую экспортную базу в марте
1932 г., был следующий: вес самцов D) из Новосибирска 341,2, самок D) 327,5 а, из Аба-
БОРОДАТАЯ КУРОПАТКА
245
кана самцов B9) 315,9, самок B0) 315,3 г, из Иркутска самцов B2) 297,4, самок A4 J96,2 г,
самцов из Верхнеудинска B7) 295,9, самок B8) 289 г; средний вес самцов (82) 305,6,
самок F6) 300,8 г.
Окраска. Пуховичок. Лоб, темя и затылок каштаново-рыжие с черно-бурыми
полосками; бока головы охристо-желтоватые с темнобурыми прерывистыми надглазничной
и подглазничной полосами. Шея сверху тусклая рыжевато-охристая с продольной темно-
бурой полосой. Верх туловища из сочетания каштаново-рыжих и темнобурых порций
пуха, последние образуют прерывистые продольные полосы на туловище и крыльях.
Подбородок и горло желтовато-белые; брюшко и грудь светлые, грязного желтовато*
серого цвета.
Рис. 40. Пуховые птенцы бородатой куропатки
Птицы в птенцовом наряде очень похожи по окраске оперения на
птенцов серой куропатки, но еще светлее и серее последних.
Молодые в первом годовом наряде по общему тону окраски
сходны со взрослыми птицами или несколько тусклее последних. Темя и затылок буро-
ватые со светлыми продольными пестринами. Охристое пятно на зобе развито слабее,
а черное пятно на брюхе имеет значительно меньшую площадь, чем у взрослых птиц,
или иногда лишь только намечено.
Взрослые птицы по общему тону окраски и рисунку оперения очень
сходны с серой куропаткой, отличаясь от последней лишь некоторыми, иногда не совсем
отчетливыми особенностями. Под глазом проходит узкая черная полоса; лоб, полоса над
глазом, щеки, подбородок и горло охристо-желтые без красноватого оттенка,свойственного
серой куропатке. По бокам горла, от ветвей нижней челюсти и ниже, растут удлиненные
перья, окрашенные одинаково с горлом и образующие бороду. Такого же цвета, как под-
бородок и горло, большое пятно, занимающее зоб и переднюю часть груди; иногда оно
узкой полосой, проходящей по нижней поверхности шеи, незаметно сливается с охристой
окраской горла. Большое подковообразное пятно на передней части брюха черного или
черно-бурого цвета. Поперечные полосы на боках туловища каштаново-бурые или кашта-
новые, без рыжеватого оттенка; более широкие и четкие, чем у серых куропаток. Опере-
ние верхней стороны туловища очень сходно с оперением серых куропаток; однако попе-
речные каштановые полосы на передней части спины более четко и резко выражены и рас-
положены гуще.
Отличия самок от самцов сходны с таковыми у серых куропаток.
Систематические замечания. Объединение туркестанского и даурского подвидов
бородатой куропатки в один подвид P. p. daurica диктуется следующими обстоятель-
ствами. Основные отличительные признаки этих подвидов—зобное пятно соединяется
с пятном на горле или они раздельны, цвет брюшного пятна черно-бурый или черный—
широко варьируют в пределах популяции из одного района. Примерно у половины про-
смотренных птиц из Тянь-Шаня охристая окраска груди и зоба отделена серой полосой
от такой же окраски горла, у половины—сливается; у части особей брюшное пятно отчет-
ливо чисточерного цвета. То же самое можно сказать о куропатках из-под Красноярска
и Кяхты: у части особей брюшные пятна черно-бурые, у многих охристая окраска горла
отделена серой полосой от охристого пятна на груди.
246 ОТРЯД КУРИНЫЕ
Птицы из Тибета (P. d. przewalskii) хорошо отличимы по общему светлопесочному
тону окраски; так же заметно отличается по более рыжему общему тону окраски и
P. d. suschkini.
Маньчжурская бородатая куропатка Perdix daurica suschkini
Poljakov
Perdix daurica suschkini. Поляков. Орнит. Вестник, 1915, стр. 38, Хабаровск.
Распространение. Ареал. Населяет Маньчжурию (на западе, пови-
димому, проникая в Б. Хинган) и Северный Китай, на юг до Шэньси и
Шаньси. Заходит в Советское Приморье и Приамурье; детали распростра-
нения этого подвида в СССР не выяснены (зимой добывалась у Хабаровска,
встречена в лесостепных участках южнее озера Ханка).
Характер пребывания Повидимому, оседлая птица, воз-
можно совершающая небольшие осенние кочевки.
Биотоп. Преимущественно степные и лесостепные участки ареала,
вблизи полей и зарослей кустарников.
Численность. В пределах Советского Союза (Приморье и При-
амурье) редкая птица, причем некоторые исследователи даже сомневаются
в достоверности ее добычи (Шульпин, 1936). В Маньчжурии, особенно в ее
западных степных частях,—обычная и многочисленная птица, увеличиваю-
щая свой район распространения в связи с вырубкой тайги (Яковлев, 1929).
Экология. Размножение. Не изучено. Вероятно в общих чертах
сходны с таковыми сибирской бородатой куропатки.
Линька. Не изучена.
Питание. Особенности питания и кормовых связей этого подвида
совершенно неизвестны.
Хозяйственное значение. Ввиду крайней редкости в пределах Совет-
ского Союза промыслового значения не имеет; в Маньчжурии—промысло-
вый вид, добываемый в больших количествах. Повидимому, при помощи
человека численность птицы на нашей территории и ее хозяйственное зна-
чение могут быть увеличены при применении простейших биотехнических
мероприятий (завоз достаточного количества производителей в подходящие
биотопы, временное запрещение промысла, подкормка в суровые зимы и т. п.).
Враги. Истребляется многими хищниками—птицами и зверями; в зим-
нее время остатки бородатых куропаток неоднократно встречались в желуд-
ках и зобах мохноногих канюков (Яковлев, 1929).
Описание Размеры и строение. Сходна с сибирской бородатой куро-
паткой. Длина крыла самцов C) 141—148, в среднем 144 мм; длина тела (самец) 310 мм,
размах крыльев 485 мм.
Окраска. Отличается от сибирской бородатой куропатки по общему тону
окраски: по верху тела более интенсивно развит рыжеватый оттенок (в сходной степени
с развитием рыжего цвета у P. p. perdix), рыжая окраска с горла дальше заходит на бока
головы и шеи; надглазничные полосы более широкие; на брюхе и нижних кроющих хвоста
серовато-желтоватый оттенок.
ОТРЯД ГУСЕОБРАЗНЫЕ
ANSERES или ANSERIFORMBS
Характеристика отряда
Морфология. Внешние признаки. Размеры крупные или сред-
ние. Самые крупные из гусеобразных нашей фауны—лебеди: лебедь-шипун
достигает 1,8лс в длину, 2,6 м в размахе и 13,6 кг веса; лебедь-кликун несколь-
ко меньших размеров и весом до 12,7 кг. Один из самых мелких наших
гусеобразных чирок-свистунок имеет длину тела до 0,38 м, размах до 0,64 м
и вес до 0,42 кг.
Большинство гусеобразных отличается несколько грузным, валько-
ватым телом, удлиненной шеей и ногами, отнесенными назад. Голова отно-
сительно крупная, слегка сжатая с боков или сверху. Клюв или короток
и погнут книзу, или удлинен, уплощен и снабжен по краям пластинками
или же зубчиками. Уздечка оперенная или голая. Окраска разнообразная:
снежнобелая, серовато-бурая, иногда из сочетания различных ярких с метал-
лическим отливом тонов или же сплошь матовочерная. У ряда форм на голове
хохол из удлиненных перьев; зачастую удлинены также плечевые и внутрен-
ние маховые. Окраска или одинакова у обоих полов, или различна (у боль-
шинства видов умеренного и холодного поясов). Самцы обычно крупнее самок.
Тело обильно покрыто густым пухом.
Молодые в первом перовом наряде окрашены тусклее взрослых и в ряде
случаев их окраска сходна с самочьей. Вершины их рулевых имеют голые,
слегка булавовидно утолщенные, лишенные бородок концы стержней. Приз-
нак этот сохраняется до 7—8-месячного возраста (от момента выхода из
яйца), т. е. до начала первой линьки. Птенцы покрыты густым бело-
ватым, желтоватым или бурым пухом с темными крупными, симметрично
расположенными пятнами или без них.
Крыло аквинтокубитальное, средних размеров, заостренное, с один-
надцатью первостепенными маховыми. У некоторых видов на сгибе крыла
твердые бугры, иногда шипы. Относительно длинный или умеренных раз-
меров хвост из 12—24 рулевых. Ноги средних размеров или невысокие.
Цевка умеренной длины, сетчатая кругом или только сзади. В этом послед-
248 ОТРЯД ГУСЕОБРАЗНЫЕ
нем случае передняя ее сторона покрыта поперечными пластинками, раз-
витыми иногда только в ее нижней части. Из четырех пальцев первый (зад-
ний) обычно мал и недоразвит; а второй, третий и четвертый соединены
полными или зачаточными перепонками, которые могут, однако, и отсут-
ствовать.
Анатомические признаки. Хорошо развитая, покрытая
пуховидными перышками копчиковая железа выделяет маслянистый сек-
рет, предохраняющий плотные контурные перья от намокания. Подкожный
жировой слой в зависимости от вида птицы, ее возраста, сезона и обилия
пищи может достигать значительной толщины и составлять 10—18% об-
щего веса тела.
Череп десмогнатический с хорошо развитыми вторичными базиптери-
гоидными отростками. Носовые части голоринальные. У большинства
vestibulum отличается утолщенными стенками, а также слабым развитием
гребешков между полостью преддверия и улитки, с одной стороны, и устья-
ми каналов—с другой. Fenestra ovalis округлой формы, по длине и шири-
не короче окна улитки. Aquaeductus vestibuli отличается длиной и хорошим
развитием бугорка у своего отверстия. Canales semicirculares сильно упло-
щены и широки. Fundus в meatus acusticus interims в виде плоскодонной
ямки. Отверстия для ушных нервов сближены или разделены (Туркевич,
1936). Затылочные фонтанели черепа или хорошо выражены, или зара-
стают. Нижняя челюсть с длинным, загнутым кверху processus angularis
posterior.
Шейных позвонков 16—25. Ребер 6—11. Грудная кость с вырезкой
или отверстием на каждой стороне. Вилочка V-образна и крепка. Имеются
m. ambiens и по две пары стернотрахеальных мышц: m. ypsilotracheales
и т. sternotracheales. У большинства язык широк, мясист и покрыт зуб-
цами, соответствующими пластинкам каждой стороны клюва. Хорошо раз-
виты желчный пузырь и функционирующие слепые кишки. Сонные артерии
парные, одинаково развитые на каждой стороне.
Нижняя гортань у самцов часто с парным или непарным костяным взду-
тием, так называемым барабаном, усиливающим звуки, издаваемые птицей»
В других случаях для усиления звуков служит удлиненная петля трахеи,
входящая в киль грудной кости. Легкие отличаются значительной емкостью,
что особенно хорошо выражено у ныряющих форм. В большинстве случаев
у самцов имеется особый выворачивающийся копулятивный орган.
Образ жизни. Гусеобразные населяют самые различные водоемы или,
в крайнем случае, обитают на их побережьях. Большинство прекрасно пла-
вает, но посадка их на воде различна. Одни больше погружают в воду перед-
нюю часть тела, причем хвостовая часть приподнята, а интертарзальное
сочленение выступает из воды; другие держат туловище горизонтально.
Степень погружения тела в воду при плавании различна, и при этом над
поверхностью воды может оставаться спина и шея или даже одна только
голова. Некоторые виды очень хорошо ныряют и могут оставаться в воде
до 3,5 мин., погружаясь на глубину до 40 м Передвигаются на суше
по-разному. Гуси могут хорошо ходить и даже бегать, большинство же
в связи с тем, что ноги их несколько отодвинуты назад, ходит по земле,,
переваливаясь с боку на бок.
Пища гусеобразных разнообразна. Хорошо ныряющие формы питаются
мелкой рыбой, которую ловят в воде, или моллюсками, которых достают
со дна водоемов. Многие виды, опрокидываясь в воду передней частью
тела и вытягивая шею, клювом добывают смешанную пищу из ила на мел-
ких местах. Наконец, гуси и паламедеи кормятся на суше исключительно
растительной пищей, скусывая клювом наиболее нежные части земновод-
ных или наземных растений.
ХАРАКТЕРИСТИКА ОТРЯДА 249
Гнездятся большей частью одиночно, реже устраиваясь разбросанной
колонией. Гнезда на земле или на мелководьях, на деревьях, в дуплах,
в норах глинистых вертикальных обнажений или на скалах, но всегда неда-
леко от воды. Иногда занимается и чужое гнездо. Устраиваемые на земле
гнезда или в виде ямки, выстилаемой пухом часто с примесью различных
растительных материалов, или в виде различных размеров кучи (иногда
до 1 м высотой) из стеблей трав, мха, тростника и других болотных растений
и даже веток кустарников или деревьев. Верх такой кучи представляет вда-
вленное углубление, построенное из тонких и нежных частей растений
и выстланное пухом самки, а иногда и самца.
Яиц от 2 до 14\ у большинства видов 8—10. Меньше всего яиц откла-
дывают паламедеи, больше всего некоторые гоголи и крохали. Период наси-
живания от 20 до 40 дней, у большинства около 25 дней. Средней величины
виды, гнездящиеся в дуплах, насиживают дольше—до 30—31 дня. Строя?
гнезда и насиживают самки.
По выходе из яиц птенцы в течение нескольких часов обсыхают и уже
на другой день добывают пищу вполне самостоятельно, лишь находясь
под присмотром самки или обоих родителей вместе. Рост идет медленно,
и от момента выхода из яйца до времени подъема на крыло у небольших
по размерам видов проходит два месяца, у средних—2—2,5, у крупных даже
до 4 месяцев.
Молодые в первый год жизни не достигают роста и веса взрослых и у
некоторых видов становятся половозрелыми лишь на второй, третий и даже
на четвертый год жизни.
Самцы у ряда видов не принимают никакого участия в насиживании
и выводе птенцов (хотя многие гусеобразные являются моногамами) и с мо-
мента начала насиживания яиц самками покидают последних, собираются
в стайки и отлетают в другие, иногда значительно удаленные от гнездо-
вых, места, где и линяют, а по окончании здесь линьки летят на зимовки.
У других же видов самцы не покидают мест гнездовья и остаются при вы-
водке не только в течение всего гнездового периода, но вместе с самкой и мо-
лодыми улетают на зимовки, весною же вся семья возвращается на гнездо-
вые места, где самец и самка, наконец, отделяются от молодых.
Ход линьки разнообразен и смена перьев на разных частях тела у раз-
личных гусеобразных протекает не одинаково. Для большинства видов
весьма характерна временная утрата способности к полету, связанная с тем,
что все маховые перья выпадают в очень короткие сроки, заменяющие их
новые дорастают до нормы приблизительно через три недели. При этом ход
линьки маховых, варьируя у разных групп, в каждой группе гусеобразных
протекает всегда в определенном порядке. Для лебедей и гусей характерно
быстрое выпадение первостепенных маховых, начиная с внешних первосте-
пенных, а также с самых внутренних второстепенных. Смена маховых,
таким образом, идет от краев к углу крыла. У уток, наоборот, маховые
сменяются, расходясь от угла крыла. У многих видов имеется сложно проте-
кающая сезонная предбрачная и послебрачная линька.
Половой диморфизм гусеобразных выражается в том, что самцы, во-
первых, как уже выше сказано, несколько крупнее самок, и, во-вторых,
в том, что у многих (но не у всех) форм окраска самок и самцов различ-
ная. Последние в таких случаях окрашены в яркие, иногда с металличе-
ским отливом, цвета и к тому же имеют иначе построенный голосовой ап-
парат.
1 Указания на большее число откладываемых яиц (до 19—26 у гоголей, до 21 у серо-
го гуся, также у некоторых уток и нырков) необходимо отнести за счет кладок двух или
более самок в одно и то же гнездо.
250 ОТРЯД ГУСЕОБРАЗНЫЕ
Продолжительность жизни, повидимому, значительна: в условиях не-
воли чирки вьгчивали до 41 года, гуси до 33—65 лет, лебеди до 20 и 24 с по-
ловиной лет; ih> данным кольцевания мы пока располагаем сведениями о жиз-
ни их в природе от 8 до 11 лет, однако данных этих недостаточно.
Систематика. Современные гусеобразные делятся на два подотряда:
паламедей Anchimae и пластинчатоклювых Lamellirostres. Первые яв-
ляются крупными грузными птицами с загнутым вниз, снабженным воско-
вицей крючкообразным клювом, лишенным пластинок или зубчиков. Отно-
сительно высокие их ноги покрыты шестиугольными чешуйками, захо-
дящими и на голень. Задний палец на уровне передних. Плавательных пере-
понок между пальцами нет. Птерилии и аптерии не выражены, и тело сплошь
покрыто перьями и густым пухом под ними. Хвост относительно длинный.
Кожа и скелет сильно пневматизированы и воздушные полости заходят даже
ъ фаланги пальцев. Затылочная часть черепа без фонтанелей. Шея из 19—
20 позвонков. Ребра без processus uncinati (единственный случай среди со-
временных Volantes). На первой и второй метакарпалиях мощные шипы.
Большая грудная мышца относительно мало развита. Сравнительно крупный
железистый желудок с большими полями желез, мускулистый же, наоборот,
мал. Слепая и толстая кишки сильно развиты. Воздушные мешки—saccus
cervicales и saccus claviculares—разделены перегородками на мелкие ячейки.
Копулятивный орган отсутствует.
Паламедей населяют побережья мелководных водоемов Южной Америки
и состоят из одного семейства Anchimidae с двумя родами: Chauna с двумя
видами и Anchima с одним видом.
Подотряд пластинчатоклювых характеризуется прежде
всего своим заканчивающимся твердым «ноготком» клювом, края верхней
и нижней половин которого вооружены поперечно расположенными пластин-
ками или зубчиками. Ноги относительно короткие. Задний палец выше уров-
ня передних, почти всегда снабженных плавательными перепонками. Из
аптерий не выражены только шейные. На перьях иногда добавочный руди-
ментарный стержень. Шея из 14—25 позвонков. Грудина удлинена. Ребер
9—11. Большая грудная мышца хорошо развита. Большинство отличается
сильным и быстрым полетом, но имеются почти не летающие виды (Neso-
netta auclandica) или вовсе не летающие (Tachyeres cinereus), крыло которых
редуцировано.
Пластинчатоклювые представлены одним семейством утиных Anati-
dae, объем отдельных группировок которого еще недостаточно выяснен.
Совершенно неубедительным является мнение, что подразделение Anatidae
на подсемейства несостоятельно, потому что «признаки, принимаемые в ос-
нову этого деления, являются чисто адаптивными» (Тугаринов, 1941), так
как все, что не адаптировано с условиями существования, постепенно уничто-
жается отбором. И как бы ни ухитрялся систематик, все свои таксономи-
ческие построения он обосновывает на признаках адаптивных. Исходя
из этого изложенное выше возражение необходимо отклонить. Между тем
использовать деление семейства утиных на более мелкие подразделения необ-
ходимо в целях обозрения этой многочисленной, относительно древней груп-
пы, представленной в настоящее время 40 родами, 161 видом и 230 формами
(Делакур и Майр, 1945). Число подсемейств, принимаемых современными
исследователями, различно и доходит от 2 (Делакур и Майр) до 8 (Бекер)
или 16 (Беттихер). В последнее время семейство Anatidae делят на подсемей-
ство Anserinae, состоящее из двух триб (Anserini и Dendrocygnini), и под-
семейство Anatinae, представленное семью трибами Tadornini, Anatim,
Aythyni, Cairinini, Mergini, Oxyurini и Merganettini (Делакур и Майр, 1945).
По современным палеонтологическим данным, птицы отряда Anseri-
formes являются древней группой, возникшей в нижнем эоцене или даже
ХАРАКТЕРИСТИКА ОТРЯДА 251
в верхнем мелу (если принимать Gallornis siraeleni из верхнего мела Фран-
ции за примитивного представителя утиных). Во всяком случае в верхнем
эоцене гусеобразные в виде самостоятельной группы уже существовали,
и из отложений этой эпохи известна Romainvillea stehlini, близкая к совре-
менным древесным уткам. В олигоцене найдены вымершие представители
современных родов Anas, Aythya и Anser, а кроме того, вымершие роды,
близкие к лебедям и уткам. Из нижнего и среднего эоцена известно несколь-
ко вымерших видов рода Anasf а в верхнем миоцене гусеобразные были распро-
странены по всем материкам, кроме Австралии. В эту эпоху уже предста-
влен современный род Cygnus и близкий к казаркам род Presbychen. В плио-
цене господствуют современные роды; в нижнем плиоцене найдены вымер-
шие представители современных родов Branta и Dendrocygna, а в верхнем
плиоцене Франции—остатки Anser anatoides, чрезвычайно близкого к совре-
менным мелким тропическим гусям рода Cheniscus, В плиоцене существо-
вали современные виды—широконоска Anas clypeata и красноголовый
нырок Aythya ferina. Плейстоценовые виды гусеобразных уже сходны с совре-
менными; в эту эпоху вымирают четыре вида лебедей и гигантский новозе-
ландский нелетающий гусь Cnemiornis calcitrans, близкий к австралийскому
роду Cereopsis. Однако имеются данные, что он существовал в голоцене.
Из бинагадинских отложений Азербайджанской ССР описаны Cygnus
olor bergmanni, Anser azerbaidzhanicus, там найдены остатки и современных
видов Anser fabalis, Anser anser и Anser albifrons (Серебровский, 1941).
Географическое распространение. Гусеобразные
распространены по всем материкам, кроме Антарктиды. Наиболее разнообраз-
ный видовой состав птиц этого отряда мы находим в тропических частях
Южной Америки, в Эфиопской области и в Австралийской. В Палеарктике
встречаются 60 видов, из которых 58 распространены в пределах СССР.
Некоторые из этих видов имеют кругополярные ареалы (чернозобые казарки),
другие широко распространены по Палеарктике (лебеди, гуменники),
третьи имеют весьма ограниченный ареал {Anas falcata, Rufibrenta rufi-
•collis). Большинство видов, гнездящихся в холодном и умеренном поясах,
являются перелетными.
Практическое значение. Заготовки гусеобразных в СССР
занимают после куриных второе место, однако по числу ежегодно добывае-
мых особей они, несомненно, превосходят боровую дичь. В значительном
количестве промышляются и отстреливаются речные, или благородные,
утки, затем нырковые утки, в меньшем—гуси и в сравнительно очень неболь-
шом—лебеди. Наиболее богаты гусеобразными побережья Ледовитого океа-
на, тундра, Зауральская озерная лесостепь и Западносибирская низменность,
откуда и поступает наибольшее количество водоплавающих.
Гусеобразные дают ценное мясо, высококачественное перо и пух. В не-
которых районах даже в настоящее время перо заготовляется сотнями кило-
граммов. Наиболее ценным продуктом, получаемым от гусеобразных, яв-
ляется пух, поступающий в наибольших количествах с Новой Земли и севера
Западной Сибири (Формозов, 1940). В качестве пошивочного материала
используются легкие, мягкие и нежные пуховые шкурки лебедей, но заго-
товки их в настоящее время единичны и случайны. В ряде районов соби-
раются также яйца гусеобразных, количество которых в одной только Бара-
<шнской степи в некоторые годы достигало 9 миллионов штук (Тугаринов,
1941). В некоторых районах был распространен промысел сбора яиц гоголя
и пеганки в искусственно устроенных гнездовьях, однако уже в 30-х годах
текущего столетия промысел этот почти прекратился (Шуммер, 1935). Ряд
видов гусеобразных одомашнен, что в значительной степени повышает
хозяйственную значимость отряда, однако в этом отношении пластинчато-
клювые еще далеко не использованы, так как многие из них легко приру-
252 ОТРЯД ГУСЕОБРАЗНЫЕ
чаются. Количество же гусеобразных на всем земном шаре в последнее вре-
мя катастрофически уменьшается.
Таблица для определения видов гусеобразных
1B) Вся передняя сторона цевки покрыта мелкими многоугольными щитками . . 3
2A) Или вся передняя сторона цевки, или только ее нижняя часть покрыта попе-
речными пластинками 33
3D) Крупные белые птицы с белыми же первостепенными махами. Шея почти
равна длине тела. Уздечка у старых птиц голая, у молодых оперенная 5
4C) Обшая окраска обычно не белая; если же белая, то первостепенные маховые
черные. Шея короче длины тела. Уздечка всегда оперенная 11
5F) Крайние рулевые не менее чем на 65 мм короче средних. На верхней стороне
надклювья, у его основания, обыкновенно имеется черный нарост
лебедь-шипун Cygnus olor
6E) Крайние рулевые только на 60 мм или даже менее короче средних. Чернога
нароста у основания нет 7
7(8) Клюв одноцветно черный. Перед глазом на уздечке небольшое желто-оранже-
вое или желтое пятно ...американский лебедь Cygnus columbianus
8G) Клюв из сочетания желтого и черного (или красного) цветов, уздечка жел-
тая 9
9A0) Крыло менее 555 мм. Желтое пятно по бокам клюва обычно не заходит вперед
под ноздрю малый лебедь Cygnus bewickii
10(9) Крыло более 555 мм. Желтое пятно по бокам клюва заходит вперед под
ноздрю лебедь-кликун Cygnus cygnus
11A2) Перья плечевых, верхних кроющих крыла, шеи и боков тела одноцветные
белый гусь Chen caerulescens
12A1) Перья плечевых, верхних кроющих крыла и боков тела двуцветные; ободки
перьев светлее или темнее, чем остальная часть пера \Ъ
13A4) Оперение лба, если смотреть сбоку, не выходит вперед за угол рта. Клюв
длиннее 60 мм сухонос Cygnopsis cygnoid
14A3) Оперение лба, если смотреть сбоку, доходит вперед до угла рта или же заходит
вперед за шею. Клюв короче 60 мм 15
15A6) Ноги, клюв и хвост не черные 17
16A5) Ноги, клюв и хвост черные, последний иногда с белой каймою .... 27
17A8) Участки белого цвета имеются только на лбу, у основания клюва, или же
их нет вовсе 19
18A7) Верх головы от лба до затылка, иногда и шея, белые 25
19B0) Клюв двуцветный. Ноготок клюва черный . . . . гуменник Anser jabalis
20A9) Клюв одноцветный. Ноготок клюва не черный 21
21B2) Белая полоса у основания клюва не шире 5 мм или ее нет вовсе. Задняя
часть спины и поясница светлопепельно-серые серый гусь Anser anser
22B1) Белая полоса у основания клюва ширинсю не менее, чем 10 мм, или же у осно-
вания клюва черноЕатая полоса, за которой находятся отдельные белые перья ... 23
23B4) Клюв длиннее 39 мм. Длина ноготка в длине всего клюва содержится четыре
с лишком раза белолобыйгусь Anser albijrons
24B3) Клюв короче 39 мм. Длина ноготка в длине всего клюеэ содержится менее
четырех раз (не более 3,5 раза) пискулька Anser erythropus
25B6) Общая окраска верха темная в пестринах. Затылок и зад шеи белые . . .
белошей Philacte canagica
26B5) Общая окраска верха светлосерая. На затылке черные поперечные полосы
или весь затылок черно-бурый горный гусь Eulabeia indica
27B8) Нижняя сторона шеи и зоб рыжие
краснозобая казарка Rufibrenta ruficollis
28B7) Низ шеи и зоб иной окраски 29*
29C0) Вся голова черная. Перья надхвостья и подхвостья доходят почти до вершин
рулевых
черная казарка Branta bernicla
30B9) На голове белые пятна. Перья надхвостья и подхвостья далеко не доходят
до вершин рулевых 31
31C2) Лоб белый, темя черное. . . . белощекая казарка Branta leucopsis
32C1) Верх головы черный канадскаяказарка Branta canadensis
33C4) Длина заднего пальца с когтем, взятая сверху от его основания, в 3,5—
4 раза больше его ширины (вместе с оторочкой) 35
34C3) Длина заднего пальца с когтем только в 3 или даже в 2 раза больше его ши-
рины (вместе с лопастью) 65
35C6) Крыло короче 295 мм. Края надклювья и подклювья, сложенные вместе,
образуют более или менее ровную линию 37
ХАРАКТЕРИСТИКА ОТРЯДА 253
36C5) Крыло длиннее 295 мм. Края надклювья и подклювья образуют изогнутую
вниз линию 61
37C8) Ноготок занимает весь передний край клюва. Подмышечные темнобурые
мандаринка Aix galericulata
38C7) Ноготок занимает Vs или Vi ширины передней стороны клюва. Подмышечные
иного цвета 39
39D0) Крыло короче 220 мм 41
40C9) Крыло длиннее 220 мм 47
41D2) Стержни первостепенных маховых белые. Подмышечные чистобелые . . .
чирок-трескунок Anas querquedula
42D1) Стержни первостепенных маховых бурые. Подмышечные белые, но с кра-
пинками или редкими светлобурыми пятнами 43
43D4) Вся птица в крупных округлых тусклобеловатых пятнах
мраморный чирок Anas angustirostris
44D3) Окраска иная 45
45D6) Длина среднего пальца с когтем менее 40 лип, длина средней пары рулевых
менее 75 мм чирок-свистунок Anas crecca
46D5) Длина среднего пальца с когтем более 40 мм, длина средней пары рулей
более 75 мм клоктун Anas formosa
47D8) Вершинная часть клюва вдвое шире основной части
широконоска Anas clу peat a
48D7) Клюв почти одинаковой ширины по всей своей длине 49
49E0) Зеркальце фиолетовое 51
50D9) Зеркальце иного цвета 53
51E2) Клюв двуцветный с желтой вершинной четвертью. Основная половина но-
готка желтая черная кряква Anas poecylorhyncha
52E1) Клюв одноцветный (лишь изредка с желтой или желтоватой вершиной).
Ноготок весь одноцветно желтоватый кряква Anas platyrhyrnchos
53E4) Клюв короче плюсны 55
54E3) Клюв или длиннее плюсны, или равен ей 57
55E6) Подмышечные в мелких крапинках свиязь Anas penelope
56E5) Подмышечные чистобелые или с бледнобуроватыми концами
американскаясвиязь Anasa americana
57E8) Белые подмышечные со светлобурыми продольными или поперечными поло-
сами шилохвость Anasacuta
58E7) Подмышечные чистобелые 59
59F0) Зеркальце одноцветное, густо темнозеленое с металлическим отливом; спереди
иногда окаймлено светлой полосой. Ноги темносерые или свинцово-серые
косатка Anas falcata
60E9) Зеркальце трехцветное или двуцветное; в последнем случае низ его серо-
бурый, верх беловатый серая утка Anas st re per a
61F2) На голове хохол из удлиненных перьев
хохлатая пеганка Tadorna cristata
62F1) Хохла на голове нет 63
63F4) Оперение из сочетания белого, черного и рыжего цветов. На брюхе черная
(иногда черно-бурая) полоса или брюхо белое . . . пеганка Tadorna tadorna . .
64F3) Общая окраска оперения охристо-рыжая
красная утка Tadorna ferruginea
65F6) Клюв уплощен, его края покрыты пластинками или ребрышками .... 67
66F5) Клюв узкий, удлиненный и округлый, по краям вооружен зубцами, напра-
вленными назад 107
67F8) Хвост из жестких рулевых, ступенчатый. Верхние кроющие прикрывают
только основную треть хвоста савка Oxyura leucocephala
68F7) Хвост не ступенчатый, рулевые не жесткие. Верхние кроющие хвоста выдаются
за основную половину хвоста 69
69G0) Оперение лба выдается вперед над кожей надклювья небольшой пологой
дугой или небольшим равносторонним треугольником 71
70F9) Оперение лба узким клином идет вперед по коньку надклювья 101
71G2) Ноготок занимает всю вершинную часть клюва 73
72 G1) Ноготок мал и занимает около четверти вершинной части клюва .... 81
73 G4) Цевка короче разреза клюва (т. е. расстояние от угла рта до вершины клюва)
Подмышечные белые малая гага Somateria stelleri
74G3) Цевка равна разрезу клюва или длиннее его. Подмышечные черные . . 75
75G6) Клюв к вершине слегка суживается 77
76G5) Клюв к вершине слегка расширяется 79
77G8) Брюхо и подхвостье темные . каменушка Histrionicus histrionicus
78G7) Брюхо и подхвостье белые .• морянка Clangula hyemalis
79(80) Крыло с белым зеркальцем 81
80G9) Крыло без зеркальца 83
254 ОТРЯД ГУСЕОБРАЗНЫХ
81(82) Ноздри узкие (щелевидные). При черной окраске оперения на основании
надклювья может иметься вздутие, но не крупная шишка
обыкновенный турпан Melanitta fusca
82(81) Ноздри широкие (округлые или треугольные). При черной окраске оперения
на основании надклювья высокая шишка
горбоносый турпан Melanitta deglandi
83(84) При темнобуром оперении клюв одноцветно черный. При черном оперении
в области ноздрей небольшое желтое пятно и на основании надклювья черная шишка
черная синьга Melanitta nigra
84(83) При темнобуром оперении в области ноздрей желтое пятно. При черном
оперении все основание надклювья желтое и на нем желтая шишка
американская синьга. Melanitta americana
85(86) Подмышечные сплошь черные или темнобурые 87
86(85) Подмышечные белые, но у некоторых видов с буроватыми или дымчатыми
вершинами
87(88) Крыло менее 185 мм малый гоголь Bucephala albeola
88(87) Крыло более 185 мм 89
89(90) Наибольшая ширина ноготка 4,5—5,75 мм. Лоб гладкий
обыкновенный гоголь Bucephala clangula
90(89) Наибольшая ширина ноготка 5,5—8 мм. Лоб с наростом, прощупываемым
под кожей исландскийгоголь Bucephala islandica
91(92) Верхние кроющие крыла оливково-глинистые. Ноги красноватые, оранжевые
или желтые. Крыло не менее 240 мм. . . . красноносый нырок Netta rufina
92(91) Верхние кроющие крыла иного цвета. Ноги серые или темные. Крыло
менее 240 мм 93
93(94) Зеркальце на крыле серое . к р а с н о г о л о в ы й нырок Aythya ferina
94(93) Зеркальце на крыле белое 95
95(96) На спине нет никаких следов беловатой крапчатости; если же крапчатость
имеется, то она рыжего цвета. У основания клюва на подбородке в большинстве случаев
маленькое белое треугольное пятно. Нижние кроющие хвоста большей частью белые.
Перья боков груди с темнокаштановыми боками 97
96(95) На спине иногда заметна серая или беловатая крапчатость или струйчатость.
У основания клюва на подбородке никогда не бывает маленького белого треугольного
пятна. Нижние кроющие хвоста не чистобелые и лишь изредка грязнобелые или пестрые
от смеси беловатых и бурых перьев. Перья боков груди с белыми или охристо-рыжими
каймами или без них 99
97(98) Основания внутренних опахал первостепенных маховых чистобелые. Голова
и шея темнорыжие или рыжевато-бурые . . . белоглазый нырок Aythya nyroca
98(98) Основания внутренних опахал первостепенных маховых с дымчатым налетом.
Голова и шея черные или темнобурые; в последнем случае на уздечке рыжеватое пятно
беров нырок Ayithya baeri
99A00) На голове в большинстве случаев хохол из удлиненных перьев, иногда
слабо выраженный. При однообразной буроватой окраске верха на сгибе крыла в боль-
шинстве случаев бурые перья хохлатый нырок Aythya fuligula
100A9) На голове нет и следов хохла из удлиненных перьев. При однообразной
окраске верха на сгибе крыла в большинстве случаев беловатые перья
морская чернеть Aythya marila
101A02) Клюв к вершине несколько расширен. Окраска черная или черно-бурая
красноклювый турпан Melanitta perspicillata
102A01) Клюв одной и той же ширины по всей своей длине или же к вершине не-
сколько сужен. Окраска иная 103
103A04) Клиновидный выступ оперения головы, спускаясь по надклювью, заходит
вперед за ноздри. Ноготок занимает у2—3/4 вершинной части клюва
очковая гага Somateria fischeri
104A03) Оперение головы спускается на надклювье тремя выступами—срединным
и двумя боковыми 105
105A06) Срединный выступ оперения лба короток и далеко не доходит до ноздрей
• обыкновенная гага Scmateria mollissima
106A05) Срединный выступ оперения лба длинен и почти доходит до заднего края
ноздрей гага-гребенушка Somateria spectabilis
107A08) Клюв короче цевки. Зеркальце темное луток Mergus albellus
108A09) Клюв длиннее цевки. Зеркальце белое 109
109A10) От переднего края ноздри до вершины клюва по краю надклювья не более
15 зубцов. Длина ноготка по хорде более 10 мм
большой крохаль Mergus merganser
110A09) От переднего края ноздри до вершины клюва по краю надклювья более 15 зуб-
цов. Длина ноготка по хорде менее 10 мм 111
111A12) На середине спины имеется, хотя бы слабо выраженная, белая крапчатость.
От заднего края ноздри до угла рта более 10 зубцов
ЛЕБЕДЬ-КЛИКУН
255
чешуйчатый крохаль Mergus squamatus
112A11) На спине нет никаких следов белой крапчатости. От заднего края ноздри
до угла рта менее 10 зубцов длинноносый крохаль Mergus senator
СЕМЕЙСТВО УТИНЫЕ ANATIDAE
ПОДСЕМЕЙСТВО ГУСИНЫЕ ANSERINAE1
РОД ЛЕБЕДИ СУ GNUS BECH STEIN, 1803
Тип. Cygnus olor
1. Лебедь-кликун Cygnus cygnus L.
Anas Cygnus. Li n n a e u s. Syst. Nat., изд. X, 1758, стр. 122, Швеция.
Синоним. Cygnus musicus. В е с h s t e i n. Natur. Vog. Deutschl., Ill, стр. 830,
1809, Европа.
Русское название. Лебедь—старинное русское народное название
Распространение. Ареал. От Исландии и севера Фенноскандии, лишь
местами выходя за северную границу лесной зоны, по европейской части
СССР и Сибири до Берингова моря. На юге разрозненно от Шотландии,
Карта 22. Распространение лебедя-кликуна Cygnus cygnus
1—С. с. cygnus, 2—С. с. islandicus; а—граница гнездовий, г—зпмовкп
Карело-Финской ССР, верховьев Волги, северного побережья Каспия,
Средней Сибири, Тарбагатая до Маньчжурии, Японии и Камчатки.
Характер пребывания. В большей части ареала перелет-
ная, местами оседлая птица, нормально зимующая в северной части Сре-
1 Систематика подсемейства гусиных дается в трактовке сводки Питерса «Список
птиц земного шара», т. 1, 1931 (на английском языке), за исключением выделения рода
Rufibrenta. Остальные группы приведены в согласии с последней переработкой отряда
гусеобразных (Делакур и Майр, 1945).
256 ОТРЯД ГУСЕОБРАЗНЫХ
диземного моря, в Передней Азии на Каспии, в Средней, Южной и Юго-Во-
сточной Азии.
Биотоп. Открытые, но заросшие по краям, большей частью глухие,
водоемы, местами и вблизи населенных пунктов.
Подвиды и варьирующие признаки. Два подвида, отличающиеся размерами: остров-
ной С. с. islandicus В г е h m. 1828—о-в Исландия, и материковый С. с. cygnus Li n-
u a e u s. 1758—в остальной части ареала.
Материковый лебедь-кликун Cygnus cygnus cygnus L.
Распространение. Ареал. Занимает главным образом лесную зону
Евразии, выходя на севере местами в подзону тундры, а на юге—в зону
открытых сухих .ландшафтов. Гнездится в Шотландии, на севере Евразии
до 70° с. ш. в Норвегии, до Шведской Лапландии, в СССР до бассейна Туломы
на Кольском п-ове, до тундр Канинского п-ова, низовий Печоры, до мыса
Круглого в Обской губе, пр бассейна верхнего течения Таза, где местами обы-
чен, до 66° с. ш. на Енисее, до бассейна Вилюя, полосы криволесья в верх-
нем течении Индигирки, до Верхне-Колымска, до Анадыря и до 65° с. ш.
на Чукотке. На юге в Фенноскандии и Карело-Финской ССР местами до
62° с. ш., весьма разрозненно в верховьях Волги, по Каме и в Камском
Приуралье (Воронцов, 1949), еще изредка гнездится по северо-западному
и северному побережьям Каспия, летом встречается по Сыр-Дарье, однако
гнездовье его здесь в настоящее время достоверно не установлено; гнездится
по Наурзумским озерам, далее доходит до Тарбагатая, озера Зайсана, Джун-
гарии, озер Монгольской Народной Республики (Косогол, Сангин-Далай-
нор, Буин-Нор и др.)» Прибайкалья, где гнездится на озерах у Усть-Бар-
гузина, также по Верхней Ангаре и на некоторых озерах Байкальских гор
(Штегман, 1936), до Северной Маньчжурии, Приморской обл., Южного Саха-
лина и Камчатки. В южной части ареала гнездится спорадично в европей-
ской части СССР, в Зауралье, в Барабинской и Кулундинской степях, а
также по северо-западной окраине Алтая.
Характер пребывания. Из северных частей ареала обычно
отлетает, но местами остается на незамерзающих проливах и полыньях.
Для большей части территории СССР является пролетной птицей, нормаль-
но зимующей лишь в южных районах страны. Частично зимует у берегов
Скандинавии, на Белом и Балтийском морях на незамерзающих водоемах
Горьковского края (Пузанов, 1937), Татарской АССР, Башкирской АССР,
в ряде мест Украинской ССР, на Южном Сахалине, Камчатке, Командо-
рах и Анадырском крае (Белопольский, 1934), однако зимовки эти не носят
регулярного характера. Обычен кликун на зимовках в северной части Сре-
диземноморья и Малой Азии, на Черном и Каспийском морях, в теплые
зимы встречается в Приазовье и в дельте Терека, иногда зимой залетает
на озеро Севан (Ляйстер и Соснин, 1942); найден в зимнее время в Чкалов-
ской области; зимует в ряде мест Средней Азии; на озерах Иссык-Куль
и Телецком, на Тихоокеанском побережье вплоть до Камчатки и до
Командор и Сахалина, на о-ве Хоккайдо (Ян, 1942), по берегам Китая;
редок в Индии, встречается в Иране.
Даты. С мест зимовок в Средиземноморье, Малой и Средней Азии,
с озера Иссык-Куль (Шнитников, 1949), Индии и Китая кликуны
начинают отлетать в первой половине февраля, но на крайнем западе
и востоке миграции в общем протекают значительно быстрее, чем в Казах-
стане и Центрально? Азии. Самые передвижения открываются или отдель-
ными парами (иногда одиночками), или небольшими стайками и уже
за 50-й параллелью проходят почти в зимней обстановке, когда всюду еще
лежит снег, водоемы покрыты льдом и лишь первые лужи на пригревах
ЛЕБЕДЬ-КЛИКУН 257
да закраины воды у речных берегов свидетельствуют о наступлении весны.
Поэтому иногда при весенних похолоданиях и снегопадах наблюдается
обратный лет лебедей в более южные районы. Из Малой Азии и Закавказья
кликуны окончательно исчезают в последних числах февраля и в это же время
появляются в районах Молдавской ССР. В конце первой декады марта или
в начале второй передовые показываются в Крыму, на Азовском побе-
режье в дельте Терека (Беме, 1925) и в Приморской обл. на Дальнем Востоке
(Шульпин, 1936). В середине марта передовые уже в низовьях Днепра,
в районе Аскания-Нова, в низовьях Волги, на Аму-Дарье и Сыр-Дарье,
в Монголии (на озере Сохо-Нор; Бианки, 1908), и в Уссурийском крае. В ран-
ние весны к 20 марта, в среднем же за 12 лет 31 марта, лебеди показываются
на территории Латвийской ССР (Лоудон, 1912) и на Балтийском побережье
Ленинградской обл. В середине третьей декады марта пролетают по сред-
нему течению Днепра, во второй половине марта на Аму-Дарье, и весь
март идут небольшие передвижения в Приморской обл. на Дальнем Востоке.
В середине марта отлетают с о-ва Хондо (Ян, 1942). В теплые весны к концу
марта окончательно исчезают из Средней Азии, но в холодные задерживаются
на Сыр-Дарье до 15 апреля (Спангенберг и Фейгин, 1936). В конце марта—
начале апреля передовые в бассейне Оки, по Средней Волге, на северном
побережье Арала, в Южном и Центральном Казахстане, в районе же Вла-
дивостока в некоторые годы идет валовой пролет D—9 апреля, Медведев,
1913). 30 марта—17 апреля наблюдаются в районе Чудского озера (Несте-
ров и Никандров, 1913), в конце марта—начале апреля в Московской обл.
(Поляков, 1910, Смолин, 1948), в начале апреля на Вятке (Круликовский,
1913), 2—17 апреля в Чер дынек ' районе (Митрофанов, 1913), 6—24 ап-
реля на Наурзумских озерах (Михеев, 1938). В середине апреля долетают
до Мурманского побережья, Средней Печоры A7 апреля 1934, Дормидонтов)
и Молотовской обл. В долине Тобола замечаются раньше E—10 апреля,
Словцов, 1892), 26 апреля отмечены первые в Тиманской тундре (Гладков,
1941), 7—28 апреля показываются в Анадырском крае (Портенко, 1939).
В конце апреля долетают до Обской губы, 17—27 апреля летят в Томском
крае, 29 апреля отмечен пролет в низовьях Вилюя. В горных местах поя-
вляются позже, чем в равнинных: 3—31 мая наблюдаются в разных местах
Алтая (Фолитарек и Дементьев, 1938), в начале мая в бассейне Зеи на Ябло-
невом хребте (Дорогостайский, 1915), 11—20 мая в районе Верхне-Колым-
ска. В настоящее время всюду летят лишь в небольших количествах,
валовой пролет наблюдается только в Восточной Сибири и особенно хорошо
выражен в Приморье, где он проходит на 15—30 дней позже появления пере-
довых. Летят очень высоко, днем и ночью; в начале весенних передвижений
отдельными особями, в разгар движения группами до 10 особей, выстроив-
шись углом и время от времени издавая звучные трубные крики, особенно
слышные по ночам. На весенних перелетах останавливаются на реках,
вскрывающихся ранее замкнутых водоемов. Запоздавшие окончательно
исчезают из разных мест миграционного ареала в мае, но часть негнездя-
щихся все же остается южнее своих теперешних гнездовых районов.
Осенний отлет с мест гнездовья связан е наступлением осенних холодов
и морозов, сковывающих льдом водоемы, на которых лебеди держатся до
последней возможности, и протекает различно на разных территориях СССР.
В конце сентября и первой трети октября исчезают кликуны с Крайнего
Севера (низовий Печоры—Самородов, 1948; низовий Оби, Анадыря—Пор-
тенко, 1939); 29 сентября—12 октября идет пролет в Олонецком крае(Испо-
латов, 1917) и в те же, примерно, числа по Охотскому побережью; 19 сен-
тября уже заметно первое появление осенних на Сиваше в Крыму (Ворон-
цов, 1937), в сентябре—ноябре пролетают у восточных берегов Балтийского
моря (Бианки, 1913, Раснер, 1913), 25—26 октября в Смоленской обл.
17 Птицы СССР, т. TV
258 ОТРЯД ГУСЕОБРАЗНЫЕ
(Граве, 1928), 19—20 октября в Московской (Поляков, 1910), в середине ок-
тября в Молотовской, 25 сентября—15 октября исчезают из района Сред-
ней Волги, 1—23 октября из предгорий Алтая, в конце первой декады октября
с Пенжины (Волков и Дементьев, 1948), с начала октября с Командор (Биа-
нки, 1909), 4 октября начинают появляться в заливе Посьета, 8 октября наблю-
дался пролет в Татарском проливе (Дулькейт и Шульпин, 1937). В конце
сентября исчезают с Балтийского побережья, с конца первой трети ноября
до второй половины ноября идет пролет в Приазовье (Боровиков, 1907)
и к середине сентября лебеди появляются на о-ве Хондо (Ян, 1942). Осенью
лебеди скопляются у северного и северо-западного побережья Каспия, а а
начале зимы в большом количестве держатся по мелководьям Аграханского
залива и близ о-ва Чечень. При замерзании заливов и мелководий пере-
мещаются сначала к кромке льда, а затем постепенно отлетают к югу, за-
держиваясь надолго у Дивичинского лимана. Основная же масса спускается
еще южнее и зимует у берегов Кызыл-Агачского залива (Исаков, 1949)
и юго-восточного побережья Каспия, начиная от низовий Атрека (Демен-
тьев, 1945). Осенние миграции большей частью идут по системам озер,
на которых кликуны местами задерживаются надолго.
Биотоп. Разнообразен, но всегда связан с водоемами, большейг
частью крупными открытыми озерами, сильно заросшими у берегов земно-
водной растительностью и особенно тростником. Иногда селятся на морских
побережьях, поросших тростниками с плесами открытой воды среди послед-
них; на заросших по берегам глухих лесных озерах и старицах; на озерах
среди кустарниковой тундры (Портенко, 1939). Там же, где кликунов не-
разоряют, они гнездятся и на небольших, заросших с берегов, прудах, не-
редко даже вблизи жилья (Круликовский, 1913; Сушкин, 1938).
Численность. Прежде обыкновенные на гнездовьях в разных
районах СССР, в настоящее время лебеди-кликуны значительно снизи-
лись в численности, из ряда же районов и вовсе исчезли. У северной гра-
ницы ареала, в малодоступных, заболоченных районах, численность их зна-
чительно выше, чем в южных, что вызывается главным образом уничто-
жением необходимых лебедям местообитаний.
Экология. Размножение. Половая зрелость наступает на чет-
вертом году жизни. Формирование пар у впервые приступающих к размно-
жению происходит на зимовках. Часто пары очень привязаны друг к другу
и, возможно, соединяются на всю жизнь. Косвенно об этом свидетельст-
вует соотношение полов приблизительно 1 : 1, а также и то обстоятельство,
что в своих гнездовых биотопах кликуны появляются всегда парами.
Выбранный гнездовой участок строго охраняется от вторжения дру-
гих лебедей. Вскоре после прилета и, во всяком случае, не позже 1—2 недель
после появления на родине, на сухом месте, но иногда и на мелководье,
на завалах старого тростника, устраивается довольно громоздкое гнездо
в виде кучи до 1 м в основании и высотою до 0,6—0,8 м. В некоторых, ред-
ких, случаях размеры могут быть и большими—до 3 м у основания и 0,8 м
в высоту (Формозов, 1937). Строительным материалом является тростник,
камыш, мох, иногда ветви, палки и пр. Лоток выстилается более тонким, неж-
ным материалом из сухих стеблей трав, мха, перьями и пухом, которые
самка выщипывает у себя с груди и брюха. Выщипывание начинается с мо-
мента откладки первого яйца и продолжается до откладки последнего (Бекер,
1929). Гнездо строится одной самкой. Кладка состоит из 4—6, редко 7 яиц
слегка блестящего белого или же слабо серовато-желтого цвета. Размеры
105,2—126,3x68,1—77.4 мм (Тугаринов, 1941); D) 114—119x74—75,8,
в среднем 117,25x74,5 мм (Барабинская степь—Иогансен, 1907); G5)
117—126,3x68,2—76.3. в среднем 112,8x72,6 мм (Джоурден). Вес 33,5—
36,5 г. При гибели первой вторая кладка всегда содержит меньшее коли-
ЛЕБЕДЬ-КЛИКУН 2Ш
чество яиц, иногда только одно. Время кладки приходится на май—июнь.
В Барабинской степи в окрестностях станции Каинской 4 ненасиженных
яйца найдены 22 мая 1903 г. (Иогансен, 1907), На озере Чаны 15 мая 1931 г.
отмечено гнездо с одним яйцом в урочище Кукаринском (Данилович, 1935),
на озере Ак-Суат в районе Наурзумского заповедника покинутое гнездо
с 6 ненасиженными яйцами найдено на высыхающем месте 26 мая 1935 г.
(Формозов, 1937). На Анадыре кладка в конце первой трети—в 20-х чи-
слах июня (Портенко, 1939).
Насиживает одна самка, но самец всегда находится поблизости и рев-
ностно охраняет гнездовой участок. При опасности самец подает голос,
и вслед за этим самка сначала закрывает яйца пухом, а также растительными
материалами краев гнезда, и затем улетает вслед за самцом. Возвращаясь
на гнездо, самка летит впереди самца и прежде чем сесть на яйца она пере-
ворачивает их. Продолжительность насиживания 35—40 дней. В При-
уралье гнездо с 6 сильно насиженными яйцами, зародыши которых уже окон-
чательно сформировались, найдено 15 мая (Сушкин, 1897).
В южных районах птенцы выклевываются из яиц в конце первой де-
кады мая, в Вологодском крае и Татарской АССР в июне, в Барабинской
степи в конце мая—начале июня B3 мая 1909 г., 5 июня 1909 г. на озере
Чаны, Иогансен, 1911), на Алтае в начале второй половины—начале третьей
декады июня (Сушкин, 1938), у северной границы ареала в Сибири в первой
декаде июля. Обсохнув под самкой после выхода из яиц, с первого же дня
жизни птенцы самостоятельно добывают корм около родителей. В случае
опасности эти последние активно защищают своих птенцов, не проявляя
при этом обычной своей осторожности, и в малонаселенных местах подпу-
скают лодку шагов на сто, а в некоторых случаях даже на сорок (Дмоховский,
1938). Обычно же взрослые кликуны при опасности уводят молодых в густые
заросли, сами же улетают и не скоро возвращаются обратно. Рост птенцов
идет быстро, и в конце июля молодые на Алтае уже крупные (Сушкин, 1938).
Семьями лебеди держатся долго и не разлучаются даже после того, как моло-
дые начинают летать; зачастую весь выводок вместе с родителями напра-
вляется в осенний путь.
Линька. Дважды в году: летняя—полная и осенне-зимняя—частич-
ная. Полная летняя линька у кликунов начинается со смены маховых перьев,
причем новые маховые окончательно вырастают и становятся крепкими
только через месяц. Ежедневный их прирост равен 9 мм. Вскоре после начала
смены махов в процесс линьки вовлекаются птерилии нижней стороны тела
и рулевые, затем и все остальные птерилии. Процесс этот протекает медленно.
Линька маховых у неполовозрелых второгодних птиц на северном побе-
режье Каспия начинается в 20-х числах июня, взрослые же размножающиеся
линяют несколько позже (Тугаринов, 1941). Смена их маховых на о-ве Ка-
нине проходит в первой декаде июля, и 9 июля маховые у здешних в тру-
бочках и еще совершенно не развиты (Дементьев, 1935), на Новой Земле
полная линька лебедей наблюдается в конце июля (Горбунов, 1929), на Ана-
дыре линька начинается с середины июля (Портенко, 1939), а на Шантар-
ских о-вах совершенно нелетные лебеди наблюдались в большом количе-
стве 20 августа (Шульпин и Дулькейт, 1937). К концу августа полная летняя
линька у взрослых и полувзрослых обыкновенно заканчивается.
Смена пухового наряда на перовой у молодых начинается тогда, когда
они достигают примерно половины размеров взрослых. Приблизительно
через две недели после окончания полной летней линьки начинается частич-
ная осенняя линька контурного оперения и рулевых, затягивающаяся
надолго.
Известны случаи добычи кликунов под Вологдой в последних числах
ноября с незаконченною еще линькой рулевых. В октябре у кликунов по
17*
260 ОТРЯД ГУСЕОБРАЗНЫЕ
всей территории СССР идет еще смена рулевых, но по-разному у отдельных
индивидов. Повидимому, основная масса лебедей заканчивает линьку только
к декабрю (Тугаринов, 1941), и этим, вероятно, объясняется, что кликуны
до последней возможности держатся у северного и западного побережий
Каспия, а также и на ряде других водоемов. В период линьки лебеди пере-
бираются на большие открытые водные пространства, где они находятся
в полной безопасности от всех возможных врагов, за исключением человека.
Или же в редких случаях на это время они забираются в самые глухие
и недоступные участки внутренних водоемов.
Питание. Кормом для птенцов являются различные водные личинки,
добываемые на мелководьях и у берегов. Взрослые, помимо мелких водя-
ных животных, используют и водяные растения. Погружая шею в воду
на мелких местах, лебеди подолгу роются в одном месте в илу, вырывая в нем
округлых очертаний ямку до 80 см в поперечнике и извлекая из него корне-
вища и молодые побеги водяных растений. При этом ловятся также мелкие
водяные животные, проплывающие у дна или находящиеся в илу. Корм
собирается обычно по утрам и вечерам. На зимовках, там, где их не тре-
вожат, лебеди подолгу остаются на одних и тех же мелководных местах,
даже если последние вследствие подъема воды становятся неудобными для
добывания на них корма. Нормально кормиться лебедям зачастую мешают
также непогода и зимние штормы, однако лебеди редко перелетают в другие
места, даже если им приходится голодать. Обессилев от бескормицы, они
потом некоторое время не имеют возможности подниматься в воздух, но
затем снова быстро восстанавливают свои силы. Однако при затяжных небла-
гоприятных для добывания корма условиях в некоторых случаях гибнут
от истощения (Исаков, 1940).
Промысел. Кроме человека, лебедь не имеет никаких других врагов,
под воздействием же человека за последнее столетие население кликунов
значительно уменьшилось. Прежде всего человек лишает кликуна гнездо-
вых угодий, используя их для своих нужд, и таким путем птицы эти исчезли
из ряда мест Европы и европейской части СССР, и только в Сибири, где
еще много болотистых, малодоступных пространств, кликун встречается
в некотором количестве. В громадных количествах, иногда сразу целыми
сотнями, лебеди истреблялись во время линьки путем «гонки», т. е. пресле-
дования линных лебедей на парусных лодках и затем при помощи сетей на
зимовках. В настоящее время в связи со значительным сокращением пого-
ловья кликунов «гонок» на них почти не устраивают, сетной же лов
вообще запрещен.
Используются лебеди в небольшом количестве ради их пера, пуха и шку-
рок. Мясо кликуна жестко и невкусно, и во всяком случае не ради него
охотятся за лебедями. Говорить в настоящее время о промысле на клику-
нов не приходится, и если прежде во время гонки добывалось на Каспии
на одну лодку до 300 шт. в день (Тугаринов, 1941), то теперь удается пой-
мать одного-двух. Однако в Иране попрежнему процветает хищнический
лов сетями, которыми сразу иногда покрывают до 80 шт.
Полевые признаки. Лебедь-кликун крупная снежнобелая птица с длин-
ной шеей. На воде виден хорошо еще издали. Передняя часть его тела погру-
жена в воду больше, чем задняя. Шея поднята вертикально, палкой, голова
направлена вперед, а клюв держится горизонтально. Крылья прижаты к
бокам и в профиль, поэтому спина кажется выпуклой. Плавает в спокойной
природной обстановке довольно медленно. Прежде чем подняться с воды,
кликун сначала быстро плывет, при этом долго хлопает по воде крыльями
и отталкивается от нее лапами. Тяжело взлетая, он летит затем быстро
с вытянутой вперед шеей, делая редкие, но сильные взмахи крыльями.
Лебедь-кликун очень осторожная птица, обладающая весьма зоркими гла-
ЛЕБЕДЬ-КЛИКУН
261
Рис. 41. Лебедь-кликун (взрослый и молодой)
зами. Так как он почти всегда держится вдали от берегов, на широких откры-
тых водных пространствах, где и ночует, то это в большинстве случаев дает
ему возможность избегать преследо-
вания со стороны человека. И толь-
ко в гнездовое время он занимает
окраины водоемов или острова, где
и устраивает свое гнездо. Голос при
плавании звучит как односложное,
звучное, трубное и гармоничное
«ганг», на полете же—как еще бо-
лее звучное двухсложное «ганг-го».
Рассерженный, кликун издает пис-
кливые звуки «кликликкликлик»
или шипит, как гусь.
Описание. Размеры и строе-
ние. Размах самцов и самок 2250 —
2500 мм. Длина тела самцов и самок
1550 — 1700 мм. Длина крыла самцов E)
595—660, самок D) 580—635, в среднем
635 и 611,5 мм. Длина клюва самцов G)
90,5—108,3, самок E) 93,8—111,5, в сред-
нем 107,8 и 102,1 мм. Высота нижней че- Рис. 42. Пуховой птенец кликуна
люсти самцов G) 7,5—8,8, самок E) 7,3—
8,4, в среднем 8 и 7,8 мм. Вес самцов и самок 7—10 кг, изредка 12,7 кг. Самка несколько
меньше и легче самца. Хвост из 10—11 рулевых, округлый. Крайние рулевые на 45—
60 мм короче. Верхний край ноздрей почти параллелен поверхности конька клюва.
262 ОТРЯД ГУСЕОБРАЗНЫЕ
Окраска. Пуховик. Верхняя сторона бледносерая, с более темной голо-
вою, верхней стороной шеи и нижней частью спины. Между лопатками два белых пятна,
с возрастом быстро исчезающих. Бока шеи, ее нижняя часть и низ тела белые.
Молодая птица в первом наряде дымчато-серая сверху с более темной головой,
более темными концами маховых и рулевых и узкими белыми каемками на перьях перед-
ней части спины. Бока тела светлее. Низ белый. Основание клюва буровато-красное,
причем цвет этот распространяется на большую часть надклювья до ноготка. Ноги серо-
вато-красные.
После второй осенней линьки сохраняется слегка дымчатый цвет оперения головы,
клюв уже и меньше, чем у взрослых. Белое оперение головы появляется только после
третьей осенней линьки.
Взрослые самец и самка снежнобелые. Но некоторые особи имеют на перьях
охр и сто-желтый налет, особенно развитый на верхних сторонах головы и шеи. Налет этот
случаен и зависит от внешнего окрашивания перьев. Уздечка и основная часть клюва
желтая или желто-оранжевая без шишки или вздутия. Желтый цвет по коньку клюва
обычно заходит за задние края ноздревых впадин, с боков же доходит до ноздрей и
распространяется впереди них до половины расстояния между ноздрями и коготком
клюва, а вниз идет до краев надклювья; остальные части клюва и ноготок черные.
Ноги черные. Радужина черно-бурая.
2. Тундровый лебедь Су gnus bewickii Y а г г ell.
Cygnus bewickii. У а гг el 1. Trans. Linn. Soc, XVI, стр. 453, 1830, Англия.
Синоним. Cygnus minor, Pallas. Бутурлин1, ex P a 1 1 a s. Поли, опред. птиц
СССР, т. II, стр. 73, 1935.
Русское название. Лебедь. Тундровый, или малый, лебедь—книжные.
Распространение. Ареал. Характерен для тундры Евразии и гнез-
дится в ней от севера Фенноскандии до дельты Колымы, а также на запад-
ных островах Ледовитого океана.
Подвиды и варьирующие признаки. Два подвида, различающиеся между собой раз-
мерами клюва и характером его раскраски: С. b. bewickii Y а г г е 1 1, 1830—тундры
Евразии на восток до Лены; С. b. janKowskii A I p h е г a k i, 1904—тундры от Лены
до Колымы.
В последнее время высказан взгляд (Тугаринов, 1941), что тундровый лебедь,
чрезвычайно варьирует по форме, длине, а также по ширине клюва и обладает лишь
трансгрессивной изменчивостью, постепенно увеличиваясь в своих размерах по напра-
влению с запада на восток. Одним из существенных доводов при этом считается, что
ширококлювые лебеди, характерные для северо-востока Сибири, встречаются и на Новой
Земле. Однако явления такого порядка широко распространены среди многих геогра-
фических форм, крайние индивидуальные варианты которых тем или иным признаком
иногда сходны с другими подвидами своего вида. Вместе с тем характер распростра-
нения желтого цвета на клюве у восточного подвида отличен от того, что имеет место
у западного. Комбинация же этих и некоторых других признаков позволяет хорошо
различать оба подвида даже в молодом возрасте (Бутурлин, 1911).
Ввиду некоторой неясности вопроса впредь до его разрешения на хорошем серийном
материале мы принимаем два слабо разграниченных подвида тундрового лебедя—запад-
ный и восточный, помимо указанных выше признаков (Бутурлин, 1911 и 1935), различаю-
щихся еще местами своих зимовок.
Западный тундровый лебедь Cygnus bewickii bewickii Yarrell.
Распространение. Ареал. Гнездится в тундре от района Печенги
на Кольском п-ве (Тугаринов, 1941) и Белого моря до 74° 30' с. ш. на побе-
режьях Таймыра, отсутствует в центральных частях последнего, обычен
в тундре по Енисею и далее на восток до дельты Лены, где встречается с вос-
точным подвидом, образуя с последним на пространстве от Восточного
Таймыра до Лены смешанную популяцию. Заселяет оба острова Новой Земли,
доходя в очень небольшом числе до мыса Желания (Антипин, 1938), будучи
1 На основании толкования текста Палласа (Zoogr. Ross.—Aziat., 1811, стр. 214) Бутур-
лин принимает для этого лебедя видовое название. Однако необходимо отметить, что тек-
стовые наименования Палласа не имели номенклатурного значения.
тундровый лебвдь
263
в то же время обыкновенной птицей Южного о-ва (Успенский, 1950). Гнез
дится на Колгуеве, Вайгаче, обычен в Большеземельской тундре, на побе-
режье Карского моря, п-ве Ямале и в низовьях всех сибирских рек до дельты
Лены включительно. На юг местами заходит в лесотундру, найден под
60° с. ш. на водоразделе Таза и Турухана (Шухов; Скалой и Слудский, 1936)
указания же на гнездовья и низовья Вилюя (Маак, 1861, Бутурлин, 1935)
вызывают сомнения.
Характер пребывания. Гнездится на севере СССР, во время
пролета встречается в различных районах страны1 и редок на зимовках
в южных ее частях. Нормально зимует на побережьях Северного моря,
Карта 23. Распространение тундрового лебедя Cygnus bewickii
I—С. с. bewickii, 2—С. с. janhowshii; а—граница гнездовий, в—вимовки
у берегов Британских о-вов и в других местах побережий Западной Европы.
На территории СССР в незначительном количестве и не регулярно встре-
чается зимою на незамерзающих местах Белого моря, в качестве редкой
птицы в зимнее время известен с Кавказа, где впервые, хотя и предполо-
жительно, отмечен Радде A884) и достоверно найден на озере Ах-Чала (Карам-
зин, 1912), затем наблюдался 9 февраля 1932 г. в Кызыл-Агачском заливе
и 20 февраля того же года на о-ве Сара (Исаков и Воробьев, 1940), однако
при этом лебеди не были добыты. Достоверно зимует в Средней Азии, где
добыт 7 января 1908 г. на Сардабе, близ станции Голодная степь (Зарудный,
1910), 17 января 1910 г. под Джаркентом (Шнитников, 1940). Возможно
зимует на Алтае (Сушкин, 1938).
Даты. Сроки отлета с мест зимовок неизвестны. Весной пролетает
по Украине, где добыт в Полтавской и Курской обл., встречается на весеннем
пролете в устье Хорога на Тянь-Шане (Алфераки, 1904; Шнитников, 1949),
1 Причем на пролетах в Сибири встречается чаще, чем в пределах русской равнины, но
через Латвийскую ССР пролетают иногда крупными стаями.
264 ОТРЯД ГУСЕОБРАЗНЫЕ
также на Алтае по Чулышману, пролетает близ Семипалатинска, обычен
на пролете по Енисею у Красноярска и найден около Минусинска (Сушкин,
1938). В районе Ленинграда пролетает между 23 марта и 9 апреля (Раснер,
1913) и 15—20 апреля (Бианки, 1947), на Салаире в Кузнецком кряже вес-
ной встречается в последней декаде апреля и добыт здесь 30 апреля 1894 г.
(Хахлов, 1937). В Тиманской тундре передовые отмечены 3 мая 1938 г.;
18—21 мая шел их валовой пролет (Гладков, 1951). На Новой Земле, поСкон-
нингу, первые отмечены 28 мая 1903 г.; но Успенский здесь первых наблюдал
17 мая 1948 г. и даже 26 апреля 1949 г. На Ямале появляются 23—26 мая,
в устье Енисея на Бреховских островах 22 мая, на оз. Ессей 3 мая, в устье
Хатанги 10 июня, у селения Хатангского 24 мая, а в тундре в начале июня
(Торгашев), в дельте Индигирки 3—4 июня, в дельте Колымы около 15 мая.
Прилет их в тундру связан с началом таяния снега и появлением промоин
на реках. В тундре в общей сложности проводят около 120—130 дней.
С мест гнездовий отлетает в сентябре. На Новой Земле перемещения
тундровых лебедей отмечены уже в самом начале сентября, а в Безымянной
губе 15 сентября 1949 г. наблюдался отлет на юг (Успенский); также в на-
чале сентября шел отлет небольшими группами по 5—6 особей в Карской
губе, массовый же отлет происходил здесь 13 и 15 сентября (Михель, 1937).
15 октября 1930 г. шел пролет по Вычегде (Дмоховский, 1933), 17 октября
1927 г. отмечены пролетные в бассейне Маны (Шухов, 1925) и в то же время
18 октября 1858 г. тундровый лебедь добыт далеко на юге, на море, у устья
Сыр-Дарьи (Северцов, 1873). Между 1—24 ноября пролет идет в районе Ленин-
града (Раснер, 1913). Таким образом, на северо-западе в районе Балтийского
моря весенний пролет начинается раньше, а осенний заканчивается позже,
чем в континентальных частях материка. Как и другие лебеди, тундровый
отлетает с мест гнездовья в связи с замерзанием водоемов, а весной на ме-
стах гнездовья появляется еще по снегу.
Биотоп. Гнездовой—заболоченные и травянистые низины тундры,
перемещающиеся с многочисленными открытыми, различных размеров озе-
рами; речные долины, изобилующие старицами и протоками. Миграцион-
ный—озера междуречий, реже старицы широких речных долин. Зимний—как
и у других лебедей.
Численность неравномерна. В районах интенсивного исполь-
зования его человеком стал положительно редким, вообще же более или
менее обыкновенен в зоне тундры. В некоторых местах он селится в большом
числе и, например, на Гусиной Земле (Новая Земля) в период его размно-
жения в 1950 г. в поле зрения при передвижении человека все время было
видно несколько десятков лебедей (Успенский). В 1951 г. на Паньковой
Земле (Новая Земля) в поле зрения можно было видеть не более одной гнез-
дящейся пары, и вообще одно гнездо приходилось приблизительно на 20 кмг\
в то же время их было особенно много на о-ве Междушарском (Кузякин).
Экология. Размножение. На гнездовья прилетает парами. Холо-
стые самцы и у этого вида пытаются отбивать самок у семейных. Однако дра-
ки между самцами случаются и в пролетное время. Так, в Тиманской тундре
вдали от гнездовых мест, когда всюду бывают видны остановившиеся на
отдых пролетные стада лебедей, некоторые из самцов, сплываясь, становятся
друг против друга, высоко поднимают шеи и машут крыльями. Перья их
при этом взъерошены. Затем они бросаются друг на друга, и схватка кончает-
ся тем, что один из противников улетает (Гладков, 1951). Брачные их игры
своеобразны и происходят на земле. Самец начинает ходить перед самкой,
вытягивает шею, временами приподнимает крылья, издает ими особый хло-
пающий звук и при этом звонко кричит. Через некоторое время пара переле-
тает на другое место, где все проделывается снова. Иногда два самца совер-
шают весь этот ритуал перед одной самкой (Бутурлин, 1905).
ТУНДРОВЫЙ ЛЕБЕДЬ 265
В постройке гнезда, начинающейся сразу же после прилета, принимает
участие одна только самка. Для гнезда выбирается небольшое сухое возвы-
шение, часто кочка, на совершенно открытом месте, имеющем большой круго-
зор. Иногда же гнездо располагается на кочке у берега водоема. Основным
материалом для постройки гнезда является выщипываемый тут же мох,
но верхние части гнезда и лоток строятся из стеблей прошлогодних осок,
пушицы и других мелких растений. Лоток обычно выстилается одним только
пухом, хотя иногда заменяется и перьями (Кузякин). На Южном о-ве
Новой Земли, на так называемой Гусиной Земле, где тундровые лебеди обыч-
ны, их гнезда располагаются на междуречных пространствах. Высота гнезд
50—60 см, их поперечник до 100 см, поэтому при таянии снега они показы-
ваются задолго до появления проталин, что дает возможность прилетевшим
лебедям занимать их еще в то время, когда всюду лежит снег. При этом ка-
ждая пара использует свое старое гнездо, лишь ремонтируя его (Успенский).
Кладка состоит из 2—3 яиц, но в редких случаях бывает 4—5 яиц,
а иногда только одно. На Новой Земле первые два яйца найдены 29 мая
1948 г. (Успенский) и 5 июня (Сконнинг). Два ненасиженных яйца найдены
19 июня 1901 г. у Черной губы (Бианки, 1902), возможно относятся к поздним
кладкам.
Мелкозернистые, вначале белые с легким палевым оттенком, яйца вскоре
покрываются желто-бурыми пятнами. Размеры A2) 104—114x65,1—70,1,
в среднем 104,1x66,4 мм (Джоурден); 99,1—108,5x64,6—68,0 мм (Туга-
ринов, 1941). Насиживает одна самка. Отправляясь покормиться, самка
аккуратно закрывает кладку перьями и пухом, подтягиваемым с краев
гнезда. Однако встречаются гнезда, в которых яйца ничем не прикрыты
(Кузякин). Насиживание продолжается 29—30 дней (Тугаринов, 1941).
Яйца с эмбрионами, уже покрытыми пухом, на Грибовой Земле (Новая
Земля) найдены 3 июля 1951 г. (Кузякин).
Время выклевывания здесь же в 1951 г. пришлось на 12—15 июля.
13 июля в гнезде найдена покрытая еще не засохшими кровеносными сосу-
дами скорлупа от трех яиц, из которых птенцы выклюнулись, повидимому,
накануне; в тот же день удалось найти еще гнездо с тремя яйцами; из одного
из них просунулся наружу клюв птенца, другое было покрыто радиаль-
ными трещинами, третье оставалось еще целым. 15 июля попалось гнездо
с двумя пуховичками и одним яйцом и в тот же день наблюдалась пара взрос-
лых лебедей с одним маленьким пуховичком (Кузякин).
На Новой Земле в 1903 г. пуховички найдены 16 июля (Сконнинг}
и 20 июля (Теель). Здесь же 26 июля 1928 г. в гнезде из трех яиц два оказа-
лись болтунами, одно же заключало совершенно оформившегося, покрытого
пухом птенца. 2 августа 1928 «г. найдено уже покинутое гнездо. Около
гнезда убит линяющий самец, самка же с пуховичками уплыла в море
(Горбунов, 1929). Таким образом, пуховички на Новой Земле появляются
между 12 и 26 июля. На Ямале взрослые с пуховичками близ Юрюбея
наблюдались 16 июля (Житков, 1912), а в устье Индигирки 8 августа
(Михель, 1932). В устьях Енисея молодые пуховички уже ростом с чирка
найдены 31 июля, а на Бреховских о-вах молодые в половину взрослых наб-
людались 19 августа. Молодые ростом с белолобую казарку при взрослых
на Новой Земле у Черной губы встречены 15 августа 1951 г. (Кузякин).
Повсеместно в Арктике выводки становятся летными уже в начале сен-
тября. Период роста молодых от момента их вылупления и до подъема на
крыло составляет не более 40—45 дней и, следовательно, тундровые лебеди
являются гораздо более скороспелыми, чем другие (Тугаринов, 1941).
После появления птенцов лебеди переселяются на воду и вместе с вывод-
ком держатся по озерам тундры и кормятся у их побережий, выходя иногда
и на берег. В малонаселенных районах, заметив приближающегося четверо-
266
ОТРЯД ГУСЕОБРАЗНЫЕ
ногого хищника, самец поднимается на крылья и летит к нему навстречу,
уже одним своим появлением отгоняя хищника. Такой же маневр он проде-
лывает и при приближении человека. Самка же в таких случаях уводит
птенцов от берега и залегает с ними среди растений и камней побережья
Линька. Повидимому, дважды в году, однако это точно не выяс-
нено. На Новой Земле, на Ямале и в ряде других мест полная летняя линька
взрослых начинается в последней декаде июля B6 июля на Новой Земле,
27 июля на Ямале). 15 августа 1951 г. у Черной губы на Новой Земле у пары
взрослых крылья были без маховых (Кузякин).
Линяют одновременно самец и самка, и начало линьки выражается
в выпадении первостепенных маховых. Смена маховых происходит быстрее,
Рис. 43. Тундровый лебедь
чем у других лебедей. Взрослые с выводками ^переходят в это время на откры-
тые водоемы, где и протекают дальнейшие стадии линьки в таком же порядке,
как и у других лебедей. Вскоре начинается частичная осенняя линька, не
заканчивающаяся на местах гнездовья. Как только окрепнут новые маховые,
взрослые тундровые лебеди вместе с молодыми покидают гнездовые места
и направляются в осенние миграции. У молодых в это время иногда наблю-
даются еще не доросшие до нормы маховые, у взрослых идет линька мелкого
пера, которое окончательно сменяется только на зимовках (Тугаринов, 1941).
Питание. Тундровые лебеди питаются не только различными частя-
ми земноводных растений, но и травянистой наземной растительностью,
причем трава около их гнезд бывает полностью выщипана (Успенский).
Чаще других лебедей они используют мелкую рыбешку (Бекер, 1929).
Хозяйственное значение. Из-за прочной кожи и теплого, легкого меха
тундровые лебеди в местах, где они обыкновении, служат источником про-
мысла в период их линьки. Из их кожи выделывают сумки, телогрейки
и прочие вещи домашнего обихода. Используется также и пух. Кроме
ТУНДРОВЫЙ ЛЕБЕДЬ 267
человека, и у этих лебедей нет настоящих врагов. Песец, завидя лебедя,
убегает от него (Успенский).
Полевые признаки. В полевых условиях тундровый лебедь почти неот-
личим от кликуна, но все же несколько мельче последнего. В южных райо-
нах его скорее можно узнать по времени пролета, так как весной на юге он
пролетает позже других лебедей. Внешне он похож на кликуна и так же
держит на воде во время плавания свою шею, но последняя у него несколько
толще и потому кажется более короткой, чем у кликуна. Голос его чуть
звонче и резче, чем у кликуна, и к тому же у него нет таких трубных звуков,
как у последнего; голос этот звучит, как «тонг... бунг»; иногда слышно «гук-
гук» или «курру». Издает также шипение. В стаях он шумлив и птицы между
собою чаще перекликаются характерным гуканьем. Отличительною его чер-
тою во время миграций, как и у других лебедей, является крайняя осторож-
ность, но в тундрах, где нет человека, он меньше боится его, чем другие
лебеди, и подпускает к себе в гнездовое время метров на ЗОО(Кузякин).
Описание. Размеры самцов и самок: размах 1730—2000, в среднем
1805 мм; длина тела 1070—1220, в среднем 1110 мм; длина крыла 490—540, в среднем
-520 мм; длина клюва 84—98, в среднем 82,7 мм; ширина клюва 28—32, в среднем 28,5 мм
вес 4,9—6,2 кг. Желтый цвет на клюве зачастую не распространяется до углов рта и обык-
новенно на 10 мм не доходит до заднего края ноздрей. Если же доходит ближе, то только
по ноздревым ямкам (Бутурлин, 1935).
Окраска. Пуховик. Спина беловатая. Голова, верх шеи и основание хво-
ста сверху темные. Межлопаточная область беловатая. Бока шеи и брюхо белые.
Молодая птица. Спина дымчато-серая, но верх шеи и бока головы дым-
чато-бурые. Горло, передняя сторона шеи и ее основная часть белые. Клюв одноцветный
желтоватый или буровато-красный. После первой осенней линьки несколько светлеет.
После второй осенней линьки уздечка и основание клюва становятся бледножелтыми.
Ноги того же цвета.
Взрослые самец и самка. Оперение, как и у других лебедей, белое. По бокам
основания клюва по желтому пятну, не доходящему до ноздрей. Остальные части клюва
черные. Лапы черные. Радужина черно-бурая.
Восточный тундровый лебедь Cygnus bewickii jankowskii Alpheraki E93
С у gnus bewickii jankowskii. Alpheraki. Прир. и охота, стр. 10, сент., 1904, Уссурий-
ский край.
Синоним. Уссурийский край. Cygnus davidi (пес Swinhoe) Giglioli
et S a 1 v a d о г i , 1887, залив Посьета1.
Распространение. Ареал. Гнездится в тундре и лесотундре от дельты
Лены до дельты Колымы. Сообщения о гнездовье на Пенжине, Анадыре,
Чукотке и южной части Сахалина недостоверны. Найден в низовьях Яны
и Индигирки.
Характер пребывания. На территории СССР гнездится
и пролетает, зимует в северной половине Китая у его морских побережий;
по р. Янцзы от устья доходит до озера Пу-Ян, где обыкновенен. Редко
у Фучжоу и в окрестностях Сватоу. Одиночные долетают до Индии (Бекер,
1929).
Даты. На обратном пролете к местам гнездовья в марте уже наблю-
дается на побережье Чжилийского залива и внутри страны в соседних с ним
районах. В Приморской обл. появляется в конце марта или в начале пер-
вой трети апреля. Большая часть поздно пролетающих в Приморье лебедей
относится к этому подвиду (Шульпин, 1936). На Яне в районе Верхоянска
восточные тундровые лебеди показываются 27 мая, в устье Индигирки со
1 Молодые лебеди этой формы, добытые в октябре 1879 года в заливе Посьета
ошибочно были отнесены Джильоли и Сальвадори в 1887 году к лебедю под на-
званием Cygnur davidi', описанному Свайно в 1870 г. из Китая. Позднее к этому же
не существующему в природе виду также ошибочно была отнесена молодая птица тундро-
вого лебедя, добытого в сентябре 1902 г. в районе Иркутска (Дорогостайский, 1913).
268 ОТРЯД ГУСЕОБРАЗНЫЕ
2 по 4 июня (поздние), в устье Колымы первое в середине мая. Смеет гнездо-
вья отлетают во время заморозков и выпадения первого снега. В дельте
Яны пролет идет в середине сентября, с низовий Индигирки окончательно
исчезают 22 сентября, примерно в то же время отлетают и из бассейна Колы-
мы. В Приморье пролет идет с начала второй трети октября и продолжается
еще и в ноябре. В начале декабря перемещаются в окрестностях Пекина,
причем раньше тундровые лебеди летели через самый город (Латуш).
Биотоп. Гнездится на возвышенной или на низменной тундре у окра-
ин небольших озер или в низменных заболоченных долинах лесотундры
вблизи стариц и речных рукавов.
Численность. Местами обыкновенен, особенно в низовьях рек.
Экология. Размножение. В связи с более поздним прилетом
сроки его размножения наступают несколько позднее, чем у западной фор-
мы, в остальном не отличается от последней. Яйца белые с палевым оттенком.
Размеры D) 108,6—112x69—73, в среднем 110,25x70,87 мм (Джоурден).
Питание. Сходно с таковым у С. с. bewickii.
Линька. Протекает в те же сроки, как и у западного тундрового
лебедя, и отличается растянутостью.
Полевые признаки. Сходен с кликуном, но меньше последнего и в срав-
нении с ним кажется более короткошеим и толстошеим. Держит шею так же,
как и кликун. Голос тоньше, серебристее, чем у кликуна, подается чаще
и в стае всеми особями сразу. Его можно передать, как звонкое «кук»... «кук»
или «крук»... «крук»... «ку-ку-ку-ку». Иногда слышится более тихое «вав»
или «вов».
Более доверчив, чем западный тундровый лебедь. Лодку подпускает
ближе, но на открытых местах обнаруживает свойственную всем лебедям
осторожность. Спугнутая стая, поднимаясь в воздух, дружно подает голоса,
летит беспорядочной кучей и садится обычно ближе километра от того места,
где лебеди снялись. Отпечатки лап меньше, чем у кликуна (Шульпин, 1936)
Описание. Размеры самцов и самок: размах 1187—2045, в среднем
1916 мм; длина тела 1170—1240, в среднем 1188 мм; длина крыла 490—550, в среднем
530 мм; длина клюва 90—99, в среднем 93 мм; ширина клюва 30,5—35, в среднем 31,6 мм.
Желтая окраска клюва по краю надклювья распространяется до углов рта, доходя часто
до заднего края ноздрей и даже в ряде случаев подходит под ноздри (Бутурлин, 1935).
У молодых крыло 400—425 мм. Клюв по коньку 54—57 мм. Клюв от переднего
края глаза 90—96 мм. Клюв от переднего края ноздрей 31— 32 мм. Наибольшая ширина
клюва 24,2—24,5 мм. Высота клюва у основания 27—31 мм. Средний палец с когтем
114—118 мм (Бутурлин, 1910).
Окраска. Пуховик. Спина беловато-серая. Голова, верх шеи и основание
хвоста сверху темные. Уздечка оперена. Большая часть клюва и ноги желтоватого цвета.
Молодые. В первом и втором пере, как у западной формы, но желтая окраска клюва
ярче и светлее.
Взрослые самец и самка. Оперение чисто белое. Желтый цвет на клюве развит
больше, чем у западной формы. Самый желтый цвет на клюве более оранжевого тона, чем
у западной формы, и расстояние от глаза до вершины клюва больше, чем у последней.
3. Американский лебедь Су gnus columbianus Ord
Cuznus columbianus. Ord. 1815, Gutrie Georg., 2-е изд., стр. 319, река Колумбия.
Русское название. Американский лебедь—книжное.
Распространение. Ареал. Прежде был широко распространен по
тундре и отчасти лесотундре Северной Америки от Аляски до Баффиновой
Земли на юг до о-вов Ноттингема и Соутгемптона. Зимует вдоль Тихооке-
анского побережья Северной Америки до Калифорнии и Атлантического—
до Флориды. В настоящее время крайне редок и спорадичен (Оливер, 1943).
В СССР залетает на Анадырь, Командорские о-ва и по непроверенным данные
на Чукотку. Добыт на р. Анадыре (без даты), затем в окрестностях Ново-
АМЕРИКАНСКИЙ ЛЕБЕДЬ
269
Марьинского 9 июня 1897 г. и 1 мая 1907 г. (Портенко, 1939), а также на о-ве
Беринга 3 ноября 1882 г. (Стейнегер, 1887).
Карта 24. Распространение американского лебедя Cygnus Columbians
а-граница гнездовий, е—зимовки, д—залеты
Биотоп. Открытая тундра по низменностям, богатым озерами,
также в речных долинах, обильных рукавами, старицами и озерами.
Экология. Размножение. Гнездовые места занимает в последней
декаде мая. Гнездо в низменной мохо-
вой тундре по окраинам открытых водо-
емоь. Строится гнездо из мха, верхняя
его часть из сухих стеблей тундровых
растений, лоток устилается мхом. Яиц
4—5, иногда только 2, редко 7. Разме-
ры 90—115,7x56—73 мм. Насижива-
ние 35—40 дней. Пуховички выклевы-
ваются из яиц с конца июня до сере-
дины июля.
Линька. Сроки не выяснены.
Взрослые линяют на открытых водое-
мах, куда переселяются сейчас же вслед
за появлением птенцов.
Питание. Пищей служат главным образом водяные растения,
отчасти же мелкие водяные животные.
Полевые признаки. Размерами крупнее тундрового лебедя, шея не-
сколько тоньше. В остальном сходен с тундровым лебедем. Селится в самых
глухих труднодоступных местах. Крайне осторожен.
Рис. 44. Голова взрослого американ-
ского лебедя
270 ОТРЯД ГУСЕОБРАЗНЫЕ
Описание. Размеры и строение. Взрослые самцы и самки: длина крыла
510—575 мм; хвост 152—192 см; средний палец с когтем 123—154 мм. Клюв по разрезу
95—106 мм. Клюв от переднего края глаза 109—122 мм. Клюв от переднего края ноздри
35—48 мм; ширина клюва у середины ноздрей 29—34 мм (Стейнегер). Вес 5,5—7 /сг.Клюв
одинаковой ширины по всей своей длине. Ноготок клюва крупный в виде равносторон-
него треугольника.
Окраска. Пуховик. Дымчато-серый.
Молодая птица. Спина и кроющие крыла свинцово-серые, поясница светлее—
беловатая, верхние кроющие хвоста серые. Верх и бока головы серовато-бурые. Брюш-
ная сторона белая с сероватым налетом. Клюв и лапы буровато-желтые.
Взрослая птица. Оперение чистобелое.
Систематические заметки. В последнее время на основании географического викариата
сделаны попытки объединить американского лебедя в одни вид с тундровым. Однако мор-
фологические данные не позволяют сделать этого. Американский лебедь крупнее тундро-
вого. Строение его клюва иное, чем у тундрового, что сразу же видно при сравнении
клювов обоих видов сверху. У американского лебедя голое пространство от места пере-
гиба носовых костей к лобным почти вдвое более, чем у тундрового. Самый клюв длиннее
и достигает по разрезу от угла рта до вершины не менее 95 мм, в то время как у
тундрового он не более 83,5 мм. Края надклювья на всем их протяжении параллельны,
у тундрового же лебедя клюв к вершине слегка лопатообразно расширяется. Зубцы над-
клювья менее развиты, невысокие, тонкие и сидят реже, чем у тундрового. Клюв, заь
исключением небольшого пространства, на границе с уздечкой, черный.
А. Лебедь-шипун Су gnus olor Gmel.
Anas olor. G m e 1 i n. Syst. Nat., 1, 2, 1759, стр. 501, Россия, Сибирь и т. д.
Синоним. Су gnus sibilans. Pallas. Zoogr., II, 1811, стр. 215. Россия.
Русски! названия. Лебедь—старинное русское название; лебедь-шипун—на -
звание, данное птице за особенности ее голоса.
Распространение. Ареал. В настоящее время гнездится разрозненна
и лишь местами на юге Фенноскандии и в прилегающих районах, в низовьях
Дуная, по степным и лесостепным районам СССР и от Китайской Народной
Республики до Уссурийского края. Переходя к деталям распространения,
необходимо отметить его редкое гнездованье в Южной Швеции, в Дании,
на юго-восточном побережье Балтийского моря, на о-ве Сарема, в Калинин-
градской обл. и, повидимому, местами в Польской республике, в низовьях
Дуная и прилежащих частях Болгарии, в Малой Азии и Иране. В СССР
гнездится в низовьях Днестра, в очень незначительных количествах встре-
чается в плавнях Кубани (Шапошников, 1932), Терека и Сулака (Беме,
1925 и 1935), изредка на северном побережье Каспия, но на территории
Астраханского государственного заповедника не селится. Найден на гнез-
довье в Волжско-Уральских степях (Волчанецкий, 1937), в степях Запад-
ного Приуралья в бассейнах Илека, Утвы, Хобды, Темира (Тугаринов, 1941),
в южной части Зауральского озерного района, в низовьях Иргиза и Тургая,
на озере Челкар, в районе Троицка и Кустаная (Тугаринов, 1941), но на
территории Наурзумского заповедника и близ него не гнездится (Михеев»
1938). Обитает на озере Ассау-Балык близ озера Кургальджин (Лавров,
1929), также на ряде других озер Казахстана, в частности на озере Сассык-
Ала-Куль к юго-востоку от Семипалатинска (Селевин, 1929), на озерах
Барабинской степи, включая озеро Чаны и мелкие озера, расположенные
к северо-востоку от него. Ни в Кулундинской степи, ни по среднему течению
Иртыша, ни на Зайсане шипуна нет. Отсутствует также в Сибири на про-
странстве от бассейна Оби до Забайкалья, где снова появляется в Даурии,
и, в частности, на озере Тарей-Нор, в Монголии найден на озере Далай-
Нор и далее на восток встречается около Сидеми в Приморской обл. В недав-
нем прошлом был обычен на оз. Ханка. В Китайской Народной Республике
встречен в центральной части Гоби (Седербом), на озере Куку-Hop, Цай-
даме, на озере Лоб-Нор и других озерах северной окраины Тибета. Далее-
на запад селится по глухим озерам у Куйтана и Сары-Кумея, на Ала-Куле
ЛЕБЕДЬ-ШИПУН
271
и на Балхаше (Шнитников, 1949), обычен в долинах Сырь-Дарьи и Аму-
Дарьи. Имеются требующие проверки указания на гнездованье в долинах
Теджена и Мургаба. Неполовозрелые задерживаются весной в Крыму на
Сиваше (Воронцов, 1937).
В ряде мест, лежащих к северу от сохранившихся еще районов гнездовья,
бывает только случайно. Залетает на Сиваш, в низовья Днепра, где недавно
он еще гнездился, в Киевскую, Харьковскую, Полтавскую, Воронежскую
и Курскую обл., в район Пензы и Горького. Еще в начале нынешнего сто-
летия на гнездовье доходил до Вятского края (Круликовский, 1913). В полу-
одомашненном состоянии встречается во многих районах Европы.
Карта 25. Распространение лебедя-шипуна Cygnus olor
а—места гнездовий, в—зимовки
Характер пребывания. На территории СССР гнездится, про*
летает и зимует. Как и кликун, если его не тревожат, шипун близ мест своих
гнездовий держится до последней возможности и отлетает лишь при полном
замерзании водоемов. Из низовий Днестра, когда лиманы сковываются
льдом, перебирается на Черное море, возвращаясь при первой возможности
обратно (Назаренко); с северных побережий Каспия перелетает на его запад-
ные и восточные берега и зимует у юго-западных и юго-восточных его побе-
режий. Обычен зимою на озере Иссык-Куль (Шнитников, 1949) и встречается
по другим крупным озерам Средней Азии. Однако из ряда мест лебеди-
шипуны мигрируют на зимовки далеко на юг и доходят до Северо-Восточ-
ной Африки, Средиземноморья, Малой Азии, Палестины, Аравии, Ирана,
Афганистана, Белуджистана и одиночными особями даже до Северо-Западной
Индии. В Китайской Народной Республике и ее восточной части зимою встре-
чен в районе Пекина и Чинцзяня.
Д а т ы. С мест зимовок на Каспийском море начинает отлетать со вто-
рой половины февраля, но на восточном берегу в низовьях Атрека задер-
живается до середины марта (Дементьев, 1945), на Талышинском же побе-
212 ОТРЯД ГУСЕОБРАЗНЫЕ
режье в некоторые годы после 26 февраля уже не встречается (Тугаринов,
1950). В середине первой—в конце второй декады марта движение идет
у северо-западных берегов Каспия в дельте Терека (8 марта—Радде, 1884;
19 марта 1921 г.—Беме, 1925) В конце первой декады марта или в начале
второй шипуны появляются в низовьях Днестра и Днепра, в Крыму и в не-
значительных количествах пролетают через Украину к Балтийскому морю;
большая же их часть направляется на северо-восток. С 15 марта по 15 ап-
реля идет пролет по Сыр-Дарье (Спангенберг и Фейгин, 1936), а в середине
последней декады апреля шипуны оказываются уже у крайней северо-
восточной границы их современного гнездового ареала—в Барабинской
степи, на озере Убинском B6 апреля—Рузский, 1940). Даты передвижений
в крайних восточных частях ареала остаются невыясненными, самый же весен-
ний пролет мало выражен. Летят днем и ночью, когда их можно узнать по
особому свисту крыльев.
Места гнездовий покидают только после того, как молодые поднимутся
на крылья, что в ряде районов совпадает с наступлением осенних холодов
и заморозков. Но на юге ареала, где лебеди-шипуны выводятся и линяют
раньше, они принуждены под влиянием начинающейся интенсивной охоты
отлетать задолго до наступления морозов. В некоторые годы одиночные
передовые или семьи показываются очень рано близ мест зимовок B4 сен-
тября 1898 г. на Атреке—Житников, 1900; в середине сентября на Сиваше).
В Средней Азии пролет наблюдается по Или, Сыр-Дарье, Аму-Дарье, по ни-
зовьям Теджена и Мургаба, в предгорьях Копет-Дага и вниз по Атреку
(Дементьев, 1952). Летят также вдоль восточного и западного побережий
Каспия. В 1937 г. на восточном берегу в Балханском заливе передовые отме-
чены 6 ноября. Затем наблюдались 15 ноября у о-ва Огурчинского и 16 де-
кабря у о-ва Челекена (Исаков и Воробьев, 1940). Ежегодные зимовки
наблюдаются у Гасан-Кули, в бухте Соймонова, у острова Даг-Ада и в заливе
Чадры. Количество оставшихся здесь шипунов (вместе с кликунами) после
отлета главной их массы на север к середине февраля доходило до 1000 шт.
(Лаптев, 1934). Регулярно шипуны зимуют также в низовьях Атрека и близ
его устья.
На западном побережье Каспия шипуны скопляются к началу декабря
в районе Аграханского залива, к середине же этого месяца перебираются
к устью р. Куры—главному месту своих зимовок в СССР. Но и здесь
количество их невелико. Встречаются они то одиночками, то парами или
семьями и лишь в редких случаях стаями до 150 особей (Исаков и Воро-
бьев, 1940). Южнее низовий Куры шипунов уже очень мало, но наблюдаются
они даже у Ленкорани и случайно залетают на озеро Севан (Шелковников,
1934). В области Азовского побережья пролетные шипуны появляются
впервой трети ноября (Боровиков, 1907), а при замерзании Азовского моря
и северных берегов Черного отсюда отлетают в Малую Азию и в низовья
Дуная. В начале декабря встречаются иногда в Новороссийской бухте
и ее окрестностях F декабря 1921 г.; Птушенко, 1939), откуда они исчезают
в середине декабря, отлетая южнее. Встречались зимой в районе Поти
(Радде, 1884), но с черноморских берегов Кавказа отлетают в Турцию.
Биотоп. Населяют большие стоячие, труднодоступные пресные и
горько-соленые водоемы, мелкие берега которых зарастают крупным трост-
ником, местами занимают сплавнины среди стоячих водоемов, где тростники
перемежаются с плесами чистой воды.
Численность. Всюду, за исключением бассейна Урала и некото-
рых казахстанских озер, лебедь-шипун крайне редок и в связи с этим се-
лится на громадных расстояних пара от пары, избегая близости человече-
ских поселений. Там же, где на лебедей не охотятся, пара от пары может
гнездиться близко.
ЛЕБЕДЬ-ШИПУН 273
Помимо непосредственного преследования, заметно снижающего плот-
ность населения этой птицы, косвенной причиной, лимитирующей числен-
ность поголовья шипуна, служат также ранние осенние похолодания, что
даже в Казахстане иногда ведет к гибели неспособного еще подняться на
крылья молодняка. Происходит это вследствие позднего размножения шипу-
нов. Возможно, что как противовес этим неблагоприятным климатическим
условиям, у лебедей-шипунов наблюдается увеличенное число откладывае-
мых яиц, большее, чем у других наших лебедей (Тугаринов, 1941). Гибнут
лебеди зимою от бескормицы, а также во время перелетов на мазутных лу-
жах и озерах (Исаков, 1940; Богачев, 1943).
Экология. Размножение. На местах гнездовья появляются ши-
пуны парами, но, вследствие того, что усиленное преследование со стороны
человека ведет к разбивке семей, у шипунов много холостых самцов, которые
и пытаются отбивать самок из уже сложившихся пар. В таких случаях наблю-
даются ожесточенные схватки между самцами, которые наносят друг другу
сильные удары крыльями, причем посторонний самец чаще всего изгоняется.
Инстинкт размножения начинает проявляться в трехлетнем возрасте.
Часть самок при этом строит гнездо и сносит одно яйцо или же, закончив
постройку гнезда, не несется вовсе. Настоящая половая зрелость наступает
в возрасте четырех лет.
Весьма характерны в брачный период токовые позы и поведение самца
и самки лебедей-шипунов. Птицы держатся в это время на воде друг около
друга. Самец приподнимает крылья и отводит их несколько в стороны
и вместе с самкой часто окунает голову в воду, а затем, сплывшись, опле-
тает ее шею своей. После долгих игр самка, наконец, погружается в воду
так, что остается видной только ее голова и шея. В этот момент происходит
спаривание. Через короткое время птицы всплывают, издают особые хрип-
лые звуки и прижимаются грудью друг к другу, затем купаются и пере-
бирают перья (Тугаринов, 1941).
Гнездо строится одной только самкой в малодоступных глухих зарослях
тростников, обычно на куче старого поваленного растительного хлама. Для
его возведения служат мелкий прошлогодний тростник, а также засохшие сло-
манные стебли и части различных земноводных растений. В поперечнике
у своего основания гнездо доходит до 110 см, в высоту до 75 см. В некоторых
случаях гнездо строится на сплавнине, и тогда диаметр его может достигать
4 му высота же 1 м (Тугаринов, 1950). Когда гнездо уже в общем построено,
по мере откладки яиц плоский его лоток выстилается пухом. Иногда невда-
леке от одного гнезда находится другое или даже третье. Это дает повод
говорить о колониях шипунов (Бекер, 1929). Во всяком случае, определен-
ного гнездового участка у них не имеется, и к тому же популяции лебедей
в настоящее время, как уже указывалось, чрезвычайно разрежены.
Полная кладка состоит из 7—9 яиц, но у молодых самок число яиц мень-
ше, а у впервые приступающих к стройке гнезда бывает только одно. Цвет
ненасиженных яиц зеленовато-оливковый, к концу же насиживания яйца
становятся грязножелтовато-белыми. Размеры: 105—122x70—80 мм,
в среднем 114,5x73,1 мм. Вес 35,0—37,5 г (Тугаринов, 1941).
К откладке яиц шипун приступает несколько позже кликуна. На Сыр-
Дарье полные ненасиженные кладки можно находить уже в конце первой
декады мая. На западном берегу Каспия в Аграханском заливе 3 нена-
сиженных яйца найдены 13 мая 1925 г. (Спангенберг). На Аральском море
в заливе Шашак-Куль (к северо-западу от залива Паскевича) 2 яйца, при-
крытые слоем метелок тростника, в гнезде, на котором сидел самец, найдены
17 мая 1947 г. (Кузякин). В Калмыцкой степи ненасиженное яйцо взято из
гнезда 27 мая (Иоганзен, 1923) и в Западной Сибири на оз. Сартлан найдена
полная кладка ненасиженных яиц 2 июня 1913 г. Таким образом, гнездовой
18 Птицы СССР, т. IV
274 ОТРЯД ГУСЕОБРАЗНЫЕ
цикл в северной части ареала протекает позже. В то же время наблюдаются
запоздавшие или вторичные кладки F июня 1927 г. на Сыр-Дарье найдено
гнездо с 3 слабо насиженными яйцами; Спангенберг и Фейгин, 1936).
Насиживает около 35 дней одна только самка, самец же находится
поблизости, охраняет ее, и при тревоге птицы ведут себя подобно кликунам.
Когда самка сходит, чтобы покормиться, она покрывает кладку материалом,
стаскиваемым с боков гнезда. Самец в таких случаях иногда садится на гнезда
поверх материала, покрывающего яйца. В случае гибели первой кладки
вторая содержит не более двух яиц. На Сыр-Дарье пуховички появляются
в середине июля. Запоздавшие кладки нередки, и поздних пуховиков, до-
стигавших примерно трети взрослых, в районе Тургая наблюдали в середине
августа (Тугаринов, 1941).
Птенцы находятся под присмотром обоих родителей на воде. Вся семья
плавает вместе, и самка часто обогревает маленьких птенцов, давая им воз-
можность взобраться к ней на спину (Мензбир, 1895). Ночь семья проводит
в гнезде, а когда молодые подрастут—на сплавниках или на старом заломлен-
ном тростнике.
Линька. Линька взрослых гнездящихся птиц, холостых и второгод-
ков, повидимому, дважды в году: летом, примерно с июня по август, про-
исходит полная смена всего оперения и осенью наблюдается частичная
линька контурных и рулевых с начала сентября по декабрь, иногда по ян-
варь, однако это требует еще точного выяснения. Молодые лебеди, после
того как они наденут первый наряд (гнездовой) и станут летними, вступают
в частичную осенне-зимнюю линьку.
В начале этого столетия на северо-восточном побережье Каспия на период
линьки к середине лета холостые и неполовозрелые второгодние лебеди-
шипуны собирались в большом количестве, с чем был связан и промысел
на них (Огнев, 1913). В настоящее время таких скоплений линных лебе-
дей уже не наблюдается. Взрослые гнездящиеся линяют разновременно.
Первой, еще при пуховичках, начинает линять самка. Линька у нее тянется
около полутора месяцев. К тому времени, когда линька у самки уже закан-
чивается, самец, до сих пор охранявший выводок, надолго забирается в мало-
доступные крепи и там сменяет свой наряд. У взрослых птиц процесс линьки
знаменуется прежде всего одновременным выпадением маховых и кроющих
крыла. Вслед за этим начинают выпадать рулевые, перья надхвостья и под-
хвостья. Затем идет линька контурных низа и верха тела и, наконец, сме-
няются перья шеи и головы. Линька маховых протекает медленнее, чем
у кликуна, и ежедневный прирост их у шипуна достигает только 7 мм.
Вылинявший самец присоединяется к семье. У молодых в это время еще идет
рост маховых, и как только они достигнут нормальных размеров и окрепнут,
выводок начинает подниматься на крыло, а вскоре затем вся семья трогается
в осенний путь.
Частичная осенняя линька начинается на родине и заканчивается на
зимовках.
Питание. Пища шипунов состоит из подводных частей растений,
корни, корневища и побеги которых эти лебеди отрывают на мелких местах
в воде клювом. Для этого они становятся на воде, как утки, вертикально
хвостом кверху, а головой вниз, и достают клювом до дна. В небольшом
количестве при этом поедают и мелкие водяные животные. Птенцы кормятся
тут же всплывающими оторванными частями растений. На зимовках пищею
являются различные харовые и нитчатые водоросли (Исаков и Воробьев,
1940). Штормовая погода и поднятие уровня воды иногда лишают шипунов
ВОЗМ0ЖНОСТИ добывать корм. В таких случаях они голодают и подчас обес-
силивают до того, что теряют способность к полету, но избранных пунктов
зимовья все же не оставляют, пережидая непогоду.
ЛЕБЕДЬ-ШИПУН
275
Хозяйственное значение. Разреженность популяций лебедя-шипуна и
вообще его сравнительная редкость делает промысел на него случайным
и мало рентабельным. Лебедей-шипунов в незначительном количестве добы-
вают лишь на местах линьки на восточном побережье Каспия и на Азовском
море. Ловят также зимующих, ослабевших от бескормицы. Используют этих
лебедей на перо и пуховые шкурки. Мясо же их невкусно и жестко. Добычу
лебедей-шипунов на ряд лет следует воспретить; вместе с тем необходимо раз-
водить их в одомашненном состоянии, так как они оказываются более ужив-
чивыми и продуктивными, чем другие лебеди.
Рис. 45. Лебедь-шипун
Полевые признаки. Такой же крупный и белоснежный, как кликун,
лебедь-шипун, плавая обычно изгибает свою шею в виде буквы S, а клюв
и голову держит наклонно к воде. Эти признаки, как и несколько более тол-
стая, и поэтому кажущаяся более короткой, шея легко позволяют отличить
его от кликуна на расстоянии. Помимо этого, контур спины у шипуна угло-
ватый, а не округлый, как у кликуна. На лету шипуна можно узнать, во-
первых, потому, что он не издает таких громких трубных звуков, как кликун,
во-вторых, по особому ритмичному, слышному за несколько сот метров поскри-
пыванию больших маховых. Голос шипуна не такой звонкий, как у кликуна,
более низкий, хриплый и слегка дребезжащий. Чаще голос этот слышен
в брачную пору, когда самец издает звуки «кгиюрр», а самка «киорр». Бу-
дучи раздраженным, он издает особый характерный шипящий звук, из-за
которого и получил свое название. При плавании гребет лапами попере-
менно, но может грести и обеими сразу. При этом он время от времени раскры-
вает крылья и машет ими.
Чрезвычайно осторожен и избегает гнездиться близ человеческих посе-
лений. Большую часть времени проводит на воде, но там, где его не беспокоят,
шипун временами выходит на берег. Ночует всегда в глухих местах водо-
18*
276 ОТРЯД ГУСЕОБРАЗНЫЕ
емов на сплавнинах и старых кучах тростников. К другим птицам относится
терпимо и иногда селится рядом с гнездами серых гусей.
Описание. Размеры и строение. Длина крыла самцов 548—630, самок
535—600 мм. Длина клюва от нароста самцов 76—85, самок 73—80 мм (по Шьёлеру,
с исправлениями). Вес 8—13 кг (Тугаринов, 1941) и до 22,5 кг (Занден, 1935). Верхний
край ноздрей с поверхностью конька надклювья образует острый угол. У основания
лба на клюве крупный нарост. Хвост из 11—12 пар рулевых, крайняя пара которых
на 80—120 мм короче средней.
Окраска. Пуховик. Спинная сторона буровато-серая; брюшная сторона и
горло грязновато-белые. Клюв черноватый.
Молодая птица. В первом наряде буровато-серая с более светлыми, почти белыми,
шеей и горлом. Маховые белые у оснований и сероватые у вершин. Рулевые серовато-
дымчатые с белыми вершинами. Выроста у основания клюва нет или он только намечен.
Уздечка оперена. Основание клюва черноватое, остальные части клюва буровато-серые.
После первой осенней линьки буровато-серая сверху, снизу светлее. После второй осен-
ней линьки птица белая, но голова, верхняя сторона шеи и кроющего крыла серовато-
бурые. Шишка на основании клюва становится крупнее. Клюв мясного цвета. Оконча-
тельный белый наряд надевается не ранее как после третьей или четвертой осенней
линьки.
Взрослые самец и самка. Чистобелые. Шишка (нарост), голая уздечка, все основа-
ние клюва, его конек до ноздей, впадины последних и края разреза рта черные. Осталь-
ные части клюва красные. Ноготок черный. Ноги, пальцы и перепонки тускло-черные.
Радужина бурая. У самки шишка менее развита.
РОД СУХОНОСЫ CYGNOPSIS BRANDT, 1836
Тип. Anas cygnoid
5. Сухонос Cygnopsis cygnoid L.
Anas cygnoid. Li nnaeus. Syst. Nat., изд. Х, стр. 122, 1758, Азия.
Синоним. Anas orientalis Gmeli n. Syst. Nat., 1, 2, стр. 503, 1788.
Русские названия. Сухонос—русское народное сибирское название, другое
ходовое название—китайский гусь.
Распространение. Ареал. Гнездится спорадически в Тарбагатае
и Зайсанской котловине, в Южном и Юго-Восточном Алтае, в Минусинской
котловине до 54° 31' с. ш. Далее, начиная от Южного Прибайкалья и Забай-
калья, сплошь заселяет бассейны левых притоков Амура до устья, заходя
на север по побережью до устья Уды и, повидимому, до Аяна (около 56°
36' с. ш.), где еще в недавнее время гнездился. На юг отсюда распростра-
нен по Приморской обл., Сахалину, Маньчжурии, северным провинциям
Китая до Северного Ордоса. Отсюда граница ареала направляется к северу
на Керулен и к западу по бассейнам Селенги, Тэси, Кобдо, до северных под-
ножий Гобийского Алтая, озера Орок-Нора (Китайская Джунгария),
верхнего течения Урунгу и Булгуна. Возможно гнездится в северо-западной
приморской части Кореи. На Алтае поднимается на гнездовье от 340 до
2400 м (Сушкин, 1938). В зимнее время в пределах СССР изредка встречается
в Узбекистане, перемещается же зимой главным образом в области восточного
побережья Китайской Народной Республики от залива Чжили до низовий
Янцзе, отчасти зимует также и в Корее; очень редок на зимовках в Японии,
где в некоторые годы его мало, в другие же появляется стаями до 100 особей
и остается на местах зимовок до середины апреля (Ян, 1942).
Характер пребывания. Гнездящаяся и перелетная птица,
местами нерегулярно и в небольшом количестве зимующая.
Даты. С мест зимовок отлетают в конце марта и 1—5 апреля по-
являются в районе озера Ханка (Пржевальский, 1870). Почти в то же
время пролетают сухоносы в центре Монгольской Народной Республики—
на оз. Орок-Нор, на полыньях которого они наблюдались 4 апреля (Козлова,
1930). На Амуре в окрестностях Мариинского первые отмечены 13 апреля,
под Новониколаевском 22 и 27 апреля (Шренк, 1861). Массовый пролет на
СУХОНОС
277
оз. Ханка отмечен 13—15 апреля, последние здесь пролетали 18 апреля
(Пржевальский, 1870). На Среднем Амуре в Буреинских горах первые отме-
чены 9 апреля, на Тарей-Норе 15 апреля. Значительно позже появляются
сухоносы в высокогорьях и, в частности, на плато Тунка в Юго-Западном
Забайкалье первые наблюдались только в начале второй трети мая (Радде,
1863). Общие направления весенних передвижений сухоносов на север и
северо-восток, на север и северо-запад.
Осенью наиболее рано покидают гнездовые места в высокогорьях и,
например, с плато Укок на Алтае сухоносы откочевывают уже к 20 августа,
Карта 26. Распространение сухоноса Cygnopsis cygnoid
a—граница гнездовий, г—зимовки, д—залеты
спускаясь в более низкие места. В то же время передовые стаи начинают
осенние миграции, как только вновь получат возможность летать, и в августе
их передвижения наблюдались уже в северной части Гоби (Пржевальский,
1876). Из более низко расположенных мест отлетают позже. В Буреинских
горах первые отлетающие стаи отмечены 9 сентября, последние отсюда скры-
лись 14 сентября. С озера Тарей-Нор и его окрестностей отлет передовых
наблюдался 14 сентября; 16 сентября сухоносов оставалось уже немного,
последние исчезли 19 сентября (Радде, 1863). Из бассейна озера Ханка
начинают отлетать 17 сентября, пролетные здесь же отмечены с 17 сентября
до начала октября (Маак, 1861). Отдельные стаи на Амуре наблюдались еще
27 сентября, и вообще в низовьях Амура, где в осеннее время господствуют
сравнительно мягкие климатические условия, задерживаются долго. Из
Монгольской Народной Республики осенние передвижения направлены на
юго-восток, из Уссурийского края—на юг и юго-запад. Места зимовок рас-
положены от побережий Желтого моря и его заливов до Шанхая, а также
в долинах рек Хуанхэ, Янцзе и на примыкающих к ним озерах.
278 ОТРЯД ГУСЕОБРАЗНЫЕ
Биотоп. Хотя сухонос встречается и в горном, и в равнинном
и в степном ландшафтах, но по существу всюду в самых разнообразных местах
он связан с речными и озерными водоемами. Во время же кочевок и миграций
может быть встречен даже в голых, бесплодных степях вдали от воды (напри-
мер, вдоль Калганского тракта в Монголии; Тугаринов, 1932). В горах
он занимает устья рек, широкие горные речные долины, узкие долины быстрых
горных рек с галечниковыми берегами и редкой растительностью, горные
лесные озера и болота. В степях и равнинных местах его можно найти в ши-
роких речных долинах с прибрежными зарослями тростников, в болотистых
речных долинах, на приречных и приозерных лугах и, наконец, на пресно-
водных и солоноватых озерах, поросших тростником и рогозом. После вывода
гусят держится по песчаным речным и озерным песчаным косам и на ровных
илистых топких берегах озер (Сушкин, 1938; Тугаринов, 1941).
Численность в западной части ареала незначительна, более обы-
чен в бассейне Амура, но за последнее время значительно уменьшился
в количестве и в ряде районов бассейна Амура, где прежде сухонос был
обилен, в настоящее время он стал редким или вовсе исчез (Шульпин,
1936).
Экология. Размножение. Начало формирования пар не про-
слежено. Возможно, что пары образуются еще на зимовках или во время
миграций, так как с первых же дней появления на местах гнездованья в ряде
случаев уже встречается парами. Однако в Буреинских горах гуси, держав-
шиеся парами, были отмечены только на седьмой день по прилете—16 апреля
(Радде, 1863). Гнездо устраивается в траве в небольшом углублении, выры-
ваемом самкой, и устилается сухими злаками и пухом. Первые отложенные
яйца на границе с Монгольской Народной Республикой найдены 27 апреля;
30 апреля на озере Ильдзе (Ульде) было уже много отложенных яиц. Вместе
с тем здесь же наблюдалось, что не все гуси принимали участие в гнездовании,
т. е. часть сухоносов оставалась холостой (Радде, 1863). В Монгольской
Народной Республике на озере Орок-Нор 11 мая найдены 10 гнезд с яйцами
(Козлова, 1932). В высокогорьях откладка яиц происходит позже, чем в мест-
ностях более низких.
В кладке5—8, чаще 5—6 белых яиц, размеры которых: 77—88,2x53,5—
57,4 мм (Тугаринов, 1941); B0) 76—89,5x53—59 мм, в среднем
81,6x56,6 мм. Вес 11,76—15,9, в среднем 14,23 г (Гебель, 1904).
Насиживает самка. Сидит на яйцах она крепко и особенно неохотно
покидает их к концу насиживания; самец же, пока она сидит на яйцах, нахо-
дится близ гнезда. Первые птенцы на Среднем Амуре появляются уже
к половине мая, наИртышеу устья Курчума—4 июня (Поляков, 1912), в Кир-
гизской ССР—в начале июня. На озере Орок-Нор птенцы выклевываются
еще позже—20 июня (Латуш, 1934). Рост молодых идет довольно быстро.
11 июня на озере Ханка молодые достигали уже размеров кряквы (Пржеваль-
ский, 1870). В конце июня на Амуре молодые были уже более половины раз-
меров взрослых, 2 августа, здесь же, по размерам они не отличались от пис-
кульки, а 4 августа их крылья у воспитываемых в неволе в длину имели
330 мм (Шульпин, 1936). В высокогорьях, где птенцы появляются позднее,
размеров кряквы они достигли только 27 июля (Алтай; Сушкин, 1938).
На Амуре количество молодых в выводке 5—6, иногда 3—4 и даже 2 (Шренк,
1860). Иногда несколько выводков соединяются вместе, и тогда можно ви-
деть большое количество молодых при 4—6 старых птицах. В некоторых
случаях при опасности взрослые пытаются «отводить» от своих выводков,
отлетая недалеко, а затем опускаются на землю и идут, прихрамывая или
волоча одно крыло. Преследуемые на суше молодые, оставшиеся без взрос-
лых, спасаются в густой траве и так искусно прячутся, что без собаки найти
их невозможно. В случае, если сухоносы держатся у берегов, при пресле-
СУХОНОС
279
довании они отплывают на середину водоема, а пуховички или ныряют или
распластываются на воде, вытянув шейки (Тугаринов, 1941).
Линька. Характер линьки точно не выяснен. Холостые и неполо-
возрелые сухоносы, которые в течение всего лета держатся стаями, начи-
нают линять раньше других. Такие птицы с первыми признаками линьки
отмечены уже 8 июня (Радде, 1863). Семейные сухоносы к линьке приступают
значительно позже, и 2 июля на Амуре у самок при выводках линька еще
не начиналась (Шульпин, 1936), но с середины июля самцы при выводках
уже сменяют перо. Вообще же июль и первая половина августа—время
линьки сухоносов. На Иртыше линяющие семейные самцы при выводках
встречены 12 июля. На Тарей-
Норе 13—19 августа линька бы-
ла уже закончена, и сухоносы
к 19 августа стали собираться в
предотлетные стаи (Радде, 1863).
На Амуре 27 августа линька так-
же была закончена, но у неко-
торых молодых шел еще рост ма-
ховых, которые были в пеньках
(Воробьев, 1937). Повидимому,
самцы сухоноса, в противополож-
ность другим гусям, на время
линьки не отделяются от вывод-
ка и линяют при нем (Поляков,
1912; Тугаринов, 1941).
Питание. Основой пита-
ния сухоноса является травяни-
стая растительность тех мест, где
он держится. На Амуре у десяти
особей, добытых 27 августа, пи-
ща состояла исключительно из
осоки Cares sp. (Воробьев, 1937).
Подробных данных по составу
кормов и их сезонной смене не
имеется.
Хозяйственное значение. Вследствие незначительной численности добы-
вается лишь в небольших количествах. ВПриамурьеместамиихяйца подкла-
дывают под домашнюю птицу или ловят птенцов и выдерживают их до подъема
на крыло. Сухонос является предковой формой так называемого китайского
гуся.
Полевые признаки и поведение. Сухонос в общем несколько напоминает
гуменника и на далеком расстоянии отличим от него с трудом, хотя и круп-
нее последнего. На близком расстоянии и в бинокль заметна беловатая спере-
ди и с боков и темная сзади окраска шеи. Полет кажется более тяжелым,
чем у других гусей. В осеннее время, собравшись перед отлетом в стаи,
ведет себя чрезвычайно шумно и крикливо. Время от времени такая стая
взлетает, делает несколько кругов, затем снова садится. На лету строится
углом. Весенние стаи не велики—в них не более 20—40 птиц. Осенью иногда
встречаются более крупные стаи.
Там, где сухоносов не тревожат, они не отличаются осторожностью
и, наоборот, проявляют весною даже вредное для себя любопытство. Заметив
присевшего в траву охотника или его собаку, сворачивают в эту сторону,
подлетают на ружейный выстрел и даже делают два-три круга над предме-
том, привлекшим их внимание (Пржевальский, 1870). Но стрелянные,
а также при выводках, близко к себе не подпускают. При подходе к взрос-
Рис. 46. Сухонос
280 ОТРЯД ГУСЕОБРАЗНЫЕ
лым семейным гусям при выводках они слетают шагов за 200, после чего
подолгу не возвращаются. Иногда, прежде чем улететь, взрослая птица
делает 3—4 круга и затем уже улетает. Линные гуси, бросившись в воду,
погружают в нее тело так, что видна только их голова, иногда же погру-
жаются в воду совсем и плывут в ней к какому-либо безопасному месту, на
которое незаметно выбираются (Поляков, 1912). Голос сухоноса громкий,
протяжный и его называют то трубным (Хартерт, 1920), то стонущим (Суш-
кин, 1938); звучит он, как звонкое, протяжное «гоооо».
Описание. Размеры. Размах самцов D) 1610—1620, среднее 1616 мм; размах
самок A) 1510 мм. Длина тела самцов D) 915—930, в среднем 921,6 мм; длина тела самок A>
800 мм. Длина крыла самцов D) 460—473, самок D) 437—445 мм, в среднем 467,6 и 444 мм.
Длина клюва самцов D) 89,—98,5, в среднем 94,8 мм; длина клюва самок A) 870 мм.
Высота нижней челюсти самцов D) 7,2—7,8, в среднем 7,5 мму у самок A) 6,8 мм. У самца
основание верхней челюсти несколько вздуто. Самка размерами меньше самца, у основания
верхней челюсти вздутия не имеется.
Окраска. Пуховик. Спинная сторона темная, серовато-оливковая. По бокам
спины позади крыльев по светлому пятну. От клюва через глаз идет темная полоса.
Голова сверху желтоватая, светлее спины. Брюшная сторона светлосерая, желтеющая
к шее и к хвосту. Клюв черный. Ноготок желтый. Ноги желтые.
Молодая птица. Спинная сторона серовато-бурая, тусклая. Бока без светлых
полос. У основания надклювья нет светлой полосы. Особенности оперения полувзрос-
лых не установлены.
Взрослые самец и самка. Спинная сторона и бока серовато-бурые в светлых волни-
стых полосах. Брюшная сторона булано-бурая, но брюхо белое. На лбу у основания
надклювья белая узкая полоса. Голова сверху и с боков до глаз, а также вперед до белой
полосы и шея сверху рыжевато-бурые. Бока шеи вдоль всего ее протяжения светлые,
в средних ее частях почти белые. Подбородок рыжевато-бурый. Спина и надхвостье
серовато-бурые, бока слегка темные. Перья боков со светлыми каймами. Малые кроющие
крыла пепельно-серые; средние буроватые со светлыми каймами. Большие пепельно-
серые, также с более светлыми каймами. Первостепенные маховые аспидно-серые; второ-
степенные черно-бурые. Рулевые того же цвета, но с белыми каймами и вершинами. Ниа
шеи булано-бурый. Зоб и грудь буровато-серые. Брюхо и подхвостье белые. Клюв
черный. Ноготок блестяще черный. Ноги красновато-оранжевые Радужина бурая или
красновато-бурая.
РОД ГОЛУБЫЕ ГУСИ PHIL ACTE BANNISTER, 1870
Тип. Philacte canadica
6, Белошей Philacte canadica Sevast.
Anas canadica. Sewastianoff. Nov. Acta. Petrop., XIII, стр. 346, 1800, Алеутские
о-ва.
Синоним. Anser pictus (non Bonnaterre). Pallas. Zoogr. Ross.—As., II, стр. 233,
1811, табл. 67, Алеутские о-ва.
Русские названия. Голубой гусь—книжное. Белошей—местное камчатское рус-
ское название.
Распространение. Ареал. Побережья Берингова моря в СССР от
низовий Анадыря до Берингова пролива и далее по Чукотскому п-ову на
запад до Колючинской губы. В Северной Америке на побережьях Аляски
до устья р. Кускокуим до мыса принца Уэльского, также и на о-ве Лаврен-
тия. Пролетает через Камчатку и Командоры. Здесь же проходит и север-
ная граница зимовок. Основные места зимовок на Алеутских о-вах и вдоль
Тихоокеанского побережья Северной Америки до залива Гумбольдта
в Калифорнии.
Характер пребывания. В пределах СССР гнездится,
встречается на пролетах и зимует. Залетает на о-в Врангеля. В пределах аре-
ала на Аляске появляется в начале—середине мая (Брандт, 1943). В СССР
первые весенние на Анадыре отмечены 14 мая (Слюнин), гнездовые места
занимаются здесь 20—26 мая. Осенью белошей исчезает отсюда в последней
БЕЛОШЕИ
28Г
декаде сентября, отдельные же стаи попадаются еще и в октябре. В зимнее
время встречается на Командорских о-вах, куда прилетает в октябре и откуда
отлетает в апреле.
Биотоп. В раннее весеннее время по прилете держится на освобо-
дившихся от снега участках тундры. Для гнездовья располагается на мор-
ских побережьях в низинах, заваленных выбросами моря—водорослями^
остатками растений, стволами деревьев и т. п. Гнездится также на островах
Карта 27. Распространение белошея Philacte canadica
а—граница гнездовий, г—зимовки
в устьях небольших рек, впадающих в море, отчасти по болотистым лугам
низменной тундры в нижнем течении рек. Зимой (на Командорах) держится
по каменистому морскому побережью.
Численность незначительна. За последнее время сильно умень-
шается в количестве.
Экология. Размножение. Прилетают парами и сразу по прилете
занимают гнездовые места. Следовательно, разбивка на пары проходит,
вероятно, на зимовках. В первое время по прилете самцы токуют, довольно
неуклюже расхаживая вокруг гусыни, покачивая головою и издавая при
этом особые хриплые звуки. До начала насиживания пары неразлучны,
и самец прогоняет не только других самцов, но и всякую вообще птицу,
приблизившуюся к самке (Тугаринов, 1941). Гнездо устраивается среди
выброшенного морем или рекою мусора, невдалеке от линии наивысшего
282 ОТРЯД ГУСЕОБРАЗНЫЕ
прилива, в небольшом углублении, сделанном самкой. В эту ямку почти
без всякой подстилки откладывается 1—2 яйца. По мере сноса дальнейших
яиц ямка устилается различными обломками растений и, наконец, при
откладке последних яиц—перьями и пухом. Гнездо поэтому выглядит чрез-
вычайно примитивно. Однако такое устройство гнезда имеет существенный
биологический смысл, так как гнездо не возвышается над уровнем почвы,
и когда на нем сидит самка—совершенно незаметно среди различного расти-
тельного хлама и мусора. К тому же самка, при приближении к ней, имеет
привычку распластываться на гнезде и, вытягивая шею палкой, класть
ее на землю. Это еще более увеличивает маскировку гнезда, мимо которого
можно пройти близко, совершенно его не заметив. Как только самка начи-
нает насиживать, гусак покидает ее и, соединяясь с другими самцами, кор-
мится на урезе воды вдалеке от гнезда. Сходя с гнезда, самка прикрывает
яйца перьями и пухом. Яйца откладываются в начале июня. Кладка состоит
из 3—8, чаще из 5—6 белых без всякого блеска яиц, размеры которых сле-
дующие: 70,3—86x48,3—56,2 мм (Тугаринов, 1941); A3) 72,7—89,6x48,8—
54,7 мм, в среднем 79,69x52,13 мм (Джоурден). По форме и размерам
яйца сильно изменчивы. По наблюдениям в неволе насиживание продол-
жается 24 дня.
Как только выклюнутся пуховички, к самке возвращается самец
и вместе с нею держится при выводке. Первые пуховички на берегах Ана-
дырского залива найдены 17 июня. Молодые птицы размером с крякву,
еще совсем покрытые пухом, с едва пробивающимися перьями на плечевых,
найдены 7 августа (Сокольников). Однако это—запоздавшие птенцы, так
как в ряде случаев молодые поднимаются на крыло в конце июля или в на-
чале августа.
Линька. Характер линьки неясен. Взрослые начинают линять
с момента появления выводка и, таким образом, семейные пары белошеев
линяют вместе. Полная летняя линька у них начинается со смены маховых,
и к концу июля—началу августа старые птицы, как и молодые, снова полу-
чают возможность летать. Но после подъема на крыло, в сентябре, молодые
сменяют перья головы и шеи. Вылинявшие, эти части тела сходны с тако-
выми взрослых, с той только разницей, что черный цвет у взрослых на шее
у молодых заменен буроватым. Такое своеобразие линьки позволяет считать,
что и у этих гусей имеется частичная линька.
Питание. Основной пищей белошея являются беспозвоночные, пре-
имущественно мидии—Mytilus eduliSy а также другие моллюски и ракообраз-
ные, собираемые в полосе отлива. Таким образом, у этого гуся мы находим
известную приспособленность к литорали, мало используемой другими
гусеобразными. Но в состав пищевого рациона белошея, правда, в значи-
тельно меньшем количестве, входят и растительные корма, в том числе ягоды,
собираемые в участках тундры.
Возможно, что благодаря питанию моллюсками, что, однако, точно не
установлено, мясо и, в особенности кожа этого гуся отличаются чрезвычайно
сильным, сходным с чесноком, запахом, который исчезает при снятии кожи,
особенно если зажарить белошея. Молодые пахнут меньше, чем взрослые,
чем и объясняются расхождения зоологов по вопросу, о запахе этого гуся.
Хозяйственное значение. Из-за крайней своей малочисленности белошей
не имеет никакого значения в заготовках дичи. Но все же на побережьях
Анадыря и Чукотки его ловят вместе с другими линными гусями. Ввиду того
что численность его с каждым годом падает, что промысловое его значение
ничтожно, а биологические особенности еще мало изучены, следует запретить
его отлов, местами влекущий за собою его полное исчезновение.
Полевые признаки. Очень крупный гусь с короткой, толстой шеей,
что хорошо заметно и на лету. Характерны также частые взмахи относи-
БЕЛОШЕЙ 283
тельно коротких крыльев. Передвигающаяся в воздухе стая летит очень
низко над землей или над водой, чуть ли не касаясь гребней волн. На
земле держится парами или небольшими группами по 3—4 птицы. Довольно
молчалив и редко приходится слышать его характерное двусложное пронзи-
тельное «кла-га»... «кла-га»... «кла-га». В малолюдных местах доверчив, но
скоро научается распознавать человека, и при встречах с ним проявляет,
наоборот, крайнюю степень осторожности.
Рис. 47. Белошей
Описание. Размеры и строение. Длина тела самцов и самок 650—700 мм.
Длина крыла самцов E) 392—420, в среднем 405 мм; длина крыла самок 375 мм. Длина
клюва самцов E) 37,4—50, в среднем 41,8 мм; длина клюва самок A) 37,2 мм. Высота
нижней челюсти у самцов E) 5,8—7,б,в среднем 6,7 мм; у самок AN мм. Вес самцов и са-
мок 2,25—2,5 кг. Клюв вдвое короче головы. Ноготок занимает всю вершину надклювья
и не загибается вниз. Короткие тупые зубчики краев надклювья в профиль заметны
лишь у самого его основания. Ноздревые ямки крупны, овальны и широки. Ноздри округ-
лые. Оперение лба по бокам основания надклювья образует выпуклые дуги, доходящие
до ноздревых ямок. Дугою выдается оперение лба также и на конек надклювья. Крылья
коротки и доходят до половины хвоста. Рулевых 8—9 пар.
Окраска. Пуховик. Спинная сторона темношелковисто-серая, без пятен,
брюшная сторона светлее. Ноги черноватые.
Молодая птица в первом пере. Общая окраска серая, тусклая, голубовато-
серая. Перья туловища заостренные, более узкие, чем у взрослых. Голова и шея темно-
серые, на голове отдельные мелкие белые пятна. Поперечные светлые полосы сверху и на
боках слабо выражены. Клюв темный. Ноги темные, слегка желтеющие. Молодая птица во
втором пере сходна с птицей в первом пере, но на задней стороне шеи темных пятен зна-
чительно больше. У половозрелой птицы после третьей линьки на голове появляется
примесь черно-бурых перьев. Взрослый наряд надевается после четвертой линьки.
Взрослые самец и самка. Спинная сторона голубовато-серая, брюшная
светлосерая.Сверху и с боков в поперечных черных и светлых полосах, в общем распола-
гающихся волнистым узором. Голова и шея сверху белые, иногда с ржавым налетом.
Подбородок, низ шеи и ее бока черновато-бурые. Каждое перо верха и боков туловища
голубоватопепельно-серое с черной предвершинной узкой полосой и белой вершинной
каемкой. Эти черные и белые части пера слабее развиты на боках и груди и мало выра-
жены на сером брюхе. Первостепенные маховые черно-серые со светлыми окаймлениями.
Второстепенные темносерые с ясными белыми каймами. Хвост белый в верхней половине
и сероватый в основной. Надклювье темнорозовое. Ноги красновато-желтые. Радужина
ореховая.
284 ОТРЯД ГУСЕОБРАЗНЫЕ
РОД ИНДИЙСКИЕ ГУСИ EULABEIA REICHEN BACH, 1852
Тип. Anas indica
7. Горный гусь Eulabeia indica Lath.
Anas indica. Latham. Ind. Orn. II, стр. 839, 1790, Индия.
Синоним. Anser Skorniakovi. Sewertzoff. Верт. и гор. распр. турк. жив.>
стр. 149, табл. X, ф. 3 и 4, 1873, озеро Чатыр-Куль.
Русские названия. Индийский или индейский гусь—книжные; горный гусь—
название, данное этому гусю Пржевальским.
Распространение. Ареал. Эндемичная птица Центральной Азии,
являющаяся характерным нагорно-азиатским видом, лишь местами выхо-
дящим из пределов Центральной Азии. Крайними северо-западными
пунктами ареала являются плоскогорье Укбк и Курайская степь на Алтае.
Северные пределы распространения здесь проходят по Чуйской степи и далее
в пределах Монгольской Народной Республики по озеру Урюг-Нор,
бассейну р. Тэсь, притока озера Убса-Нор, по озеру Косогол, долинам
рек Толы и Керулена (Сушкин, 1938; Тугаринов, 1941). Летом встречается
и севернее этих пределов—в Южном Прибайкалье, по р. Селенге и на озере
Тарей-Нор, но достоверных данных о гнездовье здесь не имеется. Крайним
восточным пунктом ареала являются Баргинские озера, находящиеся запад-
нее Большого Хингана в пределах Маньчжурии (Лукашкин, 1950). На юг
и юго-восток отсюда идет до Ордоса, озера Куку-Hop и по южной окраине
Тибета до Ладака, Кашмира, Памира и Алайской долины, являющимися
крайними юго-западными пунктами ареала. К северо-востоку отсюда по
Тянь-Шаню, Семиречью, где найден на высокогорных озерах Чатыр-Куле
C500 м над уровнем моря), на Сон-Куле, на горном озере Джасыл-Куле,
на озерах в верховьях оз. Зауки, в устье Куелю и на Баянколе (Шнитников,
1949). Еще далее на северо-восток гнездится в Джунгарии и Китайском
Туркестане. Внутри очерченных, таким образом, границ ареала гнездится
спорадично по высокогорным водоемам Центральной Азии. Зимует в Паки-
стане, в Пенджабе, Ассаме и северной части Индии. На юг доходит до Синда,
района Бомбея и изредка до южных провинций. Встречается зимою также
на крупных реках Северной Бирмы. Возможны также зимовки и в Белуд-
жистане (Тугаринов, 1941).
Характер пребывания. Гнездящаяся и перелетная птица.
В виде исключения встречается зимою на Пяндже в Южном Таджикистане
(Иванов, 1940).
Даты. С мест зимовок в Индии и Пакистане начинает отлетать
в марте, последние исчезают отсюда в апреле (Бекер, 1929). На Куку-Норе
появляются в начале второй декады марта—в середине марта (Пржеваль-
ский 1876). На озере Орок-Нор в Северной Гоби пролет начался 14 апреля
и продолжался еще во второй декаде мая, когда все другие гуси уже исчезли,
серые же гуси даже загнездились (Козлова, 1930). На северной границе
ареала появляется в начале мая и в Забайкалье был добыт 6 мая (Доманев-
ский, 1937). Повидимому, в эти же сроки появляется и на Алтае, прилетая
сюда, таким образом, сравнительно поздно. Хотя места своих гнездовий
горные гуси покидают окончательно только с замерзанием водоемов
(Тугаринов, 1941), тем не менее осенний отлет в ряде мест начинается рано,
доказательством чего может служить наличие хорошо выраженного про-
лета в Тянь-Шане уже в августе ст. ст. (Алфераки, 1904). С высоких мест
плоскогорья Укбк на Алтае уже к 20 августа перемещаются в более низкие,
покидая и эти окончательно к 25 августа (Сушкин, 1938). Таким образом,
из северных пределов ареала горные гуси исчезают довольно рано, в более
ГОРНЫЙ ГУСЬ
285
южных задерживаются до конца сентября и даже до октября. На Памире
на озере Ранг-Куль встречены и добыты 19—23 сентября (Тугари-
нов, 1930). Хорошо выраженный осенний пролет на озере Куку-Нор
наблюдался с 28 августа по сентябрь (Бианки, 1915). Нет никакого сомнения,
что в южных частях ареала задерживаются до самой последней возможности
не только местные, но и прилетевшие из более северных районов, так как
в Пакистане и Индии появляются только с половины октября (Бекер, 1929).
Карта 28. Распространение горного гуся Eulabeia indica
а—граница гнездовий, г—зимовки
Весенние и осенние направления движений, повидимому, не одинаковы,
так как, например, на Лоб-Норе осенью горный гусь обыкновенен на про-
лете, весною же, наоборот, редок (Пржевальский, 1873).
Биотоп. В горном ландшафте различные высокогорные водоемы—
озера, реки, открытые кочковатые болота в истоках рек и ручьев с имею-
щимися близ них скалами или крупными обрывами. На высокогорных пло-
скогорьях открытые равнинные пресноводные или соленоводные водоемы,
болота и реки с островами на них или с высокими деревьями по берегам.
Численность. В больших количествах и лишь местами—в Тибете
и Ладаке, в других частях ареала был довольно обыкновенен, за исклю-
чением Тянь-Шаня, где встречался редко. Однако за последнее время чис-
286 ОТРЯД ГУСЕОБРАЗНЫЕ
ленность горного гуся заметно снизилась, а из ряда мест на Памире и Тянь-
Шане он и вовсе исчез.
Экология. Размножение. Первое время по прилете на места
гнездовья горные гуси держатся небольшими группами или стайками, дохо-
дящими до 20 особей. Через некоторое время наблюдается в гнездовых местах
разбивка на пары, а вместе с тем и воздушные игры, состоящие в том, что
самец на лету мчится за самкой, переворачиваясь при этом в воздухе (Прже-
вальский, 1876).
Гнездится колониями или небольшими в 3—5—7 гнезд, расположенных
невдалеке одно от другого, или же довольно крупными, что наблюдалось
в Тибете. Гнезда устраиваются на скалах, на высоких деревьях или на земле
на островах, близ берегов водоемов или на открытых заболоченных простран-
ствах. На скалах гнезда помещаются на выступах одно невдалеке от другого
(Шефер, 1938). Гнезда, находящиеся на деревьях, строятся на высоте 4—б м
от земли (Козлова, 1932). Иногда гнезда горных гусей размещаются близ
гнезда центральноазиатского канюка Buteo hemilasius (Козлов, 1907). Гнезда
на деревьях и на скалах сооружаются из тонких веток, имеют неглубокий
лоток и отличаются небрежностью постройки. Гнезда, помещаемые на сухих
болотах, лредставляют углубление во мху, выстланное сухими остатками
растений и обильно покрытое пухом. На влажных болотах гнездо предста-
вляет кучу мха, перемешанную со стеблями злаков, с большим количеством
пуха в лотке и по краям гнезда. Следовательно, устройство гнезд довольно
разнообразно.
Кладка состоит из 4—8, чаще 4—6, белых с матовым оттенком и ше-
роховатой скорлупой яиц размерами 75—89x52,4—59 мм (Тугаринов,
1941); A9) 78,5—89x52,4—56,5 мм (Джоурден, 1920). Вес B) 13,92 и 14,22 г
(Гебель, 1904).
О сроках насиживания данных не имеется. Пуховички возрастом в не-
сколько дней наблюдались в районе Улан-Батора 22 июня (Тугаринов, 1941).
Пуховики, но уже ростом в половину старых под Кош-Агачем на Алтае
встречены 24 июня (Сушкин, 1938). Птенцы длиною около 260 мм, почти сплошь
еще покрытые пухом, в Семиречье найдены 3 августа (Северцов, 1873).
В связи с различными сроками откладки яиц птенцы на территории ареала
появляются разновременно. На Алтае в более низких местах птенцы выкле-
вываются раньше (Сушкин, 1938).
При выводке все время находятся и самец и самка. В случае опасности
родители переводят птенцов на воду, и семья отплывает в недоступное место»
На Куку-Норе и других больших водоемах десятки выводков соединяются
вместе. Обычно стадо пасется на сухопутье, но, заметив опасность, все гуси
бросаются к водоему и быстро отплывают подальше от берега (Пржеваль-
ский, 1876). Это же самое явление отмечается и для Алтая (Сушкин, 1938).
Линька. Имеется полная летняя линька в июне-июле. Вопрос о ча-
стичной осенней линьке не ясен. Раньше всего линька начинается у алтай-
ских горных гусей. Около середины июня на Ачит-Нуре уже шла смена махо-
вых, и большинство гусей уже не могло летать. В Чуйской степи в районе
Кош-Агача линька начинается несколько позже, и 24 июня взрослые были
в отрепанном пере, но еще не приступали к линьке. Снова поднялись на
Алтае на крыло к 15 августа и 24 августа на нижней части плоскогорья
У кок взрослые уже все летали, у добытых птиц махи оказались доросшим»
до нормы и окрепшими. Сменились все перья верхней части тела и росли
еще на нижней. Еще не закончилась линька рулевых и перьев верхней части
груди. Одновременно с линькой взрослых идет процесс вырастания перо-
вого покрова и у молодых; 13 августа маховые не доросли до нормы лишь
на 5 мм\ 15 августа все маховые были вполне развиты, и молодые поднялись
на крыло (Сушкин, 1938).
ГОРНЫЙ ГУСЬ
287
На Памире линька протекает позже. У взрослого самца, добытого 14 ав-
густа, маховые еще в пеньках, растут средние рулевые. Имеются еще старые,
не сменившиеся перья на затылке, на спине (немного), на средних кроющих
крыла, на плечевых и на боках. У самки, добытой в тот же день, маховые
уже почти выросли и старых перьев меньше. Молодой от 14 августа еще в пуху,
перья пробиваются на спине, верхних кроющих крыла и хвосте, махи не
доросли (Дементьев, 1932). Молодые в пуху у поздних выводков на Памире
Рис. 48. Горный гусь
встречаются даже в конце августа. Вполне оперенные молодые наблюдаются
не ранее 17 сентября. Первые вылинявшие взрослые поднимаются на крыло
к 24 августа, а запоздавшие еще в конце первой декады сентября не могут
летать (Северцов, 1879).
Полная летняя линька у взрослых начинается с одновременного выпа-
дения маховых. Вслед за этим выпадают верхние кроющие крыла, нижние
кроющие крыла одно время образуют его край, затем сменяются и они и
далее вместе с ростом маховых идет смена контурного оперения. Это
последнее продолжает линять еще после того, как выросли и окрепли
маховые, но весь процесс линьки протекает в более сжатые сроки, чем
у некоторых северных видов гусей.
Питание. Основной пищей горных гусей являются наземные расте-
ния, добываемые на берегах самых разнообразных водоемов или же в отда-
лении от них. В большом количестве они потребляют злаки и осоки с при-
брежных лугов, поедают семена злаков и бобовых, на зимовках кормятся
всходами хлебных злаков. Помимо этого, их часто можно видеть на побе-
288 ОТРЯД ГУСЕОБРАЗНЫЕ
режьях водоемов у волноприбойной полосы, где они, переворачивая наносы»
проедают водоросли, различных ракообразных и других беспозвоночных.
Хозяйственное значение. Промыслового значения в пределах СССР
благодаря своему общему небольшому количеству не имеют и являются
лишь предметом спортивной охоты на местах. Легко приручаются и могут
быть вовлечены в домашний оборот в горных условиях. Прирученные
встречаются на Памире (Молчанов и Зарудный, 1915).
Полевые признаки и поведение. Средней величины гуси, от других
издали отличающиеся своей светлой окраской и белой головой. Довольно
высоки на ногах, хорошо, хотя и несколько неуклюже, ходят, хорошо
и быстро бегают, в критических случаях помогая себе при этом крылвями.
По существу горные гуси являются наземными птицами, так как большую
часть своей жизни проводят на земле. Но в то же время они совершенно
свободно чувствуют себя на воде, и именно на ней во многих случаях ищут
спасения.
При перелетах перемещаются на большой высоте углом, диагонально или
двумя сходящимися линиями, причем передовой гусь через каждые 4—5 ми-
нут сменяется следующим. Низкий трубный голос своеобразен и резко отли-
чен от голоса серого гуся. На лету крики горных гусей удается услышать
раньше, чем увидеть их стаю. Опускаясь на водоем, птицы делают над ним
один-два круга, затем стая снижается и каждый отдельный гусь в некото-
рых случаях переворачивается раз или два в воздухе (Юм).
В местах, где их не ловят и не стреляют, они не только не боятся людей,
но даже иногда бродят у окраин поселений, разыскивая здесь пищу, причем
ведут дневной образ жизни (Тугаринов, 1941). Там же, где их преследуют,
отличаются крайней осторожностью и становятся ночными птицами, отдыхаю-
щими днем на недоступных местах водоемов.
Во время линьки держатся на воде и для кормежки выходят на мало
доступные отмели и косы. Но как только гуси вновь подымутся на крыло,
а молодые получат возможность летать, горные гуси собираются в стаи,
на день забираются в непролазные трущобы, на кормежку вылетают вече-
ром и пасутся ночами. Члены стаи чрезвычайно внимательны по отношению
друг к другу, и при стрельбе по стае и ранении одного из них гуси повора-
чивают, делают несколько кругов над упавшим товарищем, иногда опускаются
к нему и подчас даже уводят его с собой (Бианки, 1915).
Очень любопытны. В безлюдных местах их очень легко подманить к
себе, как делал это Пржевальский, который заметив издали подлетающих
горных гусей, ложился на землю и начинал помахивать фуражкой, что
всякий раз побуждало гусей сворачивать со своей дороги и налетать прямо
на него (Пржевальский, 1876).
Описание. Размеры и строение. Размах самцов и самок 1422—1676 мм.
Длина тела самцов и самок 698—850 мм. Длина крыла самцов F) 434—482, самок D)
406—442, в среднем 454 и 424 мм. Длина клюва самцов F) 42,2—60, самок DL1,5—48,5,
в среднем 48,2 и 45 мм. Высота нижней челюсти у самцов F) 7,2—7,8, у самок D) 5—7, в
среднем 7,5 и 6 мм. Вес самцов и самок 2—3,2 кг. Клюв короче головы, одинаковой ширины
на всем протяжении, кроме закругленной вершины. Ноготок занимает большую часть
надклювья, свисая наклонно над подклювьем. Ноздри узкие и длинные. Зубцы по краям
закрытого клюва не заметны. Оперение лба и щек дугами заходит на конек надклювья
и его бока. Подклювье тонкое. Крылья достигают вершины хвоста. Рулевых 8 пар, хвост
округлый. Цевка длинная. Лапа короткая и средний палец с когтем значительно короче
ее. Задний палец с хорошо развитой лопастью. Квадратная кость массивна, почти не
наклонена вперед и расположена под меньшим, чем у других гусей, углом к продольной
оси черепа (Красовский, 1941).
Окраска. Пуховик. Только что выклюнувшиеся пуховички яркосоломенно-
желтые с буровато-серым пятном на темени, со светлооливково-бурыми лопатками, спи-
ной, крыльями и поясницей и со светложелтым низом (Тугаринов, 1941). Более взрослые
птенцы с длиной клюва 27—27,5 мм (по коньку) резко отличны. У них спинная сторона
светлодымчато-бурая, задняя половина темени несколько светлее. Верх шеи пестрый
БЕЛЫЙ ГУСЬ
289
от примеси белых пушинок. Брюшная сторона белая; эта белая окраска низа несколько
заходит на верх позади плечей и голени. Бока и зоб белые с легким дымчатым оттенком
(Бутурлин, 1913).
Молодая птица. Одноцветно серая без поперечных полос. Лоб, бока головы, бока
и верхняя половина шеи, подбородок и горло белые. Темя, затылок, остальная часть
верха шеи темносерые. После второй линьки начинают обозначаться темные поперечные
полосы на темени и затылке. Окончательный взрослый наряд надевается, повидимому,
не раньше, как после третьей линьки.
Взрослые самцы и самки. Общая окраска светлая, буровато-серая. Голова и бока
шеи белые. На темени и затылке две черные поперечные полосы. Верх шеи буровато-
серый. Спина темнопепельно-серая в волнистых полосах. Горло белое. Низ шеи черновато-
бурый. Зоб пепельно-серый. Бока бурые. Клюв красновато-желтый с черным ноготком.
Ноги красновато-желтые. Радужина темнобурая.
РОД БЕЛЫЕ ГУСИ CHEN ВО I E, 1822
Тип. Anser hyperboreus
8. Белый гусь Chen caemlescens L.
Anas caerulescens. Li n n a e u s. Syst. Nat., изд. X, I, стр. 124, 1758, Канада.
Русские названия. Белый гусь, снежный гусь, северный гусь—книжные
названия.
Распространение. Ареал. В Восточной Сибири—о-в Врангеля и, воз-
можно, но мало вероятно, некоторые места побережья Чукотки. В Северной
Карта 29. Распространение белого гуся Chen caerulescens
1—Ch. с. caerulesiene, 2—С ft. с. ailantica; a—граница гнездовий, в—зимовки
Америке от Аляски до Гудзонова залива, Баффиновой Земли и северных
частей Западной Гренландии.
Подвиды в варьирующие признаки. Несмотря на некоторые работы, специально
посвященные вопросу о систематическом положении белого гуся (Кеннард, 1927; Сало-
19 Птицы СССР, т. IV
290 ОТРЯД ГУСЕОБРАЗНЫЕ
монсен, 1933), вопрос о его правильном наименовании и составе форм все еще остается
неясным. Дело осложняется отсутствием в наших коллекциях достаточного количества
материалов по этой птице из пределов СССР и почти полным отсутствием его из Америки,
откуда описан ряд форм. Приходится полагаться на иностранных авторов,которые сви-
детельствуют о беспрерывном ряде переходов между птицами с Аляски, идентичными
с нашими и птицами с крайнего востока Северной Америки. К тому же так называемый
голубой гусь Anser caerulescens имеет две вариации—одну белую, в которой он почти
не отличим от наших птиц, и другую голубую, зачаток которой в виде пепельно-серых
кроющих крыла можно найти также у наших белых гусей и крайняя степень развития
которого создает голубых гусей Северной Америки. В Гренландии, как и у нас, снова
остается одна только белая вариация. Не имея возможности из-за отсутствия фактиче-
ского материала разобраться во всех этих особенностях белых гусей, принимаем последнее
деление Саломонсена.
Можно принять, что вид Chen caerulescens делится на два подвида; более мелкий
Сп. с. caerulescens Li n n a e u s, 1758, распространенный от о-ва Врангеля до Северной
Гренландии, и более крупный СА. с. atlantica К е n n а г d, 1927, гнездящийся на севере
Западной Гренландии.
Малый белый гусь Chen caerulescens caerulescens L.
Синоним* Anser hyperboreus. Pallas. Spicil. Zool., VI, стр. 25, 1769, побережья
Ледовитого океана.
Распространение. Ареал. В настоящее время в СССР, повидимому,
сохранился только в одном месте—на о-ве Врангеля; относительно гнездо-
вания на ледовитоокеанском побережье Чукотки достоверных данных не
имеется. Прежде, в XVII и XVIII столетиях, повидимому, был широко рас-
пространен по ледовитоокеанскому побережью Восточной Сибири от устья
Пясины до крайнего востока Чукотки. Гнездился в низовьях больших сибир-
ских рек—Яны, Индигирки, Колымы и ее притока Алазеи, а также на
Большом Ляховском о-ве. В середине прошлого столетия еще существовал
на него особый промысел в Русском Устье. Однако в ряде мест он был чрез-
вычайно редок уже в самом начале XIX столетия (Паллас, 1811); к 1820—
1824 гг. перевелся в дельте Колымы (Врангель) и окончательно исчез к началу
XX столетия. Существенным моментом в исчезновении этого вида является
беспрерывное преследование его и у нас и в Америке на гнездовьях, во время
линьки, на пролетах и, наконец, на зимовках. Самое губительное, конечно,,
преследование гнездовых и линных птиц, что видно по тому, что на безлюд-
ном до начала XX столетия о-ве Врангеля оказались громадные тысячные
колонии этих птиц.
Зимует отчасти в Японии. Главные же места зимовок находятся вдоль
тихоокеанского побережья Северной Америки до Луизианы, Техаса и
Мексики.
Характер пребывания. Гнездящаяся и пролетная птица.
Однако хорошо выраженный весенний пролет наблюдался лишь вдоль ледо-
витоокеанского побережья Чукотки в районе мыса Шмидта (мыс Северный).
Даты. Передовые показались здесь 26 мая, 27 мая их было уже много,
в конце мая—начале июня шел валовой пролет, во время которого летела
стая за стаей, последние отмечены 10 июня. Таким образом, весенние передви-
жения здесь продолжались около полумесяца (Артоболевский, 1926).
В прежние времена, когда белые гуси гнездились и по ледовитоокеанскому
побережью Восточной Сибири, передовые в районе Нижне-Колымска в
1821 г. были отмечены в начале второй трети мая (Кибер, 1823). На о-ве Вран-
геля первое появление белых гусей отмечено 22 мая, и весь прилет заканчи-
вается в течение недели. Осенью отсюда они отлетают в конце августа и самые
поздние в начале сентября. Их осеннее передвижение не прослежено и в райо-
не мыса Шмидта не наблюдались, что дает возможность строить предполо-
жения о том, что осенние направления передвижений гусей иные, чем весною,.
БЕЛЫЙ ГУСЬ 291
и с о-ва Врангеля проходят над океаном (Тугаринов, 1941). Однако «Колыма»
с доктором Савичем, производившим зоологические наблюдения, на кото-
рых основаны эти соображения, подошла к мысу Шмидта только 18 сентября,
когда белые гуси, возможно, успели уже закончить здесь свои осенние пере-
движения. Во всяком случае разрешить этот вопрос можно лишь прямыми
наблюдениями.
Залеты белых гусей в различные части СССР отмечались неоднократно*
однако ни один из случаев встреч этих птиц, за исключением Восточной
Сибири, не подтвержден добытыми экземплярами. В качестве весьма ред-
кого залетного вида несколько раз встречен на Анадыре (Портенко, 1939),
залетал на Камчатку (Крашенинников, 1785, 1786, 1818), повидимому,
залетал на Командоры (Стейнегер, 1887). По расспросным данным был
добыт в октябре 1948 г. у устья р. Сунгари (Воробьев). Очень редок
в области Индигирки (Житков и Зензино,в, 1915), куда в настоящее
время может только залетать. Был встречен на Байкале (Тачановский,
1877 и 1893), в Забайкалье (Моллесон, 1890), в Минусинском крае (Яблон-
ский, 1891), в долине Волги (Яковлев, 1871; Бутурлин, 1901; Житков,
1914), в Чкаловской области (Зарудный, 1888, 1887), на Нижней Сыр-
Дарье (Спангенберг и Фейгин, 1936), на Каспийском море и в Закав-
казье (Радде, 1884, 1885, 1886; Сатунин, 1907, 1912), в 1921 г. в Краснодар-
ском крае (по сообщению Раскаля), в юго-восточной части Украины (Косю-
ченко, 1905, 1906), в Киевской обл. (Кесслер, 1852), в Черниговской (Арто-
болевский, 1926), Полтавской (Шар-лемань, 1928), в Курской в 1927 г. (Пту-
шенко), в Московской (Поляков, 1910), в Молотовской (Сабанеев, 1871). Чаще
чем в других местах, белые гуси отмечались в Поволжье, Вполне возможно,
что некоторые из приведенных случаев встреч в действительности относятся
не к белому гусю, а к альбиносам других видов гусей. По крайней мере с изве-
стной долей вероятия альбиносом можно считать приведенного для Волж-
ско-Уральской степи (Волчанецкий, 1937) гуся, наблюдавшегося 20 июня
1926 г.
Б и о т о п. На о-ве Врангеля сухие участки тундры близ мелких озер
и речек.
Численность. На о-ве Врангеля многочисленны (Портенко, 1937).
Обычные в XVIII столетии на крайнем севере Сибири и на Ново-Сибирских
о-вах, белые гуси в настоящее время здесь встречаются лишь залетом. В пре-
делах СССР вымирающая птица.
Экология. Размножение. Образование пар наблюдалось еще во
время миграций, на материке (район мыса Шмидта, Артоболевский, 1926).
В это время и затем позже в гнездовых колониях самцы держатся около самок
и часто ходят вокруг них, своеобразно покачивая головою и издавая особые
глухие звуки. На о-в Врангеля прилетают в стаях, где пары, однако, наме-
тились, и сейчас же приступают к размножению. Однако далеко не каждый
год оказывается благоприятным для размножения. В годы с запоздавшими
веснами, с возвратом холодов и снегопадов или вовсе не размножаются,
или же размножаются только немногие пары, количество же яиц в кладке
снижается. Гнездятся белые гуси колониями то небольшими в 15—20 пар,
то огромными в 1000 пар и более. Колония от колонии располагается в
1—2 км. В таких колониях среди гусиных гнезд попадаются гнезда белых
сов, около которых гуси вполне безопасны от нападения четвероногих хищ-
ников.
Гнездо белого гуся представляет собою небольшое углубление во мху,
выстланное сухими стебельками трав и незначительным количеством пуха.
Кладка состоит из 3—6, чаще из 3—5 яиц желтовато-белого или кремового
цвета с размерами 63,2—88 х 41,8—57,2 мм (Тугаринов, 1941). Яйца откла-
дывается в начале июня. Насиживание начинается с откладки первого яйца.
19*
292 ОТРЯД ГУСЕОБРАЗНЫЕ
Около 10 июня на о-ве Врангеля в кладках находили уже все яйца с эмбрио-
нами. Насиживает одна самка, которая, сходя с гнезда, тщательно прикрывает
их сухими стеблями травы. Благодаря ничтожной высоте гнезда яйца, при-
крытые сухой растительностью, совершенно незаметны. Гусак находится
поблизости или же стоит у гнезда. В случае, если часть кладки гибнет или
забирается, самка докладывает новые яйца. При гибели же всей кладки пов-
торно яйца не откладываются, и пара выходит из ряда размножающихся
птиц (Портенко, 1937).
В районе мыса Шмидта, начиная со 2 июня, происходила разбивка на
пары и, хотя после исчезновения почти всей массы пролетных оставались
еще отдельные пары и особи, тем не менее ни гнезд, ни яиц найдено не было
и лишь допускалась возможность размножения здесь белых гусей (Артобо-
левский, 1926).
Таким образом, в текущем столетии гнездование белых гусей нигде на
материке Сибири достоверно не установлено, хотя имеются неопределенные
указания на редкие встречи их в гнездовое время в прибрежных тундрах
к востоку от Чау некой губы (Тугаринов, 1950). Есть основания полагать,
что отдельные пары или мелкие колонии, пытающиеся обосноваться на ма-
терике, помимо человека, разоряются также и четвероногими хищниками,
что и лимитирует в настоящее время возможности гнездованья белого гуся
на побережьях Ледовитого океана. Наблюдений над временем вылупления
молодых и их ростом не имеется.
Линька. Как у других гусей, имеется полная летняя линька, по осен-
ней материалов нет. Первыми с середины июля начинают линьку холостые
и неполовозрелые, не гнездившиеся в данном сезоне птицы. На время линьки
они скопляются массами или в устье рек, или на морских побережьях. Дан-
ных о линьке взрослых семейных и молодых не имеется. Окончательный
снежнобелый наряд надевается, повидимому, на четвертый год. В наряде
третьего года частично еще сохраняются серые перья предшествующего года
(Артоболевский, 1926).
Питание. Основной пищей белых гусей является растительная;
состоит она из листьев, плодов, семян и ягод тундровых растений. Особенно
охотно поедаются ягоды водяницы Empetrum nigra (Тугаринов, 1941). Из
животных организмов для питания используются небольшие беспозвоночные
пресноводных и морских побережий, а также мелководий. На пролете и
зимовках (в Америке) кормятся на озимых полях.
Основными врагами белого гуся являются четвероногие хищники, круп-
ные чайки и поморники, уничтожающие его яйца и пуховиков.
Хозяйственное значение. Вследствие редкости экономическое значение
белого гуся ничтожно, но в местных условиях о-ва Врангеля в больших коли-
чествах собираются его яйца и линяющие птицы. Так как такое примитивное
использование белого гуся даже при всех его ограничениях здесь приведет
к тому же, что случилось на материке, т. е. к полному исчезновению его
с острова, необходимо теперь же тем или иным способом вовлечь его в до-
машний оборот, иначе при разрушении крупных колоний с мелкими оставши-
мися очень легко справятся четвероногие и пернатые хищники, а также
и ездовые собаки.
Полевые признаки. Общий гусиный облик, сравнительно некрупные
размеры, белая окраска с черными вершинами крыльев позволяют отличать
этот вид на расстоянии от других гусеобразных. Особенно четки признаки
этих гусей на лету, когда ясно видны на концах крыльев темные пятна,
образованные черными первостепенными маховыми. На лету строятся углом,
шеренгой или диагональю. Иногда на лету слышен их резкий, высокого
тона крик, который можно передать как «геув» или «кеаг», звучащими
слитно. В некоторых случаях летят вместе с белолобыми гусями или канад-
БЕЛЫЙ ГУСЬ 293
скими казарками (по наблюдениям в Америке). Определения белых гусей
на лету по встречам их в вершине угла (Кесслер, 1851) сомнительны, возможно
это были альбиносы.
В противоположность многим другим гусям при приближении наблюда-
теля к гнезду самец и самка не улетают и даже не убегают, а спокойно
отходят от гнезда, останавливаются невдалеке и начинают щипать траву,
негромко переговариваясь между собою. Стоит только отойти от гнезда,
как гуси снова возвращаются на него (Тугаринов, 1941). Эта их особенность
дает возможность истреблять и взрослых гусей и полностью забирать
Рис. 49. Белый гусь
их кладку. Но во время миграций они крайне осторожны, и нет никакой
возможности приблизиться на выстрел к пасущейся стае (Артоболевский.
1926).
Описание. Размеры и строение. У самцов и самок длина тела 584—736 мм;
длина крыла 365—455 мм; длина клюва 49,5—60 мм; вес 2—2,25 кг (Тугаринов, 1941
и 1950). Клюв короче головы, в профиль постепенно суживается к вершине. Ноготок
занимает всю вершину клюва, не свисая над подклювьем. Зубцы по краям надклювья
сбоку хорошо заметны. Длинные крылья достигают концов хвоста. Округлый хвост
состоит из 8 пар рулевых. Цевка длинная; средний палец без когтя равен 8Д длины цевки.
Окраска. Пуховик. Спинная сторона серовато-оливковая, блестящая. Брюш-
ная светлооливково-желтая. Лоб, щеки и горло бледножелтые.
Молодая птица сверху пестрая бледнорыжевато-серая. Верх головы, ее бока,
верх шеи светлосерые со светлыми каемками перьев. Верхние кроющие крыла пепельно-
серые, каждое перо с белой каймой. Первостепенные махи буровато-черные. Рулевые белые
с серым налетом. Шея снизу, зоб и бока сероватые с расплывчатыми пестринами. Брюш-
ная сторона белая. Клюв темносерый, с возрастом краснеющий, ноги зеленовато-серые
также изменяющиеся в цвете с возрастом. После каждой линьки птица становится все
более и более белой, а окончательный взрослый наряд и мясная окраска клюва приобре-
таются не ранее как после четвертой осенней линьки.
Взрослые самец и самка. Общая окраска оперения снежнобелая, но первосте-
пенные маховые черные, а их кроющие пепельно-серыв. Как уже отмечено в свое
время (Артоболевский, 1926), оперению белых гусей, как и лебедей и многих отдельных
особей водоплавающих, свойствен особый ржавый налет, возникающий от растворенных
в воде минеральных солей и не имеющий никакого отношения к диагностике этого вида.
294
ОТРЯД ГУСЕОБРАЗНЫЕ
Клюв мясного цвета. Ноготок серый. По разрезу рта идет черная полоса, суживающаяся
к углу рта и вершине клюва. Ноги мясного цвета или мясо-красные. Радужина корич-
невая.
РОД ГУСИ ANSER LINNAEUS, 1760
Тип. Anser domesticus == Anser anser L.
9. Серый гусь Anser anser L,
Anser anser. Li n n a e u s. Syst. Nat., изд. X, I, стр. 123, 758, Швеция.
Синоним. Anser cinereus. Meyer. Meyer et Wolf Taschenb. d. deutschl. Vogelk.,
II, стр. 552, 1810.
Русское название. Серый гусь—русское старинное название.
Распространение. Ареал и численность. Широко распро-
страненный прежде по Палеарктике от Атлантического до Тихого океана,
к северу доходивший до 60—50-й параллели, а к югу до Испании,
Карта 30. Распространение серого гуся Anser anser
а—граница гнездовий, г—зимовки, в—отдельные места гнездовий
Балканского п-ва, Мессопотамии и долины Желтой реки в Китае, серый
гусь во многих местах стал теперь редкой птицей или исчез совершенно. Па-
дение численности и полное исчезновение особенно заметно в западной поло-
вине его ареала. Пятнистый и прежде, в связи с приуроченностью серого гуся
к водоемам, его ареал в западной части в настоящее время представляет лишь
ряд весьма далеко отстоящих одно от другого пятен и только в Средней Сибири
и Зауральских степях он может рассматриваться как более или менее сплош-
ной. В настоящее время серый гусь гнездится в Исландии, Шотландии, по
западному побережью Скандинавии до Финмаркена, изредка в южной око-
нечности Швеции. На севере СССР гнездится на Кольском п-ве (Спангенберг,
1932), кое-где и лишь изредка на водоразделах Волги и Северной Двины.
Нигде в бассейнах левых притоков Верхней Волги его уже нет и только в не-
большом количестве еще встречается в верхней части течения Камы. За Ура-
лом на пространстве до Оби лишь изредка и местами доходит до 60—61° с. ш.
серый гусь 295
В Западной Сибири северная граница его распространения снижается до
55° с. ш., в бассейне Енисея он изредка встречается в Минусинских степях, так
же редко в лесостепи южнее Байкала, в Даурии и еще селится в небольшом
числе в бассейне озера Ханка (Шульпин, 1936).
Южная граница его распространения весьма условна. Он гнездится
в Дании, по Среднему и Нижнему Дунаю, в Месопотамии, Ираке, Иране,
Афганистане, в Средней Азии, в бассейнах Тарима и Хуан-Хэ, в Маньчжу-
рии и Приморской обл. СССР.
В пределах СССР во многих густонаселенных местах серый гусь уже
давно перестал гнездиться, но еще встречается в отдельных местах Бело-
руссии (Тугаринов, 1941), в районе озера Ильмень (Хомченко, 1928),
повидимому, стал гнездиться в северной части Московского моря (по наблю-
дениям Исакова), Гнездится по р. Каме, в Татарской АССР, в районе
Чердыни (Тугаринов, 1941), на некоторых пруаах и речках Винницкой обл.
(Храневич, 1925), пор. Орель (приток р. Сама >ы) в Днепропетровской обл.
(Стахровский, 1941), в низовьях Днестра, Днепра, в низовьях Дуная,
Кубани, Терека, Волги, по некоторым тростниковым озерам Азербай-
джана, на озере Гилли в Армении (Ляйстер и Соснин, 1942) и на озерах
Хозапин и Бугдашен в Джавахетии (Чхиквишвили, 1930). Начиная
от Заволжья, становится более обыкновенной птицей водоемов Средней
Сибири, Казахстана и Средней Азии. Гнездится по болотам и озерам При-
ерусланской степи (Волчанецкий, 1934), на озерах Балыкты, Сактыл, Жилан-
да, Старицком и Волжскр-Уральской степи (Волчанецкий, 1934), по многим
озерам Казахстана, по озерам Барабинской степи, системе озер Чаны (Дани-
лович, 1935) и Кулундннской степи, на Иртыше и Нижнем Кара-Иртыше,
на Балхаше, Ала-Куле, Сасык-Куле и по р. Или в Семиречье (Шнитников,
1949), по Нижней Сыр-Дарье и восточному побережью Аральского моря
(Спангенберг и Фейгин, 1936). По нижнему течению Аму-Дарьи (Гладков,
1932; Рустамов, 1945) и в низовьях Атрека (Дементьев, 1943). Гнездится на
западной окраине Алтая, на озере Марка-Куль и в Северо-Западной Монголии
(Сушкин, 1938), причем по вертикали на Алтае идет от 340 до 1500 м и до
2300 м в Монголии. Во многих перечисленных районах, в том числе и в При-
морской обл., серый гусь гнездится лишь в самых недоступных местах и исче-
зает в культурном ландшафте.
Зимует на Британских о-вах, на европейских и африканских побережьях
Средиземного моря, в Иране, в Афганистане, в Индии, Индокитае и южной
части Китайской Народной Республики. В СССР зимою в Крыму (не ежегод-
но), по юго-западному и юго-восточному побережью Каспия, в Азербай-
джане, в Туркменской, Узбекской и Таджикской ССР.
Характер пребывания. Гнездящийся, пролетный и зимую-
щий вид.
Д а т ы. С мест зимовок в Средней Азии передовые серые гуси начинают
сниматься с 15 февраля, но валовой пролет по Сыр-Дарье идет месяцем позже,
а все весенние передвижения в низовьях Сыр-Дарьи и на берегах Аральского
моря еще продолжаются до конца марта. Первые весенние прилетные на ме-
стах гнездовья в Семиречье и на Балхаше появляются уже к 3 марта (Шнит-
ников, 1949). В конце февраля—начале марта трогаются с мест закавказских
зимовок. К 10—20 марта долетают до дельты Терека1, Волги и Урала, в 20-х
числах марта идут передвижения в области Иргиза и Тургая, 10—20 марта
появляются в Приерусланской степи (Волчанецкий, 1934), в начале апреля
открывается пролет по Суре (Волчанецкий, 1925), в конце марта появляются
под Чкаловом, валовой же пролет здесь идет в середине апреля. На озере
Ханка в Приморье показываются в конце марта—начале апреля.
1 Местные гнездящиеся появляются в дельте Терека в конце февраля (Беме, 1935).
296 ОТРЯД ГУСИНООБРАЗНЫЕ
С мест европейских зимовок на Днестр, в Молдавскую ССР; серые гуси
прибывают в конце февраля, в Подолию 3 марта—2 апреля, в среднем около
1& марта, в районе Александрии появляются 17 марта—1 апреля, в сред-
нем же за 5 лет 26 марта (Браунер, 1894), в конце марта пролетают в Уман-
ском районе Киевской обл. Передвижения на Украине заканчиваются к
20 апреля. В районе Москвы весенние передвижения происходят 12 марта—
27 апреля, в среднем 12 марта (Паровщиков), с 1 по 10 апреля на Московском
морей, наконец, в районе Ленинграда 14—21 апреля (Бианки, 1907). Таким
образом, весенние миграции в Заволжье и Казахстане происходят раньше, чем
на Украине и в европейской части СССР.
Осенью из южных районов гнездовья отлетают раньше, чем здесь появля-
ются более северные особи. Между этими двумя моментами в общем проходит
обыкновенно несколько дней (Тугаринов, 1941). Однако осенний отлет ослож-
няется тем обстоятельством, что еще летом перед линькой у серых гусей
происходят значительные перемещения, что ясно видно по встречам околь-
цованных птиц. Холостые взрослые и молодые неполовозрелые птицы соби-
раются в стаи и отлетают иногда на значительные расстояния в удобные для
линьки места. Так, например, в низовья Волги собираются на линьку птицы
из Челябинской обл., Западного Казахстана и ряда казахстанских озер. Вы-
линявши, эти птицы, летят на зимовки, как показывают данные кольцева-
ния, в самые разные сроки (Дементьев, 1947).
Оставшиеся же на местах гнездовья семейные и молодые птицы из север*
ных районов отлетают 20—30 сентября. Д о конца октября они задерживаются
в Казахстане, до середины ноября в Южном Приуралье. В дельте Волги и на
северном побережье Каспия наблюдаются птицы самых различных возрастов.
Здесь и часть из прилетевших еще летом и прилетевшие вылинявшие на ме-
стах семейные птицы и молодые—все они задерживаются иногда до полного
замерзания воды.
Из дельты Волги часть серых гусей летит в Приазовье и отсюда в Малую
Азию. Большая часть перелетает на западное побережье Каспия и в теплые
зимы остается в дельте Терека. Но основная масса направляется южнее, и
многие из них попадаются на азербайджанских зимовках (Дементьев, 1947).
Наиболее значительными местами зимовок в Азербайджане является
Мильская степь на пространстве от озера Сары до озера Агджабедьи
В меньшем количестве серые гуси зимуют на разливах Ax-Чалы и в сте-
пях, граничащих с заливом имени Кирова, а также в ряде других мест
(Исаков, 1949).
В европейской части СССР осенние передвижения наблюдались под
Ленинградом 13—21 сентября (Бианки, 1907), на северных берегах Москов-
ского моря с 1 октября, валовой же пролет 6—10 октября (Исаков и Распо-
пов, 1949), в средней полосе европейской части СССР—во второй половине
октября (последние здесь отмечены 23 октября), в Украинской ССР в сере-
дине ноября, хотя даже у берегов Черного моря некоторые стаи появляются
значительно раньше (Браунер, 1894) и остаются здесь в теплые зимы. Пролет
на Северном Кавказе идет в октябре и ноябре (Беме, 1935).
Биотоп. Гнездовой—поймы рек, лиманы, плавни; травянистые боло-
та, мокрые луга, степные пресноводные озера, поросшие густыми тростниками.
Всюду, как уже указано, лишь в самых недоступных и непролазных местах
и притом по соседству с заболоченными травянистыми лугами. Зимний—побе-
режья лиманов, разливы рек и водохранилища.
Экология. Половая зрелость—на третьем, четвертом году жизни. Про-
явление ухаживания заметно уже на третьем году жизни. Выражается оно
в том, что самец постоянно держится около одной определенной самки, отго-
няет от нее других гусаков и, прогнав, возвращается к ней с особым триум-
фальным гоготом. Иногда отгоняются несуществующие самцы, т. е. выпол-
СЕРЫЙ ГУСЬ 297
няется определенный ритуал. Не всегда за подобным поведением следует
гнездование.
Даже на юге появляются на местах гнездовья самой ранней весной,
когда водоемы еще покрыты льдом, и снег только что стаял (Спанген-
берг и Фейгин, 1936; Шнитников, 1949). Часть птиц прилетает парами,
часть разбивается на пары на месте; по прилете сразу же занимаются гнездо-
вые участки. Там, где гусей много, а места представляют удобства, гнезда
устраиваются колониями шагах в десяти одно от другого (по р. Или; Шесто-
перов, 1929), в громадном же большинстве случаев пары селятся вдалеке одна
от другой.
Для гнезд выбираются сухие гривы, бугорки, кочки на берегу или на
воде, очень часто кучи старого тростника, сплавнины, а в Семиречье гнезда
устраиваются даже на туранговых деревьях (Шнитников, 1949). На сплав-
нинах гнезда довольно часто находят в тех местах, где бывают поздние
разливы рек и такое гнездо не заливается полыми водами (Тугаринов,
1941). Гнездо строит самка из самых различных растительных материалов.
В его основании находятся крупные стебли болотных растений, иногда ветки
и даже палки. Верхние части построены из более мелких сухих прошлогод-
них остатков болотных растений. Края гнезда и лоток выстилаются пухом.
Другие гнезда являются однородной постройкой из тростника. В некоторых
случаях гнезда представляют солидную цилиндрическую кучу, возвышаю-
щуюся над водою на 100—120 см и построенную на заломах тростника (Шесто-
перов, 1929). На оз. Чаны гнезда имели 60—80 см в основании и до 40 см
в высоту, с шириной лотка 20—25 см и его глубиной 20 см. Глубина же воды
у гнезда доходила до 0,5 м (Данилович, 1935). Первое время такие гнезда вид-
ны еще издали, но затем их прорастает тростник, и найти их чрезвычайно
трудно, хотя к ним гуси и прокладывают тропинки.
Кладка состоит из 4—10, чаще 4—5 белых гладких с легким палевым,
иногда легким зеленоватым оттенком яиц, размеры которых 80,6—94,4х
X 51,4—62 мм (Тугаринов, 1941); E1) 79,5—95,5x53,5—65,5, в среднем
82,2x60,3 мм. Вес 14,7—24,42, в среднем 20,02 г (Гебель, 1904).
Наиболее рано откладка яиц происходит в Средней Азии, где в Узбеки-
стане первые отложенные яйца можно найти около 20 марта, а в Семиречье—
4—6 апреля, но откладка яиц, как и стройка гнезд, оказывается растяну-
той примерно на 25—30 дней. Еще более растянуты сроки кладки в соседней
Кульдже, где первые отложенные яйца находят 14—17 марта и в то же время
ненасиженные кладки попадаются 10—11 мая. (Шестоперов, 1929). С другой
стороны, в более высоких местах строятся гнезда и откладываются яйца
позже, чем на низменностях.
На Сыр-Дарье стройка гнезд заканчивается в самом начале весны и наси-
живание начинается еще в то время, когда пресноводные водоемы покрыты
льдом, но гнездовой период и здесь оказывается растянутым, так как еще
ненасиженные яйца встречаются в начале второй половины апреля A7 апреля
1927 г. в окрестностях станции Кара-Узяк, 4 яйца), а слабо насиженные
можно найти даже в самом начале мая (Спангенберг и Фейгин, 1936). У север-
ного берега Аральского моря 5 ненасиженных яиц найдены 13 мая 1947 г.
(Кузякин), в устье же Урала на берегу моря обнаружены 24 апреля 1941 г.
4 яйца примерно 10-дневной насиженности. Полная ненасиженная кладка,
следовательно, здесь может быть найдена в середине апреля. В Закавказье
яйца 8-дневной насиженности найдены 17 апреля (Радде, 1884). На озере Ча-
ны найдены 9 мая 5 насиженных яиц, 14 мая 2 ненасиженных и 2 слабо наси-
женных, 18 мая—2 гнезда, каждое с 5 слабо насиженными яйцами (Данило-
вич, 1935). Таким образом, в Западной Сибири к кладке яиц серые гуси при-
ступают позже, чем в Средней Азии. В Приморской обл. полные кладки нахо-
дят около 15 апреля (Шульпин, 1936).
298 ОТРЯД ГУСЕОБРАЗНЫЕ
Насиживает самка, в течение 27—28 дней. Гусак держится поблизости,
чаще всего на каком-либо открытом месте или рядом с гнездом, но в насижи-
вании участия не принимает. При опасности гусак очень волнуется и подолгу
гогочет.
Первые пуховички в разных местах Симиречья найдены 9, 16, 19 мая
и 1июня (Шнитников, 1949). В районе Кульджи первые пуховички встречены
уже 18 апреля. На озере Кургальджин в Казахстане первые выводки наблю-
дались 27 мая (Лавров, 1930), в Наурзумском заповеднике 22 мая и в на-
чале июня (Михеев, 1938).
3—4-дневные птенцы по Кара-Иртышу найдены 23 и 26 июня. Ростом
с чирка там же 26 июня (сборы Полякова) и в Волжско-Уральской степи во
второй половине июля (Волчанецкий, 1937). Ростом с крякву в долине Сыр-
Дарьи (окр. Кара-Узяка) 23 июня (сборы Спангенберга) и по Кара-Ирты-
шу 26 июня. Таким образом, гнездовые явления в одних и тех же районах
оказываются значительно, растянутыми. Интересно отметить, что у птенцов
с Сыр-Дарьи по сравнению с кара-иртышскими рост замедлен, диференци-
ровка же оперения выражена значительно яснее.
Молодые находятся под присмотром обоих родителей, пока у послед-
них не начнется линька. Семья держится в наиболее глухих местах близ
гнезда и птенцы ночуют в нем под самкой, самец остается где-либо поблизо-
сти. Когда же самец начинает линять, выводок остается под присмотром одной
только самки.
Л и н ь к а. У молодых условно можно принять две линьки в году: полную
летнюю и частичную осеннюю. Взрослые же, повидимому, линяют один раз
в год. Как уже сообщалось, линька холостых, связанная с их отлетом, начи-
нается много раньше, чем у семейных. Первым начинает линять самец, при-
мерно в то время, когда молодые уже оперятся, но пока еще не летают, сам-
ка же сменяет оперение позже; линька взрослых семейных птиц на юге ареала
начинается с июня, на севере с 20-х чисел июля и к середине августа почти
всюду заканчивается, причем со старыми линяющими птицами держатся
и их выводки.
Первыми, начиная с внешнего и внутреннего краев крыла, выпадают
первостепенные и второстепенные маховые и через 7—8 дней на крыльях
остаются одни только малые кроющие, вообще медленно сменяющиеся. Через
месяц, когда новые маховые отрастут на две трети своей длины, гуси могут
уже подлётывать, но окончательно маховые вырастают только через два месяца
с момента начала линьки. За этот же срок сменяются и все верхние кроющие
крыла. Далее идет смена контурных низа тела, затем контурных спины. Вслед
за этим одновременно со сменой рулевых идет линька лопаточных и контурных
шеи. С конца августа—первых чисел сентября начинается частичная осенняя
линька контурного оперения и рулевых. В этой линьке принимают участие
и молодые, надевшие свой первый наряд. Их контурные перья сменяются
за период от октября до весны, так как весной уже не наблюдается птиц,
сохранивших оперение первого наряда (со сплошь белым, без темных
пятен, низом).
У птенцов сначала вытираются остевидные кончики пуха на брюхе, так
что становится видным густое основание пуха. Затем появляются зачатки
перьев на плечах, почти одновременно показываются зачатки рулевых и ма-
ховых, далее формируются перья на боках тела пониже крыльев. Вслед за
этим начинают появляться перья на середине брюха. Голова же и шея в это
время еще покрыты пухом.
На время линьки серые гуси переселяются в самые недоступные, бога-
тые кормами места и держатся здесь чрезвычайно скрытно.
Молодые становятся летными в Молдавской ССР и юго-западной части
Украины в середине июля, в районе Наурзумского заповедника 24—27 июля,
СЕРЫЙ ГУСЬ 299
в северных районах только к 20-м числам августа. Подъем на крыло и у старых
и у молодых птиц выводка этого года в одних и тех же местах происходит
одновременно. В связи с этим резко меняется образ жизни серых гусей.
Питание. С начала подъема на крыло после линьки серые гуси начи-
нают предпринимать кормовые перелеты семьями или группами из двух-трех
семей. Стаи вылетают на кормовые места на закате солнца, ночью возвра-
щаются на отмели и открытые недоступные части побережий, где спят, стоя
или опустившись на песок. Перед рассветом снова вылетают на кормежку,
а на день перелетают для отдыха в другие, также недоступные и открытые
места на каких-либо водоемах. На зимовках, если их не тревожат, серые гуси
и кормятся и отдыхают днем и ночью в одних и тех же местах. Там же, где
их преследуют, они постоянно предпринимают кормовые вылеты. Кормятся
на стерне, паровых полях, в степях или на побережьях и мелководьях.
Корм серого гуся довольно разнообразен. На местах гнездовья он пое-
дает разнообразные земноводные и наземные растения, но некоторым рас-
тениям отдает ясное предпочтение. На озерах Наурзумского заповедника
с большой жадностью он поедает плоды рдестов и, например, к 25—27 июля
1945 г. все созревшие плоды Potamogeton pectinalis здесь уже были им уничто-
жены. Кормится водяной настурцией, различными луговыми злаками, в край-
них же случаях довольствуется и листьями одного только тростника. В про-
летное время весною в районе разливов Мологи и Шексны на полях и лугах
серые гуси питались побегами ржи озимой, костра безостого, щучки, красной
овсяницы и разных других злаков. Поглощали также семена овса и гречихи
(Polygonum sp.). В других случаях в их экскрементах найдены остатки леер-
сии, селезеночника, зерна ржи, злаков и ежеголовки и также механические
примеси (Исаков). Гусята кормятся вместе со взрослыми, но свою пищу, со-
стоящую из побегов и нежных стеблей болотных и водяных растений, птенцы
добывают самостоятельно.
Во время осенних миграций на полях серые гуси подбирают самые раз-
личные семема диких и культурных растений и очень охотно кормятся всхо-
дами озимых хлебов. На зимовках могут питаться морской травой Zostera
marina, а также выкапывать из земли клубни и луковицы степных растений
<Тугаринов, 1950).
Молодые гусята охотно едят ряску, водяную овсяницу (Festuca fluitans),
корневища и побеги земноводных растений, которые добывают со дна взрослые
птицы, погружая подобно уткам переднюю часть своего туловища в воду.
В прежние времена, когда местами серые гуси были многочисленны,
они причиняли существенный вред озимым хлебам, в настоящее время они
могут вредить посевам только на зимовках, если собираются в крупные стаи.
Хозяйственное значение. В тех местах Сибири и Средней Азии, где
серые гуси еще обыкновенны, они активно используются местным населением,
и притом самым примитивным путем. Весною собирают их яйца, во время
линьки загоняют в сети и истребляют всеми способами во множестве. На
зимовках их добывают главным образом путем отстрела. Но промысловое
их значение в связи с падением численности в настоящее время невелико.
Серый гусь является родоначальником наших серых гусей—тульских,
холмогорских, шадринских, тулузских и эмденских.
Полевые признаки и поведение. Крупный серой окраски гусь с плотным
телосложением. Хорошо ходит и проворно бегает по земле. Часто стоит на
одной только ноге. Очень хорошо плавает, может нырять, но быстро
утомляется. Летает быстро, производя редкие, но сильные взмахи крыльями.
Во время миграций летит довольно высоко, выстраиваясь углом.
Вне периода размножения ведет стайный образ жизни. Чрезвычайно
чуток и осторожен, что особенно проявляется в стаях, где одна или несколько
птиц почти не кормятся и зорко осматриваются по сторонам. В случае опа-
300
ОТРЯД ГУСЕОБРАЗНЫЕ
сности такие птицы взлетают первыми, остальные же поднимаются дружно
вслед за ними. При таких перемещениях высоко не летают.
У серого гуся мало врагов и даже четвероногих хищников он заставляет
держаться в отдалении от гнезд, так как способен наносить им с большой
ловкостью весьма чувствитель-
ные удары .своими сильными
крыльями.Раненый гусак чрез-
вычайно энергично защищает-
ся (Мензбир, 1895).
Голос серого гуся доволь-
но разнообразен; громкие его
крики отличаются по особому
трубному, металлическому то-
ну и звучат, как «га-га-гаг»
или «гия-га-гаг». На лету из-
дает звучное и громкое «га-
ганг»... «гаганг»... «гаганг». Во
время миграций гуси летят
высоко над землей углом, вер-
шина, которого обращена впе-
ред, или жеьшеренгой.
Рис. 50. Серый гусь
Описание Размеры и
строение. Размах самцов F>
1515—1830, самок D) 1480—1635, в
среднем 1662 и 1569 мм. Длина тела
самцов (8) 780—920, самок (8) 780—
8Ь0, в среднем 855,3 и 825 мм. Дли-
на крыла самцов A6) 435—513, самок G) 395—470, в среднем 467,6 и 447,7 мм. Длина
клюва самцов A9) 59,2—77,7, самок (8) 47,1—72,7, в среднем 68,8 и 63,8 мм. Высота
нижней челюсти у самцов A9) 8,3—11,3, у самок (8) 8,1—9,5, в среднем 9,7 и 8,9 мм.
Вес самцов D) 3,05—4,37 кг, в исключительных случаях до 6 кг. Количество зубцов
в клюве колеблется, вернее—увеличивает-
ся с возрастом.
Окраска. Пуховик. Спинная сто-
рона оливково-бурая. Голова и шея свер-
ху, зоб и бока зеленовато-желтые. Брюш-
ная сторона желтовато-белая Клюв и ноги
тёмнокрасные. С возрастом птенец стано-
вится все более и более серым.
Молодая птица. Сверху темносерая с
бурым оттенком. Грудь и передняя часть
брюха серовато-бурые, без темных пятен,
на боках нет белых поперечных полос.
Клюв и ноги или светломясного цвета или
желтоватые. После первой осенней линьки
общая окраска сходна с таковой у птиц
в первом пере, лишь в редких случаях по-
являются единичные черные перья на брю-
хе. После второй осенней линьки, сохраняя
тот же окрас, перо становится более упру-
гим и появляется несколько черных пятен
на брюхе. Число этих перьев увеличивается
после третьей осенней линьки, клюв почти
дорастает до нормальных размеров. Окон- D „ г, п .
чательный взрослый наряд надевается толь- Рис" 5L ПУХОВОЙ птенец серого гуся
ко после четвертой осенней линьки.
Взрослые самец и самка сходны по окраске, но самка мельче. Общая окраска сверху
и снизу серая. Спина и поясница пепельно-серые, кроющие хвоста белые. Лоб часта
спереди с белой полосой. Голова, шея и пространство между лопатками серовато-бурые.
Зоб, грудь и передняя часть брюха серые, на груди и передней части брюха неправиль-
ные мелкие черные пятна. Задняя часть брюха и подхвостье белые. Бока серовато-
бурые с поперечными белыми полосами.
БЕЛОЛОБЫЙ ГУСЬ
301
10. Белолобый гусь Anser albifrons Scop
Branta albifrons. S с о р о 1 i. Annus I Histor. Nat., стр. 69, 1769, Италия?
Синоним. Anser casarca. G m e 1 i n, Reise Russ., II, стр. 179, Нижняя
Волга
Русские названия. Казарка или козарка—народные; белолобый гусь—книжное
название.
Распространение. Ареал. Северная полоса Северной Америки, Грен-
ландия, Исландия. В Евразии на островах Ледовитого океана и в тундре от
Карта 31. Распространение белолобого гуся Anser albifrons
1—A. a. albifrone, 2—А. а. gambelli; а—граница гнездовий, г—зимовки
Белого моря до Чукотки. Зимой до Мексики, Вест-Индии, Южной Европы»
Средней Азии, Индии, Китая, Кореи и Японии.
Характер пребывания. Гнездящийся и пролетный вид.
Биотоп. Зона тундры.
Численность. Один из самых многочисленных гусей.
Подвиды и варьирующие признаки. Распадается на два подвида: A. a. albifrons
Scopoli, 1769, который занимает восточный берег Гренландии, о-в Исландию,
север Евразии и Северной Америки до бассейна р. Мекензи; отличается мелкими
размерами, однако постепенно увеличивающимися у популяций по направлению от
запада на восток, и A. a. gambelli Hartlaub, 1852, который распространен по тундре
Северной Америки к востоку от бассейна Мекензи до побережья Западной Гренландии;
от предыдущего подвида отличается относительно крупными размерами, величиной
и соотношением частей клюва.
Западный белолобый гусь Anser albifrons albifrons Scop.
Распространение. Ареал. Западное побережье Гренландии, Исландия,
в крайне ограниченном количестве на севере Скандинавского п-ва в Финмар-
кене, отсутствует на Кольском п-ве, обычен на Новой Земле, Колгуеве, Вай-
гаче, редок на Новосибирских о-вах, где гнездится только на о-ве Большом
Ляховском. На материке, начиная от п-ва Канина, обыкновенен по всей евра-
зийской тундре, местами заходит в лесотундру. В Северной Америке по
туйдре от Аляски до Мекензи и на юг до Юкона. Таким образом, белолобый
гусь является кругополярной птицей.
302 ОТРЯД ГУСЕОБРАЗНЫЕ
Зимой до Средиземноморья и северного побережья Африки, единично
до Мадейры, в большом количестве до Египта, до Северной Индии, Бирмы,
Кореи, Китая и Японии. В СССР в небольшом количестве на Черном море,
обилен на Каспийском и в Закавказье, редок в Средней Азии. В Америке
до Калифорнии, Мексики и Кубы.
Характер пребывания. Гнездящаяся птица крайнего севе-
ра, пролетная для ряда территорий СССР, зимующая на юге страны. Во время
перелетов обильна в западной части Украины, в Приазовье, на северном бере-
гу Каспийского моря, в низовьях Волги, Урала и Эмбы, по Иргизу, Тургаю
и Тоболу. Отсутствует на пространстве от Западного Казахстана, за исклю-
чением указанных частей до Даурии и верховьев Амура (Тугаринов, 1941).
В небольшом количестве пролетает в европейской части СССР.
Даты. Самое раннее появление белолобого гуся на территории СССР,
указывается для окрестностей Павлограда в УССР, где первые пролетные
стаи были отмечены 7 февраля 1895 г. Однако в этом случае гуси должны были
возвратиться обратно в силу резко наступившего похолодания и значитель-
ных снегопадов. У северного берега Черного моря в некоторые годы наб-
людались 22 февраля. В эти же годы отмечены 1 марта в Аскании-Нова.
Нормально белолобые гуси показываются около 10 марта в устьях Дуная
и Днестра (Браунер, 1894), 18 марта в Приазовских плавнях Краснодарского
края и в этот же срок в Восточном Закавказье, через которое летят стаи,
зимовавшие, повидимому, в Иране или в смежных с ним территориях. Про-
лет здесь продолжается около 10 дней и последние отмечены 29 марта. В этом
движении не принимают участие гуси, находящиеся на зимовке в Закав-
казье, откуда они отлетают позже и окончательно исчезают только к 23 апреля
(Тугаринов, 1950). 17—22 марта появляются в дельте Терека (Беме, 1925);
в начале апреля они уже на северных берегах Каспия, 19 марта—10 апреля,
или в среднем за 6 лет 1 апреля, пролетают в Киевской обл.(Уманский район;
Гебель, 1879), причем задерживаются здесь иногда до 15 апреля; 27 марта
отмечены в долине Сыр-Дарьи у станции Джулек (Спангенберг и Фейгин,
1936); 1—5 апреля в Приморской обл. в районе Ханка, валовой же пролет
их здесь отмечен 15 апреля; 20—30 апреля в бассейне Оки в центральных
областях европейской части СССР. Гуси, подлетевшие к низовьям Волги
и северному берегу Каспия, на некоторое время задерживаются здесь, окон-
чательно исчезая отсюда только к 15 мая; с середины апреля до конца мая
пролетают побережьем Балтийского моря вдоль Латвийской ССР и Эстонской
ССР; 9 мая показываются передовые в районе Ленинграда (Бианки, 1912);
7—12 мая в северной части Московского моря (Спангенберг, 1949). С начала
апреля появляются в бассейне Урала в районе Чкалова, валовой же пролет
идет здесь 1—4 мая; 2 мая передовые отмечены в районе Тюмени, 1 мая
на оз. Чаны, где обильный пролет наблюдается до 20 мая (Данилович,
1935). В конце апреля—в первой декаде мая появляется в Тиманской тундре.
В 1938 г. передовые здесь появились 7 мая, 9—12 шел валовой пролет, 21 мая
белолобых гусей стало меньше, но затем 24 мая появилась новая волна про-
летных, 29 мая из долины р. Вельть гуси исчезли (Гладков, 1941); 18 мая по-
являются в бассейне Печоры (в окрестностях Усть-Ылыча; Дмоховский, 1933).
В устье Мологи отмечены 25 апреля 1940 г. С 25 апреля по 3 мая наблюдался
валовой пролет. Последние крупные стаи летели 4 мая, но часть гусей оста-
валась в низовьях Мологи и количество их значительно уменьшилось к 8 мая
(Исаков и Распопов, 1949). В ранние весны к концу апреля, в нормальные
же к 15 мая долетают до района Якутска, валовой пролет около которого отме-
чен 20—25 мая; 23 мая появляются на Шантарских о-вах (Дулькейт и Шуль-
пин, 1937), 9—30 мая в районе Охотска, где гуси подолгу останавливаются
и держатся часто в открытом море на плавающих льдинах (Харитонов, 1915);
на крайнем севере материка появляются в середине—конце мая; 15—25 мая
БЕЛОЛОБЫЙ ГУСЬ 303
прилетают в Новую Землю; 20 мая отмечены на Ямале, 23 мая на его восточ-
ном побережье в Обской губе, а 3 июня местные ямальские отмечены уже
в своих гнездовых участках, 1 июня на Гыданском п-ве (Наумов, 1931), 23—
25 мая на Таймыре, где валовой лет наблюдается 28 мая (Гизенко, 1938),
но на Таймырском озере появляются только 10 июня, 27 мая в устье Индигир-
ки, 20 мая в районе Верхоянска в долине Яны, в горах же появляется на
10—15 дней позже; 11 мая на Командорах (Бианки, 1909); около 1 мая пере-
довые на р. Гижиге (Бэкстон), 11—19 мая на Анадыре в районе Марково и 12—
20 мая на самом Анадыре (Портенко, 1939), 8 июня в низовьях Лены (Тор-
гашев), 9 июня в низовьях Яны и на большом Ляховском о-ве и только
15 июня на о-ве Котельном (Тугаринов, 1941).
Передовые в тундру прилетают еще при настоящей зиме и, продержавшись
здесь в течение ряда дней, при частых на севере возвратах холодов исчезают,
а затем через некоторое время появляются сразу в большом количестве.
Массовый их прилет совпадает с появлением промоин на реках и первых
проталин в тундре. Характерны также длительные остановки в богатых
кормами местах, что в ряде случаев дает возможность белолобым гусям поя-
виться на родине в весьма упитанном виде.
В западной части тундры белолобые гуси остаются 105—140 дней, на
Анадыре не более 103 дней.'Отлет с мест гнездовья на севере начинается рано,
и хорошо выраженный пролет наблюдался 15—31 августа на Камчатке (Би-
анки, 1910), 20—30 августа на Таймыре (Торгашев) и на Гыданском п-ве
(Наумов, 1931). К 4 сентября совершенно исчезают из бассейна Анадыря
(Портенко, 1939). Первыми начинают отлетать отдельные особи или неболь-
шие стайки, и таким ранним их отлетом объясняется крайне раннее появле-
ние отдельных передовых особей и стаек далеко на юге; 1 сентября в районе
Чкалова (Зарудный, 1897) и 1—15 сентября в Киевской обл. (Гебель, 1879).
Отлет с мест гнездовья с Новой Земли наблюдается 1—4 сентября, но часть
гусей здесь задерживается до сентября на Северном о-ве и до 7 октября на
Южном (Горбунов, 1929). Вместе с тем далеко на севере, в суровых осенних
условиях Арктики в ряде мест белолобые гуси остаются еще долго и отле-
тают поздно. Так, отлет с Ляховских о-вов отмечен 13 сентября, пролет на
Яне наблюдался 9 сентября, последние из ее низовьев отлетели 25 сентября
(Бунге, 1884), последние с Индигирки—30 сентября (Михель, 1930). До конца
сентября—начала октября идет пролет в окрестностях Якутска. 19 сентября
еще летят по Печоре и 18 сентября пролетают вдоль Охотского побережья,
в сентябре идет пролет на Шантарских о-вах (Дулькейт и Шульпин, 1937).
На континенте в Центральном Казахстане также в сентябре идет хорошо
выраженный пролет и последние здесь наблюдаются 24 октября (Сушкин,
1908). В районе Наурзумского заповедника осенний пролет начинается
с середины октября (Михеев, 1938). 29 сентября—9 октября пролетают в
Херсонской обл.; 7 октября в районе Ленинграда (Раснер, 1913). С 25 сен-
тября по 12 октября пролетают через территории Эстонской и Латвийской
ССР. С 1 по 18 октября идет пролет в низовьях Мологи (Исаков и Распопов,
1949). Довольно рано передовые появляются близ побережий Черного моря,
где в низовьях Днестра первые отмечены 10 октября 1949 г. и где в значитель-
ном количестве они скопляются к концу октября (Назаренко). К 30 октября
прибывают на места зимовок в Крыму, где остаются в теплые зимы до 30 мар-
та. К 9—10 ноября собираются на местах зимовок у юго-западного берега
Черного моря, где остаются до 31 марта. 15 октября отмечено их появление
в дельте Терека (Беме, 1925); примерно в это же время, с половины октября,
но чаще в конце этого месяца, слетаются на зимовки в Закавказье и остаются
в одних участках до середины марта (Исаков и Воробьев, 1940), в других—
до23 апреля (Тугаринов, 1941). Основным местом зимовок в Закавказье яв-
ляется пространство между средней частью долины реки Куры и северной
304 ОТРЯД ГУСЕОБРАЗНЫЕ
окраиной Талышинской низменности, где белолобые гуси занимают солян-
ковые степи и полупустынные территории. Однако определенная часть про-
летных стай направляется далее на юг, на зимовки в Иран и Месопотамию
(Исаков и Воробьев, 1940). Вместе с тем значительные стаи пролетных задер-
живаются в дельте Волги еще в течение всего ноября и только в начале декаб-
ря отлетают отсюда на места своих зимовок.
Необходимо отметить, на что указывалось уже давно (Мензбир,
1897), некоторое непостоянство направлений движения стай гусей в пределах
небольших территорий. В ряде мест ежегодно гуси летят по определенным
направлениям, затем в последующие годы отклоняются от них, избирая
новые, и уже не пролетают там, где прежде были обычны. Весьма возможно,
что такие перемены направления передвижений связаны с выбором новых,
более безопасных мест отдыха и кормежек (Птушенко, 1937). Далее необхо-
димо указать на количественную разницу пролета птиц в разные сезоны.
Так, на северном побережье Азовского моря осенью белолобый гусь чрез-
вычайно многочислен, весною же он здесь пролетает в ничтожных количе-
ствах (Алфераки, 1904). В других же местах, как например, в Рязанской
обл., наблюдается только на осеннем пролете (Хомяков, 1900).
Биотоп. Гнездовой—обильные реками и озерами районы кустарни-
ковой тундры. Гнездовые участки выбираются в более или менее сухих местах,
на возвышенных берегах рек, по склонам речных долин, на холмах вблизи
рек и озер, однако в сухой пятнистой тундре не гнездится. Зимний—степи,
поля и морские побережья.
Численность. Неравномерна. На Северном о-ве Новой Земли ре-
док, на Южном—местами обыкновенная птица. В наибольших количест-
вах встречается на Гыданском п-ве, на Таймыре и в районах низовий сибир-
ских рек. Во время миграций обилен в Западной Сибири, в Нижнем Поволжье,
осенью на Азовском побережье и в низовьях Днестра. На зимовках в боль-
шом количестве встречается в Закавказье.
Экология. Размножение. Изменения в половой системе насту-
пают уже у мигрирующих птиц, но они еще едва выражены. У самки, добы-
той из стаи в Приморской обл., яичник набух еще очень слабо, но отдель-
ные фолликулы достигали уже 3 мм в диаметре (Шульпин, 1936). Дальней-
шее развитие гонад происходит на местах гнездовья, однако довольно разно-
временно.
Вскоре после прилета взрослые птицы разбиваются на пары и присту-
пают к размножению, неполовозрелые же и холостые белолобые гуси дер-
жатся стаями и кочуют по тундре весь летний сезон.
Приступившие к стройке гнезд пары используют для их сооружения вся-
кого рода возвышенные точки микрорельефа—крупные кочки, бугорки, хол-
мики, обыкновенно поросшие травою. В некоторых же случаях гнездо распо-
лагается на глинистой почве или даже на песке. Трава, если она имеется
в избранном месте, уминается, иногда делается небольшое углубление и в него
обильно укладывается пух, надерганный из собственной хлупи. Некоторые
лары углубление это устилают прошлогодними сухими злаками, поверх ко-
торых настилают слой пуха. Таким образом, гнездо у белолобого гуся доволь-
но примитивно. Гнезда располагаются разбросанно, вдали одно от другого,
иногда же устраиваются близ гнезд тундровых хищников—сапсана или мох-
ноногого канюка.
С момента прилета до начала кладки проходит некоторый промежуток
времени: на Таймыре не менее 10 дней, на Гыданском п-ве—16 (Наумов, 1931).
Кладка состоит из 3—7, чаще из 5, белых с палевым оттенком яиц. Их
размеры: 70,0—89,6x46,7—58,0 мм (Тугаринов, 1941); B4) 76—88,5x49,5—
56,5, в среднем 81x53,9 мм\ вес B0) 9,96—12,48, в среднем 10,98 г (Ге-
бель, 1904).
БЕЛОЛОБЫЙ ГУСЬ 305
Первые отстроенные гнезда на Таймыре находили с 10 июня, массовая
стройка их отмечена с 15 июня (Гизенко, 1938). На Ямале в это время птицы
держались парами и у самки, добытой 13 июня, в яйцеводе оказалось яйцо,
одетое скорлупою (Житков, 1912). Первое отложенное яйцо найдено 12 июня
в устье Индигирки (Тугаринов, 1941) и 17 июня на Гыданском п-ове (Наумов,
1931); 4 ненасиженных яйца на Таймыре 26 июня, насиженные яйца 19 июня
(на Ямале; Житков, 1912); яйца с совершенно сформированными зародышами
на Новой Земле—13 июля 1951 г. (Кузякин) и 28 июля 1924 г. (Горбунов,
1929). Насиживает одна самка в течение 26—28 дней1.
Первые выклюнувшиеся пуховички—5 июля в устье Лены и 10 июля
в устье Индигирки, 2—3-дневные в дельте Колымы 8 июля. Массовый вывод
пуховичков 1—10 июля на Таймыре (Гизенко, 1938). Молодые ростом с чир-
ка; но еще в пуху 1 августа в Сагастыре (Тугаринов, 1941), 29 июля на Ямале
и 26 июля на Новой Земле. Уже подросшие молодые с пробивающимся пером
30 июля на Ямале (Житков, 1912); крупные пуховики 1 авгусга на Новой
Земле; молодые в новом пере 14 сентября—на Новой Земле.
Во время насиживания самец находится при гнезде или близ него. Подра-
стающий выводок все время держится при стариках. При преследовании
выводка, находящегося под присмотром родителей, старые птицы быстро
убегают или подлетывают непосредственно над землей, а вслед за ними быст-
ро мчатся и молодые. Иногда же, если семья заблаговременно заметит опас-
ность, старики и молодые крадутся, низко прижимаясь к земле или, отбе-
жав недалеко, затаиваются в траве.
Л и н ь к а. У молодых условно можно принять две линьки в году: пол-
ную летнюю от июля до 20-х чисел августа и частичную с сентября по январь.
Взрослые же, повидимому, линяют один раз в год летом и осенью.
Первыми начинают линять неполовозрелые белолобые гуси вывода пред-
шествующего года и оставшиеся холостыми птицы. К ним присоединяются
также семейные самцы от выводков. Перед началом линьки стаи этих гусей
совершают более или менее отдаленные перелеты, при которых выбираются
уединенные или малодоступные места. В это время белолобые гуси переле-
тают с Южного о-ва Новой Земли на Северный, с пространства между Леной
и Яной—на Новосибирские о-ва. Еще и в настоящее время на Большом Ляхов-
ском о-ве собираются тысячные стада линяющих белолобых гусей. В таких
громадных стаях лишь в крайне редких случаях можно найти единичные
особи других видов гусей.
Начало перемещений гусей перед линькой отмечено на Ямале 26 июля.
На Новой Земле линяющие холостые уже стали заметны с 28 июля, массовая
линька наблюдалась 7 августа, первые закончившие линьку маховых—16 ав-
густа, а 22 августа большинство уже снова поднялось на крыло (Горбунов,
1929). На Таймыре основная масса холостых белолобых гусей сменяла махо-
вые перья в период между 25 июля и 10 августа, снова белолобые гуси подня-
лись на крыло 15 августа (Гизенко, 1938). Указываются и другие сроки линь-
ки этих гусей на Таймыре: холостые начали линять 22 июля и 27 июля встре-
чались особи, которые уже утратили способность к полету. 8 августа часть
гусей еще продолжала терять перья, и 17 августа все гуси уже снова могли
летать (Миддендорф, 1854). За исключением некоторых деталей, линька
на Таймыре заняла и в том и в другом случае почти один и тот же промежуток
времени.
Самка, оставшаяся при выводке, приступает к линьке несколько позже
холостых. На Анадыре линька самок начинается с 1 августа. К концу этого
1 Однако Торгашев указывает, что в насиживании принимает участие и самец. Сооб-
щение это противоречит всем предшествующим данным.
20 птицы ссср. т. IV
306
ОТРЯД ГУСЕОБРАЗНЫЕ
месяца смена маховых у них полностью заканчивается и к этому же времени
и молодые поднимаются на крыло (поСокольникову, Портенко, 1939). Между
20—31 августа здесь встречались стаи, все гуси в которых при подходе
к ним взлетали, но некоторые из них, не будучи в силах лететь, вскоре опу-
скались, так как у них маховые или не доросли еще до нормы,или недостаточ-
но окрепли. К 1 сентября все белолобые гуси на Анадыре летали уже хорошо
(Портенко). С момента подъема на крыло выводков семьи окончательно раз-
биваются и начинают формироваться стаи.
Частичная линька контурного оперения и рулевых начинается еще на
родине и заканчивается на зимовках.
Питание. Вместе с окончанием летней линьки у белолобых гусей
связаны также изменения и в образе жизни. Каждая из вновь сложившихся
стай перед заходом солнца на-
правляется в избранное место
кормежки, и, наевшись здесь до-
сыта, летит отсюда на место во-
допоя и ночного отдыха, перед
утром снова совершает кормовой
вылет и после кормежки переле-
тает опять на место дневного от-
дыха. Такой порядок суточной
жизни белолобых гусей наблю-
дается до самого их отлета. На
зимовках, если их не беспокоят
охотники, у белолобых гусей
иное распределение дня. Они, по
существу, оказываются привя-
занными к месту кормежки, где
появляются с рассвета и где кор-
мятся до самого полудня, когда
летят на водопой. Отсюда они
снова возвращаются на место кор-
межки, располагаются на каком-
либо пункте с обширным круго-
зором, долго здесь отдыхают, лежат, чистятся и приводят свое оперение
в порядок. Потом пешком белолобые гуси идут кормиться, а перед закатом
солнца летят в избранное безопасное место ночлега.
Основной их пищей являются нежные зеленые части травяных растений.
Во время перелетов через культурные ландшафты они кормятся на жнивьях
и на озимях. В низовьях Шексны и Мологи на весеннем пролете первое время
основным их кормом являются прошлогодние зерна овса, которые они вместе
с серыми гусями собирают по стерне. По мере спада вешних вод и появления
зеленеющих растений на лугах они начинают кормиться побегами костра
и овсяницы (Исаков и Распопов, 1949).
На зимовках в степях Юго-Восточного Закавказья, где белолобые гуси
в теплые зимы собираются в огромных количествах, основным их кормом
являются зеленые злаки и всходы озимой пшеницы, которой на мокрых или
влажных пашнях они способны причинять значительный вред. Помимо этого,
в состав их корма входят зеленые побеги вероники и Ruppia maritima, а так-
же семена Bolbochaenus maritimus, Fimbristylus sp. и гороха (Исаков
и Воробьев, 1940).
Хозяйственное значение. На местах гнездовья и линьки еще и в настоящее
время белолобые гуси широко используются в промысловых целях. Ранней
весной по прилете их стреляют по проталинам из засидок на местах кормежек
или ловят здесь капканами. Так как в ряде мест совершенно разумно запре-
Рис. 52. Белолобый гусь
БЕЛОЛОБЫЙ ГУСЬ 307
щена охота на линяющих гусей, их теперь добывают летом и осенью только
с помощью огнестрельного оружия. В значительном количестве отстреливают-
ся на прикаспийских зимовках, где иногда наносят вред полям.
Враги. В тундре белолобые становятся жертвой песца и других четве-
роногих хищников; на Анадыре линяющих гусей удается ловить даже мед-
ведю (Сокольников). На местах зимовок белолобые гуси зачастую гибнут
в мазутных лужах и вследствие разлива нефти по воде (Верещагин, 1946).
Полевые признаки. Средней величины гуси, издали кажутся серыми, в би-
нокль у взрослых различимы белые «лбы» и черные пятна на брюхе. Во вре-
мя миграции белолобые гуси летят на большой высоте, выстраиваясь углом
или волнистой линией. На лету часто перестраиваются и кричат. Голос их по-
хож на крик серого гуся, но более высок и резок. Его можно передать как
двусложное «клинг-клинк»... и «кленг-клэк». Иногда слышно «коунг-клунк».
Белолобые гуси являются наземными птицами и их чаще можно увидеть на
земле, чем на воде, к которой они прилетают только для водопоя. На земле
предпочитают держаться на более сухих участках, хорошо ходят и могут
быстро бегать. На кормежках встречаются небольшими стаями по сухим
открытым травянистым местам. Взлетают с воды и садятся на воду легко.
Хорошо плавают, а при опасности и ныряют. В связи с обитанием в малона-
селенных местах менее осторожны, чем серые гуси.
Описание. Размеры. Размах самцов A4) 1250—1467, самок A0) 1382—1487,
среднем 1428,9 и 1382 мм. Длина тела самцов A8) 647—765, самок A3) 620—772, в сред-
нем 713 и 680 мм. Длина крыла самцов C1) 385—455, самок B7) 380—420, в среднем 421,4
и 405,7 мм. Длина клюва самцов C6) 39,8—56,5, самок C0) 39,8—51,5, в среднем 48,2
и 46,8 мм. Высота нижней челюсти самцов C6) 5—7,6, самок C0) 5—6, 8, в среднем
6,31 и 5,8 мм. Вес самцов A5) 2400—3200, самок A0) 2000—3000, в среднем 2670 и 2430 г.
Никакой существенной разницы в размерах между западными особями в европейской
части СССР и восточно-сибирскими (бассейн Колымы, Анадырь, Пенжина) не обна-
ружено. Средний размер крыла первых у самцов 423,1, у вторых 420 мм, средняя длина
клюва соответственно 46,5 и 48,75 мм. Можно только сказать, что длина клюва по нап-
равлению к востоку слегка увеличивается.
Окраска. Пуховик сверху серовато-бурый. Концы крыльев спереди и их сере-
дина сзади светложелтые. Бока головы и низ серовато-белые с желтым налетом.
Молодые птицы в первом пере покрыты еще рыхлым оперением, особенно на голове
и шее. У молодых птиц после первой осенней линьки ни у вершины лба, ни вдоль осно-
вания верхней челюсти нет белых перьев и, наоборот, здесь выделяются черные или
буровато-черные перья. На верхней стороне перья по сравнению с таковыми взрослых
узкие и заостренные, на нижней же на одну треть уже, чем у взрослых. Светлосерое
брюхо с правильно расположенными серыми крапинами. Клюв изжелта-серый или
серовато-мясного цвета. Ноготок сероватый.
Молодые птицы после второй осенней линьки. На голове узкая белая кайма из
перьев на границе с верхней челюстью. Черные пятна на брюхе едва выражены и раз-
бросаны единично. Клюв темносерый с легким красноватым оттенком и серовато-белым
ноготком. После третьей осенней линьки черные пятна на брюхе появляются в большем
количестве. Окончательный наряд надевается после четвертой осенней линьки. Взрослые
птицы сходны по окраске, но самка немного мельче и легче самца. Общая окраска сверху
тускловато-серая, снизу светлосерая с черными пятнами. На лбу, начиная от клюва, идет
белое пятно 15—26 мм шириною, за ним расположены темноокрашенные перья. Осталь-
ная часть головы, шея, лопаточные и пространство между ними, а также бока груди
буровато-серые со светлыми вершинными каймами перьев. Спина и поясница темно-
серые. Грудь и брюхо светлосерые с неправильными крупными и редкими или мелкими
и многочисленными черными пятнами.
Маховые черно-бурые с белыми стволами. Рулевые черно-бурые с белыми кай-
мами. Клюв двуцветный. В области ноздрей он розовато-телесный, под ними и по коньку
желтый с легким оранжевым оттенком. Ноги желто-оранжевые. Радужина темнобурая.
20*
308
ОТРЯД ГУСЕОБРАЗНЫЕ
11. Пискулька Anser erythropus L.
Anas erythropus. Li n n a e u s. Syst. Nat., изд. X, 1, стр. 123, 1758, Швеция.
Синонимы. Anser finmarchicus. Gunnerus in Leemi de Lappon. Comm. notis,
1767, стр. 264, Финмаркет— A nser rhodorhynchus. Бутурлин. Дикие гуси России, II,
1901, стр. 19, Западная Сибирь.
Русские названия. Малая казарка, малый белолобый гусь. Пискулька—зву-
коподражательное название, данное этому гусю за его своеобразный голос.
Распространение. А р е а л. По северу Евразии от Скандинавского п-ова
до Анадыря. В пределах СССР лишь местами и в незначительном количестве
выходит на побережья Кольского п-ова, Баренцова и Карского морей, на
Карта 32. Распространение пискульки Anser erythropus
а—граница гнездовий; е—зимовки
Таймыре на его западном побережье доходит до 73°30' с. ш., в центре до
72°30'—74° с. ш. ,на пространстве от Хатанги до Колымы на ледовитоокеанском
побережье бывает только во время линьки. На Колыме очень редка и немно-
гочисленна на Анадыре. На Чукотке не найдена (см. Бутурлин, 1935).
На юг от перечисленных мест становится все более и более обыкновенной,
а местами даже и обильной. Обыкновенна пискулька во внутренних частях
Кольского п-ова, в Тиманской, Канинской и Большеземельской тундрах,
в долине Оби гнездится до окрестностей Салехарда, в долине Енисея до Ду-
динки на Хатанге весьма многочисленна и встречается до Котуя и северной
части лесной зоны. Далее на восток она редка и доходит на юг до устья Адычи
на Яне, до Зашиверска на Индигирке и до Средне-Колымска на Колыме,
гнездится в бассейне Анадыря. Сведения о гнездовании на Камчатке (Бианки,
1909) требуют подтверждения. Залетает на Новую Землю. Таким образом,
пискулька занимает зону тундры, кроме ее крайней северной полосы, лесо-
тундру и на юг доходит до северных частей лесной зоны.
ПИСКУЛЬКА 309
Зимой в небольшом количестве и редко в Германии, Бельгии, Англии,
Франции, Венгрии и других местах Европы, часто встречается на Балканском
п-ове. В Африке долетает до Египта. В Азии бывает зимой до Малой Азии,
Ирана, Индии (в небольшом числе), в южной части Китайской Народной
Республики, в Корее и Японии.
Характер пребывания. На территории СССР гнездится,
пролетает и в теплые годы зимует в незначительном количестве в Крыму
и на Черном море, многочисленна на зимовках в Юго-Восточном Закавказье
и в годы больших разливов на Атреке. Известна с озера Иссык-Куль (Алфера-
ки, 1904)» но за последнее время не указывается для него (Шнитников, 1949).
Даты. Отлет с мест зимовок на Атреке начинается еще в феврале,
идет исподволь, и птицы в общем исчезают к середине марта. 16 марта их бы-
ло очень мало, 17 марта еще оставалась одна небольшая стая (Житников,
1900). Незначительный пролет на Узбое отмечен 24—25 марта и 1 апреля
(Шестоперов). С Атрека пискульки отлетают преимущественно через конти-
нент, так как вдоль восточного берега Каспия весною они встречаются в нич-
тожном количестве (Бостанжогло, 1911). Отлет из-Юго-Восточного Закав-
казья происходит в те же сроки, что и на Атреке. Пискульки, отлетающие
с Балканского п-ова и побережий Черного моря в начале апреля показываются
у его северных берегов. 10 апреля наблюдались на о-ве Джарылгач (Шарле-
мань и Шуммер, 1930). На Азовском побережье у Таганрога они отмечены
3 апреля (Боровиков, 1907). В Смоленской обл. наблюдались 21 апреля (Гра-
ве, 1926), в районе озера Ильмень 26 апреля. Во многих районах СССР пи-
скульки пролетают одновременно с белолобым гусем, недостоверных данных
об их пролете почти не имеется. На крайнем востоке, на озере Ханка в При-
морской обл., первое их появление отмечено между 27 марта л 2 апреля.
(Пржевальский, 1870), валовой же пролет между 10 и 27 апреля, последние
стаи пролетали 15 мая. На север в Тиманскую тундру прилетают в мае.
8 окрестностях Якутска первые отмечены 15 мая 1927 г., пролет шел 20—
25 мая 1927 г. (Воробьева, 1928). В районе с. Хатангского первые показыва-
ются 29 мая и пролет продолжается до 10 июня. В осеннее время в окрестно-
стях этого последнего пункта еще оставались 26 марта и с мест гнездовья
окончательно исчезли в сентябре. 15 сентября они уже отлетали с Таймыра
в южном направлении (Торгашев); Таким образом, на Таймыре пискульки
проводят только 95—125 дней. В окрестностях Новгорода пролет шел 29
и 30 сентября (Бианки, 1910), на территориях Эстонии, Литвы и Латвии
9 октября, в районе Наурзумского заповедника в середине октября (Михеев,
1938). На разливах Атрека у Чатлы первые стайки пискулек появились
в середине октября, массовый их пролет проходил в начале ноября, в Гасан-
Кули—в ноябре (Исаков и Воробьев, 1940). В Приморской обл. осенний
пролет идет в сентябре. Во время миграций в наибольшем количестве пискуль-
ка пролетает в Приазовье, где задерживается иногда до 10—13 декабря,
а также в Тургайской низменности и в Приморье. В западных частях РСФСР
и в УССР пролетает в очень небольшом количестве; между Тургайской низ-
менностью и Приморьем на пролете почти не наблюдается. Крайне редка
в западной части бассейна Сыр-Дарьи (Зарудный, 1915).
Биотоп. Гнездовой—разнообразнее, чем у белолобого гуся. Встре-
чается не только в тундре, но и в низовьях горных рек, в предгорьях на рано
оттаивающих уступах горных склонов, на горных озерах и заходит даже
в альпийскую область, где занимает скалистые обрывы, часто селится в ме-
стах, в начале весны богатых проталинами, где потом развивается обильная
травянистая или кустарниковая растительность, но гнездится и на местах,
покрытых редко разбросанными камнями. Зимний—открытые пространства
с широким кругозором и богатые кормами, а также степи и морские побе-
режья.
310 ОТРЯД ГУСЕОБРАЗНЫЕ
Численность неравномерна, в общем встречается несколько реже
белолобого гуся, местами даже редка; наиболее многочисленна в бассейне
р. Хатанги.
На Новой Земле в одних местах, например вдоль р. Шумилихи, на 4 км
пути в 1951 г. приходилось 8—9 гнездовых пар, в других—на 20 км пути
только одна гнездовая пара (Кузякин).
Экология. Размножение. Гнездо помещается то в тундре, то среди
кустарников, то на скалах в горном ландшафте и строится на прдталине вбли-
зи воды. Оно так же примитивно, как и у белолобого гуся. В гнездах с недав-
но отложенными яйцами обычно содержится мало пуха и, наоборот, в них
много мха. По мере насиживания количество пуха все более и более увели-
чивается (Кузякин). Откладываются 4—6—8 гладких слегка блестящих свет-
ложелтых яиц. Размеры: 69—84,5x43—52 мм (Тугаринов, 1941); G2) 69—
84,5x43—50,5, в среднем 74,48x87 мм. Средний вес B1) 7,375 г (Рей и Джо-
урден, 1920). В дальнейшем окраска яиц приобретает неровный охристо-
бурый цвет. На Новой Земле вдоль р. Шумилихи 20 июня 1951 г. найдены
гнезда с 2, 4, 5и6 ненасиженными яйцами. 20 июня 1951 г. в окрестностях
Лагерного становища гнездо с 5 слабо насиженными яйцами. 30 июля 1951 г.
на юге Паньковой Земли гнездо с 4 насиженными яйцами; 6 июля 1951 г. там
же два гнезда, в каждом из которых было по 6 сильно насиженных яиц; глаза
эмбрионов в них достигали более 8,5 мм в диаметре (Кузякин). На Большом
Айновом о-ве выклевывание из яиц наблюдалось 21 июня 1949 г. (Белополь-
ский). Однодневные птенцы на Паньковой Земле встречены 13 июля 1951 г.
(Кузякин). Пуховички в возрасте нескольких дней найдены на Боганиде на
Таймыре 5 июля (Миддендорф, 1854), в верховьях р. Дудинки 7 июля (Туга-
ринов, 1941), на Яне у р. Адычи 8 июля (Бунге, 1887), на Индигирке у Заши-
верска 8 июля (Михель, 1935), на Колыме в районе Средне-Колымска 2 июля
(Черский), 11 июля на Боганиде добыты птенцы, у которых начали появляться
плечевые перья и маховые (Миддендорф, 1854).
Линька. У молодых, условно, две линьки—полная летняя с июля
и частично осенняя; у взрослых, повидимому, одна, точных данных не
имеется. Достоверных данных о сроках полной летней линьки мало. На Но-
вой Земле первая пискулька со сплошь выпавшими маховыми добыта 1 июля
1951 г. Там же, у губы Безымянной в большой стае, наблюдавшейся 8 июля
1951 г., часть гусей еще свободно летала, часть уже не могла подняться на
крыло, а 24 июля 1951 г. здесь же у гуся, добытого из линной стаи более чем
в двадцать особей, отрастающие первостепенные маховые достигали поло-
вины своей нормальной длины (Кузякин). На Таймыре 19 июля встречены
две особи с совершенно выпавшими маховыми. 4 августа там же найдены
нелетные молодые и линяющие взрослые. 4 августа добыт гусь из уже летав-
шей стаи, состоящей, повидимому, из холостых (Торгашев). 14 августа 1937 г.
в низовьях р. Вельть в Тиманской тундре встречено сотенное стадо уже пере-
линявших гусей (Гладков, 1951). На время линьки собираются крупными
стадами.
Питание. По характеру питания является наземным гусем. На зи-
мовках в Закавказье вместе с белолобым гусем держится на сухих простран-
ствах солянковых степей и в полупустынных местах (Исаков и Воробьев,
1940). На Атреке день проводит и кормится на зеленых лугах левобережья
(Житников, 1900).
Весною в Приморье корм состоит преимущественно из свежей зелени
растущих на пригревах трав (Шульпин, 1936) и на Анадыре из стеблей,
молодых побегов и листьев травянистых растений (Портенко, 1939).
Полевые признаки. Пискулька похожа на белолобого гуся, но мельче
и легко благодаря этому различима в полевых условиях. На лету, помимо
более мелких размеров, бросается в глаза ее большая острокрылость.
ПИСКУЛЬКА 311
При местных перемещениях летит беспорядочной гурьбою с беспрестан-
ными криками, но при дальних перелетах выстраивается или косой, или на-
клонной линией, или углом. С другими гусями не смешивается (Алфераки,
1904). Ходит хорошо, бегает же настолько проворно, что даже молодого
или линного гуся поймать не всегда удается. На зимовках пискульки ночуют
в тростниках. В предрассветных сумерках пробуждаются с громкими кри-
ками, поднимаются все сразу и, сделав несколько кругов над местом ночев-
ки, летят к местам кормежек. Днем около 2—3 час. летят, также очень
высоко, к местам водопоя с огромной высоты стремительно по вертикали
спускаются вниз. У водоема снова взмывают слегка вверх и, сделав полу-
круг, садятся на мелких местах. Вообще очень осторожны, но в различных
местах проявляют разную степень осторожности. Там, где они не подвер-
Рис. 53. Пискулька
гаются .преследованию или где их мало вообще преследуют, проявляют
значительное любопытство, и в предрассветном тумане, например, спо-
собны легеть на всякий незнакомый предмет или даже на незнакомый
шум. Но в местах, где пискульки являются предметом постоянного и си-
стематического преследования, наоборот, осторожны до чрезвычайности.
На Новой Земле для гнездованья выбирают места, мало посещаемые чело-
веком, самые гнезда устраивают в трудно доступных уголках, и притом
очень хорошо их маскируют. Для линьки перемещаются в устья рек или на
горные озера, устраиваясь в таких местах, где их трудно ловить. В случае
преследования во время линьки на воде они рассыпаются в разные стороны,
быстро уплывают, а иногда погружаются в воду, выставляя наружу только
голову. Если линных выгнать на берег, они сначала чрезвычайно быстро
разбегаются в разные стороны, а затем очень искусно затаиваются среди
камней или кочек (Кузякин). Тонко реагируют на состояние погоды и в ясные
лунные ночи ведут себя оживленно, все время перекликаясь. В дождливые
ночи держатся молчаливо и иногда даже не летят на ночлег в тростники,
оставаясь у мест кормежек (Житников, 1900). Голос пискульки не так гро-
мок, как у белолобого гуся, но по тембру выше и отличается особой пискли-
востью. Его можно передать как «клик»... «клик» «юиилью-юильо», также
«тье-тьо». Иногда слышно шипенье: «кши-а-йяк». У молодых особей писк
«ки-ки-ки».
Экономическое значение. Как промысловая птица, пискулька имеет не-
сколько меньшее значение, чем белолобый гусь, и добывается вместе с пос-
ледним главным образом путем отстрела, но местами (например, на Новой
Земле) охота на нее не упорядочена и ее добывают от прилета до отлета. На
312 ОТРЯД ГУСЕОБРАЗНЫЕ
зимовках является объектом спортивной охоты. Вобщем, экономическое
значение этой птицы не велико, во-первых, вследствие относительно малой
их численности, и, во-вторых, вследствие трудности их добычи.
Описание. Размеры. Размах самцов A) 1340, самок B) 1215 и 1257 мм.
Длина тела самцов A) 615, самок B) 595 и 615 мм. Длина крыла самцов E) 381—412,
самок 10) 350—395, в среднем 391,6 и 370,8 мм. Длина клюва самцов E) 31—35,5, самок
A0) 30—35,3, в среднем 33,04 и 32,22 мм. Наибольшая высота подклювья у самцов E>
4,2—4,7, у самок A0) 3,8—5, в среднем 4,46 и 4,2Й мм. Вес самцов и самок 1,6—2,5 кг.
Сложенные крылья сильно выдаются за концы рулевых. Вокруг глаза оголенное про-
странство.
Окраска. Пуховик. Сверху темносеро-бурый. Через глаз идет темная продоль-
ная полоса. Лоб до уровня задних краев глаз зеленовато-желтый. Низ такого же цвета.
Клюв темный серовато-бурый. Ноги грязножелтые с зеленоватым оттенком.
Молодая птица в первом пере. Тона оперения тусклые. На лбу по границе с
клювом черные перья. Низ дымчато-бурый с грязножелтоватыми концами перьев без следов
черных пятен. Клюв грязнорозовато-серый. Ноги грязножелтые с оранжевым оттенком.
После первой осенней линьки на лбу начинают появляться отдельные белые перья (Суш-
кин). После второй осенней линьки белая лобная полоса на границе с верхней челюстью
получает примесь черных перышек. У вершины подбородка появляется небольшое белое
пятно. На брюхе небольшие черные пятна. Вокруг глаза бледножелтоватое вздутое
кольцо. Окончательный наряд надевается, повидимому,только после четвертой осенней
линьки.
Взрослые самец и самка. Верхняя сторона в общем буровато-серая, нижняя—
белая. Голова темнобуровато-серая, чернеющая на границах с белым оперением. Лобная
полоса у основания надклювья белая шириною 20—30 лик. Спина буровато-серая, поясница
аспидно-серая. Мелкие кроющие крыла пепельно-голубоватые с серым оттенком. Средние
такого же цвета, но вершинная половина каждого их пера с буроватым налетом. Большие—
бурые с узкими белыми вершинами. Наружные первостепенные маховые с темнеющими
вершинами. Внутренние первостепенные и внешние второстепенные маховые буровато-
черные и на сложенном крыле дают буровато-черное зеркальце. Стержни всех маховых
белые, чернеющие к вершинам. Зоб, грудь и бока тела буровато-черные со светлыми кай-
мами перьев. На груди неправильные черные пятна. Брюхо и подхвостье белые. Рулевые
черные с белыми вершинами и белыми боковыми окаймлениями.
12. Гуменник Anser fabalis Latham
Anas Fabalis. Latham. Gen. Syn.f Suppl., 1, стр. 297, 1787, Британия.
Русское название. Гуменник—русское народное название для всех форм этого
вида, связанное с появлением этих гусей во время пролета у гумен и на пашнях.
Распространение. Ареал. От Гренландии до Анадырского края; в ря-
де мест на север доходит до побережий и островов Ледовитого океана, на юг
в западной части ареала примерно до 60° с. ш., в восточной—до южной грани-
цы тайги. Зимою у берегов Европы, в Средней Азии, Китайской Народной
Республики и Японии.
Биотоп. Травянистые участки тундры, открытые пространства лесо-
тундры или глухие таежные места около водоемов.
Подвиды и варьирующие признаки. Наиболее существенными признаками для раз-
личения подвидов являются форма и соотношения частей клюва. Большинство же дру-
гих признаков—размеры различных частей тела, окраска клюва, окраска лап и т. п.*
принимавшихся прежними орнитологами, описавшими многочисленный ряд форм, пред-
ставляют собой признаки неустойчивые, проявляющиеся в разных местах в пределах
ареала всего вида, и поэтому не могут быть приняты во внимание. Из большого числа
описанных форм после критического анализа всего накопившегося материала (Дементьев,
1936) можно выделить только четыре1: 1) А. /. brachyrhynchus В a i 1 1 о п, 1833, наиболее
диференцированную форму, занимающую острова крайней западной части ареала, отли-
чающуюся мелкими размерами и розовато-мясной окраской середины надклювья;
1 Описанного из Гренландии короткоклювого гуменника Anser fabalis flavirostris
Dalgety et scott 1948 мы склонны толковать» как уклонение формы Anser fabalis
brachyrhynchvs, у которого неоднократно отмечалось пожелтение цветной перевязи клюва.
ГУМЕННИК
313.
2) А. /. fabalis Latham, 1787, довольно неустойчивую по своим признакам, более
крупную форму, распространенную от Кольского п-ова до Хатангско-Ленского водораз-
дела; 3) А. /. serrirostris Swi n h о е, 1871, крупную хорошо выраженную форму
тундры от Хатангско-Ленского водораздела до Анадыря и несколько южнее его по побе-
режью; 4) А. /. sibiricus А 1 р h e r a k i, 1904, крупную устойчивую форму тайгй Восточ-
ной Сибири.
Западный тундровый гуменник Anser fabalis fabalis Latham
Синонимы. Anser Middendorfii (parti m). Северцов. Верт. и гориз. распр. Турк.
Жив. стр. 70, 149, 1873.— A riser (Melanonyx) carneirostris. Бутурлин. Пс. и Руж. Ох.,
февр.-апр., 1901, стр. 28.—Anser neglectus. S u s h k i n, Bull. Brit. Orn. CL, V, стр. VI,
1895.—Anser serrirostris rossicus. Бутурлин. Поли. опр. пт. СССР., II, 1935, стр. 89.
Распространение. А р е а л. На севере от Финмаркена на Скандинавском
п-ове (откуда длинным языком на юго-запад спускается по внутренним озерам
к болотам Швеции) по Кольскому п-ову, Канину, южному о-ву Новой Земли,
Карта 33. Распространение гуменника Anser fabalis
J— А. /. fabalis, 2—А. /. serrirostris, 3—А. /. sibiricus, 4—А. /. bra-
chyrhynchus; a—граница гнездовий, г—зимовки
Ямалу, Гыдану, и Таймыру до водораздела между Хатангой и Леной. Но
ни в Приполярном Урале, ни в районе мыса Челюскина на Таймыре на
гнездовье не обнаружен. На юге от озера Стур в Швеции, от 62° с. ш. в Фин-
ляндии и Карело-Финской ССР, по водораздельным пространствам Северной
Двины и Печоры, с одной стороны, и бассейнам Волги, с другой, до 62° с. ш.
у западного подножия Урала; гнездится в Средней Сибири, на Елогуе и ниж-
нем течении Подкаменной Тунгуски. Отсюда на север до нижнего течения
Нижней Тунгуски далее на северо-восток до устья Мойеро и по водоразделу
между Хатангой и Леной. Но граница от Елогуя до Ледовитого океана,
собственно говоря, довольно условна. С одной стороны, на Таймыр заходят
восточно-сибирские тундровые гуменники, с другой—европейские доходят
до Лены. В бассейны же Елогуя, Подкаменной и Нижней Тунгуски вклини-
ваются еще и таежные гуменники и в общем на этом пространстве встреча-
ются особи всех трех видов и ряд переходов между ними. Отдельными
островами, отъединенными от общего ареала западного тундрового гуменни-
ка, являются его нахождения на гнездовье на о-веСарема, в районе Чудского
озера (что, однако, еще требует подтверждения) и в системе озер Чаны
в Западной Сибири (Формозов, 1934).
314 ОТРЯД ГУСЕОБРАЗНЫЕ
Зимует на побережьях Англии, Северной и Южной Европы, частично
и в небольшом количестве на Черном море и в Средней Азии. Отсутствует
на зимовках в Закавказье и на Каспийском море.
Характер пребывания. На севере СССР—летняя гнездя-
щаяся птица; в большинстве районов средней полосы—пролетная, на юге—
кроме того, местами зимующая. В конце февраля—начале марта покидает
места зимовок и появляется в это время в низовьях Днестра (Остерман, 1909),
10—20 марта показывается на территории Украины и Белоруссии (Тугари-
нов, 1941), в конце марта на Волге выше Сталинграда и быстро продвигается
по Средней Волге- Гуси, зимовавшие в Средней Азии, появляются на Нижней
Сыр-Дарье 15 марта, затем пролет здесь продолжается до 15 апреля (Спанген-
берг и Фейгин, 1936); большинство же уже в конце марта отлетает из Средней
Азии. 17 марта—29 апреля, в среднем же за 15 лет 4 апреля передовые дости-
гают Чудского озера (Нестеров и Никандров, 1913), 1—5 апреля средних
областей европейской части СССР и в некоторые годы 1—5 апреля бассейна
р. Вятки (Круликовский, 1913), но в дальнейшем миграции приостанавли-
ваются, и гуменник следует ходу весны, продвигаясь вперед по мере того,
как вскрываются реки. В связи с возвратом холодов передовые стаи, слишком
продвинувшиеся вперед, нередко принуждены бывают возвращаться обрат-
но, а местами даже делать более или менее длительные остановки. В таких
случаях день они проводят на разливах, а вечерними и утренними зорями
вылетают на пастьбу на вышедшие из-под снега озимые хлеба (Мензбир, 1895).
13—17 апреля долетают гуменники до территорий Латвийской, Литовской
и Эстонской ССР (Лоудон, 1912), 14—20 апреля до Ленинградской обл. (Биан-
ки, 1907). 24 апреля передовые показываются в районе Московского моря,
а 25 апреля—3 мая здесь проходит валовой пролет (Исаков и Распопов, 1949);
в конце апреля они уже в бассейне Камы в районе Чердыни, где последние
пролетают 9—10 мая (Тугаринов, 1941). В середине апреля идет пролет по
Чкаловской обл., а 10—30 апреля по Эмбе и Ори, откуда гуменники далее
следуют Уралом. В начале мая передовые долетают до побережья Ледови-
того окена на Мурмане, после 15 мая показываются в низовьях Печоры,
16—18 мая (по наблюдениям Успенского) появляются на южном о-ве Новой
Земли. В Тиманской тундре первые отмечены 23 апреля 1938 г., до 5 мая
шел слабый пролет, 9—12 мая летели во множестве, 21 мая стало заметно
меньше, а 29 мая исчезли последние (Гладков, 1941). 16 мая отмечены на
Ямале, где 23—25 мая наблюдался валовой пролет. 13—15 мая передовые
появились на Гыданском п-ове, но 27 мая в связи с возвратом зимы наблюдался
обратный лёт, и гуси здесь снова показались лишь 5 июня (Наумов, 1931);
3—5 июня передовые показываются на Таймыре. В 20-х числах мая первые
прилетают в устье Хатанги, а в конце мая—начале июня здесь появляются
они уже массами (Тугаринов, 1941). На Новой Земле проводят 105—145 дней;
на Таймыре же остаются только 90—102 дня. Первые отлетающие и с Север-
ного о-ва Новой Земли (Тугаринов, 1941) и с Южного (по наблюдениям Успен-
ского) отмечены 1 сентября. Последние наМаточкином Шаре отмечены 18 сен-
тября и в Грибовой губе 10 октября 1949 г. (наблюдения Успенского). В начале
сентября первые отлетают с Гыданского п-ова, 15 сентября наблюдался мас-
совый отлет, 30 сентября—последние (Наумов, 1931). Последние с Таймыр-
ского озера исчезают 18 сентября, из низовьев Хатанги 22 сентября (Тугари-
нов и Толмачев, 1928). В начале сентября идет хорошо выраженный пролет
в районе Салехарда (Дерюгин, 1898), 15 сентября в большом количестве по-
являются на Мурманском побережье, затем часть отлетает, но последние за-
держиваются до 15 ноября (Тугаринов, 1941). 22 октября—14 ноября идет
пролет на взморье в районе Ленинграда (Раснер, 1913), 10—11 ноября появ-
ляются в Прибалтийских Советских Республиках (Лоудон, 1909), 1—18
октября пролетают у северного края Московского моря (Исаков и Распо-
ГУМЕННИК 315
пов, 1949), 16—20 октября появляются в бассейне Вятки (Круликовский,
1913), 28 сентября передовые в Ветлужском районе и 7 октября здесь же
валовой пролет (Воронцов, 1935). 29 сентября—23 октября идет пролет по
Московской обл. (Поляков, 1910), в конце сентября, в октябре и начале нояб-
ря пролетают по средней полосе европейской части СССР (Тугаринов, 1914);
в то же время пролет идет и по Украинской ССР, откуда последние исчезают
к 15 ноября. 10—21 октября наблюдался пролет на озере Ак-Часты-Куль
с Ори на Эмбу (Сушкин, 1918). В то время, когда основная масса европей-
ских гуменников находится еще далеко на севере, передовые уже долетают
до мест зимовок, и, например, в Крыму, где небольшая часть их остается
в ряде случаев и на зиму, первые стаи появляются 30 сентября (Воронцов,
1937). Ввиду того что детальные наблюдения ряда последних лет в Закав-
казье показали полное отсутствие гуменников и в бассейне Куры и на Кас-
пии, трудно допустить, что гуси с Эмбы летят именно на каспийские зимовки,
а весною возвращаются с них через Эмбу же, как это принималось ранее
{Сушкин, 1908). Скорее всего—это крайнее юго-восточное отклонение птиц,
движущихся из низовьев Оби в бассейны Волги и Урала. Долетая до Ори и
озера Ак-Тасты-Куль гуменники летят по Эмбе и затем через Северный Каспий
выходят к Крыму и летят далее на юго-запад. Весной же ими проделывается
обратная дорога. Однако это предположение нуждается в прямых доказатель-
ствах, которые пока отсутствуют. Часть птиц проводит зиму в бассейне Сыр-
Дарьи, где гуменники встречались неоднократно в ноябре—феврале (Заруд-
ный 1910 и 1915). Во время своих осенних и весенних передвижений, пролетая
днем и ночью гуменники следуют от одного места остановки до другого через
речные долины, водоразделы, лесные пространства, не приурочивая своих
миграций к каким-либо речным долинам, но лишь частично используя их,
если такие долины представляют им удобства для отдыха и кормежки.
Общее направление миграций в европейской части СССР осенью идет на юго-
запад, весною на северо-восток и север.
Биотоп. Гнездовой—различные ландшафты травянистой или ку-
старниковой тундры, главным образом по соседству с водоемами, хотя не
всегда в непосредственной с ними близости. В лесной полосе глухие лесные
ручьи, широкие речные долины и острова на озерах. Зимний—открытые места:
степи, поля, береговые отмели.
Численность. Хотя за последние десятилетия этот гуменник
значительно уменьшился в количестве, но в глухих и малолюдных местах
крайнего Севера еще и в наши дни является весьма обыкновенной птицей.
Экология. Размножение. Первый период жизни на гнездовых ме-
стах от прилета до времени откладки яиц выяснен еще недостаточно. На Гы-
данском птове неоднократно сейчас же после прилета замечались групповые
полеты птиц. Обычно можно было видеть трех гуменников, гоняющихся
друг за другом в воздухе, то падающих вниз, то снова взмывающих кверху.
Остается неясным, воздушные это игры (Наумов, 1931), или разбивка на па-
ры, когда два самца преследуют самку (Тугаринов, 1941), или же попытки
холостых самцов отбить самку у семейных. Во всяком случае, появившись на
местах гнездовья с первыми проталинами и первыми лужицами воды еще
на льду, гуменники быстро приступают к гнездовью. Весна в это время еще
чрезвычайно неустойчива, температура колеблется между —3° и +2° С, еще
идет пролет более северных особей (Житков, 1912), а семейная пара уже
приступает к устройству гнезда. Гнезда помещаются на сухих незаливае-
мых местах речных долин, на низменных участках моховой тундры у края
речных долин, на бугорках или холмиках среди тундры или среди низких
и редких ивнячков. Облюбованное место утаптывается или на нем делается
небольшое углубление. Гнездо представляет собою аккуратную постройку,
в его основание укладываются различные сухие растительные остатки, бока
316 ОТРЯД ГУСЕОБРАЗНЫЕ
же возводятся из перьев и пуха, выщипываемых самкою из своей хлупи.
В противоположность некоторым другим гусям, гнездо строят и самка и самец,
вместе.
Кладка состоит из 3—6, чаще из 3—4, нежнопалевых яиц со слабо-
блестящей зернистой скорлупой. Размеры яиц: B4) 74,5—87x42—59, в сред*
нем 81,9x56,2 мм. Вес 10,74—13,38, в среднем 12,54 г. Первые яйца оказы-
ваются отложенными через 15—20 дней после появления гуменников в ме-
стах гнездовья, и на Мурманском побережье их находят 1—10 июня, на Но»
вой Земле 10 июня (Горбунов, 1925). Здесь же 4 насиженных яйца были най-
дены 2 июля (Бианки, 1902) и, очевидно, запоздавшие с совершенно готовыми
эмбрионами—26 августа (Горбунов, 1925). Первые 2 отложенных яйца на Яма-
ле найдены также 10 июня (Житков, 1912), а 13—14 июня у некоторых самок
в яйцеводе были яйца, еще покрытые только белковыми оболочками. Число
яиц здесь 5—7, чаще 6. В Гыданской тундре гнездо с 6 яйцами было найдена
16 июня (Наумов, 1931). Таким образом, начало кладки на Крайнем Севере
приходится примерно на одинаковые сроки.
Насиживает одна только самка, около 25 дней. Самец постоянно нахо-
дится поблизости от гнезда и предупреждает самку об опасности. Иногда сам-
ца приходится видеть рядом с гнездом, но не на гнезде. Слетают птицы с гнез*
да и от него только в крайних случаях и предпочитают маскироваться, при-
жимаясь к земле и вытянув палками шеи.
При выклевывании птенцов самка оттаскивает скорлупу в сторону.
Гнезда с только что выклюнувшимися пуховичками найдены на Ямале
8 и 10 июня (Житков, 1913), на Гыданском п-ове—в середине июля. Первых
пуховичков на Новой Земле находили 15 июля (Горбунов, 1945), а 8—
10-дневных на КостиномШаре 19 и 20 июля. т. е. во всех этих местах прибли-
зительно в одно и то же время. В низовьях Печоры выклевывание начинается
раньше, и на Вухтуле пуховичков находили уже 26 июня, покрупнее—
7 июля, а больших, но еще нелетных птенцов у Савиногорска 21 июля (ДАю-
ховский, 1933). В устье Енисея молодые появляются с конца первой декады
июня до середины июля.
После того, как выводок обогреется и обсохнет, вся семья переселяется
на влажные травянистые луга, в речные долины или на острова, предпочитая
те места, где имеется кустарниковая растительность куда птицы спасаются
в случае опасности. Потревоженные члены семьи ведут себя по-разному.
Птенцы затаиваются в траве или кустарнике, взрослые же отлетают несколь-
ко в сторону и беспокойно бродят здесь; по миновании же опасности возвра-
щаются к выводку.
Рост птенцов идет довольно быстро. Пуховики ростом с чирка на Мур-
манском берегу наблюдались в 20-х числах июля. Значительно оперившиеся^
но еще нелетные молодые под Салехардом встречены 28 июля. Чем ближе
подходит время линьки,тем семья все чаще и чаще начинает держаться на воде.
Линька. У молодых, условно, две линьки в году: летняя полная
и частичная осенне-зимняя, протекающая медленно с сентября—октября
по март или по самое начало апреля. У взрослых, повидимому, одна линька
в году.
Молодые вступают в свою первую осеннюю линьку с конца сентября—
начала октября и заканчивают ее к апрелю. Первыми летом начинают линять
холостые старые и второгодние неполовозрелые птицы. Для линьки они пере-
летают в более или менее отдаленные безопасные места и, например, с Южного
о-ва Новой Земли ежегодно перелетают на Северный. Холостые гуси начинают
собираться в стаи в конце июня, затем улетают в места линьки и уже в начале
июля у них начинается смена маховых.
Взрослые семейные начинают линять не раньше, чем появятся пухович-
ки. К началу линьки они перебираются на водоемы и здесь в ряде случаев
ГУМЕННИК 317
соединяются по нескольку семей вместе. Семейные гуси начинают терять
маховые с 15—20 июля.
Линька начинается с одновременного выпадения маховых. Вместе с ма-
ховыми сменяются также верхние и нижние кроющие крыла. Вслед за ними
начинают линять лопаточные перья. Уже после того как начнут расти новые
маховые, происходит смена рулевых, в ходе линьки которых особых зако-
номерностей не находят (Тугаринов, 1941). Одновременно со сменой рулевых,
иногда позже их, линяют перья надхвостья. Затем сменяются контурные
перья нижней части туловища. Далее идет линька боков тела, и последними
сменяются перья шеи и головы. Примерно месяц спустя после начала линьки
маховых гуменники вновь получают возможность летать.
На Новой Земле (по наблюдениям Успенского) линька, очевидно холо-
стых, отмечена 2 июля, но, судя по находкам выпавших маховых, она нача-
лась несколько раньше. 20 июля эти птицы еще были в линьке, 12 августа
в море встречены полулетные птицы, а 19 августа все гуменники уже летали.
На Канине 16 июля 1914 г. линяли семейные гуси, при которых были
еще нелетные молодые, 21 июля семейные при пуховичках линяли на Ха-
танге (Скалой, 1937). На Ямале с конца июня и до 7—8 августа встреча-
лись отдельные стада еще на крыльях, 15 августа все сплошь были в линьке
<Житков, 1912). Для Таймыра указываются несколько иные сроки: 30 июля
половина стад гуменников летная, 4 августа в смешанных стаях почти все
лётные (Тугаринов и Толмачов, 1928). Сроки линьки оказываются различны,
и, повидимому, в разных широтах и в различные годы оказываются не оди-
наковыми.
В общем же смена первостепенных и второстепенных маховых происхо-
дит примерно в две-три недели, затем проходит некоторое время, пока ма-
ховые не окрепнут окончательно, и даже в тех местах ареала, где линька за-
канчивается поздно, гуменники к концу августа снова подымаются на крыло.
Обычно к концу линьки взрослых отрастают и крепнут маховые и у молодых.
Они также получают возможность летать, и семейные связи к этому времени
нарушаются (Тугаринов, 1941).
Вслед за полной вскоре начинается частичная осенне-зимняя линька,
во время которой сменяются контурное оперение и рулевые. Линька эта
заканчивается только на зимовках.
Питание. Основная пища европейских гуменников состоит из зеле-»
ных частей и ягод различных наземных растений. В гнездовых местах они
поедают всевозможные травянистые растения, различные ягоды и, в особен-
ности, голубику. Во время осенних перелетов часто пасутся на зеленях, на
убранных хлебных полях, а зимою на рисовых полях и местах, бывших под
огородными культурами (Алфераки, 1904). На весеннем пролете в области
Московского моря их пища состоит из побегов озимой ржи D9%), корешков
полевых растений A8%), красной овсяницы (8,1%), злаков и осок, ягод
клюквы B4%) и голубики, за которыми они летают на междуречья, из семян
ржи, овса, гречихи и минеральных примесей (Исаков и Распопов, 1949).
Пища пуховичков состоит из различных водяных и наземных насекомых,
моллюсков, ракообразных и даже из рыбьей икры (Алфераки, 1904).
Определенных данных о врагах гуменников не имеется и можно предпо-
лагать, что их кладки и птенцов уничтожают крупные чайки, поморники,
песцы, а в речных долинах и лисицы.
Хозяйственное значение. • В тундре гуменники являются предметом
интенсивного промысла. В период гнездования собирают их яйца, летом во
время линьки их промышляют путем «гусевания» (на Канине в середине июля;
Дементьев, 1935), и местное население таким путем запасает мясную пищу
и для себя и для своих ездовых собак. При удачной охоте артель из 15 человек
добывает по нескольку сот гусей в день. Гуси, очищенные от пера и пуха,
318 ОТРЯД ГУСЕОБРАЗНЫЕ
засаливаются, вялятся, иногда же замораживаются в особых ямах прямо в
грунте (в вечной мерзлоте). Однако такое варварское истребление путем так
называемого гусевания подрывает основной костяк местных стад гусей и мед-
ленно, но верно ведет к полному их истреблению. Добывают гусей также
путем отстрела с промышленными целями, а также при спортивной охоте.
Полевые признаки и поведение. Крупные серой окраски птицы, за исклю-
чением гнездового периода, держащиеся стаями. На миграциях летят как
днем, так и ночью, чаще всего углом, но иногда и волнистой линией. Летя-
щие впереди птицы беспрерывно сменяются. Во время весенних миграций
на севере иногда плывут вниз по реке, если последняя соответствует направле»
Рис. 54, Западный тундровый гуменник
нию их полета (Тугаринов, 1941). Голос гогочущий, в общем довольно разно-
образный. Он звучит как «кайяг», «каййк», также «гаг㧻, «клиррьэ».
На взлете слышно «рат-рат». На лету во время миграций глухое «гок». Предо-
стерегающий крик звучит как «трэнг», иногда как краткое ускоряющееся
«гагагага». При раздражении шипит. Отличаются крайней осторожностью,
особенно по отношению к охотнику. При стайном образе жизни всегда име-
ются сторожа, подающие при опасности голос и взлетающие первыми, за ними
следует и вся остальная стая.
Вылинявшие особи собираются в стаи, состоящие из птиц разных вы-
водков и возрастов; вслед за этим начинается более или менее продолжи-
тельный период кочевок. День птицы проводят на тихих речных отмелях
или на больших озерах, к вечеру летят на обильные кормами места и здесь
наедаются досыта. Ночью дремлют в этих же местах, перед утренней зарей
кормятся и затем летят на раз выбранное место дневного отдыха. Такой
образ жизни птицы ведут до отлета, который начинается вместе с наступле-
нием морозов, побивающих растительные корма гусей.
Описание. Размеры и строение. Размах самцов и самок 1524—1651 мм.
Длина тела самцов и самок 730—900 мм. Длина крыла самцов П5) 404—482, самок A5)
402—460, в среднем 444,2 и 430,4 мм. Длина клюва самцов C2) 51,0—67,5, самок B6)
52,5—60, в среднем 61,2 и 56,6 мм. Высота нижней челюсти у самцов C1) 5,5—11, у са-
мок B1) 6—8,5, в среднем 7,5 и 7 мм. Форма клюва чрезвычайно изменчива, имеются все
переходы от короткого высокого у основания до удлиненного, при основании относительно
низкого. Сильно меняется и изгиб нижней челюсти.
ГУМЕННИК 31 ?
Окраска. Пуховик. Сверху оливково-бурый с более светлыми краями кры-
льев и желтоватыми пятнами позади крыльев. Брюшная сторона светлая, желтоватая.
С возрастом спина темнеет, брюхо же становится светлее. )Келтые пятна за крыльям»
исчезают. Голова дольше сохраняет птенцовый характер.
Молодая птица. С буроватыми ободками перьев на верхней стороне и с буро-
ватыми кроющими крыла. Голова и шея с рыжеватым оттенком. Боковые рулевые светло-
серые. На груди беловатые каймы перьев. После первой осенне-зимней линьки общая
окраска темноватая. Каймы перьев тусклые, рыжеватые. На шее резкий рыжеватый
оттенок. На относительно коротком клюве еще мало желтого. После второй осенней
линьки окраска становится несколько светлее, а клюв желтее. Окончательный взрослый
наряд и окраска перевязи клюва приобретаются только после третьей осенне-зимней
линьки.
Взрослые самец и самка. Спина темнобурая с легким серым налетом и свет-
лыми ободками перьев, низ спины потемнее, надхвостье белое. Маховые буровато-черные
с белыми стержнями. Верх шеи темнее спины, голова чуть посветлее. Низ шеи светло-
бурый. Грудь светлобурая со светлыми ободками перьев. Бока темнее, но с такими же
ободками, дающими поперечные светлые полосы. Брюхо и подхвостье белые. Перевязь
на клюве желто-оранжевая, розоватая, телесного цвета или же красноватая. Также
варьирует по окраске и цвет ног. Радужина ореховая.
Восточносибирский тундровый гуменник Anser fabalis serrirostris
Swinhoe
Anser segetum var. serrirostris. S w i n h о e. Proc. Zool. Soc. Lond., стр. 417, 1891, Китаи
Синоним. Melanonyx segetum anadyrensis. Бутурлин. «Наша охота», 1908>
вып. XI, стр. 21—28, Анадырь.
Распространение. Ареал. Населяет крайний север Сибири от Хатанг-
ско-Ленского междуречья до Анадыря. На Чукотке доходит только до 180° в.д.
Не найден на островах Ледовитого океана в пределах Восточной Сибири.
На крайнем востоке гнездится в узкой прибрежной полосе от Анадыря до
перешейка Камчатки и далее по побережью Охотского моря до района Охот-
ска. Южная граница проходит по северной окраине лесотундры.
Зимою в очень небольшом количестве встречается в Японии (Ян, 1942),
обычен в северной части Китайской Народной Республики и на юг доходит
до Шанхая и Амоя.
Характер пребывания. Гнездится на крайнем северо-восто-
ке Сибири, пролетает через континент и вдоль тихоокеанского побережья
Сибири, в качестве крайне редко зимующей отмечен на Анадыре (Портенко,
1939).
Весной появляется в Приморской обл. между 20—31 марта, в районе
Якутска между 19 апреля—1 мая; массовый пролет здесь идет 20—23 мая
(Воробьева, 1928). Первые у Верхоянска 7—12 мая, валовой пролет 23 мая
(Ткаченко, 1937). В Коряцкой Земле первые 1 мая, валовой пролет 20—30
мая, последние 2 июня (Волков и Дементьев, 1948). Первые на Анадыре
9 мая, в Крестовой бухте 13 мая, у мыса Беринга 18 мая; валовой пролет
20 мая (Белопольский, 1933). В области тундры появляется в устье Индигир-
ки 13 мая, под Нижне-Колымском 10 мая; массовое появление приходится
на конец мая (Тугаринов, 1941). В окрестностях Охотска остаются на лето
холостые особи (Харитонов, 1915).
Отлету предшествуют местные более или менее отдаленные перекочевки.
Из тундры отлетают 1—5 сентября, последние задержавшиеся наблюдаются
15—20 сентября. В Коряцкой Земле осенний пролет отмечен 1—30 сентября
(Волков и Дементьев, 1948), в районе Якутска задерживаются до 25 сентя-
бря (Воробьева, 1928). В окрестностях Верхне-Колымска встречаются до
15 октября при температурах —20°С (Черский). В более южных областях
Сибири во время пролета делают длительные остановки на сентябрь и даже
.320
ОТРЯД ГУСЕОБРАЗНЫЕ
до начала октября. По южной окраине Сибири пролетают с конца сентября
сю конец октября.
Биотоп. Открытая тундра с озерами, речные долины с островами
я многочисленными рукавами, отдельные небольшие речки в тундре и лесо-
тундре, иногда верхнее течение рек.
Численность. Местами в тундре многочислен, вообще же обык-
новенен на крайнем севере Восточной Сибири.
Экология. Размножение. Пары заметны сейчас же по прилете.
Для гнезда выбирается небольшое сухое пространство среди низменной тунд-
ры или речной долины, на котором в небольшом естественном углублении
«строится гнездо. Последнее у впервые строящих птиц довольно примитивно
и устраивается из сухих стеблей трав,
лишайников и пуха, выщипываемого из
брюха. У взрослых птиц в основание
гнезда укладываются мох и сухие рас-
тительные материалы, лоток выстилает-
ся более нежным материалом, края же
его окружаются пухом. Самка насижи-
вает одна, самец все время до появле-
ния пуховичков находится поблизости
от гнезда. Кладка состоит из 5—6 яиц.
Первые отложенные яйца у Русского
Устья 1 июня, полная кладка из 6 яиц
в устье Лены найдена 12 июня, 4 слегка
насиженных яйца в Хатангском заливе
в устье р. Большой Балахны 7 июля
1934 г. (Спангенберг), сильно насижен-
ные яйца восточнее устья Колымы на
Большом Барановом мысу 6 июля, на
Анадыре яйца находят с 18 июня. Но
на побережье, в Коряцкой Земле, клад-
ка из 5 яиц найдена значительно рань-
ше—20 мая 1941 г. (Волков и Дементьев,
1948). Соответственно этому здесь рань-
ше появляются и пуховички—первые в
1941 г. найдены 23—24 июня. 2—3-днев-
ные пуховички на Алазее найдены 7 и 16 июля, в дельте Колымы—8 июля.
На Индигирке первые выводки появляются только в середине июля Рост
молодых идет довольно быстро. Птенцы длиною 321—324 мм в дельте Колы-
мы найдены 16 июля, длиною 375—380 мм—20 июля. В районе Индигирки
20—28 августа они почти достигают размеров взрослых. На Анадыре в заливе
Креста 19 августа найдены такие же крупные молодые. Во время роста моло-
дых старики остаются с ними и с ними же линяют.
Линька. Общий характер линьки, как и у западного тундрового
гуменника. Первыми вступают в линьку прошлогодние неполовозрелые птицы
и перед началом ее собираются на малодоступных илисто-песчаных косах,
ца берегах озер в открытой тундре, также в приморской тундре (Портенко,
1939). С окраин материка для линьки гуси перелетают на острова Ново-
сибирского архипелага. В Коряцкой Земле линька начинается в период
с 30 июня по 4 июля. Массовое выпадение маховых наблюдается 10—20 июля.
Весь период линьки крыльев продолжается около 3—4 недель и с 14 августа
уже заметны летающие стаи (Волков и Дементьев, 1948), но в дельте Инди-
гирки первые перелинявшие птицы добывались 7 августа, а на Анадыре пере-
движения перелинявших наблюдаются с 9—10 августа. Гнездившиеся взрос-
лые начинают линять только тогда, когда достаточно подрастут молодые,
Рис. 55. Клюв восточносибирского тунд-
рового гуменника (вид сбоку и сверху)
ГУМЕННИК
321
в общем только с августа. Первым вступает в линьку самец, а затем уже самка,
и линька гнездящихся птиц протекает разновременно (Портенко, 1939).
Питание. Основою питания являются различные травянистые расте-
ния, особенно в весенний период; осенью при урожае ягод последние начи-
нают преобладать в кормах восточносибирского гуменника—это голубика,
водяника и шикша (Волков и Дементьев, 1948). У молодых в желудке нахо-
дили остатки хвощей (колымские сборы Бутурлина).
Хозяйственное значение. Для местного населения Восточной Сибири
этот гусь имеет громадное значение: там, где он обилен, его заготовляют
в значительных количествах и для потребления в пищу человеком и для
корма ездовых собак. Собираются также яйца, перья и пух. Нет сомнения,
Рис. 56. Пуховой птенец восточносибирского тундрового гуменника
что сбор этого гуся необходимо регламентировать и постоянно иметь
в виду сохранение основного стада.
Полевые признаки. Те же, что и у европейского гуменника. В полевой
обстановке вместе с тем с трудом отличим от таежного гуменника, хотя все
же несколько мельче последнего и держится более крупными стаями.
Описание. Размеры и окраска. Длина тела самцов и самок 776—825 мм.
Длина крыла самцов B3) 422—524, самок A2) 422--491, в среднем 469,4 и 446,9 мм. Длина
клюва самцов B4) 59,2—71,5, самок (9) 58,5—66, в среднем 65,2 и 62,2 мм. Высота нижней
челюсти у самцов B4) 8—12,5, самок (9) 8,1—11,3, в среднем 10,3 и 9,4 мм. Подклювье
сильно изогнуто, массивно, наибольшая его толщина 8,7—13,3 мм. По своим размерам
восточносибирский гуменник несколько крупнее европейского, по окраске сходен с ним.
Что касается окраски клюва, то в некоторых случаях перевязь может быть красноватого
цвета, обычно же она желто-оранжевая. Ноги желтые или желто-оранжевые (Тугаринов,
1941).
Таежный гуменник Anser fabalis sibiricus Alpheraki
Melanonyx arvensis sibiricus. Алфераки. 1904. Гуси России, стр. 98, Be сточная Сибирь.
Синоним. Anser fabalis middendorfii. Северцов (partim). Вертикальн. и гориз.
распр. туркест. жив., 1873, стр. 70, 149.
Русские названия. Лесной гуменник—местное сибирское народное название.
Большой гуменник, рыжеголовый гуменник, таежный гуменник—книжные.
Распространение. Ар е а л. Населяет таежную зону Восточной Сибири.
На крайнем северо-западе ареала доходит до Таймыра и бассейна Хатанги,
21 Птицы СССР, т. IV
322 ОТРЯД ГУСЕОБРАЗНЫЕ
где смешивается с A. f. fabalis и Л. /. serrirostris, но здесь вместе с переход-
ными встречаются и типично выраженные особи таежного гуся (Скалой,
1937). Северная граница ареала идет вдоль лесотундры, но местами таеж-
ный гусь по долинам больших сибирских рек гнездится вплоть до их низовий,
где встречается вместе с Л. /. serrirostris. На востоке граница ареала не
доходит до Охотского моря, побережье которого занимает А. /. serrirostris,
но от Охотска до устья Уды, повидимому, распространен по побережью.
На крайнем юго-западе ареал образует выступ, от Саян до Алтая. Граница
ареала в районе западных Саян проходит по бассейнам Маны, Сисима, Тубы,
огибает Минусинскую лесостепь, охватывает Кузнецкий Ала-Тау, кроме
Горной Шории (Залесский, 1930), и отсюда к югу идет до Алтая, охватывает
его, и по истокам Кара-Иртыша и Кобдо также и Монгольский Ала-Тау.
Таежного гуменника нет в горах Танну-Ола и в Уряханской котловине. Но
далее он обычен в бассейнах Ий-Кема, Ха-Кема, Кара-Соулука, Аны, Кара-
тога и в Монголии в Хангае. Отсюда граница идет по Южному Прибайкалью,
по южным отрогам Яблоневого, Станового и Буреинского хребтов до устья
Уссури. Прежде гнездился и южнее—на оз. Ханка (Пржевальский, 1870),
но к 30-м годам текущего столетия исчез отсюда (Шульпин, 1936). От Уссури
граница проходит по бассейнам рек Селемджи и Уды до Охотского моря
(Золотарев, 1934). Таким образом, южная граница распространения таежного
гуменника совпадает отчасти с южной границей лесной зоны, отчасти с север-
ными окраинами Центрально-Азиатских степных плоскогорий В пределах
указанного ареала селится не сплошь: на Ангаре не встречен, в долине Под-
каменной Тунгуски найден только в самых ее верховьях; в долинах Ниж-
ней Тунгуски гнездится только местами, а в бассейне Хатанги только на
р. Мойеро.
Зимует в восточных провинциях Китайской Народной Республики, в.
Японии, залетает в Бирму; в позднеосенние и зимние месяцы несколько
раз был добыт в Средней Азии в Сар дабе в долине Ангрена (Зарудный, 1910).
Характер пребывания. Гнездящаяся птица тайги Восточ-
ной Сибири, на пролете встречающаяся от Байкала до Камчатки, Командор-
ских о-вов, Сахалина и северо-восточных частей Монгольской Народной
Республики. Но ни в Гоби, ни в других районах Центральной Азии на проле-
те не найдена.
Даты. Первые весенние в Даурии на Тарей-Норе отмечены 7 марта
(Радде, 1863), 13 апреля на Алтае на Телецком озере (Фолитарек и Демен-
тьев, 1938), 12—19 апреля в верховьях р. Томи (Хахлов, 1937), 15 апреля
на р. Иркут, в конце апреля в устье Амура, 5 мая на Шантарских о-вах
Щулькейт и Шульпин, 1937).
Данных об осенних миграциях мало. Из верховьев Томи отлетает 28 сен-
тября—7 октября (Хахлов, 1937). На Алтае первые передвижения отмечены
9 августа, настоящий пролет выражен только в октябре. Передовые отмечены
3 октября, массовый пролет на Телецком озере 7—12 октября, последние
25 октября (Фолитарек и Дементьев, 1938).
Биотоп. Открытые пространства среди тайги с озерами на них, обшир-
ные моховые болота, долины рек, субальпийские моренные плато с заболо-
ченной почвой, с болотами и озерами на них, заболоченные долины высоко-
горных рек с озерами, горные речки с галечниковыми или скалистыми берега-
ми, горные лесные озера. По вертикали идет до 2400 м над уровнем моря
(Сушкин, 1938).
Численность. В связи с незначительным количеством удобных
для гнездования мест среди глухой тайги численность таежного гуся не
велика.
Экология. Размножение. На места гнездовья прилетают парами
или стайками по нескольку особей. В тайге гнезда располагаются далеко
ГУМЕННИК
323
одно от другого, и вообще таежные гуси селятся здесь весьма разрозненно.
В горных странах, хотя и спорадично, располагаются л а гнездовьях чаще.
Гнезда строятся вблизи озер, болот, речек или же на островах и островках
среди водоемов. Количество яиц 5—7. Окраска их белая с легким желтоватым
оттенком или слоново-белая, но вскоре они загрязняются и покрываются
серыми или желтыми пятнами. Размеры G) 73,6—90x53,2—59 мм (Тача-
новский, 1893): C) 92,5x61,7 мм, вес в среднем 20, 58 г (Гебель, 1904);
Первые пуховички в Курайской степи найдены 5 июня (Сушкин, 1938),
на Вилюе 20 июня (Маак, 1854). Молодые с еще недоросшими маховыми в Ту-
винской автономной области встречены на р. Систикем 4 августа; достигшие
половины размеров взрослых в Во-
сточных Саянах по р. Казыр 7 авгу-
ста (Тугаринов, 1941).
Линька. У молодых, как и у
номинальной формы, две линьки:
полная летняя и частичная осенне-
зимняя. Взрослые же, повидимому,
линяют один раз в году. На Алтае
полная летняя линька начинает-
ся в конце июня. 29 июня в стае
из 50 особей на одном из высокогор-
ных плато летными оказались только
две птицы (Фолитарек и Дементьев,
1938). Таким образом, здесь гуси ли-
няют в стаях1, в тайге же, где гнез-
довья сильно разбросаны, во время
линьки птицы держатся семьями.
Как и у других подвидов гуменника,
у таежного первыми приступают к
линьке холостые птицы, семейные
начинают линять позже. У северной
границы ареала начало линьки при-
ходится на конец первой декады
июля, в конце второй декады июля
наблюдается разгар линьки. В Саянах
молодые с недоросшими маховыми найдены 4 августа, а 17 августа на Алтае
добыт молодой уже в свежем пере. Примерно в это же время вылинивают
и вырастают до нормы маховые и у взрослых птиц: 22 августа у взрослой
самки маховые выросли и окрепли, но остальное перо еще сменяется. Вся
спина, поясница и кроющие крыла, за исключением средних, уже новые.
Середина брюха уже в новом пере. Идет интенсивная смена перьев зоба.
На голове, шее и остальной части низа линька еще не начиналась
(Сушкин, 1938). Следовательно, и у этой формы линька чрезвычайно за-
тягивается и заканчивается только во время осенних миграций или даже
на зимовках.
Питание. Основной пищей являются травянистые растения. Во
время созревания ягод наблюдаются ежедневные кормовые вылеты на ягод.*
ники. Из ягод таежный гусь часто поедает голубику.
Хозяйственное значение. Вследствие чрезвычайной разбросанности
отдельных гнезд и того обстоятельства, что в большинстве случаев таежные
гуси держатся семьями, промышленного значения этот гуменник не имеет.
Добывается он только при случае.
Рис. 57. Клюв таежного гуменника (вид
сбоку и сверху)
1 Хотя возможно, что в данном случае наблюдалась стая холостых птиц, слетевшихся
в удобное для линьки место.
21*
324 ОТРЯД ГУСЕОБРАЗНЫЕ
Полевые признаки. Наиболее крупный из гуменников, чаще всего встре-
чается небольшими группами (семьями) и даже во время миграций не соби-
рается в крупные стаи (не более 20 птиц). На воде держится легко и хорошо
ныряет. Голос более хриплый чем у других гуменников. В остальном
узнается по признакам, характерным для гуменников вообще.
Описание. Равмеры и строение. Размах до 1390 мм. Длина тела самцов
и самок 780—930 мм. Длина крыла самцов (8) 440—558, самок D) 494—524, в среднекг
495,8 и 506,2 мм. Длина клюва самцов (8) 64—81, самок E) 66—79, в среднем 73,2 и 73,6 мм.
Высота нижней челюсти у самцов (8) 7—10,5, у самок E) 8,2—9,8, в среднем 8,7 и 8,9 мм.
Однако данных этих недостаточно, что видно по средним размерам самок, превосходящих
по размерам клюва данные для самцов,чего в действительности не должно быть, так как
у гусей самки в среднем мельче самцов. В литературе приводятся предельные размеры
крыла в 562 мм (Тугаринов, 1941), не наблюдавшиеся у измеренных нами экземпляров.
Изгиб нижней челюсти относительно слабый и клюв в общем длиннее и стройнее, чем
у A. f. serrirostris. Цевка 74—84 мм.
Окраска оперения в общем как и у А. /. fabalis, но голова бурее, шея и спина не-
сколько рыжеватые. Пуховик и молодые сходны с таковыми восточносибирского гумен-
ника, но у молодых птиц оперение более тусклое и перья уже, чем у последнего.
Короткоклювый гуменник Anser fabalis brachyrhynchus Bail Ion
Anser brachyrhynchus. В a i 1 1 о n. Mem. Soc. R. Abbev., стр. 74, 1833, Франция.
Распространение. Ареал. Островной вид, распространенный по восточно-
му побережью Гренландии от 80° с. ш. до крайнего юга, по побережьям внут-
ренних водоемов о-ва Исландия и на Шпицбергене. Добыт экспедицией Седова
на Земле Франца-Иосифа (на о-ве Гукера 12 июня 1914), но гнездовье здесь
точно не установлено. Зимой встречается на побережьях Скандинавии, Север-
ной Германии, Голландии, Дании, Бельгии, Северной Франции и на Бри-
танских о-вах. Однако правильно, нормально, но не в большом количестве
зимует лишь на побережьях Голландии и Англии. Известен залет в Новгород-
скую обл., где добыт 21 апреля 1912 г. в устье Ловати на озере Ильмень.
Другие случаи залета, о которых имеются сведения в нашей орнитоло-
гической литературе, не основываются на добытом или на точно опреде-
ленном материале и поэтому недостоверны. Отмечены также залеты на тер-
ритории Польши, Германии и других мест Западной Европы.
Характер пребывания и даты. В СССР случайно залет-
ный вид северо-западной части страны и Земли Франца-Иосифа, хотя допу-
скается возможность его гнездованья в последнем месте. На Шпицбергене
появляется во второй половине мая. 26 мая отмечена первая стая, 29 мая
было уже много (Бунге, 1887). Отлетает отсюда в сентябре. 26 сентября еще
попадались здесь отдельные стаи (Бианки, 1902).
Биотоп. По наблюдениям на Шпицбергене, основными биотопами
здесь являются покрытые травою возвышенные места тундры, выходы скал
в тундре, пологие каменистые склоны, и наконец, небольшие островки, т. е.
места с достаточно хорошим кругозором, с травянистой растительностью
вблизи, и притом в большинстве случаев не на побережьях, а внутри отдель-
ных островов архипелага.
Численность. На Шпицбергене встречается спорадично, неболь-
шими скоплениями и в общем численность его здесь незначительна. На Земле
Франца-Иосифа редок.
Экология. Размножение. Короткоклювый гуменник обычно рас-
полагается небольшими колониями, устраивая гнезда неподалеку одно от дру-
гого на кочках, на вершинах бугорков или на кучах валунов. Гнездо пред-
ставляет небольшое углубление в одном из таких мест и выстилается пухом.
Иногда на мшистых склонах гнездо строится в виде кучи из различных расти-
тельных материалов—кусков мха, лишайников, стебельков трав, а сдавлен-
ная ее вершина выстилается пухом и перьями вперемежку с частями расте-
ГУМЕННИК
325
ний. В некоторых же случаях в таких условиях гнезда состоят только из раз-
личных мхов; поперечник кучи у основания гнезда доходит до 30 см, а совер-
шенно правильный лоточек выстилается одним пухом (Бируля, 1910).
Кладка состоит из 3—5, чаще 3—4, иногда до 7—8 и даже до 9, желто-
вато-белых гладких или шероховатых, слегка блестящих яиц, размеры кото-
рых E0) 69—95,7x47,4—58,3, в среднем B92) 78,29x52,35 мм. Вес B92)
16,24—8,11, в среднем 11,65 г (ле-Руа). Первые яйца находят уже в середине
июня, но кладка растянута. 6 июля найдены были 4 яйца с совершенно зре-
лыми, уже оперенными эмбрионами и в тот же день в другом гнезде 3 еще
слабо насиженных яйца (Бируля, 1910). 26 июня найдена крепко насиживаю-
щая самка, а 24 июля при старых птицах были уже быстро бегающие выводки.
Самец очень привязан к самке, в период насиживания все время расхаживает
около гнезда вперед и назад и в конце концов
вытаптывает при этом даже тропинку в траве.
Птенцов выхаживают оба родителя. Однако
семья кормится на суше, при опасности же
спасается бегством. Иногда и старые птицы и
птенцы затаиваются на земле, плотно прижи-
маясь к ней и вытянув палками шеи.
Линька. Полная летняя линька (услов-
но) из гнездового наряда в первый у молодых
начинается в последней трети июля и закан-
чивается в последней трети августа—в начале
сентября. У добытых 21 августа маховые и ру-
левые доросли до нормы, но еще не окрепли
(Бунге, 1887). У добытых 29 августа маховые до
нормы еще не доросли (Бируля, 1910). Таким
образом, имеются и индивидуальные и годич-
ные колебания в сроках полной летней линьки.
Частичная осенне-зимняя линька заканчивается
в марте или в начале апреля. Точных данных
о характере линьки взрослых нет.
Питание. Местами кормежек коротко-
К|лювых гуменников на Шпицбергене являются
обширные, слегка покатые равнины с озерками
и сбегающими к морю речушками, обильно по-
крытыми различными гипновыми мхами, коврами лишайников, отчасти и
злаками (Бируля, 1910). В летнее время этот гусь питается различными
арктическими растениями: камнеломками, ложечной травой, хвощами, также
мхами, лишайниками, злаками, их корешками и пр. Часто заглатывает
мелкие камешки, а вместе с корешками и землю.
Полевые признаки. Самый мелкий из гуменников со светлосерой окрас-
кой, темной розовато-мясной перевязью надклювья и красноватыми лапами.
Голова и шея темнее остальных частей тела.
Крик звонче, чем у других гуменников, повторяется чаще, довольно
разнообразен и в некоторых случаях в нем ясно различаются звук «р» и опреде-
ленная охриплость. Голос звучит в общем как «гагьяк» или «кагга лирр§».
Издает и иные звуки вроде «гагага»... «кикикй», также громкое «хрэ-рэ-црё»
(Леруа).
Описание. Размеры и строение. Отличается наименьшими размерами
клюва и устойчивостью признаков, хотя у некоторых популяций перевязь клювя оран-
жево-желтая, в основном же розовато-мясного цвета. Длина крыла самцов F) 406—468, са-
мок D) 410—443, в среднем 439,1 и 426,5 мм. Длина клюва самцов F) 45—50, самок D) 44—
47,5, в среднем 47,о и 45,7 мм. Высота нижней челюсти у самцов E) 4,2—6,5, у самок D)
4,3—5, в среднем 5,2 и 4,6 мм (Дементьев, 1936). В общем подклювье мало изогнуто.
Вес самцов и самок 2,75—3,29 кг.
Рис. 58. Короткоклювый гу-
менник (клюв сбоку и сверху)
326 ОТРЯД ГУСЕОБРАЗНЫЕ
Окраска. Пуховик. Только что выведшиеся птенцы сверху оливково-зелено-
го цвета, иногда с желтизною, позже они становятся темноватосерыми или даже серыми.
Шея светлее. От угла рта к глазу идет темная полоса. Края крыльев желтые. Брюшная
сторона у маленьких светложелтая, у старших буро-желтая.
Молодая птица в первом пере. Все оперение тусклых тонов. Сверху буровато-
серая. Кроющие крыла не пепельно-серые, а темнобуро-сероватые. Клюв темный, почти
черный, и лишь в вершинной части надклювья у конька намечается его покраснение.
Ноги тусклосероватые с примесью красного.
Молодая птица после осенне-зимней линьки, т. е. во втором наряде, сходна со
взрослыми, но большие и средние кроющие крыла темные и размеры мельче. Повидимому,
только после третьей осенне-зимней линьки надевается взрослый наряд.
Взрослые самец и самка. По окраске сходны, но самка мельче. Общая окраска се-
рая с белым брюхом. Голова и шея бурые. Плечевые, большие кроющие крыла, предплечье
и кисть голубовато-пепельно-серые. Поясница серовато-бурая. Маховые черно-бурые.
Рулевые того же цвета, но с белыми боковыми окаймлениями и белыми, вершинами.
Горло светлое. Зоб и грудь беловато-серые. Брюхо и подхвостье белое. Перевязь на клюве
темномясо-красная. Основание клюва черное. Лапы розоватые или бледнотелесные,
радужина бурая.
После полной летней линьки выглядит слегка темнее, чем после частичной осенне-
зимней»
РОД КРАСНОЗОБЫЕ КАЗАРКИ RU FIBRE NT A BONAPARTE, 1856
Тип. Anser ruficollis
13. Краснозобая казарка Rufibrenta ruficollis Pall.
Anser ruficollis. P a 1 1 a «. Spicil. Zool., fasc. 6, стр. 21, табл. IV, 1769, низовья Оби.
Синоним. Anas torquata. G m e 1 i n. Reise d. Russland, II, стр. 179, табл. 14, 1774,
Каспийское море.
Русские названия. Морская казарка, рыжешейка—местные; краснозобая
казарка—книжное.
Распространение. Ареал. Занимает ограниченное пространство тундры
и частью лесотундры от восточного берега п-ова Ямала до рек Пясины, Дудып-
ты, Боганиды и Новой (приток Хатанги) на Таймыре. Найдена в Авамской
тундре. На север доходит до морских побережий между Обью и Енисеем,
до 73°30' с. ш. на западном побережье Таймыра в области Енисейской губы
и до 70° с. ш. на р. Боганиде. На юг до Полярного круга на 80-м меридиане,
до 68° с. ш. на Енисее. На восточном берегу Ямала встречается по рекам
Се-Яга и Томбей-Яга (Житков, 1908). Находка Пирсоном в 1896 г. красно-
зобой казарки в гнездовое время в Лапландии никем в дальнейшем подтвер-
ждена не была и является случайной.
На зимовки летит к юго-восточным, южным и юго-западным берегам
Каспия, в Закавказье, в Иран и Месопотамию. Отдельные особи залетают
в Северо-Западную Африку, Европу, Азию и известны почти для всех евро-
пейских стран. В СССР на основании добытых экземпляров залеты установ-
лены для следующих мест: для окрестностей Архангельска, для Горьков-
ской, Московской, Рязанской, Пензенской и Тульской обл., для Мордов-
ской и Татарской АССР, для Ростовской обл., Краснодарского края, Северо-
Осетинской АССР и Украинской ССР. В азиатской части СССР залеты извест-
ны для окрестностей Иркутска, в Балаганскую степь и на Чукотку. Залетает
и в другие страны Азии, в том числе в Индию, однако достоверность этих
последних сведений не подкреплена фактическим, добытым материалом.
Эти многочисленные залеты в чрезвычайно удаленные одна от другой тер-
ритории свидетельствуют о некотором распылении популяций краснозобой
казарки, необыкновенно яркая окраска которой, привлекающая внимание
охотников и любителей природы, позволяет точно устанавливать места ее
находок. Подобное явление известно и для белого гуся, однако с меньшей
степенью достоверности его встреч, и, вероятно, свойственно всем другим
видам гусей, находки которых в огромном большинстве случаев остаются
КРАСНОЗОБАЯ КАЗАРКА
327
не замеченными. Иногда такие залетные, заблудившиеся и, повидимому,
обессиленные краснозобые казарки присоединяются к домашним гусям (близ
с. Каменского Горьковской обл.; Серебровский, 1914) или даже к уткам
(Тульская обл.; Сушкин, 1898). При своих осенних и весенних передвиже-
ниях краснозобая казарка в одни районы залетает чаще, чем в другие, что
до некоторой степени напоминает явления пролета. Так, например, редко,
но более или менее постоянно весною и осенью она встречается в Приазовье
(Алфераки, 1904; Костюченко, 1925). Такая регулярность встреч дает воз-
Карта 34. Распространение краснозобой казарки Rufibrenta ruficollis
а—граница гнездовий, г—зимовки
можность предполагать, что в прошлом в Приазовье проходил определенно
выраженный пролет по направлению к Средиземноморью и что в настоящее
время это направление движения постепенно отмирает. Доказательство же на-
личия былого пролета этого гуся в области Средиземноморья видят в том, что
в Древнем Египте, где теперь ее не находят, краснозобая казарка была, пови-
димому, обычной зимующей птицей, о чем можно судить по многочисленным
ее изображениям на египетских фресках (Мейнерцхаген 1930). Однако все
эти соображения остаются лишь интересным предположением, пока еще
мало обоснованным.
Характер пребывания. Гнездящаяся, пролетная и зимую-
щая птица СССР.
Д а т ы. С мест зимовок в Иране снимается в конце января—начале фев-
раля C0 января—4 февраля 1898 г.). 3 февраля отмечена на разливах Атрека>
328 ОТРЯД ГУСЕОБРАЗНЫЕ
19 февраля наблюдался отлет отсюда на север (Житников, 1900). Из Закав-
казья исчезает 1—14 марта. На северном побережье Каспия и в западной
части Казахской ССР появляется 14—17 марта и надолго задерживается
на степных озерах Казахстана 15 апреля—3 мая пролетает в небольшом коли-
честве по Эмбе (Сушкин, 1908), 13 апреля—13 мая по Уралу (Зарудный,
1897), 25 апреля—13 мая основная масса казарок перемещается в Тургай-
ской низменности (Тугаринов, 1941).
Первые на местах гнездовий наблюдались: 9 июня в низовьях Енисея
и 14 июня на Гыданском п-ове (Наумов, 1931). Таким образом, весенний про-
лет проходит со значительными задержками в полупустынных и степных
районах, и на местах гнездовья краснозобые казарки появляются в то время,
когда тундра уже начинает зеленеть. На родине остаются 93—113 дней и уже
с 10 сентября начинают отлетать отсюда, основная же масса совершает
осенние передвижения с 15 сентября и последние из гнездового ареала исче-
зают в конце сентября. В то же время ранее отлетевшие краснозобые казарки
уже 15 сентября показываются далеко к югу—в степях по Иргизу и в вер-
ховьях Тобола (Сушкин, 1908). Пролет в этих районах продолжается еще
и в октябре. В ноябре долетают до берегов Каспия и к концу ноября дости-
гают до мест своих зимовок в Закавказье. Массовое появление их здесь наблю-
дается 1—10 декабря. Закавказские зимовки краснозобых казарок занимают
довольно ограниченное пространство и главным образом приурочены к рай-
ону залива имени Кирова, также к его окрестностям и к протокам системы
Ax-Чала и Хуршута (Исаков и Воробьев, 1940).
Биотоп. Гнездовой—наиболее сухие и возвышенные участки тундры
и лесотундры вблизи от воды. Зимний—солянковая пустыня с участками
злаковых эфемеров в понижениях близ водоемов (Исаков и Воробьев, 1940).
Численность. Неравномерна. Редка в восточной части ареала
по окраине п-ова Ямала, наиболее многочисленна в районе Енисейско-Хатанг-
ского водораздела.
Экология. Размножение. К постройке гнезд приступает сразу же
по прилете (Алфераки, 1904). Гнезда располагаются небольшими колониями,
по 4—5 пар, на крутых склонах речных долин, на крутых берегах рек и овра-
гов, поросших кустарниками и травою. Строительным материалом являются
сухие стебли злаков и других растений, из которых сооружаются основание
и стенки гнезда. Лоток устилается пухом. Иногда же гнездо представляет
выбоину или же углубление на площадке склона, где укладываются злаки
и прикрываются сверху пухом самой птицы. Гнезда помещаются открыто,
подчас заметны еще издали и в ряде случаев строятся по соседству с гнездом
сокола-сапсана (Наумов, 1931) или мохноногого канюка (Попгем, 1901).
Кладка состоит из 3—4—6, в некоторых случаях даже 7—9 (Попгем,
1901) яиц мутнозеленоватого или беловато-палевого цвета с глубоким зеле-
новатым оттенком. Размеры: 63—69,5x44,8—47,3 мм (Тугаринов, 1941).
Ненасиженные кладки по 3—4 яйца найдены 22 июня в Гыданской тундре
(Наумов, 1931), с 4 ненасиженными яйцами на озере Хассап-По
21 июня 1927 г. (Спангенберг), с 3,4 и 5 яйцами 24 июня на Енисее близ
Омулевой, с2 яйцами 1 июня на Енисее (с гнезда добыта самка; Зибом, 1880).
2—3-дневные пуховички найдены 19 июля в устье Енисея, примерно 5-днев-
ные птицы 15 июля у Омулевой. Таким образом, в низовьях Енисея выкле-
вываются пуховички между 10—16 июля. Птенцы величиной с чирка в низо-
вьях Енисея найдены 31 июля.
Насиживает одна самка, самец же находится неподалеку от нее на берегу
или на воде. Временно покидая гнездо, самка закрывает кладку растительным
материалом или пухом, снятым с краев гнезда. Как только выклюнувшиеся
пуховички подсохнут, к выводку присоединяется самец, и вся семья начи-
нает держаться на поросших свежей травою местах. При опасности взрослые
КРАСНОЗОБАЯ КАЗАРКА 329
птицы издают беспокойные громкие крики, принимают угрожающие позы
и даже делают попытки нападения на нарушителя их покоя, но в крайних
случаях все же обращаются в бегство и ищут спасения на воде.
Линька. Полная летняя линька. Наличие частичной осенне-зимней
линьки для взрослых точно не установлено. Молодые же после подъема на
крыло вступают в частичную осенне-зимнюю линьку. Перед летней линькой
краснозобые казарки собираются в стаи, и в ряде случаев не принимавшие
участия в размножении гуси вылетают за пределы гнездового ареала. Такие
стаи наблюдаются, например, в бассейне р. Балахны, где краснозобые ка-
зарки не гнездятся. Семейные же птицы соединяются в особые стаи и остаются
линять на месте. Линька начинается с 25 июля и протекает быстро. В течение
15—20 дней идет смена маховых одновременно почти у всех казарок. Около
15 августа на Гыданском п-ове уже встречались поднявшиеся на крыло стаи,
но незакончившие линьку отдельные особи попадались еще в конце сентября
(Наумов, 1931). Вообще же к 20-м числам августа огромное большинство
взрослых и молодых птиц начинает летать. С момента подъема на крыло
и до самого отлета, т. е. примерно в течение 20 дней, краснозобые казарки
ведут стайный образ жизни.
Частичная осенняя линька молодых начинается на родине, заканчи-
вается же на зимовках.
Питание. Основными кормами краснозобых казарок являются травя-
нистые растения. Поздний прилет их в тундру связан с поздним появлением
в ней травянистой растительности. Нежными побегами трав выкармливаются
выводки, побегами трав поддерживают свое существование линяющие птицы,
ими же кормятся краснозобые казарки и на зимовках, где основной их пищей
являются эфемерные злаки степных участков солянковой пустыни. В значи-
тельно меньшем количестве поедаются в это время рдесты (Potamogeton flui-
tans), семена Galium sp. и Bolbochaenus maritimus. Иногда в большом коли-
честве потребляется бурая солянка Salicornia herbacea и часто на полях
казарки кормятся всходами озимой пшеницы. Постоянно заглатывается также
речной песок (Исаков и Воробьев, 1940). Во время перелетов по Казахстану,
помимо нежной зелени трав, поедают также луковицы дикого чеснока (Суш-
кин, 1908), клубни и корневища степных растений.
Враги. Из врагов краснозобым казаркам причиняют вред чайки и по-
морники, а также четвероногие хищники, что принуждает казарок устра-
иваться близ гнезд хищных птиц, куда песцы, например, избегают проникать.
На зимовках губителен для краснозобых казарок длительный снеговой
покров, связанный с одновременным похолоданием.
Хозяйственное значение. В тундре краснозобые казарки во время их
линьки добываются местным населением для пропитания, а также для сбора
пера и пуха. На зимовках добываются стрельбой и в значительных количе-
ствах истребяются во время больших снегопадов. Весьма ценится как де-
коративная птица для содержания в зоологических садах.
Полевые признаки. Мелкие, величиною с огаря, ярко раскрашенные
гуси с белыми отметинами. Бросается в глаза рыжеватая окраска шеи и зоба,
резко контрастирующая с черным брюхом. От других гусей и казарок отли-
чаются значительно большей подвижностью. В стаях на земле они всегда
очень вертлявы и суетливы. Это заметно и на лету. Стая, прежде чем
опуститься в воду, облетает вокруг водоема несколько раз, одновременно
поворачиваясь при этом, подобно скворцам, то в одну, то в другую сторону,
растягиваясь и снова затем сбиваясь в плотную кучу (Сушкин, 1908). Эта
особенность их посадки сразу позволяет различать краснозобых казарок
среди других наших водоплавающих. Во время перелетов в течение дня почти
не делают остановок и спускаются на озера лишь с наступлением ночи. При
посадке на воду в это время суток спускаются с громкими криками. Между
330 ОТРЯД ГУСЕОБРАЗНЫЕ
собою в высокой степени общительны и на осенних перелетах и зимою дер-
жатся стаями по нескольку сот голов и более, но от других уток и гусей дер-
жатся обособленно. Весною же летят небольшими стаями. С воды крупные
стаи поднимаются с неумолчным гоготом (Житков, 1912). Отличаются быст-
рым и сильным полетом, прекрасно плавают и ныряют, что одинаково свой-
ственно и пуховикам, и молодым, и взрослым птицам. На зимовках в пред-
рассветных сумерках летят огромными стаями с мест ночлега на водопой,
пьют воду, глотают песок и часа через полтора направляются в степные
участки. Здесь они кормятся или в течение всего дня, или в полдень направ-
ляются на водопой, возвращаясь снова в облюбованные кормовые места.
С заходом солнца летят к местам ночевок, или на море, или на соленые
Рис. 59. Краснозобая казарка
озера, или на обширные озерные плесы. Но иногда ночуют и на местах
кормежек. В штормовую погоду прячутся в тростниках (Исаков и Во-
робьев, 1940).
Голос краснозобой казарки—характерный двусложный гусиный гогот,
который звукоподражательно передается как резкое, короткое и несколько
хриплое «чак-вой», откуда и происходит местное название этой казарки
у северян (Финш, 1879). Издают они также и особое шипенье. Чрезвычайно
легко и быстро приручаются и одомашниваются.
Описание. Размеры. Длина тела самцов и самок 540—560 мм. Длина крыла
самцов F) 355—390, самок C) 350—360 в среднем, 367,1 и 354,6 мм. Длина клюва самцов F)
23,8—25,7, самок C) 24,2 — 24,5, в среднем 24,6 и 24,3 мм. Наибольшая высота
клюва у самцов F) 3—3,8, у самок C) 3—3,5, в среднем 3,3 и 3,3 мм. Вес самцов и самок
около 1 кг.
Окраска. Пуховик. Спинная сторона желтовато-серая, затылок и шея несколь-
ко светлее. Брюшная сторона желтоватая. Клюв буро-рогового цвета. Ноготок белый.
Молодая птица. Лоб, голова и шея сверху, горло и грудь серовато-черные, по-
следняя с белыми пятнами. Бока головы черновато-бурые. Шея и зоб грязнокаштановые.
На больших кроющих беловатые вершины. На рулевых белые концы. Клюв и лапы
темные. По наблюдениям в неволе, окончательный взрослый яркий наряд надевается после
четвертой осенней линьки.
Взрослые самец и самка. Общая окраска пестрая из сочетания черного, рыжего
и белого цветов. Лоб, голова и задняя сторона шеи, вся спина и хвост черные, кроме
белых верхних кроющих хвоста. Через глаз от темени идет к подбородку и горлу чер-
ная полоса. Позади глаз по большому рыжему пятну, окруженному широкой белой
каймой; последняя продолжается вниз по бокам шеи. Зоб и грудь рыжие и отделены
белой полосой от черного брюха, но самая его задняя часть и подхвостье белые. Клюв
и лапы черные. Радужина темноореховая.
ЧЕРНАЯ КАЗАРКА
331
РОД КАЗАРКИ BRANT A SCOPOLI, 1769
Тип. Anas bernicla
\А. Черная казарка Branta bernicla L.
Anas Bernicla, Li nnaeus. Syst. Nat., изд. X, 1, стр. 124, 1758, Северная Европа.
Русские названия. Черная казарка—книжное; немок—местное название у
северян.
Распространение. Ареал. Кругополярный вид побережий аркти-
ческих морей и островов Ледовитого океана.
Подвиды и варьирующие признаки. Географическая изменчивость выражается
в характере общей окраски верхней и нижней сторон тела и в ее общем потемнении по
направлению от запада к востоку, а также в расположении белых пятен на боках шеи.
Карта 35. Распространение черной казарки BrantcTbernicla
J~B. b. hrota, 2—В. Ъ. bernicla, 5—В. b. orientalis, 4—В. b. nigricana; а—граница гнездовий,
г—эимовки; цифры в кружках—номера зимующих подвидов
Имеется также разница в направлении миграций, что позволяет в пределах вида выделить
две группы форм: одних, тяготеющих к северным побережьям Атлантического океана,
и других—к берегам Тихого океана. Различают четыре подвида: В. b. hrota M u 1 1 е г,
1776—острова севера Атлантики и Европы; В. Ь, bernicla Li nnaeus, 1758—побе-
режья Ледовитого океана и его островов до Таймыра и Хатанги; В, b, orientalis Tuga-
т i now, 1941—Восточная Сибирь и В. b. nigricans Lawrence, 1846—Северная
Америка и Анадырь.
Атлантическая черная казарка Branta bernicla hrota Mull.
Anas hrota. M u 1 1 e r. Zool. Dan. Prodr., стр. 14, 1776, Исландия.
Распространение. Ареал. Острова Арктического архипелага Север-
ной Америки до 100—110° з. д. и к северу до всех исследованных мест, Восточ-
ная Гренландия от пролива Скоресби до Земли Германии, Шпицберген,
Земля Франца-Иосифа, возможно (но не доказано) гнездование на Северном
о-ве Новая Земля, а достоверность гнездованья на Колгуеве и Таймыре
сомнительна, хотя в последнем месте атлантическая казарка была добыта
16 июня (Тугаринов, 1941). Таким образом, на территории СССР определенно
гнездится лишь на Земле Франца-Иосифа. На пролете встречается на Мед-
332 ОТРЯД ГУСЕОБРАЗНЫЕ
вежьем о-ве, на Фарерских островах и в Исландии. Зимою в Европе до Ла-
манша, Голландии, Дании, Германии и случайно в Северной Норвегии.
В Америке зимою на Атлантическом побережье от Нью-Джерси до Каролины,
изредка до Флориды; на Тихоокеанском побережье в небольшом числе к югу
до Британской Колумбии.
Характер пребывания. Гнездящаяся и перелетная птица.
Общее направление миграций к югу вдоль северной части Атлантического
океана, по его островам и побережьям.
Даты. На архипелаг Земли Франца-Иосифа прилетает 3—12 июня
(Зиюня 1882 г., Зиюня 1896 г.—мыс Флора; 5 июня 1796 г.—мысРихтгофена;
10 июня 1897 г.—мыс Флора; 12 июня 1914 г.—бухта Тихая). Осенний про-
лет в сентябре и не позднее конца этого месяца D сентября 1899 г.; 6 сентября
1895 г.; 22 сентября 1881 г.). Время пребывания на архипелаге Земли Франца-
Иосифа 90—117 дней.
Биотоп. В гнездовой период—низкая тундра, покрытая травянистой
растительностью с рассеянными по ней небольшими озерами. В остальное
время—морские побережья с отмелями, косами, мелкими заливами и лагу-
нами.
Численность. Многочисленна на Шпицбергене, местами у побе-
режий Гренландии. В небольшом количестве встречается на Земле Франца-
Иосифа.
Экология. Размножение. Гнездится обществами на прибрежных
островах, реже на побережьях близ озер. На Шпицбергене к гнездовью при-
ступает с начала июня или в первой половине июня. Для гнезда атланти-
ческая черная казарка выскребывает небольшую ямку, которую выклады-
вает сухими стеблями растений, ас начала насиживания—и п>хом, похожим
на гагачий. Кладка изЗ—5, изредка даже 8 яиц желтоватого цвета с гладкой
скорлупой (Джоурден, 1945). Размеры яиц B40) 63,5—78,5x41,4—50,5,
в среднем 70,32 X46,48 лш(ле-Руа). Вес 5,14—9,5, в среднем 6,69 г. Насижи-
вает самка около 28 дней, самец находится близ гнезда. Птенцы выхаживаются
обоими родителями.
Линька. Полная летняя линька начинается в июле. У самца, добы-
того 16 июля 1914 г. на о-ве Гукера, выпали все второстепенные маховые
и уже выбились из пеньков новые. Данных о частичной осенней линьке мало.
Молодые определенно проходят осеннюю линьку.
Питание. Основной пищей в гнездовой период являются травянистые
растения тундры. На Шпицбергене пища состоит из травы, стеблей и листьев
оксирии, камнеломок, а также из мхов. На зимовках поедаются морская
трава Zostera marina и в большом количестве Enteromorpha sp. (Кэмпбел).
Полевые признаки и поведение. Небольшой гусь размерами немного
крупнее кряквы, трехцветной окраски. Издали голова, шея и верх груди
кажутся черными, спина темной, а низ беловатым. Вблизи на боках шеи
заметна белая поперечная полоса.
Подвижная общественная птица, отлетающая с мест зимовок крупными
стаями, встречающаяся такими же стаями и зимою. В период размножения
держится у гнездовых мест, а после вывода молодых—около уреза воды
в самых разнообразных местах. Зимою кормится по мелким илистым морским
побережьям, богатым зарослями зостеры. Утром на местах кормежек дер-
жится часа два, затем улетает в море на водопой и отдыхает здесь до середины
дня. С полудня и до захода солнца снова кормится у побережий, ночует
в открытом море. Голос—резкое и частое «гонк», «кронк» или «ррот».
Описание. Размеры. Длина самцов и самок 596—774 мм; длина крыла самцов.
300—335, самок 325—335 мм. Длина клюва самцов 34—37,5, самок 31—35 мм (Шиолер).
Окраска. Пуховик. Верх серый, низ светлосерый. Щеки и полоса над
глазом темносерые.
ЧЕРНАЯ КАЗАРКА 333
Молодая птица в первом наряде. На плечевых и верхних кроющих крыла
выделяются белые вершинные каймы перьев. Бока тела без светлых каемок. На шее
«ет белых пятен.
Молодые птицы после первой осенне-зимней линьки,. Имеют на шее белые пятна.
На боках тела появляются светлые каймы. На верхних кроющих крыла сохраняются
белые вершинные каемки.
Взрослые самец и самка. Голова, шея, за исключением белых пятен, и пере-
дняя часть груди черные. Спинная сторона светлосеровато-бурая с легким корич-
неватым оттенком и четкими светлыми каймами перьев. Задняя часть груди, брюхо
и бока светло-сероватые с неявственными буроватыми полосами, становящимися более
светлыми ближе к брюху. Подхвостье белое. Белые пятна шеи не смыкаются на ее передней
стороне.
Евразийская черная казарка Branta bernicla bernicla L.
Распространение. Ареал. Повидимому гнездится на Новой Земле,
однако в 1951 г. здесь не найдена (Кузякин), что побуждает сомневаться
в достоверности прежних указаний на гнездовье здесь черной казарки, гнез-
дится о-ве Колгуеве, на Ямале севернее 70 ° с. ш., на Таймыре—на западном
побережье севернее 71° с. ш., на восточном—в Хатангском заливе, не южнее
74° с. ш. (Скалой, 1932), многочисленна на о-вах Сибирякова, Расторгуева,
Моржовом, доходит до Северной Земли и гнездится на о-ве Октябрьской
Революции, на о-ве Большевик и на Домашнем под 79° 30' с. ш. (Гизенко,
1938—1940). Достоверных сведений о распространении между о-вом Кол-
гуевом и Ямалом, а также между этим последним и п-овом Таймыром не
имеется. Зимою у южной и юго-западной частей Скандинавского п-ова, по
побережьям Ютландского п-ова, между последним и п-овом Бретань и в боль-
шом количестве у берегов Британских о-вов.
Характер пребывания. Гнездящаяся и пролетная птица.
С мест зимовок движется отчасти Атлантическим побережьем Скандинав-
ского п-ова и к 15—29 мая появляется на Мурманском побережье. 15—25 мая
на Новой Земле и примерно в то же время на о-ве Колгуеве. Часть казарок
летит побережьями и островами Балтийского моря. Между 26 апреля—27 мая
их наблюдают у побережий Финского залива, на Аландских о-вах и у юго-
западного побережья Ботнического залива. Птицы эти через озера Финлян-
дии и Карело-Финской ССР летят к горлу Белого моря и через восточную
оконечность Кольского п-ова. К 21—31 мая передовые показываются на Ямале,
основная же масса пролетает здесь 2—4 июня, но пролет идет и позже. 9—
10 июня отмечены на Се-Яга (Житков, 1912). Между 30 мая и 13 июня идут
весенние перемещения на Таймыре, валовой же пролет здесь приходится
на 8—10 июня. На островах Северной Земли появляется 12—16 июня.
Помимо этих направлений движения, имеются и другие, еще мало выяснен-
ные. Хорошо выраженный пролет проходит через Латвийскую, Литовскую
и Эстонскую ССР, пролетают черные казарки также через Ладожское озеро,
но далее на восток пункты, через которые перемещаются черные казарки,
остаются еще не выясненными. Весною их находили в Смоленской обл.,
на Вятке, Каме, на реке Белой, в Барабинской степи, в частности
на оз. Чаны и в районе Томска, где черные казарки в некоторые весны появ-
ляются в большом количестве и летят отсюда на север (Данилович, 1935;
Тугаринов, 1941). Наиболее восточным пунктом их весенних находок
являются окрестности Туруханска (Паллас, 1811).
Залеты евразийской черной казарки известны для Белоруссии, Смолен-
ской, Московской, Калужской, Курской, Воронежской, Архангельской,
Кировской, Молотовской, Уфимской, Черниговской, Киевской, Днепропет-
ровской обл. и указанных выше мест Западной Сибири. В большинстве этих
районов были встречены только единичные особи.
На Таймыре черные казарки проводят летом 80—95 дней и первые осен-
ние передвижения на оз. Таймырском отмечены 17 августа, массовый отлет
334 ОТРЯД ГУСЕОБРАЗНЫЕ
отсюда—23 августа и полное исчезновение—7 сентября (Тугаринов и Тол-
мачев, 1928). Примерно в те же сроки происходит отлет с западных побе-
режий Таймыра, где первые отлетающие стаи наблюдались 21 августа, а
24 августа шел массовый пролет и отлет многочисленных стай над морским
побережьем в юго-западном направлении (Бируля, 1907). Осенние переме-
щения совершаются на севере довольно быстро, и 1—16 сентября черные ка-
зарки пролетают побережьями Карской губы. 5—15 сентября идет пролег
уже на северных и восточных побережьях Балтийского моря, но здесь его
скорость замедляется и отдельные запоздавшие стаи встречаются еще в начале
октября. Осенью на побережьях Балтики пролетает в значительно больших
количествах, нежели весною.
Биотоп. Морские* побережья или низовья речных долин близ побе-
режий с низкой тундрой, поросшей травянистой растительностью и разбро-
санными всюду отдельными небольшими озерами. Во время кочевок, миграций
и на зимовках—морские побережья с песчаными косами, отмелями и неглу-
бокими заливами.
Численность. В большинстве мест встречается довольно редка
и только в отдельных районах Таймырского побережья и прилежащих к нему
островах многочисленна. Во время миграций и особенно на зимовках соби-
рается иногда в громадные стаи. Однако в последнее время нет таких несмет-
ных полчищ, которые в прошлом столетии и еще в начале нынешнего наблю-
дались на берегах Германии, Англии и Франции, «где голоса несметных стай
заглушали шум моря и где издали рои их точно дымом затемняли свет» (Hay
манн). Все это уже уничтожено безрассудной жадностью промышленников.
Экология. Размножение. Гнездовой период начинается сразу же
по прилете и в то время, когда идет еще пролет гнездящихся восточнее. Гнезда
устраиваются на сухих возвышенных участках тундры, для чего на избран-
ном месте утаптывается небольшое углубление, обкладываемое по краям вали-
ком из стеблей трав, перевитых пухом и перьями, которыми устилается
и лоточек. По существу евразийская черная казарка является колониальным
видом, так как гнезда в удобных местах располагаются по нескольку штуку
невдалеке одно от другого. Уже через 10 дней после прилета можно находить
полные кладки из 3—6 яиц сливочно-белого или светлооливково-буроватого
цвета. Полная кладка на Ямале из 6 яиц найдена 9 июня (Житков, 1912).
Размеры их: 51,0—81,1x36,5—66,0 мм (Тугаринов, 1941). Вес 6,78—7,56г,
в среднем 7,15 г (Гебель, 1904). Насиживает самка в течение 24—26 дней,
самец держится поблизости (Тугаринов, 1941). Как только птенцы обсохнут,
взрослые птицы переводят их на ближайшие пресноводные водоемы (озера,
речные рукава, устья рек) и выхаживают вместе до начала линьки, когда
самцы отделяются от выводков. Обе взрослые птицы очень заботливы по от-
ношению к своему выводку, очень беспокоятся в случае опасности, а в
критических случаях все же бросают птенцов и отлетают в сторону (неда-
леко). Предоставленные в таких случаях самим себе, птенцы затаиваются
и через некоторое время созываются родителями. Выводки при стариках
встречены 24 июля на Таймыре (Тугаринов и Толмачев, 1928).
Линька. У взрослых ежегодно полная летняя линька, по частичной
осенне-зимней линьке нет данных. Что касается молодых, то, после того
как они поднимутся на крыло, у них начинается частичная осенне-зимняя
линька. Полная летняя линька начинается в конце первой половины июля.
Иногда линяющие присоединяются к другим видам гусей, находящимся также
в линьке. На Новой Земле 15 июля 1950 г. добыты пять линных черных каза-
рок среди линяющего стада в 200 голов гуменников и белолобых гусей
(Успенский). Здесь же 6 августа линька еще продолжалась, и было добыто
несколько линных птиц (Горбунов, 1929). На Западном Таймыре начало
смены оперения отмечено 20 июля, 1—4 августа линькой были охвачены все
ЧЕРНАЯ КАЗАРКА
335
птицы, 6 августа появились первые летные, 8 августа наблюдались уже стаи
летных. Линька маховых протекает интенсивно, и около середины августа
она уже заканчивается. После того как отрастут маховые, начинается смена
контурных перьев. Первыми сменяются контурные перья низа тела, затем
контурные верхней стороны, последними сменяются перья хвоста. Благодаря
быстрым темпам линьки часть казарок успевает сменить все оперение еще
до отлета. У молодых вслед за полной линькой начинается частичная осен-
няя линька, которая заканчивается только на зимовках, где у них появля-
ются белые перья ошейника и перья со светлыми каймами на боках тела
(Тугаринов, 1941)
Рис. 60. Евразийская черная казарка
Для летней линьки евразийские казарки собираются крупными стадами,
самцы, как указано, отделяются от самок и выводков, последние же объеди-
няются, и можно находить по нескольку самок с полутора—двумя десятками
молодых, держащихся вместе. На небольших озерах крупных стай не бывает,
но на больших водоемах все птицы—и взрослые, и молодые, и холостые—
иногда объединяются в один огромный табун. По существу, в это время птицы
остаются наземными и кормятся по берегам этого же водоема, при опасности
же спасаются на воде, отплывая на середину водоема (Тугаринов, 1941).
Питание. По прилете в тундру, когда все водоемы оказываются
скованными льдом и нет еще всходов трав, пищей казаркам служат мхи
и лишайники, которые склевываются ими с камней. Затем с наступлением
весны казарки начинают скусывать молодые побеги трав. Позже, когда стебли
травянистых растений огрубеют, казарки перегрызают их, работая то одной,
то другой стороной клюва (Тугаринов, 1941).Когда казарки держатся на водо-
емах, они кормятся и водорослями, а на морских побережьях в большом коли-
честве потребляют зостеру. Наряду с растительными кормами поедаются,
правда в меньшем количестве, моллюски и мелкие ракообразные. Молодые
336 ОТРЯД ГУСЕОБРАЗНЫЕ
питаются, в сущности, теми же кормами, что и взрослые, но поедают более
нежные части растений.
Враги. Главными врагами черной казарки являются различные круп-
ные чайки и поморники, похищающие их яйца и птенцов. Несмотря на то
что свои кладки казарки, когда им приходится сходить с гнезда, тщательно
прикрывают стеблями, пухом и перьями, все же в некоторых случаях их
гнезда оказываются разоренными.
Хозяйственное значение. Для жителей Крайнего Севера черные ка-
зарки имеют весьма существенное значение в качестве запасов продоволь-
ствия, однако использование их и до сих пор ведется по старинке, так назы-
ваемым гусеванием и сбором яиц, что в корне необходимо изменить, так как
это быстро приведет к полному истреблению черных казарок, которые
на север идут далее всех прочих гусей. Правильная же постановка охот-
ничьего промысла на казарок может дать местному населению богатый запас
мясных кормов.
Полевые признаки. Мелкий гусь, на расстоянии спереди кажущийся
черным, сзади белым. На земле проворен, быстро и хорошо ходит, при опас-
ности иногда затаивается, прижавшись телом к земле и вытянув шею. Это же
самое в некоторых случаях проделывается и на воде. Свободно и легко
плавает, нырять не может, но способен подобно уткам переворачиваться
вверх хвостом, вниз головою и доставать со дна пищу. Летает с виду тяжело,
делая частые взмахи крыльями, но способен преодолевать значительные
расстояния. Во время миграций летит волнистой линией невысоко над
землей, при местных же перемещениях летит тесной кучей низко над землей
или водой. На полете издает характерный негромкий крик, сходный со зву-
ками «кронк»... «кронк»... «кронк».
Описание. Размеры и строение. Длина тела самцов и самок 584—889 мм.
Длина крыла самцов 310—351, самок 315—331 мм. Клюв самцов 29—37, самок 31—38 мм.
Вес самцов F) 1415—1730, самок A) 1202 г (Шиолер).
Окраска. Пуховик. Спинная сторона, включая голову и шею, серо-мышиного
цвета. Полоса над глазом и щеки черновато-серые. Подбородок и горло беловатые.
Брюшная сторона светлосерая. Кончик ноготка беловатый.
Молодые птицы в первом пере. Шея без светлых пятен. Грудь немного темнее
брюха и боков тела. На последних нет светлых поперечин. На вершинах кроющих крыла
светлые каемки. Молодые птицы после первой осенней линьки окрашены несколько
светлее взрослых и на каймах перьев верха у них сохраняется рыжеватый оттенок.
Окончательный взрослый наряд надевается после четвертой осенней линьки.
Взрослые самец и самка. Общая окраска темная. Спинная сторона аспидно-
серая. Каемки перьев верха нерезкие. На брюшной стороне черная окраска зоба резко
отграничена от цвета груди. Брюхо чистобелого цвета. Белые пятна на боках шеи
у отдельных особей смыкаются спереди.
Восточносибирская черная казарка Branta bernicla orientalis
Tugari now
Branta bernicla orientalis. Тугаринов. Фауна СССР, Птицы 1,4 стр. 180, 1941,
Восточная Сибирь.
Распространение. Ареал. Побережье Ледовитого океана от дельты
Лены до верховьев бассейна Анадыря и п-ова Чукотки, Новосибирские о-ва,
о-ва де-Лонга, о-в Врангеля, где возможно встречается вместе с В. b. nigricans.
Зимою у побережий Японии и Китайской Народной Республики (Тугари-
нов, 1941).
Характер пребывания. Гнездящиеся и пролетные птицы
Северо-Восточной Сибири. С мест зимовок на побережьях Желтого моря и
Чжилийского залива улетают в мае. 25—30 мая пересекают Амур, перевали-
вают через Становой хребет и попадают в бассейн Лены, но на самую Лену
ЧЕРНАЯ КАЗАРКА 337
вылетают не западнее р. Синей. В окрестностях Якутска появляются 26—
30 мая и летят здесь до 13 июня. В устье Лены показываются 29—
30 мая; отсюда часть птиц летит на Новосибирские о-ва, куда на Новую
Сибирь и на о-в Котельный прибывают 1—5 июня и в те же сроки прилетают
в устье Индигирки птицы, движущиеся на восток вдоль арктического побе-
режья. Другое направление весенних передвижений, выраженное значительно
слабее, ведет к побережьям Охотского моря, где в окрестностях Охотска
черные казарки отмечены уже 8 мая. Пролетают также вдоль побережий
Сахалина и Охотского побережья Камчатки.
С мест гнездовий отлетают рано, Новосибирские о-ва начинают покидать
с 15 августа и окончательно исчезают отсюда 28—30 августа. Осенью изредка
залетают на Командоры, где казарка эта добыта 8 сентября, и к 20 сентября
прилетают в район Амурского залива. Осенние миграции совершаются
по иным направлениям, нежели весенние, и восточносибирские черные
казарки совершенно не встречаются в окрестностях Якутска, а также
по Лене вверх по р. Синей, где весною они обильны. Осенние передвижения
на местах гнездовья и далее по материку до Охотского моря еще не выяс-
нены. Можно только предполагать, что казарки летят речными долинами
и затем, пересекая их и Становой хребет, выходят на побережье Охотского
моря, где определенно установлены их осенние передвижения. Отсюда
казарки летят к Малому Хингану через Амур, где значительный их пролет
наблюдался 25—28 сентября и далее направляются к зимовкам на берегах
Чжилийского залива и Желтого моря.
Биотоп. Низменные участки прибрежной тундры и речных долин.
Численность. Не велика, но в малолюдных районах эти казар-
ки обыкновенны.
Экология. Размножение. Полные кладки в дельте Лены найдены
в 20-х числах июня. Сильно насиженные—в устье Индигирки 13 июля, на Но-
восибирских о-вах (о-в Фадеевский) ненасиженные — 6 июля (Тугаринов,
1941). Насиживание начинается с откладкой первого яйца, что доказывается
разной степенью развития эмбрионов в яйцах одной и той же кладки (Пор-
тенко, 1937). Только что выклюнувшиеся птенцы на мысе Большом Барановом
найдены 12 июля (Тугаринов, 1941).
Л и н ь к а. У взрослых полная летняя, у молодых, кроме того, частично
осенне-зимняя. Полная летняя линька начинается почти одновременно и на
крайних северных и у южных границ гнездового ареала. На время линьки
часть казарок перелетает на крайние северные из Новосибирских о-вов и с бо-
лее южных и с материка. Судя по тому, что при самках остаются только
молодые, можно полагать, что на линьку в другие районы улетают и холостые
птицы и самцы от выводков, однако достоверно это не установлено. На-
чинается линька с 20 июля и к середине августа заканчивается. Таким обра-
зом, и для этого подвида характерны крайне сжатые сроки линьки.
Частичная осенняя линька молодых начинается на родине и заканчивается
на зимовках.
Питание, повидимому, носит тот же характер, что и у евразийской
казарки, однако данных не имеется.
Описание. Размеры сходны с В. b. bernicla.
Окраска. Пуховик. Немного темнее, чем у В. b. bernicla.
Молодая птица. Также несколько темнее, особенно в области брюха. Смена наря-
дов, как и В. 6. bernicla.
Взрослые самец и самка. Спинная сторона темная. Снизу граница между чер-
ным зобом и грудью неявственна. Брюхо на большей части своего протяжения чер-
ное, и этот цвет в некоторых случаях доходит до области клоаки. Белые пятна шеи
сходятся спереди и почти сходятся сзади. Выше этого неполного кольца на каждой
стороне шеи располагаются по 3—4 косые белые полоски. На передней стороне шеи
белые пятна.
22 Птицы СССР.т. IV
338 ОТРЯД ГУСЕОБРАЗНЫЕ
Американская черная казарка Branta bernicla nigricans Lawrence
Anser nigricans. Lawrence. Ann. Lye. N. H. New-York, IV, стр. 171, 1846,
Ш Нью-Джерси.
Распространение. Ареал. Побережье Берингова моря от Анадыря
до Берингова пролива и отсюда, повидимому, до мыса Шмидта на Чукотке.
Возможно гнездится и на о-ве Врангеля, что окончательно можно установить
только на дополнительных сборах. Вне СССР широко распространена по
арктическому побережью Северной Америки от Аляски до 100—110° з. д.
На север до о-вов Бенкса и Мельвилля. На зиму спускается по Тихоокеан-
скому побережью Северной Америки до Калифорнии. Весною на Анадырь
прилетает 22 мая (Портенко, 1939). В начале июня валовой пролет наблю-
дался в заливе Креста. 4 июня добыта на Чукотке. Вместе с тем передовые
на мысе Шмидта, находящемся далеко от устья Индигирки, показываются
уже 20 мая. Это обстоятельство позволяет предположить, что сюда напра-
вляются Branta bernicla nigricans, а не Branta bernicla orientalis, появляющие-
ся на побережье вообще позднее. Однако доказать это можно только сбором
материала из района мыса Шмидта, а также и с о-ва Врангеля, где гнездя-
щаяся форма еще точно не установлена.
Биотоп. Приморская полоса побережий, берега заливов, устья рек.
Численность. Не многочисленна.
Экология. Сходна с предыдущим подвидом, но линька, повидимому,
начинается раньше, на Чукотке 8—11 июля (Белопольский, 1934).
Описание. Размерами не отличается от В. b. bernicla. Верх и низ черно-бурые. Осно-
вание шеи и зоб черные. Брюхо все черное до области клоаки. Вокруг шеи белое кольцо*
Спереди над ним белые пятна, а на боках косые белые полоски.
15. Белощекая казарка Branta leucopsis Bechstein
Anas leucopsis. Bechstei n. Orn. Taschenb., II, стр. 424, 1803, Германия.
Русское название. Белощекая казарка—книжное.
Распространение. Ареал. Островной вид, гнездящийся на восточном
побережье Гренландии (в заливе Скоресби), на Шпицбергене, на Лофотенах
у берегов Норвегии и на Южном о-ве Новой Земли. Зимою у берегов Ирландии
и Великобритании, Северной Франции, Голландии и Дании, отчасти на
устричных банках Шлезвига. Отдельные стаи и особи доходят до Средизем-
номорья (Италия, Испания, Марокко), Азорских о-вов и Северной Америки.
Залетает в Калининградскую обл., где добыта три раза.
Характер пребывания. Гнездящийся и пролетный вид.
С мест зимовок отлетает в конце марта—середине апреля. 28 апреля—10 мая
в качестве чрезвычайно редкой пролетной птицы появляется в Латвийской
и Эстонской ССР, на Чудском озере, 21 апреля—5 мая в Ленинградской обл.
(Бианки, 1907) и около 20 мая на Новой Земле (Горбунов, 1929). С Новой
Земли отлетает в сентябре, 28 сентября—25 октября изредка пролетает
в окрестностях Ленинграда (Бианки, 1907; Раснер, 1913) и в октябре
появляется на местах зимовок.
Биотоп. Гнездовой—горный ландшафт севера, скалистые обрывы,
скалистые побережья, крутые склоны, чередующиеся с понижениями, покры-
тыми сочной травянистой растительностью, по берегам соседних со скалами
ручьев и озер. Зимний—морские побережья.
Численность. Незначительна. За последние годы местами (на Но-
вой Земле) численность катастрофически снизилась (Кузякин).
Экология. Размножение. Гнезда располагаются на выступах
скал, на площадках крутых склонов и откосов, на вершинах невысоких гор-
ных хребтов. Гнездятся разбросанными колониями, на больших расстояниях
БЕЛОЩЕКАЯ КАЗАРКА
пара от пары. Но обыкновенно сразу видно по нескольку гнезд. Строятся
они открыто, без маскировки, и заметны издали. В одной из таких колоний
у залива Селезнева на Новой Земле насчитывалось 75 гнезд, разбросанных
на большой площади (Горбунов, 1929). Самое гнездо помещается в небольшом
естественном углублении и строится из мха и лишайников, водорослей,
сухих стеблей растений, идущих в основание и отчасти на боковые стенки,
и устилается то небольшим количеством пуха, to значительно большим, чем
у гаги. Благодаря обилию пуха в некоторых случаях гнездо издали кажется
светлосерым полушаром (Успенский). Кладка из 3—6, чаще из 4—5 яиц
Карта 36. Распространение белощекой казарки Branta leucopsis
а—граница гневдовий, •—вимовки
чистобелого цвета, с насиживанием приобретающих желтый оттенок. Их
размеры: 68,7—82,7x46,4—53,6 мм (Тугаринов, 1941); A2) 70,6—80,8 X
Х48—51,1 мм\ вес 94—114,2, в среднем 102,52 г (ле-Руа по Хартерту, 1920)
16 июня 1950 г. в районе Грибовой губы на Новой Земле найдено гнездо, вы-
стланное большим количеством светлого пуха. Одна птица постоянно находи-
лась на гнезде, другая рядом. 17 июня 1949 г. у губы Безымянной на скале,
занятой прежде колонией казарок, наблюдалась другая пара, утаптывающая
снег на месте будущего гнезда. 27 июня 1949 г. на о-ве Гольце найдены 4 гнезда
с яйцами; в двух гнездах было по одному яйцу. 24 июля 1949 г. там же гнезда
уже были пусты, рядом держались 8 пуховиков и 6 взрослых птиц, 8 июля
1948 г. здесь же было 7 гнезд: 3 с насиженными яйцами и 4 с пуховиками
(Успенский). 8 июля 1951 г. на острове у губы Безымянной найдено гнездо
с сильно насиженными яйцами (Кузякин). Пуховички в возрасте нескольких
дней также на Новой Земле наблюдались 26 июля. Точных данных о времени
насиживания не имеется. Насиживает одна самка, самца же часто можно
видеть в сторожевой позе близ гнезда. Пуховички, выклюнувшиеся из яиц»
22*
340
ОТРЯД ГУСЕОБРАЗНЫЕ
обсыхают в гнезде, затем покидают его, опускаясь со скал и обрывов и зача-
стую без вреда для себя сваливаясь вниз, где их собирают родители и уводят
на кормовые места близ пресноводных водоемов.
Линька. Полная летняя у взрослых и частичная осенняя у молодых.
На Новой Земле линька начинается с 20—31 июля. Нелетающие белощекие
казарки здесь были найдены 2 августа. Самцы приступают к линьке раньше
самок. О дальнейшем ходе линьки сведений не имеется.
Питание. Корма белощекой казарки довольно разнообразны. Из тра-
вянистых растений на лугах она поедает мятлик (Роа distans), осоку (Сагех
bulbosa), клевер ползучий (Tuifolium repens), лютики (Ranunculus sp.),
далее почки, листья и сережки полярной ивы, водоросли. Наряду с этим
использует и животные корма—различных ракообразных, водных насекомых
и, возможно, моллюсков. По крайней мере в желудках белощеких казарок
Рис. 61. Белощекая казарка
находят створки морских моллюсков, хотя возможно последние заглатыва-
ются в качестве минеральных примесей вместе с грубозернистым песком.
Наблюдается определенная сезонность смены кормов в связи с тем, что
в гнездовой период белощекая казарка держится на луговых пространствах,
а в период миграций и зимовок—на морских побережьях.
Хозяйственное значение. Добывается лишь случайно и поэтому в насто-
ящее время промыслового значения не имеет. Дает ценный пух. Необходимо
наложить запрет на истребление этого вида, гнездующегося у нас лишь на
Южном о-ве Новой Земли.
Полевые признаки. При общем гусином облике в полевых условиях легко
распознается своей двуцветностью, так как белощекая казарка кажется
черной сверху, белой снизу. Резко выделяются белые бока головы, лоб и гор-
ло. Постоянно, за исключением периода насиживания, держится стадно. Гнез-
дится, как указано, колониями, линяет в стаях, которые при преследовании
их на воде сбиваются в плотные кучи, что местами влечет за собою их полное
истребление; совершает стайные весенние и осенние перелеты, а на зимовках
стаи еще более укрупняются. Бегает быстро и настолько хорошо, что иногда
спасается бегством при преследовании во время линьки. Прекрасно плавает.
Легко летает. Громкий, издалека слышный голос белощеких казарок напо-
минает хриплые звуки «кхаг»... «кхаю>... «кхап» и некоторыми сравнивается
с собачьими лаем. Иногда слышны звуки «вррэк» или «грэ-грэ-грэ». На кор-
межке белощекие казарки постоянно громко переговариваются друг с другом.
КАНАДСКАЯ КАЗАРКА 341
Кормятся или на влажных берегах или на побережьях, залитых неглубоким
слоем воды. По сравнению с другими гусями мало осторожны, а к гнезду, не уле-
тая, подпускают на 10—15 шагов (по наблюдениям Успенского). С другими
видами не смешиваются и всегда держатся обособленными стаями.
Описание. Размеры. Длина тела 596—71 м. Крыло 365—430 мм. Клюв 25—
37 мм. Цевка 62—76 мм. Вес 1,1—2,25 кг.
Окраска. Пуховик. Сверху темносерый. Голова, основание клюва, уздечка,
кольцо вокруг глаза темносерые с буроватым налетом. Бока головы и шея светлосерые.
Спина темная, буровато-серая. Зоб буровато-серый. Грудь, бока и брюхо беловатые.
Ноги и клюв черноватые (Тугаринов, 1941).
Молодая птица в первом пере. Белые места на голове и ее боках со значительной
примесью бурого. Перед белыми вершинами перьев головы, шеи и груди бурые поля.
Вершины рулевых серовато-белые. Клюв и лапы черные с красноватым оттенком.
Молодая птица после первой осенней линьки. На белых участках головы и щек
имеются черные и темнобурые отдельные перышки. Вершины лопаточных и верхних
кроющих крыла с рыжеватым оттенком. На боках туловища широкие темносерые попереч-
ные полосы. Верх головы, шеи и грудь с примесью бурого цвета. Белые вершины перьев
спины с примесью бурого.
Взрослые самец и самка. Почти сходны по окраске, но самка несколько мельче.
Общая окраска из сочетания белого и черного. Верх спины, плечевые, кроющие крыла
и внутренние второстепенные маховые черные с белыми вершинами на перьях спины.
Низ спины и середина надхвостья буровато-черные. Бока надхвостья и верхние крою-
щие хвоста белые. Первостепенные маховые и рулевые черные. Лоб, щеки и верх горла
чистобелые. Темя, уздечка и зоб черные. Грудь и брюхо белые. Крупные буровато-серые
полосы на боках у самцов развиты сильнее.
16. Канадская казарка Branta canadensis
Anas canadensis. Linnaeus. Syst. Nat., изд. Х, стр. 123, 1758, Канада.
Русское название. Канадская казаока—книжное.
Распространение. А р е а л. От крайнего сев< ра Америки приблизительно
до 40° с. ш. В пределах СССР найдены на островах Берингова моря.
Биотип. Заболоченная тундра
Подвиды и ьарьирующие признаки. В настоящее время различают пять подвидов,
систематическое значение которых еще недостаточно выяснено: В. с. canadensis L i n-
n a e u s, 1758—занимает большую часть Северной Америки от границы лесной зоны
до восточных побережий Гудзонова залива и на юг примерно до 40° с. ш.; В. с. leu-
copareia Brandt, 1836—Северная Америка от Аляски до п-ова Мельвилля и на север
до 70°; В. с. minima R i d g w а у, 1885—восточное побережье Аляски и Алеутские
о-ва; В. с. occidentalis В a i г d, 1858—западное побережье Северной Америки от бухты
принца Вильяма до Северо-Восточной Калифорнии; В. с. butchinsii Richardson,
1831—крайние восточные острова Северной Америки от Земли Эльсмира на севере до
Баффиновой Земли на юге и крайний север Гренландии. Отличаются между собою разме-
рами, окраской и частью заменяют друг друга географически. Вся эта группа требует но-
вой ревизии. Некоторые из этих форм были встречены на территории СССР.
Средняя канадская казарка Branta canadensis leucopareia Brandt
Anser leucopareius. Brandt. Bull. Se. Ac. Petrop., 1, стр. 37, 1836, Алеутские о-ва.
Распространение. Ареал. Прибрежные тундры Северной Америки от
долины Ковак на Аляскеи устья р. Мекензи до п-ова Мельвилля,также остро-
ва на север до 70° с. ш. Гнездится также на Алеутских о-вах, отмечена
в прошлом столетии на Курильских о-вах (Стейнегер, 1898). Прежде в очень
небольшом количестве гнездилась на Командорских о-вах. Зимовки отчасти
в Японии, отчасти на Тихоокеанском побережье Северной Америки на юг до
Калифорнии и Луизианы.
Характер пребывания. Гнездящаяся, частью залетная
птица. В прежнее время—в конце прошлого и начале нынешнего столетий—
гнездилась на о-ве Беринга. Была добыта здесь на гнездовье в 1882, 1883
и 1914 гг., но позднее уже не найдена (Иогансен, 1934). На о-ве Беринга
342
ОТРЯД ГУСЕОБРАЗНЫЕ
первые весною отмечены 2 мая, на Аляску прилетают 25—30 апреля. Отлетают
с Аляски с конца сентября до середины ноября.
Биотоп. Морские побережья с солоноватыми озерами; участки тун-
дры, заходящей на побережья, покрытые травой; травянистые участки тундры
близ солоноватых озер.
Численность. В СССР всюду редка.
Экология Размножение. Гнездо помещается или в густой траве
или под ветвями отдельных кустов. Строится оно из сухих стеблей трав и вы-
Карта 37. Распространение канадской казарки Branta canadensis
1—В. с. canadensis, 2—В. с. leucopareia, 3—В. с. ocddentalis, 4—В. с. minima,
5. В. с. hutschinsi; а—граница гнездовий, в—вимовки; цифры в кружках—номера ви-
мующих подвидов
стилается пухом. Откладывает 5—8 яиц сливочно-белого или желтовато-бело-
го цвета. Их размеры: E) 85—86x54,5—57 мм. Вес 13,08—14,14 г (по Гебелю
Хартерт, 1920).
Линька. Повидимому, как и у других казарок. Взрослые птицы начи-
нают линять 20 июля и становятся летными в конце августа. Молодые под-
нимаются на крыло к 20-м числам августа.
Полевые признаки. Средней величины казарка. Поведение, как и у дру-
гих гусей. Но голос своеобразен и представляет собой громкое гоготанье,
в котором отчетливо слышен звук «р». После вывода детей и подъема на крыло
держатся по морским побережьям.
Описание. Размеры. Крыло 373—450 мм. Клюв от углов рта 40—41 мм. Цевка
71—85 мм. Рулевых 14—16 (Тугаринов, 1941 и 1950).
Окраска. Пуховик. Верх желтовато-бурый; голова светлопа лево-желта я
с бурым пятном сверху. На спине позади крыльев пара желтых пятен. Низ светлопа-
лево-желтый.
Молодая птица. Сверху серовато-бурая, снизу сероватая. На боках нет поперечных
полос. По бокам головы белые пятна, на горле разделяющиеся узкой черно-бурой поло-
сой или черно-бурыми пятнами.
КАНАДСКАЯ КАЗАРКА
343
Взрослые самец и самка. Окраска спинной стороны буровато-серая, темнеющая
к хвосту. Маховые и хвост черно-бурые или черные. Верх головы, шея и зоб лоснящиеся
черные. Горло и бока головы белые. Эта белая окраска, суживаясь, в виде треугольни-
ков распространяется за глаза и немного не доходит до затылка. Передняя часть спины,
плечевые и верхние кроющие крыла буровато-серые. Задняя часть спины и середина
Рис. 62. Канадская казарка
надхвостья черные. Бока надхвостья белые. Брюшная сторона бледносеровато-бурая со
светлыми каймами перьев. Брюхо и подхвостье белые. Бока темные серовато-бурые со
светлыми каймами перьев, образующими светлые поперечные полосы. Клюв черный.
Ноги черные. Радужина темнобурая.
Малая канадская казарка Branta canadensis minima Ridgway
Branta minima. Ridgway. Proc. U. S. Nat., Mus. VIII, стр. 22, 1885, Сент-Михель,
Аляска.
Распространение Ареал. Алеутские о-ва и побережья Аляски от
залива Нартон до залива Бристоль. Зимовки в Северной Америке на про-
странстве от штата Британская Колумбия до округа Сан-Диего в Кали-
форнии.
Характер пребывания. В пределах СССР случайно залетная
птица, дважды добытая на о-ве Беринга.
Биотоп. Заболоченные низины прибрежной тундры, островки на них,
водоемы и болотистые берега.
Численность. По количеству особей местами на Аляске превос-
ходит всех других гусей.
Экология. Размножение. На юго-западном побережье Аляски
появляется в конце первой или в начале второй декады мая, уже с 20 мая
начинается разбивка на пары. Гнезда обычно строятся на сухих островках
среди болотистых низин и 5—10 июня в них можно находить полные ненаси-
344 ОТРЯД ГУСЕОБРАЗНЫЕ
женные кладки из 4—7 яиц. Насиживает самка, самец же безотлучно нахо-
дится при гнезде и вместе с самкой принимает участие в выхаживании
птенцов.
Линька. Данных нет, но местами на побережьях Юго-Западной Аляски
22 июня наблюдались значительные передвижения по направлению на север
смешанных стай малой канадской казарки вместе с голубым гусем. Очевидно
эти передвижения совершаются холостыми птицами, направляющимися
в удобные для них места линьки.
Полевые признаки. Самая мелкая из всех канадских казарок и на лету
может быть принята за крупную утку. Сверху кажется черной, снизу темно-
серой, мышиного цвета. Голос более высок и резок, чем у других канадских
гусей, почему в некоторых местах ей и дают название крикливой казарки.
Описание. Размеры и строение. Длина крыла самцов A) 406, самок B)
346 и 360 мм. Длина клюва самцов B) 31, самок B) 29 и 31 мм (Беме, 1948)
Окраска. По общей расцветке близка к другим канадским казаркам, но отли-
чается от них более темноокрашенной нижней стороной тела, имеющей оттенок мышиного,
цвета.
Темногрудая канадская казарка Branta canadensis occidentalis В a i г d
Bernicla occidentalis. В a i г d. Report Expl. et Survey R. R. Рас, IX, стр. 766. 1858,
порт Таунсенд, Вашингтон.
Распространение. Ареал. Западное побережье Северной Америки,
от пролива принца Вильяма и о-ва Миткова к югу до островов королевы
Шарлотты. Зимою от штата Вашингтона до побережий Южной Калифорнии.
Характер пребывания. Гнездящийся перелетный вид, слу-
чайно залетает на Анадырь (добыча 27 мая 1903 г. у б. поста Нсвомарин-
ского, хранится в Дарвиновском музее в Москве).
Экология. По образу жизни сходна с В. с. leucopareia.
Описание. Размеры. Одна из крупных канадских казарик, приближающаяся
по размерам к обыкновенной канадской казарке. Число рулевых 18—20. Длина крыла
411—458 мм, длина клюва 36—41,9 мм, длина цевки 77—82 мм (Хартерт). Крыло добы-
того на Анадыре экземпляра равно 463 мм (Тугаринов, 1941).
Окраска. По расцветке оперения приближается к В. с. minima, но в общем силь-
но изменчива, имеются темноокрашенные снизу особи, светлоокрашенные я переходные-
между ними.
ПОДСЕМЕЙСТВО УТКИ AN AT INAE
РОД ПЕГАНКИ ТADORN A FLEMING, I882
Тип. Anas tadorna
С 1 н о н и *. ы. asarca Bonaparte, 1838.— Vulpanser Keyserling et В 1 a-
si us, 1840—Pseudotadorna Kuroda, 1917.
17. Пеганка Tadorna tadorna L.
Anas tadorna. Li nnaeus. Syst. Nat., изд. X, 1, 1758, стр. 122, Швеция.
Синоним. Anas cornuta. G m e 1 i n. Reise d. Russl., 1774, стр. 185, Нижняя Волга.
Русские названия. Пеганка—народное, широкораспространенное, кроме того,
га агаз (Крым), хархаль (Каспийское море), бугровая утка (Южный Урал) и др.
Распространение. Гнездовой ареал. Британские острова (их
побережья и отдельные местности внутри страны, побережья Северной Фран-
ции, Бельгии, Голландии и Северной Германии; Дания, Норвегия к северу
даже до 71° с. ш., Западная Швеция, Юго-Западная Финляндия и западная
часть польского берега. В Советской Прибалтике гнездится на о-вах
Хиуме, Сарема, Вайка, Самсала и смежных с ними. В области Средиземного
моря гнездится разрозненными колониями, состояние некоторых из них
ПЕГАНКА
345
в настоящее время неизвестно, возможно есть в Южной Испании, Сардинии,
Южной Франции и Алжире (озеро Феццара, Цедлитц); гнездится в Греции
(Фессалия; Штреземан, 1920). На Черном море гнездится вдоль берегов
Болгарии и Румынии, в приморской полосе Молдавии и Украины и в степ-
ной части Крыма, где особенно обычна в районе Сиваша. Под Днепропетров-
ском довольно регулярно появляется осенью в период пролета, в Винницкую,
Киевскую, Полтавскую и Харьковскую обл. залетает лишь изредка, чаще
Карта 38. Распространение пеганки Tadorna tadorna
•—граница гнездовий, а'—районы наибольшей численное, птиц на гнев до в ье, б—не вполне
выясненная граница гнездовий, в—изолированные гнеадовые колонии, в'—места постоянные
встреч летом и спорадичного гневдования, г—зимовки, в—валеты, э#с—места скоплений хин»
ных птиц
осенью и даже зимой. Повидимому, гнездится в Приазовье и, несомненно,
в степном Предкавказье (Маныч—Кума), но детали распространения по этой
территории неясны. Обычна в Прикаспийских степях и спускается по за-
падному берегу Каспия до степей Юго-Восточного Закавказья; гнездится
в Армении в Севанской котловине (Шелковников, 1934). В Поволжье доходит
к северу до Сарпинских озер, а в Волжско-Уральских степях до отрогов
Сыртов, т. е. примерно до 50° с. ш. (Динесман), при этом она обычна на гнез-
довье вдоль линии соленых озер: Эльтона, Аще-Сая, Рыбного Сакрала, Арал-
Сора (Волчанецкий, 1937). По р. Уралу не идет севернее Калмыкова (Бое-
танжогло, 1911), далее гнездится к югу от р. Илека (Зарудный, 1897), дохо-
дит до верховьев р. Ори и верхнего течения Иргиза (Сушкин, 1908). В За-
уралье граница резко поднимается к северу, пеганка гнездится наНаурзум-
ских озерах (Кустанайская обл.), где все же редка (Формозов, 1937), более
обычна на степных озерах под Кокчетавом (Штегман, 1934) и на озерах Горь-
346 ОТРЯД ГУСЕОБРАЗНЫЕ
кой линии (Елпатьевский, 1901). В Барабинской степи нормально гнездится
до южного берега озера Чаны, севернее же лишь спорадично и не регулярно
(Иогаисен, 1907; Карачи, Рузский, 1932 и др.) восточнее Оби ее нет; гнездит-
ся в Кулундинской степи по западной степной окраине Алтая до Усть-Ка-
меногорска (Сушкин, 1938), спорадично в Зайсанской котловине, по Кур-
чуму и Черному Иртышу. Далее граница огибает с юга Монгольский Алтай
и затем выдается к северу через озеро Барга-Нор до Убса-Нурской кот-
ловины (Сушкин, 1938; Ячушевич, 1948). Севернее известны две изолирован-
ные гнездовые территории: одна в Тувинской котловине и вторая от Абакан-
ской степи до степных озер Ачинского района (Широ, Учум и др.), т. е. при-
мерно до 55° с. ш. (Сушкин, 1914). В Хангзе пеганка отсутствует, но восточ-
нее гнездится, повидимому, по всей Монголии, заходя в пределы западной
степной Маньчжурии. В этой части ареала она известна на гнездовье в пре-
делах Союза по р. Чикой (приток Селенги, Моллесон), в Даурии на озере
Барун-Тарей и в степях по Аргуни, севернее же—в район Читы она лишь
залетает. Южная граница совсем не ясна. В Северной Гоби пеганка одна из
самых обыкновенных птиц солоноватых озер, предположительно гнездится
и в Центральной Гоби на озере Гашун-Нор (вблизи Сого-Нор) (Козлова,
1930). Предположение о гнездовании ее на Лоб-Норе, в Са-Чжоу, на озере
Куку-Hop, в южной части Алашанья и Ордосе (Сушкин, 1938) пока не до-
стоверно, так как все встречи птиц в этих местах приходятся на период проле-
тов. В низменном Семиречье пеганка гнездится всюду, но более обильна
в Восточном Прибалхашье (Шнитников, 1949). По Сыр-Дарье обычна всюду
до ее гористой части, на Аму-Дарье много реже, и от Чарджуя вверх по те-
чению на гнездовье не встречена (Зарудный, 1896). В небольшом числе гнез-
дится в низменных частях Таджикистана (Иванов, 1940), достоверных сведе-
ний о гнездованье на Памире нет. Распространена повсюду в подходящих
местах в Туркмении (прикаспийские местности, Западный Узбой) и в Иране
на юг до Сеистанских озер, возможно в Ираке и Малой Азии (Уайзерби).
Залеты пеганок за пределы гнездового ареала — явление довольно ча-
стое. Чаще всего они приходятся на сезоны пролетов, особенно на осень
и начало зимы.
Основныезимовки находятся на Британских о-вах, в Северной
и Западной Франции, Португалии и восточной части Средиземного моря как
на европейском, так и на африканском берегу. Меньше на побережьях Адриа-
тического и Эгейского морей, чаще в Малой Азии и Египте. На Черном море
пеганки зимуют у берегов Болгарии и Румынии, на Сиваше и в горле Азов-
ского моря. Большие зимовки находятся на юго-западном берегу Каспия
и на озерах Муганской степи, в массе зимуют они и на Туркменском берегу
к югу от Челекена. Зимуют в Ираке, на побережьях Ирана и в Сеистане,
в Пакистане и Индии от Синда до Ассама, но как птица преимуще-
ственно морских берегов—там редка. В Японии зимой бывает редко и не-
регулярно, чаще встречается только на о-вах Кюсю (Ян, 1942); в Корее
встречается лишь на пролетах и редко; на побережье Южного Китая зимой
обычна (Голдуэл, 1931); залетают зимой до Вьетнама и о-ва Лусона (Филип-
пины).
Скопления линных птицу пеганок выражены слабее, чем
у других видов уток, так как самцы у них некоторое время заняты заботами
о выводке и не совершают дальних миграций на линьку; стаи линных птиц
собираются на Сиваше и в некоторых других местах.
Характер пребывания. Повсюду в Европе перелетная пти-
ца, кроме Британских о-вов и средиземноморских гнездовий, где оседла.
В Азии оседлы, повидимому, гнездящиеся в Туркмении и Иране.
Сезонные миграции и их даты. Птица, скольцованная
на зимовке в Га^ан-Кули, добыта летом в Алакульском районе Алма-Атин-
ПЕГАНКА 347
ской обл. Других сведений о возврате колец на территории Союза нет.
В Прибалтике пеганки появляются в последних числах марта или в первых
числах апреля. В северозападной части Черного моря они прилетают в пос-
ледней декаде февраля, реже и в начале марта (Клименко, 1950); на Си-
ваше появляются в марте, но перелет продолжается и в апреле, в это время
они скапливаются в заливах тысячными стаями (Киселев, 1941). С берегов
Аджарии они исчезают к началу апреля (Вильконский). В Гасан-Кули
(Юго-Восточный Каспий) интенсивный пролет их происходит в марте;
в низовьях Атрека в марте число их заметно возрастает за счет стай, подлетаю-
щих с юга, а в последних числах месяца оно резко, сокращается (Житни-
ков, 1900). На озерах Узбоя пролет их начался в первой половине февраля
и продолжался в течение всего марта (Шестоперов), на о-ве Кулалы (Се-
верный Каспий) первые птицы появились 23 марта (Смирнов, 1916), а в дель-
те Волги 28 марта. В нижнем течении Сыр-Дарьи (Джусалы—Чили) нор-
мальный весенний пролет пеганок начинается в первых числах марта и про-
должается до конца месяца, но в некоторые годы птицы попадаются уже
в конце февраля (Спангенберг и Фейгин, 1936). На о-ве Барса-Кельмес
(Аральское море) прилетают, в зависимости от весны 4—22 марта (Исмаги-
лов и Васенко, 1950). В степях Западного Казахстана пеганки появляются
рано. Между Темиром и Эмбой они были замечены 6 апреля, когда
в степи еще оставалось много снега, а запоздалой весной они появились
в низовьях Иргиза 12 апреля. В последнем месте пролет бывает сильным,
птицы летят стаями до 50 шт.; заканчивается пролет в конце мая (Сушкин,
1908). В низовьях р. Или прилет пеганок отмечался 17 и 21 марта, а очень
ранней весной A948) даже 4 марта; у северного предела ареала, на Барабе
(Татарский район Новосибирской обл.), первые птицы наблюдались 13 апреля
(Слудский). В Северной Гоби, на озере Орок-Нор прилет наблюдался
очень поздно—23 апреля, после чего небольшие стайки наблюдались до кон-
ца мая (Козлова, 1930); на р. Чикой (Юго-Западное Забайкалье) пролетная-
птица добыта 12 апреля (Моллесон), а на озере Тарей-Нор (Даурия) появ-
ляется в середине апреля (Штегман, 1928). Осенью пеганки исчезают из
Даурии к середине сентября; в долине р. Толы (Монголия) пролетные птицы
наблюдались во второй половине августа, а последняя пара птиц замечена
в Юго-Восточном Хангае 25 сентября (Козлова, 1930). Отлет пеганок из
верховьев Тобола (озеро Мокрое) во второй половине сентября, самая пос-
ледняя встреча там 25 сентября; у северной окраины Больших Барсуков
наблюдались еще 19 октября (Сушкин, 1908); среднее течение Сыр-Дарьи
покидают в середине ноября. Пеганки, гнездящиеся на озерах Узбоя, отле-
тают уже в сентябре, а в октябре на этих водоемах появляются пролетные
стаи; в Красноводеком заливе первые стаи в 1937 г. замечены 10 ноября,
в ноябре же происходит наиболее интенсивный пролет в Гасан-Кули. У бе-
регов Аджарии птицы появляются в середине октября, а в северо-западной
части Черного моря отлет заканчивается во второй половине сентября, но
изредка в годы с теплой и затянувшейся осенью пеганки задерживаются
там до конца ноября.
Биотоп. Во всех частях ареала пеганка придерживается солоновато-
водных озер, наличие их зачастую определяет границу распространения
этого вида. Пеганки явно предпочитают те соленые водоемы, поблизости от
которых имеются песчаные бугры, удобные для постройки нор, или каменные
осыпи с пригодными для этой цели расщелинами. На горах, расположен-
ных среди обширных равнин с плотной глинистой почвой, пеганки держатся
летом в значительно меньшем числе, а иногда и совсем отсутствуют. Лишь
в Прибалтике и на севере Западной Европы они гнездятся также непосред-
ственно у морских заливов. На реках и пресных озерах пеганки появляются
лишь во время пролета. На зимовках пеганки скапливаются обычно огром-
348 ОТРЯД ГУСЕОБРАЗНЫЕ
ными стаями в мелководных морских заливах или на обширных, залитых
водой солончаках.
Численность. Пеганки обычны, а местами обильны и распростра-
нены наиболее равномерно в основной части своего ареала от Прикаспийских
и особенно Приаральских степей на северо-западе до озер Северного Казах-
стана (Горькая линия) на севере, аральских Кара-Кумов и Прибалхашья
на юге. Повидимому, столь же обычны они и для Монголии.
Экология. Размножение. Половая зрелость наступает на втором
году жизни. На места гнездовья птицы прилетают уже парами, пары не раз-
биваются вплоть до появления выводков; в отличие от большинства других
уток самцы принимают участие в заботе о потомстве. Возможно—пары по-
стоянные, но достоверных данных, подтверждающих это, пока не имеется.
Токование самца довольно однообразно. Сидя на берегу около самки, он
подымает кверху голову, растопыривая при этом перья на шее и приот-
крывая клюв (Формозов, 1937); иногда подпрыгивает на месте и обычно»
издает свистящий звук.
Спаривание происходит обычно на воде, но иногда и на суше.
Гнезда пеганка помещает, как правило, в норах, и притом глубоких до-
3—4 м длины; при невозможности найти или вырыть нору гнездо строится
на поверхности земли. В Барабинской степи и в Крыму находили гнезда
в скирдах прошлогодней соломы. Для гнездования используются гнезда
лис, корсаков и барсуков или (Кустанайские степи; Формозов, 1937)
старые норы сурков; на Западном Узбое—норы дикобразов (Дементьев,.
1952); используются также казахские могильники, в особенности построен-
ные из саманного кирпича, старые казахские зимовки, где (Семиречье) гнезда
устраиваются в остатках печей. Устраиваются норы и в береговых обрывах,
а открытые гнезда располагаются обычно среди кустиков, солянок (в северо-
западной части Черного моря; Клименко, 1950) и в зарослях чахлого трост-
ника. В Семиречье возможно используются норы больших песчанок, вход
в которые птицы расширяют (Шнитников, 1949). В Даурии (озеро Тарей-
Нор) пеганки поселяются в норах тарбаганов. В Волжско-Уральских пес-
ках норы пеганок помещаются обычно на невысоких песчаных береговых обры-
вах, а в казахских могильниках (мазарах) они здесь не наблюдались (Вол-
чанецкий, 1937). Норы в скирдах бывают очень глубокими, они опускаются
примерно на 1,3 jw, а затем еще около метра идут в сторону. Наконец, в Арме-
нии около озера Севан (Гилли) гнезда пеганок находили в лисьих норах
и в расщелинах скал (Шелковников, 1934). На островах Советской Эстонии,
так же, как и в Северо-Западной Польше, отмечено гнездование в лисьих
и в самостоятельно вырытых норах (в песчаных дюнах), на чердаках неза-
селенных зданий, стоящих близко к морю, и в каменных оградах около них,,
а также на земле в кустах чертополоха (Гомейер, 1887; Кох, 1911). Птицы
не избегают близкого соседства друг друга, и в некоторых сложных лисьих
норах гнездятся одновременно от 2 до 5 самок (Сиваш). Расстояние гнезд
от воды зависит от местных условий и иногда бывает более километра. От-
мечено, что некоторые норы занимаются несколько лет подряд. Гнездо обильна
выстилается пухом.
Обычные размеры полной кладки установить трудно, так как повиди-
мому, нередко случаи откладки яиц, несколькими самками в одно гнездо*
Гнезда на Барса-Кельмесе содержали по 9—12 яиц; все три кладки, изве-
стные из дельты Или—по 8 яиц; законченные кладки на озере Кургальджин
два по 8, одно 17 и два по 18; на Сиваше 12—15; в Эстонии 8—10 яиц.
Нередко из-за недостатка нор в однвм гнезде несутся по нескольку самок,
и бывает, что в гнездах скапливается до 50 яиц (Сиваш; Киселев, 1941).
Цвет яиц сливочно-белый, иногда с легким оливковым оттенком. Их
размеры 64—68,9x46,8—50 мм (Кургальджин; Лавров); 64—70x42—51 мм
ПЕГАНКА 349
(Сиваш; Киселев, 1941); обычно вес около 85 г. (Хейнрот), 76,5 г (по Нитхам-
меру). Яйца откладываются ежедневно; насиживает одна самка, 27—29 дней
(по разным наблюдениям, с откладки последнего яйца). Последние двое
суток самки с яиц не сходят. Обычно, покидая гнездо, самка закрывает яй-
ца пухом даже в тех случаях, когда они находятся в глубокой норе. Весь
период насиживания самец держится поблизости и в случае появления хищ-
ника начинает с криком летать у гнезда. Выходя на кормежку, самка не-
которое расстояние проходит пешком и лишь после этого поднимается на
крыло, самец сопровождает ее. Вес молодых в день вывода 55 г. Из гнезда
молодые выходят самостоятельно, даже выведшиеся на скалах сами спры-
гивают вниз. Родители переводят выводок на водоем,при этом птенцы следуют
за матерью, а самец держится в стороне или следует сзади. Переход менее
километра занимает около получаса. Молодые птенцы умеют нырять, но в
случае опасности стараются укрыться у берега, родители же в это время
с криком летают поблизости.
Характерной особенностью всех пеганок является объединение несколь-
ких выводков птенцов при одной паре родителей. Это происходит в резуль-
тате преследования самцами других взрослых птиц, появляющихся с вы-
водками около уже занятого самкой района. Отставшие от изгнанных
родителей птенцы присоединяются к выводку захватившей данный участок
водоема пары.
Полное развитие молодых происходит примерно за 2 месяца. Самец
покидает выводок значительно раньше самки,
В низовьях Или массовая кладка яиц начинается около 10 мая, гнездо,
найденное 14 мая, содержало уже 8 свежих яиц, но еще 30 мая добыта самка
с яйцами в яйцеводе. Только что выведшиеся пуховики наблюдались 13 июня,
а летный выводок 30 июля (Слудский). На озере Тили-Куль (Кзыл-Ордин-
екая обл.) выводки наблюдались 18 июня, а ниже по Сыр-Дарье с конца мая
по середину июня. На Теджене только что вылупившийся птенец добыт 18 мая
(Зарудный, 1896). На о-ве Барса-Кельмес пеганки гнездятся в середине мая,
появление выводков на водоеме отмечено 29 июня и 2 июля; в верховьях
Ори выводок мелких пуховиков встречен 29 июня, а в верховьях Илека в се-
редине июля встречаются как только что выведшиеся пуховики, так и уже
летние выводки. В Волжско-Уральских песках пуховики встречаются с се-
редины июня до первых чисел июля, а в начале августа уже нередко появ-
ляются летные молодые.
На озере Кургальджин кладка из 5 свежих яиц найдена 19 мая, из
8 свежих 27 мая, а из 17 и 18 сильно насиженных 2—15 июня; в Барабин-
ской степи гнездо с единственным яйцом найдено 21 мая; на озерах Горькой
линии пуховой птенец добыт 14 июля (Слудский). В Наурзумском заповеднике
(Кустанайская обл.) пуховые птенцы обнаружены 12 июня, но 11 июня
было еще найдено гнездо с яйцами, а в первой половине августа выводки уже
поднялись на крыло (Михеев, 1937). На островах Джарылгачского залива
(Черное море) в первой половине мая в гнездах уже встречается по 1—2 яй-
ца, но откладка свежих яиц отмечена и 21 июня (Шарлемань и Шуммер,
1930), а в начале второй декады июня уже появляются выводки (Клименко,
1950). В Эстонии откладка яиц приходится на май.
Линька. Как и у всех уток, двукратная, но летняя почти непосред-
ственно переходит в предбрачную, и смена оперения на ряде участков тела
происходит, очевидно, один раз в год. Самцы, повидимому, начинают
линять несколько раньше самок, но не на много, так как при выводках они
остаются очень долго.
Смена контурного оперения начинается еще в период жизни взрослых
птиц при выводке, обычно в июне. Во второй половине июня пеганки
начинают собираться в стаи, а в конце июля и в августе на Сиваше,
350 ОТРЯД ГУСЕОБРАЗНЫЕ
куда на линьку собираются пеганки со всего приморья, уже держатся
стаи их в несколько тысяч штук. Сперва выпадают перья нижней стороны
тела, отчего исчезает каштановый пояс на груди и черная полоса вдоль
брюха. Затем сменяются кроющие хвоста, часть перьев мантии и плечевых,
но не спина. Позже, уже в начале августа, линяют крылья и продолжается
смена рулевых. На Сиваше маховые отрастают у них лишь в сентябре. Пере-
линяв, они продолжают держаться там стаями по нескольку сот и даже
тысяч штук (Киселев, 1941). Следующая линька происходит с августа до
декабря, когда снова сменяется большая часть контурных перьев. Линька
у взрослых самок проходит так же, как и у самцов. Молодые птицы начинают
свою первую линьку в августе или в начале сентября и заканчивают ее
в марте. У них сменяются почти полностью все контурные перья, кроме
небольшого количества их, сохраняющихся на брюхе и груди. Перелинивает
хвост и часть внутренних второстепенных маховых и их кроющих, а иногда
также часть больших средних кроющих крыла.
Питание пеганок изучено еще слабо. Несомненно, что это—животнояд-
ная утка, кормящаяся нежным кормом: ракообразными и личинками насе-
комых, но в некоторые сезоны потребляет и растительную пищу. Весной,
по прилетев дельту Или, пеганка наряду с другими утками кормится в основ-
ном семенами лебедовых, которыми изобилуют мелководные озера на солон-
чаках (Штегман, 1949). В летнее время намечаются три основных корма:
рачки—Artemia salina (и их икра), которые иногда в массе населяют соленые
озера, личинки хирономид, существующие в озерах с довольно высокой соле-
ностью, и одноклеточные водоросли (возможно Aphanoteca salina, весьма
обычная для соленых озер). Так желудки и пищеводы у восьми крупных
птенцов, добытых 31 июля, содержали красных личинок хирономид (Слуд-
ский); по наблюдениям Зарудного A888), в степях к югу от Илека подняв-
шиеся на крыло выводки кормятся на озерах тиной и какими-то красными
личинками (несомненно, хирономидами), кроме того, бегая по берегам,
они собирают «различных насекомых, песок с перегнившими органическими
веществами, споры водорослей (вероятно, одноклеточные водоросли) и семена
солончаковых растений. Часто не только желудок, но и пищевод во всю
свою длину бывает набит этими спорами» (водорослями). Формозов A937)
наблюдал на Наурзумских озерах энергичное поедание пеганками саран-
чевых насекомых. Пять молодых пеганок, добытых 4 июля, все без
исключения имели в желудках остатки пруса (Calliptamus italicus), причем
у наиболее крупного птенца их оказалось 17 экземпляров. Во время летней
линьки (озера Горькой линии) птицы держатся по середине озер и кормятся
рачками и водорослями, перелинявшие же перемещаются к вечеру на кор-
межку к берегам (Елпатьевский, 1901). Желудок взрослого самца, добытого
19 июля на озере Сары-Муин (Наурзумские озера), был наполнен много-
численными Artemia salina (Деливрон, 1937); по Сушкину A908), в Централь-
ном Казахстане летнее питание пеганок состоит главным образом из яиц
этих же рачков, которыми кишат некоторые соры; иногда весьма помести-
тельный зоб пеганки бывает набит только яйцами их. В конце лета пеганки
дважды в сутки летают кормиться на поля и возвращаются на озеро около
8 час. утра и 5 час. вечера (не за прямокрылыми ли они летают?).
Основными кормами пеганок в Волжско-Уральской степи служат
A. salina и мухи, выводящиеся в соленой грязи (Волчанецкий, 1937). Пище-
вой рацион пеганок на Сиваше состоит в летнее время из трех основных видов
корма: рачков—Artemia salina, их яиц и мелких личинок комаров-толкун-
цов (Chironomidae). Среднее число рачков в одном желудке 1800, макси-
мальное 19 800, их яиц 10150 и доходит до 36 800, личинок комаров в сред-
нем 5970 и до 63 880 шт. Кроме того, каждый желудок содержит около 2,4 г
кашицы из деформированных организмов тех же видов, что соответствует
ПЕГАНКА
351
еще 7—12 тысячам экземпляров. Этот корм находится во взвешенном со-
стоянии в воде Сиваша, и массовые скопления его перегоняются ветром от
одних берегов к другим. Огромные стада пеганок, состоящие из линяющих
птиц и выводков молодых, перемещаются по Сивашу вслед за скоплениями
планктона (Андреев).
На соленых озерах Узбоя птицы кормились, повидимому, ракообраз-
ными, но в дни сильного лёта муравьев зобы и желудки пеганок были
набиты исключительно ими. Повидимому, они все нуждаются в пресной
воде» так как постоянно прилетают к тем местам, где в соленые озера при-
Рис. 63. Пеганка
текает пресная вода (Узбой, Сиваш, Кара-Богаз-Гол). Наконец, на зимов-
ках в заливе им. Кирова и в Муганской степи основу питания пеганок со-
ставляет растительная пища. Содержимое желудков птиц, добытых в фев-
рале, состояло лишь на 8,2% (по объему) из мелких моллюсков (Hydrobia
Theodoxus), на 16,1% из мелких семян, преимущественно руппии и камы-
шей, а в остальном из трухи вегетативных частей растений, главным образом
водорослей, и частично трухи семян.
Хозяйственное значение пеганки, как объекта промысла, невелико глав-
ным образом из-за невысокой численности этого вида. Мясо ее в некоторые
сезоны обладает неприятным привкусом (лето), но когда птица жиреет (на-
чало зимы, местами и весна) качество ее мяса повышается. Промысловая
добыча имеет место лишь на каспийских зимовках. Пеганка охотно занимает
устроенные для нее гнездовые сооружения. Так, на островах Сиваша ее
полностью перевели в искусственные, деревянные норы, гнездовые камеры
которых сообщаются крышкой с поверхностью. Норы с одним ходом и не-
сколькими гнездовыми камерами занимались иногда несколькими самками
(Киселев, 1941). Удачный опыт создания искусственных гнездовий для
352 ОТРЯД ГУСЕОБРАЗНЫЕ
пеганки был проведен в Эстонии на о-ве Вайка, где для этой цели использо-
вались случайные материалы: старые бочки, пустые мины и обломки лодок
{Транзее, 1927), местами в Северной Германии устройство искусственных
нор практикуется с целью сбора пуха, не уступающего по качеству гагачьему,
и яиц. Путем постепенного выбора яиц из гнезд кладку одной самки доводят
до 20—30 шт. и более. Пеганки легко привыкают к соседству человека,
и там, где их не преследуют, гнездятся невдалеке от жилья. Проведение
несложных биотехнических мероприятий (создание гнездовий) может зна-
чительно повысить численность этой утки и обеспечить возможность полу-
чения ценного дополнительного продукта —пуха.
Полевые признаки. Крупная утка гусеобразного типа, по земле бегает
легко и держит туловище горизонтально. Окраска пестрая, на основном
чистобелом фоне выделяется черная голова и коричнево-красная перевязь
от груди к лопаткам. Верхняя сторона крыльев белая, что хорошо видно
на полете. Клюв красный. Хорошо плавает, но ныряют только птенцы. Часто
скапливается большими стаями. Полет гусиного типа с редкими взмахами
крыльев; летает всегда молча. На перелетах летит клином или развернутым
фронтом; голос, обычно кряканье, глухое и довольно мягкое, передаваемое
слогами «га-га-га»; самец в брачное время издает довольно чистый свист вроде
«вхи-оо» или «цвие-цвье».
Описание. Строение и размеры. Крупная утка. Клюв широкий, плоский,
с округлой вершиной и несколько изогнутым кверху надклювьем; коготок узкий и малень-
кий. Роговые пластинки по краю надклювья довольно редкие и грубые, а на подклювье
они, наоборот, очень частые и тонкие. У основания надклювья у самца большой кожный
нарост, развивающийся к весне, когда он достигает 200 мм в высоту, и опадающий к осе-
ни (в декабре он достигает около 1,5 мм). Крылья довольно длинные, широкие, в сложен-
ном виде достигают конца крыла. Лапа некрупная, цевка также; она равна длине средне-
го пальца с когтем. Рулевых 14. Длина крыла самцов 310—360, самок 280—330 мм; цев-
ка самцов 48—58, самок 45—53 мм; клюв (от угла рта) у самцов 55—58, у самок 53—
56 мм.
Вес. Взрослые самцы, добытые на весеннем пролете на озерах Западного Узбоя,
весили 1100—1500 г (Шестоперов), весной же в дельте Или они бывают неплохо упитаны
и весят обычно около 1300—1350 г (Штегман), очень худые птицы около 1200 г (Слуд-
ский); там же летом вес их снижается до 1000—1150 г. В феврале на ленкоранских зимов-
ках средний вес самцов (включая и молодняк) 1180 г (830—1500 г), в Гасан-Кули 1150—
1250 г. Самки весной на Западном Узбое 900—1100 г, в июне на озере Кургальджин 1000 г,
на зимовках в феврале 813 г E62—1085 г.) В дельте Или молодые птицы в последних
числах июля и в первых числах августа весят от 625 до 1100 г. (Сл уде кий). Предельный
вес селезня 1650 г. (Сиваш, Киселев, 1941).
Окраска. Пуховой птенец. Верх головы (кроме лба), задняя сторона шеи
и спина до середины, над вестья и край крыльев серовато-бурые, все остальное белое.
Молодые птицы в первом наряде. Верх головы и шея сзади черновато-бурые, но
лоб и бока головы у основания клюва, горло и передняя сторона шеи белые. Плечевые
перья серо-бурые, остальной верх туловища и низ его белые, иногда с мелкими серова-
тыми пятнами на брюхе. Первостепенные маховые черные, второстепенные почти без
блеска, с белыми каемками на вершинах (настоящего зеркала нет). Мелкие и средние
верхние кроющие крыла беловатые, большие серые. Рулевые белые с черновато-бурыми
каймами и вершинами. Клюв и ноги розово-телесного цвета; радужина бурая.
Взрослая с а м к а. Окрашена сходно с самцом, но голова черно-бурая, лоб, щеки
и горло обыкновенно беловатые, полоса вдоль брюха темнобурая с белыми пестринами;
последние есть и на каштановом поясе груди. Плечевые перья с мелким черным крапом.
Клюв красный, но несколько бледнее, чем у самца, с черным кончиком; ноги розовые;
радужина бурая.
Самка после летней линьки похожа на самца в том же наряде, но голова более корич-
невая; центральная часть груди и брюхо—белые.
Годовалый самец после первой линьки. Окрашен, как взрослый, но зеленоватый
отлив на голове и шее тусклее; среди плечевых перьев сохраняются юношеские серые
(у взрослых белые); каштановая перевязь на груди с примесью иначе окрашенных перьев;
черная полоса вдоль брюха уже и тусклее, чем у взрослых. Крылья, в основном, как
у молодых, но внутренние второстепенные маховые и часть кроющих крыла, как у взро-
слых.
Взрослый самец в брачном наряде. Голова и шея черные с сине-зеленым отли-
вом, основание шеи и зоб белые. Через верхнюю часть груди и бока перед основанием
ОГАРЬ
353
крыльев проходит довольно широкий густокаштановый пояс, который соединяется в верх-
ней части спины. Плечевые перья черные, кроме самых внутренних белых. Вся остальная
спина, надхвостья и бока тела белые. Вдоль брюха от средней части груди проходит широ-
кая черная полоса; подхвостье светлокаштановое. Первостепенные маховые черные,
второстепенные с темнозелеными наружными опахалами, образующими зеркало, внутрен-
ние второстепенные с белыми внутренними остатками и ржаво-коричневыми наружными,
узкая средняя часть перьев черная. Все верхние кроющие крылья образуют сплошное
белое поле; подмышечные также белые. Рулевые белые с черными вершинами. Клюв
и кожистый нарост на надклювье яркокрасные, с темным ноготком; ноги розовые; раду-
жина краснобурая.
Самец в летнем наряде. Голова и шея не чисточерные, а буроватые за счет бурых,
а снизу головы беловатых каемок перьев; широкие основания перьев светлые. Каш-
тановый пояс не сплошной, на груди он прерывается белыми перьями с бурыми
каемками. Черная полоса вдоль брюха почти полностью исчезает*
18* Огарь Tadorna ferruginea Pall.
Anas ferruginea. Pallas, (аноним). Vroeg's Cat. Adumbr., 1764, стр. 5, сТатария».
Синонимы. Anas Casarca. Li nnaeus. Syst. Nat., Ill, App., Anim., 1768, стр. 224,
Астрахань.— Anas rutila. Pallas Nov. Comm. Petrop., XIV, I, 1770, стр. 579.
Русские названия. Народное—красная утка (широко распространенное), вар-
нава (Алтай, Кулунда), турпан (Байкал, Забайкалье, Северная Монголия), отайка (Сред-
ня Азия, местами)*
Распространение. Гнездовой ареал. Занимает степную и пусты н
ную зоны Палеарктики от Средиземного моря до Приамурья. Огарь гнез-
Карта 39. Распространение огаря Tadorna ferruginea
а—граница гнездовий, а'—районы наибольшей численности птиц на гнездовье, б—не вполне
выясненная граница гнездовий, в—изолированные гнеэдовые колонии, •'—места постоянных
встреч летом и спорадического гнеадованья, а—основные зимовки, д—валеты, в—встречи про-
летных птиц весною, севернее гнездовых мест, ж—места скоплений птиц на линьку, а—ме-
ста, в которых птица исчезла за два последних столетия
дится в Южной Испании, где редок, в Марокко, Алжире, возможно в Тунисе
и в Египте, а также в Малой Азии, Трансиордании (Давыдов, 1898) и Ираке.
На Балканском п-ове огарь гнездится в Греции, Турции, Болгарии и Румы-
нии. В двух последних странах он редок, и вообще на берегах Эгейского
23 птицы СССР, т. IV
354 ОТРЯД ГУСЕОБРАЗНЫЕ
и Мраморного морей чаще, чем на черноморских. В Молдавии огарь гнез-
дится на побережье вдоль Прута и на Днестровском лимане (Радаков, 1879;
Остерман 1912 и др.); на юге Украины он исчез почти совершенно. В 90-х го-
дах прошлого столетия огарь гнездился на Днепровском и некоторых соле-
ных лиманах (Браунер, 1894), но уже в начале 900-х годов он стал в низовьях
Днепра редкой залетной птицей (Пачосский, 1911), и единственный случай
гнездования его на о-ве Джарылгач отмечен в 1923 г. (две пары, Шарлемань
и Шуммер, 1930). За последние годы гнездящихся птиц на этой территории
не отмечалось (Клименко, 1950). Бесследно исчез огарь в Северном При-
азовье (Костюченко, 1926) и почти полностью в степном Крыму, где теперь
в районе Сиваша гнездятся лишь отдельные пары (Снигиревский, 1923;
Воронцов, 1937). Небольшая гнездовая колония сохраняется в полувольном
состоянии на прудах Аскания-Нова. Она происходит от завезенных, аккли-
матизированных птиц. Граница ареала этого вида между Доном и Волгой
не выяснена, но, повидимому, огарь гнездится по Куме, Манычу и, возможно,
в дельте Кубани. В дельте Волги он на гнездовье обычен и достоверно гнез-
дится на Сарпинских озерах, но дальнейшее его распространение к северу
не ясно. Повидимому, гнездится под Камышином (Гладков); есть указания
на летние встречи пар этих уток в степи по р. Еруслану (Волчанецкий и
Яльцев, 1934), а со слов охотников указывалась в качестве нередкой гнез-
дящейся птицы для южного берега Самарской Луки (Житков и Бутурлин,
1906). В Волжско-Уральских степях редок и гнездится, вероятно, только-
на юге; в среднем течении Урала (г. Чкалов) очень редок, по Хобде и в вер-
ховьях Илека обычен (Зарудный, 1897); восточнее Мугоджар огарь проникает
к северу примерно до Аман-Карагая E2,5° с. ш.), а на весеннем пролете
регулярно встречается и севернее, под Кустанаем и на Убаган-Тенизе.
Самое северное нахождение в Ишимской степи—озеро Чеглы (Советский
район Северо-Казахстанской обл.), где, однако, птицы добываются не каждый
год (Слудский), регулярно гнездится в Кулундинской степи, но в Барабин-
ской очень редок, встречается спорадично (Татарский и Купинский районы
Новосибирской обл.) и гнездование его там не доказано, столь же редок
и у Барнаула. Широко распространен огарь в Центральном Алтае, в высокой
части Западного Алтая и далее к югу до Монголии. Отсутствует в Северо-
Западном Алтае (западнее Семчинского хребта), на Иртыше в пределах Алтая
и в лесной части Северо-Восточного Алтая (к северу от Телецкого озера
и в бассейне Абакана; Сушкин, 1938). По Абаканской лесостепи поднимается
к северу до окраин Ачинского района (Ужура и озера Учум; Федюшин,
1925), где гнездится по окраине тайги. Восточнее Енисея обычен в Мину-
синской степи, но в горах в бассейне Тубы бывает лишь в качестве залетной
птицы, зато в пределах Тувинской автономной области всюду обычен. На
Байкале это обычная гнездящаяся птица к северу, по крайней мере до устья
Баргузина и острова Ольхон (Туров, 1923; Штегман, 1936). В Забайкалье наи-
более северные районы гнездованья известны на Витимском плато (озера
Арахлей, Иван; Штегман, 1928) и под Сретенском на Шилке; по Амуру он
найден лишь в Зейско-Буреинской низменности, под Благовещенском (Штег-
ман, 1928). Гнездится по всей Монголии и в Западной Маньчжурии (Ялам-Тэ;
Ямасина, 1939), но распространение по остальной территории Китая неясно.
Встречается в Ордосе, а на юге гнездится, судя по летним встречам, в Юннане,
в верховьях Янцзы (Лицзянь; Рили, 1926), в Ладаке (Оберхользер, 1900)
и определенно на озерах Западного Тибета, доходя до Гималаев (Столичка,
1868). Многочислен на Восточном Памире, но на Западном отсутствует,
гнездится в Ала-Шане и Кашгарии; заселяет большую часть Афганистана
и почти всю территорию Ирана, кроме приморских областей.
Основные зимовки огаря находятся в Алжире, Тунисе,.
Египте и особенно на Белом Ниле; зимует огарь также в Южной Аравии,.
огарь 355
в Эгейском море, Сирии, Трансиордании и Ираке. В Иране зимует почти по
всей стране, кроме ее северо-западной части и Хорасана. Особенно много
огарей в Сеистане (Зарудный, 1911). В Пакистане бывает регулярно, но не
в большом числе; зимует в Индии, не доходя до Малабарского побережья
в Ассаме, Манипуре и в Бирме в долине Иравадди. Держатся птицы здесь
парами и небольшими стайками, чаще всего по рекам. Проводят зиму и на
незамерзающих озерах Южного Тибета; Вьетнама достигают лишь одиночки.
В Китае огарь обычен зимой как по долинам рек, так и вдоль побережья
к югу от Шаньдунского п-ова, также в Корее, особенно у восточного и южного
берегов (Остин, 1948). На территории Советского Союза огари зимуют на
низменности Юго-Восточного Закавказья от Муганской и Сальянской сте-
пей до Кахетии. Но количество их здесь не велико. В низовьях Атрека, по
наблюдениям Житникова A900), зимуют в большом числе; зимуют и в других
частях Туркмении (Зарудный, 1896, Дементьев, 1952). В незначительном
числе зимует в Южном Таджикистане (Иванов, 1940).
Скопления линных птиц не бывают столь массовыми, как
у настоящих уток и нырков, так как оба родителя долго остаются при вы-
водке. Однако большие скопления линных огарей все же наблюдаются на
Балхаше, в его западной части (Слудский), на озерах Памира (особенно на
Сассык-Куле и Сары-Куле).
Характер пребывания. Повсюду перелетная птица, кроме
Средиземноморья, Северного Ирака, Сеистана и, возможно, Южного Тибета;
в СССР вероятно частично оседла в Туркмении.
Сезонные миграции и их даты. Кольцевание огаря
проводилось у настолько на юге Украины, в Аскании-Нова. Одна из околь-
цованных птиц добыта в Смирне (Малая Азия), а две на месте кольцевания
через 2 и 3 года (Шарлемань, 1933).
В Нижнем Египте отдельные мелкие стайки зимующих огарей задер-
живаются до мая, по всей вероятности они состоят из птиц-первогодков
(Хейглин, 1856). В северо-западной части Черного моря красные утки появ-
ляются в конце февраля—начале марта. В это время они часто встречаются
у Кинбурнесской косы, о-ва Долгого и в Днепро-Бугском лимане (Клименко,
1950). В Аскании-Нова возврат улетавших на зиму птиц отмечен 16 марта
(Шарлемань, 1924). В Пакистане и Северном Пенджабе встречается до марта.
Бывает на пролете в верхней части долины Куры и по Алазани; в дельте
Волги появляется в разные годы Э —26 марта. В низовьях Атрека интенсив-
ный пролет отмечен в конце первой половины марта, птицы летели на северо-
восток. В последних числах апреля, на взморье у Кара-Богаз-Гола они уже
встречались парами; на озерах Узбоя птицы наблюдались во второй поло-
вине марта. На Памир огари прилетают довольно рано, когда озера еще не
вскрылись, а болота покрыты льдом, между 15 и 20 марта, на Аличуре первое
появление отмечено 1 апреля, после чего птицы стали попадаться постоянно,
но обычно небольшими группами, по 4—5 особей (Мекленбурцев, 1946).
На Нижней Сыр-Дарье появляется весной в начале марта (Спангенберг),
но о-ве Барса-Кельмес (Аральское море) около 15 марта, а в годы с теплой
весной уже 25 февраля (Исмагилов и Васенко, 1950). В дельте Или красная
утка прилетает во второй половине марта (Штегман, 1949), первые птицы
появляются там еще до вскрытия реки, на полыньях, в разные годы от 7 до
15 марта (Слудский). На Темире их прилет был отмечен 17 марта, в низовьях
Иргиза массовый пролет наблюдался 29 марта, а в год с очень поздней весной
они появились там лишь 22 апреля; с самого начала пролета эти утки летят
парами (Сушкин, 1903). Прилет на Наурзумских озерах замечен 12 апреля
(Михеев, 1937), а под Семипалатинском 18 марта (Селевин, 1927). С южно-
китайских зимовок птицы отлетают в марте, пролет через Чжили продол-
жается с марта по май (Колдуэлл, 1931). Пролет красной утки в Монголии,
23*
356 ОТРЯД ГУСЕОБРАЗНЫЕ
в Северной Гоби и под Улан-Батором дважды отмечен 29 марта (Козлова,
1930), а в Забайкалье и на р. Чикой 14 марта (Моллесон, 1897).
В последних числах августа и в начале сентября красные утки начинают
собираться в большие стаи на соленых озерах, это предшествует их отлету.
Из области верхнего течения Иргиза они исчезают в середине или во второй
половине сентября; на Нижней Эмбе последняя встреча зарегистрирована
27 сентября (Сушкин, 1908). На взморье против дельты Волги красные утки
появляются в августе и до конца сентября держатся небольшими стайками
или выводками. В октябре они объединяются в крупные стаи и отлетают
в середине этого месяца. На озерах Атрека первая пролетная стая появилась
24 октября, в ночь на 29 октября отмечен сильный пролет, а 31 октября на
озерах они были уже массовой уткой и держались стаями до 30 шт. (Жит-
ников, 1900); на взморье у Гасан-Кули пролетные стаи наблюдались в пер-
вой половине ноября. На прудах Аскания-Нова пролетные птицы отмеча-
лись с 4 по 29 сентября, чаще в конце месяца. С о-ва Барса-Кельмес огари
отлетают в конце сентября, но отдельные птицы встречаются в течение всего
октября, в низовьях Сыр-Дарьи отлетают в конце ноября. В Северной Мон-
голии отлет красных уток начинается в 10-х числах сентября, так в Кентэе
(р. Хара) пролетная стая наблюдалась 17 сентября. Пролет через Чиили
отмечен в октябре и ноябре, а в последнем месяце огари достигают и южных
провинций Китая.
Биотоп. Весьма разнообразен. В степях Казахстана озера и реки,
обычно в холмистой местности; заросших водоемов огарь избегает, особенно
охотно гнездится на соленых и солоноватых озерах. Однако соленость водое-
ма для огаря не обязательна, и местами (Северная Монголия) он гнездится
около озер с пресной водой. В горах (Алтай) не избегает берегов с пышной
травой и даже слегка заболоченных (Байкал), если они перемежаются с уча-
стками твердой, солонцеватой или щебнистой почвы. Он гнездится от низ-
менностей Средней Азии до плоскогорий Памира и альпийских озер Алтая
в поясе полярной березки. В Гималаях достигает 3100 м, в Семиречье 4000 м,
а на Памире 4300—4500 м. Огарь заселяет местами окрайки лесов, но
сплошной тайги избегает Осенью стаи птиц группируются на озерах (обычно
соленых) с открытыми плесами. Зимует на низменностях, по широким раз-
ливам рек и озер.
Численность. Вполне обычен во всем Центральном Казахстане
и Семиречье, в Тувинской автономной области и Забайкалье. Многочислен
в Северной Монголии, весьма многочислен на Восточном Памире. Обычен
в Южной Туркмении и на значительной территории Ирана, в Южном Закав-
казье (Армения), весьма обычен в степях от Терека до Волги и несколько
западнее. На остальном протяжении ареала огарь встречается реже и распро-
странен менее равномерно.
Численность огаря в западной части ареала чрезвычайно сократилась
за сравнительно недавнее время, в Крыму он удержался лишь единицами,
в Приазовье исчез совсем, тогда как в середине прошлого столетия скопле-
ния их там достигали нескольких тысяч штук (Радде, 1854). Около 180 лет
назад огари были довольно многочисленны в Харьковской обл. (Сомов,
1897), исчезновение их там шло параллельно с исчезновением байбаков,
норами которых эти птицы пользовались для своих гнезд. В Воронежской
обл. они гнездились еще около 100 лет назад (Северцов, 1855), а в Среднем
Поволжье (Жигули) были известны еще в начале нашего века (Житков
и Бутурлин, 1906). Можно предполагать, что причиной сокращения ареала
послужило быстрое хозяйственное освоение территории; в Казахстане и За-
байкалье подобного явления не наблюдается.
Экология. Размножение. Половая зрелость, как правило, насту-
пает на втором году жизни. На места гнездовья птицы прилетают уже парами,
ОГАРЬ 357
которые у красных уток сохраняются, вероятно, в течение нескольких лет.
Брачные игры несложны. Самка ведет себя очень возбужденно, она с раскры-
тым клювом плавает вокруг самца, постоянно кричит и не подпускает близко
ни одной водяной птицы. Самка принимает позу защиты с вытянутой вперед
шеей и пригнутой к земле головой даже и при отсутствии поблизости врагов.
Судя по наблюдениям в неволе, повидимому, не самец выбирает самку, а самка
отыскивает себе самца. Самец же обычно стоит, высоко вытянув шею или
ходит около самки, наклоняя голову вперед и снова поднимая ее вверх.
Спаривание происходит чаще на земле, но иногда и на воде; в последнем
случае самец после спаривания некоторое время плавает около самки с
крыльями, высоко приподнятыми в локтях (как это делают гуси). Иногда
у огаря наблюдаются воздушные игры. Самец, поднявшись на большую вы-
соту, несется вниз по наклонной линии, сложив особенным образом крылья
и издавая ими свистящий звук. Затем птица поворачивается в воздухе, летит
в другую сторону и опускается на воду, где присоединяется к своей паре
(Козлова, 1930).
Гнезда помещаются в норах степных животных (лисиц, степных кошек,
барсуков), в трещинах, расселинах берегов и очень часто в старых казахских
могильниках, в заброшенных постройках, иногда в сараях и на чердаках
жилых построек. В дельте Волги, огарь роет норы в глинистых, круговых
обрывах. Норы эти, повидимому, очень длинны, иногда более 4 м (Бостан-
жогло, 1911). На о-ве Барса-Кельмес норы располагаются в невысоких
обрывчиках и у основания старых казахских землянок, два года одна пара
гнездилась в сарае у нежилого дома (Исмагилов и Васенко, 1950). В низовьях
Сыр-Дарьи наблюдалось однажды и открытое гнездо, построенное на большой
высоте, на карнизе огромного мазара (Спангенберг и Фейгин, 1936); в горном
Семиречье гнезда устраиваются в старых сурочьих норах (Шнитников, 1949).
В горах огарь ведет себя как настоящая горная птица. Иногда он делает
там гнезда в низких каменных буграх или грядах среди обычного для него
степного ландшафта, но чаще селится на обрывах, возвышающихся на много
десятков метров над дном долины, рядом с клушицами и горными вьюрками.
В самой северной части ареала, на границе Ачинского и Минусинского
районов, огарь гнездится в глубоких дуплах, преимущественно в дуплах
лиственниц, на высоте от 4 до 10 м. Деревья с гнездами находятся обычно
на опушках небольших полян и лесных ключей, иногда в нескольких кило-
метрах от озера. Возможно, что гнездование в дуплах иногда имеет место
также в Тувинской автономной области, на Южном Байкале и в Забайкалье.
На Среднем Байкале гнезда находятся иногда открыто на уступах скал. Нако-
нец, в парке Аскания-Нова (а также в Московском зоопарке) огари гнездятся
в ящиках, установленных для них на деревьях. Одни и те же гнезда и, пови-
димому, одними и теми же птицами нередко занимаются несколько лет подряд
(Овод, 1928; Насимович, 1951). Есть наблюдения, что красная утка успешно
выводила птенцов в одном из отнорков жилой норы лисы. Так как потре-
воженная насиживающая самка издает громкое шипение, напоминающее
шипение крупной змеи, возможно эти звуки спасают птицу, отпугивая лисиц
(Спангенберг, 1948).
Количество яиц в полной кладке обычно 8—12 (по Хейнроту 8—9), но
иногда и больше. Гнездо в дельте Или содержало 9 яиц, на Сыр-Дарье—9,
на Мангышлаке 15, в низовьях Иргиза 16, в Туркмении (Кара-Бенд) обнару-
жен выводок из 11 только что вылупившихся птенцов, а на о-ве Барса-Кель-
мес—из 12 молодых. Яйца не чистобелые, а цвета слоновой кости. Размеры
их 60,2—70,2x46—50 мм; A5) 64,9—69,2x44,9-46,9, в среднем 68,3х
X 46,23 мм (Глазков и Залетаев) вес яиц 75—85 г. Яйца откладываются по
одному в день. Срок насиживания 27—29 дней (по Уайзерби); насиживает
только самка, но, по наблюдениям Спангенберга A936), в насиживании прини-
358 ОТРЧД ГУСЕОБРАЗНЫЕ
мает участие и самец. Насиживание начинается после откладки последних
яиц. Пока кладка яиц продолжается, селезень провожает самку до гнезда,
позже он перестает подходить к гнезду, но наблюдает за его окрестностями.
При появлении опасности самец налетает на пришельца и криком дает знать
самке о грозящей опасности. Вес молодых при выводе около 52 г, в 10-дневном
возрасте 180 г, через 18 дней—430 г, в месячном возрасте—930 г и в 2-ме-
сячном—1100 г. К последнему сроку молодые вырастают полностью. Предпо-
ложение о переносе птенцов из высоко расположенных гнезд на воду самкой
неверно. Молодые выпрыгивают из гнезд сами. Острые коготки утят позво-
ляют им забираться по вертикальной дощатой стенке на высоту до метра.
При падении птенцы помогают себе, быстро махая короткими крылышками,
падают несколько по наклонной линии. Они очень легки и пушисты, что
ослабляет силу удара (Дергунов, 1926). Утята прекрасно бегают и хорошо
приспособлены к местным перекочевкам по суше. Даже взрослые птенцы
нередко убегают от наблюдателя по солонцу, ловко и быстро шмыгая между
кустиками травы. Пуховые птенцы прекрасно плавают и хорошо ныряют.
Обе взрослые птицы остаются при выводке до того времени, пока молодые
не поднимутся на крыло. Они оберегают своих утят более ревностно, чем
другие утки, иногда налетают на человека так близко, что едва не задевают
крыльями. При этом самка обычно остается у выводка, отводя молодых
в безопасное место, а самец налетает на врага, привлекая внимание на себя.
Красные утки энергично отгоняют пернатых хищников, нападая иногда даже
на орлана-белохвоста.
Огари, как и пеганки, весьма склонны объединять около себя птенцов
из чужих выводков, которых они отбивают, сражаясь с их родителями.
Однако «усыновляются» только молодые птенцы, не старше недельного воз-
раста, а более крупных птенцов чужие родители гоняют и бьют, как взрос*
лых птиц. Молодые добывают себе корм на мелководье, но столь же часто они
бродят с этой целью по солонцам. На ночь выводок собирается на берегу
среди тростников или рогоза.
На самом юге страны—в Туркмении (Кара-Бенд)—только что вывед-
шиеся птенцы встречены 8 мая (Радде и Вальтер), в Таджикистане (озеро
Кара-Куль) в конце июня встречены очень мелкие птенцы, а 8 июля они
уже были в половину матки (Иванов, 1940), на плоскогорье Памира гнездя-
щиеся птицы наблюдались с конца первой декады мая, а первые птенцы
встречены 26 июня (на высоте 4300—4500 м). В низовьях Сыр-Дарьи 16 мая
найдено гнездо с проклюнувшимися яйцами, через несколько дней уже
встречались пуховики. Но в другом гнезде самка продолжала насиживать
еще 27 мая. На о-ве Барса-Кельмес 7 апреля найдено гнездо со свежими
яйцами, а 22 апреля наблюдался выводок пуховиков, которых родители
переводили к морю. На Мангышлаке выводок, переходящий к морю, встречен
в конце мая, а 31 мая найдено гнездо, из которого птенцы вывелись через
неделю (Гладков). В верховьях Иргиза гнездо с довольно сильно насижен-
ными яйцами найдено 30 мая, а в конце июля встречались уже хорошо летаю-
щре молодые, хотя еще и с не совсем доросшими маховыми; к югу от Илека
летние молодые отмечены в середине июля. В Карагандинской обл. на озере
Соры выводки пуховиков встречены И и 13 июня (Слудский); на Наурзум-
ских озерах 21 июля наблюдался выводок мелких пуховых птенцов (Делив-
рон). В низовьях р. Или погоня селезней за утками наблюдалась 25—27 марта;
12 мая найдена кладка свежих яиц, 2 июня гнездо со слегка насиженными
яйцами, а 2 августа уже выводок поднявшихся на крыло молодых; на Ак-Су
пуховики размером с горлицу встречены 7 февраля (Слудский). На юге
Ачинского района в Красноярском крае кладка яиц у огаря начинается во
второй половине апреля и в начале мая (Федюшин, 1925); в Тункинской
долине птенцы весом 110 г (т. е. менее 10 дней) встречены 25 июня (Слудский);
ОГАРЬ 359
в Даурии (озеро Тарей-Нор) выводок найден в середине июня. На Алтае
сроки размножения очень различны в зависимости от высоты местности.
В котловине озера Марка-Куль только что выклюнувшиеся птенцы наблю-
дались 7 июня, 29 июня и даже 1 июля, а на высоком плато Укбке выводки
их 7 августа еще не летали, но к 23 августа уже все поднялись на крыло
(Сушкин, 1938).
Линька. Дважды в году и у обоих полов проходит, повидимому,
одинаково и примерно в одни сроки, хотя, возможно, у самцов немного
раньше. Начинается летняя линька поздно, в большинстве случаев в первых
числах августа, когда молодые птицы приобретают почти полную самосто-
ятельность. Так, самец, добытый 21 июля на Наурзумских озерах, еще не
имел признаков линьки. Летняя линька, повидимому, полная, но иногда
она еще продолжается до момента начала брачной. Смена маховых перьев
начинается вскоре после начала смены мелкого оперения, иногда почти
одновременно с ней, но есть индивидуальные различия. К середине сентября
маховые успевают вполне смениться (Иргиз, Тургай; Сушкин, 1908), но
линька мелкого оперения и после этого продолжается весьма энергично.
Позже всего заканчивают сменяться рули, последнее происходит в октябре
или, чаще, в ноябре. С декабря по апрель проходит частичная предбрачная
линька, во время которой сменяются большая часть контурного оперения,
средняя пара рулевых, внутренние второстепенные маховые и их кроющие.
Первая линька молодых птиц проходит, повидимому, так же, как пред-
брачная линька взрослых птиц. Начинается она с августа. Черное кольцо
на шее приобретается в феврале.
Питание. Огари кормятся как на воде, так и на суше, причем на
суше чаще. Объектами их питания служат животные и растительные корма.
Весной по прилете огари часто встречаются на зеленых лужайках, между
барханами, где кормятся преимущественно молодыми зелеными всходами
солянок и злаков (Кара-Кумы, Барса-Кельмес). Летом выводки часто кор-
мятся на солонцах, собирая насекомых, преимущественно кобылок (Северо-
Западный Казахстан); в других местах (Памир) существенным кормом слу-
жат в это время, повидимому, рачки—Artemia salina. В сентябре, когда
молодые птицы поднимаются на крыло, они начинают совершать регуляр-
ные перелеты на поля, главным образом просяные, где кормятся утром и ве-
чером, а на озера возвращаются около полудня и после заката солнца, в тем-
ноте (Тамань, Иргиз, Кургальджин, Ачинск). Радде A854) находил в пище-
воде летевших с кормежки уток по 40—65 г зерен проса. Где нет просяных
полей, огари кормятся на солонцах и мелководьях у берегов. Желудки
птиц, добытых на взморье (дельта Волги, во время пролета), содержали
остатки ракообразных и нежные вегетативные части растений. В Аскания-
Нова огари полностью переходят осенью на питание зерном (пшеница),
которое собирают на дорогах, токах и убранных полях. На этом корме они
быстро жиреют (Фортунатов, 1926). Зимой в долинах Кахетии стаи красных
уто^с держатся на озимых полях и кормятся зеленью всходов (Чхиквишвили,
1930). Кроме того, огари иногда кормятся отбросами около жилья, а Заруд-
ный A888) несколько раз замечал их на падали, которую они поедали вместе
с воронами, сарычами и коршунами.
Хозяйственное значение. Красная утка нигде не имеет большого про-
мыслового значения, но как объект охоты безусловно представляет интерес.
Мясо огаря в период питания преимущественно животной пищей имеет непри-
ятный запах, но с осени, после перехода на растительные корма, приобретает
прекрасные вкусовые качества. Птенцы легко вырастают в неволе и быстро
ручнеют, взрослая птица, где ее не преследуют, очень доверчива. Поэтому
красная утка может быть использована для заселения новых водохранилищ
и парковых водоемов. По Дунаю( в Болгарии) огаря разводили местами как
360
ОТРЯД ГУСЕОБРАЗНЫЕ
домашнюю птицу (Христович, 1890). Огарь очень вынослив; он может быть
использован для гибридизации с домашними утками.
Полевые признаки. Крупная утка; окраска одноцветная рыжая, бока
головы светлые. Ноги, клюв, хвост и маховые черные, на крыльях сверху
белые пятна. Легко передвигается по земле, держа туловище горизонтально;
плавает погружая переднюю часть туловища глубже задней и держа шею
вытянутой прямо вверх. Ныряет очень редко, обычно лишь раненая или во
время линьки. Кормится на воде, опрокидываясь головой вниз, или на суше.
Летает быстро, редко и сильно взмахивая крыльями. Птица крикливая,
Рис. 64. Красная утка, или огарь (самец и самка)
на полете постоянно слышен ее громкий гортанный крик «анг»..., «аанг*
причем у самца он звучит несколько более на «о», а у самки на «а». Кроме
того, обоим полам свойственны хриплые трескучие звуки, выражающие
призывы и возбуждение. Большую часть года птицы держатся парами, соби-
раясь в большие стаи только осенью и зимой.
Описание. Строение и размеры. Крупная утка с довольно длинными нога-
ми, расположенными несколько ближе к переду, чем у обычных речных уток и нырков,
Клюв короче головы, одинаковой ширины у основания и у вершины, коготок широкий.
занимает всю среднюю часть надклювья. Пластинки по краям челюстей крупные, тонкие
и нечастые. Цевка длинная, лапа короткая. Средний палец равен длине иевки, кожистая
оторочка заднего пальца узкая. Хвост довольно длинный, слабо закругленный из 14 руле-
вых. Длина крыла самцов 355—395, самок 315—365 мм; цевка самцов 54—63, самок 54—
61 мм; клюв самцов 44—48, самок 43—45 мм.
Вес. Взрослые самцы в апреле в среднем весят 1320 г A200—1500) (дельта Волги),
1600 г (озеро Кургальджин); в сентябре 1250 г (низовья р. Или в сентябре—октябре
самцы A5) 1200—1600 г в среднем 1360 г, а взрослая самка 1100 г (дельта Волги).
ХОХЛАТАЯ ПЕГАНКА
361
Окраска. Пуховой птенец. В основном чистобелый, но темя, полоса, проходя-
щая от него по задней стороне шеи, и спина бурые. Лапы и клюв темные.
Молодые птицы. Окрашены сходно со взрослой самкой, но несколько тусклее
ее. Отличием служит окраска верхних кроющих крыла. Большие кроющие серо-
ватые, с более светлым, иногда белым, наружным опахалом, средние роющие также
обычно слегка сероватые и с буроватой каймой. Этот признак до первой летней линьки
позволяет отличать годовалых птиц от взрослых.
Взрослая самка. Сходна с самцом, но общий тон окраски головы и шеи светлее,
а бока головы почти чистобелые; черный ошейник у нее отсутствует.
Самец в брачном наряде. В основном коричнево-рыжий. На шее окраска не-
сколько светлеет, а на верхней стороне головы переходит в светлоохристую, на боках
ее в еще более светлый цвет. В нижней части шеи черное кольцо. Задняя часть спины не-
сколько менее яркая и испещрена мелкой поперечной струйчатостью; подхвостье и хвост
черные с зеленоватым отливом. Первостепенные маховые черные, второстепенные с бле-
стящими зелеными наружными опахалами, образующими большое зеркало, внутренние
второстепенные с рыже-ко'ричневыми наружными опахалами и сероватыми внутренними.
Все верхние кроющие крыла чистобелые; они образуют широкое белое поле. Клюв к лапы
черноватые, радужина темнобурая.
Самец в летнем наряде. Характеризуется отсутствием черного ошейника.
19. Хохлатая пеганка Tadorna cristata Kuroda
Pseudotadorna cristata. Kuroda. Tori, 1, 1917, стр. 5, 1918, Корея.
Синоним. Tadorna casarcax Querquedula falcata. S с 1 a t e r. Proc. Zool. Soc. Lond.,
1890, стр. lf Владивосток.
Распространение. Ареал. В коллекциях имеются всего 4 экземпляра
этой редкой утки. Взрослая самка добыта под Владивостоком в апреле 1877 г.,
остальные в Корее, самка в
декабре 1913 г. на р. Кун-
Киан, самецв декабре 1916г.
около Пусана (Фузана); на-
конец, летом 1924 г. еще
один самец добыт вблизи от
Сеула в устье р. Кун-Киан
(Курода, 1924). Кроме того
(по сообщению Курода) в
марте 1916 г. добыты три та-
кие утки из стайки в 6 шт.
у северо-западного берега
Кореи. Экземпляры эти, к
сожалению, не сохранились.
В старых японских книгах
есть описания и изображе-
ния хохлатых пеганок с ука-
занием, что они нередко за-
\
Карта 40. Распространение хохлатой пеганки Ta-
dorna cristata
а—места, где птицы были встречены и добыты
возились в прежнее время
в Японию из Кореи. Изо-
бражения их встречаются
также и на старинных китайских картинах и гобеленах. Все это говорит о том,
что хохлатая пеганка—очень малочисленный, вероятно вымирающий, вид'
распространенный в Корее и, возможно, в южных частях Советского Приморья'
Никаких сведений об экологии нет.
Описание. Строение и размеры. Крупная утка, высоко стоящая на ногах.
Клюв широкий, прямой. Перья затылка образуют длинный хохол. Размеры (самец и сам-
ка): крыло 300—320 мм, цевка 47—49,5 мм, клюв (от лба) 41 5 45 мм
Окрас ка. Взрослая самка Основной фон сверху, снизу (от зоба до брюха вклю-
чительно) и на боках серо-бурый с более светлыми волнистыми поперечными полосками,
более крупными на боках. Оперение вокруг клюва, лоб, нижняя часть щек и бока шеи бе-
ловатые. Темя, затылок и хохол черные, со слабым зеленоватым отливом Такого же цвета
остальная часть боков головы, на фоне которых резко выделяется белое пятно вокруг
362
ОТРЯД ГУСЕОБРАЗНЫЕ
глаз, суживающееся к затылку. Подхвостье желтоватое. Крылья сверху белые (кроющие)
с ярко зеленым зеркалом, ограниченным спереди черной полоской, а от основания крыла
буро-каштановыми опахалами третьестепенных маховых. Первостепенные маховые чер-
ные. Клюв желтоватый, ноги буровато-желтые.
Взрослый самец сверху буровато-серый с буро-каштановыми окончаниями плече-
вых перьев. Низ спины, надхвостье и хвост черные. Верх и бока головы, включая область
глаз, хохол и подбородок черные с зеленым отливом. Такого же цвета нижняя часть шеи,
Рис. 65. Хохлатая пеганка
зоб и верх груди. Остальная часть головы и шеи серая. Грудь и брюшко темносерые в мел-
ких поперечных черноватых полосках. Подхвостье охристо-рыжее, а по бокам у основа-
ния его черное пятно. Крылья такие, как у самки. Клюв желтый с красноватым основа-
нием надклювья, ноги бледнооранжевые, радужина темнобурая.
Другие сезонные и возрастные наряды неизвестны.
РОД НАСТОЯЩИЕ или РЕЧНЫЕ УТКИ ANAS LINNAEUS, 1758
Тип. Anas platyrhynchos.
Синонимы. Spatula В о i e, 1822.—Dafila Stephens, 1824. —Mareca Stephens,
1824.— Querquedula S t e p h en s, 1824—Nettion Каир, 1829.—Сhaulalesmus Bona-
parte, \838.—Marmaronetta Reichenbach, 1852.— Eunetta Bonaparte,
1856.— Polionetta О a t e s, 1899.
20. Узконосый чирок Anas angustirostris Men.
Anas angustirostris. Menetries. Catalogue raisonne des objets de Zoologie rec. dans un
voyage an Caucase, 1832, стр. 58, Ленкорань.
Русское название. Узконосый, или мраморный, чирок—книжное.
Распространение. Ареал. В основном прибрежные части Средизем-
ного моря, а именно: юг Пиренейского п-ова, Канарские о-ва, все побережье
УЗКОНОСЫЙ ЧИРОК
363
Северной Африки, Кипр, Крит, Сирию, Палестину и Ирак. Одиночно эти
утки появляются на островах Средиземного и Адриатического морей, в Гре-
ции, Албании, Южной Австрии и Венгрии. В пределах Советского Союза
гнездятся по всей долине Куры, начиная от Кировабада, и в Ленкоранской
низменности. В весеннее время отмечен как обычная птица в Армении
<в б. Эриванской обл.), но на гнездовье не зарегистрирован (Бобринский, 1916).
Севернее узконосый чирок встречается кое-где в приморской полосе запад-
ного берега Каспия (устье Терека, Ильмени к западу от дельты Волги), но
повсюду там очень редок. В Заволжье доходит на север до Камыш-Самар-
Карта 41. Распространение узконосого чирка Anas angustirostrts
а—граница гнездовий, г—эимовки
ежих озер и встречается около Урды (Волчанецкий, 1937), но на восток не
достигает р. Урала. В Туркмении узконосый чирок местами наиболее рас-
пространенная утка. Он обычен в нижнем течении Атрека, по Теджену и
Мургабу. Гнездится на пресных озерах Узбоя (Шестоперов) и весьма обычен
по нижнему и среднему течению Аму-Дарьи. В верховьях Аму-Дарьи,
в Северном и Западном Таджикистане (ст. Мельниково, Кировабад, Пар-
хар) встречен на гнездовье, но редок (Иванов, 1940). Отсутствует в Кызыл-
Кумах и на восточном берегу Аральского моря, но обычен в нижнем и сред-
нем течении Сыр-Дарьи. Южнее узконосый чирок гнездится в Иране у юж-
ных берегов Каспийского моря, в бассейне Горгана и Геррируда и на озерах
Сеистанской котловины (Зарудный, 1911), повидимому, гнездится в неболь-
шом числе по низменностям Белуджистана и, возможно, Синда (Паки-
стан) .
364 ОТРЯД ГУСЕОБРАЗНЫЕ
Основные зимовки находятся в странах Северной Африки,
.не южнее Атласа и Нижнего Египта, в Сирии, Палестине и Ираке, зимует
у южных берегов Каспийского моря к северу до Ленкорани (в незначитель-
ном числе) и долины Атрека (огромными стаями), на Западном Узбое по
Мургабу и Теджену, вдоль побережья Оманского залива в бассейнах Герри-
руда и в Сеистанской котловине, всюду в низменных частях Пакистана и в се-
верной половине Индии (обычнее всего в бассейне Инда).
Характер пребывания. В северной половине своего ареала,
в том числе повсюду в нашей стране, кроме южных районов Каспийского
моря, перелетная птица. На юге (Северная Африка, Иран, Ирак, Пакистан)
ведет оседлую жизнь. Летних перелетов и скоплений селезней в местах
линьки не отмечено.
Сезонные миграции и их даты. Весенний пролет этих
чирков проходит поздно. На пресных озерах Узбоя первые птицы отмечены
31 марта (Шестоперов), а валовой пролет на Мургабе наблюдался 7—26 ап-
реля (Радде и Вальтер, 1889). На о-ве Сара (юго-западный берег Каспия
у залива им. Кирова) первые птицы были добыты 2 мая, а затем до 9 мая
наблюдалось продвижение пролетных стай (Тугаринов, 1950). Их осенний
пролет по Узбою происходит в конце сентября и в первых числах октября.
В низовьях Атрека первые пролетные птицы появляются 1 ноября, а к концу
месяца они там же встречаются в массе. Однако в середине зимы иногда исче-
зают там и откочевывают дальше на юг (Житников, 1910).
Биотоп. В период гнездованья узконосые чирки держатся обычно
на небольших озерках с илистым дном и тростниковыми зарослями. Зимой
они проводят дневное время на мелководьях открытого взморья или на пле-
сах озер, а для кормежки перелетают на разливы рек и затопленные рисовые
поля.
Чи елейность. Узконосый чирок обычен в Закавказье к югу от
долины Куры, а в равнинной части Туркмении от долины Атрека до сред-
него течения Аму-Дарьи он является наиболее многочисленной уткой. Пови-
димому, столь же обычен он и в пределах Ирана.
Экология. Размножение. Время наступления половой зрелости
неизвестно. Весенние пролетные стаи чирков состоят уже из оформившихся
пар. Брачные игры походят на таковые у шилохвостей, но несколько менее
сложны и менее разнообразны. Гнезда устраиваются на земле под прикры-
тием невысоких кустов или сухой и жесткой травы. Иногда они распола-
гаются у берегов озер, а иногда у временных водоемов на заболоченных
солончаках. Гнездо представляет собой ямку, выстланную сухой травой,
и имеет небольшое количество пуха. Есть сведения, что в восточной части
Волжской дельты, где эти утки нередки, они зачастую гнездятся в старых
вороньих гнездах (Воробьев, 1936). Число яиц в полной кладке от 7 до 12
(9—14, Белуджистан; Тайсхерст, 1927). Их цвет слегка палевый или бурова-
тый, форма широкая, размер 42,4—50,6x31,5—36 мм. Длительность наси-
живания 25—27 дней. На озере Тели-Куль и в низовьях Сары-Су (Нижняя
Сыр-Дарья) эти чирки 20—23 мая держались как стайками, так и парами,.
а у добытой самки оказалось готовое к сносу яйцо (Спангенберг и Фейгин,
1936). В дельте Волги самка с готовым к откладке яйцом добыта 15 июня
(Воробьев, 1936). В Западном Пакистане гнезда со свежими и насиженными
яйцами встречались с 14 июня по 16 июля. Период гнездования всегда растя-
нут. В течение первой и второй декады июля на Аму-Дарье одновременно
встречались и только что вылупившиеся и уже летные молодые (Зарудный>
1890). Самцы покидают самок, когда те садятся на яйца, сбиваются в неболь-
шие стайки и переходят к бродячему образу жизни.
Линька. Имеющиеся в настоящее время материалы говорят о том,
что узконосые чирки (оба пола) линяют дважды в год. У селезня от 3 октября
УЗКОНОСЫЙ ЧИРОК
365
маховые свежие, но интенсивно линяет контурное оперение (низ тела, шеи,
поясница, надхвостье) и рули, крайние из которых едва появились из пень-
ков. Повидимому, в этот период послебрачная линька закончилась, и селезни
проходят неполную предбрачную линьку. В декабре никаких признаков
линьки нет. У майских же птиц (самцов и самок) заканчивается линька кон-
турного пера на зобе, зашейке и лопатках. Либо предбрачная линька временно
прекращается зимой и на-
чинается вновь во время ве-
сеннего пролета, либо сро-
ки ее чрезвычайно варьи-
руют у разных особей и стай.
Особого летнего наряда у
селезней нет.
Питание. Кормятся
птицы обычно на мелководье
или даже на берегу, роясь
в иле и собирая корм среди
водной растительности. Со-
став питания совсем не изу-
чен, известно лишь, что в
него входят личинки насе-
комых, моллюски, вегета-
тивные части растений и се-
мена.
Полевые признаки. Мел-
кие утки, у которых оба по-
ла имеют одинаковое серое
со светлыми пятнышками
оперение. Тихие, сравни-
тельно малоподвижные и до-
верчивые птицы. Плавают,
погрузив довольно глубоко
переднюю часть тела и приподняв кверху хвост. Полет быстрый и прямолиней-
ный. Часто отдыхают, сидя на ветвях прибрежных кустов, нависших над водой.
Описание. Строение и размеры. Утки размером с чирка-трескунка. Клюв
длинный, но не длинней головы. Ноготок также длинный, размером около трети длины
надклювья и загнутый под прямым углом. Оперение выдается вперед по хребту клюва ост-
рым углом, но по бокам надклювья не выступает. Перья затылка у селезня удлинены,
образуя короткий хохолок, у самки его почти нет. Рулевых 7 пар с тупыми вершинами.
Форма хвоста клиновидная, но слегка закругленная. Надхвостье короткое, едва закры-
вает более половины длины рулевых, подхвостье доходит до их конца. Размеры: крыло
(самец и самка) 189—212 мм, цевка 35— 37 мм, клюв 40—49 мм. Вес: самка, добытая в ок-
тябре на Узбое (жирная), весила 400 г, птицы в низовьях Атрека (без разделения пола)
в декабре весили 420—500 г, в среднем 477 г.
Окраска. Пуховой птенец. На спинной стороне довольно темнобурый с бело-
ватой пятнистостью на плечах, по сторонам спины, надхвостья и бедрам. На боках головы
надбровная желтая полоска и бурая полоса от клюва через глаз. Брюшная сторона
светлая, буровато-желтая.
Молодые птицы в первом наряде. Походят на взрослых, но общий тон окраски
тусклее, пятна менее четки, брюшная сторона грязнобелая.
Взрослая самка. В отличие от селезня имеет около глаз светлобурое пятно
и меньше размером.
Взрослый самец. На спинной стороне серо-соловый; вершины перьев с круг-
лыми светлыми пятнами, создающими мраморный рисунок оперения. Так же окрашены
и бока тела. Голова буровато-серая с рыжеватым налетом сверху. Вокруг глаз буро-
ватое пятно, щеки и шея в мелких бурых крапинах. Брюшная сторона беловатая
с более темными пятнами на зобе и подхвостье. Первостепенные маховые буроватые
с серыми наружными опахалами, второстепенные маховые и верхние кроющие крыла
серо-соловые, последние со светлыми вершинами. Рулевые серые. Клюв серый, лапы
бурые, радужина бурая.
Рис. 66. Узконосый чирок
366
ОТРЯД ГУСЕОБРАЗНЫЕ
21. Шилохвость Anas acuta L.
Anas acuta. Li nnaeus, Syst. Nat., изд. X, I, 1758, стр. 126, Швеция.
Русские названия. Шилохвость, вострохвост—народные, обязаны форме хвоста.
Распространение. Гнездовой ареал. Северные и умеренные
части Европы, Азии и Северной Америки. Населяет Исландию, Шотландию
и Оркнейские о-ва, нерегулярно гнездится в северных районах Англии
и Ирландии. В Дании довольно редка, в Германии же гнездится регулярно,
в ее северо-западной части, но местами гнездится не каждый год, а лишь
в многоводные годы (Штеттин; Робин, 1923). Так же довольно спорадично-
населяет Центральную и Северную Венгрию и Верхнюю Австрию. Довольно
Карта 42. Распространение шилохвости Anas acuta
1—A. a. acuta, 2—А. о. eytoni; а—граница гнездовий, а1—районы наибольшей численности птиц на
гнездовье, б—не вполне выясненная граница гнездовий, в—обособленные гнездовые колонии, в'—
места постоянных встреч летом и спорадического гнездования, а—основные зимовки, д—валюты,
ж—места скоплений птиц на линьку
регулярно встречается летом в дельте Гвадалквивира, местами на юге Фран-
ции (Юра, дельта Роны), на озере Скутари (Албания) и кое-где в Болгарии.
Вплоть до июня шилохвость отмечали в Египте (Фаюме, Хартум). Вероятно,
в основном это молодые птицы, задержавшиеся на местах зимовки, но неко-
торые из них при известных обстоятельствах остаются там и на гнездовье.
Гнездится повсюду в Польше, Скандинавских странах и Финляндии, хотя
на юге последней редка. В пределах Советского Союза северная граница
гнездованья шилохвостей от Мурмана до Байдарацкой губы (Карское море)
следует по морским берегам. Они отсутствуют на о-ве Колгуев, но встречаются
летом на Вайгаче и вероятно гнездятся там. На Ямале достигают примерно
до 71° с. ш. (Житков, 1912), на о-ве Белом не гнездятся, но большие стаи их
отмечались в августе (Тюлин, 1938). По Енисею доходят почти до устья
G2°30' с. ш.), встречаются на северо-западном побережье Таймыра и у Тай-
мырского озера. По Лене, Индигирке и Колыме обычны до самых устьев,
но редки в Приморских тундрах. Встречаются повсеместно в Анадырском
крае, но в восточной части Чукотского п-ова на гнездовье не зарегистриро-
ваны. Обычны на Олюторском побережье, по всей Камчатке, на Командор-
ШИЛОХВОСТЬ 367
ских о-вах и вдоль берегов Охотского моря. В европейской части Советского
Союза шилохвость обычна на гнездовье к югу до Белоруссии, Калининской,
Ярославской, северной половины Горьковской и Кировской обл. включи-
тельно. В Киевской, Брянской, Орловской и Рязанской обл., а также в Та-
тарской АССР она гнездится регулярно, но не повсеместно и в малом числе.
Еще южнее, в Харьковской и Полтавской обл., под Запорожьем (Вальх,
1899), в Причерноморских степях (Аскания-Нова; Шарлемань, 1924) и на
некоторых островах Черного моря (о-в Орлов, о-в Джарылгач; Клименко,
1950) шилохвости гнездятся спорадично, лишь в отдельных пунктах и далеко
не каждый год. То же самое относится к Курской, Воронежской, Пензенской,
Ульяновской и Саратовской обл. В Заволжье шилохвость обычна всюду
в равнинной части Башкирии, но очень редка в гористой. На юг она дости-
гает Камыш-Самарских озер, где, однако, гнездится далеко не регулярно.
Изолированными районами постоянного гнездованья служат озера Ахал-
калакского плато в Джавахетии (Чхиквишвили, 1930) и озеро Севан в Арме-
нии. В некоторые годы шилохвости задерживаются на лето в ряде пунктов
Закавказья и Предкавказья и возможно, что иногда гнездятся там (Ленкоран-
ская низменность, Маныч). По долине Урала шилохвость на гнездовье дохо-
дит до нижнего течения, но на степных озерах она там очень редка. В Казах-
стане границей регулярного и обычного гнездованья служат южные пределы
лесостепи. В зоне южных степей и полупустынь она встречается не повсе-
местно и численность ее резко колеблется в разные годы, в зависимости от
степени их многоводности. Эти утки гнездятся на озерах между Эмбой и Му-
годжарами (Сушкин, 1908) и бывают обычны в некоторые годы на Наур-
зумских озерах, озере Кургальджин и на юге Кулундинской степи. В зону
пустынь шилохвости не проникают, и встречи их там носят случайный хара-
ктер. Птицы, проводившие зиму у южных берегов Каспия, нередко задер-
живаются вблизи от мест зимовки, но данных о гнездованье их там нет.
Негнездящиеся птицы встречаются летом в нижнем течении Сыр-Дарьи,
а отдельные пары иногда и гнездятся там (Спангенберг, Фейгин, 1936). То же
отмечается и для различных частей Семиречья (Баскан, Ала-Куль, Прибал-
хашье), где в некоторые годы отдельные пары остаются на лето и гнездятся
(Шнитников, 1949). Известны летние встречи шилохвостей в Кашгарии,
Северо-Западной Монголии, Южном Забайкалье и даже в долине Хуан-хэ.
На Алтае шилохвость довольно редка. Она гнездится к югу до Чуйской
степи и плато Укбк, но совершенно отсутствует в северо-восточной части
к северу от бассейна р. Лебеди (Сушкин, 1938). Регулярно, хотя и нечасто,
гнездится в Минусинской и Тувинской котловинах, в Среднем и Северном
Прибайкалье, Даурии и в бассейне р. Аргуни. Обычна местами в Северной
Маньчжурии, всюду в Уссурийском крае и на Сахалине.
На американском континенте область гнездованья шилохвости захва-
тывает значительную часть материка и выходит к морскому побережью от
залива Коронации до Аляски на севере и от последней до Лос-Анжелоса
на западе. Восточная граница проходит к западной части Гудзонова залива
и оттуда к области Великих озер. Пределом распространения к югу служат
озера Гурон и Мичиган, устье р. Миссури и верховья р. Колорадо.
Районы скоплений на время линьки располагаются
в степной, лесостепной и лесотундровой зонах. Основными местами массового
скопления линных птиц служат: дельта Волги и озера Сталинградской обл.,
среднего течения Урала, низовьев Илека и местность между Мугоджарами
и Эмбой, озерные территории Зауралья, Кустанайской обл., озеро Челкар-
Тениз, Кургальджинский озерный бассейн, Ишимская, Барабинская и Ку-
лундинская степи. Менее значительные скопления птиц на юге известны
на озерах Минусинской котловины и в Тибете на границе Сиккима и Ладака
(Штреземан, 1942). На севере линные шилохвости в значительных количе-
.368 ОТРЯД ГУСЕОБРАЗНЫЕ
ствах скопляются в низовьях Оби и на Южном Ямале, а также кое-где в ни-
зовьях Лены, Колымы и Анадыря.
Основные зимовки шилохвостей находятся у берегов Южной
Норвегии, Британских о-вов и вдоль побережий Западной Европы на запад
от Средней Германии. В неменьшем числе они зимуют в средиземноморских
странах Европы и на берегах Северной Африки, в Малой Азии, Сирии, Пале-
стине, всюду по долине Нила, включая верхнее течение реки, и кое-где на
берегах Красного моря (Аден). В незначительном числе они проводят зиму
на Черном море у берегов Колхиды и Крыма и в несколько большем на побе-
режьях Румынии и Болгарии. Массовые зимовки шилохвостей находятся
в южной части Каспийского моря (от долины Куры до Энзелийского залива
и от о. Челекена до Атрека, Гюргена, Кара-Су и Астрабадского залива).
В меньшем числе они зимуют в Средней Азии на Теджене, Мургабе и по
Верхней Аму-Дарье (до Чубека). Постоянными местами зимовки служат
Ирак, южное побережье Ирана, Сеистанская котловина, Пакистан, почти
вся территория Индии с Цейлоном, Ассам и Бирма, а также озерное плато
Южного Тибета (Ледлоу, 1950). На Дальнем Востоке зимовки шилохвостей
располагаются на Японских о-вах к северу до 40° с. ш., вдоль всех берегов
Кореи, в прибрежных местностях Китая, к югу от Чжилийского залива,
в низовьях р. Хуан-хэ, в долинах рек Янцзы и Мин, в Северном Вьетнаме]
на Тайване и на Филиппинских о-вах (о-в Минданао). На Американском
материке шилохвости зимуют всюду вдоль обоих морских побережий от
архипелага Александра и Филадельфии до Панамского перешейка, в бассейне
Миссисипи южнее впадения Миссури, на всех внутренних водоемах к югу
от Мексиканской границы, на Кубе и некоторых смежных островах.
Характер пребывания. Почти на всем протяжении своего
огромного ареала шилохвость перелетная птица, совершающая дальние
сезонные миграции. В Евразии исключением являются только шилохвости,
населяющие Британские о-ва, и быть может южнонорвежские птицы. В Аме-
рике, на значительном протяжении Тихоокеанского побережья, территории
гнездованья и зимовки совпадают; можно предполагать, что птицы, насе-
ляющие эту зону, оседлы. Как видно из описания ареала, для распростра-
нения шилохвости характерны две особенности. Первая—это резкие изме-
нения численности ее у южной границы распространения в зависимости
от климатических условий года и пульсация самой южной границы. Так,
например, в пределах Харьковской и Полтавской обл. шилохвость была
довольно обычна на гнездовье в 1893 и 1894 гг., затем полностью отсутство-
вала в 1895 и 1897 гг., но стала опять довольно обычной в 1905 г. (Зарудный,
1911). В пойме Нижней Камы в 1934 и 1935 гг. было отмечено столь резкое
увеличение количества шилохвостей, что по численности они почти не усту-
пали крякве (Першаков, 1937). На степных озерах Наурзумского заповед-
ника (Кустанайская обл.) в годы наполнения озер количество этих уток
на гнездовье бывает велико, а в годы высыхания водоемов резко сокращается
Второй характерной чертой распространения шилохвости служат ежегодно
наблюдающиеся случаи летования некоторого количества птиц во многих
пунктах вблизи от мест их зимовок и случаи спорадичного гнездованья их
Ь этих пунктах далеко за пределами ареала вида. Обычно это имеет место
при сочетании трех условий: многоводности года на юге, тяжелых условий
прошедшей зимовки и длительной задержки начала весны в районах, по
которым следуют птицы с мест их зимовки. Так было в низовьях р. Или
в 1949 г. (Слудскии), под Алма-Ата в 1936 г. (Шнитников, 1949), в Аскании-
Нова (Причерноморские степи) в 1907 и 1911 гг. (Соколов, 1926) и т д Воз-
можно, что такого рода задержкам обязано возникновение ряда постоянных
и обособленных гнездовых колоний шилохвости на горных плато Закав-
казья, на о-ве Орлове в Черном море, в устьях Роны, Гвадалквивира и др"
ШИЛОХВОСТЬ 369
Сезонные миграции. Довольно значительные материалы, полу-
ченные в результате кольцевания шилохвостей, рисуют следующую картину
их сезонных миграций и распределения по местам зимовок. Птицы, населяю-
щие северную зону европейской части Союза к востоку до Урала, а именно
западные и северо-западные области, север Кировской обл., Тиманскую
тундру и нижнее течение Печоры, отлетают на зимовку в страны Западной
Европы (на Британские о-ва, в Голландию и Францию). Судя по данным
кольцевания шилохвостей на пути их пролета (о-в Фуне в Дании), часть этих
птиц спускается в течение зимы по континентальным водоемам Европы на юг,
достигая центральных и западных районов Франции, Италии и даже Испа-
нии (Мортенсен, 1914). В меньшей степени к этой области зимовок тяготеют
птицы центральных областей европейской части Союза (Ивановская обл.,
Черниговщина), Урала и Прииртышской Сибири (Нижний Тагил, устье
Иртыша). Там же проводят зиму птицы, населяющие Скандинавию и Ислан-
дию (Томсон, 1941). Из Приуралья и Западной Сибири (бассейн Тобола,
среднее течение Иртыша и низовья Оби) шилохвости отлетают в юго-запад-
ном направлении. Через Уральские степи они достигают северного побе-
режья Каспия, где значительная часть взрослых птиц проводит время линьки.
Далее часть их спускается к южным берегам Каспия, где и зимует или про-
летает еще южнее—в Иран и Ирак. Другая, не менее значительная, часть птиц
пролетает вдоль Черного моря и остается зимовать на Балканском п-ове и в Се-
веро-Восточной Италии. В меньшем числе они достигают Египта, Северной
Африки, Южной Франции и в еще меньшем появляются у северных берегов
Европы (Дания, Голландия). Того же основного направления в сезонных
перелетах придерживается часть птиц, населяющих Тиманскую тундру,
северные части республик Коми, среднее течение Оби и Западный Алтай.
Шилохвости из южных частей Приобской и Прииртышской Сибири отле-
тают непосредственно к югу вдоль озерных областей Северного Казахстана
и долины Сыр-Дарьи, следуя, по всей вероятности, на места зимовки в Север-
ной Индии. О направлении пролета у восточносибирских птиц почти ничего
неизвестно. По Ангаре и ее притокам все утиные, в том числе и шилохвости,
пролетают весной вниз по течению, что говорит о том, что правобережье
Енисея населяется утками, зимующими в Восточной Азии. На Чукотке 20 июня
добыта шилохвость, окольцованная зимой C января 1936 г.) в Калифорнии
(Лос-Баннос). Это говорит о том, что птицы, населяющие крайний северо-
восток Сибири, посещают зимой Тихоокеанское побережье Америки (Вуче-
тич и Тугаринов, 1937; Михеев, 1949). В ряде случаев шилохвости начинают
свой осенний перелет в направлении, противоположном их дальнейшему
пути пролета. Так, молодая самка, окольцованная на Наурзумских озерах
(Кустанайская обл.) во время линьки, через месяц была добыта около Петро-
павловска, примерно в 600 км к северо-востоку от места кольцевания. В ни-
зовьях Оби и у берегов Обской губы в июле наблюдается ясно выражен-
ный пролет шилохвостей, стаями до 100 птиц, прямо на север к озерам
Среднего и Северного Ямала (Морды-Тамбей). В августе стаи вероятно уже
перелинявших птиц были отмечены на о-ве Белом. Некоторые же птицы,
наоборот, начиная свой путь, спускаются к югу до дельты Волги, где прово-
дят линьку, а по окончании ее отлетают на северо-запад к берегам Балтий-
ского и Северного морей. Данные кольцевания говорят о том, что скорость
продвижения шилохвостей во время их перелета составляет в некоторых
случаях более 90—100 км в день.
Даты. Гнездовой ареал шилохвости простирается на большое расстоя-
ние с юга на север и захватывает весьма различные ландшафтные зоны.
В связи с этим сроки отлета этой утки с мест зимовки и пролета её на юге
весьма растянуты. Зачастую от времени появления первых пролетных стай
до исчезновения последних проходит более двух месяцев. С западноевропей-
24 птицы СССР, т. TV
370 ОТРЯД ГУСЕОБРАЗНЫЕ
ских зимовок шилохвости начинают отлетать уже в марте, но интенсивный
пролет их у берегов Северной Германии продолжается еще в середине апреля.
В Прибалтийских советских республиках они появляются в первых числах
апреля, и в первой декаде этого месяца достигают Ленинградской обл. В се-
верной части Рыбинского водохранилища средняя дата появления первых
птиц 16 апреля A2—21 апреля), а на следующий день они уже прилетают
в массе. В Тиманской тундре (низовья р. Вельть) первые шилохвости наблю-
дались 8 мая, а 10—13 мая они появились стаями (Гладков, 1951)* на Кольском
п-ове (озеро Имандра) их прилет приходится на 17 мая A0—25 мая). Печоры
(в устье Ылыча) первые шилохвости достигают 29 апреля B0 апреля—6 мая),
валовой пролет начинается около 4 мая и продолжается в среднем 20 дней.
Южно-каспийские зимовки шилохвости покидают в марте. К концу этого
месяца заканчивается в основном пролет их в районе Гасан-Кули, хотя
некоторое количество птиц задерживается на разливах р. Атрека до конца
мая. В конце февраля или в первых числах марта начинается массовый про-
лет шилохвостей у Ленкорани: появление пролетных стай из Ирана и отлет
к северу птиц, проводивших зиму на месте. Пролет продолжается в течение
всего марта. В то же время отлетают к северу птицы, зимующие в Джарыл-
гачском заливе Черного моря, но последние из них отлетают 6—10 апреля
(Клименко, 1950). Около 15 марта (9—26 марта) шилохвости появляются
в Аскании-Нова (Соколов, 1926), а 10 марта начинается массовый пролет
их у Обиточенской косы Азовского моря (Костюченко, 1926). В низовьях
Днепра шилохвости появляются во второй половине февраля, а главная
масса их проходит там в конце марта, у Днепропетровска со второй поло-
вины марта до конца апреля, а под Киевом 19 марта E—30 марта). Под Калу-
гой шилохвости появляются с ледоходом в первых числах или в середине
апреля, а под Москвой в последней декаде апреля. В дельту Волги они при-
летают 22 февраля A4 февраля—5 марта). Через Башкирию они летят огром-
ными стаями в середине апреля (Сушкин, 1897), а под Бугурусланом валовой
пролет приходится на конец этого месяца. В верхнем течении Эмбы первые
птицы были отмечены 8 апреля, с 10 по 23 апреля шел валовой пролет, но от-
дельные стаи встречались до 20 мая (Сушкин, 1908). На Наурзумских озерах
они появляются около 12 апреля, пролет под Тобольском 21 апреля A2—
26 апреля), а валовой пролет их в последних числах апреля и первых числах
мая (Тарунин, 1929). На Южном Ямале первые птицы отмечались 30 мая—
3 июня (Осмоловская). Из Пакистана последние шилохвости исчезают весной
не позднее апреля, а из Индии обычно в марте (Уистлер, 1935). В среднем
и нижнем течении Сыр-Дарьи (Чиили, Кара-Узяк) первые стайки появля-
ются в ранние весны уже 25—29 февраля, с середины до конца марта там уже
проходит массовый пролет, а в начале апреля они исчезают оттуда; запоз-
далой же весной первые стайки были отмечены 13 марта (Спангенберг, 1936).
В низовьях р. Или шилохвости появляются, примерно, с 10 марта, а закан-
чивается их пролет в первой половине апреля (Штегман, 1949). В Барабин-
ской степи шилохвости прилетают в конце марта, массовый их прилет при-
ходится на середину и конец апреля, а под Семипалатинском он был отмечен
Ъ апреля (Селевин, 1927).
Отлет шилохвостей с китайских зимовок заканчивается у берегов Чжи-
лийского залива лишь в апреле. Через Среднюю Корею (Сеул) они летят
с начала марта до второй недели апреля (Вольфе, 1950) Советского При-
морья первые птицы достигают уже в начале марта, но пролет их продол-
жается в течение всего апреля и даже иногда в начале мая (Воробьев). На
восточном побережье Камчатки пролет начинается 4—6 мая и бывает весьма
интенсивным до конца месяца, но в июне пролетных птиц уже не бывает
(Аверин, 1948). На Командорских о-вах они появляются в конце апреля.
В верховьях Колымы пролет шилохвостей происходит в 20-х числах маят
ШИЛОХВОСТЬ 371
а 22 мая уже были отмечены первые птицы под Нижне-Колымском. Время
прилета на Анадыре—середина или вторая половина мая. На р. Илиме
(приток Ангары) около 1 мая ломается лед и сразу же появляются стаи шило-
хвостей. Под Якутском они отмечены с 27 апреля, у Верхоянска 15 мая, а на
Верхоянском хребте в верховьях Яны (Кен-Юрях) 25 мая. На Индигирке
(устье р. Момы) первые птицы наблюдались 17 мая, а массовый пролет на-
чался 23 мая. Ниже по реке у с. Русское Устье первые пары их отмечены
8 июня (Михель, 1935). Основная масса шилохвостей покидает Анадырь
уже к началу сентября, но отдельные стаи и группы их встречаются в тече-
ние всего сентября, а одиночки даже еще в октябре (Портенко, 1939). С Коман-
дорских о-вов они отлетают во второй половине октября. У восточного берега
Камчатки большая часть их пролетает в сентябре и первой половине октября,
последние же отмечены 26 октября (Аверин, 1948). В Приморье пролет начи-
нается во второй половине сентября, продолжается весь октябрь и закан-
чивается в начале ноября (Воробьев). В Корее шилохвости очень многочис-
ленны весь ноябрь, на местах зимовок в Китае они появляются в Чжили
с сентября, в Западном Сычуане в октябре и в Южном Китае с декабря—
января (Колдуэлл, 1931). Из Барабинской степи шилохвости отлетают в конце
октября, с 8 октября начинается пролет на Наурзумских озерах, с первых
чисел сентября до конца октября он идет в Семиречье, где отдельные стаи
пролетают и в начале ноября. По сравнению с весной он бывает весьма
незначительным. В Гыдаянской лесотундре (долина р. Юрибея) интенсивный
осенний пролет наблюдался с 20 августа до середины сентября (Наумов,
1931), в то же время он заканчивается и под Салехардом. Осенний пролет
на Печоре начинается около 16 сентября B4 августа—8 октября) и длится
около 12 дней, последние же птицы наблюдаются еще в октябре (Теплов,
1948). В районе Петсамо задерживаются до первых чисел октября, из-под
Архангельска основная масса отлетает 20— 30 октября. Под Кировом встре-
чаются еще в начале октября (последние 12 октября). Пролетные стаи появ-
ляются на Рыбинском водохранилище в конце августа или в начале сентября,
к середине октября заканчивается массовый пролет, но одиночные птицы
встречаются до конца месяца. Из Эстонии отлетает в сентябре, последние
встречи под Москвой относятся к середине октября, но количество шило-
хвостей на осеннем пролете там сравнительно невелико. Пролет под Калугой
начинается с первых чисел сентября и продолжается до глубокой осени; на
Волге, в Ульяновской обл., он приходится на конец сентября, причем шило-
хности там в это время обычны. На Днепре под Запорожьем осенний пролет
начинается в первых числах октября и продолжается до начала ноября;
в Тендровском заливе Черного моря начало пролета обычно приходится
на середину сентября, но в некоторые годы птицы появляются уже 1 сентября.
В дельте Волги первые пролетные стаи появляются осенью около 25 сентября,
массовый пролет продолжается обычно с 5 по 12 октября, а массовый отлет
падает на время между 27 октября и 2 ноября. В Ленкоранской низменности
основной пролет шилохвости происходит в октябре и ноябре. Часть птиц
оседает там на зиму, но значительная часть стай проходит на большой высоте
и следует в Иран и Ирак. В последнем месте первые птицы отмечены 13 сен-
тября. Под Красноводском пролет начинается в октябре и идет несколькими
волнами, соответствующими волнам холода, проходящим через Северный
Каспий. Такие периоды появления большого количества пролетных стай
бывают в ноябре и даже в середине декабря. На озерах Атрека интенсивный
лет шилохвости наблюдался с 25 октября, и к 5 ноября она уже была там пре-
обладающей по численности уткой. Однако с 22 декабря число ее резко со-
кратилось, так как птицы, повидимому, продвинулись к югу на разливы
Гюргена (Житников, 1900). Во время своих сезонных миграций шилохвости
летят самостоятельными стайками, обычно от 10 до 60 шт., и на значительно
24*
372 ОТРЯД ГУСЕОБРАЗНЫЕ
большей высоте, чем другие утки. На юге же они иногда летят огромными—
многосотенными и даже тысячными—стаями. Перелет совершается чаще
всего по ночам. Для шилохвостей особенно характерно свойственное многим
уткам разделение полов и возрастов на зимовке. Чем суровее зима, тем
далее к югу откочевывают самки и молодняк птицы и процент взрослых сам-
цов на наших южнокаспийских зимовках бывает больше. Холодной зимой
1932 г. соотношение самцов и самок в Ленкорани было 4 : 1, а более мягкой
зимой 1934 г.—2,1 : 1. Изменяется это соотношение и в течение зимы, так
как в начале ее заметно преобладают самки и молодняк, которые в наиболее
холодном месяце—январе—обычно продвигаются к югу. Частично разде-
ление полов сохраняется у шилохвостей и во время весеннего пролета, осо-
бенно на юге. Вероятно эта особенность и послужила причиной ошибочного
представления о том, что у шилохвостей самцы численно резко преобладают
над самками (Алфераки, 1900; Данилович, 1928; Тугаринов, 1941).
Биотоп. Шилохвость—обитательница преимущественно открытых
пространств. Наиболее многочисленна она в зоне лесотундры Шилохвость
селится на внутренних водоемах самого различного типа, но обязательным
условием служит наличие водной и прибрежной растительности. В тундре
она встречается на мелководных озерках, в поймах рек с зарослями осоки
у берегов. В лесной полосе эти утки чаще всего заселяют луговые поймы
более крупных рек, а в лесостепи они наиболее обычны в луговых займищах
рек и озер. На время линьки шилохвости собираются на обширных, заросших
травой сорах, пойменных водоемах в низовьях северных рек, на больших
тростниковых озерах лесостепи и степи, а также в обширных дельтах рек
на юге страны. На пролетах их можно встретить буквально повсюду, а зимой
шилохвости обычно проводят день на тихих плесах морских заливов или
крупных озер, а по ночам вылетают кормиться на мелководные разливы
и затопленные рисовые поля.
Численность. Шилохвость чрезвычайно широко распространен-
ная и местами весьма многочисленная утка. В лесотундре и в южных райо-
нах тундры она численно преобладает над другими видами настоящих уток.
Территория, на которой она вполне обычна в качестве гнездящейся птицы,
занимает лесотундровую и лесную зону европейской части Союза, Фин-
ляндии и Скандинавии, а также северные районы Польши и Германии, всю
Западно-Сибирскую низменность, нижнее течение всех сибирских рек, низ-
менную часть бассейна Лены и Анадыря, Охотское побережье, Уссурийский
край, бассейн Сунгари, северную часть Сахалина и Командорские о-ва.
Экология. Размножение. Половая зрелость наступает, повидимому,
в годовалом возрасте. Но частые летние встречи птиц вне пределов их гнездо-
вого ареала заставляют предполагать, что часть молодняка приступает к раз-
множению через год. В гнездовое время шилохвости держатся парами, и поли-
гамии у них, как и у других уток, не наблюдается. Где и когда оформляются
пары—пока еще не совсем ясно, так как на местах зимовки наряду со сме-
шанными стаями всегда можно встретить и стайки, состоящие из одних селез-
ней, на местах же гнездовья эти утки уже появляются парами (Рыбинское
водохранилище). По всей вероятности, пары образуются или в конце зимы,
перед самым отлетом птиц с зимовки, или даже в пути. Токовые игры у шило-
хвостей при содержании их в качестве парковой птицы начинаются зимой или
даже осенью, но наблюдать их на местах зимовки нам не приходилось.
Брачные игры их заключаются в том, что селезень плавает или ходит
по берегу с вытянутой кверху шеей и издает время от времени свист. Самка
находится около него, она часто поворачивает голову назад через плечо,
как бы отпугивая мнимого врага. Иногда селезень останавливается против
самки, приподнимает одно крыло так, что расправляется его зеркальце,
и закинув голову на спину, быстро и резко проводит клювом по очину
шилохвость
373
махового пера, издавая этим негромкий дребезжащий звук. Значительно реже
чем у кряквы наблюдается выбрасывание головы вперед и вверх. Зато часто
повторяется другой, весьма схожий прием токования. Голова, имеющая
«плоскую прическу», с плотно прижатыми перьями по бокам ее и взъеро-
шенными на лбу и затылке, резким движением выталкивается вверх. Одно-
временно с этим издается звук «геееее-геееее», к которому иногда присое-
диняется высокий свист. Иногда селезень, плавающий в спокойной позе,
Рис. 67. Позы тока шилохвости.
внезапно поднимается почти вертикально над водой, в то время как клюв
его остается погруженным в воду, затем резким движением он вздергивает
голову вверх, издавая при этом свист. Часто на какую-нибудь долю
секунды самец расправляет и вздергивает вертикально кверху хвост, при-
жимает шею к спине и поворачивается головой к своей самке. Наконец,
одной из поз во время тока бывает поворот к своей самке затылком,
при котором демонстрируются яркобелые полосы на темнокоричневом фоне
(Лоренц, 1942). Шилохвости, как и некоторым другим уткам (например,
чирку-свистунку), свойствен особый токовый полет. Стайки, состоящие из
самцов и самок числом около десятка птиц, периодически взлетают с места
отдыха и кружатся на высоте 5—7 м над водой. На полете птицы держат шею
374 ОТРЯД ГУСЕОБРАЗНЫЕ
не вытянутой, как обычно, а изогнутой ввиде буквы S. Их полет бывает
неровным, и после более резких взмахов крыльями птицы как бы оседают
на хвост, отчего тело их принимает круто наклонное положение. Самцы уча-
ствуют совместно с самкой в подыскании мест для гнезда, причем поиски
происходят ранним утром.
Шилохвость гнездится всегда на земле, обычно на открытых местах.
В тундре чаще всего в понижениях рельефа, поросших осокой или кустами,
вблизи от пойменных озер. В лесной полосе она гнездится на лугах с невы-
сокой травой, стернях посевов, а иногда и на островках с почти совсем ого-
ленной почвой. В степной полосе устраивает гнезда чаще всего на пойменных
лугах. Гнезда зачастую располагаются на расстоянии более 100 м от водоема.
На Южном Ямале гнезда находили в травянистых низинах среди редких
кустов тальника, а одно из них, найденное в пятнистой тундре, было устроено
в трещине почвы под прикрытием кустиков ерника—полярной березки. На
Рыбинском водохранилище более 60% осмотренных гнезд шилохвости поме-
щалось на участках почти без растительности (стерни, вырубки с песчаным
грунтом) и около 30% на лугах с низкой травой. В первом случае гнезда
устраиваются в небольших куртинках вейника, среди редких кустиков овсян-
ницы, в кустиках вереска или даже просто на ровном месте, без всяких укры-
тий. На лугах гнезда располагаются обычно в небольших куртинках более
густой и высокой травы, среди кочек или же на кочках осоки. Лишь один
раз гнездо было обнаружено там в редком кусте ивняка. Гнездо под низко-
рослым дубком на болоте обнаружено близ Киева (Данилович, 1928). В Бара-
бинской степи гнезда строятся чаще всего среди осоковых кочек или среди
бурьянов у края озер. Гнездо представляет собой аккуратно сделанную
ямку, почти без выстилки, в которой по мере окончания кладки появляется
пух, валиком окружающий яйца и полностью закрывающий их сверху в то
время, когда самка уходит с гнезда. Размеры гнезда: диаметр около 22 см,
высота его бортов до 11—13 см, ширина лотка 19—20 и глубина его 9—11 см.
Полная кладка содержит обычно 8 яиц, в Тиманской тундре 6—9, в пойме
Мологи A940) и теперь на Рыбинском водохранилище 8 F—11), под Киевом
6—10, на островах Черного моря 8, под Томском, в Барабе и на озере Кур-
гальджин 7—9, на Южном Ямале 6—8, в Южном Приморье 9 F—10). Яйца
слегка вытянутой формы, белые с легким желтоватым или оливковым от-
тенком. Размеры яиц одной кладки довольно постоянны. Разница в длине по
большому диаметру обычно менее 5 мм, но яйца из разных кладок сильно
различаются по величине. Предельные размеры таковы: 48,9—61 х35—42 мм.
Средние размеры для отдельных кладок: 51,8x37,3 мм; 58,5x38,5 мм и т. д.
Кладки с крупными и мелкими яйцами встречаются в одних и тех же геогра-
фических пунктах. Средний вес ненасиженных яиц (для Германии) 40 г
C5—43 г), но отдельные яйца достигают 47 г (Хейнрот, 1928). Вес яиц из
Барабы (по одной кладке) 47 г D0—50,5 г). Под Киевом полные кладки появ-
ляются в последней декаде мая, на о-ве Орлов (Черное море) кладка с сильно
насиженными яйцами найдена 5 июня. В нижнем течении р. Мологи (Верхняя
Волга) первое гнездо с 2 яйцами найдено 3 мая, когда пролет их в этом месте
еще не совсем закончился. В северной части Рыбинского водохранилища на-
чало кладки у шилохвости отмечается 1—8 мая, В Эстонии гнездится с сере-
дины мая до начала июня. Под Кировом первые яйца появляются в конце
апреля, а в начале мая находят уже полные кладки по 9 яиц (Плесский,
1948). Полные кладки в Тиманской тундре отмечены в первых числах июня.
На озере Кургальджин в последней декаде мая наряду с сильно насиженными
кладками попадались и свежие; в Барабинской степи период откладки яиц
продолжается с первой декады до конца мая (Янушевич, 1947). Полные
кладки под Тобольском появляются около 12 мая, а на Южном Ямале в на-
чале июня. Эти птицы начинают гнездиться там еще до ледохода, так, 15 июня
ШИЛОХВОСТЬ 375
яа р. Щучьей были найдены гнезда с 4, 6 и 8 яйцами, а ледоход на реке прошел
только 11 июня (Дунаева и Кучерук). В районе Якутска (р. Лунхе) гнездо
с 6 яйцами обнаружено в середине июня, на Вилюе с 8 яйцами 27 июня,
а под Верхоянском 8 свежих яиц 30 мая. На Командорских о-вах откладка
яиц происходит в первых числах июня, в Приморье первые полные кладки
яиц появляются в последней декаде апреля. Шилохвость крепко сидит на
яйцах, особенно во второй половине насиживания, когда подпускает наблю-
дателя почти вплотную к своему открыто расположенному гнезду. В это
время самка, спугнутая человеком с гнезда, как правило, обливает яйца
жидким зеленым пометом, чего не бывает в начале насиживания (это же
наблюдается и у многих других уток: кряквы, широконоски, гоголя). Воз-
можно, что это является результатом очень редких в это время отлучек самки
с гнезда. Всю первую половину насиживания селезень держится вблизи
от гнезда и встречает самку, слетающую на отдых или кормежку. При появле-
нии человека он поднимается и летает кругами над тем местом, где располо-
жено гнездо, до тех пор, пока самка не слетает с него. Продолжительность
насиживания 22—23 дня. Только что выведшиеся пуховички весят 29—31 г
(по выводку из Салехарда), они быстро бегают по земле, хорошо плавают,
но ныряют хуже, чем птенцы других речных уток. Самка энергично защищает
выводок, бегает и плавает рядом, волоча крыло и стараясь отвлечь на себя
внимание врага. Выводки держатся обычно на мелководьях с богатой расти-
тельностью, склевывая с листьев и стеблей насекомых, которые первое время
составляют их основной корм. Растут молодые быстро. Их маховые перья
в период сильного роста увеличиваются на 7 мм в день. Примерно в возрасте
42 дней они приобретают способность к полету, а к 50 дням формирование их
оперения полностью заканчивается, и летают они уже хорошо.
Выводки пуховиков под Киевом встречаются уже 1 июля, а в середине
месяца они обычны. В пойме Мологи первые птенцы были обнаружены
6 июня, на Рыбинском водохранилище они появляются тоже в июне, а в се-
редине июля A3—19 июля) выводки уже поднимаются на крыло. В Тиман-
ской тундре выводки встречались с 30 июля, в середине июля еще встреча-
лись совсем молодые пуховики, а к 29 июля многие утята были уже пол-
ностью оперены, но еше не летали; нелетающие выводки встречались там
до 10 августа (Гладков, 1951). В устье р. Полуя (Салехард) только что вывед-
шиеся пуховики встречены 16 июля, севернее же на Ямале 23 июля обна-
ружен выводок подлетков, а 13 августа молодые уже хорошо летали, хотя
16 августа был обнаружен и один нелетный выводок. На Енисее под 70° с. ш.
2 августа встречен выводок только что выведшихся молодых (Тугаринов,
1908). У Березова только что вылупившиеся молодые обнаружены 24 июня,
а под Тобольском они начинают летать около 31 июля B2 июля—6 авгу-
ста). В Барабинской степи вывод птенцов продолжается с 20 мая по 20 июня,
а за время с 20 июля по 10 августа выводки поднимаются на крыло.
На озере Кургальджин 27 мая встречались одновременно и кладки со све-
жими яйцами и уже выведшиеся птенцы, а на Наурзумских озерах 18 июня
наблюдались уже почти полностью оперенные утята, к середине июля они
поднялись на крыло. На Анадыре первые выведшиеся молодые отмечались
9 и 14 июля, выводки пуховиков встречались и в первых числах августа, но
во второй декаде августа часть молодых уже поднялась на крыло. Еще плохо
летающие молодые добыты в Чаунской губе 27 августа (Тайер и Бэнгс, 1914),
а на Командорских о-вах летные выводки попадаются уже в конце июля.
Гибель кладок у шилохвости велика. Ее гнезда зачастую строятся
совершенно открыто, а к тому же на лугах с редкой травой, которые обычно
используются под выгоны для скота. В годы с высокими паводками гнезда
их часто затопляются. Это особенно часто случается на реках Сибири,
376 ОТРЯД ГУСЕОБРАЗНЫЕ
имеющих очень поздние паводки. Возможно, что по этой причине в неко-
торые годы одновременно с самцами линяет огромное количество самок.
Гибель пуховых птенцов также велика, в пойме Мологи она составляла
до момента подъема на крыло около 37%.
Линька. Как и все речные утки, шилохвость линяет дважды в год:
после сезона размножения полностью и перед началом нового частично.
Последовательность смены оперения во время полной линьки сходна у самцов
и у самок. Первыми сменяются перья на боках груди и плечевые, прикры-
вающие собой крыло, находящееся в сложенном состоянии. Одновременно
начинает линять надхвостье (его длинная центральная пара перьев). Выпа-
дает средняя пара рулей, а вскоре затем ряд других рулевых, появляются
новые перья надхвостья и подхвостья. К тому времени, когда средняя пара
рулей дорастает, почти полностью обновляется оперение всей нижней
стороны тела и верхней стороны головы. Частично перелинивают бока
головы, плечевые, средняя часть спины и надхвостье. Неперелинявшими
остаются только передняя часть спины, нижняя сторона головы и зоб.
Маховые выпадают через 3—4 недели после начала линьки. К этому вре-
мени полностью дорастают по меньшей мере две средние пары рулевых
и у самок заканчивается смена контурного оперения, в то время как у сам-
цов она заканчивается лишь тогда, когда их маховые уже достигают при-
мерно 3Д своей полной длины. От выпадения маховых до их полного отра-
стания проходит 30—35 дней. Все перья крыла, кроме третьестепенных
и длинных кроющих кисти, линяют одновременно с маховыми. Смена рулей
имеет такую последовательность. Первой сменяется средняя пара, за ней
вторая, а когда обе они уже отрастают, то последовательно четвертая,
шестая, седьмая, а затем третья, пятая и восьмая. Смена трех-четырех
внешних пар проходит различно. У самцов, закончивших линьку в лет-
ний наряд, на зобу остается большее или меньшее число перьев брачного
наряда. На тех же местах дольше всего сохраняются и остатки ювенального
оперения.
Вскоре после того как самки крепко сядут на яйца, селезни покидают
их, объединяются в стаи и начинают линьку. В средней полосе (Рыбинское
водохранилище) это происходит в середине июня. Как правило, шилохво-
сти не линяют на месте размножения, а собираются для этой цели в опре-
деленных местах, совершая нередко весьма значительные перелеты, как,
например, из устья Иртыша в дельту Волги. Кое-где—в Северном Казах-
стане, в низовьях Оби и в других районах—передвижение самцов к местам,
линьки принимает характер ясно выраженного сезонного перелета, когда
в течение определенного времени крупные стаи их следуют одна за другой.
В устье Оби стаи шилохвостей числом до 100 шт., летящие к северу, наблю-
даются в первых числах июля. С 19 по 25 июля на р. Паюте (Южный Ямал)
наблюдались стайки линных самцов, числом примерно по 20 птиц, поте-
рявших маховые перья и неспособных к полету. Первые 4 селезня, приоб-
ретшие способность к полету, встречены там 1 августа (Кучерук, 1948).
В низовьях р. Вельть (Тиманская тундра) самцы, перелинявшие в летнее
перо, встречаются с конца июля, начала августа, но наряду с этим 4 авгу-
ста был добыт селезень, который еще не линял (Гладков, 1951). В низовьях
р. Елогуя линяющие селезни поднялись на крыло к 16 августа (Слудский).
На Анадыре 4 июля уже встречались стаи холостых селезней. На мелковод-
ных озерах, заросших осокой и злаками, в устье р. Танюрера 7 августа
встречены стаи линных самцов и самок, причем у добытого селезня махо-
вые перья отросли на половину их длины. Там же 9 и 10 августа многие
селезни уже взлетали, а вторая декада августа была временем массового
выхода из линьки (Портенко, 1939). На озерах Барабинской степи массо-
вая линька шилохвостей (период их нелетного состояния) продолжается»
ШИЛОХВОСТЬ 377
примерно, с 5 по 25 июля и с последней даты начинается подъем на крыло.
Массовый выход из линьки происходит в начале августа (Яйушевич и Золо-
тарева, 1947). На озере Кургальджин (Акмолинская обл.), где шилохвость
составляет основу среди линных уток, большие стаи селезней были встре-
чены 19 июня. Они еще не потеряли способности к полету, но линька кон-
турного пера у них шла интенсивно (Слудский). Первые перелинявшие
и поднявшиеся на крыло птицы отмечены там 7 августа (Меженный и Соко-
лов). На Наурзумских озерах линных селезней отлавливали 22 июля.
Значительная часть местных птиц в некоторые годы (маловодные) отле-
тает линять куда-то южнее (Формозов, 1937). Еще южнее, в бассейне Чел-
кар-Тениза, на мелководных соленых озерах сотенные стаи шилохвостей
появляются уже в середине июня (Сушкин, 1908). В дельте Волги большие
стаи шилохвостей, прилетавших на линьку, появляются в первой поло-
вине июня A2 июня 1946 г.). Уже в начале июля часть селезней теряет
способность к полету, но для основной массы их этот период наступает
позже, а именно в конце июля, и даже в первых числах августа. В это время
шилохвости являются преобладающим видом в стаях линяющих уток.
Изменяется по времени и соотношение полов. В середине июля самки соста-
вляют незначительней процент, а в конце месяца они иногда составляют
до 30—40%. На одном из озерков у границы Тибета и Сиккима 16 августа
добыта серия линяющих шилохвостей. По всей вероятности, там прово-
дят линьку птицы, населяющие север Средней Сибири. Изучение этой сери»
шкурок показало, что, несмотря на столь позднее время, птицы находились
там в весьма различных стадиях линьки — от самых начальных вплоть
до конечных (Штреземан, 1940).
Линька самок, имевших выводки, начинается много позже, чем у селез-
ней и самок, потерявших кладку. Смена контурного пера начинается еще
в период их жизни при выводке, но выпадение маховых обычно совпадает
с тем моментом, когда молодые уже поднимаются на крыло и переходят
к самостоятельной жизни. На Рыбинском водохранилище самка, еще не сме-
нявшая маховых, добыта 25 августа. На Наурзумских озерах две самки
с выпавшими маховыми и, несомненно, имевшие выводки в этом году
(сильно пораненные от ходьбы лапы) пойманы 31 июля. В Тиманской тундре
самки с выпавшими маховыми перьями были встречены 10 августа при
выводках с еще нелетными молодыми (Гладков).
Следующая, неполная, линька селезней в брачное оперение начинается
уже тогда, когда маховые и 2—3 крайние пары рулевых еще не закончили
роста. В это время появляются новые перья по бокам груди у основания
крыльев и в партиях плечевых. Затем происходит линька нижней стороны
тела от груди к брюху. Позднее линяет спина и позже всего дорастают перья
на голове, шее и плечах, отдельные рулевые и третьестепенные маховые.
Последовательность смены оперения у обоих полов, а также у взрослых
и молодых птиц одинакова. Взрослые селезни заканчивают эту линьку
обычно уже на зимовках, но ее сроки у разных особей и стай весьма различны.
В октябре у взрослых селезней шилохвости, летящих через дельту Волги,
в большинстве случаев недолинявшими остаются плечевые партии, спина
и кроющие хвоста, кроме того, много пеньков нового оперения на голове
и шее. У молодых селезней в это время начинается смена рулей и оперения
на нижней стороне тела, но у многих из них, так же как и у молодых самок,
линька совсем не начиналась. Большая часть селезней заканчивает линьку
в середине зимы, но у некоторых она затягивается еще дольше. Так, селезни,
добытые в середине января во Вьетнаме, имели еще много линяющего опере-
ния. Один из них, пойманный живым, не получил полного брачного оперения
до начала марта (Делакур и Жабуйль, 1925). Возможно, что так ведут себя
годовалые птицы с крайнего северо-востока Сибири.
.378 ОТРЯД ГУСЕОБРАЗНЫЕ
Линька взрослых самок происходит на протяжении с ноября по апрель.
Следы ее еще можно обнаружить весной у пролетных птиц.
Питание шилохвости имеет смешанный характер. В северных ча-
стях ареала преобладающее значение имеют животные корма, в то время как
на юге растительные. Набор кормовых объектов шилохвости определяется
способом добывания корма. Эти утки кормятся не ныряя, а лишь опроки-
дываясь вертикально, и склевывают те корма, которые могут достать со дна
или с растений. Благодаря длинной шее, шилохвости способны доставать корм
с глубины, несколько большей, чем другие речные утки. Первое время по
прилете, еще до ледохода (март) шилохвости в пойме Мологи кормились
исключительно зелеными частями растений, перезимовавшей ряской, побе-
гами элодеи, злаками и др. Сразу же после вскрытия рек в их питании в
довольно значительном количестве появились личинки хирсномид,—весьма
характерный для шилохвости корм. На Рыбинском водохранилище в апреле
животные корма по их объему составляют 60% от всего содержимого желуд-
ков. Основное значение среди них имеют личинки ручейников (Phryganea
grandisy 30%) и хирономиды (ЮЛ%), моллюски в это время отсутствуют
вовсе. Много поедают они и зеленых частей растений* злаков B5%) и рого-
листника, доля же, приходящаяся на семена, не велика. На весеннем про-
лете в дельте Волги (февраль-март) основу питания шилохвостей составляют
зеленые части растений D1.5 %), из них валлиснерия и в меньшей степени
гребенчатый рдест и нитчатые водоросли. Большой процент по объему в со-
держимом желудков составляют и семена D0,6%) камышей, валлиснерии,
рдестов и др. Однако часть из них не является пищей, а служит лишь в каче-
стве гастролитов (ежеголовник, рдесты). Животных кормов в это время шило-
хвости получают мало A7,9 %), так как мелководья дельты бедны ими ранней
весной. Чаще всего поедаются моллюски и из них лунки (Theodoxus), кото-
рых утки склевывают со дна при добывании корневищ валлиснерии. Весной
на пролете шилохвости часто кормятся на убранных полях, частично затоп-
ленных, собирая там зерна хлебных злаков. В низовьях Или они чаще всего
посещают просяные поля, а в пойме Мологи овсяные жнивья. В мае—июне
на Рыбинском водохранилище потребление животных кормов еще возрастает
{70,5%). Особенно большое значение приобретают личинки хирономид
C0%), появляются в пище моллюски A5%) и совсем исчезают зеленые части
растений. В июле—августе количество хирономид в содержимом желудков
попрежнему остается большим, значение других насекомых уменьшается,
но зато сильно возрастает роль моллюсков. Последние представлены в основ-
ном мелкими видами катушек, которых утки склевывают вместе с листьями
водных растений, развивающихся к этому времени на мелководьях.
Тогда же утки начинают питаться и клубеньками гребенчатого рдеста
A1%). В пойме Мологи летнее питание взрослых птиц состояло также на 70%
из животных кормов: личинок хирономид A5%), стрекоз B0%), других
насекомых E%) и моллюсков C0%). Вегетативные части растений, злаки,
элодея, ряска составляли 25% от общего состава пищи и семена всего 5%.
Питание пуховых птенцов в основном сходно с таковыми взрослых птиц.
Из числа животных кормов (80 %) основу составляют моллюски B0 %), личин-
ки стрекоз B0%) и хирономид A5%), а также мелкие ракообразные A5%):
веслоногие, листоногие и ракушковые рачки. Совершенно иной характер
имеет питание шилохвостей в период их линьки в дельте Волги. На 50,3%
оно состоит из зеленых частей растений, главным образом резухи (Najas
marina) и семян D8%) преимущественно того же растения. Огромные стаи
шилохвостей, которые на длительный срок лишаются возможности к полету,
кормятся вечерами на подводных лугах из резухи и валлиснерии. Почти
единственным кормом их тогда служат ветки этих растений с недозревшими
семенами. Количество животных кормов там очень невелико и в содержимом
шилохвость
379
желудков они составляют всего 1,7% (моллюски затворки и др., личинки
мух-львинок, стрекоз и некоторые другие насекомые). Обычным кормом ши-
лохвости в тайге и в тундре служат ягоды. Весной на Ямале они часто кор-
мятся перезимовавшей вороникой (Empetrum nigrum), а летом поедают мо-
рошку и бруснику. В тайге под Архангельском в сезон созревания черники
выводки шилохвостей вместе с другими утками часто пасутся на ягодниках.
В степной полосе в период массового развития прямокрылых шилохвости
наряду с другими утками энергично поедают их, причем они ловят не только
тех насекомых, которые падают в воду, но охотятся за ними и на суше. На
озерах Наурзумского заповедника в течение некоторого периода таким
кормом служит npyc(Calliptamus italicus) (Формозов, 1937). В сентябре—октя-
бре на осеннем пролете через Рыбинское водохранилище пища шилохвостей
Рис. 68. Шилохвость (самец)
попрежнему остается преимущественно животной F8%). Основными кор-
мами в это время служат личинки хирономид C9%) и ручейников B0,5%),
которых уткам становится легче добывать со дна в связи с осенним спадом
воды. Кроме того, несколько возрастает значение семян. В то же время
{октябрь) в дельте Волги основной пищей шилохвости становятся вегетатив-
ные части растений E3,4%) и, в частности, луковички стрелолиста, которые
они отрывают на мелководье, а также листья и побеги валлиснерии. В боль-
шом числе поедаются и семена (резуха, рдесты), но животных кормов в их
рационе попрежнему мало. Зимой птицы, живущие на взморье у юго-восточ-
ного берега Каспия, кормятся мелкими моллюсками и водорослями. На
азербайджанских зимовках основу пищевого рациона шилохвостей состав-
ляют семена G4,6% по весу): руппий, различных камышей, гречишек и не-
которых солончаковых растений. На долю зеленых частей растений прихо-
дится 23,7%, а на животные корма всего 1,7% (моллюски, жуки).
Хозяйственное значение. Шилохвость—одна из наиболее широко рас-
пространенных и многочисленных речных уток. Значение ее как промысло-
вого объекта очень велико. В заготовках дичи на территории Западной
Сибири на долю шилохвости приходится около 16,8% от всей суммы добы-
тых настоящих уток, а на зимовках в Закавказье она составляет около 7%
уток, добываемых за сезон. Большое значение шилохвость имеет для насе-
ления нашего севера, особенно севера Якутии, где существует массовый
промысел линных птиц и заготовка их впрок. Чистый вес мясной тушки
шилохвости составляет около 65% от ее живого веса, т. е. примерно 520—580 г
380 ОТРЯД ГУСЕОБРАЗНЫЕ
Полевые признаки. Утка среднего размера с длинной шеей и острым,
игловидным хвостом, форма которого хорошо видна и у летящей и у пла-
вающей птицы. Окраска самки однообразно серая, у самца коричневая
голова, белая грудь, передняя сторона шеи и узкие полосы по ее бокам,
доходящие до головы сзади ушей. У летящей птицы видна одна белая полоска
по краю зеркала. На лету шилохвость часто изгибает шею вниз и в стороны.
Ныряет плохо и неохотно, даже будучи ранена. Голос самки очень сходен
с голосом кряквы. Голос самца — довольно тихий и мелодичный свист,
довольно хорошо передаваемый слогами «тью-и-дишь», «тью-и-дишь».
Описание. Строение и размеры. Утка среднего размера с вытянутым туло-
вищем, длинной шеей и длинным клювом. Два средних рулевых узкие и сильно удлинен-
ные. Их длина у селезня до 200 мм, у утки до 100—120 мм. Остальные рулевые тоже
заостренные. Шилохвость хороший летун, по размеру крыльев и весу летательной муску-
латуры занимает второе место среди настоящих уток (следующее после кряквы).
Размеры: крыло самцов 260—290, самок 240—266 мм; цевка самцов 39—44, самок 37—
42 мм; клюв самцов 48—53, самок 42—50 мм.
Вес шилохвостей весной на пролете таков: в дельте Волги в феврале самцы 1000 г
(800—1150 г); в марте самцы 923 г G50—1150 г), самки 700 г; в апреле самцы—773 г G60—
800 г). Снижение веса от февраля к апрелю обязано тому, что раньше прилетают старые
птицы, а позже молодежь. К тому же условия кормежки в дельте ранней весной плохие,
и живущие в ней утки худеют. В северной части Рыбинского водохранилища в апреле
самцы весят 917 г (800—1020 г), самки 860 г; в мае самцы 888 г G75—950 г),самки 800 г.
В Барабинской степи в конце мая вес самцов 880 г G65—1035 г), самок 935 г. В Карелии
в мае самцы 856 г, самки 910 г; на Печоре в мае самцы 950 г (840—1060 г), самки 740 г
и на Ямале самцы в мае 930 г (900—970 г).
В течение периода размножения вес у обоих полов снижается. На Рыбинском водо-
хранилище он в июне у самцов 897 г (828—964 г), а у самок в июне 738 г, в июле 700 г,
в августе 775 г F70—880 г). Самки из Северного Казахстана в июле 750—850 г. Вес лин-
ных птиц (в период потери маховых) в дельте Волги: самцы в июле 944 г (800—1150 г),
в августа 929 г G00—1150 г); самки в июле 800 г F50—1050 г), в августе 848 г F00—
1200 г). Там же взрослые птицы на осеннем пролете: самцы в августе-сентябре 1081 г
A000—1150г), в октябре 1057г (900—1300г); самки в августе-сентябре 675г G50—805 г),
в октябре 870 г (800—900 г). Как видно, осенью вес взрослых селезней достигает своего-
максимума. Изменение веса молодых птиц происходит так (Рыбинское водохранилище): во
второй половине июля, наряду с пуховиками, встречаются уже молодые самцы, достигаю-
щие 830 г, и самки до 765 г; вес их в августе—самцы 180—850 г, самки 572—825 г; в
сентябре уже большая часть молодняка догоняет по весу взрослых птиц, самцы весят
810—1045 г, самки 625—830 г; в октябре самец 959 г, самка 700—799 г. Вгс молодых
птиц во время осеннего пролета на Печоре: самцы 750 г G00—800 г), самки 600 г E50—
700 г), а в Астрахани в сентябре самцы 910 г (800—975 г), самки 800 г G00—875 г) и в
октябре самцы 842 г G00—950 г), а самки 731 г F00—850 г).
За период зимовки, особенно во второй его половине вес птиц заметно снижается.
В ноябре у Гасан-Кули самцы 750—1100 г, самки 728 г D60—860 г), возраст птиц зимой
не учитывался. На ленкоранских зимовках вес шилохвостей A932) был: в январе самцы
820 г G50—1000 г), самки 650 г F00—700 г); в феврале самцы 732 г E50—930 г),самки 586 г
D00—700 г). В некоторые зимы A934) средний вес падает еще сильнее.
Окраска. Пуховой птенец. На спинной стороне темный, серовато-бурый.
На голове темная окраска ограничена лишь довольно узкой полосой. Бока головы бело-
ватые, от клюва через глаз и далее к затылку проходит узкая темная полоса, на ухе
небольшое пятно. Брюшная сторона серовато-белая. По бокам спины от основания
крыльев до поясницы узкие белые полоски или ряд пятнышек. Вдоль переднего края
кисти и заднего края предплечья кайма из белого пуха. Клюв сверху темнобурый, ного-
гок и подклювье светлые, рыжеватые. Ноги оливково-бурые.
Молодая самка в первом наряде. Общая окраска спинной стороны серо-бурая
со светлыми каемками перьев и ржавчато-бурыми неполными поперечными полосками
на некоторых из них. Голова несколько бурее и в мелких продольных черных штрихах.
Брюшная сторона сероватая, сильно испещренная продольными темными штрихами. Руле-
вые бурые обычно с поперечными беловато-охристыми полосками. Средняя пара их почти
не удлинена Верхние кроющие крыла серо-бурые или темнобурые с беловатыми, а неко-
торые со слегка охристыми каемками и с такими же поперечными пятнами по бокам пера
в его верхней части. Первостепенные маховые темнобурые, второстепенные бурые с более
светлым мраморным рисунком на наружных опахалах и белыми вершинами. Они обра-
зуют рыжеватое с бурыми пятнами зеркало, совершенно без блеска, с белыми каемками
сверху и снизу. Плечевые черно-бурые с более светлыми каймами. Клюв и лапы темно-
серые, радужина коричневая или темнокоричневая.
шилохвость 381
Молодые самцы в том же возрасте. Бывают двух типов окраски: или такой же
как самки или же менее бурой, черно-серого основного тона с беловатыми краями
перьев и буроватым крапом на них. Крылья их весьма сходны с таковыми у взрослых
•селезней. Верхние кроющие их оливково-серые; ржаво-каштановые вершины больших
кроющих образуют кайму с передней стороны зеркала крыла. Последнее образуется
бронзово-зелеными наружными опахалами второстепенных маховых и ограничено сзади
черной и белой каймой из вершин и предвершинных полосок тех же перьев. Второсте-
пенное маховое, замыкающее, зеркало, с внутренней стороны имеет широкий черный край
наружного опахала. Однако в отличие от взрослых селезней его окраска неровная
и местами переходит в аспидно-серую, На том же опахале вдоль ствола проходит
пепельно-серая полоса. Плечевые бурые со светлым крапом. Окраска голых частей и ра-
дужины та же, что и у самки.
Взрослая самка. Окраска верхней стороны головы и туловища та же, что и у моло-
дой птицы. Брюшная сторона без штрихового рисунка, сероватая с мелкой темной рябью.
Рулевые бурые с поперечными буровато-охристыми полосками. Средняя пара их заметно
удлинена. Крылья, как у молодых самок, только рыжеватое зеркало их несколько шире.
Клюв темносерый, более черный по коньку, ноги свинцово-серые; радужина бурая,
у старых желтовато-бурая.
Взрослый самец в брачном наряде. Голова и часть шеи коричневые, причем
верх головы более темный, затылок почти черный. По бокам головы, начиная от глаз
к затылку, хорошо заметный бронзово-зеленый отлив. Нижняя часть шеи сзади, весь
верх туловища и бока серые с темной струйчатостью на перьях. В задней части спины
струйчатость бывает только у старых птиц. Перья надхвостья с темными центрами,
кроющие хвоста черные, причем верхние с белыми внутренними опахалами. Грудь, зоб
и передняя часть шеи белые. От них по бокам шеи поднимается кверху довольно узкая
белая полоса, достигающая боков головы сзади ушей. Брюшко белое с серой рябью, бока
поясницы палевые. Средние удлиненные рулевые черные с металлическим отливом.
Крыло такое же, как и у селезня в первом наряде, но не бывает светлых пестрин на верх-
них кроющих, а крайнее второстепенное маховое, замыкающее зеркало, имеет ровную бар-
хатисто-черную внешнюю часть наружного опахала и почти белую полосу вдоль ствола.
Плечевые перья черные с буровато-серыми краями. Клюв голубовато-серый, лапы свинцо-
во-серые, радужина буро-желтая, а у старых желтая.
Молодой самец в первом брачном наряде. Отличается, как описано выше, по
окраске крайнего второстепенного махового. Кроме того, окраска головы у него не
коричневая, а темная серовато-бурая. Струйчатость на перьях спины выражена менее
четко, особенно в ее нижней части. Радужина бурая.
Самец в летнем наряде. Сходен по расцветке с самкой. Как и у молодых селезней
в первом пере, окраска их бывает двух типов—серо-черная и более бурая. На перьях
зачастую бывает струйчатый буроватый рисунок или крап. У самцов черно-серого основ-
ного тона рулевые одноцветные, темносерые, а у более бурых—со светлыми краями
и неправильными косыми полосками. Брюшная сторона у всех белая с большим или
меньшим числом округлых серо-бурых пятен. Оба типа окраски самцов встречаются в од-
них и тех же географических пунктах. Верным отличием полов в любом из нарядов слу-
жит зеленое зеркало на крыле у селезней и наличие мелких темных пятнышек на клюве
у самок. Зачастую белое брюхо у шилохвостей бывает окрашено в ржаво-красный цвет. Это
наблюдается местами в Казахстане и на зимовках в Вьетнаме и обязано пребыванию птиц
шилохвостей на болотах, вода которых богата железом.
Систематические замечания. Американские шилохвости выделяются в особый под-
вид A. acuta tzitzihoa V i e i 1 1 о t, 1816, отличием которого служит несколько более ши-
рокое зеркало с пурпурным отливом и более удлиненные рулевые. Просмотр имеющихся
в наших музеях коллекционных материалов не подтверждает этих различий. Оба различи-
тельных признака у птиц, добытых в пределах СССР, подвержены весьма значительной
изменчивости, полностью перекрывающей те показатели, которые характеризуют амери-
канскую форму. Американские орнитологи, различая эти две формы, считают, что запад-
ная часть Аляски населена A. a. acuta (Бейли, 1948), а также регистрируют случаи залета
A. a. tzitzihoa в Восточный Китай. Однако пока нет оснований для выделения американ-
ской шилохвости в качестве самостоятельного подвида.
В южной части Индийского океана, на о-ве Кергелен (около 50° ю. ш.) гнездится осо-
бая форма шилохвостей, близкая по строению и окраске с голоактическим шилохвостям.
Она описана как самостоятельный вид Anas eytoni ьпагре* 1875. Однако возможно, что ее
следует считать лишь подвидом A. acuta (Делакур и Майр, 1945). Эта форма известна так-
же с о-вов Крозет и мыса Доброй Надежды (находку в последнем пункте вероятно следу-
ет считать залетом).
382
ОТРЯД ГУСЕОБРАЗНЫЕ
22. Чирок-свистунок Anas crecca L.
Anas Crecca. Li n n a e u s, Syst. Nat., изд. X, 1, 1758, стр. 125, Швеция.
Русские названия. Чирок-свистунок, свистун, чирок-хрипунок—обязаны
весеннему крику самцов; чиренок, чирок малый, чирок половой.
Распространение. Ареал. Гнездовой ареал захватывает почти всю
Западную Европу, Советский Союз, кроме северных частей тундровой зоны,
а также Южного Казахстана и большей части территории Среднеазиатских
республик. На юг идет до Северного Ирана, Северо-Западной Монголии
и Маньчжурии. Населяет северную половину Японии и западную часть
Северной Америки на север до залива Коцебу и дельты р. Мекензи. На восток
Карта 43. Распространение чирка-свистунка Anas crecca
1—А. с. crecca, 2—А. с. саго linen sis; a—граница гнездовий, в—места постоянных встреч летом и
спорадического гнездования, г—основные зимовки, д—залеты, ж—места скопления птиц на линьку
до Медвежьего и Большого Невольничьего озер, устья рек Черчилль и Нель-
сон на берегу Гудзонова залива и западных окраин Великих озер. К югу
до низовьев Миссури и Лос-Анжелоса. Кроме того, обособленная гнездовая
популяция занимает озера Эри, Онтарио и верхнее течение р. св. Лаврен-
тия. Места зимовок опоясывают всю приморскую зону Европы от Южной
Норвегии на севере до Черного моря на юге. Они занимают также Северо-
Западную Африку, бассейн Нила, Красное море, Ирак, Иран, Индию,
северную часть Индо-Китая, значительную часть Китая, Корею и Японию.
В Америке зимний ареал свистунка простирается от о-вов королевы Шар-
лотты и Онтарио на севере до Мексики и Антильских о-вов (включительно)
на юге.
Характер пребывания. На протяжении большой части
ареала перелетная птица. Живет оседло только на Британских о-вах, в Ислан-
дии, в средиземноморских странах и юго-восточной части ареала на Амери-
канском материке.
Биотоп. В гнездовое время исключительно пресные внутренние водоемы
преимущественно мелкие озера, ручьи и речки. Зимой и во время пролета
ЧИРОК-СВИСТУНОК 383-
не избегает морских побережий. Встречается от лесотундры и тундры до
пустынь, но особенно характерен для водоемов в лесном ландшафте.
Подвиды и варьирующие признаки. Европейско-азиатская часть ареала занята типи-
чной формой А. с. crecca L., 1758; Америку заселяет подвид А. с. carolinensls G me li n,
1789, хорошо отличающийся тем, что зеленые и рыжие пятна на боках головы не разделя-
ются белыми полосками. Неясная светлая линия намечается только от клюва к брови. Зата
по бокам зоба перед крылом широкая белая поперечная полоса. Струйчатость мантии более
тонкая и частая, отчего птица выглядит сверху более темной. Плечевые перья без черных
наружных опахал. Самки американского подвида по внешним признакам не отличаются
от птиц восточного полушария. Часто залетает на юго-западное побережье Гренландии;
возможны залеты на северо-восток Сибири.
Чирок-свистунок Anas crecca crecca
Распространение. Ареал. В Западной Европе гнездится повсюду,,
включая Исландию, но отсутствует на Пиренейском п-ове, в Южной Италии,
Сицилии и Греции. Часто, иногда стайками, залетает весной на юго-восточ-
ный берег Гренландии, изредка на Шпицберген и Ян-Майен. На территории
Советского Союза на восток почти до устья Печоры, северная граница гне-
здованья свистунка достигает морского побережья. Восточнее он не про-
никает в зону Приморской арктической тундры, но спорадически отдель-
ными парами гнездится на юге Новой Земли (Успенский). На Ямале гне-
здится к северу за 70° с. ш., по Енисею до устья; гнездится на Таймырском
озере. Проникает до Хатангского залива и до южной части дельты Лены.
По Колыме гнездится до устья Анюя. Населяет весь бассейн Анадыря,
встречается летом в Анадырском хребте, но гнездованье там не доказано.
Южнее обычен на Камчатке, Командорских о-вах и Курильской гряде.
Южная граница гнездованья проходит по берегам Черного моря, отсекая
Крым. За последнее время нет достоверных сведений и о гнездованьи в Бол-
гарии; места, для которых он указывался прежде, в настоящее время осу-
шены (Патев, 1950). Заселяет Кавказ и Закавказье. По лесистому южному
берегу Каспия проникает в долины Гюргена и Атрека (но не в СССР). Вос-
точнее граница гнездованья чирка-свистунка в Средней Азии проходит
от северного побережья Каспийского к северному побережью Аральского
морей; не подходя вплотную к пустынным берегам последнего, спускается
в дельту Сыр-Дарьи и «языком» вдается к югу по Аму-Дарье. Однако гне-
здованье в долине Аму-Дарьи не вполне достоверно. Если эта территория
действительно входит в гнездовой ареал свистунка, то лишь как район спо-
радичного гнездования и регулярных летних встреч бродячих особей. К той
же категории можно отнести долины рек Теджена, Мургаба и пресных озер
Западного Узбоя, которые указываются в качестве обособленных гнездовий
(Зарудный, 1896).
Восточнее южная граница проходит по самым северным частям Казах-
стана, обходя все пустынные и полпустынные области. Свистунок обычен
на гнездовье на озере Чеглы Советского района Северо-Казахстанской обл.,
но в низовьях рек Тургая и Иргиза, на озерах Кургальджин Тенизской
группы и во всем Прибалхашье отсутствует (Слудский). Указания Шнит-
никова A949) о гнездованьи в горах и особенно в равнинах Семиречья
нуждаются в серьезной проверке. Параллельно Иртышу граница гнездо-
вого ареала спускается к югу и, повидимому, захватывает северо-западную
озерную область Монгольской Народной Республики. Далее пределом гне-
здовой территории служат Северо-Восточная Монголия и Маньчжурия.
Встречается летом в Северном Китае (Чжили, Хэбэй), но гнездованье его
там достоверно не доказано. В Японии заселяет остров Хоккайдо и северную
половину Хондо, где вместе с кряквой гнездится в горах на плато Ками-
кухи (Ян, 1941).
384 ОТРЯД ГУСЕОБРАЗНЫЕ
Во многих местностях южнее очерченной нами территории свистунки
встречаются летом довольно регулярно как стайками, так и одиночками.
Это относится к Трансиордании, Южной Месопотамии, Сеистанской котло-
вине, верховьям Аму-Дарьи и нижнему течению Сыр-Дарьи, бассейну Или,
Ала-Кулю, верхнему течению Хуанхэ (Синин, Нинся) и ряду других мест.
В большинстве случаев это бродячие особи, задержавшиеся вблизи от мест
зимовки, но в тех же пунктах возможны и случаи их спорадичного гнездо-
ванья. Возможно, что именно так следует расценивать неоднократные лет-
ние встречи свистунков в низовьях р. Или и добычу там в конце июля—начале
августа 1942 г. молодых с еще недоросшими маховыми (Штегман, 1949).
Области скоплений линных птиц. Основная область
массовых скоплений селезней в период линьки лежит в степной и лесостеп-
ной полосе юго-востока Европейской России и Юго-Западной Сибири. Она
простирается от дельты Волги через среднее течение Урала, бассейна Илека,
Кустанайскую и Кургальджинскую озерные области до озер Южного За-
уралья и Барабинской степи. Других пунктов столь же массового скопле-
ния линных свистунков неизвестно. Однако в некотором количестве они
линяют и в других местах. Так, свистунки встречаются в стаях линяющих
уток в бухте Мацаль-Вик (Эстония) и в затопленных лесах на Рыбинском
водохранилище. На обширных полях в низовьях Оби линные свистунки
встречаются единицами среди тысячных стай шилохвости, одиночками же
они линяют в поймах таежных и лесотундровых рек левобережья Енисея.
Наконец, незначительные скопления линных стай известны в бассейнах
Лены и Анадыря.
Основные зимовки. Массовые зимовки свистунков находятся
на побережьях Британских о-вов, а также у берегов континента от Северной
Франции до Дании и Южной Швеции. Исландские чирки частично зимуют
на месте. В большом числе чирки проводят зиму в Испании, Южной Фран-
ции, Италии и на прилежащих островах. В массе зимуют они всюду на Бал-
канском п-ове. Южными пределами регулярных зимовок служат Канарские
и Азорские о-ва, побережье Северо-Западной Африки, бассейн Нила и берега
Красного моря. В большом числе свистунки зимуют на Азовском море,
а также у берегов Южного Каспия, в Юго-Восточном Закавказье и Юго-За-
падной Туркмении. В меньшем числе они проводят зиму на Мургабе, Теджене,
Верхней Аму-Дарье и Или. В огромном числе в Ираке, Иране (Сеистане
и на побережье Оманского залива), Белуджистане и повсюду в Индии.
Зимует в Северном Индокитае и на Филиппинах. Многочислен зимой в при-
морской полосе Китая, по долинам крупных рек страны, на озерах Южного
Тибета, в Корее и всюду в Японии.
Характер пребывания. На Британских о-вах, частично
в Исландии и у Средиземного моря чирок-свистунок оседлая птица. Так же
и в Японии. На территории СССР чирок повсюду отлетает на зиму к югу.
Частичную оседлость можно предположить только в отношении Закавказья
и Камчатки, но в последнем месте он остается на зиму в весьма небольшом
числе вблизи от горячих источников.
Сезонные миграции. Данные кольцевания позволяют гово-
рить об основных направлениях сезонных миграций у различных геогра-
фических популяций чирка-свистунка.
Чирки, населяющие Северную Германию, Бельгию, Голландию, Данию,
Скандинавские страны, Финляндию, Прибалтийские советские республики,
Польшу, северную зону европейской части Союза на восток до рек Индиги
и Печоры, а на юг до Московской и Ивановской обл. зимуют на северо-
западе Европы в большом количестве на Британских о-вах. На места зимовок
они следуют в основном вдоль берегов Балтийского и Северного морей через
Северную Германию, Данию и Голландию. Туда же спускаются зимой
ЧИРОК-СВИСТУНОК 385
и чирки, гнездящиеся в Исландии. Однако некоторая, хотя и незначитель-
ная, часть птиц из этих популяций проникает значительно южнее—в Среди-
земное море и встречается зимой вплоть до Южной Франции, Италии и Испа-
нии. Сюда же отлетают и птицы, населяющие Центральную Европу. Часть
свистунков, обитающих на европейском севере к востоку от Северной Двины,
большая часть чирков из центральных областей европейской России, а также
птицы с Поволжья, Урала, из бассейна Оби и Иртыша и частично из Север-
ного Казахстана отлетают к южным берегам Каспийского моря в Закав-
казье и далее в Западный Иран, Ирак, а также в Балканские и другие страны
восточной половины Средиземного моря, на запад до Апеннинского п-ова
и на юг до Египта. Основное направление их сезонных миграций юго-запад-
ное. Птицы следуют по долинам Оби, Иртыша, Тобола, Камы, Урала, Волги
и Дона и далее к югу побережьями Черного и Каспийского морей. Огром-
ное количество селезней из пределов очерченного гнездового ареала про-
водит летнюю линьку в дельте Волги.
На вр*мя летней линьки по озерам Барабинской степи, насколько
можно судить в настоящее время, собираются птицы из Западной Сибири
(бассейн Оби и Предуралья—Молотовская обл.), преимущественно из лесо-
степных территорий. Осенняя миграция этих птиц направлена к югу и основ-
ные места зимовки находятся в Северной Индии, куда птицы следуют через
Южный Казахстан (бассейн Сыр-Дарьи) и Киргизию. Перелеты в восточной
части материка изучены слабо, достоверно известно лишь то, что свистунки,
населяющие Камчатку, проводят зиму в Японии.
Даты. Весенний отлет свистунков с их зимовки на Британских о-вах
начинается в феврале, когда они покидают западное побережье острова,
пересекая его по долине р. Темзы. С южнокаспийских зимовок отлет начи-
нается в середине февраля, и основная масса чирков исчезает к началу марта.
Однако пролет на этом полностью не заканчивается и продолжается вдоль
берегов Азербайджана еще в апреле. Очевидно, в это время отлетают птицы
из наиболее северных гнездовых популяций вида. В дельте Волги первые
свистунки появляются обычно около 25 февраля B1 февраля—7 марта),
в нижнем течении Днепра (Запорожье) в начале марта, а в середине месяца
там проходит их основной пролет тысячными стаями, под Киевом он бывает
23—30 марта. В среднем течении Урала появление первых приходится на
последнюю декаду марта и первые числа апреля, а валовой пролет на 14—
28 апреля. Под Москвой чирки прилетают в первой половине апреля (9—
15 апреля) и летят до конца месяца, обычно небольшими стайками по 7—
10 шт. В устье р. Мологи в 1940 г. пролет прошел очень дружно. Первые
появились 20 апреля, а 21—23 апреля шел валовой пролет, последние про-
летные стайки встречены 1 мая. В это же время свистунки появляются и под
Ленинградом. Под Архангельском (озеро Слободское) средняя дата при-
лета 7 мая A3 апреля—13 мая), в Тиманской тундре первые отмечены 14 мая,
а с 27 мая стали встречаться значительно чаще (Гладков, 1951). На Коль-
ском п-ове (озеро Имандра) чирки прилетают обычно к 20 мая (8 мая—3 июня);
(Владимирская, 1948), в Кировской обл. они появляются в середине апреля,
а на Печоре, в устье Ылыча, 27 апреля B2 апреля—5 мая). Пролет их там
продолжается обычно 28 дней, птицы держатся стайками по 10—15 шт.
Средняя дата прилета под Тобольском 26 апреля B0—30 апреля), а разгар
пролета 5 мая B8 апреля—14 мая). Из Белуджистана и с Мекранского побе-
режья свистунки улетают в апреле, задерживаясь там до конца этого месяца,
а некоторые даже до 9 мая (Тайсхерст, 1927). В Гиссарской долине (Таджи-
кистан) полет наблюдается в апреле и в начале мая. В низовьях же Сыр-
Дарьи первые чирки в теплые весны появляются уже в конце февраля,
а с 13 марта бывает уже валовой пролет, наибольшая интенсивность кото-
рого отмечена 25 марта (Спангенберг, 1936). В низовья р. Или они приле-
25 птицы СССР, т. IV
386 ОТРЯД ГУСЕОБРАЗНЫЕ
тают 11—19 марта (Слудский), основной же пролет проходит позже, уже
после вскрытия рек. На Эмбе валовой пролет наблюдался с 10 апреля до
конца этого месяца, а в низовьях Иргиза пролет начался с 12 апреля (Суш-
кин, 1908). Отлет свистунков, зимующих в Южном Китае (Хубэй, Сычуань),
происходит в марте, но затягивается до апреля, массовой пролет через Чжили
в марте-апреле (Колдуэлл, 1931). На Японских о-вах чирки задерживаются
еще много дольше, до начала мая, когда они объединяются в огромные стаи,
которые пролетают к северу над морем вдоль берегов (Ян, 1942); судя по
данным кольцевания, это, вероятно, птицы с Камчатки. В Южном При-
морье свистунки появляются 21—25 марта, валовой пролет их приходится
на 2—13 апреля и бывает очень интенсивен, размеры отдыхающих стай дохо-
дят до 200—400 шт. (Шульпин, 1936). Камчатки-свистунки достигают
26 апреля—5 мая, а валовой пролет их проходит обычно 10—20 мая (Аверин,
1948), на Командорских о-вах первые появляются 2 апреля, а основная
масса в конце апреля—начале мая. В районе Аяна прилет отмечен в послед-
ней трети мая (Снигиревский), а на Анадыре в разные годы с 11 по 30 мая,
а с 5 июня птицы уже держались парами (Портенко, 1939). На Индигирке,
в устье р. Момы, прилет свистунков отмечен 21 мая 1929 г. (Михель, 1935).
Осеннему пролету, как и у других уток, предшествует летняя миграция
селезней. Основная же масса отлетает с Анадыря в сентябре, с Командор-
ских о-вов в октябре, с Камчатки в сентябре-октябре. Однако в последнем
месте в районе горячих ключей, чирки задерживаются много дольше, а не-
которая часть их даже зимует. Осенний пролет на юг Приморья начинается
в конце сентября и в то же время чирки достигают мест зимовки в Японии,
а в октябре уже в массе появляются в Южном Китае. Однако, в небольшом
числе они отмечались в Квантуне уже с 30 августа. Из окрестностей Якутска
они отлетают в конце сентября. Под Салехардом сильный пролет отмечен
в начале сентября, а к 14 сентября чирков там уже не было. На Верхней
Печоре осенние пролетные стаи появляются около 23 августа A8 августа—
1 сентября), пролет продолжается в среднем 31 день и последние встречи
относятся обычно к 4 октября Из-под Архангельска основная масса отлетает
15—20 сентября, а с Кольского п-ова около 3 сентября B5 августа—9 сен-
тября. На р. Мологе откочевка к югу отмечена во второй половине сен-
тября, интенсивный пролет проходил в конце этого месяца, а последние
стайки встречались в устье до 21 октября. Под Москвой пролет идет в тече-
ние всего октября, причем пролетные стаи бывают довольно велики. В Чка-
ловской обл. осенний пролет очень длителен, он заметен уже с 18 августа,
к 26 августа уже настолько силен, что стаи достигают 100—200 птиц, но осо
бенно велика его интенсивность в конце сентября и начале октября. В дельте
Волги первые пролетные стаи появляются 25 сентября, время массового
пролета падает обычно на 10—20 октября, а массовой отлет на последние
числа месяца. В нижнем течении Или осенний пролет зарегистрирован
30 сентября—3 октября (Штегман, 1949). В верхнем течении Аму Дарьи
количество их постепенно возрастает уже с августа, в последних числах
этого месяца первые свистунки появляются в Пакистане, основная же масса
их прилетает туда в начале сентября. Наиболее интенсивный поток свистун-
ков через Голландию проходит во второй половине сентября, через Данию
в сентябре—октябре, в начале октября они достигают уже мест зимовок
в Южном Уэльсе (Англия). Численность их в последнем месте постепенно
увеличивается до середины декабря.
Биотоп. В период гнездованья чирки-свистунки заселяют внутрен-
ние водоемы самого разнообразного типа—от речек в тундре до степных
и полупустынных озер. Явное предпочтение они отдают в это время мелким
водоемам перед крупными, лесистым местностям—перед открытыми. В север-
ной части ареала свистунки характерны для равнин и редки в горах; наоборот
ЧИРОК-СВИСТУНОК 387
у южных пределов они населяют преимущественно или даже исключительно
озерные плато в горах (Закавказье, южный берег Каспия, Северо-Западная
Монголия, о-в Хондо в Японии).
На время линьки значительная часть селезней выселяется в степные
и полупустынные местности, на водоемы с богатой надводной раститель-
ностью. Зиму чирки зачастую проводят в приморских местностях, где стаи
их отдыхают днем на морских заливах, а по зорям кормятся на мелководьях
самых разнообразных внутренних водоемов, не избегая при этом и соленых.
Численность. Один из наиболее многочисленных видов уток,
во многих местах превосходящий по численности крякву, с которой имеет
весьма сходный по характеру ареал. Весьма обычен и многочислен чирок-
свистунок по всей лесной и лесостепной полосе, становясь более редким
в лесотундре и редким в тундре. Значительно реже он и в открытых степях,
а в пустынные области проникает лишь спорадично.
Экология. Размножение. Половой зрелости чирки-свистунки
достигают обычно в возрасте одного года, но некоторая часть птиц этого
вида на первый год в размно-
жении, повидимому, не участ-
вует. Именно к этой катего-
рии, вероятно, относятся птицы,
встречающиеся летом за преде-
лами своего основного гнездо-
вого ареала в Туркмении, на
Сыр-Дарье, в Семиречье, Мон- Рис# 69. позы тока чирка-свистунка
голии, Ираке, Сеистане и дру-
гих местах. Свистунка совер-
шенно ошибочно считают типичным полигамом (Тугаринов, 1941). Причи-
ной этому служит то обстоятельство, что повсюду в средней лесной полосе
чирок-свистунок наиболее многочисленная утка, которая гнездится в не-
посредственной близости пары от пары, отчего на призыв самки, сошедшей
с гнезда, зачастую слетаются селезни из соседних пар. Однако пары у сви-
стунков выражены очень четко, не менее ясно, чем у крякв. Образуются
они позже, чем у последнего вида. Почти всю зиму чирки держатся круп-
ными стаями, и только в феврале, перед отлетом с азербайджанских зимо-
вок, часть птиц внутри этих стай обособляется в пары. Дальнейшая раз-
бивка на пары происходит в пути во время весеннего перелета. По наблю-
дениям на р. Мологе, не менее 20% чирков в пролетных стаях уже раз-
бились на пары.
Токование чирка-свистунка весьма сходно с токованием кряквы.
Селезни так же плавают вокруг самок с втянутой в плечи головой, оперение
которой распушается и на щеках красуются меднокрасные и зеленые
пятна. При этом они часто резким движением выталкивают голову кверху,
издавая звонкий металлический звук «хрлюк». Утка, плавая около
выбранного ею самца, отпугивает мнимого врага клювом через плечо и по-
крякивает скороговоркой. Часто селезни расправляют крыловое зеркало,
закладывают голову за крыло и издают дребезжащий звук, проводя ногот-
ком клюва по нижней поверхности очина махового пера. Они опускают в воду
клюв, а затем быстро, почти вертикально, поднимают все туловище и вздер-
гивают голову вверх, поднимая фонтан брызг и издавая резкий свист. Как
и кряквы, они часто на одно мгновение вздергивают зад с расправленным
хвостом, приподнимают локти крыльев и прижимают голову к спине, демон-
стрируя при этом яркое крыловое зеркало, матовочерное оперение под-
хвостья со светлыми пятнами по бокам и черные каплевидные пятна
на зобу и груди. Вслед за этим селезень плавает вокруг самки, распла-
ставшись на воде с вытянутой по поверхности шеей. Затем, переходя на
25*
388 ОТРЯД ГУСЕОБРАЗНЫЕ
обычное плавание, селезень поднимает голову на вытянутой шее и все время
поворачивает ее в сторону своей самки ярко раскрашенными красно-зеле-
ными щеками. Часто стайки их кружатся над водой, летая особым токовым
полетом, описанным нами для шилохвости.
Для гнездованья свистунки выбирают места, обладающие хорошими
укрытиями, под которыми и прячут свои гнезда. В тундре они устраивают
их в зарослях тальников, на осоковых кочках около маленьких речек или
на сухих ярах под кустами кедрового стланца (Анадырь). В таежной и сред-
ней лесной полосе более 50% гнезд этого чирка помещается в лесу около
узких лесных ручьев или в лесистых гривах среди пойм рек или около круп-
ных озер и водохранилищ с лесистыми берегами. В сосновых и еловых гри-
вах они устраивают гнезда под укрытием папоротников или же в густых зарос-
лях брусники и черники, а в мелколесье и на опушках под мелкими сосен-
ками и елками. На вырубках свистунки зачастую гнездятся под кучами
валежника и хвороста. На пойменных лугах чирки охотно поселяются около
мелких озеринок с ольховыми зарослями и в кустарниках притеррасной
части поймы; редко, гнезда свистунка встречаются и на безлесных луговых
участках. В таких случаях они бывают спрятаны или в густых осоковых
кочках, или в куртинах высоких злаков и бурьянов, выделяющихся среди
основного, более низкого травостоя. Из 33 гнезд, найденных в поймах рек
Шексны и Мологи, 18 были устроены среди ивовых и ольховых кустов,
2 под елочками в лесу, 2 под кучами хвороста на торфяном болоте и 9 на осо-
ковых кочках среди болотистых лугов. 65 % гнезд, обнаруженных у берегов
Рыбинского водохранилища, было устроено под молодыми деревьями, кустами
и валежником, 28% в куртинах высокой травы и лишь 7% на земле без
хороших укрытий. В лесостепи (Барабинская степь) свистунки гнездятся
или в березовых колках или в зарослях бурьянов, реже среди осоковых кочек
по займищам. На юге своего ареала они чаще всего используют для гнездо-
вания лесные уремы в поймах рек и заросли бурьянов у мельничных прудов.
Гнезда свистунка располагаются обычно вблизи от воды, но нередко
и на значительном расстоянии (до 500 м) от нее. Последнее часто происходит
оттого, что рано приступающие к гнездованию свистунки устраивают гнезда
по окраине поймы у временных весенних луж, которые после спада воды
высыхают. Ямку для гнезда чирок подготавливает, вырывая клювом корни
растений и выравнивает ее вращательными движениями груди. За один
день самка делает часто несколько ямок, одну из которых и приспосабли-
вает для гнезда. Выстилка гнезда состоит из небольшого количества сухих
тонких злаков, а к моменту насиживанья по краям его появляются также
валики из темного пуха, которым утка покрывает все гнездо, сходя с него
на отдых. Диаметр гнезда 14—18 см, высота его бортов над землей 7—9 см>
ширина лотка 12—15 см и его глубина около 10 см.
К откладке яиц чирки приступают вскоре же после прилета. Так, на
р. Мологе в 1940 г. свистунки появились 20 апреля, 28 апреля уже были
обнаружены первые гнезда, а 11 мая они содержали полные кладки. Под
Москвой начало гнездованья приходится на первую треть апреля (9—
15 апреля). В Эстонии и в Кировской обл. время гнездованья приходится на
середину мая (Кох, 1911; Плесский, 1937). Под Тобольском полные кладки
появляются в среднем 24 мая, на Елогуе гнездо с полной кладкой найдено
6 июня, а под Красноярском 15 июня (эмбрионы в пуху) (Юдин), на Вилюе
27 июня, а на Анадыре ненасиженная кладка обнаружена 5 июля. На Ко-
мандорских о-вах яйца в гнездах появляются в первой половине июня.
Наиболее ранняя кладка в Приморье зарегистрирована 13 мая, а на Альпий-
ском плато Камимухи, на о-ве Хондо (Япония) гнездо со свежими яйцами
найдено 24 мая (Ян, 1942). Обычное число яиц в полной кладке в пойме
Мологи, в Эстонии (Кох, 1911) и в Германии (Нитхаммер, 1938) 8—10, на
ЧИРОК-СВИСТУНОК 389
Рыбинском водохранилище 9—10 (от 8 до 10), в Большеземельской тундре
9—11 (Гладков, 1951). Гнездо, найденное на Анадыре, содержало 8 яиц
(Портенко, 1939), а на Елогуе и около Красноярска по 9 (Слудский, Юдин).
Большое число яиц в кладке (до 15) бывает вследствие того, что в одно гнездо
несутся две самки, гнездо у одной из которых разорено.
Яйца белые, слегка желтоватые, разница между острым и тупым концом
яйца незначительна. Размер яиц: 41—49,5x30,5—35,2, в среднем 45,6 х
Х31,6 мм. То же для Германии—45,3x32,7 мм. Вес ненасиженных яиц
26—30 г, а для Германии в среднем 27,7 г (Нитхаммер, 1938).
Первые выводки пуховиков появляются на Кольском п-ове 24—25 июня,
среднее число утят обычно 5 (от 3 до 8), на крыло они поднимаются 6—
12 августа (Владимирская, 1948). В Тиманской тундре пуховики впервые
встречены 5—14 июля и до 5 августа основная масса птенцов еще не летала,
хотя некоторые уже доросли до размеров взрослых (Гладков). Под Архан-
гельском они появляются около 27 июня A7 июня—16 июля; Паровщиков),
на Верхней Печоре в среднем 21 июня A8 июня—2 июля), а подъем их на
крыло происходит между 31 июля и 15 августа (Теплов, 1948). Под Тоболь-
ском появление пуховиков, по многолетним данным, приходится на 10 июня,
а подъем молодых на крыло 5 августа A0 июля—19 августа, Тарунин, 1929).
На р. Елогуе первые выводки отмечены с 30 июня, а к 6 августа они под-
нялись на крыло (Слудский). На Вилюе не совсем оперившиеся птенцы
обнаружены 30 июля (Воробьева, 1928), а на Индигирке, в устье р. Момы
8 августа встречены еще пуховики (Михель, 1935). На Командорских о-вах
птенцы бывают в июле, а в середине августа они уже начинают летать (Иоган-
сен, 1934). На р. Мологе первые птенцы были встречены 23 мая 1940 г., а
25 июня уже летные выводки объединялись в стаи. В Чкаловской обл.
летные молодые появляются во второй декаде июня (Зарудный, 1897).
Пуховые птенцы в день вывода весят 26—31 г (р. Молога, 16 взвеши-
ваний). Они значительно самостоятельнее пуховиков кряквы и других реч-
ных уток. С первого же дня жизни птенцы прекрасно ныряют в случае опас-
ности и много лучше других утят бегают по земле. Примерно до 5-дневного
возраста выводок собирается по вечерам к своему гнезду, если оно нахо-
дится вблизи от воды, и проводит ночь, забираясь под самку. В случае опас-
ности самка предупреждает выводок тревожным криком, после которого
молодые разбегаются по земле и затаиваются в траве, распластавшись и вытя-
нув голову и шею или же заныривают в прибрежные заросли водной расти-
тельности. По мере роста птенцов их способность к нырянью становится
хуже. Они уже не могут подолгу оставаться под водой, и обнаружить вспуг-
нутый выводок становится много легче. На крыло птенцы поднимаются
примерно в месячном возрасте, и с этого времени выводок начинает совер-
шать перелеты с водоема на водоем. Вскоре вслед за тем начинается табу-
нение выводков и концентрация их на крупных водоемах. В средней полосе
европейской части РСФСР уже в конце июля начинаются местные передвижки
чирков-свистунков, которые, вероятно, можно рассматривать, как про-
межуточный перелет. Явление это наблюдалось нами в пойме р. Мологи.
Ночью и на рассвете огромные стаи чирков-свистунков, по нескольку сот
штук пролетали вниз по течению реки и опускались на ее широкие плесы,
а утром разлетались по озерам междуречья.
Гнезда чирков-свистунков, располагающиеся укрыто, но в легко доступ-
ных местах, в большом числе погибают от разорения четвероногими и реже
пернатыми хищниками. Одним из наиболее тяжелых периодов в жизни
птенцов бывает время вынужденного переселения выводков с мелких высы-
хающих водоемов на более крупные. Для свистунка этот период проходит
тяжелее, чем для многих других речных уток, так как он охотнее всех устраи-
вается на гнездовье около самых мелких по размеру водоемов. Поэтому
390 ОТРЯД ГУСЕОБРАЗНЫЕ
нередко в летнее время на бочагах пересохших ручьев или на небольших
озерках не более 100 м2 собирается по нескольку выводков чирка.
Линька. Чирки-свистунки линяют два раза в год. Летом, после
окончания периода размножения, их оперение сменяется полностью и вто-
рой раз осенью частично. Последовательность смены оперения на различных
частях тела во время летней линьки в общих чертах та же, что и у кряквы.
Существенные отличия намечаются, повидимому, лишь в смене рулевых,
первые шесть из которых сменяются подряд. Они закладываются еще до
выпадения старых и по мере потери последних замещают их. Маховые перья
выпадают сразу, к тому времени, когда линька в летний наряд бывает уже
почти закончена. До восстановления способности к полету проходит около
28 дней (Хейнрот, 1947). Полная послебрачная линька наступает у селезней
в конце или даже в середине июня, когда они начинают терять отдельные
контурные перья.
Селезни свистунка, так же как и других речных уток, по окончании
брачного периода покидают места гнездовья и откочевывают на линьку.
Группы пролетных селезней по нескольку штук появляются под Ленин-
градом уже в середине июня. Районы массовых скоплений линных птиц
этого вида указаны нами выше. В дельте Волги свистунок является наи-
более поздно линяющим видом уток. Период его массового залегания на
линьку (потеря маховых) приходится там на первую декаду августа. Соот-
ветственно этому количество свистунков в отловах линных уток (Астра-
ханский заповедник) составляет в начале июля от 0 до 5%, а в начале
августа доходит до 65%. Примерно в те же сроки, т. е. в самых последних
числах июля и в начале августа, свистунки линяют и на озерах Кустанай-
ской обл. (Наурзумский заповедник; Формозов, 1949). На Рыбинском водо-
хранилище все селезни, добытые в июле, имели пеньки контурных перьев
по всему телу, а у самца от 23 июля 1950 г. полностью отсутствовали махо-
вые. В Лапландии не откочевавшие на линьку селезни проходят ее в июле.
Так, 2 августа в районе озера Имандра добыты два селезня уже в новом
пере (Владимирская, 1948). В нижнем течении Оби у Пельвожа F6°15' с. ш.)
6 августа добыт нелетный самец (Дерюгин, 1898), а на р. Паюте (Южный
Ямал) 31 июля наблюдалась стайка селезней, уже надевших новый наряд,
но с еще неотросшими маховыми (Кучерук, 1949). На р. Елогуе к 6 августа
1932 г. линные селезни уже поднялись на крыло (Слудский). На Анадыре
в противоположность южным районам линьки свистунки линяют несколько
раньше шилохвостей и свиязей.
Стаи линяющих чирков-свистунков держатся в дельте Волги среди
густых зарослей рогоза и ежеголовки, на мелководьях взморья, на озерах
Северного Казахстана в тростниковых чащах, чередующихся с небольшими
плесами, густо заросшими подводной растительностью. В бухте Мацаль-
Вик (Эстония) они концентрируются в густых зарослях камышей и трост-
ников, а в нижнем течении Оби на обширных, открытых, но богатых над-
водной растительностью полоях. До потери способности к полету свистунки
объединяются в огромные стаи численностью нередко в несколько сот голов.
В это время они кормятся на мелководьях с иловатым дном или же отдыхают
на открытых отмелях. После выпадения маховых чирки уходят в густые
заросли надводных растений, в которых и проводят все дневное время.
Там они отдыхают на заломах тростника или рогоза, которые в местах пре-
бывания утиных стай бывают покрыты слоем помета и перьев. По вечерам
птицы кормятся на расположенных поблизости водных плесах. Одиночные
селезни, не покидающие на время линьки своих гнездовых территорий, про-
водят ее в таежной зоне на очень маленьких, богатых растительностью лес-
ных водоемах (Лапландия, Енисей), а в степях на заросших мельничных
прудах (Украина).
ЧИРОК-СВИСТУНОК 391
Одновременно с селезнями линяет и некоторое количество самок из
числа потерявших кладки или выводки. Самки же, находящиеся при вывод-
ках, линяют несколько позднее. В июле у птиц этой группы обычно даже
почти не бывает пеньков контурных перьев. Где проводят они время смены
маховых—на местах ли гнездовья или на пути к зимовкам, до сих пор еще
недостаточно ясно. Окончание линьки затягивается надолго, так, в октябре
у пролетных самок в дельте Волги смена оперения еще не всегда бывает
закончена.
Вторая, неполная, линька начинается у взрослых самцов непосред-
ственно после окончания первой, т. е. обычно в августе. Ее последователь-
ность не отличается существенно от того, что описано для кряквы. В сере-
дине октября на пролете в дельте Волги уже встречаются селезни, полностью
надевшие нарядное зимнее перо, но основная масса еще находится в стадии
линьки, причем некоторые экземпляры только начинают ее. Полностью
линька заканчивается у селезней только к январю, уже на местах зимовки.
У самок предбрачная линька начинается лишь зимой, а заканчивается
весной, по дороге к местам гнездовья, в апреле-мае. Одновременно со взрос-
лыми селезнями начинают линять молодые птицы обоих полов, меняющие
свое юношеское оперение на первый взрослый наряд. Молодые самцы почти
не отстают в смене оперения от взрослых. Молодые самки линяют одновре-
менно с холостыми взрослыми, зимой линька у них затухает и вновь ожи-
вляется весной, когда заканчивается смена контурного оперения и дорастает
гнездовой пух.
Питание. Питание чирка-свистунка имеет смешанный характер,
летом в нем преобладают животные корма, а зимой растительные, преиму-
щественно семена. Кормятся свистунки на самых мелких местах, там, где
они могут доставать корм со дна, перевертываясь, но не ныряя. Зачастую
они при этом даже не плавают, а расхаживают на ногах, собирая корм на
вязком дне или на обнажившихся грязевых отмелях. Ранней весной, в фев-
рале и марте, по наблюдениям в дельте Волги, чирки питаются преимуще-
ственно растительными кормами: семенами F9% содержимого желудков
по объему) резухи, рдестов, ежеголовника и других, а также зелеными
частями этих растений C0%). Животная жизнь на мелководьях в это время
еще не пробудилась и неныряющим за кормом чиркам удается добывать
лишь небольшое количество моллюсков (катушек) и водяных жуков. В сред-
ней лесной полосе Союза (р. Молога) в апреле-мае животные корма встре-
чаются уже в 77 % исследованных желудков и составляют по объему около
35% от всего содержимого. Основой служат мелкие моллюски, жуки-пла-
вунцы и личинки хирономид. Остальную часть содержимого желудков
составляют зеленые части перезимовавших растений. На Рыбинском водо-
хранилище (Немцев) весеннее питание свистунков на 84% в апреле и на 74%
в мае-июне состоит также из животных кормов, в основном из личинок
комаров-толкунцов (хирономид), личинок ручейников (Phryganea grandis) и
в меньшей степени клопов-гребляков (Corixa). В мае-июне свистунки почти
целиком переходят там на питание личинками хирономид F5% по объему).
Содержимое желудков нескольких чирков-свистунков, добытых в мае
на юге Приморья, состояло из множества мелких мушек, густо покрывав-
ших в эти дни водяную растительность и поверхность воды, а также из жел-
тых семян, напоминающих просо (Шульпин, 1936).
В гнездовое время животные корма сохраняют столь же большое зна-
чение, встречаясь в 97,5% исследованных желудков и составляя около
70% содержимого по объему (р. Молога). Состав их в это время разнообра-
зен: водяные жуки, личинки стрекоз и хирономид, моллюски, ручейники,
листоногие рачки и водные олигохеты. Примерно то же можно сказать о пита-
нии пуховиков и оперяющихся птенцов, у которых объемный процент содер-
392 ОТРЯД ГУСЕОБРАЗНЫЕ
жания животных кормов составляет 85 и 75%. Наибольшее значение в это
время имеют мелкие моллюски C0%), которых птенцы склевывают с водя-
ных растений, личинки стрекоз, хирономид и кровососущих комаров (8%),
а также водяные жуки (плавунцы). В июле-августе количество животных
кормов у взрослых птиц снижается до 60% за счет появления луковиц стре-
лолиста E%), клубеньков гребенчатого рдеста A0%) и созревающих к этому
времени семян B5%). На Рыбинском водохранилище летнее питание моло-
дых и взрослых (июль-август) состоит на 76% из животных кормов, главным
образом из личинок хирономид C7%) и в меньшей степени из моллюсков
водяных жуков и клопов. 12,3% составляют вегетативные части растений:
клубеньки рдеста и зеленые листья. В питании птенцов свистунка на Кам-
чатке (Кроноки) зарегистрированы личинки хирономид, семена и зеленые
части растений (Аверин, 1948). Во время линьки (дельта Волги) питание
чирков однообразно, так как птицы долгое время держатся на весьма огра-
ниченном участке. Животные корма (личинки хирономид) составляют всего
около 2% содержимого, основой питания в это время служат зеленые части
растений B8%) резухи, валлиснерии и других и семена G0%), главным
образом незрелые семена резухи. Осенью, в период пролета, свистунки все
более и более переходят на растительную пищу. На Рыбинском водохрани-
лище и в это время объемный процент животных кормов не падает ниже
67,3% исключительно за счет поедания личинок хирономид F0%), которых
чирки собирают на обнажающихся отмелях. На р. Мологе значение живот-
ных кормов сокращалось в сентябре-октябре до 15% (жуки, личинки мух,
моллюски), 15% составляли луковицы стрелолиста и 70% семена. В дельте
Волги животные корма у пролетных чирков (личинки хирономид и моллюски)
составляют в августе-сентябре—4%, в октябре 5,2%, в ноябре 11,7%. На
долю вегетативных частей растений падает от 14 до 35% за счет верхушек
роголистника, зимних почек сусака и ряски. Основой же пищи являются
семена E3,3—85,5%) резухи, рдестов и камыша. Примерно тот же характер
имеет осеннее питание свистунков в устье Камы (Тихвинский, 1931). Отме-
чено поедание чирком в это время спорокарпий сальвинии в дельте Волги
и Днепра (Пачосский, 1911). На местах зимовки чирки почти целиком пере-
ходят на растительные корма. В октябре-ноябре в районе Красноводска
около 45% пищи еще составляют морские моллюски (Theodoxus pallasi),
но позже (в декабре-феврале) на низменностях Азербайджана животные
корма составляют лишь доли процента от общего содержимого; это личинки
хирономид, мелкие моллюски и ракушковые рачки. Основой же зимней
пищи служат семена: на взморье—рупии и резухи, на рисовых полях—
куриного проса, некоторых злаков и камышей, а на временных степных
водоемах—лебеды и солянок. Сезонные переходы с животной пищи на расти-
тельную влекут за собой соответствующие изменения веса птиц.
Хозяйственное значение. Промысловое значение чирков-свистунков,
чрезвычайно велико. Их сравнительно небольшие размеры компенсиру-
ются прекрасными вкусовыми качествами мяса и очень хорошей упи-
танностью в некоторые сезоны. Средний вес их тушки в промысловое
время составляет 70% от живого веса, т. е. около 250 г, причем до 53 г при-
ходится на жировые отложения. Свистунок наиболее широко и повсеместно
распространенная утка. В средней полосе и особенно в населенных местах
он является наиболее многочисленным видом, так как довольствуется даже
самыми мелкими водоемами. В низовьях Днепра свистунок составляет 33%
(т. е. треть) всей добычи спортивной утиной охоты (Пачосский, 1911), а в цен-
тральных областях и еще большую часть. На закавказских зимовках добыча
чирков-свистунков составляет около 11,3% всего количества заготовляемых
уток. Десятками тысяч ловят их промысловыми ловушками на путях пролета
и в местах зимовок в Дании, Голландии и Англии.
ЧИРОК-СВИСТУНОК
393
Полевые признаки. Мелкая утка. Сидящая на воде самка равномерно
серая без ярких отметин, для селезня характерна темная голова и светлая
полоса вдоль сложенного крыла. На полете снизу заметны белое брюхо
и подбой крыльев и темная голова, сверху птица целиком темная, включая
и крылья. Полет очень быстрый и бесшумный. Голос у обоих полов—кря-
канье, более тихое и высокое, чем у кряквы. Весенний брачный крик самца—
звонкий, металлический, трудно передаваемый буквами, дребезжащий свист
«храюк» .
Рис. 70. Чирок-свистунок (самец и самка)
Чирок-свистунок имеет наиболее узкие и острые крылья среди всех реч-
ных уток и соответственно этому наиболее быстрый полет (Штегман, 1950).
Он может взлетать и с воды и с суши почти вертикально вверх, что дает
ему возможность посещать даже самые маленькие бочажки среди тростни-
ковых зарослей и кормиться в затопленных лесах во время половодья и на
водохранилищах.
Описание. Размеры и строение. Самая мелкая из наших уток. Клюв
среднего размера, несколько короче головы, одинаковой ширины от основания до верши-
ны A0—12 мм), с узким коготком. Ноздри овально вытянутые, лежат у основания надклю-
вья. Цевка короткая, короче среднего пальца. Крылья значительно не достигают конца
хвоста. Рулевых 14—16, у самцов иногда 18, с округлой вершиной. Размеры: общая дли-
на самцов 333—384, самок 305—367 мм; размах самцов 593—640 мм; крыло селезней 173—
193, самок 165—180 мм; цевка самцов 29—33, самки 25—32; клюв селезней 34—39,
самок 30—36 мм.
Вес взрослого, т. е. перезимовавшего, чирка-свистунка колеблется в пределах от
250 до 450 г у селезня и от 200 до 380 г у самки. Столь значительное различие в весе у птиц
одного вида зависит как от разницы в размерах (изменяются с возрастом и старше одного
года), так и, главным образом, от степени упитанности (ожирения), которая меняется по
394
ОТРЯД ГУСЕОБРАЗНЫЕ
Рис. 71. Клювы чирка-
свистунка (вверху моло-
дая самка, внизу старая
самка). Видно различие в
количестве черных пятны-
шек
сезонам. Количество жира у свистунка может доходить до 18% от общего веса птицы. Вес-
ной, по прилете, в марте-апреле, селезни, добытые в низовьях р. Или, весят 275—375 г,
в дельте Волги 325 г C00—350 г), на Рыбинском водохранилище 330 C00—355 г), на верх-
ней Печоре в апреле-мае 360 г C20—400 г). Самки там же 320 г C00—380 г), на Южном
Ямале в мае 410 г C50—450 г; Осмоловская), а на Камчатке в мае 302 г B67—376 г),
в июне 341 г C20—370 г; Аверин, 1948). Вес значительно
снижается за период размножения и вновь восстанавливает-
ся ко времени линьки. Селезни, добытые в августе на Рыбин-
ском водохранилище, в начале линьки весят 334 г B95—
365 г), линные в дельте Волги тогда же 334 г B50—450 г).
Самки в августе в первом месте 323 г C00—355 г), во втором
325 г C00—360 г). Особенно хорошо упитаны бывают взрос-
лые селезни свистунков и молодые птицы осенью. Так, в
Астрахани в сентябре—ноябре селезни весят 352 г C00—
450 г), а на Рыбинском водохранилище молодые селезни
в сентябре 378 г C35—420 г), а молодые утки 325 г B75—
390 г). За время осеннего перелета и пребывания на зимов-
ках свистунки теряют в весе, но не так сильно, как боль-
шинство других настоящих уток. Вес селезней в ноябре-
декабре в Гасан-Кули (Туркмения) 313 г B50—390 г) и са-
мок 272 г B00—300 г). На азербайджанских зимовках у юго-
западных берегов Каспия средний вес селезней в январе
276 г B00—350 г), в феврале 290 г B00—350 г) и в начале
марта 258 г B00—300 г), а уток соответственно в январе
23/ г B00—300 г), в феврале 250 г. Весной, как видно из
приведенных выше цифр, вес быстро восстанавливается,
причем к моменту появления на местах гнездовья птицы из
северной части ареала весят несколько более южных.
Окраска. Пуховой птенец. На спинной стороне
оливково-бурый с несколько более темным верхом головы,
снизу сернисто-желтый. Темное пятнышко на ухе, темная
полоска через глаз и светлая над глазом.
Молодые птицы. В первом пере сходны по окрас-
ке с взрослой самкой, но на верхних кроющих крыла светлые коричневатые
каемки. Хорошим отличием самцов от самок уже в этом возрасте (осенью) служат мел*
кие темные пятнышки, расположенные у последних без всякой системы по краям верх-
ней челюсти и дугам нижней.
Взрослая самка. На спинной стороне
темнобурая со светлокоричневыми краями перь-
ев. Голова в узких продольных пестринах,
на щеках светлее. Горло чистобелое. Брюшная
сторона с бурыми пятнами, более частыми на зобе
и боках. Крыло, как и у самца с зеленым зеркаль-
цем, окаймленным снаружи белой полоской, но
без черной каемки с внутренней стороны, третье-
степенные маховые и плечевые перья одного цвета
с верхом. Клюв черный с оранжево-желтыми края-
ми и с мелкими черными пятнышками на них.
Ноги оливковосерые; радужина бурая.
Самец в брачном наряде. Голова и верх-
няя часть шеи спереди рыжевато-коричневые,
подбородок и верхняя часть горла черные. По
бокам головы за глазами широкие синевато-
зеленые полосы с меднокрасным отливом, соеди-
няющиеся между собой на затылке. Зеленое пятно
окаймлено узкой белой полоской, которая продол-
жае ся от глаза к разрезу клюва. Лопатки, мантия,
бока, передняя часть спины и подхвостье светло-
серые в узких черных струйчатых полосках. Осталь-
ная спина и надхвостье буровато-серые. Самые
длинные кроющие хвоста черные; с обеих сторон над-
хвостья вниз проходит бархатисто-черная полоска,
соединяющаяся с черными же нижними кроющими хвоста. Ниже полоски соловое пятно.
Брюшная сторона бледноохристая, на груди и зобу каплевидные черные пятна. Подхвостье
черное с бледноохристыми пятнами по бокам. Плечевые светлосерые с черными струй-
ками, крайние наружные из них белые с черными каймами. При согнутом крыле они
образуют светлую продольную полосу, отделяющую крыло от туловища. Наружная
часть зеркала крыла бархатисто-черная, внутренняя яркая металлически-зеленая, гра-
ница двух цветов проходит по диагонали зеркала. Снаружи зеркало окаймлено белой
Рис. 72. Рулевые перья чирка-сви-
стунка (справа молодая птица,
слева взрослая птица)
ЧИРОК-КЛОКТУН
395
полоской, обычно с ржавым налетом. Верхние кроющие крыла лапы оливково-серые,
буровато-серые, подбой крыла белый. Рулевые буровато-серой окраски. Клюв черный,
радужина бурая.
23. Чирок-клоктун Anas formosa Georgi.
Anas formosa, Georgi. Bemerkungen Reise Russl., 1775, стр. 168, Байкал.
Синонимы. Anas glocitans. Pallas. Acta Stokkholm., XL, 1779, стр. 26, Восточная
Сибирь.—Anas cucullaia. Fi scher. Bull. Soc. Nat. Moscou, III, 1831, стр. 279, Камчатка.
Русские названия. Клоктун—книжное; моклок, гаганок обязаны голосу, мара-
душка, чирок—народные.
Распространение. Ареал. Восточносибирский вид. Западная граница
его распространения намечается следующими крайними пунктами: среднее
течение Ангары, нижнее течение Нижней Тунгуски, тундра между Енисеем
Карта 44. Распространение клоктуна Anas formosa
а—граница гнездовий, а'—районы наибольшей численности птиц на гнездовье,
б—не вполне выясненные границы гнездовий, в—места постоянных встреч ле-
том (гнездованье не доказано), г—зимовки, а—валеты, е—встречи пролетных
птиц весной в стороне от гнездового ареала
и Пясиной. Известно несколько летних встреч самцов под Туруханском
и летние встречи на р. Елогуе. В последнем месте самец добыт 21 июня 1931 г.,
кроме того, у детей эвенков и хантэ часто встречаются куклы, сделанные
из шкурок красивых селезней клоктуна (Слудски ). Весной замечается про-
лет клоктунов вверх по р. Турухану, значительное количество шкурок
селезней этого вида было заготовлено на фактории Ратта, в верхнем течении
Таза (Юдин). Эти факты говорят о возможном гнездовании клоктуна в области
рек Елогуя, Турухана и Таза, но полной уверенности в этом нет, так как
селезни многих видов уток нередко встречаются в летнее время далеко вне
пределов своего гнездового ареала. Северная граница распространения
396 ОТРЯД ГУСЕОБРАЗНЫЕ
клоктуна достигает в бассейне Енисея 70° с. ш. и проходит далее на восток,
повидимому, по границе редколесья и чистой тундры. Он обычен на Бога-
ниде, а на Юго-Восточном Таймыре заходит в зону южной тундры и нередок
в устье Хатанги по р. Большая Балахна и у озера Портнягино, но в послед-
них трех пунктах известен лишь в послегнездовое время (Тугаринов, 1941).
По Лене доходит до самых низовий, но в дельту лишь залетает. Зато на Яне,
Индигирке и Колыме он достигает устья. Встречен летом на о-ве Большом
Ляховском, но вряд ли гнездится там. В небольшом числе гнездится в запад-
ной части бассейна Анадыря, на север до южных склонов Анадырского
хребта, а также по р. Великой (Портенко, 1939). Известны неоднократные
летние встречи на Чукотке, о-ве Лаврентия, п-ове Сюард (Северная Аме-
рика), Командорских о-вах и Сахалине. Все эти пункты следует расценивать
как область частых летних залетов, в некоторых из них возможны
спорадичные случаи гнездованья. Отмечен летом для Камчатки и, по неко-
торым данным, гнездится там (Бианки, 1909; Бергман, 1935). Однако Аве-
риным A948) за ряд лет наблюдений в районе Кронок он не встречен там
ни разу. Встречается по Пенжиной и Гижиге, но не часто, под Охотском
одна из наиболее обыкновенных гнездящихся уток (Харитонов, 1915). Пови-
димому, гнездится в районе Аяна (Снигиревский), найден в устье Уды.
Достоверных сведений о гнездовании в бассейне Амура нет. Есть указание
на то, что яйца клоктуна были привезены Дыбовским с р. Дарасуна (Тача-
новский, 1873), но последующими наблюдениями в Южном Забайкалье
гнездованье там клоктуна не подтверждается. Встречи этих чирков в июле-
августе между Ононом и Борзей (Долгушин, 1941) еще не говорят об их гне-
здовье там. Несомненно гнездится клоктун у северного берега Байкала
в низовьях р. Верхней Ангары (Слудский). Неоднократно отмечались залеты
его под Томск и в Барабинскую степь.
Основные зимовки находятся на Японских о-вах, на о-ве
Хондо и далее к югу. Однако по сравнению с другими утками клоктун встре-
чается там в незначительном числе. В огромных утиных стаях численностью
от 500 до 1500 птиц клоктунов бывает всего от 5 до 50 (Ян, 1942). В Корее
бывает только на пролете (Остин, 1948). В большом количестве зимует в при-
морской зоне Среднего Китая, но в Южном очень редок, хотя доходит до
Тайвана и Хайнаня. В Индии представляет исключительную редкость,
появляется только в холодные зимы, когда встречи его зарегистрированы
в Бенгалии, Раджпутане, Саруне и Гуджерате (Финн, 1901).
Сезонные миграции и их даты. Весной в Приморье
клоктун одна из наиболее многочисленных пролетных уток. Появляется
он уже в последних числах марта. На озере Ханка интенсивный пролет
наблюдается в первой половине апреля. Клоктуны летят огромными стаями
до 100—500 шт., а некоторые стаи их на отдыхе достигают нескольких тысяч
особей. В дни массового пролета стаи следуют одна за другой круглые сутки.
Основное направление пролета прямо на север. Однако в некоторые годы
A895, 1909) клоктун, по неизвестным причинам, почти полностью отсут-
ствовал на весеннем пролете в Приморье. На пролете стаи клоктунов обычно
выстраиваются в длинные волнистые линии, вытянутые по фронту. Интен-
сивный пролет нередко продолжается около месяца, а отдельные стаи задер-
живаются до 15 и даже 20 мая (Шульпин, 1936; Воробьев). ПодМариин-
ском (на Амуре) первые были замечены 23 апреля (Шренк, 1861). У Охотска
клоктун одна из основных пролетных уток (Харитонов, 1915), а в верховьях
р. Колымы, вдоль которой они следуют на север, самая многочисленная
после свиязи. Основной пролет проходит там в 20-х числах мая (Дементьев
и Шохин, 1939), а под Нижне-Колымском прилет отмечен 31 мая (Тайер
и Бэнгс, 1914). Весенние добычи на Пенжине 25 мая и 3 июня 1941 г. (Волков
и Дементьев, 1948); появляется в небольшом количестве на Анадыре в конце
ЧИРОК-КЛОКТУН 397
первой и в начале второй декады июня (Портенко, 1939). На Индигирке
у впадения р. Момы первые 15 мая, в дельте реки у сел. Русское устье лишь
12—20 июня (Зензинов, 1915; Михель, 1935). В Даурии (озеро Тарей-Нор)
первые отмечены 8 апреля, а на Буреинском хребте 10 апреля (Радде, 1863).
Весной под Якутском пролетает по Лене в большом количестве, появляется
1—3 мая (Воробьева, 1931). В очень большом числе бывает во время пролета
на озере Далай-Нор с начала и до второй половины апреля. Западнее же
нигде не встречен (Пржевальский, 1876). Весеннее появление под Иркут-
ском 22 апреля (среднее за 6 лет), последние же экземпляры держатся до
1—15 июня. Начало валового пролета пор. Илиму, правому притоку Ангары,
18 мая (Косыгин, 1926).
Сведения об осеннем пролете клоктуна весьма отрывочны. На Индигирке
у Русского устья, последний раз клоктуны наблюдались 21 октября на
маленьком озерке, у берегов которого уже был лед, а на Анадыре они попа-
дались до конца августа. В тех местах, где весной он встречается в массе
(Приморье, Амур, Лена), осенью его обычно совсем не замечают. Возможно
потому, что на осеннем пролете клоктуны следуют не по речным долинам,
как весной, а сперва на север по течению рек, а затем круговым путем вдоль
морских побережий на восток и на юг (Тугаринов, 1930, 1941). Однако факти-
ческих материалов, подтверждающих это предложение, нет, за исключением
осенних встреч молодых птиц и линяющих селезней севернее пределов обыч-
ного гнездового ареала. Но такой промежуточный перелет свойствен мно-
гим видам уток. С другой стороны, клоктуны все же встречаются осенью,
хотя и редко, на Байкале (самец, добытый 3 сентября в районе Баргузин-
ского залива; Туров, 1923), а по Ангаре под Иркутском- наблюдается ясно
выраженный осенний пролет этого вида. В последних числах августа и в пер-
вых числах сентября B2 августа—5 сентября в разные годы) клоктуны про-
летают вниз по Ангаре, причем в некоторые дни стайками по нескольку
десятков птиц в каждой. Заметный пролет под Иркутском продол-
жается очень короткий срок, всего 1—1,5 дня, после чего встречаются
лишь одиночные случайные птицы. Пролетные стаи не делают длитель-
ных остановок в пути (Вотинцев, 1947). Известен ряд случаев добычи
клоктунов в августе и сентябре: в верховьях Амура и в низовьях его, под
Николаевском (Шренк, 1861), а также в различных частях Приморья (залив
Ольги, р. Ботчи, озеро Ханка). Несколько пролетных стай этого вида встре-
чено 14 сентября 1940 г. на р. Лефу в Приморье (Вотинцев, 1947). На зимов-
ках в Японии появляется несколько позднее других речных уток. Возможно,
что редкость чирка-клоктуна на осеннем пролете и скудость сведений по
этому вопросу объясняются тем, что пролет его происходит дружно и без
длительных остановок в пути, а большинство охотников не умеет отличать
клоктуна в его осеннем наряде, от других чирков, а своего характерного
голоса на осеннем пролете селезни не подают. Летних перелетов самцов и
массовых скоплений на линьку у этого вида неизвестно.
Биотоп. Самые разнообразные континентальные водоемы. Зимой
и на весеннем пролете рисовые поля, разливы рек и пойменные озера. Летом
мелкие лесные водоемы и острова в дельтах рек.
Численность. В пределах своего ареала клоктун обычен и даже
многочислен в верхнем течении Хатанги, в нижнем и среднем течении
Лены и ее притоков (кроме Вилюя, по которому редок), в бассейнах Яны,
Индигирки, Колымы и на морском побережье в районе Охотска.
Экология. Размножение. Пары у клоктунов оформляются уже
во время пролета. Часть клоктунов прилетает и держится парами уже в
апреле и начале мая на юге Приморья. В верховьях Колымы (около 25 мая)
селезни из пролетных стай в утренние и вечерние часы гонялись за самками
и спаривались с ними. На Индигирку они прилетают, уже разбившись на
398 ОТРЯД ГУСЕОБРАЗНЫЕ
пары. Во время ухаживания селезень поднимает и почти запрокидывает
назад голову, а затем снова опускает ее. При этом оперение на боках головы
прилегает плотно, четко обрисовывая яркую раскраску щек, а по срединной
линии оно распушено и образует как бы продольный гребень. Токование
сопровождается характерным клокотанием селезня. Игры происходят чаще
на суше и реже на воде. Условия гнездованья почти не изучены. На Вилюе
гнездо было обнаружено на открытом месте, под кустом можжевельника.
На Индигирке клоктун гнездится около совсем мелких лесных водоемов
(площадью до 4 м2). Гнездо, найденное у Нижне-Колымска, помещалось
в ивняках, а в дельте Колымы на открытой моховой тундре в 100 м от водоема.
Гнезда устраиваются на земле в вырытой птицей ямке и выстилаются сухой
осокой и другой травой или же ягелем и сухими листьями. С момента наси-
живания в нем появляется темный пух.
К размножению клоктуны приступают сразу же по прилете. Гнездо
с 9 яйцами (полная кладка) найдено на Вилюе 2 июня (Маак, 1886), под
Верхоянском первые 2 яйца обнаружены 28 мая (Тугаринов, 1932). На
Индигирке (под 69°50' с. ш.) 18 июня гнездо содержало 2 насиженных яйца
(Михель, 1935), под Нижне-Колымском 7 июня добыта самка с готовым
к откладке яйцом, 19 июня обнаружена утка, насиживавшая 6 яиц, и еще
одно гнездо обнаружено 22 июня (Тайер и Бэнгс, 1914). В дельте Колымы
гнездо, найденное 9 июля, содержало 7 яиц, насиживаемых уже 5—6 дней
(Рили, 1918). Таким образом, известные нам насиженные кладки содержали
от 6 до 9 яиц. Цвет яиц бледный, зеленовато-серый, схожий с таковыми
у кряквы. Их размеры: 46,2—47,7x33,7—35,7, среднее 47,1x34,4. Про-
должительность насиживания неизвестна.
В низовьях р. Верхней Ангары выводок из 8 молодых размером с поло-
вину матки встречен 31 июля 1933 г. Он держался в кочках осоки у берега,
а самка летала с криком над головой наблюдателя (Слудский). На озерке
в верхнем течении Яны (р. Берелях, 66° с. ш.) 16 августа встречена самка
с 10 птенцами размером с перепела. Ко 2 августа молодые еще не поднялись
на крыло (Ткаченко, 1932). На Индигирке первый выводок из 4 молодых
замечен 11 июля (у с. Майор Крест), близ с. Абыя выводок из 6 еще нелет-
ных молодых встречен 21 июля, а 17 июля выводок из 5 птенцов, уже достиг-
ших размеров взрослой птицы, но с еще недоросшими маховыми (Михель,
1935). На Анадыре к 10 августа молодые еще лишь наполовину оперились.
Маховые у них были в пеньках, низ туловища и плечи в перьях, а голова
еще покрыта пухом (Портенко, 1939).
Линька. Последовательность смены оперения во время летней,
послебрачной, линьки, по Тугаринову A941), такова. Сначала выпадают
перья на брюшной стороне тела, затем на лопатках, спине, а потом на
шее и голове. Когда почти все контурное перо уже сменено, кроме отдель-
ных участков на голове и спине, линяют крылья и начинается смена руле-
вых. Часть самцов к началу линьки появляется за северными пределами
гнездового ареала (Хатангский залив, озеро Портнягино на Юго-Восточ-
ном Таймыре, о-в Большой Ляховский), но основная масса, повидимому,
линяет на местах гнездовья. У северного побережья Байкала линяющий
самец с выпавшими маховыми добыт 14 августа на небольшом озерке среди
обширных сырых лугов, богатых водоемами, поросшими у берегов тростни-
ком. Среди многочисленных чирков-свистунков здесь встречен единствен-
ный селезень клоктуна (Слудский). На Лене близ Якутска линяющий селе-
зень без маховых встречен 11 июля (Иванов, 1929). На Яне под 66° с. ш.
2 августа селезни находились в периоде линьки. Только что окончившие
линьку селезни добывались на Таймыре (озеро Портнягино) 15 авгусга
(Тугаринов, 1941). На Индигирке около устья р. Момы 14 июня встречена
группа самцов, еще летавших, но уже собравшихся на линьку в глухой
ЧИРОК-КЛОКТУН 399-
зарастающей старице. 9 августа у с. Ожогина наблюдались уже перели-
нявшие и получившие способность к полету самцы. В устье Танюрера (Ана-
дырь) линный селезнь добыт 7 августа. Линька самок, находящихся при
выводках, обычно наступает несколько позже, чем у самцов, но некоторые
линяют одновременно с ними. Так, из трех самок с выводками, встречен-
ных на Индигирке в июле, в состоянии интенсивной линьки находилась
одна. Маховые ее 17 июля находились в пеньках.
Новая линька наступает у самцов поздно осенью и заканчивается
к январю. Таким образом, на осеннем пролете они бывают еще в летнем
наряде. У недавно перелинявшей птицы черное темя имеет буроватый тон,
Рис. 73. Чирок-клоктун (самец и самка)
соловые щеки пестрят бурыми пятнышками, а каплевидные пятна на зобу
и груди закрыты винно-желтым фоном за счет соответствующих окончаний
перьев свежего оперения. К весне каемки этих перьев обтрепываются и все
части оперения принимают свою основную окраску. Смена оперения у самок
затягивается до весны и остатки ее почти по всему телу можно отметить
в конце апреля.
Питание клоктуна почти совсем не изучено. В желудках птиц,
добытых весной в Приморье, находили много семян, похожих на мак, чумизу
и просо (Шульпин, 1936). Часто они совершают кормовые перелеты на рисо-
вые поля и пашни. Поднимали их по ночам с проезжих дорог, на которых
они кормились просыпанными при перевозке бобами сои. Повидимому,
этот корм играет заметную роль в весеннем питании клоктунов (Воробьев).
На весеннем пролете под Хабаровском клоктуны охотно кормятся желу-
дями, разыскивая их в опавшей листве, нередко удаляясь при этом за 700—
800 м от воды. В их пищеводах удавалось находить до 32 желудей (Слепцов).
Пищевод у селезня, добытого 7 августа на Анадыре, был заполнен семенами
осоки (Портенко, 1939). В желудке самки, добытой 31 июля на озере Ин-
горга (низовья Верхней Ангары), обнаружен планктон.
Хозяйственное значение. Ружейный промысел клоктуна на весеннем
пролете широко развит во всем Приморье, на Лене и местами на Колыме,
400 ОТРЯД ГУСЕОБРАЗНЫЕ
где эта птица является одной из наиболее многочисленных уток. Однако
в силу малой населенности этих мест не приобретает больших размеров.
Манера клоктунов держаться плотной стаей позволяет сбивать с одного
выстрела по нескольку штук. Птицы эти весной в Приморье бывают чрез-
вычайно жирны в противоположность другим видам пролетных уток в это
время.
Полевые признаки. Внешне очень напоминает чирка-свистунка, но
выглядит гораздо плотнее и грубее его из-за более короткой и толстой шеи.
На боках головы яркие зеленые и светлоохристые пятна, окаймленные
белыми полосками, и черная поперечная полоса. На воде сидит довольно
глубоко, летает обычно невысоко и со страшной быстротой. Голос селезней—
мягкие глухие, равномерно и довольно часто повторяющиеся звуки «кло»,
«кло», «кло», «кло», «кло». Он слышен далеко. Весной самцы кричат все
время—и на полете и сидя. Голос самки похож на кряканье свистунка,
только несколько более частое.
Описание. Строение и размеры. Весьма близок к чирку-свистунку, но
отличается относительно более короткой шеей. Клюв по сравнению с другими видами чир-
ков очень крупный. Рулей 7 пар. Размеры: крыло самцов 200—220, самок 180—210 мм;
цевка самцов 33—35, самок 30—35 мм; клюв самцов 35—38, самок 33—35 мм. Вес 500—
600 г.
Окраска. Пуховой птенец. Сходен с таковым чирка-свистунка, но отли-
чается очень светлыми желтоватыми пятнами по бокам головы, у основания клюва и свет-
лой полосой через глаз.
Молодые птицы. В первом наряде имеют оперение взрослой самки, но окрас-
ка верха тусклая и одноцветная, каемки перьев серо-бурые. Голова также серо-бурая
без продольных пестрин, зоб и бока тела буроватые без ржавых тонов, спина и над-
хвостье серо-дымчатые с темными наствольями.
Годовалые птицы. Судя по коллекционным материалам, весной наряду с селезнями
в полном брачном наряде есть много самцов, у которых спина имеет бурую окраску
с очень малым числом поперечных полосок, создающих струйчатость пера, или совсем
без них. По аналогии с другими утками, их можно считать годовалыми птицами. Значи-
тельный процент самок клоктуна имеет в оперении спинной части тела (мантия, спина,
надхвостье) отдельные перья юношеского наряда. Так как это наблюдается не у всех
птиц, то можно предполагать, что наличие юношеских перьев служит признаком самок-
первогодок.
Взрослая самка. На спинной стороне темнобурая с рыжеватыми каемками перь-
ев. Щеки и бока шеи светлее основного тона, с бурыми крапинками на середине
перьев. По бокам у основания клюва округлые светлые пятнышки размером с горошинку,
за глазом отчетливая темная полоска, а над ней узкая светлая. Зоб, верхняя часть груди и
бока тела тускло-охристые с мелкими округлыми бурыми пятнами, которые переходят
на боках в более крупные продольные. Остальная брюшная сторона буланая, с неясными
пятнами. Крылья, как у самца, но зеркало тусклее, плечевые короче и одного цвета со
спиной.
Взрослый самец в летнем наряде. Сходен с самкой, но верх головы очень
темный и без штрихов, лопатки оливково-бурые без светлых каемок, зоб и бока корич-
нево-рыжие, брюшко и подхвостье в более частых, четких и округлых, чем у самки,
пятнах.
Самец в брачном наряде. Верх головы, подбородок и горло черные. От глаза к гор-
лу черная полоса. От глаз к задней части шеи и вниз по сторонам ее располагаются
в виде широких полулуний яркие металлически-зеленые пятна, соединяющиеся ниже за-
тылка между собой. Они окаймлены сверху узкой, а по бокам более широкими белыми по-
лосками. Пространство между клювом и черной вертикальной полосой, щеки и примыка-
ющая часть шеи тускло-охристые. Основание шеи сзади и лопатки аспидные с мелким
струйчатым рисунком, спина и надхвостье у старых селезней темные, пепельно-серые,
струйчатые. Бока голубовато-аспидные с мельчайшими черными струйками, у основания
крыльев белая поперечная полоса. Зоб и верхняя часть груди розовато-винного цвета с
полукруглыми черными пятнышками. Брюхо белое, подхвостье бархатисто-черное а по
бокам у основания его белые поперечные полоски. Хвост серо-бурый. Верхние кроющие
крыла буровато-оливковые, а самые длинные из них, обрамляющие зеркало, каштаново-
охристые. Зеркало металлически-зеленое на черном фоне, окаймленное снаружи белой
полоской. Первостепенные маховые серо-бурые, третьестепенные бурые, частично с чер-
ными наружными опахалами. Клюв темный, голубовато-серый, ноги буровато-оливковые,
радужина бурая.
КАСАТКА
401
24. Касатка Anas falcata Georgi.
Anas falcata. Georgi. Bemerk. Reise Russl., 1, 1775, стр. 167. сАзиатская Россия».
Синоним. Anas falcaria. Pallas. Reise d. versch. Prov. d. Russ. Reichs, III, 1776,
стр. 701, Даурия.
Русские названия: Касатка, косатый селезень—по серповидным плечевым
перьям, народное.
Распространение. Гнездовой ареал. Восточносибирский вид.
Западным пределом распространения служит, повидимому, Енисей, в вер-
ховьях которого касатка обнаружена Янушевичем A948, 1950) в Тувинской
Карта 45. Распространение касатки Anas falcata
о—граница гнездовий, б—не вполне выясненная граница гнездовий, •—места по-
стоянных летних встреч (гнездованье не доказано), г—зимовки, е—встречи пролет-
ных птиц весной в стороне от гнездового ареала
котловине и на Восточно-Тувинском нагорье, но в этих местах она очень
редка, так как прежние исследователи края (Сушкин, Нестеров) ее там не
встретили. Всюду к северу по долине Енисея (Минусинская котловина,
Красноярск, низовья Енисея) она известна лишь как залетная птица. Касатка
редка в бассейне Ангары, встречается на Подкаменной Тунгуске, примерно
до устья р. Чуни (Тугаринов, 1923) и в верхнем течении Нижней Тунгуски
(Ткаченко, 1937), где она даже обыкновенна. В бассейне Лены нормально,
хотя и не часто, гнездится в районе Якутска, а по Вилюю попадается лишь
спорадически (Тугаринов, 1934). По Яне встречена на север до устья р. Адычи
26 Птицы СССР, т. IV
402 ОТРЯД ГУСЕОБРАЗНЫЕ
(но неизвестно, гнездится ли там). В бассейне Индигирки и Колымы неизве-
стна и вероятно отсутствует там. Встречена на Гижиге, где вряд ли гнездится
(Дементьев, 1940), но в районе Аяна, повидимому, редко гнездящаяся птицам
(Снигиревский). На Камчатке гнездится, но изредка и не повсеместно,
в восточной части полуострова (Кроноки) бывает только на весенних про-
летах, когда весьма редка, малочисленна и встречается только на море
(Аверин, 1948). На Командоры лишь залетает, но в южной части Куриль-
ской гряды обычна. Населяет весь бассейн Амура, Приморье, Сахалин и о-в
Хоккайдо в Японии. В Корее бывает на пролете и несомненно гнездится
в северной части. Обычна в северной, лесистой части Маньчжурии, но в пре-
делах Китая на гнездовье неизвестна. В Чжили летом добывались отдель-
ные бродячие особи, то же вероятно относится и к верхнему течению Хуанхэ
Гнездится в восточной Монголии (Тугаринов) и по всему Забайкалью, вклю-
чая его юго-западную Селенгинскую часть, и на Байкале. В Северной Мон-
голии бывает лишь на пролете.
Основные зимовки находятся в Японии (о-в Хоккайдо и южнее)
и в Южном Китае (Хубэй, Цзянсу, Чжецзянь, Гуандунь, Фудзянь). Регу-
лярно бывает в Северном и Центральном Вьетнаме, но в малом числе (Дела-
кур и Жабуйль, 1931). В Индии и Верхней Бирме появляется лишь слу-
чайно, но в холодные зимы бывает даже обычна в Бенгалии и служит объек-
том промысла. Интересно, что этих южных пределов зимовки достигают
преимущественно самки (Финн, 1901, 1915).
Характер пребывания. Перелетная птица.
Сезонные миграции и их даты. На зимовках в про-
винции Хубэй (Китай) касатки бывают в январе-феврале, в Цзянсу они
задерживаются до апреля, а у Чжилийского залива пролет продолжается
с марта по май (Колдуэлл, 1931). С японских зимовок отлетают в середине
апреля (Ян, 1942). В Южном Приморье появляются во второй половине
марта, а валовой пролет падает на середину апреля. На оз. Ханка в это время
касатки занимают по численности следующее место после клоктуна (наряду
со свистунком и хохлатой чернетью). Во время пролета держатся стайками
по 5—10 шт., часто присоединяясь к стаям других уток (Воробьев). Берегов
Восточной Камчатки они достигают в мае, встречи их в районе Кронок еди-
ничны и приходятся на 5—23 мая (Аверин, 1948). На Командорские о-ва
залетают только самцы, они отмечались там с 17 апреля по 4 июня (Иоган-
сен, 1934). Часть птиц летит через Монголию, по рекам Тола и Хара (Юго-
Западный Кентэй), и пересекает Северную Гоби (озеро Орок-Нор, 17 апреля;
Козлова, 1930); на р. Чикой (притокСеленги) отмечена 16 апреля (Моллесон,
1897), в Даурию прилетает во второй половине апреля (Тачановский, 1873).
На р. Илиме (приток Ангары) появляется в первой декаде мая, сразу вслед
за вскрытием реки (Косыгин, 1926), под Якутском прилет их приходится
на конец апреля—Начало мая (Иванов, 1926), под Верхоянском на 26 мая.
На осеннем пролете обычна по р. Баргузину и на озере Рангатуй
у северного берега Байкала (Туров, 1923). В Тункинской долине пролет
их начался 5 сентября, когда утром появились косяки касатых уток. Не-
большими табунами они летели еще 24 сентября, останавливаясь для кор-
межки на открытых плесах (Слудский), на Дарасуне держались до 27 сен-
тября. На р. Хара (Кентэй) пролетная птица, державшаяся в стайке сви-
язей, добыта 20 сентября. В нижнем течении Амура (Николаевск) около
1 октября касатки часто встречались мелкими стайками по 3—5 шт.; весь
сентябрь они держатся стайками в районе Ханки (Шульпин, 1936). Обычна
на пролете в Корее, на японские зимовки прибывает несколько позже других
видов, а именно с начала до середины сентября.
Биотоп. На весеннем пролете касатки придерживаются рек, озер
и их разливов, на морском побережье они встречаются очень редко. В период.
КАСАТКА 403
гнездованья эти утки держатся на мелких озерах и речных старицах
с открытыми, так и с поросшими лесом берегами. Селезни, начавшие линьку,
собираются на луговых озерах. На зимовках касатки кормятся на внутрен-
них водоемах низменностей и рисовых полях. День большие стаи их отдыхают
на открытых плесах озер и в море у берегов.
Численность. В южных частях этой территории (Приморье,
Амур, Забайкалье, Маньчжурия, о-в Хоккайдо) касатка обычна, причем
в Южном Приморье она принадлежит к числу наиболее обыкновенных и
характерных птиц. В северной половине ареала (Лена, Яна, Охотское
побережье, Камчатка) редка.
Экология. Размножение. Формирование пар заканчивается
у касаток к середине декабря, еще на местах зимовок (Ян, 1942). На места
гнездовья в Советское Приморье они прибывают стайками, состоящими из
парных птиц. Брачные игры довольно однообразны, они выражаются в по-
клонах головы, которые сочетаются с общим наклоном передней части кор-
пуса и сменяются позой, при которой голова и шея плотно прижаты к туло-
вищу. Оперение головы при этом распушается и последняя кажется неесте-
ственно большой, но красивые серповидные перья самца в это время никогда
не растопыриваются. Самки во время брачных игр принимают те же позы,
что и самцы. На юге Приморья всю весну, вплоть до конца июня, встре-
чаются пары касаток, не имеющих гнезд. Одиночные птицы отмечаются
и за южными пределами гнездового ареала (Чжили), возможно, что это
особи, не достигшие половой зрелости на первом году жизни.
Гнезда помещаются на земле. В приморье их находили по берегам озер
среди густой высокой травы и мелкого кустарника, на расстоянии до 80 м
от воды (Пржевальский; Спангенберг, 1936). Гнездо, найденное на р. Тунке
(Забайкалье), было устроено на осоковой кочке, яйца были укрыты серым
пухом (Слудский), на Дарасуне гнезда помещались в болотистых кустарни-
ках (Тачановский, 1873). Обычное число яиц в полной кладке 8. Окраска
скорлупы желтоватого тона, что служит хорошим отличием от яиц кряквы.
Средние размеры (по одной кладке) 55,8x37,8 мм, предельные (по несколь-
ким кладкам) 53—58,5x37,6—41,5 мм. Срок насиживания (по наблюдениям
в зоологическом парке) 24 дня. Первую половину срока насиживания селезень
держится около гнезда и сразу же присоединяется к слетевшей с гнезда самке.
Откладка яиц начинается позднее, чем у кряквы. В нижнем течении р. Имана
(Приморье) гнездо с 6 ненасиженными яйцами найдено 5 июня. 19 июня пой-
маны два совсем маленьких утенка, а 27 июня еще четыре величиной со
скворца. Воспитанные в зоопарке, они к концу июля стали подниматься
на крылья (Спангенберг, 1936). На р. Лефу гнездо с 5 довольно сильно наси-
женными яйцами найдено 26 июня, но там же 29 июня встречены уже опе-
рившиеся молодые, размером в три четверти старой птицы (Пржеваль-
ский). Гнездо с 8 насиженными яйцами найдено на р. Тунке 26 июня, выводки
утят размером с половину матки E и 10 птенцов) встречены там 5 и 10 июля.
В низовьях р. Верхней Ангары пуховые утята обнаружены 7 августа, а моло-
дые размером с кедровку 14 августа (Слудский).
Линька. В Южном Приморье самцы начинают линять в половине
июня. В среднем течении р. Лефу по луговым озерам в конце июня держа-
лось уже множество селезней, часто стаями по 25—30 шт. (Пржевальский).
На Имане касатки наблюдались парами до 10 июня, а позднее селезни встре-
чались уже без уток. Первый самец, приступивший к линьке, добыт 23 июня,
яо до конца месяца селезни не теряли способности к полету (Спангенберг, 1936).
У некоторых особей линька затягивается долго. Самец, добытый на озере
Ханка 25 августа, вылинял еще только наполовину, маховые перья, рулевые
и кроющие крыла еще кровоточили (Маак, 1861). В Тункинской долине
селезни собрались в табунки по 10—15 шт. к 28 июня, а в середине июля
26*
404
ОТРЯД ГУСЕОБРАЗНЫЕ
они перестали попадаться на глаза (Слудский). Молодые птицы обоего пола
линяют на зимовке в январе, сменяя контурное перо и рулевые.
Питание касатки не изучено. В желудках птиц, добытых в мае
на р. Имане, обнаружены зеленые побеги растений и песок.
Хозяйственное значение касатки на Дальнем Востоке довольно велико,
так как на весеннем пролете в Приморье она служит одним из основных про-
мысловых объектов. На Ханке из числа добытых охотниками уток занимает
третье место.
Полевые признаки. Плавающий селезень узнается на далеком расстоя-
нии по яркой зеленой голове, оперение которой в возбужденном состоянии
Рис. 74. Касатка (самец и самка)
распушается, отчего голова кажется очень большой. От кряквы отличается
грудью и желто-белым двойным пятном на подбородке и зобу. Голос самки
сходен с голосом кряквы, самец же обладает очень мелодичным высоким
свистом, несколько напоминающим крик кроншнепа. Его можно передать
слогами «тю-тю-вить»... «тю-вить»... «тю-тю-вить». Этот крик издает и сидя-
щая и летящая птица. Касатка очень осторожная утка.
Описание. Строение и размеры. Из особенностей строения касатки следу-
ет отметить удлиненные перья затылка и шеи, образующие широкий хохол и третьестепен-
ные маховые, которые длиннее больших маховых и серповидно изогнуты. Крыло самцов
230—255, самок от 225 мм; цевка самцов 35—40 мм, клюв самцов 35—40 мм. Вес селезней
около 750 г, самок 640—660 г.
Окраска. Пуховой птенец. Похож на такового у свиязи, но не имеет по-
лоски через глаз и пятна в области уха.
Молодые птицы. В первом пере напоминают взрослую самку, но темя почти
черное с зеленоватым отливом, спинная сторона тела матовочерная с узкими темно-
охристыми каемками на отдельных перьях, зашеек и лопатки почти одноцветны.
КРЯКВА 405
Самцы в этом возрасте узнаются по неясным охристым полоскам на перьях лопаток
и зашейка, которые у самок одноцветны.
Взрослая самка. На спинной стороне в общем черно-бурая. Голова в светло-
бурых продольных пестрянках, весь верх, в том числе и плечевые, с рыжими каемками и
неправильными пятнами. Брюшная сторона охристо-рыжая, с неясными черными пятнами,
особенно обильными на зобу. Крылья как у самца, но третьестепенные маховые лишь
слегка изогнуты наружу и не образуют косиц. Клюв черный, лапы серые с очень темными
перепонками, радужина темнобурая.
Самец в летнем наряде. Отличается от самки полным отсутствием косичек треть-
естепенных маховых.
Самец в брачном наряде. Верх головы от лба до затылка густокоричневый,
почти черный, бока головы и шея сзади темнозеленые с бронзовым отливом. Лопатки и
большинство плечевых серые стемноструйчатым рисунком Спина темнодымчатая, у старых
экземпляров почти вся струйчатая, основание надхвостья также струйчатое, остальная
часть черная. Подбородок, горло и неполный ошейник белые, в нижней части шеи черное
с зеленоватым отливом кольцо. Зоб и грудь стемносерым чешуйчатым рисунком, бока тела
и брюшко с тончайшим струйчатым рисунком. Подхвостье бархатисто-черное с соловыми
пятнами по бокам. Верхние кроющие крыла дымчато-серые, самые длинные из них светлее.
Первостепенные маховые серо-бурые, второстепенные образуют серо-зеленое зеркало,
удлиненные третьестепенные бархатисто-синие с очень узкой светлой каемкой по наружному
опахалу и иногда серым струйчатым рисунком внутреннего опахала. Наружные плечевые
перья черные, хвост серый, подмышечные чистобелые. Клюв, лапы и радужина, как
у самки.
25. Кряква Anas platyrhynchos L.
Anas platyrhynchos. Li n n a e u s. Syst. Nat., изд. X, 1758, стр. 125, Швеция.
Синоним. Anas boschas. Li n n a e u s. Там -же, стр. 127, 1758.
Русские названия. Кряква, крякуша, крыжень—народные, вероятно звукопод-
ражательные; селезень, матерая утка—также народные.
Распространение. Ареал. Вся Европа, кроме Пиренейского п-ова,
северной части Мурмана и тундровой зоны европейской части РСФСР.
В Сибири на север до Салехарда, Туруханска, среднего течения Нижней
Тунгуски, устья Вилюя, п-ова Тайгонос на Охотском море и Северной Кам-
чатки. На юг до Малой Азии включительно, Ирана, Гималаев, Кашмира
и Чжилийского залива в Китае. Курильские о-ва и Япония на юг до средней
части о-ва Хонсю, Марианские, Гавайские и Сандвичевы о-ва, Северная
Америка до Полярного круга, устья р. Мекензи, Гудзонова залива и Вели-
ких озер. На юг до Калифорнии и среднего течения Миссисипи. Южная
часть Гренландии на западе до 73° с. ш., на востоке до Полярного круга.
Таким образом, ареал кряквы широкой полосой опоясывает север Европы,
Азии, Северной Америки, прерываясь только в восточной части Северной
Америки.
Характер пребывания. В Англии, частично Исландии
и Гренландии и в южных частях ареала оседла. На остальной территории
перелетная птица. Основные зимовки расположены на побережьях Балтий-
ского, Северного, Средиземного, Черного и Каспийского морей, в Иране,
Ираке, Северной Индии, в приморских частях Китая, Японии, на Тихо-
океанском побережье Северной Америки, в Мексиканском заливе и в долине
Миссисипи.
Биотоп. Водоемы самого разнообразного типа, в гнездовое время
преимущественно континентальные, пресноводные, а зимой столь же часто
и тихие морские заливы. Отсутствует в зоне чистой тундры (кроме Грен-
ландии) и избегает альпийской зоны гор.
Подвиды и варьирующие признаки. Основная часть ареала вида занята номиналь-
ной формой А. р. platyrhynchos Li n n a e u s, 1758, чрезвычайно пластичной в экологиче-
ском отношении и весьма постоянной по своим морфологическим признакам на всей обшир-
ной территории ее распространения. Гренландию населяет А. р. conboschas С h г. L. В г е h in,
1831, несколько более крупная форма, которая отличается строением клюва, ьзцутого
у основания в результате сильного развития носовых желез. Последнее является ядапта
цией к жизни преимущественно на морских водах. Окраска спинной стороны более серая,
406
ОТРЯД ГУСЕОБРАЗНЫЕ
бока, грудь и брюхо сильнее испещрены. А. p. oustaleti Salvador i, 1894—на о-вах
Гуам, Сайпан и Тиниан из группы Марианских о-вов. А. p. wyvilliana S с 1 a t e г, 1878—на
Гавайских о-вах и А. р. laysanensis R о t h s h i I d, 1880, с о-ва Лайзана Гавайской груп-
пы. Наиболее характерным свойством всех трех южных островных форм служит отсутствие
брачного наряда у селезней, которые имеют оперение, сходное с летним нарядом селезней
исходной формы.
Карта 46. Распространение кряквы Anas plaiyrhynchos
I—A. p. platyrhynchos, 2—A. p. conboschas, 3—A. p. wyvilliana, 4—A. p. iai/sanensie, 5— A. p.
oustaleti; a—граница гнездовий, a'—районы наибольшей численности птип на гнездовье* в—обособ-
ленные гневдовые колонии, в'—места постоянных встреч летом (гнездование возможно, но не дока-
вано), е—8имовки, а—валеты, э#с—места скоплений птиц на линьку
Кряква Anas platyrhynchos platyrhynchos L.
Распространение. Ареал. Западная Европа на север до Исландии
включительно, кроме северной части Остфинмаркена. На Кольском п-ове
и далее на восток до границы настоящей тундры, на Печоре до устья р. Усы,
на восточном склоне Урала до долины р. Сосьвы, а по Оби спускается до
Салехарда, по Енисею до Туруханска, на Нижней Тунгуске до 61° с. ш.
В бассейне Лены гнездится при устье Вилюя и по долине Алдана на север
примерно до 64° с. ш. О гнездовании на Яне, Индигирке и Колыме поло-
жительных сведений нет. По Охотскому побережью доходит до п-ова Тай-
гонос, обыкновенна по всей Камчатке, включая о-в Карагинский, гнездится
на о-ве Беринга и Курильских о-вах. Местами севернее этой границы кряквы
довольно регулярно встречаются в летнее время, но это бывают негнездя-
щиеся бродячие особи. Таковы случаи встреч их в Финмаркене, Тиманской
тундре (Гладков, 1941), по р. Щучьей на Южном Ямале (Кучерук) и у Дудинки
на Енисее (Тугаринов). Нерегулярные залеты крякв известны: на юго-
восточном берегу Гренландии, на реках Пенжине и Анадыре. В Америке
северная граница проходит от долины Юкона на устье р. Мекензи (Бейли,
1948). На п-ове Сюард кряква сейчас очень редка и, повидимому, не гне-
здится, хотя по археологическим данным 1,5—2 тысячи лет назад она слу-
жила там обычной охотничьей добычей местного населения (Фридман).
Восточнее граница распространения кояквы резко падает к югу, через Мед-
кряква 407
вежье и Невольничье озера к берегам Гудзонова залива. Южная граница
проходит по Пиренеям, захватывает все острова Средиземного моря и Малую
Азию. Кряква встречается в летнее время в Северо-Западной Африке. Изоли-
рованная гнездовая колония известна в Алжире на горном озере Сенар
(Нитхаммер, 1938) и возможно гнездится в Агельмане (Марокко) на озере
Сиди-Али в горах Среднего Атласа на высоте более 2 000 м (Хартерт, 1926).
Она населяет большую часть Ирана, включая Сеистанскую котловину,
но отсутствует в южной приморской зоне. Гнездится в Гималаях и Кашмире
(Финн, 1901), но неизвестна в Тибете и Центральном Китае. Гнездится
в Кашгарии и Заалтайской Гоби, редка в провинции Чжили. Встречается
в летнее время (гнездится?) в верхнем течении р. Хуанхэ, Сачжоу и Шеньси
(Латуш, 1931). Отсутствует в Корее, но обычна в Маньчжурии и Советском
Приморье, однако южнее залива Посьет отсутствует и замещается черной
кряквой (Воробьев). В Японии населяет о-в Хоккайдо и встречается на юг до
средней части о-ва Хондо. Изолированная гнездовая колония находится
на о-ве Бородино из группы Риу-Киу, возникшая, повидимому, за счет
птиц, завезенных туда из Японии и разводившихся на прудах (Курода, 1925),
В Америке южная граница распространения проходит от Калифорнийского
п-ова C0° с. ш.) через устье Миссури, примерно до озера Эри, захватывает
почти всю область Великих озер и поворачивает на север к берегам Гудзо-
нова залива.
Кряква акклиматизирована на Новой Зеландии. Она полностью нату-
рализовалась там и расселилась почти по всей территории острова. Она
живет в одних стациях с местной желтоносой кряквой (A. poecilorhyncha
superciliosa) и скрещивается с ней, вытесняя и поглощая последнюю
(Коттам, 1951).
Районы скоплений на линьку. Основная область
массовых скоплений линных птиц расположена в степной и лесостепной
зонах. Она начинается с дельты Волги и идет через степи среднего течения
Урала, Илека и Зауральские степные озера к Кургальджин-Тенизской
озерной системе и Барабинской лесостепи. Другие, несомненно существую-
щие, районы скопления линных птиц до сих пор не описаны. Это явление
известно еще лишь для Рыбинского водохранилища (на Верхней Волге)
и для бухты Мацаль-Вик на Эстонском побережье Финского залива. С Бри-
танских о-вов селезни улетают линять в Голландию и Германию.
Основные зимовки. Единичные особи и небольшие группы
крякв зимуют почти повсюду в пределах своего гнездового ареала, там,
где сохраняются незамерзающие водоемы. Однако это ни в коей мере не опре-
деляет зимнее размещение вида. Основные районы скоплений зимующих
птиц таковы: западная часть Балтийского моря, побережье Южной Швеции,
Норвегии, Дании и далее на запад вплоть до Испании включительно. Массо-
вые зимовки расположены у Британских о-вов, особенно вдоль юго-восточ-
ного берега. Зимуют кряквы на Фарерских о-вах и в Исландии, но в незна-
чительном количестве. Многочисленны они по всему европейскому берегу
Средиземного моря, в Италии и на Балканах, в Алжире, Тунисе, Марокко
и долине Нила. Зимуют также на Канарских и Азорских о-вах. На Черном
море кряквы зимуют в его западной части, а также у восточного берега
Азовского моря. Многочисленны они на Южном Каспии, в Восточном Закав-
казье, Восточном Иране (Сеистанские озера), на морском побережье и в Ме-
сопотамии. Кряква одна из наиболее многочисленных зимних птиц Север-
ной и Средней Индии, Ассама и приморской зоны Китая. В массе зимует
кряква на Японских о-вах, в небольшом числе у теплых ключей на Кам-
чатке, на о-ве Беринга и вдоль Алеутской гряды На Американском конти-
ненте зона зимовок располагается вдоль побережий от Аляски и Лабрадора
до Калифорнии и берегов Мексиканского аалива включительно.
408 ОТРЯД ГУСЕОБРАЗНЫЕ
Характер пребывания. Кряква оседла в Англии, частично
в Исландии и возможно в средиземноморских странах. В Гималаях она
совершает сезонные кочевки с гор на низменности и обратно, оседла в Япо-
нии и частично на Камчатке. На всей остальной обширной территории кряква
перелетная птица.
Миграции. Результаты кольцевания позволяют наметить основ-
ные пути пролета и районы зимовки для различных географических попу-
ляций этого вида. Кряквы из Швеции, Норвегии, Финляндии, Дании, Совет-
ских Прибалтийских Республик, Северной Польши и Германии зимуют
в Англии. Туда же переселяется и часть птиц из Исландии. Кряквы, насе-
ляющие северную половину европейской части Союза, на восток до Печоры
и на юг до Верхней и Средней Волги, отлетают преимущественно в западном
направлении через Прибалтийские страны и зимуют в приморской зоне
Германии, Дании и Голландии, доходя на запад до Англии и Северной
Франции. Незначительная часть их пролетает на юго-запад через Бело-
руссию и Украину в Придунайские страны на Балканы и даже в Северную
Италию. Кряквы, населяющие восточную половину европейской части
Союза: бассейн Дона, Средней и Нижней Волги, Урала и Печоры, а также
Обь-Иртышский бассейн, озера Западной Сибири и Северного Казахстана,
составляют другую—европейско-сибирскую популяцию. Селезни этой попу-
ляции проводят линьку в лесостепной и степной полосе от Барабы на востоке
до дельты Волги на западе. Основная масса их зимует у южных побережий
Каспийского моря и южнее в Иране, Ираке, Турции и Сирии. В меньшем
числе они проводят зиму в Предкавказье на восточном побережье Азовского
и Черного морей. Значительная часть их улетает дальше, на Балканский
п-ов, в Грецию, а некоторые достигают Италии и Африканских берегов. Туда
же отлетают кряквы, населяющие Украину и центральные районы европей-
ской части РСФСР. Несколько восточнее, у Средиземноморских берегов.
Франции и Северной Италии, зимуют кряквы, гнездившиеся в Центральной
Европе (Германия). Часть крякв из Юго-Западной Сибири отлетает непосред-
ственно в южном направлении в Северную Индию и Пакистан. О распреде-
лении на зимовках восточносибирских крякв до сих пор почти ничего не-
известно, кроме находок на Северном Сахалине и на Уссури птиц, околь-
цованных зимой в Японии.
На всем протяжении своего обширного ареала кряква обнаруживает
значительное постоянство строения, размеров и окраски. Нигде на мате-
риках Евразии и Северной Америки она не образует местных форм. Это
обстоятельство принято объяснять тем, что якобы кряквы, а также и мно-
гие другие утки, слабо связаны с местами своего гнездовья. Так как раз-
бивка на пары происходит у них на зимовках, то зачастую в одну пару соеди-
няются селезень и утка уроженцы различных географических мест, в резуль-
тате чего одна из птиц, следуя за своим партнером, не возвращается на родину*
а переселяется на новое место («абмиграция»—Томсон, 1930).
Отдельные случаи, подтверждающие возможность такого происше-
ствия, зарегистрированы путем кольцевания. Однако случаи эти столь же
единичны, как и у других видов птиц. В то же время накопилось уже огром-
ное количество фактов, говорящих о чрезвычайно строгой привязанности
уток к местам их вывода, гнездовья, линьки и зимовки. Более чем 50%
крякв, закольцованных птенцами на Карельском перешейке, встречены затем
на гнездовье в том же месте (Вялинкянгас, 1933). Селезень с весьма своеобраз-
ным брачным свистом в течение нескольких лет появлялся весной на одном
и том же болоте (Спангенберг). Значительно более сотни крякв, закольцо-
ванных на линьке в дельте Волги (Астраханский заповедник), пойманы
в последующие годы там же. При этом они встречались чаще всего в тех же
самых мелководных култуках, что и при первой их поимке, в те же самые
КРЯКВА 409
сроки и нередко одними и теми же группами особей (Исаков, 1949). Наконец,
накопилось уже немало данных о том, что на зимовках одни и те же особи
этого вида появляются в одни и те же сроки, в одних и тех же местах (Дю-
пон, 1937—1940).
Одновременно существуют наблюдения и о наличии у крякв местных
географических популяций. Еще Северцов A850) обратил внимание на
различия в размерах и весе между воронежскими и московскими кряк-
вами. По его наблюдениям, первые заметно крупнее и тяжелее вторых.
По наблюдениям Куражсковского в Тульской обл., местные гнездящиеся
утки (самки) несколько темнее тех, которые бывают в области лишь на про-
лете. Тульские утки тяжелее, чем вологодские. Интересные паразитологи-
ческие наблюдения Дубинина говорят о том, что красноярские кряквы
хорошо отличаются по составу фауны перьевых клещей от птиц, прилетаю-
щих осенью с низовьев Енисея. Исходя из всех этих данных предполагать
у кряквы частую смену мест гнездовья нет оснований. Несомненно, что
в результате тщательного изучения размеров и веса крякв из разных геогра-
фических пунктов (при учете сезона и возраста птицы) выявятся морфоло-
гические различия у разных популяций этого вида.
Даты. Массовый отлет на север с азербайджанских зимовок проис-
ходит в середине февраля и продолжается 5—10 дней, но не заканчивается
и до конца марта (Верещагин, 1950). У Пехлеви, на Иранском побережье
Каспия, их число уменьшается уже в начале марта. В то же время начинается
отлет и на юго-восточном берегу Каспийского моря. Индийские зимовки
(Северный Пенджаб) кряквы покидают в феврале (Уистлер, 1920), но в
Белуджистане они задерживаются до конца марта. Юго-восточные про-
винции Китая и Ганьсу кряквы покидают в марте.
Валовой пролет по долине Узбоя в первых числах марта (Шестоперов),
а по Нижней Сыр-Дарье пролет начинается в конце февраля и становится
валовым в первой декаде и середине марта. В конце февраля кряквы по-
являются на о-ве Барса-Кельмес в Аральском море, в дельтах Днестра, Днеп-
ра, Дона и Волги, а также в степной части Крыма. В первой декаде марта
кряквы появляются в низовьях Кубани и под Казалинском, а в южной
части Приморья на озере Ханка проходит массовый пролет их, затем интен-
сивность его ослабевает, и новая волна валового пролета наблюдается там
в середине апреля. В середине марта кряквы появляются в среднем течении
Днепра у Киева и около Минска. На вторую декаду марта падает прилет
их на Урале под Чкаловом, на Хопре, под Харьковом и около Пскова.
Несколькими днями позже они появляются в Башкирии, а в самом
конце марта под Ленинградом. В последних числах этого месяца или
в первых числах апреля начинается пролет на Алатыре, в устье Камы и
на озерах Барабинской степи (Янушевич). В первой декаде апреля кряквы
прилетают в Прибалтику, Смоленскую обл., в Наурзумские степи Север-
ного Казахстана, под Челябинск, Иркутск и на Командорские о-ва.
В конце этой декады и начале следующей они появляются под Москвой.
На середину апреля приходится валовой пролет крякв в районе Верх-
ней Волги (устье р. Мологи) и появление их под Чердынью, Свердловском
и Молотовом на Урале, на Иртыше под Тобольском, в низовьях Амура и
в приморской зоне Камчатки. Во внутренних частях этого полуострова
кряквы появляются почти на 10 дней позже. В конце второй декады апреля
и в начале последней кряквы прилетают под Архангельском (Паровщиков),
Вологдой, в верхнем течении Печоры, на Средней Ангаре, у Енисейска и под
Якутском на Лене. Едва ли не позже всего, в середине мая, появляются кряк-
вы на Кольском п-ове в районе озера Имандра (Владимирская) и на Енисее
под Полярным кругом (Тугаринов). Указанные нами средние сроки весьма
значительно колеблются по годам. Весеннее появление крякв бывает приуро-
410 ОТРЯД ГУСЕОБРАЗНЫЕ
чено ко времени образования самых первых заберегов, полыней в местах
впадения мелких речек или даже просто луж талой воды на лугах. Изопип-
тезы весеннего появления крякв весьма несхожи между собой в различные
годы. Их широтное положение и форма зависят от взаимного расположения
и циркуляции воздушных масс весной данного года (Динесман).
Говорить о сроках отлета или осеннего пролета уток очень трудно, так
как откочевка селезней с мест гнездовья во многих местах происходит уже
в мае и июне. Поэтому под осенним отлетом мы будем подразумевать время
осенней миграции основной части популяции того или иного вида уток,
состоящей главным образом из молодежи и взрослых самок.
Для того чтобы показать всю сложность этого явления, надо описать
последовательность летних и осенних миграций крякв в дельте Волги.
Уже в последних числах мая (обычно 18—21 мая) здесь появляются стайки
взрослых самцов, слетающихся на летнюю линьку с весьма обширных и уда-
ленных территорий. Особенно интенсивно подлет их происходит в июне,
но продолжается он и в июле. В середине сентября A6—18 сентября),
когда дельту Волги еще не покинули огромные стаи крякв, линявших здесь,
появляются первые стайки пролетных, главным образом молодых, птиц.
Интенсивность пролета постепенно возрастает, и в течение первой половины
октября он принимает массовый характер. Основная масса птиц отлетает
с Волжского взморья 22—26 октября. Последние же утки исчезают там только
в ноябре, а иногда и в декабре. Таким образом, стаи пролетных крякв сме-
няют одна другую в дельте Волги в продолжение почти шести месяцев,
но интенсивность их движения затихает в июне и августе.
В верхнем течении Печоры пролет начинается уже в августе и продол-
жается до середины октября (Теплов). С сентября по ноябрь проходит про-
лет на Камчатке. В середине сентября он начинается в Уссурийском крае,
но продолжается весь октябрь и заканчивается в Южном Приморье лишь
в ноябре. Во второй декаде сентября начинают появляться кряквы на местах
зимовки у Обиточенской косы Азовского моря. Подлет с севера продол-
жается там до декабря (Костюченко). В конце сентября исчезают послед-
ние кряквы под Якутском, и пролет начинается на Верхней Волге, под
Москвой и на Украине; он продолжается там до ноября. В первых числах ок-
тября начинается массовый отлет из-под Тобольска и Архангельска, в первой
половине месяца исчезает основная масса птиц из Башкирии, начинается
пролет в Наурзуме, в массе скапливаются кряквы на р. Или. Валовой про-
лет их в среднем течении Урала, под Чкаловом приходится на середину
октября. В это же время кряквы появляются на местах зимовки: на Алеут-
ской гряде (Уналашка) и в Северном Китае (Чжили). Во второй половине
октября кряквы отлетают с Телецкого озера, полностью исчезают на Ала-
тыре, наблюдается валовой пролет в низовье Днепра, а в последних числах
этого месяца—по долине Узбоя. В начале ноября они исчезают на Хопре,
под Ленинградом и в основной массе отлетают из Прибалтийских респуб-
лик. В ноябре они достигают мест зимовки в Южном Китае (Хубэй, Цзяньсу).
В конце ноября отлетают с Командорских о-вов, а в конце декабря исчезают
в устье Дона. На азербайджанских зимовках кряквы появляются уже в кон-
це августа, и число их там непрерывно возрастает до ноября, когда наблю-
дается массовый прилет. В Гасан-Кули (Туркмения) заметный пролет начи-
нается в октябре и длится в октябре-ноябре. В Южном Таджикистане про-
летные кряквы наблюдаются уже в начале сентября, в этом же месяце они
достигают Белуджистана, а в октябре появляются уже в Северном Пенджабе.
Биотоп. Места обитания крякв весьма разнообразны в разных
ландшафтных зонах и в разные сезоны года. Однако можно отметить, что
в гнездовое время кряквы определенно предпочитают места лесистые или
с высоким травостоем, что соответствует их привычке устраивать хорошо
КРЯКВА 41!
укрытые гнезда. Для кормежки они предпочитают водоемы с богатой расти-
тельностью. В лесотундре кряквы придерживаются лесистых участков близ
рек, в лесной полосе они особенно охотно заселяют озера-старицы в леси-
стых поймах, но избегают узких лесных ручьев. В лесостепи придерживаются
осоковых болот и водоемов среди березовых колков, а в степях заселяют
речные уремы и мельничные пруды. В полосе пустынь кряквы редки и чаще
встречаются в заболоченных местностях или там, где есть тугаи. В горах они
поднимаются высоко, до 2250 м (Алтай), заселяя озерные плато, но избе-
гают горных рек с быстрым течением. У крайних южных пределов своего
ареала кряква гнездится только в горах, отсутствуя летом на низменности:
есть в Среднем Атласе в Северной Африке B000 ж), Гималаях A300лс),
в Пенджабе и Кашмире, плато Камикуси A400 м) в Японии.
Сказанное выше говорит лишь о предпочтении кряквой водоемов того
или иного типа, вообще же как вид, чрезвычайно широко приспособленный,
она встречается в самых разнообразных местах. Так, например, в Москве
она гнездится на прудах зоопарка, где живет оседло, в Западной Европе
обычна в некоторых больших городах; гнездится в Лондоне, Берлине, Мюнхе-
не, Лейпциге, причем не только в парках, но и на крышах домов. На крыше
одного пятиэтажного дома в центральной части Берлина кряква устраивала
гнездо 3 года подряд (Хейнрот, 1940).
Численное ть. Один из наиболее широко распространенных
и многочисленных видов настоящих уток. Однако далеко не повсюду кряква
встречается одинаково часто. Она обычна в средней лесной зоне и лесостепи,
становясь редкой у северных пределов лесной растительности, в горах
и в большей части пустынь.
Экология. Размножение. Размножаться кряквы начинают в воз-
расте одного года. Однако некоторая, хотя и незначительная, часть популя-
ции на первом году жизни в размножении, повидимому, не участвует. На
мысль об этом наводит наличие бродячих особей в течение всего гнездового
сезона в южных частях ареала вида, а также постоянное присутствие холо-
стых самцов.
Широко распространенное представление о полигамии уток (в том числе
и крякв) не соответствует действительности. В течение всего периода спари-
вания и откладки яиц у крякв существуют совершенно определенные пары.
У оседлых популяций этого вида формирование пар начинается еще осенью.
В этих местах уже в конце сентября можно наблюдать отдельные пары, хотя
многие самцы к этому времени еще не вполне заканчивают линьку в брачное
перо, а их половые железы чрезвычайно малы. У перелетных популяций
разбивка на пары происходит несколько позже—на местах зимовки. Так,
первую половину зимы в Юго-Восточном Закавказье кряквы держатся
стайками, разнообразными по количеству и половому составу особей, но
уже в начале декабря в стаях намечаются отдельные пары этих птиц, кото-
рые держатся совместно на кормежке и отдыхе. К концу месяца количество
оформившихся пар возрастает, а в январе почти все стаи расчленяются на
пары. Однако это не влечет за собой распада стай, которые сохраняют свою
целостность до конца весеннего перелета.
Период разбивки на пары характеризуется весьма своеобразным токо-
вым поведением птиц, так называемыми «общественными» играми, когда
птицы собираются небольшими группами и ведут себя возбужденно и шумно.
Оставшиеся еще без пары самки стараются привлечь к себе пролетающих
селезней громкими «куак»... «куак»... «куак»... «куа-куак». По большей части
в этих играх принимают участие оба пола, но нередко они происходят и при
отсутствии самок.
Таким образом, на места гнездовья кряквы прилетают уже окончательно
установившимися парами, а нередко наблюдающиеся весной драки между
412 ОТРЯД ГУСЕОБРАЗНЫЕ
селезнями обязаны или наличию холостых птиц, или охране спаренными пти-
цами своей гнездовой территории. Настоящие токовые позы крякв можно-
наблюдать уже зимой в ясные, солнечные дни, иногда они даже заканчиваются
попытками к спариванию. Однако во всем своем разнообразии брачные игры
проявляются лишь весной, во время перелета и уже на местах гнездовья.
У крякв они достигают наибольшего развития и разнообразия среди утиных
птиц, почему остановимся на их описании несколько подробнее.
Обычно селезни сидят на воде в обществе других селезней или медленна
плавают вокруг своей самки с головой, втянутой в плечи, слегка опущенным
клювом и постоянно вертя хвостом. Оперение головы у них при этом распу-
шается и имеет темную матовую расцветку. Утка кружится около избранного
ею супруга, поворачивает голову назад и делает через плечо отрывистые
угрожающие движения клювом, как бы отпугивая врага, которого по бли-
зости может и не быть. При этом она издает особый звук из очень часто повто-
ряющихся слогов: «куегг-егг-егг-егг-егг-егг-егг». Утка сидит высоко на воде
так как перья нижней поверхности туловища у нее распушаются.
Спокойное плавание в описанной позе нарушается нервозным движением
головы вперед и вверх. Это происходит обычно трижды в течение нескольких
секунд, причем раз от раза интенсивность движения возрастает. При послед-
нем толчке селезень даже привстает над водой. Иногда это движение прини-
мает еще более стремительный характер. Селезень погружает в воду клюв
и затем с такой быстротой выбрасывает голову высоко вверх, что поднимает
при этом фонтан брызг. В этот момент, когда корпус поднимается почти
вертикально над водой, а опущенная вниз шея сильно сдавливает трахею,
селезень издает громкий резкий свист, за которым следует низкий хрюкаю-
щий звук, издаваемый тогда, когда голова снова поднимается, а туловище
опускается на воду. Время от времени селезень останавливается против вы-
бранной им самки и заворачивает голову на спину за слегка приподнятое
крыло, зеркало которого при этом бывает полностью расправлено. Вслед
за тем резким и отрывистым движением он проводит ноготком клюва вдоль
очина махового пера, чем издает довольно громкий, слышный на много*
метров, дребезжащий звук. Движение это настолько коротко, что до сих пор
неясно—второстепенными или первостепенными маховыми при этом поль-
зуется селезень; сила звука говорит скорей о последнем.
Часто селезни на очень короткое время принимают весьма оригинальную
позу. Они вскидывают голову и отводят ее назад. Одновременно вздергивается
кверху зад, черное матовое оперение которого распушается и демонстри-
руется во всей красе. Хвост расправляется, а локти поднимаются вверх,
отчего широко раскрывается зеркальце крыла и четко обрисовываются косицы
надхвостья. Птица в этот момент кажется очень короткой и высокой. Фаза
эта длится только доли секунды, вслед за чем корпус принимает нормальное
положение и голова у парного самца поворачивается в сторону самки. Затем
он вытягивает шею и, распластавшись на поверхности воды, делает круги
вокруг самки. Последняя часто плавает тем же манером. Перья на шее у них
при этом взъерошены. Постепенно селезень переходит на обычное плавание,
причем голова его бывает высоко поднята на взъерошенной шее, но повер-
нута теперь к своей самке затылком. По сторонам головы перья плотно при-
жаты к коже и образуют на щеках яркие зеленые блики, зато по медиальной
линии от лба до самого низа затылка они растопырены и образуют матово-
черную продольную полосу, которая и демонстрируется селезнем при соот-
ветствующих поворотах шеи. Голова при этой своеобразной «прическе» при-
нимает форму плоского диска.
Вслед за описанным иногда следует еще одно движение. Селезень с мол-
ниеносной быстротой опускает в воду клюв, а в следующий момент поднимает
голову вверх, не только не выпрямляя груди, а наоборот, опуская ее еще-
КРЯКВА
413
глубже в воду. В тот момент, когда голова принимает наивысшее положе-
ние, а грудь уходит наиболее глубоко, т. е. при наибольшем растяжении
и сжатии трахеи, издается свист. При резком подъеме головы часто над
водой появляется небольшой фонтан брызг.
Рис. 75. Позы тока кряквы
Как и у всех настоящих речных уток, у обоих полов спариванию пред-
шествует подергиванье головы снизу вверх, причем клюв при своем нижнем
положении касается воды, все время оставаясь почти горизонтальным.
В заключение самка вытягивает шею, распластываясь на воде, и самец топ-
чет ее.
Весной парные селезни, обосновавшиеся на местах гнездовья, начинают
настойчиво преследовать чужих самок. Однако последние всеми силами ста-
раются избавиться от преследователей. Во время отсутствия своего селезня
414 ОТРЯД ГУСЕОБРАЗНЫЕ
самки затаиваются от чужих, своего же узнают еще издалека по голосу
и присоединяются к нему в воздухе. Причины этого преследования еще
до сих пор не совсем ясны, но во всяком случае не имеют ничего общего
с настоящей полигамией, так как отношение селезней к своим и посторонним
самкам совершенно различное. По наблюдениям Куражсковского, селезни
с особенным азартом преследуют самок, сошедших с гнезда, слетаясь на
особый крик, который утки издают, покидая гнездо.
По некоторым наблюдениям, селезень участвует в выборе места для
гнезда (Лоренц, 1935), а некоторые исследователи приписывают ему в этом
отношении даже основную роль. В устройстве гнезда самец, как правило,
никакого участия не принимает, но часто провожает самку до гнезда, когда
она направляется для откладки очередного яйца. Покидая ее на это время<
и в течение первого периода насиживания селезень держится вблизи от гнезда,
поджидая самку, слетающую к нему на время отдыха и кормежки. При при-
ближении опасности он кружится над тем местом, где расположено гнездо,
и особым тревожным криком предупреждает подругу.
Однако вскоре у селезней начинается линька, и в большинстве случаев
они покидают гнездовую территорию. Исходя из наблюдений за утками
в парках Германии, Хейнрот A928) считает, что пары у крякв разлучаются
только на тот период, пока самец линяет, а самка занята воспитанием птен-
цов. Осенью же и селезни и утки появляются в одних и тех же местах, и пары
соединяются вновь, отчего одни и те же пары могут сохраняться несколько
лет. На территории Союза селезни совершают огромные перелеты на линьку
и вслед за тем на зимовку, где только они и могут встретиться с самками.
Однако, зная исключительно четкую привязанность уток к местам их гнез-
довья, зимовки и линьки (Исаков, 1949), не исключена возможность что это
нередко происходит и в наших условиях.
Помимо постоянства пар, кряквы, повидимому, и физиологически не
способны к полигамии. В тех случаях, когда при содержании на прудах
селезней бывает меньше, чем уток, последние начинают откладывать жировые
яйца. Так, 14 уток, живших на прудах Ростовского зоопарка при 8 селез-
нях, отложили 140 яиц, но 42% из них были неоплодотворенными. Иными
словами, 8 селезней смогли обслужить только 8 самок (Денисов, 1933).
Гнезда кряквы устраивают в самых разнообразных местах, но, как
правило, поблизости от воды. Однако нередко гнезда строятся около вре-
менных лесных луж или у границы весенних разливов, отчего после спада
воды и усыхания водоемов оказываются от них на значительном расстоянии.
К гнездованью кряквы приступают рано, когда большая часть речных пойм
находится еще под водой, а степные озера и пруды переполнены талой водой
и выходят из берегов. Поэтому основная масса их гнезд размещается на пери-
ферии поймы, на коренных берегах или же на наиболее высоких гривах.
Как правило, гнезда строятся укрыто под деревьями, кустами, бурело-
мом, валежником или в куртинах высокой травы. У северной границы распро-
странения, в лесотундре (Салехард), гнезда помещаются под отдельно стоя-
щими низкорослыми елями или корявыми лиственницами с низко спускаю-
щимися ветвями, среди моховых кочкарников, иногда довольно далеко от
воды. В северно-таежной зоне кряква поселяется в долинах крупных рек,
редко на глухих ручьях и совсем отсутствует на водоразделах. Гнезда ее
помещаются чаще всего на высоких лесистых гривах или в тайге на корен-
ных берегах. Они устраиваются у стволов деревьев, под буреломом и валеж-
ником. Примерно в тех же условиях гнездится кряква в средней лесной по-
лосе и в лесостепи, но гнезда ее располагаются там чаще всего под кустами
или под низкими кронами молоденьких елочек. Однако столь же часто кряк-
ва гнездится в куртинах высоких бурьянов, или в зарослях вейника на сухо-
долах, или на больших осоковых кочках среди кочкарного болота. В пой-
кряква 415
мах, длительно заливаемых полыми водами, кряквы часто, а иногда и почти
исключительно (дельта Волги), гнездятся на деревьях в старых гнездах
ворон, цапель и других крупных птиц. То же самое наблюдается в затоплен*
ных лесах водохранилищ. Гнездятся кряквы и в дуплах с открытыми широ-
кими входами. Чаще всего это бывают дупла в сломанных стволах или же
прикорневые дупла. В условиях Рыбинского водохранилища они занимают
повешенные над водой или установленные на открытых островах гнездовые
ящики соответствующего размера, а на Московском море охотно заселяют
старые охотничьи «скрадки» и шалаши.
Гнезда представляют собой постройку различного размера. В сухих
местах это довольно глубокие и ровные ямки, лишь слегка выстланные мяг«
кой и сухой травой. Ямку птица углубляет клювом, вырывая корни расте-
ний и разрыхляя этим почву, и выравнивает ее грудью, долго крутясь на
одном месте. Материал для выстилки далеко не носят, а берут по большей
части тот, который можно достать клювом не сходя с гнезда. Такая гнездо-
вая ямка строится за один день, а если первое выбранное место оказывается
неудобным, то в тот же день утка роет одну или несколько новых ямок.
В сырых местах кряква натаскивает большую кучу строительного мате-
риала (бурьяна, травы, тростника и др.) и уже в ней устраивает гнездовую
ямку. В скворечниках с опилками на дне бывает только ямка, совсем без
гнездового материала. Обычные размеры гнезд таковы: средняя ширина лотка
16 см A5—20 см) и глубина в зависимости от места от 4 до 13 см. Обычный
диаметр самого гнезда 27—28 см B0—29 см) и высота бортов над землей
от 4 до 14,5 см. Первые яйца откладываются в еще не совсем достроенное
гнездо, и по мере увеличения кладки выстилка гнезда улучшается. Когда
откладка уже близится к концу, в гнезде появляется первый пух, который
к этому времени начинает выпадать с груди у птицы (особый гнездовой пух,
темной окраски). Количество пуха возрастает непрерывно в течение почти
всего периода насиживания. Пух утка укладывает кольцом по периферии
гнездового лотка, отчего вокруг последнего образуются довольно высокие
бортики, прикрывающие с боков насиживающую птицу. Уходя с гнезда,
самка тщательно закрывает яйца пухом, что способствует сохранению ими
тепла за время отсутствия наседки и делает гнездо мало заметным.
Откладка яиц у крякв начинается очень рано: на Нижней Сыр-Дарье
с начала апреля, в Приморье в первой половине апреля, и продолжается
до второй декады мая, а первые птенцы появляются в конце мая (Воробьев).
У Киева и в среднем течении Хопра откладка происходит в мае, в Чкалов-
ской обл. и Барабинской степи она начинается в середине апреля, под Москвой
и Казанью в последней декаде апреля, на р. Мологе в первых числах мая
A1 мая 1940 г. первая полная кладка), под Тобольском полные кладки в сред-
нем за много лет появляются 7 мая (птенцы обычно с 1 июня), в Архангель-
ской обл. около 11 мая самки уже сидят на яйцах (Паровщиков). Примерно
в то же время это происходит и в верхнем течении Печоры (устье Ылыча),
так как первые выводки появляются там в разные годы от9 июня до 5 июля.
Наконец, на Камчатке находка гнезда кряквы с яйцами зарегистрирована
14 мая (Аверин, 1948). Обращает на себя внимание то обстоятельство, что
сроки начала откладки яиц у кряквы на севере и на юге ее ареала разли-
чаются значительно меньше, чем сроки прилета ее в тех же районах. Так,
например, если кряквы под Архангельском и у Тобольска появляются
в среднем на 30—40 дней позже, чем на юге Советского Приморья, то соот-
ветствующая разница в сроках начала откладки яиц составляет всего 10—
12 дней. Причина этого заключается, повидимому, в том, что на севере кряквы
приступают к размножению сразу же по прилете, а на юге лишь через неко-
торый срок после появления (Ларионов, 1951). Обычное число яиц в кладке
от 8 до 11. Установить его можно, лишь выяснив наиболее часто повторяю-
416 ОТРЯД ГУСЕОБРАЗНЫЕ
щееся количество яиц в результате просмотра большого числа полных кла-
док. Ни в коем случае не следует определять размер кладки, как среднее
арифметическое из ряда наблюдений, так как у кряквы, как и у многих
уток, часто наблюдается потеря кладок, которая влечет за собой дополни-
тельные уменьшенные кладки или же откладку яиц в чужие гнезда. Средний
размер кладки различен в разных географических пунктах, а возможно, что
непостоянен и по годам. На юге число яиц как будто несколько больше.
В Архангельской обл., под Салехардом и Ханты-Мансийском кладки содер-
жат обычно по 7—8 яиц, на Рыбинском водохранилище 8 F—9), но на
р. Мологе в 1940 г. в средней кладке насчитывалось 11 (8—12) яиц, в Туль-
ской обл. 8 (Куражсковский). В Ульяновской обл. обычное число яиц в гнез-
дах Ю—12, в среднем течении Хопра 11 G—16), Барабинской степи 9 (8—11),
гнездо на озере Кургальджин имело 8 яиц, в Наурзумском заповеднике
(Кустанайская обл.) 11 и на Нижней Сыр-Дарье также 11. В Советском При-
морье средний размер кладки 10—11 G—12) яиц. В Германии от 8 до 14,
обычно 9—13 яиц.
Яйца кряквы откладывают ежедневно по одному, и при этом обычно
вечером. С каждым яйцом утка задерживается на гнезде все дольше и дольше,
с откладкой последнего яйца начинается настоящее насиживание. В резуль-
тате первые яйца ко времени окончания кладки имеют уже хорошо разли-
чимый зародышевый диск. Насиживает кряква 26 дней, причем все птенцы
выводятся почти одновременно на протяжении не более 10, редко 14 часов.
Таким образом, яйца, отложенные последними, проходят цикл своего раз-
вития за несколько более короткий срок, чем предыдущие. Возможно, это
обязано тому, что, будучи уже оплодотворенными, они несколько более
долгий срок остаются в организме самки. В первом периоде насиживания
кряква сходит с гнезда для кормежки и отдыха утром и вечером, но к концу
насиживания сидит уже очень плотно, неохотно покидая гнездо даже в слу-
чае грозящей опасности. По некоторым наблюдениям (Мантейфель), у наси-
живающей самки кряквы прекращается выделение секрета копчиковой
железы. Это имеет большое значение для сохранности кладки, так как от
постоянного соприкосновения с обильно смазанным жиром пером могут*
закупориваться поры скорлупы и затруднится газообмен яйца. К тому же
секрет железы, имеющий сильный запах, привлекал бы к гнезду наземных
хищников.
Яйца крякв имеют обычную яйцеобразную форму и одноцветно белую,
слегка зеленоватую, скорлупу. Зеленовато-оливковый оттенок за время
насиживания обычно исчезает. Яйца из одной кладки имеют сходные раз-
меры, но разные кладки нередко сильно отличаются одна от другой как
по величине, так и по форме яиц. Размеры яиц 49,5—66,8x37—46, среднее
53,5x39 мм. Наиболее обычный вес ненасиженных яиц около 46 г D0—52 г).
Хейнрот A928) средним весом считает 50—55 г, но, возможно, это обязано
тому, что он работал с утками в условиях зоопарка.
Вес только что выведшихся птенцов в среднем 28—28,6 г (от 25 до 31 г)
в Салехарде и Ханты-Мансийске (Исаков), 31—38 г—в Германии (Хейнрот,
1928). На обсыхание выведшегося птенца уходит два-три часа, но гнездо выво-
док покидает обычно часов через 12—16 после вылупления первого птенца.
К этому времени молодые уже способны быстро передвигаться по суше, пла-
вать и нырять. Ныряют птенцы хорошо и постоянно пользуются этим прие-
мом, спасаясь от преследования, что взрослая птица делает только будучи
раненой. Из высокорасположенных гнезд птенцы самостоятельно спрыгивают
вниз. При этом самка находится на воде или на земле под гнездом и беспре-
станно кричит, призывая к себе молодых. Благодаря очень малому весу птенцов
прыжок с высоты нескольких метров проходит для них благополучно. Самка
отводит птенцов в наиболее густо заросшую водяными и прибрежными расте-
КРЯКВА 417
ниями часть водоема, где выводок и держится довольно постоянно. Днем
утята зачастую собираются под кустами на берегу, которые отстоят от воды
иногда на расстоянии до 50 ле, и молодые птенцы греются около самки под ее
крыльями и распушенным оперением груди. Первое время они греются
не реже чем раз в два часа. От соприкосновения с перьями самки они полу-
чают необходимую жировую смазку, спасающую их пуховой наряд от быст-
рого намокания при плавании. В первые же часы своей жизни только что обсох-
шие птенцы уже пытаются схватывать корм. Вначале они хватают только
мелких насекомых и пауков, не обращая внимания на неподвижные пред-
меты. При этом они склевывают корм преимущественно с растений или друг
с друга и лишь позже начинают собирать его с земли и с воды. Первое время
птенцы ведут совершенно дневной образ жизни, но несколько позднее они
начинают кормиться и вечером. Целиком на вечерний режим кормежки
птенцы переходят, когда у них развиваются роговые пластинки на клюве
и они приобретают способность добывать себе корм процеживанием, в тем-
ноте. С первых же дней птенцы научаются узнавать друг друга и отгоняют
присоединяющихся к ним птенцов из других выводков. Так же поступает
и самка.
Голос пуховых птенцов—высокий и звонкий писк, но уже в самом ран-
нем возрасте бывает возможно определить пол птенца, так как самочки издают
двухтактный свист, в то время как у самцов он ординарный. В возрасте около
пяти недель самки начинают крякать. Примерно на 23-й день жизни у птенца
начинают разворачиваться контурные перья его первого наряда, тогда же
начинают расти и маховые перья. В 28 дней грудь и брюхо, особенно необ-
ходимые для предохранения птенца от намокания, уже бывают в пере, а еще
через 10 дней неоперенными остаются только задняя сторона шеи, часть
спины и бока тела, прикрытые крыльями. Примерно в 50-дневном возрасте
птенцы начинают уже взлетать, а в 56 дней летают хорошо, хотя первосте-
пенные маховые бывают к этому времени еще не совсем окрепшими. В воз-
расте 10 дней птенцы весят около 100 г, 20 дней—320 г, 30 дней—550—600 г
и к 60 дням (ко времени взлета)—около 800—900 г.
Большое число яиц кряквы погибает в результате разорения гнезд
четвероногими и пернатыми хищниками. Из первых наиболее существенный
урон наносят лисицы и енотовидные собаки, а из вторых—вороны и камыше-
вые луни. Устройство гнезд на островах и на деревьях обеспечивает их луч-
шую сохранность, но еще лучше сохраняются кладки в полудуплах и в спе-
циальных гнездовых ящиках. В поймах рек и по берегам водохранилищ гнезда
часто гибнут от затопления их вешними водами, что в годы высоких павод-
ков бывает иногда очень губительным для крякв. Столь же серьезным бед-
ствием для них служат пожары и практиковавшиеся до недавнего времени
весенние палы степных бурьянов и тростников, в результате которых поги-
бает иногда огромное количество гнезд. Самки, потерявшие кладку еще до
начала насиживания, обычно продолжают откладку яиц, снося их в одно
из расположенных поблизости утиных гнезд, вне зависимости от того, какому
виду оно принадлежит, или даже в гнезда других птиц, например фазана.
Иногда же, особенно в случае потери только что начатой или, наоборот,
уже законченной кладки, кряква строит новое гнездо и откладывает в него
яйца, но, как правило, повторная кладка бывает меньше обычной. Частые
случаи гибели гнезд и запоздалых повторных кладок обусловливают чрезвы-
чайную растянутость сроков размножения у кряквы и разновозрастность
птенцов, в результате которой повсюду к началу осенней охоты наряду
со вполне взматерелыми птицами имеются нелетные и даже не совсем опе-
рившиеся птенцы. Как говорилось выше, гнезда крякв, построенные у края
весенних разливов или около временных луж, оказываются летом на большом
расстоянии от воды, отчего птенцы должны совершать путешествие по суше
27 Птицы СССР, т. IV
418 ОТРЯД ГУСЕОБРАЗНЫЕ
иногда более чем за километр. При этом им приходится пересекать и совер-
шенно открытые пространства и даже окраины населенных пунктов. Осо-
бенно далекие переходы выводки крякв совершают в засушливых степных
областях. Эти переходы вызывают гибель значительного числа птенцов,
которые становятся добычей хищных птиц, а местами даже собак и кошек.
Постепенное сокращение среднего размера выводка крякв, по нашим наб-
людениям в поймах рек Шексны и Мологи (в 1940 г.), таково: яиц в кладке 11,
птенцов в выводке пуховых 9, крупных в августе до начала охоты 5, в сен-
тябре 4, в октябре ко времени объединения в стаи 3—4. По наблюдениям
в подмосковных охотхозяйствах, средний размер выводка в июне 6, а в июле
5 утят.
Линька. Кряквам свойственны две линьки: полная послебрачная
и неполная предбрачная. Полная смена оперения начинается у самцов с того
момента, когда самки уже окончательно садятся на яйца, а у самок—когда
их выводки поднимаются на крыло. Последовательность смены оперения
в общих чертах такова. Первыми выпадают черные косицы—закрученные
рулевые селезней. Затем начинается выпадение контурных перьев и появля-
ются пеньки нового оперения на шее, груди, брюшке, голове и подхвостье.
Потом линька захватывает верхнюю часть спины. Следующим этапом
бывает дружное выпадение маховых перьев. К этому моменту брюхо имеет
уже свежее оперение, в верхней части спины и на боках зимних перьев остает-
ся немного, появляются пеньки в нижней части спины и на подхвостье.
К тому времени, когда подрастают пеньки новых маховых, выпадают верх-
ние и нижние кроющие крыла. Когда новые кроющие крыла уже образуют
зеркало, линька головы и нижней половины тела почти полностью закан-
чивается, но по всей спине продолжается еще энергичная смена перьев.
Вслед за тем дорастают и разворачиваются маховые перья, в результате
чего утка вновь получает способность к полету. Заканчивается линька дора-
станием перьев спины и рулевых. Смена последних начинается на первых
этапах линьки и растягивается на длительный срок. Общая продолжи-
тельность линьки около двух месяцев. Период, в течение которого птица
теряет способность к полету в результате дружного выпадения маховых,
длится у кряквы 20—25 дней, в то время как период роста и полного раз-
вертывания этих перьев занимает 30—35 дней. Первый этап линьки до появ-
ления пеньков маховых проходит медленнее, чем дальнейший подрост
оперения.
Начиная с конца мая отбивающиеся от уток селезни соединяются в стайки
и большей частью покидают места гнездованья, отлетая на линьку. Линять
на месте остается очень незначительная часть селезней, да к тому же и далеко
не повсюду. Это наблюдается на Юго-Западной Украине (Киевщина, Пол-
тавщина), где, хотя и в небольшом числе, но постоянно линные селезни попа-
даются на заросших мельничных прудах и в поймах рек с ольховыми топями
и коблами (Гебель, Портенко). То же отмечается кое-где в болотистых пой-
мах рек Европейской России и Белорусского Полесья. Однако отовсюду,
в том числе и из перечисленных выше мест, основная масса селезней исче-
зает к моменту начала линьки и отлетает к местам массового скопления лин-
ных птиц. Районы эти, перечисленные выше, лежат обычно на путях мигра-
ции крякв к местам их зимовки.
Отлет селезней и дальнейшее продвижение их к районам линьки начи-
нается в южных частях ареала раньше, чем в северных. Так, на Рыбинском
водохранилище стайки селезней, пролетающих к месту линьки, наблюдаются
в первых числах июня. На местах отдыха они оставляют много рыжих перьев
с груди, косицы и разрисованные плечевые перья. На озерах Барабинской
степи кряквы начинают линять (терять маховые) в середине июня, массовая
линька наблюдается между 25 июня и 10 июля. С 15 июля первые птицы уже-
КРЯКВА 419
вновь поднимаются на крыло, а массовое окончание линьки приходится
на срок с 20 июля по 5 августа (Янушевич, 1947). В Кустанайской обл. (Фор-
мозов, 1937) селезни уже 25 мая теряют косицы, 30 мая у них проходит интен-
сивная линька контурных перьев, а с 9 июня начинается выпадение маховых.
Наконец, в дельте Волги по нашим наблюдениям первые стаи прилетных селе-
зней появляются уже 18—21 мая, а в начале июня они сбиваются в большие
табуны. В середине июня A6 июня) появляются первые нелетные птицы.
Массовая линька маховых наступает около 6 июля B4 июня—18 июля).
Первые птицы поднимаются на крыло около 22 июля, а в начале августа
наблюдается массовый выход из линьки, хотя отдельные нелетные птицы встре-
чаются и много позже (наиболее поздняя встреча 27 сентября 1938 г.). Срок
наступления каждого из этих биологических этапов чрезвычайно растянут,
что объясняется тем, что на линьку в дельте Волги собираются кряквы из
весьма различных географических мест, от низовий Оби и Печоры до степей
Зауралья и Западной Сибири.
Летние перелеты селезни совершают небольшими стайками по 10—15 птиц
но по прибытии на место стайки вскоре объединяются. К концу мая стаи
селезней в дельте Волги состоят уже из многих десятков птиц, а к середине
июня превращаются в многосотенные табуны. В первом периоде линьки,
когда сменяется лишь контурное оперение, огромные стаи крякв проводят
день на отмелях, а вечером они разбиваются на небольшие группы и разле-
таются на кормежку, с которой возвращаются только утром. Корма в это
время обильны, благодаря чему птицы быстро восстанавливают свой вес,
снизившийся за брачный период, и накапливают отложения жира.
Выпадение маховых перьев происходит дружно. Часто бывает так, что
у вспугнутой и взлетевшей утки после нескольких резких взмахов крыла
перья из него высыпаются, и птица падает на воду. С того момента, когда
селезни теряют способность к полету, они, как говорят, «залегают на линьку»,
а именно—уходят в хорошо укрытые места, в которых находят себе спасение
от наземных и пернатых хищников. По водоемам линные птицы распреде-
ляются весьма неравномерно. Они образуют массовые скопления далеко
не на всех озерах Барабинской степи, а из года в год на одних и тех же, так
же как и в строго определенных ильменях и култуках Волжской дельты.
Скопления линных птиц бывают иной раз настолько велики, что в расстав-
ленные сети удается загонять одновременно более 2000 уток. В дельте Волги
кряквы скапливаются чаще всего в зарослях рогоза, растущего в култуках
при глубине 15—40 см, а также в куртинах тростника, ежеголовки
или в затопленных тальниках в устьях рек. На озерах Западной Сибири
и Северного Казахстана убежищем для линяющих крякв служат трудно про-
ходимые заросли тростников. Весь день линяющие птицы проводят под защи-
той зарослей, отдыхая на заломах тростника и рогоза. Вечером они выплывают
кормиться на участки открытой воды с зарослями подводной растительности.
Так как скопления птицы бывают зачастую очень велики, то корма на бли-
жайшем расстоянии от мест дневки уток бывают ими в значительной мере
уничтожены. Редеют заросли валлиснерии, полностью бывают вырыты все
кусты стрелолиста, сострижены молодые листья частухи и других растений
со свежей зеленью. Утки начинают испытывать недостаток в полноценных
кормах, потребность в которых особенно велика от того, что разреженное
линькой оперение служит плохой термоизоляцией и потеря тепла организ-
мом возрастает. В результате этого утки за время второго периода линьки
снижают свой вес и иногда значительно (на 10% и более).
Холостые самки начинают линять почти одновременно с селезнями,
а затем к ним присоединяется некоторое количество уток, потерявших кладки
или выводки. В результате этого в районах линьки первое время скапливаются
только селезни, но потом постепенно увеличивается количество самок,
27*
420 ОТРЯД ГУСЕОБРАЗНЫЕ
которые к концу периода линьки уже составляют значительный процент.
Самки, находящиеся при выродках, линяют на месте и линька их начинается
позже. Контурное перо у них начинает выпадать только тогда, когда молодые
достигают уже размера 7з взрослой птицы, Так, на Наурзумских озерах
пеньки на брюхе у уток при выводках обнаружены 23 июня. В данном
случае мы не имеем в виду гнездовой пух, который выпадает у несущихся
самок в период откладки ими яиц.
Оперение, которое кряквы надевают в результате летней линьки, они
носят очень недолго—не более двух недель. Следующая, неполная, линька
начинается у селезней уже в августе или даже в конце июля. В октябре
большая часть селезней уже надевает новое брачное оперение. По некоторым
наблюдениям (Тугаринов, 1941), птицы, населяющие северные части ареала,
заканчивают осеннюю линьку раньше, чем кряквы, живущие на юге. Однако
делать этот вывод пока преждевременно, так как селезни из северных попу-
ляций откочевывают к югу еще летом, отчего судить о происхождении селез-
ней, добытых в осеннее время на юге страны, очень трудно. Последователь-
ность смены оперения довольно сильно варьирует у отдельных особей, но
в общих чертах она такова. Линька начинается с брюха, откуда она распро-
страняется в одну сторону на грудь и бока, а в другую на подхвостье. Вслед
за тем линяет шея и надхвостье, появляются пеньки на голове, ногах и в ниж-
ней части спины. Последними линяют передняя часть спины и плечевые пар-
тии перьев. Четкой приуроченности смены рулей к определенным этапам
линьки нет. Крылья осенью не линяют совсем. Взрослые самки начинают
линять позже, чем селезни, обычно не раньше начала сентября. К ноябрю
у них заканчивается смена контурного оперения и рулевых, но гнездовой
пух, значительно более темный, чем обычный, отрастает только в марте-
апреле, и, наконец, в апреле-мае дорастают плечевые перья. Точно так же
линяют и молодые птицы вывода данного года, меняющие свое ювенальное
перо на оперение взрослой птицы. Только сроки начала и окончания линьки
у них несколько запаздывают по сравнению со взрослыми особями того же
пола. Эту линьку молодые селезни крякв в южных частях ареала заканчи-
вают большей частью еще до отлета, а в более северных—на пути к зимовкам.
Смена оперения у самок, как говорилось выше, затягивается до зимы и после
некоторого перерыва продолжается ранней весной.
Питание. Питание крякв состоит в равной мере как из растительных,
так и из животных кормов. Пищу свою они добывают чаще всего на мелководь-
ях с глубиной до 30—35 см, где могут склевывать ее со дна, переворачиваясь
при этом вертикально вниз головой, но не ныряя. Особенно охотно кряквы
посещают для кормежки места, богатые водной растительностью, а в неко-
торых случаях выбираются и на сушу.
Ранней весной, когда лед еще не сошел, кряквы держатся на полыньях
в устьях мелких речек, и пищу их в это время составляют перезимовавшие
зеленые части водяных растений: элодеи, рясок, роголистника и осок. На
озерах Барабинской степи почти единственным кормом их первое время
по прилете служат семена и всходы Atrip lex pedunculata, Salicornia, Suedaa
corniculata, которые они собирают на разливах и лужах, наполненных
талой водой. Быстрый весенний подъем воды затрудняет кормежку растени-
ями, но зато обильные корма кряквы находят у уреза воды, где широкой
полосой скапливается смытый с затопленной территории мусор: семена раз-
личных растений, насекомые, их коконы и моллюски. Полузатопленные
пойменные гривы в средней полосе зачастую бывают заняты полями яровых.
На них кряквы собирают опавшие зерна хлебных злаков, чаще всего овса,
которые и составляют местами существенную часть их кормового рациона.
С первыми же теплыми днями, когда пробуждается животная жизнь
на мелководьях разливов и во временных лужах, животные корма начинают
КРЯКВА 421
преобладать в питании кряквы (от 60 до 97% по объему). Сперва птицы кор-
мятся листоногими рачками (жаброноги и щитни) и водяными жуками,
добывая их на временных лужах вне поймы, а затем перемещаются
на луговые разливы, где основу их рациона составляют личинки
стрекоз, комаров-толкунов (хирономид), жуки и моллюски, а на лесистых
озерах личинки ручейников. В Барабинской степи животные корма встре-
чаются в 65,6% желудков, причем первое место среди них принадлежит
насекомым, а из них личинкам хирономид. На втором месте стоят легочные
моллюски, затем ручейники и жуки. Из последних особую роль играют радуж-
ницы (Donacia), которых кряквы поедают как во взрослой стадии, так и, осо-
бенно, в стадии личинок (Федюшин, 1939). Очень важно значение радужниц
в питании кряквы и в других местах: в пойме р. Мологи и особенно в дельте
Волги. Такой разнообразный состав питания сохраняется у крякв в течение
почти всего теплого времени. В средней полосе ведущими кормами служат
личинки стрекоз, моллюски и разные группы жуков, на Рыбинском водо-
хранилище и Барабинских озерах личинки хирономид, а в дельте Волги
личинки радужниц, которых утки добывают из пазух листьев водяных расте-
ний. В озерных районах Северного Казахстана кряквы часто летом появляют-
ся в степи, где охотятся за многочисленными в это время прямокрылыми
(Формозов, 1937). Однако немалую роль в летнем питании кряквы, особенно
на юге, играют зеленые части растений: верхушки роголистника, ряска (пре-
имущественно малая), листья частухи, сусака, рдестов и многих других.
Селезни в тот период линьки, когда они не способны к полету и ведут скучен-
ный образ жизни, кормятся почти исключительно растительной пищей:
в дельте Волги побегами резухи, листьями валлиснерии и рдестов, а также
семенами этих растений; в Барабинской степи преимущественно рдестами,
местами и молодыми всходами тростника.
Пища пуховых птенцов на 83,4% состоит из животных кормов. Чаще
всего они склевывают корм с растений, возвышающихся над водой, и реже
с поверхности воды. В силу этого 35% (по объему) составляют личинки стре-
коз, взлезающие перед вылетом на стебли растений, 8% легочные моллюски
и 15% планктонные рачки. В конце июля и в августе в средней полосе, а на юге
в сентябре растительные корма начинают преобладать над животными.
Большое значение приобретают формирующиеся к этому времени зимующие
части растений: луковички стрелолиста, клубеньки гребенчатого рдеста
и сочные почки лягушатника, а местами и пузырчатки. Они составляют зача-
стую более 40% всего рациона. Тогда же созревают семена у рдестов, камы-
шей и осок, которые охотно и в большом количестве поедаются кряквами.
В некоторых местах они составляют осенью до 45% содержимого желудков.
Повсюду с началом созревания хлебов кряквы начинают регулярно летать
для кормежки на хлебные поля. Вылетая с вечера, они зачастую кормятся
целую ночь, возвращаясь на озера только к утру. Так посещают они поля
пшеницы, ржи, овса и проса, а на юге Приморья также посевы риса. Особен-
ное предпочтение они отдают, кажется, двум последним культурам и нередко
наносят им существенный ущерб на участках, расположенных поблизости
от водоемов. Иногда кряквы поедают мелких позвоночных животных, рыбью
молодь и мелких лягушек. Это, например, довольно регулярно имеет место
осенью во время пролета крякв через дельту Волги, а также на азербайджан-
ских зимовках, но корма эти для них не характерны и должны рассматри-
ваться как случайные.
Зимой в питании кряквы резко сокращается количество животных кор-
мов. В первой половине зимы на озерах Юго-Восточного Закавказья эти
утки кормятся преимущественно побегами водяных растений роголистника
и урути, а также семенами, среди которых большое значение имеют опавшие
семена риса (Верещагин, 1950). В результате зимнего недостатка раститель-
422
ОТРЯД ГУСЕОБРАЗНЫЕ
ности и выедания ее утками на местах их массовых зимних скоплений во вто-
рой половине зимы зеленые части растений в утиных желудках встречаются
редко. Основу питания (около 70% по весу) в это трудное для крякв время
составляют семена. На разливах в степи это преимущественно семена и всходы
различных солянок, а на озерах и приморских лиманах—семена куриного
проса, руппии, роголистника, гречишек, камышей и ежеголовок (последние,
однако, служат скорее в качестве жерновов при перетирании пищи, чем самой
пищей).
Хозяйственное значение кряквы велико. На большей части своего
ареала она служит одним из основных объектов спортивной, а местами
Рис. 76. Кряква (самец и самка)
и промысловой охоты. Последнее относится к области озерной лесостепи,
дельтам южных рек и районам зимовок. Так, на верхней Печоре кряква
составляет всего 5,3 % от общей добычи водоплавающих птиц, а на зимовках
в долине Куры от24,4 до 61,5% (Верещагин, 1950). Заготовки дичи в Ленкоран-
ском районе дают около 10 000 крякв за сезон, а в Западной Сибири—около
33 600, что составляет в первом месте около 37,2% от числа всех заготовлен-
ных уток, во втором 21,6%, а в дельте Волги в разные годы от 11,5 до 32,6%.
Более чем из 4000 уток, добытых в низовьях Днепра за 13 лет спортивной
охоты, кряквы составляют 10,1% (Пачосский, 1911). Чистый вес мясной
тушки кряквы составляет 69% от живого веса у селезня и 65% у утки, что
составляет для периода промысла 835 и 730 г.
О вреде, причиняемом кряквами в некоторых местах хлебным полям,
упоминалось выше. Наряду с этим они приносят пользу посевам уничтоже-
КРЯКВА 423
нием вредителей из числа прямокрылых и массовым поеданием семян вред-
ных сорняков риса. Дикая кряква—родоначальник разнообразных пород
уток. Очень легко она поддается одомашнению и в настоящее время. Нередки
случаи, когда дикие селезни живут в паре с домашними подсадными утками.
В Дарвинском заповеднике на Рыбинском водохранилище два диких селезня
за один сезон настолько привыкли к людям, что не только подпускали чело-
века на близкое расстояние, но и сами подплывали на зов для получения
корма (Куражсковский). Кряква охотно заселяет гнездовые ящики и во мно-
гих населенных пунктах становится полудомашней птицей (пруды заповед-
ники Аскания-Нова, пруды Московского зоопарка).
Полевые признаки. Крупная утка. У летящей птицы четко выде-
ляются белые полоски на крыле, окаймляющие зеркало. На полете
резко выделяется белый подбой крыльев. Самка как сверху, так и
снизу однотонно темная. У селезня темные голова, грудь и зад. У плыву-
щего селезня выделяются темнозеленая голова и белый ошейник, а у самки
оранжевый клюв и беловатый хвост. Полет очень шумный. Взмахи крыльев
сопровождаются частыми звонкими звуками «вить-вить-вить-вить», по кото-
рым крякву можно отличить даже не видя летящей птицы. Обычный голос—
тихое, слегка картавое крякание. При испуге оно более протяжно, а перед
взлетом тихое и торопливое. Осенний и зимний голос утки, подзывающей
самца,—громкое «куак-куак-куак-куак-куак». Во время ухаживания за
самками селезни издают высокий, но хриплый свист с помощью костного
барабана, который находится у них в нижней части трахеи (см. выше).
Описание. Размеры и строение. Клюв плоский, широкий с хорошо раз-
витыми гребнями роговых пластинок по краям. Одна или две пары средних рулевых, так
называемые косицы, имеют тонкие и мягкие стволы и загнуты вверх, образуя полукольцо.
Рулевых обыкновенно 9 пар. Длина самцов 575—630, самок 490—575 мм. Размах крыльев
самцов 925—1040 мм. Крыло самцов 265—295, самок 235—275 мм> цевка самцов 38—
50, самок 36—45 мм, клюв самцов 50-^58, самок 48—52 мм.
Рост кряквы продолжается, по всей вероятности, не один год, возрастные же отличия
у птиц старше одного года выражены слабо. Имеются, повидимому, и географические раз-
личия в размерах. Все это делает средние показатели мало достоверными.
Вес кряквы сильно изменяется по сезонам. Охарактеризовать его можно следующими
цифрами. Весной, по прилете на места гнездованья—в марте, апреле, начале мая,—селезень
весит в среднем 1150 г" A000—1410 г), а утка 1140 г (980—1242 г). За период размножения
кряквы заметно теряют в весе, особенно самки, общий вес яиц, снесенных ими, составляет
почти половину веса их тела. Средний вес селезня в конце мая и в июне 1100 г, утки 1050 г.
К началу линьки вес несколько возрастает, но в разгаре ее (конец июля—начало августа)
опять снижается в среднем до И 50 г A115—1450г) у селезней и 1020 г (951—1300 г) у уток. По
окончании линьки вес неуклонно возрастает и становится наивысшим в конце октября—
ноябре у селезней в среднем 1330 г A000—1700 г), а у уток 1240 г A000—1400 г). Зимой
кряквы, остающиеся на южнокаспийских зимовках, заметно падают в весе. Средний вес
селезней в декабре около 1150 г, в январе он падает до 950 г, а в феврале еще ниже; взрос-
лые утки соответственно весят в декабре 980 г, в январе 840 г и в феврале 805 г. В октябре
молодые птицы вывода данного года достигают веса взрослых. Сравнение среднего веса
крякв, добытых в дельте Волги и на Рыбинском водохранилище, показывает, что в одни
и те же месяцы как самцы, так и самки, как взрослые птицы, так и сеголетки в первом
пункте весят всегда больше, чем во втором. Весной эта разница равна 60 г, а с июня до
октября составляет 150—2иО г.
Окраска. Пуховой птенец. На спинной стороне темнооливкового цвета
с двумя парами желтовато-белых пятен на спине сзади крыла и с обеих сторон пояс-
ницы. Окраска брюшной стороны серо-желтая, переходит с возрастом в желто-пале-
вую, щеки более рыжего цвета. От верхней части клюва через глаз до затылка проходит
темная узкая полоска, в области уха темное пятно. Лапы оливково-серые, такого же
цвета и клюв с розоватым ноготком. Радужина темнобурая.
Молодые в первом пере. Очень схожи со взрослой самкой, но наствольные
пятна их перьев менее четки и образуют вдоль тела продольные полосы, в то время
как у старых птиц эти пятна более ограничены и округлы. Самцы отличаются, кроме
того, наличием струйчатого рисунка на некоторых кроющих второстепенных маховых
в области локтевого сгиба, в то время как у самок он заменен неправильными или попе-
речными буроватыми пятнами или полосками. Ноги грязнотелесного цвета или желто-
оранжевые с черноватыми перепонками.
424 ОТРЯД ГУСЕОБРАЗНЫЕ
У взрослой самки верх головы и затылок черноватые с узкими светлыми пе-
стринами, остальной верх, кроме крыльев, черно-бурый, причем каждое перо имеет
ржаво-бурую кайму и параллельные ей полосы, при выцветании светлеющие. Над глазом
проходит более светлая полоса, а через глаз темнобурая. Брюшная сторона однообразно
серовато-бурая с неясными продольными темнобурыми полосами или пятнышками. Горло
и шея спереди охристо - соловые без пятен. Рулевые беловатые с наствольными пят-
нами и крупными косыми бурыми полосами. Крылья сверху серо-бурые с блестящим фио-
летовым зеркалом, образованным наружными опахалами второстепенных маховых. Спере-
ди и сзади оно окаймлено черными и белыми полосами за счет черной предвершинной и бе-
лой вершинной полосок тех же перьев и обратного расположения их у больших кроющих
крыла. Внутренние второстепенные с буро-коричневыми наружными опахалами, третье-
степенные и большие маховые серо-бурые. Клюв оливковый, по бокам более желтый. На
нем всегда можно заметить несколько мелких черных пятнышек, в размещении которых
нет никакой закономерности. Эти пятнышки служат наиболее легким и достоверным приз-
наком для различения пола у птиц в летнем наряде и в периоде линьки. Ноги грязнооран-
жевые, радужина бурая.
Селезень в летнем пере. Схож по оперению с самкой, но отличается окра-
ской рулевых—светлосерых с широкими белыми каймами, иногда с небольшим числом
буроватых пятен. Обычно также зоб его более темен с коричневым налетом.
Взрослый селезень в брачном наряде. Голова и шея блестящие темнозеленые.
На шее они окаймляются белым ошейником, не смыкающимся сзади. Зоб и передняя
часть груди темнокоричневые. Остальная брюшная сторона серовато-белая с мел-
кими поперечными струйчатыми полосками. Передняя часть спины, лопатки и задняя
сторона шеи буровато-серые с мелкими еще более светлыми струйчатыми полосками.
Задняя часть спины черно-бурая, переходящая в сине-черное надхвостье, которое соеди-
няется с бархатисто-черным подхвостьем. Одна или две пары средних рулевых имеют мяг-
кие стержни и загнуты кверху в полукольцо (косицы). Остальные рулевые светлосерые
с широкими каймами наружных опахал. Плечевые беловатые со струйчатыми полосками.
Крыло с фиолетовым зеркалом, окаймленным спереди и сзади внутренней черной и наруж-
ной белой полосами, как это описано у самки. Размер зеркальца, повидимому, увели-
чивается с возрастом как в длину, так и в ширину. Клюв обычно зеленовато-оливковый,
с возрастом становится более желтым и приобретает даже оранжевый тон. Радужина
бурая. Ноги у вполне взрослого селезня кораллово-красные, у селезня, по первому году
живущего в паре самкой, киноварно-красные, у первогодков, не имеющих пары, они
оранжево-красные с темными (почти черными) перепонками. Другим отличием первогод-
ков служит черная окраска кожи между дужками нижней челюсти.
26. Желтоносая кряква Anas poecilorhyncha Forst.
Anas poecilorhyncha. F о г s t e г. Zoolog. Ind., 1781, стр. 23, Цейлон.
Русские названия. Желтоносая кряква—книжное, черная кряква—местное
в Уссурийском крае.
Распространение. Ареал. Крайний юг Советского Приморья, южная
часть Сахалина и южные острова Курильской гряды. Даурия, Маньчжурия,
Корея, все Японские о-ва, Западный, Южный и Центральный Китай, цен-
тральная и северная часть Индо-Китая, Бирма и Индия, кроме Северного
Кашмира, Филиппинские о-ва, Малые Зондские, Целебес, Ява и южная
часть Суматры. О-ва Фиджи, Соломоновы и Самоа. Весь материк Австралии,
Тасмания и Новая Гвинея.
Характер пребывания. К северу от Южной Кореи и Чжилий-
гкого залива—перелетная птица, всюду южнее оседлая.
Биотоп. Континентальные, пресноводные водоемы, обычно в низмен-
ных местах, богатые надводной растительностью. В горы поднимается весьма
неохотно, но в Южном Китае (Юннань) достигает высоты 2 700 м (Рили,
1926). Моря избегает даже зимой.
Подвиды и варьирующие признаки. Основная форма А. /?. poecilorhyncha F о г s t e г,
1781, населяет всю Индию, отсутствуя только в Западном Пакистане, Гималаях и Манипу-
ре. Доходит на восток до Западного Ассама, а на юг до Цейлона, где довольно редка.
Характеризуется большими желто-оранжевыми пятнами в основании надклювья и зеле-
ным крыловым зеркальцем; A. p. harringtoni О a t e s, 1907, отличается отсутствием жел-
тых пятен на надклювье, более светлым и менее испещренным низом тела. Крыловое зер-
кальце зеленое. Населяет Восточный Ассам, Бирму и Индо-Китай к югу, примерно до 17°
с. ш. На север доходит до Южного Китая (Юннань, верхнее течение Ян-Цзы), где граничит
ЖЕЛТОНОСАЯ КРЯКВА
425
со следующим подвидом. А. р. zonorhyncha SwI nhoe, 1866, отличается темным клювом
с желтой вершиной, сине-фиолетовым зеркальцем и сильно испещренным оперением. За-
селяет Советское Приморье, Корею, Японию и Китай. А. р. luzonica F г a s e г, 1839,
характеризуется одноцветной серо-коричневой окраской с черным верхом головы и поло-
сой, идущей через глаз. Зеркальце пурпурно-синее, клюв свинцово-серый. Форма эндемич-
ная для Филиппинских о-вов, где, однако, далеко не обычна. А. р. superciliosa G m e 1 i n,
1789, наиболее светлый и однородно окрашенный подвид. Населяет Австралию с прилегаю-
щими большими островами, Океанию и Зондские о-ва, кроме Борнео и северной части Сума-
Карта 47. Распространение желтоносой кряквы Anas poecilorhyncha
I—А. р. zonorhyncha, 2—A. p. harringtoni, 3—A. p. poecilorhyncha, 4—A. p.
luzonica, 6—A. p. superciliosa; a—граница гневдовий, б—не вполне выясненные
границы гневдовий, в—места изолированных гнездовий, в—зимовки (номиналь-
ного подвида), а—валеты номинального подвида
тры. На этой обширной территории возможно, однако, существование четырех подвидов:
собственно superciliosa G m e 1 i п на Новой Зеландии, А. р. rogersi M a t n e w s, 1912,
в Австралии и Тасмании, А.р. perenna R i 1 е у, 1919, на Целебесе и Яве и А. р. pelewensis
Hartlaub et Finsch, 1872, на Новой Гвинее и в Океании.
Черная кряква Anas poecilorhyncha zonorhyncha S w i n h о е
Anas zonorhyncha. S w i nhoe. The Jbis, 1866, стр. 394, Нингпо, Китай.
Распространение. А реал. Юг Советского Приморья; обыкновенна
на гнездовье в районе залива Посьет, где является одной из наиболее харак-
терных уток, не редка весной на озере Ханка, но в гнездовое время там мало-
численна, довольно редка и в бассейне Суйфуна. Севернее, в долине Уссури,
черная кряква совсем не гнездится, но встречена летом в среднем течении
Амура на Зейско-буреинской низменности (Радде, 1863; Штегман, 1928).
Встречается летом также в Даурии на Аргуни, Ононе и Дарасуне, хотя и до-
вольно редко. Там были даже добыты пуховые птенцы, что заставляет счи-
426 ОТРЯД ГУСЕОБРАЗНЫЕ
тать черную крякву гнездящейся птицей Даурии. К сожалению, эти наблю-
дения относятся к концу прошлого столетия A873—1876), а более новых,
подтверждающих их данных нет. Пролетных уток этого вида Пржевальский
наблюдал на озере Далай-Нор, встречены они в районе Буир-Нура (Иванов,
Банников). Таким образом, Восточная Монголия, повидимому, входит в ее
гнездовой ареал, а на юго-востоке этой страны она даже не редка. Наиболее
западные находки: р. Чикой 14 мая 1896 г., р. БураЗОавгуста 1914 г. (Юго-
Западное Забайкалье) и озеро Богоносор в бассейне Селенги 15 июня (Северо-
Западная Монголия), не следует рассматривать как гнездовые. Южнее известны
три находки из северо-восточного Гансу в июне и сентябре и с северного
склона Нань-Шаня, между Сачжоу и Ганьчжоу A4 апреля 1890). Эти пункты
показывают крайние западные нахождения вида в южной части его ареала.
Обычная гнездящаяся птица Маньчжурии, в значительном числе гнездится
в Восточном Китае от Чжили до Хубэя, Шаньси, Гуаньдуня и на Тайване.
Единственная гнездящаяся утка Кореи и обычнейшая утка всех Японских
о-вов до Риу-Киу на юге, причем в северной части архипелага она обычней,
чем в южной. Гнездится в южной части Сахалина, а по японским данным—
и на южных островах Курильской группы.
Характер пребывания. В северной части Японских о-вов,
Северной Корее, Маньчжурии и в Приморье черная кряква—перелетная
птица, а всюду южнее ведет оседлый образ жизни. Сезонные миграции в основ-
ном не выходят за пределы гнездового ареала подвида, только отдельные
птицы достигают зимой Филиппин и даже долины Ганга, что, однако, скорее
относится к категории залетов, как и находки под Томском и на Олекме
(в бассейне Лены).
Даты. В Советском Приморье появляется несколько позже других
уток, обычно в конце первой половины апреля (Воробьев). На озере Далай-
Нор, вМонголии, первые были встречены 14апреля (Пржевальский), а в Япо-
нии на местах гнездовья появляется уже в феврале и держится до ноября.
Биотоп. Наиболее характерны для черной кряквы открытые озера
низменности с обилием водяной растительности и наличием тростника. Гор-
ных местностей и лесов, повидимому, избегает, но не боится населенных
пунктов там, где ее не трогают. Так, на прудах в одном из парков Токио
зимует вместе с другими утками около сотни черных крякв.
Численность. Обычная, а местами и многочисленная утка низ-
менностей в районе залива Посьет, всей Японии, Кореи, равнинной Мань-
чжурии и западной части Китая. На всей остальной территории черная кряк-
ва на гнездовье редка и встречается спорадично.
Экология. Размножение. С момента прилета птицы всегда встре-
чаются парами, так что, повидимому, оформление пар происходит зимой,
а по Хейнроту—уже осенью. Брачные игры во всем схожи с таковыми
у обычной кряквы. Единственное отличие состоит в том, что оперение головы
не принимает при этом столь плоскую«прическу», как у кряквы (Лоренц, 1942).
Селезни этого вида весной столь же настойчиво преследуют чужих самок,
как и селезни кряквы. Сезон гнездованья в Южном Китае с середины апреля
и весь май (Годвэл). В Японии в конце мая птицы с птенцами, так что гнездо-
вое время падает на апрель. В Маньчжурии у Харбина 3 полные кладки
обнаружены 26 мая—2 июня 1935 (Ямасина). В Советском Приморье вы-
водки появляются в середине июля (Воробьев), под Владивостоком пуховики
отмечены 17 июля (Медведев).
На нашей территории гнезда до сих пор не найдены и не описаны. Гнездо,
найденное на р. Сунгари в районе Рыбачьих озер около Харбина, находилось
в болотистой местности, среди больших и густо заросших травой кочек на бе-
регу озера. Оно было свито очень искусно из вейника (Calamagrostis Langs-
dorfi) и выстлано пухом. Наружный диаметр гнезда 25—27 см, лоток 15—16,
ЖЕЛТОНОСАЯ КРЯКВА
427
его глубина 8 см> 16 июня 1925 г. в гнезде было 7 сильно насиженных яиц—про-
клевышей (Яковлев, 1927). В других случаях насиженные кладки под Харби-
ном содержали 7,10 и 12 яиц (Ямасина, 1939). Размеры яиц: 52,2—58,2X
Х39—44 мм. Окраска сходна с таковой у кряквы, с голубоватым оттенком;
несколько резче, чем у названного вида, заостряются к острому концу (по эк-
земпляру Зоологического музея Московского государственного университета).
Срок насиживания 24 дня (Остин, 1948). В Японии в подходящих местах
охотно гнездятся совместно—колониями. Так, на маленьком болотистом тро-
Рис. 77. Черная кряква
стниковом островке в одном из парков Токио гнездилось совместно много
дюжин пар. Одиночками же встречаются всюду на заросших тростником
ручьях между рисовыми полями (Ян, 1942).
Никаких сведений о линькеио составе питания нет.
Полевые признаки. По общему облику и манере держаться напоминают
крякву, но селезни сразу отличаются отсутствием яркого брачного оперения.
Для обоих полов характерны белые пятна на крыльях, образованные кон-
цами третьестепенных маховых, видимые и у плавающей птицы и во время
полета.
Описание. Размеры и строение. Внешне напоминает крякву, но несколько
меньше размером и имеет относительно более слабый клюв. Вес 0,75—1,5 кг. Крыло сам-
цов 256—293, самок 240—265 мм; цевка самцов 40—46, самок 38—43 мм; клюв самцов
48—60, самок 47—53 мм.
Окраска. Пуховой птенец. На спинной стороне оливково-бурый с дву-
мя парами желто-бурых пятен, расположенных (как и у кряквы) у основания
крыльев и у поясницы. Того же цвета бока головы и шея. Полоска от основания клюва
через глаз буроватая, грудь серовато-бурая, остальной низ грязно охристо-желтый.
Взрослая самка. По общему характеру оперения напоминает самку обычной
кряквы. Перья головы рыжевато-бурые на затылке с широкими, а на щеках с узкими
черными стержневыми и концевыми пятнами, отчего верх головы более темный. Такая же
широкая полоса от основания надклювья к глазу, а над ней светлый штрих. Низ головы
палево-белый. Вся спинная сторона темнобурая, как у кряквы; бока несколько светлее,
428
ОТРЯД ГУСЕОБРАЗНЫЕ
надхвостье черноватое, низ грязнобелый с серо-бурыми пятнами, более крупными на
груди. Крыло сверху серо-бурое с фиолетово-синим зеркальцем, ограниченным и спереди
и сзади черными внутренними и белыми наружными каемками. Третьестепенные маховые
серо-бурые с белыми каймами по наружному опахалу. Рулевые грязнобелые с серо-
бурыми широкими поперечными полосами. Ноги оранжевые, радужина коричневая.
Клюв черный с желтой вершиной и темным ноготком. Иногда две светлые полоски сверху
вниз у основания клюва.
Взрослый самец. Характер окраски тот же, что и у самки. Зоб тускло бело-
вато-охристый с темными пестринами. Грудь к брюху постепенно темнеет, брюхо корич-
нево-бурое, подхвостье черно-бурое. Подбой крыла и подмышечные белые (как и у самки).
Крыло, как у самки, но белые каемки выражены слабее, обычно только задняя. Особого
летнего наряда самец не имеет.
Молодые птицы. Имеют более светлую окраску. Белая передняя кайма зерка-
ла выражена всегда хорошо.
27. Серая утка Anas strepera L.
Anas strepera. Li nnaeus. Syst. Nat., изд. X, I, 1758, стр. 125, Швеция.
Синоним. Anas cinerea. G m e 1 i n. Reise d. Russl., II, 1/74, стр. 184, Южная
Россия.
Русские названия. Народное, кроме того, серуха, серушка (широко распро-
страненное), полукряква полуматерая (Европейская Россия), семенуха (местами, Повол-
жье и Каспий), нерезень (Херсонская обл.).
Распространение. Гнездовой ареал. Занимает Европу и Азию
от Британских о-вов до Приамурья, к северу до Рыбинского водохранилища,
Карта 48. Распространение серой утки Anas strepera
1—Л. s. strepera, 2—А. а. couesi; a—граница гнездовий, а'—районы наибольшей численности птиц
на гнездовье, б—не вполне выясненная граница гнездовий, в—обособленные гнездовые колонии,
«/—места постоянных встреч летом и возможного гнездованья, в—зимовки, д—валеты; е—встречи
пролетных птиц весною севернее гневдовых мест, ж—места скоплений птиц на линьку
устья Иртыша, Красноярска и 60 °с. ш. в бассейне Лены, к югу до Кашгарии,
Сеистана и Дарданелл. Северная Америка от Тихоокеанского побережья
до Великих озер. Острова группы Фаннинг в Тихом океане.
Основные зимовки находятся на побережьях Северного и Сре-
диземного морей, в Египте, Иране, Индии и Южном Китае, а также в
южной части Северной Америки.
Биотоп. В гнездовое время внутренние водоемы со стоячей или мед-
ленно текучей водой, чаще всего богатые растительностью. Во время мигра-
СЕРАЯ УТКА 429
ций и на зимовках не избегают и морских побережий, где держатся,
однако, только в заливах и лагунах.
Характер пребывания. Перелетная птица, кроме нескольких
участков на юге ареала и о-вов Фаннинг, где оседла.
Подвиды и варьирующие признаки. Существует два подвида серой утки, считавшие-
ся обычно самостоятельными видами. Номинальный подвид A. s. strepera Linnaeus
1758, заселяет всю Голарктическую часть ареала; Л. s. couesi Streets, 1876—принадле-
жит островам группы Фаннинг (о-в Нью-Йорк и о-в Вашингтон), находящимся в Тихом
океане под 6° с. ш. и 160° в. д. В настоящее время этот подвид, повидимому, следует счи-
тать вымершим.
Серая утка Anas strepera strepera L.
Распространение. Гнездовой ареал. Исландия, Юго-Восточ-
ная Англия, Голландия (очень редка), Дания, Швеция, Венгрия, северная
часть Югославии, Болгарии и Румыния. Кроме того, изолированные гнездо-
вые колонии известны в Южной Испании (Гвадалквивир), Южной Франции
(Лионский залив, устье Роны) и на озере Феццара в Алжире (Цедлитц, 1914).
В пределах Советского Союза северная граница ареала определяется следую-
щими пунктами. В Прибалтийские республики серая утка лишь залетает,
и возможно отдельные пары гнездятся на Эстонских о-вах (Гроссе и Транзее,
1929), то же относится и к Псковскому озеру, где однажды A1 мая) добыта
самка с готовым к сносу яйцом (Зарудный, 1910). Достоверных сведений
о гнездовье в Белоруссии мало, повидимому серуха там очень редка и распро-
странена спорадично. В южной части Витебской обл. она известна местным
охотникам (Федюшин, 1926), под Гомелем и Речицей изредка гнездится
(выводок подРославлем;Станчинский, 1929); спорадично и очень редко гнезди-
тся не юге Смоленской обл. (Станчинский, 1915), очень редка под Москвой
в Калужской, Тульской, Рязанской и Ивановской обл. Единично гнездится
на Рыбинском водохранилище (Дарвиновский заповедник); из Горьковской
обл. точных сведений нет, но гнездится около Малмыжа на р. Вятке (Плес-
ский, 1927). На Урале около Свердловска редка, но в Зауралье и Западной
Сибири ее ареал далеко выдается на север. Она гнездится под Тюменью,
Тобольском и доходит в небольшом числе до устья Иртыша. Далее граница
спускается до северных пределов лесостепи, т. е. до Венгеровского и Куй-
бышевского районов Барабы, Ачинской и Абаканской степи; по Чулышу
не встречена, под Красноярском бывает нерегулярно, и гнездованье там не
установлено (Шухов, 1925). Восточнее .граница не выяснена, так как на Анга-
ре, в верховьях Нижней Тунгуски и в Тункинской долине серая утка не встре-
чена, но она не редка на гнездовье в средней и северной части Байкала
(Котельниково, Усть-Баргузин, устье Верхней Ангары; Туров, 1923; Штег-
ман, 1936). Моллесон A895) отмечает эту утку для Юго-Западного Забайкалья
гнездится она в Даурской степи и по Аргуни (Штегман, 1928). Из Приамурья
достоверных сведений нет; встречена Миддендорфом на Становом хребте
под 56° с. ш., добыта под Олекминском, обнаружена летом (в 20-х числах
августа) в нижнем течении Амура на озере Эворон (Воробьев) и летом же
на Камчатке, однако эти данные не доказывают гнездованья серой утки на
Дальнем Востоке. Для Приморья она редкая залетная птица. В Маньчжурии
она, хотя и редко, но несомненно гнездится, находки ее гнезд известны на во-
сток до Харбина (Яковлев, 1939). В Монголии гнездится в долине Толы, на
юго-восточной окраине Ханганя и, повидимому, на озере Орок-Нор (Козлова,
1930), в Убса-Нурской котловине, на озере Ачит-Нур и возможно в Кашга-
рии, но на Лоб-Норе отсутствует. Западнее границей регулярного гнездова-
ния служит Горное Семиречье и Памир, на котором серая утка появляется
лишь во время пролетов (Мекленбурцев, 1946). Гнездится в низменных
430 ОТРЯД ГУСЕОБРАЗНЫЕ
частях Таджикистана (Иванов, 1940) и далее к западу всюду до Гиндукуша,
обычна в бассейне Геррируда и редка на разливах Гильменда в Сеистан-
ской котловине. Гнездится по южному берегу Касция и повсюду на Кавказе
и в Закавказье, как на низменностях, так и на озерных плато. Достоверных
сведений относительно Малой Азии нет.
На Американском континенте северная граница ареала проходит от о-ва
Ванкувер по водоразделу рек Атабаска и Саскачеван к озеру Манитоба; далее
к югу до западного берега озера Верхнего, нижнего течения Миссури и вер-
ховьев Красной реки, затем граница огибает с севера плато Колорадо и выхо-
дит к вершине Калифорнийского залива. На западе серая утка гнездится
до побережья.
Основные зимовки. В большом числе серые утки зимуют
у Британских о-вов и у берегов Северного моря от Голландии, также в Бий-
ском заливе. Еще большее значение имеют средиземноморские зимовки,
особенно многочисленные в Адриатическом море от Венецианского залива
до Балканского п-ова. В Африке они зимуют на побережье к северу от Атласа
и в Египте и залетают единично до Абиссинии, Судана и Красного моря
(Гроте). Зимуют в Сирии, Трансиордании и Ираке, причем в последнем это
наиболее обычная зимующая утка (Мейнерцхаген, 1924). В Иране она обычна
зимой у южного берега Каспия и в Сеистане, в других же частях страны
довольно редка. Обычна в подходящих местах Пакистана, но избегает
горных частей страны (Тайсхерст, 1927); обычна и в Индии, не доходит до
Майсура и Тенасерима и Верхней Бирмы. Массовые зимовки серой утки в
пределах Советского Союза находятся на Каспийском море, в Юго-Восточном
Закавказье. Талынская низменность и Прикуринские степи у западных бере-
гов Туркмении и в низовьях Атрека. Меньше их зимует на Азовском море,
у устья Кубани и на побережье Аджарии. В значительном числе серые утки
проводят зиму в долине Мургаба, Теджена и меньше по верхнему течению
Аму-Дарьи и на озере Иссык-Куль. На востоке Азии серые утки зимуют
в Китае к югу от устья Хуанхэ. В Америке зимовки расположены в примор-
ской зоне от о-ва Ванкувер и мыса Гаттерас к югу до Центральной Мексики
и в долине нижнего течения Миссисипи.
Скопления линных птиц характерны для озерных районов
степи и лесостепи. Они известны в дельте Волги, на озерах к югу от
р. Илека, в системе Иргиза и Тургая до озера Челкар-Тениз, на озерах Кур-
гальджин-Тенизской группы, на Наурзумских озерах, в Курганской обл.
(Макушино), в Барабинской и Кулундинской степях. Кроме того, линные
птицы добывались в дельте Или. За последние годы серые утки стали соби-
раться на линьку на Рыбинском водохранилище у северного предела ареа-
ла вида.
Характер пребывания. Повсюду в северных частях ареала.
Но перелетна в Англии, в Средиземноморье и, повидимому, в Иране оседла.
Сезонные миграции и их даты. Как показали резуль-
таты кольцевания, на Британских о-вах зимуют исландские утки, а по всей
вероятности также птицы, населяющие Германию, Польшу и северо-запад-
ную часть Советского Союза. Однако основная масса населяющих нашу тер-
риторию серых уток имеет тяготение к южным зимовкам. На линьку в дельте
Волги собираются селезни этого вида с широкой территории от Наурзумских
и Челябинских озер на востоке до Надеждинска Свердловской обл. на севере,
Чистополя (Татария), Самарской луки и Ульяновска на западе. Линяющие
в дельте Волги утки зимуют преимущественно на юго-западном берегу Кас-
пийского моря и в меньшем числе на его восточном берегу, в Иране, Малой
Азии, на Черном море и на Балканах (Болгария).
Серые утки из Барабинской степи (Купино, Татарка), Северного Казах-
стана (Наурзумские озера) и быть может Челябинской обл. летят прямо
СЕРАЯ УТКА 431
к югу и проводят зиму в низовьях Инда (Пакистан) и у Сринагара (Кашмир).
Пролет их идет через Аральское море, низовья Сыр-Дарьи и Бухару.
С мест зимовки на Британских о-вах серые утки отлетают с марта до кон-
ца апреля (Уайзерби); пролет в низовьях Мологи наблюдался 22—24 апреля
(Исаков и Распопов, 1949); у Малмыжа (р. Вятка) появляются во второй
половине апреля (Круликовский, 1913). В северо-западной части Черного
моря появляются в первой декаде марта (Клименко, 1950), в низовьях Днепра
первые отмечены 8 марта (Пачосский, 1911), в Аскании-Нова пролетные наб-
людались в разные годы 24 февраля—31 марта (Шарлемань, 1924). Прилет
под Днепропетровском во второй половине марта (Вальх, 1899). Из Паки-
стана зимующие утки этого вида исчезают к началу апреля, но немногие задер-
живаются и до середины мая; довольно сильный пролет по Геррируду
проходил еще 20—23 апреля, но одновременно там держались и местные
особи, уже расположившиеся для гнездования. Начало пролета в Юж-
ной Туркмении (Дорт-Кую) отмечено 14 марта, а около Учаджи он про-
должался с начала и до второй половины марта (Зарудный, 1896);
в нижнем течении Сыр-Дарьи серые утки появляются в первой декаде
марта, к концу месяца они становятся очень многочисленны, а к сере-
дине апреля остаются только гнездящиеся особи (Спангенберг). На озерах
Атрека к 6 апреля эти утки остаются лишь одиночками (Житников, 1900),
по Западному Узбою заметный пролет проходит во второй половине марта
(Шестоперов). В дельте Волги появление их наблюдалось в разные годы
24 февраля—14 марта (Исаков). Под Гурьевом первые отмечены в половине
марта, на Аральском море пролет стаями по 20—30 птиц наблюдался еще
в конце апреля (Бостанжогло, 1911). Под Чкаловом первые появляются
в конце марта, а пролет продолжается до конца второй декады апреля (Заруд-
ный, 1895); на Эмбе первые отмечены 17 апреля, а в низовьях Иргиза 16 ап-
реля (Сушкин, 1908). В Барабе начало пролета приходится на апрель, а мас-
совый пролет на середину мая (Янушевич). На озере Орок-Нор (Северная
Гоби) большие стаи серых уток наблюдались с 4 апреля до последних чисел
мая (Козлова, 1930); в низовьях р. Или прилет их наблюдается обычно
около 25 марта A2 марта—7 апреля; Слудский).
Осенний пролет серых уток проходит обычно малозаметно, нередко
без образования больших стай. В верховьях Тобола и Иргиза они исчезают
в начале октября, хотя запоздалые птицы наблюдаются и много позже. Под
Чкаловом большая часть местных птиц отлетает уже в середине сентября.
В дельте Волги первые пролетные стаи появляются около 20 сентября, мас-
совый пролет проходит 5—20 октября и отлет основной массы птиц 24—27 ок-
тября; под Днепропетровском они становятся очень редкими после 23 октября,
на Аджарское побережье они прилетают в начале декабря и держатся там
до начала марта (Вильконский, 1896). На азербайджанских зимовках мас-
совое появление серых уток приходится на ноябрь; пролет по Западному
Узбою был наиболее заметен в первых числах октября; в низовьях Атрека
утки были многочисленны уже в первой половине октября, но сильный лет
их по ночам продолжался с 25 октября по 21 ноября. В низовьях Сыр-Дарьи
число этих уток начинает возрастать уже с конца августа за счет птиц, подле-
тающих с севера; держатся они там до середины ноября; в Пакистане появля-
ются уже в половине сентября и в октябре. В низовьях Или отлет наблюдался
15—16 октября (Слудский).
Б и о топ. Предпочтительно водоемы, богатые водной растительностью;
лиманы по долинам рек и неглубокие, камышистые степные озера. Пресные
водоемы явно предпочитаются соленым; последние заселяются, если вдоль
берегов имеется густая трава и тростники. Гнездится часто в сухой степи
в отдалении от водоемов, а также вылетает в степь на кормежку. Зимует в за-
болоченных низменностях, особенно охотно на затопленных участках степи
432 ОТРЯД ГУСЕОБРАЗНЫЕ
Численность. Территория, на которой эта утка обычна в каче-
стве гнездящейся птицы, довольно четко ограничена пределами лесостепи
и степи—от Венгрии до низменного Семиречья и нижнего течения Сыр-Дарьи.
Кроме того, серая утка обычна в бассейнах Мургаба и Теджена, вдоль южного
берега Каспия и в Закавказье. На остальном пространстве ареала она зна-
чительно реже, а в горных лесистых частях Алтая отсутствует вовсе.
Во многих пунктах северней основного ареала вида серая утка регулярно
встречается во время пролетов. Кое-где в этой зоне она задерживается на лето,
а отдельные пары в некоторые годы гнездятся (Эстонские о-ва, север Белорус-
сии, нижнее течение Оби, р. Сым в Туруханском крае, Становой хребет^
низовья Амура, Камчатка). Нерегулярные залеты известны и много дальше.
Численность серой утки в различных районах сильно меняется по годам.
Так, в 20-х годах нашего столетия она резко сократилась на севере Киров-
ской обл. (Плесский, 1927); в Винницкой же обл. она в первом десятилетии
нашего века настолько возросла в числе, что стала обычнее кряквы (Герх-
нер, 1928); в низовьях Днепра серая утка обычно очень редка, но в некоторые
годы появляется в большом количестве (Пачосский, 1911). Изменение чис-
ленности у границ ареала связывается с еще более резкими изменениями ее,
происходящими на основной гнездовой территории вида. Можно предполо-
жить, что периоды сильного усыхания степных озер вызывают передвижения
серых уток к окраинам ареала и за его пределы, что иногда приводит к рас-
ширению ареала. Расселение по территории Швеции произошло после мас-
сового залета в 70-х годах (Цедлитц, 1925), в Англии гнездится с 1850 г.,
ряд местностей там заселился в 1912—1918 гг., а другие в 1936—1938 гг.
(Уайзерби, 1941).
Экология. Размножение. Половая зрелость наступает, вероятно,
в годовалом возрасте. Однако большое число птиц, задерживающихся на пути
с зимовья, говорит о том, что некоторая часть уток в размножении не уча-
ствует, скорее всего это годовалые особи. На места гнездовья птицы прилетают
уже в парах, но в брачных играх около каждой самки зачастую принимает
участие несколько самцов. Токовые позы самцов заключаются главным
образом в плавании с вытянутой кверху шеей и подергивании головой вверх
и вниз. Движения эти сопровождаются своеобразным каркающим криком
и вздергиванием поднятого и развернутого веером хвоста, при котором демон-
стрируется бархатно-черное оперение подхвостья. Весьма обычны у серых уток
воздушные погони двух или трех самцов за самкой, во время которых селезни
иногда летают особым токовым полетом (описанным нами для некоторых
других уток), при котором они время от времени на одно мгновение пере-
стают работать крыльями, отчего корпус их принимает наклонное положение.
Спаривание происходит на воде.
Гнезда, как правило, строятся на земле, причем нередко далеко от воды.
Так, на Нижнем Дону (ст. Курмаярская) найдено было гнездо в типчаковой
степи не менее чем в 800 м от водоема, а Слудский в полынно-злаковой степи
(Кургальджин) нашел гнездо не менее чем в 2 км от воды. Гнезда строятся
или под кустами, часто колючими (боярышник, терн, джингиль), или в кусти-
ках полыни и бурьянов, или же совсем открыто. Открытые гнезда нередко
находятся около гнезд чаек, крачек, кречеток и других куликов. В Барабин-
ской степи два гнезда были найдены на затопленном остатке стога, они нахо-
дились в 34 см одно от другого среди 13 гнезд обыкновенных крачек. На самом
севере ареала, на Рыбинском водохранилище, гнезда серой утки находились
на открытых островах под прикрытием спускающихся низко ветвей малень-
ких елочек. Наконец, в дельте Волги, вернее в ее приморском участке,
где высокие паводки не позволяют строить наземные гнезда, редкая
там серая утка поселяется в заломах тростника или изредка в расположен-
ных низко над водой гнездах кваквы.
СЕРАЯ УТКА
433
Как правило, все гнезда устраиваются на сухой почве и представляют
собой аккуратную ямку глубиной 11—15 см. Ямку утка роет самостоятельно
и выкладывает сухой травой тщательнее, чем другие утки, так что иногда
появляются валики, возвышающиеся над почвой. Гнездо обильно выклады-
вается пухом, который размещается по его стенкам, а во время отсутствия
самки используется ею для укрытия яиц. Диаметр гнезда 20—30 см, а лотка
его 18—20 см.
Количество яиц в полной кладке обычно 9—11, но колеблется от 6 до 12
и даже 14 (не сдвоенные ли кладки?); в Барабинской степи 9—12, в дельте
Или 11, на Кургальджине 7—9, на о-ве Орлове (Черное море) 9—Н(Кузякин).
под Кишиневом, на Хопре и под Москвой найдены гнезда с 9 яйцами, на Ры-
Рис. 78. Позы тока серой утки
бинском водохранилище с 6—7. Откладывается ежедневно по одному яйцу.
Яйца белые, слегка желтоватого или оливкового оттенка. Размеры их 51—59 X
Х36—42,2, среднее54,6 X39,2 мм (Барабинская степь); измерения яиц из дру-
гих пунктов укладываются в эти цифры; вес ненасиженных яиц 39,8—43,8,
среднее 42,1 г. Насиживание начинается с откладки последнего яйца, про-
должительность 27—28 дней. Самки сидят очень крепко, особенно в конце
насиживания. Иногда они подпускают на расстояние 1 м (те, у которых гнезда
под кустами). Вес только что выведшихся молодых около 32—33 г. В гнезде
птенцы остаются менее суток, а затем совершают переход к воде, иногда
очень длинный (до 1 км\ Янушевич). Выводки нередко объединяются, наблю-
дались группы до 35 утят при трех сопровождающих их самках (Формозов,
1937). Выводки, особенно первое время, терпят большой урон от пернатых
хищников: Камышевых луней и крупных чаек.
С мая—июня по первые дни августа погибает около 20 % молодняка (Миас-
ское озеро; Ушков, 1947). Способность к полету молодые получают в возра-
сте около 55 дней (Хейнрот).
В Барабинской степи откладка яиц происходит в основном с 15 мая по
10 июня, после 15 июня ненасиженные кладки попадаются уже редко. Время
появления молодых приходится на 15 июня—6 июля, а подъем молодых
на крыло на 5 августа—1 сентября (Янушевич и Золотарева, 1947). НаНаур-
зумских озерах 25 мая селезни гонялись за самками, среди которых были
еще и одиночные. В начале июня кряканье селезней стало реже, и последний
раз оно было слышно 12 июня. Гнездо с еще неполной кладкой G яиц) найдено
28 мая, а первый выводок еще очень мелких утят отмечен 12 июня (Формозов,
1937). На озере Кургальджин насиженная кладка найдена 12 июня, вывод
в одном гнезде наблюдался 4 июня; еще неполные кладки A—5 яиц)
продолжали встречаться 19—24 июня, вероятно повторные (Лавров,
28 Птицы СССР, т. IV
434 ОТРЯД ГУСЕОБРАЗНЫЕ
1930). В низовьях р. Или неполные кладки встречались в разные годы 8—
14 мая, а уже насиженные с 13 апреля, первые встречи выводков относятся
к первым числам июля, но на ряду с пуховичками встречались и крупные
птенцы размером с горлицу (Слудский). На Сыр-Дарье кладка, найденная
12 июня, была еще ненасиженной (Спангенберг); на о-ве Орлове (Черное море)
16—17 июня одна самка уже плавала с выводком пуховиков, но остальные
еще сидели на яйцах (Кузякин), под Кишиневом вывод из одного гнезда
наблюдался 27 июня (Остерман), а на Рыбинском водохранилище—11 июля,
другое же гнездо с сильно насиженными яйцами найдено уже 28 мая.
Линька. Дважды в год: летом полностью, осенью и зимой частично.
Селезни и самки, не имеющие выводков, начинают линять рано, сразу же
вслед за кряквами, одновременно счирком-трескунком. В дельте Волги неко-
торые стаи их теряют способность к полету уже в конце июня, и в некоторые
годы в первой половине июля A2 июля в 1935 г.) можно встретить селезней,
полностью надевших летнее перо. Примерно те же сроки наблюдаются и в
Барабинской степи. Там начало линьки (выпадение маховых) приходится на
третью декаду июня, массовая линька этого вида проходит между 20 июля
и 20 августа (Янушевич и Золотарева, 1947). На Кургальджине к 8 августа
среди линных птиц они были уже единичны (Меженный и Соколов). На Наур-
зумских озерах табуны линных селезней наблюдались 11 июня и 2 июля,
самец, добытый 7 июля, только что начал линьку контурного оперения, 17
и 22 июля ловились селезни без маховых (Формозов, 1937). Линька начинаете»
со смены оперения на груди особенно на боках ее, у основания крыльев,
плечевых и надхвостья. Спина линяет позже, маховые также в конце периода
линьки, рулевые же сменяются весьма постепенно и подрост их продолжае-
тся у перелинявших (в основном) птиц. Самки с выводками начинают
линять много позже: самка, добытая на Рыбинском водохранилище
27 июня, еще не имела пеньков нового наряда, а 13 июля расправляющиеся
новые перья обнаружены только на груди и зобу. Линька самок, повиди-
мому, бывает менее полной.
Следующая, предбрачная, линька начинается у селезней в августе,
а в середине октября некоторые из них имеют уже в основном зимний наряд
(дельта Волги, Кургальджин). Однако полностью кончается подрост опере-
ния в ноябре, а у некоторых особей затягивается и дальше. У взрослых самок
смена оперения, почти затихающая зимой, вновь активизируется в марте.
В это время сменяется часть покровных перьев (во всяком случае непол-
ностью на спине), часть рулевых (иногда лишь средняя пара), внутрен-
ние второстепенные маховые и их кроющие. Весной же отрастает гнездо-
вой пух. Молодые птицы начинают свою первую линьку в конце сентября,
а некоторые лишь в октябре, и заканчивают ее к марту. У селезней сменяются
часть линных перьев, рулевые и внутренние второстепенные маховые. У моло-
дых самок линька проходит в основном так же, но внутренние второстепен-
ные маховые и рули сменяются часто неполностью.
Линяющие серые утки обычно не образуют столь больших табунов, как.
свиязи и шилохвости, и, утратив способность к полету, ведут скрытный образ
жизни: день они проводят в зарослях тростников или рогоза и только по вече-
рам выплывают кормиться на ближние плесы.
Питание. Главным образом зеленые корма, в основном листья и побеги
растений, чаще всего водяных, но также и нежные злаки. Животные корма
могут приобретать существенное значение только летом, но не везде.
На весеннем пролете в устье Мологи серые >1ки кормились побегами элодеи,
на Рыбинском водохранилище во все сезоны они питаются только зелеными,
частями растений (99,5—99,9%). В мае очень большое значение играет трех-
дольная ряска, в июне—августе нежные, стелящиеся по воде листья простой
ежеголовки F6,6%), а в октябре листья блестящего рдеста. Животные корма
СЕРАЯ УТКА
436
(мелкие плавунцы и мелкие ручейники) не имеют какого-либо значения и,
вероятно, поедаются вместе с листьями, на которых они сидят. В дельте
Волги весной (март-апрель) зеленые части растений составляют 88,4% по объе-
му всей пищи. В основном это корневища и нежные листья валлиснерии
B5,3%), побеги роголистника A6,3%) и нитчатые водоросли A5,6%). Семена
составляют 11,3%, это главным образом резуха водяная и гречиха, а живот-
ные корма всего 0,3% (плавунцы). Во время линьки кормится исключительно
зеленью, в основном ветки резухи E0,0%) и листья валлиснерии C5,7%).
Рис. 79. Серая утка
По наблюдениям в Наурзумском заповеднике (Северный Казахстан), огром-
ное значение в летнем питании многих уток, а том числе и серых, имеют саран-
чевые насекомые. Утки поедают попавших в воду прямокрылых и вылетают
стайками в степь за 1,5—2 км от озера, где охотятся за кобылками. Даже
птенцы весом от 26 до 150 г имеют в желудках остатки саранчевых, главным
образом пруса Calliptamus italicus (Формозов, 1937). В низовьях Или (в нача-
ле июля) наблюдались регулярные вылеты селезней (стаями до 50—60 птиц)
в степь на участки, пораженные саранчей (Слудский). Ко времени созревания
посевов пшеницы, гречихи и особенно проса, а местами также и риса, стаи
серых уток начинают регулярно посещать поля и кормятся там от зари до
зари. Излюбленным кормом серых уток в начале осени служат клубеньки
гребенчатого рдеста и нежные луковички стрелолиста. На мелководье утки
достают их из грунта, отчего на местах кормежки образуются довольно
28*
436 ОТРЯД ГУСЕОБРАЗНЫЕ
большие воронки (Барабинская степь; Формозов, 1934). На зимовках (Лен-
корань—Муганская степь) во второй половине зимы серые утки иногда ощу-
щают недостаток в зеленых кормах и поедают большое количество семян
(81,4%по весу), преимущественно злаков, атакжеруппии, рдестов и камышей.
На зимовках в Северной Индии серые утки часто пасутся на луговинах, соби-
рая многочисленных там в это время прямокрылых и других насекомых.
Хозяйственное значение серой утки относительно не велико, так как
достаточно многочисленна она лишь на ограниченной территории нашего
Союза. Однако кое-где на юге она имеет немалый удельный вес в добыче
ружейных охотников. В дельте Волги в разные годы она составляет от
7 до 21% от общего числа добываемых уток. Качество мяса их очень
высокое.
Полевые признаки. Утки среднего размера; окраска головы и туловища
на расстоянии кажется однотонной. Хорошо видно белое зеркало как налету,
так и у сидячей птицы. У селезня черный зад. Полет легкий, с воды может
взлетать прямо вверх. Плавая, держит низко грудь, а зад приподнимает
над водой. «Говорливая» утка. На полете ее легко отличить по частому покря-
киванию, полет шумный, но не настолько, как у кряквы. Голос самки тре-
скучее, и часто повторяемое кряканье несколько более высокое и резкое,
чем у кряквы. Голос самца несколько напоминает крик ворона. Крупных
стай обычно не образуют.
Описание. Строение и размеры. Клюв небольшой, несколько короче голо-
вы, одинаковой ширины от основания до вершины. Хвост из 16 рулевых, обрез хвоста зак-
ругленный. Длина крыла самцов 250—280, самок 228—255 мм; цевка самцов 38—43,
самок 35—41 мм; клюв самцов 40—45, самок 37—42 мм.
Вес взрослых самцов на весеннем пролете в марте-апреле 807 г F05—1000 ш)
(дельта Волги), 665—825 г (дельта} Или; упитанность бывает здесь ниже средней), 797 г
G30—910 г) (Барабинская степь), 800 г (Рыбинское водохранилище). После брачного
периода в июне /00 г (Ала-Куль; начавший линьку, но еще летный). Линные в начале
августа 913 г (800—1000 г) (дельта Волги). На осеннем пролете в сентябре 907 г (900—
920 г), в октябре 958 г (900—1300 г) (дельта Волги), 800 г (Узбой). Взрослые самки весной
в марте-апреле 716 г F20—786 г) (дельта Волги), 750 г (дельта Или), в мае 770 г (Бара-
бинская степь); после откладки яиц в июне 500 г, в июле 717 г (Рыбинское водохра-
нилище). В начале линьки в июне 900 г (Кургальджин), линные в августе 854 г
(800—1000 г) (дельта Волги), перелинявшая 780 г (Архангельск). На осеннем пролете в сен-
тябре 850 г, в октябре 869 г (800—1000 г) (дельта Волги). Молодые птицы: в августе самец
745 г, в сентябре самка 800 г (Рыбинское водохранилище), в сентябре самцы 797 г E00—
900 г), самки 792 г G00—1000 г), в октябре самцы 895 г G00—1000 г), самки 823 г G00—
900 г) (дельта Волги). На зимовках (без учета возраста птицы) в январе очень суровой зи-
мой самцы 500—600—830 г (Азербайджан, озеро Кара-Су), в феврале самцы 800 г B)
(Красноводский залив), самцы 707 г D70—860 г), самки 663 г E62—780 г); в начале марта
самцы 655 г E60—800 г), самки 562 г D70—620 г).
Окраска. Пуховой птенец. Чрезвычайно похож на одновозрастного
птенца кряквы (см. описание). Некоторыми отличиями служат темное пятно на голове
у разреза клюва и более четкая бурая полоска через глаз, а также более желтый оттенок
пуха на щеках и горле.
Молодые птицы в первом наряде. В общем похожи на взрослую самку, но отличаются
от нее более сильно испещренной нижней стороной тела (кроме структуры рулевых). Раз-
личия у разных полов намечаются уже в этом наряде. У самцов темя почти чисточерное
с легким зеленоватым отливом; тон окраски верха темнее, основание шеи и передние пле-
чевые в узких поперечных охристых пестринах, перед спины и надхвостья матово-черные
без каемок. У самки темя черноватое, испещренное мелкими штрихами. Окраска верха
тела светлее, перья спины, лопаток и основания шеи с охристыми каемками, но без
поперечных полосок. Развитие каштанового цвета на кроющих крыла сильно варьирует
индивидуально.
Самка взрослая. Окраска спинной стороны темнобурая с ржавыми (при
выцветании белеющими) каемками и поперечными полосками на перьях. Передняя часть
груди и бока более ржавые в продольных пестринах; остальная брюшная сторона с неяс-
ными бурыми пестринами, более частыми на брюшке. Окраска крыльев в общем та же, что
и у самца, но меньше ржаво-каштановых кроющих и белого на маховых. Первые нередко
даже совсем отсутствуют. Клюв желтоватый, иногда с ржавым оттенком и черный по
коньку; по краям надклювья всегда имеются мелкие черные точки, расположенные
в беспорядке. Лапы грязножелтые с темными перепонками; радужина бурая.
СВИЯЗЬ 437
Самец годовалый. Отличается от взрослого более тусклой буроватой окрас-
кой спины, серой окраской на некоторых белых перьях зеркала и бурыми вершинами
некоторых перьев брюха.
Взрослый самец в брачном наряде. Голова буровато-серая в мелких темных
крапинках, более темная сверху. Зашеек и бока тела серые с черным струйчатым
рисунком. Спина серовато-бурая, темнеющая к надхвостью. Последнее так же, как и под-
хвостье, бархатисто-черное. Подбородок и шея с винным налетом, переходящим на шее
в рыжеватый. Зоб и верх груди черновато-серые с чешуйчатым рисунком, который соз-
дается скобовидными черными и белыми каймами на перьях. В задней части груди рису-
нок утрачивается, она почти белая с мелкой поперечной струйчатостью. Первостепенные
маховые серые, из второстепенных же первые 4 пепельно-серые, следующие с черной кай-
мой по наружнему опахалу, затем чистобелые и самые внутренние серые; подмышечные
белые; верхние кроющие вдоль сгиба крыла сероватые, средние ржаво-каштановые, боль-
шие бархатисто-черные. Рулевые серые с белыми вершинами. Клюв аспидно-серый, лапы
желтые с темными перепонками; радужина бурая.
У самца в летнем наряде зашеек, бока тела и нижняя часть груди без струй-
чатого рисунка, а бурые с более светлыми каймами перьев, как у самки. Плечевые
бурые, нижняя часть спины и поясница остаются матовочерными, низ тела сильно
испещрен округлыми пятнами.
28. Свиязь Anas penelope L.
Anas penelope. Li nnaeus. Syst. Nat., изд. X, 1, 1758, стр. 126, Швеция.
Синоним. Anas cogolea. G m e 1 i n. Reise d. Russl., 1, 1770, стр. 70, Южная Россия.
Русское название. Свиязь—народное, кроме того, широко распространенные
свияга, свистун по голосу селезня.
Распространение. Гнездовой ареал. Исландия, Фарерские
острова (?), Шотландия к северу от Глазго и местами Англия, где гнездится,
однако, не регулярно. В Дании изредка отдельными парами, в Голландии
лишь случайно. В Северной Германии гнездится регулярно в Мекленбурге
и непостоянно в Бранденбурге, а также в Северной Польше, по всей Нор-
вегии, Финляндии и в Швеции, кроме наиболее южных провинций. В пре-
делах Советского Союза гнездится на север до Кольского п-ова (Хибины,
р. Тулома), но для Восточного Мурмана пока не отмечена. Под Архан-
гельском одна из самых обычных гнездящихся уток (Паровщиков), гнез-
дится на Канине (Дементьев, 1935), в Тиманской тундре (Гладков, 1951)
и изредка на Колгуеве. По Печоре доходит до устья, редка на Вайгаче, на
Ямале прослежена до Юрибея F9° с. ш.), где гнездится (Дунаева и Осмолов-
ская), но возможно, что идет и дальше; встречается в устье Таза (Бутурлин,
1934), доходит до Енисейского залива. На Таймыре гнездится как в лесистой,
так и в тундровой зоне, по Лене и Хатанге за пределы лесов не выходит,
по Яне не известна севернее устья Адычи, южнее же по реке обычна. На Инди-
гирке самая редкая из всех настоящих уток, встречается до границы леса
(Михель, 1935), так же и по Колыме, где в лесной полосе обычна (Тайер
и Бэнгс, 1914). Гнездится по всему Анадырскому краю к северу до Анадыр-
ского хребта (Портенко, 1939) и залива Креста (Белопольский, 1933), причем
здесь она может считаться обыкновенной, а местами даже многочисленной
уткой. Гнездится по всей Камчатке, на о-ве Карагинском и Гижиге. Обычна
в верховьях Колымы (Дементьев и Шохин, 1939) и в районе Охотска (Хари-
тонов, 1915). На Командорских о-вах бывает лишь на пролете. Повидимому,
гнездится в северной части Сахалина, но достоверных данных о гнездовании
в Приморье нет. Есть яйца якобы этого вида из-под Мариинска (Шренк,
1861); указание Маака A861) о встречах в июне с птенцами на Уссури.
Южную границу ареала последить еще сложнее. Гнездится, хотя и редко
в Калининградской обл.; нов Прибалтийских Советских Республиках на гне-
здовье отсутствует, если не считать нескольких случайных гнезд на Эстон-
ских о-вах. В небольшом числе гнездится в Псковской (Зарудный, 1910)
и Новгородской обл. (Бианки, 1910). Достоверных данных о гнездовании
в Белоруссии нет; очень редко и спорадично гнездится в Смоленской обл.
438
ОТРЯД ГУСЕОБРАЗНЫЕ
(Станчинский, 1915, 1924), очень редко под Москвой, указывается для Туль-
ской обл. (Мензбир, 1879), но гнездованье там лишь случайное; редка и спо-
радична в Ивановской обл., в Горьковской известна лишь одна гнездовая
находка (Формозов, 1935). Гнездится в Татарии, но ниже по Волге очень
редка, доходя до Пензенской и Ульяновской обл. Указывалась для дельты
Волги (Яковлев, 1874; Хлебников, 1928, 1930), но в более позднее время све-
дения эти не подтверждены. Гнездится в области Сыртов, по Волжско-Ураль-
ской степи не найдена (Бостанжогло, 1910; Волчанецкий, 1937). Есть указа-
ние на гнездование в Курской обл. (Рязанцев, 1928), в степной части Крыма
(Шатилов, 1860; Молчанов, 1906) и на о-ве Бирючьем (Воронцов, 1937);
однако эти районы можно рассматривать только как пункты случайного гне-
Карта 49. Распространение свиязи Anas репе lope
а—граница гнездовий, а'—районы наибольшей численности птиц на гнездовье, б—не вполне выяс-
ненная граница гнездовий, в—места постоянных встреч летом и спорадичного гнездованья, г—зи-
мовки, в—валеты, е—встречи на пролете за пределами ареала, э*с—места скоплений птиц на линьку
здованья задержавшихся на пролете стай. Обыкновенна в среднем течении
Урала от Уральска до Орска, а также по Илену, Хобде и многим степным
озерам (Зарудный, 1888, 1896); далее гнездится до среднего течения Эмбы
и низовьев Тургая. Обычна под Семипалатинском, в Западном и Южном Ал-
тае, в Юго-Восточном Алтае очень редка, а в Северо-Восточном не найдена
совершенно (Сушкин, 1938). На Зайсане, повидимому, очень редка; гнездится
в Северо-Западной Монголии, в районе озера Ачит-Нур, в Тувинской авт. обл.
и в котловине Убса-Нур. Найдена на гнездовье в Тункинской долине (Слуд-
ский), в Забайкалье по р. Чикою (Моллесон, 1895) и на Байкале (Туров,
1923), в Даурии известна лишь на пролетах. Из Приамурья никаких сведений
нет, на этом участке южная граница распространения совсем не ясна. Изо-
лированным районом гнездования является южное Закавказье. Здесь свиязь
гнездится в Армении в районе озера Севан, в Грузии—на озерах Ахалкалак-
ского плато (Чхиквишвили, 1930) и, повидимому, единична в горах Аджарии
(Вильконский, 1896). Во многих пунктах южнее очерченного нами ареала
летом удается встречать группы или пары эгих птиц, а в отдельных случаях
даже выводки их. Однако явление это следует расценивать как случаи задерж-
СВИЯЗЬ 439
ки птиц на местах их зимовки или на пути пролета, иногда заканчивающиеся
гнездованьем отдельных пар. Сюда мы относим летние встречи и гнездованье
свиязей в Голландии, Венеции, Южной Франции, Крыму, Литве, Курской
обл., под Тулой и Пензой, на Сыр-Дарье, в Северном Пакистане, Кашгарии,
в Маньчжурии и быть может на Нижнем Амуре.
Основные зимовки находятся в Северном море от берегов Англии
до Южной Швеции, Дании и Франции. Несколько меньше птиц проводит
зиму на внутренних водоемах Юго-Западной Европы. В массе зимуют свиязи
на побережьях Средиземного моря как на европейском, так и на африкан-
ском берегах. На юге эти утки достигают Мадейры, Канарских и Азорских
о-вов, северо-западного побережья Африки, Хартума, Белого Нила, Абис-
синии и Южной Аравии, но южнее тропиков залетают только немногие экзем-
пляры. В значительном числе зимуют в Малой Азии, Сирии, Ираке, причем
в последнем месте многочисленна; повсюду в Иране, кроме центральной,
пустынной, части, но особенно многочисленна у берегов Каспийского моря и
в Сеистане. Обычны свиязи зимой в Пакистане вплоть до Мекранского бере-
га; довольно обычны в Индии, но не южнее Майсура, а также Бирме, Ассаме
и Тенасериме. Обычны в зимние месяцы в Тонкине и Центральном Аннаме
{Делакур и Жабуйль, 1931); регулярно зимуют на Филиппинах (Делакур
и Майр, 1946); в большом числе зимуют всюду вдоль побережья Юго-Вос-
точного Китая и особенно на крупных водоемах внутри страны от Хэбэя
к югу до Хайнаня; в Японии обычнейшая зимняя утка, скапливается более
всего на рисовых площадях о-ва Хондо (Ян, 1942). В пределах Союза основные
и наиболее массовые зимовки находятся в Юго-Восточном Закавказье, на во-
доемах Муганской степи и в Туркмении, на Юго-Восточном Каспии в долине
Атрека. Довольно значительные зимовки находятся также на Азовском море
и относительно малочисленные на Мургабе, Теджене и в верховьях Аму-
Дарьи.
Места скопления птиц на линьку находятся глав-
ным образом в озерной зоне лесостепи и степи от устья Волги на Западе до
Барабинской степи на востоке. Кроме того, свиязи линяют на Зауральских
озерах (Курганская обл.) и возможно Уральских (Каслинские, Кыштымские);
на Кургальджине они составляют основную массу линяющих уток; линяют
в массе на Наурзумских озерах (Кустанайская обл.) и на озерах в системе
Иргиза и Тургая. За последние годы стаи свиязей линяют на Рыбинском
водохранилище; отлавливали линных птиц в верховьях Печоры (Ныробский
район Молотовской обл.), но велики ли там скопления их—неизвестно; ли-
няют они в устьях Оби как примесь к стаям шилохвостей; бывают скопления
линных свиязей в озерном районе Якутии и по р. Турухану. Значительные
скопления известны в бассейне Анадыря и Пенжины. Кроме того, некоторое
количество селезней линяет на местах гнездовья в одиночку или маленькими
стаями (р. Сура, р. Сым в Туруханском крае и др.).
Характер пребывания. Всюду, кроме Британских о-вов,
правильно перелетная птица.
Сезонные миграции и их даты. В результате кольцевания
свиязей известно, что на западноевропейские зимовки в Данию, Голландию
и на Британские о-ва отлетают утки, населяющие северо-запад, север, а частью
и центральные районы европейской части Союза (верховья Днепра, Дона
и Волги). В тех же местах проводят зиму и некоторые из птиц, гнездящихся
на Урале (Свердловская обл.) и в нижнем течении Оби (Ханты-Мансийск).
На зимовки эти утки летят как непосредственно с мест гнездовья, так и из
района линьки их в дельте Волги. Таким образом, осенний прилет их имеет
западное, а местами ясно выраженное северо-западное направление, что хо-
рошо подтверждается непосредственными наблюдениями (Рыбинское водо-
хранилище). Однако часть птиц, помеченных во время их миграций в Голлан-
440 ОТРЯД ГУСЕОБРАЗНЫЕ
дии, не остается зимовать на севере Европы, а откочевывает затем к юго-западу
во Францию и Испанию до Гвадалквивира.
Основная масса свиязей, гнездящихся в Западной и частично в Средней
Сибири (до Иркутской обл.), а также часть птиц из Северного Казахстана
зимует на Каспийском, Азовском и Средиземном морях. В последнем месте
они встречаются чаще всего на северо-восточном побережье, в Болгарии,
Греции и Северной Италии (Венеция) и не западнее Лионского залива. На
зимовки они следуют чаще всего через дельту Волги, где селезни проводят
время линьки. Наконец, часть птиц из Обь-Иртышской Сибири (Северная
Бараба, Тобольск, р. Кеть), а вероятно и Казахстана отлетает прямо к югу
и зимует в долине Инда и Северной Индии, в Мадхьебхарате. Относительно
западно-европейских популяций свиязи интересно отметить, что птицы, насе-
ляющие Британские о-ва, в своей основной массе оседлы, но часть их (пре-
имущественно молодые) осенью вылетает на континент Европы, проникая
к югу до Португалии и Венецианского залива. В нескольких случаях околь-
цованные молодыми на Британских о-вах обнаружены затем в весеннее гнез-
довое время на территории Советского Союза (под Уральском, на р. Вятке
и в Кировской обл. и на р. Ижме в Коми АССР). В еще большей степени это
относится к исландским птицам, которые встречены у нас под Ленингра-
дом, в Архангельской обл., под Каргополем и в Чешской губе, под Тулой,
Воронежем, в Свердловской обл. и в устье Иртыша. Основные зимовки их
находятся на Британских о-вах и на территории Западной Европы, но неко-
торые птицы достигают Американского побережья (от Новой Шотландии
до Северной Каролины) (Вучетич, 1939; Михеев, 1949 и др.).
Отлет зимующих свиязей из Англии и Шотландии происходит с середины
марта до конца апреля, но в большинстве случаев прилетные птицы наблю-
даются там до середины мая, а случайно даже и в июне (Уайзерби, 1945).
В Эстонии свиязь наблюдается на пролете в апреле, но численность ее в раз-
ные годы бывает весьма различна (Кох, 1911). На Псковском озере появляется
в начале или в середине апреля, а валовой пролет проходит обычно со второй
половины апреля до первых чисел мая, однако, в некоторые годы она еще про-
должает встречаться довольно часто до конца мая. Некоторые стаи содержат
до тысячи штук (Зарудный, 1910). Под Москвой свиязь весной малочислен-
на, зато на Верхней Волге в устье Мологи пролет интенсивен. Первые птицы
отмечены там 20 апреля, а валовой пролет наблюдался 22 и 23 апреля; в север-
ной части Рыбинского водохранилища первые встречи отмечены 15 и 21 апре-
ля; массовое появление 16—26 апреля. Прилет под Архангельском (озеро
Слободское) 9 мая B4 апреля—17 мая) (Паровщиков), в Тиманскую тундру
25 мая (Гладков), на Кольский п-в (озеро Имандра)—26 мая B апреля—31 мая)
(Владимирская, 1948). По Печоре (устье Ылыча, где свиязи пролетают в массе)
первое появление приходится обычно на 2 мая B8 апреля—8 мая), а начало
валового пролета на 4 мая, средняя продолжительность около 26 дней (Теп-
лов, 1948).
Отлет с черноморских зимовок (Джарылгачский и Егорлыцкий заливы)
заканчивается обычно в половине марта, но иногда затягивается почти до кон-
ца апреля (Клименко, 1950). Начало валового пролета под Киевом отмечено
30 марта (Данилович, 1930), по Суре и Алатырю в начале апреля (Житков
и Бутурлин, 1906), а в Татарии 25 апреля. Иран свяизи покидают в начале
марта, в Пакистане же задерживаются до апреля (отдельные птицы встречались
в июне; Тайсхерст, 1922, 1927). В Азербайджане пролет свиязей начинается
в первой половине марта; в дельте Волги появление их отмечалось 10—13 мар-
та. В низовьях Атрека с 1 марта число свиязей стало возрастать за счет про-
летных с юга птиц, через два дня они уже встречались в массе, в ночь
на 18 марта наблюдался лет их к северу, а к 6 апреля этих уток осталось всего
несколько стай (Житников, 1900). По Узбою свиязь пролетает небольшими
СВИЯЗЬ 44]
стайками во второй половине марта (Шестоперов). В Таджикистане пролетает
в начале апреля (Иванов, 1940); на Памире весной очень редка и наблюдалась
только 4—6 мая (Мекленбурцев, 1946). Пролет через Семиречье слаб, в сред-
нем течении Или ее наблюдали парами и мелкими стаями с 13 марта по 2 мая
и только один раз стаей в 50 шт. (Кореев и Зарудный, 1906); в низовьях Или
много свиязей наблюдалось 25 апреля (Слудский). В долине Нижней Сыр-
Дарьи свиязь на весеннем пролете встречается в значительном количестве;
запоздалой весной первые птицы появились 13 марта, а валовой пролет про-
шел с 20 по 24 марта, бродячие стайки попадались и летом (Спангенберг и Фей-
гин, 1936). На Аральском море в заливе Сары-Чеганак пролет проходил в кон-
це апреля (Бостанжогло, 1910); в среднем течении Эмбы первые стайки заме-
чены 8 апреля, а валовой пролет шел в середине апреля и прекратился в конце
месяца (Сушкин, 1908); по Уралу под Чкаловом валовой пролет идет в пер-
вой половине апреля, а запоздалые стайки задерживаются до середины мая
(Зарудный). На Наурзумских озерах массовый пролет наблюдался 22—27 ап-
реля (Михеев, 1937); пролет под Тобольском 25 апреля A8 апреля—1 мая)
(Тарукин, 1929). На р. Елогуе (приток Енисея, 63° с. ш.) первые свиязи отме-
чены 15 мая (Слудский).
В Индо-Китае свиязи встречаются до марта; примерно тогда же они отле-
тают и с южнокитайских зимовок, в Чиили на пролете редки и бывают в апреле.
Через Корею пролетают в значительном числе (Остин, 1948); в Приморье
редкости весной не представляет, но все же довольно малочисленны в течение
всего апреля и в первых числах мая (Воробьев). На Восточной Камчатке
(Кроноки) на весеннем пролете очень многочисленны; первые встречи заре-
гистрированы в разные годы 21 апреля*—4 мая, а массовый пролет с первой
декады мая до конца месяца (Аверин, 1948); весеннее появление на Анадыре
17—18 мая (Сокольников). В верховьях Колымы интенсивный пролет свиязи
наблюдался около 25 мая (Дементьев и Шохин, 1939), прилет под Нижне-
Колымском отмечен 5 июня (Тайер и Бэнгс, 1914), а на Индигирке в устье
Момы 1—2 июня (Михель, 1935). В Северной Гоби (озеро Орок-Нор) свиязи
встречены 4 апреля, а в Забайкалье нар. Чикое добыта 25 апреля, в Даурии
наблюдались в мае. Начало пролета на р. Илиме (приток Ангары)—апрель,
с ледоходом (Косыгин, 1926); в Якутии прилет отмечен 5 мая на Амге (Мид-
дендорф) и 11 мая под Якутском (Воробьева, 1931).
На Анадыре свиязи исчезают в самом конце сентября; осенний пролет
на Восточной Камчатке идет в сентябре и заканчивается в середине октября.
Через Приморье пролетает во второй половине сентября и в октябре. По-
следняя встреча под Якутском 21 сентября. Массовый пролет в Тункинской
долине наблюдался в последних числах сентября и в первых числах октября
(Слудский), на р. Ононе в октябре начало пролета в бассейне р. Толы (юго-
восточная окраина Хангая) в середине августа и конец его в начале октября.
Зимовок в южном Китае (Западный Сы-Чуань) достигают в октябре-ноябре,
а в Индо-Китае в ноябре—декабре. На р. Сосьве (левом притоке Оби) встреча-
лись до 20 сентября (Портенко, 1937); на Верхней Печоре (устье Ылыча)
осенний пролет начинается около 30 августа B5 августа—3 сентября),
продолжается около 33 дней, причем иногда птицы летят стаями до 200—
300 шт.; последние птицы отмечались 10—22 октября; под г. Кировом свиязи
задерживаются примерно до 20 октября (Плесский, 1933); под Архангель-
ском (озеро Слободское) осеннее табунение начинается около 9 сентября
A—15 сентября), а массовый отлет приходится на 15—25 сентября. Начало
пролета на Рыбинском водохранилище отмечалось 19 августа—31 сентября,
конец массового пролета 25—28 сентября, а последние птицы 26—29 октября
(Немцев). На Псковском озере она показывается в значительном числе иногда
уже в середине августа, в первых числах сентября делается обыкновенной;
особенно многочисленна она в конце сентября и до середины октября; про
442 ОТРЯД ГУСЕОБРАЗНЫЕ
лет через Эстонию происходит в сентябре. На британские зимовки свиязи
прибывают с конца августа до конца ноября.
Осенний пролет под Чкаловом идет с середины августа и затягивается
до конца октября; в верховьях Иргиза он падает на вторую половину сен-
тября и продолжается в октябре. На озерах Узбоя одиночные птицы проле-
тают в течение всего октября; на Атреке они появляются около 20 октября,
к концу месяца уже бывают в массе, но интенсивный лет их продолжается
до конца ноября; в январе их на зимовках становится меньше, так как часть
птиц отлетает к югу (Житников, 1900); у Гасан-Кули они появляются в на-
чале октября; в Северном Пакистане в сентябре, а на Мекранском берегу
первые птицы встречены 3 октября.
По Хопру на пролете обычна, начинается пролет 15 сентября, а стано-
вится массовым в октябре (Измайлов, 1940); в дельте Волги первые пролетные
стаи появляются около 28 сентября, массовый пролет приходится на 10—
15 октября, а отлет основной массы птиц на 21—26 октября.
На азербайджанских зимовках свиязи собираются в массе в ноябре, но
первые стаи появляются рано, и в августе уже достигают Ирака. Под Киевом
начало пролета наблюдалось 16 сентября, а валовой пролет 22 сентября;
в низовьях Днепра единичные птицы появляются в первых числах сентября,
в более значительном количестве в первой же половине этого месяца и оста-
ются до глубокой осени (Пачосский, 1911); наконец, на черноморских зимов-
ках (северо-западные заливы) первые стаи оседают в начале сентября.
Биотоп. В гнездовое время озера, богатые водной растительностью
и небольшими свободными плесами. Как чересчур заросших тростниками
озер, так и озер с обширными плесами свиязи избегают; в степи местами
бывают и на соленых водоемах. Избегают гор, особенно лесистых. На зимов-
ках концентрируются в обширных заболоченных низменностях, день про-
водят на взморье или больших озерах, а по ночам кормятся на залитых
водой лугах, полях или мелководных водоемах.
Численность. Распределение свиязи внутри ареала крайне нерав-
номерно, места обилия ее чередуются с территориями, на которых вид редок.
Свиязь вполне обычна на гнездовье в Средней и Северной Финляндии, лес-
ной части Кольского п-ова, набольшей части Карелии, в северной части бас-
сейна Северной Двины и Печоры. Обычна свиязь в верховьях Камы и ее
притоках—Шишере и др. Основная территория относительного обилия
свиязи находится в Обь-Иртышской низине, на север, от тундры на юг до
северных частей Сибирской лесостепи, степных озер Зауралья и Актюбин-
ской обл., на запад до Урала, на восток через бассейн Таза к ниж-
нему течению Енисея, и, повидимому, захватывает лесотундровую полосу
Таймыра. Более чем в других местах обычна свиязь в озерной лесостепи
Ачинск—Красноярск; в северной половине Байкала и верхнем течении
Нижней Тунгуски, в озерной низменности Якутии; повидимому, на Колы-
ме, Камчатке и Анадыре. Объяснить такое распределение районов обилия
вида трудно. Можно отметить лишь, что на низменностях они обильнее,
чем в горных районах, избегают сплошной темной тайги, предпочитая ред-
колесье и криволесье (исключением является Обский бассейн).
Экология. Размножение. Время наступления половой зрелости
точно неизвестно. Вероятно, что часть особей гнездится на первом году жиз-
ни, но длительные задержки целых стаек на местах зимовок и в пути указы-
вают, что часть популяции в размножении участия не принимает; следует
предполагать, что это молодые птицы. На места гнездовья птицы прилетают
уже парами, которые несколько более стойки, чем у других уток. Селезни
свиязи чужих самок не преследуют. Их брачные игры довольно однообразны.
Селезни плавают вокруг своих самок с распушенным оперением головы
и взъерошенными удлиненными перьями лопаток. Голова у них несколько
СВИЯЗЬ 443
откинута назад при горизонтальном положении клюва. Время от времени
селезень вскидывает голову кверху и издает громкий и резкий свист, пере-
даваемый слогами «вви-у». При наличии свободных самцов, еще не имеющих
пары, между селезнями иногда бывают драки.
Гнезда устраиваются обычно около небольших озерков и речек, с заро-
слями кустов по берегам, прибрежной растительностью, небольшими откры-
тыми плесами. При отсутствии подобных мест селится и у водоемов иного
характера. В тундре и лесотундре свиязь строит гнезда близ берегов озер
и речек, под кустами ивняков на опушках их зарослей. Гнезда обычно хорошо
укрыты, особенно, если помещаются среди прошлогодней травы, нависшей
на ветках кустов (Тиманская тундра), или же находятся среди ольховников
под прикрытием их обломанных сучьев с засохшими листьями (р. Щучья;
Южный Ямал). В зоне северной темнохвойной тайги гнезда свиязей находили
в елово-кедровом лесу, недалеко от
воды (Печора); в сухом ягельном бору
под молодыми елочками и еловыми
кустами (Кольский п-ов) и в тальни-
ках у озера под ивовым кустом (Бере-
зов на Оби). В средней лесной полосе
{Рыбинское водохранилище) из 8 осмо-
тренных гнезд 2 помещались в лесу Рис. 80. Позы тока свиязи
суборив 100—250 м отводы. Они были
устроены у комлей крупных елок и скрыты под еловыми лапами. Одно было
устроено на берегу под молодой сосенкой с низко опущенными ветками.
Пять на лугах. Из числа последних 2 среди осочников в густых осоковых
кочках и три в куртинах более высокой травы, иногда совершенно закры-
вающей гнезда сверху. Наконец, в лесостепи (Бараба) гнезда свиязей нахо-
дят в бурьянах на опушках березовых колков, близко подходящих к воде.
Гнездо представляет собой ямку глубиной 5—7 см, обычно с очень скудной
растительной выстилкой или совсем без нее и значительным количеством
пуха, который укладывается высокими валиками по краям гнезда и закры-
вает яйца на время отсутствия самки. 10
Повсеместно обычное число яиц в полной кладке 9, реже 10 или 7—8
{последнее наблюдалось у нас на севере: Тиманская тундра, Кольский п-ов,
а на Британских о-вах это наиболее обычный размер кладки). Ненасиженные
яйца чистобелые, совсем без зеленоватого оттенка. Размеры 50—59,9X
Х33,2—40,7 мм, вес 41—47 г. Насиживание начинается с откладки послед-
«его яйца; срок 24—25 дней (Уайзерби, 1945), а по Хейнроту 22—23 дня.
Возможно, что столь короткий период инкубации является биологическим
приспособлением, облегчающим этой утке существование в условиях край-
него севера. Высиживает только самка, но самец первое время находится
вблизи от гнезда. Существуют отдельные наблюдения о встрече селезней
(уже в смешанном пере) при выводках молодых (Житков и Бутурлин, 1906)
и даже над участием их в защите выводка от луня (Сушкин, 1908). Однако
скорее всего наблюдения эти обязаны случайному совпадению. Вес вывед-
шихся птенцов 20—21 г. В гнезде молодые остаются менее суток, после чего
мать переводит их на водоем. Пуховые птенцы быстро передвигаются по
суше и хорошо ныряют. Способность к полету они приобретают довольно
рано, в возрасте примерно 45 дней.
В северной части Рыбинского водохранилища откладка яиц начинается
около 27 мая, в первой декаде июня встречаются полные, но не насиженные
кладки, позднее же все кладки насижены. Под Архангельском (озеро Сло-
бодское) первые встречи выводков отмечаются около 8 июля A7 июня—
29 июля); в Тиманской тундре 8 июня и 7 июля найдены гнезда с полными,
но ненасиженными кладками, а 15 и 19 июня со слегка насиженными;
444 ОТРЯД ГУСЕОБРАЗНЫЕ
19 августа встречен выводок молодых, по размеру уже сравнявшихся со взрос-
лыми. На Кольском п-ве (озеро Имандра) гнезда с полными кладками найдены
1 и 16 июня, к 27 июня в некоторых гнездах уже происходит вывод птенцов,
а оперившиеся, но еще нелетные молодые встречаются с 20-х чисел июля
до первых чисел августа; обычное число птенцов в выводке 6 C—8).
На Печоре, в устье Ылыча первые выводки появляются около 2 июля
A8 июня—15 июля; Теплов), в дельте реки они обычны в июле, но запоздавшие
бывают и много позже, так 7 августа один из выводков, встреченных в устье
Усы, состоял из пуховых птенцов (Дмоховский, 1933). На Щугоре (Север-
ный Урал) маленькие пуховики встречены 15 июля (Портенко, 1937); гнездо
с еще неполной кладкой найдено под Березовом около 22 июня, а в низовьях
Оби (Питляр) 27 июля молодые уже начинают летать (Дерюгин, 1898);
на р. Щучьей (Южный Ямал) гнездо с насиженными яйцами 26 июня, а
птенцы-хлопунцы 13 августа. На р. Елогуе (Туруханский край) 3 июня
самцы свиязей токовали и дрались между собой, некоторые самки уже
несли яйца, а 4 июля встречены первые выводки, 6 августа они поднялись
на крыло (Слудский). ВТункинской долине недавно выведшиеся пуховики
обнаружены 13 июля, а 23 июля уже встречались молодые весом от 400
до 500 г (Слудский).
В Якутии близ устья Вилюя (р. Лунхе) гнездо было найдено 2 июня
(Маас, 1886), а около Вилюйска самка с выводками встречена 11 августа
(Воробьева, 1928). На о-ве Карагинском (Камчатка) 30 июня найдено гнездо
с 7 свежими яйцами, но еще 25 июля добыта самка с яйцом в яйцеводе (Бело-
польский и Рогова, 1947), наконец, в Анадырском заливе в конце августа
молодые уже начинают летать (Белопольский, 1933).
Линька. Двукратная, полная летняя и частичная осенне-зимняя, за-
тягивающаяся у самок и молодых птиц до весны. Взрослые самцы, отделившие-
ся от выводка, начинают сперва терять контурное оперение, смена рулевых
и маховых происходит на втором этапе линьки. Позднее всего заканчивается
линька спины, которая не бывает полной. У всех самцов, полностью закон-
чивших летнюю линьку (Рыбинское водохранилище), спина на 50—80%
бывает покрыта серыми струйчатыми перьями, которые частью еще зимние,
не сменившиеся, а частью свежие, выросшие во время послебрачной линьки.
Кроме того, часть перьев спины имеет все переходы от летнего пера самочьего
типа до самцового струйчатого. Можно предполагать, что часть перьев спины
сменяется у селезней один раз в год. На Рыбинском водохранилище самец
с линяющей грудью добыт 9 июня, у самцов от 1 июля в основном уже закон-
чилась линька и только на спине сохранились развертывающиеся пеньки,
но у одного селезня от 12 августа первостепенные маховые развернулись
лишь наполовину, а пеньки имеются всюду как на верхней, так и на нижней
стороне тела. В Барабинской лесостепи линька свиязей начинается (смена
махов) в середине июня, время массовой линьки приходится на июль
A июля—1 августа), с 1 августа уже начинается подъем на крыло, а в начале
этого месяца происходит массовый выход из линьки (Янушевич и Золотарева,
1947). На Наурзумских озерах сбор на линьку селезней и холостых самок
начался в первой половине июня, а к концу месяца они были уже многочис-
ленны: отдельные табуны их достигали 300—400 голов. Селезень, потерявший
маховые перья, пойман 11 июля, в дальнейшем линные селезни встречались
до 31 июля (Формозов, 1937). На озере Кургальджин выход селезней из
линьки наблюдался во второй половине августа, там они заканчивают линьку
одновременно с шилохвостями (Меженный и Соколов); в Марыйском оа-
зисе (Туркмения) линяющий селезень добыт 17 июня (Зарудный, 1896).
В верховьях Печоры (устье Ылыча) взрослые самцы объединяются в стайки
по 10—15 экземпляров и отлетают на линьку в половине июня (Теплов);
несколько выше по течению на р. Пожеч линные селезни отлавливались для
свиязь 445
кольцевания 3 августа. На Оби, ниже устья Иртыша, стада селезней, груп-
пирующихся на линьку по 50—60 шт., наблюдались 17 июня (Дерюгин),
на р. Сым в Туруханском крае селезни в разгаре линьки (смена махов) добы-
вались 24 июля—6 августа (Исаков); под Якутском самец, начавший линьку
в брачный наряд, встречен 25 мая (Иванов, 1929), и, наконец, на Анадыре
огромные стаи свиязей (преимущественно самцов), собравшихся на линьку,
наблюдались в конце первой декады июля (Портенко, 1939).
Следующая линька селезней, из летнего в зимний наряд, начинается
вскоре после окончания предыдущей—в конце августа или в начале сен-
тября. Во время нее сменяется контурное оперение почти полностью, кроме
спины, средняя пара рулевых и внутренние второстепенные маховые. На
Рыбинском водохранилище I—3 сентября пролетные селезни на 75% уже
начинают линять на груди и боках, а часть их на подхвостье. Заканчивается
эта линька поздно, на каспийские зимовки все селезни прилетают еще,
не долинявшими; в Гасан-Кули первые птицы в брачном наряде (полном?)
отмечены 22 ноября (Дементьев, 1945), но многие из них заканчивают линьку
только к январю. На Гижиге селезень, сменяющий плечевые перья, добыт
19 сентября (Дементьев, 1940).
Холостые самки или потерявшие кладку линяют вместе с самцами, соста-
вляя незначительный процент в их стаях. Так, энергично сменяющая мелкое
оперение самка добыта на Наурзумских озерах 24 июня (Формозов). На Сыр-
Дарье, около Соло-Тюбе, 16 июня (Спангенберг), а самка, добытая на озере
Токуше в Акмолинской обл. в начале августа, была уже в новом пере
(Аверин, 1911). Линька самок, имеющих выводки, начинается позже, обыч-
но во второй половине июля, и без перерыва переходит в предбрачную линьку.
И та и другая линька, повидимому, не бывают полной, и оперение на части
птерилий сменяется единожды в год. Смена мелкого оперения начинается
еще в то время, когда самка находится при выводке. Линька быстро охваты-
вает все птерилий, и вскоре после ее начала происходит смена рулевых и ма-
ховых. В октябре летнее оперение начинает выпадать, и к декабрю-январю
самки имеют уже в основном брачный наряд, в январе-феврале сменяется
часть рулевых и только в апреле-мае заканчивается смена внутренних второ-
степенных маховых и их кроющих. Весной же отрастает гнездовой пух.
В начале сентября и в октябре на Рыбинском водохранилище у самок были
уже свежие маховые перья, но у некоторых еще проходила смена рулевых
и контурного оперения по всему туловищу. У самки, добытой 26 июля,
линька только что начиналась, и пеньки у нее появились лишь на груди;
под Якутском самка, меняющая маховые перья, добыта 24 августа (Ива-
нов, 1929),
Молодые самцы начинают линять в конце октября; к февралю сменяется
большая часть контурных перьев, рулевые (иногда все, а иногда лишь две
их средние пары), иногда часть внутренних второстепенных маховых и в
виде исключения отдельные средние кроющие крыла. Затем в марте и апреле
часто сменяются перья спины, зада и другие сохранившиеся юношеские кон-
турные перья. Линька молодых самок происходит примерно по той же схеме,
но несколько медленнее. Многие молодые птицы, особенно самки, не закан-
чивают линьку до возвращения на родину и имеют по прилете немало
перьев от юношеского наряда.
Питание. Свиязь по преимуществу растительноядная утка, кормя-
щаяся зелеными частями растений и корневищами. Семена и животные корма
имеют меньшее значение, но в некоторые сезоны свиязи их поедают охотно.
Весной, больше чем в другое время, проявляется у свиязей тяга к зеленым
кормам, недостаток которых они ощущают зимой. В марте и апреле, пролетая
через дельту Волги, свиязи кормятся главным образом листьями и побегами
водяных растений (92%): валлиснерией (Vallisneria spiralis\ 23,3%), побе-
446 ОТРЯД ГУСЕОБРАЗНЫЕ
гами нимфейника (Limnanthemum nymphoides; 24,5%), нитчатыми водо-
рослями A0%) и листьями сусака (Butomus umbellatus; 7,5%). Как под
Астраханью, так и повсюду, повидимому, в степной полосе (Бараба, дельта
Или) свиязи в весеннее время часто вылетают кормиться на затопленные талой
водой солончаки, где они кормятся молодыми всходами лебеды (Atriplex)
и солянок. На р. Мологе в конце апреля свиязи кормились исключительно-
зелеными частями растений, а именно: малой ряской G0%), побегами элодеи
A0%) и всходами щучки (Deschampsia caspitosa, 20%). На северных реках
во время весеннего пролета, который во многих местах совпадает с ледохо-
дом, свиязи любят кормиться среди несомого по течению мусора. Они на-
ходят в нем обрывки зеленых частей растений и выдранные льдинами их
корневища. Основным кормом их в это время на реках севера служат корне-
вища рдеста, которые печорские охотники даже называют «утинными кореш-
ками». Обилие плавающих корневищ рдеста бывает в разные годы различным,
что определяет обилие задерживающихся на пролете свиязей (Печора, Теп-
лов, Илим, Косыгин). Желудок свиязи, добытой в мае на Южном Ямале,
содержал почки ив и песок (Осмоловская). Летом в июне и июле на Рыбинском
водохранилище они кормятся также почти исключительно листвой и побе-
гами водяных растений (99,7%) в том числе листьями полевицы (Agrostis-
stolonizans; 35%), простой ежеголовки (Sparganium simplex; 25%) и лугового
сердечника (Cardaminae pratensis\ 25%). На севере Актюбинской обл. глав-
ную пищу свиязей составляют прямокрылые—кобылки и отчасти «тина»
(водоросли?) (Зарудный, 1888). Питание прямокрылыми в сезон их массового
развития—широко распространенное явление в степях Казахстана. Так,
на Наурзумских озерах во второй половине июня и в июле большие табуны
свиязей жили на этом корме. В период сильного развития пруса (Callipta-
mus italicus) утки подбирали саранчуков, упавших в воду, и охотились за
ними на суше (Формозов, 1937). Во время линьки в дельте Волги свиязи пита-
ются зелеными частями растений F7,5%); валлиснерией A7,5%) и главным
образом резухой (Najas marina; 50,0%), а также незрелыми семенами послед-
ней C2,5%), которые они срывали вместе с побегами. В составе летне-осен-
него питания свиязей повсюду большое значение имеют луковицы стрело-
листа (Sagittaria) и клубеньки гребенчатого рдеста (Potamogeton pectina-
tus), как это уже отмечалось для Барабинских озер (Формозов, 1934). На
осеннем пролете (Рыбинское водохранилище) свиязь попрежнему кормится
листьями водяных растений (98%), главным образом листьями блестящего
рдеста (Pot. lucens; 65%) и нитчатками A2%). В то же время в дельте Волги
вегетативные части растений составляют лишь 52,1 % рациона (главным обра-
зом валлиснерия 12,1%, резуха 16,3% и луковички стрелолиста 9,6%); 45,4%
приходится на долю семян, более всего частухи (Alisma; 32,3%) и 2,5%
составляют моллюски. На севере свиязи нередко кормятся ягодами. Так,
под Архангельском (озеро Слободское) выводки их собирают чернику на
ягодниках у берегов озер (Паровщиков), а в желудке селезня, добытого
2 октября на озере Чуна (Кольский п-ов), найдены ягоды вороники (Влади-
мирская, 1948).
На зимовках свиязи ощущают недостаток в зеленых кормах и едят глав-
ным образом семена. В Талышинской низменности и на Мугани свиязи кор-
мятся в январе-феврале на залитых водой участках степи и рисовых полях.
Здесь они находят зеленые листья некоторых злаков (Роа bulbosa и Paccine-
Ilia), осок и сусака (всего в зависимости от суровости зимы от 39,4 до 75,8%),.
остальную часть их питания составляют семена B4,2—60,6%), главным обра-
зом сорняков рисовых посевов—морского камыша (Balboschoemus maritimus),
злака (Fimbristylus) и куриного проса (Eschinochloa cms galli). На восточном
берегу Каспия (о-в Челекен) свиязи собирают зимой зеленые водоросли
(Cladophora) и ловят в зоне заплеска бокоплавов.
свиязь
447
Хозяйственное значение как промыслового объекта очень велико. Мас-
совые скопления этой утки на местах зимовок обеспечивают возможность
ее промысловой добычи. В заготовках дичи на ленкоранских зимовках она
составляет 5,8% от всего числа заготовленных уток, но в период применения
там сетного лова удельный вес ее в заготовках был гораздо больше. В уло-
вах уток «коечными» ловушками на местах зимовки свиязи в Англии эти
утки составляют в разных пунктах до 32—45%, а в Дании еще больше.
По качеству мяса свиязь одна из лучших уток; чистый вес мясной тушки се-
лезня в промысловый сезон около 475 г. Вполне возможно организовать пра-
вильный промысел этого вида на местах скопления его во время осеннего
пролета.
Рис. 81. Свиязь (самец и самка)
Полевые признаки. Утка среднего размера с относительно небольшой
головой и клювом. Селезень выделяется своей каштановой головой и шеей,
значительно более темными, чем окраска спины, и яркобелыми пятнами на
крыльях. Последние хорошо видны как у сидящей, так и летящей птицы.
Для самки (на полете) характерны светлые серые пятна на крыльях, хорошо
контрастирующие с темной спиной и маховыми. Брюхо сплошь белое.
Голос самца—громкий свист вроде «вхии-у» или «вви-у», у самки резкий
вроде «рерр...». Полет быстрый и маневренный; свиязь может подниматься
с воды почти вертикально. По земле передвигается хорошо; ныряет только
раненая.
Описание. Строение и размеры. Клюв очень короткий, короче цевкиг
к вершине чуть заметно сужается. Хвост несколько клиновидный, короткий; рулевых 14.
Плечевые перья удлинены и несколько заострены, а в летнем наряде закруглены. Размеры
крыла самцов 255—270, самок 232—268 мм; цевка самцов 37—41, самок 34—40 мм; клюв
самцов 33—38, самок 30—34 мм.
В е с. Взрослые самцы на весеннем пролете в дельте Волги в марте—апреле 650 г F00—
700 г), в апреле 570—810 г (Рыбинское море); в мае 701 г F30—765 г) (Бараба), 760—
780 г (Архангельск), 760 г G40—840 г) (Печора), 775 г G00—850 г) (Ямал). За брачный
период вес падает, в июне он 740 г (Архангельск), 681 г (Рыбинское водохранилище). Во
448 ОТРЯД ГУСЕОБРАЗНЫЕ
время линьки в августе: 838 г G00—1000 г) (дельта Волги), 700—785 г (Рыбинское водо-
хранилище), 600 г (Архангельск), 620 г (р.Сым, Туруханский край), 755 г (Ямал). Осенью
во время пролета свиязи опять прибавляют в весе. В сентябре 835 г G20—980 г) (Рыбин-
ское водохранилище), в октябре 925 г (850—1000 г) (дельта Волги). Взрослые самки на про-
лете в мае 720 г (Заонежье), 650 г (Рыбинское водохранилище), 690 г F20—770 г) (Печора),
750 г (Ямал), 650 г (Камчатка). На местах гнездовья в июне 530 г (от выводка), в июле
650 г (холостая, Архангельск), 695 г (холостая, Ямал). В различном состоянии линьки: в
июне 590 г (Наурзум), 715 г (Рыбинское водохранилище), 590 г E30—650 г) (Архангельск),
615 г F00—650 г) (р. Сым), 720 г (Ямал). На осеннем пролете в сентябре 650 г (р. Елогуй),
752 г F90—830 г), в октябре 748 г G25—770 г) (Рыбинское водохранилище), 710 г F00—
850 г) (дельта Волги). Молодые на Рыбинском водохранилище в июле до 625 г, в августе
самцы 650 г, самки 670—725 г; в сентябре самцы 848 г G20—900 г), самки 686 г F45—750 г);
в октябре самцы 831 г G60—900 г), самки 712 г F00—820 г). На зимовках без учета возра-
ста в октябре самки 650—720 г (Красноводск); в ноябре самцы 697 г E40—820 г), самки
465 г D00—590 г) (Гасан-Кули); в январе самцы 625 г F80—750 г), самки 650 г E60—670 г);
в феврале самцы 637 г E60—700 г), самки 567 г E05—620 г); в начале марта самцы 647 г
D00—800 г), самки 590 г D50—780 г).
Окраска. Пуховой птенец. Хорошо отличим от птенцов других уток. На
спинной стороне он чернобурый, иногда с легким оливковым оттенком. Бока головы
ржаво-рыжие, каемка по заднему краю крыльев и пятна по бокам спины желтовато-бурые.
Брюшная сторона беловато-желтая.
Молодая самка в первом наряде почти не отличима от старой. Только верх-
ние кроющие крыла у нее более одноцветны, а на зобу имеются неясные пестрины.
Зеркало более тусклое и часто более мраморное, а следующие за ним маховые с меньшим
количеством белого на внешних опахалах.
Взрослая самка. Голова и шея рыже-бурые, в частых темных пестринах,
особенно густых на темени, отчего верх головы кажется более темным и имеет зеленый
блеск. Спинная сторона тела серовато-бурая со светлыми краями перьев; плечевые перья
и кроющие хвоста с рыже-бурыми каймами. Зоб, верхняя часть груди, бока и подхвостье
рыже-бурые с более темной серединой перьев и светлыми вершинами; остальная брюшная
сторона белая. Первостепенные маховые бурые, четыре внешних второстепенных
аспидно-серого цвета, слегка мраморные с белыми вершинами, у остальных же нижние
опахала черноватые с белыми вершинами и зеленым блеском или совсем без блеска, верхние
кроющие крыла буровато-серые со светлыми каймами, самые длинные с белыми вершина-
ми. Хвост серо-бурый. Клюв матово-серый с черным концом, лапы свинцово-серые;
радужина бурая.
Молодой самец в первом наряде. Напоминает самку, но легко отличается по
окраске крыла, которое уже имеет зеленое зеркало, а верхние кроющие с более
широкими белыми каймами и вершинами. Окраска спинной стороны туловища бывает двух
типов: одноцветно дымчатая с сероватыми вершинами перьев или черно-бурая с рыже-охри-
стыми каймами и такого же цвета узкими поперечными полосками на перьях, некоторые
из перьев имеют, кроме того, мелкий край. Оба типа окраски встречаются примерно оди-
наково часто.
Годовалый самец. Весной, после первой линьки, имеет в основном наряд взрос-
лой птицы. Отличается серой расцветкой верхних кроющих крыла, а также черно-
дымчатой спиной и надхвостьем, не имеющими струйчатого рисунка.
Взрослый самец в брачном наряде. Лоб и верх головы желтовато-белые;
остальная голова и шея ржаво-каштановые с мелкими черными крапинками, осо-
бенно на затылке, вокруг глаз и на подбородке. Задняя часть шеи, плечевые, вся спина
и поясница серые с мелкой темной поперечной струйчатостью. Надхвостье белое по сере-
дине и черное с боков. Зоб и бока его серовато-винного цвета; остальная брюшная сторона
чистобелая, нижние кроющие хвоста черные, подмышечные белые с мелкой серой рябью.
Первостепенные маховые серо-бурые, второстепенные с узкими белыми вершинами, а у
двух первых из них внешние опахала в основной части металлически-зеленые, а в дисталь-
ной части черные (они образуют зеркало), следующие с черными внешними опахалами,
окаймленными белым. Верхние кроющие крыла вдоль сгиба серые, все остальные белые
образующие сплошное белое пятно; самые длинные с черными вершинами образуют поло-
ску, отграничивающую зеркало. Рулевые серые. Клюв свинцово-серый с темным когот-
ком, лапы также свинцово-серые с темными перепонками. Радужина бурая.
Взрослый самец в летнем пере. Вся голова и шея ржаво-коричневые в мел-
ких черных пестринках, более густых на темени и затылке. За глазами и над ними
пурпурный отлив, на затылке он слегка зеленоватый. Зашеек и мантия темнобурые с бело-
охристыми поперечными полосками и рыже-бурыми каемками. Спина и надхвостье серо-
вато-бурые со светлыми каймами перьев и поперечными полосками или пятнами. Зоб
и бока каштаново-рыжие с более темными поперечными полосками, четкими на зобу и по-
степенно исчезающими на боках. Нижняя часть груди и живот белые. Перья подхвостья
бурые с белыми каймами и поперечными полосами. Крыло, клюв и ноги, как в брачном
наряде.
АМЕРИКАНСКАЯ СВИЯЗЬ
449
29. Американская свиязь Anas americana Gm.
Anas americana, G m e 1 i n. Syst. Nat., 1, 2, 1789, стр. 526, Луизиана.
Распространение. Гнездовой ареал. Североамериканский вид,
населяющий северо-западную и центральную части материка. Северная гра-
ница ареала проходит от залива Коцебу к устью р. Мекензи и М. Батерст.
Восточным пределом служит цепь крупных озер: Медвежье, Невольничье,
Виннипег и южный берег Мичигана. Крайними южными точками являются
плато Небраска, верховья рек Ар Канзаса и Колорадо. На западе ареал свиязи
Карта 50. Распространение американской свиязи Anas americana
а—граница гнездовий, в—зимовки, д—валеты, е—регулярные валеты
не выходит к морю, а достигает восточных склонов Каскадных гор (Орегон),
севернее же границей его служат Скалистые горы, истоки Юкона и водораз-
дел между ним и р. Ку скоку им. Таким образом, весь ареал этого вида нахо-
дится в области Центральноамериканской возвышенности и естественными
границами его служат с одной стороны низменности, а с другой территории
с настоящим горным ландшафтом.
Основные зимовки американской свиязи находятся вдоль Тихо-
океанского побережья от Ванкувера на севере до Никарагуа на юге. Регу-
лярно посещают они и Гавайские о-ва. Северным пределом их на материке
служат низовье р. Колорадо и устье Миссури. На Атлантическом побережье
свиязь зимует от Филадельфии до берегов Мексиканского залива (отсутствуя
лишь на юге Флориды), а также на о-вах Ямайке и Кубе.
Залеты. 1 мая 1883 г. на о-ве Беринга найдена погибшая самка этого
вида. До настоящего времени это единственный известный случай залета
американской свиязи в СССР, но повторные встречи возможны, так как из-
вестны залеты в Японию, а для Алеутских о-вов они нередки. Залетает
изредка в Англию и на Гебридские о-ва.
Характер пребывания. Перелетная птица. В СССР—слу»
чайно залетная.
29 Птицы СССР, т. IV
450 ОТРЯД ГУСЕОБРАЗНЫЕ
Биотоп. Континентальные водоемы. Предпочтительно озера с бога-
той водной и прибрежной растительностью, зимой—реки и озера, также
морское побережье.
Численность. На своей родине многочисленный промысловый
вид уток.
Экология. Размножение. Брачные игры напоминают такие же
у европейской свиязи. Самцы плавают вокруг самок с распушенными перья-
ми головы и лопаток и, вздергивая резким движением голову вверх, издают
свой громкий брачный свист. Гнездится около озер и тихих рек, часто на
островках. Расстояние от гнезда до воды бывает различно. Часто гнезда
помещается под деревьями или кустами, среди сухой травы, но иногда и на
Рис. 82. Американская свиязь
совсем открытых местах. Гнездовая ямка вырывается на сухом месте, имеет
скудную выстилку из растительных материалов и большое количество до-
вольно светлого пуха. Яиц в полной кладке сбычно 9—11 F—13), по форме
и окраске они не отличимы от яиц обычной свиязи. Размер их 50,5—60х
Х36—42, среднее 53,9 X 38,3 лш. Время гнездования июнь. Срок насиживания
24—25 дней; период развития птенцов до взлета 45—50 дней.
Линька. Последовательность смены нарядов та же, что и у обык-
новенной свиязи. Самцы объединяются в стайки в конце июня, а в июле
уже начинают линять. Характерная для нашей свиязи неполная смена опе-
рения на спине для этого вида пока не отмечена.
Питание. Американская свиязь растительноядная утка, животные
же корма составляют в ее рационе только 6,8%; это главным образом мол-
люски и в меньшем числе личинки насекомых. Из растений летом основу
составляют рдесты D2,9%), их листья и корневища; весной поедает много
злаков (зеленые части) и осок (семена); зимой, за время пребывания на
взморье, большую роль играют водоросли. Из числа последних особенное
значение имеют Ulva lactuca и Enteromorpha intestinalis, встречающиеся почтит
ЧИРОК-ТРЕСКУНОК 451
во всех желудках и нередко составляющие до 100% их содержимого (Линч,
1935). Кроме них, свиязи в это время кормятся на море зелеными частями
руппии и Zannichelia, а на внутренних водоемах рдестами, роголистником
и резухой.
Экономическое значение. Довольно велико, так как на большом простран-
стве материка эта свиязь служит постоянным объектом спортивной охоты.
Качество ее мяса ценится очень высоко.
Полевые признаки. Утка среднего размера, посадка на воде характерна*
для настоящих речных уток. Как у сидящего на воде, так и у летящего селез-
ня обращает внимание светлый, почти белый, верх головы, ограниченный
по бокам ее широкой темной зоной, проходящей через глаз. Основная поло-
вина крыла сверху белая, зеркало зеленое. У самок верх крыла светлый,
но не белый. Голова у самок в общем серая, туловище коричневое. Спина,
грудь и подхвостье темные, но брюхо, а у самцов и поясница, сверху и с
боков белые. Довольно молчаливая птица, свист самца менее громкий, чем
у европейской свиязи.
Описание. По строению и размерам весьма сходна с обыкновенной свиязью.
Окраска. Пуховой птенец. Не отличим от такового обычной свиязи.
Молодые самцы в первом наряде. Похожи на одновозрастных европейских сви-
язей, но передняя часть головы до середины темени кремового цвета или светло-
бурая с ясными темнокоричневымн штрихами, а перья задней части темени темнокорич-
невые с белыми концами и скобовидными полосами. Бока и низ головы и шеи светлые,
обычно кремовые. Нижние кроющие крыла белые, зеркало зеленое, но менее яркое, чем
у взрослых птиц.
Молодые самки отличаются от самок обычной свиязи тем, что оперение головы,
шеи, большие верхние и нижние кроющие крыла такие же, как у молодого самца.
Взрослая самка очень похожа на европейскую свиязь, но голова несколько более
светлая, сероватая. Подмышечные и нижние кроющие крыла белые.
Взрослый самец в брачном наряде. Верх головы (лоб и темя) сливочно-белые; во-
круг глаза черное пятно, вытянутое к затылку, с зеленоватым блеском в своем
задней части. Остальная голова и шея серо-бурые с мелкими черными с металлическим бле-
ском, штрихами и пятнами. Лопатки и плечевые перья серые с винным оттенком и черной
струйчатостью. Спина и надхвостье серые струйчатые. Зоб и бока более темные, серовато-
винные, средняя часть груди, брюхо и бока поясницы белые; подхвостье и боковые крою-
щие хвоста черные. Верхние кроющие по краю крыла серые, остальные белые, самые длин-
ные с черными вершинами. Первостепенные маховые серо-бурые, второстепенные с черны-
ми, а в средней части с блестящими зелеными наружными опахалами и белыми вершина-
ми. Они образуют зеркало, окаймленное сзади белой полоской. Удлиненные третьестепен-
ные маховые серые или серо-бурые с черной центральной частью; подмышечные чисто-
белые. Рулевые серые. Клюв и лапы серо-голубые, радужина бурая.
Самец летом окрашен в общем, как самка, но тона окраски более интенсивные
и более ржавчатые. От самок хорошо отличим по оперению крыльев, а от самцов обычной
свиязи более серой головой.
30. Чирок-трескунок Anas querquedula L.
Anas querquedula. Li n n a e u s. Syst. Nat., изд. X, 1, 1758, стр. 126, Швеция.
Синоним. Anas circia. Li n n a e u s. Syst. Nat., 1, 1766, стр. 204.
Русские названия. Чирок-трескунок (широко распространенное), храпунок
(Украина), ширкунок (Ильмень) —по голосу селезня, большой чирок, сизокрылый чирок
(Астрахань)—народные.
Распространение. Ареал гнездовой занимает большую часть
европейского континента, на север до Юго-Восточной Англии (Исландии?,
по Уайзерби, 1945), Средней Швеции F1° или 64° с. ш.) и Северной Финлян-
дии F5° с. ш.). На западе трескунок заселяет Францию (кроме Бретани
и берегов Бискайского залива), доходит до Пиренеев, на юге достигает Среди-
земноморского побережья, Средней Италии, Албании, Болгарии и, возмож-
но, Северной Греции. Изолированные места гнездованья этого вида известны:
на юге Испании в устье Гвадалквивира, в Сардинии, на о-ве Наксос в Эгей-
ском море и на Кипре (Нитхаммер, 1938).
29*
452
ОТРЯД ГУСЕОБРАЗНЫЕ
В пределах Советского Союза трескунок гнездится к северу до средней
части Карелии, Соловецких о-вов (Поляков, 1929) и Архангельска. Известны
летние встречи его на Канине, но гнездованье там не доказано (Дементьев,
1940); в Тиманской и Большеземельской тундре отсутствует. В бассейне
ЯПечоры гнездится лишь в самых верховьях (Теплоухов, 1911), а уже около
устья р. Ылыча бывает ежегодно лишь весной, в период пролета, но не гнез-
дится. На Урале повсюду редок, даже в средней части его (верховья Туры).
Под Тобольском обычен, а как редкая гнездящаяся птица несомненно дохо-
дит до устья Иртыша; летом, а чаще весной, встречается под Березовом
(Малая Сосьва), единичные встречи известны и под Салехардом, но гнездо-
ванье в этих пунктах не доказано. Встречен в сентябре на водораздельных.
Карта 51. Распространение чирка-трескунка Anas querquedula
а—граница гнездовий, а'—районы наибольшей численности птиц на гнездовье, б—не
вполне выясненная граница гнездовий, п—обособленные гнездовые колонии, в'—места
постоянных встреч летом и спорадичного гнездованья, з—зимовки, в—залеты, е—встре-
чи пролетных птиц весной севернее гнездовых мест, ж—места скоплений птиц на
линьку
озерах в верховьях Таза, обнаружен на гнездовье по Елогую (Юдин). Встре-
чается в верхнем течении Нижней Тунгуски; в долине Лены гнездится к
северу до Якутска, где очень редок (Иванов, 1929), для Вилюя уже не ука-
зывается. Восточнее граница распространения не выяснена, но, повидимому,
она резко снижается к югу. Трескунок несомненно гнездится в долине Амура
и на Сахалине, хотя и редок там; отмечен летом в устье р. Уды и возможно
гнездится в районе Аяна (Снигиревский). Повидимому гнездится изредка
в южной части Камчатки и на Курильских о-вах; залетает на восточный
берег Камчатки и Командоры. Нередко чирок-трескунок встречается зна-
чительно севернее пределов своего гнездового ареала, причем в некоторых
пунктах его можно наблюдать ежегодно, особенно весной, в период пролета
(Печора, низовья Оби и др.). На юг этот вид гнездится до Черного моря,
встречаясь летом и по южному его берегу, где гнездованье, однако, не заре-
гистрировано. Заселяет Кавказ, Закавказье и южный берег Каспия, где
молодой, только что поднявшийся на крыло, был добыт 14 июля около Бен-
ЧИРОК-ТРЕСКУНОК 453
дер-Гязь (Зарудный, 1896). В Туркмении встречается летом на Мургабе
и Теджене и возможно изредка гнездится там (Зарудный, 1896), хотя Шесто-
перов A936), обследовавший последний райои, трескунка не обнаружил. Пови-
димому,. гнездится, хотя и очень редко, в долине Аму-Дарьи (Рустамов,
1945), но под Ташаузом (Шестоперов, 1936) и в Сарыкамышской котловине
не найден, да и в дельте, по последним данным, бывает лишь на пролете
(Салихбаев, 1950). По Зарудному A900), держится летом и изредка гнездится
в Иране по Геррируду и в Сеистанской котловине, однако можно предпола-
гать, что большинство этих сомнительных сведений относится к задержавшим-
ся на пролете особям, которые спорадически оседают на гнездовье в ряде
пунктов южнее их основного ареала. Гнездится по восточному берегу Араль-
ского моря и в дельте Сыр-Дарьи (Зарудный, 1916), но в нижнем течении
этой реки и прилегающих районах бывает лишь на пролетах (Спангенберг
и Фейгин, 1936). В Таджикистане редок и бывает лишь на пролетах, летом
встречен только на Искандер-Куле, но гнездованье там не доказано (Иванов,
1940); на Памире не гнездится, так же как и в гористой части Семиречья
к югу от Киргизского хребта, Заилийского Алатау и Кетменских гор,
на низменности же распространен повсеместно (Шнитников, 1949). Гнездится
в Синь-цзяне (Кульджа), известны летние встречи в Кашгарии (Дивно-
горский), в Северном Пакистане (до мая и даже июня, Тайсхерст, 1927),
в Тибете и в бассейне Желтой реки. В большинстве случаев это, несомненно,
птицы, задержавшиеся на пролете с зимовок, но возможны там и случаи
спорадичного гнездованья, так же как известны, например, два случая гнез-
дованья в Северной Индии (Уд и Маулейнин; Финн, 1901). Широко распро-
странен трескунок в пределах Монголии (Банников, 1946), гнездится по
всей Маньчжурии; Уссурийскому краю и на о-ве Хоккайдо.
Основные зимовки этого вида находятся в области Среди-
земного моря но Франции, Италии (особенно Северной) и Испании; меньше
птиц зимует в Югославии, Греции и Западной Турции. Многочисленны тре-
скунки на африканских зимовках, где они встречаются в большем числе
и заходят далее к югу, чем другие палеарктические виды уток. Они зимуют
всюду в Северной Африке, а в ее западной части спускаются до Северной
Нигерии, Камеруна и Багирми, далеко переходя за тропик. Обычны они
всюду в Египте, Судане и Абиссинии, нередки в Кении, Уганде и у Рувензори,
доходя на юге до озер Танганайки, Ньясса и до Сейшелльских о-вов (Гроте,
1930, 1931 и др.). Зимуют трескунки в Южной Аравии и на берегах Оман-
ского залива, но Ирак посещают лишь на пролете (Тайсхерст, 1923). В очень
большом числе зимуют повсюду в Индии и Пакистане. Обычны в Бирме,
Индокитае, на Филиппинских о-вах и в Китае до Гуандуня, на Хайнане и Тай-
ване. Наиболее северные зимовки на Тихом океане известны в южной
части о-ва Хонсю (Япония). На юге они встречаются изредка на Зондских
и смежных с ними островах. Залетают на Новую Гвинею. В пределах
СССР трескунок, как правило, не зимует. Лишь единичные птицы и неболь-
шие стайки самцов задерживаются на зиму в южных частях Каспийского
моря, на Азовском море против дельты Кубани (Михайлов, 1937), на Сиваше,
в заливах северо-западной части Черного моря и кое-где в Средней Азии.
Однако и это случается не каждый год, а только в более теплые зимы.
Характер пребывания. Повсюду трескунок перелетная
утка, совершающая более дальние сезонные миграции, чем большинство
других уток.
Сезонные миграции. Как показали результаты кольцевания,
трескунки, населяющие северо-запад и север европейской части Союза, отле-
тают на запад и встречаются осенью в Польше, Голландии, Франции и Швей-
царии. Зимой они вместе с местными птицами перемещаются к югу на побе-
режье Средиземного моря. В этом же направлении отлетает некоторая часть
454 ОТРЯД ГУСЕОБРАЗНЫЕ
птиц из центральных областей и даже из Западной Сибири (Омск), но основ-
ная масса их пролетает на юго-запад через дельту Волги (где птицы задер-
живаются на время линьки) или через Южную Украину и достигает запад-
ных частей Средиземного моря (Северная Италия), а частью и Африки.
Птицы из Средней Сибири, пролетая через Казахстан, достигают Индии
(Вучетич, 1939; Михеев, 1949), а из Восточной Сибири отлетают в Китай
и Индокитай.
Даты. Весной трескунки появляются поздно, одними из последних
среди настоящих уток. Их отлет со средиземноморских зимовок происходит
в начале марта, но часть птиц задерживается в тропической Африке до сере-
дины и даже конца апреля. В северо-западной части Черного моря (Джарыл-
гачский залив) трескунки появляются в первой декаде марта, а валовой
пролет их проходит обычно с 1 по 7 апреля (Клименко, 1950). Появление
их на Днестре приходится на конец марта (Остерман, 1912); в Аскании-Нова
первые птицы отмечаются около 16 марта B8 февраля—4 апреля), валовой
пролет приходится на 28 марта, а запоздалые особи встречались еще 28 апре-
ля; таким образом, пролет продолжается (как и везде на юге) очень долго,
около 57 дней, из которых 42 он бывает довольно интенсивным (Соколов, 1926;
Дергунов, 1924). Под Запорожьем первые птицы отмечались в середине марта
(Вальх, 1899), под Киевом 7—9 марта, а валовой пролет 23—31 марта (Дани-
лович, 1930). На пресных озерах Западного Узбоя первая стайка встречена
26 марта (Шестоперов); в Азербайджане пролет происходит в апреле (Вере-
щагин, 1950). На Хопре трескунки появляются 19—25 марта (Измайлов,
1940); на Суре в годы с ранней весной в первых числах апреля (Житков и
Бутурлин, 1906); под Москвой в середине апреля, причем пролет их продол-
жается до конца месяца (Поляков, 1910; Смолин, 1948); в устье Мологи
валовой пролет начался 26 апреля и продолжался всего 2 дня, после чего
до 8 мая наблюдались лишь отдельные небольшие стайки; в северной части
Рыбинского водохранилища первые птицы отмечались в разные годы 17—
30 апреля. В Эстонии трескунки появляются в конце марта—начале апреля
(Кох, 1911), под Ленинградом в первой декаде апреля (Бианки, 1907); на
Верхней Печоре 11 мая C—19 мая) и встречаются в среднем около 12 дней
(от 1 до 30), после чего исчезают. В среднем течении Урала трескунки проле-
тают в большом числе; передовые птицы появляются обычно в середине апре-
ля, а валовой пролет их тянется больше, чем у всех других уток—со второй
декады апреля до середины мая (Зарудный, 1897); на Эмбе первые птицы
были отмечены 20 апреля и пролет длился до начала мая, а в низовьях Ир-
гиза первые трескунки показались 27 апреля (Сушкин, 1908). На Наурзум-
ских озерах (Кустанайская обл.) первые особи отмечены 21 апреля, с 24 по
27 апреля наблюдался их массовый пролет, а стайки птиц, неразбившихся
на пары, встречались еще в 20-х числах мая (Михегв, 1938). На Барабе начало
прилета приходится на конец апреля, а массовый прилет на начало и сере-
дину мая (Янушевич и Золотарева, 1947); под Тобольском трескунки появля-
ются 2 мая B2 апреля—9 мая) (Тарунин, 1929). Пролет на Аму-Дарье и
Сыр-Дарье весьма незначителен, зато по Чу и Или интенсивен и проходит
в конце марта—начале апреля (Шнитников, 1949). На окраинах Алтая
у Новенского появляются в первой половине мая (Сушкин, 1938). На китай-
ских зимовках они задерживаются до апреля, а некоторые особи даже до
мая, а через Чжили пролетают с марта по май (Колдуэлл, 1931). На юге
Советского Приморья первые особи появляются в начале апреля, а валовой
пролет проходит в первой половине мая (Воробьев). На озере Орок-Нор
в Северной Гоби прилет отмечен 21 апреля (Козлова, 1930), на р. Чикой
в Юго-Западном Забайкалье 3 мая (Моллесон, 1897). На восточной Кам-
чатке (Кроноки) наблюдался 10 мая (Аверин, 1948); в верховьях Колымы
добыт 28 мая. На р. Илиме (приток Ангары) ошечен 20 мая (Косыгин, 1926),
ЧИРОК-ТРЕСКУНОК 455
.а прилет под Якутском приходится на середину мая (Воробьева, 1931).
Осенний отлет трескунков происходит очень рано, обычно раньше, чем у
всех других уток. На Верхней Печоре (устье р. Ылыча) осенний пролет
начинается около 10 сентября B0 августа—24 сентября) и продолжается
в среднем 7 дней. Птицы летят стайками по 20—30 шт. На Рыбинском
водохранилище осенний пролет начинается около 1 сентября, а последние
птицы в обычные годы отмечаются не позднее 25 сентября. Под Москвой
пролет продолжается с середины августа до конца сентября; в Эстонии
в середине и конце августа; в Ульяновской обл. они начинают пропа-
дать с середины сентября. В верховьях Иргиза и Тобола пролет про-
ходит с начала до конца сентября, последние экземпляры в районе Больших
Барсуков встречены 19 октября. В дельте Волги первые пролетные стаи
появляются около 1 сентября, массовый пролет проходит 10—20 сентября,
а к 1 октября основная масса трескунков отлетает. Пролет их у берегов
Апшеронского п-ова начинается в середине августа. По Днепру в сентябре
пролетает иногда огромными стаями. В низовьях Днепра количество тре-
скунков начинает сокращаться с середины сентября и в начале октября их
уже не бывает видно (Пачосский, 1911); в Джарылгачском заливе Черного
моря пролет начинается с конца августа, в начале сентября они бывают
в значительном числе. Первые встречи в Африке отмечены 24 августа.
В Северном Китае они появляются в августе (Чжили), а Южного достигают
в сентябре (Цзяньсу).
Места скоплений на линьку. Селезни чирка-трескунка
и оставшиеся по тем или иным причинам без выводков самки ко времени насту-
пления линьки сбиваются в стаи и отлетают иногда на значительные расстоя-
ния. Летняя миграция у этого вида выражена совершенно четко, но наряду с
этим часть селезней-трескунков остается местами линять вблизи or мест сво-
его гнездованья. Так, например, линные селезни как одиночками, так и
стайками до десятка голов встречаются в пойме Оки (Калужская обл.; Фила-
тов, 1915), по Суре и Алатырю (Житков и Бутурлин, 1906), в Воронеж-
ской обл., на Донце, кое-где на Украине и в других местах. Массовые
концентрации линных птиц известны в Барабинской лесостепи, на За-
уральских озерах, в Кустанайской обл., на Кургальджине, на озерах
Чкаловской обл. и в дельте Волги. Кроме того, они собираются на линьку
в Матцальской бухте (Эстония) и возможно на озере Ильмень.
Биотоп. Трескунок преимущественно озерная утка. Он предпочитает
водоемы открытого ландшафта и явно избегает гор. Наиболее характерен
он для широких речных пойм и для луговых степей, причем поселяется
нередко у весьма незначительных по размеру водоемов и даже у осоковых
болот. В качестве мест кормежки предпочитает водоемы, богатые водной
растительностью, как пресные, так и соленые. На взморье и крупных озерах
обычно не кормится, а только проводит время отдыха.
Численность его в разных частях ареала не равномерна. Тре-
окунок весьма обычная и даже многочисленная птица степи, лесостепи и зоны
смешанных лесов, но он редок в тайге, в районах с резко выраженным горным
ландшафтом и совсем не проникает в тундру. В пустынях он распространен
неравномерно, но местами в поймах рек весьма обычен. В Западной Европе
трескунок обычен всюду, кроме крайнего севера и средиземноморских стран.
В Витебской, Ивановской, на севере Горьковской и в Кировской обл. он
обычен, но встречается реже, чем биологически викариирующий с ним чирок-
свистунок. В Московской, Рязанской, Калужской и Тульской обл. в раз-
ные годы преобладает то тот, то другой вид. В гористых частях Урала
и Кавказа трескунок редок. В Сибири он еще обычен в устье Тобола, редок
в Нарымском и Туруханском краях и в гористой Восточной Сибири. В Уссу-
рийском крае также довольно редок и распространен спорадически.
456
ОТРЯД ГУСЕОБРАЗНЫЕ
Экология. Размножение. Время наступления половой зрелости
точно неизвестно. Возможно, что она наступает на первом году, но молодые
птицы еще сильно отличаются в этом возрасте от взрослых по оперению крыла.
Постоянные задержки значительного числа особей на юге вне пределов гнез-
дового ареала вида говорят о том, что какая-то часть популяции не уча-
ствует в размножении, и по всей вероятности это годовалые птицы. На места
гнездованья трескунки прибывают уже разбившись на пары и вскоре при-
ступают к размножению. Их брачные игры можно наблюдать весной, они
очень схожи с таковыми у широконоски и состоят в следующем. Одна или
несколько пар плавают кругами—самец за самкой, голова к хвосту, с клю-
вом, погруженным в воду. Длинные плечевые перья у них оттопыриваются,
а оперение головы распушается. Плавание это сменяется ритмическим подер-
гиванием головы вверх и вниз, с горизонтально направленным клювом.
Время от времени селезень запрокидывает голову на спину так, что лоб почти
касается надхвостья, и издает
трескучий звук, а затем резким
движением приводит голову в
прежнее положение. Иногда он
несколько расправляет крыло
и загибает шею назад, но так,
что голова приходится не сзади
крыла, а снаружи его, и резко
контрастирует на голубоватом
Ф°не КРОЮ1ДИХ перьев. Харак-
терным токовым движением для
Рис. 83. Позы тока чирка-трескунка
трескунка служит встряхива-
ние крыльев, когда селезень
привстает на воде и резко взма-
хивает крыльями. Плавая во-
круг самки, самец обычно пово-
рачивает к ней голову наиболее
ярко разрисованной боковой стороной (Лоренц, 1941). Наконец, трескункам
свойствен токовый полет, когда небольшие стайки их кружатся невысоко
над водой. Птицы летают не ровным полетом, а делая время от времени силь-
ные взмахи, при которых тело принимает наклонное положение и они как
бы оседают на хвост. Шея у них при этом бывает не вытянута, а слегка изо-
гнута. Пары трескунков в период размножения выражены очень четко, но
тем не менее преследование самцами чужих самок, а иногда даже самок дру-
гого вида (крякв), у них весьма обычное явление. На севере в лесной полосе
трескунки гнездятся обычно в открытых поймах рек около озер, озеринок
или осоковых болот. Чаще всего гнезда помещаются в заболоченной притер-
расной части поймы в непосредственной близости от воды, под кустами ивня-
ков и ольхи. Иногда гнезда находятся на открытых осоковых и злаковых
лугах в 100—150 м от водоема. Особенно охотно трескунки устраивают гнезда
в густых и высоких зарослях манника (Clyceria) и ситника (Heleocharis),
В степях Поволжья трескунки охотнее гнездятся около мелких речек и степ-
ных прудов, чем в долинах крупных рек, причем нередко они поселяются
вблизи от населенных пунктов. В плавнях Днепра они гнездятся на лугах
под кустами ив. В Винницкой обл. гнезда их находили на сухих лугах, под
кустами бурьянов, иногда далеко от воды (Герхнер, 1928); в Барабинской
степи на лугах и пашнях у берегов озер после освобождения их от талой воды
(Лавров, 1929; Янушевич, 1947). Наконец, в Семиречье они гнездятся на
луговинах и сухих участках болот вблизи от воды, а в Уссурийском крае
только около открытых луговых водоемов (Шульпин, 1936). Гнезда повсюду
устраиваются, как правило, на сухом месте и представляют собою довольна
ЧИРОК-ТРЕСКУНОК 457
глубокую ямку, которую птица роет самостоятельно или углубляет, исполь-
зуя естественное углубление почвы. При этом самка работает клювом и урав-
нивает ямку вращательными движениями груди. Гнездо выстилается сухими
травинками, а по мере откладки яиц—и пухом, который образует высокий
валик вокруг яиц и служит для укрытия их на время отсутствия самки.
По расцветке пуха и особенно примешанных к нему перьев гнезда трескунков
легко отличить от гнезд чирков-свистунков. Перья первых из них мельче
и имеют четко обрисованные бурые пятна в их центральной части на очень
светлом, почти белом, фоне.
Количество яиц в полной кладке обычно 8—10 с колебаниями от 7 до
12 и даже 15, хотя последняя цифра вероятно обязана сдвоенным кладкам.
В 21 гнезде трескунка, найденном на Мологе, полные кладки чаще всего
содержали 9—10 яиц (8—12), гнезда, под Бежецком 9 и 12, в Смоленской обл.
7—14 (Плющенков, 1950), в Эстонии 7—10, в Винницкой обл. (Литин)
7—11, на Хопре 10—15 G гнезд), под Днепропетровском 10, на о-ве Орлове
(Черное море) 6 и 8, в Крыму 7, в Семиречье обычно 8 G—8), в Уссурий-
ском крае 9, в Маньчжурии 7 и 10 (Ямасина, 1939), в Монголии на
р. Толе 7 (Козлова, 1930). Яйца удлиненно-овальные, несколько более
вытянутые, чем у свистунка. От светлопалевой до светлой желтовато-бурой
окраски, иногда с оливковым оттенком. Размер 43,5—49,2x29,4—35,6 мм,
среднее 45,3—33,2 мм. Средний размер для Барабы—46,2x33,1 мм. Вес
ненасиженных яиц 22,9—28,8 г, в среднем 27 г. Яйца откладываются
ежедневно. Самка сидит на гнезде очень крепко, иногда подпускает
наблюдателя вплотную к гнезду, а были случаи, когда даже удавалось
поймать ее на гнезде руками. Срок насиживания около 23 дней (по раз-
ным наблюдениям 21—24 дня). Самец весь период откладки яиц и первые
дни после начала насиживания их держится вблизи от гнезда. Первая пол-
ная кладка в Кельце (Польша) отмечена 6 мая (Катин, 1912), в Эсто-
нии откладка яиц продолжается с мая и до начала июня (Кох,
1911); под Бежецком вывод в двух гнездах отмечен 24 и 25 июня
(Тюлин, 1914); на Мологе первые яйца обнаружены 22 мая, а пер-
вые пуховые птенцы около 10 июня. На Хопре первые выводки бы-
ли встречены уже в конце мая, а самая поздняя встреча пуховиков при-
ходится на 8 августа (Измайлов, 1940); в Крыму гнездо с яйцами в послед-
ней стадии насиживания найдено 29 мая (Браунер, 1898). В Барабинской
степи ненасиженные кладки чаще всего встречаются в третьей декаде мая>
весь же период кладки продолжается примерно с 5 мая по 10 июня; под Змеино-
горском пуховые птенцы встречены в конце июня (Аверин и Лавров, 1912);
на Наурзумских озерах первые выводки 25 июня, а в середине июля A6—
18 июля) уже встречаются летные молодые; в Семиречье полные, но еще све-
жие кладки находили 15—21 мая; в долине Уссури 5 июня, но 19 июня там
же встречен выводок молодых. Вес пуховых птенцов при выводе около 21 г,
в 24-дневном возрасте 250 г. Летом, когда усыхают мелкие пойменные и вре-
менные степные водоемы, около которых часто гнездятся чирки-трескунки,
выводки их принуждены переселяться на соседние озера. Перекочевки эти
бывают иногда довольно дальними и во время их погибает большое количе-
ство утят, несмотря на то, что они значительно самостоятельнее и проворнее
одновозрастных птенцов кряквы. Главным образом в результате этого сред-
нее число чирят в выводке на р. Мологе сокращалось к августу с 9 до 6. Самка
очень горячо защищает свой выводок, перелетая поблизости или бегая по
земле и волоча крыло. В конце июля выводки трескунков зачастую начинают
объединяться в стайки. С августа начинаются их вечерние перелеты с одних
водоемов на другие. Предотлетные стаи трескунков местами достигают не-
скольких сотен голов (Ока—Павлов, 1879; Кургальджин—Меженный и
Соколов).
458 ОТРЯД ГУСЕОБРАЗНЫЕ
Линька. Самцы линяют дважцы в год, причем между полной и пред-
¦брачной линьками существуют перерыв, иногда довольно длительный. После
брачная линька, повидимому, полная, сперва сменяется контурное оперение
туловища и головы, а затем линяют хвост и крылья. Она начинается в июне
и длится до конца июля—августа. На Оке, под Калугой, линные селезни
(повидимому, утерявшие способность к полету) отмечались в конце июля
и 4 августа. Они держались в густых зарослях хвоща группами по 8—10 шт.
(Филатов, 1915). В Ульяновской обл. селезни в конце июля уже поднимаются
на крыло, примерно с 20 июля, на вечерних перелетах уже добывают взрос-
лых самцов, еще не полностаго закончивших линьку, у которых продолжается
линька хвоста, а маховые на 1,5—2 см от основания еще не потеряли чехлов
(Житков и Бутурлин, 1906). В среднем течении Донца (Ростовская обл.)
смена маховых начинается во второй половине июня (Аверин, 1911).
В дельте Волги трескунки линяют сразу же вслед за кряквой, раньше
других настоящих уток. Стайки их появляются там уже в первых числах
июня E июня), а к концу первой декады они скапливаются на полоях
в тысячные табуны. В 1946 г. уже 29 июля на взморье появились
большие стаи трескунков, закончивших линьку. В первый период линьки
трескунки совершают регулярные перелеты для кормежки на мелководные,
богатые животными кормами водоемы, а день проводят в стаях на откры-
тых песчаных отмелях или на воде около них. На время смены маховых
стаи скрываются в зарослях водной растительности: в Средней России в хво-
щах, в Эстонии и Западной Сибири в тростниках, а в дельте Волги в заро-
слях рогоза и ежеголовки. Из этих мест они только по ночам выплывают
кормиться на ближайшие плесы.
Следующая линька начинается у селезней поздно, не ранее октября.
Признаков ее не удалось отметить у них до самого отлета: на Рыбинском водо-
хранилище до середины сентября, а в дельте Волги до 18 октября. При этой
линьке сменяются контурное оперение на туловище и голове, внутренние
из второстепенных маховых и, очевидно, рулевые. Линька заканчивается
обычно в феврале, но в виде исключения затягивается до марта. Первая
линька молодых птиц обоего пола имеет примерно тот же характер. Она
начинается в октябре—ноябре и продолжается до весны. У них сменяются
контурное оперение, рулевые, но, повидимому, не все, а лишь несколько
пар, и внутренние второстепенные маховые (а по Уайзерби A945) также
и кроющие?). У взрослых самок, находящихся при выводках, обе линьки
заходят одна за другую, и подразделить их трудно. Пока не совсем ясно,
какие части оперения сменяются дважды и какие один раз в год. С августа
до января перелинивает контурное оперение, крылья и хвост. Самки с еще
несмененными маховыми добывались на Рыбинском водохранилище в первой
половине августа. Вновь линька начинается в марте и продолжается до мая.
Во время нее сменяется большая часть контурных перьев, главным образом
на верхней стороне тела, пух, обычно (но не всегда) все рулевые и внутренние
второстепенные маховые (и кроющие?). Весной же отрастает гнездовой пух.
Самки, не имевшие выводка в данном году (негнездившиеся или потерявшие
кладки), линяют одновременно с самцами и в одних стаях с ними.
Питание. Трескунок типичная животноядная утка, причем особенно
большую роль в его пищевом рационе составляют моллюски. На весеннем
пролете в дельте Вэлги (март—начало апреля) животные корма составляют
всего около 26,7% (по объему) от всего содержимого желудков. Вода на
взморье в это время еще не прогрелась, а богатые обычно животной жизнью
полой еще стоят без воды. На 18,3 % питание трескунков в это время состоит из
моллюсков, 6,7% приходится на водяных клопов корикс, 1,7% на ракообраз-
ных (Corophium), 23,3% на вегетативные части растений (листья валлиснерии
и ее корневища) и 50% на семена, которые частично служат в качестве гастро-
ЧИРОК-ТРЕСКУНОК 459
литов. В средней полосе первое время по прилете трескунки проводят день
на разливах рек, а по вечерам вылетают кормиться на небольшие временные
лужи, расположенные на периферии поймы (р. Молога). Там они добывают
различные животные корма (95% по объему), особенное значение среди
которых имеют листоногие ракообразные и ракушковые рачки F0%): Apus>
Branchipus, Esteria, Limandiay Ostracoda и др., которыми кишат в это время
некоторые лужи; много находят они тогда также моллюсков B7%), особенно
битиний; насекомые же (хирономиды и др.) играют в их питании малую
роль (8%). На Рыбинском водохранилище животные корма составляют вес-
ной 68%, из них: моллюски 34% (как и всегда, у трескунка, только легоч-
ные), личинки хиромид 7, мелкие плавунцы и клопы гребляки (Corixa)
27%. Остальную часть рациона составляют листья гребенчатого рдеста B%)
и семена—30% (главным образом ежеголовка, семена которой служат в
качестве гастролитов). Питание утят в июле и августе (там же) уже на 90%
состоит из животных кормов: на 54% из моллюсков, на 26% из личинок
водяных насекомых, главным образом Olyptotendipes gripenoveni и на 10%
из других насекомых. На Мологе питание утят имело несколько иной харак-
тер. Животные корма также составляли 90%, но основой C2%) являлись
личинки двукрылых, главным образом хирономид. На долю моллюсков разно-
образного видового состава приходилось 27% и 25% составляли личинки
и взрослые стрекозы. Период линьки (потери маховых)—трудное время для
трескунков. Длительный срок стаи их держатся на очень ограниченном
участке и питаются тем, что удается найти. В дельте Волги животные корма
в их рационе составляют в это время всего 8,1 %, основу питания составляют
зеленые части растений B4,9%), главным образом веточки резухи (Najas
marina) и семена F7%) преимущественно той же резухи (мягкие, недозрелые).
По выходе из линьки птицы начинают откармливаться на богатых в это вре-
мя животных кормах. В августе—сентябре (дельта Волги) содержание жи-
вотных кормов доходит до 41,2%: моллюсков 34,6% (преимущественно за-
творка Valvatapiscinalis) и насекомых 6,6%, главным образом личинок льви-
нок (Stratyomija) и тлей, сидящих на листьях растений. Вегетативные части
растений—побеги роголистника, ветки валлиснерии, резухи и рдестов—
составляют 19,1%, а семена 39,7%. Там же в октябре содержание живот-
ных кормов сокращается до 27,5% за счет большего потребления зеленых
частей растений C7,5%). Осеннее питание трескунков в средней полосе
(р. Молога) весьма разнообразно. Животные корма F0%) представлены
главным образом моллюсками D0%) восьми видов и насекомыми A9,5%),
из вегетативных частей растений A0%) чаще всего поедаются нитчатые водо-
росли и луковицы стрелолиста; семена составляют 30%. В сентябре на Ры-
бинском водохранилище на долю животных кормов приходится 77%; это
моллюски C5,5%), преимущественно прудовики, личинки водяных насеко-
мых—хирономиды B1 %) и ручейников A7%) и взрослые насекомые C,5%);
семена составляют 23%, значительная часть которых служит в качестве
гастролитов. В желудках трескунков, добытых под Киевом (Шарлемань),
обнаружены остатки жуков, главным образом плавунцов и водолюбов,
личинок водяных насекомых, в том числе стрекоз, щитня, моллюсков:
Vivipara, Bythinia, Planorbis, Valvata, Pisidium, Sphaerlum, а также семеня
и другие растительные остатки.
Хозяйственное значение. Значение трескунка, как объекта спортивной
охоты местами велико, но удельный вес его в промысловых заготовках
невелик, так как этот чирок зимует вне наших пределов и отлетает осенью
очень рано. Больше всего его добывают на юге европейской части Союза,
например в низовьях Днепра, где трескунок составляет около 30 % всей
добычи ружейных охотников, и обилие его выводков определяет успешность
охоты в том или ином году. Вкусовые качества мяса очень высоки. Вес
460
ОТРЯД ГУСЕОБРАЗНЫЕ
мясной тушки составляет 68% от живого веса, т. е. около 300 г, считая за
средний вес осеннего селезня 445 г.
Полевые признаки. Довольно мелкая утка. У селезня яркобелая поло-
са проходит от глаза назад к шее, она четко обрисовывается на общем корич-
невом фоне головы. Верхние кроющие крыла у него голубовато-серые,
на полете они выделяются, как более светлые пятна. Грудь темная в отли-
чие от близкого к нему по размерам свистунка, что хорошо заметно у летя-
щей птицы. Полет быстрый. Голос самки —кряканье, но более высокое,
чем у кряквы. Голос селезня в период весеннего возбуждения—трескучее
«крер-креррерр». Этот крик он издает как на лету, так и сидя на воде.
Осенью зачастую держится большими стаями.
Рис. 84. Чирок-трескунок (самец и самка)
Описание. Строение и размеры. Мелкая утка. Крылья значительно не
достигают конца хвоста. Плечевые перья удлинены и образуют заостренные концы. Клюв
узкий и длинный, примерно равен длине головы, одинаковой ширины по всей его длине,
с узким ноготком. Ноздри вытянутые, лежат в основной части надклювья. Размеры: кры-
ло селезней 187—198, самок 175—194 мм; цевка 25—32 мм: клюв самцов 36,5—42, самок
34—39 мм.
Вес взрослых самцов по прилете в марте 400 г (озера Узбоя), в апреле 450 г (дельта
Волги), 400 г в мае 350 г (р. Или), в апреле 320 г, в мае 346 г C00—375 г) (Рыбинское во-
дохранилище), 348 г C20—390 г) (Бараба). После окончания гнездового периода в июне
375 г C58—403 г) (водохранилище). Перед началом линьки 450 г (Кургальджин), линя-
ющие селезни в августе 367 г B50—450 г), в августе же закончившие линьку 463 г C70—
500г) в сентябре 491 г D80—505 г) (дельта Волги), 550 г (водохранилище), 500 г (Печора);
в октябре 500 г D00—600 г) (дельта Волги). Взрослые самки в августе: добытые от вывод-
ка, не начавшие линьку 332 г B85—360 г) (водохранилище), линяющие в стаях 360 г
B50—550 г), закончившие линьку 441 г D27—460 г) (дельта Волги); в сентябре 520 г (во-
дохранилище), 480 г C90—450 г), в октябре 450 г (дельта Волги). Молодые птицы в сере-
дине июня 150—176 г в июле 250 г, в августе 297 г B70—358 г), в сентябре самцы 415 г
{380—450 г), самки 452 г D00—520 г), т. е. достигается вес взрослых птиц (Рыбинское водо-
ШИРОКОНОСКА 461
хранилище). В дельте Волги это происходит несколько раньше. Молодые самцы там весят
в августе 388 г C50—460 г), в сентябре 396 г C50—425 г), в октябре 386 г C00—400 г),
а самки в августе 374 г C11—411 г), в сентябре 377 г C00—450 г) и в октябре 350 г C00—
400 г).
Окраска. Пуховой птенец. На спинной стороне черновато-бурый с более тем-
ным затылком. У основания крыльев и на пояснице мелкие светлые пятнышки. Брюш-
ная сторона желтая с более темной грудкой. Через глаз проходит темная полоска, а над
глазом светлая, на ухе темное пятно. От свистунка в том же возрасте отличается отсут-
ствием светлого кольца вокруг глаза и более узкой темной полоской на щеке.
Молодые птицы в первом наряде. Похожи на взрослую самку в летнем наряде,
яо грудь и бока несколько рыжее. Каймы на перьях большинства птерилий уже. Они
отличны также пятнистым или исчерченным низом туловища и более тусклыми крою-
щими крыла. По оперению крыла в этом возрасте полы хорошо различимы, так как крыло
имеет в общих чертах ту же расцветку, что и у взрослых. У молодых самцов кроющие
крыла бурые со светлыми оливково-сизыми каймами. У самок зеркало обычно палево-
коричневое без зеленого отлива вли с очень слабым. Кроющие бурые или аспидно-бурые
с узкими более светлыми каймами. Клюв розового цвета, ноги бурые.
Годовалая самка первым летом отличается от взрослой оперением крыла, окрашен-
ным как в первом наряде молодых.
Взрослая самка. На спинной стороне темнобурая с более бледными краями перьев.
Бока головы более светлые, у основания надклювья белые пятнышки; от клюва
через глаз проходит буроватая полоска. Подбородок и горло беловатые. Зоб и бока тела
рыжеватые с темнобурыми центрами перьев. Грудь и брюхо беловатые с мелкими неясными
пятнами, более темными на подхвостье. Кроющие крыла довольно светлые, буровато-
серые. Маховые серо-бурые, у первостепенных стержни белые; зеркало отличается тусклым
зеленоватым отливом, спереди и сзади оно окаймлено белыми полосками. Хвост бурова-
тый. Клюв очень темносерый, несколько зеленоватый, лапы серые, радужина бурая.
Самка после летней линьки имеет более узкие каймы на перьях темени, мантии,
спины и лопаток, а также на внутренних из второстепенных маховых.
Годовалый самец летом сохраняет оперение крыла, свойственное молодым птицам.
Внутренние второстепенные маховые оливково-бурые, с черновато-зелеными внешними
опахалами и беловатой каймой. Иногда часть перьев ювенального наряда сохраняется до
летней линьки.
Взрослый самец в брачном наряде. Голова сверху темнобурая с мелкими белыми
штрихами на лбу; бока головы и шея шоколадно-бурые с белыми стержневыми штри-
хами, подбородок черный. От глаза к затылку, загибаясь на шею, проходит широкая
белая подбровная полоса. Лопатки, спина и надхвостье темные серо-оливковые с буро-
ватыми каемками перьев; плечевые в продопьных черных, белых и голубовато-сизых
полосах. Задняя сторона шеи, зоб и перед груди буровато-коричневые, на зобу более тем-
ный чешуйчатый рисунок, переходящий на груди в поперечную полосатость. Бока тела
сизые с черным струйчатым рисунком, в задней части их широкая белая, а за ней голубо-
вато-серая полоса. Грудь и брюхо тела белые с буроватыми пестринками на брюхе и под-
хвостье. Верхние кроющие крыла сизо-голубые; нижний ряд их с белыми вершинами,
образующими полоску вдоль зеркала. Первостепенные маховые серо-бурые с белыми
стержнями; второстепенные с блестящими наружными опахалами, образующими светлое
зеркало зеленовато-стального цвета, белые вершины их создают полоску, окаймляющую
зеркало сзади, третьестепенные серо-бурые, подмышечные чистобелые. Хвост буро-
ватый, клюв черный; лапы серые, радужина бурая.
Самец в летнем наряде похож на самку, но сразу отличается по расцветке крыльев,
которая одинакова круглый год, а от молодых самцов третьестепенными маховыми и широ-
кими белыми концами больших кроющих крыла.
31. Широконоска Anas clypeata L.
Anas clypeata. Linnaeus. Syst. Nat., изд. X, 1758, стр. 134, Швеция.
Русское название. Широконоска, лопонос (Днепр)—народное,
по форме клюва; соксун (Сибирь)—быть может звукоподражательное.
Распространение. Гнездовой ареал. Британские, Гебридские
и Оркнейские о-ва, причем на этой территории широконоска заметно рас-
селилась и увеличилась в числе за последние 50 лет. Гнездится по всей
Польше, Голландии и Германии, в последней она более обычна в ее северо-
восточной части. Распространение во Франции спорадично—гнездится в Соло-
ни, Бренн и Домб, в устье Роны в ла-Камарг, так же на о-ве Корсика (Майо,
1936), в качестве изолированных мест гнездованья указывается Южная
Испания. Гнездится в Чехословакии, Венгрии, Австрии, Югославии, Маке-
462
ОТРЯД ГУСЕОБРАЗНЫК
доний, Румынии и Болгарии. Кроме того в Малой Азии и на о-ве Кипр.
Населяет Южную Норвегию и Швецию, Финляндию до 63° с. ш., где доходит
изредка до Улеаборга и Торнео. В пределах Советского Союза северная гра-
ница проходит через Карелию. Широконоска гнездится под Беломорском,
но отсутствует на Соловецких о-вах. Под Архангельском гнездится, хотя
и не часто; довольно обычна в Тиманской тундре до самого побережья.
Известны кладки из устья Печоры (Гебель), но по всей реке на гнездовье
редка и встречается не ежегодно (Теплов, 1948). По Оби доходит до Сале-
харда, гнездится в дельте Таза (Бутурлин, 1934); по Нижней Тунгуске гнез-
дится до устья, но в Приенисейских тундрах не наблюдалась; встречается
Карта 52. Распространение широконоски Anas clypeata
а—граница пкмдоынй, а'—районы наибольшей численности птиц на гнездовье, б—не вполне
выясненная граница гнездовий, в—места постоянных встреч летом и спорадичного гнездованья»
г—зимопки, д—залеты, е—встречи пролетных птиц весной севернее гнездовых мест, э*с—места скоп-
лений птиц на линьку
в лесной зоне Таймыра, отмечена на р. Б. Балахне в юго-восточной части.
Гнездится под Якутском и в бассейне Вилюя, но, повидимому, не выходит
на север за пределы лесной полосы. По Яне известна до устья Адычи F8° с. ш.),
для Индигирки пока не отмечена, но встречается в лесистой части низовьеЕ
Колымы на север до Нижне-Колымска F8,5°). На Анадыре, повидимому,
не гнездится, но появляется в некоторые годы на весеннем пролете (Пор-
тенко, 1939), добыта у мыса Наварина; гнездится, повидимому, по всей
Камчатке, но редка там; Командорские о-ва посещает во время обоих проле-
тов (Иогансен, 1934). Известны летние встречи в районе Гижиги, в верховьях
Колымы не встречена совершенно; под Охотском очень редка, как, пови-
димому, и на всем этом гористом побережье. Гнездится на Сахалине, доволь-
но обычна в Уссурийском крае, причем в южной части встречается чаще,
чем в северной. На юге широконоска доходит до берегов Черного моря,
гнездится на Кавказе и в Закавказье к югу до Ленкорани. Для Ирана в ка-
честве гнездящейся птицы не отмечалась. Обычна в Арало-Каспийских сте-
пях и полупустынях, но отсутствует, собственно, в пустынях. Гнездится
ШИРОКОНОСКА
в Марыйском оазисе и возможно в низовьях Теджена (Зарудный, 1896)*
но в последнем месте, видимо, нерегулярно, так как Шестоперовым A936)
не обнаружена. То же относится и к Аму-Дарье, для которой она указывается
Рустамовым A945) в качестве гнездящейся птицы, но рядом других исследова-
телей для нижнего и среднего течения не упоминается. По Нижней Сыр-
Дарье и в прилегающих районах на гнездовье очень редка, зато в течение
всего лета встречается много негнездящихся птиц, ведущих кочующий образ
жизни (Спангенберг, 1936). Гнездится в низменных частях Таджикистана
(Иванов, 1940) и повсюду в Семиречье, кроме его южных гористых окраин
(Шнитииков, 1949). Восточнее гнездится на Черном Иртыше, а возможно
и в смежных частях Синьцзяня, Тувинской авт. обл. (Убса-Нур) и в Северо-
Западной Монголии на Орок-Норе (Козлова, 1934). Редко гнездится в Запад-
ном Забайкалье и в Даурии, обычна в Маньчжурии и на о-ве Хоккайдо.
Летние встречи широконосок известны в Сеистанской котловине в восточном
Иране, Кашгарии, Кашмире и в ряде пунктов Центральной Азии. Харак-
тер пребывания их там не ясен, в большинстве случаев это вероятно не-
гнездящиеся бродячие птицы, но возможны и спорадичные случаи гнездова-
ния, как это зарегистрировано, например, для Цейлона (Уэйт, 1931) и Егип-
та (Гроте, 1930). Гнездовой ареал широконоски в Северной Америке ограни-
чен на севере линией, соединяющей залив Коцебу и устье р. Мекензи; на
востоке его граница проходит через озера Б. Медвежье и Невольничье
к западным окраинам Виннипега и Мичигана. На юге крайними пунктами
служат Чикаго, Канзас-Сити, долина р. Колорадо и ее устье. На западе
широконоска доходит до морского побережья.
Основные зимовки находятся на Британских о-вах и в области
Средиземного моря. В последнем широконоски проводят зиму как на евро-
пейском, так и на африканском берегах, доходя к югу до Азорских и Канар-
ских о-вов, Сенегамбии и Северной Нигерии на западе, Белого и Голубого
Нила, берегов Красного моря и Уганды на востоке. Имеются зимовки на
Азовском море против устьев Кубани (Михайлов, 1937) и в причерномор-
ских районах Болгарии и Румынии. Большие зимовки находятся в Юго-
Восточном Закавказье, по южному берегу Каспия и в низовьях Атрека.
В Средней Азии эти утки зимуют, кроме того, на Теджене, Мургабе и в верхо-
вьях Аму-Дарьи. Много их зимует в Иране по Гюргену на Геррируде, вСеистане
и вдоль морских побережий, особенно обычны зимой всюду в Ираке. Зимуют
повсюду в Индии и Бирме, но многочисленны лишь в северной части. В
Китае проводят зиму в долине Янцзе, в приморских частях страны, к югу
от устья этой реки и в северной части Индокитая, а также на Хонсю и более
южных Японских о-вах, Тайване и Филиппинах.
На Американском континенте северная граница зимовок проходит от
о-ва Ванкувер к верховьям р. Колорадо, затем резко спускается к югу,
но по долине Миссисипи вновь поднимается к северу до устья Миссури; на
Атлантическом берегу зона зимовок начинается примерно от Чисарикского
залива. Южным пределом служит перешеек Тегуантепек.
Скопление ли иных птиц известны в СССР в дельте Волги,
на озерах Чкаловской обл., степного Зауралья (Челябинская и Курганская
обл.), на Наурзумских озерах (Кустанайская обл.), в Барабинской степи
и на озере Кургальджин. В некотором количестве они линяют, повидимому,
на озере Ильмень и в Матцальской бухте (Эстония).
Сезонные миграции. Широконоски, населяющие Скандинав-
ский п-ов, Данию, Северную Германию, северо-запад и север европейской
части Союза, отлетают на запад и зимуют на Британских о-вах, в Голландии
и Северной Франции. Эти зимовки посещают, например, птицы с озера Иль-
мень и из Калининской обл. Птицы из восточных районов европейской части
Союза, Зауралья и Западной Сибири в значительном числе проводят линьку
464 ОТРЯД ГУСЕОБРАЗНЫЕ
в дельте Волги и на озерах Кустанайской обл., откуда они следуют на зи-
мовки в юго-западную часть Каспийского моря, на Азовское и Черное моря
и в средиземноморские страны: Болгарию, Румынию, Грецию, Северную
Италию, Египет, Ливию и Тунис. Однако некоторая, незначительная, часть
птиц этих гнездовых популяций отлетает зимовать к северным берегам Запад-
ной Европы (Голландия). Часть широконосок из Западной и Средней Сибири
линяет в Барабинской степи и отлетает зимовать как к юго-восточному
Туркменскому берегу Каспия, так и на Аму-Дарью, Иссык-Куль и в
Индию.
Характер пребывания. В пределах всего ареала перелет-
ная птица. Районы зимовки совпадают с гнездовым ареалом лишь на Бри-
танских о-вах, но и в отношении этого места известно, что по крайней мере
часть местных птиц на зиму отлетает.
Даты. Зимовки на Британских о-вах птицы начинают покидать в
конце февраля. Основной отлет происходит в марте и заканчивается он толь-
ко в конце апреля. Наших Прибалтийских республик они достигают в начале
апреля, а в середине его появляются под Псковом и Ленинградом. В устье
Мологи валовой пролет наблюдался 23—24 апреля, в северной части Рыбин-
ского водохранилища первые птицы появляются 18—26 апреля. В верхнем
течении Печоры (устье Ылыча) прилет первых птиц приходится на 7 мая
E—10 мая), а начало пролета в среднем на 13 мая (Теплов, 1948). Тиман-
ской тундры широконоски достигают 25 мая, причем пролет их заканчи-
вается довольно скоро (Гладков, 1951). Тропические зимовки широконоски
покидают поздно и часто встречаются в Африке еще в апреле. В северо-запад-
ной части Черного моря они появляются в конце марта, но вало-
вой пролет их проходит в первой декаде апреля (Клименко, 1950), массовый
пролет в низовьях Днепра в начале и середине апреля, причем большая
часть птиц держится в это время не в плавнях, а в степных водоемах (Пачос-
ский, 1911). В Аскания-Нова первые птицы отмечались 9 марта, а послед-
ние 30 апреля (Дергунов, 1926). Интенсивный пролет с закавказских зимо-
вок наблюдается на Апшероне в апреле; на Мангышлаке первые птицы отме-
чены 25 марта (Смирнов, 1916), но южнее в море у Кара-Богаз-Гола они
были еще многочисленны 28 апреля—3 мая. Появление в дельте Волги
5—16 марта; в устье Урала во второй декаде марта, а с конца месяца до
конца апреля они там весьма многочисленны. В среднем течении Урала и
по Сакмаре валовой пролет проходит в середине апреля. В верхнем течении
Оки широконоски появляются в начале апреля (Ефимов, 1915), под Москвой
пролет их продолжается с середины и до конца месяца. Валовой пролет на
Мургабе наблюдался с 7 по 26 апреля (Радде и Вальтер, 1889); на Памире
первые птицы отмечены 15 апреля, а 6 мая они были многочисленны (Меклен-
бурцев, 1946). В северной части Аральского моря пролет проходит во второй
половине апреля и длится около недели, наиболее интенсивен он был 28—
31 апреля (Бостанжогло, 1911). В низовьях Или широконоски появляются
во второй половине марта, пролет идет весь апрель и даже первую декаду
мая (Шнитников, 1949). На Эмбе первые птицы отмечены 11 апреля, а про-
лет продолжался до 3 мая; на Наурзумских озерах они появились 28 апреля.
Начало пролета на Барабе—в конце апреля, а массовый пролет в начале
мая (Янушевич, 1947); появление первых птиц под Тобольском 4 мая
B6 апреля—15 мая) и начало валового пролета 10 мая C—18 мая) (Тарунин,
1929). Отлет с южно-китайских зимовок начинается в марте, пролет в Чжили
продолжается до мая; в Уссурийском крае появляются в половине марта,
а валовой пролет приходится на первую половину апреля. На озере Орок-
Нор (Северо-Западная Монголия) широконоски прилетают в начале апреля;
«ачало пролета по Илиму (правый приток Ангары), прилет под Олекминском
и Якутском отмечены 14—15 мая; на Командорах появляются в конце апре-
ШИРОКОНОСКА 465
ля; первые встречи на Восточной Камчатке 26 апреля—4 мая (до первой
декады июля); встречи на Анадыре в конце мая—первой половине июня.
Осенний пролет в верховьях Печоры начинается около 20 сентября
(8—30 сентября) и длится в среднем 5 дней, но последние птицы встречаются
до 4 октября B3 сентября—10 октября) (Теплов, 1948). В северной части
Рыбинского водохранилища первые пролетные стаи появляются 2—9 сен-
тября, около 26 сентября основной пролет заканчивается, но отдельные
птицы задерживаются до середины октября; примерно в те же сроки прохо-
дит пролет и под Москвой, где заканчивается в начале октября. На места
зимовки у Британских О'вов они прибывают основной своей массой в октябре.
На Хопре пролетные широконоски появляются в середине сентября (Измай-
лов, 1940), в дельте Волги около 20 сентября, массовый пролет приходится
в среднем на 5—19 октября, а время массового отлета на 24 октября. Пролет
через Аскания-Нова отмечался с 30 августа по 30 сентября (Шарлемань,
1924), а в низовьях Днепра—с конца сентября до середины октября; отдель-
ные же птицы попадаются в ноябре (Пачосский, 1911); в январе и феврале
значительное количество широконосок скапливается в бухтах южного берега
Крыма (Дмитриев, 1935). Пролет у берегов Апшерона проходит в сентябре,
на пресных озерах Узбоя в первой половине сентября (Шестоперов),
в низовьях Атрека во второй трети этого месяца (Житников, 1900). На
Теджене их бывает много уже в середине августа, очевидно за счет подлета
недалеко гнездящихся или линяющих птиц. На Памире (озеро Яшиль-
Куль) широконоски наблюдались с 16 сентября (Мекленбурцев, 1946).
Отлет под Змеиногорском в предгорьях Алтая в начале октября; у Соловь-
евска (в среднем течении Амура) их было еще много в последней трети сен-
тября (Тугаринов, 1932); в Гобийском Алтае, на р. Шангин-Гол эти утки
отмечены 25 сентября. Наибольшее скопление птиц на китайских зимовках
(Хэбэй, Цзяньсу) происходит в январе и феврале.
Биотоп. Водоемы в открытых ландшафтах—лесостепи и степи,
а в лесной зоне заселяет широкие и открытые поймы. Таежных рек и озер,
у которых лес подступает к берегам, широконоска определенно избегает. В гор-
ных районах широконоски живут только на озерных плато. Для кормежки
выбирают водоемы с растительностью, но не сплошь заросшие ею, богатые
животными кормами. На соленых озерах бывают столь же обычны, как
и на пресных. Во втором периоде линьки (смена маховых) скрываются в
зарослях надводных растений. Зимой день проводят на море или открытых
плесах крупных озер. Кормятся на мелких внутренних водоемах (зимние
разливы рек, лужи в степи, рисовые поля). На гнездовье мелководные
водоемы широконоски предпочитают большим и глубоким.
Численность. Широконоска обычная, а местами и многочислен-
ная на гнездовье утка на значительном пространстве лесостепной и степной
зоны европейской части Союза, Зауралья, Западной Сибири, Северного
Казахстана и низменного Семиречья (см. карту). Как утка открытого ланд-
шафта в средней лесной полосе она значительно реже, в тайге еще более ред-
кая птица, а в тундру заходит только в северо-восточной Европе.
Экология. Размножение. Время наступления половой зрелости
точно не известно, повидимому, на первый год часть популяции не гнездится.
Об этом говорит значительное количество птиц, ведущих летом бродячий
образ жизни в южных частях ареала вида и за его пределами. На места гнез-
довья широконоски прилетают уже разбившись на пары. Нередко, однако,
можно наблюдать случаи преследования самцами чужих самок. Токовые
позы у широконосок примерно те же, что и у чирков-трескунков, но более
однообразны. Они ограничиваются плаванием одной или нескольких пар
друг за другом по кругу, с клювом, погруженным в воду. Кроме того, как
самец, так и самка покачивают головой вверх и вниз на вытянутой вперед шее,
30 Птица СССР, т. IV
466 ОТРЯД ГУСЕОБРАЗНЫЕ
причем клюв все время сохраняет горизонтальное положение. Движение
это сопровождается низким и тихим покрякиванием. Гнезда устраиваются
на земле, обычно на открытых местах и не очень далеко от воды. Гнездо,
найденное в Тиманской тундре, помещалось на небольшой кочке среди травы
(Гладков, 1951). В средней лесной полосе гнезда широконоски наиболее
обычны в широких и открытых поймах рек, устраиваясь на гнездовье чаще
всего у берегов мелких речек, протекающих по лугам и у луговых озер,
мелких озерков или даже просто временных луж, переходящих летом
в осоковые болота. Столь же охотно гнездятся широконоски и на неболь-
ших открытых островках среди рек и водохранилищ. Гнезда устраи-
вают как на участках с весьма разреженной растительностью, вырубках
и паровых полях, так и на сенокосных лугах. Они избегают, однако, ров-
ного и высокого травостоя, само же гнездо помещают или в куртинках более
высокой травы (часто бурьянов), или в бордюрах из вейника, или среди ред-
ких кустиков осок и злаков. Иногда гнезда находятся под прикрытием невы-
соких и одиночных кустов ивняка, ольхи и шиповника. Основой всегда слу-
жит глубокая ямка, вырытая на сухом месте, с выстилкой из сухих стеблей
и пуха, который появляется в конце откладки яиц и в начале насиживания
их. Обычные размеры гнезда: ширина 20—23 см (до 27), ширина лотка 15—
20 см, глубина 9—11 см. В Среднем Поволжье широконоски гнездятся в доли-
нах более или менее значительных рек, редки на мелких степных водоемах
и совсем отсутствуют на лесных озерах (Житков и Бутурлин, 1906); в сред-
нем течении Донца гнезда их встречали в ольховых зарослях у берегов глу-
хих озер (Аверин, 1911). В степной части Крыма (Тархан-Сунак) гнезда
устраиваются почти исключительно в тростниках (Сеницкий, 1898). В Чка-
ловской обл. любым местом гнездованья служат берега мелких лиманов,
поросших кугой, и тихих, неглубоких плесов степных речек; гнездятся они
и вблизи соленых водоемов. Гнезда устраиваются почти всегда около воды
(Зарудный, 1888). В Барабинской степи гнезда встречались нам на сухих
травянистых гривах и мысах близ озер, на скошенных сенокосах и на паро-
вых полях. Как правило, они помещаются под укрытием более высоких кур-
тинок травы, обычно не скошенных и не потравленных скотом бурьянов.
Только одно гнездо обнаружено в заломах старого тростника, в 800 м от
воды. На юге Барабы широконоски часто гнездятся на гривах с редкой тра-
вяной растительностью среди прибрежных солончаков и на кочковатых
осоковых болотах. В сходных условиях гнездятся они и на Наурзумских
озерах. У широконосок, как и у некоторых других уток, гнезда которых
сильно страдают от разорения воронами и лунями, ясно заметна концентра-
ция гнезд около колоний крачек, гнездовий сизых чаек и крупных куликоа
(озерная лесостепь, Рыбинское водохранилище и другие места). Наблюде-
ния показали, что расположение отдельных гнезд широконоски довольно
постоянно, а некоторые гнездовые ямки занимаются несколько лет подряд.
В тех случаях, когда условия для гнездованья ограничены, широконоски
заселяют устроенные для них искусственные гнездовья—ямки под воткну-
тыми маленькими елочками (Рыбинское водохранилище).
Число яиц в полной кладке для средней полосы 7—11, чаще всего 10*
(Рыбинское водохранилище), но иногда средний размер кладки доходит до-
12, а максимальное число яиц до 13 (р. Молога). Кладки с большим числом
яиц, до 19—20, несомненно снесены двумя самками. Количество яиц у одной
и той же самки, повидимому, постоянно год от года. Нередко встречаются
объединенные кладки с хохлатой чернетью, гнездящейся в тех же условиях
D из 27 на Рыбинском водохранилище). Размер кладки в Эстонии 8—11
(Кох, 1911), гнездо в Тиманской тундре содержало 8 яиц, на р. Лунхе
(в Якутии) 11, кладки в Барабинской степи 9—11 (по Янушевичу, 1947), 5—8
в среднем 6 (?). На Наурзумских озерах 8—9, кладка на Сыр-Дарье 8.
ШИРОКОНОСКА 467
Яйца очень светлые с легким желтоватым или желтовато-оливковым
оттенком. Их размер 48—58x34—39 мм, средний вес яйца 40 г. Ежедневно
откладывается по одному яйцу. Срок насиживания 22—23 дня. В начале
насиживания самка часто и подолгу отлучается с гнезда, тщательно закры-
вая яйца пухом. В то же время самец держится поблизости от гнезда и
встречает отлучившуются от яиц самку. Во второй половине насиживания
самка сидит очень крепко, почти не покидает гнезда и слетает с него почти
из-под самых ног наблюдателя. В это время она часто обрызгивает яйца при
взлете жидким зеленым калом; если яйца после этого не обтерты, утка часто
бросает гнездо. Гибель гнезд широконоски от разорения хищниками и осо-
бенно серой вороной чрезвычайно велика. Гнезда, хорошо укрытые кустами
и густой травой, сохраняются лучше. Средний вес однодневных пуховиков,
по Хейнроту, 28 г, по нашим данным, от 45 до 50 г (р. Молога). От птенцов
кряквы они отличаются мягкими краями надклювья. Утята значительно
охотнее и чаще ныряют, чем взрослые птицы.
В Эстонии кладки яиц происходят в мае. В Тиманской тундре гнездо
со слабо насиженными яйцами найдено 2 июня, 19 и 30 июля; там же встре-
чены выводки с полуоперившимися птенцами (Гладков, 1951). Ненасиженные
яйца под Березовом найдены 20 июня, а еще ниже по Оби 18 и 28 июля на-
блюдались выводки начинающих оперяться птенцов (Дерюгин, 1898); в рай-
оне Якутии гнездо с полной кладкой найдено 19 июля (Маак). В северной
части Рыбинского водохранилища откладка яиц начинается в первой декаде
мая, а в конце месяца бывает уже много как свежих, так и слегка насиженных
кладок; последнее гнездо с яйцами встречено 11 июля. Под Москвой присту-
пают к откладке в начале мая, срок гнездованья у этого вида очень растянут.
В Барабинской степи период откладки яиц с 5 по 30 мая (по Янушевичу,
Золотаревой, 1947), но в некоторые годы в последней декаде мая и первой
декаде июня преобладают еще неполные кладки (Слудский). На Наурзум-
ских озерах пуховики появляются в последних числах июня, с 9 июля
наблюдались почти совсем оперившиеся, а с 17 июля летные молодые (Михеев,
1936). На озере Селете-Денгиз (Сев. Казахстан) пуховик добыт 27 июля,
а на озере Кургальджин встречено 6 крупных пуховиков; в низовьях
р. Или самка с яйцом в яйцеводе добыта 24 мая (Слудский).
Линьки. Широконоски линяют дважды в год: летом почти полностью,
а осенью и зимой частично. Первые признаки смены контурного оперения
у селезней проявляются рано. На Рыбинском водохранилище птицы с пень-
ками на зобу, груди и шее отмечались 7—18 июня. На Алтае (плато Иштык-
Коль) 5 июня добыт селезень с отдельными перьями летнего периода на гру-
ди, боках и плечевых (Сушкин, 1938). В степной полосе первые летние перья
появляются уже в мае, с этим моментом совпадает процесс табунения самцов
и скопления их на местах линьки. На Барабе табуны самцов отмечены
25 мая, на озерах Кургальджинской группы 19—20 июня перелетали большие
стаи селезней, теряющих много пера, на озерах Тели-Куль (Кзыл-Ординской
обл.) они наблюдались 17 июня (Слудский).
Первыми сменяются перья на нижней стороне туловища, голове, плече-
вые и средняя пара рулевых, затем оперение спины и другие перья хвоста.
Крылья линяют последними, примерно через месяц после начала линьки.
Этот этап проходят широконоски довольно поздно, позже чем кряквы, чирки-
трескунки и серые утки. В дельте Волги первые нелетные птицы встречаются
с средины июля A7 июля); в Ульяновской обл. нелетные птицы встречались
в последних числах июля и в начале августа (Житков и Бутурлин, 1906);
в среднем течении Донца смена маховых отмечена в последних числах июня
и в начале июля (Аверин, 1911). В Барабинской степи (район озера Чаны)
начало смены махов приходится на третью декаду июня, массовая линька
наблюдается с 5 июля по 1 августа, начало подъема на крыло 25 июля и мас-
30*
468 ОТРЯД ГУСЕОБРАЗНЫЕ
совый выход из линьки в начале августа (Янушевич, 1947). На озере Кур-
гальджин к началу августа линька широконосок в основном закончилась,
и 8 августа в загоне линных уток их было всего несколько штук (Меженный
и Соколов). На озерах Наурзумского заповедника широконоски начинают
сбиваться в стаи уже в конце мая. К 1 июля отмечались табуны из 500—600
птиц, в которых самок было не более 30—40. Перья груди, брюшка и боков,
а также серповидные плечевые выпадали уже в первых числах июня, а 11
и 22 июля были пойманы селезни с наполовину отросшими маховыми, од-
нако в то же время встречались и птицы, еще не потерявшие маховых (Фор-
мозов, 1937). Первый период линьки широконоски проводят на открытых,
но богатых кормами водоемах и большими стаями отдыхают на песчаных
отмелях. К моменту потери маховых сотенные стаи их переселяются на
водоемы, богатые тростником и рогозом, в зарослях которых они проводят
день, скрываясь от хищников.
Летний наряд, который на спине не всегда бывает полным, носится очень
недолго, так как уже в августе начинается новая смена оперения на брач-
ный наряд. На осеннем пролете в октябре многие селезни имеют уже в основ-
ном брачный наряд. Линька эта заканчивается в декабре, во время нее сме-
няются контурное оперение, внутренние второстепенные маховые и средняя
пара рулевых.
Линька взрослых самок, почему-либо не имеющих выводка, начинается
рано. Они линяют в одних стаях с самцами, составляя около 13,1% от об-
щего числа линяющих широконосок на Барабе и в среднем 10,9% в дельте
Волги. Самки при выводках начинают линять поздно—в июле или даже
в августе, но зато промежуточный срок между началом линьки и сменой
маховых у них много короче. Летом сменяются контурное оперение
(вероятно не всё), рулевые и оперение крыльев. Летний наряд, приобретае-
мый в сентябре, начинает сменяться вновь по всей вероятности в ноябре и пол-
ностью перелинивает только к апрелю. За этот период вновь линяют контур-
ное оперение (вероятно всё), часть рулевых и внутренние второстепенные
маховые. Первая линька у молодых птиц начинается обычно в августе и
продолжается до марта (и дольше). Первый зимний (брачный) наряд молодые
селезни надевают гораздо позднее, чем взрослые, линька молодых и взрос-
лых самок заканчивается одновременно. У молодых селезней осенью сме-
няется контурное оперение, но весьма не полно, на спине и подхвостье, а
также часть рулевых. Зимой долинивает контурное оперение и рулевые,
а также сменяются внутренние второстепенные маховые и их кроющие.
У некоторых особей (нередко) эта линька заканчивается только к началу
летней, отчего наряду с первыми перьями летнего наряда сохраняются еще
юношеские перья на зобу и в нижней части груди. У молодых самок к янва-
рю сменяется контурное оперение, но не все, и изредка внутренние второ-
степенные маховые. Весной они линяют, как взрослые.
Питание. Широконоска животноядная утка. Корм она добывает
с поверхности воды, процеживая ее через прекрасно развитый фильтрующий
аппарат клюва и склевывая корм с растений или дна на той глубине, где
это возможно сделать не ныряя. Основным объектом питания служат мелкие
моллюски и планктонные рачки. В период массового развития дафний широ-
коноски часами вертятся на одном месте, вылавливая ракообразных, скопив-
шихся у поверхности воды (Мантейфель). Нередко они добывают ракообраз-
ных, фильтруя воду, богатую нитчатыми водорослями. В эти периоды их
кал состоит из непереварившейся зеленой нитчатки и остатков Copepoda,
Diaptomus и Ostracoda, а также коловраток (Наурзумские озера; Формозов,
1937). В желудках широконосок, добытых осенью в устье Камы, обнаружено
12 видов моллюсков, а встречаемость их в желудках достигала 94,7% (Тих-
винский, 1931). Весьма быстрая переваримость основных кормов широко
ШИРОКОНОСКА 469
носки очень затрудняет анализ содержимого их желудков, наиболее точные
данные собраны для Рыбинского водохранилища (Немцев). В мае, пока
моллюски составляют там 47% по объему от всего кормового рациона, это
главным образом катушки Planorbis contortus и др. На долю ракушковых
рачков (Ostracoda) приходится 17%, на личинок водяных насекомых 25,2%
(хирономиды, стрекозы и реже ручейники), на взрослых насекомых 6%
и на семена 4,8%. В июле—августе значение моллюсков возрастает G4,8%),
видовой состав их увеличивается до 7 видов и большую роль, наряду с ка-
тушками, приобретают также лужанки (Viviparus viviparus). Ракообразные,
насекомые и чх личинки поедаются несколько реже; семена составляют 13%.
Рис* 85. Широконоска (самец и самка)
Осенью, в сентябре—октябре, значение моллюсков остается весьма большим
F2%), личинки водных насекомых поедаются чаще C1 %), и из них особенно
часто ручейники B6%); семена растений и в это время заметной роли не
играют F,5%). По наблюдениям на р. Мологе A940), ранней весной, во время
половодья, животные корма (моллюски, хирономиды) составляли 30%, а осталь-
ное приходилось на долю зеленых частей растений (ряска трехраздельная,
элодея, роголистник), что говорит о явном недостатке основных необходи-
мых кормов. Пуховые птенцы в этом месте кормились планктонными рако-
образными (Daphne два вида, Cyclops, Diaptomus, Esteria, Limnadia, Ostra-
coda), составлявшими 75% содержимого желудков. Основной пищей птиц
на осеннем пролете служили моллюски (89,5% не менее 15 видов), более
всего лужанки. На волжском взморье, относительно бедном планктоном,
весной основным кормом служат моллюски (особенно Valvata), в августе—
сентябре они же, но в значительном числе поедаются также ракообразные
F%), водяные насекомые и их личинки 5,3% (главным образом тли, сидя-
щие на листьях чилима и нимфейника) и в малом числе личинки рыб (уклеи
и сома) 0,6%. В Барабинской степи моллюски встречены в 71,4% желудков,
жуки и их личинки в 35,7%, неопределенные насекомые в 35,7% и пауко-
образные в 7,1% (Янушевич и Золотарева, 1947). Наконец, зимой в Закав-
казье (Ленкорань) относительное значение животных кормов становится
меньше за счет значительного поедания семян, часть из которых служит кор-
мом, а часть является гастролитами. Больше всего зимой поедается моллюс-
470 ОТРЯД ГУСЕОБРАЗНЫЕ
ков (Dreissena, Planorbis, Hydrobia, Limnaea и реже другие); из насекомых
чаще других служат их пищей водяные жуки.
Хозяйственное значение. Широконоска — существенный объект спортив-
ной и промысловой охоты. Она несколько менее пуглива, чем другие виды
настоящих уток и относительно часто попадает под выстрел. Так, в низовьях
Днепра широконоски составляют около 9% от всего количества уток, добыва-
емых там охотниками (Пачосский, 1911). В промысловых заготовках уток
на территории Западной Сибири приходится 5,5% от числа настоящих уток,
а на закавказских зимовках 6,9% от общего количества уток. Больший
удельный вес в заготовках они имеют на юге своего ареала. Качество мяса
очень хорошо, чистый вес мясной тушки составляет 69% от живого веса
птицы, т. е. около 385 г для селезней в промысловый сезон.
Полевые признаки. В фигуре широконосок характерна относительно
короткая шея, маленькая голова и большой клюв, что особенно отчетливо
видно на полете. Окраска селезня пестрая: темная с металлическим отливом
голова, белые полосы по бокам спины, голубовато-сизый верх крыльев,
темное брюхо и белая грудь. Самка бурая со светлым верхом крыльев. На
воде сидит глубоко и голову держит на втянутой шее. Летает небыстро, изда-
вая (особенно при взлете) крыльями шум, похожий на шум полета кряквы.
Довольно молчаливая утка. Голос самцов—глухое и негромкое «кхо-кхо-
кхо», которое они издают чаще весной; самка крякает однообразно и доволь-
но тихо.
Описание. Строение и размеры. Средней величины утка, клюв длиннее
головы, у вершины он в два раза шире, чем около основания. Ноготок узкий, роговые пла-
стинки очень длинные. Первостепенные маховые узкие и заостренные на конце. Внутренние
второстепенные маховые и плечевые удлиненные и с острыми вершинами только в летнем
наряде, последние шире и в большинстве с закругленным концом. Рулей 14, они с заост-
ренными вершинами, форма хвоста округла. Длина крыла самцов 225—260, самок 205—
230 мм; цевка самцов 32—38, самок 30—35 мм; клюв самцов 62—70, самок 59—65 мм.
Вес взрослых самцов сразу же по прилете в апреле 589 г E50—620 г) (дельта Вол-
ги), в мае 578 г E35—610 г) (Рыбинское водохранилище), 568 г D90—645 г) (Бараба),
650 г (устье Оби), в мае 640 г (Восточная Камчатка). В сезон размножения в июне 600 г
E50—650 г), в июле 635 г F20—650 г) (Рыбинское водохранилище). Линные селезни в авгу-
сте 850 г F50—1100 г), закончившие линьку в августе—сентябре 717 г F50—800 г) (дельта
Волги), 655 г (Рыбинское водохранилище). На осеннем пролете в октябре 692 г E60—
800 г) (дельта Волги). Взрослые самки в апреле 610 г (Астрахань), 500 г (низовья р. Или);
в мае 688 г (Восточная Камчатка), в среднем 550 г, в июне 692 г F11—774 г), в июле 576 г
D77—700 г) (Рыбинское водохранилище). Линные в августе 642 г F00—700 г), закончившие
линьку в сентябре 680 г, на пролете в октябре 664 г F00—800 г) (дельта Волги). Молодые
птицы на Рыбинском водохранилище в середине июня 60—76 г, в июле от 490 до 500 г;
в августе самцы 546 г D50—648 г), самки 522 г D58—650 г); в сентябре самцы 520 г E00—
540 г), самки 533 г D40—640 г). На пролете в дельте Волги в сентябре самцы 592 г E00—
680 г), самки 555 г E20—600 г); в октябре самцы 618гE00—700 г), самки 582 г D80—770 г),
тогда же самки на озерах Узбоя 400 г. На каспийских зимовках в ноябре молодые селезни
563 г E00—660 г), самки 490 г D00— 590 г) (Гасан-Кули); в январе разные годы самцы
418—510 г D00—620 г), самки 441—478 г C12—469 г); в феврале самцы 434—441 г C37—
505 г), самки 387—435 г C00—450 г); в первой половине марта самцы 395 г C00—505 г)
(Ленкорань).
Окраска. Пуховой птенец. Похож на птенца кряквы (клюв расширен
очень мало, но края надклювья мягкие). Верхняя сторона головы и туловища обычно
темнее. Желтые пятнышки на верхней стороне светлее, щеки и полоса через глаз несколько
более коричневатые.
Молодые птицы. В первом наряде похожи на взрослую самку после летней
линьки. Верхние кроющие крыла у них более тусклые, а белая полоска, образованная вер-
шинами больших кроющих, уже. Эти признаки отличают молодых птиц до их первой пол-
ной линьки. Молодые самцы весной сохраняют часто на зобу и груди отдельные перья
юношеского наряда. Полы в первом наряде различимы по окраске верхних кроющих
крыла, которые у самцов значительно голубее, а также рисунком оперения на боках над-
хвостья (у самцов такой же, как у взрослых самцов летом).
Взрослая самка. Голова и спинная сторона туловища коричнево-бурые, на
голове узкие продольные рыжеватые штрихи, на спине того же цвета каемки перьев, а на
надхвостье также бурые поперечные пестрины. Низ тела охристо-соловый в мелких бурых
пестринах, по бокам надхвостья и поясницы бурые округлые пятна. Крылья такие же,
КРАСНОНОСЫЙ НЫРОК 471
-как у взрослого самца, но голубое пятно, образуемое верхними кроющими, имеет более
грязную окраску, а зеленое зеркало очень бледно. Внутренние второстепенные маховые
и плечевые одного цвета со спиной. Хвост как у самца, но более испещрен. Клюв буро-олив-
ковый, у краев более светлый, желтоватый, всегда с небольшим числом мелких темных
крапинок; лапы оранжевые; радужина желто-ореховая.
Взрослый самец в брачном н а р я де. Имеет черную голову и шею с пур-
пурово-синим отливом по бокам. Межлопаточная область серо-бурая; передние плечевые
белые или белые с бурыми отметинами, остальные черно-бурые с белыми центрами, самые
длинные из них голубоватые по наружному опахалу и черные по внутреннему с белой поло-
сой вдоль стержня. Спина, надхвостье и подхвостье черные с сине-зеленым отливом. Зоб
белый, грудь, брюхо и бока тела светлокоричневые, задние перья боков охристо-сероватые
с темным крапом. По сторонам надхвостья белые пятна. Маховые черно-бурые, второстепен-
ные с зеленым блестящим наружным опахалом (образуют зеркало). Большие кроющие кры-
ла бурые с белыми верхушками, создающими кайму пе|.:ед зеркалом. Мелкие кроющие обра-
зуют большое сизо-голубое поле на верхней стороне крыла. Средняя пара рулевых бурая,
остальные почти белые с неправильными буроватыми пятнами. Клюв черный, лапы оран-
жево-красные, радужина от желтой до красновато-оранжевой.
Взрослый самец летом. Похож на самку, но отличается окраской крыла,
черными без пестрин спиной и надхвостьем, а также оперением боков надхвостья, которые
имеют не белую, а соловую окраску с мелкой поперечной рябью.
РОД КРАСНОНОСЫЕ НЫРКИ NETTA КАИР, 1829
Тип. Anas rufina.
32. Красноносый нырок Netta rufina Pall,
Anas rufina. Pallas. Reise d. versch. Prov. Russ., II, 1773, стр. 713, Каспийское море.
Синоним. Anas cinerea. Gmeli n. Reise Russl., II, 1774, стр. 184, Астрахань.
Русское название. Народное, кроме того, красный нырок.
Распространение. Гнездовой ареал располагается в зоне степей
и пустынь от Средиземного моря на западе до Кашгарии на востоке, только
в Западной Европе далеко выдается на север. На западе гнездованья крас-
ноносого нырка распределены спорадично и часто представляют собой изо-
лированные, далеко отстоящие друг от друга колонии. Красноносый нырок
гнездится в Восточной Испании, в Сардинии, Сицилии и некоторых озерах
Алжира (Феццара и Цана). Известно гнездованье на юге Франции (относитель-
но Италии указаний нет), в Швейцарии (Боденское озеро), местами в Гер-
мании, где впервые отмечен в 1822 г. и с тех пор увеличился в числе (Баден,
Мекленбург, о-в Фемарн в Кильской бухте), а с 1940 г. загнездился в Дании
на о-ве Лоланд, откуда понемногу расселяется в другие места (Иесперсен,
1946). Восточнее случаи изолированного гнездованья отмечены в Южной
Венгрии, Чехословакии, Македонии (озеро Ардзан) и кое-где в очень неболь-
шом числе в Румынии и Болгарии (Силистрия, Патев, 1950); известны лет-
ние встречи на севере Малой Азии. По некоторым литературным данным
(Тугаринов, 1941; Тугаринов и Козлова, 1951), гнездится в Молдавии и на
юге Украины. Однако факты, подтверждающие эти сведения, нам неизвест-
ны, и если гнездованье красноносого нырка на этой территории имеет место,
то лишь в немногих пунктах и спорадично, так как в Крыму и в низовьях
Днепра эта утка бывает лишь на пролете и в весьма небольшом числе. Гнез-
дится в низовьях Кубани, на Северном Кавказе по Куме и Тереку (Богданов,
1879). Достоверных сведений о гнездованьи в Азербайджане нет, но найден
гнездящимся на озерах Ахалкалакского плато в Джавахетии (Чхиквишвили)
и на озере Севан в Армении. Несомненно гнездится на Сарпинских озерах
повсюду в Волжско-Уральских песках, очень редок в среднем течении Урала
по р. Илек, изолированные гнездовья отмечены в Башкирии между Уфой
и Белебеем (озера Шунгак-Куль и Аслы-Куль; Сушкин, 1897). Восточнее
красноносый нырок гнездится в небольшом числе на озерах Наурзумской
лесостепи и в Барабинской степи к северу до озера Чаны включительно.
472
ОТРЯД ГУСЕОБРАЗНЫЕ
Далее граница, не доходя до Оби, опускается к югу и проходит вдоль запад-
ных предгорий Алтая к озеру Зайсану и низовьям Черного Иртыша. Восточ-
нее Монгольского Алтая граница распространения вновь поднимается к
северу и захватывает озерную котловину Убса-Нур (Янушевич, 1948). Рас-
пространение в пределах Центральной Азии выяснено еще недостаточно
полно, известно лишь, что красноносый нырок гнездится на озере Орок-Нор
в Северной Гоби, в Кашгарии (Судиловская, 1937), особенно многочислен
на Лоб-Норе и возможно доходит до Гашун-Нора (Сушкин, 1938). Кроме
юго, пара встречена Козловым в Ордосе на р. Хуанхэ 10 мая, однако это
могли быть и пролетные птицы (Бианки, 1915). В Средней Азии распри-
Карта 53. Распространение красноносого нырка Netta rufina
а—граница гнездовий, а'—районы наибольшей численности птиц на гнездовье, б—не вполне вы-
ясненная граница гнездовий, в—обособленные гнездовые колонии, в'—места постоянных встреч
летом и спорадичного гнездованья, г—зимовки, а—залеты, ж—места скоплений птиц на лины<>
странен на юг до Памира; гнездится в верховьях Аму-Дарьи, Пянджа и на
озерах в северной части Таджикистана (Иванов, 1940) В Туркмении обычен
по Мургабу, Теджену и Атреку; в Центральных Кара-Кумах и на Усть-Урте
отсутствует. Гнездится в долине Геррируда, Сеистанской котловине, и,
невидимому, у северо-западного берега Оманского залива (Зарудный, 1911).
Нередко красноносый нырок проникает севернее своего ареала. В некоторых
пунктах Западной Сибири он довольно регулярно встречается на пролетах
(например, на севере Кустанайской обл.); в пролетное же время весной и
осенью залетает на север до Гомеля, Полтавы, Москвы, Горького и Тары.
Основные зимовкив пределах Союза находятся на озере Иссык-
Куль, где красноносый нырок скапливается тысячными стаями (Шнитников,
1949), менее значительные на Аму-Дарье, Пяндже и Вахше, большие на Мур-
габе и Теджене и массовые в юго-восточной части Каспийского моря (от
Красноводского залива до Гасан-Кули и в низовьях Атрека). В большом
числе зимует в Юго-Восточном Закавказье, особенно в Муганской степи и на
озерах Кара-Су (Спангенберг и Туров, 1950), в значительном числе у юж-
ного берега Азовского моря, а также на озере Севан. Южнее большие зи-
мовки этого вида находятся в Иране, у южного берега Каспия, в Сеистане
и в менее значительных у берегов Оманского залива. В некоторые зимы
КРАСНОНОСЫЙ НЫРОК 473
обычен в Пакистане (Тайсхерст, 1927), зимует всюду в Северной и Централь-
ной Индии (местами тысячными стаями), но редок в Декане и далее к юг\,
встречен зимой в Ассаме и Манипуре (Финн, 1921). Европейские птицы зи-
муют у берегов восточной части Средиземного моря: в Италии, на Балканском
п-ове (Скутари), на Кипре, в западной части Малой Азии и в приморских
частях Северной Африки, не залетая далее к югу.
Массовые скопления на линьку наблюдаются на озе-
рах Кургальджинского бассейна, на Челкар-Тенизе (при устье Тургая).
Вторым районом массовой линьки служит Прибалхашье (р. Чу, дельта Или).
Большие стаи селезней и негнездящихся самок проводят лето на озерах
Нижней Сыр-Дарьи и в дельте Аму-Дарьи, по всей вероятности там
они и линяют. В небольшом числе линяют селезни в Кулундинской степи
и единично в дельте Волги.
Характер пребывания. На большей части ареала перелет-
ная утка. Оседлы, повидимому, птицы, населяющие острова Средиземного
моря, Сеистан и быть может Теджен и Мургаб. Селезни совершают летние
перелеты на линьку.
Сезонные миграции и их даты. Большая часть птиц,
населяющих нашу территорию, отлетает зимовать на Каспий, в южные райо-
ны Средней Азии и далее в Ирак и Индию, но существует и юго-западное
направление пролета, о котором говорят зимовки на Азовском море, незна-
чительный, но все же регулярный пролет в северо-западной части Черного
моря, и добыча зимой в Египте нырка, окольцованного на нашей территории
(Михеев, 1949). Из Пакистана красноносые нырки отлетают в марте (но от-
дельные особи задерживаются на местах зимовки и встречались даже 28 июня);
в северо-западной части Черного моря весенний пролет заканчивается к концу
марта—началу апреля (Клименко, 1950); начало отлета с азербайджан-
ских зимовок отмечено 3 марта, последние птицы отлетают в апреле; на
озерах у Атрека пролет наблюдался 29 марта; на пресных озерах Узбоя
первые птицы появились 27 февраля и до начала апреля пролет не прекра-
щался (Шестоперов). В Красноводском заливе последняя стая отмечена
15 мая (Радде и Вальтер, 1889); сильный пролет по Геррируду наблюдался
20—23 апреля (Зарудный, 1903), на Мургабе стаи нырков отмечены 21 —
28 марта. На местах гнездовья красноносые нырки появляются поздно, позд-
нее всех других уток: в дельте Аму-Дарьи в середине апреля (Салихбаев,
1950), в нижнем течении Сыр-Дарьи первые стайки в середине апреля,
а обыкновенными они становятся в конце апреля и в начале мая (Спанген-
берг, 1936); в низовьях Или обычны в марте, но даты первого появления очень
различны в разные годы—27 февраля, 8 марта, 25 марта, 7 апреля (Слудский);
первые птицы в среднем течении Эмбы наблюдались 16 апреля и 26 апреля
они уже были обычны, в низовьях Иргиза первая пара 26 апреля, а вало-
вой пролет 29 апреля —9 мая (Сушкин, 1908); самой северной точки ареала—
Башкирии—достигают в начале мая (Сушкин, 1897).
Осеннее движение этих нырков начинается в сентябре, когда по озе-
рам начинают кочевать большие стаи поднявшегося на крыло молодняка.
В Кустанайской обл. (озеро Мокрое) пролетные птицы наблюдались 2 октяб-
ря; в Семиречье у г. Панфилова они появились в небольшом числе 21—22 сен-
тября, а в массе наблюдались 5—8 октября и 2 —4 ноября (Зарудный и Ко-
реев, 1906); на озере Ала-Куль местные птицы исчезли уже к 15 сентября,
пролетные же появляются позже и встречаются массами на юге Семи-
речья еще в начале ноября (Шнитников, 1949); на озере Иссык-Куль стаи
зимующих птиц собираются в ноябре. В это же время огромное количество
их скапливается в дельте Аму-Дарьи, где на отдельных озерах их насчиты-
вали до 4500 шт., здесь они задерживаются до морозов (Салихбаев, 1950).
На места зимовок у юго-восточного берега Каспия красноносые нырки при-
474 ОТРЯД ГУСЕОБРАЗНЫЕ
летают поздно, в Красноводском заливе первая стайка замечена 2 ноября,
вскоре нырки появились в массе, что совпало с волной холодов, а в поло-
вине января количество их заметно уменьшилось, так как значительная
часть птиц отлетела далее к югу. В Гасан-Кули они появляются также
в начале ноября, а в низовьях Атрека наблюдались большими стаями 26 ок-
тября. В Закавказье нырки прилетают в октябре, но в массе собираются
они на озерах Кара-Су и Шильян во второй половине декабря (Верещагин,
1950); в октябре птицы достигают Пакистана. На северо-западе Чер-
ного моря эти нырки пролетают в небольшом числе, начиная с середины
сентября.
Б и о т о п. В гнездовое время озера, поросшие тростником, с участками
чистой и глубокой воды. Время линьки селезни проводят на озерах с боль-
шими открытыми пляжами, а зимуют красноносые нырки на морских зали-
вах и озерах, богатых подводной растительностью.
Численность. Эта одна из наиболее обычных, а местами и мно-
гочисленных уток водоемов низменной части Средней Азии, равнинного
Семиречья, низовий Иргиза и Тургая, Кургальджинской озерной системы
и в некоторых частях Кулундинской степи (Чистоозерский район), а также,
повидимому, озеро Лоб-Нор и в Сеистане. Столь же обычен он на гнездовье
в Приуральской береговой зоне Каспия. В остальной части ареала красно-
носый нырок значительно более редок.
Экология. Размножение. К гнездованью приступают, повиди-
мому, на втором году жизни (но по Нитхаммеру, на первом году). Большин-
ство птиц прилетает на места гнездованья уже парами, но наряду с ними
присутствуют и холостые самцы, что служит причиной драк, происходящих
как на воде, так и на лету. В брачное время самцы часто летают, распушив
оперение головы, которая кажется от этого очень большой, или расхаживают
около своих самок, сидящих на берегу. На воде токование более оживленно,
селезни часто привстают, выпячивают грудь, закидывают шею на спину,
а клюв опускают книзу и прижимают к шее. При этом они расправляют
оперение головы и издают низкий свист. Период ухаживания и драк между
самцами продолжается в течение месяца.
Гнездятся красноносые нырки в довольно разнообразных условиях»
но чаще всего на озерах с густыми зарослями тростников. Гнезда их в таком
случае устраиваются в тростниках на сплавинах—купаках в центре озера,
около небольших открытых плесов или на завалах старого тростника у бере-
гов. Иногда основанием для гнезда служит куча тростника, сооруженная на
мелководье самой птицей. Особенно охотно гнезда располагаются у самой
воды, на выдающихся мысах и кочках. В долине Сыр-Дарьи гнезда их иногда
находили колониями на одних купаках вместе с белоглазыми нырками,
там же на протоке Чиили они располагались в тростниках, растущих по
берегам, а на Тели-Кульских озерах на сухих островах среди кустарников
джингиля и сухой прошлогодней травы (Спангенберг, 1936). В Туркмении,
недалеко от Мары (Дорт-Кую), найдено гнездо в кусте гребенщука на лугу,
затопленном оросительным каналом. Высота его была 32 см, ширина 24 см>
глубина лотка 12 см (Зарудный, 1896). Гнездо сооружается из стеблей
и листьев тростника и рогоза, по бокам яйца окружаются валиком пуха, кото-
рым закрывается кладка на время отсутствия самки. Число яиц в полной
кладке для Семиречья 6—9 (пуховиков наблюдалось и 10), гнездо на
озере Чаны содержало 7 яиц (в этом же гнезде было 21 яйцо красноголового
нырка; Лавров, 1929). Окраска яиц серовато- или буровато-оливковая, раз-
мер 53,8—59,8x41,8—43,7 мм (Сыр-Дарья), 58—62,2x42—44,4 мм (Ба-
раба). Вес яиц около 60 г. Срок насиживания 28 дней. Птенцы в день вывода
весят 33—39 г (Хейнрот, 1928). Кладки яиц в разной степени насиженности
находились в низовьях Или 17 мая, маленькие пуховые птенцы отмечались
КРАСНОНОСЫЙ НЫРОК 475
там 20—27 мая, а выводки подросших птенцов—в течение всего июня, один
раз и 27 августа (Слудский); в других местах Семиречья гнезда с яйцами
находились 16 мая—2 июня, а 8—9 июня на Балхаше много выводков раз-
личного возраста, 10—12 июля птенцы были уже крупными, но еще нелет-
ными (Шнитников, 1949). На нижней Сыр-Дарье гнезда находили в мае,
а в южной Туркмении 22 мая с ненасиженными яйцами. На озерах А улье-
атинского района 20 мая еще продолжалась кладка яиц, 27 мая все самки
уже сидели на яйцах, а 3 июня обнаружены первые выводки (Кашкаров,
J928). В устье Черного Иртыша самки с совсем еще маленькими пуховиками
попадались 6—10 июня (Поляков, 1914); на Эмбе и Нижнем Иргизе пер-
вые самки с яйцами в яйцеводе добыты 11 мая, гнездо с яйцами на озере Чаны
найдено 30 мая. В Закавказье (Джавахетия) самка с 5 пуховиками встре-
чена 1 августа. В случае опасности самки трещанием сзывают к себе моло-
дых, кормящихся на открытых плесах, и уводят их в заросли тростников.
На 4—5-м месяце жизни одновременно с линькой мелкого оперения темные
глаза становятся красными, а селезни приобретают трескучий голос.
Линька. Два раза в году: полная летом и частичная осенью и зимой.
Послебрачная линька селезней начинается сразу же после того, как самки
сядут на яйца; в низовьях Или первые табунки самцов наблюдались уже 24 мая,
а у самца, добытого 18 июня на Ала-Куле, интенсивно линяли грудь,
брюхо и плечевые перья. 12 июля в дельте Или нелетных (линяющих) селез-
ней было много. Их удавалось встречать до сотни за день (Слудский). В июне
же нелетных селезней встречал там и Штегман A949). В области Нижнего
Иргиза и Тургая селезни сбиваются в стаи в последних числах мая—в начале
июня. У самца, добытого там 16 мая, контурное перо сменилось более чем
на две трети (Сушкин, 1908). Вначале сменяются оперение на нижней поверх-
ности тела и рулевые, начиная со средних, спина линяет позже. Маховые
выпадают примерно через месяц после линьки. К концу июля они уже обыч-
но отрастают настолько, что птица начинает летать, но смена плечевых, перьев
шеи и рулевых еще продолжается. Полностью линька заканчивается обычно
к концу августа. Летний наряд носится менее 10 дней, и уже в первой декаде
сентября начинается линька в брачное оперение. Как и летом, сперва линяет
низ тела, а затем голова и спина, смена рулевых растягивается на долгий
срок. Эта линька застает птиц в пути к их зимовкам и заканчивается обычно
к началу ноября, но у птиц-первогодков затягивается очень долго и нередко
продолжается еще в декабре.
Смена оперения у самок происходит в том же порядке, но начинается она
дней на 20—25 позже, чем у самцов. Перерыва между летней и осенней линь-
кой нет, и часть оперения спины сменяется один раз в год. В дельте Или две
линяющие, уже нелетные самки, державшиеся поодиночке, встречены 13 ав-
густа, причем одна из них совсем не имела маховых, в то время как у добы-
того одновременно самца новые маховые уже отросли. У самки, добытой
26 июня, на Зайсане, началась смена плечевых.
Питание. Красноносый нырок растительноядная утка, питающаяся
зелеными частями растений. Летнее питание его изучено очень слабо, изве-
стно лишь, что на озерах Барабинской степи он кормится преимущественно
листьями рдестов и верхушечными побегами роголистника и урути. Зимой
в Красноводском заливе нырки питаются почти исключительно харой-лучи-
цей, которую достают, ныряя на глубину 2—4 м. Кроме этого основного
корма, составляющего 100 % содержимого полных желудков, в них встре-
чаются мелкие моллюски (Hydrobia, Theodoxus), склеванные, вероятно, вместе
с растениями. На ленкоранских зимовках основу рациона составляют также
зеленые части растений, главным образом хары и роголистника (89,7%),
но потребляются также и семена A0,3%). На озере Иссык-Куль зимующие
птицы питаются также харой и мелкими водными организмами (Фолитарек,
476
ОТРЯД ГУСЕОБРАЗНЫЕ
1945). Наиболее интенсивно кормятся нырки обычно по зорям, и желудки
птиц, добытых в дневное время, бывают обычно пустыми.
Хозяйственное значение. Благодаря своему большому весу (мясная
тушка селезня в промысловый сезон около 775 г) и неплохому качеству мяса
красноносый нырок представляет несомненный интерес как объект охоты.
Первостепенное значение в охотничьем промысле он имеет осенью в дельте
Аму-Дарьи, на Нижней Сыр-Дарье, в Прибалхашье, а также на зимовках
в Красноводском заливе и на Мугани. Вид этот быстро заселяет водоемы,
появляющиеся в южных степях и полупустынях (Алихановский канал,
разливы Кумы и Терека). Распространение и обилие его на юге нашей страны
Рис. 86. Красноносый нырок (самец и самка)
в значительной мере определяются наличием подходящих водоемов. Поэтому
строительство больших и малых водохранилищ и обводнение пустыни быстра
поднимает численность этого ценного вида.
Полевые признаки. Крупная массивная утка. У селезня большая ярко-
рыжая голова, красные клюв и лапы, черные грудь и брюхо. Самка бурова-
тая сверху со светлым низом туловища и щеками. Самец молчалив, и голос
его чаще можно услышать только весной—это своеобразный негромкий
свист. Голос самки громкий, трескучий вроде «кёрр-кёрр-кёрр». Ныряют
хорошо и на значительную глубину, но нередко кормятся и без нырянья,
переворачиваясь вверх ногами, как настоящие утки. На берег выходят часто
и ходят по земле лучше, чем другие нырки, держа тело более горизонтально.
С воды поднимается с трудом, отталкиваясь лапами; полет быстрый, по шуму
крыльев напоминает красноголового нырка. Пролетает обычно парами или
небольшими стайками, а на зимовках держится огромными стаями.
Описание. Строение и размеры. По ряду морфологических признаков
занимает промежуточное положение между речными настоящими утками и нырками. Ноги
и крылья отставлены несколько далее назад, чем у настоящих уток. Размеры лапы велики
средний палец в 1,5 раза больше цевки; задний палец имеет широкую лопасть (шириной!
КРАСНОНОСЫЙ НЫРОК 477
в 0,5 своей длины). Клюв длинный, не короче головы, постепенно суживающийся к верши-
не по всей ого длине. Ноготок узкий, не более половины ширины надклювья. Роговые пла-
стинки по краю надклювья грубые и невысокие. Перья темени и затылка у обоих полов
удлиненные и образуют густой хохол. Рулевых 16, вершины их тупые; форма хвоста слег-
ка округлая. Длина крыла самцов 250—278, самок 243—260 мм\ цевка самцов 42—45,
самок 39—44 мм\ клюв 44—54 мм.
Вес. Сведений о весе красноносых нырков мало. По Хейнроту, обычный вес селезня
около 1000 г, жирного 1200, а самки 875 г. В марте самец весил 1200, а самка 1100—1300 г
{озера Узбоя), в конце апреля самка 1000 г (дельта Или); в мае селезни 1112 г A020—1265 г),
самки 1110 г A050—1210 г) (Бараба, озеро Чаны). Осенью на озере Кургальджин (без
указания пола) 820—1500 г (Меженной и Соколов), селезни в сентябре 1075—1347 г
<дельта Или); в ноябре—декабре птицы бывают очень жирными, слой жира достигает тол-
щины около 1 см, жиром же покрыты все внутренние органы. Взрослые самцы 1500—1550 г,
молодые селезни 1350—1500 г, молодые самки 1000—1200 г, в январе взрослый селезень
1210 г (Закавказье, Карасу); в феврале селезни 1135 г (900—1170 г), самки 967 г (830—
1320 г) (Муганская степь, Ленкорань); зимой на Вахше 1038 г, на Иссык-Куле (без указа-
ния пола) 1170—1431 г.
Окраска. Пуховой птенец. На спинной стороне темный оливково-серый, у осно-
вания крыльев желтоватые пятна. Через глаз проходит темносерая полоска, а выше
ее над глазом—желтая. Брюшная сторона желтая. Клюв красновато-бурый с белым но-
готком, ноги серые с зеленоватым оттенком.
Молодые птицы в первом наряде. Имеют окраску взрослой самки, но светлее
ее. Горло и шея с сероватой рябью; по всей нижней поверхности тела неясные серо-бурые
пятна, которые на зобу и брюшке почти сливаются между собой.
Взрослая самка. Весь верх головы, задняя сторона шеи, спина глинисто-бурые,
плечевые перья со светлыми каймами. Надхвостье серое. Бока головы и шеи светлые пе-
пельно-серые. Низ грязнобелый с неясными более темными центрами перьев. Крылья такие
же, как у самца, но зеркало не белое, а пепельно-серое, окраска верхних кроющих более
тусклая, глинистая. Клюв темный, красноватый со светлой вершиной; лапы красновато-
бурые с черноватыми перепонками; радужина красная. Самки по первому году отличаются
красновато-бурой радужиной.
Самец в первом брачном наряде. Имеет ту же схему окраски, что и взрослый,
но брюхо его не черное, а буроватое, подхвостье не чисточерное и без отлива; хохол
значительно короче и более одноцветно рыжий. Радужина красновато-бурая. Клюв более
бледный, чем у взрослых самцов.
Самец взрослый в брачном наряде. Вся голова, кроме темени и затылка, а
также верхняя часть шеи буро-каштановые с винным налетом на щеках. Удлиненные
перья затылка и темени бледноохристые. Верх спины и плечевые глинистые с оливковым
отливом, остальная спина и надхвостье черные, последнее с зеленым отливом. Над
плечами белые пятна; перья боков, прикрывающие сгиб крыла, черно-бурые с белым
струйчатым рисунком; остальные боков туловища белые с буро-каштановыми верхами
опахал. Шея и вся брюшная сторона до подхвостья черные. Верхние кроющие крыла
глинисто-оливковые, сгиб крыла белый; первостепенные маховые с буроватыми, наружными
опахалами, белыми внутренними и темными вершинами; второстепенные маховые образуют
белое зеркало, а вершины их пепельно-серую кайму: внутренние второстепенные серо-гли-
нистые. В свежем брачном наряде все белые перья имеют бледнорозовый оттенок, что,
вероятно, обязано оранжевому цвету жира самца (у самок окраска жира значительно
светлее). Рулевые темносерые с белыми вершинами. Клюв яркокрасный с беловатым ногот-
ком. Только у очень старых самцов клюв совсем не имеет темных отметин, обыкновенно
же кожа между ветвями подклювья испещрена неправильными черными пятнами, а у бо-
лее молодых они есть и на ветвях подклювья. Лапы красные. Радужина красная.
Самец в летнем наряде. Имеет оперение, сходное с оперением самки, но
окраска спинной стороны темнее и более коричневая. Брюшная сторона более темная
с примесью серо-бурых перьев, особенно на зобу и брюхе, бока рыжеватые. Хорошим от-
личием от самки служит чистобелое зеркало и окраска верхних кроющих крыла.
478
ОТРЯД ГУСЕОБРАЗНЫЕ
РОД ЧЕРНЕТИ AYTHYA ВО IE, 1822
Тип. Anas ferina
Синонимы. Nyroca. Fleming, 1822.—Fuligula. Stephens, 1824.—Fulix. Sun-
d e v a 11, 1835.— Marila. Reichenbach, 1852.—Aristonetta. В a i r d, 1858.
33. Красноголовый нырок Aythya ferina L.
Anas ferina. Li nnaeus. Syst. Nat., 1, изд. X, 1758, стр. 126. Швеция.
Синоним. Anasrufa. G m e 1 i n. Syst. Nat., 1, 2, 1789, стр. 545, nomen praeoccupatum.
Русские названия. Голубая чернеть (широко распространенное), сиваш (на
Черноморском побережье), краснобаш (Средняя Азия) — народные.
Распространение. Гнездовой ареал. В Евразии от Британ-
ских о-вов до Байкала; до границы леса доходит только по Северной Двине.
На юг до Черного и Каспийского морей, дельты Аму-Дарьи и гор Семиречья»
Карта 54. Распространение красноголового нырка Aythya ferina
1—A. f. ferina, 2—А. /. americana; a—граница гнездовий, а'—районы наибольшей численности
птил на гневдовье, б—не вполне выясненная граница гнездовий, в—обособленные гневдопые коло-
нии, в'—места постоянных встреч летом и спорадичного гнездования, е—вимовки, д—валеты, в—
встречи пролетных птиц весной севернее гнездовых мест, ж—места скоплений птиц на линьку
Южным пределом служит зона настоящих пустынь. В Северной Америке
гнездится до озера Атабаски на севере, озер Буффало, Манитоба и устья Не-
браски на востоке, на юг до низовий, р. Колорадо и к западу до гор Сьер-
ра-Невада.
Зимует на Балтийском, Северном и Средиземном морях, на Черном море
и Каспии, в Иране, Индии и у Японских о-вов, в Северной Америке к югу
от о-ва Ванкувера, устья Миссури, озера Онтарио и Лонг-Айленда до
тропика.
Характер пребывания. Всюду перелетная птица, только
на Британских о-вах эти нырки ведут оседлый образ жизни. Селезни совер-
шают регулярные перелеты на линьку.
Биотоп. В гнездовой сезон внутренние водоемы, как правило,
с зарослями тростников; на зимовках обычно большие озера и морские заливы.
Подвиды и варьирующие признаки. Существует две хорошо географически изолиро-
ванные формы, которые часто принимают за самостоятельные виды. Евразию населяет
КРАСНОГОЛОВЫЙ НЫРОК 479
основная форма A. /. ferina Li n n a e u s, 1758, а Северную Америку A. f. americana
E у t о n, 1838, которая отличается меднокрасным отливом на шее, чистобелым брюхом
(у самца и у самки) и голубовато-серым основанием клюва.
Красноголовый нырок Aythya ferina ferina L.
Распространение. Гнездовой ареал. Регулярно гнездится в Ан-
глии и Шотландии, спорадично в Уэльсе; в Ирландии отсутствует. Встре-
чается всюду в Германии и в северных частях ее, обычен также и в Поль-
ше. Гнездится в Дании, Южной и Средней Швеции, Южной Норвегии
и Финляндии. Южнее гнездится в Чехословакии, Восточной Австрии и реже
на Балканах (Югославия, Румыния, Болгария). Западнее и южнее этой
зоны красноголовый нырок указывается для Голландии, Бельгии, Южной
Франции, Южной Испании, Алжира и Туниса (Уайзерби). Неясно, следует
ли включать эти территории в гнездовой ареал вида или считать их изоли-
рованными и не всегда постоянными районами гнездованья. В пределах
Советского Союза обычен в Прибалтийских республиках, гнездится в Ле-
нинградской и Псковской обл.; единичен на Онежском озере и в верхней
части бассейна Северной Двины, известны случаи гнездованья под Архан-
гельском (озеро Слободское, Паровщиков), летние встречи на Соловецких
о-вах (Поляков, 1924) и на юге Удмуртии. Гнездится в нижнем течении
Камы, в Башкирии к северу до низовьев Белой (Сушкин, 1897), по Оби до
Ханты-Мансийска (устье Иртыша, Исаков). По Енисею регулярно до Мину-
синских степей, но отдельные случаи гнездованья известны и севернее (90 км
ниже Красноярска, Юдин; р. Сым, 61° с. ш., Исаков). Встречается в верхнем
течении Ангары, Нижней Тунгуски (до 62° с. ш., Ткаченко, 1927) и на Байка-
ле (Штегман, 1936). Залетает к Якутску (Иванов, 1929), встречен Бергма-
ном A935) в мае и июне на Камчатке, причем некоторые птицы держались
парами; отмечен на Командорских о-вах и о-вах Курильской гряды; в Южном
Приморье известен как регулярно залетная птица (в сезоны пролетов). Гнез-
дится в Тункинской долине (Слудский), в Тувинской и Убсанорской котло-
винах (Янушевич, 1948) и в небольшом числе на Орок-Норе (Козлова, 1930).
Полувзрослый птенец добыт в Восточном Цайдаме (Иргицык, Бианки, 1907);
известны летние встречи в Южном Тибете и Маньчжурии (Мейзе, 1943). На
Алтае распространен спорадично в местностях полупустынного характера
с нерезким рельефом и невысокого уровня, полностью отсутствуя в высоко-
горных лесистых частях его (Сушкин, 1938). Гнездится редко на Марке-Куле
по Черному Иртышу, в равнинном Семиречье, где более обычен в восточной
части (Щнитников, 1949), в низовьях Или очень редок (Слудский), а по р. Чу
на гнездовье пока не встречен. Спорадично гнездится под Ташкентом, регу-
лярно в низовьях Сыр-Дарьи и Аму-Дарьи и на восточном берегу Араль-
ского моря, выше по этим рекам встречается только на пролетах. В Арало-
Каспийских степях южным пределом ареала служит граница распростра-
нения полупресных озер, т. е. примерно 47° с. ш. (Бостанжогло, 1911), гнез-
дится на Камыш-Самарских озерах, а по долине Волги доходит до моря;
повсюду в степях Предкавказья, на юге Украины (в Крыму отсутствует)
и в Молдавии. Изолированно гнездится в Закавказье на озерах Джавахетии
(Чхиквишвили) и в районе Севана. Иногда задерживается до лета на Мур-
габе, летует и возможно спорадично гнездится в Сеистане (Зарудный, 1903)
и Западном Пакистане (Тайсхерст, 1927).
Основные зимовки на севере находятся у берегов Англии
и на самих островах; сотенные стаи красноголовых нырков держатся зимой
у берегов Северной Германии, Бельгии и Франции. В меньшем числе они
зимуют в Южной Норвегии и в датских водах. Массовые зимовки находятся
в средиземноморских странах как в Европе, так и в Африке, причем здесь
птицы в большем числе, чем на севере, посещают внутренние водоемы. На
480 ОТРЯД ГУСЕОБРАЗНЫЕ
Африканском материке эти нырки обычны на юг до Зеленого Мыса и Хар-
тума, а также в Южной Аравии (Гроте, 1930). Зимуют в Сирии, Ираке, в при-
морских частях Ирана и Сеистане, в Пакистане и в большом числе в Север-
ной Индии (не доходя до Майсура), но нередко в Манипуре и Ассаме (Финн,
1901). В Китае зимой бывает единично, так же как и во Вьетнаме; посещает
Филиппины; в Японии нередок на больших озерах и на море, так, на озере
Бива встречался в тысячных стаях совместно с хохлатыми чернетями (Ян,
1942). В пределах Союза большие зимовки находятся на Черном море: в зали-
вах его северо-западной и западной части (Клименко, 1950), в бухтах Южного
Крыма (Федоровский, 1888) и меньше по численности у берегов Аджарии
(Вильконский, 1896), а также на Азовском море у Керчи, Обиточной косы
и устья Кубани. Численность нырка у Обиточной косы очень сильно колеб-
лется по годам. Массовые зимовки находятся в Юго-Восточном Закавказье
как на морском побережье к югу от Махач-Калы, так и на озерах низмен-
ностей Азербайджана; в большом числе зимуют эти нырки и у Туркменских
берегов Каспия к югу от Красноводского залива, также в низовьях Атрека,
по Мургабу и на озере Иссык-Куль.
Скопления на линьку селезней наблюдаются на всем протя-
жении озерной лесостепи и степи от озера Чаны, через Кулундинскую степь,
бассейн Тениза, до Челкар-Тениза, верховьев Эмбы, Наурзумских озер и озер
Зауралья. Птицы собираются в большие стаи, иногда по нескольку сотен
штук, из года в год на одних и тех же определенных озерах. Данные кольце-
вания показали, что очень большой процент селезней возвращается в после-
дующие годы линять на те же самые озера, на которых они были окольцо-
ваны во время линьки. Наряду с этим некоторые самцы линяют и на местах
гнездовья (в степной зоне) маленькими группами или даже в одиночку.
Так, повидимому, они линяют кое-где в Башкирии. Известны встречи лин-
ных птиц в дельте Волги, под Сталинградом и на Каме под Мензелинском
(Першаков). В лесной полосе они линяют в Мацальской бухте Финского
залива и в небольшом числе на пойменном лесном водоеме близ устья Иртыша
(Исаков).
Характер пребывания. Всюду, кроме Британских о-вов и сре-
диземноморских гнездовых колоний, перелетная утка. Кроме того, самцы
совершают летние перелеты на линьку, не всегда совпадающие с общим на-
правлением их пути на зимовку. Характерной особенностью этого нырка
являются частые, а иногда даже регулярные встречи его во время пролетов
(особенно весенних) далеко за пределами ареала (Белое море, Печора, ни-
зовья Оби, Красноярск, Приморье и др.). Эти регулярные вылеты, иногда мас-
совые (как под Красноярском; Шухов, 1925), нередко приводят к случаям
гнездованья отдельных пар (озеро Слободское Архангельской обл., р. Вятка,
Кондинск на Оби, р. Сым на Енисее, Маньчжурия, Камчатка и др.). При-
мерно тот же характер носит и пребывание его в северных частях ареала,
заселенных этим видом недавно и расселение в которых, повидимому, про-
должается. Так, под Москвой он стал гнездиться только с 1900 г. (озере Се-
неж, Дуров), тогда же на озере Тростенском и с 1907 г. в ряде других
мест (Поляков, 1910), а. сейчас не представляет особой редкости. Совсем не-
давно он заселил устье Иртыша, а под Тобольском стал столь же обычен,
как хохлатая чернеть и гоголь, хотя раньше был много их реже. До поло-
вины прошлого столетия красноголовый нырок был для Швеции редким
видом, а к 1902 г. заселил значительную часть страны. Одновременно засе-
лялись Финляндия, Дания, Шотландия, Оркнейские и Гебридские о-ва
(Цедлитц, 1925). Южнее основного ареала имеется ряд изолированных гнез-
довых колоний и районов, для которых зарегистрировано спорадичное гнез-
дованье этого нырка. Возможно, что неопределенность и неустойчивость
границ ареала есть результат того, что водоемы южных степей, в зоне его
КРАСНОГОЛОВЫЙ НЫРОК 481
основного обитания, весьма непостоянны, и в годы усыхания их птицы ли-
шаются мест своего гнездованья и линьки.
Сезонные миграции и их даты. Сведения о перелетах
этих нырков, полученные путем кольцевания, говорят о следующем. У Бри-
танских о-вов зимуют птицы из Северной Германии и Финляндии, а вероятно
также из Прибалтийских Советских Республик и северо-запада России.
На озерах Курганской обл. и Северного Казахстана (Кокчетавские озера)
собираются на линьку птицы, населяющие Зауралье и Западную Сибирь от
Челябинской до Новосибирской обл. и к северу до Ханты-Мансийского окру-
га. Окольцованные там селезни в основной своей массе отлетают зимовать
на Средиземное море, где добывались на протяжении от Южной Франции
до Греции, чаще всего в Северной Италии, а также в Египте (Александрия).
Они следуют на зимовки через Южный Урал, низовья Дона и юг Украины,
оставаясь частично на Черном море. Значительная часть птиц зимует на
Каспийском море как в Азербайджане, так и в Туркмении. Нырки из Казах-
стана отлетают к югу и юго-востоку. Так, нырок, окольцованный в июле
на озере Горьком, добыт в январе около Алма-Аты, а добытый на весеннем
пролете (в апреле) около Кзыл-Орды был окольцован в январе в Пакистане
на озере Хали (Михеев, 1949).
Отлет с британских зимовок начинается в половине марта и продол-
жается до конца апреля (Уайзерби); под Ленинградом эти нырки появляются
в конце первой декады апреля (Бианки, 1907), примерно тогда же на Псков-
ском озере, где наиболее обычны в конце месяца (Зарудный, 1920); в устье
Мологи первые стаи были отмечены 20 апреля и следующие два дня шел их
валовой пролет. Из Албании с озера Скутари отлетают в марте (Рейзер,
1896). Места зимовки в северо-западной части Черного моря они покидают
в первой половине апреля (Клименко, 1950), а берега Аджарии со второй
половины марта (Вильконский, 1896). Из Ирака отлетают в марте; в начале
этого месяца пролетные стайки появляются под Ленкоранью, но последние
стаи отлетают из Азербайджана в апреле. На озерах Западного Узбоя появи-
лись с начала февраля, но интенсивно летели во второй половине марта
(Шестоперов). В большом количестве пролетают вдоль Мангышлака, огибая
полуостров над морем (Смирнов, 1916). На места гнездования красноголо-
вый нырок прилетает поздно. В Среднем Поволжье он появляется в середине
апреля, но до самого конца мая еще встречаются небольшие стайки (Житков
и Бутурлин, 1906). Массовый пролет через Татарию проходит в последних
числах апреля, а в годы с поздней весной и позже (Попов, 1949). Под Чкало-
вом первые птицы появляются в начале апреля, а валовой пролет, стаями
до 150 шт., проходит в конце месяца (Зарудный, 1897); по Иргизу и Эмбе
пролет интенсивен, в верховьях последней первые стайки отмечены 9 апреля,
валовой пролет начался 18 апреля и закончился к концу месяца (Сушкин,
1908). В Барабу прилетают в конце апреля, а массовый пролет происходит
в начале мая (Янушевич и Золотарева, 1947); под Тобольском появляются
около 10 мая B8 апреля—12 мая), массовый пролет бывает 15 мая (Тарунин,
1929). На Памире (Аличур) первый селезень добыт 10 марта (на осеннем про-
лете совсем не встречается; Мекленбурцев, 1946); в низовьях Сыр-Дарьи
первые стайки наблюдались 15 марта, валовой же пролет длился с 26 марта
по 6 апреля, после чего птицы стали редки и совершенно исчезли в середине
месяца. Пролетные птицы были многочисленны, и скученные стаи их густо
покрывали воду озер (Спангенберг и Фейгин, 1936). Первые птицы в низовьях
Или отмечались 6—8 марта (Слудский), пролет через Семиречье в районе
Заилийского Алатау в середине марта, а севернее в Аягузе он наблюдался
23—25 апреля и был довольно Силен (Шнитников, 1949). Наконец, зимовки
на Японских о-вах красноголовые нырки покидают во второй половине
апреля (Ян, 1942).
31 Птицы СССР, т. IV
482 ОТРЯД ГУСЕОБРАЗНЫЕ
Появление первых осенних стай на Рыбинском водохранилище (Дар-
виновский заповедник) наблюдалось 14 августа—19 сентября, время основ-
ного пролета более постоянно, а именно с 18—22 сентября по 22—27 сен-
тября, самая поздняя встреча отмечена 1 октября. Первыми летят взрослые
селезни и только примерно через две недели после них появляются самки
и молодые птицы (Немцев); на Псковском озере эти нырки бывают особенно
многочисленны в последних числах сентября и начале октября, попадаясь
изредка до середины месяца; на места зимовки к берегам Англии они прибы-
вают с начала сентября до середины ноября; особенно интенсивный прилет
идет во второй половине октября. Основная масса селезней, линяющих на
Курганских озерах, покидает эти места в сентябрей не совершает (как пока-
зало кольцевание) кочевок перед отлетом на зимовки. Однако часть птиц
улетает очень рано и рано достигает мест зимовки; так, в Карагандинской
обл. одна птица добыта 5 августа, другая в Красноводском заливе 13 августа
и третья 10 сентября в Италии. В верховьях Тобола разгар пролета был
отмечен в первых числах октября, в верхнем течении Иргиза он начался
в первой декаде сентября, а под Чкаловом основная масса их пролетает
в сентябре. Полет по Узбою наблюдался в течение всего октября, причем
наиболее интенсивным был в последних числах месяца; на озерах Атрека
к 15 октября их было уже много (Житников, 1900); в Красноводском заливе
в 1937 г. в октябре он численно преобладал над всеми прочими утками, но
к середине ноября количество его резко сократилось, что совпало с периодом
холодов. На ленкоранские зимовки красноголовый нырок прибывает уже
й октябре, но количество еще долго продолжает возрастать, и на озерах
Кара-Су и Шильян он появляется в массе только во второй половине декабря,
т. е. тогда, когда замерзает Северный Каспий. В Московской обл. осенний
пролет проходит с середины сентября до начала ноября. Под Днепропетров-
ском в конце октября красноголовые нырки становятся очень редкими (Вальх,
1899); в заливах северозападной части Черного моря пролет, хорошо замет-
ный по увеличению числа птиц, начинается в конце августа, валовой пролет
идет в середине октября (Клименко, 1950); в Югославии они появляются
в октябре, а Северной Италии и Южной Франции (по данным кольцева-
ния) большая часть наших птиц достигает с середины ноября. На Сыр-
Дарье осенний пролет начинается с октября, но бывает много слабее весен-
него; в низовьях Или и на Ала-Куле начало пролета в первых числах
августа, а разгар с середины сентября до половины октября. До Северного
Пакистана они долетают в конце сентября, а в феврале бывают там обыч-
нейшей уткой.
Биотоп. Довольно глубокие озера в открытой местности обычно
с зарослями тростников по берегам и открытыми плесами. Обычно заселяет
йизменности, но вЦайдамеи в Тибете живет на высоте 2600 м. Во время про-
лета и зимовок большие плесы озер и морских заливов служат местами от-
дыха для крупных стай. Для кормежки же эти нырки посещают богатые вод-
йой растительностью водоемы, иногда совсем небольшие. Солоноватых озер,
если они кормны, не избегают.
Численность. На огромном протяжении ареала красноголового
йырка численность его весьма различна. Территории, на которых он вполне
обычен в качестве гнездящейся птицы, относительно невелики. Это примор-
ская зона Советских Прибалтийских Республик; Южная Украина от Дне-
пропетровской обл. до Черниговской; Поволжье от низовьев Камы до Сар-
пинских озер, юг Башкирии, лесостепная и степная зоны Зауралья, Запад-
ной Сибири й Казахстана на север до Тюмени, на восток до Барабинской
и Кулундинской степей, на юг до Челкар-Тениза; Восточное Прибалхашье
и Тункинская долина. На остальном пространстве ареала красноголовый
нырок значительно более редок, распространен неравномерно и гнез-
КРАСНОГОЛОВЫЙ НЫРОК 483
дится не везде регулярно. Так, например, он до сих пор достоверно не найден
на гнездовье в Горьковской, Ивановской, Калининской, Курской, Пензен-
ской обл. и в Белоруссии.
Экология. Размножение. Время наступления половой зрелости
неизвестно. Судя по тому, что стайки этих нырков очень долго задержи-
ваются на местах зимовки (в Пакистане нередки в мае и июне, а встречались
и в июле; Тайсхерст, 1927), можно предполагать, что по крайней мере часть
популяции на первый год в размножении не участвует. Брачные игры их
наблюдаются в пролетных стаях и продолжаются по прибытии на места
гнездовья. В первом случае около одной самки обычно присутствует по не-
скольку самцов, самка плавает среди них кругами с клювом, погруженным
в воду, и издает время от времени хриплые каркающие звуки. Селезни
откидывают голову назад так, что она почти прикасается к спине, а откры-
тый клюв направлен кверху. Шея при этом бывает раздута воздухом.
Затем резким движением голова выбрасывается вперед на вытянутой гори-
зонтально шее. Момент закидывания головы сопровождается тихим свистом,
а при вытягивании шеи вперед выдыхаемый воздух вызывает хриплый, каш-
ляющий звук вроде «икк-икк-коо». Плавая перед самкой и другими самцами,
селезни поднимают голову так высоко, как только это позволяет полностью
вытянутая шея. Поза эта сменяется плаванием с вытянутой горизонтально
над водой шеей. Пары у этих нырков сохраняются вплоть до начала наси-
живания яиц.
В зоне своего основного обитания—в полосе озерной степи и лесостепи
подавляющая масса гнезд находится на тростниковых сплавниках—лабзах,
в заломах тростника и среди прибрежных зарослей его. Из 74 гнезд, осмот-
ренных нами на озере Чаны A941), 12 помещались на сплавинах, иногда очень
небольших—всего около 1 м2, 25—в заломах тростника и 37—среди тростни-
ков, растущих на мелководье. В последнем случае гнезда строились обычно
на глубине до б см, реже до 10, и самая большая глубина, на которой гнездо
возвышалось от грунта, была 30 см. Несколько гнезд найдено на глубине
36 см; они имели скорее характер пловучих построек, сходный с таковым у лы-
сух, но обычно все же были укреплены на подводных корневищах тростника.
Все они были построены из стеблей и листьев тростника. Как правило, гнезда
строятся совсем у воды, не более чем в 2—3 м от нее. В маловодные годы основ-
ная масса гнезд располагается у главного плеса озера или у небольших
лагун среди тростников, а в годы наполнения степных озер значительная
часть гнезд перемещается на внешний край тростникового барьера, грани-
чащий с затопленными осоковыми лугами. Гнезда на заломах помещаются
обычно у края их и даже в этих местах бывают плохо укрыты сверху, те же,
которые находятся среди растущих тростников, хотя и приурочены к более
высоким и густым куртинам, всегда хорошо заметны. Высота гнезд в зави-
симости от постройки их в воде или на сухом месте колеблется от 9—10 до
31—36 см, а диаметр их от 21 до 40 см при обычном размере 27—34 см. Обыч-
ная ширина лотка 15—18 см, а глубина его 6—10 см (от 3 до 12). В особенно
многоводные годы, когда бывает мало сплавин, а тростники глубоко залиты
водой, красноголовые нырки помещают гнезда на осоковых кочках заболо-
ченных озер, в залитых водой кочкарниках и березовых колках, под кустами
на почках (Барабинская степь, Слудский). В эти годы выводков бывает мало.
Вне основной гнездовой территории этого вида (Тункинская долина, плавни
Днепра под Днепропетровском, озере Киёво под Москвой) гнездованье на
осоковых кочках в заводях рек и озер обычное явление. Для поймы Волги
и Камы в Татарии (Попов, 1949) характерны пловучие гнезда среди трост-
ников. В Чкаловской обл. как на тростниковых водоемах (иногда солонова-
тых), так и в затопленных пойменных лесах на кочках у основания ольх
(Зарудный, 1888); в сходных условиях у подножья деревьев на уединенных
31*
484 ОТРЯД ГУСЕОБРАЗНЫЕ
лесных озерах иногда гнездится нырок в Черниговской обл. (Остерский
район, Емельяненко, 1917). Гнездо продолжает достраиваться во время
кладки яиц, и нередко первые яйца лежат на мокром лотке или даже в воде.
Материалом для постройки, как правило, служит тростник, реже сухая
осока и злаки, к началу насиживания откладывается темный пух, укрываю-
щий яйца с боков, а во время отсутствия самки и сверху.
Полная кладка содержит обычно 8 F—15) яиц в Барабинской степи, до
12 (если судить по выводкам) на озере Кургальджин, кладка в 8 яиц найдена
в Тункинской долине, выводок в 8 птенцов в низовьях Или (Слудский),
кладки под Чкаловом 6—14 яиц, в Татарии и в Полтавской обл. до 12 яиц,
в Эстонии—6—10 (Кох, 1911). На Чанах 41% найденных мной гнезд содер-
жал скорлупы расклеванных яиц. В результате гибели кладки часть самок
продолжает нестись в новом гнезде, но кладка их содержит уже меньшее
количество яиц, некоторые же откладывают яйца в гнезда других самок,
в результате чего встречаются гнезда с огромным количеством яиц 22, 24, 27;
в одном гнезде найдено 21 яйцо красноголового и 7 яиц красноносого нырка.
Нередко гнезда бросаются обеими несущимися самками, которые не в состоя-
нии прогреть такие количества яиц, или часть их выкатывается из гнезда
и находится в воде рядом с ним; в одном случае наблюдалось, как в гнезде
одновременно насиживали две самки (Арбузов, 1926).
Яйца крупные, свежеснесенные светлые, зеленовато-голубой окраски,
но скоро они становятся грязнооливковыми. Размер их для Барабы EM5—
69x41—49, среднее 60,6x46 мм. Вес ненасиженных яиц 64,6 г E9—69 г)
(Исаков), для той же местности (озеро Чаны, по Лаврову, 1929) B2) 50,4—
64,4x40,6—45,2, среднее 57x48,5 мм. Насиживать самка начинает после
откладки последнего яйца и сидит довольно крепко. Срок насиживания 24—
26 дней. (Уайзерби). Только что выведшиеся молодые весят около 43—45 г
(до 37 г, по Хейнроту). День после вывода птенцы остаются в гнезде, после
чего самка сводит их на воду. На второй-третий день птенцы уже ныряют
и склевывают насекомых с листьев растений. Оперяются примерно в месяч-
ном возрасте, а в двухмесячном хорошо летают. Вес 57-дневного птенца
800 г. Выводки кормятся обычно вдоль края тростников или на подводных
лугах роголистника и рдестов. В случае опасности они скрываются в тростни-
ках или в другой прибрежной растительности. Подросшие птенцы объеди-
няются в стаи, которые переходят к кочевому образу жизни.
Откладка яиц в Барабинской степи происходит обычно с 5 до 25 мая,
к 15 июня гнезда со свежими яйцами уже отсутствуют, да и с насиженными
их остается очень мало. Вывод молодых продолжается с 5 до 30 июня, а
подъем на крыло происходит 25 июля—25 августа (Янушевич и Золотарева,
1947). В год с очень поздней весной A941) 2 июня большинство гнезд содер-
жало еще неполные кладки, а 8 июня 50% кладок уже насиживалось. Так же
затянулась откладка яиц и в очень многоводном 1930 г. На озере Кургальд-
жин выводок из крупных пуховиков встречен 28 июня; в низовьях Или моло-
дые, достигшие уже веса 500 г, добыты 18 июля, в Тункинской долине (Ингор-
га) 25—26 мая были как свежие, так и сильно насиженные кладки (Слуд-
ский). Под Тобольском большинство выводков красноголового нырка уже
летает к 29 августа B5 августа—6 сентября). В районе Иргиза и Тургая
к середине августа нелетных молодых остается мало. В юго-восточной части
Куйбышевской обл. самка с яйцом добыта 21 мая, а 19 июня самки уже
сидели на яйцах (Карамзин, 1901); в плавнях Днепра у Днепропетровска
кладка свежих яиц найдена 27 июня (Слудский), а во второй половине июля
большинство молодых уже достигает размера перепела или даже чирка
(Вальх, 1899); в Черниговской обл. (Остерский район) выводки бывают в на-
чале июня; под Рязанью в конце июля были уже крупные птенцы (Павлов,
1879).
КРАСНОГОЛОВЫЙ НЫРОК 485
Линька двукратная—летняя полная послебрачная и осенняя пред-
брачная. У самок они несколько заходят одна на другую. В Барабе самцы
начинают сбиваться в стайки и терять перья брачного наряда к началу июня.
Терять маховые птицы начинают в третьей декаде июня; время с 10 июля по
10 августа бывает обычно периодом линьки, в котором основная масса птиц
лишается способности к полету, с 1 августа первые селезни уже поднимаются
на крыло, а начало и середина августа—время массового выхода из линьки
(Янушевич и Золотарева, 1947). Однако смена рулевых, продолжающаяся
еще и тогда, когда птица вновь свободно летает, затягивается у некоторых
самцов до сентября, в этом же месяце появляются и первые перья брачного
наряда (Исаков). Нелетных линяющих селезней отлавливали для кольце-
вания на озерах Курганской обл. и Северного Казахстана в конце июля и в
первой половине августа. В Наурзумском заповеднике (Кустанайская обл.)
12 и 20 июля наблюдалось много линных селезней, но в том же году 11 июля
пойманы два самца уже в свежем летном пере и уже с почти отросшими ма-
ховыми. Линяющие красноголовые нырки держались на больших плесах
в огромных табунах вместе с другими нырками (хохлатыми селезнями,
савками, Формозов, 1937). В Барабинской степи (озеро Чаны) и на Заураль-
ских озерах они проводят день в больших объединенных стаях на обширных
плесах, а на зорях перемещаются к берегам, где и кормятся на мелководьях.
Следует отметить, что линные птицы ныряют менее охотно. На Челкар-
Тенизе в середине июня было много стай селезней, которые группировались
на линьку на Соленых озерах (Сушкин, 1908); на озере Кургальджин боль-
шие стаи перелинявших и уже летных птиц наблюдались 23 августа (Межен-
ный и Соколов). Под Мензелинском (Татария) 17 августа добыты линяющий
и уже перелинявший селезни (Першаков, 1930). Осенняя линька селезней
продолжается обычно с начала сентября до ноября. За это время у них сме-
няется контурное оперение и внутренние второстепенные маховые. Под
Москвой стайка селезней, уже заметно перелинявших в зимний наряд, наблю-
далась 14 сентября (Поляков, 1900), но из числа селезней, добытых в сентябре
наРыбинскомводохранилище, только у двух имелись пеньки нового оперения
на боках, остальные же были полностью в летнем наряде. В Гасан-Кули (Турк-
мения) селезни в полном зимнем наряде добыты 29 ноября (Дементьев, 1945).
Взрослые самки, не имеющие выводков в данном году, линяют в одних
стаях с самцами. На озере Чаны в 1941 г. они составляли 16,1% от числа
отловленных гонами уток (Янушевич). Самки с выводками линяют позже.
Линька их проходит в более сжатые сроки, контурное перо сменяется сразу
на многих птерилиях, и вскоре после начала линьки происходит смена махо-
вых и рулей. Возможно, что часть оперения у них при этой линьке не выпа-
дает. Наиболее интенсивно она проходит во второй половине августа, но
полностью прекращается лишь к ноябрю. У самок, добытых на Рыбинском
водохранилище 3 и 22 сентября, линяли брюхо, грудь и шея. С февраля по
май самки снова линяют. Весной у них сменяются контурные перья (не все),
рулевые (часть) и часть внутренних из второстепенных маховых.
Первая линька молодых птиц начинается в сентябре. Во время нее
сменяется часть контурного оперения, рулевые (у самок иногда не все)
и у некоторых птиц часть внутренних второстепенных маховых. Часть моло-
дых заканчивает свою линьку уже в конце октября, но другая часть сохра-
няет юношеское оперение на груди и брюхе до января и даже до конца фев-
раля. Столь резкая разница в сроках обязана, вероятно, возрасту птиц (из
ранних и поздних кладок), и, конечно, происхождению их из географически
разных мест. Из молодых самцов, добытых в последней декаде сентября из
пролетных стай на Рыбинском водохранилище, только некоторые имели кое-
где пеньки нового наряда и, в частности, средней пары рулей. В конце де-
кабря на ленкоранских зимовках многие молодые селезни еще линяли.
486 ОТРЯД ГУСЕОБРАЗНЫЕ
Питание. Красноголовый нырок питается как растительной, так
и животной пищей, причем соотношение частей меняется в зависимости от
сезона и места. Во время весеннего пролета по р. Мологе нырки питались
зелеными частями растений A2% по объему), побегами элодеи и ряской,
а также семенами (88%), главным образом рдестов. Желудок нырка, добы-
того в мае йа Рыбинском водохранилище, был наполнен личинками ручей-
ников (Phryganea grandis) и хирономид (Tendipes plumoses). Оперяющиеся
утята (в августе) питаются там почти исключительно крупными личинками
хирономид (99,8%). Зато в сентябре, во время осеннего пролета, характер
питания в корне изменяется. Значение животных кормов сокращается до
11,4%. Основу в это время составляют корневища блестящего рдеста (Potae-
mogeton lucetiSy 88,6%), которые птицы достают с глубины 2—3 м, в незначи-
тельном количестве поедаются семена растений (Немцев). На озерах Бара-
бинской степи красноголовый нырок первое время по прилете кормится
преимущественно растительной пищей. Особенное значение в это время
имеют молодые всходы солероса (Salicornia) и стебельчатой лебеды (A trip lex
pedunculatus), которые они, наряду со многими другими утками, собирают
на затопленных солончаках. Однако уже и в это время многие желудки бы-
вают заполнены личинками хирономид, которые в животных кормах зани-
мают на Барабе первое место (Попова). Другие животные корма—жуки и их
личинки, ручейники, пауки, моллюски и пиявки—имеют гораздо меньшее зна-
чение. В южных районах Барабы кормом служат также бокоплавы (Формо-
зов, 1934). Из числа растений наибольшую роль играют рдесты, особенно
осенью. Со времени созревания клубеньков гребенчатого рдеста (Potamogeton
pectinatus) красноголовые нырки энергично питаются этим кормом. Летом
они поедают листья рдестов (нескольких видов) и роголистника, в меньшей
мере урути и пузырчатки. Очень большое значение в кормовом рационе
имеют и семена, особенно свежие семена рдестов вскоре после их созревания
(встречаемость в желудках 92%) и семена резухи (Najas marina). Кроме того,
в значительном количестве поедаются семена камышей (Scirpus) и гречишек
(Polygonum). Зимой в Красноводском заливе пища нырков преимущественно
растительная, 60% рациона составляют харовые водоросли (Chara sp.), 8,5%
семена руппии (Ruppia mar it i та). Однако наряду с этим поедаются и мол-
люски, которые составляют 28% (основа—Cardium edule), и другие беспоз-
воночные 3,5%. На азербайджанских зимовках характер питания нырков
весьма различен в зависимости от того, где они проводят зиму. Птицы,
живущие в крупных стаях на море, питаются животной пищей. В заливе
им. Кирова и около о-ва Сара в 1932 г. содержимое желудков на 100% состояло
из моллюсков (Cardium edule и в качестве дополнения Theodoxus pallasi).
На внутренних водоемах их питание смешанное, основой его служат в пер-
вой половине зимы вегетативные части растений (роголистник, уруть, рде-
сты), а во второй половине разнообразные семена; значение животных кормов
невелико.
Хозяйственное значение красноголового нырка как объекта промысловой
охоты очень велико. По всей Западной Сибири в целом он составляет, около
23,3% от числа всех добываемых уток; в 1931 г. его было заготовлено более
36 000 шт. Основу дичной продукции Тарского округа (Западная Сибирь)
составляют утки, а из них в весеннем сезоне красноголовый нырок (Ушаков,
1925). В еще большей степени это относится к барабинской степи—одному из
основных районов по промыслу водоплавающих птиц. Продукцию весеннего
промысла уток на 86,3% там составляет красноголовый нырок. Качество мяса
его, особенно зимой и поздней осенью, хорошее. Имеются возможности орга-
низовать в это время правильный промысел его безружейными методами.
Полевые признаки. Довольно крупная утка. Голова селезня каштаново-
красная, спина и бока светлосерые с мелкими крапом, грудь и зад черные-
КРАСНОГОЛОВЫЙ НЫРОК
487
У самки передняя часть тела буро-коричневая со светлыми участками около
клюва и на горле. На полете видны белое брюхо, светлая спина и подбой
крыльев, темная грудь и голова. На воде сидит довольно глубоко, хвост
полупогружен в воду. Ныряет хорошо, находясь обычно под водой 25-—
28 секунд. Взлетает очень тяжело, но летит довольно быстро, издавая крылья-
ми резкий звук. По земле ходит неуклюже, высоко поднимая переднюю
часть туловища. На кормовых перелетах летает низко плотными стайками,
на пролете высоко, часто строясь линией или клином. На зимовках держатся
большими стаями. Голос самки хриплое карканье «кэрр-р-р», у самца, обычно
молчаливого, тоже, но в период брачных игр также и свистящий звук.
Рис. 87. Красноголовыи нырок (самец и самка)
Описание. Строение и размеры. Утка среднего размера с плотным туло-
вищем и довольно короткой шеей. Ноги отнесены довольно далеко назад, отчего туловище
у стоящей птицы имеет сильно наклонное положение; лапы крупные. Клюв примерно
равен по длине голове, слегка расширяется у вершины; ноготок узкий. В хвосте 14 руле-
вых. Плечевые со слегка округлыми вершинами. Размеры крыла самцов 213—224, самок
200—215 мм; цевка самцов 36—40, самок 43—47 мм.
Вес взрослых самцов на весеннем пролете (февраль-март) на пресных озерах Узбоя
670—900 г (Шестоперов), в мае на Рыбинском водохранилище 860 г, на озере Чаны 881 г
G20—1085 г) D22). Во время линьки на озерах Зауралья 790—800 г (Курганская обл.
Цецевинский). На пролете в августе 907 г (870—1050 г), в сентябре 998 г (930—1100 г)
(Рыбинское водохранилище). Взрослые самки в феврале-марте 770—820 г (Узбой),
в мае 1015 г (960—1110 г) (озеро Чаны), 850 г (Тункинская долина). В июле от выводка
900 г (р. Или, Слудский). Линные птицы в июле-августе: в среднем 730 г (Курганская обл.),
на пролете в сентябре 947 г (900—995 г) (Рыбинское водохранилище). Осенью на озере
Кургальджин (без указания пола) 740—1050 г (Меженный и Соколов). Молодые птицы
в августе: самцы 355 г, самки 281 г B65—298 г), в сентябре самцы 847 г G90—900 г), сам-
ки 700 г (Рыбинское водохранилище). Преимущественно молодые в октябре: самцы 600—
750 г, самки 700 г (Узбой); в ноябре самцы 678 г F00—800 г), самки 772 г F90—900 г)
(Гасан-Кули); на ленкоранских зимовках в январе-феврале птицы из крупных стай на
море: самцы 1095 г (900—1300 г), самки 905 (850—920 г); птицы с внутренних водоемов
488 ОТРЯД ГУСЕОБРАЗНЫЕ
Ленкоранской низменности и Муганской степи (средние данные для разных лет): самцы
737 и 645 г E90—953 г), самки 631 и 660 г E00—900 г). В первой половине зимы птицы жире-
ют, а зимующие на море остаются жирными до отлета.
Окраска. Пуховой птенец. На спинной стороне бурый со слегка оливковым от-
тенком, бока головы более желтоватые. Низ тела желтовато-серый.
Молодые птицы в первом наряде. Похожи на старую самку, но спинная сторона
у них более одноцветная, серо-бурая, с очень малым числом крапчатых перьев, особенно
у молодых самок. Окраска брюшной стороны однотонная сероватая с очень неясной
мелкой пятнистостью. Бока серо-бурые; зоб более темный, несколько рыжеватый.
Взрослая самка. Голова рыжевато-бурая, более светлая по бокам у основания
клюва и на горле. Лоб и темя темнобурые. Плечи и спина серо-бурые с поперечными
струйками; нижняя часть спины и надхвостье черно-бурые. Шея, зоб и бока тела ржаво-
рыжие, на груди эта окраска постепенно переходит в грязнобелую окраску брюха; подхво-
стье буроватое. Крылья и хвост окрашены, как у самца. Клюв свинцово-голубой, темный
у основания и вершины, лапы серые. У годовалого самца после первой пред-
брачной линьки голова, шея, лопатки и весь верх, как у старого самца, но черный грудной
пластрон очень неполный, пестрый. Струйчатость на перьях выражена очень слабо,
и она не черная, атемнобурая. На брюшной стороне тела обычно остается много перьев
юношеского наряда. Радужина бурая или желтовато-бурая.
Взрослый самец в брачном наряде. Голова и шея ржаво-рыжие, часто с
красно-фиолетовым отливом; зоб, передняя часть груди и плечи черные; спинная сторо-
на пепельно-серая с мельчайшей поперечной струйчатостью, задняя часть спины тем-
нее, надхвостье черное. Бока и задняя часть груди сероватые с неясной поперечной рябью,
брюшко более темное, подхвостье черное. Хвост серо-бурый. Первостепенные маховые
темносерые, второстепенные пепельно-серые с темными крапчатыми вершинами, внутрен-
ние из них серые. Верхние кроющие крыла пепельно-серые с рябью, которая с возрастом
становится все более тонкой и чистосерой. Клюв голубоватый, темнеющий к вершине.
Лапы серые, радужина красновато-бурая.
Самец в летнем наряде. Верх головы и затылок темные, почти чернобурые; бока
головы и шея светлокаштановые, более светлые на горле и шее. Спина и плечевые
буроватые, последние с темным струйчатым рисунком, но темнее, чем в брачном
наряде. Зоб и передняя часть груди серо-бурые с беловатыми каемками перьев; остальная
грудь грязнобелая, крапчатая; брюшко и подхвостье темносерые с черным крапом.
34. Белоглазый нырок Aythya nyroca G iild.
Anas nyroca. Guldenstaedt. Nov. Comm. Ac. Sc. Petrop, XIV, 1, 1769, стр. 403,
Южная Россия.
Синоним. Anas rufa. Li n n a e u s. Fauna Suec, 1761, стр. 47, Швеция.
Русские названия. Белоглазый нырок—книжное; народное—белоглазка, чер-
нушка (широко распространенные).
Распространение. Гнездовой ареал на западе достигает Испа-
нии, занимает Восточную Францию и значительную часть Германии до Бран-
денбурга и Померании включительно. Известны случаи гнездованья отдель-
ных пар в Голландии и Бельгии. Обычная птица Польши, особенно южной
ее половины, в северо-западной части страны доходит до Щецина (Робин,
1923). На юге до берегов Средиземного моря; известен на гнездовье в Марок-
ко, Алжире и Северной Сирии; гнездится в Албании и Румынии. В Болгарии
численность его и распространенность по территории за последнее время
сократились (Патев, 1949). В Прибалтийских Советских Республиках бело-
глазый нырок гнездится настолько редко и спорадично, что включать их
территорию в нормальный гнездовой ареал вида нет оснований. В районе
Псковского озера с последних лет прошлого столетия стал на гнездовье нере-
док (Зарудный, 1910); в Белоруссии же, повидимому, чрезвычайно редок,
так как вполне достоверных сведений о гнездованье его там нет. Одно гнездо
найдено около Дольска, бассейн Припяти (Цедлитц, 1921). Для северной
половины Черниговской обл. не указывается, но в южной гнездится, так же
как под Полтавой и Харьковом; под Киевом гнездится, но немногочислен.
В Орловской, Тульской, Рязанской, Московской обл. появляется во время
пролетов, но не гнездится. То же указывается в отношении Тамбовской,
Курской и Пензенской обл. Как регулярно гнездящаяся птица известен для
БЕЛОГЛАЗЫЙ НЫРОК
489
Воронежской обл. (Северцов, 1855), для долины Суры и Средней Волги (Жит-
ков и Бутурлин, 1906) и как очень редкая для Куйбышевской (Доброхотов,
1937); указания на гнездовье в лесостепной части Татарии не вполне досто-
верны. Севернее этой линии белоглазый нырок указан Генглером и Каве-
линым A909) для Калужской обл. как птица, в некоторые годы обычная
на гнездовье, но последующими исследованиями он там не обнаружен вовсе.
Отмечены случаи гнездованья его под Смоленском (Станчинский, 1915,
1924), под Ярославлем (Кузнецов, 1928) и два года подряд на Рыбинском водо-
хранилище (Немцев). Кроме того, две молодые самки были добыты однажды
под Егорьевском (Рязанская обл., Поляков, 1910). Факты эти следует рас-
Карта 55. Распространение белоглазого нырка Aythya nyroca
а—граница гнездовий, а'—районы наибольшей численности птиц на гнездовье, б-—не вполне
выясненная граница гнездовий, в—места летних встреч птиц и спорадичного гнездованья, г—зи-
мовки, д—залеты, е—встречи ^ролетных птиц весной севернее гнездовых мест
сматривать, как случайное соединение птиц, залетевших севернее границ
своего ареала, иногда единичное, иногда групповое; такие залеты, как видно
на карте, весьма свойственны данному виду. Проводить границу нормаль-
ного ареала по этим крайним точкам нет оснований.
В Низовьях Волги белоглазый нырок обычен, в Башкирии не гнездится,
и выводок (предположительно этого вида) обнаружен под Уфой всего один
раз (Сушкин, 1897). В Чкаловской обл. очень редок; с 1889—1891 гг. стал
гнездиться в небольшом числе по нижнему течению Сакмары (вероятно после
одного из массовых залетов, какой, например, имел место в 1885 г., Зарудный,
1895, 1897). Не обнаружен Сушкиным A908) в верховьях Эмбы, Ори и в сред-
нем течении Иргиза. Замечен Сабанеевым A874) на пролете на Каслинских
озерах и по собранным им сведениям гнездится там, однако после никем в
Зауралье на гнездовье не найден. Очень редко и нерегулярно гнездится на
юге Кустанайской обл., несколько обычнее на Кокчетавских озерах (Штег-
ман, 1934), очень редок на Барабе и лишь однажды был найден гнездящимся
на Чанах (Лавров, 1929). Изредка, обычно в пролетное время, проникает до
Тюмени, Новосибирска, Колывани и даже Нарымского края. До Оби гнездо-
вой ареал, повидимому, не доходит, и границы его спускаются к югу вдоль
западных окраин Алтая. Гнездится по Иртышу выше устья Бухтармы, на
Черном Иртыше, в низовьях р. Урунчу и далее в Синьцзяне и Китайской
490 ОТРЯД ГУСЕОБРАЗНЫЕ
Джунгарии до Юлдуса, Яркенда, и возможно Лоб-Нора. Регулярно гнез-
дится в Кашмире и Южном Тибете (Лхасса, озеро Храмцзо) и доходит на
восток до верховий Голубой реки. В гористой части Семиречья и на Памире
отсутствует. В Таджикистане гнездится по Аму-Дарье, Пянджу (от Термеза
до Кировабада) и в долинах других рек (Иванов, 1940). Гнездящаяся птица
Сеистанской котловины, долины Геррируда (реже), Северного и Северо-Запад-
ного Ирана (до Загроша), Северной Сирии и Малой Азии (?). Отсутствует
на Усть-Урте и в горных частях Кавказа. В последнем белоглазый нырок
обычен в бассейнах Кубани и Терека, гнездится вдоль Черноморского побе-
режья, в долинах рек Аджарии, на Ахалкалакском плато (Чхиквишвили)
и на Севане в Армении.
Основные зимовки. Страны Средиземного моря, где основная
масса зимующих птиц не пролетает южнее морских побережий Африки и Ниж-
него Египта. Сравнительно немногие птицы спускаются к югу до Зеленого
Мыса, Северной Нигерии, Хартума, Абиссинии и о-ва Сокотра. В значитель-
ном числе зимует в Трансиордании, Ираке и Иране, реже в Пакистане, но
весьма обычен всюду на равнинах Северной Индии. На восток область зимо-
вок доходит до Бенгалии, Северной Бирмы, Ассама и Манипура. На терри-
тории Советского Союза крупные зимовки находятся: в Восточном Закав-
казье как в Ленкоранской низменности, так и, особенно, на озерах Кара-Су
и Шильян; на юго-восточном берегу Каспия они многочисленны на взморье
и в низовьях Атрека. Зимует на Теджене, Мургабе и в верховьях Аму-
Дарьи. Кроме того, известны случаи зимовок этих нырков на незамерзающих
водоемах и в более северных пунктах.
Относительно скоплений на линьку и летних миграций
самцов ничего не известно. Повидимому, в основном они линяют в одиночку
на местах гнездовья, что известно для бассейна Иргиза, Тургая, р. Суры
и дельты Волги.
Характер пребывания. На большей части ареала правильно
перелетная птица. Частичная оседлость возможна лишь у птиц, населяющих
юг ареала—средиземноморские страны, озеро Севан, Сеистан и юг Средней
Азии, хотя и в этих местах часть особей совершает сезонные миграции.
Численность. На нашей территории белоглазый нырок одна из
наиболее многочисленных и ландшафтных уток Туркмении, восточного
берега Аральского моря, среднего течения Сыр-Дарьи (Чиили—Джусалы)
и низменного Семиречья (Чу, Прибалхашье), а также нижнего течения Дне-
пра. За рубежом он, повидимому, столь же обычен в Синцзяне, Сеистане и ме-
стами в Средиземноморье. В остальной части ареала этот нырок встречается
значительно реже, а у северных пределов и совсем редок, причем гнездится
разрозненно и нерегулярно. Характерными чертами распространения бело-
глазого нырка служат: неравномерность распределения его по территории,
сильные изменения численности по годам и регулярные, иногда массовые,
вылеты за пределы ареала с оседанием некоторых пар на гнездовье. Все
это делает границы ареала неопределенными, причиной чего служит крайняя
неустойчивость режима полупустынных и пустынных водоемов в зоне основ-
ного обитания этого вида.
Сезонные миграции и их даты. В северо-западной части
Черного моря весенний прилет и пролет начинаются в конце февраля—
начале марта (Клименко, 1950). В низовьях Днепра самая ранняя встреча
8 марта, а валовой пролет проходит во второй половине месяца (Пачосский,
1911), в Аскании-Нова пролетные птицы отмечались с 9 по 28 марта (Шарле-
мань, 1924), под Киевом первые 10 марта, у Черноморского побережья Кав-
каза птицы держатся стайками в течение всего апреля (Вильконский, 1897).
Под Ленкоранью подлет стаек с юга начинается в половине февраля, а отлет
на север в начале марта. Валовой пролет на Мургабе наблюдался с 5 по 26
БЕЛОГЛАЗЫЙ НЫРОК 491
апреля (Радде и Вальтер, 1889). В верховьях Эмбы белоглазые нырки отме-
чены в начале апреля, 20 апреля начался валовой пролет, закончившийся
2 мая. В низовьях Иргиза валовой пролет наблюдался с 27 апреля до конца
первой декады мая (Сушкин, 1908). В нижнем течении Сыр-Дарьи (Джулек)
первые стаи появились 6 марта, а пролет продолжался весь месяц и даже
в начале апреля (Спангенберг и Фейгин, 1936). В дельте Или появляется
обычно во второй половине марта (самая ранняя дата И марта) (Слудский)
и становится обычным к апрелю (Штегман, 1949). Осенний пролет в Семи-
речье начинается во второй половине сентября, он отмечался: на Чу—
13 сентября, на Ала-Куле 16 сентября, по Ко-Су 24 сентября, продолжается
до начала ноября (Шнитников, 1949); под Кульджей первые стаи отмечены
21 сентября, после чего они встречались в течение всего октября (Шесто-
перов, 1929). Осенью в верховьях Иргиза белоглазые нырки летели с 23 сен-
тября почти до середины октября, причем интенсивность пролета постепенно
нарастала. Тогда же наблюдался пролет и в верховьях Тобола, а в нижнем
течении Тургая он продолжался до 25 октября. В низовьях Атрека прилет-
ные с севера стаи появляются в начале октября, а 15 октября они уже встре-
чаются в массе (Житников, 1900); на ленкоранские зимовки они прибывают
в ноябре; на Черноморском побережье Кавказа стаи их наблюдаются с начала
сентября до начала октября. Пролет по Днепру и особенно по Днестру
интенсивен. В низовьях Днепра он продолжается до середины октября
и заканчивается только после начала морозов. Под Днепропетровском бело-
глазые нырки прилетают и отлетают большими стаями. На лиманах Днепра
и Днестра они скапливаются огромными табунками и держатся там до замер-
зания мелководий. В Аскания-Нова пролетные птицы наблюдались с ^ок-
тября до 15 января.
Биотоп белоглазого нырка—достаточно глубокие озера, богатые
водной растительностью и имеющие заросли тростников. Этот нырок явно
предпочитает водоемы низменностей и особенно широкие поймы южных рек,
но встречается иногда и на горных озерах (Семиречье), а в Южном Тибете
и Кашмире даже обычен. Не избегает и лесных водоемов (Киевская обл.).
Зимой проводит день на открытых плесах озер и морских заливов, а кормится
на мелких пресных водоемах.
Экология. Размножение. Время наступления половой зрелости
неизвестно. Сразу же по прибытии на места гнездовий птицы держатся па-
рочками, но наряду с ними присутствуют и холостые самцы, что служит при-
чиной частых драк и преследований между ними. Токующие селезни пла-
вают, держа шею под углом 75° к туловищу; приближаясь к самке, они
резким движением вскидывают голову вверх и назад, но не так далеко к спине,
как красноголовый нырок. В момент наибольшего натяжения трахеи они
издают низкий, хриплый и очень недалеко слышный звук. Кроме того,
они плавают, вытянув шею над поверхностью воды, приподняв и распустив
веером хвост. Для гнездовья выбираются водоемы, обязательно имеющие
заросли тростника. В дельте Или это обычно небольшие и среднего размера
озерки, но не слишком мелководные. Однако иногда гнезда и выводки встре-
чаются там и на совсем мелких озерках между песчаных барханов. На каж-
дом озерке гнездится по 1—2 пары белоглазых нырков (Слудский). На Ниж-
ней Сыр-Дарье они гнездятся небольшими колониями на озерах (обычно глу-
боководных), изобилующих сплавинами и заломами тростников. Гнездовые
колонии состоят или из особей только этого вида, или из сообщества их
с красноносым нырком. Гнезда помещаются обычно на пловучих купаках
среди отмерших стеблей тростника и другой растительности. Гнезда иногда
располагаются совсем близко одно от другого; в одном случае птицы имели
даже один проход к воде в виде узкой водяной дорожки (Спангенберг и Фей-
гин, 1936). В дельте Аму-Дарьи гнезда помещаются как на возвышающихся
492 ОТРЯД ГУСЕОБРАЗНЫЕ
из воды островках и кочках, так и в прибрежных тростниках (Салихбаев,
1950). В средней полосе эти нырки гнездятся на пойменных озерах, богатых
зарослями тростников (низовья Днепра, Умань Киевской обл.), и устраивают
гнезда охотнее всего в тростниках на сплавинах. На Рыбинском водохра-
нилище белоглазый нырок гнездился один раз на опушке затопленного леса,
на поставленном для уток плоту с навесом, а на другой год рядом в корнях
полузатопленного выворотня, причем гнездо находилось в глубокой нише,
над самой водой. Наконец, на самом севере ареала, под Щецином (Польша)
он гнездится на торфяных прудах (Робин, 1923). Повсюду, даже в районах,
где белоглазый нырок очень обычен, он гнездится далеко не повсеместно,
а обильно населяет одни озера и совсем отсутствует на других, смежных
с ними. Гнезда находятся всегда близко от воды, основание их сооружено
из растительного материала и обильно выстлано пухом.
Число яиц в полной кладке обычно 7—11, до 12 и даже 14. Большое
число яиц, безусловно, обязано откладке двух самок в одно гнездо. Кладки
в Семиречье 7—8 яиц, в дельте Аму-Дарьи 10, в Киевской обл. (Умань) 10;
на Рыбинском водохранилище 8. Свежеснесенные яйца светлые с зеленоватым
налетом, но постепенно они становятся светлобурыми. Размер яиц для Сыр-
Дарьи A4) 44,8—47,9x35,3—37 мм (Спангенберг); средний размер (по Уай-
зерби) 52,3x38,2 мм, видимо, западноевропейская популяция этого вида
имеет более крупные яйца. Все яйца около 40 г (Хейнрот). Насиживание
начинается с момента откладки последнего яйца, продолжительность его 28
(по Хейнроту), 25—27 дней (по Уайзерби). Выведшиеся молодые весят 26,5—
31 г. В месячном возрасте у них оформляется оперение на нижней поверх-
ности туловища и на боках; примерно на 35-м дне развиваются пеньки махо-
вых, и только в возрасте около двух месяцев они становятся вполне летными.
Несмотря на ранний прилет, к гнездованью белоглазый нырок присту-
пает сравнительно поздно. В низовье Или самки с яйцами в яйцеводе добы-
вались 16 и 18 мая, хотя сперма в семенниках селезней уже в начале апреля;
28 мая там обнаружена кладка из свежих яиц, только что выведшиеся пухо-
вики встречены 6 июня; на Ала-Куле выводок молодых размером с горлицу
21 июня (Слудский). В нижнем течении Сыр-Дарьи начало откладки яиц
падает на первую декаду мая, но даже 8 июня приходилось находить кладки,
содержащие свежие яйца; первые птенцы наблюдались там в середине июня
(Спангенберг и Фейгин, 1936). На Черном Иртыше две самки с маленькими
пуховиками встречены 11 и 13 июля (Поляков, 1914); в низовьях Иргиза
ухаживание селезней продолжалось до конца мая, а в середине августа
многие птенцы еще плохо летали (Сушкин, 1908). На озере Чаны кладка из
4 яиц найдена 26 мая (Лавров, 1929). В устье Днепра откладка яиц в первых
числах мая, в конце июня,в начале июля уже встречаются выводки взрослых
птенцов, но нелетные молодые бывают нередки даже в августе (Клименко,
1950; Браунер, 1894). В Бендерах выводок трехдневных молодых зареги-
стрирован 15 мая (Остерман, 1912); первая полная кладка в Кельце 20 мая.
Линька селезней двукратная: полная летом и частичная осенью
и зимой. У самок, имевших выводки, обе эти линьки сливаются в одну. Смена
контурного оперения у селезней начинается с первых чисел июля. К середине
июля (по Тугаринову, 1941) перелинивает голова, шея, лопатки и мантия,
начинается линька груди и брюшка. В августе сменяются маховые и начи-
нают выпадать рулевые. Смена последних идет медленно и заканчивается
к октябрю. Уже в сентябре отрастают первые перЪя брачного наряда в пле-
чевых партиях и на мантии в это время еще только заканчивается линька
верхней части груди, которая, возможно, линяет раз в год. Ровный корич-
невый фон оперения этой части туловища появляется в результате обновле-
ния светлых каемок перьев. Полный брачный наряд старые селезни наде-
вают в ноябре.
БЕЛОГЛАЗЫЙ НЫРОК
493
Некоторые самки, вероятно не имеющие выводков, начинают линять
одновременно с самцами, самки с выводками значительно позже их, отчего
одновременно попадаются птицы в весьма различной стадии линьки. Так,
на Черном Иртыше 6 июня добыта самка, у которой уже сменилось 9 руле-
вых, у добытой 7 июля сменилось только 3, а у самки от 13 июля—4 (Поля-
ков, 1914). Линька самок заканчивается, повидимому, зимой. Какие части
оперения линяют у них дважды в год—неясно.
Молодые самцы начинают линять в октябре и до февраля сменяют опе-
рение на голове, шее и частично на туловище, рулевые и часть внутренних
второстепенных маховых.
Рис. 88. Белоглазый нырок (самец и самка)
Питание. Белоглазый нырок преимущественно растительноядная
утка, состав питания его до сих пор изучен плохо. Он поедает листву и ко-
решки некоторых водяных растений (валлиснерии, рдестов, особенно гре-
бенчатого роголистника, лягушатника, ряски и др.) и семена. В меньшем
количестве он потребляет животные корма (личинки хирономид, стрекоз,
ручейников, водяные жуки и реже моллюски). На ленкоранских зимовках
основу пищевого рациона составляют семена главным образом камышей
{Balboschonus maritimnus) и куриного проса (Echinochloa crus-galli), в мень-
шем количестве в желудках встречаются вегетативные части растений и остат-
ки насекомых. На кормежках он ведет себя сходно с настоящими утками,
т. е. вылетает на жировку с вечера и возвращается отдыхать на открытые
плесы утром.
Хозяйственное значение белоглазого нырка относительно велико. Мясо
его очень высокого качества, но существенное значение в промысле он имеет,
пожалуй, только в дельте Аму-Дарьи и в Прибалхашье. В значительном
количестве его добывают и на зимовках.
Полевые признаки. Некрупная темноокрашенная утка с рыжевато-
каштановой головой и белым глазом (самец). На полете характерна темная
окраска брюха у самца. Голос самки грубый, каркающий, как у других ныр-
494 ОТРЯД ГУСЕОБРАЗНЫЕ
ков, голос самца удается слышать редко. Прекрасно ныряет, с воды подни-
мается довольно тяжело.
Описание. Строен и е и размеры. Клюв короче головы, неширокий A7— 20мм)
и более или менее одинаковой ширины от основания до вершины. Ноготок очень узкий.
Ноздри лежат в основной половине надклювья. Лопасть заднего пальца равна половине
его длины. Рулевых 14. Длина крыла самцов 177—193, самок 170—185 мм; цевка самцов
29—31 мм; клюв самцов (от оперения лба) 38,5—43,2, самок 36—39,5 мм.
Вес. Сведений о весе белоглазых нырков очень мало. На весеннем пролете в дель-
те Или птицы (без указания пола) весят 525—689 г (Штегман, 1949); нырок, добытый там
же 15 ноября (пол?) 450 г (Слудский). На ленкоранских зимовках в январе-феврале самцы
583 г E00—650 г), самки 520 г D10—500 г); очень суровой зимой в январе на озерах Кара-
Су средний вес (без указания пола) равнялся 400—480 г, а максимальный доходил до
530 г A2 экз.), причем птицы были хорошо упитаны и имели подкожный жир (Спангенберг
и Туров, 1950).
Окраска. Пуховой птенец. На спинной стороне темный, почти черный с отдель-
ными длинными золотистыми перышками. Бока головы желтоватые, горло и подбородок
беловатые, брюшко и бока такие же темные, как спина, остальной низ серо-желтый.
Молодые птенцы в первом наряде. Сходны со взрослой самкой, но голова их
темнобурая без рыжих тонов, с более светлыми буро-каштановыми боками. Спинная
сторона также светлее, охристо-бурая. Зоб серовато-бурый, ниже к груди окраска
светлеет. Белые части груди и подхвостья испещрены серо-бурыми пятнами. Крылья, как
у взрослых. Клюв шиферный голубовато-черный. Радужина бурая, к маю становится
жемчужно-серой.
Взрослая самка. Спинная сторона темнобурая, голова рыжевато-каштановая, но
несколько более бледная, чем у самца; кольцо у основания шеи серо-бурое. Зоб,
передняя часть груди и бока рыже-бурые, остальная брюшная сторона серовато-бурая,
грудь в темных бурых пестринках. Крылья и хвост, как у самца. Клюв очень темный,
черновато-свинцовый, ноги зеленовато-серые, радужина коричневая. Молодой самец
в первую же зиму надевает наряд, не отличимый от взрослого.
Взрослый самец в брачном наряде. Голова, часть шеи, зоб, передняя часть
груди рыжевато-каштановые с легким фиолетовым отливом; у основания шеи черное
кольцо. Задняя сторона шеи, плечи, спина и надхвостье черные. На подбородке
белое пятно, середина груди и подхвостье белые. Брюшко черновато-рыжее с мелким бе-
лым крапом, бока ржаво-бурые. Кроющие крыла черно-бурые, первостепенные маховые
бурые с беловатыми внутренними опахалами; второстепенные маховые белые с черно-бу-
рыми вершинами образуют зеркало, внутренние второстепенные черные с зеленоватым
отливом. Рулевые черно-бурые. Клюв синевато-черный, лапы свинцово-серые, радужина
белая.
Самец в летнем наряде. Голова и шея более бледного каштанового цвета, кольцо
у основания шеи бурое. Спинная сторона туловища темнобурая со светлокоричневыми кай-
мами перьев. Каштановые перья груди с широкими белыми каймами; на брюшке много
серо-бурых пятен.
35. Нырок Бэра Aythya baeri Radde
Anas (Futigula) baeri. R a d d e. Reise im Suden von Ost-Sibirien, II, 1863, стр. 376, Амур.
Русские названия. Нырок Бэра, восточный белоглазый нырок—книжные
Распространение. Ареал. Гнездится в бассейне Уссури, Ханкайской
низменности и на озерах Посьетского района, однако вполне обычен лишь
в последнем месте. Севернее же, например в низовьях Имана, этот нырок
хотя и довольно обычен, но встречается реже всех других уток. Судя по
экземплярам, добытым в летнее время, доходит на север до Амура, где встре-
чен в устье Уссури, вниз по Амуру до Мариинска и вверх до устья Зеи.
В последнем месте встречается не единично (Радде, 1863). Кроме того, гнез-
дится в Южной Даурии по Аргуни под 50° с. ш. (Тачановский, 1874), встре-
чен на Шилке под Сретенском и нередок на гнездовье и в Маньчжурии.
Залетный экземпляр добыт на Камчатке. В Корее попадается на пролете,
но редок и малоизвестен, есть экземпляры, добытые в ноябре и декабре.
Так же редок и в Японии, где встречается лишь зимой и во время пролета.
Зимует в Китае на юг до Цзяньсу и Фудзяня, где, однако, редок. Есть
экземпляр, добытый в сентябре на о-ве Малый Кукунор в Алашане (Прже-
вальский). Достигает в зимнее время Бирмы и Бенгалии. В последней поя-
НЫРОК БЭРА
495
вляется в большем или меньшем количестве каждую зиму, причем в некото-
рые годы бывает очень обычен (в холодные зимы). В Бенгалии добыто несколь-
ко экземпляров, по характеру оперения промежуточных между восточным
и обычным белоглазым нырком (Финн, 1900).
Характер пребывания. Перелетная птица, повидимому,
на всем протяжении гнездового ареала.
Даты. На местах зимовки по берегам Чжилийского залива задержи-
вается до конца марта. На юге Уссурийского края появляется в 20-х числах
Карта 56. Распространение нырка бэра Aythya baeri
а—граница гнеэдовий, б—не вполне выясненная граница гнездовий,
г—зимовки, д—валеты
марта, а валовой пролет происходит в первой половине апреля (Воробьев).
На осеннем пролете в Корее добывались в октябре и декабре (Остин, 1948).
В Фудзяне (Южный Китай) появляются в ноябре—декабре, в Цзяньсу в январе
(Колдуэлл, 1931).
Биотоп. В гнездовое время—небольшие озера низменности, пред-
почтительно с богатой водной растительностью и наличием тростников.
Повидимому, безлесные местности заселяются обильнее, чем озера с ле-
систыми берегами. Во время пролета не избегают и рек с быстрым
течением.
Численность. Обычная и характерная гнездящаяся птица на юге
Приморья и в восточной половине Маньчжурии. В остальной части ареала
довольно редок. На зимовке обычен в средней части Приморского Китая
до устья Янцзы, южнее которого редок.
Экология. Размножение. Весной у этих нырков иногда наблю-
даются своеобразные игры в воздухе, имеющие значение токового полета.
Пржевальский описывает их в своем дневнике Уссурийской экспедиции
496 ОТРЯД ГУСЕОБРАЗНЫЕ
ции следующим образом: «В конце марта (ст. ст.) я часто видел, стоя где-нибудь
в скрытом месте, как эти утки, небольшими A0—12 экз.) стаями с особым
тихим криком долго и невысоко летали около одного места, описывая круги.
Вероятно, самцы гонялись здесь за самками».
По наблюдениям на Аргуни (Тачановский, 1893) этот нырок прилетает
уже парами; однако в Уссурийском крае он довольно долгое время по при*
лете держится стайками. Так, в низовьях Имана, стайки и небольшие группы
встречались в 1938 г. до 14 мая, но в последних числах мая и в начале июня
нырки держались уже парами и одиночками. 9 июня 1938 г. там было най-
дено гнездо этого вида с 10 сильно насиженными яйцами. Гнездо обнаружено
на маленьком озерке в низменной части долины Имана. Оно было устроено
в густой осоке метрах в 7 от воды и обильно выстлано пухом самки. С 14 июня
птицы этого вида на глаза больше там не попадались, повидимому, к этому
времени все местные утки сели на яйца, а селезни, вероятно, откочевали на
линьку (Спангенберг, 1940). В Забайкалье гнезда находили на сухих местах
в траве по берегам озер и речек.
Яйца по окраске и форме похожи на яйца белоглазого нырка, но отли-
чаются крупными размерами. Их окраска желтовато-серая, такая же и на
просвет. Размеры: из Забайкалья 51—55x38—39 мм; из Приморья (по одной
кладке) 50,8—53x37,8—39,8 мм. 21—22 июня 1927 г. на озерах Посьет-
ского района встречены утки небольшими группами—холостые или потеряв-
шие выводки. Это говорит как будто бы о том, что по крайней мере часть птиц
не гнездится в годовалом возрасте.
Линька. О линьке этого вида ничего не известно. Самец, встречен-
ный 5 августа 1926 г. на озере Ханка, был в брачном пере—с коричневой
грудью и темной головой (Шульпин, 1936).
Питание. Состав питания неизвестен. Весной (по Дулькейту)
посещает на кормежке луговые прудки и лужи, уничтожая в большом числе
лягушат. Местами на зимовках в Индии считается почти несъедобной уткой
из-за рыбного привкуса мяса.
Полевые признаки. Характерна темная окраска и большое белое зеркало
на крыле, хорошо заметное издали. Посадка на воде, как у хохлатой чернети;
белое пятно на крыле заметно и у плавающих птиц.
Описание. Размеры и строение. Морфологически весьма схож с белогла-
зым нырком. Череп отличается относительной массивностью, по сравнению с другими пред-
ставителями этого рода. Мелкая утка; длина крыла самцов 200—240, самок 190—215 мм;
цевка самцов 32—36, самок от 30 мм; длина клюва самцов 44, самок 43 мм.
Окраска. Пуховой птенец неизвестен.
Молодые птицы в первом пере. Голова светлобурая с более темным теменем;
спинная сторона однообразно глинисто-бурая; зоб и передняя часть груди бурые;
задняя часть груди беловатая с серо-бурыми пятнами; брюшко рыже-бурое; подхвостье
белое. Крылья, как у взрослых птиц. Клюв темносерый, глаза бурые, лапы серые с
черноватыми перепонками.
Самка взрослая. Темя и затылок черные с легким зеленоватым блеском, бока
головы темные рыжевато-бурые. Плечи и мантия серо-бурые с рыжеватыми каймами
перьев, спина и надхвостье черно-бурые. На горле и шее спереди примесь белого, зоб
и верх груди рыжевато-бурые. Остальная часть груди и подхвостье белые с черными
пятнами, брюхо черно-бурое с большей или меньшей примесью белого. Бока ржаво-
бурые с охристыми вершинами перьев. Крылья, как у самца, только внутренние опахала
больших маховых не белые, а светлосерые. Глаза обыкновенно бурые.
Самец в брачном наряде. Голова и вся шея черные с ярким зеленым отливом.
На подбородке белое пятнышко. Спина и надхвостье матово-черные. Мантия черно-
бурая с очень мелким рыжим крапом. Зоб и передняя часть груди блестящие
рыже-каштановые, задняя часть груди и подхвостье белые, брюшко серо-бурое перехо-
дит в рыже-каштановые бока. Хвост черно-бурый. Наружные первостепенные маховые
черно-бурые с белыми внутренними опахалами, у внутренних первостепенных маховых
оба опахала белые, темной остается вершина. Второстепенные маховые белые с широкой
черной вершиной образуют белое зеркало крыла. Верхние кроющие крыла черно-бурые
с оливковым отливом. Клюв свинцово-серый с черным основанием и ноготком. Лапы также
свинцово-серые. Радужина белая или светложелтая.
ХОХЛАТАЯ ЧЕРНЕТЬ
497
36. Хохлатая чернеть Aythya fuligula L.
Anas Fuligula. Li n n a e u s. Syst. Nat., изд. X, 1, 1758, стр. 128, Швеция.
Синоним. Anas colymbis. Pallas. Zoogr. Rosso-Asiat., II, 1811, стр. 266, вся Рос-
сия и Сибирь.
Русские названия. Чернь, чернушка, белобок (обязано резкому контрасту
окраски боков и остального туловища)—широко распространенные, народные; травник
(Печора), морская чернь (Посьет, Приморье)—по занимаемым биотопам.
Распространение. Гнездовой ареал. Регулярно гнездится на
Британских о-вах, случайно в Исландии, на Фарерах и возможно на Мед-
вежьем о-ве; в Норвегии до 69,5° с. ш. (в Ост-Финмаркене бывает регулярно,
Карта 57. Распространение хохлатой чернети Aythya fuligula
а—граница гневдовий, а'—районы наибольшей численности птиц на гнездовье, б—не вполне выяс-
ненная грапица гнездовий, в—обособленные гнездовые колонии, в'—места постоянных встреч летом
и спорадичного гнездованья, г—зимовки, д—залеты, е—встречи пролетных птиц весной севернее
гнездовых мест, э#с—места скоплений птиц на линьку
но гнездованье не доказано). Повсюду в Швеции и Финляндии. Очень редко
гнездится в Голландии, Дании в северной части Германии (Бранденбург,
Померания), на севере Силезии (Щецин, Робин, 1923), в других частях Поль-
ши бывает лишь на пролетах. Есть сведения о гнездовании в Чехословакии,
Австрии, Албании, в Югославии, Румынии и Болгарии, на Кипре и в
Сирии на Антиохийском озере. Однако все эти данные не вполне достоверны
и не обрисовывают границу ареала вида, а показывают лишь пункты изоли-
рованных гнездовий. В течение последнего столетия хохлатая чернеть рас-
ширяет свой ареал на северо-западе. В 1835 г. она занимала небольшие гнез-
довые площади в Шведской Лапландии, в 1844 г. появилась на Готланде,
где к 1867 г. стала гнездиться регулярно, в конце прошлого века заселила
всю Среднюю и Южную Швецию. Расселение в Англии началось с 1850 г.,
в 1875 г. появилась в Шотландии, в 1877 г. в Ирландии, в 1903 г. на Оркней-
ских о-вах. В Дании впервые обнаружена в 1905 г. и в настоящее время рас-
селилась в Западной Исландии и по островам (Иесперсен, 1946), заметно уве-
личивается ее численность в Северной Германии. В пределах Советского
Союза обычна по всей Карелии и островам Белого моря до Соловецих вклю-
чительно. На Кольском п-ове добыта на реках Коле и Туломе, но точных дан-
ных о гнездовании нет; в районе озера Имандра (Лапландский заповедник)
32 Птицы СССР, т. IV
498 ОТРЯД ГУСЕОБРАЗНЫЕ
не встречена. Под Архангельском обычна (Паровщиков), добыта на Канине
у границы криволесья и тундры (Дементьев 1935); редка в Тиманской тундре
(низовья р. Вельть),но, возможно там гнездится (Гладков, 1951). На Печоре
доходит примерно до Полярного круга (Дмоховский, 1933). На Полярном
Урале гнездится на р. Щугор (Портенко, 1937), в устье Оби, на Полуе и из-
редка по Щучьей, по Енисею до с. Носок (Тугаринов, 1941). На Таймыре
по Хатанге и по Лене гнездится всюду в лесной и лесотундровой зонах;
по Яне прослежена до 70° с. ш., на Индигирке до устья р. Момы, на Колыме
до 69° с. ш. На Анадыре отмечена всего несколько раз в западной части края
и только во время пролетов (Портенко, 1939); на Камчатке гнездится, но
не повсюду и в небольшом числе, хотя на пролете весьма многочисленна;
на Командорских о-вах, повидимому, гнездится, но очень редка (Иогансен,
1934). Гнездится на Курильских о-вах и на Сахалине. На Гижиге очень редка,
но под Охотском обычна. В Советской Прибалтике на гнездовье редка,
так же и в Псковской обл. В отношении Белоруссии достоверных данных нет,
но предположительно гнездится под Оршей, Бобруйском, Речицей и Витеб-
ском (Федюшин). Гнездится, хотя и редко, в Украинском полесье, не выходя
в лесостепь. Она известна на юге Черниговской обл., под Киевом, Переяс-
лавль-Хмельницким и на границе Харьковской и Полтавской обл. (р. Орчик,
Зарудный, 1892). Южнее гнездится в плавнях Днепра, в Днепропетровской
обл. (Вальх, 1899) и найдена однажды на о-ве Джарылгач у устья Днепра
(Шарлемань и Шуммер, 1930); однако последний случай следует рассмат-
ривать, как задержку на гнездовье за пределами ареала. Очень редка вСмо-
ленской, Московской, Ивановской и Тульской обл. (в последней только
по Упе); в Рязанской известна лишь в северной, заокской части (Окский
заповедник); в Горьковской обычна лишь в Заволжье. В Калужской, Орлов-
ской, Курской и Тамбовской обл. известна лишь на пролете, но для Воро-
нежской (Битюг, Хопер), Пензенской и Ульяновской может считаться гнез-
дящейся, причем в последнем месте не особенно редкой (Житков и Бутур-
лин, 1906). Гнездится в устье Камы, под Куйбышевом и вниз по Волге до
Сарпинских озер. В Южном Заволжье найдена на гнездовье у Эльтона,
Баскунчака и Камыш-Самарских озер. Есть указания на случаи задержки
птиц и возможного гнездованья в устье Дона (Сарандинаки, 1907); в значи-
тельном числе гнездится в Закавказье на озерах Ахалкалакского плато
(Чхиквишвили), в Армении на Севане (Шелковников, 1934); наблюдалась
и добыта в гнездовое время по р. Алазани (Серебровский, 1925), хотя позд-
нее там не обнаружена (Чхиквишвили, 1930). Хохлатая чернеть обычна
в среднем течении Урала, Эмбы, на Челкар-Тенизе и близ Карсакпая (Слуд-
ский), ее гнездовой ареал близко подходит к северным берегам Аральского
моря. На Наурзумских озерах (Кустанайская обл.) хохлатая чернеть на
гнездовье обычна, так же и под Кокчетавом; гнездится на озерах Кургальд-
жин-Тенизской группы (Меженный и Соколов) и на озерах Горькой линии.
В Барабинской степи редка, в Кулундинской доходит до 53° с. ш., населяет
Алтай, хотя и распространена по нему спорадично (Сушкин 1938). Пови-
димому, гнездится на озере Марка-Куль (Поляков, 1914) и южнее в пределах
Монголии на озерах Ачит-Нор (Сушкин, 1938), Убса-Нур (Янушевич, 1948),
Орок-Нор, и, вероятно, в долинеТолы (Козлова, 1930). Гнездится в Юго-Запад-
ном Забайкалье (Моллесон, 1895), в небольшом числе на Аргуни (Тачанов-
ский, 1893) и, вероятно, на Верхнем Амуре, но о последнем никаких данных
нет. Указывается в качестве гнездящейся птицы для лесной части Маньчжу-
рии (Яковлев, 1929). Гнездится в северной части о-ва Хоккайдо (Ян, 1942),
но в Уссурийском крае не зарегистрирована.
Зимовки. В огромном количестве хохлатые чернети скапливаются
зимой у Британских о-вов, Северной Франции, Бельгии, Голландии и Север-
ной Германии (у берегов Бранденбурга стаи их часто достигают 400 голов).
ХОХЛАТАЯ ЧВРНЕТЬ 499
Меньше их зимует у берегов Дании, Южной Швеции и Норвегии. Зимуют
чернети и на всех незамерзающих внутренних озерах, но столь больших
скоплений, как на море, обычно не образуют. Обычны они у всех
берегов Средиземного моря; в Алжире, Марокко и Нижнем Египте много-
численны, в меньшем числе достигают Абиссинии и в еще меньшем—Судана,
малочисленны у берегов Аравии. В большом числе проводят зиму на Черном
море у берегов Румынии и Болгарии, бухтах Южного Крыма и в незамер-
зающих частях Азовского моря. Массовые зимовки этого вида находятся
на Каспии к югу от Махач-Кала и Кара-Богаз-Гола, где этот вид является
едва ли не наиболее многочисленным: стаи его достигают местами 4—5 тысяч
особей. Гораздо меньшее значение имеют зимовки на внутренних водоемах
Азербайджана, озере Севан, реках Туркмении, в Таджикистане на Кызыл-Су,
верхнем течении Аму-Дарьи и озере Кара-Куль (Иванов, 1940), а также на
Иссык-Куле. Зимуют хохлатые чернети и в Ираке, в Сеистанской котловине
(Иран) и у берегов Индии; в Индо-Гангской равнине, в Деккане, на плато
Чота-Нагпур, очень многочисленны в Манипуре. Зимуют в Бирме и Вьет-
наме. Редки у берегов Китая и обычны в водах Кореи. Встречаются зимой
на Северных Филиппинах, многочисленны всюду в Японии (как на море,
так и на больших озерах), проводят зиму на Южных Курильских о-вах
и у южных берегов Сахалина.
Скопления линных птиц, повидимому, бывают у хохла-
той чернети не столь большими, как у некоторых других нырков. Значитель-
ная часть их линяет небольшими группами, часто присоединяясь к стаям
других видов. Как районы линьки известны: озерные бассейны Иргиза
и Тургая, возможно Кургальджин-Тениза, озера Кустанайской, Челябин-
ской и Курганской обл., Барабинская и Кулундинская степи. Восточнее
стайки самцов наблюдались в Тункинской долине (Слудский). Линные
птицы добыты на озерах, расположенных на границе Сиккима и Тибета (Штре-
земан, 1940). Большие стаи держатся летом на море вблизи Охотска и, пови-
димому, там линяют (Харитонов, 1915).
Характер пребывания. Перелетная птица. Оседлы лишь
птицы, населяющие Британские о-ва, а возможно также Южную Швецию
и Сахалин.
Сезонные миграции и их даты. Результаты кольцевания
хохлатой чернети (пока незначительные) говорят о том, что на Британских
о-вах, кроме местных птиц, зимуют чернети из Исландии, Северной Фран-
ции, Дании, Финляндии и северных районов европейской части Союза.
О северо-западном же направлении пролета говорит добыча в августе в Куй-
бышевской обл. чернети, окольцованной 3 июня, за 4 года перед этим, в Ка-
лининградской обл. Птицы из средней полосы европейской части Союза
и Западной Сибири зимуют в области Каспийского и Средиземного морей.
Так, чернеть с Московского моря добыта зимой в Греции (Фракия), чернеть,
линявшая в Барабинской степи, добыта в Египте; птицы с Южного Урала
(озеро Миасово) встречены зимой в Италии (о-в Капри) и на Каспийском
море (Апшерон); наконец, птицы, линявшие в степном Зауралье (озеро
Щучье), добыты зимой на Каспии (Баку), на весеннем пролете в Актюбин-
ской обл. и одна около Алма-Ата. Последняя находка говорит о том, что
часть зауральских птиц отлетает к юго-востоку на Иссык-Куль, а возможно
и в Индию.
Отлет с мест зимовок у берегов Британии начинается с конца февраля
и заканчивается к середине мая. В Эстонию прилетают в апреле, на Псков-
ское озеро и в Ленинградскую обл. в конце первой декады этого месяца,
а валовой пролет проходит с конца апреля до первых чисел мая (Зарудный,
1910) В устье Мологи первые птицы отмечены 22 апреля и пролет продолжался
до 1 мая; в северной части Рыбинского водохранилища они появляются
32*
500 ОТРЯД ГУСЕОБРАЗНЫЕ
15—21 апреля. Прилет под Архангельском в среднем 2 мая B2 апреля—
11 мая) (Паровщиков); в Тиманской тунДре первая пара отмечена 1 июня
(вероятно, появились раньше, но не были замечены среди морских чернетей;
Гладков, 1951). На Печоре при устье Ылыча первое появление приходится
на 9 мая F—12 мая), начало валового пролета на 18 мая, а продолжительность
его бывает в среднем около 12 дней. Здесь хохлатая чернеть летит неболь-
шими стаями по 15—20 шт. (редко до 50—70), как днем так и ночью, в боль-
шинстве случаев на высоте нескольких десятков метров (Теплов, 1948).
С заливов северо-западной части Черного моря чернети отлетают в конце
марта—начале апреля (Клименко, 1950), зимовки на Азовском море они поки-
дают в середине марта (Костюченко, 1926), весенний отлет у Черноморских
берегов Кавказа начинается уже с половины февраля, но затягивается он
до конца марта (Вильконский, 1896).
Из Ирака чернети отлетают в марте (Тайсхерст, 1922), в Пакистане
исчезают к апрелю, но часть особей (вероятно, первогодков) задерживается
в Кандагаре до середины мая (Тайсхерст, 1927). Из Энзелийского залива
(Иран) основная масса птиц отлетает к концу февраля, но отдельные стаи
хохлатых чернетей остаются в течение всего марта и последняя отмечена
26 апреля (Бекстон, 1921). У азербайджанских берегов Каспия продвижение
к югу начинается с конца февраля, но отлет чрезвычайно затягивается,
и еще в середине апреля хохлатые чернети бывают там многочисленны (Вере-
щагин, 1950). То же наблюдается у Туркменского берега моря, где у пролива
Кара-Богаз-Гол встречались и в начале апреля и в начале мая.Большая растя-
нутость сроков пролета может объясняться и задержкой на местах зимовки
части первогодков, и тем, что пролетают разные, гнездящиеся на боль-
шом протяжении с севера на юг, популяции. Последними, вероятно, отле-
тают птицы из северных гнездовых популяций. Первое появление их в дельте
Волги отмечено 18 февраля; под Днепропетровском оно приходится часто
на 5—10 марта (Вальх, 1899), а под Киевом валовой пролет наблюдался
22 марта (Данилович, 1930). На Урале под Чкаловом первые птицы приле-
тают в первых числах апреля, а валовой пролет проходит во второй поло-
вине апреля (Зарудный, 1897), под Уфой прилет в середине апреля и пролет
до середины мая, в это время чернеть встречается громадными стаями (Суш-
кин, 1897). На Верхней Эмбе первая стайка замечена 14 апреля, после чего
более месяца чернеть летела стайками по 50 экземпляров и более; в низовьях
Иргиза она показалась лишь к 25 апреля и летела в течение полумесяца (Суш-
кин 1908). У Тобольска первые чернети прилетают 4 мая B8 апреля—11 мая),
а массовый пролет приходится в среднем на 15 мая (Тарунин, 1929). На
полыньях р. Елогуя они отмечены 5 мая. В дельте Или прилетают впервой
половине марта, становятся обычными после вскрытия реки, а в первой
половине апреля уже исчезают (Штегман, 1949). На озере Орок-Нор чернети
наблюдались с 4 апреля, на р. Толе с 12 апреля (Козлова, 1930), а в Юго-
Западном Забайкалье 29 апреля (Моллесон, 1897).
На озере Тарей-Нор они появляются в последних числах апреля (Радде,
1863). На притоке Ангары Илиме массовый пролет чернетей начинается
20 мая (Косыгин, 1926), под Якутском появляются в середине мая (Воробье-
ва, 1931), под Верхоянском около 1 июня. Из Южного Китая (Хубэй) отле-
тают в феврале, пролет через Чиили происходит с марта по май (Колдуэлл,
1931); на некоторых горных озерах Японии задерживаются очень долго
и бывают многочисленны еще в середине мая (Ян, 1942). В Советском При-
морье многочисленны во время пролетов как на морском побережье, так
и внутри страны. Появляются обычно в конце марта, пролет продолжается
весь апрель и начало мая. У Охотска появляются весной огромными стаями;
в верховьях Колымы на пролете реже, чем другие нырки, но все же обычны
(Дементьев и Шохин, 1939), в конце апреля—в мае это самая многочисленная
ХОХЛАТАЯ ЧЕРНЕТЬ 501
утка на море, много ее бывает до половины июня; заканчивается пролет
к концу месяца (Аверин, 1948).
Осенний пролет происходит на Камчатке в половине сентября—октябре;
в Южном Приморье с начала октября и весь месяц, встречаясь изредка
и в ноябре; в Северном Китае (Чжили) в сентябре—октябре, на юге (Хубэй)
появляется с ноября. Пролет в Тункинской долине наблюдался перед ледо-
ставом, в последних числах сентября—первых числах октября (Слудский),
под Салехардом в начале сентября (Дерюгин, 1898), на Печоре начинается
обычно 20 сентября A2—20 сентября) и продолжается около 24 дней, послед-
ние птицы отмечались 9—23 октября. Летят птицы осенью низко, обычно чуть
выше верхушек деревьев прибрежного леса (Теплов, 1948). В северной части
Рыбинского водохранилища первые пролетные стаи появляются 4—9 сен-
тября, через полмесяца или месяц начинается основной пролет, продолжаю-
щийся 15—20 дней; последние птицы наблюдались 18—24 октября. На Псков-
ском озере чернети появляются так же в начале сентября и летят до конца
октября, в то же время они пролетают через Эстонию, а на места зимовки
у Британских о-вов прибывают с середины сентября до середины ноября.
В Среднем Поволжье хохлатые чернети наиболее многочисленные пролет-
ные нырковые утки, валовой пролет проходит в начале и середине сентября,
когда они встречаются на Волге огромными стаями (Житков и Бутурлин,
1906). В начале октября чернети уже покидают Башкирию, в среднем тече-
нии Урала наиболее оживленный пролет проходит в сентябре и первой поло-
вине октября. На зимовки в Гасан-Кули прилет начинается с октября и про-
должается до декабря, каждое похолодание на севере Каспия вызывает новую
волну прилета; на азербайджанских зимовках массовое появление прихо-
дится на начало декабря, так как долгое время птицы задерживаются у бере-
гов Дагестана, где часть их проводит всю зиму. Пакистана первые чернети
достигают в сентябре (самая ранняя встреча даже 14 августа), а Ирака не
ранее октября. У берегов Аджарии они появляются в конце октября; под-
лет их на зимовки в Азовском море заканчивается к декабрю, а основной
пролет их в северо-западной части Черного моря начинается с половины
октября. Пролетные стаи чернетей повсюду не особенно крупны, они состоят
обычно из 20—30 птиц, редко более. Но на местах остановок в подходящих
местах они объединяются в огромные стаи, весьма плотные во время отдыха
и широко разбредающиеся на кормежке.
Биотоп. Широкие поймы рек и озер на открытых равнинах. В таеж-
ной зоне бывают только в широких поймах рек и озер. На пролетах стаи
придерживаются больших открытых плесов, основные скопления в зимнее вре-
мя наблюдаются у морских берегов, или в заливах, или у каменистых берегов.
Численность. Хохлатая чернеть вполне обычна, а местами и мно-
гочисленна на следующих, богатых озерами территориях: в лесостепной
и степной полосе Среднего Заволжья, Башкирии, Северного Казахстана,
Зауралья и Западной Сибири, заходя в последней и в лесную зону. Менее
обычна она на европейском севере от Рыбинского водохранилища, на восток
до Северной Двины и на запад до Южной и Средней Финляндии включительно.
Наконец, обычна она также в озерной низменности Якутии.
Экология. Размножение. Время наступления половой зрелости
вероятно в годовом возрасте, но реализуют половой цикл в этом возрасте
далеко не все особи, о чем говорит очень длительная задержка (до мая и даже
июня) целых стай на местах их зимовки. Пары образуются еще на зимовках.
Брачные игры у них такие же, как и у других нырков. Селезни поднимают
голову, затем опускают ее и поднимают вновь, сопровождая это движение
свистящим звуком. Время от времени они закидывают голову на спину
(как белоглазый нырок), издавая при этом тот же негромкий звук. Кроме
того, иногда селезни приподнимаются на воде, вытягивают кверху голову
502 ОТРЯД ГУСЕОВРАЗНЫВ
и шею и взмахивают крыльями, самка же в виде приглашения к спариванию
опускает клюв в воду. Спаривание происходит на воде.
Гнезда устраиваются чаще всего на маленьких островках среди боль-
ших озер. В широкой пойме р. Мологи A940) из 10 найденных гнезд 8 нахо-
дились у больших и малых луговых озер и только 2 у озер с лесистыми бере-
гами. В Эстонии чернеть гнездится на заросших тростником озерах (Кох,
1911), в Башкирии она также охотнее всего селится по камышистым озерам,
реже по старицам, обросшим кустами, или мелким озерам на заливных лугах
(Сушкин, 1908); у Северного Байкала преимущественно на мелких озерах,
расположенных не выше 800 м над уровнем моря (Штегман, 1936). У север-
ной части Рыбинского водохранилища 22 гнезда из 24 описанных находились
на островках, 3 на пловучих, а остальные на открытых песчаных. Почти все
гнезда были хорошо укрыты, но, как правило, помещались не в сплошных
зарослях травы, а в отдельных куртинах ее, так что насиживающая утка,
будучи укрыта от хищников, сама имеет возможность заранее видеть опас-
ность. Одно гнездо находилось под кустом березы, 7 в зарослях высокого
вейника, 8 среди кочек осоки, 7 среди дерновины щучки и в бурьянах. Наи-
более открыто расположенные гнезда нередко помещались у пней или бре-
вен плавника, что до некоторой степени предохраняло от выдувания пуха
из них ветром. В Барабинской степи гнезда хохлатой чернети чаще всего
находятся на тростниковых сплавинах или даже небольших кучах плавающего
тростника; гнездо, найденное на Б. Узене. (Волжско-Уральские пески),
находилось на лугу у берегового обрыва (Волчанецкий, 1937). Как правило,
гнезда строятся недалеко от воды, обычно не далее чем в 4 м. Если место,
выбранное для гнезда, сухое, то утка роет для него глубокую ямку, такую,
что кладка, укрытая пухом, не выдается над поверхностью почвы. На влаж-
ном месте гнездо представляет собой большую постройку в виде чаши с бор-
тами высотой до 9—10 см. Оно строится обычно из сухой прошлогодней
травы, но иногда, там где ее нет, из свежих зеленых стеблей и листьев (осока,
череда, белокрыльник). Пуха много, он темносерый и появляется после
окончания кладки, некоторые поздние, уже насиживаемые кладки, вероятно
повторные, почти не имели пуха в гнездах, видимо, он очень полно исполь-
зуется самкой в начале насиживания. Средний размер гнезд: ширина 20—
23 см, поперечник лотка 18—20 см, глубина лотка около 8 см, высота гнезда
от земли 0—10 см. Имеется ряд указаний, что, наряду с обычным для этого
вида открытым гнездованием, для хохлатой чернети свойственно и гнездо-
вание в дуплах. Достоверность этих сведений остается под большим сомне-
нием, однако чернети занимают поставленные на земле гнездовые ящики
(устье Камы, Попов); по старым сведениям Озерецковского A812, Ладож-
ское озеро), они занимают и ящики, повешенные на деревьях.
Число яиц в полной кладке: для Рыбинского водохранилища 'в среднем
11 (9—13) и до 7 в повторных кладках; для Эстонии указывается б—9; клад-
ка на Б. Узене 12. Яйца очень крупные грязнооливкового цвета. Средний
размер яиц, добытых на Рыбинском водохранилище, 57,1x41,4 мм\ пределы
для вида 53—65,9x37,7—47,2 мм (Тугаринов), вес ненасиженных 52,3 —
61 г. Срок высиживания 25^-26 дней (по Хейнроту), обычно 23—25 дней.
Самка сидит очень крепко, во втором периоде насиживания она иногда под-
пускает наблюдателя почти вплотную. Молодые проводят в гнезде менее
суток и почти сразу же по спуске на воду способны нырять. При опасности
самка отводит выводок от берега на открытую воду, а сама летает, издавая
низкое и грубое кряканье, отвлекая внимание врага на себя. Выводок дер-
жится вместе до того времени, пока молодые не оперятся. Самцы покидают
самок вскоре после начала насиживания.
Гнезда чернети нередко гибнут от затопления в случае подъема уровня
воды (от дождей, нагонных ветров и т. д.); много яиц погибает от пернатых
ХОХЛАТАЯ ЧЕРНКТЬ 503
хищников (ворон и Камышевых луней). Поэтому средний размер выводка
молодых пуховиков, как правило, меньше среднего числа яиц в кладке. Так,
на Мологе нормальное число яиц 11, а мелких пуховых птенцов 8. Зато ги-
бель птенцов несколько меньше, чем у многих других уток. На Мологе,
к началу охоты размер выводка сокращается до 6 (на 25%), а в условиях
заповедности (Миасское озеро) за это же время с 6,6 до 6,3 (всего на 4%),
Гнездится хохлатая чернеть поздно. В Эстонии время гнездования при-
ходится на июнь; в Татарии кладка начинается в конце мая, но под Мензе-
линском около 20 августа было еще много нелетных молодых (Першаков,
1930). На Рыбинском водохранилище первые полные кладки появляются
около 28 мая, но откладка яиц продолжается и всю первую декаду июня;
гнезда со свежими яйцами, найденные в начале июля, следует считать пов-
торными кладками в результате гибели первых. В Башкирии в первой де-
каде июля птенцы еще очень мелки; в Волжско-Уральских песках (Большой
Узень) 22 июня найдено гнездо с проклюнувшимися яйцами и выводок
пуховиков; на Кавказе (Джавахетия) самка с пуховиками встречена 31 июля;
в Южном Зауралье 6 июня добыта самка с готовым к откладке яйцом,
а птенцы появились в начале июля; пуховые птенцы добыты на Кургальд-
жине 25 июля, в Чуйской степи на Алтае в середине августа было много
утят, еще не начавших оперяться, а некоторые из них не достигали 20 см
в длину; гнездо с 5 яйцами найдено на озере Чаны 12 июня, у Березова две
ненасиженные кладки. 20 июня на р. Щугоре (Северный Урал) встречен
выводок молодых размером с чирка; на Елогуе утята встречены 16 июля,
а поднявшиеся на крыло молодые замечены 6 августа, хотя одновременно
встречались и птенцы весом всего 100 г. В Тункинской долине 26 июня было
найдено гнездо с 6 ненасиженными яйцами, а 24 июля вывелись птенцы
(Слудский); на Северном Байкале 7—8 августа в большей части выводков
птенцы были еще пуховыми (Штегман, 1936); около Якутска 14 июня
найдено гнездо с 4 яйцами, а в конце августа выводки уже объединялись в
стайки (Иванов, 1929).
Линька. Селезни линяют дважды в год: полностью летом и частично
осенью; самки так же, но летняя линька их менее полная. Табунки самцов,
покинувших самок, наблюдались в Тункинской долине 24 июня и 6 июля;
в низовьях Тургая линька проходит в июле, но у многих селезней еще в
середине августа маховые окончательно не дорастают (Сушкин, 1908). На
Наурзумских озерах нелетные селезни встречались 11—25 июля, но одно-
временно с ними A7 июля) наблюдались нернети, поднимающиеся на крыло,
хотя и с трудом (Формозов, 1937). В Зауралье (Макушино) линные селезни
отлавливались в начале августа, на озере Джулу-Коль в Прителецком участке
Алтая селезень, не способный к полету, встречен 22 июля (Фолитарек и
Дементьев, 1938). Наконец, в Юго-Восточном Тибете 16 августа были встре-
чены селезни как с дорастающими маховыми, так и в начальных стадиях
линьки, видимо одни из них начали линять примерно на 1,5 месяца раньше
других. Столь большая разница в состоянии линьки несомненно говорит
о том, что в одних и тех же местах линяют селезни из весьма удаленных друг
от друга географических пунктов.
Вначале сменяется контурное оперение на нижней стороне тела и голове,
затем линька захватывает спину, внутренние второстепенные маховые и
хвост. Рулевые сменяются постепенно, начиная со средней пары. Маховые
выпадают примерно через месяц после начала линьки, к этому времени птицы
уже имеют в основном летнее оперение. Хохол редеет, но нового не отрастает,
повидимому, он линяет один раз в году. До полного окончания летней линь-
ки (до конца августа) никаких признаков следующей (предбрачной) линьки
не отмечается. Линька начинается зачастую уже на местах зимовки и заканчи-
вается лишь к декабрю (в полном брачном наряде 27 ноября в Гасан-Кули;
504 ОТРЯД ГУСЕОБРАЗНЫЕ
Дементьев, 1945). За это время сменяется контурное оперение и отрастает
хохол.
Самки начинают линять позже не менее чем на 20 дней и заканчивают
смену оперения к ноябрю. Смена маховых происходит у них не на последнем
этапе линьки, как у самцов, а примерно в середине ее. После зимнего пере-
рыва линька возобновляется в марте и продолжается до мая, а у молодых
птиц иногда до июня. Весной линяет контурное оперение, хвост, внутренние
из второстепенных маховых, а также отрастает гнездовой пух. Молодые
птицы отлетают на зимовки еще до начала их первой линьки, которая про-
ходит у них с октября по май. За это время сменяется часть контурных перьев,
обычно хвост и иногда внутренние второстепенные маховые. Часть юноше-
ских перьев на брюхе и в нижней части груди сохраняется до летней линьки
и даже после нее.
Питание. Хохлатая чернеть животноядная утка, растительные кор-
ма употребляет редко; переход на них обычно говорит о недостатке нормаль-
ной пищи. Основой питания служат моллюски, которых чернети добывают
нырянием на глубину обычно не более 2—3 м. На Печоре (весной и осенью)
животная пища встречается в 50% желудков; в основном это моллюски (бо-
лее всего Limnaea), реже насекомые (личинки ручейников и жуки) и рыба
(гольяны). Более чем в половине желудков содержатся вегетативные части
растений, преимущественно листья рдестов и корни (Теплов, 1948). Весной
(в апреле) на Рыбинском водохранилище моллюски составляют 32% от содер-
жимого желудков по объему. Это главным образом Unio, реже Pisidium
и Valvata, которых чернети добывают в русловых частях водоема. Столько
же C2%) составляют крупные личинки ручейников (Phryganea grandis),
16% мелкая рыба и 10% семена (играющие, вероятно, роль гастролитов).
Пуховые птенцы (р. Молога, 1940) в июне кормятся преимущественно мол-
люсками G0%): мелкими лужанками (Vivipara duboisiana) и физами (Physa),
которых они склевывают с растений на мелководьях, и личинками хироно-
мид E%). Более крупные утята в июле—августе (Рыбинское водохранилище)
охотятся также в зарослях подводной растительности и поедают моллюсков
D4%), личинок, крупных ручейников B0%) и хирономид C3%); раститель-
ной пищи они практически не потребляют. На осеннем пролете (там же)
в сентябре—октябре значение отдельных групп кормов сохраняется тем же:
моллюски 44% (разнообразного видового состава, но преимущественно
Valvata), личинки ручейников 26%, хирономид 10%, личинки стрекоз и
плавунцы 5%, зеленые части растений 5%, семена 10%. Кормятся они в это
время среди затопленных кустарников, склевывая пищу с ветвей и висящих
на них прядей нитчатых водорослей. В октябре—ноябре на р. Мологе мол-
люски составляли 60% содержимого желудков, и основным кормом служили
перловицы (Unio), так как пролетные стаи отдыхают на руслах рек и плесах
озер. Остальное приходится на долю семян, играющих, вероятно, роль гаст-
ролитов. На севере своего ареала, Кандалакшской губе Белого моря, чер-
нети кормятся моллюсками (прудовики), рыбой колюшкой и растительной
пищей (Виноградов, 1950). В ноябре 1937 г. у Туркменского берега Каспия
основная масса чернетей, живущих в крупных стаях на море, питалась почти
исключительно моллюском Cardium edule. На этом же корме жили птицы
в крупных стаях у юго-западного берега Каспия (о-в Сара, залив им. Кирова)
в 1930—1934 гг. Более мелкие стайки чернетей, держащиеся во внутренних
частях заливов (у обоих берегов моря), кормились более разнообразной
пищей. Основу их рациона F9%) составляли мелкие моллюски (Theodoxus,
Hydrobia и Dreissena), 22% С. edule, 8% креветка (Leander) и бокоплавы
(Pontogammarus robustoides). На внутренних водоемах 26% рациона прихо-
дится на ракообразных, 7% на насекомых, 14% на моллюсков (пресновод-
ных), 47% на семена и 5% на вегетативные части растений. За последние
ХОХЛАТАЯ ЧЕРНЕТЬ 505
годы берега Каспия заселил моллюск Mytilaster lineatus, который стал
особенно многочислен у подводных каменных кос. Эти части побережья
стали основными местами зимовки хохлатых чернетей, кормящихся здесь
исключительно митиластером (Северный Азербайджан и южнее, Верещагин,
1950). Птицы кормятся обычно у вертикальных обрывов каменных гряд, где
им легче отрывать от скал друзы моллюсков. Птицы ныряют вертикально
на глубину 3—4 м (максимально до 12—14 м)> бывая под водой 30—40 се-
кунд.
Рис. 89. Чернеть хохлатая (самец и самка)
Хозяйственное значение хохлатой чернети как объекта охоты довольно
велико несмотря на слабо развитый промысел. На озере Ханка во время
весеннего пролета хохлатая чернеть составляет более 11% от всего числа
добываемых уток (Воробьев). В Северном Азербайджане (Килязи, Зорат)
вылавливают ежегодно сетями 12—16 тысяч этих нырков (Верещагин, 1950).
Массовые скопления их на местах зимовки и пролетов обеспечивают развитие
эффективного, организованного промысла. Вес мясной тушки зимой 600—
700 г. Хохлатая чернеть не боится соседства человека и нередко гнездится
на территории населенных пунктов. Один дикий селезень этого вида провел
два летних сезона на прудах Московского зоопарка; то же наблюдалось в те-
чение нескольких лет на прудах Аскания-Нова (Соколов, 1926); они гнез-
дятся на прудах в центре Лондона. Этой красивой уткой можно населить
водоемы пригородной зоны больших городов.
Полевые признаки. Утка среднего размера, плотного сложения. У пла-
вающего селезня снежнобелые бока ярко контрастируют с черной окраской
506 ОТРЯД ГУСЕОБРАЗНЫЕ
верха. На голове хохол. У летящей птицы видна четкая граница между тем-
ной верхней частью груди и белым остальным низом. Прекрасно плавает
и ныряет, под водой находится 30—40 секунд. Взлетает с воды тяжело.
Довольно молчаливая птица. Голос самки грубое и хриплое карканье «кэрр»...
«кэррр». Голос самца слышен редко и только вблизи, он напоминает слитно
и быстро произносимые звуки вроде «глю-глю» или «курлюк-рлюк». Вне
сезона размножения часто держится очень крупными стаями на открытых
плесах.
Описание. Строение и размеры. Клюв немного короче головы, широкий,
уплощенный, несколько расширяющийся к вершине, с узким ноготком. Перья темени удли-
нены и образуют хохол-косицу—у взрослого селезня длиной около 3—4 см, у взрослой сам-
ки много короче; у молодых птиц косиц нет совсем. Рулевых 14. Размер крыла самцов
189—210, самок 185—203 мм. Цевка самцов 32—37, самок 30—35 мм; клюв самцов 38—
43, самок 36—41 мм.
Вес. Взрослые самцы по прилете в апреле 782гF30—895г), в мае 715 г (Рыбинское
водохранилище), 788 г G60—840 г) (Архангельск), 742 г E95—900 г) (Бараба), 600—722 г,
в июне 952 г (Камчатка). После сезона размножения в июле 710 г (Тункинская долина),
750 г (Северный Казахстан); в августе 690 г F60—710 г), в сентябре 745 г G25—770 г)
(Рыбинское водохранилище), 970г(Кургальджин). На зимовках же птицы, которые дер-
жатся на море в больших стаях, откармливаются и прибывают в весе, более слабые дер-
жатся в глубине заливов и на внутренних водоемах. В январе птицы, пойманные в море,
весят 882 г G50—1175 г), у берегов в среднем 717 г, на пресных водоемах 550 г (Азербайд-
жан, залив им. Кирова); в феврале птицы, живущие в море, 1166 г A000—1400 г) (Зорат,
Северный Азербайждан). Взрослые самки до начала размножения: в апреле 747 г G20—
775 г) (Рыбинское водохранилище), в мае 840 г G95—955 г) (Бараба). За период размноже-
ния они сильно теряют в весе. Самки при выводках: в июле 750 г (Северный Казахстан,
эзеро Чеглы), 540 г, в августе 550 г. (р. Сым, Туруханский край), 650 г (Тункинская долина),
635 г F10—660 г), в сентябре 642 г F10—670 г), в октябре 637 г F15—660 г) (Рыбинское
водохранилище), 970 г сентябрь (Кургальджин). На зимовках в январе в больших стаях
на море в среднем 768 г, а на внутренних водоемах 530 г (залив им. Кирова), в феврале
птицы из больших стай на море 1050 г A000—1150 г). Молодые птицы на Рыбинском
водохранилище в июле до 280 г, в августе 150—538 г (с 340 г уже хлопунцы); в сентябре
самцы 725 г F50—825 г), самки 659 г E30—928 г); в ноябре самка 840 г.
Окраска. Пуховой птенец. На спинной стороне темный оливково-бу-
рый с несколько более рыжим оттенком на лбу и золотистыми концами пуха по краю
крыла. Темная окраска заходит на бока головы и туловища, захватывая бедра. Горло и
грудь рыжевато-белые с широкой темной перевязью на зобу; середина брюха буровато-
серая. Клюв темный оливково-бурый сверху и рыжий на подклювье; лапы буровато-олив-
ковые.
Молодые птицы в первом наряде. Похожи на взрослую самку.
У самцов более темные черноватые голова, шея и зоб, а у самок они коричнево-бурые,
ресколько более светлые, чем у взрослых. На задней части груди и на брюшке нерезкая
пятнистость, в области бедер более четкие буровато-серые пятна. Радужина грязножелтая.
Взрослая самка. Голова, верх туловища, шея с зобом и верх груди темно-
бурые. На спине не ясно выраженный мелкий крап. Бока тела бурые. Брюшная сторона
туловища (кроме подхвостья) белая, с большей или меньшей примесью бурых перьев.
Основной фон окраски брюшной стороны низа сильно варьирует от почти чистобелого до
почти сплошь бурого. Нередко у основания надклювья белые перышки. Клюв шиферно-
серый. Ноги светлосерые с черными перепонками, глаз желтый.
Взрослый самец в брачном наряде. Голова черная с сине-фиолето-
вым отливом. Вся верхняя сторона тела, шея, зоб, верх груди и подхвостье черные. Брюш-
ная сторона тела и бока белые. Спина, плечевые и верхние кроющие крыла с мельчайшим
белым крапом. Первостепенные маховые черно-бурые, второстепенные образуют белое
зеркало с черной каемкой, внутренние второстепенные черно-бурые с сине-зеленым от-
ливом; подмышечные белые. Клюв серо-голубой с черным ноготком; лапы свинцово-серые
с черными перепонками; радужина желтая.
Самец летом. Спинная сторона туловища черная с мелким крапом: голова, шея
и передняя часть груди буровато-черные без отлива (хохол короче). На груди, в нижней
части ее темной зоны, много белых пестрин, длинные перья боков серовато-бурые
с мелким крапом.
МОРСКАЯ ЧЕРНЕТЬ
507
37. Морская чернеть Aythya marila L.
Anas marila. Li n n a e u s. Faunae Suec, 1761, стр. 39, Лапландия.
Русские названия. Морская чернеть—книжное; народные: чернь, белобок,
плесовка; последнее по манере держаться на открытых плесах.
Распространение. Гнездовой ареал. Исландия и Оркнейские
о-ва; весь Скандинавский п-ов, кроме крайнего юга Швеции; Северная Фин-
ляндия. Случаев гнездованья в Дании не доказано, хотя оно вполне возмож-
но (Иесперсен, 1946), по наблюдениям ряда авторов, эти чернети гнездятся
иногда на о-ве Фемарн против входа в Кильскую бухту (Нитхаммер, 1938);
Карта 58. Распространение морской чернети Aythya marila
а—граница гнездовий, а'—районы наибольшей численности птиц на гнездовье, б—не вполне вы-
ясненная граница гнездовий, в—места постоянных летних встреч и спорадичного гнездованья,
г—зимовки, 0—валеты, э*с—места скоплений птиц на линьку
известен один случай гнездованья на островке в устье Одера (Робин, 1923).
В пределах СССР гнездится на Мурмане и в Северной Карелии. Далее вплоть
до Урала северная граница ареала следует по морскому побережью. Морская
чернеть весьма обычная птица Тиманской тундры и устья Печоры, гнездит-
ся и на о-ве Колгуев. На Ямале известна примерно до 70° с. ш., гнездится
в Енисейском заливе, населяет тундры Таймыра, доходя до устья Хатанги,
но, повидимому, отсутствует в арктической тундре на крайнем севере полу-
острова. Сведений из бассейна Лены нет; на Яне известна под Верхоянском;
на Индигирке обычна как в лесной полосе, так и в тундре, но в самой дельте
отсутствует (Михель, 1935), по Колыме же обычна до устья (Бутурлин,
1906), уступая в числе только морянке. Гнездится по всему Анадырскому
краю, где обыкновенна, хотя и немногочисленна; более редка у Анадырского
хребта и в Приморской полосе. Самец встречен 16 июня у мыса Северного
на Чукотке, но гнездование там этого вида мало вероятно (Артоболевский,
1926). Обычная гнездящаяся птица Камчатки, Командорских и северной
части Курильских о-вов и редкая для Коряцкого побережья, где выводок
этого вида добыт в районе Гижиги (Аллен, 1903). Указывается в качестве
поздно прилетающей птицы для района Охотска, но характер ее пребывания
там неясен (Харитонов, 1915),—весьма возможно, что это летующие в стаях
и линяющие на море птицы. Южная граница ареала выяснена еще слабее.
508 ОТРЯД ГУСЕОБРАЗНЫЕ
В небольшом числе морская чернеть гнездится в Эстонии на о-ве Сарема
и мелких островках в бухтах Хапсалу и Мацаль (Кох, 1911; Гроссе иТран-
зее, 1929). Предположения о гнездовании ее в Псковской и Новгородской
обл. (Зарудный, 1910) не основаны на достоверных фактах и мало вероятны.
Летование небольшого числа птиц на больших озерах края, как это имеет
место на финском заливе, несомненно. Сколь далеко к югу спускается мор-
ская чернеть в бассейне Северной Двины—неизвестно; на Печоре она харак-
терна для тундровой и лесотундровой зоны, но единичные случаи гнездо-
ванья известны в верхнем течении реки при устье Ылыча (Печорский запо-
ведник, Теплов, 1948). На Оби встречается почти до Березова (Устрям,
Дерюгин, 1898), а на Енисее до Туруханска; в Якутии, повидимому, не спу-
скается южнее Полярного круга.
На Американском материке граница ареала проходит вдоль побережья
Аляски от Бристольского залива до мыса Ледового G0° с. ш.). Затем она сле-
дует к устью р. Мекензи, западному берегу Большого Медвежьего озера
вплоть до озера Атабаска. Эта местность служит наиболее южным пунктом
ее ареала E9° с. ш.), откуда граница следует к Бристольскому заливу, ми-
нуя п-ов Аляску.
Основные зимовки находятся в западной части Балтийского
моря, на побережье Северного моря (до 62° с. ш.) и далее к западу до Ламан-
ша, у Британских и смежных островов. Особенно велики зимние скопления
этого вида у берегов Северной Англии, а также Голландии и Франции.
На Средиземном море зимует регулярно, но в значительно меньшем числе,
причем преимущественно молодые птицы и самки. Несколько обычнее в
восточной части моря (Балканский п-ов); Африки достигает редко. Регу-
лярно и в значительном числе зимует на Азовском море в восточной его
части (Байсугский лиман, Михайлов, 1937), Керченском проливе (Макле-
цов, 1916), а в теплые зимы и у Обиточенской косы (Костюченко, 1926).
Зимует она и на Черном море, особенно в его западной части у берегов Укра-
ины, Румынии и Болгарии. На каспийских зимовках морская чернеть бы-
вает в значительно меньшем числе, но вдоль западного берега, особенно вбли-
зи каменистых кос, встречается регулярно, хотя и не особенно большими ста-
ями. У восточного побережья она зимой редка. Никаких положительных
данных о зимовке ее на водоемах Средней Азии нет; в небольшом числе встре-
чается зимой в Иране (Геррируд, Сеистанская котловина) и совсем редка у
берегов Оманского залива (Зарудный, 1911); известны единичные встречи
в Северной Индии. На Тихоокеанском побережье регулярные зимовки рас-
положены у Южного Сахалина, например у о-ва Монерон (Гизенко). Обычна
на зимовке у Японских о-вов вплоть до Риу-Киу, держась большими стаями
на море; у берегов Кореи обычна к югу от Сеула (Остин, 1948) и вдоль бере-
гов Китая, примерно до Фудзяня, также у Тайвана, южнее которых очень
редка. На Американском материке морские чернети зимуют вдоль Алеут-
ской гряды и п-ова Аляски, у материкового побережья к югу до мексиканской
границы. На атлантическом побережье от южных частей Новой Шотландии
до берегов Мексиканского залива, а также на озерах Эри и Онтарио.
Места скопления линных птиц изучены плохо. На
Южном Ямале (р. Паюта) они встречаются стайками по 10—15 шт. (Куче-
ру к, 1948); линяют на лесных поемных водоемах в устье Иртыша; кое-где
на севере Якутии, в бассейне Лены и в массе на Камчатке (Каменское и
Курарочное озера). В последнем месте скопления бывают столь велики, что
за один загон в сети удавалось добывать более тысячи птиц (Шмидт, 1916).
Неясен характер их пребывания в районе Охотска, возможно, что там
проводят лето и линяют неполовозрелые птицы.
Характер пребывания. Почти на всем протяжении ареала
морская чернеть правильно перелетная утка. Исключением являются быть
МОРСКАЯ ЧЕРНЕТЬ 509
может только популяции, населяющие Южную Норвегию и Шотландию,
поскольку эти местности служат не только местами гнездовья, но и зимовок.
Сезонные миграции и их даты. Соответственно с распре-
делением основных мест зимовки вида главные направления пролета ведут
на запад к северным берегам Европы и на юго-запад к Азовскому, Черному
и Ионическому морям, а в Восточной Сибири на юг и юго-восток в Китай
и Японию. Отлет с английских зимовок начинается в середине марта, а закан-
чивается к началу мая (поздно отлетают стаи первогодков). В те же сроки
наблюдается пролет их у берегов Германии и Калининградской обл. СССР.
Первые птицы в районе Псковского озера показываются обычно в конце
второй декады апреля, а валовой пролет проходит в последней декаде этого
месяца и начале мая. Иногда птицы задерживаются там до второй половины
мая (Зарудный, 1910). В Ленинградской обл. на взморье Финского залива
во второй половине апреля морские чернети скапливаются тысячными ста-
ями. На фарватере вдоль отмелей они образуют сплошные широкие ленты
в несколько сот метров длиной (Алфераки, 1900). В устье Мологи наблюда-
лись первый раз 19 апреля, а 22—24 апреля были днями массового пролета.
Прилет в Тиманской тундре приходится на середину мая (Гладков, 1941),
первая пара встречена там 14 мая, но число птиц продолжало возрастать
до 26 мая; пролетные стаи в устье Мезени еще наблюдались 9 июня (Банни-
ков, 1934). В верхнем течении Печоры первые птицы появляются обычно
17 мая, а около 20 мая A6—26 мая) начинается их пролет, который продол-
жается в среднем 11 дней. Птицы летят небольшими стайками, состоящими
из 10—15 экземпляров как самцов, так и самок. Количество птиц на про-
лете в разные годы бывает чрезвычайно различно. Подсчеты показали, что
в одни годы их пролетает в 50 раз больше, чем в другие (Теплов, 1948). Отлет
с зимовок с северо-западной части Черного моря (Украина) заканчивается
в начале апреля, но отдельные стайки встречались там до 19 апреля (Кли-
менко, 1950). Первые птицы под Киевом отмечаются 14—16 марта (Дани-
лович, 1930). В Московской обл. на пролетах обычна, но интенсивность
пролета весьма колеблется по годам. На Урале под Чкаловом в некоторые
годы в середине апреля довольно многочисленна; довольно обычна на Сак-
маре во второй половине этого месяца; регулярный пролет наблюдается
вдоль русла Камы. Довольно обильный пролет по Эмбе стайками по 20—50 шт.
наблюдался с 16 апреля по 18 мая (Сушкин, 1908) ;обычна весной на Наур-
зумских озерах (Кустанайская обл.), на Тоболе и Иртыше. Появление пер-
вых птиц этого вида на Южном Ямале отмечено 11 и 16 июня (Осмоловская).
В Средней Азии на пролете бывает нерегулярно и в малом числе, в нижнем
течении Сыр-Дарьи (Чиили, Джулек) наблюдалось несколько стаек и отдель-
ных экземпляров в начале и середине апреля (Спангенберг и Фейгин, 1936).
На пролете в Корее встречается до последних чисел апреля; в Советском
Приморье очень многочисленна в конце апреля и первых числах мая вдоль
побережья, но внутри страны довольно редка. На Командорских о-вах появ-
ляется в начале мая; тогда же и у восточного берега Камчатки, где в тече-
ние всего месяца встречается в большом количестве на море; в начале
июня белобоков уже бывает немного (Аверин, 1948). Первые даты встреч
на Анадыре 13—16 мая и 3 июня (Портенко, 1939). В устье Индигирки
наблюдалась 18 июня (Михель, 1935). На первый год жизни птицы, повиди-
мому, часто не достигают своих родных мест; стайки их регулярно летуют
у берегов Англии, Голландии, Калининградской обл. СССР, в Финском
заливе и под Охотском (?), а одиночные особи нередко задерживаются на
внутренних водоемах.
Осенний пролет на Анадыре проходит незаметно, 24 сентября еще встре-
чена стайка, а 13 октября запоздалая пара. Отлет с Командорских о-вов при-
ходится на октябрь; пролет на Восточной Камчатке на сентябрь—начале
510 ОТРЯД ГУСЕОБРАЗНЫЕ
октября (последняя встреча 26 октября); у берегов Приморья пролет про-
ходит в октябре—ноябре. Осенний пролет на Юрибее (Гыданский п-ов) на-
чался в половине сентября (Наумов, 1931), а под Чкаловом он происходит
с половины сентября до половины октября, причем последние птицы встре-
чаются в конце месяца. Самая ранняя добыча в устье Днепра 19 сентября;
в половине сентября появляется и на Черном море (Украина), где валовой
пролет приходится на 7—16 октября. Начало осеннего пролета на Верхней
Печоре приходится на 22 сентября F сентября—7 октября), его продолжи-
тельность до месяца; дата последних встреч 7 октября A—10 октября);
на Рыбинском водохранилище он проходит с 3—17 сентября по 20—
26 октября; под Ленинградом с половины августа до второй половины ноября
(Бианки, 1907); в Псковской обл. появляется с середины сентября, валовой
пролет с начала до второй половины октября, последние до замерзания вод.
В Калининградской обл. первые в сентябре и иногда даже в начале его;
на местах зимовки в Северной Германии в середине октября и начале ноября;
у Британских о-вов появляется со второй половины сентября и продолжает
увеличиваться в числе до середины ноября.
Биотоп морской чернети в гнездовое время—проточные, богатые
растительностью озера тундры, лесотундры и северной редкоствольной зоны
тайги, иногда совсем небольшого размера. На время линьки птицы выбирают
озера с более крупными открытыми плесами; на пролетах они придерживают-
ся плесов рек, озер, а также морских побережий. Зиму, в основной своей
массе проводят на море, заходя в заливы и устья рек и лишь в небольшом
числе посещая удаленные от моря озера.
Численность. Обычная, а местами даже многочисленная гнез-
дящаяся утка тундры. Обилие ее на гнездовье отмечается для тундр европей-
ской части Союза, Ямала, низовьев Енисея, лесной полосы Индигирки,
низовьев Колымы (к югу до 67° с. ш.), Камчатки и Командорских о-вов.
В этих местах морская чернеть уступает по численности только морянке.
Экология. Размножение. Срок наступления половой зрелости неиз-
вестен, но, повидимому, это происходит на втором году жизни, так как на
первый год птицы еще не надевают взрослого наряда. Разбивка на пары про-
исходит, повидимому, еще на зимовках, так как весенние пролетные стаи
состоят из уже хорошо определившихся пар. В это время, как правило,
возле убитой самки задерживается, отставая от стаи, самец. Токование
селезней наблюдается однако, лишь на местах гнездовья. Самцы плавают
возле самок, распушив оперение горла и шеи, время от времени они заки-
дывают голову и шею на спину подобно другим ныркам и издают при этом
низкие воркующие, несколько трескучие звуки вроде «крр-крр-крр».
Гнезда устраиваются обычно (Ямал, Тиманская тундра) в сырых осоковых
низинах около озер и речек. Они помещаются, как правило, недалеко от
воды. Для гнезда обычно выбирается сухое местечко, но это не всегда воз-
можно, и основание гнезда иногда находится прямо в воде. Гнезда бывают
скрыты старой или свежей осокой, зачастую же они строятся в ивняках
и находятся под нависшими ветвями кустарников. Гнезда, построенные
на мокром месте, представляют собой довольно высокую постройку из прошло-
годней травы, так что лоток их бывает всегда сухим. Диаметр гнезда 25—
30 см, вышина его до 17 еле и больше. Пуха значительное количество,он темный
с небольшими светлыми центрами. Обычно число яиц в полной кладке 7—9
(Ямал, Тиманская тундра), 6—9 (Эстония). Откладка ежедневно по одному
яйцу. Цвет скорлупы серовато-оливковый, средний размер яиц 62,4x41,5 мм
(для Ямала), пределы для вида 54,5—68,1x40,7—48 мм\ вес яиц 57—72,5,
среднее 60,7 г (Ямал; Осмоловская). Откладка яиц происходит в Эсто-
нии в мае (Кох, 1911), в Тиманской тундре начинается в первых числах июня
и продолжается до конца месяца (Гладков); на Ямале полные кладки со
МОРСКАЯ ЧЕРНЕТЬ 511
свежими яйцами появляются в первой декаде июня (Дунаева, Осмоловская);
на Командорских о-вах в первых числах июня (Бианки, 1909). Насиживать
самка начинает еще до откладки последнего яйца, но вывод проходит очень
дружно, за одни сутки. Срок насиживания с момента откладки последнего
яйца в разных гнездах 26—28 дней (Гладков, 1951). Выводок в районе
озера Имандра (Кольский п-ов) отмечен 16 июня (Владимирская, 1948);
в Тиманской тундре первый 7—8 июля, а массовый вывод в конце июля
B1—31 июля); на Ямале первая встреча 30 июля, но встречались еще малые
птенцы и 10 августа (Осмоловская), под Салехардом начинающие оперяться
молодые 4 августа (Дерюгин, 1898), на Нижнем Енисее двухнедельные
птенцы 10 августа (Тугаринов, 1941); в устье Печоры еще не полностью
оперившиеся выводки наблюдались 4—9 сентября (Дмоховский, 1933),
выводок пуховиков под Гижигой 19 июля (Дементьев, 1940). Молодые под-
нимаются на крыло в возрасте 35—40 дней. Наиболее значительная гибель
молодняка происходит первые 15 дней жизни выводка, когда погибает до
30% общего числа молодых. Основной причиной гибели служит деятельность
хищников, главным образом крупных чаек; наиболее трудный период жизни
выводков—время перехода их с мест гнездованья (озер) к рекам. В дальней-
шем, вплоть до момента разделения семей, число молодых в выводках не
уменьшается (Кучерук, 1948).
Линька. Полная летняя линька самцов проходит обычно с июля по
конец августа. Сперва сменяется контурное оперение, а затем уже линяют
крылья и хвост. Оперение спины и поясницы сменяется с весьма различной
полнотой у разных особей. На Южном Ямале (р. Паюта) первые стайки
линных чернетей, среди которых были особи, потерявшие способность к
полету, встречены 19 июля. Стайки содержали по 10—20 птиц, примерно
в равном количестве самцов и самок. Нелетные селезни (маховые разверну-
лись на половину) встречались еще 25 августа (Кучерук, 1948). Линька в
брачный наряд начинается у селезней непосредственно после окончания лет-
ней, уже в сентябре, и заканчивается в декабре или даже в начале февраля,
но в основном брачный наряд они получают уже в ноябре. За это время
сменяется все контурное оперение и внутренние второстепенные маховые.
У взрослых самок, имеющих выводки (у холостых?), обе линьки сливаются
в одну, при которой оперение на некоторых частях тела сменяется дваж-
ды, а на других один раз. Линька эта начинается в августе и идет до декабря,
затем затихает на зиму и возобновляется вновь в апреле и мае. У некоторых
начало линьки затягивается до сентября. Первая линька молодых птиц
проходит с сентября или октября по апрель. За это время сменяется контур-
ное оперение, кроме перьев спины, поясницы, нижней части груди и брюха.
Линяет также хвост и внутренние второстепенные маховые, которые сменя-
ются последними иногда уже в апреле.
Питание морской чернети носит смешанный характер, почти в рав-
ной мере она потребляет и животные и растительные корма. На Печоре
(сезон?) личинки стрекоз встречены в 3 из 10 исследованных желудков,
личинки ручейников, моллюски и рыба (гольян) каждый в одном случае,
а хлорофиллоносные части растений (преимущественно листья рдестов) во
всех 10 случаях (Теплов, 1948). В октябре на Рыбинском водохранилище
31,5% по объему от всего содержимого желудков составляют моллюски,
из которых более всего встречается затворок (Valvata; 16,0%), затем перло-
виц (Unio) и беззубок (Anodonta). Личинки водяных насекомых составляют
24,0%, в основном это ручейники A4%, Phryganea grandis) и хирономиды
(8%, Tendipes plumosus). Очень большую роль играют в их рационе корне-
вища блестящего рдеста C3%, Potamogeton lucens), которые чернети обры-
вают кусками по 3—5 см длиной, ныряя на глубину около 3 м. Наконец,
10,5% приходится на долю семян (Немцев). Желудки чернетей, добытых
512 ОТРЯД ГУСЕОБРАЗНЫЕ
в октябре и ноябре на Украине (Киевская, Черниговская обл.), содержали
преимущественно моллюсков: шаровок (Sphaerium), живородок (Vivipara)
и битиний (Bithynia), нескольких клопов гладышей (Notonecta) и небольшое
число растительных остатков (Шарлемань). На каспийских зимовках в рай-
оне Ленкорани чернети живут целиком за счет моллюска Cardium edule,
который составляет 100% содержимого исследованных желудков. Каждый
желудок содержит в среднем 45 моллюсков (до 65). Птицы, зимующие несколь-
ко севернее, у каменистых берегов Азербайджана, существуют за счет дру-
гого моллюска (Mytilaster lineatus). На зимовках в Северном море их пищу
составляют также почти исключительно моллюски, особенно Litorina,
Mytilus, Nassa, Cerithium, а в заливах Dreissensia и лишь случайно ракооб-
разные, черви и семена Zostera marina (Нитхаммер, 1938).
Рис. 90. Чернеть морская (самец и самка)
Экономическое значение морской чернети как объекта промысловой
охоты велико. В значительном числе добывают ее на весеннем пролете в
Южном Приморье. Она служит главным объектом промысла на взморье под
Ленинградом, где за удачный день добывали до 60 шт. на ружье (Алфераки,
1900). Тысячами вылавливают их на зимовке в Балтийском море сетями,
растянутыми горизонтально под водой, на обычных местах кормежки стай.
Кроме того, еще совсем недавно существовал массовый промысел линных
уток, особое значение в котором морская чернеть имела на Камчатке. Мясо
морской чернети очень высокого качества.
Полевые признаки. Довольно крупная утка. У самца голова, грудь
и зад черные, спина светлосерая, брюхо и бока белые. Самка темнобурая,
у основания клюва белое кольцо шириной в палец. У летящей птицы обра-
щает внимание широкая белая полоса, проходящая вдоль заднего края кры-
ла. Прекрасно ныряет, полет быстрый, с воды слетает легко. Обычный голос
громкое и глубокое «карр».
Описание. Строение и размеры. Утка с плотным широким туловищем и не
длинной шеей. Клюв широкий и несколько расширяющийся к вершине, с узким ноготком,
снимающим только середину надклювья. Хвост короткий, округлый, из 14 рулевых. Хо-
хол на голове отсутствует. Длина крыла самцов 200—235, самок 183—225 мм; цевка сам-
пог* 34—41, самок 28—-35 мм, клюв самцов 40—47, самок 38—45 мм.
МОРСКАЯ ЧЕРНЕТЬ 513
Вес. Взрослые самцы: на зимовке в начале марта 1100 г (Ленкорань), май 990 г
(Печора), июнь 1000 г (905—1050 г), август, линный 1015 г (Ямал), сентябрь 890 г
(840—935 г), октябрь 935 г, ноябрь 805 г (Рыбинское водохранилище). Взрослые самки:
июнь 910—1100 г, июль 825—880 г (от выводка); из 6 линных самок (без выводка) 3 весили
более 1000 г; август 1000 г (Ямал), тогда же на пролете 807 г G50—900 г), сентябрь 806 г
F Ю—1025 г), октябрь 845—857 г. Молодые самцы в октябре 871 г G15—1220 г), в ноябре
980 г. Молодые самки в октябре 842 г G60—925 г) (Рыбинское водохранилище).
Окраска. Пуховой птенец. На спинной стороне темнобурый с бурым
же ошейником на зобу; снизу беловато или желтовато-серый с более темной грудкай
и брюшком; иногда по бокам спины неясные коричневато-желтые пятна.
Молодые птицы в первом наряде. Очень похожи на взрослую самку.
Отличием их, кроме юношеских рулевых, служат отсутствие белого лба и бурые перья на
других белых частях лица. На боках перья без светлых вершин. У молодых самцов на
спинной стороне тела белый крап распространен шире, чем у взрослых самок.
Взрослая самка. Голова, шея, верхняя часть спины и верхние кроющие
хвоста бурые, поясница более темная черно-бурая. Вокруг клюва белое кольцо шириной
не менее 20 мм; в области ушей беловатое пятно, соединяющееся узкой полоской с задним
краем глаз. Грудь в основном белая с большей или меньшей примесью бурых перьев, на
брюхе эта примесь больше, подхвостье бурое; бока груди буро-каштановые; бока тела
светлокоричневые с серыми вершинами перьев. Верхние кроющие крыла темно-бурые
обычно с мелким белым крапом. Маховые перья, как у самца; рулевые черно-бурые. Клюв
свинцово-серый, более темный по хребту; ноги такие же; радужина яркожелтая.
Самка впереходном летнем наряде. Белое кольцо у клюва
почти отсутствует на лбу и подбородке, на ушах светлых пятен нет. Верх головы
и плечевые более рыжие, с желто-коричневыми каймами перьев и иногда белым крапом.
Молодой самец после первой осени е-л етней линьки.
Имеет смешанное оперение Спинная сторона тела, как у взрослых, но в средней части
мантии обычно остаются юношеские перья; спина и поясница более коричневые, чем
у взрослых. Брюшная поверхность тела имеет оперение, как у взрослых, но перья юно-
шеского наряда остаются в верхней части груди, на боках и подхвостье, нижняя часть
груди и брюхо сохраняют юношеский наряд. Внутренние второстепенные маховые, как
у взрослых.
Взрослый самец в брачном наряде. Голова, шея, зоб, верхняя
часть груди и лопатки черные; голова с зеленоватым или фиолетовым отливом. Плече-
вые и спина очень светлые, почти белые благодаря густому тонкоструйчатому рисунку
перьев. Поясница и надхвостье черные. Задняя часть груди белая, брюшко беловатое;
подхвостье и нижние кроющие хвоста черно-бурые с белым крапом; бока частью белые,
частью буроватые в бглых струйчатых полосках. Верхние кроющие крыла струйчатые,
как и спина; первостепенные маховые серо-бурые, второстепенные белые с черной вер-
шиной, третьестепенные черные с зеленоватым отливом, вутренние из них с белой рябью.
Хвост черно-бурый. Клюв свинцово-серый, лапы такие же с черными перепонками;
радужина яркожелтая.
Самец в летнем наряде. Голова черноватая, без блеска, без белого или
иногда лишь с отдельными белыми пятнышками по бокам надклювья. Шея черно-бурая,
область лопаток коричнево-бурая с серым крапом. Бока серо-бурые с белой струйчатостью,
средняя часть груди черно-бурая с беловатыми концами перьев. Остальное оперение, как
а брачном наряде.
Систематические замечания. Описаны четыре формы морских чернетей: номинальная
А. т. таг На L., свойственная всей Европе и Сибири; дальневосточная А. т. mariloides
Vi go rs, 1839, ареалом который считается Камчатка и частично Анадырь и Командор-
ские о-ва; американская A. m. nearctica S t e j n e g e г, 1885, населяющая северо-запад-
ную часть Америки и указываемая (Портенко, 1939) для Командорских о-вов. Наконец,
четвертая форма, описанная как вид A. affinis Е у t о n, 1838, гнездится также в северо-
западной части Америки, распространяясь там несколько восточнее предыдущей формы.
Реальность трех первых форм как подвидов сомнительна, так как степень вариации их
различительных признаков (размеры, рисунок мантии, отлив головы) велика и зачастую
переходит за критерий данной формы. Поэтому я вполне согласен с Тугариновым A941),
не признающим за подвиды формы mariloides и nearctica. Однако форму af finis правильнее
считать не подвидом, а самостоятельным видом, за что говорят ее заметные отличия в раз-
мерах (крыло самца 185—210), в форме головы, которая напоминает хохлатую чернеть-
а также в некоторых деталях окраски. Весьма существенно и то обстоятельство, что раз-
личия эти очень стойки несмотря на то, что большая часть ареала A. affinis совпадает
с американской частью ареала А. таг На. Алфераки A900) вводит A. affinis в списки на-
шей фауны на основании добычи взрослого селезня (Вознесенским) на Камчатке у с. Ело-
вка. К сожалению, в коллекциях Зоологического института Академии наук СССР, где хра-
нятся сборы Вознесенского, взрослого селезня нет. Имеющийся же экземпляр самца с
р. Еловки (сентябрь 1845 г.)—молодая птица, судить о видовой принадлежности которой
трудно.
33 Птицы СССР, т. IV
514
ОТРЯД ГУСЕОБРАЗНЫЕ
РОД МАНДАРИНКИ AIX ВО IE, 1828
Тип. Aix galericulata
38. Мандаринка Aix galericulata L.
Anas galericulata. Li n n a e u s. Syst. Nat., изд. X, 1, 1758, стр. 128, Китай.
Русские названия. Мандаринка, японка, дупловка (последнее по манере
гнездованья).
Распространение. Ареал. Среднее и нижнее течение Амура, на
север до устья этой реки и восточных склонов Большого Хингана, Буреин-
ские горы(Радде, 1863), устье р. Зеи (Штегман, 1928), где, однако, мандаринка
встречается редко и вероят-
но гнездится далеко не по-
всюду. Весь Уссурийски!!
край (не гнездится в настоя-
щее время у залива Посьет
и в долине р. Лефу) и леси-
стая часть Северной Маньч-
журии (бассейн Сунгари и
Большой Хинган). Известны
летние встречи на Сахалине,
но гнездованье там не отме-
чено. Гнездится всюду на
Японских о-вах, Тайване, и
возможно на самых южных
из Курильских. В Корее на
гнездовье отсутствует, за ис-
ключением быть может са-
мых северных окраин стра-
ны (р. Хам-Муры). Харак-
тер ее пребывания в Китае
не ясен. Повидимому, гнез-
дится в лесистых районах,
прилегающих к Чжилийско-
му заливу, и местами в при-
морских частях Восточного
Китая. Залетные экземпля-
ры обнаружены в летнее
время на озерах Далай-Нор
и Буир-Нор.
Характер пре-
бывания. В южных ча-
Карта 59. Распространение мандаринки Aix galeri-
culata
а—граница гнездовий, а'—районы наибольшей численности
птиц на гневдовье, г—зимовки, д—галеты СТЯХ ареала, Начиная С ЮЖ-
НОЙ половины о-ва Хоккай-
до, мандаринки ведут оседлую жизнь, за исключением того, что птицы*
населяющие горные озера, спускаются зимой на низменности. В северных
частях ареала они совершают регулярные сезонные миграции. Летних отко-
чевок селезней на период линьки не отмечено.
Основные зимовки находятся на Японских о-вах к югу от
44° с. ш., в Южном Китае от нижнего течения Хуанхэ до р. Мин и на о-ве
Тайван. Известны зимние залеты в Юго-западное Ганьсу и в Ассам.
Сезонные миграции и их даты. Отлет с китайских зимо-
вок в Гуаньдуне заканчивается в марте, а пролет в Цзяньсу и Чжили затя-
гивается до апреля. В апреле идет пролет через Корею (окрестности Сеула),
МАНДАРИНКА 515
в середине этого месяца покидают и места зимовки в Японии. На юге При-
морья весенний пролет выражен довольно слабо. Местные гнездящиеся
особи появляются еще в конце марта, а пролетные несколько позже,—в
апреле и продолжают встречаться в течение всего этого месяца. Для нижнего
течения Амура (р. Хунгари) наиболее ранняя встреча 18 мая (Воробьев).
Первые пролетные стаи в Ханкайской низменности отмечались 19 августа,
в начале сентября там бывают уже большие стаи, содержащие как взрослых
так и молодых птиц. Пролет продолжается весь сентябрь и до середины ок-
тября, но одиночные птицы попадаются еще в начале ноября (Шульпин,
1936). Пролет через Корею наблюдается в октябре-ноябре (Остин, 1948),
в Чжили и Цзяньсу они появляются уже в сентябре и продолжают лететь
до ноября, когда достигают Гуаьндуня. На местах зимовки в Японии птицы
собираются с октября и проводят зиму, объединяясь в огромные многосотен-
ные стаи (Ян, 1942).
Биотоп. Долины рек с островками, поросшими лесом, протоками,
заливами и лесными озерками с зарослями лозы по берегам. Такая обстанов-
ка свойственна рекам предгорий или нижней зоны гор Уссурийского края.
Выше, в таежной зоне, мандаринки очень редки и держатся лишь в поймах
крупных рек, избегая маленьких таежных речек, а на озерах низменности
они не встречаются вовсе. На юге своего ареала, в Японии, гнездятся на всех
лесных водоемах, в том числе и на чистых горных озерах (до 1500 м н. у. м.).
Зимой мандаринки избегают приморских низменностей и проводят зиму
на лесных и горных озерах. Там, где ее не преследуют, мандаринка даже не
избегает людей и гнездится, например, в одном из парков в центре Токио,
(Ян, 1942).
Численность. Мандаринка весьма обычна, а местами и много-
численна в бассейнах рек Уссури и Сунгари и на низменности южной излу-
чины Амура, а также на всем Японском архипелаге (особенно в его север-
ной части) и на о-ве Тайван.
Экология. Размножение. На местах гнездовья птицы появля-
ются уже парами и почти сразу приступают к гнездованью. Токующий се-
лезень выпячивает грудь, распускает хохол и закидывает голову далеко
назад. При этом он несколько приподнимает крылья и раскрывает их в лок-
тевом сгибе, отчего изогнутые третьестепенные маховые становятся верти-
кально. Одновременно распушается оперение, особенно на шее, благодоря чему
фигура птицы становится короткой и высокой. Яркость и пестрота его окрасок
проявляются очень резко. Селезень делает резкие движения головой вверх
и вниз и издает мягкий мелодичный свист вроде слогов «вэк-вэк-вэк». Иног-
да селезень прижимает клюв к груди, распуская при этом хохол и удли-
ненные перья шеи или опускает клюв в воду, а затем резко вздергивает
голову вверх, сопровождая это движение глухим свистом. Нередко он заки-
дывает голову на спину, как это делается при чистке оперения, и тогда синие,
зеленые и черные цвета третьестепенных маховых резко констрастируют
с золоти сто-рыжей окраской головы. Весной во время полета утка беспре-
рывно подает голос, сильно отличающийся от кряканья других уток, селе-
зень же обычно следует за ней молча. В мае и в самом начале июня ман-
даринки часто сидят на боковых ветвях тополей и дубов. В период поисков
места для гнезд они тщательно осматривают дупла, причем самец следует
всюду за самкой. Птицы усаживаются на дуплистых деревьях, самка зале-
зает в отверстие, а селезень, вытянув шею, заглядывает в дупло. Эти утки
гнездятся обычно близ воды в сильно захламленных участках леса у берегов,
с которых свешиваются над водой стволы и корни деревьев, перевитые ло-
зами дикого винограда. Гнезда устраиваются, как правило, в дуплах, но
весьма различной высоты. Иногда они находятся не менее чем в 10 м от зем-
ли, а иногда у самой почвы в прогнивших пнях (последнее главным образом
33*
516
ОТРЯД ГУСЕОБРАЗНЫЕ
на вырубках). Значительно реже гнезда строятся прямо на земле, в углу-
блениях почвы, среди густой растительности, под прикрытием пеньков,
ветвей упавших деревьев и отмершей растительности (Спангенберг, 1940).
Количество яиц в полной кладке 9—12, которые имеют окраску от чисто-
белой до цвета слоновой кости. Размер яиц 52,6—55x40,2—41 мм. Наси-
живает только самка, причем сидит настолько крепко, что не покидает дупла
даже после ударов топором по стволу дерева. Срок насиживания 28—30
дней. В низовьях р. Имана первые полные кладки появляются в третьей
декаде апреля. Но после этого откладка яиц продолжается еще долго. Так,
самки с яйцами, готовыми к сносу, добыты 2 и 12 мая, а 22 и 31 мая еще наблю-
дались парочки около гнезд, у которых кладка, повидимому, еще не закон-
чилась (Спангенберг, 1936). Появление полных кладок на самом юге При-
морья падает также на конец апреля.
Выведшиеся птенцы самостоятельно выпрыгивают из гнезда. Они чрез-
вычайно проворны и прекрасно ныряют. Обычно утром и вечером, а во время
дождя и днем, выводок с
маткой кормится на откры-
тых плесах заводей. В
случае опасности птенцы
ныряют и затаиваются где-
либо у берега под навис-
шими кустами или ствола-
ми деревьев. Маленькие пу-
ховички B—5 днер), засти-
гнутые на берегу, прячутся
под корнями деревьев, а
подросшие птенцы стара-
ются уйти в воду. Первый
выводок пуховиков на Има-
не отмечен 28 мая, и вплоть
до 30 июня там продол-
жали попадаться выводки
только что вылупившихся
птенцов. Наиболее поздний срок встречи только что появившегося на свет
выводка на юге Приморья 20 июля (р. Лефу; Пржевальский). Поднимаю-
щиеся на крыло молодые были отмечены в устье Уссури уже 8 июня
(Деррис, 1883), но там же 8 сентября добыт молодой, крыло которого еще
на 3 см не достигало размера крыла взрослых птиц.
Линька. Полная послебрачная линька бывает у селезней в июне-
июле. Первый самец, начавший линьку, встречен на Имане 24 мая, у него
сыпалось контурное перо и отсутствовали кроющие крыла, составляющие
зеркало. Там же 4 июня обнаружен селезень, взлетевший уже с большим
трудом. В Ханкайской низменности селезень, добытый 18 июня, сильно линял,
а селезни от21 июля и 3 августа имели уже летний наряд, но махи их еще не
доросли, а еще один 4 августа уже окончил линьку. К моменту линьки
селезни собираются там стайками по 10 шт. и более и держатся по откосам
речных берегов, спасаясь в случае опасности в зарослях лозы. На Имане
линные селезни держатся в одиночку. До конца июня все самки как при
выводках, так и холостые, еще прекрасно летают. Перья брачного наряда
начинают появляться у селезней в конце августа, тогда приступают к линьке
и молодые птицы. Селезень, добытый 20 августа, уже имел новые перья на
плечах и фигурные маховые. Как у взрослых, так и у молодых заканчивается
эта линька на местах зимовки.
Питание носит смешанный характер, состоит из животных и расти-
тельных кормов. Весной в мае состав пищи весьма разнообразен. Вскрытия
Рис. 91. Позы тока мандаринки
МАНДАРИНКА 517
желудков обнаружили в них одинаково часто остатки насекомых (жуков),
улиток (наземных?), семена дикого винограда, большое количество хвоща,
жолуди (которые утки собирают, роясь клювом в опавшей листве), зерна
риса и побеги злаков (возможно хлебных, так как мандаринки посещают
в это время зеленеющие поля озимых) (Иман; Спангенберг, 1940). Кроме
того, они поедают весной и рыбу; в одном из желудков был обнаружен голь-
ян и личинка миноги (Воробьев), а в некоторых местах питание погибшей
икрой порой приводит к тому, что мясо мандаринок становится почти не
Рис. 92. Мандаринка (самец и самка)
пригодным в пищу (Деррис, 1883). Утята в июне кормятся насекомыми,
мелкой рыбкой и растительной пищей. Очень большое значение в питании
этих уток ранней осенью имеют наземные моллюски. В августе и сентябре
стайки мандаринок начинают совершать перелеты на поля гречихи и риса,
семенами которых они в это время питаются. В сентябре и начале октября
основной пищей их становятся жолуди. Выводки поднявшихся на крыло
молодых сбиваются в стайки и собираются в заводях рек, по берегам которых
растут дубы. Здесь же появляются и взрослые птицы. Они собирают пада-
ющие в воду жолуди, отчего особенно много уток скапливается в этих заво-
дях в ветреные дни. Однако утки не довольствуются одними упавшими желу-
дями, и приходилось наблюдать их обирающими жолуди непосредственно
с деревьев. В большинстве случаев желудки мандаринок, добытых в это
время, содержат только жолуди.
Хозяйственное значение мандаринки сравнительно невелико. В При-
морье ее добывают, но в незначительном количестве. В Китае и Японии она
одомашнена и разводится как декоративная птица.
518 ОТРЯД ГУСЕОБРАЗНЫЕ
Полевые признаки. Окраска селезня очень ярка и состоит из сочетания
желтого, металлически-зелейого, пурпурного и красного цветов. Самое
внутреннее маховое изогнуто и при сложенных крыльях образует угол,
бозвышающийся над спиной. На голове огромный хохол. При плавании
птицы высоко сидят на воде с несколько поднятым хвостом. Ныряют редко,
только в случае преследования раненой птицы. Полет быстрый и маневрен-
ный, легко поднимается с места, может взлетать почти прямо вверх. Часто
можно видеть сидящей на ветвях деревьев или на прибрежных скалах.
В обычное время молчаливые утки.
Описание. Строение и размеры. Небольшая утка с коротким телом и корот-
кой шеей. Крылья довольно длинные и лишь немного не достигают конца хвоста. Хвост
длинный, округлый, рулей 8 пар. Цевка короткая, пальцы длинные, задний без лопасти.
Клюв короткий, довольно высокий, с широким ноготком. У птиц обоего пола на голове
хохол. Последнее второстепенное маховое сильно изогнуто, а его внутреннее опахало рас-
ширено в 3—4 раза. При сложенном крыле оно выдается из оперения и возвышается над
спиной. Наружные плечевые перья также изогнуты кверху. Перья на шее, особенно на
боках ее, удлинены и образуют пышную епанчу. Длина крыла (самец и самка) 210—
240 мм, цевки 35—38 мм, клюва 28—32 мм.
Вес взрослых птиц в декабре: самцы 571—693 г, самки 428—608 г, среднее 628
и 512 г.
Окраска. Пуховой птенец. Сверху оливково-бурый с желтоватым налетом.
На крыльях и по бокам поясницы светложелтые пятна. Горло и бока головы светло-
желтые с буроватой полоской за глазом, низ тела желтовато-белый.
Молодые птицы в первом наряде. По окраске похожи на взрослую самку, но грудь
у них в менее ясных пятнах. Многие перья имеют штриховые пятна.
Самка взрослая. Верх головы серо-аспидный, а ее бока и шея более светлосерые.
По бокам у основания клюва белое пятно, вокруг глаза кольцо и от его основания
к затылку полоска того же цвета. Вся спинная сторона тела оливково-бурая. Зоб, перед-
няя часть и бока тела бурые с овальными беловатыми пятнами, остальная брюшная сто-
рона белая. Крылья оливково-бурые с блестящим зеленым зеркалом, окаймленным белой
полоской за счет наружных опахал части второстепенных маховых и их белых вершин.
Последнее из второстепенных маховых имеет, кроме того, белую предвершинную полоску,
которой не бывает у самцов. Хвост оливковый. Клюв буроватый с оранжевым ноготком,
лапы грязножелтые.
Взрослый самец в брачном наряде. Имеет зеленый зоб и темя с пурпурным
блеском и медно-красный затылок. Хохол сверху также медно-красный, а в остальной
части блестящий сине-зеленый. Передняя часть головы перед глазом палево-рыжая,
а щеки, подбородок и шея яркорыжие. От глаза к затылку проходит постепенно
суживающаяся белая полоса. Верх тела темнооливкового цвета, более зеленый в об-
ласти мантии и более бурый на спине и надхвостье. Передняя часть шеи снизу и зоб
медно-красные с блеском. Оперение боков груди, прикрывающее сгиб крыла, образует
3 черные и 3 белые дугообразные полоски; бока туловища серо-зеленые с черной и серо-
вато-белой поперечной струйчатостью. Брюшная сторона тела белая. Самые наружные
из плечевых перьев белые с черными каймами по наружному опахалу; кроющие крыла
и первостепенные маховые оливково-бурые. Последние с серебристыми ободками по
наружному краю и блестящими зелеными вершинами внутренних опахал. Второстепенные
маховые оливковые с белыми вершинными каймами, внутренние из них с блестящими
зелеными наружными опахалами, образующими зеркало. Крайнее третьестепенное в осно-
вании блестяще-синее, по наружному опахалу оливково-зеленое, а по внутреннему рыже-
каштановое, сзади с черной, а спереди с белой каймой. Клюв яркокрасный, лапы жел-
тые, радужина темнобурая.
Взрослый самец в летнем наряде. Имеет оперение весьма сходное с самкой, но
у него не бывает белого пятна у основания клюва, а сам клюв и лапы сохраняют ту же
окраску, что и в брачном наряде, но более тусклы.
ГАГА ОБЫКНОВЕННАЯ
619
РОД ГАГИ SOMATERIA LEACH, 1819
Тип. Anas mollissima
Синонимы. Polysticta Е у t о n, 1836.— Eniconetta Gray, 1840.—Lampronetta
Brandt, \847.—Arctonetta Gray, lS55.—Erionetta Coues, 1884.
39. Гага обыкновенная Somateria mollissima L.
Anas mollissima. Li n n a e u s. Syst. Nat., изд. X, I, стр. 124, 1758, о-в Готланд.
Синоним. Anas cutberti. Pallas. Zoographia
Rosso-Asiatica, II, стр. 235, 1811.
Русские названия. Гага, гагун (самец), гагка, гавка, близки к саамскому
названию havda и финскому hagka (происхождение названия вероятно обязано брачному
крику самцов).
Распространение. Ареал. Северные морские побережья Европы от
Исландии до Новой Земли, восточной Сибири, побережья и острова Берин-
гова и местами Охотского моря, полярные побережья Америки и Гренландии
с прилежащими островами.
Карта 60. Распространение обыкновенной гаги Somateria mollissima
IS. m. mollissima, 2—S. m. borealis, 3—S. m. dresseri, 4—S. m. v-nigrum; а-граница гнездовий,
г—зимовки, а—залеты; цифры в кружках—номера зимующих подвидов
Основные зимовки у берегов Скандинавии, Исландии, Бри-
танских о-вов и на Мурмане, в южной части Берингова моря до южной
оконечности Камчатки и Алеутских о-вов, а у берегов Америки к югу до
Ванкувера и Филадельфии.
Характер пребывания. Частью перелетная, частью кочую-
щая птица. Местами гага совершает дальние сезонные миграции (Северо-
Восточная Сибирь), а местами живет оседло, лишь откочевывая зимой
к незамерзающим участкам моря (Западная Европа).
Биотоп. Гаги—обитательницы морских побережий, обычно высоких
и каменистых. Вне сезона размножения держится на море, или вблизи от
берегов и мелководий, или у кромки льдов»
Подвиды и варьирующие признаки. Описано значительное число подвидов обыкновен-
ной гаги, из которых не внушает сомнения реальность следующих четырех: S. т. mollis-
520 ОТРЯД ГУСЕОБРАЗНЫЕ
sima Li nnaeus, 1758,—заселяет европейские побережья и острова на восток до Новой
Земли; S. т. borealisChr L. Breh m, 1824—северный подвид гаги, занимающий Гренлан-
дию, Баффинову Землю с прилежащими островами, Шпицберген и Землю Франца-Иосифа;
S* т. dresseri S h а г р е, 1871—восточноамериканская форма с Восточного Лабрадора,.
Ньюфаундленда, южной и восточной части Гудзонова залива; 5. m. v-nigrum Gray,
1855—тихоокеанский подвид, которому иногда приписывают значение вида. Населяет
Северо-Восточную Сибирь, острова Берингова моря и берега Америки от п-ова Аляски на-
•зпаде до залива Коронации на востоке.
Обыкновенная гага Somateria mollissima mollissima L.
Русские названия. Кроме обычйых гага, гавка, гагун, местами неполовозре-
лые и линные птицы называются неправильно турпанами (о-в Колгуев).
Распространение. Гнездовой ареал занимает побережья Ислан-
дии и большей части Шотландии. Спорадически гнездится на севере Ирлан-
дии, населяет Шетландские, Гебридские и Оркнейские о-ва. Гнездится в се-
верной части Ютландского п-ова и на ближних островах вдоль всех берегов
Скандинавии как со стороны Норвежского моря, так и вдоль Балтийского
побережья. В Финляндии достигает района Хельсинки. В пределах Совет-
ского Союза обычна на островах Эстонии (Сарема, Хиуме и др.) и гнездится
кое-где на побережье Латвии. Обычна гага на Мурмане, причем особенно
многочисленна на островах Айновых, группе Семь Островов и др. Почти столь
же обычна она и у берегов Белого моря в его Кандалакшской губе, но реже
в Онежской. На этой территории гаги селятся главным образом на больших
и мелких островах. Восточнее вдоль побережья гага отсутствует и в качестве
гнездящейся птицы встречается быть может лишь в северной части п-ова
Канина (Дементьев, 1940). Наиболее восточным гнездовым районом служит
западное побережье обоих островов Новой Земли и о-ва Вайгач. Встречу
гаги в Енисейском заливе следует считать лишь случаем залета, таким же,.
как находки ее под Ленинградом.
Основные зимовки гаг находятся вдоль побережий Британ-
ских о-вов, Франции, Германии, Дании, Южной Исландии и Скандинавского
п-ова. Зимуют гаги в Баренцовом море (на Мурмане) в наибольшем числе
в губе Печенгской, в меньшем в губе Мотовской и в еще меньшем у Териберки
и Семи Островов; в небольшом числе у Южного о-ва Новой Земли и в Белом
море, там, где остаются незамерзающие пространства воды.
Характер пребывания. Гаги, населяющие Новую Землю
и берега Белого моря, совершают дальние откочевки и проводят зиму на
открытых участках моря. Часть из них зимует на Мурмане. Норвежские,,
шотландские, а частично и мурманские гаги оседлы.
Сезонные миграции и их даты. Гаги, гнездящиеся на Бе*
лом море, в основной своей массе отлетают зимовать на север, к обогреваемым
Гольфштримом западным берегам Мурмана. С восточного побережья Мур-
мана (Семь Островов) большая часть гаг отлетает осенью на Западный Мурман
и в Северную Норвегию. Продвижение назад начинается уже в середине
января. Оно совершается весьма постепенно, возможно, что часть пути гаги
преодолевают вплавь. В марте уже скапливается большое количество их
в бухтах Восточного Мурмана. Между 18 апреля и И мая каждый год наблю-
дается пролет гаг в горле Белого моря. Небольшие стайки их по 8—15 шт.
пролетают к югу одна за другой, следуя над разводьями или вдоль кромки
льда (Смирнов, 1926). Птицы с Новой Земли также отлетают зимовать на
запад и возвращаются лишь в апреле, когда появляются большие стаи их
с преобладанием самцов (Успенский). Весьма интересно, что весенний пролет
гаг отмечен в Карском море у о-ва Белого (Тюлин, 1938), где они пролетали
с юго-юго-запада на северо-северо-восток. Первая стая из 80 шт. была
замечена 30 апреля, последние же пролетали с 18 мая. Куда направлялись
ГАГА ОБЫКНОВЕННАЯ 521,
эти птицы—не ясно. К сожалению, добытых там экземпляров в коллекциях
нет, отчего наблюдения эти остаются под сомнением. Из Кандалакшского
залива гаги отлетают только с ледоставом, в ноябре (Дубровский, 1936).
В Кольском заливе зимние стаи появляются уже в октябре (Спасский, 1925),
а у берегов Северной Германии в ноябре, где и задерживаются до марта.
Районы линьки селезней с беломорских и мурманских
гнездовий расположены у Терского берега Белого моря примерно в районе
становищ Тетрино-Полоньга, а быть может и несколько западнее. Новозе-
мельские гагуны линяют возможно у о-ва Колгуева, но стайки их встречают-
ся и в Карском море.
Биотоп. Гага—морская прибрежная птица, круглый год она прово-
дит в литоральной зоне и не удаляется от берегов более 3—5 км. В период
размножения она населяет морские острова и побережья. Любимые место-
обитания гаги небольшие, отдаленные от материка каменистые островки,
голые на севере и лесистые на юге ее ареала. У песчаных побережий гаги не
встречаются. На материке они гнездятся крайне редко. Гаги избегают ветре-
ных пробойных мест, население литорали в которых бывает очень бедным.
Поэтому на островах они охотнее держатся со стороны проливов или укры-
тых от ветра заливах и бухтах.
Зимой они не выходят на берег и держатся не в литоральной полосе, а на
открытых плесах проливов, заходя иногда в устья рек до 2—3 км от моря
(Кафтановский, 1941). Судя по результатам обора гагачьего пуха и степени
развития промысла, количество гнезд на этой территории превышает 90000»
Хищническое отношение к гаге (массовый сбор яиц), имевшее место в течение
длительного времени, привело местами (Мурман, страны Западной Европы)
к резкому оскудению ее численности. Введение охранных мероприятий и,
в частности, создание специальных заповедников положило конец этому
процессу. В настоящее время почти повсеместно, особенно в пределах Совет-
ского Союза, численность гаги неуклонно растет. Зимой у архипелага Семь
Островов гага самая многочисленная зимующая птица. Она держится как
одиночками, так и стайками по 50—70 шт. На каждый километр берега при-
ходится в среднем 30—33 гаги, так что всего у островов зимует около 1500 шт.
Из них взрослых самцов в зимнем наряде не более 11,5%, основную же массу
составляют самки и молодые.
Экология. Размножение. Половая зрелость.у гаг наступает в воз-
расте двух или, скорее, трех лет, так как полный брачный наряд самцы наде-
вают только на 28—30-м месяце жизни: В гнездовых колониях встречается
некоторое количество неполовозрелых самок, которые еще не несут яиц, но
выстилают своим пухом чужие гнезда и стремятся насиживать яйца (Успен-
ский). На пары гаги разбиваются уже на зимовке или в период их продвиже-
ния к местам гнездовья. В конце апреля и в мае в стаях гаг хорошо заметны
пары, однако самцы в это время количественно преобладают и между ними
нередко возникают драки. Уже с осени в ясные дни начинают иногда раз-
даваться крики гагунов, которые они издают в период ухаживания. Чем бли-
же к весне, тем все чаще можно видеть и слышать токующих селезней. Токуя,
самец-гагун привстает на воде, поднимая туловище почти вертикально, де-
монстрируя свое бархатно-черное брюхо, которое представляет резкий
контраст с белоснежной окраской верха. При этом он взмахивает крыльями,
Клюв^у токующего селезня бывает сперва прижат к груди, а затем он вздер-
гивается вертикально вверх при вытянутой кверху же шее и в этот момент
издается глухой, но далеко слышимый, воркующий крик «агууу-агуууа».
Весной, наряду с обычными парами, часто приходится видеть по две-три самки
около одного самца и, наоборот, по два-три самца около одной самки. Обычное
же соотношение полов у гаги в период спаривания 1 : 1 (Белопольский).
Гаги заселяют острова неравномерно, они скопляются на некоторых островах
Г>22 ОТРЯД ГУСЕОБРАЗНЫЕ
создавая как бы гнездовые колонии в несколько сот, а иногда и до 2000 пар.
Плотность распределения гнезд бывает иногда очень велика; так на о-ве
Крестоватике (Новая Земля) площадью около 300 м2 бывает ежегодно более
400 гнезд (Успенский). Подмечено, что в местах, богатых укрытиями для
гнезд, гаги гнездятся довольно разрозненно, плотные же колонии они обра-
зуют лишь на совершенно открытых островах. Гаги выбирают преимуще-
ственно такие острова и луды, которые хорошо изолированы от материка
даже во время отлива и недоступны для наземных хищников—лисицы и пес-
ца. На Мурмане и Новой Земле они гнездятся на открытых островах, а на
Белом море охотно занимают лесистые, забираясь иногда довольно далеко
в лес. Расстояние гнезд от воды бывает весьма различным: одни из них рас-
полагаются непосредственно за зоной заплеска, а другие не менее чем в пол-
километре от моря. Большая часть гнезд приурочена все же к береговой поло-
се, а в центре островов их бывает сравнительно мало. Высота гнезд над уров-
нем моря различна, она зависит от высоты островов. Однако почти во всех
случаях путь от гнезда к воде проходит по отлогим ложбинкам, гаги избегают
гнездиться у обрывистых берегов и на самих обрывах. На отбытых островах
Восточного Мурмана1, спасая гнезда от сильных ветров, гаги используют осо-
бенности микрорельефа. На архипелаге Семь Островов из 468 найденных
260 помещались под камнями, в расселинах между ними, в пещерках или
под навесами торфа, 142 около камней, скал, кочек и на уступах обрывов,
2 под кустами ивняка и 64 без укрытий. Половина гнезд находится там
среди дерновин злака E4,5%), а другая половина среди вороничников,
на участках открытого торфа или среди камней. Злаки около гнезд—явле-
ние вторичное, результат удобрения почвы пометом; они указывают на
длительность существования того или иного гнезда. На Айновых о-вах
большая часть гнезд помещается под навесами или в пещерках высоких
кочек торфа (в разрушенных норках тупиков), на Новой Земле охотнее
всего около стелящихся кустов ивняка, а на островах Белого моря (от Канда-
лакшской до Онежской губы) преимущественно в лесу под спускающимися
к земле ветвями елей и их поваленными стволами; под некоторыми елями
с ветвями, спускающимися до земли, бывает по 2—3 гнезда. Одни и те же
гнезда занимают гаги в течение ряда лет, что позволяет довольно легко
переводить гаг в искусственные гнездовья (Эстония, Новая Земля, Ислан-
дия). Из 568 гнезд на о-ве Б. Лицком (Восточный Мурман) 560, т. е. 98,6%,
было старых и только 8 построенных заново. Из числа старых гнезд одна треть
была прошлогодних гагачьих, а две трети принадлежали прежде крупным
чайкам, яйца которых были выбраны перед началом гнездованья гаг.
Наряду с этим есть наблюдения, что гаги избегают гнезд с остатками прошло-
годнего пуха, а устраивают новый лоток рядом с прежним. Биологический
смысл выселения из таких гнезд может заключаться в том, что в них
перезимовывает большое количество блох.
Гнездо представляет собой ямку, вырытую в торфе, шириной 20—25 см
глубиной около 10 еж. Выстилки очень мало и состоит она из материалов, на-
ходящихся около гнезда. Пух появляется в гнезде с откладкой четвертого
яйца, и за 4—6 час. после начала выщипывания он уже образует валик
вокруг яиц. Начало гнездованья на о-ве Сарема (Эстония) приходится на
конец мая—начало июня, на Восточном Мурмане первые кладки 25 мая
(9 мая—1 июня), а на Новой Земле 2—14 июня. Разгар откладки яиц бывает
в середине июня, а в конце месяца появляются лишь единичные новые
кладки, вероятно повторные в результате гибели первых. В годы, когда
прибрежные части моря долго не освобождаются ото льда, откладка
1 Основные сведения по экологии гаги на Восточном Мурмане взяты из материалов
заповедника «Семь Островов».
ГАГА ОБЫКНОВЕННАЯ 523
яиц задерживается. Обычное количество яиц в полной кладке в Кандалакш-
ском заливе 5—6, нередко до 7; на Восточном Мурмане (Семь Островов)
4, нередко до 6; на Новой Земле 3—4 (до 6); на Айновых о-вах 4; на островах
Эстонии 5—6; в Дании (о-в Эртхольм) 5 D—6). Повидимому, размеры кладки
зависят от возраста птиц и от климатических условий года. Меньшее коли-
чество яиц бывает у молодых самок и в годы с затяжной весной. В случае
потери яиц в начале гнездового сезона гаги зачастую несутся вторично.
Исследование яичников гаг обнаружило в них от 4—6 до 13—15 рубцов,
следовательно, некоторые самки могут нестись дважды или даже трижды
в год (Слепцов). В результате частичного отбора яиц из гнезд не удается
достигнуть увеличения размера кладки. Известны кладки с 11—13 и даже
19 яйцами, но, несомненно, они принадлежат нескольким самкам, несшимся
в одно гнездо. Иногда гаги откладывают отдельные яйца в гнезда других
птиц: турпана и длинноносого крохаля (Эстония), длинноносого крохаля и
чайки клуши (о-в Эртхольм), серебристой чайки (Восточный Мурман).
Яйца очень крупные, правильной яйцевидной формы, зеленовато-серого цве-
та. Размер их 69—83x47—53,5, среднее 77,9x51,3 мм, и вес 86,7—127 г,
среднее 109, 1 г (Восточный Мурман). Средние данные для других пунктов
весьма близки. Самка откладывает ежедневно по одному яйцу, но в неко-
торых случаях интервалы между двумя яйцами сокращаются до 12—15 час.
Первые яйца почти не насиживаются, плотно самки садятся лишь после
откладки четвертого яйца, а после окончания кладки они гнезда, повидимому,
не покидают совсем. К концу насиживания гаги сидят настолько плотно, что
иногда разрешают брать себя руками. За это время они, повидимому, совсем
не принимают пищи и живут за счет запасов жира. В результате вес их за
период насиживания падает примерно на 39%, а кишечник чрезвычайно истон-
чается и спадается настолько, что в нем бывает трудно отыскать просвет.
В результате выщипывания гнездового пуха на брюхе и в нижней части груди
у самок образуется наседное пятно размером около 200 см2, которое начинает
зарастать лишь к концу насиживания. Срок насиживания (для Восточного
Мурмана) в разных гнездах 24—27 дней, из которых последние 0,5—2 суток
приходятся на процесс вылупления птенца с момента надклева яйца. За время
инкубации яйца теряют от 10 до 16% своего веса. В конце июня или в начале
июля появляются птенцы. Вес их при вылуплении около 69 г E6,5—85). Птен-
цы выводятся более или менее одновременно, в гнезде они проводят 1 —
2 суток, после чего самка сводит выводок на воду. Дорога зачастую далека
и трудна, и выводок делает в пути остановки на отдых. Спуск к воде обычно
отлогий, но иногда птенцам приходится прыгать в море с 10-метровой высоты.
Птенцы большей частью сразу же попадают на море, но иногда они до-
вольно длительное время проводят на внутренних водоемах. На Семи Остро-
вах первые птенцы отмечены 25 июня (9 июня—2 июля). С первых же
дней птенцы хорошо плавают и ныряют в бурной полосе прибоя. Выводки
собираются в тихих, неглубоких бухтах, где собирают корм с наполовину
обнажающихся камней. Гаги в противоположность многим видам уток не
проявляют враждебности к птенцам из чужих выводков, в результате чего
выводки, даже различающиеся по возрасту, часто объединяются. К ним часто
присоединяются молодые и холостые самцы, образующие вместе большую
и пеструю по составу стаю. Двухмесячные птенцы почти достигают размера
взрослых птиц; к этому времени выводок распадается, и молодые переходят
к самостоятельной жизни.
Линька. Гаги линяют дважды в год: после сезона размножения пол-
ностью или почти полностью и перед началом размножения частично (кроме
крыльев). После того как самки садятся на яйца, селезни объединяются в стай-
ки и откочевывают с мест гнездовья на линьку. На Восточном Мурмане это
начинается уже в середине июня. В конце июня—начале июля стайки
524 ОТРЯД ГУСЕОБРАЗНЫЕ
самцов летят вдоль берегов на восток (отдельные стайки на запад). С этого-
времени начинается линька, и 22—24 июня появляются первые пятнистые
гагуны, а в августе у них уже выпадают маховые перья, и птицы теряют
способность к полету. У береговЭстонии линька самцов начинается в середине
июня (Кох, 1911), а на Новой Земле 24 июня часть самцов уже находится в со-
стоянии линьки (Успенский). Сперва линяет оперение туловища, кроме ниж-
ней части груди, живота и спины. Эти части тела, крылья и хвост линяют
несколько позже. Последние стадии послебрачной линьки совпадают по вре-
мени с началом следующей линьки. Иногда оперение нижней части груди
и живота так и не сменяется до начала осенней линьки, линяя; таким образом,
один раз в году. Заканчивается полная смена оперения у самцов обычно-
в августе. У взрослых самок она проходит с августа по сентябрь. У них вна-
чале сменяется оперение головы, шеи, верхней стороны туловища, верхней
части груди и подхвостья. Крылья, хвост и остальные части тела линяют
позже. Предбрачная линька проходит у селезней с августа по декабрь, а у
самок с сентября по март. Первая линька у молодых птиц чрезвычайно растя-
нута: начинаясь в сентябре, она кончается лишь в апреле. Линька эта бывает
далеко неполной, причем полнота ее весьма различна у разных особей. Так,
сменяется большая часть (но не всегда все) перьев мантии; иногда все, а иног-
да лишь некоторые перья спины, надхвостья и подхвостья; не всегда перели-
нивают низ груди и живот; рулевые сменяются у некоторых все, а у некото-
рых лишь центральная пара. Следующие линьки неполовозрелых птиц про-
ходят более или менее одновременно со взрослыми. Стаи линяющих гаг дер-
жатся в море, обычно в тихих бухтах около мелководий или подводных ри-
фов, но на большом расстоянии от берегов.
Питание. Гаги кормятся преимущественно животной пищей, к ко-
торой растительные корма служат лишь незначительным добавлением. Так,
на Восточном Мурмане основу питания взрослых птиц составляют моллюски,
и из них мидии (Mytilus edulis—встречаемость в желудках 70,3%) и лито-
рины (Litorina sp.—24,3%). В большом числе поедаются также усоногие рач-
ки балянусы (Balanus balanoides—40,5%), реже другие ракообразные, мор-
ские звезды и ежи. Весной гаги нередко кормятся перезимовавшими ягодами
вороники и зелеными частями растений. С конца июня основное значение
в питании молодых птенцов и держащихся с ними самок приобретают мол-
люски литорины, которых птицы собирают на прибрежных мелководьях,
рачки бокоплавы и поспевающие к тому времени ягоды вороники (Успенский).
Питание гаг в Онежской губе Белого моря (наши данные) имеет тот же харак-
тер, что и на Мурмане, но больший удельный вес в нем занимают литорины,
бокоплавы и черви полихеты (Nereis). В Кандалакшской губе моллюски (ми-
дии и литорины) встречаются в 100% желудков, бокоплавы в 37,5%, рыбы-
колюшки в 12,5%, насекомые в 12,5%, водоросли фукусы в 37,5% и ягоды
вороники в 25% (Виноградов, 1950). Корм гаги, как взрослые, так и птенцы,
добывают нырянием, обычно на глубину до 2—4 м. Их крепкий, массивный
клюв позволяет отрывать моллюсков, прикрепленных биссусом к подводным
камням, и разрушать известковые раковинки балянусов. Сильно наполненные
желудки содержат около 76 г корма; учитывая, что гаги кормятся 4 раза в
сутки (в периоды отлива и прилива), их суточный рацион можно принять,
за 300 г.
Хозяйственное значение гаги в экономике севера весьма существенно.
Большую ценность представляет гагачий пух, очень мягкий, эластичный
и обладающий исключительно малой теплопроводностью. Он незаменим для
изготовления зимней одежды в тех случаях, когда теплота должна сочетаться
с легкостью. Поэтому гагачий пух широко используется в авиации при изго-
товлении костюмов для летчиков. Пух, собранный из гнезд, имеет совершенна
другие товарные качества, чем пух, ощипанный с птицы. Дело в том, что для
ГАГА ОБЫКНОВЕННАЯ 525
выстилки гнезда гаги (как и другие утки) используют особый, так называемый
гнездовой пух. Он отрастает в нижней части груди и на брюхе и отличается
от пуха с других частей тела большим количеством извитых бородок первого
и второго порядка, а также несколько большей общей длиной. Благодаря
этому отдельные пушинки хорошо сцепляются между собой, образуя эластич-
ную массу. Среднее количество пуха, которое дает гагачье гнездо, 17—19 г
для Восточного Мурмана, 18—20 г для Кандалакши и 14,5—16,5 г в Ислан-
дии (Леппентин, 1936). Дополнительная откладка пуха птицей, после сбора
его из гнезд, если и происходит, то в очень небольшом количестве. Промысел
гагачьего пуха существует в России с давних времен. В документах XVII века
Рис. 93. Обыкновенная гага (самец и самка)
«птичий пух» упоминается в числе товаров, продававшихся голландским
купцам, но несомненно, что сбор пуха существовал и значительно раньше.
Масштабы промысла можно охарактеризовать тем, что в 1930 г. только на Но-
вой Земле было заготовлено 452 кг этого ценного продукта. Сбор пуха играет
заметную роль в бюджете новоземельских промышленников. Чрезвычайно
вредным явлением следует считать широко практикуемый местами еще до
сих пор свор гагачьих яиц и стрельбу самих птиц. В тех местах, где гаги
находятся под охраной, число гнездящихся птиц быстро возрастает, а боязнь
человека пропадает настолько, что они становятся полудомашними птицами.
В дореволюционное время охрана гаги с целью эксплуатации пуха проводи-
лась на Айновых и Соловецких о-вах и давала высокий хозяйственный эффект.
У нас опытные работы в этом направлении проводятся заповедниками Семь
Островов и Новая Земля. Годовой прирост поголовья в результате охраны
гнезд и борьбы с хищниками достигает 60%. Плотность населения гаг на ост-
ровах путем их всемерной охраны и устройства искусственных гнездовий
526 ОТРЯД ГУСЕОБРАЗНЫЕ
можно сделать очень высокой (до 13 000 гнезд на 1 га) и достигнуть этим повы-
шения доходности хозяйств населения крайнего севера.
Полевые признаки. Гага крупная утка, причем самец-гагун имеет бросаю-
щийся в глаза пестрый наряд. Окраска верха тела ослепительно белая, на те
мени черная «шапочка», затылок зеленый, грудь оранжево-розовая, брюхо
густо черное. Утка и молодые птицы коричнево-бурые с многочисленными
темными пестринами. Гага прекрасно ныряет, используя крылья для пере-
движения под водой. Летает низко, держась над самыми гребнями волн,
равномерно взмахивая крыльями. Скорость полета около 50—55 км в час.
Голос самца «агууу-агууу», самки низкое «корр-крр».
Описание. Строение и размеры. Крупная утка. Клюв массивный; опе-
рение по бокам надклювья выдается далеко вперед и у самцов доходит до уровня ноздрей.
Перья затылка у самцов удлинены и образуют гриву. Внутренние из второстепенных махо-
вых также удлинены и серповидно изогнуты—у самцов сильнее, у самок слабее. У гаг,
как это свойственно многим обитателям моря, очень сильно развиты нссоЕые железы, что
обусловливает высокий профиль лицевой части готовы. В случае длительного пребывания,
птиц (более года) на пресных водоемах носовые железы уменьшаются настолько, что это
бывает отчетливо заметно по контурам головы. Длина крыла самцов 269—328, самок 258—
295 мм; цевка самцов 45—55, самок 45—53 мм\ клюв от оперения надклювья самцов 49—
61, самок 44—55 мм, от конца оголенных висков у самцов 68—80 мм.
Вес взрослых селезней весной перед сезоном размножения около 2250—2600 *,
взрослых самок 2365—2895 г. В это время гаги чрезвычайно жирны. Жиром заплывают все
внутренности, и слой жира до 1,5 см залегает под кожей. За сезон размножения селезнн-
теряют всего 200—300 г, а самки около 1000 г. Вес самок перед концом насижиЕания1555—
1637 г. (Восточный Мурман). Неполовозрелые селезни в конце мая—начале июня около
2250 г B075—2485 г) (Белое море; наши данные).
По Белопольскому, вес мурманской гаги за летний период для взрослых самцов 2218 г
A384—2800 г), по месяцам: в мае 2370 г, в июне 2110 г; для взрослых самок за летний
сезон 1915 г A192—2895 г), по месяцам: в мае 2370 г, в июне 2081 г, в июле 1629 г,
в августе 1928 г.
Окраска. Пуховой птенец. На спинной стороне буровато-серый, бока головы
того же цвета, но над глазами проходит широкая светлосерая полоска. Передняя-
часть шеи и зоб дымчато-серые, грудь и брюшко серые. От птенцов других видов гаг хоро-
шо отличаются тем, что у них, как и у взрослых птиц, оперение по бокам надклювья*
далеко выдается вперед.
Молодые птицы. В общем похожи на взрослых самок, но спинная сторона туло-
вища у них темнее и с более узкими каймами перьев. Брюшная сторона туловища
серая с беловатыми или ржавыми каемками. У самца внутренние второстепенные махо-
вые темнее, сильнее изогнуты, чем у самок; второстепенные махи и большие кроющие
крыла имеют неясные белые или коричневатые вершины. Кроме того, у молодых самок
оперение спины и зада часто с более широкими бурыми окончаниями.
Самка в первом зимнем наряде.По оперению спинной стороны тела напоминает
взрослую самку в наряде после полной летней линьки. Отличается она и более
темной окраской темени и темным пепельно-коричневым затылком. Мантия, плечевые
перья и бока тела цвета сепии с более узкими буроватыми каемками, чем у взрослых.
Окраска спины, как в первом наряде. Верхние кроющие хвоста коричнево-черные
с охристыми предвершинными полосками; перья по бокам поясницы, как у взрослых. Щекн
и бока шеи коричнево-бурые, значительно сильнее чем у взрослых птиц исчерченные
темнокоричневыми полосками. Подбородок и горло пепельно-белые с узкими темнокорич-
невыми полосками, остальное оперение брюшной стороны, как у взрослых.
Самка после первой полной линьки. Имеет окраску взрослой птицы в том же наря-
де, но отличается окраской крыльев (как у молодых) и наличием отдельных перьев
ювеналылуго наряда на туловище
Самка во втором зимнем наряде. Отличима от взрослой только более узкими*
белыми вершинами второстепенных маховых и больших кроющих крыла. Во всех
последующих нарядах самки уже неотличимы от взрослых птиц.
Взрослая самка в брачном (весеннем) наряде. Однообразно буровато-коричневая,
более темная сверху. Верх головы черноватый в частых мелких продольных пестринах,
щеки и горло светлые, но также в черных штрихах. Спинная сторона тела черно-бурая
с рыжеватыми каймами перьев, такой же чешуйчатый рисунок имеют бока и почти
весь низ тела, кроме нижней части груди, которая однообразно чернобурая. Крылья
и хвост того же цвета, второстепенные маховые и большие кроющие имеют белые верши-
ны, которые образуют полоски, окаймляющие место, на котором обычно находится
зеркало. Клюв зеленовато-оливковый, темнее, чем у самца; лапы оливкоЕые.
Самка взрослая в летнем наряде (после полной линьки). Имеет оперение на
спинной стороне туловища и особенно на голове значительно более темное, чем вес-
ГАГА ОБЫКНОВЕННАЯ 527
ной. Темя коричнево-черное, затылок пепельно-коричневый. Оперение спинной стороны
и груди с более светлыми и узими краями, чем в обычном наряде. Щеки и шея с боков
с толстыми черно-коричневыми штрихами. Подбородок и горло пепельно-белые или бурова-
тые с узкими черно-коричневыми полосками. Бока темные пепельно-коричневые, перья
подхвостья светлее, с бурыми вершинами и темнокоричневыми предвершинными
пятнами.
Самец в первом зимнем наряде. Лоб и затылок табачно-коричневого цвета с
черными, более или менее четкими продольными штрихами. Нижняя часть темени
и затылок черные с голубоватым блеском, а задняя часть шеи черно-коричневая. Через
глаз проходит светлобурая или беловатая полоса; остальное оперение боков головы и шеи
коричнево-черное. Горло, шея спереди и плечевые перья черно-коричневые. На остальных
частях тела сохраняется ювенальное оперение. Весной первого года жизни, еще до полной
линьки, у самцов появляется частично белое оперение, свойственное взрослым птицам,
на затылке, щеках и шее, в мантии и на плечах. Передняя часть груди приобретает белую
или бледную розовато-кремовую окраску. В задней части темени и иногда по бокам шеи от-
растают белые перья с изумрудным налетом. Хвост, как у взрослых птиц; оперение брюшной
стороны сохраняет окраску, свойственную первому наряду. Описанный наряд имеет весь-
ма значительные индивидуальные вариации по степени развития белого оперения.
Самец после первой летней линьки. Отличается от взрослого в том же наряде
сохраняющимися перьями ювенальной окраски и оперением крыльев, присущим молодым
птицам.
Самец во втором зимнем наряде. В основном сходен со взрослым, но перья
головы и шеи с темными и желто-бурыми каемками, а перья мантии и плечевые ча-
стично с черными каемками. Средняя и задняя часть спины обычно черно-коричневые, бока
поясницы более или менее белые или с примесью белых перьев. По бокам подхвостья боль-
шие белые пятна, часть перьев на которых с черноватыми окончаниями. Остальное опере-
ние, как у взрослых, но часть перьев на горле, передней части шеи и в верхней половине
груди с узкими черноватыми полосками. Некоторые второстепенные маховые с узкими
белыми окончаниями и иногда с отметинами, но никогда не имеют столько белого, как у
взрослых. Внутренние из них ^же, чем у взрослых, только слегка изогнуты, с внешними
или с обоими белыми опахалами. Средние кроющие крыла белые с черно-коричневыми
концами и пятнами; большие кроющие с белыми окончаниями; некоторые из малых крою-
щих с белыми пятнышками. В остальном крылья как у взрослых. Количество белого на
крыле бывает весьма различно.
Самец после второй полной линьки Отличается от взрослых и годовалых птиц
в том же наряде оперением крыльев. Внутренние второстепенные маховые черновато-
коричневые с более или менее развитыми белыми отметинами, средние кроющие с черно-
ватыми концами, а малые кроющие бледнокоричневые.
Самец в третьем зимнем наряде. Имеет в основном оперение взрослой птииы,
но спина окрашена так же, как у двухгодовалых самцов; средняя часть поясницы с
примесью беловатых перьев с серо-коричневыми основаниями. Белые внутренние вто-
ростепенные маховые и кроющие с коричневатыми концами, некоторые внутренние боль-
шие и средние кроющие с коричневатыми пятнышками.
Самец после третьей полной линьки. Уже неотличим от более взрослых.
Взрослый самец в брачном наряде. Лоб, темя и верхняя половина боков головы
черные; от затылка вперед над глазами вдаются неширокие белые полосы. Затылок
и пространство позади щек с изумрудно-зеленым налетом по белому фону. Остальная
часть головы, плечи, верхняя часть спины, два пятна по сторонам поясницы и кроющие
крыла белые. Зоб и передняя часть груди также белые, но с нежным розовато-
кремовым налетом. Остальная часть брюшной стороны туловища, задняя часть спины и
надхвостье черные. Первостепенные маховые черно-бурые; второстепенные черные, внут-
ренние из них изогнутые, белые. Рулевые черно-бурые. Надклювье черное, в основной
половине зелено-оливковое, ноготок розовый; лапы оливковые; радужина черно-бурая.
Взрослый са*ец летом. Вся голова, шея, мантия и грудь грязнобурые с узкими
черными поперечными полосками. Над глазами черные и белые пестрины. Зоб и передняя
часть груди белые, иногда розоватые, с черными концами перьев. Остальные части
брюшной стороны черные; белые удлиненные перья по бокам поясницы отсутствуют.
Гренландская гага Somateria mollissima borealis В г е h m
Platypus borealis. В г е h m. Lehrb. Naturg. all. europ. Vcgel, 11, стр. 813, 1824, Баффинов
залив, Девисов пролив и западный берег Гренландии.
Распространение. Гнездовой ареал. О-в Банкса, северо-восточ-
ная часть о-ва Виктории, южные острова архипелага Парри, о-в принца Уэль-
ского, западный берег Баффиновой Земли, п-ва Бутия и Мельвилля, северная
часть Гудзонова залива и Лабрадор к югу до устья р. Гамильтон. Побережья
528 ОТРЯД ГУСЕОБРАЗНЫК
Гренландии до северных частей пролива Робсона и траверза пика Петермана;
о-в Ян-Майен, Шпицберген и Медвежий о-в. В наших пределах гренландская
гага заселяет, повидимому, южные острова Земли Франца-Иосифа. К сожале-
нию, в коллекциях до сих пор отсутствуют самцы, добытые в этом районе,
отчего окончательное решение о их подвидовой принадлежности пока не-
возможно. Основные зимовки располагаются вдоль южных берегов Гренлан-
дии, у Атлантического побережья Америки и к югу до Массачусетса, а также
в Баренцовом море.
Характер пребывания. Гаги этого наиболее северного под-
вида совершают регулярные и дальние сезонные миграции. Относительно осед-
лы лишь птицы, населяющие южную часть Гренландии и Лабрадора.
Сезонные миграции и их даты. Птица, окольцованная
летом на Шпицбергене, была добыта в ноябре на севере Норвегии (Нитхам-
мер, 1938); есть указания, что зимой они залетают на Мурман (Пальмен).
Весеннее появление гаг на Земле Франца-Иосифа приходится обычно на конец
мая—начало июня. Даты первых наблюдений 28 мая—9 июня (Горбунов,
1932). Даты прилета гаг связаны с временем появления первых промоин
и поэтому весьма различны в разные годы. Так, в 1931 г. первые гаги на о-ве.
Гукера появились уже 10 апреля, когда далеко на западе показалось «водяное
небо» (Демме, 1934). Прилет птиц на Шпицберген происходит в первых числах
июня. Время отлета с Земли Франца-Иосифа зависит от состояния льдов;
22 сентября они были там уже редки, а в конце месяца встречались лишь
единицами.
Биотоп. Период размножения эти гаги проводят на каменистых
островках и побережьях крайнего севера, а всю остальную часть года дер-
жатся в море.
Численность. На Шпицбергене, в Гренландии и на Лабрадоре
гнездует в массе. На Земле Франца-Иосифа обычна, но не многочисленна.
Экология. Размножение. На места гнездовья прилетает парами.
Огкладка яиц на Земле Франца-Иосифа происходит во второй половине
июня. Первое гнездо с 4 яйцами было найдено 21 июня. Вылупление птенцов
начинается в конце июля. Первые птенцы наблюдались 19—22 июля, но мас-
совое вылупление продолжалось в некоторые годы до 7 августа. Сроки гнез-
дованья, так же как и все остальные, бывают весьма различны в зависи-
мости от состояния льдов, в разные годы и в разных,даже весьма близких,
районах. Гнездится гренландская гага,большими обществами на высоких и су-
хих частях островов, зачастую среди камней и обычно довольно далеко от бе-
рега B00—300 м). Гнезда их иногда помещаются настолько близко одно от
другого, что бывает трудно найти место для того, чтобы поставить ногу.
Устраиваются гнезда зачастую на кочках злаков Роа arctica и Kochlearia,
причем одни и те же лотки занимаются птицами в течение многих лет (Кэм-
лин, 1879). Количество яиц в кладке для Земли Франца-Иосифа 4—6, для
Гренландии 4 C—5), редко 6—8 (Саломонсен, 1950). Их размеры там 69,9—
81,6x47,2—52,6, среднее 75,4x50,7 мм. Есть наблюдения, что самцы не сра-
зу покидают насиживающих самок и держатся некоторое время вблизи от
гнезда. Птенцы, покидая гнездо, сходят в море.
Линька. Первые стайки селезней отмечались на Земле Франца-
Иосифа 7 июля, но еще и 4 августа встречались селезни в брачном пере.
П и т а н и е. За время пребывания на море гренландские гаги кормятся
преимущественно моллюсками G0,3%), а из них мидиями (Mytilus edulis)
и другими двустворчатыми и в меньшем числе брюхоногими (Buccinum).
Ракообразные A4,3%), главным образом бокоплавы, имеют меньшее значе-
ние, зато более заметную роль, чем у других гаг, играют рыбы (8,9%), бычки,
преимущественно Myoxocephalus. Питание пуховых птенцов состоит в основ-
ном из рыбы (Cottidae) (Гренландия и Лабрадор; Коттам, 1938, 1939). Желуд-
ГАГА ОБЫКНОВЕННАЯ 629
ки гаг со Шпицбергена содержат исключительно моллюсков (Муа, Neptunia,
Bela) (Бианки, 1902).
Хозяйственное значение гренландской гаги велико. Гнездованье боль-
шими колониями облегчает сбор пуха. Промысел его на Шпицбергене
(Груманте) развит с очень давнего времени, причем уже с XVI в. в нем прини-
мали участие поморы нашего Мурмана.
Описание. Размеры. Гренландская гага несколько мельче, чем номинальная форма.
Клюв немного короче, размер его от конца голых «висков» у самцов обычно 65—70 F2—74),
а у самок 55—67 мм. Крыло короче, чем у основной формы примерно на 10 мм. Серпо-
видные же внутренние второстепенные маховые, как правило, несколько длиннее.
Вес. Наиболее жирными гаги бывают в конце июня, когда вес некоторых из них
превышает 2300 г.
Окраска. Селезень отличается от европейской гаги расцветкой клюва, кото-
рый у старых птиц, особенно в брачное время, яркий оранжево-желтый. Оперение самок
несколько более рыжее с такого же тона рыжими щеками.
Тихоокеанская гага Somateria mollissima v-nigrum Gray
Somateria v-nigrum. Gray. Proc. Zool. Soc. of Lond., стр. 212, 1855, Северная Аляска.
Русские названия. Гага, на Олюторском побережье—баул (вероятно звуко-
подражательное).
Распространение. Гнездовой ареал. Северное побережье Во-
сточной Сибири, от Чаунской губы до мыса Дежнева, о-в Врангеля и далее
к югу по берегу Берингова моря до Олюторского залива и Карагинского
о-ва. По последним данным гнездится на Новосибирских о-вах. Она обычна
у берегов Чукотки, к западу до мыса Ванкарем, но еще западнее редка и у
мыса Учкарпий наблюдается лишь единично (Тайер и Бэнгс, 1914). На Коман-
дорских о-вах гнездится в небольшом числе только на о-ве Медном. На Кам-
чатке гнездится только в северо-восточной части (Белопольский и Рогова,
1947), на Пенжине одна из самых обычных уток, причем встречается там на
расстояии до 300 км от моря (устье р. Слаутной; Волков и Дементьев, 1948).
Нередка в Гижиге. Самое южное нахождение бухта Шкиперова в Тауйской
губе (Ола), где гаги гнездятся колонией в 2—3 тысячи пар (Тугаринов, 1941);
залетают они и еще южнее, в район Аяна (Снигиревский). Вне наших преде-
лов гнездится на всех островах Берингова моря к югу от Алеутской гряды
и о-ва Кадьяка, а на Американском материке от основания полуострова
Аляски к северу, а затем к востоку до залива Франклина и устья Медной реки.
Наиболее восточным местом гнездовья служит юго-восточный берег о-ва
Виктории.
Характер пребывания. В течение всего года гаги держатся
у Командорских о-вов, Алеутской гряды и п-ва Аляски. Птицы, населяющие
другие части ареала, или откочевывают на зиму в море к границе ледяных
полен. (побережья Охотского и Берингова морей), или же совершают дальние
сезонные миграции (побережья Чукотского моря и моря Бофорта).
Основные зимовки расположены в Беринговом море. Тысяч-
ные стаи их встречаются зимой в проливе около о-ва Диомида: огромные
скопления гаг бывают в средней части моря у границы ледовых полей. Много
птиц зимует у Командорских и Алеутских о-вов, а также вдоль американского
побережья к югу до Ванкувера.
Сезонные миграции и их даты. Гаги с берегов Ледо-
витого океана, как из Сибири, так и из Америки, проводят зиму в Беринговом
море, совершая регулярные сезонные миграции. Ежегодно в первой декаде
июня происходит массовый пролет гаг к северу через Берингов пролив. За
один день там можно наблюдать в это время тысячи пролетающих птиц.
На Пенжине в устье р. Слаутной первые птицы отмечены 3 мая, на северо-
востоке Камчатки, в бухте Карагинской 23 апреля, в Анадырском заливе они
34 Птицы СССР, т. IV
530
ОТРЯД ГУСЕОБРАЗНЫЕ
появляются в последней декаде апреля, а у полярных берегов Сибири в конце
мая. Птицы прилетают парами или стайками из нескольких пар. Летом, в се-
редине июля A8 июля), проходит миграция селезней гаг в обратном направ-
лении—через Берингов пролив к югу, к местам их линьки (Бейли, 1948). Иа
Гижиги гаги исчезают во второй половине сентября, последнее наблюдение
в Карагинской бухте относится к 14 ноября, а скопление пролетных птиц
отмечено там 21 октября (Белопольский и Рогова, 1947).
Биотоп. Тихоокеанская гага в отличие от европейской гнездится
обычно в приморских тундрах.
Рис. 94. Тихоокеанская гага
Численность. Весьма обычная птица, гнездящаяся более разроз-
ненно, чем европейская форма и не образующая столь плотных гнездовых
скоплений.
Экология. Размножение. На места гнездовья гаги прилетают уже
разбившись на пары. Их токовые позы и голос такие же, как и у европейской
гаги. Гнездятся около мелких тундровых озерков. Гнезда устраиваются обыч-
но среди тундровых кустарничков, а на о-ве Карагинском, как правило,
в зарослях кедрового стланца. Диаметр гнезда около 18—24 см, а лотка
12—18 см, глубина лотка 4—9 см. Яиц обычно 5 C—6). На Командорских
о-вах в середине июня самки сидят на яйцах, а в июле появляются птенцы.
На о-ве Карагинском откладка яиц происходит в конце июня C0 июня нена-
сиженные кладки и самка с яйцом в яйцеводе). В Анадырском заливе выводок
мелких пуховиков встречен 8 июля, а полу оперившиеся птенцы 22 августа;
в бухте Провидения молодые с едва отросшими крыльями добыты 24 сен-
тября. В бассейне Пенжины птенцы появляются уже в конце июня—
начале июля, но молодые пуховики продолжают встречаться на Охотском по-
бережье до конца второй декады июля. Птенец, добытый 24 августа на п-ове
Тайгонос, был в пуху, и перья на крыльях у него только что начали проби-
ваться. Выводки долгое время живут на тундровых озерках и в устьях рек,
а на море переселяются лишь тогда, когда уже подрастут и возмужают.
Линька. Как и европейский подвид, линяет дважды в год, последо-
вательность смены оперения специально не изучалась. Послебрачная линь-
ГАГА-ГРББЕНУШКА 531
ка селезней наступает в начале или середине июля, когда они собираются в
стайки и переселяются на море. В некоторых местах перед началом линьки
наблюдается перелет самцов, но места концентрации линных птиц пока не-
известны. У берегов Аляски еще нелетные селезни, но уже надевшие новое
перо, встречены 11 августа. Самки линяют позже, уже после того, как вывод-
ки перемещаются на море.
Питание. Животные корма составляют в рационе тихоокеанской
гаги около 95,3% по объему от всего содержимого желудков. Основное зна-
чение имеют моллюски D6%) и среди них: мидии (Mytilus edulis), Seliqua,
ТеШпа, а также Litorina и другие Gastropoda. В отличие от европейской
тихоокеанская гага поедает значительное количество ракообразных C0,7 %),
за время пребывания на море это различные креветки и крабы, а на пресных
водоемах равноногие раки и бо коп лавы. В бЬльшом числе поедаются игло-
кожие A4,4%): Echinorachnis, Strongilocentrotus и Asteroidea (Аляска; Коттам,
1939).
Хозяйственное значение этой гаги невелико, особенно по сравнению
с европейской. Рассеянное гнездованье ее и отсутствие гнездовых колоний
делает сбор ее пуха нерентабельным. Добыча взрослых птиц и сбор их яиц
не имеют промыслового значения.
Полевые признаки те же, что и у европейской формы, но клюв оранжево-
желтый, а на горле у самца черное пятно в виде угла.
Описание» Размеры несколько крупнее, чем европейской формы. Крыло
самцов 260—330 мм, но обычно больше 295 мму у самок длиннее 275 мм; цевка 50—55 мм,
клюв 45—55 мм.
Вес взрослых самцов в мае и июне 2617 г B298—2809 г), а самок, добытых с мая
до середины июля, 2406 г B127—2894 г) (Брандт, 1943).
Окраска. Взрослый селезень. Отличается от номинальной формы черным
пятном на горле в виде буквы V, обращенным углом к подбородку. Зеленая окраска
распространяется от затылка до передней части щек под глазами. Клюв оранжево-желтый
с красноватым основанием и молочным ноготком; ноги грязнооранжевые или желтоватые.
Взрослая самка. Несколько светлее, чем самка европейской гаги, особенно
голова, и более охристая сверху. Горло и шея с винным налетом. Клюв и лапы оливково-
бурые.
Систематические замечания. Экземпляр самца из Тауйской губы (Охотское море)
не имеет горлового пятна, хотя по другим признакам (размеры, окраска щек и клюва)
он типичен для 5. m. v-nigrum. Есть указания, что отсутствие горлового пятна харак-
терно для гаг из указанной местности. Следует отметить, что среди гаг, населяющих
Чукотский п-ов, встречаются единичные птицы без горлового пятна (Портенко). Учиты-
вая, что слабо выраженное V-образное пятно есть и у подвида 5. m. dresseri, становится
невозможным выделение тихоокеанской гаги в самостоятельный вид, как это делают
Бутурлин A935), Тугаринов A941) и некоторые другие.
40. Гага-гребенушка Somateria spectabilis L.
Anas spectabilis. Li nnaeus. Syst. Nat., I, изд. X, 1758, стр. 123, Канада, Швеция
Синоним. Anas Beringii. G m e 1 i n. Syst. Nat., 1, 2, 1789, стр. 509, о-в Беринга.
Русские названия. Гребенушка—народное. Кроме того, горная гага (Новая
Земля) обязано гнездованию ее в стороне от побережья; туркан (самец), большая серая
утка (самка) в Якутии; баул, багул (Анадырь) звукоподражательные; турпан (непра-
вильно) неполовозрелые и линные птицы (Колгуев).
Распространение. Гнездовой ареал кругополярный. На Шпиц-
бергене гнездится в восточной части острова, но редка (Бируля, 1910; Кениг,
1911), держится в море у Медвежьего о-ва, но не гнездится.На Мурмане бывает
зимой и во время пролета, но иногда задерживается на восточном берегу до
20-х чисел июня. Как исключительный случай можно отметить встречу
двух выводков гребенушки 22 и 30 августа 1946 г. у Соловецких о-вов
(Паровщиков). Гнездится на п-ове Канин, о-ве Колгуеве и в Тиманской
тундре, как и везде, придерживаясь лишь приморской тундры и не удаляясь,
далеко от побережья. Обычная гнездящаяся птица Новой Земли, о-ва Белый
34*
532
ОТРЯД ГУСЕОБРАЗНЫЕ
и побережий материка. На Ямале доходит к югу до центральной озерной
котловины (р. Морды, 70° с. ш.; Житков, 1913), на Гыданском п-ве пример-
но до 71° с. ш. (Наумов, 1931). В Енисейском заливе идет несколько южнее
72° 15' с. ш., на Таймыре обычна и гнездится повсюду до южных берегов Таймы-
ровского озера и 72° с. ш. в долине Хатанги. Гнездится на Новосибирских о-вах,
о-ве Врангеля и обычна в материковых приморских тундрах к востоку до
Чукотского п-ва, вдоль берегов которого спускается к югу до залива Прови-
дения, бухты Креста (Белопольский, 1933) и возможно Анадырского побе-
режья. Известны летние встречи взрослых птиц на Камчатке, в Пенжинской
губе и на п-ове Тайгонос, но, по всей вероятности, в этих пунктах гребенушка
Карта 61. Распространение гаги-гребенушки Somateria spectabilis
а—граница гнездовий, в—места летних встреч птиц и спорадичного гнездования, г—зимовки,
э#с—места скоплений птиц на линьку
не гнездует. На Американском материке гнездится всюду вдоль полярного
побережья от Сюарда на западе до мыса Унгава (Лабрадор) на востоке,
отсутствуя лишь в районе залива Коронации; заселяет острова, кроме север-
ной части архипелага Парри и земли Элсмира. В Гренландии гнездится на
западном и восточном берегах, но отсутствует на юге и крайнем севере.
Основные зимовки в западном секторе Арктики находятся
на Мурмане и у северных берегов Норвегии, в море у севера Британских
о-вов, на юге Исландии и Гренландии и у американского побережья от Нью-
фаундленда до мыса Код. Наряду с этим гребенушки перезимовывают в неболь-
шом числе на незамерзающих участках моря у о-ва Колгуева юго-западных бе-
регов Новой Земли и даже в Карском море у Гыданского п-ова. На востоке
основные скопления зимующих птиц бывают у кромки льдов в средней части
Берингова моря; обычными местами зимовки служат восточное побережье
Камчатки, Алеутская гряда, п-ов Аляска. Кроме того, в некотором количестве
зимует в Анадырском заливе, у мыса Беринга и в Беринговом проливе у
о-ва Диомида.
Массовые скопления неполовозрелых и лин-
н ы х птиц наблюдаются в море около о-ва Колгуева, менее значительные
у Вайгача и вдоль западного побережья Новой Земли от Карских ворот до
южной части Северного о-ва. Около Колгуева неполовозрелые птицы держат-
ся в течение круглого года стадами по нескольку десятков тысяч штук. Туда
ГАГА-ГРЕБЕНУШКА 533
же стекаются на линьку и взрослые селезни. Огромные стаи неполово-
зрелых птиц, поражающие своей численностью, проводят лето у берегов
Чукотского п-ва и у побережья Аляски от мыса Барроу примерно до о-ва
Нунивок.
Характер пребывания. Во всех частях ареала перелетная
птица. Гребенушка совершает регулярные и иногда дальние сезонные мигра-
ции к местам зимовки, а взрослые самцы также в районы линьки.
Сезонные миграции гребенушки совершают вдоль побережий
или прямо над морем, но нигде их пролетные пути не проходят над мате-
риком. Новоземельские птицы линяют частично на месте, а частично, навер-
ное, у Колгуева, зимуют же они на Мурмане. Восточносибирские птицы со-
вершают дальний путь на восток и, только обогнув мыс Дежнева, поворачи-
вают к югу. Гребенушки, которые массами проводят зиму у берегов Финмар-
кена, исчезают оттуда в середине апреля (Смирнов, 1926), в Кольском заливе
последние птицы наблюдались 27 апреля, в большом количестве скапливаются
пролетные гребенушки весной у берегов Восточного Мурмана. Залетные пти-
цы добывались весной в шхерах Южной Финляндии (Нордман, 1860). Оди-
ночные пролетные птицы севернее о-ва Канин наблюдались 3 мая. Время
появления гаг-гребенушек у берегов Новой Земли бывает весьма различным
в зависимости от ледовых условий года: в 1950 г. первые стаи (неполовозрелых
самцов) появились 30 марта, в 1948 г.—14 апреля, а в тяжелом по состоянию
льдов 1949 г. только 21 мая (Успенский). В Карском море они появляются
поздно. Громадные стаи гребенушек встречены 3 июня в море у ледового при-
пая, к западу от Ямала, а у о-ва Белого пролет продолжался с 18 по 21 июня.
Птицы летели парочками, небольшими стаями, а иногда и стаями до 80 шт.,
чаще всего со стороны Обской губы (Тюлин, 1938). На западном берегу Таймы-
ра первые птицы были встречены 23 июня (Вальтер, 1902). Довольно обильный
весенний пролет у берегов Восточной Камчатки происходит в мае—начале
июня, примерно в то же время эти птицы появляются на Анадырском побе-
режье. Весенняя миграция гребенушек около мыса Дежнева начинается зна-
чительно раньше. Старые, половозрелые птицы появляются уже в апреле,
массовое продвижение их к гнездовым местам происходит в середине мая,
а в последней декаде его уже затихает. В первой половине июня вновь появ-
ляются пролетные гребенушки, и в середине месяца происходит массовое
движение их стай. Это бывают неполовозрелые птицы второгодки и трех-
летки (Портенко, 1947). Во время валового пролета гребенушек через Берин-
гов пролив D—19 мая) за день наблюдалось до 75 000 летящих птиц этого ви-
да. Новосибирских о-вов гребенушки достигают очень рано, что вероятно
обязано наличию полыней и разводий к северу от островов. Около о-ва Ко-
тельного первая стая была отмечена 3 мая, а затем до первой половины июня
наблюдались пролетные группы и стаи. Птицы, появляющиеся там в первой
половине мая, совершают дальний и тяжелый перелет над сплошными льдами
Восточно-Сибирского моря при 10—15° мороза, отчего нередко приходится на-
ходить на льду совершенно обессилевших или погибших птиц (Бируля,
1907). В дельте Индигирки первые гребенушки отмечены 4 июня (Михель,
1935), в Хатангском заливе 5 июня, а массовый пролет там стаями до 100 шт.
начался 10 июня (Скалой, 1935), наконец, на Восточном Таймыре 8 июня был
встречен одиночный самец, а 11 июня первая стайка (Тугаринов и Толмачев,
1934). Последние встречи этих гаг у Новосибирских о-вов (в разные годы)
17 сентября—10 октября, а у Таймырского озера 24 сентября; на Восточной
Камчатке появляется осенью в конце сентября. С о-ва Белого отлет происхо-
дит постепенно, небольшими группами, последние особи наблюдались 28 сен-
тября. У юго-западного берега Новой Земли последние стаи гребенушек наб-
людались в 1947 г. 3 октября, в 1948 г.—7—10 ноября. В октябре стаи гребе-
нушек уже появляются на Западном Мурмане (Кольский залив, Варангер-
534 ОТРЯД ГУСЕОБРАЗНЫЕ
фьорд), а у восточного берега они становятся многочисленны значительно поз-
же, в феврале-марте. Обилие их на зимовке у берегов Кольского п-ва весьма
различно в разные годы и, повидимому, зависит от гидрологического режима
года (Кафтановский, 1941).
Б и о т о п. В гнездовое время гребенушки заселяют приморские тундры
полярных островов и материкового побережья. Остальное время года, а не
половозрелые птицы и весь год, они держатся на море. Летом гребенушки
предпочитают иловатые мелководья и преду стьевые пространства рек, зиму же
проводят в открытом море, зачастую у кромки льдов.
Численность. Гребенушка, наряду с морянкой, наиболее обычная
утка приморских тундр, особенно в восточном секторе Арктики. По ориенти-
ровочным подсчетам , в летнее время на о-ве Белом держится около 4000 птиц
(Тюлин, 1938), а стаи линных селезней и неполовозрелых птиц достигают ме-
стами нескольких тысяч голов.
Экология. Размножение. Судя по смене возрастных нарядов
гребенушек, можно предполагать, что половая зрелость у них наступает на
третьем году жизни. Неполовозрелые птицы ведут стайный образ жизни и
круглый год держатся в море, зачастую далеко от своих родных мест. К местам
гнездовья гребенушки прилетают уже оформившимися парами, но так как
в это время тундра бывает еще покрыта снегом, то они принуждены иногда
долгое время держаться стаями на море у ледяного припая. Брачное поведе-
ние гребенушек таково же, как и у обыкновенной гаги. Самцы приподымают-
ся на воде, взмахивают крыльями, вздергивают кверху клюв и издают трое-
кратный громкий крик «арр»... «арр.»... «крр». Нередко наблюдаются ссоры и
драки между самцами, но никакой полигамии у них незаметно. Каждая
пара занимает определенный гнездовой участок, которого и придерживается.
Гнезда устраиваются в низинной моховой тундре около небольших озерков
и речек как непосредственно вблизи от моря, так и в стороне от него; иногда
(Новая Земля) преимущественно в сухой травянистой тундре, а иногда часть
гнезд находится в высокой полигональной тундре (о-в Белый). Гнезда обычно
разбросаны по тундре, но на некоторых речных островках, защищенных от
нападения песцов, гребенушки поселяются группами вблизи друг от друга,
часто вместе с гусями и чайками. Ямка для гнезда устраивается на сухом ме-
сте, но обычно поблизости от воды. Гнезда неглубокие, диаметром около
25 см и тщательно выстланные большим количеством довольно грубого
на ощупь пуха. Нормальный размер кладки 4—7 яиц, чаще всего 5 (Новая
Земля). Размер яиц 61,3—79,5 х 41,5—52,0, среднее 67 х 44 мм, цвет светло-
оливковый. На Новой Земле гребенушки держатся парами с начала июня и до
конца месяца. Первые гнезда с полными кладками обнаружены там 28 июня
(Успенский). В приморской части Тиманской тундры гнездо с 8 яйцами найдено
21 июня (Гладков, 1951), первое гнездо на о-ве Белом 3 июля, насиженные
кладки на Ямале 6 июля. На Новосибирских о-вах первая полная кладка
обнаружена 30 июня, а гнездо в дельте Колымы 26 июня. Самка сидит на гнез-
де очень крепко, но не так, как обыкновенная гага. Срок насиживания неиз-
вестен. Птенцы попадают сначала на пресные водоемы и только постепенно
перебираются к морю, где скапливаются зачастую в устьях рек, на местах, бо-
гатых моллюсками и другими животными кормами. Самки гребенушек охотно
принимают чужих птенцов, потерявших родителей, отчего нередко встреча-
ются выводки, состоящие из 15—18 молодых. Первые выводки в Тиманской
тундре наблюдались 23 июля, на о-ве Колгуеве и на Новой Земле они появ-
ляются во второй половине июля, а на Новосибирских о-вах 19 июля. Время
гнездованья бывает весьма растянуто, так, например, на Новой Земле выводки
мелких пуховиков встречались с июля до середины сентября (Горбунов, 1929).
Можно предполагать, что молодые из наиболее поздних выводков не успе-
вают развиться до наступления зимы и погибают.
ГАГА-ГРЕБЕНУШКА 535
Линька. Гребенушки линяют дважды в год, причем разграничить во
времени летнюю и осеннее-зимнюю линьку очень трудно, так как вторая
начинается тогда, когда еще не закончилась первая. В результате этого
оперение на многих частях тела или полностью или частично сменяется лишь
один раз в году. Особенно сильно проявляется это частичное выпадение отдель-
ных этапов линьки у неполовозрелых птиц, которые приобретают полный
взрослый наряд только на третьем году жизни. Взрослые самцы гребенушки
линяют в июле и августе. Первые стайки селезней, отбившихся от самок,
появляются в начале июля B июля в Хатангском заливе, 9 июля в устье
Индигирки). В Анадырском заливе линька самцов начинается в середине
июля. На Новой Земле 24 июля самцы уже держатся в море большими стая-
ми, теряют брачный наряд, но еще способны к полету; 10 августа в громадных
стаях линных гребенушек были уже нелетные птицы. В то же время самцы,
надевшие летнее перо, встречаются и на Новосибирских о-вах. Массовая
добыча линных птиц о-ва Колгуева происходит около 14 августа (Федосов,
1926). По характеру линьки они сходны с обычной гагой. Иногда в верхней
части мантии и на груди сохраняются перья зимнего наряда, сменяющиеся
уже при последующей предбрачной линьке. Эта последняя наступает обычно
в сентябре и продолжается до января. Взрослые самки линяют позже сам-
цов—в августе и сентябре. Вначале сменяется оперение головы, шеи, мантии
и лопаток, верхней части груди и боков, а также часть перьев спины, поясни-
цы и надхвостья. После этого линяют хвост и крылья. Смена оперения на
остальных частях тела оттягивается до начала следующей линьки. Полная
линька у молодых самцов начинается в августе, когда линяет часть оперения
головы, частично мантия, плечевые перья, бока поясницы, надхвостье, боль-
шая часть перьев в верхней части груди, бока тела и иногда подхвостье. Линька
остального оперения происходит в сентябре, когда уже начинается вторая
предбрачная линька, заканчивающаяся в январе. Первая линька молодых
птиц начинается в ноябре, когда у самцов сменяются перья на голове и шее,
а иногда также на боках груди и плечах. Такой промежуточный наряд птицы
носят зимой. С ноября по май продолжается частичная смена оперения туло-
вища, небольшого числа перьев в нижней части груди, перьев брюха, части
ттерьев мантии, плечевых и средней пары рулевых. Таким образом, эта линьки
бывает весьма неполной: кроме крыльев, совсем не линяют спина и зад, а так-
же сохраняется много ювенальных перьев на других частях тела. Линька
молодых самок еще менее полная; у них за это время сменяются лишь часть
перьев мантии и поясницы, отдельные перья спины, часть кроющих хвоста
и брюха и не всегда хвост (иногда лишь центральная пара рулевых).
Питание гребенушек по относительному значению основных кормов
распадается на два сезонных периода. Весной и летом, во время пребывания
на пресных тундровых водоемах, гребенушки кормятся преимущественно
личинками водяных насекомых и рачками-бокоплавами. Желудки самок гре-
бенушек, добытых в мае и июне на Таймыре и Новосибирских о-вах, были
наполнены красными личинками комаров-хирономид, которыми изобилуют
тундровые озера (Бируля, 1907). Самцы ранней весной почти не кормились
и имели обычно пустые желудки. С момента перехода на море состав питания
гребенушек резмо меняется. Основу его на Новой Земле составляют моллю-
ски, в большинстве случаев двустворчатые (Mytilus, Saxicava) и в меньшей
степени крабы. Однако иногда птицы переходят и на другие корма. В июле
желудки гаг, добытых в губе Безымянной (Новая Земля), содержали
почти исключительно крупных равноногих раков Jdothaea entomon и лишь
отдельных моллюсков. Там же, против Б. Карманкульского залива, в 15—
20 км от берега, 14 августа наблюдались стаи самцов гребенушек, кормив-
шихся в верхнем слое воды, вылавливая, повидимому, крылоногих, планк-
тонных моллюсков Clio borealis и Limacina limacina, которыми кишела вода
536 ОТРЯД ГУСЕОБРАЗНЫЕ
(Белопольский). На американском севере моллюски составляют 45,8% пище-
вого рациона гребенушек (в основном Mytilus edulis и Modiolaria), рако-
образные 18,6% (крабы бокоплавы, балянусы), иглокожие 17,2% (Echinora-
china, Strongilocentrotus), личинки ручейников 5,2%.
Экономическое значение гребенушек как объекта охоты велико только
в некоторых частях ареала. Так, например, на о-ве Колгуеве существует
регулярный промысел линных гребенушек, дающий 12—14 тысяч птиц в год.
Сбор пуха вследствие гнездованья этой гаги отдельными, не образующими
скоплений, парами не практикуется.
Полевые признаки. Крупная утка тяжелого склада. Как у плавающего,
так и у летящего селезня передняя часть туловища кажется белой, а задняя
черной, кроме белого верха спины. Изогнутые внутренние второстепенные
маховые при сложенном крыле выступают над спиной у плавающей птицы.
На полете видны белые пятна в основании крыльев и по бокам поясницы.
Рис. 95. Гага-гребенушка
У селезня в основании надклювья оранжевый вырост. Самка сходна с обык-
новенной гагой, однообразно бурая с темными пестринами. По земле передви-
гается тяжело, плавает и ныряет прекрасно, помогая себе под водой крылья-
ми. Весенний голос самца трехкратное «арр» или «урр», самки «кроо-кроо»
и обычное для гаг «гак-гак-гак».
Описание. Строение и размеры. Крупная утка с сильным клювом,
на основании которого у взрослых самцов имеется большой кожистый вырост. Он разви-
вается с февраля и существует в течение гнездового сезона, но после летней линьки исче-
зает. По сторонам надклювья оперение далеко не доходит до уровня ноздрей, а по коньку
клюва оно достигает их заднего края. Перья затылка у самца несколько удлинены и обра-
зуют гриву. Перья мантии и плечевые имеют у самок закругленные окончания, а не квад-
ратные, как у обыкновенной гаги. Внутренние второстепенные маховые серповидно изог-
нуты у обоих полов. Размеры: крылья самцов 270—290, самок 250—282 мм; цевки самцов
45—50, самок 45—48 мм; клюв (от оперения на хребте надклювья) самцов 28—34, самок
30—35 мм.
Вес селезней в мае 1831 г A490—2015 г), самок 1748г A500—1870г), самка в сен-
тябре 1400 г.
Окраска. Пуховой птенец. Теменная сторона головы, спинная сторона туло-
вища и бока его серовато-коричневые, но светлее, чем у птенцов обыкновенной га-
ги. Щеки светлее; светлая линия, проходящая над глазом, заметна слабо. Брюшная
сторона тела, горло и подбородок бледные пепельно-белые; пух на верхней части груди
темнокоричневыи у основания.
ГАГА-ГРЕБЕНУШКА 537
Молодые птицы в первом наряде. Темя черно-коричневое с темным штри-
ховым рисунком и светлобурыми каймами перьев. Затылок серо-коричневый, спинная сто-
рона туловища темнокоричневая с желто-бурыми каймами у большинства перьев. Перья
подбородка, горла, шеи спереди и с боков пепельно-белые с темными пятнами и штрихами.
Перья передней части груди, боков, туловища и подхвостья серовато-белые с темными
предвершинными полосками и бурыми концами. Остальная часть брюшной стороны тела
серо-коричневая, большей частью с темными полосками или вершинами перьев. Крылья
и хвост, как у взрослой самки, но рулевые с рыжими окончаниями перьев; второстепенные
маховые черно-коричневые с бледными внешними опахалами и частично с желто-бурыми
концами; часть больших кроющих с буровато-белыми концами. Самки в этом наряде
отличаются от самцов тем, что перья, образующие зеркало, светлее, а на спине и заду
с более широкими бурыми каймами.
Самки в первом весеннем наряде. Хорошо отличимы от взрослых не
только ювенальным оперением крыльев, но и остатками юношеского оперения на мно-
гих других частях тела.
Самки-второгодки. Сходны со взрослыми, но на сериях отличаются более
узкими белыми каемками больших кроющих и второстепенных маховых.
Взрослая самка. Темно-, почти черно-бурая. Перья головы с узкими черными
штрихами, а на туловище с черными центрами и ржавыми каймами (последние постепенно
обтрепываются). Брюшко и грудь одноцветные, коричнево-бурые. Второстепенные махо-
вые и их кроющие имеют белые вершины, которые образуют две белые полоски, огра-
ничивающие пространство, занятое у других видов зеркалом. Внутренние изогнутые
маховые с ржавыми каемками по наружному опахалу. Клюв и ноги зеленовато-серые,
радужина грязножелтая.
Взрослая самка после летней линьки. Оперение верха светлее, чем
зимой. Верх головы в частых черно-коричневых штрихах, затылок пепельно-коричне-
вый, мантия и плечевые перья с желто-бурыми концами. Свежие перья на верхней стороне
туловища с узкими черноватыми окончаниями и неправильными предвершинными по-
лосками.
Молодой самец в первую зиму. Темя и передняя часть затылка орехово-
или табачно-коричневые с черноватыми концами перьев и полосками на них. Уздечка и
щеки черно-коричневые с более светлыми полосками; остальная часть затылка, подбо-
родок, горло и шея черноватые; черное V-образное пятно видно более или менее заметно
только на подбородке и горле. Часть перьев на боках груди и плечах с буро-коричневыми
концами. Клюв светлый, зеленовато-серый, лапы грязножелтые; радужина тускложелтая.
Самцы-первогодки летом после окончания частичной юно-
шеской линьки. На темени и передней части затылка к зимнему оперению приме-
шиваются перламутрово-серые перья, остальной затылок становится белым. На щеках
примешиваются беловатые перья, перед глазами иногда появляется зеленый налет, а под
глазами черные пятна. Передняя часть мантии белая, остальная часть мантии, плечевые
перфя и надхвостье черные, как у взрослых птиц, а спина сохраняет юношеское опе-
рение; на боках поясницы примесь белых перьев. Подбородок, горло и передняя часть
шеи беловатые с не всегда ясно выраженным коричнево-черным V-образным пятном. Пе-
редняя часть груди буровато-кремовая. Бока груди, туловища и подхвостье черные, как
у взрослых, иногда черные перья появляются частично и на брюхе. Часто средняя пара
рулей окрашена, как у взрослых птиц. Все вновь появившиеся белые перья имеют чер-
новатые вершины. В этом возрасте птицы сохраняют много перьев первого юношеского-
наряда.
Самец после первой полной линьки. Имеет оперение, схожее с лет-
ним нарядом взрослых самцов, но в нем сохраняется некоторое количество ювенальных
перьев на туловище, особенно на его брюшной стороне. Затылок его несколько светлее,
чем у взрослого, а перья щек, лопаток и боков тела с более узкими темными окончаниями.
Самец после второй предбрачной линьки. По раскраске оперения
сходен со взрослым самцом, но серые перья затылка с темнокоричневыми концами; щеки,
бока шеи, передняя часть груди и мантия с черноватыми концами перьев. На крыльях зна-
чительно меньше белого, чем у взрослых: малые кроющие коричнево-черные, средние редко
белые, в большинстве с черноватыми концами, а иногда и одноцветные серо-бурые. Два
или три из них с белыми пятнами. Количество белого на кроющих варьирует индивиду-
ально. Самцы во втором летнем наряде и третьем зимнем уже неотличимы от взрослых.
Взрослый самец в брачном наряде. Темя и затылок голубовато-серые;
остальная часть головы, шея, верхняя часть спины, зоб и пятна по бокам надхвостья белые.
На щеках под глазами зеленый налет, а на зобу палевый. Вздутая часть надклювья огра-
ничена черной каемкой, черное пятнышко примыкает к задней части глаза, на горле черное
пятно в виде буквы V, обращенной углом к подбородку. Плечевые перья, задняя часть
спины, надхвостье, грудь и брюхо черные. Маховые черные, второстепенные обыкновенно-
с белыми кончиками; на крыле сверху белое пятно, образованное малыми и средними
кроющими, большие кроющие черные, перья по сгибу крыла черно-бурые. Хвост черно-
бурый. Нарост у основания надклювья оранжевый, остальной клюв мясного цвета; лапы
грязнооранжевые с темными перепонками; радужина желтая.
538
ОТРЯД ГУСЕОБРАЗНЫЕ
Взрослый самец летом. После полной линьки белое оперение сменяется
черно-бурым, белые перья сохраняются в небольшом числе только на зобу и конечно на
крыльях. Горло серо-бурое, щеки с мелким коричневым крапом.
41. Очковая гага Somateria fischerl Brandt
Fuligula (Lampronetta) Fischeri. Brandt. Novam AviumSpeciem..., 1847, стр. 18, Аляска.
Русские названия. Очковая гага—книжное. Народные: в Якутии дулкиян,
средняя серая утка (самка), турпан (самец—неправильно, так как это название в основ-
ном применяется к самцам гребенушки).
Распространение. Гнездовой ареал занимает узкую береговую
полосу северо-востока арктической Сибири и Аляски. В небольшом числе гнез-
дится к востоку от устья Яны и, видимо, в самом устье. Западнее отдельные
Карта 62. Распространение очковой гаги Somateria fischeri
а—места гнездовий, б—места точно не доказанных гневдовий, г—эимовки, д—валеты
встречи известны вплоть до устья Лены, где, повидимому, спорадично гнез-
дится. На Новосибирских о-вах летом держится парами, но всегда на взморье
и, вероятно, не гнездится (Бируля, 1907). Распространена по всей дельте
Индигирки и является там наиболее обычным видом гаг (Михель, 1935);
то же относится и к дельте Колымы (Бутурлин, 1906). На восток прослежена
на гнездовье до Колючинской губы Чукотского п-ва. Повидимому, залетает
и к югу вдоль побережья, так как известна эскимосам из залива Креста (Бе-
лопольский, 1933). В Америке населяет северо-западное побережье Аляски
от залива Портон до мыса Барроу.
Основные зимовки располагаются в Беринговом море у юж-
ной кромки льдов. У Командорских о-вов и азиатского побережья они даже
зимой не появляются, но бывают зимой на Уналашке (Далл, 1873), Прибыло-
вых и Алеутских о-вах. Известны единичные случаи зимних залетов в Европу:
в Норвегию у Вардб (Ионсен, 1937) и на нашу территорию в район Печенги
{Мейнертцхаген, 1938).
Характер пребывания. Гнездящаяся перелетная и в значи-
тельной мере зимующая в СССР птица.
Сезонные миграции их проходят через Берингов пролив
и далее вдоль арктического побережья Сибири и Америки. С Алеутских
о-вов отлетает в начале мая. На Новосибирских о-вах первая пара встречена
10 июня (Бируля, 1907), а в дельте Индигирки 16 июня (Михель, 1935). Отлет
с мест гнездовья происходит в середине сентября.
ОЧКОВАЯ ГАГА 539
Б и о т о п. В период гнездованья эти гаги держатся на мелких озерках
в приморской тундре. Время линьки, зиму, а неполовозрелые птицы, вероят-
но, и весь год они проводят на море иногда далеко от берегов, зимой обычно
у кромки льдов.
Численность. В дельте Индигирки и к востоку от нее вплоть
до Чукотки очковая гага вполне обычный, а местами даже малочисленный
вид. Однако, принимая во внимание, что ее ареалом служит лишь очень
узкая прибрежная полоса, можно считать, что общая численность вида
невелика. Местами (устье Индигирки) происходит промысловый сбор яиц
этого вида и численность его, по словам местного населения, сокращается.
Экология. Размножение, так же как и другие стороны экологии
очковой гаги, изучены очень слабо. Время наступления половой зрелости
неизвестно, но в аналогии с другими гагами можно предполагать, что она на-
ступает на втором или третьем году жизни (после двух или трех зимовок).
С момента прилета и до периода высиживания яиц птицы держатся парами,
причем самец сопровождает самку до гнезда. Наблюдений над токовыми играми
их нет. Очковая гага не гнездится колониями, а каждая пара занимает обычно
отдельное маленькое озерко с низкими болотистыми берегами. Только на бо-
лее крупных озерках с изрезанной береговой линией держится по нескольку
пар, невдалеке одна от другой.Гнездо представляет собой ямку во мху или
в грунте, иногда с небольшим количеством сухой травы. Пух появляется в нем
по мере завершения кладки и добавляется с началом насиживания. Обыч-
ное число яиц 5—6, но бывает и до 9. Скорлупа гладкая, светлая, по окраске
варьирует от буровато-оливковой до зеленовато- или желтовато-серой. Их
размеры 59,5—73,0x40,5—47,8 мм, в среднем 65,4x44,6 мм. В устье Инди-
гирки самка, добытая из пары 16 июня, еще не имела в яичнике развиваю-
щихся яиц. Откладка яиц начинается там в последних числах июня, а в на-
чале июля самки уже сидят на гнездах и держатся отдельно от самцов. Про-
должительность насиживания неизвестна. Самки, спугнутые с гнезда еще до
начала насиживания, часто возвращаются к нему и летают в сопровождении
самца. На полных кладках самки сидят очень плотно. Пуховые птенцы имеют
то же характерное оперение клюва, как и взрослые птицы. Молодые долгое
время держатся на тех же озерках, где они вывелись, и переселяются на
взморье уже будучи размером в половину взрослых или почти сравнявшись
с ними. В дельте Индигирки 30—31 августа молодые были размером почти
с самку, и хотя еще не могли летать, но уже подлетали на несколько метров.
В устье Колымы еще нелетные молодые были встречены 12 сентября.
Линька. Гага линяет дважды в год. Полная линька селезней начи-
нается, повидимому, в начале июля В устье Индигирки самец в брачном
наряде добыт последний раз 1 июля. Селезни, сбившись в большие стаи,ухо-
дят на море, где надевают летний наряд. Постепенно они откочевывают к ме-
стам зимовки, и на тундровых озерах появляются вновь лишь следующей ве-
сной. Точные сроки полной линьки установить трудно, но можно предпо-
лагать, что у селезней она заканчивается в середине июля. Так, 12—15 июля
в устье Колымы (Тайер и Бэнгс, 1934), а с 18 июля по 8 августа на взморье
у Индигирки (Житкова и Зензинов, 1915) наблюдались крупные стаи этих
гаг (иногда до 100 шт.) в наряде самок. Это могли быть частью перелинявшие
селезни, частью неполовозрелые, годовалые молодые. Сроки предбрачной
линьки, которую селезни проводят, вероятно, уже на зимовках, неизвестны.
Линька самок начинается с момента переселения выводков на море.
Полный взрослый наряд молодые птицы надевают, повидимому, на тре-
тьем году жизни.
Питание носит смешанный характер. Летом, за время жизни на
тундровых озерках, основным кормом служат водяные насекомые E1,1%)
л из них личинки ручейников D2,4%) и хирономид (8,3%), брюхоногие мол-
540
ОТРЯД ГУСЕОБРАЗНЫЕ
люски (9,4%) и ракообразные B,6%). Тогда же очковые гаги потребляют
порядочно растительной пищи C7%). Это главным образом листья и клубень-
ки гребенчатого рдеста A0,2%), зелень злаков, осок, водяной сосенки и дру-
гих растений, а также ягоды вороники (Empetrum nigrum). За период жизни
на море очковая гага переходит почти целиком на животную пищу, в основ-
ном на моллюсков (95,8%). Больше всего цз них поедаются морские плече-
ногие (92 %) и из них Siliqua (Коттам, 1939).
Хозяйственное значение этой гаги невелико. Промысел ее имеет чисто
местное значение. В приморской полосе Северной Якутии она добывается на
мясо наравне с другими утками.
Полевые признаки. Утка среднего размера с довольно короткой шеей
и массивной головой. Самка однообразно бурая, кроме более светлых пятен
Рис. 96. Очковая гага
около глаз и белых перьев в основании крыла, заметных и у сидячей и у ле-
тящей птицы. Самец в основном белый, с ярко белыми «очками» около глаз,
зелеными пятнами на щеках и затылке и оранжевым клювом.
Описание. Строение и размеры. Утка среднего размера с нешироким,
но довольно высоким и сильным клювом. Оперение лба широким клином выступает по
хребту надклювья и спускается по нему далее ноздрей. Оперение вокруг глаз пушистое,
образует как бы очки. Перья затылка удлинены и у самца составляют нечто вроде гривы.
Удлиненные плечевые и третьестепенные маховые перья слегка изогнуты вниз. Рулевые
на концах закругленные. Длина крыла самцов 255-267, самок 240—250 лык; цевка сам-
цов 45—50, самок 46—48 мм; клюв (от оперения надклювья) у самцов 21—23, у самок
ок 28 мм
О к р а с к а. Пуховой птенец. На спинной стороне серовато-бурый, на брюшной
светлый, почти белый. Характер оперения надклювья тот же, что и у взрослых птиц.
Вокруг глаз коричневатые пятна на месте будущих «очков», а на этом фоне за глазом тем-
ная полоска. г
Молодые птицы в первом наряде. В общем похожи на взрослую самку, спинная
сторона рыжевато-бурая, но тусклее, чем у самок и с менее выраженной поперечной
полосатостью; спина и надхвостье черноватые, каемки перьев ржавчатые. «Очки» наме-
чаются более короткими и светлыми перьями. Брюшная сторона тела довольно светлая
серовато-бурая, с неясной более темной штриховкой, продольной на зобу и поперечной
СИБИРСКАЯ ГАГА 541
на боках и груди. Плечевые и верхние кроющие крыла черноватые с бурыми краями.
Клюв буро-оливковый, лапы желтоватые.
Самка взрослая. Имеет оперение в общем рыжевато-бурое в мелких черных по-
перечных пестринах. Темя и затылок более каштановые с мелкими продольными штри-
хами. Спинная сторона, бока тела, груди и подхвостье имеют поперечно-полосатый рисунок
за счет рыжих и почти черных полосок на перьях. Вокруг глаз пятно из более светлых
серовато-бурых перьев, а перед ними на щеках и лбу более темное пятно из мелких черно-
вато-бурых перышек. Середина груди и брюшко дымчато-бурые. Большие кроющие
крыла буроватые, маховые серо-бурые, так же как и рулевые. Клюв голубоватый, лапы
желто-бурые.
Взрослый самец в брачном наряде. Раскрашен ярко, на голове вокруг глаз
округлое белое пятно (сочки») в поперечнике около 30 мм. С боков и сверху оно окай-
млено узкой черной полоской. Оперение у основания надклювья бархатисто-белое, пере-
ходящее на лбу, темени и затылке в зелено-желтый цвет; грива на затылке зеленая.
Щеки ниже «очков» и затылка бархатистые, густо зеленые. Горло, шея, мантия, плечевые
и большая часть спины белые. Задняя часть спины и надхвостье дымчато-черные с двумя
белыми пятнами по бокам поясницы. Зоб, верхняя часть груди и бока серо-стальные
с синеватым отливом. Задняя часть груди, брюхо и подхвостье дымчато-черные. Верх-
ние кроющие крыла серо-бурые, кроме малых, образующих белое пятно. Первостепенные
маховые и часть второстепенных черно-бурые, внутренние второстепенные и третьесте-
пенные чистобелые. Рулевые серовато-бурые. Клюв оранжевый, лапы буро-оливковые,
радужина молочно-белая с голубоватым кольцом.
Взрослый самец летом. Вся голова однообразно серая, и «очки* заметны лишь
в виде серого пятна. Зеленые участки (затылок, щеки и др.) заменены беловатыми.
Лопаточные перья серые. Грудь буроватая, брюхо серое с черными вершинами перьев.
42. Сибирская гага Somateria stelleri Pall.
Anas stelleri. Pallas. Spicil. Zool., VI, стр. 35, 1769, Камчатка.
Русские названия. Книжные—сибирская, стеллерова или малая гага; народ-
ное—биргиньях (Якутия).
Распространение. Гнездовой ареал, занимает узкую примор-
скую полосу в восточном секторе Арктики, но выяснен недостаточно. Наи-
более западным пунктом единичного гнездованья можно считать Новую
Землю. Там была встречена пара взрослых птиц в июне на Безымянной губе
Южного о-ва в брачном наряде, а в сентябре у Маточкиного Шара выводок
уже летных молодых (Успенский). В качестве случайно летующей птицы
известна также для п-ова Канина и дляМурмана. Возможно, что в последнем
месте спорадично гнездится, так как выводок был встречен однажды в райо-
не Варангер-фьорда (Болам, 1926), а по старым сведениям, собранным Плеске
A887), бывает летом в районе Печенги, на Айновых о-вах и на о-ве Сосновце.
Указывается для прибрежных тундр северной части Ямала, но на о-ве Белом
не отмечена (Тюлин 1938). На Гыданском п-ве и в устье Енисея не встречена
ни разу. В северо-западной части Таймыра и в Хатангском заливе наблюда-
лись единичные птицы и гнездовье не зарегистрировано (Скалой, 1935). На
Новосибирских о-вах летом нередка и возможно гнездится на ближних к ма-
терику (Бируля, 1907). В дельте Лены обычна, но в дельте Индигирки очень
редка, известна лишь по отдельным экземплярам, и вопрос о гнездовании
ее там пока не ясен (Михель, 1935). Очень редка и в устье Колымы, но к вос-
стоку от нее становится обычной и встречается повсюду до крайнего севера
Чукотки. Залетает на о-ва Врангеля и Геральд. На Анадырском побережье
сибирская гага известна как пролетная и местами летующая птица. Так,
в заливе Креста она держится все лето в большом лисле (Белопольский,
1933). На Восточной Камчатке и на Гижиге известны лишь единичные летние
встречи явно негнездящихся особей. На американском побережье гнездится
от бухты Нортон до мыса Барроу. Кроме того, есть указание, что очковая
гага оседла на Уналашке (Далл, 1873), но возможно, что оно основано на ре-
гулярном летнем пребывании неполовозрелых птиц.
Основные зимовки располагаются в южной части Берингова
моря у Командорских, Алеутских, Прибыловых о-вов и вдоль п-ова Аляски.
542
ОТРЯД ГУСЕОБРАЗНЫЕ
В меньшем числе, но регулярно эти гаги зимуют у восточного берега Камчат-
ки и на севере Курильской гряды. Залетают зимой на Сахалин и к берегам
Приморья. Ежегодно встречаются зимой в Остфинмаркене (Сконнинг, 1907)
и в Варангер-фьорде с сентября до весны (Плеске, 1887), а единичные
зимние встречи известны и на Балтийском море: в Польше, Латвии, под
Ленинградом.
Характер пребывания. Гнездящаяся перелетная и в значи-
тельной степени зимующая в СССР птица.
Сезонные миграции и их даты. Пролет сибирских гаг
проходит над морем или вдоль морских побережий. Встречи их во внутренних
частях страны (среднее течение Анадыря) случайны. С мест зимовки у о-ва
Беринга они полностью исчезают к началу мая. В мае пролетают у восточных
Карта 63. Распространение сибирской гаги Somateria stelleri
о—места гнездовий, б—места точно не доказанных гнездовий, в—места постоянных встреч
летом (возможно гнездованье), г—зимовки, д—залеты
берегов Камчатки, причем в некоторые годы бывают многочисленны (Аверин,
1948). Весенние встречи этих гаг на Анадыре зарегистрированы 19 мая—
7 июня. Массовый пролет их через Берингов пролив наблюдается у мыса
Стюарта с конца мая до середины июня (Бейли, 1948). У северных берегов
Сибири появляется в начале июня. Первые встречи их: под Нижне-Колым-
ском 9 июня, на Новосибирских о-вах 15 июня, в Хатангском заливе 18 июня.
8 середине августа они держатся уже большими стаями, постепенно продви-
гающимися к местам зимовки. В начале сентября появляются у Командор-
ских о-вов, а в середине этого месяца у Восточной Камчатки, где в октябре
становятся уже многочисленными.
Биотоп. На гнездовье придерживается приморской тундры, где
поселяется у мелких озерков. Все остальное время года проводит в море.
Численность сибирской гаги на местах гнездовья повсюду неве-
лика. На пролетах же и в местах зимовки местами она собирается в большом
числе. Численность ее за последнее время, повидимому, заметно снижается.
Так, в начале нашего столетия у Командорских о-вов эти гаги зимовали огром-
ными стаями, а в 30-х годах они стали там уже лишь обычным, но не мас-
совым видом.
СИБИРСКАЯ ГАГА 543
Экология. Размножение. Наступление половой зрелости, повиди-
мому, в возрасте 2—3 лет. В период откладки яиц и в начале насиживания
держатся парами. Токование селезня довольно просто и заключается
в вытягивании шеи кверху или закидывании головы на спину. Последнее
движение сопровождается негромким гортанным криком. Однако по сравне-
нию с другими утками ток у этих гаг проходит очень тихо. На Уналашке стаи
распадаются на пары уже в начале мая (Далл, 1873). Местом гнездованья
служат озерные понижения в моховой тундре. Гнездо представляет ямку,
выстланную мохом или сухой травой, взятыми здесь же, поблизости, и имеет
значительное количество темнсбурого пуха. Яиц обычно 6—7, изредка до 10.
Скорлупа гладкая, желтовато- или буровато-оливкового цвета. Вариации
в размерах велики: 55,5—70,5x37,0—47,1 мм. Среднее 61,4 х 42 мм. Срок
насиживания неизвестен. Пуховики хорошо отличимы от птенцов других ви-
дов гаг по оперению у основания клюва. Ненасиженная кладка на Таймыре
найдена 25 июля (Миддендорф, 1853), гнездо с яйцами в дельте Лены—4 июля
(Бунге, 1887).
Линька. Как и другие утки, гага линяет дважды в год: после сезона
размножения полностью и перед началом его частично. Последовательность
смены оперения во время летней линьки у них несколько своеобразна. Сна-
чала сменяются перья головы и шеи, а затем зоба, плеч, лопаток и боков тела.
При этом на лопатках отрастают частично темнобурые летние перья, а частич-
но сразу же черные перья брачного наряда. При осенней линьке эти участки
черных перьев уже не сменяются, а лишь дополняются новыми. Таким обра-
зом, часть оперения на некоторых участках тела (кроме крыльев) сменяется
только один раз в году. Рули сменяются поздно, уже тогда, когда линька
контурного пера зашла далеко. Вслед за ними, уже к концу летней линьки,
сменяются маховые. Селезни покидают самок с самого начала насиживания.
Линька их начинается в последней декаде июля, выпадение же маховых
приходится на конец августа. В середине сентября у селезней начинается пред-
брачная линька. Сперва перелинивают перья груди и брюха, затем спина
и плечи. В середине октября линяет голова, шея и отрастают удлиненные
плечевые перья. Сменяются ли вторично рули—неизвестно. В начале декабря
селезни уже надевают полный брачный наряд (Тугаринов, 1941).
Питание. Сибирская гага преимущественно животноядная утка.
За время ее пребывания в море она кормится почти исключительно животной
пищей и при этом в значительной мере моллюсками. В летнем же рационе
гаг, живущих на тундровых озерах, животные корма составляют около 87,1 %.
Основу составляют ракообразные, главным образом различные бокоплавы
D5,2%); в большом числе поедаются водные насекомые A3%) (личинки хиро-
номид и ручейников), моллюски A9,3%), главным образом плеченогие, в ма-
лом числе черви-полихеты и мелкие рыбки. Из растительных кормов A2,9%)
чаще всего поедаются гагами рдесты, морская трава—зостера, ягоды воро-
ники и водоросли (Коттам, 1939). В желудке селезня, добытого на Анадыре
7 июня, обнаружено большое число личинок веснянок, водяных жуков
и 1 клоп Сопха (Портенко, 1939).
Хозяйственное значение сравнительно невелико. Сбор пуха сибирской
гаги производится попутно со сбором его у других гаг, но ввиду малочислен-
ности птицы промышленного интереса он не представляет. Как объект охот-
ничьего промысла имеет некоторое значение лишь в быту кочевого населе-
ния крайнего северо-востока.
Полевые признаки. Утка среднего размера. Голова самца белая с зеле-
ным пятном на затылке. Самка рыжевато-бурая. У обоих полов имеется
блестящее крыловое зеркало с синим отливом. Довольно молчаливые утки.
8 противоположность другим видам гаг летают довольно высоко —30—50 м
над водой и не часто спускаются к волнам.
ОТРЯД ГУСЕОБРАЗНЫЕ
Описание. Строение и размеры. Утка среднего размера, несколько
мельче остальных гаг. Клюв несколько менее мощный, чем у других видов этого рода.
Оперение лба почти не выдается вперед от основания лба. Этот признак служит хорошей
отличительной чертой пуховиков данного вида от других видов гаг. Внутренние второ-
степенные маховые серповидные, изогнуты наружу. Рулевые с заостренными вершинами.
Длина крыла (самцы и самки) 208—226 мм, цевки 34—40 мм, клюва 37—43 мм.
Век самцов в мае 850 г, в октябре 500—800 г, в ноябре 1000 г; самок в июле 720 г,
в сентябре 720 г (Камчатка; Аверин 1948).
Окраска. Пуховой птенец. Темнобурый на спинной стороне с желтоватобурой
полоской за глазом, более светлым лбом и щеками. Брюшная сторона светлее верха,
буровато-серая.
Молодые птицы в первом наряде. Сходны со взрослой самкой. Спинная сторо-
на рыжевато-бурая со светлыми вершинами перьев. Брюшная сторона рыжеватосерая в
узких черных поперечных полосках. У самцов на второстепенных маховых есть слабо
выраженное синее зеркальце, у самок его нет. Большие кроющие крыла и второсте-
пенные маховые имеют белые вершины, которые образуют две узкие полоски вдоль крыла.
Рис. 97. Сибирская гага
Взрослая самка. Общий тон окраски рыжевато-бурый. На голове и шее мелкие
рыжеватые поперечные пестрины, то же на спине и надхвостье. Перья мантии со
светлобурыми каемками. Передняя часть груди более ржавого цвета, брюшко черно-
бурое. Большие кроющие крыла с белыми вершинами; первостепенные маховые черно-
бурые, второстепенные с белыми вершинами, а внутренние из них с синими наружными
опахалами. Образованное ими зеркальце имеет синий блеск и окаймлено двумя белыми
полосами. Рулевые черно-бурые. Клюв и лапы синевато-серые, радужина бурая.
Молодой самец после первой предбрачной линьки (весной по первому году).
Имеет большую часть оперения, сходную с первым нарядом. Часть плечевых перьев
получает синие наружные и белые внутренние опахала; горло черноватое и черноватый
ошейник у основания шеи.
Взрослый самец в брачном наряде. Голова сверху и с боков белая с зелено-
ватым налетом на лбу между глазами и густо зеленым поперечным пятном на затылке,
которое заканчивается по бокам черными пятнышками. Горло, шея, зашеек и вся спина
до хвоста включительно черные с синим отливом. Грудь и бока рыже-каштановые.
Середина груди, брюхо и подхвостье черные. Черные небольшие пятна на боках груди
у сгиба крыла. Кроющие крыла белые, первостепенные маховые бурые, второстепенные
с сине-фиолетовыми наружными опахалами и бурыми вершинами. Удлиненные плечевые
с блестящими синеватыми наружными опахалами и узкими белыми внутренними. Клюв
и лапы сине-серые, радужина красновато-бурая.
СИНЬГА
545
Взрослый самец летом. Вся голова, шея и лопатки темнобурые с буроохристыми
вершинами перьев. Спина вся до хвоста и плечевые того же цвета, но без каемок.
Зоб и грудь темнобурые с более рыжими каемками перьев; брюшко и подхвостье черные.
Плечевые мало удлинены.
РОД ТУРПАНЫ MELANITTA ВО IE, 1822
Тип. Anas fusca
С и н о> и м. Oidemia Fleming, 1822.— Polionetta. Каир, 1829.
43, Синьга Melanitta nlgra L.
Anas nigra. Li n n a e u s. Syst. Nat., изд. X, I, 1758, стр. 123, Лапландия.
О и н о н и м. Anas atra. Pallas. Zoographia Rosso-Asiatica, II, 1811, стр. 247, Каспий.
Русские названия. Синьга, местами чернеть, обязано окраске.
Распространение. Гнездовой ареал. Захватывает всю Ислан-
дию и Оркнейские о-ва, север Шотландии и Ирландии, большую часть Скан-
Карта 64. Распространение синьги Melanitta nigra
а—граница гнездовий, б—не вполне выясненная граница гнездовий, в—обособленные гнездовые
колонии, в*—места постоянных встреч летом и спорадичного гнездования, г—зимовки, ж—места
скоплений птиц на линьку
динавского п-ова, кроме наиболее южных областей Норвегии и Швеции, а так-
же Финляндию до широты Ваза. Встречается летом в южной части Шпиц-
бергена, на Медвежьем о-ве и гнездится там (Кениг, 1911). В пределах Совет-
ского Союза северная граница от Мурмана до Байдарацкой губы Карского
моря, проходит вдоль морского побережья, захватывая о-ва Колгуев и Вай-
гач, на которых птицы регулярно встречаются летом, но неизвестно—гнез-
дятся ли. На Южном о-ве Новой Земли синьга встречается летом и, повиди-
мому, гнездится на о-ве Междушарском (Успенский). На Ямале доходит до
границы между типичной и арктической тундрой и следует этому рубежу
35 Птицы СССР, т. IV
546 ОТРЯД ГУСЕОБРАЗНЫЕ
далее на восток до устья Хатанги, где вновь почти подходит к морскому
побережью. Восточным пределом распространения европейской синьги слу-
жит водораздел Хатанги и Лены. Она встречается в верховьях р. Оленека и в
западной части Вилюйского бассейна, хотя в последнем месте и редка (Туга-
ринов, 1934). Гнездованье на Лене не установлено, но есть экземпляры, добы-
тые в устье реки (Бунге, 1887). Гнездится в верховьях Хатанги на р. Мойеро,
известна в верхнем течении Подкаменной Тунгуски, но отсутствует в бас-
сейне Ангары. На Енисее о0ычна под Туруханском и севернее его. Встречает-
ся летом (самцы) на р. Елогуе, но вероятно не гнездится там (Слудский).
На Оби граница гнездованья синьги проходит около Березова, хотя негнез-
дящиеся птицы попадаются в летнее время и в устье Иртыша. На Печоре
она гнездится примерно до устья Усы, всюду в Тиманской тундре, вдоль
всех берегов Белого моря, в северной части Карелии и на Мурмане. Спора-
дично гнездится на островах (Сарема и Хиума) в Балтийском море (Гроссе
и Транзее, 1929).
Районы линьки. Места скопления линных птиц до сих пор почти
неизвестны. Вероятно, некоторая часть селезней линяет на тундровых озерах
в местах гнездовья, часть, хотя и очень небольшая, появляется в это время
на озерах лесостепи Зауралья (Свердловская обл.; Сабанеев, 1874; Матушино
Курганской обл.; Богачев) и, наконец, значительная часть проводит линьку
где-то у берегов Балтийского моря. О последнем говорит сильный пролет
селезней перед началом линьки через Ладожское озеро и Финский залив.
Основные зимовки синьги расположены в Северном и Бал-
тийском морях. Огромные стаи их проводят зиму у Британских о-вов, у бере-
гов Голландии, Бельгии и Франции (Пикардия). В меньшем числе они встре-
чаются зимой у берегов Пиренейского п-ова и достигают Азорских о-вов. Зи-
мует синьга также у Скандинавских берегов на север до Лофотенских о-вов
и всюду в незамерзающих частях Балтийского моря до побережий Литвы и
Латвии (Лиепая) включительно. Внутрь материка она зимой не проникает
и появляется на реках и озерах (Рейн, Боденское озеро и т. д.) только во вре-
мя сильных штормов на море. Регулярных зимовок синьги на южных морях
(Каспийское, Черное, Средиземное) нет, встречи ее там единичны и составляют
исключительное явление.
Характер пребывания. У северных побережий Англии и
Норвегии синьгу можно встретить круглый год, но во всех других частях
ареала это перелетная птица.
Сезонные миграции и их даты. Исландские птицы
спускаются зимой к Британским о-вам и далее к югу. Одна синьга, окольцо-
ванная летом в Исландии, добыта зимой у берегов Португалии, а другая на
Азорских о-вах. Птицы, населяющие север европейской части Союза, летят
на Запад вдоль морских побережий, а также в массе через озера Онежское и
Ладожское к Финскому заливу и далее над морем к берегам Западной Европы.
В верхнем течении Печоры, по Ылычу и Колве наблюдается интенсивный
пролет в широтном направлении; весной на восток и северо-восток, а осенью
на запад (Теплоухов, 1911; Теплов, 1948). Этим путем, повидимому, следуют
птицы, населяющие Урал и Зауралье. По долинам Вычегды и Северной Двины
они достигают Белого моря, а часть двигается дальше на запад. Некоторые
из них попадают на Рыбинское водохранилище и, следуя вверх по течению
Мологи, летят в сторону Балтики. По Иртышу, Оби и Енисею, вне пределов
гнездового ареала, нормального пролета нет. Вероятно из Западной Сибири
птицы осенью отлетают частью прямо на запад, частью к морю и продвигают-
ся дальше морскими путями. Ежегодно в осеннее время одиночные птицы
и их небольшие стайки появляются повсюду в средних и кое-где в южных
областях европейской части Союза (Московская, Тульская, Ульяновская
обл., Башкирия и др.) и изредка кое-где в Средней Азии. Как правило, это
синьга 547
бывают молодые птицы, много реже взрослые самки и крайне редко самцы.
Весной никакого пролета в этих местах не наблюдается. Осеннее появление
синьги на юге нельзя считать пролетом этого вида, так как на южных
зимовках его не бывает. Явление это правильнее расценивать, как пример
регулярных и массовых залетов в результате отклонения молодых птиц от
основного пролетного направления во время их первого осеннего пути.
Отклонение это для них, как и для большинства залетных птиц, кончается
гибелью.
Западноевропейские зимовки синьги покидают в марте, в начале апредя
они появляются у берегов Эстонии, где держатся большими и малыми стаями
в море, обычно далеко от берегов (Кох, 1911). Пролет их продолжается там
весь май, а под Ленинградом большие стайки их держатся в заливе с 28 мая
до 4 июня, конец же пролета затягивается до середины последнего месяца.
Кольского п-ова (озера Имандра) они достигают около 28 мая (9 мая—7 июня),
причем появляются там как парами, так и стайками от 3—4 до 25—30 шт.
смешанного полового состава (Владимирская, 1948). В Тиманской тундре пер-
вые птицы встречены 3 июня, а в верхнем течении Печоры 20 мая A4—23 мая),
валовой же пролет начинается там 25 мая и продолжается от 3 до 11 дней.
В последнем месте синьга летит весной как ночью, так и днем большими
стаями, иногда по нескольку сотен штук. Валовой пролет, как правило, про-
ходит двумя волнами, промежуток между которыми не превышает 5 дней.
Первая волна бывает более мощной. Интенсивность пролета в разные годы
чрезвычайно различна (Теплов, 1948). На Южном Ямале первая пара была
отмечена 19 июня (Осмоловская). Осенняя миграция начинается отлетом
взрослых самцов к местам их линьки. Только после значительного перерыва
появляются пролетные молодые птицы и самки. На Ямале синьга наблюдалась
еще 24 сентября, когда других уток там уже не было. На Верхней Печоре про-
летные стайки взрослых самцов числом от 5 до 9 экземпляров отмечены
в разные годы 14—24 августа, а начало осеннего пролета молодых приходится
на 27 сентября B1 сентября—3 октября). Его продолжительность составляет
обычно 14 дней A1—17), а последние птицы встречаются до 28 октября
A7 октября—1 ноября) (Теплов, 1948). На Рыбинское водохранилище попа-
дают только молодые и это бывает с 28 сентября по 14 октября, последние птицы
задерживаются до 31 октября—6 ноября. Под Архангельском (озеро Слобод-
ское) массовый отлет отмечается 5—15 октября, а в Лапландии (озеро Иманд-
ра) в конце октября перед самым ледоставом. В середине и во второй половине
июля в Карелии проходит интенсивный пролет селезней синьги. Стаи их чис-
лом до нескольких сотен штук, вытянувшись в длинную линию по фронту,
летят на значительной высоте к юго-западу. Возможно, что их же наблюдал
17—24 июля A943—1944) Семенов Тян-шанский A947), принявший их по сви-
сту крыльев за стаи гоголей. Ежегодно, примерно с 12 июля по 2 августа, про-
лет селезней громадными стаями проходит между Ладожским озером и Фин-
ским заливом и в то же время под Выборгом. Летят исключительно одни
взрослые самцы в сильно поношенном пере (Алфераки, 1900). В середине сен-
тября там же начинается пролет молодых птиц, которые в конце октября
держатся у берегов залива стаями по нескольку тысяч голов. Задерживается
там синьга до начала ноября. Те же сроки пролета для селезней и молодых
птиц указываются для Эстонии (Кох, 1911). На зимовках у берегов Фран-
ции они скапливаются в большом количестве уже в ноябре, когда море места-
ми бывает покрыто их тысячными стаями.
Биотоп. В гнездовое время открытые озера тундры и лесотундры
или же моховые болота в тайге. Во время пролетов озера и реки с обширны-
ми плесами, зимой неглубокие морские заливы и бухты.
Численность. На этой территории синьга обычна всюду в зоне се-
верной лесотундры и в тундре. В лесную зону заходит дальше всего в области
35*
548 ОТРЯД ГУСЕОБРАЗНЫЕ
Средне-Сибирского нагорья, но в ней повсюду редка. Многочисленной на гнез-
довье ее нельзя назвать нигде.
Экология. Размножение. Половой зрелости птицы достигают, ве-
роятно, в двухлетнем возрасте (после двух зимовок). Разбивка на пары
проходит на местах зимовки, и на пролете возле сбитой из стаи птицы задер-
живается ее пара. Токующие селезни запрокидывают на спину голову с
открытым клювом, расправляют и вздергивают кверху хвост. При этом в мо-
мент максимального растяжения и сжатия трахеи они издают звуки вроде
слогов «тека-тека-тека» (есть, однако, предположение, что звуки эти произво-
дятся крыльями). Это движение чередуется с плаванием с вытянутой по поверх-
ности воды шеей. Гнезда синьги находятся обычно у тундровых озер или
речек с медленным течением и открытыми берегами. Они устраиваются на
земле, обычно у самой воды, под прикрытием кустиков низкорослых ив, бере-
зок или сухой травы. Выстилка состоит из сухих растительных остатков,
имеющихся на месте. В гнездах с полной кладкой яиц много пуха, который
образует валик вокруг насиживающей самки и закрывает яйца на время
ее отсутствия. Обычное число яиц в тундрах европейской части Союза 6—8
(в Исландии 9—10; Науманн, 1910). Их окраска бледная зеленовато-бурая,
размер их колеблется в широких пределах 59—72x42,0—47,5 мм\ вес 60—
74 г. Насиживание начинается, повидимому, еще до откладки последнего
яйца, которое и остается в некоторых случаях невыведенным. Продолжи-
тельность насиживания 30—31 день (Гладков, 1951). Полные кладки в Тиман-
ской тундре появляются 15 июня—3 июля, выводки пуховиков с 17 июля.
На Кольском п-ове (Чунозеро) начало гнездованья приходится на середину
и конец июня. У самки, добытой 1 июня, было яйцо в яичнике размером 30 мм.
Первые выводки появляются в середине июля, подъем на крыло обычно 10—
23 августа (Владимирская, 1948). На Оби, близ Салехарда, 1 августа встречено
несколько выводков, большинство из которых было еще совсем в пуху. Там же
один нелетный выводок был отмечен 28 августа. Повидимому, некоторые
выводки не успевают подняться на крыло до замерзания водоемов и поги-
бают (Дерюгин, 1898).
Линька. Полная послебрачная линька начинается у взрослых сам-
цов в самом конце июля и проходит в августе— сентябре, а иногда и позже, уже
на местах зимовки. Выпадение маховых происходит в середине линьки.
Прямых наблюдений над ней очень мало. Как сказано выше, селезни прово-
дят линьку в море, где держатся большими стаями, зачастую вместе с годо-
валыми неполовозрелыми птицами. Линька самок начинается с момента подъ-
ема на крыло птенцов. Значительных изменений в расцветке в результате
линьки не происходит. Вторая, предбрачная, линька у птиц обоего пола
(сменяются перья рулевые и контурные) приходится на март—апрель.
Питание европейской синьги на местах гнездованья не изучено.
Известно лишь, что они кормятся моллюсками, насекомыми и корневищами
какого-то водяного растения. Корни они добывают нырянием обычно на
глубину от 1 до 3 м. Во время пролета на Верхней Печоре синьга кормится
преимущественно личинками ручейников и стрекоз, меньше моллюсками и еще
меньше рыбой (гольян). Растительная пища, хотя и попадается в желудках,
но имеет совсем второстепенное значение (Теплов, 1948). Осенью на Рыбин-
ском водохранилище синьга поедает преимущественно двустворчатых мол-
люсков E0% по объему), личинок ручейников A6,3%) и личинок хирономид
B0%). В море основную пищу зимующих стай составляют моллюски, и из них
особенно охотно поедаются различные мидии (Mytilus).
Хозяйственное значение. На севере повсюду добывается наряду с дру-
гими нырковыми утками. На Печоре составляет 10% от числа общего
количества добываемой охотниками водоплавающей дичи. В промысловых
масштабах добывается на Балтийском море, в том числе у берегов Фин-
СИНЬГА
549
ского залива. В массовом количестве отлавливается сетями на местах про-
лета и зимовок.
Полевые признаки. Утка среднего размера. Самец, сидящий на воде,
сплошь черный, надклювье у основания оранжевое. Самка темная со светлы-
ми щеками и зобом. На полете сверху вся птица темная, снизу у селезня
туловище и подбой крыльев черные, маховые образуют светлую кайму на
крыльях; у самки светлое брюхо и зоб. Птицы прекрасно ныряют, проводя
под водой обычно около 30 сек. (до 45). Ходят неуклюже, держат туловище
сильно приподнятым, так как ноги у них отодвинуты далеко назад. Голос
самки хриплое карканье «ре-ре-ре-ре», у самца мелодичный, звонкий—
«стрюк-люк». Взрослые селезни издают крыльями звон, который хорошо
слышен на полете. У молодых самцов и у самок полет беззвучный.
Рис. 98. Синьга (самец и самка)
Описание. Строение и размеры. Утка средней величины. Клюв очень
широкий с коротким коготком, занимающим всю ширину надклювья. У основания над-
клювья костное вздутие, глубоко раздвоенное вдоль. Оно велико у взрослых самцов
и меньше по размеру у самок, увеличивается с возрастом. Ноздри округлые, от их перед-
него края до конца клюва 23—25 мм. Оперение уздечки не доходит до заднего края
ноздрей значительно более чем на 1 см. Первое маховое у взрослых самцов резко сужено
на внутреннем опахале на протяжении около 70 мм от вершины. Эти перья издают на по-
лете особый высокий, звенящий звук. Размеры: крыло у самцов 228—242, самок 214—
230 мм; цевка самцов 44—54, самок 42—46 мм; клюв самцов 45—50, самок 40—45 мм.
Столь большой диапазон в размерах зависит от значительной возрастной изменчивости
их у птиц в первые 2—3 года жизни.
Вес. Сведений мало. Взрослые селезни, добытые весной (май) на Онежском озере,
весили в среднем 1306 г A215—1610 г); близ Архангельска в мае 1240—1270 г, в июне
960—1150 г. Самки там же в мае 990 г (Паровщиков); на Южном Ямале взрослые самки
в июне 1000 г, июле 915 г, в августе 915 г; самцы в июле 975—1100 г (Осмоловская).
Молодые птицы на осеннем пролете через Рыбинское водохранилище: самцы в октябре
778 г G10—818 г), в ноябре 1013 г, самки в октябре 735 г F00—850 г), в ноябре 852 г
G08—1022 г) (Немцев); на р. Печоре осенью (без указания пола) 760 г G00—820 г)
(Теплов, 1948).
Окраска. Пуховой птенец. На спинной стороне темнобурый с того же
цвета перевязью на зобу, на брюшной светлее—серовато-бурый со слегка сероватой задней
частью щек, серым горлом и шеей и белым пятнышком под глазом.
Молодые птицы в первом наряде. Имеют черно-бурое темя, спинная сторона,
передняя часть груди, бока и подхвостье темнобурые. Брюшная сторона тела более
светлая с неясными серо-бурыми пятнышками. Клюв матовочерный, лапы черноватые.
Самка взрослая. В основном темнобурая. Перья спинной стороны тела со свет-
лыми вершинными каемками, отчего создается неясная полосатость. Щеки, горло и
550
ОТРЯД ГУСЕОБРАЗНЫЕ
низ тела, особенно грудь, более светлые. У основания надклювья небольшое, но бро-
сающееся в глаза вздутие. Клюв и лапы черные, радужина бурая.
Самец годовалый летом. Вся спинная сторона тела, бока, шея, зоб и прилежащая
часть груди, подхвостье черные; бока головы и горло бурые. Остальная брюшная
сторона тела серо-бурая с отдельными черными перьями. Вздутия у основания надклювья
нет или оно едва заметно.
Взрослый самец в брачном наряде. Сплошь черный с синевато-зеленым отли-
вом на голове, верхней части тела, зобу и шее. Клюв черный, но хребет надклювья
от ноздрей до коготка яркожелтый. Лапы черные, радужина темнобурая.
Взрослый самец после полной линьки. Отличается лишь отсутствием сине-
ватого отлива на черном оперении и более буроватой брюшной стороной.
44. Тихоокеанская синьга Melanitta americana Swains.
Oidemia americana, Swainson in Swainson and Ri chardson, Fauna Bor.
Amer., II, 1832, стр. 450, Гудзонов залив.
Русские названия. Синьга—народное; кроме того, черная утка, свистун (на
Анадыре), последнее обязано голосу самцов или их звенящему полету.
Распространение. Гнездовой ареал занимает Северо-Восточ-
ную Сибирь и крайний северо-запад Америки. Западным пределом распро-
странения служит Ленско-Янский водораздел. Обычна по лесным озерам
Карта 65. Распространение тихоокеанской синьги Melanitta americana
а—граница гнездовий, б—не вполне выясненная граница гнездовий, в—места по-
стоянных летних встреч (гнездованье не доказано), г—зимовки
бассейна Яны, обнаружена под Верхоянском (гнездится ли там—неизвестно),
встречается до самых низовий реки. По Индигирке синьга распространена по
озерам лесной зоны, заходит в южные части тундры, одиночные птицы посе-
щают и дельту (Михель, 1935). В бассейне Колымы встречается в ее среднем
и нижнем течении, доходя до устья реки, но весьма немногочисленна там. На
Анадыре достигает Анадырского хребта. Гнездится по всей Камчатке и Коряц-
кой земле до Таунской губы включительно, а также на северных островах
Курильской гряды. Бродячие, повидимому неполовозрелые, особи попадают-
ся летом у берегов Японского моря. Установить более точно южную грани-
цу гнездовой территории не представляется возможным. На Американском
континенте заселяет все побережье Аляски на восток до залива Кука и на се-
вер до мыса Франклин. Проводит лето в восточной части Алеутской гряды
и возможно гнездится там.
О местах линьки этой синьги ничего неизвестно.
Основные зимовки. Зимой синьги нередки у восточного берега
Камчатки и в небольшом числе зимуют у Командорских о-вов, держась вда-
ТИХООКЕАНСКАЯ СИНЬГА 551
ли от берегов. Обычны они вдоль Курильской гряды и большими стаями про-
водят зиму в море у берегов Северной и Средней Японии, внутренние же озера
посещают в это время очень редко. Встречаются зимой у северо-восточных бе-
регов Кореи и сравнительно редко на западе этой страны, изредка достигают
берегов Китая. В Америке зимуют у Алеутских о-вов, восточной части п-ова
Аляски и вдоль Тихоокеанского побережья от Ванкувера до мексиканской
границы. Часть птиц пересекает материк и зимует на атлантическом берегу от
Ньюфаундленда до мыса Гаттерас.
Характер пребывания. В большей части своего ареала ти-
хоокеанская синьга перелетная птица, совершающая регулярные, довольно
дальние миграции. Лишь для Восточной Камчатки, Курильской гряды
и Аляски можно допустить, что часть популяции оседла.
Сезонные миграции и их даты. Пути пролета этой
утки еще не прослежены, но, повидимому, с мест зимовки птицы следуют вдоль
морских берегов до Камчатки и Охотского побережья, лишь оттуда проникая
в глубь страны. На юге Приморья (залив Посьет) стайки синьги отмечались
^ще в последних числах апреля. На восточном берегу Камчатки (Кроноки)
весенний пролет начинается с половины апреля, длится весь май и, видимо,
начало июня (Аверин, 1948). Многочисленна синьга во время пролета по
Пенжине и занимает среди уток второе место по численности (после морян-
ки). На Анадыре она появляется несколько позднее других уток, а именно
25—30 мая; после вскрытия реки 9 июня они там уже обычны (Портенко, 1939).
На Индигирку синьга прилетает вместе с турпаном в начале июня, осенью
же последняя, видимо задержавшаяся с отлетом, птица встречена в Русском
Устье 22 сентября, когда озера уже были затянуты льдом (Михель, 1935).
На Анадыре, в устье Танюрера, они наблюдались последний раз 13 сентября,
а в верховьях реки отмечались до 2 и даже 11 октября. Валового осеннего
пролета на этой реке не наблюдалось (Портенко, 1939). На восточном берегу
Камчатки осенний пролет проходит в октябре.
Биотоп. В гнездовое время—озера преимущественно лесной и лесо-
тундровой зоны, на плесах рек останавливаются только во время пролетов.
Зиму, а неполовозрелые годовалые птицы и круглый год, проводят на море
и иногда довольно далеко от берегов.
Экология. Размножение. Половая зрелость достигается на втором
году жизни. На места гнездовья синьги прилетают стайками, в которых птицы
уже разбиты на пары. Весной во время брачных игр селезень издает протяж-
ный, звонкий свист. Описаний гнезд из Восточной Сибири пока не сделано,
на Аляске они строятся на земле, поблизости от воды, под укрытием травы
или чахлых кустарников. Основной выстилкой служит пух самки. Обычный
размер кладки 8, реже 10 яиц. Окраска скорлупы одноцветная бледножелто-
ватая. Размер яиц из разных кладок колеблется весьма значительно, а именно
53,0—72,5x33,6—46,2 мм.
Выводки, содержащие мелких пуховых птенцов, попадаются на Ана-
дыре с середины августа (самый первый 3 августа), а в конце месяца их уже мно-
го, и птенцы довольно велики размером, хотя еще и одеты в пух. На Пен-
жине утенок добыт 28 июля (Портенко, 1939). На Камчатке (Кроноки) во
второй половине сентября на ступенчатых озерах в верховьях рек встречались
выводки птенцов, уже близких по размеру к взрослым A7 сентября)—от
730 до 813 г (Аверин, 1948). Число птенцов в выводках на Камчатке было 5,
а на Анадыре обычно 2—3, но иногда доходило до 7.
Линька. Особого летнего наряда селезни не имеют, но линяют дваж-
ды в год. Полная линька в августе—сентябре, неполная (кроме крыльев) в нача-
ле весны. К концу первой декады июля на Анадыре взрослые самцы уже
собираются в большие стаи. Во второй декаде августа количество холостых
самцов заметно увеличивается, и вскоре вслед за этим они откочевывают
552 ОТРЯД ГУСЕОБРАЗНЫЕ
на линьку. Но единичные селезни были встречены там еще 24—25 сентября.
По данным американских орнитологов, самки линяют всего один раз в год—
осенью, что, вероятно, неверно.
Питание. В июне на Анадыре основным кормом синьги служили
личинки ручейников, которые встречались у всех добытых птиц и в количестве
до 250—300 шт. на желудок (Портенко, 1939). У летних птенцов, добытых
на Камчатке 17 сентября, желудки содержали насекомых. Кормовой рацион
синьги на Американском континенте на
89,7% состоит из животных кормов.
Среди них основу составляют моллюс-
ки F5,2%; морские двустворчатые) и
ракообразные A7,3%; балянусы и де-
сятиногие раки). Насекомые (личинки
ручейников) составляют лишь 3,2%
(Коттам, 1939).
Хозяйственное значение. Благодаря
Рис. 99. Голова тихоокеанского синьги своим размерам и многочисленности
синьга служит одним из наиболее важ-
ных промысловых объектов на крайнем северо-востоке Сибири. Чаще всего
добывается стрельбой на весеннем перелете, когда в добыче анадырских
охотников она составляет около 16%.
Описание. Строение и размеры. Отличием от европейской синьги слу*
жит форма надклювья. Костная шишка у его основания выступает менее резко, округла
и не раздвоена посередине. Ноздри вытянутые, расстояние от их переднего края до конца
клюва обычно 25—28 мм. Ноготок клюва более выпуклый и загнутый, чем у европейского
вида. Эти отличия характерны как для самцов, так и для самок.
Вес самки, добытой в сентябре на Камчатке от выводка, 957 г. Данных о весе
самцов нет.
Окраска. Пуховой птенец. На спинной стороне буровато-серый с более тем-
ной головкой, белым горлом и пятнами на боках головы около глаз и на щеках.
Окраска брюшной стороны сероватая.
Молодые птицы. В основном темнобурые с еще более темным теменем и более
светлой брюшной стороной тела, в нерезких пятнышках. Бока головы, горло и часть шеи
серые. Клюв черный, матовый, лапы черноватые.
Молодой самец после первой брачной линьки. Схож с таковым у европейской
синьги, но в основании надклювья иногда появляется желтое пятнышко.
Взрослая самка. Утка темнобурая, но общий тон окраски более сероватый, чем у
европейской синьги. Беловато-серые щеки до линии глаз довольно резко отграничи-
ваются от темнобурого верха головы. Клюв темный, почти черный, в области ноздрей
желтое пятно, ноги черные.
Взрослый самец в брачном наряде. Весь черный, но тон окраски несколько-
буроватый и металлический отлив на голове и передней части туловища выражен слабо.
Надклювье от его основания до переднего края ноздрей яркожелтое, остальная часть
клюва черная. Ноги черные.
Взрослый самец после послебрачной линьки. Имеет более матовый тон опе-
рения и несколько буроватую брюшную сторону.
45. Пестроносый турпан Melanitta perspicillata L.
Anas perspicillata. Li nnaeus. Syst. Nat., изд. X, 1, 1758, стр. 125, Канада.
Русские названия. Книжные: пестроносый турпан, белолобая синьга.
Распространение. Гнездовой ареал вытянут узкой полосой с се-
вера на юг в области больших озер Канады от залива Мекензи до озера
Оленьего. Кроме того, негнездящиеся птицы весьма обычны летом в ряде
пунктов Аляски: у берегов моря Бофорта на границе с Канадой, у мыса Барроу
и в заливе Коцебу. В СССР известен на Командорских о-вах и Чукотском
п-ове, но не гнездится там.
Скопление линных птиц отмечено на взморье у устья Мекен-
зи (Бейли, 1948).
ПЕСТРОНОСЫЙ ТУРПАН
553
Основные зимовки находятся на Алеутской гряде, на Тихо-
океанском берегу от границы Аляски до Калифорнии, на озерах Верхнем,
Гурон и Мичиган и на атлантическом берегу от Новой Шотландии почти до
Флориды.
Залеты. Неоднократно добывался на Командорских о-вах в мае, июне
и октябре (взрослые и молодые самцы). На о-ве Беринга 10 мая 1931 г., в устье
р. Гаванской отмечена стайка в 15 шт. (Иогансен, 1934). Один раз A августа
1881 г.) встречен на Чукотском п-ове у мыса Ванкарем. Данных, говорящих
о гнездованье на азиатском берегу этого турпана, нет, и встречи его следует
считать за случаи залета с расположенных поблизости мест зимовок и пребы-
вания неполовозрелых птиц. Нередко залетает в Европу, где неоднократно
Карта 66. Распространение пестроносого турпана Melamlla perspicillafa
а—граница гнездовий, в—места постоянных встреч летом негнездящихся птиц,
г—зимовки, д—залеты» ж—места скоплений птиц на линьку
добывался в Финляндии, Швеции и Норвегии, вблизи от нашей границы
(Мальме, Энаре, Каресуандо, Аландские о-ва); возможны залеты наМурман
и в Советскую Прибалтику.
Экология. Размножение. Оформление пар происходит еще на зи-
мовках и к местам гнездовья турпаны прилетают в парах. Однако нередко
около одной утки токует по нескольку самцов. Токовые движения заключают-
ся в том, что селезни то вытягивают шею кверху и поднимают голову, то опу-
скают ее в воду, издавая своеобразные глухие звуки. Гнезда строятся близко
от воды, обычно под кустами или под нависшими ветвями деревьев. Они пред-
ставляют собой ямку, окруженную валиком из сухого мусора, и имеют боль-
шое количество пуха. Яиц обычно 5—7. Острый конец у них притуплён,
скорлупа желтоватого оттенка. Их средний размер 61,6 х 43 мм. Самка сидит
очень крепко. Осенью у пестроносых турпанов наблюдаются своеобразные
игры самцов, напоминающие их весеннее токование.
Линька. Число линек и сроки их такие же, как и у других турпанов.
Селезни, вскоре после того, как самки сядут на яйца, объединяются в стаи
и откочевывают к морю или на большие озера, где и проводят время полной
линьки.
Питание на 87,9% состоит из животных кормов. Основу составляют
моллюски F0,8%), и из них мидии B8,7%). На долю ракообразных прихо-
дится 10,3%, насекомых (ручейники, стрекозы, плавунцы) 9,6% и рыб всего
3,4%. Из растительных кормов поедаются семена и зеленые части рдестов,
зостеры и руппии. Молодые птицы за время их пребывания на внутренних
водоемах кормятся преимущественно насекомыми F1%); водяными жуками
554
ОТРЯД ГУСЕОБРАЗНЫЕ
A6,7%), клопами A2,6%), личинками поденок A4%) и ручейников F,7%).
Моллюски в их рационе составляют 18,6%, а растительные корма 20,4%.
Из последних наибольшее значение имеют ягоды вороники A0,3%) (Коттам,
1939).
Хозяйственное значение этого, относительно малочисленного, турпана
невелико даже у него на родине. В ОХР же он залетает лишь единицами.
Полевые признаки. Сидящие на воде птицы сплошь черные, но у селезня
два белых пятнана лбу и на затылке, а у самки у основания клюва и на ушах.
На полете обе птицы сверху темные (кроме пятен на голове), а снизу брюхо
у самки светлое. Прекрасно ныряют и при плавании под водой пользуются
крыльями. Летают обычно на высоте 40 м и выше, а в шторм низко над водой.
Голос—грубые горловые звуки, похожие на голос крохаля.
Рис. 100. Пестроносый турпан (самец и самка)
Описание. Строение и размеры. По фигуре похож на обыкновенного
турпана, но заметно меньше его. Рулевых 7 пар. Клюв широкий и у основания высокий,
но не имеет резко ограниченного вздутия в основании надклювья. Ноздри округлые.
У птиц всех возрастов оперение лба далеко выдается вперед по коньку надклювья и дохо-
дит до уровня ноздрей. В области уздечек оно выступов не образует.
Окраска. Пуховой птенец не известен.
Молодые птицы в первом наряде. Похожи на взрослую самку. На спинной сторо-
не они однообразно буроватые, на брюшной, особенно на груди, более светлые с мелкими
нерезкими пятнышками, более частыми на боках и брюшке. Белые пятна на уздечке и
ушах у них выделяются яснее, а у самцов на затылке есть беловатое пятно (последнее
скорее после первой предбрачной линьки). Радужина бурая.
Взрослая самка. Однообразно бурая с беловатой пятнистостью на груди. На
уздечке и ушах намечаются светлые пятна и еще одно намечается на затылке, на месте
белого пятна у самца. Клюв зеленовато-черный, на боках надклювья в его основной части
более или менее четко выраженное более темное пятно. Лапы желто-оранжевые с темными
перепонками, радужина красно-бурая.
Молодой самец после первой предбрачной линьки. Матовочерный, без глянца
на голове. Брюшная сторона коричневато-черная. Клюв скорее желтый, чем красный,
надклювье по краям черное. Лапы бледнокрасные, радужина серая.
Взрослый самец в брачном наряде. Весь черный, на голове, спине и зобу с зеле-
новатым отливом. На голове два белых пятна: на лбу и на затылке, последнее имеет
треугольную форму и обращено острым углом вниз. Клюв красно-оранжевый, ноготок
желтый. По бокам надклювья в его основной части большие угловатые черные
пятна, пространство от них до ноздрей беловатое. Лапы оранжево-красные, радужина
белая. У некоторых особей лобного пятна не бывает, возможно, что это неполовозрелые
птицы.
Самец после полной летней линьки. Остается черным, но не имеет металличе-
ского отлива, а грудь становится беловатой.
ЧЕРНЫЙ ТУРПАН
555
46. Черный турпан Melanitta fusca L.
Anas fusca. Li nnaeus, Syst. Nat., изд. X, 1, 1758, стр. 123, Швеция.
Синоним. Anas Carbo. Pallas. Zoographia Rosso-Asiatica, II, 1811, стр. 244.
Европа и Азия.
Русские названия. Турпан—народное, кроме того, свирок, тюльпан (Ленин-
градская обл.), чернуха, во многих местах—черная утка (Красноводск), головня (Запад-
ная Сибирь).
Распространение. Гнездовой ареал. Занимает весь Скандинав-
ский п-ов, кроме самых южных частей Норвегии и Швеции, всю Финляндию
и Аландские о-ва (обычен в шхерах около Хельсинки; Бергман, 1935). Гнез-
Карта 67. Распространение черного турпана Melanitta fusca
а—граница гнездовий, б—не вполне выясненная граница гнездовий, в—обособленные
гневдовые колонии, в'—места постоянных летних встреч и спорадичного гнездованья,
г—зимовки, Э4с—места скоплений птиц на линьку
дится на Оркнейских и Гебридских о-вах, есть данные о спорадичном гнездо-
ваньи в Шотландии и в южной части Шпицбергена. В пределах Советского
Союза гнездится на побережье Эстонии, причем на о-вах Сарема и Хиума
обычен (Гроссе и Транзее, 1929), отмечен летом у берегов Литвы, но гнездо-
ванье там не доказано, так как в течение всего лета у берегов Балтики дер-
жатся стайки неполовозрелых птиц, а во второй его половине также взрос-
лых самцов. Встречается всюду на Мурмане, где обычен лишь в лесной
части, а в северной, тундровой, возможно и не гнездится, однако ежегодно
гнездится на Айновых о-вах. Обычен в Северной Карелии, примерно до
556 ОТРЯД ГУСЕОБРАЗНЫЕ
широты Кеми, повидимому гнездится в северной части Ладожского озера.
Довольно обычен под Архангельском, гнездится на п-ове Канине, встречается
в Тиманской тундре и возможно гнездится там (Гладков, 1951). На Печоре
южнее 65° с. ш. летом не встречен (Дмоховский, 1933). На о-вах Колгуеве,
Вайгаче и Южном о-ве Новой Земли летом держатся стайки взрослых самцов
и молодых птиц, но гнездованье там не зарегистрировано. На западных скло-
нах Среднего Урала турпан на гнездовье достоверно не был отмечен, указания
на встречи его в верховьях Камы вероятно относятся к бродячим особям
(Воронцов, 1949). На Ямале его северная граница проходит примерно под
70 ° с. ш., в озерной котловине Юрибея F8—69° с. ш.) он еще обычен (Осмо-
ловская). Встречен в устье Таза, а восточнее, по Тугаринову A941), дости-
гает границы леса на Енисее, встречается на Южном Таймыре и в устье Хатан-
ги. В Зауралье и в Западной Сибири южная граница распространения тур-
пана продвигается далеко на юг. Он гнездится на Урале в бассейнах Павды
и Сосьвы, где особенно обычен на Княспинском озере (Сабанеев, 1874) и обна-
ружен на гнездовье в районе Свердловска (гнездо на озере Тыгиш в 60 км
к юго-востоку; Данилов). Гнездится, хотя и не часто, на больших степных
озерах в местности между верховьями Белой, Сакмарой и Иком (Кириков,
1935), многочислен летом на степных озерах в Челябинской обл., под
Троицком (Сабанеев) и в Курганской обл. Однако в этих пунктах несомненно
проводят лето неполовозрелые птицы и стайки взрослых самцов, гнездованье
же там прямыми наблюдениями не доказано. Восточнее турпан гнездится под
Тюменью (за р. Турой, Ларионов), повидимому, на больших озерах в север-
ной части Ишимского и Тюкалинского районов (Шухов, 1930) и несомненно
в северных районах Барабы, там, где степные участки чередуются с лесными
массивами подтаежного типа (северные части Венгеровского и Куйбышевско-
го районов; Янушевич; курорт Карачи; Рузский, 1932). Обычен на гнездовье
под Тарой (Ушаков, 1913) и на больших озерах под Тобольском (Тарунин).
Юго-восточная граница ареала пока неясна. Значительно южнее основного
ареала обнаружены случаи гнездованья турпана в ленточных борах Кулун-
динской степи на озере Горьком в 220 км юго-западнее Барнаула (Велижанин,
1928), между Карагандой и Баян-Лулом, в Кокчетавских борах (курорт Бо-
ровое; Штегман, 1934) и на озере Кургальджин в Акмолинской обл. (добыта
самка с яйцом, готовым к сносу; Лавров, 1930). Повидимому, некоторые из
этих пунктов не являются постоянными гнездовыми колониями и гнездованье
в них имеет место лишь спорадично (Кургальджин, Баян-Аул). Наконец,
совершенно изолированными районами гнездованья служат горное озеро
Севан в Армении и Ахалкалакское плато в Грузии, где турпаны регулярно
гнездятся на многих крупных озерах: Хозапинском, Мадопотинском*
Табисцхури и др.
Скопления линных и неполовозрелых птиц отме-
чены на озерах Павдинского и Сосьвинского Урала, где они с прилета и до
отлета держатся большими и мелкими стайками. Еще многочисленнее они
летом на степных озерах Челябинской и Курганской обл., где при отлове
в сети линных уток попадают и самцы турпанов (Сабанеев, 1874; Цецевин-
ский, 1931). Однако на линьку в Зауралье попадает лишь незначительная
часть птиц; где проводит этот период их основная масса—неизвестно, но
вероятно на обширных тундровых и горных озерах или в море, уже в рай-
онах зимовки. О последнем говорит то обстоятельство, что большие стаи
по 60—100 шт., состоящие из одних селезней, появляются в Северной Гер-
мании у о-ва Рюген уже в июле (Гомейер, 1889).
Основные зимовки находятся у берегов Западной Европы,
от горла Балтийского моря до побережий Англии и Германии, причем в по-
следнем месте (Померания и др.) турпан по численности кое-где превосходит
синьгу. В меньшем числе он зимует в фьордах Норвегии к северу до Ло-
ЧЕРНЫЙ ТУРПАН 557
фотенских о-вов, а небольшими стайками и у нас на Мурмане. К югу он встре-
чается у берегов Франции и Северной Испании. Чаще, чем синьга, турпан
заходит зимой на материковые водоемы, но никогда не бывает многочислен
на них. Встречи его в Средиземном море представляют собой случайное
•явление, а на Черном море единичны. Птицы из закавказских гнездовых ко-
лоний в основном зимуют на месте. Изолированным районом регулярной
зимовки турпана служит Красноводский залив Каспийского моря и смежные
территории. У западных берегов Каспия турпан попадается зимой единица-
ми, так же как и на водоемах Средней Азии.
Характер пребывания. Гнездящаяся перелетная птица,
в СССР в небольшом числе зимует.
Сезонные миграции и их даты. Отлет птиц с западно-
европейских зимовок начинается очень рано—в марте—апреле там остаются
лишь единицы. Птицы летят большей частью ночью, а если они продолжают
путь днем, то поднимаются очень высоко и выдерживают строй линией, раз-
вернутой по фронту. Турпаны обычны на пролете вдоль берегов Советской
Прибалтики, где они появляются уже в середине марта, и в Ленинградской
обл. с конца апреля. На Онежском озере пролет проходит в начале и в сере-
дине мая, Кольского п-ова (озера Имандра) турпаны достигают в конце мая
и в первых числах июня B9 мая—12 июня) (Владимирская, 1948). В Тиман-
ской тундре (низовья р. Вельть) первые птицы были замечены 14 мая и до
1 июня встречались небольшие стайки, летящие в восточном направлении
{Гладков, 1951).
На Печоре в устье Ылыча пролетные турпаны весной одиночны и проле-
тают вместе с синьгой, в верховьях этой реки (Колва, Сосьва) они также
немногочисленны и летят небольшими стайками вверх по рекам к Уралу
{Теплоухов, 1911). Под Тобольском эти нырки появляются позже всех дру-
гих уток, около 24 мая B0—28 мая) (Тарунин, 1929), на Южном Ямале пер-
вая стая их на появившихся полыньях озер отмечена 16 июня (Осмоловская).
Регулярного весеннего пролета турпанов в средней полосе и на юге европей-
ской части Союза не бывает. Отдельные птицы и небольшие стайки их встре-
чаются в мае во многих пунктах, при этом чем южнее, тем реже. Направле-
ние их полета, там, где оно прослежено, бывает восточное (по течению Оки
в Калужской обл.; Филатов, 1915). Как правило, это бывают неполовозре-
лые птицы и изредка взрослые самки. Можно предполагать, что турпаны,
появляющиеся в средней полосе, это птицы, уклонившиеся от основного
русла пролета этого вида. Осенью они появляются там же значительно
чаще, и местами (Рыбинское водохранилище) в октябре имеет место настоящий
пролет, хотя и очень слабый, на юге же они бывают крайне редки и в это
время. Отсутствие турпанов на обратном пути вероятно обязано почти по-
головной гибели молодых птиц, отклонившихся во время своего первого пу-
ти от основного, свойственного виду, пути пролета. Осенний пролет на
Печоре (устье Ылыча) бывает немногочисленным, он начинается около 2 ок-
тября B7 сентября—9 октября) и продолжается примерно 10 дней, последние
же птицы задерживаются до 13 октября (8—18 октября). Птицы летят стай-
ками по 5—10, изредка до 20—30 шт. (Теплов, 1948). Из-под Архангельска
они отлетают 5—15 октября, а с Кольского п-ова (озеро Имандра) в конце
октября, перед самым ледоставом. В Ленинградской обл. и в Прибалтике
пролет заканчивается в ноябре. Тогда же турпаны достигают мест своих зи-
мовок у берегов Германии.
Биотоп. Летом турпаны живут на озерах, в тундре на неглубоких
озерах с заросшими хвощом и осокой берегами и заводями, в тайге они не
встречаются в поймах, а заселяют лесные озера, в лесостепи молодые и линя-
ющие птицы держатся на открытых озерах. У берегов Балтики турпаны про-
водят лето на поросших травой побережьях и островках. Зиму они проводят
558
ОТРЯД ГУСЕОБРАЗНЫЕ
на море, но поблизости от берегов в заливах и бухтах, где кормятся на мел-
ководьях и отдыхают на глубоких открытых местах.
Численность турпанов на гнездовье всюду весьма невелика. По-
жалуй, наиболее обычны они в озерных районах лесотундры Западной Сиби-
ри (Ямале) и на водораздельных озерах южной зоны тайги Прииртышской
Сибири (Тобольск, Тара).
Экология. Размножение. Половой зрелости достигают на вто-
ром году жизни. Молодые птицы проводят первый год в стаях на море, вбли-
зи от мест зимовки, на озерах в южной части их ареала и на озерах тундры.
Повидимому, значительная часть их мест гнездовья в этом возрасте не дости-
гает. Разбивка на пары происходит еще на местах зимовки, и пролетные стаи
состоят уже из парных птиц. Однако стаи распадаются не сразу по при-
Рис. 101. Черный турпан
лете на место, так как гнездовые стации турпанов—прибрежные низины
озер—очень поздно освобождаются от снега, и к размножению эти утки при-
ступают позже всех других. На озерах Кольского п-ова стаи наблюдались
еще 23 июня, а на Ямале часть из них стала держаться парами лишь с 26 ию-
ня (Осмоловская). Брачные игры начинаются у них еще на зимовках, они
выражаются в плавании вокруг самки с поднятой вертикально головой
и полураскрытыми дрожащими крыльями и сопровождаются нырянием и
всплесками воды.
Гнезда турпанов помещаются на земле. На Ямале их находили на влаж-
ных низинах среди осоки и другой травяной растительности, около самой
воды озер, на Айновых о-вах в нишах торфяных кочек или между ними, но
не в кустах. На побережье Эстонии они гнездятся вблизи от моря на лугах
и пастбищах с низкой травой, устраивая гнезда под кустами ольхи или под
нависшими камнями (Кох, 1911). На Урале турпан гнездится в мелколесье,
в совершенно сухих местах, иногда далеко от озер—на расстоянии до 2—3 км
(Сабанеев, 1874). Так же в березовых колках в стороне от воды гнезда его
находили в северных районах Барабы. На о-ве Сарема (Эстония) турпаны
нередко занимают гнездовые ящики, поставленные для гаг и пеганок, при-
ЧЕРНЫЙ ТУРПАН 559
чем иногда несутся с ними одновременно в одно и то же гнездо (Транзее,
1927). В Северной Финляндии ящики для них выставляют специально с це-
лью сбора яиц. Выстилка гнезда состоит из сухой травы и большого количе-
ства пуха, прикрывающего яйца с боков и сверху. Число яиц в полной клад-
ке 6—10, как исключение до 14 (Тугаринов, 1941), в Эстонии обычно 5—8.
Яйца сливочно-белого цвета, очень крупные: 64,3—76,5x44,8—51,5 мм, сред-
нее 70,8x47,9 мм. Срок насиживания около месяца.
На Мурмане полные кладки бывают во второй половине июня (а начало
их приходится на первую декаду этого месяца—Айновы о-ва; Белопольский),
в низовьях Печоры в начале июля и до конца июня на Урале (Сабанеев).
В Хибинах (озеро Имандра) два выводка наблюдались 2 августа, они держа-
лись на мелком озерке среди сфагнового болота и состояли из 5 и 6 птенцов
(Владимирская, 1948). Под Салехардом 1 августа выводки состояли из
пуховиков и в большинстве содержали по 3—4 птенца (Дерюгин, 1898).
Выводки нередко объединяются между собой, и одна утка водит утят двух
или трех различных возрастов, иногда число птенцов бывает вдвое больше,
чем обычно. Держатся выводки на озерах. Развитие птенцов идет довольно
медленно и летать они начинают не ранее середины сентября.
Линька двукратная, как и у родственных видов. Полная послебрач-
ная проходит у взрослых селезней в августе, в результате ее наряд изменяется
лишь незначительно. У самок она начинается несколько позже, но, повиди-
мому, еще в период нахождения при выводке. Предбрачная линька селезней
происходит ранней весной и заканчивается во время пролета, у неполовозре-
лых птиц примерно в то же время.
Питание. Сведений о летнем питании турпана нет. В желудках
этих птиц, добытых во время осеннего пролета на Печоре, часто встречались
моллюски (прудовики), реже личинки ручейников и всегда остатки растений,
очевидно листья рдестов (Теплов, 1948). В октябре на Рыбинском водохра-
нилище пища их на 99,5% состоит из животных кормов. Основу среди них
составляют моллюски F2,5%; беззубки, перловица и др.), меньше встречает-
ся личинок ручейников C0%) и еще меньше мелкой рыбы G%). На зимовках
турпаны почти целиком переходят на моллюсков. На Каспийском море
(Красноводский залив) они кормятся Cardium edule, в Северном и Балтийском
морях этим же моллюском, а также мидиями и другими видами морских
двустворчатых.
Хозяйственное значение невелико, так как в силу относительной мало-
численности турпана специальная охота на него проводится редко.
Полевые признаки. Крупная черная утка. У плавающего селезня видно
небольшое белое пятно на крыле и белое пятно около глаза, а у самки на
крыле, у основания клюва и в области уха. Летящие птицы черные и сверху
и снизу, отличительным признаком служит белое зеркало. Вблизи видно
оранжевое надклювье селезня и красные ноги обеих птиц. Большую часть
времени проводит на воде, прекрасно ныряет, поднимается с воды с трудом
и летит низко над ней. Голос самца и самки—глухое, грубое карканье, вроде
*краа-краа-кра».
Описание. Строение и размеры. Крупная утка с массивным туловищем,
относительно короткой шеей и большими лапами. Хвост клиновидный, рулей 7 пар, клюв
широкий с широким ноготком. В основании небольшое вздутие, ноздри продолговатые
щелевидные. По бокам надклювья оперение почти не выдается вперед. Крыло самцов 269—
293, самок 250—265 мм; цевка самцов 48—52, самок 46—48 мм; клюв самцов (от лба)
37—50, самок 38—43 мм.
Вес. Данных о весе турпана очень мало. Вес взрослого селезня может доходить
до 1500—1750 г, самец годовалый в мае (Карелия) весил 1090 г, полуторалетний в январе
(Красноводск) 1200 г, молодой в октябре 1025 г (Рыбинское водохранилище). Взрослая
самка в июле (Ямал) 1100 г, в августе (Архангельск) 1230 г, молодые в сентябре (Рыбин-
ское море) 995 г (860—1210 г). Молодые птицы в октябре на Печоре (без указания пола)
1000—1050 г.
560 ОТРЯД ГУСЕОБРАЗНЫЕ
Окраска. Пуховой птенец. На спинной стороне темнобурый с широкой пере-
вязью того же цвета на зобу и груди, на брюшной стороне светлосерый, почти
белый до задней части щек и боков верхней части шеи. Светлых пятен на темном
фоне нет.
Молодые птицы в первом наряде. Сходны со взрослой самкой, но светлее ее, а
грудь и брюшко сильнее испещрены белыми поперечными полосками и неправиль-
ными пятнами. Клюв темный. Взрослая самка одноцветная темнобурая, брюхо с
беловатыми кончиками перьев. По бокам головы два светлых пятна: одно между клювом
и глазом, другое в области уха. Зеркало белое. Клюв бурый, лапы грязнокрасного
цвета, радужина бурая.
Самец годовалый после первой предбрачной линьки. Голова, шея, мантия, плечи
и зоб чисточерные. Спина, верхние кроющие крыла и остальная брюшная сторона бурые
с беловатыми пятнами. Белое пятно под глазом имеется всегда, но невелико. Клюв
темный, иногда с небольшим светлооранжевым пятнышком на хребте, лапы светлокрас-
ные, радужина светлосерая.
Самец взрослый в брачном наряде. Сплошь черный с зеленовато-фиолетовым
отливом, который особенно силен на голове и зобе. Под глазом белое пятнышко,
зеркало белое за счет окончаний второстепенных маховых и больших кроющих крыла.
Надклювье от основания до переднего края ноздрей и в углах рта черное, осталь-
ная часть его яркооранжевая. Лапы красные с черноватыми перепонками. Радужина
голубовато-белая.
Самец после полной линьки. Не приобретает особого наряда, но на некото-
рых частях тела, а именно на зобу передней части груди и среди кроющих крыла
появляются отдельные бурые перья такого же цвета, как у самок.
47. Горбоносый турпан Melanitta deglandi В p.
Oidemia deglandi. Bonaparte. Rev. Crit. Orn. Eur., 1850, стр. 208, Северная Америка.
Синоним. Anas Carbo. Pallas, Zoographia Rosso-Asiatica, II, 1811, стр. 244 (Пал-
лос смешивал черного и восточносибирского турпанов, отчего нельзя присвоить данное
им имя этому виду).
Русские названия. Турпан, неводчик—народные.
Распространение. Ареал. Восточная Сибирь на запад до Алтая и сред-
него течения Енисея, к северу до границы древесной растительности, к вос-
току до Тихого океана, к югу до Витимского нагорья и хребта Танну-Ола.
В Америке от Полярного моря до Плато Прерий, от Скалистых гор и Сред-
него Юкона до озер Невольничьего и Виннипег. Таким образом, ареал со-
стоит из двух разорванных между собой частей.
Характер пребывани я—перелетная птица, частично оседлы
быть может только особи, населяющие Камчатку. Сибирская популяция зи-
мует у морских берегов от Камчатки до Японии и Кореи. Американские пти-
цы проводят зиму на Тихоокеанском берегу от Аляски до Калифорнии, на
Атлантическом от Ньюфаундленда до мыса Гаттерас и в области Великих озер.
Б и о топ—озера лесотундровой зоны, а в лесной полосе—озера альпий-
ского пояса гор, примерно до высоты 2300 м. Зимой мелководные части моря
вблизи от берегов.
Подвиды и варьирующие) признаки. Распадается на три подвида, из которых два,
занимающие изолированные ареалы, имеют ярко выраженные различия в пластических
признаках и незначительные в окраске. Третья форма переходная. М. d. deglandi Bona-
parte, 1850—Северная Америка от устья Мекензи и Верхнего Юкона до Северной
Дакоты; М. d. dixoni Brooks, 1915—внутренние области Северной Америки от
Северной Дакоты до верховьев Миссури; М. d. stejnegeri Ri d g w а у, 1887—Восточ-
ная Сибирь и восточная часть Алтая.
Американский турпан Melanitta deglandi deglandi В p.
Распространение. Ареал. Северные и центральные районы Америки
от среднего течения Юкона (форт Юкон) и залива Мекензи до Северной
Дакоты и от восточных склонов Скалистых гор до озер Медвежьего, Неволь-
ничьего и Виннипег.
ГОРБОНОСЫЙ ТУРПАН
561
Карта 68. Распространение горбоносого турпана Melanitta deglandi
1—М. d. stefnegeri, 2—М. d. deQlandi; а—граница гнездовий, б—не вполне выясненная граница
гнездовий, «—места постоянных встреч летом (гневдованье не доказано), а—зимовки, а—валеты,
<—весенние встречи птиц в стороне от гнездового ареала, э#с—места скоплений птиц на линьку;
цифры в кружках—номера зимующих подвидов
Основные зи-
мовки вдоль Тихоокеан-
ского побережья от п-ова
Аляски и Алеутских о-вов
до Мексиканской грани-
цы, в области Великих
озер и на Атлантическом
берегу от Ньюфаундленда
до мыса Гаттерас.
Залеты. Несколько
раз залетал на Командор-
ские о-ва, где добывался
в апреле, мае и ноябре.
Взрослый самец добыт
2 мая 1944 г. в Южном При-
морье на взморье у р. Та-
чингоуза.
Характер пре-
бывания в СССР.
Залетная птица.
Сезонные миг-
рации и их даты.
Свои зимовки у берегов
Аляски птицы покидают в
мае и вновь собираются там
в октябре. Неполовозрелые, годовалые птицы
мест гнездовья.
Биотоп. Как и другие турпаны, это обитатель озер. Область его рас-
пространения целиком находится в зоне хвойных лесов.
Численность В СССР единичные находки.
36 Птипы СССР, т. IV
Рис. 102. Американский горбоносый турпан (самец
и самка)
повидимому, не достигают
562 ОТРЯД ГУСЕОБРАЗНЫЕ
Экология. Размножение. К размножению в годовалом возрасте
не приступают. Гнезда устраиваются на земле, часто далеко от водоема и бы-
вают скрыты растительностью. Выстилка гнезда плохая, но много черно-бу-
рого пуха. Число яиц в кладке 9—14. Скорлупа их гладкая, цвет грязно-
охристый, размер в среднем 65,3x45,7 мм. Кладка заканчивается в конце
июня. Срок насиживания около месяца.
Линька. Полная, послебрачная линька проходит в конце лета,
предбрачная (контурное оперение и рулевые) ранней весной. Молодые частич-
но сменяют свой первый наряд в середине зимы, после чего молодые самцы
становятся отличимы от самок. Первая полная линька у них наступает летом.
Питание на 94,1 % состоит из животной пищи, для добывания кото-
рой эти птицы ныряют зачастую на большую глубину. Наибольшее значение
имеют моллюски G5,3%), главным образом двустворчатые (устрицы, мидии,
Protothala и др.). Меньшую роль играют ракообразные A3,2%; десятиногие,
балянусы и др.).
Полевые признаки те же, что и у черного турпана, но на близком рас-
стоянии у самца заметно вздутие у основания подклювья.
Описание. Строение и размеры сходны с таковыми у восточносибирского подвида.
Отличие в форме клюва и характере оперения у его основания. Оперение клиновид-
ным мысом выдается вперед по коньку надклювья, обычно достигая уровня заднего края*
ноздрей, а по бокам клюва оно выступает не дугообразно, а также углом, и немного не
доходит до заднего края ноздри, что хорошо видно у птиц обоего пола и во всяком воз-
расте. Ноздри округлые. У самца при основании клюва имеется резко выраженное взду-
тие, но передняя сторона его несколько поката назад.
Окраска. Пуховой птенец. На спинной стороне коричнево-бурый, снизу сере-
бристо-белый. Под глазом белое пятнышко.
Взрослая самка. Весьма схожа с самкой обыкновенного турпана, но бока у нее
несколько более светлобурые.
Взрослый самец. По окраске оперения почти неотличим от обыкновенного, но-
плечевые с несколько буроватым, а бока тела оливковым оттенком. Клюв в основном
оранжевый, бока надклювья красноватые, черное только вздутие и черноватые угль»
рта (черных полосок по бокам надклювья нет). Особого летнего наряда у селезней нет.
Восточносибирский турпан Melanitta deglandi stejn geri Ridgw.
Oidemia stejnegeri. R i d g w a y. Man. Nord. Amer. Birds, 1887, стр. 112, Камчатка.
Русские названия. Турпан повсеме тно, неводчик на Алтае
Распространение. Гнездовой ареал. Восточносибирский под-
вид, западным пределом распространения которого служит бассейн Енисея и
Восточный Алтай. Гнездится в альпийской зоне у Телецкогоозера(Фолитарек
и Дементьев, 1938), на Чулышманском плато и на юг до верховьев Бухтармы
(Рахмановские ключи) (Сушкин, 1938), севернее встречен на озерах Ачин-
ского района, в бассейне Подкаменной Тунгуски, в верхнем течении Нижней
Тунгуски, на Вилюе и в верхнем течении Хатанги. Отдельные встречи сибир-
ского турпана зарегистрированы в низовьях Енисея (до Дудинки), но гнездо-
ванье его там сомнительно. Восточнее пределом распространения служит
северная граница древесной растительности, так как из лесотундры в типич-
ную тундру турпан почти не выходит. Он обычен на горных озерах Верхоян-
ского хребта, а в низовьях Индигирки (сел. Абый) это основная дичь местных
охотников (Михель, 1935). На Колыме доходит почти до устья, но в низовьях
малочислен (Бутурлин, 1906). Распространен по всему Анадырскому краю,
кроме узкой приморской полосы (Портенко, 1939), "обыкновенен, но не-
многочислен там. К северу за пределы леса далеко не выходит и на Чукотке,
повидимому, не гнездится. Относительно Олюторского побережья сведений
нет, по Бергману A935), гнездится по всей Камчатке, но в районе Кронок
он встречается только на пролетах и зимовке (Аверин, 1948). Обычен по Пен-
жине и Гижиге, встречается летом в районе Охотска, обычен на гнездовье
ГОРБОНОСЫЙ ТУРПАН 563
в районе Аяна (Снигиревский), но, судя по летным встречам, возможно гцез-
дованье его на Сахалине (Чайва) и на озерах в низовье Амура. Бродячие
особи часто встречаются летом в Приморье (до бухты Кит). Южная граница
распространения совсем неясна.
Возможно, что она достигает Яблоневого хребта. В Забайкалье обнару-
жен на гнездовье на водоразделе Витима и Селенги (Еравнинские озера;
Вотинцев, 1947). Обычен на Северном и Среднем Байкале до о-ва Ольхон
включительно (Штегман, 1936). Известны летние находки на озерах левых
притоков Селенги (июнь—июль, озера Гусиное и Таглей), но возможно, что
они относятся к молодым бродячим птицам. Далее турпан гнездится в Восточ-
ных Саянах на Канско-Манском белогорье (Тугаринов) и в верховьях Ени-
сея, где занимает Минусинскую, Тувинскую и Убса-Нурскую котловины,
гнездится на Танну-Ола и Восточно-Тувинском нагорье (Янушевич, 1948,
1950). Кроме того, восточносибирский турпан был добыт под Томском и на
севере Барабинской степи (Куйбышевский район; Лавров, 1929), но эти слу-
чаи можно расценивать лишь как залеты.
Массовые скопления линных селезней и неполово-
зрелых птиц наблюдаются в море вдоль Охотского побережья. Большими
стаями они проводят лето на море в районе Охотска и в других местах. В мень-
шем числе турпаны линяют на озерах Камчатки и на Сахалине; много их
бывает в это время в низовьях Колымы (Майдель, 1894) и на озерах в низо-
вьях Вилюя; стайки селезней штук по 10 наблюдались на альпийских озерах
в верховьях Маны (Юдин). В основном это птенцы и самки, так как взрослые
самцы откочевывают оттуда на линьку в другие места.
Основные зимовки находятся у побережий дальневосточных
морей. В колоссальном количестве турпаны зимуют у побережий Южной
и Юго-Восточной Камчатки, около Командорских о-вов они в это время немно-
гочисленны. Зимуют вдоль Курильской гряды и в небольшом числе у бере-
гов Приморья. Очень обычен у обоих берегов Кореи, встречается у Японских
о-вов и у Китайского побережья к югу до Фучжоу, но в южных частях зимне-
го ареала редок.
Характер пребывания. На всем протяжении ареала пере-
летная птица и быть молгет оседла лишь на Восточной Камчатке.
Сезонные миграции и их даты. Весьма обычен на про-
лете в Уссурийском крае и в апреле часто встречается на взморье большими
стаями. В районе Охотска к 8 мая турпанов бывает уже много (Харитонов,
1915). В верховьях Колымы пролетная стая наблюдалась 9 июня. Птицы,
зимующие у Командорских о-вов, исчезают в середине мая. На зимовках
у Восточной Камчатки они приходят в движение уже в конце марта, когда
разбросанные вдоль побережья табунки начинают объединяться в большие
стаи. Первая пролетная стая в 1500—2000 шт. отмечена 1 апреля (Кроноки),
в половине апреля начинается валовой пролет. С этого времени и почти до
конца мая турпаны там уступают по численности только морской чернети.
В течение ряда дней бухты бывают усеяны ими. Последняя дата наблюдения
пролета 16 июня, но возможно, что они летят и до конца месяца. Летят цепоч-
кой, углом или шеренгой (Аверин, 1948). На Анадыре турпаны появляются
весной позже всех других уток, в разные годы 27 мая—5 июня. Турпаны не-
редки на пролете через Маньчжурию, встречаются в это время всюду в Юго-
Восточном Забайкалье. На Алтае, в Чуйской степи они появляются, повиди-
мому, в первой половине июня (Сушкин, 1938). На пролете под Якутском
с 30 мая по 12 июня они бывают в огромном числе. Основная масса их проле-
тает там весной с востока, вероятно с Охотского побережья, придерживаясь
горных плато, изобилующих большими озерами (Иванов, 1926). Прилет под
Верхоянском 1 июня, массовое появление на Индигирке (у сел. Абый) около
7 июня. Во многих частях ареала уже после окончания пролета продолжают
36*
564 ОТРЯД ГУСЕОБРАЗНЫЕ
встречаться небольшие стайки турпанов. Они состоят из неполовозрелых птиц,
совершающих свой перелет последними.
Осенний пролет на Лене, южнее Якутска, отмечен еще 5 октября.
Отлет их с Анадыря происходит в конце октября, последняя стайка
встречена 2 октября. У Командорских о-вов они появляются в сентябре,
во второй половине этого же месяца проходит их пролет на Камчатке,
причем птицы встречаются не только на море, но и на некоторых озе-
рах. Под Охотском их на осеннем пролете совсем не видно, очевидно,
пролет проходит над морем в стороне от берегов. В Приморье встречается
осенью в октябре как на морском побережье, так и внутри страны, но в
небольшом числе.
Биотоп. Летним местопребыванием восточносибирского турпана
служат озера тундры, лесотундры и альпийской зоны гор. В лесной полосе
эти утки встречаются исключительно на горных озерах. В Минусинской и Ту-
винской котловинах турпан гнездится на степных озерах. Часть самцов оста-
ется и линять на озерах, но основная масса их проводит этот период в море
у берегов. Там же расположены все места зимовки турпанов и проводят пер-
вый год жизни молодые птицы.
Численность турпана на этой территории весьма неравномерна.
Наиболее обычен он, повндимому, в Якутии, а именно в ее лесной озерной
части: на Лене северо-западнее Якутска, в лесной зоне Индигирки и Колы-
мы. На территориях, не имеющих крупных озер (например, южная часть
Ленско-Амгинского водораздела, гористые верховья Индигирки), турпан от-
сутствует почти совершенно.
Экология. Размножение. Приступает к размножению не раньше,
чем на втором году жизни. Весенние пролетные стаи состоят из птиц, уже
разбившихся на пары. Учитывая это, якутские охотники выбивают из отды-
хающей стаи в первую очередь самок, так как около них обязательно задер-
живаются самцы. Об условиях гнездованья этого турпана обстоятельные
сведения имеются только в отношении Алтая, где он наблюдался Сушкиным
A938). Гнезда там помещаются у открытых берегов озер, обычно у самой
воды. На Анадыре же турпаны гнездятся на высоких берегах рек и озер, пред-
почтительно в кустах. Гнезда представляют собой ямку диаметром около 30 см
и бывают хорошо скрыты кустами или травой. Выстилкой служат те расти-
тельные материалы (мох, сухая трава), которые разгребаются при рытье ямки
и образуют валики вокруг нее. В гнездах бывает много буровато-серого пуха.
Число яиц в кладке 8—9 (Тугаринов, 1941), на Ямале 6—8. Поверхность скор-
лупы матовая, окраска бледная палево-охристая. Размеры сильно колеблются
в разных кладках, а именно 55,4—72,5x35,7—49 мм. Утка сидит очень плот-
но, подпуская к себе иногда почти вплотную.
Сроки размножения различны и зависят как от географического положе-
ния, так и от абсолютной высоты места. В низкогорной зоне Алтая (озеро Бу-
ланду-Куль) 20 июля встречены утята длиной около 12 см, а на Чулышманском
плато B300 м) 16 и 20 июля обнаружены гнезда со слабо насиженными яй-
цами. Оперившиеся птенцы с несколько недоросшими маховыми встречались
примерно там же с 21 августа по 9 сентября (Сушкин, 1938). В истоках Маны
и Кана, на озерах в горной моховой тундре, выводки молодых размером со
свистунка наблюдались 30 июля—12 августа (Юдин). На северо-западном
берегу Байкала маленькие стайки уже летных молодых встречаются в сере-
дине августа (Штегман, 1936). В низовьях Колымы пуховички встречены
еще 2 августа, на Анадыре пуховой птенец (размером с бекаса)—1 августа,
а молодые с начинающими отрастать маховыми и с остатками пуха на голове
и спине 26 августа. Пуховые птенцы хорошо бегают и прячутся в траве,
но ныряют неохотно. Самка находится при выводке до подъема молодых
на крыло.
ГОРБОНОСЫЙ ТУРПАН 565
Линька, вероятно, как и у близкого вида, двукратная. Однако
наряд, надеваемый после основной, полной линьки, почти неотличим от брач-
ного. Послебрачная линька происходит у взрослых самцов близ Охотска
уже в конце июля, когда там и проводятся охоты на нелетных птиц (Харито-
нов, 1915). На Сахалине линяющие самцы добывались с середины августа.
В истоках Маны в первой декаде августа селезни еще были летными, а самки
только начали терять мелкое перо (Юдин). На Лене, в устье Алдана, стаи
отбившихся от самок селезней отмечены уже 29 июня, а на Алтае (Чулышман-
ское плато) они были многочисленны в середине июля. С Алтая, так же как
и из ряда других мест, селезни полностью отлетают до начала линьки. Рай-
оны массовой линьки отмечены выше. Самки линяют позже самцов, в боль-
шинстве случаев на местах гнездовья, зачастую при выводках. Первая пред-
брачная линька молодых птиц происходит весной, вероятно одновременно со
взрослыми. Она заканчивается в мае. Взрослый наряд самцы получают ле-
том, после первой полной линьки.
Питание почти не изучено.
В июне на Анадыре оно состояло
почти целиком из личинок ручейни-
ков и веснянок (из семейства Per-
lidae).
Хозяйственное значение восточ-
носибирского турпана как промыс-
ловой птицы весьма ограничено. Ре-
гулярный весенний промысел его раз-
вит в Якутии. Охота с чучелами Рис. 103. Голова восточносибирского гор-
(Лена, Индигирка) бывает весьма до- боносого турпана
бычлива. В ряде мест до недавнего
времени кочевым населением, севера проводились массовые отловы линных
птиц (Вилюй, Колыма, Камчатка, Охотск); стаи нелетных птиц загоняли
на лодках в устья рек и задерживали до отлива, когда утки оставались на
обсохших лайдах. Такие охоты давали до 10—14 тысяч турпанов (Слюнин,
1903; Харитонов, 1915). В меньшем числе турпан промышлялся на озерах
Камчатки (Шмидт, 1916). Добыча линных птиц известна на Сахалине. Ловля
в сети нелетных турпанов развита на севере Якутии. Тысячами добывали
их в низовьях Колымы (Майдель, 1894) и в большом числе на озерах в ни-
зовьях Вилюя. В последнем месте охоты давали до 300—400 птиц.
Полевые признаки те же, что и у черного турпана, но у селезня обраща-
ет внимание сильно развитый нарост в основании надклювья.
Описание. Строение и размеры сходны с таковыми у обыкновенного
турпана; существенное отличие—в строении клюва и характере оперения около него.
У взрослых селезней у основания надклювья имеется резко выраженное костное вздутие
и кожный нарост на нем, образующий высокую шишку, несколько нависающую вперед.
Оперение лба не выдается мысом вперед, а образует лишь дугообразный выступ, который
далеко не достигает уровня заднего края ноздрей. Клюв в среднем несколько короче,
чем у обыкновенного турпана.
Вес самцов весной (май) 1020—1437 г, в ноябре 1300 г, самок в октябре 1030 г
(Камчатка; Аверин, 1948).
Окраска. Пуховой птенец. На спинной стороне дымчато-бурый с такого
же цвета широкой перевязью поперек зоба. Подбородок, горло, щеки и часть груди
белые. Брюшко и бока сероватые. Клюв черноватый с бурым ноготком.
Взрослая самка. Окрашена несколько темнее, чем предыдущий вид.
Взрослый самец. Весь совершенно черный, в том числе бока и плечи. Ноготок
оранжевый, бока надклювья красные. Шишка, хребет клюва и основания надклювья
черные. От краев ноготка к ноздрям проходят две узкие черные полоски. Наряд, наде-
ваемый после полной линьки, не отличается от брачного.
Молодые птицы и самцы по первому году. Сходны с таковыми у обыкновенного
турпана.
566
ОТРЯД ГУСЕОБРАЗНЫЕ
РОД КАМЕНУШКИ HISTRIONICUS LESSON, 1828
Тип. Anas histrionica.
Синоним. Cosmonessa. Каир, 1829; Phlyaconetta Brandt, 1847.
48. Каменушка Histrionicas histrionicus L.
Anas histrionica. L n пае us Syst. Nat., изд. X, lt 1758, стр.127, Ньюфаундленд.
Русские названия. Народное и книжное—каменушка—связано с характером
обитания птицы на горных реках и у каменистых морских берегов; пестрошейка—на
Анадыре.
Распространение. Гнездовой ареал разорванный, голарктиче-
ский. Северо-Восточная Сибирь на запад до бассейна р. Лены и Байкала, на
Карта 69. Распространение каменушки Histrionicus histrionicus
1—Я. 7i. paci/icue, 2—Н. h. hiatrionicus; a-граница—гнездовий, б—не вполне выясненная граница
гнездовий, в—обособленные гнездовые колонии, в'—места постоянных встреч летом (гнездованье не
доказано), г—зимовки, в—залеты, сне—места скоплений птиц на линьку
север примерно до Полярного круга, на юг до Приморья, на восток до Кам-
чатки и Командорских о-вов. Изолированным районом гнездованья служит
о-в Аскольд в Японском море. На Американском континенте северная
часть Тихоокеанского побережья, область Кордильеров и Скалистых гор
и крайняя северо-восточная часть Лабрадора. Побережья Гренландии и
Исландия.
Характер пребывания. На большей части ареала—перелет-
ная птица, но на тихоокеанских побережьях обоих материков в Южной Грен-
ландии и в Исландии часть птиц оседла.
Биотоп. В летнее время каменушки обычные обитатели горных рек
и озер, а зиму они проводят на море у каменистых берегов.
Подвиды и варьирующие признаки. Выделяют два подвида ка мен ушек: Н. h. histrio-
nicus Linnaeus, населяющий Исландию, Гренландию и Лабрадор, и Н. h. pacificus
Brooks, 1915в Северо-Восточной Сибири и западной части Америки.
КАМЕНУШКА 567
Каменушка сибирская Histrionicus histrionicus pacificus Brooks
Histrionicus histrionicus pacificus. Brooks. Bull. Mus. Сотр. Zool., LIX, 1915,
стр. 393, Камчатка.
Распространение. Гнездовой ареал. Западным пределом рас-
пространения каменушки в Сибири служит, вероятно, северо-западное побе-
режье Байкала. Взрослые птицы многократно наблюдались летом в Бай-
кальском хребте (верховья р. Горемыки). Местность эта для гнездованья ка-
менушек весьма подходящая и возможно, что они гнездятся там (Штегман,
1936). Наблюдались они летом и у южного побережья Байкала (Радде, 1863),
но эти встречи следует скорее отнести за счет летующих неполовозрелых или
взрослых линяющих птиц. От Байкала граница гнездованья каменушки про-
ходит по северо-западным отрогам Станового хребта (в бассейне Олекмы до
75° с. ш.) и вдоль западного склона Верхоянского хребта, по которому она
прослежена примерно до Верхоянска (устье Адычи; Тугаринов, 1934). На
низменности Якутии, в бассейне Лены, каменушки встречаются только на
пролете, и притом крайне редко (Иванов, 1926). Как проходит северная гра-
ница распространения—неясно, так как для всего бассейна Индигирки
они не отмечены (Михель, 1935), отсутствуют в нижнем течении Колымы, но
зато очень обычны в ее верховьях, всюду вверх от устья р. Нюречи (Демен-
тьев и Шохин, 1939). В Анадырском крае обитает спорадично, главным образом
в горных частях его. Местами не редка, но многочисленна только западней
Маркова. На север достигает Анадырского хребта, где редка (Портенко, 1939).
Гнездится по Корякскому хребту, по Пенжине пролетает весной в большом
числе, но остается на гнездовье отдельными парами, нередка в Гижиге и на
п-ове Тайгонос (Дементьев, 1940). Гнездится в небольшом числе по всей Кам-
чатке, на Командорских о-вах (восточный берег о-ва Беринга), а на пролетах
там весьма многочисленна. Встречается летом на островах Курильской гря-
ды, гнездованье там не доказано, но в северной части возможно. Распростра-
нена по всему Охотскому побережью, где в районе Аяна обычна (Снигирев-
ский) и доходит до низовьев Амура (Марийск, река Горин) включительно.
Известны летние находки взрослых птиц на р. Кур и озере Болен-Оджал, но
они еще не говорят о гнездовании там. Гнездится в северной части Сихотэ-
Алиня от залива Декастри до рек Ботчи и Кочи (Емельянов, 1929; Шульпин,
1936) и, вероятно, верховьев Бикина. Гнездится также на каменистых остро-
вах в заливе Петра Великого (о-в Аскольд, а в недавнем прошлом и другие)
и возможно на Сахалине. Южная граница ареала совсем неясна. Населяет
Яблоневый хребет, где найдена в верховьях Зеи (Окононская котловина;
Дорогостайский, 1915), но на Аргуни, Ононе, а также между Читой и Нер-
чинском встречается лишь на пролете (Штегман, 1928).
В Америке заселяет Береговой хребет, Кордильеры и Скалистые горы
от п-ова Кенан и Клондайка на севере до Ванкувера на побережье и верхо-
вьев Колорадо на юге (Филлипс, 1925). Кроме того, в небольшом числе веро-
ятно гнездится на о-ве Уналашка и некоторых других островах Алеутской
гряды (Далл, 1873).
Скопления линных селезней и неполовозрелых годова-
лых птиц отмечены: незначительные в южной части Байкала и в море у юж-
ных берегов Сахалина (Гизенко), большие вдоль всей Курильской гряды, на
Охотском побережье в районе Охотска (Харитонов, 1915) (вероятно и в
других местах), на Командорских о-вах и вдоль всех побережий Анадырского
залива от мыса Наварина до Мечигменской губы (Портенко), а также у вос-
точных берегов Берингова пролива у п-ова Сюард от залива Нортон до зали-
ва Коцебу и кое-где у Алеутской гряды.
Основные зимовки находятся у Командорских и Алеутских
о-вов, вдоль юго-восточных берегов Камчатки и Курильской гряды. Зимует
568 ОТРЯД ГУСВОБРАЗНЫЕ
у берегов Японского моря в южной части Советского Приморья, вдоль восточ-
ного берега Кореи на юг до Фузана (Остин, 1948) и, повидимому, у берегов
Северной и Средней Японии. Обычны зимой у Тихоокеанских берегов Амери-
ки от Аляски почти до Лос-Анжелоса.
Характер пребывания. Гнездящаяся перелетная, частично
зимующая в СССР птица.
Сезонные миграции и их даты. На юге Приморья в за-
ливе Петра Великого появляется в конце мая и долго держится там больши-
ми стаями (Тачановский, 1893), в низовьях Амура первые птицы были
встречены под Николаевском 26 мая, а под Мариинском 2 июня еще шел про-
лет (Шренк, 1861). В верховьях Колымы, где идет интенсивный пролет боль-
шими стаями, срок валового пролета весной очень короток, всего 4—5дней.
К середине июня птицы держатся уже преимущественно парочками (Демен-
тьев и Шохин, 1939). Нередка на весеннем пролете вдоль Пенжины, в среднем
ее течении (устье р. Слаутной) добыта 25 мая (Волков и Дементьев, 1948).
На восточном берегу Камчатки весенний пролет в мае и до половины июня
(Аверин, 1948). В заливе Креста в конце мая—начале июня наблюдались
стайки по 40—60 шт., пересекавшие залив (Белопольский, 1933). В низовьях
Анадыря прилетают 26—27 мая, а под Марковом отмечены 3 июня, когда на
реке появились большие полыньи, 6 июня они уже были нередки и держались
еще несколько дней, но с наступлением половодья исчезли (Портенко, 1939).
Об осенних миграциях известно очень мало. Они начинаются с того, что моло-
дые птицы покидают места гнездовья и спускаются к морю. На о-ве Беринга
уже в середине августа выводки исчезают в верховьях рек и спускаются
в устья (р. Половина и др.), где встречаются в это время во множестве, а с по-
ловины сентября они уже появляются всюду в прибрежной полосе (Иоган-
сен, 1934). В горах Приморья каменушки спускаются к морю (устья рек
Ботчи и Коппи) также в половине августа и сентября. В Анадырском заливе
(бухта св. Павла) последняя дата добычи 10 сентября, на Камчатке основной
пролет происходит в сентябре. Первые пролетные стаи близ устья Уссури
отмечены в самом начале октября (Маак, 1861), в низовьях Амура они добыва-
лись до 4 ноября, когда на реке были уже большие ледяные забереги (Шренк,
1860), на р. Хор встречаются в октябре, а в первой половине ноября
уже собираются на местах зимовки у о-ва Аскольд (Воробьев). Основной
пролет происходит над морем, а на внутренних водоемах каменушки бывают
редко.
Биотоп. Летом каменушки обитают в высокогорных местностях, пре-
имущественно на реках гольцовой зоны. Зимой они держатся стаями в море
у каменистых берегов, там же проводят лето стаи неполовозрелых птиц.
Численность каменушки повсюду невелика. На Азиатском мате-
рике она наиболее обычная в верховьях Колымы, где является самым распро-
страненным нырком как на пролете, так и летом, на Охотском побережье
и в западной части Анадырского края.
Экология. Размножение. Половая зрелость у каменушек насту-
пает не ранее, чем на втором году жизни (после двух зимовок), а полный брач-
ный наряд селезни надевают лишь на третьем году жизни. Первый год птицы
проводят в стаях у морских берегов. На места гнездованья они прилетают
уже разбившись на пары. На Анадыре они встречаются парами в первые же
дни по прилете E—6 июня), хотя при некоторых самках и держится по 2 сам-
ца. К убитой на охоте птице, какого бы пола она ни была, обычно всегда воз-
вращается ее пара. Токующие селезни плавают с выпяченной грудью, с слег-
ка расправленными и опущенными крыльями. Они держат голову запроки-
нутой на спину с раскрытым клювом, а затем рывком выбрасывают ее вперед,
издавая при этом громкий крик вроде «ги-эк». Самки отвечают примерно та-
ким же тоном «ги-ак».
КАМЕНУШКА
569
Гнездятся каменушки в верховьях горных речек с быстрым течением,
перекатами и галечными отмелями, на Камчатке до 400—500 м н. у. м.
В Сибири в южных частях ареала кладка начинается в первой половине июня,
а пуховой птенец добыт около устья Амура 18 июля (Алфераки, 1900). Биоло-
гия гнездованья на нашей территории почти совсем неизвестна. На Амери-
канском континенте они строят гнезда обычно близко от воды, в неровностях
почвы, часто среди камней или под прикрытием травы и кустов. Яиц в полной
кладке 3—8, цвета они слоновой кости и размером 53—62x38,7—44,5 мм.
Длительность высиживания неизвестна, но, повидимому, около месяца. На-
блюдений над жизнью выводков почти нет. Известно, что на Камчатке некото-
Рис. 104. Каменушка (самец и самка)
рые из них поднимаются на крыло очень поздно. Так, 28 сентября в низовье
р. Оленьей встречены 5 выводков с шестью, четырьмя и тремя молодыми, ве-
сом от 300 до-353 г, которые еще не взлетали (Аверин, 1948).
Л и н ь к а. В конце июня взрослые селезни исчезают из гнездовых райо-
нов и появляются на море, где сбиваются в стайки, объединяясь иногда со
стаями годовалых птиц. В конце июля и в августе они линяют. Взрослые
самки начинают линять много позже, лишь ко времени подъема молодых птиц
на крыло. Линька в брачный наряд начинается у селезней поздно и происхо-
дит на местах их зимовки. Одновременно линяют и молодые птицы. Следующая
линька происходит у них летом в то же время, как и у взрослых самцов. Осе-
нью второго года жизни молодые селезни надевают наряд, уже близкий
к взрослому, но окончательный они получают лишь осенью на третий год.
Питание каменушек составляют животные корма: насекомые, рако-
образные, моллюски. Почти все желудки птиц, добытых в первых числах июня
на Анадыре, содержали личинок ручейников до 250 шт. в одном желудке. Кро-
ме них, были обнаружены личинки веснянок (Perlidae), водные жуки, кло-
пы и остатки мелких рыбок (Портенко, 1939). Во всех желудках птенцов,
добытых в сентябре н« Камчатке, были только остатки насекомых и, как
всегда, камешки и песок. За время пребывания на море (осень—зима) состав
питания несколько иной. Основу его составляют ракообразные E7,1%),
в основном десятиногие раки B7,6%; Hemigrapsus, Pagurus и др.), амфиподы
570 ОТРЯД ГУСЕОБРАЗНЫЕ
и балянусы. Большое значение имеют моллюски B4,7%): Chiton, Lacuna,
Litorina, Margarites, Mytilus и др. Насекомые составляют всего 10,2 %
иглокожие и рыбы по 2,4% (Коттам, 1939).
Хозяйственное значение как промысловая птица имеет только местами:
в верховьях Колымы, где каменушки наиболее многочисленный вид нырко-
вых уток, под Охотском, где промышляют линяющих у берега птиц, и на
Командорских о-вах? где они служат существенным подспорьем в питании
зимой, когда других птиц около островов бывает мало.
Полевые признаки. Птица высоко сидит на воде и прекрасно ныряет,
даже в полосе бурного прибоя. Селезень темный с ржавыми боками,белым
полулунным пятном перед глазом, по бокам головы, белым ошейником и еще
несколькими белыми пятнами и полосками. Самка темная с тремя белыми
пятнышками на голове. На полете при взгляде сверху обращает внимание
маленький клюв, целиком темные крылья, темный зашеек и белые продоль-
ные полосы (у самца) на спине. Снизу селезень темный с белым лбом и поло-
сками на плечах и шее. Птицы очень непугливы. Держась в стаях, они пода-
ют голос «эк-эк-эк-эк», а селезни также хриплое «хе-хе».
Описание. Строение и размеры. Небольшая утка со сравнительно корот-
кими шеей и клювом. Ноготок во всю ширину надклювья, ноздри в середине клюва,
несколько ближе к его основанию. Рулевых 14, хвост сильно заостренный, ступенчатый.
Крайние рулевые не превышают половины длины средних. Перепончатая лапа велика,
средний палец в полтора раза длиннее цевки. Длина крыла самцов 198—210, самок 180—
200 мм; цевка самцов 34—39, самок 33—36 мм. Клюв самцов (от оперения лба) 27—30,
самок 23—30 мм, несколько более сильный, чем у атлантического подвида.
Вес каменушек 500—800 г. Взрослые селезни, добытые на Камчатке,
месили: в мае 714 г, октябре 650 г, ноябре 670 г, январе 675 г; самки в сентябре от выводка
510 г, в октябре 500 г (Аверин, 1948).
Окраска. Пуховой птенец. Сверху весь темный, серовато-коричневый.
Голова темная до уровня глаз. Передними сверху с каждой стороны маленькое округлое
белое пятнышко. По бокам головы у основания клюва также беловатые пятна, отделяю-
щиеся от белого пространства под глазами темной полоской, спускающейся сверху перед
глазом. Вся брюшная сторона беловатая с коричневатым налетом на зобу и передней
части груди. Ноги и клюв черноватые, последний с более светлым концом.
Молодые птицы в первом пере. Очень походят на взрослую самку и пол их по
оперению неразличим. Клюв у них темносвинцовый.
Взрослая самка. На спинной стороне темнобурая с оливковым отливом, такого
же цвета шея, зоб и бока тела. На боках головы три белых пятна: у основания
клюва, над передним краем глаза и сзади уха. Брюшная сторона тела беловатая с мел-
кими неясными бурыми пестринками. Крылья и хвост однотонные черно-бурые. Клюв
и лапы буровато-серые.
Самец весной после первой предбрачной линьки. По окраске головы близок к на-
ряду взрослого селезня, но белое пятно перед глазом нечеткое, особенно на темени,
а остальные белые пятна малы. Черная полоса вдоль темени лишь намечается (каштановых
полос вдоль нее нет совсем). Бока шеи и лопатки сине-аспидные, а на спине и зобу
отдельные перья того же цвета. На боках тела ржаво-каштановые перья, на боках
намечается белая полоса, остальное оперение, как в юношеском наряде. Крыло без
зеркала и белых пятен, плечевые и спина сплошь бурые.
После второй предбрачной линьки черное и белое оперение головы приобретает
четкие контуры, появляются белый ошейник и каштановые полосы по бокам темени.
Брюшная сторона тела остается пятнистой, крыло не имеет зеркала.
Взрослый самец в брачном наряде. Голова и шея черные, матовые, по бокам го-
ловы от основания клюва почти до самого глаза вытянутое в вертикальном направле-
нии белое пятно в виде узкой полоски, продолжающееся назад вдоль черного темени.
Ниже нее продольная ржаво-каштановая полоса. Она менее отчетлива и меньше по раз-
меру, чем у атлантической каменушки. Небольшое белое пятно за глазом и длинное от
уха вниз по бокам шеи. У основания шеи белое ожерелье, окаймленное снизу узким чер-
ным кольцом. Спина и надхвостье черные, мантия, верх и бока груди сине-аспидные,
задняя часть груди серая, брюхо черно-бурое, подхвостье черноватое с маленькими
белыми пятнами по бокам. На боках груди, у сгиба крыла белые поперечные пятна, окай-
мленные черным, бока тела каштановые. Плечевые белые, образуют при сложенном крыле
белую полоску вдоль спины. Кроющие крыла серо-аспидные, зеркало блестящее черно-
синее, образуется наружными опахалами второстепенных маховых, окаймленное спереди
белой полоской за счет вершин больших кроющих. Средние кроющие образуют белое
КАМЕНУШКА 571
пятнышко. Третьестепенные маховые с белыми наружными опахалами. Хвост черный.
Клюв темнооливковый с более светлым коготком. Лапы серо-бурые с черноватыми
перепонками, радужина бурая.
Самец летом. В основном черновато-бурый, но межлопаточная область сине-
аспидная. У основания клюва и за ухом белые пятна.
Каменушка атлантическая Histrionicus histrionicus histrionicus L.
Распространение. Ареал. Вся Исландия. Побережье Гренландии
примерно до 70° с. ш. на востоке и до северной части Баффинова залива, а на
Американском континенте северный мыс Лабрадора у западного берега залива
Унгава. В СССР—залетом на Балтийском и Белом морях.
Основные зимовки исландских птиц в море у южных берегов
того же острова. В южной части Гренландии, примерно до Ангмагсалика
на восточном берегу и до Полярного круга на западном. Часть каменушек
с этого острова достигает зимой Северо-Восточной Америки, где они зимуют
в море от Ньюфаундленда и залива, св. Лаврентия на севере до Бостона
на юге.
Залеты. Многократно зарегистрированы случаи залетов атлантиче-
ских каменушек на территории, расположенные много южнее и восточнее их
гнездового ареала (Англия, разные пункты Германии, Швеция, Финляндия,
Шпицберген). В пределах СССР эти птицы добывались на Куришской косе близ
Калининграда, причем там они появляются регулярно каждый год в ноябре
небольшими стайками и только молодые птицы (Флерике, 1894). Известны
единичные случаи появления каменушек на Белом море (Кемь, Архан-
гельск). Последний раз селезень добыт там в Онежском заливе у с. Лямцы
6 августа 1935 г. (Паровщиков).
Характер пребывания в СССР—залетная птица.
Сезонные миграции и их даты. В Исландии каменуш-
ки оседлы. Они совершают только местные перекочевки с внутренних водое-
мов к морю и от северных берегов острова к южным. Откочевка к югу про-
исходит в октябре, а в апреле птицы уже снова появляются в бухтах север-
ного берега. В Гренландии птицы смещаются вдоль побережий к югу, а часть
спускается к берегам Северной Америки.
Биотоп. В летнее время быстрые неширокие реки, преимущественно
со скалистыми берегами, а зимой каменистые морские побережья.
Численное ть. В СССР—единичные нахождения и изредка неболь-
шие стайки.
Экология. Размножение. В Исландии гнезда устраиваются под кар-
ликовыми березками, ивами и можжевельниками или в нишах берега под на-
висшей травой, зачастую на расстоянии менее 1 м от бурлящего потока. Они
почти не имеют выстилки, кроме незначительного количества пуха. Гнездятся
каменушки позже всех других уток. Первые яйца появляются с середины
июня, но еще 18 июля можно встретить свежую кладку. Яиц в кладке обыч-
но 5—7. Их раамер 55,3—59x40,2—44 мм.
Линька. Сбор взрослых селезней в стаи и уход их на линьку проис-
ходят в середине июля.
Питание. Двустворчатые моллюски, ракообразные и насекомые.
Описание. Строение и размеры. От сибирской формы отличается несколько
меньшей длиной крыла и более слабым клювом. Крыло самца обычно немного более 190 мм,
клюв селезня 25—29 мм.
Окраска, как у сибирской каменушки. Отличием этой формы от восточной
служит более крупная и яркая ржаво-каштановая полоса вдоль темени у взрослых
селезней.
572
ОТРЯД ГУСЕОБРАЗНЫЕ
РОД МОРЯНКИ CLANGULA LEACH., 1819
Тип. Anas hyemalis.
Синоним. Harelda Stephens, 1824.
49. Морянка Clangula hyemalis L.
Anas hyemalis. Li n n a e u s. Syst. Nat., изд. X, 1758, стр. 126, Швеция.
Синоним. Anas glacialis. Li n n a e u s. Syst. Nat.t 1, 1766, стр. 203.
Русские названия, Народные: морянка, аулейка (авлейка), саук, савка—
последние три звукоподражательные по голосу самцов.
Распространение. Гнездовой ареал кругополярный. Морянка
гнездится в Исландии, спорадично на Оркнейских о-вах, регулярно на о-ве
Медвежьем, в южной и западной части Шпицбергена, в Северной Швеции,
Карта 70. Распространение морянки Clangula Xiyemalis
а—граница гневдовий, в—места постоянных встреч летом негневдящихся птиц, г—зимовки, сие—ме-
ста скоплений птиц на линьку
Норвегии и Финляндии к югу примерно до 71° с. ш. В Дании встречается ле-
том, но гнездованье ее там неизвестно. На Кольском п-ове, примерно до устья
Поноя, на п-ове Канине, о-ве Колгуеве и Новой Земле. На материковом берегу
заселяет всю тундровую зону европейской части СССР и Сибири до границы
лесной растительности. Гнездится на всех островах Новосибирского архипе-
лага (Тугаринов, 1934, 1941), но отсутствует на о-ве Врангеля. На крайнем
северо-востоке морянка заселяет весь Чукотский п-ов, Анадырский край
к востоку от устья Танюрера, бассейны рек Великой и Пенжины (а возможно
и Гижиги), Олюторское побережье, о-в Карагинский и о-в Беринга. Изве-
стны летние встречи морянок на Северном Сахалине, в Окононской котловине
(Становой хребет), в северной части Байкала и на озерах Зауралья, однако,
нет никаких оснований предполагать гнездованье их в этих пунктах. Гнездит-
ся морянка и на всех островах Берингова моря до Алеутской гряды включи-
тельно, на п-ове Аляске и далее к востоку по всей тундровой полосе до Се-
верного Лабрадора включительно, а также на островах полярного побе-
режья, кроме северной части Земли Элсмира. В Гренландии она заселяет
оба берега и отсутствует только в самой северной части острова.
МОРЯНКА 573
Основные зимовки в западном секторе Арктики лежат у бе-
регов Западного Мурмана, вдоль берегов Швеции и Норвегии к северу до
Лофотенских о-вов, а в Балтийском море до горла Ботнического залива и по-
бережий Латвии. В массе зимуют морянки у берегов Дании и Северной Гер-
мании, реже Бельгии и Северной Франции (многочисленны в холодные зимы),
у восточного и северного побережий Британских о-вов, в южной части Ислан-
дии и Гренландии. На атлантическом берегу Америки морянки зимуют от
Ньюфаундленда на севере и далее чем до мыса Гаттерас на юге, а также в об-
ласти Великих озер внутри страны. В небольшом числе, но регулярно, морян-
ки проводят зиму на полыньях с юго-западной стороны Новой Земли (Ус-
пенский). Птицы, населяющие восточный сектор Арктики, в основной своей
массе зимуют в Беринговом море у кромки льдов, вдоль Алеутской гряды,
у Командорских о-вов, восточного и южного побережья Камчатки и у Куриль-
ской гряды; в меньшем числе в море у о-ва Хоккайдо и у северо-восточного
побережья Кореи. Значительные стаи морянок держатся в течение всей зимы
в северной части Анадырского залива и в Беринговом проливе. В менее зна-
чительном числе они зимуют на море в районе Охотска (Харитонов, 1915),
у берегов Южного Приморья и в истоках Ангары (Третьяков, 1940). Вдоль
американского побережья морянки зимуют от залива Нортон на Аляске
к югу почти до Калифорнии.
Большие скопления линных и неполовозрелых
птиц известны у о-ва Колгуева, западного берега Южного о-ва Новой Зем-
ли, в районе Югорского Шара и около Вайгача, у о-ва Белого и на озерах
северной части Ямала. Линяют морянки в низовьях Яны (с. Казачье и дру-
гие места), на озерах у св. Носа, в огромном количестве в дельте Индигирки
(с. Русское Устье и другие места), на озерах по р. Б. Чукочьей в низовьях
Колымы и Анадыря. Летние скопления морянок, главным образом неполово-
зрелых птиц, известны у Новосибирских о-вов и у мыса Шмидта. У о-ва Вран-
геля, где морянки не гнездятся, на время линьки собирается много тысяч
самцов (Портенко, 1947). Летуют неполовозрелые птицы на Анадыре,
у берегов Камчатки. На Американском континенте наиболее известный рай-
он массового скопления линных птиц находится в море против устья р. Мекен-
зи. Наконец, далеко в стороне от гнездового ареала морянки скопления лин-
ных птиц находятся на степных озерах Зауралья (Макушинский район, Кур-
ганской обл.; Цецевинский). Единичные встречи на Илеке (Зарудный, 1888).
Легуют молодые птицы и в Балтийском море у берегов Эстонии и Южной Фин-
ляндии.
Характер пребывания. На протяжении большей части сво-
его ареала морянка перелетная птица; только в Исландии, Средней Норвегии,
на Северном Мурмане, у островов южной части Берингова моря, вдоль его
восточного берега и на юге Гренландии места зимовок совпадают с местами
гнездовья и можно предполагать частичную оседлость местных птиц. Для
морянок характерны дальние сезонные миграции к местам зимовок и линь-
ки, однако во многих районах часть популяции не отлетает, а лишь откочевы-
вает к незамерзающим участкам моря.
Сезонные миграции морянок лучше изучены в западной
части ареала этого вида. Данные кольцевания показали, что основная
масса птиц, населяющих Исландию, зимует в Юго-Западной Гренландии
<Саломонсен, 1950). Птицы, окольцованные в Северной Швеции, добыты зи-
мой—одна на западном берегу Норвегии и одна на Черном море. Морянки,
линяющие на Северном Ямале, зимуют на Балтийском море от берегов Эсто-
нии (о-в Сарема) до Южной Швеции, Норвегии и Северной Германии (бухта
Травемюнде). Есть некоторые основания предполагать, что на Запад
летят и морянки, населяющие Восточный Таймыр. Основная трасса пролета
проходит вдоль морских побережий, причем часть птиц огибает Скандинав-
574 ОТРЯД ГУСЕОБРАЗНЫЕ
ский п-ов, а часть следует от Белого моря к Балтийскому, над озерами Каре-
лии и Финляндии (Михеев, 1947). Как осенью, так и весной пролет по этой
трассе имеет массовый характер. Через Ладожское озеро и вдоль берегов
Финского залива морянки летят огромными стаями; осенний промысел их
в этих местах дает по нескольку сотен птиц на ружье. У берегов Эстонии морян-
ка—наиболее массовый вид из числа пролетных уток, а в проливах Балтий-
ского моря и в Северной Польше (Штеттин) развит промысловый лов их став-
ными сетями. Наряду с этим в сезоны перелетов морянок можно встретить поч-
ти на всех реках и крупных озерах европейской части СССР, Сибири и Казах-
стана. В некоторых местах они появляются регулярно каждый год, но в боль-
шинстве случаев явление это нельзя считать нормальным пролетом. В средней
полосе Союза, как правило, появляются только молодые птицы данного года
вывода, взрослые же среди них бывают очень редки. В северной половине
лесной полосы, примерно до широты Вычегды, Сухоны и Рыбинского водо-
хранилища, морянки пролетают осенью в северо-западном направлении,
т. е. к Балтийскому морю, и пролет их можно рассматривать как движение
стай, несколько отклонившихся от основного пролетного пути. Южнее морян-
ки появляются только одиночками или, изредка, мелкими группами, причем
только осенью, совершенно или почти совершенно не встречаясь весной.
Более регулярный пролет морянок внутри страны наблюдается в Западной
Сибири и в Казахстане. Ежегодно, хотя и в небольшом числе, они проле-
тают по Иртышу и Тоболу, а затем через озера Кустанайской степи следуют
к верховьям Иртыша и далее к среднему течению Урала. Интересно отметить,
что по этому пути морянки пролетают и весной и осенью. Куда следуют пти-
цы дальше—неясно, так как в дельту Волги они не попадают, а зимовки их на
южных морях до сих пор нигде не отмечены. Исключением служат лишь
наблюдения в Джарылгачском заливе Черного моря, где пролетные морянки,
хотя и в небольшом числе, наблюдаются в оба сезона (Клименко, 1950).
Можно предполагать, что большая часть морянок, следующая этим путем,
огибает с юга Урал и летит затем на северо-запад, как это делают некоторые
другие нырки, например гоголи. Остальные же рассеиваются по южным мо-
рям, но попадают там в несвойственные им условия существования и, живя
в одиночку, без стаи, в основной своей массе погибают на первом же году
жизни. Таким образом, регулярное появление морянок осенью внутри стра-
ны следует расценивать, как массовое отклонение молодых птиц от основного
пути пролета. Чаще всего это происходит в центре материка, в бассейне Оби-
Иртыша, и по пути следования к Балтийскому морю, в бассейне Днепра.
Морянки, гнездящиеся в Восточной Сибири, пролетают также в основной
своей массе вдоль морских берегов, но летят и через Байкал, где задержи-
ваются до поздней осени и частично зимуют.
Даты. Отлет с места зимовки у Британских о-вов начинается в марте,
а из Северной Германии в середине марта—начале апреля, но пролет круп-
ными стаями продолжается в этих местах до середины мая. У берегов Эсто-
нии морянки становятся многочисленными уже в середине апреля и до нача-
ла мая численность их нарастает, а с 10—15 мая начинается отлет на север
(Кох, 1911). В Кольском заливе зимующие стаи встречаются еще 21—22 мая
(Спасский, 1925), под Архангельском первые птицы появляются около 17 мая
F—28 мая, Паровщиков), а в Тиманской тундре отмечены 26 мая, причем
27 мая они были там уже многочисленны (Гладков, 1951). В центре Кольского
п-ова на озере Имандра морянки появляются позднее других уток—только
около 6 июня B5 мая—13 июня, Владимирская, 1948), а в Кандалакшскую
губу Белого моря стаи морянок прилетают в середине июня. Эту вторую
волну пролета составляют неполовозрелые птицы, которые большую часть
года, и иногда и весь год, проводят вне мест своего гнездованья. Около о-ва
Колгуева морянки появляются в середине мая, когда пресные водоемы нахо-
МОРЯНКА 575
дятся еще подо льдом, и стаи их долгое время принуждены держаться в море
у ледяного припая (Подэкрат, 1936). На больших полыньях у юго-западных
берегов Новой Земли морянки держатся в течение всей зимы стайками до
15 шт., а в конце апреля и в мае, с отходом льдов они прилетают огромными
стаями—по 300—400 птиц (Успенский). У о-ва Белого стаи до 100 птиц и бо-
лее появились 20 мая и пролет их продолжался до 3 июня (Тюлин, 1938). На
Ямале первые 12 морянок встречены на полыньях р. Юрибея 5 июня (Осмо-
ловская), на Гыданском п-ове 9 июня, а в середине месяца они были там уже
многочисленны (Наумов, 1931). Западного побережья Таймыра они дости-
гают 5—13 июня (Вальтер, 1902; Бируля, 1907).
Встречи морянок у берегов Кореи известны весной до 27 апреля (Курода,
1918); стаи, зимующие в заливах Посьет и Петра Великого, отлетают в апре-
ле; на о-ве Манерон они бывают многочисленны до марта (Гизенко). Коман-
дорские о-ва зимующие птицы покидают в мае, у берегов Восточной Камчат-
ки с начала мая число морянок заметно возрастает, пролет их длится весь
май и июнь (Аверин, 1948). На Пенжине птицы добыты 25 мая; на Анадыре
и в Анадырском заливе они появляются 18—27 мая (Портенко, 1939). В устье
Индигирки первая стая из 200 птиц отмечена 4 июня, а массовый прилет 9 июня
(Михель, 1935); у о-ва Котельного (Новосибирские о-ва) одиночки появи-
лись уже 11 мая, но до середины июня они держались на полыньях и в тун-
дру переселились только 22 июня (Бируля, 1907). В Хатангском заливе
появление большого числа морянок отмечено 2 июня (Скалой, 1935), а на
восточном берегу Таймыра первые зарегистрированы 10 июня (Тугаринов
и Толмачев, 1934). Весенний пролет морянок в Западной части Черного моря
наблюдается с конца марта до половины апреля (Клименко, 1950), под Чка-
ловом с начала апреля до второй его половины, а на Наурзумских озерах эти
птицы задерживаются до последних чисел мая (Михеев, 1937). Пролет их
в среднем течении Печоры начинается около 27 мая B3—30 мая) и продол-
жается от 1 до 12 дней (Теплов, 1948).
Осенью морянки исчезают с мест своего гнездованья только с началом ле-
достава. На Новосибирских о-вах птицы встречаются до 14—30 сентября, пос-
ледние три особи наблюдались 10 октября, а в Хатангском заливе 24 сентября.
Из Анадырского залива морянки откочевывают в октябре, у восточных
берегов Камчатки появляются во второй половине сентября, пролет в бухте
Карагинской наблюдался с 20 октября по 14 ноября, массовым он был 30 ок*
тября, когда отмечены большие стаи общей численностью около 1500 шт.
(Белопольский и Рогова, 1947). У Командорских о-вов пролет происходит
в октябре, вдоль Курильской гряды он начинается в последних числах
сентября, а берегов Кореи птицы достигают, повидимому, к январю.
8 Гыданской тундре начало осеннего пролета приходится на первые
числа сентября, а последние птицы задерживаются еще целый месяц.
У о-ва Белого первая пролетная стая появилась 24 августа, но последние
птицы отмечены 17 октября. Отлет их под Архангельском происходит 5—
15 октября, а во внутренних частях Кольского п-ова (озеро Имандра) 27 октября
B0—29 октября). В Балтийском море у берегов Эстонии морянки появляют-
ся в конце сентября и задерживаются там до декабря, но бывают менее
обычны, чем весной. Мест зимовки у Британских о-вов они достигают в кон-
це сентября—в октябре, а Северной Германии в середине октября—в ноябре.
Даты пролета по внутренним водным путям таковы: на Средней Печоре (устье
Ылыча) с 7 октября B9 сентября—13 октября) в течение 6—13 дней, в верхо-
вьях Иргиза с 9 по 25 октября, в верховьях Тобола и на озере Боровом 11 —
19 октября, на озерах Наурзумской степи с начала октября, под Чкаловом
с первых чисел октября до замерзания, на Рыбинском водохранилище с 6—
9 по 15—17 октября, последние птицы до 18—21 ноября, а в западной части
Черного моря с половины октября, но в январе их там уже не бывает.
576 ОТРЯД ГУСЕОБРАЗНЫЕ
Биотоп морянди—внутренние водоемы тундровой зоны. На Коль-
ском п-ове и в Скандинавии морянка гнездится на горных озерах в области
ивняков и полярной березы, а на низменности только в самых северных частях
этой территории. На Ямале она охотнее всего заселяет озерные котлорины
тундры и реже встречается на реках. Зиму, зачастую время линьки, а неполо-
возрелые птицы весь год проводят на море.
Численность. Наиболее обычная и в большинстве случаев много-
численная утка тундровых водоемов, особенно на севере Сибири. Хорошее
представление об обилии морянок дают скопления неполовозрелых и линяю-
щих птиц в море и на тундровых озерах, достигающие десятков тысяч штук.
Экология. Размножение. Размножаться начинают, повидимому,
на втором году жизни. Брачное оживление у них наступает еще на местах
зимовок, на Мурмане в марте, а на Восточной Камчатке во второй полоэине
апреля. Повидимому, в это время происходит и разбивка на пары. Самцы
оживленно плавают вокруг самок; вздергивают кверху свой длинный хвост,
вытягивают шею и откидывают голову на спину, издавая при этом свой гром-
кий многосложный крик. Он довольно удачно передается слогами «кау-кау»...
«скауэ». Голоса стаи морянок сливаются в общий гул, напоминающий отдален-
ный лай собак. Голос самки слышится реже и звучит как «вед» или «воод-воод».
На места гнездовья птицы прилетают стаями* в которых нередко преоблада-
ют самцы, но пары морянок бывают к этому времени уже оформлены. Морян-
ки часто прилетают задолго до вскрытия пресных водоемов тундры и поэтому
около месяца они держатся в море и не могут приступить к гнездованью (о-в
Колгуев, Новосибирские о-ва и др.). Гнездится чаще всего в тундре около
озерков, ручьев или даже просто котловин, наполненных водой. На Среднем
Ямале это наиболее характерный, а местами и единственный вид уток, насе-
ляюцщх водораздельные озера тундры. Гнездо устраивается на сухом месте,
как правило, вблизи от водоема. Оно помещается чаще всего под укрытием
кустов полярной березки или низкорослого ивняка, но нередко и на совсем
открытом месте среди осоки. Гнездо представляет собой довольно глубокую
ямку небольшого размера с очень скудной выстилкой из растительной трухи,
собранной рядом, или даже совсем без нее. К концу откладки яиц в гнезде
появляется значительное количество темнокоричневого пуха со светлыми
центрами. Диаметр гнезда около 10 см, лотка 13 см, а глубина его 8 см. Наи-
более обычное число яиц в полной кладке 6—7 (о-в Колгуев, Тиманская тун-
дра, Ямал, Индигирка), но нередко и 8, а бывают гнезда даже с 10—12 яй-
цами. Возможно, что последнее относится к сдвоенным кладкам, так как число
рубцов на яичнике у насиживающих самок обычно 6—7. Яйца овальные или
удлиненно-овальные с гладкой скорлупой и оливково-бурым оттенком. Раз-
мер 48—60 х 35,5—40 мм, среднее—53 х 37 мм, по данным для Ямала 54,5 х
X 36,5 мм, вес38,7 г C8—40). Откладка яиц на северо-западе Мурмана (Варан-
гер-фьорд) происходит в середине июня, обычно до 1 июля; начало ее в
Лапландии (Каресуандо) 9 июля. На Восточном Мурмане (ст. Харловка)
гнездо с 10 свежими яйцами найдено 24 июня (Спангенберг, 1941). Пер-
вые полные кладки в Тиманской тундре обнаружены 7—10 июня (Гладков,
1951); на Новой Земле гнезда с яйцами и отводящие от гнезд самки встреча-
лись в конце июня и начале июля. На Южном Ямале время откладки яиц (по
добыче самок с яйцами) 10—24 июня, но еще 27 июня там наблюдались самцы,
гонявшиеся за самками, а гнезда с неполным числом яиц встречались до
5 июля (Дунаева, Осмоловская). В дельте Индигирки полные, но еще слабо
насиженные кладки найдены 3 и 8 июля, на Анадыре гнездо с 12 свежими яй-
цами 7 июля, а другое уже с сильно насиженными 12 июля; на о-ве Беринга
птицы поселяются парами на озерах в конце мая, а в июне происходит отклад-
ка яиц. Срок насиживания в природных условиях около 23—24 дней, яйца,
взятые для инкубации, выводятся за 24—25 дней (Уайзерби и др., 1945).
морянка 577
Самка сидит очень крепко, особенно в конце периода насиживания. Известны
случаи, когда ее удавалось брать на гнезде руками (Анадырь; Портенко,
1939). Существуют наблюдения, что птенцы первые несколько дней держатся
под опекой матери у гнезда и на воду не сходят. Плавают и ныряют они первое
времй плохо й держатся плотной стайкой около самки, которая сзывает их
голосом. При опасности самка уводит птенцов не на берег, а на воду (Туга-
ринов, 1941). Выводки держатся долгое время на пресных мелководных озер-
ках в тундре, предпочтительно на тех из них, которые имеют заросли осоки
вдоль берегов. На крупные водоемы и на море они переселяются обычно лишь
ко времени подъема на крыло. Большое количество яиц морянки и ев мелких
птенцов Гибнет от нападений поморников и крупных чаек. По наблюдениям
Кучерука A948), гибель эта не вызывает уменьшения среднего размера вывод-
ка, а идет за счет полного уничтожения некоторых, более слабых из них.
Присоединение к выводку чужих птенцов и полное объединение выводков
(до 19 птенцбв при двух самках) обычное явление у морянок. На Мурмане
вывод птенцов происходите конце июля (пуховикиоколо Ура-губы 31 июля).
На о-ве Колгуеве птенцы, еще не покинувшие гнезда, найдены 27 июля,
14 августа там же добыты оперенные, но еще нелетные молодые (Толмачев,
1927), а на о-ве Кильдине нелетный выводок встречен 28 августа. В Тиманской
тундре первый Выводок отмечен 12 июля, на Южном Ямале 14 июля. На Новой
Земле в июле самки с полу оперившимися птенцами уже появились в море,
в августе птенцы достигли полного роста, но еще не летали. В устье Енисея
птенцы наблюдались 27 июля, на Таймырском озере 24 июля, на Анадыре
31 июля—3 августа, а на Командорских о-вах в июле и в августе. Повсюду на-
ряду с нормальными выводками встречаются и очень поздние, появившиеся
только в августе. Можно предполагать, что большая часть их не успевает
вырасти и погибает, так как период полного развития птенцов занимает около
5 недель.
Линька. Сезонная смена нарядов у морянок несколько отличается
от общей схемы, которая свойственна большинству видов нырковых уток.
В недавнее время линька морянок на большом материале была изучена Сало-
монсеном A941). Однако мы не можем согласиться с рядом выводов этого
автора, и в дальнейшем изложении, используя его богатый фактический мате-
риал, даем несколько другую интерпретацию вопроса. Полная линька со
сменой маховых и рулевых перьев начинается у селезней из западного сек-
тора Арктики (Исландия, Гренландия) в конце июня—начале июля, а в на-
ших пределах несколько позже. Первым признаком ее служит отрастание серо-
коричневых перьев на боках перед основанием ног. Затем частично или пол-
ностью выпадают удлиненные плечевые перья и начинают отрастать новые,
одновременно выпадают и все белые перья, сохранившиеся на передней части
тела от зимнего наряда, заменяясь черно-коричневыми. Следующим этапом бы-
вает линька крыльев с выпадением маховых и кроющих крыла, а также смена
некоторых или большинства рулевых. Рост новых плечевых и длинных перьев
на боках груди, прикрывающих крыло, на этой стадии бывает завершен.
Линька хвоста начинается с выпадения трех крайних пар рулевых. Вслед за-
тем начинается интенсивная смена оперений на туловище сзади от передней
части груди, т. е. на самой груди, спине, брюхе, над- и подхвостье. Одновре-
менно выпадают несменившиеся до тех пор рулевые и последними из них
длинные рулевые средней пары. На этой стадии линька несколько затихает,
но вскоре возобновляется снова быстрой сменой контурного оперения перед-
ней части тела (к переду от линии груди), т. е. на голове, шее и в верхней ча-
сти груди. К этому же времени завершается обычно и рост средней, наиболее
длинной пары рулевых, что происходит в конце сентября или начале октяб-
ря. Однако задолго до окончания этой линьки уже начинается следующая,
сменяется оперение махов и плечевые перья, полученные в процессе летней
37 Птицы СССР, т. IV
578
ОТРЯД ГУСЕОБРАЗНЫЕ
линьки. Вслед затем начинает линять голова и верхняя часть шеи. Новое опе-
рение отрастает повсюду на боках головы, подбородке, горле и по бокам шеи,
в верхней ее части. Смена оперения проходит довольно вяло и заканчивается
в конце ноября или в декабре. Соответствующая линька самок начинается
много позже, лишь тогда, когда подрастают молодые. Последовательность
Рис. 105. Схема сезонных изменений наряда морянки: а—голова самца
в брачном наряде, б—голова самца в летнем наряде, в—голова самца
в осеннем наряде, г—плечевое перо в брачном наряде, д—е—то же, в
летнем наряде
смены оперения та же, но дополнительная линька головы и шеи не отмечена.
Зимний наряд носится дольше других—несколько более трех месяцев, сме-
на его начинается только в апреле. Первыми появляются черные перья на
темени и щеках. Вслед затем выпадают удлиненные плечевые перья зимнего
наряда и начинается смена перьев на боках головы и шеи, горле, зобу и пле-
чах. Эта линька захватывает всю переднюю часть тела до линии груди. По-
следним линяет затылок с прилегающей частью шеи. У самок предбрачная
линька, которая проходит в конце апреля и в мае, захватывает большую пло-
МОРЯНКА 579
щадь, чем у самцов. Кроме головы, шеи и зоба, вылинивает также передняя
часть груди, плечи, спина и зад.
Таким образом, для взрослых самцов морянки в их годовом цикле можно
наметить три основных сезонных наряда: зимний, брачный и летний. Описа-
ние их довольно трудно, так как в некоторых случаях новая линька начинает-
ся раньше, чем заканчивается предыдущая. Оперение туловища полностью
сменяется только один раз в год—летом и лишь частично, далеко не всегда,
весной. Голова и шея линяют трижды: два раза полностью и один частично
(осенью). Хвост линяет один раз в год, а плечевые перья сменяются три раза,
причем во всех нарядах они различны по форме, размерам и раскраске.
Первая линька молодых птиц начинается в октябре—ноябре. У самцов
в это время перелинивает голова, шея, часть перьев затылка и зоба, маховые
и плечевые перья. У некоторых особей линька заходит дальше, появляются
новые перья в передней части груди и сменяется средняя пара рулевых.
С декабря до начала апреля линька полностью затихает, и птицы носят пест-
рый, весьма различный у отдельных особей наряд. С середины апреля до кон-
ца мая линька молодых возобновляется и проходит так же, как и у взрослых
самцов. Первая смена оперения у молодых самок проходит примерно в те же
сроки и захватывает те же части тела. Первая полная летне-осенняя линька
у неполовозрелых птиц имеет ту же последовательность, как и у взрослых.
Наиболее тяжелым для уток периодом линьки бывает одновременная смена
маховых, в течение которой они на длительное время теряют способность
к полету. В разных районах это происходит в различные сроки. На р. Илеке
в Чкаловской обл. (Зарудный, 1888) линный селезень был добыт 21 июня, в
Курганской обл. отлов нелетных линяющих морянок проводился 30 июля,
а на озерах Северного Ямала с 29 июля по 13 августа (Цецевинский).
В южной части Ямала, в бассейне р. Щучьей, неохотно поднимающиеся с воды
птицы отмечались с 19 июля, 24 июля были встречены первые морян-
ки с выпавшими маховыми, к 17 августа все птицы уже поднялись на
крыло (Кучерук, 1948). На Новой Земле стаи линных самцов отмечались
в первой декаде августа. В проливе Малыгина тысячные стада линяю-
щих морянок держатся с 15 июля по 15 августа (Тюлин, 1938). В дельте
Индигирки линька маховых начинается у морянок 23—25 июля, и в первой
декаде августа огромные стаи их держатся на озерах. Местом скопления ли-
няющих птиц служат водоемы различного типа: морские мелководья у скали-
стых берегов (Новая Земля, о-в Белый), небольшие и мелководные озера
тундры (Северный Ямал, р. Чукочья), мелководные озера дельты (р. Индигир-
ка), реки тундры (Южный Ямал) и, наконец, степные мелководные озера
(Курганская обл.). В небольших стайках на реках (до 20—25 шт.) линяют
обычно самки, самцы же собираются большими стаями на озерах и частично
на море. Неполовозрелые птицы проводят первую линьку обычно в море.
Какая половозрастная категория птиц попадает на степные озера—неясно.
Питание. Пищу морянок составляют животные корма. За время
пребывания на местах гнездовья в тундре они питаются ракообразными и ли-
чинками насекомых. Ранней весной на Таймыре желудки их бывают наполне-
ны красными личинками комаров-хирономид (Бируля, 1907). На Ямале
морянки поедают личинок ручейников, а также, повидимому, и ракооб-
разных (Ostracoda, Apus, Branchipus), которыми кишат мелководные во-
доемы тундры (Осмоловская). Желудки птиц, добытых весной на Анадыре,
содержали личинок веснянок (Perlidae), иногда не менее 100 на один желу-
док, и личинок ручейников, а также остатки мелкой рыбы, икринки и над-
крылья жуков (Портенко, 1939). По наблюдениям Сокольникова, там же
весной морянки питались прошлогодней рыбой, оставшейся на берегах пос-
ле ее осеннего нерестового хода На Печоре во время осеннего пролета они
кормятся преимущественно насекомыми, а на Рыбинском водохранилище
37*
580
ОТРЯД ГУСЕОБРАЗНЫЕ
мелкой рыбой G3%): плотвой E2%) и окунем B1 %), а также личинками на1
секомых, преимущественно хирономид B0% Tendipes plutnosus-^redudus,
Glyptotendipes gripeicoueni). Меньшее значение имеют личинки ручейников
и моллюски-беззубки (Немцев). Местами кормежки морянок во время проле-
та служат открытые песчаные мелководья. На местах зимовки у Восточного
Мурмана в желудках обнаружены бокоплавы (Кафтановский, 1941), а на Кам-
чатке главным образом остатки моллюсков (Аверин, 1948). По данным для
Лабрадора и Аляски A90 желудков взрослых птиц), основу питания моря-
нок составляют ракообразные D8,2%), главным образом бокоплавы, но так-
же крабы, креветки, мизиды и др. Из моллюсков A5,7%) чаще поедаются
двустворчатые, особенно мидии и реже брюхоногие (Litorina, Lacuna).
Рис. 103. Морянка (самец и самка в брачном наряде)
Довольно большое значение имеют насекомые A0,8%), личинки ручейников
и хирономид, а также рыба (9,7%): колюшки, сельдевые, бычки. Раститель-
ные корма (зеленые части злаков и других растений) следует рассматривать
как случайную примесь. Пищу птенцов C6 желудков) составляют главным
образом ракообразные G5,6%) и из них: листоногие (Branchinecta и др.
33%), дафнии G,8%), мизиды и бокоплавы. Поедаются птенцами и раститель-
ные корма B2,6%): водяная сосенка, вороника и др. (Коттам, 1938, 1939).
Хозяйственное значение. Почти повсюду в тундровой зоне нашего Союза
морянка, по ее значению в промысле, твердо занимает первое место среди
уток. Так, на Анадыре, где существует интенсивный весенний промысел ныр-
ковых уток, дающий в год более 5500 птиц, морянка составляет около 70%
всей продукции. На Колгуеве их добывают ежегодно около 1000 шт. Огромное
значение для населения крайнего севера имел в свое время промысел линных
морянок. В дельте Индигирки, где морянки занимали первое место в промысле
линных птиц, артель из 5 человек добывала около 2 500 шт. (Житков и Зен-
зинов, 1915). Добывают морянок повсюду предпочтительно ранней весной,
так как с момента переселения на море мясо их приобретает сильный и непри-
ятный запах ворвани.
Полевые признаки. Утка среднего размера с небольшой головой и длин-
ным хвостом с нитевидными рулевыми. Зимой у птиц обоего пола голова
и шея белые с темными пятнами на щеках; у обоих также белое брюхо, а у сам-
МОРЯНКА 681
ца и бока. Крылья с обеих сторон, спина и грудь (у самца шире, у самки уже)
темные. В поздне-летнем наряде верх и передняя часть туловища темнокорич-
невые; светлые только бока головы и низ. Крылья узкие, заостренные, по-
лет быстрый; прекрасно ныряет. Часто держится очень большими стаями.
Очень шумливая утка» особенно в период весеннего возбуждения.
Описание. Строение н размеры. Некрупная утка с относительно малень-
кой головой и коротким клювом (короче головы). Ноготок во всю ширину надклювья
и сильно изогнут над нижней челюстью. Зубцы по краям челюстей острые. Хвост из
14 рулевых, в основном округлый, но две средние пары рулевых нитевидно вытянуты,
у взрослого самца они достигают 200 мм и более. Плюсна короткая, средний палец
с ногтем в 1,6 раза длиннее ее. Внутренние плечевые перья удлинены и заострены. Их
размер у взрослых самцов в зимнем наряде 149 мм A29—176 мм), в гнездовом 112,5 мм
(92—129 мм) и после летней линьки 89,4 мм (83—96 мм); у молодых самцов в первом
зимнем наряде 86—125jmjm. Зимой они вытянуты ланцетовидно, в гнездовое время так же
заострены, но короче и шире, а летом еще шире и несколько округлее. Длина крыла:
самцов 210—240, самок 180—215 мм; цевка самцов 33—38, самок 32—35 мм, клюв сам-
цов 26—29, самок 24—26 мм. Хвост самцов до 250, самок 55—70 мм.
Вес. К местам гнездовья морянки прилетают, как правило, очень жирными, но сни-
жают свой вес за период размножения. Вес взрослых самцов в мае 859 а G75—920 г)
(Архангельск; Паровщиков), 680 а (Бараба), 815—930 а и самка в апреле 690 а (Камчатка;
Аверин, 1948); В июне самцы 765 а F38—890 а), самки 685 а F13—780 а); в июле самцы
781 г F66—800 г), самки 624 а E16—730 а) (Ямал; Осмоловская), в то же время самки
590—600 г (Тиманская тундра; Гладков). Со второй половины лета и к осени вес птиц
вновь нарастает. Самки в августе 638 а E50—800 а), причем самки с выводками 550—
600 г, а холостые самки, начинающие линьку, 700—800 а, в сентябре 650 а (Ямал),
800 г (Архангельск); в октябре самец 750 а, самка 645 а (Рыбинекое водохранилище);
в ноябре самка 720 г, в январе 635 а (Камчатка). Молодые птицы в середине августа
A0—12 августа) 317—537 а, 6 сентября 570 а (Ямал); в октябре самцы 616 а E00—660 а),
самки 603 а E50—650 а), в ноябре самец 670 а, самка 805 а (Рыбинское водохранилище).
Окраска. Пуховой птенец. На спинной стороне темнобурый с еще более
темной головой и поясницей. Пятно у основания клюва над глазом и полоска позади его
беловатые; бока шеи и область уха сероватые, горло и брюшко белые, зоб темнобурый.
Молодые птицы в первом наряде. На спинной стороне черноватые, все перья
без охристых и сероватых каемок. Уздечка, область глаз и ушей беловатые; плечевые
перья одноцветные оливково-коричневые. Горло, зоб, шея с боков и оперение ног серо-
коричневые; нижняя сторона туловища белая. Кроющие крыла матово-коричневые. Хвост
однообразно темносерыи. Клюв одноцветный голубовато-серый.
Молодая самка первой зимой. В основном сохраняет свой первый наряд, но голова
и шея получают оперение взрослой птицы. Перья мантии и плечевые черно-коричне-
вые со светлосерыми каймами. Новые перья на верхней части груди и на боках тела
в основном белые.
Самка летом первого года жизни. Отличается от взрослой сохранившимися
ювенальвыми перьями туловища, хвоста и крыльев.
Самка во втором зимнем наряде. Имеет в основном оперение взрослой птицы,
но отличается тем, что плечевые перья ее имеют по большей части серые или зем-
листо-бурые края. Перья спинной стороны черные с узкой коричневой каемкой или
совсем без нее.
Взрослая самка зимой. Голова и шея в основном белые, но верхняя сторона голо-
вы и нижняя часть щек черно-коричневые. Спинная сторона туловища черно-коричне-
вая с широкими красно-коричневыми каймами перьев, особенно плечевых. Поперек зоба
Проходит довольно узкая ржаво-коричневая перевязь. Брюшная сторона тела белая.
Маховые бурые, второстепенные с коричневатыми внешними опахалами, верхние крою-
щие с охристо-бурыми вершинами. Гнездовой пух черно-коричневый. Клюв аспидно-серого
цвета, лапы черно-оливковые, радужина желтая.
Взрослая самка в гнездовом наряде (май—июль). Голова и шея черноватые;
пространство перед глазами серое; одно белое пятно проходит от глаза назад через
область уха, а другое, большое, находится рядом с ним на шее. Перья спинной стороны
тела, которые сменяются лишь частично, черноватые с узкими светлыми каемками.
Плечевые с широкими оливково-коричневыми краями. Зоб серовато-бурый.
Самка после летней линьки. Одноцветно темная на спинной стороне с серо-
коричневыми боками. Плечевые оливково-коричневые без светлых каемок.
Молодой селезень первой зимой. Имеет наряд с весьма различным количе-
ством новых перьев у разных особей. Уздечка и щеки серо-коричневые, а пятно на
боках шеи темнокоричневое, как у взрослых. Большие пространства на голове и шее
заняты белыми перьями. Плечевые голубовато-белые, но короче, чем у взрослых. Клюв
с розовой перевязью перед вершиной.
582 ОТРЯД ГУСЕОБРАЗНЫЕ
Молодой самец первым летом до летней линьки. В основном сходен со взрослым,
но хорошо отличается более узкой коричневой частью груди, ювенальными перьями
крыльев (коричневые, а не черные кроющие), более короткими плечевыми и рулевыми.
Молодой самец на втором году жизни. Отличается от вполне взрослых только
несколько более короткими средними рулевыми.
Взрослый самец в зимнем пере. Верх головы, кольцо вокруг глаза, подбородок,
горло и шея белые; бока головы от основания клюва до передней части щек дымчато-
серые; в области ушей и прилегающей части шеи большое черно-коричневое пятно, пере-
ходящее в своей нижней части в каштановое. Спина и надхвостье черные. Плечевые
перья голубовато-серые, внутренние из них сильно удлинены; бока тела тоже серые.
Передняя половина груди черно-бурая, весь остальной низ тела чистобелый. Все маховые
и кроющие крыла темно-, почти черно-бурые. Две средние пары рулевых черно-бурые,
остальные с узкой светлой каемкой, а крайние в большей или меньшей степени серые
или белые. Клюв черный с розовой или желто-оранжевой перевязью у вершины. Лапы
голубовато-серые, радужина красная.
Брачный наряд, или вернее, гнездовой (носимый с конца мая до конца июня—
начала июля) отличается от зимнего только оперением передней части тела и плеч.
От глаз по направлению к затылку тянутся довольно длинные глянцевитые белые
пятна. Область уздечки, оперение над глазами, перед и под ними серовато-коричневое,
несколько краснее, чем в зимнем наряде. Все остальные части головы, горло, шея
и зоб темнокоричневые. Перья зашейка и удлиненные плечевые имеют черную среднюю
часть и широкие каймы каштанового цвета. Цвет этих каемок постепенно выцветает и из
красно-коричневого становится желто-белым.
Осенне-летний наряд (надеваемый после полной линьки). Отличается формой
и окраской плечевых, а также оперением головы и шеи. Удлиненные плечевые с ши-
рокой, довольно светлой оливково-коричневой или землисто-коричневой каймой, которая
менее ярко, чем в предыдущем наряде, контрастирует с темнокоричневой серединой пера.
Все оперение спереди от линии груди, т. е. голова и шея, блестяще белое с оранжевым
налетом на задней части головы и затылке и с небольшими серо-коричневыми (испещрен-
ными белым) пятнами по бокам шеи в ее верхней части. Наряд этот сразу же переходит
в зимний, так что ко времени окончания линьки головы и шеи (в начале октября) часть
плечевых уже заменяется зимними.
РОД ГОГОЛИ BU СЕРН ALA BAIRD, 1860
Тип. Bucephala albeola.
Синонимы. Clangula F1 е m m i n g, 1822; Glaucion, Каир, 1829; Glaucionetta S t e j-
n e g e r, 1885; Charitonetta S t e j n e g e r, 1885.
50. Гоголь обыкновенный Bucephala claugula L.
Anas Clangula. Li nnaeus. Syst. Nal., изд. X, 1758, стр. 125, Швеция.
Синоним. Anas hyemalis. Pallas. Zoographia Rosso-Asiatica, II, 1811, стр. 270,
nee. L.
Русские названия. Гоголь—народное, вероятно из-за нарядного оперения;
местами дупленка, благодаря гнездованию в дуплах. Название известно по крайней
мере с XIII в.
Систематические замечания. Гоголь, населяющий Американский континент, выде-
ляется в особый подвид В. с. americana Bonaparte, 1838. Никаких различий в окраске
оперения этот подвид не имеет и основанием для его выделения послужили только
несколько более крупные размеры, определимые к тому же лишь при сравнении значитель-
ных серий. Имеющиеся в наших музеях экземпляры американских гоголей неотличимы
по своим размерам от гоголей Палеарктики. Мы не видим достаточных оснований для
выделения этой формы.
Распространение. Гнездовой ареал. Область гнездованья зани-
мает большую часть лесной зоны Европы, Азии и Северной Америки. В Нор-
вегии всюду, кроме крайних южных провинций, до Остфинмаркена (Варан-
гер-фьорд) на севере; по всей территории Швеции и Финляндии; в северо-за-
падной части Германии, примерно до Эльбы (известен из Мекленбурга, Бран-
денбурга и Саксонии), и в весьма незначительном количестве в Северной
Польше. В Дании пока определенно не зарегистрирован, но гнездованье его
там весьма вероятно (Иесперсен, 1946). Обнаружен на гнездовье в Шотландии
Карта 71. Распространение гоголя Bucephala clangula
а—граница гнездовий, в—обособленные гнездовые колонии, в'—места постоянных встреч летом негнездящихся птиц, з—-зимовки, д—валеты, эк*—места
скоплений птиц на линьку, з—места, в которых гоголь перестал гнездиться в последнее время
584 ОТРЯД ГУСЕОБРАЗНЫЕ
(Чешир) в 1931—1932 гг. (Уайзерби, 1939). Есть указания на наличие таких
же изолированных гнездовых колоний гоголя в Швейцарии (Бутурлин, 1935),
на горных озерах Черногории (Рейзер, 1896) и по нижнему течению Дуная
в Болгарии (Беттихер, 1927). Однако имеющиеся данные недостаточно
достоверны, и больше оснований к тому, чтобы считать эти пункты местами
чистого летования особей и стаек, в которых возможны и случаи спорадич-
ного гнездованья.
В пределах СССР северная граница гнездовой территории гоголя обус-
ловлена северным пределом высокоствольного леса. На Кольском п-ове го-
голь весьма обычная птица. У Белого моря он всюду доходит до побережья,
а устами встречается и на островах. В обеих этих местностях гоголь
является наиболее многочисленной гнездящейся уткой. По Мезени он дохо-
дит почти до устья, а на Печоре примерно до Усть-Усы F0° с. ш.), где
встречается уже редко, хотя несколько выше, в среднем течении реки, это
самый обычный нырок. В долине Оби гоголь гнездится почти до Салехардаг
т. е. достигает там Полярного круга. По Енисею он встречается даже не-
сколько севернее, так как обычен на гнездовье по всему бассейну Нижней
Тунгуски, а под Туруханском принадлежит к числу наиболее многочислен-
ных птиц. В долине Лены гоголь переходит также за Полярный круг, а по
Колыме спускается до устья Анюя. На северо-востоке гоголь достигает
верховьев Анадыря, но очень редок там на гнездовье, а в среднем и ниж-
нем течении реки даже совсем не встречается (Портенко, 1939). Гнез-
дится по р. Пенжине, почти на всей Камчатке и, вероятно, на Командор-
ских о-вах.
Южная граница гнездованья гоголя менее точно совпадает с границей
лесных массивов. Он гнездится в Литве, довольно редко в Эстонии и Латвии,
а в Белоруссии найден только на крайнем ее северо-востоке, отсутствуя в ос-
тальной части республики. В незначительном числе гнездится в Псковской
и Новгородской обл., а также в восточной части Калининской, но совер-
шенно не найден пока ни в Смоленской, ни в Московской обл. Относи-
тельно последней зарегистрирован лишь одцн случай, когда пара гоголей
в гнездовое время постоянно держалась в одном месте на р. Шерне
(Немцев). Обычны гоголи в Ярославской и в лесистой, заволжской, части
Горьксзвской обл. Наиболее южная находка сделана в бассейне р. Оки
(район Рязанской Мещеры; Бекштрем, 1928). Изолированные гнездовые
колонии гоголя совсем недавно существовали в лесистой части Воронеж-
ской и Тамбовской обл. (Северцов, 1855; Резцов, 1910), а колония
в низовьях Днепра (Ново-Воронцовский район Николаевской обл.; Данило-
вич, 1939) сохранилась идо сих пор. Не так давно число этих изолированных
колоний было, видимо, значительно больше. Так, в конце прошлого века
гоголи гнездились в дельте Волги (Яковлев, 1873), а в начале нашего века—
в Козельском районе Калужской обл. (Генглер и Кавелин, 1909). Заселение
территории и рубка старых дуплистых деревьев оттесняет эту птицу из южных
частей ее гнездового ареала. По долине Волги граница спускается до южных
пределов Татарии (Попов). Есть не вполне достоверное указание на слу-
чаи гнездованья гоголя по р. Кинеле (Карамзин, 1909). Нет никаких точных
сведений о гнездовье гоголей по нижнему течению Волги (вопреки утверж-
дению основных орнитологических сводок). Восточнее гоголь гнездится
по долинам Камы и Белой, а по предгорьям Урала спускается далеко
на юг и встречается, хотя и редко, по рекам Сакмаре и Илеку в районе Чка-
лова (Зарудный, 1888). Затем граница гнездового ареала резко поднимается
к северу и проходит по северной части Тобольской и Ишимской лесостепи.
По вклинивающимся на юг лесным участкам гоголь проникает в Барабиц-
скую стфпь до озера Чаны, а по ленточным борам заходит кое-где в Кулундиц-
скую степь (Иогансен, 1907) и даже в окрестности Семипалатинска. Еще южнее
ГОГОЛЬ ОБЫКНОВЕННЫЙ $85
известна изолированная колония гоголей в Зайсанской котловине и по Чер-
ному Иртышу. По верхнему течению Оби гоголь проникает на Алтай, гнез-
дится в его восточной части, в Саянах и в пределах Тувинской обл. На Бай-
кале гоголь обычен всюду, кроме его юго-восточного берега, где пока не
зарегистрирован. Есть указание на гнездованье гоголя э районе Кяхты (Мол-
лесон? 1895), никем, однако, впоследствии не подтвержденное. В Даурской
лесостепи гоголя нет, но уже где-то около Нерчинска южная граница его
гнездовой области пересекает Амур, проходит по северной лесистой части
Маньчжурии и захватывает Уссурийский край, где в недавнее время гоголь
обнаружен поИману и Бикину (Воробьев).. На Сахалине гоголь известен толь-
ко в северной половине острова. В Северной Америке северная граница рас-
пространения гоголя проходит от залива Нортон на Аляске к устью р. Мекен-
зи, западному берегу Б. Медвежьего озера, на озеро Невольничье и форт
Черчилль. Далее граница следует берегом Гудзонова залива, пересекает Лаб-
радор и рыходит к устью р, Гамильтон. Южная граница начинается от о-ва
Лонг-Айленд, рассекает область Великих озер и следует к верховьям Мис-
сури, откуда поднимается к северу по восточным склонам Скалистых гор
и выходит к Тихому океану в Аляскинском заливе
Наличие существующих в настоящее время и недавно исчезнувших изо-
лированных мест гнездованья гоголя южнее границ его основного ареала,
приуроченных к небольшим островам леса в степи, говорит о том, что ареал
этого вида еще недавно был значительно шире. Он, повидимсму, сильно
сократился за историческое время в результате уничтожения лесов на юге
нашей страны и в Западной Европе.
Районы линьки. Негнездящиеся и неполовозрелые птицы,
а также и линяющие самцы местами проводят лето вне пределов гнездовой
территории вида. Нередко они встречаются в это время на Восточном Мурмане
(Териберка) и в Енисейско-Тазовской лесотундре, обычно небольшими груп-
пами или одиночками, но в Кандалакшской губе Белого моря они скапли-
ваются на линьку тысячными стаями (Благосклонов, 1951). Основной тер-
риторией, на которой наблюдаются массовые скопления линяющих птиц,
служат лесостепи Западной Сибири (Бараба и Кулунда), Зауралья (Курган-
ская и Кустанайская обл.) и Южного Урала (Челябинская и меньше
Актюбинская обл.).
Зимовки. Основные места массовых зимовок гоголя в Европе: мор-
ские побережья Великобритании, Северной Франции, Голландии, Германии
и Дании. В значительно меньшем числе они проводят зиму на материковых
водоемах этих стран. В Балтийском море на незамерзающих водах гоголи
зимуют регулярно у берегов Польши и Прибалтийских советских респуб-
лик, но массовых скоплений не образуют. В небольшом числе они бывают
зимой на Мурмане в районе Варангер-фьорда. Массовые зимовки находятся
у обоих берегов Южного Каспия и менее многочисленные на морях Черном,
Адриатическом и Средиземном. В совсем небольшом числе гоголи достигают
зимой североафриканских берегов. В Азии гоголь в значительном количестве
зимует на Иссык-Куле, в Иране; совсем редко в Индии, где регулярно посещает
только долину Инда. В небольшом числе он бывает зимой в Белуджистане»
Кашгарии, в Северной и Западной Монголии. Зимует в истоках Ангары на
Оноие, у Командорских о-вов, берегов Камчатки и Южного Приморья. Южнее
гоголи зимуют в большом количестве у побережья Кореи, Японских о-вов
и в Китае на юг до Тайвана как на море, так и на внутренних водах. Кроме
того, небольшие группы гоголей или одиночные птицы проводят зиму на
незамерзающих водоемах в разных пунктах Западной Европы, на Волыни,
около Киева, на Южном Урале, Алтае и еще в ряде мест. Но эти зимние
встречи, часто отмечающиеся в литературе, должны быть отнесены к разряду
случайных зимовок,
586 ОТРЯД ГУСЕОБРАЗНЫЕ
Зимовки гоголя на Американском континенте располагаются на Алеут-
ской гряде, вдоль Тихоокеанского побережья от 60° с. ш. на севере до Ка-
лифорнии на юге, и у берегов Атлантического океана от Ньюфаундленда
до Мексиканского залива включительно. Кроме того, гоголи проводят зиму
в области Великих озер и по нижнему течению Миссисипи.
Характер пребывания. Гнездящаяся, перелетная и в зна-
чительном числе зимующая в СССР птица.
Сезонные миграции. О направлениях сезонных миграций
гоголя известно следующее. Линять на Зауральские озера (Курганская обл.),
как показали данные кольцевания, собираются птицы с обширной тер-
ритории Западной Сибири, из бассейнов Иртыша, Оби, частью с Енисея и, ве-
роятно, с Печоры. Отлет их на зимовку из районов линьки проходит по
нескольким основным направлениям. Значительная часть птиц летит сперва
на запад, а затем, обогнув или пересекши Урал, поворачивает на северо-
запад через Коми АССР к Белому морю и Мурману. Можно предполагать,
что, следуя морским побережьем, эти птицы достигают берегов Западной
Европы. Другая, не менее значительная, часть птиц направляется на юго-
запад к среднему течению р. Урала и далее на каспийские зимовки.
Некоторое количество птиц этого потока продвигается далее к западу и по
всей вероятности попадает на Черное море. Кроме того, намечается еще
одно, южное, направление отлета через Кустанайскую и Акмолинскую обл.,
вероятно к восточному берегу Каспия (Михеев, 1949). Один из птенцов, околь-
цованных в Печорском заповеднике, был убит на осеннем пролете близ Сык-
тывкара, т. е. на пути в северо-западном пролетном направлении, а другой
встречен в сентябре через пять лет после кольцевания в Курганской обл.,
еде он вероятно был на линьке. Наконец, гоголь с кольцом, надетым в конце
июля в Финляндии у Хельсинки, добыт через два года зимой на о-ве Джарыл-
гач в Черном море (Шарлемань, 1933).
Даты. Отлет гоголей с южных зимовок начинается в феврале. Из пре-
делов Белуджистана они полностью исчезают к началу марта (Тайсхерст,
1927), так же как и с берегов Адриатического моря (Рейзер и Фюрер, 1896).
Из района Ленкорани основная масса птиц этого вида отлетает в конце
февраля. Первыми, по наблюдениям Радде A885), исчезают самки и молодые
птицы, а взрослые селезни еще продолжают встречаться и в марте. Значительно
дольше задерживаются гоголи у каменистых берегов Дагестана и Северного
Азербайджана, а на восточном берегу Каспийского моря, у пролива Кара-
Богаз-Гол, они были еще довольно многочисленны в первых числах мая
{Исаков). Много гоголей держалось в море между Баку и Красноводском еще
18-апреля 1892 г., а несколько старых селезней замечено в конце мая 1889 г.
около форта Шевченко (Зарудный, 1896). Небольшая, видимо запоздалая,
пролетная стайка отмечена на Мангышлаке 3 мая (Долгушин, 1948). Валовой
пролет под Чкаловом проходит с начала апреля до конца второй декады
этого месяца. На Эмбе 9 апреля 1894 г. пролет был в разгаре, он закончился
к 18 мая. В низовьях Иргиза первые стаи отмечены только 25 апреля 1898 г.
(поздняя весна), а последние 9 мая. По количеству гоголи преобладали там
«а весеннем пролете над всеми другими видами нырковых уток. На пресных
озерах Узбоя первые стайки гоголей появляются во второй половине фев-
раля, а на Аральском море, в заливе Сары-Чеганак валовой пролет прихо-
дится на середину апреля (Бостанжогло, 1911). На Нижней Сыр-Дарье они
летят в конце марта и в начале апреля (Спангенберг, 1936). В Кустанайской
обл. на озерах Наурзумского заповедника массовый пролет отмечен 22—
27 апреля, после чего до конца мая на больших озерах наблюдались крупные,
иногда сотенные стаи гоголей. Позднее гоголи сократились в числе, но неболь-
шие стайки негнездящихся птиц встречались в течение всего лета (Михеев,
1938).
ГОГОЛЬ ОБЫКНОВЕННЫЙ 587
В Советской Прибалтике гоголи появляются в конце марта, пролет их
s Эстонии продолжается с марта до середины мая. Держатся они в это время
«арами или группами по 6—10 шт., преимущественно на море и на больших
озерах (Кох, 1911). Примерно в то же время появляются гоголи и под Ленин-
градом (Бианки, 1907). Под Киевом первые гоголи появляются еще до вскры-
тия Днепра, около 14 марта B—22 марта), а валовой пролет, который про-
ходит там интенсивно, приходится на30марта—1 апреля (Данилович, 1930).
В Московской обл. пролет продолжается в течение всего апреля. На Волге,
около устья Мологи, первые гоголи появились 19 апреля, до 21 апреля шел
их валовой пролет, а отдельные стаи по 20—30 шт. продолжали подлетать
до 28 апреля. В среднем течении Мологи пролетные стаи держались до 5 мая.
В северной части Рыбинского водохранилища первые гоголи появляются
в разные годы 11—21 апреля, а через 3—4 дня начинается массовый пролет.
Средняя дата прилета под Архангельском 22 апреля A3 апреля—4 мая),
а на озере Имандра (Лапландский заповедник) 3 мая B3 апреля—6 мая).
Реки и озера в это время находятся еще подо льдом, и гоголи держатся парами
на порожистых, незамерзающих участках рек. Со вскрытием тихих рек гоголи
исчезают с порогов (Владимирская, 1948). В верхнем течении Печоры (устье
р. Ылыча) гоголи появляются около 17 апреля A2—24 апреля). Массовый
пролет начинается 20 апреля A7—25 апреля). Интенсивность его в разные
годы колеблется весьма значительно (Теплов, 1948). Под Тобольском появле-
ние первых гоголей приходится на 15 апреля (9—20 апреля), а валовой про-
лет их на последние числа этого месяца и начало мая (Тарунин, 1929).
Пролет гоголей с китайских зимовок заканчивается в Чжили в апреле.
В конце марта первые гоголи появляются уже в Советском Приморье. На озере
Ханка они обычны в течение всего апреля. Камчатки же они достигают не
раньше 5 мая, последние же пролетные птицы встречаются там до 23 мая
(Аверин, 1948). Даты первых встреч на Анадыре: 10 мая 1901 г., 17 мая 1900 г.,
6 мая 1932 г. На р. Лене под Якутском появляется в первой трети мая
(Воробьева, 1931).
Осенний пролет почти повсюду проходит менее заметно, чем весенний.
На Камчатке он проходит во второй половине сентября, самое же позднее
нахождение одиночной птицы 3 октября. Под Салехардом сильный пролет
в начале сентября (Дерюгин, 1898). На Печоре (устье Ылыча) пролет начи-
нается появлением взрослых самцов, полностью отсутствующих в этих местах
во время линьки. Это происходит около 15 августа. В сентябре и октябре
пролет идет вяло. Стаи состоят не более чем из 10 и лишь очень в редких
случаях из 20—30 птиц. Последние птицы задерживаются обычно до 20 октяб-
ря G октября—1 ноября), но изредка одиночные птицы держатся на незамер-
зающих полыньях в ноябре и декабре (Теплов, 1948). С Кольского п-ова
гоголи (самки и молодые) отлетают в среднем 13 октября E—30 октября),
а из-под Архангельска 25 сентября—15 октября. На Рыбинском водохра-
нилище интенсивный пролет старых селезней, линявших где-то на севере,
бывает между 20 сентября и 5 октября. В эти дни пролетает много ходовых
стаек гоголей по 30, 40 и до 50 птиц в каждой. После этого пролет затихает,
и в течение 1,5 месяца гоголи летят редко, и совсем мелкими стайками от 2 до 5
и редко до 10 птиц. Это молодые вывода текущего года и взрослые самки, отле-
тающие, повидимому, выводками. Наконец, за несколько дней перед ледоста-
вом (около 20 ноября) пролет вновь оживает, причем в это время опять появля-
ются селезни в брачном пере (Немцев). Пролет под Киевом начинается около
8 сентября, в Запорожье он проходит с конца сентября до середины ноября,
а в устье Дона с октября до морозов, в октябре гоголи уже достигают мест
зимовки на Азовском море (Обиточенская коса), а на ноябрь падает разгар
их прилета туда (Кистяковский, 1926). В декабре птицы, задержавшиеся
в низовьях Днепра, сбиваются в косяки и отлетают оттуда (Браунер, 1894).
588 ОТРЯД ГУСЕОБРАЗНЫЕ
В верховьях Иргиза одиночные птицы были отмечены с 24 августа, весь сен-
тябрь пролет шел очень тихо, а с 9 октября после первых морозов начале**
валовой пролет, который шел без ослабления до конца октября (Суцжин.
1908). Под Бугурусланом пролет идет с сентября, а в Башкирии с начала
октября до замерзания водоемов. На зимовках в районе Гасан-Кули (юго-
восточный берег Каспия) первые самки и молодые птицы появляются в конце
октября, взрослые же самцы появляются в значительном количестве только
во второй декаде декабря. У берегов Азербайджана массовое появление гого-
лей приходится на первую половину декабря. Белуджистана гоголи дости-
гают к декабрю, а на местах зимовок в Китае (Чжили) прилет гоголей начи-
нается в октябре и заканчивается к декабрю. Летят перелетные стаи преиму-
щественно по ночам и на зорях, а день отдыхают на плесах озер.
Биотоп. По прилете, если реки еще не вскрылись, гоголей можно
видеть на полыньях и лужах надледной воды. Обычно во время пролета они
держатся на открытых плесах рек, озер и их разливах, но избегают времен-
ных луж на лугах. В гнездовое время для гоголя наиболее характерны тихие
таежные реки и озера поймы с лесистыми берегами. На зимовках основная
масса этих нырков проводит время на море, по большей части у входов в за-
ливы или у каменистых берегов.
Численность. Не во всех частях своего обширного гнездового
ареала гоголь одинаково характерная и многочисленная птица. Он является
преобладающим видом в таежной полосе европейской части Союза и Запад-
ной Сибири (Кольский п-ов, Архангельский север, Печора, Средний Урал,
Обь, Енисей). Повсюду южнее и восточнее этой зоны численность гоголей
н$ Европейско-Азиатском континенте заметно меньше.
Экология. Размножение. Половой зрелости гоголи достигают
в двухлетнем возрасте. Первый год своей жизни птицы живут небольшими
стайками и группами как на местах гнездовья, так и за пределами гнездо*
вого ареала (Наурзумские озера, озера Зауралья, Барабинская степь и др.).
Весь сезон размножения гоголи держатся парами, которые создаются ранней,
весной перед отлетом с зимовок или же в самом начале весеннего пролета.
По наблюдениям на Рыбинском водохранилище, основная масса гоголей появ-
ляется уже парами. Только самые первые самцы-одиночки, присоединившиеся
к стаям крякв, прилетают без самок, но первые же прилетные стайки этих
нырков состоят из птиц, уже разбившихся на пары. То же самое наблюдалось
и на р. Мологе, где, однако, наряду со взрослыми старыми птицами
появлялись весной неполовозрелые молодые, стайки которых задерживались
в пойме до 20 мая. На пролете через Эстонию гоголи держатся преимуществен*
но парами, а в Московской обл. наряду с парными птицами, хотя и изредка,
отмечались табунки из одних селезней (Поляков, 1910). Парами же появля-
ются гоголи и в Лапландском заповеднике. Несколько иную картину рисует
Сушкин A908) по своим наблюдениям на Эмбе. В первых пролетных стаях
там резко преобладали самцы, и очень многие стаи состояли из одних селезней.
К концу пролета количество самок возросло, и последние 15 дней летели
только самки. Такое же разновременное появление самцов и самок отме-
чается для Печорского заповедника. Селезни прилетают там на 3—4 дна
раньше самок, и разбивка на пары якобы происходит по прилете последних.
Токовать гоголи начинают рано, в первые же солнечные весенние дни.
Обычно это бывает в самых первых числах марта, когда все селезни пол-
ностью заканчивают линьку. Токование гоголя выражается в том, что селе-
зень распускает хвост и откидывает голову с вытянутым клювом на спину,
а затем резким движением выбрасывает ее вперед и вверх, толкая одновре-
менно ногами весь корпус вперед и поднимая ими фонтан брызг. При этом
он издает ээонкий, дребезжащий звук, который достигается сильным растя-
жением трахеи р момент закладывания шеи на спину. Оперение головы в это
ГОГОЛЬ ОБЫКНОВЕННЫЙ
589
время распушается, отчего особенно резко выступает контраст между иссиня-
черным фоном ее и яркобелыми пятнами на щеках. После этого селезень
обычно плавает вокруг самки с вытянутой шеей. Пары у гоголей очень
прочны, попытки преследования чужих самок очень редки, так же как
й драки между селезнями. Гнезда гоголи устраивают в дуплах деревьев
у берегов водоема. В долинах больших рек они заселяют обычно лесистые
гривы, расположенные у пойменных озер, а на таежных реках, у которых
лес подходит к самой воде, гнездятся у речного плеса. На севере гнезда чаще
всего бывают в дуплах осин и много реже в елях и осинах. В средней лесной
полосе Также в осинах, но, кроме того, и в дубах (р. Молога), а еще южнее
Pgc. 107. Позы тока гоголя
в осокорях (Воронежская обл.), каштанах, дубах, липах и буках (Южная
Германия). Охотно заселяют они дуплянки и гнездовые ящики, повешей-
ные на деревьях или установленные на шестах. При этом гоголи ийбгДа
поселяются почти в самых поселках, гнездятся во дворах (Кулунда) или
яа деревьях, окаймляющих автострады (Саксония). Гнездовые дупла в оси-
нах и соснах помещаются обычно высоко, в дубах же значительно ниже.
Как правило, гоголи не гнездятся нигде, кроме дупел, но, будучи вынуждены
к тому, они устраивают гнезда и в несовсем обычных условиях. Так, в 1940 г.,
когда в пойме Мологи проходила сплошная выруока леса для расчистки
ложа будущего водохранилища, из 53 осмотренных гоголиных гнезд 32 по-
мещались в обычных дуплах, 5 в прикорневых, т. е. на земле, 8 в прогнив-
ших с торца и лежащих на земле бревнах, 3 в полудуплах пней и сломанных
стволов, 2 в штабелях бревен и 3 в стогах сена.
Подстилкой для гнезда служит имеющаяся в дупле труха, в которой
самка делает аккуратную ямку. После откладки первых же яиц в гнезде
появляется белый пух, количество которого к моменту насиживания бывает
велико—около 60 г. Некоторые утиные дупла, заселяются гоголями до
30 лет подряд. Путем кольцевания установлено, что одни и те же самки по-
селяются несколько лет подряд в одних и тех же дуплах (Теплов, 1948).
590 ОТРЯД ГУСЕОБРАЗНЫЕ
Нередко количество дупел бывает недостаточным и между гоголихами про-
исходят драки из-за обладания ими. Зачастую в одно дупло несутся две самки
гоголя, или гоголь и луток, или, реже, гоголь и кряква, или большой крохаль.
Мы нашли гнездо с 30 яйцами, располагавшимися в три слоя, из которых наси-
живающая самка прогревала только верхний, в результате чего 18 яиц оста-
лись не выведшимися. Диаметр гнезда обычно не меньше 25—30 см, а расстоя-
ние его от входного отверстия нередко более 1 м (до220см). На р. Мологе пер-
вое гнездо с полной кладкой яиц было найдено 21 мая, причем яйца уже были
насижены. Первые яйца на Рыбинском водохранилище появляются около-
23 апреля A5 апреля—7 мая), полные кладки 7 мая A—20 мая), время же
массового появления полных кладок падает на середину мая. На Кольском
п-ове (Лапландский заповедник) наиболее ранняя полная кладка найдена
15 мая, но бывают там и очень запоздалые кладки. На р. Вилюе гнездо с яйца-
ми было найдено 2 июня (Маак, 1861). В Тункинской долине (Бурято-Мон-
гольская АССР) гнездо в высокой лиственнице найдено 9 мая, но спаривание
гоголей продолжалось до 19 мая (Слудский). В Саксонии, под Дрезденом,
первые яйца у гоголей появляются в начале апреля, а заканчивается боль-
шинство кладок в конце апреля или в начале мая (Бернхардт, 1940).
Среднее число яиц в полной кладке различно в разных географических точ-
ках, а возможно, что оно изменяется и год от года. На Печоре обычный размер
кладки 6 яиц (от 5 до 7), на Кольском п-ове 7 D—12), а найденные там же клад-
ки из 14—18 яиц, повидимому, были отложены двумя самками. В одном из-
таких гнезд ежедневно прибавлялось по 2 яйца. Наиболее обычное число-
яиц по наблюдениям в пойме р. Мологи равнялось 12. На Рыбинском водо-
хранилище количество яиц в насиживаемых кладках колеблется в пределах
от 5 до 24. В 1949 г. средний размер полной кладки был 7 яиц D—12). В 1950
и 1951 гг. картина оказалась много сложнее: 23,3% гнезд C9 из 168) содержали
8—10 яиц, в среднем 9, а 31,5% E3 гнезда) от 12 до 14, в среднем 13. Можно
предположить, что наличие двух различных показателей обычного размера
кладки обязано наличию двух возрастных групп самок: размножающихся
впервые и более взрослых. Более мелкие кладки, принадлежащие, повиди-
мому, молодым, появляются позже. Наблюдения за откладкой яиц в отдель-
ных гнездах показали, что считать кладки из 12—14 яиц сдвоенными нет
оснований. Обычный размер выводков в Тункинской долине 12 птенцов
(Слудский). В Саксонии число яиц в кладке доходит до 10 (Бернхардт, 1940),
а в качестве среднего размера кладки для Германии в целом указывается»
12 яиц (Хейнрот, 1928). Яйца крупные по сравнению с размером самки, скор-
лупа крепкая, глянцевитая, голубовато-зеленого цвета. Тон и интенсивность
окраски варьируют, но они постоянны для всех яиц каждой кладки. Вес яиц
на Верхней Волге (Рыбинское водохранилище) от 61 до 77 г, среднее 68 г,
размеры 53—68x40—47 мм, среднее 58,4x42,5 мм.
Вес яиц для Лапландского заповедника 52—67 г, среднее 59 г, для Гер-
мании 55—65 г, среднее 60 г (Хейнрот, 1928). Форма яиц довольно постоянна,
но наряду с яйцами обычной для уток формы попадаются и очень вытянутые
F8,0x41,2 мм) и овально-круглые E9,5x46,5 мм). Обычно каждый день
откладывается по одному яйцу, но нередко в конце кладки бывают и про-
пуски D яйца за 6 дней, 3 за 5). Насиживать самка начинает с момента окон-
чания кладки, но первое время она по утрам и вечерам надолго покидает
гнездо, находясь в нем лишь днем и ночью. Благодаря большому коли-
честву пуха, которым укрываются яйца, они остаются теплыми даже после
нескольких часов отсутствия самки. В солнечные дни, когда в дупле стано-
вится жарко, самка отсутствует там весь день. Но последние 10 дней перед
выводом она при любых условиях сидит очень плотно. В это время самку
можно поймать руками, а посаженная обратно в гнездо, она даже не всегда
вылетает из него. Срок насиживания 30 дней. Весь период откладки яиц
ГОГОЛЬ ОБЫКНОВЕННЫЙ 591
и первую половину периода насиживания селезень держится вблизи от гнез-
да. По самцам можно отыскивать занятые утками дупла. При приближении
опасности селезень шумно и со звоном крыльев поднимается с воды, что слу-
жит сигналом для утки, которая покидает дупло и с тревожным карканьем
опускается где-либо поблизости на воду.
Птенцы выводятся дружно на протяжении всего 2—3 часов, но после
этого они почти целые сутки остаются в гнезде, обсыхая под матерью и сма-
зывая жиром свой пуховой наряд под ее оперением. Вес птенцов в день вы-
вода 33,5—40 г, среднее 36,5 г. Их очень острые и сильно загнутые коготки
и жесткий хвост, служащий опорой при лазании, позволяют им без труда
выбираться из дупел любой глубины и даже из дощатых гнездовых ящиков.
Выход из гнезда совершается так. Самка, находящаяся под гнездом на суше
или на воде, подзывает молодых своим хриплым карканьем. Птенцы на голос
матери вылезают из дупла и прыгают вниз. Благодаря их очень малому весу
падение с большой высоты (до 10 л* и более) обходится для них благополучно.
Достигнув земли, гоголенок сразу же способен бежать. Когда весь выводок
оказывается внизу, утка собирает его около себя и отводит в места, имеющие
хорошие укрытия от хищников. Даже самые маленькие гоголята прекрасно
ныряют и могут находится под водой до 1,5—2 мин. Контурное оперение раз-
вивается полностью в возрасте 35 дней. К этому времени совершенно исче-
зают белые пятна по бокам головы, но ноги еще остаются темными. Способ-
ность к полету птенцы приобретают примерно к 57 дням, хотя маховые к это-
му времени еще отрастают неполностью. На Печоре молодняк поднимается на
крыло через 60—70 дней после появления первого выводка. Первые выводки
в Лапландском заповеднике отмечались 15 июня, под Архангельском в сред-
нем 11 июня B—28 июня), на Печоре (устье Ылыча) также 11 июня A—16 ию-
ня), а летные выводки появляются там около 14 августа. На Северном Урале
у границы леса недавно выведшиеся молодые встречены 7 июля (Портенко,
1937), на Елогуе (Туруханский край) первые птенцы были встречены 6 июля,
к 14 июля их было уже много, но до 24 августа еще встречались и не вполне
оперившиеся (Слудский). На Мологе первые пуховички были замечены
12 июня, а на Рыбинском водохранилище они появляются 9—12 июня, подъем
же на крыло приходится на 20 июля. Самки водят молодых не более
двух-трех недель, после чего сами объединяются в стаи.
Кладки гоголя находятся в гораздо большей безопасности, чем кладки
уток, гнездящихся открыто. Известны случаи, когда гнезда разоряла куница
(Робин, 1923), но это, конечно, редкость. Наблюдалось, что иногда сама
самка гоголя выносит из гнезда в клюве яйца и затем расклевывает их (Нем-
цев). Однако в то время, как разоренные кладки довольно редки, брошенные
в некоторые годы составляют огромный процент, особенно в гнездовых ящиках
и дуплянках. Основной причиной бросания гнезд служит замокание их от до-
ждей. Птица не в состоянии прогреть мокрую подстилку гнезда, которая
не просыхает в закрытом дупле очень долго. В 1951 г., очень дождливом,
на Рыбинском водохранилище было брошено около 50% всех гнезд, причем
яйца в них находились в самых различных стадиях насиживания вплоть
до уже крупных птенцов. Гибель птенцов в раннем возрасте сравнительно
невелика. Среднее число птенцов в выводке на Печоре 5 (яиц в кладке 6),
на Мологе средний выводок пуховых птенцов 9—10 (яиц 12), в августе до на-
чала охоты 8—9, в сентябре 6, в октябре-ноябре перед отлетом 4.
Л и н ь к а. К тому времени, когда самки уже крепко садятся на яйца»
т. е. в середине периода насиживания, селезни объединяются в группы и отле-
тают к местам линьки, которые находятся зачастую очень далеко от мест
гнездовья. С Печоры селезни отлетают 19 июня A6—22 июня), с озера Иманд-
ра (Кольский п-ов) в конце июня—начале июля. На Рыбинском водохранили-
ще в конце мая они собираются в небольшие стайки и около 1 июня исчезают.
э92
ОТРЯД ГУСЕОБРАЗНЫЕ
На озерах Северной Эстонии к 11 июня селезни уже отсутствуют (Кумари,
1941). В Саксонии последний самец отмечен 7 июня.
Сперва линяет контурное оперение, а в последней декаде июля сменяются
перья крыла и рулевые. Как и у других уток, послебрачная линька бывает
полной. Взрослые самки линяют в августе C0 июля немного пеньков на
спине). Летнее перо селезни носят довольно долго, но в сентябре старые птицы
начинают вторую, неполную, линьку. В конце сентября и начале октября на
Рыбинском водохранилище появляются старые селезни гоголя из числа прово-
Рис.
ad
108. Схема окраски крыла гоголя: а—молодой самец, б—самец в возрасте
одного года, в—взрослый самец, г—молодая самка, д—взрослая самка
давших летнюю линьку где-то на севере, многие из которых имеют уже почти
полный зимний наряд. Молодые полуторагодовалые селезни, надевающие свой
первый брачный наряд, линяют позднее. В конце ноября они находятся в со-
стоянии интенсивной линьки, и очень немногие из них имеют к этому времени
белые гуски. Заканчивается у них эта линька на зимовках в декабре-январе,
а у некоторых затягивается и до февраля. Вторичная линька самок проходит
зимой. Молодняк надевает первый зимний наряд в октябре-ноябре. В апреле
появляются перья первого брачного наряда, при этом заменяется лишь незна-
чительная часть оперения, втом числе и рули. Полную линьку молодые птицы
проходят вместе со старыми самцами в июле. Время линьки селезни проводят
на больших открытых озерах степной зоны как пресных, так и соленых.
На некоторых, каждый год одних и тех же, озерах они собираются многосо-
тенными стаями.
Питание. Гоголи добывают свой корм на дне водоема, ныряя для этой
цели на глубину до 4 м, но обычно менее глубоко. Обследуя значительные пло-
щадки дна, они собирают свой корм с поверхности его (моллюски), или скле-
вывают с затопленных кустов и бревен (личинки ручейников), или, совсем
ГОГОЛЬ ОБЫКНОВЕННЫЙ
593
изредка, охотятся в толще воды (рыба). По составу пищи гоголь животноядная
утка. Растительные же корма в его рационе в большинстве случаев следует
рассматривать, как вынужденные или как случайную примесь. Ранней вес-
ной на р. Мологе 60% рациона по объему составляла мелкая рыба, которую
гоголи добывали на полыньях и в лужах надледной воды, а 20% приходи-
лось на долю личинок хирономид. На Рыбинском водохранилище в апреле
основу питания составляют два вида корма: личинки ручейников (Phryganea
grandis и др.) 46,5% по объему и личинки хирономид 32%. Пищу пуховых птен-
цов там составляют те же самые два вида корма, а на озерах Мологской поймы
основную роль играли водяные жуки и клопы C2,5%) и личинки стрекоз
C0%), меньшее значение имели личинки хирономид, ручейников и водяные
Рис. 109. Гоголь (самец и самка)
ослики. Осенью, в сентябре—октябре, на водохранилище решающее значение
имеют те же личинки ручейников G5%), а в Мологской пойме личинки стре-
коз D5%). Кроме того, известное значение приобретают некоторые расти-
тельные корма: луковицы стрелолиста B0%) и клубеньки гребенчатого рде-
ста A3%). На Верхней Печоре наиболее обычными кормами гоголя служат
личинки ручейников, наземные насекомые, преимущественно жуки, в мень-
шей степени рыба (гольян), а также листья рдестов (Теплов, 1948). В же-
лудке гоголей, добытых в июне на Анадыре, обнаружены личинки ручейни-
ков, остатки мелкой рыбы и жук-водолюб (Портенко, 1939). На южно-кас-
пийских зимовках гоголи, живущие у каменистых туркменских берегов,
кормятся преимущественно ракообразными {Gammaridae, 46%, Potamobius,
45%). В Закавказье основу зимней пищи составляют моллюски (80—100%),
в Кизил-Агачском заливе это Cardium edule F2,1%), а у каменистых кос
Северного Азербайджана и Дагестана Mytilaster lineatus A00%).
Хозяйственное значение. Гоголь широко распространенный и местами
многочисленный вид утки, но промыслом на территории Союза он исполь-
зуется очень слабо. В массовых же количествах вылавливается гоголь
за рубежом, в Балтийском море, во время его сезонных миграций. Хорошую
производительность и высококачественную продукцию дает сетной лов его
на море у берегов Северного Азербайджана. Но результаты этого промысла
не превышают 3—4 тысяч гоголей за сезон. Местами проводится сбор яиц
38 Птицы СССР, т. IV
594 ОТРЯД ГУСЕОБРАЗНЫЕ
и пуха гоголя в поставленных для этой цели дуплянках (Ладожское
озеро, Лапландия, Печора, р. Демьянка—приток Иртыша, Барабинская
степь).
Полевые признаки. У сидящего на воде селезня обращают внимание
ярко белые грудь и бока, а также округлые белые пятна на крупной черной
голове. Туловище утки равномерно серое, голова черная, отграниченная
белым ошейником. На полете низ туловища белый, голова темная. Особенно
характерны для птиц обоего пола очень темный подбой крыла и белое пятно,
образованное второстепенными маховыми. При взгляде на летящую птицу
сверху бросаются в глаза белые зеркала на крыльях, короткая шея и боль-
шая черная голова. На полете селезень издает крыльями высокий, звенящий
звук, по которому гоголя можно отличить даже в темноте. Голос—хриплое
карканье.
Описание. Строение и размеры. Клюв короткий и высокий у основания,
значительно короче головы. Ноготок узкий, ноздри лежат ближе к вершине, чем к осно-
ванию клюва. Рулевых 16, крайние примерно вдвое короче средних. У самцов перья
затылка и темени удлинены, но не образуют ни хохла, ни гребня. У взрослого селезня
самые внешние первостепенные маховые очень узки; вибрируя во время полета, они
издают звенящий звук. Соотг-етствующие перья у самок и селезней в возрасте до года
значительно шире и на полете не звучат. Лобная часть черепа вздута. Крыло у селезней
210—231, у самок 187—215 мм; цевка самцов 35—41, самок 33—36 мм, клюв самцоа
32—41, самок 28—35 мм. Размеры американских птиц несколько крупнее: крыло селез-
ней 223—240, самок 200—215 мм (обыкновенно длиннее 205 мм). У пухового птенца
клюв довольно узкий, коготки на лапах очень острые, сильно изогнутые, ювенальные
перья на хвосте с очень жесткими стержнями. Общая длина 151 мм, цевка 26 мм,
клюв 11 мм.
Вес. Весенний вес селезней сразу по их прилете одинаков в удаленных друг от друга
географических пунктах. Под Архангельском в мае 986 г (950—1060 г), на Печоре 955 г
(925—985 г), в Карелии (Заонежье) 951 г (820—1110 г), на Рыбинском водохранилище
в апреле 956 г (900—1150 г). За время брачного периода и линьки вес селезней снижается.
Средний вес линяющих на озерах Зауралья в июле—августе 824 г, в сентябре на Рыбин-
ском водохранилище 880 г, а в октябре 912 г (850—1010 г). На зимовках (Южный Каспий)
основная часть гоголей, живущая в море у каменных кос, сильно жиреет, и вес селезней
доходит до 1400г. Птицы, живущие на мелководьях у самых берегов и на озерах, тощают,
вес их 739 г E00—875 г). Весной они задерживаются на юге почти на два месяца после
отлета основной массы гоголей.
Весенний вес самок на Печоре в апреле—мае 635 г F05—670 г), под Архангельском
в мае 708 г F45—810 г,) в Заонежье в мае 806 г G65—882 г). За время откладки яиц
и насиживания утка очень теряет в весе; так, птица с гнезда за день до вывода птенцов
(июнь) весила 400 г, а утка с маленькими птенцами (июль)—600 г. Вес их в октябре
на пролете—731 г F50—810 г), а в феврале на море в крупных стаях 885 г (850—900 г),
в то же время у берегов 541 г D50—685 г). Молодые птицы достигают веса взрослых уже
в сентябре.
Окраска. Пуховой птенец. На спинной стороне очень темный, почти черный.
На голове темная окраска спускается до основания клюва, захватывает глаз и большую
часть щек. На темном фоне выделяются два пятна белого пуха на плечевых суставах
крыла и два на крестце. На передней части груди довольно широкий бурый ошейник,
вся же остальная брюшная поверхность белая. Белый пух имеет бурые основания. Клюв
темный оливково-бурый, лапы оливковые.
Молодые птицы в первом наряде. Голова и вся шея коричневато-бурые, на зобу,
груди и боках окраска более светлая и серая, остальная брюшная сторона белая.
Только верхний ряд верхних кроющих крыла белый, остальные жетемнобурые, из кото-
рых лишь часть с неясными белыми вершинными пятнами. У самцов даже в этом воз-
расте белого несколько больше. Глаза темные.
Молодой самец после первой осенней линьки. Похож на взрослую самку, но
спинная сторона тела бурее и одноцветнее, так как перья не имеют светлых каемок. Свет-
лый ошейник менее четкий. Основным отличительным признаком остаются крылья.
Весной на некоторых частях тела у них появляется оперение взрослых птиц. Появ-
ляется чистобелое пятнышко на щеках с примесью лишь отдельных бурых перьев,
основание шеи становится чистобелым, но серая перевязь поперек зоба сохраняется.
На голове отрастают отдельные черные перышки с зеленоватым отливом. На мантии
часть перьев сменяется черными, а плечевые—белыми с черными опахалами. Глаза светло-
желтые, ноги шафраново-желтые.
Взрослая самка. Голова и часть шеи темнокоричневые. Передняя часть спины
темноаспидная со светлыми каймами перьев, остальная же спина и надхвостье черно-
ИСЛАНДСКИЙ ГОГОЛЬ
595
дымчатые. Зоб и бока аспидно-серые, грудь, брюхо и подхвостье белые. Зеркало крыла
создается белыми второстепенными маховыми и лежащими над ними белыми кроющими
с черной вершиной. Выше них два ряда перьев образуют чистобелое (у вполне взрослой
самки) пятно, отделенное рядом черных перьев. Еще выше весь верх крыла темноаспид-
ный. Первостепенные маховые и их кроющие буро-черные. На исподе крыла мелкие крою-
щие темноаспидные, а остальные перья стального цвета. Рулевые буро-черные. Клюв
черный, включая ноготок, но у многих старых самок близ вершины его бывает оранжево-
желтая перевязь. Лапы желтые с черноватыми перепонками. Радужина желтая.
Самец в летнем наряде. Походит на взрослую самку и отличается только по-
крыльям.
Самец после второй осенней линьки. Получает перо взрослой птицы, но белые
пятна на крыльях, образуемые верхними кроющими, меньше и менее чистые за счет
бурых пятен на перьях, ноги оранжево-желтые, радужина желтая.
Взрослый самец в брачном наряде. Голова черная с металлическим зеленым
отливом. У основания клюва по бокам головы округлые белые пятна. Вся спина,
бока и надхвостье, шея, грудь, брюхо и подхвостье чистобелые, кроме узкой черной
перемычки в области крестца. Белые второстепенные маховые образуют зеркало,
выше его идет около 5 рядов белых перьев, образующих чистобелое пятно, которое на
сложенном крыле смыкается с зеркалом. Остальное крыло сверху черно-бурое. Снизу
мелкие кроющие черно-бурые, а остальные перья стального цвета. Рулевые буро-черные.
Клюв черный, лапы оранжевые, радужина красновато-желтая. У некоторых (быть
может старых) ноги оранжево-красные, глаза почти огненно-красные.
51. Исландский гоголь Bucephala islandica Gm.
Anas isiandica. Gmelin, Syst. Nat., 1789, стр. 541, Исландия.
Распространение. Гнездовой ареал. Занимает всю Исландию,
юго-западный берег Гренландии до 68,5° с. ш., а в Америке северный
Карта 72. Распространение исландского гоголя Bucephala islandica
а—граница гнездовий, г—зимовки, в—залеты
конец Лабрадора (восточный берег залива Унгава), область Скалистых гор
и Тихоокеанское побережье от Ванкувера до залива Кука на Аляске.
Основные зимовки. Исландские и гренландские птицы зи-
муют на месте, передвигаясь лишь в южные части островов. Лабрадор-
ские гоголи проводят зиму на морском побережье от залива Св. Лаврен-
тия до Нью-Йорка, тихоокеанские—от архипелага Александра до Сан-
Франциско, заходя в глубь материка на плато Колорадо и в южную часть
Скалистых гор.
Залеты. Исландский гоголь нередко залетает весной и осенью на ма-
терик Европы в пункты, близкие от наших границ, дважды зарегистрирован
он и на нашей территории. В 10-х годах текущего столетия один экземпляр
38*
596 ОТРЯД ГУСЕОБРАЗНЫЕ
был добыт во время пролета (какого?) на озере Ильмень (Бутурлин, 1935).
Взрослый селезень добыт в мае 1935 г. на Шуньгском п-ове Онежского
озера из стайки обыкновенных гоголей (Исаков). Неоднократно залетал
в Северную Норвегию (Остфинмаркен) в районе Варангер-фьорда (Леппен-
тин, 1911), весьма возможны его залеты в район Печенги и на Айно-
вы о-ва.
Характер пребывания. У нас случайно залетная птица.
Даты. Гоголи, проводившие зиму в больших стаях на море далеко
от берегов, в марте возвращаются к берегам Исландии, а в конце месяца
и начале апреля появляются на местах гнездовья во внутренней части
острова. В октябре—ноябре они объединяются в стаи на море и удаляются
от острова.
Б и о т о п. На большей части ареала безлесные, тундровые территории.
Летом птицы держатся на внутренних водоемах, больших и малых озерах.
Осенью собираются в узких морских заливах, а зимой откочевывают в море
и лишь малая часть задерживается у теплых источников.
Численность. Случайный залет отдельных особей.
Экология. Размножение. На местах гнездованья появляются
в большинстве случаев уже в парах. Гнездятся около внутренних озер, иногда
далеко от моря. Гнездо устраивают на берегу вблизи от воды или на совсем
маленьких низких островках. Оно бывает скрыто под мелкими кустиками
карликовых ив и березок, под нависшими камнями, в пещерках между
камнями, а нередко в стенах овчарен, построенных из земли и торфа, но всегда
в более или менее глубокой норке. В гнезде выстилка из травинок и моха
и много пуха. Обычная кладка содержит 12—14 яиц, по форме и цвету
очень схожих с яйцами обыкновенного гоголя. Их размер несколько круп-
нее: 56—65x42,5—46,5 мм, среднее 61,2x45,2 мм. На севере Исландии
5 июля птенцов еще не было.
Л и н ь к а. В Исландии в конце июня—начале июля самцы уже собира-
ются в стаи для прохождения линьки, но некоторые из них в это время еще
держатся с самками.
Питание. Пища гоголя на 78% состоит из животных кормов
и на 22% из растительных. Основу составляют зимой моллюски A9,1%),
главным образом мидии и литорины, и ракообразные A7,7 %), а летом насе-
комые C6,4%): личинки стрекоз, ручейников, водяные клопы и жуки. Из ра-
стительной пищи чаще других объектов встречаются рдесты и валлиснерия
(Коттам, 1939).
Полевые признаки. У сидячей птицы меньшую площадь тела, чем у обык-
новенного гоголя, занимают белые бока, хорошо заметно полулунное белое
пятно по бокам головы у основания клюва. Звук, издаваемый при поле-
те, менее сильный и не такой металлический, как у обыкновенного гоголя.
Весенний крик селезня «ге-ге-ге-ге-геер», голос самки—карканье.
Описание. Строение и размеры. Весьма сходен по строению с обыкновен-
ным гоголем, но отличается более массивным клювом, а главное широким ноготком. Ши-
рина последнего у селезней 6—8 мм, у уток 5,5 мм (у обыкновенного гоголя соответственно
5 до 5,7 и 4,5—4,7). Длина клюва селезней 35 мм. Другие размеры также несколько
крупнее: крыло селезней 235—247 мм, самок 210—225 мм; цевка самцов 47 мм.
Окраска. Самка весьма похожа на самку обыкновенного гоголя, но голова
потемнее, белый ошейник шире, серая грудь темнее, зеркало на крыле меньше. Ноги
желтые с черноватыми перепонками.
Взрослый селезень в брачном наряде. Голова и прилегающая часть шеи черные
с сине-фиолетовым отливом. По бокам головы у основания клюва белое полулунное
пятно. Спинная сторона тела черная, часть плечевых перьев с большими белыми
пятнами в средней их части. Вся брюшная сторона тела чистобелая, на боках часть
перьев с черными опахалами. Зеркало большое, белое, отделенное черной полосой
от белого пятна, образованного средними кроющими крыла. Остальное оперение крыла
сверху темное. Ноги желто-красные.
ГОГОЛЬ-ГОЛОВАСТИК
597
52* Гоголь-головастик Bucephala albeola L.
Anas albeola. Li n n a e u s. Syst. Nat., изд. X, 1758, стр. 124, Ньюфаундленд.
Русские названия. Книжные—го голь-головастик, американский гоголь.
Распространение. Гнездовой ареал захватывает северо-восточ-
ную и центральную часть Северной Америки. Северная граница проходит от
залива Нортон к устью р. Мекензи через Невольничье озеро к Гудзонову
заливу у впадения р. Черчилль. На юг гнездится до южной оконечности
Карта 73. Распространение гоголя-головастика Bucephala albeola
а—граница гнездовий, а—зимовки, а—валеты
Гудзонова залива и низовьев р. Колумбии. Восточной границей почти по
всей линии служит Тихоокеанское побережье, но на островах и на п-ове
Аляске он не гнездует.
Основные зимовки располагаются вдоль тихоокеанского
берега от восточной части Алеутских о-вов и Бристольского залива на севере
до Калифорнийского залива на юге, а на Атлантическом берегу от Новой
Шотландии до берегов Мексиканского залива. Кроме того, встречается зимой
и внутри страны в бассейнах Колорадо, Рио-Гранде-дель-Норте и Миссисипи.
В пределы СССР этот гоголь залетает на о-в Беринга (Командорские о-ва),
где неоднократно добывался зимой (ноябрь —январь).
Характер пребывания. Перелетная птица, в СССР-
залетная.
Сезонные миграции и их даты. В наиболее северные
части ареала гоголь-головастик прилетает в начале мая. В бухтах Аляски
держится стайками до середины марта, а на о-ве Адак наблюдался еще
11 апреля (Лайн, 1925).
Биотоп. В летнее время лесные озера, зимой и на пролетах как
взморье, так и внутренние водоемы.
Численность. В СССР залетная птица.
598 ОТРЯД ГУСЕОБРАЗНЫЕ
Экология. Размножение. Токуя, птицы выпячивают грудь, распу-
шают перья головы и резким толчком ног скользят по воде, издавая при
этом каркающие звуки. Гнездятся в дуплах деревьев поблизости от воды.
Есть указания, что на крайнем севере при отсутствии подходящих деревьев
иногда занимают брошеные норы или гнездятся в расщелинах скал. Выстилки
обычно не бывает никакой, кроме собственного пуха с примесью перьев.
Число яиц 9—12, обычно не более 10, цвета слоновой кости или зеленоватые,
средний размер 48,5x34,7 мм.
Питание этого гоголя примерно на 79% состоит из животных кор-
мов. Среди них основу составляют насекомые D0,7%): личинки ручейников,
стрекоз, водяные жуки и клопы; ракообразные 16,7%: десятиногие раки
и амфиподы, моллюски A5,7%) более всего пресноводные брюхоногие, в очень
незначительном количестве поедается рыба G,9%) и в небольшом числе
растительные корма B1%) (Коттам, 1939).
Полевые признаки. Маленькая утка величиной с чирка. У селезня обра-
щают внимание очень широкие белые бока, грудь и большое белое пятно
поперек головы. Окраска самки в общем темная, на фоне которой выделяется
белое пятно в области уха. На полете при взгляде сверху видны широкие
белые пятна на крыльях и голове, при взгляде снизу напоминает обыкновен-
ного гоголя, но мельче его, основания крыльев светлые с двумя темными про-
дольными полосами. Голос—короткое карканье, слабее, чем у обычного гоголя.
Описание. Строение и размеры. Утка размером с крупного чирка, но
более плотного сложения. «Клюв короткий, высокий при основании заметно суживается
к концу, с узким ноготком. Хвост округло клиновидный, крайние рулевые более половины
длины средних, перья темени, затылка и верхней части шеи удлинены и образуют как бы
гребень, отчего голова кажется очень большой. Крыло самцов 165—177, самок 150—
160 мм; цевка самцов 32—35, самок 27—31 мм; клюв самцов 27—30, самок 23—27 мм.
Окраска. Пуховой птенец. Очень похож на пуховика обыкновенного гоголя.
Молодые птицы в первом наряде. Имеют окраску взрослой самки, но несколько
бледнее ее, а горло сероватого тона.
Взрослая caMjta. Голова и шея черно-бурые с белыми пятнами под глазом и
в области уха. Вся спинная сторона черно-бурая. Часть зоба и бока сероватые, грудь
и брюшко белые, подхвостье темносерое. Четыре второстепенных маховых и большие
кроющие крыла образуют широкое белое зеркало.
Самец в летнем наряде. Походит на взрослую самку, но отличается от послед-
ней серым затылком и большим размером белого пятна на боках головы.
Самец в брачном наряде. Лоб, темя, горло, бока и задняя сторона шеи черные
с зеленовато-пурпурным отливом. Бока головы, начиная от глаза, занимает большое
белое пятно, которое соединяется на затылке с таким же пятном другой стороны.
Спина, плечевые и поясница черные, верхние кроющие хвоста пепельно-серые. Мантия
и весь низ тела белые, на боках перья с узкими черными каемками по наружному
опахалу. Маховые черные, но часть второстепенных с белыми наружными опахалами,
образующими зеркало. Средние кроющие крыла белые, образуют пятно, отделенное чер-
ной полоской от зеркала, остальные кроющие темные. Рули аспидно-серые. Клюв голу-
бовато-черный, лапы телесного цвета с темными перепонками; радужина бурая.
РОД КРОХАЛИ MERGUS LINNAEUS, 1758
Тип. Mergus merganser
Синонимы. Merganser В г i s s о n, 1760, тип. М. merganser,—Mergellus S e 1 b у,
1840, тип М. albellus.—Lophodytes. Reichenbach, 1852, тип. М. cucullatus.
53. Луток Mergus albellus L.
Mergus albellus. Li n n a e u s. Syst. Nat., изд. X, 1, 1758, стр. 129, Смирна.
Русские названия. Луток,—народное широко распространенное; острота
(в низовьях Днепра).
Распространение. Гнездовой ареал занимает лесную зону
европейской части РСФСР, Сибири и Дальнего Востока, но границы его
во многих местах неясны. Луток гнездится в Северной Швеции и Норвегии
ЛУТОК
599
на север до Остфинмаркена (Варангер) и в Северной Финляндии. Заселяет
лесистую часть Кольского п-ова и всю Карелию. В Архангельской обл.
и на Печоре доходит до северного предела лесов, но на гнездовье довольно
редок и в тундру лишь залетает. По Оби доходит примерно до Салехарда,
изредка встречается весной по р. Щучьей, но там не гнездится. На Енисее
прослежен до границы тундры, т. е. несколько севернее 69° с. ш.; на Таймыре
проникает в лесотундру, нередок под Якутском, гнездится на Вилюе
Карта 74. Распространение лутка Mergus albellu
а—граница гнездовий, б—не вполне выясненная граница гнездовий, в—обособленные
гнездовые колонии, в1—места постоянного летнего пребывания птиц (гнездованье не
доказано), в—зимовки, е—весенние встречи птиц севернее гнездовых мест, э#с—места
скоплений птиц на линьку, з—места гнездованья птиц в недавнем прошлом
(Воробьева, 1928), известен из лесной зоны бассейна Яны. В бассейнах Инди-
гирки и Колымы на гнездовье пока не обнаружен, но осенью появляется
в небольшом числе в дельте последней (Бутурлин, 1906). На крайнем северо-
востоке луток встречается в западной части Анадырского края, где нередок
на гнездовье в лесистых частях верхнего течения Анадыря. Встречен в гнез-
довое время в северной части Камчатки (Бианки, 1908; Бергман, 1935), но
в районе Кронок известен лишь как пролетная птица (Аверин, 1948); самка
добыта 3 мая в Гижиге (Аллен, 1905); ни под Охотском (Харитонов, 1915),
ни в верховьях Колымы (Шохин) луток не отмечен, но гнездится в районе
Аяна (Снигиревский). Гнездится на Сахалине и Шантарских о-вах, поэтому
возможно нахождение его и в нижнем течении Амура. Южная граница рас-
600 ОТРЯД ГУСЕОБРАЗНЫЕ
пространения лутка выяснена еще менее полно. Есть указание на гнездовье
его в Новгородской обл., в районе Ильменя (Бианки, 1910), но ни для Ленин-
градской, ни для Псковской обл. он не указывается. Обычен на Онежском
озере, известно гнездованье в пойме Мологи и на Рыбинском водохранилище,
но далее граница его совсем неопределенна. Для Горьковской, Костромской
и Кировской обл. прямых данных о гнездованье нет, но в северных частях
двух последних оно вполне возможно. Для Татарии ПершаковA929) считает
лутка редко гнездящейся птицей, но основанием к этому, повидимому, слу-
жат лишь летние встречи птиц в нижнем течении Камы. Совершенно изоли-
рованное гнездованье лутка обнаружено недавно в низовьях Днепра, в Ново-
воронцовском районе Николаевской обл. (Данилович, 1939). Гнездится луток
в Башкирии, доходя на юг до верховьев Сакмары и Ика, но отсутствует или
почти отсутствует в горных частях страны (Зарудный, 1897; Сушкин, 1897).
На восточном склоне Урала обычен в районе Миасского озера (Ушков, 1947),
а восточнее—под Тюменью, Тобольском и Тарой. Проникает в северные
лесистые районы Барабы, гнездится в таежных частях Томской обл. (Нарым,
р. Чулым) и по левобережью Енисея (р. Сым, Енисейск), однако под Красно-
ярском он уже отсутствует, а гнездованье его в бассейне Ангары не доказано.
Южнее этой зоны известны неоднократные летние встречи лутка на Барабе
(озеро Карачи; Рузский, 1932), в районе ленточных боров Кулунды, под
Семипалатинском (Хахлов и Селевин, 1928) и на Черном Иртыше (Селевин,
1929), причем в последнем месте обнаружены даже пуховики, повидимому,
луток в этих пунктах спорадично гнездится. Луток известен в верхнем тече-
нии Южной Тунгуски (Ткаченко, 1937), найден на гнездовье в низовьях
Верхней Ангары (Слудский), но для самого Байкала в качестве гнездящейся
птицы не отмечался. По старым и в дальнейшем не подтвержденным данным
Тачановского A893), луток гнездится в довольно большом количестве в Дау-
рии. В Якутии указывается как редко гнездящаяся птица ее центральной
части, на юге Якутии известна лишь летняя встреча на р. Учуре (Мидден-
дорф). Гнездится, повидимому, в верховьях р. Зеи (Окононская котловина;
Дорогостайский, 1915) и возможно в районе северной излучины Амура (Алба-
зин, Джалинда); из районов, лежащих восточнее, никаких сведений о лутке
не имеется. Несмотря на крайнюю неполноту наших сведений о распростра-
нении лутка в Восточной Сибири, обращают на себя внимание два факта,
несомненно связанных между собой; чрезвычайно малая общая численность
этого вида и полное отсутствие его в наиболее гористых частях страны (хребты
Колымский, Черского, Верхоянский, Джугджур). В этом отношении луток
является прямой противоположностью другим крохалям. Ареал лутка,
повидимому, сократился за историческое время, причиной чему послужило
исчезновение его гнездовых стаций—крупноствольных приречных лесов.
Об этом говорит наличие изолированных гнездовых колоний лутка (Ильмень,
низовья Днепра) и гнездованье его в недавнем прошлом в ряде других пун-
ктов: в Добрудже (Румыния; Домбровский, 1904), в северной части Туль-
ской обл. на Оке и Упе (Мензбир, 1879), в Воронежской обл., где он был отме-
чен в 1849 и 1852 гг., хотя и тогда уже был очень редок(Северцов, 1855),
и в дельте Волги (Яковлев, 1873; Хлебников, 1930).
Основные зимовки находятся у берегов и в прибрежной полосе
Балтийского и Северного морей на запад от о-ва Готланда и на юготТронд-
хейма в Норвегии. У Британских о-вов в зимнее время немногочислен, а у
западного берега Англии в Ирландии даже редок. Обычен в зимнее время
повсюду в Западной Европе, кроме Португалии и Западной Испании, причем
встречается не только на Северном и Средиземноморском побережьях, но
и на незамерзающих водоемах внутри страны. Обилие птиц на юге нахо-
дится в прямой зависимости от степени суровости зимы. Молодые птицы
и самки отлетают далее к югу и проникают далее в глубь страны,
ЛУТОК 601
чем взрослые самцы. В небольшом числе они достигают африканского бере-
га, бывая иногда даже в Нижнем Египте. Регулярно лутки задерживаются
на зиму на реках Южной Украины, а в западной части Черного моря и на
Азовском море зимуют в большом числе. Многочислен луток у даге-
станского берега Каспия, где большие стаи его скапливаются у кромки льдов.
Южнее и на восточном берегу он бывает в меньшем числе, так же как в пре-
делах Ирана и Пакистана. В Средней Азии лутки зимуют регулярно на Сыр-
Дарье (район Джулека; Спангенберг, 1936), в верховьях Аму-Дарьи и в зна-
чительном числе на Иссык-Куле. В Северной Индии луток довольно обычен
в Пенджабе, встречается в Синде и северо-западных провинциях, доходя
изредка до Бенгалии и Чота-Начпур; изредка в Ассаме (Бекер, 1929). В вос-
точной части ареала луток довольно обычный зимний гость Японских о-вов,
где встречается как на море, так и на больших озерах; в Корее он обычен
на пролетах, но зимует лишь в небольшом числе, на юге полуострова; в Китае
встречается зимой на юг до долины Яньцзе регулярно, южнее редко, на край-
нем юго-западе в Юннане не бывает (Латуш, 1934), в северной приморской
части страны и в долинах крупных рек очень обычен.
Места скоплений линных птиц до настоящего времени
почти не выяснены. Однако откочевка селезней на время линьки, свойствен-
ная большинству видов уток, повидимому, имеет место и у лутков. Стайки
линных селезней встречались на озерах севернее Якутска; нам известно место
линьки лутков на одном из лесных пойменных озер близ устья Иртыша (Репо-
лово), на котором линяло более 200 этих птиц, преимущественно самцов при
небольшом числе самок. Кроме того, лутки регулярно линяют на озерах
Барабинской степи (Янушевич, 1947). Стайки первогодних, неполовозрелых
птиц поздно продвигаются к родным местам, а иногда и не достигают их, про-
водя лето за пределами своего гнездового ареала (Голландия, Калининград-
ская обл., Сеистанские озера).
Характер пребывания. По всему ареалу правильно пере-
летная птица, если не считать небольшого количества особей, остающихся
зимовать кое-где на незамерзающих участках рек.
Сезонные миграции и их даты. Отлет лутков с зимо-
вок у Британских о-вов происходит с середины марта до середины мая. Из
внутренних частей Германии они исчезают в конце февраля (Брауншвейг),
в марте (Баден); пролет через Северную Германию начинается в конце марта
и идет весь апрель до начала мая. В Польше луток обычен на пролете с марта
до конца апреля (Доманевский), в Латвии и Эстонии бывает в апреле, но
редок. На Псковском озере весной довольно обычен, появляется в начале
апреля, а валовой пролет приходится на вторую половину месяца, причем
некоторые стаи содержат до 50 птиц (Зарудный, 1910). На Рыбинском водо-
хранилище первые птицы прилетают 11—14 апреля, а 15—17 апреля уже
начинается их основной пролет (Немцев). Прилет под Архангельском (озеро
Слободское) около 9 мая B4 апреля—23 мая), на Кольском п-ове, в районе
озера Имандра, 26 мая A2 мая—8 июня), а в среднем течении Печоры около
устья Ылыча 3 мая B9 апреля—7 мая). С зимовок в западном Пакистане лутки
отлетают в марте, но отдельные небольшие стайки их задерживаются
долго; известны случаи добычи их там 19 и 24 мая (Тайсхерст, 1927). В ниж-
нем течении Сыр-Дарьи пролет начинается в середине марта и продолжается
до конца апреля (Спангенберг и Фейгин, 1936), а на озерах Кустанайской обл.
он проходит в 20-х числах апреля (Михеев, 1937). Из Энзели (Иран) лутки
отлетают весной одними из самых последних среди водоплавающих птиц,
а в Мильской степи (Аджи-Кабульское озеро) они были многочисленны еще
17—18 марта (Сатунин, 1907). Однако значительная часть лутков проводит
зиму на Каспийском море непосредственно у кромки льдов и потому в дельте
Волги первые птицы появляются очень рано, сразу же вслед за образованием
602 ОТРЯД ГУСЕОБРАЗНЫЕ
первых полыней—11—24 февраля, а на пресных озерах Западного Узбоя
26—28 февраля. С Днепровского лимана лутки отлетают в конце марта,
а с Азовского моря (Обиточенская коса) даже в первых днях этого месяца.
В Аскания-Нова первые птицы часто прилетают уже в начале февраля, а под
Киевом в разные годы 2—25 марта. На Урале под Чкаловом появляются
в последних числах марта, а валовой пролет наблюдается в середине этого
месяца и его второй половине; на Эмбе пролет наблюдался с 17 по 22 апреля,
причем наибольшее количество птиц пролетело в последний день; в год с позд-
ней весной в низовьях Иргиза лутки летели с 27 апреля до половины мая
(Сушкин, 1908). Пролет под Тобольском приходится на 28 апреля B3 апреля—
1 мая) (Тарунин, 1929). Из Чжилийского залива (Китай) лутки отлетают
в апреле. На юге Уссурийского края появляются очень рано, первые
стайки уже в середине марта, валовой же пролет бывает в конце этого
месяца и в первой половине апреля. Лутки пролетают здесь обычно парами
и небольшими стайками (только однажды Пржевальский наблюдал 2 апре-
ля 1869 г. стаю в 300—400 птиц). Отдельные пары в Приморье задерживаются
до последних чисел апреля. На Аяне прилетает в последней декаде мая.
На восточном берегу Камчатки встречаются в мае немногочисленными парами
и стайками (Аверин, 1948), а на Командорских о-вах, где бывает лишь на весен-
нем пролете, с конца апреля до середины мая (Бианки, 1909). Под Якутском
появляется в середине мая (Воробьева 1931).
Осенний пролет на Камчатке наблюдается в октябре, в Приморье с конца
сентября и весь октябрь; на китайских зимовках (Чжили, Хубэй) птицы
появляются в ноябре. В среднем течении Печоры осеннего пролета не бывает
заметно, птицы отлетают незаметно, выводками; последние встречи их при-
ходятся на 5—10 октября. На озере Кургальджин8—12 октября луток встре-
чался громадными стаями до 150 шт. (Меженный и Соколов). На Наурзум-
ских озерах (Кустанайская обл.) пролет начинается в первых числах октября;
на Урале под Чкаловом первые птицы появляются в конце августа, а главная
масса летит с начала октября до замерзания рек; пролет по Иргизу наблю-
дался с 9 по 20 октября. В Уфимской обл. осеннего пролета незаметно, но позже
начала сентября они там уже не встречаются. В низовьях А трека лутки появи-
лись во множестве с 22 ноября (Житников, 1900). Около Ленкорани самки и мо-
лодые птицы появляются в декабре, а взрослые самцы в январе. На Азовское
море (Обиточенская коса) прилетают поздно и заканчивается пролет в декаб-
ре, а в Днепровском лимане лутки пролетают в половине октября и задер-
живаются на зиму в море. На р. Мологе первые пролетные стаи отмечены
28 августа, а последние 26 октября, а на Рыбинском водохранилище пролет
продолжается с 20—25 сентября по 21—23 ноября и бывает на своем послед-
нем этапе весьма интенсивным. Под Псковом луток пролетает осенью в мень-
шем числе, чем весной, с первых чисел сентября до ноября; в Польше первые
особи появляются в половине сентября, но основной пролет проходит в нояб-
ре; на места зимовок в Германии прибывают с конца октября. Летят лутки по
ночам или на утренних зорях, весной небольшими группами или парами,
а осенью иногда большими стаями. В полете стая строится гуськом или вол-
нистой линией.
Биотоп. В гнездовое время—лесные реки и озера, но горных рек
с особенно быстрым течением луток определенно избегает, предпочитая
богатые животными кормами озера в поймах рек. В противоположность дру-
гому дуплогнезднику—гоголю—луток избегает узких таежных речек, а пред-
почитает селиться в поймах более крупных рек, в островах леса на гривах
или в затопляемых приречных уремах. На пролетах лутки посещают водо-
емы самого разнообразного типа, а зимой предпочитают держаться на мор-
ских мелководьях у кромки льдов, а также в устьях и предустьевых про-
странствах рек.
ЛУТОК 603
Численность. На гнездовье луток нигде не бывает многочислен,
но он довольно обычен на большом пространстве низинной тайги европей-
ской части СССР и Западной Сибири. Общая численность вида велика, о чем
свидетельствуют сотенные стаи, которыми он пролетает осенью, в разгар
валового пролета.
Экология. Размножение. Достоверных данных о времени наступ-
ления половой зрелости нет, но, повидимому, птицы начинают размножаться
на втором году жизни. Весной птицы летят уже оформившимися парами,
по крайней мере вблизи от мест их гнездованья. Токовые игры их начинаются
-еще зимой, а в условиях паркового содержания иногда даже с ноября. Самец
плавает около самки, распушив оперение на горле и лопатках и гребнем рас-
правив хохол. Он делает движения головой и издает хриплые, каркающие
звуки. Прямых наблюдений над гнездованьем лутка очень мало. Известно,
что, как правило, он гнездится в дуплах деревьев: на Мологе чаще всего
в дубах, на Иртыше под Тобольском в ветлах и осинах. Дупла его в последнем
месте помещаются обычно совсем невысоко, и нередко их можно осматривать
даже не влезая на дерево. Охотно занимают лутки и гнездовые ящики, пове-
шенные на деревьях (Ладожское озеро, Рыбинское водохранилище). Под-
стилки, как правило, никакой не бывает, если она не принесена прежним
хозяином дупла, например скворцом. Гнездового пуха много, он светлой,
почти чистобелой расцветки. Яиц в полной кладке обычно 8, реже 10—11
на Рыбинском водохранилище, 8—10 в дельте Волги в недавнем прошлом
(по Яковлеву). Яйца белые, размер их 47,7—58x34—40,5 мм, среднее52,4х
X 37,5 мм. Нередко встречаются объединенные кладки с гоголем, причем выси-
живает обыкновенно луток, который выводит и своих птенцов и гоголят.
Срок инкубации около месяца, но точно не прослежен. Во второй половине
насиживания самка сидит так плотно, что позволяет брать себя руками и са-
жать обратно в гнездовой ящик. Весь период откладки яиц и некоторое время
после того как самка сядет на яйца, селезень неотлучно держится около
гнезда, так что наблюдая за селезнями, можно находить занятые гнезда лутка.
Позднее самцы откочевывают с мест гнездовья к местам линьки. На Рыбин-
ском водохранилище полные кладки свежих яиц найдены 17 мая, а насижен-
ные отмечались 9—13 июня. На Печоре гнезда с насиженными яйцами
находили в половине июня; на севере Финляндии F9° с. ш.) яйца най-
дены 26 июня. Выводки лутков были отмечены: на Кольском п-ове (озеро
Имандра) 26 июня и 27 июля; на Онежском озере 29 июля (Кесслер,
1868); первые пуховики на Средней Печоре 7 июля; на р. Елогуе птенцы
уже Jb пере, но с еще неразвитыми крыльями, встречены 5 августа
(Юдин); в низовьях Верхней Ангары летные молодые 13 августа, а опе-
рившиеся, но нелетные 20 августа (Слудский); пуховики на Шантар-
ских о-вах 3 августа и на озере Зайсан 23 июня. Некоторые выводки
бывают очень поздними, так, на Печоре, где обычно в начале августа
молодые бывают в полном пере (Теплоухов, 1911), 6 августа были встре-
чены и пуховики (Дмоховский, 1933); в Уфимской обл. даже в первых числах
сентября встречаются молодые, у которых маховые перья еще в пеньках
{Сушкин, 1897).
Линька. Как и другие утки, луток линяет дважды—летом почти пол-
ностью, осенью и зимой частично. Взрослые самцы начинают линять в июне.
Сначала происходит смена контурного оперения и лишь в конце ее линяют
хвост и перья. Селезни в периоде смены маховых добывались в Якутии еще
15 августа. В Уфимской обл. к концу августа отрастание маховых у селезней
заканчивается, но на темени и кое-где на спине сохраняются перья старого
наряда. Между летней и осенней линькой существует довольно большой пере-
рыв, чем луток, повидимому, отличается от других крохалей. Первые особи
с пеньками нового наряда начинают попадаться в пролетных стаях в начале
604
ОТРЯД ГУСЕОБРАЗНЫЕ
октября, а чаще в середине этого месяца. Во время этой линьки сменяется
контурное оперение и внутренние второстепенные маховые. Линька закан-
чивается в декабре, но некоторые особи пролетают через Рыбинское водохра-
нилище в ноябре, имея уже почти полный зимний наряд. У взрослых самок
линька начинается значительно позже, только в августе, и незаметно перехо-
дит в осеннюю, которую нельзя отграничить от летней. Специального летнего
наряда у самки нет, и какие части оперения линяют у нее дважды в год—
неясно.
Первая линька молодых птиц начинается у некоторых особей уже в сен-
тябре, в то время как у других только в середине октября. Начинается она
появлением пеньков на нижней поверхности тела и сменой средней пары
Рис. 110. Луток
рулей. За время этой линьки сменяется контурное оперение (спина и поясни-
ца?), но на груди выпадают не все старые перья. Сменяются также рулевые
перья и часть внутренних из второстепенных маховых. Когда заканчивается
эта линька—неизвестно.
Питание. Лутки, в отличие от других крохалей, питаются в боль-
шей степени водными насекомыми, нежели рыбой. Ранней весной (в апреле)
на р. Мологе 60% содержимого желудков этих уток составляла рыба,
в основном плотва (Rutilus rutilus), остальное приходилось на долю жуков
(плавунцы—Macrodytes marginalis и др.) и в меньшей степени на личинок
хирономид. На втором этапе пролета, когда вскрываются озера поймы
и лутки начинают кормиться на них, рыба в их рационе исчезает. Основу
питания их в это время (май) составляли жуки F5%—плавунцы, водолюбы
и др.) и личинки стрекоз C5%). На Рыбинском водохранилище в апреле
68% составляли личинки ручейников (Phryganea grandis), 6% жуки-плавунцы
и 26% рыба, в основном ерши и в малом числе плотва и окуни (все годовики);
там же в мае 100% содержимого составляли плавунцы Macrodytes margi-
nalis. В Уфимской обл. в желудках птиц, добытых в весеннее время, встре-
чались преимущественно личинки стрекоз (Сушкин, 1897). Желудок лутка,
добытого на Анадыре 6 июня, содержал в большом числе личинок ручей-
ников и в меньшем жуков-плавунцов и других (Портенко, 1939). В дельте
Волги весной (март) питание лутков состоит на 76% из рыбы, главным обра-
ЛУТОК 605
зом прошлогодних сазанчиков, водяных клопов (Corixa—14%) и в малой
доле из жуков, личинок ручейников, стрекоз, хирономид и ракообразных.
Тот же характер питания сохраняется и на Печоре (сезон неизвестен), где
рыба (гольян, сорога) и водяные насекомые (жуки-плавунцы, личинки стре-
коз и ручейников) встречаются примерно одинаково часто (Теплов, 1948).
На первом этапе осеннего пролета питание лутков остается таким же. На
Рыбинском водохранилище в сентябре—октябре на 77,1% оно состоит из
личинок ручейников, на 6,4% из других насекомых и на 16% из рыбы,
в ноябре картина резко меняется: первое место в их рационе занимает рыба
(86,5%), главным образом ерши-годовики F4%), а на долю ручейников при-
ходится лишь 13%. Эта перемена обязана похолоданию воды, в результате
которого водные беспозвоночные мигрируют в глубину и становятся трудно
доступными. Наконец, в зимнее время исследованные нами желудки лутков
содержали исключительно рыбу, в Ленкорани главным образом годовиков
сазана (Cyprinus carpio, до 10 см) и меньше уклеек, а в устье р. Сулака
исключительно только быстрянок, которых птицы вылавливали в ледовых
полыньях.
Экономическое значение лутка как охотничьего объекта невелико.
Специального промысла не существует, хотя во время валового осеннего
пролета вполне возможна промысловая добыча с чучелами. Охотятся на
лутков, попутно с охотой на другие виды нырков, местами на Каспийском
и" Азовском морях.
Полевые признаки. Мелкая утка. Самец в брачном наряде почти цели-
ком белый, с черным рисунком. У летящей птицы обращают внимание чер-
ные сверху крылья с белым пятном на месте крылового зеркала. У самки
верх головы коричневый, туловища—серый, вся нижняя поверхность белая.
Хорошо ныряет; плавает, не поднимая хвоста над водой. Голос—короткий
и трескучий звук вроде «экк» или «крр».
Описание. Строение и размеры. По величине немного крупнее чирка-
свистунка. Клюв небольшой, короче головы, суживается от основания к вершине; ноготок
во всю ширину надклювья. По краям обеих челюстей мелкие, острые и часто сидящие
зубцы. Лапа крупная, средний палец (с когтем) в полтора раза длиннее цевки. Хвост
округлый, рулевых 8 пар. Перья затылка удлинены и образуют широкий хохол—боль-
шой у самца в брачном наряде и небольшой у самки и у самца в летнем наряде. Размер
крыла у самцов 192—205, самок 178—190 мм; цевки самцов 30—34, самок 30—32 мм;
клюв самцов 28—30, самок 25—29 мм.
Вес. На весеннем пролете лутки имеют хорошую упитанность и отложения жира.
Вес их в это время таков: в апреле взрослые самцы 667 г E50—750 г) , самка 670 г (дельта
Волги), самцы 700 г E90—795 г), самки 510—670 г (Рыбинское водохранилище); в мае
самцы 702 г F15—755 г) (Бараба), 622 г E65—650 г) (Рыбинское водохранилище), 718 г
F15—860 г) (Заонежье), в среднем 750 г (Салехард); в июле 655 г (Архангельск). На осен-
нем пролете, особенно в конце его, птицы бывают упитаны хорошо и очень жирны.
Вес взрослых самцов в октябре 652 г E40—825 г), в ноябре 814 г G20—935 г), взрос-
лых самок в октябре 568 г E15—630 г), в ноябре 572 г E50—650 г). Молодые птицы
в октябре почти догоняют по весу взрослых, а в ноябре бывают даже несколько тяжелее
их. Вес молодых самцов в октябре 630 г E00—760 г), в ноябре 822 г F45—920 г), молодых
самок в октябре 556 г E00—680 г), в ноябре 588 г E35—670 г) (Рыбинское водохранилище).
Окраска. Пуховой птенец. На спинной стороне черно-бурый с особенно темной
головой и задней частью спины. Под глазами и у основания крыльев белые пятна.
Шея с боков рыжеватая, грудь и бока сероватые, остальной низ белый.
Молодые птицы в первом наряде. Похожи на взрослую самку (хохол короче),
но зоб и бока поясницы почти сплошь темносерые. Область уздечки не темнее, чем верх
головы. Средние верхние кроющие крыла с коричневатыми каемками. Самец несколько
крупнее одновозрастной самки, задние маховые у него более светлосерые.
Взрослая самка. Верх головы и задняя сторона шеи рыже-бурые. Спинная сторона
туловища темносерая, плечевые несколько буроватые. Перед глазом обилие черных
перышек образует темное пятно. Горло, передняя часть шеи и остальной низ белые, но
на зобу, боках и брюшке неясные серо-бурые пестринки. Большие маховые черно-серые,
второстепенные черные с белыми вершинами, самые внутренние из них серые. Верхние
кроющие крыла по сгибу темносерые, средние белые, большие черные с белыми концами.
Хвост серый. Клюв свинцовый; лапы серые; радужина бурая.
606 ОТРЯД ГУСЕОБРАЗНЫЕ
Молодой самец после первой зимней линьки. Имеет в основном окраску взрос-
лой самки, но на темени имеется уже некоторое количество белых перьев, а пятно перед
глазом и перевязь, идущая от крыльев поперек груди, черные; плечевые также белые,
но с серо-бурыми центрами или концами.
Взрослый самец в брачном наряде. Имеет белые голову и шею. На боках головы
между клювом и глазом бархатисто-черное пятно с зеленым отливом; такое же пятно
на затылке, но оно прикрыто хохлом и кажется имеющим вид буквы V. Спина чер-
ная. Плечевые белые с черной каймой по внешнему опахалу. Остальная спина и над-
хвостье темносерые; бока тела с тонким серым струйчатым рисунком. Вся брюшная
сторона, начиная от горла, чистобелая. Крыло такое же, как описано у самки, но белое
пятно на нем несколько больше. Хвост серый. Клюв свинцово-серый; лапы серые с тем-
ными перепонками; радужина мутнобелая.
Самец в летнем наряде. Весьма похож на самку, но пространство перед глазом
у него бурое, а не черное, а на темени и спине, как правило, сохраняются отдель-
ные перья брачного наряда.
54. Длинноносый, или средний, крохаль
Mergus serrator L.
Mergus serrator. Li n n a e u s. Syst. Nat., 1, изд. X, 1758, стр. 129, Швеция.
Русские названия. Длинноносый крохаль—книжное. Народные и широко
распространенные—крохаль, малый [крохаль.
з «а п п п. длпппипи^от i\pu/
распространенные—крохаль.
Распространение. Ареал. В гнездовое время населяет Исландию, Фае-
ры, север Шотландии и острова Гебридские, Оркнейские и Шотландские
и северо-западную часть Ирландии; гнездится на Ютландском п-ове и Дат-
ских о-вах; в Северной Германии занимает прибрежную зону, доходя на запад
до Шлезвига и Голштинии. Нередок в Северной Польше (Мазурские боло-
та), довольно обычен в приморской полосе Литвы и Латвии, а в Эстонии и на
прилежащих островах вполне обычен. Населяет весь Скандинавский п-ов,
Мурман и Карелию, в районе Белого моря всюду доходит до побережья и
гнездится на островах (Соловецкие о-ва). Гнездится на Канине, обычен в Ти-
манской тундре, по долине Печоры доходит до устья, отсутствуя лишь в при-
морской тундре. На о-ве Колгуеве и обоих островах Новой Земли неоднократ-
но встречался летом, но гнездованье там не доказано. Обычен на всех реках
Полярного Урала; на Ямале гнездится в долине р. Юрибея, но севернее
вероятно отсутствует. Отмечен летом на Гыданском п-ове, по Енисею подходит
к морю; северная граница на Таймыре неизвестна, но во всяком случае
средний крохаль заселяет всю лесотундровую зону полуострова и гнездится
в тундре. По долинам Лены и Яны спускается до морского берега, по Колыме
также гнездится до устья, но в тундровой части дельты Индигирки отсут-
ствует (Михель, 1935). Не гнездится на Новосибирских о-вах и о-ве Вранге-
ля; встречается по южным склонам Анадырского хребта, по всему течению
Анадыря и гнездится на ближних к берегу морских островах (о-в Алюмка)
(Портенко, 1939). Южнее гнездится на Олюторском побережье, Командор-
ских о-вах, повсюду на Камчатке, на северных островах Курильской гряды,
в бассейне р. Пенжины, на п-ове Тайгонос, в районе Охотска, Аяна и устьях
Уды. Южная граница гнездовой территории совсем неясна. В Ленинградской
обл. нередко летует на Финском заливе и Ладожском озере, но фактов, гово-
рящих о гнездованье, нет, хотя в шхерах Южной Финляндии несомненно
гнездится.Восточнее гнездованье его известно наОнежском озере (Повенецская
губа, Шуньгский п-ов) и у Архангельска. В Предуралье граница ареала резко
спускается к югу. Средний крохаль гнездится по всему бассейну Печоры до
самых верховий (причем особенно обычен на реках, текущих с Урала;
Дмоховский, 1933), на притоках Камы (реки Колва и Вишера; Теплоухов,
1911) и в верховьях Тавды (р. Лозьва; Резцов, 1904). Обычен по рекам вос-
точного склона Урала, но в долине Оби встречается только в лесотундровой
зоне, ниже Березова. На левобережье Енисея гнездится, повидимому, к югу
ДЛИННОНОСЫЙ, ИЛИ СРЕДНИЙ, КРОХАЛЬ
607
до р. Турухана (на р. Елогуе уже не встречен), по правому же, гористому, бе-
регу спускается до 60° с. ш. (по Тугаринову, 1941) и заселяет весь бассейн
р. Нижней Тунгуски до ее верховьев (Ткаченко, 1937); известны летние
встречи из бассейна Ангары. Гнездится на Байкале, а в его северной части
нередок (Туров, 1923). Восточнее граница совсем неясна. По некоторым лите-
ратурным данным, средний крохаль гнездится в Юго-Западном Забайкалье—
между Читой и Нерчинском, по Онону и Аргуни (Тачаьовский, 1893;Молле-
сон, 1907); в верхнем течении Амура до 53° с. ш. и может быть на Буреин-
ском хребте (Шульпин, 1936). По нижнему течению Амура и на Сихотэ-Али-
не не встречен. Однако и в пределах очерченного ареала местами этот кро-
Карта 75, Распространение длинноносого крохаля Mergus senator
*—граница гнездовий, б—не вполне выясненная граница 'гнездовий, в—обособленная гнеэдоваг
колония, •'—места постоянных встреч летом негневдящихся птиц, э—вимовки, а—залеты, otc—мест;
скоплений птиц на линьку
халь не встречен совсем, например, по среднему течению Лены, около Якут-
ска между Вилюем, Леной и Алданом. На основании нескольких летних
встреч предполагается гнездованье его на р. Мане в Восточных Саянах (Шу-
хов, 1925) и на севере Барабинской степи (озеро Карачи; Рузский, 1932).
Однако последнее из этих предположений мало вероятно. Обособленно от
основного ареала находятся две гнездовые колонии среднего крохаля: в Кер-
кинитском заливе Черного моря на островах Сары-Булатских (Киселев, 1950),
Джарылгач, Орлов и смежных с ними (Шарлемань и Шуммер, 1930; Климен-
ко, 1950) и в Армении на озере Севан. На Американском материке северная
граница гнездового ареала проходит от бухты Нортон к устью р. Мекензи,
через восточные окраины Медвежьего и Невольничьего озер до устья р. Чер-
чилль. Затем вдоль берегов Гудзонова залива до устья Большой Китовой реки,
а оттуда к восточному берегу залива Унгава на Лабрадоре. Далее границей
служит морское побережье, включая и о-в Ньюфаундленд, на юг до мыса
Код. Крайними южными точками служат область Великих озер, озеро Рин-
нипег и остров королевы Шарлотты. Кроме того, эти крохали населяют Кум-
берленд, южную часть Баффиновой Земли; западный берег Гренландии, на
север до Умнака и Упернивика, даже до 74°39' с. ш.; на восточном берегу
Гренландии до Скоресби-зунда отдельные находки и севернее (Саломон-
сен, 1950).
608 ОТРЯД ГУСЕОБРАЗНЫЕ
Основные зимовки в Европе расположены в Северном море
у берегов Дании, Германии и Британских о-вов; в Балтийском море до Аланд-
ских о-вов и в Норвегии до Полярного круга. В значительно меньшем числе
зимуют крохали на Западном Мурмане, в Исландии и Северной Франции.
Еще меньше птиц проводит зиму у берегов Пиренейского п-ова, Южной Фран-
ции и Италии. Единицы зимуют в Северной Африке от Гибралтара до Туниса.
Обильнее крохали бывают зимой у берегов Адриатического моря и в западной
части Черного. У берегов Малой Азии, Крыма и Кавказа крохали редки. Зи-
муют они на озере Севан, у обоих берегов Южного Каспия, в Сеистане и у бе-
регов Персидского залива. Однако в Иране крохали редки, а в Индии и Паки-
стане известны лишь залеты. У азиатских берегов крохали в небольшом чис-
ле зимуют на Восточной Камчатке и у Командорских о-вов. Обильнее они на
Курильской гряде, Японских о-вах и особенно в Корее, реже в Восточном
Китае на юг до низовьев Яньцзе, Фудзяня и Гуаньдуня, так же на Тайване.
В малом числе они проводят зиму в ряде пунктов внутри страны, в том числе
в истоках Ангары. В Америке они зимуют на Тихоокеанском побережье от
о-ва Ванкувера до Калифорнии включительно, а на Атлантическом побережье
от Новой Шотландии до устья Рио-Гранде-дель-Норте. Внутри материка
средние крохали зимуют в долине Миссисипи и на озерах Гурон и Мичиган.
Зимуют они и на крайнем юге Гренландии.
Районы скопления птиц на линьку до сих пор почти
неизвестны. Возможно, что в большинстве случаев птицы не совершают лет-
них миграций, а линяют на месте. Однако значительные стаи их в это время
наблюдаются в море у юго-западных берегов Новой Земли, в Кандалакшском
заливе, на реках Ямала и около о-ва Карагинского B8 июня).
Характер пребывания. Почти повсюду средний крохаль
правильно перелетная утка, совершающая дальние миграции. Частично оседл
он в Исландии, на Британских и смежных островах, в горле Балтийского
моря, у берегов Дании и Швеции. В еще меньшей степени он зимует на месте
у берегов Мурмана, Камчатки и Командорских о-вов. На Атлантическом бе-
регу Америки и в Юго-Западной Гренландии зимовки совпадают с гнездовой
территорией на очень малом протяжении.
Сезонные миграции и их даты. В европейской части
ареала осеннее движение птиц имеет два основных направления: на запад
и, реже, к югу. Это подтверждается немногочисленными пока данными коль-
цевания. Птицы из разных частей Швеции встречены поздней осенью и зимой
у западного берега Норвегии, в проливах Балтийского моря (о-в Борнхольм)
и в Силезии. Крохали, окольцованные молодыми в Исландии, зимовали
в Шотландии B находки) и в Голландии A находка), наконец, крохаль с о-ва
Фор (Дания) добыт зимой в Верхней Италии. Более дальние миграции со-
вершают молодые птицы и самки, самцы же относительно более оседлы и пе-
редвижения их зачастую ограничиваются лишь перекочевкой с пресных водо-
емов к морским побережьям. Отлет с мест зимовок у Британских о-вов на-
чинается в середине марта и заканчивается только в середине мая. Как и
у других уток, позднее пролетают неполовозрелые птицы, не достигающие
на первый год своей родины. Птицы, зимующие в Центральной Германии
(Баден, Брауншвейг), отлетают еще раньше, до начала марта. Пролетают они
мелкими стайками по ночам. На местах гнездовья в Северо-Восточной Гер-
мании появляются в середине апреля—начале мая; пролет через Польшу
наблюдается в марте. В Прибалтийских Советских Республиках средние кро-
хали появляются весной около 1 апреля, а в первой декаде этого месяца под
Ленинградом (Бианки, 1907). Однако пролет затягивается очень долго, по-
видимому за счет передвижения стай годовалых птиц; так, в Эстонии (Мацаль-
Вик) пролетная стая наблюдалась еще 27 мая (Лоудон и Бутурлин, 1908).
На озерах Псковской обл. появляется в первой половине апреля, валовой
ДЛИННОНОСЫЙ, ИЛИ СРЕДНИЙ, КРОХАЛЬ 609
пролет его стаями до 70 шт. проходит там во второй половине месяца, а от-
дельные стаи задерживаются до первых чисел мая (Зарудный, 1910). В устье
Мологи пролет их был отмечен 23—24 апреля. На Кольский п-ов (Лапланд-
ский заповедник) средние крохали прилетают около 14 мая F—26 мая)
(Владимирская, 1948), в то же время в середине мая они появляются и в Ти-
манской тундре (р. Вельть). Первые крохали на Печоре (устье р. Ылыча)
появляются около 12 мая G—17 мая), а массовый пролет начинается 14—
26 мая, причем крупных стай весной не встречается (Теплов, 1948).
На крайнем юге зимнего своего ареала—в Западном Пакистане длинно-
носый крохаль встречается небольшими стайками в декабре-январе, но
отдельные птицы задерживаются до первых чисел апреля (Тайсхерст, 1927).
В северо-западной части Черного моря (о-в Джарылгач) пролет проходит
в первой половине апреля, а зимовки в Азовском море (Обиточенская коса)
они покидают даже в начале марта. Пролет по Уралу и Эмбе чрезвычайно
слаб; первый экземпляр под Чкаловом добыт 31 марта, а на Эмбе пролет наблю-
дался 19—22 апреля (Сушкин, 1908). По Сыр-Дарье, Или, Тургаю и через
сибирские степи пролета этого крохаля не бывает. Отлет с китайских зимо-
вок (Гуаньдунь) задерживается до конца апреля, последняя дата добычи в
Корее 26 апреля (Курода, 1918). Через Приморье пролетает несколько позже
других крохалей; первая встреча 19 апреля, частые встречи 22 апреля—7 мая,
а в низовьях Амура нередок еще 1 июня (Шульпин, 1936). Первая добыча
под Охотском 19 мая; прилет на Командорские о-ва в середине мая; появле-
ние на Анадыре 6—26 мая (вид крохаля неизвестен), а первая добыча заведо-
мо длинноносого крохаля (под Марковом) 3 июня; прилет на Индигирке
(устье Момы) 18 мая. Последние встречи на Индигирке 27—29 сентября;
в этой местности крохали редки, но ежегодно появляются большими стаями
перед самой шугой (Михель, 1935). На Анадыре во второй половине сентября
количество их резко сокращается, а последние птицы встречены 9 октября
(Портенко, 1939). С о-ва Беринга крохали отлетают в октябре, и на зиму там
остаются лишь единицы (Иогансен, 1934). В нижнем течении Амура (озеро
Кизи) первая птица добыта 22 августа, на юге Приморья пролетные крохали
появляются во второй половине сентября и встречаются в течение всего октя-
бря; на зимовки в Южном Китае (Гуаньдунь) прибывают в ноябре. Пролет
в верховьях Тобола наблюдался 16—18 Iоктября, а в верховьях Иргиза
18—20 октября (Сушкин, 1908). Осенний отлет со среднего течения Печоры
18 октября A0—25 октября); пролетают птицы стайками по 8—12, редко по
20—30 шт. На Рыбинском водохранилище пролет наблюдался с 1 по 5 октя-
бря, а последние птицы наблюдались 17 октября. С Кольского п-ова эти кро-
хали отлетают 27 октября A2 октября—7Гноября), отдельные птицы зимуют
на порожистых речках. На Онежском озере (Повенец) пролетная птица добы-
та И октября; в Псковской обл. осенью встречается в течение всего октября,
но реже, чем весной. Из-под Ленинграда и из Прибалтики исчезает в конце
октября—в первых числах ноября. Пролет через Польшу происходит в ноя-
бре; в конце сентября—в октябре птицы уже появляются на зимовках у
Британских о-вов. Под Красноводском ранняя встреча—1 ноября; на
Азовском море у Обиточенской косы пролет среднего крохаля происходит со
второй половины октября до середины декабря, несколько раньше, чем у боль-
шого крохаля; табунки его задерживаются там на зимовку (Костюченко,
1926); в Джарылгачском заливе Черного моря пролетные птицы появляются
в начале октября.
Биотоп. В гнездовой период средние крохали заселяют весьма раз-
личные биотопы: шхеры северных морских побережий и приморские озера,
лагуны, речки и проточные озера тундровой зоны, реки гор и предгорий
северной лесной полосы и, наконец, открытые песчаные острова южных
морей. На пролетах они придерживаются открытых плесов рек и озер, а на
39 птицы СССР, т. IV
610 ОТРЯД ГУСЕОБРАЗНЫЕ
зимовке населяют преимущественно взморье и особенно предустьевые про-
странства рек.
Численность. Обычная птица на большей части своего ареала.
Особенно обычны длинноносые крохали в местностях с гористым ландшафтом:
в Скандинавии и на Кольском п-ове, на Урале и в Предуралье, на правобережье
Енисея и в верховьях Колымы. Именно в горных частях страны гнездовой
ареал их далеко выдается к югу, зато на обширных низменностях они, как
правило, отсутствуют.
Экология. Размножение. Половая зрелость достигается на втором
году жизни; на первый год птицы зачастую не долетают до тех мест, где они
вывелись, и живут стайками на море или больших озерах. На места гнез-
довья птицы прилетают уже парами. Брачные игры у длинноносого крохаля
проходят более бурно, чем у лутков и больших крохалей. Они заключаются
в плавании с глубоко погруженной передней частью тела и приподнятым задом;
шея при этом вытянута под углом около 75° к поверхности и делает поклоны
вперед, хвост поднят и раскрыт веером. Часто шея и голова быстро подни-
маются вертикально вверх, клюв в это время широко раскрыт и демонстри-
руется яркокрасная полость рта; хохол прижат к шее. Затем столь же быст-
ро голова опускается вниз. Птица часто делает движения, как во время купа-
ния, забрызгивая крыльями воду себе на спину. Игры сопровождаются хло-
паньем крыльев, погоней друг за другом, а иногда и драками. Движения самок
примерно те же, что и у самцов, но менее сложны. Даже в это время селезни
довольно молчаливы, но в момент вскидывания головы они издают довольно-
долгий и глухой двухтактный звук, трудно передаваемый слогами, вроде
сда-аа». Игры крохалей всегда наблюдаются в пролетных стаях, но продол-
жаются и по црилете на место.
Характер гнездованья различен в разных частях ареала. В шхерах и на
морских голых островах северных морских побережий (Эстония, Финляндия,
Белое море) гнезда чаще всего устраиваются в полупещерках или в щелях
между камней, но не далее 40 см от входа; как правило, вблизи от воды.
В тундре (Ямал, Тиманская тундра) гнезда помещаются под густыми кустами
в зарослях ивняков по берегам рек. В лесной части Кольского п-ова (озеро
Имандра) средние крохали гнездятся около порожистых речек под елями или
кустами можжевельника. В Печорской тайге гнезда строятся или в расщели-
нах скал, или среди подмытых водою корней деревьев, или в трещинах обва-
лов глинистых берегов (Теплов, 1948). В Анадырском крае средний крохаль
гнездится как по берегам рек по соседству с перекатами, так и на взморье;
в обоих случаях на островках с густой и высокой травой. Найденные там гне-
зда помещались среди травы, были полностью, как сводом, закрыты ею свер-
ху и имели боковой вход (Портенко, 1939). В Прибалтийских республиках
(Эстония, Латвия) этот крохаль часто гнездится около крупных озер с заро-
слями тростников. Гнезда его в этих условиях строятся открыто среди трост-
ников и высокой травы. Наконец, на юге Украины, в северо-западной части
Черного моря, длинноносый крохаль гнездится на открытых песчаных остро-
вах и устраивает свои гнезда среди чахлых тростников на прибрежных валах,
зачастую среди выбросов морской травы «камки» (Киселев, 1950). Устройство
гнезда во всех случаях очень примитивно. Это неглубокая ямка, выстланная
небольшим количеством растительного хлама и с большим количеством пуха,
который появляется по мере окончания кладки. Гнездовой пух у открыто гнез-
дящегося среднего крохаля, в отличие от дуплогнездника большого крохаля,
темный, хорошо гармонирующий по окраске с почвой и отмершей раститель-
ностью, среди которой строятся гнезда. Нередко эти крохали занимают одно
и то же гнездо несколько лет. В Лапландском заповеднике (озеро Чуна) одно-
гнездо, устроенное под елью, занималось в 1931,1932 и 1935 гг., а дру-
гое два года подряд (Владимирская, 1948).
ДЛИННОНОСЫЙ, ИЛИ СРЕДНИЙ, КРОХАЛЬ
Число яиц в полной кладке: в Эстонии 8—10, в Тиманской тундре 9
A гнездо), на Кольском п-ове (озеро Чуна) 8—10 C), на Ямале 9 B), на Анады-
ре 7 B), на Черном море 9—12. Кладки с числом яиц более 12 A5 и даже 20),
на которые имеются указания в литературе, следует, по всей вероятности,
считать сдвоенными. Скорлупа яиц гладкая, цвет бледнсоливковый. Размер
около 65,2x45 мм E6,5—70,7x40,3—47,6 мм)\ вес 70,7 г F4,5—80 г).
Откладка яиц на Черном море в мае—июне A6—17 июня яйца в половине
насиживания), в Эстонии в конце мая и начале июня, на Кольском п-ове (озере
Чуна) самки в периоде откладки яиц добыты 1—9 июня, но еще 28 июля най-
дено гнездо с сильно насиженными яйцами. В Тиманской тундре (р. Вельть)
кладка с сидящей на гнезде самкой встречена 7 июля (Гладков, 1951). На
Ямале 3 самки, добытые 11 июля, находились в периоде откладки яиц, самка
16 июля закончила кладку (Осмоловская), а 2 августа найдено гнездо с силь-
но насиженными яйцами (Житков, 1913); сильно насиженные кладки на Ана-
дыре обнаружены 16 июля и 1 августа. Насиживает одна самка; срок насижи-
вания 31—32 дня (Хейнрот). Вес птенцов в день вывода 44—49 г. Оперяться
они начинают примерно с 16-го дня, и подъем на крыло происходит очень
поздно. Средний размер выводка в Латвии 8—10 птенцов, на Кольском п-ове 6,
на Печоре 6,6. В последнем месте размер выводка заметно различается по
годам. Малое число птенцов по сравнению с размером кладки говорит о ги-
бели молодых, нередко из-за паразитарных болезней. Птенцы прекрасно
плавают, ныряют, так же хорошо бегают по земле. Зачастую два-три выводка
объединяются вместе, и стайка до 16—24 птенцов плавает в сопровождении не-
скольких самок. Иногда они объединяются и с утятами других видов, напри-
мер гоголя (озеро Имандра). На Иртыше Сабанеев наблюдал стаю из более 100
крохалят различного возраста, сплывавшую по реке в сопровождении взрос-
лых птиц. Спуск по течению еще не летных, а иногда даже не оперившихся
выводков отмечался на многих реках: текущих с Урала притоках Камы,
Печоры, Оби и Тобола. Биологическое значение этого явления велико, так
как выводки крохалей зачастую не поднимаются на крыло почти до самого
ледостава. Этой осенней откочевкой вплавь можно объяснить некоторые встре-
чи нелетных молодых за пределами гнездового ареала (Кама, Белая,
Иртыш). На Черном море (Ягорлыцкий залив) выводок пуховиков встречен
22 июня (Шарлемань и Шуммер, 1930), а одна очень ранняя кладка проклюну-
лась 6 июня (Клименко, 1950). В Северо-Восточной Латвии выводки появ-
ляются в середине июля (Бутурлин, 1900); на Кольском п-ове сроки вывода
чрезвычайно растянуты, пуховики встречаются с 8 июля по 30 августа, опе-
рившиеся, но еще не летные молодые с 20 июля по 12 сентября, а первые лет-
ные выводки с 20 августа (Владимирская, 1948). В Кольском заливе малень-
кий пуховичок добыт 11 августа, а на Онежском озере (Повенец) пуховой
птенец размером с галку 12 августа (Исполатов, 1916). На Печоре первые
выводки появляются около 5 июля B7 июня—11 июля), а массовый вывод
происходит в середине июля, на крыло они поднимаются в середине сентя-
бря—12 сентября F—19 сентября). Совсем маленькие пуховики встречены на
р. Вельть 17 июля (Тиманская тундра; Гладков, 1951). На р. Лозьве (при-
ток Тавды) 7 августа добыт птенец, у которого перья появились толь-
ко на плечах и боках, а маховые только что начали выбиваться из пень-
ков (Резцов, 1904). В низовье Енисея пуховики были добыты 25 ав-
густа, а 7-дневный птенец под Верхоянском 3 августа. На Охотском побере-
жье (Пенжина) пуховой птенец добыт 22 августа, а молодые с растущими
маховыми и остатками пуха (п-ов Тайгонос) 28—29 агуста (Дементьев, 1940).
На Командорских о-вах нелетные птенцы 7—11 августа; на Карагинском о-ве
нелетные выводки наблюдались 2 и 7 сентября; на Анадыре выводок разме-
ром с чирка 16 августа, тот же выводок 4 сентября еще не поднялся на
крыло.
39*
612
ОТРЯД ГУСЕОБРАЗНЫЕ
Линька. Взрослые самцы линяют дважды: почти полностью после
сезона размножения и частично осенью и зимой. Линька самцов начинается
с конца июня, когда они покидают занятых насиживанием самок и объеди-
няются в стайки. Одновременно с ними линяют и годовалые птицы. Осо-
бенно больших скоплений линных птиц этого вида неизвестно, но на Ямале
(р. Щучья и севернее) встречались стаи более 100 голов (Кучерук, 1948),
большие стаи наблюдаются и в море около южного берега Новой Земли (Мол-
чанов, 1908). Вначале происходит смена контурного оперения на голове
и туловище птицы, кроме нижней части груди и части спины, позднее ли-
няют хвост и крылья. Смена маховых происходит в июле и в августе, в конце
августа линька обычно заканчивается B4 августа на р. Щучьей были еще
не летные селезни). Следующая линька самцов (в брачный наряд) начинается
Рис. 111. Длинноносый крохаль (самец и самка)
в сентябре и продолжается до января. За это время сменяется оперение тела,
хвост (с октября) и часть внутренних второстепенных маховых. Взрослые
самки начинают линять не ранее августа. У самки, добытой 9 августа от
выводка (р. Лозьва), новые перья появились только среди плечевых. Лет-
няя линька у них переходит без перерыва в осенне-зимнюю, которая закан-
чивается в феврале. Для описания ее нет материалов. Первая линька моло-
дых проходит с октября по февраль. За это время частично сменяется опе-
рение на голове и туловище, хвосте и часть внутренних второстепенных
маховых, но спина и зад почти полностью сохраняют птенцовый наряд до
первой летней линьки.
Питание. Длинноносый крохаль—типичная животноядная утка.
На кормежке крохаль обычно плывет, погрузив в воду клюв и часть головы,
высматривая рыбу, а заметив ее, преследует под водой, помогая себе при
этом крыльями. Нередко крохали устраивают совместные охоты выводками
или стайками, двигаясь при этом по фронту изогнутой линией. Основной
пищей служит рыба, но немалую роль играют также водяные насекомые.
В двух желудках крохалей, добытых в мае на Белом море (Онежская губа),
обнаружены корюшки размером около б см: На Печоре (устье Ылыча) рыба
ДЛИННОНОСЫЙ, ИЛИ СРЕДНИЙ, КРОХАЛЬ 613
встречена в 63% исследованных желудков, причем в основном это был гольян,
в меньшем числе бычок-подкаменщик (Coitus sibiricus) и в еще меньшем ха-
риус. В одном случае обнаружена лягушка и почти в 50% желудков водяные
насекомые: личинки ручейников и жуки-плавунцы (Теплов, 1948). У трех
крохалей, добытых во время осеннего пролета на Рыбинском водохранили-
ще, желудки были заполнены ручейниками {Phryganea grandis). На каспий-
ских зимовках (Ленкорань) в желудках крохалей обнаружена только рыба»
чаще всего молодые сазаны E—15 см) и быстрянки {Alburnoides). У птицы,
добытой 8 декабря в море около Сухуми, пищевод был наполнен мелкими
рыбками Blennius (Сатунин, 1913).
Экономическое значение как объекта охоты мало. Специального промысла
этого вида нигде не существует. Так как длинноносый крохаль служит окон-
чательным хозяином лигулы (Ligula intestinal is) и зараженность птиц этим
паразитом почти поголовная, то местами они могут играть роль в распро-
странении лигулеза среди рыб.
Полевые признаки. Крупная утка с длинным и узким клювом. У самца
голова и верхняя часть шеи черная, у самки коричневая; на затылке двой-
ной хохол. Белая передняя сторона шеи самца четко выделяется между
темной головой и ржавым зобом, так же как белые перья сложенного крыла
между черной спиной и серыми боками. У самки голова примерно того же
цвета, что и спина. Клюв яркокрасный. У летящего самца спина темная,
основная часть крыльев сверху белая; снизу виден ржавый зоб. Прекрасно
плавает, глубоко погружая туловище в воду; прекрасно ныряет, обычно пре-
бывая под водой около 20—25 сек. (с паузами 15—20 сек.). Шум от полета
слегка свистящий. Голос самца, слышный только в брачное время, тихий
и глухой, самка же издает долгое и грубое «кар-р-р».
Описание. Строение и размеры. Довольно крупная утка с удлиненным
телом, длинной и- узкой шеей и далеко отставленными назад ногами. Клюв необычной
для уток формы, сжатый с боков, с узким ноготком, загнутым в виде тупого крючка.
По краям обеих челюстей острые, направленные назад роговые зубцы. В отличие от боль-
шого крохаля, в верхней челюсти от переднего края ноздрей до вершины их 18 или более
(у пуховых птенцов 16—19); от заднего края ноздрей до углов рта обычно менее 11 зубцов.
Ноздри лежат целиком в основной трети надклювья. На затылке у обоих полов двойной
хохол из узких и довольно редких, но длинных перьев. Рулевых 9 пар, крайние значи-
тельно короче средних.
Вес птиц в феврале на зимовках в Юго-Восточном Закавказье: взрослый самец
1350 г, самка 1150 г, молодые самки 688 г F44—760 г); взрослые самцы в мае 1100 г
(Ямал), 1082 г (970—1120 г) (Заонежье), в июне 1265 г (о-в Медвежий, Белое море). Гнез-
дящиеся птицы в июле: взрослые самцы 1100 г A000—1200 г), самка 860 г (Ямал); в сен-
тябре взрослая самка 870 г (Заонежье), в октябре 780—930 г и молодой самец 1050 г
(Рыбинское водохранилище).
Окраска. Пуховой птенец. Голова сверху буровато-каштановая, а остальной
верх темный оливково-бурый, несколько более коричневый, чем у большого крохаля.
У основания крыльев и по бокам поясницы белые пятна. Темная полоска, отходящая
от разреза рта, выражена очень слабо, кремового пятна под глазом, как у большого кро-
халя, нет, на щеках больше коричневого и меньше белого цвета. Низ тела серебристо-
белый.
Молодые птицы в первом наряде. В основном сходны со взрослой самкой, но окраска
верха более одноцветная, темносерая, пятна на зобу более мелкие, на подхвостье
значительная примесь серого цвета. Кроме того, хохол на голове у них много короче.
Клюв красновато-рогового цвета, лапы коричневато-желтые, радужина желтая.
Самка после первой зимней линьки. Имеет оперение взрослой, но у нее нет
черных пятнышек над глазом, как у последней. Хохол на затылке короче. После
первой линьки неотличима от взрослой.
Взрослая самка. Шея и голова рыже-каштановые, последняя с буровато-серым
верхом. Туловище сверху серовато-бурое с более светлыми, серыми концами перьев.
Зоб и бока буроватые, также со светлыми вершинами перьев; остальная брюшная сто-
рона тела белая, подхвостье с небольшой серо-бурой пятнистостью. Большая часть
кроющих крыла той же расцветки, что и спина, кроме больших кроющих, у которых*
вершинная половина белая. Второстепенные маховые черные в основной половине и белые
в верхней. Белое пятно на крыле меньше, чем у самца, и с одной черной полоской. Перво-
614 ОТРЯД ГУСЕОБРАЗНЫЕ
степенные маховые черно-бурые, третьестепенные буро-серые. Хвост темносерый. Клюв
карминово-красный, но более тусклый; лапы красно-оранжевые, радужина красная.
Самец после первой осенне-зимней линьки. Голова и шея до основания буро-
каштановые, но с большим числом черных перьев взрослого наряда, особенно на верху
головы. Частично черные спина и плечи, но окраска новых перьев буроватая. Лапы
оранжево-красные, радужина оранжево-желтая.
Взрослый самец в брачном наряде. Голова черная с сине-зеленым отливом,
шея белая, но по задней стороне ее проходит узкая черная полоса. Плечевые перья
и верхняя часть спины черные; нижняя часть спины, бока тела, поясница и надхвостье
темносерые с черной струйчатостью. Зоб ржавого цвета, на боках его и у осно-
вания шеи черно-бурые пятна. Остальная брюшная сторона белая, часто с розовато-
оранжевым налетом Мелкие верхние кроющие крыла черные, средние белые, большие
черные с белыми вершинами (составляющими кайму перед зеркалом), второстепенные
маховые черные в основной и белые в верхней половине. В результате в основной части
крыла сверху образуется большое белое пятно, разделяемое двумя черными поперечными
полосками. Первостепенные маховые черные, третьестепенные белые с черной наружной
каймой. Хвост серо-бурый. Клюв карминово-красный с темным хребтом и вершиной;
лапы оранжево-красные, радужина красная.
Самец в летнем наряде. Окрашен, как самка, но спина несколько темнее. Крыло,
как в брачном наряде, но внутренние второстепенные маховые черно-бурые вместо
черных. Хохол короче, чем зимой.
Систематические замечания. Весьма однородный вид как по окраске, так и по раз-
мерам на всем протяжении его ареала. Некоторые авторы в качестве особого подвида
выделяют лишь форму, населяющую Гренландию: М. s. schioleri Salomonsen, 1950.
Основанием к этому служат несколько более крупные размеры ее, однако разница эта
мала и выявляется лишь на сериях [размеры крыла самцов 257 мм B48—265 мм), в то
время как у типичной формы 248,2 мм B38—260 мм)]. Ввиду отсутствия отличий
в расцветке оперения и в биологии выделение этого подвида приходится считать неосно-
вательным.
55. Чешуйчатый крохаль Мег gas squamatus Gould
Mergus squamatus. Gould. Proc. Zool. Soc. Lond., 1864, стр. 184, Китай.
Русское название. Чешуйчатый крохаль—книжное, по окраске груди и боков.
Распространение. Ареал. Сведения о летнем ареале чешуйчатого
крохаля очень отрывочны. До недавнего времени коллекционные материалы
по этому виду ограничивались следующими: добыт 13 августа 1910 г. на р.
Кур в 80 км к северо-западу от Хабаровска, полуразложившаяся самка
найдена 20 сентября 1925 г. на берегу бухты Наталья на Коряцском побе-
режье Берингова моря F1° с. ш.), 2 взрослых селезня 4 июня и самец на вто-
ром году жизни 19 июня 1911 г. на о-ве Медном (Командорские о-ва), годо-
валая самка в июле (августе) 1930 г. на р. Мотай, притоке Хора D7,5° с. ш.)
и самец 24 апреля 1933 г. в нижнем течении Имана D5,7° с. ш.). За послед-
нее время наши сведения об этом крохале несколько пополнились. Спанген-
берг нашел его довольно обычным в летнее время в среднем течении р. Имана,
а по наблюдениям сотрудников Сихотэ-Алиньского заповедника (сообщенным
нам Бромлеем) эти крохали живут на реках восточного склона хребта:
Сице, Белимбэ, Такеме и др. Кроме того, этот вид встречен в Маньчжурии,
где один экземпляр добыт 25 августа 1925 г. около ст. Яблоня на юго-востоке
от Харбина, повидимому на весеннем пролете, и в значительном количестве
обнаружен осенью 1931 г. на озере Цинбоху в Гиринской провинции.
В последнем месте добыты 4 самца и, по словам рыбаков, там этот крохаль
гнездится. Таким образом, если расценивать пункты крайних северных нахо-
док чешуйчатых крохалей как места их залетов, то можно предполагать,
что областью гнездованья этого вида служат средняя и южная части горной
страны Сихотэ-Алиня и гористая часть Северо-Восточной Маньчжурии.
В зимнее время чешуйчатые крохали были добыты в Корее, Китае (Тянь-цзин,
Сычуань, Юго-Западный Юннань, Хубэй и Фудзянь), в Тонкине и Бирме.
Во всех этих пунктах крохаль известен не как регулярно зимующая птица,
а лишь в виде единичных, случайных находок. Повидимому, значительная
ЧЕШУЙЧАТЫЙ КРОХАЛЬ
615
часть птиц проводит зиму на местах своего гнездовья. Так, они регулярно
зимуют на незамерзающих речках Приморья в районе бухты Терней и на
р. Тачингоуза в Судзухинском заповеднике.
Характер пребывания. Не выяснен, возможно оседлая
птица.
Биотоп. В гнездовое время держатся в среднем течении нешироких
горных рек, в средней и верхней таежной зоне. Так, на реке Сице, текущей
с восточного склона Сихотэ-Алиня, и ее притоках, в 35—60 км от моря, их
КартаO6. Распространение чешуйчатого крохаля Mergus squamatus
а—иввестные места гнеедованья, а—весенние иХдетние встречи, в'—осенние и вившие
встречи
выводки нередки, а в 75 км уже редки. Они избегают слишком узких прито-
ков, в которых трудно нырять, и совсем не встречаются там, где берега реки
не покрыты лесом. Зимой крохали держатся в нижнем течении рек (на р. Сице
в 9—19 км от моря), там, где они не замерзают. В Маньчжурии на озере Цин-
боху эти крохали гнездились, по словам местных рыбаков, на его скалистых
островках (Лукашкин, 1934).
Численность. Редкий вид с весьма ограниченным ареалом распро-
странения.
Экология. Размножение. С 20 апреля 1941 г. в течение 10—12
дней Бромлей наблюдал ежедневно с восходом солнца перелеты группы
крохалей этого вида: 1 самец и 2 самки вдоль рек Бичана и Фаты, притоков
р. Сицы. В середине мая селезень наблюдался здесь уже один, без самок
616 ОТРЯД ГУСЕОБРАЗНЫВ
всегда на определенных плесах, повидимому наиболее рыбных. Бромлей
описывает два дупла на р. Сице и ее притоке Фате, в которых наблюдателем
охраны заповедника были найдены гнезда чешуйчатого крохаля. Одно дупло
находилось в торце гнилого, сломанного и нависшего над рекой ствола,
в 1,5 м над водой. Другое находилось в толстой липе A5—18 см в диаметре),
склонившейся над рекой, на нижней поверхности ее ствола, в 3—3,5 м над
водой. Под дуплом был большой нарост, на который выходила утка, поки-
дая гнездо.
По размеру выводка чешуйчатый крохаль, повидимому, близок к длинно-
носому. Так, в июле 1940 г. Бромлей и Салмин встретили на р. Сице выводок
из 10—12 довольно взрослых крохалей уже в перьях, которые с шумом,
прихлопывая крыльями, поплыли вверх по реке. В конце августа 1940 г.
ими добыт крупный птенец этого вида, уже в полном оперении. В верхнем
течении р. Сицы G5 км от устья) держались две семьи крохалей в 8—10
и 10—12 шт. В сентябре выводки держатся еще вблизи от мест своего гнездо-
вья в среднем течении р. Сицы, и их неоднократно наблюдали (в 57 км от
устья) проплывавшими вверх и вниз по течению. Взрослый самец с черной
головой (с переливами) держался отдельно и встречен за лето всего 4—5 раз.
Наблюдения говорят о том, что этот крохаль вообще довольно редка
летает и по большей части передвигается вплавь. Повидимому, это относится
и к его сезонным кочевкам из среднего течения рек в их низовья и
обратно. Можно высказать предположение, что значительная оседлость
этой птицы, ограниченный и труднодоступный район ее обитания
служат той причиной, по которой ее биология до сих пор почти неизвестна.
Линька. О линьке этого вида почти ничего неизвестно. Оперяются
птенцы в июле-августе, способность к полету получают в конце августа.
Молодой селезень, начавший линять из первого брачного в летнее перо,
был добыт 9 июня, а взрослый самец в летнем пере—13 августа.
Питание. В желудке и пищеводе крохаленка, добытого в августе
на р. Сице, обнаружены две рыбки 12—15 см длины, одна из них мальма
(Salvelinus malma cur ill us и Oncorynchus masu).
Полевые признаки. Средний по размерам, сходный с длинноносым
крохалем. Самец в брачном наряде отличим по очень длинному хохлу и оба
пола по чешуйчатому рисунку боков туловища.
Описание. Размеры и строение. Клюв узкий обычного крохалиного
типа с острыми зубцами по краям. От конца клюва до ноздри их 18—20 и от заднего края
ноздри до угла рта 11—12. Перья темени и затылка образуют узкий и длинный, до ПО мм
у самца и 62 мм у самки, двойной хохол. Длина крыла самцов 250—265, самок 240—
250 мм; цевки самцов 46—48, самок около 45 мм; клюв самцов 52—57, самок 43—48 мм.
Окраска. Пуховые птенцы неизвестны.
Молодая самка. Бока головы и шеи рыжие, низ шеи бледнее, горло беловатое.
Спинная сторона чистосерая, окраска головы и шеи темнобурые со слегка рыжеватым от-
тенком на шее. Брюшная сторона белая. Третьестепенные маховые с широким черноватым
наружным краем. Рулевые буроватые с серым налетом.
Самка взрослая. Голова с хохлом и шея оливково-бурые. Бока их и нижняя
сторона шеи несколько рыжее, горло белое. Мантия, мелкие и средние кроющие крыла
чисто пепельно-серые с чуть голубоватым оттенком, перья остальной части спины и верхней
части надхвостья с белыми и серыми дугообразными полосками, длинные кроющие хво-
ста серые. Настволья перьев верхней стороны темные. Брюшная сторона тела, зоб, испод
крыла и подхвостье белые с легким лиловым оттенком. Перья на боках зоба, груди, брюха
и поясницы, а также на узкой полоске поперек задней части брюха (в области клоаки)
имеют резко очерченные, довольно широкие серые дугообразные полосы вдоль их краев,
создавая чешуйчатый рисунок на этих частях тела. Прекрасным отличием птиц обоего
пола от длинноносого крохаля служит окраска пуха, чистобелая у настоящего вида
и серо-бурая у последнего. Зеркало крыла белое, разделено на две части черными основа-
ниями второстепенных маховых. Хохол довольно густой из очень узких длинных перьев.
Осенний и зимние экземпляры уток чешуйчатого рисунка не имеют (очевидно, в резуль-
тате послебрачной линьки он пропадает), на заду и боках имеется лишь пепельно-серый,
налет (Бутурлин, 1935).
БОЛЬШОЙ КРОХАЛЬ
617
Селезень в брачном пере. Голова и шея черные с зеленым отливом, двойной
хохол из сильно удлиненных узких перьев. Зашеек лопатки и плечевые перья черные.
Спина и поясница серые с черным чешуйчатым рисунком от полукруглых каемок перьев.
Надхвостье пепельно-серое с черноватой рябью. Весь низ тела, включая зоб, белый с лег-
ким семужным налетом. На боках тела крупный черный, а на подхвостье неясный чешуй-
чатый рисунок. Верхние кроющие крыла вдоль сгиба черные, средние белые и нижние
черные с белой верхней половиной. Первостепенные маховые и наружные второстепенные
черные, внутренние второстепенные с белой верхней частью, третьестепенные с белым
наружным опахалом и черной каймой по краю. Хвост серо-бурый. Клюв у обоих полов
красный с темными хребтом и концом. Лапы яркокрасные, радужина сероватая или
белая.
Рис. 112. Чешуйчатый крохаль (самец и самка)
Самец в летнем пере. Верх головы и задняя часть шеи оливково-бурые. Бока
головы и шеи светлые охристо-рыжеватые, подбородок и шея спереди еще светлее. На
затылке пучок удлиненных перьев. Шея и зоб с серовато-белыми и чешуйчатыми перьями.
Плечевые и спина не чисточерные, а с большой примесью аспидных перьев, остальное
оперение, как в брачном наряде. Клюв и лапы, как весной.
56. Большой крохаль Мег gas merganser L.
Mergus merganser. Li n n a e u s. Syst. Nat., изд. X, 1, 1758, стр. 129, Швеция.
Синоним. Mergus asiaticus. G m e 1 i n. Reise. d. Russl. II, 1774, стр. 188, устье Волги.
Русские названия. Народное—крохаль большой, баклан-утка (Южный Кас-
пий); краснобрюхий зубарь (Азовское море); большая острота (низовья Днепра).
Распространение. Ареал. Исландия, Шотландия, северная часть
Европы от Дании на западе и изолированно в Швейцарии и Южной Германии.
В пределах Союза от западных границ до Тихого океана, на север до границы
лесной растительности, но пока не найден к северо-востоку от Верхоянского
хребта, в бассейнах Индигирки и Колымы. На юг до р. Мологи, южных пре-
618
ОТРЯД ГУСЕОБРАЗНЫЕ
делов бассейна Северной Двины и южных предгорий Урала. Изолированная
колония в Армении на озере Севан. Гнездится на Алтае, в Северо-Западной
Монголии, Забайкалье, Приамурье, Уссурийском крае и Северной Маньчжу-
рии, на Сахалине, Курильских о-вах и Камчатке. По горным хребтам Сред-
ней Азии достигает Памира, населяет Гималаи, Тибет, Юго-Западные Ганьсу
и западнее Юннани. В Америке гнездится от Тихоокеанского до Атланти-
ческого побережья на север до архипелага Александра, Невольничьего
озера, южного берега залива Джемса, устья реки св. Лаврентия и Нью-
фаундленда. Южная граница проходит через штат Мек, озера Гурон и Мичи-
ган, среднее течение Миссури, спускается к югу по южным окраинам Ска-
листых гор, огибает с севера пустыню Невада и вновь опускается к югу по
береговым хребтам до устья р. Сакраменто.
Карта 77. Распространение большого крохаля Mergus merganser
1—М. т. merganser, 2—M. m. orientalist*—M. m. americanus; а—граница гнездовий, а'—районы
наибольшей численности птиц на гнездовье, в—обособленная гнев до в ая колония, в*—места постоян-
ных встреч летом (гнездованье не докавано), в—зимовки; цифры в кружках-номера вимующих
подвидов
Основные зимовки находятся на побережьях и в приморских
местностях Балтийского и Северного морей, в Исландии, на Британских
о-вах, во Франции и других странах Средиземного моря до Северной Африки
включительно. На Черном и Каспийском морях, в Средней Азии от Таджи-
кистана до Северной Индии, повсюду в Китае, Корее и Японии. На тихоо-
кеанском берегу Америки от Ванкувера до Калифорнии, на атлантическом
от Новой Шотландии до Мексиканского залива включительно, в бассейне
Миссисипи и в южной части Великих озер.
Характер пребывания. На большей части ареала пере-
летная птица. На юге и в приморских местностях большая часть популяции
оседла; в горах Средней и Центральной Азии эти птицы совершают верти-
кальные миграции.
Биотоп. Большой крохаль—обитатель рек и чистых незаросших
озер. Местности с горным ландшафтом он явно предпочитает равнинным,
а на юге селится только в горах. На море крохали встречаются сравнительно
редко, преимущественно неполовозрелые и линные птицы.
Численность. Обычными на гнездовье большие крохали бывают
только в горах и предгорьях. Большие скопления по нескольку сот голов они
БОЛЬШОЙ КРОХАЛЬ 619
иногда образуют на местах линьки, осеннем пролете и на зимовке. Однако
это происходит редко, только местами и преимущественно в северной части
ареала вида.
Подвиды и варьирующие признаки. Большой крохаль образует три довольно четко
отличимых друг от друга подвида. Номинальная форма М. т. merganser Linnaeus, 1758,
населяет Европу, бореальную и частично арктическую зону Азии; М. т. orientalis Gould,
1845, отличающийся размерами и деталями окраски, живет в горах Средней и Централь-
ной Азии; М. т. americanus С a s si n, 1853, североамериканская форма, отличается от
типичной формы более коротким крючком на конце надклювья и тем, что на белом крыло-
вом пятне селезня имеется черная поперечная полоса (шириной 7,5 мм), образованная про-
свечивающими основаниями больших кроющих крыла; самка темнее обычной, передняя
часть спины темноаспидная с серыми краями перьев. Этот крохаль регулярно залетает на
Уналашку и Прибыловские о-ва; вполне возможны залеты его на Командоры, Камчатку
« Курильскую гряду. Один экземпляр с черной поперечной полоской на зеркале, но более
узкой, чем у американских птиц, был добыт на Командорских о-вах (Хартерт, 1915).
Обыкновенный, большой, крохаль Mergus merganser merganser L.
Распространение. Ареал. Исландия, северная часть Шотландии,
весь Скандинавский п-ов, южная и средняя части Ютландского п-ова, Северо-
Западная Германия (до Мекленбурга, Бранденбурга и Шлезвига) и Север-
ная Польша. Изолированно от основного ареала большие крохали гнездятся
в южной части Баварии (на притоках Дуная), в Швейцарии и на южном
берегу Женевского озера. Кроме того, есть указания на гнездованье их
в Боснии и Добрудже (Нитхаммер, 1938), Распространение этого вида в пре-
делах Советского Союза изучено еще недостаточно. На Кольском п-ове он
встречается всюду, кроме самой северной тундровой части, где хотя и попа-
дается летом, но на гнездовье не обнаружен. Населяет всю Карелию, доходя
до Беломорского побережья. К востоку от Северной Двины северная гра-
ница распространения этого крохаля примерно совпадает с пределом древес-
ной растительности. На Канине, в Тиманской и Болыыеземельской тундрах
он отсутствует. На Северном Урале, повидимому, переходит за Полярный
круг, в долине Оби достигает Салехарда, населяет лесную часть бассейна
Таза, а на Енисее доходит до устья р. Курейки. На Таймыре не отмечен,
в бассейне Вилюя встречен под 66° с. ш., на Лене, в устье р. Лунхи F4° с. ш.).
В бассейнах Индигирки и Колымы до сих пор не добыт, но в Анадырском
крае нередок и встречается от верхнего течения реки до низовий (факт гнез-
дованья в нижнем течении реки не подтвержден). Гнездится по всей Кам-
чатке, единично на о-ве Беринга и, повидимому, на островах Курильской
гряды. На Олюторском берегу и в северной части Охотского побережья
пока не добыт, но в лесистой части последнего по всей вероятности будет
найден, так как под Охотском встречается. Южная граница намечается еще
менее четко. Большой крохаль гнездится в Прибалтийских Советских Рес-
публиках, хотя и немногочислен там. О Псковской обл. достоверных све-
дений нет, в Новгородской возможно гнездится (Бианки, 1910), известны
единичные случаи гнездованья около Невеля, в Великолукской обл. (Федю-
шин, 1926). В Смоленской, Московской и Калининской обл. бывает лишь на
пролетах, но гнездился в небольшом числе в нижнем течении р. Мологи,
до образования там Рыбинского водохранилища. Несомненно заселяет
бассейн Северной Двины, но южнее его в Центральной России на гнездовье
никем не отмечен. Для Горьковской обл. пролетная птица, но для Волги
и Камы в пределах Татарии Першаков A929) считает его редко гнездящимся.
Основанием этого служат летние встречи стаек и находка одного гнезда
около Тетюшей. Повидимому, если крохаль и гнездится здесь, то единично
и нерегулярно, отчего включать эту территорию в основной ареал вида
достаточных оснований нет. На Урале и в Предуралье крохали весьма обычны,
и в этой области их гнездовой ареал далеко выдвигается к югу. Крайними
620 ОТРЯД ГУСЕОБРАЗНЫЕ
южными точками служат верховья Белой и Уфы, а также реки Ика, Сак-
мары и район Верхнеуральска (Зарудный, 1888, 1897). В долинах Иртыша
и Оби крохаль редок, но весьма обычен на их левых притоках, берущих
начало с Урала. Под Тюменью (Ларионов, 1927), Тарой (Ушаков, 1913)
и Тобольском (Тарунин, 1928) он отмечен лишь на пролетах; более обычным
на Оби он становится лишь от широты Березова, хотя по всей вероятности
гнездится и на севере Томской обл. Гнездованье крохаля, на основании
летних встреч, указывается для некоторых пунктов северной части Барабы
(озеро Карачи; Рузский, 1932), Ишимской лесостепи и Кулундинских лен-
точных боров (Залесский, 1931). По правобережью Оби граница резко спу-
скается к югу. Крохаль гнездится в Кузнецком Ала-Тау и повсюду на Алтае,
кроме его степной части, проникает в Монгольский Алтай, а возможно ив Го-
бийский, так как самка добыта у гнезда на р. Урунгу в Китайской Джунга-
рии (Сушкин, 1938). Южнее крохаль гнездится в Тарбагатае (Аягуз) и изо-
лированно, часто небольшими колониями, в нескольких местах Джунгар-
ского Ала-Тау (реки Тентек, Аганакатты, Каратал, Акичке, Коксу и Усек),
Киргизского хребта (Карабалта) и Кунгей-Ала-Тау, Чолпан-Ата, М. Аксу
(Шнитников, 1949). Далее на восток крохаль заселяет Западные и Восточ-
ные Саяны и Северо-Западную Монголию до южных и восточных склонов
Хангая (Сушкин, 1938). В Северной Монголии встречается в бассейне Толы
и в Юго-Западном Кентэе (Козлова, 1930). Обычная гнездящаяся птица
Байкала, Забайкалья (Чикой, Онон, Аргунь) и левобережья Амура. Гнез-
дится на Сахалине, во всяком случае в его северной и средней части, обычен
повсюду в Приморье и встречается в гор но-лесистой части Маньчжурии
(Хинган, Сунгари). Совершенно изолированная гнездовая колония нахо-
дится на озере Севан в Армении.
Основные места зимовок этого крохаля находятся на
побережьях Северного и в незамерзающих частях Балтийского морей. На
Британских о-вах, в Исландии и в Северной Германии, на озерах Бранден-
бурга в январе встречаются стаи по 300—400 птиц, причем не менее третьей
части их составляют взрослые самцы (Гессе, 1920). Внутри страны крохали
также нередки, например на Рейне и Боденском озере, а в холодные зимы
появляются также на мелких притоках. Однако там молодые птицы и самки
значительно преобладают над самцами. В меньшем числе крохали проводят
зиму во Франции и Северной Испании. В западной части Средиземного моря
крохали редки и появляются только в холодные зимы, причем Северной Аф-
рики достигают лишь единицами. В этом районе зимовок преобладают самки
и особенно молодняк. В несколько большем числе зимуют они на Балкан-
ском п-ове, а в западной части Черного моря (Бургас, Дунай, Ягорлыцкий
и Тендровский заливы) крохали в зимнее время обычны. В большом числе
зимуют на Азовском море (Обиточенская коса), где встречаются табунами
(Костюченко, 1926). На Каспийском море зимуют одиночками и неболь-
шими группами, редки зимой в Иране. Есть указания на зимние встречи
в верховьях Аму-Дарьи, зимует на полыньях в низовьях Сыр-Дарьи (Джу-
лек); обычны зимой в низовьях Или и в большом числе проводят зиму на
Иссык-Куле (Шнитников, 1949). В небольшом числе зимует на Камчатке,
о-вах Курильской гряды у берегов Приморья. Обычны зимой на боль-
ших озерах Японии, но несколько реже встречаются на реках (Ян, 1942);
зимуют в Южной Корее и повсюду в Китае как в приморской его полосе на
юг до границ Вьетнама, так и по долинам рек. Местами в юго-западных
и южных районах Китая проводят зиму совместно обе формы крохаля—и ти-
пичная и гималайская (Латуш, 1925).
Места скопления линных птиц известны очень плохо.
Огромные стаи их по нескольку сотен птиц линяют у берегов Новой Земли
(Маточкин Шар, Сконнинг; Гусиная Земля, Успенский). Очень большая
БОЛЬШОЙ КРОХАЛЬ 621
стая линных крохалей встречена в Ачитнурской котловине Северо-Запад-
ной Монголии (озеро Цаган-Нур; Сушкин, 1938). Часть птиц линяет на ме-
стах гнездовья, держась в одиночку или объединяясь в небольшие группы
(Телецкое озеро, Укбк на Алтае, притоки Печоры).
Характер пребывания. Почти по всему ареалу правильно
перелетная птица. Частично оседлы лишь крохали, населяющие Исландию,
Шотландию, области Северного и Балтийского морей, озеро Севан, Кам-
чатку, Курилы и Приморье. Кроме того, почти во всех частях ареала не-
большое число птиц остается зимовать на незамерзающих участках рек
{Карелия, Эстония, Украина, Московская обл., Урал, Енисей, Байкал,
Тола и Орхон в Северной Монголии и др.).
Сезонные миграции и их даты. На территории Союза
намечаются три основных направления, по которым проходят сезонные
миграции больших крохалей: в европейской части на запад; с Урала, Запад-
ной Сибири и Северо-Западного Казахстана на юго-запад; в Юго-Восточном
Казахстане к югу; в Восточной Сибири и на Дальнем Востоке на юг и юго-
восток. Данные кольцевания показывают, что птицы из Финляндии и со
Скандинавского п-ова обычно зимуют в Англии и частью у берегов Германии.
В последнем месте зимой также встречаются птицы, гнездящиеся в Исландии
и бывающие на осеннем пролете, в октябре, на Днепре (Шюц и Вейгольд,
1931). Отлет крохалей с мест зимовки у Британских о-вов происходит с конца
марта до начала мая; с Балтийского побережья Польши и из Центральной
Германии в марте. В Эстонии на незамерзающих участках моря они появ-
ляются уже в конце февраля, стайками по 5—20 шт. Появление первых
птиц внутри страны совпадает обычно с образованием полыней, а время ва-
лового пролета—с началом ледохода или с последними днями перед ним.
В Ленинградской обл. прилет их приходится на вторую половину марта,
на р. Мологе и Рыбинском водохранилище 12—19 апреля, встречи в Киров-
ской обл. 25—26 апреля, в Варангер-фьорде в апреле, в Финской Лапландии
6—10 мая, в центре Кольского п-ова (озеро Имандра) около середины мая.
В среднем течении Печоры первые крохали появляются около 15 апреля
G—22 апреля), а валовой пролет приходится на 17—23 апреля (Теплов,
1948). В западной части Черного моря отлет зимующих крохалей заканчи-
вается в первой декаде апреля, но поступательное движение их начинается
много раньше; так, на полыньях Днепра у Херсона они появляются уже
в конце февраля (Клименко, 1950). Азовское море они покидают в начале
марта; первые встречи в Аскания-Нова приходятся на 28 февраля F февраля
21 марта) (Соколов, 1926), а под Киевом в разные годы на 9—27 марта (Дани-
лович, 1930). Самая первая встреча в дельте Волги 13 февраля, но последние
задерживаются до конца апреля. По среднему течению Урала и Эмбе пролет
крохалей проходит интенсивно. Далее основная масса их следует по рекам
Ори и Сакмаре, а меньшая попадает в верховья Иргиза. Обильны они на
пролете также по Тоболу, озерам между ним и Улаганом и по Иртышу. Под
Чкаловом на пролете с конца марта до конца апреля, валовой пролет по
Эмбе 15—22 апреля, по Сакмаре в последней декаде апреля или несколько
раньше. Первая встреча на р. Обве (приток Камы) 3 мая, добыча на озере
Чусовском 8 мая. Пролетные стайки на Сыр-Дарье появляются в первой
декаде марта; в среднем течении Или первые с 25 февраля—2 марта, обычны
весь март и исчезают к 4 апреля (Слудский). Весенний пролет на Южном
Алтае в середине марта; встреча на Елогуе 3 июня, у Туруханска 1 июня.
В Японии часть птиц задерживается до середины апреля; на юге Приморья
крохали появляются 17 марта—2 апреля; а в первой или во второй половине
апреля происходит их валовой пролет, к 10 мая пролетные особи исчезают.
На Орок-Норе (Северо-Западная Монголия) небольшие стайки встречены
уже 4 апреля, а в Юго-Западном Забайкалье (р. Чикой)—5 апреля (Мол-
622 ОТРЯД ГУСЕОБРАЗНЫЕ
лесон). На о-ве Беринга пролетные птицы появляются в первой половине
апреля, на Камчатке 11—21 апреля, первая встреча у Охотска 6 мая, а на
Анадыре 3 мая.
Осенний валовой пролет у крохалей происходит поздно, обычно уже
перед самым ледоставом. С Анадыря они исчезают в начале октября, послед-
ние птицы отмечены 11 октября, Южное Приморье покидают в конце этого
месяца с появлением льда на реках. Среднего Китая (долина Янцзе) дости-
гают в ноябре, а в Южном встречаются в декабре-январе. На Печоре пос-
ледние отмечаются около 22 октября A3—30 октября). Осенний отлет их
оттуда происходит постепенно, в первую очередь исчезают выводки моло-
дых и позже всех взрослые самцы. В Кировской обл. крохали добывались
до 28 октября, на Рыбинском водохранилище первые пролетные птицы
появляются в конце сентября, основной пролет начинается с первых чисел
октября и продолжается до 26 октября—12 ноября, последние же птицы
встречаются до 23—25 ноября. Из Ленинградской обл. и Прибалтики отле-
тают в начале ноября. Обильный пролет в ноябре через бассейн Припяти
(Цедлитц, 1920); на местах зимовки (в Англии) появляются с конца сентября
и в октябре. В верховьях Иргиза первые отмечены 9 октября сразу же после
первого мороза. Осенний пролет в низовьях Или примерно с 20 октября по
20 ноября; пролет под Чкаловом в начале октября. Первые крохали на озе-
рах Атрека отмечены 22 ноября; прилет на зимовки в Азовском море с конца
октября до середины декабря; пролет через Аскания-Нова в течение всего
ноября и до начала декабря; в западной части Черного моря появляются
в середине октября. Весной крохали летят обычно по утренним зорям, а день
отдыхают и кормятся. Стайки их в весеннее время невелики, обычно по
4—6, реже 10—15 шт. и редко до 30 шт. Осенью же, наоборот, стаи бывают
очень велики, нередко из нескольких сотен птиц, особенно на местах их от-
дыха. На полете такие стаи часто выстраиваются углом или «ключом» по-
добно журавлям.
Биотоп большого крохаля—озера и реки, весьма различные по хара-
ктеру, но обязательно богатые рыбой и с прозрачной водой. В горных мест-
ностях крохали гораздо обычней, чем на равнинах, и именно по горным
хребтам ареал их выдвигается к югу. На море крохали встречаются обычно
лишь в зимнее время и иногда в периоде линьки. Наиболее характерная для
них зимняя стация—устья рек и их предустьевые пространства. Лишь
кое-где по берегам Балтийского (Эстония) и Северного (Шотландия) морей
они живут и летом на морских побережьях, поселяясь даже во фьордах и на
небольших островках. В основной части своего ареала этот крохаль—оби-
татель лесного ландшафта, что связано с характером его гнездованья,
и в безлесных местностях его можно встретить лишь в горах на юге
страны. Рубка лесов и уничтожение старых дуплистых деревьев вызвали
за последнее время исчезновение крохалей на гнездовье в ряде пунктов
и сокращение их ареала (Дания; Иесперсен, 1946; б. Сувалкская губ.
в Польше; Тачановский, 1888; повидимому, в дельте Волги; Хлебников,
1930; низовья Дуная).
Численность. Ареал большого крохаля велик, но в пределах
его он далеко не везде может считаться обычной птицей. Обычен и даже
довольно многочислен он на реках предгорий и в горных местностях: Коль-
ский п-ов, Урал и Предуралье, Алтай, Саяны, Байкал и Сихотэ-Алинь.
В Хибинах оба крохаля, взятые вместе, самые многочисленные виды уток,
на Печоре выводки больших крохалей составляют 24% от всего числа
встреченных утиных выводков. Общая численность, вида велика; осенью,
в дни валового пролета через Рыбинское водохранилище, за день наблю-
дается до 1500 птиц; пролетные и зимующие стаи часто достигают несколь-
ких сотен голов.
БОЛЬШОЙ КРОХАЛЬ 623
Экология. Размножение. Половой зрелости достигают, повиди-
мому, на втором году жизни. На места гнездованья они прилетают уже офор-
мившись парами, но токовые игры, а иногда и драки между самцами продол-
жаются и по прибытии на место. Основные токовые позы крохаля таковы.
Птицы плавают, погрузив в воду грудь настолько, что зашеек их находится
вровень с поверхностью, в то время как зад бывает приподнят над водойг
хвост вздернут вверх и расправлен веером, а крылья слегка расставлены,
демонстрируя белые плечевые перья. При этом птицы часто плещутся, как
во время купания, а голова на вытянутой шее делает поклоны вперед. Вне-
запно и быстро шея и голова с раскрытым клювом вытягиваются перпен-
дикулярно вверх, а затем столь же быстро принимают обычное положение.
Иногда селезень привстает на воде так, что тело его принимает почти верти-
кальное положение, причем клюв в это время касается груди. Движение
это часто сопровождается взмахом крыльев. Резким спазматическим ударом
обеих ног туловище выбрасывается вперед на 30—60 см, г сзади над поверх-
ностью поднимается струя воды. В тех случаях, когда игры происходят
в стае, плеск воды и глухие каркающие крики самцов бывают слышны изда-
лека. Кроме того, крохалям свойствен токовый полет, когда птицы летают,
делая круги на значительной высоте. Самки также принимают участие в иг-
рах, но их движения менее сложны и обычно выражаются в приподнимании
корпуса над водой, толкании его вперед ударом ног и плескании крыльями.
Брачные игры начинаются уже в ноябре, во время осеннего пролета. Спари-
вание также наблюдается иногда уже на зимовках; оно происходит на воде,
и самец при этом полностью топит самку.
Гнездятся крохали вблизи от водоемов, отдавая предпочтение рыбным
речкам с быстрым течением. Чаще всего они устраивают гнезда в дуплах
старых деревьев. Так они поступают в Уссурийском крае, на Печоре,
в Поволжье, Новгородской обл., Эстонии, Германии и Дании. Крохали-
ные дупла известны в осинах, вязах, ивах и ольхах как в прибрежных
лесах, так и в старых парках. Высота дупел от земли различна, от 1 до
18 ле; часто используются дупла, сделанные черным дятлом. Иногда они
гнездятся в старых постройках, сараях и развалинах каменных зданий
(Эстония). Устраиваются гнезда и в трещинах прибрежных скал, изредка
в Эстонии (Кох, 1911), и, повидимому, как правило, на Алтае (Сушкин,
1938). В последнем месте крохали даже отсутствуют на тех реках и озерах,
к берегам которых не подходят отвесные скалы. Там, где крохали на
гнездовье обычны, они охотно занимают гнездовые ящики как повешен-
ные на деревьях (Печора, Ладожское озеро), так и установленные на земле
(о-ва Вайка в Эстонии, Транзее, 1927). В некоторых случаях гнездятся они
и открыто на земле, под кустами. В таких случаях выстилка из травы,
веточек и перьев бывает гораздо обильней (Юго-Западный Кентэй; Коз-
лова, 1933). Выстилка из растительных остатков в гнездах почти или совер-
шенно отсутствует, но зато бывает много пуха очень светлого, почти
белого цвета. Обычное число яиц в полной кладке 8—9 до 13 на Печоре,
15 в Эстонии. Цвет яиц почти белый или сливочный, скорлупа гладкая,
размер 55,4—74,5x37—50 мм, средний вес 84 г F9—97,6 г), по Хейнроту
75 г. Ежедневно откладывается по одному яйцу. Насиживание начинается
со времени откладки последнего яйца; срок его 32 дня (Хейнрот, 1928).
Ни в подыскании места для гнезда, ни в устройстве его самец участия не при-
нимает, но некоторое время он держится около гнезда и в случае опасности
старается отвлечь внимание на себя. Данных о сроках гнездованья мало.
В Эстонии откладка яиц приходится на середину мая, но затягивается и до
последних чисел его; в Щецине (Польша) кладка яиц обнаружена 20 апреля
(Робин, 1923); в Северной Монголии (Юго-Западный Кентэй) гнездо с 8 яйца-
ми найдено 13 июля.
624 ОТРЯД ГУСЕОБРАЗНЫЕ
Птенцы сутки или двое находятся в гнезде. Из дупла они выпрыгивают
самостоятельно на голос матери, находящейся под гнездом. Каким образом
птенцы благополучно достигают воды на речках Алтая, где птицы гнездятся
на отвесных обрывах,—неясно. По некоторым наблюдениям, самка перено-
сит молодых на воду в клюве, однако по отношению ни к одной утке эти
наблюдения до сих пор не подтверждены. Хорошо нырять молодые научают-
ся примерно через 8 дней, но способность к полету они приобретают очень
поздно, повидимому в возрасте 60 (Нитхаммер) или 70 дней (Теплов). Раньше
всего у молодых отрастают плечевые и рулевые перья. Затем оперяются
надхвостье, грудь, брюшко и бока тела. Потом голова и шея, вслед за ними
спина и одновременно появляются второстепенные и третьестепенные махо-
вые. Большие маховые отрастают последними. Молодые птенцы часто отды-
хают на спине у самки. Плавают они быстро, легко справляясь с сильным
течением. Спасаясь от преследования, часто бегут по воде, хлопая крылыш-
ками, причем долго не утомляются. Известны случаи, когда птенцы, спасаясь
от лодки, поднимались по реке более чем на 8 км. Средний размер выводка
пуховиков на Печоре 7—8 птенцов по Теплову A940), не более б по Дмохов-
скому A933); на Южном Урале (Миасское озеро) 5—9 (Ушков, 1947), на
р. Елогуе в среднем 7,2 F—12) (Слудский); выводки, наблюдавшиеся на
Мологе, в Кентэе и на Имане, содержали по 6 птенцов. Основным врагом кро-
халят на Печоре служит выдра. Часто несколько выводков объединяется,
и тогда стайка до 30—35 птенцов живет при одной или нескольких самках.
Во многих местах выводки еще не оперившихся молодых сплывают с самками
вниз по течению к более крупным рекам или озерам. На Алтае это может
быть объяснено слишком сильным течением мелких речек, с которым птен-
цам трудно бороться, а на реках, текущих с Урала, лучшей кормностью
и лучшими условиями защитности крупных рек. Выводок пуховиков в Вели-
колукской обл. (Невель, озеро Иван) обнаружен в конце июня, а на р. Мо-
логе 22 июня. Появление первых выводков на Печоре приходится в среднем
на 23 июня (9 июня—4 июля), в середине августа B0 июля—25 августа)
они объединяются между собой и в массе появляются на магистральных
реках; в августе летные выводки там почти никогда не встречаются, часто
попадаются нелегные молодые и в первой половине сентября. Начавшие
оперяться птенцы наблюдались в низовьях Оби (Парават) 1 августа, а совсем
мелкие пуховички на Имане 31 мая (Спангенберг, 1940). На Восточной Кам-
чатке нелетные выводки обнаружены еще 27 сентября и 4 октября (Аверин,
1948). В более низких частях Северо-Восточного Алтая молодые появляются
около половины июня. На Абакане несколькодневные птенцы 24 июня, на
притоках Абакана и Лебеди с 30 июня по 5 июля молодые размером с чирка,
на Телецком озере птенцы того же размера в середине июля. Выше в горах
выводки бывают более поздними; так, на Верхнем Уйгуте и в истоках Кара-
Аллаха молодые размером в две трети самки 16 и 28 августа (Сушкин, 1938).
Первый выводок на р. Толе (Северная Монголия) 22 июня; в Джунгарском
Ала-Тау (Усек) молодые размером немного мельче взрослых 9 июля, а 30 июня
летный выводок (Борохоро).
Линька. Как и другие утки, крохали линяют дважды в год: летом
почти полностью, осенью и зимой частично (кроме крыльев и хвоста). У сам-
цов окончание первой линьки несколько заходит за начало второй, у самок
они слиты еще более. Взрослые самцы начинают линять с того времени, как
самки сядут на яйца, т. е. в Прибалтике с конца мая, на Печоре и Алтае
в июне. Сперва сменяется контурное оперение головы и плечевые перья,
затем перья шеи и туловища, кроме части перьев спины, надхвостья и под-
хвостья, а также частично нижней части груди и брюха. К середине июля
самцы бывают уже в полном летнем наряде. Вслед за тем линяют крылья
и начинается линька хвоста, которая затягивается иногда почти до зимы.
БОЛЬШОЙ КРОХАЛЬ 526
Маховые выпадают во второй половине июля или в начале августа. К концу
этого месяца самцы уже обычно приобретают вновь способность к полету.
В августе же появляются первые перья брачного наряда на голове и в верхней
части спины. Некоторая часть старого брачного наряда на перечисленных
вып№ частях тела, особенно на спине, сохраняется до начала осенней линьки
и сменяется непосредственно перьями нового брачного наряда. Более энер-
гично предбрачная линька начинается в сентябре и заканчивается лишь
в декабре. Во время нее сменяется все контурное оперение и некоторые из
внутренних второстепенных маховых.
Линька взрослых самок прослежена значительно хуже. У самок, живу-
щих при выводках, она начинается много позже, чем у самцов, а именно
в середине июля (Алтай). Летняя и осенне-зимняя линьки у них сливаются
в одну общую затяжную линьку, почему смена оперения на многих частях
тела бывает у них однократной. Едва ли не раньше всего начинает линять
спина, вскоре затем выпадают маховые, а вслед происходит смена рулевых
и контурного оперения. Можно считать, что у самок почти полностью выпа-
дает летняя линька, так как соответствующий ей летний наряд отсутствует.
Полностью заканчивается смена оперения обычно к январю, но у отдельных
особей растущие перья на туловище можно встретить до марта.
Первая линька у молодых самцов начинается поздней, чем у взрослых,
и бывает весьма неполной. У некоторых птиц, добытых в октябре на Рыбин-
ском водохранилище, ее еще нет. У них сменяются рулевые, часть внутрен-
них второстепенных маховых и большая часть перьев туловища. Голова
и шея не перелинивают. Линька у них затихает в январе, но, видимо, вновь
возобновляется весной. Два молодых самца, добытых в апреле на Рыбин-
ском водохранилище, имели в надхвостье развертывающиеся перья, а на
основании некоторых рулевых еще не спавшие чехлики B0 апреля). Первая
полная линька молодых птиц проходит так же, как у взрослых.
Взрослые самцы к началу линьки собираются в стайки и зачастую отле-
тают с мест гнездованья на Алтае к большим открытым озерам, на севере
частично к морю. Но значительная часть самцов линяет на месте одиночками
и небольшими группами.
Питание. Пищу крохалей почти целиком составляет рыба; водяные
насекомые и моллюски являются лишь примесью. В Печорском заповед-
нике рыба обнаружена в 70% исследованных желудков большого крохаля,
причем чаще всего встречены гольян и хариус, кроме того, обнаружены семга
A0 см), бычок-подкаменщик и плотва. Нередко A0,9%) желудки содержали
травяных лягушек и в одном случае найдена землеройка. В 16,4% желудков
были остатки водяных насекомых (жуков-плавунцов, личинок стрекоз
и ручейников) и в 2,7% моллюски. Растительные остатки, встреченные
в желудках, должны расцениваться как случайная примесь (Теплов, 1948).
На р. Мологе во время весеннего пролета 75% пищи крохаля составляла
рыба—голавли и окуни (однолетки), а остальное приходилось на долю
моллюсков-горошинок (Pisidium amnicum). На Рыбинском водохранилище
весной рыба составляет 100% содержимого желудков, а осенью 97%. Весной
это плотва и налимчики, осенью же основу составляет плотва F8%), затем
идет ерш A8%) и в меньшем числе поедаются окуни и снетки (Osmerus грет-
lanus). 3% содержимого по объему составляют личинки ручейников (Phry-
ganea grandis). Все встреченные здесь рыбы были годовички, но в литературе
есть указания, что крохали заглатывают рыб до 18 см длиной и даже таких,
которые остаются некоторое время торчать у них изо рта. В Семиречье,
кроме рыб-османов (Diptychus), крохали едят также водяных ужей, кото-
рые местами очень многочисленны в горных потоках. В одном случае из зоба
у подстреленного крохаля выползла еще живая, видимо недавно заглочен-
ная, змея (Шнитников, 1949). В дельте Волги на осеннем пролете большие
40 Птицы СССР, т. IV
,626
ОТРЯД ГУСЕОБРАЗНЫЕ
крохали .кормятся только рыбой (вобла, молодь сазана и щуки, разная
мелочь). Зимой у каменистых берегов Туркмении (Каспийское море) они
питаются преимущественно бычками {Gobiidae) и атеринками. Большие кро-
хали активно преследуют добычу под водой, ныряя обычно на глубину до
2—4 м. Осенью, когда крохали собираются на пролетах сотенными стаями,
можно наблюдать их коллективные охоты за рыбой. Стая их быстро плывет
в каком-либо направлении, развернувшись широким фронтом. Птицы дер-
жат клюв и часть головы подводой, высматривая добычу и то и дело ныряя
за ней. Отстающие догоняют стаю по воздуху и опускаются в ее передний
ряд. Обычно такую охотящуюся стаю сопровождают чайки, которые, пи-
кируя на воду, схватывают рыбок, выпугнутых на поверхность крохалями.
Рис. 113. Большой крохаль (самец и самка)
Экономическое значение. Специального промысла на больших кроха-
лей не существует. Их добывают попутно с другими утками, в более значи-
тельном числе на Печоре (около 15% от всей добычи водоплавающих птиц),
в истоках Ангары и на Азовском море. Мясо их имеет в некоторые периоды
неприятный запах. Крохаль—рыбоядная утка, но горные речки не осваи-
ваются рыбным промыслом, почему он и не наносит вреда поеданием рыбы.
Однако крохали бывают сильно заражены лентецом Ligula, и в тех местах,
где эти птицы обильны, они могут способствовать размножению и расселе-
нию этого паразита. Шкурка крохаля с выщипанным пером после соответ-
ствующей выделки идет на изготовление детских шапок и отделку шуб.
Полевые признаки. Очень крупная утка с длинной шеей и узким клю-
вом.У самца голова и верхняя часть шеи черные с металлическим блеском;
остальная часть шеи, бока и низ тела белые. Последний иногда с оранжево-
розовым налетом. У летящей птицы, при взгляде на нее сверху, обращает
внимание целиком белая основная половина крыла. Клюв яркокрасный.
У самки рыжая голова с двойным широким хохлом. Горло и зоб белые (в отли-
БОЛЬШОЙ КРОХАЛЬ , 627
чие от самки длинноносого крохаля). Птицы прекрасно ныряют, при пла-
вании туловище глубоко погружается в воду. Шум крыльев на полете
слегка свистящий. Голос самца глухой «ба-а-баб*, у самки трескучее
«карр-карр».
Описание. Строение и размеры. Клюв узкий, ноготок круто загнут вниз;
по краям челюстей острые роговые зубцы, слегка загнутые назад. В верхней челюсти от
переднего края ноздрей до вершины 13—15 зубцов, столько же их бывает и у пуховых
птенцов. У селезня голова без хохла, хотя перья затылка несколько удлинены, а самка
с двойным, довольно широким хохлом. Размеры крыла у самцов (циркулем) 262—290,
у самок 235—272 мм; цевка самцов 45—54, самок более 43 мм; клюв самцов 51—60,
самок 40—52 мм.
Вес. Взвешиваний большого крохаля проведено немного. Весной птицы прилета-
ют обычно хорошо упитанными и имеют отложения жира. Взрослые селезни в марте-апре-
ле весят 1800—2060 г (дельта Волги), в апреле 1700—1785 г (р. Или; Штегман; 1949),
1200—1640 г (Печора; не первогодки ли?). Взрослые самки: в апреле 1400 г (Печора), в
октябре 1700 г (Рыбинское водохранилище), 1150 г (дельта Волги). Молодые птицы в поло-
вине августа и в сентябре 1000 г G50—1200 г) (Печора), в сентябре самцы 1550 г, в октябре
1390 г A185—1535 г); самки 1385—1550 г; в ноябре самец 1970 г, самка 1490 г. Самцы-
прошлогодки в апреле 1650—1850 г (Рыбинское водохранилище). Зимой крохали очень
жиреют, к декабрю они бывают буквально облиты жиром, и мясо их к этому времени
теряет неприятный запах (Азовское море; Костюченко, 1926).
Окраска. Пуховой птенец. На спинной стороне темный оливково-бурый, верх
головы буро-каштановый. От надклювья через глаз и от разреза рта под глазом про-
ходят две узкие черновато-коричневые полоски, соединяющиеся за глазом с такой же
окраской боков головы. Они ограничивают узкое вытянутое пятно кремового цвета. Сред-
няя часть щек и бока шеи с рыжеватым налетом. Подбородок, горло, нижняя часть щек и
остальной низ тела серебристо-белые. У основания крыльев и по бокам поясницы по одно-
му белому пятну с каждой стороны.
Молодые птицы. Окрашены в общем, как взрослые самки (хохол на затылке коро-
че). Спинная сторона коричнево-серая, перья на лбу и темени с темными штрихами,
на передней стороне шеи вниз от горла проходит узкая беловатая полоса и область
уздечки также беловатая. Бока тела одноцветные серо-дымчатые. Клюв темный крас-
новато-бурый; лапы грязножелтые, радужина темнокоричневая.
Перезимовавшая самка. Иногда отличается светлыми перьями на уздечке и перед-
ней стороне шеи.
Взрослая самка. Голова и шея рыжевато-бурые, темя более серое, горло беловатое.
Задняя сторона шеи у ее основания, бока зоба и весь верх туловища и хвост аспидно-
серые. Зоб и бока тела светлосерые остальная брюшная сторона белая. На белом брюхе
иногда бывает неяркий оранжевый налет. Большие и наружные второстепенные маховые
черно-бурые, внутренние же белые, а третьестепенные темносерые, одноцветные со спи-
ной. Малые и средние верхние кроющие крыла серые; большие кроющие белые и обра-
зуют сплошное белое пятно с второстепенными маховыми. Гнездовой пух очень светлосе-
рый. Клюв красный, но менее яркий, чем у самца; лапы оранжевые с темными перепон-
ками; радужина темнокоричневая, желтоватая по краям.
Молодой самец после первой осенне-зимней линьки. Голова и шея раскрашены
как у самки (хохол короче), они не перелинивают. На основании шеи сзади большая
примесь белых перьев. Спина серо-аспидная с отдельными черными перьями; бока тела
с серым поперечным рисунком.
Самец после первой летней линьки. Имеет оперение взрослого, но верхние
кроющие крыла аспидно-серые.
Взрослый самец в брачном наряде. Голова и верх шеи черные с зеленым отли-
вом, остальная часть шеи белая. Внутренние плечевые перья и верхняя часть спины
черные, остальная спина, надхвостье и хвост аспидно-серые. Бока поясницы и перья
голени с мелким серым крапом. Зашеек, зоб и весь низ тела белые с розовато-оранже-
вым налетом. Большие маховые и внешние второстепенные черно-бурые, внутренние же
белые, третьестепенные и наружные плечевые белые с черной каймой по наружному
опахалу (ширина которой не более 1,2 мм). Мелкие верхние кроющие крыла серые,
остальные же белые и образуют вместе с второстепенными маховыми большое белое пятно
поперек всего крыла. Розоватый (семужый) налет на груди и брюхе весьма нестоек; тон
и интенсивность его различны у разных особей; на снятых шкурках он быстро исчезает, то
же происходит обычно и при содержании в неволе. Клюв красный с черной вершиной;
лапы красно-оранжевые, радужина красная.
Самец после летней линьки. Имеет оперение схожее с самкой. Спина его окрашена
несколько темнее и на ней всегда сохраняется некоторое количество перьев брачного
наряда, которые в том или ином числе сохраняются и на других частях тела (грудь,
поясница). Легко отличим от самки и по окраске крыла; внутренние маховые короче,
чем в зимнем наряде, их внешние опахала светлосерые, внутренние темные.
40*
628 ОТРЯД ГУСЕОБРАЗНЫЕ
Гималайский большой крохаль Mergus merganser orientalis Gould
Mergus orientalis. Gould. Proc. Zool. Soc. Lond., 1, 1845, Амой в Южном Китае.
Русские названия. Южный большой крохаль, азиатский большой крохаль,
центральноазиатский крохаль, гималайский крохаль. Все названия книжные.
Распространение. Гнездовой ареал в пределах СССР занимает
большую часть Памира, где этот крохаль известен на озерах Ранг-Куль
и Яшиль-Куль, на реках Памире, Ак-Су и Мургабе. Обычен он также в
Рушане (Пасхув, Кала-и-Вамар) и Шугнане (Шах-Дара; Иванов, 1940).
Повидимому, к этому же подвиду должны быть отнесены крохали, гнездя-
щиеся на озере Искандер-Куль (северо-западный склон Гиссарского хребта;
Плеске, 1888). Севернее стайка крохалей этого подвида встречена в первой
половине сентября в Северо-Западной Монголии на р. Буянту недалеко
от Кобдо (добытый там экземпляр был неправильно определен Бианки как
М. senator, Сушкин, 1938). Учитывая значительную оседлость этой формы
и раннюю дату встречи, можно предполагать гнездованье ее там. В таком
случае Монгольский Алтай можно считать местом соприкосновения двух
подвидов большого крохаля. Основной ареал этой формы захватывает
Гималаи, Куэнь-Лунь, значительную часть Тибета (Бэнгс и Питере, 1928)
и доходит до Юго-Западного Ганьсу на северо-востоке и верховьев Хуанхэ
и Янцзе в провинции Юннань на востоке. Детали ареала очень неясны,
недостаточно ясен вопрос и о подвидовой принадлежности птиц, насе-
ляющих горные хребты Семиречья.
Зимовки. На Западном Памире крохали зимуют по всем крупным
рекам, так как последние никогда не замерзают благодаря своему быстрому
течению. В восточной части этой территории местами зимовки служат те
участки рек и озер, которые не покрываются льдом из-за обилия теплых под-
земных вод (Аличур, Булун-Куль; Мекленбурцев, 1946). Какая форма встре-
чается зимой в верхнем течении Аму-Дарьи—неизвестно. В Индии птицы
зимуют в южных предгорьях Гималаев, в местности между Гангом и Года-
вари и на холмах Ассама; встречаются они также в Северной Бирме и дости-
гают иногда Бомбейской бухты. Держатся в зимнее время в среднем течении
р. Ялу, а в Фудзяне добывались птицы, по своим признакам переходные
между двумя формами крохаля. Указания на то, что осенние птицы из
Маньчжурии (Ямасина, 1939) и все птицы, проводящие зиму в Корее (Остин,
1948), принадлежат к гималайскому подвиду, явно ошибочны.
Характер пребывания. Птицы, населяющие Памир и Гиссар-
ский хребет, оседлы; в Гималаях они совершают регулярные вертикальные
миграции, несколько расширяя зимой свой ареал; в Юго-Западном Ганьсу
оседлы, но птицы, населяющие Восточной Тибет и Юннань, лишь частично
зимуют на месте, в большей же своей части, повидимому, перелетны.
Даты сезонных миграций. На местах зимовки в северной
части Пенджаба эти птицы бывают обычны в январе и феврале (Уайстлер,
1926). Под Сталинабадом они встречены в декабре и в конце января (Иванов,
1949), а на Восточном Памире еще в середине февраля они не покидают
своих мест зимовки (на Булун-Куле).
Биотоп. Для гималайского крохаля характерен горный ландшафт.
Большую часть года проводит на горных реках и озерах, спускаясь в долины
лишь на короткое время, да и то далеко не везде и не всегда. В Восточном
Тибете он встречается на высоте 3300 м.
Численность. Обычная, но немногочисленная птица, даже на
пролетах и во время линьки не встречающаяся столь большими стаями, как
типичная форма.
Экология. О размножении этого крохаля известно очень мало.
Гнездится он по берегам рек и озер со сравнительно спокойным течением.
САВКА 629
Гнезда свои в большинстве случаев делает в скалах, иногда довольно далеко
от воды и на громадной высоте от днища долины. Два гнезда, найденные
Мекленбурцевым A946), помещались в глубоких нишах отвесных утесов
высотой до нескольких десятков метров. Ни к одному из них добраться не
удалось. Количество яиц и время их откладки неизвестны. На Памире птен-
цы появляются в начале июня, и матери сейчас же уводят их на речки, где
выводки ведут скрытую жизнь, прячась под нависающими берегами. Неко-
торые выводки держатся на озерках (Мекленбурцев, 1946). На озере Искан-
дер-Куль выводок из 9 птенцов встречен 25 июня, а 8 июля молодые уже
достигали там размера в половину самки. Нелетные, но уже крупные птен-
цы добыты в Рушане 28 июля и 7 сентября (Иванов, 1940). Молодые, сравняв-
шиеся размером со взрослыми, но еще не поднявшиеся на крыло, встречались
на Яшиль-Куле вплоть до 14 сентября (Мекленбурцев, 1946).
О линьке этой формы крохаля ничего неизвестно. На Памире перед
началом линьки большая часть селезней, покинувших своих самок, соби-
рается на озерках. Самец во вполне развитом зимнем наряде добыт 19 сен-
тября (Кала-и -Вамар).
Питание. В период гнездованья на Памире кормится рыбой, кото-
рая в это время в громадном количестве поднимается для нереста по рекам
и ручьям.
Экономического значения этот крохаль практически не имеет, так как
численность его невелика, а живет он в местах по большей части совсем не
населенных.
Описание. Строение и размеры. От типичной формы отличается несколь-
ко меньшими размерами тела и клюва. Крыло самца обыкновенно короче 280 мм, а у сам-
ки всегда короче 260 мм, обыкновенно же меньше 255 мм. Клюв самца 47,5—57 мм, обы-
кновенно короче 55 мм; у самки 40—47 мм.
Окраска. Взрослый селезень. Отличается от типичной формы более бледной
серой поясницей, гуще испещренной белой струйчатостью. Больше черного на плечевых
перьях и третьестепенных маховых. Ширина черных каемок на плечевых доходит
до 2,2 мм.
Взрослая самка. Отличается более светлой буро-каштановой окраской головы и шеи.
РОД САВКИ OXYURA BONAPARTF, 1828
Тип. Anas jamajcensis
Синоним. Erismatura, Bonaparte, 1832.
57. Савка Oxyara leucocephala S с о р о I i
Anas leucocephala. S с о р о 1 i. Ann. I, Hist Nat., 1768, стр. 65, Италия.
Синоним. Anas mersa. Pallas. Reised. versch. Prov. Russ. Reichs. II, 1773, стр. 713,
Зауралье.
Русские названия. Савка—народное; также синеносая утка; белоголовая утка.
Распространение. Ареал. На гнездовьях состоит из нескольких изо-
лированных друг от друга участков и лежит целиком в области засушливых
степей и пустынь. Первый участок находится в западной половине Среди-
земного моря. Он занимает юг Испании, Сардинию, Сицилию, самый юг
Италии, Западную Грецию и Албанию (озеро Скутари), а также побережье
Северной Африки от Марокко до Туниса. Кроме того, гнездованье савки
отмечено на озере Феленче в степной части Венгрии. Изолированным пунктом
гнездованья служит Горная Армения в районе озера Севан, где савка очень
обыкновенна на мелких озерах около Шорджи (Шелковников, 1934). У бере-
гов Черного и Азовского морей встречается крайне редко, лишь как залетная
птица. В низовьях Днепра известен всего один случай добычи (Шарлемань,
1924). Есть указание, что савка обычна на пролете под Павлоградом и ветре-
630
ОТРЯД ГУСЕОБРАЗНЫЕ
чается там изредка летом. В Александровских плавнях Днепра были добыты
молодые (Вальх, 1899). В дальнейшем ни в этих, ни в смежных местах савка
на гнездовье не отмечалась. Ее основной ареал начинается в степях При-
каспия и Нижнего Поволжья (Сарпинские озера), доходит на север примерно
до Сталинграда, занимает Волжско-Уральские пески и следует среднему
течению Урала. Севернее, в пределах Заволжья и Башкирии, зарегистриро-
ваны лишь отдельные точки, в которых савки, в числе нескольких пар, гнез-
дятся по нескольку лет подряд, но непостоянно. Это пруд у с. Языково
G0 км к западу от Уфы; Сушкин, 1897,1901) и пруд по р. Чембулатке (в Бу-
гурусланском районе; Карамзин, 1901). В Зауралье граница распростране-
Карта 78. Распространение савки Oxyura leucocephala
а—граница гнездовий, а'—районы наибольшей численности птиц, в—обособленные гневдо-
вые колонии, в'—места постоянных встреч летом (гнездованье не доказано), в—зимовки
ния поднимается к северу, и савка обычна на степных озерах в Кустанай-
ской и Челябинской обл. Далее северным пределом служат озера Горькой
линии в Ишимской степи (озеро Чаглы; Слудский), степные озера Тюкалин-
ского района и Барабинская степь, где савка обычна в южной части Чанов,
но лишь спорадично встречается севернее их (в мае на озерах Карачи; Руз-
ский, 1932). Гнездится всюду в Кулундинской степи, доходя до окрестностей
Барнаула, Приалейской степи, Семипалатинска и Змеиногорска. Естествен-
ным пределом ее распространения на восток служат северо-западные и запад-
ные предгорья Алтая. Добыта в сентябре на озере Зайсане, но вряд ли там
гнездится (Селевин, 1929). Далее граница ареала проходит на Ала-Куль
и следует на юго-запад вдоль предгорий Джунгарского Алатау и хребтов
Семиречья. Обычна по Нижней и Средней Сыр-Дарье, гнездится в Туркме-
нии по Аму-Дарье, Теджену и Мургабу, быть может в низовьях Атрека и по
Западному Узбою, но по восточному берегу Каспия и в пустынях между
нижним течением Урала и Аральским морем отсутствует. Гнездится на озерах
низменностей Северного и Юго-Западного Таджикистана (ст. Мельниково
в Ферганской долине, низовья Вахша. Пархар; Иванов, 1940). Наиболее
САВКА 63}
южным районом гнездованья служит Сеистанская озерная котловина на
границе Ирана и Афганистана (Зарудный, 1900). Изолированная от основ-
ного ареала гнездовая область находится в верховьях Енисея. Там савка
найдена на гнездовье, хотя и в небольшом числе, в Тувинской и Убса-
Нурской котловинах (Янушевич, 1948). Добывалась летом в озерной части
Ачинского района, но гнездованье ее там не доказано.
Районы линьки савки очертить трудно. Стаи селезней, поки-
нувших самок, встречаются на крупных озерах всех водных систем, на кото-
рых гнездятся эти птицы, но случаи добычи линных самцов, утерявших
способность к полету, пока неизвестна.
Основные зимовки в пределах Союза находятся у юго-вос-
точного побережья Каспия (Красноводский залив—Гасан-Кули), в то время
как у берегов Завкавказья савки являются лишь залетными птицами.
Кроме того, они обычны зимой на Мургабе и Теджене, а также в пределах
Ирана на Геррируде и в Сеистанской котловине, а в Ираке в низовьях Тигра
и Евфрата. Зимуют савки всюду в Северной Индии, спускаясь иногда до
Калькутты и Дели. Зимуют савки на северном побережье Африки и появля-
ются в это время в Нижнем Египте.
Характер пребывания. На побережьях Средиземного моря,
в Южной Туркмении и Сеистане савка оседлая птица. В остальных частях
ареала перелетная.
Сезонные миграции и их даты. Со средиземноморских
зимовок птицы отлетают рано, и уже в конце марта появляются в Венгрии
на озере Феленче, но на территории Союза савка одна из наиболее поздно
прилетающих уток, что говорит о том, что основная область зимовок наших
птиц лежит восточнее Средиземноморского бассейна. О том же свидетель-
ствует и отсутствие сколько-нибудь выраженного пролета савок у берегов
Азовского и Черного морей; в Болгарии редка (Патев, 1950), на Украине
пролет отмечен лишь для Запорожья 15—25 апреля (Вальх, 1899). Весенний
отлет их из Белуджистана начинается в феврале, но отдельные птицы задер-
живаются до апреля; в Ираке, где савки довольно редки, они отмечались до
21 апреля (Тайсхерст, 1922, 1927). В Красноводском заливе Каспийского
моря небольшие стайки савок попадаются еще в конце апреля, но в боль-
шинстве случаев так долго задерживается неполовозрелый молодняк. На
пресных озерах Западного Узбоя, первые птицы появляются в конце
марта, в низовьях р. Или тогда же, а в начале апреля они там уже
довольно обыкновенны (Штегман, 1949), однако пролет их там продолжается
и в конце апреля. По Уралу (под Чкаловом) савки пролетают весной едини-
цами, по Эмбе они не встречены вовсе, но восточнее Мугоджар в низовьях
р. Иргиза обычны. Первые отмечались с 26 апреля, причем летели они одиноч-
ками и парами, повидимому это были местные птицы. Валовой пролет стай-
ками до 30 шт. начался 29 апреля и закончился около 5 мая (Сушкин, 1908).
Во время пролета савки не пересекают горных хребтов, а огибают их, следуя
вдоль предгорий. На озере Кургальджин савки отмечались до 9 октября,
«а в низовьях Или последняя встреча зарегистрирована 16 октября. На озе-
рах Узбоя пролетные птицы встречались с конца сентября до второй поло-
вины октября, на местах зимовки у юго-восточных берегов Каспия они скапли-
ваются в большом числе уже к середине октября, а в начале ноября коли-
чество их еще возрастает. Белуджистана савки достигают в ноябре. Зиму
на море савки проводят в крупных стаях до 400—500 голов, причем взрослые
птицы держатся дальше от берегов, а молодые ближе к ним. Сильные штормы
заставляют их концентрироваться в заливах и бухтах.
Биотоп. Летом савки держатся на степных озерах, предпочтительнее
на солоноватых, чем на пресных. Озера эти обязательно имеют заросли
тростников и открытые плесы с подводной растительностью. В тростниках
632 ОТРЯД ГУСЕОБРАЗНЫЕ
птицы гнездится и держатся с молодыми птенцами, а остальное время прово-
дят на открытых плесах. Зимуют савки на больших открытых водоемах,
озерах или в морских заливах, чаще с каменистыми берегами. Во время
пролетов встречаются на водоемах различного типа, вплоть до рек, протекаю-
щих в скалистых берегах, но обычно на открытых степных озерах.
Численность. Савки весьма обычны на Кустанайских озерах
в системе Тургая и на озере Челкар-Тениз, в Кургальджин-Тенизском
озерном бассейне, в Кулундинской степи, в Прибалхашье и в нижнем тече-
нии Сыр-Дарьи, но распространены в этих районах весьма спорадично.
Численность савок в северных частях ареала сильно изменяется по годам.
Она резко повышается в периоды усыхания степных озер, сопровождающиеся
осолонением их, и сильно сокращается в многоводные годы (Формозов,
1949). В наиболее северных точках ареала савки то появляются на гнез-
довье, то исчезают совершенно, в результате чего северная граница их рас-
пространения пульсирует в разные годы. Массовым видом савку нельзя
считать нигде.
Экология. Размножение. Время наступления половой зрелости
неизвестно. На места гнездовья птицы появляются уже в парах. Токовые
позы селезней состоят в том, что они плавают вокруг самки с вздернутым
кверху и расправленным веером хвостом, раздувают грудь, вытягивают шею
и двигают ею взад и вперед, хлопая клювом по раздутой груди. Иногда
они прижимают клюв к груди и опускают его в воду, а затем сильным уда-
ром обеих ног толкают туловище вперед, поднимая при этом фонтан брызг.
Самки плавают с раскрытым клювом и время от времени вытягивают шею.
Гнезда савка устраивает пловучие, среди тростников, часто используя
для этого сплавины из старых стеблей тростника или старые гнезда других
птиц (лысухи, хохлатой чернети, белоглазого нырка). Гнездо представляет
собой постройку из обломков тростника и собранного поблизости раститель-
ного мусора, а лоток его бывает выложен пухом. Гнезда помещаются под
укрытием свисающих стеблей тростника или в глубине тростниковых зало-
мов и располагаются всегда непосредственно около пространства открытой
воды, хотя бы и небольшого. Последнее необходимо савке ввиду того, что
в случае тревоги она всегда уходит с гнезда, проныривая некоторое расстоя-
ние под водой. Самец держится поблизости от гнезда и спугнутая с гнезда
самка присоединяется к нему (Спангенберг и Фейгин, 1936).
Яиц в полной кладке обыкновенно 6 (Сыр-Дарья), в некоторых случаях
до 13 (по Генке), но весьма возможно, что это сдвоенные кладки. Каждый
день откладывается по 1 яйцу. Яйца очень велики и имеют форму широкого
эллипса. Скорлупа грубая, крупнозернистая, слегка шероховатая, грязно-
белого цвета. Их размеры (по трем кладкам с Нижней Сыр-Дарьи) 66,6—69,9 х
Х47,9—51 мм. Насиживающую самку никогда не удается застать в гнезде,
что объясняется обычно особенной осторожностью ее в это время. Однако-
можно высказать и другое предположение, что самка проводит на гнезде
мало времени, легко и надолго покидая его без вреда для яиц. Насиженные
яйца савки, взятые из гнезда где-то под Астраханью и находившиеся в ком-
натах без всякого подогрева, развивались нормально, всегда оставались теп-
лыми и вывелись через неделю (Генке). Можно предположить, что очень круп-
ные яйца савки нуждаются в постоянном согревании лишь недолгое время,
и развивающиеся в них эмбрионы очень скоро получают способность к само-
стоятельной терморегуляции (Мейер и Штреземан, 1928). Пуховые птенцы
имеют жесткие рулевые перья и держат свой хвост поднятым вверх, как это
делают взрослые птицы. Выводки с самками держатся у края тростников
и скрываются в них в случае опасности. Для кормежки они выплывают на
более открытые места только по вечерам. Самцы покидают самок, когда те
заняты насиживанием яиц. Участия селезней в заботах о выводке, как это
САВКА
633
указывается для близкого вида О. jamaicensis, у белоголовой савки не
наблюдалось.
Период размножения у савки проходит очень поздно, на Наурзумских
озерах брачные игры наблюдались еще 18 июня, а как исключение даже
16 июля. Начало гнездованья приходится там на середину июня. У самки,
добытой 6 мая в низовьях р. Или, яичник содержал еще мелкие желтки. 28 мая
найдено 7 гнезд, содержавших от 2 до 6 яиц, 8 июня гнездо с сильно насижен-
ными яйцами, а 9 июня гнезда с 1 и 2 яйцами (Спангенберг и Фейгин, 1936).
Выводок некрупных пуховиков встречен на озере Ала-Куль в Семиречье
11 июля (Шнитников, 1949), птенцы весом 125—167 г в Советском районе
Северо-Казахстанской обл. 24 июля (Слудский) и начавший оперяться уте-
нок в Наурзумском заповеднике 11 августа.
Рис. 114. Савка (самец и самка)
Л и н ь к а савок почти не изучена. Наличие двух различных сезонных
нарядов заставляет считать,что линька происходит, как у всех других утокг
дважды в год. Уже в начале июня на озерах в низовьях Иргиза самцы соби-
раются в большие и малые стаи, которые держатся на открытых плесах тех же
озер, где они провели гнездовой период, или на смежных. Когда именно про-
ходит полная послебрачная линька селезней—неясно, так как случаи добычи
их с выпавшими маховыми перьями неизвестны. Наряду с этим очень интере-
сен факт добычи в январе в Северной Индии линяющего селезня, неспособ-
ного к полету (Финн, 1901). Как расценивать этот случай—сказать трудно,
но следует иметь в виду, что у близкого вида О. jamaicensis выпадение махо-
вых происходит, повидимому, в октябре, а рост их заканчивается в ноябре,
уже на местах зимовки. Линька в брачный наряд, судя по коллекционным
материалам, происходит в апреле.
Питание савки состоит преимущественно из растительных кормов:
листьев, рдестов, валлиснерии, зостеры и хары, семян рдестов, руппии, ка-
мышей и резухи, а также из насекомых, преимущественно личинок хиро-
номид, небольшого количества моллюсков и ракообразных. Птицы, добытые
зимой на Каспийском море, имели в желудках икру какого-то беспозво-
634 ОТРЯД ГУСЕОБРАЗНЫЕ
ночного, моллюсков Hydrobia и других, а также водоросли Chara и Poly-
siphonia и семена Ruppia maritima.
Хозяйственное значение савки как промысловой птицы мало, так
как численность ее нигде не бывает высокой.
Полевые признаки. Спокойно плавающая птица сидит на воде довольно
высоко, но, испуганная, погружается так, что спина у нее скрывается под
водой и над поверхностью возвышаются только голова и поднятый кверху
хвост. У взрослого самца издалека видна белая голова, а вблизи различим
также лазорево-синий клюв. Молодые птицы однообразно буро-серые. Савок
почти никогда не удается увидеть на земле, так как ходят они с трудом. Все
время проводят на воде, плавают и ныряют прекрасно. Взлетают крайне
неохотно. От опасности обычно стараются спастись ныряньем и скрыться
в тростниках. Птицы в обычное время весьма молчаливые, но в период уха-
живания за самками селезни издают пискливые и гогочущие звуки.
Описание. Строение и размеры. Утка среднего размера с округлым и
компактным телом, с короткой и очень толстой шеей и большой головой. Надклювье широ-
кое, вздутое у основания, образует седлообразный прогиб в средней части. Ноготок неболь-
шой и крючкообразно изогнутый. Хвост длинный из 9 пар узких и чрезвычайно жестких
рулевых. Он имеет ступенчатую форму, так как крайние перья равны обычно половине
длины средних. Перья надхвостья и подхвостья очень короткие и едва прикрывают основа-
ние хвоста. Савка наиболее приспособленный к нырянию вид среди всей группы уток.
Соответственно этому она имеет самые короткие крылья, которые в сложенном виде полно-
стью закрываются покровным оперением и не затрудняют благодаря этому передвижение
под водой. Ее ноги отнесены очень далеко назад, лапы очень велики, а мускулатура
задних конечностей обладает необыкновенной мощностью, что делает савку наиболее ис-
кусным и неутомимым пловцом. Крылышки савки несоразмерно малы по сравнению с боль-
шим и тяжелым телом этой утки. Размах ее довольно узких крыльев превышает длину
тела всего в 3,3 раза, несомненно, что савка вообще одна из самых малокрылых птиц, спо-
собных летать. Полет у савки стремительный с очень быстрыми ударами крыльев. Она
неспособна делать резкие повороты, значительно изменять скорость полета или быстро
взмывать вверх. Для взлета савке необходим постепенный разбег по воде, переходящий
в глиссирование. При посадке она также некоторое время стремительно скользит по воде.
Таким образом, взлетать савка может только с воды и опускаться лишь на воду. В обычное
время савки летают очень редко, хотя и способны совершать большие перелеты во время
их сезонных миграций (Штегман, 1950). Длина крыла (самцов и самок) 147—160 мм,
цевки 34—37 мм, клюва 46—50 мм; хвост самцов 110—120 мм, самок до 95 мм.
Вес. Данных о весе савок мало. Самки весной, до начала кладки, в апреле 720 г,
в мае 900 г; самцы в апреле 755 г G20—800 г) (низовья р. Или; Слудский); там же весной
(без указания пола) 750—760г (Штегман, 1949); в сентябре-октябре на озере Кургальджин
(без указания пола) 505—820 г, (Меженный и Соколов). В октябре в пресных озерах Узбоя,
вероятно молодые, самец 650 г, самки 400—500 г (Шестоперов), в ноябре в Красноводском
заливе молодые самки 600—650 г.
Окраска. Пуховой птенец. Верх и бока головы почти черные, а остальная спин-
ная сторона темнобурая, под глазом проходит четко выделяющаяся светлая полоска.
Подбородок белый, шея и грудь буроватые, остальная брюшная сторона светлосерая.
Молодые птицы в первом наряде. Похожи на самку, но тон окраски спины и боков
менее рыжий. На спинной стороне тела неправильный черный струйчатый рисунок. Полы
у них различать трудно, но у молодых самцов на голове и шее рисунок более ровный, а на
зобе ржаво-коричневый налет, отсутствующий у самок. Клюв светлосвинцового цвета,
лапы светлокоричневые, радужина серовато-белая.
Взрослая самка. Верх головы и щеки темнобурые; от клюва под глазом прохо-
дит широкая светлая полоса, пестрящая мелкими бурыми перышками. Вся спинная
сторона светлая желтовато-ржаво-коричневая с черной поперечной полосатостью. Под-
бородок и верхняя часть горла почти белые, а бока шеи беловатые. Брюшная сторона
грязно-беловато-желтая с серыми пятнышками и поперечными полосками. Клюв темный
свинцового цвета; лапы свинцово-серые со слегка красноватым оттенком на цевке и паль-
цах. Радужина светложелтая.
Взрослый самец в брачном наряде. Имеет белую голову с черной шапочкой на
темени и черный ошейник, более широкий сзади,а спереди достигающий подбородка.
Вся спина и бока тела серовато-ржавчато-бурые с мелким черным крапом, верхние
кроющие хвоста густо ржаво-бурые с фиолетовым отливом. Низ шеи, зоб и лопатки рыже-
коричневые с черным крапом. Грудь и брюшко серо-коричневые с серыми поперечными
полосами, подхвостье светлоохристое. Хвост почти черный. Зеркала на крыльях нет. Кро-
ющие крыла и маховые бурые с белокрапчатыми наружными опахалами, первостепенные
САВКА
635
маховые без крапа и с темными концами. Клюв небесно-голубого цвета, особено яркий в
период размножения. Лапы красновато-серые с черноватыми перепонками и сустава-
ми. Радужина яркожелтая или даже померанцевого цвета.
Взрослый самец после полной линьки (зимой). Имеет чистобелую голову с черным
теменем и буровато-серую шею с более темным пятном на затылке. Зоб серовато-бурый,
низ сильно испещрен темносерыми пятнами. Рули темносерые. (Тугаринов, 1940).
У некоторых июньских и некоторых весенних экземпляров голова и шея почти целиком
черные с некоторым количеством белых перьев на щеках, боках шеи и горле. Следует ли
этот наряд считать сезонным для взрослых самцов или возрастным для неполовозре-
лых—сказать пока нельзя.
Дополнение к тому третьему
ОТРЯД КУЛИКИ
РОД ПЕРЕПОНЧАТОПАЛЫЕ ПЕСОЧНИКИ EREUNETES, ILLIGER, 1811
Тип. Tringa pusilla
Перепончатопалый песочник Ereunetes maurii С ab.
Ereunetes maurii. C a b a n i s. Journ. f. Orn., 1856 A857), стр. 410, Куба.
Распространение. Птица эта основной частью ареала принадлежит
прибрежным тундрам Аляски, где гнездится от дельты Юкона до мыса Бар
роу, гнездится и на о-ве Нунивок. Согласно Нельсону A887), она найдена
в некоторых местах северо-восточного берега Сибири, но так как ряд
указаний Нельсона о нахождении им птиц на территории Сибири ока-
зался в дальнейшем не подтвержденным, то это свидетельство также оста-
валось под сомнением, и перепончатопалый песочник в состав фауны птиц
СССР не включался. Теперь исследованиями Портенко выяснено, что птица
эта несомненно гнездится на Чукотском п-ве. Зимовки расположены
от штатов Северная Каролина и Вашингтон на севере до Венецуэлы и Перу
на юге. Прилет на места гнездовья на Аляске происходит в средних числах
мая. Гнездовое местообитание довольно разнообразно—от сильно увлажнен-
ных тундр до скалистых, покрытых мохом склонов гор.
Экология. В кладке 4 яйца. Полные кладки у бухты Хупера обычно
26—30 мая. Длительность насиживания 18—19 дней, пуховые птенцы поя-
вляются в середине июня. Размер яиц A20J8,7—32,5x21,1—21,8 мм,
среднее 30,7x21,8 мм (Брандт, 1943).
Полевые признаки. Ввиду большого внешнего сходства этой птицы
с песочниками полагаться в определении ее на глаз невозможно; взятая
в руки узнается по наличию при основании пальцев хорошо развитой пере-
понки.
Описание. Длина крыла 90—99,5 мм (Портенко и Штегман, 1951). В летнем наряде
имеет рыжеватую шапочку, верхнюю сторону тела охристо-серую с черными, а на плечах:
с рыжими пятнами. Грудь бледнобуроватая с черно-бурыми пятнышками.
ОГЛАВЛЕНИЕ
Отряд куриные
Стр.
Характеристика отряда (Г. 77. Дементьев) 2
Таблица для определения видов куриных птиц СССР . . : 8
Семейство тетеревиные (А. В. Михеев) 10
Род белые куропатки 10
1. Белая куропатка 10
Лапландская белая куропатка 11
Среднерусская белая куропатка 31
Северосибирская белая куропатка 33
Большая белая куропатка 35
Малоклювая белая куропатка 39
Восточносибирская белая куропатка 40
Сахалинская белая куропатка 41
2. Тундреная куропатка 42
Лапландская тундреная куропатка 43
Сибирская тундреная куропатка 47
Алтайская тундреная куропатка 49
Командорская тундреная куропатка 51
3. Тетерев 52
Североевропейский тетерев 52
Степной тетерев 62
Енисейский тетерев 71
Тяньшаньский тетерев 72
Байкальский тетерев 72
Уссурийский тетерев 76
4. Кавказский тетерев 78
Род глухари (С. В. Кириков) 84
5. Глухарь 84
Белобрюхий глухарь 87
Таежный темный глухарь 96
Чернобрюхий западноевропейский глухарь 100
6. Каменный глухарь 103
Камчатский каменный глухарь 104
Темный каменный глухарь 106
7. Дикуша (А. В. Михеев) 107
8. Рябчик 112
Восточноевропейский рябчик 113
Сибирский рябчик 118
Амурский рябчик 127
Семейство фазановые 133
Род перепелы (#. И. Картсаиев) 133
9. Обыкновенный перепел 133
Обыкновенный перепел 134
Немой, или восточносибирский перепел 146
638 ОГЛАВЛЕНИЕ
Род пустынные куропатки 149
10. Пустынная куропатка 149
Туркестанская пустынная куропатка 149
Род каменные куропатки 154
11. Кеклик, или каменная куропатка 154
Кавказский кеклик 155
Копетдагский кеклик 160
Западнотуркменский равнинный кеклик 162
Восточноиранский кеклик 164
Кызылкумский кеклик 165
Тяньшаньский кеклик 167
Кара-Корумский кеклик 171
Джунгарский кеклик 172
Кашгарский кеклик 173
Род Турач 174
12. турачи 174
Европейский турач : 174
Род Улары 180
13. Кавказский улар 180
14. Каспийский улар 184
Армянский каспийский улар 184
Южнокаспийский улар 187
15. Гималайский, или темнобрюхий, улар 188
Туркестанский темнобрюхий (или гималайский) улар 188
16. Алтайский улар 193
17. Тибетский улар 197
Западный тибетский улар 198
Род фазаны (Я. А. Гладков) 199
18. Обыкновенный фазан 199
Закавказский фазан 201
Северокавказский фазан 204
Талышский фазан 206
Персидский фазан 207
Мургабский фазан 207
Аму-Дарьинский фазан 209
Таджикистане кий фазан 211
Хивинский фазан 216
Зеравшанский фазан 216
Сыр-Дарьинский фазан 218
Семиреченский фазан 220
Маньчжурский фазан 223
Род куропатки (Я. Я. Карташов) 226
19. ?ерая куропатка 227
Среднеевропейская серая куропатка 227
Среднерусская серая куропатка 230
Закавказская серая куропатка 235
Сибирская серая куропатка 237
20. Бородатая куропатка 241
Сибирская бородатая куропатка 242
Маньчжурская бородатая куропатка 246
Отряд гусеобразгые
Характеристика отряда (Е. С. Птушенко) 247
Таблица для определения видов гусеообразных 252
Семейство утиные 255
Подсемейство гусиные 255
Род лебеди ч 255
1. Лебедь-кликун 255
Материковый лебедь-кликун 256
2. Тундровый лебедь 262
Западный тундровый лебедь 262
Восточный тундровый лебедь 267
3. Американский лебедь 268
4. Лебедь-шипун 270
ОГЛАВЛЕНИЕ
Род сухоносы 276
б.Сухонос 276
Род голубые гуси 280
6. Белошей 280
Род индийские гуси 284
7. Горный гусь 284
Род белый гусь 289
8. Белые гуси 289
Малый белый гусь 290
Род гуси . 294
9. Серый гусь ; . 294
10. Белолобый гусь 301
Западный белолобый гусь •. . . . 301
11. Пискулька 308
12. Гуменник 312
Западный тундровый гуменник 313
Восточносибирский тундровый гуменник . 319
Таежный гуменник 321
Короткоклювый гуменни< ... 324
Род краснозобые казарки 326
13. Краснозобая казарка 326
Род казарки 331
14. Черная казарка 331
Атлантическая черная казарка 331
Евразийская черная казарка 333
Восточносибирская черная казарка 338
Американская черная казарка 338
15. Белощекая казарка 341
16. Канадская казарка 341
Средняя канадская казарка ... 343
Малая канадская казарка 344
Темногрудая канадская казарка • 344
Подсемейство утки (Ю. А. Исаков) 344
Род пеганки 344
17. Пеганка 344
18. Огарь 353
19. Хохлатая пеганка 361
Род настоящие или речные утки 362
20. Узконосый чирок 362
21. Шилохвость 366
Чирок-свистунок 383
22. Чирок-свистунок 382
23. Чирок-клоктун 395
24. Касатка 401
25. Кряква 405
Кряква 406
26. Желтоносая кряква 424
Черная кряква 425
27. Серая утка 428
Серая утка 429
28. Свиязь 437
29. Американская свиязь 449
30. Чирок-треску нок 451
31. Широконоска 461
Род красноносые нырки * 471
32. Красноносый нырок 471
Род чернети 478
33. Красноголовый нырок 478
Красноголовый нырок 479
34. Белоглазый нырок 488
35. Нырок Бэра 494
36. Хохлатая чернеть 497
37. Морская чернеть 507
Род мандаринки 514
38. Мандаринка 514
640 ОГЛАВЛЕНИЕ
Род гаги 519
39. Гага обыкновенная 519
Обыкновенная гага 520
Гренландская гага 527
Тихоокеанская гага 529
40. Гага-гребенушка 531
41. Очковая гага 538
42. Сибирская гага 541
Род турпаны 545
43. Синьга 545
44. Тихоокеанская синьга 550
45. Пестроносый турпан 552
46. Черный турпан 555
47. Горбоносый турпан 560
Американский турпан 560
Восточносибирский турпан 562
Род каменушки 566
48. Каменушка 567
Каменушка сибирская 571
Каменушка атлантическая 572
Род морянки 572
49. Морянка 572
Род гоголи 582
50. Гоголь обыкновенный 582
51. Исландский гоголь 595
52» Гоголь-головастик 597
Род крохали 598
53. Луток 598
54. Длинноносый, или средний крохаль 606
55. Чешуйчатый крохаль 614
56. Большой крохаль 617
Обыкновенный большой крохаль ." 619
Гималайский большой крохаль 628
Род савки 629
57. Савка 629
Дополнение к тому третьему: отряд кулики (Я. А. Гладков) 636
Род перепончатопалые песочники 636
Перепончатопалый песочник 636
Редактор В. И. Сидорова Технич. редактор Л. И. Стрельникова
Т 07940 Подп. к печати 30.IX.52 г. Ти.раж 5000 экз.
Бумага 70 X 108Vm =¦ 20 бум. л. = 40 печ. л. + 6 цв. вклейки. = 62 уч. изд. л.
Цена в переплете 32 руб. 50 коп.
Изд. № 57. Заказ 788
Набрано в 16-й типографии Главполиграфиздата при Совете Министров СССР
Москва, Трехпрудный пер., 9.
Отпечатано во 2-й типографии Издательства Академии Наук СССР
Москва, Шубинский пер., д. 10.
Том 4-й содержит описание двух отрядов птиц: куриных и
гусеобразных. Отряд куриных описан Н. А. Гладковым,
Н. Н. Карташевым, С. В. Кириковым, А. В. Михеевым; гусе-
образных — Ю. А. Исаковым, Е. С. Птушенко.