/















































































































































































































































































































































































































































































































































































































































































Автор: Дементьев Г.П. Спангенберг Е.П. Гладков Н.А. Птушенко Е.С. Судиловская А.М.
Теги: птицы фауна ссср орнитология птицы советского союза
Год: 1951




Текст
Пти цы
СОВЕТСКОГО
СОЮЗА
Г. П. ДЕМЕНТЬЕВ, Н. А. ГЛАДКОВ,
Е. С. ПТУШЕНКО, Е. П. ПАНГЕНБЕРГ,
А. М. СУДИЛОВСКАЯ
ПТИЦЫ
СОВЕТСКОГО
СОЮЗА
Том I
Под общей редакцией
Г. П. Дементьева и Н. А. Гладкова
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО
„СОВЕТСКАЯ НАУКА"
Москва—1 951
Том 1 содержит описание девяти отридол птиц.
Отряды хищных и со» описаны Г. П. Дементьев ы м, дятлов — Н. А. Гладко-
в ы м, стрижей — Е. С. П т у ш с и К о, коаодоен — Е. П. С п а и г е и б е р г о м,
веслоногих, кукушек, ракш и удодов — Л. М, С у д и л о в с к о й.
Отп. редактор //. И. Кузнецов
Рисунки художников: А. И. Комарова, Н. Н. Кондакова и В. А. Ватагина.
Ведущий редактор Л. П. Костина. Художественная редакция К. И. Журинской.
Технический редактор А. И. Прояева.
^^4^ГГ1одпис7шо и печать 23 ЛХ.50 гГ'Тйраж 10.000 (1 —5000) экз. Объем 403/4 п. л. 4цветн.
вклейки. Учгавт. 04,0. Формат 70 х ЮНУю. Цена в переплете 35 руб. Заказ № 679.
2-я типография «Печатный Двор» им. А. М. Горького Главполиграфиздата при Совете
Министров СССР. Ленинград, Гатчинская, 26.
ПРЕДИСЛОВИЕ
Птицы — обширный класс позвоночных животных, состоящий более чем
из 8000 видов, из которых на территории Советского Союза до настоящего
времени известно около 700 видов. По числу обитающих в нашей стране особей
это, несомненно, наиболее многочисленная группа среди наземных
позвоночных. Распространены птицы чрезвычайно широко и встречаются во всех
районах нашей страны — от островов и побережий Северного Ледовитого океана
до жарких пустынь и высоких гор крайнего юга Союза. Естественно поэтому,
что жизнь птиц представляет для нас не только и не столько теоретический
интерес (конечно, весьма большой), но имеет и огромное практическое значение.
Это значение весьма многообразно, прежде всего, в сельском и лесном,
хозяйстве. Без преувеличения можно сказать, что роль птиц в деле борьбы
с вредителями,, в деле защиты урожая и защиты наших лесов, огромна.
Биологические методы борьбы с вредителями хозяйства имеют большое значение,
а самым дешевым и вероятно одним из самых эффективных методов такой
борьбы с вредителями служит использование их естественных врагов.
Особенную важность этот вопрос приобретает теперь в связи с историческим
постановлением партии и правительства о полезащитных насаждениях и
о других мероприятиях, обеспечивающих высокие и устойчивые урожаи.
Не следует, конечно, думать, что польза птиц ограничивается истреблением
или снижением численности вредных насекомых и грызунов. Например,
зерноядные птицы могут приносить и фактически приносят пользу уничтожением
семян сорняков. Птицы имеют значение и как истребители переносчиков
заразных болезней.
Общеизвестна польза, приносимая охотничье-промысловыми птицами,
добываемыми для пищи> получения пуха, пера и т. п. Стоимость товарной
продукции дичи в дореволюционной России — при отсутствии достаточно
полной статистики — исчислялась миллионами рублей. Надо при этом иметь
в виду, что значение охотничьих птиц в качестве местного источника пищевых
продуктов особенно велико и до сих пор. К тому же охота на птиц — один из
лучших видов спорта, воспитывающий выносливых, наблюдательных и метких
стрелков и разведчиков.
Домашние птицы издавна занимают значительное место в нашем хозяйстве.
Нельзя недооценивать и эстетически-воспитательных сторон изучения птиц,
в частности — любительского содержания певчих птиц. Оно несомненно
способствует развитию интереса и любви к природе нашей великой Родины.
Само собою разумеется, что широкое и разностороннее использование
птиц невозможно без знания состава и распределения авифауны Советского
4
ПРЕДИСЛОВИЕ
Союза, а в особенности образа жизни отдельных видов в конкретных условиях
различных частей страны. Тем более, что птицы, как мы уже отметили,
встречаются в самых разнообразных условиях и представляют собою яркий пример
и важный предмет изучения пригнанное™ животного организма к условиям
существования.
Однако разрешение указанных выше вопросов в значительной степени
затрудняется отсутствием обобщающих сводок материалов о птицах.
Орнитологическая литература, касающаяся авифауны Советского Союза, весьма
обширна и насчитывает много тысяч названий. В особенности значительный
размах орнитологические исследования приняли после Великой Октябрьской
революции. Но литература эта, разбросанная по множеству отдельных изданий,
трудно доступна даже для специалиста.
После выхода в свет в 1895 году известной книги М. А. Мензбира
«Птицы России», на которой воспитался ряд поколений наших зоологов и
любителей природы, в литературе не появлялось более доведенных до конца сводок
сведений о птицах нашей страны. За истекшие полвека изучение животного
мира получило большое развитие. Естественно поэтому, что превосходная
книга М. А. Мензбира уже не может удовлетворять наших запросов,
тем более, что она относится только к части территории Союза (европейской
России и Кавказу).
Авторы и поставили перед собою задачу дать читателю сводку всех
имеющихся сведений о птицах нашей страны. Это тем более важно, если учесть то
обстоятельство, что сводка всех разбросанные в обширной литературе и
накопившихся лично у них сведений по биологии птиц может наконец выяснить
особенности образа жизни птиц в разных климатических и иных условиях,
способствовать разрешению чрезвычайно важного вопроса о «географических»
изменениях экологии видов, главным образом, широко распространенных,
сопоставить это явление с определенными морфологическими особенностями и т. п.
Такого рода обобщающие работы имеют всегда и существенное практическое
значение.
Поэтому при составлении книги авторы главное внимание уделяли
вопросам географического распространения ив особенности образа жизни птиц.
При описании распространения авторы отошли от обычного в подобного рода
монографиях шаблона, характеризуя его не только ареалом, но и биотопами
и особенностями сезонного размещения. Использованы накопившиеся за
последние двадцать лет результаты кольцевания птиц в Союзе.
Главное внимание обращено на выяснение всех подробностей
распространения вида в пределах Советского Союза. Распространение видов вне нашей
страны дается главным образом по лучшим и новейшим работам справочного
характера, без претензии на критический разбор всей относящейся к
зарубежным странам фаунистической литературы.
При описании образа жизни птиц полностью использованы все
доступные авторам материалы и литература. При этом круг вопросов неизбежно
пришлось ограничить важнейшими сторонами биологии видов, в
особенности периодическими явлениями. В этом отношении авторы следовали
примеру основоположника современной экологии Н. А. Северцову:из
периодических явлений с возможной подробностью рассматриваются вопросы
размножения, сезонного размещения и линьки. Вопросам питания в книге
уделяется несколько меньшее место. При всем стремлении обобщить материал
авторы старались приводить его с предельно возможной конкретностью,
исходя из указанной выше теоретической задачи исследования и чисто
практических целей.
В основе разбора вопросов, относящихся к систематике, положено
главным образом изучение коллекций Зоологического музея Московского ордена
ПРЕДИСЛОВИЕ
5
18.
19.
20.
21.
22.
23.
24.
25.
26.
Хищные
Совы
Кукушки
Козодои
Ракши
Удоды
Дятлы
Длиннокрылые
Воробьиные
Ленина государственного университета имени Ломоносова. Однако по
возможности, в особенности в отношении мало изученных и редких в коллекциях
видов, изучались материалы и других собраний.
Много времени и труда было затрачено на составление карт
распространения птиц фауны Советского Союза, тем более, что авторы поставили перед
собою задачу изобразить ареалы видов в целом, а не только в пределах СССР,
отразив при этом сезонное размещение, и географическую изменчивость.
Весь материал располагается в шести томах, включающих описание 26
отрядов птиц, представленных в авифауне СССР. Сюда входят отряды:
1. Куриные 14. Г у с и н ы е или пластин-
2. Трехперстки чатоклювые
3. Голуби 15. Веслоногие
4. Рябки 16. Голенастые
5. П а с т у ш к и 17. Краснокрылы'
6. Журавли
7. Дрофы
8. Кулики
9. Чайки
10. Чистики
11. Гагары
12. Поганки
13. Трубконос ые
Последовательность описания отрядов в книге не соответствует этой
системе. В первом томе помещено описание отрядов веслоногих, хищных, сов,
кукушек, козодоев, ракш, удодов, длиннокрылых и дятлов. Во втором —
голубей, рябков, трехперсток, журавлей, дроф, чистиков, гагар, поганок, трубко-
носых, голенастых и краснокрылов. В третьем томе описаны отряды куликов,
чаек и пастушков. Остальные отряды, кроме воробьиных, будут описаны
в четвертом томе. Описание воробьиных будет составлять содержание пятого
и шестого томов.
При описании каждого отряда дается определительная таблица его
представителей в СССР, а также общая характеристика отряда. Пояснение
анатомической терминологии, которой авторы при этом пользовались, легко можно
найти в соответствующих зоологических руководствах (например, в шестом
томе «Руководства по зоологии», 1940, или в «Общей орнитологии» Шульпина,
1940).
Описанию отдельных отрядов предшествует общая определительная
таблица, позволяющая отнести птицу к тому или иному отряду. Определительные
таблицы построены по так называемому дихотомическому способу. То-есть
признаки, по которым производится определение, скомбинированы во взаимно
исключающие друг друга пары. Определяющий должен сравнивать
интересующую его птицу с признаками, отмеченными в таблице, начиная с признака,
отмеченного первым номером, и сравнивать то, что он видит на птице, как
с признаком, отмеченным в первом номере, так и с противоположным ему.
Чтобы легче находить противоположный признак, его номер ставится в
скобках рядом с номером основного признака. Подходящий признак или приводит
к названию птицы, или к другому номеру (признаку), который надо опять
сравнить и т. д. Определение продолжается до тех пор, пока не дойдет до
названия птицы. После этого необходимо проверить определение по имеющемуся
в тексте описанию птицы.
Следует иметь в виду, что определительные таблицы рассчитаны только
на виды птиц, встречающиеся в СССР (то-есть на тех птиц, о которых идет речь
6
ПРЕДИСЛОВИЕ
в книге), и ограничиваются теми возрастами и нарядами, относительно которых
пользование таблицей может быть вполне надежным. В некоторых случаях
составить определительную таблицу, по которой можно определить птицу
в любом возрасте и наряде, еще не представляется возможным.
Для того, чтобы легче было уяснить терминологию отдельных частей тела
птицы, мы даем соответствующий рисунок — схему (рис. 1).
Рис. 1. Название частей тела и оперения птицы (из С. А. Бутурлина, 1934).
А — спинная, дорзальная или верхняя сторона тела; В — брюшная, вентральная или нижняя сторона
тела; 7 — лоб; 2 — темя; 3 — затылок; 4 — уздечка; 5 — бровь; 6 — щека; 7 — кроющие уха, ухо; 8 —
верхняя челюсть или надклювье; 9 — нижняя челюсть или подклювье; 10 — конек или хребет
надклювья—линия от середины основания клюва у лба до вершины (конца) клюва; 7/ — края челюсти
или разрез рта; 12 — угол рта; 13 — горло; 14 — передняя или нижняя часть шеи и зоб; 15 — грудь;
16 — брюхо; 17 — верхняя или передняя часть спины; 18 — нижняя или задняя часть спины; 19 —
плечевые или лопаточные перья; 20 — первостепенные маховые. Маховые считаются по порядку от
наружного края крыла внутрь, ближайшие к краю крыла маховые называются передними или наружными>
противоположные им, т. е. расположенные ближе к туловищу — задними или внутренними; 21 —
второстепенные маховые; 22 — задние второстепенные или третьестепенные маховые; * 23 — малые верхние
кроющие крыла или малые кроющие; первые расположены на мясистой передней части крыла,
вторые — на передней летательной перепонке, так называемой патагиальной или патагиуме; 24 — средние
верхние кроющие крыла или средние кроющие; 25 — большие верхние кроющие предплечья или
большие кроющие; 26 — большие верхние кроющие кисти или кроющие кисти (нижние кроющие
крыла все вместе называются обычно подкрыльем); 27 — крылышко, твердые перья, прикрепленные к
большому пальцу; 28 — надхвостье или верхние кроющие хвоста; 29 — рулевые или правильные перья
(рулевые перья считают от боковой или крайней пары к середине хвоста); 30 — подхвостье, нижние
кроющие хвоста или хлупь; 31 — цевка или плюсна; 32 — задний или первый палец; 33 — внутренний
или второй палец; 34 — средний или третий палец; 35 — наружный или четвертый палец; 36 — пятка;
37 — передний или кистевой сгиб крыла; 38 — голень; 39 — бок (передняя часть его является собственно
боком груди, а задняя — боком брюха); 40 — поясница; 41 —- подбородок (расположен между ветвями
нижней челюсти); 42 — верхняя сторона шеи; 43 — бока шеи.
Общая длина птицы измеряется по спине—от конца клюва до конца хвоста,
размах крыльев — по спине между крайними точками максимально
растянутых крыльев. Длина крыла измеряется от кистевого сгиба крыла до конца
самого длинного махового пера. Длина хвоста измеряется от точки, где средняя
пара хвостовых (рулевых) перьев входит в кожу, до конца самого длинного
из этих перьев. Напомним, что при измерении крыла и хвоста надо следить,
доросло ли перо до своего полного развития (недоросшие перья одеты обычно
у основания «пеньками», роговым чехликом), и не оббит ли в результате изна-
ПРЕДИСЛОВИЕ
7
шивания его конец. Цевка (неправильно называемая у птиц плюсной)
измеряется от впадины сустава у задней поверхности ее верхнего конца до
соединения ее передней поверхности с основанием среднего пальца. Клюв
измеряется по прямой линии от места окончания лба, то-есть от места схождения
оперения с голым покровом хребта клюва, до самого конца верхней челюсти.
Представление об этих способах измерения дают прилагаемые рисунки (рис. 2);
Рис. 2. Способы измерения частей тела птицы.
а — крыло, б — хвост (от основания средней пары перьев до конца самого длинного рулевого), в — из*
мерение цевки, г — длина клюва от оперения лба до вершины.
Данные по размерам приводятся нами в книге следующим образом. В
скобках дается число промеренных экземпляров, затем дается минимальный и
максимальный размер, затем среднее. При этом сначала даются крайние
величины размеров для каждого пола отдельно и лишь после этого — средние для
самцов и для самок. Если измерения даются по литературным источникам, и
имеется несколько таких источников, мы даем соответственно несколько серий
измерений, указывая после каждой серии литературный источник (в скобках).
Работа, предлагаемая вниманию читателей, представляет собою в основном
труд зоологов Московского университета, но к участию в ней привлечены и
зоологи других научных учреждений Советского Союза. Имена авторов
соответствующих разделов указаны на титульном листе и в оглавлении.
Г Дементьев
Н. Гладков
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ОТРЯДОВ ПТИЦ
1 (2). Задний палец соединен полной перепонкой со внутренним пальцем (рис. 3)
Отр. Веслоногие — Steganopodes.
2 (1). Задний палец не соединен полной перепонкой с внутренним пальцем 3
3 (4). Ноздри в одной или в двух трубочках (рис. 4). Ротовые щетинки отсутствуют
Отр. Трубконос ые — Tubinares:
4 (3). Ноздри не в трубочках. Если же ноздри открываются в трубкообразных
образованиях, то имеются ротовые щетинки . 5
5 (б). Края надклювья и подклювья образуют многочисленные поперечные или косые
пластинки или же ряд хорошо развитых зубцов (рис. 5) 7
6 (5). Края надклювья и подклювья совершенно гладкие, во всяком случае не
образуют никаких пластинок или зубцов 9
7 (8). Обнаженная часть голени занимает пространство не более 40 мм
Отр. Гусиные — Anseres.
8 (7). Обнаженная часть голени занимает пространство не менее, чем в 150 мм. . . .
Отр. Краснокрылы — Phoenicopteri.
9 (10). Ноги далеко отнесены назад (рис. 6, //). Хвост укороченный. Три передних
пальца соединены полными перепонками или же каждый палец несет отдельную лопасть
Хрис.7 и 8) • 11
10 (9). Ноги отходят приблизительно у середины туловища (рис. 6, /). Хвост из
удлиненных рулевых. Если же хвост укороченный, то три передних пальца не соединены
полными или выемчатыми перепонками (рис. 9) или же, в крайнем случае, каждая фаланга
пальцев несет отдельную лопасть 15
11 (12). Заднего пальца нет Отр. Чистики — Alcae.
12 (11). Задний палец имеется 13
13 (14). Три передних пальца одеты каждый в отдельности особой лопастью (рис. 7)
Отр. П о га н к и — Colymbi.
14 (13). Три передних пальца соединены вместе перепонками (рис. 8)
Отр. Гагары — Gaviae.
15 (16). Передние пальцы соединены перепонками почти на всем своем протяжении
или хотя бы до половины (рис. 9) Otp. Чайки — Lari.
16 (15). Передние пальцы соединены перепонками лишь у своих оснований или же
свободны до самого основания * 17
17 (18). Нижняя часть голени не оперена (за исключением малой выпи lxobrychus
minutus, у которой уздечка голая, клюв длинный и прямой, голова плоская) 19
18 (17). Нижняя часть голени оперена 27
19 (20). Уздечка или пространство вокруг глаза или и то и другое вместе лишены
перьев (рис. 10) Отр. Голенастые — Gressores.
20 (19). Уздечка и пространство вокруг глаза полностью оперены 21
21 (22). Видимое 1-е маховое длиннее 2-го или равно 2-му (в некоторых случаях, как
например, у чибиса — Vanellus, кречетки — Chettusia иавдотки Burhinus, оно
короче 2-го). Настоящее первое маховое узкое, острое и скрыто кроющими крыла (рис. 11)
Отр. К у л и к и — Limicolae.
22 (21). Видимое 1-е маховое короче 2-го. Настоящее 1-е маховое совершенно
рудиментарно и неразличимо см. 23
23 (24). Заднего пальца нет Отр. Дрофы — Of ides.
24 (23). Задний палец имеется 25
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ОТРЯДОВ ПТИЦ
9
25 (26). Крупные птицы. Клюв от угла рта не короче 60 мм
Отр. Журавли — Grues.
26 (25). Средней величины и мелкие птицы. Клюв от угла рта не длиннее 45 мм. . .
Отр. Пастушки — RallL
27 (28). Ноздри открываются в голой коже (Еосковице) у основной части надклювья,
не на клюве (рис. 12) 29
28 (27). Ноздри открываются непосредственно на клюве 33
29 (30). Когти не острые, клюв без острого крючка на конце и без острых режущих
краев («не хищный») Отр. Голуби — Columbae.
30 (29). Когти острые или заостренные, клюв с острым крючком на конце, с острыми
режущими краями 31
31 (32). Глаза крупные, направленные прямо вперед (рис. 13). Пальцы обычно покрыты
жесткими перышками или волосиками, хотя бы у оснований .... Отр. Совы — Striges.
32 (33). Глаза умеренной величины, направлены вбок. Пальцы всегда обнаженные
Отр. Дневные хищники — Accipitres.
33 (32). Заднего пальца нет или он зачаточный 35
34 (33). Задний палец вполне развитый 37
35 (36). Ноги сплошь оперены до самых когтей или плюсна оперена на своей передней
поверхности.
36 (35). Ноги совершенно голые. Отр. Трехперстки — Tarnices.
37 (38). Рулевых более 12 (за исключением перепела Coturnix, у которого хвост
прикрыт кроющими и первостепенное маховое узкое и закругленное). Куриные — Rasores.
38 (37). Рулевых не более 12 39
39 (40). Рулевых 12 41
40 (39). Рулевых 10 -. . . . 43
41 (42). Из 12 рулевых хорошо развиты только 10; два же боковых небольшие и
нежные (рис. 14). Клюв долотообразный. Два пальца направлены вперед, два или один — назад
(рис. 15) Отр. Дятлы— Picariae.
42 (41). Все 12 рулевых развиты одинаково или же крыло более 60 мм. Клюв не
долотообразный. Расположение пальцев иное 53
43 (44). Крыло не более 60 мм
Отр. Воробьиные — Passeres (сем. Славки — Sylviidae).
44 (43). Крыло более 80 мм
45 (46). На голове большой пестрый хохол (рис. 16) Отр. Удоды — Upupae.
46 (45). На голове хохла нет 47
47 (48). Два пальца направлены впе|ред и два назад (рис. 17)
Отр. Кукушки — Cuculi.
48 (47). Расположение пальцев иное 49
49 (50). Все четыре пальца направлены вперед и несколько вбок (рис. 18)
Отр. Длиннокрылые — Macrochires,
50 (49). Три пальца направлены вперед, один назад 51
51 (52). Средний палец значительно длиннее внешнего и внутреннего; есть коготь,
гребенчатый (рис. 19). Хвост мягкий и стержни не выдаются наружу иглообразно
Отр. Козодои — Caprimulgi.
52 (51). Средний палец приблизительно той же величины, что внешний и
внутренний; его когти без зазубрин. Рулевые жесткие и их стержни выдаются наружу наподобие
игол г . . . Отр. Длиннокрылые — Macrochires.
53 (54). Окраска передней части спины и надхвостья голубовато-зеленая,
синевато-зеленая или яркозеленая. В последнем случае не менее 145 лш. . . Отр. Ракши — Coraciae.
54 (53). Окраска спины иная. При зеленоватой окраске спины размеры крыла не более
75 мм Отр. Воробьиные — Passeres.
Рис. 3. Лапа большого баклана.
Рис. 4. Клюв альбатроса.
Рис, 5. Клюв серого гуся.
Рис. б. Положение ног по отношению к туловищу
/ — у обыкновенной чайки; U — у большой поганки.
Рис. 7. Лапа большой поганки.
Рис. 9. Лапа обыкновенной чайки
(слева) и лапа белокрылой крачки
(справа).
Рис. 8. Лапа краснозобой гагары.
Рис. 10. Голова серой цапли.
Рис. 11. Крыло чибиса. Настоящее первое маховое показано стрелкой.
Рис. 12. Клюв голубя.
Рис. 13. Голова длиннохвостой неясыти.
Рис. 14. Хвост большого пестрого дятла.
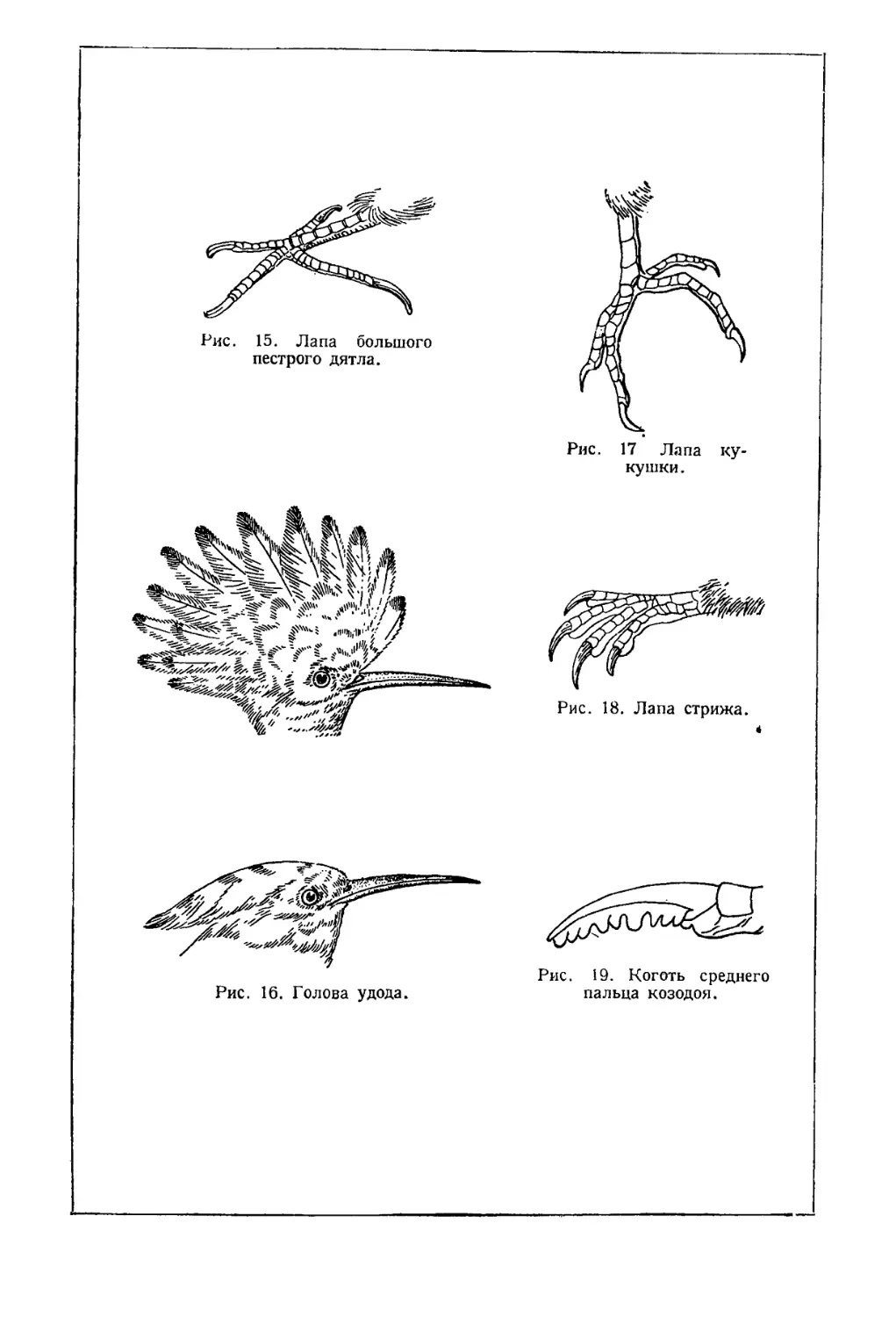
Рис. 15. Лапа большого
пестрого дятла.
Рис. 17 Лапа
кукушки .
'\*^vr ^ffy
Рис. 18. Лапа стрижа.
Рис. 16. Голова удода.
Рис. 19. Коготь среднего
пальца козодоя.
ОТРЯД ВЕСЛОНОГИЕ
STECANOPODES или PELECANIFORMES
Характеристика отряда
Морфология. Внешние признаки. Отряд веслоногих составляют
птицы крупных и средних размеров с разнообразной формой тела. Наибольших
размеров достигают пеликаны. Вес кудрявого пеликана доходит до 13 кг, длина
тела — до 1800 Аш, размах крыльев — до 3000 мм, длина крыла — до 770 мм.
Наименьшие размеры у фаэтонов (не представлены в нашей фауне), которые по
величине равны сизой чайке. В нашей фауне самым мелким представителем
веслоногих является малый баклан. Вес его 800 г, длина тела достигает 550 мм,
длина крыла — до 200 мм. Самцы крупнее самок, в особенности у пеликанов.
Клюв у одних птиц (пеликаны) очень длинный, в 4—5 раз превышающий
длину головы, сильно уплощенный, особенно в вершинной половине, и
оканчивающийся резко загнутым вниз крючком. У других (бакланы, фрегаты) —
клюв длинный, но не превышающий двойной длины головы и не плоский.
Надклювье вогнуто в срединной части и оканчивается, как и у пеликанов,
резко загнутым вниз крючком. У остальных веслоногих—олуш, фаэтонов,
змеешеек — клюв конический, слегка загнутый вниз в вершинной половине,
но без вершинного крючка. Ноздри несквозные (исключение составляют
фаэтоны). У пеликанов и фаэтонов наружные отверстия ноздрей развиты
нормально; у бакланов и фрегатов эти отверстия сильно редуцированы, а у олуш
они совсем отсутствуют. У всех представителей отряда сложная рамфотека,
состоящая на надклювье из 3—4 пластинок.
Оперение у веслоногих густое (за исключением пеликанов), жесткое и
плотно прилегающее к телу. Побочного ствола перьев обычно нет (только
у фрегатов имеется слабо развитый побочный ствол). Пух равномерно
покрывает все тело, распределяясь как по птерилиям, так и по аптериям. Аптерии
слабо развиты: спинная и брюшная представлены узкими полосами, исключая
пеликанов, у которых развита одна спинная аптерия. Копчиковая железа хорошо
развита и оперена. На голове у многих видов имеются удлиненные украшающие
14
ОТРЯД ВЕСЛОНОГИЕ
перья, образующие хохлы, которые развиваются преимущественно в брачный
период. Появляются они в декабре — январе и исчезают в июне — июле. У
некоторых птиц, например, у кудрявого пеликана, удлиненные перья имеются
не только на голове, но и вдоль верхней стороны шеи, образуя «гриву», а у
змеешеек такие перья бывают и на спине. На голове, шее, а у бакланов иногда
и на теле, появляются в брачный период белые ланцетовидные или каплевидные
перья. Лицевые части в разной степени оголены и только у фаэтонов оперены
полностью. Имеется горловой мешок, особенно растяжимый и достигающий
громадных размеров у пеликанов. Этот мешок, исключая фаэтонов, голый
или частично оперен.
Окраска оперения обычно темная, часто черная с металлическим блеском,
или белая с розовым или серым налетом. Первостепенные маховые перья всегда
темные.
Полового диморфизма в окраске оперения нет, за исключением фрегатов;
но имеются большие сезонные и возрастные изменения. Некоторые виды,
например, бакланы, носят брачный наряд очень долго — до полугода и более.
Молодые птицы только на 3-й или 4-й год жизни надевают полный взрослый
наряд. Пуховой наряд птенца у некоторых видов темный, у других — светлый.
У взрослых птиц бывают обычно две линьки в году: частичная (предбрачная) и
полная (послебрачная). Смена маховых перьев идет постепенно, так что птицы
не теряют способности к полету, за исключением змеешеек, у которых сразу
выпадают все маховые перья, и птицы некоторое время не могут летать.
Первостепенных маховых имеется 11 и только у олуш—10. Крыло
аквинтокубитальное. Формула крыла очень разнообразна.
Хвост состоит из 12—24 рулевых перьев разной формы и длины. У
пеликанов хвост короткий, округлый, мягкий и состоит из 20—24 рулевых перьев;
у бакланов и змеешеек хвост длинный, ступенчатый и состоит из 12—14жестких
рулевых перьев; у олуш —длинный, клиновидный и образован из 12—18
рулету фрегатов — вильчатый, крайняя пара рулевых сильно удлинена и имеет
всего 12 перьев; у фаэтонов —также 12 рулевых, но сильно удлинена одна
средняя пара.
Цевка короткая, особенно у фрегатов и фаэтонов, и только у фрегатов
оперена, у остальных представителей отряда голая, обычно сетчатая. Задний палец
находится на одном уровне с другими пальцами и направлен вперед. Все четыре
пальца соединены плавательной перепонкой.
Ноги у видов хорошо ныряющих и плавающих (бакланы, змеешейки)
смещены далеко назад, вследствие чего при ходьбе и сиденьи их туловище
занимает почти вертикальное положение. У пеликанов и олуш ноги
расположены ближе к средней части тела. Ноги фрегата настолько слабы, коротки
и с недоразвитыми плавательными перепонками, что он передвигается с
трудом по земле и по воде. У фаэтонов ноги слабые и короткие, но с полной
плавательной перепонкой; они с трудом передвигаются по суше, но могут плавать.
Анатомические признаки. Череп голоринальный,
переходного типа между схизогнатическим и десмогнатическим, за исключением
фаэтонов; базиптеригоидные отростки не развиты. Сошник имеется только у
фаэтонов и фрегатов, у остальных же представителей отряда не развит. Череп
характеризуется сильным развитием гребней, служащих для прикрепления хорошо
развитой жевательной и шейной мускулатуры. Число шейных позвонков
колеблется у разных представителей отряда. У фрегатов и фаэтонов 14—15,
у олуш и пеликанов — 16, у бакланов — 12, у змеешеек — 19—20. Шейные
позвонки у большинства видов (олуши, змеешейки, бакланы и др.)
приспособлены вследствие особого устройства их сочленовных поверхностей для резкого
толчка вперед при схватывании добычи. Так, передний отдел шеи может
изгибаться только вперед, срединный — назад и последний опять вперед. Таким
ХАРАКТЕРИСТИКА ОТРЯДА
15
образом, при спокойном состоянии птицы шея принимает S-образное положение.
Форма грудных позвонков различна: у бакланов и олуш опистоцельная, у
фаэтонов и фрегатов — гетероцельная. Грудина у бакланов, пеликанов,
фрегатов широкая, почти квадратная. У пеликанов в ширину несколько больше,
чем в длину; у олуш довольно узкая и сильно удлиненная. У хорошо
ныряющих птиц, например, у бакланов, олуш и других, угол соединения ребер с
грудиной острее, чем у неныряющих или плохо ныряющих птиц. Такое соединение
ребер с грудиной позволяет хорошо ныряющим птицам свободнее дышать под
водой. Соединение грудины с вилочкой различно: у пеликанов и фрегатов
происходит полное сращение между этими костями, причем у фрегатов вилочка
неподвижно срастается с коракоидными костями. У других видов имеется
подвижное соединение ключицы с грудиной при помощи соединительнотканных
связок. Таз у видов хорошо ныряющих, например, олуш, сильно вытянут,
у пеликанов, фрегатов и фаэтонов — короткий и широкий, у бакланов —
средней длины. ф
Скелет пневматичен, особенно развита пневматичность скелета у
пеликанов, олуш и фрегатов, у которых воздухоносные полости имеются почти во
всех костях. У бакланов и у других ныряющих и плавающих под водою видов
пневматичность скелета выражена слабо: воздухоносные полости имеются
только в немногих костях. У пеликанов, олуш и фаэтонов хорошо развита
сеть подкожных разветвлений воздушных мешков, которые образуют
воздухоносный слой, особенно сильно выраженный на брюшной поверхности тела.
Носовые железы развиты слабо, слюнные совсем отсутствуют. Язык
рудиментарный. Пищевод, железистый и мускульный желудок легко растягиваются,
что позволяет птице заглатывать крупную добычу. В железистом желудке
имеется очень большое количество пищеварительных желез, а также пилориче-
ский отдел. Кишечник длинный, слепые кишки обычно рудиментарны, только
у пеликанов они достигают 50 мм длины. Переваривание и всасывание пищи
идет быстро.
Сонная артерия у бакланов, фрегатов и фаэтонов парная, у пеликанов и
змеешеек — только одна левая; у олуш — правая. Обводящая мышца имеется
у бакланов, олуш и фрегатов; у пеликанов и фаэтонов она отсутствует.
Большая грудная мышца состоит обычно из двух слоев. Исключение составляют
бакланы и фаэтоны, у которых эта мышца состоит только из одного слоя.
Температура тела от 39,7° до 42,2°/
Образ жизни. Веслоногие — дневные птицы, тесно связанные с водою,
преимущественно с морями и океанами, и в меньшей степени с внутренними
водоемами. Селятся преимущественно на побережьях: либо скалистых, либо
покрытых деревьями или тростниками.
Большинство видов превосходно летает, а некоторые способны парить
(пеликаны, олуши, фрегаты); у фаэтонов и бакланов полет активный, гребной.
Все они хорошо плавают, за исключением фрегата. Некоторые виды хорошо
ныряют и плавают под водой, например, бакланы. Олуши, фаэтоны и бурый
пеликан P. occidentalis ныряют в воду с разлета; пеликаны и фрегаты совсем
не ныряют и с ровной поверхности с трудом поднимаются в воздух.
Пищу веслоногих составляет, главным образом, рыба; змеешейки питаются
и водными беспозвоночными. Способы добывания пищи весьма разнообразны:
пеликаны, за исключением бурого пеликана, захватывают рыбу клювом, как
черпаком, плавая на мелководье; бакланы — ныряя с поверхности воды и
плавая под водой; олуши, фаэтоны и бурый пеликан — бросаясь с разлета
в воду и погружаясь при этом на довольно большую глубину. Фрегаты
схватывают рыб, плавающих близко к поверхности воды, или летучих рыб; иногда
они отнимают добычу у других птиц. Пеликаны и бакланы чаще ловят рыбу
стаями, иногда самостоятельными, а иногда соединяясь в общие стаи.
16
ОТРЯД ВЕСЛОНОГИЕ
Веслоногие гнездятся обычно колониями и даже вне периода
размножения часто держатся стаями. Колонии помещаются всегда близ воды. Гнезда
устраивают на деревьях, на кустарниках, на скалах, на земле среди зарослей
тростника, на плову чих островах и т. д. Даже у одного и того же вида
наблюдается большое разнообразие в месторасположении гнезд. Одни и те
же гнезда птицы занимают в течение ряда лет. Период размножения —
высиживания и выкармливания птенцов продолжителен и занимает 3—
4 месяца.
Устройство гнезда очень простое. Строительным материалом служат
сучья, ветви, водоросли, камыши и т. д. Гнездо мелкое, с небольшим
количеством мягкой выстилки в лотке. Некоторые виды олуш и фаэтоны совсем не
строят гнезд, а откладывают яйца прямо на земле без подстилки. В местах, где
расположены гнезда веслоногих птиц, накапливается большое количество
птичьего помета. Высыхая на поверхности, помет цементирует гнезда.
Яиц в кладке у пеликанов обычно 2—4, у бакланов и змеешеек 3—6, у
фрегатов и фаэтонов 1 и у олуш 1—2. Яйца, относительно размеров птиц, мелкие,
однотонные голубоватые или зеленоватые, покрытые густым известковым слоем.
У фаэтонов яйца пестрые. В постройке гнезда, в высиживании и
выкармливании птенцов участвуют оба родителя.
Срок высиживания у бакланов 28—30 дней, у пеликанов — 33—40 дней.
Птенцы вылупляются совершенно беспомощными — голыми и слепыми. Глаза
открываются на 3—5-й день, на 5—8-й день начинает появляться пух, который
через несколько дней густо покрывает все тело птенца. Первое время
родители кормят птенцов полупереваренной пищей, которую они отрыгивают им
прямо в рот. Приносимая родителями вода также выливается в рот птенца.
Птенцы кормятся 2—3 раза в сутки, быстро прибавляют в весе и обычно
к концу выкармливания достигают веса родителей и даже его превышают.
Период выкармливания у бакланов продолжается 40—45 дней, у пеликанов и
олуш — 50—60 дней. К этому времени птенцы надевают уже почти полностью
первый наряд и способны летать. Некоторые же риды, например, олуши,
бросают кормить птенцов задолго до их поднятия на крыло.
После окончания гнездового периода молодые и старые птицы собираются
вместе и кочуют, иногда удаляясь от гнездовья на большие расстояния. Но
настоящими перелетными птицами можно считать лишь пеликанов. Бакланы
частично отлетают, частично откочевывают от мест своих гнездований в районы
незамерзающих вод, где возможна добыча рыбы. Северные виды олуш, также,
как и бакланы, полуперелетные, полукочующие птицы. Большинство же видов
веслоногих птиц оседлы.
Систематика. Отряд состоит из пяти резко различающихся между собою
семейств: бакланы Phalacrocoracidae, пеликаны Pelecanidae, олуши Sulidae,
фрегаты Fregatidae и фаэтоны Phaethontidae.
Семейство фрегаты Fregatidae составляют птицы крупных размеров — весом
около 1500 г, с коротким телом и шеей, с очень длинным вильчатым хвостом,
с длинными и острыми крыльями; вершину крыла образует первое
первостепенное маховое. Киль грудины высокий, короткий, не доходящий до половины
ее длины. В локтевом сочленении крыла имеется сесамоидная кость,
помогающая удерживать крыло в раскрытом положении при парящем полете. Кости
цевки, как у пингвинов, не имеют во взрослом состоянии полного сращения
между собою. Пальцы длинные, особенно средний палец; когти круто загнуты
и сжаты с боков; внутренняя сторона когтя среднего пальца зазубрена.
Плавательная перепонка развита слабо и сильно вырезана, далеко не доходит до
ногтевых фаланг. Подбородок, горло и зоб —голые, окрашенные у самцов
в яркокрасный цвет. Окраска оперения черная, с металлическим блеском на
голове и спине, а у некоторых видов с белыми пятнами на брюшной стороне тела*
ХАРАКТЕРИСТИКА ОТРЯДА
17
Фрегаты превосходные летуны, не уступающие в быстроте и неутомимости
полета альбатросам.
Семейство состоит из одного рода — фрегаты Fregata — с пятью видами,
которые населяют тропические воды всего земного шара и гнездятся на мелких
океанических островах.
Семейство олуши Sulidae составляют птицы средних и крупных
размеров. Тело удлиненное, шея короткая, крылья длинные, острые; вершину
крыла составляет первое первостепенное маховое. Хвост клиновидный с сильно
удлиненной парой средних рулей. Края клюва зазубрены и помогают
удерживать скользкую живую добычу. Разрез рта большой, заходящий за глаз.
Внутренние ноздревые камеры между собою не соединены, наружных
отверстий ноздрей нет. Наружный палец равен среднему или несколько длиннее
его; наружный и средний пальцы длиннее внутреннего. Когти широкие
и плоские; коготь среднего пальца по внутренней стороне зазубрен. Кости
нижней челюсти в области os spleniale имеют неполное сращение и в связи
с особым строением сочленения с квадратной костью позволяют широко
открывать рот при заглатывании крупной добычи. Киль грудины заходит несколько
далее половины ее длины. Копчиковая железа открывается пятью выводными
отверстиями. Хорошо развитый в особенности на брюшной стороне
подкожный пневматический слой предохраняет птицу от повреждений при нырянии
с разлета, смягчая удар о воду. Лицо, подбородок и середина горла голые.
Оперение голени несколько не доходит до пяточного сочленения. Окраска
оперения взрослых птиц преимущественно белая с черными маховыми; у
молодых птиц оперение пестрое.
Хорошие летуны, могут в течение продолжительного времени парить в
воздухе. Обычно придерживаются побережий и далеко в открытых океанах
не встречаются.
Это семейство состоит из одного рода — олуши Sula -г- с семью видами,
распространенными преимущественно в субтропиках и тропиках всего земного
шара, только один вид — северная олуша Sula bassana — проникает на
гнездовье далеко к северу до Исландии.
Семейство фаэтоны Phaetontidae состоит из птиц средней величины
с короткой шеей и ногами, по внешнему виду несколько сходных с олушами,
но меньше. Края клюва тонко зазубрены, Ноздри сквозные, наружные носовые
отверстия щелевидны и расположены у основания клюва. Коготь среднего
пальца по своей внутренней стороне гладкий. Язык по сравнению с другими
представителями отряда веслоногих птиц довольно хорошо развит. Пилориче-
ский отдел желудка небольших размеров. М. iliofibularis не проходит через
сухожильную петлю. Крыло острое, но не длинное: средняя пара рулей сильно
удлинена — в среднем до 50 см. Лицо и горловой мешок оперены, последний
очень небольших размеров и слабо растягивается. Окраска оперения
преимущественно белая.
Это семейство содержит один род — фаэтоны Phaeton — с тремя видами,
распространенными в субтропиках и тропиках.
Семейство пеликаны Pelecanidae составляют огромной величины птицы
с длинной шеей и клювом и с большим горловым мешком, легко
растягивающимся. Этот мешок начинается от вершины подклювья и тянется по ветвям
подклювья до основания клюва. Межглазничная перегородка полностью
окостеневает и имеет только одну фонтанель. Киль грудины длинный и идет почти
вдоль всей грудины. Имеется os dorsale. Мускулы нижней гортани не развиты.
Зоб отсутствует. Копчиковая железа с 6—8 выводными отверстиями. Ноги
короткие, цевка сжата с боков. Средний палец самый длинный, внутренний,
край его когтя гладкий. Крылья длинные, но не острые, вершина крыла
образована 2-м и 3-м первостепенными маховыми. Лицо и горловой мешок голые*
2 Птицы СССР, т. I
18
ОТРЯД ВЕСЛОНОГИЕ
Окраска оперения наших пеликанов белая с розовым или сероватым налетом,
но среди других видов имеются птицы, окрашенные в серый цвет с черными и
белыми партиями перьев. Сильно пневматичный подкожный слой уменьшает
удельный вес птицы и помогает ей легко держать на воде и в воздухе свое
огромное тело.
Семейство состоит из одного рода — пеликаны Pelecanus. Этот род
распадается на восемь видов, широко распространенных по всем частям света, но
населяющих преимущественно тропики и субтропики; только ограниченное
количество видов доходит до стран с умеренным климатом.
Семейство бакланы Phalacrocoracidae составляют птицы крупных и
средних размеров с валькообразным телом и с длинной шеей. Края клюва
гладкие, исключая змеешеек, у которых на клюве есть зазубрины. Крылья
тупые и недлинные, вершина крыла образована 2-м и 3-м первостепенными
маховыми. Ноги сильные; наружный палец самый длинный. Когти сжаты с боков
и круто загнуты вниз; коготь среднего пальца по внутренней стороне
зазубрен. Наружные отверстия ноздрей у бакланов во взрослом состоянии
зарастают, а у молодых имеются; у змеешеек и во взрослом состоянии развиты в виде
узких щелей. Череп с резко развитыми гребнями сильно уплощен сверху вниз
и вытянут в длину. В затылочной области бакланов имеется особая сесамоидная
кость os retrooccipitale, образованная окостенениями соединительной ткани.
Мышцы, сгибающие голову, прикрепляются к сесамоиду, а не к шейным
позвонкам. Образование сесамоидной кости, сильное развитие сгибающих мышц
головы и прикрепление их к сесамоиду обусловливает крепкое сжимание клюва
и помогает птице удерживать живую добычу. Обводящая мышца у бакланов
проходит сквозь коленную чашечку, а у змеешеек лежит в желобке над
коленной чашечкой. Зоба нет, но имеется расширение пищевода, заменяющего,
повидимому, зоб. Копчиковая железа открывается двумя отверстиями. Глаз
в связи с плаванием и добыванием пищи под водой имеет сильно развитую
аккомодацию, достигающую у бакланов 20—45 диоптрий. В результате
большого давления, которое под водой испытывает глаз, роговица глаза сильно
утолщена. Окраска оперения черная с металлическим блеском, но среди
тропических птиц есть виды, имеющие темносерую или черную с белым окраску-
На верхней стороне тела развит чешуйчатый рисунок, который у тропических
видов заменяется крапчатостью.
Семейство это делится на 2 подсемейства. Первое подсемейство — бакланы
Phalacrocoracinae содержит 2 рода: первый род — бакланы Phalacrocorax с 29
видами, широко распространенными почти по всему свету, и второй —
нелетающий галапогосский баклан Nannopterum с одним видом N. harrisii, населяющий
Галапогосские о-ва; второе подсемейство — змеешейки или анхинги Anchin-
ginae — с одним родом — анхинги или змеешейки Anchingae — и 4 видами,
обитающими в тропиках Восточного и Западного полушария.
Ископаемые остатки (их известно 47 видов) веслоногих
находят в Европе, Азии, Северной и Южной Америке, Австралии и на островах
Суматра и Маскаренских. Близкие к современным пеликанам виды описаны
из верхнего олигоцена Франции, эоцена Англии, мела и эоцена Югославии.
В СССР из окрестностей Одессы из нижнего плиоцена описан вымерший
пеликан Pelecanus odessanus. Представители фаэтонов, фрегатов, олуш, змеешеек
и бакланов, близкие уже к современным видам, известны из плейстоцена. Род
олуш выделился в олигоцене и наиболее богато представлен в миоцене Южной
Америки. Род бакланов известен с начала плиоцена. Змеешейка найдена в
ископаемом состоянии в плиоцене Венгрии — Anchinga pannonica. Из СССР
описан род Pliocarbo, близкий к бакланам, найденный в раскопках под
Одессой в понтических известняках нижнего плиоцена. В середине прошлого
столетия вымер стеллеров баклан Phalacrocorax perspicillatus, живший оседло
ХАРАКТЕРИСТИКА ОТРЯДА
19
на о. Беринге. В 1741 г., по описанию Стеллера, впервые открывшего этого
баклана, он на острове был многочисленен, а в 1883 г. здесь не было уже ни
одной птицы. Вероятно, истребление человеком, а также, возможно,
возникновение сильной эпизоотии, послужило гибелью этих птиц. Стеллеров баклан
был много крупнее большого баклана: длина клюва достигала 95 мм; размеры
коротких, слабо развитых крыльев не превышали 360 мм. Последнее
обстоятельство дает возможность предположить, что стеллеров баклан летал хуже
ныне живущих бакланов.
Веслоногие птицы близки с одной стороны к трубконосым> с другой —
к голенастым.
Географическое распространение. Веслоногие птицы населяют морские
и океанические побережья и острова всех частей света; в небольшом количестве
видов обитают на внутренних пресноводных водоемах и реках. Наибольшее
число видов встречается в субтропиках и тропиках, лишь отдельные виды —
в Арктике и Антарктике. Среди веслоногих птиц имеется большое количество
видов с узким распространением. Виды с широким распространением обычно
имеют громадные разрывы в ареале.
Из 54 видов веслоногих в СССР встречается 11, из которых 8 гнездящихся
и 3 случайно залетных, всего 20,4%- Гнездящиеся у нас виды принадлежат
к семейству пеликанов (2 вида) и к семейству бакланов (6 видов); и залетные —
2 к семейству олуш (северная олуша и красноногая олуша) и 1 к семейству
фрегатов (большой фрегат). Семейство фаэтоны совсем не представлено в нашей
фауне.
Распределение веслоногих на гнездовье в нашей стране следующее:
I — зона Арктика, подзона побережья — Phalacrocorax pelagicus, Phalacro-
corax aristotelis aristotelis, Phalacrocoraxcarbo carbon Phalacrocorax mile
выходит за пределы подзоны (Командорские о-ва); II — зона открытых сухих
пространств, подзона степей (озерная часть Зап. Сибири и Казахстана) и
подзона полупустыни и пустыни (водоемы) — Phalacrocorax carbo sinensis,
Phalacrocorax pygmaeus, Phalacrocorax aristotelis desmarestii, Pelecanus crispus
и Pelecanus onocrotalus и III — горная зона, высокогорная бореоальпийская
подзона — Phalacrocorax carbo sinensis.
Практическое значение. В некоторых местах веслоногие птицы приносят
вред рыбному хозяйству, поедая имеющих промысловое значение рыб,
преимущественно среднего размера. В особенности вредны птицы, населяющие
пресноводные водоемы.
Мясо веслоногих в свежем виде не вкусно, но в консервированном — вполне
пригодно к употреблению в пищу. Шкурки их выделываются и идут на рынок,
как «птичий мех», пригодный для мелких теплых вещей (воротники, шапки,
шляпы и т. д.).
Большие скопления птиц на суше в период размножения в условиях сухого
климата способствуют накоплению гуано, имеющего большое значение как
удобрение. Громадные скопления гуано находятся на побережье Перу между
5° и 19° ю. ш., в Патогонии, по берегам Караибского моря и на островах близ
западного берега Южной Африки. Скопления гуано принадлежат
преимущественно американскому бурому пеликану Pelecanus occidentalis, баклану
Phalacrocorax bougainvillei и олуше Sula variegata.
Таблица для определения видов веслоногих
1 (2). Хвост закругленный или клинообразный, крайние рулевые короче средних;
цевка не оперена; плавательные перепонки доходят до конца пальцев 3
2 (1). Хвост вильчато-вырезанный, средние рулевые короче крайних; цевка оперена до
пальцев; плавательные перепонки не доходят до половины длины пальцев
Большой фрегат Fregata minor.
3 (4). Клюв конический, без вершинного коготка, 1-е маховое длиннее 2-го 5
*
Рис. 20. Голова уссурийского баклана.
Рис. 21. Голова большого баклана.
Рис. 22. Головы бакланов (вверху — бериигов баклан,
внизу -— краснолицый баклан).
Рис. 23. Голова розового пеликана (слева) и кудрявого пеликана (справа).
БОЛЬШОЙ БАКЛАН
21
4 (3). Клюв плоский или округлый с вершинным коготком, резко загнутым вниз,
1-е маховое короче 2-го 7
5 (6). Рулевых б пар; голая часть подбородка в виде узкой полосы глубоко вдается
вдоль срединной части горла и основания шеи Севернаяолуша Sula bassana.
6 (5). Рулевых 7 пар; голая часть подбородка не распространяется в виде узкой ленты
пдоль горла, а граничит с оперенной областью подбородка на уровне разреза рта по прямой
поперечной линии Красноногая олуша Sula sula.
7 (8). Клюв длинный, не короче 300 мм) уплощенный; оперение светлое; горловой мешок
больших размеров 9
8 (7). Клюв короче 100 мм, не уплощенный; оперение темное; горловой мешок мало
заметен 11
9 (10). Стержни первостепенных маховых темные; лоб оперен, но в срединной части
разделен глубоко входящим голым хребтом надклювья; ноги серые
Кудрявый пеликан Pelecanus crispus.
10 (9): Стержни первостепенных маховых светлые; оперение головы в виде постепенно
суживающейся полосы заходит на голый лоб; ноги розовые или желтые
Розовый пеликан Pelecanus onocrotalus.
11 (12). Хвост из 14 рулевых, крыло более 300 мм 13
12 (11). Хвост из 12 рулевых, крыло менее 300 мм 15
13 (14). Оперение на боках нижней челюсти идет вперед и достигает или даже заходит
за линию, проведенную вертикально через угол рта
Уссурийский баклан Phalacrocorax filamentosus.
14 (13). Оперение на боках нижней челюсти идет назад или прямо вниз и не доходит
до линии, проведенной вертикально через угол рта
Обыкновенный баклан Phalacrocorax carbo.
15 (16). Крыло менее 225, клюв короче 35 мм. Малыйбаклан Phalacrocorax pygmaeus.
16 (15). Крыло более 240, клюв более 40 мм 17
17 (18). Длина клюва 58—70 мму высота у основания 16—17 мм; в брачном наряде на
голове один хохол Длинноносый баклан Phalacrocorax aristotelis.
18 (17). Длина клюва 49—55 мм, высота 8—13 мм; в брачном наряде 2 хохла на голове
19
19 (22). Оперение лба доходит до ноздрей; на боках нижней челюсти оперение доходит
до заднего края глаз Берингов баклан Phalacrocorax pelagicus.
20 (19). Оперение лба не доходит до ноздрей; на боках нижней челюсти оперение не
доходит до заднего края глаз Краснолицый баклан Phalacrocorax urHe.
СЕМЕЙСТВО БАКЛАНОВЫЕ PHALACROCORACIDAE
РОД БАКЛАНЫ PHALACROCORAX BRISSON 1760
Тип Pelecanus carbo
\. Большой баклан Phalacrocorax carbo L.
Pelecanus carbo. Li nnaeus. Syst. Nat., изд. X, 1758, стр. 133, Швеция.
Русское название.* Баклан — народное, впервые упоминаемое в
зоологической литературе у Палласа (1811), от тюркского слова бакла.
Распространение. Ареал. Европа — от Исландии, Фарерских и
Британских островов на западе до Кольского п-ова, на юг до Средиземного и
Черного, морей и Закавказья; Азия — южная и Средняя, на север—до
сев. Казахстана, Байкала и Приморья; Африка, кроме Сахары и большей
части южного побережья Средиземного моря, острова Индо-Австралийского
архипелага, Австралия, Тасмания и Новая Зеландия; в Сев. Америке
теперь только в Гренландии; ранее большой баклан гнездился на северо-
востоке до северного побережья залива св. Лаврентия и о, Магдалины, быть
может местами остался и в настоящее время.
Характер пребывания в большей части ареала оседлый, на
севере перелетный. Биотоп: побережья пресных и морских водоемов.
Численност ь неравномерна, зависит от обилия кормовых ресурсов
Карта 1. Распространение большого баклана Phalacrocorax carbo
7— Ph. с. carbo, 2 -- Ph. с. sinensis, 3 — Ph. с. maroccanus, 4— Ph. с Iucidus, 5 — Ph. с lugubris, 6 — Ph. с novaehollandiae 7 — Ph с steaoi-
« - граница гнездовой области, 6 - не вполне выясненная граница распространения, г - область зимовок (цифры в кружках - местонахо^ениё
соответствующего подвида на зимовке).
БОЛЬШОЙ БАКЛАН
23
(рыбы). Местами (Азия, Африка) весьма обычен. В Зап. Европе и в Сев.
Америке во многих местах исчез в XIX--XX веке.
Подвиды и варьирующие признаки. Имеется семь подвидов, различающихся между
собою морфологическими особенностями (окраска и размеры) и деталями экологии: Ph. с.
carbo L., 1758 — Гренландия и сев. Европа от Исландии до Норвегии и зап. побережья
Кольского п-ова; Ph. с. sinensis Shaw, 1801 —от средней и южной Европы до Китая и
Японии, Индо-Австралийский архипелаг; Ph. с. maroccanus Н а г t е г t, 1906 — западное
побережье Марокко и от Могадора до мыса Бланко; Ph. с. lucidus Lichtenstein, 1823 —
от островов Зеленого Мыса и Сенегала на западном побережье Африки до Капской Земли
на юге и до реки Тана на восточном побережье Африки; Ph. с. lugubris R ii р р е 1 1,1845 — от
дельты Нила и Суэцкого канала до озера Виктория-Ньянца и Тангаиайки; Ph. с. novaehollan-
diae Stephens, 1826 — Австралия, за исключением ее центральных областей, Тасмания;
Ph. с. steadi Mathews et Iredale, 1913 — Новая Зеландия.
Материковый большой баклан Phalacrocorax carbo sinensis Shaw et Nodder
Pelecahus sinensis. Shaw et Nodder, Nat. Misc., XIII, табл. 529, 1801, Китай.
Синонимы: Carbo subcormoranus. С h r. L. В r e h m. Ornis, I, 1824, стр. 42, Голландия —
Phalacrocorax carbo medius. N i I s s о n. Scand. Fauna Fuglarna II, 1835, стр. 478, Балтийское
море. Carbo phalacrocorax. Северцов. Верт. и Гориз. Распространение Туркестанских
Животных, 1873, стр. 70.
Распространение. Ареал. Литва, местами по южному берегу
Балтийского моря к востоку от Эльбы, у Рюгена, на Цитхензее в б. зап. Пруссии,
в Калининградской обл.; Голландия, Бельгия, юго-западная Франция,
вероятно сев. Испания, Италия, Балканский п-ов за исключением Греции,
на север до Добруджи и Придунайской низменности, острова Средиземного
моря (Балеарские, Сардиния, Сицилия, Крит); Малая Азия.
В СССР, кроме указанных мест в Прибалтике, материковый большой
баклан гнездится в южной части страны: Днестровский и Днепровский лиманы,
прилежащие части Черноморского побережья и по Днепру приблизительно до
48° с. ш. в заповеднике Конча-Заспа (Шарлемань, 1930); Крым, в особенности
по южному берегу (Пузанов, 1933), раньше регулярно гнездился и в степной
части; побережье Азовского моря; плавни Кубани и от устья Терека и Кизляра
(Беме, 1926) по Каспийскому побережью до дельты Волги, где очень много-
численен. На север по Волге доходит почти до Сталинграда. Южнее в
центральном Кавказе и почти по всему Черноморскому побережью Кавказа баклана
на гнездовье нет. В Закавказье гнездится местами в Грузии по Черноморскому
побережью к югу от Батуми и быть может от Поти; в Армении — в бассейне
озера Севан и др. (Ляйстер и Соснин, 1942). На восток в Азербайджане до Лен-
коранской низменности и берегов Каспия, на север до о. Глиняного близ
ст. Алят (Баньковский, 1912) и вероятно до Баку.
В Волжско-Уральских степях к северу доходит до Камыш-Самарских озер,
то-есть приблизительно до 49° с. ш. (Волчанецкий, 1937); вероятно, что баклан
поднимается по крупным озерам и выше к северу, до 50—бО1^0 с. ш.
Гнездится на северном и северо-восточном побережье Каспия с устьями
рек Урала и Эмбы, к югу до северных частей Усть-Урта. По Эмбе ранее
доходил почти до верховьев реки (у Кок-Джиды); по Уралу поднимается от
дельты до устья Илека и вероятно несколько выше, но до Чкалова не
доходит (Зарудный, 1888); по Илеку дальше низовьев реки не идет.
В сев. Казахстане и в южных областях Зап. Сибири распространен еще
с большей спорадичностью: в низовьях Иргиза и по близлежащим озерам,
в низовьях Тургая к северу до озера Сары-копа и Майкарау и в верховьях
Тобола; до Кустаная не доходит, в Наурзуме ранее гнездился, но в настоящее
время баклана там нет (Михеев, 1938); на озере Боровом и Мокром (Сушкин,
1908), в Кулундинской степи на озере Бакланьем (Иогансен, 1907), в Барабин-
ской степи на озере Чанах (Янушевич и Золотарева, 1947). Также
спорадично распространен и южней, где отмечен в окрестностях Семипалатинска, на
24
ОТРЯД ВЕСЛОНОГИЕ
Иртыше собственно на Алтае, на Кара-иртыше и Зайсане, быть может на Марка-
куле; в Семиречье на озерах Ала-куль, Сасык-куль, Балхаш и на Или.
В Тянь-Шане на озере Иссык-куль — на Карабулуне и на горном озере Тюп
(Шнитников, 1949) и на других горных озерах.
К востоку от Алтая — на озере Джигетай-куль в Танну-Тувинской обл.,
в с.-з. Монголии на озере Урюг-нур и Ачит-нури, на озере Даин-гол в
Монгольском Алтае.
В Туркмении баклан распространен на юго-восточном Каспии к северу
до Кара-богаза, на Мургабе, Теджене и Аму-Дарье; на западном Узбое в
настоящее время вероятно не гнездится, во всяком случае регулярно (Рустамов,
1947). Гнездится в юго-западном Таджикистане и на Памире (Иванов, 1940).
На Аральском море: по юго-восточному, восточному и северо-восточному
побережьям, с прилежащими островами, по островам у западного побережья и
быть может местами по западному побережью; в дельте и на нижнем течении
Сыр-Дарьи, в нижнем течении Сары-су, на озере Тели-куль (Спангенберг и
Фейгин, 1936), может быть и верхнем течении Сыр-Дарьи и в низовьях Чу
(Долгушин, 1939).
На Саянах- и в Минусинском крае отсутствует, но распространен от
Байкала и далее на восток до Приморья. На зап. берегу Байкала идет к северу
до Малого моря, на восточном — до Святого носа и Чивыркуйского залива
(Туров, 1923), к югу от Байкала будто бы гнездится на р. Селенге, Чикою,
Орхону и нижнему течению Кирана (Моллесон, 1896), что сомнительно.
Далее — на Аргуни, верхнем течении Амура, озере Далай-нор по нижнему
течению Амура, на Уссури от устья до верховьев. Гнездится ли в
остальных частях Приморья и по морскому его побережью, — неясно; по берегу
Приморья гнездится уссурийский баклан Ph. filamentosus, трудно отличимый
в природных условиях от большого баклана, и наблюдения в этих районах
очень противоречивы. Некоторые исследователи считают, что в Приморье
большой баклан — птица внутренних пресноводных водоемов, а
уссурийский— морских побережий.
Вне СССР к югу баклан распространен в,Иране, М. Азии, в Японии,
Корее, но в Монголии и Манчжурии, исключая упомянутых выше озер
северных частей, повидимому, не гнездится. В/Центральной Азии: Кашгария с Лоб-
нором и Куку-нор с прилежащими озерами и реками, не доходит до верхней
Хуан-хе, юго-восточной Монголии и Тибета. В вост. Китае, но как далеко идет
внутрь страны неясно, так как там бакланы содержатся в полудомашнем
состоянии; в Индо-Китае, Индии и на островах Индо-Австралийского архипелага.
Характер пребывания. Большой баклан — гнездящаяся,
перелетная и в южных частях ареала оседлая птица. У нас зимует обычно в
местах гнездовий только в самой южной части страны, там, где имеются богатые
рыбой незамерзающие водоемы. В большом количестве зимует в Крыму,
преимущественно по южному побережью и изредка в степной части. По кавказскому
побережью Черного моря наблюдается даже несколько северней мест
гнездовий, например, у сочинского берега. Встречен на озерах Армении, причем
на озере Гилли гнездятся, а зимуют, по сообщению Соснихиной, на
озере Севане, у Ленкорани и в Кызыл-Агачском заливе, где они особенно
многочисленны (Тугаринов и Козлова, 1935, 1938). Максимум зимующих птиц в
Ленкорани наблюдается в декабре (Исаков и Воробьев, 1940). По
юго-восточному побережью Каспия, в устье Атрека, у Гассан-Кули, у Чикишляра,
на озерах Делили большой баклан многочисленен осенью, зимовать же
остается там в ограниченном количестве. К северу от Гассан-Кули по
восточному берегу Каспия наблюдаются зимой до Балханского залива, где они
довольно многочисленны, но в Красноводский залив большой баклан зимой
уже не заходит. В большом количестве зимует в Иране по южному берегу
БОЛЬШОЙ БАКЛАН
25
Каспия, особенно в Астрабадском заливе. В небольшом количестве — на Мур-
габе и на Теджене в Туркмении и в южном Таджикистане.
Вне нашей страны в небольшом количестве зимует на Азорских о-вах,
Мадейре, Канарских и Марокко. Обычен на зимовках на островах
Средиземного моря (Сицилия, Корсика, Мальта, Кипр), в Греции, Алжире, Тунисе,
Египте, Синае, Палестине, Малой Азии, Месопотамии, Аравии, в южном Иране,
Белуджистане и Афганистане.
В юго-восточной части своего ареала: в Индии, Индо-Китае, Южном
и восточном Китае, на Индо-Австралийском архипелаге и т. д. большой
баклан оседл.
Весною в дельте Волги бакланы появляются в первой половине марта,
чаще между 7 и 12, иногда в связи с ранним временным потеплением бакланы
прилетают и раньше; например в 1926 г. они прилетели 22 февраля, но обычно
при возврате холодов бакланы отлетают к югу и возвращаются обратно в конце
первой половины марта. В Туркмении, преимущественно на Мургабе, пролет
наблюдается с конца марта и только изредка — ранее, например у Имам-баба
12 марта (Лоудон, 1910) и в феврале под Ашхабадом. У Репетека и Анненкова
пролетные птицы отмечены 23 и 29 марта, в Уч-аджи — в феврале, марте.
В зап. Туркмении пролетные птицы наблюдаются в начале марта, на озере
Делили и в Гассан-Кули, но до 13—19 марта бакланы редки (Дементьев, 1945).
В юго-вост. части Кызыл-Кумов (ур. Алабне) пролетные бакланы встречались
12 апреля (Спангенберг и Фейгин, 1936); судя по. очень поздней дате, это,
повидимому, были холостые, не гнездившиеся в этом году птицы. На нижней
Сыр-Дарье пролет начинается в конце первой трети марта и продолжается до
начала апреля. На нижнем течении Иргиза первые птицы отмечены 25 апреля
(Сушкин, 1908); в Семиречье первые пролетные птицы наблюдаются в
середине второй половины марта; в Кулунде — в начале мая, в с. Новенское — 4 мая
и 12 июня (вероятно птицы, встреченные 12 июня, были не пролетные, а
летние кочующие). В Читинской обл. на р. Чикой первые птицы появились
24 марта, валовый пролет наблюдался 10 апреля. В Приморье на Сунгачи
первые стаи прилетали 14—18 марта (температура утром доходила до 20°С, было
много снега) и валовый пролет шел или до середины первой половины апреля,
или до середины того же месяца (Пржевальский, 1870).
По прилете на места гнездовий бакланы первые дни малоподвижны, затем,
они начинают довольно оживленно передвигаться с места на место в поисках
пищи, которой обычно в первое время очень мало. Особенное оживление и
интенсивная ловля рыбы наблюдается после освобождения водоемов от льда.
Осенью бакланы небольшими стаями долгое время кочуют, обычно у мест
гнездовий, но иногда и довольно далеко от них. Перед самым отлетом мелкие
стаи сбиваются в большие. На Иртыше пролетные стаи бакланов появились
13 сентября, у Семипалатинска пролет наблюдался 15—16 сентября. В
Семиречье, начиная с 20 августа, по течениям рек встречаются кочующие птицы,
повидимому, постепенно передвигающиеся к югу; но отдельные птицы остаются
на местах гнездовий до первых чисел ноября, хотя и не зимуют даже на Иссык-
куле. С Иргиза бакланы начинают отлетать обычно в середине сентября.
С дельты Волги главная масса птиц отлетает в течение октября, собираясь
в тысячные стаи, но одиночки наблюдаются даже еще в первых числах декабря
(Воробьев, 1936), в Кизляре бакланы встречались еще в октябре (Беме, 1925).
На Сыр-Дарье в окрестностях Джулека и Чиили в середине ноября птицы
были еще обычны, на Калган-Дарье встречались в конце сентября, в южной
части Аральского моря между Куван-Дарьей и Джаны-Дарьей стайки
бакланов наблюдались 28 ноября на пловучей льдине. На юго-восточном берегу
Каспия у Чикишляра стаи бакланов держатся еще в половине декабря. Но
пролетные бакланы в Туркмении наблюдаются в первых числах сентября;.
26
ОТРЯД ВЕСЛОНОГИЕ
так, у Узуп-ада большие стаи бакланов летели 5—б сентября. На Байкале в
Чивыркуйском заливе отлет идет повидимому в начале третьей декады
сентября (17 сентября птицы еще наблюдались в заливе, а 25 сентября их уже
не было, Туров, 1923). В Приморском крае в середине октября бакланы еще
обыкновенно встречаются.
Результаты кольцевания. Бакланы кольцевались на озере
Ала-куль и на о. Комсомольском на Аральском море. Из окольцованных
на Ала-куле 4. VI1. 1939 г. птиц одна была обнаружена там же в 1940 г., другая
на кочевке 30.X 1.1939 г. на р. Или ниже Баканаса. На о. Комсомольском
бакланы кольцевались птенцами в конце мая — начале июня 1948 г. В августе
1948 г. две из них добыты на озере Узун-каир на Аральском море; три птицы
добыты 1,13 и 16. IX. 1948 г. в районе озера Уялы на Аральском море; осенью
того же года три баклана встречены в низовьях Сыр-Дарьи в Казалинском
районе; в дальнейшем птицы далеко откочевывают на юго-восток: одна добыта
на озере Джили-куль в ю.-з. Таджикистане; три — на Мургабе в Туркмении,
а именно 8.Х 1.1948 г у Иолотани, 2. IV. 1949 г. на Коушут-бентском
водохранилище в Туркменкалинском районе, 9. IV. 1949 г. — у Ташкепри.
Залеты. Одиночки или небольшие стаи птиц залетают (главным
образом осенью) внутрь страны далеко от мест гнездований. На Украине
и Кавказе залетные птицы встречались весною и осенью, например в Подо-
лии, на Днепре и в окрестностях Киева, на сев.-вост. Кавказе — весною в марте
и апреле, в горной части Кавказа осенью в августе — сентябре, в сентябре на
р. Киши и осенью на р. Белой. Только осенние залеты отмечены в Курской
обл., в Московской, в Смоленской в Ельнинском районе, в Оршанский
район в вост. Белоруссии, в Прибалтику залетали даже зимой в январе,
в Ленинградскую обл. —в сентябре, октябре и в первой половине ноября
в Кировской обл.— окрестности г. Кирова — добыт 27 октября, в Нагорском
районе в ноябре. В сев. Казахстане и в южной части Зап. Сибири бакланы
встречаются в разное время года. Там, повидимому, наблюдаются не только
залеты, а главным образом кочевки холостых стай бакланов, которые ведут
в течение летне-осеннего сезона с апреля по октябрь кочевой образ жизни.
Бакланы встречаются в окрестностях г. Ишима, на Иртыше в окрестностях
Павлодара, в окрестностях г. Тюмени, под Омском, близ Каинска, на озере
Карачи, в Атбасарском районе, в Наурзумском заповеднике, в горной Шории,
в Новосибирском районе и других местах.
Биотоп. Богатые рыбой водоемы с медленным течением и чистой водой,
В гнездовое время — скалистые высокие берега морских водоемов и обычно
низкие, поросшие тростником и древесной растительностью, берега
внутренних водоемов. В горных районах поднимается на Алтае и Иссык-куле не выше
1600 м9 в с.-з. Монголии — на озере Даин-гол гнездится на высоте 2200 м,
на Памире —до 3500 м и в Закавказье в Армении в бассейне озера Севан
до 2000 м.
Численность.В устье Волги, в Ленкоранской низменности, по
нижнему течению Куры и Сыр-Дарьи, на Аральском море, в низовьях Аму-Дарьи,
на Балхаше и в некоторые годы на Сары-су большой баклан водится в большом
количестве. По побережью Черного моря его довольно мало, но местами,
например, по южному побережью Крыма, он более многочисленен. По южному
и юго-восточному берегу Азовского моря баклана несколько больше, чем на
Черноморском побережье. Довольно редок баклан по р. Уралу, более обычен-
у устья реки. Редок в низовьях р. Илека, более обычен в сев.-зап. Казахстане
и в Зайсанской котловине. Редок в юго-западной части Таджикистана
и обычен на высокогорном Памире. Редок в Туркмении, за
исключением юго-вост, побережья Каспия. В Приморье малочисленен, на Байкале,
обычен.
БОЛЬШОЙ БАКЛАН
27
Иа зимовках водится в большом количестве только в Ленкоранской
низменности.
Экология. Размножение. Половая зрелость у больших бакланов
наступает в возрасте около трех лет. Моногамы. На места гнездовий бакланы
прилетают уже в брачном наряде и в парах, которые повидимому постоянны.
Первые дни по прилете бакланы держатся близ старых гнезд, но к постройке
и ремонту гнезд приступают несколько позднее. Гнездятся колониями,
иногда достигающими громадных размеров — на небольшом о. Комсомольском,
Аральское море, более 1000 гнезд (Гладков, 1949), в низовьях Сыр-Дарьи на
отдельных озерах наблюдается до 2—3 тысяч гнездящихся бакланов; местами
впрочем бакланы гнездятся отдельными парами или небольшими группами.
Места расположения колоний очень постоянны, и птицы гнездятся в них из года
в год даже при постоянном преследовании, разорении гнезд, истреблении
птенцов и т. д. Обычно в колонии бакланов гнездятся и другие птицы — в Армении:
каравайки, рыжая и египетская цапли, кваквы, колпицы, чайки-хохотуньи,
серые вороны, грачи, усатые синицы и т. д.; у Ленкорани — малые бакланы,
кваквы, малая белая цапля, желтые цапли и грачи, по окраинам колонии —
черные аисты, коршуны, тювики,,серые вороны; в дельте Волги — серые цапли,
малые белые цапли, каравайки, реже желтые цапли и серые вороны; на
о. Комсомольском, Аральское море — серебристые чайки, черноголовые
хохотуны, в низовьях Сыр-Дарьи — пеликаны, малые бакланы, колпицы, каравайки,
кваквы, желтые цапли. Колонии, состоящие только из гнезд бакланов,
встречаются редко (дельта Волги, низовья Сыр-Дарьи у Соло-тюбе). Это,
повидимому, новые, только что образовавшиеся колонии, к которым еще не успели
присоединиться другие птицы. Позднее колония заселяется разными видами
птиц. Первыми поселяются серые цапли (Ленкорань), гнездящиеся так же,
как и бакланы, в верхнем ярусе деревьев, затем и другие птицы. Чем старше
колония, тем больше число видов, из которых она состоит.
Расположены колонии либо среди тростника, либо на деревьях, либо на
голых скалистых побережьях. Размеры гнезд и материалы, из которых они
сделаны, различны. В Армении встречаются гнезда конусообразной формы из
старого тростника. Они тесно сгруппированы и иногда даже соприкасаются
друг с другом. Помещаются гнезда среди воды в густых зарослях тростника или
на земле, или на заломленных стеблях старого тростника на высоте до 50 см над
землею. Высота гнезда бывает от 0,75 см до 1 м, ширина у основания около 1 м,
с лотком диаметром в 30—40 см. Гнезда выстланы листьями тростника. В
Ленкорани гнезда делаются из сучьев и ветвей, расположены они на вершинах
крупных деревьев (дуб, карагач, клен и др.). Здесь, как и в дельте Волги,
на одном дереве обычно размещается несколько гнезд. Причем деревья в
Ленкорани обычно отстоят от воды на 5—8 км в глубине широколиственного леса.
В дельте Волги гнезда устраиваются на деревьях, растущих у самой воды.
Диаметр гнезд 40—50 см, лоток не глубокий, выстлан сухой травой и листьями
тростника. На о. Комсомольском гнезда высотой до 0,5 м находятся на земле.
Сделаны они из веточек саксаула, тамариска, тростника. Часть гнезд
устраивается на саксауловых деревьях; иногда гнезда расположены по одиночке,
иногда группируются по 15—30 штук, образуя своеобразный бугор. В
низовьях Сыр-Дарьи гнездовые колонии находятся на островках среди заросших
тростником глубоководных озер и проток. На каждом таком озере, размером
приблизительно в Ькм*, помещается 2—3 тысячи бакланов. Обычно гнезда
лежат по краю острова, с одной стороны открытого к чистой воде, а с другой
стороны к нему примыкает тростник. Иногда они разбросаны по всему острову.
Гнезда высотой до 1 м построены из сухого тростника с неглубоким лотком.
Иногда гнезда расположены рядами, причем так плотно, что почти невозможно
отделить одно гнездо от другого. В ряде мест — на озере Сасык-куль и озере
28
ОТРЯД ВЕСЛОНОГИЕ
Ала-куль (Шнитников, 1949), на Черном море, западном побережье Арала,
туркменском побережье Каспия — гнезда построены из ветвей и тростника
на скалах. Во всех случаях гнезда плотно сцементированы пометом бакланов,
загрязняющим и все место колонии.
Период размножения большого баклана сильно растянут: в одной колонии
одновременно можно встретить ненасиженные и насиженные яйца и различного
возраста птенцов. Явление это может быть объясйено разорением бакланьих
кладок, главным образом, воронами и чайками и вызванными вследствие
этого вторичными кладками. Кладка у бакланов начинается рано, но точных
сведений о сроке начала ее очень мало. Можно считать, что начало кладки
падает на первую половину апреля. В Днепровском лимане у самого моря
в половине мая найдены гнезда с птенцами (5—10 дней возраста) и с
насиженными и ненасйженными яйцами (Браунер, 1891); в дельте Волги кладка яиц
начинается с 8—10 апреля (Огнев, 1913); в устье Урала в середине апреля (по
старому стилю) птицы сидели на яйцах (Бостанжогло, 1911), на нижней Сыр-
Дарье в середине апреля можно найти еще незаконченные кладки (Спанген-
берг и Фейгин, 1936).
В кладке обычно 5 яиц, голубого или бледнозеленого цвета, покрытых
сверху густым известковым слоем, сквозь который местами просвечивает основной
тон. Яйца овально удлиненной формы с несколько заостренным тупым концом.
Размеры:51— 61 х 34—38 мм (дельта Волги, Дюнин, 1936); (23) 58—66 х 37—42,6,
в среднем 62,3 х 38,6 мм (восточный берег Аральского моря, Зарудный, 1916);
62,4x39,1 мм и 66,9x39,9 мм (нижняя Сыр-Дарья, Спангенберг, 1936);
(4) 61—70 х 36—45 мм, в среднем 65,02 х 40,7 мм (устье Урала, Бостанжогло,
1911); (16)63,5x38,7 мм (озеро Севан, Соснихина). Вес 37—50 г (дельта
Волги, Дюнин, 1936); (5) 42,7—49,4, в среднем 46,2 г (Армения, Соснихина).
Срок насиживания 28—30 дней (дельта Волги, Ромашова). Насиживают
яйца оба родителя, начиная с откладки первого яйца, поэтому птенцы в одном
гнезде разновозрастны. В Армении 8 июля вывод птенцов уже закончился
во всех гнездах (Ляйстер и Соснин, 1942); в Днепровском лимане в середине
мая старого стиля в некоторых гнездах были птенцы в возрасте от 5 до 10 дней;
в дельте Волги средняя дата вылупления птенцов 6—8 мая; на берегу
Аральского моря (о. Комсомольский) 21 мая во многих гнездах были птенцы разного
возраста, 5 июня птенцы еще продолжали выводиться (Гладков, 1949); в устье
Урала в последней трети апреля старого стиля начинают вылупляться птенцы
(Бостанжогло, 1911). На нижней Сыр-Дарье 25 мая во многих гнездах были
уже оперившиеся птенцы (Спангенберг и Фейгин, 1936). Как правило, почти
в каждой кладке, в особенности содержащей 5—6 яиц, одно яйцо остается
не оплодотворенным. Птенцы появляются голыми и слепыми (глаза
открываются на 3—4-й день).
В возрасте приблизительно 2 недель птенцы покрываются густым пухом,
и у них начинают появляться маховые и рулевые перья. Младшие слабые
птенцы нередко погибают; процент гибели птенцов довольно велик. Так, на
о. Комсомольском (Аральское море) 25 мая в одной колонии был 51 птенец,
а 30 мая в ней осталось всего 15. В возрасте 3 недель птенцы имеют довольно
хорошо развитые плечевые перья, маховые, средние и большие кроющие крыла
и рулевые. В 4-недельном возрасте птенец почти весь одет пером, но маховые
и рулевые перья еще короткие; вес птенца в этом возрасте превосходит вес
рзрослой птицы, достигая иногда до 2500 г. Приблизительно в 7-недельном
возрасте птенец полностью надевает наряд молодой птицы; маховые и рулевые
перья достигают нормальных размеров. Родители выкармливают птенцов
отрыгиваемой рыбой, в более раннем возрасте эта рыба — полупереварена.
Прилетевшая с кормом взрослая птица усаживается на край гнезда и издает
характерный глухой крик. Птенцы начинают пищать и приближаться к краю гнезда.
БОЛЬШОЙ БАКЛАН
29
Принесшая пищу птица открывает рот, делает несколько конвульсивных
движений головой и шеей и, захватывая голову птенца в рот, отрыгивает пищу.
В один прилет взрослая птица старается накормить всех птенцов, но более
сильные отталкивают слабых и иногда получают двойную порцию.
Кроме рыбы, родители приносят птенцам иногда и растительную пищу,
главным образом водоросли. Приносят они и воду, которую дают птенцу таким
же приемом, как и пищу. Пища доставляется птенцам обычно 3, реже 2 или 5 раз
в день. В возрасте 1*/2 недель птенцы уже самостоятельно заглатывают
принесенную родителями целую рыбу. В один прием в общей сложности все птенцы в
гнезде в возрасте 2—3 недель получают 500—600 г рыбы, позднее—до 700 г.
Птенцы в колониях на деревьях приблизительно в возрасте 3 недель
начинают карабкаться по ветвям, в 4-недельном возрасте уже хорошо лазают по
деревьям, цепляясь лапами и помогая крыльями, шеей и клювом. В возрасте
около 7 недель птенцы приобретают способность перепархивать, а в 8а/2 недель
оставляют гнездо. Птенцы, выведшиеся в колониях, устроенных в камышах
на земле, также очень рано начинают лазить по гнездам и, еще не достигнув
способности к полету, уже начинают оставлять на некоторое время свои гнезда.
Способность к плаванию и нырянию они приобретают рано и обычно при
малейшей опасности птенцы стремятся добраться до воды и нырнуть.
О сроках вылета молодых птиц имеется очень мало сведений. В Армении
на озере Гйлли 2 августа молодые бакланы почти все плавали, но еще не
летали; в гнездах их оставалось мало; 11 августа они уже летали. В дельте
Волги массовый вылет бакланов обычно происходит в первых числах июля.
На нижней Сыр-Дарье 25 мая некоторые птенцы были уже оперены, хорошо
лазали по гнездам и могли нырять, 11 июня большинство было хорошо
оперено и ныряло на большие расстояния, а в начале июля большая часть
птенцов летала. В большинстве случаев с момента вылета молодых из гнезда, т. е.
с момента приобретения способности к полету, забота взрослых птиц о них
прекращается, и только в редких случаях родители еще подкармливают более
слабых молодых птиц.
Линька. Последовательность смены нарядов: пуховой —гнездовый —
первый брачный — первый зимний — второй брачный — второй зимний
и т. д. Птицы надевают окончательный наряд — третий брачный и третий
зимний — на 4-й год жизни. У молодых птиц две линьки в году: частичная в
августе — сентябре 'и полная с апреля по август. Гнездовый наряд молодые
птицы надевают в августе, в этом наряде они зимуют, а в апреле начинается
линька рулевых, кроющих крыла и затем маховых перьев. Линька
заканчивается к июлю. У взрослых птиц две линьки в году. Первая неполная, предбрач-
ная начинается в декабре и заканчивается в январе. Птицы надевают брачный
наряд и носят его до мая или июня. Вторая линька полная, послебрачная,
сильно растянута в связи с растянутостью периода размножения и начинается
с мая — июня. При этом птицы сначала теряют белые украшающие перья
брачного наряда, затем начинается линька остального мелкого пера,
заканчивающаяся в сентябре — октябре. В августе идет смена рулевых и в сентябре
или с конца августа и до середины октября происходит линька маховых перьев.
Линька рулевых происходит от центральных перьев к крайним, маховых —
от внутренних к наружным.
Питание. Бакланы питаются преимущественно рыбой размером,
обычно, в 20—25 см, редко — крупнее. Какого-либо предпочтения отдельным
видам рыб, повидимому, бакланы не отдают, поедая тех, которыми изобилует в
настоящий момент водоем. В течение суток птицы дважды вылетают за кормом.
Взрослая птица за сутки поедает примерно до 1 кг, в среднем 700 — 750 г.
Кроме взрослых рыб баклан в очень незначительном количестве поедает
мальков и случайно моллюсков, насекомых, земноводных и растения.
30 ОТРЯД ВЕСЛОНОГИЕ
Добывают рыбу бакланы на глубине не более 4 м, так что в море за.кормом
залетают не далее 50 км от берега (Каспийское море). Осенью, когда рыба в
море уходит на большую глубину, и бакланы достать ее не могут, они обычно
заметно теряют в весе. Охотясь за рыбой, бакланы плавают низко опустив
голову и высматривая добычу; увидав рыбу, ныряют и бьют ее сильным ударом
в бок, чем вероятно замедляют ее ход, а затем захватывают рыбу клювом;
вынырнув на поверхность, баклан умерщвляет рыбу, повреждая ей клювом
жаберную полость и резким движением ирловы подбрасывает ее в воздух,
чтобы затем подхватить ее в более удобном положении для заглатывания.
Заглатывает птица рыбу с головы.
Бакланы охотятся в одиночку или стаями, достигающими больших
размеров (на Аральском море, например, стая насчитывает до 250 птиц и даже
более), иногда совместно с пеликанами (Аральское море, Зарудный, 1916).
Рано утром, около 6—7 часов, бакланы вылетают с мест ночевок
небольшими стаями по 40—50 птиц и отправляются на ловлю рыбы. В дельте Волги
они обычно летят в предустьевую часть Каспийского моря, на западном берегу
Аральского моря (о. Комсомольский) — в море, на нижнем течении Сыр-Дарьи
птицы обычно летят на мелководные пространства и разливы рек, изредка и на
глубокие водоемы, но избегают соленых озер и мутных вод Сыр-Дарьи. Через
1х/2—2 часа, наевшись, бакланы возвращаются на берег к местам ночевок.
Птицы, имеющие птенцов, приступают к их кормежке, не имеющие —
располагаются на отдых и просушку своего оперения. Если на берегу растут деревья,
то бакланы на них рассаживаются группами (дельта Волги), если деревьев
нет или деревья далеко (Ленкорань, о. Комсомольский), то птицы усаживаются
на земле у самой линии воды. Часа в 4 дня бакланы вновь улетают за кормом
и к 6 часам вечера возвращаются на ночлег (дельта Волги); на о.
Комсомольском бакланы прилетают на ночлег только к 7—8 часам вечера.
Вред, приносимый бакланами рыбному хозяйству, несомненен в том
случае, когда численность птиц достаточно велика. Представление о вреде,
приносимом бакланами в местах их летнего пребывания, можно составить на
основании данных учета, произведенного Сушкиной (1932) и Дюниным (1936)
в дельте Волги. Количество бакланов осенью в дельте Волги исчисляется
приблизительно от 16 до 18 тысяч, из них около 8 тысяч взрослых и около
10 тысяч молодых. На местах гнездовий бакланы проводят приблизительно
8 месяцев. Взрослая птица за сутки поедает в среднем 700 г, так что
количество рыбы, поедаемой взрослыми бакланами за время летнего пребывания,
равняется 1344 т. Кроме того, птенцы поедают за летнее пребывание до 1050 т.
В общем, в дельте Волги бакланы съедают около 2394 т рыбы. На местах
охоты бакланов и около гнезд наблюдается много рыбы, убитой бакланами,
но не съеденной. Количество этой убитой, но не использованной рыбы по-
видимому превышает количество съеденной на 20—25%. Если даже считать
эти количества равными, то вред, приносимый бакланами рыбному хозяйству
в течение их летнего пребывания в дельте Волги, исчисляется в 4788 т*
В бассейне озера Севан вред, приносимый бакланами в течение 2 летних
месяцев, определяется в количестве 411 ц рыбы (Соснихина).
Охотясь за косяками рыб (дельта» Волги), бакланы часто отпугивают их
от захода в реки и тем самым препятствуют нересту. Во многих районах
бакланы поедают наиболее ценные промысловые виды рыб, например форель в
бассейне озера Севан.
Кроме того, селясь на деревьях целыми колониями птицы своими
экскрементами способствуют образованию сухостоя и гибели леса.
На Каспии и в Приморье до 1914 г. баклана добывали, используя шкурки
как мех и украшение и частично экспортируя за границу. В настоящее время
этот вид промысла совершенно заглох. В Приморье однако баклан иногда
БОЛЬШОЙ БАКЛАН 31
потребляется в пищу в свежем виде. В Астраханском заповеднике пробовали
использовать в пищу молодых готовых к вылету из гнезда птиц. Их особым
способом потрошили, затем солили и коптили, мясо получалось вполне
пригодное для' еды. Добывали молодых птиц от 5 до 10 тысяч штук в год, но
в настоящее время их не заготовляют. В Китае бакланов держат в
полудомашнем состоянии и используют для ловли рыбы.
Полевые признаки. Бакланы в гнездовое и внегнездовое время
держатся стаями. Очень осторожны и при приближении опасности обычно
стараются улететь. Полет сильный, но несколько тяжелый, с довольно частыми
взмахами крыльев; только изредка баклан на короткое время останавливается
в воздухе, как бы парит. Ноги и шея при полете вытянуты горизонтально,
по прямой линии. На воздух подымается с трудом с разбега или с какого-
либо возвышения. При перелетах обычно летит по прямой линии. Перемещаясь
на короткие расстояния, держится низко над водою, во время сезонных
перелетов летит на большой высоте.
По земле баклан ходит плохо, медленно, держа туловище почти
вертикально. На отдыхе бакланы собираются на берегу в небольшие стаи и усаживаются
рядами на земле, на скалах, или группами на вершинах деревьев. На отдыхе
птицы просушивают оперение, помахивая крыльями. Плавают они очень
хорошо. Ныряют на глубину не более 4 метров и под водой держатся до 40 секунд.
С воды поднимаются с разбега, работая в это время крыльями и ногами на
протяжении около 100 и более метров, и только затем взлетают в воздух. Баклан
может лазать по деревьям, цепляясь ногами и помогая крыльями; особенно
хорошо лазают птенцы, начиная с 4-недельного возраста.
32
ОТРЯД ВЕСЛОНОГИЕ
Баклан довольно молчаливая птица, но у гнезд, в особенности во время
кормления, они обычно шумливы и крикливы. Голос их похож на глухое
кряканье или карканье.
Описание. Размеры истроение. Длина тела самцов (4) 880—910, в среднем
864 мм; самок (4) 800—900, в среднем 824 мм.Размах крыльев у самцов (4) 1270—1560, в
среднем 1395 мм; самок(4) 1250—1500, в среднем 1382 мм. Длина крыла у самцов (20) 333—383лш,
в среднем 353 мм; у самок (24) 325—365 мм, в среднем 341 мм. Длина клюва у самцов (20)
59—75, в среднем 66,5 мм; самок (24) 52—62, в среднем 62,1 мм.
Вес сильно колеблется: от 1 800 г до 3 000 г; в среднем у самцов (55) 2529 г, самок (25)
2025 г. Кроме того, состояние упитанности птиц в разные месяцы года, даже в одном и том
же районе, весьма различно; например, в дельте Волги средний вес самцов в марте — 2369 г,
в апреле 2650 г, в мае — 2421 г, в сентябре — 2570 г, в октябре — 2670 г; средний вес самок
в марте —1999 г, в апреле — 2158 г, в мае —2112 г, в июне —1928 г, в сентябре — 1882 г
(вычислено по данным Дюнина, 1936). Первостепенных маховых перьев 11. Формула крыла
2>3>1>4... или 2=3>1=4>5... Вершины передних первостепенных маховых
приблизительно до половины их длины сужены на внутренних опахалах. Хвост округлый,
умеренной длины, состоит из 14 жестких рулевых перьев. Оперение густое, плотно прилегающее
к телу Уздечка, кольцо вокруг глаза и основания нижней челюсти у взрослых и молодых
Рис. 25. Птенцы большого баклана.
птиц голые; голые части заходят на нижней челюсти до угла разреза рта, так что линия,
проведенная перпендикулярно от угла рта по нижней челюсти, проходит по голой коже.
Окраска. Вылупившийся птенец совершенно голый. Кожа окрашена в черный цвет
с несколько розоватым тоном на голове и точечным рисунком на местах будущих птерилий.
Пуховой птенец. Буровато-черный с беловатым оттенком в средней части груди и брюха
и по переднему краю крыла. Голая кожа горлового мешка светлая, клюв светлосерый.
Гнездовый наряд. Голова, шея и верхняя часть зоба буровато-серые с мелкими
белыми крапинами, которые особенно многочисленны на нижней стороне шеи.
Полукольцо, охватывающее нижнюю челюсть до глаз, грязнобелое с мелкими темными
пестринами. Брюшная сторона тела белая или белая с редкими крупными бурыми пятнами. Бока
тела, спина и надхвостье темнобурые без блеска с более светлыми окаймлениями перьев.
Верхние кроющие крыла, плечевые и лопаточные буровато-серые с более темными
окаймлениями отдельных перьев. Радужина бурая. Клюв темнобурый, несколько светлый на под,-
клювье. Голая кожа горлового мешка желтая.
Первый брачный наряд. Оперение головы и шеи сходно с гнездовым нарядом —
буровато-серое с мелкими белыми крапинами. Белое полукольцо у основания подклювья грязно-
белое, без темных пестрин. Брюшная сторона тела белая с темнобурыми продольными
крупными пятнами, в особенности многочисленными на груди и на боках груди и брюха. Под-
хвостье буровато-черное с редкими белыми пестринами. Спинная сторона, за исключением
головы и шеи, похожа на взрослую птицу, но с очень слабым блеском.
Первый послебрачный наряд. Верх головы и шеи темнобурый с сероватым налетом
и редкими мелкими беловато-бурыми пестринами. На нижней стороне шеи и ее боках пёстрин
больше и они несколько крупнее. Брюхо темнобурое с редкими крупными продольными
пестринами. Спинная сторона тела похожа на взрослую птицу, но с меньшим блеском.
Второй брачный наряд сходен со взрослым. Оперение имеет хорошо развитый
металлический блеск. На бедрах во второй половине зимы появляется незначительное белое
БОЛЬШОЙ БАКЛАН
33
пятно. На голове хохла из черных перьев еще нет, но кое-где на голове и шее начинают
появляться белые узкие перья.
Взрослая птица в зимнем (послебрачном) наряде. Оперение черное с
металлическим зеленрвато-фиолетовым блеском и темными окаймлениями отдельных перьев
спины. Плечевые, лопаточные, верхние кроющие крыла и второстепенные маховые перья
зеленовато-бурые с бронзовым отливом и с широкими темносиними окаймлениями каждого
пера, придающими верхней стороне тела чешуйчатый рисунок. Первостепенные маховые и
рулевые перья буровато-черные со слабым металлическим блеском на рулевых. Широкое белое
полукольцо охватывает голову снизу, доходя до глаз и заходя треугольником на нижнюю
челюсть, соприкасаясь передним своим краем с неоперенными частями головы. Радужина,
светлозеленая, клюв буровато-черный, голые части головы желтые, голое кольцо вокруг
глаза зеленовато-бурое, ноги черные.
Брачный наряд (взрослые самцы и самки). На затылке удлиненные черные
перья образуют небольшой хохол. На голове и шее появляются узкие, удлиненные
тонкие белые перья, покрывающие, в особенности густо у старых особей, почти всю
голову и шею, только хохол на затылке остается черным. На наружной стороне бедра и голени
развиваются удлиненные белые перья, образующие большое белое пятно.
Атлантический большой баклан Phalacrocorax carbo carbo L.
Распространение. Ареал. Южн. и зап. Гренландия, на север до 69° с. ш.
и до Годхавн на о. Диско, Исландия, Фарерские о-ва, Великобританские о-ва,
Норвегия ис.-з. часть Кольского п-ова до Кольского залива, нерегулярно до
заповедника Семи островов (гнездился на о. Кувшин, Модестов, Горчаковская).
Вопрос о гнездовании в Сев. Америке, где бакланы встречаются от восточного
побережья Баффиновой Земли (Кумберленд) до Лабрадора, Новой Шотландии
и Ньюфаундленда неясен, повидимому, баклан здесь истреблен.
Характер пребывания. Гнездящаяся, оседлая и частично
кочующая птица. В Англии — частью оседлы, частью кочуют, доходя к югу
до берегов Пиренейского п-ова, достигая с.-з. Африки и Канарских о-в.
В небольшом числе может быть встречен и зимой вдоль Мурманского
побережья и в Кольском заливе. От берегов Гренландии кочуют вдоль
Атлантического побережья Сев. Америки, доходя к югу до Каролины.
У заповедника Семи островов появляются весною в середине марта, но
первое время по прилете держатся в море и мало заметны (Горчаковская). Часть
птиц, повидимому, не гнездящихся, кочует в небольшом количестве и летом
у берегов Семи островов.
Биотоп. В северных частях распространения скалистые
побережья морей, заливов и рек, мелкие скалистые острова и отдельно стоящие
скалы; в южных частях своего ареала и побережья внутренних водоемов —
заросшие древесной и тростниковой растительностью.
Численность. На Мурманском побережье в конце прошлого
столетия был повидимому обыкновенным и даже многочисленным видом; еще в начале
нынешнего столетия местами был обычен, в настоящее время редок и кое-где
совсем исчез (Федюшин, 1925). На Семи островах с 1938—1941 г. большого
баклана было меньше, чем длинноносого баклана, но численность его все эти
годы не изменялась. Гнездился атлантический баклан только на о. Кувшине,
где было около 50 гнезд (Модестов, Успенский, 1941). В 1947 г. ни на одном из
островов заповедника «Семи островов» баклан, по сообщению Карташева, не
только не гнездился, но даже и не залетал.
Экология. Размножение. Данные о размножении по Мурманскому
побережью почти полностью отсутствуют. Гнездятся атлантические бакланы
небольшими колониями, иногда совместно с другими птицами, так на о.
Кувшине в заповеднике «Семь островов» совместно с длинноносым бакланом и кайрами.
Гнезда устраивают на высоких отвесных скалах, на больших открытых
карнизах скал. Расстояние между гнездами 1—2 метра (Горчаковская).
Птицы занимают одно и то же гнездо в течение нескольких лет, только из
года в год подновляя его. Строят гнездо самец и самка; строительный мате-
3 Птицы СССР, т. I
34
ОТРЯД ВЕСЛОНОГИЕ
риал — ветви, сучья, водоросли и травы — собирают на побережье близ
колоний. Все гнездо, как и окружающие его скалы или деревья, бывают густо
запачканы пометом.
Количество яиц в кладке по Мурманскому побережью не выяснено. В
Великобритании 3—4, иногда 5 и очень редко 6 яиц (Уайзерби, 1945). В Сев.
Америке обычно 4—5, редко 6 яиц (Бент, 1922). Яйца овальной формы, светло-
голубые с известковым поверхностным слоем, похожие на яйца материкового
большого баклана. Размеры яиц (100) 57,5—74 х 38—42, в среднем 65,8 х 40,7 мм
(Англия, Уайзерби, 1940); (42)61—70,5x38,5—43,5, в среднем 64,8x40,9 мм
(Сев. Америка, Бент, 1922).
Насиживают яйца самец и самка; период насиживания около 28 дней. Одна
кладка в году. Время кладки у нас не выяснено. В Шотландии — во второй
половине мая и до начала июня, в южных частях Британских о-вов — во
второй половине апреля и в начале мая. У нас в Кольском заливе 16. VI. 1902 г.
в Пола-Губе был добыт пуховой птенец из гнезда (Бианки, 1902), а 6. VIII.
1925 г. у Сеть-Наволока добыты 3 птенца с подрастающими маховыми и
рулевыми перьями (Спасский, 1925).
Линька. Как у материкового подвида.
Питание. Атлантические большие бакланы питаются рыбой мелких
и средних размеров, в очень ограниченном количестве ракообразными, а
в период размножения и растениями. Добывают рыбу так же, как и
материковый баклан.
Описание. Размеры и строение. Длина крыла мурманских самцов и
самок (4) 335—360 мм, в среднем 350 мм; длина клюва самцов и самок (4) 62—74 мм,
в среднем 65 мм. Длина крыла самцов 330—370 мм, клюва от оперения 66—75 мм; длина
крыла самок 325—365 мм, клюва 65—75 мм (Англия, Уайзерби, 1945).
Длина тела (Мурман) самки 850 мм, размах крыла 1320. Вес 3017 г.
Окраска. Пуховой птенец. Пух буровато-черный, похожий на пух птенца
материкового баклана. Возрастные изменения такие же, как у материкового баклана. Взрослая
птица отличается от материкового баклана фиолетово-синим металлическим блеском
оперения. Брачный наряд такой же, как и у материкового баклана.
2. Уссурийский баклан Phalacrocorax filamentosas
Те mm. et Schleg.
Carbo filameritosus. Temminck et Schlegel. Fauna Jap., Aves, 1850, стр 129,
Япония.
Синоним: Carbo capillatus. Temminck et Schlegel. Fauna Jap., Aves,
1850, табл. 83, Япония.
Распространение* Ареал. Побережье южн. Приморья и прилежащие
острова (о. Аскольд, о. Коврижка, о. Русский и др.), к северу достоверно
найден до устья р. Тетюхе (Шульпин, 1936), может быть и несколько северней — до
45° с. ш.; Корея; Япония, о. Хондо (Ян, 1942); вост. Китай к югу до Амоя.
Характер пребывания. В Приморье гнездящаяся, и там, где
море не замерзает, частично зимующая птица. Наблюдалась в большом
количестве в Амурском заливе в первых числах ноября; в конце ноября близ устья
р. Сучана встречалась уже редко, а 21 декабря отмечены только две птицы в
заливе Ольга (Пржевальский, 1870). Добыта в январе 1926 г. в устье р^. Самарги.
В Японии и Китае оседла. Весною у мест гнездовий появляются во jJ-й половине
апреля. Осенью долго держится в море близ своих гнездовий. Зимой к югу
доходит до Южн. Китая.
Биотоп. Преимущественно скалистые морские побережья, острова
и отдельные скалы в море.
Численность. По берегам Приморья уссурийский баклан
распределен неравномерно, но в общем довольно обыкновенен, только у северной гра-
УССУРИЙСКИЙ БАКЛАН
35
ницы распространения (у р. Тетюхе) встречается редко. Большие колонии по
побережью отмечены на мысе Гамова и в заливе Славянка. В большом
количестве водится на некоторых островах южн. Приморья. Так, большие колонии
имеются на о. Аскольд, в заливе Абрек, на о. Коврижка в заливе Посьет, на
о. Фуругельм и на о. Русском. В
прошлом столетии большая
колония существовала на скалах,
расположенных между берегом и
о. Аскольд, в настоящее время
этой колонии нет(Шульпин, 1936).
На Хондо обыкновенен, большие
колонии имеются близ Токио и у
Нагоя.
Экология* Размножение.
Моногамы. В конце апреля птицы
встречаются в парах у мест
гнездований. Обычно гнездятся
колониями различных размеров, но
иногда — чаще у северной границы
своего распространения — всего
ЛИШЬ двумя ИЛИ тремя парами ИЛИ Карта 2. Распространение Уссурийского бак-
даже отдельными парами. В При- лана Phalacrocorax filamentosus
МОрье КОЛОНИИ уССуриЙСКОГО бакла- « — граница гнездовий, г — область зимовок.
на населены только этой птицей.
В Японии имеются смешанные колонии — с кваквами и белыми цаплями.
Кроме того близ Токио уссурийский баклан гнездится с обыкновенным
бакланом в общей колонии.
Гнезда обычно устраиваются на уединенных малодоступных скалах,
расположенных у самого берега или выступающих в море, но в Японии, кроме
того, встречаются гнезда, устроенные на соснах и других хвойных деревьях.
Рис. 26. Уссурийский баклан.
В этих случаях колонии бывают часто расположены в 2—3 км от берега (Ян,
1942). Время начала кладки и вылуплёния птенцов не выяснено. В кладке
4 яйца, похожих по окраске и форме на яйца большого баклана.
Размеры :(10) 60—63,8 х37,8—42,8, в среднем 62,5 х 40,3 лш (Тачановский, 1893).
На яйцах птицы сидят плотно. Спугнутые с гнезд птицы сейчас же, не
обращая внимания на присутствие человека, возвращаются на свои гнезда. Молодые
з*
36
ОТРЯД ВЕСЛОНОГИЕ
самостоятельные птицы обычно наблюдаются на побережье Приморья (бухта
Тетюхе) в первых числах июля. В Японии (Хондо) в начале апреля в гнездах
находили нелетающих птенцов, величиною с половину взрослой птицы (Ян,
1942). Линька не изучена.
Питание. Уссурийский баклан питается преимущественно рыбой,
добываемой им, повидимому, только в море. Бакланов, кормящихся на пресной
воде, даже в приморских частях пресных рек не наблюдалось; в тех случаях,
когда колонии расположены относительно далеко от морского берега (Хондо),
птицы за пищей летают на море. '
Полевые признаки. В поле уссурийский баклан трудно отличим от
большого баклана, их повадки сходны.
Описание. Размеры и строение. Величиною с обыкновенного баклана. Длина
тела (1) 840 мм, длина крыла (3) 320—332 мм, длина клюва (3) 61—67 мм.
Хвост округлый, состоит из 14 рулевых перьев. Первостепенных маховых 11, формула
крыла 2>3>4>1>5...Уздечка, кольцо вокруг глаз, пространство за глазом и боковые
стороны у основания нижней челюсти голые. Пространство, занимаемое голой кожей, на нижней
челюсти меньше, чем у большого баклана; оперение идет вниз и вперед, так что линия,
проведенная перпендикулярно от угла рта, проходит до оперенной части нижней челюсти.
Окраска. Пуховой птенец не описан. Молодые птицы. Повидимому, похожи
на молодых птиц большого баклана. Гнездовый наряд. Верхняя сторона тела
серовато-бурая без металлического блеска. Перья спины, лопаточные и верхние
кроющие крыла с темными предвершинными и беловато-серыми вершинными окаймлениями.
Брюшная сторона тела белая с легким буроватым налетом, подхвостье бурое. На
боках тела бурые, как бы размытые, крупные пятна. Ноги черные. Клюв
желтовато-грязный с темной полосой посредине надклювья. Взрослая птица похожа на
обыкновенного баклана, но отличается большим развитием зеленого блеска в оперении.
Голова и шея темнозеленовато-синие. Плечевые, верхние кроющие крыла бронзово-зеленые
с темнозелеными окаймлениями. Белое полукольцо, которое охватывает снизу голову и
примыкает к ее неоперенной части, густо покрыто зеленовато-черными пестринами. Ноги
черные, радужина желтая, голые части лицевых частей желто-оранжевые.
В брачном наряде на голове и шее имеются белые перья, но в отличие от большого
баклана они длиннее и реже расположены, так что не закрывают темного оперения головы
и шеи. На бедрах имеется по пучку белых удлиненных перьев.
Географическая изменчивость повидимому не выражена.
3. Берингов баклан Phalacrocorax pelagicus Pall.
Phalacrocorax pelagicus. Pallas. Zoogr. Rosso—Asiatica, 1811, стр. 303, вост. Камчатка.
Распространение. Ареал. Тихоокеанские побережья: от о. Врангеля
и Чукотского п-ова до Курильских о-вов, Сахалина, сев. Японии —
в Азии и от Аляски и Алеутских о-вов до Нижней Калифорнии в Сев.
Америке.
Характер пребывания. На севере перелетная, на юге
оседлая птица.
Биотоп. Скалистые морские побережья материков и островов.
Подвиды и варьирующие признаки. Два подвида, различающихся между собою
размерами и образом жизни: Ph. p, pelagicus Pall., 1811 — азиатское побережье с прилежащими
о-вами, Аляска и Алеутские о-ва; Ph. p. resplendens А и g u b о п, 1838— Сев. Америка от
Вашингтона до м. Сан-Лукас в Нижней Калифорнии и Мазатлана в сев. Мексике.
Западный берингов баклан Phalacrocorax pelagicus pelagicus Pallas
Синонимы: Phalacrocorax aeolus. Swinhoe. The Ibis, 1867, стр. 395, Амой.—
Phalacrocorax pelagicus robustus. Ridgway, Brewer etRidgway. Water — Birds
N. Amer. 11, 1884, стр. 160, Аляска.
Распространение. Ареал. По азиатскому материку распространен от
о. Врангеля и м. Северный (м. Рыркайпий) на Чукотском п-ове по побережью*
и прилежащим островам до Камчатки; сев. и сев.-западные берега Охотского
моря — Пенжинская и Гижигинская губа, Шантары, может быть в северной
БЕРИНГОВ БАКЛДН 37
части Приморья (встречены и добыты с мая по август, Шульпин, 1936); Команд
доры, Курилы, Сахалин. Вне СССР встречается в Японии — Хоккайдо и
северная часть Хондо (зал. Аомори). В Сев. Америке — Аляска от залива
Нортон и может быть даже северней, от залива Коцебу (встречен в июле). При-
быловы о-ва, о. св. Лаврентия и другие острова в Беринговом море,
Алеутские о-ва, о. Кадьяк и тихоокеанское побережье Сев. Америки к югу до
о. Ванкувера.
Характер пребывания. В северных частях своего ареала
перелетная, в южных — оседлая и зимующая птица.
Карта 3. Распространение берингова баклана Phalacrосотах pelagicas
а — граница гнездовий, г — область зимовок.
Осенью птицы держатся у мест гнездовий очень долго — до полного
покрытия льдом морей (21 сентябрянаблюдалисьум. Северного, Чукотка,
Артоболевский, 1926). Отлет у побережья Чукотки происходит в самом конце сентября
и может быть в начале октября (26 сентября в небольшом количестве птицы
наблюдались у м. Северного). Анадырский край они покидают около середины
октября. У Охотского побережья птицы встречаются в течение сентября —
октября (о. Рейнеке, м. Мухтеля," Дулькейт и Шульпин, 1937). В северной
части Приморья и в Татарском проливе птицы наблюдаются с сентября до конца
ноября (Шульпин, 1936, Воробьев, 1938). На Камчатке, вдоль восточного
побережья у Кроноцкого п-ова — наблюдаются в небольшом количестве
и зимой, но большинство бакланов, начиная с сентября, отлетают к югу. У
Командорских о-вов встречается в течение всей зимы, но в несколько
меньшем количестве, чем летом. Зимою у Командорских о-вов птицы собираются
в большие стаи и держатся преимущественно на восточной стороне островов,
38
ОТРЯД ВЕСЛОНОГИЕ
например, на Тонком мысу о. Беринга. Зимой бакланы встречаются вдоль
берегов южн. Приморья, Японии (до южной окраины Хондо), Кореи и до Южн.
Китая.
Весною к местам гнездовий птицы прилетают как только начинается
движение льдов и появляются открытые водные пространства. Так, на Чукотке у м.
Северного 7 мая птицы уже прилетели. На Анадыре в заливе Креста первые птицы
появились 1-го мая и 18-го их было уже много(Белопольский, 1934), впрочем
по данным Сокольникова (Портенко, 1939), бакланы прилетают в Анадырский
край только в первых числах июня (по старому стилю), что весьма сомнительно;
у Гижиги — в конце первой недели мая. На Камчатке первые перелетные
бакланы появляются с первых чисел марта (5.III), интенсивный прилет и пролет идет
с конца марта до начала апреля, последние одиночные пролетные птицы
встречаются до конца первой декады мая (Аверин, 1948).
Биотоп. Крутые скалистые морские побережья материков и островов,
рифы и скалы, выступающие из воды.
Численность. Баклан встречается в очень большом количестве по
северному побережью Чукотки ум. Северного; на Анадыре обычен и местами мно-
гочисленен, например между Анадырским лиманом и заливом Креста. Обыкно-
венен на Камчатке, у Гижиги многочисленен, но на Б. Шантарах и в Приморье
бакланов мало. Многочисленен в настоящее время на Командорах. Особенно
многочисленен был там до 1876 г., но зимой 1876—77 г. птицы гибли тысячами,
вследствие какой-то эпизоотии. На следующее лето птиц было мало, но через
5 лет они вновь стали уже обыкновенными. На Алеутской и Курильской гряде
обыкновенен и на некоторых островах многочисленен. В* Японии обычен.
Экология. Размножение. Половая зрелость наступает в возрасте
около 2 лет, когда птицы не надели еще окончательного наряда. Обычно
гнездятся колониями, изредка отдельными парами среди колоний других птиц
(Командоры, Иогансен, 1934). Колонии различных размеров от нескольких пар
до нескольких сотен и тысяч. Большие колонии имеются на м. Северном
(Артоболевский, 1926), в бухте Ребкена, на побережье между заливом Креста и
Анадырским лиманом (Белопольский, 1934) в Гижигинской губе, на Командорских
о-вах (Иогансен, 1934), на Камчатке и Курильских о-вах. Не все птицы,
прилетевшие к местам гнездовий, приступают к размножению. Негнездящиеся
холостые птицы держатся в гнездовом сезоне близ колоний, проводя весь день
в море и прилетая на скалы только для отдыха и ночлега.
Гнезда устраиваются в расщелинах, на карнизах и уступах скал и утесов,
довольно высоко от основания скалы, так что волны не могут достать их. Гнездо
больших размеров, широкое; сделано из водорослей, мха, травы и других
растений, внутри выстилается более нежными частями растений. Обычно гнезда
служат в течение нескольких лет. Все места, где расположены гнезда,
испачканы экскрементами птиц; по этому признаку гнездовья легко обнаружить.
Кладка яиц начинается на Командорах в первой половине мая или в конце
мая, иногда и в начале июня, но еще в июле встречаются и ненасиженные яйца;
на Камчатке в начале 2-й половины июня (19.VI) в гнездах были яйца. На
основании нахождения на Командорах ненасиженных яиц в мае, июне и июле
было высказано Стейнегером (1885) предположение, что у Берингова баклана
на Командорах 2 кладки в году, связанные между собой птенцами переходного
возраста: 1-я с мая по июль и 2-я с июля по сентябрь, но это предположение
сомнительно. Растянутость периода кладки в колонии бакланов.объясняется,
вероятно, наличием дополнительных кладок взамен погибших и разновозра-
стностью состава птиц в колонии.
В кладке обычно 3—4 яйца, реже 2—5, но иногда будто бы 6—7 (Командоры,
Гижига). Яйца продолговато-овальной формы, голубого цвета, который хорошо
виден на только что отложенных яйцах; позже скорлупа обычно- покрыта сверху
БЕРИНГОВ БАКЛАН 39
белым известковым слоем. Размеры: (41) 53,3—63x35—41, в среднем 58,3 х
Х.37,4 мм (Бент, 1922). Продолжительность насиживания 26 дней, в
насиживании принимает участие и самец. Время вылупления и время оставления гнезда
птенцами точно не выяснено. Птенцы вылупляются голые и покрываются пухом
в возрасте нескольких дней. На Камчатке в августе найдены в гнездах птенцы
больших размеров (Бергман, 1935 ), на Командорах летные птенцы наблюдались
с конца июля и по сентябрь включительно (Бианки, 1909), на Анадыре 21—22.
VIII. 1931 г. в гнездах еще были нелетающие птенцы (Белопольский, 1934),
на м. Северном (Чукотка) 8 июня птицы еще сидели на яйцах
(Артоболевский, 1926).
Рис. 27. Берингов баклан (взрослая птица и молодая).
Линька. Последовательность смены нарядов, — как у большого
баклана. Две линьки в году: частичная предбрачная — в декабре —январе
и полная послебрачная в июле — октябре и вероятно до ноября. Начало после-
брачной линьки у различных особей различное, и можно встретить птиц почти
в одной стадии линьки и в июле и в октябре. Например, у птицы, добытой
29.VI 1.1948 г. на Итурупе, Курильские о-ва, 3 внутренних маховых свежие,
остальные маховые старые, внутренняя пара рулевых свежая, но еще короткая,
остальные старые; белых перьев в оперении уже нет, на голове остатки хохлов.
У птицы, добытой 8.Х. 1947 г. на Сикотане, Курильские о-ва, 1-е маховое
старое, 2-е свежее, но недоросшее; остальные маховые свежие; рулевые все
сменились, крайняя пара еще не достигла своих нормальных размеров. Молодые
птицы надевают взрослое оперение только на 3-м году жизни. Линька маховых
перьев идет постепенно от внутренних к наружным, рулевые сменяются от
центральных к крайним.
Питание. Пищу берингова баклана составляет рыба.
Полевые признаки* В природных условиях плохо различим от
краснолицего баклана, с которым сходен по общим размерам. Повадки — как
у большого баклана. Мало осторожен.
40
ОТРЯД ВЕСЛОНОГИЕ
Описание. Размеры и строение. Длина тела самок (2) 720—770 мм. Длина
крыла самцов и самок(15) 266—291, в среднем 284, 2 мм. Длина клюва (15) 48—56, в
среднем 51,7 мм. Вес около 1500—1600 г.
Первостепенных маховых 11, формула крыла 2>3>4>5=1>6... Хвост округлый,
состоит из 12 рулевых перьев. Клюв тоньше, чем у большого и краснолицого баклана.
Голые участки кожи по бокам головы незначительных размеров. Оперение на лицевых
частях доходит до ноздрей и верхнего края глаз, так что образуется обычно голое
полукольцо, охватывающее глаз снизу; только у старых птиц имеется вокруг глаза узкое голое
кольцо. На боках нижней челюсти голое пространство, также очень незначительно развитое.
Окраска. Пуховой птенец темносерый. Гнездовый наряд. Все оперение бурое,
светлее на голове и шее и с едва заметным зеленоватым металлическим блеском на спинной
стороне тела, за исключением головы и шеи. Перья тела с узкими темными предвершинными
и светлыми вершинными окаймлениями. Клюв и ноги буроватые.
Первый брачный наряд. Темнобурый со слабым металлическим зеленым блеском на
спинной и брюшной сторонах тела. Голова и шея бурые, без блеска, с мелкими белыми
крапинками, образуемыми белыми вершинами отдельных перьев. Первый зимний наряд.
Металлический блеск развит сильней, чем у птиц в первом брачном наряде, но еще не достигает
такого развития, как у взрослых птиц. На перьях спины имеются более темные окаймления
перьев. Голова и шея темнобурые со слабым синеватым блеском, кое-где на голове и шее
имеются еще оставшиеся перья с белыми вершинами.
Взрослая птица в послебрачном наряде. Оперение черное с сильным
металлическим блеском. Голова, нижняя часть шеи, спина, поясница, зоб и вся брюшная
сторона тела — с зеленым блеском, верхняя часть шеи — с синевато-фиолетовым,-кроющие
крыла с зеленовато-бронзовым и с темными каймами отдельных перьев. Маховые,
рулевые и подхвостье черные со слабым, едва заметным зеленым блеском. Клюв черный, только
у основания нижней челюсти желтый. Ноги черные. Радужина зеленая. Голая кожа
вокруг основания клюва и глаза черная, испещренная желтовато-красными бородавками.
Взрослая птица в брачном наряде имеет на шее, боках, зобе, на нижней части
спины, пояснице и на бедрах удлиненные узкие белые перья. На голове (на затылке и
темени) хорошо заметны Z хохла из черных перьев.
4. Краснолицый баклан Phalacrocorax urile G m e 1 i n
Pelecanus Urile. Gme 1 i n- Syst. Nat., изд. XIII, 1789, стр. 575, Камчатка.
Синоним: Phalacrocorax bicristatus. Pallas. Zoogr. Rosso — As., II, 1811, стр. 301,
Камчатка.
Распространение. Ареал. Командорские о-ва, но в настоящее время
встречаются, повидимому, только на о. Медном; старыми авторами приводится
для Камчатки, но в последние годы никем там не наблюдался (Бергман, 1935,
Аверин, 1948 и др.). К северу от Камчатки по азиатскому побережью и на
прилежащих островах краснолицый баклан не отмечен. Приводился ранее для
Курильских о-вов, в настоящее время встречен осенью только на одном из
южных островов — на о. Сикотане (добыл Гизенко 8.Х.1947 г., коллекция
Зоол. музея Моск. гос. университета). Вне нашей страны на Прибыловых
о-вах и может быть на западных островах Алеутской гряды (Бент, 1922).
Характер пребывания. Краснолицый баклан оседлая и
частично перелетная птица. На Командорских о-вах оседла, на Прибыловых —
перелетна. Зимой наблюдается у Алеутских о-вов, Командор, Курильской
гряды, у южн. Сахалина (добыт в феврале Шульпиным, 1936), у берегов
Японии и южней, до Формозы. Время прилета весной к местам гнездовий и
начало осенних кочевок неизвестно, но 8 октября добыта, повидимому, уже
кочующая 'птица на о. Сикотан. Молодые птицы, еще не размножающиеся,
дольше задерживаются весною на зимовках, а часть из них проводит там
даже все лето.
Биотоп. Скалистые, каменистые побережья островов и отдельные скалы,
стоящие в море близ берегов.
Численность. На Командорских о-вах во второй половине
прошлого столетия и в начале нынешнего краснолицый баклан был обыкно-
венен, но на о. Медном его было больше, чем на о. Беринга. В последние
годы, повидимому, на о. Беринга совсем не гнездится. На Курильских о-вах
КРАСНОЛИЦЫЙ БАКЛАН 41
и в прежние времена был редок, а в настоящее время, вероятно, и совсем н$
гнездится.
Если считать, что сведения старых авторов правильны, то можно придти
к выводу, что количество краснолицых бакланов постепенно уменьшается, а
ареал их сокращается.
Экология. Размножение. Половая зрелость наступает на третьзм
году жизни (Пальмер, 1899). Гнездятся колониями на плоских открытых
выступах скал и утесов. Гнездо построено главным образом из водорослей, которые
Карта 4. Распространение краснолицого баклана Phalacrocorax urile
а — граница гнездовий, г — область зимовок.
птицы собирают у берега и вылавливают, ныряя в море. Выстилка из нежных;
частей растений и перьев чаек. Размеры гнезда — в среднем 40—50 см в
диаметре и 15 см высотою; оно по всей вероятности служит птицам в течение
нескольких лет (Прибыловы о-ва, Бент, 1922).
Количество яиц в кладке точно не известно. Размеры: в среднем (3) 63,5 х
38,66 мм (о. Медный, Стейнегер, 1885); 61 х39мм(о. Беринга, Стейнегер, 1885);
(7) 60—63x38,7, в среднем 61,61 х40,4 мм (Тачановский, 1893); (47) 53,2—
65,5 х33—41,5, в среднем 60,3 х37,6 мм (Бент, 1922). Яйца
удлиненно-овальной формы, бледноголубого цвета, покрыты сверху белым известковым слоем..
Насиживание продолжается около 3 недель (Прибыловы о-ва).
Откладывание яиц начинается, повидимому, не одновременно у всех птиц колоний, как и у
некоторых других видов веслоногих. Одна кладка в году. На о-ве Медном
яйца находили в конце мая, в июне и ненасиженные яйца встречались даже в
июле. В июле в это же время во многих гнездах были уже птенцы, величиной
,42
ОТРЯД ВЕСЛОНОГИЕ
с половину взрослой птицы. Летные птенцы наблюдались с конца июля по
сентябрь включительно. На Прибыл овых о-вах насиженные яйца находили
1 июня, первых птенцов —13 июня; но и в начале июля в некоторых гнездах были
птенцы разного возраста, а в некоторых — насиженные яйца, повидимому,
отложенные в середине июня или даже позднее. Вероятно, птенцы этих поздних
кладок покидают гнезда только в конце августа.
Весь период размножения в гнездовой колонии краснолицого баклана
занимает 4—4,5 месяца. Птенцы вылупляются голыми, слепыми, темного багрово-
коричневого цвета. Растут они быстро, как и птенцы других видов бакланов.
Через несколько дней появляется пух вдоль срединной части спины и вскоре
все тело птенца покрывается
пухом. Через полтора месяца
птенцы достигают размеров
взрослой птицы, но тело их
еще покрыто пухом.
Контурные перья только что
пробиваются, но рулевые и маховые
перья развиты и достигают
почти нормальных размеров.
В гнездовом оперении
краснолицые бакланы
отличимы от других видов
молодых птиц, за исключением бе-
рингова баклана, только
слабым развитием голых лицевых
частей и линией оперения,
окружающей голую лицевую
часть головы, и почти не
отличимы по этим признакам от бе-
рингова баклана, который
отличается только менее
массивным клювом.
Линька. Точно не
изучена, в общем как у
других видов бакланов.
Питание. Пищу
краснолицого баклана составляет
преимущественно рыба. Птенцы выкармливаются родителями первое время
полуперевариваемой пищей, затем целой мелкой рыбой'и рачками.
Полевые признаки. В природе не отличим от берингова баклана, с
которым сходен и по повадкам.
Описание. Размеры и строение. Средних размеров птица, приблизительно
таких же, как берингов баклан, но с несколько более массивным клювом. Вес около
2 кг. Длина крыла (3) 270—285, в среднем 275 'мм, длина клюва (3) 53—57, в среднем 55 мм,
высота клюва у основания (3) 13—14,5, в среднем 13,8 мм.
Первостепенных маховых 11, формула крыла 2>3>4>1>5... Хвост закругленный
из 12 рулевых перьев. Передняя часть лба у взрослых птиц голая, вокруг глаз широкое
голое кольцо. У молодых птиц лоб оперен, но оперение не доходит до ноздрей; голое
кольцо вокруг глаз узкое и иногда выражено только в виде полукольца под глазом. На
боках нижней челюсти и головы голое пространство заходит глубоко за задний край глаза.
Окраска. Пуховой птенец темносерый, испещренный на брюшной стороне
белыми крапинами.
Гнездовый наряд черно-бурый, со слабым пурпуровым блеском на плечевых и
кроющих крыла. Голова и верхняя часть шеи дымчато-серые.
Взрослая птица в зимнем наряде. Оперение черное с металлическим блеском. Голова
и прилегающая часть шеи с зеленовато-синим блеском, надхвостье и брюшная сторона
Рис. 28. Краснолицый баклан.
ДЛИННОНОСЫЙ БАКЛАН
43
тела с бронзово-зеленым отливом.; спина и плечевые зеленовато-лиловые. Темных каемок
на перьях мантии нет.
Голая кожа вокруг основания клюва и глаз бугорчатая оранжево-красная. Голая
кожа горла синяя. Радужная—-бурая, ноги черные.
Брачный наряд. На темени и затылке расположено два черных хохла с
бронзово-зеленым блеском. На бедрах — по пучку белых удлиненных узких перьев. У
основания шеи и на подхвостье имеются редкие узкие белые перья, иногда
отсутствующие на надхвостье. Промежуточные — между гнездовым и окончательным (вторым
брачным, вторым зимним) — наряды не описаны.
5. Длинноносый баклан Phalacrocorax aristotelis L.
Pelecanus Aristotelis Linnaeus. Fn. Svec, Faunula, 1761, стр. 5, № 146, Швеция.
Синонимы: Pelecanus Graculus. Linnaeus. Syst. Nat. 1766, стр. 217, Швеция.
Распространение. Ареал. Зап. Палеарктика от Мурмана и
Норвегии до Исландии, Фарерских о-вов, Англии, Ирландии и по западному
берегу Европы до Пиренейского п-ова; побережье и острова Средиземного
и Черного морей.
Характер пребывания — оседлая и кочующая птица. Биотоп.
Скалистые побережья, острова и отдельно стоящие в море скалы.
Численность. Менее многочисленен и обыкновенен, чем большой баклан.
Распространение спорадично.
Подвиды и варьирующие признаки. Длинноносый баклан имеет три подвида,
различающихся пропорциями клюва, окраской голых частей головы и перепонок лап, а также
деталями экологии: РЛ. ar, aristotelis Linnaeus, 1761 — от Мурманского побережья и
Исландии до Испании и Португалии; РЛ. ar. desmarestii Payraudeau, 1924 — острова
и северное побережье Средиземного моря и сев.-западного побережья Черного моря и
РЛ. ar. riggenbachi H a r t e r t, 1923 — западный берег Марокко от Могадора до м. Бланко.
Атлантический длинноносый баклан Phalacrocorax aristotelis aristotelis L.
Распространение. Ареал. Атлантическое побережье Португалии,
Испании, Франции, Бельгии, Голландии, Британские о-ва (южное и восточное
побережье Англии, Уэльс, восточное побережье Шотландии, ю.-з. и зап.
побережье Ирландии), Исландия, Фарерские о-ва, Норвегия и Кольский п-ов, на
восток по Мурманскому побережью и по прилежащим островам до Иоканги.
Характер пребывания. На Мурмане гнездится в небольшом
числе, зимует и кочует. На Британских о-вах и южней оседлый. Немногие
встречи окольцованных на вост. Мурмане, о. Харлов, птиц показывают, что
они откочевывают вдоль берегов Скандинавии в ю.-з. направлении, достигая
Норвегии; некоторые остаются у берегов Мурмана и зимою. Баклан,
окольцованный 11.VII. 1938 г., добыт 17. XI того же года у Фолксблада в Норвегии,
около 65° с.ш.; окольцованный 29.VII.1940 г. —31. XII.1940 г. в Гамвике,
Финмаркен; окольцованный 10.VII.1939 г. добыт 30.1.1940 г. у г. Полярного
на Мурмане; окольцованный 29.VI.1939 г. встречен 28.11.1940 г. близ Гар-
стада, Стагнес, Норвегия.
Биотоп. Высокие, обрывистые скалы с глубокими расщелинами,
нишами, уступами, лежащие в море или на морском побережье материков и
островов. С сушей связаны только в период размножения, остальное время
проводят в море близ берегов.
Численность. По побережью Кольского п-ова обыкновенен.
В последние годы повидимому стал увеличиваться в количестве и начал заселять
некоторые из островов по Мурманскому побережью. Ранее был обыкновенен
на гнездовье к востоку лишь до Варангер-фиорда. До 1929 г. совсем не отмечен
для «Семи островов», в 1929 г. встречены на о. Харлове единичные не
гнездившиеся птицы, в 1932 г. найдена одна гнездящаяся пара, в 1935 г. на о. Харлове
44
ОТРЯД ВЕСЛОНОГИЕ
был довольно многочисленен, но на других островах не отмечен. В 1938 и 1939 гг.
гнездился на двух островах: па о. Харлопе и о. Кувшине. В 1938 г. на о.
Харлове зарегистрировано 39 гнезд, на о. Кувшине до 10; в 1939 г. на о. Харлове
найдено 44 гнезда, па о. Кувшине — не более 15. Атлантический длинноносый
баклан численно преобладал над большим бакланом. В последующие годы до
1947 г., вследствие консервации заповедника «Семи островов», сведений о коли-
Карта 5. Распространение длинноносого баклана Phalacrocorax aristotelis
1 — Ph. a. aristotelis, 2 — Ph. a. desmarestli, 3 — Ph. a. riggenbachi; a —граница
гнездовий, г — область зимовок (цифры в кружках — местонахождение соответствующего подвида
на зимовке).
честве бакланов там нет. Но в 1947 г. на о. Харлове, согласно сообщению
Карташева, гнездилось всего 2 пары, на о. Кувшине не найдено ни одного
гнезда, зато на о. Вешняке обнаружены новые гнездовые колонии, не
отмеченные ранее. Всего найдено там 3 колонии, повидимому, переселившиеся сюда
с о. Кувшина и состоящие из 13 гнезд (6, 4 и 3).
Экология. Размножение. К местам гнездовья на «Семь островов»
прилетает рано — в середине марта, но держится в море по одиночке или
небольшими стаями. В 1939 г. в конце апреля (с 29. IV) на о. Харлове часто
уже наблюдались бакланы, сидящие стаями на плоских камнях у самого моря
близ своих колоний. 1.V.1939 г. птицы держались в парах на местах колоний,
но гнезд не строили (Модестов).
Колонии на «Семи островах» обычно смешанные, совместно с большим
бакланом, кайрами, гагарками, моевками и другими птицами; иногда
встречаются колонии, состоящие из одних только длинноносых бакланов. В колонии
ДЛИННОНОСЫЙ БАКЛАН
45
бывает от 10 до 15 гнезд, помещающихся на скалах с глубокими трещинами,
нишами и т. д.
Гнезда устраиваются укрыто — в горизонтальных и вертикальных
расщелинах скал', в карнизах, под большими глыбами, в нишах и т. п., так что не
только не видно гнезда, но и вся колония укрыта. Изредка встречаются и
открытые гнезда. Например, на о. Харлове в 1938 г. из 39 гнезд два были устроены
открыто; впрочем, на о. Кувшине большинство встречавшихся гнезд устроены
открыто на широких уступах, которые, повидимому, ранее занимались кайрами,
вытесненными длинноносым бакланом. В 1939 г. 5—6.V началась постройка
и поправка гнезд. Строительный материал птицы собирали поблизости на
берегу моря. Гнездо на «Семи островах» строится из ветвей ивы, в небольшом
количестве идут на постройку гнезд ветви можжевельника, слоевища фукуса
и т. д. Лоток выстлан кустиками вороницы и сухой прошлогодней травой.
Гнезда неправильно округленной формы, в диаметре до 45—55 смУ высота до
\Ъсм (Успенский, 1941). До 15—20 мая шла постройка гнезд во всех колониях.
На устройство гнезда у отдельной пары уходило в среднем 2—3 дня. Одни и
те же гнезда птицы занимают в течение нескольких лет. Кладка яиц начинается
у нас на «Семи островах» с середины мая; так, на о. Харлове в 1932 г. в кладках
20, 23 и 24 мая было по 3 свежих яйца, а 6 июня найдена кладка с сильно
насиженными яйцами (Спангенберг). В 1939 г. 20. V в двух гнездах было уже по
2 яйца,. 22.V в этих же гнездах — по 3 яйца. На Британских о-вах кладка
яиц начинается в конце марта или в начале апреля. Количество яиц в кладке
неравномерно: на «Семи островах» обычно 3, редко меньше, в Норвегии будто
бы до 8 яиц, на Британских о-вах нормально 3, но довольно часто
встречается 4 и 5 яиц и как исключение б, но иногда встречаются кладки в 7 и 11 яиц,
отложенные, повидимому, двумя самками в одно гнездо. Яйца светлоголубого
основного тона, покрытые снаружи белым известковым слоем, продолговато-
овальной или овальной формы, похожие на яйца большого баклана, но меньших
размеров. Размеры: (18)60,3—69, 9x35,5—39, в среднем 61,9x37,3 мм
(о. Харлов, Модестов); (100) 56,6—74,6 х 34,9—41,7, в среднем 62—87 х 38,4 мм
(Британские о-ва, Уайзерби, 1940). Вес: (17), в среднем 42,6 г,
максимальный 50,2 г, минимальный 36,2 г (о. Харлов).
Перерывы между откладываемыми яйцами от 1 до 5 дней (о. Харлов,
Модестов). В году бывает одна кладка, но обычно при гибели первой кладки
бывает вторая — дополнительная. В случае, если кладка уничтожена в
самом начале насиживания или еще даже не окончена, птицы сейчас же
приступают ко второй кладке (на Британских о-вах в мае — июне). Если же кладка
уничтожена в конце насиживания, то вторая кладка наступает через большой
промежуток времени; на Британских о-вах — в июле и августе.
Насиживание начинается с откладки первого яйца, насиживают оба родителя, на
яйцах сидят плотно, в особенности при приближении срока вылупления.
Период насиживания 30—32 дня (о. Харлов, Модестов); для Британских о-вов
по старым данным указывается 24—28 дней, но вероятно здесь вкралась
ошибка. Птенцы разновозрастны.
Первые птенцы вылупляются в середине июня (на о. Харлове — 14.VI.
1939 г.). Птенцы вылупляются голыми, слепыми, с белым яйцевым бугорком
на надклювье, с темноокрашенной, влажной кожей и непропорционально
длинной шеей. Глаза на девятый день намечаются в виде узких щелей, на
двенадцатый день открываются полностью. Вес вылупившегося птенца от
30,5 до 31,5 г; первые 3 дня вес прибавляется медленно, и на 3-й день птенец
весит 35 г, на 8-й — 75 г. Затем вес резко начинает увеличиваться и
десятидневный птенец весит 125 г, 13-дневный — 250 г, 15-дневный 375 г, 42-дневный—
1760 г. После 42-дневного возраста увеличение веса идет очень медленно,
часто совсем не происходит, а иногда даже уменьшается. На 60-й день к
46
ОТРЯД ВЕСЛОНОГИЕ .
моменту вылета птенцы весят от 1700—1800 г. Развитие оперения идет
медленно: первые редкие пушинки появляются на 8—9-й день, 13-дневный
птенец покрыт еще неравномерно и не вполне развитым пухом. На 20-й день
весь птенец в пуху, на крыльях имеются 2 ряда пробивающихся маховых
перьев, и начинают появляться рулевые перья. У Зб-дневного птенца на концах
маховых и рулевых еще держатся пушинки. На 50—52-й день птенец одет
полностью в гнездовый наряд и на 60-й день, т. е. в середине августа, первые
птенцы на о. Харлове покидают гнездо, полет их еще неуклюж (Модестов).
Выкармливают птенцов, оба родителя, первое время — отрыгиваемой
полупереваренной пищей, причем взрослая птица захватывает голову птенца
глубоко в клюв. Первые 2 недели старые птицы отлучаются от гнезда
поочередно: одна сидит на гнезде, другая — добывает корм.
Рис. 29. Длинноносый баклан (взрослая птица и молодая).
Птенцы очень чувствительны к холоду, в особенности первые две недели
жизни, и при температуре +Ю°С, спустя 8—10 минут после того, как взрослая
птица оставила гнездо, начинают холодеть, движения их замедляются и через
короткое время они умирают (Модестов).
Линька. Последовательность смены нарядов —как у большого
баклана. У взрослых птиц 2 линьки в году: частичная предбрачная в декабре и
январе, в течение которой происходит смена кроющих крыла, мелкого пера
и вырастает хохол, и полная послебрачная с июня по октябрь, начинающаяся
потерей перьев хохла, частичной сменой мелкого оперения и в сентябре —
октябре линькой маховых и рулевых перьев. В Англии линька маховых и
рулевых продолжается до января.
Питание. Пищу составляет преимущественно морская рыба.
У Мурманского побережья: треска, сельдь, песчанка, бычки и отмечен только
один случай, когда в желудке было обнаружено два краба. У Британских
о-вов также почти исключительно морская рыба: угри, сарганы, сельдь,
губаны и, как исключение, моллюски и ракообразные. Охотятся обычно
одиночно, ныряя за кормом, задерживаются под водой до 3—4 минут («Семь
островов», Модестов).
ДЛИННОНОСЫЙ БАКЛАН
47
Полевые признаки. Исключительно морская птица. Похожа на большого
баклана, но несколько меньших размеров, с более тонким и длинным клювом,
более тонким телом и с оливково-зеленым тоном в оперении. В брачном наряде
нет белых перьев на голове, шее и бедре, но имеется хохол на голове из темных
перьев, вершинами загнутых вперед.
В повадках, в манере летать отличий от большого баклана нет. Осторожен,
хорошо плавает и ныряет. Под водою проплывает большие расстояния и ныряет
на глубину до 4 м. Летает довольно тяжело и долго в воздухе не держится.
Взлетает с трудом, обычно со скалы или с уступа, с воды — с разбега.
Атлантический баклан очень молчаливая птица; издает низкие хрипящие
звуки только чем-либо потревоженная на гнездовьях или при кормлении
птенцов.
Описание. Размеры и строение. Длина тела самцов и самок (4) 690—720,
в среднем 709 мм. Размах крыльев (3) 900—940, в среднем 927 мм. Длина крыла (7) 265—295,
в среднем 279,3 мм. Длина клюва (б) 53—60, в среднем 57,2 мм; высота клюва у основания
(6)14—17, в среднем \6 мм. Вес в среднем 1865 г.
Первостепенных маховых 11, формула крыла 2>3>1>4... Хвост закругленный,-
состоит из 12 жестких рулевых. Оперение плотно прилегающее к телу, довольно жесткое.
Окраска. Пуховой птенец имеет черновато-бурый пух, несколько более светлый
на голове и шее. Голая кожа вокруг глаз темная, у угла разреза рта и на нижней челюсти
грязного желтовато-оранжевого цвета. Радужина светлобурая. Ноги буровато-черные.
Молодая птица. Гнездовый наряд. Верх головы, шея, спина, поясница, лопаточные и
плечевые перья черные с едва заметным зеленым металлическим блеском и — за исключением
верха головы, шеи, спины и подхвостья — с широкими темными неблестящими окаймлениями
отдельных перьев. Подбородок грязнобелый, брюшная сторона шеи и ее бока бурые с мелкими,,
частыми продольными пестринами, несколько более темными на боках шеи. Грудь, брюхо,
бока тела грязнобелые с редкими светлобурыми, как бы размытыми пятнами на груди и боках;
брюха. Подхвостья и перья голени бурые. Маховые и рулевые темнобурые без блеска.
Верхние кроющие крыла бурые со светлобурыми широкими окаймлениями перьев. Ног»
грязнорозовые. Клюв бурый с более темной полоской вдоль конька надклювья.
Первый брачный наряд. Верхняя сторона головы, шея и спина с более сильным зе->.
леным металлическим блеском, чем у птиц в гнездовом наряде. Мелкие кроющие крыла,
второстепенные и первостепенные маховые темнобурые с металлическим блеском на
кроющих крыла и на наружных опахалах второстепенных и первостепенных маховых. Нижняя-
сторона тела бурая без пестрин и блеска, светлеющая на шее к подбородку. Подбородок
грязнобелый. Ноги бурые.
Первый зимний наряд. Похож на взрослый, но низ тела темнобурый с очень слабым,
металлическим блеском, подбородок бурый с редкими мелкими белыми пестринами. Ноги
буровато-черные.
Взрослые птицы, самцы и самки, в послебрачном (зимнем) наряде. Оперение
черное с зеленым металлическим блеском, особенно сильно выражающимся%а верхней стороне
головы и шеи. Перья передней части спины, лопаточные, плечевые и кроющие крыла с
зеленовато-бронзовым металлическим блеском и с блестящими .бархатно-черными окаймлениями
отдельных перьев, придающих чешуйчатый рисунок этим частям тела.
Ноги черные. Клюв черный, бледнеющий на вершине и желтоватый у основания ниж^
ней челюсти. Голая кожа вокруг глаз и у основания клюва желтая, испещренная вокруг
клюва черными пестринками. Радужина яркозеленая.
Взрослые птицы, брачный наряд. На передней части головы имеется небольшой
черный хохол, перья которого на вершине загнуты вперед.
Средиземноморский длинноносый баклан Phalacrocorax aristotelis desmareStii
Payr.
Carbo desmarestii. Payradeau. Ann. Sci. Nat. (1), VIII, 1826, стр. 464, Сардиния.
Распространение. Ареал. Северное побережье Средиземного моря и
прилежащие острова, средиземноморское побережье Малой Азии, западная
часть северного берега Черного моря (Днепровский лиман —о. Березин) и
берега Крымского п-ова (Ак-Мечеть, Гурзуф, Балаклава и др., Мензбир,1918)..
Сведения старых авторов о встречах этой птицы на Каспийском, море
исследованиями последних лет не подтверждаются.
48
ОТРЯД ВЕСЛОНОГИЕ
Характер пребывания. Как у предыдущего. Во время своих
осенних и зимних кочевок держится у берегов, обычно близ мест гнездовий.
Иногда откочевывает и несколько далее — встречается осенью в Азовском
море, а зимой в Средиземном море у берегов африканского материка.
У Крымского п-ова наблюдается в течение всей зимы, в особенности
в большом количестве в некоторых районах южного берега, например, у
Балаклавы. Направление кочевок в зимнее и осеннее время в Черном море
определяется скоплением и миграциями стай рыб: хамсы и др.
Биотоп. Прибрежные высокие обрывистые скалы с пещерами и с
глубокими расщелинами, нишами и уступами, мелкие скалистые острова и отдельно
стоящие в море скалы.
Численность. В Днепровском лимане малочисленен, но по
побережью Крымского п-ова обыкновенен и местами даже в большем
количестве, чем большой баклан, например, у Ак-Мечети, Балаклавы (Мензбир, 1918).
Экология. Размножение. Данные о размножении длинноносого
баклана на Крымском побережье у нас почти отсутствуют, имеются лишь
некоторые сведения, собранные корреспондентами М. А. Мензбира (1918).
Гнездятся колониями, чаще самостоятельными, но иногда вместе с другими птицами,
преимущественно с большим бакланом. Гнездо устраивает в трудно доступных
местах, высоко над водою — метрах в 50—60, в щелях, нишах скал, в пещерах,
на уступах скал и т. д. У птиц, гнездящихся по европейскому побережью
Средиземного моря, гнездо массивно, построено из водорослей и внутри выстлано
травой. Одни и те же гнезда служат птицам в течение нескольких лет. Вся
скала, как и само гнездо, густо испачканы пометом и хорошо заметны издали.
В кладке бывает от 3 до 5 бледноголубых яиц, покрытых сверху толстым
известковым слоем. Яйца овальной формы, несколько более удлиненной, и
меньших размеров, чем яйца атлантического подвида. Размеры (30) 54,2—
69,7 х 34,3—40, в среднем 60,84 х 37,14 мм (Хартерт, 1915).
Точная дата начала кладки в СССР не выяснена, но в конце марта
начинается насиживание яиц. На европейском побережье Средиземного моря
кладка начинается в начале марта, и с середины месяца птицы сидят на яйцах.
В конце апреля выклевываются уже птенцы. Данных о времени вылупления
птенцов, а также об их росте, нет, но постэмбриоцальное развитие, повидимому,
очень медленное, так как молодые выводки этого года наблюдаются в море
только с половины июня. До осени молодые птицы вместе с взрослыми
держатся у мест своих гнездовий (Мензбир, 1918).
Линька не изучена. В начале января начинает появляться хохол
брачного наряда, который к концу апреля исчезает.
Питание. Пищу составляет рыба, повидимому, преимущественно
ведущая стайный образ жизни и совершающая сезонные миграции, так как
осенние и зимние кочевки длинноносого баклана в Черном море связаны
с передвижением стай хамсы и другой рыбы (Мензбир, 1918). Детали не изучены.
Описание. Слабо отличается от атлантического длинноносого баклана. Размеры такие
же. Только клюв несколько длиннее — от 60—71 мм, а хохол на голове короче.Юкраска как
и у атлантического подвида, только голые части головы с большим развитием желтого цвета
и перепонки лап — буроватого тона.
6. Малый баклан Phalacrocorax pygmaeas Pall.
Pelecanus pygmaeus. Pallas. Reise durch verschied. Prov. d. Russ. Reichs, 11, 1773,
Каспийское море.
Распространение. Ареал. Южн. Европа, Передняя и Средняя Азия;
еще недавно гнездился на озере Феццара в Алжире, указания на
гнездования в Египте не подтверждаются. В Европе на юго-востоке от Венгрии,
Югославии (в Словении, в Обедска-Бара) и Румынии (низовья и дельта Дуная),
МАЛЫЙ БАКЛАН,
49
далее на Балканском п-ове, кроме Греции; Крым, быть может, по юго-вост.
побережью Азовского моря (у Бейсугского лимана 26.VI добыта самка с сильно
развитыми фолликулами, Птушенко); быть может дельта Волги (Хлебников,
1928), вероятно, на Кавказе на северо-востоке, на Темирсоевских озерах между
Хасав-юртом и Махач-Кала, южней в Армении и Азербайджане — в районе
Ленкорани; в Средней Азии в Туркмении, вероятно, у ю.-в. Каспия и в
низовьях Теджена (Зарудный, 1896); на Аральском море на вост. побережье,
на Куван-Дарье, на озере Камышлы-баш; в низовьях Сыр-Дарьи вверх до
Джулека и Чиили (Спангенберг, 1936) и в сев. и ю.-з. Таджикистане (Иванов,
1940); вне пределов СССР в Малой Азии, Сирии, может быть, в Палестине,
Ираке и Иране на восток до Сеистана (но в Хорассане отсутствует).
Характер пребывания. Оседлая, кочующая и перелетная
птица. Область зимовок — в Средиземном море у с.-з. Африки, в Адриатическом
и Эгейском морях, у побережья Греции, вероятно, по южн. побережью Черного
Карта б. Распространение малого баклана Phalacrocorax pygmaeus
а — граница гнездовий, б — не вполне выясненная граница распространения,
г — область зимовок.
моря, в Палестине, Иране, Сеистане, Хорассане, по южн. побережью Каспия.
У нас зимует в Закавказье и Средней Азии. Даты: весной на пролете — в
Туркмении на Теджене, и в пустыне между Мары и Аму-Дарьей встречен в марте
(3 и 26) и в начале апреля. Первые птицы на Сыр-Дарью прилетают в начале
марта, а иногда и раньше — в конце февраля, валовый прилет идет в конце
марта. Осенний отлет начинается очень поздно и мало заметно, так как часть
птиц остается зимовать на местах гнездовий. Зимуют в Ленкорани (Тугаринов
и Козлова, 1938), на Сыр-Дарье, причем здесь количество зимующих птиц
сильно колеблется в зависимости от температуры зимы — в мягкие зимы
встречаются в довольно большом количестве, в суровые — лишь одиночные
особи. В Туркмении — в ю.-в. Каспии (Атрек, озеро Делили) пролет
наблюдается в первых числах ноября, в декабре часть птиц улетает к югу,
а часть остается зимовать (Дементьев, Воробьев и Исаков, 1940). На Теджене —
пролет в ноябре, в небольшом количестве, повидимому, зимуют у Мары(28.ХП
добыт, Зарудный, 1896). В Таджикистане зимует в бассейне озера Митель-куль,
под Ленинабадом и вероятно на юге по Пянджу.
Во время кочевок, в особенности осенних, малый баклан наблюдается
довольно далеко от своих гнездовий — встречается на сев.-западном берегу
4 Птицы СССР, т. I
50
ОТРЯД 1*7,СЛ0П0ГМЕ
Черного моря, на Азовском морс. Залетает случайно на Украину в бывшую
Екатеринославскую губ., и окрестности Краснодара (Птушенко), в
центральное Закавказье — и Закаталы, и Осетию (конец октября) и т. д. Упоминание
о встрече малого баклана на Черном Иртыше 1.VI.1876 (Финш, 1879),
сомнительно, повидимому, здесь вкралась ошибка. Случайные залеты одиночных
птиц наблюдались в разные страны средней Европы, на север до Польши,
Германии и Франции.
Б и о т о п. Густо заросшие тростниковой или древесной растительностью
побережья как пресноводных, так и морских водоемов, богатых рыбой, с
прозрачной водою. В горных районах (Армения, Таджикистан) по водоемам,
расположенным в низменных частях страны, в остальном условия
существования те же.
Численность. В дельте Волги, в Армении по вост. побережью
Аральского моря и в Туркмении редкая, малочисленная птица, в
Таджикистане малый баклан несколько более обыкновенен. В Ленкорани обычен и
даже многочисленен, в особенности зимою. В дельте и по нижнему течению
Сыр-Дарьи и на прилежащих озерах обыкновенен, и местами многочисленен.
Экология. Размножение. Моногам. Гнездится малый баклан
колониально, обычно совместно с другими видами птиц: большим бакланом, кол-
пиком, каравайкой, кваквой и др. Колонии на Сыр-Дарье' помещаются или
среди камышей или на деревьях и кустарниках; особенно излюбленные места —
небольшие пловучие островки из отмершей растительности, поросшие мелким
камышем, на глубоководных Камышевых озерах, в рукавах и протоках реки
(Спангенберг, 1936). В Ленкорани малый баклан селится в смешанных
колониях с другими видами птиц. Эти колонии, по сообщению Спангенберга,
размещаются среди большого высокоствольного леса, отстоящего от морского
побережья на 3—5 км.
Гнезда строят на нижней Сыр-Дарье (Соло-тюбе) среди тростника на
заломленных стеблях этого растения, на высоте 1—Р/2 метра от воды. На
деревьях гнезда строятся в среднем ярусе кроны, иногда и на вершинах
кустарников, совместно с кваквами (Ленкорань, Сыр-Дарья). Гнезда сделаны из
сучьев, камыша и т. д., приносимых обеими птицами. Лоток выстлан листьями
и нежными частями тростника, сильно углублен и плотно утоптан, так что
насиживающая птица почти не видна в гнезде. Форма гнезд
полушарообразная, размеры в диаметре до 25 см (Зарудный, 1916, Спангенберг, 1936).
Одни и те же гнезда занимаются малыми бакланами в течение ряда лет и
каждое гнездо в колонии в тростниках приобретает вид пирамиды.
На Сыр-Дарье к размножению малый баклан приступает значительно
позже больших бакланов: обычно в гнездах больших бакланов бывают уже
хорошо оперившиеся птенцы, а в гнездах малого баклана —еще не насиженные
и слабо насиженные яйца.
Кладка на Сыр-Дарье начинается в начале мая (1—3 мая найдены гнезда
с только что отложенными яйцами), но в некоторые годы и позже, например,
в 1925 г. 24—28.V яйца в гнездах были еще слабо насижены, в 1926 г. 10.VI
встречались одновременно гнезда с ненасиженными кладками и насиженные.
По вост. берегу Аральского моря также отмечена неодновременность кладки
в одной и той же колонии (Зарудный, 1916). В Ленкорани размножение
начинается с конца апреля или с начала мая (найдены кладки с 5 свежими
яйцами 2 мая, с 5 насиженными яйцами 2 июня и 3 насиженными — 15 июня).
В кладке обычно 4—6, редко 3 или 7 яиц, они похожи по форме на яйца
большого баклана, но меньших размеров. Известковый поверхностный слой
тбньше.
Размер: (40) 42,4—53,5 х 28,7—31,3 мм (нижнее течение Сыр-Дарьи,
Спангенберг, 1936); (16)45—47,3 х 28—34 мм (дельта Сыр-Дарьи, Зар>дный, 1916).
малый баклан
51
Насиживают оба родителя с первого яйца (птенцы разновозрастны).
Продолжительность насиживания неизвестна—повидимому, несколько менее месяца*
Первые птенцы на Сыр-Дарье вылупляются в середине июня. Вылупляются
они голыми, но опушаются и растут быстро — в первых числах июля у
пуховиков появляются маховые, плечевые и рулевые перья; в начале 2-й половины
июля пух у птенцов остается только на голове, шее, местами на боках тела,
бедрах, на надхвостье и подхвостье (птенец, добытый 20.VII. 1926 г., Кара-
Узяк, Сыр-Дарья; Зоологический музей Моск. университета). К концу июля
птенцы надевают гнездовый наряд. Еще с недоросшими маховыми и не умея
летать, птенцы покидают гнезда и передвигаются по площади колонии на
гнездовых островках или по берегу (Спангенберг, 1936). Вначале сентября
молодые уже хорошо летают, но
держатся вместе со старыми птицами.
Выкармливают птенцов оба
родителя — вначале
полупереваренной пищей, которую они
отрыгивают в рот птенца, затем рыбой,
сначала мелкой и только в конце
гнездового периода более крупной,
достигающей 10—12 см. Первое
время, пока не окрепнут птенцы,
взрослые птицы летают за
кормом по очереди — одна из них
остается в гнезде, согревая птенцов.
Линька.
Последовательность смены нарядов — как у
других бакланов. У взрослых птип
2 линьки в году: частичная пред-
брачная в декабре— январе, во
время которой птица надевает
брачный наряд и носит его
приблизительно до июня, и полная после-
брачнай" с июня по октябрь —
ноябрь. Во время полной линьки
происходит смена рулевых,
маховых и мелкого пера, причем смена всех указанных перьев начинается не
одновременно у разных особей. Заканчивается линька первостепенных маховых
и рулевых к августу — сентябрю, мелкого пера к октябрю — ноябрю. Смена
первостепенных маховых и рулевых идет от внутренних перьев к наружным.
Питание. Пищу малого баклана составляет мелкая рыба — до 12—
15 см: красноперка, сазан, щука, вобла и т. д.
В Ленкорани малый баклан в поисках кормных мест совершает небольшие
кочевки на морские побережья, пресноводные водоемы и даже рисовые поля.
Если в найденном месте оказывается достаточно пищи, бакланы проводят там
долгое время. Обычно они дважды в сутки вылетают на ловлю рыбы. Как
только в водоеме рыба убывает, бакланы начинают вновь кочевать
(Тугаринов и Козлова, 1938).
Полевые признаки. Почти в два раза меньше большого баклана, в брачном
наряде белые перья расположены по всему телу и имеют каплевидную форму.
Мало осторожен. Превосходно плавает и ныряет. Довольно легкий полет со
спокойными редкими взмахиваниями крыльев, иногда птица на короткое
время совсем не работает ими. Обычно держатся одиночно или парами, только
осенью встречаются небольшими стаями, состоящими чаще из одного выводка.
Отдыхает, сидя на ветвях деревьев, на камыше или на каких-либо возвышениях
Рис
Малый баклан (взрослый и молодой).
4*
52
ОТРЯД ВЕСЛОНОГИЕ
близ воды. Сидит прямо, почти вертикально, изогнув S-образно шею, обхватив
лапой ветку или камыш и опираясь слегка на хвост. В это время бакланы
обычно просушивают свое оперение, раскрыв крылья и изредка
помахивая ими.
Малый баклан молчаливая птица, только в период размножения
становится крикливой, издавая звуки, подобные кряканью большого баклана.
Описание. Размеры и строение. Длина тела около 550 мм. Длина крыла у
самцов (10) 193—208, в среднем 198,7 мм; у самок (9) 185—205, в среднем 193,4 мм. Вес около
800 г. Первостепенных маховых 11, с вырезкой на внутреннем опахале. Формула крыла
2>3>4>1>5... Хвост закругленный, из 12 жестких рулевых.
Окраска. Пуховой птенец имеет однотонный, темнобурый пух.
Гнездовый наряд. Верх головы и шеи бурый, подбородок грязнобеловатый, нижняя
сторона шеи светлобурая. Спина и бока тела темнобурые, перья верхней части спины со
светлобурыми окаймлениями. Плечевые и верхние кроющие крыла с сероватым налетом
и с темными каймами. Брюшная сторона тела буровато-белая, в особенности посредине
брюха, с рыжевато-бурыми размытыми пятнами на груди и зобе, на задней части брюшной
стороны тела пятна черновато-бурые, продолговатой формы, переходящие постепенно в тем-
нобурое подхвостье. Уже в этом наряде в январе появляются на боках зоба несколько
(3—5) белых каплевидных перышек.
Первый брачный наряд. Верх головы и шеи буровато-коричневый, нижняя сторона
шеи несколько светлей, подбородок светлобурый. Спина и надхвостье черные с зеленоватым
металлическим блеском. Кроющие крыла, плечевые и второстепенные маховые с сероватым
налетом, с темными предвершинными и бурыми вершинными каймами. Низ тела темный,
коричнево-бурый с крупными нерезкими, более светлыми пятнами, образуемыми широкими
светлыми каймами перьев. Бока тела и подхвостье черновато-бурые. В январе появляются
белые каплевидные перья на боках зоба, шеи и на бедре в несколько большем количестве,
чем в гнездовом наряде.
Второй брачный наряд. Верх головы, шеи и вся верхняя сторона тела окрашены как
у взрослой птицы, нижняя сторона тела несколько светлей. Подбородок•»светлобурый, низ
тела однообразно буровато-черный без блеска. Белые каплевидные перья", появившиеся в
январе, еще очень редкие, но уже разбросанные по всему телу.
Второй зимний (послебрачный) наряд почти не отличим от взрослой птицы. Нижняя
сторона с металлическим блеском, но более слабым, чем у взрослой птицы. Кроме того, на
нижней части брюха еще имеются отдельные более светлые перья. Этот наряд носится, по-
видимому, до января.
У взрослой птицы в послебрачном (зимнем) наряде голова и верхняя половина шеи
темнокоричневые, почти черные с буроватым налетом; кольцо у основания клюва почти
черное. Остальное оперение черное с зеленым металлическим блеском на спине и нижней
стороне тела. Плечевые, лопаточные и верхние кроющие крыла черные с сероватым
оттенком и с черными каймами отдельных перьев, образующих чешуйчатый рисунок. Клюв
черновато-бурый, ноги черные, радужина бурая.
Взрослые в брачном наряде имеют в оперении спинной и в особенности
брюшной стороны тела и на боках редкие мелкие каплевидные белые перья. Такие же перья,
но еще меньших размеров, расположены вокруг основания клюва и глаза. Самцы и самки
окрашены одинаково.
СЕМЕЙСТВО ПЕЛИКАНОВЫЕ PELECANIDAE
РОД ПЕЛИКАНЫ PELECANUS LINNAEUS, 1758
Тип Pelecanus onocrotalus
7. Кудрявый пеликан Pelecanus crispas Bruch
Pelecanus crispus. Bruch. Isis. 1832, стр. 1109, табл. V, рис. 1, Далмация*
Русское название. Пеликан — книжное, от латинского pelecanus, как и в
других европейских языках. Народное на юге — баба, баба-птица.
Распространение. Ареал. Места гнездований расположены
спорадично в ю.-в. Европе, Передней, Средней и Центральной Азии. Еще недавно
гнездился в Венгрии и Далмации, в историческое время — в Австрии и
Германии, в настоящее время в Европе вне пределов СССР — местами на
Балканском п-ове — в низовьях Дуная в Румынии и Болгарии, у рек Ка-
ламас, Аскропотамос и Вардар в Греции, по лагунам Албании, на озерах
КУДРЯВЫЙ ПЕЛИКАН IVf
Охрадском, Скутари и др. в Югославии. В Европейской части СССР — и
низовьях Днепра, быть может в плавнях у Никополя (Тугаринов, 1947);
в Крыму на Сиваше; на побережьях Азовского моря, в устьях Дона и Ку*
банских плавнях; у Каспийского моря — в дельте Волги (Астраханский
заповедник), на зап. берегу Каспия от устья Сулака и быть может от Махач-Кала
до устья Терека; в Закавказье — по горным озерам Армении (Соснин и Ляй-
стер, 1942), на озере Чалдир-гель (Нестеров, 1911), по Араксу (Бобринский,
1916), а затем на крайнем юго-востоке Закавказья от порта Ильича, быть может
северней до устья Куры, внутри Азербайджана, по озерам до Кюрдамира; на
вост. берегу Каспия от устьев Урала (но не в устьях Эмбы), до Мертвого Кул-
тука. В Заволжье на больших Камыш-Самарских озерах, быть может на озере
Карта 7. Распространение кудрявого пеликана.Реlecanus crispus.
а — граница гнездовий, г — область зимовок.
Джумарт-куль и Чалкар-тениз у сев. окраины Б. Барсуков (Бостанжогло,
1911), но это требует подтверждения. Гнездование в Туркмении и
Таджикистане не доказано. На Аральском море по островам и побережьям, кроме
скалистых обрывистых участков; в низовьях Сыр-Дарьи от дельты до г.
Туркестана, повидимому на озере Тели-куль и на нижней Сары-су; наконец на
нижнем течении и в дельте Или. К югу от СССР в Азии — по южн. берегу
Каспия, в Сеистане, у Персидского залива, в сев. Сирии, вероятно в Ираке.
Распространение в Центральной Азии неясно; обычен в Синьцзяне на р. Хай-
дык-гол и на озере Лоб-норе, в начале сентября найден в низовьях Кобдо
и на озере Хара-усу; отдельные особи наблюдались на озере Далай-нор и
на верхней Хуан-хе; в вост. Китае не встречен.
Характер пребывания. Кудрявый пеликан — гнездящаяся
перелетная, в небольшом числе зимующая у нас птица.
Их стаи на пролете иногда достигают больших размеров — до 300 и более
особей, но обычно к местам гнездовий прилетают небольшими группами или
даже одиночками. Во время пролета пеликаны летят шеренгой, вытянутой
54
ОТРЯД ВЕСЛОНОГИЕ
.прямой или несколько волнистой линией, держась близко друг к другу. Часто
наблюдаются стаи, смешанные с розовым пеликаном. Можно наметить
несколько направлений пролета, в особенности ясно выраженных при движении весной:
вдоль берегов Черноморского побережья Кавказа к Азовскому морю, по
зап. побережью Каспия — к дельте Волги и Урала; по ю.-в. и вост.
побережью Каспия; но зап. окраине пустыни Кизыл-Кумов и по Теджену,
Мургабу и Аму-Дарье. Кроме того, повидимому, летят в Центральной Азии,
так как наблюдались в конце апреля (29.IV) по долине р. Дзенкан между
Кобдо и Улясутаем (Березовский, 1877).
Прилетают весной очень рано, в среднем в середине марта. В Закавказье
у Батуми вдоль Черноморского побережья пролетные птицы наблюдаются
уже в конце февраля или в начале марта, в зависимости от весны. В б. Ареш-
ском уезде 26.11. 1894 (Мензбир, 1918) и у Ленкорани по сообщению Спанген-
берга первые птицы появляются в конце февраля. В дельту Волги прилетают
в середине марта. В Туркмении на пролете наблюдается нерегулярно: на
Мургабе — во второй половине марта (21.111), но иногда пролетные стаи
встречаются и раньше, в конце февраля (Лоудон, 1902), в долине Кушки — во
второй половине марта, у Чемен-и-бида 20 марта наблюдалась стая в 200—250
особей кудрявых и розовых пеликанов (Шестоперов, 1935). На Сыр-Дарье
прилет и пролет обычно идете середины марта (15. III. 1927) и до конца апреля
(Спангенберг и Фейгин, 1936).
Осенний отлет начинается поздно; птицы, главным образом, молодые,
задерживаются иногда на незамерзающих водоемах до начала зимы и в
незначительном числе наблюдаются даже зимой. Дельту Волги покидают с конца
октября. В Армении на озере Гилли одиночные экземпляры наблюдались еще
в начале ноября (Ляйстер и Соснин, 1942). На Сыр-Дарье отлет
начинается с середины октября и продолжается до конца ноября, но в небольшом
количестве встречается и в начале зимы. На ю.-в. Каспии (Гассан-кули,
Чикишляр) пролетные птицы наблюдаются в конце ноября и начале декабря
(Дементьев, 1950).
Кудрявые пеликаны кольцевались в Казахстане в августе 1938 на озере
Сассык-куль и в июне 1939 г. у Арал-тюбе. Из сассык-кульских птиц три
были добыты в марте 1939 г. на озере Ала-куль; одна 15.Х. 1938 г. на озере
Балхаш; другая 19. V. 1939 г. на Балхаше в низовьях Или; одна в 1939 г.
у ст. Джалагаш в низовьях Сыр-Дарьи; одна в 1940 г. у г. Кагана в зап.
Узбекистане; наконец, 4.VI.1939r. в Алтайском крае в Толмачевском районе
у с. Зайцево. Основное направление кочевок молодых пеликанов, таким
образом, западное и юго-западное. Однако имеют место и кочевки на восток
(Алтайский край). Судя по встрече пеликана у Кагана, представляется вероятным
движение по крайней мере части казахстанских пеликанов в направлении
к каспийским зимовкам, где, как упоминалось, этот вид пролетает регулярно.
Арал-тюбинские пеликаны встречены: один 22.IXЛ939 г. на Каратале, другой
26.IX. 1939 на озере Сассык-куль.
Как правило, кудрявый пеликан у нас не зимует, но одиночные
экземпляры, иногда и небольшие стаи наблюдаются у нас зимой. Встречены в
декабре— январе 1924—25 гг. на Тереке у с. Коби (Военно-Грузинская дорога) и
на р.Комбилеевке(Беме,1926; при этом морозы доходили до 25°); в сев.-западной
горной части Кавказа на горе Дудугуш (высота 1200 м над уровнем моря)
встречены в феврале (Аверин и Насимович, 1928). В ю.-з. Закавказье — в Колхиде
зимует более или менее регулярно в довольно большом количестве
(Нестеров, 1911); в небольшом числе каждую зиму встречается в Ленкорани (конец
января и февраль, Спангенберг); в устье Куры (2 экземпляра 20.XII.1916,
Зоол. музей Моск. гос. университета); встречается и по юго-вост. побережью
Каспия (залив Михайловский, около 20 экземпляров, Зарудный, 1896).
КУДРЯВЫЙ ПЕЛИКАН
55
Главные места зимовок расположены вне нашей страны: в ю.-в. Китае
к югу от Голубой реки; в с.-з. Индии, в Белуджистане, по берегам
Персидского залива, в Сеистане, в Хорассане, в Загроше, по южнокаспийскому
побережью, в Ираке и в Египте.
Залет ы. Негнездящиеся птицы — молодые и взрослые — держатся
обычно близ своих гнездорых колоний; но иногда откочевывают от них на
довольно большие расстояния. Кроме того, можно отметить случайные залеты
единичных экземпляров, иногда небольших стай. Залеты эти бывают
преимущественно в осеннее время года, реже в летнее или весеннее; Крым —. Ялта,
Полтавская обл., окрестности Киева на Днепре, Польша, Дзауджикау
(Владикавказ), окрестности Тагила, окрестности г. Кириллова б. Череповецкой губ.
Наурзум — озеро Ак-суат, окрестности Омска, окрестности Семипалатинска,
южн. части б. Тобольской губ. и Красноярский край.
Биотоп. Глубокие, богатые водной растительностью водоемы — озера,
низовья и дельты рек и т. п.
Численность. Численность кудрявого пеликана подвержена
сильным колебаниям. Это особенно наглядно видно на гнездовьях в дельте Волги.
Во 2-й половине прошлого столетия кудрявый пеликан был весьма много-
численен на гнездовье не только в дельте Волги, но и в нижнем ее течении, на
Сарпинских озерах, в небольшом количестве доходил до Самары, а одиночные
гнездовые пары отмечены под Казанью, в б. Белебеевском уезде Уфимской
губ. и в устье Илека. В начале нынешнего столетия почти во всех
вышеуказанных местах он не гнездился, за исключением устьев Волги и Урала, но там был
еще многочисленен. Затем численность кудрявого пеликана стала резко падать
и в 1927—1929 гг. в дельте Волги и на прилежащем взморье гнездовья
пеликанов совсем не отмечались; в 1930 г. птицы опять начали гнездиться, и в дельте
Волги и на взморье найдено около 60 гнезд (Воробьев, 1936). В последующие
годы там началось увеличение количества гнездящихся кудрявых пеликанов.
В настоящее время — в 1949 г., в дельте Волги и на взморье гнездится
приблизительно около 300 пар, а может быть и более. Надо иметь в виду, что —
эти сведения относятся к территории Астраханского заповедника.
В нагорной Армении пеликан редок, но в начале XX века был обыкно-
венен и даже многочисленен. Обычен на Азовском море в ю.-в. Закавказье, по
с.-з. и сев. побережьям Каспия с прилегающими островами и устьями рек, на
Аральском море и в дельте Сыр-Дарьи. Многочисленен в низовьях Сыр-Дарьи.
Экология. Р а.&м И о ж е н и е. Пары, повидимому, постоянны; птицы
парами держатся во всяком случае с прилета. Половая зрелость, вероятно,
наступает на 3-м году жизни. Гнездятся колониями, обычно из нескольких
десятков пар или меньше — в 4—5 пар; редко изолированными парами (Сыр-
Дарья, Спангенберг, 1936). Гнездовья располагаются очень уединенно и,
как правило, отдельно от колоний других птиц; только изредка совместно
с другими птицами, например, розовым пеликаном (Сыр-Дарья, дельта Волги),
большим бакланом (Аральское море, о. Комсомольский, Гладков, 1949). В таком
случае обычно кудрявые пеликаны располагаются по краю колонии,
обособленно и на некотором расстоянии от гнездовий других видов птиц.
Вскоре по прилете самки (дельта Волги) размещаются по гнездовьям.
В это же время начинаются брачные игры и спаривание; самец, раскрыв крылья,
ходит вокруг самки, то приближаясь, то отходя, бросается в воду, отплывает
на недалекое расстояние и опять возвращается к самке, трется грудью и шеей
о самку, перебирает клювом ее перья. После этого происходит спаривание;
все это проходит в разное время дня, иногда и ночью, с промежутками в 10—
15 минут. Во время спаривания самец поднимает крылья и бьет ими.
В этот же период происходит и постройка гнезда. К устройству гнезд
кудрявый пеликан приступает на 10—15 дней раньше, чем розовый пеликан;
56
ОТРЯД ВЕСЛОНОГИЕ
в дельте Волги — по сообщению Ромашевой, в начале апреля, но может быть
раньше, в конце марта; на Сыр-Дарье — в конце апреля и в первой половине
мая. Гнездо строят только самки, но весь строительный материал подносят
самцы. Самец в клюве приносит траву, камни, сучья и даже палки — до метра
длины и 5—7 см ширины; часто наблюдаются драки между самцами из-за
строительного материала. За сутки самец успевает 25—40 раз поднести к гнезду
материал. Строится гнездо 3—4, редко 5 дней.
Гнездо представляет собою кучу небрежно набросанного, утоптанного
сухого камыша, покрытого пометом, придающим большую прочность постройке.
Диаметр гнезда от 1 до l1^ Щ высота 1—1х/2 м над уровнем воды. По мере
подрастания птенцов гнездо сильно оседает и поднимается только на 1/2 м над
водой, даже опускаясь до ее уровня. Помещается гнездо на пловучем островке
близ чистой воды или прямо среди густых зарослей камыша в некотором
отдалении от воды (Сыр-Дарья). Иногда на одном островке помещается до
30 пар пеликанов (б. Арешский уезд, Шелковников, по Мензбиру, 1918).
Встречаются гнезда и на совершенно плоских низменных островках, лишенных
почти всякой растительности и лежащих среди лагун (Аральское море, о.
Комсомольский, Гладков, 1949).
Кладка яиц происходит неодновременно во всех гнездах одной колонии.
В дельте Волги первые яйца откладываются в 20 числах апреля, но еще в конце
мая (22 и 24) можно найти в некоторых гнездах свежие яйца. На Сыр-Дарье
кладка начинается в конце апреля и идет до середины мая, но еще в середине
июля встречаются гнезда со слабо насиженными яйцами, тогда как в
других гнездах имеются уже совсем оперившиеся птенцы. На Аральском море
31 мая найдена кладка свежих яиц (Спангенберг). В дельте Волги в кладке
обычно 2,3, реже 4 яйца, на Сыр-Дарье обычно 3—4, реже 5 и, как исключение,
даже 6 яиц.
Яйца белые, неравномерно покрытые известковым слоем, придающим
скорлупе шероховатую наружную поверхность. Размеры: 86—102 х 53—65 мм;
вес 143—195 г (дельта Волги, Ромашева). Как правило бывает одна кладка
в году, но то, что в июле найдены слабо насиженные яйца (Сыр-Дарья), можно
объяснить только вторичными кладками.
Продолжительность насиживания 39—40 дней. Насиживание начинается
по всей вероятности с откладки первого яйца, так как в одном гнезде
встречаются птенцы различного возраста. Насиживает преимущественно самка,
самец сидит на яйцах только утром и вечером на время кормежки самки.
Птица на гнезде сидит очень крепко. Гнездо с яйцами хорошо скрыто и найти
его среди густых зарослей камыша трудно; только после вылупления
птенцов, по их голосам, найти гнездо становится значительно проще.
Вылупляются птенцы голыми, слепыми, с кожей, окрашенной в розовый
цвет, с светлым подклювьем и более темным надклювьем. Птенец быстро
прибавляет в весе и достигает размеров взрослой птицы раньше, чем наденет
полностью покров из перьев. Вес только что вылупившегося птенца ПО г,
на 2-й день — 202 г (дельта Волги), на 6-й день — 480 г, на 12-й — 1950,
на 20-й — 3500, в возрасте около 30 дней — 6930 г и в возрасте 2 месяцев —
9200, т. е. равен и даже превышает вес взрослой птицы. Первые редкие
пушинки, равномерно покрывающие все тело птенца, появляются на 5-й день
(дельта Волги), на 9-й день птенец покрыт густым пухом. В возрасте 20 дней
имеются маховые перья, кроющие крыла и начинают пробиваться перья
на нижней стороне тела; дольше всего пух держится на спине — почти
до 2-месячного возраста. Приблизительно через 2х/2 месяца птенец становится
летным. Но плавать птенцы уже могут в возрасте приблизительно 7—8 дней,
еще одетые редким пухом (дельта Волги). Вспугнутые птенцы торопятся
выбраться из гнезда и, тяжело шлепая всей ступней по земле, опираясь на заднюю
КУДРЯВЫЙ ПЕЛИКАН
bf
часть туловища, стараются добраться до воды и отплыть (Аральское море,
о. Комсомольский).
Первое время родители выкармливают птенцов отрыгиваемой
полупереваренной рыбой, при этом они глубоко захватывают в рот голову птенцов;
так же в клюве приносят птенцам и воду. Молодые птицы свободно
проглатывают рыбу в 800—1200 г. За кормом обычно пеликаны далеко не летают, а ловят
рыбу близ гнездовой колонии. Подлетев к гнездовой колонии, птица опускается
на воду и плывет к гнезду; если гнездо расположено на краю острова у воды,
взбирается на берег почти непосредственно у самого гнезда. В тех же случаях,
когда гнездо находится глубоко в густых камышах, птица опускается на чистую
воду и подплывает к камышам, и уже по ним продвигается к гнезду, протаптывая
широкие тропы (Сыр-Дарья). Птенцы встречают родителей глухим ревом и
стараются пробраться вперед, толкая друг друга. Спугнутые с гнезда взрослые
удаляются от него или «пешком» по тропе в камышах и, дойдя до чистой воды,
взлетают почти вертикально (Сыр-Дарья), или, если вокруг колонии чистая вода,
непосредственно поднимаясь с воды в воздух (Закавказье, б. Арешский уезд).
После того, как молодые птенцы станут на крыло, ойи вместе с родителями
удаляются с мест гнездовых колоний на глубокие озера и морские берега. Здесь
они, сбившись в стаи, ведут до отлета кочевой образ жизни.
Линька. Полная послебрачная линька начинается после конца периода
размножения и идет с июля по октябрь и может быть ноябрь. Линька сильно
растянута и начинается неодновременно у птиц в разных районах (у птицы,
добытой 11 августа, Дагестан, первых 3 первостепенных маховых нет,
остальные свежие, не доросшие до нормальных размеров; экземпляр от 14 июля,
дельта Или, не имеет даже следов начала линьки, а у птицы от 16 августа,
дельта Или, идет сильная линька маховых и мелкого пера). Это, по всей
вероятности, связано с отмеченными особенностями размножения. В декабре все
птицы в свежем наряде. Смена первостепенных маховых идет от внутренних
перьев к наружным.
Молодые птицы надевают взрослый наряд на 3-й год жизни.
Последовательность смены нарядов неясна, в частности — в отношении наличия пред-
брачной линьки.
Питание. Пищу кудрявого пеликана составляет средних размеров рыба—
2,5—Зкг и молодь. В отличие от розового пеликана, кудрявый добывает рыбу
не только на мелководье, но и на глубокой воде; медленно плавая, кудрявый
пеликан высматривает подплывающую к поверхности рыбу и быстрым
движением схватывает ее (Сыр-Дарья). Часто, в особенности осенью, после поднятия
молодых на крыло, наблюдается ловля рыбы семьями, или даже несколько
выводков пеликанов соединяются вместе; иногда к ним присоединяются розовые
пеликаны и бакланы. Вылетая на ловлю рыбы, стая пеликанов, снявшись
с земли, кружится некоторое время в воздухе, затем выстраивается по прямой
линии или под углом, и летит на море или озеро. Опустившись на воду, птицы
вытягиваются в линию, держась близко друг к другу и, хлопая крыльями,
начинают медленно двигаться к берегу. Присоединившиеся к ним бакланы
плывут вместе с ними, то и дело ныряя и выплывая в разных направлениях.
Такие стаи нередко сопровождают чайки, которые носятся в воздухе, время
от времени стремительно бросаясь за рыбой. Вся стая поднимает большой шум,
пугая рыбу и направляя ее на мелководье, где с ней уже легко справиться.
Пеликаны при отсутствии корма могут переносить длительную голодовку;
3—4 дня голодовки совершенно не отражаются на общем состоянии здоровья,
но более длительные голодовки — до 2 недель — сильно истощают птицу
(Спангенберг, 1936).
Кудрявый пеликан в известных условиях приносит несомненный вред
рыбному хозяйству, питаясь сазаном, лещем, окунем, воблой, густерой, куту-
58
ОТРЯД ИКСЛОИОГИК
мом, сельдью и т. д. Пара пирослмх птиц м 2 птенца (Ромашева) поедают, в
среднем, за 8 месяцев пребыпания и дельте Волги 1080 кг рыбы. Впрочем, учитывая
немногочисленность кудрявого пеликана, спорадичность гнездового ареала,
едва ли правильно ставить вопрос об уничтожении.
Полевые признаки. Фигуры пеликана очень 'характерна. Похож он — из
птиц фауны СССР —только на розового пеликана, но крупней; на темени и
затылке «кудрявые» перья; горловой мешок оранжевый, лапы темносерые,
радужина беловатая. Хорошо летает, нередко парит. Во время полета шея
изогнута и голова положена на спину, так, что только клюв несколько выдается
вперед. Ноги вытянуты назад. Плавает превосходно, на воде сидит высоко,
i олову и шею во время отдыха на воде держит, как и на полете, а при быстром
плавании вытягивает шею кверху, почти прямо, опустив клюв несколько вниз
Рис. 31. Кудрявый пеликан.
так, что между шеей и головой образуется острый угол. Не ныряет: добывает
рыбу, погружая в воду только голову, шею и переднюю часть туловища, причем
в воде голова птицы остается лишь очень короткое время. На воде проводит
много времени не только во время рыбной ловли, но и на отдыхе. Перья
пеликанов сильно намокают и птица, даже плавая в воде, «отжимает» клювом
перья, захватывая их у основания и постепенно передвигая клюв к вершине
пера. Кроме того, на воде пеликан высоко поднимает крылья, стараясь
предохранить их от намокания. С воды поднимается довольно легко, отталкиваясь от нее
одновременно обеими лапами, но когда горловой мешок переполнен добычей,
поднимается с трудом. По земле передвигается медленно, вперевалку. С земли
поднимается с трудом, при помощи нескольких прыжков, во время которых
отталкивается от земли, как и во время поднятия с воды — обеими лапами
сразу. Присаживается на деревья, причем также, как и бакланы, обхватывает
сучок, направляя три передних пальца вперед, а задний назад.
Описание. Размеры и строение. Длина тела около 1800 мм, размах около
2 000 мм, длина крыла самцов (7) 725—770, в среднем 750 мм, самок (2) 685—700 мм. Вес
около 9 кг; по Бутурлину (1935) — до 12 кг; по Тугаринову (1947) — до 13 кг.
На голове и верхней стороне шеи имеются удлиненные, закрученные «курчавые» перья,
образующие подобие гривы. Голое пространство, занимающее уздечку и кольцо вокруг глаза
РОЗОВЫЙ ПЕЛИКАН
59
и вдающееся острым мысом на лоб, очень мало, лоб оперенный. Первостепенных маховых —
11, формула крыла 4>3>2>5>1>6; хвост короткий, почти прямой, из 22 мягких рулевых.
Окраска. Пуховой птенец — беловатый пух, клюв буровато-черный.
Гнездовый наряд грязнобелый, с сильно развитым буровато-охристым и серым
налетом. Голова и шея грязносерые, спина беловатая с охристым налетом, кроющие
крыла охристые, плечевые буровато-черные с серыми вершинами. Первостепенные маховые
бурые. Рулевые светлосерые с почти белыми окаймлениями. Брюшная сторона тела грязно-
белая, бока тела светлобурые. Удлиненные перья на голове и верхней стороне тела почти
неразвиты.
Второй наряд, похож на взрослую птицу, но имеются еще серые тона на голове
и шее и буровато-охристые на кроющих крыла и плечевых.
Взрослая птица имеет оперение в основном белое. Спина, плечевые и верхние
кроющие крыла светлосерые с более темными стволами перьев и с белыми вершинами.
Первостепенные маховые и их кроющие буровато-черные с темными стержнями,
второстепенные маховые — серые с белыми каймами по краю внутренних опахал. Горловой мешок и
прилежащие голые части «лица» желтые. Клюв серо-роговой с оранжево-красным коготком.
Ноги серые. Радужина желтовато-белая.
В брачном на ряде горловой мешок становится красновато-оранжевым, на зобе желтое или
рыжеватое пятно. Самцы и самки во всех возрастах окрашены одинаково.
8. Розовый пеликан Pelecanus onocrotalus L.
Pelecanus onocrotalus. Linnaeus. Syst, Nat. изд., X, 1758, стр. 132, Африка.
Синоним. Pelecanus minor Ruppell. Mus. Senckenb. 11, 1837, стр. 186, Молдава
и Египет.
Русское название. Розовый пеликан — книжное, народное на юге — баба-
бабу ра, баба-птица.
Распространение. Ареал. Ю.-в. Европа, Африка, Передняя, Средняя
и ю.-з. Азия. В Европе еще недавно гнездился в Венгрии и повидимому в Че-
Карта 8. Распространение розового пеликана Pelecanus onocrotalus
а — граница гнездовий, г — область зимовок.
хословакии; в настоящее время вне пределов СССР в низовьях Дуная и в До-
брудже. В СССР — в Молдавской республике в Днестровских плавнях
60
ОТРЯД ВЕСЛОНОГИЕ
(Измайловская обл.), в Днепровском лимане, на о. Бабине в Каркинит-
ском заливе, быть может в Тендровском лимане у Одессы и по островам
в ю.-в. части Азовского моря между Ейском и Кривой косой; в дельте
!Золги; в Крыму и на Кавказе не гнездится. В Азии в пределах СССР
быть может на с.-в. побережье Каспия, к югу до Мертвого Калтука; на
Аральском море от о-вов Чушка-бас до о. Толмачева и от дельты Сыр-Дарьи
до Аму-Дарьи, возможно и на нижнем течении этой реки и даже на о-вах
выше Керков в Туркмении (Зарудный, 1896, 1916); по Сыр-Дарье и на
прилежащих озерах от дельты до ст. Туркестана, быть может и несколько
выше; в низовьях Чу, на Балхаше, главным образом в области устьев Или,
Каратала, Аксу и др.; на озере Сассык-куль (Шнитников, 1949), в Зайсанской
котловине (Хахлов, 1928); в Джунгарии (р. Кран, Сушкин, 1938). К югу от
СССР в Иране на озере Урмия (Радугин, 1917); в южн. части Хорассана,
у Персидского залива, быть может, в Сеистане, в Месопотамии, Сирии (озеро
Антиохия)ив с.-з. Индии до Синда. В вост. Азии распространение неясно, быть
может, гнездится где-то в Центральной Азии; в Китае не гнездится. В
Африке от Сенегала до озера Ньясса. В негнездовое время у Черного и
Каспийского морей, в Иране, Белуджистане, Месопотамии, на Балканах, в Египте.
Характер пребывания. Розовый пеликан — гнездящаяся
кочующая и перелетная птица. В пределах СССР на зимовках отмечен в
Днепровском лимане, у южного побережья Крыма в юго-вост. углу Черного моря
у Кавказа, на Каспии у Ленкорани, иногда — на юго-вост. у Гассан-кули.
Даты. Сведения о пролете и отлете этих птиц отрывочны. У Одессы
прилетает в конце февраля или в начале марта, в Днепровский лиман —
в конце марта, иногда даже в начале апреля. В дельту Волги — в первых
числах марта. В Туркмении пролет идет сначала марта до начала апреля (у
Фараба на Аму-Дарье 31 марта, Лоудон, 1902; у Кушки 20 марта,
Шестоперов, 1936; в подгорной долине у Геок-тепе 4 апреля, Радде и Вальтер, 1889;
у Гассан-кули в начале марта и до 5 апреля, Дементьев, 1950). В низовьях
Сыр-Дары^— со второй половины марта. Первые птицы на озере Балхаш —-
11 марта (Долгушин, 1934).
Осенью перед отлетом птицы кочуют в разных направлениях. Стаи
пеликанов наблюдались в низовьях Днепра, изредка вверх до Днепропетровска;
на Азовском море, в низовьях р. Урал до Инде рекой и редко до Калмыкова.
Осенний пролет начинается поздно, обычно после заморозков. На сев.
побережье Каспия и в устьях Урала, на нижней Сыр-Дарье держатся до начала
ноября, в ю.-в. Туркмении — до начала декабря. На перелетах пеликаны
летят обычно «клином», на большой высоте. Отмечено, что они пересекают
высокие горы (Сурамский перевал, Радде, 1885), но как правило летят вдоль
речных долин и морских побережий.
Залеты. Холостые, вероятно, главным образом, неполовозрелые
пеликаны встречаются иногда далеко от мест гнездований. Так, они
наблюдались на Днестре, в низовьях Днепра, под Мариуполем, в Волжско-Ураль-
ских степях (озеро Ак-сор), на вост. Каспии (Узун-ада), у Ленкорани,
на Аму-Дарье, на озере Ала-куль и т. д. Отдельные залеты отмечены и
северней — в Черниговской обл., в Харьковской, в Татарской республике,
в Пензенской обл., в Котельническом районе Кировской обл., близ Можайска
на р. Москва, в Ельнинском и Вяземском районах Смоленской обл., в
Ленинградской обл., в Прибалтике; в Сибири в Приалтайских степях и у
Семипалатинска.
Биотоп. Мелководные внутренние водоемы с заросшими водной
растительностью берегами — озера, реки, включая их дельты.
Численность. В СССР розовый пеликан встречается в меньшем
количестве, чем кудрявый пеликан: относительно много розового пеликана
РОЗОВЫЙ ПЕЛИКАН (II
в низовьях Сыр-Дарьи, на Балхаше и на озере Сассык-куль. Численность
этого вида за последние десятилетия сокращается. Это наблюдается в
частности на Аральском море, где в конце прошлого — начале нынешнего века
пеликан был относительно обыкновенен; такое же положение было и в дельте Волги,
"но в последние годы, после организации заповедника, розовый пеликан стал
там обычен.
Экология* Размножение. К местам гнездовья розовые
пеликаны прилетают стаями, но вскоре начинают держатся парами. В это время
наблюдаются брачные игры — птицы, с бормотанием, издавая низкие глухие
звуки, подняв крылья, подпрыгивают на месте, поднимаются в воздух и кружат
в нем, затем, опустившись на землю, трутся клювами друг об друга. В это
время наблюдается и «символическая» постройка гнезд — самцы срывают и
собирают траву, носят ее, но затем бросают. Моногамы.
Гнездятся, в отличие от кудрявых пеликанов, только колониями и, как
исключение, отдельными парами. Колонии эти достигают больших размеров
и обычно располагаются (нижняя Сыр-Дарья) в центре обмелевшего озера,
сильно засоренного различной растительностью, на совершенно вытоптанном
камыше, образующем ряд отдельных небольших плотин. Эти плотины почти
целиком покрыты неглубоким слоем воды, доходящим местами сантиметров
до 15. Они свободно выдерживают несколько громадных рыхлых гнезд
пеликанов и самих птиц. Постройка гнезд начинается в дельте Волги около 20 апреля.
Гнездо строит самка. Самец доставляет строительный материал, главным
образом, траву. Постройка гнезд идет очень быстро, и через 2—3 дня гнездо уже
готово. В это же время наблюдается и спаривание. Оно происходит в течение
всего дня с промежутками в 10—15 минут; в момент спаривания самец опускает
крылья, как бы прикрывая самку. Гнезда в колонии располагаются тесно
друг около друга.
Кладка начинается в дельте Волги 20 апреля (Ромашева), на Сыр-Дарье
в конце апреля или в начале мая (Спангенберг, 1936). В кладке обычно
2, реже 1 или 3 яйца белого цвета с толстым известковым налетом, овальной
формы. Размер: 80—112 х 50—75 мм (дельта Волги); 90—106,3 х 58,9—60,5 мм
(Сыр-Дарья, Спангенберг, 1936). Вес яиц 155—195 г (дельта Волги).
Одна кладка в году, но в случае гибели кладки в начале насиживания
(не позже 10 дней после его начала) птица вторично несется. Перерывы между
откладыванием яиц 1—2 дня.
Насиживание начинается с 1-го яйца и продолжается 33 дня. Насиживает
преимущественно самка, самец лишь сменяет ее на короткий срок утром
и вечером.
Птенцы выводятся почти одновременно во всей колонии. В дельте Волги
вылупление падает на последнюю треть мая (Ромашева), на Сыр-Дарье на
конец мая —начало июня (Спангенберг, 1936), на Сассык-Куле — в конце мая
были маленькие, но уже одетые пухом птенцы. Птенцы вылупляются
слепыми, голыми и совершенно беспомощными. Голая кожа только что
вылупившегося птенца бледнорозовая, но через несколько часов приобретает сероватую,
а затем черно-бурую окраску. Очень скоро начинает появляться пух, и на
8—10-й день вылупления птенцы равномерно покрыты густым пухом.
Первые дни по вылуплению птенцов кормят полупереваренной пищей,
(грамм 30—40 в одну кормежку). Родители корм отрыгивают непосредственно
птенцам в рот. В день обычно старики кормят молодых дважды. 5-дневный
птенец уже получает в пищу мелкую свежую рыбу, 10-дневный птенец
съедает от 200—250 г, месячный — около 1 кг. Птенцы, завидев летящих
с кормом стариков, поднимают страшный крик, толкая друг друга, стараются
передвинуться к краю гнезда. Пуховые птенцы могут уже плавать и в случае
опасности стараются уйти скорее в воду. Не оперившиеся еще полностью
62
ОТРЯД ВЕСЛОНОГИЕ
птенцы переходят на воду до поднятия на крыло, так что родители
кормят их на воде. В возрасте около 21/2 месяцев птенцы становятся
летными: в дельте Волги — в начале августа; на Сыр-Дарье 9 августа
значительная часть птенцов уже поднялась на крыло. На о. Меньшикова,
Аральское море, 18—19 июля птенцы еще были нелетны (Зарудный, 1916).
Первое время после вылета молодых старые птицы их подкармливают,
затем, к концу августа, начинают отгонять от себя. Имеется «абортивный»
осенний половой цикл, в августе у самцов вновь замечается стремление к
спариванию.
Рис. 32. Розовый пеликан (взрослый и птенцы).
Линька. Плохо изучена. Начинается в конце июля (у пеликана,
добытого 22 июня в дельте Волги, идет линька первостепенных маховых), время
окончания не выяснено, у экземпляра, убитого в начале января в Казалинске,
линька уже закончена и все оперение свежее. Есть ли предбрачная линька — не
выяснено. Смена первостепенных маховых и рулевых идет от внутренних перьев
к наружным. Молодые птицы надевают взрослый наряд на 3-м году жизни.
Питание. Пищу розового пеликана составляет различная рыба,
преимущественно средних размеров, которую пеликан вылавливает на
мелководье клювом, сильно растягивая при этом горловой мешок наподобие черпака,
и подхватывая им добычу. Вместе с рыбой в горловой мешок попадает вода,
которую птицы, прежде чем проглотить добычу, выпускают изо рта. В настоящее
время, повидимОму, почти уже нигде не сохранилась совместная охота
собравшихся в огромные стаи пеликанов и присоединившихся к ним бакланов.
Кормящиеся пеликаны выстраиваются ломаной линией и медленно передвигаются
по воде, поджидая рыбу.
РОЗОВЫЙ ПЕЛИКАН
63
Розовый пеликан приносит известный вред рыбному хозяйству, но область
его распространения незначительна, численность'низка.
Полевые признаки. Розовый пеликан сходен с" кудрявым пеликаном, но
отличается от него даже на довольно далеком расстоянии отсутствием гривы
и деталями окраски. Кроме того, манера держаться на воде несколько другая:
никогда не держит шею вертикально вверх или наклонив ее вниз, а всегда
закидывает шею и голову назад на спину. Голос не отличим от голоса кудрявого
пеликана.
Описание. Размеры и строение. Длина крыла самца (5) 700—720, в среднем
716 мм, самок (2) 640—690 мм. Вес самцов достигает И кг, самок — 10 кг (Бутурлин, 1935).
Клюв очень длинный, уплощенный, оканчивающийся резко загнутым вниз крючком.
Шея довольно длинная, ноги короткие. Большой, сильно растягивающийся горловой мешок.
Лоб, уздечка, кольцо вокруг глаза, пространство за глазом и основание нижней челюсти
не оперены. Оперение головы острым мысом вдается на голый лоб. На голове имеется хохол
из удлиненных, заостренных перьев.
Первостепенных маховых 11, формула крыла 2=3>4>1=5>6... Хвост почти
прямой, из 24 рулевых. Оперение плотно прилегает к телу, но редкое.
Окраска. Пуховой птенец коричневый. Ноги и клюв черноватые, горловой
мешок темносвинцового цвета.
Гнездовый наряд. Голова и шея серовато-бурые, светлеющие к" спине. Спина светлого-
лубая; плечевые, средние и большие кроющие крыла серобурые с более светлыми
вершинами, малые кроющие крыла светлобурые с охристым оттенком. Маховые черновато-бурые
с более светлыми окаймлениями на первостепенных маховых и с серебристо-серым налетом
на второстепенных. Рулевые светлосерые. Брюшная сторона тела белая с бурым налетом.
При последующих линьках бурые тона постепенно исчезают и птица после 3-й линьки,
т. е. на 3-м году жизни, приобретает взрослый наряд.
Взрослая птица. Оперение белое с бледнорозовым оттенком, несколько более
интенсивно развитым на брюшной стороне тела. На груди охристо-желтое пятно.
Маховые черные с легким буроватым налетом, с белыми стержнями и пепельно-серебристыми
наружными опахалами второстепенных маховых. Внутренние второстепенные маховые
светлей наружных. Голые участки вокруг глаз желтые, горловой мешок желтоватый с
просвечивающими красными кровеносными сосудами. Надклювье серовато-голубое с
красноватыми пятнами, розовыми краями и беловатым вершинным крючком, подклювье у основания
серовато-голубое постепенно желтеющее к вершине клюва. Радужина светлок()асная. Ноги
желтые, на сгибах оранжевые.
В брачном наряде передняя, неоперенная часть лба образует вздутие. Голые части
окрашены более ярко — в красный цвет с желтоватым оттенком; горловой мешок
желто-охристый. Радужина тёмнокрасная. Ноги желтовато-красные. Окраска клюва также становится
ярче. Самцы и самки окрашены одинаково.
Систематические замечания. Систематические взаимоотношения розового пеликана
с ближайшими его родственниками и внутри вида до сих пор неясны и сложны, а в связи
с этим неясны границы его распространения. Многие авторы: Мензбир (1918), Склетер (1924),
Питере (1931) и др. склонны считать различные формы розовых пеликанов'за отдельные виды,
ряд других авторов: Бутурлин (1935), Хартерт (1912—21) и т. д. принимали их за подвиды
одного вида.
Некоторые авторы рассматривали птиц, населяющих нашу страну, как
принадлежащих к типичному розовому пеликану и к малому розовому пеликану, причем одни считали,
что последний является особым видом Pelecanus minor (Мензбир, 1918), другие — подвидом
Pelecantts onocrotalus roseus(Бутурлин, 1935), сводя minor в синоним Р. о. onocrotalus.
Исследования последних лет и просмотренный нами коллекционный материал позволяют отнести
всех розовых пеликанов, населяющих нашу страну, к Pelecanus onocrotalus, не
образующему у нас подвидов; все имеющиеся различия считать индивидуальной и возрастной
изменчивостью. Признание существования у нас двух подвидов: Р. о. onocrotalus и Р. о.
roseus совершенно невозможно, так как они почти на всем своем распространении
перекрывают друг друга, и у них существует полный ряд переходных особей.
Однако, как далеко выходит за пределы нашей страны P. onocrotalus и являются ли
P. roseus и P. philippensis подвидами или самостоятельными видами, — не имея большого
коллекционного материала и при существующем разногласии различных авторов — решить
невозможно. Если считать P. onocrotalus, P. roseusw P. philippensis за самостоятельные виды,
то распространение их будет следующее: P. onocrotalus вне нашей страны указано выше;
распространение Pelecanus roseus G m e 1 i n, 1789: Вост. Азия (в Китае розовых
пеликанов нет, Латуш, 1934), Белый Нил, Ангола, Замбези, побережье Мазамбика и на озере Нгами
и Pelecanus philippensis G me 1 i ri, 1789: Индия, за исключением северо-западной, Цейлон,
Бирма, Малайский п-ов, Кохинхина, Сиам, может быть Южн. Китай и о-ва Хайнань,
Ява, Люцон и Минданао.
ОТРЯД ВЕСЛОНОГИЕ
СЕМЕЙСТВО ОЛУШИ SUUDAE
РОД ОЛУША SULA BRISSON, 1760
Тип Pelecanus leucogcester
9. Олуша Sula bassana L.
Pelecanus Bassanus. Linnaeus, Syst. Nat. изд. X, 1758, стр. 133, Шотландия.
Русское название — олуша, олуша-глупыш.
Распространение. Ареал. Сев. Атлантика, Великобритания, Фарерские
о-ва, Исландия; Сев. Америка — острова в заливе св. Лаврентия,
Ньюфаундленд и Новая Шотландия; острова и побережья южн. Африки (Капская
колония), юго-вост. берег Австралии, острова в Бассовом проливе; Тасмания и
северный о-в Новой Зеландии.
Характер пребывания. Олуша — гнездящаяся и кочующая
птица. Частично в негнездовое время держится у мест гнездовий, частично
доходит до Азорских и Канарских о-вов, до сев. и зап. Африки (Марокко и
Сенегамбии); к северу в небольшом количестве до Норвегии и Прибалтики;
в Сев. Америке до Гренландии; по берегам Сев. Америки спускается к югу
до Мексиканского залива.
Подвиды и варьирующие признаки. 4 подвида, различающиеся между собою
морфологическими особенностями оперения и экологией: S. Ъ. bassana L, 1758 — Англия, Исландия,
Сев. Америка; S. Ь. capensis Lichtenstein, 1823 — побережье и прилежащие острова
Капской Земли; S. b. senator Gray, 1843 — острова в Бассовом проливе, юго-вост.
побережье Австралии (к югу от Брисбена) и Тасмания; S. Ь. rex Mathews et Ire dale,
1921—сев. о-в Новой Зеландии.
Атлантическая олуша или северная Sula bassana bassana L.
Распространение. Ареал. Ю.-з. Ирландия, Англия (Ленди и Бемп-
тон), Уэльс и прилежащие острова у берегов Шотландии; Фарерские о-ва,
Исландия; Сев. Америка — острова в заливе св. Лаврентия, Ньюфаундленд
и Новая Шотландия. В СССР редкая, случайно залетная птица у берегов
Мурмана.
Вопрос о залетах северной олуши к берегам Кольского п-ова неясен.
Все сведения, касающиеся залетов северной олуши, относятся, главным образом,
к наблюдениям, сделанным в прошлом столетии, и никто ее не добывал: Лил-
льеборг (1852) встречал олушу во время поездки на судне между Нордкапом
и Щурецкой; Мальм (1845) — по берегу моря, в холодное время года; Кол-
летт (1876) — до русской границы и Гебель (1903) упоминает о встрече олуши
у Мурманского побережья.
Характер пребывания. В Великобритании и Сев. Америке
северная олуша — гнездящаяся и частично оседлая и кочующая птица, причем
в Англии кочевки хорошо выражены только у молодых птиц, взрослые птицы
далее побережья Великобритании не откочевывают.
Биотоп. Гнездовья — обрывистые, крутые скалы побережий материков
и островов. В негнездовое время — море.
Численность. У нас единичные и случайные встречи. На местах
гнездовий весьма многочисленны.
Экология. Размножение. Моногамы. Северные олуши гнездятся
колониями. Гнезда строят на скалах; места гнездовий очень постоянны (на
о. Магдалины, Сев. Америка, известны с 1534 г.). Строительным материалом
служат преимущественно различные морские водоросли и в небольшом
количестве наземные растения.
ОЛУША
65
Одна кладка в году, и она начинается неодновременно у всех членов
колонии. У берегов Великобритании кладка обычно в апреле, реже в мае. В Сев.
Америке первые кладки находят в- середине мая и в конце июня, встречаются
одновременно гнезда с ненасиженными яйцами и с не вполне оперившимися
птенцами. В кладке 1, редко 2 яйца, округло-овальной формы, светлоголубого
цвета, покрытых сверху густым известковым слоем.
Размеры: (100) 62,5 — 87,5 х 43,1— 53,7, в среднем 78,06 X 53,7 мм
(Уайзерби, 1940); (44) 53,5 — 86,5 х 37—47, в среднем 77,6 X 47 мм (Сев.
Америка, Бент, 1922).
Насиживают самец и самка, период насиживания для атлантического
подвида около 43 — 45 дней (Бент, 1922, Уайзерби, 1940). Вылупляются
птенцы голыми, слепыми и совершенно беспомощными, но вскоре начинает
Рис. 33. Северная олуша (взрослая птица и молодая).
появляться пух, и в возрасте около 10 дней он весь одет пухом. В возрасте
около 3 недель вырастают маховые и рулевые перья и к 2 месяцам птенец
надевает полный перовой (гнездовый) наряд, а в 9—10 недель становится летным.
Вылупившихся птенцов оба родителя сначала кормят отрыгиваемой
полупереваренной пищей, позже — целой мелкой рыбой. В Англии птенцы покидают
гнезда с первых чисел августа, основная же масса птенцов уходит на воду
в сентябре.
Линька. У взрослых птиц 2 линьки в году: частичная предбрачная
в марте—апреле и может быть до мая, во время которой сменяются перья головы,
и полная послебрачная с августа — ноября и может быть до января.
Питание. Пищу северной олуши составляет мелкая рыба.
Полевые признаки. Средних размеров птицы с удлиненным телом и
сильным коническим, длинным клювом, слегка изогнутым в вершинной части
надклювья. Шея и ноги короткие. С сушей связаны только в период размножения,
причем держатся только на побережьях, внутри материков, встречаются
лишь случайно.
Олуши — прекрасные летуны, в течение продолжительного времени могут
парить в воздухе, делая изредка несколько резких сильных ударов крыльями.
5 Птицы СССР, т. I
66
ОТРЯД ВЕСЛОНОГИЕ
Хорошо плавают, гребя одновременно обеими ногами, но спят или отдыхают
обычно на воде, не работая ногами. Ныряют северные олуши, добывая рыбу,
только с разлета, иногда с очень большой высоты, причем в момент падения
складывают крылья и летят вниз головой, силой тяжести тела разрезая при таком
падении воду на большую глубину. По земле передвигается без труда. Это —
молчаливые, тихие птицы, только в гнездовый период крикливы и шумливы.
Описание. Размеры и строение. Длина крыла самцов 475—510, самок 475—
505 мм; длина клюва от оперения лба 95—100 мм (Уайзерби, 1940). Вес около 3 500 г.
{Первостепенныхмаховых 10. Формула крыла 1>2>3>4>5... Внутреннее опахало 1-го и 2-го
'маховых резко сужено в вершинной части, третьестепенные маховые хорошо развиты. Хвост
клиновидный, из 12 перьев, средняя пара сильно удлинена. По краям надклювья и под-
клювья имеются зазубрины, служащие для удержания живой добычи. Наружные
отверстия ноздрей отсутствуют. Лицевые части и кольцо вокруг глаз не оперены. Ноги короткие,
цевка не оперена, покрыта многогранными пластинками. Оперение густое, плотное.
Окраска. Пуховой птенец белый, лицевые части голые, черные. Гнездовый наряд:
голова, шея, спина, крылья и хвост темнобурые с мелкими белыми пестринами на голове,
шее и спине. Брюшная сторона тела и надхвостье грязнобелые с буровато-серыми пятнами.
Ноги серовато-черные. Первый брачный наряд: голова, шея и брюшная сторона тела белые
(с редкими темнобурыми пятнами. Верхние кроющие крыла и плечевые темнобурые с
белыми отдельными перьями.
Первый зимний (послебрачный) наряд: на голове, шее и нижней части тела нет
темных пестрин. Малые верхние кроющие крыла белые с редкими бурыми крапинами. Рулевые
белые.
Второй брачный наряд. Молодая птица похожа на взрослую, только кое-где на
больших кроющих крыла еще имеются бурые пестрины.
Взрослая птица: оперение белое с кремовым налетом на голове и шее, за
исключением первостепенных маховых, их кроющих и крылышка, которые буровато-черные. Голая
кожа лицевых частей и полоса на горле черноватые. Ноги серовато-черные с зеленоватыми
полосами на передней стороне цевки и на наружной стороне пальцев. Радужина желтовато-
белая. В брачном наряде голова и шея ярче — желтовато-коричневые, в остальном
оперение без изменения. Самцы и самки окрашены одинаково.
10. Красноногая олуша Sula sula L.
Pelecanus Sula. Li nnaeus. Syst. Nat. изд. XII, 1766, стр. 218, о. Вознесения.
Распространение. Ареал. Галапагосские о-ва, Вест-Индия, побережье
Венецуэлы, о. Вознесения, о. Южный Тринидад, острова в Индийском океане
и острова в зап. и центр, части тропической области Тихого океана (точно
гнездовья для ряда островов Тихого и Индийского океанов не установлены).
Характер пребывания. Красноногая олуша — оседлая птица.
Подвиды и варьирующие признаки. 5. 5. sula L., 1766 — острова Вест-Индии (о. Порто-
Рико, о. Доминика, о. Батавия, о. Кик-ем-Джени и др.), о-ва и скалы у побережья
Венецуэлы, о. Вознесения и о. Южный Тринидад; 5. s. websteri Rothschi Id, 1898 —
Галапагосские о-ва и о. Ревилья-Гигедо; S. s. rubripes Gould, 1833 — о-ва в Индийском
океане и острова в зап. и центр, части тропической области Тихого океана.
Восточная красноногая олуша Sula sula rubripes Gould
Sula rubripes. Gould. Syn. Bds. Aust. pt. 4, 1838, app. стр. 7, Южн. Уэльс.
Распространение. Ареал. Острова в Индийском океане (о. Родригес,
о. Реюньон и др.) и в зап. и центр, частях тропической области Тихого
океана (Филиппины, о. Лейзан, о. Феннинг, о. Кристмес, о. Пальмира и др.).
В СССР молодая птица добыта в Татарском проливе в бухте де-Кастри
(Финш, 1872). Подвид точно не установлен.
Биотоп. Океанические о-ва, поросшие густыми зарослями
кустарников и деревьев.
Численность. На местах гнездовий образует многочисленные
колонии.
Экология. Размножение. Моногамы. Гнездится восточная
красноногая олуша колониями. Гнезда на кустах и деревьях» растущих вблизи лагун.
КРАСНОНОГАЯ ОЛУША
07
Гнездо плоское, открытое, сделано из сучьев и ветвей, внутри выстлано травой.
В кладке 1, редко 2 яйца светлоголубого цвета, покрытые сверху густым
известковым слоем. Форма яиц сильно варьирует: от продолговато-овальной
до округло-овальной. Размеры (38) 59 — 72 х 35 — 48,5, в среднем 62,7 х
X 41 мм (Бент, 1922).
На о-ве Пальмира в декабре в гнездах были не вполне оперившиеся птенцы,
на о-ве Кристмес в январе все птицы сидели еще на яйцах. Насиживают и самец
и самка, период насиживания около 45 дней. Птенцы вылупляются голыми,
слепыми и совершенно беспомощными, но вскоре открываются глаза и тело
покрывается густым пухом. Первые перья показываются на крыльях и хвосте,
а приблизительно через 2 месяца птенцы, достигнув размеров взрослой птицы
и надев перо, покидают гнезда. Птенцы выкармливаются родителями в начале
полупереваренной отрыгиваемой пищей, затем цельной рыбой.
Рис. 34. Красноногая олуша.
Линька. Слабо изучена. Линяющие птицы встречены в июне и в августе,
повидимому, в этот период бывает предбрачная линька. Окончательный
наряд на 4-м году жизни.
Питание. Пищу восточной красноногой олуши составляет
преимущественно различная рыба и в небольшом количестве рачки. Рыбу добывает как и
обыкновенная олуша, стремительно бросаясь с воздуха с большой высоты в
воду. При этом птица складывает крылья и падает вниз головой.
Описание. Размер ыистроени е. Длина крыла 380—420 мм, длина клюва 88—
08 мм. Первостепенных маховых 10, внутренние опахала у 1-го и 2-го первостепенных
маховых в вершинной части пера резко сужены;формула крыла 1>2>3>4... Рулевых б пар,
средняя пара длиннее других, остальные постепенно уменьшаются по направлению к крайним.
Лицевые части и кольцо вокруг глаз голые. Края надклювья и подклювья зазубрены.
Оперение густое и плотное.
Окраска. Пуховой птенец белый. Гнездовый наряд серовато-бурый, более светлый
на брюшной стороне. Маховые и большие верхние кроющие крыла темносерые. Голая кожа
лицевых частей свинцово-голубая, на горле телесного цвета. В дальнейшем — в первом
брачном и первом зимнем нарядах в оперении постепенно развивается белый цвет. В первую
очередь белеет голова, шея и нижняя часть тела. При линьке в первый послебрачный
появляются отдельные белые перья на спине. Окончательный наряд надевают на 4-м году
жизни.
Взрослая птица имеет белое оперение, за исключением первостепенных
маховых, наружных опахал и вершин второстепенных маховых и больших верхних кроющих
крыла, которые темносерые с серебристым налетом. При обнашивании пера они буреют.
Голая кожа у основания клюва и кольцо вокруг глаз черноватые. Ноги красные, разных
оттенков. Клюв голубоватый, несколько краснеющий к основанию. Радужина серая.
68
ОТРЯД ВЕСЛОНОГИЕ
СЕМЕЙСТВО ФРЕГАТОВ FREGATIDAE
РОД ФРЕГАТЫ FREG AT A LACEPEDE, 1790
Тип Pelecanus minor
11. Большой фрегат Fregata minor Gm.
Pelecanus minor. G me 1 i n. Syst. Nat. 1 (2), стр. 572, 1789, Ямайка.
Русское название — фрегат дано птице в связи с ее превосходным полетом,
напоминающим быстроходные корабли — фрегаты.
Распространение. Ареал. О. Лейзан, Гавайские о-ва, Каролинские,
Маршальские, к югу до Новой Гвинеи, Квинсленда (Австралия) и Новой
Зеландии; о. Кристмес (близ Явы), риф Парацельс (Китайское море), южн.
Тринидад (Атлантический океан) и Галапагосские о-ва.
Подвиды и варьирующие признаки. 6 подвидов, морфологически слабо различающихся
между собою. Fr. т. minor Gmelin, 1789 — о, Кристмес, риф Парацельс, кочует в
восточной части Индийского океана; Fr.m. aldabrensis Mathews, 1914—Сейшельские
о-ва, о. Альдабра, кочует в зап. части Индийского океана; Fr. m. peninsulae Mathews,
1924 — Каролинские, Маршальские о-ва, к югу до Новой Гвинеи и Квинсленда; Fr. m.
palmerstoni Gmelin, 1789 — о. Лейзан, Гавайские о-ва, к югу до Новой Зеландии; Fr.
т. ridgwayi Mathews, 1914 — Галапагосские о-ва; Fr. т. nicolli Mathews, 1914,—
южный Тринидад, кочует в Атлантическом океане.
БОЛЬШОЙ ФРЕГАТ
69
Гавайский фрегат Fregata minor palmerstoni Gm.
PelecanusPalmerstonu G me 1 i n. Syst. Nat. 1 pt. 2, 1789, стр. 573, о. Пальмерстопа.
Распространение* Ареал. О. Лейзан, Гавайские о-ва, к югу до Новой
Зеландии.
Залетный экземпляр, повидимому этого подвида, добыт в 5 км от
Хабаровска. 1. VI. 1926 г. птица летела со стороны Амура. Добытый экземпляр
хранится в Хабаровском музее (Кардаков, 1928).
Экология. Размножение. Моногамы. Гавайские фрегаты гнездятся
колониями, которые иногда расположены рядом с колониями других птиц,
например, олуш. Гнезда устраивают из сучьев и веток на вершинах густо
растущих кустарников или на деревьях; внутри лоток выстилают стеблями и
листьями. Время гнездовья на Гавайских о-вах с мая по июль. В кладке 1,
редко 2 белых яйца. Продолжительность насиживания точно не установлена.
Высиживают и самец и самка. Птенцы вылупляются голыми и совершенно
беспомощными, но вскоре покрываются густым пухом. Выкармливаются
птенцы полупереваренной пищей, отрыгиваемой родителями в рот птенцам.
Питание. Питаются рыбой, которую добывают сами, или отнимают
у других птиц (олуш, пеликанов, бакланов, чаек и т. д.), заставляя их бросить
схваченную или даже отрыгнуть проглоченную пищу.
Полевые признаки. Океанические птицы больших размеров с клювом
сильным и длинным, несколько вогнутым по коньку надклювья и
оканчивающимся крючком, загнутым вниз. Встречаются во время кочевок далеко от
берега в открытом океане. Прекрасные неутомимые летуны, имеющие
способность целыми часами парить в воздухе. Плавает и передвигается по земле
очень плохо. Совсем не ныряет. Подняться с ровной поверхности в воздух не
может и поэтому обычно на суше опускается лишь на деревья или кустарники.
Описание. Размеры и строение. Самки несколько крупней самцов: длина
крыла самцов 585—603 мм, самок 605—621 мм, длина клюва самцов 116—130 мм, самок
130—150 мм.
Лицевые части головы оперены, но подбородок, горло и зоб голые, причем у самок
голое пространство несколько меньше, чему самцов. Крылья длинные, острые; вершину крыла
образует 1-е первостепенное маховое. Первостепенных маховых— 11. Хвост сильно
вильчатый и состоит из 12 рулевых, из которых особенно сильно удлинена крайняя пара рулевых.
Ноги очень короткие и оперены до пальцев. Пальцев 4, соединенных между собою слабо
развитой, сильно выемчатой перепонкой, доходящей до половины длины пальцев. Особенно
длинен средний палец.
Оперение густое и плотное. Половой диморфизм имеется не только в размерах, но и
в окраске оперения.
Окраска. Пуховой птенец покрыт белым пухом. В гнездовом наряде верх тела
бурый, низ белый; голова, шея и зоб белые с буровато-охристым налетом, переходящим в бурый
тон на груди. Верхние кроющие крыла серые. Молодые птицы надевают взрослый наряд
повидимому только на 4-й год жизни.
Взрослый самец имеет оперение черное с коричневатым оттенком на нижней
стороне тела. Перья головы и спины удлинены, заострены и с зеленовато-бронзовым
металлическим блеском. Малые верхние кроющие крыла и третьестепенные маховые черновато-
бурые. Голая кожа на подбородке, горле и зобе кармино-красная. Клюв темносерый. Ноги
черные. Радужина коричневая. У самки голова и шея черные,спина
черно-бурая,заостренные перья головы и спины с легким зеленовато-бронзовым отливом. Зоб, грудь и кольцо,
охватывающее основание шеи, белые. Брюхо и подхвостье черно-бурые. Малые верхние
кроющие крыла с широкими серыми окаймлениями. Горловой мешок неяркого красного
цвета. Клюв голубовато-серый.
ACCIPITRES или FALCONIFORMES
Характеристика отряда
Морфология. Внешние признаки. Размеры средние и крупные
(мелки только тропические соколы-карлики). Самые большие хищные, птицы
нашей фауны — грифы: серый гриф имеет более метра в длину, почти три
метра в размахе крыльев и около 6—7,5 кг веса. Вес камчатского орлана
достигает 8,9 кг. Одна из самых мелких — степная пустельга имеет длину
в 30—35 ему с размахом крыльев в 65—75 см и весит 150—200 г. Хищники
средних размеров имеют около 120—150 см в размахе крыльев и весят около
800—1000 г.
Перья жесткие, плотно прилегающие к телу. Не оперены пальцы ног,
цевка (у отдельных видов на разном протяжении), мягкая кожа у основания
надклювья (восковица), кольцо вокруг глаза и складка у разреза клюва.
У некоторых видов, питающихся падалью, голова и шея голые или покрыты
только пухом. Оперение неяркое и обычно представляет собой сочетание
бурого, серого, черного и белого цветов. У большинства видов в окончательном
(взрослом) наряде рисунок поперечный, в гнездовом — продольный. Во всех
возрастах поперечный рисунок хорошо развит обычно на крыльях (маховые
перья) и на хвосте (рулевые). Иногда в окраске проявляется половой
диморфизм. Неоперенные участки кожи (лапы, восковица, кольцо вокруг глаз)
окрашены чаще в желтый или синеватый цвет.
Кроющие перья обычно имеют закругленные концы, но иногда (например,
на затылке и на зашейке у орлов) и заостренные. У основания клюва и на
уздечке — между клювом и глазом — перья образуют щетинки. На голени
перья удлинены и имеют форму так называемых «штанов». Маховые и рулевые
очень жесткие, крылья длинные, острые, или закругленные. На
первостепенных маховых имеются вырезки и выемки, число первостепенных маховых
обычно 10—11, рулевых 12 (иногда — у грифов — 14).
Клюв у хищных птиц сравнительно короткий с загибающимся книзу
крючком и острыми режущими краями. На пальцах острые загнутые когти,
служащие для захвата и умерщвления добычи; на внутренней стороне пальцев имеются
ХАРАКТЕРИСТИКА ОТРЯДА
71
подушечки, помогающие удерживать схваченную добычу; первый (задний)
палец всегда имеется; вперед обращены три пальца (только у скопы внутренний
палец оборотный). Цевка — за исключением птицы-секретаря — короткая,,
покрыта поперечными щитками или сетчатыми пластинками. Глаза большие/
обращенные не только в стороны, но несколько вперед, что увеличивает поле
бинокулярного зрения. Радужина чаще всего темнобурая или желтая.
Анатомические признаки. Череп десмогнатический (реже.,
как например, у дымчатых коршунов, схизогнатический), довольно большого
размера, закругленный в затылочной области; затылочное отверстие широкое'
и направлено прямо вниз; базиптеригоидные отростки у взрослых птиц обычно,
отсутствуют. Переднелобная кость имеет надглазничный отросток, обычно
обращенный назад и прикрывающий орбиту сверху; иногда он имеет особое
продолжение—так называемую надглазничную кость osorbitale.Шея умеренной
длины, из 14—15—17 позвонков.. Опорный скелет туловища — короткий и
прочный. Грудные позвонки то сливаются в так называемую os dorsale, то
остаются свободными. Грудина длинная и широкая, с высоким гребнем, что
связано с сильным развитием двигающей крыло мускулатуры; вороньи кости —
толстые и широкие, вилочка также широкая и соединяется с грудиной связкой.'
Таз широкий. Сильно развиты крючковидные отростки ребер. Скелет
конечностей мощный. Как и у всех хорошо летающих форм, самый короткий отдел{
в крыле — плечо; боковой гребень его больших размеров. Самый длинный
отдел крыла — предплечье, иногда кисть имеет одинаковую с ним длину-
Ноги умеренной длины, пальцы длинные. Самая длинная фаланга пальцев ног —
четвертая (на третьем и четвертом пальцах), что связано с захватом и
закалыванием добычи. Мускулатура ног хорошо развита, в особенности мышцы,
сгибающие ногу и пальцы; приспособление сухожилия длинного глубокого сгибателя
пальцев, фиксирующее согнутые пальцы, сильно развито. Имеется обводящая
мышца. Мозг относительно большой, окципито-темпорального типа. Хищные
птицы обладают очень острым зрением, нервные окончания в сетчатке хорошо
развиты и расположены главным образом в верхнем ее секторе. Слух, тонкий,
отверстие наружного уха большое (в особенности у лесных видов), с кожистой
складкой; кроющие уха с расщепленными опахалами очень подвижны и
образуют замечательный звукоуловитель. Воздушные мешки большие. В скелете
грудины, плечевых костях, позвоночнике, черепе имеются пневматические
полости. Сердце относительно велико. Зоб и железистый желудок хорошо
развиты, слепые кишки рудиментарны, иногда же вообще отсутствуют. Часто
имеется не только левый, но и правый функционирующий яичник.Перья с
побочным стволом (его нет только у американских грифов, а у скопы — на брюшной
стороне тела). Пух расположен и по птерилиям, и по аптериям. Копчиковая
железа слабо развита (исключение — скопа) и оперена.
Образ жизни. Деятельны хищные птицы преимущественно днем. Встретить
их можно повсеместно, в самых разнообразных ландшафтах от Арктики до
тропиков, от альпийской зоны до равнин, лежащих ниже уровня моря. Часть
видов оседла, часть кочует по сезонам, некоторые совершают правильные
перелеты. В отношении характера сезонного размещения есть и внутривидовые
отличия между отдельными подвидами. Пища за редкими исключениями
животная (африканский грифовый орлан, гвианская каракара — плодоядны).
Кормовой режим, равно как и способы добывания пищи, весьма разнообразны.
В отношении кормового режима более или менее различаются такие группы:
орнитофаги (питающиеся птицами), миофаги (питающиеся мелкими
млекопитающими, главным образом, грызунами), ихтиофаги (питающиеся рыбой),
герпетофаги (питающиеся гадами), энтомофаги (питающиеся насекомыми),
саирофаги (питающиеся падалью). Значительна группа полифагов или пан-
тофагов, т. е. группа кормящихся разнообразной животной пищей.
72
ОТРЯД ХИЩНЫЕ ПТИЦЫ
В качестве примера орнитофагов можно привести сокола или кречета,
ястребов перепелятника и тетеревятника. Примером миофага служат сарычи.
Ихтиофаг — скопа, герпетофаг — змееяд, энтомофаг — осоед, сапрофаг —
гриф. К пантофагам относятся орлы, луни. Впрочем, группы эти условны и даже
такие специализированные орнитофаги, как настоящий сокол, весной в тундре
кормится леммингами и полевками, а на юге в Туркмении или Месопотамии
нападает по вечерним зорям на летучих мышей. В питании сарычей известное
место занимают и птицы. Орлы-подорлики в значительной мере кормятся гадами,
но также зверями, птицами и т. п. Более исключительны в отношении питания
сапрофаги, не промышляющие «живой» добычей.
В пределах указанных групп у отдельных особей и быть может популяций
наблюдается и индивидуальная или групповая специализация, вероятно даже
наследственная. Она выражается в предпочтении, оказываемом птицей
определенному виду добычи. Так, например, среди настоящих соколов есть птицы,
систематически нападающие на домашних голубей, у других главное место
занимают в питании чайки, у третьих — утки, у четвертых — грачи или
галки и т. д.
Сезонные различия в кормовом режиме хищных птиц в общем
незначительны, в особенности у перелетных форм, сезонное размещение которых в общем
более или менее совпадает с сезонным размещением главных видов добычи.
У оседлых птиц разница более значительна и, например, у ястребов и соколов
средних широт Европы в зимнее время главное место в питании занимают
птицы культурного ландшафта и поселений человека. Сезонные изменения
кормового режима хищных птиц связываются и с общим ходом годового цикла
их жизни (в особенности с выкармливанием выводка).
Кормовой режим и специализация в области питания находятся в связи
с определенными морфо-функциональными свойствами данного вида птиц.
В этом отношении перечисленные выше группы — орнитофаги, миофаги,
сапрофаги и т. д. — могут рассматриваться и как определенные «жизненные
формы».
Для грифов-сапрофагов характерно отсутствие контурного оперения на
голове, длинная и часто неоперенная шея с воротником из удлиненных перьев
у ее основания, длинные и широкие крылья, относительно длинный и мощный
клюв, тупые и малозагнутые когти. Форма клюва и длина шеи связаны с тем,
что грифы по преимуществу питаются- трупами павших животных большой
величины, отрывая большие куски и начиная расчленение трупа с извлечения
внутренностей их полости тела. Воротник при этом предохраняет от загрязнения
оперение туловища. Трупы больших животных попадаются не часто. В связи
с этим грифы обладают весьма экономным в смысле траты энергии способом
полета, очень полно используют найденные трупы, и, наконец, могут подолгу
оставаться без пищи.
Обычный тип полета грифа — парение, и его широкие крылья весьма к нему
приспособлены. У грифов удлинен и скелет крыла, и маховые перья, но форма
крыла — тупая, причем расходящиеся вершины маховых, снабженные
вырезками, напоминают раскрытую кисть руки. Для ускорения полета грифу
достаточно подогнуть пальцы и сложить таким образом передние маховые, не сгибая
даже кистевого отдела крыла; к сгибанию последнего грифы прибегают изредка
для достижения быстрого скольжения, в частности, для спуска. Для этой
группы хищных птиц характерна большая длина и ширина опахал маховых
и рулевых перьев молодых птиц, и это имеет для них большое значение, так как
мускулатура у них развита слабо. Увеличенная несущая поверхность большую
роль играет при парении.
Противоположный тип строения у сокола, ловящего птиц на лету.
Разыскивая и нападая на добычу, соколы не прибегают к парению. Весовая
ХАРАКТЕРИСТИКА ОТРЯДА
73
нагрузка крыла у соколов, в отличие от грифов, велика, вес тела
относительно площади крыльев высокий; крылья узкие и длинные, по длине кисть
примерно равна предплечью, тогда как у грифов предплечье значительно
длиннее кисти; хвост короткий, скелет туловища очень мощный грудная
мускулатура хорошо развита. Узость крыльев уменьшает лобовое
сопротивление, а большая тяжесть тела способствует увеличению скорости
движения вперед при активном скользящем полете. Сокола поэтому по быстроте
полета можно сравнить со стрижами и ласточками (которых они и ловят).
Нападая на добычу, сокол взлетает над нею, подбирает крылья так, что
первостепенные маховые располагаются параллельно продольной оси тела,
складывает рулевые, втягивает голову «в плечи» и бросается на добычу, делая так
называемую «ставку». Приблизившись к добыче, сокол несколько шире
расправляет крылья и наносит ей удар когтями задних пальцев или схватывает ее*
В соответствии с этим у соколов цевки короткие и толстые, пальцы длинные
и тонкие (ими соколы подхватывают добычу).
Орлы, сарычи, коршуны разыскивают добычу, состоящую, за редким
исключением, из находящихся на земле животных. При этом они, пользуясь
парением, как грифы, или медленным скользящим полетом, бросаются на
добычу, быстро скользя вниз и сильно подгибая при этом кистевой отдел
крыла; при таком движении относительно длинный хвост служит для быстрого
торможения. Летательный аппарат всех этих птиц более напоминает грифов,
чем соколов.
Особым типом строения крыльев и хвоста и особым способом нападения на
добычу обладают те виды хищных птиц, которые в отличие от соколов, сарычей,
орлов ловят добычу не в воздухе и не на открытых пространствах, но среди
деревьев, кустарников и других зарослей (как например, ястребы в европейской
фауне). Управляющая полетом мускулатура развита у них не менее, чем у
соколов, но крылья короткие, закругленные и хвост длинный. Быстроте полета у
ястребов способствует незначительная несущая поверхность крыла и
заостренная форма передних первостепенных маховых, образующих при полусложенном
крыле сплошную поверхность; длинный хвост обеспечивает им быстрое
торможение и поворотливость. Схватывание находящейся в укрытии (среди куста
и т. п.) добычи облегчается длиной цевки и способностью быстро и далеко
выбрасывать вперед лапу.
Среди хищных птиц есть и такие виды (правда, немногочисленные),
которые ловят добычу не на лету, а ходя по земле; таковы например, секретарь,
который ловит таким образом змей в африканских саваннах; малый подорлик,
ловящий лягушек; осоед, разрывающий гнезда
общественных'перепончатокрылых и т. д. Некоторые виды подстерегают добычу, сидя на земле (степной
орел, курганник и т. п.).
Умерщвляется добыча обычно лапой, причем хищная птица сдавливает
ее когтями (главным образом когтями заднего и внутреннего пальцев, которые
в связи с этим и по размерам больше, длинней и толще, чем когти на других
пальцах). При этом когтевые фаланги сгибаются и вдавливаются в добычу.
Соколы умерщвляют пойманных животных, ломая им шею клювом или
повреждая укусом затылочную область.
Отыскиванию добычи в большей мере способствует прекрасно развитое
•у хищных птиц зрение; некоторым птицам помогает также и слух
(лесным видам, например ястребам), а американским грифам еще и обоняние.
Пойманная добыча придерживается лапами и расчленяется и измельчается
клювом.
Пищеварение хищных птиц протекает весьма энергично, в железистом
желудке перевариваются частично и кости. Непереваренные же остатки —
кости, зубы, шерсть, хитин,— выбрасываются обратно через рот(так называемые
74
ОТРЯД ХИЩНЫЕ ПТИЦЫ
погадки). Суточная потребность в пище составляет около 10% веса птицы.
Хищные птицы едят не всегда ежедневно, но при возможности съедают много(яст*
реб-тетеревятник, например, может за сутки съесть утку, немного лишь
уступающую ему по весу). Поэтому-то хищные птицы способны и подолгу голодать:
орлан-белохвост — до 45 дней, черный коршун — до 3 недель. Нормально
хищные птицы едят и охотятся один-два раза в день, вылетая за добычей утром
и под вечер. Значительно чаще им приходится охотиться во время вывода
птенцов. В литературе часто встречаются утверждения, будто бы молодые птицы
обучаются «охотничьему искусству» у родителей. Это неправильно: способы
нападения на добычу, процедура питания и т. п. у хищных птиц строго
наследственны.
Хищные птицы — моногамы и образуют более или менее постоянные
пары. Вне периода размножения самец и самка держатся однако на некотором
расстоянии, и каждая птица охотится «за себя*. Но имеется и излишек или
резерв холостых птиц, главным образом молодых годовалых, которые при гибели
одного из партнеров гнездовой пары заменяют выбывшего. Начало периода
размножения находится в известном соответствии с кормовыми возможностями.
Поздно гнездятся виды, кормящиеся гадами, так как последние становятся
активными сравнительно поздно, а также те, которые выкармливают птенцов
молодыми птицами мелких видов (перепелятник, малый перепелятник, чеглок)
или насекомыми (кобчик, степная пустельга, осоед). Еще позднее гнездятся
некоторые виды, выкармливающие птенцов пролетными мелкими птицами
(средиземноморский чеглок). Рано гнездятся хищные птицы, кормящиеся
оседлыми видами птиц, даже на далеком Севере (например, кречеты) и те, птенцы
которых развиваются в очень длительный срок, как бородач, грифы и т. п.
В общем сроки размножения падают на раннюю весну, но довольно
значительно варьируют географически и в пределах вида у различных его
подвидов. Тем самым, при разнице в сроках весеннего развития половых желез
на 2—3 недели, между подвидами создается физиологический барьер,
исключающий возможность синхронизации их полового цикла.
Гнездятся хищные птицы отдельными парами, только некоторые (главным
образом грифы, а также мелкие соколы, коршуны) небольшими колониями.
Гнезда располагаются разнообразно: на скалах, на деревьях, на земле, в
человеческих сооружениях и т. п. В различных условиях птицы одного и того же
вида гнездятся по-разному. В этом иногда выражаются экологические
различия между подвидами: например, соколы в тундре гнездятся на земле, в
лесной зоне — на деревьях или выходах скал, в Туркестане — на скалах или
обрывах; беркут в лесной зоне гнездится на деревьях; в Туркестане, на
Кавказе и в Крыму — на скалах и т. д.
Наличие удобных мест для устройства гнезда в известной мере определяет
и географическое распространение вида. Искусных гнездостроителей среди
хищных птиц нет. Большинство видов пользуется брошенными гнездами
других птиц или отбивает чужие гнезда (например, у воронов). Устраиваемые
самими хищными птицами гнезда очень простого строения. Гнезда используются
полного лет и достигают иногда огромных размеров (у орлов и орланов).
У одной пары обычно несколько (2—3) гнезд, занимаемых более или менее
последовательно в разные годы. Гнезда эти расположены одно близ другого
в пределах гнездового участка пары. Смена гнезд объясняется вероятно тем,
что в конце периода размножения гнездо часто бывает заражено паразитами.
В устройстве гнезда участвуют оба пола. Гнездовые участки довольно велики —
у средних и крупных видов не менее 2—3 км2.
Посторонние особи того же вида, в частности, молодые, прошлогоднего
вывода, из гнездового участка в период размножения удаляются. Но держатся
они обычно вблизи, образуя частично резерв холостых особей, или расселяются
ХАРАКТЕРИСТИКА ОТРЯДА
75
на не очень далекое расстояние от места вывода (например, в одном случае
сокол, окольцованный в Финляндии птенцом под 62°8' с. ш. и 22°32' в. д., был
через 15 лет добыт на гнездовье в Финляндии же, но под 64°8' с. ш. и 25°22'
в. д., т. е. в 225 км к северу от места рождения).
Начало периода размножения характеризуется брачным полетом:
самцы и самки с громкими криками проделывают воздушные эволюции
над гнездовым участком (иногда такой брачный полет наблюдается и после
вывода птенцов в начале осени — в этом аналогия с осенним пением или
токованием других видов; такой повторный половой цикл у хищных птиц,
в отличие от некоторых других групп, бывает всегда неполным). Спаривание
происходит на земле или на дереве, в пределах гнездового участка, а иногда
на гнезде.
У всех видов бывает одна кладка в году. Число яиц в кладке различно:
крупные виды кладут 1—2 яйца, мелкие 6—7 и даже более. Число яиц в кладке
как правило варьирует географически, и у живущих на севере подвидов их
бывает более, чем у обитающих на юге. Как у некоторых других птиц, у ряда
видов хищных птиц количество яиц в кладке может меняться в зависимости
от кормовых (трофических) условий текущего года — это относится в первую
очередь к миофагам, главная пища которых — грызуны — подвержена
периодическим колебаниям численности. В неблагоприятные годы некоторые
хищные птицы повидимому вовсе не приступают к размножению.
Эмбриональное и постэмбриональное развитие у хищных птиц происходит
довольно медленно. С эволюционной точки зрения эту особенность нужно
рассматривать в связи с относительной безопасностью условий роста птенцов
(немногочисленность врагов, активная защита со стороны родителей и т. п.).
Другая характерная особенность — значительное совпадение сроков
насиживания (эмбрионального развития) у близких по систематическому положению
видов, вне прямой зависимости от разницы в размерах взрослых особей, иногда
весьма значительной (например, кречет и пустельга, ястреб-тетеревятник и
перепелятник).
Насиживание начинается с откладки первого яйца. Недавно высказано
предположение (Вагн Хольстейн, 1942), что эмбриональное развитие
начинается через большой промежуток времени после начала насиживания (у ястреба-
тетеревятника только после 16—18 дней), но предположение это требует
подтверждения. При наличии нескольких яиц в кладке и при промежутке
в откладывании яиц по крайней мере в 48, а иногда в 72 часа — в возрасте
старших и младших птенцов получается большое различие. Это обстоятельство
связано с характерным для хищных птиц авторегулированием популяции
в соответствии с кормовыми (и вероятно иными) условиями и приводит к
повышенной смертности приплода при неблагоприятных обстоятельствах; в
частности к гибели младшего птенца или птенцов в выводке (почти всегда у
бородача и беркута, часто у тетеревятника, обычно у миофагов в годы плохого
«урожая» или «неурожая» грызунов).
Насиживают у большинства видов хищных птиц оба родителя, но главным
образом самка, причем самец сменяет ее на короткое время утром, около
полудня или под вечер. Равномерное участие родителей в высиживании установлено
для осоеда; у ястреба-перепелятника и повидимому полевого луня самец
сменяет насиживающую самку только в крайне редких случаях. Наседных пятен
у соколов три (два на груди и одно на брюшной стороне), у других видов два
(на груди), Наседные пятна имеются и у самцов, но они слабо выражены. В
период насиживания корм для самки носит самец, он же ловит добычу для
выводка в первый период жизни птенцов. Насиживание длится от 4 недель
(у соколов) до 50—55 дней (грифы), продолжаясь в среднем около 5 недель.
Вес только что вылупившегося из яйца птенца составляет около % веса
76
ОТРЯД ХИЩНЫЕ ПТИЦЫ
свежего яйца. По типу постэмбрионального развития хищные птицы в
известной мере промежуточны между птенцовыми и выводковыми: они выходят из
яйца с открытыми глазами и опушенными, хотя еще нуждаются в обогревании
родителями. Пуховых нарядов два. Первый состоит частью из вершин
окончательного пуха, а частью из вершин контурных перьев; оба эти образования
отпадают в возрасте около 10—20 дней, когда появляется второй пуховой наряд,
более густой и плотный и состоящий из окончательного пуха. Во втором
пуховом наряде птенец уже обладает самостоятельной постоянной температурой и
не нуждается в обогревании взрослой птицей. Выкармливание птенцов
начинается с первого дня их жизни. В начале птенцового периода самка приносит
в гнездо переданную ей самцом добычу ощипанной или ободранной,
обезглавленной и часто без конечностей; при этом самка оделяет птенцов
мелкими кусочками мяса по очереди. Когда птенцы подрастут, добыча кладется
им в гнездо и они расчленяют ее сами. Выкармливание выводка, даже при
обилии пищи, требует от родителей большего напряжения, и самец охотится
в это время с рассвета до заката.
Вылетают молодые хищные птицы в возрасте от одного (мелкие и средние
виды) до трех месяцев (крупные). Покидают гнездо они уже тогда, когда еще
не умеют хорошо летать и когда крылья и хвост у них еще не доросли до
полной длины. Вес их в это время достигает максимума. Летным птенцам родители
носят пищу только изредка, нерегулярно, поэтому слетки быстро худеют.
Существенно, что рефлекс нападения на живую добычу у хищных птиц
появляется повидимому только тогда, когда двигательный их аппарат, в частности,
первостепенные маховые, достигает полного развития.
Возрастные изменения перового покрова продолжаются и после
наступления половой зрелости, которая у мелких, средних и у части крупных' видов
наступает обычно в возрасте несколько менее года.
Первый годовой или гнездовый наряд сменяется окончательным после
первой же линьки у большинства видов. Исключение в этом отношении составляют
только самые крупные виды, которые надевают окончательный наряд только
после трех или даже четырех линек. Линька полная годовая. Однако у
крупных видов (грифы, орлы, орланы) после линяния остается часть несмененного
оперения — мелких перьев и даже крупных (маховые, рулевые). Начало линьки
совпадает со второй половиной периода размножения, с угасанием полового
цикла, а именно с нахождением в гнезде птенцов во втором пуховом наряде:
в это время у старых птиц начинают сменяться средние или задние
первостепенные маховые. Первостепенные маховые линяют или от средних (5—6—7-е
перо) к внутреннему и наружному краю крыла или от заднего края к
переднему, или, наконец, полицентрически, из трех центров в каждом отделе крыла
(у крупных видов — орлов и грифов). Рулевые линяют центробежно, но
последними сменяются не крайние, а вторая от края пара рулевых. Весь процесс
занимает около 4—5 месяцев, т. е. идет очень медленно, что обеспечивает
хищным птицам сохранение высоких летных качеств в период смены пера. Схема
возрастных изменений наряда хищных птиц такова: первый пуховой наряд —
второй пуховой наряд — первый годовой (гнездовый) наряд — (полная
годовая линька) — второй годовой наряд и т. д. Рост хищных птиц весьма
интенсивен, в особенности до формирования второго пухового наряда; период
интенсивного роста заканчивается с формированием крупного оперения, когда
идет образование опахал маховых и рулевых. Половой диморфизм у хищных
птиц выражается обычно в том, что самки больше самцов, иногда же и
в окраске. Различия в размерах связаны вероятно с тем, что самке
принадлежит главная роль в насиживании и обогревании потомства, а самцу —
в выкармливании выводка, по крайней мере, в первый период его жизни;
при этом маленький самец более приспособлен к ловле мелкой добычи. Про-
ХАРАКТЕРИСТИКА ОТРЯДА
77
должительность жизни хищных птиц сравнительно велика. У видов
средней и мелкой величины она исчисляется десятилетиями. Охотничьи
кречеты жили более 25 лет, для беркута в условиях неволи указывается 46
и даже более 80 лет и т. д. (но известное сообщение, в Gentleman's
Magazine, 1793, что у мыса Доброй Надежды в начале сентября 1792 года был
пойман сокол с кольцом, указывающим на принадлежность его в 1610 году королю
Якову I Стюарту, невероятно).
Систематика. Современные хищные птицы делятся на два подотряда:
американских грифов Cathartae и настоящих или нормальных хищных птиц FaU
cones. Первые отличаются рядом резко выраженных морфологических
особенностей: у них имеются базиптеригоидные отростки, ноздри сквозные,
бронхиальные кольца сплошные, и они лишены голоса, побочных стволов в
оперении нет, нет слепых кишок, копчиковая железа оперена; первый палец ноги
развит слабо и прикреплен к цевке выше остальных. Этот подотряд объединяет
трупоядных птиц, принадлежащих к пяти родам — Vultur, Gymnogyps, Sarco-
ramphus, Coragyps, Cathartes, распространенным по всему Зап. полушарию,
кроме северных частей американского материка; всего 6 видов.
Нормальные дневные хищники, характеризующиеся по сравнению с
американскими грифами противоположными признаками, объединяют 264 вида,
относящихся к четырем семействам — африканским секретарям Sagitthariidae,
состоящим всего из одного вида; ястребиным Accipitridae с 205 видами;
соколиным Falconidae с 58 видами и скопинымРапЛял/Ч/яесодним, широко
распространенным в обоих полушариях видом. В палеарктической и нашей фауне
представлены три последних семейства.
Морфологические различия соколиных от ястребиных таковы: у
соколиных на твердом небе имеется продольный гребневидный выступ, на верхней
челюсти и надклювье резко выраженный зубец, ноздри круглые с возвышением
посредине, граница оперения по бокам нижней челюсти прямая, шейных
позвонков 15, спинная кость os dorsale имеется, membrana tympaniformis externa
хорошо развита, hemipterygoideum отсутствует. Кроме того, различна и
экология, в частности, способы питания, детали репродуктивного цикла, ход
линяния (например, первостепенные маховые сменяются у соколов центробежно —
от 6-го или 7-го и кончая 2-м, 1-м и 10-м, у ястребиных — от заднего края к
переднему, т. е. начиная с 10-го и кончая 1-м, или — у орлов и грифов — поли-
центрически).
Соколиные птицы принадлежат к родам Falco, Ieracidea, Microhierax,
Poliohierax и к ним причисляют еще уклоняющиеся американские группы
Polyborus, Milvago, Daptrius, Herpetotheres, Micrastur.
Ястребиные не имеют зубца на надклювье, возвышения посредине
ноздри, без os dorsale, без membrana tympaniformis externa, без продольного
гребневидного выступа твердого неба, с 14—17 шейными позвонками, с
хорошим развитием hemipterygoideum, с выступающей вперед острым углом
границей оперения на нижней челюсти. К этому семейству относятся роды:
Accipiter, Haliaeetus, Milvus, Buieo, Aquila, Elanus, Elanoides, Gypaetus,
Aegypius, Gyps, Neophron, Torgos, Morphnus, Harpyia, Pithecophaga, Circus,
Haliastur, Machaeramphus, Buiastur, Circaetus и другие.
Скопиные, будучи сходными с ястребиными по устройству нижней гортани,
оперенной копчиковой железе, отсутствию базиптеригоидных отростков, имеют
оборотный наружный палец ноги, побочный ствол—только на перьях брюшной
стороны, в пояснице и межлопаточной области. Многие особенности птерило-
зиса, а также строение сгибателей пальцев ноги, сближают скоп с
американскими грифами (Комптон, 1938). Все это заставляет выделить скопу в особое
семейство, тем более, что она крайне специализирована и в экологическом
оггношении.
?8
ОТРЯД ХИЩНЫЕ ПТИЦЫ
В морфологическом и таксономическом отношении хищных птиц обычно
сближают с голенастыми и с веслоногими. Однако реальность этих связей
не ясна, во всяком случае обособление хищных птиц относится к очень давнему
времени, по крайней мере верхнему эоцену (Lithornis vulturinus, найденный
в Англии, очень своеобразный и далекий от современной фауны вид, в
известной степени напоминающий грифов); современные роды хищных птиц известны
главным образом с миоцена. В плейстоцене, за редкими исключениями,
ископаемые формы уже довольно близки к современным и идентичны с ними, как
правило, в родовом отношении. Вопрос о близости хищных птиц, к совам требует
еще выяснения, но во всяком случае едва ли можно согласиться с широко
распространенным объяснением всех общих черт этих отрядов приспособлением
к сходному типу питания. В высшей степени сомнительно, чтобы совы в
отношении генеалогии стояли ближе к ракшеобразным, а не к дневным хищным.
Географическое распространение. Отряд хищных птиц распространен по
всему свету, за исключением Антарктики и некоторых океанических островов.
Наибольшее число видов распространено в жарком поясе. В палеарктиче-
ской области живут только представители подотряда нормальных хищных
птиц. Общее число видов хищных птиц фауны СССР — 44, подвидов и
монотипических, т.-е. не распадающихся на подвиды, видов — 97; из этого числа
9 видов и 33 формы относятся к семейству соколиных, а 34 вида и 63 формы —
к ястребиным; кроме того, 1 вид и 1 форму в фауне СССР составляет скопа —
единственный представитель семейства скопиных.
Процент числа видов хищных птиц в отношении общей численности этого
отряда в мировой фауне относительно высокий и равняется 16,9%.
Распределение хищных птиц фауны СССР по жизненным зонам может
быть схематически представлено таким образом (все относится только к
гнездящимся птицам). Характерные формы подчеркнуты. В числителе — число
видов, в знаменателе — подвидов и монотипических видов.
I. Арктика (зона)
Falco gyrfalco, F. peregrinus, F. columbarius, Accipiter gentilis, Buteo lagopus, Haliaeetus
albicUla, H. pelagicus 7
а. Побережье (подзона)
F. gyrfalco (gyrfalco, intermedius, grebnitzkii), F. peregrinus (leucogenys), H.
albicUla. 3/5
б. Тундра (подзона)
F. peregrinus (leucogenys), F. columbarius (ae$al*L insignis, pacificus), B. lagopus
(lagopus, menzbieri), H. albicUla (albicilla) ** 4/8
в. Лесотундра (подзона)
F. gyrfalco (intermedius, grebnitzkii), F. columbarius (aesalon, regulus, insignis,pacificus),
A. gentUis (buteoides, albidus), B. lagopus (lagopus, menzbieri, kamtschatkensis), H. albicUla
(albicUla), H. pelagicus 6/15
П. Лесная зона
Falco peregrinus, F. subbuteo, F columbarius, F. cherrug, F. tinnunculus, F. naumanni,
F. vespertinus, Accipiter gentilis, A. nisus, A. badius, A. virgatus, Miluus milvus, M. korschun,
Haliaeetus albicilla, H. pelagicus, Circus aeruginosus, C. cyaneus, C7pygargus,AquUa chrysaetos,
A. heliaca, A. pennata, A. clanga, A. pomarina, Buteo buteo, Circaetusferox, Butastur indicus,
Pernis apivoruSf P. ptilorhynchus, Pandion haliaetus 29
г. Тайга (подзона)
F. peregrinus (brevirostris, kleinschmidti, pleskei), F. subbuteo (jakutensis), F. columbarius
Yaesalon, insjgnis, pacificus), F, tinnunculus (tinnunculus, doerriesi), A. gentUis (gentilis,
buteoides, albidus, schvedowi, fujiyamae), A. virgatus (gular is), A. nisus (nisus, nisosimiiis, pal-
lens), M. korschun (korschun, lineatus), H. albicilla (albicilla), H. pelagicus, A. chrysaetus
„(chrysaetus, canadensis), A. clanga (clanga), B. buteo (vulpinus, burmanicus), P. apivorus,
P. ptilorhynchos (orientalis), P. haliaetus (haliaetus), С cyaneus (cyaneus) 17/33
ХАРАКТЕРИСТИКА ОТРЯДА
79
д. Смешанный и лиственный лес (подзона)
F. peregrinus (brevirostris), F cherrug (danubialis, cherrug), F. subbuteo (subbuteo), F.
tinnunculus (tinnunculus, dotrriesi), A. gentilis (gentilis, suschkini, fufyamae), A. nisus (nisus,
nisosimilis), A. virgatus (gularis), A. badius (brevipes), M. milvus (milvus), M. korschun
(korschun, lineatus), H. albicilla (albicilla), A. chrysaetus (chrysaetus, canadensis), A. heliaca
(heliaca), A. clanga (clanga), A. pomarina (pomarina), A. pennata(pennata), B. buteo (buteo,
uulpinus, burmanicus, Сferox (ferox), P. apivorus, P. ptilorhynchus (orientalis), P. haliaetus
(haliaetus), B. indicus, C. cyaneus (cyaneus), C. aeruginosus (aeruginosus, spilonotus), C. pyga-
rgus 25/36
е. Лесостепь (подзона)
F. peregrinus (brevirostris), F. cherrug (danubialis,cherrug, milvipes), F. subbuteo (subbuteo),
F. tinnunculus (tinnunculus, doerriesi), F. naumanni, F. vespertinus (vespertinus, amurensis),
A. gentilis (suschkini), A. badius (brevipes), C. pygargus,C. cyaneus (cyaneus), C. aeruginosus
(aeruginosus, spilonotus), M. korschun (korschun, lineatus), A. heliaca (heliaca), H, albicilla
(albicilla), C\ ferox (ferox) 14/21
III. Зона открытых сухих ландшафтов
Falco cherrug, F. columbarius, F. tinnunculus, F. naumanni, F. vespertinus, Circus pygargus,
C. macrourus, C. melanoleucus, C. cyaneus, C. aeruginosus, Hatiaeetus leucoryphus, Aquila ra-
pax, A. chrysaetus, Buteo rufinus, B. hemilasius, Circaetus ferox, Neophron percnopierus 17
ж. Степь (подзона)
F. cherrug (danubialis, cherrug, milvipes), F. columbarius (christiani-ludovici), F.
tinnunculus (tinnunculus, doerriesi), F. naumanni, F. vespertinus (vespertinus, amurensis), C. pygar-
gus, C. macrourus, C. melanoleucus, C. aeruginosus (aeruginosus, spilonotus), H. leucoryphus,
A. rapax (nipalensis, orientalis), B. rufinus (rufinus), B. hemilasius 14/20
з. Полупустыни и пустыни (подзона)
F. naumanni, F. tinnunculus (tinnunculus), H. leucorhyphus, A. chrysaetus (fulva),
N percnopterus (percnopierus), B. rufinus (rufinus), C. ferox (heptneri) 7/8
IV. Горы (зона)
F. peregrinus, F. gyrfalco, F. cherrug, F. columbarius, F. subbuteo, F biarmicus, F.
tinnunculus, F. naumanni, A. gentilis, A. nisus, C. cyaneus, N. percnopterus, Aegypius monachus,
Gypaetus barbatus, gyps fuivus, A. chrysaetus, A. pomarina, A. fasciata, A. pennata, B. buteo,
B. hemilasius 21
и. В ы с о к о г о р н а я б о р е о а л ь п и й с к а я подзона
F gyrfalco (altaicus), G. fuivus (himalayensis), A. chrysaetus (daphanea), G. barbatus
(Qureus, hemachalanus) 4/5
к. Горно-лесйая подзона
F. subbuteo (subbuteo), F. columbarius (lymani), A. gentilis(schvedowi), A. nisus (nisus,
nisosimilis, melaschistus), A. monachus (monachus), A. chrysaetus (daphanea, canadensis), A.
pomarina (pomarina), A. fasciata (fasciata), A. pennata (albipectus), B. buteo (menet-
riesi) 9/12
л. Горно-степная и пустынная подзона
F. peregrinus (babylonicus, caucasicus), F. biarmicus (feldeggii), F cherrug (saceroides,
cherrug, hendersoni), F. columbarius (lymani), F. tinnunculus (tinnunculus), F. naumanni,
С cyaneus (cyaneus), N. percnopterus (percnopterus), G. fuivus (fuivus), A. monachus
(monachus), A. chrysaetus (daphanea, canadensis), G. barbatus (aureus, hemachalanus), B.
hemilasius 13/18
Из списка, носящего несколько схематический характер, видно
значительное преобладание видов, распространенных в местностях с древесной
растительностью, относительная бедность открытых ландшафтов (тундра, степь,
полупустыня и пустыня), интразональный характер горной фауны.
Значительная часть видов имеет широкое распространение. Зональный характер
проявлений географической изменчивости, географического изоморфизма также
выражен хорошо, причем именно в географических подвидах отражается эндемизм.
По характеру пребывания в фауне хищных птиц арктической зоны один
оседлый вид — кречет F. gyrfalco', три кочующих и частично перелетных —
зимняк В. lagopus, тетеревятник A. gentilis и тихоокеанский орлан
Н. pelagicus, последний только в подзоне лесотундры; три перелетных — дерб-
ник F. columbarius, настоящий сокол F. peregrinus и белохвост Н albicilla.
В фауне птиц лесной зоны оседлы или полуоседлы 3 вида (F. peregrinus,
A. gentilis, A. chrysaetus), кочующие или частично перелетны — 2 (Н. albicilla,
80
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Н. pelagicus), перелетны 24 (F. subbuteo, F. colambarius, F. cherrug, F. tinnun-
cuius, F. naumanni, F. vespertinus, A. nisus, A. badius, A. virgatus, M. milvus,
M. korschun, С aeruginosus, С cyaneus, С pygargus, A. heliaca, A. pennata,
A. clanga, A. pomarina, B. buteo, C. ferox, B. indicus, P. apivorus, P. ptilorhyn-
chus, P. habiaetus).
В зоне открытых пространств и сухих открытых ландшафтов оседлых и
полуоседлых — 6 видов (Н. leucoryphus, A. chrysaetus, В. hemilasius, В. rufi-
nus, С. aeruginosus, быть может, N. percnopterus), перелетных 11 (F. cherrug,
F. columbarius, F. tinnunculus, F. naumanni, F. vespertinus, С pygargus, C.
macrourus, C. melanoleucus, C. cyaneus, A. rapax, С ferox).
Наконец, в горной зоне оседлы или полуоседлы 13 видов (F. peregrinus,
F. gyrfalco, F. cherrug, F. columbarius, F. biarmicus, A. gentilis, G. barbatus, G.
fulvus, A. monachus, A. chrysaetus, A. fasciata, B. hemilasius, B. buteo), а
перелетны 8 (F. subbuteo, F. tinnunculus, F. naumanni, A. nisus, C. cyaneus, A.
pomarina, A. pennata, N. percnopterus).
Практическое значение. На протяжении долгих столетий, в
продолжение которых люди занимались охотой и интересовались
живой природой, отношение к хищным птицам менялось несколько
раз. В самые древние времена оно было нейтральным, безразличным. В средние
века, когда начался рост, а потом и расцвет соколиной охоты, хищные птицы —
кречеты, соколы, ястребы, орлы-беркуты — высоко ценились и охранялись.
В частности, у нас места гнездовий кречетов в Двинской земле были заповедны
как государева «заповедь», повидимому, уже с XIII столетия, и эти «кречатьи
седьбища» представляют собою одни из древнейших заповедников на земном
шаре (к ним относится в частности и ныне существующий государственный
заповедник «Семь островов» на вост. Мурмане).
Охрана хищных птиц продолжалась до XVI11 столетия, когда в моду вошла
ружейная охота. В это время в Западной Европе охотничье хозяйство приняло
парковый или полупарковый характер, что в значительной мере было
обусловлено истреблением лесов в период социальных кризисов конца XVIII века.
При этом темпы уничтожения дичи при применении огнестрельного оружия
и концентрации животных на небольших относительно пространствах
значительно возросли, и охотник стал видеть в четвероногих и пернатых хищниках
опасных конкурентов.
Лишенное какой-либо научной основы мнение о вреде хищных птиц
получило в Европе с конца XVIII и до конца XIX столетия, а местами и позже,
широкое распространение. Преследование хищных птиц продолжалось почти
сто лет. Их стало значительно меньше, число же дичи, однако, также не
увеличилось. В новый, современный, период отношение человека к хищным птицам
опирается на строго научные основы. Можно считать теперь вполне
доказанным, что удельный вес гибели животных от хищных птиц (и зверей) в общей
естественной их убыли незначителен. К тому же установлен факт известного
«саморегулирования» численности популяции хищных птиц: в годы с
неблагоприятными кормовыми условиями они размножаются слабее. Биологическое
значение этого явления понятно. Далее, хищные птицы нигде в сущности
не бывают многочисленны в сравнении с их «жертвами». Несколько больше
хищных птиц в необжитых местах.
«Конкуренция» с человеком могла бы иметь значение только в наиболее
населенных местах, где баланс природы напряжен. Между тем, хищные птицы
избегают сильно измененных человеком природных условий. Поэтому в общем
и могущие быть в известных обстоятельствах вредными для охотничьего или
сельского хозяйства хищники в обжитой полосе нашей страны редки и хотя
бы потому не могут принести большого ущерба, а в тех районах, где хищных
птиц больше (например на далеком Севере), дичные богатства вообще едва
ХАРАКТЕРИСТИКА ОТРЯДА
81
использованы и говорить о реальном вреде от хищников просто несерьезно.
Скорее для плотных многочисленных популяций «жертв» хищникам может
принадлежать известная положительная роль в качестве орудия естественного
отбора. К тому же следует помнить, что огромное большинство видов
хищников питается преимущественно нежелательными или прямо вредными в
сельскохозяйственном отношении грызунами или насекомыми.
Наконец, культурно-эстетическое и научное значение хищных птиц очень
велико и в отношении их, как и любого памятника природы, все вопросы нельзя
решать с позиций примитивного утилитаризма. Неразборчивая борьба с
хищными птицами есть в сущности плод более или менее злостного невежества.
Все сказанное разумеется не означает, что нельзя застрелить в
налаженном охотничьем хозяйстве ястреба-тетеревятника, разгоняющего тетеревиные
тока, или на птицеферме — перепелятника, таскающего цыплят, и т. п.
Научных и практических предпосылок для истребления того или иного вида хищных
птиц нет ни вообще, ни в наших условиях. Вопрос может итти только об
известном контроле количества; некоторых видов в особых условиях и в населенных
районах. К этим видам могут быть отнесены ястребы — тетеревятник и
перепелятник и камышевый лунь.
Таблица для определения видов хищных птиц
1 (2). На верхней челюсти резкий выступ в виде зубца; в отверстии ноздри посредине
виден бугорок; граница оперения нижней челюсти (подклювья) образует сбоку угол ... 3
2 (1). Зубца на верхней челюсти нет; отверстие ноздри без бугорка посредине;
граница оперения нижней челюсти (подклювья) сбоку угла не образует 21
3 (4). Крупней, крыло от 300 мм и более, а если менее, то средний палец длиннее
40 мм ; 5
4 (3). Меньше, крыло всегда короче 300 мму средний палец короче 35 мм . . .'. . .13
5 (6). Наружный палец заметно длиннее внутреннего, а средний палец длинней цевки;
1-е маховое длиннее 3-го сокол Falco peregrinus
6 (5). Наружный и внутренний пальцы примерно равной длины; средний палец короче
цевки; 1-е маховое короче 3-го 7
7 (8). Цевка оперена более чем на половину длины, обычно на 2/8; в окраске обычно
(исключение — бурая фаза алтайского подвида) рыжего цвета нет, общий тон ее беловатый,
сероватый или бурый . '. кречет Falco gyrfalco
8 (7). Цевка оперена на половину или несколько менее; в окраске заметна обычно
примесь рыжего или рыжеватого цвета 9
9 (10). Темя рыжее, окаймленное темными пятнами; первостепенные маховые с
правильным светлым поперечным рисунком на внутренних опахалах
средиземноморский сокол Falco biarmicus
10 (9). Темя иной окраски, охристое или рыжеватое с более или менее
равномерными темными наствольями; первостепенные маховые с более или менее сливающимся
беловатым полем на внутреннем опахале 11
11 (12). Меньше, крыло самки короче 375 мм, самца 340 мм; цевка оперена спереди
примерно на 7з длины; оперение голени обычно одноцветное бурое
• лаггар Falco jtigger
12 (11). Больше, крыло самки длинней 375, самца 340 мм; цевка спереди оперена более
чем на 7з длины; оперение тюлени обычно пестрое балобан Falco cherrug
13 (14). Когти черные 15
14 (13). Когти светлые (бледнороговые или беловатые) 19
15 (16). На спинной стороне тела преобладает рыжий цвет; хвост резко ступенчатый;
разница длины боковых и средних рулевых более 20 мм. . .пустельга Falco tinnunculus
16 (15). На спинной стороне тела преобладает бурый или серый цвет; хвост слегка
закруглен, разница в длине боковых и средних рулевых менее 20 мм 17
17 (18). Крыло обычно длинней 240 мм; внутреннее опахало 2-го махового без вырезки;
средние рулевые без поперечного рисунка чеглок Falco subbuteo
18 (17). Крыло обычно короче 240 мм; внутреннее опахало 2-го махового с вырезкой;
средние рулевые обычно с поперечным рисунком, хотя бы в виде вершинной полосы. . .
дербник Falco columbarius.
19 (20). Первое маховое короче 3-го; окраска спинной стороны с преобладанием
рыжего цвета степная пустельга Falco naumanni
20 (19). Первое маховое длиннее 3-го; окраска спинной стороны с преобладанием
бурого или серого тона кобчик Falco vespertinus
6 Птицы СССР, т. I
82
ОТРЯД ХИЩНЫЕ ПТИЦЫ
21 (22)» Голова голая или только опушена 23
22 (21). Голова покрыта перьями 29
23 (24). Меньше, крыло короче 500 мм; клюв низкий и короче (при измерении от
переднего края восковицы) 40 мм; затылок и передняя сторона шеи оперены
стервятник Neophron percnopterus
24 (23). Больше, крыло длиннее 530 мм; клюв высокий и длинный, более 40 мм;
затылок и передняя сторона шеи покрыты пухом 25
25 (26). Крыло короче 610 мм; окраска черная с белым (взрослые) или шоколадно-бу-
рая с беловатыми пестринами (молодые)
индийский гриф Pseudogyps bengalensis
26 (25). Крыло длиннее 650 мм; окраска иная 27
27 (28). Ноздря щелевидная, шея покрыта беловатым пухом, общая окраска светло-
бурая или рыжевато-бурая белоголовый сип Gyps fulvus
28 (27). Ноздря округлая, шея голая, общая окраска темнобурая или черноватая
бурый гриф Aegypius monachus
29 (30). Хвост с резкой выемкой по
заднему краю, вилообразный 31
30 (29). Хвост без выемки по заднему
краю 35
31 (32). Крыло короче 300 мм; самое
длинное маховое 2-е; цевка спереди
сетчатая; щетинки на уздечке очень развиты и
прикрывают частично восковицу
дымчатый коршун Elanus coeruleus
32 (31). Крыло длиннее 400 мм;самые
длинные маховые 3-е и 4-е: щетинки на
уздечке менее развиты и не заходят на
восковицу 33
33 (34). Зоб и плечевые красно-рыжие;
средние рулевые без поперечного рисунка
...красный коршун Milvus milvus
34 (33). Зоб и плечевые без красно-
рыжего цвета; средние рулевые
поперечнополосаты
. . . черный коршун Milvus korschun
35 (36). Ноздри прикрыты густыми
черными щетинками; под клювом черная
борода из щетинок, обращенных вперед;
крыло длиннее 750 мм
........бородач Gypaetus barbatus
36 (35). Ноздри не прикрыты
щетинками, бороды нет, крыло короче 750 мм
37
Цевка оперена до пальцев 39
Цевка не оперена в нижней половине 55
Задняя поверхность цевки голая 41
Задняя поверхность цевки оперена 43
Цевка короче 80 мм; на конце хвоста широкая черная полоса, заметно шире,
чем другие полосы зимняк Buteo lagopus
42(41). Цевка длиннее 80 мм; конечная полоса на хвосте не шире, чем другие
на нем полосы, или хвост одноцветный
,. среднеазиатский канюк Buteo hemilasius
43 (44). Ноздри круглые (рис. 36) 45
44 (43). Ноздри щелевидные или бобовидные (рис. 36) 47
45 (46). Наружное опахало 7-го махового резко вырезано, высота надклювья
перед восковицей не менее 17 мм
малый подорлик, орел-крикун Aquila pomarina
46 (45). Наружное опахало 7-го махового сужено, без резкой вырезки; высота
надклювья перед восковицей менее 17 мм . . . . большой подорлик Aquila clanga
47 (48). Коготь заднего пальца длиннее самого пальца и длиннее надклювья
(измеренного от восковицы); наружное опахало 7-го махового без вырезок и сужения .... 49
48 (47). Коготь заднего пальца короче самого пальца и надклювья; наружное опахало
7-го махового сужено 53
49 (50). Крупней, крыло длиннее 600 мм; перья затылка и задней стороны шеи
заострены беркут Aquila chrysaetus
50 (49). Меньше, крыло меньше 550 мм; перья затылка и задней стороны шеи
закруглены . . " • 51
Рис. 36. Форма ноздрей орлов. Вверху —
степной орел, внизу — подорлик.
37 (38).
38 (37).
39 (40).
40 (39).
41 (42).
ХАРАКТЕРИСТИКА ОТРЯДА
83
51 (52). Хвост длиннее 250 мм; крыло длиннее 450 мм
ястребиный орел Aquila fasciata
52 (51) Хвост короче 250 мм; крыло (за редкими исключениями) короче 450 мм. . .
. орел-карлик Aquila pennata
53 (54). Спинная сторона однообразно бурая или только со светлыми вершинными
пятнами на кроющих крыла и надхвостье степной орел Aquila тарах
54 (53). Спинная сторона черно-бурая с белыми плечами или светлобурая с
охристым продольным рисунком могильник Aquila heliaca
55 (56). Уздечка покрыта мелкими чешуевидными перышками, когти почти прямые
или во всяком случае мало изогнуты 57
56 (55). Уздечка без чешуевидных
перышек, когти заметно изогнуты 59
57 (58). Средний палец длиннее 45 мм;
наружное опахало 6-го махового с
вырезкой, 2-е маховое приблизительно равно по
длине 6-му
. . .хохлатый осоед Pernis ptilorhynchus
58 (57). Средний палец короче 45 мм;'
наружное опахало 6-го махового без
вырезки; 2-е маховое заметно короче шестого \\ \ ||
. .обыкновенный осоед Pernis apivorus
59 (60). Оперение голени не удлинено и
не образует «штанов»; цевка покрыта мелкими \\ \^ J[
сетчатыми щитками 61
60 (59). Оперение голени удлинено;
цевка хотя бы в нижней части покрыта
поперечными пластинками 65
61 (62). Внутренняя (нижняя)
поверхность пальцев покрыта шипиками; нижняя
поверхность когтей закругленная
скопа Pandion haliaetus
62 (61). Внутренняя (нижняя) поверхность
пальцев без шипов; нижняя поверхность
пальцев плоская или вогнутая с режущими краями
63
63 (64). Крыло длиннее 400 мм;
внутреннее опахало 5-го махового с вырезкой;
окраска брюха с преобладанием белого цвета.
зм,ееяд Circaetus ferox
64 (63). Крыло короче 350 мм;
внутреннее опахало 5-го махового крыла без
вырезки ; окраска брюха с преобладанием бурого
цвета
..ястребиный канюк Butastur indicus
65 (66). Крупнее, крыло длиннее 600 мм
67
66 (65). Меньше, крыло короче 550 мм. .
73
67 (68). Хвост из 14 рулевых перьев; клюв очень высокий, надклювье выше 80 мм
• • . тихоокеанский орлан Haliaeetus pelagicus
68 (67). Хвост из 12 рулевых перьев; клюв ниже, надклювье ниже 80 мм. ..... 69
69 (70). Первое мдховое длиннее 7-го; ноздри овальные; если хвост не белый,
то оперение щек и уха резко выделяется черным цветом
долгохвост Haliaeetus leucoryphus
70(69). Первое маховое короче 7-го; ноздри щелевидные; если хвост не белый,
то оперение щек и уха не выделяется своей темной окраской 71
71 (72). У старых птиц оперение головы, шеи и поясницы не белое (при белом хвосте);
у молодых (с темным хвостом) горло буроватое. . . белохвост Haliaeetus alb ic ill a
72 (71). У взрослых птиц (с белым хвостом) оперение головы, шеи, поясницы белое;
у молодых (с темным хвостом) горло беловатое
белоголовый орлан Haliaeetus I otocephalus
73 (74). Жесткие перья вокруг лица образуют ошейник, напоминающий лицевой диск
сов; 2-е маховое длиннее 5-го \ ^ . . 75
74 (73). Ошейника или ожерелья нет; второе маховое короче шестого ...... ИЗ
75 (76). Надхвостье без белого или почти без белого; оперение голени темнобурое;
длина клюва (от восковицы) более 20 мм; цевка не короче 80 мм ........ .. i . .
•• болотный луч ь Circus aeruginosas
Рис. 37. Маховые перья степного луня
(вверху слева), лугового луня (вверху
справа) и пегого луня (внизу).
в*
84
ОТРЯД ХИЩНЫЕ ПТИЦЫ
76 (75). Надхвостье с заметной примесью белого цвета; оперение голени не бурое;
клюв короче 20 мм; цевка короче 80 мм 77
77 (78). Наружное опахало 5-го махового и внутреннее 4-го с вырезками ... .79
78 (77). Наружное опахало 5-го махового и внутреннее 4-го без вырезок .... 81
79 (80). В надхвостье имеется обычно поперечный рисунок; вырезки наружного
опахала 2-го махового и внутреннего опахала 1-го махового достигают конца кроющих
кисти степной лунь Circus macrourus
80 (79). В надхвостье нет поперечного рисунка; вырезка наружного опахала 2-го
махового и внутреннего опахала 1-го махового выходят за вершины кроющих кисти на
полтора—три см луговой лунь CircuspygarguS
81 (82). Второе маховое длиннее 5-го махового . пегий лунь Circus melanoleucu*
82 (81). Второе маховое короче 5-го махового . . полевой лунь Circus cyaneus
83 (84). Длина хвоста больше, чем половина длины крыла; передняя сторона цевки
в крупных поперечных пластинках; внутреннее опахало 5-го махового с вырезкой. . .85
84 (83). Хвост короче, чем половина длины крыла; цевка спереди не покрыта крупными
поперечными пластинками; внутреннее опахало 5-го махового без вырезки. . . см. 9!
85 (86). Крыло длиннее 300 мм
большой ястреб, тетеревятник Accipiter gentilis
86 (85). Крыло короче 300 мм 87
87 (88). Наружное опахало 6-го махового с вырезкой; горло без резкой темной
продольной черты .малый ястреб, перепелятник Accipiter nisus
88 (87). Наружное опахало 6-го махового без вырезки; на горле заметная
продольная темная черта 89
89 (90). Крыло длиннее 200 мм, хвост длиннее 140 мм . . . тюв и к Accipiter badiui
90 (89). Крыло короче 200 мм, хвост короче 140 мм
южноазиатский малый ястреб Accipiter virgatus
91 (92). Цевка короче 75 мм; ноздри расположены параллельно разрезу рта; размеры
меньше (крыло самок короче 440 мм, самцов 400 мм)
обыкновенный сарыч Buteo buteo
92 (91). Цевка длиннее 75 мм, ноздри расположены косо (под углом) в отношении
линии разреза рта; размеры крупнее (крыло самок больше 450 мм, самцов 410 мм) . . . .
курганник Buteo rufinia
СЕМЕЙСТВО СОКОЛИНЫЕ FALCONIDAE
РОД СОКОЛЫ FALCO LINNAEUS
Тип F. subbuteo,.
Важнейшие синонимы. Hierofalco С u v i е г, 1817, тип Falco candicans. —
Tinnunculus V i e i 1 1 о t, 1807, тип Falco sparverius. — Cerchneis Boie, 1826, тип Falc9
rupicola. — Hypotriorchis Boie, 1826, тип Falco subbuteo. — Erythropus С h r. L. В г е h m,
1828, тип Falco rufipes. — Aesalon Каир, 1829, тип Falco aesalon. — Gennaja Каир, 1847,
тип Falco lanarius
1. Сокол Falco peregrinus Tunst.
Falco peregrinus. T и n s t a 1 I. Ornith. Brit. 1771, стр. I, Англия.
Русское название. В древнем русском языке и у охотников именно этому
виду дано было название «сокол», которое теперь носит родовой характер. В русской
зоологической литературе сокола обычно называют — начиная со второй половины XIX века —
калмыцким словом «сапсан», что конечно не оправдано.
Распространение. Ареал. Распространен почти по всему земному
шару: Европа, Азия, Африка, Сев. Америка (включая Гренландию, где
достигает 70° с. ш. на западном берегу); Южная Америка, Австралия и Индо-Австра-
лийский архипелаг. Отсутствует лишь в ровных открытых ландшафтах —
например, в степях ю.-в.„ Европы и ю.-з. Сибири, в пампе Южной Америки,
в центральной Австралии и т. п.
Характер пребывания сокола весьма разнообразен. Часть
его подвидов оседла, часть перелетна, часть совершает кочевки.
Б и от о п. Для охоты сокол, нападающий почти исключительно на
летящую добычу, нуждается в открытых пространствах. Гнездится он или в
перемежающихся с открытыми пространствами (культурный ландшафт, луга,
сокол
85
водоемы и т. д.) лесах, или на скалах, в тундре — по высоким речным
обрывам («ярам»), хотя иногда и на кочках в ровной тундре; изредка он гнездится
и на земле в моховых болотах средней Европы и даже в старых каменных
сооружениях: башнях, колокольнях, развалинах. Соседства человека сокол
не избегает, хотя конечно, не может вынести неумеренного преследования,
обычного в окрестностях больших поселений. Избегает сокол больших
сомкнутых лесных массивов и открытых степей, но в последних встречается на
пролете и зимою. В горах более чем на 27«—3 км высоты он едва ли поднимается.
Подвиды и варьирующие признаки. Подвидов у сокола много. В тундрах Европы и
Азии F. p. leucogenys, Chr.L. Brehm, 1854; в лесной полосе Вост. Европы и Сибири F. р.
brevirostris, Me nz bier, 1882; в Якутии (кроме крайнего Севера) F. p. kleinschmidti,
Dementiev, 1934; на Сахалине, Шантарских о-вах и быть может на Камчатке F. p. pleskei,
Dementiev, 1933; в средней Европе от Швеции до Польши и Англии F. p. peregrinus,
Т u n s t а 11,1774; в Западной Европе от Рейна до Франции F. p. rhenanus Kleinschmidt,
1912; в Средиземноморье F. p. brookei Sh ar ре, 1894; на Кавказе и вероятно Малой Азии
Карта 9. Распространение сокола Falco peregrinus
1 — F. p. anatum, 2 — F. p. cassini, 3 — F. p. kreyerborgi, 4— F. p, perconfusus, 5 — F. p. radama, 6 —
F. p. arabicus, 7 — F. p. pelegrinoides, 8 — F. p. brookei, 9 — F. p. caucasicus, 10 — F. p. macropus, 11 —
F. p. peregrinator, 12 — F. p. ernesti, 13 — F. p. peregrinus, (14 — F. p. germanicus), 15 — F. p.
brevirostris, 16 — F. p. kleinschmidti, 17 — F. p. pealei, 18 — F. p. pleskei, 19 — F. p. leucogenys, 20 — F. p.
babylonicus, 21 — F. p. frujtii, 22 — F. p. nesiotes; a — граница гнездовий, г — зимовки.
близкий к нему F. p. caucasicus, Kleinschmidt, 1907; в тропической Африке F. p. per-
confusus, Н а г t e r t, 1927; на Мадагаскаре F. p. radama H a r 11 a u b, 1861; в Индии и
ю.-в. Азии F. p. peregrinator Sundeval, 1837; на Филиппинских и Зондских о-вах,
Н. Гвинее F. p. ernesti S h а г р е, 1894; на островах между Н. Гвинеей и Австралией (Ново-
Гебридские, Лойальти, Новая Каледония) F. p. nesiotes M а у е г, 1941; в Австралии и
Тасмании F. p. macropus Swains on, 1837; в Сев. Америке F. p. anatum Bonaparte,
1938; на островах северной части Тихого океана F. p. pealei Ridgway, 1873; на о-вах
Вулкана F. p. frujtii М о m i у a m а; в центральном ЧилиР. p. cassini S h а г р е, 1873;
в южном Чили F. p. kreyenborgi, Kleinschmidt, 1927; наконец, в палеарктических
пустынях в Африке и Аравии F. p. pelegrinoides Temminck, 1829 (вероятный синоним —.
F. p. arabicus Е г 1 a n g е г, 1903); в зап. Азии F. p. babylonicus S с 1 a t е г, 1861. Быть может
заслуживает выделения и восточногерманский F. p. germanicus Er longer, 1903. При
обширном распространении и большом различии условий существования географически
варьирует у соколов много признаков — и морфологических и экологических. Из
первых—размеры (общие, длина крыла, пропорции); из вторых — характер пребывания,
оседлость и перелетность, биотопическое распределение, календарь (сроки) годовых
периодических явлений, особенности размножения (плодовитость), трофические связи, отношение
86 ОТРЯД ХИЩНЫЕ ПТИЦЫ
к температуре (перелетные подвиды относительно стенотермны, оседлые эвритермны).3амеча-
телен факт с одной стороны наличия «зональной» географической изменчивости, с широкими
ареалами подвидов, с другой — нескольких узко распространенных островных подвидов.
Тундровый или белощекий сокол Falco peregrinus leacogenys В г е h m
Falco leucogenys. Chr. L. Brehm. Naumannia 1854, стр. 51, тип — из долины Заалы
в Тюрингии, добыт 28. Х.1825
Синонимы. Falco peregrinus harterti Бутурлин «Псовая и ружейная охота»,
XIII, 1907, стр. 100, низовья Колымы.— Falco peregrinus ussuriensis. Бутурлин, там же, по
пролетным птицам из Уссурийского края; — Falco peregrinus coeruleiceps Stegmanrr
journ. f. Orn. 1934, стр. 227, тип. местн. Гыданский полуостров. — Falco peregrinus cali-
dus многих авторов.
Местное название. У ненцев и народов Зап. Сибири «ханавей», у якутов «мок-
сокол», у народов Средней Азии «бахрын», «бахарин».
Распространение. Ареал. Гнездится в тундрах и на островах Северного
Ледовитого океана в Восточном полушарии, в северной части лесной зоны вдоль
речных долин. На севере встречен на Канинском п-ове на Камне (67°),
на Колгуеве (за 68°), Вайгаче, на Новой Земле от Маточкина Шара (самый
северный пункт — земля Гемскерка — 78°) и в других местах, на полярном Урале
(Ладахей, за 68°), в низовьях Оби, на Ямале (за 70°), на Гыдакском п-ове
у р. Хассейн-то (за 71°); в низовьях Енисея около 71—72°; на о. Диксон и
в прилежащих частях Пясинской тундры (около 73°); на земле Маркгама
к северу от Таймыра (75° 30'); и даже еще северней (у 76° с. ш.) на о-ве
Сталинец около мыса Тилло, на Хатанге (Крестовые Яры, 72°4б'); в дельте Лены
(ок.72°), в низовьях Индигирки и Колымы (ок. 68°30'). Есть указания о соколах
на о. Белом к северу от Ямала, на о. Фаддеевском и Беннета. Южная граница
неясна. Нормальная область гнездовья повидимому расположена к северу
от 64—65° (р. Казым, Малая Сосьва в Зап. Сибири, Туруханский край, но
отдельные находки и южней — на Сев. Урале еще под 61°). Область зимовок —
от восточных частей Средиземного моря, Черного моря, Каспийского моря до
Средней Азии (Семиречье, Фергана, Ташкентский оазис, Туркмения), Ирана,
Месопотамии, сев. Индии и вероятно в ю.-в. Азии. Северные точки зимнего
ареала у нас — 46° в Аскания-Нова и около.43° в Семиречье, нормальные
зимовки расположены между 30°—40°. Наибольшие скопления тундровых
соколов на зимовке отмечены в области массовых зимовок водоплавающих
на южном Каспии от Ленкорани до Гюргеня и Атрека. Миграционный ареал
связывает далеко разобщенные области гнездовья и зимовки и покрывает всю
территорию нашей страны от Украины (Полтавской обл.; Аскания-Нова) до
Приморья.
Характер пребывания. Гнездящаяся перелетная птица:
ареал резко разобщен — зимовки отделены от мест гнездовья огромными
пространствами (около 30° по широте).
Даты. Тундровый сокол прилетает в тундру весною в разные числа мая,
в среднем во второй трети этого месяца; отлет его с мест зимовок относится
к началу апреля или концу марта. Места гнездовья он покидает в начале
сентября и появляется на местах зимовок в октябре. Движение, следовательно,
довольно медленное, около месяца.
Биотоп. Открытые пространства — летом тундры, главным образом
долины рек с высокими обрывистыми берегами (ярами), реже водораздельная
тундра; на Сев. Урале и на северо-востоке Сибири — прибрежные скалы:
зимовки в ровных тундроподобных ландшафтах, обильных водою и птицами
(Нильская долина, ю.-в. Закавказье, ю.-з. Туркмения и т. и.), пролет главным
образом приурочен к долинам рек.
Численность. При несколько спорадическом распространении
местами в тундре, где имеются речные долины с ярами, нередок (южн. Ямал,
низовья Индигирки и Колымы, низовья Енисея).
сокол
87
Экология* Размножение. Сроки размножения, по сравнению
с южными подвидами, поздние. Через среднюю Россию тундровые соколы летят
с неразвитыми гонадами весною, тогда, когда у местовых соколов цикл
размножения в полном разгаре. По всей вероятности пары у этих соколов постоянные,
так как они не разбиваются и на зимовках. Привязанность к гнездовой
территории весьма велика (в семидесятых годах XIX столетия в низовьях Печоры
Сибом нашел гнездо сокола в том же месте, где оно было найдено еще в XVII
веке; на Ямале у Тир-седе гнездо, найденное Житковым в 1908 г., было занято
и в 1942). Первый раз молодые птицы гнездятся в возрасте около одного года,
едва начав смену гнездового наряда- Но нужно отметить, что не все они
гнездятся в этом возрасте (возможно из-за отсутствия гнездовой территории).
У соколов имеется, повидимому, известный резерв холостых птиц, так как
выбывшие из пары птицы заменяются быстро, особенно самки. Брачные игры,
по которым присутствие соколов легко обнаружить, заключаются в
стремительном полете в воздухе: сокол, складывая крылья, падает вниз, переворачивается
в воздухе, опять поднимается ввысь и т. д., издавая при этом громкий крик
«кееек-кееек-кееек». Брачный полет наблюдается во второй половине мая.
Гнездовые территории (участки) у тундровых соколов бывают различной
величины, но вообще говоря, по сравнению с другими подвидами сокола, невелики.
Гнездясь на земле, сокол нуждается в месте, сравнительно защищенном от
непогоды, относительно рано освобождающемся от снега и с хорошим
кругозором. Этим требованиям удовлетворяют высокие обрывистые берега рек
в тундре: «яры» и каменистые утесы. На Колыме, южн. Ямале расстояние между
гнездами не превышает иногда 3—5 км по прямой, обычно же гнезда
тундрового сокола расположены на 15—20 км одно от другого (Тиманская тундра,
Индигирка). Настоящих гнезд соколы не строят и кладут яйца прямо на землю
(но на мягкую почву); подстилка случайная, из остатков пищи, перьев,
травяных стеблей или вовсе отсутствует.
Кладка происходит в конце мая — начале июня. Число яиц в кладке
в среднем больше, чем у других подвидов. Обычно 4, редко 3 яйца.
Полная кладка происходит в начале июня (свежие законченные кладки на
южн. Ямале были 1,0 июня). Одно из яиц, как и у других соколов, чаще
бывает неоплодотворенным. Насиживание начинается с откладки первого
яйца (Осмоловская). Насиживают оба родителя, но главным образом самка.
Период насиживания продолжается по всей вероятности 28 дней (прямых
наблюдений, относящихся к этой форме, нет). Терморегуляция у птенцов
устанавливается на 10—12-й день по вылуплении, при надевании второго пухового
наряда. Выклевываются из яиц птенцы в начале июля (на Ямале в 1941—42 гг.
вылупление наблюдалось между 5—22 июля). Летные молодые появляются
в тундре в разные числа августа; на крайнем севере позднее; так под 75°30' с. ш.
на земле Маркгама 23 августа птенцы были еще во втором пуховом наряде
с только что пробивающимися рулевыми и маховыми; два птенца такого же
возраста в конце августа наблюдались Розановым под 76° с. ш. на о. Сталинец.
Следовательно, гнездовый период продолжается 35—40р дней. В конце
августа и начале сентября выводки еще не разбиваются и держатся у мест
гнездовья. Смертность молодых в гнездовый период невелика, во всяком
случае по сравнению с мышеядными хищниками тундры, так как старые птицы
повидимому всегда в состоянии добыть для выводка достаточно пищи, а врагов
у соколов очень мало. Не бывает, несмотря на разницу в возрасте птенцов,
и каннибализма. Выводок и самку на гнезде кормит самец, который в период
насиживания и роста птенцов ловит добычу и передает ее самке.
Линька. Смена первостепенных маховых у старых соколов начинается
в то время, когда птенцы в гнезде находятся во втором пуховом наряде.
Следовательно, тундровые соколы линяют значительно позднее других подвидов
88
ОТРЯД ХИЩНЫЕ ПТИЦЫ
того же вида. Начинается линька в середине июля, кончается в декабре.
Порядок линьки такой же, как у других крупных соколов: маховые сменяются
в последовательности 7—6—5—8—4—3—9—2—10—1 (или 6—7—5 или 5—6—7
и далее как в первом случае). Линька второстепенных маховых и плечевых
кончается сменой дистальных перьев. Последовательность смены нарядов
такова: первый пуховой наряд (первые 10—12 дней жизни); второй пуховой
наряд; первый годовой илигнездовый наряд, который в возрасте около 17
месяцев после полной годовой линьки сменяется вторым годовым или
окончательным нарядом.
Питание. Тундровый сокол, как и другие подвиды этого вида,
путается — за крайне редкими исключениями — птицами, которых ловит на лету.
Для тундры в качестве его пищи указаны также лемминги и полевки, а для
области зимовок — летучие мыши (Месопотамия, Тайсхерст, 1922). Морфология,
экология и поведение сокола обнаруживают большую степень кормовой,
специализации. Основную пищу тундрового сокола составляют птицы средней
величины: от серебристой' до обыкновенной чайки, от кряквы и шилохвости
до чирка. Но охотничьи соколы напускаются и на цапель, гусей, а иногда и
журавлей, обычно — попарно. Отдельные соколиные пары оказывают
предпочтение определенным видам птиц. Кормовой режим меняется и по сезонам: в
период выкорма птенцов, пока последние еще малы, самец охотится главным
образом за мелкими птицами. Но непосредственно вблизи от гнезда (радиусом метров
в 200) он не охотится. Этим и объясняется «мирное» гнездованье соколов в
тундре рядом с водоплавающими (гусями): в гнездовом участке, пока птенцы
в гнезде, инстинкт защиты потомства подавляет у соколов, как, повидимому,
и у других хищных птиц, инстинкт нападения на добычу.
Непосредственными новыми наблюдениями для тундровых соколов
установлены такие кормовые объекты: утки-чирки, аулейка, свиязи, шилохвости,
морская чернеть и др., белая куропатка, длиннохвостый поморник, кулики
(бекас, турухтан, золотистая ржанка, плавунчик, щеголь и др.), полярная
крачка, лапландский подорожник, варакушка, краснозобый конек, болотная
сова и т. д. Молодые в первом пуховом наряде выкармливаются
преимущественно воробьиными и мелкими куликами, в кормовом режиме взрослых
соколов преобладают утки и кулики. Белых куропаток, в противоположность
кречету, сокол в тундре преследует немного, главным образом весною, когда
другие птицы еще не все на месте. Трофические связи сокола в тундре, в отличие
от кречета, направлены главным образом в сторону только гнездящихся и
перелетных в тундре видов. Отмечено определенное соответствие в сроках и
направлениях перелетов тундровых соколов и их кормовых объектов (в
частности, весенний прилет сокола в Тиманскую и Большеземельскую тундры
более или менее совпадает с прилетом туда шилохвости). Эта связь сохраняется
и на зимовке: на южном Каспии соколы держатся главным образом в области
зимовок северных водоплавающих птиц; у Гассан-кули в ю.-з. Туркмении
зимующие соколы кормятся утками (хохлатой чернетью, чирками,
шилохвостью, свиязью, красноголовыми нырками), чайками (серебристой,
обыкновенной, морским голубком и т. д.). Можно, однако, добавить, что на крайнем севере
отношения несколько меняются: на Новой Земле соколы в значительной мере
питаются чистиковыми у птичьих базаров, в других частях Арктики —
куликами и т. д.
Рефлекс нападения вызывается у сокола видом улетающей или
пролетающей птицы; на неподвижно сидящих на земле животных он не нападает. Сокол,
преследуя добычу, делает так называемую ставку; оглушенная им добыча или
падает на землю, куда к ней спускается сокол, или подхватывается им в
воздухе. Если удар неудачен, сокол опять идет вверх и повторяет ставку. Иногда
это повторяется несколько раз, пока птица не будет убита или пока сокол не
сокол
89
устанет. Сбитой на земле птице сокол разрывает шею, продевая зубец
надклювья в место сочленения позвонков. Скорость бросающегося на добычу
сокола очень велика (по данным, относящимся к другому подвиду сокола,
он бросается на добычу на расстоянии до 1 */2 км и при ставке развивает
скорость при угле падения до 25° — в75 м/сек, а при более крутом — идо ЮОм/сек.).
Естественно, что бросающегося на добычу сокола плохо видно, часто
видна только проносящаяся в воздухе тень и слышен звук рассекаемого
воздуха.
Часы охоты сокола совпадают с активностью других птиц и именно тех,
которые служат для данной особи главной добычей. В период выкармливания
сокол занят почти все светлое
время дня, с некоторым
перерывом около полудня. Так, на
южн. Ямале при незаходящем
солнце соколы начинали кормить
молодых около 3 часов
пополуночи и до 9—10часов утра, затем
следовал перерыв до 15 часов,
после которого снова начинали
и кончали уже часов в 7—8
вечера; с конца июля активность
старых птиц снижается
(Осмоловская). На зимовках охота
тундровых соколов наблюдалась
и после захода солнца, в лунные
ночи. Потребность в пище сокола
составляет в сутки, по
наблюдениям сокольников, около 125—
150 г (это, конечно, не предел),
для выкорма выводка в летний
период соколам нужно повиди-
мому до 300 птиц среднего
размера. Птенцам сначала добыча
приносится ощипанной, часто без
головы и конечностей, кормит их
самка, разрывая добычу на
мелкие части; позднее ощипанная
птица кладется молодым в
гнездо, и они разрывают ее сами.
Для ощипывания и «отделки»
добычи служит определенное место
в пределах гнездового участка.
Несведёнными соколы оставляют
обычно крылья с частью или более или менее целым плечевым поясом.
Полевые признаки. Окраска на расстоянии или контрастная—темный
верх и беловатый низ (взрослые), или буроватая сверху и снизу
(молодые); хорошо заметны черные «усы» по сторонам светлого горла; сидит птица
прямо («столбиком»), летает очень быстро, с более или менее подогнутыми
в кисти длинными острыми крыльями (первостепенные маховые никогда не
отстоят одно от другого в вершинной части, а касаются друг друга — в этом
отличие от кречетов и балобанов). Хвост короткий. По размерам тундровый
сокол крупней голубя. Голос «кееек-кееек-кееек» или «кьяк-кьяк-кьяк»,
отрывистый и резкий. Не парит, держится в открытых пространствах, нападает
на летящую добычу.
Рис. 38. Соколы (взрослые самец и самка и
молодой самец).
90
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Описание. Размеры и строение. Тундровый сокол — самый крупный из
всех палеарктических подвидов сокола, крупнее его только северо-тихоокеанский подвид
F. p. pealei. Сложение мощное, грудь широкая (очень большая грудина с высоким килем,
большие грудные мышцы), крылья узкие и длинные, с длинными и очень твердыми
маховыми; вырезка на внутреннем опахале 1-го и наружном опахале 2-го махового; формула крыла
2>1>3; хвост короткий, слегка закругленный; цевка короткая, оперенная до половины;
пальцы длинные и тонкие, с сильно развитыми подушечками на внутренней (нижней)
стороне фаланг, что способствует хватанию добычи в воздухе; средний (третий) палец длиннее
цевки, наружный (второй) палец заметно длиннее внутреннего. Клюв короткий, круто
загнутый, предвершинный зубец надклювья резко выражен. Глаза большие, несколько выпук^
лые, надбровная часть орбиты выдается не резко, меньше, чем у кречета, что увеличивает
кругозор; голое кольцо вокруг глаза широкое. Длина самцов (4)410—430, самок (14) 475—
505, в среднем 419,25 и 490мм; размах крыльев самцов (4) 850—970, самок (12) 1070—1170,
в среднем 910 и 1141, 1 мм. Вес самцов (4) 678—740, самок (4) 1002—1300, в среднем 709,5
и 1133 г. Крыло самцов (28) 305—330, самок (51) 350—378 мм} в среднем 314,4 и 357,1 мм.
Окраска. Это самый светлый и бледный из подвидов соколов, с отсутствием рыжих
тонов во всяком возрасте. Первый пуховой наряд — снежнобелый, второй — менее чистого
белого цвета. В первом годовом гнездовом наряде общий тон верхней стороны тела бледно-
бурый со светлыми охристыми каймами перьев; лоб, а часто и темя — бледноохристые,
с более или менее развитыми темными бурыми серединами перьев; маховые с беловатыми
поперечными пятнами на внутреннем опахале, рулевые — с поперечным
охристо-рыжеватым рисунком из пятен или полос; нижняя сторона тела — охристо-белая с бледнобурым
продольным рисунком на груди и брюхе; темные пятна над глазами («усы») узкие,
но длинные, кроющие уха чуть буроваты. Восковица и голые кольца вокруг глаза
синеватые, лапы бледнржелтые, радужина темнобурая, клюв синеватый, когти черные. В
окончательном наряде общая окраска верха — серовато-бурая с более или менее резким сизо-серым
поперечным рисунком; голова обычно серовато-сизая, не темнее или слегка темнее спины;
маховые — черновато-бурые с беловатым поперечным рисунком на внутренних опахалах;
рулевые черновато-бурые или серовато-бурые с бледносизыми поперечными полосками;
длинные серовато-черноватые усы; темные пятна под глазами развиты слабо или отсутствуют;
общая окраска низа белая, с хорошо заметным в свежем пере охристым оттенком, с тонким
поперечным черновато-серым рисунком на боках, голени, подхвостье. У самок о§щий тон
окраски верха темней, а полосы и пестрины на нижней стороне тела более развиты, чем
у самцов. Значительны и индивидуальные вариации окраски, выражающиеся как в общем
тоне окраски верха, так и в развитии темного рисунка на нижней стороне тела.
Птицы во втором годовом наряде отличаются относительно темной, менее сизой
окраской верхней стороны тела и большим развитием темных пятен и полос на нижней стороне
тела. На груди у них обычно продольные штрихи или пятна. Восковица, голое кольцо вокруг
глаз, лапы — яркожелтые, остальные неоперенные части окрашены как у молодых.
Якутский сокол Falco peregrinus kleinschmidti Dem.
Falco peregrinus kleinschmidti. Dementiev. L/Oiseau et la Revue Frang. d'Orn. 1934,
стр. 480, Олекминск.
Местные названия. Якутское — «моксокол».
Распространение. Ареал. Якутский сокол распространен в
Восточной Сибири, кроме крайнего севера и юга, — в среднем течении Лены, в
бассейне Олекмы, Вилюя, Алдана, на Охотском побережье и по крайней мере
в южных частях Коряцкой Земли. Точные границы его распространения неясны.
Популяция со смешанными признаками на Анадыре, Енисее; сходные особи
встречаются и среди F. p. leucogenys с Яны, Индигирки, Колымы. Быть может
к этой расе относятся и соколы с Камчатки, откуда впрочем нет до настоящего
времени в коллекциях достоверно гнездовых экземпляров; по крайней мере
часть пролетных на востоке Камчатки соколов относится к этому подвиду.
Птицы из Забайкалья и Приамурья относятся к другому подвиду и в общем
весьма близки к европейским. Зимний ареал неясен: одна птица такого типа
окраски добыта в Ассаме; несомненно, что к этой расе должны относиться часть
зимующих в Китае и ю.-в. Азии соколов, но материала оттуда у нас не было;
возможно, что сюда же относятся те из зимующих в Индии птиц, которые
определяются разными авторами как «F. /?. peregrinus»; то же вероятно в отношении
части зимующих в Уссурийском крае соколов.
сокол
91
Характер пребывания. Якутский сокол — перелетная
птица; в СССР гнездится и пролетает. Прилетает в разные числа второй половины
апреля (у Якутска 22 апреля по Миддендорфу, по Воробьевой 29 апреля),
отлетает — в сентябре. По крайней мере часть этого подвида пролетает по
Камчатке с половины сентября — в октябре.
Биотоп. Якутский сокол встречается в зоне тайги, и лишь в
Корячкой Земле заходит в южные пределы тундры; для охоты он нуждается в
открытых пространствах, а для гнездованья — в скалах (до сих пор гнездованье его
на деревьях не установлено, и все известные места гнездовья расположены
или по обрывам речных долин, или по морским побережьям). Пролет, по
крайней мере частично, морскими побережьями — Камчатка, Курильские и
Командорские о-ва. Встречается сравнительно нередко.
Экология. В общих чертах сходна с образом жизни тундрового подвида —
как в отношении характера пребывания (перелетная бинареальная птица), так
и по позднему годовому циклу периодических явлений и т. п. Впрочем,
экологические сведения весьма неполны.
Размножение. Пары постоянны, т. е. уже с прилета самец и самка
держатся вместе. Гнезда, как и у других соколов, в сущности отсутствуют.
В одном и том же месте пара соколов гнездится много лет. Расположены эти
гнезда исключительно на скалах (иногда в непосредственном соседстве с
ласточками и стрижами, воронами), по карнизам, в щелях и т. п. В одном случае
гнездо найдено прямо на земле у реки между сваленными деревьями
(Воробьева, 1928). Судя по тому, что в выводках обычно бывает 3, иногда же 4 птенца
(отмечен впрочем и один), число яиц в кладке в среднем 4, т. е. как у
тундровой формы. Точных сведений о времени кладки нет, вероятно она падает на
начало июня, так как птенцы появляются около начала июля. Так, 15 июля
у Ботоми на левом берегу Вилюя в гнезде было 3 пуховых птенца; 20 июля
у устья Мархи птенцы оперялись, однако есть сведения о встрече подлетка
уже 7 июля там же, на Вилюе; на п-ове Тайгонрс в южн. части Коряц-
кой Земли полуоперившийся птенец с остатками пуха и короткими крыльями
и хвостом добыт 4 августа. Летные молодые, вполне уже развитые, добывались
в середине августа. Можно таким образом полагать, что брачный период и цикл
размножения проходят, как у тундрового подвида. То же можно сказать о
линьке — как в отношении последовательности, постоянной для вида вообще,
так и о сроках (около середины июля птицы сменяют средние первостепенные
маховые — 6-е и 7-е, сильная линька идет в конце августа — сентябре).
Питание. В связи с распространением, в частности — с
биотопическим размещением, в кормовом режиме якутского сокола особенное место
принадлежит разным водным птицам — уткам и куликам. В частности, из уток
отмечены шилохвости, свиязи, чирки (Якутск, Олекминск, Коряцкая Земля,
Охотское побережье), чистиковые и чайки птичьих базаров (п-ов Тай-
гонос), плавунчики,' песочники, ржанки (Охотское побережье). Вероятно
к этому же подвиду относятся соколы, нападающие на Камчатке на качурку
Oceanodroma fareata и на золотистую ржанку Charadrius dominicus, охотящиеся
на южном Сахалине в октябре — ноябре за утками, темным дроздом, черной
и большеголовой воронами, молодой серебристой чайкой, чернохвостой чайкой.
Относительно зимующих в Индии, а также в вост. Китае соколов, часть из
которых по всей вероятности относится к якутским, известно
преимущественное питание утками. Кормовые повадки, способы нападения на добычу и т. д. —
как у других подвидов сокола (см. описание тундровой формы). Однако у
молодых якутских соколов в гнезде на Вилюе найдены «мыши» (Маак, 1886).
Описание. Размеры и строение. Якутские соколы — крупная форма, сходная
по величине с тундровой. Крыло самцов (17) 296—325, самок (12) 350—372, в среднем 312,9
и 360,9 мм.
92
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Окраска очень темная, насыщенная и яркая, напоминающая американских соколов
F. p. anatum и в то же время представляющая как бы переход к еще более темнымF. р. ре-
alei. Птицы в первом годовом (гнездовом) наряде сверху густо черновато-буры, в свежем пере
аспидно-черноваты, темя обычно бурое, часто одноцветное, реже со светлыми каемками;
темный продольный рисунок низа не очень широкий, но густого бурого цвета, от европейских
соколов отличается охристым, но не рыжим, цветом светлых каемок перьев. Во втором
годовом и последующих нарядах голова и верхняя часть спины — шиферно-серовато-черноватые;
нижняя часть спины и кроющие крыла сизые с темнобурым поперечным рисунком, темней,
чем у тундровых соколов, но посветлей, чем у среднерусских F. p. breuirostrus; вершины
рулевых обычно черноваты; усы черные, длинные и широкие, но щеки посветлей, чему
европейских соколов, приближаясь в этом отношении к тундровой расе; окраска низа в свежем
пере у самок нередко рыжевата на груди, с сероватыми боками; у самцов сероватый налет
на нижней стороне тела, особенно на боках, голенях, подхвостье очень развит; темные
пестрины низа — продольные у самок на груди, поперечные у самок и самцов на боках,
голени, подхвостье — густо черноватого тона, более широкие и правильные, чем у
тундровых соколов. Эти птицы очень сходны с американскими F. p. anatum и повидимому
идентичны с ними. Последний вопрос требует еще выяснения.
Алеутский или черный сокол Falco peregrinus pealei Ridgway
Falco communis var. Pealei. Ridgway. Bull. Essex Inst. V 1873, стр. 201, по зимней птице
из Орегона.
Русское название. «Черный ястреб» (Командорские о-ва).
Распространение. Ареал. Область гнездования весьма ограничена и
расположена на островах северной части Тихого океана примерно между 50—
55° с. ш.: Алеутских, Королевы Шарлотты и Командорских (Беринга и Медном).
На Курильских о-вах встречаются будто бы соколы переходные между
сибирскими («calidus») и черными (Ямасина, 1931). Во внегнездовое время
попадается на побережье Сев. Америки от Британской Колумбии (р. Скина)
и Калифорнии до Орегона, делает залеты в Японию (Хакодате), в
Манчжурию (?), в Приморье и на Камчатку; в Приморье по всей вероятности
регулярно, отдельные особи там попадаются и зимой.
Характер пребывания. Оседлый, часть особей кочует.
Биотоп. Скалистые морские побережья, часто у птичьих базаров.
Численность. В общем малочисленен, вследствие небольшой
области распространения, но внутри ее нередок. На о-ве Королевы Шарлотты в с.-з.
углу — 13 гнезд, на о-ве Лонсара той же группы по паре на каждые 20 кв. миль;
иногда гнездо от гнезда только в 2 милях (Грин, 1916).
Экология. Сведения отрывочны.
Размножение. Даты неясны. Молодые со следами пуха и
недоразвитыми маховыми и рулевыми добыты на о-ве Медном довольно рано — 30 июня.
Это позволяет предположить, что кладка происходит в первой трети мая,
окончательный вылет из гнезда можно отнести к середине июля (считая гнездо-
вый период в 35, а насиживанье в 28 дней, как у других соколов).
Следовательно, брачный период начинается раньше, чем' у тундровой и якутской расы.
Гнезда алеутского сокола — примитивны, помещаются на скалах (иногда легко
доступных), а нередко и среди птичьих колоний — гаг (Тернер, 1886),
поморников и беринговых бакланов, причем гнездо бакланов может находиться в
1,2 м от соколиного. В кладке 4 яйца, и в этом — сходство с северными
подвидами. Размеры (34) яиц в среднем 53,3x41,4, предельные 58x43; 48,5х
Х41; 52,9 х 39 мм (Грин, 1916). Отмечены повторные кладки: при этом,
если отнять у птиц кладки до начала насиживания, вторая кладка произойдет
в течение 10дней; если же насиживание началось, то только через 3 недели.
Одиночные молодые из разбившихся уже выводков наблюдались в августе
(Командорские о-ва, Иогансен, 1934).
Питание. Питается алеутский сокол почти исключительно морскими
птицами, причем за ними сокол летает довольно далеко от берегов (например,
сокол
93
имеются наблюдения, как черный сокол поймал песочника Calidris minutilla
в 50 милях от берега) и поедает на лету. Отмечается нападение этого сокола
на белых куропаток, на поганку, чайку, уток, среди которых попадаются
шилохвость и синьга, разных куликов, но главным образом на чистиков
—топорика Lunda cirrhata, кайр, люриков — старика Synthliboramphus antiquus,
пестрого рыжика Brachyramphus marmoratus и др. В Сев. Америке в качестве
пищи этого сокола указываются главным образом чистики Synthliboramphus
antiquus, Ptychoramphus aleuticus, Cerorhinca monocerata, Brachyramphus
marmoratus, также кулики, реже белые куропатки (Бонд, 1946). Местами соколы
кормятся едва ли не исключительно гнездящимися в колониях стариками
(с.-з. часть о-ва Королевы Шарлотты). В Приморье у пойманного алеутского
сокола в желудке была найдена конюга; наблюдалось нападение сокола на
оляпку. У берегов Командорских о-вов и Камчатки осенью соколы кормятся
мелкими чистиками, например стариками, а также утка,ми и глупышами.
Описание. Размеры и строение. Алеутский сокол— самый крупный из соколов:
крыло самцов (5) 318—322, самок (7) 365—387 мм, в среднем 326 и 376,85 мм. Хвост
относительно длинней, чем у других рас: отношение длины среднего пальца к длине хвоста 0,320—
0,263, в среднем 0,287, тогда как у тундровых и среднерусских соколов 0,345—0,290, в
среднем 0,318 (Штегман, 1929). Вес молодой самки (1) 1179 г.
Окраска. Самый темный подвид сокола нашей фауны. В первом (гнездовом) наряде
верхняя сторона почти одноцветно темнобурая, светлых каемок перьев нет или они чуть
заметны; нижняя сторона также бурая с белым подбородком и горлом и беловатыми краями
перьев; на боках — беловатые поперечные пестрины. В окончательном наряде сверху
похожи на якутских соколов, с черноватой головой и межлопаточной областью, снизу —
беловатые, без рыжеватого оттенка, с очень грубым и густым темным рисунком, продольным на
зобу и груди, поперечным на боках, голенях, подхвостье. Самцы несколько светлей самок.
Сахалинский или малый сокол Falco peregrinus pleskei D e m.
Falco peregrinus pleskei. Dementiev. Falco 1933, стр. 13, о. Б. Шантар.
Распространение. Ареал. Сахалинский сокол найден в северных
частях Тихого океана и в Южн. Китае, где в разное время были добыты
несколько соколов необычайно малых размеров. В гнездовую пору такие птицы
найдены на Сахалине. Точные местонахождения таковы: о. Б. Шантар, р. Як-
шина; устье р. Коль, Сахалинский залив; бухта Ольги — Камчатка; Охотское
море в 150 км от Сахалина (Бутурлин, 1911); Сахалин, Кайбато и Кайтиото
(Такахаси, 1938; Курода, 1935), Гуань-Чжоу-ван, южная часть провинции
Гуань-Дун, ю.-в. Китай (Жабуйль, 1935). Эти мелкие соколы гнездятся пови-
димому на Сахалине и возможно на Шантарских о-вах.
Характер пребывания. Судя по находкам в Гуань-Дуне, по
крайней мере часть этих птиц находится на зимовке в ю.-в. Азии. Однако на
Сахалине соколы встречены и в ноябре (Мунстеръельм, 1921), так что там
птицы может быть и оседлы (в данном случае наблюдения могут относиться
и к особям северных пролетных или зимующих подвидов).
Биотоп. На Сахалине и Шантарских о-вах, на Камчатке, на Охотском
побережье эти соколы встречены на скалистых берегах.
Численность. Численность сахалинского сокола точно неизвестна,
но во всяком случае это редкая птица.
Экология. Сведений, определенно относящихся к этому подвиду, почти
нет.
Питание. В гнездовое время на Сахалине наблюдалось нападение
сокола на фифи 16 мая, на самку кряквы 30 мая, в мае — июне на куликов и
уток.
Описание. Р азмеры истроение. Диагностический признак этого подвида—малые
размеры: самки равны самцам северных рас. Длина крыла у приведенных выше
экземпляров (в порядке перечисления): 9313,9312, 9302,9318,9315,Q320,<J305,9311,9323,9315 мм.
94
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Вес самки с Камчатки — только 500 г. Заслуживает внимания и относительно
незначительный размах полового диморфизма; это явление отмечено и для другого мелкого подвида
сокола — южноафриканского F, p. perconfusus. Строение — как у других подвидов.
Окраска — довольно темная, в общем сходна с окраской якутского подвида.
Систематические заметки. Реальность существования подвида F. p. pleskei
вызывает возражения у некоторых авторов. В частности, сомневались в правильности
определения пола перечисленных выше самок и — с зоогеографической точки зрения —
считался невероятным узкий ареал соколиной расы.Однако трудно допустить ошибку в
определении пола, неоднократно повторяемую зоологами: она вообще говоря возможна, но не
очень вероятна. Весьма ограниченные гнездовые ареалы известны и для других подвидов
соколов^, p. pealei(см. выше), F. p. frujtii. Пока, следовательно, для отрицания F. p.
pleskei нет достаточных оснований. Окончательное решение вопроса требует, конечно,
получения дополнительного материала.
Русский сокол Falco peregrinus brevirostris M e n z b i e r
Falco peregrinus brevirostris. Мензбир. Орнитол. География, 1888, стр. 276, Москва.
Синоним. Falco peregrinus riphaeus. Бутурлин. Псов, и Руж. Охота, 1907, №7,
стр. 99 (Урал).
Распространение. Ареал. Русский сокол занимает лесную зону
Европы и Азии на западе, примерно, от средних частей Белоруссии и Прибалтики,
Зап. Украины, на востоке до Тихого океана (низовья Амура и Уссури); с севера
на юг около 60—50° с. ш. Птицы из Зап. Белоруссии и украинские не вполне
типичны и представляют собой как бы переход к германо-скандинавским
соколам. К форме brevirostris относятся осмотренные нами соколы, гнездящиеся
в областях Ленинградской, Новгородской, Калининской, Смоленской,
Московской, Ярославской, Вологодской, Рязанской, Калужской, Тульской,
Курской, Воронежской, Куйбышевской (Жигули), в Приуралье, в Башкирии,
Свердловской обл., у Миаса, в районе Томска, Тюмени, в Салаире и
Кузнецкой степи, на Алтае, в Минусинском крае, в южных частях Красноярского края,
в Прибайкалье (Тунка, окрестности Читы, Александровский завод), в сев.
Монголии и Приморье. На литературные данные положиться нелегко из-за
известной ненадежности подвидовых определений, но вероятно сюда же относятся
соколы из Манчжурии.
Характер пребывания. Часть особей оседла, часть пови-
димому кочует, двигаясь несколько к югу и появляясь тогда в степных и
лесостепных местностях, где не гнездятся (например, на Украине, в низовьях Дона,
в Приволжских степях ю.-з. Сибири и т. п.). Впрочем, разрыва между
гнездовым и зимним ареалом не получается. Вероятно, часть особей, двигаясь на
3. Ю. 3., зимой может проникнуть до средней Европы, но экспериментально
путем кольцевания это не доказано, хотя установлено, что во Франции зимою
бывают соколы из Германии, Скандинавии и Финляндии. Откочевка этих
соколов не носит такого определенного характера, как пролет тундровых и
якутских. Известное движение их осенью замечается уже с августа и до октября:
весною—в марте и.даже в начале апреля:
Биотоп. Этот подвид обитает в лесной зоне и гнездится главным
образом на высоких деревьях, но для охоты, как и все настоящие соколы, нуждается
в открытых пространствах. Однако в тех районах, где есть скалы, например,
на Урале, в Забайкалье, в Салаире и Алтае, — он держится в скалистом
ландшафте. В зимнее время, с обеднением лесной фауны, происходит перемещение
большинства соколов в другие биотопы, связанные с культурным ландшафтом,
например в города или области, где долго держится водяная птица.
Численность. Русский сокол немногочисленная птица. Однако
местами, например, в предгорьях Салаира, в Салтымановском хребте на
р. Томь, у Красноярска — нередок. Для Салаира отмечены пары на расстоянии
4—5 км одна от другой, в Салтымановском хребте даже в 2—3 км. Для Жигуг
лей на Волге приводятся 3 пары на протяжении 50—55 км; для Ильменских
сокол
95
гор, Южн. Урал, 2 пары при плотности пара на 200 км2 (Ушков). Число
соколов более или менее стабильно (если не считать изменений его под влиянием
антропического фактора).
Экология. По сравнению с северными подвидами — якутскими и
тундровыми — существенные различия. Русские соколы оседлы и полуоседлы,
северные — перелетны. С другой стороны, северные соколы тундр весь год
пребывают в экологически-сходном (конечно, не идентичном географически)
биотопе, а формы лесной зоны в холодное время года меняют биотоп,
передвигаясь в культурный ландшафт. Различное отношение к температуре:
тундровые соколы круглый год проводят в условиях средних положительных
температур (летом примерно +8, +12° С, зимой от +4° до +8°), годовая амплитуда
8?, следовательно стенотермны. Русские соколы в гнездовое время держатся
в зоне с температурой в среднем +16° или +20°, зимой там же при температуре
от —12° до —20°, годовая амплитуда до 40°, птица явно эвритермная. Годовые
циклы проходят иначе: размножение, а следовательно и линька, начинаются
на месяц раньше, чем у северных. Тем самым, между русскими и тундровыми
соколами создается физиологическая изоляция, развитие половых желез у них
идет несинхронно, и спаривание тундровых птиц с русскими соколами,
пролетающими на север через их гнездовую область, невозможно. В апреле, когда
обе популяции могут быть встречены в одной области, у тундровых соколов
гонады не развиты, а у русских — сезонное развитие половой системы достигло
высшей точки, и птицы готовы к откладке яиц. Есть различия и в выборе мест
для гнезд. Плодовитость у северных соколов более высокая. Различен
кормовой режим и его сезонные изменения, характер трофических связей. Наконец,
есть основания предполагать наличие глубоких физиологических особенностей
(конституциональных), особенностей обмена окислительных процессов и т. п.
Размножение. Сходны с другими подвидами соколов следующие
элементы цикла: образование постоянных пар, брачные церемонии (полет),
продолжительность насиживания (28—29 дней), начало насиживания с
откладки первого яйца, степень участия полов в насиживании, число и расположение
наседлых пятен, разделение функций в заботе о потомстве, продолжительность
постэмбрионального развития и роста птенцов (около 35 дней), наступление
половой зрелости в возрасте около одного года; соотношение между
размножением и линькой (смена крупного пера, в частности 7-го и 6-го первостепенного
маховых, совпадает с получением птенцами второго пухового наряда).
Брачный полет в средней полосе России происходит вскоре после
появления птиц у гнезд, около середины апреля. Пары, как говорилось, постоянные,
гнездовые участки занимаются в течение долгих лет; в пределах участка обычно
бывает более одного гнезда, принадлежащего одной и той же паре. В Москве
на колокольне Симонова монастыря соколы гнездились по крайней мере
с начала XIX столетия. В Погонно-Лосином Острове под Москвой гнездо-
ванье соколов известно очень давно, при этом с 1925 по 1934 г. соколы
гнездились в одном гнезде, а после гибели дерева — отняли гнездо у ворона в 8 км
от первого, где и стали ежегодно гнездиться.
Кроме гнездящихся птиц, повидимому, всегда имеется небольшое число
холостых, главным образом, молодых, прошлогоднего выводка. Эти птицы
быстро замещают погибших по тем или иным причинам соколов. Наличие этих
птиц показывает, что и молодые соколы возвращаются в общем после первой
зимы в район гнездовья и рождения. В выводках отношение полов различно:
например, у Красноярска в одном выводке 3 самца и 1 самка, в другом — 1
самец и 2 самки. При двух птенцах в выводке чаще бывают 1 самец и 1 самка.
Гнезда — на деревьях (обычно на больших соснах, высоко от земли) и на
скалах, где таковые имеются; в горных местностях (Алтай, Салаир, Урал)
определенное предпочтение оказывается скалам. Собственно гнезд соколы не строят,
96
ОТРЯД ХИЩНЫЕ ПТИЦЫ
и гнезда на деревьях отнимаются у других птиц, чаще всего у ворона. Нередко
гнезда располагаются близ колоний разных птиц — грачей, цапель, галок.
В качестве редкого случая отмечено гнездование на земле на моховых болотах
(Прибалтика, Смоленская и Рязанская обл. и др.) и на высоких зданиях в
городах (Троицкая башня московского Кремля; Псков; Калуга). Гнездование
в дуплах невероятно. Число яиц в кладке 2—4, чаще всего 3. Кладка
происходит в половине апреля — начале мая (в окрестностях Москвы), у
Красноярска — 8 мая неполная кладка из 2 яиц, 12 мая свежая полная кладка,
однако в Ильменских горах между 16—25 апреля полная. Отмечены случаи
эмбриональной гибели (у Красноярска Юдин 12 августа нашел в гнезде
летного птенца задохлика в яйце и неоплодотворенное яйцо), а иногда по
невыясненным причинам —гибнут птенцы. Размеры яиц (5) 51,0—52,1 х 41,0—42Д
в среднем 51,46 х 41,46 мм; вес (5) 49,65—49,7 г, в среднем 49,68 г.
Птенцы проклевываются в первую половину июня (Москва — около 1 июня,
Салаир — 7 июня, 18 июня —Алтай, в Ильменских горах будто бы около
середины мая). Второй пуховой наряд надевают в конце июня, через 10—12 дней после
вывода (Москва, 11—18 июня, Красноярск — 22 июня). Число птенцов в выводке
обычно меньше, чем количество яиц в кладке. В гнезде в Погонно-Лосином
Острове, Москва, отмечались в 1938 г. — 1, в 1939 г. —2, в 1940 г. — 1, в
1941 г.—2 птенца, во всех случаях при кладке в 3 яйца. Для Южн. Урала
отмечено 2—3, чаще 2 птенца, для Алтая 3, для Красноярска 1—4, обычно 3 птенца,
для Забайкалья 2, для Уссури 2—3. Обычно у русских соколов бывает 2 птенца
(хотя Сушкин, 1897, писал, что видел в равнинной части б. Уфимской губ.
два выводка соколов, в которых было до 5 птенцов). Летными птенцы
становятся обычно во второй половине июля. Таковы наблюдения в Москве (Дементьев),
в южн. Зауралье (Снегиревский), у Телецкого озера (Сушкин, 1938), у
Красноярска (Юдин), для Приамурья (разные числа июля, Шульпин, 1936 и др.).
Для южн. Забайкалья наблюдения над летным выводком приводятся в начале
августа, но не ясно, действительно ли это были даты вылета. Впрочем, повиди-
мому, есть основания считать, что в различные годы бывает у одной и той же
пары соколов разница в сроке вывода птенцов дней в 10. Выводки держатся
вместе еще в августе.
Линька. Общее течение линьки то же, что и у других подвидов.
Начинается она в июне, кончается в октябре.
Питание. Как и другие подвиды русские соколы питаются главным
образом птицами средней величины, изредка млекопитающими (например, Micro-
tus ungurensis в ю.-в. Забайкалье). У них, в отличие от тундрового сокола,
кормовой режим довольно значительно меняется по сезонам. Объясняется это тем,
что зимние откочевки многих видов птиц умеренной зоны нарушают для
оседлого или полуоседлого сокола его весенне-летние кормовые связи. В общем,
в период размножения в питании соколов преобладают птицы водные, лесные,
горные, в холодное время года — птицы культурного ландшафта. Отмечена
и резко выраженная индивидуальная специализация. В окрестностях Москвы
в Погонно-Лосином Острове соколы одной пары из года в год кормились
главным образом обыкновенными чайками из большой колонии их на озере Киево.
Кормовой режим их за месяц в 1941 г. следующий: обыкновенная чайка — 54,
грач —5, домашний голубь — 2, скворец — 2, чибис — 2, сизоворонка, сорока,
кукушка, вальдшнеп — 1. В Серебряном Бору под Москвой гнездившиеся там
до 1924 г. соколы кормились серыми воронами, грачами, обыкновенными
чайками, скворцами, изредка голубями. Гнездящиеся близ грачевника у г.
Алексина в Тульской обл. соколы кормились главным образом грачами, кроме
того в гнезде найдены остатки двух вальдшнепов, чирка-трескунка и
пустельги. Для горных частей Южн. Урала в гнездовое время в питании соколов
отмечены утки, кулики, крачки, чайки, в равнинном Приуралье галки, вороны,
сокол
97
голуби, сойки, рябчики. Гнездящиеся в Жигулях соколы питаются главным
образом чайками. Нападают соколы и на мелких птиц, в особенности, когда
9 птенцы малы. Зимней пищей служат в культурном ландшафте голуби, галки,
' вороны (Москва, Ленинград и т. д.), а также в местах пролета и у
незамерзающих водоемов разные утки и кулики.
Описание. Размеры и строение — как у тундровых птиц. Крыло самцов
(23) 294—334, самок (21) 343—375, в среднем 313 и 356,6 мм.
Окраска. Русские соколы — форма темноокрашенная. От тундрового подвида
хорошо отличается более темным общим цветом верхней стороны тела и большим развитием
черноватого рисунка у взрослых птиц. Более темным общим бурым тоном верхней стороны
тела, меньшими размерами на ней светлых каемок перьев, а также более широкими и
темными продольными полосами на нижней стороне тела — у птиц в первом годовом наряде.
Окраска щек темнее, усы сливаются с темными кроющими уха и подглазными полосами. По
сравнению с якутскими соколами — менее ярки, окраска головы и верхней части спины
буровато-аспидная, менее черновата; светлосизые партии менее чистого тона, обычно с
буроватым оттенком; рисунок на нижней стороне тела менее яркий и грубый, но тоньше; это —
у взрослых. При сравнении молодых птиц русские соколы отличаются от якутских более
узким, менее «мазаным» темным рисунком низа и более широкими усами и полосами под
глазами. Общий тон окраски верхней стороны тела русских соколов в окончательном наряде
темный шиферно-серый, голова и верхняя часть спины темней остальных частей; сизый
поперечный рисунок на спине и крыльях с буроватым оттенком; у самок в этом возрасте грудь
всегда несет продольные штрихи или черточки, чего нет у тундровых птиц (за исключением
личных отклонений, а также второго годового наряда).
Второй наряд у русских соколов нередко резко меланистичен (cornicum у Мензбира,
1882): черноватый, с малым развитием сизых тонов сверху, снизу ярко рыжеватый на зобе
и груди, грубо и ярко испещренный, причем черные полосы в общем занимают такое же
пространство как и светлый фон. Значительно развита, как и у других подвидов, личная
изменчивость, выражающаяся как в развитии поперечного рисунка на верхней и нижней стороне
тела, так и в более или менее светлой или темной общей окраске. Молодые птицы обычно
с темной головой, с рыжеватыми (не охристыми, как у тундровых) светлыми каемками темно-
бурых перьев спины и крыльев, с продольным бурым рисунком низа, занимающим во всяком
случае больше места, чем светлые края перьев.
Следует отметить, что в Сибири у Томска, в Кузнецкой степи, по Енисею попадаются
особи и более или менее типичной для F. p. brevirostris окраски, и другие, более светлые,
с более светлым серо-сизым тоном верха и более тонким черноватым рисунком низа,
приближающиеся в этом отношении к тундровым. Среди соколов существуют, повидимому, кроме
географических и параллельных им личных изменений окраски, также и групповые, в
пределах отдельных замкнутых популяций. Так например, все осмотренные нами соколы из
среднего и южного Приуралья очень темны (им было дано название F, p. riphaeus; такие
примеры известны и для других стран, например, светлые соколы с о-ва Ленди у зап.
Англии).
Кавказский сокол Falco peregrinus caucasicus Kleinschmidt
Falco peregrinus caucasicus. Kleinschmidt. Falco HI, 1907, стр. 62, Владикавказ.
Синоним. Falco peregrinator atriceps Hume у Мензбира, 1916.
Распространение. Ареал. Гнездится кавказский сокол на южном
побережье Крыма (между Балаклавой и Феодосией) и на Кавказе, откуда
имеются добытые в разное время года экземпляры из Махач-Кала, Дзауджикау,
Кизляра, Киши, из Хосты, Батуми, р. Чорох, с озера Гиллии Севан в
Армении и т. д. Залетный (повидимому, молодой) самец добыт 17.11. 1911уАска-
ния-Нова.
Характер пребывания: оседлая птица, во внегнездовое время
и за добычей спускающаяся на равину, но некоторые особи и зимуют в горах.
Биотоп. Гнездится в нижнем и среднем поясе гор, в Армении около 1800 м9
в Сванетии однако до 3355 м.
Численность. Кавказский сокол встречается редко.
Экология. Сведения совершенно недостаточны.
Размножение. Гнезда на скалах, весьма постоянные. Брачный
крик и полет до апреля. Старые птицы у гнезд появились в Закатальском
округе уже в конце февраля. Птенцы вполне оперены и летают уже в конце мая,
7 Птицы СССР, т. I
98
ОТРЯД ХИЩНЫЕ ПТИЦЫ
что позволяет предположить, что они выводятся в конце апреля и что кладка
падает на конец марта *. В выводке, вынутом из гнезда в Закатальском округе,
было 3 птенца, 2 самца и 1 самка. Чаще, повидимому, выводок из 2 птенцов.
Неразбившиеся выводки наблюдались еще в августе.
Линька. В соответствии с ранним циклом размножения линька также
начинается рано. У самки с большими наседными пятнами, добытой у
Балаклавы 8 мая, начали сменяться первостепенные маховые (7-е перья свежие).
Линька, следовательно, кончается у взрослых птиц к октябрю.
Питание. В качестве добычи кавказских соколов называются дикие
голуби, галки, в зимнее время утки; отмечены нападения весною на домашних
голубей, а также на дикого гуся (7 марта 1901, Алчайское ущелье).
Описание. Размеры и строение. По величине кавказский сокол уступает
соколам северной группы: длина крыла самцов (8) 288—312, самок (13) 320—355, в среднем
294,9 и 335,9 мм.
Окраска кавказского сокола очень насыщенная и яркая, со значительным
развитием рыжеватых тонов в виде пестрин на затылке и на нижней стороне тела. Взрослые
птицы темные, с черноватой головой с узким светлым сизым поперечным рисунком на спине
и крыльях; на затылке и задней стороне шеи рыжеватые пестрины; иногда в свежем пере
рыжеватые края мелких перьев спины и крыльев (как у пустынной туркестанской формы),
грудь рыжеватая, бока сероватые, темные полосы на них расположены густо. В первом
годовом наряде — темные и яркие, обычно с сильно развитыми рыжими каймами перьев, часто
с поперечными сизоватыми или рыжеватыми поперечными пятнами на плечевых, больших
кроющих крыла, рулевых, нередко с сизым оттенком оперения (так сказать «прогрессивный»
тип окраски, несколько приближающийся к окончательному наряду и часто встречающийся—
особенно у самцов — у пустынных среднеазиатских соколов; эта окраска названа Бремом
Старшим и Клейншмидтом «paradoxus»); на затылке наствольные пятна; нижняя сторона
в свежем пере также сильно рыжеватая и с широким бурым рисунком (на боках нередко
принимающим, особенно у самцов, поперечный характер и тоже относящийся к «прогрессивному»
типу «paradoxus»). Личная изменчивость у кавказских соколов вообще довольно
значительна, хотя от'других форм вост. Палеарктики они отличаются резко.
Систематические замечания. Кавказский сапсан очень близок и быть может
идентичен со средиземноморским подвидом F. p. brookei, с которым сходен и по величине, и по
яркой, окраске; эти формы представляют собой в известной мере как бы переход от серых
северных крупных подвидов к рыжевато-серым мелким пустынным подвидам юга (F. p. pelt-
grinoides, F. p. babylonicus).
Рыжеголовый или пустынный сокол, шахин Falco peregrinus babylonicus S с 1 a t e r
Falco babylonicus! Sclater. The Ibis. 1861, стр. 218, табл. VII, Уд, в Индии, зимующая птица.
Синонимы. Falco tscherniaievi Северцов, Верт. и Гор. Распр. Турк. Жив.,
1873, стр. 114, Аулье-Ата — Falco peregrinus gobicus Stegmann. Journ. i., 1934, стр. 235,
Лукчун, вост. Туркестан.
Местные названия. «Лачин» у туркестанских сокольников, «шахин» в Индии
и в Иране.
Распространение. Ареал. Этот подвид сокола распространен в
Средней и Центральной Азии от монгольского Алтая (бассейн Кобдо, к западу от
озера Орок-нор), Са-Чжоу в южн. Гоби, быть может юго-вост. частей русского
Алтая, вост. Туркестана (от Хами и Турфана до Кашгара); в Белуджистане
{Келат), в Афганистане (Кандагар); на западе в Иране в восточных и
центральных частях (Хорассан, Керман и Кугистан, Зарудный, 1911), на запад до Заг-
роша, но видимо не в Каспийских его провинциях, где пока не найден; в
Туркмении до зап. Копет-Дага (Сонгу-Даг, Кызыл-Арват), Бадхыза (Кзыл-Джар,
Гязь-гедык, Фисташковый хребет у Кушки) и Кугитанга, в Таджикистане
(Бабатаг, Гиссар), в Узбекистане (Нура-Тау), на север до Кызыл-Кумов
на западном берегу Аральского моря (Уч-Коку, Зарудный, 1915), Таласского
Ала-Тау и его западного продолжения — хребта Казгурт (Долгушин); на
с.-в. до Александровского хребта (Ала-Медин; окрестности Фрунзе) и Цен-
* С другой стороны, подлеток с не вполне доросшими крыльями и хвостом был добыт
в Крыму у Судака только 15 июля.
Рыжеголовый сокол
сокол
99
трального Тянь-Шаня (Нарын-кол, Пржевальск, Тургень). Кроме того, зимою
встречаются в сев. Индии, к югу до Нарбадда, на восток до Уда, а по данным
некоторых авторов гнездится в Кашмире и в горах с.-з. Пенджаба*. Залеты
на Памир,
Характер пребывания. Часть особей оседла, кроме, повиди-
мому, самых северных частей ареала (Тянь-Шань), где на зиму остаются лишь
одиночные экземпляры. Заметный пролет одновременно с белобрюхими
рябками в Туркмении в конце октября — начале ноября. В Пенджабе рыжеголовые
соколы появляются очень рано (29 августа, 14 сентября и остаются до
февраля, Уистлер, 1922).
Биотоп. Гнездовый — горы или обрывы холмов в пустынных
местностях, зимой на равнинах (примерно с августа до марта; впрочем, кормовые
вылеты взрослых птиц на равнины бывают и в летнее время).
Численность. Птица малочисленная, встречается редко.
Экология. Сведения весьма отрывочны.
Размножение. Гнездовые участки, размер которых трудно устано-
вить, так как птица очень редкая, весьма постоянны (з зап. Копет-Даге, к югу от
Кызыл-Арвата, гнездовье соколов в одном и том же месте было обнаружено и
в 1916 и в 1942 гг.). Половая зрелость, как у других соколов, наступает еще
в первом годовом наряде. Спаривание и брачные игры происходят в самом
начале апреля. Гнезда помещаются в нишах, на карнизах скал, в стенах
глинистых обрывов и обычно трудно доступны. Кладка начинается в первой трети
мая (может быть с конца апреля), из 2—3 яиц. Число птенцов в выводке обычно 2,
реже 1 или даже 3. Выводки не разбиваются еще в августе, хотя в это время
вылетают уже на равнины и отдаляются от мест гнездовья. Около середины
июня птенцы начинают оперяться (24 июня в гнезде у обрывов Уч-Коку,
Аральское море, были найдены 2 полуоперившихся птенца). Летные птенцы
обнаружены около середины июля, а самостоятельные — в августе. Птицы одной пары
держатся вместе круглый год (Дементьев, 1948).
Линька. Последовательность смены нарядов, порядок смены оперения,
общая продолжительность линьки (полной годовой) — как у других подвидов
сокола. Начало линяния взрослых птиц относится примерно к концу июня,
конец же его — к концу ноября — началу декабря. У птиц в первом годовом
наряде отдельные перья начинают сменяться уже осенью на первом году жизни,
в середине мая мелкое перо сменяется уже наполовину, вырастают новые
средние (6-е и 7-е) первостепенные маховые, выпадают средние рулевые. Однако
в дальнейшем в связи с размножением линька этих птиц задерживается, и конец
ее падает примерно на то же время, что и у старых птиц, т. е. на осень. Взрослые
птицы с недоросшим старым 1-м маховым добывались в ноябре.
Питание. В гнездовый период отмечены нападения рыжеголового
сокола на кекликов,. щурок, рябков (центральный Копет-Даг), на сизых
голубей, галок, хохлатых жаворонков (зап. Копет-Даг), на белобрюхих
стрижей (Кызыл-Джар в Бадхызе), воробьев (Кушка); в негнездовое время
в Туркмении соколы питаются белобрюхими рябками, стрепетами, чирками-
свистунками, сизыми голубями, хохлатыми, белокрылыми и полевыми
жаворонками, каменками. В Монголии соколы питаются бульдуруками,
куропатками, чирками; в Иране — кекликами. Есть наблюдения над нападением
рыжеголового сокола поздно вечером на летучих мышей (Ашхабад, Моргунов-
ское на Кушке, Басра и Багдад в Месопотамии).
Описание. Размер ы,и строение. Рыжеголовый сокол — мелкий подвид: крыло
самцов (14) 274—302 (по литературным данным до 310), самок (23) 312—338 мм (по литера-
* Сведения о находках в с.-в. Африке основаны скорее всего на неточном определении
подвида (в Аравии встречен *юка только pelegrinoides).
7*
100
ОТРЯД ХИЩНЫЕ ПТИЦЫ
турным данным до 343); в среднем соответственно 283,7 и 323,1 мм. Сравнительно
короткохвостый и длиннокрылый. Длина самцов (5) 354—365, в среднем 359 мм, размах (5) 835—
883, в среднем 860;6 мм, те же величины для самок (5) 409,6—435; в среднем 421,6 и (5)
960—1005,2, в среднем 987,5 см (Юм, 1887). Вес: молодые самцы из Туркмении — 330 и
350, молодая самка 615 г, взрослая —765 г.
Окраска светлая и бледная. У взрослых птиц на лбу, темени, зашейке перья
красновато-рыжие, верх бледносерый, слегка дымчатый с более темным буроватым поперечным
рисунком, иногда выраженным лишь в межлопаточной области и на плечевых, часто перья
окаймлены рыжеватыми каемками; низ рыжий в разных оттенках со слабым развитием
темного рисунка. Молодые птицы в первом годовом наряде иногда несколько напоминают
тундровых по бледной и светлой окраске, но хорошо отличаются от них меньшими
размерами, обычно сильной рыжеватостью свежего оперения, правильно поперечнополосатым
хвостом, сизым на нем налетом, более узкими продольными полосами на брюшной стороне,
глинистым оттенком перьев спинной стороны, меньшими размерами «усов». Очень часто
(у самцов повидимому всегда) большие перья плеч и крыла несут и в первом наряде
поперечный светлый сизоватоохристый рисунок («paradoxus»), У молодых соколов в первую осень
восковица, кольцо вокруг глаз и ноги — бледные, беловато-желтые, на следующую весну
по достижении половой зрелости становятся яркожелтыми.
2. Балобан Falco cherrug Gray
Falco cherrug Gray, in Hardwicke's Illustr. of Ind. Zool., 1833—1834, табл. 25,
Индия. Русское название. Балобан или балобан — слово, до XVIII столетия не
известное. Происходит от персидского названия этой птицы. Иранские сокольники называют
«балабан» пролетных и зимующих соколов этого вида, а гнездарей называют «шарг» (близко
к индийскому их имени «cherrug», перешедшему и в научную номенклатуру). Народы
Средней Азии называют этого сокола «ительги» или «ительге». Из славянских названий
балобана цадо еще упомянуть о польском «rarog», несомненно заимствованным от венгерского
«raroh».
Распространение. Ареал. Балобан распространен в степной и
лесостепной полосе Вост. Европы и сев. Азии и в Центральной Азии. Гнездовый ареал
от Богемии, Венгрии, Прикарпатья до Балкан, средней полосы Европейской
части СССР, далее в южных частях Зап. и средней Сибири, Забайкалья,
Монголии, к югу до вост. Ирана, Средней Азии, вероятно Джунгарии, Синь-
цзяня, Нань-Шаня, Сычуаня и Тибета до Гималаев. Гнездованье в Палестине
сомнительно. На Балканах встречен на гнездовье лишь в бассейне Дуная —
в Румынии, Добрудже, Болгарии; в других же местах повидимому замещен
F. biarmicus, что имеет место и в Малой Азии. На Кавказе, несмотря на
указания многих авторов, не гнездится. Вне гнездового периода встречен в
небольшом числе в сев. Африке (Египет, Судан, Абиссиния, также в Алжире и Тунисе);
в Алжире будто бы гнездится в небольшом числе (Лош, 1867), но указание это
требует подтверждения; на Балканах, в Малой Азии, на Кавказе, в Иране,
Месопотамии, Палестине, в Афганистане, Белуджистане, сев. Индии (главным
образом в Пенджабе), Китае. Залеты отмечены в средней Европе (Германия,
Франция) и в Средиземноморье.
Характер пребывания. Часть особей не покидает гнездовой
области (кроме северных ее частей) и зимою; некоторые особи (тибетская форма)
местами строго оседлы, другие совершают миграции главным образом
вертикальные; наконец, часть популяций перелетна.
Биотоп. Лиственные и смешанные леса, окруженные открытыми
пространствами или пересеченные ими (речные долины, поля и степь); скалы в
сухих и безлесных или слабо лесистых местностях, в горах (Средняя Азия, Алтай
и т. д.), или в степи (Забайкалье, Монголия); высокогорная степь (Средняя
Азия, Монголия, Тибет). Вне периода размножения балобаны могут быть
встречены и в других ландшафтах, однако сплошных лесных массивов избегают.
Подвиды и варьирующие признаки. В систематическом отношении балобаны весьма
близки к кречетам и в сущности в морфологическом отношении все формы этих соколов от
западных (европейских) балобанов через центральноазиатских (монгольских и тибетских) до
арктических кречетов представляют один непрерывный ряд вариаций окраски и размеров,
БАЛОБАН
101
причем промежуточным звеном между кречетами и балобанами служит алтайский кречет.
Весьма сходны у этих птиц и экологические особенности. Поэтому в сущности было бы вполне
справедливым считать этих соколов за один вид, если бы не тот факт, что ареал
алтайского кречета по крайней мере частично совпадает с ареалом одного из
подвидов балобанов. Можно, таким образом,
видеть в этих двух группах случай,
пограничный между категориями вид и
подвид. Общая схема географической
изменчивости у балобанов сводится к
постепенному и «прогрессивному» развитию
в оперении верхней стороны тела
поперечного светлого рисунка и сизых тонов
(в особенности в надхвостье и на
рулевых), в приобретении взрослыми
птицами черноватого (а не буроватого)
рисунка низа, в развитии поперечного
рисунка на перьях голени, боков, под-
хвостья. Все эти признаки постепенно
(в географическом отношении)
проявляются в направлении с запада на восток,
и обитающие на крайнем востоке
балобаны (монгольские, китайские и
тибетские) в ряде отношений сходны с
северными кречетами. Наоборот, западная
(европейская и западносибирская)группа,
«примитивна», без поперечного рисунка
и сизых тонов. К тому же на западе
возрастной диморфизм—различие в окраске
первого годового (гнездового) и
окончательного (второго годового) нарядов —
невелик, а на востоке — значителен (как
у северных кречетов). Экологически
балобаны устойчивая группа, но известные различия между образом жизни отдельных форм
имеются (менее значительные, чем у F. peregrinus). Балобаны распадаются на следующие формы:
в западной части ареала, к востоку до Волги — F. ch. danubialis Kleinschmidt
1939; к востоку от него в ю.-з. Сибири, Казахстане F. ch. cherrug Gray, 1833—34; в горах
средней Сибири F. ch. saceroides В i a n с h i, 1907; в Средней Азии F. ch. coatsi Demen-
t i e v, 1945; в Забайкалье и МонголииF. ch. milvipesje r d о n, 1871; наконец, на юге —
в Нань-Шане, Тибете, вероятно в Ладаке, на Памире F. ch. hendersoni, Hume, 1871.
Остаются не вполне ясными систематические отношения между собственно балобанами
и индийским лаггаром F. jugger, а также между балобаном и мексиканским соколом F.
mexicanus. Эти птицы, несомненно, крайне близки, но пока, впредь до более детального
морфологического и экологического изучения, их так же как и F. biarmkus, приходится
считать за отдельные виды.
Карта 10. Распространение балабана Falco
cherrug
1 — F. с. danubialis, 2 — F. с. cherrug, 3 — F. с.
saceroides, 4 — F. с coatsi, 5 — F. с milvipes,
6— F. с hendersoni, 7 — F. mexicanus; a — граница
гнездовий, г — область зимовок и кочевок.
102
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Европейский балобан Falco cherrug danubialis Kleinschmidt
Falco hierofalco danubialis. Kleinschmidt. Falco XXXV, 1939, стр. 27, Румыния.
Распространение. Ареал. Этот подвид балобана распространен в Европе
от Богемии до центральных областей Европейской части СССР, в Венгрии,
Бескидах и Карпатах, в прикарпатских частях Зап. Украины, в Болгарии и
Добрудже; у нас в степной и лесостепной полосе на юг до Крыма, на восток до
бассейна Дона (к этой форме относятся балобаны из Воронежской, Пензенской,
Рязанской областей), к северу до южных частей Московской обл., Тульской
"обл., на западе несколько южней—до области Черниговской, Киевской
и Подольской. Зимует в Венгрии, на Балканах, в Крыму, на Кавказе, местами
на Украине, и к этой же форме, повидимому, относятся соколы, проникающие
в холодное время года в Африку.
Характер пребывания. Главным образом кочующий и
перелетный. Оседла, повидимому, крымская популяция. Впрочем, возможно, что
впечатление об оседлости здесь балобанов получается в результате появления
птиц других (более северных) популяций, в то время как местовые откочевывают.
В Венгрии улетает только в декабре, появляясь обратно уже в феврале; в
северной части ареала (Тульская обл.) прилетает во второй трети или начале
последней трети марта; на Украине (Подол ия, Киев) в разные числа марта —
от начала месяца до конца (однако последние даты, как и наблюдения над
появлением балобанов в апреле, могут скорее относиться к пролетным
северным или холостым особям), исчезает в первой трети октября, часть остается до
конца этого месяца.
Такие же примерно сроки отмечены для Харьковской обл., причем первое
появление иногда бывает при снеге и морозах (Сомов, 1897).
Биотоп. Лиственные и смешанные леса гор, предгорий или речных
долин, окруженные открытыми пространствами — степями и т. п. Деревья
необходимы балобану для гнездованья, открытые пространства — для охоты.
При этих условиях балобаны встречаются как в глубине леса, так и на опушках
и окраинах. Балобаны держатся в старых высокоствольных лесах, так как они
гнездятся на высоких деревьях.
Численность. Балобаны местами положительно нередки (у нас
например, в б. Новосильском уезде Тульской обл., в Воронежской обл. в Хре-
новском бору, на Северном Донце между Изюмом и Волчанском, в Кишевском
лесничестве Подольский обл.), но распространение их спорадично, как и
других крупных соколов. Это объясняется, с одной стороны, значительным
постоянством птицы в выборе местообитаний, а с другой — влиянием человека.
Относительно первой можно упомянуть, что в Богемии к северу от Праги балобаны
исчезли (вследствие истребления) в 1843 г., но в 1934 г. опять появились и
стали гнездиться в прежних местах.
Из практики современных соколиных охотников известно, что пойманные
в средней Европе (главным образом Венгрии) в холодное время года балобаны,
ввозившиеся в Германию, в гнездовое время обычно улетали от своих
владельцев на восток, к местам гнездовья (один такой сокол «Мардук», улетевший в
в апреле 1934 г. в Оберштадионв Вюртемберге, в феврале 1935 г. был найден
в Рязанской обл. в 15 км на северо-восток от Михайлова; держался там до
20 апреля, когда погиб).
Мнение, что с последней четверти прошлого столетия и начале XX века
балобан расселялся к северу в европейской России, вытесняя при этом
настоящего сокола, едва ли верно. В лесостепи и в степной полосе, где балобаны
гнездятся, соколы отсутствуют или редки не в силу конкуренции, а за
отсутствием подходящих биотопов. Самое же появление балобанов в местах, где их
прежде не было, в сущности остается точно не установленным. Балобан в зна-
БАЛОБАН
103
чительной мере миофаг, и можно предположить, что число гнездящихся пар
этого сокола и плодовитость их в той или иной местности может колебаться
в зависимости от «урожая» или «неурожая» грызунов. Так, на Северном Донце
с 1927 по 1933г. балобаны были редки, в 1935 и 1936—часты (Рудинский и
Горленко, 1937).
Колебания численности объясняются, вероятно, гнездованием выведшихся
в предшествующем подъему году молодых балобанов, в иных обстоятельствах
остающихся холостыми или гибнущими, а также относительно высокой
плодовитостью этих птиц в благоприятные по кормовым условиям годы. Наличие
холостых молодых особей, потенциально половозрелых, следует из того, что
в случае гибели в начале периода размножения одного из соколов в паре, он
замещается довольно быстро другой птицей и при этом обычно прошлогоднего
вывода, еще в первом годовом (гнездовом) наряде. Впрочем, этот вопрос
требует дальнейшего изучения.
Экология. Размножение. Половозрел ость, как указано, наступает
в возрасте несколько менее года. Пары, повидимому, постоянные, хотя вне
брачного периода самец и самка держатся на некотором расстоянии друг от
друга и охотятся врозь.
Однако весной в гнездовой области птицы в паре держатся уже с прилета
и уже в начале марта находятся в гнездовом участке, поправляя гнездо
в конце этого месяца — в первой трети апреля. В это же время происходят
и брачные игры (полет), птицы нередко присаживаются на гнездо, происходит
символический» принос гнездовых материалов.
Гнездовые участки и места гнездовья весьма постоянны и занимаются из
года в год, хотя у одной пары нередко бывает 2—3 гнезда, расположенных одно
близ другого, занимаемых поочередно. Расположены гнезда на деревьях (сосна,
дуб, ясень, старая ольха, береза, обычно высоко, в 15—20 м от земли), или —
что очень редко — на скалах (Богемия).
Собственные гнезда сокол повидимому строит редко и занимает чаще
всего готовое гнездо ворона, причем птенцы последнего выбрасываются
(Тульская обл.), или других хищных птиц, иногда грачей или цапель. Само гнездо
походит на воронье: это грубая постройка из сучьев около 70—75 см в
диаметре с неглубоким лотком, без подстилки, или со скудной подстилкой из
тонких веточек или сухой травы. Нередко гнезда балобанов находили в
колониях серых цапель.
Кладка имеет место в апреле, от начала до середины месяца, в редких
случаях уже в последней трети марта (22—27 марта, в Киевской обл. по Гебелю,
1879). Число яиц в кладке 3—5 (редко) чаще, повидимому, 4 (редко 6). Птенцов
из яиц выводится меньше, обычно от 1 до 3, но изредка бывает 4 и даже 5 (Огнев
и Воробьев, 1923). Яйца обычного соколиного типа, овально-закругленные, с
матовой мелкозернистой скорлупой с шероховатой поверхностью. Фон окраски
охристый или бледнобуроватый, но он мало проступает, так как за редкими
исключениями прикрыт темными пятнами различной формы и оттенка; цвет этих
пятен варьирует от красноватого или фиолетово-бурого до рыжевато-бурого;
иногда имеются черные или черноватые штрихи. Длина яиц: 47,5—60, изредка
62, ширина 39,6,— 45, изредка 36 и 48 мм (Сомов, 1896, Гебель, 1879,
Рудницкий и Горленко, 1937).
Насиживание начинается с первого яйца, так как в одном гнезде птенцы
бывают разновозрастными. Молодые вылупляются около середины мая и,
следовательно, насиживание продолжается около месяца (вернее 28 дней).
Насиживают оба родителя, но главным образом самка, слетающая с гнезда только утром,
около 12 часов дня и под вечер. В это время ее сменяет самец. В случае гибели
кладки она заменяется новой примерно в месячный срок. К концу мая птенцы
сменяют первый пуховой наряд на второй и начинают оперяться; к середине
104
ОТРЯД ХИЩНЫЕ ПТИЦЫ
июня перьями покрыто уже все тело, остатки пуха заметны на голове и горле,
маховые и рулевые короткие. К концу июня птенцы начинают вылезать из
гнезда на соседние с ним ветви, часто машут крыльями и встряхиваются (Суш-
кин, 1894). Летные птенцы встречены в начале июля. Гнездовый период длится
таким образом 40—45 дней.
Старые птицы и выводки держатся у гнезд довольно долго, во всяком
случае весь июль и август, причем старики еще приносят и передают молодым
добычу, хотя последние уже ловят
и сами. Осенью у гнезд отмечены
игры и крики стариков («осеннее
токование», Гавриленко, 1929).
Линька протекает так же,
как у кречетов. Когда половой
цикл затихает и в гнезде находятся
птенцы во втором пуховом наряде,
начинается смена первостепенных
маховых. Начинается она со смены
средних перьев — 7-го и б-го и
кончается передними — 2-м, 1-м и 10-м.
Рулевые линяют центробежно, но
вторая с краю пара сменяется
последней. В общем, линька
начинается во второй трети мая,
заканчиваясь во второй половине
сентября, когда первое и второр
маховые достигают полного развития.
Продолжительность линьки,
следовательно, около 5 месяцев, линька
полная годовая.
Последовательность смены нарядов —как у
обыкновенного сокола. Часто
встречающееся в литературе утверждение,
что балобаны надевают
окончательный наряд после нескольких
годовых линек, неверно: окончательный
наряд —второй годовой, надевается
в возрасте около 17 месяцев после
первой годовой линьки.
Питание. Балобан менее
специализирован в отношении
питания — способов нападения и кор-
Рис. 39. Балобан (взрослая и молодая птица), мового режима, — чем кречет и в
особенности настоящий сокол (с
этим связаны и некоторые особенности скелета, определяющие летные
качества). Мелкие звери занимают в питании балобана значительное место,
и он нередко берет добычу с земли. Впрочем, на земле балобан не хватает добычи,
как ястреб или сарыч, но, по наблюдениям соколиных охотников, бьет ее
«по-соколиному» (кроме, вероятно, мелких зверьков). На птиц балобан нападает
обычным соколиным способом—«ставкой». Кормовые остатки, погадки и т. д. -—
как у сокола и кречета. Охотничий участок, по крайней мере в гнездовый период,
велик — до 20 км от гнезда. Часы охоты — утро и под вечер, около полудня
птица обычно отдыхает. Пищей балобану служат степные и лесостепные
млекопитающие — главным образом суслики, хомяки, полевки и мыши, затем птиць*
средней величины: галки, грачи, вороны, голуби, сизоворонки, на пролете —
БАЛОБАН
105
утки. В период выкармливания молодых, пока птенцы малы, самец ловит много
мелких птиц, главным образом жаворонков.
Самая крупная добыча балобана в природных условиях — серая цапля.
Впрочем, охотничьи балобаны приучаются брать и гусей, дроф, стрепетов,
черных коршунов, зайцев и газелей. В период пролета и ранней весной балобан
кормится главным образом птицами. По некоторым отдельным сведениям
кормом служат главным образом суслики, иногда голуби, хомяки, полевки, мыши,
полевой жаворонок, галки, кваква, вяхирь, горлица, сойка, сизоворонка,
грач, полевки и водяные крысы.
Полевые признаки. Балобан сходен с кречетом, но кажется менее
массивным как на полете, так и при сиденьи. На расстоянии молодые кажутся
однообразно бурыми, старые — бурыми сверху, беловатыми снизу. На полете от
сокола отличаются большими размерами, более широкими крыльями и
длинным хвостом. Голос — соколиное «кьяк, кьяк-кьяк» или «кееек-кееек-кееею»,
или трель (в брачный период).
Описание. Размеры и строение. По длине крыла близок к кречету, но
меньше; по весу самец балобана больше самца настоящего сокола, но самка балобана
уступает в весе самке последнего. Длина тела у самцов (15) 425—500, самок (10) 520—591,
в среднем 473 и 555 мм; размах самцов (14) 1040—1140, самок (9) 1200—1290 мм, в среднем
1095 и 1260 мм. Вес самцов (4) 820—890, самок (3) 970—1130 г (эти измерения относятся
частично и к следующему подвиду). Крыло самцов (17)345—368, самок (19) 383—415, в
среднем 356,3 и 397,6 мм. Особенности строения — как у кречета, но цевка оперена
менее, только до половины (есть и некоторые различия в анатомических деталях).
Окраска. Первый пуховой наряд шелковистый, желтовато-белый, второй —
матово серовато-белый. Первый годовой (гнездовый) на спинной стороне бурый с
рыжеватыми или охристо-рыжеватыми каймами перьев спины и крыльев, иногда с небольшими
пестринами на плечевых, кроющих крыла и второстепенных маховых; голова бурая
или бурая со светлыми беловатыми краями перьев, беловатой «бровью» и пятнами на
затылке; «усы» выражены слабо, маховые темнобурые со светлыми каемками и широкими
беловатыми полями на внутренних опахалах; средняя пара рулевых обычно темнобурая,
остальные темнобурые с охристым поперечным рисунком, больше развитым на наружных
опахалах; низ — белый с бурыми продольными полосами, большие перья боков бурые
со светлыми охристыми пестринами. Радужина темнобурая, почти черная, клюв синевато-
роговой; лапы, восковица, кольцо вокруг глаза —синеватые. Взрослые (во втором годовом
наряде и следующих) в общем сходны с молодыми, но общий тон верхней стороны тела с
сероватым оттенком, посветлей, рыжеватые каймы перьев шире, а на брюшной стороне, по
крайней мере на груди и брюхе, продольные полосы заменены более или менее правильным
рисунком из вершинных пятен, иногда — особенно у самцов — слабо развитым и на зобе и
груди вовсе отсутствующим. Лапы, восковица и кольцо вокруг глаза становятся желтыми.
Обыкновенный балобан Falco cherrug cherrug Gray
Синонимы. Falco sacer gurneyi. M e n z b i e r. Ornithol. du Turkestan, I, 1888,
стр. 286, по экземплярам из разных местностей, в качестве типичной местности считаем
«Киргизские степи». — Falco Hierofalco aralocaspius Kleinschmidt. Falco XXXV,
1939, стр. 28, по зимней птице из Ленкорани.
Распространение. Ареал. Обыкновенный балобан распространен к
востоку от предыдущего подвида, начиная от бассейна Волги и Южн. Урала
в казахских и южносибирских степях, на восток до Кулундинских и
Минусинских степей. Северная граница — на Волге в бассейне Суры и в Закамье
у Чистополя (Рузский, 1892), в Западной Сибири у Тюмени и Красноярска (где
встречается повидимому переходная популяция в сторону saceroides); добытые
у гнезд на скалах соколы относятся к saceroides, а из гнезда на дереве близ
деревни Дадоновки 26.V. 1926 г. была добыта взрослая самка формы cherrug;
сюда же относится повидимому птица, наблюдавшаяся Котсом у гнезда на
лиственнице у озера Сайбаш в Абаканской степи. Южная граница — в
Казахстане в лесах Наурзума, Терсек, Сыпсын, далее — в восточноказахской склад-
чатой стране и окраине Алая, далее — у Ачинска и Минусинска. На зимовке
этот сокол встречен на Кавказе — в восточных его частях (Дагестан, Азербай-
106
ОТРЯД ХИЩНЫЕ ПТИЦЫ
джан, где особенно многочисленен в районе Ленкорани, на местах зимовок
водоплавающих), вероятно в сев. Иране, несомненно в ю.-з. Туркмении и
других частях Средней Азии; наконец, в Индии. По всей вероятности к этой же
форме относятся балобаны, зимующие в Месопотамии и других странах
Передней Азии. Зимуют балобаны уже в Предкавказье и в Астраханской обл.,
изредка в сев. Казахстане, есть особи, добытые зимою в Зап. Сибири.
Характер пребывания такой же, как и у предыдущего подвида.
Часть особей оседла, часть кочует, но основная часть популяций, во всяком
случае в северных частях гнездового ареала, перелетна. Прилет к местам
гнездовья — в разные числа марта, главным образом во второй его половине
(Куйбышевская и Ульяновская обл., сев. Казахстан, леса Наурзум, Терсек, Сыпсын).
То же показали и наблюдения в Дагестане: здесь под Махач-Кала балобаны
летят уже в середине февраля, и пролет их продолжается с месяц. Соколы летят
и по берегу моря, и между горами и морем, и через горы, обычно одиночками или
парами; в это время летят и утки. Пролет в низовьях Сыр-Дарьи наблюдался
в конце марта. Отлет падает на сентябрь —начало октября. На зимовках в
Туркмении появляется в октябре и остается до февраля—марта.
Биотоп. Островные леса в лесостепной полосе, в бассейне Суры и
в б. Семипалатинской обл. — в борах; в степи, например на Урале, по Эмбе —
в уремах речных долин; для безлесных местностей в период размножения
отмечен лишь в виде исключения в северных Мугоджарах. Во внегнездовое
время — в самых разнообразных открытых ландшафтах: у морских побережий
(Ленкорань), в речных долинах, в степях и пустынях (ю.-в. Туркмения).
Численность та же, что у предыдущего подвида. Для крупного
сокола птица относительно многочисленна. Карамзин (1901) определяет
плотность заселения балобаном некоторых мест б. Бугурусланского уезда
Самарской губ. в 3—4 гнезда на 100 десятин. В Наурзумском заповеднике в 1938 г.
был учтен 21 выводок балобана (на самом деле соколов там было повидимому
больше). В качестве мест, где балобаны положительно многочисленны, можно
назвать в гнездовое время долину р. Урала; леса Наурзум, Терсек и Сыпсын
в с.-з. Казахстане; Кузнецкую степь; в холодное время года ю.-в. Закавказье,
а также Пенджаб. Колебания численности не изучены, но судя по некоторым
сведениям о разной плодовитости в разные годы, имеют место.
Экология. Годовой цикл очень сходен с циклом danubialis и представляет
в основном те же черты, что у северных кречетов.
Размножение. Брачный период наступает рано, впрочем, на одну—
полторы недели поздней, чем у danubialis, так что кладка (Наурзум) падает на
последнюю треть апреля. Как и F. p. danubialis, F. ch. cherrug, в отличие от
других подвидов балобанов, гнездится на деревьях: на березах в
североказахских и западносибирских колках, на осокорях и ольхах в поймах и уремах
рек, на соснах в присурских и семипалатинских борах. Однако на Ергенях,
по Общему Сырту и на Мугоджарах гнезда расположены по обрывам и скалам.
Кладка из 3—5 яиц, чаще из 4, изредка 6 (Наурзум, Гибет). Размеры яиц и
окраска — как у danubialis.
Птенцы выходят из яиц в последней трети мая — начале июня. Так,
например, в Наурзумском заповеднике в 1939 г. птенцы в первом пуховом наряде
были найдены 2 июня, начавшие оперяться — 19 июня (при большой уже
величине, вес самца 890 г), 25 июня птенцы были вполне оперены, но маховые и
рулевые не вполне доросли. В конце июня птенцы уже могут перепархивать.
В это время птенцы, защищаясь от врага, не пробуют улетать, а падают на
спину, обороняясь вытянутыми лапами. Содержавшиеся в неволе (Дементьев,
1938) балобаны достигли полного веса к 20 июля. К 10 июля рулевые у них
были уже нормальной длины, но первостепенные маховые не доросли. Это
-отставание роста первостепенных маховых объясняется вероятно тем, что
БАЛОБАН
107
активный полет появляется только после достижения птицей всех необходимых
для воздушного передвижения качеств (развитие мускулатуры, ориентации,
достаточная площадь несущей поверхности в связи с ростом рулевых и
второстепенных маховых и т. д.). После того, как все это имеется налицо, рост
первостепенных маховых идет быстро — по 5—6 и даже 7 мм в день. К 20 июля эти
соколы уже свободно летали, а около середины июля у них появился рефлекс
нападения на живую (улетающую или убегающую) добычу.
Способность нападения на добычу развивается, таким образом, параллельно
со способностью к полету и вне всякой зависимости от научения со стороны
родителей. Число птенцов в выводке весьма различно, даже в одном и том же
году и в одной и той же местности: в гнездах в Наурзумском заповеднике
в 1938 г. было б выводков с одним птенцом, б — с двумя, 5 — с тремя и 4 —
с четырьмя. Кочевки молодых начинаются во второй половине августа, когда
разбиваются выводки. При этом до осенних миграций молодые разлетаются
повидимому в разных направлениях (окольцованный 19 июня 1938 г. в лесу
Терсек, Наурзумский заповедник, молодой балобан был найден 21 августа
того же хода у Кустаная).
Линька. Линька взрослых начинается в середине мая, когда сменяются
средние первостепенные маховые, кончается около середины сентября, когда
у взрослых птиц первое и второе первостепенные маховые еще не достигают
полной длины. Последовательность смены маховых:7—б—5—8—4—3—9—2—
—10—1. Рулевые сменяются как у других крупных соколов. Продолжается
линька около 5 месяцев. Имеются небольшие личные колебания как в
отношении последовательности смены перьев, так и начала линьки. Впрочем,
взрослые птицы, добытые в ноябре и апреле, не несут следов линьки.
Последовательность смены нарядов и т. д. — общая для вида (см. F. ch. danubialis).
Питание. Подробные наблюдения по питанию балобана проведены
были в Наурзумском заповеднике Волошиным (1949). Им в 128 погадках
найдены : во всех случаях млекопитающие (на первом месте — серый суслик,
около 60 экземпляров, также рыжеватый суслик, большой тушканчик,
горностай, хомяк, стадная ..полевка, степная пеструшка, обыкновенная полевка,
ушастый еж), в 68 погадках — птицы (стрепет, белая куропатка, кряква»
розовый скворец, плешанка, полевой жаворонок, сизовронка, сорока, грач,
крачка, кулик, пустельга), в 3 погадках — прыткая ящерица, в 3 погадках—
жуки. Кормовые остатки у гнезд в 47 случаях содержали степных сусликов,
серых сусликов, большого тушканчика, степную пеструшку, зайцев (молодых);
из птиц — стрепета, серого гуся, крякву, красноголовую чернеть, розового
скворца, полевого жаворонка, сороку, грача, пустельгу. В присланных
Дементьеву из Наурзумского заповедника остатках пищи балобана были обнаружены
части сусликов и полевок, белых куропаток, тетеревов, чирков, широконосок,
серух, кречетки, тиркушки, кроншнепа, черного и белокрылого жаворонка.
В общем, количественно преобладают мелкопитающие и птицы средней
величины; многие птицы добываются главным образом в период выкорма птенцов.
На зимовке в Индии балобаны в значительной мере кормятся водяной птицей,
так же как у Ленкорани. Однако в Туркмении зимующие в пустынях у ю.-в.
угла Каспийского моря балобаны кормятся главным образом пустынными и
степными птицами от джека и стрепета до жаворонков и зверями от песчаного
суслика до землероек; кроме того, как пища сокола отмечены дрозды и утки*
Описание* Размеры и строение как у danubialis. Крыло самцов (59) 343—370,
самок (69) 380—423, в среднем 357,14 и 393,5 мм.
Окраска оперения хорошо отличается от danubialis тем, что и старые и молодые птицы
бледнее и светлее. У молодых в первом годовом наряде при этом более развиты рыжие каймы
перьев на спине и крыльях; у взрослых пестрины на брюшной стороне развиты меньше.
Бурый тон менее насыщенный, боле,е сероват. Кроме того, в качестве индивидуальных
отклонений, на плечевых, больших кроющих крыла, второстепенных маховых нередко имеются
108
ОТРЯД ХИЩНЫЕ ПТИЦЫ
охристые пятна, расположенные более или менее поперечно, что сближает этих балобанов
с подвидом балобана из средней Сибири и с туркестанскими подвидами.
Сибирский балобан Falco cherrug saceroides Bianchi (ex Menzbier)
Gennaja saceroides «Menzbier». Бианки. Монголия и Нам. Птицы. 1907, стр. 204,
по зимним птицам из ю.-з. Сибири.
Распространение. Ареал. Средняя Сибирь — Алтай (ю.-в. Алтай,
прилежащие части центрального Алтая, окраина западного Алтая, Сайлюгем),
Саян, Тарбагатай, с.-з. Монголия и ответвления среднесибирских гор в бассейне
Енисея; Танну-тува, Минусинская и Абаканская степи, окрестности
Красноярска (Базаиха). Впрочем, особи из последних мест не все типичны и наряду
с «настоящими» F. ch. saceroides попадаются и птицы, окрашенные как F. ch.
cherrug (см. выше), так что здесь можно видеть переходную популяцию. Зимою
эти птицы попадаются в ю.-з. Сибири (к сожалению, экземпляры неточно
датированы), в Зайсанской котловине, в Средней Азии, в Индии.
Характер пребывания. По характеру пребывания сибирский
балобан сходен с другими подвидами балобанов. Зимою многочисленен уже
в Тарбагатае; возможно,# отдельные особи зимуют и в ю.-з. Сибири. Даты, как
и всего годового цикла, те же, что и у F. ch. cherrug.
Биотоп. В отличие от предыдущих подвидов, гнездится, насколько
известно, только на скалах. Типичный гнездовый ландшафт — сухие горы,
безлесные или бедные лесом, открытые площади с выходом скал (Алтай,
Тарбагатай) или стены с отвесными выходами скал (Абакан, Минусинск), или,
наконец, большие выходы скал в лесной зоне (Красноярск, р. Базаиха). На Алтае
эта птица в гнездовое время встречается на высоте 1700—3000м (вместе с Phoeni-
curus erythrogaster, Montifringilla nivalis), в Тарбагатае — в верхнем поясе,
но в Минусинской степи гнездо на скале у Божьего озера найдено только на
высоте 360 м; впрочем, тут обитала и клушица. Зимою встречается и в нижнем
поясе гор, и на равнинах.
Численность. Местами, повидимому, нередок; зимой
многочисленен в Зайсанской котловине; в Индии, судя по коллекционному материалу,
на зимовке значительно уступает F. ch. cherrug.
Экология. Размножение. Точные наблюдения произведены на
Алтае Сушкиным (1938), у Красноярска —Юдиным и у Божьего озера в
Минусинском районе — Котсом. Гнезда расположены на уступах скал,
защищенных стеной или навесом, или в нишах. Высота расположения гнезд различна,
на Алтае от 8 м до 50 от подножья скалы. Вблизи гнезда на Базаихе у
Красноярска выводятся пустельги, но воронов балабаны из гнездового участка
выгоняют. Гнездо сибирского балобана сделано из крупных прутьев, постройка
его небрежна, часто в нем обнаруживаются остатки пищи. В одном случае
Сушкин нашел в гнезде обрывки веревки и ребра барана. За выступом в 100 лс
было расположено гнездо Buteo hemilasius, что дает основания полагать, что
сокол занял одно из гнезд этой птицы, имеющей обыкновение использовать
в гнезде всякого рода отбросы у стоянок казахов и монголов. У Красноярска
балобан пользовался гнездом, построенным вороном.
Птенцов 3—5 (Алтай); 2—4 (Красноярск); 2 (Божье озеро); они, как и
следует ожидать, разновозрастны. Так, в ю.-в. Алтае 20 июня старший птенец
оперился и был с остатками пуха, а у младшего только начали пробиваться
перья и образовалась «маска» на лице, что соответствует возрасту около 20 дней.
У взятых из гнезда птенцов 8 июля крылья сантиметра на 3 не доросли; 10 июля
выводки вылетели, а у добытой самки наседные пятна зарастали (Сушкин, 1938).
У Красноярска 24 июня найдены в гнезде 1 самец и 3 самки; рулевые и
маховые у них не доразвились. В 1944 г. в том же гнезде 15 июля было три
летных птицы — 2 самки и 1 самец, у которых недоразвито было лишь первое
БАЛОБАН
109
маховое. Вылет птенцов из этого же гнезда наблюдался 11 июля 1943 г. В
другом гнезде у Красноярска 2 подлетка было 22.VI. 1928. Прямых наблюдений
над временем откладки нет, но судя по датам вылета птенцов, она происходит
в последней трети апреля (самка насиживала у Базаихи, близ Красноярска,
24 апреля 1942 г. и 30 апреля 1943 г.; 30 мая в том же гнезде насиживал самец).
У гнезда выводки держались еще в августе. Упоминаемое гнездо
под Красноярском находилось под наблюдением с 1928 по 1944 год;
из него неоднократно добывались молодые, в 1939 г. были убиты обе
старые птицы; в 1943 г. старая самка была найдена мертвой. Несмотря на это,
соколы гнездились ежегодно. По наблюдениям Юдина, во время
насиживания пищу добывает самец, передающий добычу самке где-нибудь
неподалеку от гнезда.
Линька. Полных наблюдений за линькой нет, общее течение линяния
как у F. ch. cherrug и F. ch. danubialis. Начало линьки относится к маю (смена
средних первостепенных маховых, средних рулевых). Во второй годовой наряд,
являющийся окончательным, линька начинается иногда уже зимой, как у
других крупных соколов.
Питание. В отношении питания сибирские балобаны в общем не
отличаются от других балобанов. В гнездовое время они питаются главным образом
сусликами, причем у гнезд соколы делают «запасы»: 24 июня были найдены под
гнездом 30 штук полусгнивших и чуть расклеванных сусликов. В одном случае
в погадке была найдена узкочерепная полевка (Юдин). Летным молодым старики
приносят части сусликов, которых передают на лету. На Алтае в желудках
балобанов находили главным образом грызунов. С этим связан не совсем
обычный для соколов способ охоты: балобан караулит грызунов, сидя где-нибудь
на камне и на бугре, и бросается за ними «в угон». Кроме того, на Алтае соколы
ловят мелких птиц (для выкорма птенцов), у гнезда найдены остатки саджи.
В холодное время года соколы питаются повидимому главным образом
птицами: у Красноярска на Каче 14.XI.1924 было отмечено нападение балобана на
домашнего голубя, В Минусинском районе в октябре — на тетеревов.
Описание. Размеры и строение — как у предыдущего вида. Крыло самцов (14)
352—372, самок (12) 375—412, в среднем 364,46 и 400,1 мм.
Окраска. Окраска сибирского балобана представляет собой переход между
окраской «примитивных» западных подвидов балобана и «прогрессивных» восточных
его подвидов с сильно развитым поперечным рисунком. От западных форм сибирский
балобан отличается большим развитием поперечного рисунка на верхней стороне тела и
большей редукцией темных пестрин на брюшной стороне у взрослых птиц. Спина, плечи,
кроющие крыла и рулевые с охристыми поперечными пятнами, особенно развитыми на
крупных перьях; надхвостье у самцов сизоватое, нередко сизый цвет распространяется и на
заднюю часть спины (поясницу), хвост — бурый с поперечным рисунком и охристо-рыжеватыми
пятнами, образующими на внутренних опахалах боковых пар рулевых более или менее
правильные полосы; светлые каемки перьев спины и крыльев уже; голова обычно светлая,
охристая, с темнобурыми наствольями, иногда бурая, со светлыми надбровными полосами; низ
беловато-охристый, с каплевидными пятнами на брюхе, часто без темного рисунка на груди
(у самцов); на боках и «штанах» зачаток поперечного рисунка в виде охристых симметрично
расположенных пятен. У самок светлые пестрины верха развиты слабо, надхвостье не серое,
низ более испещрен. Обычно начиная со второго годового наряда, у сибирских балобанов на
мантии у вершины пера появляются супротивные охристые пятнышки, разделенные стволом
пера, тогда как у предыдущих подвидов эти перья имеют сплошные рыжеватые каемки.
Молодые походят на cherrug, но у них чаще бывают беловато-охристые пестрины на крупных
перьях спины и крыльев.
Личные отклонения окраски — по большему или меньшему развитию поперечных
пестрин—значительны. Известны случаи сходных с сибирским балобаном личных отклонений
среди западных балобанов: одна такая птица добыта на Кавказе (в Зоол..институте
Академии наук), в Австрии (I. VIII. 1840 у Энцендорфа, в Венском музее); в Венгрии (в комитате
Тибару Перез-Пушта 18. XI. 1929, в Венгерск. орнит. институте); наконец, в Казахстане
из трех птенцов, взятых из одного гнезда в 1937 г. в лесу Сыпсын, один оказался не
отличимым otF. ch. saceroides, два — типичные F. ch. cherrug. Птицы из Танну-тувинской области
как бы переходны к монгольским балобанам.
no
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Туркестанский балобан Falco cherrug coatsi D e m.
Falco cherrug coatsi. Дементьев. Учен. зап. Моск. гос. ун-та, 83, 1945, стр. 93, Кугитанг.
Распространение. Ареал. Туркестанский балобан распространен в горах
Средней Азии от центрального и восточного Тянь-Шаня (достоверные
местонахождения впрочем только к югу от Иссык-куля, в системе Терскея, а также
в Малом Юлдусе), Кара-Тау до Нура-Тау, горного Таджикистана, на юге —
Кугитанга, отрогов Паропамиза и Копет-Дага. Быть может сюда же
относятся балобаны с Усть-Урта и Б. Балханов. Вне границ СССР этот балобан
гнездится в восточном Иране, к югу —до Сеистана, к западу —шовидимому до
Керман-шаха (самка добыта 27 мая, поУайзерби, 1910), в Афганистане. Зимой
туркестанский балобан был найден в Средней Азии. По всей вероятности к этому
подвиду относятся и добытые вне гнездового времени птицы из Синьцзяня,
Джунгарии (Баграч-куль), Афганистана, Ирана, Месопотамии. Такая же птица
добыта 5 ноября 1916 г. у Хамур-Санджа в Эрзерумском вилайете в М. Азии.
Характер пребывания. Этот подвид совершает, повидимому,
менее правильные миграции, чем европейские и сибирские птицы. Сезонные
перемещения ограничиваются вертикальными передвижениями из гор на
равнины и относительно небольшими кочевками. В этом отношении туркестанский
балобан приближается к оседлым и полуоседлым тибетским балобанам.
Биотоп. В гнездовое время — горы или хотя бы высокие глинистые
холмы и обрывы (Бадхыз, Карабиль), в Тянь-Шане примерно на высоте 2600—
2925 м, в Кугитанге — в ущельях нижней зоны гор. В 1903 г. был отмечен
редкий случай гнездованья в с.-в. Иране и на равнине. Во внегнездовое время
туркестанский балобан распространен в предгорьях, полупустынях, в речных
долинах. На равнинах эта птица попадается с августа и сентября по март, в горах
(Копет-Даг) и зимой, но в небольшом количестве.
Численность. Туркестанский балобан — птица редкая,
немногочисленная.
Экология* Сведения по экологии туркестанского балобана весьма
отрывочны.
Размножение. Цикл более ранний, чем у европейских и сибирских
птиц. Вылет птенцов в долине Кафирнигана наблюдался 4 июня (Иванов, 1940);
летные молодые у Ирдыка, к югу от Иссык-куля, были добыты 16 июня (Шалов,
1908); 20 марта в Фисташковом хребте у Кушки добыта от гнезда самка с
большими наседными пятнами, а 2 мая в Карабиле —птенец в возрасте около 3
недель. Поэтому можно считать, что кладка происходит уже в половине марта (при
40—45 днях гнездового периода и 28 днях насиживания). Гнездится
туркестанский балобан на скалах или по глинистым холмам и обрывам, есть
сведения, что иногда даже в стенах засыпанных колодцев (Радде и Вальтер, 1889).
Как и другие соколы, туркестанский балобан постоянно придерживается своего
гнездового района (в 1945 г. из гор Ислим на р. Егригек птенцы вынуты
повидимому из того гнезда, где, по Вальтеру, соколы гнездились в 1887 г.). Гнезда
обычно трудно доступны, в нишах или под навесом. В Карабиле гнездо найдено
в старом гнезде стервятника. В сев.-вост. Иране гнездо было найдено на
саксауле, но поскольку птицы добыты не были, то и уверенности в
принадлежности гнезда именно балобану нет. В долине Кафирнигана между Ханака-
Дарьей и Дюшамбе-Дарьей в 1934 г. 24 апреля наблюдалась взрослая птица
в гнезде, 27 апреля там было 2 пуховых птенца, 13 мая они стали оперяться,
20 мая у одною из птенцов были остатки пуха, у другого пух еще преобладал.
Они уже выходили из гнездовой ниши. В 1935 г. балобаны опять гнездились
там же, но в другом гнезде, так как ниша была размыта. В середине мая там
было 3 птенца во втором пуховом наряде, уже начавшие оперяться; 4 апреля
БАЛОБАН , 111
эти птенцы были уже на взлете (Иванов, 1940). В Бадхызе у Егригека 5 мая*
1887 были найдены 2 оперявшихся, в Карабиле 2 мая у колодца Дос-бай в гнезде
один птенец во втором пуховом наряде с пробивающимися плечевыми,
маховыми и рулевыми. Летный молодой добыт у Джулека на Сыр-Дарье уже 7 июня
1930. Число птенцов, таким образом, 2—3, иногда 1; следовательно яиц в кладке
обычно 3—4. Яйца не описаны.
Линька. Общий ход линьки тот же, что и у других балобанов, но
происходит в более ранние сроки, в соответствии с более ранним циклом
размножения. Позднеосенние экземпляры (октябрь) — в свежем пере.
Питание. В отличие от других подвидов балобана, у этого «подвида
птицы занимают в кормовом режиме относительно большее место, во всяком
случае в тех местностях, где мало грызунов, например, в горном Таджикистане.
Там в качестве добычи балобана указываются сизые голуби, щурки,
сизоворонки, удоды и т. д. В Туркмении балобаны в гнездовое время нападают на
голубей, кекликов, сизоворонок, жаворонков; у Кушки кормятся
тонкопалыми сусликами и песчанками (Радде и Вальтер, 1889), в Карабиле — бело-
спинным чеканом, голубями, щурками. Осенью и зимою в Закаспийском крае
балобаны кормятся жаворонками, большими песчанками, сусликами, по
рассказам даже дрофами-джеками. На склонах Туркестанского хребта
наблюдались нападения гнездовавших там балобанов на живущих неподалеку
розовых скворцов, причем в период выкармливания птенцов соколы ловили до
10 скворцов в день.
Описание. Размеры и строение. Как у предыдущих. Крыло самцов (14)
336—363, самок (17) 375—410, в среднем соответственно 352,6 и 393,4 мм. Длина (2) 530—
580, размах (2) 1245—1255 мм. Вес взрослой самки 950 г.
Окраска. По окраске туркестанский балобан близок к F. ch. saceroides, но
более ярок. Темя у взрослых с кирпично-рыжеватым или красновато-винным налетом (в
свежем пере); спинная сторона темней, чему сибирских птиц—темнобурая и с большим
развитием сероватого налета; светлые пестрины на спинной стороне ярче, не
охристо-рыжеватые, а красновато-рыжие; поперечный рисунок на спине, плечах, крыльях — как у
saceroides) пятна на брюшной стороне темнобурые, более насыщенного тона, чем у saceroides.
Молодые птицы обоих этих подвидов по окраске не различимы.
Монгольский балобан Falco cherrug milvipes J е г d о n
Falco milvipes. Jerdon. The Ibis 1871, стр.420, описание взрослой самки, добытой
в феврале 1867 г. у Умбалла в Пенджабе.
Синоним. Falco cherrug progressus. Ш т е г м а н, Доклады Акад. наук, 1925, стр. 62,.
Урга в с.-з. Монголии.
Распространение. Ареал. Монгольский балобан распространен в южн.
Забайкалье (Даурская степь, к югу от Аги, вероятно на север до Нерчинска),
в вост. части Монголии, на запад до Хангая, на юг до Алашаня, восточного
Нань-Шаня; Манчжурия на вост. до Б. Хингана (?). В негнездовое время от
южн. Монголии и Сев. Китая до вост. Туркестана, Индии.
Характер пребывания тот же, что и у F. ch. cherrug и F. ch.
saceroides: большинство особей перелетны или кочуют на довольно большие
расстояния, часть остается в гнездовой области и зимою, даже на севере ареала
(Забайкалье, Ага).
Биотоп. Встречается в предгорьях, в низких безлесных горах, главным
образом в степных местностях с холмами или выходами скал. В холодное время
года в самых разнообразных открытых ландшафтах.
Численность. В Забайкалье на гнездовье не част, повидимому из-за
недостатка подходящих для гнездования мест (скалы и холмы). В Монголии
местами многочисленен (на Калганском тракте в июне в течение 3 часов добыто
14 птиц, Козлова, 1930).
Экология. Размножение. Годовой цикл как у F. ch. saceroides hF, ch.
cherrug9 периодические явления начинаются позже, чем yF. ch. coat si. У гнезд
112
ОТРЯД ХИЩНЫЕ ПТИЦЫ
в Забайкалье балобаны появляются около середины апреля. Наблюдения над
размножением в Забайкалье сделаны Штегманом (1929) и Павловым (1935).
Гнезда на холмах и выходах скал, часто невысоко (у Соктуя на 5—10 м от
земли), в углублениях или под защитой стены. Иногда на деревьях (Ононские
боры; на вязе, растущем на склоне бархана у озера Холбо-нур в Монголии).
Гнездо монгольского балобана представляет собой небрежную постройку из
тонких ветвей, выстилкой служат шерсть tf перья, т. е. остатки пищи. Одно и
то же гнездо используется неоднократно, в период размножения оно
подновляется, отчего в нем образуются годовые слои (у Соктуя 5-^-6). Откладка яиц
происходит около середины апреля, число их 3—5 и даже б (Насимович). Трех
пуховиков, повидимому, только что вылупившихся, нашли у озера Холбо-нур
12 мая. В одном из гнезд у Соктуя 15 июня было 5 птенцов различного возраста,
младший — пуховик, а старший уже значительно оперился; в другом гнезде
там было 4 птенца, из них 3 самца и 1 самка. В Кентее у Арангату 10 июня
в гнезде было 3 пуховых птенца; наконец, в южн. Забайкалье 14 июня 1931—
3 оперяющихся птенца. Летные птенцы около середины июля, полное развитие
маховых отмечено 20 июля. Пищу птенцам приносит самец, передавая ее самке,
которая расчленяет добычу и оделяет птенцов; приносится добыча уже
ощипанной; в случае гибели самки выкармливание выводка продолжает самец
(Козлова, 1930). В гнездах много остатков пищи, привлекающих мух, и гнезда
вообще содержатся соколами грязно. Повидимому, именно в связи с этим
пара, гнездясь постоянно в одном районе, имеет обычно 2—3 гнезда,
которыми пользуется поочередно. Пустующие гнезда нередко занимаются Buteo
hemilasius.
Линька. Полных наблюдений нет, но в общем она проходит как у F. ch.
cherrug. У взрослых с середины мая до середины сентября, у птиц в первом
годовом наряде начинается несколько ранее (смена мелкого пера), но затем
в связи с гнездовым периодом задерживается, заканчиваясь в те же сроки,
как и у старых птиц. Эта нерегулярность линяния приводит, — как и у других
хищных птиц,— к тому, что при линьке из первого годового наряда во второй
известное число перьев (мелкое перо, второстепенные маховые) остается непе-
релинявшим. В сев. Монголии у взрослого самца в конце мая сменились, но
не доросли 5-е и 6-е первостепенные маховые; у добытых там же 20 и 26 сентября
в свежем пере линька закончилась. У взрослой самки 26 августа линька почти
закончена, остались старые средние (5) рулевые и часть второстепенных
маховых. У добытой 2 июня самки появились новые средние рулевые, сменялись
средние первостепенные маховые. У другой самки, добытой в тот же день и
в том же месте (Калганский тракт), были свежие недоросшие 5-е и 6-е маховые,
4-е пробивались, уже росли средние рулевые, сменялись плечевые и мелкое
оперение. У третьей самки 2 июня первый годовой наряд сменялся на второй,
начиная с 5-го линяли маховые, росли средние рулевые. У самца 2 мая росли
5-е и 6-е маховые, свежие перья в надхвостье и-среди второстепенных маховых.
Два самца в гнездовом (первом годовом наряде), добытые 2 июня, линяли:
сменялись рулевые, начиная от средней пары, маховые, мелкое перо; у одной
птицы по сравнению с другой линька несколько отставала (очевидно по причине
разницы в возрасте). На спинной стороне мелкое перо сменяется ранее, чем на
брюшной (Козлова, 1930).
Питание. В сев. Монголии кормовой режим состоит в значительной
степени из мышевидных грызунов-полевок и т. п., в Забайкалье—из
сусликов, пищух, молодых сурков и зайцев-толаев, разных птиц: от
каменки-плясуньи, полевого жаворонка, рюма, желтой плиски до уток (чирков—свистунка
и клоктуна, касатки), болотной совы и даже болотного луня; кроме того, саджи,
скапистого голубя, чибиса, удода. Мелкие птицы и грызуны идут повидимому
на выкармливание птенцов.
БАЛОВАН 113
Описание. Размеры и строение. Крыло самцов (13) 348—380, самок (11)
387—411, в среднем 358,6 и 398,4 мм. Окраска отличается отF. ch. saceroides и F. ch. coatsi
тем, что у взрослых поперечный рисунок развит больше и правильнее. У взрослых самцов
на спине, плечевых, больших кроющих крыла этот рисунок принимает иногда характер
полос; окраска этих поперечных пятен и полос в свежем оперении ярче, чем у saceroides,
не охристая, а красновато-рыжая; больше развит у самцов монгольского балобана и серый
оттенок на пояснице и в надхвостье, на рулевых, которые часто сероваты с буроватыми
поперечными полосами; рисунок нижней стороны тела развит слабо, однако на боках нередко
бывают поперечно-вытянутые пятна или даже полосы; также на перьях голени. У самок
поперечный рисунок не принимает характера полосатости и выражен слабее, хотя и больше,
чем у saceroides; во всяком случае, полосы на спине, хвосте, надхвостье у них не бывают
сквозными и правильными, а хвост не серый, но бурый в разных оттенках; поперечный рисунок
на боках у самок только намечен. Индивидуальные вариации окраски значительны. Окраска
взрослых в общем как бы переходная от saceroides и coatsi к тибетским hendersoni. Молодые
птицы в первом годовом наряде не отличимы от F. ch. saceroides.
Тибетский балобан Falco cherrug hendersoni Hume
Falco hendersoni. Hume. The Ibis 1871, стр. 471, тип. местн. Кичикийлак к с.-з. от
перевала Санджу, Кашгария.
Синоним: F. ch. milvipes у разных авторов.
Распространение. Ареал. Тибетский балобан распространен в
восточной части высокой Центральной Азии от Нань-Шаня, Ганьсу и Куку-нора на
севере по Тибету до Гималаев (птенец добыт в мае 1875 г. на границе Тибета и
Сиккима, в кол л. Британского музея); в вост. Туркестане (Яркенд, Алтын-таг);
Ладаке и повидимому на Памире, так как взрослая самка 10.VIII.1878 добыта
Северцовым у Кызыл-арта в Алайском хребте (единственным экземпляр этого
сокола из СССР); может быть к этой форме (но скорее к предыдущей) относятся
балобаны, встреченные в Корее и у Пекина.
Характер пребывания. В отличие от предыдущего, тибетский
балобан — оседлая птица, более или менее случайно вылетающая за пределы
гнездовой области (Шефер, 1938), на Памире также встречен и зимой (Меклен-
бурцев, 1946). Сезонные перемещения ограничены вертикальными миграциями
из высокогорья в нижний пояс гор или даже на равнины (в Кашгарии).
Биотоп. В гнездовое время в Тибете на высотах свыше 4000 м (от 4000
до 4700 м), где особенно многочисленны пищухи; реже — в зоне кустарников
от 3000 до 3800 м. В альпийской и лесной зоне не гнездится. Весьма вынослив
к суровым условиям тибетской зимы, что при малоснежии и круглогодичной
активности основного вида добычи (пищуха), вероятно, определяет оседлость
этой птицы.
Численность. Распространен спорадически, местами многочисле-
нен: в степях Тибета в местах, где были обильны пищухи Ochotona melanostoma,
попадались до 20 птиц в день (Шефер). Зимою тибетский балобан обычен у Куку-
нора и в земле Цахаров (Пржевальский, 1876). В западной части ареала во
всяком случае очень редок и, кроме упомянутого экземпляра с Алая, в Средней
Азии никем не добывался. Зарудный упоминает, что видел эту птицу у
местных сокольников, но ни определение подвида, ни происхождение птиц не
могут считаться достаточно установленными.
Экология. Размножение. Цикл относительно ранний, как у
туркестанского балобана. Спаривание наблюдалось в Тибете 14 марта, птицы
держались на скале (Шефер). Кладка происходит повидимому в конце марта (в
верховьях Хуан-хе у Синина близ Дандер-тина 22 марта 1880 в гнезде было
обнаружено одно яйцо). Полная кладка из 3 яиц отмечалась на южных склонах
Кукунорского хребта 26 апреля 1884, когда самка уже насиживала
(Пржевальский). Самки с развитыми фолликулами добывались 30 марта у Мулукуцзе
в Ганьсу (Штреземан, Мейзе, Шенветтер, 1938). Гнезда расположены обычно на
скалах, по обрывам, иногда же в сообществе с горным гусем, отайкой, филином.
8 Птицы СССР, т. I
114
ОТРЯД ХИЩНЫЕ ПТИЦЫ
скалистыми голубями, бородачами и центральноазиатскими курганниками.
Птенцы выходят из яиц, повидимому, в конце апреля — начале мая, так как в
мае на границе Тибета с Сиккимом добыт птенец уже оперившийся, но с
остатками пуха и с короткими крыльями и хвостом. Два пуховика, но уже подросших,
в гнезде на конгломератовых обрывах южн. склона хребта Бурхан-будда
найдены 13 июня. Три птенца в гнезде на предгорьях хребта Амнен-кор в верховьях
Желтой реки 23 июня уже оперились, но крылья и хвост не доразвились, и
имелись остатки пуха. Нелетные птенцы были обнаружены в гнезде у Мулукуцзе
22 июня. Таким образом вылет птенцов тибетского балобана из гнезд
происходит в начале июля. По причинам, которые пока не выяснены, бывают годы,
когда соколы не откладывают яиц: в 1938 г. в середине марта наблюдалось
спаривание, брачный полет; в апреле добывались самки с развитыми до размеров
голубиного яйца фолликулами, и было найдено 30 гнезд, но ни одной кладки.
Пищух было много, так что связывать «неразмножение» с кормовыми условиями
пока нет оснований. Яйца но величине несколько крупнее, чем у других
балобанов: средние размеры 55,97 х 42,23 (Хартерт, 1913); 53,6—57,6 X 41 —
44, в среднем 55,6 х 42,5 мм (Бекер, 1928).
Линька. Линька проходит так же как и у других балобанов. Период
линьки май—сентябрь или начало октября; интенсивная смена мелкого пера
происходит в июне—июле. Второй годовой наряд — окончательный.
Питание. О значении пищух в кормовом режиме тибетского сокола
уже говорилось. В местах, где пищух много, охотничьи вылеты сокола
происходят на небольшом расстоянии от гнезда — в радиусе 1 —2 км (Шефер).
Охотясь, сокол летит низко над землею и иногда трепещет крыльями как
пустельга. Из других млекопитающих в качестве добычи балобана упоминается
заяц, которого птица била «по-соколиному», но не хватала, и полевки.
Тибетский балобан охотится также на птиц: садж, уток, даже гусей и кроме того на
мелкие виды. На Памире в декабре наблюдалось нападение сокола на вьюрков
Leucosticte brandti и ему же, повидимому, принадлежали найденные в Аличур-
ской долине остатки чирков, лысух, горлиц и голубей (Мекленбурцев).
Описание. Размеры и строение. В среднем тибетский балобан несколько
больше других балобанов. Длина крыла самцов (12) 351—367, самок (16) 386—412 и даже
425 мм, в среднем 358,25 и 402,2 мм.
Окраска. Пуховой наряд не описан. В первом годовом (гнездовом) наряде в
среднем темнее других подвидов балобанов, с хорошо выраженными темными «усами», со
светлыми пестринами на больших кроющих крыла и плечевых и с поперечными пятнами на
рулевых (что у других балобанов бывает не часто). Взрослые — со второго годового наряда и
в последующих — резко поперечно полосаты, напоминая в этом отношении северных
кречетов; но светлый поперечный рисунок не охристый или дымчато-сизый (на спинной стороне),
а кирпично-рыжий. Темя темнобурое с рыжеватыми надглазными полосами («бровями»),
надхвостье сизое с сизо-буроватым поперечным рисунком; рулевые серовато-буры со сквозными
серо-сизыми поперечными полосами; низ белый, на зобе и груди обычно без пестрин, брюхо
с черными каплевидными пятнами, бока, голени и подхвостье с черным поперечным
рисунком из полос. Таким образом, разница между первым годовым и вторым, возрастный
диморфизм весьма значительны по сравнению с F. ch. danubialis, F. ch. cherrug и даже
F ch. saceroides и F. ch. coatsi; в некоторых отношениях окраска приближается к
северным кречетам.
3. Лаггар Falco jugger Gray
Falco jugger. Gray in H a r d w i с k e* s Illustr. of Ind. Zool. 1833—1834, табл. 2,
Индия.
Русское название. Лаггар—индийское имя этого сокола, принятое в европейских
языках. У Мензбира (1916) — «индийский балобан».
Распространение. Ареал. Лаггар распространен в Индии, к северу до
Афганистана, к востоку до Ассама и Бирмы. Зарудный (1911) указывал*
что лаггар в Туркестане встречен у Гульчи в Фергане, по среднему течению
р. Ангрен, в горах Кара-Тау. .Он же упоминает, что в кишлаке Варух видел
ЛАГГАР
115
ловчую птицу этого вида, вынутую из гнезда в горах Ахун-Тау и в 1910 г. двух
ловчих птиц, будто бы пойманных в один день в конце октября 1908 г. у Бальд-
жуана. К выводу о гнездовании лаггара в Кара-Тау Зарудный пришел, как он
пишет, на основании добычи молодой птицы 11 августа 1909 г. в Хантагском
ущелье. К сожалению, все эти сведения не были подтверждены ни экземплярами
в коллекции Зарудного, ни позднейшими находками. Приходится сомневаться
или в точности определения, или в правильности рассказов владельцев птиц За-
рудному об их происхождении. Однако Руссов 26 апреля 1878 г. у Чиназа добыл
самку лаггара, хранящуюся ныне в академической коллекции в Ленинграде.
Другая взрослая самка была добыта 14 апреля
1946 г. в 25 км западней Чаача (южн.
Туркмения)—очевидно, залетная (Дементьев, 1947).
Карта 11. Распространение лаггара Falco
jugger
а — граница распространения; д — залеты.
Рис. 40. Лаггар.
Характер пребывания. Лаггар — оседлая птица.
Экология. Относительно Средней Азии сведений почти нет, и все
наблюдения сделаны в Индии.
Размножение. Гнездится в южн. Индии в январе, северней — в
феврале, у Гималаев —в марте и даже апреле.,У добытой 14 марта в
Туркмении взрослой самки половые железы были неразвиты. Самка из Чиназа —
с зарастающими наседными пятнами. Гнезда размещаются на деревьях,
скалах, на пагодах и других сооружениях. Кладка состоит из 3—5 яиц.
Детали гнездового периода известны мало.
Линька. Обе добытые весной в Туркестане птицы сменяют средние
маховые и частично мелкое перо.
Питание. В Индии кормится птицами средней величины (голуби,
сизоворонки и др.), мелкими зверьками, пресмыкающимися (в особенности
повидимому Uromastyx hardwlckii). Кормовые остатки добытого в Туркмении
лаггара состояли из большой песчанки.
8*
116
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Описание. Размеры и строение. Сложением лаггар похож на балобана
но меньше по размерам и легче. Крыло самцов 305—330, самок 358—370 мм. Вес взрослой
самки из Туркмении — 755 г. Наружный палец лапы относительно длиннее, цевка оперена
только в верхней трети; внутреннее опахало второго махового обычно без вырезки.
Окраска. Взрослые на спинной стороне серовато-буры, на темени с беловатыми
каймами; нижняя сторона белая с бурыми боками и перьями голени, иногда с бурыми
пестринами на груди и брюшной стороне; средние рулевые одноцветно-бурые, остальные
бурые с беловатыми поперечными полосами на внутренних опахалах. Молодые птицы
в первом годовом наряде сверху бурые с рыжими каймами перьев, снизу
охристо-беловатые с широкими бурыми продольными полосами. Неоперенные части окрашены как
у балобана.
Систематические замечания. Крайне близок к балобанам, некоторые авторы (Клейн-
шмидт) считают за подвид последнего.
4. Средиземноморский сокол Falco biarmicus Temm.
Falco biarmicus. Temminck. Planches Coloriees livr. 55, №324, 1825, земля Каффров.
Русское название. В сущности, русского названия не имеется. У Галлера
(1883) — балобан, что неправильно. Название «средиземноморский сокол» нами
предложено потому, что многие формы вида встречаются у Средиземного моря, и что отсюда он
стал известен средневековым сокольникам. У последних он носил имя lanier, lanner.
Распространение. Ареал. Средиземноморский сокол замещает балобана
в Африке и в странах по сев. побережью Средиземного моря и в Малой Азии.
В Африке он гнездится от Туниса, Ливийской пустыни, Египта на севере до
Карта 12. Распространение средиземноморского сокола
7 — F. Ь. feJdeggii, 2—F. b. eriangeri, 3—F. b. tanypterus, 4 — F. b.
abyssinicus, 5—F. b. biarmicus; a — граница распространения.
СРЕДИЗЕМНОМОРСКИЙ СОКОЛ
117
Капской Земли на юге; кроме того, в Аравии и Месопотамии (Фао); в Европе
в Италии, Далмации, на Балканском п-ове. Недавно найден в Малой Азии
(у Кастамуну) и в Закавказье (Армения).
Биотоп. Скалистые местности —как у морских побережий, так и внутри
материка. По характеру пребывания оседлая птица. По
численности — редок.
Подвиды и варьирующие признаки. Географическая изменчивость выражается в
большей или меньшей насыщенности окраски верхней стороны тела и в развитии пестрин на
нижней стороне. Экологические различия вероятны, но точно не установлены. Отличают подвиды
F Ь. feldeggii S с h 1 е g e 1, 1844, в европейской части
ареала и Малой Азии;F. b. tanypterus Sсh 1 е gel,
1844, из Египта (к югу до Хартума), Синая, Аравии,
Месопотамии; F. b. erlangeri Kleinschmid t,
1901, из с.-з. Африки, к востоку до оазиса Куфра, к
югу до Ахаггара; F. b. abyssinicus Neumann, 1904,
из Нигерии, к югу до Уганды и области озер, к
востоку до Египетского Судана, Абиссинии, Эритреи,
Сомали; наконец, F. b. biarmicus Те тт., 1825, из
южн. Африки, к северу до Анголы и Кении.
Европейский средиземноморский сокол Fa [со
btarmicus feldeggii S с h 1 е g e 1
Faico feldeggii. Schlegel in Schlegel und
SusemihI. Vogel Europa's, табл. 8-a, 1843—1844,
Далмация.
Распространение. Ареал. Область
распространения европейского
средиземноморского сокола такова: Италия к
северу до Этрусских гор, Далмация;
Балканский п-ов от Боснии и Герцоговины до
Черногории, Болгарии, Албании, Греции;
М. Азия; может бьгг>, Испания. 21 июля
1940 г. молодая птица в гнездовом наряде с
немного недоросшими маховыми, т. е.
выведшаяся где-нибудь поблизости, добыта у
Еревана в Армении. К этому же виду по всей
вероятности относятся соколы, приводимые Ляй-
стером и Сосниным (1942) в сведениях о
встречах В Армении «балобанов» В гнездовое время, Рис. 41. Средиземноморский сокол,
так как балобан на Кавказе не гнездится (в
августе 1928 г. у озера Гилли; в июле 1935 г. на перевале горы Кюки-Даг
в Даралагезе; в августе 1938 г. у села Туджур в Красносельском
районе; в мае 1939 г. в хребте Сарай-булаг; наконец, в гнездовое время
у Норазбуза в Новобаязетском районе). Остальные указания на встречи
этого сокола в России (Гебель, 1879 г., Галлер, 1883, Хартерт, 1913) основаны на
недоразумениях. В частности, Гебель указывает на кладки этой птицы из
Уманского уезда Киевской губернии. Эти сведения, как и приписываемые
средиземноморскому соколу яйца «с Волги», относятся к балобану. Хартерт
сообщил, что взрослая самка была добыта у Сарепты, но тут по всей
вероятности датировка экземпляра была неверная (тем более, что птица определена
Хартертом как F. b. tanypterus). Нет этой птицы, вопреки указанию некоторых
авторов (Бленфорд), и в Иране.
Биотоп, характер пребывания, численность-
см. выше.
Экология* Сведений из нашей страны нет.
118
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Размножение. Гнездится
на скалах, кладка в южн. Европе
в конце марта. В кладке 3—4 яйца.
Вылет из гнезда, судя по
упомянутому экземпляру из Армении,
в последней трети июля. П и-
т а е т с я европейский
средиземноморский сокол птицей средней
величины, мелкими млекопитающими,
пресмыкающимися. Степень
кормовой специализации в отношении
питания птицами — как у
балобанов, этот сокол также свободно
берет добычу с земли, хотя и менее
охотно, чем в воздухе.
Линька не изучена, общий
ход процесса кйк у других видов
крупных соколов.
Полевые признаки. По
особенностям полета, голосу,
поведению похож на балобана, но меньше
и более легкого сложения.
Описание. Раэмеры и
строение. Длина около 440—500 мм (Эн-
гельманн, 1928), крыло самцов 307—344,
самок 345—374 мм (по Клейншмидту).
Крыло самки из Армении 352 и 358 мм
(асимметрия). Цевка оперена слабо,
только в верхней трети; средний палец
короче цевки, наружный палец немного
(на один поперечный щиток) длинней
внутреннего; хвост относительно
длинный; формула крыла 2>1>3>4.
Окраска. Молодые, до первой
смены оперения, в первом годовом
(гнездовом) наряде: сверху темнобурые с
рыжеватыми каемками, темя светлорыжее,
окаймленное черноватым; низстемнобу-
рым продольным рисунком, окаймленным
узкими беловатыми каймами. Взрослые
птицы (во втором • годовом наряде и
последующих) : верхняя сторона сйзо-серая
с более темным серовато-бурым
поперечным рисунком; темя розовато-рыжее с
более или менее заметными темными
штрихами, окаймленное черным; хвост
поперечно-полосатый, из серых, более
светлых и буро-серых, более темных
полос ; низ рыжеватый или беловатый с пе-
стринками черновато-бурого цвета на
груди и поперечными полосами того же
цвета на боках; иногда низ почти
одноцветный, темный рисунок лишь намечен.
Радужина темнобурая, когти черные. Во-
сковица, кольцо вокруг глаза, ноги
желтые у взрослых, синеватые — у молодых
в первом годовом наряде.
о
о.
с
ед a
ев
&
ЕГ
а>
CL
О.
I
5S
ЯР
а s
5*
&'
в 2
х х
. v а
ja«
м У т, т.
а.
с ~
ев
а. =
N
СО
is
СО ьд
* ЕГ *»
; х о 2
*Ф
111
:Ц.
л» x g
• ** н
a « х
о х о
*Н
сса°
о а> l.
X о
асе
н а
oi
О х
9» ев
й о
* at
КРЕЧЕТ
119
5. Кречет Falco gyrfalco L.
Falco Gyrfalco Linnaeus. Syst. Nat. изд. X, 1758, стр. 91, Швеция.
Синоним. Falco rusticolus <<L i n n a e u s» у многих авторов-
Русское название. Слово «кречет» встречается по крайней мере с XII века
(в «Слове о полку Игореве»); в настоящее время сохранилось в живом народном языке
на севере европейской России в местах бывшей деятельности помытчиков (Печора,
Тиман). Происходит слово по всей вероятности от старовенгерского «керечет» или «ке-
речен» и относится, следовательно, ко времени пребывания прамадьяр в Югорской
Земле. У сокольников «кречетом» называлась самка, самец назывался «кречатьим чел игом»
или «чегликом». Слово «челиг» с прилагательным (соколий, ястребий, кречетий и т. п.)
обозначает самца хищных птиц и происходит от тюркского «чаулю» того же значения.
Распространение. Ареал. Бореоальпийского типа. Область
распространения — арктическая и субарктическая зона Европы, Азии и Сев.
Америки; обособленный подвид имеется на Алтае, Саяне, центральном (вероятно
восточном) Тянь-Шане. Самые северные пункты —в Гренландии под 82°15' с. ш.
и 83°45'; самые южные, кроме горно-азиатского подвида—средняя
Скандинавия, Командорские о-ва (о. Беринга, около 55° с. ш.). На кочевках в
холодное время года примерно до бСРс. ш. в Сев. Америке, Азии, Европе,
отдельные особи и южнее.
Характер пребывания. Некоторые особи оседлы (зимние
находки в Гренландии, Лапландии, на Таймыре и т. д.), другие в течение
осени и зимы откочевывают к югу, сосредоточиваясь главным образом в
лесотундре, отчасти и в лесной полосе. Кроме того имеются вертикальные
миграции (горный среднеазиатский подвид спускается из альпийской зоны в долины).
Подвиды и варьирующие признаки. Кречет имеет несколько подвидов: в Гренландии
F. g. candicans G me 1., 1788, на севере, F. g. holbolii S h а г р е, 1887, на юге; в Сев.
Америке F. g.alascanus S w a n n, 1922, в Аляске, F. g. obsoletus Gmel., 1788в восточных
частях к югу до Лабрадора; F. g. islandus В г й n n i с h, 1764 в Исландии; F. g. gyrfalco
L., 1758, в сев. Скандинавии и сев.-зап. частях европейской России до Канина; далее на
Восток до Лены F. g. intermedius G I о g е г, 1834, на с.-в. Сибири F. g. grebnitzkii S e v e г t-
z о w, 1885, в горах средней Сибири и Тянь-Шаня, Хангае F. g. alt aims Menzb., 1892.
Географически варьирует величина: размеры постепенно увеличиваются от северной
Атлантики по направлению к с.-в. Азии и северу Гренландии; центральноазиатская форма—
при сходных линейных размерах, значительно уступает северным по величине. Другой
варьирующий признак — окраска: соответственно увеличению размеров она светлеет» при
этом наряду с посветлением окраски появляются и более или менее сильно депигментирован-
ные птицы («белые» кречеты, составляющие около половины популяций этих птиц в с.-в.
Сибири и почти всю популяцию в сев. Гренландии). Экологически все формы кречета
чрезвычайно сходны, включая горную азиатскую форму; северная группа — типичные «эваркты».
Центральноазиатская форма морфологически резко обособлена.
Лапландский кречет Falco gyrfalco gyrfalco L.
Русское название. Сокол (местное на Мурмане); «норвежский кречет»
(книжное).
Распространение. Ареал. Гнездовый — сев. Скандинавия от Лофо-
тенских и Вестерольских о-вов на западе, до Нордкапа и Вадсе на севере, на
юге до Лангфьельда и Доврефьельда в Норвегии (т. е. до 609 с. ш., лишь в
альпийской зоне гор), Иемтланда в Швеции; в верховьях Питеа, Торнеа, Муонио
в Шведской Лапландии, у Печенги (Петсамо), Утсиоки, Нуорлиярви и Энон-
текис; на Мурмане (Еретики, Тулома, Иоканга), местами во внутренней
Лапландии (Нявка — тундра), на Кильдине и Семи островах (Харлов), на п-ове
Канине. В августе взрослый кречет наблюдался в Кандалакшском
заповеднике — вероятно птица гнездится в районе Кандалакшской губы.
Миграционный ареал — сев. и сев.-зап. части Европейской части
Советского Союза, области Архангельская, Новгородская, Псковская, отдельные
залеты до Аскания-Нова и б. Ананьевского уезда Херсонской обл.; в Западной
Европе — в южн. Скандинавии, изредка в Дании, Германии, Голландии, воз-
120
ОТРЯД ХИЩНЫЕ ПТИЦЫ,
можно (определение подвида ненадежно), — во Франции, Англии, Германии.
Регулярные миграции ограничиваются Скандинавией и сев.-зап. областями
СССР, едва ли выходя южнее 60Р. Часть особей зимует в гнездовой области
(верховья Муонио, р. Кенкямя, в Лапландии; Семь островов на Myрмане).
Миграционный ареал, таким образом, частично лишь выходит за пределы
гнездового и географически от него не оторван. Повидиому, лапландский кречет
остается зимой и у мест гнездовья (главным образом старые птицы).
Даты. Начинаются кочевки уже в конце июля, причем дальнейшее
движение кречетов к югу не носит характера пролета, а постепенного
продвижения к югу, вероятно в связи с миграциями белых куропаток. Весной у гнезд
кречеты появляются в апреле.
Биотоп. Скалистые морские берега, главным образом у птичьих гор;
альпийская зона гор; речные долины с обрывами и т. д. в лапландских тундрах;
лесотундра; частично, повидимому, проникает в лесную зону (Лапландия).
На кочевках — в лесотундрах, в речных долинах и т. д., даже в городах (Псков,
Ленинград).
Численность. Лапландский кречет — редкая птица.
Экология. Размножение. Пары, повидимому, постоянные, хотя
прямых наблюдений (кроме Исландии) нет. Весьма постоянны и места гнездовья,
используемые в течение столетий. Так, еще в XVII веке (а вероятно и
значительно ранее) помытчики знали гнездовье лапландского кречета на о. Харлове,
вост. Мурман; там же гнездо было найдено в 1938 г. Пары гнездятся на большом
расстоянии одна от другой. У каждой пары повидимому 2—3 гнезда,
используемые в разные годы посменно (вероятно для освобождения гнезд от паразитов).
Гнезд кречеты обычно не строят, а пользуются часто гнездами воронов
или канюков. Соколы только подновляют эти гнезда, причем в этом участвуют.
и самка и самец. Расположены гнезда на скалах, в расселинах или нишах,
чаще на карнизах, прикрытых выступом или навесом, но иногда и на
открытых склонах. В лесотундре гнездится и на деревьях (высокая сосна, в 7 м
над землей, Ель-лухт, Русская Лапландия; в Шведской Лапландии, по Уоллею,
1864). Гнездовый биотоп—выходы скал и обрывистые берега лесных
долин, морские побережья, лесотундра. Гнездо примитивное, с грубой
подстилкой из перьев, мха, ивовых плодов или сухой травы, с плоским лотком
(размеры 1 м в поперечнике при 0,5 м высоты, Ель-лухт, Владимирская, 1948).
Брачные игры — как у других соколов, выражаются в брачном полете: птицы
высоко поднимаются в воздух, круто бросаются вниз и т. д., много кричат.
Начинается период размножения и спаривания тогда, когда ландшафт
на родине кречета носит еще вполне зимний характер, температура ниже нуля
и почва покрыта глубоким слоем снега. Кладка происходит в апреле (вторая
половина месяца) — начале мая. Число яиц в кладке обычно 3—4, иногда от
1 до 5 (и даже 7—9 у Каутокейно в Финмаркене, в 1906 г.). Число яиц
повидимому колеблется в зависимости от кормовых условий года (от «урожая»
леммингов и белых куропаток). Промежуток между откладыванием яиц точно
неизвестен, по некоторым наблюдениям он доходит до 3 дней.
Насиживание начинается с первого яйца. Участвуют в насиживании оба
родителя, но преимущественно самка (у которой три больших наседных пятна—
два по бокам и одно несколько выше на середине брюха; небольшие наседные
пятна есть и у самцов). Продолжительность насиживания вероятно 28—29 дней.
Окраска яиц буроватая или красноватая с темными красновато-бурыми
крапинами различной величины, светлей, чем яйца обыкновенного сокола, и
крупней. Размеры средние 58,69x46,65, наибольшие 62,6 х 46 и 57,3 х 48,5,
наименьшие 55x45,7 и 56x43 мм (Хартерт, 1913).
Птенцы выходят из гнезда в мае, во второй половине этого месяца надевают
второй пуховой наряд, становятся на крылья в июле. Продолжительность
КРЕЧЕТ
121
пребывания в гнезде точно неизвестна (повидимому около 2 месяцев).
Насиживающей самке, а также птенцам, по крайней мере первое время, корм носит
самец, передающий добычу самке, а последняя оделяет птенцов.
Уже в конце июля — начале августа начинается откочевка кречетов с мест
гнездовий (молодой у Холмогор добыт 25 июля). В середине июля наблюдаются
неразбившиеся выводки (Лофотенские о-ва, Ребуссен, 1933). Половозрелыми
молодые кречеты становятся на следующую весну своей жизни, и еще далеко
не закончив линяния из гнездового наряда во второй.
Линька. Последовательность смены нарядов: первый пуховой — второй
пуховой, — первый годовой или гнездовый (полная годовая линька) — второй
годовой или окончательный (полная годовая линька) — третий годовой и т. д.
Линька взрослых продолжается около 5 месяцев, начинается в середине мая,
кончается около середины октября. Начинается смена маховых в то время,
когда птенцы еще во втором пуховом наряде. Последовательность смены
первостепенных маховых такова: 7—6—5—8—4—3—9—2—10—1 (бывают
незначительные отклонения, но центробежный характер линяния и окончания его
сменой 1-го, 2-го и 10-го маховых — постоянное явление). Второстепенные
маховые линяют от наружного края крыла к внутреннему; рулевые — центробежно,
от средней пары к краю хвоста, с тем, однако, исключением, что крайняя пара
сменяется предпоследней, а соседняя с ней — с последней. Интенсивная смена
мелкого пера происходит в июле. Первая годовая линька происходит несколько
менее регулярно, отдельные перья начинают сменяться зимою, но крупные
перья сменяются в те же сроки, что и у старых птиц. Кроме того, обычно после
первой годовой линьки в оперении остается несколько несмененных перьев
первого годового (гнездового) наряда.
Питание. Кормовые объекты лапландского кречета — птицы средней
величины, в меньшем количестве млекопитающие (лемминги). Из птиц главным
образом белые куропатки, с количеством и перемещением которых связана
плодовитость и сезонные миграции кречетов, также морские птицы, живущие
на птичьих базарах, и т. д. Существует определенная разница между
кречетами, живущими близ птичьих базаров и тем#, которые гнездятся вдали от моря.
В питании первых (о. Харлов) найдены чистиковые птицы (гагарка, кайры,
тупики, чистики) морянка, чайки (трехпалая, сизая, серебристая), гаги
(обыкновенная и сибирская), кулики (в том числе морской песочник), пуночка и другие
воробьиные, белая куропатка, лемминги, горностай. Основное место занимают
чистики, за коими следуют чайки (моевка). У гнездящихся в Лапландской тайге
(Ель-лухт) кречетов пищевой режим состоял из разных лесных птиц — белых
куропаток, тетеревов, ястребиной совы, мохноногого и воробьиного сыча, кукш
и других воробьиных; чирка; леммингов и рыжих полевок. При этом главную
пищу составляли белые куропатки. Для Скандинавии отмечен такой же состав
кормового режима — чистики и чайки, населяющие птичьи базары; утиные,
кулики; из млекопитающих —зайцы беляки, лемминги, белки. На кочевках
кречеты нападают на тетеревов, уток, домашних голубей. Сезонные изменения
питания сводятся к тому, что зимою в лесотундре белые куропатки становятся
еще более преобладающей пищей кречетов, точно так же, как оседлые у
незамерзающих вод кречеты кормятся исключительно водяными птицами.
Ежедневная потребность кречета в пище составляет около 200 г; в
естественных условиях кречет может съесть, конечно, и больше. Если пищи много,
кречет объедает у пойманной птицы лишь крупные мышцы. Ощипывает и ест
добычу кречет на определенном месте в районе гнезда или зимовки. Здесь
находят остатки пищи, и погадки из костей, перьев и шерсти. Когда птенцы малы,
добычу для них ловит самец, а самка ощипывает ее и отрывает голову и
конечности. Это делается вне гнезда, поэтому перьев в гнезде не бывает. Нападает
кречет на добычу по-соколиному, подлетая к ней сверху и, складывая крылья,
122
ОТРЯД ХИЩНЫЕ ПТИЦЫ
хватает лапами. Ловит преимущественно летающих птиц. Умерщвляет
пойманную добычу клювом, ломая ей шею или прокусывая затылок. Вне времени
размножения кречеты одной пары, как и другие соколы, охотятся порознь,
но держатся, повидимому, в одном охотничьем участке.
Полевые признаки. Типичный сокол. На лету бросаются в глаза длинные
острые крылья; полет быстрый, после нескольких взмахов птица быстро несется
вперед, не парит. Сидящий кречет держится прямо. На расстоянии верх кажется
темным, низ беловатым (взрослый), темным и сверху и снизу (молодой). Походит
на обыкновенного сокола, но
отличается от него крупной
величиной и относительно длинным
хвостом. Голос «кьяк-кьяк-кьяк»
или «кееек-кееек-кееек», похож
на соколиный крик, но погрубей
и пониже. В брачный период
кречет издает довольно тихую
высокую трель.
Описание. Размеры и
строение. Крупные соколы, с
размахом крыльев около 120—ХЪЪсм
при общей длине около 55—60 см.
Вес самца несколько более 1000 г
(1192 г), самки — около 1500—2000 г
(1400 г, 1550 г, 1686 г, 1980 г). Самка
таким образом заметно больше самца;
крыло самцов (17) 342—372, самок (29)
380—407 мму в среднем у первых 358,
у вторых 395,8 мм. Телосложение
массивное, цевки оперены на 2/8
длины, крылья длинные и острые,
формула крыла обычно 2>3>1, хвост
относительно длинный; в отличие от
настоящих соколов, длина наружного
и внутреннего пальцев примерно
одинаковая, а средний палец равен
цевке.
Окраска. Первый пуховой
наряд снежно-белый, второй пуховой
наряд гуще и плотней,
серовато-беловатый. Гнездовый (первый годовой)
наряд таков: верхняя сторона тела
темнобурая со слабозаметными
светлыми каймами перьев; голова темная,
редко со светлыми краями перьев,
обычно лишь с беловатыми пятнами на задней стороне шеи; маховые и рулевые
с беловатым поперечно вытянутым рисунком; усы темнобурые, широкие, сливающиеся
с темными щеками; низ белый с широкими бурыми продольными полосами,
занимающими больше места, чем светлые края перьев. Клюв темный синевато-роговой, неопе-
ренные части ног синеватые, когти черные; радужина темнобурая; восковица синеватая,
голое кольцо вокруг глаза бледное синевато-беловатое, радужина темнобурая.
После первой линьки, кончающейся в возрасте около 15 месяцев, надевается
окончательный наряд. Птица на верхней стороне темная серовато-бурая, с таким же теменем, со
светлыми пятнами на затылке, на теле и крыльях более светлые дымчато-сизые поперечные
пятна и полосы; на внутренних опахалах первостепенных маховых поперечно вытянутые
беловатые пятна; рулевые светлей спины, буровато-серые с поперечными сизыми полосками.
Брюшная сторона более или менее чистого белого цвета с темным бурым рисунком —
продольным на зобе и груди, поперечным на боках, перьях голени, в подхвостье; размеры этих
пятен и полос очень изменчивы. Лапы, восковица, кольцо вокруг глаза более или менее
яркого желтоватого цвета. В окраске кречетов, как и других крупных соколов, имеется
таким образом половой мономорфизм, а возрастный диморфизм, если не считать пуховых
нарядов, ограничивается в сущности указанными выше изменениями.
Рис. 42. Лапландский кречет (молодая и старая
самки).
КРЕЧЕТ
123
Сибирский кречет Fatco gyrfalco intermedins G1 о g e г
Falco gyrfalco var. intermedia. Gloger. Vollstand. Handb. d. Naturg. d. Vdgel Eur.,
1834, стр. 42, Урал
Синоним. Hierofalco uralensis, M e н з б и р, Орнитологическая география. Ученые
записки Имп. Моск. университета, отд. естеств.-историческ., вып.2—3, 1882, стр.288,
описан по птицам из Владимирской губернии.
Распространение. Ареал. Сибирский кречет распространен к востоку от
предыдущего подвида, от Болынеземельской тундры до бассейна Енисея и
Лены. Достоверные находки гнездящихся птиц у нижней Печоры, на полярном
Урале, на Тазовском п-ове, в Туруханском крае, на Нижней Тунгуске (устье
Нижней Качомы, 62°30' с. ш.), на нижней Лене. Самое северное место, где
наблюдался этот кречет, — о. Бэра, 75°30' (в августе, Миддендорф).
Миграционный ареал — северная полоса Вост. Европы и Зап. Сибири,
причем часть особей зимует в гнездовой области (в бассейне Хатанги за
71—72° с.ш. встречен в январе и декабре, Скалой, 1938). Самые южные находки
на кочевке —Украина (Киевская, Черниговская, Полтавская, Херсонская
области), среднее течение Волги, низовья Дона и Маныч (Торне, 1941), сев.
части Средней Азии (Сузак, Талас и др.), Тарбагатай. Общее направление
кочевок Ю и ЮЮЗ. Нормально кречеты редко выходят зимою за 60° с. ш.
к югу, держась главным образом в лесотундре.
Даты кочевок. Начало уже в июле и августе, но обычно не ранее
сентября—октября. В мае кречеты уже у гнезд, так что обратное движение
на север происходит не позже апреля.
Биотоп. Как у предыдущего подвида, однако достоверных находок
гнездовий западносибирских кречетов на гнездовье на морских побережьях
нет (что быть может связано с отсутствием у берегов Зап. Сибири птичьих
базаров). В ровной тундре кречеты не гнездятся и бывают там лишь вне времени
гнездования. Гнездовый биотоп связан или со скалами или обрывами речных
долин, или с лесотундрой, поскольку для очень рано гнездящегося кречета
необходимы более иди менее защищенные скалы, или деревья.
Численность. Невелика, гнездовья разбросаны, повидимому,
спорадично и на значительном расстоянии одно от другого.
Экология* Размножение. В основном как у предыдущего, прямых
наблюдений мало. Все найденные за последние годы гнезда сибирского кречета
располагались на лиственницах в лесотундре (р. Хадуттей, Тазовский п-ов;
Толстый Нос в низовьях Енисея; р. Ямная, в 37 км от Дудинки,
Туруханский край). Гнездо представляет собой грубую постройку из сучьев, почти
без лотка. Число яиц в кладке 3—4, число птенцов в выводке 2—3, чаще 2
(3 — Сургутский район, август 1937; 2 — р. Ямная, Туруханский край, июль
1916, Тугаринов; 2 — Н. Качома, приток Н. Тунгуски, июль 1918 г., Ткаченко,
1922). Насиживает главным образом самка, но также и самец, судя по
присутствию наседных пятен (Таз; низовья Лены). Яйца не описаны. Нет сведений и
о продолжительности насиживания (вероятно, как у других кречетов 28—
29 дней). Вылет птенцов из гнезда в половине июля (подлетки добыты 8—
17 июля в бассейне Енисея). Откочевка молодых от гнездовий начинается в
августе, может быть в конце июля. Неразбившиеся выводки наблюдались в августе
и сентябре.
Линька и смена нарядов — как у лапландского кречета;
годовая линька происходит между маем и октябрем.
Питание. Основной пищей сибирских кречетов в лесотундре служат
белые куропатки. Зависимость сезонного размещения сибирских кречетов от
куропаток установлена, вероятно наличие и других зависимостей (детали
распространения, плодовитость). В гнезде, найденном 7 июля 1916 г. у р. Ямной
124
ОТРЯД ХИЩНЫЕ ПТИЦЫ
в Туруханском крае, были остатки около 10 куропаток, и самка передавала
находившемуся в нем птенцу принесенную самцом белую куропатку, без головы
и шеи, ощипанную, без внутренностей. Связь кочевок и зимнего размещения
кречетов с белыми куропатками указывалась для Печоры, Тимана. Кочующие
кречеты охотятся и за водяной птицей.
Зимой кречеты у поселений нападают на галок, ворон, голубей, в
лесной местности — на тетеревов и зайцев. Способы нападения на добычу — как
у предыдущего.
Описание* Размеры и строение. Как у лапландского кречета, но величина
чуть больше. Крыло самцов (49) 343—372, самок (75) 381—415 мм, в среднем у первых 360,2
и 397,5 мм. Вес молодых самок 1500 и 1588 г; взрослой самки 1940 г. Длина самцов 51,25—
53,8, у самок 51— 62,7 см, размах соответственно 114—115 и 124,5—126,6 еж (цифры не
предельны, по немногим экземплярам).
Окраска. Первый и второй пуховой наряд не описаны. Как молодые в гнездовом
оперении, так и взрослые (во втором годовом наряде и последующих)отличаются от
лапландских кречетов более светлой окраской. Весьма изменчив. Имеется не только серый, но и
белый тип окраски, с резким преобладанием белого цвета. Немногие особи сходны по окраске
с лапландскими, большинство окрашено так: серая фаза, первый годовой (гнездовий)
наряд — голова охристо-белая с темнобурыми наствольными чертами на темени и затылке,
резко отделяется от серо-бурой спины и крыльев; по сравнению с лапландским кречетом,
верхняя сторона тела менее бурая, светлей и с большим развитием светлых (беловатых)
каемок и пестрин; рулевые с хорошо развитым беловатым поперечным рисунком.
Во втором и последующих нарядах окраска головы светлей спины, темя и затылок
беловатые с серовато^бурыми продольными полосками, спина и крылья, хвост серо-дымчато-
бурые, светлей, чем у лапландского кречета и с более светлым и обычно более правильным
поперечным рисунком (охристо-сероватым или беловатым); темные полосы у разреза рта—
так называемые «усы» — менее выражены; темный рисунок на нижней стороне тела —
продольный на груди, поперечный на боках, подхвостье, голени — менее развит, иногда имеется
в виде остатков, так что на расстоянии птица кажется белой; общий тон брюшной стороны
тела более чистый, белый. Неоперенные части окрашены у всех кречетов одинаково.
Кречеты белой фазы (среди intermediusnx мало, в изученном нами материале 6 из 124 экз.,
т. е. немногим более 4%) отличаются тем, что белый цвет на брюшной стороне у них резко
преобладает, темный рисунок очень незначителен или отсутствует; на верхней стороне тела
у взрослых темный рисунок в виде тонких поперечных полос или пятен (сердцевидных,
поперечно вытянутых), буровато-серого тона, по белому фону, заметно преобладающему; у
молодых спинная сторона белая с бурыми центральными частями перьев и с более или менее
правильным поперечным рисунком на второстепенных маховых и рулевых; первостепенные
маховые — белые с бурыми вершинными частями. У некоторых особей клюв и когти светло-
рогового, желтоватого цвета.
Восточносибирский или камчатский кречет Falco gyrfalco grebnitzkti S e v.
Hierofalco Grebnitzkii. Severtzow. Nouv. Memoires de la Soc. Imp.des Naturalistes
de Moscou, XV, 3, 1885, стр. 69, о. Беринга.
Распространение. Ареал. Гнездовий — с.-в. Сибирь к востоку от дельты
Лены до Анадырского бассейна, Коряцкой Земли, вероятно Камчатки и
Командорских о-вов (о. Беринга), но встречен в нёгнездовое время и на о. Медном.
Миграционный — Вост. Сибирь от Байкала до Приамурья, Курильских
о-вов, у Сахалина, самые южные находки в Японии (Хоккайдо), в
Манчжурии и у Пекина. На севере найден на о. Врангеля, но гнездованье там не
доказано. Часть особей зимует в гнездовой области (Коряцкая Земля, декабрь;
Чукотская Земля, февраль), часть откочевывает на юг. В Америке на
кочевках на Алеутской гряде и Прибыловых о-вах. Начинаются кочевки
по крайней мере в сентябре (вероятно и ранее), у мест гнездовья кречеты
появляются в апреле (дельта Лены, Чубукулах; Русский хребет, Анадырь).
На Камчатке кречеты часто попадаются между сентябрем и первой половиной
апреля; в гнездовое время — в мае — июле добыты пока в сущности только
три птицы — у Старого Острога, в Кроноках и в районе Петропавловска, но
в небольшом числе кречет в Кроноках держится и в августе (Аверин, 1948).
Кам чатскии кречет
КРЕЧЕТ
125
Указания японских авторов на гнездование на о. Матсува и других Курильских
по ряду причин невероятно.
Биотоп. Лесотундра (Русский хребет, Анадырь), прибрежные скалы
у морских берегов (о. Беринга, Коряцкая Земля) и в долинах рек (Утесики,
Анадырь); в 1946 г. в Кроноках в горах внутри полуострова, на высоте около
1800 м. На миграциях у морских побережий, в лесотундре, по долинам рек
и даже в лесах (Буреинскиегоры). На Камчатке в Кроноках установлено, что
кречет из года в год держится на зимовке в определенном участке местности.
Численность. В южн. частях Вост. Сибири (Иркутск, Чита и т. д.)
кречет зимою не очень редок; это позволяет предполагать, что и на
гнездовье на с.-в. Сибири он относительно обычен. Впрочем, прямых наблюдений
над кречетами там очень мало.
Экология* Сведения по экологии весьма недостаточны.
Размножение. Развитие половых желез относится к концу апреля.
Гнезда размещаются на деревьях (Populus suaveolens, лиственница) или на
скалах. Кладка, яйца и т. д. — неизвестны. Молодые на крыле наблюдались
в конце июля; кочевки молодых в сентябре (когда они появляются уже у
Иркутска, в Приамурье, на Курильских и Прибыловых о-вах).
Линька, судя по неполному бывшему у нас материалу, протекает так
же как и у лапландского кречета. Смена нарядов у всех кречетов одинакова.
Питание. Кормовые объекты — белые куропатки, на зимних
кочевках, кроме белых куропаток, глухарки, разные воробьиные (Прибыловы о-ва),
водоплавающие птицы, чайки, турпаны, морянки, каменушки, серокрылая
чайка (Камчатка), из млекопитающих— полевки и землеройки (Sorex macropyg-
maeus —Камчатка) и белки. Основные корма зимующих на вост. Камчатке
кречетов — утки, чайки и вороны у моря, внутри страны — куропатки. Способы
добывания и умерщвления добычи — как у всех других кречетов.
Описание. Размеры и строение. Кречеты с.-в. Сибири несколько больше
западносибирских. Крыло самцов (12) 360—382, самок (40) 390—418, средняя соответственно
370,3—410,3 мм. Вес самок 1450, 1500, 1503, 1900, 1900 и 2100 г.
Окраска. Белый тип окраски встречается часто, во всяком случае не менее часто,
чем серый (50% среди изученных нами экземпляров). У серых птиц в первом годовом
(гнездовом) наряде бурые пестрины на бркшшой стороне тела очень слабо развиты и не образуют
продольного сплошного рисунка. Эти особенности, в связи с несколько более значительными
размерами, заставляют выделять восточносибирских кречетов в качестве отдельной
географической формы.
Алтайский кречет Falco gyrfalco altaicus M e n z.
Hierofalco altaicus. Menibier. Orn. du Turkestan 1891, стр. 272, Уймон в Центр.
Алтае.
Синоним. Hierofalco lorenzi M e n z b i е г. Bull, of the British Orn. Club v. XI n°
LXXXIV, 1900, стр. 3, тип происходит из Енисейской губернии.
Местные названия.У азиатских народов шумкар, сункар (как и для
северных кречетов).
Распространение. Ареал. Область распространения алтайского кречета
оторвана от области северных кречетов (вид имеет таким образом так
называемое «бореоальпийское» распространение). Гнездовая область —
высокогорье Алтая, Саяна, центр. Тянь-Шаня, Хангая, быть может Тарбагатай.
Во внегнездовое время в разных частях Зап. и средней Сибири, в Тянь-
Шане, в воет, Туркестане, в Монголии (Куку-нор, Улан-Батор, округ Та-
Цзинь-Шань в провинции Суй-Юань). Миграции носят и горизонтальный и
вертикальный характер. Часть птиц остается и зимою в гнездовой области.
Начинаются кочевки в августе, в конце июля выводки еще у гнезд, зимой
(январь —февраль) кречеты наблюдались у Зайсана, в центр, Тянь-Шане,
126
ОТРЯД ХИЩНЫЕ ПТИЦЫ.
в центр. Алтае, в Тарбагатае. У гнезд кречеты появляются в мае, быть может
в июне.
Биотоп. Гнездовый напоминает ландшафты, населяемые северными
кречетами — это открытые пространства высокогорья (не ниже 1500—2 000 м,
в Хангае около 2500 м), окруженные скалами. На кочевках кречеты
спускаются в лесную зону гор, в предгорья и в равнины.
Численность. Очень редкая птица (известно немногим более
50 экз.).
Экология. Размножение. Период размножения начинается повиди-
мому в апреле. Гнезда их — до сих пор найдено только 5 — одно у Лепсинска
в Джунгарском Ала-Тау, одно в Терскей-Ала-Тау, одно в ю.-в. Алтае и два в Сай-
люгеме, —на скалах, из ветвей (в одном случае карликовой березы), очень
грубой постройки, без выстилки, которую заменяют остатки добычи — перья,
шерсть и т. п. Размер гнезда в ю.-в. Алтае — 100 х 75 см. Места гнездовий весьма
постоянны; по наблюдениям Сушкина (1938), гнездо, найденное в ю.-в. Алтае
на р. Кушка-нур в 1914 г., несомненно занималось многие годы; в 1915 г. оно
было опять занято, хотя в предыдущем году самец был убит, а птенцы вынуты.
Кладка из 2—5 яиц. Размеры 56—56,5 х 42—42,3 мм\ цвет —типичный кре-
четиный, с мелкими красновато-бурыми пестринами по бледному охристо ржав-
чато-буроватому тону. Даты развития: 18 мая — яйца (Джергалан,
Терскей-Ала-Тау), 10—15 мая — вылупление (Хангай), 9 июня —птенцы во 2-м
пуховом наряде (ю.-в. Алтай), июль — птенец с недоразвитыми маховыми, но
вполне оперившийся (Саяны), 10 июля молодые вылетели из гнезда, 25 июля —
неразбившийся выводок держался близ гнезда (Сайлюгем); 31 июля уже вполне
развитые птенцы (Хангай). Насиживание начинается с первого яйца, так как
в кладке из Терскей-Ала-Тау одно яйцо совершенно свежее, другое — е
начавшим развиваться, примерно трехдневным, зародышем (П. П. Дементьев).
Промежуток в кладке яиц, следовательно, 3 суток. В добытом Сушкиным на
р. Кушка-нур в ю.-в. Алтае выводке было 5 птенцов и все самки, в гнезде на
южн. Хангае Тарасовым взяты птенцы—самец и самка.
Линька— как у других кречетов, в течение 5 месяцев, с апреля по
октябрь. В начале ноября кречеты — в свежем* пере (сев. Монголия).
Питание. Питается алтайский кречет птицами и млекопитающими.
Отмечены — клушица, горная куропатка, водоплавающие птицы; грызуны.
Полевые признаки. Большинство особей, будучи сходны с северными
кречетами, отличаются от них однообразной темной окраской.
Описание. Размеры (линейные) и строение—как у северных кречетов. Длина
тела самцов 50,1—54, самок 54,6—58,5 см, размах соответственно 109,8—118 и 124,5—
126,6 мм, крыло самцов (19) 348—377, самок (33) 385—415 лш, в среднем 366,75 и 403,0 мм.
Но вес (т. е. реальные размеры) меньше. Вес самцов (3) 800—850, самок (7) 990—1627,
в среднем 818,6 и 1169,6 г.
Окраска. Пуховые наряды не описаны. В первом гнездовом наряде однообразно
бурого цвета, бурый цвет резко преобладает над светлыми пестринами и каймами перьев;
перья верхней стороны без светлых ободков, обычно и без пятен, средние рулевые чаще —
одноцветно-бурые. Кроме таких темных «меланистических» особей изредка встречаются и
более светлые, со светлой охристой головой (с бурыми наствольями), с рыжеватыми
каемками перьев спины и крыльев (что сближает их с балобанами).
После первой линьки алтайские кречеты бывают окрашены различно. Некоторые птицы
на спинной стороне темнобурые с чуть намеченным сероватым поперечным рисунком и бурые
с небольшими светлыми каймами перьев на брюшной стороне (перья боков и подкрылья бурые
с беловатыми поперечными пятнами), или они более приближаются к северным кречетам (но
менее серы, с рыжеватым поперечным рисунком на спине и крыльях), или наконец, дымчато-
черновато-буры на спине с рыжим поперечным рисунком, напоминающие балобанов, с сизым
поперечно-полосатым хвостом. Однако, они могут быть отличены от центральноазиатских
балобанов соответствующего возраста сильным развитием пестрин на нижней стороне тела,
черноватым тоном темных частей перьев спины и крыльев, меньшим развитием светлого поля
внутреннего опахала первостепенных маховых и, наконец, большим оперением цевки.
ЧЕГЛОК
127
6. Чеглок Falco subbuteo L.
Falco subbuteo Linnaeus, Syst. Nat. изд. X, 1758, стр. 89, Швеция.
Русское название. В научной литературе, а отчасти и в живом народном
языке, для этого вида установилось имя чеглбк. Чеглок, чеглик, челиг означало ранее
самца всех употреблявшихся для соколиной охоты птиц и соответствовало
западноевропейским словам tiercelet, tiercel, Terzel; таким образом, соколом звали самку F. рете-
grinus, а самца этого вида сокольим челигом, кречетом — самку F. gyrfalco, самца же кре-
чатьим челигом и т. п. Этимология слова челиг (чеглок) — от тюркского «чаули»,
охотничьего термина того же значения. У русских сокольников видовым названием для F. subbuteo
было кобец или кобчик (см. напр. Урядник Сокольничья Пути), перешедшее затем на F. ves-
pertinus. Местами к этой птице применяют название белогорлик.
Распространение. Ареал. Область распространения чеглока— умеренная
полоса Азии, Европы и сев. Африка; от Англии (но не в Шотландии и Ир-
Карта 14. Распространение чеглока Falco subbuteo
/ —F. s. subbuteo, 2—F. s. jakutensls, 3 — F. s. jugurtha, 4— F. s. streichi; а —граница гнездовий,
г — область зимовок.
ландии) и Португалии на западе до бассейна Колымы, Охотского моря,
Камчатки и Приамурья на востоке; к югу до Малой Азии, Ирана, Гималаев, в ю.-в.
Азии почти до тропиков (Сватоу, Шаньские государства). Северная граница
совпадает примерно с границей высокоствольного леса и только местами
(Лапландия, Зап. Сибирь, Якутия) достигает полярного круга. Зимовки в
тропической Африке и Азии. Чеглок — птица перелетная, поздно появляющаяся на
местах гнездовья и рано отлетающая. Оседла повидимому в ю.-в. Китае.
128
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Биотоп. Лиственные и смешанные леса, перемежающиеся с открытыми
пространствами; встречается и в тайге, и в-лесостепи; в первой немногочисленен,
во второй обычен; в Туркестане и на Кавказе — птица горных лесов,
поднимающаяся в вертикальном отношении до 2 000 и даже 3 000 м.
Подвиды и варьирующие признаки. В таежной полосе от Карелии до Якутии с запада
на восток, к югу до Вологодской обл., среднего течения Печоры и Оби, сев. Байкала —
F. S. jakutensis В u t., 1911; к югу от него F. s. subbuteo L., заменяемый в сев. Африке
(Марокко, Алжире, Тунисе и Триполи) F. s. jugurtha Hartert et Neumann, 1907, а в
Среднем и Южн. Китае отЦинлинских гор до Юннани и Шаньских государств F. s. streichi
Hartert et Neumann, 1907. Различия географических рас выражаются в окраске
и размерах; при этом северный подвид окрашен в кажущемся противоречии с правилами
географического изоморфизма темнее; африканский и китайский подвиды мелкие.
Экологические черты вида довольно постоянны и не дают резких географических отклонений.
Впрочем, по крайней мере часть популяций F. s. streichi оседлы, тогда как остальные чеглоки
перелетны.
Обыкновенный чеглок Falco subbuteo subbuteo L.
Синонимы. Falco subbuteo cyanescens. Lounberg. Ark. f. Zool, 1905, ctp. 6,
Баим-гол в центральном Тянь-Шане.—F. s. centralasiae. Бутурлин Орнитол. Вестник,
1911, стр. 175, нов. имя для предыдущего.— Hypotriorchis subbuteo irkutensis. I o h a n s e n Orn.
Iahrbuch, 1914, № 3, стр. 83. Омолон в Киренскомуезде.—Hypotriorchis subbuteo distinguendus.
Портенко, Известия Акад. наук, о. ф. м. 1930, стр. 303, Воронежская губ. — Hypotriorchis
subbuteo planicola Портенко, цит. соч., стр. 308, Кашгария.—Fa/ca subbuteo ussuriensis. Do-
maniewski, C.-R. Soc. Sciences Varsovie, X, 1917, № 3, стр. 260, Сидеми.
Распространение. Ареал. Обыкновенный чеглок распространен в лесной
зоне СССР, кроме северных участков тайги, от Прибалтики, Белоруссии и
Украины до Дальнего Востока. На юге — до Закавказья, Средней Азии (Ко-
пет-Даг, южн. Таджикистан), бассейна Уссури (Ханка). Северная граница
определяется южной границей следующего подвида. Вне наших цределов
распространен в Зап. Европе, кроме того — в М. Азии, Иране, Синьцзяне,
Монголии, на юг до Ладака и с.-з. Индии (Кашмир, Симла) в Азии, в Европе
до Испании, Италии, Греции. Бинареальная птица, зимовки в Африке к югу
от Кении и Виктория-Нианца, главным образом в Родезии и Катанге, реже
на крайнем юге; отсутствует в лесной тропической зоне зап. Африки,
а также в южн. Азии от Индии до юга Китая:
Характер пребывания— перелетная гнездящаяся птица.
Даты. Прилет поздний, что связано с поздним циклом годовых
периодических явлений, в частности с размножением. Последнее в свою очередь
находится повидимому в зависимости от относительно позднего наступления
благоприятных условий выкармливания выводка — массовым появлением насекомых
и подлетков мелких птиц. Появление весной отмечено на юге (Бессарабия,
Украина, Кавказ, Средняя Азия) с начала апреля до первых чисел мая,
отдельные особи несколько раньше и позднее, с конца марта (Харьковская обл.) и
до середины мая (Сыр-Дарья). В средней полосе европейской России — Псков,
Смоленск, Москва, Тамбов, Орел, Курск, Казань, Куйбышев — главным
образом во второй половине апреля и в начале мая (однако для Пскова начало
пролета отмечено уже 20 марта; для Тамбовской обл. 3 апреля); для Урала
и Зауралья валовой пролет в конце апреля — начале мая, Башкирия
26 апреля; Чкалов, валовой пролет в середине апреля по ст. ст., первые в конце
марта по ст. ст., последние — 14 мая; для Зап. и средней Сибири и сев.
Казахстана — Эмба, Иргиз, Наурзум, Бараба, Кулунда, западные окраины
Алтая, Минусинск, Красноярск — во второй половине апреля — первой
половине мая (в Каменском округе Новосибирской обл. 17.V, в быв. Змеино-
горском уезде 18 апреля — 1 мая; у Красноярска во второй половине апреля
по ст. ст.; у Минусинска 12 мая, в центр. Алтае у Онгудая первые будто бы лишь
26 мая, но на Чулышмане уже 26 апреля). На Дальнем Востоке пролет отмечен
только в первой неделе мая (Пикан, Зея).
ЧЕГЛОК
129
Известная разница между северными и южными частями ареала таким
образом имеется, хотя установить, цакие именно данные относятся к прилету
местовых, а какие к пролету северных птиц, — трудно. Растянутость сроков*
пролета объясняется участием в нем разных популяций. Равным образом
ошибочен вывод о том, что между прилетом чеглоков и началом их гнездования
проходит большой промежуток времени. По тем же причинам растянут и период
отлета и осеннего пролета, отмеченный в разных местах со второй половины
августа до конца сентября. Валовой пролет падает на первую половину
сентября, но отдельные особи задерживаются до первых чисел октября (в низовьях
Днепра добыт 4 октября; последний в Харьковской обл. 2 октября; у
Астрахани будто бы даже регулярно в начале октября, последний 11 октября; в Крыму
в начале октября нередки, в Наурзуме в сев. Казахстане добыт 5 октября).
На юге — в Крыму, на Кавказе и в Туркестане — пролет падает в основном
на вторую половину сентября (например, у Феодосии 16 октября, у Сочи 23 —
29 сентября, а у Дзауджикау и 7—11 октября; в Фергане последние, по Се-
верцову, 16 октября; у Фрунзе 23—26 сентября, одиночный добыт 17 октября;
у Ашхабада первые пролетные добывались 15 сентября, одиночный у Теджена
22 октября). В виде редкого исключения отдельные особи остаются зимой
в Средней Азии: кроме наблюдения у Красноводска 16.1.1938 (к сожалению,
добыт не был), чеглок был добыт 14.1.1941 г. на озере Джили-куль в
Таджикистане. Также нерегулярно встречается зимою чеглок в Центральной Азии
(Алашань). На зимовках чеглок появляется поздно, в октябре — ноябре.
Биотоп. Охотясь за добычей в воздухе, чеглок держится в светлых
негустых лесах, перемежающихся с открытыми пространствами — степью,
полями, лугами, речными долинами и т. п. предпочитаемый тип леса —старые
высокие сосняки или смешанные леса, только на юге — лиственные леса
(березовые колки ю.-з. Сибири и сев. Казахстана, тугаи туркестанских речных
долин, лиственные леса гор Средней Азии и т. п.). На пролете чеглок встречается
более или менее повсеместно, залетая даже в пустыни (Кара-Кумы), но, по
сравнению с настоящим соколом, менее связан с побережьями и с культурным
ландшафтом городов и селений. В вертикальном отношении распространение чеглока
ограничено пределами -высокоствольной древесной растительности.
Гнездование чеглока отмечено на Кавказе в Армении у озера Севан на
высоте около 2000 м (Соснин и Ляйстер, 1942), в Хевсуретии до 3050 м (Чхикви-
швили, 1943), в Средней Азии в Копет-Даге до пределов арчи, т. е. примерно
до 2000 м, в Таджикистане до 2500 и даже 2950 м (Даль, 1936, Иванов, 1940),
но чеглока нет в безлесном вост. Памире, в Тянь-Шане, Киргизском
хребте, Заилийском, Кунгей- и Терскей-Ала-Тау до 2000—2700 м; на Урале
у Иремели на юге и Тельпосизе на севере, т. е. до 1000—1600 м\ на Алтае
до 2000 м (Сушкин, 1938).
Численность. По сравнению с крупными соколами, нередок;
однако, в отличие от .пустельги, не образует гнездовых колоний и даже во
время пролета держится одиночками и парами (однако Зарудный, 1888,
сообщает, что осенью на пролете под Оренбургом чеглоки встречались
стаями до 50 птиц).
Экология. Размножение. Половая зрелость наступает в возрасте
около 10 месяцев, когда птица находится еще в первом годовом (гнездовом)
наряде; однако, не ясно, все ли особи приступают к размножению в этом
возрасте и не образуют ли они частично резерв холостых птиц, несомненно
имеющийся у чеглоков, так как убитая или погибшая из гнездовой пары птица
обычно быстро, в том же*сезоне, заменяется новой. Пары повидимому
постоянные. Цикл размножения поздний.
Спариванье происходит на деревьях у гнезд (конец апреля, Туркмения).
Гнездовые участки постоянны, гнезда занимаются несколько лет подряд, но
9 Птицы СССР, т. I
130
ОТРЯД ХИЩНЫЕ ПТИЦЫ
может быть, что каждая пара в одном участке имеет несколько (два—три)
последовательно занимаемых гнезда. Размер гнездового участка не установлен-
Указание о гнездовых колониях чеглоков (4—5 пар близ Киева, Шарлемавь,
1915) сомнительно. К другим птицам в гнездовом участке чеглок относится
терпимо (например, Зарудный, 1888, нашел в б. Оренбургском крае на одном
дереве занятые гнезда чеглока, пустельги, черного коршуна, вороны, черноло-
бого сорокопута).
Своих гнезд чеглоки повидимому не строят, а пользуются главным образом
постройками или ворон, или других хищных птиц, или сорок. Гнезда
помещаются высоко от земли, чаще всего в 10—20 му на соснах, елях, лиственницах,
дубах, тополях, карагачах, ивах. Очень редко в виде исключения можно
встретить гнезда на скалах (Киргизский хребет). Существенное условие в выборе
места для постройки гнезда—возможность свободного и широкого обзора
с гнезда окружающей местности.
Кладка происходит в конце мая — начале июня. Например, полные
свежие кладки найдены на Или 20 мая, на Зайсане 31 мая, 9 июня в Белоруссии
у Дольска, 2 июня в Тамбовской обл., в конце мая в Киевской обл. и в
Харьковской обл., в конце мая в Чкаловской обл., 23 мая на Имане в Приморье,
в начале июня у Красноярска. Имеется ряд указаний на еще более поздние
кладки, но здесь возникает вопрос, который пока еще остается неясным: нет ли
тут вторичной кладки взамен утерянной, ибо свежие кладки в низовьях Сыр-
Дарьи были найдены 20 июля; на озере Севан в Армении 26 июня, на озере
Искандер-куль в Таджикистане 6 июля и т. д. Число яиц в кладке обычно 2—4,
изредка 5 (напр. в Днепропетровской обл., Вальх, 1899) и даже 6 (Наурзум).
Географических различий в величине кладки не установлено. О/^но яйцо
часто бывает неоплодотворенным, и птенцов в выводке чаще всего два.
Колебаний плодовитости в зависимости от кормовых условий того или иного года
также не установлено. Изредка в выводках встречается до 4 птенцов
(Свердловская обл., Волчанецкий, 1927; Котон-Карагай,южн. Алтай, Сушкин, 1938).
Размеры яиц (Харьковская обл., Сомов, 1897, Гебель, 1879) 39,4—44,5X31,1 —
34 мм. Окраска яиц: фон охристый, густо покрыт глинисто-буроватыми
и красновато-бурыми пятнами и точками, иногда сливающимися в большие
бурые поля.
У чеглоков, как и у других соколов, 3 наседных пятна —два на брюшной
стороне и одно на груди. Процесс насиживания у нас не прослежен; судя по
разнице в возрасте птенцов при двухдневном промежутке между откладкой яиц,
насиживанье начинается с первого (но возможно, что и со второго) яйца.
Продолжительность насиживания 28 дней, причем некоторое участие в
высиживании принимает вероятно и самец (у самцов естьнаседные пятна), который
ловит и приносит добычу и для выводка и для насиживающей самки. Пуховые
птенцы в разных местностях (Витебск, Харьков, Москва, Куйбышев, Наурзум,
Сыр-Дарья, Минусинск) отмечены с последних чисел июня и в разных числах
июля. Появление птенцов иногда запаздывает (от вторых «дополнительных»
кладок?). Гнездовый период длится около месяца.
В месячном возрасте птенцы выходят из гнезда и начинают перепархивать.
Второй пуховой наряд надевается, судя по данным западноевропейских
авторов, в возрасте около 8 дней. Вопрос о птенцовой смертности и о каннибализме
у молодых чеглоков не изучен, но повидимому гибели птенцов от недостатка
пищи не бывает (единственный случай каннибализма в совершенно необычных
условиях для Минусинской степи описан Зверевым, 1928; это случай, когда
2 птенца чеглока выкармливались известный период кобчиком и перед вылетом
старший съел младшего). Подлетки наблюдались с конца июля — середины
августа (6—11 августа, Воронеж; 16—20 августа, Наурзум; 18 августа,
Минусинск; 2 августа, Новгород). Однако птицы с не вполне доросшими маховыми
ЧЕГЛОК
131
и рулевыми добывались и позднее: 28 августа у Музарта в центральном Тянь-
Шане, 5 сентября в Наурзуме и даже 11 сентября в Аман-Карагае в сев.
Казахстане.
Неразбившиеся выводки держатся вместе повидимому до отлета, так они
наблюдались в разных местностях — средней и южной полосе Советского
Союза, в Средней Азии, в Сибири в разные числа сентября. Следует отметить
наблюдение Соснина (1942) о летнем (вторичном) спаривании чеглоков в
Армении 3 июля 1930 г.
Линька. Линька у чеглоков протекает довольно своеобразно; чеглок
линяет на зимовке, и сроки линьки (начала) находятся в менее строгом
соответствии с окончанием периода размножения, чем у настоящего сокола,
балобана и кречета. Наблюдаются и значительные индивидуальные колебания в
сроках линьки. Птицы в первом годовом наряде начинают сменять мелкое оперение
весною в марте—мае в возрасте около 8—9 месяцев, причем смена мелкого
пера у них продолжается до наступления активного периода размножения
(например, у одного самца — Курская обл. — еще 20 мая; у другого — из
Воронежской обл. — даже 12 июня) затем линька приостанавливается,
возобновляясь осенью.
Взрослые птицы (во 2-м годовом наряде и последующих) прилетают к нам
в свежем пере, закончив линьку (начинающуюся главным образом в сентябре)
в марте, иногда в апреле. Однако, иногда бывает, что смена внутренних
первостепенных маховых (9-го и 10-го, изредка даже 8-го) затягивается и кончается
только в конце апреля и даже начале мая (самки, Байрам-али, 24 апреля;
Фрунзе, 9 мая; Махтум-Кала, 3 мая; самец, Багир, 10 мая).
Нелиняющие взрослые птицы в разных частях СССР добывались между
концом апреля и разными числами сентября (поздние даты в средней полосе
Советского Союза 5 сентября, на юге ее 7 сентября, в Башкирии и Приуралье
11 сентября; в сев. Казахстане 18 сентября, в средней Сибири 10 сентября,
в Средней Азии даже 24 сентября; на Кавказе 23 сентября). Однако большинство
чеглоков начинает линьку у нас, причем мелкое перо сменяется сначала на
плечевых, на зобе и брюшной стороне. Смена маховых идет в
последовательности 7—6—5—8 (иногда б—7 и т. д.) и этим до отлета ограничиваются; 5—8-е
маховые сменились только у одного изученного нами самца 8. IX, Телецкое
озеро, Алтай; при этом у него сменилось и почти все мелкое оперение; лишь
в трех случаях встречены птицы, сменившие до отлета 5-е — 7-е маховые
между 18 августа и 16 сентября. В очень редких случаях на гнездовье
сменяется средняя пара рулевых. Обычно же линька чеглоков в СССР захватывает
часть мелкого оперения и 6-е и 7-е первостепенные маховые: птицы в разных
стадиях такой линьки встречаются уже с последних чисел июня (взрослая
самка, Фрунзе 17.VI) и разными числами сентября (взрослый самец, оттуда
же, 26 сентября; взрослая самка, Курск 14. IX), причем линька проходит у них
очень разновременно, но без определенного отношения к географической
широте. Обращает на себя внимание длительность периода линяния — около
7 месяцев (тогда как у крупных соколов только около 5*/г месяцев).
Линька — полная годовая, последовательность смены нарядов
обычная для соколов: первый пуховой наряд — второй пуховой наряд — первый
годовой (гнездовый) наряд — второй годовой (окончательный или взрослый
наряд) — третий годовой наряд и т. д.
Питание. Чеглок в отношении способов нападения на добычу и
кормового режима повидимому еще более специализирован, чем настоящий сокол.
За редкими исключениями, чеглоки берут добычу на лету в воздухе и она,
естественно, состоит либо из птиц, либо из насекомых. Из птиц чеглоки
предпочитают летающие стайками в открытых пространствах виды. Насекомых также
хватают лапами, причем нередко и едят их в воздухе. Быстрота полета чеглока
9*
132
ОТРЯД ХИЩНЫЕ ПТИЦЫ
очень велика (до 150 км в час, Тинберген, 1936, а, вероятно и больше), бьет
добычу он по-соколиному, не делая впрочем таких длинных ставок, как
F. peregrinus.
Потребность в пище взрослой птицы относительно невелика — 25—30 г
мяса в день. В качестве корма чеглока в нашей стране указываются ласт
точки, в особенности береговые и деревенские (по Сомову, 1897, существует
определенная зависимость между пролетом чеглока и ласточек);
жаворонки, дрозды, скворцы, овсянки (например, обыкновенная и Ё. fucata),
коньки (среди них А.
richardi), желтые и белые
трясогузки,
черноголовые чекканы, реполовы,
щеглы, зяблики,
чечевицы, воробьи, также
кулики, перепела, по
некоторым данным
молодые тетерева и серые
куропатки. В качестве
самой крупной добычи
указываются чирок,
тетерка, витютень,
молодые утки, ореховка.
На земле чеглоки
иногда хватают
жаворонков, а из
млекопитающих — грызунов. В
качестве добычи у
чеглоков на Сев. Урале
указывается пищуха, а в
Дагестане — летучие мыши,
которых они ловят по
зорям. Из насекомых
чеглок ловит главным
образом стрекоз и
крупных жуков (хрущи,
навозники, бронзовки,
усачи), отчасти саранчевых,
кобылок, даже муравьев.
Несомненно, что
некоторые насекомые также
собираются чеглоком на земле (в желудках, находил и муравьев, гусениц; есть
наблюдения как чеглоки собирали с травы гусениц Vanessa polychloros).
Как у многих других хищных птиц, у чеглоков имеется индивидуальная
специализация в питании. В частности, одни особи (и семьи) предпочитают
питаться насекомыми, другие—птицами. Впрочем, эти различия носят отчасти
сезонный характер: в период валового лета насекомых главную пищу
составляют насекомые, в период массового вылета из гнезд молодых мелких птиц—
подлетки. Индивидуальная специализация отдельных птиц выражается и при
выкармливании ими своего выводка, хотя повидимому как правило в раннем
возрасте птенцы выкармливаются преимущественно птицами. Насекомых
чеглоки поедают на лету, птиц ощипывают всегда где-либо на дереве. Кормовые
остатки — типичные соколиные поеди из костей плечевого пояса и больших
перьев, а также погадки (размеры последних около 3X1,5 ел*). Охотятся
чеглоки сравнительно поздно, утром после того,, как воздух обогреется и
Рис. 43. Чеглок (самка, молодая птица, самец).
ЧЕГЛОК
133
начнется лет насекомых, а по вечерам до полной темноты (иногда даже при
полной луне ночью, Цедлитц, 1920).
Полевые признаки. На полете чеглок похож на обыкновенного сокола,
но крылья уже и длиннее; еще более похож он на стрижа, но он больше
размером, примерно с голубя. Полет очень быстрый, взмахи крыла чередуются со
скольжением, не парит. Хорошо выделяется по контрасту белое горло («бело-
горлик»), темные полосы вокруг горла и идущее книзу темное пятно на ушных
партиях, образующее как бы двойные «усы». Голос звонкое «кли-кли-кли» или
«кик-кик^кию». Встречается чеглок в местностях, где древесная растительность
перемежается с открытыми пространствами.
Описание. Размеры и строение. В общем напоминает уменьшенного
настоящего сокола, но склад более легкий и крылья относительно длинней. Вес самцов (3) 165,2,
190,208 г, самок (3) 237,5,242,247г.Длина самцов (12)320—350,самок (10)330—368 см; в
среднем 334 и 350,7 мм; размах крыльев самцов (12)720—810, у самок (10)780—840мм,
всреднем 757,25 и 820,5 мм; длина крыла самцов (119)247—275, самок(110)258—296 мм, всреднем
259,8 и 275,6 мм. Крылья у чеглока длинные и узкие, в сложенном виде выходят обычно за
конец хвоста. Формула крыла 2>1>3. Клюв относительно слабый и короткий. Цевки
короткие и относительно толстые, оперены в верхней половине; пальцы очень длинные и тонкие,
средний палец длиннее цевки, внешний (второй) палец заметно длинней внутреннего
(четвертого).
Окраска. Первый пуховой наряд у чеглока — чисто белый — носится в течение
первых 8—14 дней жизни; второй пуховой наряд — серовато-белый с охристым брюшком. Первый
годовой (гнездовый) наряд надевается в возрасте около месяца. Он сходен у обоих полов.
Общий тон спинной стороны темнобурый, в свежем пере с сероватым налетом, с охристыми
светлыми каемками, особенно развитыми на голове и больших перьях — плечевых, больших
кроющих крыла; низ беловато-охристый с продольным бурым рисунком; маховые — темно-
бурые со светлыми каемками и охристыми поперечными пятнами на внутренних опахалах;
рулевые черно-бурые с охристой вершинной полосой и рыжевато-бурыми поперечными
пятнами, отсутствующими на средней паре. Радужина темнобурая, клюв черновато-серый,
голубоватый у основания; восковица и кольцо вокруг глаза голубоватые, лапы бледножелтые,
когти черные. Взрослые птицы на спинной стороне бурые с более или менее развитым сизым
оттенком (особенно у самцов); впрочем этот признак изменчив и в обношенном пере
пропадает; на задней части шеи по сторонам имеются белые пестрины; полосы под глазом, усы и
кроющие уха черноваты; подхвостье иногда с темным поперечным рисунком; грудь, бока,
брюшная сторона — белые, с широкими бурыми продольными пятнами; нижняя (задняя)
часть брюшной стороны, перья голени и подхвостья у самцов яркорыжие, у самок —
охристые или рыжеватые с бурыми пятнами; маховые черно-бурые с рыжеватыми поперечными
пятнами на внутренних опахалах; рулевые бурые, средние из них одноцветны, остальные
с рыжевато-охристыми поперечными пятнами, особенно развитыми на внутренних опахалах.
Неоперенные части — как у молодых, но восковица и кольцо вокруг глаз желтые, а ноги
яркожелтые. Во втором годовом наряде окраска спинной стороны тела бурее, без сизых
оттенков, а подхвостье и голени у самцов окрашены как у старых самок.
Северный чеглок Falco subbuteo jakutensis But.
Falco saturates Buturlin. J. f. Orn. 1908, стр. 283, Абый на Индигирке, nomen pracoc-
cupatum. —Falco subbuteo jakutensis. Бутурлин, Наша Охота, 1911, стр. 71, новое имя взамен
предыдущего.
Распространение. Ареал. Северная часть области распространения вида
в Европе и Азии (к северу приблизительно до границы высокоствольного леса).
Чеглок не отмечен достоверно в Русской Лапландии, в Финляндии не выходит
(за исключением быть может отдельных случаев) за 65° с. ш.; в области Белого
моря на Соловецких о-вах и у Неноксы около 65° с. ш.; по этой же
параллели идет граница и восточней (Архангельск, Пин era), несколько повышаясь
к северу в бассейне Печоры (Усть-уса, 66°); наблюдался даже у Великовисоч-
ного за 67°; на Оби примерно до 66—67° (м. Круглый на Ямале), в Туруханском
крае примерно до 63° (р. Б. Елагуй), далее к востоку по Енисею граница резко
понижается — до 58° (Енисейск), достигая однако в низовьях Нижней
Тунгуски 65°; в сев. Якутии чеглок рарпространен спорадично, но он
достоверно найден на Яне ниже Верхоянска (Ченики, Эттях), в нижнем течении
Г34
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Индигирки (Абый, около 68°) и Колымы (Бутурлин, 1908), Последнее однако
требует подтверждения, так как самая северная точка, откуда имеются
экземпляры,—Сеимчан, около 63? с. ш. На крайнем с.-в, Сибири в бассейне Анадыря
северный чеглок не найден, но он гнездится по северному побережью Охотского
моря (экз. из Наяхана) и на Камчатке, хотя и редко. Южная граница —
ввиду постепенных переходов к номинальной расе — может быть определена
лишь условно и за нее принимается Обонежье, Вологодская обл. (Яренск,
бассейн Вычегды, примерно 62° с. ш.), средняя Печора, район Тюмени в Зап.
Сибири, Байкал, северный берег Охотского моря и вероятно Камчатка. Область
зимовок неясна, так как большинство авторов не отличают этой формы от
других: темные чеглоки вероятно этой формы найдены в Ладаке ив Ассаме.
Пролетные птицы изучены из Сахалина, Приморья, Прибайкалья, из вост.
Туркмении (Кушка).
Характер пребывания и биотоп— как у предыдущей
формы. На севере встречается только в тайге, пересеченной, конечно,
открытыми пространствами.
Даты. Прилетает северный чеглок в разные числа мая — несколько
позже, чем F. s. subbuteo: около середины мая у Архангельска, на севере
Вологодской обл. и в сев. Приуралье, в начале мая на Олекме (5 мая), уже
в конце апреля на Шантарских о-вах (Шульпин и Дулькейт, 1937), в конце
апреля на Камчатке (Аверин, 1948). Отлет его происходит в период от конца
августа до середины сентября (Карелия, средняя Печора, Тюмень, Олекма);
на Камчатке же (по Бергману, 1935) только в начале октября, последние
наблюдались 5 октября, хотя на северном побережье Охотского моря, по Мидден-
дорфу, 1857, уже 13 сентября, а в Кроноках, на вост. Камчатке (по Аверину,
1948), последние 26 сентября. Пролетные в Приморье появляются в конце
августа (22.VIII р. Хунгари; 25-VIIIо. Лангр; 27.VIII —Иман, авеснойтамже
встречены еще в середине мая).
Численность. В северной тайге чеглок распространен спорадически
и в общем должен считаться редким.
Экология. Сведения очень скудны.
Размножение. Сильно насиженные яйца были найдены на р. Обве
(сев. Приуралье) 24 мая (Резцов, 1904). Выводки на Печоре были обнаружены
в конце августа, однако у Обдорска летные птенцы попадались будто бы 1
августа.
Линька. Нелиняющие взрослые птицы добывались между 7 мая и 9
сентября. Начало линяния—смена б-го и 7-го первостепенных маховых
—отмечено у взрослых птиц в августе, у некоторых в конце этого месяца сменяется
и 8-е маховое.
Питание. В качестве добычи северного чеглока отмечены: стрекозы
и мелкие птицы (в частности зеленые коньки), и белые трясогузки,
обыкновенная чайка, подлетки рябчиков и белых куропаток. В общем, кормовой
режим сходен с питанием номинальной формы, но птицы занимают у
северного чеглока в питании большое место.
Описание. Размеры и строение. Максимальные (и средние) размеры северных
чеглоков уступают размерам южных. Крыло самцов(35) 243—270, самок( 11)262—285, в
среднем 256 и 272,3 мм. Вес самцов 131, 200, 204, 208 г; самок 141, 245, 258, 275 и 325 г.
Окраска. По сравнению с номинальной формой окраска северного чеглока очень
темная, сизого цвета на верхней стороне нет даже в свежем пере; общий тон спинной стороны —
аспидно-бурый, с черноватой головой; темный рисунок на брюшной стороне очень широкий
и черноватый, рыжий цвет голеней и подхвостья яркий и насыщенный. Молодые также очень
темные, ободки перьев спины и крыльев интенсивно ржавчато-рыжие, общий тон окраски
спинной стороны черноватый.
Систематические замечания. Географическая изменчивость чеглоков в сев. и Вост.
Европе и в сев. Азии не носит ярко выраженного характера и проявляется в постепенном
потемнении окраски в направлении с юга на север. Самые светлые из изученных нами чеглоков
ДЕРБНИК
135
происходят из Закавказья и Туркестана, салые темные — из северной тайш от Архангельска
до Якутии. Переходы между темными северными п светлыми южными популяциями носят
вполне постепенный характер — и если принимать крайние популяции за подвиды — гранима
между последними может быть проведена лишь условно. С формально-номенклатурной точки
зрения существенно то, что название subbuteo относится именно к промежуточной, а не к одной
из крайних популяций. Все же среднеевропейские птицы, гнездящиеся в хвойных,
смешанных и лиственных лесах, ближе к южным, чем к северным, таежным. Поэтому удобнее
выделить последних, тем более, что они выделяются и экологически (несколько более поздний
цикл годовых периодических явлений; биотопическое распределение; почти исключительная
орнитофагия и т. п.). Все же разделение европейских и североазиатских чеглоков на две
географические формы может быть только условным.
7. Дербник Falco colambarius L.
Falco columbarius. Linnaeus. Syst. Nat. Изд. X, 1758, стр. 90, Каролина.
Русское название. Старинное слово, быть может имеющее общий корень со
словом «деряба» или названием ястреба в некоторых местностях — «дербень». У Алексея
Михайловича в «Уряднике Сокольничья Пути» — дермлиг. Впрочем, слово «дремлик» для
обозначения этой птицы для западно-славянских языков известно у вендов.
Распространение. Ареал. В область распространения дербника входят:
Сев. Америка, Европа и сев. Азия; в Америке от Аляски, Юкона и Мекензи
до Лабрадора, Ньюфаундленда, к югу до южн. Квебека, штатов Мен, Айова,
Колорадо, ю.-з. Уайоминга, сев. Юта, сев. Невада, южн. Орегона (на гнездовье),
на зимовках — к югу до южных Штатов, Мексики, северо-западных частей
Южн. Америки; в Европе от Исландии, сев. Скандинавии до Прибалтики,
средней полосы Европейской части СССР (на гнездовье), в Азии и лесной полосе
от Зауралья до Приамурья и Сахалина, на севере до Лапландии, Канина,
Тиманской и Большеземельской тундры, Колгуева, на Оби до 67° с. ш., на
Енисее до 72°, на Хатанге до 74°, на Колыме до 68°, до Анадыря; кроме того,
в степях южн. Зауралья и в горах центрального Тянь-Шаня, Алтая и
Монголии; зимовки в средней и южн. Европе, сев. Африке, М. Азии, Туркестане,
Индии, в Японии и Китае.
Характер пребывания. Дербник главным образом перелетная
птица, во всяком случае в пределах СССР только перелетная, за исключением
быть может гор Алтая и Средней Азии. Пролет ранний, в Зап. Европе уже
в марте, у нас — в зависимости от широты — от мая на севере до апреля на
юге. Отлет с северных гнездовий происходит в августе, с южных в сентябре
и даже октябре.
Биотоп. Открытые места, в лесной зоне, например, моховые болота
среди сосняка и т. п.; тундра и лесотундра; степи и лесостепи ю.-з. Сибири
и сев. Казахстана; сырты азиатских гор. В холодное время года в самых
разнообразных открытых ландшафтах или в угодьях, где древесная растительность
перемежается с открытыми пространствами.
Численность. Умеренная, периодических колебаний не
установлено. Известное скопление дербников замечается только на зимовках в
Закавказье и Средней Азии.
Подвиды и варьирующие признаки. В Америке имеется три формы дербника — бледный
F. с. rkhardsoni R i d gway, 1871 из степной зоны штатов Альберта, Саскачевана, Монтана
и Дакота; темный F. с. suckleyi R i d g w a y, 1873, на северо-западе ареала и, наконец,
промежуточный по окраске F. с. columbarius L. на востоке ареала; в Восточном полушарии
различаются очень темный F. s. subaesalon В г е h m (1827) из Исландии; F. с. aesalon T u n s t.
(1771) несколько посветлее и с более длинными крыльями в с.-в. Европе, на Сев. Урале и
в Зап. Сибири; еще более бледный F. с. insignis Clark (1907) в бассейне Енисея, Лены,
Индигирки, Колымы; наконец, более темный, близкий к американским F. с. pacificus S te g-
mann (1929) на с.-в. Сибири в бассейне Анадыря и на Охотском побережье; обособленный
ареал на юге от степей Зауралья и ю.-з. Сибири занимает бледная форма F. с. chrisiiani —
ludovici Kleinschmidt (19(7), другая светлая форма в горах Средней Азии, в
Монголии и Забайкалье, на Алтае — F. с, lymani Bangs (1912). Кроме окраски, варьирующей
в оттенках, изменчивы и размеры: длина крыла дербника, распространенного на европейско-
136
ОТРЯД ХИЩНЫЕ ПТИЦЫ
азиатском материке, возрастает в направлении с запада на восток; самые
длиннокрылые формы — южные степной и горный подвиды.
Карта 15. Распространение дербника
Falco, columbarius
1—F. с. aesalon, 2— F. с. christiani-ludovici. 3 —
F.c.insignis, 4 — F. c. pacificus,5 — F. c. lymani.
Ареалы американских подвидов не обозначены.
а — граница гнездовий, б — неясные границы
распространения, г — область зимовок.
Обыкновенный дербник Falco
columbarius Т u n s t.
Falco aesalon.
1771, стр.
T u ns ta 1 1
1, Франция,
птицы.
Orn. Brit.,
зимующие
Синонимы: Falco regulus Pa lias.
Reise d. versch. Prov. d. Russ. Reichs,
11,1773, стр.707, Зап. Сибирь —Falco
sibiricus Shaw. Gen. Zool. VII, 1809,
стр.* 207, Сибирь— Falco columbarius
alaunicus. Федю шин, Докл. Акад.
наук, А. 1927, стр. 71, вост. Россия.
Распространение. Ареал.
Обыкновенный дербник
распространен в сев. Европе и Зап. Сибири —
от Фарерских о-вов, Ирландии,
Шотландии и Англии (горы Дербишира, сев. Йоркшира) до Скандинавии
и Финляндии (до 62° с. ш.) и Польши (Беловежская пуща). В СССР—от
Прибалтики, Кольского п-ова (Кильдин), на Мезени, Канине, в Тиман-
ской тундре, на Колгуеве, на Печоре у Усть-цыльмы (65° 26'), в низовьях
Оби (67° 50'), на южн. Ямале (Ямсале, Находка, за 66°), на Тазовском
п-ове, в низовьях Енисея под 72° (Бреховские о-ва в дельте, Попгейм, 1898);
ДЕРБНИК
137
залеты отмечены у Шпицбергена (Мунстерьельм, 1910) под 73? 30'и 74°, на
Вайгаче (Хейглин, 1872), на Новой Земле (где быть может гнездится), насе-
верном берегу Ямала (о. Белый), на р. Рене из архипелага Кьелльмана (Кол-
летт и Нансен).
Южная граница гнездовой области проходит в Эстонии, у Пскова, Минска,
Смоленска, Рязани, Москвы, Ярославля, Горького, Казани, горных частей
Южн. Урала, т. е. примерно по 55° с. ш.; в Зап. Сибири немного севернее, около
56—57° (Нарым—Томск), далее в лесистых частях с.-в. Алтая (Телецкое озеро)
и южных склонов зап. Саяна. Восточную границу в области Енисея установить
трудно, но неотличимые от aesalon темные особи добывались на Хатанге (71° 49'
с. ш., Скалой, 1938) и у "Байкала (Култук, Баргузин). За Енисеем
встречаются однако и светлые особи (у Туруханска, на Таймыре и т. д.), так что здесь
распространена вероятно «смешанная» популяция.
Имеются указания на гнездование в Тульской и Тамбовской обл.,
но указания эти требуют подтверждения. Что же касается сведений о
гнездовании на Украине (на границе Полтавской и Харьковской обл., в
Черниговской обл. и на Дону), то это какое-то недоразумение. На пролете и зимой дерб-
ник обнаружен в Зап. и южн. Европе, в с.-з. Африке (Атлас), в Египте,в средней
и южной полосе Европейской части СССР, в Крыму, Закавказье, в Малой Азии,
Иране, в Туркестане от Туркмении до Киргизии и ю.-в. Казахстана, в Синьц-
зяне и южн. Забайкалье. Область зимовок, где дербник встречается регулярно,
расположена в Зап. и южн. Европе, в СССР от южной полосы в европейской
части до Крыма и Закавказья, главным образом восточного, и в Туркестане.
Севернее (Псков, Смоленск, Калуга, Москва, Ярославль, Владимир, Рязань,
Казань, Курск, также Прибалтика) — в мягкие малоснежные зимы зимуют
отдельные особи.
Характер пребывания. Дербник перелетная птица, но может
быть, что некоторые особи в южной полосе гнездовой области оседлы, так как
там дербники встречаются и зимою. Возможно, что это прикочевавшие с севера
особи, так как растянутость сроков пролета дербника в лесной зоне указывает,
что они оттуда откочевывают по всей вероятности к югу. В Англии дербники
оседлы, или, если совершают кочевки, то незначительные.
Даты. Прилет на северные места гнездовья происходит в мае: в
Лапландии и Тиманской тундре около середины этого месяца (в Лапландском
заповеднике в среднем 18 мая, крайние сроки за 5 лет наблюдений 12 мая и 27 мая,
Владимирская, 1948), между 5 и 28 мая на нижней Печоре (Зибом, 1876),
на Новой Земле только 8 июня (Сконнинг, 1903); так как южнее прилет
проходит повидимому одновременно с пролетом, то даты первого установить трудно.
Весенний пролет в зап. Закавказье отмечен в конце марта (вероятно, начинается
и ранее), продолжаясь весь апрель; в Туркмении в конце февраля и первой
половине марта, в северных частях Средней Азии в марте, главным образом,
во второй его половине, а последние пролетают в начале апреля; в начале
апреля последние дербники отмечались и в Крыму, в конце марта на юге
Украины; в северных частях Украины пролет и отлет идет с конца февраля и
до конца марта (Харьков); запоздалые особи попадались еще в начале апреля;
в средней полосе (Псков, Калинин, Москва) пролетают в апреле от начала и до
конца месяца; в Прибалтике пролет отмечен во второй половине марта — первой
половине апреля. Осеннее движение на севере начинается уже в. августе,
главным образом в начале сентября. На Новой Земле последние дербники
отмечались 16 сентября, на Колгуеве — 17 сентября; пролет на южн. Ямале 18
августа — 2 сентября (Кучерук и Дунаева, 1940), на реке Таз 9 сентября; у Яму-
тарида на Таймыре — 13 сентября (Тугаринов и Толмачев, 1934), в
Лапландском заповеднике —между 27 августа и 16 сентября, в среднем 3 сентября;
у Варангер-фьорда даже в начале октября (Плеске, 1886). В средней полосе
138
ОТРЯД ХИЩНЫЕ ПТИЦЫ
пролет идет с конца августа до сентября (на средней Печоре еще 28—30 сентября,
у Тюмени 23 сентября), отдельные особи и позднее (например, у Кирова 12
октября). Часть птиц задерживается, и в зависимости от кормовых и
температурных условий движение к югу продолжается и в сентябре, например, заметный
пролет в Каменском округе, Зап. Сибирь, отмечен до двадцатых чисел октября;
у Телецкого озера на Алтае до двадцатых чисел ноября. На местах зимовок
дербники появляются в разное время — в Закавказье с конца сентября до
разных чисел ноября, в Туркмении в конце октября и позднее, на Аральском
море с первой половины октября (Зарудный, 1916), на Сыр-Дарье с конца
сентября (Спангенберг, 1936), тогда же в "Семиречье. По некоторым наблюдениям,
осенью сначала летят молодые птицы, а старые, в особенности самцы, иногда
задерживаются и на зиму в средней полосе (Сомов, 1897). В холодные и
снежные зимы дербники продвигаются на юг далее обычного. В известной мерс
период пролета совпадает с пролетом разных воробьиных — дрозда белобровика
и рябинника (сев. Казахстан), жаворонков (Сыр-Дарья), пуночек (Печора).
Биотоп. Тундра, лесотундра и участки тайги, перемежающиеся с
моховыми болотами и другими открытыми пространствами; глухой тайги избегает. На
зимовке в открытых ландшафтах, малоснежных, вместе с мелкими птицами,
главным образом, жаворонками. В горы идет невысоко, на Алтае до 900 м9
однако в Армении на зимовке отмечен на высоте 1550 м (Даль, 1948).
Численность. Относительно редкая птица. Как и у других птице-
ядных хищников, периодических колебаний численности повидимому не
бывает. Местами на зимовке дербники собираются в значительных количествах
(ю.-в. Закавказье, Туркмения, предгорья Тянь-Шаня).
Экология. Размножение. Половая зрелость наступает на
следующую весну после рождения, т. е. в возрасте около года, когда птица находится
еще в первом годовом (гнездовом) наряде. Пары повидимому постоянные, так
как самец и самка держатся вместе и зимою (Сомов, 1897). Начало цикла
размножения наступает позднее, чем у северных подвидов настоящего сокола
(перелетная птица). Гнезда размещаются- на скалах (Лапландия), на земле —
на кочках, в болотах, среди сосняка в лесной, полосе, реже на деревьях:
лиственнице — в лесотундрах, соснах и елях — в лесах; наконец, среди
кустарниковых зарослей по береговым обрывам в тундре. Одно и то же гнездо
используется несколько лет подряд. Гнезда на деревьях устраивают вероятно сами
дербники, иногда же они отнимают гнезда у других птиц. Расстояние между
гнездовыми парами невелико —в благоприятных условиях не более Щг км.
Кладка происходит в мае, полные находят в конце этого месяца (свежая кладка
на Кольском п-ве отмечена 1 июня, Пирсон, 1904; в Шведской Лапландии
22 мая, Уоллей, 1864; неполная кладка в Серебряном Бору близ Москвы 20 мая;
насиженная кладка 9 мая, Псковская обл.). Число яиц в кладке 3—4, иногда
2 или 5, в Шведской Лапландии даже б и 7. Окраска яиц охристая с более или
менее крупными и густо расположенными рыжевато-бурыми или красновато-
бурыми крапинами. Размеры (32) 37—42,3 х 29,1—33,5 мм.
Число птенцов в выводке чаще 3, нередко 4, изредка 5 (Ямал, Лапландия).
Птенцы вылупляются в начале июля (Лапландия), но вероятно обычно с конца
июня. Насиживание, следовательно, продолжается около месяца (точно у нас
не прослежено). Насиживают и самка и самец (у первой, как обычно у соколов,
три наседных пятна). Около середины июля птенцы уже во втором пуховом наряде
с началом развития перьев (встречены в тундре 14 июля, Ямал); птенец во
втором пуховом наряде без появления перьев добыт на Канине 28 июня. Впрочем,
такая разница в возрасте может быть отнесена* за счет насиживания с первого
яйца. Из гнезда молодые выбираются еще нелетными, с недоросшйми крыльями
и хвостом, в конце июля и в разные числа августа становятся на крыло (птенцы
с не вполне доросшими крыльями встречены в Витебщине 21 июля; Федюшин,
ЛЕРБНйК 139
1926; в Лапландия 26 июля, на Канине — 27—31 июля, на Печоре еще 4
августа, на Ямале 5 августа, на реке Таз у Хальмер-ееде еще 14 августа). С
первой половины августа попадаются вполне развитые молодые дербники
(экземпляры от 4 августа до 19 августа, Соловецкие о-ва, Карелия, Лапландия,
Тиман, Печора, Ямал, Ивановская обл., Москва). Весь август выводки
держатся вместе. Гнездовый период жизни птенцов таким образом длится около
месяца.
В период насиживания и выкармливания выводка ловит и приносит
добычу самец, а оделяет птенцов (и обогревает их на гнезде) самка. В начале
добыча приносится ощицанной, причем обработка ее производится вне гнезда.
Гнездо содержится в чистоте: самка выносит из него остатки пищи. Только что
покинувшие гнездо птенцы мало летают и сидят почти неподвижно; пищу им
носят родители, но уже не ощипанную, а целых птиц (Ямал).
Линька. Несмотря на большую широтную разницу мест обитания
отдельных популяций дербника, у всех них линяние (как и размножение)
протекает приблизительно в одно время. Линька бывает раз в год, полная (на нашем
материале не подтверждается указание Уайзерби, 1924, о том, что у взрослых
дербников, кроме полной годовой, имеется еще частичная весенняя линька
между февралем—апрелем, охватывающая мелкое перо). Птицы в первом
годовом наряде начинают линьку ранее взрослых: смена мелкого оперения заметна
уже в начале апреля (2 апреля, Балаклава), к концу этого месяца сменяется
средняя пара рулевых; линька продолжается до конца мая (22 мая,
Лапландия, смена мелкого пера, нет средних рулевых, 6-е маховые растут,
а 5-е выпали); затем повидимому у гнездящихся птиц линька прерывается,
возобновляясь после вывода птенцов и заканчиваясь в то же время, что и у старых
птиц. Можно предполагать, что у оставшихся холостыми и лишь потенциально
половозрелых прошлогодних молодых линяние продолжается равномерно,
но идет медленно, так как ранее начала сентября птиц в свежем (втором
годовом) наряде не встречено. Линяние взрослых проходит между серединой июня
н концом августа или началом сентября. В первой половине сентября
встречаются уже совсем перелинявшие птицы. Общая продолжительность линьки,
следовательно, равна приблизительно 2г[2 месяцам. Линька у самок начинается
несколько ранее, чем у самцов. Причина, повидимому, заключается в
разделении труда при выводе потомства, как и у других соколов. У самок линяние
идет на 2 маховых пера впереди по сравнению с самцами. В некоторых случаях—
по неясным причинам—линяниесущественно запаздывает (у самки от 28сентября,
Усть-Ылыч, на средней Печоре, 2-е маховое еще не доросло, 1-е и 10-е
отсутствуют; может быть, что это вызвано второй «дополнительной» кладкой).
Последовательность смены первостепенных маховых: 6—7—5 (или 7—6—5 или даже
5—6—7) — 8—4—3—9—2—10—1. Рулевые сменяются от средней пары к краю
партии с тем, однако, что смена 6-й крайней пары предшествует смене 5-й.
Последовательность ■ смены нарядов: первый пуховой — второй пуховой
наряд — первый годовой (гнездовый) наряд — второй годовой (окончательный)
наряд и т. д.
Питание. Основную пищу дербника составляют, главным образом
мелкие птицы, и также мелкие зверьки, и судя по относящимся к другим
странам сведениям — насекомые (Шведская Лапландия). Добычу дербник берет
главным образом на полете, но также и с земли. Способ охоты: низкий
бреющий полет среди кустарника, над землею и т. п., реже4«ставки» в открытых
пространствах; иногда дербник подкарауливает добычу, сидя на земле и бросаясь
на приближающихся птиц и зверьков (Ямал). В негнездовое время дербники
иногда охотятся и парами, причем одна из птиц летит низом, а другая
метров на 10—20 выше (Сомов, 1897). Отмечено нападение дербников на
выпугиваемых другими хищниками птиц.
140
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Основная добыча — птицы открытых ландшафтов: коньки, трясогузки* ов-
сянки, жаворонки и т. п., в гнездовыи период в значительной мере подлетки;
как и другие хищники, дербник в пределах гнездового участка не охотится.
Еис. 44. Дербник (7 —взрослый самец, ^—-взрослая самка, 3 — молодой самец,
4 —молодой самец восточносибирского-дербника).
ДЕРБНИК
141,
Вылет молодых воробьиных.совпадает на севере со временем выкармлива*
ния дербниками молодых — в августе. Полет дербника менее быстр, чем у
чеглока, но все же и он может ловить ласточек (Мензбир, 1895). В гнездовое время
в качестве кормов дербника на севере отмечены из птиц пуночки, лапландские
подорожники, трясогузки, краснозобые коньки, в других условиях —
лапландские подорожники (главным образом), также краснозобые коньки, овсянки-
крошки, чечетки, каменки, рябинник, юрок—на южн. Ямале; там же какой-то
нырок, чирок-свистунок, белая куропатка, дупель, хрустан и золотистая ржанка,
белохвостый песочник; кормовой режим отдельных пар довольно значительно
разнится. В Кузнецкой степи и Салаирском кряже в качестве корма отмечены
домовые воробьи и скворцы. На пролете в зимовках дербники кормятся
овсянками, жаворонками, воробьями (Москва), дроздами (сев. Казахстан), рогатыми
жаворонками, обыкновенной и степной овсянками, пуночкой, воробьями, даже
домашними голубями (Харьков), жаворонками, болотными овсянками, бекасами
и гаршнепами (Сыр-Дарья); жаворонками разных видов, особенно хохлатыми
и серыми, также скворцами (Туркмения); птицами, размерами до бекаса и чирка
(Ленкорань). Кроме того, ЭверсманЦ (1866) указывает в качестве добычи
дербника молодых тетеревов и серых куропаток, а на средней Печоре наблюдалось
нападение дербника на глухарят. Из млекопитающих большую, хотя
второстепенную роль в питании дербника играют лемминги. Для Европы это
относится к норвежскому лелшингу, для Ямала к копытному. Для Лапландии даже
отмечается возрастание плотности размещения гнезд дербников и увеличение
плодовитости до 4—6 и даже 7 яиц вместо 2 в «урожайные» по леммингу годы.
Из других видов в качестве корма дербника указывается полевка Stenocranius
major (Ямал), полевка Словцова (Славгород), кроме того землеройки и т. д.
(Лапландия).
Полевые признаки. Дербник —мелкий сокол с относительно короткими
крыльями и длинным хвостом; размеры сходны с самцом перепелятника, но
крылья острее, вся фигура плотнее. Самец — характерной окраски, сизой на
спине и беловатой или рыжеватой с продольным темным рисунком на брюшной
стороне; молодые и самки на расстоянии кажутся сверху бурыми, снизу —
бурыми со светлыми краями перьев. На полете крылья обычно серпообразно
изогнуты, движения быстры и поворотливы, птица летит обычно «низом»,
не трясется и не парит. Голос «ки-ки-ки» или «кек-кек-кекк
Описание. Размеры и строение. По сложению дербник несколько напоминает
миниатюрного кречета: хвост относительно длинный, крылья в сложенном виде не доходят
до его конца. Формула крыла 2>3> 1 или 3>2>1. Имеются вырезки на внутренних опахалах
1-го и 2-го маховых. Цевка относительно длинная, оперена до половины. Пальцы длинные
и тонкие, наружный длиннее внутреннего. Надклювье сжато с боков, с узким гребнем.
Дпина тела самцов (5) 276—311, самок (8)315—343, в среднем 296 и 322 мм. Размах самцов
(4)562—625, самок (6) 635—691, в среднем 595,5 и 663 мм. Крыло самцов (44) 192—208, самок
(47) 211—224, в среднем 197,8 и 223,7 мм. Вес самцов (4) 170—215, самок (4) 218—255,
в среднем 186,5 и 231,5 г.
Окраска. Первый пуховой наряд чисто белый из редких длинных пушинок.
Второй пуховой наряд гуще и плотнее, серовато-белый на спинной стороне, охристо-беловатый
на брюшной. Первый годовой (гнездовый) наряд сходен у обоих полов, но в дальнейшем
окраска самца и самки различна. В гнездовом наряде спинная сторона темнобурая с узкими
ржавыми каемками перьев, со светлыми пестринами на затылке и черноватыми наствольями
на темени; маховые темнобурые с охристым поперечным рисунком из пятен на внутренних
опахалах, рулевые темнобурые со светлой вершинной каймой и светлыми охристыми
поперечными полосами; брюшная сторона охристая с бурыми продольными пятнами, перья
боков бурые с беловатыми поперечновытянутыми пятнами. Иногда на больших перьях
(плечевых, кроющих крыла) ржаво-буроватые поперечные пятна; такие птицы напоминают
восточносибирских дербников.
Взрослые самки (во втором годовом и последующих нарядах) сходны в общем с
молодыми, но посветлее по общему тону, с большим развитием поперечных пестрин на верхней
стороне, с сизоватым (а не бурым) надхвостьем; иногда попадаются особи, по всей вероятности
старые, с сизым налетом на верхней стороне тела, особенно на пояснице и надхвостье^ с серо-:
142
ОТРЯД ХИЩНЫЕ ПТИЦЫ
ветыми (поперечнополосатыми) рулевыми, со слаба испещренной брюшной стороной — такие
птицы в известной мере приближаются к самцрвой окраске.
Взрослые самцы на спинной стороне темно сизо-серые, с черными стволами перьев;
надхвостье светлее спины, голова цвета спины с широкими черными наствольями и светлыми
краями перьев; задняя сторона шеи рыжевата с темными пестринами; усы слабо заметны;
маховые аспидно-бурые с беловатым поперечным рисунком на внутренних опахалах; рулевые
сизые с широкой черной предвершинной полосой и с черноватым поперечным рисунком,
нередко, впрочем, отсутствующим или очень ослабленным; брюшная сторона
охристо-беловатая, с темным продольным рисунком из пятен, иногда — лишь с наствольными
штрихами: «штаны» более или менее рыжеваты. Радужина темнобурая, клюв сине-роговой,
чернеющий к вершине; когти черные, восковица, кольцо вокруг глаза, ноги — желтые
(во всех возрастах).
Систематические замечания. Эта раса дербника отличается от прочих темной
окраской, слабым развитием поперечного рисунка у самок и молодых и относительно мелкими
размерами. В северной части ареала эти свойства постепенно изменяются — величина
дербников возрастает к востоку, поперечный рисунок получает большее выражение, общий
тон окраски светлеет. Только на крайнем северо-востоке Азии окраска дербников опять
темнеет, приближаясь к окраске американских форм этого вида. Поскольку все такие
изменения проходят постепенно (в пространстве), выделение подвидов по таким признакам может
быть лишь условным. Поэтому к крайним вариантам — темному североевропейскому и
светлому из заенисейской Сибири — едва ли можно прибавлять еще какие-либо промежуточные
формы, тем более, что экологических различий между всеми этими популяциями нет (иное
дело южная степная и нагорноазиатская формы дербника). Таким образом отделение подт
вида F. с. regulus от западного подвида F. с. aesalon не может быть принято.
Восточносибирский дербник Falco columbarius insignis Clark
Aesalon regulus insignis. Clark. Proceed. U. S. Nat. Mus. XXXII, 1907, стр. 470, Фузан
в Корее.
Распространение. Ареал. Восточносибирский дербник распространен
к востоку от предыдущей формы в заенисейской Сибири. Как указывалось
выше, в области между Енисеем и Леной от Туруханского края до Таймыра
и Хатанги встречаются особи, сходные с F. с. aesalon, и особи, сходные с F. с.
insignis (смешанная популяция). В общем, область распространения
восточносибирского подвида простирается на восток от бассейна Лены, но точные
границы неясны. К северу на Лене он отмечен до устья (73° 26' Бунге, 1883, но
указание это требует проверки), в бассейне Яны до 70° 30'; на Индигирке и
Колыме распространение неясно, но по некоторым сведениям идет здесь до
68° 30'. Южная граница вероятно проходит по средней Лене (Вилюй, Якутск),
на Алдане и в Становом хребте (истоки Зеи, Штегман, 1928). В бассейне
Анадыря, Коряцкой Земле и на Охотском побережье заменен другой расой.Область
зимовок восточносибирского дербника на западе достигает южного побережья
Каспия (ю.-в. Закавказье от Армении до Талыша, ю.-з. Туркмения и сев. Иран),
на юге Индии (Кандагар, Пешин, Гильгит, Синд и Пенджаб), на востоке — ю.-в.
Китая; северные пределы области зимовок —ю.-в. Закавказье, Туркестан от
Туркмении до Киргизии (предгорья центрального Тянь-Шаня), Синьцзянь,
южн. Монголия и зап. Китай, Чжили, южн. Приморье, Корея и Япония.
Характер пребывания. Восточносибирский дербник —
перелетная птица, среди европейско-азиатских дербников этот подвид совершает
наиболее дальние миграции и имеет наиболее обширную область зимовок.
Даты выяснены плохо. Весеннее появление на местах гнездовья повиди-
мому происходит в конце апреля — первой половине мая. На зимовке в
Средней Азии дербники эти встречаются еще в марте (14—19 марта отмечается
пролет в ю.-з. Туркмении; в разные числа марта и севернее). Еще в апреле
добывались в ю.-в. Китае (Фу-чжоу). В Даурии весенний пролет отмечался 21 апреля,
на Зее у Пикана 3 мая, на Уссури в мае, на Алдане в первой половине мая
(конец апреля ст. ст.), у Якутска 29 апреля, в верховьях Колымы еще 15 мая.
Осенний пролет отмечен в сентябре—октябре, в Прибайкалье — с первой
половины сентября, на Алтае в конце сентября — начале октября; в Средней
Азии дербники появляются в октябре — ноябре.
ДЕРБНИК
143
Биотоп. Как у предыдущего; в южн. Якутии невидимому,
главны»образом, в горах.
Численность. Редок в с.-в. Якутии, повидимому и в других местах.
На зимовке в Средней Азии собирается в большом количестве.
Экология- Размножение. Не изучено, вероятно сходно в основных
чертах с размножением предыдущего.
Линька. Птицы, в первом годовом наряде начинающие смену мелкого
пера, добывались на зимовках в апреле; в конце июня сменяются средние
(6-е и 7-е) маховые и средняя пара рулевых у гнездящихся птиц. Взрослые
птицы в мае не линяют, добытые в первой половине сентября птицы были в
свежем пере.
Питание. По крайней мере часть из приведенных выше наблюдений
над питанием дербников на туркестанских зимовках должна относиться к
восточносибирскому подвиду. В Забайкалье пролетные дербники нападают на
овсянок.
Описание. Размеры и строение. Отличается от F. с. aesalon несколько
большею величиной: крыло самцов (21) 190—208, самок (10) 220—231, в среднем соответственно
199,7 и 225,1 мм; по Штегману (1929) у самцов (45) 193—208, у самок (26) 213—232, в среднем
202 и 221 мм. Длина самцов (2) 285—313, самок (2) 325—334 мм. Размах самцов (2) 589—600,
самок (2) 623—625 мм. Вес самца (1) 190 г.
Окраска. Восточносибирский дербник светлее предыдущего во всяком наряде:
старые самцы на спинной стороне бледносизые, лоб беловатый, средние рулевые обычно
без темных полос, кроме вершинной; брюшная сторона беловата или рыжевата с оч^нь
узкими продольными пестринами; беловатый поперечный рисунок на внутренних опахалах
первостепенных маховых очень развит и часто сливается у ствола в сплошное поле. У самок
и молодых общий бурый тон спинной стороны и полос на брюшной стороне значительно
светлее, чем у aesalon, с глинистым оттенком; на спинной стороне у них хорошо развит ржавчато-
бурый светлый поперечный рисунок; поперечные светлые полосы на рулевых правильные и
широкие.
Тихоокеанский дербник Falco columbarius pacificus Stegmann
Aesalon columbarius pacificus. Stegmann. Изв. Акад. наук СССР, 1929, стр. 591,
Разбойник, Охотское море.
Распространение. Ареал. В область распространения тихоокеанского
дербника входят прилежащие к Тихому океану части с.-в. Сибири от бассейна
Анадыря до Коряцкой Земли и Охотского побережья на запад до Аяна и Удского
острога (однако, здесь встречаются и не отличимые от F. с. insignis особи),
на восток до Сахалина; сюда же вероятно относятся дербники с Шантарских
о-вов и может быть добытые у Омолона и — позднею весной 25.V.1912 —
у залива де Кастри в Приморье (вероятно на гнездовье). Пролетные с
Курильских о-вов, из Японии, Манчжурии, из южных частей Приморья. Залетает
к Байкалу. Зимние экземпляры известны из Японии и Фу-чжоу в Китае.
Характер пребывания. Перелетная птица. Появление на
Анадыре относится к кокцу мая (Портенко, 1939), на Охотском побережье
появляется в разные числа этого месяца (6 мая, Шантарские о-ва; 21—22 мая
у Охотска). В Приморье пролетает в мае (Шульпин, 1936), вероятно, и в апреле,
судя по экземплярам. Осенний отлет на Анадыре происходит 9—18 сентября.
На зимовке в Приморье дербники появляются в октябре, быть может ранее.
Биотоп. На Анадыре и в Коряцкой Земле, главным образом в речных
долинах среди тальниковых и олыианых кустарников, в тундровом и
лесотундровом ландшафте.
Численность тихоокеанского дербника невелика. Это довольно
редкая птица.
Экология. Размножение. Календарь годового цикла периодических
явлений —размножения и линяния— сходен с другими северными подвидами
дербников (в частности F. с. aesalon). Молодые с чуть недоразвитыми крыльями
ОТРЯД ХИЩНЫЕ ПТИЦЫ
добыты23—27августа; вполне развитые29августа (Анадырь), 21 августа (Пен-
жина), 28 августа (река Айган, Тайгонос). Начало периода размножения,
следовательно, падает на первые числа июня, а вылупление птенцов — на начало
июля. Выводки вместе держатся до отлета (наблюдались 2—4 сентября).
Линька. Взрослые птицы в конце мая не линяют, а к концу августа,
т. е. ко времени окончания роста молодых, линька у стариков почти закончена:
21 августа у самца из Краснина на Анадыре остались старые 1-е, 2-е и 10-е маховые
и росла 5-я (вторая с края) пара рулевых, остальное оперение было уже свежее.
Сентябрьские дербники уже надели свежее перо, линяние у них уже кончено..
Питание. Вероятно, как у других дербников, главным образом,
мелкие птицы (в частности, крачки на Анадыре; осенью в долине Гижиги
трясогузки и подорожники, Аллен, 1905).
Описание. Размеры и строение. Относительно крупный дербник: крыло
самцов (8) 200—209, самок (6) 214—226, в среднем 205 и 219,4 мм. По Штегману (1929)
у самцов (7) 202—209, самок (6) 218—226, в среднем 205 и 223 мм.
Окраска. В общем сходная с окраской F. s. aesalon, темная, но может быть более
контрастная, с большим развитием пестрин на спинной стороне.От aesalon отличается главным
образом размерами. По окраске pacificus в известной мере приближается к американским
дербникам. Таким образом особенности распространения и окраски этого сокола
представляют интересную аналогию с зимняками (темная западная раса, светлая
восточносибирская, темная сходная с американской в Коряцкой Земле, на Охотском побережье и на
Камчатке). Личная изменчивость у тихоокеанских дербников развита значительно, и среди них
попадаются особи, не отличимые от F. с. insignis по светлому тону окраски и развитию
светлых пестрин.
Центральноазиатский дербник Falco columbarius lymani Bangs
Falco aesalon lymani. Bangs. Bull. Mus Compar. Zool. Harvard Coll. 1912, стр. 465,' Чеган-
бургази, Алтай.
Распространение. Ареал. Область распространения центральноазиат-
ского дербника такова: северная часть нагорной Азии от Джунгарского Ала-
Тау, центрального и вост. Тянь-Шаня, на западе, по крайней мере, от
Нарына и восточных частей Киргизского хребта (долина реки Иссык-ата),
по Северцову — будто бы на Алае; на восток до Турфана, в юго-восточных
частях русского Алтая (Чуйская степь), в Сайлюгеме, в Гобийском Алтае (до
Ихебогдо), в восточном Саяне (хребет Мунку-сардык), на Хангае, к северу до
Косогола, повидимому в горах ю.-в. Забайкалья. В негнездовое время
распространен в вост. Туркестане, редко в Цайдаме. Залетные особи добыты в с.-в.
Алтае (Яйлю, 20.XI. 1933), в низовьях Каратала (20.VIII.1935), у Иркутска
(7. IX. 1935, Скалой, 1935).
Характер пребывания. Большинство зимних экземпляров
добыты в гнездовой области, птица почти оседлая и ее сезонные перемещения
сводятся, повидимому, главным образом к вертикальным миграциям.
Передвижение дербников наблюдалось в Усекских горйх близ Джаркента в ноябре—
декабре, в это время они были часты; в январе—феврале птиц стало
меньше, в марте опять больше (Зарудый, 1906). В равнинных частях зап.
Туркестана не добывался зимою, следовательно, если и бывает там, то лишь
случайно.
Биотоп. Альпийская и субальпийская зоны гор, повидшиому и лесная
зона. На Алтае в гнездовое время от 1750 до 2300 м, в центральном Тянь-Шане
от 1800 до 3000 м.
Численность невелика.
Экология. Размножение. Календарь размножения — как у
северных форм дербника. Гнезда располагаются на деревьях: на лиственницах
в колках Чуйской степи в ю.-в. Алтае (Бенго, 1913; Сушкин, 1928), в ельниках
в Тянь-Шане, вероятно впрочем на склонах и на земле. Кладка из 3—4 яиц.
ДЕРБНИК
145
У Нарына кладка в 4 яйца найдена 2 июня, размеры их 39,7—-42,1X30—32 мм
(Бекер, 1928; Суанн, 1936). Яйца отличаются очень бледной окраской —
красновато-бурой с шоколадного цвета крапинами. Птенцы вылупляются в
конце июня — начале июля (у Алма-Ата найдены 29 июня). В середине июля
птенцы надевают второй пуховой наряд. В гнезде их бывает иногда только 2.
Оперяются молодые к концу июля, но маховые дорастают у них только
20 августа.
Линька. Смена пера у взрослых начинается в июле, кончается — в
начале сентября (Алтай, Сушкин, 1938; Хангай, Козлова, 1932). Детали линяния
не изучены.
Питание. Как у других подвидов дербника, главным образом,
воробьиные, живущие в открытом ландшафте (жаворонки, луговые коньки);
дербник нередко подстерегает их, сидя на земле.
Описание. Размеры и строение. Отличается от других подвидов длиннокры-
лостью (но не общей величиной): крыло самцов (9) 221—235, самок (7) 248—258, в среднем
229,6 и 252,2 мм.
Окраска бледная, напоминающая окраску восточносибирских дербников, но с
менее резким поперечным рисунком у молодых и у самок и с глинистым оттенком бурого тона
на верхней стороне тела (в этом отношении наблюдается сходство с последующим подвидом).
Сходство с последующим подвидом заключается и в том, что у старых самцов на плечевых
и кроющих крыла имеются узкие бледно рыжеватые каемки.
Степной дербник Falco columbarius christiani-ludovici Kleinschmidt
Falco christiani-ludovici. Kleinschmidt. Falco 13, 1917, стр. 10, новое имя для:
Lithofalco aesalon pallidas S u s с h k i n, Bull. Br. Orn. Club. II, 1900, стр. 5, Средняя
Киргизская степь.
Распространение. Ареал. Область распространения степного дербника
охватывает степи Казахстана и ю.-з. Сибири, примерно между 49° и 54—55°с. ш.
и 55—80° в. д. (от Гринвича). Гнездовая область расположена таким образом
в области левых притоков Урала у Чкалова, по Илеку, Хобдо, в Мугоджарах,
на севере у Орска, Верхнеуральска, Троицка (54° с. ш.), далее у Омска и озера
Чаны в Барабинской степи (55°), быть может до Барнаула (молодая птица
добыта 21.VIII. 1913), откуда к Югу по Кулундинской степи (бассейн Алея,
западные предгорья Алтая —ус. Новенского) и Семипалатинска; южная граница
не вполне ясна — она проходит примерно между Мугоджар по степям к
северу от Аральского моря у озера Чалкар-тениз, у р. Улькояк, около 40°
с. ш., подымаясь далее к 50° Залеты до Уфы. Зимою на Кавказе от Дзауд-
жикау до Колхиды и Ленкорани, главным образом в ю.-в. Закавказье; также
в М. Азии (Трапезунд), в Палестине и Месопотамии, в Аравии; вероятно в
Иране; в Средней Азии от Киргизии до Туркмении; в с.-з. Индии (Пешавар,
Кохат, Гильгит, Синд), наконец, в бассейне Тарима и вероятно в Тибете
(Гианцзе).
Характер пребывания. Обильные снегопады и суровость зим
в гнездовой области степного дербника делают ее непригодной для
большинства воробьиных в зимнее время и тем самым для дербника; поэтому, в
отличие от горного подвида дербников (F. с. lymani), степной
дербник—настоящая перелетная птица. После размножения в июле—августе дербник
перемещается к степным озерам и сорам, где в это время скапливаются мелкие
птицы.
Даты. Осенний пролет и отлет ранний, в сентябре, когда линька
взрослых подходит к концу. Около середины сентября дербники появляются уже
в северных частях Туркестана у Аральского моря, на Сыр-Дарье, Чу, в
предгорьях Тянь-Шаня, где часть птиц задерживается, а другие еще в ноябре
продолжают движение к югу. В зап. Закавказье добыт уже 19 сентября. В октябре
дербники этого подвида отмечены на зимовках в Туркмении, где весною остаются
10 Птицы СССР, т. I
146
ОТРЯД ХИЩНЫЕ ПТИЦЫ
по крайней мере до марта (также на Сыр-Дарье, в Дагестане), но пролет
оканчивается вероятно и позднее, в начале апреля (в это время добывались и в
Пенджабе, Уистлер, 1922). Прилет степных дербников в гнездовую область не
прослежен.
Биотоп. Сухие степи, преимущественно с солянками и полынью, с
плотной солончаковой почвой, у солоноватых озер; долины степных рек с
зарослями таволги и березы и т. п.
Численность степных дербников невысокая.
Экология. Размножение. Цикл несколько более ранний, чем у
других подвидов — кладка происходит во второй половине мая. Имеется повиди-
мому «резерв» холостых особей (два самца добыты Спангенбергом у одного
гнезда у Ак-булака, Чкаловская обл.). Гнезда располагаются на кустах
ветлы, таволги и березы в 1*/2 — 5 л* от земли, на земле и, в виде исключения,
на старом стоге (Зарудный, 1886). Начало кладки у Жулдуза близ Ак-булака
18 июня, промежуток между откладкой яиц — 48 часов (Спангенберг). Число
яиц в кладке 2—6. 3 насиженных яйца были обнаружены у Омска 14 июня.
Размер яиц (3) 42—43x32 мм (Шухов, 1928). Насиживают оба родителя:
у обоих имеются наседные пятна, самцы добывались на гнезде, 4 птенца
пуховика добыты в Майчеку в горах Семей-Тау у Семипалатинска 21 июня (Залес-
ский, 1931), в районе Кургальджина 18 июля (Слудский, 1948), однако в ту же
дату в уреме долины Урала «птенцы в полроста» старых (Зарудный, 1892);
4 пуховика в Наурзуме 21 июня. Вполне развитые молодые добывались уже
в июле (9 июля, Байназар-копа; 19 июля, Наурзум; 30 июля Каратерень),
т. е. несколько ранее, чем у северных подвидов.
Линька. В апреле —маевзрослые птицы не линяют. У птиц в первом
годовом (гнездовом) наряде смена мелкого пера начинается уже в конце марта,
линька продолжается до конца мая, затем приостанавливается. У взрослых
начало линяния относится к середине июня, как обычно, со смены средних
первостепенных маховых (7-го и 6-го) и средней пары рулевых. Интенсивная
линька имеет место в июле, когда птенцы оперяются. В это время обычно
старые — 1-е, 2-е и 10-е маховые, иногда 3-е и 8-е., остальные свежие. В конце
августа сменяются 1-е, 2-е и 10-е маховые и 5-я (вторая от края) пара рулевых.
Последовательность смены маховых как у F. с. aesalon: 7—6—5—8—4—3—9—
—2—10—1. Продолжительность линяния около 21/* месяцев. В первых числах
сентября птицы уже в свежем пере, но у некоторых особей линька затягивается
до середины сентября. Смена мелкого оперения на спинной стороне идет
несколько ранее, чем на брюшной. Перед линькой перо старых птиц очень
обношено и выцвело, более, чем у северных дербников.
Питание. Как и другие подвиды дербника, степной дербник питается
главным образом мелкими птицами открытых ландшафтов — жаворонками,
коньками, мелкими куликами и т. п., особенно белокрылыми жаворонками
(гоняется и за ласточками-касатками, Формозов, 1942). И этот подвид не только
ловит птиц на лету, но и подстерегает их, лежа или сидя среди кустов солянок
и т. д. Дербники также хватают нелетных птенцов.
Кормовой режим в Наурзуме: полевой и белокрылый жаворонки, степной
конек, обыкновенная каменка,черноголовый чеккан, бормотушка, серая славка,
перепел, полевка Microtus arvalis, пеструшка, мышевка Sicista subtilis* прыткая
ящерица, стрекозы, кобылки, чернотелки, жужелицы и т. д.; основная пища —
воробьиные, в особенности жаворонки (Осмоловская).
Описание. Размеры и строени е. Крупный подвид с длинными крыльями: у
самцов (19) 202—214, у самок (12) 226—235, в среднем 207,7 и 230,6 мм. По Штегману (1929)
у самцов (19) 200—215, у самок (11) 220—232, в среднем 206 и 227 мм. Длина самцов (3)
293—310, самок (2) 320—332 мм. Вес самцов (3) 168, 180, 188 г, самок (4) 224, 235, 238 и
261 г, в среднем 178 и 232,3 г.
ПУСТЕЛЬГА
147
Окраска. Степной дербник самый бледный из европейско-азиатских подвидов дерб-
ника во всех нарядах. Пуховые птенцы менее сероваты, чем у aesalon. Взрослые самцы на
спинной стороне бледносизые, с охристо-рыжеватыми каемками перьев на спине, плечах и
кроющих крыла; голова глинисто-рыжеватая, шея бледнорыжая; темные настволья на
спинной стороне тела и пестрины на брюшной очень узки; первостепенные маховые с резким
преобладанием белого цвета на внутренних опахалах; рулевые с узкой темной вершинной
полосой; основной цвет брюшной стороны варьирует от бледно глинисто-охристого до белого,
темный рисунок развит слабо, часто только в виде узких наствольных черт. Старые самки
и молодые птицы на спинной стороне бледные, общего песочного или глинисто-рыжеватого
тона, с нерезко выделяющимися (из-за бледности основного тона) широкими поперечными
пестринами; на внутренних первостепенных маховых светлый поперечный рисунок
преобладает; светлые полосы на рулевых шире темных; бурые^пестрины на брюшной стороне бледные
и сравнительно узкие.
8. Обыкновенная пустельга Falco tinnunculus L.
Falco tinnunculus. Linnaeus. Syst., Nat. изд. X, 1758, стр. 90, Швеция.
Русское название. Пустельга — от слова «пустой», вероятно потому, что птица
непригодна для соколиной охоты.
Распространение. Ареал. Область распространения обыкновенной
пустельги — Европа и Азия, кроме крайнего Севера; Африка.
Характер пребывания. В северных частях ареала перелетная
птица, в субтропическом и тропическом поясах оседлая.
Биотоп. Убиквист: леса (кроме только больших массивов сомкнутого
леса, предпочитает опушки, разреженные насаждения и т. п.), лесостепь,
культурный ландшафт, горы, пустыни и т. д.
Подвиды и варьирующие признаки. В северо-западной части ареала от бассейна
Енисея на востоке и сев. Африки и Передней Азии на юге F. /. tinnunculus L., 1758; в Вост.
Сибири от бассейна Лены F. t. perpallidus Clark, 1907; в Китае, Японии, Гималаях F.
t. interstinctus И о г s f i e 1 d, 1840; в Юннани, вероятно в Бирме F t. saturates В ly th,
1859; в южн. Индии и Цейлоне F. t. objurgatus Baker, 1927; в южн. Аравии и Адене
F. /. buryi Grant et Mack wor th-Praed, 1933; в Египте, Нубии и сев. Сомали
F. t. rupicolaeformis С h г. L. В г е h m, 1855; на западных Канарских о-вах и Мадейре
F. t. canariensis К б n i g, 1890; на восточных Канарских о-вах F. t. dacotiae H a r t e r t,
1913; на о-вах Зеленого Мыса F. Л neglectus S с h lege l, 1873; в Судане и Абиссинии,
в тропической Африке в области озер/7. /. carlo Hartertet Neumann, 1907; в
Сенегале, Камеруне и Бельгийском Конго F. t. rufescens S w a i n s о n, 1843; на о-вах Сокотра,
в Сомали, к югу до Кении и Ламу F. t. archeri Hartertet Neumann, 1932; в ю.-з.
Африке F. t. rhodesi Finsh-Davies, 1920; в южн. Африке F. /. rupicolus D a u d i n,
1800*.
Варьирующие признаки: по величине и окраске южные формы в общем мельче и ярче
северных (но южноафриканский подвид крупней европейского и с большим развитием
«прогрессивного» серого цвета в окраске). Географические изменения экологии изучены
недостаточно, они касаются характера пребывания (некоторые формы оседлы, другие кочуют,
третьи перелетны), календаря периодических явлений и, повидимому, также особенностей
цикла размножения и трофических связей.
Обыкновенная пустельга Falco tinnunculus tinnunculus L.
Синонимы: Falco tinnunculus ultratinnunculus. Kleinschmidt. Falco, 1929,
стр. 35, Владикавказ. —Falco tinnunculus stegmanni. Portenko. Mitteil. aus d. Zool.
Mus. Berlin, XVII, 1931, стр. 415, Кашгария.
Распространение. Ареал. Европа от 71° в Скандинавии, 68° с. ш. в
Финляндии (обычно до 65°), к югу до сев. Африки от Марокко до Триполи;
Передняя и Средняя Азия от М. Азии до Ирана, Средней Азии, вост. Туркестана
и с.-з. Монголии. В СССР северная граница проходит у Онежского озера,
Архангельска (однако возможно, что гнездится в «лемминговые» годы и
северней, до Лапландского заповедника, Владимирская, 1948), далее примерно по
* Таксономия среднеафриканских и восточноазиатских пустельг требует еще
выяснения.
148
ОТРЯД ХИЩНЫЕ ПТИЦЫ
64° с. ш. до верхней Печоры, на Сев. Урале у Няыси, у Березова, обходя с юга
в Зап. Сибири таежную полосу по линии Тюмень,Тобольск, низовья р. Томь—
Алтай; на Енисее регулярно встречается только до 60°, восточная граница
в Сибири определяется повидимому бассейном Верхней Тунгуски. Зимует в
южных частях СССР — на юге Украины, в Крыму, на Кавказе, в Средней Азии от
Киргизии (и быть может от Зайсана) до Туркмении; кроме того, в зап. и южн.
Европе, в Африке до Золотого берега, Нигерии, Камеруна, Конго, Танганайки,
Карта 16. Распространение пустельги Falco tinnunculus
1 — F. t. tinnunculus, 2 — F. t. perpailidus, 3 — F. t. interstinctus, 4— F. t. saturatus, 5 — F. t. objurgatus,
6 — F. t. buryi, 7 — F. t. rupicolaeformis, 8— F. t. archeri, 9 — F. t. canadensis, 70 — F. t. dacotiae,
11 — F. t. neglectus, 12— F. t. rufescens, 13— F. t. carlo, 14 — F. t. rhodesi, 15 — F. t. rupicolus;
a — граница гнездовий4, б—не вполне выясненная граница, г — область зимовок.
в Передней Азии и с.-з. Индии. Залеты к северу от гнездовой области —на
Канин, в Туруханский край к р. Елагуй; отмечены трансатлантические
залеты — в США (Массачузетс) и в Гренландию.
Характер пребывания. В северных частях ареала в СССР
пустельга — перелетная птица; в южных частях (см. выше) зимует, но не вполне
ясно, находятся ли там гнездовые или только прикочевавшие с севера птиды,
или и те и другие; в Англии и средней Европе, южн. Европе —встречается
круглый год. Оседлость пустельги в Закавказье и южных частях Средней Азии
вероятна.
Даты. На местах зимовок в Африке пустельги встречаются с начала
октября до конца марта, в Индии с сентября — октября до конца марта —
начала апреля. Кочевки молодых начинаются рано, в августе; с середины
сентября в Зап. Европе идет правильный пролет, продолжающийся до октября,
ПУСТЕЛЬГА
149
весенний прилет там начинается с конца марта, В южной полосе СССР пролет
пустельги весной отмечен со второй половины марта и до начала апреля (северные
популяции), иногда же движение начинается с конца февраля (Крым, Кавказ,
Украина — Киев, Мариуполь, Аскания-Нова, Умань, Харьков и т. д.; Средняя
Азия — Туркмения, Сыр-Дарья, предгорья Тянь-Шаня); в средней полосе
пустельги появляются с конца марта и до конца апреля, валовой пролет
надает на первую половину апреля (Москва, Тула, Рязань, Калинин, Псков,
Смоленск, Минск, Киров, Бугуруслан, Чкалов, Казань, Куйбышев и т. д.), на
севере (северная часть б. Пермской губ., Архангельск) и в Сибири (Новенское
Змеиногорского района; Красноярск; Минусинск, Томск) с половины апреля,
главным образом, к концу этого месяца.
Отлет происходит в разные числа сентября, на севере уже с начала этого
месяца, продолжаясь до начала октября; еще в октябре отдельные особи
встречаются в средней полосе европейской России и сев. Казахстане, но валовой
пролет падает на середину сентября. Внешние условия повидимому мало влияют
на ход пролета пустельги, так как прилет ее происходит в средней полосе еще
при зимнем ландшафте, до таяния снега и вскрытия рек. У некоторых
популяций перелеты носят характер кочевок, причем течение их находится в зависи-t
мости от кормовых условий; в частности, пустельги задерживаются в местах
массового размножения грызунов (Радде, 1895). Резкие похолодания вызывают
местные перемещения и в области зимовок. В местностях, где пустельги оседлы,
они совершают все же сезонные вертикальные перемещения. Пролет
разрозненный — парами и одиночками. Как и следовало ожидать, большое постоянство
местообитаний: в Аскании-Нова окольцованные там пустельги найдены в
марте — апреле и июне через год и через три года после кольцевания (Фальц-
Фейн, 1915).
Биотоп. Убиквист. В Палеарктике встречается всюду, кроме тундры,
лесотундры и глухой тайги: в области последней, главным образом, в долинах
рек и культурном ландшафте. Однако, в лесостепи встречается преимущественно
там, где имеются деревья или хотя бы кустарники, так как эта растительность
предпочитается для гнездования. На пролете и зимовке — преимущественно
в открытом ландшафте.
В вертикальном отношении от равнин до высокогорья: на Урале до
верхней границы леса (950 м в Яман-Тау, 1200 м на Иремели); на Кавказе
до 3000—3500 м (Насимович и Аверин, 1938; Соснин и Ляйстер, 1942), на
Алтае до 2400 м; в Копет-Даге до 2200 м, в Зеравшанском хребте обычна между
1600 и 2000 мУ доходя до 3150 м (Даль, 1936), в Фергане около 2000 м (Каш-
каров, 1927), в центральном Тянь-Шане гнездовья расположены главным
образом между 1000—1500 м, кормовые вылеты и кочевки —до 3100 лс.
Численность. Самый многочисленный из соколов нашей фауны.
Несмотря на то, что в обычное время главную пищу пустельги составляют
грызуны, колебаний численности не установлено, что" указывает вероятно на
большую пластичность трофических связей вида.
Экология. Размножение. Пары вероятно постоянные, так как самцы
и самки держатся вместе уже на пролете. Половая зрелость наступает в
возрасте около года, на следующую же весну после рождения (у гнезд добывались
неоднократно птицы в первом годовом наряде). Имеется резерв холостых
особей, главным образом, первогодков. Цикл размножения почти одновременный
во всем ареале, поэтому имеется разница в промежутке между пролетом и
размножением у южных и северных популяций. Совокупление происходит
не только в начале периода размножения (конец апреля), но и в конце его.
Расположение гнезд разнообразно; в обрывах речных берегов— глинистых и
скалистых, в сооружениях (дома и т. п.), в нишах скал, в норах (сизоворонки
и др.), на земле, в дуплах, на деревьях. В последнем случае занимаются по-
150
ОТРЯД ХИЩНЫЕ ПТИЦЫ
стройки грачей, серых и черных ворон и других птиц. Несмотря на указание
некоторых авторов, мало вероятно, что пустельга строит собственное гнездо;
гнездостроение у нее ограничивается устройством внутренней выстилки
гнезда — из тонких веточек, стеблей травы, луба, шерсти, кошмы, погадок. Иногда
гнезда расположены близко одно от другого (Псков), изредка даже в виде
колонии (Закавказье, Баньковский, 1913). Гнезда на деревьях, расположены
обычно высоко от земли (10—15 м), но в степных и полупустынных местностях
низко, на саксаульниках и песчаных акациях, кустах джузгуна в 1 —2 м высоты.
Кладка имеет место в конце апреля — первой половине мая; причины
колебаний срока, у нас не географические, неясны (в Англии однако кладка
производится с середины апреля и иногда даже с конца марта). Число яиц
в кладке от 3 до 8, обычно же 4—5. В случае гибели первой кладки бывает
дополнительная. Число яиц в ней меньше, чем в нормальной, не более 2 (Гебель,
1879). Промежуток между откладкой яиц 24 и 48 часов (Туркмения, Птушенко).
Часть яиц в кладке обычно не оплодотворена.
Яйца окрашены в охристый цвет, по которому густо расположены ржав-
чато-бурые пестрины и темнобурые пятна и точки. Размеры яиц: (283) 36—43 х
X 29—34, в среднем 38,9 х 31,5 лм* (Гебель, 1879); (24)32—41,4 х 30—36,3 мм
(Спангенберг, 1936).
Срок насиживания у нас точно не установлен (по западноевропейским
данным 28 дней); начинается насиживание с первого яйца. Насиживают
невидимому оба родителя, судя по наблюдениям и наличию у самцов наседных
пятен. Гнездовый период длится около 30 дней. В начале июня птенцы надевают
второй пуховой наряд, в это время пробиваются маховые. Оперившиеся птенцы
с недоразвитыми крыльями и хвостом попадаются в конце июня — первой
половине июля*/ в конце июля — начале августа молодые в неразбившихся
выводках уже вполне вырастают. Разделение труда при выкармливании
птенцов обычное для соколов: самец ловит добычу и передает ее самке, а последняя
оделяет птенцов, по крайней мере первое время их жизни в гнезде. Выводки
не разбиваются до отлета.
Линька. Общий ход линяния — как у всех соколов. Линька у
взрослых начинается во второй фазе периода размножения, т. е. с первой половины
июня. Последовательность смены первостепенных маховых 7—6 (или 6—7) —
5—8 —4 (или 4—8) 3—9—2—10—1. Рулевые сменяются, начиная со средней
пары и кончая второй от края хвоста. Конец линяния взрослых относится к
первой половине сентября; общая продолжительность линьки, следовательно,
около трех месяцев; у некоторых особей (возможно после второй
дополнительной кладки) конец линяния затягивается до начала октября, но, как правило,
взрослые птицы между сентябрем и маем не линяют. У птиц в первом годовом
наряде смена мелкого пера начинается с весны, в марте —апреле; некоторые
особи начинают линяние постепенно уже с декабря, но в период размножения
линька у гнездящихся птиц приостанавливается-в мае — июне, а потом
протекает как у взрослых. Тип линьки: полная годовая. Последовательность смены
нарядов: первый пуховой наряд — второй пуховой наряд — первый годовой
(гнездовый) наряд—второй годовой наряд—третий годовой наряд и т.д.
Питание. В отличие от «благородных» соколов —настоящего сокола,
кречета, чеглока и других, — пустельга берет добычу главным образом с земли.
Способ охоты: птица летает невысоко над землею и высматривает добычу,
«трясясь» в воздухе над одним местом; иногда подстерегает добычу как дербник,
сидя на земле. Охотится в открытом ландшафте. Охотничий участок около
1—3 км от гнезда, иногда больше (средняя Россия, Туркестан). Птиц на лету
пустельга ловит с трудом. Умерщвление добычи производит по-соколиному,
ломая шею или пробивая добыче затылок клювом. Потребность взрослой
птицы в пище около а/5 ее живого веса, т. е. около 40—50 г. Имеются сезонные
ПУСТЕЛЬГА.
151
изменения кормового режима: зимой пустельга кормится главным образом
грызунами и отчасти мелкими птицами, летом — насекомыми, рептилиями и
грызунами.
В качестве добычи пустельги в СССР отмечаются из млекопитающих
разные полевки и мыши Microtus arualis, Apodemus agrarius, Stenocranius gregalis
и др., суслики, например, Citellus undulatus, мышевки Sicista, степная
пеструшка" Lagurus lagurus,
водяная крыса Arvicola fer-
restris, хомячок Cricetulus
migratorius, тушканчик Di-
pus sagitta, в виде
исключения ласка; из
птиц—главным образом птенцы и
подлетки воробьиных, также
взрослые жаворонки, кроме
того отмечены
обыкновенная чечевица,
обыкновенная каменка, домовые
воробьи, перепела,
чернозобый дрозд; ящерицы;
разные насекомые, в
особенности саранчевые, также
жуки, стрекозы и т. д.
Колебаний численности в связи
с кормовыми условиями не
отмечено, но скопление вне
гнездового времени в
местах массового размножения
грызунов наблюдается.
Полевые признаки.
Преобладание рыжего цвета на
верхней стороне тела;
длинный ступенчатый хвост;
крылья длинные, несколько
более закругленные, чем у
других соколов. Часто
«трясется» на одном месте в
воздухе с несколько
опущенным хвостом и поднятыми
крыльями. Голос — обычно
звонкое «КЛИ-КЛИ», выше, Рис. 45. Обыкновенная пустельга (самец и самка),
чем у чеглоков.
Описание. Размеры и строение. Первое маховое более или менее равно
4-му, в отличие от степной пустельги, у которой 1-е маховое длиннее 4-го. Оперение
довольно мягкое. Крыло не очень острое, его формула: 2 немного более 3>4>5. Вырезки на
наружных опахалах 2-го и 3-го маховых и на внутренних опахалах 1-го и 2-го маховых
(в наличии вырезки внутреннего опахала 2-го махового — отличительный признак от
степной пустельги). Цевка длинная, пальцы короткие, средний палец не более 2/з длины
цевки, наружный и внутренний пальцы примерно равны. Длина самцов (33)310—355, самок
(40) 330—380, в среднем 336 и 367 мм. Размах крыльев самцов (24) 690—800, самок (34)
731— 820, в среднем 774и805лм/. Вес самцов (9) 159—200, самок (10)197—240,5, в среднем
180,9 и 212,9 г. Крыло взрослых самцов (62) 230—253, самок (64) 242—275, в среднем
242,45 и 254,4 мм.
Окраска. Первый пуховой наряд белый; второй пуховой наряд, надеваемый в
возрасте около 10 дней, серовато-белый на спинной стороне, белый на брюшной.
Первый годовой (гнездовый) наряд, сходен у обоих полов: на спинной стороне ржаво-
рыжий с продольными полосами на голове и поперечными темнобурого цвета на спине^
152
ОТРЯД ХИЩНЫЕ ПТИЦЫ
плечах, крыльях и хвосте; первостепенные маховые черновато-бурые с беловатыми
внутренними опахалами и с широкими охристыми каймами перьев; брюшная сторона охристая
с бурым продольным рисунком; подкрылья беловатые с темнобурым поперечным рисунком
в виде пятен или полос.
В окончательном (третьем годовом) наряде хорошо выражен половой диморфизм.
Самцы на спинной стороне тела рыжие с темнобурыми поперечными пестринами на спине и
крыльях; маховые темнобурые с зигзагообразными белыми пятнами на внутренних
опахалах; рулевые серые с черной предвершинной полосой и белой вершинной каймою; брюшная
сторона охристая с бурым продольным рисунком, более или менее развитым. Самка на
спинной стороне ржаво-бурая с широким темнобурым поперечным рисунком на спине, плечах и
хвосте, с продольными полосами на голове, с сероватым надхвостьем; подкрылья и у самцов,
и у самок беловатые с бурыми поперечными крапинами. У некоторых особей хвост серый
с поперечным бурым рисунком, иногда и темя серое. Первый годовой наряд сходен, таким
образом, с самочьим, но бурый поперечный рисунок спинной стороны шире, а рулевые лишь
редко несут сизый оттенок. Во втором годовом (промежуточном) наряде у самца голова обычно
бурая с продольным темнобурым рисунком, пестрины на спинной и брюшной стороне более
развиты; на рулевых более или менее развитый поперечный рисунок. Самки во втором
годовом наряде неотличимы от старых птиц. Кольцо вокруг глаза, восковица, лапы
желтые; радужина темнобурая, клюв сине-роговой, чернеющий у вершины; когти черные.
Восточная пустельга Falco tinnunculus perpallidus Clark.
Cerchneis petpallida Clark. Proceed. U. S. Nat. Museum XXXII, 1907, ст. 470, Корея.
Синоним. Cerchneis tinnunculus doerrisi. Swann. Synopt. List of Accipitres, 1920, стр. 146.
Распространение. Ареал. От бассейна Лены до Дальнего Востока.
На Яне до 66°30; на Вилюе около 65°, далее достоверно только у Якутска;
однако гнездится и на Индигирке (за полярным кругом, Васьковский), и на
Колыме под 68°; на Анадыре отсутствует; гнездится на северном побережье
Охотского моря; к югу до Приамурья, Забайкалья и вероятно вост. Монголии.
В холодное время года в СССР изредка встречается в с.-в. Казахстане
(Караганда, Жарык, 25.1.1944) и в центральном Тянь-Шане (Кашкасу, X. 1910).
Зимою повидимому в Индии, Китае, к югу до о-ва Формоза и Филиппин.
Залет на крайнем с.-в. Сибири —около 70° между мысами Якан и
Биллингса.
Характер пребывания. Перелетная птица.
Дат ы. Прилет у Якутска 26 апреля — 6 мая, в среднем 2 мая
(Воробьева, 1934), 22 апреля у Пикана на Зее (Дорогостайский, 1915); пролет
в Приморье отмечен в первой половине апреля (Шульпин, 1936). Осенью на
местах гнездовья и Забайкалье пустельга встречается еще в первой половине
сентября (Чита, 10. IX). В Забайкалье и Приморье встречается и зимою.
Биотоп как у номинальной формы.
Численность. В Вост. Сибири пустельга, повидимому, редка.
Экология. Размножение. У Якутска 29 мая в гнезде на лиственнице
были обнаружены 5 слегка насиженных яиц; гнезда в Забайкалье располагаются
на скалах. В Приморье в кладке находили 4—7 яиц, подлетки были добыты
8 августа (Шульпин, 1936). В вост. Монголии выводки разбились в середине
августа, почти доросшие подлетки встречены 18 июля (Тугаринов, 1932); в ю.-в.
Забайкалье 5 подлетков наблюдались 14 июля (Скалой, 1936), вполне развитые
молодые у Олекминска 5 июля.
Л и н ь к а. До конца мая линяния у взрослых нет. Начало линьки
относится к концу июня (Олекминск, 21.VI: 7-е маховые свежие, 6-е выпали,
8-е растут). Кончается линька в первой половине сентября. Общее течение
процесса — как у номинальной расы.
Питание. Питается, главным образом, грызунами — в Забайкалье
Microtus mongolicus, М. michnoi, Stenocranius gregalis» Micromys minutus; из
птиц — мелкими видами, в том числе Calandrella brachydactyla, овсянками;
ящерицами Lacerta uivipara; насекомыми, главным образом, саранчевыми,
СТЕПНАЯ ПУСТЕЛЬГА
153
также кобылками и кузнечиками, жуками (сем. усачей) и т. п. (Скалой, 1935,
1936).
Описание. Размер ыистроение. Крыло самцов (5) 235—255, самок (10) 250—
275, в среднем 247 и 262,9 мм. Величина, вопреки указаниям Суанна (1936), сходна с
таковой у номинальной формы.
Окраска. Птицы из Вост. Сибири заметно бледнее европейских, западносибирских
и среднеазиатских, в особенности по рыжему тону спинной стороны тела.
Систематические замечания. Так как диагностические признаки пустельги, описанной
Кларком из Кореи, совпадают с особенностями восточносибирских птиц (бледная окраска),
а японские пустельги этими особенностями не обладают, название «doerrisi» приходится
считать синонимом perpallidus. Для окончательного решения вопроса требуется впрочем
непосредственное сравнение типов.
9. Степная пустельга Falco naumanni Fleischer
Falco naumanni. Fleischer. Sylvan, ein Jahrbuch fur Forstmanner auf
die Jahre 1817 und 1818, стр. 174, южн. Германия.
Синонимы. Falco cenchris «F г i s с h», N a u m a n n. Naturgeschichted. Vogel Deutsch-
lands 1, 1820, стр. 318, Италия. —Falco cenchris var. pekinensis Swinhoe. Proceed. Zool.
Soc.of London, 1870, стр.442, Пекин.—Cercheis naumanni turkestanicusf Зарудный,
Орнитологический Вестник, 1912, стр. 114, Туркестан. — Cerchneis naumanni sarmatieus. D u n a-
jewski Comptes Rendus de la Sosieti des Sciences de Varsovi, X, 1917, стр. 1046, Люблин.
Распространение. Ареал. Область распространения степной пустельги —
сев. Африка —Марокко, Алжир, Тунис; южн. Испания и южн. Португалия,
Карта 17. Распространение степной пустельги Falco naumanni
а — граница гнездовий, г — граница области зимовок.
южн. и средняя Италия, о-ва Балеарские, Сицилия, Сардиния, Крит и Кипр,
Балканский п-ов к северу до Добруджи; кроме того, Тироль, Каринтия,
южн. Штирия, низовья Дуная и Трансильвания, южн. Польша (Люблин);
Украина к северу примерно до 49° с. ш. (Умань — Полтава — Старобельск),
Крым, низовья Дона, Предкавказье и Кавказ, на Волге примерно до Пуга-
154
ОТРЯД ХИЩНЫЕ ПТИЦЫ
чевска и Бузулука, т. е. около 52°—52°30' с. ш.; изолированная колония
значительно севернее, у Кук-мора, 56°15' с. ш. (Першаков, 1929); в Зауралье
до Миаса, около 55° с. ш.; затем граница идет по южным частям Курганского
округа, а восточнее несколько спускается к югу, проходя по Кокчетавской
горно-сопочной стране, не доходя до Омска и проходя по Кулундинской и
Бельагачской степи (Чистозерное, Ключевое, Семиярское, по Залесскому, 1930)
к Алтаю; далее у Красноярска и в Минусинской и Абаканской лесостепи, в
Танну-туве, в с.-з. Монголии (спорадически) — отдельные находки сделаны
в ю.-в. Забайкалье (Кяхта, Дзаргучей, 23—26 мая, Леннберг, 1909; Кяхта,
Скалой, 1936); затем, после перерыва, в вост. Монголии (Удэ), в
Манчжурии и в Сев. Китае (Чжили и Жехол), где подробности распространения
неясны. К югу до М. Азии, Сирии, Палестины, Ирана, Средней Азии (от
Киргизии до Туркмении, но отсутствует на Памире и в Больших и Малых Балха-
нах), Джунгарии. Залеты к Нижнеудинску (в мае, Штегман, 1929), к Омску,
в Хреновскую степь в б. Воронежскую губ. (Северцов), к Алатырю (Волчанец-
кий, 1926) и даже к Пскову (Зарудный, 1910), в Германию, южн. и вост.
Францию и Англию. Зимовки в Африке от Судана и Абиссинии до Капской Земли,
главным образом в саваннах ю.-з. и ю.-в. Африки, в Аравии, в Белуджистане
и в Индии; расположение азиатских зимовок впрочем известно недостаточно.
Характер пребывания. Перелетная птица с типичным бина-
реальным распространением и резкой сезонной сменой ареала.
Даты. Прилет поздний, отлет ранний, при сравнении с обыкновенной
пустельгой. На африканских зимовках появляется с конца сентября и
держится до конца марта, осенью появляются сначала молодые птицы. В Зап.
Европе прилет начинается с конца марта (в южн. Испании, по Ирб^, 1895,
уже в конце февраля) и продолжается до начала мая, отлет происходит в
августе — сентябре. Пролет в Туркмении весною отмечен с конца марта и в
первой половине апреля, осенью — в сентябре и до конца октября. На Сыр-Дарье
и Аральском море весенний прилет и пролет отмечается с последних чисел
марта, главным образом, в апреле, отлет — в августе, когда к югу начинают
двигаться молодые птицы, и в сентябре. В это же время — с конца марта и в
апреле, в августе — сентябре пролетают пустельги на Кавказе и в европейской
части ареала, только на самом севере у Чкалова и в сев. Казахстане прилетают
позднее, с половины апреля и регулярно исчезают осенью ранее, во второй
половине августа. Как редкое исключение, пустельга найдена на зимовке в зап.
Кара-Кумах у Джебела (Рустамов).
Биотоп. Гнездовый — лесостепь, степи с выходами скал и оврагами,
сухие долины рек, полупустыни, горы. В вертикальном отношении — до
2200 м в Копет-Даге, до 1300—1500 м в Таджикистане, до 3000, а на
пролете даже до 3600 м в Тянь-Шане (Зарудный и Кореев, 1906), на Алтае
до 1750 м (Сушкин, 1938). В горах предпочтение все же оказывается нижнему
поясу и всюду — открытому ландшафту. На пролете повсеместно в открытых
местностях, на зимовке, главным образом, в степеподобных саваннах и т. п.
ландшафтах. Распространение несколько спорадично, так как связано и с
наличием удобных мест для гнездовья — обрывов, скал, камней, развалин, и с
присутствием достаточного количества прямокрылых насекомых,
составляющих главную пищу этого сокола (в Африке установлены случаи перекочевок
зимующих там степных пустельг в места появления саранчи и т. п.).
Численность. На Украине, в Крыму, на Кавказе, в Средней Азии,
кроме пустынь ^обыкновенна; в северной части ареала спорадична (см.
выше).
Экология. Размножение. Начинается поздно (при массовом
появлении насекомых). В парах у гнезд держатся с прилета, но спаривание и брачный
полет происходит в конце мая и даже в начале июня (в это время были добыты
СТЕПНАЯ ПУСТЕЛЬГА
155
самки с яйцом в яйцеводе). Часть из птиц первогодков вероятно .образует
известный резерв холостых, хотя другие размножаются и в этом возрасте
(Першаков, 1929). Гнезда располагаются в камнях, скалах, развалинах и даже
под крышами, в речных глинистых обрывах, в дуплах, в норах (сизоворонки
и т. п.). Чаще всего они расположены колониями от нескольких пар до
нескольких десятков (в дуплах больших карагачей в Измаильском районе,
Закавказье). Встречаются и одиночные пары. Нередко колонии бывают смешанные
с обыкновенной пустельгой. При этом недавно в сев. Казахстане (Наурзум) на
гнездовье в таких смешанных колониях обнаружены особи гибриды между этими
видами (в Зоологическом музее Московского ун-та, сборы Осмоловской). Кладки
со второй половины мая и до начала июня. В Армении 14 мая были обнаружены
пять насиженных яиц и три свежих; 14 мая была найдена кладка в Закавказье:
27.V —пять яиц, у Нарына в Тянь-Шане; в конце мая у Чкалова; 28.IV —
13. V у Мариуполя, 17.V полная кладка в Аскания-Нова; 14.V сильно
насиженные яйца у Тышкана, Джунгарский Ала-Тау; 21— 25. IV в Копет-Даге у Су-
люкли; 11—14.VI насиженные яйца в Минусинской степи; 27.V первые кладки
в сев. Казахстане.
Число яиц в кладке 3—7, чаще 4—5. Они сходны с яйцами обыкновенной
пустельги по окраске: желтовато-красные с более темными кирпично-красными
пестринами или охристые с красно-бурым рисунком. Размеры: (80) 31,6—
37,5 х 26—31, в среднем 34,78x28,66 мм (Хартерт, 1913).
Насиживают оба родителя, самец сменяет самку в полуденные часы.
Длительность насиживания, вероятно, 28 дней. Гнездовый период немногим
более месяца. Птенцы во втором пуховом наряде, чуть начавшие оперяться,
появляются в июле (Котон-Карагай, 15.VII, быть может запоздавшие); в конце
июля большинство птенцов уже на крыле, но еще держатся близ гнезд (18—
23.VII, Харьковская обл.; Ростов-на-Дону; 10.VII, Дагестан; конец июля,
сев. Казахстан; Джаркент). На крайнем юге подлетки встречены и в начале
июля (5.VII, Туркмения; 2.VII, Таджикистан). В гнездовый период добычу,
несмотря на насекомоядность, ловит только самец, а оделяет выводок самка.
Вполне развитые птенцы обнаружены в выводках в разные числа августа,
когда уже образовались стайки и начинаются откочевки. Число птенцов в
выводке 3—5, редко 2. Указания на две кладки в лето (Волчанецкий, 1937)
основаны повидимому на недоразумении (быть может, вторая кладка имеет
место взамен утерянной).
Линька. Птицы в гнездовом наряде линяют весной (начало мая),
сменяя мелкое оперение. Мнение Штреземанна о том, что зимою молодые линяют
полностью, на нашем материале не подтверждается, но возможно, что смена
мелкого оперения на зимовке начинается у молодых уже с ноября и тянется
до марта. Линька взрослых — полная годовая — длится с середины июля до
середины сентября. Смена маховых, как у других соколов, начинается со
средних (7-е, 6-е, 5-е); одновременно со сменой средних маховых сменяется и средняя
пара рулевых. Заканчивается линька взрослых сменой 2-го, 10-го и 1-го
первостепенных маховых, а также задних плечевых, передних второстепенных и
маховых пятой (второй от края) пары рулевых. Последовательность смены нарядов:
первый пуховой наряд — второй пуховой наряд—первый годовой (гнездовый)
наряд — второй годовой наряд — третий годовой (окончательный) наряд и т. д.
Питание. В основном энтомофаг: прямокрылые (саранча, кобылки);
кроме того, жуки и стрекозы; указываются также фаланги, ящерицы, мелкие
грызуны (степная пеструшка и т. д.); изредка мелкие птицы (пеночки,
горихвостки на пролете; Северцов и Мензбир, 1894). Есть требующие подтверждения
указания на преобладание в корме степной пустельги в степях грызунов Micro-
tus arvalis и Lagurus lagurus, в пустыне ящериц Eremias arguta и Phrynocepholus
guttatus (Шварц, 1947).
156
ОТРЯД ХИЩНЫЕ ПТИЦЫ
О связи распространения и сезонных первхмещений со скоплениями саран-
чевых уже упоминалось.
Полевые признаки. Походит на обыкновенную пустельгу, но полет легче,
не трясется; размером значительно меньше; хвост шире и круглее; голос
протяжнее и выше, вроде «клии-клии-клии».
Описание. Размеры и строение. Сходна с обыкновенной пустельгой, но
строение крыла иное: второе маховое без вырезки на внутреннем опахале, первое маховое обычно
длинней, чем третье. Лапы меньше (цевка обычно менее 35 мм); общие размеры также
мельче. Длина самцов и самок (9) 310—335, размах (9) 702—745, в среднем 322,7 и 728,2 мм.
Рис. 46. Степная пустельга (самец и самка).
Крыло самцов (56) 223—245, самок (20) 228—246 (255) мм, в среднем соответственно 233,7
и 237,5 мм. Разница в размерах полов, следовательно, незначительна.
Окраска. Первый пуховой наряд не описан. Второй пуховой наряд серовато-
белый на спинной стороне, белый на брюшной. Гнездовый наряд — как у обыкновенной
пустельги, но поперечные полосы на спинной стороне правильнее и уже.
Взрослые самки рыжие, с продольными темнобурыми пестринами на голове и нижней
стороне тела, с сероватыми (обычно) рулевыми, поперечнополосатыми или с размытым
поперечным рисунком; маховые темнобурые с широким беловатым поперечным рисунком
на внутренних опахалах, иногда сливающимся в сплошное поперечное поле.
Взрослые самцы с серой головой, кирпично-рыжей не испещренной спиной, плечами и
кроющими крыла, сизым надхвостьем; первостепенные маховые темнобурые с белым полем
на внутреннем опахале; второстепенные маховые серые, внутренние — с рыжими каймами;
подкрылья охристые с черными пятнами, рулевые серые с черной предвершинной полосой
и белой вершинной каймой; брюшная сторона охристая с более или менее развитым бурым
продольным рисунком. Малые кроющие, а иногда и средние и часть больших кроющих часто
бывают сизыми, изредка встречаются особи с сильным распространением сизого на гтлечах
и крыльях. Во втором годовом наряде у самцов темя рыжеватое с продольными пестринами;
КОБЧИК
157
на спине иногда имеются бурые пестрины, брюшная сторона в продольных бурых пятнах.
Во всех нарядах восковица, кольцо вокруг глаза и лапы желтые, клюв сине-роговой,
темнеющий у вершины, радужина темнобурая, а когти беловатые (в чем хорошо отличается от
обыкновенной пустельги).
Систематические замечания. Все степные пустельги из СССР сходны по окраске,
и всюду встречаются особи с сизой расцветкой кроющих крыла, хотя число их быть может
несколько выше на востоке ареала. Так как этот признак принимается главным образом за
диагностический при выделении восточной формы pekinensis, а вместе с тем материал по
степным пустельгам из крайних восточных частей ареала вида очень мал, и к тому же
самое распространение там этого сокола выяснено плохо, от признания существования
двух подвидов степной пустельги приходится воздержаться (индивидуальная изменчивость
велика и географически не локализована).
10. Кобчик Falco vespertinus L.
Falco vespertinus. Linnaeus. Syst. Nat., изд. XII, I, 1766, стр. 129, тип. местн. Ингрия,
т. е. б. СПБ губ.
Русское название. Кобчик, кобец, копец — старинное русское имя для всех
мелких соколов. У соколиных охотников (см. <<Урядник Сокольничья Пути») кобцом
назывался чеглок. Как книжное название, имя кобец или кобчик закрепилось за F. vespertinus
с конца XVIII столетия, со времен Палласа. Любопытно, что слово «кобец», «Kobez» перешло
в качестве названия этой птицы во французский язык.
Распространение* Ареал. Область распространения кобчика
простирается от с.-з. частей СССР — Литвы, Латвии и Эстонии, Белоруссии, а также
Румынии, Венгрии, южн. Болгарии, Югославии на восток до Предкавказья, сев.
Казахстана, Алтая, Монголии, Манчжурии и с.-в. Китая. Зимой в южн. Азии,
от Китая до Индии, но главным образом в тропической и южн. Африке.
Характер пребывания. Перелетная птица с разобщенными
областями гнездовий и зимовок.
Биотоп. Степи и лесостепи, открытые участки культурного ландшафта.
Подвиды и варьирующие признаки. В западной части ареала — в Европе и зап. и
Средней Азии F. v. vespertinus L.; в восточной части от ю-в. Забайкалья и с.-в. Монголии до
Манчжурии и с.-в. Китая F. у. amurensis R a d d e, 1863. Отличия в окраске характерны
главным образом для самок, у самцов они незначительные; средние размеры восточного
подвида меньше, различны относительное расположение гнездового и зимнего ареалов;
экологически оба подвида весьма близки.
Западный или обыкновенный кобчик Falco vespertinus vespertinus Linnaeus
Синонимы. Falco rufipes. В e s e с к е. Beytrage f, Naturg. Vog. Kurlands, 1792,
стр. 20, табл. 4, Курляндия. — Falco vespertinus* obscurus. Tschusi Orn. lahrb. XV,
1904, стр. 229, Томск. — Erythropus vespertinus transriphaeus, Бутурлин, Орнитологич.
Вестник, 1915, стр. 127, Змеиногорск. — Falo pyrrhogaster Reichenow. Ornithol.
Monatsber., 1915, стр. 5, Бозум в вост. Камеруне.
Распространение. Ареал. Область обитания западного кобчика в
Польше неясна; нерегулярно гнездится в Прибалтике (Лоудон, 1913, Гроссе
и фон-Транзее, 1929), так же как и в Псковской обл.; обычен в Подолии и на
Волыни, в Венгрии и Румынии, доходя до южных частей Болгарии и
Югославии, Крыма и степной части Сев. Кавказа (но сведения о гнездовании в
Закавказье, Соснин и Ляйстер, 1942, наверное ошибочны), сев. частей Малой
Азии; восточная граница в области правых притоков Енисея и верхней Лены
до реки Марха в верховьях Нижней Тунгуски, в Балаганской степи до
Иркутска и Тунки, где спорадичен. Северная граница определяется находками в
Обонежье, у Архангельска, Усть-Сысольска, на средней Печоре под 62° (Усть*
унья, Пажгино); по Оби более или менее спорадичен на Малой Сосьве (Шух-
тункурт, в верховьях Конды, т. е. около 61—62° с. ш.), восточнее граница
спускается на юг, проходя примерно у Тюмени, Тары, Тобольска, быть может
Нарына, Томска и Красноярска, далее на Алтае (кроме высоких и лесистых
158
ОТРЯД ХИЩНЫЕ ПТИЦЫ
частей) до Марка-куля; за пределами Алтая на юге на Урунгу и в Китайской
Джунгарии; обычен в Канской и Минусинской лесостепи, но в с.-з. Монголии
и далее на запад у Зайсана не гнездится; южная граница в Зап. Сибири идет
около 51°—51°30' с. ш., а восточнее — у Семипалатинска. Зимовки от Судана
(где редок), в Анголе и Дамараленде; южнее малочисленен, будучи заменен
восточносибирской формой. Повидимому в связи с кормовыми условиями
кобчик иногда встречается далеко от гнездовой области на залетах — в Лаплан-
Карта 18. Распространение кобчика Falco vespertinus
•^— F. v. vespertinus, 2 — F. v. amurensis; a — граница гнездовий, г — граница зимовок (стрелка
показывает направление пролета восточноазиатского подвида).
дии (Мурманск), Норвегии, Дании, Англии, Швейцарии, Франции,
Голландии, Бельгии, Испании, с.-з. Африке. Имеются требующие подтверждения
указания о случайном гнездовании в ю.-в. Моравии, Лаузице и Силезии, в
Финляндии и вост. Швеции; о существовании в середине прошлого столетия
гнездовой колонии в Алжире у Джельфы. Возможны единичные случаи зимовки
к северу от нормальной области зимовок: в Крыму, в Турции, в Македонии.
Даты. Общее направление перелета в Сибири — западное осенью и
восточное весной; затем у Каспийского моря соответственно на юго-запад или
на юг и на северо-восток или на север, огибая при этом Туркестан. В Среднем
Азии кобчик отмечен только случайно весною в приаральских Кара-Кумах;
изредка у Ташкента в конце апреля — начале мая; сведения Руссова о про-
КОБЧИК
15У
лете у Искандер-куля и тем более Зарудного (1896) о гнездовании в
Туркмении ошибочны. Весенний пролет начинается на юге, на Украине около
середины апреля, в Бессарабии будто бы с конца марта; в средней полосе
кобчики летят в конце апреля и начале мая; у северной границы ареала —
в первой половине мая и даже во второй половине этого месяца (Молотовская
обл.). Так как сведения эти относятся к движению различных популяций, то
весь период пролета кобчиков кажется растянутым примерно на месяц. Еще
на севере Африки пролет кобчиков продолжается в конце апреля и начале мая
(в Тунисе; в Киренаике; в Египте). Поздно появляется кобчик и на сибирских
гнездовьях: уже в Зауралье только в начале мая, тогда же у Тюмени, в Барабе
и Кулунде, у Красноярска. Впрочем, и тут, несмотря на небольшое число
наблюдений, имеются, повидимому, колебания в сроках прилета (например в
Минусинской степи прилет отмечен 21 апреля, Зверев, 1930). В разные годы валовой
пролет проходит, повидимому, в различные сроки. Известная нерегулярность
установлена и для осеннего пролета, носящего в Западной Европе иногда
массовый характер, а иногда незначительный. Образование стай и отлет
начинается в августе, недели через две после вылета молодых из гнезд, проходя
главным образом в первой половине сентября и затягиваясь иногда до конца
этого месяца. Чем далее к югу, тем более значителен состав стай. Пролетные
кобчики наблюдаются в средней Европе от Венгрии до Балкан, на Украине,
в М. Азии, в с.-з. и южно-каспийских провинциях Ирана, но в Закавказье —
в малом количестве; затем на Синайском п-ове и в Африке.
Биотоп. Открытые равнины — степи и лесостепи, культурный
ландшафт с садами, парками и рощами (при этих условиях даже в городах,
например в Полтаве), уремы речных долин. Глухих лесов избегает и этим,
повидимому, а не историческими причинами (вопреки предположению Станчинского,
1922) объясняется редкость и спорадичность кобчика в центральной полосе
европейской России к северу от Украины, Курской, Орловской и
Воронежской областей и среднего Поволжья. Здесь, как и на севере, кобчик приурочен
либо к культурному ландшафту, либо к лесному, к гарям и вырубкам, или
обширным богатым стрекозами болотам и т. п. В горы вовсе не поднимается:
поэтому на Кавказе ограничен полосой предгорий по Кубани и Тереку; на
Алтае, впрочем, поднимается до 1000—1100 (Сушкин, 1938), местами, в виде
исключения, до 1500 м.
Численность. Распространение кобчика несколько спорадично,
так как зависит от комбинации условий, не часто встречающихся — групп
высоких деревьев среди открытого ландшафта. В таких местах кобчики
образуют колонии, иногда по нескольку десятков пар. В отличие от пустельги,
кобчик и на пролете держится обычно стаями, особенно значительными
на юге; в стаях встречается и на зимовках. По годам численность кобчика
неравномерна, как и меняется его размещение, что связано, повидимому, с
«урожаем» или «неурожаем» прямокрылых насекомых, составляющих основной корм
этого сокола в степях. Колонии кобчиков на севере — в лесной полосе —
меньше, чем в лесостепи и степи.
Экология. Размножение. Календарь размножения поздний, время
выкорма птенцов более или менее совпадает с массовым появлением насекомых.
Вместе с тем, период размножения растянут, что отчасти объясняется
индивидуальными колебаниями сроков размножения у отдельных пар, отчасти
наличием дополнительных кладок взамен утраченных, отчасти
метеорологическими условиями (например в Наурзуме, сев. Казахстан, в неблагоприятном
по кормовым условиям 1936 году кобчики гнездились на две недели позже, чем
в 1937). Половозрелость в возрасте около года, в следующую весну после
рождения. Судя по линянию некоторых особей, не все птицы в этом возрасте
размножаются (резерв холостых особей).
160
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Спариванье начинается с прилета и сопровождается брачным полетом
над местом гнездовья; самка часто присаживается на гнездо, са*мец, а иногда
и оба партнера, летают широкими кругами со своеобразными глубокими
взмахами крыльев. Спариванье происходит на деревьях (29 апреля,
Воронежская обл., Северцов, 1855). Есть указания, что кобчики иногда сами строят свои
гнезда, что вполне вероятно, так как для колониально гнездящейся птицы в
степной полосе не всюду можно найти подходящие гнезда других птиц; в таком
случае гнезда будто бы очень малы. Впрочем, постройка кобчиками своих
гнезд требует подтверждения (см., в частности, Гебель, 1879). Обычно кобчики
пользуются гнездами других птиц — в особенности грачей, а также ворон,
сорок, коршунов. Иногда на одном дереве размещается сразу несколько гнезд
(до 9, Зарудный, 1888). Кроме того, кобчики гнездятся в дуплах (на севере,
повидимому, преимущественно), на кустах (карагана и ива в ю.-з. Сибири),
а изредка даже в норах и обрывах или на земле среди кустов чилиги и
бобовника. Одиночные пары гнездятся редко, обычно гнезда расположены
колониями. Располагаются колонии по опушкам, в отдельно стоящих группах
деревьев, в садах и парках, у болот, вырубок и гарей, но не в глубине сплошных
лесных массивов. Кладка производится в конце мая — начале июня (на юге
Украины быть может уже в середине мая).
В кладке 4—5, обычно же 3—4 яйца, редко 1—2 или даже 6. Вероятно,
что размер кладки колеблется по годам в зависимости от кормовых условий.
Окраска яиц сходна с таковой у пустельги, но обычно поярче, а самые яйца
короче и легче. Размеры (7) 35—42 х 29—32 (Харьков), 34,5—37,6 х 25,5—30,8
(б. Енисейская губ.); (108) 32—41, 5 х 25, 5—31, в среднем 36, 5 х 29,4 (Умань).
Насиживают оба родителя, но ночью и в конце инкубационного периода —
только самка. Птенцы появляются в конце мая — начале июля, в некоторых
случаях позднее, во второй половине июля (20 июля Наурзум, Осмоловская,
1940). Насиживание производится с первого яйца, длительность его около
одного месяца (повидимому 28 дней). В кладке бывают обычно и неоплодо-
творенные яйца (одно — два), так как число птенцов в выводке чаще
всего 3. При плохих кормовых условиях плодовитость ниже: в Наурзуме в
1936 году в среднем на гнездо 2,6 птенца, а- 1937—3,2. Разница в возрасте
птенцов 3—4 дня, вес новорожденного 10, 1—13,5 г при вылуплении, 15,5—17
на следующий день; птенцы из последних в кладке яиц бывают меньших
размеров. Нередко в первые дни жизни младшие птенцы гибнут — это объясняется
тем, что при выкорме птенцов кобчики, как и другие насекомоядные птицы,
несколько менее тщательно оделяют птенцов пищей, чем виды, кормящиеся
более крупной добычей. За кормом птенцам сначала летает только самец,
передающий добычу самке, которая оделяет птенцов. Когда птенцы в 10 —
12-дневном возрасте надевают второй пуховой наряд, начинает летать за кормом
и самка; двухнедельным птенцам родители приносят насекомых до 57 раз в день
(Минусинская степь). С середины июля и до начала августа происходит вылет
птенцов, есть впрочем указания и на более ранние и на более поздние сроки
(22 июня, Харьковская обл., Сомов 1897; конец июня, Оренбургский край,
Зарудный, 1888; 16 —22 августа, Минусинск, Су шкин, 1913; 5 августа, Канск—не
доросшие птенцы; 14 августа Канск —подлетки, в Зоологическом музее Московского
университета и т. д.). Неразбившиеся выводки держатся у гнезд около двух
недель, после чего во второй половине августа начинается предотлетное стаенйе.
Линька. Изучена недостаточно. У птиц в гнездовом наряде смена
мелкого пера начинается ранней весной и с прилета такие птицы попадаются
только с остатками гнездового наряда — обычно это задние плечевые, рулевые
кроме средней пары, всегда первостепенные маховые (по крайней мере в мае —
начале июня). Затем у некоторых (вероятно, размножающихся) особей линяние
приостанавливается и только в июне начинается смена маховых. С другой сто-
КОБЧИК
161
роны, имеются особи, у которых в первой половине июня уже сменились 6-е
и 8-е первостепенные маховые, 5-е растут (быть может у холостых), у таких
особей линька кончается в сентябре, еще перед отлетом. Линька взрослых
начинается с половины июля, хотя отдельные нелиняющие особи встречаются
даже в начале августа (повидимому, от поздних выводков).
Рис. 47. Обыкновенный кобчик (самец, самка, молодая птица).
Конец линяния взрослых наступает на зимних квартирах.
Последовательность смены маховых — как и рулевых — сходна с тем, что известно для
других соколов (первостепенные маховые сменяются от середины, б-е или 7-е, затем
5-е, 7-е, 4-е, 3-е, 9-е, 2-е, 10-е и 1-е). Рулевые сменяются от средней пары,
но последней сменяется вторая пара от края хвоста, а крайняя —
предпоследней. Последовательность смены нарядов: первый пуховой наряд — второй
пуховой наряд — первый годовой (гнездовый) наряд — второй годовой
наряд—третий годовой (окончательный) наряд и т.д. По Уайзерби (1938),
линька в гнездовом наряде начинается с февраля и, захватывая мелкое перо и
средние рулевые, продолжается до мая; обычно с июля линька возобновляется,
распространяясь на остальные рулевые, кроющие и маховые, и заканчивается
к октябрю; взрослые птицы, по тому же автору, линяют с июля до октября»
11 Птицы СССР, т. I
162
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Питание. Основной корм — насекомые, которых кобчик ловит лапами
на лету или собирает с земли: прямокрылые —саранча и кобылки;
стрекозы; пилильщики; жуки-навозники, усачи, хрущи и др.; мелкие
млекопитающие — мыши и полевки, среди них Mas musculus, Stenocranius gregalis, Lagurus
lagurus; молодые суслики Citellus pygmaeu$y землеройки Sorex araneus; ящерицы
Lacerta agilis, Eremias arguta; птицы повидимому в виде редкого исключения.
С прилета большое место в кормовом режиме занимают грызуны (Житков и
Бутурлин, 1906), в период размножения и на зимовке —насекомые.
Полевые признаки* Кобчик — мелкий сокол с длинными, крыльями,
напоминающий в общем чеглока; полет менее стремительный, чем у последнего;
старые самцы хорошо отличаются от всех других наших соколов по темной
окраске брюшной сторвны, молодые —от.чеглока —резко поперечнополосатым
хвостом и светлым беловатым лбом и щеками. Мало осторожен, в особенности
у гнезд. Держится часто группами или в стаях. Голос — «ки-ки-ки», как у
пустельги, но протяжнее. Кобчики крикливы, особенно в период размножения.
Описание. Размеры и строение. Крылья длинные, формула крыла 2>1>3.
Клюв относительно слабый, как и пальцы, короткие и довольно толстые. Длина самцов
(19) 284—315, самок (19) 278—332, в среднем 301,0 и 309,2 мм. Размах самцов (16) 690—750,
самок (16) 714—770, в среднем 718,0 и 746,0 мм. Вес самцов (5) 130—164, самок (5) 162,5—
197,3, в среднем 149,4 и 182 24 г. Крыло взрослых самцов (50) 224—253, самок (52) 232—255,
в среднем 239,2 и 255 мм.
Окраска. Первый пуховой наряд снежнобелый, второй серовато-белый.
Взрослые самцы сизо-серые с серебристым налетом на маховых, с черноватой
головой и рулевыми; оттенки серого цвета варьируют индивидуально; подбой крыла
черновато-серый; задняя часть брюха, голени и подхвостье рыжие.
Взрослые самки серые с темносерыми поперечными полосами на спине, крыльях,
хвосте; темя рыжее или охристое с темными наствольями; у глаза черное пятно; горло
беловатое, брюшная сторона ржавчато-рыжая или охристая, иногда с продольными темными
штрихами; маховые серые с белыми поперечными пятнами на внутренних опахалах. Во
втором годовом наряде у большинства самцов рыжие пятна на затылке, брюшная сторона
частично рыжеватая или серая с черноватыми наствольями; у самок в этом возрасте
брюшная сторона менее рыжая, беловатая и обычно с большим развитием темных продольных
полос. Гнездовый наряд на спинной стороне темнобурый с охристыми каймами перьев и
поперечнополосатым хвостом, с беловатым лбом и широкими светлыми каймами перьев
темени; уздечка, пятно вокруг глаза и усы черноватые;'брюшная сторона охристо-беловатая
с бурым продольным рисунком. Радужина темнобурая; когти белые; клюв голубоватый,
чернеющий к вершине; ноги, кольцо вокруг глаза и восковица у молодых желтые, у взрослых
самок оранжевые* у самцов красные.
Восточный или амурский кобчик Falco vespertinus amurensis R a d d e
Falco vespertinus var. amurensis. R a d d e. Reisen im Siiden von Ostsibirien, II, 1863,
стр. 102, табл. I, фиг. 2, 3-я, Приамурье.
Синоним, Falco Raddei. Finsch und H a r 11 a u b. Vogel Ost. Afrikas, 1870,
стр. 74, новое имя для предыдущего.
Распространение. Ареал. Восточный кобчик распространен к востоку
от предыдущего: в ю.-в. Забайкалье, Приморье, Манчжурии, с.-з. Монголии,
Корее, Сев. Китае к югу до Циньлинских гор и Цзяньсу. В Забайкалье по
Аргуни и Онону, у Кяхты и Нижнеудинска; на Амуре вверх по течению до
Кумары, к северу до устья реки Горин; в вост. Приморье отсутствует.
Отдельные гнездовые находки — в Центральной Азии (Дынь-юань-инь) и
в Бирме.Зимовки в небольшом числе в Индии до Ассама, в Южн. Китае, глав-
ным же образом в южн. Африке. При этом область зимовок западного подвида
лежит к северу от области зимовок амурского кобчика, последний зимует
южнее земель Дамара и Ниасса. Пролет идет, повидимому, через сев.
Монголию, Индию и Афганистан и далее, вероятно, от с.-з. Индии через Индийский
океан в вост. Африку. Залеты в Коряцкую Землю (Дементьев, 1941), к
верховьям Лены (Закаменское 8.VI), на Цейлон (Уейт, 1931).
Характер пребывания. Перелетная птица.
КОБЧИК
163
Даты. В Африке на зимовке появляется только около конца декабря,
отлетая в начале марта. В Индии осенью в сентябре — октябре, весной в
марте — апреле. Пролет —. громадными стаями. На местах гнездовья
появляется поздно, в начале мая (но на Сунгачи будто бы в начале апреля, что
впрочем сомнительно). В с.-в. Китае пролетает весною в апреле —мае (Уайл-
дер и Хеббард, 1924), прилетает в Сев. Китай в апреле (Латуш, 1932). Осенью
на севере пролетает с середины до конца сентября (сев. Монголия, Приморье).
Биотоп. Лесостепь, речные долины с уремой, широколиственный лес
среди влажных болотистых мест и т. п. Избегает безлесной степи и сомкнутого
леса. На зимовке в Африке — в саваннах.
Рис. 48. Амурский кобчик (самец, самка).
Численность. На гнездовье спорадичен, на зимовке образует
большие скопления.
Экология. Размножение. Половая зрелость наступает, когда птица
находится еще в первом годовом наряде, в возрасте около 10 месяцев. Цикл
размножения поздний, повидимому, как и у номинальной формы, в
соответствии со временем массового появления насекомых, в частности саранчевых.
Начало развития, гонад в начале мая, вскоре после прилета. В отличие от
западного подвида, не образует больших колоний. Расположены гнезда в
Приморье— на лиственных деревьях, в с.-в. Монголии—на березах, в
Забайкалье — на ели, на соснах и лиственницах. Возможно, что кобчик не только
занимает чужие, в частности вороньи (в Китае сорочьи) гнезда, но строит и
свои (Шульпин, 1936).
Кладка в начале июня, т. е. примерно через месяц после прилета,
Неполные кладки из 3 яиц найдены Спангенбергом на Имане еще 5—6.VI, но там
11*
164
ОТРЯД ХИЩНЫЕ ПТИЦЫ
же найдено было только одно яйцо в гнезде 13.VI; нормально полная кладка
производится около середины июня. Промежуток между откладкой отдельных
яиц — 48 часов. Число яиц в кладке от 3 до 5, очень редко до б (Бекер, 1928).
Окраска как у европейского кобчика. Размеры (10) 34,4—37,4 х 28—29,9
(Спангенберг); (50) 33—38,9 х 27,1—32, в среднем 35,8 х 28,9 мм (Бекер,
1928), (29) 33—38,5 х 26—29,5 мм (Латуш, 1932). Насиживание продолжается
около месяца, так как пуховиков в Забайкалье находили во второй половине
июля. Вполне развитые летные молодые появляются в августе.
Линька. Птицы в первом годовом наряде линяют с прилета, к началу
июня у них уже свежие мелкие перья, а также часть рулевых; линька,
следовательно, начинается уже на зимовке; во второй половине июля у этих птиц,
как и у взрослых, начинается смена маховых (с б-го или 7-го) и рулевых, начиная
со средней пары; во второй половине августа (экз. от 19.VIII) от гнездового
наряда остаются только 2-^3 передних первостепенных маховых. Взрослые
птицы в окончательном наряде в июне еще не линяют, а во второй половине
августа у них только сменяются средние маховые (5-е, 6-е и 7-е). Эти
различия в сроках линяния позволяют предполагать, что рано линяющие молодые
не гнездятся и образуют резерв холостых потенциально половозрелых особей.
Кончается линька у взрослых на зимовке.
Описание* Размеры и строение. Длина крыла самцов (12) 220—230, самок
(4) 230—235, вс реднем соответственно 224,2 и 232,3 мм, т. е. меньше, чем у западной расы.
Вес самцов (2) 114,5—140 г.
Окраска. Пуховые наряды неизвестны. Первый годовой наряд — как у западного
подвида, но на темени охристые и рыжеватые оттенки менее развиты, каемки на спинной
стороне тела обычно поуже и бледнее. Самцы в окончательном наряде как у номинальной
расы, но с белыми подкрыльями. Самки очень своеобразны, без рыжих тонов в окраске, на
спинной стороне серые с шиферно-серым темным поперечным рисунком, с серой головой и
хорошо выраженными черноватыми усами; брюшная сторона у них белая с продольным
темным серо-бурым рисунком, переходящим на боках в неправильные поперечные полосы;
перья голени и подхвостья беловаты, без темного рисунка. Переходный наряд (второй
годовой) с теми же особенностями, как у западного подвида.
СЕМЕЙСТВО ЯСТРЕБИНЫЕ ACCIPITRIDAE
РОД ЯСТРЕБЫ ACCIPITER BRISSON, 1760
Тип A. ntsus
С и и о и и м: Astur Lacepede, 1769, тип A. palumbarius.
\\. Большой ястреб, ястреб тетеревятник Accipitergentilis L.
Falco gentilis, Linnaeus. Syst. Nat., изд. X, 1758, стр. 89 Делекарлия.
Синоним. Falco Palumbarius, Linnaeus. Syst. Nat. Изд. X, 1758, стр. 91, Швеция.
Русское название. Ястреб — старинное русское слово. Ястреб в качестве
ловчей птицы упоминается уже в завещании Владимира Мономаха, т. е. около 1125 года.
Этимологически оно близко к польскому имени этой птицы и быть может к средневековому
латинскому astur, откуда французское auto ur, итальянское astore и т. д. В XIX веке эту
птйцу.называли иногда утятником — по объяснению Аксакова потому, что «им обыкновенно
травят уток». Тетеревятник — название книжное и мало удачное, так как создает у охотника
не натуралиста ложное представление об односторонности питания этой птицы. В XVII
веке, судя по «Уряднику Сокольничья Пути», птицу эту звали просто ястребом, а соколиные
охотники в XVIII—XIX столетиях—большим ястребом.
Распространение. Ареал. Область распространения ястреба
тетеревятника: лесная зона Сев. Америки, Европы, северной и Средней Азии; в
Африке только в сев. Марокко; к северу до лесотундры, к югу до Италии, южн.
Испании, Малой Азии и Палестины, сев. Ирана, далее — юго-западной Сибири,
Алтая, зап. Китая, Тибета, Японии. Вероятно к этому же виду относится
мадагаскарский A hensti.
Карта 19. Распространение тетеревятника Accipiter gentilis
1 — A. g. atricapillus, 2 — A. g. striatulus, 3 —A. g. arrigonii, 4— A. (g.) hensti, 5 — A. g. khamensis, 6 — A. g. gentilis, 7 — A. g. gallinarum, 8 — A. g. marginatus,
9—A. g. caucasicus, 10 — A. g. buteoides, 11—A. g. suschkini, 12 — A. g. albidus, 13 — A. g. schvedowi, 14 — A. g. fujiyamae; a—граница гнездовий,
г —область кочевок; цыфры в кружках—местонахождения соответствующих подвидов зимой.
166
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Характер пребывания. Отчасти оседлый, но на севере в
лесотундре и северной полосе тайги перелетный, хотя часть птиц остается там и
зимою. Впрочем, сезонные перемещения носят только характер рассеивания
и некоторого смещения популяции к югу; ареал же, в отличие от настоящих
перелетных форм, остается непрерывным, но площадь его несколько
возрастает в южном направлении, достигая Южн. Китая (Юннань), Бирмы и сев.
Индии, Средней Азии и Ирана. Большой ястреб, будучи связан с древесной
растительностью, не выходит за ее пределы и зимою. Миграции в Америке
носят аналогичный характер —зимою ястреб доходит в ней до сев. Мексики,
Техаса, Миссури, Индианы и Виргинии.
Численность высокая, за исключением близких к границе ареала
о бластей, а также густо населенных.
Подвиды и варьирующие признаки. Как и большинство оседлых и полуоседлых видов,
тетеревятник образует большое число географических рас. Гнездовые области их таковы.
В сев. Европе от Скандинавии на западе до центральной полосы Европейской части
СССР — A. g. gentilis L., 1758; в центральной и вероятно в Западной Европе A. g. gallinarum
С h г. L. В г е h m, 1831; в Сардинии, Корсике, Испании и Марокко A. g. arrigonii Klein-
schmidt, 1903; в Крыму, на Кавказе и в сев. Иране A. g. caucasicus Kleinschmidt,
1923; в вост. Средиземноморье — на Балканах от Славонии и Боснии до Греции A. g. таг-
ginatus PilleretMitterpacher, 1783 (сюда быть может и итальянские?); в северной
полосе европейской тайги и в таежной полосе Западной Сибири A. g. bateoides M e n z b i e г,
1882; к югу от него в лесах Приуралья и ю.-з. Сибири A. g. suschkini Dementiev,
1940; в Восточной Сибири от Алтая и Енисея до Забайкалья и Лены A. g. schvedowi M e n z b.,
1882; на крайнем с.-в. Сибири от бассейна Индигирки и Колымы до Анадыря, Коряцкой
Земли и Камчатки A. g. albidus Me n z b., 1882; на Дальнем Востоке — в Приморье и
Японии — A. g. fujiyamae Н а г t. et S w a n n, 1923; в ю.-в. Тибете, ю.-з. Ганьсу и сев. Юннани,
на север до сев. склонов хребта Гумбольдта A. g. khamensis В i a n с h i, 1906; в Сев. Америке
от с.-з. Аляски, с.-з. Мекензи, сев. Манитоба, ю.-в. Онтарио, Унгава и Ньюфаундленде до
внутренних частей Британской Колумбии, Мичигана, сев. Нью-Йорка, севера Новой Англии,
Массачузетса, Пенсильвании и зап. Мериленда A. g. atricapillus Wilson, 1812; в западных
прилегающих к Тихому океану частях Сев. Америки от юга Аляски до Иосемита, Новой
Мексики, Чихуахуа A. g. striatulus R i d g w а у, 1874; наконец, на Мадагаскаре A. (g) henstl
S с 1 е g e 1, 1873. Морфологически варьирующие признаки: размеры, постепенно
уменьшающиеся с севера на юг, окраска более светлая у северных форм и более насыщенная на юге;
детали морфологии, например размеры сердца. Подвиды крайнего севера (albidus, buteoides)
при этом диморфны и кроме обычной «серой» фазы имеют и белую (аналогия с северными
кречетами).
Экологически варьирующие признаки касаются характера пребывания в ареале
(большая или меньшая степень оседлости и т. п.), календаря периодических явлений, повидимому
также особенностей размножения и пищевых связей. Морфологически резко обособлены от
европейско-азиатских американские ястребы и в особенности мадагаскарский, многими
считаемый за отдельный вид и занимающий обособленную область распространения;
остальные формы распадаются на южную группу marginatus, arrigonii, caucasicus, также schvedowi,
khamensis, fujiyamae и северную albidus, buteoides, gent His. Подвиды gullinarum и sushkini
занимают промежуточное положение.
Среднеевропейский тетеревятник Accipiter gentilis gentilis L.
Синоним. Astur gentilis moscoviae. Sushkin. Proceed. Boston Soc. Nat. History,
1928, стр. 8, Рязанская губ., Михайлов.
Распространение. Ареал. Область распространения среднеевропейского
тетеревятника — Скандинавия, Финляндия, кроме, повидимому, крайнего
севера этих стран; у нас — в европейской части страны, за исключением
северной полосы тайги — областей Вологодской и Архангельской, Молотовскои и
сев. части Кировской, возможно — Обонежья; однако отдельные экземпляры
с Кольского п-ова не отличимы от номинальной формы; на восток
распространен до Волги (включительно), на юг — до Украины (Харьков, Полтава,
Киев); в западной части ареала —на Украине уже от Полтавы, в
Белоруссии и Литве — встречаются и особи переходного к западноевропейской форме
gallinarum типа, более темной окраски; на севере и востоке — примерно
уже от Москвы и Рязани — в популяции заметна примесь крови светлой
БОЛЬШОЙ ЯСТРЕБ, ЯСТРЕБ ТЕТЕРЕВЯТНИК
167
северо-таежной расы buteoides; такие же особи обнаружены в Финляндии.
Среднерусская популяция морфологически сходна — при всем своем*
разнообразии — со шведскими ястребами, которые в сущности тоже представляют собой
форму, переходную между западно и среднеевропейскими и таежными. В
негнездовое время залетает на запад до южн. Германии и Франции (главным
образом молодые особи).
Характер пребывания. Оседлая птица, совершающая однако
перекочевки в негнездовое время. Ястреб, окольцованный 28.VI.1940 у Шток-
мансгофа в Латвии, был добыт в декабре 1940 в Починковском районе
Смоленской обл. Другой, окольцованный 29.Х И. 1932 у Цейссена, добыт 15. IV. 1942 в
Могилевском районе. Птенец, окольцованный 18. VI. 1948 г. в Тульских Засеках,
был добыт неподалеку 23.Х 1.1948 в Теплоогаревском районе Тульской обл.
Результаты кольцевания (Людерс, 1938) показывают, что эта форма ястреба
в отношении ориентации не приспособлена к дальним передвижениям, так как
завезенные на расстояния более 100 км обратно не возвращались. Имеется
известная смена биотопов — в зимнее время года ястребы перемещаются из лесистых
местностей в культурный ландшафт. С этим, повидимому, связаны, как у F. р.
brevirostris, сезонные изменения кормового режима.
Биотоп. Леса разного типа —хвойные, смешанные, лиственные,
старого и среднего возраста (гнезда размещаются всегда на высоких деревьях);
избегает однако сплошных глухих массивов. Охотно держится среди
культурного ландшафта, даже в окрестностях больших городов. Охотничьи участки
ястребов в гнездовое время около 4—б км от гнезда (окрестности Москвы),
в зимнее время более обширны. В СССР эта форма встречается на равнинах,
кроме лесистых частей Карпат.
Численность. Обыкновенен. Имеются некоторые колебания
численности популяций (плодовитость, особенно смертность молодых в выводках),
отражающие кормовые условия того или иного года (Уттендерфер, 1939).
Экология. Размножение. Как у соколов и других хищных птиц,,
потенциальная половая зрелость наступает на первую весну после вывода в
возрасте около одного года, когда птица находится еще в гнездовом наряде;
но к гнездованию приступают в этом возрасте далеко не все. Пары постоянные,
так как старые ястребы держатся вместе и вне периода размножения, но
охотятся отдельно и на некотором расстоянии один от другого (до начала марта);
молодые первогодки держатся в первую зиму одиночками. Цикл размножения
ранний. Брачный полет начинается уже с конца февраля —начала марта
(Белоруссия, Федюшин, 1926; окрестности Москвы, Дементьев).
У каждой пары имеется несколько гнезд в одном постоянно занимаемом
участке; используются гнезда по годам посменно. Гнезда располагаются на
деревьях, иногда они устраиваются самим ястребом, иногда используются
чужие; обычно расположены высоко над землей, от 8 до 20 м. Молодые птицы,
повидимому, строят гнезда и ниже. На севере гнезда размещаются чаще на
хвойных деревьях — елях, соснах, на юге на дубах, липах, грабах, березе.
К периоду размножения гнездо подновляется, на лоток кладутся свежие
зеленые ветви. Обычные размеры гнезда около 600—750 мм в диаметре, около
500 мм в высоту; лоток неглубокий, выстилка слабая из тонких веточек, на
крае гнезда и на лотке обычно настилаются зеленые ветви. Расположены
гнезда в развилке или на ветви близко от ствола, нередко у вершины дерева;
чаще у просек, опушек или вблизи от них. В занятом гнезде или под ним
обычно обнаруживаются перья линяющей самки, часто остатки пищи.
Кладка из 3—4 яиц, редко до 5 (Шнитников, 1913) или только 2.
Кладка — в разные числа апреля, на юге, повидимому, уже в первой половине
<Умань, Самара, Карамзин, 1901; Харьков, Сомов, 1897). В случае гибели
кладки производится повторная, меньшая: вторая из 2—3 яиц, третья из 1—2
168
ОТРЯД ХИЩНЫЕ ПТИЦЫ
(Гебель, 1879), при этом птица кладет яйца в новое или другое гнездо (Хару-
зин, 1926). Окраска яиц зеленовато-белая, без блеска, чаще всего однотонная
или со слабо заметными зеленоватыми, реже буроватыми, пятнами. Размеры:
(133) 51,5—63 х 40—49,5, в среднем 57 х 44,5 (Умань, Киевская обл., Гебель,
1879); (6) 55,9—61 х 44,1— 45,5 (Харьковская обл., Сомов, 1897); 55— 62 х
X 43—46 (Минск, Шнитников, 1913).
Насиживание начинается с первого яйца, участвуют в нем оба родителя
(у самцов имеются небольшие наседные пятна), но главным образом самка.
Самец приносит корм насиживающей самке. Продолжительность насиживания
у нас точно не прослежена (по западноевропейским данным, Нитхаммер, 1938,
35 дней). Вылупление птенцов начинается со второй половины мая (ранний
выводок уже 22. IV) и в начале июня. Вылет молодых происходит во второй
половине июля (24.VII, Подолия; 14.VII, Киев; 13.VII, вост. Белоруссия;
26—27.VII, Москва; в июле в Поволжье, в Тульской обл., в Калужской обл.).
Гнездовый период, следовательно, продолжается около полутора месяцев.
Число птенцов в выводке обычно меньше количества яиц в кладке, так как
одно из яиц нередко неоплодотворено, а в годы с неблагоприятными
кормовыми условиями младшие птенцы, как правило, гибнут и нередко съедаются
старшими (Псков, Зарудный, 1910 и др.).
Первое время после вылета из гнезда птенцы держатся около него (не
умеющие летать птенцы выбираются обычно из гнезда и сидят на ветвях гнездового
дерева). В августе выводки разбиваются, птенцам в это время около 50 дней.
Линька. У молодых ястребов в первом годовом (гнездовом) наряде
отдельные перья второго годового наряда начинают появляться уже в первую
осень, особенно на брюшной стороне тела и на голени (на последней быть может
взамен случайно утраченных). Однако смена крупного оперения — маховых
и рулевых — начинается и у годовалых и у старых птиц во второй половине
гнездового периода. Последовательность смены маховых у ястребов, как и у
других видов сем. ястребиных и в отличие от семейства соколиных, от
внутреннего края крыла к внешнему, от 10-го к 1-му. Линяние начинается в последних
числах апреля, к началу мая сменяются 9-е и 10-е маховые, в начале июня
линька маховых наполовину закончена; около половины сентября линяние
заканчивается, продолжительность его, следовательно, равняется 4х/2 месяцам*
Рулевые сменяются от средней пары к краю хвоста, центробежно, но последней
сменяется вторая от края пара, а крайняя — предпоследней. Линька бывает
раз в году, полная. Последовательность смены нарядов: первый пуховой
наряд — второй пуховой наряд — первый годовой (гнездовый) наряд —
второй годовой наряд — третий годовой наряд и т. д. Второй годовой наряд —
окончательный, но отличается от последующих некоторыми незначительными
особенностями (см. ниже).
Питание. Большой ястреб — главным образом орнитофаг, но в
отличие от соколов охотится преимущественно в пересеченной местности. Он может
ловить летящую добычу, но не бьет ее на высоком расстоянии от земли, а
стремится «прижать» преследуемую птицу к земле, строениям и т. п., а затем
схватить ее лапами. Еще чаще схватывает ястреб добычу, сидящую на земле или
на деревьях или добычу, бегущую по земле, будь это птица или
млекопитающее. Способы нападения также отличны от соколиных и характеризуются, как
и у других ястребов, следующими чертами. Издалека на добычу, за редкими
исключениями, ястреб не бросается, —он начинает нападение с расстояния
около 100—150 м, неожиданно вылетая из-за прикрытия —группы деревьев
и т. п.; нередко ястреб сидит в засаде, бросаясь затем на приближающееся
животное. Охотясь на открытых пространствах, обычно летает «низом» и
выпугивает добычу. Однако при нападениях на домашних голубей ястреб летит
рысоко, стараясь приблизиться к ним сверху, а затем стремится отбить одну
БОЛЬШОЙ ЯСТРЕЕ, ЯСТРЕБ ТЕТЕРЕВЯТНИК
169
из птиц из стаи и вынудить ее снизиться. Преследует добычу ястреб «в угон»,
не делая соколиных ставок; в крайнем случае для быстроты нападения на
летящую птицу ястреб проделывает в воздухе «сальтомортале», чем ускоряет спуск
и приближение к жертве.
Полет ястреба очень быстрый и увертливый, короткие крылья позволяют
ему летать между деревьями и среди кустов, сильное развитие крылышка —
круто тормозить, длинный хвост — быстро менять направления полета. На
птиц в воздухе ястреб может поэтому бросаться не только сверху, но и сбоку
и снизу. Быстрота полета обеспечивается, несмотря на небольшую длину
крыльев, большим развитием грудной мускулатуры (отношение ее веса к общему
весу тела примерно такое же, как у крупных соколов, около 8% у самок и
9—9,5% у самцов; при этом у молодых птиц мышцы эти достигают своего
полного развития уже осенью). Добычу ястреб умерщвляет лапой, сжимая ее
когтями.
В период насиживания ловит добычу главным образом самец и передает
ее самке; когда вылупятся птенцы, первое время кормит выводок самец,
передавая пойманную добычу самке для о деления молодых. Первое время добыча
приносится «очищенной», т. е. ощипанной, без перьев или шерсти, часто без головы
или с оторванными конечностями. Для такой разделки имеется нередко особое
место (в холодное время года это производится на дереве или деревьях,
служащих для ночлега). Вблизи занятого гнезда ястребы, как правило, не охотятся.
Кормовые остатки — перья и шерсть добычи на месте ее разделки, также
погадки (продолговато-овальной формы, около 3,5—6 см в длину и около
2,5—3 см в ширину, обыкновенно из перьев и шерсти и обломков
полупереваренных костей) и поеди — части крупных костей или отдельные кости.
Потребность в пище взрослого ястреба около 200 г мяса в день (судя по
опыту соколиных охотников).
Несмотря на разнообразие кормового режима ястреба, у него
наблюдается известное «саморегулирование» численности, главным образом в
результате гибели части птенцов как в раннем возрасте, так и на взлете. Как уже
указывалось, при 3^4 яйцах в кладке число гнездарей 2—3, иногда и менее. Общее
число ястребов в одной местности остается таким образом более или менее
постоянным (если исключить конечно прямое преследование со стороны
человека), но при резком изменении ландшафта — вырубке леса — ястреб исчезает.
В качестве корма ястреба в европейской России указываются главным
образом птицы средней величины — лесные голуби, рябчики, тетерева, серые
и белые куропатки, молодые и изредка даже взрослые глухари, дрозды, скворцы,
сойки, вороновые, дятлы, совы (ушастая), другие хищные птицы —
перепелятник, чеглок, относительно редко утки, из домашней птицы куры и голуби
(включая почтовых, которых ястребы могут даже догнать при полете в
воздухе); из млекопитающих —зайцы, главным образом беляки, чаще молодые,
но и взрослые; белки, реже мелкие грызуны. У отдельных особей (пар) имеется
определенная специализация кормового режима.
Сезонные изменения питания отражают обеднение лесной фауны в
холодное время года: с октября по конец февраля — начало марта ястребы главным
образом переселяются в культурный ландшафт и кормятся воронами, галками,
сороками, голубями; в Лапландии в период гнездования питаются
преимущественно воробьиными средних размеров, как кукши и дрозды; зимой
куриными — белыми куропатками, рябчиками, даже глухарями.
Полевые признаки. Старые птицы на спинной стороне тела бурые, на
брюшной беловатые с поперечным рисунком; молодые бурые на спинной стороне,
охристые или беловатые с бурым продольным рисунком на брюшной стороне;
самцы заметно меньше самок. Крылья короткие, на лету обычно с
расставленными передними маховыми, хвост длинный. Полет — см. выше. На открытых
170
ОТРЯД ХИЩНЫЕ ПТИЦЫ
местах появляется редко, тогда быстро летит либо верхом, на большой высоте,
либо низом, по прямому направлению. Появление ястреба вызывает резкую
реакцию испуга (бегство, затаивание) у других птиц. Голос — «кьяк-кьяк»
или «гик-гик-гик» или «киррк». Не избегая соседства человека, осторожен,
даже в гнездовый период.
Описание. Размеры и строение. Телосложение плотное, плоская голова с круто
из огнутым клювом (на краю верхней челюсти выступ); длинная и сильная, оперенная почти до
половины цевка; длинные и крепкие пальцы с большими когтями, особенно на заднем и
внутреннем пальцах. Формула крыла: 4>3>5>6>2>7>8>1 ...Вырезки на внутренних опахалах
1-го — 5-го и на внешних опахалах 2-го—б-го маховых; сильно развитое крылышко. Хвост
длинный, слегка закругленный. Длина самцов (13) 520—560, самок (6) 600—645, в среднем
соответственно 582,2 и 619,7 мм. Размах самцов (12) 960—1150, самок (6) 1090—1212, в
среднем 1061 и 1174 мм. Вес самцов (5) 838—960, самок (3) 1251—1477, в среднем 885,6 и 1377,7.
Крыло самцов (71) 303—342, самок (63) 350—385, в среднем 322,74 и 360,47 мм. Самцы,
следовательно, заметно меньше самок.
Окраска. Первый пуховой наряд охристо-белый, лапы розовые, радужина
черноватая. Второй пуховой наряд, надеваемый в возрасте между 7—14 днями, гуще, плотнее,
сероватый на спинной стороне, охристый на брюшной, лапы желтые, радужина серая.
Первый годовой наряд на спинной стороне бурый с более светлыми основаниями перьев,
с неправильными охристыми пестринами на плечевых и кроющих крыла, с охристыми каемками
перьев; второстепенные маховые у вершины серые, у основания бурые с поперечным
рисунком; первостепенные маховые темнобурые с бледными охристыми поперечными полосами
на внутренних опахалах; рулевые бурые с темнобурыми поперечными полосами,
окаймленными охристым, боковые рулевые с беловатыми мраморными пестринами; на затылке
охристые или рыжеватые пятна, надхвостье и поясница с широкими охристыми пестринами;
брюшная сторона охристая или рыжеватая с черновато-бурыми продольными пестринами, перья
голени иногда одноцветно охристые, иногда с бурыми пятнами.
Только что оперившиеся птенцы окрашены ярче, светлые части их оперения рыжеваты;
осенью к началу зимы они выцветают и бледнеют, доходя до белесоватых тонов. Кроме того,
яркость охристо-рыжеватых тонов подвержена большим индивидуальным колебаниям. На
западе области распространения встречаются чаще более темно окрашенные рыжеватые
с грубой пестриной на брюшной стороне и с насыщенным бурым тоном спинной стороны —
переходная или вернее смешанная с A. g. gallinarum популяция.
Взрослые ястребы на спинной стороне серовато-бурые, самцы несколько светлее и серее
самок; над глазом и на затылке беловатые пестрины; лоб светлый, темя черновато-бурое;
маховые темнобурые со светлыми полосами на внутренних опахалах; рулевые бурые с
поперечным рисунком, часто неправильным у старых особей; брюшная сторона беловата с
бурым поперечным рисунком из полос и темными стволами перьев горла, зоба, груди, верхней
части боков и брюха; подхвостье белое, на некоторых его перьях иногда бурые поперечные
полосы; подкрылья цвета боков тела.
Индивидуальные вариации выражаются в большем или меньшем развитии серого
тона на спинной стороне и в различной ширине темного рисунка на брюшной стороне; при
этом на западе ареала — на Украине, в Белоруссии — встречаются особи очень темнобурые
на спинной стороне и грубо разрисованные на брюшной стороне (популяции переходные
к среднеевропейской форме), тогда как в центральной России такие особи очень редки.
Зато там встречаются — и чем севернее и восточнее, тем чаще — относительно светлые, а в
первом годовом наряде сильно пестрые, особенно на крыле, ястребы, приближающиеся
к A. g. buteoides. Очень редка беловатая вариация (Нитхаммер, 1938).
С возрастом ширина поперечных полос на брюшной стороне, повидимому, сокращается
(по наблюдениям над содержащимися в неволе птицами). Радужина у молодых бледножел-
тая, у взрослых оранжевая или даже красноватая (у самцов); восковица бледножелтая,
разрез рта и ноги восково-желтые, клюв и когти черные.
Западносибирский тетеревятник Accipiter gent His buteoides Menz.
Astur palumbarius buteoides. Мензбир. Орнитологическая География 1882, стр. 440,
описан по зимним экз. из Владимирской губ.
Синоним. Astur gent il is poecilopterus. Lonnberg. Fauna och Flora 1922, стр. 228,
Архангельск.
Распространение. Ареал. Север лесной полосы Восточной Европы и
с.-з. Сибири — от Обонежья на западе до бассейна Енисея и Лены на востоке;
на Яне —^ переходные популяции к восточносибирскому подвиду; южная
граница не вполне ясна —к этому подвиду относятся ястребы, добытые у Онежского
БОЛЬШОЙ ЯСТРЕБ, ЯСТРЕБ ТЕТЕРЕВЯТНИК
171
озера, в северных частях Вологодской обл. и республики Коми (б. Яренский
у., следовательно, около 62° с. ш.), спускается далее южнее —до Кирова (где
встречаются и особи переходные к номинальному подвиду) и к среднему
Уралу у Свердловска; в Западной Сибири граница идет примерно по южной
окраине тайги, по линии Тюмень — Томск, т.-е. около 56—57° с. ш., на Енисее
граница неясна (возможно от Н. Тунгуски). Северная граница определяется,
невидимому, пределами лесотундры — отмечен у Мезени, под Архангельском,
на залете в Тиманской тундре (в марте), в гнездовое время в кустарниковой
тундре в низовьях Печоры по р. Куя* на Оби по Надыму около 65° с. ш., на
Ямале у м. Круглого (белая вариация), на Тазовском п-ове на р. Хадут-
тей, на Гыданском п-ове под 70°1Г с. ш., в нижнем течении Енисея
у Игарки (белая вариация), на южн. Таймыре у границы леса (Скалой, 1937).
Б зимнее время в средней полосе европейской России, в Финляндии (Войпио,
1946), в Лапландии под 68—70° с. ш.; единичные особи в Голландии, в вост.
Германии, Дании, Швеции, Польше, Венгрии, также в южн. Украине (Аскания-
Нова), в Крыму (Тамак), на Сев. Кавказе (Пятигорск), у Ростова на Дону,
в низовьях Волги у Астрахани, в Казахстане (Гурьев, Наурзум, низовья
Сыр-Дарьи, Аулье-ата), в Киргизии (в окрестностях Фрунзе, в Киргизском
хребте, по Хартерту, 1913 —на Иссык-куле), в Туркмении на ю.-з. (Гассан-
кули), в Сибири на Алтае, у Минусинска и в Забайкалье, в Центральной Азии
в южн. Синьцзяне у Саджоу (Сушкин, 1928).
Характер пребывания. Часть особей, как видно из
предыдущего, совершают довольно значительные перелеты; другие кочуют, некоторые
остаются на севере и зимою.
Даты. Пролет северных ястребов в средней полосе начинается с сентября
<Псков; экземпляры из окрестностей Загорска; экземпляр из района озера
Чаны в ю.-з. Сибири), но главным образом в октябре — ноябре. В Туркестане
««белые» ястребы чаще появляются в холодные зимы, что показывает на
характерную для кочующих нерегулярно птиц зависимость большего или
меньшего удаления из гнездовой области от метеорологических условий того
или иного сезона. Весной на местах зимовки остается до марта и первой
половины апреля.
Биотоп. Как у номинальной формы, но также лесотундра.
Численность. Обычен, кроме северной окраины области
распространения. В средней полосе европейской России (например под Москвой) к этой форме
в некоторые годы принадлежит подавляющее большинство зимующих ястребов.
Экология изучена слабо, имеются лишь весьма отрывочные сведения,
указывающие однако на большие различия с A. g. gentilis (перелетность,
календарь периодических явлений и т. п.).
Размножение. Цикл проходит значительно, почти на месяц, позднее,
чем у предыдущей формы. Кладки в Лапландии имеют место около середины
мая, из 3—4 яиц; в то же время на севере Вологодской обл. и в
Архангельской обл., также в Кировской обл. Вполне развитые молодые
подлетки в самом конце июля и в начале августа (25.VII у Надыма, бассейн
Оби; 31.VII в низовьях Печоры; только 5.VIII на средней Печоре; в начале
августа в Кировской обл. и т. д.). Есть указания, будто бы эта форма
иногда гнездится и на скалах (Плеске, 1886).
Л и н ь к а. У птиц в первом годовом наряде мелкое перо начинает
понемногу сменяться уже в январе — феврале, т. е. в возрасте около полугода,
когда появляются отдельные новые перья на голенях и брюхе, но смена больших
перьев не начинается по крайней мере до второй половины мая. Смена половины
первостепенных маховых — от 10-го до 5-го — у птиц в первом годовом наряде
заканчивается в первой половине июля (Болыыеземельская тундра, холостые
особи?), у взрослых гнездящихся птиц протекает на месяц позднее. С конца сен-
172
ОТРЯД ХИЩНЫЕ ПТИЦЫ
тября встречаются уже взрослые особи в свежем наряде. Последовательность
смены нарядов и смены крупного оперения — как у предыдущего.
Питание в общем сходно с кормовым режимом номинальной формы:
главным образом птицы средней и крупной величины, например на Печоре
глухари, тетерева, белые куропатки; также зайцы беляки (при этом отмечается
известное соответствие между численностью ястреба и урожаем зайцев, факт,
требующий, впрочем, проверки). На кочевках у Москвы зимой эти ястребы
успешно нападают на голубей, даже почтовых, кроме того на ворон, галок и т. д.
У одной птицы в зобу найдено 480 г мяса (Мылва, республика Коми).
Описание, Размеры и строение. Крупней номинальной формы. Длина самцов
(б) 520—615, самок (16) 607—680, в среднем 558,6 и 640,7 мм. Размах самцов (3) 970—1070,
самок (10) 1086—1270, в среднем 1031,3 и 1162,7 мм. Вес самцов (10) 870—1170, самок (10)
1185—1509, в среднем 1016,5 и 1354,7 г. Крыло самцов (29) 311—345, самок (71) 352—383,
БОЛЬШОЙ ЯСТРЕБ, ЯСТРЕБ ТЕТЕРЕВЯТНИК
173
в среднем 323,3 и 368,7 мм. Кроме того, в сравнении с южными и среднеевропейскими
ястребами, описываемая форма ястреба отличается относительно большими размерами сердца
и большей грудиной (это отражает быть может перелетность расы).
Окраска заметно светлее, чем у номинальной формы. Взрослые бледные, самцы
на спине голубоваты, самки сероваты; темя относительно бледное, иногда с преобладанием
белого цвета на лбу и затылке; на задней части шеи обычно, а на передней части спины—реже,
имеется поперечный рисунок из серо-бурых полос по белому фону, напоминающий по
характеру окраску брюшной стороны у взрослых тетеревятников; широкая белая бровь, слабо
испещренная темными пятнами; на кроющих крыла нередко имеются беловатые каймы, а на
больших кроющих и белые пятна; поперечный рисунок брюшной стороны бледный и узкий,
с сероватым оттенком, полосы на ней расположены довольно редко, темные настволья
выражены слабо; середина брюха часто без темного рисунка; на рулевых темные поперечные
полосы окаймлены обычно белым, на боковых парах значительная примесь белого цвета.У самок
все эти признаки выражены менее резко, чем у самцов.
Некоторые особи отличаются крайней бледностью бурого тона спинной стороны, голова
у них беловатая с немногими темными пестринами, затылок и шея белые с бурым поперечным
рисунком, крылья с крупными белыми пестринами, брюшная сторона с узкими, редкими и
очень бледными поперечными полосками — это как бы переход к белой фазе или вариации.
В первом годовом наряде голова беловатая с относительно узкимитемными наствольями,
на плечевых и кроющих крыла крупные беловатые пестрины, светлые каемки перьев
широкие, рулевые сероватые с относительно узкими широко окаймленными белым поперечными
бурыми полосками; брюшная сторона с узкими темными продольными полосками; при этом
основной тон окраски светлых частей оперения бледноохристый (отнюдь не рыжеватый)
даже в свежем пере, белый с ранней осени; общий тон бурых частей спинной стороны
светлый, сероватый. Кроме того, в отличие от номинальной формы, белая фаза у которой крайне
редкое исключение, у buteoides белые птицы встречаются среди популяций более или менее
регулярно.
Окраска этих белых птиц варьирует от чисто белой со слабыми следами бледносерых
настволий и размытого темного рисунка на верхней стороне тела до белой с темнобурыми
более или менее поперечными на спинной стороне и продольными на брюшной стороне
пятнами — в первом наряде; у взрослых птиц имеется довольно явственный бледный
«размытый» остаток бурого рисунка на белом фоне. По окраске такие птицы не отличимы от
восточносибирских белых ястребов подвида albidus. На севере они найдены пока на
нижней Оби и у Надыма под 65° с. ш. (выводок из двух птиц, из них одна белая, а другая
темная), в бассейне Конды, на севере Сургутского района, на Ямале (мыс Круглый, в
августе), в низовьях Енисея у Игарки под 69° с. ш. и у Дудинки, В негнездовое время белые
ястребы встречались в Башкирии, в Туркестане (Сыр-Дарья, предгорья Киргизского хребта,
Иссык-куль), у Ростова на Дону (экземпляры в Дарвинском музее в Москве), также в
Финляндии, но главным образом в Западной Сибири (Тобольск, Тара, Омск, Томск, Чаны).
Возможно, что распространение на гнездовье особей белой фазы ограничено северной
частью ареала, вероятно лишь на окраине тайги и в лесотундре. Характерно, что только там
встречаются, повидимому, и наиболее светлые особи «серой» фазы с развитыми светлыми
пестринами на спинной стороне тела.
Камчатский большой ястреб Accipiter gentilis albidus Menz.
Astur palumbarius albidus. Мензбир. Орнитологическая География, 1882, стр. 438,
Камчатка (новое имя для «varietas Kamtschatica alba» ястреба, описанной Палласом, 1811).
Синонимы. Astur candidissimus. Dybowski. Bull. Soc. Zool. de France 1883,
стр. 353, Камчатка. — Astur caesius. Бутурлин, Псовая и Ружейная Охота, 1907, VII,
стр. 9, Среднеколымск.
Распространение. А реа л. Область распространения камчатского ястреба—
с.-в. Сибирь от Индигирки, Колымы, Анадыря, Коряцкой Земли до Камчатки;
к северу до дельты Индигирки и Колымы, т. е. примерно до 70° с. ш.; к югу
вероятно до Верхоянского хребта; западная граница не вполне ясна (с Яны
имеются птицы, весьма сходные с buteoides, нотакже и настоящие—по окраске—
albidus, так что тут распространена повидимому смешанная популяция). В
негнездовое время встречен на Командорских о-вах, в Японии, в Приморье
и Уссурийском крае, в Даурии на Ононе, на западе до Лены (Жиганск)
и может быть Ачинска (взрослый самец добыт в октябре у с. Балахтинского,
быть может однако особь белой фазы buteoides).
Характер пребывания. Часть особей оседла (на Колыме даже
в области дельты; в низовьях Индигирки в конце февраля; на Анадыре ветре-
174
ОТРЯД ХИЩНЫЕ ПТИЦЫ
чается круглый год, Портенко, 1930). Другие откочевывают на юг, достигая
ю.-в. Сибири. Кочевки начинаются уже в первой половине сентября; весной
в Приамурье белые ястребы встречаются еще в начале апреля (9. IV), движение
на север идет, повидимому, в марте — начале апреля.
Биотоп. Гнездовый —. тайга, лесотундра, повидимому, чаще в
последней. В тундре бывает только залетом или в холодное время года; притом редко.
Зимой — в лесах (например в Буреинском хребте), а также среди равнинных
кустарников как в глубине материка, так и на морском побережье.
Численность. Камчатский ястреб — редкая птица. Может быть
представление о малочисленности усиливается вследствие большой
осторожности этого ястреба.
Экология. Размножение. Гнезда в лесотундре на Колыме
помещаются на тальниковых кустах или лиственницах (Бутурлин); по Анадырю
на тальниках (Портенко, 1939); на Камчатке будто бы даже на скалах (Тачанов-
ский, 1891). Из гнезда «на скале» Дыбовскому были доставлены камчадалами
яйца беловато-охристой окраски с маленькими буроватыми пятнами; длина их
58—59, ширина 45—45,5 мм (Тачановский). Единственные более или менее
детальные наблюдения над размножением белого ястреба произведены на
вост. Камчатке Авериным (1948) на территории Кроноцкого заповедника i
пара белых ястребов гнездится здесь из года в год в долине реки Богачевки.
По крайней мере с 1943 по 1946 год занималось одно и то же гнездо,
расположенное метрах в 7 от земли в развилке на старой березе; построено оно самими
ястребами из довольно тонких березовых веточек, подстилка из бересты;
диаметр гнезда около 70—75 см, так что сидящая в нем птица почти не видна.
Оба родителя были белой окраски, и все птенцы рождались также белые. 20.V
самка насиживала; 16.VI в гнезде были обнаружены 3 сильно насиненных
яйца; вылупление произошло в конце июля (около 25.VI). 23.VII в гнезде
сидели три почти оперившихся птенца с короткими маховыми и рулевыми,
У гнезд ястребы очень осторожны, только в конце насиживания самка сидит
очень плотно. Насиживает главным образом самка, самец в это время носит
ей корм (белых куропаток и т. п.). Вполне выросшие самостоятельные молодые
из разбившихся уже выводков наблюдались и добывались в конце августа.
Питание. На Камчатке —. зайцы, утки, глухари, куропатки, между
размещением которых и распространением ястребов* замечается, повидимому,
некоторая зависимость. Зимою в Малом Хингане нападает на белок. На
Индигирке поедает попавшихся в силки белых куропаток (а во второй
половине зимы его самого ловит местное население в различного рода снасти
и употребляет для прокорма ездовых собак). На Колыме в лесотундре ястреб
кормится главным образом белыми куропатками и утками, в таежной полосе —
беляками, глухарями, рябчиками, белыми куропатками, белками, но также и
мелкой птицей.
Описание. Размер ыистроение. Камчатский ястреб — самая крупная форма
большого ястреба: крыло самцов (10) 333—342; самок(8) 370—388 мм (по Сушкину, 1928,
до 395 мм). Средние величины соответственно 336,5 и 379,3 мм. Даже самцы весят более 1200 г
(три фунта, по Палласу, 1811); самка слеток с недоросшими крыльями и хвостом весила
1390 г у летная молодая 1320 г, молодой самец 894 г (Кроноцкий заповедник).
Окраска. Самая светлая форма, при этом имеются налицо серая и белая фазы
или вариации. Белые птицы встречены на Анадыре,в Коряцкой Земле и на Камчатке.
Взрослые ястребы этого типа окраски белые с незначительной примесью пестрин, обычно на
голове, кроющих уха, на мантии, на маховых и на брюшной стороне, на спинной стороне эти
следы пигментации сизоваты, на брюшной серовато-буры, здесь они носят характер наст-
вольных черт на зобу и груди, иногда и слабо намеченных остатков поперечных полос на тех
же местах. Некоторые особи почти чисто белы.
В первом годовом наряде у белых птиц имеется более или менее заметный бледнобурый
рисунок на голове, шее, зобу, груди, спине и следы поперечной полосатости на маховых и
рулевых; самые светлые особи имеют только следы бледнобурых штрихов на голове, шее,
передней части спины.
БОЛЬШОЙ ЯСТРЕБ, ЯСТРЕБ ТЕТЕРЕВЯТНИК
175
Птицы темной фазы в первом годовом наряде отличаются очень значительным
развитием белых пестрин (всовершенно свежем пере слегка охристых) наспиннойстороне, особенно
на крыльях; очень бледным тоном бурого цвета; узким темным рисунком на брюшно#
Рис. 50. Камчатский тетеревятник (взрослый самец темноц фазы,
молодой самец темной фазы, самка белой фазы).
стороне; белый цвет сильно развит и на маховых, и на рулевых перьях. Взррслые птицц
на спинной стороне очень бледного сизого цвета, темя бледносерое, лоб и брови белые, на
перьях затылка, шеи, нередко на верхней части спины и кроющих крыла поперечный
♦ультравентральный» рисунок, темные полосы на маховых и рулевых еда,бо развиты, на.
176
ОТРЯД ХИЩНЫЕ ПТИЦЫ
боковых рулевых—заметный белый крап; брюшная сторона белая с очень узкими и
бледными поперечными полосками и темными наствольями на зобу и груди. Нередко сизый
цвет на спинной стороне на шее, передней части спины, кроющих крыла становится как бы
размытым или струйчатым, в виде мраморно-серых пестрин.
От buteoides восточносибирская форма ястреба отличается относительно большим
количеством в популяциях белых особей, а также деталями окраски.Серые птицы в окончательном
наряде еще светлее и сизее,чем большинство buteoidesyc более слабым рисунком на брюшной
стороне. В первом годовом наряде восточносибирские ястребы светлей западносибирских,
бурый тон у них бледнее, темный рисунок на брюшной стороне тела уже и светлее, больше
беловатых пестрин на спине и крыльях. Белые птицы по окраске неразличимы (мнение Суш-
кина, 1928, и других о том, что белая вариация buteoides никогда не достигает такой степени
депигментации, как albidus, вызывает большие сомнения). Кроме того, albidus несколько
крупнее и с более оперенными — до половины длины — цевками.
Систематические замечания. Размещение по ареалу белой и темной фазы ястребов
представляет интерес. Отметим, что у большого ястреба оно дает любопытную аналогию с
кречетом. Что касается восточносибирской фазы большого ястреба, то птицы белой фазы в
гнездовое время не добывались в западной части ареала, по крайней мере они неизвестны из
бассейна Индигирки и Колымы. На Анадыре и Камчатке белые ястребы, повидимому,
численно преобладают. Среди изученного материала по восточносибирскому ястребу
(Зоологический музей Московского университета, Зоологический институт Академии наук,
Дарвиновский музей в Москве) было 13 белых и 25 темных птицы. Белые ястребы
происходили с Камчатки (4 молодых птицы), из Коряцкой Земли (3 молодых птицы), из бассейна
Анадыря (2 взрослых и 2 молодых), с о. Медного (молодая птица), из окрестностей Ачинска
(взрослая птица).Темные ястребы добыты на Яне (взрослая и молодая птица), на Индигирке (4 взрос-
лых и 2 молодых), на Колыме (взрослая и 3 молодых), на Анадыре (взрослая и 2 молодых),
в Коряцкой Земле (молодая птица), на Камчатке (взрослая и 3 молодых), на Лене у Жиганет
(молодая птица), на Дальнем Востоке (3 молодых из Удского Острога, с низовьев Амура,
Сидеми) и Даурии (Онон, молодая птица).
Таким образом, по аналогии с кречетами, на крайнем северо-востоке Сибири белые
ястребы составляют вероятно около 50% в популяциях, но с достоверностью не найдены ни
на Колыме, ни на Индигирке. Последнее обстоятельство может в известной мере оправдать
выделение особого для этих местностей подвида A. g. caesius.
Южноуральский большой ястреб Accipiter gentilis suschkint D e m.
Accipiter gentilis suschkini. Дементьев. Бюлл. Моск. Общества испыт» природы,
отд. биолог., 1940, стр, 21, Бурзянский район, Башкирия.
Распространение. Ареал. Юго-западная Сибирь от Южн. Урала (до
Зилаира и вероятно нижнего течения Сакмары) до бассейна Тобола (Тюмень)
и Иртыша (Омск), Детали распространения не вполне ясны.
Характер пребывания. Часть особей оседла, другие с
сентября до начала апреля совершают более или менее отдаленные кочевки.
В негнездовое время ястребы этого подвида добывались у Уржума (февраль),
в окрестностях Тары (зимой), у Томска (март) на севере; у Куйбышева (октябрь)
на западе; у Краснодара (декабрь), в Туркмении у Багира под Ашхабадом
(ноябрь), в Киргизии у Фрунзе (апрель) на юге.
Биотоп. Леса в лесостепной полосе; на Урале и в хвойных, и в
лиственных, и в смешанных, но в горно-таежных районах редок.
Численность. В связи с особенностями расположения в ареале
лесной растительности, форма эта размещена спорадично, но в подходящих
биотопах птица нередка.
Экология. Размножение. Наблюдения производились Кириковым,
Ушковым, Крюковой. У гнезд на Южн. Урале тетеревятник появляется в
начале — середине апреля и начинает подправлять гнездо. Одно и то же гнездо
занимается несколько лет подряд, но чаще у пары бывает 3—4 сменных гнезда.
Расположены гнезда на березах и соснах, метрах в 5—10 от земли, часто вблизи
от глухариных токов, обильных водяными птицами водоемов или у гарей и
вырубок, богатых куриными и т. д. Расстояние между гнездящимися парами
от 6 до 16 км (Ильменский заповедник, Ушков, 1949). Кладка поздняя—■ в
первой половине мая (в Башкирском заповеднике начало 5—7 мая, в Ильменском
заповеднике в 1940 году 11—13 мая).
БОЛЬШОЙ ЯСТРЕБ, ЯСТРЕБ ТЕТЕРЕВЯТНИК
177
Число яиц в кладке 2—3, редко 4; птенцов в выводке 2—3, иногда 1.
Насиживание начинается с первого яйца, в течение 35 дней. Надклюнутые яйца
в Башкирском заповеднике 2^6 июня, в это время и только что
вылупившиеся птенцы.
Летными птенцы становятся в возрасте 37 дней. Летные птенцы вне гнезда
попадались во второй половине июля (например, 22.VII, Ильменский
заповедник), но подлетки ранее (8—11.VII, Башкирия, Сушкин, 1904). Нередко
в гнезде один из птенцов погибает. Выкармливает птенцов самка, которой
самец приносит добычу; последний повидимому не может оделять птенцов,
пока те малы, и в случае гибели самки выводок гибнет с голоду, хотя самец
продолжает приносить ему добычу и бросает ее на гнездо.
Л и н ь к а. У добытых у гнезд взрослых самцов в конце мая (25.V)
линяния нет, в начале июня (6.VI) идет смена мелкого пера и первостепенных
маховых (9-го и 10-го), а также средних рулевых. У взрослых самок в конце мая
(20.V) выпадают 7-е, 8-е и 9-е маховые, 10-е свежее; в начале июня (6.VI) 8-е,
9-е и 10-е маховые у самки уже свежие, 7-е растет, 6-е выпало, меняется мелкое
перо; в таком же состоянии находится линька и у других добытых на гнездах
вначале июня (7.VI—10.VI) самок. У всех этих птиц сменяются средние пары
рулевых. Последовательность смены маховых, следовательно, от 10-го к 1-му.
В октябре линяние заканчивается (24.Х добыты птицы в свежем пере), общая
его продолжительность следовательно несколько менее 5 месяцев. Первый
пуховой наряд сменяется на второй на 10-й день жизни и тогда же чуть
начинают пробиваться маховые, а немного позднее и рулевые.
Питание. В Башкирском заповеднике кормовой режим этого ястреба
составляют главным образом разные птицы — глухари, в особенности молодые,
тетерева и другие лесные птицы; в одном гнезде найдены 4 глухаренка, 3
молодых сойки, молодой гоголь, дрозд и пустельга, в другом 3 глухаренка, 3 молодых
дрозда, 2 кукушки, 3 сойки, 1 козодой, 1 луговой чеккан, 2 бельченка. В
желудках добытых птиц обнаружены глухарята, птенцы воробьиных, бурундук,
полевки Clethrionomys rut Has и Microtus arvalis; в местах кормежки взрослых
птиц остатки 17 рябчиков, 5 тетеревов, 3 соек, глухарки, вальдшнепа, 2
кукушек, 3 воробьиных птиц, полевки.
В Ильменском заповеднике в 150 погадках и 200 кормовых остатках найдены
5 видов зверей и 41 вид птиц. Из зверей ястреб ест белку, беляка, хомяка, водяную
крысу, полевку Clethrionomysglareolus; из птиц кормом служит белая куропатка,
тетерев, рябчик, серая куропатка, перепел, домашняя курица, вяхирь, горлица,
лысуха, чибис, черныш, вальдшнеп, сизая чайка, обыкновенная крачка, чирок
трескунок, чирок свистунок, кряква, серуха, пустельга, перепелятник, лунь,
черный коршун, рыжий канюк, тетеревятник (каннибализм), седой дятел,
кукушка, желна, большой пестрый дятел, серая ворона, грач, сорока, галка,
скворец, иволга, зяблик, обыкновенная овсянка, дрозды и т. д.; насекомые
(возможно из желудков жертв). Из птиц основным кормом являются утиные (39,3 %)х
куриные (46,6%) и воробьиные (64% в период выкорма птенцов), из зверей
мышевидные грызуны (32% встреч.). У одного гнезда найдены несколько
скелетов глухарей, в том числе взрослых и почти целых — как под деревом, так
и в гнезде; у другого гнезда 6 скелетов тетеревят и глухарят, 3 скелета крякв,
2 чирка, ноги 2 крупных зайчат (Ушков, 1949).
Описание. Размеры и строение. По величине несколько больше, чем
восточносибирский A. g. schvedowi, но меньше, чем buteoides. Длина самцов (4) 506—550, самок (3)
570—604, в среднем 533,7 и 592,7 мм. Размах крыльев самца (1) 1030 мм. Вес самцов (3)
688, 860, 900 г, самки (1) 1287 г. Крыло самцов (17) 3G5—335, самок (13) 343—380 мм, в
среднем у самцов 316, у самок 360,7 мм.
Окраска. В окраске взрослых птиц на спинной стороне сильно развит сизый цвет,
не только у самцов, но и у самок; в свежем пере у самцов края перьев мантии образуют
характерный чешуйчатый рисунок. Окраска взрослых в общем близка к окраске восточно-
12 Птицы СССР, т. I.
178
ОТРЯД ХИЩНЫЕ ПТИЦЫ
сибирских schvedowi, но побледнее; темя и кроющие уха черноваты, широкая белая бровь;
полосы на рулевых развиты слабо, поперечный рисунок на брюшной стороне тонкий,
правильный и бледный, иногда как бы выцветший. Птицы в первом годовом (гнездовом) наряде,
при значительной индивидуальной изменчивости, на спинной стороне довольно светлы и
пестры, напоминая в этом отношении buteoides; на брюшной стороне темные с сильно
развитым бурым продольным рисунком.
Среднесибирский большой ястреб Accipiter geniilis schvedowi М е n z.
Astur palumbariusschvedowi. Мензбир, Орнитологическая география, 1882, стр. 439,
Да расу н в Забайкалье.
Распространение. Ареал. Средняя Сибирь, кроме самых северных ее
частей; Алтай, южные части бассейна Енисея, бассейн Лены до Мархи и
Якутска, на восток до низовьев Алдана; Танну-тува; верховья Н. Тунгуски;
Прибайкалье, на восток до ю.-в. Забайкалья; сев. Монголия —Монгольский
Алтай, Хангай, Кентей. В холодное время года в Приамурье, Туркестане
(Тянь-Шань, Фергана, Ташкентский оазис), вероятно к этому подвиду
принадлежат и ястребы, бывающие зимою в Синьцзяне; на юг до сев. Индии
(Бекер, 1928).
Характер пребывания. По крайней мере в северных частях
ареала перелетная птица, зимующая, как указывалось, в северных частях
Средней Азии, * на юге средней Сибири (Алтай, Красноярск, Прибайкалье).
Указания на гнездование в Туркестане (ельники центр. Тянь-Шаня, Шульпин,
1939; горы Били-тау в бассейне Или, Шестоперов, 1929) требуют
подтверждения. В Семиречье зимуют главным образом молодые птицы.
Даты. Осенью в Киргизии ястребы появляются в сентябре, но движение
их продолжается и в октябре—ноябре—декабре; на Сыр-Дарье главный
пролет начинается с ноября и продолжается до середины декабря. Те же даты
указываются и для Забайкалья и для осеннего пролета на с.-в. Алтае.
Весенний пролет в Забайкалье отмечен около половины апреля, в Средней Азии
ястребы начинают исчезать во второй половине февраля и в марте, отдельные
задерживаются до апреля. На севере у Иркутска прилет 13. IV, отлет 30. IX.
Биотоп. Хвойные леса — лиственничники, кедрачи и т. п. На Алтае
до 1600 м.
Численность. В Восточной Сибири ястребы немногочисленны.
Экология. Сведения отрывочны. Размножение. Цикл поздний,
кладка в начале мая (2 яйца у Дарасуна, Тачановский, 1891), пуховые
птенцы наблюдались в конце июня (2 птенца 20.VI, Салаир, Иогансен,
1935; 3 птенца 4.VII, Барнаул; 2 птенца 11.VII, Танну-ола, Тугаринов, 1916).
Гнезда помещаются на лиственницах и соснах. Подлетки встречены
в начале августа; летные, вполне выросшие молодые в разные числа
августа (10.VIII, Алтай; 21.VIII, Минусинск; 19—26.VIII, сев. Байкал).
В выводке обычно 2, реже 3 птенца; плодовитость следовательно ниже, чем
у северных подвидов (gentilis, buteoides) и сходна с таковой у suschkini.
Линька. Взрослые птицы в сентябре — октябре надевают свежий
наряд; молодые в первом годовом наряде уже в декабре начинают сменять
отдельные мелкие перья на брюшной стороне. Ход линяния в общем как у
других форм.
Питание. В качестве добычи восточносибирского ястреба указываются
в зимнее время белки (Хинган), утки, фазаны, кеклики, зайцы (Туркестан);
* Значительное большинство зимующих в Туркестане больших ястребов—A.g. schvedowi.
Кроме того, в меньшем количестве встречаются suschkini и как редкость buteoides. Для всех
этих ястребов у туркестанских сокольников имеются особые имена: редкая белая вариация
buteoides, высоко ценимая, называется «туйгун», светлые и пестрые сибирские buteoides-— «тунд-
жур»; schvedowi и suschkini— «куш».
БОЛЬШОЙ ЯСТРЕБ, ЯСТРЕБ ТЕТЕРЕВЯТНИК 179
в гнездовой области из млекопитающих бурундуки, пищухи, суслики Citellus
undulatus (Скалой, 1935, Штегман, 1936 и др.), в других местах также белки,
например, на р. Марха, притоке Вилюя и т. д. У гнезда с двумя пуховыми
птенцами в Салаире найдены б галок, 2 больших горлицы, большой пестрый
дятел, бурундук и летяга (Иогансен, 1935). Бедность восточносибирской
тайги птицами увеличивает значение млекопитающих в кормовом режиме
ястреба.
Описание. Размер ыистроение. Относится к южной, темной и некрупной по
росту группе ястребов, с пропорционально слабыми лапами; цевка в среднем на 5 мм короче,
чему северных ястребов; такая же разница в длине среднего пальца, измеренного без когтя.
Крыло самцов (9) 300—322, самок (16) 326—361, в среднем 309,4 и 346,5 мм (Зоологический
музей Московского университета); крыло самцов 310—333, самок 345—358 лш(Сушкин, 1928).
Вес самцов (2) 556, 600, самки (1) 1000 г.
Окраска. Взрослые птицы на спинной стороне темные, с сильным развитием
сизого тона, темя и кроющие уха у самцов черные, у самок темноаспидные, с резко
выделяющейся белой бровью; поперечный рисунок рулевых развит слабо, на средней паре часто
только в виде пятен или их остатков. Поперечный рисунок на брюшной стороне выражен
довольно слабо—полосы серовато-бурые и узкие, впрочем расположены довольно часто; темные
настволья на зобе и груди чуть намечены или отсутствуют. Окраска молодых в первом
годовом наряде довольно значительно варьирует; общая окраска их темная, на спинной
стороне черноватая или темнобурая; на груди и брюхе сильно желтоватая, с резким темным
продольным рисунком;светлые пестрины на плечах и крыльях развиты слабо. Белой фазы нет.
Японский большой ястреб Accipiter gentilis fujiyamae Swann et Hartert
Astur gentHis fujiyamae. Swann and Hartert. Bull. Brit. Ornith. Club, XLIII, 1923,
стр, 170, Сагами нокуни, Япония.
Распространение. Ареал. Япония (Хоккайдо, Хондо), вероятно
Курильские о-ва, Корея и Формоза; в СССР на Сахалине и в вост.
Приморье. Быть может сюда же (или к schvedowi) относятся ястребы, спорадически
гнездящиеся в горах Северного и Среднего Китая.
Характер пребывания неясен, часть особей оседла, но во всяком
случае отлет тетеревятника отмечен с Уссури и озера Ханка, осенняя
миграция в сентябре — октябре. Впрочем, эти сведения, как и данные о зимовках
ястреба в Северном и Среднем Китае, могут относиться и к другим подвидам,
гнездящимся севернее и западнее.
Биотоп. Леса разного типа — как манчжурского, так и охотского. В
вертикальном отношении в Японии прослежен до верхней границы леса в горах,
но обычно на высоте около 1000 м, зимой на равнине (Ян, 1942).
Численность. На Дальнем Востоке редок, повидимому из-за
густоты леса и подседа, затрудняющих охоту, быть может и из-за
малочисленности куриных птиц.
Экология. Размножение. В Японии гнезда на соснах, около
5—15 л* от земли. Кладки в Японии состоят из 2—4 яиц (Ян, 1942). 1. VI на
горе Фуджи была найдена будто бы свежая кладка; размеры яиц
51—54,5x43—43,5 мм (Суанн, 1925). 15.VI в гнезде были обнаружены птенцы
в возрасте 2—3 недель, 20. VII —слеток (Ян, 1942). Летные молодые с недо-
росшими рулевыми — 2 молодых самки в одном гнезде, на реке Тачингоуза,
Судзухе, добыты 6. VIII. В половине августа вполне выросшие летные молодые
в Приамурье.
Линька. У взрослой самки, добытой 21. VII, мелкое оперение уже
сменилось, перелиняли средние рулевые, задние — от 10-го до б-го —
первостепенные маховые; росло 5-е маховое; линяние, следовательно, наполовину
завершенное. У взрослой самки 21. IX линяние закончено, но не доросли две
боковых пары рулевых.
Питание. В качестве добычи японского большого ястреба на
Дальнем Востоке отмечены куриные — тетерева и фазаны; в Японии главным
12*
180
ОТРЯД ХИЩНЫЕ ПТИЦЫ
образом разные птицы—дрозды, сойки, голуби, из зверей белки; там же обыкно.
венный и медный фазан.
Описание. Размер ыистроение. Размеры менее, чем у A. g. schvedowi: крыло
взрослых самок 337, 340, 348 мм, молодых 333,5, 337, 338, 340, 345, молодого самца 302 мм
(Зоологический музей Московского университета); крыло самцов 280—300, самок323—339иси
(Суанн, 1926); крыло молодых самцов (2) 295—299, самок 352—353 мм (Сушкин, 1928).
Окраска. Взрослые птицы бурее по основному тону, темнее на спинной стороне, с
более грубым и ярким поперечным рисунком на брюшной стороне, чем schvedowi; в первом
годовом наряде эти ястребы также очень ярки, рыжеваты на брюшной стороне и темны.
Систематические замечания. Реальность отличий этой формы, от A. g. schvedowi
многими авторами подвергается сомнению (см. в частности «A Hand—list of the Japanese
Birds», 1932; Хартерти Штейнбахер, 1936). Неясно и распространение: обычно его
ограничивают Японией и Сахалином. Бывший у нас в руках небольшой материал из Приамурья и
Приморья (3 взрослых и б молодых птиц) все же заметно отличается от большой серии
Рис. 51. Кавказский тетеревятник (взрослая и молодая самки).
настоящих schvedowi указанными выше признаками (малый рост и несколько более темная
окраска). Едва ли можно объединять этих ястребов с подвидом A. g. khamensis (ю.-в. Тибет,
Ганьсу, повидимому сев. Юннань), также близкими к schvedowi и сходными с ним по размерам
(крыло самцов 308—332, самок 342—376 мм), но несколько более темными по окраске.
Кавказский большой ястреб Accipiter gentilis caucasicus Kleinschmidt
Falco palumbarius caucasicus. Kleinschmidt. Berajah, Zoographia Infinita, 1923,
стр. 10, Владикавказ.
Распространение. Ареал. Южное побережье Крыма, Кавказ и
Закавказье. Сюда же повидимому относятся ястребы, населяющие южнокаспийские
провинции Ирана (Гилян, Мазандеран, повидимому Астрабад), а также птица,
добытая 28.VII. 1884 у Гермаба в Копет-Даге, в пределах Туркмении.
Относящийся несомненно к этому подвиду молодой самец добыт 16. VIII. 1942 г.
у Челаюка в ю.-з. Туркмении.
тювик
181
Характер пребывания оседлый, но в холодное время года
ястреб совершает вертикальные перемещения; в это время отдельные особи^
уже с августа — залетают в сев. Предкавказье к Краснодару и даже
Таганрогу.
Биотоп. Лиственные и хвойные леса, в вертикальном отношении
поднимающиеся до верхней границы леса — 1800 (Сатунин, 1912), 1980 (Даль, 1948)
и даже 2500 м (Чхиквишвили, 1930).
Численность кавказского большого ястреба невелика.
Экология не изучена, имеются только отрывочные сведения.
Размножение. Цикл, 'повидимому, поздний, судя по линянию
взрослых особей и потому, что вполне развитые молодые попадаются только
в начале августа, т. е. недели на три позднее, чем в европейской России.
Линька. У взрослых самцов в начале августа линька маховых уже
наполовину завершена (I. VIII первые 5 маховых старые, б-е выпало, 7-е,
8-е перелиняли; 8. VIII первые 4 старые, 5-е выпало, б-е растет, 7-е—-10-е —
новые). Взрослые, окончившие линяние, добывались в октябре.
Описание. Размеры и строение. Мелкая раса: крыло самцов (5) 308—310,
самок (9) 336—353, в среднем 309,3 и 340,6 мм. По Сушкину (1928) крыло самцов 308—314,
самок 339—364.' Ноги относительно сильные.
Окраска. Темная, но в отличие от schvedowi, без сизых тонов в окраске взрослых
птиц и с темным и широким поперечным рисунком на брюшной стороне; общий тон спинной
стороны взрослых темный аспидно-бурый с черновато-бурой головой; белые надбровные
полосы узкие и сильно испещренные, мало выделяющиеся; поперечный рисунок рулевых
хорошо развит и правильный, поперечные полосы на груди, боках, брюхе темные, черновато-
буроватые и широкие, на груди, зобе, горле хорошо развиты темные настволья.
Птицы в первом годовом наряде темные и яркие, брюшная сторона у них ржавчато-
рыжая в свежем пере, охристо-рыжеватая зимой; темя черновато-бурое, бровь слабо
выделяется, пестрины на крыле и плечах мало заметны, темный продольный рисунок черноватый,
широкий, продольные отметины имеются и на оперении голени.
12. Тювик Accipiter badius Gmelin.
Falco badius. Gmelin. Syst. Nat., изд. XIII, 1788, стр. 280, Цейлон.
Русское название. Упоминается впервые у Левшина (Совершенный Егерь,
1774); еще в середине прошлого столетия Северцов нашел название «тювик» употребительным
у воронежских ястребятников. Теперь это книжное слово. Этимология повидимому
звукоподражательная, по голосу птицы.
Распространение. Ареал. Ю.-в. Европа от Балк^ и южн. Украины
до Поволжья и Кавказа; Азия от Анатолии, Ирана, Туркестана до Кашмира,
Южн. Китая, Индии, Цейлона, Никобарских о-вов, Индо-Китая, Малайи;
Африка, кроме палеарктических частей — от Сенегала, Золотого берега и
южн. Нигерии на западе до Эритреи (и ю-з. Аравии) на востоке, к югу до
Оранжевой реки.
Характер пребывания в тропиках и субтропиках Азии и
Африки оседлый, северные формы — европейская и среднеазиатская — перелетны.
Биотоп. Леса как естественные, так и в культурном ландшафте (гнездится
только на деревьях); этим объясняется отсутствие тювика в сев. Африке. В
горах поднимается невысоко, на гнездовье обнаружен в нижнем поясе (в
Гималаях до 1500 м).
Подвиды и варьирующие признаки. Тювик в южн. Азии образует несколько очень
близких географических форм, к ним же относится среднеазиатский подвид и подвид
тропической Африки; на периферии ареала вида живут резко отличные подвиды—европейский и
южноафриканский; возможно, кроме того, что к этому же виду относится распространенный
в Китае от Кореи и Чжили до Гуань-дуня A.solocnsls Н о г s f i e Id (птица также перелетная,
зимующая на Малайском п-ове и в Ост-Индии до Вайгиу). Кроме того, на
Никобарских о-вах живут две формы с изолированным узким распространением, близкие к ин-
182
ОТРЯД ХИЩНЫЕ ПТИЦЫ
дийским тювикам. Подвиды: Л. b. brevipes Severtzow, 1850, в ю.-в. Европе, на Кавказе
и в сев. Иране; A. b, cenchroides Severtzow, 1872, в Туркестане, Иране, Белуджистане,
Афганистане,быть может,Синде; A. b. dussumieriT e m m i n с к, 1824, в Индии от Кашмира
и с.-з.провинций до Сиккима и Бенгалии, к югу доТраванкора (исключительно); A. b. badius
G m e 1 i n, 1788, в южн. Индии от Траванкора до Цейлона; A. b. poliopsis Hume, 1874,
в Ассаме, Бирме, на юге до Тенассерима, в Южн. Китае до Гуаньси и Хайнаня, в Индо-Ки-
тае в сев. частях Малайского п-ова; A. b. sphenurus R u р р е 1 1, 1836, в сев. части
африканского ареала на юг до Камеруна, Уганды и Кении; A. b. polyzonoides Smith,
1838, к югу от предыдущего, от Танганайки, сев. Родезии, Ниасса до Оранжевой реки;
A. b. butleri G u г п е у, 1889, на о. Кар-никобар; A. b. obsoletus Richmond, 1902, на
о. Качал, южн. Никобары.
Карта 20. Распространение тювика Accipiter badius
1 — A. b. brevipes, 2 — ЛЬ. cenchroides, 3 — A. b. dussumieri, 4 — А. Ь. badius, 5 — A. b. butleri, 6 —
A. b. obsoletus, 7 — A. b. poliopsis, 8 — A. b. sphenurus, 9 — A. b. polyzonoides; a — граница гнездовий,
г — область зимовок, б — ареал викарирующего вида Accipiter soloensis.
Морфологические варьирующие признаки: окраска, величина, формула крыла,
окраска яиц (пестрая у A. b. polyzonoides, как у перепелятника; у остальных более или менее
однотонная). A. b. badius — самый темный и мелкий, близки к нему Никобарские формы, но
у них почти отсутствует поперечный рисунок на маховых и рулевых; poliopsis темный как
badius, но крупней, крыло длинней в среднем на 10 мм; dussumieri по размерам сходен с
poliopsis, но бледней; cenchroides еще посветлей, чем dussumieri; все эти подвиды весьма близки,
несколько более обособлены Никобарские формы. Brevipes темный и яркий, с особенностями
в строении крыла и с более резким, чем у других, половым диморфизмом в окраске, крупный,
резко обособленный; sphenurus очень близок к основной южноазиатской группе, но
потемней, с более развитым темным рисунком на брюшной стороне, мелкий; polyzonoides более
обособленный, очень мелкий, мельче чем badius, с иной крыловой формулой и большим
числом полос на хвосте, кроме других особенностей в окраске.
В соответствии с правилом Бергманна, на севере в умеренной зоне тювики больше
(brevipes, cenchroides), чем на юге; окраска также отражает экологический изоморфизм —
наиболее темные формы во влажных тропических лесах, а также обитающие в относительно
холодном европейском климате, самая бледная раса — в ксерофильном ландшафте Средней
и Передней Азии.
тювик
183
Туркестанский тювик Accipiter badius cenchroides Severtzow
Astur cenchroides. Северцов. Вертикальное и горизонтальное распространение
Туркестанских животных, 1873 (1872), стр. 113, Чимкент.
Синоним. Accipiter badius chorassanicus. H e p t n e r und S t а с h a n о w. J. f.
Orn. 1930, стр. 514, Махтум-Кала, Туркмения.
Распространение. Ареал. Равнинные части Туркестана к югу от
Аральского моря (дельта Аму-Дарьи — Казалинск); от зап. Копет-Дага (Сонгу-
Даг, бассейн* Сумбара), Теджена, Кушки, Мургаба на юго-западе до зап.
Тянь-Шаня (Аулье-Ата), Ферганы на востоке; в бассейне Аму-Дарьи наПяндже
до Пархара; на Памире отсутствует; изолированная колония имеется в
низовьях Или и Каратала; в Иране распространен от южнокаспийских
провинций по Хорассану и Паропамизу до Кермана; Афганистан и Белуджистан,
до южн. Вазиристана; быть может в Синде. Из с.-з. Ирана проникает в ю.-в.
Закавказье, где у сел. Вель близ Ленкорани добыты в июне три птицы, одна
у гнезда. Изредка залетает в центр. Тянь-Шань (Нарын 6. V.1910 на высоте
2100 м, Лаубманн, 1913), на ю-з. Туркмении (Атрек, Чатлы 22. IX; Молла-
Кара; Кошаба у Карабогаза). Зимует в южн. Индии, Белуджистане и сев.
Индии от Синда до Пенджаба.
Характер пребывания. В сев. части ареала перелетный,
на юге Ирана и в Белуджистане повидимому оседлый, так как встречается там
и зимою. Предположение о зимовке в Туркмении не подтверждается.
Даты. Прилет поздний, отлет ранний. В Туркмении первые прилетают
обычно в начале апреля в (1942 г. у Ашхабада 1-8.IV; у Гермаба уже 16.III);
валовой пролет имеет место в начале второй половины апреля, запоздавшие
прилетают в мае (Дементьев, 1950). На среднем и нижнем течении Сыр-Дарьи
прилет относится к концу апреля. Отлет происходит в разные числа сентября,
отдельные особи в Туркмении остаются в начале октября, на Сыр-Дарье —
также в начале октября.
Биотоп. Тювик —»птица лесистых или богатых деревьями
культурных ландшафтов. В Туркестане живет в приречных тугаях (Туркмения, Сыр-
Дарья, Или и Каратал) и в оазисах, в садах, рощах у кишлаков, на кладбищах,
в городах и т. п. На юге довольно высоко подымается в горы (подвид dussu-
mieri в Гималаях до 1500 м), но на севере ареала в Средней Азии за редкими
исключениями живет на равнинах, подымаясь в горы лишь там, где имеется
достаточно развитый культурный ландшафт (у Фирюзы в Копет-Даге; у
озера Искандер-куль в Таджикистане, но там быть может на пролете,
нормально в Таджикистане до 1000—1200 м9 Иванов, 1940). На охоте и после
вывода залетает и в пустыню (Репетек в Кара-Кумах).
Численность. В Туркестане обычен, но распространение — в связи
с характером биотопов —спорадично.
Экология. Размножение. Половая зрелость наступает в возрасте
несколько менее года, птицы размножаются будучи еще в гнездовом (первом
годовом) наряде; причем, судя по ходу линяния некоторых добытых весною
молодых птиц, часть их в этом возрасте остается холостой. Гнезда помещаются
только на деревьях, часто они строятся самим ястребом, иногда же ястребы
используют чужие гнезда, которые в этом случае перестраиваются (в Копет-
Даге и на Сыр-Дарье, главным образом, сорочьи гнезда). Гнездо имеет вид
опрокинутого вершиной вниз конуса, рыхлой постройки малых размеров.
Расположены гнезда на ветлах, карагачах, урюке, турангах, гледичии,
тутовнике и т. п., обычно метрах в 5—8, иногда и до 12, от земли. По крайней мере
иногда пара располагает двумя-тремя последовательно используемыми
гнездами. У гнезд птицы появляются с начала мая, в это время и позднее
совершается брачный полет. Самец летает над гнездовым участком, поднимаясь
184
ОТРЯД ХИЩНЫЕ ПТИЦЫ
и опускаясь, с характерными небыстрыми и глубокими взмахами крыльев,
часто крича; иногда к нему присоединяется и самка.
Кладка происходит в последней трети мая, обычно из 3—4 яиц, по Заруд-
ному, изредка из 2 и до 7. В случае гибели первой кладки она повторяется, но яиц
бывает не более 5; при гибели второй кладки иногда бывает третья из 2 яиц.
Яйца молочно-белого цвета с зеленоватым оттенком. Размеры (38) 37,2—42,9 х
Х28,7—33,9 (Спангенберг, 1936); (60) 36,8—41,2x29,1—31,3, в среднем
39 хЗО,2(Бекер, 1928). Насиживает самка, в течение 33—35 дней (Туркмения,
Александров). Летные птенцы встречаются в начале августа». Соотношение
полов в выводке точно неизвестно (у Ислама на Аму-Дарье 2 самца и 2 самки,
6. VIII. 1941). Цикл размножения таким образом поздний, позже чем у
европейского тювика, с одной стороны, и чем у тювиков в Индии — с другой (по
Бекеру, 1928, период гнездования там в марте и апреле).
Линька полная годовая. Начало линьки у взрослых гнездящихся птиц
совпадает с регрессией полового аппарата и нахождением в гнезде птенцов
во втором пуховом наряде, начинающих оперяться. Молодые из первого
годового наряда начинают линять ранее, но в первой фазе периода размножения
линька у них приостанавливается, возобновляясь одновременно с началом
линьки взрослых и оканчиваясь одновременно с концом линьки у последних.
Во всех частях среднеазиатского ареала тювики линяют одновременно.
Взрослые самки начинают линять несколько ранее, чем самцы, с июня; около
10.VI у них выпадают задние первостепенные маховые — 9-е и 10-е; примерно
в то же время выпадают средние рулевые и начинается линяние мелкого
оперения. У самцов линьки в июне нет, что связано с очень четким разделением
функций в заботе о потомстве, так как самец ловит добычу и снабжает ею
насиживающую самку, а потом и птенцов, не участвуя в насиживании и оделении
кормом молодых.
Линька взрослых самцов начинается с начала июля, продолжаясь весьма
интенсивно, так что в августе самцы в отношении линьки сравниваются с
самками. В начале сентября у тювиков остаются неперелинявшими 3—4 передних
первостепенных маховых и, повидимому, в сентябре, к отлету, линяние
заканчивается, продолжаясь у самок около 4, а у самцов только 3 месяца.
Птицы в первом годовом наряде начинают сменять мелкое перо уже в мае,
иногда к концу этого месяца теряя среднюю пару рулевых; некоторые молодые
самки в это время по линьке значительно отстают, что связано вероятно с тем,
что они размножаются, а интенсивно линяют птицы, составляющие резерв
холостых особей; большинство молодых птиц в первом годовом наряде сменяют
маховые одновременно со взрослыми.
Последовательность смены первостепенных маховых — как у других
ястребов, от внутреннего края к наружному, т. е. от 10-го к 1-му. Рулевые
сменяются начиная со средней пары. Смена мелкого пера кончается скорее на
брюшной стороне, чем на спинной. Последовательность смены нарядов:
первый пуховой наряд — второй пуховой наряд — первый годовой (гнездовый)
наряд — второй годовой (окончательный) наряд и т. д.
Питание. Тювик охотится главным образом в открытых местах по
окраинам культурного ландшафта, берет добычу с земли и сравнительно редко
преследует птиц в угон на лету. В соответствии с этим пищу его составляют
главным образом пресмыкающиеся (например гекконы Gymnodactylus russowi,
сцинки Ablepharus deserti на Сыр-Дарье, Спангенберг, 1936); Мелкие зверьки,
крупные насекомые —прямокрылые и жесткокрылые; из птиц отмечены воробьи,
щурки, также молодые кеклики, пустынные куропатки и фазанята.
Охотничий участок около 3—4 км от гнезда (Зарудный, 1890). Охотится и в сумерках.
Полевые признаки. Птица в общем походит на перепелятника, но крылья
более длинные, хвост относительно короче, самцы — с белым подбоем крыла
тювик
185
и брюхом; молодые птицы с продольным рисунком на брюшной стороне. Нередко
парит, полет менее поворотливый и быстрый, чем у перепелятника, с глубокими
взмахами крыльев. Голос — мелодичный растянутый посвист «тююю-ви-тююю-
вю. По сравнению с другими ястребами, мало осторожен.
Описание. Размеры и строение. Тювик принадлежит к группе ястребов,
промежуточной между тетеревятниками и перепелятниками. Клюв у него сильный, лапы
с относительно короткой цевкой и пальцами: средний палец менее длинный, внутренний
палец без когтя доходит до середины второй фаланги среднего пальца (у перепелятника до
Рис. 52. Тювик (взрослые самец и самка и молодой самец).
^первой фаланги), внешний палец заходит за конец второй фаланги среднего пальца (у
перепелятника доходит до этого конца); общая длина среднего пальца менее 30 мм, а у самки
и даже самца перепелятника обычно более 30 мм. Крылья относительно длинные, формула
jcpbuia 3>4>5>2>6>7 и т.д. (у перепелятника 4>5>3>6>2 >*7), вырезки на
наружных опахалах первых пяти маховых (у перепелятника и на б-м), на 5-м слабо
выраженная; внутреннее опахало 4-го махового резко вырезано. Длина самцов (2) 300—330,
самок (1) 380 мм. Размах самцов (2) 615—630, самок (1) 690 мм. Вес самца (1) 193, самки (1)
266г. Крыло самцов (39) 182—196, самок (29) 205—230, в среднем 190,5 и 217,03 мм.
Окраска. Половой диморфизм выражен, но слабо; возрастный — резко. Первый
пуховой наряд и второй пуховой наряд охристы. Первый годовой (гнездовый) наряд на
спинной стороне бледный серовато-бурый, с широкими охристыми каймами перьев на спине,
плечевых и крыльях, с поперечными беловатыми (прикрытыми) пестринами у оснований
перьев (больших); надхвостье беловатое с бурым поперечным рисунком; брюшная сторона
беловатая или охристая с темнобурой продольной полосой на горле и с бурым рисунком из
продольно вытянутых пятен на груди и брюхе, из поперечно вытянутых — на боках. Махо-
186
ОТРЯД ХИЩНЫЕ ПТИЦЫ
вые темнобурые, рулевые серо-бурые с темнобурым поперечным рисунком; основания
внутренних опахал первостепенных охристые.
Взрослые самцы (со второго годового наряда) на спинной стороне бурые с сизоватым
оттенком, рыжеватые на зашейке,с белыми основаниями перьев;маховые темнобурые с
неясными бурыми поперечными полосами; рулевые бурые, средние одноцветные или с остаткам^
темного поперечного рисунка у стволов, боковые с узким темным поперечным рисунком,
хорошо заметным на внутренних опахалах; брюшная сторона рыжеватая с
охристо-беловатым узким поперечным рисунком, продольной бурой полосой на горле и белым подхвостьем;
подкрылья, брюхо, подхвостье, голени — почти чистобелые. Самки на спинной стороне
бурые, без сизого тона; окраска их тусклее и бурее, чем у самцов, поперечный рисунок
брюшной стороны более правильный, тон его бурей, чем у самцов. Личная изменчивость (общий
тон, поперечный рисунок) довольно значительна. Радужина у молодых бурая, у старых
красновато-бурая, оранжевая у зрачка; клюв черновато-роговой, светлее у основания; когти
черные; восковица, разрез рта и лапы желтые.
Систематические замечания. Очень близок к индийской расе тювика A. b. dus-
sumieri, но несколько бледнее по окраске. В ю.-в. части ареала очень редко
встречаются особи с признаками, переходными к А. b. brevipes(экземпляр из Кошаба, Карабогаз,
в Зоологическом музее Моск. университета; о другом из окрестностей Ашхабада упоминает
Штегман, 1937).
Европейский тювик Accipiter badius brevipes Sev.
Astur brevipes. Severtzoff. Bull. Soc. Imp. Naturatistes Moscou, XXIII, 1850,
стр. 234, табл. 1—3, Воронежская губ.
Распространение. Ареал. Ю.-в. Европа от Балканского п-ова до
нижнего течения Волги и Кавказа; Малая Азия. Сев. граница в Европе
—низовья Дуная, Бессарабия (?), низовья Днепра, далее у Полтавы, Харькова,
в Стрелецкой степи в Курской обл., в Воронежской обл,, на Волге —у
Вольска; в степном Заволжье отсутствует, на пролете добыт в низовьях реки Урал;
в Крыму повидимому только на южн. берегу; на Кавказе отсутствует в
лесистых зап. частях, встречаясь от Дагестана до Ленкорани и Армении; юго-вост.
граница не вполне ясна. Зимою и на пролете отмечен на Синайском п-ове, в
Аравии, Месопотамии, Иране (от с.-з. провинций), отдельные особи —
в Египте. Область зимовок впрочем выяснена плохо и зимних особей в
коллекциях очень немного. Залетает европейский тювик в Венгрию и в
Среднюю Азию (Казалинск, Зарудный, 1908; Челаюк на Атреке, 13. IX. 1941,
Дарганата на Аму-Дарье 25. V. 1941; есть и другие указания, но они
недостоверны).
Характер пребывания. Гнездящаяся перелетная птица.
Даты. Как и туркестанский тювик, прилетает весною поздно —с конца
апреля до начала мая (например, в Аскания-Нова только 9.V). Отлет начинается
с первых чисел сентября, последние улетают в конце этого месяца и даже в
начале октября, из Крыма еще позднее, по наступлении морозов (23.X 1.1890,
Сомов, 1892). Пролет в Малой Азии наблюдался в начале апреля (10. IV, Крюпер,
1869), но еще в конце апреля тювики добывались в Египте (Мейнертцхаген,
1930). Летят тювики одиночками, стаями и небольшими группами.
Биотоп. Главным образом ольховые и смешанные леса речных долин"
(европейская Россия), рощи и перелески, сады (Кавказ, Крым). В горы
поднимается невысоко — в Крыму на высоту около 500 м, на Кавказе обычно
до 1000 му но в Армении отмечен и на 1980 м (Даль, 1948).
Численность в СССР умеренная.
Экология. Размножение. Цикл поздний, но более ранний, чем у
туркестанского подвида. Половая зрелость наступает на первом году, когда
тювик еще в гнездовом пере; имеется резерв холостых особей в этом возрасте,
убитый у гнезда член пары заменяется. Гнезда располагаются на деревьях,
в Харьковской и Воронежской обл. главным образом на ольхах (отсюда
старое охотничье название этой птицы «ольшаник», «ольшаный ястреб»), реже
на осинах, дубах и других деревьях; расположены обычно у ствола, высоко
тювик
187
от земли, метрах в 6—12. Обычно гнезда устраиваются самим ястребом,
возможно, что используются и чужие (это установлено в Малой Азии).
Гнездо — как у туркестанского тювика, небольшое и рыхлое, выстланное
внутри тонкими веточками и изредка сухими листьями, форма гнезда —
опрокинутый конус.
Кладка производится в мае (самая ранняя в Харьковской обл. — 12.V,
обычно же там во второй половине мая; у Еревана 5 слабо насиженных яиц
были найдены 21.V). Яйца откладываются через день, возможно, что иногда
и ежедневно. В случае гибели первой кладки производится вторая. Число яиц
в кладке 4—5, иногда 3 (во вторых, дополнительных, 2—3). Свежеснесенные
яйца матовые голубовато-зеленые с мелкими нерезкими зеленоватыми пятнами;
по мере насиживания—яйца становятся грязно серовато-белыми. Размеры:
(9)39—44x32—35 (Сомов, 1897); (58)40, 18x31, 74, в среднем (Джоурден, по
Суанну, 1925). Яйца несколько крупнее, чем у туркестанского тювика; шире,
но короче, чем у перепелятника.
Насиживает только самка, вероятно — судя по разновозрастности
птенцов — с первого яйца (указание Сомова, 1892,'что самка начинает насиживать
после снесения последнего яйца, ошибочно). Самец в период насиживания
кормит самку, а позднее и выводок. Срок насиживания точно не прослежен (по
Сомову, 1892, будто бы немного более 3 недель, но он считал от откладки
последнего яйца, что неверно; на самом деле около месяца, вероятней всего
35 дней). Число птенцов 2—5 (имеются колебания по годам, причины
которых не вполне ясны). Вылупление начинается со второй половины июня,
чаще в конце его. Половой состав выводков: в нескольких гнездах по 1 самцу
и 1 самке, в одном случае 2 самца, кроме того 2 самца и 1 самка, 3 самца и
1 самка, 3 самца и 2 самки. Пуховики в гнездах на юге Украины наблюдались
15.VII (Аскания-Нова); выбравшиеся из гнезда, но еще нелетные —28.VII
(Митякинская, Донская обл, Аверин, 1911), вполне развитые молодые
в конце июля и начале августа (указание Сомова, 1892, на вылет птенцов в
конце июня —начале июля ст. ст., вероятно, ошибочно). Покидают гнездо птенцы
еще нелетными и иногда ловят насекомых, бегая по земле. Гнездовый период
длится, таким образом, около 45 дней; у гнезд выводки держатся недели две
после того, как их покинут.
Линька. Протекает как у предыдущего, но, в соответствии с более
ранним размножением, начинается и кончается соответственно ранее. К началу
июля у самок остаются неперелинявшими половина первостепенных маховых
(передние), у самцов смена этих перьев в это время также приближается к
половине.
Питание.В гнездовый период охотится самец, осенью тювики охотятся
парами, постепенно облетая опушки лесов, приречные луга, края поселков и
рощ и т. п. Летит при этом обычно невысоко —метров в 6—10 над
землей.Охотится и в сумерках. Молодые птицы кормятся главным образом ящерицами,
также крупными насекомыми (кузнечики, жуки). Взрослые охотятся на
ящериц (Lacerta viridis, L. agilis и др.), на насекомых (жуки, в частности, жук-
носорог; также саранча, кузнечики и др.), но кроме того на птиц (воробьи,
Штегман, 1937; зяблики, Ляйстер и Соснин, 1942; щурки и даже ласточки
касатки), и мелких зверьков (Apodemus agrarius, Огнев и Воробьев, 1924;
Arvicola terrestris, Sylvimus sylvaticus; в Малой Азии даже летучие мыши,
Крюпер, 1872).
Описание. Размеры и строение. Крыло острее, чем у туркестанского подвида
тювика: 3-е маховое почти равно 4-му и заметно длиннее 5-го, тогда как у туркестанского
тювика перья эти приблизительно равной длины; 2-е маховое значительно длиннее б-го;
наружное опахало 5-го махового без сужения или вырезки. Размеры крупнее: длина крыла
самцов (15) 208—228, самок (10) 224—236; в среднем 215,8 и 230,3 мм.
188
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Окраска. Темней и ярче, чем туркестанский подвид, с более резкими различиями
в окраске полов. Взрослый самец на спинной стороне аспидно-сизый (как взрослый
перепелятник), без рыжего ожерелья на задней стороне шеи, общий тон серей и темней, чем у
туркестанских птиц; полосы на рулевых, даже боковых парах, только намечены; брюшная сторона
более насыщенного ржаво-рыжего цвета, с более контрастным белым поперечным рисунком;
рыжие полосы более заметны и на брюхе и на голени; щеки серые, а не серо-бурые. Самка
на спинной стороне темнобурая, на брюшной охристая с резким бурым, а не ржаво-рыжим
(как у туркестанского) поперечным рисунком, часто распространяющимся на подхвостье и
всегда хорошо заметным на подкрыльях; щеки темные, того же оттенка, как темя (у
туркестанских щеки бледно серовато-бурые, заметно светлее темени); все цвета по сравнению
с туркестанским тюбиком темнее. Молодые в первом годовом (гнездовом) наряде на спинной
стороне темнобурые, рыжеватые каемки перьев у них чуть намечены, брюшная сторона белая
с резким темнобурым рисунком, продольным на зобе, сердцевидным на груди, брюхе,
передней части боков и имеющим характер поперечно вытянутых пятен на задней части боков,
голенях, подхвостье; у туркестанских тюбиков в этом возрасте, при более бледном тоне
темного рисунка на брюшной стороне, грудь в продольных полосах, а не в сердцевидных пятнах;
поперечный характер рисунка менее выражен на голенях и на перьях подхвостья. Неоперен-
ные части окрашены, как у предыдущего. Указывается, что у птенцов в гнезде радужина
буровато-серая, а у подлетков серая.
13. Малый перепелятник Accipiter virgatus Те mm.
Falco virgatus. Temminck. Planches Coloriees livr. 19, 1822, табл. 129, Ява.
Распространение. Ареал. Вост. Азия от Кузнецкого Ала-Тау,
Алтая, Красноярска до Курильских о-вов, Японии, Филиппин, Индо-Ки-
тая, Малайского
архипелага, зап. Гималаев.
Характер
пребывания. Северные
популяции перелетны, южные,
повидимому, — оседлы — в
холодное время года
встречается в Южн. Китае и на
Зондских о-вах.
Биотоп. Как и
другие ястребы малый
перепелятник — лесная птица.
Подвиды и варьирующие
признаки.В зап. Гималаях, ю.-з.
Китае, Ассаме и сев. Бирме
A. v. affinis Hodgson, 1836;
в южн. Индии и на Цейлоне
A., v. besra J e r d о п, 1839; на
Суматре, Яве, Борнео А. у.
virgatus T e m m i n с к, 1822; на
Филиппинах A. v. contusus,
Hartert, 1910; в Сев.
Китае, Японии (Риу-Киу), ю.-в.
Сибири A. v. gularis
Temminck et Sch lege I, 1843;
в Южн. Китае, на
Андаманских о-вах, в Малайе A. v.
nisoides Blyth, 1847; на
Целебесе и Банггаи A. v. rhodoga-
ster S с h 1 е g e I, 1862; на о-вах
Сула А. и. sulaensis S с h 1 е-
g e 1, 1866. Экологические
различия мало изучены, но на
юге, как указывалось, малые ястребы оседлы, на севере перелетны; существенные
различия имеются в календаре периодических явлений. Морфологические различия в
окраске и размерах, абсолютных и относительных: в частности, вершина крыла у
совершающего дальние перелеты подвида gularis 40—60 мм, у оседлых virgatus и confusus 25—29 мм.
Карта 21.
Распространение малого перепелятника
Accipiter virgatus
1 — A. v. gularis, 2 — A. v. affinis, 3 — A. v. besra, 4 — A. v.
nisoides, 5 — A. v. confusus, 6 — A. v. virgatus, 7 — A. v. rhodo-
gaster, 8 — A. v. sulaensis; a — граница гнездовий, б —
невыясненная граница распространения, г — область зимовок.
МАЛЫЙ ПЕРЕПЕЛЯТНИК
189
Сибирский малый перепелятник Accipiter vigratus gularis Temm. et Schleg.
AstUT gularis. Temminck et Schlegel. Fauna Japonica, Aves 1844, стр. 5, табл. 2,
Япония.
Распространение. Ареал. Южная полоса средней и Вост. Сибири
от Томска (Семилужское) и Кузнецкого Ала-Тау в горной Шории (бассейн Мрас-
су), с.-в. Алтая (верховья
Абакана, Телецкое озеро),
зап. Саяна, Красноярска,
южн. Байкала (Иркутск,
Нижнеудинск, Култук),
Забайкалья (примерно на
широте Читы), Приамурья (на
север до озера Кизи),
Сахалина, Курильских о-вов
(Уруп) и Японии (но
достоверно только на Риу-Киу,
Ян, 1942), к югу до
Манчжурии, Кореи, Сев.
Китая; на западе в сев.
Монголии (Кентей, Гобийский
Алтай). Зимовки в Японии
на о. Риу-Киу, в Южн.
Китае, на Малайском п-ове, в
Индо-Китае, на Б.
Зондских о-вах, на пролете в
Китае и вост. Индии,
залеты на Филиппины.
Характер
пребывания. Перелетная
птица, появляющаяся на
севере в Уссурийском крае
поздно (на о. Аскольд в
1878 году только 15.V) и
отлетающая в сентябре и
начале октября
(Лиственничное, ю.-з. Байкал; о.
Аскольд, 6.Х, Тачанов-
ский, 1891).
Биотоп. Леса — хвойные,. смешанные, лиственные — последние по-
видимому предпочитаются. В горах поднимается до 1800 м (Кентей, Козлова,
1932).
Численность. В южном Приморье обычен, северней и западней
очень редок.
Экология. Размножение. Цикл поздний, во второй половине мая
кладки еще нет. Гнезда малы и компактны, располагаются на деревьях
довольно высоко от земли (например, на Имане на бересте UlmiiSy в 6—»13 м,
из веточек вербы с зеленой листвой, наружный слой из сухих веток, Спан-
генберг, 1940; у Красноярска на сосне в 6—7 м от земли). Гнезд у каждой
пары повидимому несколько, используются они по годам посменно, у
Красноярска Крутовской найдено 2 запасных. У добытой 10.VI самки обнаружены
зрелые яйца в яичнике (Шульпин, 1936). Незаконченная кладка из 4 яиц
была найдена 4.VI. Полная кладка состоит из 5 яиц. Яйца походят на яйца
перепелятника, но меньше (максимальные размеры 36 х 28,8, минимальные
Рис. 53. Малый перепелятник (взрослые самка и
самец, молодая самка).
190
ОТРЯД ХИЩНЫЕ ПТИЦЫ
34,9 х 29,Спангенберг, 1940; Тачановский, 1891—39 х 29 и 36 х 29 мм). Пуховые
птенцы найдены 12.VII (Столбы у Красноярска). Вылет молодых
происходит в первой половине августа: 12.VIII выводок — самка и 2 молодых с
недоразвитыми крыльями и хвостом —добыт в хребте Таскыл в зап. Саяне;
в Приамурье на р. Хунгари недоросший молодой был добыт 22.VIII, в
Манчжурии— б.УПНМейзе, 1934); самка с недоросшими крыльями под
Красноярском—13.VIIL
Линька. Как у перепелятника, в сроках линяния заметен половой
диморфизм: самка линяет раньше, чем самец, повидимому в связи с разделением
функций в периоде размножения. У взрослой самки, добытой 13.VII (Телецкое
озеро), линька кончалась, а передние маховые в трубках еще не доросли;
взрослая самка с р. Судзухе (Приморье), добытая 28.VII, была уже в свежем пере:
у взрослого самца 23.VIU (Иман), часть старых рулевых и первые 5 маховых,
6-е выпало, 7-е—10-е перелиняли; у самца во втором годовом наряде 13.VIII
(Судзухе, сопка Туманная) свежее перо с незначительными остатками первого
наряда среди перьев шеи, плечевых и кроющих крыла. Смена маховых—как
у других ястребиных, от 10-го к 1-му.
Питание. Мелкие птицы, главным образом воробьиные, например
седоголовые и др. овсянки, воробьи (Спангенберг, 1940), синицы-московки
(Шульпин, 1936); японская совка (Воробьев) и т. д. Малый перепелятник —
энергичная ловчая птица, употребляющаяся для охоты в Индии, Японии,
Китае.
Описание, Размеры и строение. Меньше обыкновенного перепелятника,
лапы несколько короче, в особенности средний палец;крыло острее, формула его 4>3>
5>2>6. Вес самца (2) 142—102, самки (1) 192,5 г. Крыло самцов (б) 155,5—170, самок
(4) 170,5—202 лш, в среднем 161,8 и 187,6 мм.
Окраска. Первый годовой (гнездовый) наряд: спинная сторона темнобурая с
рыжеватыми каймами, рулевые и маховые с поперечным рисунком; брюшная сторона — белая
с бурым продольным рисунком на груди, сердцевидными или поперечными пятнами на боках
и брюхе; на горле более или менее явственная продольная полоса. Второй (окончательный)
и последующие наряды диморфны: самец шиферно-сизый на спинной стороне, с беловатыми
основаниями плечевых, с поперечнополосатым хвостом и маховыми, на брюшной стороне
бледнорыжеватый с более или менее развитым поперечным беловатым рисунком, с белым под-
хвостьем; самка на спинной стороне темнобурая или серовато-бурая, на брюшной стороне
белая с правильным бурым поперечным рисунком на зобе, груди, брюхе, боках, подкрыльях.
Восковица и ноги желтые, клюв черноватый, когти черные, радужина желтая. Пуховые
наряды — как первый, так и второй — неизвестны.
14. Перепелятник, малый ястреб Accipiter nisus L.
Falco Nisus. Linnaeus. Syst. Nat. изд. X, 1758, стр. 92, Швеция.
Русское название. Происходит оттого, что птицей этой издавна, а местами
и ныне, травят перепелов. У охотников XIX века назывался и малым ястребом.
Распространение. Ареал. В область распространения перепелятника
входит вся территория Палеарктики (кроме безлесных тундр, степей и
пустынь), протяженностью на север примерно до границы леса, на юг—до Марокко,
Алжира, Туниса, Малой Азии, Ирана, Гималаев, сев. Бирмы, Юннани,
Среднего Китая (до Голубой реки) и Японии. Зимой в Африке —- до Судана,
Абиссинии, Аравии и южн. Азии до Индии и Южн. Китая (кроме южных частей
гнездового ареала).
Характер пребывания. В северной части гнездовой области,
примерно до 50° с. ш., перелетный; южней —оседлый.
Биотоп. Лесистые, пересеченные открытыми пространствами
местности, в особенности светлые леса и рощи, обильно населенные мелкими птицами.
В негнездовое время перепелятник обнаружен также и в культурном
ландшафте. От равнин до верхнего пояса лесной растительности в горах.
Численность. Обыкновенная птица.
ПЕРЕПЕЛЯТНИК, МАЛЫЙ ЯСТРЕБ
191
Подвиды и варьирующие признаки. Перепелятник или малый ястреб объединяет
довольно много подвидов: в европейской части ареала и Зап. Сибири, кроме Средиземноморья,
А. л. nisus L. 1758; в Марокко, Алжире, Тунисе и может быть Испании А. п. punicus
Е г 1 a n g е г, 1897; на Канарских о-вах А. п. teneriffae Laubmann, 1912; на
Мадейре А. п. granti S h а г р е, 1890; на Корсике, в Сардинии А. л. wolterstorfii Klein-
schmidt, 1901; в вост. Азии от средней Сибири до Японии и Сев. Китая А. л.
nisosimilis T i с к е 1 1, 1833; на Камчатке А. п. pallens Stejneger, 1893; в горах
Центральной Азии от Таджикистана и Гималаев до Нань-Шаня, Ганьсу и Сычуани A. n.mela-
schistos Hyme, 1869. Подвиды эти обладают рядом морфологически варьирующих
признаков: величина, окраска, строение скелета. Крупные подвиды распространены на севере,
причем величина крыла постепенно возрастает с запада на' восток, от Европы к Вост.
Карта 22. Распространение перепелятника Accipiter nisus
7 —А. п. nisus, 2 — А. п. nisosimilis, 3 — А. п. pallens, 4 — А. п. melaschistus, 5 — А. п. punlcus,
6 — А. п. wolterstorffi, 7 — А. п. granti, 8 — А. п. teneriffae; а — граница гнездовий, г — область зимовок.
Сибири; крупные формы кроме того населяют горы Центральной Азии, также Африку;
мелкие же подвиды распространены в Средиземноморье. Помимо различия в величине,
южные формы, кроме бледного марокканско-тунисского подвида, из ксерофильных лесов, —
окрашены ярче и интенсивнее, чем северные. Характерно наличие ряда островных
оседлых рас с узкими ареалами. Экологические различия сводятся к характеру пребывания
в ареале, календарю периодических явлений и особенностям хода последних и т.д. В
морфологии (окраска) имеются любопытные типы личной изменчивости, параллельные в
различных расах и свойственные всем возрастным нарядам.
Обыкновенный малый ястреб или перепелятник Accipiter nisus nisus L.
Синоним. Accipiter nisus peregrinoides. К lei nschmid t in Grote. Aus d.
Orn. Literatur Russlands. №-3, 1921, стр. 56, Росситтен, на пролете.
Распространение. Ареал. Обыкновенный малый ястреб распространен
в Европе от Великобритании и Ирландии (на Корсике, Сардинии и вероятно
в южн. Испании—другие формы); в Передней Азии от Кавказа и Малой Азии
до зап. Ирана, в горных его частях к югу до Загроша и быть может Ширазл
192
ОТРЯД ХИЩНЫЕ ПТИЦЫ
(около 30° с. ш.); в Зап. Сибири примерно до бассейна Енисея. Зимою в
Передней и Средней Азии, в Иране, Ираке, Палестине, Синае, в Африке от
Египта до южн. Аравии, Судана, Абиссинии, б. Германской Вост. Африки
(между Меру и Килиманджаро, Шиллинге, 1906). Сев. граница проходит
в Лапландии примерно у 69° с. ш., у Архангельска, на Печоре и Оби
приблизительно у полярного круга (Усть-уса, Дмоховский, 1933; Обдорск,
Дерюгин, 1899), далее в с.-з. Сибири неясна. На юге перепелятник гнездится
всюду, за исключением безлесных местностей, отсутствуя в степной части
Казахстана и Туркестана; в последней он пока найден только в зап. Туркмении
в арчевых лесах гор Б. Балхан и гнездится быть может в Копет-Даге, где две
молодых птицы добыты 15.VIII на Душаке.
Характер пребывания. Начиная от средней полосы, примерно
от 55—56° с. ш., иногда и еще далее к северу, перепелятники встречаются
в средней России и зимою; встречаются они зимою и в сев. Европе
(Скандинавия, Финляндия, в СССР изредка до Кировской обл., Плесский, 1948),
но главным образом в средней и южной, у нас в Крыму, на Кавказе и в
Туркестане на юго-западе, до низовья Сыр-Дарьи и предгорий центр. Тянь-Шаня
на севере (впрочем, относительно последних—из-за неразличимости
отдельных особей — принадлежность к подвиду nisasy а не nisosimilis не может
в сущности быть установлена). В Африке — до Судана и Абиссинии. В Зап.
Сибири и сев. Казахстане не зимует. Таким образом, в северных и восточных
частях ареала перепелятник — определенно перелетный; за отсутствием
полученных путем кольцевания точных данных трудно судить, является ли
он в средней полосе хотя бы частично оседлым или там на зиму задерживаются
птицы с севера, а местные подаются на юг. Судя по количеству встреч
перепелятника зимою в средней России, большинство особей оттуда откочевывает.
Наоборот, на юге — в Крыму, на Кавказе перепелятники зимою становятся
многочисленными, а в Средней Азии и сопредельных странах встречаются почти
только зимой.
Даты. На пролете и вне времени гнездования самцы держатся повиди-
мому отдельно от самок. Пролетают одиночками, обычно в связи с пролетом
воробьиных птиц. Время пролета весьма растянуто (неодновременное движение
разных популяций). На севере гнездового ареала (Архангельск; Печора; М. Со-
сьва в сев. Зауралье) ястребы еще могут чбыть встречены в конце сентября
(20—27. IX). В то же время в средней полосе заметный пролет идет уже с середины
сентября, продолжаясь в октябре и ослабевая заметно во второй половине этого
месяца. На юге области зимовок в СССР перепелятники появляются в октябре-
ноябре (Талыш, Туркмения, Сыр-Дарья). Весенний пролет начинается в апреле
(хотя некоторое движение заметно уже в марте — как на юге, так и в средней
полосе), причем в Лапландию перепелятники прилетают в середине апреля —
начале мая (16. IV—9.V). Значительный пролет перепелятников в европейской
части нашей страны идет в первой половине апреля, а в конце этого месяца —
начале мая перепелятники появляются у гнезд. Направление пролета ю. и ю.-з.
Биотоп. Пересеченные открытыми пространствами лесистые
местности, лесные опушки, рощи и перелески, реже — большие лесные массивы.
Зимою — в культурном ландшафте, даже в больших городах. В горах — в'
пределах лесной зоны, до 2000 и даже 2700 м (Чорох, Дерюгин, 1900).
Численность. Обычная птица.
Экология. Размножение. Половая зрелость наступает в первом
годовом наряде, в возрасте несколько менее одного года. Гнездовые участки
весьма постоянны и используются много лет. Возможно, что у каждой пары
имеется несколько гнезд в участке. О постоянстве пар точных сведений нет.
Цикл размножения поздний, начало кладки с конца апреля или в первой
половине мая. Гнезда строятся самим ястребом, чаще всего на молодых или сред-
ПЕРЕПЕЛЯТНИК, МАЛЫЙ ЯСТРЕБ
193
невозрастных хвойных деревьях — елях и соснах, реже на лиственных (на
юге); в Зауралье на березах; обычно невысоко, метрах в 2—4, реже до 8.
Гнездо—небольшая (около 35—40 см в диаметре), рыхлая постройка из ветвей,
обычно близ ствола, реже в развилке; лоток довольно глубокий, так как края
гнезда приподняты; выстилка — тонкие веточки, хвоя й т. п., — небрежная.
В отличие от большого ястреба, в гнезде свежих зеленых ветвей не бывает.
Кладка из 3—6 яиц, редко из 2 и до 7 (Кесслер, 1851), или 1, обычно же из
4^5. Промежуток между откладкой 48 часов (Зарудный, 1910). Яйца овально
закругленные, матово-белые с буроватыми от охристых до темнобурых пятнами
и крапинами, различных размеров. Величина яиц (12) 37,2—43 х 30,1—-33
(Сомов, 1897); (5) 37—42 х 30, 5—33, в среднем 40 х 32 (Гебель, 1879); 36^42,5 X
30—33,5 (Минск, Шнитников, 1913). В случае гибели кладки, она
повторяется, при этом иногда и в дополнительной кладке бывает до б яиц
(Зарудный, 1910).
Насиживает только самка. Срок насиживания весьма длительный,—32 дня
(Познанин, 1946); по данным других европейских стран — немного более месяца,
31—33 дня или 35 дней. Насиживание начинается повидимому с откладки
первого яйца, так как птенцы разновозрастны. Вылупление происходит в конце
июня — начале июля. Соотношение полов в выводке не установлено. Птенцов
в гнезде 2—6, в неблагоприятные по кормовым условиям годы обычно 3 или
2 — это связано и с неоплодотворением яиц, и со смертностью молодых.Таким
образом у перепелятников, как и у многих других хищных птиц, имеет место
известное саморегулирование популяции в зависимости от внешних условий.
Постэмбриональное развитие проходит очень быстро, около месяца, и в разные
числа июля птенцы уже оперяются и с недоросшими маховыми и рулевыми
вылезают из гнезда; вполне летными становятся они в возрасте 5 недель, весь
период размножения тянется таким образом около 3 месяцев. Полного веса
молодые достигают уже в трехнедельном возрасте, при этом развитие самок
идет несколько быстрее, чем самцов. Птенцы запоздалых выводков (повидимому
вторых «дополнительных» кладок) или младшие в больших выводках имеют
еще недоросшие крылья около середины августа, тогда как старшие выводки
вполне вырастают на месяц ранее (Московская, Калужская, Горьковская
область — особи от 8.VII и 17.VIII). У гнезд слетки держатся около двух
недель, после чего становятся самостоятельными.
В период насиживания и в первом периоде роста птенцов охотится только
самец, передающий пищу самке. Последняя оделяет птенцов и в случае гибели
самки, хотя самец продолжает носить добычу и бросать ее в гнездо, птенцы,
не умеющие расклевывать добычу самостоятельно, обычно умирают с голоду.
Линька. Взрослые птицы начинают линять в июне, при этом самки —
в соответствии с разделением функций во второй фазе цикла размножения —
раньше, чем самцы. Вскоре после окончания кладки и в период насиживания
(конец мая — начало июня) самка начинает смену маховых, у самцов этот
процесс начинается с половины июня. Последовательность смены
первостепенных маховых — как у других ястребиных, от 10*го к 1-му. Рулевые линяют,
начиная от средней пары и кончая второй от края. К началу августа—в
последних числах июля — остаются неперелинявшими 2—3 передних первостепенных
маховых и к концу августа или в первой трети сентября линяние заканчивается.
Взрослые самцы линяют в такой же последовательности, как самки, но
несколько скорее, так как линька у них начинается позже, а заканчивается одновременно
с самками. Общая продолжительность процесса линьки у самок 3 месяца, у
самцов 21/* месяца. Линька полная годовая. Как у других видов, птицы в первом
годовом наряде начинают линять раньше взрослых, смена мелкого пера у
самцов начинается в мае, затем линяние у размножающихся особей
приостанавливается; есть ли остановка в линянии у холостых —. неизвестно. Последова-
13 Птицы СССР, т. I
194
ОТРЯД ХИЩНЫЕ ПТИЦЫ
тельность смены нарядов: первый пуховой наряд — второй пуховой наряд —
первый годовой (гнездовый) наряд — второй годовой (окончательный) наряд
и т. д.
Питание. В отношении общих приспособлений к питанию
перепелятник сходен с большим ястребом — это орнитофаг, охотящийся в
пересеченной лесистой местности. Способы нападения на добычу у этих видов сходны:
перепелятник сидит на дереве или в каком-либо другом укрытии у опушки,
Рис, 54. Перепелятник (взрослый и молодой самцы и молодая самка).
дороги и т. п. и бросается на появляющуюся- вблизи добычу, или летает над
вершинами кустов, среди деревьев, хватая сидящую или взлетающую добычу.
За летящей птицей перепелятник гонится в угон, причем особенно часто
хватает ее несколько сбоку. Потребность в пище —за сутки 2—3 птицы, величиной
с воробья для самца, 3—4 таких птицы для самки (у охотничьих птиц).
Потребность выводка значительно больше — в зависимости от количества птенцов —
12—15 птиц в день. Как максимум указывается 2 птенца рябчика, 4 певчих
дрозда, 1 королек, 1 крапивник, 1 малый пестрый дятел, 1 лесной конек, 2
зяблика, 4 славки и 4 пеночки, 6 мышей — всего 26 штук (Плеске и Бихнер, 1888).
Потребность выводка в пище так велика, что ко времени надевания птенцами
второго пухового наряда и установления у них автономной терморегуляции,
в возрасте 8—10 дней, начинает охотиться и самка. Общее количество
мелких птиц, необходимых для выкармливания одного молодого ястреба с вы-
ПЕРЕПЕЛЯТНИК, МАЛЫЙ ЯСТРЕБ
195
лупления до вылета, т. е. примерно в течение 45 дней, составляет 100 штук
(Уттендерфер, 1939). В кормовом режиме молодых большое значение имеют
подлетки воробьиных — быть может этим объясняется позднее гнездование
перепелятника. Надо отметить, что в период размножения перепелятник
охотится на расстоянии до 4—5 км от гнезда; в непосредственной близости от гнезда
перепелятник в это время не охотится.
Самцы перепелятника охотятся главным образом за мелкими воробьиными,
размером с зяблика, самки — за более крупной добычей, размерами с дрозда
и до галки, вороны и голубя. Иногда ястреб нападает на птицу, превышающую
его размерами — если видит, что она ослаблена или нездорова (Кесслер, 1851).
Главной пищей ястребу служат домовые воробьи, зяблики, полевые
жаворонки, обыкновенные овсянки, большие синицы, деревенские ласточки,
певчие и черные дрозды. Наблюдается известная специализация в кормовом
режиме, связанная с определенными биотопами (различные типы леса,
культурный ландшафт, пригороды и т. п.). В холодное время года перепелятники
передвигаются в культурный ландшафт и кормятся главным образом воробьями—
полевыми и домашними, галками, голубями (первыми — самцы, последними—
самки). Из млекопитающих в качестве пищи перепелятника у нас отмечены
полевки (в частности Microtus arvalis), лемминги, мыши и суслики.
Указывается связь гнездования перепелятника в Лапландии с размножением
леммингов. Кормовые остатки — погадки (средние их размеры 3 х 2 см, состав —
остатки костей и перья), поеди и перья — обычно находятся где-либо под
деревом, на котором ястреб более или менее постоянно обрабатывает или поедает
добычу.
Полевые признаки. Короткокрылая и ширококрылая длиннохвостая
птица, бурая на спинной стороне, поперечнополосатая — набрюшной. Летает или
быстро взмахивая крыльями, или скользя, обычно низко над землею, над
кустами, среди деревьев, вдоль изгородей и т. д. Не парит. Осторожен и
молчалив. Голос у гнезда «кик-кик-кик», иногда при испуге отрывистое «кирк»,
весной в период брачных игр «кью-кью-кьюю».
Описание. Размеры и строение. Похож на большого ястреба, но значительно
меньше и с более длинной цевкой и пальцами. Крыло короткое и широкое, формула: 4>5>
3>6>2>7>8>9> 10^:1. Вырезки на наружных опахалах 3-го — 6-го и на внутренних
опахалах 2-го — 5-го маховых. Длинная цевка оперена только на протяжении верхней
четверти, пальцы длинные и тонкие, особенно средний. Длина самцов (15) 313—340, самок
(16) 370—410, в среднем 325,8 и 388,5 мм. Размах самцов (15) 615—653, самок (16) 703—780,
в среднем 632,4 и 742,4 мм. Вес самцов (3) 124, 153, 175 г; самок (4) 232, 244, 257 и 278 г.
Крыло самцов (96) 186—208, самок (120) 223—248, в среднем соответственно 198,3 и 236,35 мм.
Окраска. Первый пуховой наряд снежнобелый с коротким пухом на темени
(у большого ястреба в этом возрасте — удлиненный пух образует там «шапочку» с темным
кольцом вокруг глаза). Второй пуховой наряд белый с желтовато-охристым налетом на
спинной стороне.
Первый годовой наряд на спинной стороне бурый с рыжеватыми или
охристо-желтоватыми каемками перьев, с беловатыми основаниями перьев на плечах, спине, крыльях, с белой
бровью и белыми пестринами на затылке; маховые бурые со светлыми основаниями
внутренних опахал и с темными поперечными полосами; рулевые бледнобурые с поперечными
перевязями; брюшная сторона белая с бурым сердцевидным рисунком на зобе и поперечными
полосами на груди, боках, брюхе, подкрыльях. Весьма значительна личная изменчивость:
в развитии и правильности поперечного рисунка на брюшной стороне (тут иногда имеются
только сердцевидные пятна, иногда правильные и густо расположенные поперечные
полосы); в ширине светлых каемок на спинной стороне; в интенсивности бурого тона; в
развитии охристого оттенка на брюшной стороне (некоторые особи рыжеваты,
противоположный вариант — чисто белая с поперечным бледнобурым рисунком брюшная сторона).
Взрослые самки темнобурые с более или менее развитым сизоватым оттенком на спинной
стороне, с поперечнополосатым хвостоми первостепенными маховыми; на затылке белые пятна„
над глазом белая бровь; брюшная сторона с правильным поперечным рисунком из бурых до<-
лос, подхвостье белое. Личная изменчивость состоит в оттенках бурого цвета, в большем или
меньшем развитии сизого налета на спинной стороне и в характере полосатости на брюшной.
Взрослые самцы существенно иной окраски (таким образом, у перепелятника половой
диморфизм в размерах выражен во всяком возрасте, а окраска мономорфна только в гнездсь
196
ОТРЯД ХИЩНЫЕ ПТИЦЫ
вом наряде). На спинной стороне самцы шиферно-сизые или серо-сизые, на затылке белые
основания перьев, основания плечевых с белыми пестринами, прикрытыми серыми
вершинами; маховые буровато-серые с беловатыми основаниями и черноватыми поперечными
полосками, рулевые сизые с 4 нерезкими поперечными темнобурыми полосами и белой
вершинной каемкой; брюшная сторона беловатая с рыжими поперечными полосками, более
или менее сливающимися на боках, с белым подхвостьем. Личная изменчивость выражается
в более или менее темном тоне сизого цвета, в различных оттенках темного рисунка брюшной
стороны — от яркорыжего до тусклого темнобурого, а также в распространении этого
рисунка. Клюв серо-роговой, черноватый у вершины; радужина у взрослых оранжевая или
яркожелтая, у молодых бледножелтая, у пуховиков сероватая; когти черные; восковица,
разрез рта и лапы лимонно-желтые.
Систематические замечания. При большой личной изменчивости отграничить в
континентальной Европе и зап. Азии подвиды перепелятников невозможно, хотя среди них
наблюдается правильное возрастание величины (длина крыла, размеры грудины) с запада на северо-
восток. Об отграничении восточносибирской формы см. ниже.
Восточносибирский малый ястреб или перепелятник Accipiter nisus
\nisosimilis Tickell
Falco nisosimilis. Tickell. Journ. of the Asiatic Soc. of Bengal II 1833—1834, стр. 571,
Марха, Борабхум, Индия.
Распространение. Ареал. Сев. Азия к востоку от области
номинальной формы — Сибирь от ее средних частей (Алтай) на западе до бассейна
Колымы, Охотского побережья (Охотск, Шантарские о-ва), Японии и Сев.
Китая; гнездование в Приморье не доказано (быть может на севере, Шульпин,
1936). За Колымой на Анадыре и в Коряцкой Земле не найден. На севере по
Енисею граница не прослежена, на Яне в Верхоянском округе еще несколько
северней 65° с. ш. (Ткаченко, 1932), на Индигирке даже до 69°50' (Хаягастах,
Михель, 1935), на Колыме до 69° с. ш. (Бутурлин, 1908). На юг до Зайсана,
восточного и быть может зап. Тянь-Шаня (Заилийский Ала-Тау, отдельные
летние находки даже в Таласском Ала-Тау), сев. Монголии, Сев. и вост. Китая
(возможно до Янцзыцзяна). Зимою от Туркестана (от вост. Туркмении, долина
Кушки) на западе и южн. частей Приморья до Южн. Китая (Хайнань),
Бирмы, Индии, Цейлона и Индо-Китая. Отдельные особи зимуют уже в
средней Сибири (центр. Алтай, Сушкин, 1938; Томск, Залесский, 1915)'
Характер пребывания. В Сибири перелетный гнездящийся;
относительно Китая вопрос неясен.
Даты. Прилет поздний, в самом конце апреля (сначала самцы, сев.
Монголия), но главным образом в начале мая; во второй трети мая наблюдался уже
в низовьях Индигирки, также на Охотском побережье. Отлет с зимовок и
пролет разных популяций растянут: в Средней Азии с конца марта до конца
апреля (Киргизия), отдельные особи в мае; в Приморье с начала апреля и
до начала мая, в это время проходит пролет к северу и в Сев. Китае.
На индийских зимовках перепелятник находится с октября до марта. Осенью
движение тянется медленно, начинается оно с августа (Приморье), главным же
образом проходя в сентябре. Около середины сентября перепелятники
исчезают в северных частях ареала (в Якутии — в начале сентября последний
наблюдался на Вилюе, Маак, 1886; на Алдане за 63° с. ш. наблюдался еще
16. IX, Ткаченко, 1932), в средней Сибири задерживается до ноября (Каменский
округ на Оби, Велижанин, 1931), у Байкала по крайней мере до конца октября.
На нижней Сыр-Дарье пролет отмечен Н.Х—20.Х (Спангенберг, 1936).
Биотоп. Типичный для вида. В горах на Алтае распространен до 2000,
редко до 2300 м.
В холодное время года и осенью в Тянь-Шане выходит за пределы лесной
зоны, появляясь в степной и субальпийской, хотя обычно в это время года
придерживается там лесной зоны гор, а также предгорий и культурного
ландшафта равнин.
ПЕРЕПЕЛЯТНИК, МАЛЫЙ ЯСТРЕБ
197
Численность. По сравнению с европейским подвидом численность
перепелятника ограниченная, на востоке и северо-востоке ареала в Сибири —
в Якутии — редок.
Экология* Размножение. Половая зрелость наступает в первом
годовом (гнездовом) наряде (Сушкин, 1938). Одно и то же гнездо используется
много лет (Тачановский, 1891 и др.). Размножение позднее: кладка
происходит в конце мая (еще 30. V у самки было обнаружено яйцо в яйцеводе,
Алтай); яйца в гнезде на Индигирке были найдены 17. VI; пуховики с
пробивающимися маховыми встречаются еще в середине июля (II. VII, Алтай;
16. VII в бассейне Или, Шестоперов, 1929). Подлетки с короткими крыльями
найдены в гнезде в начале августа (б. VIII, Якутск), вполне развитые
молодые со второй половины и в конце августа (Алтай, Байкал); в это время
ястребы еще держатся выводками. Число птенцов в гнезде 2—4; яиц в кладке
4—5, изредка б и даже 7 (Тачановский, 1891). Размеры яиц (20) 35,5—44 х 30—
35 (Даурия, Тачановский).
Линька. Материал ограничен, общее течение как у европейского
перепелятника. У взрослых самок в конце июля сменяются задние 5 маховых
(Байкал); у самцов в конце июля сменяются только 3 задних маховых (там же),
а в конце августа остались неперелинявшими 3 передних маховых (23. VIII,
Яблоновый хребет). Добытые в конце мая и во второй половине сентября
взрослые птицы не линяют. У птиц в первом годовом наряде смена мелкого оперения
начинается до периода размножения, в мае, а затем вероятно
приостанавливается, если птица размножается.
Питание. В качестве пищи восточносибирского перепелятника
отмечены главным образом мелкие птицы — воробьи, синицы, овсянки,
трясогузки, лесные коньки, также дрозды; из птиц средней величины —чибисы
(Приморье), краснозобый песочник (Большие Шантарские о-ва), кеклики и
фазаны (Алма-Ата), молодые рябчики (Забайкалье), даже самка кряква
(Барнаул). Наблюдались нападения перепелятника на стрижа (Приморье,
Пржевальский). Из млекопитающих ястреб ловит полевок (Индигирка,
Шантарские о-ва). Потребность ястреба в пище определяется примерно в 80 и
не более 120 г, т. е. 3—4 воробья в сутки (Зверев, 1930, наблюдения на Абакане).
Описание. Размеры и строение. В среднем несколько больше номинальной
формы, хотя отдельные особи и неотличимы. На территории Европы и сев. Азии размеры
ястребов постепенно и непрерывно возрастают с запада на восток. Размеры птиц, относимых нами
здесь к nisosimilis, таковы: крыло самцов (24) 200—213, самок (24) 240—260, в среднем 206,3
и 245,7 мм. Окраска. Как у номинальной расы, с той же амплитудой личной
изменчивости .
Систематические замечания. Разделение европейско-сибирских ястребов на два
подвида носит условный характер; еще более условный характер носит установление
географической границы между подвидами. Принимая описанные дьа подвида, мы следуем
в сущности установившейся практике.
Камчатский малый ястреб или перепелятник Accipiter nisuspallens S t ej n e ge r
Accipiter pallens. Stejneger. Proceedings of the U.S.A. Nat. Museum, XVI, 1893,
стр. 625, Хитачи, Япония.
Распространение. Ареал. Эта форма описана по пролетной птице —
самке из Японии, причем Стейнегером высказано предположение о гнездовании
этого ястреба на Камчатке. На Камчатке перепелятник очень редок;
Бергман (1935) не встретил его вовсе за несколько лет пребывания; однако он
встречался неоднократно в Кроноцком заповеднике в 1941—1946 годах
как в гнездовое время (22. VI. 1941, устье р. Столбовой, Кроноцкий залив),
так и на осеннем пролете (5—19. IX. 1940) и зимою (устье р. Медвежьей,
21. I. 1943). В высокогорье — до 1000 м в лесном поясе, осенью на морском но-
198
ОТРЯД ХИЩНЫЕ ПТИЦЫ
бережье (Аверин, 1948). В Японии таких ястребов также очень мало (нередок
-4. п. nisosimilis), и они очень ценятся сокольниками*
Быть может камчатские перепелятники все же представляют собой особую
географическую расу, так как изученные мною в Зоологическом институте
Академии наук молодые самцы в первом годовом наряде от 18. IX. 1846 с реки
Камчатки и от 25. VIII. 1919 из Усть-Камчатска (также молодая самка из
Иокогамы) действительно отличаются особенностями окраски, указанными Стей-
негером. Тачановский (1891) также отмечает, что взрослый и молодой самец
с Камчатки необычайно бледны на спинной стороне тела. Очень бледно окрашен
и взрослый самец, добытый 21. V. 1930 у Громовской протоки близ Охотска
(Зоологический музей Московского университета). В вост. Камчатке биотоп —
горные и равнинные леса, в горах вертикально до зоны кустарников. Оседлый.
Экология* Сведений нет (на пролете в горах в Кроноцком заповеднике
5. IX. 1940 охотился за пуночками и горными коньками, Аверин, 1948).
Описание. Размеры и строение. Крупный перепелятник — крыло
упомянутых 2 самцов с Камчатки 212 и 224 мм; самца с Охотского побережья 208 мм> молодой
самки из Иокогамы 258 мм. По Тачановскому (1891) крыло самцов с Камчатки 217 и 222 мм.
Окраска. В первом годовом наряде спинная сторона бледнобурая, с сизым
оттенком, на голове широкие светлые каймы и «брови»; поперечный рисунок рулевых слабый,
вокруг темных полос беловатые пестрины; светлые каймы перьев спины и крыльев беловаты
или чуть охристы; темный рисунок на брюшной стороне мало насыщенный.
Взрослый самец из Охотска бледный, серовато-сизый на спинной стороне, белый с бледнорыжим
поперечным рисунком на брюшной стороне. Взрослая самка, по Стейнегеру, на спинной
стороне светлей и серей, чем номинальная раса, с резко выделяющимися темными на-
ствольями, слабо выраженными темными полосами и широкой белой вершинной каймой
на хвосте, с сероватыми поперечными полосами на брюшной стороне.
Систематические замечания. Материал по этой форме очень мал, и самостоятельность
ее требует еще подтверждения (быть может крайний вариант индивидуальных отклонений
у А. п. nisomilis).
Гималайский малый ястреб или перепелятник Accipiternisus melaschistos Hume
Accipiter melaschistos. Hume. Rough Notes, 1869, стр. 128, Гималаи.
Синонимы. Accipiter melanoschisius, Hume, The Ibis, 1869, стр. 356, Симла. —
Accipiter nisus ladygini. Bianchi. Bull. B. Orn Club, 1906, стр. 69, Кам.
Распространение. Ареал. Горы высокой Центральной Азии от
Кашмира на западе до вост. Ассама, Юннани, вост. Тибета (Кам), хребта Минь-Шань
на границе Ганьсу и Сычуани, к югу до сев. Бирмы; на север до
Белуджистана, Афганистана и Таджикистана на западе, до Ладака, Тетунгских гор
и хребта Рихтгофена в Нань-Шане на востоке. В Туркестане найден достоверно
на озере Искандер-куль (взрослый самец 7. IX; молодая самка 6. IX; молодая
самка с устья р. Серима 15.VIII. 1936, сборы Бибикова в Зоологическом музее
Московского университета). К этой же форме вероятно относились ястребы,
наблюдавшиеся в Зеравшанском хребте 17—29. VI (Даль, 1936), а также
молодой экземпляр с озера Сары-чилек в Фергане (Кашкаров, 1927). В
последнем случае, область распространения гималайского перепелятника в
Туркестане идет довольно далеко на северо-восток.
Общее распространение перепелятника в Туркестане выяснилось недавно
и представляет большой интерес. На крайнем северо-востоке — в восточном и
быть может центральном Тянь-Шане — обитает А. л. nisosimilis; на крайнем
западе в Б. Балханах, Туркмения, гнездится изредка А. п. nisus; он же
спорадически, повидимому, в высоких частях центр. Копет-Дага (Хейрабад); на
юге в горах, указанных выше, встречается А. л. melaschistos. При этом
гнездовые области этих трех подвидов на территории Туркестана разобщены.
Характер пребывания. Оседлая птица. Зимой в Гималаях
совершает повидимому вертикальные перемещения.
ПОЛЕВОЙ ЛУНЬ
199
Биотоп, Высокогорные, главным образом, хвойные леса и кустарники
Центральной Азии — ельники и арчевники и т. д. В Туркестане на высотах
около 2500—3000 м, в вост. Тибете примерно между 3 000—4 000 м (Биан-
ки, 1907); в Ладаке около 3000 м (Мейнерцхаген, 1927), в Гималаях от 1200 до
3000 м, главным образом между 1800 и 2400 м (Бекер, 1928).
Экология* По Туркестану наблюдений нет.
Размножение. Добытый в устье р. Серима в Таджикистане
молодой перепелятник 15. VIII во вполне развитом наряде, с доросшими крыльями
и хвостом. В Тибете в гнезде на карликовой иве насиженная кладка из 3 яиц
найдена 8 июня; размеры яиц 44,1 х34,8, 45,5x34,1, 46x34, т. е. больше, чем
у европейских перепелятников (Шефер, 1938). В Гималаях кладки из 4-^5, редко
из 3 или б яиц, размеры их: (60) 36,0--40,3 х29—33, в среднем 39,1 х32,6ш*
(Бекер, 1928). У гнезд очень неосторожен. В Индии гнезда строит сам ястреб или
отнимает гнезда других птиц; в виде исключения гнездится будто бы на скалах.
Линька. Добытый 7 сентября в Таджикистане самец в свежем пере
с двумя недоросшими передними маховыми и старой второй от края парой
рулевых. В Кашмире сильно линяющие самцы добывались 30 августа
(Мейнерцхаген, 1927); еще незакончившие линяния самцы в Тибете добыты 1 октября
(Бианки, 1907).
Питание. Мелкие птицы лесов и кустарников, в Тибете например разные
тимелии Garrulax, кроме того чечевицы Ervthrina thura и Е. eos, дрозды Turdus
kessleri и Т rubrocanus и др. В Индии главным образом мелкие птицы, воробьи
и т. п., но также и средних размеров, величиною до голубя; кроме того, мелкие
звери, рептилии и насекомые.
Описание. Размеры и строение. Крупная форма — крыло самцов 202—213,
самок 248—260 (Хартерт), у тибетских самцов (6) 205—215, в среднем 210,5 (Бианки), у гань-
суйского самца (1) 210, самок (3) 241—246, в среднем 244 (Штреземанн, Мейзе, Шенветтер,
1938), у гималайских самцов 212—219, самок 245—260 (Бекер), у самцов из Таджикистана
201 и 208, у самки 236 мм.
Окраска. Очень темная раса. Взрослые самцы с черноватой головой и черноватой
спиной, с густорыжим поперечным рисунком на брюшной стороне. Самка также окрашена
темнее (взрослые экземпляры из СССР пока неизвестны). Молодые птицы в первом годовом
наряде на спинной стороне черновато-бурые с густо рыжими каемками на крыльях и плечах,
с широким черновато-бурым рисунком на брюшной стороне.
РОД ЛУНИ CIRCUS LACEPEDE, 1799
Тип С. aeruginosas
15* Полевой лунь Circus cyaneus L. 1796]
Falco Cyaneus. Linnaeus. Syst. Nat. изд. XII, 1766, стр. 126, окрестности Лондона.
Русское название. Лунь — старинное русское название, обозначающее
беловатых хищных птиц, но не соколов и не ястребов. В первую очередь оно относится к
беловатым или бледносизым самцам полевого, лугового или степного луня, но местами на севере
лунем зовут белую сову. Видовые названия луней — полевой, луговой и т. д. — книжные.
Распространение. Ареал. Европа и Азия от границ тундры до
средиземноморских стран, М. Азии, Ирана, Туркестана, Джунгарии, Монголии и
Сев. Китая; также в Сев. Америке от севера Канады до южн. Соединенных
Штатов. Зимовки в южн. и Зап. Европе, в сев. Африке, в Индии и Китае, в
Передней Азии (до Туркмении на севере); североамериканские луни зимуют
от южн. штатов и далее до Колумбии, Кубы и Багамских о-вов.
Характер пребывания. В южной части ареала оседлая или
кочующая птица, на севере, в частности в СССР — перелетная.
Биотоп. Открытые ландшафты — поля, луга речных долин, лесные
моховые болота, степь и лесостепь; как на равнинах, так и в горах.
"^UU
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Подвиды и варьирующие признаки. Два подвида — в Старом СветеТ. с. cyaneusL.,\T(&,
в Сев. Америке С. с. hudsonius L., 1766. Отличия в окраске (у американского подвида самцы
Карта 23. Распространение полевого луня Circus cyaneus
7 — С. с. hudsonius, 2 — С. с. cyaneus; а — границы гнездовий, г — область зимовок.
с пестринами на брюшной стороне, молодые птицы окрашены темней, чем европейско-азиат-
ские; резко выраженная у самцов географическая изменчивость окраски — явление
характерное для некоторых видов луней). Экологические различия невелики.
Обыкновенный полевой лунь Circus cyaneus cyaneus L.
Синонимы. Circus iaissiae. В u t u r 1 i n J. f. Orn., 1908, стр. 283, Среднеко-
лымск. — Circus cyaneus cernuus. Thayer and.
Bangs. Proceed. N. Engl. Zool. Club. V, 1914, стр. 32, низовья Колымы.
Распространение. Ареал. Сев. Европа и Азия. В Норвегии до 69°30\
в Швеции до 68°30' с. ш., в южной части Тиманской тундры (Гладков, 1941),
в Большеземельской тундре (р. Черная), но регулярно на Печоре только до
65°, на Оби по крайней мере до 67°, по Енисею до Полярного круга и вероятно
севернее, в бассейне Вилюя по крайней мере до 65° (Маак, 1886), на Индигирке,
еще у 67°30' (Михель, 1937), на Колыме у Нижнеколымска (Тайер и Бангс,
1914); отмечен в среднем течении Анадыря, также на Камчатке, на Охотском
побережье, Шантарских о-вах, Сахалине, вероятно на Курильских о-вах,
на Хоккайдо. Южная граница — в средних частях Португалии, сев.
Испании, сев. Италии, Югославии (Сербия), Албании, Румынии, в Крыму, на
Волге у Бузулука и Бугуруслана, в Закавказье, М. Азии, Иране, Туркмении
(Копет-Даг), далее в Джунгарии, Монголии (до южн. Алашайя), в южн.
Манчжурии и Корее. К югу от среднего течения Урала, в Казахстане и Туркестане
не гнездится (луня нет на гнездовье уже северней озера Ак-тасты-куль и у
верховьев Илека, Сушкин, 1908), на Алтае гнездится, повидимому, по окраинам.
Добыт в негнездовое время на Командорских о-вах —в феврале на о-ве Беринга
(Шульпин, 1936). Зимою в сев. Африке (к северу до Сахары, быть может в Кор-
дофане, Нубии, Сомали, Египте), Передней Азии, в Иране до Загроша, Сеиста-
на, в Белуджистане, в Индии, Бирме и в Китае (до Юннани и низовьев Яньцзе),
также в Зап. и южн. Европе. В СССР зимою добыт в Туркмении, на Сыр-
Дарье и в других частях Туркестана, повидимому в низовьях Волги; в
Закавказье (Армения), в Крыму, в южн. Приморье; в мягкие малоснежные зимы —
на Украине (Умань), даже севернее, в Белоруссии (Полесье, Цедлитц, 1920;
Шнитников, 1913)/
Характер пребывания. В зап. и южн. Европе оседлый и
кочующий, на востоке и севере ареала перелетный, хотя и там (см. выше)
некоторые особи зимуют в гнездовой области.
ПОЛЕВОЙ ЛУНЬ
201
Даты. Миграции нолевого луня протекают весной раньше, чем у
лугового и степного. В Средней Азии движение к северу отмечено уже в феврале
(Сыр-Дарья, Туркмения), заметный пролет там в марте, но тянется и в апреле
(повидимому северные популяции). В конце марта — начале апреля
протекает весенний пролет и на Украине, в средней полосе европейской России он
падает главным образом на первую половину апреля, а на севере ее и в Сибири
полевой лунь появляется в конце апреля и в начале мая (Салаир, прилет 25. IV;
у Красноярска в конце апреля —начале мая ст. ст.; на Вилюе 29.V, Маак,
1886; у Якутска 28. IV—1.V, по Воробьевой, 1931, 25—28. IV, по Иванову,
1929; на Шантарских о-вах 5. V, Дулькейт и Шульпин, 1937; в верховьях
Колымы в начале мая); однако, в ю.-в. Сибири— в Забайкалье и Приморье —
уже в начале апреля. Осенний отлет и пролет на севере и в средней полосе
начинается уже в августе, но главным образом в сентябре; в средней полосе он
продолжается и в октябре, заканчиваясь около середины этого месяца; в
Средней Азии пролет и отлет тянется и в ноябре (Сыр-Дарья; Туркмения). На
весеннем пролете луни летят преимущественно поодиночке, ближе к местам
гнездовий и парами, осенью — одиночками, парами и небольшими группами. Есть
наблюдения, что весною сначала—недели на две или хотя бы одну ранее самок-—
появляются самцы (Киевская обл.).
Биотоп. Открытые местности — поля, луга, речные долины, даже
лесные моховые болота и гари, тундра (кустарниковая, Колыма; Тиман), степи и
лесостепи. На пролете встречается и в пустынях. В вертикальном отношении
главным образом на равнинах и в предгорьях, но также и в горной степи
(Кавказ, Копет-Даг, Эльбурс, Тянь-Шань и т. д.). На Кавказе до 2700 м (Сатунин,
1912), на Алтае до 2300 м (Сушкин, 1938), в Средней Азии местами до
альпийской зоны (Тянь-Шань).
Численность. Биотопические условия делают распространение
полевого луня несколько спорадическим: он поэтому не гнездится
повидимому в Таджикистане, в с.-з. Казахстане на юг от бора Аман-карагай и озера
Ак-тасты-куль (Сушкин, 1908), в южн. части Приморья, в зоне сплошных
лесов и среди тайги,,-в пустыне и т. п. В культурном и полукультурном
ландшафте средней полосы европейской России, местами в Зап. Сибири — лунь
весьма обычен. В Зап. Европе число полевых луней уменьшилось; этой птицы
нет на гнездовье в Дании, ее стало очень мало в Голландии, Бельгии, Англии
и Шотландии (но она еще регулярно гнездится на Гебридских о-вах и в
Ирландии). Периодических колебаний численности точно не установлено (как и
у других луней).
Экология. Размножение. Половая зрелость наступает до
достижения окончательного наряда. Спаривание начинается с прилета, в средней
полосе с середины апреля, когда наблюдается и брачный полет. Гнезда
размещаются на земле, на полях, лугах, сухих болотах. Средние их размеры около
50 см, редко до 80 см при высоте в 30 см (Умань, Гебель,1879). Гнезда из травы,
мелких сучьев и т. п. с неглубоким лотком, очень чистые, так как самка очищает
гнездо, вынося из него остатки пищи, погадки, и т. п. Кладка производится
около середины мая, причем географических различий между популяциями
в отношении сроков кладки установить не удалось. В кладке 3—5 яиц, редко
6 (Зарудный, 1888). Неизвестно, есть ли колебания плодовитости в
зависимости от кормовых условий сезона размножения. Окраска яиц чаще всего белая
одноцветная, реже слегка испещренная мелкими коричневыми крапинами.
Размеры яиц (4) 42,5—44x35—37, в среднем 43,2 х 36,5 мм (Умань, Гебель,
1889); (100) 40—52,1 х 32—38, в среднем 46,23 х 36,13 мм (Уайзерби, 1939).
Насиживает только самка, с первого яйца, так как птенцы весьма
разновозрастны. Между родителями — строгое разделение функций заботы о
потомстве. Самка согревает птенцов и сидит у гнезда, оделяет их кормом, самец
202
ОТРЯД ХИЩНЫЕ ПТИЦЫ
охотится и приносит пищу; по мере роста птенцов в добывании для них пищи
начинает принимать участие и самка. Пуховые птенцы появляются с начала
июня, насиживание, следовательно, длится около одного месяца; летные
молодые обнаружены во второй половине или середине июля; гнездовый период
продолжается приблизительно 6 недель.
Линька. Изучена недостаточно и протекает в значительной своей части
на зимовке. Птицы с прилета в свежем оперении (апрель), но молодые в первом
годовом (гнездовом) наряде только с примесью свежих перьев. Взрослые
линяют с июля до октября, но нередко линяние затягивается позднее и в общем
идет у некоторых особей менее регулярно, чем обычно у других хищных птиц.
Последовательность смены первостепенных маховых — типичная для
семейства, от заднего края партии к переднему, т. е. от 10-го к 1-му. Рулевые
линяют центробежно, от средней пары; крайние пары быть может без строгой
последовательности. Последовательность смены нарядов: первый пуховой
наряд — второй пуховой наряд — первый годовой (гнездовый) наряд — второй
годовой наряд (у самцов переходный; у самок возможно окончательный) —-
третий годовой (окончательный) наряд и т. д. Линька полная годовая.
Питание. Строение луня отражает применяемые им способы охоты —
розыск добычи на земле в открытых пространствах. Вес луня относительно
небольшой, размах крыльев значителен, несущая поверхность крыльев весьма
большая, хвост длинный и широкий. Цевки длинные, напоминающие
ястребиные (схватывание добычи среди травы и т. п.), но пальцы короткие со слабыми,
хотя и острыми когтями (нападения только на мелких животных). В
соответствии со всем этим находятся и способы охоты луней: они не парят, но легко и
тихо летают на небольшой высоте над землею, следуя за рельефом почвы и
неожиданно появляясь над самой добычей. Подлетев к ней, лунь бросается с
вытянутыми лапами, часто делая при этом ловкие повороты.
Пищу луня составляют в первую очередь мелкие млекопитающие, но
также — особенно в период пролета и гнездовья — гнездящиеся на земле птицы,
их птенцы и яйца; кроме того, ящерицы, насекомые и иногда падаль. Как
объекты питания луня в СССР указываются: из зверей суслики, мыши, полевки,
пеструшки (Ерусланская степь), мыши (Вилюй), различные грызуны, например,
Stenocranius gregalis, Microius ungurensis (Забайкалье); из птиц, — птенцы
береговых ласточек (Кайский район, Горьковская обл.), домашний
голубь (Дзауджикау), жаворонки, коньки, утята (Чкалов), чечетки и рюмы на
пролете (Вилюй), коньки Anthus richardi и A. campestris, жаворонки,
трясогузки, перепел (Забайкалье); ящерицы; насекомые, главным образом,
прямокрылые, также жуки и муравьи.
Полевые признаки. Размеры средние, сложение легкое; длинные крылья
и длинный прямо усеченный хвост. Самец бледносизый на спинной стороне,
беловатый на брюшной, с черными вершинами, крыльев, самка буроватая на
спинной стороне, беловатая с бурыми продольными пятнами на брюшной
стороне. Белое надхвостье отличает полевого луня от других луней сходной
окраски. Полет легкий и плавный, с медленными взмахами крыльев —
вершины первостепенных маховых при этом слегка расставлены и загнуты кверху.
Голос «ги-ги-ги» или дребезжащий звук вроде «пииррь», как у других луней.
Описание. Размеры и строение. По величине уступает из наших луней
только болотному, крупнее степного и лугового, с относительно короткими крыльями,
не достигающими вершины хвоста. Формула крыла 4^3>2>5>6>1>7. Вырезки на
наружных опахалах 2-го — 5-го маховых, при этом вырезка на 2-м маховом прикрыта
кроющими кисти. На внутренних опахалах маховых вырезки на 1-м — 4-м перьях. Длина
самцов (20) 445—515, самок (20) 490—564, в среднем 475,2 и 531,1 мм. Размах самцов (20)
990—1060, самок (16) 1100—1215, в среднем 1025 и 1146,7 мм. Вес самцов (4) 300, 350, 360,
400, самок (3) 480, 500, 600 г. Крыло самцов (50) 330—358, самок (50) 355—396, в среднем
341,54 и 372,8 мм.
ЛУГОВОЙ ЛУНЬ
203
Окраска. Первый пуховой наряд белый с серовато-охристым налетом и темным
пятном у глаза. Второй пуховой наряд охристо-сероватый с рыжеватым налетом на спинной
стороне и груди.
Первый годовой наряд у обоих полов одинаков: на спинной стороне темнобурый с
охристыми каймами перьев; на затылке беловатые пятна; надхвостье белое; маховые темнобурые
с беловатыми основаниями внутренних опахал и темным поперечным рисунком; средние
рулевые серо-бурые с темным поперечным рисунком, боковые пары охристые с бурыми
поперечными полосами; брюшная сторона рыжевато-охристая с бурым продольным рисунком.
Самки во втором годовом наряде и последующих — как молодые птицы, но каймы
перьев на спинной стороне менее развиты, а общий тон брюшной стороны беловатый, без
рыжеватого оттенка. Самцы во втором годовом (переходном) наряде отличаются сильным
развитием продольных пятен на брюшной стороне и общим сероватым тоном спинной стороны.
Самцы в третьем годовом (окончательном) наряде на спинной стороне серо-сизые с
серебристым налетом, с белым надхвостьем; передние первостепенные маховые с черной
вершиной, остальные сизые, все с беловатыми основаниями внутренних опахал; средние рулевые
сизые, боковые сизые на наружных опахалах, белые с буроватыми пестринами на
внутренних опахалах; брюшная сторона белая, только горло и зоб, а иногда и грудь, сизые; изредка
на брюхе и боках мелкие пестрины. Радужина у молодых бурая, у старых желтая, восковица
и ноги желтые; клюв черный с синеватым оттенком; когти черные. Заслуживает внимания
тот факт, что ход возрастных изменений проходит быстрей, чем у степного и лугового луня.
16. Луговой лунь Circus pygargus L.
Falco Pygargus. Linnaeus. Syst. Nat., изд. X, 1, 1758, стр. 89, Англия.
Синоним. Circus cineraccus abdullae. Floericke. Orn. Monatsber., 1896, стр. 156,
Закаспийская обл.
Распространение. Ареал. В Европе от Англии, Голландии и Германии,
Дании и южн. Швеции до Прибалтики, средней полосы Европейской части
204 ОТРЯД ХИЩНЫЕ ПТИЦЫ
СССР, к северу, примерно, до линии Псков —Москва — Ярославль — Казань,
поднимаясь восточней до Молотова, Свердловска, а в Зап. Сибири до Тюмени,
Тары, Красноярска; на восток до предгорий Алтая и Минусинской лесостепи;
к югу до Португалии, Испании, средней Франции, сев. Италии, Венгрии,
Румынии, Крыма, южн. Закавказья, Ирана (с.-з. части — Хорассан, быть
может Керман и Кугистан), Средней Азии (где на ю.-з. в Таджикистане
спорадичен, а в Туркмении вероятно и не гнездится), южн. Алтая (Котон-карагай)
и Зайсана (Кендерлык); кроме того, в с.-з. Африке.в Марокко, Алжире.
Карта 24. Распространение лугового луня Circus pygargus
а — границы гнездовий, г — область зимовок.
Отдельные особи зимуют уже в южн. Европе и Туркестане (в виде
исключения и в средней России, например в Тамбовской обл.); в низовьях Сыр-
Дарьи у Соло-тюбе; по Мензбиру и др. — на Украине и в Крыму. Впрочем, все
сведения требуют подтверждения. Регулярные зимовки в южн. Азии — в Индии
в области Гималаев, редко на равнинах; также в Бирме; на Цейлоне и в
Китае, но, главным образом, в Африке — в тропических и южных ее частях от
Сенегамбии, Конго на западе и Абиссинии на востоке; больше всего луговых
луней зимует в южн. Африке — бывшей германской ю.-з. Африке, Родезии,
Трансваале и Капской Земле; отдельные особи зимуют также на о-вах
Зеленого мыса й Канарских. Вероятны зимовки в Месопотамии. Залеты в СССР
отмечены в Ленинградскую обл. и к Усть-ряму на нижней Оби.
Характер пребывания. Перелетная птица с разомкнутым
ареалом гнездовья и зимовок.
ЛУГОВОЙ ЛУНЬ
205
Даты. Появление на зимовках позднее— в Африке на севере первые
появляются в конце октября, на юге в ноябре, весенний отлет с конца марта и до
середины апреля. В Туркмении весенний пролет бывает в первой половине
апреля, на Сыр-Дарье в конце марта — начале апреля (Спангенберг, 1936),
в сев. Казахстане во второй половине апреля (Сушкин, 1908), вЧкаловскойобл.
главным образом, в начале мая, но первые уже 6. IV, в Зауралье у Троицка
2. IV; на Украине уже со второй половины марта, но еще-и в течение апреля.
Осеннее движение начинается в августе, сначала летят молодые птицы;
кончается пролет и отлет в октябре, в Средней Азии (Туркмения; Сыр-Дарья) — еще
в начале ноября. На осеннем пролете луни летят одиночками, парами и
небольшими группами.
Биотоп. Открытый ландшафт, обычно увлажненный. В Средней Азии —
речные долины, в Зап. Сибири лесостепь и степь близ озер и болот, в
Европейской России и на Украине сырые луга; характерно предпочтение кустарниковых
зарослей; в виде исключения в сухих ландшафтах (барханы, с.-з. Казахстан,
Сушкин, 1908), а также в горных лугах (Армения, Соснин и Ляйстер, 1942;
Нарын в центр. Тянь-Шане; у Хорога на Пяндже, Молчанов, 1915). В горы
обычно высоко не идет: в Средней Азии примерно до 1500 м\ редко до 1800
(Северцов), на Алтае до 1000, редко до 1500 м (Сушкин, 1938).
Численность. Распространение спорадическое, в связи с
особенностями биотопического распределения — в Средней Азии, например, при
ограниченной площади открытых влажных ландшафтов весьма редок. По этой
же причине местами гнездовья носят характер колоний, в Наурзумскои степи
на площади в 3—Зх/г км2 13 жилых гнезд и еще 5 вблизи. Имеются, повидимому,
колебания численности в связи с кормовыми условиями («урожаи» грызунов).
Экология. Размножение. Гнезда на земле, простого устройства,
обычно невдалеке от воды или на сыром лугу, иногда среди камышей; из травы
или тонких веточек, небольших размеров, около 50 см в диаметре. Если гнездо
на сырой почве — в нем выстилка из сухой травы, более тщательная. Нередко
пары гнездятся близко одна от другой. Кладка в первой половине мая. Число
яиц 3—5, иногда 6. Размеры (8) 41,2—44,5x34—35 (Сомов, 1897). Окраска
яиц белая, изредка С бурыми крапинами. Одно и нередко 2 яйца бывают не
оплодотворены. Промежуток между откладкой яиц 24 часа, иногда до 3—4 дней
(Наурзум, Рябов, 1940). В случае утраты первой кладки — бывает
дополнительная. По западноевропейским данным, насиживает повидимому только
самка. Продолжительность насиживания — месяц.
В связи с обычным у луней резким разделением функций самца и самки
в период размножения, насиживающей самке и птенцам первое время их жизни
корм носит самец (Зап. Европа).
Вылупление птенцов в сев. Казахстане начинается в первой половине июня.
Вес птенца в первом пуховом наряде от 14 г, во втором пуховом наряде, в
зависимости от возраста,-100—200 г. Вылет птенцов из гнезда происходит около
конца июля, гнездовый период следовательно около 35 дней (в Наурзумском
заповеднике в 1947 г. птенцы вылетают с 9 по 20.VII). Иногда птенцы вылезают
из гнезда и разбредаются в возрасте уже 10—14 дней. В больших выводках
младшие птенцы погибают (каннибализм) и птенцов остается не более 4 даже
в благоприятные годы (Осмоловская).
Линька. В общем сходна с линянием у других луней, полная годовая.
Последовательность смены первостепенных маховых от внутреннего края крыла
к наружному, т. е. от 10-го к 1-му. Начало линьки у самок и у самцов после
периода размножения, между VII—X (детали и сроки требуют изучения).
Имеются большие личные отклонения в сроках линьки, причина которых
неясна. Последовательность смены нарядов: первый пуховой —второй
пуховой —. первый годовой (гнездовый) и т. д. Окончательный наряд самцами на-
206
ОТРЯД ХИЩНЫЕ ПТИЦЫ
девается после третьей годовой линьки на четвертом году, самками после второй
годовой линьки по третьему году.
Питание. Способы охоты—как у полевого луня. Как у других луней, пищу
составляют главным образом мелкие животные, которых лунь берет на земле—
мелкие млекопитающие, гнездящиеся на земле птицы, в особенности птенцы.
Рис. 56. Луговой лунь (самец, самка и молодая птица).
В частности, в гнездовое время в качестве корма лугового луня отмечены:
мыши, полевки, суслики; разные мелкие птицы, а с прилета —цыплята,
воробьи и т. д.; яйца птиц; ящерицы, крупные насекомые (Харьковская обл.),
мелкие птицы, в особенности жаворонки, кроме того гаршнеп (Туркмения);
яйца рябка, молодой удод (Кашкаров, 1928); млекопитающие Microtus arva-
lis, M. ratticeps, Lagarus lagarus, Apodemus sylvaticus, Cricetus cricetus, Sicista
subtilis, Scirtopoda telum\ птицы — обыкновенный и серый жаворонок, степной
конек, Hippolais caligata, славка-завирушка, черноголовый чеккан,
перепел, тетеревята, овсянка; ящерицы — Lacerta agilisf Eremias arguta;
насекомые— жужелицы, чернотелки, щелкуны, долгоносики, кузька, божья коровка,
СТЕПНОЙ ЛУНЬ
207
листоеды, притворяшки Byrrhus, саранча перелетная, стрекозы (сев. Казахстан,
Осмоловская).
Полевые признаки. От других луней луговой лунь отличается меньшей
величиной, более легким сложением и относительно длинными крыльями.
У взрослых самцов имеется черное пятно при основании второстепенных
маховых и продольный буровато-рыжий рисунок на брюхе. Полет легкий, сходный
с другими лунями. Голос — обычно дребезжащее ширрь-пиррь» или
отрывистое «пик-пик-пик», издается довольно редко. Как и другие луни, держится
на земле, на деревья не садится.
Описание. Размеры и строение. Самый мелкий из луней. Длина самцов
(17) 410—465, самок (9) 470—520, в среднем 441,3 и 487,7 мм. Размах самцов (16) 970—1120,
самок (10) 990—1150, в среднем 1043,5 и 1084,5 мм. Вес самцов (5) 258—288, в среднем 275,42,
самок (4) 340—380, в среднем 356,25 г (у старой насиживающей птицы только 290 г). Крыло
взрослых самцов (50) 320—380, самок (22> 325—390, в среднем 300,3 и 382 мм. Формула крыла
3>2=4>5>1>6>7 или 6>1>7, крыло следовательно острее, чем у других луней.
Вырезки на внешних опахалах 2-го — 4-го маховых, на внутренних опахалах 2-го — 3-го
иаховых, при этом край вырезки наружного опахала 2-го махового выдается на 15—30 мм
за вершины кроющих кисти.
Окраска. По западноевропейским данным первый пуховой наряд белый с
желтовато-серым налетом, особенно на спинной стороне; второй пуховой наряд с охристо-
рыжим налетом на спинной стороне и на груди. Первый годовой (гнездовый) наряд сходен
у обоих полов: спинная сторона темнобурая с яркими рыжевато-охристыми каймами; на
затылке беловатые пестрины; надхвостье белое с рыжими пятнами; рулевые серо-бурые,
поперечнополосатые; маховые темнобурые с нерезким темным поперечным рисунком и
мраморными пестринами в основной части внутренних опахал; брюшная сторона ярко ржаво-
рыжая с узкими черноватыми наствольями (их нет у степного луня).
Самка во втором годовом наряде отличается главным образом тем, что брюшная
сторона у нее становится беловатой с продольным рыжевато-охристым рисунком, рулевые как
в первом годовом наряде. Третий годовой наряд у самки окончательный: он отличается от
гнездового тем, что каймы перьев на спинной стороне тела узкие и охристые, иногда
отсутствуют вовсе; рулевые иной окраски, средние пары серовато-бурые, боковые беловатые с
бурым поперечным рисунком и широкой предвершинной полосой; кроющие крыла, а иногда
и маховые, с серым налетом; брюшная сторона охристо-белая с рыжеватыми продольными
полосами (как во втором годовом наряде).
Второй г.одовой наряд самцов более или менее похож на взрослых самок, но на
спинной стороне тела, а также и на зобе развит сероватый оттенок; кроющие крыла тоже сероваты;
грудь, брюхо и бока белые с рыже-бурым продольным рисунком. В третьем годовом наряде
самец еще серее, общий тон спинной стороны серый, но на голове и затылке сохраняются
ржавчатые каймы, и общий оттенок окраски спинной стороны все же буроват; продольный
рисунок груди, брюха, боков — широкий; на крыле темный рисунок — черная полоса на
второстепенных маховых и больших кроющих — не развит. В окончательном — четвертом
годовом — наряде самцы на спинной стороне аспидно-сизы, заметно темней степных луней;
надхвостье белое с бурыми или серыми пестринами; горло, зоб, грудь сизо-серые; брюхо,
бока, подхвостье белые с узкими рыжеватыми продольными полосами, подкрылья белые,
подмышечные с рыжим поперечным рисунком; на кроющих крыла черная полоса, скрытая
при сложенном крыле; передние первостепенные маховые черные, остальные, как и
второстепенные, серые с узкими черными полосами; средние рулевые серые, боковые беловатые
с рыжим поперечным рисунком. Иногда встречаются птицы с сильным развитием серого
цвета на брюшной стороне.
Клюв и когти черные, радужина у взрослых яркожелтая, у молодых бурая (у самцов
серовато-бурая); восковица желтая. Темнобурая меланистическая вариация лугового луня
из пределов СССР неизвестна.
17. Степной лунь Circus macrourus S. G. Gmelin
Accipiter macrourus. G m e 1 i n. Novi Commentarii Acad. Caes. Petropolitanae XV pro
1770, 1771, стр. 439, табл. VIII, IX, Воронеж.
Распространение. Ареал. Степная полоса ю.-в. Европы, на западе
до Добруджи, Подолии и Белоруссии (бассейн Припяти); в Азии на восток до
Джунгарии, Алтая, ю.-з. Забайкалья; северная граница проходит примерно
до Москвы, Тулы, Рязани, Казани, Кирова (гнездовье там не доказано), далее у
Уфы, затем у Свердловска, однако отмечен летом у Архангельска, в Сибири
208
ОТРЯД ХИЩНЫЕ ПТИЦЫ
у Тюмени, Омска, Красноярска; к югу до Крыма и Кавказа, Ирана (с.-з. Иран,
Хорассан, быть может Керман и Кугистан), в Туркестане (но не в зап. и ю.-з.
Туркмении и не в Таджикистане). Более или менее случайные находки
степного луня известны и из других местностей: из Швеции, Германии, у нас в
Прибалтике; по крайней мере часть этих находок несомненно гнездовые. На
пролете отмечен в с.-з. Монголии. Зимует в Индии (до Цейлона) и Бирме, в
Месопотамии и Иране; в Африке всюду, где нет густых тропических лесов, но
главным образом к югу от Сахары. Залеты в Китай. Одиночные особи зимуют
Карта 25. Распространена степного луня Circus macrourus
а — границы гнездовий г — область зимовок.
в южной полосе СССР: в Крыму (Сеницкий), на с.-з. Кавказе (Насимович
и Аверин, 1938), в низовьях Волги (Воробьев, 1938), в Арало-Каспийских
степях (Бостанжогло, 1911).
Даты. Перелетают примерно в то же время, что и луговой лунь, весной
в среднем немного ранее. На зимовки в Месопотамии прилетают в сентябре —
октябре, отлет в марте—апреле. В Африке появляются осенью уже в конце
сентября, но главным образом в начале октября (в тропиках), движение к
северу из южн. Африки начинается уже в январе — феврале, а отлет — в
марте—апреле; некоторые особи летуют. Прилет и пролет в южн. частях
СССР —на Кавказе, Украине, в южн. Поволжье, Средней Азии — происходит
в половине или конце марта, но главным образом в начале апреля, севернее —.
в конце апреля. Осенний отлет на зимовки начинается уже с середины августа
(молодые птицы) и проходит в сентябре, отдельные особи в европейской части
страны задерживаются до октября. В Туркестане и на Кавказе осенью движе-
ние в октябре носит еще регулярный характер, и в ноябре здесь степные луни
еще встречаются.
Биотоп. Открытый ландшафт, более сухой, чем тот, где обычно
встречается луговой^ лунь. Особенно характерны сухие степи, хотя степной лунь
может быть встречен и в долинах рек, по окраинам степных балок и т. п.
В гнездовое время на равнине или невысоко в горах: на Кавказе до 1725 м
(Армения), на Алтае до 1000 м\ в Средней Азии примерно до 1350 м (Северцов
по Мензбиру, 1891). Вне периода гнездования поднимается и выше —на Алтае
до 2300 м, на Памире до 2750 м (озеро Шор-куль, Тугаринов, 1930), в Африке
до 3300 м.
Численность. Обычная птица в подходящих биотопах (сухая
степь), но в других ландшафтах — лесостепи, влажной степи, культурной поло-
Рис. 57. Степной лунь (самец).
се — встречается более или менее спорадично. Вырубка лесов и распашка
земель способствуют, повидимому, расселению степного луня на север в
средней полосе (Московская, Тульская обл.). В некоторые годы в Зап. Европе
на пролете отмечено массовое появление степного луня осенью, носящее в
известной мере инвазионный характер.
Экология. Размножение. В парах встречается уже на весеннем
пролете. Цикл начинается недели на две раньше, чем у лугового луня. Брачный
полет и игры начинаются с прилета, в конце апреля; птицы взмывают в
воздух, переворачиваются, самец «преследует» самку; после начала кладки брачный
«фигурный» полет продолжает один самец. Гнездо весьма простого
устройства, небольшого размера (около ЪОсм в диаметре при диаметре лотка в 15—20л*)
с неглубоким лотком, иногда это только ямка, окруженная сухой травой; часто
оно располагается на кочке или небольшом возвышении среди бурьяна,
зарослей чилиги или бобовника и т. п., реже среди хлебов или на влажных луговых,
даже заболоченных, участках с осокой, таволгой и т. д. (Бараба, Зверев, 1930).
Кладка происходит в разные числа мая, на юге с конца апреля (Сыр-Дарья,
Спангенберг, 1936); возможно, что сроки кладки находятся в зависимости от
широты местности. Число яиц в кладке 3—6, обычно же 3—5. Окраска яиц
белая, изредка с небольшими бурыми пестринами. Размеры (80) 40,1—50 х
Х32,6—37, в среднем 44,77x34,77 мм (Уайзерби, 1939). В случае гибели
кладки — бывает вторая, дополнительная (Наурзум, Осмоловская). Насижи-
14 Птицы СССР, т. I
210 ОТРЯД ХИЩНЫЕ ПТИЦЫ
вание начинается с откладки первого яйца (разновозрастные птенцы),
насиживает только самка (Карамзин, 1900). Срок насиживания около месяца.
Вылупляются птенцы в конце июня — начале июля; летные птенцы
появляются в середине июля, выводки держатся вместе до августа.
Продолжительность гнездового периода, следовательно, около 40—45 дней. Насиживающую
самку и птенцов в первое время их жизни (когда они в первом пуховом наряде)
кормит самец, позднее начинает охотиться и самка.
Линька. Как у лугового луня — полная годовая. Последовательность
смены маховых от 10-го к 1-му; рулевых—от середины хвоста к краю. Сильно
линяющие молодые в первом годовом наряде встречаются и летом (возможно
холостые особи). Последовательность смены нарядов — как и у лугового луня.
Питание. Степной лунь, как и другие луни, охотится за двигающейся
или сидящей на земле добычей. Главное место в его кормовом режиме
занимают мелкие млекопитающие, но когда мышей мало, он переходит на
питание ящерицами, гнездящимися на земле птицами т. п. В качестве пищи
степного луня в СССР указываются разные мыши и полевки, в частности
Sienocranius gregalis, S. slowzowi, Microtus arvalis, M. oeconomust Micromys
minutus, Arvicola terrestris, Apodemus sylvaticus; пеструшка Lagurus lagurus,
хомяк Cricetus cricetus, суслики, среди них Citellas erythrogenys и С. pygmaeus,
землеройка Sorex araneus; из птиц — степной конек, жаворонки и их птенцы,
славковые, перепел, тетеревята, болотная сова, кулики, широконоска, утята;
на Алтае молодые белые куропатки и ящерицы; разные крупные насекомые —
жуки, саранча, кобылки, кузнечики, стрекозы и т. д.
Охотничьи участки у степных луней небольшие. Как и другие луни, они
облетают их по определенному маршруту, летя низко над землею медленным
полетом, и быстро снижаются на добычу, тормозя распущенным хвостом, занося
крылья с далеко отставленным (длинным) крылышком и далеко выбрасывая
вперед лапы.
Полевые признаки. Некрупный лунь, меньше полевого, с более узкими
и острыми крыльями, Самец бледно сизый, брюшная сторона белая, вершины
ПЕГИЙ ЛУНЬ
211
крыльев черные, черная полоса на второстепенных маховых, и кроющих крыла
отсутствует. Самки похожи на полевого луня, молодые снизу ржавчато-рыжие,
лод глазом белое пятно. Осторожен. Голос: «гик-гик-гик»или характерное
для луней «пиррь-пиррь».
Описание. Размеры и строение. Формула крыла 3>4>2>5> 6>1.
Вырезки на наружных опахалах 2-го — 4-го маховых (не на 5-м), на внутренних опахалах
4-го—3-го;конец вырезки внутреннего опахала1-го махового едва выдается за вершину
кроющих кисти; вырезка наружного опахала 2-го махового прикрыта кроющими кисти. Длина
самцов (30) 435—480, самок (16) 480—525, в среднем 453,7 и 504 мм. Размах крыльев самцов
(26)990—1100, самок (12) 1080—1170, в среднем 1040,1 и 1134,3 мм. Вес самцов (4)311—374,
в среднем 331,5, самок (17) 402—550, в среднем 445,2 г, в том числе 427 у молодых и 461,9
у старых. Крыло самцов (50) 310—350, самок (50) 345—375, в среднем 333,4 и 359,7 мм.
Окраска. Первый пуховой наряд не описан. Второй пуховой наряд белый с охристо-
ржавчатым оттенком на спине и груди. Первый годовой (гнездовий) наряд темнобурый на
спинной стороне, темя рыжее с темными наствольями, остальные перья с
рыжевато-охристыми каймами; маховые темнобурые у вершины, с белым полем у основания внутреннего
опахала и с поперечным рисунком; хвост бурый с широкими темнобурыми полосами,
боковые рулевые бледноохристые с бурым поперечным рисунком; кроющие хвоста белые с
неясными темными пестринами; брюшная сторона одноцветная, охристо-рыжеватая в разных
оттенках; под глазом белое пятно. В этом наряде оба пола сходны.
Взрослый самец на спинной стороне сизо-серый; маховые серые с беловатыми
основаниями внутренних опахал, 2-е — 6-е с черной вершиной"; кроющие хвоста белые с серым
поперечным рисунком; средние рулевые серые, боковые белые с серыми полосами; брюшная
сторона белая с более или менее заметным сизым налетом на горле и зобе. Этот наряд
надевается после трех линек на 4-м году жизни. Второй и третий годовые наряды самцов пере-
ходны — серовато-бурые на спинной стороне, на голове с рыжими каемками, на брюшной
стороне белые с рыжеватыми наствольями, рулевые с поперечным рисунком. При этом птицы
в третьем наряде серее и с меньшим развитием пестрин на брюшной стороне.
Взрослые самки походят на полевых луней, на спинной стороне бурые с охристыми
каймами, на брюшной — беловатые с рыжеватым продольным рисунком, с темнобурым
поперечнополосатым хвостом, но надхвостье у них с охристо-буроватым рисунком, нередко
в виде поперечных пятен. Второй годовой наряд самок отличается от окончательного
(третьего) несколько более темным оттенком бурого цвета и большим развитием рыжевато-
охристых каемок перьев на спинной стороне. Клюв черный, когти черные, радужина ярко-
желтая у взрослых птиц, бурая — у молодых, восковица и ноги желтые.
18. Пегий лунь Circus melanoleucus Penn.
Falco melanoleucus. Pennant. Indian Zoology, 1769, стр. 2, табл. II, Цейлон (в холодное
время года).
Распространение. Ареал. Вост. Азия — от Приамурья и
Забайкалья (Даурия) до Манчжурии. Северная граница за Зее у Пикана; на Амуре
к западу до Кумары (т. е. около 250 км выше Благовещенска); восточнее Сихо-
те-Алиня отсутствует; спорадически в ю.-в. Забайкалье (Аргунь, Борзя, к
западу от озера Тарейнор и Акши на Ононе, Штегман, 1937); сев. граница
на Амуре неясна. Зимовка в ю.-в. Азии: на Филиппинах, в Индо-Китае,
Малайе, на Борнео, в Ассаме, Бирме; в Индии, главным образом на
востоке, редко до с.-з. провинций, к югу до Цейлона, но не в центральных частях
и не на западе полуострова.
Характер пребывания. Перелетная птица с разомкнутым
гнездовым и зимним ареалами.
Даты. Весенний пролет поздний, позже, чем у полевого луня. В первом
периоде пролета в конце марта и начале апреля появляются самцы (23. III..-*
5. IV, Никольск-Уссурийск, Дулькейт, 1928); в апреле, примерно в десятых
числах, — самки. В Забайкалье луни прилетают около середины апреля.
Осенние перемещения, носящие характер кочевок, начинаются в сентябре,
отлет в октябре, в южн. и ю.-з. направлении. Некоторые особи
задерживаются и возможно даже зимуют на крайнем юге Приморья в годы с благоприятъ
14*
212
ОТРЯД ХИЩНЫЕ ПТИЦЫ
ными кормовыми условиями (обилие грызунов); в пограничных частях
Манчжурии молодые птицы отмечены до декабря; самка у Сидеми добыта Калинов-
ским 13. XI. 1885. В Индии держится с октября по апрель, весенний пролет
в Китае — с начала сентября до начала мая, осенью с конца августа до
середины октября.
Биотоп. Как и у других луней, открытый ландшафт. Распределение
в известной мере напоминает распределение лугового луня: луговые долины
рек, сопки среди полей, полустенной ландшафт; определенное предпочтение
при этом оказывается влажным местам. В сев. Приамурье — по болотам,
поросшим карликовой березкой.
Численность. Ограниченный ареал предопределяет умеренную
численность вида в целом. Распространение носит при этом в известной мере
спорадический характер. Обычен в
бассейне Уссури, у озера Ханка и по
рекам, впадающим в залив Петра
Великого. Редок в сев. Приамурье и
Забайкалье.
Экология. Размножение.
Изучено удовлетворительно (Дуль-
кейт, 1928; Спангенберг, 1940). В
парах луни бывают с середины апреля,
вскоре после прилета. Брачные игры
происходят с начала мая и до начала
июня. В это время, главным образом
по утрам, птицы проделывают
воздушные эволюции над гнездовым
участком, летая по кругу, взвиваются,
падают и т. п., самец кричит «кий-ви»
(вроде крика чибиса), самка часто
отзывается коротким «кий-кий-кий».
Спаривание происходит сначала на
Карта 26. Распространение пегого луня земле (при этом самка складывает за-
Circus melanoleucus несенные над спиной крылья), позже,
а —границы гнездовий, г —область зимовок. ПОСЛе ПОСТрОЙКИ ГНвЗДа —- На ГНеЗДС
Имеется, повидимому, резерв
холостых (вероятно, прошлогодних) особей. Гнезда строятся в середине мая,
обычно в траве, чаще на болотах или лугах с кустарником. Расстояние гнезда
от гнезда около 1 км (Иман). Размер гнезда в диаметре около 40—50см, лоток
17—18 см в диаметре и 4,5—5 см глубины. Гнезда используются по несколько
лет.
Кладка со второй половины мая (12.V первое яйцо), промежуток между
откладкой около 48 часов (16.V было обнаружено одно яйцо, 20.V —два, 25.
V. — четыре; в другом гнезде 29.V — одно яйцо, 1. VI — два). Полная кладка
из 4—5 яиц. Окраска яиц белая или чуть зеленоватая, иногда с немногими
буроватыми пестринами..Размеры: (5) 43—44x32—35 (Дулькейт); (10) 43,6—
46,7 х 33,4-35,6 (Спангенберг); (28) 40,5—45 х 32—36, в среднем 43,6 х 34,5
(Бекер, 1928).
Насиживание с первого яйца, участвуют в насиживании повидимому оба
родителя, но главным образом самка. В случае гибели самки насиживает самец.
Вылупление повидимому происходит в начале июня (по Дулькейту, иногда даже
во второй половине июня); срок насиживания, следовательно, немного более
месяца. Кормят птенцов оба родителя (носят главным образом полевок, реже
полевых мышей; также прямокрылых, жуков и мелких птиц). Подлетки
встречаются в первой половине августа. В это время молодые сидят у гнезд и их
ПЕГИЙ ЛУНЬ
213
кормят старые птицы; с конца августа молодые охотятся самостоятельно.
Линька. Цикл как у полевого луня. Точные даты неизвестны.
Питание. На охоте летает плавно и низко в 1 —2 м от земли, в
открытых местностях, иногда покачиваясь, а затем бросаясь в траву и хватая добычу.
Характерны очень длинные цевки (высокотравие?). Весной и летом
охотятся попарно, самец и самка. На лету птиц не ловит. Весною пищу
Рис. 59. Пегий лунь (самец, самка и молодая птица).
составляют главным образом грызуны, реже землеройки, лягушки, из птиц
перепела, коньки Anthus richardi, жаворонки, желтые плиски, даже
фазанки; в период выкармливания молодых — воробьи. В погадках полевки,
полевые мыши, мелкие птицы, жуки, прямокрылые. В Забайкалье в желудке
пегого луня найдены полевки Microtas ungurensis и Stenocranius gregalis. На
пролете в Китае нередко лягушки.
Полевые признаки. Размеры — с лугового луня. Взрослые самцы хорошо
отличаются пестрой двуцветной «сорочьей» окраской. Полет медленный и
плавный, как у других луней.
Описание. Размеры и строение. Вырезки на первостепенных маховых —
как у полевого луня. Формула крыла: 3>4>2>5>1=6>7. Цевки очень длинные, более
214
ОТРЯД ХИЩНЫЕ ПТИЦЫ
75 мм у самцов и более 80 мм у самок. Длина самцов (2) 415—435, самок (3) 440—463,
мах крыльев самцов (2) 1038—1065, самок (1) 1155 мм. Вес самца (1) 270, самки (1) 4!
Крыло самцов (12) 345—366, самок (5) 355—373, в среднем 352,2 и 365,8 мм.
Окраска. Пуховые наряды не описаны. Первый годовой наряд (у обоих по
на спинной стороне темнобурый с рыжеватым теменем и черноватыми пестринам
белыми основаниями перьев на затылке; надхвостье охристо-рыжеватое с темным прод
ным рисунком; брюшная сторона буровато-рыжая, иногда с беловатыми каймами пер
маховые черно-бурые с серовато-рыжими поперечными полосами на внутренних опаха.
средние рулевые темнобурые, боковые рыже-бурые с черновато-бурым поперечным ри
ком. Второй годовой наряд с беловатой брюшной стороной и продольным по ней рыжева
рисунком.
Окончательный наряд — третий, с резким половым диморфизмом. Самка на спш
стороне темнобурая с узкими светлыми краями перьев, с белыми малыми кроющими кр!
надхвостье белое с продольными рыжеватыми отметинами; брюшная сторона белая с о:
стыми продольными полосами, маховые темнобурые с поперечным рисунком в основной ч
пера; рулевые серовато-бурые с темным поперечным рисунком. У старого самца чер
голова, спина, средние кроющие крыла и первостепенные маховые; большие кроющие Kpi
второстепенные маховые, рулевые — серые; малые кроющие крыла и часть перьев надхва
белые; горло и грудь черные, остальная брюшная сторона белая. Клюв черноватый, во
вица и лапы желтые, когти черные; радужина у молодых бурая, у старых желтая.
19. Болотный или камышевый лунь Circus aeruginosas I
Falco aeruginosas Linnaeus. Syst. Nat. изд. X, 1758, стр. 91, Швеция.
Распространение. Ареал. Вост. полушарие — Палеарктическая о*
Африка, Мадагаскар, Австралия и Тасмания, Новая Зеландия, также Но
Каледония и Фиджи. Зимой в южн, Азии и тропической Африке.
Карта 27. Распространение болотного или камышевого луня Circus aeruginosm
1 —С. a. aeruginosus, 2 — С. a. spilonotus, 3 — С a. harterti, 4 — С. a. aequatorialis, 5 — С. a. rani
0 — С. a. macrosceles, 7 — С a. maillardi, 8 — С a. gouldi, 9 — С. a. wolfi, 10 — С. а. арргох
а — границы гнездовий, б — не вполне выясненная граница распространения; цифры в кружке —
нахождения соответствующего подвида на зимовке.
Характер пребывания. На севере перелетная птица, на ю;
оседлая.
БОЛОТНЫЙ ИЛИ КАМЫШЕВЫЙ ЛУНЬ
215
Биотоп. Безлесные влажные местности — болота, камышевые заросли
прибрежий рек и озер и т. п.
Подвиды и варьирующие признаки. С. a. aeruginosas L., 1758, в Зап., средней и Вост.
Европе, в Зап. Сибири до Енисея, в Передней и Средней Азии; С. a. harterti Z e d i i t z,
i914, в Средиземноморье от южн. Испании до Марокко и сев. Алжира, вероятно к этой же
форме луни из Туниса и Египта; С. a. aequatorialis Stresemann, 1924, в вост.
Африке от Уганды до Ниасса; С. a. ranivorus D a u d i n, 1800, в южн. Африке к северу до
Трансвааля; С. a. spilonotus Каир, 1847, в вост. Азии от Забайкалья и Монголии,
Приморья и Сев. Китая до Тибета; С. a. maillardi Vег гeaux, 1862, на о. Реюньон; С. a. macro-
scetes N е u t о n, 1863, на Мадагаскаре и Коморских о-вах; С. a. approximans Р е а 1 е,
1848, на о. Фиджи; С. a. wolfi Gould, 1866, на о. Новая Каледония и быть может
Новогебридских о-вах; С. a. gouldi Bonaparte, 1850, на ю.-з. Новой Гвинеи, в Австралии,
Тасмании и Нов. Зеландии. Варьирующие морфологические признаки—окраска и размеры.
При этом камышевые луни подразделяются на четыре' группы естественно близких форм:
aeruginosus (aeruginosus + harterti)y ranivoras (ranivorus -{-aequatorialis), approximans
(approximans -f- wolfi + gouldi )y spilonotus, maillardi (maillardi + macro sceles), которых
многие авторы считают за виды, но правильнее объединять их в один политипический вид.
Имеются различия между отдельными формами и в проявлении полового диморфизма,
возрастных изменений и в других деталях экологии. Географические вариации окраски
выражены главным образом только у взрослых самцов.
Обыкновенный болотный лунь Circus aeruginosus aeruginosus L.
Синонимы. Circus aeruginosus var. unicolor. R a d d e. Ornis Caucasica, 1884,
стр. 106, табл. HI, Ленкорань.
Распространение. Ареал. Европа, кроме северной полосы—примерно
до Англии, южн. Швеции и южн. Финляндии, далее до Карелии,
Архангельска, около 58° с. ш., в Приуралье (но отсутствует в Кировской обл.), в
Зап. Сибири до Тюмени, Тобольска, Томска, Красноярска, к востоку до Енисея,
Минусинской лесостепи, зап. части Таннутувинской обл., с.-з. Монголии
(р. Тесь, Ачит-нур, Улюнгур, по Сушкину, 1938); на юг до Балеарских о-вов,
Корсики, Сардинии, Сицилии, севера Малой Азии, Закавказья, Ирана (до
Сеистана), Месопотамии, Средней Азии, Джунгарии (Лоб-нор) и Синьцзяня.
Зимовки кроме того в Африке, в основном к югу от Сахары, на юг до б.
германской вост. Африки и Анголы, также в Конго; в передней Азии к югу от
Закавказья, в Туркестане, на юг до сев. Индии. Сведения о зимовках в Южн.
Китае очевидно относятся к молодым птицам формы spilonotus.
Характер пребывания. В сев. части ареала перелетная
птица; отдельные особи зимуют уже в средней Европе и южн. полосе
Европейской России (впрочем, птица, окольцованная в низовьях Волги, у
Астрахани 27. VI. 1939, добыта на зимовке 5. II. 1947 у Кюрдамира); в частности,
повидимому, встречается зимою в низовьях Волги, в декабре встречена в Крыму
(Никольский, 1892). В Закавказье вероятно, по крайней мере частично,
оседлый (Армения, Азербайджан). Несомненно зимует в Туркестане, на юго-западе
его ^регулярно. Отмечен, например в декабре—январе в Фергане, 19—20.
XII на Куванджарме;вТуркмении встречен зимою в подгорной полосе,наУзбое,
на Атреке и т. д. Однако часть луней из Туркестана, по крайней мере из ее
сев. частей, отлетает, например, с низовья Сыр-Дарьи, с Или.
Даты. Прилет на африканские зимовки протекает с конца сентября
до начала ноября, отлет — в марте и апреле, отдельные особи еще в маечи даже
летуют.
Сначала летят молодые птицы. Весенний пролет —одиночками на разной
высоте; есть указания, что самцы прилетают несколько ранее самок (Сушкин,
1908, Бостанжогло, 1911). Область гнездовья занимается постепенно,
появление луней связывается с таянием водоемов и спадом полой воды, но гнездование
во всех частях ареала идет почти одновременно. На Украине, в Бессарабии,
в Крыму пролет и прилет луней происходит уже с середины марта, севернее,
216
ОТРЯД ХИЩНЫЕ ПТИЦЫ
в Харьковской обл., валовой пролет в начале апреля, первых—*в середине марта;
в Среднюю Азию прилетает с середины марта. На Кавказе прилет отмечен
в начале апреля. В средней полосе лунь появляется главным образом во второй
половине апреля, но иногда уже в начале этого месяца. Отлет луней начинается
исподволь с августа, проходя главным образом в сентябре и кончаясь в начале
октября. Довольно значительные колебания сроков пролета луней по
отдельным годам и местностям связываются вероятно с гидрологическим режимом
(таяние и замерзание, изменение уровня вод).
Биотоп, Болотистые, заросшие тростником и камышем местности,
долины рек, побережья озер и т. д. В негнездовое время в культурном
ландшафте и других открытых местностях, но всегда вблизи воды. На равнинах,
в горы поднимается невысоко (на Алтае, по Сушкину, до 1400 м; в Средней
Азии, по Северцову, примерно до 2000 м).
Численность. При наличии подходящих биотопов многочисленен;
гнезда нередко располагаются близко одно от другого, даже на расстоянии
до 2 м (Тюмень, Словцов, 1892), иногда небольшими колониями; плотность
при этом значительная —до 10 гнезд на 1 км2 (Бараба, Зверев, 1930) или
1 гнездо на 7—13 гектар (Бараба, Жданович, 1931). Нормальная область
распространения болотного луня связана с зонами открытых ландшафтов — степи
и лесостепи и т. п. В лесной зоне он редок и спорадичен.
Экология. Размножение. Половая зрелость наступает на
следующий год после рождения. Имеется резерв прошлогодних холостых особей,
вероятно потенциально половозрелых. Гнездовые участки ограничены,
постоянство их связано с состоянием тростниковых зарослей. Спаривание
начинается с прилета (в конце апреля). Брачный полет продолжаете^ около
месяца (Сушкин, 1908). При этом обе птицы летают на довольно
значительной высоте кругами, затем круто снижаются («падают») с характерным
пискливым криком. Гнезда среди камышей и тростников, хорошо укрытые; реже—
среди прибрежных кустарников. Обычно это большие кучи тростника, куги
и т. п., размеры гнезда около 80—100 см в диаметре, высота 25—50 см, лрток
15—20 см. Строят гнездо оба родителя. Кладка состоит из 2—6, обычно из
4—5 яиц. Промежуток между откладкой яиц до 3 дней, судя по разнице в
возрасте птенцов. Время кладки — со второй половины апреля, главным образом,
в начале мая; широтных различий не установлено. В случае утраты кладки
гнездо иногда бросается, чаще однако кладка повторяется, причем в ней
бывает до 5 яиц (Полесье, б. Орловский уезД, Барабинская степь, Воронежская
обл.). В таких случаях свежие яйца можно найти даже в конце июня (22. VI).
Окраска яиц белая с легким зеленоватым оттенком, иногда с охристыми
пестринами. Размеры: (14) 45,8—51,9 х 35,3—39,1 (Сыр-Дарья, Спангенберг,
1936); (14) 47—56 х 37—41, в среднем 51,3 х 39 (Умань, Гебель, 1879);
(11)48,5—52 х37—40,5 (Харьковская обл., Сомов, 1897); 47,6—51,6 х 36,3—
39,7 (Воронежская обл., Огнев и Воробьев," 1924); 46—53 х 37— 40
(Полесье, Шнитников, 1913).
Насиживание происходит с первого яйца, птенцы разновозрастны. На
гнездах добывались только самки, так что и у этого луня, как и у других,
разделение функций партнеров в брачном цикле строгое (исключение повидимому
составляет лишь пегий лунь). Самец приносит корм и насиживающей самке,
и выводку; при этом иногда передает добычу самке, а иногда бросает ее на
гнездо с воздуха. Продолжительность насиживания около месяца, повидимому
немногими днями более. Пуховики в разных местностях отмечены около
середины июня; по западноевропейским данным длительность насиживания 32—-
33 дня или даже 36 дней. Новорожденный птенец весит около 30 г (Бараба,
Зверев, 1931). С конца июля и в начале августа встречаются летные молодые
(от вторых дополнительных кладок позднее). Гнездовый период продолжается,.
БОЛОТНЫЙ ИЛИ КАМЫШЕВЫЙ ЛУНЬ
217
следовательно, несколько менее двух месяцев. Молодых в выводке обычно
меньше, чем яиц в кладке — от 2 до 4 (это вызвано смертностью птенцов при
больших кладках). Задолго перед вылетом птенцы, как обычно у гнездящихся
на земле птиц, начинают вылезать из гнезда и прятаться в окружающих
камышах, укрываясь от жары. Разбиваются выводки около середины августа.
Линька. Как у дру-
гих луней, полная годовая.
Птицы в первом годовом
наряде начинают линять
уже весной (мелкое
оперение) с прилета и повиди-
мому уже на зимовке; затем
линька у них замедляется
или приостанавливается,
крупные перья начинают
сменяться летом, и к
отлету весь процесс
заканчивается. Взрослые начинают
линять во второй половине
периода размножения — в
июне и кончают линяние к
отлету в сентябре. Иногда
линька протекает с
отклонениями от указанной
нормы (у неразмножающихся
особей или у птиц с
затянувшимся периодом
размножения в результате
второй дополнительной
откладки яиц).
Последовательность
смены крупного оперения —
как и у других видов луней:
смена первостепенных
маховых от заднего, 10-го,
к переднему, 1-му; смена
рулевых центробежная.
Последовательность смены
нарядов : первый пуховой
наряд — второй пуховой
наряд — первый годовой
(гнездовый) наряд —
второй годовой наряд (переходный) ■— третий годовой (окончательный) наряд.
Так у самок; при этом возрастные изменения у них незначительны. Самцы
надевают окончательный наряд только после третьей годовой линьки, на
четвертом году жизни, и разница между гнездовым и окончательным нарядом у
них значительная. Как у других луней, половой диморфизм отмечается после
второй годовой линьки, причем до этого времени самцы носят «самочий» наряд»
но уже способны к размножению.
Питание. Камышевый лунь берет добычу с земли, низко летая над
поверхностью последней и схватывая жертву лапами; еще чаще охотится он
над водою или над Камышевыми зарослями; в редких случаях ловит добычу
над сухой степью вдали от воды. Вылетает за добычей главным образом с
рассвета, ранним утром; в жаркое время дня не вылетает и возобновляет охоту
Рис. 60. Обыкновенный болотный лунь (самец и самка).
218
ОТРЯД ХИЩНЫЕ ПТИЦЫ
только под вечер. Охотничий участок относительно невелик и определяется
площадью того водоема и окружающих его непосредственно камышей, где лунь
гнездится. На пролете и зимовке положение несколько изменяется, хотя и тут
луни более или менее постоянно придерживаются определенных участков.
Пищу болотного луня в значительной мере составляют птицы, в особенности
насиживающие самки, птенцы и яйца водных видов; в меньшей степени
грызуны; кроме того, ящерицы и насекомые. Весной с прилета лунь ест и падаль.
В качестве добычи луня в гнездовое время в с.-з. Казахстане (Наурзум) отмечены:
из птиц кряква, серая утка, чирки трескунок и свистунок, широконоска,
красноголовая чернеть, шилохвость, лысуха, ушастая поганка, разные кулики,
в том числе круглоносый плавунчик, веретенник, грач и мелкие воробьиные;
прыткая ящерица; насекомые, главным образом саранчевые; из зверей
тушканчики Alactaga jaculus, ушастый еж Hemiechinus auritus, водяная крыса Arvi-
cola amphibius, серый суслик Citellus pygmaeus, пеструшка Lagurus lagurus,
полевка Stenocranius gregalis. Утки, не только птенцы, но и взрослые,
главным образом линные; нередко яйца. Главное место (66—80%) —
занимают птицы, а среди них утки; из млекопитающих водяная крыса (40%). Для
других местностей в гнездовом времени в качестве добычи луня указываются
нырки, поганки и т. д. (Чкаловская обл.), крачка, тиркушка, жаворонки,
желтые плиски (Бараба), удод (Волжско-Уральская степь), дупели и бекасы
(б. Симбирская губ.),чибисы (Горьковская обл.), коростели, перепела
(Орловская обл.) и т. д.; из млекопитающих крапчатый суслик—Spermophilus guttatus
(Харьковская обл.), хомяк Cricetus cricetus, суслик Citellus erythrogenyst
землеройка Sorex araneus(Бараба); жабы и лягушки (Заволжье) и т. д. На зимовке в
качестве добычи луня в Талыше приводится падаль, птицы подранки, также
разные мелкие и средней величины здоровые птицы, например пастушки и
кулики; рыбы и мелкие грызуны; в ю.-з. Туркмении — главным образом
водяная птица-кряква, свиязь, красноголовый нырок, чирок свистунок,
лысуха, кроме того турач, пастушок, мелкие грызуны, ящерицы (агама) и вобла.
Полевые признаки. Крупный лунь. Самка и молодые кажутся однообразно
темными со светлой головой. Старые самцы сверху бурые, со светлым сизым
хвостом и пятнами на крыльях. Полет — как у других луней. Осторожен, за
исключением гнездового времени. Голос в брачный период — гнусавый писк
вроде «пиюю-пиюю», иногда легкий свист или звонкий крик «ке-ке-кее». У
самца еще громкое «куак-куак».
Описание. Размеры и строение. От других луней нашей фауны отличается
крупной величиной и относительно сильными лапами. Формула крыла: 3>2>4>5>1=6...
Вырезки на наружных опахалах первых пяти маховых; вырезка внутреннего опахала 1-го
махового выдается за вершины кроющих кисти. Длина самцов (37) 490—543, самок (40)
521—600, в среднем 526,75 и 560,92 мм. Размах самцов (32) 1110—1308, самок (31) 1230—
1415, в среднем 1177,9 и 1322,58 мм. Вес самцов (10) 405—667, в среднем 509,6 г. Самок, по
Нитхаммеру (1938) 620—740, в среднем 684 г. Крыло взрослых самцов (44) 360—413, самок
(35) 383—432, в среднем 390,3 и 410,7 мм.
Окраска. Первый пуховой наряд желтоватый с белой головой. Второй пуховой
наряд такой же, но с темным пятном у глаза.
Первый годовой (гнездовый) наряд однообразно темнобурый с охристым теменем и
такими же пятнами на горле; иногда на темени черноватые настволья; на спине нередко
бледноохристые каймы, на груди охристые пятна. Иногда птицы однообразно темнобурые,
без пятен (так называемые unicolor, описанные Радде). Эта вариация встречается
относительно нередко и без определенной географической локализации (по Зарудному, 1911,
в Полтавской обл. в одном выводке болотных луней было 3 unicolor и один нормально
окрашенный птенец, в другом 3 нормальных птенца и один unicolor).
Самки в окончательном—третьем годовом—наряде: на темени и затылке перья
охристые с буроватыми наствольями; спина одноцветная темнобурая или чаще темно-
бурая со светлыми охристыми каймами перьев; хвост темнобурый с ржавчатыми пестринами
на боковых перьях; маховые бурые со светлыми пестринами на основной части внутренних
опахал; брюшная сторона бурая, горло охристое; на больших кроющих нередко серо-сизый
налет. Таким образом, взрослые самки очень сходны с птицами в первом годовом наряде.
БОЛОТНЫЙ ИЛИ КАМЫШЕВЫМ ЛУНЬ
219
Сходно окрашены и самки во втором годовом наряде, но у них не бывает серого налета на
крыле.
Взрослые самцы (в 4-м годовом наряде) с охристой головой, испещренной темными на-
■ствольями; спина у них темнобурая, в свежем пере обычно с ржавчатыми каймами;
плечевые темнобурые с поперечным серым рисунком у оснований; малые кроющие крыла
охристые с темными наствольями, остальные кроющие темнобурые с охристыми каймами,
некоторые из больших кроющих крыла сизо-серые с беловатыми основаниями и каймами;
надхвостье бурое с сероватыми пестринами; маховые черноваты у вершин, с белым
основанием внутреннего опахала передних перьев; внутренние первостепенные и второстепенные
•серо-сизые с беловатым рисунком; рулевые сизые; горло беловатое, остальная брюшная
сторона рыжая в разных оттенках, с темными наствольными пятнами; подкрылья белые. Этот
•наряд надевается только после трех линек; во втором наряде самцы сходны со взрослыми
•самками, но не имеют сизого налета на крыльях; в третьем наряде у самца хорошо
заметен сизый налет на-крыльях и хвосте, но брюшная сторона еще бурая.
Радужина у взрослых желтая, у молодых бурая; клюв черный с синеватым
основанием; когти черные; восковина, разрез рта и лапы желтые.
У болотных луней, кроме описанной выше вариации unicolor, значительно развиты и
иные проявления индивидуальной изменчивости окраски, главным образом в отношении
распространения сизо-серого цвета в оперении. Некоторые самки и молодые птицы имеют
поперечнополосатый рисунок на хвосте, вариант, параллельный восточноазиатским и
африканским расам.
Восточный болотный лунь Circus aeruginosas spilonotus Каир
Circus spilonotus. Каир injardine's Contrib. Orn. for 1850, стр. 59, Азия.
Распространение. Ареал. От южн. Якутии (Якутск, Олекминск)
и Витимского нагорья до южн. Забайкалья и Приамурья, Уссурийского
края, Сахалина; вне наших пределов в сев. и вост. Монголии до Кентея и Толы
(быть может до Гоби и Орок-нора), в Манчжурии. Залеты отмечены в Японии.
Зимою — в разных частях Среднего и Южн. Китая, в Индо-Китае,
Бирме, Малакке, на Борнео и Филиппинах, Формозе и Риу-Киу. Добыт на
Сахалине (Шульпин, 1936), в сев. Прибайкалье (Тунка, Скалой, 1935), у
Минусинска.
Даты. Прилет в Забайкалье и Приморье во второй половине апреля,
тогда же примерно у Якутска и Олекминска; но на Витимском нагорье
пролетные встречаются еще в начале мая (Павлов, 1948). Отлет начинается с
середины сентября и продолжается до октября.
Биотоп. Как у номинальной формы, обширные камышевые и
тростниковые болота или берега озер; после вывода также луга и другие открытые
ландшафты.
Численность. Распространение несколько спорадично и в общем
лунь редок.
Экология. Размножение. Половая зрелость наступает, вероятно,
как у номинальной формы. Брачные игры начинаются с прилета, в конце апреля
и в начале мая. Самец поднимается в воздух к гнусавым «квааа», складывает
крылья и бросается вниз, самка летает несколько ниже, издавая крик вроде
«пии» или «иих» (Шульпин, 1936), иногда оба партнера высоко взлетают на
кругах. Гнезда размещаются на земле среди камышей, редко на кустах; сделаны
они главным образом из тростниковых стеблей, без подстилки. Свежие кладки
обнаружены в последней трети мая (Приморье, Забайкалье). Число яиц в кладке
3—4, редко 5. Размеры 47,2—60 х 38—39 мм (Тачановский, 1891). Окраска как
у номинальной расы. Летные молодые появляются в конце июля—начале августа.
Линька. Как у западной расы. Взрослые птицы начинают линять
в начале или первой половине июня, когда сменяются задние маховые
(9-е — 10-е); к середине августа (19. VIII, Борзинский район) линяние
проходит на 50%: 6-е — 10-е маховые новые, 5-е выпало, сменяется мелкое
оперение. Кончается линяние на зимовке. Возрастные изменения —• как у
западного камышевого луня.
220
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Питание. Главным образом птицы, их яйца и птенцы; также
млекопитающие. Последние все же местами (Забайкалье) преобладают. Из
птиц в качестве пищи луня указываются утки — шилохвости (линные); полевой
жаворонок; коньки, в том числе Anthusrichardi) овсянки, среди них дубровник;
камнешарка и др., кулики и т. д.; из зверей суслик Citellus eversmanni,
Рис. 61. Восточный болотный лунь (самец и самка).
мышевки Sicista, полевки Stenocranius gregalisy Microtus michnoi, M. ungurensis,
хомячки; из насекомых кобылки и т. п.
Описание. Размеры и строение. Крупный лунь, размеры несколько больше
чем у номинальной формы. Крыло самцов (12) 378—410, самок (8) 395—423, в среднем 392,5
и 409,4 мм. Вес самцов (2) 580, 610 г, самки (1) 780 г.
Окраска. Пуховые наряды не описаны.
Первый годовой наряд в общем сходен с таковым западной формы, но с явственным
поперечнополосатым хвостом (у большинства особей); охристый цвет занимает большее
пространство на голове и спине, а также на зобе. Взрослые самки сходны с ломинальной
формой, но также с полосатым хвостом. Взрослые самцы резко отличны. Голова и шея у них
беловатые с черными наствольями, щеки и кроющие уха черные с сероватыми каймами перьев
и пестринами; передние 5 первостепенных маховых белые у основания, черные у вершины,
с серебристо-серым налетом на внешних опахалах; остальные маховые серые с черным
поперечным рисунком и белыми полями внутренних опахал; второстепенные маховые
ЧЕРНЫЙ КОРШУН
221
серые с белыми полями внутренних опахал; рулевые серые; брюшная сторона белая с
черными наствольями на зобе и груди. Этот наряд самцы надевают, повидимому, на четвертый
год жизни, после третьей годовой линьки. Во втором годовом наряде рулевые серые, но
общий тон спинной стороны еще буроватый; в третьем наряде второстепенные маховые
серые, как и рулевые, на спинной стороне птица буровата с охристыми пятнами, брюшная
сторона беловатая с узкими бурыми наствольями.
Радужина у взрослых желтая, у молодых бурая; клюв и когти черные; ноги и вос-
ковица желтые.
Систематические замечания. Как видно из приведенного описания, самки и молодые
восточного болотного луня почти идентичны с западной формой, самцы же резко различаются.
Эта особенность характерна для всей группы Камышевых луней в широком понимании вида.
Определение отдельных особей (кроме старых самцов) может встретить поэтому затруднения.
Резко выраженный признак окраски восточной формы — поперечный рисунок на рулевых —
также не имеет абсолютного значения. С одной стороны, имеются самки и молодые spilonotus'
без поперечных полос на рулевых. С другой стороны, в качестве индивидуальных вариаций
особи с поперечнополосатым хвостом попадаются и среди западных луней: такие птицы
неоднократно добывались в Средней Азии, в частности молодая особь — в Аламединской щели
в Киргизском хребте, линные самцы — на Мургабе и у Фирюзы в центральном Копет-Даге
в Туркмении и т. д. Наличие таких птиц и дало повод к утверждению многими авторами
(начиная с Северцова и Мензбира) о появлении восточной формы болотного луня в
Туркестане. Особи с признаками, до известной степени «переходными» между aeruginosas и
spilonotus найдены и в других местностях: в Балаганской степи (Скалой, 1935), в с.-з. Монголии
на озере Хараусу, в Синьцзяне у Яркенда и Мазар-тага (Штегман,1937). Поэтому рассматри-
» вать западного и восточного болотных луней в качестве отдельных видов, как это делают
Суанн (1925) и Питере (1931), нет никаких оснований.
РОД КОРШУН MILVUS LACEPEDE, 1799
Тип Falco milvus
20. Черный коршун Milvus korschun Gmelin
Accipiter korschun. S.G. Gmelin. Novi Commentarii Academiae Caesareae Petropoli-
tanae pro 1770, 1771, стр. 444, табл. Х1-а, Россия.
Русское название. Коршун — старинное русское слово; этимология неясна.
В XVII веке, судя по переписке царя Алексея Михайловича, в соколиной охоте эту птицу
звали коршаком и травили ее кречетами.
Распространение. Ареал. Европа и Азия, кроме северной части тайги
и тундры; Африка и Мадагаскар; Индо-австралийский архипелаг и Австралия.
Характер пребывания. В Палеарктической области перелетный,
в других частях ареала оседлый.
Биотоп. Лесистые местности близ воды.
Подвиды и варьирующие признаки. В Европе, с.-з. Африке, Передней Азии М. к.
korschun; в Египте М. к. aegyptius Gmelin, 1788; в Африке к югу от Сахары, на Мадагаскаре
М. к. parasitus D a u d i п, 1880; в южн. Аравии и Сомали М. к. arabicus Swann, 1922;
в Индии и Бирме к югу до Цейлона и Малайского п-ова М. к. govinda S у k e s, 1832;
на М. Зондских о-вах, Тиморе, Целебесег на Н. Гвинее, о. Бисмарка и в Австралии М. к.
affinis Gould, 1838; в Сибири, Японии, Средней и Центральной Азии М. к. lineatus
Gray, 1831; на Формозе М. к. formosanus Kuroda, 1920.
Морфологически варьирующие признаки: окраска (в частности клюва и подкрыльев),
размеры. Экологические различия между формами касаются характера пребывания,
особенностей размножения и т. п.
Европейский черный коршун Milvus korschun korschun Gmel.
Синоним. Milvus korschun rufiventer Бутурлин. Наша Охота, 1908, сентябрь,
стр. б, Мургаб в Туркмении.
Распространение, Ареал. С.-з. Африка к югу до центр. Сахары, Европа
на юге от Пиренейского п-ова, средних частей Франции и Бельгии до Италии,
Балканского п-ова и Румынии; на север до южн. Швеции; в европейской
России на север до Обонежья, Архангельска, южн. части Вологодской обл.,
средней Печоры (Усть-Илыч, Якша, Аранец); вост. граница неопределенна,
222
ОТРЯД ХИЩНЫЕ ПТИЦЫ
так как начиная от Прикамья и Заволжья на западе и далее в сев. Казахстане
и Тянь-Шане до Памира встречается популяция переходного характера, среди
которой большинство особей обладают признаками, промежуточными между
европейской и сибирской формами. При этом в Приуралье, Заволжье и в сев.
Карта 28. Распространение черного коршуна Milvus korschun
1 — M. k. korschun, 2 —M. к. lineatus, 3 — M. к. aegyptius, 4 —M. к. parasitus, 5 — M. к. arabicus, 6 —
М. к. govinda, 7 — M. к. formosanus, 8 — M. к. affinis; а — границы гнездовий, г —область зимовок»
Казахстане преобладают птицы более или менее близкие к korschun, а в
горном Туркестане — к lineatus. Данные У истлера о том, что в Гималаях —в Кулу
и Курраме — наряду с lineatus встречается на гнездовье и korschun,
указывают, что эта гибридная популяция возможно доходит и до Гималайской
горной страны. На Кавказе и в Закавказье, в М. Азии и с.Иране, в Туркмении
и Таджикистане (кроме Памиро-Алая), в Афганистане распространен
европейский черный коршун. Зимою в Африке, главным образом в тропической,
к югу до Овамбо и Замбези, редко южней, и на Мадагаскаре. Кроме того,
в Передней Азии — в Месопотамии, Иране, Белуджистане, на север до Сеи-
стана, Хорассана, Загроша и южнокаспийских провинций, в Белуджистане;
у нас в небольшом числе и более или менее случайно на Атреке. Указание на
ЧЕРНЫЙ КОРШУН
223
зимовки этой формы в Индии требует проверки — оно скорее относится к особям
смешанных популяций.
Характер пребывания. Перелетная птица, как и другие
виды умеренной полосы, связанные с водоемами. Оседлы ли южные (иранские)
популяции или они заменяются там зимою прилетевшими с севера, а сами
передвигаются к югу — не ясно. Весеннее движение коршунов в гнездовую
область медленное и связано с освобождением водоемов от ледового покрова.
Осенью перелет растянутый, так как разные популяции, как и весною,
пролетают в разное время. Уже в начале марта коршуны появляются на юге
Средней Азии, в разные числа марта идет пролет на Кавказе, в Бессарабии,
вюжн. Украине (одиночки еще ранее —23.11 у Полтавы, Гавриленко, 1929).
Северные популяции, однако, летят значительно позднее, и коршуны в Африке
и Месопотамии встречаются еще в апреле (отдельные особии летуют). В среднее
полосе коршуны появляются около середины апреля, на севере—во второй
половине этого месяца (18—22. IV, окрестности Кирова, у Молотова с 14. IV, там же
25. IV; на Лозьве под 60° 20. IV, у Чердыни 6.V.; у Миаса в начале мая). Позднее
появление в Зауралье связано с тем, что водоемы там оттаивают поздно.
Отдельные особи задерживаются на пролете весь май. Летят коршуны весною на юге
небольшими стаями и одиночно.
Осенний пролет также разновременный: первые птицы в Месопотамии
появляются уже в конце июля, но в большом количестве коршуны по*
являются там в октябре. На севере и в средней полосе отлет коршунов
происходит в конце августа — начале сентября, южнее птицы задерживаются
дольше, до начала октября (Днепропетровск; Бессарабия; дельта Волги;
Таджикистан; Туркмения). В ю.-в. Закавказье коршуны держатся до ноября,,
но в декабре отлетают к югу (после 20. XII попадаются немногие самцы.
Тугаринов и Козлова, 1938). На отлете коршуны держатся стайками и чем
далее к югу, тем многочисленнее становятся эти стаи (иногда громадные —
в Мухранской долине в Закавказье в тысячу особей, Баньковский, 1907;
в Дагестане — до 800, Беме, 1926).
Биотоп. Коршун нуждается в деревьях для гнездования с одной
стороны, и в воде, обитатели которой имеют большое значение в питании детей
птицы, — с другой. Местами синантроп, хотя в меньшей степени, чем
восточный подвид. Встречается как на равнинах, так и в лесном поясе гор (на Урале
поднимается на высоту до 750 м, на Кавказе до 3300 м, но обычно ниже; в
Таджикистане до 1600—1800 м; в Копет-Даге до 2200 м, в Б. Балханах —*
до 1618 м, в Зеравшанском хребте от 1160 до 3300 м).
Численность. В силу особенностей биотопического
распространения, несколько спорадичен. Наиболее обычен в пойменных долинах больших
рек средней и южной полосы. У сев. границы ареала редок (примерно
от 54—55°), так как вообще избегает глухой тайги. Местами в средней
полосе число коршунов уменьшилось в результате вырубки подходящих
для гнездовий деревьев или быстрого роста городских поселений (так,
около 15 лет тому назад прекратила свое существование гнездовая колония
коршунов в Сокольниках под Москвой). У коршуна, как и у других полифагов,
периодических колебаний численности не установлено.
Экология. Размножение. Летом холостые особи, обычно в стайках,
заключающих до 150 особей, встречаются и вне гнездовой области, при этом
среди таких стай есть и взрослые особи, и птицы, еще не надевшие окончатель-*
ного наряда (Самородов, 1935). Впрочем достоверных случаев встречи у гнезд
птиц в первом годовом наряде неизвестно. Можно во всяком случае полагать,
что по крайней мере большинство птиц в неокончательном наряде не
размножается. Брачные игры начинаются с прилета (фигурный полет обоих партнеров
и характерная трель самца).
224
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Гнезда нередко располагаются группами, гнездовые участки пар слабо
ограничены или даже совсем не ограничены; иногда гнезда можно встретить
среди колоний цапель (вероятно, в связи с обилием пищи в виде дохлых
птенцов, отбросов и т. д.). Расстояние между гнездами иногда метров 100(Средний
Урал). Для Поволжья плотность указывается от 0,05 до 0,8 гнезд на гектар
(первая цифра для Пензенской обл., Артоболевский, 1924, вторая—для
Куйбышевской, Карамзин, 1901). В местах, бедных древесной
растительностью,— несколько гнезд размещаются на одном дереве (Туркмения). Гнезда
строит обычно сам коршун, значительно реже он пользуется старыми
постройками других птиц (сарычей, серых ворон). К постройке или исправлению
старого гнезда коршуны приступают в конце апреля, с прилета. Одно и то же
гнездо служит несколько лет (до б годовых слоев подстилки, Чкаловская
обл., Зарудный).
Расположены гнезда чаще всего на деревьях, близ опушек или «речных
долин; на юге изредка на скалах (например, у Ходжента; в Туркмении, в
вост. Иране). На севере коршун обычно гнездится на соснах, на юге на
дубах, тополях, березах, карагачах, ивах, чаще всего на средней высоте
около 10м (но и до 21 м), иногда впрочем низко (3 м—дельта Волги; 4 м^
Башкирия, 6 м — Зауралье). Размеры гнезд различны, в среднем около 50—
70 см в диаметре и около 30—40 см высотой, снеглубоким лотком, выстланном
тряпками, шерстью, остатками пищи, навозом, кошмой, клочками бумаги, сухой
травой и т. п. В стенках гнезд коршуна иногда поселяются полевые воробьи.
В последней трети апреля —начале мая происходит кладка; разница в
сроках кладки в зависимости от широты не установлена. В Бессарабии полная
кладка бывает в начале мая, у Умани — в конце апреля и начале мая;
в 20-х числах апреля в дельте Волги; слабо насиженные яйца найдены около
середины мая на вост. Кавказе; в низовьях Сыр-Дарьи полная кладка
обнаружена 31. IV; 15—18.IV — в ю.-в. Туркмении; в начале мая — у Рязани,
в Зауралье, Куйбышевской, Казанской, Московской и Тамбовской обл.
и т. д. В эти же сроки —в Средней и Зап..Европе. В случае гибели кладки
происходит дополнительная. Это производит впечатление растянутости периода
размножения, чего на самом деле нет. Яйца с бурыми пятнами и черточками по
белому фону. Размеры: (294) 49,5—60x39,5—47, в среднем 54,7 х 43,4 мм
(Умань, Гебель, 1879); (12) 51—58,6 х41, 5— 47 (Харьков, Сомов, 1897);
54—59 х 42,6— 44 мм (Сыр-Дарья, Спангенберг, 1936). Число яиц в кладке 2—4,
редко 1 или 5. Промежуток между откладкой яиц повидимому 2—3 дня, судя
по разнице в возрасте птенцов. Насиживание начинается с откладки
первого яйца.
Участие самца в насиживании твердо не установлено, но во всяком случае
невелико. Период насиживания около 30 дней. Птенцы появляются в конце
мая —начале июня. Одно из яиц в кладке обычно болтун. Проклевывающемуся
птенцу помогает освободиться от скорлупы самка. Разница в вылуплении
2—3 дня. Смертность птенцов значительна: чаще гибнет младший птенец,
иногда в результате каннибализма. Птенцы оперяются в возрасте 25 дней и
покидают гнездо в возрасте около 6 недель, держатся первые дни близ гнезда.
Летные молодые встречаются в разные числа июля. Выводки держатся вместе
до отлета, когда образуются стаи. Осеннее возбуждение у самцов — брачный
крик и т. п., т. е. неполный повторный половой цикл—отмечен в ноябре в ю.-в.
Закавказье (Тугаринов и Козлова, 1938).
Линька. Птицы в первом годовом наряде начинают линять весной
с прилета, в апреле у них сменяется мелкое оперение. Линька взрослых
начинается в конце гнездового периода в июне — июле, кончается
повидимому после отлета, на зимовке. Прилетные весной старые птицы находятся
уже в свежем пере и не имеют следов линяния.
ЧЕРНЫЙ КОРШУН
225
Линька полная годовая. Последовательность смены маховых от заднего
края партии (10-го) к переднему (1-му). Смена нарядов: первый пуховой
наряд — второй пуховой наряд — первый годовой (гнездовый) наряд — второй
годовой (промежуточный) наряд —третий годовой (окончательный) наряд и т. д.
Питание. Коршун — полифаг, с известной наклонностью к
паразитизму: охотно питается падалью, всякого рода отбросами, а также иногда
отнимает добычу у других птиц (например, в СССР указывается это относительно
балобана). Этим же вероятно объясняется его синантропия, впрочем особенно
сильно выраженная у других подвидов в Индии, Китае и Африке. Близ
поселений коршун кормится отбросами на неблагоустроенных бойнях и на свалках,
Рис. 62. Черный коршун (взрослая птица).
в местах массового выпаса скота (например, в Туркмении) падалью, последами
объягнившихся овец и другими отбросами скотоводства. Возможно, что по
этим же причинам распространение коршуна в значительной мере связывается
с водоемами, где обычно имеются выбросы, снулая рыба и т. д. Так,
указывается, что небольшая гнездовая колония в Башкирии образовалась там, где
рыбаки выбрасывали рыбу—отход ловли (Сушкин, 1897). Активное
нападение на добычу для коршуна мало доступно — на это указывают его слабые
лапы с короткими пальцами: он в состоянии схватить только небольших
животных.
Способы полета и розыска добычи сближают коршуна в известной мере
с настоящими падальниками — грифами: коршун хорошо парит, обладая при
относительно легком весе очень длинными и широкими крыльями и большим
хвостом. Охотится коршун в открытых местах, медленно облетая большие
участки, на высоте примерно около 70—100 м. Добычу хватает лапами, причем
за рыбой иногда ныряет с разлета по-скопиному.
Охотятся коршуны иногда далеко от гнезда, километров за 5—6, причем
у отдельных пар охотничьи участки не ограничены так строго, как у других
хищных птиц. Вылетает на охоту вскоре после рассвета, в середине дня коршун
отдыхает, а под вечер с 16—17 часов вновь охотится.
15 Птицы СССР, т. I
226
ОТРЯД ХИЩНЫЕ ПТИЦЫ
В качестве кормов коршунов в СССР указываются: различные грызуны
(роль их повышается в годы массового размножения), в частности суслики,
тушканчики, слепушонка, степная пеструшка, полевки, водяная крыса, хомяк,
мыши, черная и серая крысы; землеройки, ласка и горностай (падаль?),
ушастый еж; разные молодые птицы, или мелкие виды, держащиеся и
кормящиеся на земле: жаворонки, овсянки (дубровник, обыкновенная, садовая,
камышевая), чечевица, зяблик, дрозды, грачата, кукушки, горлица,
перевозчик, чибис, кроншнеп, молодые чайки и крачки; птенцы цапель, утята,
в том числе кряквы и
чирки; цыплята;
пустельга, чеглок; в
значительной мере эти
* птицы—птенцы или
подлетки или падаль, или
отняты от других
хищников; рыба небольших
размеров, главным
образом снулая (или сильно
пораженная лигулезом);
различные
пресмыкающиеся и земноводные —
прыткая ящерица,
водяной уж, лягушки и
головастики; из
беспозвоночных —моллюски,
прудовики и беззубки; из
насекомых—саранчевые,
водяные жуки и др. В
качестве корма коршуна
на юге (Туркмения, вост.
Иран) указываются,
кроме падали и отбросов,
молодые водяные птицы,
молодые кеклики и
пустынные куропатки,
голубята, мелкие зверьки;
ящерицы, рыбы,
насекомые, в том числе саранча;
речной краб (в Конет-Даге). Полифагия коршуна выступает достаточно ясно,
но при этом следует иметь в виду его предпочтение падали и всякого рода
отбросам. С**4 -
Полевые признаки/ Темнобурая средней величины хищная птица с
длинными и широкими крыльями и длинным вырезанным по заднему краю хвостом.
Часто парит и летает на кругах. Голос — мелодичный свист, трель, также
частое «ки—ки—ки».
Описание. Размеры и строение. Крылья длинные, вырезки на наружных
2-го — 4-го маховых и внешних опахалах 1-го — 5-го маховых; формула крыла: 4^3>
5>2>б>7> 1. Хвост длинный и выемчатый. Клюв с длинным крючком. Ноздря косо
расположенная, щелевидная. Плюсна короткая, оперенная примерно до половины, покрытая
спереди поперечными щитками; пальцы короткие, когти острые. Длина крыла самцов (45)
410—4С0, самок (44) 437—505, в среднем 453,6 и 464,6 мм.
Окраска. Первый пуховой наряд рыжевато-бурый, с желтоватым брюшком, с
темным пятном у глаза; на темени пух удлинен. Второй пуховой наряд сероватый с рыжеватым
оттенком на брюшной стороне.
Первый годовой наряд темнобурый с охристыми пятнами на вершинах перьев, черно-
бурыми, окаймленными охристыми поперечнополосатыми рулевыми; маховые черно-бурые
Рис. 63. Черный коршун (молодая птица).
ЧЕРНЫЙ КОРШУН
227
со светлыми сероватыми основаниями внутренних опахал. Промежуточный (второй
годовой) наряд с остатками охристых пятен. В окончательном (третьем годовом) наряде
общая окраска темнобурая, с более светлой бледнобурой (в обношенном пере беловатой)
головой, с темными на ней наствольями; горло беловатое, грудь бурая; на брюхе, штанах,
подхвостье обычно рыжеватый оттенок; спинная сторона тела темнобурая, маховые темно-
бурые со светлыми сероватыми полями на основаниях внутренних опахал; рулевые бурые
с темным поперечным рисунком; вся брюшная сторона с темными наствольями. Оба пола
во всех возрастах окрашены одинаково.
Радужина бледнобурая или желто-бурая; клюв и когти черные; восковица, разрез
рта и лапы желтые.
' Сибирский черный коршун Milvus korschun lineatus Gray
Haliaetus lineatus Gray in Hardwicke's Illustr. of Indian Zoology v. I, 1832, стр. I,
табл. 18, Китай.
Синонимы. Milvus melanotis. Temminck et Schlegel. Fauna Japonica,
Aves, 18-44, стр. 14, табл. V, V-B, Япония. — Milvus melanotis ferghanensis
Бутурлин, Наша Охота, 1908 (май), стр. 69, Лянгар к югу от Гульчи. — Milvus korschun tiansha-
nieus. Бутурлин. Определитель видов птиц СССР, вып. I, 1928, стр. 73, Нарын.
Распространение. Ареал. К востоку от предыдущей формы—в Сибири от
бассейна Оби и Иртыша до Приморья; Японии, Манчжурии и Китая (к югу
до Фучжоу и Гуаньдуня), сев. Тибета и сев. Гималаев (Кашмир), Ладака,
Каракорума; на западе в Кашгарии и Тянь-Шане (см. ниже). Северная граница
в бассейне Оби у Тобольска, на Енисее под 61° у Енисейска, на Вилюе
до 65°, на Лене у Якутска, на Алдане до 63°30' с. ш., далее к востоку на Шан-
тарских о-вах (Дулькейт и Шульпин, 1937). Один раз добыт на Камчатке
(Шульпин, 1936), встречен на Сахалине и на Курильских о-вах (гнездится
на Кунашири). Зимовки в южн. Азии—в Индии к югу примерно до 18°
с. ш., в Бирме, Южн. Китае, на п-ове Хайнань, Индо-Китае и о. Формоза.
Характер пребывания. Перелетная птица (возможно, что на
юге ареала делает вертикальные миграции).
Даты. Период пролета растянут, как у европейского подвида. В южн.
Приморье и Туркестан прилетает уже во второй половине марта; в
Кашгарии первые птицы появляются уже в конце февраля (23.11), валовой пролет
падает на середину марта. Пролет северных особей идет позднее и на юге, до
последней трети апреля. В разные числа апреля коршуны появляются у
северной границы гнездового ареала (у Якутска 8—29. IV, в Зап. Сибири 2. IV —
27. IV и даже I.V; на Шантарских о-вах только в начале мая). Отлет
начинается уже в августе и тянется до конца сентября на севере (последние у
Якутска встречены еще З.Х; на Дальнем Востоке на Бурее с 22. IX по 1.Х,
последние еще 17.X; в Приморье на р. Самарга добыт 14.XI; на Курильских
о-вах попадается до конца ноября). По некоторым наблюдениям, южные
популяции отлетают .раньше, северные пролетные задерживаются в тех же
местах позднее (в бассейне р. Или отлет местовых с 17.VIII, пролет северных
с 25.VIII и до середины сентября, Шестоперов, 1929). Залеты в зап. части
Средней Азии (Туркмения). *
Биотоп. Как и для европейской формы характерные^ биотопические
условия —сочетание древесной растительности и водоемов; впрочем в горах
на юге в Средней и Центральной Азии связан и с горно-степным ландшафтом.
В горах подымается в Тянь-Шане до 3150 м (Семиречье), на Алтае до 2000 мУ
в Тибете от 600 до 4700 м.
Численность. В южной части ареала обычен, на севере редок.
Распространение в связи с биотопическими условиями спорадично, так как
в глухой тайге коршуна нет. Отмечено негнездование, несмотря на обилие
пищи (Танну-тува, Сушкин, 1913), или оставление родителями кладки (вост.
Китай, Кольтхофф, 1932).
15*
228
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Экология. Размножение. В отличие от европейского коршуна,
имеется разница в сроках гнездования северных и южных популяций. На юге
ареала кладка отмечена уже в конце апреля или даже несколько ранее (в вост.
Китае, в Кашмире). В Средней и Центральной Азии коршун несет яйца
позднее, во второй половине апреля. Наконец, на севере кладка производится
в конце апреля и начале мая (в Сибири, в частности, у Якутска в первой
половине мая, Иванов, 1929, Воробьев, 1931, тогда же у Томска, Залесский, 1915).
Гнезда — как у номинальной формы; по наблюдениям в Китае строит их
самка, самец приносит строительный материал. Расположены гнезда главным
образом на деревьях (ель, лиственница, сосна, тальник, береза, дуб и др.),
на различной высоте от земли, от 6 м и выше, обычно на 10—12 м от земли;
размеры гнезд различны — от 30 см в диаметре (Семиречье) до метра и несколько
более (Монголия, Китай). Выстилка в Туркестане —тряпки, кошма, конский
навоз и т. п. В горных местностях гнездится и на скалах (Алтай, Тибет).
i Кладки состоят из 2^3 яиц, редко из 4 (Якутск, Воробьева, 1931). Размеры
яиц крупнее, чем у европейского черного коршуна: 56,3—64 х 45—47,5
(Вост. Сибирь, Тачановский, 1891), 53—61 х 44—46 (Забайкалье, Радде, 1863),
54—57 х44—43 (Семиречье, Зарудный и Кореев, 1906), 55—63 х41— 48
(зап. Китай, Штреземанн, 1938), 65—66 х 44,5—45,9 мм (Тибет, Шефер, 1938).
Насиживание длится около месяца, так как пуховики в Сибири встречены
в начале июня (Якутск 10.V — неполная кладка, 12.VI вылупились птенцы;
у Томска 18.V. 2 яйца, 12.VI первый пуховик; в зап. Китае соответственно
ранее, первые птенцы вылупились 14.VI). Летные птенцы и оперенные
гнездари встречаются со второй половины июля, запоздавшие и в первой
половине августа.
Линька. Изучена недостаточно, общее ее течение как у европейских птиц.
Питание. Полифаг, кормовой режим в общем как у европейского
коршуна. В Центральной Азии и Китае — типичный синантроп, гнездящийся
в городах и кормящийся главным образом отбросами, в Монголии даже
человеческими трупами. Падаль, снулая рыба — едва ли не главная пища коршуна
в Сибири. С другой стороны, он, повидимому, более энергичный хищник, чем
европейский подвид. Из птиц в качестве добычи коршуна в Сибири
указываются мелкие наземные виды — коньки и жаворонки (Танну-тува), молодые
черные вороны (Семиречье), домашняя птица, также чайки, бакланы, старик,
чистик Cepphus carbo и др. (Приморье); кроме того, мелкие звери —хомяки,
зайчата, мыши; змеи и лягушки; крупные насекомые — саранчевые и
жуки и т. д.
Полевые признаки. От европейского коршуна отличается большей
величиной и белым пятном у оснований маховых в подкрыльях, бросающимся
в глаза на лету. Крик как будто тише и короче, чем у европейских птиц
(Тугаринов и Бутурлин, 1911).
Описание. Размеры и строение. Крупней европейской расы. Крыло самцов
(28) 450—505, самок (20) 475—515, в среднем 476,7 и 489,1 мм. Крыло несколько тупее;
2-е маховое обычно короче его, редко равно ему, а 1-е маховое ближе к 8-му, чем к 7-му
(у номинальной формы соотношения обратные).
Окраска. Старые птицы более равномерного и темного бурого цвета, чем
европейские, голова бурая, лишь немного светлей спины; брюшная сторона без рыжеватого оттенка,
у основания внутренних опахал первостепенных маховых белое или белое с темными
«мраморными» пестринами поле, образующее при полете на нижней поверхности развернутого
крыла белое зеркальце; кроющие уха черноваты. В первом годовом наряде
охристо-беловатые каймы перьев очень развиты, а брюшная сторона с бледноохристыми в свежем пере и
беловатыми в обношенном пере наствольями. В этом наряде восковица и ноги голубоваты или
беловаты. Второй годовой (переходный) наряд лучше выражен, чем у европейского
подвида, — светлый продольный рисунок на брюшной стороне резкий.
Систематические замечания. Коршуны из Тянь-Шаня и прилежащих местностей к
западу до Ферганы и Памира, как отмечалось выше, образуют повидимому смешанную
популяцию с признаками, промежуточными между европейскими и восточно- и центрально-
КРАСНЫЙ КОРШУН
229
азиатскими птицами. Промежуточны как размеры, так и окраска. Наравне с такими
особями попадаются и птицы, внешне типичные lineatus и очень близкие или идентичные с
korschun.
21. Красный коршун Milvus milvus L.
Falco milvus. Linnaeus.
Распространение. Ареал.
Скандинавии (примерно до 60—61°
М. Азия и сев. Иран; с.-з. Африка,
о-ва Зеленого Мыса и Канарские.
Характер пребывания.
На севере перелетная птица. У
Средиземного моря оседлая. Зимует в
Испании, Италии, Греции, в
Африке к северу от Сахары. Биотоп.
Высокоствольные леса, охотно
вблизи культурного ландшафта.
Главным образом на равнинах.
Подвиды и варьирующие признаки.
Два подвида: на о-вах Зеленого Мыса
М. т. fasciicauda H a r t e r t, 1914, в
остальной части ареала М. /п. milvus
L. Морфологические различия в
развитии поперечного рисунка на хвосте, в
меньшей его выемке, в менее ярком тоне
ржаво-красного цвета на спинной и
брюшной стороне у fasciicauda. Эта раса,
таким образом, в известной мере сближает
красного и черного коршуна.
Экологические отличия номинальной формы и
fasciicauda слабо выяснены (первая главным
вторая оседлая, гнездится на скалах и т.
Sys. Nat. изд. X, v. I, стр. 89, Швеция.
Средняя и южн. Европа к северу до южн.
с. ш.), к востоку до Украины; Кавказ,
II v .i
да^ш
$ж&/1
?С^я
^У Л1
1 P^—h 1
Ш г
Карта 29. Распространение красного коршуна
Milvus milvus
7 — М. m. milvus, 2 — M. m. fasciicauda; a — границы
гнездовий, г — область зимовок.
образом
д.).
перелетная, гнездится" на деревьях,
Обыкновенный красный коршун Milvus milvus milvus L.
Синоним. Milvus milvus caucasicus. Бутурлин. Сист. заметки о птицах Сев.
Кавказа, 1929, стр. 13, Сочи.
Распространение. Ареал. В Зап. Европе спорадичен в Англии (Уэльс),
гнездится в Испании и Португалии, на Балеарских о-вах, в вост. и южн.
Франции, на Корсике, Сицилии и Сардинии, в Италии, Швейцарии,
Германии, в Швеции до 60—61°, в Польше, Чехословакии, Югославии, на
Балканах (Сербия, Македония, Румыния), в М.Азии и быть может в Палестине;
в Африке в Марокко, Тунисе, Алжире, на Канарских о-вах. В СССР
в Латвии и возможно в Эстонии на севере, в Белоруссии отмечен
в Беловеже; на Украине в Подолии и Киевской обл., может быть в южных
частях Черниговского Полесья (Шарлемань, 1937); кроме того, на Кавказе,
где на западе встречается на юг от Сочинского района (Хоста), к северу по
крайней мере до Телава (Чхиквишвили, 1930), к востоку до Шуши. Залеты
отмечены у Пскова и Изборска (июнь — июль), в Харьковскую и
Полтавскую обл. (июль), по требующим подтверждения и весьма- сомнительным
старым сведениям в центральной части страны до Тулы и Орла. Залеты отмечены
также на Кавказе —до Дзауджикау на сев.-востоке. Наконец, залетает в
низовья Атрека в Туркмении (Зарудный, 1911), очевидно из Астрабадской
провинции Ирана, где гнездится и находит восточный предел
распространения.
Характер пребывания. В СССР перелетный, оседлый в Англии
и у Средиземного моря, где расположены зимовки. Вероятно оседлый в Закав-
230
отряд хищные птицы
казье (добыт зимой у Ленкорани; в долине Риона, в Телаве). Редкие случаи
зимовок приводятся и для средней Европы.
Даты. Прилет ранний, отлет поздний, в Зап. Европе в феврале — марте
и в сентябре—октябре. В Беловеже добыт 30. IV, 2—З.У(Цедлитц, 1920). В
Киевской обл. прилетает в конце марта, отлетая оттуда в начале октября.
Биотоп. В СССР в лиственных или смешанных старых лесах,
перемежающихся с культурным ландшафтом или открытыми пространствами.
В Закавказье в горах на высоте до 2100— 2400 м.
Численность. В Зап. Европе численность заметно надает: почти
исчез в Англии, не гнездится более в Голландии и Дании. Для Франции однако
отмечается некоторое увеличение числа коршунов за последние два десяти-
Рис. 64. Красный коршун.
летая (анкета в «Alauda» 1932—1937). Коршун нередок в Испании, где местами
образует большие гнездовые колонии. В СССР очень редок.
Экология. Размножение. Гнезда в Киевской обл. располагаются
на опушках леса —«на дубах и липах или соснах, обычно на горизонтальном
суку, реже в развилке. Строятся гнезда из сучьев, с выстилкой, как обычно
у коршунов, из шерсти, тряпок, бумаги и т. п. Занимаются несколько лет подряд
(до 10) и иногда строятся самим коршуном, иногда используется постройка
других птиц (ворона, канюка). Размер гнезда небольшой, от 400 до 850 мм
в диаметре, высотой 200—500 мм, с лотком в 200 мм (Шарлемань). Яиц 2—3,
чаще 2. Свежие кладки найдены во второй половине апреля, при гибели
одного яйца оно заменяется. Окраска яиц беловатая с более или менее
развитым бурым рисунком, в общем сходная с цветом яиц сарыча. Размеры (34)
51,5—61,5x43—57,4 мм (Умань, Гебель, 1879). Вылупление происходит во
второй половине мая (19. V у Киева обнаружены 2 пуховика и яйцо с готовым
к вылуплению птенцом), насиживание, следовательно, длится около месяца.
Впрочем гнездовый период изучен слабо. Выводки из 4 птиц наблюдались
в августе (Владимир-Волынский).
КРАСНЫЙ КОРШУН
231
Линька. Повидимому, поздняя (с октября по февраль, Нитхаммер,
1938; из СССР материалов нет).
Питание. В общем, сходно с питанием черного коршуна, но менее
связано с водоемами: падаль, беспозвоночные, мелкие позвоночные. На
Кавказе в период пролета ловит перепелов и коростелей. У гнезда в окрестностях
Киева найдены остатки полевки Clethrionomys glareolus, вяхиря и уклеи.
Полевые признаки. Похож на черного коршуна, но крупней, с более
глубоко вырезанным хвостом и более светлой красновато-рыжей окраской, голова
на расстоянии кажется беловатой. Голос «хияя-хияя-хияя» или трель «хи-хи-
хи-хии».
Описание. Размеры и строение. Крупней черного коршуна; размеры полов
примерно сходны. Крыло у птиц из СССР (4) 480—520, в среднем 500 мм. У
западноевропейских коршунов (по Нитхаммеру, 1938) длина крыла самцов 475—500, самок 480—530 мм.
Выемка хвоста у кавказских 71—95 мм (Штегман, 1937). Формула крыла 3^2>5>6>1>7;
вырезки на наружных опахалах 2-го — 5-го маховых, на внутренних опахалах 1-го —
4-го маховых, внутреннее опахало 5-го махового сужено.
Окраска. Пуховой наряд из СССР неизвестен. Западноевропейские в первом
наряде рыжевато-белые на спинной стороне, с удлиненным пухом на темени, с черным пятном
у глаза, охристо-белые на брюшной стороне; во втором пуховом наряде белые с более или
менее рыжеватым налетом на спине.
Первый годовой наряд: темя бурое с буроватыми наствольями и беловатыми крапинами
у вершин; спинная сторона красно-бурая с бледноохристыми пятнами и каймами у вершин
перьев; брюшная сторона бледнорыжая с охристо-беловатыми наствольями; первостепенные
маховые черные с большим белым полем у основания внутренних опахал; рулевые рыжие,
боковые пары черноваты у вершин, все перья с более или менее заметным темнобурым
поперечным рисунком.
Окончательный наряд: темя беловатое, в свежем пере рыжеватое, с черными
наствольями, перья затылка удлинены и заострены; спинная сторона темнобурая с черными
стволами и рыжими краями перьев, особенно широкими в межлопаточной области и на
кроющих крыла; лоб белый, щеки и горло беловатые с черными продольными штрихами,
остальная брюшная сторона ржавчато-рыжая с темными наствольями, на груди широкими,
далее суживающимися; маховые и рулевые как у птиц в первом наряде, но белое поле у
основания первостепенных маховых менее развито. Этот наряд быть может связан с первым
годовым промежуточным, выраженным не резко и отличающимся от окончательного менее
беловатой головой.
Клюв у взрослых ..желтоватый, у птиц в первом годовом наряде черный; радужина
белая или бледножелтая; восковица и лапы желтые; когти черные.
РОД ЧЕРНОКРЫЛЫЕ КОРШУНЫ ELANUS SAVIGNY, 1809
Тип Falco caeruleus Desfontaines
22. Чернокрылый коршун Elanus caeruleus Desfjonjtaines
Falco caeruleus. Desfontaines. Hist. (Mem.) Acad. Royale d. Sciences (Paris),
1787 (1789), стр. 503, табл. XV, Алжир.
Распространение. Ареал. Африка от Марокко и Алжира, Египта и
Капской провинции; Мадагаскар; Португалия, Индия, Цейлон, ю.-з. Китай
(Юннань) и Индо-Китай, Филиппины, Большие Зондские о-ва, архипелаг
Сула. Известны залеты в Европу (до Бельгии и Германии), в Туркестан.
Характер пребывания. Оседлая птица.
Биотоп. Местности с древесной или кустарниковой растительностью,
культурный ландшафт.
Подвиды и варьирующие признаки. В Африке Е. с. caeruleus Desfontaines;
в Индии до Бирмы, Ассама ю.-з. Китая, на Цейлоне Е. с. vociferus L a 11 h a m, 1790; на
Филиппинских, Зондских о-вах, на архипелаге Сула Е. с. hypoleucus Goal d, 1859.
Варьирующие признаки — величина и окраска.
Индийский чернокрылый коршун Elanus caeruleus vociferus Lath.
Falco vociferus. Lattham. Index Orn. 1, 1790, стр. 46, Коромандельский берег, Индия.
Распространение. Ареал. Индия от Гималаев до Цейлона и
Лаккадивских о-вов; на восток до Бирмы, Индо-Китая, Юннани; в апреле 1872 добыт
232
ОТРЯД ХИЩНЫЕ ПТИЦЫ
у гнезда в пров.Чжецзянь в вост. Китае (Латуш, 1932). Взрослый самец, по-
видимому залетный, добыт 19. IV 1929 г. у Термеза на Аму-Дарье (Иванов,
1940). Характер пребывания, биотоп — см. выше.
Карта 30. Распространение чернокрылого коршуна Elanus caeruleus
7 — Е. с. vociferus,r2— Е. с. hypoleucus, 3 — Е. с. caeruleus,
а — граница гнездовий.
Численность подвержена колебаниям, так как в Индии отмечаются
случаи массового появления чернокрылого коршуна в таких местах, где
обычно эта птица редка: например,
в 1877—1879 г. в Пуна, в 1902-^
1904 г. в Гуджерате, в 1899 г. в
Бехаре (Бекер, 1928).
Экология. Размножение.
По наблюдениям в Индии
возможно, что цикл размножения
типичный тропический, календарно не
фиксированный, так как кладки
были найдены в различные времена
года: в разных местностях в
ноябре — январе, в Раджпутане в
августе, в Гарвале с апреля до июня
(Юм)*. Гнезда на невысоких
деревьях, близ вершин; кладка
состоит из 3—5 яиц, с красно-бурыми
пестринами по беловатому фону;
средние размеры их 39,3 х 30,9 мм
(Бекер).
Линька не изучена.
Питание. Насекомые,
грызуны, крабы, мелкие птицы. В
общем полифаг, сходный в этом
отношении с настоящими коршунами.
Описание. Размеры и строе-
Рис. 65. Чернокрылый коршун. ни е. Некрупная птица, с самку пере-
ОРЛАН БЕЛОХВОСТ
233
пелятника или пустельгу, но крылья длинней: у самцов 255—268, у самок 262—268 мм
и до 272 мм (Бекер, 1928, Суани, 1935). Самое длинное маховое — второе. Клюв круто
загнут, с длинным крючком на вершине. Лапа короткая с короткими пальцами, покрытая
мелкими сетчатыми пластинами. Хвост короткий со слабо выраженной выемкой на вершине.
Окраска. Сходна у обоих полов. Первый годовой наряд: на спинной стороне
<:еро-бурый со светлыми охристо-рыжеватыми каемками, брюшная сторона охристая, часто
с продольными полосами на груди и серовато-бурым налетом. Взрослые птицы
пепельно-серые на спинной стороне тела, с черным пятном на уздечке и под глазом, с черными
малыми и средними кроющими крыла; маховые с буроватым оттенком на внутренних
опахалах и у вершин; средние рулевые серые, боковые белые, промежуточные пары серы
на наружных опахалах. Брюшная сторона белая, иногда со слабым сероватым оттенком
на груди и боках. Радужина красная у взрослых, желтоватая у молодых; восковица и
лапы желтые, когти и клюв черные.
РОД ОРЛАНЫ HALIAEETUS SAVIGNY, 1809.
Тип haliaeetus nisus=Falco alb kill a.
Синонимы. Thalassoaetus Каир, 1844, тип Т pelagicus; Cuncuma Hodgson,
1837, тип С. albipes = H. leucoryphus.
23. Орлан белохвост Haliaeetus albicilla L.
Falco albicilla. Linnaeus. Syst. Nat. изд. X, 1758, стр. 89, Швеция.
Русское название. Орлан — слово, повидимому, книжное. В народном
языке орланы едва ли различаются от орлов. Белохвост, как имя этой птицы, впервые
встречается в XVIII веке у Палласа, но взято из живого языка. Эпитет белохвост, белохвостый,
прилагается к этой птице и в других европейских языках. Слово орлак, встречающееся в
русских памятниках XVII века («Урядник Сокольничья Пути»), относится повидимому
к малым видам орлов (подорликам). У народов Туркестана «су-буркут», т. е. водяной орел.
Распространение. Ареал. Сев. Азия от тундр до Японии, Китая (до
-Цзяньсу, 28 с. ш.), Монголии, низовьев реки Или, Аральского и Каспий-
Карта 31. Распространение белохвоста Haliaeetus albicilla
7 —НЛа. albicilla, 2 — Н. a. groenlandicus; а—границы гнездовий, б—-невыясненная граница
распространения, г — область зимовок и кочевок, ж — местности, где орлан исчез в XIX—XX веке.
ского моря, сев. Ирана, Малой Азии. Европа от крайнего севера (Финмаркен,
Лапландия) до Румынии, Балкан, Венгрии, побережий Балтийского моря;
Корсика и Сардиния, Гебридские и Шетландские о-ва; Исландия и
Гренландия. Зимою за пределами гнездовой области в разных европейских странах, в
Азии к югу до с.-з. Индии и Китая, также и в сев. Африке (где ранее
гнездился в Египте и у берегов Красного моря).
Характер пребывания. Оседлая и кочующая птица, на севере
перелетная. Биотоп. Побережья рек, озер, морей, главным образом там,
234
ОТРЯД ХИЩНЫЕ ПТИЦЫ
где есть высокие деревья, удобные для гнездования, или скалы (в
частности на островах севера).
Подвиды и варьирующие признаки. Орлан белохвост образует два подвида: в
Гренландии на зап. берегу от м. Фаруэлл до Диско (однажды гнездо найдено на земле Кембер-
ленд, Девисов пролив, Кемлин, 1879) — Н. a. groenlandicus С h г. L. В г е h m, 1831 и Я. а
albicilla L. из других частей ареала. Первый подвид крупней. Экологических различий
между этими формами повидимому нет. Очень близок к белохвосту американский
белоголовый орлан.
Обыкновенный орлан белохвост Haliaeetus albicilla albicilla L.
Распространение. Ареал. Область распространения вида, кроме
Гренландии. В СССР на севере встречается регулярно в лесотундре, но и в тундре
там, где по долинам рек имеются высокие деревья: в Лапландии до 70° с. ш.
(западная Норвегия), в Тиманской тундре (кочевки?), на Новой Земле и на
Вайгаче, на Оби у устья, в низовьях Таза, на Енисее по крайней мере до 72°,
на ю.-в. Таймыре около 71—72°, в низовьях Лены, Индигирки и Колымы, на
Анадыре, в Коряцкой Земле и на Камчатке; в средней Сибири быть может еще
севернее, до 75° (Сев. Таймыр). На юг область распространения простирается до
Украины (заповедник Конча-заспа в низовьях Днепра), Крыма, низовьев
Волги, южн. Закавказья (Армения, Ленкорань); южнокаспийских провинций
Ирана; в Ираке; в Туркестане в низовьях Аму-Дарьи, на нижней Или; далее у
Зайсана, на Ачит-нуре, по Теси, в ю.-з. Забайкалье, по Аргуни, у озера
Далай-нор, в области Хингана и на Амуре (на севере у озера Кизи), на
Сахалине и на о. Сикотан. В гнездовое время встречается и в разных странах
Западной и средней Европы, в Передней (М. Азия, Закавказье, Иран) и
Средней Азии, в Японии, Китае. Зимой обычен на Каспийских побережьях, у
Иссык-куля, в степной полосе Европейской части СССР. Кочуют в основном
молодые (неполовозрелые) птицы.
Характер пребывания. В северной части ареала перелетный,
южней оседлый или кочующий.
Даты. Прилет ранний, отлет поздний. Заметный пролет бывает в
Средней Азии в марте (12—20. III на Сыр-Дарье; в разных числах марта на Чу и
у Иссык-куля); в Приморье уже во второй половине февраля и начале марта,
в средней России в марте — апреле. Появление на крайнем севере довольно
разновременно, но главным образом в апреле (2. IV, 29. IV, в среднем 19. IV,
Лапландия; 13. IV, Ямал; 7. IV на Индигирке, под 70°29'с. ш.; у Хатангского
под 71°48' с. ш. будто бы только 25.V; на Анадыре в первой половине апреля).
На Шантарских о-вах отмечен 20. III, в устьях Лунхи на Вилюе только 2.V.
Отлет на крайнем севере проходит преимущественно в октябре, но движение
молодых начинается и ранее. В Гыданской тундре отлетают после исчезновения
гусей (Наумов, 1931). Первые пролетные в средней полосе отмечены около
середины сентября (Чкалов). В конце октября — начале ноября белохвосты
становятся обычными на южнокаспийских зимовках как на западе, в
Азербайджане, так и на востоке, в Туркмении, и держатся тут примерно до конца
марта — начала апреля. Движение белохвостов носит, таким образом, медленный
и растянутый характер, так как в нем участвуют птицы различных популяций
и разных возрастов. Отмечено, что в негнездовый период птицы держатся не
только одиночками и парами, но и группами (до 30—40 птиц, низовья Дона,
Алфераки, 1910, Полтавская обл., Гавриленко, 1929); впрочем такие
скопления имеют место только там, где имеется обилие пищи, например, в местах
массового пролета водоплавающих птиц, у падали при падеже скота и т. п.
Связи кочевок белохвоста со вскрытием и замерзанием водоемов (как это
имеет место, например, у скопы) не установлено, возможно из-за полифагии
орлана. Однако замерзание водоемов влияет повидимому на сезонную смену
ОРЛАН БЕЛОХВОСТ
235
биотопов — переселение орланов с побережий в степь (отмечено еще
Богдановым, 1871). Относительно дальности кочевок орланов дает известное
представление добыча окольцованной у Росситгена на Куришской косе птицы
близ Пскова.
Биотоп. В гнездовый период связан главным образом с близостью
водоемов (морские побережья, речные долины, озера) и наличием подходящих
для гнездования высоких деревьев. В негнездовое время иногда и далеко от
воды, в степи, но не в лесистом ландшафте. В горах на Алтае поднимается
до 1400 м, случайно до 2300 м, в Закавказье до 2000 м (Севан), в центр. Тянь-
Шане до 1500 и*.
Численность. Как и другие крупные хищники, орлан легко
вытесняется человеком и в Европе местами истреблен (Дания, Шотландия, Англия,
Фарерские о-ва, также в сев. Африке в Египте у озера Мензалех), местами стал
очень редок, например в Швеции (всего до 20 пар, Цедлитц, 1922). Реже
местами стал он и у нас на Украине (быть может, только две пары в низовьях
Днепра, Рудинский и Горленко, 1937), в Поволжье. Птица еще
многочисленна на севере в тундре и лесотундре по долинам больших рек (Обь, Енисей
и его притоки, Колыма). На с.-в. Сибири к востоку от Колымы малочисленен,
но южней, на Камчатке, а также в Приамурье—обычен (впрочем и там
численность орлана в последнее время повидимому уменьшается, Шульпин, 1936).
Экология. Размножение. Пары постоянные, взрослые держатся
вместе и на зимовках, но молодые первогодки поодиночке (Туркмения).
Гнездовые участки весьма постоянны, известны случаи, когда одно гнездо
занималось по крайней мере 30 лет подряд, даже на севере — при обилии подходящих
угодий—26 лет (южн. Ямал, Кучерук и Дунаев, 1941). У каждой пары обычно
имеется по нескольку гнезд, используемых поочередно (в низовьях Сунгачи,
Приморье, из 10 гнезд было занято 2). Размер гнездового участка значителен,
хотя иногда, при ограниченности подходящих биотопов, одна пара
располагается близко от другой (на Енисее были обнаружены 5 жилых гнезд на
протяжении 80 км, Тугаринов и Бутурлин, 1911; в Харьковской обл. 8 пар на 400 км2,
Аверин, 1910).
Гнезда располагаются на деревьях, в лесотундре — на лиственницах или
тополях, в других местностях — на соснах, дубах, вязах, осокорях, ветлах,
ольхах, березах и др. Гнездо находится обычно высоко от земли, на верхушке
дерева, в развилке или на большом боковом суку. В основе гнезда —■ толстые
плотно уложенные сучья, лоток небольшой, выстилка из веточек — сухих
или зеленых, коры, перьев и т. п. От земли гнездо обычно в 15—20 и до 25 м,
в лесотундре и ниже, в б—1м; гнездо на иве, расположенное низко от земли, —
в б м — найдено в Приморье. В тундре (Енисей) и на северных островах
повидимому гнездится и на скалах, что установлено и для Зап. Европы.
Размер гнезда значителен, но варьирует в зависимости от
продолжительности использования; старые и столетние гнезда очень велики, до 1,5—2 м
в диаметре и до 1-7—1,5 л« в высоту; новые постройки меньше, около 1 м
в диаметре.
В стенках гнезд иногда гнездятся полевые воробьи. На юге птицы
появляются у гнезд уже в феврале; в это же время наблюдается и спаривание (Херсон,
Браунер, 1894). Наступление периода размножения варьирует повидимому
в различных местностях, так как в то время, когда на юге у орланов
происходят кладки, зимующие в тех же местностях северные птицы не обнаруживают
проявлений (в поведении) полового цикла, а самый их отлет на север к местам
гнездовья начинается поздней (Каспийское море). Соответственно этому начало
кладки значительно варьирует географически.
На юге кладка происходит в первой половине марта, даже в конце февраля
(26.11, низовья Волги), в марте кладки производятся на Украине (Умань,
236
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Харьковская обл.), на Сев. Кавказе и в Туркестане (Илийский край,
Шестоперов, 1929). Тогда же, судя по находкам птенцов, происходят кладки и в
других местах юга Европы и в Приморье. В средней полосе России кладка
бывает в разные числа апреля. В лесотундре и тундре, судя по находкам птенцов
и датам прилета туда орланов, кладка бывает только в конце апреля и
возможно даже в начале мая. Полные кладки на о. Сикотан, Курильский
архипелаг, —в конце марта и начале апреля (Кобайяси). Яиц в кладке— 2,
редко 1 или 3 (т. е. больше, чем у орлов и других крупных хищников).
Окраска яиц белая, иногда со
слабо заметными охристыми пятнами.
Размеры: (4) 75,1—81 х 58—59,&
(Харьковская обл., Сомов, 1897), (39)
69,5—77,5 х 57—62 мм (Умань, Ге-
бель, 1879). В случае гибели кладки
производится дополнительная, в
течение 4 недель (Гебель).
Судя по разнице в возрасте
птенцов насиживание начинается с
откладки первого яйца; участвуют в нем
оба родителя, но доля самки
значительно выше. Продолжительность
насиживания точно не установлена, но
повидимому невелика для такой
крупной птицы, составляя немногим более
месяца (вылупление на юге
происходит в разных числах апреля,
северней — в Поволжье, в Зап. и средней
Сибири— в мае). Число птенцов
обычно 2 и 1, редко 3. Гибель
младших птенцов не носит такого
регулярного характера, как у орлов или
бородача. Оперившиеся птенцы с не-
доросшими маховыми и рулевыми
встречены на крайнем юге в конце
июня (Бендершах, южн. Каспий;
заповедник Конча Заспа в низовьях
Днепра; в это время и в
Воронежской, и в Харьковской обл.).
Северней — с середины и во второй
половине июля. В начале августа всюду уже появляются летные молодые (но не
все птенцы доживают до вылета, иногда они вываливаются из гнезда и
становятся добычей хищных зверей, например, в Лапландии, по Мантейфелю,
и в Харьковской обл., по Сомову). Продолжительность гнездового периода
таким образом около 272 месяцев.
Линька полная годовая; впрочем, часть старых перьев остается несме-
ненной, даже маховые и рулевые, так что смена всего оперения происходит
в сущности за 2 года. У молодых в первом годовом наряде линяние начинается
в феврале, у взрослых протекает с апреля до октября, причем крупные перья
сменяются между серединой мая и концом сентября (Московский зоопарк).
Следует ожидать, что начало и конец линяния у разных популяций белохвоста
неодинаковы в связи с разницей в сроках размножения. Общая
продолжительность линяния около 5 месяцев (хотя отдельные неперелинявшие перья остаются
и дольше, поэтому Хортлинг, 1931, определяет продолжительность линьки
в 7 месяцев). Последовательность смены первостепенных маховых не вполне
Рис. 66. Орлан-Белохвост.
ОРЛАН БЕЛОХВОСТ
237
ясна, по типу линька — полицентрическая, но начинается с внутреннего края
партии (10-го махового). Последовательность смены нарядов: первый пуховой
наряд — второй пуховой наряд —первый годовой (гнездовый) наряд — второй
годовой (переходный) наряд—третий годовой (переходный)
наряд—четвертый годовой (окончательный) наряд и т. д.
Питание. Орлан — полифаг. Связь его распространения с водоемами
указывает на то, что в его кормовом режиме значительное место занимает рыба.
В частности, отмечена известная связь распространения орлана с численностью
красной рыбы в Приморье, с нерестовым ходом рыбы на Каспии и т. п. Орлан
кормится млекопитающими средней величины — зайцами, сусликами,
хомяками, водяными крысами, ондатрой и т. п., на севере леммингами и молодыми
оленями; из домашних животных нападает зимой на собак и кошек. Из птиц
орланы нападают на гусей как домашних, так и диких, на уток, лысух,
цапель (кваква и др.), гаг, чаек, белых куропаток, тетеревов, глухарей,
стрепетов и даже дроф, лебедей и фламинго. Из рыб кормятся щуками, налимами,
сазанами, бешенкой, воблой, лососевыми. Охотно едят падаль и отбросы (на
юге отбросы рыбного промысла и боен; на Канине отбросы белушьего
промысла, Банников, 1934). В весеннее и летнее время значительное место в
питании орланов и в выкармливании ими выводка занимает рыба, на осеннем
пролете и зимовках водяная птица, на зимовке в степях зайцы. Орлан ловит
добычу на земле, птицу также в воздухе близко над поверхностью земли или
воды (и на воде), рыбу — на мелководье лапами или бросаясь в воду с
разлета. Разыскивает добычу паря более или менее высоко в воздухе. Иногда
отбивает добычу у других птиц, например у скопы. Охотничий район пары
значителен —например в Лапландии 15—16 км от гнезда (Владимирская, 1948).
Полевые признаки. Крупный орел с широкими и длинными крыльями около
2—2х/2 м в размахе, однообразно бурой окраски; хвост короткий, клиновидный,
у старых птиц белый. Маховые на полете пальцеобразно расставлены, как у
настоящих орлов и грифов; крылья при этом обычно не согнуты в кистевых
суставах. Осторожен. Голос — чаще всего лающее или каркающее «кра-кра-
кра» или реже «кии-кии-кии».
Описание. Размеры и строение. Клюв массивный, ноздри длинные, щеле-
видные и косо поставленные. Цевка короткая, покрытая спереди поперечными щитками,
оперена — в отличие от орлов — только до половины. Пальцы сильные, покрыты на нижней
поверхности заостренными шипиками. Крылья широкие и длинные; формула крыла
3>4>4>5>2>6>7>1>8; вырезки на наружных опахалах 2-го — 7-го и на внутренних
опахалах 1-го — 5-го маховых. Рулевых 12, они короткие и широкие, крайние пары короче
средних. Белохвост, при приблизительно равной длине крыла, массивней и тяжелей беркута:
вес самцов (2) 3075—3800, самок (3) 4900, 4920, 6560 г; длина самцов (3) 755, 855, 900, самок
(6) 855—980, в среднем 939 мм; размах самцов (3) 1992, 2025, 2250, самок (6) 2020—2278,
в среднем 2135,5 мм. Крыло самцов (23) 575—625, самок (17) 635—690, в среднем 594,8 и
653 мм. Крылья и хвост молодых птиц в первом годовом наряде длинней, чем у старых,
так как рулевые и маховые у них шире и длиннее (увеличение несущей поверхности,
компенсирующее относительно слабое развитие мускулатуры).
Окраска. Первый пуховой наряд — из длинных редких пушинок серовато-бурого
цвета, на голове пух особенно длинный и чисто белый, с темным пятном у глаза; лапы и воско-
вица розовые; клюв черный. Второй пуховой наряд надевается в возрасте сколо 3 недель—
он густой и короткий, темный буровато-серый, между пухом пробиваются белые пушистые
вершины перьев; лапы и восковица бледножелтые, клюв черный.
Первый годовой наряд с черновато-бурой головой, на спинной стороне бурый со
светлым охристо-бурым рисунком из пятен и каемок перьев и с белыми (прикрытыми)
основаниями перьев; края перьев иногда беловаты; маховые темнобурые, рулевые черно-бурые
с белым мраморным рисунком, иногда грязнобелые с темными пестринами, образующими
каймы перьев; брюшная сторона светлобурая с темнобурыми пятнами и белыми
основаниями перьев. Радужина бурая, клюв черноватый, восковица и лапы бледножелтые,
когти черные.
Взрослые птицы (в четвертом и последующих годовых нарядах) однообразно бурые,
в свежем пере темные с фиолетовым отливом, в обношенном серовато-бурые; оттенки бурого
цвета повидимому варьируют и индивидуально; шея и голова светлые, в обношенном пере
238
ОТРЯД ХИЩНЫЕ ПТИЦЫ
грязно охристо-белые; перья спины, зоба, груди, брюха и боков с нерезкими бледными
краями; маховые черно-бурые; длинные перья надхвостья белые, обычно с черными
вершинными отметинами; рулевые белые. Радужина от бледной орехово-бурой до желтой,
клюв желто-роговой, лапы и восковица яркожелтые, когти черные. Этот наряд надевается
после трех годовых линек и связан с первым годовым промежуточными.
Второй годовой наряд похож на первый, но голова светлей, на спинной стороне еще
заметны белые основания перьев, надхвостье с примесью белого, хвост белый с черноватыми
пестринами. В третьем годовом наряде и возможно в четвертом голова окрашена несколько
темней, чем у взрослых, белые основания перьев на груди еще сохраняются в той или иной
степени, остатки темных пестрин имеются на боковых парах рулевых. Оба пола окрашены
одинаково.
Географической изменчивости окраски и размеров среди белохвостое из разных частей
нашей страны не наблюдается.
24. Белоголовый орлан Haliaeetus leucocephalus L.
Falco leucocephalus. Linnaeus. Syst. Nat. изд. XII, 1766, стр. 124, Южная Каролина.
Распространение. Ареал. В область распространения белоголового
орлана входит Сев. Америка от Аляски (кроме ее крайнего севера), се-
Карта 32. Распространение белоголового орлана Haliaeetus leucocephalus
7 — Н. 1. washingtoniensis, 2 — Н. 1. leucocephalus; a — граница гнездовий.
верных частей Мекензи (до побережья моря Бофорта), Кьюотина (к югу от
полярного круга), Лабрадора и Ньюфаундленда на севере, до Калифорнии,
Мексики, Техаса, Луизианы и Флориды на юге; отсутствует в пустынных
районах США и местами истреблен в наиболее населенных частях этой страны.
Характер пребывания. В большей части ареала оседлый, но
на севере по крайней мере большая часть птиц откочевывает к югу.
Биотоп. Как у орлана белохвоста — побережья рек, больших озер,
приморские местности.
БЕЛОГОЛОВЫЙ ОРЛАН
239
Подвиды и варьирующие признаки. На севере ареала, к югу до Британской Колумбии
и с.-в. США—Я. /. waschingtoniensis Audubon, 1827; в южных Штатах и Мексике
Н. 1. leucocephalus L. Северные птицы крупней южных, возрастание размеров с юга
на север идет постепенно и разграничение этих подвидов весьма условно.
Систематические заме-
чания. Некоторыми авторами
белоголовые орланы
объединяются в один вид с бело-
хвостами. Вопрос о видовом
тождестве этих весьма
различных в окончательном
наряде орланов требует
выяснения. В пользу объединения
говорит то, что молодые
птицы в первом годовом наряде
практически неразличимы,
что географически обе птицы
строго викарируют и что
экология их одинакова.
Северный белоголовый
орлан
Haliaeetus leuco-
cephaluswaschingtoniensis
Audu bon.
Falco waschingtoniensis. A u-
dubon. Birds of America,
1827, табл. П, Кентуки.
Синонимы. Haliaetus
leucocephalus alascanus
С. Н. Townsend.
Proceed. Biol. Soc. Waschington,
11, 1897, стр. 145, Уналаш-
ка.—Haliaetus hypoleucus,
R i d g w а у. Proceed. U.
S. Nation Mus. 1883, стр.
90, о. Беринга.
Распространение.
Ареал. В Сев.
Америке —см. выше.
Гнездился до недавнего времени
Yпоследние наблюдения
имели место, повиди-
мому, в 1882—1884, Стей-
негер, 1885) на о-ве
Беринга. Сомнительные —
в смысле верности
определения—залеты к Ниж-
неколымску и на вост.
побережье Камчатки (Карага). Отмечен один случай залета в Европу (Швеция,
около 1850). На о-ве Беринга впервые найден Стелл ером (1774). В последнее
время на Командорских о-вах не найден, наблюдения, проведенные у бухты
Лисенкова в южн. части о-ва Беринга, весьма сомнительны (Иогансен, 1934).
Характер пребывания. Во времена Стейнегера белоголовый
орлан был редок и встречался лишь в скалистых южных частях о-ва Беринга;
добыты птицы в негнездовое время в октябре, декабре, феврале и марте, одна
молодая самка—15.V.
Экология. Размножение. Гнездо, найденное Стейнегером на о-ве
Беринга, было расположено на скале и устроено из ветвей; кладка «около на-
Рис. 67. Белоголовый орлан.
240
ОТРЯД ХИЩНЫЕ ПТИЦЫ
чала июня» из 2 яиц. Однако Стейнегер указывает, что в начале июня в гнезде
были птенцы в белом пуху» Старые птицы были очень осторожны.
В Сев. Америке эта птица гнездится на скалах и на деревьях, причем
гнезда используются десятилетиями. Яйца по окраске, как у белохвоста,
размеры 50 экземпляров из Аляски и сев. Канады: 69,4—84,3 х 53,1—63,4, в
среднем 74,4 х 57,1 мм. Насиживание 34—35 дней, гнездовый период длится
около 75 (72—74) дней. Птенцы разновозрастны, насиживание начинается
с первого яйца. Пищу птенцам носят оба родителя (и оба насиживают).
Линька. Смена маховых происходит в июле — сентябре у южного
подвида.
Питание. Главным образом, рыба, также мелкие млекопитающие,птицы,
падаль. Из птиц —чистиковые, утки, кулики, также мелкие воробьиные (все
сведения по экологии американских орланов по Бенту, 1937, и Мею, 1935).
Полевые признаки. Типичный орлан, весьма сходный с белохвостом;
взрослые отличаются от последнего белой головой и темной черноватой окраской;
молодые практически неразличимы.
Описание. Размеры и строение» Крупный орлан. Вес птиц с о-ва Беринга
(по Стейнегеру, 1882): молодого самца 4100, самки 5840 г. Длина крыла у самцов 600,635,
648, 650 мм; самок 705, 705 мм (у типа hypoleucus молодой самки, по тому же автору,
только 630 мм); у птиц, пол которых не был определен, 677, 680, 690, 700, 715 мм.
Окраска. Взрослые птицы темнобурые с белой головой, шеей, кроющими хвоста
и рулевыми. Радужина желтовато-белая (Стейнегер), клюв желтоватый, когти черные,
восковица и ноги желтые. Молодые в первом годовом наряде, как белохвосты: темнобурые
со светлыми беловатыми пестринами, причем белый цвет обычно более развит, чем у
белохвоста. Клюв у молодых бурый или черноватый, восковица оливковая (Стейнегер). Полный
взрослый наряд надевается после 4 линек (Бент, 1937).
25. Орлан долгохвост Haliaeetus leucoryphus Pallas
Russ. Reichs, 1, 1771,
Natur. Moscou, XXV,
d.
Aquila leucorypha. Pallas. Reise d. verschied. Pro v.
стр. 454, низовья р. Урал.
Синонимы. Aquila deserticola. Eversmann. Bull. Soc
1852, стр. 545, табл. 8.
Распространение. Ареал. Ю.-в. Европа от Волжско-Уральских
степей (у р. Еруслан, по Камыш-Самарским озерам) и среднего течения р. Урала;
ю.-з. Сибирь и сев.
Казахстан до Эмбы (48°30')
и Тургая, далее у
Аральского моря (и на о. Бар-
са-кельмес) и низовьев
Сыр-Дарьи, Балхаша,
Сергиополя, Аягуза, Зай-
санаиМарка-куля; южн.
и ю.-в. Алтай, Тан-
ну-тува, на Байкале до
Усть-Баргузина
(вероятно гнездится, Туров,
1924); ю.-в. Забайкалье,
зап. Манчжурия
(возможно гнездится, Мейзе,
1934, Ямасина, 1935);
Монголия (до Ордоса),
зап. Китай (Ганьсу,
Шаньси), Тибет, вост.
Туркестан, к югу до сев. Индии и сев. Бирмы, возможно в Белуджистане (Тайс-
херст, 1927), В Средней Азии долгохвост повидимому не гнездится в Туркмении
к западу от Аму-Дарьи (но и в бассейне этой реки, в нижнем течении гнездовье
Карта 33. Распространение долгохвоста Haliaeetus
leucoryphus
а — граница гнездовий, б — невыясненная граница распространения,
г — область зимовок.
орлан долгохвост
241
не доказано; летние находки сделаны у южного чинка Усть-У рта, в нижнем
течении Аму-Дарьи; изредка на Мургабе; сообщение Зарудного, 1896, о гнездовании
у Гермаба, Фирюзы, на Чандыре и Атреке безусловно ошибочно); в
Таджикистане гнездится лишь на вост. Памире. Залетает в Приазовье (но не добыт), на
вост. Кавказ, на нижнюю Волгу (самый северный пункт — река Самара у
Бузулука), к Каспийскому морю — в Туркмению и ю.-в. Закавказье, в Зап.
Сибирь у Белокаменного и Ямышевского (ок. 52° с. ш.), на Амур. Сведения
0 гнездовании в Крыму на р. Каче ошибочны, впрочем, Дрессер упоминает об
одной добытой там птице.
Характер пребывания. Только немногие особи остаются на
зиму в северных частях ареала (Средняя Азия), большинство отлетает в Индию,
где птица в холодное время года многочисленна.
Даты. Прилет весной поздний—в конце апреля: в сев. Казахстане
у Кок-Джиды первые птицы обнаружены 23. IV, в низовьях Иргиза —20. IV;
на Памире первые птицы отмечены 23. IV, много птиц только в мае; тогда
же в ю.-в. Забайкалье. В Тибете пролет отмечен 10.V, но и 17.VI пролетные
стаи по 5—7 птиц встечались у Иекундо (Шефер, 1938). Эти поздние даты едва ли
однако относятся к прилету гнездовых; так как размножение долгохвостое
начинается не позже марта, приводимые наблюдения скорее указывают на
перекочевки холостых птиц, тем более, например, что начало появления
долгохвоста на Сыр-Дарье относится уже к февралю.
Появление долгохвостов весной связано повидимому со вскрытием
водоемов; где есть незамерзающие водные пространства, орланы зимуют (даже
в высокогорье, например, в Кашмире, Осместон, 1927); на Памире с прилета
орлан держится у сазов, теплых ручьев и незамерзающих ключей и т. п.
Осенью отлет начинается уже в сентябре (Сыр-Дарья), но проследить его
трудно, так как часть птиц зимует в гнездовой области.
Биотоп. Открытые полупустынные местности — как на равнинах,
так и в горах, чаще вблизи водоемов — речные долины, плато с озерами и т. д.
В вертикальном отношении на Алтае поднимается от 400 до 2400 м, в Тянь-
Шане примерно до 3200 м, на Памире до озера Зор-куль, Яшиль-куль и др.,
в Тибете между 4000 и 4700 м, в Ладаке до 3600 и даже 4500 м. Древесная
растительность не мешает, но и не необходима.
Численность. В СССР немногочисленен и спорадичен.
Экология. Размножение. Имеется известное количество неразмно-
жающихся птиц, так как еще в середине мая попадаются особи с неразвитыми
половыми железами и ожиревшие (на р. Урале в сев. Казахстане). Гнездо-
ванье в Туркестане раннее, на Сыр-Дарье яйца повидимому уже в конце марта,
при морозах. Гнезда на кучах тростника, из тростника, ветвей джидовника
и т. д. (Сыр-Дарья, Спангенберг), также на деревьях, например у Джаркента
на карагаче в 24 м от земли; размеры последнего гнезда в диаметре около 100,
в высоту около 65 см, диаметр лотка 45, глубина его 20 см; выстилка была из
тонких ветвей, клочков бараньей шерсти и сухого конского навоза (Зарудный
и Кореев, 1906). Описанное гнездо принадлежало повидимому ранее ворону
(впрочем, для Индии указывается, что долгохвост гнездится на деревьях и
всегда сам строит гнездо, не занимая чужих).
Кладка состоит из 2 яиц (Сыр-Дарья, Джаркент) в Средней Азии; в
Индии из 2—4. Свежие кладки найдены 10. III. на Сыр-Дарье (Спангенберг),
26. III. у Джаркента (Зарудный и Кореев). Яйца как у белохвоста, чисто
белые. Размеры 65,5—68x55—55,5 (Джаркент, Зарудный и Кореев),
средние (40) 69,7 х 55,1, предельные 64,3—76,8 х 52,3 — 57,9 мм (Индия,
Бекер, 1928). Птенцы у озера Марка-куль найдены 8.VI., неразбившийся
выводок в ю.-в. Алтае —-21—-22.VIII. Вполне развитые молодые добывались
1 .VII. (Сыр-Дарья, Соло-тюбе), в середине июля у Лбищенска на реке Урал.
16 Птицы СССР, т. I
242
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Линька. Наличие холостых особей затрудняет изучение линьки.
Взрослые (гнездовые) птицы меняют средние маховые (5-е, б-е и 7-е) и начинают смену
рулевых в разные числа июля; с другой стороны, некоторые особи не линяют
в "середине и даже в конце июля, а другие находятся в состоянии линьки в конце
мая. Около середины
августа большинство старых
птиц сменяет большую часть
мелкого пера, а в сентябре
встречаются уже совсем
перелинявшие особи.
Окончательный наряд
надевается повидимому после 3
или 4 линек. Линька
полная годовая, хотя как у
других крупных хищных
птиц обычно остается
некоторое число старых перьев
(даже маховых).
Последовательность смены нарядов:
первый пуховой — второй
пуховой — первый годовой
(гнездовый) наряд —второй
годовой (переходный)
наряд — третий годовой
(переходный) наряд —
четвертый годовой
(окончательный) наряд и т. д.
Питание. Как у
белохвоста: рыба — сазан,
плотва, усач и т. д.;
мелкие млекопитающие
—суслики (Казахстан, Алтай),
сеноставки (Забайкалье;
Ochotona melanostoma в
Тибете, Шефер, 1938); разная,
главным образом, водяная
птица от серого гуся
(Индия), горного гуся (Ладак)
и сухоноса (Забайкалье) до
погоныша (Волжско-У раль-
ская степь); особенно
страдают от долгохвоста
молодые и линные утки и
лысухи; указывается даже такая крупная добыча, как кудрявый пеликан
(Сыр-Дарья* Спангенберг, 1936); кроме того, земноводные (Индия). Долгохвост
охотно ест и падаль, в особенности снулую рыбу, причем в местах замора
наблюдались скопления этого орлана (Соло-тюбе у Сыр-Дарьи). Рыбу
долгохвост ловит повидимому только на мелководье и не нападает на хорошо
летающих птиц. Иногда отбивает добычу от других птиц (с этой целью на
Алтае гоняется за Buteo hemilasius).
Полевые признаки. Сходен с белохвостом — крупная птица, в размахе
крыльев около 2 м, но в общем легче и стройнее, старые птицы не серо-бурые,
а черно-бурые, молодые однообразно бурые без пестрин. Мало осторожен.
Голос как у других орланов, громкое «квок-квок-квок».
fir
4
Рис. 68. Орлан-долгохвост.
ТИХООКЕАНСКИЙ ОРЛАН
243
Описание. Размеры и строение. Цевка оперена слабо, в верхней половине;
«штаны» короткие; хвост слабо закруглен, из 12 рулевых; имеются вырезки на внутренних
опахалах 1-го — 5-го, на наружных опахалах 2-го — 7-го маховых: 1-е маховое длинней
7-го; ноздри закругленные. Длина самцов (4) 610—785, самок (10) 830—880, в среднем 721,2
и 846,5 мм. Размах самцов (3) 2020—2070, самок (8) 2050—2480, в среднем 2035 и 2130 мм.
Вес самцов (3) 2620, 2900, 3278 г; самки (1) 3272 г. Крыло самцов (18) 545—585, самок (20)
593—623, в среднем 572,6 и 605,9 мм.
Окраска. Пуховые наряды не описаны. Окончательный (4-годовой?) наряд: общая
окраска черновато-бурая, в свежем пере с фиолетовым оттенком; грудь и верхняя часть
спины, подхвостье и штаны коричневые; бока, плечевые, кроющие крыла и маховые черные,
голова охристо-буроватая, в изношенном пере беловатая, горло беловатое; рулевые белые
с черными основаниями и широкой черной вершинной полосой.
Первый годовой (гнездовый) наряд на спинной стороне светлобурый с
охристо-беловатыми каймами перьев, поясница и надхвостье беловатые с бурыми пестринами; под глазом
и на кроющих уха широкая темнобурая полоса; брюшная сторона бурая; маховые черные
с белыми пестринами на основной части внутренних опахал; рулевые черноватые с более
или менее развитыми мелкими белыми пестринами. Во втором годовом наряде больше белых
пятен на рулевых, в третьем общий бурый тон спинной
стороны ровнее и темнее и еще больше белого на
рулевых. В четвертом наряде птица — как взрослая, но на
рулевых имеются еще черноватые пестрины в средней
их части. Последовательность смены этих
промежуточных нарядов требует еще дополнительного
изучения и учета личной изменчивости, повидимому
значительной. Хорошо установлена только характеристика
первого годового и окончательного наряда. Радужина
орехово-бурая, клюв синевато-роговый (у молодых
буровато-роговой), восковица серовато-синеватая, лапы
грязножелтые с черными когтями.
26. Тихоокеанский орлан Haliaeetus
pelagicus Pallas
Aquila pelagica. Pallas. Zoographia Rosso-Asiatica
1, 1811, стр. 343, Камчатка.
Синоним. Thalassaetus macrurus. M e n z b i e г.
Bull. Brit. Orn. Club XI, 1900, стр. 4, Якутск.
Русское назва ни е. У местного населения
Камчатки «морской орел»; «орел»; в Приморье —
«пестрый морской орел». Книжные — «камчатский орлан»,
«белоплечий орлан» — оба неправильны, так как
область обитания этой птицы не ограничивается
Камчаткой и оперение плеч этого орлана не белое.
Распространение. Ареал. Гнездовая
область — побережья Берингова и Охотского
морей, на севере от Коряцкой Земли (среднее
течение р. Апуки, примерно в 150 км от ее
впадения в Олюторский залив); северные части
Приамурья (озеро Кизи); Шантарские о-ва;
Сахалин (р. Тымь); Камчатка (бассейны рек
Карага, Авача, Паратунка, Большая, Тигил,
Богачевка, Тихая и т. д.). Область зимовок
отчасти совпадает с гнездовой, но
значительная часть птиц передвигается на юг,
достигая южн. Приамурья и Уссурийского края,
Кореи и Японии (о-ва Хоккайдо, Хондо,
Сикоку, Квельпарт, Амами-осима). Отмечены
залеты на Прибыловы о-ва (св. Павла),
к Якутску (два раза), на Алеутские о-ва
к Пекину.
16*
Карта 34. Распространение
тихоокеанского орлана Haliaeetus
pelagicus
а — граница гнездовий, в — случайные
залеты, г — область зимовок.
на Анадырь (один случай),
(Кадьяк), Риу-Киу; в 1865 г.
244
ОТРЯД ХИЩНЫЕ ПТ^ЦЫ
Характер пребывания. Часть особей оседла (близ
незамерзающих водоемов), остальные откочевывают к югу, относительно недалеко.
Даты. Весеннее движение на север в южн. Приморье начинается уже
15.11, прилет на Шантарские о-ва отмечен в последней трети марта; в это
же время орланы появляются у гнезд на Камчатке. Осенью на южн. берегу
Охотского моря орланы стали редкими в августе, пролет на юг шел около 10.Х
(Миддендорф, 1853).
Биотоп. Гнездовый — долины низовьев рек с высокими деревьями;
скалистые морские побережья. Зимою побережья водоемов, главным образом
в незамерзающих местах.
Численность. Нередок; однако, при узкой области распространения
абсолютная численность невысока.
Экология* Размножение. Пара орланов из года в год пользуется
определенным гнездовым участком. Брачные игры бывают в марте. Спаривание
происходит на гнезде (25. III. 1941 г., Кроноцкий заповедник, вост.
Камчатка, по Аверину, 1948 г.). Гнездо — огромное сооружение, используемое
много лет и ежегодно подновляемое; диаметр его до 272 м, постройка из
больших сучьев, высоко на дереве (тополь Populus suaveolens, береза Betula ermanni,
лиственница Larix daurica). Наличие подходящих для гнездованья деревьев и
определяет распространение орлана в речных долинах. Реже гнезда
расположены на скалах.
Кладка состоит из 1 —3, обычно 2 яиц, производится она в конце апреля
(по Дыбовскому в апреле; по Бергману найдена в мае, что вероятно относится не
к свежей, а к насиженной уже кладке *; б мая в долине р. Русак на с.-в.
Камчатки в гнезде на высокой березе обнаружены 2 яйца (Белопольский и Рогова, 1947).
Окраска яиц беловатая со слабым фисташково-зеленоватым оттенком.
Размеры: 65—58 х 85—78 (Тачановский, Хартерт), вес 149 г.
Птенцы вылупляются в первые дни июня, начинают оперяться в июле,
птенец в полроста найден на Камчатке у Вилючинской бухты 12.VII (Бергман,
1935), летные молодые появляются в августе. В выводке у Ольской губы, сев.
побережье Охотского моря, было 2 самки и 1 самец.
Линька. Полная годовая (но часть перьев все же остается старыми,
как у белохвоста), проходит с середины мая, т. е. второй фазы периода
размножения, до конца сентября — начала октября; продолжительность процесса
около 5 месяцев. Мелкое перо сменяется одновременно с крупным,
первостепенные маховые начинают линять несколько ранее второстепенных; смена
рулевых происходит между июнем и сентябрем. Последовательность смены
первостепенных маховых от периферии к середине, начиная с десятого и кончая
четвертым и пятым; второстепенные маховые линяют от наружного края к
внутреннему; рулевые сменяются от крайней пары к средней, асимметрично
(установлено наблюдениями по жившим в Московском зоопарке птицам).
Последовательность смены нарядов такова: первый пуховой наряд — второй пуховой
наряд — гнездовый или первый годовой наряд — второй годовой
(промежуточный) наряд — третий годовой (промежуточный) наряд — четвертый
годовой (окончательный) наряд и т. д.
Питание. Крупная рыба, птицы крупной и средней величины
(глухари, белые куропатки), звери (зайцы, молодые тюлени, песцы); по Бергману
будто бы соболя, морские выбросы и падаль (отбросы зверобойного промысла,
* Упоминание Стейнегера (1885) о добыче на Камчатке у Петропавловска 21 и
25. V. 1883 самцов с небольшими семенниками и 25. V. 1883 самки со слабо развитыми
яичниками отнюдь не доказывает позднего размножения — вопреки мнению Шульпина
<1936).Зто птицы, закончившие первую фазу размножения —спариванье и кладку,у которых
лоловые железы начали регрессировать; правильность такого взгляда подтверждает и
ешличие у самки больших наседных пятен.
ТИХООКЕАНСКИЙ ОРЛАН 245
отравленные лисицы, гнилая рыба), также беспозвоночные (двухстворчатые
моллюски, головоногие, крабы, по Аверину). Отмечено неудачное нападение на
лису (Белопольский и Рогова, 1947). Красная рыба, как это отмечено еще
старыми авторами (Киттлиц), имеет для орлана большое значение, в местах ее хода
и собираются орланы. С
наличием снулой красной рыбы
связана и зимовка тихоокеанских
орланов у незамерзших
водоемов—в таких местах (в
частности, по Спангенбергу, осенью
и зимой 1938—1939 гг. на р. Иман)
орланы сидят и на скалах, и на
деревьях, и на льду, причем
можно одновременно видеть по
нескольку птиц сразу;
откочевка орланов из таких мест
совпадает с исчезновением падали.
Полевые признаки. Крупная
величина, белые «плечи»,
большой желтый клюв хорошо
отличают сидящую взрослую птицу;
на полете при общем сходстве с
белохвостом отличается от него
белым резко клиновидным
хвостом, яркожелтым клювом;
маховые обычно загнуты и
пальцеобразно расставлены. Молодые
сходны с белохвостами, но клюв
сильней и вся птица кажется
массивней. Размах крыльев
около 2—27з м. Повадки, как у
других орланов.
Осторожная птица, даже
вблизи от гнезд. Голос—клекот,
как у белохвоста, но громче и
ниже.
Описание. Размеры истрое-
н и е. Вес молодых самцов 5—б кг,
взрослого(1) 7,5 кг; вес взрослых самок
6,8 и8,97кг. Длина крыла570—680 лш,
хвост 320—345 мм у взрослых птиц,
у молодых в первом наряде (как и Рис. 69. Тихоокеанский орлан.
у других орланов) маховые и рулевые
длиннее. Самцы заметно меньше самок. Клюв массивный, как бы вздутый, плюсна спереди
покрыта сетчатыми мелкими щитками; хвост из 14 рулевых, резко клинообразный.
Окраска. Первый пуховой наряд шелковисто-белый, второй — в отличие от
белохвоста — дымчатый буровато-серый (Стейнегер). Первый годовой (гнездовий) наряд
однообразный темнобурый с белыми основаниями перьев и светлыми пестринами на рулевых.
Радужина темнобурая, ноги беловатые, когти черные, клюв черновато-бурый. Второй
годовой наряд: белые основания перьев туловища меньше, на хвосте и больших перьях
крыла белых пестрин больше, на боковых рулевых бурый цвет занимает вершину пера и
небольшое поле у основания и по краю наружного опахала; радужина и клюв, как в
гнездовом наряде. Третий годовой наряд: общая окраска туловища и крыльев, головы и шеи
темнобурая с бронзовым отливом; на лбу иногда белые пестрины; рулевые белые с
неправильным черноватым крапом у краев пера; кроющие хвоста белые с бурыми пестринами;
часть мелких и средних кроющих крыла белые; радужина темнобурая; клюв желтый
с небольшим темным пятном на конце надклювья. Четвертый годовой (окончательный)
наряд: общая окраска головы, шеи, туловища, крыльев — бурая с бронзовым отливом в
246
ОТРЯД ХИЩНЫЕ ПТИЦЫ
свежем пере; лоб, малые передние кроющие крыла, рулевые, кроющие хвоста, перья
голени — белые. Лапы и клюв яркожелтые; радужина светлая орехово-бурая.
Систематические замечания. Из Кореи описан в качестве особого вида Haliaetus
niger H e u d e, 1887 (синоним Haliaetus ВгапккИ Taczanowski, 1888) орлан, в общем
очень сходный с тихоокеанским, но без белого лба, без белых перьев на крыле и более
тусклого и темного, черноватого тона в окончательном наряде. Молодые, судя по имеющимся
описаниям, неотличимы. По литературным данным такие орланы будто бы гнездятся в Корее
(Хейаинандо, Кочендо, Кейкидо, по Hand—list of the Japanese Birds, 1932), откуда
происходит тип (Цзен-пьен, между Сеулом и Владивостоком) и где наблюдалось и добывалось
большинство особей этой редкой птицы. Указания Годлевского о наблюдениях наОноне в Даурии
ненадежны. Один взрослый самец добыт 25.11.1902 в Приморье у Сидеми Янковским, будто
бы видевшим там еще двух птиц. Этого орлана теперь обычно рассматривают в качестве
подвида тихоокеанского орлана, но скорее всего это цветовая вариация, быть может и
локализованная в Сев. Корее, но не географическая форма и во всяком случае не вид. Всего этих
черных орланов добыто 18 экземпляров (Остин, 1948).
РОД СТЕРВЯТНИКИ NEOPHRON SAVIGNY, 1809
Тип Valter percnopterus
27. Стервятник Neophron percnopterus L.
Vultur Percnopterus. Linnaeus. Syst. Nat. ed. X, 1758, стр. 37, Египет.
Распространение. Ареал. В область распространения стервятника
входят: Африка, южн. Европа, Передняя Азия, М. Азия, Палестина,
Месопотамия, Иран, Туркестан,
Афганистан, Индия.
Характер
пребывания. В северной части ареала
перелетный (за исключением быть
может отдельных особей), на юге
оседлый.
Биотоп. Пустынные и
полупустынные, реже степные местности
вблизи гор, с кручами, обрывами
и т. п.
Подвиды и варьирующие признаки.
В Европе, Африке, Передней Азии N. р.
percnopterus L. (вероятный синоним N. р.
rubripersonatus Zarudny etHaerms,
1902, из Персидского Белуджистана); в
Индии этот подвид распространен в
Кашмире, Синде, Пенджабе, с.-з.
провинциях; N. p. ginginianus La tha m, 1790,
в Индии к югу от предыдущего (но в
Соединенных провинциях, Раджпутане и
Куче распространена смешанная
популяция, где наряду с птицами с
признаками одного или другого подвида
встречаются и «переходные») особи). Морфологические различия выражаются в размерах и окраске:
индийский подвид меньше и с желтым, а не черным клювом. Экологические различия
подвидов заключаются в цикле периодических явлений: северный подвид преимущественно
перелетный, южный оседлый, имеется повидимому и разница в календарных сроках
размножения и т. п.
Обыкновенный стервятник Neophron percnopterus percnopterus L.
Синонимы. Vultur Meleagris. Pallas. Zoographia Rosso-Asiatica 1, 1811, стр.377,
Крым*.— Neophron percnopterus rubripersonatus Zarudny. Haerms.Orn. Monatsber, 1902,
стр. 52, Персидский Белуджистан.
Распространение. Ареал. Обыкновенный стервятник распространен в
Европе: Португалия, Испания (в северной реже), Пиренеи, во Франции — Цен-
* Предположение Эверсмана, 1866, что К. percnopterus у Палласа есть «К. kolbii»,
несомненно ошибочно.
^Къ^ОГ^^- 11
W^
шЛ
Карта 35. Распространение стервятника
Neophron percnepterus
7 — N. p. percnopterus, 2 — N. p. ginginianus; a —
граница гнездовий, г — граница зимовок.
СТЕРВЯТНИК
247
тральный массив, Севенны и Альпы (до Мон-Салех в Швейцарии), спорадически
в Италии; Балеарские о-ва, Сардиния и Сицилия; на Балканах к северу
до Добруджи и далее до Буковины; изолированная колония —в Подолии на
Днестре; Крым, Кавказ. В Африке, кроме западной лесной зоны, в южн,
Африке спорадичен; о-ва Зеленого Мыса и Канарские. В Азии—М. Азия,
Палестина, Месопотамия, Иран, Афганистан и сев. Индия; Туркестан, к северу
быть может до Усть-Урта (во всяком случае, до Шах-Кадама на берегу
Каспийского моря), Кара-Тау и центрального Тянь-Шаня, не доходя до Синьцзяня.
Залеты в СССР в низовья Волги, на Общий Сырт, в Башкирию (Сушкин,
1897), в Воронежскую обл. (Северцов, 1855), к Полтаве (Гавриленко, 1929)
и Мариуполю (Голицинский, 1937), в Зап. Сибири и Таре и даже на р. Конду
у Шухтункурта, около 61°30' с.ш., где был отмечен Раевским в июне 1944 г.
В Европе отмечены залеты в Англию, в Германию (до Силезии), в Данию
и т. д.
Характер пребывания. В Европе в северных частях ареала —
перелетная птица, на юге ареала —оседла. Есть указания Нордманна (1840)
и Никольского (1892) на оседлость стервятника в Крыму (по указаниям
Никольского, зимою вылетает в степи) и на то, что отдельные особи остаются
зимовать на Кавказе (Радде, 1885). Но это все же сомнительно. В Европе, М. Азии
и даже сев. Африке стервятник — перелетный, он зимует в Судане у южной
границы Сахары; причем молодые —до наступления половой зрелости
—держатся главным образом в Африке (Штреземанн, 1944). В Туркестане на зиму
отлетает (Туркмения, Кизыл-Кумы, центр. Тянь-Шань и т. д.), повидимому
в Иран, Месопотамию и возможно в Белуджистан. Более вероятно, что и на
Кавказе стервятника зимою нет, как это указывается рядом авторов (Телав,
Грузия; даже на юге в Армении). Летят стервятники одиночками и парами.
Даты: в южн. Европе (Испания) появляется с африканских зимовок
в марте. В Армении — первые весною в конце марта (29. III). В Туркмению
прилетают в марте, первые — уже в начале этого месяца, но обычным становится
к началу апреля. Отлет в Туркмении начинается в первой половине октября.
Биотоп. В гнездовое время — предгорья и горы среди пустынного и
степного ландшафта, во всяком случае местности с неровным рельефом,
обрывами, холмами и т. п. В негнездовое время и на равнинах, куда регулярно
прилетает кормиться в гнездовом периоде. В вертикальном отношении на
Кавказе поднимаются до 2100 м в Грузии (Чхиквишвили, 1930) и до 3600 м в
Армении (Соснин и Ляйстер, 1942); в Зеравшанском хребте от 1600 до 3000 м
(Даль, 1936), в Таджикистане обычно до 1200—1300 м, реже до 1600—1700 м
(Иванов, 1940); встречен и на Памире, но в восточных его частях редок, на
зап. Памире поднимается до среднего течения р, Гунт (Мекленбурцев); в Тянь-
Шане обычно до 1200—1500 л*, изредка до 2100 (верховья Нарына, Северцов);
в Индии до 2500 м.
Численность. Обычная птица в южных частях Туркестана —в
Туркмении (но не в Кара-Кумах), *в Таджикистане, в юж. Узбекистане, в зап.
Тянь-Шане и в Фергане в пределах Киргизии; также в Закавказье. В центр.
Тянь-Шане -и в Крыму редок, на вост. Памире более или менее случаен, в
Подолии только несколько пар. Численность и распространение стервятника
находятся повидимому в известной зависимости от деятельности человека,
доставляющего птице пищу в виде отбросов, падали на выпасах и т. д. В
Туркмении отмечается зависимость распределения стервятника от перемещения
стад. В отличие от других грифов, стервятники обычно держатся парами и
одиночками, редко (весной) образуя небольшие стаи, еще реже гнездясь
группами или колониями. Временные скопления стервятников (как и других
грифов) замечаются в местах падежа скота (подобное же явление отмечено в
Севастопольскую кампанию на южн. берегу Крыма, Ирби). Колебаний численности
248
ОТРЯД ХИЩНЫЕ ПТИЦЫ
за счет изменений плодовитости, повышения смертности — не установлено,
как и для других трупоядных видов.
Экология. Размножение. Значительное развитие половых желез
обнаруживается уже с прилета, в марте (8. III, Атрек, ю.-з. Туркмения); в конце
марта — начале апреля птицы уже в парах. Кладка происходит во второй по-
Рис. 70. Стервятник (старая и молодая птица).
ловине апреля (полная свежая кладка с чуть насиженным первым яйцом
обнаружена 25. IV, Фисташковый хребет). Число яиц в кладке обычно 2 (в Индии
изредка 1). Судя по разнице в развитии эмбрионов, промежуток между
откладкой яиц около 3 суток (Туркмения). Яйца покрыты густо буровато-красными
пятнами различных размеров, фон окраски бледный, желтовато-белый. Размеры
(100) 58,2—76,4x43—56,1, в среднем 66,18 х 50 мм (Уайзерби, 1939). Гнезда
на Кавказе и в Средней Азии расположены на скалах, склонах холмов, по
обрывам в стенках оврагов, иногда в развалинах (например, мечеть Султан-
санджар в Старом Мерве); в Индии—и на деревьях. Чаще всего гнездо
устраивается в нишах, под карнизом, так, чтобы на него не падали лучи
полуденного солнца. Строится гнездо из ветвей, костей и т. п., лоток выстлан
СТЕРВЯТНИК
249
шерстью и разными отбросами; диаметр гнезда около 700—1000мм (с.-з. Кавказ;
Туркмения). Насиживание начинается с откладки первого яйца, участвуют в
нем оба родителя (Туркмения). Продолжительность насиживания повидимому
около 40 дней. Пуховые птенцы найдены около половины июня (14.VI на
р. Учкулак к югу от Баталпашинска); однако в южн. Туркмении (р. Егригек,
Бадхыз) Гептнером 19.VI добыты молодые, начавшие оперяться. Летные
молодые встречаются в июле (в Дагестане в начале июля; 18.VII плохо
летающий птенец найден у Реданта в Дагестане, Беме, 1926; подлеток в
Таджикистане добыт даже 27.V и в начале июня, Иванов, 1940; в Туркмении —в июле;
7.VI—в сев. Иране, Зарудный, 1900). Разница в сроке вылета объясняется
вероятно различиями в возрасте птенцов, но быть может отчасти и неточностью
наблюдений. Возможно, что период размножения затягивается ввиду того,
что утраченные кладки заменяются стервятником новыми (у Кеши под
Ашхабадом яйца найдены 20.V1).
Л и н ь к а. На материале из СССР не изучена. По Уайзерби (1939) птицы
из первого годового наряда начинают линять в марте, кончая линьку к зиме,
взрослые линяют с мая до зимы. Линька — полная годовая, но, как у других
крупных хищных птиц, часть перьев обычно остается неперелинявшей.
Ход возрастных изменений сложен — окончательный наряд надевается,
повидимому, только после 5 линек, на 6-м году жизни.
П*и т а н и е. Главным образом падаль и отбросы, экскременты. Охотно
держатся поэтому у боен, выпасов, свалок и т. п., в Туркестане —в городах
и поселениях. В гнезде стервятника в ю.-в. Туркмении найдены задняя нога
зайца, нога корсака, клочья шерсти, кроме того крупный варан и агама; в
другом в хребте Гязь-гедык в Туркмении—главным образом щиты черепах, в гнезде
на с.-з. Кавказе—кости, гнилое мясо, остатки змеи; в желудках найдена
падаль (Кавказ, Средняя Азия и т, д.). Из предыдущего видно, что стервятник
кормится и живой добычей (пресмыкающимися). Стервятники охотно пьют воду
и слетаются к рекам, где и купаются.
Полевые признаки. Взрослый стервятник характерной окраски, белый
с черными крыльями: бросается в глаза желтое лицо и слабый «нехищный»
клюв. На полете голова втянута, крылья широко развернуты, хвост кажется
клиновидным. Полет легкий и довольно быстрый, парит по сравнению с
другими грифами реже и чаще пользуется в поисках пищи активным полетом.
Хорошо двигается по земле. Мало осторожен. Очень молчалив (голос
напоминает звон колокольчика, Туркмения).
Описание. Размеры и строение. Небольшой гриф с узким, слабым, сильно
вытянутым клювом, длинной восковицей и параллельными разрезу рта ноздрями; лицо,
темя; горло покрыты пуховидными волосками, часто отсутствующими; перья затылка и шеи
удлиннены и заострены, образуя«воротник»; на зобе голое пятно; крылья длинные, самое
длинное маховое 3-е, 1-е маховое между 5-м и б-м. Хвост из 14 рулевых, клиновидный. Цевка
длинная, пальцы довольно тонкие с тупыми когтями, основания 3-го и 4-го пальцев
соединены заметной перепонкой. Длина самцов и самок (4) 630—745, в среднем 681 мм> размах
(3) 1630—1706, в среднем 1679 мм. Вес (2) 2lt)0—2400 г. Крыло (15) 468—520, в среднем
487,4 мм у в том числе у самцов 497—520, самок 493—515 мм, у птиц неизвестного пола 468—
508 мм; различий в размерах между полами нет.
Окраска. Первый пуховой наряд темный шоколадно-бурый, второй — серовато-
бурый.
Первый годовой наряд темнобурый с охристо-рыжеватыми пестринами на вершинах
перьев шеи и спины; плечевые и кроющие крыла с охристыми каемками; поясница, нижние
и верхние кроющие хвоста рыжевато-охристые.
Второй годовой наряд без светлых каемок или с менее развитыми каемками, общий
бурый тон его бледный, сероватый. В двух последующих нарядах (но на нашем материале
это не выяснено точно) окраска становится еще более бледной сероватой с беловатыми
вершинами перьев, в 5-м птица почти белая с сероватым налетом на больших кроющих.
Окончательный наряд: оперение белое, иногда с желтоватым и (редко) розоватым
оттенком, маховые черные с сероватым блестящим налетом на наружных опахалах передних
маховых и на обоих опахалах задних. Полового диморфизма в окраске нет. Радужина у взрос-
250
ОТРЯД ХИЩНЫЕ ПТИЦЫ
лых желтоватая, у молодых бурая; клюв взрослых черноватый, у молодых сероватый; лапы
голубовато-серые или красноватые; восковица и голая кожа лица оранжевые у взрослых,
бледножелтые у молодых.
РОД СИПЫ GYPS SAVIGNY, 1809
Тип Gyps fulvus
28. Белоголовый сип Gyps fulvus Н а Ы i z 1
Vultur fulvus. H a b 1 i z 1. Neue Nordische Beytraege, IV, 1788, стр. 58, Гилян.
Русское название. Происхождение слова «сип» неясно, но во всяком случае
оно относится уже к XVIII веку. Паллас (1811) считает это название родовым для всех
грифов («rossice generico nomine»). Белоголовый сип — книжное название. У некоторых
авторов — «бурый гриф», что может дать повод к недоразумениям — смешению с Aegypius
monachus.
Распространение. Ареал. Сев. Африка от Алжира, Марокко и Египта,
к югу до Абиссинии и Судана, на берегу Красного моря до 16° с. ш.; Аравия
Карта 36. Распространение белоголового сипа Gyps fulvus
1—G. f. fulvus, 2 — G. f. himalayensis, 3 — G. f. fulvescens; a — граница
распространения (цифры в кружках показывают местонахождение подвидов на
зимовке и кочевках).
(Синай); Европа — Пиренейский п-ов, южн. Франция (Пиренеи и Косе),
Сицилия и Сардиния, Крит и Кипр, Югославия до Славонии и Далмации,
Балканский п-ов, Бессарабия, ю.-з. Украина (?), Крым, Кавказ, Южн.
Урал (?) и Мугоджары (?), ю.-в. Алтай (?); горы Туркестана; Центральная
Азия до Ганьсу, Нань-Шаня, Тибета, Гималаев; на западе и юге в Малой Азии,
Палестине, Иране, Афганистане, Белуджистане, с.-з. Индии.
Характер пребывания. Оседлая птица, совершающая
нерегулярные кочевки далеко от гнездовой области — до средней Европы и Зап.
Сибири.
Биотоп. Сухие открытые ландшафты как в горах, так и на равнинах;
для гнездования сипу необходимы скалы, обрывы, в крайнем случае хотя бы
неровности почвы.
Подвиды и варьирующие признаки. В сев. Африке, Европе и Передней Азии на восток
до Туркестана и Ирана G. /. fulvus H a b 1 i z 1, 1788; в Белуджистане и с.-з. Индии G. /.
fulvescens Hume, 1869; различия между этими птицами незначительны и выражаются
в окраске. Экологические отличия не установлены. В высокой Центральной Азии —
снежный гриф G. /. himalayensis Hume, 1869, который однако на крайнем западе ареала (Кир-
БЕЛОГОЛОВЫЙ СИП
251
гизский хребет) встречается вместе с белоголовым сипом, хотя экологически викарирует,
занимая более высокие области гор. Отличия в первом годовом наряде между снежным грифом
и номинальной формой весьма значительны. Систематические отношения тут не вполне ясны.
Обыкновенный белоголовый сип Gyps fulvus fulvus H a b 1 i z 1
Синонимы Vultur persicus. Pallas. Zoographia Rosso-Asia tica 1, 1811, стр. 377,
Персия.—Vultur Percnopterus. Pallas, там же, стр. 375, Персия. — Gyps rutilans. S e v e г t-
z о w. J. f. Orn. 1876, стр. 169, Туркестан.
Распространение* Ареал. Западная часть области распространения
вида — в Африке, Европе и Передней Азии, на восток до Тарбагатая и быть
может в ю.-в. Алтае (Гессе, 1913), Сайлюгеме (Сушкин*, 1938), в Туркестане
до зап. Тянь-Шаня (Таласский Ала-Тау, Александровский хребет), Чаткала,
Алая, зап. Памира (в долине Пянджа между Кала-и-Хумб и Шиц), в зап. и
среднем Иране до Кугистана и Кермана; на север до Мугоджар, Кара-Тау и гор
Ак-Тау и Тохта-Тау в вост. Кызыл-Кумах (Зарудный, 1915). В центр. Тянь-Шане
и вост. Туркестане заменен снежным грифом. Сведения о распространении
в Индии неясны, возможно встречается в Гималаях на восток до Сиккима и
даже Бутана, но гнездится ли там или только залетает и нет ли смешения при
определении с G. /. fulvescens или G. /. himalayensis — неясно.
Залеты далеко от гнездовой области, на севере в Ирландию, Голландию,
Бельгию, Германию, Данию, Финляндию, Польшу; в СССР в Прибалтику,
более или менее регулярно на Украину (Подолия, Волынь, Полтава, Киев,
реже до Харьковской обл.; на Черном море у Мариуполя и т. д.), на Сев.
Кавказ, в нижнее Поволжье и Приуралье; редкие появления отмечены в
средней полосе—в Белоруссии (Витебщина), у Кирсанова в Тамбовской обл.
в Московской обл. (в Зоол. музее Московского университета), в Рязанской обл.,
далее в Прикамье (устье Белой, низовья Свияги), у Уфы, в Кировской обл., на
Урале (Павда) и т. д. Более или менее регулярно встречается в сев. Казахстане,
реже в Зап. Сибири (Тобольск 15.VI. 1940, Барнаул, Рубцово и т. д.). При этом
грифы появляются и в холодное время года, и летом (июнь—^июль), держась
подолгу в одной и той же местности (у устья Свияги почти два года, Першаков, 1937).
Залетают и молодые, и старые птицы, но преимущественно первые. Возможно,
что при преимущественно пассивном способе полета грифа (парение), залеты
их представляют собой в сущности занос неблагоприятными ветрами и т. п.
(это относится и к другим видам грифов — бурому и стервятнику). Все эти
обстоятельства существенно затемняют вопрос о реальном существовании
небольших рассеянных гнездовых колоний сипов вдали от гнездовой области,
в частности о гнездовании их у Днепра, на Мугоджарах и в особенности в с.-з.
Казахстане и по Южн. Уралу. Относительно Южн. Урала еще Эверсманн
(1866) сообщал, что грифы не гнездились там до 20-х годов XIX века, а потом
появились; Зарудный (1888) писал о гнездовании сипа в сев. Казахстане в
горах Биш-Тау и по словам казахов — в Джарык-Тау между Темиром и Илеком;
Сабанеев (1874) приводит даже данные для Каслинского Урала. Все эти
указания верней всего ошибочны, так как позже никто сипов на гнездовье в этих
местах не находил.
Широкие кочевки грифов и появление их вне гнездовой области связаны,
кроме уже отмеченной выше внешней причины, и с нахождением среди них
холостых особей, а быть может и с тем, что некоторые грифы гнездятся не
ежегодно. Несомненно, далее, что эти кочевки имеют отношение к кормовым
условиям — массовому падежу скота (годы «джута» в сев. Казахстане; в
Рубцовском округе летом в июле — много сипов появилось в связи с эпизоотией
сибирской язвы), войне (например в Туркестанском походе после сражения у Ак-
булака в июле 1864; после штурма Чимкента; в Севастопольскую кампанию
зимой 1654—1855, в особенности у Балаклавы). Последний значительный
252
ОТРЯД ХИЩНЫЕ ПТИЦЫ
налет к северу был отмечен в 1946 году, когда холостые, судя по состоянию
гонад, долетели до Ярославской и
Кировской обл.,
в июне
где были добыты
и июле.
Характер
пребывания. Оседлая
птица, совершающая, однако,
нерегулярные
передвижения за пределы гнездовой
области (см. выше).
Биотоп. Сухие
открытые ландшафты от
равнин до гор, при условии
наличия подходящего для
гнездования рельефа
(скалы, обрывы, холмы и т. п.),
горы все же
предпочитаются; впрочем, гнезда
найдены даже на холмах в
пустыне (Кара-Кумы, Лоудон,
1915). В горах Кавказа
поднимаются до 2750 м и выше
(Армения), в Туркестане
обычно от 900 до 1 500 м,
во всяком случае ниже
2000м; выше в Тянь-Шане
гнездится уже кумай. На
кормовых вылетах и выше,
до 3300 м (Даль, 1936). В
холодное время года и за
кормом обычно опускается
в долины. В районах
большого скотоводства
наблюдается перемещение сипов
за стадами скота как
осенью, так и весной
(Таджикистан, зап. и центр.
Тянь-Шань, Туркмения).
Численность. Из
крупных грифов
относительно самый
многочисленный, хотя распространен
спорадически, а гнездовые
колонии в общем невелики.
Экология. Размножение. Сведения недостаточны. Длительность
эмбрионального и постэмбрионального развития определяет очень раннее
начало полового цикла, обеспечивающего вылет молодых из гнезда в наиболее
благоприятных для них условиях (конец лета — начало осени). Несомненно
имеются холостые особи (молодые?). Спаривание происходит зимой, в январе
(Московский зоопарк). Совокупление — на земле, в течение около 1 минуты.
Птицы при этом издают хриплые каркающие звуки. Самец перед спариванием
ходит близ самки, держась горизонтально, приподняв хвост и пол у распустив
крылья. Период спаривания тянется до апреля (Северцов, по Мензбиру, 1891).
Гнезда делаются из сучьев, с подстилкой из веточек или сухих трав, иногда
прямо на земле, на крутых обрывах и выемках — скалистых, конгломератовых,
Рис. 71. Белоголовый сип.
БЕЛОГОЛОВЫЙ СИП
253
глинистых. Расположены гнезда группами или небольшими колониями (от 2—3
и до 20 пар, Туркестан, Кавказ). Иногда две птицы откладывают яйца в одном
гнезде (Испания).
В кладке 1 яйцо, происходит она повидимому в конце февраля — начале
марта. Окраска яиц белая, изредка с буроватыми пестринами. Размер (60)
81,5—101,2 х 64,5—75, в среднем 92 х70,1 мм (Уайзерби, 1939).
Насиживают, судя по наблюдениям в неволе, оба родителя, сменяя один
другого через 24—48 часов. Срок насиживания около 50 дней (Суанн, 1924).
В начале июня в Туркестане в гнездах обнаружены птенцы, начинающие one-
ряться. Летными птенцы становятся, повидимому, с конца июня (Северцов,
1891) —в начале июля (Крым, Штегман> 1937).
Линька. Мало известна. В соответствии с большими размерами
птицы, весь процесс длительный. В начале зимы взрослые сипы в более или
менее равномерном свежем наряде. Интенсивная линька начинается со второй
половины периода размножения, т. е. в апреле, но особенно в мае — июне.
Последовательность смены крупных перьев (маховых и рулевых) не
установлена. Последовательность смены нарядов: первый пуховой наряд — второй
пуховой наряд — первый годовой (гнездовый) наряд — второй годовой
(переходный) наряд и т. д.; окончательный наряд надевается повидимому на 5-м году
(точно не установлено). Линька —полная годовая; утверждение старых
авторов, что грифы линяют не ежегодно и как бы постоянно исподволь сменяют
оперение,—неверно и основано повидимому на том, что у сипов, как и у других
крупных хищников, часть оперения, в том числе маховые и рулевые, и после
линяния остается от старого наряда.
Питание. Пища —трупы крупных животных, главным образом более
или менее разложившиеся, но и свежие. В первую очередь сип поедает
внутренности падали, разрывая своим крепким клювом кожу брюшной полости и
извлекая внутренние органы; в дальнейшем падаль поедается изнутри, и кожа,
сухожилия, скелет оставляются нетронутыми. В связи с этим сип погадок не
сбрасывает. Как и другие крупные падальники сип наедается доотвала и после
этого садится на отдых неподалеку от места кормежки, иногда даже не будучи
в состоянии взлететь- не отрыгнув пищи. Птенцы выкармливаются
отрыгиваемой родителями (обоими) пищей. На живую добычу сип не нападает. Может
подолгу голодать. Как и другие падальники, охотно пьет.
Полевые признаки. Это одна из самых крупных птиц фауны СССР,
превышающая по размерам орлов. Общая окраска бледнобурая с длинной беловатой,
несколько втянутой на полете, шеей. Крылья длинные и широкие, на полете
характерны широко расставленные пальцеобразные передние маховые.
Подымается с земли с известным трудом, с довольно длительного разбега на ровном
месте. Главный способ полета —парение. По земле ходит легко и может даже
бежать. Обычно встречается группами — как на добыче, так и у гнезд и на
кочевках. Довольно.осторожен. Голос —свист или шипенье, также кряхтенье
или карканье.
Описание. Размеры и строение. Небольшая покрытая пухом голова,
вытянутый и сжатый с боков клюв с поперечно поставленными щелевидными ноздрями. Цевка
короткая, короче среднего пальца (у бурого грифа несколько длинней); когти тупые. Формула
крыла: 3>4>5>6>1... Второстепенные маховые только немного уступают в длине
первостепенным. Хвост слегка закругленный, из 14 рулевых (у бурого грифа из 12 рулевых).
Полового диморфизма в размерах нет, или быть может самцы несколько крупней самок.
Длина (2) 990—1125, размах (2) 2400—2557 мм. Крыло самцов (4) 700, 720, 720, 735 мм,
самок (5) 690, 700, 700, 710, 725 мм.
Окраска. Первый пуховой наряд белый, второй (в возрасте около месяца) охристо-
белый (Хейнрот, 1924). Первый годовой (гнездовый) наряд с беловатым пухом на
голове; зоб черный; общий тон спинной стороны рыжевато-бурый с более или менее
заметными светлыми охристыми полосами вдоль стволов перьев; брюшная сторона
рыжевато-бурая; маховые и рулевые темнобурые. Взрослый (окончательный) наряд светлый, бланжево-
254
ОТРЯД ХИЩНЫЕ ПТИЦЫ
бурый; голова в пуху, беловатая;ожерелье не из рассученных заостренных перьев, как
у молодых птиц, но из короткого густого пуха; по сторонам головы и шеи голые пятна;
брюшная сторона несколько рыжевата, темней спинной, которая серовато-бурая; зоб бурый;
на брюшной стороне светлые наствольные полосы; первостепенные маховые и рулевые
черновато-бурые.
Окончательный наряд связан с первым годовым переходным. Во втором годовом и
последующих переходных (число которых не установлено, вероятно два или три) нарядах
окраска несколько темней, и ошейник состоит из рассученных перьев, а не из пуха. Птицы
с ошейником из перьев уже способны к размножению.
Радужина ореховая или желтовато-бурая; клюв буровато-роговой, желтоватый у
вершины и основания; восковица сероватая; голые пятна на голове и шее голубоватые;
ноги свинцово-серые с буроватыми пальцами; когти черноваты. Полового диморфизма
в окраске нет.
Индийский белоголовый сип Gyps fulvus fulvescens Hume
Gyps fulvescens. Hume. The Ibis, 1869, стр. 356, Гургаон, Пенджаб.
Распространение. Ареал. Сев.-зап. Индия, к югу до Хандеша и Деккана,
быть может в Афганистане. По определению П. П. Сушкина, к этой форме
относится экземпляр, убитый в ноябре на Памире (Шугнан).
Характер пребывания. Оседлая птица, совершающая
нерегулярные кочевки.
Биотоп. Как в горах, так и на равнинах, где только имеются обрывы,
холмы и т. п. В вертикальном отношении в Индии поднимается до 1800 м
(Бекер, 1928).
Экология. Размножение. Период размножения в Индии длится
с февраля по апрель в Кашмире, с ноября до начала марта на равнинах. Гнезда
на обрывах скал, из сучьев, используются много лет подряд, пока не будут
сброшены муссонами. В гнездовых колониях отмечено 6—20 пар. Кладка
состоит из одного яйца, белого цвета, иногда испещренного бледно-красноватым.
Размеры: (25) 83,8—97 х 65—73, в среднем 90,7 х 70,2 мм (Бекер).
Питание как и у других сипов, — трупы крупных животных.
Описание. Размеры. Длина крыла 675—740 мм (Бекер, 1928). Окраска.
Отличается от номинальной формы более бледной и рыжей окраской, зоб не темнобурый, а
коричневый.
Систематические замечания. При незначительности различий индийского
белоголового сипа от обыкновенного, нет полной уверенности в том, что упомянутый выше
экземпляр с Памира, из колл. Мензбира, хранящейся ныне в Зоологическом институте
Академии наук, есть действительно fulvescens, а не индивидуальная вариация G.f.fulvus
1907, в сторону fulvescens; тем более, что последний подвид — оседлый.
Снежный гриф или кумай Gyps fulvus himalayensis Hume
Gyps himalayensis. H u m e. My Scrap Book or Roygh Notes, 1869, стр. 12, Гималаи.
Синонимы. Gyps nivicola Северцов. Вертикальное и горизонтальное
распространение туркестанских животных, 1873, стр. 111, высокогорная зона Тянь-Шаня; Семире-
ченский Ала-Тау, верховья Нарына. — Gyps cinnamomeus. Reichenow. Orn. Monatsber.
1907, стр. 30, Нарын.
Русское название. Снежный гриф — книжное название, введенное Северцовым
и Пржевальским. Кумай или гуммай — киргизское название этой птицы в Туркестане. На
Памире эта птица носит название ак-кальджир, т. е. белый гриф.
Распространение. Ареал. От Тянь-Шаня (центр, части Киргизского
хребта), вост. Ферганы (несколько восточнее Гульчи) и Памира на западе до
Нань-Шаня, Ганьсу, Тибета на востоке; Гималаев (до зап. Ассама) на юге.
Указывается для Тарбагатая, что сомнительно. Есть указания на нахождение
отдельных особей в Туркестанском хребте у верховьев Исфары, у Искандер-
куля, в вост. Бухаре, но они требуют подтверждения. Зимой будто бы
залетает в долину Пянджа у Иола и Хирманджоу.
Характер пребывания. Оседлая птица, совершающая вылеты
за пищей в подгорные долины.
БЕЛОГОЛОВЫЙ СИП
255
Биотоп. Гнездится в высокогорье от 2000 до 5200 м, в
субальпийской и альпийской зонах, выше верхней границы леса.
Численность. Относительно многочисленен в зап. Китае, Тибете
и Гималаях, у зап. границы ареала в Туркестане редок.
Рис. 72. Кумай.
Экология. Размножение. Весьма раннее спаривание происходит
в конце января (Московский зоопарк) — в это время у птиц уже сильно
развиты половые железы. В Гималаях период размножения длится с января до
марта. Кладка происходит в феврале (Кашмир), в ней одно яйцо зеленовато-
белого цвета, изредка с буроватыми крапинами; размеры яиц (5) 87—95,8 х
X 65—74 (Кашмир, Хартерт, 1913); (50) 89,2—103,6 х 65—74, в среднем
94,8 х 70 мм (Бекер, 1928). Из пределов СССР яйца не описаны. Гнездится
отдельными парами или колониями, небольшими, из 4—5 пар (Киргизский
хребет, Спангенберг, 1935; Памир, Мекленбурцев). Гнезда расположены в
нишах или по обрывам скал, занимаются по много лет подряд (Гималаи). В начале
256
ОТРЯД ХИЩНЫЕ ПТИЦЫ
июля молодые уже оперяются, но крылья и хвост у них еще короткие (5.VII,
Усекские горы у Джаркента, Зарудный и Кореев, 1806), слетки в начале августа
(Иркештам, Мензбир, 1888). Период гнездования тянется следовательно около
6х/2—7 месяцев.
Л и н ь к а не изучена, как и последовательность смены нарядов. Первый
годовой наряд связывается с окончательным промежуточным, но число
последних неизвестно.
Питание. Как и другие грифы, питается трупами. Вылетает на
кормежку относительно поздно, когда от нагретой поверхности земли потекут
восходящие струи воздуха и рассеется туман. На кормовых вылетах в стаях
иногда встречается по нескольку десятков особей (Тибет, Памир). Массовое
скопление кумаев и других грифов в Тянь-Шане и Памире бывает при джуте,
когда гибнет не только домашний скот, но и архары. В Тибете, где местами
человеческие трупы оставляются открыто для своеобразных похорон, кумай
кормится и ими. Как и другие грифы, кумай в падали проделывает дыру в брюхе
и пожирает сначала внутренности, выворачивая затем иногда кожу (Тибет,
Шефер, 1938). Птица крайне прожорлива; несколько грифов съедают в Тибете
труп человека за полчаса, объедают до скелета труп яка за 2 часа. Завидев
падаль, кумай опускается на нее с полусложенными крыльями, с характерным
дребезжащим шумом. На падали весьма агрессивен, разгоняя всех прочих
птиц, включая#бородачей. Наевшись, или отлетает, или отбегает к ближайшим
холмам, где потом долгое время отдыхает. В поисках корма парит очень
высоко, часто вне поля зрения стоящего на земле человека; основной способ
передвижения —парение.
Полевые признаки. Похож на белоголового сипа, но отличается от него
белыми подкрыльями и светлым тоном окраски (у взрослых птиц). Весьма
осторожен. Голос как у белоголового сипа, кряхтенье или шипенье (Московский
зоопарк).
Описание. Размеры и строение. Очень крупный гриф, не уступающий по
величине бурому. Вес взрослых самцов 10—12 кг (Пржевальский, 1876), 8—10 кг (Шефер)
у птиц из Тибета; у старой птицы из центрального Тянь-Шаня 11 кг, молодой —10,5 кг. Длина
самца 1243 (Пржевальский), самки 1497 мм (Северцов), размах крыльев соответственно 2616
и 3063 мм. Крыло самцов и самок 765—839 мм (Штегман, 1937).
Окраска. Пуховые наряды не описаны. Первый годовой (гнездовый) наряд с
буроватым пухом на голове, брюшная сторона темнобурая с охристо-белыми наствольями,
спинная сторона темнобурая со светлобурыми продольными полосами; маховые и рулевые
черные; характерна белая окраска подкрыльев; восковицане черноватая, как у номинальной
формы, а бледнобурая. В окончательном наряде похожа на белоголового сипа, но заметно
бледней, спинная сторона не серовато-рыжеватая, а бледнопалево-серая, пятно на зобе не темно-
бурое, а бледнобурое; брюшная сторона беловато-охристого общего тона; подкрылья белые;
ожерелье не пуховое, а из рассученных заостренных перьев, охристо-буроватых со светлыми
наствольями (в отношении ожерелья G. /. fulvescens занимает промежуточное положение
между обыкновенным белоголовым сипом и кумаем, так как пуховое ожерелье у него или
слабо развито или вовсе отсутствует даже у взрослых птиц).
Клюв желтовато-роговой, радужина бледнобурая или желтоватая, ноги серовато-
белые, когти черные. Внешних признаков полового диморфизма нет.
РОД PSEUDOGYPS SHARPE 1873
Тип P. bengalensis
29. Индийский гриф Pseudogyps bengalensis Gmelin
Vultur bengalensis. Gmelin Syst. Nat. 1, 1788, стр. 245, Бенгалия.
Распространение. Ареал. Южная Азия от Сиама и Кохинхины на
востоке, Малайского архипелага на юге, и Индии (Пенджаб, Синд) на севере;
повидимому в с.-з. Белуджистане. В СССР в теплый сезон 1939 года взрослый
индийский гриф
257
экземпляр был добыт в окрестностях Ростова-на-Дону; птица в 1940 году
поступила в Зоологический музей Московского университета. К сожалению,
подробности этого исключительного залета остались не установленными.
Рис. 73. Индийский гриф.
Карта 37. Распространение индийского грифа Pseudogyps benga-
lensis
а — граница гнездовий (кружок — место залета в СССР).
17 Птицы СССР, т. I
258
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Характер пребывания. Индийский гриф — оседлая птица.
Главным образом синантроп, живущий в окрестностях поселений. В горах
встречается только в нижнем поясе.
Экология. Размножение. По сведениям новейших авторов (Бекер,
1928, Суанн, 1924), индийский гриф гнездится с октября до февраля,
большинство кладок в декабре — январе. Гнезда расположены на деревьях,
преимущественно колониями, обычно в культурном ландшафте. В кладке
бывает одно яйцо, белого цвета, часто с зеленоватым оттенком, изредка
с красновато-бурым крапом.
Средние их размеры 85,8 х 64,2 мм. Насиживают оба родителя, пови-
димому в течение 45 дней.
Питание. Пищу индийского грифа составляют падаль и отбросы.
Описание. Размеры и строение. Крыло самцов и самок 535—578(Бекерг
1928), 557—608 мм (Суанн, 1924). Окраска взрослых птиц черная с белой спиной и
поясницей, с белым пуховым ошейником; наружные опахала второстепенных маховых серовато-
бурые; подкрылья белые. Первый годовой наряд буро-шоколадный, некоторые перья со
светлыми ободками, нижняя часть спины и поясница бледней, со светлыми наствольями;
голова и шея покрыты беловатым пухом, ошейник не пуховой, а из заостренных перьев
белого цвета, окаймленных бурым; брюшная сторона бледнобурая со светлыми наствольями;
подкрылья бурые. Клюв буровато-серый, голые участки кожи на голове свиндово-серые,
радужина бурая, ноги черные. Полового диморфизма нет. Линька и возрастные' изменения
не изучены.
РОД СЕРЫЕ ГРИФЫ AEGYPIUS SAVIQNY, 1809
Тип Viltur. monachus"
30. Черный или серый гриф Aegypius monachus L.
Vultur monachus. Linnaeus. Syst. Nat. ed. XII, 1766, стр. 122, Аравия.
Русское название. Название «гриф* (латинское gryphus от греческого 7Р°4')
относится у древних к легендарной птице с крючковатым клювом, орлиными крыльями и
львиным телом. В современном понимании оно было впервые применено к этой птице
французскими академиками в 1666 году, от них перешло к Бюффону, а от него к позднейшим
авторам. В русском языке слово гриф, как имя легендарного животного, встречается очень
давно; гриф в качестве геральдической эмблемы известен у нас по крайней мере с XVI века,
а возможно и ранее. Черный гриф—книжное название, птица в сущности не черная, а
серовато-бурая (только молодые в первом годовом наряде черноватые, впрочем в полете на
светлом фоне неба гриф кажется черным).
Распространение. Ареал. Южная Европа — Испания и быть может
Португалия, Балеары (Майорка), Сардиния и Сицилия, Балканы к северу до
Славонии и Румынии (Добруджа), Трансильвания; до недавнего времени повиди-
мому в с.-з. Африке и быть может в Египте; Кипр, М. Азия, Палестина, сев.
Аравия и Аден, Иран, Афганистан, Белуджистан (горы), зап. Гималаи;
Монголия, возможно с.-з. Манчжурия и Сев. Китай (Чжили). В СССР в Бессарабии
(лес Корнешти в б. Белецком уезде, Остерман, 1916, сохранился ли?), Крым,
Кавказ, Туркестан от Копет-Дага и Б. Балханов до Тарбагатая; в южн. и
быть может в юго-вост. Алтае, в хребте Кокон в зап. Алтае, у Горно-Алтайска
(Улала), в центр. Алтае. Залетают черные грифы в сев. Францию, Германию,
Данию, Польшу, нередко в разные страны южн. Европы. В СССР в
Прибалтику, на Украину (Проскуров, Черниговская обл.; Аскания-Нова,
Бердянск и т. д.), к Пскову; в Белоруссию (Полесье); в Поволжье (Арзамас);
в Приуралье (Молотов, Кунгур, Кизелово; самый северный пункт —у устья
Подчерема под 64° с. ш.), регулярно бывают в южн. Приуралье и сев.
Казахстане; в ю.-з. Сибири у Барнаула, Петропавловска, Колывани; более или
менее регулярно в южн. Приморье, в вост. Китае. Эти залеты бывают и летом
(очевидно, холостые птицы), встречаются и одиночные птицы, и группы (осенью
и весною). В последнем случае залеты связаны с падежом скота в результате
черный или серый гриф
259
зимней бескормицы, в Китае — однажды с наличием множества набранных
трупов после восстания тайпингов в долине Яньцзе (Стайен, 1891).
Характер пребывания. В Крыму и на Кавказе черный гриф —
оседлая птица, в Туркестане в холодное время года отлетает на юг, даже из
Копет-Дага. На севере Туркестана прилетает к местам гнездовья только в
начале апреля, в Туркмении в феврале, исчезая оттуда в конце октября—ноября.
Откочевка в южн. Алтае в верховьях реки Сары-джаматы наблюдалась в начале
августа (Сушкин, 1938). После вывода грифы совершают и вертикальные
миграции, а холостые особи во всякое время года залетают за пределы гнездовой
области..
Биотоп. Главным образом горы, однако местами и равнины. Возможно, что
на равнинах птицы гнездятся при наступлении особо благоприятных кормовых
Карта 38. Распространение черного грифа Aegypius monachus
а — граница гнездовий, г -— область зимовок и кочевок.
условий — наличии большого количества падали (Сыр-Дарья, Спангенберг).
В горах в Крыму на высоте 800—1000 л*(Акимов, 1940), на Кавказе до 3 000 м,
на Алтае выше 2000 м, в Тянь-Шане до 3 000 м (Аксай), на Памире до 3 500—
3 600 м, в Тибете даже до 4 500 м. В субальпийскую и альпийскую зону гриф
только залетает, гнездится же в Средней Азии, главным образом в зоне арчи
на высотах около 500—2 000 м.
Численность. Ограниченная, гнездовые колонии состоят из
немногих пар.
Экология. Размножение. Судя по тому, что и летом холостые особи
встречаются вне гнездовой области, в том числе и молодые в первом годовом
наряде, способными к размножению грифы становятся в возрасте более года
(или позже); противоположное мнение высказывалось однако Рейзером. Цикл
размножения ранний. У гнезд в Крыму грифы появляются уже в конце февраля
(21 —26 февраля, Акимов), когда старые постройки ремонтируются. Впрочем
иногда грифы держатся у гнезд и зимою. Гнезда — в отличие от сипов —
расположены на деревьях: на соснах (Крым), на арче (Копет-Даг, Тянь-Шань),
на фисташке (Бадхыз); в виде очень редкого исключения на скалах (ю. Алтай,
Сушкин), на саксауле (Сыр-Дарья, Спангенберг). Гнезда используются много
лет, но вероятно попеременно, так как имеются и незанятые. Гнездовые участки
невелики — расстояние одного гнезда от другого 750—1 500 м (Крым). Размеры
новых гнезд в Крыму 130—140 см в диаметре, при высоте в 60 см; старых—до
200 см и в толщину более 100 см. Старые гнезда в Тянь-Шане так велики, что
17*
260
ОТРЯД ХИЩНЫЕ ПТИЦЫ
в них, шуэассказам киргизов, иногда отдыхают барсы. Грифы нередко
устраивают гнезда невысоко от земли (Крым). Выстилка из мелких веток, коры, травы,
клочков кошмы, шерсти и т. п. Спариванье происходит на гнезде, при этом
грифы издают кряхтенье и шипенье.
Кладка бывает в начале марта (Крым), из одного яйца. Яйца грязновато-
белые с более или менее заметными красновато-бурыми штрихами и пятнами.
Размеры: 97—84 х 72—64 мм (Крым, Акимов).
Насиживают оба родителя, 55 дней (Акимов). Вылупление наступает в
начале мая (Крым). Птенцов родители кормят отрыгиваемой пищей. Рост птенцов
Рис. 74. Черный гриф.
медленный— в месячном возрасте только начинается развитие контурных
перьев и маховые достигают 2—3 см в длину; в возрасте около 2 месяцев,
в начале июля контурное оперение вполне развито, но имеются остатки пуха,
хвост и крылья короткие (Крым); однако уже в начале августа попадаются
выросшие летные молодые. Гнездовый период таким образом продолжается
около 3— Зх/г месяцев. Возможно, что в Туркестане цикл размножения
несколько более ранний, чем в Крыму, так как выросшие молодые добывались
уже в конце июля (28 июля Сары-чилек, Фергана, Кашкаров, 1927 и т. д.).
Еще полтора — два месяца после вылета птенцы остаются в районе гнезда,
сбиваясь в сентябре — октябре в стаи, после чего начинаются откочевки.
Линька. Плохо выяснена. Заметная линька у взрослых происходит
в июне—июле (Московский зоопарк).
Линька полная годовая, хотя в оперении всегда остаются неперелинявшие
мелкие и крупные перья. Последовательность смены нарядов: первый пуховой
БОРОДАЧ ИЛИ ЯГНЯТНИК
261
наряд — второй пуховой наряд — первый годовой (гнездовый) наряд —
второй годовой наряд и т. д. Представляет ли собой второй годовой наряд
окончательный или переходный — неясно, но во всяком случае возрастные
изменения после смены первого годового наряда незначительны и твердо не
установлены.
Питание. Пищу черного грифа составляет падаль, главным образом
трупы крупных животных, которых он разыскивает, паря на большой высоте.
В отличие от сипов, кормится преимущественно мясом, отчасти костями
трупов, проглатывая также кожу и шерсть и выбрасывает их затем в виде погадок.
Изредка нападает на живую добычу: мелких млекопитающих (суслики),
пресмыкающихся (ящерицы и черепахи), домашних животных (ягнята, Велижа-
нин, 1928).
Полевые признаки. Одна из самых больших птиц, с размахом крыльев около
22/2 м, черный гриф имеет темную однообразную окраску; крылья на полете
длинные и широкие, при пареньи маховые расставлены пальцеобразно; хвост
короткий и клинообразный; голова втянута. Характерен очень быстрый и
сопровождаемый шумом спуск с высоты с подогнутыми крыльями,
приподнятым хвостом и вытянутыми лапами. Держится стаями и одиночками, часто
парит; кормовые вылеты бывают в теплое время суток, с восхождением от земли
воздушных потоков. Голос —хриплое кряканье или шипенье.
Описание. Размеры и строение. Самая большая хищная птица нашей фауны.
Голова большая, покрытая пухом; клюв массивный, высокий, сжатый с боков, надклювья
с длинными острым крючком; ноздри широкие и округлые. Ноги короткие и толстые, пальцы
длинные с тупыми и мало загнутыми когтями. Крылья длинные и широкие, предплечье очень
длинное. Хвост клинообразный, из 12 рулевых. Вес (2) 6310, 7657 г (у 20 самцов из Румынии
7 000—11 500 г, у 21 самки оттуда же 7 500—12 500 г, Домбровский, 1912). Крыло
самцов (5) 715—825, самок (5) 705—813, в среднем 764,6 мм. Следовательно диморфизм в
размерах выражен незначительно.
Окраска. Первый пуховой наряд однообразный желтовато-серый. Второй пуховой
наряд с желтоватой головой, голубыми голыми участками кожи под глазами и на зобе,
общего дымчато-серого тона (Акимов, 1948). Первый годовой (гнездовый) наряд: пух на
голове жесткий, черноватого цвета, зоб черный; общая окраска черноватая со слабым
блеском. Окончательный наряд: пух на голове бледнобурый или беловатый, перья «ожерелья»
бледнобурые, общая окраска темнобурая с более темными маховыми и перьями голени со
светлыми наствольными чертами на брюшной стороне; нередко среди кроющих крыла и на
груди отдельные белые перья. Радужина темнобурая или золотисто-бурая, клюв темный
серо-роговой, когти черные; ноги, голое пятно на шее и восковица у молодых белые, у старых
восковица и пятно на шее голубоватые, ноги беловато-буроватые.
Систематические замечания» Восточноазиатские грифы, по некоторым авторам,
будто бы крупней западных и могут в таком случае быть отделены под названием А. /п.
chincou (Vulturchincou Da u d i n. Traite cTOrnith. 11, 1800, стр. 12, Житай»). У нас не было
достаточного материала для решения этого вопроса.
РОД БОРОДАЧИ GYPAETUS STORR 1784
Тип V. barb at us
31. Бородач или ягнятник Gypae'tus barbatus L.
Vultur barbatus. Linnaeus. Syst. Nat. ed. X, 1758, стр. 87, Санта-Круц, Оран.
Русское название. «Ягнятник» — перевод западноевропейского названия этой
птицы Lammergeyer, но смысл этого названия неправилен, так как ягнятник не ест овец,
а- кормится падалью. Имя бородач — от пучка жестких перьев или щетинок под клювом,
образующих бородку; в русском языке оба названия книжные.
Распространение. Ареал. Горные местности южн. ,Европы, Африки,
Передней и Центральной Азии.
Характер пребывания. Оседлая птица.
Биотоп. Скалистые участки гор, открытые, расположенные выше
границы древесной растительности, в альпийской и субальпийской зонах.
262
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Подвиды и варьирующие признаки» В северо-западной Африке — Атласских горах от
Марокко до Туниса G. Ь. barbatus L.; на Пиренейском п-ове, по гористым островам
Средиземного моря, на Балканах, в Трансильвании, на Кавказе, в Передней Азии, в Аравии,
в Малой Азии, в Иране и Туркмении G. b. aureus H a b 1 i z 1, 1783; в Центральной Азии от
зап. Китая до Гималаев, Тянь-Шаня и Памира G. b. hemachalanus, H u 11 о п, 1838; в
Абиссинии и южн. Африке G. b. meridionalis Keyserling et Blasiu s,1840.
Морфологические различия в деталях окраски, в оперении цевки, в размерах (южноафриканские
бородачи самые мелкие, ярко окрашенные на брюшной стороне, с обнаженной нижней половиной
Карта 39„ Распространение бородача Gypaetus barbatus
J —G. b. aureus, 2 —О. b. hemachalanus, J—G. b. barbatus, 4 — O, b.
meridionalis; a — граница гнездовий, е — прежний ареал.
цевки; у других подвидов цевка оперена до пальцев; из них подвид из сев.-зап. Африки ярко
окрашен на брюшной стороне и меньших размеров, примерно как G. b. meridionalis; передне-
азиатский подвид крупнее, с меньшим развитием ржавого оттенка на груди и брюхе; цен-
тральноазиатские птицы достигают наибольших размеров, для них характерно развитие
темных пестрин на зобе). Экологические различия не выяснены.
Переднеазиатский бородач Gypaetus barbatus aureus HablizI
Falco aureus. HablizI. Neue Nordische Beytraege, IV, 1788, стр. 64, Самамизские горы,
сев. Иран.
Распространение. Ареал. Горы Испании, Пиренеи, Сицилия, Сардиния,
Корсика, Крит, Балканский п-ов от Греции до Трансильванских Альп,
М. Азия (Тавр), Аравия от Синая до Йемена, Палестина, Иран; возможно к этой
же форме относятся белуджистанские бородачи. В СССР на Кавказе и в
Туркмении от Б. Балханов до Копет-Дага (в зап. Копет-Даге повидимому
отсутствует), также в Таджикистане (Баба-таг, Гузар, Джандак). Точная восточная
граница в Туркестане неясна (быть может сюда же относятся птицы из Чат-
кальских гор —- в центр. Тянь-Шане следующий подвид).
Характер пребывания. Оседлая птица, в холодное время года
не регулярно совершающая незначительные вертикальные перемещения.
Шестоперов сообщает (1937) будто бы из Туркмении бородачи отлетают на зиму,
но это не подтверждается. Сообщение о появлении бородача в Муганской степи,
БОРОДАЧ ИЛИ ЯГНЯТНИК
263
где он будто бы охотится на ягнят и молодых джейранов, — явная
несообразность.
Биотоп. См. выше относительно вида. На Кавказе на с.-з. летом не ниже
2000—2200 (Кавказский заповедник), в Армении на высоте 3300 м, отмечен
и выше — до 4800 м (Радде, 1884), в Копет-Даге обычно около 1500—2000 м,
в Таджикистане от 1670 до 3100 м. Распространение бородача находится в
известной зависимости от наличия крупных копытных, в особенности козлов.
Численность. На Кавказе и в Средней Азии нередок. В Европе
местами исчез (Форарльберг, Каринтия, Швейцария, по крайней мере с 80-х
годов прошлого века; последний бо]родач в" Швейцарии убит в Граубюндене
повидимому в 1887 году; однако пара будто бы наблюдалась в августе
1929 года, Уоллис, 1929).
Экология. Размножение. В СССР не изучено, основные сведения,
по наблюдениям в неволе, таковы (Софийский зоопарк, Шуманн, 1928, Петере,
1940): цикл размножения очень ранний, зимою, что связано с
длительным ростом и развитием молодых —тем самым первые месяцы их жизни
падают на климатически благоприятное время года. Первые спаривания
происходят уже в начале ноября, затем продолжаются до февраля (Софийский
зоопарк); в ноябре — декабре (Московский зоопарк). Гнезда расположены на
скалах, построены из сучьев и прикрыты сухой травой или мхом; иногда
в них бывают сухие кости. В некоторых случаях яйца откладываются прямо
на камни. Иногда гнезда расположены близко одно от другого. Используются
гнезда по многа лет.
Кладка происходит в конце декабря — январе (то же на Кавказе,
Богданов, 1879), с довольно значительными (индивидуальными?) колебаниями. Яиц
в кладке 2, редко 1. Промежуток между откладкой яиц 4—5 суток. Насиживают
оба родителя, но преимущественно около 2/з всего периода — самка, у которой
более развиты наседные пятна.
Насиживание продолжается около 55—60 дней. Вылупление наступает
соответственно с кладкой, от второй трети февраля до первой трети марта
(птенцы в марте находились на Кавказе, по опросным данным, Мензбир, 1895).
Процесс вылупления продолжается около 50 часов. Окраска яиц охристая или
беловатая с коричневыми или серо-фиолетовыми пестринами, размеры их
(71) 75—92,2 х 56—70, в среднем 87,77 х 64,58 мм (Хартерт, 1914). Если
оба яйца оказываются оплодотворенными, то младший птенец как правило
погибает в первые дни жизни; его убивают и пожирают родители. Развитие
птенцов медленное, первый пуховой наряд сменяется вторым в возрасте около
3 недель, а после 35 дней начинается процесс оперения. В июле в Дагестане
наблюдались выросшие молодые, а в конце июля вполне развитые молодые
добывались и в Туркмении.
Линька. Не изучена. Окончательный наряд — в возрасте не менее
5 лет (Петере, 1940)-. В начале зимы взрослые особи бывают в свежем пере,
обычно с незначительной примесью старых перьев (начало декабря).
Питание. Уже строение летательного аппарата бородача, несмотря
на ряд особенностей, указывает на его сходство в отношении способов питания
с грифами. Бородач — птица с большой несущей поверхностью (длинные
крылья и хвост), превосходно приспособленная к длительному полету в
поисках падали крупных животных. Тупые когти и относительно короткие пальцы
делают лапы бородача непригодными для умерщвления живой добычи. Однако,
в отличие от грифов, бородач иногда носит пищу в лапах (при этом даже целую
лошадиную ногу, Армения). Старый спор об опасности или безопасности
бородача для крупных млекопитающих и даже человека решается отрицательно.
Точных доказательств нападений бородачей на крупную живую добычу нет
(хотя Гиртаннер, 1870, писал о нападении этой птицы на 14-летнего мальчика).
264
ОТРЯД ХИЩНЫЕ ПТИЦЫ
С другой стороны, известная зависимость распределения бородача от наличия
копытных—диких или домашних—несомненна. Повидимому однако это
объясняется тем, что бородачи питаются павшими животными, предпочитая,
в отличие от грифов, свежее мясо
'разложившемуся. Впрочем, не
исключается и ловля бородачем мелких
животных, быть может даже
умерщвление новорожденных ягнят и
козлят. Бородач будто бы умерщвляет
степных черепах, бросая их с воздуха
на землю (Туркмения, Зарудный,
1896), ловит кекликов (Филлогг, 1908)*
Основное место в питании бородача
занимают все же кости, которые он
«обрабатывает», разбивая о землю
(см. ниже вцентральноазиатскойрасе);
с питанием костями связано
чрезвычайно значительное развитие желез в
пищеварительном тракте. В желудке
убитого на с.-з. Кавказе бородача
найдено: часть черепа и рог серны, часть
ноги серны, нога лисицы, лапы
голубя вяхиря и т. п. (Насимович и
Аверин, 1938).
Полевые признаки. На лету
бородач несколько похож на огромного
сокола — крылья длинные и узкие,
хвост также длинный. Полет быстрый
и поворотливый. При этом в отличие
от грифов, разыскивающих падаль,
паря на большой высоте, бородач
быстро облетает свой охотничий
участок, следуя за очертанием гор и
низко —на высоте 50—100 л* —летя
над обрывами. При этом крылья
летящего бородача с необычайно
длинными маховыми, с твердыми и
толстыми их стволами издают своеобразный
дребезжащий шум.. Характерно, что
реакция горных зверей (и птиц —
кеклики, улары) на появление
бородача иная, чем при виде летящего
грифа: на последнего они не
обращают внимания, но при виде бородача
проявляют беспокойство. Обычно пара
бородачей — самец и самка — летают на небольшом расстоянии один от
другого. Увидев корм (падаль) бородачи сначала долго кружат над нею в
воздухе, потом опускаются ниже и, наконец, сев на землю в некотором
отдалении, идут к добыче «пешком». Утром бородачи облетают свои участки после
того как рассеется туман и облачность, часа через 2 после восхода солнца.
Около 16—17 часов дня бородачи становятся менее активными, но в общем
могут быть встречены на крыле до захода солнца. Во внешности бородача
бросается в глаза темная черноватая окраска спинной стороны и беловатая
окраска брюшной стороны у старых птиц; молодые темнобурые; под клювом
Рис. 75. Бородач.
БОРОДАЧ ИЛИ ЯГНЯТНИК
265
издали заметный пучок перьев — «борода». Осторожен. Голос — свист или
мяукающий звук.
Описание. Размеры и строение. Голова и шея в отличие от настоящих грифов
сплошь оперены, что указывает на меньшую специализацию в питании падалью; перья
затылка и шеи удлинены и заострены, вокруг клюва находятся густые и жесткие обращенные
вперед щетинкообразные перышки, образующие на подбородке бороду. Клюв удлиненный,
сжатый с боков и резко загибающийся на конце,менее массивный, чему грифов. Цевка
короткая, оперена до пальцев, когти тупые, но круто изогнутые. Маховые длинные, формула крыла
3>2>4>5>1>6, вырезки на внутренних опахалах 1-го—4-го маховых. Второстепенные
маховые короткие. Рулевых 12, хвост резко ступенчат. Крыло самцов и самок (11) 720—810,
в среднем 776 мм. Крыло кавказских экземпляров по Штегману (1937) 780—832, в среднем
808 мм. Вес (10) 5000—6750, в среднем 5680 г (европейские особи, Петере, 1940).
Окраска.У взрослых голова белая, либо желтоватая или ржавчато-рыжая с черными
пятнами на темени и с черной «бородой»; спинная сторона черновато-бурая с серебристо-
серым оттенком на больших перьях и в надхвостье, с белыми стволами перьев и наствольями;
маховые и рулевые серые с серебристым налетом и с бурыми стволами; горло и зоб белые или
охристые с отдельными черными крапинами, не образующими — в отличие от других
азиатских рас — ожерелья, хотя бы и неполного; остальная брюшная сторона белая, охристая или
рыжеватая. Свежее перо белое или беловатое, порыжение брюшной стороны зависит повиди-
мому от пропитывания пера секретом копчиковой железы.
В первом годовом наряде бородач с черной головой и шеей, буровато-сероватый на
брюшной стороне, на спинной — темнобурый с черно-бурыми маховыми и темнобурыми
рулевыми, с более или менее заметным беловатым размытым рисунком на некоторых перьях
крыла и на рулевых. Промежуточные наряды плохо выяснены, после первой годовой линьки
бородач становится несколько светлей, а на спине и шее появляются отдельные белые перья.
За этим нарядом следует еще один или два, приближающиеся к окончательному, но с
менее чистыми цветами и буроватым тоном брюшной стороны. У взрослых радужина желтая
или беловатая, окаймленная красным (видным) кольцом склеротики; клюв голубовато-
роговой, темнороговой у вершины; ноги серые, когти черные. У молодых радужина бурая
(с красным кольцом склеротики). Первый пуховой наряд охристо-белый, второй пуховой
наряд желтовато-серый.
Центральноазиатский бородач Gypaetus barbatus hemachalanus Hutton.
Gypaetus hemachalanus. Hutton. Journ. Asiatic Society of Bengal, VII, 1838, стр. 22,Симла-
Синоним. Gypaetusaltaicus. S harpe. Catal. Birds Brit. Mus. v. 1, 1874, стр. 229,Алтай.
Распространение. Ареал. Восточные части ареала вида в Азии от Тянь-
Шаня (Киргизский хребет и далее на восток) и Памира, к югу до Гималаев и
Бутана; горы Синда и Пенджаба (Соленый и Солиманов хребты); на востоке до
Нань-Шаня, Ганьсу, Сычуаня, Циньлинских гор и Тибета, возможно—до Хубея
и Юннани; на север до Джунгарского Ала-Тау и ю.-в. Алтая (редко в центр.
Алтае, сев. предел—Курайский хребет и Катунские альпы, западный Уймон,
Сушкин, 1938). Быть может в южн. Забайкалье: Тачановский(1891) сообЪцал
о гнездовании в Тункинских горах. Еще Палласом центральноазиатский
бородач наблюдался в 1772 году в горах Адунчалон между Чикоем и Аргунью, но
видимо он исчез оттуда после истребления там диких баранов (Радде, 1863),
хотя изредка попадался там и позднее. Добыт в 1907 г. на р. Мензе, притоке
Чикоя (Штегман, 1937). Залеты отмечены на водораздел между Леной и
Нижней Тунгуской, к сев. от Киренска ^ добыт осенью 1902 г. (экз. в
Зоологическом музее Московского университета), в сев. Манчжурию (Барга), в Порт-
Артур, в Сев. Китай, изредка в равнины Туркестана (Кызыл-Кумы). Сведения
о залете на Таймыр к Пясинскому озеру (Миддендорф) по всей вероятности
ошибочны.
Характер пребывания. Оседлая птица, на равнинах появляется
только случайно, хотя на охоте спускается до нижнего пояса гор. Имеются
указания о редкости птицы на вост. Памире в январе — феврале и о
возвращении туда в марте (Мекленбурцев, 1946).
Биотоп. Альпийская и субальпийская зона гор с осыпями, ущельями
и т. п. В вертикальном распространении одна из наиболее далеко идущих вверх
птиц. На Алтае бывает не ниже 2400 м, в Тянь-Шане выше 2000 м, обычно
266
ОТРЯД ХИЩНЫЕ ПТИЦЫ
около 3000 м и до 4270 м, на Памире на высоте 3500—3600 м, в Ладаке
до 4200 и даже 5000 м, в Тибете по крайней мере до такой же высоты,
чаще на 4000 м. Бородач, наконец, наблюдался над Эверестом в Гималаях на
уровне 7310 м.
Численность. Распространение несколько спорадично, но птица
в подходящих местах нередка.
Экология. Размножени е.. Большинство наблюдений сделаны за
пределами нашей страны. Цикл в общем сходный с циклом у переднеазиатского
бородача, начинается рано и тянется долго. Спаривание происходит на земле,
уже в середине января; брачный полет бывает—при этом птица на лету иногда
опрокидывается спиною вниз —в конце января —начале февраля (вост. Тибет,
Дза-чю в бассейне Меконга, Козлов, по Бианки, 1907). Спаривание и
брачный полет продолжаются и в период кладки и даже при насиживании:
в феврале —марте, а второй и в мае (Тибет, Шефер, 1938).
Кладка из 2 яиц, реже из 1 (Гималаи), изредка даже 3 (Тибет, Бенгт,
Берг, по Петерсу, 1940). Размеры: (60) в среднем 76,5 х 91,55 мм (Тибет и
Гималаи, Бекер, 1928). Две кладки из центрального Тянь-Шаня, найденные
18 и 19 марта, состояли одна из двух, другая из одного яйца; размеры их 80,8^
92x62—67,2 мм (Бамберг, 1908).
Гнезда расположены на скалах в более или менее глубоких нишах; сделаны
из сучьев с выстилкой из шерсти, веточек и т. п., иногда громадных размеров;
используются птицами много лет подряд. Пуховики бывают в апреле (Дза-чю,
Тибет, Козлов, 1907). Вылупление вероятно происходит уже в марте, так как
в конце июня в гнездах бывают крупные птенцы ростом со взрослых (25 июня
Киргизский хребет, Спангенберг, 1936), а в конце июля — начале августа
встречены летные молодые (29 июля, Тибет, Шефер, 1938; начало августа, верховья
р. Шахдара, Памир, Мекленбурцев, 1936; 20 июля, Киргизский хребет).
На возможность запоздалых по какой-то причине выводков указывает добыча
18 августа в Могол-Тау в Таджикистане молодого с недоросшими крыльями
(Иванов, 1940). Старые, в отличие от грифов, не кормят птенцов отрыжкой,
а приносят им корм в гнездо (Гималаи).
Линька. Не изучена, по всей вероятности сходна с линянием
предыдущей формы. В декабре старые птицы уже в свежем пере. Смена оперения
у птиц, не надевших еще окончательного наряда, начинается уже в марте —
апреле. Птица в 3-м — 4-м наряде, почти закончившая линяние, добыта в
Тянь-Шане 25 июля.
Питание. Относительно питания центральноазиатского бородача
существуют такие же противоречивые сведения, как об европейском. Несомненно
все же, что распространение этой птицы связано с наличием крупных копытных,
в особенности горного козла. Дикие звери (копытные) и даже птицы (кеклики,
улары) тревожно реагируют на появление бородача. Бородач может повиди-
мому переносить в лапах довольно значительные тяжести: в Гималаях в гнезде
найден целый труп козленка, полноги быка и т. п. (Бенгт, Берг). Бородач
предпочитает свежие трупы, но главным образом кости.
Относительно нападений бородача на баранов при перегонах их с горных
пастбищ Тянь-Шаня в долины, особенно при неблагоприятных
обстоятельствах — раннем выпадении снега и т. п. — сообщают киргизские пастухи:
птица при этом будто бы пытается сбросить одного из теснящихся у края обрыва
баранов, в особенности если он упадет или поскользнется. О таком же
способе нападения на взрослого горного козла в Джунгарском Ала-Тау писал
Карелин (1840).
Тому же наблюдателю принадлежит известный рассказ о нападении
бородача в Джунгарском Ала-Тау на казака при весьма опасных для последнего
обстоятельствах. Казак оступился и, не решаясь сам встать на краю обрыва,
БЕРКУТ
267
держался в ожидании помощи за куст. Налетевший бородач стал бить казака
крыльями и был отогнан только стрельбою. Надо полагать, что в этом случае
бородач принял казака за мертвого, так как на лету птица эта— без особых
враждебных намерений — нередко приближается к неподвижно сидящему и
в особенности лежащему человеку.
Вообще в Центральной Азии бородачи менее осторожны, чем в Европе
{Тибет, по Козлову и Шеферу). Найденные в желудках бородачей кеклики
(Афганистан), зайцы (Бекер), фазаны (Тибет, Козлов), быть может и свежая
голова козленка теке (Тянь-Шань) свидетельствуют о нападении этих птиц
на мелких живых животных. Есть указания на нападение бородача в Тянь-Шане
на сурков, в Тибете на сурков и зайцев (Пржевальский); на козляттека в Заи-
лийском Ала-Тау (Шульпин, 1929), будто бы даже на джейранов (Северцов).
Можно повидимому считать установленным фактом, что при благоприятных
обстоятельствах бородачи в Центральной Азии ловят живую добычу. Но место
ее в кормовом режиме бородача все же весьма незначительно. Главное значение
в пище этой птицы имеют падаль и кости {Памир). При этом бородач иногда
раздробляет кости, но в противоположность широко распространенному
мнению не бросает их для этой цели с высоты, а с силой ударяет о камни при
быстром спуске (Бенгт, Берг, 1931).
Описание. Размер ыистроени е. Крупней предыдущего: крыло (13) 800—930,
в одном случае 795, в среднем 850 мм. По Штегману (1937) крыло 813—893, в среднем 860 мм.
Окраска.У птиц в окончательном наряде темные пестрины по бокам зоба образуют
ожерелье, более или менее смыкающееся посередине зоба.
Систематические замечания. Как видно из предыдущего, морфологические различия
между G. b. aureus и hemachalanus реальны, хотя и незначительны. Замечательно, что
экология повидимому совершенно одинакова. Увеличение размеров с запада на восток в Азии
идет постепенно, и граница между подвидами проводится нами условно. Таджикиста некие
птицы принадлежат все же к западной, меньшей по размерам, форме. Крыло у них, по Ме-
кленбурцеву, 750—800 мм (птицы из Бабатага, Гузара, Джандака); у б птиц из Даштиджун-
ского района в Зоологическом музее Московского университета крыло 720—780 мм. У па-
мирских бородачей, по Мекленбурцеву, крыло 840—890 мм. У 9 птиц с Тянь-Шаня
(Киргизский хребет, Терскей Ала-Тау, Аксай, Сарыджас), хранящихся в Зоологическом музее
Московского университета, крыло 812—930 лш, у 10 птиц с Тянь-Шаня в Дарвиновском музее
в Москве крыло 845—912 мм. Средняя длина крыла у таджикистанских (7) 763, у тяныиан-
ских (19) 866 мм,
РОД ОРЛЫ AQUILA BRISSON, 1760
Тип Aquila mogilnik
Синонимы: Aetos Severtzow, 1885. — Hieraaetus Каир, 1814.
32 Беркут. Орел-беркут Aquila chrysae'tus L.
Falco chrysaetus. Linnaeus. Syst. Nat. ed. X, 1758, стр. 88, Швеция.
Русское название. Слово орел — старинное русское, сходное с названием этой
птицы у других славянских народов и возможно близкое к древне-германскому Ааг, Аго
(скандинавское От, шотландское Erne). Названия у русских писателей XVIII века, в
частности у В. Левшина, «каменный» и «золотой» орел — вероятно, перевод с немецкого (Stein-
adler, Goldadler). Имя «камский орел», по замечанию того же автора, объясняется тем,
что беркутов башкиры ловили и вынимали из гнезд на Каме для продажи в Киргизской степи.
Со времени Па л ласа многие русские авторы, включая Северцова и Мензбира,
принимали для орлов два видовых имени: беркут и халзан; при этом беркутом называли молодых,
а халзаном старых птиц. Название беркут (буркут, бюркют), тюркское и широко
распространено у народов Средней Азии, где с этой птицей много охотятся и теперь. У киргизских
сокольников — беркут (но не карагуш, как пишут некоторые). Любопытно, что у жителей
Уэльса для крупных хищных птиц существует имя «баргуд» («bargud»), сходство которого
со словом беркут, по справедливому замечанию А. Ньютона, заслуживает внимания.
Распространение. Ареал. В Европе несколько спорадичен —* в
Шотландии, южн. Франции (Ардеш, Косе), на Пиренейском п-ове, в Альпах,
Апеннинах, на Балканском п-ове, на Карпатах, в Скандинавии и на
востоке от Финляндии до границы леса на севере СССР; на островах Средизем-
268
ОТРЯД ХИЩНЫЕ ПТИЦЫ
ного моря — Балеарах, Корсике и Сардинии; в сев. Африке — Египте, Тунисе,
Алжире, Марокко; на Синайском п-ове и в Передней Азии — в М. Азии,
Сирии, Иране; в Средней и Центральной Азии до зап. Китая и Гималаев; по
всему лесному поясу Сибири до Камчатки; в Японии; в Сев. Америке к югу до
Мексики.
Карта 40. Распространение
беркута Aquila chrysaetus
7 — А. с. fulva, 2 — А. с.
chrysaetus, 3 — А. с. kamtschatica, 4 — А.
с. daphanea, 5 — А. с. homeyeri,
6 — А. с. canadensis; a — граница
гнездовий, б — невыясненные
границы распространения; цифры в
кружке соответствуют номеру
подвида и показывают зимние находки их.
Характер пребывания. Оседлая птица в большей части ареала,
совершающая однако зимой кочевки -^ вертикальные (из гор на равнины) и
горизонтальные — последние более или менее регулярно в Сибири»
Биотоп. В южной части ареала —за редкими исключениями — гористые
местности; в Вост. Европе и в Сибири —на равнинах, в высокоствольных лесах
и в горах.
Подвиды и варьирующие признаки. Изучены недостаточно, и описанные формы во вся*
ком случае крайне близки и в морфологическом, и в экологическом отношении.
В западной части ареала, в Зап. Европе, на юг до М. Азии, Кавказа, Ирана и Туркмении
A. ch.fulva L., 1758; в сев. и вост. Европе от Скандинавии до Зап. Сибири A. ch. chrysaetus
L., 1758; в ю.-з. Европе на Пиренейском п-ове и в сев. Африке A. ch. homeyeri Sever t-
zo w, 1888; в средней и Восточной Сибири A. ch. kamtschatica Severtzow, 1888; в
Центральной Азии A. ch. daphanea M e n z b i е г 1888; в Сев. Америке A. ch. canadensis L.,
1758. Морфологически варьируют размеры и окраска: южные подвиды темной окраски и
небольших относительно размеров, за исключением только горной расы daphanea, очень
крупной; северные подвиды — chrysaetus, kamtschatica, canadensis темнеют с запада на восток;
БЕРКУТ
269
величина из них наименьшая у chrysaetus, наибольшая у kamtschatica и canadensis. Изменения
©краски и размеров соответствуют в общем так называемым правилам Глогера и Бергманна.
Экологические различия незначительны: биотопическое размещение и детали годичного
цикла выяснены недостаточно.
Североевропейский орел-беркут Aquila chrysaetus chrysaetus L.
Синоним. Aquila nobilis. Pallas. Zoographia Rosso-Asiatica, 1811, стр. 338, Урал,
Распространение. Ареал. Скандинавия (в Норвегии от 70° на севере до
Кристианзанда и Хамара на юге, в Швеции в северных частях до Даларне и
Херьедален на юге); Финляндия; Европейская часть СССР на север до границы
высокоствольного леса—около 69°—70° с. ш. в Лапландии, у Архангельска,
на Печоре у полярного круга; в Сибири на восток до Енисея; к югу до
Прибалтики и Полесья; Подолия, Черниговской и Харьковской обл., Тамбовской
обл., на Волге по крайней мере до Бузулукского района в Куйбышевской
обл.; на Южн. Урале примерно до 52° с. ш. (Зилаир); спорадичен и южней
(б. Эристеневский улус, урочище Цаган-нур-улун, Россиков, 1916); быть
может на Общем Сырте и сев. склоне Мугоджар; в борах и колках юго-зап.
Сибири, к востоку до предгорий Алтая. В Туркестане и на Кавказе—другой
подвид.
Характер пребывания. Оседлая птица, совершающая в
негнездовое время кочевки (особенно молодые птицы). В это время беркуты более
или менее регулярно появляются в различных странах Западной Европы,
в степях таврических, северокавказских и донских, заволжских и зауральских,
в ю.-з. Сибири и Казахстане. Поэтому в южной полосе орлов больше зимою,
чем летом. Откочевки и зимние кочевки орлов отмечены в сев. и средней
полосе (Псков, Вологодская обл.; Поволжье —б. Симбирская губ.), с другой
стороны и на севере (Вологодская обл., Лапландия и т. д.) беркуты
встречаются и зимой.
Биотоп. Необходимая для беркута комбинация гнездовых и кормовых
условий сводится к; наличию скал или деревьев, где он гнездится, и достаточно
широких открытых пространств, так как размах крыльев затрудняет ему охоту
между деревьями в лесу. В тайге беркут встречается поэтому в тех местах, где
лесные массивы перемежаются с открытыми местами — речными долинами,
болотами и т. п. Населенных районов беркут избегает. Например, в Западной и
средней Европе, где он когда-то был распространен и в равнинных лесах,
теперь встречается только в малодоступных скалистых горных участках.
Численность. Число орлов в Европе значительно сократилось —
как в результате вырубки старых лесов, так и из-за прямого преследования,
продолжавшегося с середины XVIII по конец XIX столетия. В Германии
беркуты повидимому вовсе перестали гнездиться, в Швеции их осталось 40—
47 пар (Цедлитц, 1922). Плотность распространения внутри ареала невелика,
повидимому, даже при наличии подходящих биотопов, так как гнездовые и
охотничьи участки отдельных пар значительны. Относительно многочисленен
беркут на северо-востоке европейской части нашей страны. Периодические
колебания численности не отмечены, что вероятно отражает разнообразие кормового
режима.
Экология. Размножение. Так как у гнезд добывались и
наблюдались и молодые особи-птицы с белым основанием рулевых, то очевидно орлы
размножаются до достижения полного взрослого наряда. Гнездовые участки
весьма велики и постоянны (расстояние пары от пары около 10 км,
Ульяновская обл., Житков и Бутурлин, 1906). У каждой пары 2—4 гнезда,
используемых по годам постепенно, но иногда и подряд (Лапландия). Расположены
гнезда на больших деревьях—соснах, дубах, грабах, реже на скалах или
обрывах (Калмыцкая степь, Россиков, 1916), обычно высоко от земли, в 12— 20 м
270
ОТ РЯД ХИЩНЫЕ ПТИЦЫ
и более. Строится гнездо из толстых сучьев, причем, подновляясь, с годами
достигает огромных размеров: до 200 см высоты и столько же в диаметре
(Мензелинск). Подстилка из тонких частей растений, шерсти, волоса и т. д.
Сезон размножения ранний, пары, судя по другим подвидам, постоянные и дер»
жатся вместе круглый год. Брачный полет уже в марте.
Кладка в разные числа первой половины апреля (полная кладка в
Лапландии 25 апреля, Плеске, 1886; 14—18 апреля, Харьковская обл., Горленко и
Рудинский,1937; 23 апреля, Куйбышевская обл., Карамзин, 1901; начало апреля
старого стиля, б. Шацкий у. Тамбовской губ.; 21 апреля, Черниговская обл.).
Число яиц в кладке колеблется от одного до трех, чаще бывает два. Окраска
яиц грязнобелая с бурыми пестринами и крапинами, скорлупа грубая.
Размеры (6) 73,5—78 х 56,7—59,2 мм (Горленко и Рудинский, 1937). В
случае гибели яиц бывает дополнительная кладка. Промежуток между откладкой
яиц, судя по разнице в возрасте птенцов, 3—5 дней.
Насиживание происходит с первого яйца. Участие родителей у нас не
прослежено, но по западноевропейским данным насиживают самец и самка:
продолжительность насиживания повидимому, около 45 дней. Пуховики — два
или один — находятся в гнезде в конце мая, подлетки близ гнезд в конце
июля — начале августа. После вывода птенцы долго держатся у гнезд и только
к концу августа становятся самостоятельными. Весь гнездовый период
продолжается таким образом около 75 дней.
Разбиваются выводки в конце сентября — начале октября, когда молодые
орлы держатся отдельно, а старые птицы держатся в парах весь год. Кормит
птенцов, пока они малы, самка, которой самец передает добычу. При этом близ
гнезда орлы не охотятся. Нередкое явление —гибель младшего птенца еще
в раннем возрасте, причем происходит она повидимому или в результате
агрессивности старшего птенца (к которой родители относятся безразлично)
или из-за недокорма. Нередко впрочем вырастают и оба птенца (Полесье, Шнит-
пиков, 1913; Мензелинск). Отмечено, что благополучное выращивание двух
нтенцов происходит в годы, обильные кормом (по.Россикову, в мышиные годы
орлы выкармливают птенцов главным образом полевками и мышами). Старые
орлы у гнезд осторожны и, потревоженные неоднократно, бросают даже
птенцов (Лапландия).
Линька изучена недостаточно. Полная годовая. Общая
продолжительность приблизительно с апреля до октября. Последовательность смены крупного
оперения не выяснена. Как и другие крупные хищники, беркуты даже после
окончания линяния имеют некоторое число старых выцветших перьев среди
оперения (в том числе маховых и рулевых). Последовательность смены нарядов:
первый пуховой наряд — второй пуховой наряд — первый годовой
(гнездовый) наряд—второй годовой (промежуточный) наряд и т.д., пока после
четвертой линьки не надевается окончательной — пятый годовой наряд.
Мнение многих старых авторов, будто бы орлы линяют не каждый год и что
линяние чередуется у них по годам с размножением, —неосновательно.
Питание. Беркут наиболее энергичный из орлов, питающийся поэтому
относительно крупной добычей. Мало специализирован в отношении кормового
режима и нападает поэтому и на зверей, и на птиц, но не может однако ловить
последних на лету. Главное место в питании занимают звери средней величины
от лисицы до зайца, и куриные птицы, начиная от самцов глухарей. В виде
исключения орлы нападают и на более крупную добычу (телята северных
оленей, козуль и т. п.), и на более мелкую. Выношенные птицы берут антилоп
(джейраны), волков, даже куланов (Северцов). Имеются сведения о
безрезультатном нападении орла на проходившую в лесу женщину зимой 1938—1939
близ г. Уржума (Плесский), а также о том, что охотничий орел убил казаха,
вцепившись в его лисий малахай (Зарудный, 1888).
БЕРКУТ
271
В качестве пищи беркутов в СССР указываются зайцы, суслики, хомяки,
новорожденные северные олени, козули, лесные куницы, горностай, белка,
лисица, ежи, разные полевки и мыши; из птиц глухари, тетерева, белые
куропатки, перепела, утки, цапли, домашние гуси, быть может даже дрофы и жу-*
равли, галки, сороки, сарыч, филин, длиннохвостая неясыть и т. д. Зимой пишд
орлов более однообразна, состоит главным образом из зайцев и куриных птиц,
В этот период орлы,
особенно молодые,
нападают и на домашних
животных— птиц и собак,
а также охотно клюют
падаль.
Потребность в пище
взрослого орла около
1 000 г в день (судя по
опыту соколиных
охотников). Без пищи орел
может прожить долго
(до 5 недель). Вне
периода размножения орлы
охотятся обычно парами.
На охоте беркут парит,
то высоко, то низко над
землею, но часто
пользуется и активным
полетом и, завидев добычу,
стремительно бросается
на нее. При нападении
беркут весьма быстр и
поворотлив. Захват и
умерщвление добычи
происходит по-разному.
Чаще всего орел хватает
зверя одной лапой за
голову, а другой за
спину, стараясь
переломить ему позвоночник,
или разрывает
схваченному животному
крупные шейные сосуды
клювом. Сидя на крупной
сопротивляющейся
добыче, орел не складывает крыльев, а балансирует ими, сгибая то одно, та
другое и нанося удары твердыми маховыми. Добыча меньших размеров
умерщвляется по-ястребиному, когтями.
Полевые признаки. Крупная птица, около 2 м в размахе, темнобурой, на
расстоянии однообразной, окраски; у молодых резко двуцветный хвост (белый
с черным концом). На полете, как и другие орлы, беркут пальцеобразно
расставляет передние маховые; крылья довольно узкие; хвост слегка закругленный
и относительно длинный. Очень осторожен. Голос «кьек-кьек-кьек», так
называемый клекот, походящий на лай; кроме того,мелодичная трель и иные звуки
Описание, Размеры и строение. Крупный орел гс относительно узкими
крыльями и длинным слегка закругленным хвостом из 12 рулевых. Клюв высокий, довольна
сжатый с боков, с узкой вертикальной ноздрей. Лапы сильные, с очень мощными когтями,
Рис
76. Беркут (слева хвост молодой птицы).
272
ОТРЯД ХИЩНЫЕ ПТИЦЫ
особенно велики когти заднего и внутреннего пальцев, остальные меньше (лапа типа
«ястреба»). Формула крыла: 3>5>6>2>7>8>1>9>10; вырезки на наружных опахалах 2-го —
6-го маховых, на внутренних опахалах 1-го — 5-го маховых. Длина самцов (2) 820—850,
самок (3) 910—928 лш, размах самцов (2) 1880—2015 мм. Вес самцов (2) 2840—3150 г, самок
(1) 3840. Крыло самцов (13) 595—635, самок (16) 642—708, в среднем 614,8 и 668,8 мм.
Окраска- Первый пуховой наряд белый с сероватым налетом, второй чисто белый.
Первый годовой наряд темнобурый с беловатыми основаниями перьев, иногда выступающими
на брюшной стороне, но обычно прикрытыми темной вершиной; задние первостепенные
маховые и второстепенные с белыми основаниями перьев, выступающими в виде белого пятна на
нижней поверхности развернутого крыла; оперение цевки беловато; задняя часть головы
и шея светлоохристые; маховые черновато-буры, рулевые белые с широкой черной вершинной
полосой.
В окончательном (вероятно 5-м годовом) наряде общая окраска спинной стороны темно-
бурая, в свежем пере с фиолетовым отливом, перья с прикрытыми белыми основаниями; перья
затылка и зашейка заострены, ржавчато-желтоватые, лоб и темя темнобурые(но не черноватые
и не образуют резкой «шапочки», как у других рас). Брюшная сторона несколько светлей
спинной, обычно с золотисто-рыжими пятнами на зобе, груди (иногда на брюхе) и голени; эта
«золотистая» вариация не встречается у других подвидов беркута в СССР; подхвостье
рыжеватое ; маховые черноватые с серым размытым мраморным рисунком у оснований внутренних
опахал; рулевые с черноватой вершиной и серыми, испещренными неправильным поперечным
рисунком, основаниями. Второй и третий годовой наряды отличаются от окончательного
тем, что основание хвоста беловатое или сероватое с немногими и нерезкими пестринами;
рыже-золотистые пятна на брюшной стороне развиты слабо или совсем отсутствуют.
Четвертый наряд повидимому отличается от окончательного тем, что самые основания рулевых и
маховых еще беловатые. Оба пола окрашены одинаково. Клюв черноватый, синевато-серый
у основания; восковица, разрез рта и лапы желтые; когти черные; радужина золотисто-
буроватая или ореховая.
Восточносибирский орел-беркут Aquila chrysaetus kamtschitica S e v.
Aquila fulva kamtschatica. Severtzow Nov. Mem. Soc. Imp. Natural. Moscou livr.
5, v. XV, 1888, стр, 180, Камчатка.
Синонимы. Aquila chrysaetus obscurior. Сушкин. Список и распределение птиц
Русского Алтая, 1925, стр. 59, Алтай. — Aquila fulva japonica. Severtzow. Nouv. Mem.
Soc. Imp. Natural. Moscou, v. XV, 1888, стр. 182, Япония.
Распространение. Ареал. В Азии к востоку от предыдущего в заени-
сейской Сибири, на севере в низовьях Яны, на Индигирке до 69°60' с. ш. (Хаяга-
стах, Михель, 1935); на Колыме повидимому всюду в горных районах; на
Охотском побережье; на Анадыре в лесной области; на восток до Коряцкой Земли и
Камчатки; быть может гнездится в Приморье; на юг до Алтая, Абаканской и
Минусинской лесостепи, ю.-в. Забайкалья, а также сев. Монголии (Гобий-
ский Алтай, Ханген и Кентей). Повидимому к этой же форме относятся
беркуты из Японии (Хондо), Курильских о-вов, гор Манчжурии и Сев. Китая.
Характер пребывания. В северной части ареала, в Якутии, —
перелетная птица, но на Анадыре и в Коряцкой Земле уже оседлая; в низовьях
Яны отмечен в конце сентября (Бунге, 1887). У Якутска прилет отмечен в конце
марта —начале апреля, отлет в начале октября (Воробьева, 1931; впрочем,
добыт еще 1 ноября в 100 км северней, Иванов, 1929).
Биотоп. На с.-в. Сибири пока найден лишь в лесистых местностях,
а гнезда лишь на деревьях; южнее встречается и в горах, где гнездится на
скалах (Алтай, Забайкалье, Монголия).
Численность невысока. Расстояние между гнездовыми участками
в районе Верхоянска около 10 км (Ткаченко, 1932 г.); в ю.-в. Забайкалье 20—
25 км (Штегман, 1928). На Камчатке, в Коряцкой Земле и на Анадыре редок.
Экология. Размножение. Сроки размножения повидимому те же,
что у A. chrysaetus. Гнезда в Якутии расположены на деревьях (главным
образом на лиственнице); в Минусинской лесостепи — на березе; в с*-в. Сибири
также на тополе, часто невысоко от земли, в 4—8 м; в горах на скалах, обычно
на труднодоступных утесах. Одно и то же гнездо используется много лет,
хотя вероятно у каждой пары есть и «запасные» гнезда. У Верхоянска
БЕРКУТ
273
24 мая в яйце наблюдался почти сформировавшийся зародыш; начинающий
оперяться птенец на Яне — 5 июля. Готовый к вылуплению зародыш обнаружен
ниже Верхоянска 26 мая. Подлетки с недоросшими крыльями встречаются
с половины мая (Якутск; Забайкалье; Яна; Минусинская лесостепь; Алтай)
и до начала августа. В выводке обычно отмечен только один птенец; однако в
зап. Саяне, устье р. Тупсен-кем, за 4 года у пары беркутов вывелось 7 птенцов.
Линька. Изучена недостаточно, общий ход протекает как у
номинальной расы.
Питание. В качестве добычи восточносибирского беркута приводятся:
зайцы, суслики, тарбаганы, пищухи, белки, попавшие в капкан звери, в том
числе соболи; лисицы, возможно молодые телята северного оленя; молодые
сибирские горные козлы, домашние козлята; из птиц утки, улары и другие
куриные и т. п.
Описание. Размер ыистроение. Несколько крупней североевропейской расы:
крыло самцов (4) 620—683, самок (4) 650—700, в среднем 640,7 и 674,3 мм. Вес самца 3450,
самки 4220 г (Кроноцкий заповедник, Камчатка).
Окраска. Заметно темней, чем номинальная раса; у взрослых на темени хорошо
заметная темная черноватая «шапочка*, доходящая до бровей; заостренные перья затылка
и прилегающих частей шеи густо рыжие, а не охристые; перья эти уже; общий бурый тон
темней; охристо-ржавых пестрин на брюшной стороне нет или они слабо развиты; темный
рисунок на рулевых преобладает над светлым. Молодые однообразной черноватой окраски,
с относительно слабым распространением белого цвета на основаниях перьев; оперение цевкн
буроватое.
Систематические замечания. В коллекциях имеется недостаточно материала по
восточносибирским беркутам для решения вопроса об отношении их к американским
и для точного разграничения отдельных форм в вост. Азии. Относительно японских
орлов указывается, что они, как и манчжурские и северокитайские, меньше по
размерам, чем восточносибирские. Птицы с крайнего востока Сибири — от Якутска до границы
леса на севере, Анадыря на востоке — отождествляются нередко с американскими. При
ограниченности материала и большой индивидуальной изменчивости у этого вида кажется
правильней объединять всех восточноазиатских орлов в одну форму, оставив открытым
вопрос об отношении их к A. ch. candensis.
Южноевропейский орел-беркут Aquila chrysaetus fulva L.
Falco fulva. Linnaeus. Syst. Nat. ed. X, 1758, стр. 88, Англия,
Синоним: Aquila fulva intermedia. Северцов. Вертикальное и горизонтальное
распространение туркестанских животных, 1873, стр. 112, равнинный Туркестан.
Распространение. Ареал. Горы Зап. и южной Европы от Шотландии до
Франции, Пиренеев и Альп; Балканский п-ов; вероятно к этому же
подвиду орлы из Карпат. Кроме того на Кавказе (в Крыму гнездованье не доказано,
но оно весьма вероятно в горной части; в равнинной части зимою там
встречается номинальная раса), а также в Туркмении от Больших Балханов и
Куба-Дага до вост. Копет-Дага. Сюда же относятся и гнездящиеся в Кара-
Кумах и Кызыл-Кумах беркуты, а также птицы с Усть-Урта. В Азии вне СССР,
в М. Азии и прилежащих частях Ирана.
Характер пребывания. Оседлый.
Биотоп. Скалистые участки гор; в Передней Азии и на равнине в пустыне,
где есть выходы скал или более или менее^высокие саксаульники, или наконец
развалины (например крепость Шах-сенем в вост. Кара-Кумах). На Кавказе
от 1200 до 3100 (Армения, центр. Закавказье), в Туркмении до вершин Б.
Балханов и Копет-Дага, т. е. несколько более 2000 м.
Численность. Редок.
Экология. Размножение. Пары постоянные, самцы и самки
держатся вместе целый год (наблюдения в Закавказье — в январе —феврале, Цвет-*
ков; в декабре —феврале в Копет-Даге, Дементьев, 1950). Брачный полет
начинается уже в феврале (центр. Копет-Даг). Гнезда расположены на скалах,
в горах, в пустынях на останцевых холмах, выходах каменистых пород, развали-
18 Птицы СССР, т. I
274
ОТРЯД ХИЩНЫЕ ПТИЦЫ
нах сооружений, на саксауловых деревьях (черный саксаул), редко на кандыме
или джидовнике; в сев. Иране на фисташковых деревьях (Зарудный, 1903);
Гнезда устроены из ветвей со скудной выстилкой из сухой травы.
Число яиц в кладке 1—2. Размеры их 72,5—80x56,5—57,2 (Кызыл-
Кумы, Зарудный, 1915). Сроки кладки не выяснены, но повидимому ранние—
в первой половине марта и в таком случае раньше, чем у номинальной расы (по
сведениям местных охотников казахов в Кызыл-Кумах кладка начинается уже
в феврале по старому стилю). Взамен утраченных кладок бывают вторые.
Пуховые, начинавшие оперяться птенцы добывались в мае (Армения,
Челекен). В конце июня находятся уже вполне оперенные или имеющие
незначительные остатки пуха орлята с недоросшими крыльями и хвостом (22 июня
на Аму-Дарье ниже Турт-куля; 22 июня в Кызыл-Кумах; 28 июня в Копет-Даге
у Чули; с другой стороны, молодой с преобладанием пуха на брюшной стороне
у Самура, Дагестан, еще 2 июля). Хорошо летающие молодые встречаются при
стариках в Туркмении и на Кавказе в разные числа июля, при этом отмечается
обычно один только птенец (редко два, Дарваза, в центр. Кара-Кумах, Каш-
каров и Коровин, 1929). Неразбившиеся выводки в Закавказье бывают еще
в середине октября.
Линька. В деталях не изучена. В соответствии с более ранним
размножением, у переднеазиатских орлов линяние происходит несколько раньше, чем
у номинальной расы.
Питание. В Туркмении пищу южноевропейского беркута составляют
зайцы, суслики, молодые джейраны, молодые горные бараны и козлы, кек-
лики; на пролете водоплавающие птицы; кроме того, падаль. В вост. Иране—
джеки, зайцы, суслики и песчанки в долинах; зайцы и кеклики в горах, нередко
молодые джейраны, даже кулан жеребенок (Зарудный, 1903). В Кызыл-Кумах
молодой архар, домашний козленок, джейран, черепаха, еж, суслики, зайцы,
песчанки,джеки, кеклики. В Закавказье— зайцы, улары,кеклики(Закаталы).
Описание.Р а з ме р ы. Сходны с номинальной расой; крыло кавказских и копет-
дагских самцов (12) 600—643, самок (5) 635—684, в среднем соответственно 622,5 и 662 мм.
Окраска. В среднем посветлей сибирских беркутов, гнездовое оперение не
черноватое, а темнобурое; перья зашейка и затылка бледнобурые, а не темнорыжие; у
взрослых пятна на груди, подхвостье, перья цевки бледнобурые, не ржавчато-рыжие, как
у номинальной расы. «Шапочка» на голове, хорошо заметна.
Центральноазиатский беркут Aquila chrysaetus daphanea Menzbier
Aquila fulva daphanea. Menzbier. Ornithologie du Turkestan, 1888, стр. 75, Непал.
Синонимы. Aquila chrysaetus hodgsoni Ticehurst. Bull. Brit. Orn. Club LI I,
1931, стр. 25, Хазара, в с.-з. Индии.
Распространение. Ареал. Высокогорная центральная Азия от горных
частей Китая (Ганьсу, Сы-Чуань, Нань-Шань, Чжили, Шеньси, Хубей,Юннайь),
на юге до Гималаев, включая Ассам; на западе до Куэн-луня в Синцзяне,
Белуджистана и Афганистана, в СССР в Тяньшанской и Памиро-Алтайской
горной стране, на западе до Таджикистана (вероятно к этой форме и беркуты
из Кугитанга). К северу быть может до Тарбагатая. Зимой в Среднем Китае,
быть может сюда же орлы из Манчжурии (Хартерт и Штейнбахер, 1937).
Характер пребывания. Оседлая птица, совершающая однако
зимой кочевки и спускающаяся с гор в долины (часть особей остается в
высокогорье и зимою).
Биотоп. Скалистые участки гор, как в альпийской зоне, так и в зоне
хвойного леса й арчевника на гнездовье; зимой встречается и ниже.
В вертикальном отношении в Гималаях примерно от 2000 до 3000 л/, в
Тибете от 2600 до 4000, в Тянь-Шане от 2000 до 3000 м, на Памире — до 3700 м
(озеро Ранг-куль, Тугаринов, 1930).
ОРЕЛ-МОГИЛЬНИК
275
Численность. Относительно нередок, хотя и немногочисленен.
Экология. Размножение. Брачный полет происходит уже в январе
(Памир). Пары постоянные, птицы держатся вместе и зимой. Цикл
размножения ранний, но повидимому начинается поздней, чем у A. ch. fulva.
Кладки в Гималаях бывают в феврале—марте, в Средней Азии — в сере^
дине марта. Число яиц 1—2 (Тянь-Шань; Фергана; Ганьсу; Нань-Шань).
Размеры яиц из Туркестана 70,8—71,6 х 57,1—58,7; из Тянь-Шаня 78 х 61,4
и 74 х 56,8 мм (Бамберг, 1906); из Ганьсу 78,5—79,9x62,8—64,3 (Штрезе-
манн, 1938), из Гималаев в среднем (5) 78,1-60,4 мм (Бекер, 1928).
Гнезда расположены главным образом на скалах, но иногда и на деревьях
(сосна, в 15—18 м от земли, Ганьсу; ели в Тянь-Шане, по сведениям киргизских
сокольников). Вывод птенцов наступает в начале мая. Относительно нередко
бывает два птенца (Или, Нань-Шань). Во второй половине июня птенцы
оперены и в начале июля — еще с недоразвитыми маховыми —покидают гнездо.
Вполне выросшие молодые — в начале августа.
Линька. Как у других беркутов.
Питание. В качестве кормов этого орла указываются кеклики и сурки
{Киргизский хребет), молодые копытные—козлы и бараны, сурки, зайцы,
лисицы, кеклики и улары (центр. Тянь-Шань), зайцы и улары, падаль
(Памир), сурки, молодые копытные, улары, ушастые фазаны HjT. д. (Тибет).
Этот подвид беркута часто употребляется в Средней Азии для охоты —
выношенные орлы берут джейранов, лисиц и даже волков.
Описание* Крупный орел, размерами с восточносибирского: крыло около 640—725мм.
Вес самцов (2) 4000—4100, самки 6350 г (центр. Тянь-Шань). По окраске близок к fulva и kamt-
schatica, но в общем темнее (в свежем пере черноватый), с резкой черноватой «шапочкой»
на темени, с рыже-коричневым затылком и зашейком; на груди и голени заметны ржавчато-
бурые пятна, никогда не достигающие впрочем такого развития, как у A. ch. chrysaitus.
Светлые основания перьев слабо развиты уже и в первом наряде; корневая белизна
рулевых исчезает у взрослых полностью и темный рисунок на них достигает наибольшего
развития среди беркутов.
33. Орел-могильник Aquila heliaca Savigny
Aquila heliaca. Savigny. Description cTEgypte, 1809, стр. 82, табл. 12, Египет.
Русское название. Название могильник происходит оттого, что птица эта обычно
встречается в степях на курганах и могильных холмах. У азиатских народов «каракуш»,
т. е. черная птица — в отличие от беркута.
Распространение. Ареал, Южная Европа от Пиренеев и Венгрии,
южная полоса Европейской части СССР и до Передней .Азии; с.-з. Индия,
южн. Сибирь и Монголия.
Характер пребывания: на севере перелетная, на юге — в
Передней Азии и Индии — оседлая птица.
Биотоп. Лесостепь и полоса островных лесов,
Подвиды и варьирующие признаки. В Испании и возможно в Марокко A. h, adalberti
В г е h m, 1861, в остальной части ареала—А. Л. heliaca Sa vi g ny. Различия в окраске,
экологические особенности подвидов не выяснены.
Обыкновенный орел-могильник Aquila heliaca heliaca Savigny
Распространение. Ареал. От Югославии (Славония, Сербия) по
Балканам до Добруджи; Кипр, М. Азия, Иран, Белуджистан, с.-з. Индия; кроме того,
в Джунгарии и сев. Монголии до Хангая и быть может Сев. Китая. В СССР
встречается в южн. половине Европейской части страны, к северу до сев. частей
Украины (Киевская, Харьковская обл., южн. часть Черниговской),
бассейна Оки и Камы, в Зауралье и Зап. Сибири на север примерно до 56° с. ш;
(Ялуторовск, Салаир, Томск, Красноярск), к югу до Крыма, Кавказа (но там рас^
is*
276
ОТРЯД ХИЩНЫЕ ПТИЦЫ
пространение неясно, в Армении, например, на гнездовье достоверно не найден),
Туркестана, где спорадически гнездится в низовьях Сыр-Дарьи, в Джунгарском
Ала-Тау и в вост. Тянь-Шане (Или), спорадически в Кызыл-Кумах и быть
может в Туркмении; далее на Алтае, в Минусинской лесостепи и у Ачинска, затем
спорадически в Балаганской степи, у Иркутска, Троицкосавска и Аги. Залеты
отмечены к северу до Псковской, Московской, Тульской обл., в Литву и
Прибалтику; в Зап. Европе в Австрию, Богемию, Германию, Голландию, южн. Францию,
Италию и даже в Швецию и Лапландию, на востоке до Японии, Кореи и разных
частей Китая от севера до Фучжоу. Зимою в Закавказье, Туркестане, Иране,
Монголии, Индии (до вост. Бенгалиии Деккана), Китае (отЧжили и Шаньси до
Карта 41. Распространение могильника Aquila heliaca
1 — A. h. adalbertl, 2 — A. h. heliaca; a — границы гнездовий, г — область зимовок.
Фуцзяня и быть может центральных провинций), в Лаосе и с.-в. Тонкине
(Делакур, 1931), в Африке от южн. Аравии, Египта до Эритреи, Судана, Нубии
и Абиссинии.
Характер пребывания. На севере, в частности в СССР,
перелетная птица; часть особей впрочем зимует уже в Закавказье и Туркестане;
южнее оседлая или кочующая. Даты, Прилет —в первой половине апреля,
отлет в сентябре и октябре, последние особи — не позже первой половины этого
месяца. Пролет и появление в Туркестане начинается уже с конца февраля
(Кызыл-Кумы) и в марте. Весной летят парами и одиночками или небольшими
группами; осенью в стаях, иногда по несколько сот штук, часто вместе со
степными орлами (Зарудный, 1888). Движение к гнездовьям весной медленное,
с задержками и остановками в пути (например пролетные орлы несколько
дней держались у пос. Джамбейта Уральской обл., близ большой грачиной
колонии и кормились грачами, Павлов, 1927).
Биотоп. В лесостепной и степной полосе — поросшие древесной
растительностью участки, в Туркестане и в пустыне. Предпочитает смешанные и
лиственные леса; в Сибири и в сев. Казахстане встречается и в борах. В горах на
гнездовье до 1300—1400 м; на охоте и выше, например, на Алтае до 2000 м,
в Кашмире до 2100 м.
орел-могильник 277
Численность. В лесостепной полосе и в лесах юга европейской
России относительно многочисленен, в Туркестане и к востоку от Енисея редок
и спорадичен. Редок и у северной границы ареала, где гнездится нерегулярно
(например, в южн. Приуралье в Ильменском заповеднике залетал в 1936—-
1939 г., стал гнездиться с 1940 г.). В богатых пищей лесах гнезда иногда
находятся на расстоянии только до l1^ км (Воронежская обл., Огнев и Воробьев,
1924); в Башкирии примерно 4 пары на 10 км2 (Сушкин, 1897). Полифагия
могильников служит
вероятной причиной их
высокой
плодовитости; по той же
причине (в отличие от
других орлов)
младшие птенцы в
выводках обычно
выживают.
Экология*
Размножение.
Половая зрелость
наступает до приобретения
окончательного
наряда. Гнездовые участки
весьма постоянны,
различных размеров—
в зависимости от
биотопических условий,
иногда в глубине
леса, иногда на
опушке. Одно и то же
гнездо используется
много лет подряд»
причем у пары орлов
есть обычно 2—3
гнезда, которые орлы
занимают посменно.
Размеры гнезда
варьируют в зависимости от
его возраста, но уже в
первый год оно
достигает 1 /2 м В диаметре Рис. 77. Могильник (старая и молодая птицы).
и 70 м в высоте
(Зауралье, Ильменский заповедник, Ушков). Гнезда расположены на различных
деревьях — на соснах, дубах, тополях, в Туркестане и на саксауле и джидов-
нике, в безлесных местностях на отдельно стоящих деревьях и в
исключительно редких случаях по обрывам (сев. Казахстан, Сушкин, 1908). Гнезда
сделаны из толстых сучьев, строятся обоими родителями, примерно 10 дней.
Высота расположения гнезда от земли различна, иногда только в 2 м, но
обычно выше, от 10 до 25 и более метров (в зависимости от наличия подходящих
деревьев). Лоток выстлан сухой травой, веточками, иногда обрывками кошмы
или пометом. Обычно гнездо, а также птенцы при отсутствии родителей
закрываются зелеными ветвями. Спариванье наступает в конце апреля после
прилета; в этот период наблюдается характерный брачный полет над гнездовым
участком, с громким клекотом. Совокупление происходит на дереве близ
гнезда.
278
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Кладка из 2—3 яиц, причем период ее довольно растянут и повидимому
варьирует географически. В Средней Азии слабо насиженные кладки находили
в первой половине апреля (11 апреля, Кызыл-Кумы; 12 апреля у Илийска;
даже 26 марта р. Или). Северней — во второй половине апреля. Яйца белые,
размеры их (5) 70—74,4x55,7—58,2 (Харьковская обл., Горленко и Рудин-
ский, 1937); 72—78x58—60 (Умань, Гебель, 1879); (3) 72,3—75x56,4—58,2
(Красноярск, Тугаринов и Бутурлин, 1911); (4) 70,1—78,5 х 56,9—62 (р. Или,
Шестоперов); (2) 70,8—71 х 55,2—56,3 (Семиречье, Зарудныйи Кореев).
Насиживанье с первого яйца и продолжается 43 дня (Ушков) при
участии обоих родителей. Птенцы вылупляются от конца мая до начала июня
(в Кызыл-Кумах уже 18 мая). Над выкармливанием птенцов подробные
наблюдения произведены в Ильменском заповеднике Ушковым. Выкармливает
птенцов самка, а самец ловит и приносит добычу. Если птенцов два, то
одного из них она кормит утром, а другого вечером (при достаточно крупной
добыче; если добыча мелкая, то кормление происходит 5—6 раз в день).
Гибель младшего птенца наблюдается, но не имеет столь регулярного
характера, как у беркута; выживают и два, и даже три птенца. В возрасте
около 2 недель у птенцов начинает пробиваться оперение—маховые, рулевые
и плечевые; в 35—40 дней пух остается только на голове и шее, в 55 дней
процесс оперения заканчивается и примерно в 2-месячном возрасте птенцы
покидают, гнездо, летают, хотя долго еще ночуют в гнезде.
Линька. Изучена недостаточно (с теми же оговорками, как относительно
других орлов). Полная годовая, очень медленная, происходит между маем
и ноябрем—декабрем. Последовательность смены нарядов: первый пуховой—
второй пуховой — первый годовой (гнездовый) — второй годовой
(промежуточный) и т. д.; только в возрасте 4 или 5 лет — окончательный наряд.
Питание. Могильник — менее энергичный хищник, чем беркут, на
что указывает и более слабый клюв и лапы. Главная пища могильника —
средней и мелкой величины млекопитающие, в особенности суслики (имеется
некоторое соответствие между численностью сусликов и распространением
могильника), также зайцы. Летающих птиц могильник не ловит. Отмечена и известная
индивидуальная специализация отдельных орлов в связи с обилием в их
охотничьем участке определенных видов добычи. Потребность в пище в среднем
около 400—600 г в день, но съедается и 200 и до 1200 г (Ушков). В качестве корма
могильников в СССР указываются зайцы песчаники, русаки и беляки, суслики,
тушканчики, водяная крыса, хомяк, полевки, сурки, ежи; из птиц белая и
серая куропатка, тетерев, перепел, глухарь, молодые дрофы и журавли (Заруд-
ный, 1880),голуби, галки, грачи, серые вороны, сороки, утки, в том числе кряква
и хохлатая чернеть; гусь (домашний), коростель, чомга, сизая чайка, болотная
сова, полевой и болотный лунь и т. д. Могильник охотно клюет падаль.
Полевые признаки. Крупный орел, но несколько мельче беркута, с
относительно более длинными крыльями и коротким хвостом. Без белого на хвосте.
Молодые продольно полосатые с примесью охристо-рыжего цвета, старые тем-
нобурые, иногда с белыми пятнами на плечах. Голос похож на лай собаки,
вроде «тьяф-тьяф-тьяф» (кроме того, и другие звуки). Полет менее быстрый;
чем у беркута.
Описание. Размеры и строение. Ноздри вытянутые, а не круглые, как у
подорликов. Лапы довольно слабые, в особенности по сравнению с беркутом—коготь
внутреннего пальца. Формула крыла как у беркута, наружное опахало 7-го махового сужено. Длина
самцов (14) 726—830, самок (4) 810—835, в среднем 777 и 827,5лш. Размах крыльев
самцов (4) 1900—2010, самок 2050—2110, в среднем 1954 и 2071,7 мм. Вес самки (1) 3160 г.
Крыло самцов (18) 540—600, самок (20) 580—645, в среднем 542,5 и 605,3 мм.
Окраска. Первый и второй пуховые наряды белые. Первый годовой наряд бурый
с широкими охристыми продольными полосами, расширяющимися к вершине пера,
на голове, спине, плечевых, кроющих крыла, брюшной стороне; голени и кроющие хвоста
СТЕПНОЙ ОРЕЛ
279
охристые; первостепенные маховые черно-бурые с сероватым поперечным рисунком на
основаниях внутренних опахал; на второстепенных маховых еще и светлые пятна на вершинах;
рулевые черновато-бурые с сероватым поперечным рисунком на внутренних опахалах и
светлыми охристыми вершинами. Во втором годовом наряде светлые охристые пестрины на
спинной стороне несколько сокращаются, и цвет перьев голени и надхвостья темнеет.В третьем
годовом наряде светлых пятен еще менее, цевка и надхвостье с бурыми пятнами, темя и
затылок равномерно охристые.
В четвертом наряде орел уже темнобурый, на брюхе, в надхвостье и на голенях —
светлые каемки. В окончательном наряде общая окраска брюшной и спинной стороны
черно-бурая; задняя часть темени, затылок, «брови», бледножелтовато-охристые; часть плечевых
обычно (не всегда) белые и образуют «эполеты»; надхвостье светлобурое; нижняя часть цевки
беловата; мелкие перья с белыми основаниями, прикрытыми темнобурой вершиной;
маховые черно-бурые с серым поперечным рисунком на основаниях внутренних опахал; рулевые
темносерые с темнобурым поперечным рисунком из 5—7 полос и с широкой черновато-бурой
вершинной каймой.
Радужина желтая или орехово-бурая, у молодых сероватая, клюв темный черновато-
роговой у вершины, синеватый у основания; когти черные; восковица, разрез рта и лапы
желтые.
34. Степной орел Aquila rapax Temminck
Falco rapax. Temminck. Planches Coloriees livr. 76, pi. 455, 1828, ю. Африка.
Распространение. Ареал. Ю.-в. Европа, ю.-з. Сибирь, Передняя и
Средняя Азия от Туркестана до с.-з. Китая; Индия; Аравия; Африка. X а р а к-
Карта 42. Распространение степного орла Aquila rapax
1 — А. г. nipalensis, 2 — А. г. orientalis, 3 — А. г. vindhiana, 4 — А. г. belisarius, 5 — А. г.
raptor, 6 — А. г. rapax; а -~ граница гнездовий (цифры в кружках соответствуют номеру
подвидов, показывают местонахождение на зимовке и кочевках).
тер пребывания. На севере перелетная, на юге оседлая птица. Б и о-
т о п. Открытые сухие равнины степного и полупустынного характера.
Подвиды и варьирующие признаки. В Добрудже, ю.-в. полосе Европейской части СССР,
в сев. Казахстане и ю.-з. Сибири А. г. orientalis С a b a n i s, 1857; в Передней и Средней
Азии А, г. nipalensis Hodgson, 1833; в Индии от Персидского Белуджистана и Гималаев
А. г. vindhiana F г a n k 1 i n, 1831; в с.-в. Африке от Абиссинии и Эритреи, в Сомали и Кор-
дофане, в ю.-з. Аравии, изредка в Египте (Мейнерцхаген, 1931) и в Сирии А. г. raptor
В г е h m, 1856; в Марокко, Алжире, Тунисе А. г. belisarius L e v a i 1 1 a n t, 1850; в южн.
280
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Африке от Кении и Анголы до Капской колонии А. г. г арах Т е m m i n с к, 1828.
Морфологические отличия в размерах и окраске; в частности, северные подвиды nipalensisw orienta*
lis отличаются от остальных рас положением ноздри, более короткими пальцами и иным
ходом возрастных изменений, а также относительно большей величиной. Экологические
различия в характере пребывания (северные подвиды перелетны), в деталях биотопического
распределения и в ходе периодических явлений.
Европейский степной орел Aquila rapax orientalis С a b a n i s
Aquila orientalis. Cabanis. J. f. Orn., 1857, стр. 369, Сарепта.
Синонимы. Aquila Pallasii «A. Breh m». С h r. L. Brehm. Der vol 1st. Vogelfang,.
1855, стр. 413, низовья Волги. — Aquila Glitschii Severtzow. Stray Feathers III,
1875, стр. 422, nom. nud.—-M ензбир. Орнитологическая География, 1882, стр. 387, южн.
Россия.
Распространение. Ареал. Добруджа (низовья Дуная), Бессарабия,
Украина на север до Днепропетровской, Николаевской и Одесской областей,
возможно до южн. части Киевской, ранее спорадически до Харьковской (Сомов,
1897); далее на восток в степях Донских, Ставропольских и Терских, к северу
доХреновской степи; на Волге и в Заволжье примерно до линии Бузулук—
Чкалов; в арало-каспийских и североказахстанских степях на север до
верховьев Тобола, широты Акмолинска и Павлодара (устье реки Селеты),
далее на восток повидимому до Тарбагатая и сев. Балхаша, на юг да
Аральского моря; не встречен однако к востоку от Иртыша в степях
ю.-в. Сибири — в Кулундинской и Барабинской. Детали распространения
неясны особенно в Туркестане, и подвидовое положение гнездящихся там
орлов неясно. Какие-то степные орлы в небольшом числе гнездятся у
Аральского моря, в Кызыл-Кумах и нижнем течении Сыр-Дарьи, но экземпляров
оттуда в изученных коллекциях нет и к какой форме они относятся —
неизвестно. Вне пределов СССР в Иране (подвид?). Быть может в Туркмении
(Дорт-кую, Зарудный, 1896). Наиболее вероятным представляется, что
восточная граница распространения этого орла проходит в с.-з. Казахстане до
среднего течения Урала, дальше у Аральского моря и в Кызыл-Кумах, а на ю.-в.—в
Иране. В ю.-в. Казахстане уже следующая форма — область гнездовья подвидов
быть может прервана. На пролете встречается спорадически в разных частях
Туркестана, регулярно у Каспийского моря в ю.-з. Туркмении и на Кавказе.
Область зимовок не вполне ясна —добытые в Африке и Дарфуре и Кордофане
(Лайне) птицы относятся к этой форме; сюда же относятся повидимому орлы,
добытые зимой в Месопотамии и Палестине, в Судане, Дамараленде. Залеты
к устьям Уя и на Иртыш около 160 км выше Семипалатинска (Северцов).
Характер пребывания. Перелетная птица.
Даты. Прилет относительно поздний, недели на две позже, чем у
могильника — весной не ранее начала апреля и до второй половины месяца,
осенью с конца августа и в разные числа сентября, последние в начале октября
(наблюдения в Заволжье и с.-з. Казахстане, Зарудный, 1888, Сушкин, 1908);
в Туркмении осенью еще около середины октября (Чатлы), на Сев. Кавказе
весной уже около середины марта (Беме, 1925), тогда же и в Крыму (Сиваш).
Биотоп. Открытый широко обозреваемый ровный ландшафт —
целинные степи, с ковыльной и полынной растительностью; полупустыни; при
наличии таких площадей встречается и в пустынях — приаральских Кара-Кумах
и Кызых-Кумах. Возможно наличие связи распространения с обилием грызунов
(в частности, с мугоджарским сусликом). В горы не идет, но охотно держится
в всхолмленных местностях (отроги Мугоджар и т. п.).
Численность. В Европейской части СССР весьма сократилась в
историческое время в результате распашки целинной степи. В Заволжье и степях
сев. Казахстана, на Сев. Кавказе нередок; южней, в области Аральского моря
и далее в Средней Азии спорадичен и малочисленен.
СТЕПНОЙ ОРЕЛ
281
Экология. Размножение. Начало периода размножения относится
к середине апреля, когда строятся новые или ремонтируются старые гнезда»
В парах держатся с прилета, когда начинаются и брачные игры. Гнезда
чаще всего расположены на земле, иногда на старых сооружениях,
например, мазарах (Казахстан) на
склонах оврагов среди кустов
(Актюбинск), на песчаных островах и
старых скирдах сена (Крым, Брау-
нер) и т. п., редко на низких
отдельно стоящих деревьях,
например, на тополе в 2 лют земли (Урда,
Волчанецкий, 1937) или саксауле,
в 1 м от земли (Сушкин, 1908).
Построены гнезда из сучьев,
бурьяна, иногда в стенки
помещены ребра домашних животных.
Размеры гнезд различны, в среднем
около 700—1000 мм в диаметре.
Выстилка из шерсти, обрывков
кошмы, сухого помета, с
прибавлением линных перьев старых
орлов. Занимаются гнезда
длительное время, но не каждый год
подряд (у каждой пары бывает повиди-
мому 2 или 3 гнезда). В Добрудже,
где степных орлов сильно
преследуют, и степи распаханы, орлы
стали гнездиться главным образом
на деревьях.
Кладка во второй половине
апреля (ранние 13^—-15 апреля,
Сталинграду Сиваша уже 8 апреля;
поздние в начале мая, Волжско-
Уральская степь). Число яиц в
кладке 1—3, чаще 2. Яйца белого
цвета с буроватыми крапинами, по
размерам меньше, чем у
могильника. Размеры 66—68 х 48—54
(Аральское море, Бостанжогло,
1911); (30) 62,5—74 х 50,8—57,1,
в среднем 69,2 х 54,35 мм (Джо-
урден, по Хартерту, 1913).
Насиживанье—с первого яйца,
продолжается около 45 дней; вылу-
пление происходит в начале июня (1 июня, сев. Казахстан; 1 июня первый
птенец, 3 июня—второй, Актюбинск, Карамзин, 1917; в первой неделе июня,
Чкалов, Зарудный, 1897 и т. д.). Так как яйца находят и поздней (23 июня,
дельта Волги, Воробьев, 1938), то при нередкой гибели кладок из доступных
гнезд, повидимому, бывают дополнительные. Птенцов в гнезде один — два,
редко три. В ловле добычи участвуют оба родителя, но оделяет птенцов самка.
Известны случаи гибели младшего птенца в гнезде. Выводка родители не
защищают. В начале августа птенцы вполне оперены и становятся летными
(Чкалов; в августе, Волжско-Уральская степь; 12 августа, Аральское море).
Гнездовый период, следовательно, продолжается около 60 дней.
Рис. 78. Степной орел.
282
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Линька. Начало смены крупного пера во второй половине периода
размножения, после вывода птенцов. Последовательность смены маховых и общая
продолжительность линьки точно неизвестны. Последовательность смены
нарядов: первый пуховой наряд — второй пуховой наряд — первый годовой (гнез-
довый) наряд —второй годовой (промежуточный) наряд —третий годовой
(промежуточный) наряд и т. д. Окончательный наряд надевается повидимому
после 4 линек.
Питание. Пищу европейского степного орла составляют главным
образом средней величины грызуны — самое появление орла на местах гнездовья
связано быть может с пробуждением от спячки сусликов. Суточный цикл также
совпадает с активностью грызунов — вылеты на охоту в период наибольшей
деятельности последних (примерно с 7 часов утра до 10, затем под вечер,
Сушкин, 1895). Охотится паря и затем бросаясь на замеченную добычу;
подстерегает ее сидя у нор или, наконец, ходит по земле, как подорлик. В качестве
пищи степного орла указываются суслики, зайцы и мелкие грызуны, ушастый
еж, иногда подлетки или молодые птицы (дрофенок, стрепет, серая куропатка,
чибис, перепел), также змеи и, наконец, падаль (Крым, с.-з. Казахстан, Волж-
ско-Уральские степи и т. д.).
Полевые признаки. По размерам уступает могильнику; более светлой
окраски. Менее осторожен, чем другие крупные орлы. Довольно молчалив —*
голос напоминает тявканье небольшой собачки или глухое карканье или
кряхтенье (Аргиропуло, 1928). Характерна манера охотиться на земле пешком
или сидя на земле и бросаясь коротким взлетом на приблизившуюся добычу.
Описание. Размеры и строение. Клюв высокий и крепкий, заметно
сильней, чем у подорликов; ноздря поперечно вытянутая, а не круглая; лапы относительно
слабые и с короткими когтями. Крылья широкие и длинные; формула как у других
орлов.
На наружном опахале 7-го первостепенного махового есть вырезка. Длина самцов
668,5—740, самок (4) 755—800, в среднем 717,5 и 780. Размах самцов (6) 1745—1912,
самок (4) 1920—2 000, в среднем 1838,7 и 1965 мм. Вес самца 2700, самки 4850 г. Крыло
самцов (17) 510—560, самок (18) 525—600, в среднем 527,2 и 556,8 мм.
Окраска. Первый пуховой наряд чисто белый, второй пуховой наряд
серовато-белый. Первый годовой (гнездовый) наряд бледный глинисто-бурый, плечевые
с серовато-бурым с поперечным рисунком, кроме того на плечевых, средних и больших
кроющих крыла бледноохристые вершинные пятна; надхвостье охристое; на затылке
иногда рыжеватое пятно; маховые темнобурые с охристыми каймами на
второстепенных; рулевые бурые с охристыми каемками; брюшная сторона бледнобурая, иногда со
светлыми пятнами. В окончательном, повидимому, пятом, годовом наряде птица темно-
бурая, более бледная в обношенном пере; надхвостье с неясным светлым поперечным
рисунком; маховые темнобурые с сероватыми поперечными пестринами у оснований
внутренних опахал; рулевые темнобурые с серыми поперечными полосами; брюшная
сторона однообразно бурая; очень редко на затылке светлое рыжеватое пятно. Оба пола
окрашены одинаково.
Радужина орехово-бурая, клюв черноватый у вершины, сероватый у основания;
восковицы, разрез рта и ноги желтые. Окончательный наряд отделен от первого
годового двумя и быть может тремя промежуточными, которые характеризуются
постепенным потемнением окраски и сокращением размеров, а потом исчезновением охристых
пестрин.
ВосточныйХстепной орел Aqulla тарах nipalensis Hodgson
Circaetus nipalensis. Hodgson. Asiatic Researches XVIII (2), 1833, стр. 13, Непал.
Синоним. Aquila amurensis. Swinho'e. Proceed. Zool. Soc. of London, 1871,
стр. 338, Амур.
Распространение. Ареал. От восточного Алтая до ю.-в. Забайкалья,
Монголии, повидимому, Сев. Китая (Чжили) и Средн. Китая, сев. Тибета,
Джунгарии. Будто бы гнездится в с.-з. Индии (Бекер, 1928), что вызывает
большие сомнения, как и сведения о гнездовании этой формы в Туркестане —
вероятно гнездящиеся там орлы относятся к европейской расе. Зимой в централь-
СТЕПНОЙ ОРЕЛ
283
ном Китае, ю.-в. Монголии (Куку-нор, Пржевальский, 1876), Индии, в вост.
Африке на юг до Абиссинии, Дамараленда и даже области Уелле (Чепин, 1932),
в Передней Азии, Месопотамии, в Иране (Сеистан, Зарудный, 1911). В СССР
на зимовке встречен только на крайнем юге в области отрогов Паропамиза
(в декабре—феврале добыты у Кушки). Залеты в Туркестан от с.-з.
Казахстана (Ак-шушан), до Кугитанга и Шугнана, в Сибири до Красноярска,
Минусинска и Иркутска.
Характер пребывай, и я. Перелетная птица, но быть может не
по всему ареалу. Даты. Осеннее скопление орлов в стаи и передвижения их
наблюдались в августе в вост. Монголии. В Тибете появление с юга уже в марте
{Шефер, 1938). В сев. Монголии экземпляры наблюдались и добывались между
концом апреля и концом октября (Козлова, 1930; экземпляры в Зоол. музее
Московского университета).
Биотоп. В отличие от западного подвида, гнездящегося в ровных
степных и полупустынных ландшафтах, восточный степной орел предпочитает
холмистые местности с выходами камней, а не плоские долины. В горах встречается
довольно высоко — на Алтае до 2300 м, в Хангае до 2000 м; в сев. Монголии
вообще не ниже 1600 м (Сушкин, 1938).
Численность. В* ю.-в. Забайкалье и Монголии (кроме повидимому
центральных частей Гоби) многочисленен. Гнездовые участки обширны —в Бор-
зинском районе одно гнездо на 110—185 км2 (Тарасов, 1944). Плотность
населения восточного степного орла находится в известном соответствии с
распространением грызунов—тарбаганов и сусликов в Забайкалье (и пищухи Ochotona
4aurica в Монголии и Тибете).
Экология. Размножение. Гнезда расположены на скалах (Алтай,
Хангай, ю.-в. Монголия), среди камней на вершинах бугров или холмов, иногда
даже на тарбаганьем бутане (Павлов, 1935). Построены гнезда из сучьев или
камыша; выстланы шерстью, обрывками шкур и т. п. (Тарасов, 1944). Кладка
бывает в середине мая, из 2 яиц белого цвета с бурыми пестринами. Размеры
(2)71,2-^76 х51— 55 лш (Тачановский, 1891); (3) 69,5-^71,7 X 51,7— 54 мм
(Хартерт, 1913).
Насиживанье происходит с первого яйца, птенцы разновозрастны.
Длительность насиживанья повидимому около 45 дней, так как птенцы появляются
около конца июня (во второй трети этого месяца, Тарасов, 1944). Смертность
птенцов значительна, вследствие разорения гнезд хищниками (гнезда не
защищены, птенцы видны издалека, запах разлагающейся пищи привлекает к гнезду
хищников—лис и т. п., —поэтому в Борзинском районе в одном участке из
11 гнезд погибли 8, т. е. 72%, а в другом участке из 10 гнезд все, т. е. 100%) и
гибели одного из птенцов, младшего, как это нередко бывает у орлов (Павлов,
1935). Оперяются птенцы в разные числа июля (пуховики с пробивающимися
лерьями на крыле, Алтай; 18 июля в сев. Гоби и т. д.). Гнездовый период
продолжается около 55 дней, летные появляются с конца августа (первые летные
на Алтае 24 августа) до первой трети сентября (Борзинский район; Алтай)*
Линька. Не изучена.
Питание. Пищу степного орла составляют главным образом
грызуны. Охотится, паря довольно высоко над степью и бросаясь на добычу с
высоты 150—200 м (Штегман, 1937), или сидя в засаде на бугре у норы
грызунов. В Забайкалье в значительной мере кормится тарбаганами (откуда
местное название «тарбажин»), главным образом молодыми, но вероятно и
взрослыми, а также их трупами. При этом часто ест чумных зверей, но чумой не
заражается (Павлов). Из других грызунов в качестве пищи степного орла
отмечены зайцы, даурские суслики, даурские пищухи, тушканчики Alactaga saltator,
полевки Microtus raddei, из зверей других отрядов — степной хорь, солонгой,
даурский еж (Алтай, Забайкалье). Зимою в ю.-в. Туркмении в долине Кушки
284
ОТРЯД ХИЩНЫЕ ПТИЦЫ
питается главным образом большими песчанками. Из птиц — куропатки, один
раз в желудке степного орла был найден молодой филин (Пржевальский, 1876).
Описание. Размеры и строение. Заметно крупней европейского подвида:
крыло самцов (10) 540—590, самок (4) 618—650 мм, в среднем 577,5 и 632 мм. Длина самцов
(2) 752,820, самок (3) 782, 830,870. Размах самцов (3)1920—2000, самок (3) 2020,2080,2600 мм.
Вес самца (1)3500, самок (4) 3800—4850 г. По Штегману (1937) крыло у самцов 565—600,
у самок 597—638 мм.
Окраска.В свежем пере немного темней, чем европейская раса; на затылке бывает
обычно светлое охристо-рыжеватое пятно, иногда значительных размеров*.
35. Большой подорлик или орел-крикун Aquila clanga Pallas
Aquila clanga. Pallas. Zoographia Rosso-Asiatica, 1, 1811, стр. 351, Европейская Россия.
Русское название. Употреблявшееся в XVII веке сокольниками название
«орлак» (на орлаков напускали кречетов), вероятно, относится к этому виду. Из других
книжных названий наиболее распространено два: большой подорлик и орел-крикун. Второе
быть может удобней для различения от следующего вида. С другой стороны, оно не отражает
систематической близости A. clanga и A. pomarina.
Распространение. Ареал. Вост. Европа, на запад до Калининградской
обл., Венгрии, Югославии (Кроатия), на восток до Приморья и с.-в. Китая;
Карта 43. Распространение большого подорлика Aquila clanga
а — граница гнездовий, г — область зимовок.
к югу до Македонии, Добруджи, сев. Украины (Харьковская и Черниговская
обл.), нижней Волги (Куйбышевской обл.), южн. Зауралья (Троицк), далее в
лесостепной полосе ю.-з. Сибири, в Танну-туве и сев. Монголии (Кентей),
Манчжурии (Ямасина, 1935) и Приморье, наконец, в Китае (долина Яньцзе, быть
может в Чжили, Латуш, 1932). На север до Обонежья, Холмогор (взрослый самец
добыт Паровщиковым 20.VI.1941), быть может средней Печоры (Дмоховский,
1935), в Зауралье до р. Няыси и устьев Конды (примерно 60° с. ш.), Сургута,
Томска, Енисейска; на Алтае, в Зайсанской котловине (Хахлов, 1928); у Ки-
ренска, в Балаганском округе, у Иркутска и Тунки; в ю.-в. Забайкалье
примерно до Читы и Сретенска; в Приамурье у озера Иван, устье Кумары и устье
Уссури. Границы распространения как в Европе, так и в Азии плохо известны.
Отмечен для Крыма (Никольский, 1892); будто бы в гнездовое время на
Кавказе (Джавахетия, Чхиквишвили; Дагестан —вероятно все же не этот вид.
БОЛЬШОЙ ПОДОРЛИК ИЛИ ОРЕЛ-КРИКУН
285
a A. pomarlna), в Туркестане (старые сведения, Северцов; 24.VII.1935
встречен в долине Санзара в Туркестанском хребте, Даль, 1936; летом 1926 года у
Соло-тюбе в низовьях Сыр-Дарьи, Спангенберг, 1936; в Зоологическом институте
Академии наук есть экземпляры, добытые в летнее время в Кара-Тау, а в
Зоологическом музее Московского университета — в центральном Тянь-Шане),
возможно в сев. Иране (Зарудный, 1911; однако там добывался A. pomarlna).
Кроме того, гнездится быть может в с.-з. Индии. Зимойвю.-в. Закавказье,Иране
(Сеистан),Месопотамии, Индии, Бирме, Южн. Китае, Кохинхине,также в Египте.
Отдельные особи встречаются и северней, в частности в Туркмении у Кушки.
Залеты в среднюю и ю.-з. Европу от Англии и Ирландии, Бельгии и Голландии,
Дании и Швеции до Франции, Испании и Италии.
Характер пребывания. Перелетная птица (кроме Индии), но
область зимовок, кроме быть может ю.-в. Азии, лежит близко от гнездовой.
Даты. Весенний прилет ранний, отлет поздний. Появление в гнездовой
области в разные числа апреля, ранние особи — с конца марта: в Прибалтике
6 апреля — 24 апреля (Лоудон, 1913), в Кировской обл. 20 апреля; в
Поволжье первые появляются 8—9 апреля, валовой прилет 19—22 апреля,
запоздавшие птицы прилетают 7 мая; в Харьковской области 22 марта — 28 апреля;
у Киева пролет 5 апреля; в б. Воронежской губ. 11 апреля; у Чкалова и
в Зауралье в апреле; у Зайсана 27 апреля —2 мая (Хахлов, 1914); в Вост.
Сибири в начале апреля; пролет в области Аральского моря в начале
апреля, на Сыр-Дарье в апреле — начале мая, у Ташкента 5 апреля—2 мая;
в Туркмении на Мургабе будто бы уже 8—9 марта, у Кушки—29 марта—
11 апреля, в Киргизии 21 марта—6 апреля (по экземплярам). Осенний
отлет происходит в разных числах сентября и до начала октября: 3 сентября,
28 октября в Харьковской обл. (Сомов), в низовьях Днепра даже 4 ноября,
в сентябре в Воронежской обл., с 20 сентября до 10 октября в
Куйбышевской обл., с 12 до 30 сентября у Бугуруслана, в сентябре—октябре у
Чкалова; у Красноярска в сентябре, у Абакана еще 24 сентября, в Забайкалье
в долине Аргуни последний 23 сентября, в Приморье между 11—25 сентября
и до конца октября (22 октября у Посьета); в Туркестане на Сыр-Дарье у Джу-
лека с конца сентября и в октябре (экземпляры коллекций), в предгорьях центр.
Тянь-Шаня встречается еще 1 ноября, в Туркмении от сентября и до конца
ноября (22 ноября, Аннау). Пролетают парами, одиночками и небольшими
стайками по 2—5 птиц.
Биотоп. Высокоствольные леса, негустые, обычно близ водоемов —в
долинах рек и на болотах; в Сибири главным образом в лесостепи; в тайге живет
только по речным долинам. Предпочитает равнины, но в невысоких горах
(Южн. Урал) доходит до верхней границы леса; на Алтае на гнездовье не
выше 500 м, на охоте и выше —до 1000 и редко даже до 1700 м\ в Средней
Азии в предгорьях и по речным долинам.
Численность умеренная. В Сибири редок и спорадичен. Малочисле-
нен на пролете в Туркестане. Местами значительная плотность гнездовий:
4 гнезда на 6 десятинах (Чкалов, Зарудный, 1888), одно гнездо находится на
расстоянии 400 м от другого (Башкирия, Сушкин, 1897). Колебаний
численности не установлено (полифаг).
Экология* Размножение. Брачный полет наблюдается в первой
половине апреля (Башкирия). Гнездовые участки весьма постоянны, и гнезда
используются птицами по много лет (Сомов, 1897, и др.). Иногда занимается и
перестраивается чужое гнездо, иногда строится собственное. Гнезда расположены на
соснах, дубах, ольхах, ивах на высоте 8—12, иногда и выше до 25 м
(Харьковская обл., Рудинский и Горленко, 1937); в Зап. Сибири в Барабинской степи
на тальнике в 3 м от земли (Коте, .1910). Размеры различны —примерно от
700 до 1100 л« в диаметре (Рудинский и Горленко). Лоток плоский, около 50 мм
286
ОТРЯД ХИЩНЫЕ ПТИЦЫ
глубиной; выстилка примитивна, главным образом кора и тонкие веточки;
в гнезде обычно свежие зеленые ветви.
Кладка бывает в мае с первых чисел и до середины месяца (одно яйцо
1 мая, среднее течение Сев. Донца; 8 мая полная кладка из 2 свежих яиц,
Харьковская обл.; тоже 10 мая, Бугуруслан; одно яйцо там же 6 мая; слегка,
насиженные яйца 14—16 мая, Куйбышевская обл.; 10 мая свежее яйцо,
Башкирия; 9—10 мая полная кладка, Иман, Приморье). Число яиц в кладке
обычно 2, иногда 1 (по некоторым авторам—Эверсманн, 1866, Зарудный, 1888*
Рис. 79. Большой подорлик.
Сомов, 1897, до 3). В Индии чаще 1, редко 2 (Бекер, 1928). Окраска яиц белая с
фиолетовыми и буроватыми пестринами, обычно немногочисленными. Размеры:
(3) 66—72x51,1—54 (Сомов); (4) 68,3—71 х52,9—57,9 (Иман, Спангенберг);
(56) 61,2 —74,6 х 51 —58, в среднем 68,2 х 54,2 мм (разного происхождения,
Хартерт, 1913). В случае гибели кладки бывает вторая дополнительная (Север-
цов, 1855).
Насиживанье происходит с первого яйца, срок точно не прослежен, судя
по находкам в гнездах пуховиков (15 июня пуховики и проклевыши, вост.
Белоруссия, Федюшин, 1928, и др.) —около 6 недель. Число птенцов в выводке
обычно 1, так как младший птенец чаще всего погибает. Однако бывают случаи,
когда благополучно выживают оба птенца (б. Александровский уезд
Владимирской губ.; Бобровский р. Воронежской обл.). В конце июля—начале авгу-
БОЛЬШОЙ ПОДОРЛИК ИЛИ ОРЕЛ-КРИКУН
287
ста птенцы бывают уже вполне оперенными, но с недоросшими маховыми. Около
середины августа покидает гнездо сначала старший птенец, а через несколько
дней и младший. Запоздавшие особи возможно от повторных кладок вылетают
в последней трети августа. Гнездовый период тянется таким образом более 7
недель. Выводки у гнезд держатся весь август, а потом до отлета кочуют в
окрестностях, перемещаясь иногда из лесов в степь.
Линька. Полная годовая, начинается со второй половины гнездового
периода (с конца июля — начала августа). Конец линяния бывает на зимовке.
Маховые сменяются от заднего края к переднему (не всегда в строгой
последовательности, как и у других орлов). Последовательность смены нарядов:
первый пуховой наряд — второй пуховой наряд — первый годовой (гнездовый)
наряд — второй годовой (промежуточный) наряд, — третий годовой
(промежуточный) наряд — четвертый годовой (окончательный) наряд.
Питание. Охотится на лету, высматривая добычу, или сидит на земле
и подстерегает ее, или, наконец, ходит по земле и хватает встреченных
животных. Питается средней и мелкой величины позвоночными, крупными
насекомыми, падалью. Из млекопитающих —тарбаганом (Забайкалье, Радде, 1863),
лесчанками, тушканчиками, пищухами, хомячками, сусликами (Вост. Сибирь,
Тачановский, 1891), кроме того полевками Microtus ungurensis, M. michnoi и
землеройками (Забайкалье, Скалой, 1936); зверями от зайца до белки и мыши
(Чкаловская область, Эверсманн); на Украине поедает разных
грызунов—крапчатого суслика, хомяка, мышей и полевок (Сомов). Из птиц питается утками и
их яйцами, тетеревами, грачами, воронами, сороками, болотными совами,
домашней птицей (разные авторы), овсянками (Скалой), нелетными птицами
жаворонков, которых ловит «пешком» (Кокчетав, Штегман, 1937); кроме того ест
рыбу (Чкаловская обл., Эверсманн; Приморье, Маак, 1855, Джулек на
Сыр-Дарье, Грачев, 1939); поедает в значительном числе лягушек и жаб,
ящериц, крупных насекомых (кузнечики, саранча, жуки и бабочки, Забайкалье,
Скалой). Наконец, охотно съедает разную падаль.
Полевые признаки* Некрупный орел, обычно однообразно бурой окраски;
молодые черновато-бурые со светлыми пестринами, надхвостье беловатое.
Полет быстрый и ловкий, кроме паренья часто пользуется и активным
передвижением по воздуху. Хорошо ходит по земле. Осторожен. Голос обычно
звонкое «кьяк-кьяк-кьяк».
Описание. Размеры и строение. Клюв и лапы относительно слабые, цевка
длинная. Формула крыла 4>5^6>3>2>7>8>1>9>10. Вырезка на наружном опахале
7-го махового. Ноздря округлая, высота надклювья обычно 16—19 мм (перед восковицей).
Длина самцов (8) 650—702, самок (8) 670—730, в среднем 675 и 700 мм. Размах самцов (8)
1585—1700, самок (7) 1670—1820, в среднем 1668,5 и 1747,6 мм. Вес самцов (3) 1600, 1600,
2000, самок (4) 2150, 2260,3100, 3200 г. Крылр самцов (41) 490—520, самок (27) 520—545,
в среднем 503,8 и 529,4 мм.
Окраска. Первый пуховой наряд буровато-серый, второй пуховой наряд
(надевается в возрасте около-3 недель) серовато-белый. Первый годовой наряд темный
черно-бурый с фиолетовым оттенком в свежем пере, со светлыми-буроватыми или охристыми наст-
вольями разной ширины, особенно развитыми на задней части спины; надхвостье светлей
спины, часто беловатое; брюшная сторона черновата с широкими бледнобурыми или
рыжеватыми или охристыми продольными полосами, подхвостье охристое, оперение ног беловато-
охристое; маховые черновато-буры, рулевые черноваты. Во втором и третьем годовом
нарядах светлые пестрины постепенно сокращаются, а основной бурый цвет бледнеет. При этом
в третьем наряде пестрины обычно сохраняются лишь в надхвостье. Взрослая птица темно-
бурая с несколько более светлым затылком и зашейком, беловатыми длинными верхними
хвоста; маховые черноваты, бледнеют к основанию пера; хвост темнобурый, иногда с более
или менее заметным поперечным рисунком. Самцы и хамки окрашены одинаково.
Радужина бурая; клюв синевато-роговой у основания, чернеющий к вершине; воско-
вица и лапы желтые; когти черные.
У молодых птиц развитие и цвет светлых пестрин очень варьирует: от особей с немногими
светлыми отметинами до таких, у которых светлая окраска резки преобладает на брюшной
стороне и образует большие пятна на спинной. Крайний вариант с общим преобладанием
в окраске охристо-золотистого или охристо-беловатого тона — относительно редок и счи-
288
ОТРЯД ХИЩНЫЕ ПТИЦЫ
та лея за особый вид «желтый подорлик» Aquila fulvescens Gray. Эта светлая вариация
встречается и в окончательном наряде, причем бурые тона заменены в таком случае золотисто-
желтыми (с темными наствольями на голове и зобе и с темными центрами больших кроющих
и задними плечевыми; с бурыми маховыми и рулевыми). Однако эти особи связаны с
«типичными» полным рядом переходов и могут быть встречены в одном и том же выводке с птицами
обычной окраски (например в Курганском районе, в Зоологическом музее Московского
университета). В Бузулукском районе, Куйбышевской обл. орлята «fulvescens» были
добыты из гнезда, где и старые птицы были желтыми (Мензбир, 1913). В Калининской обл.
в гнезде оба птенца были желтыми, в б. Владимирской губ. один птенец был желтый,
другой промежуточный. Географически распространение светлой вариации не локализовано.
В СССР, в частности, такие птицы добывались в Прибайкалье, на пролете на Сыр-Дарье
(Зарудный, 1911; экземпляры в Зоологическом музее Московского университета);
в б. Курганском округе (см. выше); в низовьях Днепра у Корсунского Монастыря, также из
окрестностей Чимкента, Бухары, Минусинска, Томска, в Абаканской степи; у Иркутска;
в районе Владимира, Бузулука, Астрахани, у Гатчины. Вне пределов СССР из Албании,
Пруссии и т. д.
36. Малый подорлик Aquila pomarina В г е h m
Aquila pomarina. С h г. L.Brehm. Handb. d. Naturgesch. aller Vbgel Deutschl., 1831,
стр. 27, Померания.
Распространение. Ареал. Разъединенный: средняя Европа от Вост.
Германии и Австрии до средней России, Балкан, М. Азии, Кавказа и сев. Ирана
с одной стороны; с
другой—Индия, кроме
Синда, сев. Бирма до Пегу,
Ассам. Зимовки повиди-
мому от южн. Европы
(Греция) и Передней
Азии до Африки.
Массовые зимовки в Африке,
главным образом, на
юге, в сев. Родезии и в б.
Германской ю.-з.
Африке (Штреземанн, 1935).
Характер
пребывания.
Европейские особи перелетны,
также повидимому и
пере днеазиатские.
Индийские оседлы.
Биотоп. В
гнездовой области леса,
главным образом смешанные
и лиственные, в сырых
местностях вблизи
заливных лугов, болот, в
речных долинах и т. п.
Подвиды и
варьирующие признаки. В Европе и
Передней Азии А. р.
pomarina Brehm, 1831, в Индии
А. р. hastata Lesson,
1834. Варьирующие
морфологические признаки:
окраска, особенно у молодых
птиц в первом годовом наряде; пропорции (более слабая лапа у индийского
подорлика), размеры сходны. Экологические различия — в характере пребывания, в ходе
периодических явлений (в частности, у индийского подвида в кладке чаще 1 яйцо, а у
европейского обычно 2).
Карта
44. Распространение малого подорлика Aquila
pomarina
1 — Ан. p. pomarina, 2 — A. p. haetata; a — границы гнездовий (цифры
в кружке соответствуют номеру подвидов и показывают область их
зимовок).
МАЛЫЙ ПОДОРЛИК
289
Систематические замечания. Малый подорлик чрезвычайно близок к орлу-крикуну и
может считаться западным и южным представителем последнего. Сходство распространяется
как на морфологию, так и на экологию. Однако области гнездования этих птиц
перекрываются на западе Европейской части СССР и далее в Венгрии и Югославии (Хорватии).
С другой стороньГхарактерно, что в Индии орел-крикун встречается на северо-западе
повидимому на гнездовье, и что в этой части полуострова^/ш/а^а повидимому не гнездится. Точно
так же викарируют эти^птицы в Закавказье. Относительно общая площадь ареала
орла-крикуна очень мала, и факт этот представляет собой по всей вероятности случай вторичного
(послеледникового) расселения одной из этих двух форм. Систематические взаимоотношения
этих двух орлов — один из примеров категорий, пограничных между понятиями вид и
подвид, хотя характер ареала говорит скорее в пользу объединения орла-крикуна и
подорлика в политипический вид.
Европейский малый подорлик Aquila pomarina pomarina Brehm
Распространение. Ареал. В Европе от долготы Эльбы, озера Нейнзид-
лер и Хорватии до Балканского п-ова (Македония); М. Азия и сев. Иран на
восток до Астрабада; в СССР западные
части страны до Ленинграда, Пскова,
Новгорода, Смоленска, западных
частей Московской обл. (Можайск),
Полтавской и Киевской обл.; Кавказ
от Кубанских лесов и Дагестана на
севере, до Армении на юге. Имеется
повидимому ошибочное указание на
гнездование в б. Костромской губ.
(Кирпичников, 1915). Залеты в Зап.
Европе от Франции и Испании, в южн.
Швецию и Данию, в Финляндию; у
нас в Крым (Тотакой), в Воронежскую
и Харьковскую обл.; к Серахсу в
Туркмению (но не все случаи
достоверны). Зимовки главным образом в
Африке от Египта yi Кении до ю.-з.
материка и сев. Родезии; также быть
может в Греции и Передней Азии, у
нас будто бы в Бессарабии и
Закавказье.
Характер пребывания.
Перелетная птица.
Даты. Отлет с африканских
зимовок в марте, осенний пролет в
Средиземноморье в октябре—ноябре.
Прилет в апреле, в разные числа (в
Прибалтике от 6 до 25; у Пскова
6 апреля, 4 апреля — 8 мая, в б.
УманСКОМ у. Киевской Обл. будто бы Рис. 80. Малый подорлик.
уже в конце марта, Гебель, 1879; к
Можайску в первой половине апреля старого стиля, Огнев, 1911). Отлет
происходит в сентябре и до начала октября (Умань; Псков; Можайск; пролет
в Колхиде 14 сентября). На пролете осенью встречаются одиночки, пары и
небольшие стайки по 3—5 птиц. Пролет в вост. Средиземноморье в ю.-в.
направлении, поэтому на пролете в Зап. и ю.-з. Европе бывают только
случайно. Результат кольцевания: окольцованный в Курляндии
орел добыт в сентябре в южн. Болгарии. Биотоп. См. относительно вида.
В горах Кавказа живет на высоте около 1500—1800 л* в кленово-буковых
лесах (Караклис) и до 2200 м (Помбак, Даль, 1943).
19 Птицы СССР, т. I
290
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Численность. В Зап. Европе спорадичен; в Полесье, Псковской
обл., на Украине обычен. Гнездовые и охотничьи участки относительно
невелики — в Псковской обл. расстояние между гнездовыми парами иногда
немного более 1 км, чаще около 3—5 км. Подорлик избегает культурных
разреженных лесных участков и поэтому постепенно исчезает в Зап. Европе
(средняя Германия). Так как подорлик всеяден, то периодических колебаний
численности, характерных для хищников миофагов, у него неизвестно; однако
смертность птенцов в раннем возрасте, особенно младшего, при недокорме
родителями — нередкое явление.
Экология. Размножение. Гнездовые участки и гнезда используются
птицами весьма постоянно (гнездо занималось 12 лет подряд). В связи с этим
размеры гнезд весьма различны: новые постройки меньше, старые больше
(диаметром 555—900 мм, 300—500 мм высотой, Гебель, 1879; от 710 до 1700 мм
диаметром и от 400 до 1200 мм высотой, с лотком в диаметре 480—700 мм и 60—
85 мм глубиной, Зарудный, 1911). Подорлики строят гнезда повидимому и
самостоятельно и только исправляют гнезда других видов. Обычно гнездо
расположено у опушки, высоко от земли, в 6—18л«, на разных деревьях —на севере
на березах и елях, реже на соснах (Псков, Можайск), на юге на дубах или
липах (Умань, Подолия), на буках (Армения). Гнездо как правило обложено
свежими ветвями (сосновыми, Можайск, Огнев; дубовыми, Подолия, Портенко,
1928). Брачный полет у гнезд начинается в конце сентября. Имеется
повидимому резерв холостых особей (два самца у гнезда, Бобруйск, Федюшин, 1928).
Кладка во второй половине апреля или в первых числах мая. Число яиц
обычно 2, редко 1 или 3 (две кладки из 10 по 3 яйца, Полесье, Шнитников,
1913; только иногда 3 яйца, Псков, Зарудный, 1911). При 3 яйцах одно обычно
не оплодотворено. Окраска яиц охристая с бурыми различной яркости пестри-
нами, обычно более многочисленными, чем у орла-крикуна. Размеры яицменыне,
чем у орла-крикуна: (43)56,5—72 х 45—55, в среднем 65 х 51,7 мм (Умань,
Гебель, 1879); 60-69 х41— 53мм (Полесье, Шнитников, 1913); 60,5-^65,8 х 49—
—51,5 мм (Псков); (2) 61—62,3 х 49,8 — 51,7 (Армения, Спангенберг). В
случае гибели кладки бывает дополнительная.
Насиживают оба родителя, самка сидит очень крепко; при гибели самки
самец иногда бросает гнездо. Птенец обычно только один (однако в
исключительных случаях даже 3, Зарудный, 1911). Вылупление происходит во второй
половине мая, обычно к концу этого месяца; насиживание, следовательно, длится
около 45 дней (по Зиверту, 1932, в Германии 43 дня, с откладки первого яйца).
Около середины июля птенцы оперяются и в самом конце июля — начале
августа вылетают (29 июля, вылет, Подолия; на крыле в конце июля, Псков;
выводки в августе, Смоленск; вылет в начале августа, запоздавших 17 августа,
Можайск). Гнездовый период продолжается около 8 недель.
Линька. Полная годовая, с конца июля — начала августа. У самок
начинается несколько ранее, чем у самцов. Маховые сменяются от заднего края
к переднему. Рулевые — от средней пары к краю хвоста. Последовательность
смены крупных перьев иногда частично нарушается. Конец линяния зимой
(в середине августа сменяются только 4 задних первостепенных маховых).
Последовательность смены нарядов: первый пуховой наряд —второй пуховой
наряд — первый годовой (гнездовый) наряд — второй годовой (переходный)
наряд — третий годовой (переходный) наряд — четвертый годовой
(окончательный) наряд и т. д.
Питание. Способы охоты и кормовой режим подорлика сходен в основных
чертах с питанием орла-крикуна. Способы охоты — медленный полет с
остановками в воздухе. Заметив добычу, орел «трясется», как пустельга, с вытянутыми
лапами; иногда подстерегает добычу сидя; очень часто разыскивает добычу
пешком, ходя по земле — с этим связана быть может относительно большая
ЯСТРЕБИНЫЙ ИЛИ ДЛИННОХВОСТЫЙ ОРЕЛ
291
длина цевки. Малый подорлик всеяден, но не ловит птиц на лету и не берет
крупной добычи. Повидимому, у отдельных птиц (пар?) имеется известная
специализация кормового режима — чаще в сторону питания пресмыкающимися
и земноводными, реже в сторону мышеядности (Уттендерфер, 1939). В качестве
корма, приносимого подорликами для птенцов, указываются полевка Clethrio-
namys glareolus, лесная мышь, крупные жуки и лягушки (Можайск). В гнездо-
вый период — молодой рябчик (Белоруссия), зайченок, хомяк, суслик, крысы,
ежи, цыпленок (Подолия), мыши, полевки, крысы, белки — в одном случае
до двух десятков хвостов в гнезде, зайчата, молодые птицы — коньки, дрозды,
жаворонки, тетерева, белая куропатка; лягушка, гадюки; майские жуки и
навозники (Псков), лягушки (Алазань). На пролете в с.-з. Закавказье подорлик
питается иногда кузнечиками (Домбровский, 1913). Главное место все же
занимают лягушки, ящерицы и змеи, а за ними мышеядные грызуны.
Полевые признаки. Небольшой орел, более стройного и легкого сложения,
чем крикун. Полет легкий; чаще, чем крупные орлы, пользуется активным
скользящим полетом. Хорошо ходит по земле. При полете маховые обычно
«пальцеобразно» расставлены. Окраска бледнобурая, однообразная, у
молодых со светлым пятном на затылке и бледными перевязями на крыльях;
без пестрин на брюшной стороне. Голос похож на тявканье небольшой собаки,
вроде «кьек-кьек-кьек».
Описание. Размеры и строение.В отличие от орла-крикуна, в крыле 7-е махо-
вое£относительно короткое и короче 4-го махового более чем на 60 мм. Наружное опахало
7-го махового без резкой вырезки. Размеры меньше, чем у крикуна. Цевка относительно
длинней, клюв слабей — высота надклювья перед восковицей менее 17 мм, у крикуна обычно
более. Формула крыла 4>5>3>6>2>7>1>8. Наружные опахала 2-го —б-го, внутренние
2-го—7-го с вырезками. Длина самца (1) 625, самок (3) 630—650, средняя 643 мм. Размах
самца (1) 1455, самки (1) 1593 мм. Крыло самцов (8) 444—480, самок (11) 475—505, в
среднем 467 и 492,3 мм.
Окраска. Первый пуховой наряд серый с резко выступающим темным теменем,
второй пуховой чисто белый. Первый годовой (гнездовый) наряд: бледнобурый на спинной
и брюшной стороне, светлей, чему орла-крикуна и менее пестрый; на затылке светлое
охристое пятно; на кроющих крыла охристые пятна, образующие полосы, такие же пятна на
плечевых и в надхвостье; последнее все же в основном бурое (не белое, как у орла-крикуна);
на брюшной стороне нередко беловатые настволья, менее развитые, чем у орла-крикуна,
маховые и рулевые темнобурые. Изредка встречаются особи с преобладанием
охристо-золотистого тона в окраске,напоминающие так называемых << Aquila fulvescens»: такие экземпляры
однако из пределов СССР неизвестны.
Окончательный наряд бледно серовато-бурый, светлей, чем у орла-крикуна и без
фиолетового оттенка; голова бледней и серей спины, маховые темнобурые, рулевые бурые,
иногда с темным поперечным рисунком. Этот наряд надевается по всей вероятности после
нескольких (2 или 3) линек, при которых последовательно уменьшаются светлые охристые
пестрины, причем уже со 2-го годового наряда эти пятна исчезают на спине и брюшной
стороне.
Радужина у взрослых бледная желто-бурая,у молодых серовато-бурая; клюв синевато-
черный: восковица, разрез рта и лапы желтые, когти черные.
37, Ястребиный или длиннохвостый орел Aquila fasciata
Vieillot
Aquila fasciata. Vieillot. Mem. Soc. Linn., Paris 11 (2), 1822, стр. 152, Монпелье.
Распространение. Ареал. Южная Европа от Испании и о-ва Майорка,
южн. Франции, Корсики, Сардинии, Сицилии, Кипра, Крита (в самой Италии
вероятно не гнездится), до Балканского п-ова; Азия от Йемена, М. Азии,
Палестины, Ирака, Ирана, Туркестана, Афганистана до Индии (от Гималаев)
и Бирмы, Среднего и Южного Китая (от Чжили до вост. Фуцзяня и сев. Гуань-
дуня, в долине Яньцзе от Чжецзяня до Хубея); Африка от с.-з. частей до Анголы,
Ниасса, Родезии, изредка до Капской Земли; наконец на о-вах Сумбава,
Флорес, Тимор.
19*
292
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Характер пребывания. Оседлая птица. В холодное время года
совершает кочевки как на север, так и на юг, появляясь тогда вне гнездовой
области — в Египте, средней и северной Франции, Бельгии, Италии, Венгрии,
Богемии и возможно в Германии, но нерегулярно.
Биотоп. Ксерофильные участки гор и предгорий, на кочевках и на
равнинах. В Индии лесистые местности как в горах, так и на плоскости, в
последних реже и обычно во влажных участках.
Подвиды и варьирующие признаки. В южной Европе, сев. Африке и Азии до Китая и
Индии А. /. fasciata V i e i 1 I о t, 1822; в средней и южн. Африке от сев: Нигерии, Эритреи
и1Абиссинии A./, spilogaster В р., 1850; на М. Зондских о-вах A./. renschiS tresemann,
1932. По мнению Суанна иУитмора (1933) более крупные размеры и темная окраска отличают
Карта 45. Распространение ястребиного орла Aquila fasciata
1 — A. f. fasciata, 2 — A. f. renschl, 3 — A. f. spilogaster; a — границы гнездовий (цифры в кружках
соответствуют номеру подвида и показывают местонахождение их в зимнее время).
индийских длиннохвостых орлов от прочих — в таком случае они должны называться А. /.
grandis Hodgson, 1844. Отличия между расами в размерах и окраске, экологические
особенности у них также имеются, но они плохо выражены (в отношении биотопического
распределения, в календаре периодических явлений и т. п.).
Северный ястребиный или длиннохвостый орел Aquila fasciata fasciata
Vieillot
Распространение. Ареал. Южная Европа, Передняя и Средняя Азия,
Индия, Китай, с.-з. Африка (Марокко, Алжир, Тунис). В СССР приводится
в литературе для б. Уманского уезда Киевской губ., Одессы (Нордман, 1840),
Воронежской обл. (Северцов), Сарепты (Яковлев, 1872) в европейской части
страны. Все эти случаи сомнительны и не подтверждены добытыми экземплярами.
Несомненно гнездится в южных частях Средней Азии—в Туркмении от Сумбара
ЯСТРЕБИНЫЙ ИЛИ ДЛИННОХВОСТЫЙ ОРЕЛ
293
до центральных и вероятно восточных частей Копет-Дага, а также быть может
и в Бадхызе и Карабиле. В гнездовое время (в апреле) добыт у Курган-тюбе
в Таджикистане, добывался повидимому у Бальджуана. Зарудный наблюдал
в Кугитанге с 24 по 31.V. 1910, на Сурхан-Дарье, на Пяндже у Чубека и Саята
и считает, что этот орел в гнездовое время встречается на Аму-Дарье между
Келифом и Кировабадом. Нередок в горах, окаймляющих Гиссарскую долину
(Иванов, 1948). По Зарудному (1911) добыт летом и севернее, у Шах-и-мардана
в Фергане, а 4. Х.1906 у Кок-булака к ю.-з. от Ташкента. Отмечен в Нура-Тау
(Мекленбурцев, 1937). Встречен в Кызыл-Кумах, в горах Аристан-бель-тау,
Ак-Тау и Тохта-Тау в июне 1912 (Зарудный, 1915). Наконец, добыт в Кара-
Тау (Мекленбурцев).
Рис. 81. Ястребиный орел.
Биотоп. Полупустынные и пустынные горы, предпочтительно с
кустарниковой или древесной растительностью (отсутствует поэтому во влажных лесах
«европейского» типа Закавказья и сев. Ирана).
Численность. В Средней Азии весьма редок.
Экология. В СССР не изучена. Размножение. Гнезда в Кызыл-
Кумах расположены на скалах, в Туркмении на скалах и на деревьях
(Зарудный, 1896). Летный молодой добыт на Сум баре в начале августа, В Таджикистане
на левом берегу Кафирнигана 30.III. 1946 орлы строили гнездо из ветвей на
невысоком уступе (Иванов, 1948). В сопредельных местностях (с.-в. Иран)
пара довольно крупных птенцов в гнезде на круче наблюдалась в хребте Зуль-
пенай 4 мая, а 3 августа наблюдался выводок из двух темных молодых птиц.
В Испании ястребиный орел имеет 2 или даже 3 гнезда в одном участке, занимая
их по годам попеременно; кладка из 2 яиц, начало ее в феврале, вылупление
в середине марта, насиживание следовательно около 40 дней, гнезда
расположены также на скалах и деревьях. Однако в Белуджистане у Кветты
кладка из 2 яиц найдена 26 мая, в Кашмире у Сринагара 15 мая. В Индии эта
птица гнездится с ноября по февраль. В Китае кладка в марте. В кладке 2 яйца,
реже 1, однажды даже 3; насиживают оба родителя (Юм, 1869; Бекер, 1928).
Яйца белые или беловатые, чаще всего с бледными буроватыми крапинами;
размеры (40) 65-71,2 х 51,2-^56,9 мм (Индия, Бекер, 1928).
294
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Линька. Не изучена, как и последовательность смены нарядов. После
обычных двух пуховых нарядов надевается первый годовой — гнездовый —
отличный от окончательного. Число и характер промежуточных нарядов
неизвестны.
Питание. В кормовом режиме большое место занимают птицы, в
частности в Кызыл-Кумах кеклики. Для сев. Ирана приводятся зайцы, однажды
пластинчатозубая крыса; Для Индии водяные птицы, также куропатки и
голуби; для Андалузии горные красноногие куропатки, для Палестины — зайцы,
дрофа красотка, также рыбы и рептилии (Мейнерцхаген, 1931).
Полевые признаки. Полет этого орла напоминает ястребиный, он быстрей
и поворотливей, чем у других орлов. Парит редко.
Описание. Размеры и строение. От других орлов ястребиный орел, как и орел-
карлик, отличается более стройным и легким сложением, длинным хвостом, относительно
короткими крыльями и коротким клювом с резким выступом по краю надклювья, с длинными
цевками и длинными пальцами, в особенности средним; когти очень длинные и острые. Вес
самца из Индии несколько более 1500 г, самки около 2500 г. Длина крыла особей из СССР:
самцов 473, 507 (Курган-тюбе), 465 (Сумбар), самок 478 (Тутлы, зап. Копет-Даг), 510
(Ташкент), 510 (Кара-Тау), 485 (Кашка-Дарья); птиц неизвестного пола 495 (Оби-Гарм, Бальд-
жуан), 483 (Тигровая Балка, Таджикистан). Для других местностей крыло самцов 460—
490, самок 490—520 мм (Хартерт), у индийских самцов 482—520, самок 530—550 мм (Бекер).
Окраска. Первый пуховой наряд не описан. Второй пуховой наряд судя по
оперяющимся птенцам, серовато-белый (Дрессер). Первый годовой наряд на спинной стороне темно-
бурый, на голове и шее рыжеватые каемки или вершинные крапины, хвост серовато-бурый
с тонкими черноватыми полосами; брюшная сторона охристо-рыжеватая или рыже-бурая
(два типа окраски—темный и светлый), с наствольными чертами на зобе и груди. Старые
птицы на спинной стороне черновато-бурые с серым поперечнополосатым хвостом, на
брюшной стороне белые или охристые с черными продольными штрихами и поперечными полосками
на штанах и в подхвостье. Клюв синеватый чернеющий к вершине; восковица и разрез рта
желтые, лапы беловато-желтые, когти черные; радужина у взрослых желтая — от
оранжевой до бледножелтой (Дрессер), у молодых бледнобурая.
Карта 46. Распространение орла-карлика Aquila pennata
1 — A. p. pennata, 2 — А. р. milvoides, 3 — A. p. ayresil, 4 — A. p. weiskel, 5 — A. p. morphnoides;
/ — A. kienerl kieneri, II— A. k. formosanus; a — границы гнездовий (цифры в кружках соответствуют
номеру подвидов и показывают область их зимовок).
ОРЕЛ-КАРЛИК
295
38. Орел-карлик Aquila pennata Gmel.
Falco pennatus. Gmel in, Syst. Nat. ed. XIII, 1788, стр. 272, Франция.
Распространение. Ареал. Южн. Европа, Африка, Средняя и южная
Азия, Новая Гвинея, Австралия.
Характер пребывания. Палеарктические птицы перелетны,
остальные оседлы.
Биотоп. Лесистые местности как на равнинах, так и в горах.
Подвиды и варьирующие признаки. В Европе, сев. Африке, Передней Азии А. р.
pennata Gmel., 1788; в Туркестане от Таджикистана до Казахстана и Киргизии, в Вост. Сибири
и Приморье, сев. Монголии и быть может спорадично в Индии A. p. milvoides, J е г d о п,
1839; в восточной и западной Австралии А. р. morphnoides Gould, 1841; на Новой Гвинее
А. р. weiskei Reichenow, 1900; в тропической Африке от Того и Сомали через
Бельгийское Конго и зап. Кению до Ниасса, Делагоа, Наталя А. р. ayresii G u r n e у, 1862.
Варьирующие признаки: величина (южные формы в общем мельче, северные крупней) и окраска
(как различие оттенков, так и наличие или отсутствие полиморфизма; последний характерен
для палеарктических подвидов), наличие или отсутствие хохла и некоторые особенности
строения. Имеются некоторые географические различия в экологии, но они выяснены
недостаточно.
Систематические замечания. К этому же виду Штреземанн (1926) относит еще
две формы: A. kienerii kienerii Geoffroy, 1835, которая распространена в
восточных Гималаях от Непала до вост. Ассама, вост. Бенгалии и по ю.-з. побережью Индии от
Бомбейской провинции, также на Цейлоне; близкий к прыдыдущему подвид А. к. formosanus
Stresemann, 1924, живущий на Суматре, Яве, Борнео, Целебесе, на М. Зондских
о-вах, до Сумбавы и на Филиппинских о-вах. Будучи очень близкими к A. pennata,
эти орлы отличаются от него некоторыми особенностями строения (длиннопалые и
длинноногие) ; к тому же неясно, не перекрываются ли их области распространения в Индии, где
орел-карлик встречается в разных местах в холодное время года, но имеются указания и на
гнездование, правда спорадическое и редкое.
Европейский орел-карлик Aquila pennata pennata Gmel.
Распространение. Ареал. В Европе область распространения разом-
кнута в области Альп: с одной стороны, Франция (кроме севера; спорадично»
главным образом на юге), на Пиренейском п-ове и Балеарских о-вах;
в сев. Африке ^Алжире, Марокко, вероятно в Тунисе; с другой стороны —
на юго-востоке от Балканского п-ова до Хорватии, Венгрии, Зап. Украины
(Галичина), быть может случайно до Полесья и Беловежской пущи
(Грассман, 1918); на севере до Киевской, Черниговской и Харьковской обл^
далее до южных частей Московской обл. и сопредельных частей Калужской
обл. (Боровский район), Тульской обл. (Новосиль), Тамбовской, Воронежской;
в Крыму (гнездование не доказано), на Кавказе, в Туркмении от Больших Бал-
ханов по Копет-Дагу до долин Теджена, Мургаба, Аму-Дарьи и Кугитанга;
кроме того, в Сирии и М. Азии, Иране. Зимой в Передней Азии и Индии,
некоторые особи быть может уже в Туркмении и Таджикистане (Зарудный и Биль-
кевич, 1918); также в Африке к югу от Сахары, где пролетает в центральных
частях пустыни; африканские зимовки расположены главным образом в
восточных частях материка — Ниасса, Узамбара, Уганда, редко в Натале. Залеты
в Германию, Италию, сев. Богемию; в СССР в Татарскую АССР, на границе
с Куйбышевской обл. (Бугульминский у.), в Новоторжский район Калининской
обл. к Пскову, к Чкалову, к Гурьеву.
Не все эти сведения достоверны, а сообщение о гнездовании на среднем
Урале (Сабанеев, 1876) несомненно ошибочно.
Характер пребывания. Перелетная птица, кроме самых южных
частей ареала.
Даты. Прилет в конце марта — первой половине апреля; 1 апреля ■—
19 апреля в Туркмении; в середине апреля Бессарабия; к Киеву 7 апреля;
22 марта—3 апреля,Черниговская обл.; 26 марта—»25 апреля, Харьковскаяобл.
296
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Осенью отлет наступает с середины сентября и длится до начала октября.
Отлет отдельных особей бывает уже с конца августа: 30 августа — 2 октября,
Харьковская обл.; середина сентября, Чернигов и т. д. Весной летят
одиночками, осенью иногда небольшими стаями.
Биотоп. Смешанные, но чаще лиственные высокоствольные леса, как
на равнинах, так и в горах; на Кавказе примерно до высоты 1500 м и даже
1980 м (Армения).
Численность. В местах, где сохранились старые леса, численность
достигает высокой плотности. Гнездо от гнезда находится на расстоянии
200 м, 75—100 особей на
2489 десятин (Подолия,
Портенко, 1929), 5 гнезд
на 4 км2 (Харьков,
Сомов, 1897). Изменение
биотопических условий
вызывает быстрое
падение численности орлов-
карликов: на Сев.
Донце в Харьковской обл.
на площади, где в 1934 г.
было 4 занятых гнезда,
в 1936 г. было только
одно жилое (Горленко и
Рудинский, 1937). Это
при низкой
плодовитости вида приводит к
быстрому общему
снижению популяции (как это
имело место во Франции,
где ареал этой птицы
значительно сократился в
XIX—начале XX
столетия, Майо, 1937).
Экология.
Размножение. Брачный
полет начинается с конца
апреля, в парах птицы
держатся уже с середины
апреля, т. е. с прилета,
В это время птицы со звонким криком носятся над лесом и проделывают
воздушные эволюции, часто в виде мертвых петель. Гнездовые участки невелики
(см. выше), энергично защищаются. Гнезда расположены на деревьях (в
Туркмении будто бы на скалах, Радде и Вальтер, 1889) разных пород: соснах, дубах,
буках, липах, ясенях, вязах, ольхах, в Туркмении на разнолистных тополях;
обычно в верхней трети дерева, в европейской России метрах в 12—22 от земли,
в Туркмении и ниже— до 4 м. Нередко орлы строят гнезда в колонии цапель
(Умань, Харьков, Новосиль). Обычно орел использует чужое гнездо
—канюка, коршуна, подорлика, балобана, белохвоста, цапли, иногда строит свое:
в этом случае в постройке гнезда участвуют оба родителя, но больше самка.
Постройка или подновление гнезда занимает около 2 недель (Фененко, 1909).
Постройка в 60—70 см в диаметре, с плоским лотком. Используются гнезда
несколько лет.
Кладка обычно из 2 яиц, иногда из 3 (Зарудный, 1896, Портенко, 1929),
откладываемых с промежутками в 2—4 дня. Окраска яиц зеленовато-белая,
Рис. 82. Орел-карлик (самец и самка).
ОРЕЛ-КАР ЛИК
297
иногда с немногими красноватыми пестринами. Размеры: (97) 52—64 х 42,5—
48,5, в среднем 56,4x45,6 мм (Гебель, 1879); (7) 53,2—58,4 х 43—47,2 мм
(Горленко и Рудинский, 1937); (7) 52,7—61,6x44,2—49,1 мм (Сомов, 1897);
53,4—56 х 43,6—44,7 мм (Харузин, 1926). Полные кладки 8—10 мая (Тула),
с конца апреля до середины мая (Харьков, Сомов), в начале мая (Чернигов,
Фененко).
Насиживанье начинается с первого яйца, в течение приблизительно месяца
(Харузин); сидит главным образом самка. В начале июня происходит вылупле-
ние (31 мая два пуховика, Бессарабия; около 10 июня, Тула; в начале июня
Чернигов). Птенцы разновозрастны. Самец носит корм и насиживающей самке
и птенцам, по крайней мере пока те малы. В конце июля — начале августа
птенцы оперены и покидают гнездо, хотя еще держатся близ него (вылет
в конце июля в Туркмении; 30 июля Воронеж, около 5—20 августа Тула).
Гнездовый период, следовательно, длится около 9 недель. Выводки держатся
вместе, по крайней мере до конца августа.
Линька. Слабо изучена. Насиживающая самка начинает сменять
маховые, к концу периода размножения остаются несмененными передние
первостепенные маховые. Конец линяния не прослежен (на зимовке?).
Последовательность смены нарядов: первый пуховой наряд — второй пуховой
наряд —первый годовой (гнездовый) наряд — второй годовой (окончательный)
наряд и т. д.
Питание. По строению и кормовым повадкам орел-карлик напоминает
ястребиного орла. Лапы его чрезвычайно мощны, с длинными пальцами и
большими когтями, крепкой и длинной цевкой. Крылья относительно короткие и
узкие, хвост длинный. Орел-карлик поэтому отлично справляется с
преследованием в лесу довольно крупной добычи и ловит даже летающих птиц, летая
быстро и поворотливо и напоминая в этом отношении ястреба. Подобно
последнему, он или бросается на добычу из засады, или выпугивает ее, быстро летая
среди деревьев; реже парит по-орлиному, высматривая добычу. Пища
—разные птицы средней и мелкой величины — от лесных голубей, горлиц и сорок
до жаворонков, скворцов, чернолобого сорокопута, коростеля, лесного конька,
желтой плиски (Вильконский, 1892, Портенко, 1928, Сомов, 1897); из зверей
суслики и другие грызуны.
Полевые признаки. Небольшой орел размерами с сарыча, с ловким полетом,
очень подвижный. Крылья большие и довольно узкие, хвост длинный. Полет
разнообразен — и паренье, и быстрое скольжение с подогнутыми крыльями.
В брачный период очень заметен, как при частых воздушных играх, так и из-за
часто подаваемого голоса, вроде мелодичного «ки-ки-ки» или «кю-кю-кю», также
двухсложного «тервить-тервиты». Менее осторожен, чем крупные виды орлов.
Описание. Размеры и строение. См. выше. Клюв относительно короткий и
круто загнутый. Цевкаоколо 60 мм длины, оперена до пальцев. Длина самца (1) 480, размах
1210 мм. Крыло самцов (18) 352—378, самок (10) 375—403, в среднем 361,2 и 391,5 мм.
Окраска. Первый пуховой наряд желтовато-белый с синеватой радужиной и бледно-
желтой восковицей и ногами; второй пуховой наряд — сероватый, надевается в возрасте
около 2 недель. Первый годовой наряд с черноватым лбом, светлобурым затылком и шеей
с темными стволами, бледнобурыми с широкими более светлыми каймами передними
плечевыми, большими средними кроющими крыла и надхвостьем: темнобурыми спиной, задними
плечевыми и малыми кроющими крыла (у некоторых птиц часть последней партии белая);
маховые черно-бурые с сероватым рисунком на внутренних опахалах; рулевые черно-бурые
со слабо выраженным рисунком; брюшная сторона или охристая с более или менее
развитыми наствольными чертами (светлая «фаза») или темнобурая с охристым оттенком (темная
«фаза» или вариация,так называемая «minuta» старых авторов) или, наконец промежуточного
типа —. охристая с бледным буроватым рисунком или бурая с охристыми мелкими округлыми
пятнами, образующими поперечный рисунок.
Взрослые птицы светлой вариации (во втором годовом и последующих нарядах)
отличаются большей дифференциацией рисунка брюшной стороны — развитием настволий и
пестрин, а также тем, что светлая «вариация» на брюхе белая, а не охристая. Возрастные
298
ОТРЯД ХИЩНЫЕ ПТИЦЫ
изменения окраски, таким образэм, ничтожны. Оба пола окрашены одинаково. Клюв
голубоватый у основания, с черноватой вершиной; восковица и пальцы желтые, когти
черные ; радужина бурая у молодых, буровато-желтая у взрослых.
Описанные вариации окраски носят вполне индивидуальный характер. Тенденция
к гомогамии у птиц одной вариации окраски не наблюдается — смешанные пары встречаются
быть может чаще, чем пары однотипных птиц. Например, смешанные пары найдены в
Приазовье (Алфераки, 1910), в Харьковской обл. (Сомов), в Подолии (Портенко), в Тульской обл.
(Харузин), в Воронежской обл. Состав пар, найденных в Тульской обл., таков (Харузин,
1926): из 16 пар смешанных 9, в них одна птица светлой, а другая темной вариации; 2 пары
из светлых птиц, 5 пар — из темных птиц. При этом у пары из темного самца и светлой самки
оба птенца темных, у светлой пары — один темный птенец. Соотношение особей светлой и
темной вариации весьма изменчиво в пределах узких популяций и не дает картины
определенной географической локализации. Среди изученного материала с Кавказа 50% светлых
и 50% темных орлов; из б. Новосильского уезда Тульской губ. 70% темных и 30% светлых;
в б. Бобровском уезде Воронежской губ. 25% светлых, 75% темных; в Туркмении 80%
темных и 20% светлых.
Среднеазиатский орел-карлик Aquila pennata milvoides Jerdon
Spizaetus milvoides. Jerdon. Madras Journ. Lit. and Sc. X, 1839, стр. 75, Трихинополи,
Индия.
Синонимы. Aquila albipectus, Северцов. Вертикальное и горизонтальное
распространение туркестанских животных, 1873, стр. 111, Верное. — Hieraetas pennatas harterti.
Stegmann. Orn. Monatsber. 1935, стр. 151, «Южная Сибирь и северная Монголия».
Распространение. Ареал. Алтай, Танну-тува, Прибайкалье от Тунки и
Иркутска до верхней Лены (Жигалова, около 55° с. ш.), Сретенска и Аргуни на
севере; сев. Монголия (Хангай) на юге; Туркестан от Тянь-Шаня (восточные
и центральные части) на севере до Кара-Тау, на ю.-з. до южн. Таджикистана.
Зимой в Индии, откуда, как и из сев. Белуджистана, имеются впрочем и не
вполне определенные указания о редких находках в гнездовое время (Юм,
1869, и др.).
Характер пребывания. Перелетная птица. На севере
Средней Азии —. прилет от начала апреля, отлет в конце сентября.
Биотоп: лиственные леса гор, чаще всего от 900 до 1800 м, изредка
до 2400 м (Северцов и Мензбир, 1891).
Численность, Редкая птица.
Экология. Размножение. Гнезда расположены как на деревьях
(карагачах, тополях и т. п.), так и на скалах (Нура-Тау, Мекленбурцев, 1937).
Кладки из 1—2 яиц (по Тачановскому, 1891, в ю.-в. Сибири даже 3), в Тянь-
Шане в начале мая (3 мая у Алма-Ата, Северцов). Оперяющиеся птенцы
встречены в конце июля, летные с остатками пуха в середине августа (Хангай,
Козлова, 1932). Птенцов в выводке 1^2.
Линька. Как у предыдущего. Нелиняющие птицы добывались в конце
мая, птицы в свежем пере, почти закончившие линяние, — в конце августа.
Питание. Пищу составляют главным образом птицы —. голуби,
горлинки, кеклики, жаворонки (Киргизия, Узбекистан): белошапочная овсянка,
сойка (Забайкалье, Скалой, 1936); мелкие звери —»суслики (Хангай),
Описание Размеры. От номинальной формы отличается более крупными
размерами — величина у европейско-азиатских орлов-карликов возрастает с запада на восток,
.достигая максимума у прибайкальских особей. Крыло туркестанских самцов (7) 362—376,
самок (5) 392—408, в среднем 369,3 и 399 мм; у забайкальских самцов 380—395, у самок
405—435,-в среднем 388 и 420 мм (Штегман, 1937). Размеры западных — см. выше. Крупные
орлы встречаются и на зимовке в Индии.
Окраска. Как у европейских, встречаются обе вариации, причем в направлении
на восток начинает повидимому возрастать относительное число темных особей (см. выше).
В Средней Азии темные птицы в изученном материале составляют около 50%. В одном и том
же выводке встречаются иногда разноокрашенные птенцы — один светлый, другой темный
(Или, Шестоперов, 1929) или оба темных (Хангай, Козлова) или только светлые (Фергана).
Светлые птицы у гнезд наблюдались и в Прибайкалье (Тунка, Скалой, 1936).
САРЫЧ ИЛИ КАНЮК
299
РОД САРЫЧИ BUTEO LACEPEDE 1799
Тип. В. buteo
Синоним: Archibuteo Brehm, 1828
39. Сарыч или канюк Buteo buteo L.
Falco Buteo. Linnaeus. Syst. Nat. ed. X, t. I, 1758, стр. 90, Швеция.
Русское название. Слова сарыч и «канюк» повидимому и в народном языке
синонимы. Этимология первого не ясна, быть может связывается с тюркским «сары» —
желтый — из-за окраски^птицы. С другой стороны у этой птицы на польском языке название
«сарнь», так что слово сарыч имеет быть может и славянское происхождение. Канюк
(у старых авторов иногда неправильно «конюх», у Даля канюга) — от канючить, жалобно
кричать, клянчить, связывается с жалобным криком птицы. Интересно, что аналогичное имя
птица имеет на германских языках—buzzard, Bussard (от старонемецкого Bus-aro, что значит
«мяукающий орел»).
Распространение. Лесная и лесостепная полоса Европы и Азии, в Европе
к югу примерно от Шотландии, средней Скандинавии и Финляндии (около
Карта 47. Распространение обыкновенного сарыча Buteo buteo
1 —В. b. buteo, 2 — В. b. vulpinus, 3 — В. b. burmanicus, 4—'В. b. menetriesh
5 — B. b. tokoshimai, 6 — B. b. arrigonii, 7 — B. b. harterti, 8 — B. b. insularum,
9 — B. b. bannermanl, 10 — B. b. refectus; a — границы гнездовий (цифры в
кружках соответствуют номеру подвидов и показывают область их зимовок).
65° с.ш.), далее до Обонежья и Архангельска, в Зап. Сибири около 60° с. ш.,
восточней в Якутии около 63°30', затем — на Охотском побережье; к востоку до
Японии и о. Бонин; к югу до Средиземноморья (Испания, Италия, Балканы;
Корсика и Сардиния); о-ва Азорские, Канарские, Мадейра и Зеленого мыса,
далее до Кавказа, М. Азии, сев. Ирана, Гималаев, Тибета и зап. Китая. Зимой,
кроме того, в Средней Азии, сев.-зап. Индии, Бирме, Южн. Китае, повидимому
в Месопотамии (Фао), Аравии, в Африке к югу от Сахары.
Характер пребывания. Северные птицы — из Восточной
Европы, а также сибирские и восточноазиатские — перелетны, остальные —
оседлы.
Биотоп. Лесистые местности, покрытые хвойным, смешанным или
лиственным лесом, на равнинах и в горах (Азия). Внегнездовое время и в открытом
ландшафте.
300
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Подвиды и варьирующие признаки. В Зап., средней и южн. Европе В. b. buteo L.f
1758; в Вост. Европе и Зап. Сибири В. b. vulpinus G 1 о ge г, 1833; на Кавказе, в сев. Иране
и вероятно М. Азии В. b. menetriesi Bogdanow, 1879; в Вост. Сибири и Японии, Корее,
Манчжурии и сев. Монголии В. Ъ. burmanicus О a t e s, 1875; в горах зап. Китая, Тибете и
Гималаях B.b.refectusPo г te n k о, 1935(7);на о.Бонин В. b. toyshimai M om i у ата,1927(?);
на Азорских о-вах В. b. rothschildi Swann, 1919; на Мадейре В. b. harterti Swann, 1919;
на Канарских о-вах В. b. insularum F 1 о е г i с к е, 1903; на о. Зеленого Мыса В. b. banner-
maniS w a n n, 1919; на Корсике и Сардинии В. b. arrigoniiP i с с h i, 1903. Морфологически
варьируют окраска, относительная численность разных цветовых вариаций, размеры и
пропорции. Экологические различия касаются характера пребывания, особенностей хода
периодических явлений.
Малый или рыжий сарыч Buteo buteo vulpinus Gloger
Falco vulpinus. Gloger. Das Abandern d. Vbgel durch Einfluss d. Klimas, 1833, зимние
птицы из южн. Африки.
Синонимы. Buteo zimmermannae. E h m с к е. J. f. Orn., 1893, стр. 117, Гумбиннен
в В. Пруссии.— Buteo vulpinus ruficaudus s. typicus. M e n z b i e r. Ornithologie du Turkestan,
1888, стр. 195. — Buteo vulpinus intermedius B. V fusco-ater s. fuliginosus M e n z b i e r
о. с, стр. 197, Россия.
Распространение* Ареал. Лесная полоса Вост. Европы и^ Зап.
Сибири, на запад до Финляндии, Швеции, Литвы, Польши, Зап. Украины,
Румынии и Болгарии, на восток до Зап. Саяна и Танну-тувы, Алтая, Енисея
(Красноярск), быть может до низовьев Ангары и даже левых притоков Вилюя
(Марха). К северу в Финляндии до 65° с. ш., далее к востоку до Обонежья
(Медвежья Гора), Архангельска, на Печоре до 66° с. ш. (Дмоховский, 1935),
далее на М. Сосьве около 62° с. ш. и затем южней (точная граница не ясна)
около 60° с. ш.; на Енисее у Красноярска; южная граница в СССР в
Бессарабии, Подолии и Волыни, на Северном Кавказе (у Дзауджикау повидимому
вместе с 5. b. menetriesi)*, далее в лесостепи ю.-з. Сибири, в сев. и центр.
Казахстане (Кокчетау, Баян-аул, Каркаралинск, Долгушин, 1938), в восточном и
центральном Тянь-Шане (Джаркент, Алма-Ата, Иссык-куль, горы между Аксу
и Борохудзиром, Портенко, 1929). Зимой изредка в ю.-в. Европе (Хорватия,
Болгария), в виде исключения в Европейской части СССР, более или менее
регулярно у Ленкорани в ю.-в. Закавказье и в Туркмении, изредка в Киргизии
(декабрь—январь, окрестности Фрунзе), кроме того в с.-з. Индии, Аравии, но
главным образом в Африке от Судана, Абиссинии, Анголы и Конго до Капской
Земли. Быть может в юго-вост. Азии (Малаккский п-ов, Штреземанн, по
Хартерту, 1913).
Характер пребывания. Перелетная птица с разомкнутыми
гнездовым и зимним ареалами (в отличие от оседлых западноевропейской и
островных африканских рас).
Д а т ы. В Африке появляется в октябре, исчезая оттуда в марте—апреле,
отдельные особи встречаются еще в мае. Весенний- пролет на юге Средней Азии
в Туркмении в апреле—мае. Прилет на гнездовье в разные числа апреля, на
юге отдельные особи уже в марте (Украина), северней в конце марта, но в
основном около середины апреля; на севере ареала и в Сибири во второй половине
апреля (Киров 22—24 апреля; север Молотовской обл. 27—29 апреля; Томск
23 апреля; Минусинская лесостепь 22 апреля). Осенью отлет и пролет протекают
в сентябре; на севере птицы исчезают уже около середины этого месяца.
Отдельные особи задерживаются до октября (Псковская обл.; Харьковская обл.;
экземпляры из окрестностей Бердянска, с Кавказа и из Туркестана и т. д.).
Весенний пролет совершается одиночками и парами, осенний одиночками,
небольшими стаями и группами, иногда до нескольких десятков птиц.
* По Портенко, малый сарыч встречен на гнездовье в Старом Крыму и в Мазандеране
в Сев. Иране. Однако автору из Крыма известны пока только гнездовые В. b. menetriesi,
и к этой же расе принадлежат все осмотренные канюки из Астрабадской провинции.
САРЫЧ ИЛИ КАНЮК
301
Биотоп. Лесистые местности —. хвойные, смешанные и лиственные
леса; лесостепь (колки ю.-з. Сибири). На пролете и в открытом ландшафте.
В горах до границы леса (в Тянь-Шане в верхней части пояса лиственных лесов
и в ельниках; на Алтае до 2200 м, на Урале до Иремели).
Численность. Один из самых обычных видов хищных птиц с плотным
распространением (в благоприятных условиях гнездовый участок 2—5 км2)»
Колебания численности не изучены, но повидимому имеются; после суровой
зимы 1939—-1940 года в Европейской части СССР местами (например в
Московской и Калининской обл.) число гнездовых пар сарычей сократилось и в
выводках было не более 2 молодых.
Экология. Размножение. Половая зрелость наступает в возрасте
несколько менее года, в первом годовом (гнездовом) наряде. Имеется резерв
холостых особей, так как погибший партнер заменяется другой птицей. Гнезда
на деревьях, как хвойных (сосна, ель), так и лиственных (береза, дуб, ива,
ясень, граб), расположены ближе к опушкам. Гнездастроит сам сарыч, но иногда
(Харьковская обл., Сомов, 1897; Бессарабия, Остерман, 1916) занимается или
надстраивается чужое (воронье) гнездо. В центральном Казахстане найдено
гнездо на скале (Долгушин, 1938). Гнезда из ветвей, размер их около 50—
80 см в диаметре, высота около 20—50 см; лоток плоский, выстланный корой,
мхом, пухом; гнездо обычно прикрыто зелеными ветвями; расположено чаще на
средней высоте, в 8—-12 м, реже ниже, в 4—5 (Томск, Белышев, 1929). Брачный
полет — с прилета, со второй половины апреля. Кладка с конца апреля
до середины мая (на юге, повидимому, раньше, в б. Минской губ. 15—20 апреля,
22 апреля в Уманском уезде Киевской обл.; 19 апреля в Подолии, 22 апреля
в б. Екатеринославской губ.; два яйца 19 апреля, Псков; в Сибири в мае, у
Томска 17 мая чуть насижены; 11 мая в Барабинской степи, в первой трети мая
на Южном Урале).
В кладке обычно 2—3 яйца, редко 4 или даже 5 (Минусинск, Зверев, 1930);
взамен утраченной кладки очень редко встречается одно яйцо (Сомов, 1897).
Число яиц в кладках колеблется по годам, что вероятно связано с кормовыми
условиями; шышиньде» годы дают большие кладки, как это установлено и
в Зап. Европе, хотя у перелетного vulpinus такое влияние кормовых условий
должно бы быть менее резким, чем у оседлого В. b. buleo. Окраска яиц беловато-
зеленоватая, с поверхностными каштаново-бурыми и глубокими фиолетово-
бурыми крапинами.
Размеры (8) 54—58,3 х 41,5—46 (Харьковская обл., Сомов, 1897), (10)
50—53x40,5—41,5, в среднем 50,9 х 41,2 для «vulpinus» и (24) 50—61 X
х 40,5—48, в среднем 55,1 X 41,2 мм для «buteo* (Умань, Гебель, 1879);
58,5—50 х 41,5—45 мм (Полесье, Шнитников, 1913). В случае частых
беспокойств, сарычи иногда бросают кладку, но в других случаях взамен
утраченной птицы снова откладывают яйца (Томск, Иогансен, 1906).
Насиживание происходит с первого яйца, участвуют в нем оба родителя, но
главным образом самка (Подолия, Московская обл.). Длительность
насиживания около месяца (28 дней, Бессарабия, Остерман, 1916). Начало кладки в
Башкирии 15 мая, вылупление там начинается с 15 июня. Насиживающей самке,
так же как и выводку, во всяком случае в первое время, корм носит самец,
самка же оделяет птенцов. Когда птенцы несколько подрастут, наденут второй
пуховой наряд — начинают охотиться оба родителя. Они бросают птенцам
добычу в гнездо. Отмечена борьба между птенцами из-за корма (Белышев,
1929). Смертность птенцов довольно высока: в Башкирии «отмечена
эмбриональная смертность до 50%, постэмбриональная до 17,5% (Крюкова). Только что
вылупившиеся птенцы наблюдались в разные числа июня (в б. Минской губ.
уже 20 мая, в Минусинской лесостепи 12 июня и даже в конце июня, у
Чкалова 4 июня). Число птенцов в выводке обычно 2—3, редко до 4 (Поволжье;
302
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Псков; Минск; Минусинск). Птенцы начинают летать с конца июля, гнездовый
период продолжается следовательно около 40 дней (41—42 дня, Москва,
Дементьев). Запоздавшие птенцы (от вторых кладок?) вылетают в начале августа и
примерно до конца этого месяца держатся в гнездовом участке (средняя полоса).
Выводки держатся вместе до
отлета.
Линька. Полная
годовая. Взрослые птицы
начинают линять в первой
половине июня (в конце мая сарычи
еще не линяют). Самцы
линяют несколько поздней, чем
самки, быть может в связи
с малым участием в
насиживании. В первой половине
сентября попадаются сарычи
в свежем пере, к отлету
линька заканчивается. Общая
продолжительность процесса
таким образом около Зх/2 ме*
сяцев, т. е. значительно менее,
чем у соколов и повидимому,
чем у оседлого В. b. buteo. У
отдельных особей линька
запаздывает.
Последовательность смены маховых от 10-го
к 1-му; рулевые линяют цен-
тробежно от середины к краям,
но не всегда в строгой
последовательности. Птицы в
первом годовом наряде начинают
линять несколько ранее, чем
старые. Последовательность
смены нарядов: первый
пуховой наряд — второй пуховой
наряд—первый годовой
(гнездовый) наряд — второй
годовой (окончательный) наряд
и т. д.
Питание. Сарыч-по-
.лифаг. В кормовом его
режиме в СССР отмечены
млекопитающие мелкой
величины: белки, водяные крысы, полевки (в том числе Clethrionomys rutllus,
С. glareolus, Microtus oeconomus, M. arvalis, Arvicola amphibius); суслики,
мыши Silvymus sylvaticus, изредка М. minutus и др.; землеройки и
быть может выхухоль; молодые птицы —дрозды, тетерева, белые куропатки,
большой пестрый дятел; жаворонки, перепела, голуби, молодые домашние
птицы — цыплята и утята и т. д.; пресмыкающиеся —змеи, ящерицы;
земноводные — полосатый тритон Triton taeniatus, лягушки, жабы, разные
насекомые — жуки, гусеницы, саранчевые и др.
Все же наибольшее значение в питании сарыча имеют грызуны, и в Зап.
Европе установлена зависимость между размножением сарычей и «урожаем»
мышевидных грызунов.
Рис. 83. Обыкновенный сарыч (взрослый самец и
молодая птица).
САРЫЧ ИЛИ КАНЮК
303
Среди сарычей имеется впрочем известная индивидуальная специализация
кормового режима (в частности, для Печоры отмечаются сарычи —
специалисты по белым куропаткам, Донауров). С другой стороны, в годы, бедные
мышами, сарычи довольно быстро переключаются на питание и другой добычей и
в средней полосе выкармливают тогда молодых главным образом птицами-
подлетками, птенцами и т. д. Дневная потребность сарыча в пище
составляет около 100—120 г, т. е. относительно невысока (наблюдения в
Московском зоопарке). Сравнительно слабые лапы, короткие пальцы и относительно
тихий полет сарыча не позволяют ему нападать на летающих птиц. Сарыч
охотится паря над открытыми пространствами, изредка трепещет в воздухе на
месте, как пустельга, иногда же довольно быстро летает скользящим полетом.
Другой способ — подстерегание добычи сидя на возвышенном месте — дереве,
телеграфной проволоке, столбе, стоге и т. п. Во время массового размножения
саранчевых Pachytilusn CalliptamusB б. Симбирской губ. в 1899 г. было
замечено, что сарычи охотились на саранчу, ходя «пешком» (Житков и
Бутурлин, 1906).
Сарыч иногда ест и падаль и, наконец, отнимает добычу у мелких хищников
(перепелятника, дербника и др.), хотя сам нередко становится жертвой других
птиц (тетеревятника, филина и сокола).
Полевые признаки. Хищная птица средней величины с" широкими и
довольно короткими крыльями, коротким и закругленным хвостом. Окраска
бурая, на спинной стороне однообразная, на брюшной иногда с преобладанием
беловато-охристого цвета. Полет сравнительно медленный и мало поворотливый.
Осторожен. Голос — несколько напоминающий мяуканье ши-пия» |или
посвист.
Описание. Размеры и строение. Оперение рыхлое и мягкое, крылья с
длинным предплечьем и относительно короткой кистью. Хвост из 12 рулевых, слегка
закругленный. Формула крыла 4^3>5>6>2>7>8>1 или 7>1>8. Вырезки на наружных
опахалах первых пяти, на внутренних опахалах первых четырех первостепенных маховых.
Клюв резко сжатый с боков, слабый, но с длинным крючком надклювья. Цевка оперена
только в верхней половине, спереди покрыта большими поперечными щитками; пальцы
короткие, когти менее острые и изогнутые, чем у соколов и ястребов. Длина самцов (10)
460—492, самок (16) 495—530, в среднем соответственно 480 и 513,8 мм. Размах самцов (10)
1018—1200; самок (17) 1176—1277, в среднем 1133,3 и 1227,5 мм. Вес самцов (4) 600—675,5,
самок (5) 710—1175, в среднем 627,4 и 851 г. Крыло самцов (82) 342—372, самок (77) 360—386,
в среднем соответственно 357,1 и 373 мм.
Окраска. Полиморфная: три типа окраски, различимые во всех возрастах;
однообразный темнобурый (названный Мензбиром «fuscoater»), бурый, с примесью рыжего цвета
и с пестрой брюшной стороной («ruficauda») и наконец бурый с примесью охристого цвета
на брюшной стороне («intermedius»). Преобладает второй тип окраски, наиболее редок
первый, составляющий в изученном материале 7%; в отдельных местностях это количество
меняется, составляя на нашем материале в средней России 4%; в Поволжье и Приуралье13%;
среди пролетных и зимующих на Кавказе птиц 13%; на севере от Карелии до Печоры и
бассейна левых притоков Оби «fuscoater* не встречается. Темные птицы вариации «fuscoater»BCipe-
чены и в Сибири (Омск,-Томск, Алтай, зап. Саяны, Минусинск; зимою в Семиречье). Форма
«intermedius» по отсутствию рыжего цвета как бы промежуточна между В. Ь. buteo и В. Ь.
vulpinus; она встречается также по всему ареалу первого подвида, на восток до Енисея и
Алтая, но численно заметно уступает вариации с примесью рыжего; более многочисленна
она в западной части ареала, приблизительно до долготы Москвы. Однако соотношение
числа особей различных цветовых вариаций не характеризует определенных местностей
и не отражает каких-либо географических закономерностей.
Первый пуховой наряд буровато-серый, из длинных, особенно на голове, но редких
пушинок. Второй пуховой наряд густой и короткий, беловато-сероватый, разных оттенков.
Окончательный наряд вариации «ruficauda»: голова, спина, кроющие крыла серовато-бурые,
с ржаво-рыжими каймами, причем иногда эти каймы так широки, что преобладают над
основным тоном; стволы перьев черноваты; маховые бурые с нерезким темным поперечным
рисунком и беловатым основанием внутренних опахал; рулевые ржавчато-бурые с узкими
черноватыми поперечными полосами и черной предвершинной широкой перевязью; у некоторых
птиц хвост рыжий с черной вершинной полосой; горло охристое с буроватыми или
рыжеватыми наствольями, грудь ржаво-бурая с охристо-беловатыми пятнами, брюхо и бока рыжие
с белым поперечным рисунком. Вариация «intermedius» отличается отсутствием ржавчато-
304
ОТРЯД ХИЩНЫЕ ПТИЦЫ
рыжих тонов или они едва заметны на спине и крыльях, в виде оттенка на хвосте
(последний обычно сероватый с темнобурым поперечным рисунком). Птицы «fuseoater»
темнобурые с серовато-бурым хвостом, покрытым темнобурыми поперечными полосами,
основания внутренних опахал первостепенных маховых беловатые. Птицы в первом
годовом наряде отличаются от старых более слабым развитием ржавчатых тонов — они
заменены у них главным образом охристыми. Кроме того, поперечный рисунок на брюшной
стороне тела заменен продольным. Птицы темной вариации «fuseoater» в первом годовом
наряде обычно имеют узкие рыжевато-охристые каемки перьев. Оба пола окрашены
одинаково, хотя среди самок редко встречаются столь интенсивно рыжие особи, как среди самцов.
Радужина бурая разных оттенков, клюв синевато-роговой, у вершины черноватый, когти
черные, восковица и лапы желгые.
Систематические замечания. Большинство авторов указывает, что в западной
части нашей страны, в Прибалтике, Бессарабии, на Украине в Подолии и на Волыни,
в Днепропетровской обл., в зап. частях Киевской губ, (Умань), быть может в
Харьковской обл. — встречается и западноевропейская раса сарыча В. b. buteo, отличающаяся
огтВ.Ь. uulpinus, в среднем более темной окраской и большими размерами, отсутствием рыжих
тонов, иной амплитудой индивидуальной изменчивости (нет рыжей вариации, но кроме
темнобурой и бурой с пестрым низом, есть еще беловатая вариация), а также рядом
экологических отличий: оседлостью, повадками и т. п. Однако тут же встречаются и переходные
особи (по ним описаны В. b. zimmermannoe), и настоящие uulpinus. Вероятно правильней
считать, что по всей этой переходной полосе, занимающей около 400 км (Штреземанн, 1925),
встречается смешанная популяция (buteo x uulpinus) и что среди этой популяции могут
встречаться особи, внешне не отличимые от исходных форм. Тем более, что имеются указания,
на наличие (редкое?) пар, в которых одна из птиц «buteo», а другая «vulpinus» (например,
Подолия, Портенко, 1928). Заслуживает внимания то обстоятельство, что в данном случае
скрещиваются две расы, из которых одна перелетная, а другая оседлая: однако сроки
размножения у них совпадают (кладка у Bfo. buteo в Германии обычно во второй и последней трети
апреля). *
В литературе (Радде, 1885, Портенко, 1929) есть указания на нахождение В. b. buteo
в негнездовое время на западном Кавказе. Остается неясным, гнездится ли в пределах
СССР настоящий В. b. buteo в б. Галиции (Штреземанн, 1925), откуда мы пока не имеем
достаточных материалов. При таком положении вещей конечно невозможно считать buteo
и vulpinus за разные виды, тем более, что кавказские и североиранские сарычи составляют
между ними «переход» (размеры крупные, как в buteo, а окраска с рыжими тонами). С
эволюционной точки зрения интересны некоторые особенности географической изменчивости
сарычей buteo и vulpinus.
Птицы, гнездящиеся в более суровом климате, меньше по размерам и более яркой
рыжеватой окраски, что кажется исключением из обычных закономерностей. На самом деле
это исключение кажущееся, так как vulpinus перелетный, a buteo оседлый и более эври-
термный, чем первый.
Кавказский сарыч Buteo buteo menetriesl Bogdanow
Buteo Menetriesi. Богданов. Птицы Кавказа. Тр. Общ. естествоисп. при Имп«
Казанском унив., VIII, 1879, стр. 45, Кавказ.
Распространение. Ареал. Кавказ от бассейна Кубани на западе и
Дагестана на востоке, на севере, к югу в Закавказье и южнокаспийских провинциях
Ирана, на восток до Астрабада; горный Крым, вероятно, Малая Азия (вост.
части?). Приводится для Добруджи (Портенко, J929). Особи с признаками,
переходными между В. b. buteo и menetriesl (крупные, с сильным развитием
ржавчатых тонов), отмечены для Балкан (Чузи, 1890, Рейзер, 1896) и Венгрии
(Чергей, 1906).
Характер пребывания. Кавказский сарыч оседлая птица, но
в негнездовое время часть особей кочует, спускаясь с гор и достигая низовьев
Дона (Ростов) и Добруджи (Портенко, 1929).
Биотоп. Пояс горных лесов, до 1800 м (Армения, Даль, 1948), на охоте
и на альпийских и субальпийских лугах, вне гнездового времени и на
равнинах.
Численность. Обычен на Кавказе, в Крыму редок. Есть указания
на колебания численности в связи с «урожаем» мышевидных грызунов (на
с.-з. Кавказе в 1934 году более многочисленен, чем в 1933, Насимович и
Аверин, 1938).
САРЫЧ ИЛИ КАНЮК
305
Экология. Размножение. Половой цикл относительно ранний.
Гнезда расположены на деревьях в лесах у опушек. Число яиц 2—4, чаще 3
(Беме, 1926). Птенец иногда только 1 (Крым, Молчанов, 1906). Вылет птенцов
начинается уже в конце июня —* начале июля, так что кладка вероятно в конце
марта — начале апреля (в конце июня вылет в Дагестане, летные молодые
13 июля в ю.-з. Закавказье; вполне выросшие молодые в Закатальском округе
5 июля, в Армении покинутое птенцами гнездо найдено 13 июля).
Линька. Общий ход — как у предыдущего подвида. Самки, сменившие
больше половины первостепенных маховых, добывались во второй половине
июля (остались старые 4—5 первостепенных маховых), у самцов в это время
сменились только 2—3 первостепенных маховых. Сарычи, кончившие линьку,
добывались в конце августа — в начале сентября (у некоторых особей линька
повидимому затягивается и позднее).
Питание. В Крыму в качестве корма сарыча отмечены сойка, мелкие
птицы—снегирь, зеленушка, поползень; ящерицы (Молчанов, 1906), на Кавказе
средние и мелкие по размерам птицы, желна, сойка и т. д., звери — зайченок,
полевки, лесные мыши и другие мышевидные грызуны; пресмыкающиеся —
ящерицы, ужи; насекомые —жуки (жужелицы и др.), кузнечики и кобылки,
саранча и т. п. При этом указывается (Стахровский, 1941), что сарыч ловит
цикад «по слуху», сидя и слушая их стрекотание. Главное место в кормовом
режиме занимают мышевидные грызуны (75% встреч летом 1936 в Кавказском
заповеднике, Жарков, 1938).
Описание. Размеры и строение. Величина крупная, как у номинальной
расы: крыло самцов (15) 370—385, самок (12) 390—413, в среднем 377,4 и 396,3 мм.
Окраска. Отличается от номинальной расы присутствием ржавчато-рыжих тонов
в окраске, в этом сходство с vulpinus. Спинная сторона бурая с широкими
рыжевато-охристыми или красновато-охристыми каемками; брюшная сторона рыжая с беловатым (у старых
птиц поперечным) рисунком. У птиц в первом годовом наряде брюшная сторона охристая с
рыжим продольным рисунком. Рулевые с преобладанием рыжего цвета и с более или менее
развитым темным поперечным рисунком. Некоторые особи с резким преобладанием красно-
рыжего цвета.
Имеется незначительное число одноцветно-бурых или почти одноцветно-бурых
особей {«fuscoater»).
Восточноазиатский сарыч Butco buteo burmanicus Oates
Buteo burmanicus. Oates. Stray Feathers v. Ill, 1875, стр. 60, Тайетмио, Бирма.
Синоним. Falco buteo japonicus. Temminck et Schlegel. Fauna Japonica
Aves, 1844—1845, стр. 16, табл. VI—VI, Япония, nom. praeoccupatum.
Распространение. Ареал. Вост. Сибирь, на запад до Чоны и Муры,
верховьев Н. Тунгуски, Нижнеудинска, на север до Вилюя и Якутска (63э30'),
к востоку до сев. побережья Охотского мор'я (р. Половинная, Аян, бухта Герт-
нера, Магадан), Шантарские о-ва; южные Курилы, Сахалин; на Камчатке
отсутствует; к югу до Японии (Хоккайдо, Хондо, Изу, Сикоку), южных частей
Приморья, Манчжурии и Кореи, сев. Монголии (Кентей и Хангай). Зимовки
в Средней Азии — южн. Казахстане и Киргизии (центр. Тянь-Шань у Джар-
кента, Алма-Ата, Фрунзе, к западу до Джелалабада, Оша, Андижана, Аулье-ата,
Ташкента); по Зарудному и Билькевичу (1918), до верхнего течения Аму-Дарьи
(нами осмотрен один экземпляр из Репетека в вост. Кара-Кумах от 22.1.1917, и
другой из окрестностей Ашхабада от 8. V 1948); в Синьцзяне, Китае (до
Хайнаня), в Бирме и Индии; в Индо-Китае; на севере — в южн. Японии и
Корее, в долине Янцзы в Китае, на Формозе.
Характер пребывания. Перелетная птица, отлетающая с мест
гнездовья в сентябре и появляющаяся на гнездовье в апреле. На зимовке в
Туркестане сарычи встречаются с начала ноября и находятся там до середины
20 Птицы СССР, т. I
306
ОТРЯД ХИЩНЫЕ ПТИЦЫ
апреля, запоздавшие особи встречаются еще в конце этого месяца (30. IV. 1928,
окрестности Фрунзе). Пролет в Сев. Китае в Чжили в феврале-мае.
Пролет на с.-в. Алтае уже с конца августа, но главным образом в сентябре
(Яйлю, Дементьев иФолитарек, 1938). В Приморье пролет уже с конца марта
и тянется около месяца (Шульпин, 1936), там же осенью до начала октября
(запоздалый сарыч на о. Аскольд добыт еще 8 ноября). Прилет на места
гнездовья отмечен в Забайкалье в ю.-в. частях Витимского нагорья в разных
числах апреля (Павлов, 1948), на Шантарских о-вах в конце апреля
(Шульпин и Дулькейт, 1937), северней, у Олекминска 12—18 апреля
(Воробьева, 1928).
Биотоп. Леса — редкие или пересеченные открытыми пространствами,
с редким подседом (что связано с охотой на грызунов); глухой тайги избегает.
На зимовке в Средней Азии главным образом в подгорном культурном
ландшафте.
Численность. Повидимому, ограниченная, хотя в Киргизии и
южн. Казахстане на зимовке нередок, а на пролете значительно преобладает
над vulpinus.
Экология. Размножение. Сроки размножения совпадают с таковыми
В. b. vulpinus. Гнезда расположены на деревьях, чаще всего на лиственницах,
также на березах (Борзинский район, Скалой, 1936), сосне (Чита, Штегман,
1928); в верхней части дерева, обычно у вершины, иногда впрочем низко, в Зм
от земли (Скалой, 1936). В кладке 2—4 яйца;размеры (9) 52,3—59 х 42,6—47 мм
(Тачановский, 1891).
Кладка в начале мая (Витимское нагорье). Насиживанье продолжается
около месяца, появление птенцов в начале июня (6 июня только что
вылупившиеся птенцы и наклюнутое яйцо, Витимское нагорье, Павлов). Число
птенцов 2—3. Летные молодые птенцы в конце июля или начале августа, но
маховые у них еще не доросли, и даже в конце августа — начале сентября попадаются
молодые с недоросшими маховыми (1 сентября, Хангай, Козлова, 1932; 23
августа, Кентей, Козлова, 1933).
Линька. Как у vulpinus.
Питание. Полифаг, однако, грызуны занимают главное место в
кормовом режиме — с наличием их (и отсутствием густой травы и подседа) связано
и распределение канюка в Вост. Сибири. В качестве пищи восточноазиатского
сарыча указываются главным образом грызуны-бурундуки, полевки Micro-
tus ungurensis, Clethrionomys rutilus, M. michnol, Stenocranius gregalts, также
пищухи; лесной лемминг Myopus schisticolor; разные птицы, в частности воробьи
(Скалой, 1936; Некипелов, 1935; Штегман, 1928; Павлов, 1948). Указывается,
что сарыч вечерами охотится на бурундуков, сидя на земле и бросаясь на
их свист.
Описание. Размеры и строение. Цевка обычно оперена несколько более,
чем на половину, в чем отличие от европейских и кавказских сарычей; вырезка наружного
опахала 4-го махового обычно располагается между вершинами 9-го и 10-го маховых (у
vulpinus и menetriesi между вершинами 8-го и 9-го маховых). Размеры близки к кавказским
сарычам и больше, чем у vulpinus. Длина крыла самцов (14) 365—388, самок (20) 378—
426, в среднем соответственно 377,9 и 399,2 мм. Длина самцов (б) 507—532, самок (10)
530—574, в среднем 521,9 и 548,7 мм. Размах крыльев самцов (6) 1104—1220, самок (10)
1200—1315.
Окраска. Более однообразная, чем у В. Ь. vulpinus, с меньшим развитием ярко-
рыжих тонов; взрослые птицы на спинной стороне темнобурые или серовато-бурые
с охристо-рыжеватыми или беловатыми краями перьев; серовато-бурый поперечнополосатый
хвост; брюшная сторона с беловатыми или охристыми горлом и грудью, иногда более или
менее испещренными продольными бурыми полосками; брюхо и бока ржавчато-бурые с
беловатым поперечным рисунком, оперение голени бурое с узкими охристыми поперечными
полосами. Молодые в первом годовом (гнездовом) наряде менее рыжеваты, имеют охристые
каймы на перьях спинной стороны, брюшная сторона без поперечного рисунка. Бурая
вариация в пределах СССР не найдена, но встречается в Центральной Азии.
МОХНОНОГИЙ КАНЮК ИЛИ ЗИМНЯК
307
40. Мохноногий канюк
или зимняк Buteo
lagopus Briinn.
Falco Lagopus BrQnnich Or-
nithologia Borealis, 1764, стр.4,о.
Борнгольм.
Русское название.
Мохноногий канюк — книжное имя,
общее по смыслу почти всем
европейским языкам. На севере
местное название этой птицы —
мышеловка или мышелов. Зимняком ее
называют местами в средней
полосе, где она появляется в
холодное время года.
Распространение. Ареал.
Север Европы, Азии и Сев.
Америки, в зоне тундр и
лесотундр; в холодное время года
и на пролете в средней Европе
к югу до Пиренеев и Балкан,
Крыма, Сев. Кавказа,
Туркестана, Монголии, Манчжурии,
сев. Японии, южных частей
Соединенных Штатов до
Калифорнии, Техаса, Луизианы и
сев. Каролины.
Характер
пребывания. Перелетная птица.
Биотоп. Открытые
ландшафты — тундры и
лесотундры в гнездовое время, но
на Камчатке и в лесу; на
зимовке в степном и культурном
ландшафте, сплошных лесов
избегает.
Подвиды и варьирующие
признаки. Четыре географических
расы. В сев. Европе и Азии от
Скандинавии до Оби В. Ь. lagopus
Brunnich; в заенисейской
Вост. Сибири В, I. menzbieri (см.
ниже); на Охотском побережье и
на Камчатке В. I. kamtschatken-
sis D e m e n t i e v, 1931; в Сев.
Америке В. /. sancti- iohann is
Gtnelin, 1788. Различия — в
окраске; американский подвид
диморфен (кроме обычного типа
окраски попадаются меланистиче-
ские однообразно окрашенные тем-
нобурые птицы); общая окраска
американских птиц темная; близок
к американским, но без мелани-
стической вариации, камчатский
подвид; восточносибирские птицы
самые светлые и пестрые;
европейские и западносибирские — бурые
20*
iff
X u
* I
Я I
S о
CO in
» i
= §
X js
Q> О
X S3
M
!«?
* 4
CO „f
« s
§■*
i
J*
И
I
ffl
ffl
308
ОТРЯД ХИЩНЫЕ ПТИЦЫ
и темные, менее контрастно окрашенные, чем американские и камчатские. Американские
птицы по величине несколько меньше сибирских и камчатских. Экологическая географиче^
екая изменчивость изучена недостаточно и к тому же выражена слабо.
Западный или европейский мохноногий канюк Bute о lagopus lagopus Brunn.
Распространение. Ареал. Север Европы и Азии за пределами сомкнутого
высокоствольного леса — от Лапландии до Ямала и Таза. Южная граница не
вполне ясна, в Норвегии еще у 59°, в Швеции до 60°; в Финляндии несколько
северней 65° (в финской Карелии самый южный пункт — Куохатти, Хортлинг,
1931), в советской Карелии у Сегозера, немного северней 63° (гнездится ли — не
выяснено), далее на Мезени, до южной части Канина, в Тиманской и Больше-
земельской тундре; на полярном Урале добыт несколько южней полярного
круга (на горе Пирбью, Гофман); в Зап. Сибири граница не прослежена. Едва
ли выходит на гнездовье за пределы криволесья, хотя имеется ряд
литературных указаний о гнездовании на севере лесной полосы и даже в средних частях
европейской России и Сибири (по Руссову, 1880, гнездо найдено под Ригой;
Хлебников, по Бианки, 1910, добыл будто бы 18.VII. 1882 в б. Новгородской губ.;
по Воронцову, 1935, будто бы гнездится в Ветлужском районе; есть указания
на летние находки в Петербургской и Владимирской губ.; Богданов и Эверсманн
приводят для Поволжья, Зарудный будто бы в середине июля 1883 нашел
канюка в Киргизских степях в урочище Джирень-копа; по Сабанееву, 1874,
будто бы гнездится в б. Екатеринбургском и Шадринском уезде Пермской губ.;
далее, Залесский, 1935, указывает, что зимняк гнездится в Нарымском крае,
а также в Барабинской степи у озера Сартлан и в Тарском уезде, где будто бы
найдены гнезда; Ермолаев, 1921, приводит канюка в качестве гнездящейся
птицы окрестностей Гурьевского завода в б. Кузнецком уезде). Однако все эти
сведения пока приходится считать недостоверными: они основаны или на неверных
определениях на расстоянии, или на встречах поздних пролетных особей, или,
наконец, на находках задержавшихся почему-либо вне гнездовой области птиц.
Экземплярами они не подтверждены (как между прочим и не подтверждено
гнездование/ хотя бы случайное, зимняка в сев. Германии). Из достоверных
находок зимняков на юге можно только привести добычу его 9. VII. 1897 у Сул-
тангулова в Самарской губ. (Карамзин, 1901) и о встрече его у горы Шаман
в с.-в. Алтае 19июня(Сушкин, 1938). На севере канюк распространен до
прибрежных частей тундры, найден на о-вах Колгуев и Вайгач, но не на Новой
Земле, где встречается повидимому только случайно (Горбунов, 1929, Антипин,
1938). Область нормальных зимовок в средней Европе от южн. Скандинавии,
равнинных частей сев. Германии, в Украинских и Таврических степях, на
Сев. Кавказе, в Казахстане, в северных частях Туркестана от центрального
Тянь-Шаня до нижнего и среднего течения Сыр-Дарьи, быть может в ю.-з.
Сибири. Нерегулярно зимою встречается в Англии, Франции (к югу до Пиренеев),
на Балканском п-ове, в средней России, в Закавказье (до Ленкорани);
залетает в Италию, Швейцарию, М. Азию, южн. части Туркестана (Туркмения),
в Египет.
Даты. Как сроки, так и общий ход перелетов зимняков довольно
разнообразны и подвержены значительным изменениям в разные годы. В иные годы
зимняки появляются в большем, в иные в меньшем количестве. Большинство
птиц зимует в открытых относительно малоснежных степных участках, в
лесной же полосе лишь нерегулярно, вероятно при наличии особо благоприятных
кормовых условий (или при неблагоприятных условиях — глубоком снеге —
в области нормальных зимовок на юге?). Многие авторы отмечают связь между
численностью пролетающих и зимующих канюков и обилием грызунов. В
бедные леммингами годы откочевки канюков из тундр носят более широкий
характер и быть может начинаются ранее, в «лемминговые» годы часть птиц даже зиму-
мохноногий канюк или зимняк 309
ет (например, в Скандинавии, где добывались зимою у Карлсботтен и у Пол^
мака за 70°с. ш.).Годы массового появления зимняков вХарьковской обл. 1888—
1889 (Сомов, 1897), в б.Минской губ. 1899—1900 и 1903—1904 (Шнитников, 1913);
в средней полосе множество в 1931—1932, 1935—1936 и в 1938—1939
годах. Отмечено и обратное явление. Так, в Лапландии, по Веслену (цитируется
по Уттендерферу, 1939) наблюдается иногда внезапное вымирание леммингов,
бывших многочисленными еще в начале периода размножения канюков;
последние в таком случае в большинстве покидают кладки и начинают кочевать.
Именно этим обстоятельством, а не переменой места гнездованья, объясняется
по всей вероятности добыча летом у Омы в Чешской губе окольцованного
птенцом в сев. Норвегии зимняка; это было в нелемминговый год, и зимняк на
Белом море вероятно не гнездился.
Всюду на пролете и зимою молодые птицы в первом годовом наряде
преобладают численно над старыми. Отлет с мест гнездовья отмечен в сентябре
(в низовьях Енисея на Луковой протоке уже в августе, Тугаринов и Бутурлин,
1911; однако в Лапландии птицы задерживаются и в октябре (отлет в
Лапландском заповеднике 3 октября—17 ноября, в среднем 19 октября, Владимирская,
1948). В сентябре зимняки появляются в лесной полосе, причем движение их
продолжается и в октябре. Южных зимовок канюки достигают с конца октября
(первые особи), но главное появление бывает только в ноябре (например на Сыр-
Дарье 20 ноября, Спангенберг, 1936; в ноябре и на юге Украины, на Дону и
Сев. Кавказе; 3 ноября в Армении, Спангенберг, 1948). Осенний пролет
проходит довольно быстро, но иногда и медленно. Отлет с мест зимовок начинается
уже в марте, главным образом в апреле.
Весеннее движение на север медленное и быть может связано с таянием
снега и увеличением активности грызунов в средней полосе. На севере лесной
полосы пролетающие зимняки встречаются еще в мае (Псков—2 мая; в Молотовской
обл.; экземпляры из Плеса, б. Костромской губ. 13 мая; из Богородского района
Московской обл. 16 мая и т. д.). Массовый отлет на южных зимовках
заканчивается в первой половине апреля. В самом конце апреля — начале мая
канюки появляются на севере (30 апреля, 2—3 мая, в среднем 7 мая, в Лапландском
заповеднике, Владимирская, 1948; на южном Ямале у Ярро-то в конце апреля,
но у озера Нейтто под 70° с. ш. 11 мая, Житков, 1913; в Тиманской тундре
первые встречены 30 апреля, Гладков, 1941; в низовьях Енисея —в начале мая,
Тугаринов и Бутурлин, 1911).
Биотоп. Тундра и лесотундра в гнездовое время, зимою — открытые
пространства; на пролете в лесной зоне, главным образом в речных долинах и
в культурном ландшафте.
Численное тф. Как у всех тесно связанных в питании с
мышевидными грызунами хищных птиц, численность зимняков и в гнездовой области,
и на пролетах, и зимовках сильно меняется по годам. Это объясняется
усиленным размножением в лемминговые годы, а также повышением смертности и
негнездованием в годы неурожая грызунов. Вообще в тундре на гнездовье
нередок, но в неблагоприятные годы число гнездящихся пар чрезвычайно
сокращается (в 15 раз, Лапландия, Ларсон, 1935).
Экология. Размножение. Зимняки держатся парами уже с прилета,
пары вероятно постоянны. Гнездовый период — в конце мая, когда тундра
почти свободна от снега, но земля не протаяла; поэтому в гнездах всегда
толстая подстилка из сухой травы (около 6—7 см, Осмоловская, 1943). Размеры
гнезд различны — в зависимости от биотопа: более крупные, около 70—80 см
в диаметре, на обрывах; меньшие, около 50-60 см на мокрой ровной тундре.
На размеры гнезда влияет и длительность его использования. Устроено гнездо
из довольно толстых сучьев, хорошо защищающих кладку и насиживающую
птицу от ветра и снега. В редких случаях гнезда располагаются на деревьях
310
OTP ЯД ХИЩНЫЕ ПТИЦЫ
или скалах (Лапландия). Гнездовый биотоп —лесотундра речных долин, мокрая
ровная тундра, сухая водораздельная тундра, обрывы и яры побережий-
тундровых рек и т. д. Располагаются гнезда предпочтительно в местах с удобным и
широким обзором. У каждой пары по нескольку гнезд, занимаемых
последовательно в разные соды. Спаривание, брачные игры (полет и характерный свист),
починка гнезд происходят в последних числах мая, тогда же или в первых
числах июня — кладка. Гнездовые участки различной величины (в Тиманской
тундре пара от пары находятся на расстоянии 2—3 км, площадь участка 4—
9 км2, Гладков/1941; на южн. Ямале расстояния между гнездами 2—3 км,
Осмоловская). Защищаются участки неэнергично.
Число яиц в кладке сильно колеблется, повидимому в связи с кормовыми
условиями: обычно 3—4, в «хорошие» годы до 7, в «плохие» 2—3. Несомненно
негнездование зимняков на севере при отсутствии леммингов (Ларсон, 1935,
считал, что в Лапландии число гнездовых пар в год «неурожая» грызунов в 15
раз менее, чем в благоприятные годы; на Кольском п-ове в местах, где
в 1895 г. гнезда канюков встречались ежедневно, в 1901 г. не было ни одного
занятого гнезда, Пирсон, 1904; в Лапландии при внезапной эпизоотии у
леммингов из 67 занятых канюками гнезд выводки (при том только из одного птенца)
остались лишь в пяти гнездах, остальные были брошены родителями, Веслен,
и т. д. Размеры яиц (110)48—59x40,5—46,5 в среднем 54,97x43,54 мм
(Хартерт, 1913).
Насиживание начинается, судя по разнице в возрасте птенцов, с откладки
первого яйца, насиживают оба родителя (по наблюдениям в Сев. Америке,
Берне, 1915, в течение 28 дней). Появление птенцов с конца июня и в первой
половине июля, в зависимости от срока кладки (по наблюдениям Осмоловской
на Ямале разница в сроке кладки определяется в известной мере
местоположением гнезда: на высоких обрывах гнезда занимаются несколько раньше, чем
в мокрой низкой тундре). Птенец при выходе из яйца весит 34,5—45 г,
двухдневный около 50 г (Ямал, Осмоловская). Только что вылупившиеся птенцы
найдены в Лапландии 22 июня и 5 июля (Пирсон, 1899,1904), в Тиманской тундре
24 июня— 10 июля (Гладков, 1941), на о. Вайгач 27 июня (Пирсон, 1899), на южн.
Ямале 9—16 июля. Уже на 3—4 день после вылупления у птенцов начинает
пробиваться второй пуховой наряд, причем смена первого пухового наряда,
кроме головы, заканчивается в возрасте 7 дней, а к 10 дням второй пуховый
наряд развивается полностью; в 12—14 дней начинают пробиваться пеньки
плечевых, маховых, затем рулевых; в 28 дней у птенца остается только немного
пуха, в 35 дней молодые оперены полностью, но маховые у них еще не достигли
полной длины; в это время они находятся вне гнезда. Летные молодые в
Лапландии найдены 17 августа, около середины этого месяца и в Тиманской тундре;
на Ямале 13—20 августа. Вес летных молодых почти достигает в это время веса
взрослых: у самцов900—1000, у самок 1100—1200г (Осмоловская). Смертность
птенцов в неблагоприятные по кормовым условиям годы очень высока: в 1942 г.
на южн. Ямале при кладке в 4 яйца в выводках было в среднем 2,7
молодых (Осмоловская.) Младшие птенцы при этом погибают обычно в возрасте
10—14 дней, когда самка перестает оделять выводок, эстолько кладет в гнездо
добычу (Кучерук и Дунаева, 1941). Иногда младшие птенцы съедаются
старшими, иногда гибнут от голода. Выводки канюков кроме того страдают от
песцов, ездовых собак и быть может от белой совы. По наблюдениям в
Тиманской тундре яйца канюков похищают иногда поморники (Гладков, 1941).
Сильно Досаждают птенцам комары и гнус (Финш, 1879).
Л и н ь к а. Изучена недостаточно. У птиц в первом годовом наряде
она начЦтется ранней весной и даже в конце зимовки: смена мелкого
оперения на^ плечевых, спине, кроющих крыла, зобе в начале мая; затем
линяние^-в связи повидимому с размножением — приостанавливается, возобно-
мохноногий канюк или зимняк 311
вляясь в июле (?). Начало линяния взрослых—в конце периода размножения
(на Канине, в Тиманской и Большеземельской тундре, на Ямале в первой
половине июля). Поздние осенние экземпляры (встречаются в конце октября—
ноября) уже в свежем пере. К отлету в августе у старых птиц несменив-
шимися остаются передние 3—4 первостепенных маховых. Последовательность
смены маховых не вполне ясна — быть может в отличие от других видов
семейства начинается с середины, от 7-го махового и идет к краям—внешнему
и внутреннему. Рулевые линяют от средней пары, последней сменяется вторая
от края хвоста пара.
В ходе линяния
имеются довольно
значительные личные
уклонения.
Последовательность смены нарядов
обычная: первый
пуховой наряд—второй
пуховой наряд —
первый годовой (гнездо-
вый) наряд — второй
годовой
(окончательный) наряд и т. д.
Питание. В
области гнездовья
основную пищу
канюков составляют
лемминги, на западе Lem-
mus lemmus, на
востоке L. obensis и в
особенности Dichrostonyx
torquatus. Зависимость
плодовитости,
смертности, сезонного
размещения от «урожая»
леммингов указана
выше. Охотится
зимняк или скрадом,
сидя на земле и ожидая
приближающуюся
добычу, ИЛИ на лету, Рис. 85. Зимняк.
летая медленно и
низко, околов—\0м над землею, и временами «трясясь» на месте, как пустельга.
Охотничий участок около lVs^3 км (Ямал). Если леммингов много, то
сезонные изменения питания невелики. Но в неблагоприятные годы в
кормовом режиме большую роль начинают играть птицы, особенно белые
куропатки, хотя таким образом нормальная потребность молодых в пище не
может быть обеспечена.
В гнездовый период в качестве корма зимняка на Ямале отмечена, кроме
леммингов, полевка Миддендорфа, узкочерепная полевка, водяная крыса,
зайчата беляки, горностай, ласка —из млекопитающих; из птиц белая
куропатка, молодая синьга, гусята гуменники, один раз взрослый гусь этого вида,
лапландский подорожник и т. п. (Ямал), на Вайгаче — пуночка; в неурожайный
по леммингам 1938 год в Тиманской тундре отмечено предпочтительное питание
куропатками; в таких же условиях в Лапландии — дрозд белобровик,
береговой конек и другие птицы. На зимовках в европейской России канюки кормят-
312
ОТРЯД ХИЩНЫЕ ПТИЦЫ
ся разными мышевидными грызунами; а также другими млекопитающими
(упоминается ласка, для Германии даже заяц) и птицами, в том числе серыми
куропатками (Боровиков, 1907, Сомов, 1897 и др.). Имеются требующие
подтверждения сведения о питании канюков отбросами белужьего и рыбного промыслов
(первое — на Канине, Банников, 1934; второе — в низовьях Енисея, Тугаринов
и Бутурлин, 1911).
Полевые признаки» Типичный сарыч, на полете кажется крупнее
обыкновенного канюка. Охотнее, чем последний, садится на землю. На лету характерно
белое основание хвоста и белое зеркало — основания маховых — на нижней
поверхности крыла. Голос напоминает мяуканье или протяжный свист.
Описание. Размеры и строение. Длинные крылья и хвост, относительно
пышное оперение, небольшой клюв с косорасположенной ноздрей, длинная оперенная
спереди и с боков (но не сзади) цевка, короткие пальцы. Формула крыла: 3>4>5>6>
2>7>1>8. Вырезки на внутренних опахалах 1-го — 4-го и на наружных опахалах 2-го —
5-го маховых. Вес самцов (5) 800—930, самок (5) 990—1280, в среднем 846 и 1151,4 г. Длина
самцов (5) 530—545, самок (И) 555—608, в среднем 539 и 582 мм. Размах самцов (7) 1991—
1382, самок (10) 1310—1442, в среднем 1332,3 и 1388,8 мм. Крыло самцов (42) 385—432, самок
(49) 425—473, в среднем 414,5 и 438,1 мм.
Окраска. Первый пуховой наряд сероватый на спинной стороне, беловатый на брюхе;
второй пуховой наряд буровато-серый; характерно, что цевка оперена уже у пуховиков.
Первый годовой (гнездовый) наряд с охристо-беловатой, испещренной продольными
черноватыми или темнобурыми полосками головой, с темнобурой окаймленной охристыми
каймами спиной, плечевыми и кроющими крыла; брюшная сторона охристая с бурыми более или
менее сливающимися в темное поле продольными пестринами на груди (темный «пластрон»),
с бурыми боками и брюхом; оперение голеней беловато-охристое с продольными или только
слегка расширяющимися в поперечном направлении пестринами; подхвостье беловатое;
надхвостье с бурым продольным рисунком; маховые темнобурые с белыми основаниями;
рулевые белые с охристым оттенком и широкой бурой вершинной полосой. В окончательном
наряде бурый цвет более насыщенный, черноватый; на плечевых, брюхе, надхвостье, голенях
рисунок более или менее правильный поперечный; перья спинной стороны тела окаймлены
сероватыми каемками; маховые с сероватым налетом и нерезким поперечным рисунком,
с белыми полями у оснований внутренних опахал; рулевые белые с черными поперечными
полосами и широкой предвершинной каймой.
Во втором годовом наряде, в общем сходном с окончательным, окраска несколько менее
контрастная, поперечный рисунок на брюхе и голенях менее тонкий и правильный, менее
развита поперечная полосатость рулевых. Впрочем, эти'особенности встречаются повиди-
мому и у старых птиц в виде индивидуальной вариации, как встречаются среди европейских
зимняков особи со значительной примесью белого цвета в оперении, напоминающие в этом
отношении восточносибирских птиц. Радужина у молодых серовато-бурая, или бледнобурая,
у старых темнобурая или желтовато-бурая; клюв черноватый, светлеющий у основания;
когти черные; восковица, разрез рта и лапы желтые.
Сибирский мохноногий канюк ВШео lagopus menzbieri, nomen novum.
emend, pro
Archibuteo pallidus. Menzbier Ornithologie du Turkestan, 1888, стр. 163, описано
по зимующим в Туркестане особям; за типичную местность здесь предлагается принять сев.
Якутию — низовья Колымы (см. ниже систематические замечания), nom. praeoccup. (пес
Buteo pallidus Lesson. Traite d'Orn. I. 2, 1830, стр. 82).
С и н о н и мы. Archibuteo lagopus var. Slbirca S e ve r t zо w. J. f. Orn., 1875, стр. 170,
nom. nudum.
Распространение. Ареал. Северная Сибирь, на восток от предыдущей
расы. На Ямале и э области нижнего Енисея распространена популяция с
переходными признаками, в которой немногие особи сходны с lagopus, другие — с
североякутскими, большинство же более или менее промежуточны. Типичные
menzbieri начинают встречаться примерно в бассейне Хатанги и Нижней
Тунгуски. С этими оговорками, областью распространения сибирского зимняка
представляется полоса тундр и лесотундр Восточной Сибири: бассейн Енисея
к югу до Туруханска (65° с. ш.) и даже устья притока Н. Тунгуски, Кочечумо
(под 62° с. ш., в гнездовое время добыт Н. П. Наумовым); Таймыр, бассейны
Хатанги, Оленека, Анабары, Яны, Индигирки, Колымы и Анадыря
мохноногий канюк или зимняк
313
(приморская часть). На севере в низовьях Енисея, на о-вах Сибирякова
и Расторгуева, на материке у Омулевой (73° 30' с. ш.), на Хатанге у Сандаска,
(Торгашев, Яковлев, под73°30' с. ш.), на Таймыре у Боганиды, но не в северной
части полуострова, далее к востоку до побережья океана. Указание на
Новосибирские о-ва (Большой Ляховский) сомнительно. Южная граница точно не
установлена, но едва ли выходит за пределы лесотундры (во всяком случае,
в относительно хорошо изученных в орнитологическом отношении долинах
Индигирки и Колымы, по наблюдениям Михеля и Бутурлина). Сведения о
гнездовании у Якутска не подтверждаются. В зимнее время встречен в вост.
частях Туркестана (Семиречье), у Зайсан-нора, в Прибайкалье, Приамурье и
бассейне Уссури, по литературным данным в сев. Монголии и Манчжурии,
сев. Японии, Корее, быть может в Сев. Китае. Светлоокрашенные особи и птицы
промежуточного типа, происходящие вероятно с севера средней Сибири, в
небольшом числе попадаются на пролете и зимой в европейской части нашей
страны (области Московская, Воронежская, Поволжье, Сарепта и б. Самарский
уезд, Азовское побережье у Бердянска, окрестности Ростова на Дону, Сев.
Кавказ от Кубани до Дзауджикау), в Зап. Сибири (Тара, окраины Алтайской
горной страны) и составляют большинство зимующих в Туркестане канюков.
Биотоп и характер пребывания. Как у
предыдущего подвида.
Даты. Прилет на северные места гнездовья несколько позднее, чем у
западного подвида. В Енисейско-Пясинской тундре отмечен в начале мая, на
Хатанге у Хатангского под 71°48' с. ш. 8 мая, у мыса Б. Карго под 73° 30'
13 мая, на Индигирке у Русского устья 6 и 20 мая, в дельте Колымы
у Сухарного 16 мая, на Анадыре 10—21 мая. Отлет наблюдался у Хатангского
29 сентября (последние, по Торгашеву), 19 сентября на Алазее, в начале
сентября и не позднее 13 сентября на Индигирке, в первой половине сентября
последние птицы еще в конце этого месяца — на Анадыре. Пролет у Якутска
наблюдался осенью с конца сентября, весной в первой половине мая, в
Забайкалье в конце сентября — начале октября, весною до начала мая, в Приморье
с конца октября и весною с начала апреля до первой половины мая (12 мая);
в Семиречье с ноября до марта (по Шестоперову, 1929, на Или до начала апреля).
Численность. Колебания численности на зимовке отмечены в
Приморье, и Средней Азии, они имеются конечно и в гнездовой области, но не
изучены. В тундрах Якутии зимняк многочисленен.
Экология. Сведения отрывочны, основные черты повидимому сходны с
номинальной расой.
Размножение. Гнезда расположены на деревьях (лиственницы —
Н. Тунгуска, Хатанга, Таймыр, Колыма), на скалах и береговых обрывах
и холмах (преимущественно). Предпочитаются речные долины. В парах птицы
держатся с прилета (на Индигирке с середины мая, Житков и Зензинов, 1913).
В кладках 2—5 яиц, чаще 3—4, изредка больше (в выводке, добытом 3 июля
Бутурлиным у м. Каменного, 7 птенцов). В начале июля пуховые птенцы
разных возрастов: в первом пуховом наряде, в линьке из первого пухового
наряда во второй, во втором пуховом наряде с чуть пробивающимися
плечевыми и маховыми (Таймыр, Хатанга, Лена, Индигирка, Алазея, Колыма).
Вылупление происходит таким образом в последних числах июня — начале
июля, а кладка в конце мая — начале июня. Летные молодые в конце августа
(Колыма).
Линька. Как у номинальной расы. Начало смены маховых (обычно по-
видимому с 7-го или 6-го) и средних рулевых у взрослых птиц в первой половине
июля, около середины августа несмененными остаются передние два и заднее —
десятое — маховое: добытые на пролете и зимовке в конце октября —ноября
зимняки уже закончили линяние.
314
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Питание. В основном питаются мелкими грызунами, леммингами и
полевками; кроме того, приводятся зайцы, вероятно молодые (Индигирка; на
зимовке по р. Или); землеройки, полевки Microtus oeconomus, M. major
(Забайкалье), молодые птицы; также белые куропатки.
Описание. Размеры и строение. Немного крупней предыдущего. Длина тела
самцов (7) 530—570, самок (7) 575—605 мм, в среднем 552,9 и 584,4 мм, размах самцов (5)
1290—1377, самок (б) 1239—1430 мм, в среднем 1344 и 1391,7 мм. Длина крыла самцов (21)
408—442, самок (16) 434—470, в среднем 421,5 и 451 мм.
Окраска. Светлей, чем у европейского зимняка. В первом годовом наряде общий
бурый тон бледный, темные полосы на голове узкие, светлые каймы перьев спинной стороны
тела развиты больше, а темные бурые пестрины на груди слабо, на ней преобладает белый
цвет; темные пятна по сторонам брюха узкие, уже и темная вершинная полоса на рулевых.
У взрослых птиц больше белого на спинной стороне, рисунок на плечевых и больших
кроющих крыла часто носит характер светлых (сероватых в свежем пере, белых в обношенном)
полос; на зобе только остатки темного «пластрона», поперечный рисунок боков, брюха, голени,
хвоста развит слабей; белый цвет на брюшной стороне в общем значительно преобладает.
Систематические замечания. Средний тип окраски восточносибирских канюков в общем
хорошо отличим от европейских. О промежуточной полосе уже упоминалось. Вопрос о
систематическом их положении все же осложняется тем, что типами для описания pallidus
послужили Мензбиру зимние птицы из Туркестана, относящиеся повидимому как раз к
промежуточной популяции. Далее, между этими формами нет, повидимому, экологических различий,
так что они не вполне быть может подходят к понятию «подвид», отражающему проявления
географического (экологического) изоморфизма (возможно, однако суровый климат
Восточной Сибири вызывает у зимняков физиологические к нему приспособления, выражающиеся
в особенностях обмена, коррелятом которых служит особенность их окраски). Надо
добавить, что окрашенные как menzbieri птицы известны и из гнездовой области lagopus (например
из Тиманской тундры), а темноокрашенные зимняки в небольшом числе встречены и на
востоке, например, на пролете у Якутска, на гнездовье в низовьях Колымы.
Камчатский мохноногий канюк Buteo lagopus kamtschatkensis D e m.
Buteo lagopus kamtschatkensis. Dementiev. Orn. Monatsberichte, 1931, стр. 53,
Камчатка.
Распространение. Ареал. К этой форме относятся зимняки, гнездящиеся
на Камчатке и по сев. побережью Охотского моря (п-ов Тайгонос). Вероятно
с ними тождественны птицы из западных частей Охотского побережья
(Удекой Острог; Аян, Шантарские о-ва), а также с Курильских о-вов. Как
проходит граница этого подвида и предыдущего на севере Коряцкой Земли
(в области Олюторского залива?) —неясно. В негнездовое время камчатские
канюки появляются вероятно в Аляске (судя по описаниям добытых там и
отнесенных к восточносибирскому подвиду зимняков, Фридманн, 1934). В
холодное время года этот подвид встречен, наконец, в Приморье (Вяземская, 17. X.
1910; Сиваковка, -1-4. X. 1932), вероятно доходя до Манчжурии и Японии.
Б и о т о п. В отличие от других рас зимняков, встречается главным
образом в лесах (Камчатка, Бергман, 1935, Аверин, 1948; о. Большой Шантар,
Шульпин и Дулькейт, 1937).
Характер пребывания. Из области Охотского моря улетает,
появляясь там и на Камчатке в конце апреля (Шантарские о-ва; Коряцкая
земля); на Камчатке встречается в небольшом числе и зимой, хотя там
отмечается и пролет (птицы из с.-в. Сибири?). Численность. Повидимому, редок
(Камчатка).
Экология* Сведения крайне неполны. В гнезде у Кроноцкого озера, вост.
Камчатка, на р. Татьяна 27 мая найдены 2 свежих яйца. Однако, на о. Большой
Шантар еще в последней трети июля кладка из 6 насиженных яиц. Окраска
яиц типичная для вида, с крупными и мелкими темнобурыми пятнами по
охристому фону.
Гнезда расположены на деревьях — тополях, лиственницах и др.
(Камчатка); на п-ове Тайгонос гнездо на скале (Дементьев, 1941). Не вылетевший
МОХНОНОГИЙ КУРГАННИК
315
еще из гнезда выводок из 5 оперенных молодых наблюдался 13 августа
(Бергман). В гнезде на п-ове Тайгонос 4 августа было два птенца с короткими
крыльями и остатками пуха. Вполне развитые летные молодые на Камчатке
наблюдались в августе.
Линька. У взрослого самца 4 августа сменились 7-е и 8-е маховые,
"6-е выпало, растут средние рулевые и свежее мелкое перо. Около середины
октября добыты взрослые, закончившие линяние.
Питание, как у других рас, главным образом мелкие грызуны. На
Камчатке отмечены полевки, землеройки, молодые зайцы, на о. Большой Шан-
тар красная полевка Clethrionomys rutilus, в Коряцкой Земле лемминги и
полевки; из птиц—белая куропатка (Аверин, 1948). Отмечена зависимость между
численностью зимняков и полевок.
Описание. Размеры и строение. Как у других рас зимняка восточного
полушария, быть может немного крупней. Вес самца S00, самок (2) 1050, 1209 г. Крыло
самцов (4) 420—445, самок (4) 447—483, в среднем 435 и 468,3 мм.
Окраска. Хорошо отличается от восточносибирского канюка более темным цветом,
интенсивно черноватым с большим развитием темного рисунка у взрослых, меньшим
развитием белого цвета у молодых. В этом отношении походит на европейский подвид, но все же
пестрей его и с большим развитием поперечного рисунка «(сероватого в свежем оперении) на
спинной стороне тела. По окраске таким образом камчатский канюк приближается в
известной степени к североамериканскому подвиду В. /. sancti-iohannis, также окрашенному
темней и ярче, чем B.l. menzbieri.Однако у камчатского канюка нет свойственной американскому
темнобурой меланистической вариации или фазы окраски, и к тому же американские зимняки
меньше по размерам азиатских, чего о камчатских сказать нельзя.
41. Мохноногий курганник Buteo hemilasius Temminck et
Schlegel
Buteo hemilasius. Temminck et Schlegel. Fauna Japonica. Aves, 1844, стр. 81,
табл. VII, Япония.
Распространение. Ареал. Центральноазиатское нагорье и его окраины —
от центрального Тянь-Шаня (где гнездование впрочем еще не доказано), Тар-
багатая, ю.-в. Алтая и прилежащих к
нему частей центрального Алтая, быть
может Танну-тувы — хребта Танну-ола,
ю.-в. Забайкалья (до Нерчинска и
Борзи), зап. Манчжурии, Монголии (но
не Гоби), к югу до Тибета и Непала, быть
может в Гималаях — Кашмир и Кумаон.
В негнездовое время, кроме того, в
различных частях Туркестана до Ферганы
и Ташкентского оазиса (случайно даже
по средней Аму-Дарье между Чарджоу
и Келифом, Зарудный и Билькевич,
J 918), в Гоби, в Кашгарии, в Китае до
Шанхая и Хубея, залетая к Фуцзяни и
Сватоу в сев. Гуаньдуне (Латуш, 1932),
в Бирме и с.-з. Индии. Залеты отмечены
в Приморье (Сидеми, Тачановский, 1891; Карта 49. Распространение мохноногого
Аскольд, Болау, 1880) и в сев. Японии курганника вщео hemilasius
(Нагасаки). о. — границы гнездовий, г — область зимовок и
Характер пребывания. кочевок.
Выяснен недостаточно. Вероятно часть птиц зимует в гнездовой области, где
точных наблюдений за прилетом нет; в Манчжурии (Соверби, 1923) оседлая
птица, но вместе с тем, в холодное время года канюки появляются в местно-
316
ОТРЯД ХИЩНЫЕ ПТИЦЫ
стях, отстоящих более или менее далеко от гнездовой области; в горах по-
видимому имеют место вертикальные кочевки. На равнинах Семиречья
появляется в ноябре и остается до февраля, у гнезд появляется в конце апреля
(Забайкалье, с.-з. Монголия, Тибет). В Чжили в Сев. Китае пролет
наблюдался в марте—апреле и в октябре — ноябре (Латуш, 1920). Для
Забайкалья прилет указывался в конце марта, позже 16 сентября не наблюдался
(Радде, 1863).
Биотоп. Сухие степи с выходами скал или хотя бы всхолмленные,
обычно в горах или предгорьях, реже на равнинах; широкие горные долины
с мягким рельефом; в общем биотоп сходен с таковым обыкновенного курган-
ника, но последний предпочитает равнины, а мохноногий курганник — горы.
Вертикальное распространение на Алтае от 1500 до 2300м; в Тибете до 4500 м\
в Забайкалье встречен на равнинах и в гнездовое время.
Численность. Распространен несколько спорадично, что
определяется наличием удобных для гнездованья мест (скалы), и кормовыми
условиями (обилие грызунов, в Тибете в частности пищухи Ochotona melanostomar
по Шеферу, 1938). В ю.-в. Забайкалье в Кулусутаевском районе одно гнезда
приходится на 42,5 км2, в фктуй-Молозайском только на 186 км2 (Тарасов,
1944). В Тибете местами многочисленен — уСешупоЗО— 40 птиц за 2—3 часа
экскурсии, 3 гнезда расположены на протяжении 200 км, причем иногда одно
гнездо от другого отстоит на 50 м (Шефер, 1938); гнезда иногда размещаются
на разных склонах одного и того же холма (Забайкалье, Скалой, 1936). В
Туркестане редок и в холодное время года, не говоря уже о гнездовье. Численность
мохноногого курганника, как и других мышеядных хищников, подвержена по-
видимому периодическим колебаниям в зависимости от кормовых условий,
слагающихся из увеличения или уменьшения плодовитости, смертности и т. д.
Закономерности этих колебаний не изучены. Зимой птицы держатся иногда
группами по несколько особей (Монголия, Бианки, 1915).
Экология. Размножение. Цикл в общем сходен с таковым у
обыкновенного курганника, но начинается поздней почти на месяц (горная птица).
Кладка в начале мая и в конце апреля (Забайкалье, Алтай, в это же время
в Тибете). Спариванье в Тибете наблюдалось в конце марта, на земле (Козлов,
по Бианки, 1907) и 20 апреля (Шефер, 1938). Гнезда главным образом
расположены на скалах, хотя бы на небольших каменистых выходах в степи, обычно под
защитой кустарников; реже на холмах, сурчинах тарбаганов (бутанах), в виде
исключения даже на Слогах сена (Тарасов, 1944). У пары имеется повидимому
2 гнезда, занимаемых поочередно в разные годы. Гнезда небольшого размера,
построены из ветвей; лоток плоский, выстланный тряпками, сухой травой,
шерстью, отбросами; старые гнезда достигают до 1 м в диаметре (Алтай, Суш-
кин, 1938).
В случае утраты кладки возможно ее повторение, но число яиц меньше
(Бекер, 1928). В кладке 2—4 яйца (изредка 5, Бианки; 1907). Окраска яиц
напоминает цвет яиц зимняка: по охристо-желтоватому фону разбросаны мелкие
крапины и пятна красновато-бурого цвета, более многочисленные на узком
конце яйца. Размеры яиц из Забайкалья: (10) 59—65,5 х 48—50,3 (Тача-
новский, 1893); 70—61x47—52 (Радде), из других мест (14) 56,1—62,7 х
44,5—47,7 (Суанн, 1926), 53,5— 64 х 42,6—47,9 мм (Бекер, 1928).
Птенцы появляются в первых числах июня, насиживанье следовательно
продолжается около месяца. Молодые птенцы вылетают из гнезд в середине
июля (Забайкалье), в начале августа (Тибет), гнездовый период тянется 45 дней
(Забайкалье, Тарасов, 1944). Птенцов в выводке 1—4, чаще 2. В неблагоприят:
ные годы отмечен каннибализм: старшие птенцы поедают младших
(Скалой, 1936). Неразбившиеся выводки встречаются еще в конце августа
(Алтай).
мохноногий курганник
317
Линька. Взрослые начинают линять в первой половине июня, т. е.
в конце гнездового периода. Последовательность смены нарядов —обычная:
первый и второй пуховые, первый годовой (гнездовый), сменяющиеся затем
вторым годовым, окончательным, нарядом.
Питание. Мохноногий курганник — типичный миофаг, как и другие
•европейско-азиатские сарычи. Для Манчжурии в качестве пищи курганника
указываются зайцы, вероятно, молодые; песчанки, суслики; в Тибете и
Монголии установлена определенная
зависимость между распространением
сеноставцев и курганника. Для Алтая главным
кормом являются полевки, суслики,
сеноставцы (Сушкин, 1938). В Забайкалье
из грызунов Citellus dauricus, Stenocrani-
us gregalis, Microtus mongolicus, из птиц
жаворонки Alauda arvensis, Calandrella
cinerea; из насекомых жужелицы,
щелкуны, навозники, кобылки, муравьи
и т. д. (Скалой, 1936); молодые
тарбаганы, даурские суслики, сеноставки,
полевки, из птиц полевые жаворонки,
рюм, каменный воробей Anthus richardi,
перепелка; кроме того, полоз и жаба
Bufo raddei (Тарасов, 1944); полевки,
суслики, сеноставки, тушки ободранных
охотниками тарбаганов (Павлов, 1935);
во всех случаях количественно
преобладают грызуны. Для вост. Алтая (озеро
Джулю-куль) указывается питание
белыми куропатками. На охоте
мохноногий курганник или выслеживает добычу,
сидя на земле, или,-летает.
Полевые признаки. Типичный сарыч
крупной величины. На полете кажется
стройным и длиннокрылым, полет
легкий, нередко парит и «трясется» в
воздухе на месте. Птицы светлой окраски
на полете отличаются рыжеватыми
подкрыльями со светлым беловатым пятном
у кистевого сгиба и полосатым хвостом.
Птицы темной вариации — однообразной
темнобурой окраски. Осторожен. Голос напоминает обыкновенного сарыча
или подорлика.
Описание. Размеры и 'строение. От ближайшего вида — курганника —
отличается тем, что цевка в верхней части покрыта не поперечными крупными пластинками,
а сетчатыми щитками; кроме того, обычно цевка более оперена — нередко до основания
пальцев или до 2/8 длины, но иногда и менее (причем оперение бывает иногда асимметрично
расположено). Размеры крупные. Длина самца (1) 610, самки (1) 720 мм, размах самки (1)
1580 мм. Вес только что начавшего летать самца 2000 г. Крыло самцов (4) 445—480, самок
(7) 490—504, в среднем 467,5 и 498,3 мм. #
Окраска. В первом пуховом наряде птенцы беловато-серые (Бианки, 1907),
второй пуховой наряд бледносерый (Портенко, 1929). Как у многих других сарычей, окраска
диморфна. Птицы темной вариации или фазы более или менее насыщенного бурого цвета
с полосатым хвостом, иногда с рыжеватыми светлыми каемками на перьях груди. Сарычи
этой вариации окрашены сходно и в первом годовом, и в последующих нарядах. Птицы
светлой вариации в первом годовом наряде бурые на верхней стороне тела, с более или менее
развитыми светлыми каймами перьев — беловатыми или рыжеватыми: голова белая или
Рис. 84. Мохноногий курганник.
318
ОТРЯД ХИЩНЫЕ ПТИЦЫ
охристая с темными наствольями; маховые темнобурые с белыми основаниями,
образующими на развернутом крыле «зеркальце», рулевые бурые, часто беловатые у стволов, с
неправильным темнобурым поперечным рисунком (иногда столь «размытым», что от него остаются
только продольные наствольные пятна); брюшная сторона охристая с бурыми пятнами на
зобу, груди, брюхе и боках; бока иногда сплошь темнобурые; голени охристые с бурым
продольным рисунком. У птиц во втором годовом и последующих нарядах на боках, брюхе,
штанах более или менее правильный бурый поперечный рисунок. Других возрастных
изменений нет, как нет и полового диморфизма в окраске. Иногда в одном выводке птенцы
бывают один светлой, а другой темной вариации (Штегман, 1928). Географическая вариация
локализации цветовых фаз не изучена, но в Тибете численно преобладает повидимому темная
вариация, а в Забайкалье светлая. Радужина светлокаряя или желтоватая, клюв черный,
когти черные, восковица зеленовато-желтая, лапы желтые.
Систематические замечания* Хартерт (1933) считает Buteo hemilasiusea географическую
форму обыкновенного курганника. В пользу этого взгляда говорит большое сходство
морфологии этих птиц, сходство экологии, одинаковая амплитуда вариаций окраски. Оба —
птицы открытых пространств, главным образом сухих, но rufinus равнинный, a hemiiasius
горный. На стыке ареалов имеются повидимому особи с переходными признаками (Портенко,
1929). Однако области распространения этих сарычей перекрываются на значительном
пространстве и их лучше считать за отдельные виды.
42. Степной сарыч, курганник Buteo rufinus Cretzschmar
Falco rufinus. Cretzschmar. Atlas zu der Reise im nQrdlichen Africa von E* Ruppell,
1826, стр. 40—41, табл. 27, сев. Африка.
Синонимы. Buteo ferox auctorum, nee G m e 1 i n, 1771.
Распространение. Ареал. Сев. Африка к югу до Ахаггарского плато
в Сахаре; ю.-в. Европа (Греция, Заволжье, Кавказ); Передняя и Средняя
Карта 50. Распространение курганника Buteo rufinus
1 — В. г. rufinus, 2 — В. г. cirtensis; а — границы гнездовий, г — область зимовок.
Азия от М. Азии, Ирана, Месопотамии, Палестины, Туркестана, Афганистана,
Белуджистана, на восток до Синьцзяня, Даурии (Тарейнор), Монголии (от
Кяхты и Улан-Батора на восток до Цайдама и Алашаня), к югу до с.-з.
Индии (до Кашмира и Гарвал; на ю.-з. до Ноушера). В негнездовое время
на Ниле до Судана и Абиссинии, в Аравии, Ираке, в сев. Индии, а также в
Закавказье (Армения, Азербайджан), в Туркестане в Семиречье, Фергане,
Ташкентском оазисе, Таджикистане и Туркмении, в Синьцзяне и Монголии, За-
СТЕПНОЙ САРЫЧ, КУРГАННИК
319
леты в Зап. Европу — Балканские страны, Венгрию, Богемию, Германию,
Швейцарию, Италию, Голландию, Францию, на Украину.
Характер пребывания. В сев. части ареала перелетная птица,
в южн. — оседлая. Однако неясно, не происходит ли откочевка части особей
южных популяций.
Биотоп. Открытый сухой ландшафт — пустыни и полупустыни,
степи, горные долины (сырты); в вертикальном отношении в Кашмире доходит
повидимому почти до 4000 м (нет ли смешения с мохноногими курганниками?);
на Памире почти до 4000 м у озера Ранг-куль, в Тань-Шане до 3000 м, в
Закавказье до 2000 м (Армения).
Подвиды и варьирующие признаки. В сев. Африке от Марокко, Алжира, Туниса,
Киренаики, к югу до Сахары, на восток до Египта и Синайского п-ова, вероятно в южн.
Палестине (Мейнертцхаген, 1931) В. г. cirtensis Levai 1 1 a n t, 1867, отличающийся
мелкими размерами и отсутствием темнобурой вариации окраски; в остальной части ареала
вида В. г. rufinus. Образ жизни обеих рас сходен.
Восточный курганник Buteo rufinus rufinus С г ёЧ z s с h m a r.
Синонимы. Buteo leucurus. N a u m a n п. Naumannia, 1853, стр. 256, Сарепта. —
Buteo nigricans Северцов. Вертикальное и горизонтальное распространение туркест.
животных, 1873/1872, стр. 112. Туркестан. — Buteo ferox raddei. Loudon Verhandl. V,
Intern. Orn. Kongr. Berlin, 1910, 1911 (1912), стр. 356, Кумбаши, Азербайджан.
Распространение. Ареал. В гнездовое время в европейской и азиатской
части ареала вида. В СССР от низовьев Дона, Ергеней и Кумоманычской
впадины на западе, в Заволжье примерно до 52—52°30#с. ш. (Новоузенский район;
Чкалов); на Сев. Кавказе в равнинных безлесных местностях; Закавказье
до Муганской степи и Армянского нагорья; в Зауралье наблюдался в верховьях
р. Уй (Рузский, 1895) и добыт у Троицка, т. е. около 54° с. ш.; восточнее
в сев. Казахстане найден в верховьях Тургая, под 52°—52°30', и далее до при-
алтайских степей граница идет примерно по этой широте (у западных предгорий
Алтая у Рубцова), спускаясь затем к Тарбагатаю и Зайсану; в Алтайских горах
и в Забайкалье не гнездится. Кроме того, распространен по всему Туркестану
от Киргизии и Казахстана до Памира и Туркмении. Залеты в европейскую
Россию к северу от гнездового ареала в Воронежскую обл. (Северцов, 1855),
к Ростову-на-Дону, в Полтавскую обл. (Гавриленко, 1929), Крым (Мензбир,
1895), в Азии к Омску, на Алтай (Укек, Сушкин, 1938), к Иркутску (Пор-
тенко, 1929).
Характер пребывания. В сев. части ареала большинство
особей перелетные, например в Казахстане, кроме его ю.-в. частей, на зиму
остаются немногие. В южной части ареала — в Таджикистане, Узбекистане,
Киргизии, также в Туркмении и в Закавказье встречается круглый год. Однако
и здесь происходят вертикальные миграции, и на равнинах Семиречья и
Ферганы зимою собирается большое количество курганников — как прилетных с
севера, так и спустившихся с гор. В значительном числе курганники зимуют
в сев. Индии (с.-з. провинции, Раджпутана, Пенджаб, Синд, редко южней) и
появляются в холодное время в с.-в. Африке.
Даты. Появление кочующих курганников на равнинах Средней Азии
наблюдается уже с половины сентября — начала октября; во второй половине
марта число их здесь уменьшается. В ю.-в. России (Чкаловская обл.) прилет
отмечен весной в первой половине апреля, тогда же в сев. Казахстане,
однако на Эмбе пролет затягивается и до 3 мая. На нижней и средней Сыр-
Дарье прилет и пролет в середине марта —начале апреля, пролет и отлет в
ноябре и до середины декабря; у Чимкента пролет с конца февраля и в марте.
В горах на перевале Санташ, Киргизский хребет, значительный пролет — за
день до сотни птиц —наблюдался 14 сентября (Д. П. Дементьев). Для ю.-в.
320
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Забайкалья пролет отмечен в конце марта, осенью не наблюдались позже 14
сентября (Радде, 1863, но нет уверенности, не относятся ли эти данные к
мохноногому курганнику). В Египте в дельте Нила и в Файюмском оазисе курганники
наблюдаются с ноября по март, на Синайском п-ове с ноября по февраль (Мей-
нертцхаген, 1931).
Биотоп. См. выше, относительно вида: сухие степи на равнинах и в
горах,, полупустыни — наиболее типичные местообитания. В вертикальном
отношении на Тянь-Шане до 2000—2500 м, на Памире у озера Ранг-куль свыше
4000^ (кочевка?).
Численность. Распространение курганника находится в
известной зависимости от распространения грызунов, составляющих главный его
корм (курганник, например, редок в ю.-з. Туркмении и Таджикистане,
весьма обилен в Волжско-Уральской степи и на равнинах предгорий зап. Тянь-
Шаня, равно как и на сыртах центрального Тянь-Шаня). Значительные
скопления курганников зимою отмечены в Ташкентском оазисе, в Фергане, в
подгорной полосе Туркмении у центрального и вост. Копет-Дага, по Тед*
жену. Как у других миофагов, имеются колебания численности, но они не
изучены. Для Туркмении указывается, что в благоприятный год 10 жилых гнезд
курганника найдено на протяжении несколько большем, чем 3 км (Зарудный,
1896); у ст. Анненково Закаспийской ж. д. в 1901 г. найдено 5—6 гнезд с
кладками из 5, лишь в одном случае из 4 яиц; там же в 1903 г. лишь одно гнездо
с кладкой в 3 яйца (Лоудон, 1910).
Экология* Размножение. На юге гнездовый период начинается
с конца марта (Туркмения), но в большинстве других местностей в Туркестане—
в разные числа апреля; в апреле находили полные кладки и в низовьях Волги.
Так как курганники гнездятся в относительно легко доступных местах и
нередко теряют поэтому кладку, взамен которой несутся вторично, то разница
в сроках размножения, отмеченная рядом авторов, вероятно не географическая,
а объясняется именно наличием этих вторичных более поздних кладок. В пользу
этого говорит и то обстоятельство, что в сходных географических условиях
птенцы одного примерно возраста добывались и всередине июня и месяцем
поздней (например в Туркмении); подлетков в Гиссаре в то же время встречали уже
27 мая (Иванов, 1940); трехнедельная разница в возрасте молодых наблюдалась
и в Закавказье (Ардаган, Ольта, Нестеров, 1911).
Гнезда располагаются или на скалах или на глинистых обрывах и
холмах, в горах иногда на деревьях (ель, центр. Тянь-Шань, Зарудный и Ко-
реев, 1906; Шестоперов, 1929); в пустынях Туркестана — Кара-Кумах и
Кызыл-Кумах — главным образом на деревьях или кустах саксаула, джингиля,
кандыма, реже на бархане среди кустов. При этом гнездо располагается так,
чтобы в жаркое время дня оно было по возможности затенено (Зарудный,
1916). Постройка из веток кустарников, полыни, сухой травы: размеры гнезд
около 70 см в диаметре и 35—50 см в высоту.
Число яиц в кладке 2—5, чаще всего повидимому 4, в благоприятные по
кормовым условиям годы больше, в плохие меньше. Окраска яиц
разнообразна — по беловатому фону они испещрены то бледножелтоватыми, то более
или менее темнобурыми пятнами весьма различных размеров. Величина яиц:
54,9—56,4 х43,1—-45 мм (Сыр-Дарья, Спангенберг, 1936).
Ход насиживания не выяснен. В мае в гнездах находятся пуховики, а в
конце этого месяца и в июне птицы оперяются и выходят из гнезда; к концу
июня молодые летают. Ввиду недостаточной изученности периода
размножения у курганников, приводим важнейшие даты: Армения — 23 июня летный
молодой (Соснин и Ляйстер, 1942): Кара-Тау, верховья рек Машат, почти
летные птенцы 25 мая (Северцов); низовья Сыр-Дарьи 22 мая пуховой птенец,
22 июня молодые на взлете (Спангенберг, 1936); Кызыл-Кумы—21 июня
СТЕПНОЙ САРЫЧ, КУРГАННИК 321
Летные птенцы (Зарудный, 1915); Гиссар —12 мая значительно оперившиеся
молодые, 27 мая покинули гнездо (Иванов, 1946); Туркмения — летные на
Мургабе в июне (Зарудный, 1896), летные в июне и в конце мая (Лоудон, 1910),
молодые с хорошо развитым оперением, но недоросшими крыльями и хвостом
17 июня, 14 и 16 июля, вполне развитый 5 июня (Дементьев, 1950).
Семиречье — пуховики 19 мая, птенцы 27 мая, летные молодые 8 июля (Зарудный
и Кореев, 1906, Шестоперов, 1929); ю,-з.
Сибирь, Рубцово, вполне развитые молодые
25—30 июля (Велижанин, 1930); сев.
Казахстан—вылет в начале июля (Зарудный,
1910); Чкаловская обл. —летные в июле
(Сушкин, 1908); Ергени — запоздалый не-
разбившийся выводок отмечен 30 июля.
Линька. Выяснена недостаточно.
Линяние проходит повидимому
сравнительно быстро, так как с одной стороны
взрослые птицы, начавшие смену оперения,
добывались в конце мая, а с другой —
большинство особей в конце сентября — начале
октября одеты уже в свежее перо.
Последовательность смены нарядов: первый
пуховой — второй пуховой—первый годовой
(гнездовый) —второй годовой
(окончательный) наряд и т. д.
Питание. Пищу составляют
главным образом грызуны и другие мелкие
млекопитающие: молодые зайцы, суслики,
в том числе тонкопалый, песчанки — в
особенности большая, хомячки, мыши,
тушканчики; ежи (ушастый). В Заволжье, сев.
Казахстане и ю.-з. Сибири особенно
большое значение в питании курганника имеют
суслики (Сушкин, 1908, предполагает
наличие кормовой конкуренции в этом
отношении между курганником и степным орлом),
в Кара-Кумах и Кызыл-Кумах песчанки;
связь распространения курганника и
песчанок отмечена в Туркмении, в Синьцяне
(Шестоперов, 1929), наконец в Индии (с Рис. 86. Курганник.
краснохвостой песчанкой, Юм, 1869). Кроме
млекопитающих курганник кормится ящерицами, змеями (гадюка), лягушками,
жабами; молодыми птицами, яйцами, иногда и взрослыми, например,
жаворонками (Туркмения). Иногда курганник нападает и на более крупных птиц—
на фазанку (Зарудный, 1896), на болотную сову (Тугаринов и Козлова, 1935),
даже на журавля, на последнего без успеха (Зарудный, 1888), на кур (сев*
Казахстан, Мензбир, 1895). Ест и падаль: в Египте снулую рыбу, мясо
дохлого буйвола и т. п. (Мейнертцхаген, 1931).
Охотится курганник, подстерегая зверей (сусликов, песчанок) у нор,
сидя где-либо поблизости, высматривая добычу на лету. Кормовые вылеты
начинаются на рассвете, в жаркое время дня птица сидит где-либо в тени или
у воды. Повидимому, курганник нуждается в питьевой воде, так как в
туркестанских пустынях если не ежедневно, то регулярно собирается у рек в тех
местах, где нет других источников поверхностных вод и при этом не только пьет,
но и заходит в воду (Зарудный, 1896, на Мургабе у Султанбента наблюдал на
21 Птицы СССР, т. I
322
ОТРЯД ХИЩНЫЕ ПТИЦЫ
водопое одновременно 48 курганников; после вывода молодых курганники
к воде прилетают группами и стайками).
Полевые признаки. Крупный длинноногий и длиннокрылый сарыч. От
мелких орлов хорошо отличается на полете светлым пятном у основания
маховых. Полет легкий, часто парит. Осторожен. Садится обычно на землю или
скалы, но редко на деревья. Голос подает редко, при этом крик более напоминает
подорликов (клекот), а не «мяуканье» сарычей.
Описание. Размеры1 и строение. Размеры относительно крупные, как у мохно-
.ногого курганника, с которым сходен в общем и по окраске (та же амплитуда вариаций от
бурой с охристыми пестринами и краями перьев до почти одноцветной темнобурой). Крылья
относительно длинные, цевки длинные (вдвое длинней среднего пальца, измеренного без
когтя). Оперена цевка только в верхней трети ее длины: передняя ее сторона покрыта — в
отличие от мохноногого курганника — крупными поперечными пластинками. Длина самцов
(5) 573—595, в среднем 582, самок (4) 600—637, в среднем 612,5 мм. Размах самцов (5) 1262—
1430, самок (4) 1405—1480, в среднем соответственно 1355,4 и 1445 мм. Крыло самцов (24)
405—438, самок (24) 442—475, в среднем соответственно 426,8 и 456,3 мм. По литературным
данным, у самок крыло до 525 мм (Штегман, 1937), у самцов до 400 мм (Портенко, 1929).
В нашем материале имеются птицы, определенные сборщиками как самцы с крылом в 443,
444 и 446 мм, и самки с крылом в 432 и 435 мм. Во всех этих случаях правильность
определения пола вызывает сомнения.
Окраска. Первый пуховой наряд не описан. Второй пуховой наряд белый
с охристым оттенком на спинной стороне. В дальнейшем развивается два типа
окраски: светлый и темный. Темные птицы однообразно бурые в различных оттенках
с серовато-бурым поперечнополосатым хвостом; изредка на мелких перьях бывают
беловатые пестрины или рыжеватые края. Возрастный диморфизм у сарычей темной вариации
окраски не выражен. Светлый тип окраски: голова, шея, грудь, каймы перьев спины и
плеч, кроющих крыла бледноохристые или беловатые или (реже) ржавчато-рыжие; на голове,
шее, груди — темные продольные полосы, брюхо бурое или беловатое с бурым рисунком,
оперение голени с бурыми поперечными полосками; этот поперечный рисунок
распространяется иногда на брюхо и бока; иногда брюшная сторона почти одноцветная беловатая с
немногими продольными бурыми пестринами; маховые темнобурые, первостепенные —
с белыми основаниями; рулевые охристо-беловатые или ржавчато-рыжие с белыми
стволами, часто с бурыми поперечными полосами. В первом годовом (гнездовом) наряде
поперечный рисунок более развит на хвосте (птиц без полос на хвосте не бывает), но отсутствует
на брюшной стороне тела. Оба пола окрашены одинаково. Радужина желтовато-бурая или
бледнобурая, клюв черноватый, когти черные, восковица и ноги желтые.
Систематические замечания. Курганник близок к В. hemilasius и объединяется с ним
некоторыми авторами в один вид. Экологически первый— птица главным образом равнинная,
а второй — горная. Однако hemilasius в Монголии и Забайкалье встречен на равнинах, а
rufinus в Туркестане —в горах. Ареалы этих птиц на востоке частично перекрываются.
Пока, до детального изучения распространения этих птиц в Центральной Азии, их лучше
считать за различные виды. Требует изучения и географическая локализация особей темной
и светлой вариации: первая более редка, но в небольшом числе встречена и в сев. Африке,
и в Иране, и в Туркестане и Заволжье. Она однако численно преобладает в высокогорье
центр. Тянь-Шаня и в Гималаях.
РОД ОСОЕДЫ PERN IS С U V I E R 1817
Тип. P. apivorus
43. Хохлатый осоед Pernis ptilorhynchus T e m m i n с к
Falco ptilorhynchus. Temminck. Planches Coloriees livr., 8, 1821, табл. 44, Ява.
Распространение. A p e а л. Ю.-в. и вост. Азия от Вост. Сибири и Приморья;
Япония, Индия к северу до предгорьев Гималаев (на восток до сев. Юннани,
в ю.-з. Китае); Сиам; Минданао, Цебу, Люсон, Палаван на Филиппинском
архипелаге, Малайский п-ов; Зондские о-ва — Ява, Борнео, Суматра.
Биотоп. Лесистые местности.
Характер пребывания. На севере перелетная, на юге отчасти
кочующая, отчасти оседлая птица.
ХОХЛАТЫЙ ОСОЕД
323
Подвиды и варьирующие признаки. В Вост. Сибири и Приморье, вероятно в
Манчжурии, Корее и Сев. Китае P. p. orientalis Taczanows k i, 1897; в Японии на Иессо и
Хондо P. p. japonicus К и г о d a, 1925 (?); в Индии от Гималаев до Ассама, Бирмы и
в сев. Юннани, на юг до Цейлона P. p. mficollis Lesson, 1831; зимою в нижней Бирме
P. p. gurneyi Stresemann, 1940 (гнездится в Сиаме?); на Филиппинских о-вах
P. p. philippensis, Мауг, 1939; на Малайском п-ове и южн. Сиаме, вероятно в южн.
Тенассериме, на Суматре и Борнео P. p. torquatus Lesson, 1831 ;■ на-Лалаване P. p. pala-
wanensis Stresemann, 1940; на Яве P. p. ptilorhynchus Temminck. Варьирующие
морфологические признаки: окраска (рулевые); развитие хохла на голове;'
пропорции длины крыла (вершины) и хвоста;
относительные размеры клюва; общая величина.
Экологические географические изменения плохо
выяснены (в частности, некоторые расы оседлы-
другие перелетны, что выражено в относитель,
ной длине вершины крыла). Относительно
«примитивен» P. p. philippensis (самый
примитивный из осоедов вообще), наиболее
специализированы северные P.p. orientalis и P. p.
japonicus.
Восточносибирский осоед Pernis
ptilorhynchus orientalis Taczanowski
Pernis apivorus orientalis. Taczanowski.
Faune Ornith. de la Siberie Orientale, 1891,
стр. 50, низовья Уссури.
Распространение* Ареал.
Восточная Сибирь от Байкала до Приморья и
Сахалина; вероятно сюда же относятся
птицы из Манчжурии (Мейзе, 1934),
Кореи и Сев. Китая (где распространение
еще неясно). Западная граница
распространения не вполне ясна: к этой
форме относятся осоеды, добытые у
Красноярска, у Котон-карагая в южн. Алтае,
на Н. Тунгуске у Подволочной (Штегман,
1937). Однако область регулярного
гнездовья едва ли выходит за пределы
Прибайкалья и Приморья. Залетный добыт у Чимкента (взрослая самка 29. IX.
1866, Северцов), у Алма-Ата (Шульпин, 1936), в Аксу -^ Джебаглы в
Таласском Ала-Тау (самец 15. IV. 1937), у Джулека (8. IV. 1948). Самка,
«отложившая яйца», добыта будто бы на р. Итокар близ Намангана (Зарудный,
1911), но случай этот вызывает сомнения. Зимою на Зондских о-вах, Яве,
Биллитоне, Борнео, Суматре, на Малайском п-ове, в Индо-Китае, быть может
в Сиаме и Бирме (Штреземанн, 1940).
Характер" пребывания. Перелетная птица.
Даты. Прилет поздний. У Бомнака на Зее 29 мая (Дорогостайский,
1915), у Байкала осенью молодые еще встречены около середины сентября.
В Приморье добыты еще в начале октября (молодая птица у м. Гамова, залив
Петра Великого, 8. X. 1946; на о. Аскольд даже 17 ноября). На пролете в вост.
Китае от 12 мая до 13 июня и с 21 сентября до 7 октября (о. Шавейшан,
Латуш, 1932), в Хубее с марта до мая и с сентября до октября (Чоу, 1936).
Биотоп: лиственные леса.
Численность. На гнездовье редок, быть может за исключением
южных частей Приморья. На пролете в вост. Китае и зимою на Зондских о-вах
собирается в значительном количестве (Латуш наблюдал 4. X. 1910 на Ша~
вейшане до 40 пролетавших с севера птиц).
Экология. Размножение. Не изучено.
21*
Карта 51. Распространение хохлатого
осоеда Pernis ptilorhynchus
1 — P. p. orientalis, 2 — P. p. Japonicus, 3— P.
p. riificollis, 4 — P. p. gurneyi, 5 ■— P. p.
torquatus, 6 — P. p. palawanensia, 7 — P. p.
ptilorhynchus, 8 — P. p. philippensis; и — граница
гнездовий (цифры в кружке соответствуют
номеру подвидов и показывают область их
зимовок).
324
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Питание. В желудке добытых на Яве птиц найдены только пчелы, осы
и их личинки. Перепончатокрылые найдены и в желудках добытых в Приморье
осоедов.
Рис. 87. Хохлатый осоед (внизу—старый самец, сверху — молодая самка).
Линька. Как у других осоедов, смена маховых начинается с 10-го и
идет к переднему краю крыла. Начало линяния, повидимому, в соответствии
с поздним календарем размножения, в июле. Вполне выросшие молодые
отмечены около середины августа.
Описание. Размеры и строение. Заметно крупнее, чем европейский осоед,
и с иной формулой крыла. Наружное опахало сужено на первых 6 первостепенных маховых,
*огда как у европейского осоеда только на 5. 2-е короче или только немного длиннее 6-го
(у европейского осоеда заметно короче б-го). Лапы больше,-средний палец без когтя обычно
длинней 45 мм (у европейского осоеда не более 45 мм). Вес также больше: у молодого самца
из Приморья 819 г, у другого взрослого из Казахстана 1282г;у 9 птиц 750—1490, в среднем
1066 (Штреземанн, 1940). Общая длина самцов (2) 629—637, размах их 1414 и 1394 мм (Тача-
новский, 1891). Крыло самцов (2) 450—475 мм (Уссури, Байкал, Тачановский); самцов и
ЕВРОПЕЙСКИЙ ОСОЕД
325
самок (9) 425—465 и даже .495 мм (Вост. Сибирь, Штегман, 1937); самцов и самок (8) 420—
463 мм (Байкал, Аксу-Джебаглы, Джулек, Красноярский край, Приморье, по экз. в Зоол.
музее Моск. университета). У зимующих и пролетных самцов (9) 414—448, самок (9) 413—
467, в среднем соответственно 437 и 444 мм, у птиц, пол которых неизвестен (6) 405—430,
в среднем 423 мм; общая средняя 436 мм (Штреземанн, 1940). Подвидовьш отличия:
относительная длина крыла и хвоста (длина хвоста, умноженная на сто и разделенная на длину
крыла) меньше 65, т. е. хвост относительно короткий. Вершина крыла длинная, более 1\0мм.
На затылке часто более или менее длинный хохол.
Окраска. В общем сходна с окраской европейских осоедов, но у старых самцов
обычно 2, а не 3, темных поперечных полосы на хвосте: на зобе часто подковообразное
темное пятно, а на горле продольная полоса, идущая к подклювью. Общая окраска спинной
стороны бурая, брюшная сторона чаще всего белая с более или менее развитым бурым
поперечным рисунком. У старых самцов темя иногда серовато. Имеется и однотонная темнобурая
вариация. Молодые птицы чаще с белой чуть испещренной бурыми наствольями и
поперечными полосами брюшной стороной, на боках неясные поперечные полоски; иногда
брюшная сторона рыжевато-бурая; на хвосте узкие и частые поперечные полосы; светлые
вершинные пятна на перьях затылка, шеи, спины. Частая поперечная полосатость на хвосте
удерживается повидимому и у взрослых самок (Латуш, 1932, Бартельс по Штреземанну,
1940). Радужина красноватая (как у южноазиатских осоедов, в отличие от европейского
вида); восковица у взрослых черноватая, у молодых желтая; ноги желтые, клюв черноватый
на конце; буроватый у основания; когти черные.
Систематические замечания. Этот осоед большинством современных авторов относится
к виду P. apivorus, а Питере (1931) считает и весь род Pernis монотипическим. Недавно,
однако, Штреземанн (1940) установил определенные морфологические отличия группы вос-
точноазиатских осоедов (P. ptilorhynchus и P. celebensis), причем эти признаки выходят
за пределы вариаций у P. apivorus. Размеры (длина крыла, величина лап) трансгрессируют.
Особи переходных размеров между европейскими и восточносибирскими найдены в Зап.
Сибири и даже в с.-в. России (самцы, Печора: с крылом в 416 и 420 мм; М. Сосьва, Конда,
Лобсынья—405, 415, 423 мм; Чумыш, Барнаульский округ — 420 мм), а также в сев.
Иране в Астрабадской провинции (самка с крылом в 447 мм), следовательно все эти находки
расположены у восточных границ ареала европейского осоеда. Это обстоятельство было
отмечено Птушенко (1938). Однако строение маховых у всех этих птиц и рисунок хвоста
отличны от восточноазиатских. К тому же в Котон-карагае, откуда приводится добытый
26. IX.1926 молодой orientalis (Штегман, 1937), добыт был и настоящий apivorus (самец с
крылом в 395 мм и типичными пластическими признаками в Зоол. музее Моск.
университета). Таким образом не исключается и совместное распространение или частичное
совпадение ареалов этих двух осоедов в средней Сибири, что также говорит в пользу видового
обособления apivorus и orientalis.
44. Европейский осоед Pernis apivorus L.
Falco apivorus. Linnaeus. Syst. Nat. ed. X, 1756, I, стр. 91, Швеция
Синоним. Accipiter lacertarius. Pallas. Zoographia Rosso-Asiatica, t. I, 1811,
стр. 359, ■„«Россия».
Распространение. Ареал. Европа, кроме крайнего севера; Передняя
Азия на восток до Астрабадской (Горганской) провинции Ирана; Зап. Сибирь
до Алтая. В Европе к северу до Англии (редок), в сев. Франции, Бельгии,
Голландии, Дании, южн. Норвегии (до 62° с. ш.), в Швеции до Лапландии (64°),
в Финляндии до бб°30' с. ш.; на юг до сев. Испании, средней Франции, средней
Италии, Греции. В Передней Азии в Анатолии (Куммерлеве и Нитхаммер,
1935), также в южнокаспийских провинциях Ирана, на восток до Астрабада
(Терсакан). В СССР на севере до Обонежья, Архангельска, Соловецких о-вов
(Паровщиков), в области Коми (Удора, 63° 20'), у верхней Печоры (около 62°),
на востоке по ту сторону Урала около 62^63° с. ш. (Конда, Лобсынья, М.
Сосьва). Далее на Востоке в Сибири встречается спорадично: имеются указания
относительно рек Васюгана и Кети в Нарымском крае, Пыжмы в б. Тобольской губ.,
Тобольска, Каменского округа, Салаира, верховьев р. Чумыш, Томска,
Новосибирска, Барнаула, Котон-карагая в южн. Алтае (экз. в Зоол. музее
Московского университета). На юге —в Крыму, на Кавказе (Б. Карачай, Кизлярский
округ, Дзауджикау и другие местности на с.-в. Кавказе; бассейн Кубани на
с.-з. Кавказе; бассейн Алазани и Караклис в Закавказье). Зимовки в
тропических лесах Африки, в особенности в Либерии, на Гвинейском берегу, в Ан-
326
ОТРЯД ХИЩНЫЕ ПТИЦЫ
голе, Камеруне, Конго, реже в Кении и Уганде, в вост. Африке и Натале. На
пролете от Босфора до Гибралтара, в СССР главным образом в южной части
европейской России, в Крыму и на Кавказе; в Туркестане редок (отдельные
находки осенью, август — сентябрь, в предгорьях Кара-Тау и на Сыр-Дарье,
Зарудный, 1911; в Туркмении у Гоудана. Возможно сюда же относится осоед
из Ганьсу (Леннберг, 1924) и добытый на пролете в Синьцзяне в верховьях
Яркенд-Дарьи (Шарп, 1891).
Характер пребывания. Перелетная птица с широким разрывом
между областями гнездовья и зимовки. Прилет поздний (по Штреземанну 1941,
в связи со сроками
размножения
общественных ос). На
зимовках с октября до
апреля. Прилет в
СССР наблюдался в
конце апреля —
первой половине мая
(однако для
Бессарабии будто бы в конце
марта, тогда же и в
Харьковской обл.; для
Смоленска
указывается в середине
апреля — 14 апреля, 18
апреля). Для севера и
Приуралья
приводятся более поздние даты
(вторая половина мая
в Молотовской обл.,
у Уфы будто бы
только 26 мая; однако
в Каменском округе
в Зап. Сибири 10
апреля). Пролетные
особи встречаются пови-
димому и поздней, до
последней трети мая
(22 мая,
Куйбышевская обл., 21 мая
Ростов-на-Дону) .
Осеннее движение
наблюдается уже с августа, но еще в конце этого месяца осоеды добывались и у
Холмогор, а в середине — на средней Печоре. Валовой пролет бывает в
разные числа сентября, в конце сентября птиц меньше (у Москвы 25 сентября,
в Курской обл. 27 сентября), последние в октябре (Кавказ, Крым, у
Полтавы еще 23 октября). Пролет одиночками, парами и стаями от 5 до 50 птиц
(Чкаловская обл., Зарудный, 1892).
Биотоп. Высокоствольные леса, обычно смешанные или лиственные,
режб —на севере — хвойные, в таком случае —сосняки. Распространение
связано с наличием перепончатокрылых (осы), поэтому предпочитается сочетание
леса с открытыми пространствами. Отдельные пары на значительном
расстоянии одна от другой. Главным образом на равнине, реже в предгорьях; однако
на Кавказе до 1200—1300 м (Птушенко, 1938) и даже 1980 м (Даль,
1948).
Карта 52. Распространение европейского осоеда Pernis api-
vorus
а — границы гнездовий, г — область зимовок.
ЕВРОПЕЙСКИЙ ОСОЕД
327
Численность. Умеренная, нигде не может быть назван
многочисленным (на пролете обычен местами — в Крыму и на Кавказе), к северу от
55—56° и в Сибири весьма редок. Отмечены колебания численности,
приписываемые обилию или малочисленности составляющих главную пищу осоеда
насекомых (в Харьковской обл., по Сомову, 1897, редок в начале XIX века,
позже обычен; множество летом 1892 г. при большом размножении ос; по
Хомякову, 1900, в Рязанской обл. много в 1892 г. и мало в 1898). Есть указания
на изменения плодовитости в зависимости от кормовых условий (слабое
размножение и высокая смертность в годы с сырым дождливым летом, Уттендер-
фер, 1939; повышенное размножение в благоприятные годы, Сомов; последнее
вызывает однако сомнения).
Экология. Размножени е. Цикл поздний во всех частях ареала, без каких-
либо широтных различий (исключение — Кавказ). В конце мая — начале июня
свежие или чуть насиженные кладки отмечены в Лапландии, у Пскова,
Горького, Москвы, Калуги, в Белоруссии и на Украине (Умань). В это же время
кладки и в Зап. Европе. Впрочем, у Хангакурта на Малой Сосьве, Зап.
Сибирь, еще 3 июня в гнезде осоеда не было яиц (Скалой, 1940). На Кавказе цикл
более ранний: свежая полная кладка у Дзауджикау найдена 11 мая 1925 г.
(Беме, 1926), а 12 июня 1937 г. у Караклиса в Армении в одном гнезде найдено
сильно насиженное яйцо, а в другом—проклевывавшиеся птенцы (Спанген-
берг, 1948). Гнезда расположены в лесах, чаще ближе к опушке или близ просек,
используются несколько лет (Москва), на деревьях разных пород (береза, граб,
дуб, сосна, ель, лиственница). Иногда занимаются чужие постройки — ворбн
или канюков, иногда гнезда строятся самим осоедом. Размеры их относительно
небольшие (диаметр 600—650, высота 200—300 мм., Ленинградская обл.,Плеске
и Бихнер, 1881; диаметр 590, высота 370, Карачай, Птушенко, 1915); обычно
гнезда расположены на боковых ветвях близ развилки, чаще высоко от земли,
в 10—20 м, иногда и ниже (3—3,5 м, Малая Сосьва, Скалой). Гнезда из сучьев,
иногда с примесью сосновых лап, коры и т. д., почти без подстилки, но
«украшены» зелеными ветвями и листьями (быть может в связи с тем, что —в
отличие от большинства других хищных птиц — птенцы осоеда испражняются
в гнезде, а остатки пищи, из гнезда старыми осоедами не удаляются).
Наступление половой зрелости, постоянство пар и гнездовых участков, брачные
игры — у нас не изучены. Кладка из 2 яиц, реже из 1, в виде исключения
указываются 3 или 4 яйца (3—б. Казанская губ.; 2—3 яйца — Куйбышевская
обл., 3—4—Харьковская обл., 3—Горьковская обл., 3—4—Псковская обл.),
однако здесь возможны и недоразумения. Яйца закругленные с густым и ярким
каштановым рисунком, почти закрывающим основной фон. Размеры (8)
47—52,4x40,7—43,2 (Сомов, 1897), 47—51x38,5—41—41,5 мм (Минск,
Шнитников, 1913), (3) 51—53x41,9—43 (Армения, Спангенберг, 1948).
Птенцов в гнезде бывает 1—2 (Залесский, 1921, указывает на нахождение
у Томска 4. VII. 1917 3 пуховиков в гнезде; Хахлов, 1937, сообщает о
наблюдении 28. VI в верховьях р. Чумыш выводка из 2 старых и 3 молодых, но эта дата
невероятно ранняя). Насиживают оба родителя (у гнезда добывались и самцы),
срок насиживания около месяца: пуховые птенцы в первом пуховом наряде
в первой трети июля (10 июля Ленинградская обл., 7—8 июля вост.
Белоруссия, 11 июля окрестности Москвы и т. д.); оперяющиеся в конце июля
подлетки с не вполне развитыми крыльями около середины августа (ранние —
в первых числах этого месяца). Птенцы выбираются из гнезда с еще недорос-
шими (менее 300 мм длины) маховыми и долго держатся близ него. Гнездовый
период продолжается таким образом, недолго, около 40—45 дней. В
выкармливании птенцов участвуют оба родителя, но главная роль принадлежит самке:
отмечены случаи, когда при гибели самки самец бросал выводок (Бихнер и
Плеске, 1881).
328
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Линька. Изучена недостаточно. Как у других хищных птиц, смена
первого годового наряда начинается ранее, чем линька у старых птиц, уже с при*
лета (начало мая), причем процесс захватывает мелкое перо и задние
первостепенные маховые (9-е и 10-е), но потом повидимому приостанавливается.
Взрослые птицы начинают смену крупного пера во время смены птенцами первого
пухового наряда на второй, т. е. около середины июля, с довольно
значительными колебаниями сроков: самцы несколько позже самок, в начале августа.
Порядок смены маховых —как у других представителей семейства ястребиных,
от 10-го к 1-му; рулевых — от средней пары к краю хвоста (последними быть
может сменяются вторые от края перья, а крайняя пара — предпоследней).
Ход линяния медленный, через месяц после его начала сменяются только 3—
4 задних первостепенных маховых (обычно 10-е—-8-е), к отлету линька далеко
еще не закончена (по Уайзерби, 1938, взрослые птицы линяют до октября, но
иногда до зимы и будто бы даже весны). Линька — раз в году, полная.
Последовательность смены нарядов: первый пуховой наряд — второй пуховой
наряд — первый годовой (гнездовый) наряд — второй годовой (окончательный)
наряд и т. д.
Питание. Все осоеды узко специализированные энтомофаги. Главная
их пища — личинки общественных перепончатокрылых, в первую очередь, ос,
затем шмелей. Способ охоты: осоед, сидя на ветке дерева, или медленно летая,
следит за насекомыми, подмечает гнездо, а найдя его — разрывает лапами
(закапываясь иногда на полтуловища). Ос ест вместе с жалом. В связи с этими
особенностями питания осоеда стоит то, что у него оперение лба и уздечки —
из плотных перышек, а не из щетинок; цевка короткая, но сильная,
защищенная толстыми щитками, пальцы с длинными, но мало изогнутыми когтями,
примерно одинаково развитыми — они служат главным образом для рытья, а
не для схватывания и умерщвления добычи; слабый изгиб когтей облегчает
и передвижение по земле (ходьбу); ноздри узкие, щелевидные, защищенные
от засорения при работе в земле. «Урожай» перепончатокрылых вероятно
влияет на размножение осоеда (число яиц в кладке; размеры яиц? по Сомову^
1897). В качестве пищи осоеда в СССР указываются личинки, осы и взрослые
насекомые Vespa germanica, Polistes gallicus и др., в качестве
дополнительного питания — гусеницы, кобылки, крупные жуки, также лягушки, змеи,
мелкие птицы и грызуны. Корм с прилета и перед отлетом, повидимому, более
разнообразен, чем в период размножения, когда осоед питается почти
исключительно перепончатокрылыми (птицы носят их птенцам иногда целыми
гнездами). В связи с характером питания осоедов, они мало сбрасывают погадок
(погадки однако находятся даже в гнездах, Зарудный, 1911).
Полевые признаки. Похож на сарыча, но на лету отличается более
длинными и узкими крыльями, длинным хвостом и более узкой головой,
вытянутой вперед. Полет легкий, но довольно медленный, с редкими взмахами
крыльев; парит редко. Мало осторожен, в особенности у гнезда. Молчалив, голос —
короткий, звонкий свист «пи-и-ия».
Описание. Размеры и строение. Сложение легкое, крылья и хвост длинные,
так что птица кажется больше, чем на самом деле. Клюв низкий, без выступа на крае
надклювья (в связи с характером питания). Ноздри узкие, щелевидные, косо расположенные.
Уздечка и лоб, участки вокруг глаз покрыты чешуеподобными плотными перышками.
Формула крыла 3>4>2>5>6> 1 >7. Опахала сужены — внутренние на 1-м — 5-м,
наружные на 2-м — 5-м маховых. Хвост слегка закруглен, из 14 рулевых. Цевки оперены до
половины, покрыты мелкими щитками. Лапа по сравнению с восточноазиатскими осоедами
слабая: длина среднего пальца обычно менее 45 мм (без когтя). Вес самцов 4 (580), 600, 718,
730, самок (3) 825, 930, 950 г. Крыло самцов (46) 370—423; самок (50) 372—435, в одном
случае 447 мм, в среднем соответственно 402,2 и 409,9 мм.
Окраска. Первый пуховой наряд белый с желтоватым оттенком на спине; второй
пуховой наряд— в возрасте около 2 недель — белый с легким сероватым оттенком. Окраска
оперения полиморфна и в общем может быть сведена к четырем типам: 1) однообразно бурому
ЕВРОПЕЙСКИЙ ОСОЕД
329
на спинной и брюшной стороне, иногда несколько более светлой; 2) бурому на спинной
стороне, светлобурому с более или менее заметными белыми основаниями перьев на брюшной
стороне; 3) бурому на спинной стороне и пестрому — белому с широкими бурыми попереч-
Рис. 88. Европейский осоед (взрослый и молодой самцы).
ными полосами на груди, боках, брюхе; 4) бурому на спинной стороне и белому с немногими
темными продольными пестринами на брюшной стороне. Этот полиморфизм свойственен всем
возрастам, но особенно выражен у птиц в первом годовом наряде. В окончательном (втором
годовом) наряде осоед на спинной стороне серовато-бурый с более или менее заметными
темными наствольями, иногда расширяющимися в продольные пятна; на темени, уздечке,
щеках нередко серый налет (повидимому исключительно у самцов); маховые бурые с
черновато-бурыми вершинами, с белыми основаниями и темными поперечными полосами; рулевые
с 3—4 поперечными полосами — широкой у вершины и узкими ближе к основанию и
характерным «муаровым» рисунком; при этом вершинная полоса отделена от лежащих ближе
к основанию широким промежутком. Иногда на голове и шее развит белый цвет в виде пестрин
330
ОТРЯД ХИЩНЫЕ ПТИЦЫ
у оснований перьев или даже голова белая с темнобурыми пестринами. Брюшная сторона
бурая или белая с бурым поперечным рисунком, или бурая с белыми основаниями перьев,
или белая с немногими темными продольными штрихами.
В первом годовом наряде осоеды на спинной стороне бурые, часто с
охристо-беловатыми крапинами у вершин перьев на голове, шее, кроющих крыла; на голове серого оттенка
не бывает; чаще попадаются светлоголовые особи. Брюшная сторона как у взрослых, с
такими же вариациями, но особи с правильным бурым поперечным рисунком не встречаются.
Поперечный рисунок хвоста обычно более равномерный. Радужина у взрослых желтая или
оранжевая, у молодых серовато-желтая или сероватая; восковица у взрослых темносерая,
у молодых бледножелтая, клюв черноватый, желтеющий у основания нижней челюсти;
когти черные.
Систематические замечания. Популяции осоедов в отдельных частях ареала не вполне
однородны, как это и следовало ожидать, но не образуют географических рас.
Экологических различий между ними нет. Распределение типов окраски во всей изученной нами серии
(около 200 экземпляров) таково: светлых особей 6%, пестрых 34%, «промежуточных» между
ними и темными 26%, темных 34%. В отдельных популяциях соотношение птиц разных
типов окраски меняется: в Московской обл., например, 5% светлых, 15% пестрых,
30% «промежуточных», 50% темных. В Поволжье в б. Корсунском уезде светлых 0%,
пестрых 44%, «промежуточных» 36%, темных 20%. У восточных границ ареала в с.-з. Сибири
и Приуралье темных птиц в коллекции нет. Надо отметить, что в одном гнезде (Московская
обл.) из двух птенцов встречен один темный и один пестрый. Имеются некоторые
различия и в средних размерах крыла отдельных популяций. Они видны из таблицы.
3 1
Самки
Архангельские и Печорские.
Зап.-сибирские
Московские
Ульяновские
Московские
Ульяновские
Число особей
3
4
7
12
7
8
Колебания длины
крыла в мм
415—420
405—423
385—420
395—417
380—425
402—435
Среднее
417
414,5
402,6
404,6
403
417
Таким образом, различия между популяциями не выражают географического
изоморфизма, так как мелкие и крупные особи или популяции с преобладанием определенных
типов окраски встречаются в «перемешку». Несколько особняком стоят осоеды,
гнездящиеся в горных лесах Кавказа и сев. Ирана (Астрабад). Среди 5 известных гнездовых
особей с Кавказа и сев. Ирана (Карачай, Алазань, Караклис на Кавказе, Терсакан и сев.
Иран) — все птицы темной бурой окраски. Величина их значительна: крыло самцов (2) 412—
422, в среднем 417, самок (3) 405,420 и 447, в среднем 424 мм. К тому же эти птицы отличаются
и в некоторых экологических отношениях (ранний цикл периодических явлений).
РОД ЯСТРЕБИНЫЕ САРЫЧИ BUTASTUR HODGSON, 1843
Тип. В. teesa
45. Ястребиный сарыч Butastar indiciis Gmel.
Falco indicus. G m e 1 i n. Syst. Nat. t. I, 1788, стр. 264, Ява.
Распространение. Ареал. Вост. Азия — Япония, Сев. Китай
(провинции Жехол и Чжили), Манчжурия (?), в СССР — Приморье на запад до
Малого Хингана (Штегман), на север до устьев Бурей. Зимовки в южн. Азии
ЯСТРЕБИНЫЙ САРЫЧ
331
до Южн. Китая, с. в. Бирмы, Малайского п-ова, в Индо-Китае, Тенассериме,
Сиаме, на о-вах Филипповских, сев. Целебесе; одиночки на Новой Гвинее,
сев. Молукках Вайгиу, Салавати, Борнео и Яве.
Характер пребывания. Перелетная птица; прилет в
Уссурийском крае отмечен в начале апреля (Деррис), отлет, повидимому, в начале
сентября (массовый пролет на Филиппинах в сентябре — октябре).
Биотоп, Широколиственный или смешанный лес с близлежащими
лугами, рисовыми полями и другими открытыми пространствами, чаще близ
воды или у болотистых мест, что связано повидимому с кормовым режимом
(амфибии).
Численность. Обычно указывается, что ястребиный сарыч в
гнездовой области немногочисленен, чему однако противоречит широкая область
зимовок и относительная там
многочисленность. Местами в Приморье
обычен (на Имане и Седанке; на
Имане одна пара находилась на
расстоянии 2 км от другой, Спангенберг).
Экология. Размножение.
Гнезда расположены на деревьях в
лесу вблизи от лугов (Спангенберг)
или среди рисовых полей (Оустон).
Находятся гнезда на высоте 7—12 м,
построены из веток, камыша, осоки;
размеры гнезда относительно
невелики. Кладка в начале мая. Число
яиц обычно четыре, окраска их белая,
на просвет яркозеленая, длина 49,1 —
51, ширина 38,2—39,9 мм
(Спангенберг). Нелетные оперяющиеся птенцы
на Имане найдены 25 июля. Детали
цикла размножения' —
продолжительность насиживания, нахождения птен- Карта 53. Распространение ястребиного са-
цов в гнезде и т. п. — неизвестны. рыча Butasfur indicus
ЛИНЬКа. Изучена ПЛОХО. МО- а —граница гнездовий, г — область зимовок.
лодые в мае еще не линяют. Начало
линьки взрослых с первых чисел июня (4 июня смена 9-го и 10-го
первостепенных маховых); в первой половине августа у самца смена мелкого пера на
брюшной стороне, много свежего пера на спине и плечевых, первые два
маховых старые, 3-е растет, остальные новые, смена рулевых.
Смена нарядов: первый годовой (гнездовий) — второй годовой
(окончательный) и т. п.-
Питание. Кормовой режим —лягушки, ящерицы, змеи, грызуны
(Иман, Спангенберг). Птицы на появление ястребиного сарыча будто бы не
реагируют, хотя есть наблюдения над нападением канюка на самку фазана.
С кормовым режимом находятся в соответствии и распространение канюка, и
особенности его строения: длинные, слабо оперенные цевки, короткие, покрытые
грубыми щитками, пальцы и т. п. Способ охоты: птица сидит на дереве и
осматривает окрестности, бросаясь на добычу наискось; кормовые вылеты по утрам
и вечерам.
Полевые признаки. В период размножения высоко летает над лесом
кругами, при этом полет — с глубокими взмахами крыльев — напоминает
ястребов, а форма крыльев — сарыча. Сидят ястребиные канюки обычно на
вершинах одиноко стоящих сухих деревьев среди сырых лугов, болот или выгонов.
Сидящая птица очень похожа на сарыча, отличаясь от него несколько мень.
332
ОТРЯД ХИЩНЫЕ ПТИЦЫ
шими размерами, и более легким сложением. Не очень осторожен, нередко
охотится близ селений. Часто подает голос, напоминающий крик ястреба-тн>
вика ^громкое и протяжное «ти-виии.. ти-вии» (Спангенберг).
Рис. 89. Ястребиный сарыч (взрослая и молодая птицы).
Описание. Размеры и строение. Телосложение легкое, длинные и довольно
острые крылья (формула 3>4>5), цевки длинные, оперены только в верхней трети, покрыты
мелкими щитками; пальцы короткие, когти короткие с крутым изгибом; хвост длинный,
прямой на вершине. Крыло самцов (7) 310—330, самок (3) 327—340 мм, в среднем 315,6
и 333 мм.
Окраска. Пухов'ой наряд не описан. Первый годовой наряд на спинной стороне бурый
со светлыми краями перьев на голове, на брюшной стороне охристый с продольным бурым
рисунком на груди, с поперечновытянутыми бурыми пятнами на боках и бурыми наствольями
на штанах. Взрослая птица: сверху бурая с серовато-бурой головой; маховые с поперечным
темнобурым рисунком, на хвосте 8 поперечных темных полос; низ беловатый с темной полосой
вдоль горла и поперечными бурыми полосками на груди и брюхе. У самца кроющие крыла
более яркие рыжие. Радужина желтая; ноги и восковица яркожелтые; клюв и когти
черные.
ЗМЕЕЯД
333
РОД ЗМЕЕЯДЫ CIRCAETUS V I E I L L О Т, 1816
Тип С. Gallicus
46. Змееяд Circaetus ferox Gmelin
Accipiter ferox. S. G. G me 1 i n. Novi Commentarii Acad. Caesar. Petropolitanae pro 1770,
1771, стр. 442, табл. Х, Астрахань.
Синонимы. Falco gallicus. Gmelin. Syst. Nat. ed. XII, I, 1788, стр. 259, Фран*
ция. — Accipiter hypoleucos. Pallas. Zoographia Rosso — Asiatica, t. I, 1811, стр. 354,
южн. Россия.
Русское название. Название происходит от питания этой птицы (главным
образом пресмыкающиеся и земноводные). Происхождение другого имени этой птицы —
крачун — неясно.
Распространение. Южная и средняя Европа, ю.-з. Сибирь, Средняя Азия,
сев. Монголия, к югу до передней Индии; Африка.
Карта 54. Распространение змееяда Circaetus ferox
1 — С. f. ferox, 2 — С. f. heptneri, 3 — C. f. beaudouini, 4 — C. f. pectoralls; a —
граница гнездовий, г—область зимовок.
Характер пребывания. В сев. части ареала перелетная, в
южной оседлая птица.
Биотоп. На севере —леса, южнее — сухие местности с достаточно
развитой, но хотя бы редкой древесной растительностью (саванны). Равнины,
гор избегает. Имеется известная связь биотопического распределения и
численности пресмыкающихся.
Численность. Распространение, довольно спорадическое, нигде не
многочисленен.
334
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Подвиды и варьирующие признаки. В сев. Африке и Европе, на Кавказе, в М. Азии,,
в Зап. Сибири, вероятно в Иране С. /. ferox Gmelin, 1771; в Средней Азии и сев.
Монголии С. /. heptneri Dement., 1932; в Африке от Сенегамбии до Кордофана, к югу до
Камеруна С. /. beaudouini Verreaux et de Murs, 1862; в южн. и вост. Африке от Белого*
Нила до Земли Дамара, Родезии, Трансвааля и Капской Земли С. /. pectoral is S m i t hy
1830. Варьирующие признаки — размеры (тропический beaudouini самый мелкий;
южноафриканский pectoralis и южноевропейский ferox примерно сходны; среднеазиатский
heptneri самый крупный) и окраска, главным образом брюшной стороны тела.
Экологические проявления географической изменчивости не изучены, но ferox и heptneri перелетны,
остальные оседлы.
Европейский змееяд Circae'tas ferox ferox G m.
Распространение. Ареал. Северная граница в Европе от Пиренейского
п-ова до 48—49° с. ш. во Франции (низовья Луары, ле-Ман, Мелен, Шалон
и Нанси); Люксембург, средний Рейн; в Ганновере до 55°, далее в Меклен-
бурге, Померании, Пруссии, в СССР в Прибалтике (Эстония), до Пскова,
Ленинграда, Новгорода, в "Владимирской обл., Рыбинска, Костромы, в Горьков-
ской обл. в Ветлужском и Семеновском районах (Пузанов, 1942), в Поволжье
около 66° (Першаков, 1929), на Урале около 52°; в с.-з. Казахстане граница
неясна, в Аралокаспийских степях редок и гнездованье не доказано (Бостан-
жогло, 1911), самка с наседным пятном добыта у Чалкар-тениза (Сушкин,
1908). Далее граница идет к северу, поднимаясь до Караганды (Кызыл-рай,
49° с. ш.) и Павлодара, а в ю.-з. Сибири примерно до Барнаула (Горькое
озеро, Велижанин, 1928, примерно до 52°). Южная граница—в Сицилии,.
в сев. Африке, где распространен от Марокко до Египта; в Передней Азии до
Аравии, Палестины, сев. Ирана (Астрабад). Вероятно к этой же расе относятся
гнездящиеся в зап. Туркмении змееяды (см. ниже), и, судя по размерам (Бекер,
1928), гнездящиеся в Индии, где змееяды встречаются на восток до нижней
Бенгалии.
Характер пребывания. Перелетная птица в северных частях
ареала, в частности всюду в СССР. Зимует изредка в южн. Франции, а главным
образом в Передней Азии, начиная от Палестины (там вероятно частично осе-
дел, Тристрам, 1865), в южн. Аравии и особенно в сев. Африке; к югу от Сахары
только в Абиссинии, на Белом и Голубом Ниле и в лесах бассейна Нигера у
южной границы зап. Сахары.
Даты. На зимовках встречается с середины сентября, чаще с октября, и
до марта, отдельные особи до мая. Прилет поздний и растянутый, на юге
Европейской части СССР в первой половине апреля (Умань —начало апреля;
Полтава— 27 марта — 11 апреля; Полесье— 6 апреля, Минская обл., — начало
апреля, Прибалтика — 5 апреля), на севере быть может и поздней (23 апреля —
9 мая Псков, Зарудный, 1911). Весь пролет тянется около месяца (пролетные
змееяды у Киева встречены еще в конце апреля — начале мая, Шарлемань,.
1926; в Подолии в конце апреля, Герхнер, 1928). Отлет и осенний пролет
происходит в разные числа сентября, до конца этого месяца или до начала
октября; отдельные особи начинают отлетать еще ранее—в августе (стая из
20 птиц, Золотоноша, Гавриленко, 1929) и даже в июле (19 июля, Аска-
ния-Нова, Гроте, 1914). В Крыму змееяды встречены еще 30 октября. Летят
змееяды и одиночками, и парами, и стаями (на с.-в. Кавказе по 40—50 штук,
Беме, 1926).
Биотоп. Змееяд связан с двумя типами биотопов: на севере с влажными
лесами, перемежающимися с открытым ландшафтом, богатыми ужами и
амфибиями (впрочем, в Прибалтике и в сухих сосняках, Лоудон, 1907); на юге—
с сухими, изобилующими рептилиями угодиями. При этом гнезда устраиваются
почти исключительно на деревьях, хотя бы и невысоких. В вертикальном
отношении, из-за бедности высокогорья рептилиями—вверх далеко не подни-
ЗМЕЕЯД
335
мается, хотя и не избегает лесной зоны гор. Во Франции в горах 1200—1500 дл,
на Балканах до 1300 м в Черногории, на Кавказе до 1300 м и даже 1980 м.
Численность. Стенофагия змееяда делает его распространение
спорадичным; у северного предела ареала, а также в центральной и восточной
полосе Европейской части СССР он редок, как редок в сев. Казахстане и ю.-з.
Сибири; но на широтах между Волынью —Минском —Псковом, в богатых
ужами районах Полесья — многочисленен. Слабое размножение змееяда
приводит к быстрому его вытеснению из культурного ландшафта в Зап. Европе.
Экология. Размножение. Сведения отрывочны. Гнезда повидимому
всегда строятся самим змееядом, на соснах (Минск, Псков, Киев), также на
дубах и липах (Умань); обычно высоко от земли, около 6—15 м, изредка
низко — до 2 м. Гнезда чаще расположены невдалеке от опушки, реже в
глубине леса. Судя по разнице в размерах (у Пскова, по Зарудному, 1910, от 68
до 136 см ширины, 62—112,5 см высоты, с лотком от 35 до 50 ел* в диаметре
и от 13 до 21 см глубины; в б. Уманском уезде по Гебелю, 1879, в 45—55 см
в диаметре и 30—40 см в высоту) — гнезда используются много лет. В редких
случаях гнездо располагается на земле на кочке (Полтава), на скалах (сев.
Казахстан, Долгушин, 1938). Постройка из сучьев, без выстилки, но обычно
с зелеными свежими ветвями. Кладка из 1 яйца, редко из 2 (Джоурден по
Бекеру, 1928). Слабость размножения находится вероятно в связи с
трудностью прокормления птенца в результате стенофагии.
Окраска яиц белая (желтоватая или зеленоватая на просвет); размеры
74,3—79,1 х 54,3—59,5 (Псков, Зарудный), 75 х 60(Полесье, Шнитников, 1913),
74 х 60 (Киев, Шарлемань, 1928); у индийских птиц (30) 73,6 х 58,4, в среднем
67,7—80 х50,4—64,6 мм. Кладка в конце апреля — начале мая: ранняя 18
апреля (Умань, Гебель), свежие 6 мая (Полтава, Гавриленко); 6 мая (Киев), в
начале мая (Псков). В Индии цикл, повидимому, более ранний, на юге в январе —
марте, в Гималаях в марте — апреле. В случае гибели кладки бывает новая,
в том же гнезде, иногда через 2—3 недели (отмечена 12—15 мая).
Насиживают, судя по наседным пятнам, оба родителя (над участием самца
в насиживании прямые наблюдения у Киева, Шарлемань, 1926; в Силезии,
Цебе, 1936, во Франции, Энар и Бланш, 1937). Длительность насиживания —
несколько больше месяца (Гавриленко, 1929, Зарудный, 1911); вылупление
молодых происходит около середины июня. В конце июля птенцы оперяются
и к середине августа становятся на крыло (10 августа в б. Новосильском уезде
Тульской обл. молодые со слегка недоросшими маховыми и рулевыми, Хару-
зин, 1926; 13 августа и 23 августа подлетки в Полтавской обл., Гавриленко,
1929 и т. д.). Гнездовый период продолжительный, около 10 недель. Кормят
птенцов обе старых птицы, при этом в начале гнездового периода самка
разрывает добычу на куски, поздней змеи и другая добыча передаются птенцу
целиком.
Линька. Выяснена недостаточно. Полная годовая, между апрелем и
сентябрем, следовательно, весьма медленная и без обычной четкой корреляции
с определенными фазами репродуктивного цикла. Последовательность смены
первостепенных маховых — от 10-го к 1-му. Последовательность смены
нарядов : пуховой первый — пуховой второй — первый годовой (гнездовый) —
второй годовой (окончательный). Возрастные изменения окраски
незначительны.
Питание. Кормится змееяд главным образом пресмыкающимися и
земноводными, с наличием их связано и распространение этой птицы. Вылетает
на охоту поздно, когда земля и воздух прогреваются. Высматривает добычу,
останавливаясь на полете с вытянутыми лапами, развернутыми крыльями и
хвостом, но не «трепещет» подобно пустельге и сарычам. Реже охотится
«пешком» на земле или в мелкой воде. В качестве корма в СССР приводятся
336
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Anguls fragilis, Coronella austriaca, Coluber berus (Киев, Шарлемань, 1926);
в зобе у одного найдено 10 прытких ящериц и гадюка (Волжско-Уральская
степь, Волчанецкий, 1936); ящерицы (Полесье), в желудке одной птицы 7 Со-
luber renardi (Приазовье, Алфераки, 1910); змеи, лягушки, ящерицы, также
мелкие зверьки и
тетеревенок (Псков); змеи,
преимущественно ужи,
также гадюки, ящерицы
Lacerta и Eremias, иногда
мыши (Харьковская обл,
Сев. Кавказ); из птиц —
лесной конек, из
насекомых — саранча.
Характерно, что в
дождливое лето 1925 г. при
низкой численности гадов,
змееяды кормились в
Полтавской обл.
мышами, а также птицами —
дроздами, сойками,
горлицами, главным
образом нелетными птенцами
(Гавриленко, 1929). Есть
указания на похищение
змееядом кладки
подорлика (Зарудный, 1911).
Змеям отрывается
голова, после чего они
заглатываются с
переднего конца тела; у
летящего змееяда из клюва
нередко висит хвостовая
часть змеи, так как
змееяд не носит добычи в
лапах. Размеры
добываемых змей до 1 м (Мадон,
1929). Несмотря на то,
что змеи — едва ли не
основная пища этой
птицы, змееяд восприимчив
к действию змеиного яда.
Потребность в пище выводка (родителей и птенца) за лето определяется
примерно в 800—1000 змей (Уттендерфер, 1939) —этим хорошо объясняется
спорадичность и редкость-этой птицы на севере.
Полевые признаки. Характерна большая голова с большими желтыми
глазами и небольшим относительно клювом; окраска на спинной стороне
темная, на брюшной белая с темным зобом или почти совсем белая; маховые на
полете несколько расставлены, крылья длинные и обычно полусогнуты в
кистевом сочленении; хвост относительно длинный с небольшим числом
поперечных полос. Размеры с подорлика, заметно крупней сарыча. Голос главным
образом протяжное «хиий-о, хиий—о» или мяукающие звуки (у самки).
Описание. Размеры и строение. Оперение густое и мягкое; крылья длинные
и широкие. Формула крыла 3>4>5...., вырезки на внутренних опахалах первых пяти
первостепенных маховых. Цевки длинные, покрытые сетчатыми щитками, слабо оперенные
Рис. 90. Змееяд.
ЗМЕЕЯД
337
только в верхней части; пальцы короткие, но крепкие, покрытые толстыми щитками, когти
острые и круто загнутые. Длина самцов (3) 670—720, в среднем 700, самки (1) 720 мм.
Крыло самцов (8) 520—552, самок (16) 520—570, в среднем 532,4 и 539 мм; у одного
самца из Непала (в колл. Зоологического института Акад. наук СССР) крыло 473 мм.
Окраска. Первый пуховой наряд — белый, на голове пух удлинен; просвечивает
розоватая кожа, у орбиты — голое пягно; радужина мутно-серая, восковица бледноголубая,
клюв черно-синеватый, лапы розовые; характерна большая голова (Энар и Бланш, 1937).
Второй пуховой наряд белый, гуще и пушистее предыдущего, с темным кольцом вокруг
глаз; радужина желтоватая (Нитхаммер, 1938).
Взрослая птица (второй годовой и последующие наряды): общая окраска спинной
стороны бурая, с черноватыми стволами перьев и с беловатыми их основаниями;
маховые черно-бурые, задние беловаты с темным поперечным рисунком на внутренних
опахалах; рулевые бледнобурые с 2—4 темными поперечными полосами; брюшная сторона
белая, у некоторых особей с немногими бурыми продольными наствольями, у других с
бурым горлом и зобом и с редкими широкими бурыми поперечными пятнами на груди, брюхе
и боках или с широким бурым продольным рисунком на зобе и груди; изредка встречаются
особи с белой головой, исчерченной бурыми наствольями. Первый годовой наряд бледнее
на спинной стороне, с охристым оттенком, а темный рисунок на брюшной стороне
также бледнее и чаще не поперечный, а продольный.
Самцы и самки окрашены одинаково. Радужина яркожелтая или оранжевая;
восковица серо-голубая или желтоватая; клюв серо-роговой, голубоватый у основания; лапы
синевато-серые с черными когтями.
Туркестанский змееяд Ciraetus ferox he pt пег I Dem.
Circaetus gallicus heptneri. Dementiev. Orn. Monatsber., 1932, стр. 172, Фрунзе,
Киргизия.
Распространение. Ареал. Средняя Азия от Кара-Тау на севере до
Туркмении (Бадхыз и Карабиль) на юге, Таджикистана и Киргизии на востоке.
В сев. Иране (Астрабадская провинция близ Терсакана) в гнездовое время
добыта птица европейского подвида. К туркестанскому подвиду относятся
змееяды из Монголии (Гудзирте, Кентей; горы Харанорин-ула у Желтой
реки, Ордос).
Характер пребывания. В Средней Азии не зимует. Птицы
этой формы должны на зимовках встречаться в Индии и по всей вероятности
в Передней Азии (в Иране в Сеистане, в Белуджистане); к ним же должны
относиться сведения о залетах змееядов в ю.-в. Азию — в Сиам, на Малайский
п-ов (Бекер, 1928), на о. Тимор (И. 1829).
Даты. Пролет в сев. частях Средней Азии на р. Чу осенью с конца
августа отдельными особями и группами до 8 птиц; в Туркмении наблюдается
по крайней мере до конца сентября (добыт 29. IX. у Кушки). Весенний
пролет и прилет не прослежен; в ю.-в. Туркмении птицы на месте уже во второй
половине апреля (20—23 апреля в Бадгисе между Акар-чешме и Ак-рабатом).
Биотоп. Сухие, богатые пресмыкающимися местности с ксерофильной
растительностью. Главным образом в предгорьях и холмах и в нижнем поясе
гор, примерно до 1500 м (Гульча). По Северцову (1873) от 300 до 2400 м, но
последнее маловероятно, как невероятно указание этого автора на нахождение
змееяда у озера Чатыр-куль, если только это не был случайный залет.
Встречается и в пустынях Кызыл-Кумы и Кара-Кумы, куда птицу привлекает
обилие пресмыкающихся.
Численность. ВЗаилийском Ал а-Тау найдено на протяжении 10 км
б (занятых?) гнезд (Северцов).
Экология. Размножение. Сведения отрывочны. Даты цикла пови-
димомутеже, что и у европейского подвида. Гнезда расположены на деревьях
(в Монголии; у Сыр-Дарьи; на фисташке в 2 м от земли, Фисташковый хребет
у Кушки; на черном саксауле в Кара-Кумах, например уРепетека, Лоудон
1913 и др.) или на скалах (Нура-Тау, Тянь-Шань). Кладка в Туркмении
бывает в апреле; 30 апреля у Кушки добыт птенец в первом пуховом наряде
(Воробьев). Размер яйца 72,1X55,6 мм (Урме в Кызыл-Кумах). В кладке
22 Птицы СССР, т. I
338
ОТРЯД ХИЩНЫЕ ПТИЦЫ
1 яйцо. Выросшие молодые отмечены в середине августа. Л и н ь к а не изучена.
Питание. Почти исключительно ящерицы и змеи. В гнезде у Кушки
найдена контия и тонкопалый суслик. Пара змееядов наблюдалась у Ак-рабата
в Бадхызе 23 апреля на охоте за ящерицами.
Описание. Крупнее, чем номинальная форма: крыло самцов (12) 542—585, самок (5)
561—605, в среднем 552 и 589,3 мм. Окраска. Как у европейских змееядов.
РОД СКОПЫ PANDION SAVIGNY 1809
Тип Я. haliaetus
47. Скопа Pandion haliaetus L.
Falco Haliaetus Linnaeus, Syst. Nat. ed. X, t. I, 1758, стр. 91, Швеция.
Русское название. Слово «скопа» — старинное обозначение крупной хищной
птицы, питающейся рыбой (поэтому на севере местами зовут скопой и орлеана-белохвоста).
Этимология неясна. Предположение Палласа о происхождении «скопа» от скопить, скопец —
маловероятно и непонятно.
Распространение. Ареал. Гнездится в Европе и Азии на север до
границы высокоствольного леса; на юг до сев. Индии (Гималаи) и Филиппинских
Карта 55. Распространение скопы Pandion haliaetus
7 — P. h. haliaetus, 2 — P. h. cristatus, 3 — P. h. microhaliaetus, 4 — P. h. carolinensis,
5 — P. h. ridgwayi; a — граница гнездовий, г — область зимовок.
о-вов; в сев. Африке на юг до Сомали; на о-вах Зеленого мыса; на о-вах Индо-
австралийского архипелага и в Австралии; в Сев. и Южн. Америке. В
негнездовое время северные подвиды встречаются в тропической и южн. Африке,
в южн. Азии от Индии до Филиппинских и Зондских о-вов в восточном
полушарии и от южных Соединенных Штатов, Мексики, Вест-Индии до Перу, сев.
Аргентины и Парагвая в западном полушарии.
Характер пребывания. Северные популяции в Европе,
Азии и Америке перелетны, южные оседлы.
Биотоп. Скопа — специализированный ихтиофаг и поэтому живет
вблизи от водоемов; гнездится, за редкими исключениями, на деревьях.
Сочетание этих условий и определяет распределение скопы в биотопическом
отношении. В безлесных местностях, — тундрах, степях, высокогорьях —
отсутствует.
СКОПА
339
Подвиды и варьирующие признаки. Географическая изменчивость выражена слабо и
птицы по всему обширному ареалу в сущности весьма сходны. Описаны 5 подвидов: P. Л.
haliaitus L., 1758, в умеренной зоне Европы и Азии; P. ft. carolinensis Gme lin, 1788, в Сев.
Америке от с.-з. Аляски, с.-з. Мекензи, Унгава и Ньюфаундленда до юга Соединенных
Штатов и Мексики; Р. Л. ridgwayi May nard, 1887, оседлая форма Багамских о-вов
и побережий Юкатана и Британского Гондураса; Р. Л. cristatusV i e i 1 1 о t, 1816, на Яве,
Целебесе, М. Зондских о-вах, Молуккских, Новой Гвинее, на архипелаге Бисмарка,
Соломоновых о-вах, в Австралии и Тасмании; Р. Л. microhaliaetus В га si 1, 1916, в
Новой Каледонии. Из этих подвидов реальность ridgwayi и microhaliaetus требует
подтверждения. Различия главным образом в окраске. Экология в основных чертах сходна, но
характер пребывания и календарь периодических явлений варьируют.
Обыкновенная скопа Pandion haliaetus haliaitus L:
Распространение. Ареал. Европа от Лапландии до Средиземного моря;
отсутствует в Англии, Бельгии, Голландии, Дании, Франции, Греции,
Болгарии и Сербии; в Германии гнездится в Мекленбурге и Бранденбурге. Азия от
границы лесной растительности (по долинам рек в лесотундре) до Камчатки
и Японии на востоке, Южн. Китая (главным образом в восточных приморских
частях), Гималаев, Белуджистана и Ирана (Южнокаспийские провинции, Хо-
рассан) на юге; южная Аравия, сев. Африка (Алжир, Тунис). В СССР на севере
встречается на Кольском п-ове, на Соловецких о-вах, далее у
Архангельска, на нижней Печоре, по Оби у Обдорска (66° 30' с. ш.), в Туруханском
крае на р. Большой Елагуй и в верховьях Таза; на Нижней Тунгуске у Илим-
пеи; далее к востоку северная граница неясна — найдена на Витиме и в
верховьях Колымы, на северном побережье Охотского моря и на Камчатке. В
Средней Азии на Балхаше, Иссык-куле, возможно в низовьях Сыр-Дарьи и в
горной Фергане (озеро Сары-чилек); в Таджикистане и Туркмении не гнездится,
но обычна на ю.-в. Каспии в пределах сев. Ирана. Отсутствует в Центр.
Азии, доходя к югу до Танну-тувинской обл., сев. Монголии (с.-з. Кентей)
и Прибайкалья. Возможно спорадическое гнездование в тропической Африке
(Виктория-Ньянца; озеро Наиваша). Зимовки в области Средиземного моря»
а также в Африке к югу от сев. Нигерии и Конго на западе, Сомали и южн.
Аравии на востоке, ,.а также в Передней Азии и на юге этого материка
(Месопотамия), Иран у Персидского залива, у Каспийского моря и в Сеистане;
в Белуджистане, Индии и Бирме до Цейлона, в Индо-Китае.
Характер пребывания. Перелетная птица в северной части
ареала, в частности в СССР (зимовки у нас только на Атреке в ю.-з. Туркмении
ив мягкие зимы, по Радде, 1886, в Талыше).
Даты. На африканских зимовках отмечена с августа по апрель (Гроте).
Прилет в СССР главным образом в апреле, в течение примерно месяца
(разновременный пролет разных гнездовых популяций). В сев. части области
распространения — со второй половины апреля и в начале мая (rta Камчатке в мае;
на Шантарских о-вах примерно в середине мая; в начале мая на Байкале;
у Архангельска в начале мая). На юге в первой половине апреля и даже в конце
марта. Осенью пролет проходит с конца августа и до начала ноября, но
главным образом в сентябре. Весенний пролет связывается по всей вероятности с
таянием льдов и вскрытием рек. Летят одиночками и парами.
Биотоп. См. выше, относительно вида. Водоемы, в которых скопа
ловит рыбу, должны быть достаточно прозрачными. В вертикальном
отношении в горах до лесной зоны; в Туркестане примерно до 1800 и быть может да
2500 м; в Закавказье в Армении до 2000 м, до таких же высот на Алтае.
Численность. Неравномерна. Распределение связано с нечасто
встречающимися условиями (прозрачные, богатые рыбой водоемы и
прибрежные высокоствольные леса), поэтому скопа многочисленна только местами
(например, в южн. Закавказье и сев. Иране). Обычно гнездовые пары
располагаются на большом расстоянии друг от друга.
22*
340
ОТРЯД ХИЩНЫЕ ПТИЦЫ
Экология. Размножение. Гнездовые участки весьма постоянны и
используются много лет (в одном случае, например, 18 лет, Рязанская обл.,
Хомяков, 1900). У пары повидимому только одно гнездо. Брачный полет
начинается с прилета (в конце апреля в Средней Азии, Северцов). За редкими
исключениями гнезда расположены на высоких деревьях; как правило гнезда
находятся на деревьях с отмершей вершиной (возможно в результате
долголетнего гнездования), — на лиственницах, соснах, березах, елях; обычно высоко от
земли, от 8—-10 и до 20—25 м. Средний диаметр гнезда около 1 м, высота 50—
70 см. Постройка из толстых
ветвей, с закругленным днищем и
неглубоким лотком. В редких случаях
гнезда находятся далеко от берегов
водоемов (до 2 и даже 3 км,
Баргузин, Туров, 1923). В очень
редких случаях гнезда на скалах
(озеро Гилли, Армения, Соснин и
Ляйстер, 1942).
Кладка из 2—3 яиц (по Север-
цову и из 4), в последних числах
апреля — начале мая.
Географические различия в сроках кладки
не установлены, но наличие их
весьма вероятно. Окраска яиц: по
бледному голубоватому фону
красновато-бурые и лиловато - бурые
пестрины разных размеров; фон
окраски иногда красноватый или
бурый в различных оттенках.
Размеры: 62,5—64x46—47,6 (Вост.
Сибирь, Тачановский, 1891); 63—
66 х 43—47 мм (низовья Волги,
Бостанжогло, 1911).
Насиживание начинается с
откладки первого яйца, при участии
Рис. 91. Скопа. обоих родителей (наседные пятна
и у самцов). Срок насиживания
несколько более месяца (35 дней, Нитхаммер, 1938; в СССР прямых
наблюдений нет). Полуоперенные птенцы встречаются около середины июля(Бараба;
Новгородская обл.; 15 июля — Воронежская обл.; 25 июля — нижняя Обь).
Летные молодые встречаются со второй половины августа. Гнездовый период
следовательно около 8 недель. Семьи держатся вместе до отлета.
Линька. Требует дополнительного изучения. Птицы, добытые между
октябрем и апрелем, в свежем пере. Смена первостепенных маховых идет со
второй половины периода размножения — в июле. Последовательность смены
маховых вероятно от заднего края партии к переднему. Смена нарядов:
первый пуховой — второй пуховой — первый годовой (гнездовый) — второй
годовой (окончательный) и т. д.
Питание. Стенофаг: кормится рыбой средней величины, весом
примерно до 2—3 кг (Аверин, 1910). Рыба ловится с полета, путем ныряния —
поэтому непрозрачность воды, дождливая или туманная погода препятствуют охоте
скопы. Весной, когда на разливах вода мутна, скопе приходится кормиться и
лягушками, сусликами, мышами, пеструшками, даже галками и утками и т. п.
(Зарудный, 1888, Бергман, 1935 и др.). В это же время скопа нападает на
подранков, например, на уток, даже на серебристую чайку (Печора, Дмоховский,
СКОПА 341
1935). Указания старых авторов, будто скопа иногда гибнет, вцепившись
в крупную рыбу и будучи увлеченной ею под воду, сомнительны. В связи со
способом охоты скопы находится ряд ее морфологических особенностей:
плотное и жесткое оперение, слабая оперенность голени и цевки, большое развитие
копчиковой железы, наличие поворотного пальца, шипики на подошвенной
поверхности пальцев, длинные и крутозагнутые когти и т. д. Крупная рыба
схватывается двумя лапами, мелкая одной.
Полевые признаки. Скопа в природе хорошо отличается двухцветной
окраской — темнобурой на спинной и белой на брюшной стороне тела. На полете
по размерам напоминает подорлика по величине и длинным и широким
крыльям, но более легкого сложения. Охотится над водой, останавливаясь и
«трясясь» с вытянутыми лапами над замеченной добычей и бросаясь за ней с
разлета в воду. Голос «кай-кай-кай», у гнезда иногда <ц<и-ки-ки». Отдыхает на
деревьях. Осторожна.
Описание. Размеры и строение. Крылья длинные, формула 3>2>4>5...
Вырезки на внутренних опахалах первых 3 первостепенных маховых. Перья на брюшной
стороне без побочного ствола. Хвост короткий, прямо-усеченный, из 12 рулевых. Клюв
вытянутый, с резко загибающимся и длинным крючком. Лапы сильные и толстые, цевка
короткая, без «штанов», покрытая толстыми щитками. Пальцы развиты более или менее
равномерно, с длинными острыми когтями, сильно выпуклыми на нижней поверхности, наружный
палец оборотный; нижняя поверхность пальцев покрыта острыми шипиками. Длина самцов
(5) 560—598, самок (4) 575—615, в среднем 574 и 595 мм. Размах самцов (4) 1470—1663,
самок (3) 1540—1683, в среднем 1590,5 и 1631,7 мм. Вес самцов (3) 1300, 1356, 1600, самки
(1) 1900 г. Крыло самцов (26) 448—492, самок (17) 485—515, в среднем 470 и 498,2 мм.
Окраска. Первый пуховой наряд серовато-белый, с охристой'продольной
окаймленной бурым и сероватым полосой на спине, с охристо-буроватыми крыльями и темным
пятном у глаз (Нитхаммер, 1938). Второй пуховой наряд темносерый с беловатой полосой на
спине, брюхо беловато-сероватое.
Первый годовой наряд на спинной стороне темнобурый с беловатыми каемками
перьев, особенно развитыми на плечевых и кроющих крыла; темя белое с бурыми на-
ствольями, перья затылка удлинены и образуют хохол; маховые темнобурые со светлыми
основаниями; рулевые темнобурые с черноватым поперечным рисунком и светлыми
поперечными полосами на внутренних опахалах боковых перьев; брюшная сторона белая
с более или менее развитым темным бурым рисунком на зобе. Второй годовой
(окончательный) наряд отличается д>т описанного тем, что на спинной стороне нет охристых каемок.
Оба пола окрашены одинаково. Радужина желтая, у молодых иногда буроватая; клюв
черно-роговой, синеватый у основания; когти черные; восковица и нога свинцово-серые
или зеленовато-серые.
ОТРЯД CODbl
STRIDES или STRICIFORMES
Характеристика отряда
Морфология. Внешние признаки. Совы — птицы чаще всего
средней величины, немногие виды — крупные или мелкие. Самые большие
совы нашей фауны —филины —имеют вес у самцов около 2,5 кг, у самок
3—3,25 кг, при размахе крыльев около 150—180 см. Средние размеры сов
таковы: у ушастой совы вес самцов около 250г, самок около 300 г; у болотной
совы соответственно около 300—325 и 350—400 г; у серой неясыти вес самцов
450—500, а самок около 600—685 г. Размах крыльев у ушастых сов 90—100,
у болотных 90—110, у серой неясыти 95—105 см: Самая маленькая сова
—воробьиный сыч — имеет вес у самцов 50—60 г, у самок 70—80 г, при размахе
крыльев 35—39 см.
Оперение сов мягкое и рыхлое, только у некоторых северных видов
(белая сова, ястребиная сова) относительно жесткое и плотно прилегающее
к телу. Перья покрывают обычно не только цевку, но и пальцы ног (за
редкими исключениями, например, у рыбных сов, неоперена только воско-
вица, прикрытая густыми и длинными вибриссами, обращенными вперед,
нижняя поверхность пальцев, иногда и внутренняя, задняя, поверхность цевки).
Окраска перьев неяркая, часто «ландшафтная», из смеси серых, бурых,
охристых и черноватых тонов. Полового диморфизма в окраске нет, как нет и
возрастных отличий между первым годовым и окончательным нарядом.
Оперение лица образует так называемый «лицевой диск» из рассученных
концентрически расположенных вокруг глаз перьев, отделенный от лобных,
ушных и горловых перьев короткими и плотными перышками. Маховые и
рулевые относительно мягкие. Число первостепенных маховых — 10 (не считая
рудиментарного, скрытого под кроющими), рулевых обычно 12.
Клюв короткий, круто загибающийся на надклювье, с крючком на конце
и острыми режущими краями. Пальцы ног с очень острыми круто загнутыми
когтями, служащими для схватывания и умерщвления (закалывания) добычи;
на внутренней поверхности пальцев на подошвах имеются подушечки с
жесткими сосочками, помогающими удерживать схваченную добычу. Лапы всегда
четырехпалые, при этом четвертый палец оборотный — он может
поворачиваться назад. Цевка относительно длинная.
ХАРАКТЕРИСТИКА ОТРЯДА
343
Глаза очень большие, обращенные вперед, с относительно большим полем
бинокулярного зрения (60%), но с ограниченным тотальным (160%). Самые
глаза неподвижные, из-за сращения склеротического кольца с черепом.
Неподвижность глаз, с одной стороны, компенсируется большой подвижностью шеи,
позволяющей голове вращаться до 270°, и чрезвычайной тонкостью слуха —
с другой.
Анатомические признаки. Череп десмогнатический, голо-
ринальный. Сошник обычно имеется. Базиптеригоидные отростки
функционально развиты. Шейных позвонков 14, шея умеренной длины. Грудные позвонки
свободны и не сливаются в os dorsale. Грудина — с большим килем, на заднем
ее крае имеются две, реже одна пара вырезок. Скелет конечностей напоминает
по строению скелет дневных хищных, однако пропорции отделов иные. В
скелете крыла самый длинный отдел — предплечье, плечо длинней кисти,
последняя — самый короткий отдел. В строении фаланг пальцев ноги также большое
сходство с дневными хищными птицами, основные фаланги 3-го и 4-го пальцев
короткие. Мощно развита мускулатура лап (сгибатели). Обводящей мышцы нет.
Мозг окципито-темпорального типа, относительно большой. Отверстия
наружного уха большие, часто окруженные кожистой складкой, с сильно
развитой системой двигательных мышц. Уши (и соответственно скелет черепа)
нередко асимметричны. Язык мясистый. Зоба нет. Функционирующие слепые
кишки.
Перья — без побочного ствола, с длинными пуховыми основаниями.
Концы бородок второго разряда удлинены и не сцеплены — отсюда мягкость
оперения и бесшумность полета. Пух есть только на крыловых птерилиях.
Копчиковая железа голая или почти голая. Нижняя гортань с одной парой мышц,
часто с хорошо развитой наружной голосовой мембраной (от этого —
разнообразие издаваемых совами звуков).
Экология. Большинство сов ведет ночной или сумеречный образ жизни,
но виды, обитающие на севере, активны неизбежно и днем. Распространены
совы повсеместно, но все же большая часть видов в той или иной мере связана
с древесной растительностью.
Чаще всего совы оседлы или полуоседлы; некоторые совершают
нерегулярные откочевки в связи с наступлением благоприятных или
неблагоприятных, главным образом, кормовых условий; немногие перелетны (у н^с,
например, сплюшка, иглоногая сова и др.).
Пища сов —животная, при этом совы едят свежепойманную добычу.
В отношении кормового режима^ главное место среди сов занимают миофаги
(питающиеся мелкими млекопитающими, в первую очередь грызунами);
довольно много энтомофагов и полифагов. Как редкая специализация встречаются
ихтиофаги (род рыбные совы).
В отношении кормовой специализации и способов нападения на добычу
среди сов наблюдается мало разнообразия. Совы разыскивают добычу,
пользуясь главным образом слухом, но отчасти и зрением
Охотятся или облетая охотничий участок (относительно небольшой) по
строго определенному маршруту, или сидя на земле или на дереве и подстерегая
добычу.
Рыбные совы хватают рыбу и ракообразных (крабы), заходя «пешком»
в воду на мелких местах. Добыча схватывается и умерщвляется лапой, при этом
когтевые фаланги вдавливаются предпоследней фалангой в тело добычи, таким
образом повреждая ее внутренние органы. За исключением относительно
крупных зверей и птиц, совы не измельчают добычи клювом, а заглатывают ее
целиком. При этом — в отличие от дневных хищников — у сов в кишечном тракте
кости не перевариваются, и сбрасывание погадок у них носит более правильный
характер, чем у дневных хищных птиц.
344
ОТРЯД СОВЫ
% Весьма характерно для сов влияние кормовых условий на размножение.
При благоприятных условиях — в годы массового размножения мышевидных
грызунов, например леммингов на севере, — совы усиленно размножаются,
у них возрастает и число яиц в кладке и даже бывают повторные кладки
поздней осенью. Кроме того, в этих случаях смертность птенцов в выводках
невысока. В противоположных случаях совы с трудом переходят на питание
другими кормами, плодовитость снижается как за счет уменьшения числа яиц
в кладке, так и за счет повышенной смертности молодых (при подобных
обстоятельствах младших птенцов иногда съедают старшие птенцы в выводке или
даже родители). Кроме того, в особенности на севере, наблюдаются и случаи
негнездования сов при отсутствии мышевидных грызунов (у белой совы,
бородатой неясыти и др.), и массовые откочевки из гнездовой области. Наконец,
и у старых птиц смертность в холодном сезоне в такие «плохие» годы
повышается.
Таким образом, численность многих сов подвержена колебаниям,
параллельным с колебаниями численности основных видов их кормов.
Совы — моногамы, гнездятся отдельными парами. Пары, повидимому,
постоянны. %
Незначительность возрастных изменений позволяет предполагать, что
совы становятся способными к размножению в возрасте около года, на
первую весну после года рождения.
Места гнездовий разнообразны, но расположены гнезда —за редкими
исключениями — закрыто, в дуплах, пещерах и щелях, норах. Однако
некоторые совы кладут яйца прямо на землю, а другие используют гнезда,
устроенные на деревьях иными видами птиц. Своих гнезд совы не строят
(исключением в нашей фауне служит повидимому только болотная сова). Насиживает
как правило одна-самка.
Срок насиживания у разных видов примерно от 25 до 35 дней. Начало
насиживания с откладки первого яйца, птенцы поэтому разновозрастны. Яйца,
как правило, белые, закругленной формы.
Число яиц в кладке варьирует и по годам-(как указано выше), и у
разных видов. Крупные кладки — у видов севера — у белой совы от 3—5 до 8—9
и даже 11 яиц; у мохноногого сыча 4—6, до 7—8 и 10 яиц; у воробьиного сыча
4—6 и, до 7 яиц; у ястребиной совы 4, но и до 13 яиц. У крупных видов —
филинов и рыбных сов — в тропиках и субтропиках — число яиц 1 —2.
Среднее число яиц в кладках сов 4—5.
Тип развития молодых — птенцовый. Пвенцы вылупляются с закрытыми
глазами, без автономной температуры тела. Постэмбриональное развитие
медленное.
После первого пухового наряда надевается так называемый мезоптиль,
состоящий из мягких полупуховых перьевк кроме окончательных
маховых и рулевых (исключение—сипухи, у которых, как у дневных хищников,
имеется два пуховых наряда). В мезоптиле молодые покидают гнездо,
а затем становятся на крыло.
Гнездовый период продолжается примерно от 3 недель (у мелких видов,
как например сплюшек) до 6 недель (у крупных).
В первую осень мягкие перья мезоптиля сменяются на контурные
обычного типа, окрашенные как у старых птиц. Возрастные изменения на этом и
ограничиваются. Этот первоосенний комбинированный наряд (из новых
контурных перьев и оставшихся от мезоптиля маховых и рулевых) сменяется
полностью после первой годовой линьки во второй половине лета — осенью
следующего за рождением года жизни. В дальнейшем в то же время бывают
полные годовые линьки. Линяние в общем продолжительное, что
обеспечивает общее сохранение в период линьки летательных способностей на вы-
ХАРАКТЕРИСТИКА ОТРЯДА
345
соком уровне. Длительность линяния у крупных видов более 4 месяцев,
у мелких — около 3 месяцев.
Половой диморфизм у сов выражен в размерах; при этом в отличие от
большинства других групп птиц, но как у дневных хищников, самки
превосходят величиной самцов. Окраска обоих полов сходна.
Продолжительность жизни у сов относительно велика. В отношении
крупных видов- (филины), содержавшихся в неволе, установлено, что они жили
53 и 68 лет.
Систематика. Современные совы Делятся на два семейства: сипухи
Tytonidae и настоящие совы -^Strlgtdae.
У Tytonidae череп узкий и длинный; орбиты небольшие; вилочка непнев-
матизированная; задний край грудины с одной парой вырезок; клюв
вытянутый и относительно слабый; копчиковая железа оперена; уши симметричны;
второй и третий пальцы ног развиты одинаково; коготь среднего пальца
зазубрен на внутреннем крае; цевка длинная. В постэмбриональном развитии — два
пуховых наряда (как у дневных хищных), но нет мезоптиля. Яйца
удлиненные. Семейство имеет два рода Tyto с 10 видами, распространенными в
жарком и умеренном поясе обоих полушарий, и Phodilus с одним видом в
ю.-в. Азии.
У Strigidae череп закругленный; орбиты большие; вилочка пневматична;
на заднем крае грудины две пары вырезок; клюв сильный, короткий и круто
загнутый; копчиковая железа голая, уши нередко асимметричны, причем эта
асимметрия распространяется и на кости черепа; третий палец длинней
второго, режущие края когтя среднего пальца гладкие; цевка короткая. В
постэмбриональном развитии — один пуховой наряд, второй пуховой наряд заменен
мезоптилем. Яйца округленные. Семейство, распадается на 28 родов (Otus*
Pyrrhoglaux, Mimizuka, Jubula, Lophostrixy Bubo, Pseudoptynx, Ketupa, Scoto-
pella, Pulsatrix, Novipulsatrix, Nyctea, Surnia, Glaucldlum, Micrathene, Urog-
laux, Ninox, Gymnoglaux, Sceloglaux, Athene, Speotyto, Ciccaba, Nesaslo, Aego-
lius> Pseudoscops, Asio, Rhinoptynx, Strix) с 123 видами. Распространено оно
по всему свету.
Ископаемые совы известны с эоцена (из штата Уайоминг). Рецентные роды
найдены уже в олигоцене (Bubo, Aslo, Strix).
Сов обычно сближали с ракшеобразными птицами и в частности с
козодоями. Это однако неосновательно. Многие особенности сов сходны с
хищными птицами, а некоторые черты, общие с козодоями, явно конвергентны.
Правильней всего считать сов за отдельный отряд, притом резко диференциро-
ванный.
Географическое распространение. Совы распространены по всему земному
шару, за исключением некоторых океанических островов и Антарктики.
Наибольшее число видов — в жарком поясе восточного полушария, где, быть
может, и лежит центр возникновения и распространения отряда. Общее число
видов сов в фауне СССР 28; подвидов и монотипических, т. е. не образующих
подвидов, видов 46. Из этого числа 1 вид (в двух подвидах) относится к
семейству сипух, а 16 видов и 44 формы к семейству настоящих сов. Число видов*
сов фауны СССР от общего числа видов отряда составляет 12,68%.
Распределение сов в фауне СССР по жизненным зонам таково (в числителе
число видов, в знаменателе — подвидов и монотипических видов; характерные
формы подчеркнуты):
I. Арктика (зона). Nyctea scandiaca, Asio flammeus 2
а. Побережье (подзона) N. scandiaca 1/1
в! ^Х^тТндТа (подзона) } "• *™««* A' /• "<»~ ^
346
ОТРЯД СОВЫ
II. Лесная зона. Bubo bubo, Ketupa zeylonensis, Otus bakkamoena, O. scops, O. sunia,
Asio otus, Ninox scutulata, Aegolius fanereus, Glaucidium passerinum, Surnia ulula, Strix
nebulosa, S. uralensis, S. aluco 14
г. Тайга (подзона) В. b. bubo, ruthenus, sibiricus, yenisseensis, yakutensis, ussu-
riensis; K- z. doerrisi; A. o. otus; A. /. finereus, sibiricus, magnus; G. p. passerinum, orientate;
S. u. ulula; S. n. lapponica; S. u. liturata, uralensis, yenisseensis, buturlini, nikolskii;
S. a. siberiae 9/2Г
д. Смешанный и лиственный лес (подзона) В. b. bubo, ruthenus; О. s.
scops, pulchellus; О. s. stictonotus; A. o. otus; N. s. macroptera; S. a. aluco 6/8
е. Лесостепь (подзона) В. b. interpositus; О. s. scops, pulchellus; A. o. otus;
A. n. noctua, indigena; S. u. ulula; T. a. guttata (культурный ландшафт) 6/8
III. Зона открытых сухих ландшафтов Bubo bubo, Otus brucei, Asio flammeus, Athene
noctua 4
ж. Степь (подзона) В. b. interpositus, ussuriensis; A. /. flammeus; А. л. noctua,
indigena, plumipes 4/7
а. Полупустыни и пустыни (подзона) В. b. turcomanus, omissus; 0. brucei;
А. л. bactriana 3/4
IV. Горная зона Bubo bubo, Otus scops, Asio otus, Athene noctua, Surnia ulula, Strix
uralensis, S. aluco 8
и. Высокогорная бореоальпийекая подзона О
к. Горно-лесная подзона- В. b. interpositus, yenisseensis; О, s. pulchellus,
А. о. otus; A. /. caucasicus, pallens; S. u. tianschanica; S. u. yenisseensis, buturlini (?);•
S. a. wilkonskii, harmsi 7/12
л. Горно-степная и пустынная подзон а В. b. omissus, hemachalanus,
А. п. bactriana, orientalis *Ш
Из приведенных списков видно, несмотря на их неизбежную
схематичность, преобладание среди сов нашей фауны дендрофильных видов;
своеобразие и высокий эндемизм таежной фауны сов, зональный характер проявлений
географического изоморфизма. у
По характеру j пребывания, как уже говорилось, большинство сов
оседлы.
Хозяйственное значение. Как и относительно дневных
хищных птиц, можно отметить, что экологические исследования последних
лет ясно показали всю несостоятельность бывших широко распространенными
в прошлом столетии и в начале текущего взглядов о вреде так называемых
хищников. Совы весьма полезны, в первую очередь как истребители вредных
грызунов. В отношении крупных видов, кормящихся*и более значительной по.
размерам добычей, в частности охотничьими животными (например зайцами),
надо все же иметь в виду ряд обстоятельств. Во-первых, кормовой режим
этих сов, например филина, разнообразен и в значительной мере состоит из
видов нежелательных или вредных. Во-вторых, численность крупных видов
сов невысока, и они тем самым не могут реально влиять на сокращение
популяций своей добычи. Надо, далее, иметь в виду отмеченное выше
саморегулирование численности сов в зависимости от колебаний численности
основных видов кормов. Те же виды сов, которые у нас многочисленны, —.
по кормовому режиму главным образом миофаги — и поэтому заслуживают
охраны.
Таблица для определения видов сов
1(2). Лицевой диск резко суживается книзу и имеет сердцеобразную форму; средний
и внутренний пальцы приблизительно равны по длине; внутренний край когтя среднего
пальца в зазубринах; брюшная сторона в мелких глазчатых крапинах
сипуха Tyto alba
ХАРАКТЕРИСТИКА ОТРЯДА
347
2(1). Лицевой диск более или менее округленный; средний палец заметно длинней
внутреннего; внутренний край когтя среднего пальца без зазубрин; брюшная сторона в
полосах или пятнах 3
3(4), Крыло длинней 350 мм . ♦ 5
4(3). Крыло короче 350 мм 9
5(6). Общая окраска белая с более или менее крупными бурыми пестринами; нижние
кроющие хвоста доходят до конца рулевых перьев; «ушки» незаметны или мало заметны. . . .
белаясова Nyctea scandiaca
6(5). Окраска иная; нижние кроющие хвоста не доходят до вершины рулевых; «ушки»
хорошо заметны 7
7(8). Пальцы ног оперены; край наружных опахал передних маховых зазубрен; плечи
в резких черноватых пестринах; самое длинное маховое — третье
филин Bubo bubo
8(7). Пальцы ног не оперены; край наружных опахал передних маховых без зазубрин;
темный рисунок на плечах нерезкий; самое длинное маховое четвертое или даже пятое. . • .
рыбнаясова Ketupa zeylonensis
9(10). По сторонам темени заметны удлиненные пучки перьев («ушки») 1
10(9). По сторонам темени удлиненных пучков перьев нет 21
11(12). Крыло длинней 200 мм 13
12(11). Крыло короче 150 мм .15
13(14). «Ушки» длинные; оперение зоба и груди с поперечным рисунком
ушастая сова Asio otus
14(13). «Ушки» короткие; оперение зоба и груди без поперечного рисунка. . . . . .
болотнаясова Asio flammeus
15(16). Пальцы оперены в основной трети; на зашейке охристое или беловатое пятно;
первое маховое короче восьмого
ошейниковаясовка Otus bakkamaena
16(15). Пальцы не оперены или оперены только у самого основания; на зашейке нет
охристого или беловатого пятна; первое маховое длинней восьмого 17
17(18). Нижняя часть цевки голая; первое маховое короче седьмого, второе короче
лятого (изредка равно ему по длине)
южноазиатскаясовка Otus sunia
18(17). Нижняя часть цевки оперена; первое маховое длиннее седьмого; второе маховое
длинней пятого 19
19(20). Малые кроющие крыла без ржавчатого оттенка; настволья перьев резко
выделяются; первое маховое короче шестого пустыннаясовка Otus brucei
20(19). Малые кроющие крыла с ржавчатым оттенком; темные настволья перьев
выделяются нерезко; первое маховое длинней шестого
.-• обыкновенная совка Otus scops
21(22). Хвост резко ступенчатый и длинный (приблизительно8Д длины крыла); грудь,
<юка и брюхо с правильными темными поперечными полосами
ястребинаясова Surnia ulula
22(21). Строение хвоста и рисунок оперения груди, боков и брюха иные 23
23(24). Крыло длинней 250 мм 25
24(23). Крыло короче 250 мм 29
25(26). Крыло длинней 400 мм; темный рисунок лицевого диска из тонких правильных
концентрических кругов бородатая неясыть Strix nebulosa
26(25). Крыло короче 400 мм; рисунок лицевого диска иной . 27
27(28). Брюшная сторона тела с темным продольным рисунком; хвост длинней 225 мм;
четвертое маховое короче пятого
длиннохвостая неясыть Strix uralensis
28(27). Брюшная сторона тела с темным продольным и поперечным рисунком; хвост
короче 225 мм; четвертое маховое длинней пятого .
обыкновенная неясыть Strix aluco
29(30). Голова, задняя часть шеи и спина однообразно бурой окраски без пестрин,
«пальцы покрыты щетинками иглоногаясова Ninox scutulata
30(29). Голова иной окраски; пальцы покрыты перышками 31
31(32). Крыло короче Ш мм
воробьиныйсыч Glaucidium passerinum
32 (31). Крыло длинней 100 мм 33
33(34). Хвост длинный, равный примерно 2/з длины крыла; наружное опахало
четвертого махового без вырезки; ушные отверстия несимметричны
мохноногий сыч Aegolius funereus
34(33). Хвост короткий, равный примерно 7* длины крыла; наружное опахало
четвертого махового с вырезкой; ушные отверстия симметричны
домов ыйсыч Athene noctua
348
ОТРЯД СОВЫ
СЕМЕЙСТВО НАСТОЯЩИЕ СОВЫ STRIGIDAE
РОД БЕЛЫЕ СОВЫ NYCTEA, STEPHENS, 1826
Тип Strix nyctea
1. Белая или полярная сова Nyctea scandiaca Linn
Strix Scandiaca, Linnaeus. Sys. Nat. изд. X, 1758, стр. 92, горы Лапландии.
Русское название. Белая сова — книжное. У русских на севере «лунь»
или «улунь», общее для многих светлоокрашенных, белесых хищных птиц. В Якутии
народное название — «снежная бабушка».
Распространение. Ареал. Гнездится кругополярно на островах
Северного Ледовитого океана и в тундре. На севере в Гренландии (обычно у Ско-
ресби-зунда и о-ва Шаннон на востоке, у бухты Кена, м. Септон и пролива
Робсона на западе (быть может и северней, у Сенк-Год-харбор), на Земле
Гриннеля (82°33* с. ш.), на Новой Земле, кроме крайнего ее севера; на о-вах
Карта 56. Распространение белой совы Nyctea scandiaca
1 — граница гнездовий, г — область зимовок и кочевок.
Вайгач и Колгуев (?); на Новосибирских о-вах; на о-ве Врангеля; к югу
до южных границ тундры, в Скандинавии до Кристианзанда в Норвегии,
до Херьедален в Швеции, в Финляндии в северной горной части страны; в
Восточной Сибири до северных частей Коряцкой Земли и о-ва Беринга; в
Америке до Квебека, с.-в. Манитоба, с.-з. Кьюатина, с.-з. Мекензи и в Аляске;
гнездование на Земле Франца-Иосифа, Шпицбергене и Ян-майене, равно
как и на Камчатке и Сахалине —не доказано; возможно, что гнездится и
в Приямской тундре на северном побережье Охотского моря. Сведения старых
авторов (Эверсманн, 1866, Сабанеев, 1874, Зарудный, 1884) о гнездовании, хотя
бы случайном, к югу от полосы тундры не заслуживают внимания.
Достоверные сведения о встречах белых сов летом к югу от обычной гнездовой их
области имеются, но они не доказывают гнездования (окрестности Москвы, июль
1867; б. Халтуринский уезд Кировской обл., 12 июля; окрестности Молотова,
23 июля; устье Великой, Псков, 10 июня; Ленинградская обл., конец июля;
Нарым, Зап. Сибирь, 17 июля; озеро Кулусунах, Якутия, 15 июля и т. д.).
Характер пребывания. Белая сова отчасти оседлая, но
преимущественно кочующая птица. Кочевкинерегулярны, в различном количестве
и на разные расстояния, в зависимости от местных условий снегового покрова,
кормовых и т. п. В зимнее время отмечена на Анадыре (Портенко, 1939), в
низовьях Индигирки и Колымы, между Анабарой и Хатангой (экз. от
15. I. 1927), на Новой Земле (Пахтусовым, затем Горбуновым, 1929 и др.),
БЕЛАЯ ИЛИ ПОЛЯРНАЯ СОВА
349
на Канине, на Кольском п-ове и т. д. Оседлость белой совы на севере
связана с обилием леммингов (в обычные годы она, например, отлетает на зиму
с о-ва Врангеля, но в 1931—1932 гг. зимовала, по сведениям Минеева; тоже
на Кольском п-ове, Плеске, 1884). Обычно большинство (или все?) сов
улетает с крайнего севера на зиму, оседая в открытых ландшафтах —
лесотундре, в степной полосе, на юге в Вост. Европе и в Сибири до 50° с. ш.
Регулярные зимовки в степях Поволжья, Зап. Сибири и Казахстана, в ю.-в.
Забайкалье и Приамурье, в Манчжурии и сев. Японии.
Отдельные особи встречены и значительно южнее — до сев. Крыма
(Перекоп, Паллас, 1811), Сев. Кавказа, Каспийского моря (Мангышлак), о-ва
Тюленьего, Туркмении (Фараб, Артык), Семиречья (Верный, 22. II. 1886, Фрунзе,
1926—1927). Однажды—в сев. Индии (Мардан, Пенджаб) и в Сев. Китае
(Чжили). Кроме того, вне СССР — до Франции, Бельгии, Голландии, Англии,
Дании, Германии, Австрии, Венгрии, один раз — в Югославии (Хорватия)
и на Азорских о-вах. В Сев. Америке до Бермудских о-вов, Георгии, Техаса
и Калифорнии.
Кочевки иногда принимают характер массовых. Годы значительных
налетов белых сов в Европейскую часть СССР: 1889—1899, 1911—1912, 1915—1916,
1919—1920, 1922—1923; в особенности 1926—1927, когда область миграций
захватила территорию от Украины до Семиречья, 1932—1933, 1935—1936.
В конце прошлого столетия особенно большая кочевка белых сов наблюдалась
в 1895—1896 гг. (Украина, Сомов, 1897, Башкирия, Сушкин, 1894). Кочевки
носят местный характер и обычно не захватывают всей области распространения
(например, нет совпадений между массовыми появлениями в умеренной полосе
Европы и Сев. Америке; в последней значительные кочевки белых сов были
в 1917—1918, 1926—1927,1930—1932, 1934—1935, 1941—1942, по Шелфорду,
1945). Это явление отражает неравномерность колебаний численности
леммингов в разных частях гнездового ареала совы.
Даты. На зимовках белые совы обычно появляются в октябре и остаются
до апреля, в редких случаях ранее или позднее этих дат (в сентябре и до
первых чисел мая). Отлет на юг из тундры в разные числа сентября и по ноябрь
растянут; прилет также растягивается между апрелем и маем (например на
о-ве Врангеля 1 мая, на Новосибирских о-вах 26 марта — 11 апреля, на
Таймыре 17 апреля, на Хатанге в конце апреля, в Пясинской тундре с конца
марта, на Ямале в мае, в Тиманской тундре 10 апреля).
Биотоп. Тундра, преимущественно — на гнездовье —
водораздельная, как на равнине, так и вторах (Лапландия, Вост. Сибирь и т. п.). На
кочевках в лесотундре и открытых безлесных ландшафтах—побережья, болота,
степь, поля и т. п.
Численность. Колеблется по годам, повидимому, в первую
очередь в зависимости от кормовых условий. Падение численности происходит
на следующий или следующие после вымирания леммингов годы, как в
результате возрастания смертности, отчасти в связи с массовыми миграциями, так
и из-за уменьшения интенсивности размножения или даже его отсутствия
в особо неблагоприятные годы (например на Новой Земле в 1899, по Пирсону,
не было леммингов и было очень мало сов; в Тиманской тундре летом 1938 г.
не было леммингов, совы не гнездились и держались по одиночке; на
восточном Таймыре по тем же причинам совы, повидимому, не гнездились в 1933 г.,
на севере Сибири в местах работы русской Полярной экспедиции это
наблюдалось в 1902 г., Бируля, 1907). В благополучные же годы совы гнездятся
плотно (на о-ве Врангеля по Минееву, в 2—3 км пара от пары) и дают
большой приплод.
Экология. Размножение. Неравномерное по годам (см. выше),
как у большинства видов мышеядных хищников. Возможно, что в годы изоби-
350
ОТРЯД СОВЫ
лия леммингов имеет место и известное расширение ареала в южном
направлении (например на Ямале совы иногда гнездятся к югу от 69° с. ш.).
Постоянство пар(?) Половая зрелость(?) (вероятно, потенциальная на
первую весну после вывода, в возрасте менее года).
Гнезда расположены как в высокой, так и в низкой тундре, но
предпочтение оказывается высоким холмам и сухому грунту, так как птица
начинает кладку, когда местность обычно еще покрыта снегом. Гнезда
используются несколько лет, но не каждый год подряд. Собственно гнезда нет,
яйца кладутся в ямку на земле. Спаривание на Новой Земле отмечено 13 мая
(Сконнинг, 1903).
Кладка происходит около середины мая (18 мая, Новая Земля; 9 мая,
низовья Лены; последняя неделя мая, Новосибирские о-ва и т. д.). Кладки
варьируют от 3—5 до 8—9 яиц, даже до 11 (для Новой Земли от 4 до 9 яиц,
для Хатанги до 9 яиц, Ямала 4—8 и до 10 яиц; Сагастырь, дельта Лены, зап.
Таймыр 3—-11, низовья Колымы 3—10, о. Врангеля до 10). Промежуток между
откладкой яиц, судя по большей, до 2 недель, разновозрастности птенцов,
значительный (на Новой Земле 7 яиц за 14 дней, Сконнинг, 1903). Насиживание
с первого яйца. Срок насиживания немного более месяца, между 32 и 34 днями,
но Сконнинг (1903), указывает только 27 дней.
Яйца округлые, белые или охристо-белые. Размеры 55,2—70,2 х 42—49,3 мм
(Сконнинг, 1903), 53,7—62 х 43,5— 47,5 мм (Плеске, 1928).
Насиживает самка, самец носит ей, а затем и выводку, добычу; но в
случае гибели самки насиживать продолжает самец (Таймыр). Гнездо защищается
совами (самцом) очень активно. Птенцы вылупляются в конце июня (старшие)
и в начале июля (младшие).
Гнездовый период продолжается около месяца, отмечены подлетки в ме-
зоптиле с конца июля — начала августа (Канин), вполне выросшие молодые —
в разные числа августа. Значительная разновозрастность птенцов — старшие
оперяются, когда младшие только вылупляются — связана быть может с тем,
что при больших выводках прокормление их требует частого отсутствия обоих
родителей (по Гизенко, при птенцах в возрасте нескольких дней старые совы
улетают уже за 5 и даже 10 км); насиживание последних яиц носит весьма
нерегулярный характер и заменяется обогреванием их старшими птенцами.
Только первые дни птенцов кормит самка, вскоре же на охоту летают оба
родителя. Птенцы разбредаются из гнезда и держатся близ него задолго еще
до приобретения способности к полету. Смертность птенцов значительна. О
колебании ее, негнездовании и связанных с этим изменениях численности
см. выше*
Линька. Полная годовая. Последовательность смены первостепенных
маховых — от задних к передним. Взрослые птицы линяют со второй половины
периода размножения, между концом июня или началом июля и поздней
осенью; совы в свежем наряде в октябре —начале ноября. Первое осеннее
оперение молодых надевается в конце сентября — начале октября.
Последовательность смены нарядов: первый пуховой наряд — мезоптиль — первый годовой
наряд — второй годовой наряд и т. д. При этом первый годовой наряд —
комбинированный, так как в нем сохраняются от мезоптиля маховые и рулевые.
Питание. Пищу белой совы в гнездовое время составляют главным
образом мышевидные грызуны и в первую очередь норвежский, обский и
копытный лемминги. «Урожаи» и «неурожаи» леммингов определяют ход
основных периодических явлений в жизни белых сов — размножения, сезонного
размещения и т. п. (см. выше). На о. Беринга отмечена зависимость увеличения
численности сов от размножения завезенных туда рыжих полевок Clethriono-
mys rutilus и домовых мышей (Стейнегер, 1887). В нормальные годы — мелкие
грызуны (кроме леммингов — полевка Миддендорфа, большая узкочерепная
БЕЛАЯ ИЛИ ПОЛЯРНАЯ СОВА
35 Г
полевка и др., Осмоловская, Ямал; суслики на Анадыре, Портенко, 1939),
в период выкармливания птенцов — молодые птицы, в особенности белые ку~
-*в§
Рис. 92. Белая сова (взрослая, молодая, птенец).
ропатки, также кулики, например куличок-воробей, воробьиные —.
лапландский подорожник и др., разные чистики, молодые чайки и гагачата (Анадырь,
Белопольский, 1932). В неблагоприятные для сов по численности грызунов
352
ОТРЯД СОВЫ
годы — птицы, в особенности белые куропатки, имеют в кормовом режиме сов
большее значение.
В негнездовое время пища более разнообразна — зайцы беляки, пищухи
(Радде, 1863), прочие грызуны; горностай, фазаны, тетерева, утки величиной
до кряквы; разные мелкие птицы (подорожники). Отмечалось совпадение
массовых кочевок белых куропаток и сов в ю.-з. Сибири (Тара, 1917, Ушаков)
и в сев. Якутии (Колыма).
Охотится белая сова, главным образом, сидя на земле, предпочтительно
на возвышении, и бросаясь на приближающуюся добычу. В сумерках иногда
охотится и на лету, трепеща при этом на одном месте в воздухе, подобно
пустельге. Хотя белая сова и не строго ночная птица, все же охотничьи вылеты
обычно падают на раннее утро или вечер.
Полевые признаки. Крупная сова, величиной почти с филина; голова
круглая, небольшая, щетинистые перья у основания клюва почти полностью
его прикрывают; глаза небольшие, оперение лап густое и длинное, почти
полностью закрывающее когти; крылья относительно короткие и широкие, хвост
довольно длинный; окраска белая или белая с бурыми крапинами. Держится
обычно на земле, неохотно присаживаясь на деревья. Полет быстрый,
напоминающий дневных хищных птиц. Довольно осторожна. Крик — громкое и
резкое «кик-кик-кик>, а также звук, напоминающий карканье.
Описание* Размеры и строение. На голове небольшие «ушки», часто
незаметные, каждое состоит из 10—12 коротких перышек; лицевые перья и оперение ног очень
длинные и густые, лицевой диск выражен относительно нерезко; перья подхвостья очень
длинны и почти одинаковой длины с рулевыми. Формула крыла: 4>3>5>2^6=7
1-е маховое рудиментарно и прикрыто кроющими. 3-е, 4-е и 5-е с резкими, б-е — со слабыми
вырезками на наружном опахале. Хвост закругленный, из 12 рулевых. Клюв резко загнутый
и сжатый с боков. Когти очень острые и длинные, обычно 3 пальца обращены вперед, а
первый назад. Длина самцов (5) 560—610, размах (5) 1485—1540, в среднем 579,6 и 1494 мм.
Самка крупней: длина (1) 650, размах (1) 1610 мм. Вес самцов (5) 1350, 1530, 1600, 1595,
2000 г; самок (3) 1770, 2210, 2480 г; в среднем 1615 и 2153,3 г. Крыло самцов (86) 384—423;
самок (63) 428—462, в среднем 405,4 и 437,9 мм. Географических различий в размерах нет.
Окраска. Пуховой наряд — белый. Мезоптиль- бурый с беловатыми вершинами
(остатки пухового наряда) на брюшной стороне; спина и плечевые более жесткие, бурые
с беловатыми полосами; лицевой диск беловатый; маховые и рулевые белые с бурыми
поперечными полосами и бурым мраморным крапом. Первый годовой нар яд самок белый с широкими
бурыми поперечными полосами, занимающими на плечах, спине и крыльях примерно столько
же места, как и белый цвет; кроющие кисти с мелким бурым крапом; на голове крупные
белые пятна. У самцов бурый рисунок занимает меньше места, чему самок, и носит характер
тонких поперечных полос. Рулевые и первостепенные маховые в первом годовом наряде
остаются от мезоптиля и сменяются только при линянии во второй годовой наряд. Второй
годовой (окончательный) наряд снежно-белый, у самцов с незначительной примесью бурых
пестрин, иногда только у вершин маховых и на затылке, иногда же в виде тонких поперечных
полос или поперечно вытянутых пятен на плечевых, кроющих крыла, брюхе, маховых,
рулевых; между этими крайностями — промежуточные типы окраски. У самок окраска белая
с более или менее широким бурым поперечным рисунком на брюхе, боках, верхней стороне
тела, маховых и рулевых. Оперение лица, горла, подкрылья и ноги всегда белые. Радужина
золотисто-желтая, реже лимонно-желтая; клюв и когти черные.
РОД ФИЛИНЫ BUBO D U М Ё R I L, 1806
Тип Bubo bubo
2. Филин Bubo bubo Linn
Strix Bubo. Linnaeus. Syst. Nat. изд. X, 1758, стр. 92; Швеция.
Русское название. Филин — народное и охотничье, в литературе известно по
крайней мере с XVIII века («Совершенный Егерь» Левшина).
Распространение. Ареал. Европа и сев. Азия, к северу до границы
леса, на восток до Якутии, Охотского побережья, Сахалина, Японии; на юг
до Средиземного моря, Крыма, Кавказа, Аравии, Палестины, Месопотамии,
ФИЛИН
353
Южн. Китая; палеарктическая Африка от Марокко, Египта, Синая до
Тимбукту, Аира и Судана.
Характер пребывания. Филин — оседлая, отчасти
кочующая птица.
Биотоп. В биотопическом отношении типичный убиквист,
встречающийся в лесах, степях, пустынях, в равнинах ниже уровня моря и в горах на
высотах выше 3000 м в Тянь-Шане и даже 4700 м в Тибете.
Карта 57. Распространение филина Bubo bubo
/—В. b. ascilaphus, 2— В. b. dssertorum, 3—В. b. hispinus, 4— B. b. bubo, 5 — B. b. ruthenus, 6 —
B. b. intjrpositus, 7 — B. b. sibiricus, 8 — B. b. yenisseensis, 9 — B. b. yakutensis, 10— B. b. ussuiien-
sis, 11 — B. b. turcomanjs, 12 — B. b. omissus, 13 — B. b. nikolskii, 14 — B. b. hemachalanus, 15 — B. b.
tenuipes, 76 — B. b. kiautschensis, 17 — B. b. swinhoei; a — граница гнездовий.
Численность относительно высокая, колеблющаяся по годам
в*результате «неразмножения» или высокой смертности молодых в
неблагоприятные по кормовым (климатическим?) условиям годы. В густо населенных
местностях Европы становится редким, местами исчез.
Подвиды и варьирующие признаки. Морфологически варьирует окраска и размеры,
оперенность пальцев; экологически — детали цикла периодических явлений и трофические
связи. В сев. Африке от Марокко до Египта, на Синайском п-ове, в южн. Палестине
и Сирии В. b. ascalaphus S a v i g n у, 1809; в Сахаре к югу от предыдущего, от Туниса до
Нила, к югу до Тимбукту и Судана, в южн. Аравии В. b. desertorum Е г I a n g e г, 1897;
в зап. части европейского ареала от Скандинавии и Пиренеев до средней России и Балкан
В. b. bubo L., 1758; на Пиренейском п-ове В. b. hispanus Rothschild Hartert,
1910; в южн. России и вост. России на восток примерно до Урала В. b. ruthenus Z h i t-
kov et Buturlin, 1906; на Украине, в Крыму, в Малой Азии и на Кавказе, в Сирии
В. b. interpositus R о t s с h. et Hart., 1910; в Зап. Сибири от Зауралья до Томска
и зап. предгорий Алтая В. b. sibiricus G lo ge г, 1883; в средней Сибири от Енисея
и Алтая до зап. Забайкалья В. b. yenisseensis But., 1911; в ю.-в. Забайкалье и Приморье
В. b. ussuriensis Р о 1 j a k о w, 1910; в Якутии от Лены до Охотского моря, на Сахалине
В. b. jakutensis But., 1908; в области низовьев Урала и сев. Казахстана, к югу до Усть-
Урта В. b. turcomanus Eversmann, 1835; в горах Средней Азии на Памире и в Тянь-
Шане, в Тибете и Гималаях В. b. hemachalanus Hume, 1873; в зап. Таджикистане и Туркме-
23 Птицы СССР, т. I
ОТРЯД СОВЫ
нии В. b. omissus Dementiev, 1932; в Луристане и Арабистане, Иран В. b. nikolskii
Za ru d ny, 1905; в Манчжурии и Сев. Китае, Японии (приводится для Хоккайдо, Хондо,
Амамиосима, кроме того для южн. Курильских о-вов) В. b. tenuipes Clark, 1907
(вероятный синоним В, b. inexpectatus В a ngs); в центральном и вост. Китае от Сычуаня и
Хубея до Аньхоя, Шань-Дуна и Цзяньсу В. b. kiautschuensis Reichenow, 1903 (к этой
же форме вероятно и юннаньские филины, описанные под названием B.b. jarlandi La t о u-
che); в ю.-в. Китае — Цзяньси, Чжецзяне, Фуцзяне, Гуань-Дуне В. b. swinhoei Наг-
tert, 1913 (проникает в тропики).
Филины очень изменчивы индивидуально, что затрудняет правильное отграничение
географических форм. Наиболее обособлена южная группа ascalaphus-desertorum, однако она
примыкает к центральноазиатской и в особенности южноиранской и закаспийской расам,
бледно окрашенным.
Близкий вид, замещающий палеарктического филина — в Америке от Аляски, Мекензи
и Гудсонова залива до Магелланова пролива Bubo virginianus Gme 1., 1788, вид столь же
эвритопный и политипический, как Bubo bubo. Другая очень близкая форма гнездится в
непалеарктической Индии к югу от Кашмира и южных склонов Гималаев до Нильгири,
Синда, Мадраса и Арракана — В. bengalensis Franklin, 1831. Наконец, в Африке викари-
рующий и очень близкий вид В. capensis A. S m i t h, 1834, распространенный в трех
подвидах от Абиссинии, Кении и Танганаики до Капской провинции и Наталя. Все эти четыре
филина образуют кругосветный или почти кругосветный genus geographicum. Африканские
пустынные филины (ascolaphus и desertorum) занимают, как уже упоминалось, несколько
особое положение и повидимому могут считаться в отношении других рас В. bubo — за формы,
пограничные между категориями «вид» и «подвид». Из Келесирии и гор Абу-шух в Палестине
известны особи с признаками гибридизации между interositusu ascalaphus; крупные, близкие
к hispanus филины, встречаются в сев. Алжире близ мыса Бон в ареале В. b. ascalaphus
(Хартерт, 1921); такие же гибриды были вероятно описаны под названием В. b. aharonii
Rotschild et Hartert, 1910 из Вади-Суенит в Палестине, причем в этих горах
встречены были и птицы типа ascalophus и «interpositus», и промежуточные (Хартерт, 1912).
Среднеевропейский филин Bubo bubo bubo \Lin n.
Синонимы. Bubo europaeus Lesson, Traite cTOrnith. 1831, стр. 115, «большие леса
Венгрии, России, Германии». — Bubo septentrional is. Chr. L. Brehm. Handb. d* Na-
turgesch. Vog. Deutschland, 1831, стр. 120, «к Северу от Германии»*
Распространение. Ареал. Сев. и зап. части европейской области
распространения вида от Скандинавии (в Норвегии до полярного круга, по Вал-
ленгрену в Лапландии будто бы по безлесным прибрежным островам) до
Франции и Пиренеев (Центральный Массив), Италии, Сицилии, Греции (однако в
последней, быть может, как и вообще на Балканах в Румынии, Югославии,
Албании и Болгарии, южноевропейская форма В. b. interpositus). Отсутствует
в зап. Франции, Бельгии, Англии и Дании. В СССР — северные, западные
и центральные области Европейской части, на севере в лесах Лапландии
(около 66°22' у Имандры, залетный у Екатерининской гавани на Мурмане),
у Кандалакши на Белом море, у Пинеги (за 64°15',), Архангельска, на востоке
до бассейна Мезени (смешанная популяция — переход к В. b. ruthenus в
бассейне реки Удоры, притока Вашки, до 63°); зимние залеты в Тиманскую тундру
(низовья реки Вельть, Гладков, 1938). Южней вост. граница определяется
верхним течением Волги (Горьковская обл.), проходя далее в Тамбовскую и
Воронежскую обл., в последней найдена переходная популяция (часть
особей —как bubo, часть —как ruthenus; часть со смешанными признаками);
такая же смешанная популяция в лесных частях Украины (Харьковская обл.,
Подолия), чем определяется южная граница ареала.
Характер пребывания. Среднеевропейский филин —
оседлая птица, совершающая небольшие кочевки зимой и иногда залетающая за
пределы ареала (Англия, разные части Франции; у нас —до тундры, см. выше).
В средней полосе СССР зимой приближается к поселениям, не исключая
больших городов (например залетал в центральную часть Москвы, в Ленинград и
т. д.). В Скандинавии отмечены кочевки молодых птиц в поисках гнездовой
территории; известное расселение реакклиматизированных филинов за счет
занятия молодыми участков, лежащих за пределами гнездовых территорий (но
ФИЛИН
355
неподалеку от родителей), отмечено и в Германии. Окольцованный у
Гельсингфорса в Финляндии филин добыт 16. III. 1940 в Лохвицком районе
Калининской обл.
Биотоп. Среднеевропейский филин — типичный убиквист; избегает
лишь густо населенных мест и близости человека, во всяком случае в гнездовый
период. В Европейской части СССР — в лесах, на равнинах, но в средней
Европе главным образом в скалах (быть может в результате преследования
человеком). Основные требования к биотопу: защитность и кормность угодий.
Численность. В населенных местностях — в средней полосе и
в особенности на Украине — редок; в сев. участках тайги, примерно к
северу от 55—56° с. ш. — обычен. В Зап. Европе редок (например в
Германии 100—ПО пар). Несмотря на полифагию филинов, у них установлены
колебания численности и плодовитости в зависимости от кормовых условий
года; в неблагоприятные годы выводки страдают от каннибализма; замечены
известные корреляции между численностью зайцев и филинов (средняя полоса
Европейской части Союза, то же в отношении Туркестана).
Экология. Размножение. Половая зрелость у среднеевропейского
филина наступает, повидимому, в возрасте около года, у самок быть может на
второй год. Большое постоянство гнездовых участков, но в случае беспокойств
птицы меняют гнездо. Известны случаи, когда при гибели самца самка через
год спаривалась с другим самцом в том же гнездовом участке (Минское
Полесье, Шнитников, 1913). Цикл ранний, начало кладки в средней России еще
при снеге. Гнезда на земле, простая ямка, вытоптанная самкой, без подстилки,
в лесу, на моховых болотах среди леса и т. п. Иногда занимаются старые
гнезда хищных птиц на деревьях (подорлик, Полесье, Шнитников;
Ленинградская обл., сарыч и т. д.). Есть указания на редкое гнездование в
дуплах (Зап. Европа, Нитхаммер, 1938). Брачный крик начинается с конца
февраля, интенсивно продолжается в марте и первой половине апреля. При
токовании самец ходит по земле вокруг самки, плотно прижав оперение,
оттого фигура его кажется тонкой и высоконогой; громко кричит, раздувая
шею, причем хорошо выделяется белое горловое пятно (привлекает
внимание самки к токующему самцу?); токование, начавшись в сумерках
(Московский зоопарк), продолжалось до глубокой ночи.
Кладка в первой половине апреля (2—18 апреля, Московская обл.,
Дементьев, 1949); ранние кладки в конце марта (даже 15. III., Минская
обл., Шнитников). В случае гибели кладки бывает дополнительная, чем и
объясняется растянутость сроков нахождения свежих яиц. На севере кладка,
повидимому, позднее (в Лапландии от середины апреля). Число яиц в кладке
обычно 2—3, даже 4 (по западноевропейским данным до 5, Нитхаммер, 1938).
Колебание плодовитости находится в зависимости от кормовых условий перед
периодом размножения или в его начале.
Во время выкармливания птенцов при недостатке корма бывает
«саморегулирование» численности популяций — в результате уничтожения молодых,
которых — иногда всех, чаще же младшего — убивают и съедают старики.
Яйца круглые, белые, с грубо зернистой скорлупой. Размеры (5) 54—63 X 44,5
—49,6, в среднем 57,1 х 45,9 мм (Шнитников, 1913). Промежуток между
откладкой яиц от 2 до 4 дней, кладка ночью (Московский зоопарк).
Насиживает только самка, с первого яйца; длительность насиживания 35
дней (Дементьев, 1946; сообщение Филатова, 1915, будто бы
продолжительность насиживания у филина «три недели» — явно ошибочно). Птенцы в
средней полосе появляются в начале мая — Минск, Калуга, с промежутком до 5 и
даже 7 дней. Около середины июня они покидают гнездо, будучи одетыми в
мезоптиль, но нелетными. Хорошо летают молодые филины в возрасте около
100 дней. Число птенцов в выводке обычно менее, чем яиц в кладке. Старики;
23 *
356
ОТРЯД СОВЫ
активно защищают гнездо и молодых. Ухает филин и осенью, что указывает
на наличие у него неполного осеннего полового цикла.
Линька. Полная годовая, последовательность смены нарядов:
пуховой наряд — мезоптиль — первый годовой наряд (окончательный по окраске,
комбинированный, с маховыми и рулевыми от мезоптиля) — второй годовой
наряд и т. д. Смена первостепенных маховых от заднего края партии к
переднему, от 10-го к первому. Начало смены
крупного пера в июле, конец з декабре.
Питание. Пищу
среднеевропейского филина составляют различные
мелкие и средней величины позвоночные,
главным образом все же грызуны от
зайцев — русака и беляка — до полевки и
мыши; также насекомоядные (ежи) и
мелкие хищные. В «мышиные» годы мыши
составляют основное питание, до 90%
(Уттендерфер, 1939). Отмечено
соотношение между числом зайцев и филинов (см.
выше). Имеется успешное нападение
филина на самку козули. Списки кормов
среднеевропейских филинов
(Уттендерфер, 1939), содержат в себе 32 вида
млекопитающих и 80 видов птиц. Среди
первых: ежи, случайно летучие мыши,
хищные— от лисят до куницы, горностая,
ласки; белки, хомяки, водяные крысы,
пасюк и т. д., молодые и даже взрослые
козули. Среди птиц: хищники от
настоящего сокола, тетеревятника, сарыча,
коршуна, зимняка до чеглока, пустельги,
перепелятйика; совы—молодые филины,
болотная ушастая неясыть, сычи
воробьиный, мохноногий и домовый;
лесные и домашние голуби; глухари,
тетерева, рябчики, серые куропатки, фазаны,
перепелы, молодые цапли (из гнездовых
колоний), выпь, серебристая чайка;
утки —кряква, шилохвость и свистунок,
нырки; мелкие птицы вплоть до крапив-
Рис. 93. Филин. ника; кроме того, повидимому случайно,
лягушки и рыбы.
Филин, таким образом, настоящий полифаг, но с известной
наклонностью к миофагии. Потребность в пище около 300 г чистого мяса в сутки
(т. е. около 500 г живого веса), но, разумеется, филин может съесть и много
больше. Погадки—типичные совиные, с непереваренными костями и
шерстью, сбрасываются при ночной охоте, не на отдыхе днем, как у многих
других сов. У каждого гнезда филина —несколько столовых и иногда запасы,
которые в гнездовый период филин-самец носит насиживающей самке.
Охотится филин и по открытым пространствам, и в лесу, начиная с сумерек;
в лунные ночи летает на большой высоте. У северной границы ареала
активен и днем.
Полевые признаки. Крупная сова черновато-желтой окраски, с
большими «ушными* пучками, которые чаще направлены в стороны; голова
большая. Сидит прямо, на земле или на дерезьях. Характерный медленный и вол-
ФИЛИН
357
нистый полет, обычно низко над землей. Крылья длинные и широкие, хвост
умеренной длины. Активен главным образом в темноте. Голос —
общеизвестное «уханье», глубокое, но не очень громкое «у-ху» или «у-ху-ху», иногда «квеею».
Самка у гнезда издает своеобразное верещанье или клохтанье. Держится
одиночками или парами.
Описание. Размеры и строение, Лицевой диск неполный, длинные «ушки».
Формула крыла 3 = 4^2>1>5... Вырезки на наружных опахалах 3-го — 5-го маховых,
на 6-м — выемка. Хвост из 12 рулевых, закругленный; длинное подхвостье. Клюв сильный,
круто загибающийся почти от основания. Цевки и пальцы сильные, густо оперенные (до
концевой фаланги пальцев), когти большие и острые, с двумя режущими краями. Длина самцов
(4) 620—670, самок (4) 695—725, в среднем соответственно 647,5 и 715 мм. Размах самцов
(2) 1550—1560, самок (3) 1676—1800 мм, в среднем 1555 и 1754 мм. Вес самцов 2100, 2260,
2450, 2570, 2670,2700 г, самок 3075, 3100, 3150, 3150, 3250 и 3260 г (эти данные относятся и
к расе ruthenus, имеющей одинаковые размеры с номинальной формой). Крыло самцов (20)
430—465, самок (20) 475—520, в среднем соответственно 452,5 и 485,4 мм.
Окраска. Пуховой наряд беловато-охристый. Мезоптиль бледный рыжевато-
охристый с темными черновато-бурыми тонкими поперечными полосками. Взрослые самцы
и самки основного ржаво-желтого цвета с черными пестринами. Лоб охристо-беловатый с
мелкими поперечными черточками; темя, затылок и шея с широкими черными продольными
полосами и нерезким мелким черноватым поперечным рисунком; «уши» черные с
желтоватыми пестринами и краями (на внутренних опахалах); спина и плечевые — черные с
небольшой примесью охристо-беловатого цвета у оснований и по краям перьев; на плечевых черные
пестрины сливаются в сплошное черное поле; кроющие крыла — как плечевые, но с большей
примесью светлых пестрин; надхвостье рыжее с неправильным узким черноватым
поперечным рисунком; второстепенные маховые черноватые, желтеющие у края внутреннего опахала,
с нерезким светлым поперечным рисунком; первостепенные маховые ржаво-желтые у
основания, черновато-бурые у вершины, с неправильным темным поперечным рисунком на
внутренних опахалах. Средние рулевые черные со следами узкого поперечного рыжевато-охристого
рисунка; на остальных парах рулевых, по мере приближения к краю хвоста, этот светлый
рисунок занимает все большее и большее место, так что боковые пары рулевых охристо-
рыжеватые с узкими черными поперечными полосами и черным крапом.
Рулевые и маховые гнездового наряда — мезоптиля — окрашены как у взрослых и не
сменяются при первой осенней линьке; лицевой диск сероватый с неясным поперечным
рисунком и с резкими черными наствольями у горла, последнее — белое; остальная
брюшная сторона ржаво-рыжая с широкими черными наствольями, постепенно суживающимися
от зоба и груди к брюху и бокам с резкой, правильной поперечной полосатостью.
Подкрылья охристо-беловатые с бурыми пестринами; ноги более или менее испещрены
черновато-бурым, подхвостье с темными наствольями и поперечным рисунком.
Радужина красновато-оранжевая; клюв и когти черновато-серые.
От других рас филинов нашей страны номинальный подвид отличается яркой и темной
окраской. На голове, спине, плечевых, средних рулевых преобладает черный цвет. На груди
черный продольный рисунок весьма широк, занимает около а/8 и даже а/* ширины пера.
Темные наствольные черты идут далеко вниз по брюху. Поперечный рисунок на брюхе,
боках, голенях частый и относительно широкий. Как указывалось, на восточной и южной
окраинах ареала встречаются особи с признаками, переходными к соседним формам, менее
черноватые, с более «размытым» темным рисунком и с более сероватым тоном светлых частей
оперения.
Русский филин Bubo bubo ruthenus Zhitkow et Buturlin
Bubo ruthenus. Ж и т[к ов и Бутурлин. Известия Имп. Русского География. Общества i
XVI, 1906, стр. 271, Промзино, бассейн Суры.
Распространение. Ареал. К востоку от предыдущего и к западу от
сибирской расы: восточная полоса Европейской части СССР от бассейна Печоры
и среднего течения Волги, верхнего течения Дона (Воронежская обл.) и
до низовьев Волги и зап. Приуралья (Уфа, Чкалов). На Печоре отмечен
к северу до 66°(Усть-цыльма); самая ю.-вост. точка гнездования ареала — До-
санг в Калмыцкой степи.
Характер пребывания. Русский филин оседлая,
совершающая незначительные передвижения в холодное время года птица — в это время
зимует в дельте Волги и в Оренбургских степях.
Биотоп. Леса, речные обрывы, выходы скал, степные овраги и балки
358
ОТРЯД СОВЫ
Численность. Умеренная, более обычен в таежной полосе.
Экология. Размножение. Цикл в общем протекает сходно с циклом
у номинального подвида, однако на самом севере, повидимому, поздней (в мае на
Княсьпинском Урале, Резцов, 1904). Брачный крик в Поволжье наблюдался уже
в марте, затем до мая, особенно по лунным ночам (Карамзин, 1902, Житков и
Бутурлин, 1906). Гнезда занимаются постоянно, расположены по обрывам,
в стенах балок и оврагов, в лесу на земле или выгнивших пнях, вероятно и
в старых постройках других видов птиц (Поволжье, Богданов, 1871, Карамзин,
1902; север Молотовск. обл., Резцов, 1904; Воронежская обл., Огнев и Воробьев,
1924; Печора).
Кладка из 2—4 яиц, быть может даже до 6 (Казань, Рузский, 1891),
в апреле (Богданов, 1871 и др.), но иногда позже —в мае (свежая кладка
из 2 яиц у Ак-булака, Чкаловская обл., 17. V. 1941, Спангенберг, но быть
может дополнительная кладка взамен утерянной?). Гибель яиц и птенцов
наблюдалась при весенних заморозках (Печора, Те плов). На наличие поздних и,
по всей вероятности, дополнительных кладок указывает и нахождение птенцов—
поздняков (начинающий линяние мезоптиля добыт 3. IX. 1922, Хреновской
бор. Воронежской обл., но там же молодой того же примерно возраста 16. VII.
1922; в Башкирии крупные птенцы, линяющие из мезоптиля в контурное перо,
в середине июня и 1 августа 1897). Особи, завершающие линяние в контурное
перо первого годового наряда, встречаются в конце сентября (Укедин, Печора,
20. IX. 1935). Птенцы разновозрастны, насиживание с первого яйца. В выводках
птенцов обычно меньше, чем яиц в кладке — часто 1, обычно 2, иногда и до 4
(летные, Казань, Печора). Неполный половой цикл (токование) осенью
(Житков и Бутурлин).
Линька. Начало линяния старых происходит в конце июня —* начале
июля, птицы в свежем пере бывают в декабре. Линяние молодых см. выше.
Питание. В общем сходное с питанием номинального подвида (и
других). Отмечено, что зимою филин приближается к поселениям и даже ловит
кур в курятниках, а также делает запасы пищи, пряча в дупла или в снег
(Чкаловская обл.). В качестве кормов русского филина отмечены грачи
(Воронежская обл.), ящерицы и малые жаворонки (Заволжье), зайцы (более или менее
повсеместно, вероятна корреляция плодовитости филина и «урожая» зайцев;
особенное значение имеют для филина зайцы в снежный период года, В. и Е. Теп-
ловы, 1948), изредка соболи, белка, мышевидные грызуны, также белые и серые
куропатки, глухари, рябчики, тетерева, вороновые, даже воробьи. Потребность
в пище за сутки, повидимому, около 800 г.
Описание. Размеры и строение. Крыло самцов»(17) 430—468, самок(22)
471—515, в среднем 445,6 и 485,4 мм. Окраска. Основной фон окраски русского филина
серовато-темный рисунок, менее интенсивный, чем у номинального подвида, — не черный,
а скорее буровато-черный; на крыльях по кроющим хорошо развиты беловатые пятна, так
что птица кажется более пестрой; поперечный рисунок брюшной стороны менее частый и
правильный; на спинной стороне темный рисунок менее распространен, чему В. b. bubo,
особенно на затылке и шее; птица в общем кажется светлей и пестрей.
Южный филин Bubo bubo interposit us Rotschild et Hartert
Bubo bubo interpositus. Rotschild and Hartert. NovitatusZoologicae, 1910, стр. Ill,
Эрегли, М. Азия.
Синонимы. Bubo bubo tauricus, Бутурлин. Определитель видов птиц СССР, 1928,
стр. 9, Кара-Актачи, Крым. — Bubo bubo nativus, Гавриленко. Зборник Полт. Держ. Муз. I,
1923, стр. 279, Миргород.— Bubo bubo transcaucasicus Tschikwischwi li. Bull. Mus.
Georgie V, 1930, стр. 979, Закавказье.
Распространение. Ареал. Степные части Украины к югу от области
В. b. bubo (см. выше); Бессарабия; Крым; Кавказ; М.Азия; вероятно в Сирии.
ФИЛИН
359
Северные границы примерно в южных частях Харьковской обл., в
Киевской обл., в Подолии, Бессарабии, по Таврическим степям, в низовьях
Днепра, на Сев. Кавказе, на восток по крайней мере до Эрзерумского
вилайета, к югу до Сирии, сев. Палестины и Ирана.
Характер пребывания. Южный филин — оседлая птица;
в горах совершает вертикальные миграции, собираясь зимой в долинах. На
равнине зимой приближается к поселениям.
Биотоп. Как другие подвиды — убиквист. В горах выше пределов леса,
до 2700 м в Армении (Соснин и Л'яйстер, 1941).
Численность. На Кавказе обычен, на Украине и в Крыму
относительно редок, вероятно в результате преследования человеком.
Экология. Размножение в общем сходно с размножением других
филинов. Гнезда на земле — у пней в лесу, в трещинах обрывов и скал,
иногда прямо на земле в степи и даже на скирдах сена (Сиваш, Сеницкий, 1898),
редко на деревьях (Бессарабия, Остерман, 1916).
Кладка повидимому ранняя — с ко^ца марта (Бессарабия, быть может
с середины этого месяца, Гебель, 1879). Кладка из 2—3 яиц, для севера ареала
(Харьковская обл.) указывается преобладание первого числа, для юга —>
второго (в 11 гнездах по 3 яйца) (Остерман, 1916). Редко бывает и 4 (Сиваш).
При всех обстоятельствах полные кладки — в начале апреля.
При обилии мышей будто бы — в исключительных случаях — повторный
цикл: в начале сентября 1898 или 1894 —кладка из двух яиц в Ростовском
округе (Сарандинаки, 1909). При утрате кладки бывает дополнительная.
Размеры яиц (8) 55—63,5x43,2—53,4 мм (Сомов, 1897).
Насиживание с первого яйца, птенцы разновозрастны. Вылупление
происходит в начале мая (быть может в Бессарабии в конце апреля). В начале
июня птенцы выбираются из гнезда, хотя не летают, летными они становятся
в конце июня. Конец смены мезоптиля повидимому в сентябре.
Линька. Изучена плохо. Интенсивная смена маховых происходит
в середине июля (около 40%, Дагестан). Взрослые в свежем пере отмечены
в октябре.
Питание. Охота с сумерек до рассвета, в мае еще в 5 часов утра.
Южный филин питается преимущественно грызунами, от зайца до полевки. В
частности для Харьковской обл. отмечены зайцы и тушканчики, для Бессарабии
зайцы, суслики и т. д., для Сиваша — суслики и разные птицы, для Армении —
грызуны, главным образом полевки, также обыкновенная полевка Microtus
arvalis, кустарная полевка Pitymys majari, хомячки Cricetulus, migratorius,
снежная полевка Chionomys nivalis, водяная крыса Arvicola terrestris,
бурозубка Sorex raddei, русак, сизый голубь (Даль, 1948); для окрестностей
Ворошилова на Сев. Кавказе 67,5% вредных грызунов, среди них полевки, слепыш,
хомячок Mesocricetus nigricans, кроме того, заяц русак, кавказский крот,
землеройка белозубка Crocidura saaveolens; перепел, кузнечики Decticus,
жужелица Platisma и т. д. (Лютый, 1940). Указывается, что на Сиваше в суровую
весну при гибели пролетных пластинчатоклювых филины кормились мертвыми
утками (Радде, 1854).
Описание. Размеры и строение. Южный филин несколько меньше, чем
номинальная раса: крыло самцов (25) 428—463, самок (19) 468—502, в среднем 444,7 и
480 мм.
Окраска. Очень похож на русского филина по развитию и характеру темного
рисунка, но тон этого рисунка более насыщенный и яркий (черный, как у В. b. bubo, но у
последнего пятна крупней, менее «размыты»); основной фон окраски более желтоват и менее
сероват, чем у В. b. ruthenus, так что южный филин по сравнению с ним кажется более ярким.
Закавказские филины особенно желтоваты.
360
отряд совы
Западносибирский филин Bubo bubo sibiricus G1 о g e r
Strix sibirka. Gloger. Abandern d. Vogel d. Einfluss d. Klima's, 1833, стр. 142, Урал.
Распространение* Ареал. Зап. Сибирь от Пермского Урала (Чердынь,
где переходная к ruthcnus популяция) и Башкирии (такая же популяция,
описанная Сушкиным, как «baschkiricusb) до средней Оби (Томск) и западных при-
алтайских районов; северная граница не прослежена, повидимому она
проходит где-то в б. Березовском уезде на Оби или в бассейне Таза,
Характер пребывания. Западносибирский филин частично
оседлая, но в значительной мере кочующая птица. Этот филин совершает, более
или менее регулярно, далекие откочевки. Зимние находки на западе до
Карелии (Медвежья гора, зима 1937), бассейна верхней Мезени (р. Удора, б. Ярен-
ский уезд, зима 1932), Башкирских степей (Бирск, поздняя осень 1930), ю.-в.
Каспия (Гассан-кули 6. II. 1938), низовьев Сыр-Дарьи (Джулек 14 января,
и Алма-ата).
Биотоп. Тайга, но не «глухая»; охотно вблизи водоемов. В Уральских
горах до 1000 м (Иремель).
Численность. Невысокая — суровые условия существования,
вынуждающие эту форму к кочевкам, вызывают и колебания численности:
в районе Тюмени филинов в иные годы много (в 1883—1884, в особенности
в 1887), в иные нет совсем (Словцов, 1892); такие же колебания численности
отмечаются и для Зауралья (Ушков).
Экология. Размножение. В Ильменском заповеднике, по Ушкову,
одна пара филинов располагается от другой на расстоянии 4—5—6, в среднем
немного более 5 км. Гнезда в нишах среди камней, на южных, скорее освобо-г
ждающихся от снега склонах, используются много лет (вероятно посменно?).
Начало полового цикла очень раннее — уже в начале марта, когда все условия
носят зимний характер, слышно уханье и «хохот» филинов. В кладке 2 яйца.
Размеры(12) 58,5—62 х 50—51, в среднем 59,43 х 50,1 мм (Ушков). В 1937—
1941 гг. полные кладки 9—15 апреля (однако «свежие» кладки для района
Тюмени отмечены будто бы и 18 мая, Словцов, 1892). Насиживание происходит
с первого яйца, птенцы разновозрастны. Насиживает только самка, самец
сторожит гнездо и носит ей добычу, принося уже перед вылуплением птенцов
известные запасы (в одном случае — 9 водяных крыс). Вылупление отмечено
15—20 мая, период насиживания следовательно 35 дней. Птенцы в мезоптиле —
в возрасте 20 дней. В 30 дней птенцы выползают из гнезда и держатся близ
него; в 50 дней они становятся летными. Первые 10 дней самка все время
присутствует в гнезде и обогревает молодых, а самец приносит пищу.
Питание. Летом сибирский филин охотится главным образом вокруг
гнездового участка, зимой — нередко вблизи поселений. В кормовом режиме
сибирского филина позвоночные занимают 98,t)%, из них млекопитающие
90,4%. Среди млекопитающих на первом месте—мышевидные, 83,8%, из
них 49,3% водяная крыса, 51 % полевки, кроме того заяц беляк и русак, белка,
бурундук, лесная мышь, крот, молодые козули. Птиц встречено 24,1 %, среди
них преимущественно куриные (белая куропатка, тетерев, глухарь), утки
(хохлатая чернеть, чирки свистунок и трескунок, кряква), лысуха, чомга,
чернозобая гагара, болотный лунь, малый канюк, черный коршун, болотная
сова, горлица, клинтух, вальдшнеп, сизая чайка, седой дятел, большой
пестрый дятел, желна, кукушка, из воробьиных—главным образом вороновые
(серая ворона, сорока, галка, грач, сойка), кроме того — зяблик и дрозд
деряба; изредка —насекомые. Летом мышевидные грызуны занимают в
питании филина резко преобладающее место, зимой повышается удельный вес птиц
(вороновые, куриные) и зайцев. Потребность в пище в сутки около 650—670 г
(у взрослых и больших птенцов). Охотятся филины ночью, а зимой и в пасмурную
ФИЛИН
361
погоду — и днем. Все приведенные сведения относятся к Ильменскому
заповеднику и принадлежат Ушкову, Связь численности сибирского филина и
зайцев уже отмечалась.
Имеются указания на нападение сибирских филинов на домашних птиц
(Нарым, Ильменский заповедник).
Описание. Размеры и строение. Крупный филин с густо оперенными до
когтей пальцами ног. Крыло самцов (7) 438—465, самок (14) 472—515, в среднем 451 и 492 мм.
Длина самки (1) 732, размах 1586 мм.
Окраска. Самый светлоокрашенный подвид как по бледному основному тону
оперения, так и по слабому развитию темных пестрин. Основной тон окраски сливочно-
белый с легкой примесью охристого цвета. Темный рисунок развит незначительно,
поперечные полосы на брюхе и боках обычно неправильны, тонки и далеко стоят одна
от другой; темные продольные полосы на затылке и шее очень узки, темные пятна на
плечевых невелики и занимают лишь самую вершину пера; на плечевых и кроющих крыла
хорошо выражены крупные беловатые пестрины. При значительной индивидуальной
изменчивости сибирских филинов для них все же всегда характерен общий белесоватый тон
окраски оперения, очень мелкий «размытый» темный рисунок и неправильная поперечная по-
лосатость брюха и боков.
Енисейский филин Bubo bubo yenisseensis But.
Bubo bubo yenisseensis. Бутурлин. Орнитологический Вестник, 1911, стр. 28, Красноярск.
Синонимы. Bubo bubo zaissanensis. Хахлов. Орнитологический Вестник, 1915,
стр. 224, Саур.
Распространение. Ареал. Средняя Сибирь —от Туруханского края до
Алтая, Тарбагатая и Саура, Танну-тува, Минусинской лесостепи, зап.
Прибайкалья; здесь, как и в сев. Монголии у озера Косогол, в горах Кентея и Хан-
гая — популяция переходная к дальневосточной расе; такие же переходные
формы, но к sibiricus, у Томска. Северная граница на Енисее проходит примерно
около 58—59° с. ш. (Тугаринов и Бутурлин, 1911), однако залеты отмечены до
границы леса — в лиственничниках повидимому до Норильска и Хатангского
за 71° с. ш. (Скалой, 1938), на западе в Туруханском крае на Тазе и Б. Елагуе.
Характер пребывания. Енисейский филин — оседлая птица,
совершающая в горах вертикальные перекочевки (Алтай).
Биотоп. На равнине в тайге, особенно по окраинным ее участкам;
в горах главным образом на голых или покрытых скудной растительностью
скалах. Вертикальное распространение до 2400 м, т. е. до альпийской зоны
(Алтай). Численность высокая.
Экология. Размножение. Интенсивный брачный крик начинается
в марте — апреле (с.-в. Алтай, Дементьев и Фолитарек, 1938). Гнезда на Алтае
в скалах. Кладка? Птенцы в мезоптиле у Чаган-бургази 7 июля (Бенгс,
1913), в другом выводке в Цаган-нура начинающие смену мезоптиля птенцы
21 июля (Сушкин, 1938). В обоих этих выводках по 2 птенца. Относительно
равнинной тайги сведений нет, кроме добытых в б. Минусинском уезде подлетков
18 июня — 1 июля. Неразбившиеся выводки встречены в начале августа (Сай-
люгем, Сушкин, 1938).
Линька. Линяние начинается со второй половины лета, после
снижения репродуктивного цикла, и продолжается до осени. Остатки старого пера
на темени наблюдаются еще в начале зимы. Питание. Енисейский филин —
вылетает на охоту засветло, до заката солнца (Алтай). У гнезда на Алтае —
остатки зайцев и красных уток (Сушкин). У Красноярска — главным
образом зайцы, также куриные птицы.
Описание. Размеры и строение. Крупная раса. Длина самцов (2) 660—680,
самок (2) 708—730, размах самцов (2) 1670—1680, самки (1) 1800 мм. Крыло самцов (18)
443—468, самок (18) 473—518, в среднем 456 и 487 мм.
О к р а с к а. От западносибирского филина отличается более темной окраской; общий
тон ее сероватый, с большей или меньшей примесью охристого цвета; темный рисунок сильнр
362
ОТРЯД СОВЫ
развит, продольные полосы на груди широки и доходят до брюха; поперечный рисунок
на брюшной стороне грубый и широкий, но не очень правильный (в чем отличие от
европейских рас); спинная сторона сильно испещрена черновато-бурым, в этом отношении
енисейский филин приближается к русскому, но, как и другие азиатские филины, отличается от
него тем, что рисунок очень разбит и даже на плечевых не образует сплошных черноватых
партий.
Якутский филин Bubo bubo jakutensis But.
Bubo jakutensis, В u t u г 1 i n. Journ. f. Orn., 1908, стр. 287, Якутск.
Распространение. Ареал. С.-в. Сибирь. Якутия от южных частей
бассейна Лены и Олекмы до 63°30'— 64°с. ш., на запад по крайней мере до
Вилюя, на восток до Охотского побережья (Удекой Острог, Охотск, Магадан)
и быть может до лиственничных лесов верховьев Анадыря (в ноябре 1930 добыт
на Ерополе, по Белопольскому, 1931). Экземпляры известны из следующих
мест: окрестности Якутска; Чурапча к с.-в. от Якутска; Удской Острог;
Магадан; Эльгенская долина в верховьях Колымы. Отсутствует в сев. Якутии
за Верхоянским хребтом, в Коряцкой Земле и на Камчатке.
Характер пребывания. Якутский филин повидимому оседлая
птица, в гнездовой области добывался в декабре и январе. Биотоп. Тайга
и скалистые участки, главным образом в речных долинах.
Экология* Сведений почти нет. Размножение. Птенцы в мезоп-
тиле на утесе на Вилюе встречены 13 июня, в конце августа у Якутска и Уд-
ского Острога молодые заканчивают линяние из мезоптиля в контурное перо.
У Охотска весенний брачный крик интенсивен в мае, слабое повторение его
еще в конце сентября, что показывает на наличие неполного осеннего полового
цикла (Харитонов, 1915).
Описание. Размеры и строение. Величина крупная: крыло самцов (3) 452—
468, самок (5) 475—495, в среднем 458 и 484 мм. Окраска. Близок к среднесибирскому
(енисейскому) подвиду, но в общем темней и ярче: темные пестрины черноваты и очень
многочисленны, хотя и «размыты»; на брюхе широкие, частые и правильные поперечные
полосы; общий тон окраски ржавчато-желтоватый.
Дальневосточный филин Bubo bubo ussuriensis Р о I j a k о w
Bubo bubo ussuriensis, Поляков. Орнитологический Вестник, 1915, стр. 44, Никольск-
Уссурийский.
Синонимы. Bubo bubo dauricus. Stegmann (ex. Suschkin M. S.). Ежегодник
Зоологического музея Академии наук XXIX (1929), стр. 178, Кулуссутаевск. — В. b. boris*
sowi Hesse J. f. Ornith. 1915, стр. Збб, Сахалин.
Распространение. Ареал. Ю.-в. Сибирь до Даурии и сев.
Монголии (Улан-Батор, долина Толы, Удэ, по Тугаринову, Г932; в Кентее, Хан-
гае и у озера Косогол, в зап. Забайкалье у Янды — переходная к
среднесибирской смешанная популяция); до бассейна Уссури и низовьев Амура
(озеро Эворон, р. Хунгари ок. 51° с, ш.), Сахалина. Повидимому, сюда же
относятся филины Курильских о-вов и из пограничных лесистых частей вост.
Манчжурии.
Характер пребывания. Дальневосточный филин — оседлая
птица.
Биотоп. Тайга и степные местности с выходами скал и камне й.
Численность. Низкая (Забайкалье, Павлов, 1935).
Экология* Размножение. Половой цикл у дальневосточного
филина начинается рано: уханье в феврале, кладка уже с конца марта (Тачанов-
ский, 1891); 1 апреля самка уже насиживала у озера Тарей-нор (Радде, 1863).
В Забайкалье гнезда помещаются на скалах, в нишах, под камнями и т. п.
Филин размножается много лет в одном и том же месте, но сменяя гнездо. Число
ФИЛИН
363
яиц 2. Размеры (8) 58,3^64 х 48—51 (Тачановский), (3) 59 х 49 мм (Радде).
Корм птенцам к гнезду носит самец, иногда у гнезда складываются «запасы».
Подлетки в мезоптиле наблюдались в южн. Приморье 22 июня. Птенцы,
кончающие линяние из мезоптиля в контурное перо, добывались на среднем Амуре
15 августа, в Монголии птенцы в мезоптиле 3 мая, но и в августе (Козлова,
1930).
Линька. Взрослые птицы в конце октября бывают в свежем пере.
Питание. Пищу дальневосточного филина составляют главным
образом мелкие млекопитающие —зайцы, сеноставки, суслик Эверсманна,
тушканчики, полевки, хомячки, разные птицы, в том числе перепелятник; на
Сахалине отмечено нападение филина на черную ворону (Мунстерьельм, 1920).
Описание. Размеры и строение. Крупная форма: крыло самцов (10) 430—465,
самок (11) 470—502, в среднем 448 и 483 мм. Окраска. Общий тон спинной стороны
сероватый; темный цвет на ней мелкий и «размытый», менее развитый, чему енисейского филина;
светлых пятен на спинной стороне почти не заметно; поперечный рисунок на боках и брюхе
очень тонкий и частый; темные настволья на брюшной стороне обычно не заходят на брюхо.
Общий тон мезоптиля — интенсивно желтый.
Систематические замечания. Птицы из степной сев. Монголии и Даурии были намечены
Сушкиным к описанию в качестве особой расы «dauricus»; часть из них действительно слегка
желтей и менее сероваты, чем птицы из Приморья, но другие особи неразличимы, и самые
отличия в сущности столь невелики и нестойки, что не позволяют при ограниченности
материала считать их проявлением географической изменчивости. Такие же замечания относятся
к сахалинским птицам. Совершенно непонятно, на каких основаниях Хартерт и Штейнбахер
(1936) соединяют даурских филинов в одну расу с тянынанскими.
Казахский филин Bubo bubo tarcomanus Eversm.
Strix turcomana. Eversmann. Addenda ad celeberrimi Pallasii Zoographiane Rosso —
Asiaticam fasc. I, 1835, стр. 3, Усть-Урт. — Bubo bubo eversmanm. Dementiev. Alauda
1931, стр. 381, залив Паскевича, Аральское море.
Распространение. Ареал. Полоса степей и полупустынь между Волгой
и низовьями Урала на западе и Казахской складчатой страной на востоке (до
Семипалатинска). К северу до Мугоджар и Караганды, на юг до Усть-Урта,
Мангышлака и Карабогаза, на востоке граница неясна (повидимому, бассейн
Чу, Прибалхашье). Зимою вне гнездовой области — на Волге у Астрахани,
8 низовьях реки Урал (Гурьев), в Туркмении главным образом на Каспийском
побережье (Чикишляр, Гассан-кули, низовья Атрека), также на зап. Узбое
(озера Топиатан, Ясха) и в низовьях Аму-Дарьи (Эльджик).
Характер пребывания. Ввиду мало благоприятных условий
зимы в сев. Казахстане (низкие температуры, многоснежие) значительная часть
филинов здесь совершает откочевки на юг, главным образом на Каспийское
побережье. Там наблюдается более или менее регулярный пролет с середины
октября; отлет зимующих филинов начинается в феврале, но отдельные особи
задерживаются до конца марта и даже начала апреля. У Красноводска в конце
прошлого — начале текущего столетия существовал промысел на филинов,
добывалось ежегодно до 500—600 птиц, использовались шкурки и перья.
Численность. На Каспийских зимовках высокая, в гнездовой
области спорадичен, но местами обычен.
Экология. Размножение. Пары постоянные, птицы держатся
вместе и вне периода гнездования. Гнезда в приаральских степях (Топускен) и
в Индерских горах — среди камней, иногда с 2 выходами. В кладке 3 и
даже 4 яйца, одно обычно не оплодотворено. Размеры яиц 58 х 49 мм в
среднем. Насиживает яйца самка, —сидит очень плотно. Цикл ранний: 17 мая
найдены пуховее птенцы, в середине июля молодые уходят из гнезда, а в конце
июля уже сменяют мезоптиль (Бостанжогло, 1911). Л и н ь к а. Птицы в
свежем пере бывают в конце сентября; в начале июня у самцов линяния еще нет.
364
ОТРЯД СОВЫ
Питание. Казахский филин в гнездовое время питается
тушканчиками Dipus, Allactaga, реже сусликами (Бостанжогло, 1911). Зимою в ю.-з.
Туркмении —в значительной мере водоплавающими птицами, области зимовок
которых совпадают с зимовками казахских филинов (среди них отмечены
утки, лысухи, кроме того, коростели), из млекопитающих — пластинчатозу-
бой крысой, песчанкой, ежами.
Описание. Размеры и строение. Филин средней величины: длина самцов (2)
650—670, размах самцов (2) 1645—1685; длина самок (4) 680—725, размах (4) 1735—1790,
в среднем 701,25 и 1770 мм. Крыло самцов (20) 420—468, самок (17) 470—492, в среднем
442,5 и 481,5 мм.
Окраска. Бледно окрашенная форма, в свежем пере общий тон окраски бледный
желтовато-охристый, в обношенном — беловатый; темный рисунок на плечевых, зашейке,
брюшной стороне более разбит, чем у европейских форм (русского и южного филинов).
На плечевых черноватые пестрины занимают примерно столько же места, как и светлый фон;
темный продольный рисунок на груди не заходит на брюхо; поперечный рисунок боков и
брюха правильный и чистый, но тонкий и бледный. От горного туркестанского подвида
казахский филин отличается несколько меньшей в среднем величиною и более слабым развитием
темного рисунка.
Туркменский филин Bubo bubo omissus D e m.
Bubo bubo omissus. Dementiev. Alauda, 1932, стр. 392, Ашхабад.
Распространение* Ареал. Туркмения и прилежащие части Ирана;
ю.-з. Таджикистан. Границы ареала на юге и юго-востоке неясны. В
Туркмении от Б. и М. Балханов, низовьев Атрека по Копет-Дагу от Сонгу-Дага до
восточных его частей (Меана-Чаача), по долинам Теджена, Мургаба и
Кушки, по отрогам Паропамиза в Бадхызе и Карабиле (хребты Гязь-гедык,
Фисташковый, Кермили), в рКугитанге, на Аму-Дарье, в Кара-Кумах. Возможно
к этой форме (но также и к В. b. turcomanus?) относятся сведения о гнездовании
филинов в горах между Красноводском и Карабогазом и в Куба-Даге и даже
на Мангышлаке. В Иране по крайней мере до Астрабада и центрального Сеи-
стана. В Таджикистане гнездовые особи этой расы были добыты на Сурхан-
Дарье близ устья Кафирнигана, у Кулут-куля, Суджина, Сталинабада
(восточные особи в известной степени переходны к В. b. hemachalanus,
распространенному на Памире).
Характер пребывания. Туркменский филин — оседлая
птица, совершающая зимою в горах вертикальные перемещения и
приближающаяся в это время к поселениям.
Биотоп. Как и всюду, филин весьма эвритопен, встречаясь как на
равнинах, так и в горах (до верхнего пояса Копет-Дага у Хейрабада на высоте
в 2027 м, у Чаека, Шах-шаха, Донгузтепе и т. д.). Предпочтение оказывается
береговым обрывам речных долин (Теджен, Кушка, Аму-Дарья) и обрывам
в горах; в пустыне по саксаульникам, но также и на склонах холмов; наконгц,
в развалинах сооружений (мечеть Султан-санджар в старом Мерве;
развалины у Ашхабада, Аннау и т. д.).
Численность. В горах и речных долинах нередок; в Кара-Кумах
малочисленен. Возможно, что имеется известное соотношение между числен-,
ностью филина и кормовыми условиями; в частности количеством зайцев (в
ю.-в. Туркмении — Бадхызе и Карабиле —весной 1946 г. было мало и филинов
и зайцев).
Экология. Размножение. Уханье филинов слышится в Туркмении
с начала марта, больше всего птица кричит между закатом и полуночью. Яйца
откладываются прямо на землю, в расщелинах береговых обрывов рек, на
карнизах в нишах, в пустыне даже на склонах Холмов, иногда в развалинах.
Кладка из 1—2 яиц, в начале апреля; в выводке обычно 1—2 птенца. Птенцы в мезо-
птиле, линяющие, вне гнезда, в конце мая — начале июня (25 мая, Тахтаба-
ФИЛИН
365
зар; 9'июня у Чакашлы близ Репетека и т. д.); почти заканчивающие смену
мезоптиля отмечены в конце июня, в середине июля — только остатки мезоп-
тиля на голове. Линька. Старые птицы линяют между июнем и сентябрем;
в конце сентября птицы бывают в свежем пере.
Питание. В качестве пищи туркменского филина указываются кек-
лики, клушицы, голуби, пищухи и даже молодые козлы в Копет-Даге, на
равнине — ежи, песчанки и средней величины птицы (Зарудный, 1896). В мечети
Султан-санджар филины нападали на голубей и галок. В погадках и поедях
филина у Ашхабада зимой 1941—1942 гг. найдены жаворонки, главным
образом хохлатые, много ежей и больших песчанок. На озере Б. Делили, нижний
Атрек, в марте 1942 г. — зайцы, большие песчанки, утки, главным образом
пластинчатозубая крыса. В ю.-в. Туркмении и в Кара-Кумах зимою
—преимущественно зайцы. У Тахтабазара 23—25.V. 1942 — сольпуга, жуки носороги,
лысуха, погоныш, чирок, горлица, полевки, слепушонка, песчанки, летучие
мыши (по Фррмозову).
Описание. Размер ыистроение. Мелкий подвид, как бы переходный к
южноиранскому В. b. nikolskii (у последнего крыло у самца 378, самки 394 мм); крыло самцов
(7) 404—424, самок (15) 425—460 мм, в среднем 415 и 445 мм. Пальцы оперены слабо:
конечная фаланга их обычно голая.
Окраска. У взрослых бледная, рыжевато-желтая по общему тону в свежем пере,
белесоватая—в обношенном;темный рисунок развит слабо как на спинной, так и набрюшной
стороне, общая окраска довольно однотонная и гармонирующая с окружающим итицу
пустынным ландшафтом; на плечевых светлый фон преобладает над черноватым рисунком;
темные настволья на затылке очень узки; продольный рисунок на брюшной стороне не
доходит до брюха; поперечные полосы на брюхе и боках очень частые и правильные, но тонкие
и бледные.
Гималайский филин Bubo bubo hemachalanus Hume
Bubo Hemachalana. Hume. Stray Feathers 1, 1873, стр. 315, Кулу в Гималаях.
Синонимы. Bubo bubo auspicabilis. Dementiev. Alauda 1931, стр. 35,
Александровский хребет. — ? Bubo bubo tibetanus. В i a n с h i. Bull. Brit. Orn. Club XVI, 1906,
стр. 69, Дзы-чю, верховья Голубой Реки.
Местное назва ни е. У народов Туркестана, как и др. филины, «уку» или «укю»,
звукоподражательное.
Распространение. Ареал. Высокая нагорная Азия —от Тянь-Шаня
и Ферганы до Памира на юго-западе, Кара-Тау на севере, Белуджистана и
Гималаев на юге; сюда же повидимому птицы из Тибета. Сходные филины
населяют равнины Туркестана (за исключением областей двух предыдущих форм),
т. е. бассейн Сыр-Дарьи и Кызыл-Кумы.
Характер пребывания. Гималайский филин — оседлая,
совершающая в горах вертикальные миграции, птица. Биотоп. Главным
образом скалистый ландшафт в Туркестане, кроме того речные обрывы и останцы
в пустынях; в горных лесах отсутствует. В вертикальном отношении в Тянь-
Шане до 2500 м, даже до 3000, на Памире до 3500—4000 м, в Тибете даже до
4700 м. Численность. Обычен.
Экология. Размножение. Изучено недостаточно. Начало брачного
крика бывает уже с конца января, разгар его в феврале, когда филин кричит
и днем (Памир, Мекленбурцев, 1946; Или, Шестоперов, 1929). Кладка однако
поздняя (в горах есть ли различия в зависимости от вертикального уровня
местности —остается неясным) —в конце апреля; в с.-з. отрогах Кара-Тау и
близлежащих местностях три кладки (свежих) 21 апреля (Спангенберг, 1936).
В Тибете насиженные яйца находились еще 11 июня, что указывает, вероятно,
на дополнительную кладку в начале мая. Гнезда в Кызыл-Кумах бывают на
земле в выдувах, в горах — в стенках обрывов, нишах и т. п. В кладке от
двух до трех яиц, но чаще два. Судя по развитию наседных пятен, яйца
насиживает одна самка, хотя у гнезда держатся обе птицы (Сухой хребет в окр.
366
ОТРЯД СОВЫ
Пржевальска). Судя по возрасту птенцов, вылупление происходит во второй
половине мая (Тянь-Шань). Птенцы во вполне развитом мезоптиле добывались
в первой трети июня; в конце июня начинается линяние из мезоптиля в
первый годовой наряд. В конце августа — первой половине сентября молодые
уже в полном осеннем пере. Птенцов в выводках обычно 2.
Питание. Главную пищу гималайского филина составляют грызуны,
в летнее время полевки, зимою зайцы и пищухи (Тянь-Шань, Памир). В ка^е^
стве корма филинов указываются для Кара-Тау и Кызыл-Кумов ежи, зайцы,
кеклики (Спангенберг, 1936), для центрального Тянь-Шаня—водяная крыса,
горлица, сорока, коростель, пустельга (Шульпин, 1939), голуби, песчанки,
галки, зайцы (Шестоперов, 1929); для Памира —зайцы, молодой сурок,
хомячки, пищухи, полевки; пустельга, мелкие птицы; рыба Schyzopygopsis,
которую филин ловит во время нереста (Мекленбурцев, 1946), и т. д. В
Гималаях у Кулу отмечено нападение филина на улара (Уистлер, 1926).
Описание. Размеры и строение. Крупная форма. Длина самцов (2) 685—700,
размах их 1520—1535 мм. Длина самок (5) 715—740, размах (5) 1595—740, в среднем 722
и 1665 мм. Крыло самцов (15) 433—466, самок (16) 473—508, в среднем 450,73 и 485,5 мм.
Окраска. Близок по цвету к казахскому филину, как по бледности основного тона,
так и по слабому развитию темного рисунка. Крупней, чем Туркменский филин и с более
контрастным темным рисунком на спинной стороне.
Общий тон окраски желтоватый; на плечевых темный рисунок занимает примерно
одинаковое пространство со светлым основным фоном или даже пребладает; продольный
рисунок на брюшной стороне заходит на верхнюю часть брюха; поперечный рисунок груди,
брюха и боков более грубый и менее правильный, чем у других туркестанских форм филина —
turcomanus и omissus. В развитии темного рисунка отличия от turcomanus: у последнего на
брюшной стороне тела продольный рисунок не заходит на брюхо, а поперечный рисунок на
брюхе и боках правильный; общий тон менее желтоват. В известной мере горные
туркестанские филины приближаются к среднесибирским.
РОД РЫБНЫЕ СОВЫ KETUPA LESSON 1831
Тип Ketupa javanensis
3. Рыбная сова Ketupa zeylonensis Gmelin
Strix zeylonensis. Gm?lin. Syst. Nat. 1788, стр. 287, Цейлон.
Распространение. Ареал. Вост. и южн. Азия от Охотского
побережья и Сахалина на севере, Манчжурии и Китая на востоке, Палестины,
ю.-з. Малой Азии, Ирана на западе, к югу до Цейлона, Индии и Индо-
Китая. Биотоп. Речные долины с лесными и кустарниковыми зарослями.
Характер пребывания. Оседлая птица. Численность.
Редкая; распространение спорадично.
Подвиды и варьирующие признаки. Изучены плохо из-за недостаточности материала.
Наиболее вероятно такое подразделение вида:К. z. semenowi Z а г u d n у, 1905—Палестина,
ю.-з. Малая Азия, Месопотамия, Иран, Индия до Синда с с.-зап. провинций; К. z. zeylonensis
Gmelin, 1788 (вероятный синоним К. z. leschenault Temminck, 1820) —Индия к
востоку от предыдущей, к югу от Гималаев, Ассам, Бирма (кроме северо-востока), Сиам;
К. 2. orientalis D е 1 а с о и г, 1926 от с.-в. Бирмы и ю.-в. Китая (Гуань-Дун и Цзяньси,
вероятно Юннань) до Индо-Китая; К. z. blakistoni Seebohm, 1883,— Япония (Хоккайдо),
южные Курилы (Кунашири), возможно Сахалин; К. z. doerriesi Seebohm, 1895, от
Охотского побережья до Приморья, Кореи и Манчжурии. Варьируют размеры (северная группа
крупней южной), окраска и быть может экология.
Северная рыбная сова Ketupa zeylonensis doerriesi Seebohm
Bubo doerriesi. Seebohm. Bull. Brit. Ornith. Club 1895, стр. IV, Сидеми.
Синоним. Bubo blakistoni piscivorus. M e i s e. Ornith. Monatsberichte, XLI (6), 1933,
стр. 169, Якши, к западу от Б. Хингана, Манчжурия.
Распространение. Ареал. От Охотского моря, где обычна на сев.
побережье (Армань, Уптар, Магадан) до Приморья (р. Горин, окрестности Вла-
РЫБНАЯ СОВА
дивостока, Иман, Сидеми, повидимому Ханка), Манчжурии (Б. Хинган) и
вероятно в прилежащих частях Кореи.
Характер пребывания. Северная рыбная сова — оседлая
птица. Биотоп. Главным образом у незамерзающих ручьев и рек с
быстрым течением или хотя бы с незамерзающими участками (в Манчжурии —* по
Б. Хингану). Численность. Редка. Указание Фирсова, по Лукашкину,
1934, будто рыбная сова обычна в приморских частях бассейна Уссури,
сомнительно, однако местами у рек, например на Имане и на Охотском побережье
(Васьковский), она встречается чаще, чем обыкновенный филин.
Экология* Почти неизвестна. Размножение. У японского подвида
на о-ве Кунашири найден один птенец в апреле (Бергман, 1935). Ста-
Карта 58. Распространение рыбной совы Ketupa zeylonensis
1 — К. z. semenowi, 2 — К. z. zeylonensis, 3 — К. z. orientalls, 4 — K. z. doerriesi,
5 — К. z. blakistoni; a — граница гнездовий (в кружках — местонахождения
подвидов в зимнее время).
рое гнездо найдено на сломанном дереве; гнездится повидимому и на земле
(Спангенберг).
Питание. Пищу северной рыбной совы составляют главным образом
рыбы и раки Astacus schrenckii; сова ловит их, заходя в воду и бродя по
мелководью, или сидя у воды и бросаясь на добычу; охотится с наступлением
темноты (Манчжурия, Яковлев, 1929).
Полевые признаки. Крупная сова, строго ночная птица, по виду
в общем напоминающая филина, но с голыми пальцами и с однообразной мало
контрастной окраской. Голос «баууу», звучит протяжно и глухо. В отличие
от других сов много бродит по земле, по берегам речек даже протаптывая
хорошо заметные тропы.
Описание. Размеры и строение. Лицевой диск развит слабо; образующие
«ушки» пучки перьев широкие и длинные; маховые прямые, редко суженные к концу, на
наружных опахалах нет зазубрин, характерных для многих других сов и в частности для
филинов; формула крыла 4>5 (или5>4) >3>6>2>7>8>1>9>10. Хвост слегка
закругленный, из 12 рулевых. Цевки оперены почти до основания голых пальцев. Клюв загнут отно-.
368
ОТРЯД СОВЫ
сительно слабо. Длина самца 710, размах 1780 (Яковлев, 1929), длина самки 710, размах
1890(Тачановский, 1891).
Крыло самцов (7) 510—550, самок (9) 538—560, в среднем соответственно 530
и 548 мм.
Окраска. Пуховой наряд и мезоптиль не описаны. Первый годовой
(окончательный) наряд, с мчховыми и рулевыми от мезоптиля, впрочем сходен по цвету и рисунку
с последующими нарядами: общая серовато-охристо-буроватая в различных оттенках;на лбу,
Рис. 94. Рыбная сова.
темени и затылке темные бурые настволья и нерезкий поперечный бурый рисунок; спина
и кроющие крыла с широкими бурыми наствольями и нерезкими пестринами; маховые бурые
с бледным охристым продольным и поперечным рисунком и крапинами, рулевые охристые
с неправильным бурым поперечным рисунком, слабо развитым на крайних парах. На
затылке или темени часто несколько белых перьев, иногда они имеются и среди малых
кроющих крыла; лицевой диск бледно серовато-бурый; горло беловатое, грудь, брюхо,
бока — с узкими темнобурыми наствольями и бледнобурыми поперечными черточками у
вершин перьев; подкрылья булано-охристые с мелкими бурыми черточками; цевки беловатые.
Клюв светлый серовато-роговой, желтоватый у вершин; пальцы темносерые; когти бурые;
радужина оранжево-желтая.
Систематические замечания. Систематические отношения северной группы рыбных сов,
часто объединяемых в особый вид Bubo blakistoni, требуют выяснения, что трудно вследствие
редкости птицы. Указываемые различия — окраска — даже на небольшом материале
оказывается подверженной большим индивидуальным колебаниям. Диагноз «Bubo blakistoni
ОШЕЙНИКОВАЯ СОВКА
369
piscivorusb у Мейзе совершенно несостоятелен (как неправильны на наш взгляд и его
соображения о принадлежности рыбных сов к роду Bubo и о самостоятельности вида В. blakistoni).
Совершенно неясным остается положение описанной Курода с южн. Сахалина («Tori»
1931, стр. 41) формы Bubo blakistoni karafutonis. Птица эта будто бы отличается более
темной окраской спинной стороны тела и меньшими размерами. Для самца (единственный
экземпляр) приводится длина крыла в 501 мм. Но первый признак варьирует у рыбных сов
индивидуально в весьма значительной степени и едва ли диагностичен; то же приходится
сказать и о длине крыла (см. приведенные выше измерения приморских особей). Неясно,
относится ли «Karafutonis» к blakistoni или doerriesi. Но и последние две формы вероятнее
всего также синонимы.
РОД СОВКИ OTUS PENNANT 1769
Тип Otus bokkamoena
4. Ошейниковая совка Otus bokkamoena Pennant
Otus Ваккатоепа. Pennant. Indian Zoology, 1769, стр. З, табл. З, Цейлон.
Распространение. Ареал. Вост. и южн. Азия от Приморья,
Сахалина, южн. Курильских о-вов и Японии до Индии, Китая, Филиппинских
и Зондских о-вов. Биотоп. Леса
долин и нижнего пояса гор.
Характер
пребывания. Ошейниковая совка
оседлая птица.
Подвиды и варьирующие признаки.
Множество форм, иные с узким
островным распространением, различающихся
размерами, окраской и т. п. Описаны:
О. b. semitorques Temmincket
S с h 1 е g., 1850—Курильские о-ва (Куна-
шири), Япония (Хоккайдо, Хондо, Оси-
ма, Сикоку, Цусима, Кюсю), о. Квель-
парт; О. b. ussuriensis,B u t, 1910 —
Приморье на юг до Кореи, южн.
Манчжурия, Сахалин; О. b. pryeri G u г n e у,
1889—о-ва Риу-Киу (Хачизо и Окинава);
О. b. glabripes S w i n h о е, 1870 — Китай
от Аньхуея и возможно Сычуани до
Юннани; Тонкий, Формоза: О. b. umbra-
tilis S w i n h о е, 1870— о. Хайнань;
О. b. plumipes Hume, 1870 — с.-з.
Гималаи: Наини-таль, Муррей, Гарвал,
Котегур; О. b. deserticola Ticehurst,
1922 — Белуджистан и Синд, быть
может ю.-в. Аравия (Маскат); О. b. mar at-
hae T i с е h u r s t, 1922 — центральные
провинции Индии, на восток до ю.-з.
Бенгалии (Манбхум и Самбалпур); О. Ь.
gangeticus Ticehurst, 1922 — Радж-
путана и Соединенные провинции Индии;
О. Ь. Ьаккатоепа Pennant, 1769 —
южн, Индия и Цейлон; О. b. lettia
Н о d s о п, 1836—вост. Гималаи от
Непала до вост. Ассама; Бирма, сев.
Сиам; О. b. lempiji Н о г s f i e 1 d,
1821 —Малайский п-ов, Суматра, Банка,
Биллитон, Ява, Борнео, Бали, северная группа о-вов Натуна; О. b. kangeana M а у г,
1938—0. Кангеан (к сев. от Бали); О. b. Kondorensis К 1 о s s, 1930—о. Пуло-кондор
у устьев Меконга, быть может южные части Малайского п-ова; О. b. mentawi Chasen
et К 1 о s s, 1926 — острова Сиберут, Сипора, архипелаг Пагэ; О. b. fuliginosa S h а г р е,
1888 — Палаван; О. b. boholensis Т е. G г е g о г, 1907 — о. Бохол; О. b. everett Tweeddale,
1878 — о-ва Самар, Минданао и Базилан, Филиппины.
Карта 59. Распространение ошейниковой совки
Otus Ьаккатоепа
1 — О. b. ussuriensis, 2 — О. b. semitorques, 3 — О. Ь.
glabripes, 4 — О. b. pryeri, 5 — О. Ь. lettia, 6 — О. b.
plumipes, 7 — О. b. umbratilis, 8— О. b. deserticola,
9 ^O. b. gangeticus, 10 — O. b. marathae, 11 — O. b.
bakkamoena, 12 — O. b. lempiji, 13 — O. b. kangeana,
14 — O. b. kondorensis, 15 — O. b. mentawi, 16 — O.
b. fuliginosa, 17 — O. b. boholensis, 18 — O.'b. everet-
ti; a — граница области распространения.
24 Птицы СССР, т. I
370
ОТРЯД СОВЫ
Уссурийская ошейниковая совка Otus bakkamoena ussuriensts But.
Scops semitorques ussuriensis. Бутурлин. Орнит. Вестник 1910, стр. 119, озеро Ханка.
Распространение. Ареал. Южные части Приморья (Сидеми,
Владивосток, Славянка, бассейн р. Иман, о. Аскольд и т. д.), Сахалин, Корея,
Манчжурия.
Биотоп. Лиственные леса (Спангенберг, 1940). Характер
пребывания. Уссурийская ошейниковая совка оседлая птица (в Приморье
добывалась в декабре и в январе;
указания Воробьева, 1946, о
весеннем прилете —. повидимому
ошибочны). Численность.
Невысокая (окраина ареала).
Экология. Почти вовсе неиз:
вестна.
Размножение. В период
гнездования кричит и днем, до
июля (Спангенберг, 1940). У самки,
добытой 23 апреля, яичник
неразвит. 6 июля у реки Псуфун добыта
молодая сменяющая пух на мезоп-
тиль птица с короткими рулевыми
и маховыми; 23 июня молодая птица
во вполне развитом мезоптиле
(Зоологический институт Академии
наук). Японский подвид
semitorques, очень близкий к
уссурийскому, гнездится главным образом
в дуплах, редко старых гнездах
хищных птиц или даже под
крышами зданий (Ян, 1942).
Питание. Пищу
составляют главным образом
насекомые. В желудке птиц, добытых в
южн. Приморье, найдены остатки
грызунов, мелких птиц, гусениц-
пядениц Geometra (Воробьев, 1946).
Полевые признаки. Самая крупная из наших совок, примерно с домового
сыча. Голос двусложный «ху-кху», несколько сходный по интонации с криком
буланой совки. Главным образом ночная, отчасти сумеречная птица.
Описание. Размеры и строение. Пальцы ног оперены на V» длины от
основания. Крылья длинные, формула 4>5>3>6>2>7>8>1; на наружном опахале 5-го
махового имеется сужение. Длина самцов (2) 230—245, самки (1) 270, размах самцов (2>
605—610, самки (1)660лш (Тачановский, 1893). Крыло (5)159—176, в среднем 168,6мм,
при этому одного самца 159, у самки 176 мм.
Окраска. Спинная сторона буроватая с примесью охристо-желтоватых пестрин,
с темными бурыми наствольями и поперечными чертами; на задней стороне шеи светлое
охристое ожерелье; кроме того, на наружных опахалах плечевых резко выступающие пятна;
спина и крылья светлей головы и шеи; маховые буровато-серые, на наружных опахалах
светлые пятна, внутренние опахала темнобурые с неясным поперечным рисунком; вершины
маховых сероваты; рулевые с темными мраморными бурыми пестринами и с 7—8
неправильными тонкими поперечными полосками; лицевой диск сероватый с мелкими бурыми пестрин-
ками; середина горла и зоба беловатые, остальная брюшная сторона охристая с бурыми на-
ствольными чертами и крапом. Радужина желтая или оранжево-желтая, реже бурая; когти
светлороговые; клюв буро-роговой.
В отличие от японских ошейниковых совок, нам пока неизвестны особи из Приморья
желтой вариации окраски.
Рис. 95. Ошейниковая совка.
ПУСТЫННАЯ СОВКА
371
Систематические замечания* Уссурийская ошейниковая совка очень близка к
японскому подвиду О. b. semitorques. Длина крыла у измеренных экземпляров этого подвида
самцов и самок (9) 155—182, в среднем 167,6 мм; размеры, следовательно, идентичны.
Отличия в окраске: уссурийская совка несколько менее рыжевата и буровата; светлые
пестрины у нее бледней, с менее развитым золотистым оттенком.
5. Пустынная совка Otus brucei Hume
Ephialtes Brucei. Hume. Stray Feathers, 1 1873, стр. 8, Рахури, Бомбейская провинция.
Синонимы. Scops semenowi Sarudny und H a e г m s. Ornith. Monatsber., 1902,
стр. 49, Персидский Белуджистан.—Scops obsoleta. С a b a n i s Journ. f. Ornith.1875, стр. 126,
Бухара.
Распространение. Ареал. Передняя Азия на север до южного берега
Аральского моря (низовья Аму-Дарьи и Сыр-Дарьи), на восток до Ферганской
долины (Уч-курган,
Андижан), Кашгарии (Куэн-Лунь
и Вахан), Таджикистана (до
Памира исключительно),
Афганистана, Белуджистана,
вост. Ирана; на запад до
центрального Копет-Дага (район
Ашхабада), Месопотамии и
Палестины; отмечена на
Синайском п-ове (Мейнерцхаген,
1930). Находки в сев. Индии—
в Ахмеднагаре и Ратнагири
в Бомбейском президентстве;
в Хайдерабаде; в Умаркоти и
Чипра в Синде, в Султан-
пуре, в Уде (Бекер, 1927)—
относятся повидимому только Карта бо. распространение пустынной совки Otus
к зимующим птицам. Запад- brucei
НЫе наХОДКИ В СССР ДОСТО- д — граница гнездовий, г — область зимовок.
верны для районов Ашхабада
и Кушки, также для Мургаба. Указания Флерике (1897) о встрече у Дже-
бела и Левчука (1904) —на Челекене в зап. Туркмении сомнительны.
Характер пребывания. Пустынная совка — перелетная птица.
Прилет в конце марта — начале апреля (Фергана, Уч-курган — 22—29 марта,
сб. Северцова; на нижней Сыр-Дарье; в Туркмении). Отлет начинается в
сентябре (Туркмения) и продолжается до октября (Сыр-Дарья).
Биотоп. Приречные тугаи, но главным образом, культурный
ландшафт (сады) долин, при этом и в городах; в горы не поднимается, по крайней
мере в Туркмении и в Тянь-Шане; сообщение Даля (1936) о распространении
в Зеравшанском и Туркестанском хребтах до высот 2440—2875 м требует
подтверждения; достоверны лишь залеты в горы (в частности к верховьям реки
Аличур, Памир, Мекленбурцев, 1936).
Численность. В соответствии с характером биотопа,
неравномерная; в культурном ландшафте на Аму-Дарье и Сыр-Дарье многочисленна;
у западной границы ареала на Мургабе и Теджене, в подгорной полосе Копет-
Дага — редка.
Экология. Размножение. Половой цикл более ранний, чем у
обыкновенной совки; в начале апреля семенники самцов только начинают
развиваться (Ашхабад, низовья Сыр-Дарьи). Образование пар наступает с
прилета (28 марта, Сыр-Дарья). Гнезда — в дуплах, иногда занятых другими
птицами, например чешуйчатым дятлом на Мургабе (Радде и Вальтер); чаще всего
24*
ГГ—Г—М
Ib.L сч_
1г" ^r><^y<fe5<&
' ~) У ^ V_,> s^ ji
ш°Д
372
ОТРЯД СОВЫ
находятся на разнолистных тополях или ветлах (в Месопотамии — на
финиковых пальмах); иногда гнезда — открытые, на деревьях, даже на саксауле
(Аральское море, Зарудный, 1916)- Совка больше всего пользуется сорочьими
гнездами, сохраняя при этом их крышу (Джили-куль, Иванов, 1940). В гнездо-
вый период совки крикливы, голос их часто слышится с конца марта до июня
(Сыр-Дарья). Кладка начинается в последней трети апреля, полные встречаются
в начале мая (Туркмения, Сыр-Дарья). Число яиц 4—6. Окраска яиц белая,
величина их больше, чем у обыкновенной совки; (60) 28,6—34X26—29,4 мм
(Сыр-Дарья, Спангенберг). Насиживает повидимому только самка; сидит очень
крепко; самец приносит ей в этот период корм. Срок? (повидимому, около
4 недель). Птенцов не более 4. Летные молодые в мезоптиле с конца июня и
в июле; выводки держатся
вместе до августа.
Продолжительность гнездового периода,
следовательно, около 7
недель.
Линька. Не изучена.
Мезоптиль сменяется вероятно
с августа. Взрослые птицы во
время пребывания в гнездовой
области, между концом марта и
сентябрем, не линяют;
линяние, следовательно, проходит
на зимовке, с поздней осени.
Питание. Пищу
составляют главным образом
насекомые, также летучие
мыши и мелкие воробьиные
птицы.
Полевые признаки.
Пустынная совка похожа на
обыкновенную совку, но
светлей по окраске и с иным
голосом. Брачный крик «ух-ух»,
долго повторяемый; кроме
того, начинающийся трелью
«цирр-ва-ваа». Полет легкий, длинные крылья, хорошо заметные «ушки».
Образ жизни — строго ночной.
Описание. Размеры и строение. Отличается от обыкновенной совки иной
формулой крыла, менее заостренного (недалекие перелеты): 3>2! = 4>5>6>1 или
3>2>4>5... (иногда 1-е длинней 6-го). Нижняя часть девки оперена. Длина самцов (2)
2Ю—211, самок (2)207—210, размах самцов (2) 540, самки (1) 575 мм. Вес самца 110 г.
Крыло самцов (24) 150—165, самок (11)155—168, в среднем 158,8 и 161,1 мм; половой
диморфизм размеров, следовательно, незначителен.
Окраска. Пуховой наряд не описан. Мезоптиль бледно песочно-сероватый
(буланый) с тонким поперечным буроватым рисунком на перьях брюха, боков, головы; маховые
и рулевые, сохраняющиеся от мезоптиля и в первом годовом наряде, окрашены как у
взрослых (см. ниже). Взрослые самцы и самки (в первом годовом наряде и последующих) бледно
песочно-серые с неясным более светлым рисунком и резкими темными наствольями и пестри-
нами; на затылке и наружных опахалах плечевых светлые пятна охристого цвета; темные
настволья особенно широки на лбу, несколько ^же на спине, еще уже — на плечах и
крыльях; малые кроющие крыла без рыжеватого оттенка (в этом различие от обыкновенной
и дальневосточной сплюшек); первостепенные маховые бурые с сероватой вершиной, неясным
светлым рисунком'на внутренних опахалах и с 3—4 беловатыми пятнами на наружных
опахалах; брюшная ^сторона тела по основному тону окраски сходна со спинной, но несколько
бледней; на оперении ног обычно резкие наствольные черты; подкрылья беловаты с
небольшой примесью серовато-бурых пестрин; у основания маховых на нижней поверхности крыла
Рис. 96. Пустынная совка (взрослая и молодая).
ОБЫКНОВЕННАЯ СПЛЮШКА ИЛИ ЗОРЬКА 373
большое бурое пятно. Радужина желтая, когти темные роговые, клюв буро-роговой,
желтоватый у основания нижней челюсти.
Окраска пустынных совок довольно изменчива и по общему тону, и по развитию тем*
ного рисунка. В общем, как у многих других видов сов, намечаются два типа: более светлый,
с пребладанием или значительным развитием палево-желтоватого цвета, и более темный,
буровато-серый. При этом у птиц темного типа брюшная сторона обычно более испещрена
мелким темным крапом. Географической локализации эти типы окраски не имеют (однако,
изученные экземпляры из Таджикистана — Каракуль, Курган-тюбе, Шаартуз — все
темные).
6. Обыкновенная сплюшка или зорька Otus scops Linn.
Strix Scops. Linnaeus. Syst. Nat. изд. X, 1758, стр. 92, Италия.
Русское название. «Сплюшка» — звукоподражательное; «зорька» —
южнославянское (у Левшина, 1774, «маленький род сыча»).
Распространение. Ареал. Южн., средняя и вост. Европа, сев. Африка
от Туниса и Марокко до сев. Сахары; вост. Канарские о-ва; Кипр, Крит,
Карта 61. Распространение обыкновенной сплюшки Otus scops
7—О. s. scops, 2 — О. s. pulchellus, 3 — О. е. mallorcae, 4 — О. s. cycladum,
5 — О. s. cyprius; a — граница гнездовий, г — область зимовок.
Циклады, Малая и Передняя Азия, Туркестан; ю.-з. Сибирь до Алтая.
Зимой в сев. частях тропической Африки и ю.-з. Азии до с.-з. Индии.
Характер пребывания. Обыкновенная сплюшка — перелетная
птица, кроме некоторых местностей Средиземноморья. Биотоп. Лиственные
леса, в южной части ареала — в горах (сев. Африка, Кавказ, Туркестан).
Численность. Обычная птица Средиземноморья и южной Европы, на
востоке ареала редка.
Подвиды и варьирующие признаки» Морфологические различия между подвидами
в размерах и окраске, экологические — в оседлости и перелетности, в особенностях цикла
периодических явлений. В сев. Африке, южн. и средней Европе, до Кавказа и Туркмении на
374
ОТРЯД СОВЫ
востоке О. s. scops L., 1758; на Балеарских о-вах О. s. mallorcae J о г d a n s, 1921; на Крите
и Цикладских о-вах „-оседлая О. s. cycladum T s с h u s i, 1904; на Кипре О. s> сургШ
Madarasz, 1901; в Вост. Европе от средней России и Волги до Енисея на востоке (Тан-
ну-тува), Тянь-Шаня и Ферганы на юге О. s, pulchellus P a 11 a s, 1801.
Европейская сплюшка Otas scops scops Linn.
Синоним. Plsorhina scops turanica. Loudon. Orn. Monatsber. 1905, стр. 129,
Репетек в ю.-в. Кара-Кумах.
Распространение* Ареал, Южн. и средняя Европа, Пиренейский п-ов,
Франция, кроме севера, южные части Швейцарии (Граубюнден, Тессин,
Валлис), Италия, Корсика, Сардиния, Сицилия, Балканский п-ов, южн.
Австрия (Тироль, Штирия), Венгрия; Канарские о-ва (Ланцероте); Алжир,
Тунис, Марокко, Малая Азия, Сирия, Палестина; в СССР на Украине, в Крыму
и на Кавказе^ сюда же птицы из Туркмении и, вероятно, сев. Ирана; на
<:евер в Европейской России до Ленинградской, Псковской, Смоленской,
Московской, сев. части Рязанской областей, Ярославской (Переяславль); на
восток до Тульской и Курской (Стрелецкая степь). В бассейне Волги —
следующая раса. Зимою в Африке от Сенегамбии до Судана и до Кении и
Уганды. Залеты в Ирландию, Англию, Бельгию, Голландию, Данию и
Германию.
Характер пребывания. Европейская сплюшка — перелетная
птица. Есть требующие проверки указания на нахождение сплюшки зимой
в Крыму, будто бы изредка у Симферополя.
Даты. Прилет в южной полосе Европейской части СССР начинается
в разных числах апреля, валовой —во второй половине этого месяца, чаще
после 25-го; в средней полосе примерно в то же время или в самом начале
мая. В Туркестане около 20—25 апреля; в исключительных случаях ранее,
например 19 марта у Хейрабада в центральном Копет-Даге (скорее всего
случайный залет). В сев. Африке пролет начинается в середине — конце апреля.
Осенний пролет бывает уже с конца августа, он мало заметен и продолжается
весь сентябрь. Отдельные особи встречаются на юге до конца сентября и
даже в октябре (Туркмения; 14 октября у Джулека на нижней Сыр-Дарье).
Биотоп. Лиственные и смешанные (на севере) рощи с подседом и старыми
дуплистыми деревьями; сады, парки и уличные аллеи (в городах, в частности
в Зап. Европе). На пролете даже в пустынях (Кара-Кумы, Сахара). В горах —
до верхней границы высокоствольного леса: в Закавказье до 1700 м (Армения),
в Туркмении, где встречается на гнездовье только в горах, до 2000 м
(Хейрабад).
Численность. На юге — на Кавказе, на Украине — обычная
птица; в средней полосе редка, в Туркмении —также. Колебаний численности
и плодовитости не установлено, хотя быть может имеет место известное
расширение ареала по направлению на север (по гарям, вырубкам и т. п.).
Экология. Размножение. Брачный крик начинается с прилета, при
этом совка кричит и днем. Гнезда главным образом бывают в дуплах разных
пород деревьев (дуб, Московская обл.; осина, Минское Полесье; липа, Умань,
в диких фруктовых деревьях —яблонях, грушах и пр., Харьковская обл.);
реже — в старых гнездах других птиц; гнезда эти расположены как на деревьях
(сорочьи), так и в норках или расщелинах (сизоворонка, скалистый поползень);
иногда — в трещинах камней (Копет-Даг) и даже в постройках (за обшивкой
деревянного дома, Курская обл. Птушенко). Подстилки в гнездах нет.
ч, Кладка — в конце мая как на юге ареала, так и на севере (готовое к сносу
яйцо у самки 10 мая, б. Екатеринославская губ , полная кладка в конце
мая, Харьковская обл., незаконченная кладка у Сулюкли в Копет-Даге
29 мая, 7 июня 4 насиженных яйца в Бессарабии, 14 июня 4 насиженных
ОБЫКНОВЕННАЯ СПЛЮШКА ИЛИ ЗОРЬКА
375
яйца у Острова, в Псковской обл.). В кладке 2—5, быть может до 6 яиц. Яйца
чисто белые, блестящие и мелкозернистые. Размеры: (11) 30—33 х 27—28,
в среднем 32 х27,4 мм (Сомов, 1897); (2) 30,5—32,5 х 25,5— 27 (Гебель,
1879); (1) 31,5 х 26 мм (Шнитников, 1913). При утрате кладки бывает иногда
дополнительная.
Насиживание начинается с откладки первого яйца, чем объясняется то,
что степень насиженности отдельных яиц в кладке и возраст птенцов в одном
и том же выводке различны. Процесс насиживания у нас не изучен (по
западноевропейским данным насиживает только самка, в течение 24—25 дней;
срок пребывания в гнезде
птенцов короткий, около 3
недель (Уайзерби, 1938). Вылу-
пление — в различных числах
июня летные молодые в мезоп-
тиле с недоросшими крыльями
во второй половине июля (14
июля, Стрелецкая степь,
Курская обл. 9 июля, Тамак,
Крым; 15 июля, Мцхет; 4—
б июля Фирюза, 20 июля в
Гермабе, Копет-Даг, и т. д.).
Летные птенцы затем держатся
вместе (в выводке бывает до
5 молодых) по крайней мере
до начала августа. Отдельные
выводки запаздывают, и
молодые в мезоптиле с
недоросшими крыльями могут быть
встречены еще в августе
(Мцхет, дополнительная
кладка?).
Линька.
Последовательность смены нарядовобыч-
ная для сов: пуховой наряд—
мезоптиль — первый годовой
наряд (ПО окраске ОКОНЧа- Рис. 97. Обыкновенная сплюшка (взрослая и
тельный, но сохраняющий молодая),
от мезоптиля маховые и
рулевые) — второй годовой наряд и т. д. Мезоптиль носится до августа, во
второй половине августа —начале сентября молодые уже в контурном пере.
Взрослые птицы в СССР не линяют; линяние протекает у совок на зимовке,
продолжаясь до января — февраля.
Питание. Европейская сплюшка в значительной мере
насекомоядная птица: в качестве пищи сплюшки в СССР отмечаются главным образом
жуки (в том числе хрущи Melolontha hippocastani); ночные бабочки; гусеницы;
пауки Epeira; реже мелкие грызуны и птицы (Московская обл.; Харьковская
обл., Бессарабия, Туркмения и т. п.). Охотится сплюшка после наступления
темноты.
Полевые признаки. Одна из самых мелких сов нашей фауны, величиной
примерно с крупного дрозда, по окраске оперения напоминающая вертишейку
или козодоя. Телосложение стройное (в сравнении с сычами), заметные «ушки»,
впрочем часто прижатые. Длиннокрылая, с относительно коротким хвостом.
Мало осторожна. Ночная птица, днем сидит обычно на дереве у ствола или в
кустарнике, вытянувшись и прижав перья. Плохо видит при дневном свете.
376
ОТРЯД СОВЫ
Голос—мелодический посвист, весьма характерный, вроде «сплю-ю, сплю-ю»
или «кьюю-кьюю», кроме того — шипенье.
Описание. Р азмеры и строение. Стройная сова с небольшой головой;
наружное ухо небольшое; перьевые «ушки» широкие; лицевой диск неполный; клюв сжатый
с боков, восковица вздутая; цевки густо оперены, пальцы голые, покрытые жесткими
щетинками (схватывание насекомых). Крылья длинные и острые, формула 3>2>4>5>1>6
(не считая рудиментарного скрытого под кроющими истинного первого махового). Вырезки
наружных опахал 2-го и 3-го первостепенных маховых. Хвост прямо срезанный. Длина
самцов (3) 201—206, самок (2) 208—210. Размах самцов (1) 505,5, самок (2) 531—542 мм.
Вес? Крыло птиц Европейской части СССР у самцов (11) 143—161, в среднем 150,6 мм,
самок (11) 146—163, в среднем 154,2 мм, в Крыму у самцов (6) 145—152, в среднем 148,8,
у самок (2) 156, в среднем 156 мм; на Кавказе у самцов (18) 143—157, в среднем 154 мм;
в Туркмении у самцов (2) 153—156, в среднем 154, у самок (2) 154—163, в среднем 158,5 мм.
Средние величины по всему ареалу у самцов (37) 149,8, у самок (22) 154,7 мм.
Окраска. Пуховой птенец белый. Взрослые птицы, самцы и самки: общий тон
спинной стороны буровато-серый с большей или меньшей примесью ржавчато-рыжеватого цвета,
с темнобурыми наствольными чертами и мелким поперечным буроватым рисунком. Кроме
темного рисунка—беловатые пестрины, особенно заметные на затылке и на внешних опахалах
плечевых и средних кроющих крыла; малые кроющие крыла золотисто-желтоватые; маховые
бурые, сероватые у вершин, с дымчатым светлым поперечным рисунком на внутренних
опахалах и 4—5 беловатыми пятнами на наружных опахалах; рулевые серовато-бурые
с темными пестринами и неправильным светлым поперечным рисунком; лицевой диск
пестрый, бледносерый с темными крапинами, более многочисленными у горла и ушей; брюшная
сторона светлей спинной, с явственным беловатым поперечным рисунком и темными на-
ствольями; брюхо беловатое; оперение ног ржавчатое с резкими продольными черточками.
Индивидуальная изменчивость окраски очень велика; главным образом варьирует
развитие рыжеватого оттенка в оперении;у некоторых особей он преобладает над серым,так что
как бы намечается наличие двух типов или вариаций окраски — серого и рыжего, впрочем
менее резко выраженных, чем у О. sunia (при этом серые птицы спариваются с рыжими).
Мезоптиль отличается более светлой окраской со слабым развитием настволий на брюшной
стороне, последняя кажется поперечнополосатой. Радужина бледножелтая у молодых,
оранжевая у взрослых; клюв бурый, черноватый у вершины; восковица темнобурая; лапы
желтовато-бурые; когти желтоватые у основания, черноватые у вершины.
Сибирская сплюшка Otus scops pulchellus Pallas
Stryx pulchella. Pallas. Reise d. verschied. Provinzen d. Russ. Reichs I, 1801, стр. 456,
Поволжье.
Синонимы. Pisorhina scops zarudnyi. Tschusi. Ornith. Jahrbuch XIV, 1903,
стр. 189, Сарепта. — Pisorhina scops bascanica. I o h a n s e n. Ornith. Jahrbuch XVIII, 1904,
стр. 202, Баскан, сев. Семиречье. — Scops scops sibirica. Бутурлин. Орн. Вестник,
1910, стр. 260, Красноярск. — Scops scops irtyshensis. Бутурлин. Наша Охота, 1912,
июль, стр. 46, Тара. — Scops scops ferghanensis. Бутурлин, цит. соч., стр. 45, Ош.
Распространение. А р е а л. К востоку от предыдущей — от долины Волги,
в Приуралье, Зап. и средней Сибири и в Туркестане; она же повидимому в с.-в.
Иране. Северная граница в Европе — в Горьковской обл. и около 56° с. ш.
у Казани (Першаков, 1929), в Зап. Сибири у Тюмени, Новосибирска, Томска,
далее у Красноярска; южная граница —в низовьях Волги и вероятно Урала,
затем по границе леса (отмечена в июле в Наурзумском бору, где более не
наблюдалась) в Туркестане к югу до Таджикистана, Ферганы и вост. Тянь-Шаня;
в Тарбагатае; в южн. и центральном (но не ю.-в.) Алтае; в Танну-тувинской
обл.; самый восточный пункт—Троицкосавск. Этот подвид также зимует
в Африке, вероятно вместе с номинальной расой (до Кении и Уганды, также
в ю.-з. Аравии).
Характер пребывания. Сибирская сплюшка — перелетная
птица. Даты. Прилет и отлет происходит примерно в те же сроки, как у
номинальной расы, в конце апреля — начале мая. В Сибири, однако, повидимому,
несколько поздней, во второй половине мая (у Новенского, западная окраина
Алтая, 16 мая; у Онгудая в центральном Алтае даже 27 мая; в Кузнецкой степи
13 мая). Пролет на Урале и на Эмбе бывает в середине мая, на реке Или
В0СТ0ЧН0АЗИАТСКАЯ СОВКА
377
в Семиречье 17—25 апреля. Осенний отлет протекает в конце августа и в
сентябре. Запоздавшие пролетные птицы задерживаются иногда в октябре
(Усть-Урт, 2 октября; Новенское 15 октября). Залет в гнездовое время
к Сарапулу (Першаков, 1929). Вероятно, только залетает на вост. Памир, но
на зап. Памире гнездится. Биотоп. Боровые леса с гарями или обширными
полянами; лесостепь; горные леса Туркестана (яблонники и осинники в
центральном Тянь-Шане; арчевники — Кугитанг, Фергана; фисташники в Баба-
таге). Вертикально до 1000 м в Алтае, где залетает и до 2300 м, 1400—1800 м
в Таджикистане, 1000—1500 м в Тянь-Шане.
Численность. Редкая птица, быть может, в связи с
чувствительностью к низким температурам (отмечались случаи гибели прилетных сплюшек
от весенних заморозко.в I.V. 1884, Зарудный, 1888). С другой стороны, быть
может, намечается расселение совки на север в Прикамской тайге в связи
с лесными пожарами 1920—1921 годов и размножением на гарях насекомых.
Экология. Размножение. У самки, добытой под Томском 19 мая,
сильно развиты яичники (Залесский, 1915); птенцы в мезоптиле с совершенно
короткими крыльями отмечены 22 июня (Башкирия), подлетки в мезоптиле —
6 июля (Ташкент), 15 июля (Красноярск), 30 июля (вполне выросшая птица
из выводка, Бийск). Следовательно цикл размножения происходит вероятно
тогда же, когда у номинальной расы. Гнезда находятся в дуплах тополей и
других деревьев, но также занимаются и постройки других видов птиц,
например, сорок (Алмаатинский заповедник, Шульпин, 1929). Число яиц 2—4
(Эверсманн, 1866), вероятно бывает и более. В выводках до 4 птенцов.
Запоздавшие молодые в мезоптиле еще 17 августа (Бузулук, маховые не вполне
доросли, вероятно от вынужденной второй кладки).
Линька. Изучена недостаточно, в основных чертах, как у
номинальной расы.
Питание. Сибирская сплюшка питается главным образом
насекомыми— жуками, бабочками; изредка мелкими птицами.
Описание. Размеры и строение. Длина самцов (4) 190—207, самок (3) 207—220,
в среднем 198,2 и 216 мм. Размах самцов (4) 490—525, самок (3) 523—545, в среднем 509,7
и 531,3 мм. Вес самца (1) 80,5, самки (1) 82,5 г. Крыло самцов (45) 145—161, самок (27) 150—
167,5 в среднем соответственно 153,1 и 158,1 мм, т. е. крыло немного, примерно на 5 мм
в среднем, длинней, чем у номинальной расы (правило Сибома, в связи с более далекими
перелетами?).
Окраска. Отличия от европейских совок незначительны и сводятся к тому, что
общий тон окраски серей и бледней, темный рисунок поэтому более выделяется, а беловатые
пестрины более развиты. Рыжей вариации окраски пока не найдено.
7. Восточноазиатская совка Otus sunia Hodgson
Scops Sunia. Hodgson Asiatic. Researchs, 1836, стр. 175. Непал.
Распространение. Ареал. Вост. Азия от Приморья, Японии и
Манчжурии на севере до Индии, Цейлона, Малайи и Индо-Китая; острова у
восточного берега Азии от Риу-Киу до Филиппин; Андаманские и Никобарские о-ва;
о. Мантанани у Борнео.
Характер пребывания. Восточноазиатская совка на севере
перелетная (Япония, Манчжурия, Приморье, быть может Сев. Китай), в других
частях ареала оседлая птица. Биотоп. Леса.
Подвиды и варьирующие признаки. Морфологические различия — окраска, величина,
и оперенность ног; также имеются различия в экологии, требующие впрочем выяснения,
так как большинство рас плохо известны. Описаны: О. s. stictonota S h a r p e, 1875, из
Манчжурии, Приморья, Сев. Китая и Кореи; О. s. japonica TemmincketSchlegel, 1850,
из Японии от Хоккайдо до Кюсю; О. s. malayanus Hay, 1847, из Малайи, Ассама к югу до
Брамапутры, Южн. Китая (южн. Юннань, Цзяньси, Гуаньдунь, Фуцзянь); Тенассерима,
Сиама, Индо-Китая, Андаманских и Никобарских о-вов; О. 5. sunia Hodgson, 1836, из
378
ОТРЯД СОВЫ
сев. Индии от нижнего пояса Гималаев (Кумаон-Бутан), на юг до Пенджаба, центральных
провинций, Бенгалии; О. s. rufipennis Sharpe, 1875, из южн. Индии, к югу от Бомбея
и Мадраса; О. 5. leggei Ticehurst,
1923, с Цейлона; О, $. interpo situs К и-
г о d а с о-ва Бородино; О. s. elegans
С a s s i n, 1852, на о-вах Риу-Киу; О. $.
botelensis К и г о d a, 1928, с о-ва Ботель-
Тобаго; 0. 5. calayensis Mc. Gregor,
1904, с о-ва Калайян, Филиппины; 0. s.
longicornis О g i I v i e-G rant, 1894, с
о-ва Люсон; 0. 5. mindorensis Wh i-
te h e a d, 1899, с о-ва Миндоро; 0. s.
romblontsM с. G r e g о г, 1905,с о-вов Бан-
тон и Ромблон, Филиппины; О. s.cujen-
sis М с. Gregor, 1904, с о-ва Куйо,
Филиппины; 0. s. mantananensis S h a r-
p e, 1892, с о-ва Мантанани к северу от
Борнео. Из этих 15 форм только 4
материковых, остальные островные, из них
филиппинские (и некоторые другие) —
с узким распространением и нередкое
нарушением обычных закономерностей
географических вариаций (например
форма с Риу-Киу очень крупных
размеСистематические замечания. Эта
группа многими авторами
рассматривается в качестве .подвидов Otus scops.
Однако признаки, отличающие
восточных совок от западных — при общем
характере изменчивости у совок и' при
наличии разобщенного ареала —
заставляют нас пока присоединиться к
мнению Тайсхерста, принимающего группу
0. sunia за особый вид. Для
окончательного суждения мы не располагали
материалом по тропическим формам этой
птицы.
•** Положение совок scops и sunia
относится по всей вероятности к
категории так называемых superspecies, причем
к ним следует в этом смысле присоединить ряд африканских и американских форм
(группа senegalensis, flammeolus и др., Делакур, 1931).
Карта 62. Распространение восточноазиатской
совки Otus sunia
2 — О. 8. japonlca, 3 — О. 8. ma-
sunia, 5 — О.
7 — О. 8. stictonotus, .
Iayanus, 4 — О. s. sunia, 5 — О. s. rufipennis, б — О.
s. leggei, 7 — О. s. interpositus, 8 — O. 8. elegans, 9 —
O. 8. botelensis, 10 — O. s. calayensis, 77 — O. 8. lon-
ficomie, 12— O. s. mindorensis, 13 — O. s. romblonis,
4 — o. 8. cujensis, 15 — O. s. mentananensis; a —
граница гнездовий.
Уссурийская совка Otus sunia stictonotus Sharpe
Scops stictonotus. Sharpe. Catalogue of the Birds in the British Museum, 1875, стр.54,
табл. Ill, фиг. 2, Китай.
Распространение. Ареал. Приморье на север приблизительно до 50°
с. ш. (река Хунгари), на запад до верхнего Амура и Большого Хингана;
Сахалин, Манчжурия, Сев. Китай (распространение там не вполне ясно), гнездится
в Чжили и южной части провинции Аньхуей, быть может к этой же расе
относятся совки, оседлые в провинциях Фу-цзянь и Гуаньдунь.
Характер пребывания. Уссурийская совка перелетная птица
(однако см. выше о гуаньдунских и фуцзянских птицах). Зимовки в ю.-в.
Китае и на о-ве Формоза.
Даты. Прилет в Приморье начинается в конце апреля (25.IV. 1933,
Иман), отлет—в конце сентября (пролет на о-ве Фуругельм 26—27. IX,
1946, у залива Посьет добыта еще 1.Х.1945). Весенний пролет в вост. Китае
бывает еще в мае. Биотоп. Равнинные леса, предпочтительно
лиственные, но также и хвойная тайга, охотно в долинах рек. Также и в городских
садах (Харбин, Яковлев, 1929). Численность. Нередкая птица Приморья.
В0СТ0ЧН0АЗИАТСКАЯ СОВКА
379
Экология. Сведения совершенно отрывочны. Размножение. Брачный
крик начинается с прилета и продолжается до второй трети июня, главным
образом по ночам, иногда и днем. Гнездится уссурийская совка в дуплах высо-
ких7деревьев. Самки с сильно развитыми яичниками добывались в конце мая.
Кладка повидимому в конце мая или в начале июня. Яйца закругленные,
овальной формы, матово-белые, размеры (3) 29,7; 30; 30,7 X 24,5; 25,5; 25,7 мм
(Латуш, 1932). Летные молодые в мезоптиле, не линяющие еще в контурный
наряд, добыты 14—23 августа на реке Хунгари. Разновозрастность птенцов
указывает на
насиживание с откладки
первого яйца.
Питание.
Уссурийская совка
питается главным
образом насекомыми
(Приморье); в Манчжурии,
преимущественно
жуками, также
воробьями и мышевидными
грызунами.
Полевые
признаки. Строго ночная
сова, весьма похожая
по внешности на
обыкновенную сплюшку,
но некоторые особи
ярко рыжие. Иной
голос: двухсложное
мелодичное «ке-вюю-ке-
вюю» или громкое
«тьок-клок-клок».
Описание. Р а з м е-
р ы и строение. В
отличие от обыкновенной
сплюшки — другая
формула крыла: 4>3>5>2>
>б>7>1. Кроме того, не
только пальцы, но и
нижняя часть цевки
голая. Вес самцов 75, 86,3 г, самок 78,4, 82,8 и 95 г (Горчаковская). Длина самцов (2)
190 (Дементьев), 179,6 (Черский, 1915), 184, 194 (Тачановский, 1891), в среднем 186,9 мм,
длина самок 192,8 (Черский), 184, 195, 203 (Тачановский), в среднем 196,9 мм. Размах
самцов 505, 510 (Дементьев), 507 (Черский), 510, 517 (Тачановский), в среднем 509,7 мм;
размах самок 525 (Черский), 523, 526 (Тачановский), 497, 522, 525 (Дементьев), в среднем
524 мм. Крыло самцов^(б) 138—152, самок (14) 140—158, в среднем 145,6 и 147,9 мм.
Окраска. Пуховой наряд? Мезоптиль — как у О. scops, но наствольные черточки
на спинной стороне развиты слабо, а поперечный рисунок на груди и брюхе обычно редкий,
весь общий тон серый (у серых птиц). Взрослые самцы и самки отличаются от О. scops очень
тонким, частым и мелким поперечным рисунком (пестринками) на спинной стороне, темно-
бурые настволья там менее резки, птица кажется поэтому однотонной и менее пестрой, чем
scops. Диморфизм в окраске выражен хорошо: имеется не только серый или бурый тип
окраски, то светлый, то темный, но и рыжеватый в разных оттенках. Среди изученного
материала из Приморья серый тип окраски количественно заметно преобладает. Редко
встречаются совки ярко золотистой ржавчато-рыжей основной окраски (из 33 особей из Приморья,
изученных нами —только одна; среди 7 японских тоже только одна).
Систематические замечания. Вопрос о том, следует ли соединять или разделять
континентальных stictonotus от островных японских, все еще нельзя считать окончательно
решенным, Обе формы во всяком случае очень близки, но на небольшом материале из Японии,
бывшем у нас, кажется, что темный рисунок на спинной стороне развит у японских птиц
ж'
Hi'
Рис. 98. Восточноазиатская совка.
380
ОТРЯД СОВЫ
слабей и тоньше, чем у материковых; быть может среди японских несколько чаще встречается
ярко рыжая вариация окраски; наконец, японские птицы несколько меньше, но впрочем
незначительно; крыло самцов из Японии (3) 142—143, самок (3) 146—150 мм.
РОД УШАСТЫЕ СОВЫ ASIO SCHAEFFER
Тип Asio otus
8. Ушастая сова Asio otus Linn.
Strix Otus Linnaeus Syst. Nat. изд. X, 1758, стр. 92, Швеция.
Распространение. Ареал. Европа и сев. Азия к северу до пределов
леса, на восток до Охотского побережья и Японии (Хоккайдо); к югу до Ирана,
Туркестана, Гималаев, Ганьсу и Сычуани в Китае; сев. Африка и Абиссиния;
Сев. Америка от Британской Колумбии и Канады до Южн. Китая и с.-з. Индии.
Карта 63. Распространение ушастой' совы Asio otus
1 — А. о. otus, 2 — А. о. canadensis, 3 — А. о. abyssinicus, 4 — А. о. wilsonianus; a — граница
гнездовий, г — область зимовок.
Характер пребывания. Ушастая сова оседлая, Кочующая,,
отчасти перелетная птица. Биотоп. Высокоствольные леса гор и долин*
Численность. Высокая; как и у других мышеядов, колебания по годам.
Подвиды и варьирующие признаки. По окраске и величине, а также экологическим
признакам (в частности, календарю периодических явлений, большей или меньшей оседлости
и т. д.) различают четыре подвида, из них — один островной и три с континентальным
распространением; из последних два — с весьма широким. В Европе, сев. Африке, Азии —
А. о. otus Linn., 1758; на зап. Канарских .о-вах —Гран-Канариа, Тенериффа,
Пальма —А. о. canadensis Madarasz, 1901; в высоких горах Абиссинии, Шоа,
в горных лесах восточной части Бельгийского Конго к западу от Танганайки А. о.
abyssinicus Guerin Meneville, 1843; в Сев. Америке от средних частей Британской
Колумбии, южных частей Мекензи, севера Онтарио, юга Квебека и Ньюфаундленда до северо-
запада Нижней Калифорнии, сев. Техаса, Арканзаса и Виргинии А. о. wilsonianus
Lesson, 1830; американская форма зимою доходит до Мексики, берегов Мексиканского залива,
и Флориды.
Обыкновенная ушастая сова Asio otus otus Linn.
Синонимы. Stryx deminuta. Pallas. Reise d. verschied.. Prov. d. Russ. Reichs,
II, 1773, стр. 706, Урал. — Asio otus turcmenica Зарудный и Билькевич.
Известия Закаспийского музея, 1918, стр. 16, «Теджен и Мургаб».
Русское название. Книжное.
Распространение. Ареал. С.-з. Африка и Азорские о-ва; Европа —
в Норвегии примерно до 65°, в Швеции около 68°, в Финляндии до 63°, около
УШАСТАЯ СОВА
381
•65° в Архангельской обл., около 62° в верховьях Вычегды (Устькуломское),
на Урале около 6(Р; на юг до Средиземного моря; сев. Азия по южной полосе
тайги, в Зап. Сибири граница известна плохо, по крайней мере до Тюмени;
в бассейне Енисея она доходит примерно до 60°, на Лене несколько южней
Якутска, далее —до Охотского моря (Удекой острог; с северного побережья,
как и из области с.-в. Сибири и е Камчатки и Сахалина, неизвестна), Приамурья
и Японии (Хоккайдо, Хонд*'), к югу до Малой Азии, Палестины, Ирана,
(южнокаспийские провинции, Паропамиз), Монголии, Ганьсу и Нань-Шаня,
Циньлинских гор в Китае; на о-ве Формоза; однажды найдены на гнездовье
в Кашмире. Зимою к югу от гнездовой области в с.-з. Индии в Пенджабе,
Синде и Кече, в Южн. Китае (Гуаньдунь, Фу-цзян); на Корсике, Кипре,
Египте; в Африке не встречается однако и зимой за пределами Палеарктики.
Залеты на Курильские о-ва, на Памир.
Характер пребывания. Оседлая и кочующая птица,
перелетная у северных пределов ареала (Архангельск — Молотов, равно как и в
Сибири). Даты. Пролет и отлет поздний, как это бывает обыкновенно у
полуоседлых видов. В районе Молотова прилет — около середины апреля; у
Чкалова уже около середины марта, у Красноярска в конце марта. Осенью отлет
на севере бывает с конца сентября (Приозерный район Архангельской обл.;
Молотов, Чкалов) и до конца ноября ; в средней полосе также еще в ноябре
(Тамбов, Чкалов), причем часть особей здесь зимует.
Биотоп. Леса — как в лесной зоне, так и в лесостепи. Предпочтение
оказывается хвойным деревьям, в особенности в холодное время года (в Сибири
однако гнездится и в осиновых и березовых колках, Залесские, 1915). Вместе
с тем, ушастая сова для охоты нуждается в перелесках и открытых
пространствах. Не избегает культурного ландшафта — парков, садов, кладбищ
(Туркестан). На юге —в горах: на Алтае до 2000 л£, в центральном Тянь-Шане до
3000 м> в Закавказье до 2750 м (Армения); зимою в горных местностях —
вертикальные миграции. В открытых безлесных местах, например в Туркестане,
отсутствует (в частности у Аральского моря, в Кара-Кумах; в последних однако
на кочевках встречается даже в центральных частях, например у Дарвазы.)
Численность. Высокая. Количество особей, как у других миофа-
гов, подвержено по годам колебаниям (смертность, плодовитость). К северу
от 60° редка, как спорадична и на юге (Туркестан; в Туркмении с
достоверностью найдена на гнездовье только на Теджене, и вероятно гнездится в низовьях
Аму-Дарьи). На пролете и зимой встречаются стайки и группы до 30 птиц
(но также и одиночки)*
Экология. Размножение. Гнездится в лесах, быть может в особо
благоприятные по кормовым условиям годы и в соответствующих местах среди
открытого ландшафта (Крым, Ирби, 1856). Обычно гнездится в старых гнездах
других птиц, главным образом вороновых (сорок, ворон, грачей), также
хищных птиц (сарычей, коршунов, в Приморье ястребиного сарыча). Есть
требующие подтверждения сведения о постройке совой собственных гнезд (Вальх,
1899); реже сова гнездится в дуплах, еще реже — на земле. В
неблагоприятные годы кладок иногда не бывает; в годы массового размножения грызунов
бывает полный повторный половой цикл. Брачный крик начинается в разных
числах марта. Кладка в апреле, обычно в первой половине, быть может уже
с конца марта, так как в конце апреля попадаются пуховики (Ленинград,
Новгород, Сыр-Дарья).
Период кладки растянут, причем зависимостей от широты установить
не удается. Полные кладки найдены в апреле (слабо насиженные и частью
свежие), отмечены и в мае (начале мая, Молотов; 25 мая едва насиженные
у Красноярска). Число яиц в кладке в среднем 4—5, иногда менее (3, Еруслан;
2, Омск), в благоприятные годы яиц больше —до 7,8 и даже 9 (Ленинград),
382
ОТРЯД СОВЫ
в исключительных случаях 10 (Казань, Рузский, 1891). Промежуток между
откладкой яиц — 28 часов. При утрате кладки вторая кладка состоит из
меньшего числа яиц (3, по Спангенбергу, 1936). Яйца белые, эллиптической
формы. Размеры (26) 39—45 х 32—34,5, в среднем 41,4 х 33,3 мм (Умань,
Гебель, 1879), (12) 39—41,9 х 32,4—34 мм (Харьковская обл., Сомов, 1897),
(9) 39,6—45x33—34 мм (Забайкалье, Тачановский, 1891).
Насиживание начинается с первого яйца (разновозрастные птенцы и
зародыши); судя по добытым у гнезд птицам —насиживает только самка. Срок
насиживания около месяца (птенцы вылупляются в первой половине мая,
в разных местностях СССР; по западноевропейским данным — Уайзерби,
1938 и др.—длительность насиживания 27—28 дней). Летные молодые
встречаются около середины июня; в это время у них начинается линяние
мезоптиля. Гнездовый период продолжается следовательно около месяца (по
упомянутым уже западноевропейским источникам — 24 дня). Одно яйцо в
кладке нередко бывает неоплодотворенным, а число птенцов в выводке
меньше, чем число яиц. В неблагоприятные годы имеет место каннибализм
(Уттендерфер, 1939; для СССР прямых наблюдений нет). Носит добычу для
птенцов самец, оделяет их самка. При обилии пищи у гнезда образуются
запасы, а в исключительно благоприятные («мышиные») годы совы имеют и
вторую кладку в конце лета и даже осенью.
Линька. Изучена недостаточно. Смена нарядов обычная для сов:
пуховой наряд — мезоптиль — первый годовой наряд (с сохранением маховых
и рулевых от мезоптиля) — второй годовой наряд (по окраске не отличимый
от первого годового и окончательный) — третий годовой наряд и т. д. Линька
(за исключением смены мезоптиля) полная годовая. Смена мезоптиля на:
контурное перо происходит с середины июня, конец ее — в последних числах
августа или в начале сентября. Годовая линька (по небольшому материалу)
начинается еще у гнезда, повидимому в начале июля; в конце этого месяца
сменяются средние первостепенные маховые, к концу августа линяние маховых
продвигается на 60—70%. Последовательность сдоены маховых идет от заднего
края партии к переднему. Однако по европейским авторам линяние начинается
в сентябре и продолжается до декабря. Конец линьки вероятно наступает
зимой; в ноябре — декабре добытые в Туркмении птицы в свежем пере.
Питание. Пищу обыкновенной ушастой совы составляют, в основном,
мышевидные грызуны (например, среди исследованных птиц на с.-з. Кавказе
97,9%, Жарков, 1930). О связи между размножением сов и «урожаем» грызунов
см. выше. Потребность в пище одной совы — около 30 г в сутки, т. е. примерно
до 3 мышей. К выводку суточный принос, не считая запасов, 10—12 мышей
(Вальх, 1899). Из кормов ушастой совы в СССР указываются разные полевки,
в особенности Microtus arvalis; лесные и полевые мыши, водяные крысы;
пластинчатозубая крыса (в Туркмении, зимой); кустарниковые полевки
Pitymys та fori (с.-з. Кавказ, Жарков, 1938); пеструшки Lagurus tagurus;
мышевки Sicista (Забайкалье, Скалой, 1935); крысы домовые; тамарисковые
песчанки (Спангенберг, 1936).
Птицы занимают в кормовом режиме ушастой совы относительно
небольшое место и сова, повидимому, не может полностью перейти на орнитофагию
при «неурожае» мышевидных грызунов (для СССР указываются из птиц большой
пестрый дятел, разные виды от воробья до галки; разные виды жаворонков),
Насекомые в пище совы имеют повидимому случайное значение, как и лягушки
(Зап. Европа). Погадки похожи на погадки неясыти, размер их примерно
5 х 2 см.
Полевые признаки. Сова средней величины, с явственными перяными
«ушками». Брюшная сторона с поперечным и продольным рисунком. Образ
жизни строго ночной, днем сидит на дереве у ствола, вытянувшись и прижав
БОЛОТНАЯ СОВА
383
оперение» Брачный полет — зигзагообразный, от дерева к дереву. Голос
(в брачный период) «ху-хуу».
Описание. Размеры и строение. Размеры средние, есть длинные перьевые
ушки; лицевой диск хорошо выражен; ушные отверстия велики и симметричны, но
кожистая складка правого и левого наружных ушей
несимметрична; пальцы ног густо оперены;
крылья длинные, формула 2>3>1>4>5...,
наружное опахало сужено на 1-м маховом: 3-е
маховое с вырезкой. Хвост закругленный, из
12 рулевых. Общая длина самцов (10) 350—
375, самок (14) 371—391, в среднем 362 и
378,46 мм. Размах самцов (10) 860—930, самок
(15) 930—1010, в среднем 893,7 и 972,5 мм.
Вес самцов (3) 233—257, самок (5) 284—330,
в среднем 243,3 и 303,4 г. Крыло (самцов) (125)
276—309, самок (150) 282—320, в среднем 294,8
и 298,6 мм.
Окраска. Пухоиой наряд охристый,
удлинение «ушек» уже заметно. Мезоптиль:
перья спины, крыльев, груди, брюха серовато-
охристые с рыжеватыми вершинами (поэтому
птенец, одетый растущим мезоптилем, кажется
рыжеватым) с i емнобурым, тонким поперечным
рисунком; «лицо» почти черное; маховое и
рулевые, как у взрослых. Взрослые птицы (самцы
и самки): основной тон окраски более или
менее насыщенный охристый с темным буровато -
серым рисунком. Спинная сторона с темнобу-
рыми наствольными чертами и мелкими
буроватыми пестринками, образующими
«мраморный» рисунок; белый цвет выступает на
наружных опахалах плечевых, средних и больших
кроющих крыла; маховые с широкими бурыми
поперечными полосами и мраморными
пестринками, у стволов — желтоватые, у краев опахал
бледнее; надхвостье с мелкой
буровато-мраморной пестриной; рулевые ржавчато-охристые
с темными поперечными полосами и мелкими
пятнами; лицевой диск охристый с черным
пятном у глаз; горло беловатое; брюшная
сторона рыжеватая, охристая или даже беловатая
с широкими продольными и узкими
поперечными полосами на зобу, груди и брюхе;
подкрылья желтоватые с бурым пятном у
основания маховых и часто с бурыми наствольями;
подхвостье с резким продольным и поперечным
бурым рисунком. Радужина желтая или
оранжевая, клюв и когти черные; восковицасерая.
Систематические замечания. Окраска ушастых сов весьма изменчива как по основному
тону, так и по развитию темного рисунка. Основной тон варьирует от охристо-беловатого до
насыщенного ржавчато-рыжего; общий тон спинной стороны иногда не буроватый, а серый.
Темный рисунок различной интенсивности от серовато-бурого до темнобурого; развитие era
также различно—поперечные полосы на боках то сплошные и правильные, то прерванные и
редкие; подкрылья иногда без пятен, иногда же резко испещрены; полос на рулевых разное
число (от 4 до 7 на крайних парах). Все эти вариации носят индивидуальный характер и
географически не локализованы (среди изученных свыше 300 птиц самые бледные из
Джунгарии с Урунгу и из Синьцзяна в окрестностях Кашгара, но таких только две и они не
достоверно гнездовые местные). В Европе и Азии ушастые совы, судя по нашему материалу,
географически не варьируют.
9. Болотная сова Asio flammeus Pontoppldan
Strix Flammea. Pontoppidan. Danske Atlas, I, 1763, стр. 617, табл. 25, Дания.
Русское название. Книжное.
Распространение. Ареал. Европа от тундр до Средиземного моря; сев.
Азия от тундр на восток до Камчатки и Сахалина, на юг до Палестины, М. Азии„
Рис. 99. Ушастая сова.
384
ОТРЯД СОВЫ
Месопотамии, Туркестана и Монголии; Америка от севера Аляски и Мекензи
до юга Перу, Боливии и Бразилии, Галапагосских о-вов; кроме того на
Каролинских и Гавайских о-вах. В холодное время года от умеренной
полосы Европы и Туркестана до сев, Африки, южн. Аравии, сев. Индии,
ю.-в. Китая и Аннама, средней Америки.
Характер пребывания. Болотная сова в северных частях
ареала перелетная, в остальных кочующая и оседлая птица. Сезонные
передвижения (как и колебания численности, зависящие от «урожая» кормов), носят
иногда нерегулярный, «инвазионный» характер. Биотоп. Открытые
ландшафты—тундра, речные долины, степи,—как равнины, так и горы.
Численность. Высокая, но меняющаяся по годам.
Карта 64. Распространение болотной совы Asio flammeus
7 — A. f. flammsus, 2 — A. f. bogotensis, 3 — A. f. suindae, 4 — A. f. domingensis, 5 — A. f. portoncensis,
6 — A. f. galapagoensis, 7 — A. 1. sanfordi, 8 — A. f. sandwichensis, 9 — A. f. pokapensis; a — граница
гнездовий, б — неясная граница гнездовой области, г — область зимовок. Область — распространения
^викарирующего вида Asio capensis I. А. с. tingitanus. — П. А. с. capensis. — III. А. с. nova.
Подвиды и варьирующие признаки. Вид мало изменчивый в отношении морфологии —
все северное полушарие занято одним подвидом. Но в южном полушарии имеется несколько
различающихся по окраске и размерам островных форм с ограниченным распространением
(не вполне, быть может равноценных в таксономическом отношении с географическими
расами в строгом смысле слова). Географические различия в экологии — в характере
пребывания (оседлость — перелетность), быть может в особенностях репродуктивного цикла и
в трофических связях. Описаны подвиды: в Европе, Азии и Америке А. /. flammeus Pon-
to р р., 1763; в Южн. Америке на юг от южн. частей Боливии, Бразилии и Перу А. /.
suinda Vieillot, 1817; на Фалкландских о-вах А. /. sanfordi Bangs, 1919; на о-ве
Испаниола в группе Больших Антильских A./, domingensis P. L. S. Miiller, 1776; на о-ве
Порто-Рико в томже архипелаге A.f.portoricensis R i d g w а у, 1882(быть может уже
истреблена); на Галапагосских о-вах А. /. galapagoensis Gould, 1837; в сухих высокогорьях
Анд в Колумбии и Эквадоре А. /. bogotensis Chapman, 1915; на Гавайских о-вах А. /.
sandwichensis В I о х h a m, 1826; на о-ве Понапе из группы Каролинских A. f. ропареп-
sis M а у г, 1933.
В Африке от Марокко и сев. Алжира до крайнего юга и Мадагаскара распространен
очень близкий вид Asio capensis A. S m i t h, 1834, распадающийся на три подвида, палеарк-
тический А.с. tingitanus Loche, 1867 в южн. Испании, Алжире, Марокко, у озера Чад;
А. е. capensis S m i t h на эфиопской части материка к югу от тропических лесов; на севере
до Анголы, Кении и вероятно Абиссинии; А. с. nova Streseraann, 1922 на Мадагаскаре.
БОЛОТНАЯ СОВА
385
Обыкновенная болотная сова Asio flammeus flammeus Р о n t о р p.
Синонимы. Stryx accipitrina. Pallas. Reise d. verschied. Prov. d. Russ. Reichs
I, 1771, стр. 455, Каспийское море. — Stryx Caspia. Shaw. Gener. Zool. VII, 1809, стр. 272,
Каспийское море. — Stryx Aegolius. Pallas. Zoographia Rosso—Asiat. 1. 1811, стр. 309,
сев. Сибирь. — Otus leucopsis. С h r. L. В re h m. Der vollstandige Vogelfang, 1855, стр. 413,
Сарепта. — Asio accipitrinus pallidus. S а г u d n у und Loudon. Ornith. Monatsberichte,
1906, стр. 151, Зап. Сибирь.
Распространение. Ареал. От южной полосы тундр на севере — в
Скандинавии около 70° с. ш., в Советской Лапландии до морского побережья;
Архангельск, Канин, Тиманская и Болынеземельская тундра; на Ямале до
70° с. ш. (Житков, 1913), на Енисее по крайней мере до Дудинки, на Таймыре
до Боганиды (Миддендорф), в Вост. Сибири в бассейнах Лены, Индигирки и
Колымы — до устья этих рек; на восток до Анадыря, Охотского моря,
Камчатки, Сахалина; на Командорских, Прибыловых и Алеутских о-вах. В Европе
отсутствует в Ирландии, на Пиренейском п-ове и в Греции. Гнездится
в Европейской части СССР к югу до Черного моря (Украина, Крым), Кавказа;
в Азии до Малой Азии, Палестины, Месопотамии, сев. частей Туркестана
(низовья Аму-Дарьи у Айбугира, Молчанов, 1912, возможно несколько южней;
на юге до Аральского моря на восточном его побережье; в приаральских
Кара-Кумах, в зап. Казахстане в северной степной части), в Забайкалье и сев.
Монголии (Кентей), центральной Манчжурии, Приморья; точная граница в*
Центральной Азии не выяснена. В Сев. Америке от северных частей Аляски
(мыс Блоссом, река Кобук, мыс Барроу), сев. Мекензи, Квебека (Унгава),
ю.-в. части Баффиновой Земли до Калифорнии, Колорадо, южн. Канзаса,
Миссури, сев. Огайо, сев. Индианы, берегов Нью-Джерсея. В негнездовое
время —■ в средней и южн. Европе, главным образом в Средиземноморье
и в черноморских степях; в сев. Африке в области саванн к югу от Сахары до
южн. Нигерии, озера Чад, Сомали, Абиссинии, Судана и даже Кении; в
Азии до южн. Аравии, Ирана, Туркестана, Белуджистана, сев. Индии и
Бирмы, Цейлона, ю.-в. Китая, Аннама, Японии и Формозы; в Америке от
Британской Колумбии, Уайоминга, Миннесота, Огайо, Массачузетса, Луизианы,
Гватемалы и Кубы.
Характер пребывания. Обыкновенная болотная сова на севере —
примерно от 50° с. ш. — перелетная, южней —кочующая и быть может оседлая
птица. Отдельные особи отмечались зимой и на севере—к северу от Якутска
(27 декабря, Ткаченко, 1932), в центральном Алтае (Сушкин, 1938), у Ленинграда,
Пскова, в южн. Скандинавии. Миграции носят отчасти нерегулярный
характер и повидимому зависят от обилия основной пищи (грызуны) и ее доступности
(снеговой покров). При благоприятных условиях совы зимуют и в средней
полосе (Приуралье, 1889—1890, у Чкалова, в сев. Казахстане). В другие
годы совы в большом количестве скопляются в местах массового размножения
грызунов, например у Днепропетровска в 1894 г. (Вальх, 1894), в
новороссийских степях и южной Бессарабии при массовом размножении полевых и
домовых мышей в 1906 году (Браунер), зимой 1945—1946 г: в с.-з. Казахстане
и т. д. Массовые налеты в вост. Германию — от Силезии до Саксонии — имели
место в 1930, 1931, 1933, 1934, 1936—1937 гг. (Нитхаммер, 1938). Все же и при
таких «инвазионных» миграциях наблюдается, повидимому, определенное
направление движения на запад и на юго-запад (окольцованная у Аскания-
Нова в таврических степях сова добыта в январе у Будапешта; окольцованная
в Силезии на пролете добыта через год в Латвии и т. д.).
Даты. Весенний пролет и прилет ранний —в Туркмении (ю.-в. Каспий,
бассейн Атрека) уже в начале марта, пролет в Туркестане. Кончается около
середины апреля (северные тундровые популяции?), единицы задерживаются
и позднее. В эти же примерно числа протекает и пролет на Украине (Харь-
25 Птицы СССР, т. I
386
ОТРЯД СОВЫ
ков, Киевская обл.). В средней полосе появляется около середины апреля
(Ленинград, Москва, Смоленск, Псков, Казань), в тундре — в разных числах
мая, обычно около середины этого месяца — в это время южные
популяции уже гнездятся. Отлет — поздний и растянутый, главным образом в
сентябре (впрочем, отдельные особи задерживаются позже даже на севере,
например в низовьях реки Таз у Хальмер-седэ 26 сентября; 17 октября добыта
Сокольниковым на Анадыре; у Дудыпты на южн. Таймыре —9 октября; на
Колыме добыта Бутурлиным 19 сентября, а на Алазее им же 21 сентября).
Северные популяции в средней полосе пролетают в октябре. Движение
медленное: на юге, на Украине и в Туркестане заметный пролет еще в
ноябре.Появление на среднеазиатских зимовках отмечено главным образом в ноябре
(Таджикистан, Туркмения, Киргизия).
Биотоп. Открытый ландшафт, в гнездовое время связанный с
водоемами: речные поймы и луга, тундры, но также и сухие степи (Украина, Крым,
Сев. Кавказ, Казахстан), главным образом на равнинах, но также и в горах:
на ю.-в. Алтае до 2000—2350 м, в Армении до 2000—2350 м. Зимой охотно
в открытом культурном ландшафте. Встречается в холодное время и в пустыне
(Кызыл-Кумы, Кара-Кумы).
Численность. Высокая, подверженная периодическим колебаниям
по годам, повидимому на основе изменения плодовитости, в связи с кормовыми
условиями («урожай» или «неурожай» мышевидных грызунов). Большая
плодовитость обеспечивает быстрый подъем численности болотных сов в
благоприятные годы. В этих случаях наблюдаются и повторные кладки (см. ниже)
в местах массового скопления грызунов, где совы держатся группами и
стайками по 20—30 птиц. Размещение сов по ареалу носит несколько
спорадический характер и плотность не равномерна. Высокой численности вида не
препятствует и то обстоятельство, что болотная сова, при дневном образе жизни
и привязанности к открытому ландшафту, чаще других сов становится жертвой
разного рода хищников — в частности, крупных соколов, тетеревятников,
лисиц (даже пустельги, Мургаб, Лоудон, 1910).
Экология. Размножение. Период размножения у разных
популяций существенно варьирует в зависимости от широты. Брачный полет и
токование начинаются с прилета. Птица в это время летает над гнездовым участком
с характерным криком и хлопаньем крыльев, не только в сумерках, но и днем.
Болотная сова — единственный из видов сов нашей фауны, строящий
собственные гнезда. Расположено гнездо на земле чаще под кустом и сделано из сухих
растений. Иногда впрочем гнездо — только слегка расчищенная площадка с
вытоптанной во мху ямкой (тундры) и т. п., в относительно сухих местах.
Указание Волчанецкого (1927) об использовании болотной совой сорочьего
гнезда относится, конечно, к ушастой сове.
Кладка на юге (Украина: Харьковская обл., б. Екатеринославская
губ.) начинается около середины апреля, в Сибири поздней, —в конце
апреля — начале мая (судя по датам вылупления и роста птенцов и т. д.), в тундре
еще поздней, хотя прямых наблюдений там нет (в низовьях Лены птенцы не
вывелись еще 23 июня; вывод на южн. Ямале только 22 июля, Осмоловская,
1944). Число яиц в кладке варьирует, повидимому в связи с кормовыми
условиями: обычно 3—5, но в «мышиные» годы —до 7 (Иман, Спангенберг, 1940;
б. Екатеринославская губ.) и даже 9—10 (9 — Чкалов, 9 — Ерусланская
степь, 9— Ямал, 10 —б. Екатеринославская губ.; по Хартерту, 1914, даже
до 14 яиц). В «мышиный» год в Днепропетровской обл. в четырех гнездах по
8, в трех по 9, в одном 7, в одном —10 яиц (Вальх, 1899). Промежуток между
откладыванием яиц, судя по датам вылупления птенцов, 48 часов. Яйца
белые, менее блестящие и продолговатые, чем у ушастой совы. Размеры
38,2—42,3x31,5— 33 мм (низовья Лены, Плеске, 1928). (12)38,2—42,3x31,5—
БОЛОТНАЯ СОВА
387
33 мм (Даурия, Тачановский, 1891); (7) 38—40 х 30,5—31,6 мм (Иман, Спан-
генберг, 1940). В исключительные по кормовым условиям годы, при массовом
размножении мышей, имеются вторые поздне-осенние и даже зимние кладки;
такое явление имело место на Сев. Кавказе в 1929 году; в 1906 году, при
обилии Microtus arvalis и Mus musculus hortulaniis в Бессарабии и
Новороссийских степях вторая кладка была в начале зимы, подлетки в декабре 1906 и
в январе 1907. В 1948 г. летом в Приморье было массовое размножение
грызунов. В связи с этим — у болотных сов был второй цикл, и 19 ноября в районе
ст. Лиховцы, в 40 км к с.-з. от Ворошилова-Уссурийского в гнезде найдено
5 птенцов и 2 яйца. Численность сов была очень высокой (Воробьев).
Насиживание, судя по возрасту птенцов и зародышей, с первого яйца.
Насиживает самка, а самец носит ей в это время корм (Чкалов, Ямал). Вылупле-
ние птенцов происходит в зависимости от широты —от конца мая (Казань,
Иман и т. д.) до второй половины июля (Ямал). Вес птенца по вылуплении
13,7—14 г (Осмоловская). Птенцы в выводке весьма разновозрастны: когда
старшие начинают оперяться, младшие еще в пуху. Срок насиживания,
следовательно, около месяца (у нас точно не прослежен; по западноевропейским
данным 24—29, в среднем 27 дней), старшие птенцы несколько дольше, чем
младшие, так как в первые дни насиживание неполное (Уайзерби, 1938). Гнездовый
период короткий, в средней полосе около середины июня птенцы уже на крыле
и оперились. Самостоятельные молодые там же (например, в Наурзуме, с.-з.
Казахстан), в первой трети августа. Из гнезд птенцы выбираются очень
рано, в мезоптиле и нелетными, в возрасте 14—17 дней (Зап. Европа). При
этом птенцы просят корм у родителей шипеньем, а последние разыскивают
сидящих в разных местах вокруг гнезда птенцов по слуху. Летными молодые
болотные совы становятся в возрасте около месяца. При неблагоприятных
обстоятельствах в выводках бывают случаи каннибализма (южн. Ямал).
Требует выяснения вопрос о различиях в экологии размножения
североамериканских и европейско-азиатских болотных сов: у первых будто бы насиживают
и самец и самка, в течение трех недель (Берне, 1915). Осенью происходит
неполный вторичный половой цикл (брачный полет отмечен в июле).
Линька. Обычная для сов последовательность смены нарядов:
пуховой — мезоптиль — первый годовой (по окраске окончательный, с маховыми
и рулевыми от мезоптиля) — второй годовой наряд и т. д. Пуховой наряд
заменяется мезоптилем в возрасте около 2 недель. Смена мезоптиля контурным
первым годовым нарядом наступает в июле — августе, в сентябре она
закончена. Линька полная годовая (кроме частичной смены мезоптиля), длительная,
между второй половиной гнездового периода и отлетом, прослежена еще
недостаточно. Уже в половине сентября встречаются особи в свежем пере,
закончившие линяние (13сентября, с.-в. Алтай); с другой стороны,
западноевропейские авторы указывают, что линька болотных сов заканчивается в ноябре.
Растянутость линьки находится в соотношении с различиями в сроках
размножения разных популяций. Последовательность смены маховых — обычная
у сов, от внутреннего края крыла к наружному (от 10-го к 1-му).
Питание. Болотная сова — типичный миофаг, остальные животные
носят в ее кормовом режиме случайный характер. Типичные экологические
отношения для миофагов, отчасти отмеченные выше: авторегуляция
численности популяций путем повышения или снижения плодовитости в соответствии
с кормовыми условиями, высокая потенциальная плодовитость, нерегулярный
«инвазионный» характер сезонных перемещений и т. д. Колебания численности
сов идут параллельно с колебаниями численности грызунов (Харьковская
обл., Сомов, 1897; в 1922 у Кизляра, Беме, 1925; в 1895, в б. Екатеринослав-
ской губ., Вальх, 1899; в 1928 г. в Волжско-Уральской степи, Волчанецкий,
1937; в 1906—1907 в окрестностях Одессы, Браунер). Охотится сова на севере
25*
388
ОТРЯД СОВЫ
и днем (в тундре в июне — июле), но предпочитает сумеречные часы. На охоте
чаще всего летает над землею, «трясется» редко. Есть указания на
подстерегание совами добычи сидя на земле (Нарым). Выводку — по крайней мере
когда птенцы малы — добычу носит самец, в благоприятные годы у гнезд
большие запасы (например, у гнезда с 8 птенцами б Arvicola terrestrls, 4 Microtus
arvalls, 4 Ellobius talpinus, 2 Lagurus lagurus, 1 Locnstella fluvlatllis, 1 Porzana
Рис. 100. Болотная сова.
pusllla) у гнезда с 4 насиженными яйцами 5 Ochotona pusilla и б Lagurus
lagurus, Чкалов, Зарудный, 1892; у гнезда с 6 птенцами 7 Stenocranlus slovzoul,
Приалтайская степь, Штегман, 1926). И на севере, и на юге грызуны составляют
более 90% пищи болотных сов. Примеры типичных кормов: разные полевки
91%, в том числе Dlcrostonyx torquatus, Lemmus obensis, Microtus mtddendorffl,
Stenocranlus major, кроме того, немного воробьиных птиц и яйца (р. Щучья,
южн. Ямал Осмоловская); млекопитающих 94%, в том числе Microtus arvalis,
М. oeconomus, Clethrtonomys glareolus, Arvicola amphlblus, Apodemus agrarlus,
немного жуков, среди них Geotrupes и Hydrous (Татарская республика, Жарков
и Теплов, 1932)..
Другими авторами упоминаются в качестве корма болотной совы:
лемминги (Лапландия); летучие мыши (Ульяновская обл., Житков и Бутурлин,
БОЛОТНАЯ СОВА
389
1906), «мыши», полевки, тушканчики Alactaga jaculus, кроме того мелкие
птицы и насекомые (Харьковская обл., Сомов, 1897), полевки (дельта
Индигирки, Михель, 1935; Кизляр, Беме, 1925; Волжско-Уральская степь, Волча-
нецкий, 1937); полевые мыши, полевки, водяные крысы, хомяки, ежи,
тушканчики, иногда мелкие птицы (Поволжье, Богданов, 1871); степные пеструшки и
полевки (Приерусланские степи, Волчанецкий, 1934); жуки навозники
(Забайкалье, Скалой, 1935); водяные крысы, домашние мыши, CrocidurQ (Талыш,
Тугаринов и Козлова, 1938); водяные крысы, степные пеструшки,
тушканчики, хомяки, утята; мыши; воробьи; удод, молодые голуби—в гнездовое
время, а осенью и весною взрослые галки, утки и т. п.; иногда ночующие в
камышах ласточки (Чкаловская обл., Зарудный, 1888, 1892). Намечается,
таким образом известное изменение кормового режима по сезонам — в
негнездовое время года болотные совы менее исключительные мышеяды, чем
в гнездовое.
В Туркмении на зимовке в подгорной полосе по окраине Кара-Кумов
болотные совы охотятся главным образом по зорям на больших песчанок и
нападают на ночующие стаи хохлатых жаворонков.
Сова может съесть, очень много — в желудке одного экземпляра были
найдены 3 взрослых мыши, 5 мышат и жук (Горьковская обл., Серебров-
ский, 1918).
Полевые признаки. Одна из немногих наших сов, встречающихся в
открытом ландшафте и избегающая деревьев (не садится на них, ночует и отдыхает
на земле).
На полете при носке добычи несколько напоминает луня (длинные
крылья, летает на небольшой высоте).
Голова относительно небольшая, желтые небольшие глаза, «черное* лицо,
стройное сложение. Нередко активна и днем. Будучи сходной по размерам
с ушастой совой (средняя величина), отличается от нее короткими, обычно
прижатыми «ушками»; спинная сторона не серовато-бурая, а
охристо-буроватая, брюшная сторона только с продольным рисунком. Брачный полет —
с хлопаньем крыльев, «пение» — ряд повышающихся звуков «бу-бу-бу-буб»,
крик «кьяв-кьяв».
Описание. Размеры и строение. Перьевые ушки короткие. Крылья длинные:
формула крыла 2>3>1>5>6... (настоящее 1-е маховое рудиментарно и в счет здесь не
принимается); сужение наружного опахала 1-го махового и внутреннего 2-го махового. Длина
самцов (36) 340—415, самок (28) 340—423, в среднем 372,8 и 381,8 мм. Размах самцов
(30) 870—1090, самок (20) 926—1092, в среднем 1000,2 и 1045 мм. Крыло самцов (90) 281—
329, самок (88) 285—335, в среднем 307,65 и 310,2 мм. Вес самцов (10) 320—385, самок (4)
400—430, в среднем 350 и 410,5 г.
Окраска. Пуховой наряд охристо-беловатый. Мезоптиль рыжеватый на брюшной
стороне, бурый с рыжеватыми пятнами и каемками на спинной; лицо черное; маховые и
рулевые, как у взрослых (ем. ниже). Взрослые самцы и самки: общий тон окраски очень изменчив
и колеблется от ржавчато-желтого в разных оттенках до бледноохристого и даже белого; по
светлому фону бурый продольный рисунок, к которому на спине и крыльях присоединяются
поперечные пестринки; на плечевых и больших кроющих крыла беловатые пятна на внешних
опахалах; первостепенные маховые охристо-рыжеватые с темными стволами и слабо
развитым бурым поперечным рисунком в вершинной части пера, последняя серовато-бурая;
второстепенные маховые с правильным серовато-бурым поперечным рисунком и узкой темной
полосой по стволу пера.
Лицевой диск белый, с черным кольцом вокруг глаза и с черными пестринками
у горла; брюшная сторона тела с темными бурыми продольными наствольями,-более
широкими на зобе и суживающимися на боках и брюхе; ноги и подхвостье не испещрены;
подкрылья беловатые с немногочисленными маленькими продольными полосками и темно-
бурым пятном у основания маховых. Клюв и когти черные, радужина желтая.
Общий тон очень выцветает перед линькой. Самцы в среднем бледней самок. Впрочем,
индивидуальные вариации очень велики, и отдельные особи среди самцов бывают такими
же желтыми, как самки, а отдельные самки — почти белыми по основному фону окраски.
Географических вариаций окраски в пределах СССР нет.
390
ОТРЯД СОВЫ
РОД ИГЛОНОГИЕ СОВЫ NINOX HODGSON
10. Иглоногая сова Ninox scutulata Raffles
Тип «Ninox nipalensis>>
Strix scutulata. Raffles. Transact. Linn. Soc. of London, 1822, 13 pt. 2, стр. 280, Суматра.
Распространение. Ареал. Вост. Азия от Японии и Приморья до Южн.
Китая, Индо-Китая, Индии и Цейлона, Больших и Малых Зондских о-вов.
Характер
пребывания. На севере —. в Приморье
и Японии иглоногая сова
перелетная, а на юге — оседлая птица.
Биотоп. Лесистые местности,
главным образом на равнинах,
также — в нижнем поясе гор (в
Японии до 1000 м)\ не избегает
культурного ландшафта.
Подвиды и варьирующие признаки.
Несколько географических форм,
различающихся размерами и окраской. В
Японии, средней и южн. Корее, в Китае
до Цзяньсу и Фуцзяна N. s. scutalata
Raffles, 1822; в Приморье, сев.
Корее и с.-в. Манчжурии N. s. macroptera
В 1 a s i u s, 1888; в ю.-в. Азии от Ассама
и южн. Юннани до Сиама и Индо-Китая
N. s. burmanica Hume, 1876; на севере
Индии, к югу до центральных частей
п-ова (на востоке до Мадраса) N. s. lugu-
bris T i с k e 1 1, 1833; в южн. Индии
и на Цейлоне N. s. hirsuta Tern-
m i n с k, 1824; на Андаманских и
Никобарских о-вах N. s. obscura Hume,
1873; на Малайском п-ове, Суматре и
прилежащих островах N. s. moluccensis
Е у t о n, 1845; на зап. Яве N. s. java-
nensis Stresemann, 1928; на Бор-
Натуна N. s. borneensis Bonaparte, 1850.
Карта 65. Распространение иглоногой совы
Ninox scutulata
1 — N. s. macroptera, 2 —N. s. scutulata, 3 — N. s.
burmanica, 4 — N. s. moluccensis, 5 — N. 8. obscura,
6 — N. s. javanensis, 7 — N. s. borneensis, 8 — N. s.
ugubris, 9 — N. s. hirsuta; a — граница гнездовий,
г — места зимних встреч N. s. macroptera.
нео и северных островах архипелага
Уссурийская иглоногая сова Ninox scutalata macroptera В1 a s i u s
Ninox macroptera. Bias i us. Braunschweiger Anzeiger, 1888, № 9, стр. 86, о. Большой
Сангир к северу от Целебеса.
Синоним. Ninox scutalata ussuriensis. Бутурлин. Орнитологический Вестник, 1910
стр. 187, бассейн Уссури.
Распространение. Ареал. С.-в. Манчжурия, сев. Корея, Приморье,
на север до р. Хунгари (ок. 50° с. ш.), на запад до Хабаровска. Детали
распространения неясны. Характер пребывания. Перелетная
птица. Зимние экземпляры известны с Борнео (Бутурлин, 1910, Хартерт,
1912), с Филиппинских и Зондских о-вов (Мейзе, 1941). Даты. В
Приморье добывались в середине мая и в сентябре, даже в начале октября
(Черский, 1915). Близкий подвид в Японии появляется у Кобе в конце апреля,
северней — в начале мая и будто бы остается до ноября (Ян, 1942). Биотоп.
Леса, смешанные, лиственные, с примесью кедра и пихты (Воробьев, 1943),
также приречные заросли. Численность. В Приморье редка.
Экология. В пределах СССР неизвестна. Размножение. У самки
из Приморья наседные пятна еще не заросли 7 июля (Тачановский, 1891),
выводок 3 августа, у молодой птицы из него маховые не доросли
(Бутурлин, 1911).
мохноногий сыч
391
Питание. Пищу уссурийской иглоногой совы составляют главным
образом насекомые, охотится она в сумерках. В желудке добытой птицы
найдены остатки насекомых, в том числе водяные жуки Cybister faponicus
(Черский, 1915, Бутурлин, 1911). У других главным образом жужелицы,
навозники Qeotrupes stercorarius, ночные бабочки (Воробьев, 1948).
Полевые признаки. Внешность напоминает дневную хищную птицу,
голова небольшая, крылья длинные. Голос «ху-ху», двусложный.
Описание. Размеры и строение. Сова средней величины. Лицевой диск
выражен слабо. Ушные отверстия невелики и закруглены. Перьевых ушек нет. Оперение жесткое и
плотно прилегающее к телу. Крылья длинные и острые (формула крыла: 3>4>2>5>6> 1 >7...
не считая рудиментарного настоящего
1-го махового). Хвост относительно
короткий, слегка закругленный. Цевки
оперены до пальцев, а пальцы покрыты
жесткими щетинками (ловит лапой
насекомых). Длина самцов (2) 310—320,
в среднем 315, самки (I) 324 мм. Размах
самцов (2) 776—796, в среднем 786,
самки (1) 797 мм (Тачановский, 1891).
Крыло самцов (5) 229—245, самок (4)
222,1—240, в среднем соответственно
235,4, 230,5 мм (едва ли реально самки
меньше самцов, приведенные измерения
относятся к очень небольшому
материалу).
Окраска. Общий тон спинной
стороны темнобурый, более темный на
голове; на-плечевых белый поперечный
рисунок; маховые со светлым
поперечным рисунком и рыжеватыми
наружными опахалами; рулевые бледнобурые с
широкими черными поперечными
полосами; брюшная сторона беловатая с
бурым продольным рисунком; надхвостье
белое с бурыми наствольями; цевки
бурые с беловатыми крапинами. Радужина
яркожелтая; клюв черноватый с желтым
коньком; пальцы желтые. Первый наряд
не различается от последующих.
Пуховой наряд и мезоптиль неизвестны, по
крайней мере из пределов СССР.
Систематические замечания. Игло-
ногие совы из северных частей
материкового ареала вида — перелетные, как и
японские, и при полном сходстве с
последними в окраске имеют несколько
более длинное крыло. Кроме приведенных выше измерений, для птиц, зимующих в ю.-в.
Азии, указывается длина крыла в 239—241 мм у самок (Мейзе, 1914), для пролетных
птиц вероятно тех же популяций 229—233 мм. Крыло у 4 измеренных самцов и самок из
Японии 211,5—229, в среднем 218 мм. В описании N. macroptera у Блазиуса указывается длина
крыла в 223—240 мм .О номенклатуре этих форм см. Мейзе (1941). Различия между японскими
и приморскими материковыми совами требуют все же еще подтверждения на большем
материале.
РОД МОХНОНОГИЕ СЫЧИ AEGOLIUS KAUP 1829
Тип Strix tengmalmi
11. Мохноногий сыч Aegolius funereus Linn.
Strix funerea. Linnaeus. Systema Naturae ed X, t. 1, 1758, стр. 93, Швеция.
Распространение. Ареал. Сев. и средняя Европа, Азия от Урала до
Камчатки и Курильских о-вов; на север примерно до границы
высокоствольного леса, на юг до гор южной Европы от Пиренеев и Альп до Балкан^
Рис. 101. Иглоногая сова.
392
ОТРЯД СОВЫ
Кавказа, быть может Крыма; в Азии до Тарбагатая, Тянь-Шаня и Зеравшан-
ского хребта, сев. Монголии и Ганьсу; Сев. Америка от северной границы леса
до Британской Колумбии, Альберта, Манитоба, залива св. Лаврентия и Новой
Шотландии. Характер пребывания. Мохноногий сыч оседлая и
кочующая птица. Биотоп. Главным образом хвойные, отчасти и смешанные
леса; на юге только в горах.
Подвиды и варьирующие признаки. Морфологически варьируют окраска, также
величина (крупная и светлая форма на крайнем северо-востоке Сибири; мелкие темные формы
на Кавказе в центральной Азии; темный подвид в Сев. Америке; в таежной полосе северной
Европы и Азии постепенное побледнение окраски в направлении с запада на восток и северо-
восток). Описаны подвиды: A. f .funereus L.y 1766, из европейской части ареала, на восток до
Урала; А. /. sibiricusВ u t., 1910, из Сибири, кроме северо-востока; А. /. magnus В u t., 1907,
из северной Якутии за Верхоянским хребтом, бассейна Анадыря, Коряцкой Земли и
Камчатки; А. /. caucaskus But., 1907, из Кавказа (быть может в лесах Крыма); А. /. pallens
S с h а 1 о w, 1908, из гор Туркестана от Тянь-Шаня до Зеравшана; А. /. beickianus S t г е-
Карта 66. Распространение мохноногого сыча Aegolius funereus
7 — A. f. funereus, 2 — A. f. sibiricus, 3 — A. f. magnus, 4 — A. f. caucasicus, 5 — A. f. pallens,
6 — A. f. beickianus, 7 — A. f. richardsoni; a — граница гнездовий, б — недостаточно выясненная
граница гнездовой области, г — область зимних кочевок»
semann, 1928, из сев. Ганьсу (Тетунгские горы); А. /. richardsoni Bonaparte, 1838^
из средних частей Аляски, северного Юкона, Мекензи, северной Манитобы, Квебека и да
севера Британской Колумбии, Альберты, залива Св. Лаврентия и Нов. Шотландии. Другой
близкий вид Aegolius acadicus G m e 1. распространен в Сев. Америке несколько южней, чем
A. funereus.
Европейский мохноногий сыч Aegolius funereus funereus L.
Синоним. Cryptoglaux tengmalmi transvolgensis. Бутурлин. Наша Охота, 1910,.
ноябрь, стр. И, частично (описан из вост. России и Зап. Сибири до Красноярска, тип из Тары)
Распространение. Ареал. Европа от Сев. полярного круга в
Скандинавии, около 69° с. ш. в Русской Лапландии, Соловецких о-вов, Архангельска,
верхнего течения Печоры (Якша), на юг до б. Восточной Пруссии, Белоруссии
(Витебск, Лепель, Минск, Гомельское Полесье), Прикамья (Татарская
республика), средней Волги (Промзино в бассейне Суры), Южн. Урала;
изолированные горные колонии (реликтовые?), в Пиренеях, Французской Юре, Альпах,
в горных лесах средней Германии, Чехословакии, в Карпатах, в Боснии,
Черногории, Болгарии. В негнездовое время встречается и южней — на Украине в-
Полтавской и Харьковской обл., в б. Александровском уезде Екатеринослав-
скойгуб., в Курской обл., в низовьях Волги и Урала, в Западной Европе —
до Франции, Бельгии, Англии и Балкан.
Биотоп. Тайга, отчасти смешанные леса. В европейской России на
равнинах, в южной части ареала в изолированных горных лесах. Характер
пребывания. Европейский мохноногий сыч — оседлая, отчасти
кочующая между октябрем и февралем птица. На кочевках будто бы встречается
мохноногий сыч
393
группами до 10 птиц (Зарудный, 1892). В холодное время охотно держится •
у поселений.
Численность. При скрытном и строго ночном образе жизни кажется
более редким, чем на самом деле. В северной полосе тайги обыкновенен.
Экология. Размножение. Цикл ранний — брачный крик уже
с середины марта, иногда с конца апреля и до середины апреля — мая. Гнезда
в дуплах (осина, береза, сосна); охотно использует гнездо черного дятла.
Характерный брачный полет над вершинами деревьев.
Кладка от середины до конца апреля. Число яиц в кладке 4—6, редко
больше (по Эверсманну 2—3, но это, вероятно, неполные кладки).
Плодовитость находится в известной зависимости от кормовых условий —
численности грызунов (для Скандинавии в «урожайные» годы отмечено до 7—8,
быть может 10 яиц, а также возможно
и повторение кладки). Яйца чисто
белые, размеры (100) 29—36,5 х 23,6—
28,5, в среднем 32,3 х 26,3 мм
(Скандинавия, Нитхаммер, 1938).
В европейской России
наблюдений за насиживанием и гнездовым
периодом нет. Западноевропейские
данные таковы. Насиживает самка
(Швейцария), в течение 25 и быть
может 31 дня (Норвегия). Гнездовый
период продолжительный, до 30 дней.
В СССР вполне выросшие молодые
в мезоптиле — с начала второй трети
и в конце июня (Поволжье,
Лапландия). Выводки вместе еще до ноября
(Повенец, Карелия). Крик в июне
(Конда, Раевский) свидетельствует о
наличии вторичного неполного
полового цикла.
Линька. Изучена
недостаточно. Мезоптиль сменяется к октябрю.
Линька взрослых полная годовая,
между июнем и ноябрем. Птицы,
добытые в декабре — январе, в свежем
пере, без следов линяния.
Питание. Пищу европейского мохноногого сыча составляют в основном
мелкие грызуны и мелкие птицы. Для Зап. Европы (Швейцарская Юра)
указываются землеройки, сони, мыши, из птиц — крапивник, молодые дрозды,
синицы и т. д. Для средней полосы России — домовые воробьи, синицы,
пищухи, также грызуны. В кормовом режиме имеются и сезонные изменения:
зимой в нем больше места занимают птицы, причем сыч в дуплах устраивает
запасы. О вероятных колебаниях численности и плодовитости в зависимости
от кормовых условий — см. выше.
Полевые признаки. Размеры небольшие, с домового сыча, но отличается
от последнего тем, что голова относительно больше, оперение более рыхлое,
а крылья и хвост длинней, так что птица кажется больше. Белые пятна на
плечевых более или менее сливаются; полет быстрый и прямолинейный. Голос
в брачный период «у-пу-пу-пу», несколько напоминающий крик удода; кроме
того, трель «ва-ва-ваа», а в тревоге — свистящее «тс-иии».
Описание. Размеры и строение. Большая голова, небольшие относительно
глаза, резко выраженный лицевой диск, асимметричные ушные отверстия (и внутреннее ухо).
Рис. 102. Мохноногий сыч.
394
ОТРЯД СОВЫ
Крылья длинные, формула 3>4^2>5>6>7>1... и ли 4>3... Вырезки наружных опахал 2-го
и 3-го маховых, на внутренних опахалах 1-го и 2-го маховых, на внутреннем опахале 3-го
махового выемки нет или она мало заметна. Хвост длинный, около 2/з длины крыла. Пальцы ног
густо оперены.до когтей. Длина самца (1) 207, самок (5) 246—270, в среднем 260 мм. Размах
самца (1) 550, самок (4) 590—620, в среднем 604 мм. Вес самца (1) 116, самки (2) 177 и 197 г.
Крыло самцов (21) 154—170, самок (34) 163—181,4, в среднем 103 и 174,7 мм.
Окраска. Пуховой наряд на спинной стороне буровато-белый, на брюшной — белый.
Мезоптиль: спинная сторона темная шоколадно-бурая, со слабо развитыми светлыми пестри-
нами на голове и спине; брюшная сторона темнобурая с примесью беловато-охристого
рисунка. Оперение в отношении структуры близко к дефинитивному. Взрослые птицы (самцы
и самки): спинная сторона бурая с белой (не охристой в отличие от домового сыча)
пестриной; белые крапины на затылке мелкие и частые, на задней части шеи крупные, верхняя
(передняя) часть спины однообразно бурая, так как светлые пестрины прикрыты темными
частями перьев; плечевые с крупными поперечными белыми пятнами, сливающимися в
полосы; кроющие крыла с более или менее крупными белыми крапинами; маховые бурые с
белыми поперечными пятнами; рулевые бурые с 4—5 узкими белыми поперечными полосами;
лицевой диск белый, испещренный бурым у глаз и ушей и с черным пятном перед глазами;
брюшная сторона белая с бурым продольным рисунком. Радужина желтая (у птиц в мезо-
птиле — с красной каймой); клюв желтый; когти черные. Индивидуальные вариации окраски
взрослых птиц значительны и выражаются в том, что у одних общий тон бурой окраски
несколько рыжеватый, у других— сероватый. Поэтому разграничение этой и следующей расы
в с.-з. Сибири, на границе их областей, носит условный характер и требует выяснения. Сычи
из с.-з. частей Зауралья (бассейн Конды) ближе к европейским, чем к сибирским.
Кавказский мохноногий сыч Aegolius funereus caucasicus But.
Nyctala caticasica. Бутурлин. Псовая и Ружейная Охота, 1907, март, стр. 87, Кисловодск.
Распространение. Ареал. Кавказ от лесов бассейна Кубани и ее
притоков и Дагестана (Дзауджикау) до центрального Закавказья (Белый Ключ);
однажды встречен в Крыму (окрестности Алушты, Пузанов, 1931).
Характер пребывания. Кавказский мохноногий сыч — оседлая птица.
Биотоп. Горные леса, как хвойные, так и лиственные. Численность.
Очень редок.
Экология. Не изучена. Размножение. Молодой в мезоптиле не
линяющий в контурное перо добыт у Черкесска еще 22 августа (поздняя дата).
Питание. Пищу кавказского мохноногого сыча составляют главным
образом мелкие млекопитающие: землеройки Sorex araneus, S. minutus,
различные мышевидные грызуны, в том числе Apodemus agrarius, Sylvimus sylvaticus,
Mas musculus, Pitymvs majori; в небольшом числе птицы (с.-з. Кавказ, Жарков,
1938).
Описание. Размеры и строение. Величина мелкая: крыло самцов и самок (7)
151—159, в среднем 156,4 мм.
Окраска. Темная, общий тон спинной стороны не сероватый, а шоколадно-бурый;
светлые пятна на затылке мелкие, на спине прикрыты темными частями перьев; на плечевых
темный бурый цвет преобладает над светлыми пестринами; на рулевых 3—4 белых
поперечных полоски; брюшная сторона сильно испещрена бурым; оперение ног всегда с бурыми
крапинами.
Сибирский мохноногий сыч Aegolius funereus sibiricus В u t,
Cryptoglaux tengmalmi sibiricus. Бутурлин, Наша Охота, 1910, ноябрь, стр. 11, оз.Ханка.
Синонимы. Cryptoglaux tengmalmi transvolgensis. Бутурлин. Цит. соч., частью. —
Nyctala jakutorum В u t u г 1 i n. Journ. f. Orn. 1908, стр. 287, Якутия.
Распространение. Ареал. Сибирь от бассейна Оби до Туруханского
края (р. Сым), южной Якутии (Олекминск, река Кямпендяй, Якутск) и
северного побережья Охотского моря (бухта Нагаева), на юг до границы тайги
в Западной Сибири (Тюмень, Тара), Кокчетавских боров (Залесские, 1931),
Алтая, Минусинской тайги, в сев. Монголии до Хангая и Кентея, Манчжурии
<Хинган) и Приморья (Ханка); Сахалин. Отмечены залеты на Курильские
о-ва и на Хоккайдо.
мохноногий сыч
395
Характер пребывания. Сибирский мохноногий сыч —
оседлая птица. Биотоп. Хвойная тайга. Вертикально на Алтае до 2000 м,
в Хингане до 1200 м.
Экология. Почти неизвестна. Размножение. На реке Ем-еган в
бассейне М. Сосьвы 15 мая добыта самка, не закончившая еще кладку. С другой
стороны, молодые в мезоптиле с недоросшими крыльями добыты у Олекминска
уже 19 июня, так что период размножения повидимому растянут
(дополнительные или вторичные кладки?). Размеры яиц (3) 31—37 х 28—30,5 мм
(Тачановский, 1891). Линька. В. свежем пере без следов линяния сычи
добывались в конце октября и в середине мая. Питание. Как и европейская
форма, сибирский сыч питается главным образом грызунами (Скалой, 1935).
Описание. Размеры и строение. Крыло самцов (19) 156—173, самок (23)
166—183, в среднем 166,25 и 176,1 мм. Длина самца (1) 255, самки 270, размах соответственно
580 и 590 (Тачановский, 1891).
Окраска. Несколько серее европейской формы, менее рыжеватый (хотя рыжеватые
особи, как указывалось, встречаются в небольшом количестве как индивидуальная вариация
в Западной Сибири, в частности таков тип transvolgensis); белый рисунок часто более развит,
чем у европейских птиц.
Систематические заметки. В Восточной Сибири были добыты (устья реки Кямпендяй,
бассейн Вилюя, Хаяхсытский наслег в 200 км на ВСВ от Якутска; сходные птицы из Кара-
тузского района Минусинского округа и с Большой Речки, Баргузин, с.-в. Байкал) сычи,
отличающиеся от обычных сибирских более светлым сероватым общим тоном окраски,
меньшим развитием темных пестрин на брюшной стороне и большим распространением белого
цвета на спинной. Эти сычи приближаются таким образом к подвиду из сев .-вост. Сибири, но
отличаются от него меньшей величиной (как у сибирского сыча). Они были описаны
Бутурлиным как подвид jakutorum, но по географическим соображениям и по крайней
малочисленности подобного рода особей их целесообразней считать за индивидуальные вариации
sibiricus или за особи-гибриды между sibiricus и magnus.
Большой мохноногий сыч Aegolius fanereus magnus But.
Nyctala magna Бутурлин. Псовая и Ружейная Охота, 1907, март, стр. 87, урочище
Эмях близ Среднеколымска.
Распространение.- Ареал. Северо-восток Сибири от среднего течения
Колымы (в низовьях ее до 68°30') до бассейна Анадыря (кроме приморских
его частей), Коряцкой Земли, Камчатки. Быть может в бассейне Индигирки.
Отмечены залеты на о-в Беринга и о-в Св. Павла (Прибыловы о-ва).
Характер пребывания. Большой сыч оседлая птица.
Биотоп. Лиственничные и тополевые леса речных долин. Численность. Редок,
Экология неизвестна. Размножение. Брачный крик «ку-ку-ку»
начинается в середине мая (Анадырь, Портенко, 1939). Питание. В желудках
птиц, добытых на Колыме и в Коряцкой Земле, найдены полевки; на Анадыре —
гаички.
Описание. Размер ыистроение. Самый крупный из мохноногих сычей: крыло
самцов (6) 172—188, самок (3) 180—191,5, в среднем 180 и 187 мм. Вес (1) 109 г.
Окраска. Очень бледная, общий тон спинной стороны светлый пепельно серовато-
буроватый; белый рисунок на спинной стороне преобладает над бурым фоном; на рулевых
имеются 3 поперечно вытянутых белых пятна; бурый рисунок на брюшной стороне мало
развит, оперение ног белое без пестрин.
Туркестанский мохноногий сыч Aegolius funereus pallens S с h a 1 о w
Nyctala tengmalmi pallens. Schalow. Journ. f. Ornith. 1908, стр. 109, Кашка-су
в центр. Тянь-Шане.
Распространение. Ареал. Центральный Тянь-Шань от Алма-Ата до
Иссык-Куля; на запад, повидимому, до восточных частей Киргизского хребта
и возможно Ферганы (Падша-ата, Зарудный, 1911). В 1935 году найден на гнез-
396
ОТРЯД СОВЫ
довье у Артуча в Зеравшанском хребте (Даль, 1936). Сюда же относятся
добытые в Тарбагатае птицы, быть может кочующие. Биотоп. Горные хвойные
леса — в Тянь-Шане ельники, в Зеравшанском хребте — арчевники.
Численность. Весьма редок.
Экология. Неизвестна. Размножение. Добытый в Зеравшанском
хребте 20.VI. 1935 сыч — молодой, в мезоптиле, с недоросшими еще
крыльями.
Описание. Размеры и строение как у других сибирских рае (кроме, конечно,
magnus). Крыло самцов (3) 155,5—170, самок (3) 164—176, в среднем 165 и 167 мм.
Окраска. Относительно темно окрашенный подвид, впрочем очень похожий на
sibiricus, несколько менее сероватый по общему тону, но не так рыжеват, как большинство
европейских особей.
Систематические заметки* Самостоятельность этого подвида, очень близкого к сибирскому,
вызывает сомнения, так как по мохноногим, сычам вообще, а по туркестанским — в
особенности, известно еще слишком мало экземпляров для установления амплитуды индивидуальных
вариаций окраски. С другой стороны, гнездовая область этого сыча обособлена, что говорит
в пользу сохранения этой формы как отдельной. Надо и тут впрочем иметь в виду, что
раздельно живущие уже с ледникового времени в горах южной Европы популяции мохноногих
сычей не выделились в особые расы и ни морфологически, ни экологически не отличимы 07
номинального подвида. Вопрос остается таким образом открытым.
РОД ДОМОВЫЕ СЫЧИ ATHENE ВО IE 1822
^Тип Athene rwctua
12. Домовый сыч Athene noctua Scopoli
Stris noctua. Scopoli. Annus Historico-Naturalis I, 1769, стр. 22, Крайна.
Русское название. Книжное. Сыч — обычное народное обозначение мелких
видов сов.
Распространение. Ареал. Умеренная и южная Европа, сев. Африка,
включая Сахару, на юг до Эритреи и Судана, вост. Абиссинии и сев. частей
Карта 67. Распространение домового'сыча Athene noctua
1 — А. п. vidali, 2— А. п. sarda, 3— А. п. noctua, 4 — А. п. indigena, 5 — А. п. bactriana,
6 — A. oriental is, 7 — А. п. ludlowl, 8 — А. п. plumipes, 9 — А. п. glaux, 10 — А. п. saharae,.
77 —А. п. solitudinis, 12— А. п. epUogaster, 13 — А. п. somaliensis; a — граница гнездовий.
Британского Сомали; Передняя и Средняя Азия, на юг до Месопотамии,
Афганистана и Белуджистана, на восток до Сев. Китая (Шаньси, Чжили), Кореи н
Тибета.
домовый сыч
397
Характер пребывания. Оседлая птица. Биотоп.
Открытый ландшафт; в горах (примерно до 3000 м в альпийской зоне) и на равнинах.
На севере главным образом в культурном ландшафте, на юге преимущественно
в пустынях и полупустынях и в «сухих» горах. Численность. Высокая,
в пределах ареала всюду обычен.
Подвиды и варьирующие признаки. Варьируют размеры и окраска. Крупные птицы —
в Центральной Азии, мелкие в Передней Азии и сев. Африке; максимальные размеры в Тянь-
Шане, Ладаке и Тибете. Окраска бледная в пустынных областях, насыщенная в горах и
в средней Европе. Экологически варьирующие признаки — детали годового
цикла*периодических явлений и отчасти трофические связи. Распространение отдельных форм таково:
в Западной Европе от Пиренейского п-ова до Франции и Бельгии (интродуцирован
в Голландии и южной Англии с конца XIX века) A.n.vidali С h г. L. В r'e h m 1857; в
средней Европе от Рейна до Дании, б. восточной Пруссии, Литвы, Прибалтики, на
восток до Чкалова, к югу до Италии, Сицилии, Боснии и Венгрии А. п. noctua S с о р о 1 i,
1769; на Сардинии, возможно на Корсике, А. п. sarda Kleinschmidt, 1907; в ю.-в.
Европе на Балканах, на юге Украины, в Крыму, на Кавказе и в сев. частях Малой Азии
А. п. indigena В re h m, 1855; в оазисах Египта, в северных частях Марокко, Алжира и
Туниса, вероятно также на Кипре А. п. glaux S a v i g n у 1809; в пустынных частях Египта,
Сахаре, кроме центральных ее частей, в южн. Алжире, южн. Тунисе, в Аравии, в Палестине,
Сирии, Месопотамии, южных частях М. Азии, южн. Иране А. л. saharae Kleinschmidt,
1909 (синоним А. п. lilith H a r te r t, 1912, см. Бете, 1937); в центральной Сахаре — Аире
и Ахаггаре — А. п. solitudinis H a r t e r t, 1924; в равнинной Средней Азии от Туркмении
до бассейна Сыр-Дарьи, в Иране, Афганистане и Белуджистане А. п. bactriana Hutton,
1847; в горном Туркестане, Джунгарии, Синьцзяне, на север до Тарбагатая А. п. orientalis
Severtzow, 1872; в Северном Китае, Корее, Монголии, на западе до ю.-в. Алтая и ю.-з.
Забайкалья А. л. plumipes S w i n h о е, 1870; в Ладаке, Тибете А. п. ludtowi Baker,
1926; в тропической Африке — в Эритрее и прилегающих к Красному морю частях Судана
А. л. spilogaster H e u g I i п, 1869; в восточной Абиссинии (бассейн реки Гауаш) и северных
частях Британского Сомали А. л. somaliensis Reichenow, 1905.
Западный домовый сыч Athene noctua noctua S с о р о 1 i
Распространение. Ареал. Европа до Рейна на западе, до южной Дании
(Ютландия, Фюнен), западной Латвии, Литвы, Белоруссии, Пскова, Калуги,
Рязани, Тулы, Казани, Ульяновска на севере, до Чкалова, Куйбышева,
среднего течения Урала й Илека на востоке, на юг до Италии, Венгрии, Боснии,
Бессарабии, Киевской, Харьковской и Воронежской областей.
Характер пребывания. Западный домовый сыч — оседлая
птица, но слегка перемещается из гнездовых участков в зимнее время.
Максимальное расстояние таких перемещений, по западноевропейским данным
300 км (Саксония — Верхняя Австрия). Есть требующие подтверждения
указания на откочевку зимой с северных частей ареала (Рязанская обл.,
Хомяков, 1900). Б и о то п. Главным образом связанные с человеком ландшафты —
сады, парки, в том числе в больших городах (встречен в Пскове, Москве и т. д.).
Кроме того, в оврагах, балках и тому подобных пересеченных или открытых
местностях.
Численность. На севере ареала довольно низкая, у 55° с. ш. редок
и спорадичен, южней обычен; но и на севере повидимому увеличивается в
числе с развитием культурного ландшафта (расселение в частности отмечено
у Казани, Першаков, 1929).
Экология. Размножение. Брачный крик с конца марта, в апреле
и до мая (Поволжье), цикл относительно ранний. Пары постоянные, самец
и самка держатся вместе и зимою (Минское Полесье, Шнитников, 1913).
Гнезда — закрытые, в норах и сооружениях (амбары, чердаки и т. п.), даже
в скирдах сена, изредка повидимому в дуплах.
Кладка обычно происходит в конце апреля и начале мая (Харьков). Число
яиц в кладке 4—5, изредка до б (Карамзин, 1901), по западноевропейским
данным и до 8. Яйца белые, слабо блестящие, с мелкозернистой скорлупой. Раз-
398 отряд совы
меры: (9) 33—35 х 28—30 (Харьковская обл., Сомов, 1897). В случае утраты
кладки бывает дополнительная, до 3 яиц.
В отличие от большинства других сов, по указанию ряда авторов, начало
насиживания не с первого яйца, а быть может даже с последнего, так как
птенцы в выводках одинакового возраста. Впрочем, это требует еще
проверки. Длительность насиживания около одного месяца (28 дней, по Уай-
зерби, 1938), у нас не прослежена. Во второй половине мая появляются птенцы
(Куйбышевская обл.). Участие самца в насиживании неясно. Через месяц
после вйлупления птенцы покидают гнездо (в первой половине июля,
Харьковская обл.; около 1 июля, Куйбышевская обл.), но вполне летными они
становятся только в возрасте около 5
недель. В выводках отмечено до 5
птенцов. Неразбившиеся семьи держатся
до августа.
Линька. Как у других сов,
полная годовая за исключением смены
мезоптиля. Последний сменяется
частично, кроме больших кроющих
крыла, маховых и рулевых. Порядок
смены маховых—от заднего края к
переднему (от 10-го к 1-му),
рулевых — от середины, центробежный.
Начало линяния взрослых в июле;
птицы в свежем пере встречены в
конце октября—начале ноября. Смена
мезоптиля начинается в возрасте
около 6 недель, затягиваясь до
октября— начала ноября.
Последовательность смены нарядов: пуховой
наряд— гнездовый наряд илимезоптиль
(мало отличающийся по структуре и
окраске от окончательного) — первый
годовой (окончательный по окраске,
но комбинированный по составу, с
гнездовыми большими кроющими, ма-
Рис. 103. Домовый сыч. ховыми и рулевыми) — второй годовой
(окончательный) наряд и т. д.
Питание. При существенных географических и быть может
индивидуальных различиях в кормовом режиме домовых сычей, питание их все же
может быть охарактеризовано как мало специализированное: грызуны отнюдь
не занимают в нем исключительного места, относительно большее значение
имеют насекомые (в особенности жуки и прямокрылые); птицы занимают
меньшее место, на юге большое значение имеют пресмыкающиеся. Сезонные
изменения кормового режима в общей форме сводятся к тому, что летом в нем
значительное место принадлежит насекомым, а зимой —мышевидным
грызунам. Быть может именно поэтому, в соответствии с данными
западноевропейских авторов, в суровые и бедные мышами зимы замечается гибель домовых
сычей от голода; и быть может именно поэтому домовый сыч не распространен
на севере.
Охотится сыч и днем, нападает главным образом на сидячую, реже на
бегающую добычу. При защите гнезда нападает и на таких относительно больших
по сравнению с птицей зверей, как кошка (Сомов, 1897). Самая крупная
добыча — ласка и крыса (Зап. Европа, Уттендерфер). В СССР в качестве
кормов домового сыча отмечены: 92% мышевидных грызунов, 10% птиц, 30,4%
домовый сыч
399
насекомых (Татарская республика, Жарков и Теплов); мыши, насекомые*
реже мелкие птицы (Харьковская обл., Сомов); тушканчик Alactagulus aeon*
Hon и тарантулы (Сталинград, Агриропуло, 1928) и т.д. Есть указания,что
в бескормные суровые годы сычи откочевывают; с другой стороны, в Германии
имеются наблюдения, что в «мышиные» годы сычи кладут яйца даже в сентябре,
•но в наших условиях (северная граница ареала?) этого, повидимому, не
бывает. Погадки — типичные совиные, с костями, хитином, шерстью, обычно
3—4 х 1,5 мм.
Полевые признаки* Небольшая сова плотного сложения, с плоской голо-
вой без перяных «ушей»; крылья и хвост относительно длинные, лицевой диск
выражен нерезко; глаза небольшие. Сидит обычно не на деревьях, а на земле
или на зданиях, на камнях и т.п. При испуге—характерные «поклоны».
Активен и при свете, особенно по вечерам. Полет днем волнообразный,
напоминающий дятлов (это помогает сычу, повидимому, избегать преследования
хищников), ночью прямой, низко над землею. Голос «ку-вит-ку-вит» или «кькь
кьюю».
Описание. Размеры и строение. Ушные отверстия небольшие и симметричные..
Восковица вздутая. Формула крыла 3>4>2>5>1>6... (не считая рудиментарного,
прикрытого кроющими настоящего 1-го); на 2-м — 4-м маховых вырезки на наружных опахалах, на
5-м легкое сужение. Хвост прямо обрезанный, из 12 рулевых; подхвостье доходит до 2/з длины
хвоста. Пальцы покрыты волосообразными щетинками, три обычно обращены вперед, один
назад. Размеры относительно мелкие. Длина самцов (5) 232—250, самок (7) 243—275, в
среднем 241,6 и 252,6 мм. Размах самцов (6) 565—590, самок (8) 590—612, в среднем 575,4 и
601,7 мм. Вес самцов (1) 158, самки (1) 182 г. Крыло самцов (37) 152—169, самок (26) 158—177,
в среднем 160,2 и 168,4 мм.
Окраска. Пуховой наряд чистобелый. Взрослые птицы, самцы и самки, на спинной
стороне темнобурые со светлыми беловатыми пестринами; на брюшной стороне беловатые
с бурым продольным рисунком; на темени и затылке удлиненные белые крапины, на шее
эти крапины крупные; такие же, но еще более крупные, пестрины, отчасти прикрытые
основным бурым цветом, на плечах, спине и крыльях; маховые бурые с поперечным беловатым
рисунком и с сероватыми вершинами; рулевые бурые с 4—5 охристо-беловатыми
поперечными полосками; горло белое; зоб, грудь и брюхо беловатые с бурыми продольными пятнами,
более всего развитыми и сливающимися на боках груди; подхвостье и оперение цевки белые,
без пятен; лицевой диск^елый с бурым пятном на кроющих уха.
Мезоптиль по окраске не отличается от окончательного наряда, быть может слегка
серей, и более пушистый; разница между мезоптилем и окончательным нарядом таким
образом меньшая, чем у многих других сов.
Пальцы — в серовато-белых щетинках. Радужина яркожелтая, у молодых бледножел-
тая; клюв восково-желтый, когти черные.
Это подвид — самый темный по окраске из водящихся в СССР домовых сычей, как по
насыщенному бурому основному тону, так и по значительному развитию бурого рисунка
на брюшной стороне. К югу и востоку окраска домовых сычей постепенно светлеет (и
изменяются размеры).
Южноевропейский домовый сыч Athene noctm indigena Brehm
Athene indigena. Chr. L. Brehm. Der vollst. Vogelfang 1855, стр. 37, Аттика.
Синонимы. Athene glaux Kessleri. Семенов. Зап. Импер. Академии наук (8>
VII, 1899, стр. 74, Крым. — Carine noctua caucasica. Sarudny unci Loudon. Om.
Jahrbuch, XV, 1904, стр. 56, Баку.
Распространение. Ареал. К югу от номинальной формы —на
Балканском п-ове до Греции; в Крыму, на Кавказе и в прилежащих к северу
степных частях бассейнов Днепра и Дона (экземпляры из окрестностей
Бердянска, с низовьев Дона, с Маныча). К этой же форме относятся
немногочисленные изученные экземпляры из М. Азии и с.-з. Ирана (Курдистан и
Иранский Азербайджан). На востоке на нижней Волге у Сталинграда и даже в с.-з.
Казахстане (бассейн реки Урал) встречаются сычи, переходные к
закаспийской форме.
400
ОТРЯД СОВЫ
Характер пребывания. Оседлая птица. Биотоп. Равнины
и горы: на Кавказе до 1800—2000 м (Грузия, Армения). Обрывы скал, овраги
и балки, стены речных террас, человеческие поселения и т. п.
Численность. Обычная птица; плотность населения в СССР выше, чем у
номинальной расы.
Экология. Размножение. В общем цикл сходен с таковым у
предыдущего, но кладка начинается повидимому несколько раньше —- с середины
апреля (14—16, IV, Днепропетровск). Обычное число яиц 4—5, но указывается
и 8 (вост. Закавказье, Сатунин, 1914). Кладки в старых зданиях, в норах
(низовья Днепра, Попов, 1937), в щелях обрывов, степных балок и оврагов, в
стенах старых колодцев (Сатунин, 1914), в скирдах (сев. Крым, Сеницкий, 1898),
в зданиях—мазарах, ветряных мельницах (с.-з. Казахстан, Волжско-Ураль-
ская степь, Волчанецкий, 1937). Подлетки в мезоптиле в конце июня — начале
июля (экземпляры из разных частей Кавказа; в Днепропетровской обл.,
по Боровикову, 1907; в Волжско-Уральской степи, по Волчанецкому, 1937).
Молодые, достигшие роста взрослых, держатся в выводках по крайней мере
до начала августа (Армения).
Линька. Как у номинального подвида; взрослые сычи в свежем пере
встречаются с конца октября — начала ноября и до мая.
Питание. В низовьях Днепра в погадках найдены жуки —
жужелицы и навозник, два черепа Citellus suslika, 3 Mus musculus, 1 Crocidura leu-
codon (Попов, 1937). В Крыму кормится мышами, насекомыми и мелкими
птицами. В 45 погадках у Бердянска 97,7% грызунов, в том числе 39,8% полевок
и 24,4% мышей, особенно часты среди грызунов Mus musculus hortulanus, реже
Cricetulus migratorius, Sylvimus syluaticus, Microtusarualis, M. socialis; в 2,2%
ящерицы Lacerta agilis; в б, 6% мелкие птицы; наконец в значительной доле—
62,2% — жесткокрылые насекомые (Кучерук и Дунаева, 1938).
Описание. Размерыистроение. Величина как у номинального подвида. Крыло
самцов (27) 163—168, самок (33) 156—172, в среднем 162,4 и 164,3 мм. Вес самца (1) 160,
самки (1) 190,5 г. Пальцы в основной части обычно покрыты редкими щетинками.
Окраска. Довольно изменчива, но птицы в общем светлей и рыжеватей, чем
номинальный подвид, оставляя как бы переход к закаспийским. Оперение брюшной стороны
поэтому менее контрастно; поперечный рисунок на хвосте выражен слабей, обычно в виде
вытянутых пятен. Особенно светлы сычи в восточных частях ареала—в Волжско-Уральских
степях, в Закавказье и сев. Иране.
Каспийский или пустынный домовый сыч Athene noctua bactriana Hutton
Athene bactriana. Hutton. Journ. of the Asiatic Society of Bengal, XIV, 1847, стр.776,
Кандагар.
Распространение. Ареал. От северных берегов.Аральского моря, Усть-
Урта и зап. Туркмении (Большие Балханы, низовья Атрека и т. д.) до
северного (южнокаспийские провинции — Астрабад и т. д.) и восточного Ирана,
Афганистана и Белуджистана, низменных частей Таджикистана, Узбекистана
(Чиназ, Самарканд, Ташкент, Узгент) и нижнего среднего течения Сыр-Дарьи;
граница на северо-востоке не вполне ясна, в горном Семиречье —другая раса.
Характер пребывания. Оседлая птица. Биотоп. Песчаные
и глинистые пустыни, долины рек, пустынный ландшафт. В верхней зоне гор
отсутствует (Копет-Даг). Численность. Высокая, но подвержена
известным колебаниям, по крайней мере в отношении плодовитости
(размеры кладок).
Экология. Размножение. Цикл ранний. У самцов с половины марта
сильно развитые половые железы. В это время сычи кричат и днем. Гнезда*
расположены в самых разнообразных местах, под карнизами скал, в норах
больших песчанок и тонкопалых сусликов, в гнездах скалистых поползней
домовый сыч
401
(Зарудный, 1896), в норах, вырытых самими сычами, в стенах оврагов и .силевых
промоин, степных колодцев, в могильниках и старых зданиях, в развалинах.
Типичное гнездо имеет около 1— l1^ м хода и камеру, дно которой плотно
выстлано обломками хитина насекомых, костями и т. д.; диаметр гнездовой
камеры около 25—30 см (Туркмения, Дементьев, 1945). У каждой пары 2—3
гнезда, занимаемых посменно в разные годы; кроме того —норы для отдыха и т. п.
Гнездовые участки очень постоянны. Старые птицы держатся в них круглый
год, молодые — разлетаются (Сыр-Дарья).
Кладки быть может уже с конца марта, в начале апреля — полные. Число
яиц 4—6 (Сыр-Дарья, Туркмения) и до 7 и даже 9 (Байгакум). В середине
апреля полные кладки, в конце месяца сильно насиженные. В случае утраты
кладки бывает дополнительная, в мае (Туркмения) и даже до июля (Сыр-
Дарья). Размеры яиц (10) 30,9—34,5x26,1—28,9 мм (Сыр-Дарья, Спанген-
берг); (6) 32,2—33,6 х28,1—28,4л*л*(Бугунь, Аральское море, Зарудный, 1916).
Насиживание начинается с первого яйца (разновозрастные зародыши и
птенцы), при участии обоих родителей. Подлетки в мезоптиле с конца мая
(Туркмения) и в начале июня (Сыр-Дарья, Аральское море). В августе —брачный
крик, абортивный половой цикл (Туркмения).
Линька. Соответственно раньше, чем у европейских птиц. Птицы
в свежем пере бывают уже с конца сентября.
Питание. Охотится сыч и днем, но главный период суточной
активности — с вечера и до 2—3 часов полуночи. Кормовой режим —мелкие
грызуны и другие млекопитающие; мелкие птицы, главным образом жаворонки,
домашние и полевые воробьи; пресмыкающиеся и насекомые. Подробные
анализы погадок и кормовых остатков в сев.-зап. Казахстане (Сассык-тау):
70—79% встреч — мелкие млекопитающие (Merlones meridianus, Mas musculus,
редко Dlpas sagitta; Crocidara saaveolens, Diplomesodon palchellam); 77—80%
встреч— насекохмые, особенно летом, среди них главным образом жуки
—жужелицы, навозники, чернотелки, кроме того прямокрылые, также гусеницы,
бабочки и перепончатокрылые; 16,3—44% встреч—рептилии, среди которых
Phryhocephalas guttatus, Eremias argata; птиц мало—до 1%. Процент встреч
пресмыкающихся повышается осенью. В южной Туркмении у Рабат-кашана:
около 83% встреч — мелкие зверьки, главным образом Phaiomys afghanus,
кроме того Mas mascalas, Meriones erythroaras; землеройки, около 2% птиц,
2% рептилий, 5,4% амфибий и 13,8% насекомых (Дунаева и Кучерук, 1938).
Описание. Размеры и строение. Длина самцов (6) 230—250, самок (5) 230—
250, в среднем 241,7 и 242 мм. Размах самцов (8) 570—590, самок (5) 580—610, в среднем
582,5 и 592,5 мм. Вес самцов (4) 140,3—162 г, самки (1) 168 г. Крыло самцов (90) 153—170,
самок (50) 157—175, в среднем 162,6 и 165 мм. Пальцы обычно оперены на 2/з длины.
Окраска. Самая бледная форма среди домовых сычей фауны СССР. Общий тон
окраски спинной стороны тела двух типов: светлого серовато-песочного или рыжевато-
песочного. Светлые пестрины на спине, плечах, крыльях, задней стороне шеи очень крупны
и многочисленны; на хвосте широкие и правильно поперечные полосы; темные пятна на
брюшной стороне невелики, неясны и не резко очерчены.
Систематические замечания. От горнотуркестанских домовых сычей, с которыми
каспийские сычи объединяются многими авторами в одну форму, пустынные сычи отличаются
заметно более бледной окраской и меньшей величиной. Каспийские сычи очень сходны с
месопотамскими, палестинскими и южноиранскими сычами расы А. п. saharae, но последняя
по окраске несколько рыжеватей, с менее испещренной брюшной стороной и с неоперенными,
покрытыми только щетинками, основаниями пальцев.
Горноазиатский домовый сыч Athene noctua orientalls Severtzow
Athene oriental is. Северцов. Вертикальное и горизонтальное распространение
туркестанских животных, 1873, стр. 115, Тянь-Шань.
Распространение. Ареал. Горный Туркестан от восточных частей, Тадт
жикистана до Ферганы, Тянь-Шаня, Памира и Тарбагатая; Синьцзянь, Био-
26 Птицы СССР, т. I
402
ОТРЯД СОВЫ
т о п. В горах до 4200 м на Памире (р. Беик, сборы Грум—Гржимайло), в Тянь-
Шане до 2000 м. Глинистые обрывы, скалы, речные долины; также в
культурном ландшафте, включая человеческие поселения. Численность. Ниже,
чем у равнинного туркестанского подвида.
Экология. Сведения совершенно недостаточны. Размножение. Цикл
более поздний, чем у пустынного сыча, так как разновозрастные подлетки
встречаются в конце июня—начале июля, не сменив еще мезоптиля (Памир,
р. Беик; центр. Тянь-Шань). Гнезда главным образом расположены-в щелях
обрывов и скал, также в норах грызунов и сизоворонок и т. д. Л и н ь к а.
Линяющие взрослые наблюдались с конца июня, а птицы, кончившие линяние,—
в октябре. Питание. Как и у других подвидов пищу горноазиатского сыча
составляют насекомые, грызуны, мелкие птицы. Охотится нередко и днем.
Описание. Размеры и строение. Величина крупная, самый большой из
домовых сычей, уступающий по длине крыла только тибетско-ладакским птицам. Длина
самца (1) 260, самки (1) 275, размах самца (1) 605, самки (1) 635 мм. Крыло самцов (10) 165—
172, самок (5) 170—181, в среднем 168,1 и 175,8 мм. Пальцы ног оперены густо по крайней
мере до основания концевой фаланги.
Окраска. От пустынных сычей хорошо отличается более темной и насыщенной бурой
окраской; светлых пестрин на спинной стороне меньше; пятнала брюшной стороне крупней
и темней; хвост в поперечных полосах.
Систематические замечания. Сычи из Синьцзяня (Хами, Ния, Карашар, Ойт-ограк,
Ваш-шары, Лукчун-кыр) сходны по окраске с тяньшанскими, но меньше их: крыло
самцов (10) 152—166, самок (3) 166—169, в среднем 162 и 167,6 мм.
Монгольский домовый сыч Athene noctua plumipes S w i n h о е
Athene plumipes. S win hoe. Proceed Zool. Soc. Loudon, 1870, стр. 448, «Sha to» у
Нанькоу, Чжили.
Распространение. Ареал. Ю.-в. Алтай, Монголия, ю.-з. Забайкалье, до
Цайдама, Алашаня, Ордоса и сев. Нань-Шаня на юге; Манчжурия и Корея;
Сев. Китай (Чжили, Шаньси). Характер пребывания. Оседлая
птица. Биотоп. Сухие холмистые степи, полупустыни и пустыни; сухие
безлесные горы, примерно до высот в 2000—2800>и (Алтай). Численность.
Умеренная, распространен несколько спорадично.
Экология. Размножение. Сроки размножения — как у
горнотуркестанского подвида. Молодые в мезоптиле летают с начала июля (Алтай, 6. VII,
Сушкин, 1938; Кайластуй, Забайкалье, Скалой, 1936). В выводках отмечалось
3—4 птенца, по первому автору, в одном случае все 3 молодых были самки.
Неразбившиеся выводки на Алтае встречались в начале августа.
Линька. Начало линяния взрослых в первой неделе июля, конец — в
октябре. Конец смены мезоптиля во второй половине августа.
Описание. Размеры и строение. Крупный сыч, несколько уступающий
впрочем по величине горнотуркестанскому. Крыло самцов (23) 158—170, самок (9) 167—
178,5, в среднем 163,3 и 173,7 мм. Пальцы ног густо оперены. Окраска. Близкая к
окраске горнотуркестанских сычей, но бурый цвет менее насыщенный, серей и светлей; на
спинной стороне много белых пестрин. #
Пустынные сычи из западной Азии — А. п. bactriana — заметно серей, бледней и
мельче гобийских. Птицы из высоких частей западного Китая, из области Куку-нора и
Тибета заметно темней и рыжей монгольских и больше их по размерам.
РОД ВОРОБЬИНЫЕ СЫЧИ GLAUCIDIUM BOIE 1826
Тип Glaucidium passerinum
13. Воробьиный сыч Glaucidium passerinum Linn.
Strix passerina. Linnaeus. Syst. Nat. изд. X, 1758, стр. 93, Швеция.
Распространение. Ареал. Полоса хвойных лесов Азии и северной
Европы от Пиренеев, Французской Юры и Альп на западе до полярного круга в
ВОРОБЬИНЫЙ СЫЧ
403
Скандинавии, Архангельска, около 62°30' с. ш. на Печоре до Испании, сев.
Италии, Бескидов, Карпат, Трансильвании и Боснии, средней полосы
европейской России, на юге —в Европе; в Азии и Сибири северная граница идет около
60° с. ш., к востоку до Охотского побережья и Сахалина, к югу до Алтая и
Саяна, Монголии (Кентей), Манчжурии и Приморья.
Биотоп: высокоствольный лес, поэтому в Зап. Европе и на юге ареала
в восточной Азии спорадичен, только в горах (ледниковый реликт).
Характер пребывания — оседлая птица. Численность невысокая.
Подвиды и варьирующие признаки. Две географических расы, слабо различимые по
окраске: G. p. passerinum L., 1758— в Европе и Зап. Сибири, примерно до Енисея; G. р.
orientate Taczanowski, 1891 — в восточной части ареала. Очень близкий вид Glau-
Карта 68. Распространение воробьиного сыча Glaucidium passerinum
1 —О. p. passerinum, 2 — О; p. orientate; a — граница области распространения (оседлый), X—места
залетов.
cidium gnoma W a g l e г заменяет воробьиного сыча в Америке от Аляски и Британской
Колумбии на севере и до Центральной Америки на юге.
Западный воробьиный сыч Glaucidium passerinum passerinum L.
Синоним. Strix torquata. Fischer. Memoires de la Societe Imp. des Naturalistes de
Moscou 1812, стр. 276, Петровско-Разумовское под Москвой.
Распространение. Ареал. Европа — Швеция и Норвегия до 68°30' с. ш.,
Русская Лапландия до границы леса, Карелия, Зап. Сибирь — около 60°
в бассейне Конды и Малой Сосьвы, у Тобольска, Красноярска; на юге до
Пиренеев, Юры, Альп (Швейцария, сев. Италия), Бескид, Карпат,
Трансильванских Альп, Боснии, в Европейской части СССР до Белоруссии (Пинский район,
Беловежская пуща), Смоленской, Московской, северной части Рязанской
областей, Татарской республики (Казань), Бугуруслана; в Сибири до Тюмени,
Красноярска и северного Алтая (у Телецкого озера); на юге указания на Крым
и Кавказ — ошибочны. Некоторые авторы указывают на залеты в холодное
время года на Украину. Приводится и для Сибирской лесостепи в б.
Барнаульском уезде (дер. Плотникова, Иогансен, 1907).
Характер пребывания. Оседлая птица. В гнездовое время
совершает повидимому незначительные местные передвижения, спускаясь
с гор (Зап. Европа) или приближаясь к поселениям, где условия питания
становятся благоприятней. Вероятно в таких случаях сыч несколько выходит за
26*
404
ОТРЯД СОВЫ
южные пределы гнездовой области (см. выше; также экземпляры из
окружности Калуги; Темникова Тамбовской обл., Льгова Курской обл. —в
Зоологическом музее Московского университета). Биотоп. Леса,
преимущественно хвойные, охотно селится в глухих ельниках. В Зап. Европе в гс-
рах — до верхней границы леса.
Численность. Невелика, в Западной и средней Сибири редок и
далеко не доходит до северной границы тайги. Отмечены известные колебания
численности (например, в б. Казанской губ. Рузский, 1891).
Экология* Относящиеся к СССР сведения весьма отрывочны.
Размножение. Брачный крик в средней полосе начинается с марта и продолжается
до конца мая (на севере у Молотова — со
второй половины апреля). Гнезда расположены в
дуплах, главным образом осин, реже берез.
Кладка, по старым сведениям будто бы из 2—
3 яиц (Эверсманн, 1866), но на самом деле пови-
димому больше, из 4—6 и даже 7 яиц
(Западная Европа). Яйца белые, слегка блестящие.
Размеры (?); у западноевропейских в среднем
28,8 X 22,8/ амплитуда 27—31 х 21,7—24 мм,
(Нитхаммер). Судя по птенцам в мезоптиле,
свежие кладки у нас с конца апреля.
Насиживание продолжается 28 дней (Западная Европа).
Вполне выросшие летные птенцы в мезоптиле
встречаются в последней трети августа.
Линька. Не изучена. К началу зимы, в
октябре молодые сменяют мезоптиль, а старые
находятся в свежем наряде.
Питание. Воробьиный сыч, как и другие
таежные совы, охотится и днем, но главным
образом перед утренним рассветом и в сумерках.
Корм — мышевидные грызуны и мелкие птицы.
Из млекопитающих для Швеции указываются
лемминги и другие грызуны, летучие мыши,
полевки, лесные и домовые мыши и землеройка
Sorex araneus. Самая крупная добыча — хомяк
(Томск, Иогансен, 1912). Из птиц—для Запад-
Рис. 104. Воробьиный сыч. ной Европы—синицы, воробьи, зяблики,
зарянки. Летом повидимому, поедаются и
насекомые. Потребность в пище, по определению на содержащихся в неволе птицах,
2—3 мыши в день, погадки с костями скидываются через 2—21/2 дня после
еды; воды сыч не пьет (Промптов, 1927). Воррбьиный сыч склонен делать
кормовые запасы, в особенности зимой, помещая их в дуплах.
Полевые признаки» Самая малая из наших сов, размерами со скворца,
с небольшой головой и коротким хвостом, глаза небольшие, лицевой диск
развит слабо; клюв резко выдается вперед. Спинная сторона буроватая с
небольшими светлыми пестринами; брюшная сторона беловатая с бурыми
продольными пятнами. Сидя, часто подергивает хвостом. Очень быстр, подвижен и
неосторожен. Деятелен и днем. Голос в брачное время — свист «тю-тю-тю>—
своеобразная песнь; кроме того, «кувитт», вроде крика домового сыча.
Описание. Размеры и строение. Крыло очень закруглено, формула 4>
>3>5>2>7>8>9>10 ^ 1 (не считая настоящего 1-го махового, рудиментарного).
Вырезки на внутренних опахалах первых 5 маховых (на 5-м неглубокая). Короткий, слегка
закругленный хвост. Лапы оперены до когтей. Длина самцов (2) 152—170, размах (1) 356 мм;
длина самок (3) 174, 175, размах (2) 340 и 390 мм. Крыло самцов (29) 92—102, самок (41)
100—102, в среднем соответственно 97,8 и 103,6 мм. Различия в линейных размерах, следо-
ВОРОБЬИНЫЙ СЫЧ
405
вательно, невелики, но по весу самки заметно больше: вес самцов (2) 56 и 61,5, самок (2)
70 и 78,8 г.
Окраска. Пуховой наряд беловатый. Мезоптиль напоминает мохноногого сыча
и резко отличен от домового сыча: пуховых перьев в нем нет, и наряд состоит из несколько
более рыхлых, по сравнению с окончательным, контурных перьев. Весь тон окраски спинной
стороны шоколадно-бурый, без беловатых пестрин на голове, шее и спине (в этом отличие
от взрослых); брюшная сторона беловатая с отдельными бурыми пятнами, на боках зоба и
груди темные бурые поля (без поперечных беловатых полос, имеющихся у взрослых).
Взрослые самцы и самки: общий тон спинной стороны серовато- (изредка рыжевато-) бурый со
светлыми пестринами; брюшная сторона белая с бурым рисунком; на голове, спине, крыльях
мелкие округленные беловатые пятнышки, часто прикрытые бурым основным цветом
оперения; на задней стороне шеи эти пятна крупней и образуют светлый полуошейник; на
средних кроющих крыла большие светлые пятна у вершин наружных опахал; маховые землисто-
бурые с неясным светлым поперечным рисунком, переходящим у краев опахал в белые
пятна; рулевые с 3—4 белыми поперечными полосами и беловатой вершинной каймой;
лицевой диск и лицо беловатые с мелкими бурыми пестринами; бока зоба и груди бурые с белым
поперечным рисунком; остальные части брюшной стороны белые с бурым продольным
рисунком; оперение ног белое или белое с бурыми крапинами. Радужина желтая, клюв
желтый, когти темнобурые.
Восточный воробьиный сыч Glaucidium passerinam orientate Taczanowski
Glaucidium passerinum orientale. T а с za n о w s k i. Faune Ornithol., de la Siberie
Orientate, 1891, стр. 128, Удской Острог.
Распространение. Ареал. Восточная Сибирь от верхней Лены (Олек-
минск) и сев. Байкала до Охотского побережья (Удской Острог) и Сахалина на
востоке, на юг до Саяна, Забайкалья, сев. Монголии (Кентей) и Манчжурии,
Приморья (бассейн Уссури). Зимой добыт в с.-в. Чжили в Китае. Есть
требующие подтверждения указания на нахождение на Анадыре в устье реки Щучьей
(Портенко, 1938). X а р а кт е р пребывания, биотоп и
численность. Как у предыдущего.
Экология. Не изучена. Размножение. Судя по добыче в урочище
Крестях близ Олекминска 24.VI. 1909 птенца в мезоптиле с недоросшими
крыльями, размножение проходит примерно в то же время, как у европейской формы.
Описание. Размеры и строение. Как у предыдущего: крыло самцов (7)
94—102, самок (5)* 103—108, в среднем соответственно 98,6 и 105,8 мм. Вес самца (1) 69 г.
Окраска. Основной тон спинной стороны .серей; светлые пестрины «чище», белее; часто
эти светлые пестрины выступают резче, чем у европейских птиц.
Систематические замечания. Индивидуальная изменчивость у воробьиных сычей
велика и проявляется параллельно у восточной и западной форм: среди особей и той и другой
встречаются то более бурые, то более сероватые, хотя подавляющее большинство изученных
нами восточных птиц серей, а западных — бурей. Еще на Алтае (Телецкое озеро) встречены
особи, не отличимые от номинальной расы. С другой стороны, у Хо*могор и на М. Сосьве
в Зап. Сибири — добыты птицы, столь же сероватые, как восточносибирские.
РОД ЯСТРЕБИНЫЕ СОВЫ SURNIA DUMERIL 1806
Тип Strix hudsonica
14. Ястребиная сова Surnia ulula Linn.
Strix Ulula. Linnaeus. Systema Naturae, изд. X, 1758, стр. 93, Швеция.
Распространение. Ареал. Сев. Европа и Азия до границы леса на севере
и до средней Скандинавии, центральных частей европейской России, южной
окраины тайги в Сибири — Тюмени, Тары, Алтая; кроме того, в Тарбагатае,
Тянь-Шане, сев. Монголии, Манчжурии, Приморье; на Сахалине; в Сев.
Америке в Аляске, Канаде, Британской Колумбии. Характер
пребывания. Оседлая птица, совершающая зимой нерегулярные перекочевки и
406
ОТРЯД СОВЫ
появляющаяся тогда к югу от гнездовой области. Биотоп. Леса, главным
образом, хвойные.
Подвиды и варьирующие признаки. Европейско-сибирский подвид отличаетсяотсеверо-
американского более интенсивной и темной окраской, тяньшанский подвид отличается
большей длиной крыла. S. и. ulula L, 1758, распространена в Европе и Азии от Скандинавии до
Анадыря, Камчатки и Сахалина; S. и. tianschanica Smallbones, 1906, в
центральном и восточном Тянь-Шане; S. и. caparoch M u 1 1 е г, 1776, — в Сев. Америке от Аляски,
с.-з. Мекензи и Гудзонова залива до юга Британской Колумбии, средних частей Альберты,
Унгавы (зимой до южной Канады и более или менее случайно в северных Соединенных
Штатах).
Карта 69. Распространение ястребиной *совы Surnia ulula
1 — S. u. ulula, 2 — S. u. tianschanica, 3 — S. u. caparoch; a — граница гнездовий, г — область .зимних
кочевок.
Обыкновенная ястребиная сова Surnia ulula ulula Linn.
Синонимы. Surnia ulula pallasi. Buturlin. Ornith. Monatsber., 1907, стр. 100,
Алазея и Среднеколымск. — Surnia ulula orokensis. Stachanow. Koezag, IV, 1931, стр. 21.
Люди на Сахалине. — Strix doliata. Pallas Zoographia Rosso-Asiat. 1811, стр. 316,
новое имя для 5. ulula. — Strix uralensi Shaw. Gen Zool. v. VII, 1809, стр. 277,
табл. 35, Сибирь и Урал.
Русское название. Книжное. Местами в Сибири у местных жителей
«березовая сова».
Распространение. Ареал. В Европе —в северной Скандинавии от 70°
с. ш. до Даларне и сев. Вермланда в Швеции; в сев. Финляндии повидимому
до Куопио; в СССР от 69° с. ш. в Лапландии (граница леса), Соловецких
о-вов, Обонежья, Архангельска, Мезени, средней Печоры; в Зап. Сибири до
Обдорска, Туруханского края, южн. Таймыра (Хатангское, по наблюдениям
Скалона, 1938); в бассейне Лены и восточнее до 68—70° с. ш. (Аллаиха на
Индигирке, на Колыме по крайней мере до 68°30' с. ш.), на Анадыре, в Коряц-
кой Земле, Камчатке; к югу до Смоленска, Калинина, Москвы, Костромы,
Горьковской обл. (Макарьев, Ветлуга), Казани; в Западной Сибири до
Тюмени, Тары, Томска, Алтая, Тарбагатая и Саяна, ю.-з. Забайкалья и сев.
Монголии (Кентей и Хангай); северные части Приморья; Сахалин; Б. Хинган
в Манчжурии. В негнездовое время —к югу от гнездовой области --в
Прибалтике, южной Норвегии и Швеции, в средней России (Рязанская, Орловская,
Курская обл., средняя Волга, Башкирия; в виде исключения даже до
Киева ), в сев. Казахстане (Аман-карагай, Зерендинское); кроме того в Англии,
Дании, в вост. Германии, изредка в Австрии, Венгрии, с.-в. Франции, Бельгии,
Голландии, Швейцарии. В Азии на Курильских о-вах и т. д.
Биотоп. Тайга, главным образом хвойная или смешанная: на севере
в лесотундре—ивовые и тополевые заросли по долинам рек. В гнездовое время
в лесостепи, в березовых колках в ю.-з. Сибири, по вырубкам и гарям. Охотно
ЯСТРЕБИНАЯ СОВА
407
придерживается участков с преобладанием лиственницы. В горах — до
верхней границы леса (до 1800—2000 м на Алтае).
Характер пребывания. Оседлая и кочующая птица. В Зап.
Сибири часть сов зимует в тайге, часть откочевывает в лесостепь (см. выше).
Вообще у ястребиных сов, как у других миофагов, в негнездовое время
усиливается тяготение к открытому и обжитому ландшафту. Расстояние
кочевок довольно значительное, а направления их разнообразны: сова,
окольцованная 24.VI. 1946 у Квиккьока в Шведской Лапландии, добыта была
3.XI. 1946 у Мурманска.
Численность. В северной полосе тайги — обычная птица.
Численность, как у других миофагов, колеблется повидимому по годам, как колеблются
и плодовитость (число яиц в кладке), и кочевки — в зависимости от «урожая»
грызунов. Хотя ястребиная сова питается и птицами, но негнездование ее на
севере в годы отсутствия грызунов весьма вероятно. В зависимости от наличия
грызунов, зимнее размещение сов варьирует локально. В разных местностях
средней России она появляется не ежегодно (в Ленинградской обл., в
Псковской, где их было много в 1894—1895 годах, по Зарудному, 1910; в
Куйбышевской обл., где сов было много в 1886 г., а затем они не попадались зимой до
1897 г., Карамзин, 1901;в Татарской республике сов было много в 1887—1888 г.,
мало в 1889—1890, и где они вовсе не попадались в 1891—1892, Рузский, 1891,
не встречена вовсе в неурожайные по грызунам годы на Анадыре, Портенко, 1939).
Экология. Размножение. Брачный крик и спаривание начинается
уже с середины марта (южная Финляндия; Ленинградская обл., Камчатка).
Гнезда главным образом расположены на обломанных вершинах деревьев
(Лапландия) нередко в дуплах осин; иногда используется старое гнездо
вороновых (сорок, ворон) или хищных птиц, а также дятловые дупла. Подстилки
нет (немногие случайно выпавшие или линные перья самой совы).
Кладка около середины апреля (для СССР точных наблюдений не
опубликовано), быть может с конца марта. Однако яйца в Лапландии найдены ив
июне, что возможно указывает не на дополнительную взамен утерянной кладку,
а на наличие второй при благоприятных кормовых условиях (У ил раит, 1871).
Число яиц обычно 3—4, но иногда до 9—10 и даже 13, у нас кладки в 7 и 9 яиц
найдены в частности у Томска (В. Иогансен, 1923); большие цифры —в годы
урожая грызунов. Судя по развитию птенцов, кладка у нас происходит в
такое же время и быть может одновременно по всему ареалу. Яйца
эллиптически-закругленные, белые и блестящие. Размеры (100): в среднем 40X31,8,
колебания 36—44X29—34,4 мм (Уайзерби, 1938).
Насиживание с первого яйца, пбэтому птенцы очень разновозрастны.
Насиживает главным образом самка, но быть может при известном участии самца.
Птенцы в мезоптиле — подлетки встречаются обычно во второй половине июня
(Минусинская лесостепь, Божье озеро, 15—27 июня; 15 июня у Мажарского
озера; 29 июля у Якутска; 23 июня на Алтае; 1*5 июня на Камчатке; 22 июня
у реки Туры, Туруханский край); только что вылупившиеся ниже Верхоянска
встречены 24 мая. Летные выводки — в разные числа июля.
Линька. Полная годовая. Последовательность смены нарядов:
пуховой — мезоптиль — первый годовой — второй годовой и т. д. При смене ме-
зоптиля, как у других сов, маховые и рулевые остаются от гнездового наряда
и линяние таким образом частичное. Последовательность смены маховых от
заднего края к переднему (от 10-го к 1-му). Линька мезоптиля с июня по
август, заканчивается повидимому в сентябре, когда остатков гнездового наряда
среди мелкого оперения уже нет. Взрослые линяют со второй половины июня
и до конца сентября, в начале октября перо свежее.
Питание. Пищу обыкновенной ястребиной совы составляют главным
образом грызуны, в первую очередь различные полевки и лемминги. Отмечается
408
ОТРЯД СОВЫ
зависимость плодовитости от «урожая» грызунов (см. выше). Охотится
преимущественно в светлое время дня, рано утром и затем под вечер. Способы
охоты: сова сидит на дереве, преимущественно на сухой вершине, подстерегая
появляющуюся добычу, и бросается на нее, или низко, по-ястребиному, летает
над землей, вспугивая и хватая добычу. Отмечено нападение на летящих птиц
(кукша, Уилрайт, 1871). В качестве корма ястребиной совы в нашей стране
указываются главным образом полевки (до 80%, Зап. Сибирь, Шлезигер, 1929),
кроме того, водяные крысы, землеройки, мыши и т. д. Из птиц упоминается
о нападении даже на куропаток, на тетеревиные чучела, кроме того на воробьев,
чечеток и др.
Рис. 105. Ястребиная сова.
Полевые признаки. Сова средней величины, бурая со светлыми пестринами
на спинной стороне, на брюшной беловатая с поперечным «ястребиным»
рисунком. Небольшая голова с нерезким лицевым диском, без перовых ушей;
небольшие желтые глаза; относительно короткие крылья и длинный
ступенчатый хвост. Полет напоминает дневных хищных птиц — быстро летает, после
нескольких взмахов проносится вперед, не шевеля крыльями и т. д. Иногда
«трясется» на месте в воздухе, как пустельга. Очень подвижна, деятельна и
днем, или главным образом днем. Неосторожна, активно защищает птенцов и
гнездо. Ночует на деревьях на ветвях, но не в дуплах. Крик «ки-ки-ки» или
вроде ястребиного «кия-кия-кияяк».
Описание. Размеры и строение. Оперение жесткое, плотно прилегающее
к телу. Ушные отверстия симметричны.
Лапы оперены до когтей. Формула крыла: 3>4>2>1>6>7 (не считая истинного 1-го
рудиментарного махового). Вырезка на наружных опахалах 2-го, 3-го и 4-го маховых.
Длина самцов (10) 360—390, самок (8) 390—410, в среднем 371 и 402 мм. Размах самцов
(7)710—792, самок (5) 790—808 мм, в среднем 766,4 и 802,4 мм. Вес самцов (6) 247—375,
в среднем 314 г, самок (3) 323, 350 и 371 г, в среднем 346 г. Крыло птиц из Европы: самцов
ЯСТРЕБИНАЯ СОВА
409
(19) 222—242, в среднем 233,8 мм; самок (22) 224—244, в среднем 235,3 мм; из Зап. и
средней Сибири —самцов (25) 223—241, в среднем 232,7 мм, самок (14) 228—242, в среднем
236 мм; из Вост. Сибири самцов (22) 221—241, в среднем 233 мм, самок (28) 228—243,
в среднем 239,5 мм.
Окраска. Пуховой наряд белый с охристым оттенком. Мезоптиль темнобурый
на спинной стороне, с беловатыми пятнами; на брюшной стороне грязно-белый с тонким
поперечным рисунком. Окончательный наряд (первый годовой и последующие) — самцы и
самки: общая окраска спинной стороны бурая со светлыми белыми пестринками и полосами;
брюшная сторона белая с темнобурым поперечным рисунком; на темени и затылке мелкие
белые пятна у вершины и поперечные белые полоски на каждом пере; на задней части шеи
пятна крупней и часто почти сливаются в белое поле; верхняя (передняя) часть спины
одноцветно бурая, с немногими белыми пятнами: на плечевых бурый цвет лишь в виде пятна у
вершины пера и тонких поперечных полос; малые кроющие крыла бурые, прочие — бурые
с крупными продольными белыми пятнами; надхвостье бурое, светлей спины, с белым
поперечным рисунком; маховые бурые с беловатой каймой и поперечными белыми пятнами;
рулевые с узкими белыми полосами и белой вершинной каймой; лицевой диск белый, с темно-
бурым пятном у ушей; горло белое с бурым пятном посередине; верхняя часть груди более
или менее чисто белая с правильным темнобурым поперечным рисунком различной ширины.
Индивидуальные вариации окраски довольно значительны и выражаются в разнице
основного бурого тона спинной стороны (иногда сероватого, что впрочем часто в значительной
мере связано с выгоранием пера), в развитии белых пестрин на затылке, плечах, крыльях
(у некоторых особей эти пестрины резко преобладают над основным бурым фоном на затылке
и шее), в ширине темных полос на брюшной стороне. Самые темные особи приближаются
к американскому подвиду 5. и. caparoch, но встречаются они в самых разнообразных частях
ареала (например на Соловецких о-вах, на севере Вологодской обл., у Нижнеудинска
и т. д.). Радужина 'желтая, клюв желтый, когти черные.
Систематические признаки. Экспедиция «Вега» 22.V. 1879 г. добыла на северном
побережье Чукотского п-ова у Питтлекая сову, которую Пальмен (1887) отнес к
американскому подвиду. Мной осмотрены 3 птицы из земли Чукчей, они, как и анадырские,
не отличимы от сибирских. Принимая во внимание большук/индивидуальную изменчивость
окраски этой совы и ее характер (см. выше), весьма вероятно/что упомянутая особь — лишь
индивидуальная вариация европейско-азиатской расы ястребиной совы.
Тяньшанская ястребиная сова Surnia alula tianschanica Smallbones
Surnia ulula tianschanica. Smallbones. Ornith. Monatsber. 1906, стр. 27, Кашка-
су в Терскей-Ала-тау.
Синоним. Surnia rulula korejewi. Sarudny aud Loudon. Orn. Monatsber., 1907,
стр. 2, Джунгарский Ала-Тау.
Распространение. Ареал. Восточный и центральный Тянь-Шань—от
Джунгарского Ала-Тау на севере до Боро-хоро и долины Текеса (Хелль-
майр, 1929) на востоке, на запад до восточных частей Киргизского хребта, на
юг до южных хребтов Тянь-Шаня—Нарын, Аксай, Джергалан, Каракол.
Биотоп. Леса тяньшанской ели Picea Schrenckiana, примерно от 1500 до 3000 м.
Характер пребывания. Оседлая птица. Численность. Редка.
Экология. Размножение. По датам совпадает с размножением птиц
из северной тайги. Птенец с совершенно короткими крыльями, в мезоптиле,
добыт на Талгаре 4 июня. В Боро-хоро 25 июля добыта'птица, кончающая смену
мезоптиля на контурное перо. Судя по этим данным, кладка происходит в
апреле. Гнезда расположены на елях. Яйца не описаны. Линька. Птицы
в свежем пере добыты в конце сентября; иногда линяние затягивается до
начала октября (Шульпин, 1939), быть может в результате затянувшихся или
вторых кладок. Питание. Пищу таньшанской совы составляют главным
образом грызуны —полевки. Охотится сидя на'вершинах'елей и высматривая
появление добычи (Шульпин, 1939).
Описание. Размеры и строение. От северо-таежных птиц отличается
большей длиной крыла: длина крыла самцов (И) 238—251, самок (7) 243—252, в среднем
соответственно 243,4 и 247,8 мм. Следовательно, крыло в среднем на 1 см больше. Окраска.
Темная, но такие же птицы встречаются и в Европе, и в Сибири: бурый цвет верхней стороны
тела насыщенный; белые пестрины развиты относительно слабо; светлых вариаций
окраски — по основному тону и по развитию белых пестрин — неизвестно.
410
ОТРЯД СОВЫ
РОД НЕЯСЫТИ STRIX LINNAEUS 1758
Тип Strix aluco
15. Бородатая неясыть Strix nebulosa Forster.
Strix nebulosa. Forster. Philosophical Transactions, t. 62, 1772, стр. 424, Гудзонов
залив.
Русское название. Книжное, происходит от черного пятна под клювом.
Местное название в Сибири — «каменная сова».
Распространение. Ареал. Северные леса Европы, Азии и Сев. Америки.
В Европе и сев. Скандинавии, Финляндии, в СССР от границы леса примерно
до 52° с. ш., на юге в Европейской части, в Сибири до южной окраины тайги.
В Сев. Америке от средней Аляски, с.-з. Мекензи, Квебека, на юг до севера
Британской Колумбии, Альберта, севера Манитоба, с.-в. Онтарио, в горах до
Сьерра-Невада, в Калифорнии, Айдахо и зап. Монтана. Характер пре-
Карта 70. Распространение'бородатой неясыти Strix nebulosa
1 — S. n. lapponica, 2 — S. n. nebulosa; a — область гнездовий, г — область зимних кочевок (в кружках
местонахождение подвидов в зимнее время).
бывания. Оседлая и нерегулярно кочующая птица. Биотоп.
Высокоствольный хвойный лес как в низменности, так и в горах.
Подвиды и варьирующие признаки. В американской части ареала S. п. nebulosa
Forster, 1772; в европейской, азиатской 5. п. lapponica Thunberg, 1798. Морфологические
различия в окраске — американская раса темней; образ жизни весьма сходен,
экологические различия в трофических связях и деталях цикла периодических явлений.
Лапландская бородатая неясыть Strix nebulosa lapponica Thunberg
Strix lapponica. Thunberg. KungL Vetensk. nya Handl, 1798, стр. 184, Лапландия.
Синонимы. Strix barbata. Latham. Index Ornith. 1, 1790, стр. 62, горы Восточ-
ной Сибири. — Strix tridactyla Fischer. Memoires Soc. Imp. Naturalistes Moscou, v. 3,
1812, стр. 278, между Можайском и Смоленском. — Strix microphatalmos. Tyzenhauz
Ornich. Powszechna, 1842—1846,стр. 26, Литва.— Syrnium cinereum sakhalinense.
Бутурлин. Псовая и Ружейная Охота, февр. 1907, стр. 8, Сахалин. — Strix nebulosa elizabethae.
Stegmann. Доклады Академии наук, 1925, стр. 61, Кентей.
Распространение. Ареал. Сев. Европа и Азия, в Европе в сев.
Скандинавии — в Норвегии, Швеции и Лапландии, восточней — в
Архангельской, Вологодской, Кировской, Молотовской обл. на юг до Ленинградской,
Псковской, Смоленской обл., в Белоруссии —до Полесья, в Литве,
Ярославской обл., быть может в Татарской республике (самец с сильно развитыми
семенниками добыт в мае 1926), в Свердловской области на Среднем Урале;
в Сибири на юг до Тюмени, Тары, Алтая, Забайкалья (где доходит до Кентея
в сев. Монголии) и Приамурья (Бурея, Вира); к северу по крайней мере до
БОРОДАТАЯ НЕЯСЫТЬ
411
Обдорска, на реке Таз до 65° с. ш., в Якутии примерно до 68°, на востоке до
лесных частей бассейна Анадыря и Коряцкой Земли (Пенжина), побережья
Охотского моря и Сахалина. Залеты бывают еще далее на север — до Хатанг-
ского на Таймыре (72° с. ш.), Казачьего на Яне. Есть сведения о гнездовании
в б. Лифляндской губ. (Руссов), в Киевской обл. (Шарлемань, 1937); о
встречах на Камчатке. Все эти сведения по всей вероятности ошибочны. В
негнездовое время встречается и южней, достигая Прибалтики (Калининград),
Калининской, Московской, Орловской и Чкаловской обл.
Характер пребывания. Оседлая, отчасти кочующая птица
(нерегулярные миграции в связи с неблагоприятными кормовыми условиями
в гнездовой области?). Во всяком случае, в Лапландии в некоторые годы
довольно обычна, в другие вовсе не гнездится. Зимой встречена и далеко на
севере — на Анадыре и у Среднеколымска и т. п. Б и о т о п. Крупный старый лес,
как в горах (Лапландия, Алтай, Восточная Сибирь), так и на равнинах.
Главным образом хвойная тайга; в Восточной Сибири лиственничники. Зимою
иногда у жилья.
Численность. В Европе немногочисленна, в Восточной Сибири
(Якутия) одна из обыкновенных сов. Численность подвержена периодическим
колебаниям (Лапландия, ряд авторов, бассейн Колымы, устн. сообщение
Бутурлина).
Экология. Размножение. Цикл ранний, географических различий
в календаре размножения не установлено. Гнезда расположены на деревьях,
чаще всего со сломанными вершинами (как у ястребиной совы), открытые, не
в дуплах. В Сибири чаще всего на лиственницах (в бассейне Пенжины на
тополе), в Европейской части СССР на елях (в 6—-10 м от земли; размеры гнезда
до 1 м в диаметре, при высоте в 40 см; лоток плоский; выстлано гнездо хвоей
и еловыми веточками, Смоленская обл., Сушкин, 1917). Сама сова их пови-
димому не строит, но пользуется сооружениями других птиц. Одно гнездо
используется много лет.
Кладка начинается около середины апреля, полная -^ обычно к началу
мая (3 яйца у Охотска 1 мая; 5 яиц в Поречье, Минская обл., 27 апреля, Шнит-
ников, 1913; 4 яйца 26 апреля в Лапландии, Уоллей, 1867). Число яиц в кладке
3—5, чаще 4, но иногда одно. В неблагоприятные годы вовсе не гнездится.
Яйца продолговатой формы, длинней, чем у других неясытей, белые матовые
или со слабым блеском. Размеры (97) 54,05X42,54 мм, в среднем, колебания
49—58,1X39—44,7 мм (Хартерт, 1913).
Насиживает повидимому одна самка, с первого яйца, высоко подняв хвост
и крылья, и сидя очень крепко. Слепые только что вылупившиеся птенцы
найдены 12 мая; глаза открываются на 7-й день, в десятидневном возрасте
начинает пробиваться оперение (маховые), полный мезоптиль развивается в
возрасте несколько большем 5 недель, птенцы тогда начинают перелетывать
(Сушкин). Летные птенцы в Смоленской юбл. встречены 20 июня, на Вилюе 23 июня
(Воробьева, 1928), однако, из-за разновозрастности птенцов вылет их
растягивается до разных чисел июля (в устье Алдана еще 8 июля птенцы с выросшими
примерно на 2/3 маховыми, Иванов, 1929), на Оби у Сарай-гор подлеток 10 июля;
птица у Тары с недоразвитыми маховыми — 14 июля. В конце июля
начинается смена мезоптиля, заканчивающаяся примерно через полтора месяца, в
начале сентября (приблизившаяся к окончанию линяния из мезоптиля сова у
Сур-язы, с.-в. Алтай, добыта 24 августа). Выводки долго держатся вместе
и родители кормят вылетевших птенцов.
Линька. Полная годовая, как у других сов, за исключением смены
мезоптиля, когда от гнездового наряда остаются маховые рулевые и большие
кроющие крыла. Последовательность смены нарядов: пуховой —мезоптиль —
первый годовой (окончательный по окраске, но комбинированный по составу)—
412
ОТРЯД СОВЫ
второй годовой наряд и т. д. Детали линяния не изучены. Смена маховых идет
от заднего края партии к переднему. Птицы в свежем пере отмечены в ноябре,
у апрельских линяние еще не
начинается.
Питание. Пищу
лапландской неясыти составляют главным
образом грызуны, колебание
численности которых, как уже
указывалось, влияет на размножение
и географическое размещение. В
Лапландии — лемминги. Кроме
того —другие полевки, также
мыши (в частности, у Якутска
Stenocranius gregalis, Arvicola ter-
restris), белки (Колыма, Анадырь),
даже соболь (Кондо-Сосвенский
заповедник, Раевский 1947). Кроме
того, птицы мелкой и средней
величины (кукши, рябчики, Якутия, по
Бутурлину). Охотится
преимущественно в светлое время дня, по
крайней мере в гнездовое время.
Полевые признаки. Крупная
темноокрашенная сова с большой
головой, круговой концентрической
окраской лицевого диска, темной
«бородой» под клювом, с
небольшими желтыми глазами. Строение
легкое, длинный хвост; вытянутая
фигура. Полет плавный и легкий,
довольно медленный; голос«ху-ху-
хуу», самка у гнезда «гукает», как
клинтух, птенцы кричат, как
обыкновенные неясыти. Деятельна и
днем.
Описание. Размеры и
строение. Крупная сова с очень рыхлым и
пушистым оперением. Формула крыла
4>5>6>3>2>1... (не считая истинного 1-го рудиментарного махового). Хвост
закругленный по заднему краю. Длина самцов и самок (7) 630—660, в среднем 648,5 мм, размах
самцов и самок (7) 1310—1425, в среднем 1379 мм. Вес самцов 700, 725, 745, 810, самок
995 и 1200 <?. Крыло самцов (21) 405—436, самок (57^ 438—480 мм, в среднем соответственно
432,5 и 464,8 мм.
Окраска. Пуховой наряд сероватый на спине, чисто белый на брюшной стороне;
ноги желтоватые, радужина желтовато-серая. Мезоптиль на спинной и брюшной стороне
тела темнобурый с беловатым поперечным рисунком. Маховые и рулевые — как в
окончательном наряде. Окончательный наряд самиов и самок: спинная сторона беловатая, сильно
испещренная серовато-бурым, кажущаяся в общем сероватой; темя и затылок с охристым
оттенком, с бурым поперечным и продольным рисунком, мелким и темным; на спине и
крыльях такой темный рисунок очень развит, и белый цвет заметен только на внешних опахалах
плечевых и кроющих крыла; продольные темные полосы на спине и крыльях широкие;
маховые темнобурые с охристым поперечным светлым рисунком, более развитым на
внутренних опахалах, рулевые бурые с беловатыми мраморными пестринами, образующими
неправильный рисунок. Лицевой диск беловатый в резких черноватых концентрических кругах;
горло черновато-бурое («борода»); грудь, брюхо, бока в широких «мазаных» бурых
продольных полосах и с мелким буроватым крапом, ноги и подхвостье в мелких бурых пестринах.
Индивидуально варьирует общий тон окраски, то беловатый, то охристый, и распространение
темного рисунка. Клюв желтый, радужина яркожелтая, когти черноватые.
Рис. 106. Бородатая неясыть.
УРАЛЬСКАЯ ИЛИ ДЛИННОХВОСТАЯ НЕЯСЫТЬ
413
16. Уральская или длиннохвостая неясыть Strix uralensis Pallas
Strix uralensis, Pallas. Reise d. verschied. Prov. d. Russ. Reichs I, Anhang 1771,
стр. З, Урал.
Русское название. Неясыть — родовое обозначение для крупных серых
сов, народное. «Длиннохвостая» — по морфологии, «уральская» — перевод латинского
названия, неудачного, так как птица распространена не только на Урале; оба имени —
книжные.
Распространение. Ареал. Сев. Европа и сев. Азия, к югу до
Калининградской обл. Альп, Карпат, средней России, в Зап. Сибири примерно до
Карта 71. Распространение длиннохвостой неясыти Strix uralensis
1 — S. u. Iiturata, 2 — S. u. uralensis, 3 — S. u. yenisseensis, 4 — S. u. buturlini, 5 — S. u. nikolskli, б — S.
u. coreensii, 7 — S. u. hondoensis, 8 — S. u. momiyamae, 9 — S. u. fuscescens, 10 — S. u. davidi, И — S.
u. macrura; a —» границы гнездовий, б — недостаточно выясненные границы гнездовий, г — область
зимних кочевок (цифры в кружках обозначают зимние местонахождения разных подвидов и соответствуют
номеру подвида).
54° с. ш., далее до Алтая, Саяна, сев. Монголии, Манчжурии, Кореи,
Сахалина, Японии; в зап. Сычуани в Китае.
Характер пребывания. Оседлая и кочующая птица.
Биотоп. Хвойные и смешанные леса таежного типа, на равнинах; на юге ареала —
в горах. Численность. В таежной полосе с.-в. Европы и Сибири обычна,
у южного края ареала и в разбросанных горных колониях редка (ледниковый
реликт).
Подвиды и варьирующие признаки. Морфологически варьируют размеры и окраска
(в соответствии с так называемыми правилами Бергманна и Глогера, расы холодного и
сурового климата светлей и контрастно окрашены, с черноватым рисунком, и крупнее; в мягком
климате обратные явления). Экологические географические изменения слабо изучены. В
западной части ареала — в Скандинавии, в горах средней Европы, на восток до западной
полосы европейской России 5. и. Iiturata T e n g m a 1 m, 1793; в восточной части
европейской России и Зап. Сибири S. и. uralensis Pallas, 1771; в средней Сибири от Енисея,
Алтая до сев. Монголии (Хангай) S. и. yenisseensis В u t., 1915; в Якутии и на Охотском
побережье 5. и. buturlini Dementiev; в Забайкалье, прилежащих частях Монголии
(Кентей), на Амуре, в бассейне Уссури, в М. Хингане, на Сахалине 5. и. nikolskii В u t,
1907;вю.-в. Манчжурии, в Корее, на о-ве Хоккайдо сомнительная S. и. corensis M о m i-
414
ОТРЯД СОВЫ
у a m a, 1927; далее в Японии формы, взаимоотношения и самостоятельность которых не
ясны — на северном Хондо, к югу до 37° с. ш. S. и. hondoensis Clark, 1907; в средних
частях Хондо S. и. momiyamae Так а-Т s u к a s а, 1931; в южной и западной части
Хондо, на Авачи, Сикоку и Кюсю 5. и. fuscescensT emmincket Schlege 1,1847; наконец,
в западной Сычуани в качестве изолированной колонии S. и. dauidi S h a r p е, 1875.
Значение hondoensis и momiyamae, а быть может и coreensis, в качестве географических форм
кажется весьма сомнительным, как и отграничение первых двух от fuscescens, а третьей —
от nikolskii. Для окончательного суждения у нас, к сожалению, не было материала.
Западная длиннохвостая неясыть Strix uralensis liturata T e n g m a 1 m
Strix liturata. Tengmalm. Kongl. Vetensk. Akad. Handlingar, 14, 1793, стр. 267,
Швеция.
Распространение. Ареал. Средняя Европа от Лапландии, Швеции (на
юге до северных частей Даларне), ю.-в. Норвегии, Финляндии до 64° с. ш.,
к югу до Прибалтики, Карпат, вост. Каринтии, Крайны, Штирии и Богемского
леса, Балкан (Югославия, сев. Албания). В горах средней Европы более или
менее изолированные колонии. В СССР от Лапландии (Лапландский
заповедник), Белого моря (Кандалакша) и Архангельска к югу до южной границы
тайги, примерно 53° с. ш., в Белоруссии (Пинск), Калининградской обл.,
Смоленской, Псковской, Ленинградской, Новгородской областей, на востоке
до Костромской и Московской (в последней гнездование не доказано).
Характер пребывания. Оседлая и быть может лишь недалеко
кочующая птица. Залеты вне гнездовой области редки. Биотоп.
Высокоствольный лес — хвойный или смешанный. В негнездовое время
приближается к поселениям (парки, окрестности сел, где зимою у гумен скапливаются
грызуны и птицы; даже города, например Ленинград, в 1878 г,, по Плеске и
Бихнеру, 1881; 8 февраля добыта в Москве).
Численность. Невысокая, быть может в связи с реликтовым
характером распространения в южных и западных частях'ареала вида, но также в
связи со слабой и к тому же неравномерной по годам плодовитостью. Как и
другие миофаги, в годы «неурожая» грызунов длиннохвостая неясыть не
размножается. По наблюдениям Эйхлера в Калининградской обл. в 1931г., после
суровой зимы, сова на участке работ автора не гнездилась, а зимой там были
найдены три мертвых птицы; между тем, тут же в 1930 г. было 4 гнезда, в
которых вывелось в общем 20 молодых. В 1932 г. длиннохвостая неясыть
там опять не гнездилась и только в 1933 г. отмечалось нормальное ее
размножение. При этом и число яиц в кладке у этого вида невелико.
Экология. Размножение. Цикл размножения ранний. Брачный
крик уже в марте (Ленинградская обл.). Гнезда расположены изредка
в дуплах (крупная птица), обычно же в старых гнездах других птиц,
высоко от земли, в 10—20, но иногда и только в 4 м (Нитхаммер, 1938). В
неблагоприятные по кормовым условиям годы не размножается. Число яиц
в кладке обычно 3—4, редко 2 или до 6. Насиживает (в Скандинавии и по
наблюдениям в неволе) только самка, в течение 27—29 дней (гнездовый период
продолжается немного более месяца — около 34 дней). Птенцы в мезоптиле
у Пскова в начале июня (выводок из 3 разновозрастных птенцов, насиживание,
следовательно, с первого яйца), в конце июня птенцы подросли, но смена мезо-
птиля еще не начиналась. Вполне выросшие птенцы в контурном пере в начале
сентября.
Линька. Начало линяния из мезоптиля в июле (в возрасте около
45 дней), конец — в начале сентября. Линька взрослых — полная годовая,
вероятно с мая, в начале июня смена маховых проходит уже на 40^45%. Птицы
в свежем пере с сентября. Последовательность смены первостепенных маховых
идет от 10-го к 1-му. Смена нарядов обычного у неясытей типа.
УРАЛЬСКАЯ ИЛИ ДЛИННОХВОСТАЯ НЕЯСЫТЬ
415
Питание. Западная длиннохвостая неясыть в основном миофаг,
поэтому заметны колебания плодовитости и численности (см. выше). Летом
охотится на опушках и лесных полянах, зимою — кроме того на открытых местах
у поселений. Охотится чаще всего сидя на дереве и выслеживая появляющуюся
добычу, иногда же руководствуясь слухом. Потребность в пище за сутки около
125 г (Уттендерфер, 1939). В качестве пищи указываются полевки Clethrio-
nomys glareolus для Лапландии (Се-
менов-Тяншанский, 1937), мыши
(Череповец, Богачев, 1927), белки,
полевки, лесные и полевые мыши,
зайчата (средняя полоса), кроме того, в
особенности зимою, птицы: галки,
сороки, вороны, сойки, голуби,
рябчики, тетерева, также мелкие
воробьиные.
Полевые признаки. Крупная и
светлая, неясыть больше
обыкновенной; рисунок брюшной стороны резко
продольный по беловатому фону;
небольшие темные глаза; длинный
слегка закругленный хвост,
провисающий при полете книзу. Очень
подвижна, деятельна и днем, в
особенности в пасмурную погоду. Голосом
похожа на обыкновенную неясыть, но
крик глуше, как «ху-ху-хуу»; кроме
того, лающее «хау-хау».
Описание. Размеры и
строение. Формула крыла и вырезки — как у
других неясытей 5^4>3>2>6>7> 1... (не
считая, как всегда у сов, истинного первого
рудиментарного махового). Лапы оперены
до пальцев. Длина (?) Размах (?) Вес
самцов (4) 680—750 в среднем 720 г, самок
(5) 820—970, в среднем 888 г. Крыло
самцов (21) 342—368, самок (26) 357—382, в
среднем соответственно 354 и 366,5 мм.
Несколько крупней номинального подвида.
Окраска. Пуховой наряд белый с
охристым оттенком. Мезоптиль охристый с
бурым поперечным рисунком и с широкими
белыми краями перьев на голове, шее,
спине, груди, брюхе, боках. Сразу после
смены пухового наряда, когда выступают
главным образом белые вершины перьев,
молодая неясыть кажется беловатой, затем, по мере роста пера, — бурой с беловатым
поперечным рисунком. Окончательный наряд самцов и самок: общий тон охри<;то-
беловатый с бурым рисунком; темя, затылок и задняя сторона шеи с широкими
бурыми наствольями; на плечевых эти наствольные полосы менее правильны и расширяются
к вершине, так что плечевые, как и спина, кажутся бурыми с охристыми каймами перьев;
малые кроющие крыла бурые, средние и большие — бурые с супротивными
беловато-охристыми пятнами, особенно заметными на наружных опахалах; маховые и рулевые буровато-
охристые с темнобурым поперечным рисунком. Лицевой диск серовато-охристый с темными
стволами перьев; брюшная сторона беловато-охристая с широкими бурыми наствольными
чертами; оперение ног охристое с бурыми крапинами. Радужина темнобурая, клюв желтый,
когти черные.
Систематические замечания. Возможность выделения юго-западных — карпатских и
других — длиннохвостых неясытей в особую форму 5. и. macrura Wolf, 1810, как это
предложил А. Дунаевский (1940), кажется нам сомнительной, так как и liturata нерезко
различается от номинальной расы. Остается однако фактом наличие в Карпатах большого
Рис. 107. Длиннохвостая неясыть.
416
ОТРЯД СОВЫ
числа особей темнобурой вариации окраски — наряду с обычными серыми. Такой
вариации — аналога окраски «wilkonskii» у обыкновенной неясыти — в других частях ареала
длиннохвостой неясыти — нет, но служит ли этот сам по себе любопытный факт наличия
в Карпатах таких бурых особей основанием для признания этих популяций особой
географической формой? На это, как нам кажется, можно дать отрицательный ответ.
Западносибирская длиннохвостая неясыть Strix uralensis uralensis Pallas
Синоним. Syrnium uralense sibiritum. T s с h u s i. Ornith. Jahrbuch, XIV, 1903, стр. 166,
Томск.
Распространение. Ареал. К востоку от предыдущей: бассейн Печоры
и ее притоков, бассейн Волги — в Европейской части Союза, Урал и При-
уралье; Зап. Сибирь на восток до Томска; на юг до средней Волги (бассейн
Суры, Самары и Бузулукский бор; Пензенская обл.), Южн. Урала
(бассейн Белой, Миас), Тюмени, Ялуторовска, Тары и Каинска в Зап. Сибири.
В с.-в. Европе северная граница не вполне ясна, на самом севере Урала вероятно
отсутствует; добыта на средней Печоре (Якша) и в верхнем течении этой реки,
в б. Яренском у., ныне территория Коми АССР, в бассейне реки Удоры; за
Уралом у Шухтункурта на М. Сосьве, на Оби у Саранпаула; далее на
восток северная граница не прослежена.
Характер пребывания и биотоп. Как у предыдущей расы.
В Зап. Сибири кочевки зимой повидимому более дальние, так как к этому
подвиду относятся птицы, добытые в холодное время года у Красноярска и на
Алтае (экз. в Зоологическом музее Московского университета). На кочевках
в Европейской чисти страны до низовьев Волги и Урала, нередко в городах и
их окрестностях (Чкалов, Киров).
Численность. Выше, чем у предыдущей расы. В воет, России и
Зап. Сибири обыкновенная
Экология. Размножение. Брачный крик начинается с середины
апреля (Молотов). Кладка «как сойдет снег» (б. Сызранский у., Богданов,
1871) во второй половине марта (Татарская АССР, Рузский, 1891). Число яиц
в кладке 2—3 (Богданов, Рузский) и даже одно (Эверсманн, 1866). Яйца чисто
белого цвета, размеры (2) 45,5—45,6X38,5—39,5 мм (Рузский, 1891). Кладки
быть может иногда в дуплах, иногда же на земле между корнями деревьев
или даже в расщелинах песчаника по склонам лесных оврагов (Богданов)
или в гнездах других птиц. Птенцы вылупляются в мае, насиживание
следовательно продолжается около месяца; подлетки в мезоптиле с середины
июня, гнездовый период длится немного более 30 дней. Птенцов в выводке
бывает 2—3, они разновозрастные, следовательно, насиживание с первого яйца.
Линька. Как у предыдущей формы. Взрослые интенсивно линяют
в июне, июле и до августа, в начале сентября птицы в свежем пере. Начало
линяния, повидимому, в мае. 30 июня на р. М. Сосьве у самки сменились все
задние первостепенные маховые, 4-е выпало, передние 3 старые, линька
продвинулась на 70%. Мезоптиль сменяется на контурное оперение во второй
половине августа, сохраняясь дольше всего на затылке.
Питание. В летнее время пищу западносибирской неясыти составляют
главным образом млекопитающие —мыши и полевки, молодые зайцы; в
меньшей мере птицы, изредка насекомые. Зимою она приближается к поселкам, и
тогда в кормовом режиме большое место начинают занимать птицы: галки,
вороны, сороки, даже голуби, которых сова ловит на чердаках (Чкалов);
также мыши, домашние куры, серые куропатки и тетерева (б. Сызранский район,
Богданов; низовья Волги и Урала, Бостанжогло, 1911),
Описание* Размеры и строение. Длина самцов (2) 505—550, самок (2) 570—
585, в среднем 527,5 и 578 мм. Размах самцов (2) 1140—1150, самок (4) 1200—1251, в среднем
1145 и 1225 мм. Вес самцов (3) 560—712, в среднем 657,3 г, самки (1) 950 г. Крыло несколько
ДАЛЬНЕВОСТОЧНАЯ ДЛИННОХВОСТАЯ НЕЯСЫТЬ 417
короче, чем у liturata. Длина крыла самцов (18) 338—354 (368), самок (40) 348—368, в
среднем 347,4 и 357,7 мм.
Окраска. Наряду с якутским, самый светлый по окраске подвид длиннохвостой
неясыти. Основной фон чистобелый; темные полосы на задней стороне шеи узкие; лицевой
диск беловатый; на средних и больших кроющих крыла белые пестрины преобладают над
бурым фоном; настволья на груди, боках, брюхе узкие, оперение ног не испещрено или чуть
испещрено. В мезоптиле белый цвет чище и занимает большее пространство. Вся окраска
ярче, светлей и контрастней, чем у liturata.
Систематические замечания. Указанные особенности полностью приложимы к особям
из Приуралья и Зап. Сибири. В средней полосе Европейской части СССР — на север
примерно до Вологодской области, на юге в Московской, Рязанской и Горьковской —
встречаются птицы, объединяющие признаки liturata и uratensis и приближающиеся то к одной,
то к другой форме, по всей вероятности гибридная популяция.
Среднесибирская длиннохвостая неясыть Strix uralensis yenisseensis В u t u r 1 i n
Strix uralensis yenisseensis. Бутурлин. Орн. Вестник, 1915, стр. 133, Красноярск.
Распространение. Ареал. К востоку от номинальной формы — в
бассейне Енисея (на север быть может до полярного круга, Тугаринов и Бутурлин,
1911, но экземпляров нет), на юг до Алтая, Саяна и с.-з. Монголии, на восток
до Иркутска. В негнездовое время встречается в Забайкалье (Троицкосавск
и т. д.).
Биотоп. Светлая, в особенности лиственничная тайга; в горах до
1500—1600 м (Алтай). Как и другие расы, зимой в культурном ландшафте, даже
в городах (Барнаул; Красноярск, где наблюдалась с начала марта до начала
апреля; Иркутск). Заметный пролет в с.-в. Алтае у Телецкого озера начинается
с последней трети сентября и продолжается до начала ноября.
Экология. Размножение. Календарь, повидимому как у европейской
и западносибирской формы. Подлетки на Алтае у Топучей обнаружены 4 июля
(Сушкин, 1938).
Линька. Птицы в свежем пере бывают в середине сентября, конец
смены мезоптиля — остатки его на затылке и на спине — 18 августа (экз.
из Коноваловой, на границе Енисейской и Иркутской обл.). Нелиняющие
взрослые весной бывают по крайней мере до конца апреля. Питание. Пищу
составляют главным образом грызуны ^мыши и полевки (Алтай; б.
Енисейская губ.).
Описание. Размеры и строение. Величина некрупная: крыло самцов (17)
328—344, самок (28) 335—359, в среднем 338 и 349 мм. Окраска. Темная и
однообразная : бурые полосы на брюшной стороне тела развиты значительно, белые пестрины на
спинной стороне занимают небольшое пространство; бурый цвет пестрин не интенсивный, с
охристым оттенком; общий тон светлых частей охристый.
Якутская длиннохвостая неясыть Strix uralensis buturlini subs p. nova
Распространение. Ареал. Крайний сев ро-восток ареала вида: Якутия
и северное побережье Охотского моря. Известны экземпляры с Вилюя, где
птицы доходят на север до 66°, Оленека, с притока Вилюя р, Марха, из
окрестностей Якутска, добытые в 100 км ю.-з. Якутска; на северном побережье
Охотского моря в 100 км северней Охотска; у Удского острога.
Экология. Не изучена. Размножение. Молодые в мезоптиле добыты
у Якутска в середине августа (Иванов, 1929).
Линька. В мае у Якутска добыта нелиняющая птица, в сентябре у
озера Энердех в бассейне Мархи — тоже. Питание. Полевки, северная
пищуха (Вилюй, Маак), летяга.
Описание. Размеры. Крыло самцов и самок (4) 334—354, в среднем 342 мм,
следовательно примерно сходно со среднесибирским подвидом и несколько крупней, чем у
дальневосточного. Окраска. Вместе с S. и. uralensis, самая светлая по окраске раса, с чисто
27 Птицы СССР, т. I
418
ОТРЯД СОВЫ
белым основным тоном окраски; темный рисунок на брюшной стороне резкий и контрастный;
бурый цвет более насыщен, чем у uralensis; от yenisseensisn nikolskii отличается более
светлой окраской. Тип: самка, сентябрь 1938, озеро Энердех, река Марха, Якутия, в
Зоологическом музее Московского университета.
Дальневосточная длиннохвостая неясыть Strix uralensis nikolskii В u t u r 1 i n.
Syrnium uralensie nikolskii. В u t u г 1 i n, Journ f. Ornith., 1907, стр. 333, Приморье.
Синонимы. Strix uralensis tatibanae. M о m i у a m a, Bull. Brit. Ornith. Club»
CCCXV11, 1927, стр. 21, Сисука, Южн. Сахалин — Strix uralensis daurica. Stegmann
(ex Suschkin MS). Journal fur Ornithologie. 1930, стр. 463, ю.-в. Забайкалье (?) — Strix
uratensus coreensis. Momiyama, Journ Chosen Nat. Hist Soc. V. 4, 1927, стр. 1, Тайян-
цан, Корея.
Распространение. Ареал. Забайкалье, к северу до Баргузина и Витима;
Приамурье, Сахалин ;к югу до Кентея, Хингана и быть может Кореи(uoreensisrt).
Биотоп. Тайга как северного, так и манчжурского типа; в Забайкалье
главным образом в горах. Численность. Обычная птица.
Экология. Размножение. Брачный крик начинается с конца
февраля до апреля (ю.-в. Забайкалье, Штегман, 1928). Птенцы у гнезда на
Сахалине встречены еще 8 июля (Никольский, 1889). Молодой в мезоптиле
отмечен 11 августа (Бальзино, ю.-в. Забайкалье).
Линька. В апреле и сентябре птицы не линяют. Питание. В
Забайкалье длиннохвостая неясыть питается мелкими грызунами, зайцами
(Тачановский, 1891); кроме того, в Приамурье —полевками Microtus pelliceus
и Clethrionomys rufocanus, рябчиками; в Манчжурии мелкими грызунами и
птицами, в частности пестрым дятлом и серыми куропатками (Яковлев, 1929)
и т. д.
Описание. Размеры. Мелкие: крыло самцов (18) 318—335, самок (36) 328—355,
в среднем 329,3 и 343,4 мм. Длина самца (1) 480, самок (1) 525, размах соответственно 1100
и 1120 мм. Ьес самца (1) 630, самок (4) 608—842, в среднем 702,2 г.
Окраска. Самая темная из встречающихся в СССР длиннохвостых неясытей,
приближающаяся в этом отношении к японским. Общий тон светлых партий, особенно на
голове и на брюшной стороне, охристо-желтоватый; на'стволья там очень развиты и нерезко
очерчены, как бы «размыты» по краям; бурый тон впрочем менее насыщенный, чем у
уральского и в особенности якутского подвидов; ноги сильно испещрены.
Систематические замечания» Неясыти из западных частей ареала — из Забайкалья и
Кентея—в среднем несколько светлей, чем приморские, и были выделены как особая раса под
названием S. и. daurica; однако большинство особей неотличимы от приморских и, при
значительной индивидуальной изменчивости окраски длиннохвостых неясытей, реальность
существования daurica кажется весьма сомнительной. С биогеографической точки зрения здесь
можно видеть случай «проникновения» манчжурского фаунистического элемента в лесные и
лесостепные участки вост. Забайкалья, что имеет место и у других видов. Птицы из Сев.
Сахалина, изученные нами, в общем неотличимы от приморских, и таким образом по всей
вероятности имя tatibanai — синоним nikolskii, тем более, что в основе оригинального
описания лежит скудный материал, а указываемые в нем признаки — сомнительны; все же среди
добытых на Южн. Сахалине птиц имеются особи, несколько более светлые, чем nikolskii,
с признаками как бы переходными к предыдущей форме.
17. Обыкновенная или серая неясыть Strix aluco Linn.
Strix Aluco. Linnaeus. Syst. Nat. Изд. X, 1758, стр. 93, Швеция.
Русское название. Неясыть — народное имя сов. В XVIII веке (Левшин)
этот вид повидимому носил название «лесная сова». У Палласа (1811), Двигубского (1831)
и Эверсманна (1855) — «серая сова».
Распространение. Ареал. Европа, кроме крайнего севера, Зап. Сибирь;
Передняя и Центральная Азия, на восток до Китая и Кореи, на юг до с.-з.
Индии; с.-з. Африка.
X арактер пребывания. Оседлая, на севере кочующая
нерегулярно птица. Биотоп. Леса разного типа — лиственные, смешанные,
ОБЫКНОВЕННАЯ ИЛИ СЕРАЯ НЕЯСЫТЬ
419
хвойные, в последних реже; древесная растительность в культурном ландшафте.
Как в горах (до среднего пояса), так и на равнинах. В самой южной части
ареала —только в горах.
Численность. Многочисленна в средних частях ареала, редка у
границ, в частности в Китае, с.-в. Европе, Зап. Сибири и Туркестане. Отмечены
колебания численности в зависимости от кормовых и быть может
температурных условий (средняя Европа).
Подвиды и варьирующие признаки* Много географических форм. В Англии и Зап.
Европе до Франции, Пиренейского п-ова и быть может Голландии 5. a. sylvatica
Shaw, 1809; в Скандинавии, средней и Вост. Европе, в Средиземноморье S. a. aluco
Карта 72. Распространение обыкновенной неясыти Strix aluco
1 — S. a. sylvatica, 2 — S. a. aluco, 3 — S. a. siberiae, 4 ~ S. a. wilkonskii, 5 — S. a. sancti-nicolal, 6 —
S. a. harmsi, 7 — S. a. biddulphl, 8 — S. a. nivicola, 9 — S. a. ma, 10 — S. a. yamadae, 11 — S. a. maure-
tanica; a — граница гнездовий; цифры в кружках обозначают номер подвида и показывают его
местонахождения в зимнее время.
L, 1758;в Приуралье и Зап. Сибири S. a. siberiae D е т., 1933; на Кавказе, в южнокаспийских
провинциях Ирана и в зап. Копет-Даге (возможно в Малой Азии и Сирии) S. a. wilkonskii
МепгЬ., 1896; в с.-з. Африке — сев. Марокко, Алжир и Тунис, к северу от Атласа 5. а.
mauretanica, W i t h e r b y, 1905; в зап. и ю.-з. Иране 5. a. sancti-nicolai Zarudny,
1905; в Туркестане от Кугитанга до зап. Тянь-Шаня 5. a. harmsi Zarud n у, 1911; в с.-з.
Индии (Гильтит, Кашмир), в Белуджистане и быть может в Афганистане 5. a. biddulphi
Scully, 1881; в Гималаях от Муррея до вост. Ассама к северу от Брамапутры в Шань-
ских государствах, в горах Китая — Ганьсу, Сычуани, Юннани, Хубее, Гуаньдуне, Гуйч-
жоу S. a. nivicola В 1 у t h, 1845; в Сев. Китае в провинциях Чжили и Жехол и в Корее S. а.
та С 1 а г к, 1907; наконец, в горах на юге о-ва Формоза S. a, yamadae Jamashina,
1936. Варьирующие признаки морфологии — размеры и окраска, в частности характер
полиморфизма (процентное соотношение цветовых вариаций в популяциях, Дементьев, 1933).
По размерам и характеру окраски неясыти могут быть разделены на две группы:
северозападную (sylvatica, alucof siberiae, wilkonskii, mauretanicajn юго-восточную (остальные
подвиды), соединяемые западноиранской sancti-nicolai. Для первой группы характерен
диморфизм окраски и некоторое ее детали, затем — постепенное возрастание величины к северо-
востоку. Вторая, южная, группа — в среднем крупней, при этом размеры возрастают к
северу: самый большой подвид туркестанский; окраска повидимому мономорфная, по
крайней мере на западе. Экологические различия — в биотопическом распределении (южная
группа форм — горная, северная преимущественно равнинная), в ходе периодических
явлений, в трофических связях.
27*
ОТРЯД СОВЫ
Восточноевропейская неясыть Strix aluco aluco Linn.
Распространение. Ареал. Лесная зона, но к северу не доходит до
границы высокоствольного леса: Скандинавия до 67° с. ш. в Норвегии, 64°в
Швеции; на юг до Средиземного моря (побережья; Сицилия) и Черного моря;
в СССР приблизительно до 60° с. ш., от Ленинградской обл. до
юго-восточных частей Вологодской (Никольск) и Кировской обл. на севере, на юг до
Крыма и Предкавказья, на восток до бассейна Камы и Поволжья, Башкирии
и Чкаловской обл. Птицы из средней и вост. России переходйы к сибирскому
подвиду.
Характер пребывания. Оседлая птица, совершающая однако
нерегулярные кочевки в холодное время года (молодые птицы) и появляющаяся
в это время в лесостепном ландшафте (Чкалов, Астрахань). В гнездовое время
встречается в лесах разного типа, на севере в хвойных, но преимущественно в
лиственных и смешанных. В Зап. Европе и в горах, и на равнине. В СССР на
равнинах, но в Крыму только в горных лесах южного берега (на гнездовье).
Не избегает культурного ландшафта — садов, парков и т. д., где гнездится и
куда в большом числе прикочевывает в холодное время года (даже в больших
городах—Москва, Ленинград, Псков и т. д.). Окольцованная в октябре
у Инстербурга в Калининградской обл. сова добыта в мае у Либавы в'Латвии,
в 210 км северней; это дает известное представление о размерах и направлении
кочевок (относительно средней Европы см. Людерс, 1937 — предельное
извечное расстояние кочевки неясыти 300 км, обычно не более 10—50 км от
гнездового участка).
Численность. В Зап. Европе и в средней полосе Европейской
части СССР обычна, редка однако в таежной полосе и в Крыму. Колебания
численности установлены: отмечено и негнездование в неблагоприятные годы,
и каннибализм птенцов, и понижение числа яиц в кладке против нормы.
Общее возрастание численности популяций отмечено в Поволжье, повидимому
в связи с мышиным «урожаем», в 1889—1890 и 1891—1892 г. В Швеции в «лем-
минговые» годы в кладке до 9 яиц.
Экология* Размножение. Половой цикл ранний — токование
(крики, щелканье клювом и хлопанье крыльями) начинается с февраля (Москва,
Ленинград, Ульяновск) и даже в январе (Полесье, Цедлитц, 1920). Спаривание
в Зап. Европе происходит в феврале—марте. Гнездится главным
образом в дуплах на разной высоте от земли, примерно от 50—65 ои до 10 л* (Умань,
Гебель, 1879, Харьковская обл., Сомов, 1897), в дубах, липах, на севере в
березах (Яранск); нередко занимает чужое гнездо—сорок, ворон, воронов,
ястребов-тетеревятников и т. д.; даже в строениях на чердаках и т. п.; в виде
исключения будто бы в щелях между камнями (Поволжье). Подстилка в дупле
весьма скудная —гнилушки, шерсть, перья и т. п. Гнездовые участки очень
постоянны, и дупла занимаются много лет подряд, повидимому отчасти и
потому, что подходящих дупел для этой большой птицы немного (занятие одного
дупла прослежено в течение 10 лет подряд, Федюшин, 1926). Поэтому
расстояние между гнездами бывает иногда весьма мало, до 1 км (в Германии даже 300—
400 м, Уттендерфер, 1939).
Кладка главным образом происходит в начале апреля, однако по неясным
пока причинам, но без явной связи с географическим положенцем местности,
отмечена и ранее — в марте. Как редкое исключение — даже в феврале (13
февраля, Полтавская обл. Гавриленко, 1929; на это указывает и нахождение
в конце марта птенцов, например 28 марта в Полесье). Даты кладок: конец
марта — Днепропетровская обл. (Вальх, 1899), начало апреля — Бессарабия
(Остерман, 1916); начало кладки — одно яйцо, 16 марта, полная кладка 22
и 30 марта, Орловская обл. (Ефимов, 1907); два яйца 11 апреля, Тамбов-
ОБЫКНОВЕННАЯ ИЛИ СЕРАЯ НЕЯСЫТЬ
421
екая обл. (Предтеченский, 1928); первое яйцо 7 апреля, Поволжье
(Богданов, 1871); полная слегка насиженная кладка отмечена 19 апреля, Псков
(Зарудный, 1911); кладка с 10 марта, насиживание — с 20 марта—б.
Витебская губ, (Федюшин, 1926); полная слегка насиженная кладка 24 апреля,
Тульская обл. (Харузин, 1926); незаконченная кладка 22 апреля—2 яйца и -
4 больших желтка у самки, Яранск Кировской обл. В случае гибели яиц —
кладка дополняется до нормы, если взята вся кладка— сова кладет новую
в другом гнезде, но меньшую (из 2—3 яиц). Новая кладка взамен утерянных
отмечена в мае. Число яиц в кладке обычно 2—6, т. е. больше, чем в Зап.
Европе, где их обычно 2—4. Число яиц повидимому колеблется в зависимости
от кормовых и климатических условий, так как отмечены и 2—3 яйца (и
птенца), а с другой стороны 7—8 (Харузин, 1920). Промежуток между
откладкой яиц 48 часов, но возможно и 72 часа (Ефимов, 1907). Яйца белые,
слабо блестящие, закругленные. Размеры (77) 45,5—50x36,5—41, в среднем
47,5x39,2 мм (Гебель, 1879), (6) 46— 49,5 х 35,7 мм —38,5 мм (Сомов, 1897).
Насиживанье происходит с первого яйца, насиживает самка.
Продолжительность насиживания 30 дней (Ефимов, 1907 и др.). Старые птицы активно
защищают гнездо. Птенцов обычно 2—4, появляются они с начала мая.
Подлетки из гнезда выбираются в мезоптиле, постепенно, с промежутком в
несколько дней, старшие в начале июня или даже в конце мая, следующие —
до середины июня. Первое время для выводка добычу приносит самец; иногда
в случае гибели самки он благополучно выкармливает птенцов (Федюшин,
1926). Гнездовый период продолжается немного более месяца, приблизительно
30—35 дней, быть может затягиваясь от кормовых и метеорологических
условий. После вылета птенцы начинают сменять мезоптиль. Вылетевших
птенцов родители кормят довольно длительное время — до конца июля, в августе
выводки разбиваются (при неблагоприятных условиях ранее). Отмечен
неполный осенний половой цикл — токование, — начинается в августе.
Линька. Полное годовое линяние происходит со второй половины
периода размножения, с мая и до конца сентября, — начала октября, общей
продолжительностью около 472 месяцев. Маховые сменяются от внутреннего
края крыла к наружному, т. е. от 10-го к 1-му. Последовательность смены
нарядов: пуховой наряд — мезоптиль — первый годовой наряд — второй
годовой наряд и т. д. Первый наряд по окраске окончательный, но в нем от ме-
зоптиля остаются маховые, рулевые и часть кроющих крыла; он,
следовательно, комбинированный.
Питание. Неясыти по кормовому режиму довольно универсальны.
Хотя мышевидные грызуны и предпочитаются, но в годы «неурожая» мышей,
а также в гнездовый период, когда имеется много молодых птиц, неясыть легко
переходит на питание птицами. Охотничий участок расположен в
непосредственной близости от гнезда и совпадает таким образом с гнездовым.
Охотится неясыть по ночам (осенью будто бы и днем), ловит добычу
главным образом на земле, но также хватает птиц на ветвях, в дуплах и т. п. В
качестве кормов неясыти для СССР указываются разные мелкой и средней
величины звери, птицы, рептилии и амфибии, крупные насекомые. Среди
млекопитающих хомяки, тушканчики, молодые зайцы, белки, ласка, суслик Ci-
tellus pygmaeus, степная пеструшка, хомячок Cricetulus migratorius; разные
мыши, среди которых Sylvimus sylvaticus, Mas muscutus; полевки Clethrionomys
glureolus, Microtus arvalis и др.; землеройки (7 землероек в одном желудке,
Кострома, Леман, 1920), летучие мыши Nyctalus teisteri, большой подковонос
(при этом, так как летучие мыши в желудках неясытей встречаются единицами,
сова ловит их повидимому на лету) и т. д. Из птиц —голуби, грачата, галки,
сойки, куропатки, сизоворонки, удоды, чибисы, перепела, дрозды, синицы,
зеленушка, овсянки, зяблики, воробьи, ласточки; из рептилий ужи, из ам-
422
ОТРЯД СОВЫ
фибий лягушки, в частности чесночница; крупные насекомые —жуки и др.
Изредка сова ест свежую падаль (коростель, форель, Крым-Даль и Шерешев-
ский, 1931).
О связи кормовых условий и плодовитости см. выше. Неясыть повидимому
чувствительна и к неблагоприятным внешним, в частности температурным,
условиям, так как зимой прячется обычно в укрытия, мало активна в холодную
погоду и даже гибнет в особо суровые зимы (1939—1940, 1941—1942).
Полевые признаки.
Сова средней величины,
темной сероватой или
буроватой окраски, с большой
круглой головой без
«перяных» ушек. Большие
темные глаза. Крылья
относительно короткие и
закругленные, хвост короткий.
Держится главным образом
на деревьях, реже на
зданиях и т. п.
Преимущественно ночная, хотя
зимою и весною может быть
встречена и днем. Брачный
крик, самца «ху-ху-хууу»,
самки «кувитт»; кроме
того, иногда у гнезда «век».
Описание. Размеры
и строение. Уши
асимметричные, правое больше левого.
Лапы оперены до когтей.
Формула крыла 4>5>3>2>1>
6... (не считая истинного 1-го,
рудиментарного). Вырезки на
наружных опахалах 3-го—6-го
маховых. Рулевых 12. Длина
самцов (5) 410—435, самок (8)
435—460, в среднем 421 и 447
мм. Размах самцов (5) 910—
950, самок (8) 940—1050, в
среднем 940 и 1008,1 мм. Вес
самцов 450, 475, 490 г, самок 590,
622, 670, 685 г. Крыло самцов
(53) 268—295, самок (66) 277-
311, в среднем 283,2 и 296,4 г.
Окраска. Пуховой наряд белый с охристым оттенком. Мезоптиль — охристо-
буроватый с тонким темным поперечным рисунком и беловатыми вершинами перьев;
рулевые и маховые серовато-бурые или рыжевато-бурые, со светлым неправильным поперечным
рисунком; уже в мезоптиле выражены две вариации окраски — серая и рыжая.
Окончательный наряд (после смены мезоптиля), серая вариация или фаза: общий тон оперения спинной
стороны беловатый с широкими бурыми наствольными чертами, густо испещренный мелкими
сероватыми пестринками; белый цвет выступает в виде пятен на внешних опахалах
плечевых и средних и больших кроющих крыла; надхвостье сероватое с темным мелким
поперечным рисунком; маховые и рулевые серовато-бурые с неправильным светлым поперечным
рисунком; рулевые с беловатой вершинной каемкой. Лицевой диск серый; горло с охристыми
и бурыми пестринами и белым пятном по середине; остальная брюшная сторона белая с
широкими темнобурыми наствольями и неправильным поперечным рисунком; оперение ног
беловатое с темными крапинами. У птиц рыжей вариации или фазы беловатый цвет заменен
охристо-рыжеватым в разных оттенках, а темный рисунок бурый с рыжеватым оттенком.
Самцы и самки окрашены одинаково. Радужина темнобурая, края век красноватые, клюв
желтоватый, когти черновато-бурые. „ .
Систематические замечания. Количественные соотношения цветовых вариации или
фаз — серой и рыжей — характеризуют отдельные популяции обыкновенной неясыти (Де-
Рис. 108. Обыкновенная неясыть.
ОБЫКНОВЕННАЯ ИЛИ СЕРАЯ НЕЯСЫТЬ
423
ментьев, 1933). При этом на территории Европы и Зап. Сибири соотношения эти меняются
с запада к востоку, причем на западе преобладают особи рыжей, а на востоке серой вариаций.
Первая составляет почти 100% в Англии; в Европейской части СССР на восток до средней
полосы обе вариации представлены примерно в равном числе особей; далее к востоку в
областях Московской, Костромской, Горьковской, Кировской, Рязанской, Тульской, Воро-г.
нежской, Ульяновской, Куйбышевской, Чкаловской начинают количественно преобладать
серые птицы — в бассейне Волги рыжая вариация и переходные между рыжими и серыми
особи составляют не более 25% популяций. На северо-востоке Европейской части Союза
и в Зап. Сибири имеются только серые птицы.
Увеличение количества серых птиц идет параллельно с общим посветлением окраски
(и возрастанием размеров). Номинальный подвид относительно темный: белый «фон»
оперения спинной стороны почти весь покрыт темными пестринами; продольный темный рисунок
широкий; на мантии бурые настволья резко выделяются; брюшная сторона сильно
испещрена, темные пестрины преобладают над беловатым фоном, поперечный рисунок довольно
грубый; ноги сильно испещрены, рисунок на них вытянут поперечно.
Особи рыжей и серой вариации спариваются между собой; в популяциях нередко
встречаются «переходные» по окраске особи (в Европейской части СССР они составляют около */в
всех изученных экземпляров).
Сибирская неясыть Strix altico siberiae Dementi ev
Strix aluco siberiae. Dementiev. Alauda V, 1933, стр. 339, Тобольск.
Распространение. Ареал. Северо-восточная часть области
распространения вида в нашей стране —* от пермского Урала (Кунгур, быть может сюда
же птицы из Молотовского округа) и юго-западного угла долины Белой,
Общего Сырта и среднего течения Урала до Иртыша на востоке (Ишим), Тюмени
и Тобольска на севере. Возможно также в Троицко-Челябинских березняках
(Словцов, 1892; но по Снигиревскому, здесь не встречается). К этой же форме
повидимому относится добытая у Джаркента залетная птица (3. XII. 1906,
Зарудный, 1911).
Характер пребывания. Оседлая птица (добыта у Тюмени
в феврале и у Тобольска в январе). Биотоп. Главным образом высокое
старое чернолесье, также в городах (в Тобольске, например, гнездится регулярно
на одной старой церкви). Численность. Ограниченна, в центральных
частях Южного Урала отсутствует, распространение в Сибири несколько
спорадично.
Экология. Сведения почти отсутствуют. Размножение. Вылупле-
ние птенцов в Молотовском округе начинается в последней трети апреля
(Ушков, 1926). Подлеток в мезоптиле встречен у Тюмени 27 мая, т. е. сроки
размножения те же, что у номинального подвида.
Описание. Размеры. Крупные: крыло самцов (4) 280—300, са ок (3) 301—307'
в среднем 290,6 и 303,3 мм. Окраска. Очень светлая, общий тон бледносерый, темный
рисунок мелкий; белый цвет чистый и резко выступающий; на задней части шеи, крыльях,
плечевых много белых пятен; настволья на брюшной стороне очень узки, поперечный
рисунок только у самой вершины перьев, белый цвет на брюшной стороне преобладает. Известна
только серая вариация.
Кавказская неясыть Strix aluco witkonskli Menzbie*v
Syrnium Wilkonskii (errore). M e n z b i e r. Bull. Brit. Ornithologist's Club v. VI, 1896,
стр. VI, Закавказье.
Синоним. Strix aluco obscurata. Stegmann. Bull. Brit. Ornith. Club. V. XLVII,
1926, стр. 39, Ленкорань.
Распространение. Ареал. Кавказ от Краснодарского края и Дагестана
до Армении и Азербайджана; южнокаспийские провинции Ирана от Талыша
до Астрабада; найдена в западном Копет-Даге в Каракалинском районе у
Лидере 30. IV. 1937. Возможно к этой же форме относятся малоазиатские неясыти.
Биотоп. Высокоствольный лес как на равнине, так и в горах (Талыш; се
424
ОТРЯД СОВЫ
верный Иран). В вертикальном отношении до 2350 м (Арарат, Сардар-булаг,
Бобринский, 1916). Численность. Невысокая, редка на с.-з. Кавказе,
как и в Талыше на равнине (в последнем вероятно из-за малочисленности там
толстых дуплистых деревьев, Тугаринов и Козлова, 1938).
Экология. Размножение. Календарь — как у европейской неясыти.
Самки с развитым яичником добывались у Кутаиси 10 марта (Кобылин, 1908),
подлетки там же 10 июня. Молодые еще в дупле, подлетки, 22 июня на
Черноморском побережье (Лауниц, 1912). В сев. Иране 25 апреля яйцо и молодой
птенец, 22 июля выросшие молодые в мезоптиле. Линька как у
номинальной формы.
Питание. На с.-з. Кавказе пища кавказской неясыти разнообразна,
но преобладают грызуны, среди них — лесные мыши, полевые мыши, домовые
мыши, кустарниковая полевка, полевка Роберта, полчок, лесная соня;
насекомоядные —. еж, крот, малая бурозубка, серая белозубка Crocidura lasia;
рукокрылые — рыжая вечерница Nyctatus noctula. Из птиц отмечены дрозд,
снегирь и разные мелкие зерноядные и насекомоядные виды. Кроме того,
насекомые — жуки усачи, жужелицы, навозники. Изредка наконец рыбы. При
этом наблюдается индивидуальная специализация: обычно преобладают
грызуны, среди них лесная мышь (около 50% встреч позвоночных), у одной пары—
полчки; иногда повышается удельный вес птиц. Примерная годовая
потребность в корме составляет 624 мелких грызуна, 112 мелких птиц и 88 насекомых
(Кавказский заповедник, Жарков, 1938).
Описание. Размеры. Несколько меньше восточноевропейских неясытей: крыло
самцов (17) 266—296, самок (33) 282—305, в среднем 276,2 и 296,2 мм.
Окраска. Наиболее темно окрашенная форма неясытей. Птицы из ю.-в.
Закавказья (Куба, Талыш) и сев. Ирана со слабо выделяющимся светлым фоном; продольные
темные полосы очень широки и ярки; поперечный рисунок брюшной стороны резкий и
грубый, ноги и лицевой диск сильно испещрены. Северокавказские птицы менее темной и
однообразной окраски, но все же в среднем темней европейских. Рыжая вариация составляет на
Сев. Кавказе около V* всего количества изученных экземпляров, в Закавказье около 7г-
Кроме того, на Кавказе встречается третья — кофейно-бурая вариация, со слабо
выделяющимся темным рисунком; у таких птиц лицевой диск ,темнобурый, рулевые и маховые
почти одноцветные.
Систематические замечания. Кавказские совы, как видно из предыдущего, не все
однотипны. Самая темная окраска и одновременно высокий процент рыжей вариации в
популяциях — в ю.-в. Закавказье и сев. Иране. Этим птицам и дано было название obscurata. Птицы
на Сев. Кавказе несколько светлей, но темней европейских. Для всех популяций
характерно присутствие темнобурой вариации, послужившей для описания wilkonskii. Как
типы этой формы, Мензбир отметил темнобурых птиц из окрестностей Батуми и из Шуши.
Темнобурая вариация встречена у Майкопа, Дзауджикау, Геленджика, Хосты. В других
частях СССР такие птицы не найдены, но в качестве очень редких индивидуальных
уклонений добыты в Венгрии, у Константинополя и даже в Италии. При большой индивидуальной
изменчивости отделять особую географическую расу в ю.-в. Закавказье и сев. Иране едва
ли возможно. Размеры всех этих птиц сходны. Крыло кавказских (кроме ю.-в. Закавказья)
[266—296, в среднем 274,4 мм у 14 самцов, 282—304, в среднем 296 мм у 23 самок. У «obscurata»
из ю.-в. Закавказья, сев. Ирана и Копет-Дага крыло у Зсамцов 278—293, в среднем 284,3 мм,
у 10 самок 290—305, в среднем 297,7 мм.
Туркестанская обыкновенная неясыть Strix aluco harmsi S a r u d n у
Syrnium hclrmsi. S а г u d n y. Orn. Monatsberichte* 1911, стр. 34, Чирчик.
Распространение. Ареал. Горы Туркестана от Кугитанга и с.-з.
Таджикистана (Искандер-куль; северные склоны Зеравшанского хребта у Ургута;
Пенджакент) на западе, на север до северных склонов Таласского Ала-Тау
(Балбадерек и Джебаглы, Шульпин, 1936), на востоке до Ферганы, Сусамыра
и Терскей-Алатау (Зарудный, 1911).
Биотоп. Лиственные леса гор. В вертикальном отношении до 2000—
2275 м в западном Тянь-Шане (Гульча, Чаткал и т. д.). Характер пре-
СИПУХА
425
б ы в а н и я: оседлая птица; в холодное время года иногда появляется на
равнинах (Ташкент). Численность: редка.
Экология* Размножение. Подлетки в мезоптиле—три в выводке —
у озера Сары-чилек в Фергане 26 июня (Кашкаров, 1927). Линяющая из ме-
зоптиля молодая птица
встречена у Джелалабада 11 июля.
В Кугитанге неясыти в апреле
держались парой, гнездо
находилось в щели скалы (Тиш-
кин). Брачный крик
начинается еще в апреле. Цикл по
времени совпадает, повиди-
мому, с циклом европейской
неясыти.
Описание. Размеры.
Самая крупная форма из
обитающих в СССР неясытей. Крыло
самцов (4) 300—316, самок (5)
315—332, в среднем306,5 и 323 мм.
Окраска. Бледная:
серый тон до известной степени
напоминает сибирских птиц, но
белый цвет менее выделяется, и число
мелких серых пестрин больше,
темные настволья на брюшной
стороне узкие, а поперечный
рисунок резкий и прямолинейный,
перпендикулярный к стволу пера
(у европейских и кавказских
птиц зигзагообразный,
неправильный). Встречена только серая
вариация.
Систематические замечания.
Туркестанские неясыти отличаются Рис- 109« Туркестанская неясыть.
от гималайских S. a. ,biddulphi
бледной окраской, гималайские ярче и темней, бурые полосы на брюшной стороне
черноваты; беловатый фон у гималайских сов слабо заметен. 5. a. nivicola еще темней. Размеры
hartnsi и biddulphi, повидимому, сходны.
СЕМЕЙСТВО СИПУХИ TYTONIDAE
|РОД СИПУХИ TYTO BILLBOERG 1928
Тип Tyto alba
18. Сипуха Tyto alba Scopoli
Strixalba. Sc о р о I i, Annus Historico Naturalis. I, 1769, стр. 21, Фриуль, сев. Италия.
Русское название. Звукоподражательное, по шипящему голосу птицы.
Распространение. Ареал. Америка, кроме севера; Азия, кроме Сибири,
Средней и Центральной Азии; вся Африка с Мадагаскаром; Европа, кроме
северо-восточных частей; Индонезия и Австралия, кроме Новой Зеландии.
Характер пребывания. Оседлая или кочующая птица. Биотоп.
При почти кругосветном распространении весьма разнообразен. Первично
вероятно связан со скалами. В культурном ландшафте селится близ
человеческих поселений. Тропические подвиды обитают в весьма разнообразных
условиях, трудно поддающихся общей характеристике.
Подвиды и варьирующие признаки. Морфологические различия между подвидами в
размерах и окраске. Характерно, что в ряде случаев соседние островные расы отличаются одна
от другой очень резко. Некоторые формы — с широким, другие (главным образом островные)
426
ОТРЯД СОВЫ
с очень узким ареалом. Обычно различают: 7\ a. alba S с о р о 1 i, 1769, Британские о-ва,
Франция, Пиренейский п-ов, Италия, Средиземноморье (Алжир, Тунис, Марокко,
Египет), кроме Корсики и Сардинии; Т. a. schmitzi Hartert, 1900 — Мадейра; Г. с.
gracilirostris Hartert, 1905 — восточные Канарские о-ва — Фуэртавентура и Ланца-
роте; Т. a. ernesti К le i nsch mi d t, 1901 —Корсика и Сардиния; Т. a. guttata Chr.
L. Brehm, 1831 — средняя Европа; Т a. detorta Hartert, 1913 — о-ва Зеленого
Мыса (Сант-Яго и Сант-Винсент); Т a. affinis В 1 у t h, 1862—Африка к югу от Сенегамбии,
южн. Сахары и Судана; Т a. thomensis Н a r 11 a u b, 1852 — о-ва Сао-Томе в Гвинейском
заливе; Т a. erlangeri W Sclater, 1921— Аравия на восток по Маската, к северу
возможно до южн. Палестины и Месопотамии; Т. a. hypermetra G г о t e, 1928 — Мадагаскар
и Коморские о-ва; Т. a. stertens Hartert, 1929 — Индия, Ассам, вероятно сев. Бирма;
Цейлон; Т a. javanica G m e 1 i n, 1788 — ю.-в. Азия — Сиам, Индо-Китай (возможно
южн. Бирма), Ява, Тысяча Островов, Кангеан, Ломбок; Т. a. de-roepstorffi Hume,
1875—- Андаманские о-ва; Т. a. sumbaensis Hartert, 1897 — 0. Сумба; Т. a. everetti
Карта 73. Распространение сипухи Tyto alba
7 — Т. a. alba, 2 — Т. a. ernesti, 3 — Т. a. guttata, 4 — Т< a. affinis, 5 — Т. a. hypermetra,
6 — Т. a. detorta, 7 — Т. a. schmitzi, 8 — Т. a. gracilistris, 9 — Т. a. thomensis, 10 — Т. а.
«rlangeri, 77 —Т. a. stertens, 12— Т. a. javanica, 13 — Т. a. de-ropstorffi, 14 — Т. a. sumbaensis,
15 — Т. a. everetti, lb — Т. a. kuehni, 17 — Т. a. mecki, 18 — Т. a. delicatuia, 19 — Т. a. cras-
sirostiis, 20 — Т. a. interposita, 21 —Т. a. lulu, 22 — Т. a. pratincole, 23 ^Т. a. guatemalae,
24 — Т. a. lucayana, 25—Т. a. furcata, 26 — Т. a. glaucops, 27 — Т. a. bargei, 28 — Т.
a» conttmpta, 2У — Т. a. hellmayri, 30 — Т. a. tuidara, 31 — Т. a. punctatissima, 32 — Т. а.
nigrescens, 33 — Т. a. insularis; a — границы гнездовий.
Hartert, 1929 — о. Саву в 100 км западней Тимора; Т. a. kuehni Hartert, 1929—0.
Кисар и вероятно Малые Зондские от Флореса до Тимора; Т. a. mecki Rothschild et
Hartert, 1907 — о-ва Вулкановы и Дампьер, ю.-в. Новой Гвинеи от Коллингвудбай
на севере и до Порт-Морсби на юге; Т. a. delicatuia Gould, 1837 — Соломоновы о-ва;
Австралия; 7\ a. crassirostris M а у г, 1935 — о-в Боанг, архипелаг Бисмарка; Т. а.
interposita М а у г, 1935 — северные Ново-Гебридские" о-ва, о. Бенкса и Санта-Круц;
Т. a. lulu Р е а 1 е, 1848 — южные Ново-Гебридские о-ва, Новая Каледония, Лойальти,
Фиджи, Тонга, Самоа и острова Товарищества; Т. a. pratincola Bonaparte, 1838—
южные части Сев. Америки (к югу от сев. Калифорнии, Небраски, южн. Уисконсина,
южн. Мичигана, зап. Нью-Йорка и юга Новой Англии), центральная Америка до востока
Гватемалы и быть может Никарагуа; Т. a. guatemalae R i d g w ay, 1873 — зап.
Гватемала, Сальвадор, зап. Никарагуа, Панама; Т. a. lucayana Riley, 1913 — Багамские о-ва;
Т. a. furcata Temminck, 1827 — Куба, о. Пинос, Гран-Кайман, Кайман-брек,
Ямайка; Т. a. glaucops Каир, 1852 — о-ва Тортуга и Испаниола; Г. a. bargeiHa rtert,
1892 — о. Кюрасо; Г. а. contempta Hartert, 1898 (Г. a. subandeana Kelso, 1938?) —
Колумбия, Эквадор, Перу и Венецуэла; Т. a. hellmayri Griscom et Greenway,
1937 — Гвиана к югу до Амазонки; Т a. tuidara J. Gray, 1829 — Бразилия к югу от
Амазонки; Чили, Аргентина, до Огненной Земли; Т. a. nigrescens L a w r e n с е, 1878 —
о. Доминика (из М. Антильских); Т a. insular is P e 1 z e 1 n, 1872 — южные острова
архипелага Малых Антильских: св. Люция, св. Винсент, Бекия, Карриаку, Унион, Гренада;
Т. a. punctatissima G.Gray, 1838 — Галапагосские о-ва.
СИПУХА
427
Среднеевропейская сипуха Tyto alba guttata Brehm
Strix guttata. С h r. L. Brehm. Handbuch d. Naturgeschichte aller Vogel Deutsch-
lands, 1931, стр. 107, остров Рюген.
Распространение. Ареал. Средняя Европа на север до южн. Швеции,
Дании, Литвы; на запад до Эльзаса и Лотарингии, Голландии (в восточной
Франции и Бельгии
промежуточная
между номинальной и
среднеевропейской расами
популяция), к югу до
Альп (сев.
Швейцария, Австрия),
Югославии, Болгарии и
Румынии, на восток
до Латвии, Румынии,
Польши,
Приднепровья. В СССР в
Калининградской обл., в
западной Латвии
(достоверная гнездовая
находка у Митавы 10.
VI. 1947), далее в
Белоруссии в б. Пинском
и Бобруйском уездах,
к югу до Днепра,
местами немного
переходя на левый его
берег (отмечена в
разных местностях
Бессарабии, Подолии, , в
Киевской обл. у Ка-
нева, Конча-Заспа,
Белой Церкви; в
Полтавской обл. у Золо-
тоноши, Переяславля,
Пирятийа, Прилук,
Л у бен, Лохвицы; до
Полтавы не доходит,
Гавриленко,1929;
возможно в б.
Константинова деком у., Заруд-
ный, 1892; в
Черниговской обл. у
Нежина, Шарлемань, 1936,
у Конотопа). Сведения
о распространении в
Крыму, а также в
средней и в
особенности в восточной
полосе Европейской
части СССР невероятны,
как ошибочно, конечно, и некритическое сообщение о выводке сипух в б.
Бобровском у., Воронежской обл. (Воробьев, 1924).
Рис. 110. Сипуха.
428
ОТРЯД СОВЫ
Характер пребывания. Оседлая птица, совершающая однака
нерегулярные кочевки в негнездовое время (молодые?). Осенью 1934 года
кочевка сипух наблюдалась в средней Европе и в Эстонии (Герме, 1935).
Биотоп. Главным образом около человеческих поселений, в
особенности при наличии заброшенных или малопосещаемых зданий и сооружений,
а также дуплистых деревьев.
Численность. В СССР редкая и спорадически встречающаяся птица
(граница ареала). Отмечены колебания численности и плодовитости по годам
в связи с кормовыми условиями (урожай или неурожай грызунов и быть
может землероек). В Киевской обл. (Конча-Заспа) сипухи исчезли после
суровой зимы (1929—-1930 гг.), потом опять появились (Шарлемань, 1930, 1933)*
Экология. Относящиеся к СССР сведения крайне отрывочны.
Размножение. Гнезда расположены в старых зданиях и в дуплах
деревьев. Брачный крик начинается уже в начале марта (Киев,
Шарлемань, 1930). Кладка — повидимому в конце апреля — начале мая (свежие и
слегка насиженные отмечены б мая, Гебель, 1879, в Уманском р.; 11 мая
в Пинском р., Шнитников, 1913). Кладка из 6—8 яиц. Яйца белые,
матовые, продолговатой формы. Размеры (13) 36,5—41,5X30—30,5, в среднем
38,6X30,2 мм (Гебель). Насиживание? (по западноевропейским данным
насиживает только самка; продолжительность насиживания 30—34 дня). В
неблагоприятные годы высокая смертность птенцов и каннибализм (например
в Киевской обл. после гибели грызунов от высокого паводка в 1931 г. в
выводке из 4 птенцов один исчез 24 июня, а 4 июля оставшиеся были съедены
родителями (Шарлемань, 1933). В обильные грызунами годы плодовитость
повидимому выше и к тому же поздней осенью птицы иногда вторично
откладывают яйца: вылет молодых ЗО.Х. 1930, заповедник Конча-Заспа. Подлетки
встречены у Киева 30 июля (Шарлемань, 1933); продолжительность
гнездового периода следовательно около 8 недель.
Линька. Полная годовая; у нас не изучена. В средней Европе
происходит между июлем и ноябрем. Последовательность смены нарядов:
первый пуховой наряд -*. второй пуховой наряд — первый годовой
(окончательный) наряд и т. д.
Питание. Пищу среднеевропейской сипухи составляют главным
образом грызуны (мышевидные) и землеройки; редко плохо летающие птицы,
преимущественно молодые. В годы «неурожая» грызунов птицы имеют большее
значение в пище сипух — при таких обстоятельствах сипуха ест и лягушек
(чесночница, Шарлемань, 1935). В погадках сипух у Киева найдено 78,5%
остатков грызунов, среди них преимущественно Microtas arualis, Sylvimus sylva-
ticas> Arvicola amphibius; птиц—только 6,7% и 1,5 амфибий. При выводке
сипухи ловят за месяц 85—128 грызунов и вылавливают в среднем по 13 штук
мышей с гектара (Пидопличка, 1930).
Полевые признаки. Сова легкого сложения с большой головой, очеш»
резко выраженным лицевым диском почти треугольной формы. Крылья
длинные, хвост короткий. Образ жизни строго ночной, активность начинается с пс-
черними и прекращается с утренними сумерками. Голос — главным образом,
кряхтенье и шипенье.
Описание. Размеры и строение. Череп длинный и узкий, лицевой диск
имеет форму сердца. Глаза небольшие. Клюв длинный и слабый, слегка лишь загибающийся
к концу. Уши симметричные. Формула крыла: 2> 1>3>4>5... (не считая, как обычно
у сов, истинного 1-го махового, рудиментарного и скрытого под кроющими крыла). Хвост
короткий и закругленный, из 12 рулевых. Цевка длинная. Внутренний и средний палец
одинаковой длины. Внутренний край когтя среднего пальца в зазубринах (у взрослых).
Длина самца (1) 332, самки (1) 393, размах соответственно 903 и 925 мм. Крыло самцов и
самок (10) 274—297, в среднем 288,1 мм (вес одной самки из Полтавы 254 г; у германских
сипух вес около 300—355, минимум 241 и 265 г).
СИПУХА
429
Окраска. Первый пуховой наряд чисто белый ^носится около 2 недель). Второй
пуховой наряд — не мезоптиль, как у других наших сов — белый с желтоватым оттенком
на брюшной стороне и сероватым — на спинной. Первый и>последующие годовые наряды,
самцы и самки: общая окраска спинной стороны бледнопепельно-серая с черноватыми
мелкими пестринами и белым пятнышком на каждом пере; основания перьев рыжеватые и хорошо
выступают на больших кроющих крыла; маховые с рыжеватым поперечным рисунком;
основная окраска внутренних опахал первостепенных маховых черноватая; рулевые рыжие
с сероватой испещренной беловатым вершиной и черноватыми полосками у края пера;
брюшная сторона рыжеватая с беловатым, окаймленным бурым («глазчатым») пятнышком у
вершин перьев; лицевой диск беловатый с примесью рыжего и бурого по краям. В развитии
рыжеватого оттенка, особенно на брюшной стороне, имеется довольно значительная
индивидуальная изменчивость. Радужина темнобурая, клюв сероватый или желтовато-белый;
когти черные.
Средиземноморская сипуха Tyto alba alba Scopoli
Распространение. Ареал. Британские о-ва, зап. Франция, Бельгия,
Швейцария, Испания, Португалия, Италия, Балеарские о-ва, Мальта,
Сицилия, Кипр, М. Азия, Палестина,"Ирак, сев. Африка от Марокко до Египта.
В фауну СССР вносится на основании добычи взрослой самки 4.Х.1942 г.
в Гассан-кули на каспийском побережье Туркмении. Этот залет трудно
объяснить, так как сипуха оседлая птица; с другой стороны, она не встречена никем
в Иране. Возможно все же, что эта сова гнездится в небольшом числе в
южнокаспийских провинциях Ирана, откуда и могла залететь в Гассан-кули
упомянутая выше птица.
Экология. Размножение. Гнездится в развалинах, старых
строениях, расщелинах скал, дуплах. Начало размножения обычно в апреле или
первых числах мая, в благоприятные по кормовым условиям годы и ранее—в марте
или даже феврале. Количество яиц 4—7, но иногда их бывает до 11. Форма яиц
удлиненная, окраска — матово-белая. Размеры (100) 36,2—45x29,1—33,5,
в среднем 39,74x31,57 мм (Уайзербы, 1938). Насиживает только самка в
течение 32—34 дней. В возрасте около двух месяцев молодые оперяются и
становятся на крыло. В «мышиные» годы бывает вторая кладка в конце лета
или даже поздней осенью (в июле и сентябре, в виде исключения в декабре).
Линька. Как у предыдущего подвида, полная годовая, между июлем и
ноябрем.
Питание. Преимущественно мелкие грызуны, также птицы и
насекомые (жуки, реже ночные бабочки, Уайзерби, 1938).
Описание. Длина крыла у самки из Гассан-кули 308 мм.
Окраска. Спинная сторона светлей по общему тону, чем у среднеевропейской
сипухи, с большей примесью охристых оттенков. Брюшная сторона белая, иногда — с
охристым оттенком по сторонам груди, слабо испещренная тонкими буроватыми наствольными
пятнами на брюхе и боках.
отряд кукушки
CUCULl или CUCULIFORMES
Характеристика отряда
Морфология. Внешние признаки. Птицы средних размеров.
В фауне СССР самых крупных размеров достигает обыкновенная кукушка —.-
длина крыла у нее в среднем около 210—225 мм, при размахе крыльев около
580—-620 мм. Самых мелких размеров — малая кукушка Cuculus poliocephaliis*
у которой крыло имеет длину около 160 мм. Среди тропических представителей
отряда встречаются и более крупные птицы, например, некоторые виды турако,
из которых самый крупный Corythacola cristata имеет длину крыла в 300—
350 мм.
Оперение довольно жесткое, плотно прилегающее к телу, только на зобе
несколько мягче и пушистей. Цевка оперена на наружной стороне
приблизительно до половины длины, с внутренней стороны — не оперена. Перья
голени удлинены и образуют «штаны». Голые, не оперенные участки кожи на
голове имеются только у турако. Украшающих перьев, развивающихся в брач?
ный период, нет. У некоторых видов на голове хохол. Половой диморфизм и
окраске оперения или совсем отсутствует, или развит слабо, за исключением
некоторых самок рода Cuculus, имеющих кроме сходного с самцом серого
оперения еще и рыжий «гепатический» тип оперения. Пух распределяется по аптс-
риям. Первостепенных маховых — 10, самое длинное 3-е маховое.
Второстепенных маховых—9—10 или 13 (Scythrops) Эутаксия. Рулевых—.10 (8 у
Guira и Crotophaga). Хвост довольно длинный, ступенчатый, прямой или
закругленный.
Окраска оперения у кукушек скромная — серая, бурая, рыжая, черпая
с белым низом тела, или с поперечными белыми и темными полосами, или с
темными продольными пятнами на нижней стороне тела; у турако— яркая;
фиолетовая, пурпурно-красная, зеленая, синяя.
Ноги четырехпалые, зигодактильного типа или приближающиеся к нему,
Четвертый палец обычно может загибаться назад. Цевка короткая, покрытая
в неоперенной части щитками.
ХАРАКТЕРИСТИКА ОТРЯДА
431
Клюв средних размеров, несколько изогнутый вниз в своей вершинной
части, с ясно выраженным хребтом надклювья (кукушки), или короткий,
сильно выпуклый по хребту надклювья (турако). Края клюва обычно гладкие*
только у некоторых видов турако зазубрены. Разрез рта большой.
Анатомические признаки. Череп десмогнатический, голо-
ринальный. Базиптеригоидные отростки отсутствуют. Сошник слабо развит или
даже совсем отсутствует. Ноздри сквозные. Грудина с одной парой вырезок,
у некоторых видов зарастающих или образующих фонтанели (у взрослых птиц).
Шейных позвонков 13—14. Сонная артерия парная. Носовых желез нет.
Гортань обычно трахео-бронхиального типа, но у некоторых видов встречается и
бронхиальная. Обводящая мышца имеется. Кожа тонкая. Боковая шейная
артерия хорошо развита.
Образ жизни. Кукушки — дневные древесные и кустарниковые или
наземные птицы. Образ жизни очень разнообразен —-> от видов, всю жизнь,
проводящих среди крон деревьев и очень редко, лишь на короткое время,
спускающихся на землю, до чисто наземных, живущих в кустарниковых равнинах,.
степях и полупустынях. Хорошо летают, некоторые виды хорошо бегают по
земле. Преимущественно оседлые птицы, за исключением нескольких кукушек,.
например, некоторых видов рода Cuculus, хохлатой кукушки Clamator glan^
darius, новозеландских кукушек Chalcites lucidus и Urodynamis taitensis, аме-.
риканской кукушки Coccyzus americanus. Некоторые виды рода Cuculus, гнездя-т
щиеся в Палеарктике, как, например, обыкновенная и глухая кукушка,
находящие предел своего гнездового ареала у северной границы леса, летят на
зимовки далеко на юг — в тропическую и южн. Африку и в южн. Азию.
Новозеландская бронзовая кукушка Chalcites lucidus гнездится в Новой
Зеландии и летит на зимовки на Соломоновы и Бисмарка о-ва через
Австралию и морем по пути более 1200 миль. Новозеландская длиннохвостая
кукушка Urodynamis taitensis летит из Новой Зеландии на зимовку еще
далее —на Каролинские, Маршальские и Маркизовы о-ва. Американская
кукушка Coccyzus americanus, распространенная в Канаде, летит на зимовку
в Аргентину.
По роду пищи всех кукушек можно разделить на две резко разграничен-?
ные группы: турако питаются растительной пищей — ягодами, фруктами,
молодыми побегами и почками растений, и собственно кукушки питаются
животной пищей (за исключением рода Eudynamis, пища которого растительная).
В состав животных кормов входят преимущественно различные насекомые,
их личинки, другие беспозвоночные и в небольшом количестве птенцы и яйца
других птиц. Только Centropus, Geococcyx и Saurothera поедают рептилий и
мелких млекопитающих. Насекомых собирают с деревьев, кустов и на земле,
а также ловят их и на лету. Кукушки несомненно полезны для сельского
и лесного хозяйства, так как уничтожают большое количество насекомых и
их личинок.
Большинство видов моногамы; у некоторых видов наблюдается полигамия,
и для них характерен гнездовой паразитизм. Паразитические виды кукушек пар
не образуют и количественное соотношение полов у них очень изменчиво.
Обычно на участке можно встретить одну самку и несколько самцов, но иногда
наблюдается и обратное явление. Гнездовой территории в общепринятом
понимании у них нет. Есть территория, на которой самка отыскивает подходящие
гнезда для откладывания своих яиц —. этот участок и можно считать гнездо^
вым. Иногда на одном участке можно встретить двух самок, но в этом случае
они паразитируют на разных видах птиц. Всего видов, принадлежащих
подотряду кукушки, — 126, из них около 80 видов паразитируют полностью,
т. е. не строят гнезда,, подкидывают яйца в чужие гнезда, не высиживают их
и не воспитывают птенцов. Интересно отметить, что паразитические виды куку-.
432
ОТРЯД КУКУШКИ
шек обитают преимущественно в Восточном полушарии, а непаразитирую-
щие ^в Западном. Паразитирующие виды кукушек принадлежат к родам:
Cuculus, Cacom ntis7 Chrysococcyx, Urodynamis> Sarniculus, Chalcytes, Clamator,
Scythrops, Eudynamis, Tapera, Dromococcyx и др.
Американские кукушки рода Coccyzus обычно сами строят гнезда и
высиживают птенцов, за исключением Coccyzus melanocoryphus, которые не строят
гнезд и кладут яйца в чужие гнезда, но сами высиживают и выкармливают
птенцов. Южноамериканский вид Tapera naevia откладывает свои яйца в
чужие, еще не достроенные гнезда. У видов рода Crotophaga несколько самок
откладывают свои яйца в одно общее гнездо, сделанное самими птицами.
Представители другого американского рода Guira в период размножения образуют
пары, но гнезд не строят, а откладывают яйца в чужие гнезда, хотя затем сами
высиживают и выкармливают птенцов. И наконец, имеется целый ряд видов,
строящих гнезда, высиживающих и выкармливающих птенцов; они
принадлежат к родам Taccocua, Zanclostomus, Rhamphococcyx, Phaenicophaius, Geococcyx,
Centropus и др.
Таким образом мы имеем полный ряд переходов от непаразитирующих
видов к паразитирующим. Непаразитирующие виды строят открытые гнезда
на земле и на деревьях из ветвей, стеблей растений и т. п., причем вся постройка
очень рыхлая. Кладка состоит из двух белых, беловато-розовых или беловато-
желтых яиц (турако), или из двух-шести и даже семи белых с голубоватым и
зеленоватым налетом яиц (кукушки). У полупаразитических форм яйца
крупные и достигают 35% веса тела самки Guira, Crotophaga, у паразитических форм
яйца очень мелкие.
Паразитические виды кукушек откладывают яйца, главным образом, в
гнезда воробьиных птиц. Причем не только отдельные роды или виды, но и
разные самки имеют определенных «хозяев», в гнезда которых откладывают свои
яйца. Хохлатая кукушка Clamator glandarius несет яйца в гнезда вороновых
птиц. Ее яйца находили в гнездах сороки, африканского ворона, серой
вороны и др. Яйца ее мало изменчивы, и по форме и окраске очень похожи на яйца
вороновых птиц; здесь уже имеется узкая специализация паразита.
Кукушки, принадлежащие к роду кеглей Eudynamis, откладывают свои
яйца в гнезда вороновых птиц, иволг, разных скворцов и др.; Coccyzusmelano-
coryphus—в гнезда голубей Zenaidura auriculata; Tapera naevia —в гнезда
13 разных видов птиц. Cuculus sparverioides паразитирует на птицах,
принадлежащих к сем. мухоловок и дроздовых. Различные подвиды ширококрылой
кукушки Cuculus fugax паразитируют на разных видах птиц: так — С. /. hyperyt-
hus откладывают яйца обычно в гнезда Luscinia суапе; С. /. nisicolor в гнезда —
Proparus phayeri, Drymocataphus assamensis. Малая кукушка Cuculus poltocc»
phalus—в гнезда Parus, Saxicola, Phylloscopus. Глухая кукушка Cuculus satu-
ratus — в гнезда пеночек и коньков. Для обыкновенной кукушки С. canorus,
биология которой изучена лучше других видов кукушек, приводится 147 видом,
главным образом воробьиных птиц, в гнезда которых она откладывает спои
яйца. Но особенно часто встречаются яйца кукушек в гнездах 20—30 видов.
Причем интересно отметить, что в различных местностях обыкновенная
кукушка паразитирует на нескольких (2—3) строго определенных видах птиц, хотя
здесь имеется и целый ряд других обычных видов птиц. Каждая самка кукушки
откладывает яйца в гнезда одного определенного вида хозяина; повидимому,
в гнезда того вида, в котором сама вывелась; меняют своих хозяев кукушки
очень редко.
Обычно птицы резко реагируют на появление кукушки у гнезда и на
отложенные кукушкой яйца. Некоторые бросают гнезда с отложенным в нем
кукушечьим яйцом или прикрывают свои яйца сверху, устраивая новый лоток;
иногда выбрасывают яйцо кукушки из гнезда.
ХАРАКТЕРИСТИКА ОТРЯДА
433
Яйца обыкновенной кукушки часто бывают вполне сходны по окраске,
форме и размерам с яйцами птиц-хозяев; иногда имеются небольшие
различия, и только в редких случаях яйца кукушки сильно отличаются от яиц
хозяев.
Время размножения паразитических видов кукушки тесно связано с
временем размножения их хозяев. Яйца подкладываются кукушками обычно к
начатым или незаконченным кладкам, только как исключение к кладкам,
насиживание которых уже началось. Как правило, обыкновенная кукушка
откладывает в гнездо хозяина одно яйцо: в тех случаях, когда в гнезде встречается
два яйца кукушки, они, повидимому, принадлежат двум самкам. Хохлатая
кукушка подбрасывает несколько яиц в одно гнездо. Подбрасывание яиц
происходит различно: некоторые кукушки откладывают яйцо на землю,
затем при помощи клюва переносят его в намеченное заранее гнездо; другие
кукушки откладывают яйцо непосредственно в гнездо хозяина. Иногда
кукушки при откладывании яйца удаляют одно или несколько яиц птиц —»хозяев
гнезда. Кладка яйц обычно происходит днем, когда гнездо нередко
оставляется хозяевами без присмотра; иногда кукушка-самец отвлекает
внимание хозяев гнезда. Яйца кукушка несет через день и даже иногда каждый
день. Количество яиц, отложенных за сезон, велико, не менее 12 штук,
иногда доходит до 20 и даже 25 на одну самку. Такая большая кладка
имеется потому, что из этого числа большое количество яиц кукушки
гибнет.
У непаразитических форм насиживание идет 18—-20 дней и в нем принимают
участие оба пола. У паразитических видов эмбриональное развитие очень
быстрое —* короче, чем у многих мелких воробьиных птиц. Для обыкновенной
кукушки по последним наблюдениям оно продолжается 11,5 дней. Птенцы
и у паразитических видов и у непаразитических вылупляются голыми и
слепыми.
У турако и у кукушек рода Centropus птенцы быстро покрываются густым
пухом. Птенцы остальных видов кукушек сразу одеваются перьями
птенцового наряда. Постэмбриональное развитие у паразитических видов быстрое,
у обыкновенной кукушки птенцы дней через 20 после вылупления покидают
гнездо. У только что выведшихся птенцов обыкновенной кукушки имеются
приспособления, «рефлекс выбрасывания» яиц и птенцов птиц — хозяев гнезда.
У непаразитических видов постэмбриональное развитие идет медленно,
особенно у турако. У них лишь через 6—7 недель птенцы начинают только
перепархивать с дерева на дерево, но пуховой наряду них еще сохраняется. Птенцы
турако на первом пальце крыла имеют хорошо развитые когти, при помощи
которых они могут лазать по деревьям подобно птенцам гоацина.
Есть указания не только на сходство яиц кукушек с яйцами хозяев, но и
на сходство птенцов кукушек с птенцами птиц-хозяев, например, у коелей
Eudynamis scolapacea mindanensis, и даже на сходство голоса птенцов кукушек
с голосом птенцов хозяев.
Возникновение гнездового паразитизма не ясно. Кроме отряда кукушек*
он имеет место и в других отрядах птиц —у воробьиных (ткачи — Ploceidae,
кассики Icteridae, воловьи птицы Molofhrus, тираны Tyrannidae и т. д.), у
дятлов (медоуказчики Indicatoroidae, некоторые из бородастиков Capitonidae),
у утиных (Heteronetta atricapilla). У нормально гнездящихся птиц также
известны случаи, когда птица предпочитает селиться в гнездах других птиц
(соколы, некоторые совы и т. д.). Если для некоторых видов птиц развитие
гнездового паразитизма можно объяснить недостатком удобных для гнездовья мест,
как, например, у утки Heteronetta, то для кукушек и кассиков этим объяснить
возникновение гнездового паразитизма нельзя. Фридман (1935) объясняет это
тем, что гнездовой паразитизм развился вследствие постепенной утраты.
28 Птицы СССР, т. I
434
ОТРЯД КУКУШКИ
инстинкта гнездостроения и неравномерного численного соотношения
полов.
Систематика. Отряд кукушек делится на два подотряда — настоящих
кукушек Caculi и турако или бананоедов Turaci, или Musophagi. Подотряд
Turaci отличается от кукушек отсутствием сошника и hypocleidium, наличием
пяти пар ребер, достигающих грудины. Грудина с двумя парами вырезок.
Ключица V-образной формы. Кишечник короткий, широкий. Слепые кишки
отсутствуют. Спинная птерилия короткая. Копчиковая железа оперена. Полового
диморфизма нет. Клюв короткий с сильно выпуклым хребтом, надклювья
желтого цвета с красной вершиной. Края клюва обычно зазубрены. Носовые
отверстия щелевидные и лежат ближе к вершине и к хребту надклювья. Четвертый
палец полностью назад не обратим. Имеется побочный ствол перьев.
Эмбриональное и постэмбриональное развитие
проходит медленно. Птенцы вылупляются
голыми, но скоро покрываются пуховым
нарядом, который держится долго —
более семи недель.
Подотряд турако Turaci содержит
одно семейство — бананоеды Musopha-
gidae с 8 родами (Touraco, Gallirex, Ru-
wenzororniSy Musophaga, Corythaeola,
Crinifer, Corythaixoides, Gymnoschizornis)
и с 20 видами, распространенными в
Африке.
Подотряд кукушек характеризуется
присутствием сошника, хотя у
большинства видов сильно редуцированного.
Вырезки на грудине часто превращаются
у взрослых в фонтанели или даже совсем
зарастают. Вилочка V-образной формы,
hypocleidium имеется за исключением
рода Scythrops. Только 3—4 пары ребер
доходят до грудины. Кишечник довольно
длинный с четырьмя петлями, слепые
кишки функционируют. Клюв средней
длины, сильно изогнутый по хребту
надклювья. Края клюва гладкие (исключение Scythrops). Носовые
отверстия округлые и лежат в основной части клюва. Четвертый палец
свободно может вращаться назад. Копчиковая железа не оперена. Побочные
стволы перьев отсутствуют или слабо развиты. На голове хохла нет, за
исключением рода Clamator. Подотряд кукушек содержит одно семейство кукушек
Cuculidae, распадающееся на 40 родов (Cuculus, Clamator, Chalcites, Cacomuntls,
Chrysococcyxy ScythropSy Coccyzus, Crotophaga, Guira, Tapera, Geococcyx, Centro-
pus, Com, Piaya, Urodynamis, Saurothera, Eydynamis и др.) и на 126 видов.
Распространены очень широко и в Восточном и в Западном полушарии,
исключая собственно Арктику и Антарктику и прилежащие острова.
Ископаемые остатки кукушек известны из верхнего эоцена, нижнего олиго-
цена (Франция), из первых описан Dynamopteras velox, из среднего миоцена
(Франция) -^Necrornis palustris,^ из плейстоцена (Мадагаскар) описан Соиа
primaeva. Рецентные виды Caculus canorus и роды Crotophaga, Geococcyx, Taper a
известны из плейстоцена.
Географическое распространение. Кукушки распространены по всему
свету, за исключением Субарктики. Наибольшее разнообразие видов
встречается в тропических и субтропических странах. Общее число видов отряда
«iff
in
ш
Рис. 111. Сгиб крыла обыкновенной
кукушки (справа) и глухой кукушки
(слева).
ХАРАКТЕРИСТИКА ОТРЯДА
435
кукушек 146, из них фауне СССР свойственны 6, т. е. 4,05%, принадлежащих
2 родам подотряда, кукушек: хохлатая кукушка Clamator с 1 видом, случайно
у нас залетным, и кукушка Cuculus с 5 гнездящимися у нас видами —
обыкновенная кукушка Cuculus canorus, глухая С. saturates, малая С. poliocephalus,
индийская С. micropterus и ширококрылая С* fugax. Такое разнообразие видов
встречается только на Дальнем Востоке — в Приморье. В остальной части
СССР, и то далеко не повсюду, обитают только два вида кукушек
—обыкновенная и глухая. Обыкновенная кукушка широко распространена по всей
стране и доходит на север до предела лесной растительности. Глухая кукушка
встречается только в северо-восточных областях Европейской части СССР и
в Сибири.
Из общего числа видов (146) отряда кукушек монотипических видов, т. е.
не образующих подвидов, —. 66, из них собственно подотряду кукушек
принадлежит 60 видов. Монотипические виды в СССР, за исключением залетной
хохлатой кукушки, не представлены.
Распределение кукушек на гнездовье по зонам в нашей стране следующее
(в числителе показано число видов, в знаменателе — число подвидов и
монотипических видов, характерные формы подчеркнуты):
I. Арктика (зона) Cuculus canorus l
а) побережье (подзона)
б) тундра (подзона)
в) лесотундра (подзона) Cuculus canorus canorus ■1/1
II. Лесная зона Cuculus canorus, Cuculus sat иг at us, Cuculus fugax, Cuculus microp-
terus, Cuculus poliocephalus 5
г) тайга (подзона) С. с. canorus, С. s. horsfieldi 2/2
д) смешанный лес (подзона) С. с. canorus, С. s, horsfieldi, С. policephalis,
С. micropterus, С. fugax Б/а
е) лесостепь (подзона) С. с. canorus Vi
III. Зона открытых сухих пространств С. canorus 1
ж) степь (подзона) С. с. canorus (озерная сторона Зап. Сибири и Казахстана) */i
з) полупустыни и пустыни (подзона) С. с. subtelephonus */i
IV. Горная зона С. canorus 1
и) высокогорная бореоальпийская подзона
к) горно-степная подзона С. с. canorus. С. с. subtephonus г/2
л) горно-степная и пустынная подзона С. с. subtelephonus 1f1
Таблица для определения видов кукушек
1(2) На затылке имеется хохол. Брюшная сторона тела без темных поперечных полос
или продольных пестрин. Хвост длиннее крыла
Хохлатая кукушка Clamator glandarius.
2(1) На затылке нет хохла. На брюшной стороне тела имеются темные поперечные
полосы или продольные пестрины. Хвост короче крыла 3
3(4) Брюшная сторона тела испещрена продольными темными пестринами. На хвосте
(при серой окраске верха тела) несколько темных поперечных полос (4—5). Вершины
второстепенных маховых доходят приблизительно до 2/3 длины крыла
Ширококрылая кукушка Cuculus fugax
4(3) На брюшной стороне тела имеются поперечные темные полосы. На хвосте при
серой окраске верха или совсем нет темных поперечных полос, или имеется только одна
предвершинная полоса. Вершины второстепенных маховых доходят приблизительно до
половины длины крыла , 5
5(6) Крыло короче 170 мм Малая кукушка Cuculus poliocephalus
6(5) Крыло длиннее 175 мм 7
7(8) Спинная сторона тела серовато-бурая. На рулевых имеется одна широкая темная
предвершинная полоса Индийская кукушка Cuculus micropterus
8(7) Спинная сторона тела шиферно-серая или рыжая с темной поперечной полосато-
стью. На рулевых нет предвершинной полосы, а если рулевые поперечно полосаты, то
окраска оперения рыжая 9
9(10). Кроющие кисти, прикрывающие большие кроющие крыла, — белые с темными
поперечными полосками. Горло и зоб светлосерые
Обыкновенная кукушка Cuculus canorus:
*28
436
ОТРЯД КУКУШКИ
10(9). Кроющие кисти, прикрывающие большие кроющие крыла, — светложелтые или
беловатые, иногда с редкими темными отметинами. Горло и зоб серые
. ..... Глухая кукушка Cuculus saturates.
СЕМЕЙСТВО КУКУШКИ CUCULIDAE
РОД ХОХЛАТЫЕ КУКУШКИ CLAMATOR KAUP 1829
Тип Cuculus glandarius
Синонимы: Coccystes G 1 о g e r. 1842; Oxylophu S w a i n s о n, 1837
1. Хохлатая или пятнистая кукушка Clamator glandarius L.
Cuculus glandarius. Linnaeus. Syst. Nat., изд. Xt 1758, стр. Ill «северная Африка
и южная Европа».
Распространение. Ареал. Южн. Европа: Испания, Португалия, южн.
Франция (Эро, Арль), Кипр, вост. Греция; Малая и Передняя Азии — Сирия,
Карта 74. Распространение хохлатой кукушки Clamator glandarius
а — граница гнездовий, г — область зимовок: п — палеарктических птиц, * —
африканских, д — залеты
Палестина, с.-з. Месопотамия, ю.-з. Иран до Персидского залива; с.-з. Африка
(Марокко, Алжир, Тунис), Египет, тропическая и южн. Африка от Сиерра
Леоне на западе до Сомали на востоке, но за исключением крайнего ю.-з.
Африки. Птицы, гнездящиеся в Палеарктике, зимуют в тропической Африке,
гнездящиеся в тропической Африке — в южн. Африке.
ХОХЛАТАЯ ИЛИ ПЯТНИСТАЯ КУКУШКА 437
Характер пребывания. В СССР случайно залетная птица.
Залет в Бессарабию указан еще Нордманном (1840), позднее добыта 27.VI. 1914 г.
в б. Аккерманском у. близ с. Чобруг-Бессарабский (Парамонов, 1915).
Рис. 112. Хохлатая кукушка.
Известны залеты хохлатой кукушки на Канарские о-ва, Мадейру, Британские
о-ва, в сев. Францию, Бельгию, Данию, Германию, на Балканский п-ов, на
острова Средиземного моря и в сев. Иран. Птицы, обитающие в
Средиземноморском бассейне и в южн. Африке, перелетны, в остальных местах— оседлы.
Биотоп. Леса, перемежающиеся с полянами, лесные окраины,
кустарниковые заросли с разбросанными между ними деревьями.Ч и с л е н н о с т ь.
438
ОТРЯД КУКУШКИ
В Испании и Передней Азии обыкновения; во Франции, Греции и
Португалии — редка.
Экология. Размножение. Яйца откладывает в чужие гнезда,
главным образом, в гнезда вороновых птиц. В Испании и Португалии обычно
откладывает яйца в гнезда ворона, сороки, реже—голубой сороки; в с.-з, Африке —в
гнезда сороки, в Египте и Палестине —вороны, в Сомалии — Corvus rhipidurus,
на Кипре — сороки, в южн. Африке —вгнезда С. capensis, Amydrus тоНоищ).
Кладка яиц в Испании происходит в июне—июле, в сев. Африке —в марте—
апреле. Часты случаи нахождения в одном и том же гнезде нескольких (2, 3,
4) яиц хохлатой кукушки; однажды в гнезде найдено 8 яиц этой птицы.
Невидимому, в этих случаях несколько самок кукушек откладывают яйца в одно
гнездо'. Вскрытие яичника показало, что хохлатая кукушка может отложить
в один сезон 12—15 яиц (Мейнертцхаген, 1930). Яйца хохлатой кукушки,
найденные в гнездах вороновых птиц, похожи по окраске на вороновые —
зеленоватого цвета с коричневыми пятнами и крапинами, но несколько мельче и
с более тонкой скорлупой. Размеры (78) 28,4—35,4 х 26,3 в среднем 32,1 х 24 мм
(Уайзерби, 1938).
Линька. У взрослых одна линька в году, с декабря по март.
Питание. Преимущественно различные крупные насекомые
(прямокрылые, стрекозы, жуки и т. д.) и их личинки.
Полевые признаки. Длинный хвост, хохол, крупные белые пестрины
на кроющих крыла позволяют легко узнать птицу. По общему виду и
размерам несколько напоминает сороку.
Держится на деревьях, кустах, нередко на земле, часто подает голос;
кукование в роде—«кии-оу-кии-ау», при беспокойстве —* «карк-карк», крик
самки ~ «барру-барру».
Описание. Размеры и строение. На голове небольшой хохол. Ноздри
щелевидной формы, расположены у основания клюва. Первостепенных маховых — 10,
первое короче половины длины крыла, самые длинные — 3-е и 4-е, формула крыла 3 =
= 4>2>5>6>7>8>9>10. Хвост длинный, ступенчатый, из 12 рулевых. Размеры:
длина крыла самца (Алжир) 205, самки (Египет) 196 мм (Зоологический музей Моск.
университета); 195—220 мм (Египет); 186—224 мм (вост. Средиземноморье), 193—211 мм
(зап. Средиземноморье, Мейнерцхаген, 1930); крыло (12) самцов 191—222 мм, самок 182—
206 мм (Уайзерби, 1938).
Окраска. У взрослого самца голова серая, верх тела бурый с легким сероватым
«налетом; усиливающимся на подхвостье. Плечевые, кроющие крыла и хвоста,
второстепенные маховые — темнобурые с крупными белыми вершинными пятнами. Первостепенные
маховые темнобурые с небольшими белыми вершинными пятнами. Рулевые черно-бурые
с широкими белыми вершинами, кроме средней пары, у которой белые вершины узки.
Нижняя сторона тела белая с охристым налетом на горле, верхней части зоба и по бокам шеи.
Самка похожа на самца, но отличается меньшим хохлом, более тусклым бурым тоном
верха и менее ярким охристым налетом на боках шеи. Первостепенные маховые с рыжепа-
тыми в основных двух третях внутренними опахалами, иногда рыжий цвет около стержня
заходит на наружные опахала.
В первом осеннем наряде брюшная сторона грязносерая; на горле и
зобе и на боках шеи грязноохристый оттенок. В первом весеннем наряде самец
похож на самку, но с меньшим хохлом, с оливково-бурым верхом тела. Верх голопы тсм-
нобурый, зоб, грудь и нижние кроющие хвоста с охристым оттенком. На первостепенных
маховых рыжий цвет сильно развит.
РОД КУКУШКИ CUCULUS LINNAEUS 1758
Тип Cuculus canorus
2. Обыкновенная кукушка Cuculus canorus L.
Cuculus canorus. Linnaeus. Syst. Nat., изд. X, 1753, стр. ПО, Швеция,
Распространение. Ареал. Европа и Азия к северу до предела лесной
растительности, к югу в Европе, до Средиземного и Черного морей; в Азии —
ОБЫКНОВЕННАЯ КУКУШКА
439
до Палестины, Сирии, Малой Азии, Ирана, Туркестана, Афганистана,
Белуджистана, северной Индии, Ассама, Бирмы; на востоке в Китае, Монголии*
Манчжурии, Корее и Японии; с.-з., тропическая и южная Африка.
Зимует в тропической и -южн. Африке к югу от Сахары; в южн. Аравии,
в Индии, на Цейлоне, в Южн. Китае, на Малайском п-ове, Молуккских о-вах;
на восток до Новой Гвинеи.
Карта 75. Распространение обыкновенной кукушки Cuculus canorus
7 — G .с. canorus, 2 — С. с. kleinschmidti, 3 — С. с. bangsi, 4 — С. с. subtelephonus, 5 — С, с. bakeri, 6—
С. с. fallax, 7 — С. с. gularis.
а — граница гнездовий, 6 — не вполне выясненные границы гнездовий, г — зимовки (цифры в
кружках — зимние местонахождения соответствующих подвидов).
Характер пребывания. Гнездящаяся и перелетная птица.
Залеты отмечены в различных областях Сахары, в Аравии, в Исландии и в сев.
Аляске.
Биотоп. Весьма разнообразен, тесно связан со стационарным
распределением воробьиных птиц, в гнезда которых кукушка откладывает свои яйца,
также и с обилием пищи. В связи с этой зависимостью кукушку можно найти
в самых различных местообитаниях: на окраине тайги, в смешанных лесах,
в лесостепи, в степи, в пойменных насаждениях по рекам и озерам, в
Камышевых и тростниковых зарослях, в культурных местностях, близ человеческих
поселений, в горах и даже по окраинам пустыни.
440
ОТРЯД КУКУШКИ
Подвиды и варьирующие признаки. Семь подвидов, различающихся между собою
размерами, окраской и экологией: С. с. canorus Linn. 1758— Европа и Азия от северной
границы лесной растительности до Средиземного моря, Кавказа, Малой Азии, в Сибири до
Приморского края; Манчжурия, Монголия, Япония, Корея; С. с. kleinschmidti S с h i e b e 1,
1910 — Корсика (?), Сардиния, повидимому синоним С. с. canorus, но для окончательного
решения у нас нет коллекционного материала; С. с. bangsi Oberholser, 1919 —
Португалия, Испания, Балеарские о-ва, Марокко, Алжир и Тунис; С. с. gularis Stephens,
1815 — тропическая и южн. Африка от Сенегамбии и низовьев Нигера на западе до
Египетского Судана на востоке. К югу до Земли Дамара, Земли Бечуанов, Южноафриканского
союза (о гнездовании в Капской Земле достоверных данных нет); С, с. subtelephonus Za-
rudny, 1914 — Иран, за исключением западного; Средняя Азия, Афганистан:
Белуджистан, Кашгария; Гималаи,Тибет и Ала-Шань; С. с. bakeri H a r te r t, 1912 —Куку-нор,
ю.-з. и зап. Ганьсу, зап. Сычуань и может быть вост. Тибет, Ассам до Брамапутры, Кочин,
Юннань к югу до Бирмы; С. с. jallax Stresemann, 1930 — Китай к северу до Пекина,
к югу до Гуандуня, Гуаньси и по Яньцзы-цзяню до центральных частей Сычуаня.
Обыкновенная кукушка Cuculus canorus canorus L.
Синонимы: С. tetephonus. Heine. J. f. Orn., 1863, стр. 352, Япония, — С. с.
johanseni. T s с h u 8 i. Orn. Jahrb. XIV, 1903,стр. 165. Томск.—С. с. maximus. Neumann»
Anz. Orn. Ges. Bayern, II, 8, 1934, стр. 332, Саяны
Русское название: кукушка — народное, звукоподражательное.
Распространение. Ареал. Вся Зап. Европа (за исключением
Пиренейского п-ова, Балеарских о-вов, Корсики, может быть Сардинии, где другие
подвиды); Палестина, Сирия, Малая Азия, зап. Иран. Вся наша страна,
кроме Средней Азии, Япония, Корея, Манчжурия, Монголия. Северная
граница распространения обыкновенной кукушки в основном совпадает с границей
лесной растительности. В Скандинавии на севере идет до округа Варангер
(70° с. ш.), бассейна р. Пазы и округа Энаре (69° с. ш.); в СССР на Кольском
п-ове доходит до 68°—69° с. ш. (устья рек Туломы, Колы, Б. Тювы, бухты
Лесной, Нотозера, оз. Лов и Лявозеро, района озера Имандры, Хибин,
р. Нива, Пельозера и Пинозера, далее по реке Поной до Белого моря).
Гнездится у Кандалакшского залива, до Порье-губы и Кобды, на Соловецких о-вах
(найден птенец, Гебель, 1903; Поляков, 1929), в окрестностях Архангельска
и по всей лесной зоне Архангельской обл. до р. Кулою, по Мезени до 66°
с. ш., между Песью и Вижасом почти до 67° с. ш., (на Печоре —до
северной границы сплошного леса, т. е. до 63°30' с. ш., может быть и до полярного
круга (Сибом и Броун, 1876, встречали ее у полярного круга). На Сев.
Урале доходит до верховьев Сухой Сыньи (около 65° с. ш.), верховьев Маньи
(65° с. ш.), р. Щугора (64°30' с. ш. — 63°20' с. ш.), Ляпина. Гнездится по средней
и нижней Оби до Обдорска, р. Казым, до юрты Ильборской и Березова; по
р. Тазу до 67°'с. ш. (Скалой и Слудский, 1941), на Енисее —до Турухаиска,
в бассейне Вилюя и у Якутска, в бассейне Яны и в бассейне Индигирки до
70° 29' с.ш.,т. е. до северного предела лесной растительности (Михель, 1935);
в бассейне Колымы — до дельты (под 69°4' с. ш. найдена в кустарниках южной
части тундры); в Анадырском крае доходит до южных склонов Анадырского
хребта (озеро Красное, с. Марково и до р. Великой, в приморскую полосу не
заходит, Портенко, 1935); в Коряцкой Земле (Дементьев, 1940); на Камчатке
и Командорских о-вах (на Командорах встречена в конце мая и в июне);
Охотское побережье, на Сахалине по всей Приморской обл. За пределами
нашей страны гнездится в Японии, Корее, Манчжурии и Монголии. Южная
граница ареала в зап. Азии спускается к Каспийскому морю, повидимому,
захватывая с востока Урал и Эмбу (здесь граница между С. с. canorus и
С. с. subtelephonus еще не выяснена). Далее от Южн. Урала идет по
Казахстану, приблизительно по 47-й параллели (тут расположена северная
граница туркестанской кукушки С. с. subtelephonus); охватывает с юга Зайсан-
скую котловину, Тарбагатай и Алтай.
ОБЫКНОВЕННАЯ КУКУШКА
441
Зимует в тропической (кроме западных частей и Сахары) и в южн.
Африке, в южн. Аравии, в Индии, на Цейлоне, на Малайском п-ове, на Больших
Зондских о-вах, на восток до Новой Гвинеи.
Характер пребывания. Гнездящаяся и перелетная птица.
Даты. В Африку прилетает очень рано —во второй половине июля
(первые отдельные взрослые особи наблюдались в Египте 19.VII, в Судане —
31.VII, в Уганде — 20.VII). В августе пролет уже ясно выражен, но валовый-
проходит в Египте в начале сентября, в Египетском Судане — в середине
сентября. Зимовок в вост. и южн. Африке первые птицы достигают в октябре,
Карта 76. Изохроны первого кукования кукушки (по Д. Н. Кайгородову).
но главная масса кукушек прилетает в южн. Африку в декабре. С первых
чисел марта начинается отлет кукушек с зимовок. Массовый пролет через
с.-в. Африку, центральную Сахару и Египет протекает во второй половине
апреля и заканчивается в первой половине мая.
В СССР кукушка весной появляется, начиная с половины апреля (южные
части^страны) и до конца мая и даже начала июня (у северной границы
распространения). Сроки прилета сильно колеблются в зависимости от хода весны.
Прилетают по одиночке, в первую очередь самцы, а через 3—4 дня — самки.
ЧерезУнесколько дней (5—б) после прилета местных птиц начинается валовый
пролет птиц из лежащих севернее местностей. Во время пролета на юге
кукушки скопляются большими стаями. По Кайгородову (1907), весеннее
движение кукушек по территории Европейской части Союза в общем имеет
направление с юго-запада на северо-восток. Скорость весеннего
движения кукушек на территории Европейской части Союза составляет в среднем
около 80 км в день. При такой скорости вся гнездовая территория до самых
442
ОТРЯД КУКУШКИ
северных пределов распространения вида может быть занята птицей в течение
одного месяца. О сроках прилета кукушек имеется много наблюдений,
основанных, главным образом, на первом ее куковании. Подытоживая эти сведения
и данные Кайгородова (1907), приводим ниже только средние прилета кукушек
для разных местностей и крайние сроки особенно раннего и позднего их
появления. Кукушки в западные части Азербайджана прилетают в среднем 15. IV,
самый ранний прилет отмечен 26. III, самый поздний 3.V; в ю,-в.
Азербайджан—в среднем 1—3.V (время для прилета слишком позднее,
вероятно, отмечен пролет более северных популяций). В Тбилиси, Ленкорань,
Дербент прилетают в среднем в конце апреля 25. IV; на Черноморском
побережье Кавказа в среднем пролет отмечен 11. IV, самый ранний — 5. IV, самый
поздний—18. IV; в горной части с.-з. Кавказа — 16,17.IV; на с.-в. Кавказе во
второй половине апреля; на Кубани — в среднем 22. IV, самый ранний прилет
отмечен 6. IV, самый поздний 26. IV. В Крым —в среднем 15. IV; самый ранний
прилет 12. IV, самый поздний — 28. IV. В Бессарабию в среднем — 25. IV,
самый ранний прилет 13. IV, как исключение в 1905 г. 8. IV. На Украину —
в колце апреля (25. IV), самый ранний прилет в Харьковскую обл. и
Киевскую 15. IV (Сомов, 1897; Данилович, 1930), в Подолию — 17.IV, самый
поздний прилет в Харьковскую и Киевскую обл. 4.V, в Подолию — 30. IV.
В Астраханскую обл. и Сталинградскую прилетают в среднем 1.V, самая
ранняя дата отмечена 28. IV (как исключение —18. IV), самая поздняя
6.V. В Куйбышевскую обл. 5.V, наиболее ранний прилет —1.V, наиболее
поздний — 11.V, в восточные районы —в б. Бугурусланский у. 10.V,
самый ранний прилет 5.V, самый поздний—13. V (Карамзин, 1909). В Башкирию—
12.V, самый ранний прилет 3.V, самый поздний — 14.V. В Татарскую
республику 8.V, самый ранний — 4.V (как исключение—22. IV), самый поздний
12. V (Кайгородов, 1907)^
В центральную полосу к северу от Украины и до Московской обл.
прилетают в самом конце апреля и в начале мая, в среднем — 1.V, наиболее ранние
сроки прилета в Калужской обл.— 17. IV (как исключение 7. IV. 1906), в
Московской обл. — 15.IV.1929, в Смоленской — 20. IV, в Курской и Орловской —
24. IV, в Воронежской, Тамбовской и Тульской — 27. IV, в Пензенской — 30. IV.
Самый поздний прилет —9— 10. V. В Белоруссию прилетают в среднем—28. IV,
крайние сроки прилета — 20. IV и 3.V (Шнитников, 1913; Федюшин,
1912).
Несколько северней прилет отмечен: в среднем 3.V, самый ранний в г. Ме-
ленки Владимирской обл. — 21. IV; в Старице и Ржеве Калининской обл. —
26. IV, в Ярославской обл.—28. IV, в Галиче Костромской обл.—1 .V, самый
поздний прилет зарегистрирован —11 .V; в Ивановскую обл. самый ранний прилет —
23,IV, массовый прилет в среднем — 3—4.V (Птушенко, 1937); в Горьковский
край — 2.V, наиболее ранний прилет, как исключение в Ардатове — 28. IV,
самый поздний—8.V; в Кировскую обл.—12. V, самый ранний—4. V, самый
поздний—24. V. В Ленинградскую обл. — 4.V, самый ранний прилет 25. IV,
в Ленинград—12.V, особенно поздние даты прилета отмечены в 1853, 1857 и
1873 —19 и 20. V (Бихнер, 1884); в Прибалтику— 3.V, самый ранний прилет —
26. IV, самый поздний — 12.V. В Онежский район (64°54' с. ш.) — 22.V, на
Кемь (64°57'с. ш.)—24. V, самый ранний прилет—21. V. На Кольский п-ов
(68°Г с. ш.)—21. V, самый ранний—13. V. В Архангельскую обл.—в конце первой
половины мая, в среднем— 11.V, самый ранний прилет —5. V, самый поздний—
16.V, особенно поздно прилетают в Сыктывкар (61Q 40'с.ш.), в среднем 13.V,
самый ранний прилет— 10.V, самый поздний— 16.V. В Молотов —в
среднем— 12.V, самый ранний прилет — 30. IV, самые поздние—18. V и 21.V.
В Свердловск прилетают в среднем—20—21.V; в Верхотурье в среднем —
16-17.V.
ОБЫКНОВЕННАЯ КУКУШКА
443
В Средней Азии обыкновенная кукушка обычная пролетная птица, но
установить время ее пролета трудно, так как в поле она не отличима от обитающей
в Средней Азии туркестанской кукушки. Судя по времени прилета и началу
размножения туркестанской кукушки, обыкновенная кукушка, повидимому,
летит несколько поздней туркестанской —с половины апреля и до половины
мая. Так, у ю.-в. Каспия в районе Чикишляра пролетные обыкновенные кукушки
встречаются до половины мая (Исаков и Воробьев, 1940), под Кушкою
пролетные кукушки наблюдались 4.V. 1887, главный пролет на Теджене шел
13—14.V. 1887 (Зарудный, 1896). В большом количестве пролетают кукушки
долиною Сыр-Дарьи, окраиной Кызыл-Кумов и восточным побережьем
Аральского моря; у залива Сарычеганак пролет идет в первой половине мая. На Эмбе
первая кукушка появилась 15. IV, главная масса летит в конце апреля,
в области Иргиза большой пролет наблюдается в начале мая., В Уральске
первые кукушки наблюдались 20. IV, под Чкаловым — в конце апреля;
валовой пролет идет в начале мая, последние птицы летят до 20.V (Зарудный,
1888). В район Акмолинска 16.V кукушки уже прилетели (Карамзин, 1917),
в Кустанай, Наурзум и Омск (Шухов, 1913) прилетают в половине мая.
В Алтайский край —в половине мая, в верховьях р. Алей, на р. Сара-Чумыш
и в междуречье Бин-Катум в среднем 11.V (Залесский, 1928). В Кузнецкую
степь в среднем— 19.V (Хахлов, 1937). Под Минусинск в начале второй
половины мая —24.V кукушек было уже много (Сушкин, 1914). В
Красноярск — в первой половине мая, в среднем 6.V; в Туруханск — в конце мая,
в 1917 г. прилетели 26.V; в устье р, Курейки—23.V (Тугаринов и
Бутурлин, 1925). В Якутск в среднем 20. V (в 1844 г. прилетели очень рано ^4.V;
к Лунхе —-в конце мая, в 1854 прилетели 30.V; в Амгинскую слободу
в 1903г. —24.V, в Сулгачи—20.V; Иванов, 1929), В Индигирский край —
в конце- мая. В верховья Колымы^-в начале июня (5.VI, Дементьев,
1940), на Анадырь — в начале июня, первое кукование в среднем бывает 5.VI,
самое позднее 9.VI (Портенко, 1939). В окрестности Охотска—в первой
половине мая. На Камчатку (Кроноцкий заповедник) в среднем — 4.VI, самое
раннее появление — 2. VI (Аверин, 1948). На Сахалин прилетают в конце мая.
В Приморский край в среднем во шорой половине мая — 20.V
(Воробьев, 1938). На Зее у Бомбака первые кукушки появляются в конце мая.
В Кентее и в Гобийском Алтае — в начале второй половины мая.
Осенний отлет. Об осеннем отлете и пролете имеется мало
наблюдений. Отлет начинается рано и идет в начале медленно. В первую очередь
по одиночке или парами отлетают старые птицы, затем отлет становится
заметнее —. птицы уже летят небольшими стайками. В это же время начинают
отлетать и молодые птицы, но иногда одиночные молодые птицы задерживаются
в местах гнездовья очень долго — до конца сентября. В горных районах птицы
вначале покидают высокие местности, затем улетают и из более низких районов.
В Карело-Финской ССР, в Повенецком районе, первые отлетающие кукушки
отмечены 26—27.VII (Исполатов, 1916), в Ленинградской, Псковской и в
Новгородской обл. отлет идет с начала августа и до первой половины сентября.
Последние одиночные птицы наблюдались до конца сентября — 23. IX (Бианки,
1910; Зарудный, 1910). Прибалтику покидают в конце сентября, из Беловежья
улетают- в начале сентября. В центральных областях Европейской части Союза
отлет идет с конца июля (одиночные старые птицы), в половине августа
большинство взрослых птиц уже покидают места гнездовья, а к концу сентября
улетают и молодые птицы. В конце августа и в начале сентября в центральных
областях Союза наблюдается пролет более северных птиц. В Молотовской
обл.. отлет кукушек ясно выражен со второй половины августа и до середины
сентября; последние одиночные птицы встречаются до конца сентября; в
Башкирии отлет отмечен в конце сентября. В Поволжье — начинается с конца июля
444
ОТРЯД КУКУШКИ
идет до сентября; в конце августа в степях у Бугульмы наблюдались стаи до
40 и более экземпляров, молодых и старых птиц, в б. Вугурусланском у.
одиночные птицы начинают улетать уже в конце июля, в первой половине
августа идет валовой пролет. Под Чкаловом отдельные экземпляры начинают
улетать в конце августа, валовый пролет идет в первой половине сентября,
запоздавшие птицы встречаются до конца октября. В северных частях Украины
(Харьков) отлет идет с первых чисел августа до конца августа; с конца августа
до середины сентября наблюдается пролет северных кукушек (Сомов, 1896).
В ю.-з. Украине (Подолия) местные старые кукушки улетают в конце июли и
в начале августа, молодые покидают гнездовья в конце августа, и до конца
сентября отлетают запоздавшие одиночные местные молодые птицы, в это время
пролетают и северные. В южн. Украине (Днепропетровск, Сиваш, Приазовье)
пролет наблюдается в*течение августа и сентября, одиночные экземпляры
встречаются еще и в начале октября. Из Крыма отлетают в сентябре. На
Кавказе отлет идет в течение августа и в начале сентября, пролет северных
птиц — до конца сентября.
По осеннему отлету и пролету кукушек в Сибири и Средней Азии имеется
еще меньше наблюдений. У Красноярска отлет наблюдается в последней трети
августа. На Алтае в высоких районах отлет идет до конца, августа, в
низменных — до середины сентября, последние птицы встречаются до конца месяца.
В районе Омска в начале сентября пролетные кукушки встречаются в
большом количестве, большой пролет наблюдается у Барнаула и Семипалатинска.
В верховьях Иргиза пролет отмечается с конца августа до середины
сентября и позднее 16. IX кукушки не встречаются (Сушкин, 1908). В устьях
Урала кукушки Ца пролете появляются в начале второй половины сентября.
У Карабогаза пролет идет с первых чисел августа (начиная с 8.VIII). По
Атреку и в Астрабадской провинции в конце сентября — 24. IX
(Шестоперов, 1928). В Средней Азии под Джаркентом пролет идет в течение всего
сентября. У Гарма и на озере Яшиль-куль последние пролетные кукушки
наблюдались 8—12. IX (Иванов, 1940).
Биотоп. Как указано в очерке вида, биотоп кукушки чрезвычайно
разнообразен и тесно связан со стационарным распределением воробьиных птиц,
в гнезда которых она откладывает свои яйца. Большое значение имеет также и
наличие необходимых кормов — различных насекомых. Основная масса
кукушек приурочена к сельскохозяйственным угодьям, густо заселенным
воробьиными птицами. В лесной полосе основной биотоп — смешанные лесные
массивы, населенные в большом количестве лесным коньком и представителями
сем. славковых. Встречаются и в лиственных лесах, и в тайге, в выборе породы
леса совершенно неразборчива. Густой тайги, бедной воробьиными птицами,
кукушка избегает и обычно заселяет окраины тайги, редколесье, тальниковые
заросли, пойменные насаждения по берегам рек и опушки лесов. В
лесостепных районах Украины кукушка селится по колкам, лесным и кустарниковым
зарослям, по долинам рек и т. д., в степных районах Украины она обитает
главным образом в котловинах степных озер, поросших кустарниками и
тростниками, по оврагам, в зарослях бурьяна, но встречается и в совершенно голой
степи. Откладывая особенно часто свои яйца в гнезда белой трясогузки,
кукушка преимущественно населяет сельскохозяйственные угодья, сады, парки
и обычно встречается в непосредственной близости от жилья человека. В
горных районах Кавказа встречается повсюду — в смешанных и лиственных лесах,
в долинах рек и культурных местностях, поднимаясь в горы приблизительно до
границы лесной растительности и местами даже выше. В Грузии доходит до
Боржома, верховьев Куры —до 2500 м (Радде, 1885), в Азербайджане, по
сообщению Спангенберга —до 1800 м, в Армении—до субальпийских лугов —
до 2500 м (Ляйстер и Соснин, 1942). На Алтае распространена повсюду, за
ОБЫКНОВЕННАЯ КУКУШКА
445
исключением глухой тайги; в горы поднимается до 2200 м, заходя в область
альпийских кустарников (Сушкин, 1938). На Камчатке —по всему лесному
поясу, поднимаясь в горы до 1000 м (Аверин, 1948).
Численность.В связи с широким распространением кукушки общая
численность ее в Союзе велика, но неравномерна в различных частях ареала.
Такая неравномерность связана с плотностью гнездования воробьиных птиц и
с трофическими условиями местности. Крайне многочисленна в лесостепных
районах Украины, где есть перелески и степные пруды. Реже встречается в
степной Украине, главным образом по оврагам и озерам. В средней полосе Союза
кукушка в большом числе встречается в смешанных открытых лесах. В северной
части своего ареала — в полосе тайги — она становится положительно редкой,
встречаясь преимущественно близ окраин тайги. В горных районах численность
кукушки резко сокращается в вертикальном направлении. В предгорьях и
нижнем поясе гор ее много, но как только видовой состав воробьиных птиц по
мере поднятия в горы сокращается — численность кукушки резко падает.
Экология. Размножение. У обыкновенной кукушки особенно резко
выражен гнездовой паразитизм — она всегда откладывает свои яйца в чужие
гнезда и никогда не высиживает птенцов. Обычно наблюдается полиандрия
(два-четыре самца держатся при одной самке). Можно считать, что у
кукушек существует гнездовая территория, если за это принять район, на
котором держится самка, разыскивая гнезда мелких воробьиных птиц для
откладки в них своих яиц. Прилетевший ранее самки на место гнездования
самец занимает тот же участок, что занимал и в прошлые годы, и своим
криком привлекает пролетающих самок. Самки, повидимому, также
возвращаются на свои старые гнездовые участки, и гнездовый участок самки не всегда
совпадает с гнездовым участком самца. Размеры гнездовой территории сильно
колеблются в зависимости от количества воробьиных птиц, в гнезда которых
кукушка откладывает свои яйца, и от кормовых условий. Растительность,
ландшафт, климат и другие факторы не имеют значения. Для Ленинградской
обл., в местности, где преобладают смешанные леса, размеры гнездовой
территории исчисляются 1—1,5 кв. км (Промптов и Лукина, 1940). Драк самцов
и самок из-за территории не наблюдается, но в период откладки самкой яиц
самцы сильно возбуждены и в это время между ними происходят драки.
Куковать самцы начинают на второй — четвертый день после прилета.
Кукуют и днем и ночью, но особенно усиленно на утренней и вечерней заре.
Метеорологические условия мало влияют на кукование. В первой половине
июня кукование ослабевает и иногда совсем прекращается, затем в конце июля
вновь усиливается, может быть в это время у кукушек начинаются
дополнительные кладки. Кукует самец во время полета или сидя на каком-либо
возвышении — в лесных местностях на деревьях, причем предпочитает держаться
в верхней части кроны деревьев; в степных местностях —^на кустарнике и
в совершенно безлесных местностях — сидя на камне или на каком-либо ином
возвышении. В момент кукования самец опускает крылья, приподымает и
раскрывает хвост, пригибает голову, слегка вытянув шею, раздувает горло и издает
неоднократно повторяющийся крик.
Кукушки откладывают свои яйца преимущественно в гнезда мелких
воробьиных птиц и только как исключение в гнезда птиц других отрядов,
например, дятлов, куликов и т. д. В Ленинградской обл. откладывает яйца
преимущественно в гнезда горихвосток; в сев. Украине (район Харькова)
большинство яиц встречается в гнездах белой трясогузки; в юж. Украине
(Днепропетровск) — в гнездах дроздовидной камышевки, серой славки, реже в гнездах
жулана и серой мухоловки; в центральных областях Союза -— в гнездах белой
трясогузки, лесного конька и зарянки; в Средней Азии (район низовья Сыр-
Дарьи и Фрунзе) —в гнездах дроздовидной камышевки, черноголового чеккана
446
ОТРЯД КУКУШКИ
и несколько реже в гнездах жулана; в Туркмении в гнездах Scotocerca
inquieta; в Приморском крае —в гнездах амурского жулана итолстоклювой
камышевки Phragamaticola aedon. По подсчетам, яйца обыкновенной кукушки
найдены у 125 видов воробьиных птиц, но особенно часто яйца кукушек
откладываются в гнезда примерно 20 видов воробьиных птиц. Самка кукушка
откладывает в одной и той же местности яйца в гнезда определенных видов
воробьиных птиц и даже в гнезда той же пары, что и в прошлые годы, повидимому
в гнезда того самого вида воробьиных птиц, в которых она сама вывелась.
Иногда, очень редко, в гнездах встречаются два яйца кукушек, но тогда,
повидимому, они принадлежат двум различным самкам, гнездовые участки которых
перекрываются. Чаще всего кукушка откладывает яйца в гнезда с только что
начавшейся кладкой или с полной, но с ненасиженными яйцами. Однако,
наблюдались случаи, когда яйца подкидывались в недостроенные гнезда или
в гнезда с сильно насиженной кладкой. Самка кукушка к моменту откладки
яйца бывает в сильно возбужденном состоянии — она облетает свой гнездовый
участок, повидимому подыскивая подходящие гнезда; найдя гнездо, некоторое
время ведет наблюдение за ним и за хозяевами гнезда. Улучив момент, когда
хозяева отсутствуют, иногда самец помогает ей отвлечь внимание хозяев от
гнезда, кукушка откладывает свое яйцо, что чаще всего происходит днем.
Подкладывает свои яйца кукушка различным способом — иногда она
откладывает яйцо непосредственно в гнездо, влетая в него; иногда, когда отверстие
в гнезде мало, как это наблюдается у птиц дупляногнездников, она подлетает
к гнезду и плотно прижимает клоаку к отверстию гнезда, выбрасывая туда
яйцо. В таких случаях иногда находят разбитые кукушечьи яйца у гнезда —
кукушка не попадает в гнездо. Иногда яйца откладываются на землю близ
гнезда и при помощи клюва туда переносятся. Часто при этом кукушка
выбрасывает или уносит одно или несколько яиц хозяев гнезда. Количество яиц,
откладываемых самкою кукушки за сезон, велико —не менее 12, иногда 19—
20 и даже 25. Промежутки между откладкой каждого яйца 1—2 дня. Период
овуляции у самок продолжается 36—46 и даже 47 дней. Весь период
размножения сильно растянут — с конца апреля и начала мая до половины июля,
считая с начала кукования до его конца. Так, на Украине первое кукование
отмечено в среднем с 20. IV и продолжается до 20.VII; в Белоруссии —с 28. IV
и по 5.VII; в центральных обл. с 26. IV по 20.VII; в Поволжье с 6.V по 10.VII;
в Ленинградской обл. с 2.V по 10.VII; в северных областях Европейской
части Союза с 1.V и по 15.VII.
Яйца кукушек сравнительно с размерами птицы мелки. Размеры: (5) 23,4—
21 х 18—16,6 мм (Джаркент, Шестоперов, 1928); (100) 20—26,4 х 15,2—18,8,
в среднем 23,05 х П,23 мм (Уайзерби, 1938)
Форма яиц — разнообразная, скорлупа толстая и различно окрашена.
По сходству формы и окраски с яйцами птиц-хозяев, яйца кукушек
разбиваются на три группы: 1 — полное сходство в окраске и форме, 2 — неполное
сходство в окраске и форме и 3 — отсутствие какого-либо сходства в окраске
и форме. Повидимому, такое различие и сходство с яйцами птиц, в гнездо
которых кукушка откладывает свои яйца, показывает, как давно паразитирует
кукушка на данном виде птиц. Также различно и отношение различных видов
птиц, а иногда и отдельных пар одного вида, к появлению у гнезда кукушки и
к найденному кукушечьему яйцу. Обычно большинство птиц остро реагирует на
появление кукушки у гнезда, стараясь отогнать ее прочь, но некоторые
относятся к появлению кукушки совершенно безразлично. Поставленные опыты
с чучелом кукушки (Промптов и Лукина, 1940) в Ленинградской обл.
показали, как реагируют птицы на выставленное чучело кукушки. Так, из 25 пар
подопытных пеночек-весничек некоторые пары при виде кукушки
приходили в сильное возбуждение — поднимали крик, старались отогнать кукушку,
ОБЫКНОВЕННАЯ КУКУШКА
447
другие слабо реагировали, немногие оставались совсем спокойными. Сходная
картина наблюдалась и у пеночек-трещоток и зарянок.
Отношение-воробьиных птиц к подложенному кукушкой яйцу разнообразно. Некоторые птицы
бросают гнезда с полными уже кладками (крапивники, некоторые славки),
другие делают новую подстилку в гнезде и ею прикрывают кладку с яйцом
кукушки (различные камышевки, горихвостки); большая часть видов воробьиных
птиц выбрасывают яйцо кукушки из гнезда. Наконец некоторые из воробьиных
Рис. 113. Птенец кукушки, выбрасывающий из гнезда других птенцов.
птиц принимают и высиживают яйца кукушки, причем иногда совершенно
отличные по окраске и форме от их яиц (завирушки, зарянки, жуланы, коньки
и т. д.).
Срок развития яиц кукушки очень короткий, короче срока насиживания
яиц многих мелких воробьиных птиц — 11—13 дней (13 дней—Шолей,
1923; 12x/8—Хейнрот, 1924—26; 11а/2 — Промптов и Лукина, 1940; 11 —
Венцель, 1914; Джоурден, 1925).
Вылупляется птенец голым, темнорозового цвета. В весе прибавляет очень
быстро, так что развитие перьев отстает. На пятый день открываются у
кукушонка глаза. Вес выведшегося птенца 2,9 г. Постэмбриональное развитие
кукушонка идет быстро, особенно первые два-три дня. Он перегоняет в росте
птенцов хозяев гнезда, даже в том случае, когда яйцо было подложено в гнездо
с начавшимся уже насиживанием и кукушонок вывелся несколько позже дру-
448
ОТРЯД КУКУШКИ
гих птенцов. Через несколько часов после вылупления (10—16 часов) у
кукушонка начинает проявляться рефлекс «выбрасывания» всех соприкасающихся
со спиной предметов. На спине развиваются осязательные тельца, которые
наиболее сильно развиты на нижней части спины, в особом углублении крестца.
При прикосновении к ним у кукушонка сейчас же появляется рефлекс
«выбрасывания». Кукушонок, загнув голову и шею вниз, широко расставив ноги и
опираясь головой о дно гнезда, пытается подлезть под выбрасываемое яйцо или
Рис. 114. Птенец обыкновенной кукушки.
птенца. Несколькими толчками тела он продвигает выбрасываемый предмет
на спину в ямку в области крестца. Крепко придерживая яйцо или птенца
далеко закинутыми на спину крыльями, кукушонок старается со своей ношей
добраться до края гнезда и при помощи резких толчков тела выбрасывает
находящийся на спине предмет через край гнезда. При этом иногда выбрасывает
предметы, весом превышающие в два раза вес самого кукушонка» Рефлекс
выбрасывания продолжается три-четыре дня. Изменения внешней
температуры сильно влияют на активность кукушонка. Наиболее активен, подвижен
и энергичен кукушонок при относительно низкой температуре 15°, в то время
как находящиеся в гнезде птенцы пеночек цепенеют при такой температуре.
При температуре в 25° кукушонок становится вялым и неактивным, а птенцы
пеночек наиболее оживленны.
Пеньки махов появляются у кукушонка на четвертые сутки, первое
разрывание пеньков отмечается на 9 сутки, развертывание маховых перьев начи-
ОБЫКНОВЕННАЯ КУКУШКА
449
нается на 12 сутки. В гнезде кукушонок остается 20 дней, но покидая гнездо
кукушонок еще плохо летает и только на 25—27-й день он летает хорошо. К этому
Рис. 115. Обыкновенная кукушка (самец и самка).
времени или дня через 2—4 птенец надевает полное оперение. При кормлении
птенца кукушки приемные родители засовывают голову в широко открытый
рот кукушонка. После вылета птенца из гнезда приемные родители
продолжают кормить его еще месяц или полтора.
Долго дискутировался вопрос о том, имеет ли кукушонок голос в первые
дни по вылуплении. Наблюдения Промптова и Лукиной (1940) показали, что
кукушонок подает голос с момента вылупления, при этом писк его в первые
29 Птицы СССР, т. Т
450
ОТРЯД КУКУШКИ
дни сходен с писком птенцов большинства воробьиных птиц. На 5-й день писк
уже переходит в громкий крик.
Линька. Последовательность смены нарядов: гнездовый — первый
годовой— второй годовой и т. д. Первый годовой наряд — окончательный (впрочем
имеющий некоторые особенности в деталях окраски). Линька взрослых—на
зимовках, между ноябрем и мартом. В литературе имеются указания, что
у взрослых кукушек после окончания периода размножения — в июне—июле—
имеется еще частичная линька; на материалах СССР это не подтверждается;
если такая линька у некоторых особей и бывает, то ее вероятно надо
рассматривать как ранние фазы обычной годовой линьки, течение которых прекращается
перед отлетом и возобновляется на зимовке. Гнездовый наряд молодые кукушки
сменяют на зимовке в те же сроки, когда проходит линька взрослых особей,
между ноябрем и мартом. У большинства особей после этого линяния остается
некоторое количество ювенальных перьев — среди верхних кроющих крыла и
в особенности среди второстепенных маховых (обычно второе и третье
второстепенное, иногда ряд перьев от второго до пятого); эти перья отличаются не
только изношенностью, но и рыжей окраской. Последовательность смены
первостепенных маховых — от заднего (внутреннего) края партии к переднему.
Питание. Пища, главным образом, различные насекомые и их
личинки, в небольшом количестве ягоды, крушина и др. и птичьи яйца. В выборе
насекомых кукушка неприхотлива; различные жуки, кобылки и особенно в
большом количестве бескрылые Acridium podestra и др., личинки Smerinthus populi,
S. ocellatus, Deilephila euphorbiae и т. д. и гусеницы; в особенности в большом
количестве поедают мохнатых гусениц. В 19 вскрытых желудках птиц, добытых
в Калининской обл., оказалось 427 гусениц и личинок, 129 разных жуков и в
небольшом количестве остатки птичьих яиц.
«Волосы» мохнатых гусениц задерживаются в кутикуле стенок желудка,
которая периодически сбрасывается; хитин кукушки удаляют из кишечного
тракта в виде погадок.
Полевые признаки. Средних размеров птицы с довольно длинным хвостом,
окрашенным у самцов и некоторых самок в темносерый цвет со светлыми
пятнами и в рыжий с широкой полосатостью у самок и молодых. Сверху птица
.обычно темносерая, реже рыжеватая, снизу поперечно полосата. На раскрытых
крыльях при полете хорошо заметна поперечная полосатость нижней
поверхности крыла. По окраске и размерам несколько напоминает ястреба-
перепелятника. Крик самца — хорошо известное всем кукование «ку-ку, ку-ку».
Крик самки сильно отличается от крика самца и бывает двух родов: довольно
часто издаваемая трель, особенно при встрече с самцом, «кли-кли-кли-кли» и
редко издаваемый глухой крик, похожий как бы на приглушенный хохот.
Описание. Строение и размеры. Первостепенных маховых — 10, первое
короткое, достигает приблизительно 2/а длимы крыла; самое длинное — третье. Формула
крыла: 3>2>4>5>6>7>8>1>9>10>11>12. Рулевых — 10, хвост закругленный
ступенчатый. Клюв средних размеров, слегка загнутый у вершины. Ноздри округлые,
открытые, и расположены у основания клюва. Размеры: длина тела самцов (30) 322—395, в
среднем 360,1 мм, самок (19) 310—360, в среднем 336,5 мм. Размах крыльев самцов (27) 540—
690, в среднем 621,22 мм; самок (17) 580—635, в среднем 581,78 мм. Длина крыла самцов
(86) 211,5—239,5, в среднем 224,1 мм, самок (68) 195—227, в среднем 212,2 мм. Вес около
100—115 г (самцы в среднем тяжелее).
Окраска. Птенец вылупляется голым, приблизительно через месяц надевает
первый ювенальный наряд (пухового нет). Молодые птицы — гнездовый первый (осенний)
наряд. Независимо от пола, осенью встречаются молодые кукушки, окрашенные в серый
и рыжий цвет, но и у серых птиц имеется рыжий оттенок. Серые птицы: спинная сторона
буровато-серая с рыжими поперечными пятнами, переходящими иногда в неполные рыжие
полосы; рыжий оттенок имеется на крыльях, голове, шее и хвосте. Рыжие птицы: спинная
сторона с широкой поперечной рыжей полосатостью, надхвостье с рыжими пятнами. Серые
и рыжие птицы имеют широкие белые вершинные окаймления перьев головы, спинной
стороны, крыльев и хвоста. На затылке и шее часто встречаются белые пятна. Брюшная сторона
ОБЫКНОВЕННАЯ КУКУШКА
451
с широкой темной поперечной полосатостью, особенно сильно развитой на горле и зобе, где
белые полосы совсем узки; горло и зоб кажутся темней по сравнению с остальной брюшной
стороной. Охристый оттенок на брюшной стороне более интенсивен у рыжих особей.
Маховые темнобурые с поперечными рыжими пятнами на наружных опахалах и на вершинных
частях внутренних и белыми на остальном протяжении внутреннего опахала. Рулевые
темнобурые с белыми вершинами, белыми редкими наствольными пятнами и пятнами по краю
опахал; иногда на внутренних опахалах, образующих полосы. Радужина бурая, края век
желтые. .
Молодые птицы — второй годовой наряд. Самец. Отличается от взрослого
буроватым налетом на спинной стороне тела и охристым налетом на зобе, иногда со слабой,
едва заметной поперечной полосатостью на нем и рыжими пятнами на наружных опахалах
первостепенных маховых. У молодых самок в сером оперении охристый налет на боках зоба
развит сильнее, чем у самцов и иногда заходит на бока шеи. Среди самцов рыжих птиц
не бывает.
Взрослые птицы. У самца верх тела темносерый, светлеющий к пояснице. Крылья
серые с бурым налетом, кроющие кисти (крылышка) белые с поперечной темной полосатостью,
иногда прерывистой. Хвост темносерый. На затылке чуть заметные более темные стволы
перьев. Горло, зоб и грудь светлосерые. Остальная брюшная сторона белая с темной поперечной
полосатостью, иногда с охристым оттенком. Ширина темных полос сильно варьирует.
Нижняя часть брюха и подхвостье светлоохристого цвета различной интенсивности с редкой узкой
темной полосатостью, иногда даже в виде черточек, отдельных крапин или отсутствует
совсем. Рулевые темносерые, темнеющие к вершинной части. Вдоль стволов пера расположены
редкие продолговатой формы белые пятна, более крупные на крайних рулевых, иногда
доходящие на них до края наружного опахала. Вершины рулевых перьев белые. Маховыетемно-
серые с бурым налетом. На внутреннем опахале первостепенных маховых в основных двух
третях его имеются белые поперечные пятна. Второстепенные маховые с белыми основаниями.
Радужина и края век желтые. У самок имеется два типа окраски оперения — серый,
сходный с оперением самца, но с буроватым тоном верхней стороны тела и головы и с охристым
налетом на зобе различной интенсивности у отдельных экземпляров. Второй тип окраски —
рыжий (гепатический). Верх головы и тела ржавчато-рыжие с черными поперечными
полосами различной ширины. Маховые бурые с ржавчато-рыжими пятнами и полосами. На
внутренних опахалах первых пяти или шести первостепенных маховых имеются белые
поперечные полосы, постепенно переходящие на остальных маховых в рыжий цвет. Поясница
и верхние кроющие хвоста ржавчато-рыжие, иногда с редкими черными отметинами и
белыми вершинами перьев. Рулевые ржавчато-рыжие .с белой вершиной и с широкими
черными поперечными полосами, из которых предвершинная полоса особенно широкая. Вдоль
ствола небольшие продольные пятна, на крайней паре рулей белые пятна развиты
сильней и на внешнем опахале доходят до его наружного края, заменяя частично рыжий цвет.
Брюшная сторона белая в черно-бурых поперечных полосах, более частых на горле и зобе
и почти исчезающих на подхвостье. Горло, зоб и подхвостье с охристым оттенком, иногда
охристый тон распространяется на всю нижнюю поверхность тела. Радужина у серых —
желтая, у рыжих — ореховая. Края век желтые.
Систематические замечания* С территории нашей страны описано несколько форм
кукушек, из которых дольше других признавались две формы johanseni и telephonus, в
особенности последняя. Просмотренный большой коллекционный материал из разных
местностей нашей страны не позволяет признать существование ни johanseni, ни telephonus.
Указанные для этих форм отличительные признаки — относительно узкие полосы на
брюшной стороне, мелкие размеры и оттенки в общей окраске оперения носят возрастной и
индивидуальный характер и не могут быть рассматриваемы как подвидовые различия.
Туркестанская обыкновенная кукушка Cuculas canorus sabtelephonus Zar.
Cuculus canorus subtelephonus. За рудный. Орнитологический Вестник, 1914 (№ 2), стр.
114, Туркестан, Закаспийский край.
С и н о н и м ы: С. с. kwenlunensis. Portenko. Mitt, aus d. Zool. Berlin, Bd. 17 (3), 1931,
стр. 417, оазис Ния, Кашгария.
Распространение. Ареал. В СССР по всей Средней Азии, как низменной,
так и горной, Туркмения, Таджикистан, Узбекистан, Киргизия и прилежащие
южные части Казахстана, Западная и северная границы изучены слабо.
Гнездится от восточного побережья Каспийского моря, Арало-Каспийских степей,
но доходит ли до Эмбы иМугоджар —неясно; на Южн. Урале —уже обык-
йовенная кукушка. Далее северная граница идет по северному побережью
Аральского моря; Аральские Кара-Кумы, Сыр-Дарья с прилежащими частями
Сары-су, озеро Тели-куль, Кара-Тау, р. Чу,р. Или и южное побережье Балхаша.
29*
ОТРЯД КУКУШКИ
Какая из них населяет ближайшие северные районы —. не выяснено, но северо-
восточнее Балхаша, в Тарбагатае и Алтае обитает уже обыкновенная кукушка.
Вне нашей страны населяет Кашгарию, Ала-Шань, Тибет, Гималаи, Афганистан,
Белуджистан, южн. и вост. Иран и Месопотамию. Зимовки расположены в вост.
Африке.
Характер пребывания. Гнездящаяся и перелетная птица.
В Среднюю Азию прилетает в первой половине апреля. Так, в долине среднего
Сумбара первые прилетевшие кукушки отмечены с б апреля 1937 (Исаков и
Воробьев, 1940). По Атреку и в Астрабадской провинции 14. IV кукушки уже
куковали (Шестоперов, 1926), под Ашхабадом первые прилетевшие кукушки
наблюдались в 1886г.—14.IV (Радде и Вальтер, 1889), в 1916г.— 7. IV, в 1926—
2. IV (Шестоперов, 1928), на Мургаб у Сары-язы прилетают 22. IV, у Мары —
27—28.1У(Лоудон, 1910). В Кара-Кумах—в апреле (Лоудон, 1913); последние
даты для прилета туркестанской кукушки очень поздние, повидимому
наблюдались не прилетные местовые, а пролетные обыкновенные кукушки. В Гис-
саре первая прилетевшая кукушка отмечена 8. IV (Иванов, 1940); в окрестностях
Кзыл-Орды в 1927 г.— 4.IV, в Джулеке в 1928г. — 30.111, в 1930г. —9.1V
(Спангенберг, 1936). Сроки осеннего отлета туркестанской кукушки не
изучены, имеющиеся наблюдения над пролетом и исчезновением последних
кукушек из Средней Азии могут относиться и к обыкновенной кукушке, которая
пролетает через Среднюю Азию. Пролет идет в течение сентября, последние
птицы наблюдаются до половины октября. На Чу последние кукушки
наблюдались в первой половине сентября (Гладков и Гринберг, 1932), под Гиссаром и
на озере Яшил-куль последние пролетные кукушки встречались 8—12. IX
(Иванов, 1940), на Аму-Дарье последние кукушки наблюдались в конце
августа (Гладков, 1932), у Ашхабада пролет кукушек наблюдался с конца августа,
в окрестностях ст. Душак —в начале сентября 1886 (Радде и Вальтер, 1889),
в Ахал-текинской долине в крнце августа в начале сентября (Заруд-
ный, 1896).
Биотоп. В низменных частях Средней Азии распределение кукушки
тесно связано с распространением главным образом дроздовидной камышевки.
В связи с этим, основной биотоп туркестанской кукушки в низменных частях
Средней Азии — камыши и тростниковые заросли. В этом биотопе также
большое значение имеют и некоторые виды жесткокрылых, обитающие
в огромном количестве в камышах и тростниках и служащие пищей для
кукушки. Кроме того, она населяет оазисы, сады, древесные насаждения по
долинам рек, заросли тамариска и других кустарников. В горы поднимается
довольно высоко, так на Гиссарском и Дарвазском хребтах доходит до 2900—
3000 м (Иванов, 1940), в Семиречье —выше границы елового пояса, до пояса
верхнего можжевельника, т. е. до 3000 м (Зарудный и Кореев, 1906). Населяет
окраины пустынь, но вглубь собственно пустынь не заходит. По восточному
берегу Каспия селится по бугристым пескам, закрепленным редкой
кустарниковой растительностью.
Численность. Очень многочисленна по речным долинам,
протекающим по низменным, равнинным местностям Средней Азии. В горных районах
в большом количестве кукушка встречается только в предгорьях; с высотой
численность ее резко падает. В небольшом числе населяет окраины пустынь,
в большом — культурные ландшафты.
Экология* Размножение. Период размножения начинается с
половины апреля и продолжается до конца июня и даже начала июля. Кладка яиц
происходит с конца второй половины апреля и идет до середины мая. Яйца
встречаются особенно часто в гнездах дроздовидной камышевки Acrocephalus,
aradinaceus, реже в гнездах других камышевок, а также в гнездах горной
трясогузки Motacllla cinerea, черного чеккана Saxicola caprata, черноголовой
ГЛУХАЯ КУКУШКА
453
трясогузки Motacilla flava, соловья Luscinia luscinia и др. Вылет птенцов из
гнезд происходит обычно с середины июня, летные молодые встречались с
конца июня и до конца июля (Мургаб — 27 июня, Аму-Дарья — 27—28 июля,
Чу — б августа).
Линька. Как и у обыкновенной кукушки, линька бывает раз в году, на
зимовках.
Питание. Различные крупные насекомые и в небольшом количестве
яйца птиц.
Полевые признаки, В поле не отличима от обыкновенной кукушки. Голос
также похож на голос обыкновенной кукушки.
Описание. Размеры и строение. Размеры несколько мельче. Длина тела
самцов —(3) 335—360 мм, в среднем 354 мм, самок (3) 326—340, в среднем 331,6 мм;
размах самцов (3) 546—630, в среднем 586,3 мм, самок (3) 520—602, в среднем 561 мм; длина
крыла самцов (18) 206—222, в среднем 215,3 мм, самок (13) 191—230, в среднем 203,5 мм.
Окраска. Молодые птицы от молодых обыкновенных кукушек отличаются
более светлым общим тоном оперения.
Взрослые птицы — самец и самка отличаются от обыкновенной кукушки несколько
более светлым оперением.
Систематические замечания. Кашгарские птицы были выделены в особую форму С. с.
kwenlunensis (Портенко, 1931) из-за более крупных размеров и более светлой окраски
оперения по сравнению с sabtelephonus. Но при сравнении размеров kwenlunensis с размерами
subtelephonus выяснилось, что реальных различий нет. Длина крыла среднеазиатских самцов
и самок приведена выше. У кашгарских самцов —крыло 200—228, в среднем 216,4 мм;
у самок 180—218, в среднем 202,6 мм, реальных различий нет. Также и по окраске
кашгарские птицы не отличимы от туркестанских.
3, Глухая кукушка Cacalus saturatus Blyth
Cuculus saturatus «H о d g s о n». Blyth. Journ. As. Soc. Bengal, 1842, стр. 942. Непал,
Синонимы Cuculus optatus. Gould. Pr. Zooi. Soc. of. Lond. 1845, стр. 18, Сев.
Австралия. — Cuculus peninsulae. Stejneger. Bull. U. S. Nat. Mus. № 29, 1885, стр 227,
Камчатка.
Русское название. По голосу птицы.
Распространение* Ареал. Лесная полоса Европы и Азии. Европейская
часть СССР, Сибирь (к северу до предела лесной растительности) на восток до
Камчатки, Сахалина и Курильских о-вов; Япония (Хоккайдо, Хондо);
Корея; Манчжурия; Китай; о. Формоза; Бирма; Ассам; южные склоны
Гималаев; быть может Афганистан. Зимовки: южн. Индия, Бирма, Малайский
п-ов, Андаманские и Никобарские о-ва, Борнео, Суматра, Ява, Малые
Зондские о-ва, Целебес, Молукские о-ва, архипелаг Бисмарка, Соломоновы о-ва,
Новая Гвинея, Вайгеу и Австралия.
Характер пребывания. Гнездящаяся и перелетная птица.
Биотоп. Большие, высокоствольные, сырые леса, как хвойные, так и
смешанные, реже лиственные леса и заросли кустарников.
Подвиды и варьирующие признаки. Два подвида, различающихся размерами и
экологией: С. s. saturatus Blyth, 1842 — южн. Гималаи, Ассам, Бирма, Южн. Китай и
о. Формоза; С. 5. horsfieldi Moore, 1857 — от с.-в. Европы до Анадыря; Курильских о-вов
и Сев. Китая.
Северная глухая кукушка Cuculus saturatus horsfieldi Moore
Cuculus horsfieldi. Moore and H о rsf i e Id. Catal. Birds Hon. East Ind. Сотр. 1856—1858
(1857), стр. 703, Ява.
Распространение. Ареал. В СССР в северо-восточных областях
Европейской части, по всей Сибири на восток до Камчатки, Приамурья и Приморья;
Командоры, Сахалин и Курилы; вне пределов Союза—в сев. Японии
(Хоккайдо, Хондо), Корее, Сев. Китае, Манчжурии и сев. Монголии. Зимовки в южн.
454
ОТРЯД КУКУШКИ
Индии, на Филиппинских и Зондских о-вах, Целебесе, Молуккских о-вах,
архипелаге Бисмарка, Соломоновых о-вах, Новой Гвинее, Вайгеу и в
Австралии (впрочем, разграничение области зимовок подвидов глухой кукушки
выяснено плохо).
Карта 77. Распространение глухой кукушки Cuculus saturates
1 — С. 8. horsf ieldi, 2 — С. 8. saturatus; a — граница гнездовий, г — зимовки.
Детали распространения в Европейской части СССР таковы. На западе
отмечена между неверным побережьем Мурмана и Белым морем (вероятно
гнездится, Плеске, 1928), в Латвии у Лабана (Бутурлин, 1936); в Смоленской обл.,
(Граве, 1926), в Калининской обл. (Осташков, в июне—июле, вероятно на
гнездовье, Данилович, 1934); на севере в области Коми к югу от Сыктывкара,
на Сев. Урале (Шугор, Сев. Сосьва), в области Пермского Урала, южнее в
Кировской обл., в Ярославской (Некоузкий район, экз. Зоолог, музея Моск.
университета), Костромской, Горьковской обл., в Татарской республике, в
среднем течении Волги в Алатырском районе, в Городищенском районе Пензенской
обл. Наблюдалась в Московской обл. и добыта в Хвалынском районе
Саратовской обл., в Башкирии, на Южн. Урале до Зилаира и восточного склона
Урал-Тау (Кириков). В Сибири на севере отмечена на нижней Оби (до Боль-
ГЛУХАЯ КУКУШКА
455
ших Алтынских юрт), на Енисее почти до полярного круга (Тугаринов и
Бутурлин, 1910), далее к востоку северная граница неясна и совпадает, по-
видимому, с границей высокоствольного леса, гнездится на Вилюе, в окресг-
ностях Олекмы, на средней Лене, в Якутске, по Алдану, Курейке, на
Индигирке; на Анадыре — отмечена в западной его части к югу от р. Анадырь,
отсутствуя в приморской полосе. На-юге Сибири встречена у Златоуста и
Троицка, у Петропавловска, Омска, добыта на озере Салеты-денгиз, где быть
может гнездится (экз. в Зоолог, музее Моск. университета), у Томска, в Салаир-
ском кряже в Кузнецком районе, в борах приалтайских степей, в центральном,
с.-в. и с.-з. Алтае, быть может гнездится и в зап. Алтае, Зайсанской котловине,
на Саянах, в Прибайкалье до Даурии, далее на Амуре. На крайнем востоке —
в Коряцкой Земле (Пенжина, Дементьев, 1940), по Охотскому побережью, на
Шантарских о-вах (Дулькейт и Шульпин, 1937), на Камчатке и по всему
Приморью; на Сахалине, Курильских и Командорских о-вах.
Область распространения глухой кукушки, вследствие ее скрытного образа
жизни, изучена довольно слабо. Нахождение ее в тех местах, где она ранее не
была обнаружена, скорее всего можно отнести не за счет расширения ее ареала,
а за счет более детального изучения авифауны страны.
Характер пребывания. Гнездящаяся и перелетная птица. На
перелете единичные экземпляры встречаются далеко от мест гнездовий и
зимовок. Добытая в начале 1884 г. на Сев. Кавказе близ Кисловодска, кукушка,
которая была определена как глухая, но позднее из-за небольших ее размеров
Сатунин (1907) определил как С. poliocephalus, повидимому была залетной
глухой кукушкой. 7 июня 1907 г. кукушка была добыта на нижнем течении Урала
близ Гурьева у с. Калмыкова. Весною на Иргизе, 15.V и 4. IX, у Кустаная
18.V. 1934, близ Наурзума. В Таджикистане у Вишкента — 24. IV. 1914;
(Зоолог, музей Моск. университета), в окрестностях Сталинабада 24.V. 1935
(Иванов, 1940); в Хатаке, 15.V. 1910 г.; в Кызыл-Кумах у Дарбаза
14.V.1943 г. (Мекленбурцев). Зарудный и Билькевич (1918) указывали,
будто бы глухая кукушка гнездится и пролетает в области верхнего течения
Аму-Дарьи между Келифом и Кировабадом. Гнездование глухой кукушки в
этих местах невероятно, скорее всего вопрос идет о поздно пролетных
птицах, если только они были определены правильно.
Данных о времени появления на местах гнездований и об ее отлете почти
совсем нет, В Молотовской обл. прилет и отлет глухой кукушки совпадает
с прилетом и отлетом обыкновенной кукушки, т. е. в среднем 10—12.V (Ушков,
1927); на Сев. Урале первый крик слыхали 11.V (Портенко, 1937). В
Кузнецкую степь прилет отмечен в средине мая, в окрестностях Семипалатинска
встречается в небольшом числе регулярно на весеннем и осеннем пролете; на
Алтай прилетает поздней обыкновенной, первый крик на Онгудае услышан
29.V (Сушкин, 1939); в области Енисея также прилетает несколько позднее
обыкновенной, у Красноярска —»во второй трети мая, а у полярного круга
в первых числах июня (Тугаринов и Бутурлин, 1910). У устья Вилюя, на
Лунхе, в 1854 г. первое кукование услышано 30.V, около Иркутска — 21.V,
на Шилке 20.V (Маак, 1859). На Зее у Бомбака — 28.V (Дорогостайский,
1915). В Южно-Уссурийском крае первая встреча глухой кукушки отмечена
21.V.1938 г. (Спангенберг), в окрестностях Охотска 17.V (Харитонов, 1915),
в Анадырский край прилетела в 1912 г. 30.V (Портенко, 1939).
Биотоп. Высокоствольные участки старого леса: хвойные, еловые,
елово-пихтовые, пихтовые, сосновые, кедровые, смешанные —. с примесью
березы, осины, клена, липы, лиственницы и ильма. Изрезанные обширными
моховыми болотами, оврагами, реками пространства; невысокие лесистые
склоны предгорий и гор. Реже в чисто лиственных лесах и кустарниках. На
Южн. Урале, в Зап. Сибири и на Анадыре — по колкам, приречным таль-
456
ОТРЯД КУКУШКИ
никам и кустарниковым зарослям. В горы идет в вертикальном направлении
ниже обыкновенной кукушки, на Алтае —до 1900 м (Сушкин, 1938).
Ч и с л е н н о t т ь. Обыкновенна, но немногочисленна в Горьковском
крае; в Татарской распублике обычна, и в таежных участках мало уступает
численности обыкновенной, в лесостепи редка, но количественно и тут повидимому
стала возрастать. В Кировской обл. по численности уступает обыкновенной.
В области Пермского Урала настолько часто встречается, что трудно определить,
какая из кукушек более многочисленна; на Сев. Урале—чаще обыкновенной;
в окрестностях Омска —наравне с обыкновенной. В окрестностях Томска
глухая кукушка нередка, но летом 1928 г. наблюдалась в огромном количестве и
встречалась значительно чаще, чем обыкновенная, особенно в большом
количестве по опушкам хвойных лесов (Белышев, 1929). В Горной Шории, в
Кузнецкой степи, в Салаире обычна повсюду, но количественно уступает
обыкновенной. Для различных местностей Салаира была определена плотность
населения глухой кукушки: в сосновых борах она равняется 1 особи на
2 кв. км, в притаежной полосе вместе с тайгой 1 особь на 1 кв. км, по р. Сюхта
на расстоянии 2 кв. км отмечена 4 раза, в то время как обыкновенная — 2 раза
(Хахлов, 1937). В Забайкалье (р. Ингода и ее притоки), по свидетельству
Н. А. Гладкова, обыкновенна в высокоствольной горной тайге, тогда как в
долинах рек ее нет, в последних она заменяется обыкновенной кукушкой. На Алтае
обыкновенна в северных и северо-восточных предгорьях, там, где имеется
сплошная тайга, спорадична в северо-западном и центральном Алтае, где
лесной покров разбросан пятнами. В Красноярском районе так же обычна,
как и обыкновенная. В Якутском округе обыкновенна, но встречается
неравномерно, более обычна в глухих таежных участках и многочисленна по сырым
ельникам Алдана (Иванов, 1929). В Анадырском крае обыкновенна и
многочисленна. В Приморском крае густо населяет смешанные леса приречной
полосы, обыкновенна в широколиственных лесах сопок и редка в глухих таежных
массивах (Спангенберг).
Экология. Размножение. Данных о поведении глухой кукушки
в брачный период и о размножении ее почти нет. Выбор самкой гнездового
участка, как и у обыкновенной кукушки, зависит от наличия мелких воробьиных
птиц, в гнезда которых глухая кукушка откладывает свои яйца.
На Сев, Урале первый крик глухой кукушки услышан 11.VI, и 14.VII
она уже кончила куковать, одновременно с обыкновенной. На Алтае первое
кукование услышано на Онгудае 29.V, в Кузнецкой степи — 18.V.1928, у Бом-
бакана Зее 28.V, в устье Вилюя—30.V, в окрестностях Иркутска —21.V, на
Шилке 20.VI.
Яйца глухой кукушки найдены на Анадыре в 2 гнездах пеночки-таловки,
причем в одном гнезде 27.VI, кроме кукушечьего яйца, было 3 яйца пеночки.
Все яйца по окраске были похожи на яйца пеночки-таловки, но почти вдвое
крупнее (Портенко, 1939). В южн. Приморье, на Имане 8.VL1939
наблюдалось как глухая кукушка несколько раз подлетала к гнезду древесной
трясогузки Dendronanthus indicus, но отгонялась хозяевами гнезда (Спангенберг).
В Даурии было найдено яйцо глухой кукушки в гнезде лесного конька Anthus
trivialis (Тачановский, 1893). Наиболее часто хозяевами яиц глухой кукушки
бывают пятнистый конек Anthus hodgsoni и светлоголовая пеночка Phylloscopus
occipitalis corcnatus. В Гималаях найдены яйца в гнездах пеночек Phyltoscopus>
Orthomus, Enicurus, scfiistaceus. В Китае — в гнездах черноголового чеккана
Saxicola torquata и Pyctoris. Окраска яиц различна и часто сходна с яйцами
хозяев гнезда. В Даурии найдено яйцо — блестящего серого цвета с фиолетово-
серыми и коричневыми пятнами на широком конце (Тачановский, 1893). В
Гималаях найдены белые яйца, похожие на яйца пеночек, грязновато-зеленого
цвета и светлорозового. В Китае, кроме обычной светлой окраски, встречаются
ГЛУХАЯ КУКУШКА
457
яйца серовато-голубые с красными
пятнами и красноватые с тёмнокрасными
пятнами.
Размеры: (34) 19,7—25,4х 13—-16,4,
в среднем 21 х 13ш*(Бекер, 1927).
Линька. Проходит на зимовках,
детали неясны, смена нарядов, как у
обыкновенной кукушки.
Питание. Различные крупные
насекомые и их личинки. У птицы,
добытой на Анадыре 12.VI, в желудке найдены
части жуков, ос, двухкрылых и много
волос гусениц (Портенко, 1939).
Полевые признаки, В повадках,
поведении, полете, окраске оперения не
отличима в поле от обыкновенной
кукушки. Любит держаться в кроне верхнего
или среднего яруса крупных деревьев.
В период размножения ведет особенно
скрытный образ жизни, придерживаясь
глухих мало доступных местностей. Крик
самца похож на голос удода, но
несколько ниже тоном и глуше. Его можно
передать как глухое «ду-ду, ду-ду» или
«туту, ту-ту», повторяемое шесть — восемь
раз подряд. Этому основному крику
предшествует вступление из быстро
чередующихся четырехсложных «ду-ду,
дуду». Самец чаще кричит на утренней и вечерней заре, сидя обыкновенно на
сухом сучке высокого дерева и, реже, во время полета, при этом широко
распускает свой хвост. Голос самки похож на голос самки обыкновенной кукушки.
Рис.
116. Глухая кукушка (взрослый
самец и молодая птица).
458
ОТРЯД КУКУШКИ
Описание. Размеры и строение. Меньше обыкновенной кукушки. Первое
первостепенное короткое, немного длиннее половины крыла. Формула крыла: 3>2==
= 4>5>6>7>1>9>10. Клюв несколько массивней, выше и с более резким изгибом в
вершинной части, чем у обыкновенной кукушки. Размеры: длина тела самца (1 экз.)
340 мму самки (1) 325 мм, размах крыльев самца (1) 540 мм, самки (1) 550 мм. Длина крыла
самцов (24) 177—217, в среднем 205 мм; самок (12) 175—210, в среднем 195,2 мм.
Окраска. Гнездовый наряд также, как и у обыкновенной, бывает рыжего и серого
типа окраски. От молодых обыкновенных кукушек хорошо отличается своим темным низом
тела, особенно зоба и груди. Подбородок, зоб и грудь черноватые с редкими мало заметными
прерывистыми или белыми, или светлоохристыми полосами. Брюшная сторона с сильным
охристым налетом. Спинная сторона буровато-черная с широкими белыми окаймлениями
перьев. Радужина бурая. Первый годовой наряд несколько светлее, чем гнездовый.
Белые окаймления перьев уже имеются только на вершинной части пера. Подбородок,
зоб и грудь с широкими поперечными темными и белыми или светлоохристыми полосами.
Рыжие самки похожи на взрослых, отличаясь несколько более тусклым тоном оперения и
более темной брюшной стороной. Взрослая птица — самец похож на самца обыкновенной
кукушки, отличаясь более темной общей окраской оперения. Спинная сторона темносе-
рая, зоб и грудь более темного серого цвета, чем у обыкновенной кукушки: поперечные
полосы на брюшной стороне обычно шире, темнее и грубее, охристый тон на брюхе и под-
хвостье хорошо выражен. Кроющие кисти (крылышка) беловаты или светлоеливочны, без
поперечных темных полос, иногда с едва заметной темной крапчатостью. Нижние
кроющие крыла с слабо заметной поперечной полосатостью. Надклювье темнокоричневое,
подклювье у основания желтое, в вершинной части коричневое. Радужина темнобурая,
веки желтые. Ноги желтые. У самок, как и у самок обыкновенной кукушки, существует
два типа окраски: серый и рыжий. У рыжих самок темные полосы на верхней стороне
тела шире и общий тон темней, чем у самок обыкновенной кукушки. У серых на верхней
стороне тела развит довольно сильный бурый оттенок, в остальном они сходны с^самцами.
Рыжий тон имеется на затылке, верхней стороне шеи и по бокам зоба.
4. Малая кукушка Cuculus poliocephalus Lath,
Cuculus poliocephalus. Latham. Ind. Orn. 1, 1790, стр. 214. Индия.
Распространение. Ареал. Вост. Азия — Южно-Уссурийский (Край,
Япония, Корея, Манчжурия, Китай до юга Ганьсу, Сычуаня и Юннаня,
на запад до сев. Бирмы и Брамапутры; Гималаи, вост. Афганистан,
Большие и Малые Зондские о-ва; Мадагаскар.
Характер пребывания. На севере перелетная (зимует в ю. Азии)
на юге оседла.
Б и от о п. Высокоствольные леса.
Подвиды и варьирующие признаки. Четыре подвида, различающихся окраской,
размерами и экологией: Ср. poliocephalus Lath. 1790 — Южно-Уссурийский край, Япония,
Корея, Манчжурия, зап. и центральный Китай, сев. Бирма, Гималаи и вост. Афганистан;
С p. insulindae Н а г t e r t, 1912 — Борнео; С p. lepidus M u 11 е г, 1845 — Суматра,
Ява Бали, Ломбок, Сумбава, Флорес, Пантар, Сумба и Тимор; Ср. rochii H а г 11 a u b,
1862 — Мадагаскар (встречены молодые в Африке до Уганды и вост. Конго и на о.
Маврикия).
.Китайская малая кукушка Cuculus poliocephalus poliocephalus Latham
Распространение. Ареал. В СССР — Южно-Уссурийский 'край, где
найдена на гнездовье на Имане, в окр. с. Славянска, по нижнему течению Суй-
фуна, в районе Сучана (Шульпин, 1927), в окр. Владивостока (добыта
25.VI. 1919 Фирсовым), в Сидеми и на р. Пачихеза близ ст. Пачихеза
(Воробьев, 1948); Япония (Хондо, Сико и Кюсю); Корея, Манчжурия, зап. и
центральный Китай до южн. Ганьсу, Сычуаня и Юннаня, сев. Бирма на
запад до Брамапутры. Гималаи от вост. Ассама до Гильгита и Кашмира и
вост. Афганистан. Зимовки — Индо-Китай, южн. Индия, Цейлон, Андаманские
о-ва, о. Бонин, о. Формоза и^о-ва Риу-Киу.
МАЛАЯ КУКУШКА
459
Характер пребывания. В СССР гнездящаяся и перелетная птица.
В Приморье прилетает в конце мая, в 1938 г. первый крик малой кукушки услы-
шен на Имане 29.V (Спангенберг), близ Владивостока, ст. Пачихеза-з-в
последних числах мая (Воробьев, 1948); в горные районы Индии —в конце апреля и
в начале мая; улетает в сентябре и в начале октября.
Карта 78. Распространение малой кукушки Cuculus poliocephalus
1 — С. p. poliocephalus, 2 — С. p. insulindaee, 3 — С. p. lepidus, 4 — С. p. rochli; а
—граница гнездовий, г — зимовки (цифры в кружках — зимние местонахождения
соответствующих подвидов).
Отмечены три сомнительных случая залета: дважды — в Среднюю Азию
(Чарджоу) и один раз на Сев. Кавказ, в начале мая 1884 близ Кисловодска.
К сожалению, экземпляры не сохранились, скорее всего птицы были ошибочно
определены, так как позднее малая кукушка так далеко к западу никем
не наблюдалась.
Биотоп. В Южно-Уссурийском крае—большие светлые лиственные леса,
состоящие преимущественно из березы, граба, клена, дуба и т. д., с
разнообразным кустарником, разреженные полянами, болотами, лугами, выгонами,
оврагами и ручьями, в более южных частях ареала —»вечнозеленые леса. На
Гималаях подымается в горы до 2000 м и выше.
Численность. В Южн. Приморье местами обычна, но
распространена спорадично.
Экология. Размножение. Данных о размножении малой кукушки
в СССР почти нет. Крик ее слышен с конца мая по июль, в начале июля кричит
еще очень интенсивно, особенно в первую половину дня, иногда даже в самые
жаркие часы (Воробьев, 1948). В Индии гнездовыи сезон начинается с конца
мая и идет до конца июля. Яйца откладывает в гнезда Cettia, Horornis,
460
ОТРЯД КУКУШКИ
Tesia castaneiventris, Phylloscopus, Parus, Saxicola, Pnoepyga и др. Яйца двух
типов окраски — или белые, или от розовато-каштанового до темношоколад-
ного цвета.
Размеры: (30) 19,5—23,0х 13,5—-15,5, в среднем 21,0 х 14,2 мм (Бекер,
1927).
Линька. Не изучена.
Питание. Различные насекомые и их личинки. В желудке птицы,
добытой Воробьевым 11 июня в южном Приморье (ст. Пачихеза), найдены мелкие
гусеницы, сем. пядениц и совок (Geometridae и Noctuidae).
Рис. 117. Малая кукушка.
Полевые признаки. В поле по внешнему виду отличается от обыкновенной
кукушки более мелким размером, а также по голосу. Очень осторожная птица,
любит держаться среди листвы кроны дерева. Голос — мелодичная трель
«ле-ля-ла-лю» (Спангенберг) или «ти-тю-тю» (Воробьев, 1948), издаваемая
птицей с различными интонациями и ударениями.
!&"" Описание. Строение и размеры. Первое маховое короткое,
достигает только половины длины крыла, самое длинное—3-е. Формула крыла:
3>2:>4>5>6>7=1>8>9>10. Размеры: длина тела одного самца 272мм, длина
крыла самцов и самок (4) 156—165 мм} в среднем 160 мм; 152—171 мм (Бекер, 1927),
Окраска. Гнездовой наряд. Спинная сторона и верх головы бурые с белыми
каймами перьев; на лбу и затылке имеются пятна, образуемые белыми перьями; на боках
шеи — рыжая поперечная полосатость; горло, зоб, грудь с более охристым налетом и
узкими поперечными белыми полосами и широкими темными; остальная брюшная сторона —
с редкими широкими поперечными темными полосами; подхвостье светлоохристое.
У взрослых птиц спинная сторона тела и верх головы шиферно-серые; брюшная,
сторона — белая, с редкой узкой прерывистой черноватой поперечной полосатостью;
горло, зоб — серые с легким охристым налетом. Маховые темнобурые с сероватым нале-
ИНДИЙСКАЯ КУКУШКА
461
том. На внутренних опахалах первостепенных маховых, исключая вершинную часть пера,
белые продолговатые, поперечные пятна. Рулевые буровато-серые с редкими белыми иа-
сгвольными пятнами, которые на крайних рулевых образуют прерывистую поперечную
полосатость. Радужина коричневая, веки желтые, клюв черновато-бурый, у основания —
желтый; ноги коричнево-желтые.
5. Индийская кукушка Cuculus microptems Gould
Cuculus microptems» Gould. Pr. Zool. Soc. of London, 1837, стр. 137. Гималаи
Распространение. Ареал. Среднее течение Амура, Манчжурия, Китай,
Индо-Китай, Бирма, Индия, Цейлон, Ява, Борнео и Суматра.
Рис. 118. Индийская кукушка.
Характер пребывания. Северные птицы перелетны (зимуют на
Филиппинах, Борнео, Суматре, Яве, о. Банка и Архипелаге Тысяча Островов),
южные — оседлы. Биотоп. Большие высокоствольные сырые леса по
долинам рек.
Подвиды и варьирующие признаки. Два подвида, различающихся размерами и эколсг
гией: С. т. microptems Gould, 1837 — от среднего течения Амура, Манчжурии и Китая
до Индо-Китая, Индии и Цейлона; Ст. concretus M u 1 1 е г, 1845—Борнео, Ява и Суматра.
Обыкновенная индийская кукушка Cuculus microptems microptems Gould
Распространение. Ареал. В СССР северная граница проходит по
среднему течению Амура: взрослый экземпляр добыт 13.VI. 1857 г. на Амуре
близ Благовещенска (Радде, 1863); второй экземпляр — взрослый самец —
15.VL1928 г. по среднему течению Амура у Кумары (Штегман, 1931) и
третий — молодая птица 26.VI. 1946 г. на р. Архара, северном притоке среднего
462
ОТРЯД КУКУШКИ
течения Амура, к востоку от Бурей (Куренцовым). Встречается также в горах
Бурей и на восточных склонах Хингана. В Приморье впервые добыта Вог
робьевым у ст. Губерово 22.VI. 1949 г. и наблюдалась им 17.VI на нижнем
течении р. Бикина. Неоднократно в гнездовое время добывалась в
Манчжурии, где, повидимому, гнездится; встречена в Корее. Гнездится в Китае or
Шеньси, Чжили и Жехола до ГуандуняиГуаньси; вИндо-Китае, Бирме; в
Индии до Гималаев и Бенгалии (в сухих частях центральной Индии, в
Пенджабе, Синде и Раджпутане отсутствует) и на Цейлоне.
Характер пребывания. Особи, гнездящиеся на Амуре, в
Приморье, в Китае, в Манчжурии — перелетны; в других частях ареала — оседлы.
Биотоп. Сырые, большие высокоствольные леса. На Амуре обычна в сырых
лиственных лесах по
долинам рек, в
Приморье—в дубовых
лесах по небольшим
сопкам и в долинах
рек, в Гималаях
поднимается до 2000—
2§00 м.
Численность.
На Амуре местами
нередка, в Индии —
обычна.
Экология.
Размножение. О
размножении индийской
кукушки у нас нет
никаких данных и
вообще сведений о ее
размножении очень мало.
В Гималаях период
размножения
начинается со второй
половины мая и
заканчивается в июне; в
равнинной Индии с
апреля и
продолжается до августа
включительно. Яйца
индийской кукушки
найдены в гнездах: Trochalopterum lineatum, Hodgsonius phoenicuroldes,
Larvivora brunnea и Saxicola torqmta indica. Окраска яиц обычно голубая.
Размеры: (25) 22,8—26,0 х 17—20, в среднем 23,7 х 18,1лш (Бекер, 1927),
Линька. Не изучена. Питание. Различные насекомые и их
личинки.
Полевые признаки. Трудно отличима от обыкновенной кукушки, но по
сообщению натуралистов, слышавших ее, хорошо отличается голосом.. Голос.
Не описан.
Описание. Размеры и строение. Первое маховое немного короче длины
половины крыла, самое длинное 3-е маховое. Формула крыла 3>2 = 4>5>б>7>1>8>
>9>10. Размеры: длина крыла (2) 200—205 мм (Зоологический музей Моск. уииверси?
тета); 169—209 мм (Бекер, 1927).
Окраска. Гнездовый наряд. Темнобурый с рыжими светлеющими к наружному
краю каймами перьев. Пятна по краям рулевых рыжие. На голове и шее — белые или
охристые поперечные полосы, края перьев рыжие; брюшная сторона охристо-беловатая с широ-
КЗ г
Карта 79.
Распространение индийской кукушки Cuculus mi-
cropterus
1 — С. m. micropterus, 2—С. m. concretus; a — граница гнездовий,
г — зимовки (цифры в кружках обозначают номер зимующих подвидов),
в — изолированные находки в гнездовое время.
ШИРОКОКРЫЛАЯ КУКУШКА
463
кими темными поперечными полосами и с сильным охристым налетом. Первый годовой наряд
как у взрослых, только с рыжими окаймлениями перьев мантии и крыльев. На шее иногда
сохраняются поперечные рыжие полосы. Белые пятна на рулевых окаймлены рыжим, пятна
по краю рулевых рыжие. Взрослая птица, самец: спинная сторона тела серая с
сильным бурым налетом; только голова, шея и зоб чисто серые; брюшная сторона
охристо-беловатая с широкими темными, черновато-бурыми поперечными полосами; рулевые темнобурые
с черной предвершинной полосой. Самка: на горле, зобе и груди — охристый оттенок
выражен сильней, чем у самцов. У обоих полов радужина темнокоричневая, веки желтовато-
зеленые; клюв темнобурый, надклювье буровато-черное со светлым основанием, подклювье
зеленовато-бурое с желтоватой вершиной; ноги желтые или желтовато-оранжевые.
Карта 80. Распространение ширококрылой кукушки Cuculus fugax
Г~— С. f, hyperythrus, 2 — С. f. fugax, 3 — С. f. nlsicolor, 4 — С. f. pectoral is,
a — граница гнездовий, г — зимовки (цифры в кружках — зимовки
соответствующих подвидов).
6. Ширококрылая кукушка Cuculus fugax Horsf.
Cuculus fugax. H о r s f i e 1 d. Trans. Linn. Soc. XIII, 1821, стр. 178, Ява.
Распространение. Ареал. Приморье, Япония (Хондо), Манчжурия,
Китай, Гималаи, Индо-Китай, Малайский п-ов и острова Индо-Австралийского
архипелага.
Характер пребывания. Северная форма перелетная (зимовки
в ю.-в. Азии), южные формы оседлые или совершают более или менее
значительные кочевки.
Подвиды и варьирующие признаки. Четыре подвида, различающихся между собой
размерами, окраской и экологией: С/, fugax. Horsf, 1821 — Сиам, Малайский п-ов,
Суматра, Ява, Борнео, о-ва Бату, Сиберут, архипелаги Рис, о. Банка, о. Биллитон; о-ва
Каримата; С. /. nisicolory В 1 yth, 1843 — Гималаи от Непала и Сиккима на западе, до
Ассама, Бенгалии, Бирмы на юге; Индо-Китай, за исключением Сиама; Южн. Китай
(Гуаньси); С. /. hyperythrus Gould, 1856— Приморье, Япония (Хондо); Манчжурия, Сев.
Китай, Чжили, низовья Янцзе; С. /. pectoralis Cabanis et Heine, 1862 — о-ва Лкь
сон, Миндоро, Цебу и на Филиппинском архипелаге.
464
ОТРЯД КУКУШКИ
Манчжурская ширококрылая кукушка Cuculux fugax hyperythrus Gould
Cuculus hyperythrus. Gould — Proc. Zool Soc. Lond. 1856, стр. 96, Шанхай.
Распространение* Ареал. В СССР — в Приморье найдена на нижнем
течении Амура, на р. Яза-бира между Горин и Хунгари (добыта 14. VIII. 1855,
Маак, 1859), в с.-в. части хребта Сихоте-Алинь у устья р. Ботчи (два молодых
экземпляра 28.VIII, Емельянов, 1929); Судзухинский заповедник на р. Супу-
тинке, притока р. Суйфуна (взрослый самец 18.VI.1940 г. добыт Ивановым)
и в Супутинском заповеднике (20.IX. 1947 г., Воробьев, 1948); в верховьях
р. Курсый— Мурьи (21.VI, Янковский), на р. Имане и в Иманском районе,
(Спангенберг), Муравьева-Амурского (Воробьев, 1948). Гнездится в
Манчжурии в Сев. Китае (Чжили), на юге найдена в Гуаньси (Кольтоф, 1932), в сев.
Японии (Хондо), встречена на о-вах Сикоку, Риу-Киу ив Корее, но гнездование
там не доказано. Зимовки: Южн. Китай, Филиппины, Борнео, сев. Целебес.
Рис. 119. Ширококрылая кукушка.
Характер пребывания: гнездящаяся и перелетная птица.
Б и о т о п: в Приморье—тайга, изрезанная глубокими оврагами и реками,
состоящая преимущественно из кедра и пихты с незначительной примесью
лиственного леса и смешанные большие леса из дуба, клена, ясеня, граба, кедра,
ели и т. д.
Численность. В Южн. Приморье обыкновенна, в особенности на
среднем и верхнем Имане и у р. Супутинки (Воробьев, 1948), в остальных
частях Приморья редка и спорадична/
Экология. Размножение. Данных о размножении очень мало, из
наших пределов совсем нет. В Бирме и Сиккиме крик самца С. /. nisicolor
слышен с конца мая и до конца августа, однажды яйцо ширококрылой кукушки
найдено 14. IX (Бекер, 1927).Яйца ширококрылой кукушки найдены в гнездах
Stachyridopsisrififrons, Pellorueum, Proparas nipalensis/Dromocataphusassamensis
и особенно часто в гнездах Heteroxenicus nepalensis. В Японии чаще всего яйца
ШИРОКОКРЫЛАЯ КУКУШКА
465
ширококрылой кукушки встречаются в гнездах: Larvivora cyane. Яйца окрашены
от серовато-зеленого до желтоватого или даже светлокоричневого цвета с
красновато-коричневыми пятнами. Размеры не выяснены.
Линька. Плохо изучена. У экземпляра, добытого 20. IX, линька не
закончена —мелкие перья еще в чехликах (Воробьев, 1948).
Питание. Мелкие беспозвоночные преимущественно различные
насекомые и их личинки — бражник Smerinthas, пилильщики Tentheredinidae,
шелкопряд Dendrolimus.
Полевые признаки. Скрытная и осторожная птица, любит держаться в
кронах больших деревьев. Во время полета хорошо заметна темная поперечная
полосатость хвоста, светлый низ тела с охристым налетом на груди и зобе и
темный аспидно-серый верх тела. Голос. «Жиу-ичи, жиу-ичи» или «чиль-чи,
чиль-чи», напоминающий отчасти чириканье молодого воробья, или «пиу-чи,
пиу-чи». Этот крик с короткими паузами повторяется 8—10 раз подряд, причем
первый крик очень слабый, затем все усиливающийся и ускоряющийся,
достигнув максимального напряжения — обрывается. Вскоре птица снова начинает
кричать. Особенно часто кричит в пасмурную погоду во время мелкого
дождя или в сумерках (Воробьев, 1948).
Описание. Размеры и строение. Отличается широким и тупым крылом.
Первостепенные маховые —10; на сложенном крыле длиннее второстепенных примерно
на 11ь—11г общей длины крыла. Размеры: длина тела 312 мм (1 экз.); длина крыла (4 экз.)
194—207 мм, среднее 200,7 мм; 175 мм — 212 мм (Хартерт, 1912—21).
Окраска. Гнездовый наряд. Спинная сторона тела и верх головы бурые с
рыжеватыми окаймлениями перьев и с неясными рыжеватыми поперечными полосами; на
затылке белые перья; маховые бурые с рыжеватыми поперечными полосами, светлеющими
к краю наружного опахала; два внутренних третьестепенных маховых беловато-серые
с темными срединными пятнами; рулевые с широкими черными и рыжими (4—5)
поперечными полосами, предвершинная темная полоса шире других, подбородок и горло темно-
бурые; остальной низ тела белый с охристым налетом на груди и с темными продольными
широкими пятнами на груди и с более узкими на брюхе; радужина светлокоричневая; клюв
темнобурый, светлеющий к вершине. Первый годовой наряд. На верхней стороне тела нет
поперечных полос; на нижней стороне тела темные штрихи более узкие; от взрослого
отличается бурым налетом на верхней стороне тела и голове. Взрослые птицы — самцы и самки:
спинная сторона тела и верх головы аспидно-серые с белыми пятнами по бокам шеи;
длинные перья надхвостья серые с черноватыми вершинами; рулевые серые с четырьмя темными
поперечными полосами, вокруг полос и на вершине хвоста рыжеватые каймы; маховые тем-
носерые с зубчатым серовато-белым рисунком на внутренних опахалах, два внутренних
третьестепенных маховых беловато-охристые или беловато-серые; щеки и подбородок серые
одного цвета со спиной, горло светлосерое с чуть намечающимися более темными штрихами;
зоб белый с охристо-серым налетом; грудь и бока тела рыжевато-охристые с темными
стержнями перьев на передней части груди; брюхо и подхвостье белые, иногда с
охристо-рыжеватым налетом на брюхе, подкрылья белые или светлокремовые; радужина оранжево-красная,
клюв темный, у основания желтовато-зеленый.
30 Птицы СССР, т. I
ОТРЯД КОЗОДОИ
CAPRIMULCI или CAPRIMUWIFORMES
Характеристика отряда
Морфология. Внешние признаки. Птицы средних размеров.
Представители нашей фауны — настоящие козодои — небольшие птицы,
достигающие величины кукушки и веса несколько больше 100 г.
Оперение козодоев мягкое и рыхлое, несколько напоминает оперение сов.
Исключение составляют своеобразные крупные козодои Южн. Америки — гуахаро,
оперение которых более плотно. Маховые и рулевые мягкие. Рулевых 10,
первостепенных маховых тоже 10, только у Nyctibius и Podargus — 11;
имеется побочный ствол, кроме Steatornis; пух на аптериях. Окраска
оперения «покровительственная», напоминающая окраску древесной коры или общий
тон отмершего растительного покрова почвы. Половой диморфизм отсутствует
или слабо выражен; молодые по окраске оперения близки к взрослым самкам.
В полете козодои напоминают крупных ласточек. Хвост и крылья у
козодоев длинные. Это при относительно небольшом весе птиц позволяет им долго
летать и резко изменять направление полета. Клюв у одних видов очень мал и
слаб, у других более массивен, загнут на конце в виде крючка и обладает
зубчатым выступом на крае надклювья. Разрез рта очень широк, что связано со
способом добывания пищи. Из-за короткой цевки и слабых пальцев козодои
передвигаются по земле неуверенными прыжками, исключение — немногие
хорошо бегающие виды.
Ряд видов не способен охватывать пальцами ног древесные ветви и, садясь
на дерево, усаживаются вдоль них. Глаза всех козодоев большие, обращенные
несколько вперед.
Анатомические признаки. Череп козодоев эгитогнатический
шп /есмогнатический, голоринальный, ноздри не сквозные, исключая Steator-
ХАРАКТЕРИСТИКА. ОТРЯДА
467
nis и Podargus; шейных позвонков — 14 (кроме Steatornis и Podargus); ребер —
4 пары. Гортань трахеобронхиального или бронхиального типа. Зоб отсутствует.
Хорошо развиты слепые кишки. Имеется желчный пузырь. Все роды, кроме
Nyctibas и Podargus, имеют две сонных артерии. Нет обводящей мышцы. Кожа
очень тонкая; перья с побочным стволом, по микроструктуре напоминают
перья сов. Копчиковая железа небольшая, голая. Первостепенных маховых
10 или И, рулевых 10.
Образ жизни. Ночные или сумеречные птицы (немногие американские
виды деятельны и днем). В дневные часы укрываются на земле, на ветвях
деревьев или, как гуахаро, прячутся в пещерах. На охоту вылетают после
захода солнца. Наиболее деятельны козодои в сумерках, несколько менее
в середине ночи. Места обитания крайне разнообразны: пустыни, предгорья,
горы, местности лесостепного характера и леса. Виды тропических стран оседлы,
умеренного пояса — перелетны. Козодои, преимущественно, насекомоядные
птицы, немногие поедают сочные древесные плоды, другие — мелких
позвоночных животных. Насекомых ловят на лету, реже склевывают их с земли,
травы и листьев. Плодоядные виды (гуахаро) на лету обкусывают сочные плоды,
висящие на тонких ветвях высоких деревьев. При добывании пищи на лету
облегчает схватывание добычи широкий разрез рта, характерный для всей
группы козодоев. ;■
Козодои несомненно полезны для сельского и лесного хозяйства, так как
уничтожают большое количество насекомых.
Козодои — монргамы. Гуахаро постоянно держатся группами или стаями,
другие представители отряда после окончания гнездового периода держатся
некоторое время семьями, а позднее одиночно. У перелетных видов
осенний лёт совершается одиночками, весенний — одиночками, изредка парами
и группами.
Сроки размножения чрезвычайно разнообразны. Наши козодои
приступают к размножению сравнительно поздно. Наблюдается несовпадение сроков
гнездования отдельных пар, что вероятно зависит от возраста отдельных
особей, возможны также повторные кладки после гибели первых. За немног
гими исключениями козодои гнездятся обособленно. Колониальное гнездовье
свойственно только гуахаро.
Гнезда у большинства видов — ямки на земле. Настоящие гнезда строят
немногие виды. Гуахаро сооружают их из косточек плодов, выбрасываемых
в виде погадок; другие — из тонких веточек и мха, третьи — из перьев.
Некоторые козодои гнездятся в дуплах, на пнях и обломленных сучьях деревьев.
Гнездовой подстилкой в этих случаях служит древесная труха. Настоящие
козодои откладывают яйца прямо на землю. Размножению предшествует
своеобразный крик (песня) и брачный полет. Число яиц в кладке колеблется от
1 до 4. Настоящие козодои откладывают обычно 2 яйца. Окраска скорлупы
у видов, гнездящихся в пещерах и дуплах — белая, у видов, гнездящихся
открыто — пятнистая (мраморная), с хорошо выраженным
«покровительственным» рисунком. Эмбриональное развитие быстрое. Насиживание у видов фауны
СССР —около 17 дней. Птицы приступают к насиживанию после отложения
первого яйца. Насиживают оба родителя. Птенцы вылупляются зрячими,
у большинства видов покрытые густым коротким пухом. Их правильнее
относить к выводковым, нежели птенцовым птицам (исключение — гуахаро, птенцы
которых выклевываются голыми и лишь позднее покрываются желтым пухом).
Родители выкармливают птенцов из глотки, при этом птенец обхватывает своим
клювом клюв взрослой птицы. Подросшие птенцы обычно удаляются от местз
вывода, но еще долгое время родители проявляют о них заботу. Молодые птенцы
надевают окончательный наряд к осени и на следующую весну становятся
способными к размножению.
зо*
468
ОТРЯД КОЗОДОИ
Систематика. Три семейства: Sfeatornithidae, Podargidae и Caprimulgi-
dae. Общее число видов — 92, из них в фауне Союза представлены 3 или
3,26%. К семейству Stea-
tornithidae принадлежит
один род с единственным
видом гуахаро —*Steator-
nis caripensis,
распространенным в горных
лесах Южн. Америки от
Гвианы и Венецуэлы до
Эквадора и Перу.
Особенности: плотное
оперение; сильный,
загнутый на конце клюв с
зубчатым выступом на
крае подклювья;
сильные, оголенные до
пяточного сустава, ноги с
пальцами,
вооруженными крепкими когтями;
бронхиальная гортань;
шейных позвонков 15;
сильно развит подкож-
Рис. 120. Крыло буланого (слева) необыкновенного (справа) ный жир
козодоев- ' Птенцовые птицы,
гнездящиеся колониями
в глубоких пещерах. Своеобразно питание — древесные плоды и сооружение
гнезд из погадок. Яйца —в количестве 1—2 —белого цвета. Птенцы
выклевываются голыми, позднее одеваются темным пухом.
Семейство Podargidae — совиные козодои или белоноги. Характерные
особенности: вырезок грудины —две пары; имеется порошковый пух,
отсутствие базиптеригоидных отростков;
гортань промежуточного (между
бронхиальным и трахеальным) типа; не-
Рис. 121. ^Свост большого (справа) и обыкновенного (слева) козодоя.
парная сонная артерия; оборонный четвертый палец из пяти фаланг; сильный
клюв; тонкий язык. Включает три рода с 19 видами: Podargus, Batrachosto-
mus, Aegotheles, распространенные на Малайском архипелаге, в Папуасии,
Австралии и Тасмании. Птицы промежуточные между птенцовыми и выводко-
ОБЫКНОВЕННЫЙ КОЗОДОЙ 469
вымй: некоторые (Aegotheles) гнездятся в дуплах, другие (Batracfwstomus) —
строят гнезда на деревьях; яиц в кладке от 1 до 4.
Семейство Caprimulgidae—настоящие козодои. Признаки следующие: нога
слабая, не способная охватывать ветви; шейных позвонков 14; трахео-брон-
хиальная гортань; коготь среднего пальца по краям зазубрен; отсутствие
processus basipierygoidei; клюв слабый, короткий; толстый язык.
Семейство распадается на роды: Caprimulgus, Nyctibius, Hordeiles, Cosme-
tornis, Nyctidromus, Hydrapsalis, Macrodipterus, Scotoruis с 72 видами. Среди
них резко уклоняется Nyctibius, распространенный в средней и Южн.
Америке, у него, в отличие от других родов, IV палец имеет пять фаланг (у
прочих 4); ноготь среднего пальца не зазубрен, вырезок грудины — 2; одна
сонная артерия. Некоторые представители Caprimulgidae обладают удлиненными
рулевыми или маховыми.Семейство имеет весьма широкое распространение.
Географическое распространение. Большинство видов отряда свойственны
тропическому поясу, немногие проникают далеко к северу, но в полярных
странах отсутствуют.
В пределах Союза встречается один род Caprimulgus с 3 видами.
Из них С. europaeus широко, распространен в лесной зоне* в лесостепи и
степи, местами в пустынях. С- aegyptius ограничен только участками песчаных
кустарниковых пустынь южного типа. С. indicus гнездится в лесах —
преимущественно лиственных или смешанных южных частей Воет* Сибири. В
пределах СССР все козодои перелетны.
Таблица для определения видов козодоев
1. (2) Внутренний край опахала трех или четырех первостепенных маховых белый
с зубчатыми выступами к стержню перьев. Общая окраска оперения песчано-охристая.
Буланый козодой- Caprimulgus aegyptius.
2. (1) Зубчатые выступы белого цвета на внутренних опахалах первостепенных
маховых отсутствуют, общая окраска оперения более темная 3
3. (4) Четыре крайних пары рулей с большими белыми предвершинными пятнами,
концы рулей темные — (самец) Большой козодой Caprimulgus indicus
4. (3) На крайних рулях предвершинных пятен нет . . i * . . 5
5. (6) На двух крайних рулевых по крупному белому вершинному пятну (взрослый
самец) .Обыкновенный козодой Caprimulgus europaeus
6. (5) Белые пятна на рулевых отсутствуют ; . . 7
7. (8) Внутренние опахала первостепенных маховых на 3—4 см от основания почти
одноцветные и лишь слабо испещрены узкими охристыми полосками, длина крыла
обыкновенно более 20 см (самка) Большой козодой Caprimulgus indicus
8. (7) Внутренние опахала нескольких первостепенных маховых испещрены частыми
и ясными поперечными охристыми полосками, длина крыла обыкновенно короче 20 см
(самка). i....i.i Обыкновенный козодой^ Caprimulgus europaeus
СЕМЕЙСТВО НАСТОЯЩИЕ КОЗОДОИ CAPRIMULGIDAE
РОД КОЗОДОЙ CAPRIMULGUS LINNAEUS 1758
Тип С. europaeus
1. Обыкновенный козодой Caprimulgus europaeus L.
Caprimulgus europaeus. Linnaeus. Syst. Nat., изд. X, 1758, стр. 193; Швеция.
Русское название. Козодой — народное, как и на других европейских
языках — в силу поверья, что птица по ночам своим широким клювом выдаивает коз. Другое
народное название — полуночник, из-за ночного образа жизни птицы.
Распространение. Ареал. Европа; с.-з. Африка (Марокко); Азия на
восток до южн. Забайкалья, Вост. Монголии (р. Тола) и Гоби; Кентейские
горы; отдельные находки в Даурии. На севере до Скандинавии,
Финляндии, Карелии и до Сев. Урала, Тобольска, Томска, Енисейска, Иркутска.
470
ОТРЯД КОЗОДОИ
К югу включительно до с.-з. Африки —в Алжире, Тунисе; в Малой Азии
и ю.-з. Азии, до с.-з. Индии. " ^
Характер пребывания. В Европе и в умеренных частях Азии
перелетная, в южных частях ареала (в Африке) повидимому оседлая.
Биотоп. Леса, лесостепь, кустарниковые полупустыни и окраины
пустынь, безлесные или бедные лесом участки гор и предгорий.
. Подвиды и варьирующие признаки. Обыкновенный козодой объединяет пять подвидов,
отливающихся, главным образом, окраской оперения и размерами. В местах соприкоснове-
Карта 81. Распространение обыкновенного козодоя Caprimulgus europaeus
1 — С. е. europaeus, 2 — С. с. meridlonalis, 3 — Се. zarudnii, 4 — С. е. plimipes, 5— Се.
unwini; а—граница гнездовий, б — не вполне выясненная граница гнездовий, г — зимовки
(цифры в кружках соответствуют номеру зимующих подвидов).
ния ареалов географических форм отличия во многих случаях крайне слабы и не позволяют
разобраться в систематической принадлежности экземпляров. Лучше выражены
экологические различия (гнездовый биотоп) географических форм.
С. е. europaeus L., 1-758 — лесные области сев. и средней Европы и средней и южи.
Сибири до Кентейских гор в Монголии; С. е. meridional is Hart. 1896 — с.-з. Африка и
южные части Европы, Малая Азия; С. e.zarudnyi Hart. 1911 — от восточных берегов
Каспия и низовьев Дона до р. Толы и центральной Гоби и Кентейских гор в Монголии; С. *.
plumipes Przew, 1876 — Кашгария, Хами, ю.-в. Монголия до великой излучины Желтой
реки. Залетает в пределы Туркестана; С. е. unwini Hume 1871 — ю.-з. Иран,
Афганистан, Белуджистан до с.-з. Гималаев.
Систематические замечания. Уже после подготовки нами текста к печати вышла работа
Б. К. Штегмана «Географическая изменчивость козодоя» (1949). Этот автор пришел к ряду
новых выводов о границах распространения и характеристике подвидов козодоя, описал
подвид С. е. dementievi из сев. и Центральной Монголии и южных степных частей
Забайкалья. Мы не имели возможности сопоставить выводов Штегмана с нашими и не учли их
поэтому в нашем тексте.
ОБЫКНОВЕННЫЙ КОЗОДОЙ
471
Северный обыкновенный козодой Caprimulgas europaeus earopaeus L.
Распространение, Ареал. Сев. и средняя Европа, средняя и южн.
Сибирь. Северная граница пересекает Скандинавию на 64° с. ш., Финляндию
на 63°, южн. Карелию. Проходит по лесным районам Архангельской обл.;
на зап. Урале встречается еще под Чердынью, в вост. Зауралье прослежен до
Петропавловска включительно. По Бутурлину (1936) на Урале
распространен приблизительно до 60°. В Зап. Сибири граница проходит через район
Тобольска: южнее, в частности, под Тюменью, обыКновенен. Далее граница
идет через район Томска, проходит у Енисейска, не проникая севернее этого
города и идет к Байкалу. Неоднократно отмечался на Ангаре в районе
г. Усолье и Иркутска и обычен в районе Троицкосавска. Отдельные находки
известны в Даурии. Вероятно эта же форма обитает в лесных массивах
Кентейских гор в Монголии, где отмечена Козловой (1930). Южная
граница охватывает с юга Францию, сев. Италию, большую часть (исключая
юг) Чехословакии, Польшу, а в пределах СССР спускается до юга Полтав-
щины; в среднем Поволжье почти до Саратова; вероятно проходит вблизи
Уральска и устья Илека, направляясь отсюда к Мугоджарам. Далее на
восток граница не ясна, так как номинальная раса козодоя распространена
спорадически, отсутствуя в настоящей степной обстановке и заселяя
пятнами степные балки, заросшие древесной и кустарниковой растительностью.
Между южной границей С. е. earopaeus и северной границей С. е. zarad-
nyi в Тургайских степях существует значительный разрыв, где козодоев
в гнездовое время нет вообще. На Алтае оба подвида козодоев обитают
.рядом, причем северный живет в лесах, а другой в безлесных местностях.
Зимует, главным образом, в южн* и вост. Африке.
Характер пребывания. У нас гнездящаяся и перелетная
птица. По сравнению с другими формами козодоев, прилет относительно
ранний и происходит еще при неустановившейся весенней погоде. В южной и юго-
восточной половине ареала нормально птицы появляются в последних числах
апреля: Ростов—30 апреля (Сарандинаки, 1909), Харьков—>20 апреля
<Сомов, 1897), Смоленск —20, 30 апреля (Гржибовский, 1912), Минск 21 апреля
(Шнитников, 1913), Тула —- конец апреля, Тамбов — 30 апреля (Резцов, 1910)..
Несколько раньше — в первой трети апреля птицы появляются в районе
Чкалова. В северных областях Европейской части Союза прилет первых особей
нормально совпадает с началом мая: Псков — 4 мая (Нестеров и Никандров,
1913); Новгород — в первых числах мая, в районе Молотова — в первой трети
мая. Но и здесь в теплые и ранние вёсны первые козодои прилетают в конце
апреля: 25 апреля—.в районе Можайска (Огнев, 1913); в конце апреля—»
•в Ярославской обл. В Сибирь козодои прилетают в более поздние сроки —
обычно во второй половине мая; под Красноярском даже 23 мая (Тугаринов и
Бутурлин, 1911). Пролет сильно затягивается и валовое движение птиц
наблюдается значительно позднее появления первых особей: до 28 мая — в районе
Ростова (Сарандинаки, 1909); до конца второй трети мая — в Ярославской обл.;
в середине мая — под Чкаловом. Осенью взрослые отлетают раньше молодых.
Первые признаки осеннего движения к югу во многих пунктах ареала
отмечаются уже около половины августа: район Ленинграда в половине августа,
под Молотовом — в августе, у Красноярска — в конце августа, в районе
Чкалова — в последних числах августа, а в вост. Приазовье — в последней
трети августа. В первой половине сентября, реже в конце этого месяца, птицы
исчезают из северных частей ареала, а в более южных районах пролет часто
затягивается на продолжительное время. Отставшие пролетные особи
отмечались: в Ярославской обл. до 10 октября (Птушенко, 1937), под Рязанью —
© конце первой трети октября, под Чкаловом — до 2 ноября (Зарудный, 1888,
472
ОТРЯД КОЗОДОИ
1897), в районе Харькова — б октября (Сомов, 1897), в вост. Приазовье — до
второй трети октября. В Сибири, видимо, пролет повсеместно кончается в
последней трети сентября. Перелет совершается ночью и на зорях. Большинство
птиц летит по одиночке, но иногда и маленькими разрозненными группами.
Биотоп. Гнездовый — леса и рощи; птица явно избегает настоящей степи.
В средней полосе Европейской части Союза сосновые боры, смешанные леса и
лиственное мелколесье. Птицы предпочитают держаться близ вырубок, лесных
полян и по окраинам леса. В Казахских степях козодои избегают безлесной
степи и держатся лишь там, где есть лес и кусты, или по склонам степных балок,
заросших деревцами дикой вишни и тополем. В Сибири (под Красноярском)—
сухие боры, разреженные смешанные леса, степные колки и чаще в редкой
хвойной тайге по вершинам хребтов. В поисках пищи наиболее охотно посещает
лесные поляны, вырубки, луга и сельскохозяйственные угодья, прилегающие
к лесным массивам, или стоячие водоемы.
Численность. Обычная птица, численность которой вследствие
скрытного образа жизни учесть трудно.
Экология. Размножение. Даже при большой плотности населения
всегда гнездится отдельными парами. Гнезда не строит. Яйца откладываются
иногда на голую почву, чаще на хвое, прошлогодней листве, трухе сгнившего
дерева. Место откладки яиц обычно скрыто нависшим кустиком, папоротником,
нижними ветвями молодых хвойных деревьев и т. п. Иногда «гнездо»
помещается между корнями крупного дерева, у упавшего и полусгнившего ствола
или под прикрытием кучи сухих ветвей (Шнитников, 1913). По мере
насиживания на месте, где лежат яйца, образуется небольшое углубление.
Нормальная кладка содержит два яйца. Наличие в гнезде одного яйца
несомненно связано с гибелью первого яйца, кладка в три яйца —
исключительная редкость. Яйца имеют эллипсовидную форму с одинаково овальными
конусами. Скорлупа их гладкая и блестящая. Основной фон фарфорово-белый
или серовато-белый. По основному фону разбросаны пятна двух типов:
светлосерые глубокие с размытыми краями и поверхностные, резко очерченные
серые с буроватым оттенком. На большой серии яйца этого козодоя
отличаются от яиц других подвидов несколько большими размерами. Размеры
28,3—35,6X20,7 — 23,9 мм.
Откладка яиц у различных пар происходит не одновременно. Это связано
отчасти с потерей первых кладок и, повидимому, с более поздним гнездованием
молодых особей. Весьма вероятно наличие двух кладок. Откладке яиц
предшествует своеобразное токование самцов, весенний крик и хлопанье крыльями.
Его удается слышать вскоре по прилете птиц (в конце апреля и в мае) и до
последних чисел июля (Тугаринов и Бутурлин, 1911), а иногда (Мордовский
заповедник) до 7.VIII (Птушенко, 1938), под Калугой до 22.VIII (Филатов, 1915).
Спаривание происходит на земле. Самые ранние кладки в средней и южной
полосе Европейской части Союза обнаруживаются около 10 мая (в районе
Чернигова в конце мая найдены совершенно насиженные яйца) (Емельяненко,
1917). На севере (быв. Новгородская губ.) начинает нестись во второй половине
мая. Массовая откладка яиц совпадает с последними числами мая или
происходит в первой половине июня. С этого времени кладки со свежими яйцами удается
находить до половины июля (в районе Харькова 7.VII, Сомов, 1897), под
Рязанью 14 июля (Спангенберг). В Сибири кладка происходит несколько позднее.
Под Томском насиженные яйца найдены 21 июня (Залесский, 1915), а массовое
появление птенцов в районе Красноярска видимо совпадает с концом первой
трети июля.
Первые птенцы в Европейской части Союза появляются в.начале июня,
чаще же в более поздние сроки. Наиболее ранняя молодь приобретает
способность к полету в последней трети июня (24 июня в Воронежской обл.; Огнев
ОБЫКНОВЕННЫЙ КОЗОДОЙ 47,4
и Воробьев, 1924). Плохо летающих молодых козодоев удавалось находить
в Мордовском заповеднике даже 25 августа (Птушенко, 1938). Промежуток
времени между откладкой первого и второго яйца точно не установлен. Яйца
откладываются в ночное время. Насиживание начинается после первого отложенного
яйца, причем самка сидит на нем так же прочно, как и на полной кладке. В
насиживании принимают участие оба родителя, но самец все же меньше, чем
Рис. 122. Обыкновенный козодой.
самка. Срок насиживания — 17—18 дней. Птенцы выклевываются из яиц
зрячие и покрытые густым пухом. Вылупление птенцов происходит ночью
и неодновременно. Второй птенец вылупляется более, чем на сутки позднее
первого. При кормлении птенец охватывает своим клювом клюв родителей и
пища из глотки взрослого поступает в рот птенца. Родители продолжают
кормить птенцов, когда они уже приобретают способность летать и отличаются от
взрослых только более коротким хвостом. Вполне самостоятельными птенцы
становятся со второй трети июля в Европейской части Союза (Харьковская
обл.) и в первой трети августа в Сибири (Красноярск). От гнезд с птенцами
родители пытаются отвести врага, подражая раненой птице.
Питание. Разнообразные насекомые. Летающие ночные жуки и
бабочки. В желудке козодоя, добытого 5 августа в Мордовском заповеднике,
1 экз. Aeschna grandis и 5 экземпляров бабочек из семейства Noctaiclae. Основное
474
ОТРЯД КОЗОДОИ
количество насекомых ловится на лету, но как и близкие виды козодоев,
вероятно, берет пищу и с земли.
Линька. Взрослые птицы линяют на зимовках в декабре —феврале;
частичная смена мелкого пера после гнездового периода отмечена в июле—
сентябре. Мелкое оперение гнездового периода сменяется в сентябре—октябре.
Последовательность смены нарядов: пуховой, первый зимний, первый
годовой и т. д.
Полевые признаки и повадка. Сумеречная длиннокрылая бесшумно
летающая птица, иногда трясется в воздухе на одном месте, как это делают пустельги.
В дневные часы прочно сидит на земле в тени и, будучи спугнутой, перелетает
небольшое расстояние и усаживается на землю, реже на горизонтальную ветвь
дерева вдоль ее. Не боится человека и во время охоты за насекомыми совсем
близко подлетает к людям, влетает в селения или кружится над пасущимися
домашними животными. Голос козодоя — самца особенно часто удается слышать
в период размножения. Эта весенняя песня птиц по мнению многих
исследователей имеет нечто общее с мурлыканье^ кошки. Самец тянет свою песню до
5 минут и заканчивает резким криком, повторяющимся до 4—5 раз. Этот крик
сопровождается хлопаньем одно о другое крыльев. Особенно часто, иногда
и днем, козодои кричат от времени прилета и до начала июля. Реже крики
самцов удается слышать в августе. При взлете оба пола издают другой звук,
напоминающий приглушенное, резкое «чоканье».
Описание. Размеры и строение. Длина тела самцов и самок 260—280 мм,
размах крыльев 545—580 мм. Вес около 75 г, осенние экземпляры весят несколько больше 100 г.
Формула крыла 2>1>3>4>5>6... Вырезки на наружных опахалах 2-го и 3-го маховых.
Второстепенные маховые постепенно удлиняются к внутреннему краю партии, вершины их
закруглены. Первостепенных маховых 10, рулевых 10. Клюв слабый, короткий и широкий у
основания, слегка загнутый у вершины надклювья; разрез рта очень широк и окаймлен вдоль
верхней челюсти обращенными вперед щетинками. Цевка короткая, оперена по передней
стороне на ZU длины. Задний палец обращен внутрь, основания передних соединены
перепонкой до второй фаланги, на когте среднего пальца зазубрины, слабо развитые у молодых.
Большие глаза и наружное ушное отверстие. Крыло самцов (29) 182—200, самок (32) 177—
204, в среднем соответственно 187,9 и 188,4 мм.
Окраска. Пуховой наряд буроватый на спинной, охристо-сероватый на
брюшной стороне. Взрослые самцы: спинная сторона буровато-сероватая с тонкими более темными
пестрянками и с черноватыми продольными полосами, особенно развитыми на темени и
плечевых, а также с более или менее развитым ржавчатым оттенком; брюшная сторона охристо-
рыжеватая с темными черновато-бурыми тонкими поперечными полосками; на горле два
супротивно расположенных белых пятна; маховые темнобурые с рыжеватыми пятнами и
сероватым оттенком на вершине; на внутреннем опахале первых трех первостепенных
маховых — большое белое пятно; рулевые — средняя пара серобурая с черными поперечными
полосами; две крайних пары с длинными белыми вершинными пятнами. Самки и молодые
■обоего пола в гнездовом наряде — как самцы, но без белых пятен на маховых и боковых
рулевых. Клюв черный, ноги бурые, радужина темнобурая.
Южный обыкновенный] козодой Caprimalgus europaeus meridionalis H а г t.
Caprimalgus europaeus meridionalis. H a r te r t. Ibis 1896, стр. 370, Греция.
Распространение. Ареал. От с.-з. Африки к северу включительно,
Пиренейский, Аппенинский и Балканский п-ова, южная половина
Чехословакии, Румыния, Венгрия и прикаспийские части Ирана. У нас гнездится
б Крыму, на всем Кавказе, вероятно в Копет-Даге и проникает на Украину
до южной границы Полтавской обл. и среднего течения Дона. Основные
зимовки в вост. и южн. Африке, изредка в зап. Африке (Гвинея, Камерун).
Характер пребывания. Гнездящаяся и перелетная птица.
Весенний прилет относительно поздний. Сроки пролета мало изучены. У
Ленкорани птицы появляются около середины апреля. В половине апреля
отмечены в районе Тбилиси, в последних числах апреля — в Аджарии и Гурии.
В горных частях Армении (Делижанское ущелье) прилет происходит, вероятно,
ОБЫКНОВЕННЫЙ КОЗОДОЙ
475
те ранее мая. На Сев. Кавказе (Дзауджикау) пролет происходит в апреле.
Вероятно птицы прилетают сюда, огибая главный Кавказский хребет. В самых
последних числах апреля птицы появляются и севернее: 20 апреля в районе
Мариуполя (Боровиков, 1907), 17 апреля под Павлоградом (Вальх, 1911).
Осеннее движение к югу хорошо выражено в сентябре. На южн. Украине
птицы летят в первой декаде октября. В горной части с.-з, Кавказа последние
птицы отмечены 17 октября (Аверин и Насимович, 1938). У Дзауджикау пролет
шдет с середины сентября и по конец октября. В Армении козодой наблюдался
/до 30 сентября (Ляйстер и Соснин, 1942), а в Делижанском ущелье в половине
юктября. В Абхазии две птицы отмечены даже 7 ноября (Сатунин, 1937).
Вероятно, как и другие подвиды, они совершают перелет ночью.
Биотоп. Гнездовый. Населяет леса в горах и на равнинах. В горах
придерживается кустарниковых порослей, лесных полян и вырубок,
поднимается до верхней границы леса — на Сев. Кавказе до 2 200 м (Аверин и
Насимович, 1938), в Армении до 2 500 м (Ляйстер и Соснин, 1942). В поисках
пищи посещает открытые пространства: вырубки, луга, поля и участки,
покрытые низкорослыми кустарниковыми зарослями.
Численность. Обыкновенная, местами довольно многочисленная
птица.
Экология. Размножение. Как и прочие козодои — гнезд не строят.
Яйца откладываются на покрывающую почву, сухую, прошлогоднюю листву,
:на опавшую хвою, труху выгнившего пня или прямо на голую землю. «Гнездо»
помещается то среди редких кустарников под низким развесистьш грабом, то
на окраине крупного леса или мелколесья. Полная кладка — два яйца и очень
редко (дополнительная) — только \ яйцо. По размерам, форме и окраске
яйца южные козодои совершенно не отличимы от северного подвида. Откладка
яиц отдельных самок в одних и тех же районах сильно колеблется в сроках.
Вероятно здесь имеет значение возра&т птицы или это связано с потерей первой
кладки. Вполне возможно, как и у других козодоев, наличие двух кладок в лето.
Массовая кладка яиц имеет место, повидимому, в последних числах мая.
В районе Запорожья слабо насиженные кладки удавалось находить в конце
мая и в конце июня. Самка, добытая в Армении 12 июня, имела фоликулы
величиной с мелкую горошину (Ляйстер и Соснин, 1942)> другая, добытая в зап.
Закавказье 18 июня, оказалась с яйцом, готовым к сносу (Нестеров, 1911).
В Делижанском ущелье 2 почти свежих яйца найдены 27 мая (Спангенберг).
Вылупление птенцов из яиц происходит около середины июня. 15 июня в «гнезде»,
найденном у Дзауджикау, 1 птенец и 1 яйцо (Беме, 1926); 21 июня 2 пуховых
птенца близ Белого Ключа (Цветков, 1911); 7 июля полувзрослые птенцы в
районе Тифлиса (Баньковский, 1913). Промежуток времени между откладкой
первого и второго яйца вероятно около суток, так как второй птенец появляется
на свет в следующую ночь после первого. Самка приступает к насиживанию
после кладки первого яйца и сидит на нем очень крепко. В насиживании
принимают участие оба родителя. Срок насиживания около 16—17 дней. Кормление—
как у других козодоев. В случае опасности взрослая птица пытается отводить
ют птенцов, хлопает крыльями и, широко раскрыв клюз, издает шипящие
звуки. Летные птенцы появляются около половины июля.
Питание. В общих чертах схоже с питанием северного подвида.
Главным образом ночные насекомые. В желудке птицы, добытой в зап.
Закавказье, обнаружены: навозный жук, жужелицы, коровки и пр. Способ
добывания пищи схож с таковым у других близких видов и рас.
Линька 'не изучена.
Описание. Размеры и строение. На больших сериях южный козодой
отливается от северного подвида окраской, которая несколько светлее. Размеры мельче. Крыло
самцов (19) 173—190, самок (16) 171—187, в среднем 181 и 182,2 мм.
476
ОТРЯД КОЗОДОИ
Пустынный обыкновенный козодой Caprimulgus europaeus zarudnyi H а г t e r t
Caprimulgus europaeus zarudnyi. Hartert. Vog. Pal. Fauna II, 1912, 849 стр., 1912,
Тарбагатай.
Распространение. А р е а л. От низовьев Дона, сев. и вост. берегов Каспия
через пустыни Туркестана, Тарбагатая и Алтая до центральных частей Гоби.
Северная граница идет по песчаным степям вдоль сев. побережья Каспия
(экземпляры из дельты Волги), огибает с севера степи у Аральского моря, проходя
вблизи южной оконечности Мугоджар и несколько севернее ст. Челкар,
пересекает пустыню Аральские Кара-Кумы. Далее граница не ясна. В область
настоящих степей не проникает. На Алтае близко друг от друга, но в резко
различных биотопах, гнездятся две формы — типичный С. е. europaeus с переходными
признаками, принадлежащий сырой тайге, и пустынный, свойственный
каменистым склонам, поросшим кустарником, каменным грядам и степным буграм.
От Алтая область распространения пустынного козодоя продолжается в
Монголию, достигая долины р. Толы и центральных частей Гоби (Козлова, 1930).
Километрах в 80—100 к северу от этого места в лесистых горах Кентея обитает
уже другая форма козодоя, которую вероятно, следует отнести к номинальной.
Южная граница на западе начинается в юго-зап. углу Туркмении, проходит
вдоль северных склонов Копет-Дага, затем спускается к Кушке и следует на
восток. Далее пустынный козодой спорадически гнездится в Таджикистане,
встречается в предгориях и равнинах Ферганы, в Киргизском Алатау, во всем
Семиречье и около Кульджи, восточнее, в низовье р. Эдзин-гол в Центральной
части Гоби.
Характер пребывания. Гнездящаяся и перелетная птица.
Весенний пролет поздний и происходит в то время, когда погода вполне
установится. Сроки прилета выяснены недостаточно. Нормально прилет в Средней
Азии происходит в первой половине апреля, в Сибири — во второй половине
этого месяца. В Туркмении редкие одиночки появляются значительно раньше.
Голос козодоя удалось слышать 14 марта, а первая птица наблюдалась у Сары-
Язы на Мургабе 16 марта. Массовый прилет в устье Теджена 21—24 мая,
в районе Мары-Чарджоу — между 21 апреля и 9 мая. У ю.-в. Каспия
птицы летят в течение мая. Под Ташкентом Зарудный наблюдал этих
козодоев в середине апреля и 23 мая добыл экземпляр. На нижней Сыр-Дарье
в районе Кзыл-Орды появление первых козодоев совпадает с началом апреля,
массовый же прилет птиц происходит в середине этого месяца. В с.-з. Алтае
был добыт 25 апреля (Залесский, 1929), на Орок-норе в Монголии — 14 мая
(Козлова, 1930). Осенний отлет вероятно начинается с последних чисел
августа и безусловно с первых чисел сентября. С этого времени он
затягивается до конца октября, реже даже до начала ноября. Пролетная птица
7 сентября была добыта в Наурзумском заповеднике (Михеев, 1938). В
Семиречье в районе Джаркента пролет происходит в первой половине сентября
и тянется до половины октября. На Сыр-Дарье численность в октябре
начинает сокращаться, но отдельные особи наблюдались под Байгакумом да
8 ноября (Спангенберг 1936). В Туркмении летят в сентябре и остаются до
конца октября. Самая поздняя дата добычи —12 октября. В Таджикистане
экземпляр добыт 8 сентября (Иванов, 1940), на Памире отмечен — 11
сентября (Мекленбурцев, 1936). Перелет совершается в ночное время; птицы*
вероятно, летят одиночками и разрозненными группами.
Биотоп. Гнездовый, как и у других форм козодоев, разнообразен.
Птицы гнездятся в песчаных пустынях, в глинистых полупустынях, на
участках щебнистой почвы в безлесных горах и холмистых предгориях, а местами
в тугаях, в разреженных горных лесах, но не в лесных массивах. В Туркмении
гнездится преимущественно в горах, достигая в вертикальном направлении их.
ОБЫКНОВЕННЫЙ КОЗОДОЙ
477
верхнего пояса. Здесь придерживается родников, речных долин, поросших
кустарниками или деревьями. В равнинах заселяет тугаи, сады и пустынные
площади с саксаулом и песчаной акацией. В Бадхызе держится в фисташковых
районах (Дементьев, 1950).
В дельте Аму-Дарьи заселяют тугаи и возвышенные участки, например,
Дель-Тау, в Кызыл-Кумах пески и заросли кустарников, на восточном
побережье Аральского моря, поросшие кустарниками песчанок, глинистые и
солончаковые пространства. В области нижней Сыр-Дарьи гнездится в пухлых
солончаках, поросших редким кустарником джингиля и колючки, чередующимися
с голыми клочками солончаковой почвы. Обычен также в полынковых степях,
в песках и в пустынных горах с.-з. отрогов хребта Кара-Тау, в песках,
поросших кустарником Artemisia inodora и Elymas sabalosus. Когда птица находится
в поисках пищи, ее чаще всего можно встретить близ колодцев, аулов и
других населенных пунктов.
Численность. Обыкновенная, местами (Казахстан) многочисленная
;птица как в гнездовый период, так и во время пролетов.
Экология* Размножение. С прилета и в период размножения
удается слышать характерный весенний крик (песню), которая достигает
наибольшей интенсивности в апреле. Иногда птицы собираются обществом и среди
них происходит нечто в роде тока. Мурлыкая, козодои взлетают и кувыркаются
в воздухе. Брачный полет совершается на вечерней и утренней заре. Как и
другие козодои — настоящего гнезда не строят и откладывают яйца на голую
песчаную, глинистую, щебнистую почву или на растительную ветошь,
скопившуюся в трещинах и выбоинах земной поверхности. Большинство гнезд
располагается открыто на солнцепеке, другие несколько скрыты нависшим
кустарником. Полная кладка —2 яйца и как исключение —. 1, что вероятно
происходит вследствие гибели первого отложенного яйца. Яйца по окраске и форме
не отличимы от яиц северного и южного подвида. Средняя величина их в общем
чуть меньше. Размеры: 32,6—27,0 и 23,0—20,9 (Спангенберг, 1936; Заруд-
ный, 1896). Размножение у отдельных пар не одновременное. Несомненно
существуют две кладки в лето. Законченные кладки с едва насиженными яйцами
удается находить с середины мая и в течение всего июня. В Копет-Даге 1 яйцо
найдено 2 июня, в других частях Туркмении кладки найдены у Куркулаба —
слабо насиженные 2 июня, 2 яйца на Теджене — 19 мая. Самка с яйцом в
яйцеводе добыта на Челекене — 18 мая; летные птенцы в конце июня и в первых
числах июля. В Кызыл-Кумах—21 мая — 1 свежеснесенное яйцо, другое
в яйцеводе самки (Зарудный, 1915). На Сыр-Дарье в районе Кзыл-Орды 16 мая
.первое снесенное яйцо, а 25 мая два чуть насиженных яйца на вершине горы
Ак-тау. Первые птенцы отмечены в конце первой трети июня, летные птенцы
в июле. В дельте Волги две кладки найдены в июне, на Алтае на голой
солончаковой площади чуть насиженная кладка в 2 яйца найдена 15 июня, в горах
Бектау-ата (сев. Прибалхашье) едва насиженные яйца 4 июня (Спангенберг).
Из первого яйца птенец вылупляется более чем на сутки раньше второго.
Насиживание начинается по отложении первого яйца. Первые дни насиживает
только самка, а самец держится поблизости. Позднее и самец принимает
участие в насиживании. Продолжительность насиживания — 16—17 дней. Птенцы
вылупляются зрячими и покрыты густым пухом.
Питание. Как и у других козодоев — насекомые. При вскрытии
желудков добытых птиц обычно удается находить хитинные остатки жуков
•и бабочек.
Линька не изучена.
Описание. Размер ыистроение. Отличается от номинальной формы мелкими
размерами и общим светлым оперением. Крыло самцов (32)175—191, самок 171—184(29),
среднее 184 и 182 мм.
478
ОТРЯД КОЗОДОИ
Систематические замечания. Признаки подвидов unwini и zarudnyi не стойки.
Вследствие этого очертить ареалы более или менее точно оказывается совершенно невозможным. От
северных козодоев С. с. europaeus козодои, населяющие пустыни, отличаются двумя
основными признаками — несколько меньшими размерами и особенно светлой окраской оперения.
Других признаков, исключая гнездовых биотопов, повидимому не существует. Подвидьштиini
и zarudnyi по существу не отличаются друг от друга размерами, но будто бы отличаются
окраской оперения. У zarudnyi общий тон оперения светлосерый, у unwini с несколько
рыжеватым оттенком. Однако можно с уверенностью сказать, что в пределах русского Туркестана
нет ни одного географического района, где бы общий тон окраски оперения козодоев был
постоянен. В Туркмении значительный процент особей окрашены, как unwini, но среди
них не редки особи с окраской zarudnyi. В низовьях Сыр-Дарьи несколько чаще встречаются
козодои с окраской zarudnyi, но значительный процент обладает окраской, свойственной
unwini. Таким образом, обширная пустынная территория, от Каспия до центральной Гоби,,
оказывается заселенной мелкими светлыми козодсями с сильными колебаниями общего тона
окраски оперения.
Считая, что настоящий туркестанский козодой Се. unwini Hume, описанный из
Гималаев, не встречается у нас, мы рассматриваем наших пустынных козодоев как гибридную^
популяцию, условно называя, ее Caprimulgus europaeus zarudnyi Hart. От северных:
козодоев, распространенных в лесах Европы и лесах Сибири, эти козодои хорошо отличаются
в серии светлой окраской и несколько меньшими размерами.
Китайский обыкновенный козодой Caprimulgus europaeus plumipes Przew*
Caprimulgus plumipes, Пржевальский. Монголия и страна Тангутов II, ст.22,
1876, северная часть излучины Желтой реки.
Распространение. Ареал. Гнездовая область простирается от Кашгарии
и ю.-в. Монголии до излучины Желтой реки.
Характер пребывания. Случайно залетает к нам во время
весенних и осенних миграций. Известно два случая добычи в Туркмении: самка
1 октября 1935 г. у Ясха и самка 17 сентября 1926 г. у ст. Драгомирова
(Дементьев, 1950). Кроме того, самка 20 мая 1910 г. добыта близ г. Фергана.
Помимо этого есть указания о встречах с этим козодоем в бассейне Сыр-Дарьи
и Ташкентского базиса. Не имея в руках экземпляров, трудно что-либо
определенное сказать о их подвидовой принадлежности.
Описание. Размеры и строение. По окраске ближе всего к С е. zarudnyi p
но с иольшим развитием охристо-рыжих тонов. Подхвостье почти не полосатое.
Крыло 180, 195 мм (Бутурлин, 1936); у экземпляров Зоологического музея Моек*
университета, самок 19,45 и 18,3 мм.
2. Буланый козодой Caprimulgus aegyptias Lie hi
Caprimulgus aegyptius. Lichtenstein. Verz. Dcubl. etc., стр. 59, 1823, Верхний Египет,
Распространение. Ареал. От Марокко, Алжира, Туниса и Ливийской
пустыни через Месопотамию, южн. и вост. Иран до Афганистана, Средней
Азии. Зимует в с.-в. Африке.
Характер пребывания. В Африке оседлый (быть может
перелетный в Египте), в Азии перелетный.
Биотоп. Низменные и жаркие пустыни и полупустыни с мягкой
почвой. Гористых пространств и щебнистой почвы в гнездовой период избегает.
Местами населяет участки непосредственно, прилегающие к рекам и другим
водоемам.
Подвиды и варьирующие признаки. Буланый козодой объединяет три подвида, слабо-
отличакщиеся окраской оперения и тем, что одна форма оседла, другие перелетиы:
С. a. arenicolor Severtzow, 1875—Южн. и вост. Иран, Белуджистан, Афганистан
и пустыни Средней Азии; С. a. aegyptius Lichtenstein. Нубия, Египет, Ирак;
С. a. sahapde Е г 1 a n g е г. 1.899 — сев. Сахара — от Марокко и Алжира до западных
берегов Нила.
БУЛАНЫЙ КОЗОДОЙ
479
Северный буланый козодой Caprimulgus acgyptius arenicolor Severtz.
Caprtmulgus aegyptius arenicolor. Severtzov. The Ibis, 1875, стр. 491, Низовья Аму-Дарьи.
Распространение. Ареал. Южн. и вост. Иран, Белуджистан,
Афганистан, в СССР — Средняя Азия. В СССР распространен в пустынях и
полупустынях Туркмении от низовьев Аму-Дарьи и Кара-Богаза на севере до
Бадхыза на юге. В область распространения входят также северный и южный
Таджикистан: верхнее течение Сыр-Дарьи у ст. Мельниково, по Аму-Дарье
и Пянджу от Чарджоу до Кировабада, по Кафирнигану под Микоянабадом и у
озера Джиликуль на Вахте; пустыня Кызыл-Кум; восточное побережье
Аральского моря с островами из группы Мергень-тау; средние части пустынь
Аральские Кара-Кумы и Муюн-Кумы.
Карта 82. Распространение буланого козодоя Caprimulgus aegyptius.
1 — С. a. arenicolor, 2 — С. a. aegyptius, 3—С. a. saharae; а — граница гнездовий, г —
места зимовок (цифры в кружках обозначают номера зимующих подвидов).
Характер пребывания. В Туркестане гнездящаяся и перелетная
птица. Весенний пролет сравнительно поздний (позднее, чем у обыкновенного
козодоя) и растянутый. В вост. Иране первые птицы отмечались 18 апреля,
оживленный пролет — между 20 и 23 апреля и с этого времени тянулся до 4
мая (Зарудный, 1903). В Туркмении ясно выраженное весеннее движение
отмечено между 24 апреля и 10 мая (Зарудный, 1896). Однако первые буланые
козодои нормально появляются несколько раньше, так как их весенний крик
отмечался с середины апреля и ряд экземпляров Зоологического музея
Московского университета добыты между 10 и 12 апреля. Осенний отлет относительно
ранний, при высоких дневных температурах. Из пустынь, прилегающих к
нижнему течению Сыр-Дарьи, буланые козодои отлетают в первой половине
сентября. В восточных частях пустыни Кызыл-Кум он будто бы происходит в
сентябре (по старому стилю). В Туркмении обычно отлетают в начале сентября,
но некоторые особи задерживаются до последней трети сентября и даже позднее.
На озере Ильчик в низ. Аму-Дарьи экземпляр добыт 23 сентября, у Ашхабада
одна птица 25 сентября, а другая 24 октября (Дементьев, 1950). "В Сеистане
осеннепролетные буланые козодои в громадном числе скопляются между
23 сентября и 9 октября (Зарудный, 1903). Во время весеннего движения птицы
480
ОТРЯД КОЗОДОИ
летят одиночками, парами и группами до 10 особей. Пролет наблюдается
ранним утром, причем козодои совершают перелет на большой высоте.
Биотоп. Гнездовый, не однороден в различных частях ареала. Как
правило, птицы избегают гористых местностей и щебнистой почвы. Указания
в литературе о нахождении буланого козодоя в горах—Хорассанский участок
(Зарудный, 1896), Султан-уйздаг на Аму-Дарье (Гладков, 1832), видимо
относятся к негнездящимся птицам. В равнинах, прилегающих к нижнему течению
Сыр-Дарьи, явно избегает гнездиться в богатых водой участках, где есть тугаи
и тамариоковые заросли. Гнездовым биотопом здесь служат то безводные,
то едва всхолмленные барханные пески с бедной травянистой
растительностью и редкими деревцами саксаула и песчаной акации в котловинах.
Реже гнездится в глинистых пустынных с навеенными песками участках
(Спангенберг и Фейгин, 1936). В Закаспии населяет низкие и жаркие равнины— -
песчаные и глинистые с отдельностоящими кустами тамариска, джузгуна и
саксаула. Наиболее охотно-поселяется на участках пустыни, расположенной
вблизи пресной воды, где есть камыши и луга. В глубине пустынь, вдали от
воды, гнездятся редко и с наступлением наиболее жаркого времени
откочевывают отсюда к рекам (Зарудный, 1896).
Кормовой биотоп более разнообразен. В поисках пищи козодои посещают
тихие стоячие водоемы, ловя насекомых близ водной поверхности. Здесь же
поблизости от воды козодои иногда остаются на дневку. На Аму-Дарье в августе
удавалось встречать птиц на сухих участках почвы среди сплошных разливов.
В поисках насекомых птицы скопляются близ кустарников и колодцев. Иногда
у колодцев удается одновременно наблюдать до семи особей, ловящих
насекомых (Шестоперов, 1934).
Численность. Нередкая, а местами и многочисленная птица. Лишь
близ границы ареала, например в северных частях Аральских Кара-Кумов,
попадается редко.
Экология. Размножение. Гнезда не строит и откладывает яйца
на землю в едва заметное углубление песчаной почвы то совершенно открыто
на солнцепеке, то в тени кустика. Полная кладка содержит два яйца (как
исключение — одно). Как и у прочих козодоев, яйца имеют эллипсовидную
форму, они вытянуты и с одинаково округлыми конусами, скорлупа
мелкозернистая, гладкая и блестящая. Окраска: по основному серо-охристому фону
разбросаны глубокие светлосерые и поверхностные рыжевато-бурые то мелкие,
то более крупные пятна, образующие мраморный рисунок. Размеры: (5) 33,0—
31,3X22,0—22,9 мм. От яиц Caprimulgas europaeas отличаются общей рыжеватой
окраской и обычно более крупной величиной. Сроки размножения колеблются
как в отдельных частях ареала, так и в одной и той же местности у отдельных
пар, гнездящихся поблизости друг от друга. Часть птиц прилетает с
неразвитыми семенниками. Вполне возможно наличие двух кладов. Откладке яиц
предшествует песня самцов, которую удается слышать вскоре после прилета, в
Туркмении— в середине апреля.
Откладка яиц в низовьях Сыр-Дарьи происходит около середины мая.
19 мая в гнезде неполная кладка — 1 яйцо, другое —в яйцеводе самки; 26
мая—полные, слегка насиженные кладки (Спангенберг и Фейгин, 1936).
В Туркмении свежие яйца в гнездах обычно обнаруживаются во второй
половине мая. Насиживание начинается после откладки первого яйца, причем
самка сидит на нем так же прочно, как и на законченной кладке. Повидимому
оба родителя принимают участие в насиживании яиц. Когда самка
насиживает, самец держится поблизости от гнезда. Срок насиживания, видимо, как
и у других козодоев, —»17—18 дней. В Закаспии хорошо летающие птенцы
отмечались уже в конце первой трети июня, а на Мургабе плохо летающие —
22 июля (Зарудный, 1896). В Кызыл-Кумах 19 июня найден пуховой птенец,
БУЛАНЫЙ КОЗОДОЙ
481
в Сеистане 18 июня — яйца, из которых в тот же день должны были вывестись
птенцы и 14 июня— птенцы в возрасте недели. Птенцы выводятся зрячими
и покрытыми бледным густым пухом.
Питание. Изучено недостаточно, во всяком случае — разнообразные
насекомые. Основная масса пищи добывается во время полета. Значительно
реже с земли (в желудках козодоев найдены бескрылые насекомые). В Закас-
пии питается различными жуками, иногда довольно крупными, как, например,
Mouodon nentodoa, Orthoptera, Lepidoptera преимущественно разными Noctua
и другими насекомыми; особенно охотно преследует он Pentatoma (Заруд-
ный, 1896).
Линька. Первые признаки линьки обнаружены у птиц, добытых 22 мая
и 4 июня (нет 10 махового); 15 июня —линька 9-го и 10-го махового; 1 июля
Рис. 123. Буланый козодой.
у самца первые 4 маховых старые; 5-го нет, 6—10 — новые, нет средних рулевых;
б августа 1-е и 2-е маховые — старые, 3 растет, 4 и 10 новые, идет смена
рулевых —растут крайние. По Зарудному (1896) конец линьки взрослых птиц
совпадает с началом августа, конец линьки на наших экземплярах 29 августа.
Смена маховых идет от 10-го к 1-му; рулевые линяют от центра к крайним.
Линька молодых происходит в конце июля — начале августа. Молодая птица,
добытая на Теджене 18 августа, заканчивала линьку.
Полевые признаки. Отличается от других козодоев более крупными
размерами, но, главным образом, светлой золотисто-рыжей окраской оперения.
В дневные часы обычно прочно сидит в тени кустика или глинистого обрывчика
и слетает лишь при крайней необходимости. С заходом солнца и ночью охотно
усаживается на пыльные дороги и у арыков. Во время охоты летает бесшумно
и низко над землей, порою взмывая вверх, чтобы схватить летящих насекомых,
и вновь спускаясь к самой земле. Вечерами его часто удается видеть
летающим над тихой водой, где он занят охотой или налету пьет воду. Буланый
козодой не пуглив и близко подпускает к себе человека, охотясь среди юрт, в аулах
и среди пасущихся домашних животных. Он отличается молчаливостью. Обычно
удается слышать голоса самцов только в период размножения. В это время
самцы кричат все ночи напролет. Эти весенние крики можно передать, как
быстро следующие друг за другом слоги: «кре-кре-ке»..., причем звук
переходит порою то в ясное «у», то в «о» и звучит металлически. Во время пения ко^
31 Птицы СССР, т. I
482
ОТРЯД КОЗОДОИ
зодои подпускают человека особенно близко, умолкая в самый последний
момент (Зарудный, 1896).
Описание. Размеры и строение. Длина крыла: самцов (11) 193—212, самок
(9) 195—208 мм, среднее 209 и 208 мм.
Окраска. Птенцы покрыты светлым пухом. Молодые птицы светлее
взрослых. Общий тон оперения песчано-охристый. На груди и брюшке хорошо заметен
тонкий буроватый поперечный рисунок. Хвост птицы также разрисован бурыми поперечными
полосами, но более широкими и грубыми. Темя и спина испещрены тонкими продольными
штрихами, переходящими на верхней части крыла в крупные отметины. На горле — два
больших белых пятна. Первостепенные маховые темнобурые. На внутренних опахалах
их крупные белые волнообразные пятна. Наружные пары рулевых с охристо-белыми
концами. Самцы неотличимы от самок.
3. Большой козодой Caprimulgus indicus Lath.
Caprimulgus indicus. Latham. Ind. Orn. II, 1790, стр. 588. Индия.
Распространение. Ареал. Индия, Цейлон, Бирма, Китай, Япония,
Приамурье и ю.-в. Забайкалье. Зимой спускается до Индо-Китая и даже до Новой
Гвинеи.
Характер
пребывания. В северных частях
гнездящаяся, перелетная птица;
южные подвиды оседлы. Биотоп.
Леса, перемежающиеся с
открытыми пространствами как в
невысоких горах, так и в
низменностях.
Подвиды и варьирующие
признаки. Большой козодой объединяет
пять подвидов, отличающихся
окраской, размерами и тем, что северные
подвиды перелетны: С. i. hazarae
Whistleret Kinnear, 1935—
Гималаи от Симлы и Муссори до
Ассама и Юниани; Бирма и
Малайский п-ов; С. /. Indicus Latham
1790—-Индия к югу от Гималаев;
С. /. kelaarti Blyth, 1851 —
Цейлон; С. /. jotaka Temminck
et S с h 1 е g e I, 1847—ю.-в. Сибирь,
Амур и Уссурийский край, к югу до
Ганьсу, Манчжурии и сев. Чжили,
горы с.-з. Фудзяня, о-ва Хоккайдо,
Хондо и Риу-Киу; во время
пролета — в вост. Китае, зимою в Южн.
Китае, Индо-Китае, на Малайском п-ове, Суматре, Яве и Борнео; C.i. phalaena H а г t-
laub et Finsch, 1872 — острова Пелеу (Каролинский архипелаг).
Карта 83. Распространение большого козодоя
Caprimulgus indicus,
1 — С. i. hasarae, 2 — С. I. Indicus, •? — С. I. kelaarti, 4 —
С. i. jotaca, 5 — C. i. phalaena; a — граница гнездовий,
6 — не вполне выясненная граница гнездовий, г — зимовки
(цифры в кружках обозначают номера зимующих подвидов).
Северный большой козодой Caprimulgus indicus jotaka Temminck
et Schlegel
Caprimulgus jotaka. Temminck et Schlegel* in Siebold's Fauna Japonica, Aves
1847, стр. 37, Япония.
Распространение. Ареал. Япония (о-ва: Хоккайдо, Хондо и Риу-Киу),
Корея, Манчжурия, Китай от сев. Чжили до Гань-Су, Монголия до
подножья Хингана. В пределах Союза распространение выяснено недостаточно.
Населяет Приморье, вероятно проникая до устья Амура. Найден во всей
БОЛЬШОЙ КОЗОДОЙ
483
южной его части(Иогансен, 1927; Белопольский, 1950; Горчаковская и другие).
Обычен по среднему и нижнему Иману. На левобережье Амура гнездится близ
т>зера Болонь-оджал, обычен в бассейне р. Горин, есть экземпляр с берегов
озера Эворон в Зоологическом музее Московского университета.
На западе достигает Яблонового хребта (встречен на Зее, Дорогоетайский,
1915), населяет южные и западные части Читинской обл. (Гладков),
встречен под Троицкосавском (Моллесон, 1895). Зимует в Южном Китае, Индо-
Рис. 124. Большой козодой.
Китае, на Малайском п-ове, Суматре, Яве, Борнео, достигая даже Новой
Гвинеи.
Характер пребывания. В пределах Союза ■— гнездящаяся
и перелетная птица. Сроки прилета и отлета выяснены крайне слабо. Прилет
в Приморье около середины мая, на Имане первый отмечен — 14 мая (Спан-
генберг, 1940). Отлет начинается в сентябре и полностью заканчивается в конце
второй трети месяца. Перелет совершается вероятно одиночками и в
ночное время.
Биотоп. Гнездовый. Равнины и горные территории. Сплошных лесных
массивов и хвойной тайги избегает, поселяясь на лесных окраинах,
граничащих с лесными полянами и открытыми пространствами лугов и полей, на
вырубках и особенно на выгоревших участках лиственного леса. Реже птицы гнез-
31*
484
отряд козодои
дятся в небольших рощах, пятнами разбросанных среди
сельскохозяйственных угодий.
В поисках пищи широко посещают открытые местности речных долин,
поля, сухие и сырые луга, вейниковые болота, низкорослые кустарниковые
поросли, лесные поляны и озера с берегами, поросшими тальником, черемухой
и тополем.
Ч и с л е н~н о с т ь. Обычная, а местами в подходящих биотопах и
многочисленная птица.
Экология. Размножение. Гнездовые участки значительны. Гнезда
не строят. Яйца откладываются в небольшое углубление почвы, среди мелкого
валежника или полусгнившей прошлогодней листвы, иногда яйца лежат на
оголенной почве без всякой подстилки. Все найденные в СССР гнезда содержали
по два яйца. Яйца крупнее, чем европейского козодоя. Форма их вытянутая,
эллипсовидная, скорлупа гладкая и блестящая; окраска довольно постоянная.
По основному, слегка серовато-белому фону разбросаны светлосерые глубокие
и мелкие буроватые пятна, образующие красивый мраморный рисунок. Размер
яиц: (4) 32,6—31,0 х 23,2—21,9 мм. Данные о сроках размножения
чрезвычайно скудны. Откладке яиц предшествуют своеобразные крики самцов,
которые удается слышать в мае — июне и в первой части июля. Это позволяет
предполагать, что некоторые самки (вероятно молодые или потерявшие первые яйца)
откладывают яйца и в июле.
На Имане первые весенние голоса отмечены 27 мая, первое отложенное
яйцо — 8 июня, а законченные кладки со слабо насиженными яйцами —
13 июня. [На Зее и Яблоновом хребте голоса птиц были слышны 28 мая
(Дорогостайский, 1915).
Птица начинает насиживать, как только откладывается первое яйцо и уже
в это время слетает с гнезда лишь при крайней необходимости. У гнезд удается
находить и самца и самку. Как и у других козодоев, и самец вероятно принимает
участие в насиживании. Срок насиживания около 17 дней. На Имане совсем
маленькие птенчики найдены в гнезде 30 июня (Спангенберг, 1940), на реке
Горин 2 июля в гнезде один только что выведшийся птенец и одно яйцо
(Воробьев, 1938), всего один недавно выведшийся птенец найден 11 июля в
верховьях р. Читы (Павлов, 1948); летные молодые наблюдались у г. Имана
9 июля; летная молодая самка добыта на р. Горин 8 июля.
Питание. В желудках добытых на Имане птиц — остатки некрупных
жуков и ночных бабочек; на реке Куре и озере Болонь-оджал—♦остатки
насекомых и древесных семян (Бутурлин, 1913).
Линька. Линька происходит на зимовках. Взрослые экземпляры,
добытые с конца мая по конец августа, не имеют признаков линьки.
Полевые признаки. По внешнему виду и повадкам очень похож на других
наших козодоев -и отличается от них своей темной окраской оперения и
крупными размерами.
Не пуглив, мало боится человека и нередко охотится за насекомыми
среди селений. Голоса птиц удается слышать только в период
размножения, с вечерних сумерек и в течение всей ночи. Голос громкий, далеко
слышный и звучит как не слишком часто следуемые друг за другом слоги «вак-
вак-вак».
Описание. Размеры и строение. Длина крыла самцов (8) 202—221 мм, самок
(7) 199—217 мм, среднее 212 и 209 мм. По Бутурлину (1936) максимальный размер крыла
225 мм. Вес самца (1) 107 г, самок (2) 85 и 88 г.
Окраска. Пуховички отличаются от пуховичков европейского козодоя тем,
что брюшная сторона и бока тела имеют розовато-палевый оттенок, а на груди розовато-
палевое пятно (у европейского вида это пятно бурого цвета, Воробьев, 1938). Молодые
птицы похожи на самок, но все же отличаются от них менее ясными темными отметинами
оперения. Самки отличаются от взрослых самцов тем, что у, них на маховых и горле
БОЛЬШОЙ КОЗОДОЙ
485
белые пятна заменены охристыми, кроме того эти пятна меньших размеров, на четырех
парах рулевых. У самок также нет предвершинных белых пятен. Взрослые птицы. Общая
окраска; темная—из сочетания почти черных, темнобурых и охристых цветов. Темя и
спина покрыты продольными то мелкими, то более крупными черными пятнами. На брюшке
хорошо выражен сравнительно тонкий поперечнополосатый рисунок. Широкие поперечные
полосы хорошо видны также на хвосте птицы и на подхвостье. На горле белое большое
пятно. У самцов на первом маховом — белое пятно на внутреннем опахале; на 2-м, 3-м и
4-м маховых — поперечная белая полоса. Четыре крайние пары рулей с широкими белыми
предвершинными пятнами. Верхняя часть клюва черная, нижняя серая, ноги серые, глаз
коричневый, вокруг него желтое кольцо (Бутурлин, 1913). /
ОТРЯД СИЗОВОРОНКИ
ИЛИ РАКШИ
CORACIAE или CORACIIFORMES
Характеристика отряда
Морфология. Внешние признаки. Птицы средних и мелких
размеров; в нашей фауне самый крупный вид — сизоворонка, вес ее около 175 г,
размах крыльев около 640—660 мм и длина крыла в среднем около 160 мм.
Самый мелкий представитель отряда в нашей фауне обыкновенный зимородок:
длина крыла его около 70 мм, вес в среднем около 33 г. Среди представителей
жарких стран имеется несколько большая амплитуда колебаний в размерах.
Там наиболее крупные птицы размером с сизоворонку, но мелкие птицы
подотряда тоди, величиною приблизительно с нашего крапивника (длина
крыла 40—50 лш).
Оперение довольно жесткое, плотно прилегающее к телу. Цевка и пальцы
не оперены; цевка покрыта поперечными щитками (сизоворонки), или спереди
покрыта щитками, а сзади сетчатая (щурки). Окраска оперения яркая, иногда
с металлическим блеском, или пестрая — черная с белым. Полового
диморфизма в окраске нет или он выражен очень слабо, только у куролов Leptosomati-
dae половой диморфизм сильно развит. Возрастные изменения слабые и у
большинства видов отличия между первым гнездовым нарядом и взрослым
отсутствуют. Только у сизоворонок и зимородков первый годовой наряд отличим
от взрослого, — окончательный наряд они надевают на второй год жизни.
Крылья различной формы и величины: у зимородков и тоди они короткие,
тупые; у щурок, за исключением Nyctiornis,— острые и длинные. Эутаксия,
только у сизоворонок и у некоторых зимородков, преимущественно не
рыбоядных (Dacelo, Ceryle, Clytoceyx и др.) диастатаксия. Первостепенных маховых
обычно И, за исключением Nyctiornis, у которых 10. Рулевых—12, только у
Tanysitera — 10. У некоторых видов средняя пара рулевых удлинена; у щу-
ХАРАКТЕРИСТИКА ОТРЯДА
487
рок к вершине сужена, у момотов опахала средней пары рулевых
выщипываются самой птицей в предвершинной части пера.
Клюв обычно довольно длинный, сильный и почти прямой; у щурок
тонкий, слегка изогнутый вниз; у широкоротое —короткий большой и очень
широкий у основания; у тоди сильно уплощенный сверху вниз, с щетинкоподобными
перьями-вибриссами у основания клюва. Края клюва у сизоворонок, щурок
и зимородков гладкие, у тоди и момотов — зазубрены.
Ноги четырехпалые, обычно синдактильного типа, очень слабо выраженного
у момотов, и только у собственно сизоворонок ноги анизодактильного типа.
У большинства представителей отряда ноги слабые, но у сизоворонок, куролов
и момотов — сильные и довольно длинные. Когти острые, но слабые.
Анатомические признаки. Череп десмогнатический, голо-
ринальный. Базиптеригоидные отростки не развиты. Носовая перегородка
имеется. Сошник или рудиментарный, или отсутствует совсем. Шейных
позвонков 13—15, шея умеренной длины; но у зимородков короткая. Грудина
обычно с двумя парами вырезок (у сизоворонок чаще с одной). Гортань трахео-
бронхиального типа. Обводящая мышца отсутствует. Копчиковая железа
обычно не оперена, только у тоди и зимородков (за исключением рода Тапу-
siptera) оперена; у куролов копчиковая железа вовсе отсутствует. Пуха нет
ни у взрослых, ни у птенцов, кроме зимородков, у которых пух имеется даже
и на аптериях. Побочный ствол контурных перьев есть, но у некоторых
представителей (тоди и момоты) он рудиментарен.
Образ жизни* Дневные, древесные и наземные птицы. Большая часть видов
связана с разреженными, лиственными, смешанными лесами и водоемами,
некоторые — с большими тропическими, низменными лесами; незначительная
часть видов с открытыми степями, полупустынями и даже пустынями.
Обитающие в Палеарктике виды перелетны; за исключением обыкновенного зимородка;
преимущественно же это оседлые или кочующие птицы. На зимовках птицы
остаются короткое время, например, сизоворонки, зимующие в южной Африке
проводят на зимовках 1г/2—»2 месяца, прилетая туда в начале декабря и
отлетая в конце января или в начале февраля.
Пища преимущественно животная, в небольшом количестве растительная.
Большинство принадлежащих к отряду видов — энтомофаги, несколько меньше
ихтиофагов и еще меньше полифагов. Зимородки преимущественно рыбоядные
птицы, только небольшое число видов, принадлежащих отдельным родам —
Ceryle, Dacelo, Clytoceyx и др. питаются мелкими млекопитающими,
рептилиями и амфибиями. Обычно зимородки охотятся или облетая на небольшой
высоте свой кормовой участок, или выжидают добычу, сидя на дереве,
кустарнике или на каком-либо возвышении. Ловят и умерщвляют добычу клювом.
Рыбу ловят мелкую, при этом бросаются с разлета в воду и даже несколько
погружаются в нее, но не ныряют. Заглатывают добычу целиком. Щурки
широкороты ловят насекомых на лету, Nyctiornis собирают их с деревьев,
земляные ракши разыскивают насекомых в земле, разрывая ее при помощи
своеобразно устроенных когтей на пальцах ног. Сизоворонки и куролы ловят
насекомых и на лету, и берут их как и мелких позвоночных —* с земли.
Моногамы. У некоторых видов наблюдается колониальное гнездование.
Число пар в колонии очень различно — от двух-трех до нескольких сот.
К размножению приступают в первую весну; в возрасте около года у
некоторых видов (щурки,* широкороты) в гнездовое время наблюдаются стаи,
состоящие повидимому из холостых, прошлогодних птиц, по каким-то причинам не
приступивших к размножению. Пары постоянны. Места для гнезд разнообразны,
но обычно связаны с лесной растительностью; гнезда всегда устраиваются
закрыто — в дуплах деревьев, пней, в щелях построек, скал (сизоворонки,
куролы, широкороты, некоторые зимородки, но последние редко) и в норах
488
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
(щурки, некоторые зимородки, тоди, момоты и сизоворонки). Обычно норы
копают сами птицы — самец и самка, за исключением сизоворонки, которая
гнездится в норах, сделанных другими животными. Роют норы при помощи
клюва, ногами же лишь выбрасывают выкопанную землю. Длина норы различна
и часто зависит от твердости грунта. Гнезда занимаются птицами в течение
ряда лет. От готовности гнезда —норы сильно колеблются сроки начала
гнездования. Например, у щурок и зимородков можно найти одновременно
(у щурок даже в одной колонии) гнезда с законченной кладкой, только что
начавшейся, и еще недостроенные гнезда. На копание новой норы затрачивается
от 10 до 20 дней. Насиживает преимущественно самка, самец сменяет ее
лишь на короткое время. Срок насиживания 18—20 дней. Насиживание
начинается различно — у зимородков после окончания кладки, у
сизоворонок до окончания кладки, у щурок -— с первого отложенного яйца. В году
одна кладка, только у зимородков — две, иногда даже три. Количество яиц
в кладке очень различно у разных видов: у тоди — 3—4, редко 5, у куролов и
момотов 4—5, у сизоворонок обычно 5, иногда 4—7, у широкоротов 3—б, у
щурок б—8, реже 4—10, у Nycttornis 4—Q, у обыкновенного зимородка 6—7,
иногда 4—10, у крупных тропических зимородков 2—3. Яйца белые,
блестящие, округлой формы.
Птенцовые птицы; птенцы вылупляются слепыми и голыми.
Постэмбриональное развитие довольно медленное. Пухового наряда нет, птенцы одеваются
перьями гнездового наряда, который похож на взрослый, но отличается более
тусклым, бледным и грязноватым тоном. Первая ювенальная линька
начинается осенью и заканчивается в ноябре — декабре. Окончательный взрослый
наряд надевают обычно в первую зиму, за исключением зимородков и
сизоворонок, которые надевают полный взрослый наряд на второй год жизни. У
взрослых две линьки в году — и частичная — осенью, полная в конце зимы —
весной. Процесс линьки медленный, занимает 3—4 месяца и птицы не теряют
способность к полету.
Систематика. Отряд сизоворонок делится на 5 подотрядов. 1. Зимородки
Halcyones. 2. Тоди Todi. 3. Момоты Momoti. 4. Щурки Meropes. 5. Сизоворонки
Coraclae.
Подотряд зимородки Halcyones содержит одно семейство —зимородки
Halcyontdae. Сошник не развит. Язык рудиментарный. Сонная артерия
парная. Слепые кишки рудиментарны. 14—15 шейных позвонков. Грудина
с двумя парами вырезок. Клюв прямой, длинный, острый и сильный, края
клюва гладкие. Ноздри продолговатой формы и расположены у основания
клюва. Голова большая, шея короткая. Ноги слабые, короткие, 1-й и 2-й, 3-й
и 4-й пальцы сращены в основной своей части. Хвост короткий, у некоторых
видов короче клюва, прикрытый больше чем на половину длины верхними
кроющими перьями хвоста. Пух имеется и у взрослых и у птенцов. Окончательный
наряд надевают на второй год жизни. У взрослых полная линька с августа по
ноябрь, частичная с января — до марта. 14 родов (Alcedo, Ceryle, Chloroceryle,
Clytoceyx, Halcyon и др.) с 86 видами. Распространены в умеренных,
субтропических и тропических странах, главным образом, Восточного полушария,
только один род обитает в Северной и Южной Америке. В СССР — 3 рода:
Alcedo, Ceryle к Halcyon.
Подотряд сизоворонки Coraclae характеризуется рудиментарным
сошником, длинным языком без роговых бородок по краю языка или на его вершине.
Грудина с одной или двумя парами вырезок. Сонная артерия парная. 13—14
шейных позвонков, шея умеренной длины. Слепые кишки функционирующие.
У взрослых частичная линька проходит с июля по октябрь, полная с января
по апрель. Ноги сильные. Два семейства: собственно сизоворонки Coraciidae
UKyvom>\Leptosomatidae. Последние характеризуются бронхиальной гортанью,
ХАРАКТЕРИСТИКА ОТРЯДА
489
отсутствием копчиковой железы, развитием в поясничной области пудреток,
выделяющих порошковидный пух, 4-м оборотным пальцем, иризирующей
окраской оперения и расположением посредине клюва щелевидных ноздрей,
прикрытых роговыми покрышками. Половой диморфизм сильно развит. Ку-
ролы содержат один род куролы Leptosomus и один вид Leptosomus discolor,
обитающий на Коморских о-вах и Мадагаскаре*
Семейство сизоворонки Coraciidae содержит 5 родов (Coracias, Eurystomus,
Brachypteracias, Atelornis и Uratelornis, с 16 видами, распространенными в
умеренных и тропических странах Восточного полушария. Многими авторами роды
Brachypteraciast Atelornis и Uratelornisy обитающие на Мадагаскаре, выделяются
в самостоятельное семейство земляных ракш Brachypteraciidae, главным
образом, на основании особенностей расположения когтей на пальцах и
экологии (наземные птицы).
Подотряд щурки Meropes. Характерные особенности: рудиментарный язык,
функционирующие длинные слепые кишки, левая сонная артерия (за
исключением Nyctiornis, у которых она парная). Копчиковая железа не оперена.
Пуха нет, побочный ствол контурных перьев имеется. 14 шейных позвонков,
шея умеренной длины. Грудина с двумя парами вырезок. Ноги слабые с
довольно слабыми, но острыми когтями. Клюв тонкий несколько изогнутый
вниз, с резко выделяющимся хребтом надклювья. Края клюва не зазубрены,
ноздри округлой формы, расположены у основания клюва и несколько
прикрыты мелкими перьями лба. Крылья острые длинные (за исключением
Nyctiornis), самое длинное перо второе маховое. Одно семейство щурки Meropldae
с 7 родами (Merops, Dtcrocercus, Melittophagus, Aeropsf Nyctiornisf Moropogon
и Bombylonax) и с 23 видами, распространенными преимущественно в
тропических и субтропических странах Восточного полушария. К нам доходят только
два вида М. apiaster и М. superciliosus.
Подотряд момоты Momoti характеризуется рудиментарным сошником,
длинным языком с бахромчатыми выростами на конце; отсутствием слепых
кишок; сонная артерия парная; 15 шейных позвонков. Грудина с одной парой
вырезок. Ноги довольно сильные, слабо выраженного синдактильного типа. Пуха
нет, побочный ствол контурных перьев рудиментарный. Одно семейство момоты
Momotidae с б родами (Momotus, Baryphthengus, Eumomota, Electron, Aspatha,
Hylomanes), с 8 видами, распространенными в Средней и Южной Америке.
Подотряд тоди Todi. Самые мелкие птицы отряда. Сошник отсутствует,
15 шейных позвонков, шея умеренной длины. Язык длинный, заостренный,
без бахромчатых выростов. Межглазничная перегородка имеет большие фон-
танели. Пуха нет, побочный ствол контурных перьев рудиментарный.
Копчиковая железа оперена. Слепые кишки функционирующие. Короткие крылья
и хвост. Одно семейство Todidae с одним родом тоди Todas и с 5 видами,
распространенными на Антильских о-вах.
Ископаемые остатки сизоворонок найдены в верхне-эоценовых и нижне-
олигоценовых отложениях (Франция), по ним описан вид Geranoptems alatus.
Рецентные роды Coracias, Momotus, Alcedo и Ceryle известны из плейстоцена.
Сизоворонок соединяли вместе с совами, козодоями, удодами и кукушками
в один отряд ракшеобразных, но многие своеобразные черты морфологии и
экологии сизоворонок позволяют их выделить в самостоятельный отряд.
Географическое распространение. Сизоворонки распространены
преимущественно в тропических, субтропических и умеренных странах Восточного и
Западного полушарий. В Западном полушарии они представлены двумя
подотрядами—тоди и момоты — с небольшим количеством видов и одним родом из
подотрядов зимородков Haley ones. Представители подотрядов щурок и
сизоворонок в Западное полушарие не заходят, так же как и тоди и момоты — в
Восточное* Наибольшее разнообразие видов наблюдается в тропических и субтропи-
490
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
ческих странах Старого Света. Общее число видов в отряде сизоворонок 141,
из них в Западном полушарии обитает 14 видов, остальные 127 — в Старом
Свете. В фауне СССР гнездится 5 видов и четыре вида случайно залетают (всего,
следовательно, 5,67% общего числа видов отрядов). Они относятся к трем
подотрядам—щуркам, сизоворонкам и зимородкам.
В отряде сизоворонок наблюдается большое количество монотипических
видов — 45 от общего количества видов, в фауне СССР — один
монотипический вид.
Распределение сизоворонок на гнездовье по зонам в нашей стране таково
(в числителе показано число видов, в знаменателе —* число подвидов и
монотипических видов, характерные формы подчеркнуты):
I. Арктика (зона) —О
II. Лесная зона Coracias garrulus, Alcedo atthis, Merops apiaster, Eurystomus
orientalis— 4
а. Тайга (подзона) —О
б. Смешанный лес (подзона)Coraciasg. garrulus, Alcedo a. atthis, Alcedo a. ispida,
Alcedo a. bengalensis, Eurystomus orientalis abundus — 8/e
в. Лесостепь (подзона) Coracias g. garrulus, Coracias g. semenowii, Alcedo atthis
atthis, Alcedo atthis bengalensis, Merops apiaster —*fA
III. Зона открытых сухих пространств Coracias garrulus, Merops apiaster, Merops
superciliosus — 3
г. Степь (подзона) (озерная страна Зап. Сибири и Казахстана). Coracias g. garrulus,
Coracias g. semenovi, Alcedo a. atthis, Merops apiaster, Merops superciliosus persicus —4/4
д. Полупустыни и пустыни (подзона) Merops apiaster, Merops superciliosus
persicus, Coracias g. semenovi — 3/2
IV. Горная зона Merops apiaster, Merops superciliosus, Coracias garrulus, Alcedo
atthus — 4
е. Высокогорная бореоальпийская подзона — О
ж. Горно-лесная (подзона) Coracias g. garrulus, Coracias g. semenovi, Alcedo a.
atthis, Alcedo a. bengalensis, Merops apiaster, Merops superciliosus persicus — 4/e
з. Горностепная и пустынная (подзона) Merops apiaster, Merops
superciliosus persicus, Alcedo a. atthis, A. a. bengalensis — 3/з
Экономическое значение. Щурки в некоторых районах, поселившись близ
пасек, приносят вред пчеловодству. Остальные представители отряда —
полезные птицы.
Таблица для определения видов отряда сизоворонки
1(2) Клюв длинный, тонкий, загнутый вниз. Рулевых — 5 пар. Средняя пара рулевых
удлинена и в вершинной части сужена 3
2(1) Клюв прямой, сильный, длинный или короткий, в последнем случае очень широкий
у основания. Рулевых — б пар. Средняя пара рулевых не удлинена 5
3(4) Затылок рыжевато-коричневый, горла желтое. .
Золотистая щурка Merops apiaster
4(3) Затылок зеленый, горло рыжевато-коричневое
Зеленаящурка Merops superciliosus
5(6) Низ тела голубой или зеленый. Крыло длиннее 170 мм, цевка длиннее 19 см . .7
6(5) Окраска низа иная. Крыло короче 170 мм, цевка короче 19 мм 9
7(8) Клюв длинный, сильный, прямой. В окраске спины преобладают охристо-рыжие
тона Сизоворонка Coracias garrulus
8(7) Клюв короткий, но очень широкий у основания. Окраска спины с преобладанием
темнозеленого цвета Широкорот Eurystomus orientalis
9/10 Клюв длинный, сжатый с боков и с резко выступающим хребтом надклювья
10(9) Клюв у основания широкий, с боков не сжат, хребет надклювья округлый. .... 13
11(12) Окраска оперения голубая с рыжим. Крыло короче 90 мм. Хвост короче клюва
или равен клюву .Обыкновенный з и мор о д о к Alceda atthis
СИЗОВОРОНКА
491
12(11) Окраска оперения черная с белым. Крыло длиннее ПОлш. Хвост длиннее клюва
15
13(14) Верх головы матово-черный, спина блестяще голубая, вокруг шеи белый
ошейник О шей ни ковы й зимородок Halcyon pileata.
14(13) Верх головы и шеи и передняя часть спины коричнево-бурые. Ошейника нет
. . Белогрудый зимород'о'к Halcyon smyrnensis
15(16) Крыло короче 150 мм, спина пестрая \
• Малый пестрый зимородок Ceryle rudis.
16(15) Крыло более 160 мм, спина поперечнополосатая
Большой пегий зимородок Ceryle lugubris.
СЕМЕЙСТВО СИЗОВОРОНКОВЫЕ CORACIIDAE
РОД СИЗОВОРОНКА CORACIAS LINNAEUS 1758
Тип Coracias Garmlus
1. Сизоворонка Coracias garmlus L.
Coracias Garmlus. Linnaeus. Syst. Nat., изд. X, 1758, стр. 107, Швеция.
Русское название. Сизоворонка — название народное, от окраски птицы и
некоторого внешнего сходства ее с воронами. Приводится еще Палласом. Ракша — народное
украинское название.
Распространение. Ареал. Европа, за исключением севера; Средняя и
ю.-з. Азия и с.-з. Африка. Швеция к югу от 61°с. ш., Германия, Прибалтика,
южная часть Ленинградской обл., Калининская и Горьковская обл.,
Татарская республика и от южной оконечности Урала, Актюбинска и Омска
на восток до западного Алтая. К югу идет до Средиземного моря (Марокко,
Алжир, Тунис), Малой Азии, Палестины, Сирии, Месопотамии, Ирана, южного
Закаспия, Средней Азии, Афганистана, Белуджистана и с.-з. Индии.
В пределах очерченной области сизоворонки отсутствуют на большом
пространстве, начиная от низовьев Урала, Усть-Урта и далее к востоку до
пустыни Бек-Пак-Дала, нет ее также и в Зайсанской котловине и на Тар-
багатае.
Зимует в тропической и южной Африке к северу до Кении, Уганды,
Египетского Судана, южной Нигерии и Камеруна; к югу до Капской Земли.
Характер пребывания. Гнездящаяся и перелетная птица. Из
южной Африки сизоворонки отлетают уже в конце января и в начале февраля.
Через тропическую Африку пролетают в марте и в апреле. Главная масса летит
через Египет, Красное море и Аравию. Осенью первые птицы появляются
в Египте в двадцатых числах июля (23 июля), массовый пролет происходит
в августе и идет до конца сентября. В Судане пролет наблюдается в сентябре
и в октябре. В Кении и бывшей немецкой восточной Африке пролет идет в
начале ноября. В южную Африку (в Землю Наталь) прилетают в декабре (Гроте,
1930, Мейнерцхаген, 1930).
Залеты отмечены в Норвегии, Исландии, Ирландии, Великобритании, на
о-вах Оркнейских, Фарерских и Канарских, о. Мадейра, в Бельгии, Дании,
северной Франции, Голландии, Финляндии, на Кольском п-ове; на севере
СССР, даже на Новой Земле.
Биотоп. Лесостепь, культурный ландшафт, открытые степи с отдельно
стоящими деревьями и совершенно лишенные деревьев, но изрезанные
обрывами, оврагами, реками и т. д., полупустыни, речные долины.
Подвиды и варьирующие признаки. Два подвида, различающихся между собою окраской
оперения и экологией: С. g. garmlus Linn., 1758— Средняя и Южная Европа, с.-з. Африка
■и Азия — за исключением северных и восточных частей и области распространения
следующего подвида; С. g. semenowi Loudon etTschusi, 1902 — от Средней Азии до Ирака
и с.-з. Индии.
492
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
Обыкновенная сизоворонка Coracias garrulus garrulus L.
Синоним. Coracias garrulus caucasica. Бутурлин, Орнитологический
Вестник, 1910, стр. 190, Закавказье.
Распространение. Ареал. Западная Европа от южной Франции, Гер-
мании и южной Швеции (к северу до 61° с.ш,) к югу до Средиземного моря и его
островов (Сардинии, Сицилии и Кипра). Также с.-з. Африка—Марокко,
северный Алжир и Тунис. Малая Азия, Палестина, Сирия. В СССР сизоворонка
Карта 84. Распространение сизоворонки Coracias garrulus
1 — C.g. garrulus, 2 — С. g. semenowi; a — граница гнездовий, г — зимовки (цифры
в кружках обозначают номер зимующих подвидов).
распространена от Украины, Крыма и Кавказа, к северу до Прибалтики,
южной части Ленинградской обл.: Луги, Гдова, Пскова и Тихвина. Доходит
до Устюга Вологодской обл. Есть в Калининской обл., в окрестностях Костромы
и в Горьковской обл., в последней наблюдались весной во многих районах,
но гнездовье доказано только для районов Лукояновского и Ардатовского,
(в Ветлужском районе на гнездовье не найдена, Воронцов, 1935) и между
Муромом и Арзамасом (Кузнецов в 1938 г. на притоке р. Теша нашел 5—6
гнездящихся пар). Однако гнездится и в южной части Кировского края на север
до Яранского района (Плесский, 1933). По Средней Волге доходит на север
до Казани, есть в окрестностях Бугуруслана и Бузулука, но в б. Уфимской
губ. на гнездовье не найдена. На Урале распространена к северу до предгорий
Южного Урала — 52° с. ш. — на р. Сакмара, Илек, в Губерлинских горах и
Иремельской тайге. *
СИЗОВОРОНКА
493
К югу по среднему течению Урала гнездится до Уральска, Красноярской,
станицы, в низовьях Урала никем не встречена, хотя в низовьях Волги и в
Волжско-Уральских степях одна из самых обычных птиц на гнездовье. Далее
узкой полосой идет на восток, спускаясь к югу до Актюбинска, Мугоджар и
Семипалатинска. К северу поднимается до Наурзума, Кустаная, Кокчетавской
возвышенности, в небольшом количестве доходит до Омска; есть в
окрестностях Усть-Каменогорска, в верховьях Иртыша. Отсутствует в большей части
Барабинской степи, гнездится в Кулундинской степи, в западной окраине
Алтая, но в горы не заходит, есть в окрестностях Барнаула (с. Ключи 130 ос от
Барнаула, с. Шульбинское и с. Новенское — это самые северо-восточные
встречи сизоворонки на гнездовье). Встречена летом на У бе, но гнездование
там не доказано. Щ
Гнездится в восточной части Зайсанской котловины, в предгорьях Бар-
лыка и в Эмильской котловине, но на Тарбагатае отсутствует.
Известны залеты сизоворонки довольно далеко на север от ее гнездовой
области. Так, сизоворонка наблюдалась близ деревни Борки Петергофского
района Ленинградской обл., в б. Олонецкой губ., в Ростовском районе
Ярославской обл., в Вологодской обл., в окрестностях г. Молотова, в июне, в июле
ив августе — в Уржуме и в Прикамье, в мае — в Красно-Уфимске и под
Соликамском, в горнолесных районах — Южного Урала. Самые далекие залеты
сизоворонки отмечены в октябрей ноябре 1868г. в округе Варангер в
Финмаркете, 12 апреля 1903 г. на Новой Земле между Белушьей [губой и
Кармакулами ив 1921 г. на Маточкиншар.
В общем можно сказать, что в Европейской части нашей страны северная
граница распространения сизоворонки совпадает с северной границей
широколиственных лесов, хотя гнездовые местообитания ее здесь приурочены не
столько к широколиственным лесам, сколько к разреженным сосновым борам.
В Азиатской части северная граница повидимому совпадает с южной границей
сосновых колков лесостепной полосы. Рубки лесов и лесные пожары^способ-
ствовали проникновению сизоворонки к северу. Так, она стала обычной в
начале этого столетия в б. Псковской губернии, появившись впервые в 80-х годах
прошлого столетия, в Ленинградской обл. (окрестностях Гдова) количество
сизоворонок постепенно увеличивается: в 1903 г. гнездилось там 1—2 пары,
в 1905 и 1906 гг. уже было 5—б пар (Исполатов, 1907).
Характер пребывания. У нас гнездящаяся перелетная птица.
С мест зимовок отлетает большими стаями, но на юге нашей страны появляется
небольшими стайками в 4-^5 штук. Летят сизоворонки обычно на средней
высоте. К нам прилетают поздно. Так, в Армению прилетают в средних и
двадцатых числах апреля. К Ленкорани и в Талышскую низменность — 19—25
апреля, к Тбилиси и на среднее течение Куры несколько поздней —2—5 мая,
к Дербенту на западный берег Каспия —2 мая (Радде, 1885). На северном
Кавказе наблюдаются на пролете с середины апреля до середины мая, в среднем
в конце апреля, в горной части с.-з. Кавказа — 8—10 мая (Аверин и Наси-
мович, 1938). В Крым прилетают в конце апреля и начале мая. На Украину—^
со второй половины апреля и пролет идет до конца первой декады мая. Для
Харьковской обл. средняя дата прилета — 23 апреля, самое раннее появление
отмечено в 1890 г. ■—. 16 апреля, самое позднее в 1892—5 мая (Сомов, 1897),
для окрестностей Киева среднее прилета 22 апреля, самый ранний прилет от>-
мечен в 1924 г. — 17 апреля, самый поздний в 1921 г. —7 мая (Данилович,
1930). В Днепропетровскую обл. самый ранний прилет отмечен в 1906 г. —
25 апреля, самый поздний 10 мая 1901 г. (Боровиков, 1907). В Подолии
появляются во второй половине апреля. В Астраханский край прилетают в конце
апреля и пролет идет до конца первой декады мая, на Среднюю Волгу
прилетают с первых чисел мая и до середины мая. На Эмбе пролетные стаи сизоворо-
494
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
нок наблюдались до конца мая. В центральные области начинают появляться
с конца апреля и летят до середины второй половины мая, самый ранний
прилет наблюдался в Рязанской обл. — 18 апреля (Хомяков, 1900), самый поздний
в Тамбовской и Смоленской — 18 мая (Резцов, 1910; Огнев, 1909). В
Белоруссию — в первой половине мая и только в очень ранние вёсны — в конце
апреля. В Прибалтику—в конце апреля. В Ленинградскую обл. —в первых
числах мая и пролет идет до середины мая. В Средней Азии пролет идет с
половины апреля и до конца мая. Так, в Гассан-кули пролет наблюдался с половины
апреля и до конца мая (Лоудон, 1909), между Мары и Чарджоу —. 27 апреля
1892 г. встречались пролетные стайки, в окрестностях Караджа —. Батыр —
2 мая, у Гелле-Чешме в Карабиле —. 15 апреля (Зарудный, 1896). В Каза-
линек первые птицы появились б мая 1927 г. и 11 мая 1928 г., ранний
прилет отмечен в окрестностях Джулека — 27 апреля 1930 г. (Спангенберг,
1936). В Гиссарской долине пролет наблюдался 27 апреля.
Осенний отлет сизоворонок начинается в первых числах августа.
Вначале покидают места гнездовья взрослые птицы, затем молодые. Улетают
небольшими стайками, парами и в одиночку. Из Прибалтики последние птицы
отлетают в начале октября, из Ленинградской обл. отлет начинается с
первой половины августа и идет до конца сентября. В центральных областях
отлет начинается со второй половины августа и заканчивается в начале сентября.
Так, в Калужской обл. первые пролетные сизоворонки отмечены 18 августа
(Филатов, 1915), в Рязанской обл. последние птицы наблюдались 25 августа
1943 г. и 10 сентября 1944 г., из Тамбовской — последние птицы улетели
8 сентября 1936 г. (Птушенко, 1937), из Смоленской обл. в среднем начинают
отлетать 17 августа. Из Белоруссии отлет начинается с середины августа и
идет до начала сентября, в Минском полесье последние птицы встретились
7 сентября 1905 г. и 5 сентября 1901 г. На Украине — с конца августа и до
начала октября: в Харьковской обл, в среднем начинается 7—-8 сентября,
самый ранний отлет — 1891 г. и 1893 г. 13—15 августа, но он прекратился на
несколько дней и вновь возобновился 24—25 августа. Самый поздний отлет
в 1885 г. 22 сентября (Сомов, 1897). В Днепропетровской обл. последние
пролетные стаи встречаются в течение сентября, из Подолии последние птицы
исчезли в начале октября. С середины августа в южных частях Украины
сизоворонки начинают встречаться в большом количестве всюду и только
к началу октября число их значительно уменьшается.
На Кавказе пролет сизоворонок идет с конца августа и до октября, во
время пролета птицы поднимаются высоко в горы, иногда до перевалов (Беме,
1926). Местные птицы отлетают на Северном Кавказе с первых чисел
сентября и до конца этого месяца и даже начала октября (Северная
Осетия и Ингушетия; горная часть северо-западного Кавказа), в Закавказье
местные птицы исчезают в начале сентября, из Армении— в конце августа.
8 северном Казахстане пролет идет с первых чисел августа и до начала
октября; северное побережье Каспия покидают во второй половине сентября
(Бостанжогло, 1911).
Биотоп. Лесостепь, степь, культурный ландшафт. У северной границы
своего распространения сизоворонки предпочитают разреженные сосновые
боры и дубравы и избегают густых еловых лесов, охотно держатся по вырубкам
и гарям. В лесостепной полосе они селятся преимущественно в колковых и
долинных лесах, в степных дубравах, в садах и в ивовых насаждениях. В чисто
степных районах обитают по обрывам, оврагам, берегам рек и т. д. В Крыму
в степных и предгорных местностях. На Кавказе предпочитает селиться
в низких, равнинных местностях, поросших колючим кустарником, по широким
долинам рек с древесной растительностью и в предгорьях; в горы подымается
не выше 800--1000 м (Цветков, 1892; Радде, 1887).
СИЗОВОРОНКА
495
Численность. Многочисленна на Украине, в низменных равнинных:
местностях Крыма и Кавказа, в предгорьях уже редка. В большом количестве
встречается по Волге от устья до Куйбышева, по среднему течению Урала,
по мере приближения к северной границе распространения количество
сизоворонок уменьшается. У самой северной границы встречаются только
одиночные пары не ежегодно гнездящиеся или даже, повидимому, одиночные
холостые экземпляры. Так, в Бузулукском районе она еще весьма обычна, в Бугу-
русланском встречается уже значительно реже. В Московской обл.
сизоворонка редка на гнездовье во всех районах, за исключением
Орехово-Зуевского, где довольно обычна. В Калининской обл. также редка на гнездовье и
только в Зубцовском районе встречается в большом количестве (Давыдов, 1896).
Экология* Размножение. Возраст наступления половой зрелости:
неясен. Сейчас же после прилета у сизоворонок начинаются игры и
спаривание. К размножению приступают довольно поздно. Сизоворонки моногамы,
держатся обычно парами и устраивают гнезда на большом расстоянии друг от
друга, но иногда встречаются несколько гнезд на очень близком расстоянии
одно от другого, что повидимому объясняется трудностью нахождения
удобных для гнездовья мест.
В лесных местностях устраивают гнезда почти исключительно в дуплах,
деревьев и в пустотах пней. В степной зоне сизоворонки чаще устраиваюг
гнезда в норах, в обрывах и оврагах, в глинистых берегах рек, в трещинах,
зданий и колодцев, чем в дуплах деревьев. Так, в степных безлесных
местностях Харьковской обл. сизоворонки гнездятся в расщелинах глинистых или-
меловых оврагов и в норах. В Поволжских степях часто устраивают гнезда
в глинистых обрывах балок и речных долин. На Кавказе выкапывают
глубокие норы в глинистых обрывах, но также устраивают гнезда и в дуплах
деревьев. В Армении гнездятся в норах, по обрывам и в щелях каменных и
глинистых домов.
Дупла, занимаемые для гнезд, расположены на различной высоте, начиная:
от самой земли и до 15 метров высоты. Высота гнезда 75 мм> диаметр гнезда
120 мм, диаметр лотка 75 мм, высота 40 мм (Харьковская обл., Сомов, 1897).
Норы роются в обрывах обычно глубиною от 3—5 м, диаметр отверстия норы —
10 см, горизонтальный ход около 60 см длины, иногда перед образованием
гнездовой камеры заворачивает в сторону. Гнездовая камера в диаметре около 25 см,
высота L5 см (Предкавказье, Перекопов, 1940). Яйца в гнездах иногда лежат без
всякой подстилки, иногда сизоворонки устраивают плотную подстилку. Дно
гнезда, устроенного в дупле, обычно выстлано полусгнившими листьями и
измельченной гнилой древесиной, гнездо в норе — сухими травами с
небольшим количеством сухих листьев, лоток сделан из тех же, но более нежных,
материалов.
В кладке от 4 до 6 яиц почти правильной округлой формы, с гладкой,
плотной и блестящей белой скорлупой. Размеры: (208) 40—32x31,5-^-25,5 мм,
среднее 35,44x28,43 мм (Умань; Гебель, 1879); 37—34,4x29,6—27,5 мм,.
среднее 35,5 х 28,6 мм (Харьковская обл.; Сомов, 1897).
На Кавказе кладка яиц начинается со второй половины мая и продолжается,
до конца месяца и даже до начала июня. Так, в Закавказье (Аралых) гнезда,
с неполной кладкой от 1 до 4 яиц встречались в конце мая (Сатунин, 1907).
На Украине кладка яиц идет со второй половины мая и до середины июня:
в Днепропетровской обл. кладки встречались с 21 мая по 16 июня, а
насиженные с 31 мая (Боровиков, 1907); в Харьковской обл. — ненасиженные кладкиг.
с середины мая и до первой декады июня. В Поволжье и Заволжье — с конца
мая ц до середины июня (в бассейне р. Еруслан 16 июня найдено гнездо
с четырьмя яйцами, близкими к вылуплению; Волчанецкий и Яльцев, 1934);
в Бугурусланском районе встречено 8 июня два гнезда—в одном было 3 не-
496
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
насиженных яйца, в другом птицы сидели уже на яйцах (Карамзин, 1909).
В центральных областях кладка яиц начинается в конце мая или в начале
июня: в Смоленской обл. полные кладки встречались 17 июня (Огнев, 1909),
в окрестностях Вонлярова найдено гнездо с яйцами 3 июня (Воронцов, 1938),
в Рязанской обл. 12 июля были найдены только что вылупившиеся птенцы
(Хомяков, 1910).
В Белоруссии к кладке яиц сизоворонки приступают с конца мая и нена-
сиженные кладки яиц встречаются до второй половины июня, в Минском
Полесье самые ранние, еще неполные кладки в 3 яйца встречались с 26 мая и
самые поздние полные, но ненасиженные кладки 19 июня (Шнитников, 1913).
В окрестностях Пскова найдено гнездо с двумя ненасиженными яйцами
20 мая 1895 г. и 19 мая 1902 г. гнездо с 4 ненасиженными яйцами (Зарудный,
1910). На основании этих сведений о сроках гнездования можно сказать, что
больших различий во времени начала гнездования на юге страны и на севере
нет. В году бывает одна кладка.
Насиживание начинается с первого яйца и продолжается 18—19 дней;
на яйцах птицы сидят крепко. Вылупляются птенцы голыми; выкармливают
их самец и самка; гнездовый период длится 26—28 дней. Вылет птенцов на
Кавказе начинается со второй половины июня: близ Мцхета наблюдался вылет
птенцов в начале второй половины июня (Баньковский, 1913), в Кизлярском
окр. 5 июля 1912 г. найдены в гнезде уже оперившиеся птенцы (Беме, 1926).
На Украине вылет птенцов наблюдается с конца июня или с первых чисел
июля и продолжается до конца месяца (в Харьковской обл. самый поздний
вылет птенцов наблюдался 30 июля, Сомов, 1897; в Черниговской обл.
вылетевшие молодые встречаются в конце июня, Емельяненко, 1917; в Подолии
летные молодые птицы наблюдаются с последней трети июля, Портенко,
1928). В Поволжье вполне оперившиеся молодые встречаются около
середины августа (12 августа, Богданов, 1871). В Рязанской обл. вылет птенцов
начинается с середины июля 13—.18 июля (Хомяков, 1910). В Белоруссии
(б. Минской губ.) с 1 июля и до конца месяца (Шнитников, 1913). После вылета
птенцов из гнезда родители продолжают некоторое время кормить их, и они
держатся в это время выводками близ гнезд. Позже сизоворонки начинают
кочевать по степям и полям, если поблизости есть леса, то на ночь они
улетают в леса, в безлесных местностях ночуют в открытых местах, там, где
проводят день, В это время их можно видеть всюду: часто на телеграфных
проволоках, в одиночку и стаями, нередко в перемежку с горлицами,
сорокопутами, ласточками и даже мелкими соколами.
Линька. У взрослых бывает две линьки в году, одна — частичная -—
послебрачная с июля по ноябрь. Она начинается после вылета птенцов из
гнезд, но к моменту отлета сизоворонки не успевают сменить все мелкое
оперение и улетают на зимовки со смесью старого и нового мелкого пера. Рулевые
и маховые не сменяются. Вторая линька—полная—происходит на зимовках
с января по апрель. Частичная линька молодых происходит в ноябре —»
декабре. Предбрачная первая полная линька бывает с января по май, но
молодые птицы в первом весеннем наряде еще отличимы от взрослых.
Питание. Пищу составляют, главным образом, различные крупные
насекомые: медведки, кузнечики, жуки (жужелицы, щелкунчики), саранча.
Сизоворонки собирают добычу обычно на земле. Ловят также ящериц и в
исключительных случаях мелких грызунов, птиц и лягушек. Во вторую половину
лета, когда в поле сняты хлеба, сизоворонки в большом количестве встречаются
на полях, и тогда пища их состоит, главным образом, из прямокрылых» Осенью
с момента созревания фруктов в состав корма входят и различные фрукты;
в особенности виноград на юге страны, но большого вреда они виноградникам не
приносят. Сизоворонку можно считать полезной птицей. В питании сизоворонки
СИЗОВОРОНКА
497
в районе г. Ворошиловска, Сев. Кавказ, в июне и июле исследовано 400 экз. —
взрослых птиц; главную массу (92,7%) составляют жуки; из них Pentodon
46,2%, Gymnopleurus mopsus. 18,1%, Pitosia hungarica — 16,8%, Rhizotrogus
sp. —1,5%, Dorcadion sp. —3,3%, Copris lunaris —3,7%; прямокрылых
4,1%—2,5%, Tettigoniidae, 1,2%. Кроме того слизни Limacidae. Второстепенная
пища — позвоночные 0,5%; остальное — случайный корм. Вредных для
сельского хозяйства животных 56,7%, полезных—22,3%, остальные
нейтральные (Прекопов, 1940).
Полевые признаки; Сизоворонка легко узнается благодаря яркой
блестящей зеленовато-голубой окраске оперения, прямому, сильному клюву
и быстроте движений. Полет быстрый; по земле передвигается довольно
неуклюже и встречается обычно только во время собирания насекомых. Чаще всего
птицу можно видеть на лесных полянах, опушках лесов, сидящей на суке
дерева, копне, стоге или телеграфных проволоках, высматривающей добычу.
Увидав добычу, она стремительно бросается за ней. Ненапуганные
сизоворонки допускают человека довольно близко, но в случае преследования
становятся осторожными. Голос резкий; немного хрипловатый крик «ра-рак»
или «рэк-рэк» — птица издает, перелетая с дерева на дерево или слетая на
землю.
Описание. Размеры и строение. Птицы средних размеров, приблизительно
с галку. Клюв сильный, сжатый с боков, слегка крючковатый у вершины. Ноздри щелевидны,
расположены у основания клюва и несколько прикрыты перьями лба. У угла разреза клюва
имеются жесткие, щетиноподобные вибриссы. Ноги и пальцы довольно сильные, когти
слабые. Цевка покрыта щитками. Первостепенных маховых — 1К Формула крыла: 2>3>1>4
>5... Обрез хвоста прямой, рулевых — 12. Длина тела самцов (8) 300—343, среднее 329,2 мм,
самок (5) 320—342, среднее 330,2 мм; длина крыла самцов (44) 180—208 мм, среднее
192,7; самок (46) 181—207; среднее 190,6 мм. Размах самцов (4) 620—650, среднее 639,2 мм,
самок (2) 660—666 мм. Вес самца (1) — 183 г.
Окраска. Птенцы. Вылупляются голыми, слепыми, на 4—5 день покрываются
пеньками перьев, которые долго не раскрываются. Молодые птицы окрашены сходно со
взрослыми, но более тускло. На зобе и груди буроватый той. Горло беловатое. Лиловый цвет
на малых кроющих крыла слабо развит. Первый (годовой) наряд сходен с последую-
32 Птицы СССР, т. I
498
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
щими, отличаясь только тем, что малые кроющие крыла не имеют еще яркого лилового тона.
Взрослый самец. Голова, шея, брюшная сторона тела, большие и средние верхние
кроющие крыла, и нижние кроющие крыла зеленовато-голубые. Спина, плечевые,
третьестепенные маховые рыжевато-коричневые. Поясница и маховые темнобурые с синеватым
тоном на наружных опахалах. Малые кроющие крыла яркие лилово-синие. Рулевые
голубовато-синие, кроме средней пары, которая темнобурая с синевато-лиловым налетом.
Самка окрашена сходно с самцом. Ноги светлобурые, клюв темнобурый, радужина
светлокоричневая. В зимнем наряде на темени и зобе появляется буроватый налет.
Систематические замечания. Птицы, распространенные на Кавказе, выделены
Бутурлиным как особый подвид caucasica, отличающийся от типичных птиц более светлой спиной,,
чуть более сильным зеленоватым тоном рулевых и меньшими размерами. Просмотренный
большой коллекционный материал не подтверждает реальности существования этой формы,
хотя необходимо отметить, что на большой серии очень слабые отличия в окраске можно
уловить—чуть более светлый и мягкий тон в оперении, но размеры совершенно не отличимы.
Длина крыла у кавказских птиц: у самца (17) 184—208 мм, у самки (20) 181—204 мм,
среднее 192,1 мм и 190,2 мм. У всех остальных за исключением Кавказа: длина крыла самцов
(27) 180—205 мм, самок (26) 181—207 мм, среднее 192,7 и 189,8 мм.
Туркестанская сизоворонка Coracias garrulus semenowi Loud, et Tschusi
Coracias garrulus semenowi, Loudon et Tschusi. Orn. Jahrb. XIII, стр. 148,
1902, «Закаспийский край*.
Распространение. Ареал. Месопотамия, южн. и вост. Иран,
Афганистан, Белуджистан, с.-з. Индия (Кашмир, Пешавар), Средняя Азия от
южн. Закаспия и вост. побережья Аральского моря (в Больших Барсуках
найдена гнездящаяся пара среди зарослей джиды и саксаула, Бостанжогло,
1911) до долины Сыр-Дарьи, Чу Или и Ала-куля. Гнездится в Туркмении,
в низменном Узбекистане и повсеместно в южн. Таджикистане, в северном
обыкновенна в долине Зеравшана и прилегающих частях Зеравшанского
хребта, в долине Исфары, в Гиссарском хребте, по р. р. Ханаке, Варзобу,
Кафирнигану до Гарма и Тавил-даа; западная часть Алайской долины и в
небольшом количестве в ю.-з. Памире, возможно на Вахане (Иванов, 1940).
В низменном Семиречье гнездится на север до Балхаша и на восток до южн.
степей у Ала-куля. Зимует повидимому в Африке вместе с обыкновенной
сизоворонкой. Так как в зимнем оперении туркестанская сизоворонка трудно
отличима от типичной формы, более точно определить ее зимовки пока невозможно.
Характер пребывания. Гнездящаяся и перелетная птица.
Точных данных о прилете туркестанской сизоворонки в Среднюю Азию мало;
имеющиеся сведения о пролете касаются главным образом обыкновенной
сизоворонки, которая пролетает через Среднюю Азию. В Среднюю Азию
сизоворонка прилетает во второй половине апреля, у Гассан-кули местные птицы
в половине апреля уже прилетели, в это же время шел еще пролет северных
птиц, продолжавшийся до конца месяца. Добытые в это время на пролете птицы
принадлежали к форме garrulus. В Ашхабаде в 1942 г. появились 22 апреля
(Дементьев), на Мургабе у Сары-язы прилетели 21 апреля (Лоудон, 1910).
Осенний пролет. Перед отлетом сизоворонки собираются в
небольшие стаи в 10—15 штук и начинают кочевать. В это время их можно встретить
и в различных местностях сильно отличающихся от их основного биотопа.
Отлет в Средней Азии начинается с середины августа и заканчивается в сентябре
и быть может в начале октября. Под Джаркентом отлет идет с середины августа
и в течение всего сентября, последняя сизоворонка (Зарудный и Кореев,
1906), может быть пролетная обыкновенная, встречена там 9 октября. В Гис-
сарской долине отлет начинается в начале сентября и заканчивается в
двадцатых числах сентября; в Кулябе сизоворонки начинают убывать с конца августа
(1933 г.), в середине сентября встречались только одиночки; в Гарме (1935)
исчезают между 10—15 сентября (Иванов, 1940 г.). В низовьях Сыр-Дарьи —
в последней трети августа, между Аральском и Казалинском — в начале
сентября (Спангенберг, 1936). В Закаспии общая численность сизоворонок к на-
сизоворонка 499
чалу августа сильно уменьшается, к концу августа они становятся совсем
редки, отдельные запоздавшие особи встречаются еще и в октябре. В
Ашхабаде отлет местных птиц закончился около 1 августа, на Теджене большой
пролет отмечен в самом начале сентября. (Дементьев, 1950). Биотоп.
Степные, полупустынные и пустынные местности с обрывами, оврагами, иногда
совершенно лишенные всякой растительности, иногда покрытые редкой
древесной и кустарниковой растительностью; долины рек, арыки, человеческие
поселения. В горных районах обычно селится в низменностях и в горы
высоко не идет; так в Семиречье поднимается до 1700 м, выше встречаются только
одиночные, вероятно кочующие птицы (Зарудный и Кореев, 1906). В
Таджикистане — до 1500 м> редко и в небольшом количестве при наличии
удобных для гнездовья местностей — лессовых обрывов доходит до 3000 м (Иванов,
1940). В собственно горных лесах не встречается; только изредка, повидимому,
холостые, одиночные птицы на осеннем пролете заходят и выше лесной зоны.
Численность. Многочисленна по всем низменным местностям
Средней Азии; в предгорьях количество сизоворонок резко снижается.
Численность, повидимому, всецело зависит от наличия удобных для гнездования
мест (овраги, обрывы, глинистые сооружения человека и т. д.).
Экология. Размножение. В степной и полупустынной зонах
сизоворонка чаще всего устраивает гнезда в норах по обрывам, оврагам, арыкам,
крутым берегам рек, в отвесных стенах колодцев, а также в трещинах зданий,
под их крышами, в глинистых постройках и т. д. В местностях с лесной
растительностью предпочитает гнездиться в дуплах различных деревьев — саксаула
и др. В горных районах при отсутствии лесной растительности в норах, по
глинистым и лессовым обрывам, в трещинах скал. В обрывах по берегам рек,
иногда наблюдается колониальное гнездование, на Аму-Дарье встречено на
пространстве 20 м2 7 гнезд (Зарудный, 1896).
Точных данных о времени гнездования сизоворонки в Средней Азии очень
мало. К гнездованию, повидимому, приступают поздно — в мае. В Закаспии
у убитой 25 мая птицы в яйцеводе было готовое к сносу яйцо, но еще в мягкой
скорлупе (Вальтер и Радде, 1889). В окр. Ашхабада ненасиженные кладки
найдены 23 мая (5 яиц), 25 мая (7 яиц), в Сулюкли — 22 мая (4 яйца) и 27 мая
найдена слегка насиженная кладка (Шестоперов, 1928). В Таджикистане
приступают к гнездованию в середине мая. На нижней Сыр-Дарье и в
прилежащих районах сизоворонки 10—12 мая разбиваются на пары, кладка идет в
течение мая и июня (кладки найдены: 13 мая» 26 мая, б июня, 24 июня, 30июня,
Спангенберг, 1936 г.). Количество яиц в кладке и сроки высиживания такие
же как у обыкновенной сизоворонки. Размеры: (24) 33,0—36,0 х 25,7 — 28 мм
(Спангенберг, 1936); (50)39,0 — 33,3 х 30,0 — 26,2, среднее 35,9 х28,1ш*,
(Индия, Бекер, 1927). Вылет птенцов начинается в Таджикистане в конце
июля, и к середине августа все птенцы уже покидают гнезда (Иванов, 1940).
В Закаспии вылет птенцов наблюдается с середины июля (Зарудный, 1896).
Линька. Так же как и у обыкновенной сизоворонки имеют две линьки
в году —> частичную послебрачную и полную предбрачную. Частичная линька
мелкого пера начинается рано —. в Таджикистане линные птицы встречены
с 10—15 июля, у добытых на Памире птиц 29 июня и 6 августа мелкое перо
сменялось. Полная линька происходит на зимовках с января по апрель.
Питание. Пищу, как и у обыкновенной сизоворонки, составляют
главным образом крупные насекомые. В Таджикистане в первую половину лета,
до появления прямокрылых, сизоворонки питаются жуками (навозными,
златками, жужжелицами), моллюсками, многоножками; во вторую половину лета
прямокрылыми (саранча, богомолы, кузнечики) с очень незначительной
примесью насекомых других отрядов (Иванов, 1940). Это —»полезная для сельского
хозяйства птица.
32*
500
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
Полевые признаки. В поле от обыкновенной сизоворонки неотличима.
Голос такой же, как и у обыкновенной сизоворонки.
Описание» Строение и размеры. Отличий от номинальной формы в строении и
размерах нет. Длина тела: самцы (5) 330—352 мм, самки (2) 330—-335 мм, среднее 340,4
и 332,5 мм) длина крыла самцов (21) 182—205 мм, самок (9) 184—201 мм, среднее 191,3 и
187,5; размах крыла самцов (4) от 630—665, среднее 650 мм; самок (2) 650 и 660 мм. Длина
крыла от 194—210 мм Кашмир; 180—199 мм Месопотамия, Иран, Афганистан, Белуджистан,
Пенджаб (Бекер, 1927).
Окраска.Молодая птица похожа на молодую европейскую сизоворонку,
но малые кроющие крыла зеленовато-голубые. В з р о с л а я отличается от предыдущей формы
более светлой окраской спины, сильным развитием зеленого оттенка на частях тела,
окрашенных у обыкновенной сизоворонки в голубовато-зеленый тон и более светлым синим тоном
малых кроющих крыла.
РОД ШИРОКОРОТЫ EURYSTOMUS VIE I L L О Т, 1816
Тип. Coracias orientalis
2. Восточный широкорот Earystomus orientalis L
Coracias orientalis. Linnaeus. Syst. Nat. изд. XII, 1766, стр. 159, Индия.
Распространение. Ареал. Приморская обл. до 50° с. ш. к северу
Манчжурия, Корея, Япония (к югу от средней части Хондо и на о-вах
4
°2- ©
v-
JLL
Карта 85. Распространение восточного широкорота Eurystomus orientalis
7—Е. о. abundus, 2—Е. о. orientalis, 3—Е. о. gigans, 4 — Е. о. deignani, 5—Е. о.
connectens, 6 — Е. о. solomonensis, 7 — Е. о. crassirostris, 8 — Е. о. pacificus, 9 — Е. о.
oberholsteri, 10 — Е. о. azureus, 77— Е. о. waigionensis; а — граница гнездовий, г — зимовки
(цифры в кружках обозначают номер зимующих подвидов).
ВОСТОЧНЫЙ ШИРОКОРОТ
501
Кюсю и Риу-Киу), вост. и ю.-в. Китай, Гималаи, вост. Индия (от Бенгалии
до Мадраса) и ю.-в. Индия (Траванкор), Цейлон, Индо-Китай, Малайский
п-ов, о-ва Индо-Австралийского архипелага и Австралия.
Рис. 126. Восточный широкорот.
Характер пребывания. В северных частях ареала
гнездящаяся и перелетная птица, в южных — оседлая и частично перелетная.
Биотоп. Высокоствольные леса, особенно любит небольшие лесные
поляны, вырубки и речные долины, окруженные лесом, но не избегает и
густых лесов.
Подвиды и варьирующие признаки. Восточный
широкорот имеет И подвидов, из которых только один
abundus доходит на Дальнем Востоке до СССР, остальные
распространены вне нашей страны: Е. о, abundus R i р-
1 е у, 1942, Приморская обл. Манчжурия, Корея, Япония
вост. и ю.-в. Китай, вост. Гималаи; Е. о. orientalis
Linnaeus, 1766 (синоним calonyx S h a r p e, 1850). —
Вост. Индия от южн. Гималаев и Бенгалии до Мадраса
и зап. Индия от Траванкора до Уинаба, Цейлон, Бирма,
Ассам, Индо-Китай — до центрального и южн. Сиама,
Малайский п-ов, Большие Зондские о-ва и о-ва Талаур,
Бали и Филиппины; зимой встречается на сев.
Целебесе, о-вах Халмахера и Б. Санги; Е. о. deignani
Ripley, 1942 — северные части французского Индо-Китая,
Суматры, Ниаса, Явы, Борнео и, вероятно, на Малайском п-ове;
mann, 1913. Андаманские о-ва; Е. о. connectens Stresemann, 1913, Малые Зондские
о-ва от Ломбока и Дамара, до Дампеа, Муна, Тукан-Беси, Бонарете, Калао-туа; южн.
Целебес и Кангеан; Е. о. oberholseri J u n g e, 1936, о-ва Сималур; £. о, azureus Gray,
Рис.
127. Птенец восточного
широкорота.
сев. Сиам; зимой — до
Е. 0. gigans S t r e se-
502
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
I860 —сев. Молукские о-ва, Тернате, Гальмагейра, Тидоре и Батьян; Е. о. solomonensis
Elliott, 1890 Соломоновы о-ва; Е. о. waigionensis Elliott, 1871 зап. Папуасские
о-ва, о. Иоби, Новая Гвинея, о-ва Каркар, Рук, Тробриан, архипелаг Д' Антркасто и
архипелаг Луизиады; Е. о. pacificus Latham. 1800 — Австралия; Е. о. crassirostris S с 1 а-
te г, 1869 — архипелаг Бисмарка. Морфологические варьирующие признаки — размеры,
окраска и экология.
Восточный азиатский широкорот Eurystomus orientalis abundus Ripley
Eurystomusorientalisabundas. Ripley. Proc. Biolog. Soc. Washington, 55, 1942, стр. 170.
Нанкин.
Распространение. Ареал. Гнездится в Приморском крае — на север
до 50° с. щ., найден на гнездовье у с. Гармах, на р. Кур, на р, Хунгари, в
предгорьях Сихоте-Алиня, на озере Ханка и в бассейне озера Ханка, в
окрестностях Владивостока, в урочище Славянка, на Имане, в Посьетском районе и на
о-ве Русском. Вост. Манчжурия от ст. Пограничная, Имянпо, в 140 км
К юго-вост. от Харбина, в провинции Гирин, Мукден, порт Дальний, в горах
Чан-бо-шань. Корея. Япония, средние и южные части Хондо, Кюсю, Риу-Киу.
Встречается на о-ве Формозе, о-ве Квельпарт, о-ве Цусима, но гнездовье там
не доказано. Распространена в вост. и ю.-в. Китае от Чжили, Цзяньсу и Хубея
до Юннаня и Фу-Чжоу. На основании одного экземпляра, добытого в конце
июня, в зап. Алашане (Тянцзинский Музей), считают Алашань самым сев.-зап.
местом гнездования широкорота. Скорее всего это может быть залетом, так
как в пустынном Ордосе и в пустынном вост. Алашане широкорот вероятно
не гнездится. Гнездится в верхнем Ассаме, на восток до сев. Качара, и
в Гималаях, на запад до Кумаона.
Зимовки расположены в Индии, Южн. Китае, Индо-Китае, на
Малайском п-ове, и на о-вах Суматре, Сималуре и Борнео.
Характер пребывания, В северных частях ареала
перелетная птица, в южных — оседлая. К нам прилетает поздно — во второй
половине мая. Так, в 1931 г. появились в южн. Приморье 22 мая; в 1945 г.
(холодная поздняя весна) в Кедровую Падь — 27 мая (Воробьев, 1948). На Имане
в 1938г. прилетели 14 мая (Спангенберг); в Фу-чжоу (вост. Китай) --в конце
апреля, в Цинь-дао —. в первых числах мая. Осенний пролет наблюдался у
нас близ Амурского залива — 5 сентября» Птицы летели небольшой стайкой
в количестве 20 штук вечером перед заходом солнца (Воробьев, 1948). Из
Фу-чжоу отлетают в конце сентября или в начале октября, из Цинь-Дао в
то же время.
Биотоп. Птица высокоствольного леса, резреженного лесными поля-
цами, речными долинами и вырубами. У нас в Приморском крае предпочитает
'широколиственные леса (липа, ясень, клен), но охотно селится в лесах с
преобладанием хвойных деревьев и даже не избегает тайги, если там имеются
открытые поляны. В Сихоте-Алине поднимается до горной тайги. В Китае
гнездится в сосновых рощах и среди других лесов, состоящих из крупных деревьев,
в особенности любит леса, окружающие рисовые поля. В Японии часто селится
в больших лесах, растущих вокруг буддийских монастырей. В Гималаях —
в лесах черного дерева, поднимаясь в горы до 1000л*. В Индо-Китае
предпочитает селиться в высоких бамбуковых лесах.
Численность. В приморье довольно обыкновенная птица.
Гнездящиеся пары встречаются равномерно разбросанными на довольно большом
расстоянии друг от друга. Многочисленна только в низовье Имана, но в
верховьях этой же реки встречается уже реже. В Японии обычна повсюду, но
спорадична. В горных частях вост. Китая весьма обыкновенная птица, также
как и в остальных частях своего распространения, за исключением юго-зап.
берега Индии, где она редка.
Широкорот
восточный широкорот
503
Экология. Размножение. Весною в первую очередь прилетают
старые птицы, в парах. Несколько поздней появляются стаи холостых
(Спангенберг считает их молодыми прошлогодними птицами), которые будто бы к
размножению не приступают; такие стаи от 100 и более особей наблюдались
на р. Имане в июне. Сейчас же по прилете у широкоротов начинаются
брачные игры и выбор мест для гнездовья. В это время они шумливы, крикливы и
очень заметны, но как только сядут на яйца, становятся молчаливыми и
осторожными. Гнезда используются птицами иногда несколько лет подряд. Гнезда
устраиваются в дуплах больших деревьев, высоко над землею. В Приморье
чаще всего для гнезд выбирается дуб (Спангенберг), южнее, где уже растет
бамбук, предпочитается бамбуковое дерево. На больших ветвях этих деревьев
часто появляются болезненные наросты—вздутия до 60 см в диаметре с гнилым
центром, в котором легко образуются дупла. Они чаще всего расположены
высоко от земли до 20—30 м. Иногда широкороты устраивают гнезда в дуплах
пней, иногда гнездятся в сорочьих гнездах. Если дупло мало, то сами птицы
увеличивают его, причем трудятся и самец и самка. Отверстие в дупле
оставляют маленькое, так что едва пролезает птица. Подстилкой в гнезде служит
измельченная гнилая древесина. Число яиц в кладке в Приморье 4—6
(Спангенберг); в вост. Китае 3—4 (Латуш, 1931), в Индии и Индо-Китае — 4
(Бекер, 1927). Свежие кладки на Имане в 1940 г. и 1941 г. найдены Спан-
генбергом 14 июня; в Южно-уссурийском крае добыто несколько самок в
период от 14 до 19 июня 1945 г., у которых были сильно развиты яйцевод и
яичник, содержавший крупные фолликулы в 10 мм в диаметре (Воробьев,
1948). В вост. Китае гнезда с яйцами находили с конца мая до середины
июня. В Индии гнездовый сезон наступает с марта и тянется до мая,
исключая Траванкор, где гнездовый сезон длится с сентября по май, хотя большее
количество яиц откладывается в марте и апреле.
Яйца блестящего белого цвета, похожи на яйца сизоворонок, овальной
формы. Размеры: (4) 34,08—31,7 х 28,1—26,0 мм, среднее 33,3 х 27,5 мм
(р. Иман, Спангенберг); (29) среднее 39,5 х 26,0 мм (вост. Китай, Латуш,
1931); (25) 36,9—31,7 х26,1 ^25 мм, среднее 36,3 х 28,2 мм, (Индия,
Бекер, 1927 г.).
Вылупляются птенцы голыми. Гнездо с 6 птенцами старше
двухнедельного возраста найдено* на Имане 3 июля и второе с 4 птенцами в возрасте
четырех — пяти дней 8 июля. Неоперившиеся птенцы встречаются до середины
августа. Пока птенцы малы, родители с кормом влетают в дупло, но как
только птенцы подрастут, то они сами высовывают головы в отверстие дупла
навстречу прилетевшим родителям. Летный молодой широкорот добыт 28
августа у с. Гармахта на р. Кур (Бутурлин, 1913).
Л и н ь к а не изучена. Экзе*мпляры, добытые в Приморье в конце августа
(26 и 28), еще без следов линьки, в сильно изношенном оперении. Повиди-
мому линька происходит на зимовках.
Питание. Пища состоит главным образом из различных лесных
насекомых — жуков, клопов, цикад, кузнечиков, мух, пчел и также мелких
моллюсков (Спангенберг). В желудках широкоротов, добытых в Приморье в июне,
были найдены различные жуки: бронзовки Cetonia, хрущи Sertca,
мертвоеды, усачи Merochammus arussowi; из других отрядов — пилильщики Cimbex
(Воробьев). Пищу широкороты обычно добывают в воздухе, но иногда берут
насекомое и с дерева или с земли. Поймав насекомое, усаживаются на высокие
ветви деревьев и там поедают его. В вечерние сумерки, собираясь в стаи,
они устраивают оживленную охоту. Площадь, занимаемая стаей во время
охоты, достигает до 0,5 км2. Уничтожением вредных насекомых они приносят
пользу лесному хозяйству. Но, поселившись вблизи пасеки, в большом
количестве поедают пчел.
504
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
В течение всего дня широкороты сидят неподвижно на вершине деревьев
или на сухом сучке, погрузив голову в распушенные перья шеи и зоба, как
будто спят, только время от времени издают грубое карканье. Увидав
пролетающее насекомое, быстро слетают, чтобы поймать его, и затем возвращаются
обратно на дерево, съедают его, сидя на ветке, и снова начинают дремать.
Только в вечерние сумерки они оживлены. Птицы в погоне за насекомыми и
играя высоко летают над землею кружащим полетом.
Полевые признаки и повадки. Своими повадками несколько напоминают
сизоворонок, но менее подвижные; скорее сумеречные, нежели дневные
птицы. Обычным местом пребывания широкорота служит крона деревьев,
в особенности он любит сухие вершины деревьев. Широкорота легко узнать
по яркой окраске оперения. В полете, кроме того, хорошо заметно на
открытом крыле светлое зеркальце. Полет очень характерен: быстрый,
кружащий, несколько напоминающий полет ласточек. Голос неприятный, грубый
горловой крик «карр-карр» или, как его передает Спангенберг, «чер-чер»,
повторяющийся старыми и молодыми птицами через каждые 10—15 минут.
Описание. Размеры и строение. Клюв большой, широкий у основания, слегка
загнутый у вершины. Ноздри большие, щелевидные, слегка прикрытые перьями лба.
Вибриссы у угла клюва имеются, но развиты слабее и короче, чем у сизоворонок. Ноги
короткие, слабые. Цевка покрыта спереди щитками, сзади сетчатая. Первостепенных маховых —
11, самое длинное — 2, формула такая же, как и у сизоворонки. Рулевых — 12, обрез хвоста
прямой. Длина крыла самцов (6) 193—184 мм, самок (4) 180—186 .мл*, среднее 187,1 и 186 лш.
Крыло индийских птиц 178—202 мм (Бекер, 1927).
Окраска. Вылупившийся птенец — голый, на 4—5 день начинают появляться
темные перья на голове, крыльях и на птерилиях тела. Голые части тела желтые.
Яйцевой зуб исчезает на 5—6 день. Надклювье буровато-черное с беловатой полоской
вдоль края клюва, подклювье — светлого рогового цвета. Глаза открываются на
6—7 день.
Молодые птицы отличаются от взрослых буровато-черной головой без зеленого
тона и меньшим развитием зеленого на спине. Спина буровато-черная со слабым темно-
зеленым налетом, усиливающимся к задней части спины и к надхвостью. На горле нет
фиолетово-синего пятна. Горло, зоб и грудь — буроватые, брюхо и подхвостье —
голубовато-зеленые. На крыльях и хвосте синий цвет развит слабее. Клюв буровато-черный,
более светлый на подклювье.
Взрослый самец. Голова и спина черноватые с зеленым оттенком иа голове,
постепенно переходящим на спине в темиозеленый цвет. Хвост черный с темиосиним
основанием. Плечевые и кроющие крыла голубовато-зеленые. Маховые и кроющие
первостепенные маховые темиобурые с фиолетово-синим наружным опахалом. На крыле бледиоголу-
бое зеркальце, расположенное на внутренних и наружных опахалах первостепенных
маховых, за исключением первого, у которого зеркальце имеется только на внутреннем опахале.
Брюшная сторона темиоголубая с зеленоватым оттенком и фиолетово-синим пятном иа
горле и зобе. Клюв красный с черной вершиной. Ноги красноватые. Радужина темнокори-
чневая. Самка окрашена сходно с самцом, только верх головы без зеленого тона.
СЕМЕЙСТВО ЩУРКОВЫХ Meropidae
РОД ЩУРКИ MEROPS LINNAEU 1758
Тип. Мег ops a piaster.
3. Золотистая щурка Merops apiaster L.
Merops apiaster. Linnaeus. Syst. Nat., изд. X, 1758, стр. 117, южн. Европа.
Русское название: щурка — народное, упоминается еще Палласом. Местами
пчелоед, пчелоедка.
Распространение. Ареал. Португалия, Испания, южн. Франция?
Италия (в северной нерегулярно), о-ва Средиземного моря, Балканы,
Венгрия, Австрия, Югославия, южн. полоса европейской части СССР, сев. Африка
(Марокко, Алжир и Тунис), на юг до Сахары, южн. Африка, Сирия, Палестина,
ЗОЛОТИСТАЯ ЩУРКА
505
Месопотамия, Малая Азия, Иран, Афганистан, Белуджистан, с.-з. Индия
(от Кашмира, Пенджаба до Раджпутаны и Синда) и Средняя Азия до ю.-з.
Сибири, ю.-з. Алтая и Тарбагатая.
В СССР гнездится от Крыма, Кавказа и устьев Волги и Урала до
Бессарабии, Подолии, Житомира, Киевской и Черниговской обл. (в последней
наиболее северное местонахождение отмечено у с. Мезань) — в Волыни на.
гнездовье не отмечена. В Курской обл. на север доходит до Дмитриевского
района, гнездится в Воронежской обл. и в Тамбовской обл. к северу до
Карта 86. Распространение золотистой щурки Merops apiaster
а — граница гнездовий, г — зимовки.
Тамбова, в Пензенской обл. —к северу до Городищенского района; обычна
на гнездовье в бассейне р. Усы в ю.-в. части Самарской луки и в б. Сызранском
ив б. Симбирском уездах (указана еще Лепехиным, 1805). В Татарской
республике гнездится до Лаишева, Спасска и до устья Камы. Затем северная граница
резко спускается к югу — к Бузулуку и Бузулукским степям (коллекции
зоологического музея Моск. университета), к Уралу и р. Илеку; на Южн.
Урале проходит у р. Сакмары (приток Урала) и близ устьев Б. Ика (К и-
р и к о в); верховьев Эмбы (о гнездовании на нижней Эмбе сведений нет), устья
Темирау Кок-джидаи до Актюбинска, затем резко спускается к югу к ст.
Аральское море (в Аральских Кара-Кумах ее нет). В область
распространения этой птицы входят северное и восточное побережье Аральского моря,
долина р. Сыр-Дарьи, отроги Кара-Тау, р. Сары-Су, р. Чу и Или, озера
Балхаш, Семипалатинск, юго-зап. и южная окраина Алтая — до с. Локоть и
Змеиногорска, Барнаул* Это, повидимому, самые сев.-вост. пункты нахождения
506
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
гнездования щурки. В Барабинской и Кулундинской степях не гнездится, в
Кузнецкой степи и в предгорьях Салаирского кряжа ежегодно встречается,
но гнездование ее там не установлено (Хахлов, 1937). Гнездится в Приал-
тайской степи, в бассейне верховьев Иртыша, в Зайсанской и Эмильской
котловинах, в предгорьях Барлыка, на Тарбагатае и Ала-куле. Это самые
восточные нахождения гнездовий золотистой щурки. К югу от очерченной
границы встречается у нас повсеместно по удобным для гнездования местам,
но в центральных Кара-Кумах ее нет.
Отдельные одиночные гнездования или колонии, а иногда встречи
холостых золотистых щурок в период размножения, наблюдались много северней,
ее нормальной северной границы распространения. В Московской обл. в
течение всего лета 1880 г. наблюдалось пять щурок у дер. Мазилово, в
Можайском районе добыта 12 июня 1910 г. щурка из стайки в шесть штук, в яйцеводе
ее было обнаружено готовое к сносу яйцо; в Рязанской обл. в Окском
заповеднике наблюдалось несколько, повидимому, гнездящихся там щурок в июне
и июле, в Клязьменском заповеднике (нижнее течение Клязьмы, Муромский р-н)
найдена гнездовая колония; отмечены в июле 1885 г. у Раненбурга.
Встречались залетные щурки в южной части Тульской губ., в б. Новосильском уезде;
с 1945 по 1949 найдена гнездящейся в Крапивенском районе Тульской обл.
к югу от г. Тулы, причем в предыдущие годы (с 1935—»44 г.) ее там совсем
не встречали. В Тамбовской обл. молодая щурка добыта 8 августа под г. Мор-
шанском, в августе и сентябре наблюдалась в Темниковском районе в
Мордовском заповеднике. Однажды добыта в окр. г. Васильсурска Горьковского
края. Отмечены залеты в окрестности Пскова 19 мая 1893 и 28 мая 1902, в
Прибалтику, на Кольский п-ов у Муониониска, где была добыта 3 июня
1865 г. Ежегодно отмечены залеты щурки в Зап. Сибири в Кузнецкую
степь и Салаир. Известны залеты и далеко от мест гнездования: Фарерские
о-ва, Дания, Швеция, Норвегия, о. Гельголанд и другие страны Зап. Европы.
Северная граница распространения золотистой щурки в европейской части
нашей страны проходит по северной окраине степной зоны. В Азиатской части
страны северная граница распространения щурки идет по южной окраине
степной зоны Казахстана, т. е. значительно южней, чем в. европейской части.
Северная граница определяется, вероятно тем, что золотистая щурка плохо
переносит холода и нередко гибнет в большом количестве в своих наиболее
северных пунктах распространения в те годы, когда происходит возврат
холодов. Так, в 1904 г. наблюдалась массовая гибель щурок под Чкаловом при
возврате холодов весною. Зимует щурка в тропической и южн. Африке от
Сахары до Капской Земли, в южн. Аравии и в зап. Индии от Раджпутаны до
Бомбея.
Характер п р е б ы в а и и я. У нас гнездящаяся и перелетная птица.
На весеннем пролете щурки летят небольшими рассеянными стаями в 10—15
штук, обычно на высоте около 100 метров, но иногда и очень высоко. Во время
пролета отвлекаются от главного направления —. все время ловят насекомых.
При похолодании весной пролет прекращается до возвращения теплых дней.
Летят птицы главным образом днем, ночуют обычно на деревьях в лесу, садах,
но иногда и в камышах у озер (Спангенберг, 1936). Пролет идет довольно долго,
в продолжении 15—20 дней; в южных районах прилетевшие золотистые щурки
начинают готовиться к гнездовью, в то время, как обитающие в более северных
местах птицы еще летят. Наблюдается значительная разница в сроках прилета
щурки в одну и ту же местность в разные годы. Повидимому эта разница
зависит от характера весны. В Египте на пролете отмечен с первых чисел апреля
до половины мая.
В Закавказье пролет начинается с середины апреля. В Ленкоранском
районе первые птицы появляются около 15 числа этого месяца, в Армении
ЗОЛОТИСТАЯ ЩУРКА
507
€ 20 апреля и пролет захватывает начало мая (Ляйстер и Соснин, 1942), в
окрестности Батуми пролет в 1910 наблюдался очень поздно: 21—23 мая
(Нестеров, 1911), повидимому холодная погода задержала пролет. По
Черноморскому побережью — с конца апреля и до конца мая, валовый пролет
с 10—17 мая. На Сев. Кавказе с конца апреля и до конца мая (в
Северной Осетии и Ингушетии с конца апреля, на средней и нижней Кубани с 20
апреля, на среднем и нижнем Тереке — в мае, в окрестностях Краснодара —
в начале мая, у Ставрополя —в начале второй половины мая Радде, 1885).
В окрестностях Ростова-на-Дону первые птицы появляются 15 мая (Саран-
динаки, 1908). В Крым прилетают в конце апреля — в начале мая (5 мая).
На Украину — в конце апреля и в первой половине мая. Так, в
Днепропетровск первые птицы прилетают в последней трети апреля и в начале мая,
прилет продолжается до 20 мая (Боровиков, 1907), в Подолию —в конце
апреля, под Киев —> в первой половине мая и в средине мая, в
Харьковскую обл. в среднем — 11 мая; самый ранний прилет наблюдался в 1892 г.—>
23 апреля, самый поздний в 1879 и 1891 г.—21 мая (Сомов, 1897). В
Крапивенский район Тульской обл. прилетает в среднем 20 мая (самый ранний
прилет отмечен 14 мая, самый поздний —» 22 мая (Лихачев, 4 года наблюдений).
Под Чкаловом первые птицы появляются в первых числах мая, валовый
пролет наблюдается в середине мая и заканчивается пролёт к концу мая, в Бугу-
русланский уезд прилетают во второй декаде мая. В Средней Азии пролет
идет с середины апреля до начала мая. В Туркмению прилетают в середине
апреля, валовый пролет идет в конце апреля. В Ашхабад — в среднем
18 апреля, самый ранний прилет отмечен 12 апреля 1923 г., самый поздний —-
23 апреля 1916 г. (Шестоперов, 1926), валовый пролет в Ашхабаде
наблюдался в 1886—24 апреля (Вальтер и Радде, 1889); по Мургабу валовый
пролет — 26 марта, между Мары и Чарджоу 25^26 апреля, у Атрека 11 апреля
и в мае, Дорт-Кую—17 апреля, у Карабогаза прилет щурок отмечен 1 апреля
1939 (Воробьев и Исаков, 1940). В Арало-Каспийской низменности — с конца
апреля и до начала мая. В низовьях Сыр-Дарьи появились в 1928 г.—
25 апреля и в 1930 г. -^27 апреля, в Узбекистане щурки летят с первой
половины апреля до начала мая. В Таджикистане — в двадцатых числах апреля:
так, в Гиссарской долине прилетели 22 апреля 1924, валовый пролет отмечался
в начале мая, в Денау —в конце апреля. В Джаркенте с первых чисел апреля
(7 апреля). В Алтайский край под Усть-Каменогорск прилетают в мае.
Отлетают щурки — в конце, а иногда даже в середине августа. С конца
июля они собираются в стаи, состоящие из молодых и старых птиц. Кочующие
в поисках пищи стаи наблюдаются в это время в различных частях нашей
страны, иногда далеко от мест гнездовий; в горных районах они поднимаются
высоко в горы, вплоть до перевалов. На Кавказе встречаются в конце июля
(22—»24 июля) в области высокоствольного леса, в истоках Риона, Уцери, под
Боржомом и т. д. (Радде, ,1885). К концу августа спускаются с гор в низкие
места, так как количество насекомых в горах к этому времени сильно
уменьшается. С первых чисел августа в равнинных местностях начинают щурки
готовиться к отлету, иногда собираясь в стаи вместе с ласточками и стрижами.
Вначале улетают старые птицы, затем молодые; на их место появляются птицы из
более северных районов гнездовья. Так, например, на Кавказе местные птицы
улетают начиная с середины августа, пролет более северных птиц происходит
во второй половине сентября (с 14 по 30 сентября). Таким образом, осенний
пролет золотистых щурок тянется с августа по сентябрь, но валовый пролет
обычно идет в конце августа и в начале сентября; одиночные птицы
встречаются на пролете и в начале октября. Летят и большими стаями более ста
штук, и небольшими группами, перекликаясь между собою. Во время пролета
держатся высоко. Летят прямо через водоразделы, преимущественно по утрам
508
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
до полудня, но иногда и по ночам. Около кормовых мест образуются большие
скопления птиц. Часто пролет, тянувшийся несколько дней, прекращается на
некоторое время, затем вновь возобновляется и так повторяется несколько
раз. Золотистые щурки из северных районов своего распространения улетают
в начале августа. В Тульской обл. отлет идет с 1—6 августа и до 3—8 сентября
(5 лет наблюдений, Лихачев), в Тамбовской обл. со 2—3 августа (Предтеченский,
1928). В Воронежской обл. последние отлетающие стаи щурок наблюдались
в 1920 г. 24 сентября (Огнев и Воробьев, 1924), из Бугуруслана первые щурки
улетают в начале августа, и пролет идет до половины сентября (Карамзин,
1909). С Украины начинают улетать со второй половины августа, пролет обычно
заканчивается во второй половине сентября, хотя иногда задерживается и до
начала октября (Подолия, Храневич, 1926). В Харьковской губ. самый
ранний пролет начался 13 августа 1891 г. и 1892 г. и 14 августа 1890 г., но
закончился, как и в годы с более поздним началом, в конце сентября (23—27
сентября, Сомов, 1897). Из окрестностей Киева и Днепропетровска отлетают
с середины августа и пролет идет до половины и даже конца сентября.
В дельте Волги и в Волжско-Уральских степях — с конца августа и до начала
сентября. Из окрестностей Чкалова золотистые щурки исчезают в первых
числах сентября. С Сев. Кавказа — с половины августа и до конца сентября:
по Черноморскому побережью валовый пролет наблюдается в конце августа
и в начале сентября, причем летят золотистые щурки по нижней полосе гор
и по прибрежной низменности (Сатунин, 1907); из Сочинского района —
с конца августа и пролет тянется до середины сентября. Из Армении —
в начале августа и до конца сентября. Во время пролета на Кавказе щурки
наблюдаются высоко в горах вплоть до ледников и перевалов (17 августа
встречались у Гергетского ледника, Сушкин, 1914); 29 августа —у Боржома;
2 сентября в Мальмутах; 8 сентября—у ст. Конорбель; в ущелье Куры;
у ст. Беграхатум и на Малом Кавказе на высоте около 2 000 м (Сатунин, 1907).
В районе Семипалатинска на правом берегу Иртыша и в окрестностях Усть-Бух-
тарминской 16 сентября встречены многочисленные стаи пролетных щурок.
(Залесские, 1931). Из Семиречья начинают отлетать с первых чисел августа
и пролет идет до конца второй половины сентября (24 сентября, Зарудныи и
Кореев, 1906). В Таджикистане пролет заканчивается в середине сентября:
в Гиссарской долине отлет закончился 18—20 сентября, в Гарме — 10—12
сентября 1936г. (Иванов, 1940). Из Узбекистана отлетают с конца августа и до
конца сентября (Корелов, 1949). В низовьях Сыр-Дарьи держатся до конца
сентября. В Арало-Каспийских степях летят с половины августа и до конца
сентября. В Туркмении отлет начинается с первых чисел августа и тянется
до конца сентября, валовый пролет — в конце августа и в начале сентября,
отдельные экземпляры наблюдаются вплоть до октября: из окрестностей
Ашхабада щурки отлетают во второй половине октября, так, в 1923 г.—
встречались до 25 октября и в 1925 г. —до 16 октября (Шестоперов, 1928); из
окрестностей Репетека в 1925 г. улетали 25 августа (Снигиревский). По
нижнему течению Аму-Дарьи—с половины августа, в Закаспийском крае—с первых
чисел августа, валовый пролет — в конце августа и в начале сентября. Большие
стайки щурок, летевшие в направлении на юго-запад, замечены в конце сентября
в открытом море, у Красноводска. (Зарудныи, 1896).В Приатрекской пустыне
пролет идет в сентябре, птицы летят в юго-западном направлении* последние
птицы отмечены 12 октября (Исаков и Воробьев, 1940). В Астрабадской
провинции 5 октября наблюдался пролет золотистых щурок совместно с зеленой
(Шестоперов, 1928). В Синае и Египте от середины августа до конца октября
(Мейнерцхаген, 1930).
Биотоп. Открытые степные пространства, пересеченные обрывами,
оврагами, балками и реками с обрывистыми глинистыми берегами, поросшие ку-
ЗОЛОТИСТАЯ ЩУРКА
509
старниками, долинным лесом или даже отдельно стоящими деревьями. Не
избегает соседства с человеком, селится вблизи деревень, садов и особенно охотно
вблизи пасек.. В горных районах предпочитает равнинные, низменные
местности, в горы высоко не поднимается: на Кавказе до 1500 м (Радде, 1885), в
Закавказье — в Хевсуретии — до 2000 м в Армении не выше 2500 м (Ляйстер
и Соснин, 1940). В Семиречье доходит в горы до лиственных лесов, т. е. до
1800 м (Зарудный и Кореев, 1906). В Таджикистане найдена на гнездовье до
1800—-1900 м (Иванов, 1940). В полупустынях обычна по берегам рек, оврагов,
поросших кустарниковой растительностью, и изредка встречается в пустыне,
причем в песчаной чаще, чем в щебенистой.
Численность. В северных частях гнездового ареала золотистая
щурка встречается спорадично и общая ее численность там не велика.
В местностях, где много подходящих для гнездовий мест или где развито
пчеловодство, щурка становится весьма многочисленна. Например, в Тульской обл.
первое появление щурок на гнездовье отмечено с момента увеличения
пчеловодства, уменьшение пчеловодства в 1946 г. отразилось и на количестве
гнездящихся щурок. Увеличение числа щурок в 1949 г. (осенью наблюдались стаи
до 100 особей) совпало с развитием пчеловодства (Лихачев). В Бессарабии и в
Подолии — это редко гнездящаяся птица. Небольшими отдельными колониями
доходит в Черниговщине до с. Мезень, в Курской обл. обычна в южной части
области и редка в северной, в Воронежской обл. обычна на гнездовье, но более
многочисленные колонии встречаются в долине p.p. Битюга и Хопра и по
берегам степных прудов. Особенно многочисленна на Украине южнее Харькова
и Днепропетровска, в степной части Крыма, на Северном Кавказе до предгорий
и в восточном Закавказье, в западном уже реже. По Волге многочисленна от
устья до Сталинградской обл., местами до Куйбышева, в Сызранском районе
еще обычна, в Пензенской обл. спорадично гнездящаяся птица, причем более
обычная в степной части области. В окрестностях Чкалова обычна, а в западной
половине Казахстана, в низменных местностях Киргизии, в долинах рек и
в низменностях Узбекистана, Таджикистана и по всей Туркмении
многочисленна.
Экология. Размножение. Первое время после прилета золотистые
щурки ведут бродячий образ жизни, отыскивая старые норы или выбирая места
для новых. Вскоре затем начинают собираться около обычных мест гнездовья—
берегов, оврагов, обрывов и т. д. Гнездятся колониями, изредка парами или
в одиночку. Колонии по своей величине очень различны, от нескольких пар
до нескольких сотен пар. Норы устраиваются преимущественно в высоких
обрывах (в Предкавказье от 3 до 5 м высоты, Прекопов, 1940), совершенно
голых или поросших кустарниками; иногда роются норы и в ровной
поверхности. Грунт не играет большой роли для постройки норы, но суглинки
предпочитаются.
Роют нору сами птицы, причем в этом участвуют попеременно самец и
самка. Рытье норы происходит главным образом утром с 9—10 часов и вечером
с 17—18 ч. Копают клювом, ногами лишь отбрасывают землю пятясь из норы
к выходу. Выброшенная земля весит в среднем до 12 кг (Корелов, 1949).На
копание норы затрачивается 10—20 дней.
Разница во времени начала кладки даже в одной колонии очень
значительна, что, повидимому, вызвано готовностью гнезд. Щурки, которые по
каким-либо причинам были вынуждены рыть нору для гнезда вновь, начинают
кладку значительно поздней тех особей, у которых для гнезда нора уже
готова.
Нора роется в виде длинного почти горизонтального хода, иногда
имеющего изгиб. Длина хода достигает до 2 м, обычно 1—1,5л* (в Предкавказье
около 60 см, Прекопов, 1940). Отверстие норы и диаметр хода 8—10 см. Ход
510
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
заканчивается расширением, лежащим несколько ниже; в этом расширении
помещается собственно гнездо. На расстоянии от 30 см до 2 м от отверстия
норы роются добавочные — вторая, третья, а иногда и четвертая (Узбекистан,
Корелов, 1949). Вторая нора обычно короткая, не достигает и 50 см длины,
диаметр такой же, как у главной норы, но она не заканчивается гнездовой
камерой. Роется вторая нора одновременно с главной, но значительно медленней
(Узбекистан). Гнездовая камера шириною 15—25 см, высотой до 12—15 см.
Почва, составляющая основание гнезда, сильно разрыхлена, и, если нора
служит первый год, яйца лежат прямо на земле без всякой подстилки; в старых
норах основание гнезда усыпано слоем хитиновых остатков насекомых,
который достигает толщины в 2—4 см, много хитиновых остатков имеется и на
протяжении всего хода. Хитин лежит ровным слоем в камере, в центре его делается
небольшое углубление, в котором помещаются яйца.
Спаривание происходит после того, как готово гнездо. С этого времени до
начала кладки самка много времени проводит в гнезде или около гнезда, но на
ночь улетает на деревья. В году одна кладка. В кладке обычно 5—-6 яиц, реже
4 или 7—8, иногда и 10. Яйца белого блестящего цвета, до насиживания с
легким розоватым оттенком (от просвечивающего желтка). Скорлупа тонкая
мелкозернистая. Форма яиц почти шарообразная. Размеры: (9). 27—25x22,5—
21,5, среднее 26,11 х 21,73 мм (Харьковская обл., Сомов, 1897); 24,0—
26,3 х 20,9—22,1 мм (низ. Сыр-Дарьи, Спангенберг 1936); (96) 27,3 — 24,0 х
23,1—20,0, среднее 25,5 х 21,55 мм (Хартерт, 1912—21); (100) 28,7 —
24,1 х 23,1 — 19,9, в среднем 26,6 х 22,4 мм (Индия, Бекер, 1927).
На Кавказе шурки к устройству гнезд приступают в начале мая, к кладке
яиц в середине мая. В Закавказье в конце мая встречались гнезда с полными
кладками и гнезда, в которых кладка только начиналась. В северной Осетии и
Ингушетии полные ненасиженные кладки встречались в начале июня, а 15 июня
в десяти просмотренных гнездах были найдены яйца от 3 до 7 дней насижен-
ности (Беме, 1926). На Украине кладка яиц начинается в первых числах
июня и идет до конца месяца: в Харьковской обл.— с конца первой декады
июня и до конца месяца; в Днепропетровской — с первых чисел июня. В
низовьях Сыр-Дарьи — с конца мая и до середины июня. В Узбекистане — со
второй половины мая и до начала июля, но особи, гнездящиеся в
полупустынной и предгорной части Узбекистана, приступают к гнездованию поздней
дней на 10 (Корелов, 1949). В Туркмении период кладки начинается с середины
мая и продолжается до начала июня, ненасиженные яйца найдены 3 июня 1916
и 16 мая 1929 (Шестоперов, 1928).
В насиживании принимают участие оба пола, но, главным образом, самка,
оставляющая гнездо приблизительно на час-два в день, и на это время ее
заменяет самец. В период насиживания самец кормит самку, которая в это
время мало добывает себе пищи. На гнезде сидят крепко. Насиживание
начинается с откладки первого яйца. Промежуток времени между откладкой
каждого яйца не выяснен, но, повидимому, значителен, так как выведшиеся
в гнезде птенцы всегда оказывались различного возраста — от голых до
вполне уже оперившихся. Срок высиживания около 20 дней. В выкармливании
птенцов принимают участие оба родителя (за 10 минут приносят 5 насекомых,
главным образОхМ прямокрылых — Узбекистан, Корелов, 1949).
Растут птенцы быстро. Первое время родители с кормом влетают в нору,
но перед вылетом птенцов из гнезда, они при появлении родителей у гнезда
подходят к отверстию норы и родители уже не влетают в нору. До вылета птенцов
самка ночует в норе. Покидают птенцы гнездо на 30-ый день после вылупления.
На Кавказе птенцы начинают вылупляться в середине июня, молодые вылетают
в начале июля. На Украине вылет наблюдается в июле, а в конце июля молодые
уже хорошо летают; в Воронежской обл. 18 августа добыты молодые еще плохо
ЗОЛОТИСТАЯ ЩУРКА
511
летающие щурки (Огнев и Воробьев, 1924); в Приерусланской степи первый
вылет птенцов отмечен 15 июля, в это время в других гнездах были еще
только яйца (Волчанецкий и Яльцев, 1936); в окрестностях Бугуруслана вылет
отмечен 19 июля (Карамзин, 1909). В низовьях Сыр-Дарьи первые летные
птенцы встречаются с десятых чисел июля. В Узбекистане птенцы начинают
выклевываться во второй половине июня и первые вылетевшие птенцы
наблюдаются в середине июля. В это же время встречены молодые летные птицы
и в Туркмении. После вылета из гнезда птенцы первое время держатся вместе
с родителями, которые продолжают их кормить еще в течение 2—3 недель.
Если в период насиживания или к концу кладки гнездо разорено, то
золотистые щурки ко второй кладке не приступают. В размножении принимают
участие не все птицы; часть щурок, повидимому молодых, прошлогодних,
остаются холостыми. Они собираются в стаи и кочуют. Так, в разгар гнездового
периода щурок в Узбекистане приходилось наблюдать появление таких стай
далеко от мест гнездовий —в Ташкенте, Самарканде и т. д., причем щурки
вели себя так, как ведут они себя в послегнездовое время (Корелов, 1940).
Линька. О линьке щурки очень мало данных. У взрослых две линьки
в году — частичная и полная. Полная — зимой с января по март,
частичная— с конца июня по сентябрь; птицы улетают на зимовки, сменив только
мелкое перо» Молодые птицы улетают от нас в гнездовом наряде. Их первая
линька происходит на зимовках, и птицы возвращаются к нам весною в полном
взрослом оперении.
Питание. Различные крупные насекомые (перепончатокрылые, жуки,
бабочки, двукрылые, прямокрылые и др.). Насекомых ловят на лету. В
зависимости от особенностей местности и времени года видовой состав насекомых,
употребляемых в пищу, сильно меняется. В первое время по прилете щурки
совершают в поисках пищи большие кочевки. В это время, как правило,
пасеки ими не посещаются (птицы залетают на пасеки только в холодные и
пасмурные дни или в солнечные дни после сильного дождя, или если колония
расположена близко от пасеки). Пища в весеннее время состоит главным
образом из ос, диких пчел, жужелиц, стрекоз, плавунцов, чернотелок,
последние количественно преобладают (Корелов, 1949), так что в этот период года они
даже полезны. В гнездовое время пища состоит преимущественно из саранче-
вых, диких пчел, навозниц, стрекоз и т. д.; пасеки совсем не посещаются.
В годы появления большого количества Anisophia austriaca щурки охотно
поедают их, снимая с колосьев и ловя на лету (Харьковская обл.). Осенью во
время своих кочевок часто посещают пасеки, даже далеко отстоящие от мест
гнездовья, в это время года щурки вредны пчеловодству (Корелов, 1949).
Во время пролета, перед тем как устроиться на ночлег, и утром, перед
отлетом в пасмурные холодные дни, птицы также посещают пасеки, а в
солнечные дни только после сильного дождя.
Полевые признаки. Золотистая щурка легко узнается благодаря яркому
оперению, тонкому телосложению, заостренным крыльям, удлиненной паре
средних рулевых перьев, тонкому, удлиненному и изогнутому вниз клюву и
характерному полету, напоминающему полет ласточек. Хорошо заметна
каштановая спина, переходящая в золотистую на задней части спины, яркожел-
тое горло, черная полоса на груди и зеленовато-голубой остальной низ тела и
крылья. Молодых можно узнать по более тусклому оперению и по короткой
паре средних рулевых перьев.
Золотистые щурки часто собираются в стаи, особенно во второй половине,
лета, перед отлетом. Почти целый день проводят они в полете. Полет прямой,
слегка волнистый, птицы быстро носятся в воздухе, догоняя друг друга,
совершая различные круговые фигуры, часто останавливаются в воздухе,
трепеща крыльями. Затем снова стремительно несутся, играя или ловя насеко-
512
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
мых. Утомившись любят
отдыхать, присаживаясь
на телефонные провода,
деревья и кустарники, но
через короткий
промежуток времени опять с
громким криком
взлетают. Особенно часто их
можно видеть
отдыхающими на телеграфных
проводах и т. д. во время
пролета. К вечеру щурки
с громким криком летят
ночевать в стоящие
поблизости рощи, сады или
кустарники, растущие по
берегам рек и озер.
Голос громкий, несколько
гортанный, крик «крю-
крю-крю».... или «кюрт-
кюрт-кюрт», повторяется
несколько раз.
Описание. Размеры
и строение. Клюв
длинный, тонкий, слегка
изогнутый. Ноздри округлые и
расположены у основания
клюва, частично прикрыты ще-
тинкоподобными перьями.
Первостепенных маховых 11,
первое рудиментарное.
Формула крыла 2>3>4>5>6
>7>8>9>10>11>1 (3-е
короче 2-го на 0,4—0,7 мм).
Наружные второстепенные
маховые короче внутренних.
Ноги слабые, короткие,
плюсна покрыта спереди попереч •
ными пластинками, сзади
сетчатая. Пальцы слабые,
наружный палец соединен со
средним до последней
фаланги, внутренний со средним—
в основной фаланге. Рулевых
12, обрез хвоста слегка
округлый, средняя пара рулевых
сильно удлинена и опахала
на конце сужены. Длина тела
самцов (12) 240—300 лш,
самок (5) 230—295лш, среднее
277,6 и 269,2 мм. Размах
крыльев самцов (6) 400—450
мм у самок (4) 441—465 мм,
среднее 432,5 и 445, 2 мм.
Длина крыла самцов (54)
140—157лш, самок (29) 138—
150 мм, среднее 146 и 143 мм.
Вес (3) 50—60 г, среднее 55 г.
Окраска. Вылупившийся птенец голый, розовато-желтого цвета; края рта
желтовато-красные, без пятен; клюв прямой, короткий, серый. Молодые самцы и самки
окрашены одинаково. Похожи на взрослую самку, но бледней и тусклей. Спина окрашена
Рис. 128. Золотистая щурка (сверху — взрослая птица,
снизу — молодая птица).
ЗЕЛЕНАЯ ЩУРКА
513
в светлый каштановый тон. Большая часть темени зеленая, остальной верх головы
голубовато-зеленый. Плечевые перья голубовато-серые и иногда буровато-зеленые. Подбородок и
горло — бледножелтые, светлее чем у взрослой птицы, черная полоса на груди уже и
нерезкая, как бы с размытыми краями. Остальной низ тела бледней, чем у взрослой самки и
с грязноватым налетом. Хвост несколько темней, чем у взрослой и обычно с более голубо»
ватым оттенком. Средняя пара рулевых короткая, почти одинаковой длины с остальными
или немного длиннее. Клюв короче и изогнут слабее, чем у взрослой птицы.
У взрослого самца верх головы, шея и передняя часть спины каштановые, постепенно
светлеющие к задней части спины. Задняя часть спины, поясница — золотисто-рыжие.
Кроющие хвоста голубовато-зеленые, рулевые зеленовато-синие с темными вершинами. Лоб у
основания клюва белый, переходящий в голубовато-зеленый и затем каштановый цвет верха
головы. Уздечка, полоса через глаз и кроющие уха черные. Подбородок и горло яркожелтые,
светлеющие на боках шеи. Бровь и узкая полоса между черной полосой под глазом и желтым
горлом зеленовато-голубые. Брюшная сторона тела зеленовато-голубая, отделенная от зоба
черной полосой. Первостепенные маховые буровато-синие, темнеющие на внутренних
опахалах и на вершине. Второстепенные маховые каштановые с темнобурыми вершинами.
Третьестепенные маховые зеленоватые, голубеющие к вершине. Средние и большие кроющие
крыла каштановые, малые — буровато-зеленые. Клюв черный. Ноги буровато-черные.
Радужина красная.
Самка окрашена сходно с самцом, только верх головы, плечевые, поясница и низ
тела с легким зеленым налетом. Каштановый тон второстепенных маховых несколько
бледней. Горло чуть светлей и тусклей. Средняя пара рулевых короче, чем у самцов.
4. Зеленая щурка Merops superciliosas L.
Merops superciliosus. Linnaeus. Syst. Nat., ed. XII, 1766, стр. 183, Мадагаскар.
Распространение. Ареал. Южный Тунис, южный Алжир (быть может
Марокко), Египет, северная Сахара, восточная Африка, Мадагаскар, Пале-
Карта 87. Распространение зеленой щурки Merops superciliosus
1—ГМ. s. persicus, 2 — М. s. chrysocercus, 3 — М. s. superciliosus, 4 — M. s. phllippinus, 5 — M. s. salva—
dori; a — граница гнездовий, г — зимовки.
стина, Сирия, Месопотамия, возможно западная и центральная Аравия;
Армения, Иран, Афганистан, Белуджистан, Индия, Бирма, ю.-в. Китай,
Индокитай, северная часть Малайского п-ова, Андаманские о-ва, Новая Гвинея
и ряд мелких островов Индо-Австралийского архипелага.
33 Птицы, СССР т. I
514
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
Зимует в южной Аравии, в тропической, западной и южной Африке от
Сенегамбии, Нигерии, озера Чад, Камеруна, Конго и Сомали до Капской
Земли и на островах Индо-Австралийского архипелага.
Характер пребывания. Перелетная птица. Залетает на
Канарские о-ва, в южную Францию, в Италию, на Сицилию, Мальту и на Кавказ.
Биотоп. Степи, пустыни и полупустыни с небольшими холмами, песчаными
буграми, оврагами; речные долины с обрывистыми берегами.
Рис. 129. Зеленая щурка.
Подвиды и варьирующие признаки. Пять подвидов, различающихся между собой
окраской, размерами и экологией: M.s. superciltosus Li n n. 1766, — Мадагаскар, о. Пемба,
и в Бейра (восточная Африка);имеются единичные указания на гнездование в колонии Кения,
в Кавирондо и близ Кано, встречена на Коморских о-вах; М. s. persicus Pallas 1773—
Египет, Палестина, Сирия, может быть западная и центральная Аравия, Иран, Армения,
Средняя Азия, Афганистан, Белуджистан и северо-западная Индия; М. s. chrysocercus
С a b a n i з et Heine, I860 — южный Алжир, южный Тунис, северная Сахара, возможно
Марокко, зимует в западной Африке от Сенегамбии и Нигерии до озера Чад, Камерун,
Бельгийского Конго и на о. св. Томе; М. s. philippinus Linnaeus 1766 — Индия, за
исключением с.-з., на северо-востоке доходит до подножья Гималаев; Бирма, ю.-в. Китай
(провинции Юннань, Гуаньдун), Индо-Китай, северная часть Малайского п-ова и
Андаманские о-ва. Зимой встречается на Цейлоне, Суматре, архипелаге Рио и островах: Сималуре,
Сиберуте, Пагэ, Борнео, Ява, Бали, Кангеане, Ломбоке, Сумбаве, Флорес, Тиморе, Целебесе
и Филиппинах; М. s. salvadorii Meyer 1871—0. Новая Британия, о. Лонг,
отдельные гнездовые колонии на северном и южном берегах восточной Новой Гвинеи.
ЗЕЛЕНАЯ ЩУРКА
515
Обыкновенная зеленая шурка Merops superciliosus persicus Pall.
Merops persicus. Pallas. Reise d. versch. Prov. d. russ. Reichs. II, 1773, стр. 708, северный
берег Каспия.
Распространение. Ареал. Гнездовый ареал зеленой щурки в СССР
ограничен Средней Азией, побережьем Каспийского моря и восточным
Закавказьем, причем сведения о гнездовании ее в некоторых местах побережья
Каспийского моря и Закавказья очень неточны-и противоречивы. В Армении
зеленая щурка — залетная птица, вероятно, случайно гнездящаяся.
Гнездится в Ленкоранской низменности и по прилежащему берегу Каспия от
Иранской границы на север, вероятно, до ст. Алят, близ Баку, где встречена в
гнездовое время (Баньковский, 1912); вМуганской степи, у ст. Кумбаши; в Саль-
янах на Куре и в окрестностях Ленкорани. Еще меньше сведений имеется о
гнездовании зеленой щурки северней по западному берегу Каспийского моря
и на Северном Кавказе. В гнездовое время птицы "встречены в 1925 и 1926 г.
в Терекли-Мектеб Кизлярского района (коллекция Зоологического музея
Московского университета), найдена гнездовая колония в Ставропольской
обл. (Мориц, 1920). Старые сведения о гнездовании в низовьях Волги
подтверждены Формозовым, нашедшим в 1947 г. гнездовую колонию близ
с. Ганюшкино.
Гнездится зеленая щурка в устье Урала доходя на север до
Гурьева; отмечены встречи ее северней по Уралу и даже в б. Оренбургском крае
(Зарудный, 1888), но это вероятно случайные залеты. Что касается восточного
берега Каспия, то зеленая щурка добыта в летнее время на Мангышлаке
(коллекция Зоологического музея Московского университета), у Карабогаза,
близ Сартаса (Михель, 1941), в М. Балханах (для Б. Балханов не приводится).
Гнездится по всей Туркмении от Гассан-кули и Копет-Дага до Аму-Дарьи и
Кызыл-Кумов; в низменных и равнинных местностях Узбекистана и
Таджикистана. В южном Казахстане гнездится по северному побережью Аральского
моря, в южных частях Аральских Кара-Кумов, по нижнему и среднему
течению Сыр-Дарьи, на озере Тели-Куль, по нижнему течению Сары-су, по
р. Чу, в дельте и нижнем течении Или и в районе Ак-Дала (юго-западный
Балхаш). Таким образом, северная граница распространения зеленой щурки в
нашей стране проходит значительно южней, чем у золотистой. В зоне степи
зеленая щурка почти не гнездится и встречена на гнездовье в зоне пустыни и
полупустыни и в переходной полосе между степной и пустынной зонами.
Вне нашей страны гнездится в Афганистане, с.-з. Индии, Белуджистане,
Иране, Месопотамии, Палестине, Сирии, быть может в западной и центральной
Аравии и в Египте.* 3 и м о в к и. Отмечены в южной и тропической Африке,
к югу от Сомали, Египетского Судана, Виктории-Ньясса, в верхних и
нижних областях Конго и Анголы и в южной Аравии.
Характер пребывания. Гнездящаяся и перелетная птица.
Сроки прилета в различные части нашей страны различны. Местами в
северные районы своего распространения зеленая щурка прилетает в более ранние
сроки, нежели в южные (вероятно это связано с различными, то более близкими,
то более отдаленными местами зимовок отдельных популяций и различными
направлениями пролета). В общем, зеленые щурки прилетают в Среднюю Азию
и на Кавказ в различные числа второй половины апреля и в начале мая, лишь
иногда в холодные весны пролет захватывает и первую половину мая. В
Туркмении прилет зеленых щурок наблюдается на несколько дней позднее прилета
золотистых, но в 1942 г. под Ашхабадом они появились одновременно
(Дементьев, 1950), в Приатрекской пустыне зеленая щурка встречается на
пролете несколько поздней золотистой, обычно в конце апреля, и в меньшем
количестве, чем последняя. На нижней Сыр-Дарье, в районе Кзыл-Орды зе-
зз*
516
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
леная щурка появляется ранее золотистой, так же и в Узбекистане, где зеленые
щурки приступают к гнездованию в то время, как золотистые только
появляются и их пролет на север еще продолжается (Спангенберг, 1936; Корелов,
1949).
В Туркмении пролет наблюдался под Ашхабадом 19 апреля, в пустыне
между Мары и Аму-Дарьей большой пролет шел 9 мая, в устье Атрека
валовый пролет—13 мая (Зарудный, 1896), под Чикишляром—1.1 мая (Радде и
Вальтер, 1889), на Кушке у Кала-и-мора, на Мургабе, в Карабиле в 1946 г.—
между 24 апреля и 2 мая, отдельные пролетные птицы встречались до 8 мая.
У ст. Анненково первые прилетевшие птицы наблюдались 29 апреля, у
Артыка—4 мая (Лоудон, 1913), В Кара-Кумах поздний пролет наблюдался
в 1927 г.: у Дарваза— 10 мая, у Ербента — 4—5 мая, у бугра Чеммерли — 14
мая и у Кош-Аджи — 11 мая (Кашкаров и Курбатов, 1929). На северном
побережье Арала зеленые щурки впервые появились 17 мая; в устье Урала в
конце первой трети мая. На Кавказ они прилетают позднее золотистой. В
Ленкорани и Талышской низменности прилет первых птиц наблюдался 30 апреля
и пролет шел до конца первой декады мая; над Махач-Кала — 3—4 мая
1927 г. щурки летели на север непрерывными стаями, в Ставрополе — 7 мая
и 19 мая (Радде, 1885; Беме, 1932).
Об осеннем пролете и отлете зеленых щурок почти нет никаких данных,
сроки отлета, повидимому, тесно связаны с температурными условиями: щурки,
как и стрижи, покидают родину, как только резко сократится количество
летающих насекомых. Особенно рано щурки улетают из районов Гурьева,
Аральского моря и Семиречья, где осенью часто наблюдаются кратковременные,
но резкие похолодания. Но в 1899 г. в Семиречье близ Усека 4—5 октября было
встречено несколько стай (до 30 шт. в каждой), летящих в направлении на
запад и юго-запад (Зарудныйи Кореев, 1906). Такая поздняя встреча
пролетающих зеленых щурок вероятно связана с равномерной и теплой осенью 1899 г.
Северное побережье Арала покидают в конце августа. В южных частях
Средней Азии, где влияние с.-в. ветров не столь резко, щурки задерживаются на
местах гнездованья на длительное время. В Гиссарской долине первые
пролетные стайки появляются в первых числах сентября, последние—- в начале
октября (7—8 октября, Иванов, 1940), тогда как у золотистой щурки пролет
уже закончился в конце второй декады сентября. Пролет по Теджену
наблюдался в начале сентября, отдельные особи задерживаются в Туркмении до
октября (в Бадхызе добыта 4 октября); на Атреке у Чатлы пролет шел еще 18 октября
(Шестоперов, 1928). На Кавказе в районе Махач-Кала пролет зеленых щурок
наблюдался в двадцатых числах сентября. ♦
Биотоп. Пустыни, полупустыни, изрезанные холмами, обрывами,
оврагами, долинами рек, каналами и озерами с берегами, поросшими редкой'
древесной и кустарниковой растительностью или камышом. Оазисы,
человеческие поселения, родники, колодцы, морские побережья и т. д. Местностей,
совершенно лишенных какой-либо растительности, избегает. В горы высоко не
подымается: так в Семиречье идет до 1700 м (Зарудный и Кореев, 1906,
вероятно, это кочующие птицы), в Таджикистане до 600—700 м (Иванов, 1940).
Численность. В Ленкорани многочисленна. В Средней Азии
спорадична и местами многочисленна. В большом количестве встречается по Мур-
габу и Теджену, в дельтах Аму- и Сыр-Дарьи, в долинах нижнего и среднего
течения этих рек, но в верховьях встречается уже реже. Многочисленна в
оазисах, в низменностях горных районов, в предгорьях уже реже.
Экология. Размножение. Гнездится колониями, иногда
достигающими больших размеров, и отдельными парами. Норы роет в крутых обрывах
холмов, бугров, в берегах рек, оврагах,, в арыках, иногда норы устраиваются
на небольших возвышениях почвы, даже на совершенно ровных местах —»в
ЗЕЛЕНАЯ ЩУРКА
517
песках морского побережья, пустынях и изредка в расщелинах скал. Особенно
любит берега рек и озер, поросших какой-либо, хотя бы редкой,
растительностью. Мягкий песчаный грунт предпочитается глинистому, так как клюв ее,
более слабый по сравнению с клювом золотистых щурок, затрудняет ей
устройство норы в твердом грунте. Нора лежит неглубоко под землею и ход идет
горизонтально, почти параллельно поверхности земли. Ход короче, чем у золотистой
щурки (что также вероятно объясняется более слабым клювом), и длина его
сильно колеблется в зависимости от твердости грунта; в рыхлом она достигает
2—3 м} в твердом от полуметра до 1 м (Зарудный, 1896). Близ Ленкорани
зеленые щурки роют нору прямо в прибрежной морской полосе, где почва состоит
из измельченных раковин. Длина хода здесь около метра и гнездо расположено
на глубинеЛО см от поверхности почвы (Радде, 1885). Нора обычно служит только
один год, ее роют клювами сами птицы —. самец и самка, земля выбрасывается
наружу лапами. Собственно гнездом является расширение на конце хода.
Подстилкой в гнезде служат веточки трав, стебельки злаков, хитиновые остатки
насекомых и растертые погадки птиц. В период кладки яиц и насиживания
отверстия нор, которые находятся на ровных местностях, часто забиваются
щурками на ночь или на время оставления норы птицами землею, и таким
образом они становятся недоступными для змей, хищников и других врагов
(Кзыл-Орда, Спангенберг, 1936).
Кладка яиц в Туркмении начинается в конце мая (Зарудный, 1896), у
добытых в Туркмении 28 апреля 1946 г. птиц половые органы не развиты, 2 мая
отдельные пары у Мургаба начинают рыть норы; у добытой близ Серахса самки
яичник в начале мая был слегка гроздевидным. В низовьях Сыр-Дарьи
кладка яиц начинается также в конце мая, затягиваясь у отдельных особей
до середины июля. В Узбекистане зеленые щурки приступают к постройке
гнезда раньше золотистой и кладка начинается в середине мая, что повиди-
мому связано с более ранним прилетом по сравнению с золотистой щуркой.
В Ленкорани 25 мая 1898 г. еще не было достроено ни одной гнездовой норы:
первое гнездо с незаконченной кладкой было найдено там 17 июня 1898 г.
(Сатунин, 1907). Там же гнездо с незаконченной кладкой найдено 13 июня
(Радде, 1885); повидимому кладка яиц в Ленкорани начинается значительно
позже, чем в Средней Азии.
В году одна кладка; в кладке 4—6, редко 7 яиц. Яйца по форме и
окраске похожи на яйца золотистой щурки —. почти округлой формы,
блестящего белого цвета с мелкозернистой скорлупой. Размеры: (36) 23,6—26,1 мм х
20,0—22,0 мм (нижняя Сыр-Дарья, Спангенберг, 1936); (100) 24,1—27,1 х
20 — 22,6, в среднем 26,2 х 20,9 мм (Индия, Бекер, 1927).
Срок насиживания — около трех недель, насиживают оба пола. Вылет
молодых в низовье Сыр-Дарьи происходит приблизительно в то же время,
как и у золотистой щурки — в десятых числах июля, вылет молодых в
Туркмении— в конце июня и в начале июля. Первое время после вылета молодые
держатся с родителями, которые продолжают их кормить в течение 21/2— 3
недель.
Линька изучена слабо. Зеленая щурка, также как и золотистая, по-
видимому, имеет две линьки в году — одну частичную, с конца июля и до
сентября, во время которой сменяется только мелкое оперение, и вторую -^
полную, происходящую на зимовках с января до марта. У молодых птиц после-
гнездовая линька бывает зимой с января по февраль, когда гнездовое оперение
сменяется взрослым летним нарядом.
Питание. Зеленая щурка, как и золотистая, питается крупными
насекомыми, главным образом, перепончатокрылыми, пчелами, кузнечиками и
осами, которых ловит на лету. В некоторых случаях может приносить
значительный вред пчеловодству.
518
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
Полевые признаки. По повадкам и внешности очень похожа на золотистую,
но меньше размерами и более легкого телосложения. От золотистой отличается
кроме размеров, отсутствием на горле яркого желтого пятна (горло рыжевато-
коричневое) и зеленой спиной. Как и золотистая щурка, много времени
проводит в воздухе — играя и ловя насекомых. Легко отличается от других птиц
своим длинным тонким и слегка изогнутым клювом, удлиненной и суженной
к вершине средней парой рулевых. Молодых птиц в первое время после вылета
можно узнать по неуверенным движениям в воздухе и короткому клюву
и хвосту Голос похож на голос золотистой щурки, но более резкий и
низкий. Может быть передан как звук «кри-кри-кри», повторяемый птицами
несколько раз подряд.
Описание. Размеры и строение. Клюв длинный, тонкий, слегка загнутый книзу.
Ноздри округлой формы, лежат у основания клюва, прикрыты частично щетиноподобными
перышками. Первостепенных маховых— 11, первое рудиментарное, второстепенных— 12.
Формула крыла: 2>3>4>5>6>7>8>9>10>11>1, второе длиннее третьего на 0,9—1,1 мм.
Обрез хвоста округлый, средняя пара рулевых удлинена до 70 мм и в вершинной части
сужена, рулевых—12. Ноги слабые и короткие, плюсна спереди покрыта щитками, сзади
сетчатая. Пальцы слабые, наружный палец срастается со средним на всем протяжении—
до последней фаланги, внутренний — только с основной фалангой. Длина тела самцов
(6) 293 — 337 мм, самок (4) 295—305 мм, в среднем 322 и 299 мм. Размах крыла самцов
(6) 421 — 463 мм, в среднем 451 мм, самок (4) 447—450 мм, в среднем 448 мм. Длина
крыла самцов (32) 145—160 мм, самок (17) 158—144 мм, в среднем 153,3 и 148 мм.
Окраска. Птенец вылупляется голым, с коротким серого цвета клювом.
Молодая птица- Гнездовый наряд. Похожа на взрослую, отличаясь только
буровато-оливковым тоном оперения, светлеющим на спине к задней ее части. Белого под глазами совсем
нет и на лбу очень мало. Голубая бровь или очень узкая, или ее совсем нет.
Подбородок беловатый, зоб тусклого бледнорыжего цвета. Клюв короткий, средняя пара
рулевых или такой же длины как и остальные, или чуть длиннее. Радужина бурая, ноги
темноте лесные. Взрослые птицы. Самец и самка имеют оперение зеленое. Уздечка, полоса
через глаз и кроющие уха — черные. Лоб, бровь и полоса под глазом — голубые. Лоб
у основания клюва, узкая полоса под глазом — белые с черными и голубыми крапинами.
Подбородок яркожелтый — постепенно переходящий на зобу в рыжевато-каштановый цвет.
Рулевые и первостепенные маховые зеленовато-оливковые с темными вершинами на
внутренних опахалах первостепенных маховых. Второстепенные маховые с
буровато-оливковыми наружными опахалами и рыжеватыми темнеющими к вершине внутренними. Средние
кроющие крыла — каштановые. Кроющие хвоста и плечевые с голубыми вершинами
перьев. Клюв черный, ноги буровато-коричневые, радужина красная.
СЕМЕЙСТВО ЗИМОРОДКОВЫЕ ALCEDINIDAE
РОД ЗИМОРОДОК ALCEDO LINNAEUS 1758
Тип Alcedo ispida
5. Обыкновенный зимородок Alcedo atthis L.
Gracula Atthis. Linnaeus. Syet. Nat. I, изд. X, 1758, стр. 109, Египет.
Распространение. Ареал. Средняя и южная Европа, умеренная, южная
игпередняя Азия и северная Африка. Северная граница проходит от южной
Швеции, Прибалтики, по 58° с. ш. в зап. областях Европейской части СССР
и по 56° с. ш. в восточных; от северного Казахстана, к Барнаулу, Ачинску,
Красноярску, охватывает с севера Забайкалье, Приморскую область и
Сахалин. Гнездится в Японии, Корее, Китае, на Формозе, Хайнане, на островах
Индо-Австралийского архипелага, в Индо-Китае, Бирме, Индии, Цейлоне,
Белуджистане, Афганистане, Иране, Армении, Малой Азии, Месопотамии,
Сирии, Палестине и в северной Африке.
Характер пребывания. В северных областях ареала
перелетная птица, изредка остающаяся в теплые зимы на незамерзающих водоемах;
ОБЫКНОВЕННЫЙ ЗИМОРОДОК
519
в южных — оседлая. Биотоп. Обрывистые, покрытые кустарниковой и
древесной растительностью берега рек, ручьев, озер, канав, каналов и других
водоемов с чистой прозрачной водой и тихим течением.
Подвиды и варьирующие признаки. Семь слабо различающихся морфологически
(размеры и окраска) и экологически подвидов: A. a. ispida Linn., 1758 — Западная
Европа, на север до южных частей Скандинавии и Британских о-вов, на восток до
Прибалтики, Польши и может быть Минска (восточная граница нуждается в уточнении); к югу—
до Пиренейского п-ова, южной Франции, северной Италии, Югославии и западной
Румынии; A. a. atthis Linn., 1758 — с.-з. Африка, Египет, Сирия, Палестина, Малая Азия,
острова Средиземного моря, южная Италия, Балканы на север до нижнего и среднего
течения Дуная, южная и средняя полоса Европейской части СССР на север приблизительно
до 58° с. ш. в западной части и 56° с. ш. — в восточной, Средняя Азия к северу до Казахстана,
средняя Сибирь к северу до Красноярска, Минусинска, на восток доАлтая, подножья Саян,
Зайсанской котловины, Тарбагатая; к югу до с.-з. Индии, Белуджистана, Афганистана,
Карта 88. Распространение обыкновенного зимородка Alcedo atthis
1 — A. a. atthis, 2 — A. a. ispida, 3 — A. a. bengalensis, 4 — A. a. taprobana, 5 — A. a. flore-
sciana, 6 — A. a. hispidoides, 7 — A. a. salomonensis; а — граница гнездовий, г — зимовки.
Ирана, Закавказья и Крыма; A. a. bengalensis G m e 1 i n, 1788 — ю.-в. Сибирь от Байкала и
Даурии до Приморья; Сахалин, Япония, Формоза, Корея, Манчжурия, восточный и ю.-в.
Китай, Индо-Китай, Бирма, Индия на север до Бутана, Сиккима и Непала на юг до Орисса,
Бенгалии, р. Годавари, Бихара и Деккана; Малаккский п-ов, Андаманские и
Никобарские о-ва, Большие Зондские о-ва, Филиппины и Хайнань; A. a. taprobana Klein-
schmidt, 1894 — Цейлон, южная Индия, на север до Бомбея, Пуна, р. Годавари и
южных границ предыдущей формы; A. a. floresiana S h а г р е, 1892 —острова Бали, Сум-
бава, Флорес, Алор, Ветер, Сумва, Тимор; A. a. hispidoides Lesson, 1837 — острова
Сангир, Целебес, Бутон, группа островов Туканбеси, о-ва. Дампьер, Калао, Семиер, Пелинг,
Сула, Молуккские о-ва, западные Папуасские (Мисоль, Салавати), с.-в. Новая Гвинея,
архипелаг Д'Антрекасто, Луизиады и Бисмарка, о-ва Адмиралтейства, о-ва Скалли,
святого Матвея, Фени и Ниссан; A. a. salomonensis Rotsch. etHartcrt 1905 —
Соломоновы о-ва, от Бугэнвиля и Моно до Санта-Крус.
Русский обыкновенный зимородок Alcedo atthis atthis L.
Синонимы: Alcedo Pallasii. Reich enbach, Handb. spec. Orn. Investigatores,
Alcedineae, 1851, стр. 3, Зап. Сибирь.—Alcedo pallida 3 a p у д н ы й. Орнитолог.,
фауна Закасп. 1896, стр. 354, Закаспий, пес. Chr. L. В г е h m, 1855. — Alcedo margelanica.
Madarasz, Annal. Musei Nat. Hungar., II, 1904, стр. 86, Маргелан. — Alcedo ispida
suschkini P u s a n о w. Бюлл. Моск. Общ. Испыт. Прир., и. сер. XLII, 1933, стр. 15, Крым.
Распространение. Ареал. Северная Африка (Марокко, Алжир, Тунис,
в Египте, вероятно, не гнездится); Сирия, Палестина, с.-в. Аравия,
Месопотамия, Иран, Армения, Малая Азия, острова Средиземного моря (Кипр, Сицилия,
Сардиния, Корсика), южная Италия, Балканы на север до среднего и нижнего
520
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
течения Дуная, ю.-в. Румыния; южная и средняя полоса Европейской части
СССР от берегов Черного моря, Кавказа и низовьев Волги на север до Гдова.
окрестностей Пскова (повидимому случайное гнездование указано на озере
Сямго Онежского района Житковым и Бутурлиным, 1901), Калининской обл.,
Ростовского района Ярославской обл., Костромы, Горьковского края, Ветлуги,
Кировской обл. к северу до Малмыжа и Уржума. В Татарской республике
до Казани; гнездится в Башкирии до Мензелинского, Уфимского и Бирского
районов (Сушкин, 1897); в долине среднего Урала, в низ. Илека (Заруд-
ный, 1888); по южной оконечности Урала (Сушкин, 1897); к северу до Б. Ика
и р. Картала, близ впадения их в приток Урала — Сакмару.
В низовьях Урала зимородка на гнездовье повидимому нет; по Эверсманну
зимородок встречается в нижнем течении Урала, но до Гурьева не доходит,
позднейшими исследователями на нижнем Урале не найден, нет его на ЭмбеР
в песках Малые и Большие Барсуки, на Усть-Урте, в пустынях Кара-Кумы и
Кызыл-Кумы. Однако могут быть и здесь встречены по арыкам одиночно
гнездящиеся пары зимородков. Спорадично гнездится по водоемам Туркмении;
в низовьях Атрека его нет, гнездится по западному Узбою и по среднему
течению Атрека; также в юго-восточном углу Каспийского моря, по Аму-Дарье,
по Сурхан-Дарье; в низменных частях Узбекистана и западного Таджикистана —
в долине Исфары, на Гиссарском хребте, в долине Гиссара, по долине Кафир-
нигана, Вахша, Кызыл-Су и Пянджа (Иванов, 1940). Есть в Киргизии и
Казахстане, по всему течению Сыр-Дарьи, в Кара-тау, по долинам рек Чу и
Или и у Балхаша. Далее к северо-востоку распространен до
Усть-Каменогорска и Змеиногорска, окрестностей Семипалатинска и Барнаула. Населяет
северную, западную и южную окраины Алтая, заходя неглубоко внутрь края,
и только по долине Бухтармы идет до Котон-Карагая, найден на Телецком
озере, но гнездование его там не установлено (Сушкин, 1938). Гнездится в
верховьях Оби, в окрестностях г. Томска и в Кузнецкой степи, по р. Томи,
в окрестностях г. Кузнецка, в Барабинской степи, под Омском, по Иртышу,
на р. Имбок против устья Елагуя (Скалой и Слудский, 1941) и на р. Кеть
в районе Енисейска (Тугаринов и Бутурлин, 1925)—это самые северные
места нахождения русского зимородка. Обычен у Ачинска, Красноярска,
Каинска, в Минусинском крае, в верховьях Енисея и его притоках —
Абакану, Тубе, по нижнему течению Кебсжа, на Можарских озерах. К югу и
востоку идет до пустынных окраин Центральной Азии по подножью
северозападных Саян (в Таниу-тувииской области и в северо-западной Монголии
его нет), через Зайсан-нора, Ала-куль, Сассык-куль, Кульджу (коллекция
Зоолргического музея Московского университета) и через Среднюю Азию
(в восточном Туркестане его нет) до северо-западной Индии (Кашмир, Раджпу-
тан, Пенджаб, Синд) и берегов Персидского залива.
Характер пребывания. У нас гнездящаяся и перелетная птица;
только в Закавказье и в Туркмении живет оседло. Зимует в небольшом
количестве на юге Украины Шодолия, Херсон), по Черноморскому побережью
Кавказа, в большом количестве скопляется на зиму в низменностях
Закавказья по рекам (по нижнему течению р. Храма и Квеши; Цветков, 1892; в
низовьях Арагвы), в Ленкоранской низменности, по южному и юго-восточному
берегу Каспия, на р. Мургабе, в Пендинском оазисе (Зарудный, 1896). В
остальной области распространения у нас зимородки зимой встречаются лишь
случайно на незамерзающих водоемах, в особенности в теплые зимы
(наблюдались в Псковской обл.. 2 января 1898, Зарудный, 1910; близ Смоленска —
7 января 1914, в Гомельском полесье, Станчинский, 1929; в Минусинске,
Сушкин, 1914 и т. д.).
Весною одиночные птицы начинают появляться в южных районах нашей
страны уже в первых числах апреля и даже в конце марта. Во время пролета
ОБЫКНОВЕННЫЙ ЗИМОРОДОК _ 521
летят небольшими стайками, придерживаясь рек, низко над водою. На
Украину прилетают в среднем в середине апреля: в Днепропетровской обл. —
в среднем 15 апреля, но в некоторые годы появляются уже числа 6—7 апреля,
иногда в конце .марта (Боровиков, 1907); в окрестностях Киева — в среднем
20 апреля, но в 1921 г. прилетели 7 мая, самый ранний прилет отмечен 10
апреля 1922 г. (Данилович, 1932); в Харьковскую обл. в среднем прилетают
14 апреля; самый ранний срок зарегистрирован 6 апреля 1890, самый
поздний — 14 мая 1881 г. (Сомов, 1897).
У Чкалова и в Поволжье зимородки прилетают в среднем в середине мая;
у Бугуруслана — в Л887 г. прилетели 17 апреля (Карамзин, 1909), в
Башкирии первый прилетный зимородок встречен 27 мая (Сушкин, 1897). В
областях средней полосы прилетают во второй половине апреля и даже в начале
мая (в Тамбовской обл. однажды отмечен зимородок 3 апреля, скорее всего
был встречен зимовавший там зимородок), В низовье Сыр-Дарьи и прилежащие
районы прилетают в начале апреля (Спангенберг, 1936), в Илийский край —
30 апреля (Шестоперов, 1929), в Алтайский край —в середине мая и в
низовья Оби — в конце мая (Сушкин, 1938).
С первых чисел августа у зимородков начинаются уже кочевки, в которых
главным образом участвуют молодые птицы. Наблюдаются большие колебания
в сроках начала отлета, зависящие от наступления заморозков и покрытия
водоемов льдом. В конце августа и в начале сентября одиночные птицы
начинают уже отлетать, причем в'первую очередь улетают старые. Отлет идет в
течение всего сентября. Но в октябре встречаются еще иногда одиночки, чаще
молодые птицы. Из центральных областей Европейской части СССР зимородки
отлетают в первой половине сентября, но одиночные птицы встречаются до конца
сентября и даже начала октября (близ Пскова последний зимородок
наблюдался в середине сентября, Зарудный, 1910; в Рязанской обл. 22 сентября,
Хомяков, 1910); на Суре и ее притоках держатся до конца сентября, в б. Бу-
гурусланском уезде и до начала октября. В Чкаловской обл. отлет идет с
конца августа, и в середине сентября встречаются только одиночные экземпляры
по Уралу. На Украине пролет начинается с конца августа и идет до конца
сентября и даже начала октября. На Кавказе зимородки в конце октября и
в начале ноября спускаются с гор в низменности, скопляясь в большом
количестве по речкам. В Алтайском крае из высоких горных районов исчезают
в конце августа, вообще же Алтай покидают в первой половине сентября.
В низовьях Сыр-Дарьи и в прилегающих районах зимородки задерживаются
осенью очень долго и начинают отлетать только тогда, когда водоемы
покрываются льдом (Спангенберг, 1936). Залетный зимородок 30 августа 1939 г.
добыт Тепловой на притоке Печоры — р. Кедровке.
Биотоп. Обрывистые глинистые или песчаные, покрытые
кустарниковой или хотя бы редкой древесной растительностью берега рек, ручьев, озер,
канав, арыков и других водоемов с прозрачной водой, В горных районах
Кавказа, Средней Азии, Алтая и др. придерживается низменных местностей —
долинных рек с открытыми, но обрывистыми берегами, покрытых
растительностью. Узких скалистых ущелий, рек со стремительным течением и мутной
водой избегает. В горы распространен не выше 2000 м и там гнездится
преимущественно на озерах. На Кавказе отмечен до 1500—1800 м (Богданов,
1879, Радде, 1885), в Армении на высоте 2000 м— озера Севан и Гилли (Ляйстер
и Соснин), на Алтае до 1100 м (Сушкин, 1938).
Численность. Повсюду немногочисленен, у северной границы
своего распространения редок и спорадичен, в несколько большем количестве
встречается только в низменностях Ленкорани и Талыша, по рекам Черноморского
побережья, в низменностях Семиречья, северного Таджикистана и Копет-
Дага.
522
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
Экология. Размножение* Пары сезонные; вне периода размножения
самец и самка держатся отдельно. Гнезда устраивают в норах. Гнездящиеся
пары селятся, в зависимости от удобных для гнездовья мест, на расстоянии от
0,3 до 1 км и более. Раз поселившись, строго придерживаются выбранного
места. Норы занимаются птицами в течение ряда лет. Гнездовый и кормовой
участки строго разделены и часто кормовые участки удалены от гнезда на
километр. Норы роют в обрывистом берегу реки или какого-либо другого водоема,
покрытом кустарником и древесной растительностью. Высота откоса, на
котором помещается нора, весьма различна — от 4 до 17 м. В отношении грунта,
в котором роет нору, зимородок мало разборчив, но каменистых берегов
избегает, предпочитает глинисто-песчаные, суглинистые и глинистые почвы,
иногда роет и в черноземе и редко в меловых выходах.
Гнездо помещается чаще всего у самой воды. Входное отверстие норы
расположено обычно над водой от 75 см до 2 м, иногда до 4—7 м. Отверстие норы
в диаметре 4—б см и почти всегда скрыто ветвями деревьев, кустарниками,
корнями или пнем, но встречаются и совершенно открытые отверстия в нору.
Реже устраивают норы в отдалении от воды, иногда на расстоянии до одного
километра, в последнем случае он устраивает гнезда в плотинах, песчаных ямах,
дуплах деревьев, в отверстиях стен и т. д. (Перекрестов, 1914; Вальх, 1899).
Форма летка очень различна.
Норы имеют почти горизонтальный ход под землею и только вырытые
в черноземе или суглинке —с изгибами (Вальх, 1899). Длина хода в
зависимости от грунта колеблется от 30 см до 1 м. На конце ход постепенно
расширяется и образует довольно большую камеру, часто направленную несколько
в сторону или кверху сантиметров на 15.
Нору роют самец и самка клювом, выбрасывая лапками выкопанную
землю. На копание норы уходит от 7 до 12 и более дней, повидимому, в
зависимости от твердости грунта. Так, в Илийском крае птицы приступили 12 мая к
копанию норы, и 1 июня в этом гнезде было уже 7 яиц, под-Джаркентом птицы
с 25 апреля по 4 мая копали норы (Шестоперов, 1929). В камере помещается,
как правило, одно гнездо, но иногда и два.
Длина гнезда от 12 до 23 см, в среднем — 16,2 см; ширина гнезда от 11,5 слс
до 20 см, в среднем 17,3 см\ высота от 9 до 14 см, в среднем 11,4 см. Как все иор-
ники, настоящего гнезда не строят; в поре, вырытой в первый год, яйца лежат
или прямо на земле, или на небольшой подстилке, сделанной из сухой травы.
В старых же гнездах яйца откладываются в углубление, сделанное в подстилке
из размельченных рыбьих остатков. За период высиживания и выкармливания
птенцов в гнезде скопляется толстый слой мелких рыбьих костей, чешуи, над-
крыльев жуков и экскрементов птенцов, перемешанных и измельченных. Норы
зимородков весьма грязны, кишат личинками мух и издают зловоние от
разлагающихся мелких рыбешек.
В кладке обычно 6—7 яиц, но бывает 4—8 и редко 10. Яйца
блестящего белого цвета с тонкой мелкозернистой, но прочной скорлупой почти
шарообразной формы.
Размеры (12) 21,0—23,6 х 17,6—18,2, в среднем 28,6 х 17,9 мм
(Харьковская обл., Сомов, 1896). В году бывает одна, местами две кладки.
Насиживают оба пола, насиживание начинается после того, как кладка
закончена. Период насиживания 21 день. По вылуплению птенцов часть
скорлупы выбрасывается родителями из гнезда, часть остается в гнезде и
размельчается. Кормит птенцов только самка, самец в период выкармливания держится
близ гнездового участка, но участия в выкармливании птенцов не принимает.
Птица с кормом влетает в нору и быстро возвращается обратно. Первая кладка
обычно начинается в начале или середине мая и идет до середины или конца
июня, вторая —с конца июня и начала июля и до середины или конца августа.
ОБЫКНОВЕННЫЙ ЗИМОРОДОК
523
Гнездовый сезон сильно растянут — можно одновременно встретить гнезда
с яйцами, с неоперившимися птенцами и хорошо летающих молодых птиц.
Повидимому птицы (скорее всего старые), которые занимают готовые
прошлогодние гнезда, раньше приступают к кладке яиц.
Точных данных о нахождении у нас гнезд и кладок очень мало. На Кер-
женце 16 мая найдено гнездо с 4 ненасиженными яйцами (Серебровский,
1918); в Рязанской обл. 26 мая найдены три начатых гнезда, из которых одно
было почти закончено (Хомяков, 1910); в Башкирии встречены два гнезда 6 и
7 июля с птенцами (Штехер, 1915); в Воронежской обл. найдены в 1919 г. гнезда
21 мая с одним яйцом, 22 июля с шестью голыми птенцами и 24 июля с тремя
свежими яйцами, в 1922 г. 25 июня наблюдалось спаривание, 20 июля — гнездо
с семью сильно насиженными яйцами (Огнев и Воробьев, 1924); в
Харьковской обл. кладки яиц встречаются с середины мая до конца июня,
одновременно — гнезда с незаконченными кладками и гнезда с неоперившимися
птенцами (Сомов, 1897). В Черниговской обл. в конце июня наблюдались почти
оперившиеся птенцы (Емельяненко, 1917), в Подолии 8 июля 1926 г. добыта самка
с готовым к сносу яйцом (Портенко, 1927). На Кавказе в Северной Осетии и
Ингушетии полные кладки найдены 13 мая 1915 г., 8 июля 1915 г. и 17 мая
1917 г. (Беме, 1926); в Закавказье 19 июля 1906 г. встречено гнездо со слегка
насиженными яйцами (Баньковский, 1913). В окрестностях Томска кладки
зимородков находили в начале июня. У Джаркента кладки с ненасиженными
яйцами встречены в течение мая (19 мая—4, яйца. 22мая — 6, 22 мая—7,
29 мая — 1,1 июля — 7; Шестоперов, 1929). Вылет птенцов из гнезд
происходит на 24—26 день после вылупления. Первое время после оставления
молодыми гнезда — птицы держатся одной семьей и родители еще некоторое
время кормят молодых. Только ставши совершенно самостоятельными,
молодые разлетаются врозь и начинают вести в одиночку бродячую жизнь. В
Воронежской обл. 22 августа убит одиночный молодой зимородок (Огнев и
Воробьев, 1924), и в это же время наблюдались только что вылетевшие птенцы,
в 1920 г. 16 июня добыт молодой вполне уже оперившийся зимородок, а
30 июня выводки уже разбились, и молодые вели самостоятельную жизнь. В
Северной Осетии и Ингушетии вылетевшие молодые встречаются с половины июня.
Линька. В году две линьки: полная послебрачная — в августе —
сентябре по ноябрь и частичная предбрачная в январе — марте. В центральных
областях европейской полосы линька начинается с конца августа (у экз. от
29 августа, Тамбовская обл. Темниковский район — четыре первостепенных
маховых свежие и остальные старые), на юге линька, повидимому идет в то же
время: у экз. от 29 и 28 августа, Бердянск, и от 31 августа, Черкассы, линька
маховых уже заканчивается. В Западной Сибири линька идет с первых чисел
августа (у экз. от 17 августа, Томск, все первостепенные маховые свежие, за
исключением трех, крайняя пара рулевых свежая, перья еще в чехликах,
остальные старые), но иногда начинается и раньше, в конце июня (у экз. с
р. Кальджир-от 29 июня 4 маховых выпало на обоих крыльях). В Средней Азии
линька начинается поздней—в сентябре (экземпляры, добытые на Сыр-Дарье
31 августа в старом изношенном пере, — к линьке еще не приступали). В конце
декабря и в январе птица уже в свежем наряде (экз. от 24 декабря, Дзау-
джикау, йот 18 января, Ленкорань), но иногда линька заканчивается и раньше,
в сентябре (от 4 сентября, окрестности Батуми, и от 10 ноября, Мцхет, птицы
в свежем наряде). Линька ювенального наряда в неполный взрослый —
частичная и идет с августа по декабрь; линька молодых птиц на 2-м году жизни
с августа по ноябрь — полная. Смена маховых перьев идет от внутренних
маховых к наружным, а рулевых от крайних к внутренним.
Питание. Питаются главным образом мелкой рыбой, не более 60 мм
длины (составляет до 65% содержимого желудка), насекомыми и другими
524
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
беспозвоночными. Из насекомых преимущественно водные и их личинки, в том
числе в большом количестве личинки стрекоз. Из других беспозвоночных:
моллюски, креветки, земляные черви и т. д. Иногда поедают и мелких лягушек,
в небольшом количестве в пищу идут водные растения.
Пищу зимородки добывают главным образом в воде. Обычно они сидят на
ветке, склоненной над водою, или на каком-либо возвышении у воды и
высматривают добычу, иногда летают низко над водою, время от времени
останавливаясь в воздухе и трепеща крыльями. Увидев добычу, они стремительно
падают в воду, при этом часто сами слегка погружаются в неё, хватают добычу
клювом и уносят или в нору или возвращаются с нею на старое место, где и
поедают ее. Нырять зимородки не могут
и ловят только ту рыбу, которая
плавает близко к поверхности воды. Хотя
в питании зимородка рыба стоит на
первом месте, они большого вреда
рыбному хозяйству не приносят, так
как поедают только мелкую рыбу; к
тому же зимородки у нас
немногочисленны и селятся они на большом
расстоянии пара от пары.
Уничтожением насекомых и в том числе
личинок стрекоз, поедающих икру рыб,
приносят некоторую пользу.
Полевые признаки. Блестящий
зеленовато-голубой верх тела с
мелкими светлыми крапинами на
крыльях и затылке, рыжеватокоричне-
вый низ тела, длинный прямой
клюв, большая голова и короткие
крылья и хвост легко позволяют
узнать зимородка в природе.
Часто подолгу сидит неподвижно или
Рис. 130. Обыкновенный зимородок. "* СУЧ1<С стоящего у самой воды
кустарника или на выступающем из
поды камне и выглядывает добычу.
Также зимородка можно видеть летающим с пронзительным криком над
самой водой, иногда останавливающимся на короткое время в воздухе, быстро
и часто машущим крыльями. Голос — тонкий протяжный крик,
напоминающий свиет «пиик-пиик-пиик».
Описание. Размерыистросиие. Клюв длинный, прямой, с резко выступающим
хребтом надклювья. Ноздри щелевидной формы и расположены у самого основания клюва.
Первостепенных маховых 10,. первое рудиментарное, самое длинное 3 и 4,2 и 5 чуть короче.
Рулевых — 12. Обрез хвоста почти прямой формы, крайние рулевые очень незначительно
короче средней пары. Кроющие хвоста более чем на половину длины хвоста покрывают
его сверху. Ноги короткие, слабые, пальцы анизодактильного типа. Вес самцов 27,5—38 г.
Длина тела самцов (14) 170—187 мм, самок (13) 165—180 мм, среднее 177,8 мм и 174,5 мм,
длина молодых (8) 170—180 мм, среднее 177,15лш. Размах крыльев самцов (13) 260—283 лш,
самок (12) 260—280 мм, среднее 270,4—и 263,7 мм, молодых (6) 250—283лш, среднее 270,5 мм.
Длина крыла самцов (72) 69—75 мм, самок (34) 69—75 мм, среднее 72,7 мм и 72,6 мм,
молодых (41) 69—75 мм, среднее 72,1 мм.
Окраска. Птенец. Вылупившийся птенец голый, но вскоре покрывается перьями,
долго сохраняющими чехлики. Молодые птицы в первом осеннем наряде похожи на самок, но
с более тусклым и темным тоном оперения с более сильным развитием зеленого цвета на
плечевых, голове, боках зоба и грязноохристым тоном на зобе. Первый весенний наряд отличается
от взрослого грязноватым тоном на зобе и зеленым на верхней стороне тела, придающим
птице более темный оттенок. Ноги и клюв бурые, у некоторых экземпляров подклювье у
основания беловатое.
ОБЫКНОВЕННЫЙ ЗИМОРОДОК
525
Взрослая птица. Окончательный наряд надевается после полной линьки (зимой) на
второй год жизни. Верхняя сторона тела темноголубая. Верх головы темноголубой в
поперечных черноватых и голубых полосках. Ширина голубых предвершинных полос подвержена
сильным индивидуальным колебаниям, так что иногда верх головы не поперечно
полосатый, а крапчатый. Уздечка темнобурая, пятно на ней, полоса под глазом и кроющие уха —
охристые. От угла подклювья по бокам головы и шеи тянется голубая с темными
поперечными пестринами полоса, отграничивающая белое горло от охристой полосы под глазом.
Горло и бока шеи белые, бока зоба голубые. Остальная брюшная сторона охристо-рыжая.
Спина, надхвостье и кроющие хвоста яркого голубовато-синего цвета. Рулевые — голубые
с черными стволами перьев. Плечевые — голубые с зеленоватым налетом, постепенно
уменьшающимся с возрастом и совсем отсутствующим у очень старых птиц. Кроющие крыла
зеленовато-голубые с более светлыми блестящими вершинами перьев, причем величина
светлых вершин сильно варьирует, то они в виде крупных пятен, то в виде мелких крапин.
Маховые темнобурые с голубыми краями по наружному опахалу: Клюв черный. Ноги-
телесно-красные. Радужина бурая. Самка похожа на самца, но окрашена более тускло. На
верхней стороне тела у нее имеется легкий зеленоватый налет. Нижняя сторона тела светло-
охристая. Часто основание подклювья, а иногда и все подклювье светлорогового цвета.
Систематические замечания. В 1935 г. Тугариновым и Козловой предложено
пользоваться названиями margelanica для зимородков из северо-восточного Туркестана
(Прибалхашье, Семиречье, Фергана) и Кашгарии (в последней зимородки только зимуют).
Отличаются они от Л. a. atthis— более светлой окраской спинной стороны тела. Птиц из Гис-
сарского хребта, района Ташкента, Самарканда и Ашхабада считать за переходные.
Просмотренный нами коллекционный материал показывает, что у зимородков имеется
очень большая изменчивость — как возрастная, так в особенности и индивидуальная,
например, ширина пестрин на верхней стороне головы и на кроющих крыла. Более светло
окрашенные птицы — старые. У перезимовавших молодых окраска оперения еще довольно темная,
с зеленоватым оттенком на верхней стороне тела и с грязноватым тоном на зобе или только
боках зоба и т. д. На основании этого, мы не считаем возможным признать реальность
существования формы A, a. margelanica.
Западный обыкновенный зимородок Alcedo atthis ispida L
Alcedo ispida Linnaeus. Syst. Nat., изд. X, 1758, стр. 115, Швеция.
Распространение. Ареал. Зап. и Средняя Европа на север до
Великобритании, южных частей Скандинавии, Прибалтики, к югу до Португалии,
южной Испании, южной Франции, до северной границы Югославии и
западной Румынии. Восточная граница неясна, повидимому от Польши,
Прибалтики (у нас от Калининградской обл. до Эстонии) до западной Румынии,
может быть доходит до Ленинграда и Минска, но в Смоленской, Могилевской
Гомельской, Киевской, Подольской обл. и в Придунайской низменности —
уже предыдущий подвид.
Характер пребывания. В северных частях области
распространения — перелетная птица, в южных — оседлая. Единичные экземпляры
остаются на зиму у незамерзающих водоемов в северных частях своего
распространения, в Польше и частично в Прибалтике живут оседло.
Биотоп. Обрывистые, покрытые кустарниковой и древесной
растительностью берега различных водоемов с чистой прозрачной водой и
спокойным течением. Численность. Всюду обычен, но малочисленен.
Экология. Размножение. Отличий от русского зимородка в выборе
мест для нор, в постройке нор и в устройстве гнезд нет. Длина хода норы
колеблется от 35 см до 1 м, в среднем 56 см, высота хода от 5,3 до 9 см, в среднем
6,6 см, ширина от 4,5 до 8см (Кумари, 1939). Размеры гнездовой камеры такие
же, как и русского зимородка. Толщина слоя подстилки в гнезде доходит от
0,5 до 10,5 см, в среднем 4 см, вес ее от 3,7 до 384,4 г, в среднем --104,5 г.
Норы сильно загрязнены. В тех районах, где зимородки оседлы, наблюдаются
иногда три кладки в году, и к кладке они приступают раньше, чем русский
зимородок. В Прибалтике первая кладка обычно начинается в конце апреля и
идет до середины июня; вторая кладка — с середины июня и конца июня до
начала августа и третья кладка, встречающаяся довольно редко, —в начале
526
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
или в середине августа до середины сентября, причем часто яйца третьей кладки
откладываются во вторую кладку. В Англии кладка яиц начинается в конце'
апреля — в начале мая, но иногда встречаются кладки и в марте. Количество
яиц такое же, как и у русского зимородка.
Яйца по окраске и форме не отличимы от яиц русского зимородка. Раз-;
меры: (100) 24,2—21,2 х 19,7 — 16,7 мм, в среднем 22,64 х 18,73 мм (Англия,
Уайзерби, 1938); (10)23,1—22,2 х 19,3— 18,5 мм, в среднем 22,57х 18,85 мм.
Вес насиженных (4) яиц в среднем 4,135 г (Прибалтика, Кумари, 1939).
Срок высиживания и выкармливания такой же, как у западного зимородка.
Линька. Вопрос о количестве линек в году не совсем ясен. Повидимому
бывает две линьки в году — полная послебрачная в августе — сентябре и
частичная предбрачная в январе — марте, но может быть и одна послебрачная
полная линька с сентября по декабрь, что также весьма вероятно для
некоторых районов при наличии там двух-трех кладок в году.
Питание. В пище преобладает мелкая рыба и мальки, а также
различные водные насекомые и другие беспозвоночные.
Описание. Размеры и строение. Отличается от русского зимородка более
крупными размерами. Длина крыла самцов (5) 73—78 мм, среднее 75,5, самок (1) 73, молодых
(4)65-—68 мм, среднее 66,25. (Западная Европа); самцов 75—78мм, самки 75—80 мм
(Англия, Уайзерби, 1935).
Окраска сходна с окраской русского зимородка.
Восточный обыкновенный зимородок Alcedo atthis bengalensls Gm.
Atthis bengalensis Gmelin Syst. Nat. 1788, стр. 450, Бенгалия.
Синоним. Alcedo japonica, Bonaparte. Consp. Vol. Anis. 1854, стр. 10.
Распространение» Ареал. От Байкала, где он гнездится по всем рекам
Читинской обл., до Приморья и Сахалина; в Приморье к северу доходит до озера
Эворон, реки Эворон и низовьев Амура. К югу до северо-восточной Монголии;
Манчжурия, Япония, Формоза, Хайнань, Китай, Индо-Китай, Бирма,
северная Индия (на север до Бутана, Сиккима и Непала , на юг до Бенгалии,
Центральных провинций, Бихара, Орисса и р. Годовари), Андаманские и
Никобарские о-ва, Малаккский п-ов, Ява, Суматра, Борнео и Филиппины.
Характер пребывания. У нас гнездящаяся птица, случайно
остающаяся зимовать на незамерзающих водоемах. В южных областях своего
распространения — оседлая птица* Наблюдений о времени прилета и
отлета нет.
Биотоп. Тот же, что и у других подвидов.
Численность. В Приморье многочисленен.
Экология. Размножение. Места для нор и норы выбирает и строит
так же, как и русский зимородок. О сроках размножения восточного зимородка
у нас нет почти никаких данных. Повидимому у них так же как и у русского
зимородка начало кладки сильно зависит от готовности норы и в году так же
бывает и две кладки. К размножению приступают, повидимому, в начале мая
и период размножения тянется до конца июля и, может быть, начала августа.
Так на Имане в южном Приморье, по сообщению Спангенберга, 12 мая была
добыта самка с яйцом в яйцеводе; 30 мая найдено гнездо с шестью ненасижен-
ными яйцами; 31 июня —гнездо с четырьмя ненасиженными яйцами; 1 июня
законченная нора, но яиц в ней еще не было; 4 июня в этом гнезде оказалось
два яйца. 18 июня добыта самка с яйцом в яйцеводе и встречены молодые птицы,
хорошо летающие, 29 июня — гнездо с семью слабо насиженными яйцами.
Форма и окраска яиц такая же, как у русского зимородка. Размеры (7) 22,3—
22,7 х 18,5—18,9 мм (Иман, Приморье; Спангенберг); (40) 19,9 — 22,4 х 17,0—
19,0, в среднем 20,9 х 17,6 мм (Индия, Бекер, 1927).
ОБЫКНОВЕННЫЙ ЗИМОРОДОК
52?
Линька не изучена. У экземпляра, добытого 7 августа на р. Имане,.
4 маховых свежие, короткие, еще в чехликах, остальные —старые; у экз.
от 19 июля, озера Ханка, линька еще не начиналась.
Питание. В питании преобладает мелкая рыба; поедает также водных
насекомых других беспозвоночных, мелких лягушек и водные растения.
Описание. Размеры и строение. Отличается от русского зимородка более
мелкими размерами. Размеры: длина тела самцов (5).159—173, среднее 166,1 мм, молодых
(2 экз.) 168—170 мм. Размах крыльев самцов (5) 253—272, среднее 259,5 мм, молодых
(2) 255—263 мм. Длина крыла самцов (25) 69—73, самок (4) 69—72, молодых (12) 68—73 мм,
среднее 70,9 мм, 70,5 мм и 70,6 мм. Вес самцов (1) 30,6 г, молодых обоего пола (5)
29,4 — 35,9 г, среднее 32,5.
Окраска. Отличается от западного зимородка несколько более темным оперением..
РОД ПЕГИЙ ЗИМОРОДОК CERYLE ВО I E 1828
Тип. Ceryle rudis
6. Малый пегий зимородок Ceryle rudis L.
Alcedo rudis. Linnaeus. Syst. Nat., изд. X, 1758, стр. 116, Египет.
Распространение. Ареал. Африка — долина Нила от устья до истоков^,
на восток до Красного моря и на юг от Сахары до Капской Земли. Сирия, Па-
Карта 89. Распространение малого пегого зимородка Ceryle rudis
1 — С г. rudis, 2 —С. г. leucomelanura, 3 — С. г. Insignis; a — граница области распространения,
д — залеты.
лестина, Малая Азия, Месопотамия, берега Персидского залива, Белуджистан,.
Индия, к северу до Синда, Кашмира и Гималаев. Цейлон, Бирма, Юннань,
Южный Китай, Хайнань и Индо-Китай. Характер пребывания —
оседлая птица. Биотоп. Берега водоемов, покрытых редкой древесной
растительностью или одиночными деревьями с чистой прозрачной водой и с
медленным течением.
Подвиды и варьирующие признаки. Три подвида малого пегого зимородка,
различающихся окраской оперения, размерами тела и клюва: С. г. rudis Linnaeus, 1758—Африка,
528
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
к югу от Сахары и до Капской Земли, долина Нила (от истоков до устья), Сирия, Палестина,
Малая Азия, Месопотамия и берег Персидского залива; —С. г. leucomelanura R e i с h e п-
b а с h, 1851 — Индия на север до Синда, Кашмира и Гималаев, Цейлон, Бирма, Сиам и
Тенасеррим; — С. г. insignus H a r t e r t, 1931 — Южный Китай, Юннань, восточный
Индо-Китай и Хайнань.
Западный малый пегий зимородок Ceryle rudis rudis L
Распространение. Ареал. Африка от Сахары, дельты и долины Нила до
Капской Земли, о. Кипр, Сирия, Палестина, Малая Азия, Месопотамия, юго-
западный Иран и берег Персидского залива.
Рис. 131. Малый пегий зимородок.
Характер пребывания. Оседлая птица. У нас случайно
залетная— Рикбейль добыл в феврале 1861 г. пегого зимородка под Сарептою,
там же в марте 1869 г. встретил второго Богданов (1871); Штегман весною
1926 г. наблюдал в Крыму близ Судака. Кроме того, залеты наблюдались
в Польше,у на островах Греческого архипелага, Сицилии, Испании и в
северозападной Африке.
Биотоп.. Берега водоемов с прозрачной водой, стоячих или с тихим
спокойным течением.
Экология. Размножение. Период размножения в Египте совпадает
с понижением воды в Ниле. Гнездо устраивает в трещинах скал, в норах,
которые роют сами птицы, в отвесном берегу реки, канала или какого-либо
другого водоема. Отверстие норы расположено на различном расстоянии от воды.
Длина хода в зависимости от твердости грунта тянется от 60 до 175 см. Ход
заканчивается расширением, образующим камеру, служащую гнездом. В
большинстве случаев яйца откладываются прямо на землю, без подстилки и только
БОЛЬШОЙ ПЕГИЙ ЗИМОРОДОК
529
изредка камера бывает выстлана травой. В тех норах, в которых уже имеются
птенцы, много остатков костей рыб и экскрементов.
Яиц в кладке обычно 4, редко 3 или 6. В году бывает две кладки. Яйца
округлой формы, чисто белого цвета с блестящей, тонкой, но очень крепкой
скорлупой, похожи на яйца других зимородков. Размеры: (96) 32,2 —
25,5 х 23,9—20,7 мм, в среднем 28,9 х 22,4 мм (Хартерт, 1911); (100) 32,0—
27,9x24,0—23,8 мм, в среднем 29,9x24,1 мм (Индия, Бекер, 1927).
Линька. Происходит зимой в декабре — январе.
Питание. Различные мелкие водные животные, главным образом,
мелкая рыба, крупные водные насекомые, креветки, головастики и мелкие
лягушки. Ловит рыбу, в отличие от обыкновенного зимородка, паря в воздухе
на высоте трех-восьми метров от поверхности воды. Увидав добычу,
стремительно бросается за ней, погружаясь в воду и даже часто на короткое время
исчезая совершенно под водой.
Полевые признаки. Крупней обыкновенного зимородка, легко узнается
благодаря своей пестрой — черной с белым — окраске оперения и большому,
черному клюву. Голос — щебетание, непрерывно издаваемое во время полета
и переходящее в громкий крик, когда птица стремительно бросается в воду
за добычей.
Описание. Размеры и строение. Голова большая, клюв длинный и прямой,
крылья относительно короткие. Первостепенных маховых— 11, первое зачаточное, самые
длинные—3-е и 4-е. Формула крыла :3 = 4>5>2>6>7>8>9>10>1. Рулевых 12, обрез
хвоста прямой. Ноги короткие, пальцы слабые (анизодактильного типа). Длина крыла
(7) 136—142, в среднем 138,3 мм.
Окраска. Взрослый самец. Оперение пестрое, черное с белым. На затылке
черный хохол с белыми поперечными пестринами. От клюва к глазу и над глазом проходит
черная полоса с узкими белыми пестринами. Спина черная с белыми поперечными
полосами. Первостепенные маховые белые в основной своей части и черные в вершинной.
Второстепенные маховые белые с черным большим неправильной формы пятном в вершинной
части внешнего опахала и незначительным черным пятном на внутреннем опахале.
Надхвостье черное с белыми поперечными полосами. Рулевые в основной своей части белые,
в вершинной — черные с белыми концами, за исключением двух средних пар. Брюшная
сторона — белая с двумя черными, иногда посредине прерывистыми поперечными
полосами на груди, причем передняя более широкая, задняя более узкая. Клюв и ноги черные.
Радужина бурая. Самка окрашена сходно с самцом, отличаясь тем, что черный цветне
такой яркий — с буроватым оттенком и поперечные полосы на груди или совсем отсутствуют
или имеется только одна.
7. Большой пегий зимородок Ceryle lugubris Те mm.
Alcedo lugubris. Те mmi nc k. PI. Col., 1834, стр. 548, Япония;
Распространение. Ареал. Япония, вост. и юго-вост. Китай, сев. и вост.
Индо-Китай, Бирма и сев. Индия на запад до Кашмира.
Характер пребывания. Оседлая птица, совершающая
небольшие кочевки, обычно в вертикальном направлении и только на самом крайнем
севере своего распространения (Хоккайдо) перелетная.
Биотоп. Реки с большим количеством рыбы, протекающие по
невысоким горам и в предгориях, с берегами, поросшими кустарником и древесной
растительностью. В горы высоко не поднимается: в Гималаях — от 600 до
2100 ж. Зимой спускается ниже — обычен на высоте 150—-300. л£.
Численность. Обычен, но всюду малочисленен.
Подвиды и варьирующие признаки. Два подвида, различающиеся окраской оперения —
С. I. lugubris Те mm., окрашен более бледно; С A. lugubris Temminck, 1834—-Япония;
С. L guttulata Stejneger, 1893 — Гималаи от Кашмира до Ассама, Бирма, Тенассерим
до Амхерста; Индо-Китай (Лаос, Аннам, Тонкий) и Китай от Юнианя до Ганьсу, Цинь-линь-
шаня и Чжили.
34 Птипы СССР, т. Т
530
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
Японский большой пегий зимородок Ceryle lugubris lugubris Те mm.
Синонимы. Ceryle lugubris sikokiana. Mo mi у a ma. Annot. Orn. Orient-, I, стр. 67,
1927, Сукоку. — Ceryle lugubris jamasemi. Momiyama. Annot. Orn. Orient., I, стр. 69,
1927, Хондо. — Ceryle lugubris pallida. Momiyama. Annot. Orn. Orient., I, стр. 70у
1927, Хоккайдо.
Распространение. Ареал. У нас случайно залетный. Встречен на о-ве
Кунасири, Курильские о-ва, 18, 20 сентября и в первых числах октября
1948 г. (Портенко, 1950).
Биотоп. Реки различной величины с большим количеством рыбы,
протекающие по холмистой и горной местности.
Численность. Обычен, но немногочисленен.
Экология. Размножение. Моногамы. Гнездящиеся пары удалены
друг от друга на большое расстояние. Гнезда устраивает в норах, вырытых
в берегах рек, в оврагах и
обрывах, покрытых
кустарниковой и древесной
растительностью. Норы, независимо от
почвы, имеют короткий ход,
оканчивающийся большим
расширением —гнездовой
камерой. Дно камеры, как и у
обыкновенного зимородка,
выстлано костями и чешуей
рыб. Толщина выстилки
гнезда колеблется в зависимости
от количества лет
использования его птицами. В кладке
3—4 яйца, редко 2—5,
похожих на яйца зимородков, но
относительно размеров птицы
мелкие. Размеры: (20) 39,4—
37,3x35,0—30,1 в среднем
38,5X32,5 мм (Индия, Бекер,
1927).
В году вероятно одна
кладка. Время размножения в
Индии с марта до начала
Рис. 132. Большой пеги И мимородок.
Японии (Ян, 1942) с апреля по июнь,
мая, иногда июня (Бекер, 1927).
Линька. Не изучена.
Питание. Рыба, которую подстерегает, сидя на ветвях деревьев,
кустов, низко спускающихся к воде. За добычей стремительно бросается,
часто погружаясь в воду головой и передней частью тела.
Полевые признаки. Хорошо заметен своей пестрой — черной с белым —
окраской оперения, хохлом, большой величиною, в особенности клюва и головы,
и короткими крыльями. Летает быстро и часто взмахивая крыльями, обычно
на короткие расстояния, низко над водою, но может летать высоко в воздухе
с довольно большой скоростью и на большие расстояния. Любит тенистые
места, часто забирается в глубь кустов, но, по наблюдению Портенко (1950), на
о-ве Кунасири зимородок выбирал освещенные солнцем места. Очень
осторожная птица. Голос передается разными авторами различно, повидимому,
сходен с голосом обыкновенного зимородка, но громче «питт-птитт» (Ян, 1942).
Портенко (1950) находит, что вначале голос сходен с трещанием олянки,
затем заканчивается звуками «кеть-кеть». В брачный период издает другие
звуки — «киарарера» (Ян, 1942).
0ШЕЙНИК0ВЫЙ ЗИМОРОДОК
531
Описание. Размеры и строение: Крупных размеров зимородок, с большой
головой и клювом и короткими крыльями и хвостом. На голове хохол. Размеры: длина крыла
177—195 мм, длина клюва 55—70 мм (Бутурлин, 1936).
Первостепенных маховых 11, первое рудиментарное, самое длинное—3-е. Хвост
прямой, рулевых — 12. Ноги короткие, пальцы слабые. Существенных различий в размерах
между самцом и самкой нет.
Окраска. У самца спинная сторона тела в белых и серовато-черных поперечных
полосах. Брюшная сторона белая с черной поперечной полосатостью на зобе и на боку тела.
На зобе и щеках охристый! налет. На шее белый ошейник. По бокам шеи ряд белых и черных
пятен. Хохол черный с редкими белыми пятнами. Маховые и рулевые черные с белыми
пятнами. Верхние кроющие крыла черные с белыми вершинными пятнами. Подмышечные
белые с черными крапинами. Радужина бурая. Клюв темнобурый, почти черный.
Самка: окраска оперения сходна с самцом, отличаясь только отсутствием охристого
тона на зобе и щеках и рыжеватыми подмышечными.
Молодые птицы похожи на самок.
РОД КРАСНОНОСЫЙ ЗИМОРОДОК HALCYON SWAINSON, 1820
Тип Alcedo senegalensis
8. Ошейниковый зимородок Halcyon pileata В о d d.
Adcedo pileata. Boddaert. Tabl. PL EnL, 1783, стр. 41, Китай (рис. 10, табл. X).
Распространение. Ареал. Китай на север до вост. Ганьсу, Шань-
си, Чжили; Корея; Индия; Цейлон; Ассам; Бирма; Индо-Китай; Малайский
Карта 90. Распространение ошейникового зимородка Halcyon pileata
а — граница гнездовий, г — зимовки, д — залет.
п-ов; Андаманские и Никобарские о-ва; о-ва Риу-Киу; Хайнань; Филиппины
(о-в Палаван, о-в Балабак, о-в Барелон и о-в Тави-тави); о-в Северная Натуна,
о-в Борнео, о-в Суматра, архипелаг Рио, о-в Сималур, о-в Паге. У нас — случайно
залетная птица: добыт 24 мая 1878 Янковским на о-ве Аскольд.
34*
532
ОТРЯД СИЗОВОРОНКИ ИЛИ РАКШИ
Экология. Размножение. Гнездятся в норах, похожих на норы
красноносого зимородка Я. smyrnensis (см. дальше), которые делают в берегах рек,
морских бухт. Время размножения в Индии — май — июнь, иногда
захватывает апрель и июль. Яйца белые, блестящие, округлой формы, похожи на
яйца Я. smyrnensis. Размеры: (8) 33—35 х 28,2—29,2, в среднем 33,9 х 28,6мм
(Тачановский, 1891); (42) 28,0—31,2 х 25,3 —28,0, в среднем 29,6 х 26,3 мм
(Индия, Бекер, 1927). Л и н ь к а. Не изучена. Питание. Преимущественно
рыба и, кроме того, различные
крупные водные насекомые и
другие беспозвоночные.
Полевые признаки.
Крупней обыкновенного
зимородка, легко узнается по своей
яркой окраске оперения, ко
раллово-красному большому
клюву, коротким крыльям и
хвосту и довольно крупным
размерам тела. Полет прямой
и мягкий. Голос похож на
голос красноносого
зимородка, но более громкий,
резкий и пронзительный.
Описание. Размеры и
строение. Крылья и хвост
короткие, большая голова с длинным
клювом. Первостепенных маховых И,
первое рудиментарное 3-е, 4-е и 5-е
почти равны между собою, б>2=7.
Рулевых — 12. Обрез хвоста
прямой. Ноги короткие и слабые, но
сильнее, чем у пегого зимородка.
Размеры: длина крыла (1)125 мм
(Зоологический музей Московского
университета); 126—136 (Хартерт,
1912—1921).
Окраска. Взрослый
самец. Верх и бока головы и шеи,
передняя часть спины и верхние
кроющие крыла — черные. Вокруг
шеи светлоохристый ошейник.
Спина, плечевые, надхвостье и
рулевые кобальтово-синие с фиолетовым
оттенком. Первостепенные маховые
черные в своей вершинной части и
белые в основной, с узким синим
окаймлением по краю внешнего опахала. Второстепенные маховые с черным
внутренним опахалом и вершинной частью пера и синим внешним опахалом. Горло, зоб и
передняя часть груди— белые, грудь в остальной своей части светлоохристая. Брюхо, бока
тела и подхвостье охристые. Ноги и клюв кораллово-красные. Радужина — бурая. Самка
похожа на самца, но окрашена несколько тусклей и перья зоба и груди с узкими
черными каймами. Молодые птицы похожи на самок, но охристый налет распространен и
на зобе и на передней части груди. Перья зоба и груди, как и у самок, с темными
узкими окаймлениями.
9. Красноносый зимородок ^Halcyon smyrnensis L.
Alcedo smyrnensis. Linnaeusi Syst. Nat., изд. X, 1758, стр. 116, Малая Азия.
Распространение. Ареал. Малая Азия, Сирия, Палестина, Месопотамия,
сев. Аравия, Иран., южное побережье Каспийского моря, Белуджистан,
Афганистан, Индия, Цейлон, Андаманские и Никобарские о-ва, Малак-
кский п-ов, Индо-Китай, Южн. Китай, Хайнань и Формоза.
Рис. 133. Ошейниковый зимородок.
КРАСНОНОСЫЙ ЗИМОРОДОК
533
Карта 91. Распространение красноносого зимородка Halcyon smyrnensis
7 — н. s. smyrnensis, 2 — Н. s. fusca, 3— H. s. generosa, 4— H. s. saturatus; a — граница области
распространения (цифры в кружках обозначают номера зимующих подвидов), в — изолированные находки.
Характер -пребывания. Оседлая птица. Биотоп. Берега
рек, озер и других водоемов с чистой прозрачной водой и медленным
течением, иногда встречается далеко от воды.
Рис. 134. Красноносый зимородок.
Подвиды и варьирующие признаки. Четыре подвида: Н. s. smyrnensis Linnaeus,
1758—Малая Азия, Сирия, Палестина, сев. Аравия, Месопотамия, Иран, южное
побережье Каспия, Афганистан, Белуджистан и сев.-зап. Индия; H.s.jusca Boddaert,
1783 — Индия на северо-запад до границ предыдущей формы, на юг до Траванкоры;
Бирма, Южн. Китай, Индо-Китай, Малаккский п-ов, Формоза; Я. $. generosa M а-
d а г a z, 1904 — Цейлон; Н. s. saturatior Hume, 1874—Андаманские и Никобарские о-ва.
Морфологические варьирующие признаки — размеры и окраска оперения;
экологической изменчивости, повидимому, нет.
r534 отряд сизоворонки или ракши
Белогрудый красноносый зимородок Halcyon smyrnensis smyrnensis L.
Распространение. Ареал- Малая Азия, Сирия, Палестина, сев.
Аравия, Месопотамия, Иран, южный берег Каспия в Иране, может быть Лен-
коранская низменность, Афганистан, Белуджистан и сев.-зап. Индия (Кашмир,
Пенджаб, Синд).
Характер пребывания. Оседлая птица, в СССР только
залетает. У нас добыты в Ленкоранской низменности два экземпляра: один
в июне 1884 г. (Радде, 1885), другой 27 января, 1908 г. (Лоудон, 1910), во
втором случае птица держалась вблизи небольшого водоема в густом лесу.
Основываясь на наблюдениях Лоудона и на времени встречи этой птицы
(январь), Тугаринов и Козлова (1938) красноносого зимородка считают
редкой оседлой птицей Талыша. У северной границы своего распространения
это — редко гнездящаяся, но не оседлая птица, таким образом правильней
считать его у нас случайно залетной птицей.
Экология* Размножение. Период размножения в Индии — с
февраля по апрель, причем начало гнездового сезона совпадает с окончанием
разлива рек. Норы в берегах рек могут быть выкопаны только тогда, когда
окончится дождевой сезон и реки войдут в свои русла. Длина хода норы, в
зависимости от твердости грунта, тянется от 60 до 150 см, заканчивается
камерой, диаметром в 15—^20 см. Яйца откладываются прямо на дно камеры
без подстилки. В кладке 4—7 яиц, обычно б, округлой формы с белой
блестящей скорлупой, похожих на яйца обыкновенного зимородка. Размеры:
(8) 31,0—30,8x27,8—26,4 мм, в среднем 30,81 х 26,89 мм (Хартерт, 1911);
(30) 31,7 — 28,2 х 25,9—24,6 мм, в среднем 29,4x26,2 мм (Индия, Бекер,
1927).
Линька. Не изучена.
Питан и,е. Водные насекомые, крабы и другие беспозвоночные,
лягушки, ящерицы и в небольшом количестве рыба. "
Полевые признаки. Много крупнее обыкновенного зимородка. В полете
сходен с обыкновенным зимородком, но легко отличается благодаря своим
размерам, яркой окраске оперения и красному длинному и прямому клюву.
Крылья и хвост относительно короткие.
Описание. Размер ы и строен и с, Первостепенных маховых—11, первое
рудиментарно. 3-е, 4-е, 5-е, 0-е почти рлппы, 3-е и 4-е чуть длиннее других. Рулевых—12. Обрез
хвоста округлый, крайние рулевые короче средней пары на 20 мм. Ноги короткие, слабые,
но сильнее ног пегого и ошейпикопого зимородков. Размеры: длина крыла (1) 128 мм:
118—128 мм (Бекер, 1927), 123—130 мм (Хартерт, 1911).
Окраска. Взрослые самцы и самки. Спинная сторона тела кобальтово-синяя,
переходящая постепенно к пояснице н зеленовато-голубой тон. Верх и бока головы
и шеи, передняя часть спины, бока зоба и груди, нижние кроющие крыла, брюхо и бока
тела каштаново-коричневые. Горло, середина зоба и груди белые. Малые кроющие крыла
рыжевато-коричневые, средние кроющие крыла черные,большие кроющие крыла темноголу-
бые с зеленоватым оттенком. Первостепенные маховые светлоголубые у основания
наружного опахала, белые на внутреннем опахале основной части и черные — в вершинной.
Рулевые зеленовато-голубые сверху с черными стволами перьев и буроватые снизу. Клюв и
ноги кораллово-красные. Радужина бурая. Молодые птицы похожи, на взрослых, но верх
головы буровато-коричневый. Белые перья на горле, зобе и груди с узкими буроватыми
окаймлениями и вершинами. Клюв темнокоричиевый с красноватым тоном на подклювье.
отряд удоды
UPUPAE или UPUPIFORMES
Характеристика отряда
Морфология. Внешние признаки. Отряд удодов объединяет
небольшую группу птиц, по внешнему строению сильно различающихся между
собою — собственно удодов и птиц-носорогов. Сюда входят птицы средних и
крупных размеров, но чаще — средних. Единственный представитель отряда
в нашей фауне — удод Upupa epops, весит 70 г при длине тела около
300—310 мм. Некоторые населяющие тропические страны виды достигают
очень больших размеров (птицы-носороги). У калао Dichoceros bicornis длина
тела 1200 мм, длина крыла 500--520 мм.
Оперение довольно жесткое (особенно у птиц-носорогов), плотно
прилегающее к телу. Неоперенных, голых участков кожи на голове и шее нет.
Цевка не оперена и покрыта щитками спереди и сзади. Пуха и побочного ствола
контурных перьев нет (редуцированный побочный ствол имеют лесные удоды
Phoeniculus). Копчиковая железа не оперена. Первостепенных маховых — 10,
крыло эутаксическое, округлое, довольно широкое и тупое. Рулевых — 10,
обрез хвоста прямой или округлый, только у Phoeniculus хвост ступенчатый
и очень длинный, а у Rhinoplax с удлиненной средней парой рулевых.
Окраска оперения разнообразная, то глинисто-рыжая с черным и белым
(удоды), то черная с белым и желтым (птицы-носороги), то с металлическим
блеском (лесные удоды). Полового диморфизма в окраске оперения нет или, если
есть, то выражен очень слабо. Молодые птицы в первом осеннем наряде слабо
отличаются от взрослых. Сезонных отличий нет. Клюв по форме и величине
весьма разнообразен: то тонкий, длинный, слегка изогнутый вниз с гладкими
краями—у настоящих удодов Upupae, то огромный и сильно изогнутый вниз
с зазубренными краями челюстей (за исключением токов) и с костнороговыми
полыми выростами — у Bucerotes.
Ноги анизодактильного типа (за исключением рогатых воронов Висог-
vus abyssinicus), четырехпалые, с короткой цевкой и пальцами (только у
обыкновенного удода и рогатых воронов ноги довольно длинные).
Анатомические признаки. Череп десмогнатический, голо-
ринальный, с непрободенной носовой перегородкой. Атлас и эпистрофей у
удодов свободны, а у птиц-носорогов срастаются вместе. Грудина с одной
парой вырезок, иногда (удоды) зарастающих в фонтанели. Вилочка
536
ОТРЯД УДОДЫ
U-образной формы. Шея умеренной длины, шейных позвонков — 14.
Скелет у удодов слабо пневматичен, у птиц-носорогов сильно пневматичен.
Гортань трахео-бронхиального типа. Язык рудиментарный. Зоб, слепые
кишки и обводящая мышца отсутствуют. Сонная артерия — левая, и только
у рогатых воронов — парная.
Образ жизни. Удоды —дневные птицы, древесные и наземные. Населяют
леса низменностей (за исключением обыкновенного удода и рогатых воронов,
обитающих в степных пространствах с разбросанными по ним группами
отдельно стоящих деревьев и кустарников). В горы поднимаются до 2000—2500 м.
По земле, кроме обыкновенного удода и рогатых воронов, передвигаются плохо,
но хорошо лазают по ветвям деревьев. Некоторые виды (лесные удоды) редко
спускаются на землю, и только обыкновенный удод и рогатый ворон
—настоящие наземные птицы. Полет слабый, волнистый. Большинство птиц этого
отряда не может долго держаться в воздухе, но удод, хотя и плохо летает, все
же совершает перелеты. Это—оседлые птицы, только один подвид
обыкновенного удода U.e. epops, населяющий умеренные страны Европы и Азии,
перелетный.
Представители подотряда удодов питаются преимущественно животной
пищей — различными насекомыми и их личинками. Корм разыскивают в
мягкой, рыхлой земле, в навозных и мусорных кучах и т. п. (обыкновенный
удод), или в щелях коры деревьев (лесные удоды); при этом лесные удоды
лазают по деревьям, подобно дятлам и пищухам, опираясь на хвост. В
небольшом количестве в состав пищи у некоторых видов входит и растительный
корм. У представителей подотряда птиц-носорогов пища преимущественно
растительная (мягкие плоды и фрукты, которые птицы срывают с деревьев).
Только рогатый ворон — всеядный: питается насекомыми, их личинками,
разными позвоночными — мелкими грызунами, ящерицами и др.
Удоды — моногамы. Гнездятся отдельными парами. Пары, повидимому,
постоянны. Способными к размножению становятся в первую весну после
рождения,.^ е. в возрасте около года. Гнезда расположены, за редким
исключением, в дуплах деревьев, пней, иногда в щелях скал и построек и т. д. Гнезда
занимаются птицами в течение ряда лет (собственно гнезда они не строят).
Подстилкой служит гнилая измельченная древесина.
Насиживает, как правило, одна самка, но у обыкновенных удодов в конце
насиживания принимает участие и самец.
В кладке у удодов 5—7 яиц, реже 3—4, или 8—9 яиц, у птиц-носорогов
1—4, редко 5. Яйца или чистобелые (птицы-носороги, лесные удоды), или
беловатые, с голубоватым и светлоохристым налетом (обыкновенные удоды),
продолговатой формы. Насиживание начинается после окончания кладки.
У птиц-носорогов, за исключением рогатых воронов, самец замуровывает
самку в дупле или (Tockas bradfteldi) в* расселине скалы при помощи глины,
земли, гнилого дерева, помета, смоченных выделениями слюнных желез.
Остается только небольшая щель, в которую самка может просунуть клюв.
Замуровывание, повидимому, имеет значение для охраны яиц и птенцов от
врагов. Замурованная самка линяет в период высиживания яиц и
выкармливания птенцов, причем теряет почти сразу все перья и становится на некоторое
время неспособной к полету. Насиживают удоды 17—18 дней, у Tockas decked
самка остается замурованной 58 дней, у Bycanistes cristatus —150—175 дней.
Птенцов у удодов выкармливают и самец и самка насекомыми и их
личинками. У птиц-носорогов самец кормит замурованную самку и птенцов,
подавая самке корм прямо в просунутый в щель клюв. Корм состоит из
отрыгиваемых самцом полупереваренных плодов и фруктов, обернутых оболочкой,
образованной из выделений стенок желудка. Самец за этот период приносит
самке и птенцам до 24 тысяч фруктов и плодов и сам настолько истощается, что
ХАРАКТЕРИСТИКА ОТРЯДА
537
иногда гибнет; в этом случае заботу о самке и птенцах принимают на себя
другие самцы.
Птенцы вылупляются или одетые настоящим пухом (обыкновенный удод),
или щетиноподобным пухом — трихоптилиями (лесные удоды), или голые,
с сильно развитым и пневматизированным подкожным слоем
(птицы-носороги). В возрасте 20—22 дней удоды покидают гнездо в ювенальном наряде,
мало отличимом от взрослого; у Bycanistes cristatus птенцы при открытии
гнезда могут сразу летать, у Tockus deckeri после оставления самкой гнезда
(на 58-й день с момента замурования ее), птенцы остаются в гнезде еще около
месяца. Осенью молодые птицы линяют и приобретают наряд, не отличимый
от наряда взрослых. В дальнейшем у взрослых птиц ежегодно осенью
проходит полная линька. У птиц-носорогов самка линяет в дупле, сейчас же
после вылупления птенцов. Удоды линяют в продолжение 3—4 месяцев,
сохраняя в это время летательные способности; самки же птиц-носорогов
в период линьки совершенно теряют способность к полету.
Систематика* Отряд удодов делится на два подотряда: удоды Upupae и
птицы-носороги Bucerotes, Общее число видов — 52, из них в фауне
Советского Союза представлен 1, т. е. 1,92%. Подотряд удодов Upupae объединяет
птиц средних размеров с длинным, тонким, слегка изогнутым вниз клювом.
Края клюва гладкие. Пух имеется только у птенцов. Ресниц на верхнем веке
нет. Третий и четвертый пальцы ног срастаются только в основной фаланге,
второй и третий — свободны. Базиптеригоидные отростки отсутствуют.
Спинная птерилия заходит на шею. Подкожных разветвлений воздушных мешков
нет. Копчиковая железа выделяет коричнево-черную жидкость, имеющую
неприятный запах. Сухожилия m. flexor digitorum profundus perforans и m,
flexor hallucis longus не соединены.
Удоды объединяют два семейства: настоящие удоды Upupidae и лесные
удоды Phoeniculidae. Лесные удоды отличаются от настоящих удодов наличием
металлических тонов в окраске оперения, наличием редуцированного
побочного ствола контурных перьев, длинным ступенчатым хвостом и отсутствием
хохла на голове. Семейство удоды Upupidae содержит один род удоды Upupa
с одним видом Upupa epops, распространенным в умеренных и южных областях
Европы и Азии и в Африке. Семейство лесные удоды Phoeniculidae распадается
на два рода: Phoeniculus и Rhinomastus с б видами, распространенными в
тропической и южн. Африке.
У подотряда птиц-носорогов Bucerotes клюв больших размеров,
массивный, резко загнутый вниз, чаще с костнороговыми полыми выростами на голове
и клюве, различной формы и величины, только у Tockus выростов нет. На
верхнем веке развиты ресницы. Пух у птенцов и взрослых птиц отсутствует.
Спинная птерилия заходит на шею. Пальцы ног срастаются: второй и третий —
на половину длины основной фаланги, третий и четвертый на 2—.3 фаланги и
только у рогатых воронов пальцы не сращены. Базиптеригоидные отростки
имеются, но они не соединены с os pterygoideum. Сильно развита фаринго-
нозальная система воздушных мешков. В подкожном слое сильно развиты
ответвления воздушных мешков. Сухожилия m. flexor hallucis longus и flexor
digitorum profundus perforans соединяются в дистальной части цевки. Подотряд
имеет одно семейство птицы-носороги Bucerotidae, распадающееся на 12 родов:
Tockus, BerinicorniSy Ptilolaemus, Anorrhinus, Penelopidesy Aceros, AnthracoceroSy
Bycanistes у Ceratogytnna, Bucerosf Rhinoplax и Bucorvus с 45 видами,
распространенными в тропической и южн. Африке, в Индии, Индо-Китае, Южн. Китае
и на островах Индо-Австралийского архипелага.
Ископаемые остатки удодов известны с верхнего эоцена (Франция),
откуда описан один вид Cryptornis antiquus7 похожий несколько на род
Tockus.
538
ОТРЯД УДОДЫ
Географическое распространение. Удоды распространены преимущественно
в южных областях Европы и Азии, в Африке и на островах Индо-Австралий-
ского архипелага. Наибольшее разнообразие видов и подвидов — в
тропических и субтропических странах, где повидимому находится центр
возникновения и распространения отряда. В умеренные страны доходит всего лишь один
вид — удод Upupa epops, представленный в фауне СССР и широко
распространенный в южных частях страны.
СЕМЕЙСТВО УДОДЫ UPUPIDAE
РОД УДОДЫ UPUPA LINNAEUS, 1758
Тип Upupa epops
1. Удод Upupa epops L.
Vpupa epops. Linnaeus. Syst. Nat., изд. X, 1758, стр. 117, Швеция.
С и н о и и м. Upupa vulgaris. Pallas. Zoogr. R-As., 1, 1811, стр. 433, nom. emend.
Русское название. Удод, звукоподражательное, от весеннего призывного крика
птицы; потатуйка, пастушка, пустошка, лесной пастушок — от местообитания: пустошь.
Распространение* Ареал. Средняя и южн. Европа; Зап. Сибирь к
северу приблизительно до 55° с, ш., Алтай, но в с.-в. Алтае отсутствует,
Забайкалье, ю.-в. Сибирь, Приморская обл., к северу до 50° с. ш, К югу гнездится
до Канарских о-вов, почти по всей Африке, на Мадагаскаре, в Палестине,
Сирии, Ираке, Малой Азии, Иране, Средней и Центральной Азии, в южн.
и ю.-в. Азии, на о-ве Суматра и Хайнань.
Характер пребывания. Северные птицы перелетны, южные
оседлы.
Биотоп. Открытые местности в лесостепной зоне, степи, полупустыни
и пустыни с небольшими лесами, рощами, отдельно стоящими деревьями или
кустарниковой растительностью. В горных районах населяет преимущественно
низменности и, как исключение, в горы поднимается до 2500 м.
Подвиды и варьирующие признаки. Удоды распадаются на 8 подвидов,
различающихся между собою окраской оперсиия, размерами и экологией: U. е. epops Linn., 1758 —
европейская и сибирская часть ареала вида на посток до Приморья, Манчжурии и Китая,
на юг до Мадейры и Канарских о-поп и с.-п. Африки; Передняя Азия, Иран, Афганистан,
Белуджистан, с.-з. Индия и Средняя и Центральная Азия; U. с. major Brehm, 1855 —
Египет от Александрии до Суэцкого кнпплн и до Вади-Хальфа; U. е. senegalensis Swain-
son, 1837 — южн. Сахара, от Сенегала и Золотого берега до Асбена, Египетского Судана,
Абиссинии, Эритреи, и па юг до иерхопьев Нила, Уганды, Камеруна, Того и Кении (из
Камеруна описана форма walbeli, по самостоятельность ее подвергается сомнению;
большинство авторов считают ее синонимом senegalensis); U. е. africana В е с h s t e i n, 1811—
в южн. Африке на север до Конго, южн. Уганды, Ландана, озер Киву и Эдвард; U. е. таг-
ginata С a b a n i s, I860 — Мадагаскар; U, е. orientalis Baker, 1921 — сев. Индия от
Пенджаба до подножья Гималаев, Бихара и Бсигалии, южная граница от Бомбейской
провинции на широте Хандеша по сев. Деккану до Мадрасской провинции на востоке
(сомнительная форма, быть может синоним номинальной формы); U. е. ceylonensis Reich.,
1853 — Цейлон, южн. Индия на север до южной границы предыдущей формы; U. е. Ion-
girostris J e r d о n, 1862 — Ассам, Бирма, Юннань, Индо-Китай, Малайский п-ов, о.
Суматра и о. Хайнань.
Удод Upupa epops epops L.
Синонимы. Upupa epops loudoni. T s с h u s i. Orn. J a h r b. XII, 1902, стр. 70,
Закаспийский край. — Upupa epops saturatus. L6 nnberg. Ark. f. Zool., 1909, № 9, стр. 29,
Кяхта.
Распространение. Ареал. Зап. Европа до Англии (в Ирландии нет),
Сев. Франции, Бельгии, Голландии, Германии и южн. Швеции (на север
до Кольмара), к югу до островов Средиземного моря; с.-з. Африка;
Канарские о-ва и Мадейра; Синай, Палестина, Сирия, Месопотамия, с.-з.
удод 539
Индия, Центральная Азия и Китай, за исключением самых южных частей
его. В СССР распространен от южных государственных границ на север
до Прибалтики, южных частей Ленинградской обл. (дважды встречен в
окрестностях Петербурга весною 1876 г.), окрестностей Гдова, Пскова
(Зарудный, 1910), Новгорода (найден на р. Волхове), Калининской обл.,
до Тутаевского района в Ярославской обл., Горьковского края, Казани,
Карта 92. Распространение удода Upupa epops
— D. е. epops, 2 — U. е. major, 3 — U. е. senegalensis, 4 — U. е. africana, 5 — U. е. marginata, б —
U. е. orientalis, 7 — U. е. ceylonensis. 8 — U. е. longirostris; а — граница гнездовий, 2—■ зимовки (цифры
в кружках обозначают номера зимующих подвидов).
на север доходит до Малмыжа и устья р. Кильмези (Першаков, 1929).
Гнездится в южных частях Чистопольского района, в б. Бугульминском,
Бугурусланском и Белебеевском уездах (северней в б. Уфимской губ. на
гнездовье не найден, Сушкин, 1897). В Приуралье северная граница
распространения спускается южнее, проходя у Чкалова и по р. Сакмаре (Шайтан-
тау); восточней Урала граница опять поднимается к северу до Челябинска,
Кургана, Омска (однако, в Барабинской степи, в Петропавловском и Кокче-
тавском районах отсутствует). Есть удод в окрестностях г. Барнаула и
р. Барнаулки, в Кулундинской степи, в Колывани. По степям доходит до
Томска, Ачинска, Минусинска, Кузнецка, Салаира, по всему Алтаю, за
исключением таежного с.-в. Алтая; на Саянах его нет (Сушкин, 1938). В с.-з.
Монголии и Тувинской обл. обычен. Обычен на гнездовье в окрестностях
Иркутска, в степях Балаганска (Скалой и Слудский, 1933), в степной части
ю.-з. Забайкалья, в Баргузинских степях, по юго-восточному берегу Байкала
540
ОТРЯД УДОДЫ
до Сосновки (коллекции Зоологического музея Московского университета;,
южн. Забайкалье, в Даурских степях, вЧитинской обл. на север до юго-восточных
частей Витимского плато; верхнем и среднем течении Амура и его притоков—
Зеи и Бурей; в Приморской обл. к северу до Хунгари, с. Воскресенское, т. е.
до 50° с. ш. (Воробьев, 1938), к югу —до озера Ханка, Владивостока и
побережья Японского моря. По опросным сведениям отмечен для Б. Шантар (Дуль-
кейт и Шульпин, 1937).
Удоды, населяющие Европу, зимуют в тропической Африке, к югу до
Сомали, Египетского Судана, Виктории-Ньясса и Анголы; птицы,
гнездящиеся в Сибири, в Средней и Центральной Азии — в Южн. Китае, Бирме,
Индии, по южному берегу Каспия, в Загроше и в Месопотамии. В небольшом
количестве зимует в ю.-з. Туркмении; кроме того, добыт один экземпляр
в декабре у Мары. Некоторые особи может быть остаются зимовать в
Закавказье — в Талышской низменности, хотя позднейшие исследователи его там
зимой не находили (Тугаринов и Козлова, 1935).
Характер пребывания. Удод —.гнездящаяся и перелетная
птица, зимует только в ю.-з. Туркмении и может быть в ю.-в. Закавказье
(Талыш). Осенью первые удоды появляются в Египте в конце августа и летят
до конца сентября. В Судане пролет идет в течение всего сентября, в сев.
Сомали — с конца сентября до середины ноября. Зимовки они покидают
в начале марта и даже в феврале. В Судане весенний пролет идет в течение всего
апреля и одиночные птицы летят еще в мае. В Египте главная масса летит с се^
редины марта до середины апреля, в зап. Сахаре и Рио-де-Оро —в марте и апреле.
В СССР на юг прилетают рано, летят одиночками, парами и, реже,
небольшими группами в 8—10 штук. В Закавказье — в Ленкорань и Талышскую
низменность — первые птицы прилетают в первых числах марта (5-^6 марта,
иногда даже в конце февраля, Спангенберг), в двадцатых числах марта они там
уже обыкновенны. В горы на высоту около 1000 м прилетают в конце
марта, в Тифлис — в конце марта, в Дербент — в середине марта (Радде, 1885).
В сев. Ингушетию и Осетию прилетают в самом начале апреля (4. IV, Беме, 1926).
По Черноморскому побережью пролет идет с первых чисел апреля по первые
числа мая. На с.-з. Кавказе и на средней и нижней Кубани появляются в
середине апреля, и пролет идет до конца месяца и даже захватывает май. В вост.
Приазовье, в Крыму и на юго-западном берегу Азовского моря первые
птицы появляются в конце марта. На Украину в среднем прилетают в середине
апреля: в Подолии самый ранний прилет отмечен 1.IV. 1911 г., самый
поздний—.21. IV. 1896 (15 лет наблюдений, Брацлавский район, Бауман, 1916),
в Харьковскую обл. самый ранний прилет—21. III. 1890, самый поздний
29.IV. 1893 (14 лет наблюдений, Сомов, 1896), в окрестностях Киева самый
ранний прилет 29. III. 1925, самый поздний —20. IV-1921(11 лет наблюдений,
Данилович, 1933). В Белоруссию прилетают в среднем в начале второй половины
апреля (Минское Полесье, Шнитников, 1913). В Смоленскую обл. первые птицы
прилетают в середине апреля и пролет идет до конца месяца и даже захватывает
первые числа мая (Граве, 1926). В центральные области Европейской части СССР
удоды прилетают во второй половине апреля: в Тамбовскую обл., Моршан-
ский район — 24 апреля (Резцов, 1904), в Рязанскую обл.—29 апреля
(Хомяков, 1910), в Московскую—17 апреля (Поляков, 1910). В окрестности Бу-
гуруслана — в конце второй половины апреля, и пролет идет здесь до середины
мая. Под Гурьевом в конце апреля удоды обыкновенны. В Чкаловской обл.
первые птицы появляются во второй половине апреля, валовый пролет
наблюдается в конце апреля, и последние птицы летят в начале мая (Заруд-
ный, 1888); на Эмбе —в первой половине апреля, в низовьях Иргиза —
в среднем 14—15 апреля, но иногда прилетают рано, когда снег еще лежит
глубиной до 30 см (Сушкин, 1908); на Южн. Урале первые птицы появляются,
удод
541
в среднем, в конце апреля. Под Омском —в середине мая и даже в начале
второй половины мая, в западную степную окраину Алтая —в конце апреля (27—
28 числа) и в начале мая, внутрь Алтая —в середине мая (Сушкин, 1938).
В Средней Азии удоды появляются рано. На юго-восточном побережье
Каспия удод—обычная пролетная птица, и в низовьях Атрека первые птицы
отмечены в первой половине марта, в предгорьях центрального Копет-Дага
встречены 6 марта, на озере Б. Делили —14 марта, в Бахардене--4 апреля
(Дементьев, 1950), под Ашхабадом первые птицы прилетают в начале марта
(7 марта), в подгорной долине центрального и вост. Копёт-Дага валовый пролет
отмечен с середины апреля и до конца месяца (Радде и Вальтер, 1889). В Кара-
Кумах единичные экземпляры наблюдались 13 марта, множество 28 марта,
у Репетека—небольшие стойки —*3 марта, на Мургабе —9.марта (Лоудон,
1910). В низовьях Сыр-Дарьи —в середине марта, но в 1928г. удоды
наблюдались 27 февраля и с этого времени они то появлялись, то исчезали и только
с 19 марта стали обычными (Спангенберг, 1936). В Таджикистане—в начале
марта. У Алма-Ата — во второй половине апреля (24 апреля) и до конца
месяца были редки, под Джаркентом первая птица встретилась 21 марта (Заруд-
ный и Кореев, 1906).
На осеннем пролете летят, как и весной, или в одиночку, или
небольшими рассеянными стайками в 10—15 штук, на незначительной высоте,
преимущественно утром и вечером, иногда днем и даже ночью. Если на
пути встречаются кормные места, то птицы делают остановки, иногда на
несколько дней. Из северных районов своего распространения в Европейской
части СССР удоды улетают в начале сентября и только одиночные экземпляры
задерживаются до конца сентября. Прибалтику покидают в конце сентября,
(Лоудон, 1909). Из центральных областей улетают в конце августа, затем идет
пролет северных птиц, который продолжается до половины сентября и даже
до конца месяца. В Харьковской обл. отлет начинается в среднем в самом
конце августа —27, самый ранний отлет отмечен в 1891 г. 15 августа и
закончился он 11 сентября, самый поздний отлет — в 1882 г. —»11 сентября (Сомов,
1896). На Сев. Кавказе отлет местных птиц начинается с середины августа и
пролет идет до конца сентября. По Черноморскому побережью пролет
наблюдается с конца июля и до начала сентября. По Батумскому побережью —
даже до середины сентября (Сатунин, 1911). Пролет под Чкаловом идет
с конца августа до конца сентября. В Арал о-Каспийских степях —в середине
сентября. В низовьях Сыр-Дарьи —с середины августа до конца сентября.
На юго-восточном побережье Каспия — с первых чисел августа и идет песками
вдоль морского побережья на юг. В Таджикистане в Гиссарской долине удоды
встречаются до середины октября (Иванов, 1940). Под Джаркентом к концу
июля численность удодов сильно уменьшается, в конце августа встречаются
совсем редко и одиночные экземпляры — до середины сентября. Высокогорный
Алтай удоды покидают в самом начале сентября, более низкие местности Алтая
в конце первой половины сентября (Сушкин, 1938).
Залеты отмечены в Исландии, на Фарерских о-вах, Шпицбергене,
Норвегии, в сев. Швеции, на Кольском п-ове—в округе Энаре, на р. Тана,
Утсиоки, у Польмака в сентябре 1849 и сентябре 1876; в б. Повенецком уезде
б. Олонецкой губ., в Ростовском районе Ярославской обл., в окрестностях
г. Кирова, на р. Шакве Молотовской обл., неоднократно в верховьях Печоры
(в начале VI, 1934, 23. IX. 1934, IX. 1935 и 12. Х.1942, Теплова), в
Ивановской обл., добыт в Башкирии в восьмидесятых годах прошлого столетия
(Сушкин, 1897). Кроме того, отмечены залеты в окрестности г. Красноярска
и на Сахалине (Бутурлин, 1935).
Б и о т о п. В южных районах лесной зоны европейской части страны удоды
обитают преимущественно по разреженным сосновым и дубовым лесам, по-
542
ОТРЯД УДОДЫ
близости от полян, выгонов, лугов и т. д. В лесостепи и степи селятся по
лесным колкам, поемным лесам, в лесных садах, близ человеческих поселений,
в береговых обрывах и пр. В лесных районах Зап. Сибири встречается там;
где лес чередуется со степью, полянами и другими открытыми местностями,
а в открытых местностях — по лесным колкам, по отдельно стоящим
деревьям, по обрывам, каменным осыпям и т. д. В Средней Азии — в степных и
полупустынных местностях, в скалах, ущельях, по долинам рек, в
культурном ландшафте. В горы идет обычно не выше 2000 м, но на Алтае встречен
до альпийской зоны (Сушкин, 1938), повидимому кочующие птицы; в
Семиречье— до 1800 м (Зарудный и Кореев, 1906), местами до 2500 и более
метров, вероятно на кочевках; в Таджикистане поднимается до 4000 мУ но на
гнездовья до 2000 м (Иванов, 1940); на Кавказе обычно — до 1500 л* (Радде,
1885); в Закавказье доходит до альпийской зоны, в Армении—до 2500 м
(Ляйстер и Соснин, 1942).
Численность. У северной границы распространения удоды
встречаются в небольшом количестве, местами повидимому гнездятся не ежегодно.
Чем далее к югу, тем они становятся многочисленней. Так, в
Ленинградской обл. удод редок, в Московской и Калужской обл. редок и спорадичен,
в Тульской редок, но гнездится регулярно. В Рязанской и Орловской обл.
удод — обычная гнездящаяся, хотя и немногочисленная птица. В
Смоленской обл. гнездящаяся птица, но в северных и восточных частях области
малочисленна, в б. Вяземском уезде до 1910 г. наблюдался на гнездовье не ежегодно,
с 1910 стал обычным (Граве, 1926). На Украине, в Приазовье, в степях и
предгорьях Крыма удод — одна из самых обычных и многочисленных птиц.
Весьма обычен он по Волге до Куйбышева, но северней становится редким.
На Кавказе удод—одна из самых многочисленных птиц степей, низменностей,
долин рек и предгорий. В с.-з. Закавказье в 1897 г. удод совсем не
встречался, позднее встречался в небольшом количестве, в настоящее время обычен
на гнездовье.
В Средней Азии удод широко распространен —это одна из самых
обычных и многочисленных птиц Арало-Каспийских степей, во множестве
встречается во всех среднеазиатских республиках, кроме больших пустынных
пространств. В горных районах Средней Азии и Сибири распространен
главным образом по предгорьям.
Экология. Размножение. Удоды способны к размножению в
первую весну, т. е. в возрасте около года. Мопогамы, пары вероятно постоянны.
Вскоре после прилета начинаются брачные игры к спаривание. В разгар
брачного периода до конца июня и даже начала июля крики удодов слышны от зари
до зари. В это время самцы усиленно гоняются за самками и издают свой
крик «ду-ду».
Вскоре, разбившись на пары, удоды приступают к гнездованию. Гнезда на
севере — в лесной и лесостепной полосе —чаще всего делают в дуплах старых
деревьев (дуб, липа, вяз, ветла, фруктовые деревья и т. д.), в пустотах пней,
реже в грудах камней и мусора, в застрехах домов и т. д. В степных и пустынных
местностях устраивают гнезда в щелях и дырах глинистых построек, в
каменных укреплениях, в расщелинах скал и обрывах, в кучах кизяка, в норах,
оставленных другими животными, даже в скелетах падали, в колеях дорог и
на песчаных обрывах. Гнезда занимаются птицами в течение ряда лет.
Подстилкой служит гнилая древесина с небольшим количеством сухой травы,
конского волоса, шерсти, перьев и т. д. Гнезда, устраиваемые в горах, в щелях
скал, обычно помещаются высоко от дна ущелья, а устроенные в дуплах
—»обычно не выше 2 м над землей (Емельяненко, 1915; Измайлов, 1940; Сомов, 1896).
В кладке 5—6 яиц, реже 3—9, а иногда даже 12. Яйца матово-белые, с
сероватым, голубоватым и охристым налетом, овальной, несколько удлиненной фор-
УДОД 543
мы, с хорошо выраженными острым и тупым концами. Скорлупа тонкая,
шероховатая с заметными порами.
Размеры: (13)24,5—29,3 х 17 — 19,2, в среднем 26,83 х 18,46 мм (Харь-
ковская обл., Сомов, 1896); (6) 27,0—30,3x19,0—19,8, в среднем 28,73 х
19,3 мм (Семиречье, Шнитников, 1949); (93) 23,1— 29,3 х 16,6—19,4, в среднем
25,83 х 17,88 мм (Хартерт, 1912); (100) 23,1—28,6 х 16,3-18,5, в среднем
26,1 х 17,5 мм (Индия, Бекер, 1927).
Откладка яиц происходит с промежутком в 24 часа. Количество кладок
в году не выяснено, повидимому на юге страны — две. Период размножения
в центральных областях начинается с конца мая и продолжается до июля.
Гнезда с яйцами в Московской губ. найдены 19.VI. 1884 г. (насиженные яйца,
Поляков, 1911), в Рязанской обл. —14 июля (Хомяков, 1910), в Минском
Полесье полные кладки встречаются с середины мая (13.V), в Бугуруслан-
ском районе полные кладки находили с конца мая (с 22.V, Карамзин, 1913),
в Чкаловской обл. первые яйца находили с первых чисел мая и до середины
мая. На Украине кладка яиц начинается с первых чисел мая. Насиженные
кладки у Ростова-на-Дону находили в конце мая — 20 мая. С середины июня
в южн. Украине начинается, повидимому, вторая кладка
(Днепропетровская обл., 15. VI найдено гнездо с незаконченной кладкой в 2 яйца,
Боровиков, 1907). В Крыму кладка начинается с первой половины мая. (В 1894 г.
была холодная весна, первая кладка найдена 20. V у Симферополя). На
Кавказе — с конца апреля и начала мая и до конца июня и даже начала июля.
(В сев. Осетии и Ингушетии полные кладки встречались с первых чисел мая,
в Тбилиси гнездо с 4 насиженными яйцами найдено 6 мая.) В Туркмении
кладка яиц начинается с последних чисел марта, но кладки встречаются и
в июне, вероятно вторые. В низовьях Сыр-Дарьи—с конца марта; в гнезде,
найденном 13. IV. 1927 г., было 8 слегка насиженных яиц (Спангенберг, 1936).
В Семиречье ненасиженные полные кладки найдены 19.V, 30.V и 1.VI,
насиженные —4.V, 23.V и 30.V (Зарудный и Кореев, 1906).
Насиживание начинается по окончании кладки, но самка уже после
откладки первого яйца много времени проводит в гнезде. В начале высиживает
одна самка, самец только ее кормит, но перед концом инкубации в
высиживании принимает участие и самец. Период высиживания в среднем 17 дней (16—
18 дней). Вылупляются птенцы, покрытые редким рыжеватым с черными
вершинами волосовидным пухом. Выкармливают птенцов самец и самка
различными насекомыми и их личинками. В Семиречье вылупившиеся птенцы
встречаются уже во второй половине мая (23.V, 28.V, 22—23.V, Шнитников, 1949).
В гнезде птенцы находятся 22—24 дня. Вылет птенцов наблюдается в
центральных областях с конца июня, однако в это время в некоторых гнездах
находятся еще не оперившиеся птенцы. В б. Покровском уезде Владимирской
обл. 8—9 июля вылетели молодые, а в гнезде, найденном там же 12 июля, были
оперившиеся птенцы (Поляков, 1911); в Рязанской обл. 14 июля 5 птенцов имели
еще махи, не доросшие до половины длины; в Смоленской обл. 25 июня птенцы
только что вылупились, 9—10 июля молодые покинули гнезда (Огнев, 1909);
в Минском Полесье в конце мая в гнездах были птенцы, и с 23 июня они стали
покидать гнезда; в Тамбовской обл. в найденном 4 августа гнезде находились
взрослые птенцы, 14 августа весь выводок уже откочевал; в Воронежской обл.г
на Хопре, в гнездах — 17.VI были оперившиеся птенцы, 29.VI вылетели из
гнезда; в Бугурусланском районе молодые вылетели из гнезд 30 июня (Карамзинt
1909); в Волжско-Уральских степях летные молодые наблюдаются в двадцатых
числах июня; в Чкаловской обл. молодые покидают гнезда в середине июня.
На Украине вылет птенцов начинается со второй половины июня: в
Черниговской обл. в начале июня птенцы уже оперились, в середине июня вылетели;
в Харьковской обл. птенцы в среднем вылетают во второй половине июня (с
544
ОТРЯД УДОДЫ
с 22 июня по 4 июля, Сомов, 1896); у Ростова-на-Дону вполне оперившиеся
молодые встречались с 1 июля (Сарандинаки, 1908). На Кавказе в Сев. Осетии и
Ингушетии молодые вылетают в десятых числах июня, в южн. Закавказье
летные молодые наблюдались 15.VI (Сатунин, 1912). В Туркмении—Мары —
8 июня молодые уже вылетели из гнезд (Зарудный, 1888). В Таджикистане
молодые птицы встречаются с 21 июня по 30 августа (вероятно две кладки).
В сев. Казахстане (Наурзум) к концу июля молодые вылетели из гнезд.
В Семиречье 9 июня встречались летающие молодые удоды, а 17 августа летали
уже выводками (Шнитников, 1949). На Алтае молодые встречаются с
двадцатых чисел июля. На Зайсане хорошо летающие молодые наблюдались 14.VI
(Поляков, 1912). В ю.-з. Забайкалье в начале июня найдено гнездо с
оперившимися птенцами (Козлова, 1930). В Приморской обл. нар. Имане в гнезде
21 мая были только что выведшиеся птенцы, 11 июня они уже оперились,
18—19 июня вылетели (Спангенберг).
После вылета молодые птицы некоторое время держатся выводками вместе
со старыми, которые продолжают их кормить, но вскоре старые отделяются от
них и начинают отдельно кочевать по выгонам, лугам и т. д.
Линька. У взрослых одна линька в году после вывода птенцов,
с июля по ноябрь. В средней полосе Европейской части СССР линька маховых
начинается в конце июля, в Средней Азии — в половине июля. Смена маховых
идет от внутренних к наружным. Рулевые сменяются несколько позднее
маховых — в августе и в начале сентября. Линька рулевых идет от крайних пар
к центру; у экземпляров, добытых на Сев. Кавказе, в середине сентября линька
рулевых была уже закончена. Во время линьки птицы держатся обычно
одиночками или парами в глухих местах, если есть поблизости лес, то в лесах.
Улетают от нас, сменив махи и рулевые, мелкое оперение сменяется на
зимовках. Как происходит линька у тех удодов, которые имеют вторую кладку —
не имеется наблюдений. Линька молодых идет с августа по октябрь, рулевые
и маховые успевают смениться еще до отлета на зимовку (одна линька). Первый
зимний наряд не отличим от взрослого.
Питание. Кормом удодам служат различные насекомые и их личинки:
жуки, прямокрылые, бабочки, двукрылые, перепончатокрылые, кузнечики,
последних в особенно большом количестве поедают в степях. Пищу добывают
на земле при помощи острого и длинного клюва. Добыв насекомое, удод
умерщвляет его, подбрасывает в воздух и вновь ловит на лету, стараясь поймать так,
чтобы насекомое было направлено вдоль длины клюва.
Полевые признаки. Удода легко узнать по его длинному изогнутому клюву'
веерообразному хохлу и розовато-рыжему с черным и белым оперению. Полет
волнообразный, с быстрыми и частыми ударами крыльев. На полете хорошо
заметны белые и черные полосы крыла, длинный черный с белой серединной
полосой хвост и охристо-рыжий с черными вершинами перьев хохол. Удод
хорошо бегает по земле, может сидеть не только на земле, но и на стенках
построек и на деревьях; по деревьям может лазать подобно дятлу, но делает это
редко. В поисках пищи удод быстрым шагом передвигается с места на место,
иногда пробегая небольшое расстояние. Время от времени он то наклоняет
голову, чтобы схватить насекомое, то поднимает ее, хохол все время находится
в движении. Воды, повидимому, йе пьет.
Обычно удоды держатся в одиночку или парами, только на кочевках и
пролете можно встретить небольшие группы их. Птица осторожная и робкая,
во время опасности ложится на землю, подняв кверху почти вертикально клюв
и распустив крылья, лежит некоторое время совсем без движения. Молодые
птицы менее осторожны, чем старые. Голос удода — глухой крик «ду-ду»
или «пу-пу-пу».
УДОД
545
Описание. Размеры и строение. Клюв длинный, согнутый слегка книзу; на
голове хохол, расположенный вдоль темени от основания клюва до затылка. Ноздри
округлой формы, лежат у основания клюва и слегка прикрыты перьями лба. Ноги средних
размеров. Первостепенных маховых 10, первое маховое короткое и не достигает половины
длины клюва. Формула крыла: 4 = 5>3 >6>7>2>8>9>10>1. Рулевых 10, обрез
хвоста прямой. Длина тела самцов (21) 281—325 мм, самок (14) 280—310 мм, среднее
соответственно 311,7 мм и 298,1 мм; размах крыльев самцов (15)420—490 мм, самок
Рис. 135. Удод.
(8) 440—470 мм, среднее 466,8 и 457,5 мм; длина крыла самцов (79) 136—152 мм, самок
(54) 135—151 мм, среднее 146,8 мм и 146,5 мм. Вес 68—72 г, в среднем 70 г.
Окраска. Вылупившийся птенец покрыт редкими волосовидными рыжеватыми
с черными вершинами перьями. Вскоре на голове и на птерилиях тела начинает
появляться пух и 6—7-дневный птенец покрывается пухом: сверху — розоватым, снизу
беловатым. На 10-й день начинают появляться пеньки маховых перьев. Край ротовой полости
светлорозового цвета, без каких-либо пятен. Клюв прямой короткий и тонкий, грязноголу-
бого цвета. Птенцовый наряд. Похож на взрослых и особенно на самок. Обычно без
розового тона на зобе и груди. Клюв несколько короче, чем у взрослых и слабо изогнут.
Взрослый самец. Оперение пестро охристо-рыжее с черным и белым. Перья хохла
охристо-рыжие с черными вершинами и белыми предвершинными пятнами на задних из них.
35 Птицы СССР, тг I
546 отряд удодов
Мантия, зоб и грудь розовато-рыжие, подбородок беловатый, поясница, верхние кроющие
хвоста и нижняя часть брюшной стороны белые. На боках тела черноватые продольные
полосы. На нижней части спины две поперечные черные полосы. Хвост черный с широкой
поперечной белой полосой, расположенной приблизительно посредине хвоста. Крылья
черные с белым. В вершинной половине первостепенных маховых проходит поперечная
белая полоса, на второстепенных маховых белые пятна образуют на сложенном крыле
четыре поперечные полосы. Плечевые перья с продольными белыми полосами. Клюв черный,
ноги черно-бурые, радужина светлобурая. Различий в оперении по сезонам нет, но при
обнашивании оперение несколько темнеет. У птиц, живущих в пустынных, сухих жарких
местностях, оперение довольно сильно выцветает. Детали распределения белых пятен от
оттенков охристо-рыжего тона оперения подвергаются большим индивидуальным колебаниям,
часто некоторыми авторами принимаемым за подвидовые особенности.
Взрослая самка похожа на самца, только зоб, грудь с более коричневато-рыжим,
а не розоватым оттенком. Перья подбородка грязнобелые. Но и эти слабые
отличительные признаки часто отсутствуют, и тогда самок нельзя отличить от самцов.
ОТРЯД ДЯТЛЫ
PICARIAE или PICIFORMES
Характеристика отряда
Морфология- Отряд распадается на четыре естественных группы,
обнимающих птиц средней и мелкой величины, хорошо характеризующихся
анатомически и резко отличных одна от другой как внешним видом, так и
образом жизни. Общие признаки, объединяющие эти группы, следующие: череп
десмогнатический или эгитогнатический; строение носовых костей голориналь-
ного типа, носовая перегородка непрободенная; крыловидных отростков
носовой кости нет. Os dorsale нет; шейных позвонков 14; ребер 5 пар; на каудаль-
ном конце грудины две пары очень глубоких вырезок; каракоиды разобщены;
вилочка в виде «U»; обводящей мышцы нет, дельтовидная мышца сильно
развита; ноги зигодактильные, два пальца обращены вперед, два назад;
нижняя гортань только с одной парой трахеобронхиальных мускулов, в
оперении характерна узость птерилий и широкие аптерии, обыкновенный пух
отсутствует, крыло эвтаксическое, тупое, первостепенных маховых 10—11,
второстепенных 10—12.
Биологическая характеристика. Все представители
отряда гнездятся в дуплах или норах. Яйца откладывают одноцветные, белые.
Птенцы рождаются слепыми и голыми, без пухового наряда (он есть только у
дятла Centuras aurifrons).
Систематическое положение.. Отряд имеет много черт
сходства с воробьиными птицами (с наиболее примитивными из них — Eurylaimi и
Clamatores). Филогенетически связывается с примитивными ракшеобразными,
ведя свое происхождение от форм, близких к зимородкам и момотам.
Систематика. Отряд дятлов делится на два подотряда: якамаров или бле-
етянок Galbulae и собственно дятлов Pici. Якамары или блестянки
характеризуются десмогнатическим черепом, двумя сонными артериями, отсутствием
hypocleidium, отсутствием желчного пузыря, сильным развитием слепых кишок
и сгибателей четвертого пальца ноги, тонким и гладким языком, десятью
первостепенными маховыми, двенадцатью рулевыми, голой копчиковой железой и
тонкой кожей. К нему относятся два семейства — собственно якамары Galbulidae
с блестящей окраской, длинным и тонким клювом (роды Galbula, Jacamaralcyon
и др.) и ленивки или пуховки Bucconidae, с рыхлым и мягким оперением
35*
548
ОТРЯД ДЯТЛЫ
с коротким и массивным клювом (роды Вйссо, Monacha, Maiacoptila и др.) —
всего 15 видов, распространенных в Средней и Южн. Америке.
К подотряду дятлов относятся четыре семейства: собственно дятлов Picidae,
бородаток Capitonidae, медоуказчиков Indicator idae и туканов Rhamphastidae.
Семейство дятловых Picidae состоит из мелких и средней величины птиц
с характерным долотообразным клювом и, за редкими исключениями, жесткими
рулевыми перьями, на которые птица опирается при лазании по деревьям.
Распространены они по всему свету, исключая Нотогею и Мадагаскар. Ноги
короткие с обращенными назад двумя пальцами и с крепкими загнутыми
когтями. Крылья умеренной длины, закругленные, «лесного» типа.
Первостепенных маховых 11, рулевых б пар, но крайняя пара очень мала и скрыта
кроющими у основания второй пары. К этому семейству относятся 224 вида птиц.
Семейство бородаток Capitonidae весьма сходно с дятлами, но
принадлежащие к нему виды отличаются выгнутым по коньку надклювьем и сильным
развитием щетинок у основания клюва; череп у этих птиц промежуточного
между эгитогнатическим и десмогнатическим типом; ключицы не образуют
вилочки. Сюда относятся птицы, распространенные в тропической зоне
Средней и Южн. Америки, Азии, Африки, всего 72 вида, относящиеся к родам
Capito, Megalaema, Lybius, Pogonialus, Gymnobucco и др.
Семейство медоуказчиков Indicatoridae отличается эгитогнатическим
черепом, 9—10 первостепенными маховыми, 10—12 рулевыми, оперенной копчи-
• ковой железой. К нему относятся несколько родов в Африке, один — в Сиккиме
и один на Больших Зондских о-вах и Малакке. Всего 11 видов, относящихся
к родам Indicatir, Melichneutes, Melignomon, Protodiscus.
Туканы или перцеяды Rhamphastidae морфологически связываются с
собственно дятлами через семейство бородастиков. Они характеризуются
огромным ярко окрашенным клювом, десмогнатическим черепом, тонкостенным и
объемистым мускульным желудком, относительно коротким и широким
кишечником, отсутствием слепых кишок, короткими и не образующими вилочки
ключицами, одной (левой) сонной артерией, длинным бахромчатым по краям
и на конце языком. Первостепенных маховых 11, рулевых 10. Распространены
в Средней и Южн. Америке; всего 37 видов, относящихся к родам Rhamphastus,
Pteroglossus, Selenidera и другим.
Всего в отряде дятловых, таким образом, 359 видов. Из них в фауне
Советского Союза представлены 15, т. е. 4,17%. Эти 15 видов относятся к семейству
дятловых. Распространение их строго связано с лесами, но в общем носит
весьма широкий характер: дятлы встречаются от северной тайги до горных
лесов крайнего юга Кавказа, Средней Азии и даже в саксаульниках южных
пустынь — Кара-Кумов и Кызыл-Кумов.
Дятловые — дневные, связанные (за редким исключением) с лесом птицы.
Большинство представителей подотряда типичные лазающие птицы.
Исключение представляют некоторые американские представители рода Colaptes и
южноафриканский Geocolaptes, которые держатся на земле, гнездятся в норах
и добывают пищу, склевывая ее на земле. Гнезда, за указанным выше исключен
нием, в дуплах деревьев. Их выдалбливают сами птицы или расширяют уже
имеющиеся. Подстилки в гнезде нет. Число яиц в кладке от 3—5 до 11—12
и даже 14. Моногамные птицы, к размножению приступают в возрасте около
года, насиживают оба пола, но больше самка. В случае гибели кладки,
откладывают яйца вновь. Насиживание длится 11—13 дней.
Птенцы рождаются слепые, голые, совершенно без пуха, на интертарзаль-
ном сочленении имеют так называемую пяточную мозоль (утолщение с
шиловидными сосочками), которая, возможно, помогает им карабкаться по стенам
дупла или служит своего рода амортизатором при подпрыгивании навстречу
приносящим корм родителям. Подросшие птенцы иногда вылазят из дупла и
ХАРАКТЕРИСТИКА ОТРЯДА
549
вновь в него возвращаются. Пр вылете очень короткое время держатся имеете,
в дальнейшем выводки разбиваются и птицы держатся порознь.
Дятлы, как правило, неуживчивые птицы и вместе могут быть встречены
только при большом обилии корма. Уже в августе (в наших широтах)
начинаются кочевки. Сначала птицы выходят в другое место обитания, появляясь
в лесах, где они не гнездились из-за отсутствия там деревьев, подходящих для
долбления дупел. Зимой откочевывают к югу, но в большинстве мест гнездового
ареала встречаются круглый год, так как на место откочевавших
прикочевывают другие с севера.
Из гнезда молодые вылетают в наряде, в общем сходном с нарядом
взрослых птиц. Но отличаются более тусклым и грязноватым тоном оперения и
особенно тем, что первое (недоразвитое) маховое перо у них примерно в полтора
раза длиннее и шире, чем у взрослых. Линька в первоосеннее перо происходит
в конце лета — начале осени, красные перышки на темени задерживаются
иногда до зимы.
Взрослые дятлы линяют с конца июня по октябрь (мелкое перо начинает
сменяться раньше) раз в год, полностью. Маховые перья сменяются обычно
постепенно, перо за пером (у вертишейки быстрее). Рулевые сменяются попарно
от периферии к центру. В,зимнее время дятлы, как правило, имеют более
светлое оперение, летом белые части оперения обычно имеют буроватый налет.
Рамфотека при усердном долблении стирается, поэтому северные представители
нашей фауны имеют более широкий (стертый) клюв, а южные, которые долбят
меньше, более тонкий и длинный.
Питаются насекомыми, часто вредными для лесного хозяйства, личинками
ксилофагов, которых достают либо из-под коры, отламывая ее, либо
склевывают с поверхности, либо выдалбливают из древесины. Зимой многие поедают
семена хвойных. Для долбления шишки устраивается «станок»:
выдалбливается место, куда защемляется раздалбливаемая шишка. В поисках ^корма
лазают обычно по стволу дерева, головой вверх и опираются на жесткий хвост
при прыжках. Дерево, пораженное вредителями, обследуется довольно
тщательно, птица обычно не улетает пока не выберет всех доступных ей
насекомых.
Систематика. Семейство заключает в себе два подсемейства: вертишейки
и дятлы. Вертишейки — Jynginae отличаются мягкими и незаостренными
рулевыми и довольно слабым клювом. Кончик языка гладкий, без щетинок.
Один род Jynx: два вида в Африке и один в Евразии. Дятлы (настоящие дятлы)
РШпае имеют долотообразный клюв, кончик языка несет щетинки. В семействе
221 вид (роды: Dryocopus, Picus, Picoides, Dendrocopos, YungipiciiSy Picumnus,
Sasia, Colaptes, Geocolaptes, Melanerpes и др.). В СССР 14 видов, относящихся
к 5 родам.
Географическое распространение. Широко распространены в лесах земного
шара, за исключением Австралии, Новой Гвинеи и Мадагаскара. Особенно
много дятлов в Южн. Америке. В ископаемом состоянии известны начиная от
верхнего олигоцена (Франция).
Практическое значение. Все дятлы нашей фауны полезные птицы. Кроме
истребления вредителей леса, они оказываются чрезвычайно полезными тем,
что выдалбливают в поврежденных деревьях дупла, которые впоследствии
заселяются мелкими дуплогнездниками. Снимая с больного дерева кору,
дятел убивает его и тем самым приостанавливает дальнейшее развитие в нем
вредителей ксилофагов. Незараженных деревьев дятлы не долбят: долбление
здорового на вид дерева служит указанием, что дерево поражено. Поэтому
наблюдение за дятлами облегчает рубку ухода. Известный вред дятлы
приносят истреблением (иногда полезных) муравьев и истреблением семян
хвойных. Вопрос о соотношении вреда и пользы, приносимых дятлами, должен
550 отряд дятлы
разрешаться для каждого вида отдельно и применительно к условиям той
или иной местности. Развешиванием дуплянок можно привлекать дятлов в лес
возраст которого еще не позволяет птицам устраивать в деревьях дупла.
Таблица для определения видов дятлов
I (2) Перья хвоста мягкие, незаостренные. Окраска спинной стороны буроватая с
темными пестринами. Ноздри не прикрыты щетинками. . . . Вертишейка Jynx torquiila.
2(1) Перья хвоста жестки и заострены. Окраска иная. Ноздри прикрыты обращенными
вперед щетинками 3.
3 (4) Окраска спинной и брюшной стороны черная, крупная птица, крыло длиннее
200мм Черный дятел Dryocopus martias.
4 (3) Окраска не черная, крыло короче 200 мм 5.
5 (6) Преобладающая окраска спинной стороны тела зеленая или зеленоватая . . 7.
6 (5) Зеленого в окраске нет .11.
7 (8) Перья груди и брюха окаймлены узкой черноватой полосой, образующей чешуи-
чатый рисунок. Клюв желтый, над глазами светлая беловатая полоса
Чешуйчатый дятел Picus squamatus.
8 (7) Перья груди и брюха одноцветные или с поперечными пятнами, без темного
чешуйчатого рисунка. Клюв темный сероватый (или черноватый у вершины), над глазом белой
полосы нет 9.
9 (10) Перья затылка серые без красных отметин. Перья голени и подхвостья
одноцветны или с нерезкими и неправильными пестринами. ♦ . . Седой дятел Picus canus*
10 (9) Перья затылка не серые и несут красные отметины. Перья голени и подхвостья
с явным поперечным рисунком Зеленый дятел Picus viridis.
II (12) Число пальцев ноги—три. . . Трехпалый дяте л Picaides tridactylus.
12 (11) Число пальцев ноги — четыре 13.
13 (14) Мелкая птица, крыло короче 110 лш . .г 15.
14 (13) Крупнее, крыло длиннее 110 мм . . . .' 19.
15 (16) 2-е маховое короче 7-го. Окраска темени (у взрослых) черная или красная
Малый пестрый дятел Dendrocopos minor»
16 (15) 2-е маховое длиннее 7-го. Окраска темени (у взрослых) серовато-бурая ... 17.
17 (18). Крыло длинней 95 мм. Затылок черный, середина спины без поперечного черно-
белого рисунка. Белый рисунок на внутренних второстепенных маховых неправильный
Большой острокрылый дятел Yungipicus nanus,
18 (17) Крыло короче 95мм. Затылок серо-бурый, середина спины в правильном
поперечном черном и белом рисунке. Белый рисунок на внутренних пторостс пенных маховых
резко поперечный Малый острокрылый дятел Yungipicus kizuki.
19 (20) Брюхо и бока с сернисто-желтым оттенком
Средний пестрый лятсл Dendrocopos medius.
20 (19). Брюхо и бока без желтого 21.
21 (22) Спина белая или с примесью Со лого
Бе л ос п и к ими дятел Dendrocopos teucotos.
22 (21) Спина черная 23-
23 (24) Белые щеки отделены от бокоп шеи черной поперечной полосой, доходящей до
затылка '. Большой пестрый дятел Dendrocopos major.
24 (23) Белая окраска щек слипается с окраской шеи, поперечной черной полосы здесь
нет Сирийский дятел Dendrocopos syriacusm
РОД ЖЕЛНЫ DRYOCOPUS ВО IE, 1826
Тип Picus martius
1. Желна или черный дятел Dryocopus martius L.
Picus martius. Linnaeus. Syst. Nat., изд. X, 1758, стр. 112, Швеция.
Распространение. Ареал. От северных частей Испании и Италии и от
Греции до границы леса в Лапландии (в Англии отсутствует). Лесная полоса
Европейской части СССР к югу до Полтавы и Воронежа; Крым и Кавказ, сев.
Иран. В Сибири до 69° с. ш. (Индигирка), до верхнего течения Колымы;
Камчатка, Сахалин, Японские о-ва, Корея, Манчжурия, Сев. Китай, горы
Монголии. Осенью -^в Малой Азии. Изолированное нахождение — восточные склоны
Тибетского плоскогорья.
ЖЕЛИА ИЛИ ЧЕРНЫЙ ДЯТЕЛ
551
Характер пребывания. Оседлая птица. Во внегнездовое время
совершает небольшие кочевки, выходя за пределы гнездового ареала.
Биотоп. Высокоствольные леса таежного типа.
Численность. Довольно обыкновенная, распространенная в силу
биотопических условий — неравномерно.
Карта 93. Распространение желны Dryocopus martiuc
7 — D. m. martins, 2 — D. m. pinetorum, 3— D. m. khamensis; a — граница гнездовой области, г — места
регулярного выхода зимой за пределы гнездовой области.
Подвиды и варьирующие признаки. Вид довольно устойчивый по размерами по окраске;
распадается на 3 подвида. Номинальная форма — D. т. martius Linnaeus, 1758 —
Вост. Европа и сев. Азия, самая крупная раса; D.m. pinetorum Brehm, 1831 — Зап.
Европа, Крым, Кавказ; мельче предыдущей; D. т. khamensis В u t и г 1 i n, 1908 —
восточные склоны Тибета, имеет клюв меньших размеров и менее оперенную плюсну.
Обыкновенная желна Dryocopus martius martius L.
Синонимы: Dryocopus martius reichenowi. К о t h e. Orn. Mon. 1906, стр. 95, Уссури. —
Picus jacutomm. Buturlin. Journ. f. Orn. 1908, стр. 284, Колыма, Верхоянск. — Dryocopus
martius silvifragus. R i 1 e y. Pr. Biol. Soc. Wach, XXVIII, 1915, стр. 162, Ecco, Сахалин. —
Dryocopus martius sakhalinensis. Mo mi у a ma. Annot. Orn. Oriental., 1928, стр. 211, Южн.
Сахалин.
Распространение. Ареал. Скандинавия до бЭ^с ш., Норвегия,
Кольский п-ов, включая озеро Энаре, басе. Туломы, Имандру и Хибинские горы.
Берега Белого моря, включая и Соловецкие о-ва; р. Печора до нижнего ее
течения. В Зап. Сибири до Шеркалина на Оби (62° с. ш.), до Тарко-Сале под 64° с.
ш. на р. Пур (Вдовкин, 1941). На север до бассейна р. Таз, басе. Елагуя и
Нижней Тунгуски. Восточнее северная граница недостаточно ясна. По р.
Оленек доходит почти до 68° с. ш. На Индигирке до 69° с. ш., на Яне и Колыме
до 672/з° с. ш. В басе. Анадыря и на Пенжине отсутствует. Есть на Камчатке,
Шантарских о-вах, Сахалине, Кунашири (Курильские о-ва), о-ве Иессо и
изолированная колония — в северной части Хондо (Ян, 1942), Корее, Манчжурии,
с.-в. Китае к югу до Чжили и Шаньси. Южная граница, огибая безлесные части
Монголии, включает в себя хребты Кентей, Хангай, Тарбагатай, обходит с юга
и запада Алтай и выходит к Семипалатинску. Отсюда через Кулундинскую
552
ОТРЯД ДЯТЛЫ
степь, по лесам Казахской складчатой страны, огибая с севера степную часть
Казахстана и Зап. Сибири, южнее Тюменского округа, включая боры Аман-
Карагай и Наурзум-Карагай (51°30' с. ш.), граница идет по лесостепи Южн.
Урала в направлении Чкалова. В Европе граница тянется через устья Сак-
мары, по южной границе Бузулукского бора, на запад к Пензе (Городище)
и Тамбову, к Карачеву и от него на юг к Остру на Десне (южнее Чернигова),
отсюда севернее Овруча к Владимир-Волынскому. Западнее — другая форма.
Граница между ними может быть проведена условно по западным границам
Советского Союза к Балтийскому морю. Зимний ареал в общем совпадает с
летним, но несколько шире. На севере до 69° в бассейне Индигирки (Михель,
1935), на юге включает в себя Воронежскую и Курскую обл.; по Волге почти
до Астрахани.
Характер пребывания. Почти по всей области
распространения — оседлый. Местами откочевывает зимой к югу, выходя за пределы
гнездового арела; местами продвигается к северу и перемещается в пределах
гнездового ареала, появляясь в зимнее время там, где летом не держится.
Возвращение на гнездовые местообитания происходит в марте.
Биотоп. Большие высокоствольные, преимущественно хвойные, леса.
Местами (Саяны) отсутствие сосны означает и отсутствие дятла на гнездовье,
но, бывает, гнездится и в уреме. В некоторых местностях предпочитает
однотипному лесу смешанный. Согласно Огневу (Смоленская обл.), основной биотоп
желны — высокоствольные смешанные леса с моховыми болотами, по другим
авторам, предпочитает высокоствольные сухие и в Башкирии (Сушкин, 1897)
довольно светлые леса. В Гомельской обл. чаще в широколиственных
дубравах, в сосновых насаждениях реже. Главное для птицы — это наличие
подходящих для устройства дупел деревьев. В Сибири — сосновые боры,
лиственнички и глухие кедровики, хотя на Алтае темных однородных участков
кедровой и елово-пихтовой тайги избегает, селясь лишь по опушкам леса или
где тайга прорежена. В горы выше 1700—2000 м не поднимается (Сушкин,
1938), по среднему Амуру в лиственничных лесах селится только в случае,
если там есть ели, чистых лиственничных лесов избегает (Штегман, 1930).
Во внегнездовое время особенно охотно держится на гарях, на участках леса,
пораженных короедом, около муравьиных куч и в сосновых вырубках, где любит
сидеть на отдельных высоких деревьях, оставленных для осеменения. Во время
кочевок отмечен по всем большим и малым лесам, где есть сухие деревья.
В южной части Вятского края отмечен как частый гость берез, которыми
обсажены там с XVIII века почтовые дороги (Круликовский, 1913).
Численность. В подходящих местах обыкновенная, довольно
многочисленная птица. Местами (Якутия) почти не уступает в численности большому
пестрому и трехпалому дятлам. В других местах более редок. Отмечено, что по
мере уничтожения лесов численность черного дятла уменьшается.
Экология. Размножение. Первая весенняя «дробь» на Индигирке
отмечена 26. III (Михель, 1935). В Башкирии спаривание происходит в начале
апреля (Сушкин, 1897). В это время черный дятел становится еще крикливее
и барабанит громче других дятлов. Однажды Сушкину удалось наблюдать
у этой птицы некоторое подобие тока. 9 апреля не менее 15 самцов без самки
держались вместе, барабанили, дрались, перекликались и кроме обычного для
них голоса часто издавали крик, похожий на «любовную песню» сыча. При
этом дятлы быстро перелетали с места на место.
Гнезда устраиваются обычно на трудно доступных высоких и гладких
деревьях. Чаще всего (Европейская часть СССР) это осина, причем дупло
устраивается на высоте 8 м, чаще выше. В других случаях в качестве дерева
для гнезда отмечается сосна. Сосны избираются высотой около 30 мУ а дупла
бывают на высоте 18—20 м, однажды на высоте 27 м (Циммерман). Известны^
ЖЕЛНА ИЛИ ЧЕРНЫЙ ДЯТЕЛ
553.
впрочем, гнезда в дуплах сосны и на высоте 8—10 м. Однажды дупло было
выдолблено в сосновом столбе высоковольтной линии (Ушков). Дупло имеет
цилиндрическую форму. Входное отверстие очень узкое, прямоугольное,
подстилки в дупле нет. Яиц бывает от трех до пяти. Размер их 30—31 на 27 мм..
Длительность насиживания, судя по другому подвиду, 12—14 дней. 2.V в
Минской обл. найдены насиженные яйца (числом три). В других местах в это
время находят уже вполне оперившихся птенцов (Ленинградская обл. Бихнер,
1884). Кормящие птицы в окрестности Киева — 25.V, выводок из 5 молодых и
одной старой птицы замечен там же 24.VI (Шарлемань, 1915). В Ильменском
заповеднике в 1946 г. птенцы замечены 18 мая, а оставили дупло 4 июня
(Ушков). На Алтае (долина р. Камги) птенцы в гнезде 21—22 мая. (Фолитарек
и Дементьев, 1938). В Бузулукском бору оперяющиеся молодые найдены 7 и
14 июня (Карамзин, 1909). Вылет из гнезда в Среднем Урале в начале июня
(Уварова), в Забайкалье во второй половине июня (Павлов, 1948).
Линька. Смена маховых перьев происходит постепенно, перо за пером.
Первостепенные маховые начинают выпадать в самом начале июля или несколько
раньше. У экземпляров от б июля (из-под Звенигорода), от 4 июня (из-под
Бобруйска) и от 5 июля (Краснодарский край) нет восьмого первостепенного
махового пера. Седьмое, шестое и пятое перо сменяются в июле и августе, четвертое —»
в августе, третье — в самом конце августа, но главным образом в сентябре.
Смена второго и первого первостепенных маховых почти исключительно в
октябре. В коллекциях музея Московского университета имеется экземпляр из
Верхне-Колымска от 3 декабря, у которого отсутствуют первое, второе и третье
маховые перья. Наряду с этим экземпляры, добытые в ноябре и некоторые
в октябре, перелиняли уже полностью. Смена рулевых перьев —в июле.и
августе, частично захватывая конец июня и начало сентября. Есть
экземпляр от 11.1. 1913 г. из Владимирской обл., у которого средняя пара рулевых
выросла не более как на половину своей нормальной длины. Молодые,
меняющие мелкое перо, маховые и рули, в окрестностях Чердыни обнаруживаются
во второй половине июня (Резцов, 1905). В Башкирии линька во взрослый
наряд начинается в первых числах июля и к началу августа заканчивается
(Сушкин, 1897).
Питание. В летнее время личинками насекомых: дровосеков, короедов
(березовых заболонников в одном желудке до 300—650 штук, Шевырев, 1892),
куколками обыкновенного рогохвоста, в большом количестве муравьями.
На Алтае муравьев предпочитают всякой другой пище (Фолитарек и Дементьев,.
1938). Зимой питается в основном личинками жуков ксилофагов, частично
растительной пищей (семена хвойных). Зимой 1947 г. поврежденные короедом
деревья в Московской обл. были очищены черным дятлом от коры, начиная
от земли до вершины. Каждый квадратный дециметр содержал 10—г12 уже
пустых люлек короеда, на каждом дереве около тысячи. Неповрежденные
короедом деревья не были тронуты (Благосклонов, 1949). Разрушает иногда
дупла, извлекая оттуда пчел. Согласно Эверсманну( 1866), таскает пчел также
и из ульев. Шишек сосны и ели, вопреки указаниям в литературе, не долбит
(Формозов, Осмоловская, Благосклонов, 1950).
Полевые признаки. Легко отличается от прочих дятлов большой величиной
и черным оперением. Выглядит большеголовым с тонкой шеей. Как и у других
дятлов, бросаются в глаза закругленные крылья и как бы порхающий полет.
Свое присутствие выдает громким, весьма характерным криком, звучащим
примерно «кри-кри-кри-кри-кри»... Иногда издает заунывно-мяукающий крик
«кэээ». Мало общительная, одиночно держащаяся птица; с апреля начинает
встречаться парами.
Описание. Размеры и строение. Длина тела (25) 420—486 мм, размах крыльев
(21) 715—800 мм, вес (2) 298—308 г. Формула крыла у старых птиц преимущественна
554
ОТРЯД ДЯТЛЫ
5>6>4>7>3>8>9>2. Крыло самцов (91) 232—260 мм, самок(74) 230—254мм,
среднее 245 и 242 мм. Первое маховое недоразвито (до 50 мм длины), на конце заострено,
второе маховое тоже заострено, оба сильно сужены в вершинных частях и имеют
саблевидную форму. У молодых птиц первое маховое перо длиннее (до 70—85 мм) и имеет, как и
второе перо, широкое окончание. Вершину крыла у молодых птиц чаще всего составляет
Рис. 136. Желна.
четвертое перо, а если пятое, то четвертое почти всегда длиннее шестого. У старых птиц
клюв имеет долотовидное окончание; конец надклювья сжат с боков и круто (почти
вертикально) срезан. У молодых птиц конец надклювья оканчивается заостренно, крутого
среза нет. Длина клюва самцов в пределах 55,4—66,5, самок 53—63,5 мм.
Окраска. Молодые отличаются от взрослых буроватым оттенком оперения,
отсутствием блеска на верхней стороне тела и меньшим развитием красного цвета на голове.
Взрослый самец имеет все оперение черное (в обношенном пере он слегка буроват). Верхняя
сторона тела с некоторым блеском, тогда как на нижней стороне тела более заметен бурова-
ЖЕЛНА ИЛИ ЧЕРНЫЙ ДЯТЕЛ
555
тый оттенок. Верхняя сторона головы, т. е. лоб, темя и затылок, яркокрасные. У самок блеска
в оперении нет, и красный цвет на голове ограничен областью затылка. Клюв
серовато-роговой с примесью желтоватого на нижней челюсти, ноги темносерые, когти черноватые.
Западный черный дятел Dryocopus martias pinetorum В г е h m
Dendrocopus pinetorum. Brehm. Handb. Naturg. Vogel. Deutschl. 1831, стр. 185,
Шварцвальд.
Распространение. Ареал. Область широколиственных лесов
европейского типа. В СССР — Кавказ, Крым. В Зап. Европе соответственно указанному
выше распространению вида. Сев. Иран к востоку до Астрабада (Дементьев,
1948). Восточная граница примерно по государственной границе СССР и
Польской Народной Республики, в Скандинавии—гноминальная форма.
Характер пребывания. Оседлый, на зиму опускается с гор
в долины.
Биотоп. В Кавказском заповеднике на с.-з. Кавказе
^широколиственные и темнохвойные леса. В Кабардино-Балкарии лишь в больших
высокоствольных горных лесах до самого верхнего пояса их, не спускается вниз даже
во время кочевок (Радищев, 1926), в окрестностях Дзауджикау — буковые
и хвойныелеса, зимой спускается в равнины (Беме, 1926). В центральном
Кавказе и Закавказье — высокоствольные лиственные леса средней полосы гор,
в Батумской обл. и выше — хвойный лес до границы с субальпийской
растительностью (Сатунин, 1907). В южнокаспийских провинциях Ирана оседлый
в дубово-буковых лесах гор.
Численность. В большинстве мест довольно редок, на с.-з,
Кавказе — обычная птица, зимой в темнохвойных лесах чаще всех других дятлов
(Аверин и Насймович, 1938).
Экология. Размножение. Спаривание по наблюдениям в Зап.
Европе (Нитхаммер, 1938) в марте. Дупла устраивает в соснах, реже в елях и
пихтах, во многих местах в буках. Кроме того в ольхах, осинах, дубах, березах
и ясенях. Если подходящих деревьев достаточно, выдалбливает каждый год
новое дупло, часто рядом с прошлогодним, чаще, однако, пользуется одним
дуплом несколько лет подряд. Нередко бросает начатое дупло неоконченным и
делает другое; неоконченное служит ему как ночное убежище. Покинутые им
дупла заселяются впоследствии галками, клинтухами, удодами, сизоворонками,
гоголем и мохноногим сычем, также и мелкими птицами (поползни, скворцы,
стрижи, горихвостка). Дупло выдалбливается 10—15 дней (Нитхаммер,
1938), по другим данным до 25 дней (Доуле, 1886), преимущественно самцом.
Глубина дупла обычно 35—55 см, как исключение до метра, ширина
15—20 см. Леток овальный (в горизонтальном направлении), ширина его
примерно 8,5 см и до 10—12 см.
Яиц в кладке 4—5, редко 6, по Хартерту, чаще 3. Полные кладки с середины
апреля и в марте, как исключение раньше, иногда в середине мая, но бывают
даже и в середине июня. Яйца откладываются ежедневно, цвет их белый
блестящий, размер: (28) 31,0—37,5x22,0—27 мм, среднее 34,9x25,9 мм.
Насиживание, повидимому, с первого яйца, длительность его не более 12—
14 дней. Птенец весит 9 г (вес яйца около 12 г). Кормит птенцов преимущественно
самец. Длительность гнездового времени 24—28 дней. В дальнейшем старые
птицы держатся невдалеке от гнезда, изгоняя ставших самостоятельными
молодых из своего гнездового участка. Л и н ь к а не изучена, в ноябре птицы уже
в свежем пере.
Питание. Предпочитает (по западноевропейским данным) крупных
лесных муравьев, ест также мелких муравьев, жуков (Rhagium, Oxymiras,
Leptura, Tetropium, Cerambyx) и других насекомых как в личиночном
состоянии, так и взрослых и куколок.
556
ОТРЯД ДЯТЛЫ
Описание. Размеры. Меньше номинального подвида — длина крыла самцов (14)
228—245, самок (10) 228—240 мм, среднее 232,9 и 232,3 мм (Дементьев, 1939), по Нитхам-
меру (1938): (16) 230—240 мм, среднее 235,5 мм. Длина клюва самцов в пределах 50—60
самок 57—58 лш.Вес(6)201— 215ги до (1)250 г. Окраска сходна с окраской
номинальной формы, но южный дятел в среднем несколько бурее.
РОД ЗЕЛЕНЫЕ ДЯТЛЫ PICUS LINNAEUS, 1758
Тип Picus viridis
Синоним. Gecinus В о i e, 1831.
2. Зеленый дятел Picas viridis L.
Picus viridis. Linnaeus. Syst. Nat., изд. X, 1758, стр. 113, Швеция,
Распространение* Ареал. От Испании и Португалии, Италии и Греции
до Англии и до б9х/20 с. ш. в Норвегии. В СССР к северу до Костромы, Казани
и среднего Поволжья, к югу до Харьковской обл. и порожистой части Днепра,
на Кавказе; сев. Турция и сев. Иран к югу до Загроша. *
Характер пребывания. Встречается круглый год, но
перелетная птица. Гнездовая популяция во многих местах заменяется прилетевшей
зимующей. Биотоп. Лиственные и смешанные, более или менее
осветленные леса. Численность. Довольно обычен, но немногочисленен.
Подвиды и варьирующие признаки. Три подвида, слегка различающихся размерами
и быть может интенсивностью зеленой окраски оперения: P. v. viridis Linnaeus, 1758 —
самый крупный подвид — Вост. Европа; P. v. virescens, Brehm, 1831— мельче
предыдущего — Зап. Европа; P. v. karelini Brandt, 1841—еще мельче, Кавказ, Передним
Азия, Балканский п-ов, Италия.
Северный зеленый дятел Picus viridis viridis L.
Распространение* Ареал. От южных берегов Финского залива граница
идет на восток южным берегом Ладожского озера, огибает окрестности
Тихвина и Череповца с севера, пересекает р. Ужу выше Макарьева и направляется
к Казани, затем идет вдоль Волги и около Городища, огибая Городищенский
и Пензенский районы с юга, сворачивает на запад, идя через Воронежскую
обл. южнее Бобровского района. На Украине распространена другая форма.
Граница между ними может быть проведена только условно по южной границе
РСФСР и западной границе БССР и далее от Беловежской пущи к Штетину
в Польше. Датские острова и Скандинавия (в Норвегии до 691/*0 с. ш., в Швеции
до бЗЧа0 с. ш.).
Характер пребывания. Встречается круглый год, но не
принадлежит к оседлым птицам, так как гнездившиеся заменяются на зиму
прикочевавшими с севера. Хорошо заметное движение к югу установлено в
осеннее время в окрестностях Пскова, но и зимой там этот дятел не редок. Частичная
откочевка к югу происходит в Калужской обл. (Филатов, 1915), небольшие
перекочевки в южном направлении в сентябре в Мордовии (Птушенко, 1938).
В Пензенской обл. встречается только летом (Федорович, 1915), в соседней
Тамбовской — на гнездовье и кочёвках.
Биотоп. Почти исключительно лиственные, реже смешанные леса.
В Смоленской обл. кроме лиственных лесов, чередующихся с полянами и
хвойными рощами, посещает и боры с лиственным подседом, где много куч муравьев
(Огнёв, 1909). В окрестностях Ржева населяет почти исключительно берёзовые
пустошные леса (Давыдов, 1896), На севере Горьковского края главным
образом смешанные леса, составленные из осин, берёз и ели, причём охотно
держится на еловых деревьях. Осенью залетает в усадьбы и сады, даже далеко
расположенные от леса.
зеленый дятел
557
Численность. На большей площади ареала довольно обыкновенная
птица, но безусловно реже, чем большой пёстрый дятел, и в некоторых местах
реже ряда других дятлов. В Белоруссии реже седого дятла, а в окрестностях
Пскова чаще его. На границе распространения в Ленинградской обл. зелёный
дятел редкая птица, повидимому увеличивается в числе (Бианки, 1914). В Гдов-
ском районе и в окрестностях Петродворца появился недавно — в конце
прошлого столетия (Бихнер, 1884; Бианки, 1915). В Костромской, в Тульской,
Орловской и Харьковской областях редок.
Экология. Размножение. В начале гнездового времени очень
криклив, самец и самка кричат по очереди почти целый день. Иногда, сидя на
вершине дерева, издает
громкие крики, похожие "
на «удивленный хохот»
(Исполатов, 1903). Гнёзда
устраивает
преимущественно в гнилых деревьях,
выдалбливая в них дупла
очень правильной формы с
круглым отверстием или
овальным (Благосклонов,
1948). Чаще гнёзда
устраиваются в осине, нередко в
осокори и ивах. Гнездовый
участок не велик, но пары
гнездятся далеко друг от
друга (Федарака, 1928).
Подстилка в
гнезде—древесная труха. Высота дупла
от земли различна, иногда
оно помещается очень
высоко. В кладке —» 5—9 яиц
(Шнитников, 1913, приводит
сообщения лесников,
согласно которым зелёный
дятел откладывает 8—10
яиц). В Минской обл.
свежие яйца 19 мая,
молодые, уже подлётывающие птенцы 12 июня (Шнитников, 1913), в Псковской
обл. свежие кладки 2 июня. Насиженные кладки и кладки с частично
выведшимися уже птенцами находили в тех же местах 11 июня, в тот же день
Н. А. Зарудный добыл у гнезда самку, ещё не закончившую кладку. В
Ленинградской обл. 29 июня —3 июля птенцы ещё не летают, а 15 июля молодые
уже лётные (Бианки, 1915). В Мордовии самостоятельные молодые были
отмечены в начале июля (Птушенко, 1938). В Смоленской обл. вылет моло-:
дых происходит в начале июня (Огнёв, 1909). Кочевки в Белоруссии начинаются
уже в июле (Федарака, 1928). Осенний пролёт начинается в начале сентября
и продолжается одиночными экземплярами, изредка по несколько штук,
до середины ноября (Новгородская обл., Исполатов, 1903).
Линька. Линька крупного пера происходит во второй половине лета.
Экземпляр с недоросшими восьмым и девятым маховыми перьями добыт близ
Переславля 11 июня, восьмое короче девятого и седьмое едва растёт — 28 июня
(Костромская обл.), седьмое короче первого -—21 июля (Арзамасский район),
шестое начинает расти —15 июля и короче седьмого —23 июля (Вяземский
район), пятое короче первого— 18 июля (Москва), четвёртое короче второго —
Карта 94. Распространение зеленого дятла Picus viridis
1 — P. v, viridis, 2 — P. v. virescens, 3 — P. v. karelini; a —
граница области распространения, г — переходная полоса между
северными и другими подвидами зеленого дятла.
558
ОТРЯД ДЯТЛЫ
13 июля (Пенза), недоросшее третье перо — 2 августа (Ленинград) и 5 августа
(Ульяновская обл.). Второе перо, в той или иной степени недоросшее,
встречено в числа: 2 (Калуга), 16 (Ульяновская обл.), 19 (Мордовия), 24 (Подольский
район) сентября и 1 и 8 октября (Ульяновская обл.). Без первого пера
экземпляр от 14 сентября (Ульяновская обл.). В середине октября птицы с полностью
перелинявшим крупным оперением. В
общем линька первостепенных маховых
перьев длится с середины июня по начало
октября. Смена рулевых перьев
происходит с середины июня по середину
сентября; всё это время можно видеть птиц,
у которых средние рулевые в той или
иной степени недоросшие. У молодых
птиц смена мелкого пера на первоосен-
нее оперение начинается с конца второй
декады августа и в середине сентября,
однако, почти полностью перелинявшие
встречаются уже в самом начале
сентября. Есть один экземпляр, у которого
следы ряби на брюшной стороне и на
спине сохранились до 10 мая (Макарьев-
ский район).
Питание. В желудках взрослых
и молодых птиц находят обычно
муравьев (рыжие, лесные) и муравьиные
яйца. В поисках пищи птицы вырывают
глубокие ходы внутри муравейника.
Полевые признаки. Очень легко
узнаётся как дятел по общей зеленой
окраске. От сходного с ним седого дятла
легко отличается расцветкой головы
(красная до зашейка включительно), а
молодые, кроме того, сильной испещ-
рСмиостыо низа.
Описание. Размеры и строение.
Формула крыла преимущественно 4>5>б или
4>5>3, очень редко самое длинное перо пятое.
2-е обычно короче 8-го или равно ему, у
молодых птиц длиннее 8-го. Первое (рудиментарное)
перо у молодых более чем в полтора раза
длиннее, чем у старых птиц. Длина тела самца (12)
330—360лш,самки(12)340—37\мм. Длина
крыла взрослых самцов (15) 161—172 лш, самок (13)
164—175 мм,среднее 169и170лш. Вес 186—250г.
Окраска. Молодая птица отличается от старой более грязным цветом
(оливковым) спинной стороны тела и светлыми пестринами на нём, перья надхвостья охристые,
нижняя сторона тела белесая, иногда с охристым оттенком, и вся в пестринах от
черноватых поперечных полос на перьях. Низ и бока головы в продольных пестринах, тёмные
«усы» едва только намечаются, причём у самцов заметны и красные перышки. Красный цвет
верха головы менее интенсивен, чем у взрослой птицы.
Взрослая: общая окраска верха яркозеленая, надхвостье — блестяще желтое. В
обношенном пере окраска более блеклая. Верх головы и зашеек красные с проглядывающими
серыми основаниями перьев. Лоб,уздечка, пространство вокруг глаза—чёрные. Вдоль нижней
стороны щёк проходит красная полоса (усы), в той или иной степени окаймлённая чёрным.
Подбородок и горло, а также ушные партии, беловатые с зелёным налётом, более
интенсивным на ушных партиях. Брюшная сторона тела бледнозелёная с неясными тёмными
пестринами на задней части груди и брюха и с более ясно выраженной полосатостью на боках, голени и
подхвостье. Верхние кроющие крыла одноцветны с мантиец. Первостепенные маховые буро-
Рис. 137. Зеленый дятел.
ЗЕЛЕНЫЙ ДЯТЕЛ
559
ватые с белыми пятнами на внешних опахалах, на основной части внутренних опахал имеются
белые, поперечно расположенные полосы, которые до стержня пера не доходят.
Второстепенные маховые перья в неприкрытой части опахал зеленовато-песочного цвета, который
распространяется также и на внутренние первостепенные перья. Под этим цветом едва
проступает более светлая поперечная полосатость. Внутренние (прикрытые) части
второстепенных маховых перьев сходны по расцветке с внутренними опахалами первостепенных
маховых. Нижние кроющие крыла беловато-жёлтые с бурыми поперечными полосами, на
подмышечных поперечная полосатость едва выражена. Рулевые перья тсмнобурые с
оливковым налётом и слабо выраженной более светлой поперечной полосатостыо.
Ноги тёмносерые с зелёным оттенком. Клюв серовато-чёрный с жёлтым основанием
подклювья. Зрачок тёмнокарий, радужина беловатая с розоватым оттенком. Самка по цвету
оперения сходна с самцом, но «усы» черные без красного и брюшная сторона тела более:
блеклая, а тёмные пестрины несколько резче выражены.
Среднеевропейский зеленый дятел Pirns viridis uirescens В г е h nt
Gecinusvirescens. В г е h m. Handb. Naturg. Vog. Deutschl., 1831, стр. 199, Германия.
Распространение. Ареал. Эта слабо отличающаяся форма принадлежит
у нас Украине, на восток до Полтавской и Харьковской областей, а далее
на юге до р. Самары включительно. К западу от Днепра этой птицы нет в лесах
верховьев Ингула и Ингульца (Шевченко, 1940). Обходя Проскурой с севера,
граница выходит к Карпатам. Вне СССР этот подвид распространен в Польше
без северо-восточных её частей, на Ютландском п-ове, в Англии, Германии,
Франции и Нидерландах, в сев. Швейцарии, в Австрии и в Чехословакии.
Зимний ареал несколько обширнее летнего, встречается значительно южнее
Днепропетровска и, насколько можно полагаться на правильность подвидо-
вого определения, доходит даже до Волги (Казанский район, Бутурлин, 1936).
Характер пребывания. В пределах СССР гнездящаяся и
бродячая, вернее даже перелётная птица. В Харьковской обл. весенний пролет
происходит в середине апреля. Осенние кочёвки начинаются с середины
августа, постепенно переходя в правильный перелёт. Летят сначала старые птицы,
потом появляются молодые. Заканчивается пролёт в конце октября. Дятлы
летят в одиночку в южно,м или в юго-западном направлении (весной в
противоположном) из леса в лес, от одного дерева к другому, днем прямо через степи.
Биотоп. Лиственные и смешанные леса, растущие в лесных долинах.
В глубине больших однообразных лесов, особенно если нет лужаек и отдельно
стоящих старых деревьев, встречается очень редко, совсем редко в сосновых
борах. В послегнездовое время встречается и в нагорных дубравах, в усадьбах
и садах.
Численность. Довольно редкая птица, осенью и весной встречается
чаще, чем летом, зимой реже.
Экология. Размножение. Гнезда устраивает в дуплах старых осин,
осокорей, ив. Глубина дупла —210 мм, подстилка —древесная труха. Пары
гнездятся далеко друг от друга. В кладке от 3 до 7, как исключение, 8 яиц.
Яйца белые, блестящие. Длина их 33—34 мм,'ширина 22,5—-23 мм.
Продолжительность насиживания (по западноевропейским данным, Нитхаммер, 1938)
15—17 дней, длительность гнездового времени — 19 дней. Гнездо с двумя
птенцами и тремя сильно насиженными яйцами найдено в Изюмском районе
18 мая (Аверин, 1910).
Линька. Примерно совпадает в сроках с линькой номинальной формы,
точнее не изучена. У молодых смена оперения шапки происходит в середине
сентября. В конце сентября линька заканчивается.
Питание. Насекомые, преимущественно муравьи и их личинки. В
муравейниках добывает также личинок бронзовок. Как исключение, в желудках
цаходят древесные почки (Сомов, 1897).
560
ОТРЯД ДЯТЛЫ
Описание. Размеры и строение. Длина крыла (24) 159,5—168 мм, среднее
163,5 мм (по экз. из Баварии, Нитхаммер, 1938), т. е. меньше, чем у номинального
подвида. Вес самцов (2) 185 и 206 г, самок (2) 197 и 210 г (Нитхаммер, 1938). Окраска
такая же, как и у северного подвида.
Южный зеленый дятел Picus viridis karelini Brandt
Picus karelini. Brandt. Bull. Sc. Acad. Imp. Petersb, 1841 (на обложке журнала
указано 1842), стр. 12, Северная Персия.
Синонимы: Gecinus saundersi. Taczanowski. Journ. f. Orn. 1873, стр., 349. Ла-
годехи в басе. Алазани. — Picus viridis romaniae. Stresemann. Anz. Orn. Ges. Bayern,
1919, стр. 5, Молдавия и Валахия.
Распространение. Ареал. Область распространения этого подвида
распадается на две разделенных части: восточную и западную. На востоке ареал
доходит к северу до границы предгорных лесов Кавказа, включая в себя и
долину Терека, охватывает весь Кавказ и Закавказье, сев. Турцию и сев,
Иран, далее к югу, включая в себя Белуджистан, доходит почти до
Персидского залива. В Туркмении найден на Сумбаре (Дементьев, 1950). На
западе этот дятел встречается у нас в районе Проскурова (Шарлемань, 1933)
и в Закарпатской Украине (Страутман, 1948). Вне пределов СССР в Европе —>
Балканский п-ов, Венгрия, южн. Швейцария и Италия.
Характер пребывания. Встречается круглый год, совершает
незначительные по протяжению вертикальные миграции. Осенние кочевки
начинаются в августе. Местами оседлая птица.
Биотоп. Лесные предгорья и пойменные леса и рощи. В Грузии чисто
буковых лесов избегает (Цветков, 1892). В Армении предпочитает леса, в
которых имеются дикие плодовые деревья и держится в них, преимущественно ближе
к опушкам (Ляйстер и Соснин, 1942). Фруктовые сады. Вблизи южного берега
Каспийского моря охотно селится и в буковых лесах. В Абхазии до высоты
1400 м, у Лагодех до 3000л*. В зимнее время появляется на бульварах Еревана,
в Закаталах — по большим зарослям кустарника. Во время кочевок часто
можно видеть на отдельно стоящих диких фруктовых деревьях. Во внегнездо-
вое время встречен также и в сосновом бору (близ Дзауджикау, Беме, Красов-
ский и Чернов, 1926).
Численность. В большинстве подходящих для него мест зеленый
дятел обычная птица, в Аджарии и Гурии редок (Вильконский, 1897). Довольно
редок в Закарпатской Украине.
Экология. Р а з м и о ж е и и е. Гнезда устраивает в дуплах, используя
в долине Малки и Терека для этой цели дупла тополей (Радищев, 1926). В
Дагестане гнездятся в середине мая, молодые вылетают в конце июня —» начале июля
(Беме, 1926). В Закавказье (с. Дедели) гнездо с птенцами найдено 12 июля (Са-
тунин, 1907). Выводки в Армении встречаются в июле и позднее (например,
29 июня и 19 июля, Ляйстер и Соснин, 1942). В окрестностях Кизляра молодые,
которых старики еще кормили, встречены 5 июля (Беме, 1925).
Самостоятельных молодых можно видеть уже в конце первой декады июня (7^9 июня;
Ляйстер и Соснин, 1942).
Линька. Начинается во второй половине лета. По отдельным
экземплярам можно установить следующее: 2 июля меняется 8-е маховое; 25 июня —-
6-е; 19 июля —5-е еще короче 7-го; 4-е короче 2-го —30 августа; 3-е
короче 2-го — 16 сентября и 2-е короче 1-го в сентябре. Рулевые перья меняются
в июле—августе. Линька первостепенных маховых у молодых происходит
примерно в те же сроки. Седьмое перо меняется 9 июля и 22 июля. Смена
пестрых мелких перьев на зеленые происходит в августе — сентябре.
Есть экземпляры от 16 ноября, у которых линька мелкого пера почти
совершенно окончилась, сохранилось только несколько отдельных пестрых
перьев на спине и зобе.
ЧЕШУЙЧАТЫЙ ДЯТЕЛ
561
Питание. Летом и осенью поедает много личинок и взрослых муравьев.
В желудке самки, добытой 7 сентября 1933 г. в Кавказском заповеднике, было
найдено более 2000 муравьев. Полупереваренные остатки их заполняли также
весь кишечник (Аверин и Насимович, 1938). Долбит, как и все другие
кавказские дятлы^ мало.
Описание. Размеры и строение. Мельче номинальной формы. Длина крыла
самцов (19) 159—169 мм, самок (27) 155—169 мм, среднее 163 и 161 мм.
Окраска. Различий в окраске у разных подвидов зеленого дятла нет.
3. Чешуйчатый дятел Picas squamatus Vigors
Picas squamatus. Vigors. Proceed. Committee of
стр. 8, Гималаи.
the Zool. Soc. of London, 1831,
Распространение. Ареал. Гималаи, Афганистан, Белуджистан, вост.
Иран и южн. Туркмения (бассейн Мургаба).
Биотоп. Лиственные леса—
как горные, так и тугаи речных
долин. В Гималаях примерно от
1500 до 2700 м (Бекер, 1927).
Характер
пребывания. Оседлый.
Численность. В
Гималаях обычен, в северной
равнинной части ареала спорадичен и
редок.
Подвиды и варьирующие признаки.
В Гималаях от Гильгита и Кашмира до
Кумаона, вероятно в западном Непале—
P. s. squamatus Vigors, 1831; в вост.
Иране, Белуджистане, Афганистане и
Туркмении—P. s. gorii H a r g i 11, 1.887.
Морфологические различия в окраске —
южная гималайская раса заметно темней
на спинной стороне, с менее резким
поперечным рисунком на ней, с широким и
темным чешуйчатым рисунком на
брюшной стороне, с зеленовато-серыми зобами и грудью. Равнинная туркменско-афганская раса
бледней и желтоватей по общему тону, с заметным поперечным рисунком на мантии
(Дементьев, 1939). Экологические различия — в биотопическом распределении и, повидимому,
в особенностях цикла периодических явлений; кладка у горной формы происходит в
марте — апреле (Бекер, 1927), у равнинной в апреле и т. д.
Карта 95. Распространение чешуйчатого дятла
Picus squamatus
i — P. s. squamatus, 2 — P. s. gorii; a — граница
области распространения.
Мургабский чешуйчатый дятел Picus squamatus gorii Н а г g.
Gecinus gorii. H а г g i 11. The Ibis, 1887, стр. 74, Гильмент в Афганистане.
Синонимы. Gecinus flavirostris, «Zarudny». Menzbier. Bull. Soc. Imp. Nat.,
Moscou, 1886, стр. 440, Мургаб, nomen praeoccupatum. — Gecinus zarudnyi. Menzbier.
The Ibis, 1887, стр. 301, nomen emendatum. tta
Распространение. Ареал. Афганистан, Белуджистан (Кветта), вост.
Иран, Туркмения —по Мургабу. В 1886—1887 гг. дятел был распространён
в южн. Туркмении по пространству километров на 12 выше Сары-язы, с одной
стороны, и несколько ниже Имам-баба —с другой (Радде и Вальтер, 1889).
В настоящее время в долине Мургаба от Ташкепри до Казыклыбента и ниже
36 Птицы СССР, т. I
562
ОТРЯД ДЯТЛЫ
последнего километров на 8—10 (Формозов), но повидимому только на правом
берегу с лучше сохранившимися тугаями (Дементьев, 1950).
Характер пребывания. Строго оседлый.
Биотоп. Приречные тугаи, главным образом из разнолистного тополя,
предпочитает крупные дуплистые деревья, но за отсутствием таковых
гнездится и в тополях, сантиметров в 15 толщиной (Лоудон, 1913; Александров,
устное сообщение).
Численность. Весьма ограниченная в связи с сокращением площади
тугаев. Современный ареал чешуйчатого дятла в Туркмении носит уже
«островной» или «пятнистый» характер, и
отделён от афганского. Птица нуждается
в охране, как и ее биотоп — мургаб-
ские приречные тугаи (характерно ее
отсутствие на Теджене, где тугайные
леса сохранились лучше, чем на Мур-
габе). Больше всего дятлов
сохранилось на участке между Сары-чопом и
Казыклыбентом: число встреч за день
здесь от 1 до 10 и даже 14 (Формозов,
устное сообщение: 7 июня — 10 птиц,
8 июня—7 или 8, 10июня—5,11
июня—4, 15 июня —14 и 17 июня —1).
Экология. Р а з м н о ж с и и с.
Гнездовые участки теперь
изолированы и пары редко гнездятся иблини
одна от другой (в одном случае D тупи»
около 1 км длиной и 200 ишгоп
шириной — 2 занятых гнезда и 50 шагах
одно от другого, Лоудон, 1913).
Брачный крик — «ти-тию» или «клю-клю*
(Раддо и Вальтер, Лоудон) —
постоянно слышен и апреле. Гнезда в
дуплах разнолистного тополя иногда ни:»»
ко (от 00 и даже 30см9 Радде и Валь-
тор) и до 3,6 л* от земли. Входное
отверстие узкое, около 5—6 см, глубина
дупла около 50—60 см. Подстилки и
сущности нет. В первой половине
апреля гнезда заняты, 11 июня в гнезде
4 слегка насиженных яйца (Радде и
Вальтер); 1 апреля у Имам-бабы в
одном гнезде 5 насиженных яиц (Лоудон,
1902). О росте птенцов дают
представление следующие сведения: 21 апреля в гнезде 4 птенца (самец и три
самки), которые вылетели бы через неделю; 22 апреля в другом гнезде—3 птенца
(самец и две самки), несколько старше; 24 апреля найдено 3 гнезда —в одном
3 птенца (самец и две самки) сходного с предыдущими возраста, в других по
четыре птенца (самец и три самки), почти лётные; наконец, 27 апреля в одном
гнезде 6 птенцов, из которых 2 живых самки и 4 мёртвых (Лоудон). Лётные
молодые в птенцовом наряде наблюдались и добывались в мае (Формозов).
Кладка, следовательно, состоит из 63 — яиц. Яйца белые, блестящие, как
у зелёного дятла, Максимальные их размеры 31 х 22 мм (Радде и Вальтер,
1889).
Линька. Не изучена.
Рис. 138. Чешуйчатый дятел.
Чешуйчатый дятел
СЕДОЙ ДЯТЕЛ
563
Питание. Повадки сходны с зелёным дятлом, кормится главным
образом на земле, добывая насекомых среди тамарисковых зарослей. В гнезде
молодые кормятся какими-то зелёными гусеницами (Лоудон, 1913), старые почти
исключительно муравьиными яйцами и муравьями (Радде и Вальтер, Лоудон).
Полевые признаки — сходны с зелёным дятлом.
Описание. Размеры и строение. Как у зелёного дятла, формула крыла
4>5>6>3>7 (1-е рудиментарно). Крыло самцов (7) 156,1—160, самок (7) 149—158,2, в
среднем 157,3 и 153,5 мм. Вес самца 165 г, самки 159 г.
Окраска. Молодые несколько бледней взрослых, половой диморфизм в окраске
темени выражен и в гнездовом наряде. Взрослый самец на спинной стороне бледный
оливково-зеленый с чешуйчатым неясным рисунком на мантии, большие кроющие крыла
и задние маховые с нерезким поперечным рисунком; первостепенные маховые бурые с.
беловатым поперечным рисунком; поясница и надхвостье светлозелёные с золотистым
оттенком; рулевые бурые с белым поперечным рисунком; темя красное, щёки беловатые
с черноватыми крапинками, образующими «усы», горло охристое, зоб и грудь с зеленоватым
оттенком и черновато-бурым чешуйчатым рисунком.
У самки темя не красное, а чёрное и окраска несколько тусклей. Клюв у взрослых
желтый, у молодых сероватый; лапы сероватые с бледнороговыми когтями.
4. Седой дятел Picas cartas Gmel.
Picus canus. Gmelin. Syst. Nat., изд. XIII, 1788, стр. 434, Норвегия.
Распространение. Ареал. Широко распространённый в Палеарктике
и частично за её пределами вид. От Франции (Вогезы) и Бельгии до Сахалина
Карта 96. Распространение седого дятла Picus canus
1 — Р с. canus, 2— Р. с. biedermannl, 3— Р. с. jessoensis, 4— Р. с, zimmermanni. 5 — Р. с;
guerini, 6 — Р. с. sordidior, 7 — Р. с. cogo, S — Р. с. tancolo, 9 — Р. с. ricketti, 10 — Р. с. hai-
nanus, 11— Р. с. hessei, 12 — Р. с. gylderistolpei, 13 — P. с. sanguiniseps, 14 — P. с micror-
hynchus, 15 — P. с dedemi; a — граница области распространения, г — переходные зоны между
северными подвидами, д — изолированное нахождение в зимнее время.
и Хоккайдо. В Норвегии до 64° с. ш., в Сибири до устьев Иртыша и верховьев*.
Лены. На юг до Формозы, Малакки и Суматры, до- Непала, южн... Алтая,
Семипалатинска, Днепропетровска, Венгрии, сев. Швейцарии.
36*
564
ОТРЯД ДЯТЛЫ
Характер пребывания. Оседлая и кочующая птица.
Биотоп. Более или менее осветлённые леса как лиственные, так
частично и хвойные.
Численность. В большинстве мест относительно редкая птица.
Подвиды и варьирующие признаки. На севере Палеарктики довольно устойчив,
образуя слабо выраженные географические формы, отличающиеся различной степенью
интенсивности зелёного и серого цвета в оперении. На юго-востоке образует очень большое число
подвидов, различающихся как размерами, так и степенью развития тёмной штриховки в
оперении и большим общим потемнением окраски. Различают 15 подвидов, возможно, однако,
что число их будет в дальнейшем сокращено: Р. с. canus G m е 1 i n, 1788 — Европа и Зап.
Сибирь; Р. с. biedermanni Hesse, 1911 — Алтай, Саяны, Забайкалье, более сероват,
чем номинальная форма; Р. с. jessoensis Stejneger, 1886 —? вост. Азия до
Хоккайдо и Пекина — в обношенном пере значительно серее, чем номинальная форма;
Р. с. zimmermanni R e i с he now, 1903 — переходная к следующей расе, плохо
охарактеризованная форма, распространена от Пекина до нижнего течения Янцзы; Р. с. guerini
Ma l h e r b e, 1849 — от дельты до среднего течения р. Янцзы, темнее всех предыдущих
форм; Р. с. sordidior R i р р о п, 1906 — Юннань, темнее предыдущей и крупнее; Р. с.
kogo В i a n с h i, 1906 — от Кансу до верховьев Янцзы и до Меконга, тёмная штриховка,
уже, чем у guerinil; P. с. tancolo Gould, 1862 — Формоза, мелкая тёмнозелёная форма;
Р. с. ricketti Baker, 1919 — Фучжоу, Тонкий, еще темнее, чем guerini; P. с. hainanus,
Grant, 1899 — Хайнань, мельче чем tancolo; Р. с. hessei Gyldens stolpe, 1916 —
от сев. Бирмы до Кохинхины, в оперении больше оливково-зеленых тонов, чем у следующей
формы, и клюв сильнее; Р. с. gyldenstolpei Baker, 1918 — от вост. Непала до Ассама,
сверху с золотисто-бронзовыми тонами, снизу желтоват; Р. с. sanguiniceps Baker,
1926 — зап. Гималаи и зап. Непал; Р. с. microrhynchus Robinson et Kloss, 1919 —
юго-зап. Сиам и Малакка: Р. с. dedemi van Oort, 1911 — Суматра, имеет
красный цвет в оперении.
Обыкновенный седой дятел Picas canus canus Gmcl.
Синоним. Picus canus dzieduszyckii. Domanewski. Ann. Zool,, Mile. Pol.
Hist. Nat., 1925, стр. 75, Львов.
Распространение. Ареал. Европа и Зап. Сибирь. В Норвегии доходит
до 64° с. ш., в Финляндии до 62°. Спорадические нахождения много севернее
сплошной области распространения, например, укапан для Торнео. Зимние
экземпляры известны из окрестностей озера Слободского, невдалеке от Хол-
могор. Оставляя эти изолированные нахождения в стороне, северная
граница сплошного распространения может быть очерчена так: от Ленинграда
и местностей севернее Череповца идет на восток, огибая с севера Шарью и
Кировский край, захватывает верховья Печоры (Усть-Унья), выходя на Урале
севернее Денежкина Камня в направлении к Никито-ивделю (Резцов, 1904).
Отсюда граница проходит около устья Иртыша (с. Самаровское), далее севернее
Ангары и, пересекая верховья Нижней Тунгуски, выходит к верховьям Лены.
Далее от Байкала — другая форма, а граница номинальной формы
поворачивает на юго-запад, проходя между Томском и Красноярском, юго-восточнее
Новосибирска к Змеиногорску и Семипалатинску.* Затем драница
поднимается несколько к северу и, проходя южнее Тары и Тюмени, идёт в
направлении к Чкалову (севернее его).
Включая Башкирию и Бузулукский бор, область распространения
спускается по пойме Волги почти до устья этой реки. Западнее Пензенская обл.
видимо исключается из гнездовой области, тогда как Тамбовская и
Воронежская входят в нее. От Воронежской обл. южная граница распространения
идёт на юг к станице Митякинской на Северном Донце и, оставляя севернее
* Ранне-летние экземпляры из северной части Зап. Сибири нам неизвестны, поэтому
принадлежность седых дятлов из этих мест к номинальной форме требует ещё доказательств.
Кроме того, должны быть уточнены северные пределы распространения этого дятла в Зап.
Сибири.
СЕДОЙ ДЯТЕЛ
565
себя Изюм, идёт к Днепру, пересекая его южнее Павлограда и Днепропетровска.
На запад от Днепра граница охватывает с юга Чёрный лес (верховья Ингульца)
и обходит с севера окрестности Умани. По Днестру седой дятел гнездится до
дельты включительно. В зимнее время несколько выходит за очерченную
область к югу, вероятно залетает в Испанию и на Кавказ. На Балканах проникает
до Константинополя и Македонии, западней—в Австрию и южн. Альпы,
спорадически во Франции и в Центральной Европе.
Характер пребывания. Гнездящаяся птица. Во внегнездовое
время предпринимает довольно обширные перекочёвки. Для Псковской обл.
Зарудный (1910) приводит зелёного дятла как оседлого, под Ленинградом эта
птица добыта во внегнездовое время; в окрестностях Петродворца, согласно
Бианки, оседла. Для северной части Горьковского края, по Серебровскому
(1918), пролётная (откуда?) и быть может зимующая птица, близ Шарьи
гнездится, гнездится в Башкирии, оседла в Тамбовской обл. В Пензенской обл. —.
на кочёвках и зимою. В Воронежской и Харьковской областях оседла, оседла
близ Днепропетровска и в Подолии. По всей территории в пределах очерченного
выше ареала, где не гнездится, бывает на осенних и зимних кочёвках и в
большинстве случаев также зимой. Кроме того, во внегнездовое время встречается
к югу от гнездовой области, например, в окрестностях Чкалова в
Астраханском крае (по Хлебникову, он гнездится здесь, что однако требует проверки).
Встречается на Кавказе (два экземпляра из Сев. Осетии); для Закавказья
отмечен Нордманном и Менетрие (1833), но позднее никто седого дятла там не
наблюдал. В юго-западном углу нашей страны на пролете и] зимой во всех
подходящих для этого местах.
Биотоп. Сходен с биотопом зелёного дятла, но не однороден во всей
области распространения подвида. В западной части ареала на восток до Волги
(пойма р. Суры и Башкирия) это по большей части лиственные леса (в ряде
случаев с преобладанием широколиственных пород) и у ремы. Гнездится также
и в смешанных лесах. По Станчинскому (1929), в дубово-сосновых лесах
Смоленской обл. даже чаще, чем в других типах насаждений* Чисто хвойных лесов
в большей части своего ареала избегает даже зимой. Давыдов (1896) указывает
для Ржевского района Калининской обл. в качестве гнездовой станции почти
исключительно пустотные берёзовые леса. Портенко (1928) встретил один
выводок в «лесных садах» Подолии. На севере Татарской республики станция
этого дятла — ельники в борах и елово-пихтовые леса, также
широколиственные леса. По Эверсманну (1866), седой дятел гнездится преимущественно в
хвойных лесах, но с большими травяными полянами, кроме того, в красных лесах
и лесах побережий. В Южн. Урале как в нагорно-лесных, так и в нагорно-
лесостепных ландшафтах; в первых —- его станция преимущественно сосново-
березовые леса, реже, повреждённые пожарами и шелкопрядами
лиственничники, во вторых — дубово-липовые леса (Кириков). Во время кочевок держится
кроме того в порубах с молодой порослью, в садах и усадьбах.
Численность. Довольно редкая или, во всяком случае, редко
попадающаяся на глаза птица. В большинстве мест реже, чем зелёный дятел. У
северной и также у южной границы ареала безусловно редок. Однако в
Харьковской обл. седой дятел численно мало уступает пёстрому (Сомов, 1877).
Также довольно обыкновенен в Воронежской и Курской областях и по линии
Павлодар — Днепропетровск. Обыкновенен в Черниговской и Полтавской обл.
У ст. Митякинской (Сев. Донец) редок. На кочёвках более общителен, чем
зеленый, и встречается чаще. Сомов (1897) наблюдал осенью близ Ахтырки
за 2 часа 10 экз. (9 самок и 1 самца). В Псковской обл. наблюдали стайку в 10
штук (Зарудный, 1910).
Экология. Размножение. Гнезда устраивает преимущественно в
осинах, также в вербах и других деревьях с мягкой древесиной. Отверстие дупла
566
ОТРЯД ДЯТЛЫ
в 3—5 м от поверхности земли. Откладывает от 5 до 7 яиц (в Финляндии 6—
8, Хортлинг, 1931). Скорлупа их блестяще чистобелого цвета. Размеры яиц (по
5 экз.): длина—28,5—28,5—28,5-^29,5^29,6 мм, ширина—21,3—21,2—
21,2—21,2—21,4 мм (Аверин, 1910), Лежат на подстилке из мелких щепочек
полугнилой древесины. Яйцекладущая самка добыта в Харьковской обл.
2 мая, а ненасиженная кладка из 5 яиц— 7 мая и 11 мая. Молодые, уже
оперяющиеся, найдены под Киевом 25 мая (Шарлемань, 1915). Самостоятельно
летающие молодые в Мордовии обнаружены в первой декаде июля(Птушенко, 1938).
Рис. 130. Седой дятел.
В окрестностях Трубчевска первые лбтные молодые— 27 июня (Суслова, 1937).
В садах Подолии летающий выводок— 10 июля (Портенко, 1928). В
Ленинградской обл. выводки появляются в середине июля (Бианки, 1915).
Линька. Смена крупного пера происходит с июля по сентябрь. 6-е и
5-е маховые меняются в июле, 4-е и 3-е — в августе, 2-е и 1-е — в сентябре.
Рули сменяются в июле и августе.
Питание. Сходно с питанием других дятлов, но этот вид чаще попадается
на муравьиных кучах. Бывает, ищет насекомых в глиняной обмазке изб, стояг
щих в лесу и даже на краю селений.
Полевые признаки. Молчаливая скрытно держащаяся птица. Несколько
мельче зелёного дятла. Довольно хорошо отличается от последнего расцветкой
головы.
Описание. Размеры и строение. Формула крыла взрослой птицы большей
частью 5>4>6>3>7>2>8, иногда самое длинное перо 4-е, иногда 3-е длиннее б-го, 2-е
может быть длиннее 8-го или короче его, очень редко оно короче 9-го. Молодая птица имеет
более тупое крыло: часто 3>4>5>2>6. Первое (рудиментарное) перо у молодой птицы почти
СЕДОЙ ДЯТЕЛ
567
•■в полтора раза длиннее, чем у старой и значительно шире. Длина крыла самцов (37) 136—
150, самок (27) 141—154, среднее 143 и 146 мм. Вес самки 98 г (1 экз.).
Окраска. Молодая птица серее старой, снизу грязновато-серая с мелкими
тёмными поперечными полосами и пятнышками. У взрослых преобладающий цвет в оперении
зелёный или оливково-зелёный, в начале зимы интенсивный, насыщенный, к концу зимы и
летом более блеклый, сероватый. У самца в свежем оперении голова и шея серые, но верх
головы почти от основания клюва до середины темени^красный. Уздечка угольно-чёрная,
отделяется от красного цвета верха головы довольно широкой серой полосой.
Угольно-чёрная полоса («усы») идёт от подклювья и отделяет серые перья боков головы от более светлого
подбородка. На перьях задней части темени нередко видны более или менее резко
выраженные черноватые настволья. Кроме того, на темени и зашейке бывает иногда зеленоватый или
охристо-зеленоватый налет. Мантия зелёная с оливковым оттенком и иногда с желтоватым
налётом. Перья поясницы и надхвостья имеют широкие золотисто-жёлтые краевые полосы.
Нижняя сторона тела светлосерая с зеленоватым оттенком. Горло беловатое, нижние
кроющие хвоста с тёмной пятнистостью, не всегда ясно выраженной. Первостепенные маховые
лерья буровато-чёрные с белыми пятнами на внешних опахалах, на основной части
внутренних опахал имеются белые поперечно расположенные полосы, которые, однако, не доходят
до стержня пера. Второстепенные маховые перья в неприкрытой части опахал, зеленовато-
оливковые со слабо выраженной и не всегда заметной поперечной полосатостью.
Рулевые перья черновато-оливковые, самые кончики их нередко золотисто-жёлтого цвета
{чаще всего обломаны). По краям двух внутренних пар рулевых перьев оливково-жёлтый
налет, а внутренние опахала средней пары рулей имеют беловатые поперечные полосы,
реже подобные полосы бывают и на наружных опахалах, а также на смежной паре
рулевых перьев, но на них они менее ясно выражены. Нижние кроющие крыла с буровато-
серой поперечной полосатостью. Ноги зеленовато-серые. Клюв буро-роговой, подклювье
•ближе к основанию более светлое, желтоватое. Самка без красного на голове, иногда бывают
отдельные красные перышки. Общий тон оперения несколько более серый.
Среднесибирский седой дятел Picus canas biedermanni Hesse
Picus canus biedermanni. Hesse; Orn. Monatsb., 1911, стр. 183, Алтай
Распространение. Ареал. Алтай, Саяны, Забайкалье. На севере и северо-
востоке область распространения непосредственно смыкается с
распространением номинальной формы, по Байкалу поднимается до его северных берегов,
затем, огибая с юга Витимское плато, граница проходит на юг к Чите,
охватывает бассейн Онона и Аргуни (как по реке Чита, так и по последним двум рекам
седой дятел найден только во внегнездовое время). Во внегнездовое время
встречен на Керулене; населяет зап. Хангай и Тарбагатай. Далее область
распространения подходит вплотную к Зайсану, экземпляры, добытые на Черном
Иртыше, принадлежат к этой форме. Охватывая Алтай, граница идет на северо-
восток, причем экземпляры из села Новенского (окрестности Змеиногорска)
определены мною как Р. с. canus.
Биотоп. Лиственничные леса и уремы, часто встречается на душистых
тополях. Осенью встречается также и в чистых кедровниках. В типичной
тайге и вообще в чистых хвойных участках в гнездовое время отсутствует.
Чаще всего встречается там, где лесные насаждения более разнообразны
и нередко прерываются травянистыми яркоцветными полянами, где есть
вырубки и гари с возобновлением зарослей берёзы, а в пониженных элементах
рельефа и осины.
В горы выше 1000—1100 м не поднимается. В связи со сказанным, распре^
делён седой дятел в очерченной выше области его распространения весьма
спорадично.
Характер пребывания. Кочующая птица. Во внегнездовое время
появляется в местах, где летом его не наблюдали (например по р. Чите, по р. Ке-
рулену и в других местах).
Численность. Редкая птица, более или менее обычная только
местами, например у северной части Телецкого озера (Фолитарек и Дементьев,
1948). В осеннее время во многих местах довольно обыкновенен.
568
ОТРЯД ДЯТЛЫ
Экология* Размножение. В апреле самцы весьма активны,
характерный их крик можно слышать всюду. Часто кричащий самец сидит неподвижно
в верхушке дерева (берёзы), нередко располагаясь не вдоль, сука, как сидят
обычно* другие дятлы, а поперёк его. Покричав, он внимательно прислушивается
к ответному крику и нередко летит на него. В двадцатых числах апреля самец
и самка уже лазили в гнездо (Алтай, Фолитарек и Дементьев, 1938). В общем,
биология размножения этой формы седого дятла совершенно не изучена.
Линька. Линька происходит во второй половине лета и заканчивается
ко второй половине сентября. Добытые 13 и 16 сентября экземпляры в
Монголии (Козлова-Пушкарёва, 1933) имели свежее перо. Средние рулевые у самца
только показались из пеньков, у самки они почти достигли нормальных
размеров.
Питание. Не изучено. Два экземпляра, добытые в декабре в Тувинской
области, имели в желудке орехи кедра.
Описание. Размеры и строение. Длина крыла самцов (9) 137—150, самок
(5) 141—146, среднее 146 и 144 мм.
Окраска, Сходен с номинальной формой, но верхняя сторона не оливково-зелё-
ного тона, а серовато-зеленого цвета. Различие это хорошо видно и у осенних свежепере-
линявших птиц, к лету становится еще более ярко выраженным. Нижняя сторона тела гряз-
новатосеро-зелёная. Самые зеленые экземпляры этого подвида весьма сходны с самыми
светлыми canas и практически не различимы.
Дальневосточный седой дятел Picus canas jessoensis Stejn.
Picus canus jessoensis. S te j n e ge r. Proc. U. S. Nat. Mus., 1886, стр. 106, Хоккайдо.
Синоним. Picus canus perpallidus. Ste jneger^ там же, 1886, стр. 107, Сидеми
(Уссурийский край).
Распространение. Ареал. По всему Приморскому краю на север да
р. Хунгари (с. Вознесеновка, Воробьев, 1938). Сахалин, о-в Хоккайдо, Корея,
Манчжурия, с.-з. Китай почти до Пекина.
Биотоп. Равнинные и частью горные смешанные и лиственные леса.
Характерен для дубовых насаждений и открытых долин рек, где имеются
небольшие перелески и рощицы.
Характер пребывания. Бродячая птица. В Судзухинском
заповеднике, например, в январе и феврале отсутствует, появляется с 23—28 марта.
Численность. Сильно зависит от урожая жолудей (Белополь-
ский). Вообще говоря, по среднему течению Амура очень редок, в бассейне
р. Уссури обычен.
Экология. 13 июня в низовьях Имана найдено в дупле 4 совсем маленьких
птенца. Молодые, только что вылетевшие из гнезда, добыты в Сучанском районе
10 июня, рулевые у них короче половины нормальной длины, вес 64,2 и 71 г.
Линька. Крупное перо сменяется с начала июля по сентябрь.
Описание. Размеры и строение. Длина крыла самцов (13) 140—151 мм±
самок (9) 145—157, среднее 147 и 149 мм. Вес самцов (4) 121—144 г, самок (4) 110—138 г,,
среднее 134 и 129 г.
Окраска. В свежем оперении практически не отличим от номинальной формы, быть
может лишь несколько посветлее (посерее). В обношенном пере очень сероват, почти без
зеленого, причем общий серый тон еще сильнее выражен, чем у Р. с. biedermanni в
соответствующем оперении.
Систематические заметки. Подвидовое разграничение седых дятлов, так же как и
остальных дятловых нашей фауны, очень затруднительно. Сущкин предложил причислить
западносибирских птиц (Тюмень, Тара) к Алтайской форме biedermanni. Однако катлы из
окрестностей Тюмени, Новосибирска, Томска и даже из Змеиногорского района практически от
номинальной формы неотличимы. С другой стороны, птицы с Алтая, из Зайсана и Забайкалья,
даже в свежем пере, довольно хорошо отличаются от западносибирских сероватым цветом,
а летом это различие выражено ещё более ярко. Поэтому я считаю возможным не
причислять западносибирских дятлов к форме biedermanni, хотя для проведения границы этой
формы от Зайсана.к Байкалу требуется дополнительный (гнездовый) материал. Разграни-
ТРЕХПАЛЫЙ ДЯТЕЛ
569
чение форм по коллекционному материалу затрудняется еще тем обстоятельством, что во
внегнездовое время в области распространения biedermanni могут быть встречены
прикочевавшие с севера canus. За отсутствием материала совершенно ничего не известно о подпм-
довой принадлежности седого дятла из области Амура от Аргуни до Малого Хингана
(впрочем, нет Даже точных подтверждений, гнездятся ли там эти дятлы). Птицы с Бурей
относятся во всяком случае к уссурийской форме (Штегман, 1931). Во многих литературных
источниках указывается, что дальневосточная раса почти не отличима от европейской. Это не
совсем верно. В свежем пере дятлы из бассейна р. Уссури действительно не отличимы от
европейских, но в летнем обношенном оперении они резко от них отличаются, как более
серые, и по этому признаку становятся почти не отличимы от алтайского подвида, но еще
более резко отличаются в это время от Р. с. canus, чем Р. с. bkdermanni.
РОД ТРЕХПАЛЫЕ ДЯТЛЫ PICOIDES LACEPEDE
Тип Picus tridactulus
5. Трехпалый дятел Picoides tridactylus L.
Picus tridactylus, Linnaeus. Syst. Nat., Изд. X, 1758 стр. 114, Швеция —
Далекарлия.
Распространение. Ареал. От Альп (Шамони, Савойя на западе) да
Камчатки и Сахалина. Кроме того, Сев. Америка. На север до 70° с. ш.
Карта 97. Распространение трехпалого дятла Picoides tridactylus
1 — P. t. tridactylus, 2 — P. t. tianschanicus, 3 — P. t. alpinus, 4 — P. t* funebris, 5 — P. t. albidior, 6 —
P. t. fasciatus, 7 — P. t. dorsalis, 8 — P. t. bacatus; a — граница области распространения, г — неясные
границы между подвидами.
в Норвегии и до лесотундры на с.-в. Сибири (Индигирка). На юг до сев.
Кореи, Юннаня в Китае и еловых лесов Тянь-Шаня. В Европе по горным лесам
до Болгарии, Австрии, Швейцарии. В Сев. Америке от Аляски (где идёт
к северу до полярного круга) до Ньюфаундленда (где распространен к северу
до Лабрадора). К югу до северных границ Мексики.
Характер пребывания. Зимой кочует, продвигаясь несколько
южнее от гнездового местообитания.
Биотоп. Обширные и глухие леса северного типа (преимущественно
хвойные), на юге — горные леса.
Численность. В северных частях ареала довольно обыкновенная
птица. К югу реже.
Подвиды и варьирующие признаки. Сильно изменчивый вид, главным образом по
соотношению белого и черного цвета в оперении. Намечаются два типа оперения: белый и
черный. Образует 8 подвидов: P. t. tridactylus, Linnaeus, 1758 — сильно изменчивая по
степени развития белого цвета форма, светлее двух следующих; P.t. tianchanicus, В u t u r-
1 i n, 1907 — тёмная форма горных лесов Тянь-Шаня и Алтая; P. t. alpinus, В г е h m,
1831—тёмная форма горных лесов Европы, едва темнее предыдущей; P. t. funebris,.
Verreaux, 1870—очень тёмная форма, нижняя сторона почти чёрная. Зап. Сычуань,
ю-з. Юннань (Китай); P. t. albidior, Stejneger, 1885 — самый светлый подвид,
населяет Камчатку; P. t. fasciatus, В a i г d, 1870 —Аляска и зап. Канада, на боках и груди
ясные поперечные полосы; P. t. dorsalis, В a i r d, 1858. — США от Канады почти до границ.
Мексики, самая светлая и самая крупная в Америке форма; P. t. bacatus, Bangs, 1900 —
восточная' часть Канады и прилегающие окраины США, самая тёмная в Америке форма.
570
ОТРЯД ДЯТЛЫ
Северный трехпалый дятел Picoides tridactylus tridactylus L.
Синонимы. Apternus crissoleucus. Reichenbach. Handb. spec. Orn. Scans.
Picarinae, 1854, стр. 362, Иртыш. — Picoides uralensis. В u t u г I i n. Orn. Mon., 1907, стр. 9,
вост. Россия от Суры и Зап. 'Сибирь. — Picoides sakhalinensis. В u t u r 1 i n. Orn.
Mon., 1907, стр. 9, Сахалин и прилежащие части материка. — Picoides tridactylus
stechowi. Sachtleben. Anzeig. Orn. Ges. Bayern, 1920, стр. 20, Литва. — Picoides
tridactylus, kolymensis. Бутурлин. Орнитол. Вест. 1917, стр. 13, Среднеколымск,
Олекминск.
Распространение. Ареал. Птица северных лесных частей Евразии.
Северная граница распространения близка к северной границе леса и в некоторых
местах совпадает с ней. В Норвегии трехпалый дятел известен до 70 °с. ш.,
на Кольском п-ове к.северу до Печенги, Туломы, устья р. Колы и севернее
Хибинских гор; в лесах по берегам Белого моря (есть на Соловецких о-вах),
близ Чешской губы доходит до Верхней Пёши, на Печоре примерно
до 67° с. ш. (Сибом и Броун, 1876). Между Печорой и Уралом --на Колве
и в бассейне Адзьвы (на Харуте под 67° с. ш.). На Урале известен
в верховьях Маньи. Далее в Сибирь северная граница распространения идёт
к Берёзову и по северной границе бассейна реки Надым. Есть сведения, что
трёхпалый дятел проникает вниз по Оби до Салехарда (Дерюгин, 1898), пови-
димому гнездится в бассейне Таза и Елагуя (Скалой и Слудский, 1936) и на
Курейке. Далее есть в бассейне Нижней Тунгуски и бассейне Вилюя. О р. Яне
точных сведений нет. На Индигирке до лесотундры (примерно до селения
Аллаиха, т. е. севернее, чем до 70° с. ш.). Восточнее северная граница
направляется к Анадырю, причём верховья левых притоков Анадыря остаются
вне области распространения этого дятла. Огибая с востока Марково (не доходя
Крепости), граница сворачивает на юг и, идя вдоль гор, ограничивающих
■бассейн р. Пенжины с востока, спускается к Пенжинской губе. Далее область
распространения трёхпалого дятла такова: восточная граница — побережье
Охотского моря, Сахалин и частично Японское море, на юге — с.-в. Корея,
окраины Манчжурии, Кентей, Хангай, Танну-ола. Минуя Алтай, граница
идёт к Барнаулу, южнее Томска и Тары и к среднему течению Урала
(в Башкирском заповеднике редкая, но правильно гнездящаяся птица, Ки-
риков), отсюда к реке Суре, к Москве, Рославлю к Беловежской пуще, сев.
Польше. Зимний ареал обширнее. Встречается к югу до Калуги, Тулы,
Воронежа.
Характерпребывания. Почти во всех работах отмечен как
оседлый. На деле гнездящийся и кочующий, в зимнее время смещающийся довольно
далеко от гнездовых мест к югу.
Биотоп. Обширные и глухие леса северного типа. В Европейской части
СССР населяет хвойные и смешанные леса, где особенно охотно выбирает
тенистые, сырые и порою болотистые места, еще более охотно держится
на пожарищах и гарях, вообще на тех участках леса, где особенно много мертвого
леса. В европейской части ареала ближе к южной границе распространения
избирает более глухие и наиболее обширные участки леса, тогда как
севернее, где леса более глухи и более обширны, оказывается предпочтение тем,
где лесная глушь нарушена пожарами и имеется много мертвого леса или
где этот мёртвый лес появился в результате разработки лесных массивов
человеком.
В Сибири основное местообитание трёхпалого дятла—лиственничная тайга,
хотя он гнездится и в смешанных лесах; в низовьях Оби встречается изредка
и в ивовых зарослях (Дерюгин, 1898). В бассейне р. Анадыря часто селится
в тополевых лесах, попадается также и в березовых лесах, а в лиственничных
редок; гнездится также и в таловых зарослях, причём выбирает высокостволь-
ТРЕХПАЛЫЙ ДЯТЕЛ
571
ные заросли, у которых основания стволов достаточно толсты и корявы (Пор-
тенко, 1939). В Вост. Сибири предпочитает пожарища, и в ненаселённых
пространствах с нетронутыми лесами более редок, нежели там, где
девственный характер лесов нарушен деятельностью человека. Во внегнездовое время
чаще встречается в тальниковых зарослях, попадается и в затопленных лесах.
Численность. В
Сибири, кроме самых южных
частей ареала, очень обыкновенен,
местами многочисленен. В
Европе более обычен в северных
частях ареала.
Экология*
Размножение. Рано гнездящийся дятел.
Первая барабанная трель
отмечена на Анадыре 5 февраля.
Постукивание было слышно до
27 мая. В это время трёхпалые
дятлы очень оживлены, стреко-
чат и кричат протяжно,
несколько напоминая этим малого
пёстрого дятла. Дупла
устраивает чаще всего в лиственницах,
живых или обгоревших, и даже
в лиственничных пнях.
Устраивает гнёзда и в сухих
прогнивших стволах ели. В Башкирии
гнездо найдено в безвершинной
сухостойной березе (Кириков).
Высота дупел от земли
небольшая, иногда даже менее одного
метра, иногда до 6 м. Диаметр
дупла от б до 14 см, глубина
дупла (до подстилки) 20—30 см.
Толщина слоя подстилки (по
одному гнезду) 6 см.
Длительность насиживания неизвестна.
Откладывает от 3 до 5 и даже
€ белых яиц; размеры их 21,3—
28,3x16,8—19,3лш (Хортлинг).
В окрестностях Витебска
молодые птицы, недавно вылетевшие
,из гнезда, наблюдались 18 июля.
В Лапландии у Муониониска
:(Плеске, 1886) яйца найдены 28 июня. На Сев. Урале пять птенцов
было найдено 12 июня, и 1 июля—гнездо с тремя птенцами (два самца и
одна самка), у которых перья только начали выходить из пеньков.
Яйцекладущая самка добыта у Денежного Камня—25 мая. В Башкирии птенцы
в гнезде — 7 декабря (Кириков). На Вилюе оперившиеся птенцы
встречены 13 июля (Воробьев, 1928). На Анадыре (Марково) птенцы в гнезде
найдены 17 июня (Бианки, 1908). Там же найдены птенцы 28 июня (Пор-
тенко, 1939).
Линька. У взрослых птиц происходит в течение июля — сентября,
затягиваясь у запоздавших особей до октября. У одного июльского экземпляра
*{от 8-го) сменяется 8-е первостепенное маховое, но у большинства других осо-
Рис. 140. Трехпалый дятел.
572
ОТРЯД ДЯТЛЫ
бей в этом месяце меняется 4-е перо и 3-е, встречаются запоздавшие птицы,
у которых в это время сменяется 5-е перо. У августовских птиц меняется
преимущественно 2-е перо, но есть птицы, у которых меняется 3-е и 4-е, а у
отдельных особей и 5-е перо. У некоторых, видимо запоздавших, птиц огсмена 2-го
махового пера происходит в октябре. Линька молодых менее растянута.
В июле встречаются птицы, у которых меняется 8-е, 7-е, 4-е и даже 3-е перо.
Питание. Пищей служат древесные насекомые, муравьи, свежие
семена и ягоды (Эверсманн, 1866). В желудках птиц, добытых зимой на Анадыре,
найдены личинки и частично куколки жуков, ближе — неопределённые яйца
насекомых и представители семейства хризомелид (Портенко, 1939). В
Московской обл. основная пища зимой —личинки короедов. За неполный зимний день
одна птица может ободрать кору с крупной ели, на которой по
приблизительным подсчётам было около 10 тысяч личинок короедов. Пока не будет таким
образом обработано всё дерево, дятел не принимается за новое. Пищу добывают
почти исключительно с помощью долбления. Открыто живущих насекомых ест
не более полутора месяцев в году. В 28 желудках, собранных летом в
Костромской обл., было 1222 экз. насекомых и их личинок, 94°/0 из них добыты
долблением. К безусловно вредным принадлежали 1113 экз. (91 %), из них 72,5%
короеды и их личинки (короед полиграф — 540 экз., типограф — 164 экз., также
личинки усачей), затем личинки златок, долгоносиков, рогохвостов и др.
(Формозов, Осмоловская, Благосклонов, 1950). Птенцов в дупле (во всяком
случае подросших) выкармливает тоже личинками короедов. В желудках двух
птенцов, взятых из гнезда, Кириков обнаружил по одному большому белому
камешку.
Это одна из очень полезных птиц хвойного леса.
Полевые признаки. Несколько мельче, чем большой пёстрый дятел и
выглядит более тёмным. Нет красного цвета на голове, внимательно
приглядываясь, можно заметить желтую (у самца) и седую (у самки) шапочку на голове*
Молчаливая птица.
Описание. Размеры и строение. На ногах 3 пальца. Вершину крыла, как
у большинства пёстрых дятлов, составляют 4-е и 5-е маховые перья, затем идут
3-е и б-е перо. Реже формула крыла 4>5>3>б. Дйина крыла самцов (82) 121 мм —
132 мм, самок (79) 118—129 мм} среднее 124 и 123 мм.
Окраска. У самца верхняя сторона головы золотисто-жёлтая (каждое перо у
основания чёрное, примерно в середине имеет белую поперечную полосу, кончик его жёлтый).
По бокам и сзади жёлтого темени более или менее ясно выраженный седой налёт,
происходящий от того, что концы перьев в указанных частях головы белые. Бока головы и затылок
чёрные, но от глаза назад идёт белая полоса, которая сливается сзади с белым цветом
задней стороны шеи. Другая белая полоса проходит от основания клюва по бокам головы ниже
ушных партий: снизу ее ограничивают черные «усы». От задней стороны шеи вдоль спины
проходит довольно широкая белая полоса, которая иногда прерывается чёрными
отметинками. В остальном спинная сторона тела чёрная или черновато-бурая. Короткие верхние
кроющие хвоста имеют иногда белые окончания. Брюшная сторона тела белая с черными
поперечными полосками по бокам тела в области брюха и с продольным опятнением по бокам
передней части груди. Нижние кроющие хвоста или чисто белые, или с чёрными
поперечными полосами.
Степень развития чёрного цвета на нижней стороне тела весьма изменчива. Маховые
перья, как и у пёстрых дятлов, чёрные (или чёрно-бурые) с белыми пятнами на наружных
опахалах и с тоже белыми, но более крупными, продолговато-округлыми пятнами на
прикрытых частях внутренней стороны внутренних маховых. Верхние кроющие крыла чёрные,
нижние кроющие с белыми и чёрными полосами. Рулевые перья чёрные, но две-три
крайних пары с белыми поперечными полосами, у некоторых особей белый цвет на самой
крайней паре рулевых преобладает.
Степень развития белого и чёрного цвета подвержена сильной индивидуальной
изменчивости, так что при сопоставлении крайних форм можно говорить о существовании у
трёхпалого дятла светлого и тёмного типа окраски (Портенко, 1937). В период линьки,
когда часть белых перьев на спине выпала, обнажается тёмная пуховая часть
соседних перьев, в результате чего общая окраска спины кажется темнее, чем
обычно.
ТРЕХПАЛЫЙ ДЯТЕЛ
573
Тяньшанский трехпалый дятел Picoides tridadylus tianchanicas Buturlin
Picoides tridadylus. Buturlin. Orn. Mon., 1907, стр. 8, Тянь-Шань,
Синонимы. Picoides tridadylus altaicus. Buturlin. Orn. Mon. 1907, стр. 9.
Алтай. — Picoides dzungaricus. Buturlin. Orn. Mon. 1907, стр. 9, сев.-зап. Монголия.
Распространение. Ареал. Область еловых лесов Тянь-Шаня, Алтай.
Прщы из северо-западных склонов Тарбагатая принадлежат, вероятно, к этой
же форме.
Биотоп. Населяет глухие участки однородной тайги — елово-пихтовой,
кедровой или лиственничной. В светлых сосновых борах или в лесу со сколько-
нибудь значительной примесью лиственных пород не держится. Встречается
и на обширных гарях среди тайги. Во внегнездовое время держится выводками
в полосах тайги и на отдельных деревьях, выбегающих иногда далеко от
сплошного леса на пустынные плоскогорья. В таких случаях отдельные выводки
встречаются иногда рядом с каменкой-плешанкой. Однако необходимое
условие для такого нахождения — сравнительная близость густой тайги.
Характер пребывания. Более или менее оседлая птица.
Численность. Во многих местах Алтая это один из самых
многочисленных дятлов. Близ Телецкого озера, в долине Чулышмана, в
ю.-в. и с.-з. Алтае редок. На зиму численность их по всем данным
не увеличивается заметно, существует небольшая прикочёвка с севера
(другая форма).
Экология* Размножение. Мало изучена. На Абакане, в
северо-восточных предгорьях Алтая гнездо с уже крупными, начавшими оперяться,
птенцами найдено 24 июня, вылет молодых в северных предгорьях Алтая приходится
около половины лета. В лиственничниках Улаганского плоскогорья — метров
на 1000 выше — летающие выводки встречены недели на три позднее, 4и5ав-
густа. На горах гнездование его установлено в обгорелых и засохших на
корню стволах.
Линька. Молодые птицы заканчивают линьку к концу первой декады
сентября. Старые линяют дольше. Начинается линька у взрослых птиц еще
в июне (есть экземпляр от 7 июля, у которого 7-е маховое меняется) и тянется
до половины сентября. У экземпляров от 1 августа 4-е маховое перо
короче 2-го, у экземпляра от 14 сентября второе маховое едва только начало расти.
Питание. В желудках двух дятлов, добытых в Тувинской обл. в конце
января и в начале февраля, найдены короеды — Pityognes chaleographus (Tep-
новский, 1950).
Описание. Размеры и строение. Похож на номинальную форму, но темнее
и крупнее. Длина крыла самцов (28) 122—132 мм, самок (10) 123—127 мм в среднем 128 и
125 мм. Окраска. В общем похож на других трёхпалых дятлов, но темнее. Бока зоба
и груди, брюхо, бока туловища густо покрыты тёмными пестринами, белого на спине мало,
чёрные «усы» широкие, белые пятна на маховых мелкие, белый поперечный рисунок на двух
крайних рулевых часто уже, чем чёрные полоски. В зимнее время несколько светлее, чем
летом, но в зимнем наряде темнотой оперения сильнее отличается от других форм, чем
летом.
Камчатский трехпалый дятел Picoides tridadylus albidior S t e j n.
Picoides albidior, S t e j n e g e r. Proc. U. S. Nat. Nus. 1885, стр. 321j Камчатка.
Распространение. Леса Камчатки, оседлая птица.
Биология. Не изучена.
Описание. Самая светлая форма. Низ и крайние рулевые чисто белые, подхвостье
белое.
Систематические замечания. Трёхпалый дятел обладает большой индивидуальной из*
менчивостью. В результате незнания пределов этой изменчивости было описано много
подвидов. В настоящее время можно только сказать, что на севере (особенно на северо-восток
574
ОТРЯД ДЯТЛЫ
от Сибири) трёхпалые дятлы несколько светлее, чем на юге, но выяснение систематического
положения их затрудняется тем, что зимние экземпляры значительно светлее летних
«чёрных» птиц. Возможно, в дальнейшем камчатская форма будет слита с северной формой,,
а птицы горных лесов Тянь-Шаня с горной формой Европы.
Альпийский трехпалый дятел Picoides tridactylas olpinus Brebtn.
Picoides alpinus. В г e h m. Handb. Naturg. Vog Deutschl., 1831, стр. 194, Швейцария*
Распространение. Ареал. В СССР — Карпаты, как водораздел, так и
северные их склоны. Кроме того, горные части средней Европы: Швейцария,,
Бавария, Австрия (Тироль), Чехословакия, Югославия (Босния, к югу до
Сербии), Болгария, Румыния и южн. Польша. Возможно в южн. Франции
(Савой и Юра).
Характер пребывания. Оседлая птица. Биотоп. Хвойные
леса в горах. Численность. Довольно редок.
Экология» Не изучена. По наблюдениям в Зап. Европе спаривание —
в марте — апреле, иногда даже раньше, яйца, числом 4—5, откладываются
в конце мая — начале июня. Размеры яиц, продолжительность насиживания,
равно как и длительность всего гнездового периода, неизвестны. В
выкармливании принимают участие оба родителя, наиболее интенсивно кормят в конце
первой половины дня (в это время прилетают с кормом каждые 5—10 мин.)*
Описание. По общей расцветке очень сходен с тянынанским подвидом, быть может
даже несколько темнее. По размерам не отличим от номинальной формы.
РОД ПЕСТРЫЕ ДЯТЛЫ DENDROCOPOS KOCH, I816
Тип Picus major
Важнейшие синонимы. Dryobates. В о i e, 1826, тип Picus pubescens; Den-
rodromas. Каир, 1829, тип P. leucotos; Leuconotopicus. M a 1 h e г b e, 1845, тип Р. stric-
klandi; Dendrotypes. Cabanis et Heine, 1864, тип P. macei; Xylocopus. Caba-
nis et H e i n e, 1864, тип Р. minor; Dendrocoptes. Cabanis et Heine, 1863, тип
' P. medius.
6. Большой пестрый дятел Dendrocopos major L.
Picus major. Linnaeus. Syst. Nat, изд. X, 1758, стр. 114, Швеция.
Синоним. Picus Cissa. Pallas. Zoogr. Rosso-Asiat., 1811, стр. 412, «Россия
и Сибирь», пот. emend. •
Распространение. Ареал: от Канарских и Британских о-вов и от
Марокко до Курильских о-вов, Японии, Южн. Китая и Хайнаня. К северу
до 70° с. ш. в Скандинавии, до Канина п-ова, в Азии — бассейны Вилюя
и Нижней Тунгуски, Кавказ и Крым. Характер пребывания.
Оседлый, но совершает кочёвки. Биотоп. Леса разных типов.
Численность. Многочисленный вид почти по всей области распространения.
Подвиды и варьирующие признаки. Заметно выраженная изменчивость по форме и
величине клюва, в северной части ареала имеет индивидуальный и сезонный характер. В
основу описания географических форм положена быть не может. Южные формы, как правило,
мельче северных, имеют более стройный и иногда более короткий клюв и темнее северных
по окраске. Известно 18 подвидов: D. т. major Linnaeus, 1758 — самый крупный и
(после камчатского) самый светлый подвид. Вост. Европа и вся сев. Азия без
Камчатки; D. т. candidus Stresemann, 1919 — несколько мельче — Румыния,
возможно Венгрия, Украина; D. т. pinetorum В г е h m, 1828 — центр. Европа и
Франция, к югу до Албании, Македонии и Турции, мельче номинальной формы, клюв стройнее;
D. т. anglicus H а г t е г t, 1900 — Англия (без Ирландии), от предыдущего подвида
отличается буроватым низом; D. т. hispanus S с h 1 u t е г, 1908 — Испания и Португалия,
брюшная сторона тела ещё темнее; D. т. canadensis Кое-nig, 1889 — Канарские
о-ва, мелкая форма, брюшная сторона с буроватым налётом; D. т. mauritanus Brehm,
1855 — Марокко; D. m. numidus Mai he г be, 1842 — сев. Алжир и Тунис; D. т.
БОЛЬШОЙ ПЕСТРЫЙ ДЯТЕЛ 575
harterti А г г i g о n i, 1904 — Сардиния, очень близок к испанской форме; D. т. par-
roti Hartert, 1911—Корсика, очень близок к центральноевропейской форме;
D. т. italiae Stresem.ann, 1919 — Италия, мельче среднеевропейской формы и клюв
стройнее; D. т. praealpinus Burg, 1921—сев. Италия, сомнительная форма; D. т.
tenuirostris В u t и г 1 i п, 1906 — Крым и Кавказ, мельче номинальной формы с более
тонким стройным клювом; D. т. poelzami Bogdanow, 1879 — Ленкорань и Талыш,
мелкая темная форма; D. т. tianschankus В u t и г 1 i п., 1910 — Тянь-Шань, сомнитель-.
ная, слабо отличимая от номинальной, форма; D. т. kamtschaticus В u t и г I i п, 1883 —*
Камчатка, самый светлый подвид; D. т. faponicus Seebohm, 1883 — от Сев. Кореи
до Курильских о-вов, характеризуется супротивными пятнами на второстепенных махах;
Карта 98. Распространение большого пестрого дятла Dendrocopos major
I — D. m. major, 2— D. m. candidus, 3—D. m. pinetorum, 4— D. m. anglicus, о — D. m. hisparnus^
6 — D. m. canariensis, 7 — D. m. mauritahus, 8 — D. m. numidus, 9 — D. m. harterti, 10 — D. m. parroti,
II —. D. m. italiae, 72-— D. m. praealpinus, 73 — D. m. tenuirostris, 14—D. m. poelzami, /5— D. m.
tianshanicus, 16 — D. m. kamtschaticus, 17 — D. m. japonicus, 18 — D. m. hondoensi9, 19 — D. m. paph-.
lagoniae; a — граница области распространения, б — не вполне установленная граница, г — переходная
зона между номинальным и другими подвидами, д —места залетов.
D. т. hondoensis Kuroda, 1921 — Хондо (средняя и южная часть), Цусима, Южн. Ко^
рея; D. т. paphlagoniae, Kummerlowe etNiethammer — Малая Азия, клюв
тоньше, чем у среднеевропейского дятла, крыло длиннее. Возможно к этому же виду следует-
причислить и дятлов группы cabanisi, населяющих ю.-в- Азию.
Обыкновенный большой пестрый дятел Dendrocopus major major L.
Синонимы. Picus brevirostris. Reichenbach. Handb. d. spec. Orn. Seas., 1854,
стр. 365, Иртыш. — Dendrocopus major mongolus, Lonnberg. Ark. f. Zool., 1909,^
стр. 24, Кяхта. —Dendrocopusmajor kirghizorum. Б у т у р л и н.Орнитол. вест., 1910,стр.202,^
Семипалатинск. — Dendrocopus major sibiricus. Бутурлин, Там же, 1910, стр. 201, Олек-.
минск. — Dryobates major rossicus. Domaniewski. Ann. Zool. Mus. Pol., 1925, стр. 81,,
Саратовская губ.
Распространение. Ареал. Принадлежит Вост. Европе и сев. Азии
до её восточных пределов. На« Кольском п-ове северная граница распростра-.
нения проходит от Варангер-фиорда, примерно по северной границе леса,
однако в Хибинских горах большой пестрый дятел не обнаружен (Шиба-<
нов, 1927). Далее граница идет через Белое море, видимо по северной границе
леса (включая и Соловецкие о-ва) и на Канинском п-ове поднимается к северу-
до р. Чижи и м. Конушина (т. е. севернее 67° с. ш.). На Печоре граница.
576
ОТРЯД ДЯТЛЫ
проходит по криволесью, спускаясь на Урале несколько южнее, по
нижней Оби распространён по всей лесной области (по р. Казым его во всяком
случае много — Шухов, 1915), по Енисею до устьев Курейки. Далее к востоку
область распространения включает в себя бассейн Нижней Тунгуски и-бассейн
Вилюя. Граница проходит, видимо, по водоразделу р. Алдана и Яны
(Верхоянский хребет) и далее к р. Гижига, на юг идет берегами Охотского моря,
и по нижнему течению Амура (или несколько западнее) к Советско-Манчжурской
границе; лесной областью Забайкалья выходит в Монголию, охватывая Кентей
и Хангай. В с.-з. Монголии, так же как в южн, и ю.-в. Алтае, большого
пестрого дятла нет.
Южная граница распространения выходит на Иртыш севернее р. Бух-
тармы, лесами и рощами сев. Казахстана выходит на р. Илек, огибает
Волжско-Уральские степи с севера и по пойме Волги спускается почти до самых
устьев этой реки. Далее к западу граница распространения номинальной формы
дятла не ясна, так как южная часть Вост. Европы входит в область
распространения украинского большого пёстрого дятла D. т. Candidas*
Границу между этими двумя формами следует, повидимому, проводить от Саратова
на восток, севернее Чернигова к Карпатам, затем вдоль этих гор на северо-
запад к Балтийскому морю, так что о-в Борнхольм и Скандинавский п-ов
принадлежат к области распространения номинальной формы. Во вне-
гнездовое время дятлы появляются значительно южнее гнездового ареала,
а также и севернее. Дятлы, найденные на п-ове Канин (середина августа),
очевидно не гнездятся в этих не подходящих для них местах, а прикочевали
с юга. Залетают в Анадырский край (Марково), на совершенно безлесные
Командорские о-ва, появляются в Уссурийском крае, Манчжурии, в южн,
Алтае и на Тарбагатае, Как далеко проникают на юг в Вост. Европе —
неизвестно, так как определить к какой форме (major или Candidas) принадлежат
дятлы, добытые во внегнездовое время на Украине, невозможно.
Характер пребывания. Считается оседлой, на самом деле
гнездящаяся бродячая птица. Во внегнездовое время появляется (сначала
молодые, а затем и старые птицы) в местах, где она не гнездилась, севернее
своей гнездовой области, залетает иногда в степные местности и продвигается
довольно далеко к югу. Молодые птицы, повидимому, более подвижны, чем
старые. Постепенное передвижение к югу начинается в августе, но массовое
происходит в октябре и даже в ноябре. Возврат в старые местообитания
протекает незаметно и приходится на февраль — март.
Биотоп. В гнездовое время населяет преимущественно
высокоствольные леса и рощи, где есть деревья, пригодные для устройства дупел. Наиболее
благоприятны для него южные массивы смешанно-хвойного леса,
характеризующиеся обилием мягких лиственных пород, и в частности осины в сочетании
с елью и сосной (Формозов, Осмоловская и Благосклонов, 1950). На территории
Европейской части СССР не делает различия между лиственными и хвойными
лесами, хотя дремучих ельников избегает (Сушкин, Башкирия, 1897), а в
окрестностях Вязьмы и Ржева предпочитает сосновые боры. В центр, и Вост. Сибири
связан во время гнездования главным образом с сосновыми лесами настолько,
что местами в тайге голос его служит указанием присутствия сосны (И.
Поляков, 1873), чистых лиственничных лесов избегает (Штегман, 1930). На Алтае,
где встречается спорадично, также в сильной степени связан с распространением
сосны, а также и пихты. Возможно это связана с тем, что основные массивы
кедровых и лиственничных лесов Горно-Алтайской обл. мало поражены
вредителями, и дятел не находит здесь достаточного количества корма летом,
тогда как пихта сильно заражена вредителями, напенной гнилью и другими
болезнями (Фолитарек и Дементьев, 1938). Гнездится и в берёзовых
насаждениях. В горьгна Алтае поднимается примерно до высоты 1000, редко до 1200 м
БОЛЬШОЙ ПЕСТРЫЙ ДЯТЕЛ 577
и не доходит до верхней границы леса, тогда как в Саянах встречается
всюду равномерно, кроме верхней зоны криволесья. В ю.-з. Забайкалье и
Кентее преимущественно в нагорной тайге, изредка залетает и в долины, где
есть высокоствольная урёмная растительность. Во внегнездовое время
появляется в мелких лесах, где вовсе нет дуплистых деревьев, в речных долинах
и в фруктовых садах, даже среди больших городов. Из гор спускается в долины.
В безлесной Кулундинской степи был встречен дятел, который то прыгал
по земле, то лазал по телеге, то неуклюжими прыжками передвигался по крыше
покрытого соломой хлева (Иоганзен, 1907). В тундре (Канин)
придерживается домов и берега, где много плавника, однако его можно видеть и на
земле, и взбирающимся по ветвям ивняка.
Численность. В большинстве мест, где он встречается, это
обыкновенная на гнездовье птица и почти всюду численно превосходит других дятлов.
Ближе к северной окраине ареала становится редким. В смешанно-хвойных лесах
Финляндии — 1 пара на 1км2 (Пальмгрен, 1930), в Чернухинском районе Горь-
ковской обл. 37,5 пар на 1 км2 (Шапошников, 1938), в дубравах Харьковской обл.
(другой подвид) — 4 пары на 1 о*2(Компаниец, 1940). В горах Хангая уступает
количественно трехпалому дятлу. Во внегнездовое время численность его во
многих местах зависит от урожая семян сосны: птицы в большом количестве
скопляются в тех местах, где урожай семян сосны больше.
Экология. Размножение. С приближением гнездового времени
становится много крикливее и подвижнее. В это время часто можно слышать
«трель», состоящую из быстро следующих друг за другом ударов по сухому
дереву. После первой порции сливающихся в один сплошной треск ударов,
дятел выжидает некоторое время (2—*3 минуты), затем повторяет «трель» в том
же месте или перелетев на ближайший подходящий для того сук. Разгар
подобного тока в Калужской обл. бывает в апреле. Иногда удаётся наблюдать,
как после «трели» к самцу подлетает самка, происходит спаривание, и самец
прекращает свой треск. Начинается подобный ток в средней полосе обычно
в конце марта, в мае бывает реже, летом как исключение (Филатов, 1915).
В средней полосе СССР «трель» дятлов можно слышать уже в теплые дни
февраля и даже раньше. В Ильменском заповеднике, с 11 февраля по 2 мая
(Ушков). На Печоре «трель» до середины июля. Крики, погоня за самками и
драки в среднем Поволжье и Южн. Урале начинаются с первых чисел апреля,
в окрестностях Якутска — возбуждённые крики, указывающие на
приближение гнездового времени, отмечены с первых чисел марта (Иванов, 1929). На
Алтае (Телецкое озеро) самцы, добытые в конце марта, имели ещё небольшие
семенники (длина 3 и 4 мм), только в середине апреля семенники увеличились
до б мм. Спаривание дятлов в Полесье наблюдалось 17 апреля (Федюшин,
1927). В общем, начало брачного периода следует относить к середине апреля.
Для устройства гнёзд использует чаще всего осину, как наиболее мягкое
дерево, реже ольху и еще реже берёзу. Известны также гнезда в дуплах дуба
и лип, а также на соснах. В общем (включая и южные формы), большой
пёстрый дятел использует для устройства гнездовых дупел до 29
древесных пород (Благосклонов, 1949). Известен случай, когда дятел устроил
своё гнездо в безлесной местности (на берегу р. Хобда), выдолбив дупло
в отвесном склоне канавы вокруг стога сена (Зарудный, 1888). Для
долбления дупла дятел использует либо совершенно сухие (до 40% по
Благосклонову), либо повреждённые деревья. Лишь в виде исключения он
занимает использованное уже ранее дупло, обычно делает его заново или
расширяет естественное дупло (одно дупло у озера Миассово в течение 5 лет
занималось одним и тем же дятлом, Ушков). Основой для дупла большого пёстрого
дятла может послужить также и маленькое дупло, использованное раньше
малым пёстрым дятлом. Устраивает гнёзда и в дуплянках. Дупла делаются на разг
37 Птицы СССР, т. I
578
ОТРЯД ДЯТЛЫ
личной высоте, от нескольких десятков сантиметров (Благосклонов) до 20 м
(Циммерман), чаще всего 2—5 м. При случае, если дерево поражено
грибом, дятел устраивает входное отверстие в дупло так, чтобы оно несколько
прикрывалось им (Сушкин, 1897)» Глубина дупла 28—35 см, леток круглый,
диаметром 5—5,5 см. Подстилкой служат сильно измельченные кусочки
древесины (Ильменский заповедник, Ушков).
Согласно К. Н. Благосклонову, гнездовый район большого пёстрого
дятла довольно велик. Гнездо от гнезда располагаются не ближе 160 м. Однако
И. Б. Волчанецкий (1927) находил в южн. Зауралье по нескольку дупел
в одной старой сосне. За кормом, согласно Благосклонову, птицы
отлетают до 125 м от гнезда. Самка приносит корм обычно через каждые
три—пять минут и успевает покормить птенцов три раза, прежде чем
прилетит с кормом самец (Филатов, 1915). Яиц в кладке обычно б, реже 5, как
исключение 7. Размеры яиц одной кладки: 27x20, 27x19, 27x19, 27 х
X 17 мм. Колебания размеров от 25 до 27 мм (дайна), от 19 до 21 мм (ширина).
По Ушкову, средние размеры 25,0 х 16,0 мм. Цвет блестяще-белый.
Откладывание яиц начинается в конце апреля и продолжается в мае. Длительность
насиживания примерно 12—13 дней. В конце мая—начале июня появляются
птенцы, во второй половине июля большинство птенцов уже лётные, хотя в
отдельных гнёздах они задерживаются до августа. Более точные сведения по
разным местам следующие: в Минской обл. и в Южн. Урале первые яйца
откладываются в конце апреля, в Ленинградской обл. — в середине мая; в
Псковской обл. полные, но не насиженные ещё кладки находят в конце мая. В
десятых числах июня много гнёзд уже с птенцами, писк которых слышен за 100
шагов. Следовательно, насиживание продолжается не более 14 дней. В конце
.июня молодые вылезают уже по временам из гнезда. В Чувашии много лётных
молодых наблюдается с самых первых чисел июля, но там же найдено 1 июля
дупло с птенцами (Першаков, 1932). В бассейне реки Летки (приток Вятки)
находили нелётных птенцов в конце июня. В окрестностях Костромы можно
встретить летающих молодых дятлов в конце июня — начале июля. В бассейне
р. Суры в средних числах мая был отмечен крик птенцов в дуплах, а
самостоятельные молодые наблюдались во второй половине июня. В Орловской обл.
25 мая наблюдали дупло с совсем молодыми птенцами, а 5 июня найдено гнездо
с птенцами, которые были почти совсем оперены. Близ Калуги совершенно
оперившиеся птенцы найдены в гнезде 28 июня, а 19 и 22 июня там же найдены
гнезда с более молодыми птенцами, 20 июня добыт лётный птенец. В
окрестностях Переелавля-Залесского 20 июля птенцы большого пёстрого дятла
уже совершенно оперились, но с сильно недоросшими первостепенными
маховыми перьями, и сидели ещё в гнезде. В Белоруссии лётные птенцы
наблюдаются уже 15 июня, в это же время птенцы покидают гнёзда и в Башкирии,
тогда как в окрестностях Карачева вылет наблюдался несколько позднее —
в конце июня. Взрослые птенцы, выглядывавшие из дупел, в окрестностях
Бугуруслана, замечены 13 июня. Вылет молодых дятлов в южн. Башкирии
(д. Сосновка) отмечен 17 июня (Кириков). В Ильменском заповеднике (Южный
Урал) кладка 20—29 мая, птенцы появляются между 4—13 июня, оставляют
гнездо между 30 июня и 15 июля. В южн. Зауралье писк первых птенцов
слышен в начале июня, в начале — середине июля птенцы ведут
самостоятельный образ жизни. Однако, на р. Хобдо Н. А. Зарудным гнездо (в канаве)
с птенцами на взлёте найдено 19 июля. В Кулундинской степи гнездо с тремя
хорошо оперёнными птенцами найдено 2 июля, хотя, как правило, в
окрестностях Красноярска в конце июля молодые дятлы уже лётные. В окрестностях
Якутска 23 июня найдено пять молодых птиц, возраст которых был
приблизительно семь дней. Обычно вылет молодых происходит здесь в первой
половине июля. В Минусинском крае, 29 июня наблюдались молодые вполне
БОЛЬШОЙ ПЕСТРЫЙ ДЯТЕЛ 579
самостоятельные, а в горах, на высоте примерно 1500 м, найдены птенцы
в гнезде 11 июля. Птенцы в дупле, особенно когда они несколько подрастут,
ведут себя очень крикливо, их можно услышать шагов за сто. Старые птицы
ведут себя около гнезда при появлении человека или довольно безразлично
или встречают и провожают наблюдателя громким пиканьем, заглушающим
писк птенцов. Вылетает из дупла в присутствии человека старая птица обычно
молча, и начинает тревожно кричать лишь отлетев на несколько десятков
метров. Первые 25—30 дней по вылете из гнезда выводок держится вместе, и
старики кормят молодых (отлично уже летающих), затем молодые птицы переходят
к самостоятельному образу жизни и начинают бродячую жизнь, переселяясь
обычно туда, где корма для них больше. В осеннее время дятлы присоединяются!
иногда ^смешанным стайкам синиц и бродят по лесу с ними, представляя собой
как бы вожаков этих стаек.
Линька. Смена крупного оперения крыльев и хвоста начинается в июне,
иногда в июле и заканчивается в ноябре, иногда раньше; как исключение —г
в декабре. В Башкирии добыты экземпляры с меняющимся 4-м маховым пером,
между 15 августа и 15 сентября, 3-е перо меняется с б сентября по 14 октября
и 2-е перо с 15 сентября по 15 октября. В Московской обл. 6-е перо
меняется в августе, 4-е в октябре, 3-е и 2-е в октябре и ноябре. В Вост.
Сибири добыты птицы с меняющимся 8-м маховым в июне и июле, в те же
месяцы меняется и 7-е. У южной границы распространения подвида линька
происходит несколько раньше. А именно: известен экземпляр из Курской обл..
от 3 августа с меняющимся 2-м маховым (у другого экземпляра от 4 марта
меняется 5-е маховое), тогда как 3-е перо в окрестностях Олекмы
сменяется 9 сентября. Рулевые сменяются во второй половине лета, средняя
пара рулевых в октябре. Смена мелкого кроющего пера (по экз. из Дмитриева
Курской обл. и из Переелавля-Залесского Владимирской обл.) начинается
в марте и продолжается до декабря. В октябре и даже в ноябре можно встретить
птиц, уже закончивших линьку, а в апреле — ещё не начавших. Молодые
птицы начинают смену маховых перьев вскоре по вылете из гнезда. Седьмое
и 8-е маховые меняются в августе и даже раньше, в сентябре — октябре
линька заканчивается. В это же время исчезают и последние остатки красных
перьев на темени молодой птицы. Последовательность смены нарядов: гнез-
довый — первый годовой — второй годовой и т. д. Вопрос о линянии
рамфотеки остается неясным.
Питание. Животная (насекомые) и растительная (семена) пища. По
наблюдениям Зарудного (1888), зимой большие пёстрые дятлы едят даже падаль.
Как исключение, склевывают яйца мелких певчих птиц (Кириков). В весеннее
время, в короткий период лёта майских жуков, большой пёстрый дятел
истребляет их в большом количестве. В летнее время питается главным образом
муравьями и их «яйцами» и совершенно не долбит. В местах, мало пораженных
вредителями леса, последние составляют незначительный процент в питании
этой птицы. В лесах, пораженных подкорными вредителями, дятел истребляет
их в большом числе. Отмечено (Карамзин, 1907), например, что, питаясь гусе,-
ницами пахучего древоточца, дятел сам пропитывается их отвратительным
запахом. На стволах, покрытых сетью ходов заболонника, или где велика
плотность личинок усача, дятел отбивает кору большими участками; встречаются
стволы пораженных вредителями дубов, почти нацело очищенные дятлом, or
коры. В местах/ сравнительно слабо пораженных вредителем, очищает
отдельные деревья более тщательно, в сильно пораженных — его внимание
привлекают главным образом места наибольшого скопления вредителя. Своим
длинным языком может достать короеда (Xyloborus saxenseni) из хода,, глубиной
в 9 см, причем пробитая клювом воронка равна 2 см (Шевырев, 1892). В
сосняках большой пёстрый дятел питается преимущественно муравьями и семенами
37*
ОТРЯД ДЯТЛЫ
сосны. В зимнее время, как правило, переходит на растительную пищу. Ест
семена сосны, ели и лиственницы, семена можжевельника, орехи (лещину),
жолуди. В его желудке попадаются иногда ягоды малины, черёмухи, крушины
и рябины. Ранней весной, как только начнется весеннее сокодвижение, дятлы
систематически «кольцуют» деревья и пьют
древесный сок. Последний служит весьма
важным компонентом питания большого
пёстрого дятла, так как это время совпадает с
наиболее голодным периодом жизни этой
птицы. На кольцевание деревьев дятел
затрачивает около трети времени, которое он тратит
на добывание пищи. Изредка дятел «кольцует»
деревья и летом. Кольцуются ели, берёзы,
пихты, клёны, в общем не менее 14 пород
деревьев (Осмоловская, 1946).
«Кольцуя» дерево, дятел делает
горизонтальные ряды пробоин в его коре, ждёт
появления сока и потом поочередно
прикладывает клюв к каждому отверстию. Затем он
делает небольшой прыжок кверху и повторяет
то же самое. В результате на стволе остаются
параллельные ряды ранок, охватывающие
дерево полными или, чаще, неполными
кольцами. На одном дереве могут кормиться таким
образом две, иногда даже три птицы
одновременно. Для разбивания шишек сосны и ели
дятел устраивает «станок» или «кузницу». С
этой целью он выдалбливает в коре или сухой
вершине дерева продолговатый глубокий
желобок или ямку шириной в 3— 3,5см. Шишка
вставляется в этот «станок» и раздалбливается
в нём. На обработку каждой шишки
затрачивается от 2 мин. 40 сек., до 8 мин. 20 сек.
(Ушков).
Хозяйственное значение.
Определить хозяйственное значение большого
пёстрого дятла по характеру его питания
затруднительно. Пища его разнообразна, и в
«? зависимости от обстоятельств отдельные
компоненты её играют различную роль. Здоровых
Рис. 141. Большой пестрый дятел, деревьев большой пёстрый дятел никогда не
долбит. Долбление дятлом на вид здорового
дерева служит верным указанием, что данное дерево поражено вредителем.
Выдолбленные дятлом дупла на следующий год в значительном проценте
занимаются мелкими дуплогнездниками (синицы, мухоловки, пеструшки, скворцы,
сычи и др.) и таким образом служат для привлечения в лес этих полезных
птиц. «Кольцевание» не приносит деревьям вреда, нанесённые птицей небольшие
ранки, как правило, заживают очень хорошо. Исключение составляют
сравнительно редкие случаи, когда дятлы пьют сок на одних и тех же «излюбленных»
ими стволах деревьев. Относительно вредным большой пёстрый дятел может
быть признан лишь в тех сосновых насаждениях, всё существование которых
зависит от нормального осеменения. Во всех других случаях дятел (по
характеру питания) в большей или меньшей степени полезная птица, в худшем
случае, безразличная. Устройством дупел в деревьях он везде полезен.
БОЛЬШОЙ ПЕСТРЫЙ ДЯТЕЛ
581
Полевые признаки. Легко узнаётся как довольно крупный, пёстрой
расцветки дятел. Нижняя сторона тела у взрослых птиц без розового налёта.
Молодые имеют красное темя; самцы — красный затылок; взрослые самки
с черной головой. Как правило, большой пестрый дятел держится на
стволах деревьев, реже его можно видеть на толстых сучьях в более или
менее горизонтальном положении. Обследуя дерево, продвигается по стволу
винтообразно, прыжками, иногда почти до самого комля. Очень редко можно
увидеть дятла подвесившимся, подобно синице, к тонким концевым веточкам,
чтобы склюнуть гусеницу или сорвать шишку. Заметив наблюдателя, обычно
перемещается на противоположную сторону, постоянно наблюдая из-за
ствола за опасностью. Кричит чаще всего на лету или только сев на дерево.
Держится, как правило, одиночно, зимой присоединяется к стаям синиц*
В местах концентрации вредителей собирается небольшими группами. Муравьев
собирает в основном на земле и лишь случайно на стволах деревьев, в это время
он молчалив и почти не выдаёт своего присутствия.
Описание. Размеры и. строение. Длина тела самцов и самок (100)
225—265 мм, размах крыльев 425—470 мм. Формула крыла чаще всего 4>5>3>6>7>2,
иногда 2-е больше 7-го. Клюв в зимнее время короток и широк и только к вершине резко
заострен. Весной клюв растёт и становится длиннее, к июню — июлю принимает стройную
форму, в результате обновления узкой вершинной части. К зиме удлиненная узкая часть
клюва стирается вследствие усиленного долбления. Длина крыла самцов (261) 135—150 лш,
самок (201) 135—150 мм, среднее 143,4 мм и 142,2 мм. Длина клюва самцов и самок
21—30,5 мм. Вес самцов (7) 70—96 г, самок 79—97 г, среднее 89,7 и 89,6 г.
Окраска. Молодая птица отличается от старой несколько более тусклой и
грязноватой расцветкой оперения. На брюхе слабо выражена полосатость. Более ясно выражена
полосатость на плечевых перьях и больших (внутренних) кроющих крыла. Верх головы
красный, задняя сторона головы и затылок чёрный.
Взрослая птица, самец. Верхняя сторона головы чёрная с синеватым блеском, лоб
буровато-белый. Поперёк передней части затылка яркокрасная полоса. Бока головы
(полоса под глазом и щёки) белые. Снизу они отграничиваются широкой чёрной полосой,
идущей от подклювья к затылку. В области затылка полосы сливаются друг с другом,
ограничивая сзади красную полосу на затылке. Сверху на задней стороне шеи видны два белых
пятна. Спина и надхвостье чёрные, с синеватым блеском. Брюшная сторона тела, кроме
задней части брюха и подхвостья, зимой чистобелая, летом в той или иной степени бурая,
у отдельных экземпляров почти шоколадная. По бокам передней стороны груди от зашейка
спускается довольно широкая чёрная полоса. Задняя часть брюха и подхвостье красные.
Маховые перья чёрные с белыми пятнами на внутренних и внешних опахалах (за
исключением первого и второго маховых). Верхние кроющие крыла чёрные, но самые внутренние
из среднего и большого ряда, так же как и плечевые перья, белые. Две средние пары
рулевых перьев чёрные, обычно без металлического оттенка, смежная с ними пара чёрная с
белым кончиком и с белой предвершинной полосой, следующие (внешние) две пары у
основания чёрные на конце белые с черными поперечными полосами. Соотношение белого и
чёрного цвета на рулевых перьях сильно варьирует. Радужина красная или розовая, клюв
тёмносерый, ноги зеленовато-серые. Щетинки у основания клюва чёрные. Самка отличается
от самца отсутствием красной полосы на затылке.
Украинский большой пестрый дятел Dendrocopos major Candidas Stres.
Dryobates major candidus. Stresemann. Anz. Orn. Ges. Bayern, 1919, стр. *0,
Румыния.
Распространение* Ареал. Румыния и юго-зап. часть СССР, возможно
Венгрия. Северная граница проходит в СССР от Карпатских гор на восток
примерно к Саратову, непосредственно примыкая к области распространения
номинальной формы дятла. По пойменным лесам Волги проникает на гнездовье
почти до самых устьев реки. Между Волгой, с одной стороны, и рекой Иловлей
и нижним течением Дона, с другой, его нет; в пойме названных рек есть,
а западнее, в Донецком бассейне, доходит к югу примерно до Сталино. Включая
в себя долину р. Самары, область распространения подходит к Днепру и
спускается к югу по его пойме по крайней мере до плавней. Оставляя к югу Вла-
582
ОТРЯД ДЯТЛЫ
димирское лесничество и включая Чёрный лес, граница поднимается несколько
к северу, по Днестру спускается почти до устьев реки и, проходя водоразделом
Днестра и Прута южнее Кишинёва, выходит из пределов нашей страны.
В негнездовое время этот дятел встречается далеко к югу от гнездовой
области, доходя до Ростова и Кубани, до Мариуполя и до берегов Чёрного
моря.
Характер пребывания. Гнездящаяся и бродячая птица. В
окрестностях Камышина молодые одиночные птицы появляются в
несвойственных им ранее местах уже в конце июля. В это же время начинаются кочёвки и
в Подольской обл. В конце августа дятлы приближаются к Мариуполю,
в начале сентября появляются под Херсоном; в конце февраля и в марте
возвращаются в свою гнездовую область. ,
Биотоп. Лиственные и смешанные леса, также и сосновные боры, но
реже. На пролёте во всех степных садах и байраках, в садах городов, может
быть встречен в степи на телеграфных столбах.
Численность. В большинстве мест наиболее обыкновенный из
дятлов; в Днепропетровской обл., как и всюду ближе к южной границе ареала,
довольно редок.
Экология- Размножение. Гнёзда устраивает преимущественно
в осине, а также в берёзе, сосне, грушевом и других деревьях, иногда в ольхе
и очень редко в дубе. Высота гнезд над землей от 2 до 10 м. В качестве
исключения на одной осине можно найти несколько насиживающих пар.
Откладывает 5—6 чистобелых с мелкозернистой скорлупой яиц. Длина их от 27,5
до 28,3 мм, ширина от 18,8 до 20,1 мм (Сомов, 1897). Форма эллипсоидальная,
но встречаются то более шарообразные, то более удлиненные яйца. В
Харьковской обл. полные ненасиженные кладки бывают в начале и середине мая,
я в конце мая и в начале июня встречаются уже только что вылупившиеся
птенцы. В конце июня птенцы вылетают из гнёзд (Сомов; 1897). В окрестностях
Киева гнёзда с первым отложенным яйцом были найдены 21 мая, в тот же день
в некоторых дуплах были птенцы, птенцы в гнёздах найдены здесь и
6 июня.
Незадолго до вылета из гнезда (в Днепропетровской обл. в десятых
числах — середине июня) птенцы выходят иногда из него и ползают по коре
дерева вокруг дупла, забираясь почти до вершины дерева. Самостоятельные
молодые в Киевской обл. появляются с 23 июня. В Подольской обл. в
конце первой декады июля, чаще в последней трети этого месяца (Портенко,
1928). В Киевской обл. вылет из гнёзд происходит^ в конце июня. В
Молдавской ССР птенцов в гнёздах находили в конце июня. «Трель» дятла можно
слышать с января, чаще всего весной и изредка в июне-^июле.
Линька. Смена крупного оперения взрослых птиц наступает в конце
июля. Птенцы заканчивают линьку в первый взрослый наряд к началу сентября
(Сомов, 1897).
Питание. Питаются преимущественно насекомыми, в том числе в
большом количестве муравьями. Из растительной пищи употребляют главным
образом сосновые семена, также лесные орехи, косточки слив и даже древесные
тючки. В южных лиственных лесах продолжает питаться животной пищей и
в зимнее время (короеды, личинки златок, гусеницы древесницы въедливой),
добывая её долблением (Померанцев и Шевырёв, 1910). Для долбления сосновых
шишек устраивает специальные «кузницы».
Описание. Размеры и строение. Очень сходен с предыдущей формой, но
имеет более тонкий и стройный клюв, а также возможно несколько меньшие размеры, Длина
крыла самцов и самок (7) 139—144 мм, среднее 141,6 мм.
Окраска. Сходен с номинальной формой, но нижняя сторона почти никогда не
^бывает столь чисто белой.
БОЛЬШОЙ ПЕСТРЫЙ ДЯТЕЛ 583
Кавказский большой пестрый дятел Dendrocopos major tenuirostris But.
Dendrocopus major tenuirostris. Buturlin. The Ibis, 1906, стр. 412, Кутаиси.
Синоним. Dryobates major kurae. Laubmann. Orn. Jahrb. Bd. 26, 1915,
стр. 145, Тбилиси.
Распространение. Ареал. Широко распространен в лесах по всему
Кавказу и Закавказью, кроме Талыша; также в Крыму. На Сев. Кавказе примерно
до северной границы лесов.
Характер пребывания. Гнездящаяся и частично кочующая
птица. Далёких кочёвок не совершает, но на зиму спускается частично с гор
в долины, появляясь в конце октября и исчезая в середине марта.
Биотоп. Лиственные, смешанные и реже хвойные леса гор, а также
пойменные леса, сады и рощи. В Закавказье встречается на гнездовье до
2000—2100 м — полосы субальпийского редколесья. Но более обычен в
средней полосе гор. В Кавказском заповеднике гнездится в полосе
широколиственных и нижней зоне темнохвойных лесов. В полосе верхнего предела
лесов редок. В ю.-з. Закавказье предпочитает еловые и пихтовые леса,
в меньшей степени смешанные. В Армении —в высокоствольных лесах. Зимой
в Предкавказье держится в широколиственных лесах и в садах. В Крыму —
горные леса, откуда осенью откочевывает в сады.
Численность. Обычная птица. Почти повсеместно превосходит
численно других дятлов. Только местами в дубовых лесах уступает малому
пёстрому дятлу.
Экология. Размножение. Биология этой формы дятла изучена
недостаточно. Гнёзда устраивает в дуплах разнообразных деревьев, в
частности и в пихте. Гнёзда с 5 свежими яйцами найдены в Крыму в
окрестностях Симферополя 3 мая (Молчанов, 1906). Птенцов, готовых к вылету из
дупла, наблюдали в Аджарии 11 июня. В Сванетии на высоте 1400 м
оперившиеся птенцы в гнезде б июля. Самостоятельные одиночные молодые
встречаются с начала или с середины июля. В середине июля в Армении
наблюдали молодых дятлов, кочующих со стайками' синиц.
Линька. Смена крупного пера начинается в конце июня и заканчивается
в сентябре — октябре. Экземпляры с меняющимся 7-м маховым пером
добыты 1 и 14 июля. Шестое перо меняется 2 июля и 1 августа, 4-е —16 и 25
августа, 3-е и 2-е в середине и конце сентября.
Питание. Пища кавказского дятла разнообразная, но преимущественно
животная (различные насекомые на разных стадиях их развития).
Растительная пища состоит из различных плодов, ягод, иногда молодой кукурузы. На
Сев. Кавказе поедает в большом количестве жолуди (даже и летом). В
связи с урожаем и неурожаем жолудей в Кавказском заповеднике изменяется и
численность дятлов (Аверин и Насимович, 1938). Известен случай, что дятел
вытащил из гнезда уже оперившегося птенца малой мухоловки и начал
долбить ему череп. Уничтожает иногда яйца открыто гнездящихся птиц (зяблика).
В парках Крыма долбит зимою кипарисовые шишки. В зап. Грузии наносит
чувствительный вред бамбуковым насаждениям, выдалбливая в их стволах
значительные дыры. Повидимому это новое для дятла дерево вводит его в
заблуждение, так как звук пустоты при ударе означает для дятла повреждение
дерева вредителями (Чхиквишвили, 1941). «Кольцевание» деревьев для дятлов
Крыма и Кавказа отмечено лишь в виде исключения.
"" ^.Описание. Размеры и строение. Отличается от "номинальной формы
•более тонким и стройным клювом, который ещё стройнее, нежели у D. т. Candidas.
В среднем несколько мельче других подвидов. Длина крыла самцов (38) 131—145, самок
(21) 131—144, среднее 137 и 136 мм. Клюв самцов и самок 23—27 мм.
Окраска. В среднем брюшная сторона тела и бока головы более темные, чем у
номинальной формы. Попадаются, особенно в вост. Грузии и Азербайджане, экземпляры
с очень густо окрашенным низом, приближающиеся по этому признаку к талышской форме.
584
ОТРЯД ДЯТЛЫ
Талышский большой пестрый дятел Dendrocopos major poelzami BogcL
Picus Poelzami. Богданов. Птицы Кавказа, 1879, стр. 121, Ленкорань.
Распространение. Ареал. Южнокаспийские районы от Ленкорани и
Талыша на западе до Астрабада на востоке. Одиночное нахождение — в Кара-
каралинском районе в Туркмении (Дементьев, 1950).
Характер пребывания. Оседлый, зимой частично спускается
с гор в долины и появляется в большом количестве в садах Ленкорани.
Биотоп. Лиственные леса гор и низменностей. В горы поднимается до 2 500 м.
Численность. Обычен, даже многочисленен. Зимой в долинах чаще,
чем летом.
Экология. Размножение. В низменных местах приступает к
насиживанию в конце марта. Гнёзда устраивает в дуплах дуба, бука, вяза и др.
Л и н ь к а не изучена. П и т а н и е не изучено.
Полевые признаки. В общем те же, что и тонкоклювого дятла, но сразу
узнаётся по более тёмной окраске брюшной стороны (около Тбилиси и в Заката-
лах встречаются однако среди D. т. tenuirostris экземпляры, сильно
приближающиеся по своей расцветке к D. т. poelzami).
Описание. Размеры и строение. Мельче и темнее всех остальных форм
большого пестрого детла. Клюв тонкий, стройный. Длина крыла самцов (16) 121—130 мм,
самок (24) 120—127 мм, среднее 125 и 123,6 мм. Клюв около 21— 26 мм. Формула крыла
4>5>3 и 5>4>3.
Окраска. При общем сходстве окраски с другими большими пёстрыми дятлами
очень хорошо отличается темно окрашенным, шоколадно-бурого цвета низом. Ушные партии
и даже плечи тоже тёмнобуроватого цвета. Белый цвет на верхней стороне развит очень мало,
черные полоски на боковых рулевых широкие;
Тяньшанский большой пестрый дятел Dendrocopos major tianshamcus But.
Dendrocopus major tianshanicus. Бутурлин. Орнитол. Вест. 1910, стр.200,
Каменная р. у Джаркента.
Распространение. Ареал. Леса средней части Тянь-Шаня; Каменная река
у Джаркента, Китай-пикет на р. Кунгесе, Юлдус. Зимой до Иртыша.
Характер пребывания. Оседлая птица. Совершает кочевки, частично
выходя за пределы гнездового ареала. Биотоп. Горные леса Тянь-Шаня
(подробнее неизвестно). Численность. Сравнительно редок.
Экология не изучена.
Описание. Размеры и строение. Несколько мельче номинальной формы.
Длина крыла самцов и самок (10) 139—144 мм, среднее 141 мм, клюв 24—26 мм.
Окраска. Сходен окраской с номинальной формой. На крыле несколько больше
белого, чем приближается к следующему виду. Реальность существования подвида требует
дальнейшего подтверждения.
Камчатский большой пестрый дятел Dendrocopos major kamtschaticus Dyb.
Picus major kamtschaticus. D у b о w s k i. Bull. S. Z. Fr., 1883, стр. 368, Камчатка.
Распространение. Ареал. Гнездится на Камчатке. Залетает на
Анадырь, в окрестности Гижиги и на о-в Беринга. Характер
пребывания. Оседлая птица, на кочёвках залетает в другие местообитания. Кочёвки
начинаются в конце августа. Биотоп. Берёзовые и лиственные леса.
Высокоствольным лесом долин рек поднимается довольно высоко вверх. Зимой
залетает в пояс стланцовых кустарников. Численность. Обычная птица.
Экология не изучена. Питание: в сентябре — октябре расклёвывает
орехи кедрового стланца (Аверин, 1948).
Полевые признаки» В общем те же, что и у номинальной формы.
Описание. Размеры и строение. Длина крыла самцов 139 и 142 мм, самки
135 и 137 мм, вес самца 110г. Окраска. Самый светлый подвид. Брюшная сторона
чистобелая, много белого на хвосте и крыльях. На второстепенных маховых супротивные
белые пятна на внутренних и внешних опахалах (признак, сближающий этот подвид со
следующим). ,
БЕЛОКРЫЛЫЙ ДЯТЕЛ 585>
Уссурийский большой пестрый дятел Dendrocopos major japonicus Sccb.
Picus major japonicus. Seebohm. The Ibis, 1893, стр. 24, Хоккайдо.
Синоним. Dendrocopus japonicus tscherskii. Бутурлин. Наша охота, 1910,.
стр. 53, Ханка.
Распространение. Ареал. Бассейн Уссури, Сахалин, Курильские о-ва
(Уруппу-Кунашири), Хоккайдо, северная часть Хондо, с.-в. Корея (зимой
до центральной). Частично в
Манчжурии.
Характер пребывания.
Оседлая птица, совершает небольшие
кочёвки.
Биотоп. Широколиственные леса
речных долин и гор. В меньшей степени
тайга.
Численность. В лиственных
лесах самый многочисленный дятел, в
тайге очень редок.
Экология. Р а'з м н о ж е н и е ! не
изучено, б июня найдено гнездо с совсем
маленькими птенцами. В конце второй
трети июня попадаются семьи с хорошо
летающими молодыми (Спангенберг,
устное сообщение). Линька. У
экземпляра от 7 июля недоросшие 4-е и 8-е
маховые, нет средних рулевых. П и-
т а н и е не изучено.
Описание. Размеры и строение.
Мельче номинальной формы. Длина крыла
самцов (6) 130—142, самок 82(1)5—135, среднее
134,7 и 132 мм, клюв 23—31 мм. Вес самца (1)
76 г, самок (3) 70,7—105 г.
Окраска. Отличается от номинальной
формы наличием супротивных белых пятен на
второстепенных маховых, расположенных на
внутренних и на внешних опахалах. Белый
цвет на плечевых развит меньше. На вершинах
первостепенных маховых всегда — и у
взрослых— имеются белые пестрины;
7. Белокрылый дятел
Dendrocopos leucopterus Salvadori
Picus (Dendrocopus) leucopterus. S a 1 v a d о г i.
Atti. R. Acad. Sci., Torino, 1871, стр. 129,
Яркенд, Кашгария. Рис. 142. Уссурийский большой пестрый
Распространение. Ареал. Сред- дятел,
няя Азия от западного Узбоя и
Аральского моря до подножья Памира и высокого Тянь-Шаня, Синьцзянь до Урумчи
в южной Джунгарии и Лоб-нора; Характер пребывания. Оседлая
птица, во внегнездовое время совершает небольшие кочёвки в пределах
гнездового ареала. Биотоп. Тугайные заросли речных долин и
саксауловые заросли в пустыне.
Численность. Распространён спорадично, но в подходящих местах,
обычен.
586
ОТРЯД ДЯТЛЫ
Подвиды и варьирующие признаки. Четыре
подвида, различающихся размерами и главным
образом степенью развития белого цвета в
оперении: £). /. leucopterus Salvador i, 1871 —
восточный Туркестан, по распространению белого
цвета на крыле и хвосте занимает промежуточное
положение между двумя другими; D. I. leptor-
hynchus Severtzow, 1875 — от Кара-Тау до
Балхаша и Кульджи, к югу до Пянджа; белый цвет
на крыле развит в меньшей степени, чем у других
подвидов; D. I. albipennis В u t и г 1 i n, 1908 —
Туркмения и прилежащие к ней части Средней
Азии к северу до Кара-Тау; самая мелкая и самая
белокрылая форма (номинальная форма по
распространению на крыле белого цвета стоит ближе
к третьему подвиду — D. I. albipennis, чем к
подвиду D. I. leptorhynchus); D. I. spangenbergi
subsp. nova, окрестности озера Сары-Чилек.
Горный белокрылый дятел Dendrocopos
leucopterus leptorhynchus Severtzow
Picus leptorhynchus. Severtzow. The
Ibis, 1875, стр. 487, Ташкент.
Синонимы: Dryobates leucopterus
leptorhynchus natio korejewi. Зарудный. Изв. Турк.
отд. Русс, геогр. о-ва, 1923, стр. 95, Кульджа.—
Dryobates leucopterus leptorhynchus natio bucharen-
sis. Зарудный, там же, 1923, стр. 97, Хаз-
рет-баба, Горная Бухара.
Распространение. Ареал. От Кара-Тау до Балхаша и Суйдуна (между
Джаркентом и Кульджей). На юге огибает высокогорную область Тянь-Шаня,
включает в себя Ферганскую долину с прилежащими горами (без озера Сары-
Чилек) и Таджикистан до Пянджа, но без Памира. На юго-западе до Кугитанга
и до Самарканда.
Характер пребывания. Оседлая птица, но вне гнездового
времени совершает небольшие кочёвки, залетая в места, где летом не бывает.
Биотоп. Тугаи по берегам/ рек, кишлачные сады, реже арчевники,
*в зап. Памире —заросли ив. Зимой на равнине —заросли саксаула.
Встречается в городских садах.
Численность. Довольно обыкновенная, но всё же
немногочисленная, спорадически распространённая птица.
Экология. Размножение не изучено. Линька у старых с
середины июня до начала сентября. Интенсивная линька 18 июня и 10 июля,
кончают осеннее линяние —25 и 31 июля. 8-е перо меняется 7 июня (Кугитанг)
и 15 августа (р. Или), 5-е перо 18 августа (Искандеркуль), недоросшее 3-е —10
сентября (Айвадж). Птенцы начинают линять в первый осенний наряд
с 21 июня и позже —в конце июля, начале августа. Питание не изучено.
Описание. Размеры и строение. Длина крыла самцов (9) 124—130 мм,
самок (7) 123—129 мм, среднее 126 и 127 мм.
Окраска. Молодые похожи на взрослых, отличаются несколько большим
развитием белого на рулевых и на маховых перьях, а также тем, что красный цвет у них не на
зашейке, а на темени (у молодых самок красного цвета может и не быть). Взрослая птица
по общему характеру окраски напоминает большого пёстрого дятла, но белый цвет сильнее
развит, особенно на крыльях. Белый цвет лба шире, чем у большого пёстрого дятла, изредка
ближе ко лбу бывает глинисто-охристый налёт. Щеки почти всегда чистобелые. На маховых
перьях много белого. На наружном опахале второго, третьего и следующих маховых белого
цвета не меньше, чем чёрного. Белые пятна на второстепенных крупные и на внутренних
второстепенных сливаются в продольную окраинную полосу. Внутренние средние и
большие верхние кроющие крыла, а также плечевые — белые. Крыло снизу выглядит белым
Карта 99. Распространение
белокрылого дятла Dendrocopos leucopterus
1 —D. 1. leucopterus, 2 — D. I. leptorhynchus,
3 — D. I. albipennis, 4 — D. I.
spangenbergi; a — граница области распространения,
г — граница между горным и белокрылым
подвидами.
БЕЛОКРЫЛЫЙ ДЯТЕЛ
587
с чёрными полосами, либо полосатым равномерно. Красный цвет брюха доходит до задней
части груди. Верхняя сторона чёрная. Рулевые чёрные. На крайней паре их две или одна
белых поперечных полосы и белый кончик, ма второй паре одна полоса (или нет вовсе) и
белый кончик.
Равнинный белокрылый дятел Dendrocopos leucopteras albipennis But;
Dendrocopus albipennis. Бутурлин. «Семья охотников», 1908, «15 марта»,
стр. 108, Теджен.
Синонимы: Dendrocopus major transcaspius. Z а г u d n у. Orn. Mon. atsber. 1908,
стр. 66, вост. часть Закаспийского края» — Dryobates leucopteras albipennis natio jaxarten-
sis. Зарудный. Изв. Турк. отд. Русс, геогр. о-ва,
1923, стр. 100, низовья Сыр-Дарьи.
Распространение. Ареал. Туркмения от
западного Узбоя (Ясха) до Теджена, Мургаба и
примерно до Термеза на Аму-Дарье (в Кугитанге
уже другая форма). На север — до низовий Аму-
Дарьи, Кызыл-Кумы, о-в Барак на Аральском
море, Сыр-Дарья от низовий до предгорий Тянь-
Шаня (Кара-Тау и у Самарканда — уже другая
форма). Характер пребывания.
Оседлая птица. Во внегнездовое время совершает
небольшие кочёвки в пределах гнездового
ареала. Биотоп. Тугайные прибрежные заросли.
В пустыне — саксауловые леса.
Численность. Спорадичен в связи с
особенностями распространения в равнинной
части Средней Азии древесной растительности.
Довольно обыкновенен в восточных Кара-Кумах —
там, где сохранились черные саксаульники. По
Аму-Дарье между Чарджоу и Келифом, в связи
с вырубкой естественных насаждений, редкая
птица. С другой стороны на Сыр-Дарье между
Яны-Курганом и Джусалами обыкновенен.
Экология. Размножение. Пара от
пары селится довольно далеко, и в местах, где
леса разделены на небольшие острова, каждый
такой островок занят лишь одной парой, если
лес довольно велик — по паре с
противоположных концов и пара в середине. Иногда гнездится
на одиночно стоящем дереве и летает оттуда в
ближайший лесок, занятый другой парой
(Зарудный, 1896). Дупла выбиваются преимущест- Рис. 143. Белокрылый дятел,
венно в стволах разнолистного тополя или
осокоря и в саксауле, также в фруктовых деревьях. Высота дупел над землей от
одного до пяти метров. Близ Репетека было найдено гнездо, вырытое в откосе
сильно уплотненного песчаного бугра. Дно норы было покрыто толстым
слоем из кусочков коры и древесины, сухих листьев и стручков пустынных
растений (Зарудный, 1896).
Кладка в конце марта —> начале апреля. Число яиц 4—^6, реже 7. На
Мургабе найдены слабо насиженные яйца 28 марта и 15 апреля ещё свежая
кладка, в Джулеке 4 ненасиженных яйца найдены в середине мая (Спанген-
берг, 1936). Птенцы, готовые к вылету, найдены около Карабента на Теджене
в конце мая, около Репетека 11 июня (Зарудный, 1896) и лётные молодые из
Репетека 15 мая и из района колодца Чакашлы 9 июня (Дементьев, 1950).
Линька. Молодые линяют во взрослый наряд в июле — августе.
Молодая самка из дельты Аму-Дарьи от 27 июля имела 6 первостепенных рулевых
588
ОТРЯД ДЯТЛЫ
махов старых, остальное крупное перо новое, 7-е сильно недоросшее. Взрослые
также начинают линьку в июле и кончают её в августе. Самка от 27 июля:
5 первостепенных маховых старые, 6-е только растёт, 7-е недоросшее.
Питание не изучено.
Полевые признаки те же, что и у горной формы. Встретив что-либо
обращающее их внимание, поднимают громкий крик, барабанят и долбят по дереву.
Мало уживчивые, раздельно держащиеся птицы.
Описание. Размеры и строение. Длина крыла самцов (7 экз.) 124—131 мм,
самок (4) 124—133 мм, среднее 126 и 126 мм.
Окраска. В общем похож на предыдущую форму, но в оперении хвоста и крыльев
больше белого. Наружные опахала 4—5—б-го маховых перьев, за исключением кончиков,
почти сплошь белые. Сильно развито белое окаймление внутренних второстепенных махов,
в результате чего на сложенном крыле образуется большое белое пятно. Испод крыла
представляется на большей площади белым, иногда с 2, редко с 3 чёрными полосами.
Крайние рулевые в вершинной части либо белые с чёрными полосами или пятнами, либо с двумя
широкими белыми полосами и белым концом.
Темный белокрылый дятел Dendrocopos leucopterus spangenbergi subsp. nova
Распространение. Ареал. Очень небольшая территория по горной реке
Ходжа-ата, вытекающей из озера Сары-Чилек в южных отрогах Чаткальского
хребта (урочище Аркит).
Рис. 144. Темный белокрылый дятел.
Характер пребывания. Оседлая птица.
Биотоп. Горные широколиственные леса с преобладанием ореха,
значительной примесью фруктовых деревьев и с незначительной примесью арчи
и ели. Численность, Весьма обычная птица.
Экология. Размножение не изучено. Гнёзда устраивает видимо
в дуплах ореха. 27 июня Е. П. Спангенберг нашёл гнездо (в дупле ореха) с
готовыми к вылету птенцами. По всей вероятности это запоздалый выводок.
Линька. С конца июля по октябрь, наиболее интенсивная линька
проходит в июле — августе, у одного экземпляра, добытого 28 июля, 7-е маховое перо
СИРИЙСКИЙ ДЯТЕЛ
589
меняется, а у другого экземпляра в это же время меняются 5-е и 10-е перья*
2-е недоросшее перо обнаружено у двух экземпляров от 22 октября. Как
исключение, смена первого пера затягивается до января. Экземпляр от 28 июля
не имеет второй пары (от середины) рулевых перьев, а у другого от того же
числа не доросли вторая пара от середины и крайняя пара. У молодой птицы
от 26 июля седьмое маховое примерно 2/з нормальной длины.
Диагноз. Наиболее тёмная форма вида. Лоб тёмный глинисто-охристый,
нижняя сторона тела с глинисто-охристым налётом, особенно интенсивным
в области горла и зоба. Подобный налёт может быть на второстепенных маховых
и на рулевых. Щеки белые. Развитие белого цвета на крыле сильно варьирует.
Крайние рулевые как правило с двумя поперечными белыми полосами (одна
из них не полная) и с белыми концами. Длина крыла самцов (11) 123—128 мм,
самок (б) 123—128 мм, среднее 125 и 125 мм. Клюв тонкий, стройный.
Тип: № R —25564, самка, 26 июня 1935, урочище Аркит; котип: № R —
55584, самец, 15 октября 1937, урочище Аркит.
Исследовано 18 экземпляров от июня, июля и октября. Молодые (2 экз.)
несколько светлее старых. Называю эту формулу дятла в честь Е. П. Спан-
генберга, который сообщил также и приведенные здесь биологические сведения.
Систематические замечания. Вновь описанная форма занимает очень узкий ареал, но
как хорошо охарактеризованная заслуживает выделения в самостоятельный подвид. Вся
серия в целом и каждый её представитель хорошо отличаются от любого представителя
других подвидов.
Что касается вида в целом, то он, будучи близок к большому пёстрому дятлу,
хорошо отличается тем, что на внутренних второстепенных маховых белый цвет образует
не супротивные пятна, но продольное окаймление.
8. Сирийский дятел Dendrocopos syriacus Hempr. et Ehrenb.
Picas syriacus. HemprichetEhrenberg. Symb. Phys. Aves, Ливан.
Распространение.
казье, Иран,
Палестина, Сирия, Ирак.
Характер
пребывания.
Оседлая птица.
Биотоп.
Примерно совпадает с
биотопом южных форм
большого пестрого
дятла.
Численность.
Более или менее
обыкновенная, но не
часто встречающаяся
птица.
Ареал: Югославия, Болгария, Малая Азия, Закав-
Карта 100. Распространение сирийского
syriacus
дятла Dendrocopos
syriacus, 2 — D.s. baicaaicus. 3— D. s. transcaucasicus, 4 — D.
s. milleri; a — граница области распространения.
Подвиды и
варьирующие признаки.
Распадается на 4 слабо отли* / — D. s.
чающиеся размерами и
некоторыми
подробностями окраски подвида: D. s. syriacus Hemprich et Ehrenberg, 1838 —Сирия,
Ирак, Иран до Кирмана, Малая Азия, номинальная форма; D. s. balcanicus Gen-
gler et Stresemann, 1919 — Балканский п-ов; D. s. transcaucasicus
Бутурлин, 1910 — Закавказье; D. s. milleri Z a r u d n y, 1909 — Кух-и-туфтан (Иранский
Белуджистан), отличается более тонким клювом.
590
ОТРЯД ДЯТЛЫ
Северный сирийский дятел Dendrocopos syriacus transcaucasicus But.
Dendrocopus syriacus transcaucasicus. Бутурлин. Орнитол. Вестн., 1910, стр. 193„
вост. Закавказье.
Распространение. Ареал. Закавказье и южнокаспийские провинции
Ирана на восток до Астрабада. Детали распространения требуют выяснения.
Характер пребывания. Оседлая птица.
Биотоп. Примерно сходен с биотопом большого пестрого дятла,
держится также на ветлах, зимой переселяется в тростниковые заросли.
Численность. Довольно редок. Около Кюрдамира зимой обыкновенен.
Экология. Размножение не изучено. Известно, что южная форма
откладывает до 11 яиц, максимальный размер их 28 х 19 мм, минимальный 24,4 X
18,5лш(Хартерт, 1912). 22.V.1914 у
г. Бальха (сев. Иран) добыт молодой,
видимо, уже летный, но с недорос-
шими еще рулями и с длиной крыла
109 мм.
Линька начинается в мае.
Экземпляр от 21 июня имеет 9-е
первостепенное маховое недоросшее. У
молодых красный цвет на голове
сохраняется до середины сентября.
Питание не изучено.
Поздней осенью птицы, ночевавшие на
приусадебных деревьях (около
Кюрдамира), перелетают по восходе
солнца в тростниковые заросли и проводят
там в поисках корма целый день,
собирая крупных белых гусениц. К
вечеру возвращаются на деревья
(устное сообщение Спангенберга).
Описание* Размеры и
строение. Формула крыла 4>5^3>6>7.
Длина крыла самцов (7) 120—132, самок (10)
120—134, среднее 124 и 124 мм.
Окраска. Молодая птица
окрашена тусклее взрослой, затылок без красного,
темя красное. Часто бывает красный цвет на
зобе в виде поперечной полосы (последний
признак может быть встречен у большого
пестрого дятла лишь в виде исключения).
Распределение цветов в оперении взрослой
птицы, в общем, как у большого пестрого
дятла. Хорошо отличается тем, что черной
полоски, соединяющей позади уха черную шейную полосу с затылком, нет; поэтому белый
цвет боков головы и шеи сливается в одну сплошную белую полосу. Белый цвет на лбу
шире, чему большого пестрого дятла. Самка отличается от самца отсутствием красного цвета
на затылке.
Балканский сирийский дятел Dryvbates syriacusbaleanlcus Gengl. et Stres.
Dryobates major balcanicus. Gengler et Stresemann. Ang. Orn. Ges. Bayern,.
1919, стр. 2, Македония, Сербия, Болгария.
Распространение. Ареал. Балканский п-ов: Сербия, Македония,
Болгария и Румыния. В СССР обнаружена недавно Ф. И. Страутманом в
Закарпатской Украине. Характер пребывания. Оседлая птица.
Рис
Сирийский дятел.
БЕЛОСПИННЫЙ ДЯТЕЛ
591
Биотоп. В Закарпатской Украине найден главным образом в
фруктовых садах.
Численность. Довольно редок.
Экология. Не изучена. П и т а н и е. По сообщению Страутман^ летом в
желудках — остатки насекомых, главным образом жесткокрылых и муравьев.
Кроме того шкурки груш и семена винограда. В желудке добытой в январе
птицы была масса из раздробленных ядер миндаля. Голос более звонкий, чем,
у большого пёстрого дятла, содержит много высоких нот.
Описание. Размеры и строение. Длина крыла самцов и самок 125—135 мм--
(Хартерт, 1921). Клюв относительно слабый.
Окраска. Отличается от номинальной и от кавказской формы наличием ясно,
выраженных продольных тёмных полос на боках тела.
9. Белоспинный дятел Dendrocopos leucotos Bechst.
Picas leucotos. Bechstein. Orn. Taschenb. 1803, стр. бб. Силезия.
Распространение. Ареал. От Баварии и Австрии до Курильских о-воа
и юго-вост. Азии. К северу до 64° в Норвегии, до Елагуя, Станового хре&га
Карта 101. Распространение белоспинного дятла Dendrocopos leucotos
— D. I. leucotos, 2 — D. 1. uralensis, £ — D. 1. woznessenskfl, 4— D. I. subcirris, 5 — D. 1.
stejnegeri, 6 — D. 1. kurodae, 7 — D. 1. namiyei, 8 — D. 1. sinicus, 9 — D. 1. owstoni, 10 — D. 1.
insularis, 11 — D. 1. fohkiensis, 12 — D. I. quelpartensis, 13 — D. 1. takahashii, Ы — D. 1. carpat-
hicus, 15 — D. 1. Iilfordi; a — граница области распространения, г —переходные зоны между
подвидами, д — места нахождении во внегнездовое время.
и Камчатки. К югу — Австрия, Венгрия, Румыния, изредка Италия и Греция*
Малая Азия, Кавказ, Алтай и юго-вост. Китай.
Характер пребывания: оседлая, местами гнездящаяся ю
бродячая птица.
Биотоп. Осветлённые леса разного состава.
Численность. Довольно обыкновенная птица*
592
ОТРЯД ДЯТЛЫ
Подвиды и варьирующие признаки. Географическое значение имеют степень развития
белого цвета, степень развития продольной пятнистости и ее интенсивность и размеры.
Различают 15 подвидов: D. /. leucotos Bechstein, 1803 — северо-восточные части Европы
и Азия от Байкала до Сахалина; D. I. uralensis Ma 1 h e r b e, 1861 — от Волги до Байкала,
едва крупнее номинальной формы, белого в оперении значительно больше; D. /. wozne*
senskii В u t u r 1 i n, 1907 — Камчатка, самая светлая форма; D. L subcirris S t e j n e-
g e г, 1886 — Кунашири (Курильские о-ва) и Хоккайдо, ясный желтовато-коричневый
налет на нижней стороне тела; £>. I. stejnegeri Kuroda, 1921 — сев. и центр, часть
Хондо, темнее предыдущего; D. Lkurodae G б t z, 1926 — вост. часть Хондо и Сикоку,
форма переходная к следующей, надхвостье с широкими чёрными полосами; D. I. namiyei
S t e j n e g e r, 1886 — юго-зап. Хондо и Кюсю, надхвостье с широкими чёрными
полосами; D. /. sinicusB u t u г 1 i n, 1907 — сев.-вост. Китай (Пекин), сверху как
номинальная форма, снизу как кавказская; D.l. owstoni Ogawa, 1905 — севернее Риу-Киу,
очень тёмный, надхвостье чёрное; D.l. insular is Gould, 1862 — Формоза,
маленький, широкие темные полосы на нижней стороне тела; D. I. fohkiensls В u t и г 1 i n,
1908 — юго-вост. Китай, крупнее предыдущего; D.l. quelpartensis Kuroda et Mori,
1918 — о-в Квельпарт (к югу от Кореи), маленький и тёмный; D. I. tokqhashii Kuroda
et М о г i, 1920 — о-в Даджелет (к востоку от Кореи), клюв меньше и стройнее, белые пятна
на крыльях больше; D. I. carpathicus Бутурлин, 1907, — Буковина, Румыния,
Венгрия, темнее номинальной формы; D.l. lilfordi Sharpe et Dresser, 1871—
Балканский п-ов, Малая Азия, Закавказье, задняя часть спины в белых поперечных полосах.
Обыкновенный белоспинный дятел Dendrocopos leucotos leucotos Bechstein.
Синонимы: Dendrocopus leuconolus ussuriensis. Бутурлин. Изв. Кавк. музея. 1907,
т. Ill, стр. 47, Сицтн.—Dryobates leuconolus ussurianus. Clark. Proc. U. S. Nat. Mus, 1907,
XXXII, стр. 473, «Уссури». — Dendrodromas leuconolus clarkl. В u t u г I i п. Ежег. Зоолог,
музея А. Н., 1908, т. XIII, стр. 234, Аян, no me n e me nda turn. — Dendrodrom as leucono tus
saghalinensis. Yamashina. Tori, 1931 (VII), стр. 31, южная часть Сахалина. —
Dendrodromas leucotos stechowi. Sachtleben. Verh. Orn. Ges. Bayern, 1919, стр. 181, Гродно.
Распространение. Ареал. Северные местонахождения
—Ленинградская, Калининская, Костромская, Горьковская обл. и Татарская
республика. На Каме водится уже другой подвид. Граница между ними
начинается широкой полосой между Вязьмой и Камой, идёт вдоль нижнего течения
Камы и по Волге примерно до Сызрани; несколько южнее Сызрани
сворачивает на запад и юго-запад, огибая с юга Пензенскую и с юго-востока
Тамбовскую и Воронежскую обл., с юга Харьковскую и идя в направлении к
Днепропетровску. Западнее Днепра граница поднимается несколько севернее, огибая
с юга Подолию и Волынь. Центральная Европа от Западной Германии до
Югославии, в Скандинавии до 64° с. ш. Во Франции и Швейцарии случайно.
Другая, отделённая тысячами километров часть ареала начинается от
северной оконечности Байкала, идя на севере видимо по Становому хребту до
Аяна на Охотском море, включает в себя Сахалин, Приморье и Уссурийский
край, Корею, Манчжурию, горы к северу от Пекина, Забайкалье, Кентей.
Зимний ареал в значительной степени совпадает с летним; в Европе
расширяется к востоку за Волгу до Башкирии, к югу до лесистых балок Ергеней около
Сарепты и юго-западнее Днепропетровска. По старым данным (Гебель, 1903)
встречается на Соловецких о-вах и в южной Лапландии (Плеске, 1886).
Регулярно залетает на Шантарские о-ва.
Характер пребывания. Гнездится. Встречается круглый год,
но совершает пространственно ограниченные кочёвки. Весеннее и осеннее
увеличение'численности дятла, указывающее на пролёт, отмечено для Псковской
обл. (Зарудный, 1910). Как говорит Шнитников, он прикочёвывает на зиму
в Минскую обл. (1913). В некоторых местностях белоспинный дятел
отмечен только на кочёвках и зимой, например, в Новосильском районе Тульской
обл. (Харузин, 1926), для очень многих мест Орловской обл. отмечен
только как залётный (Горбачев, 1925), но в Трубчевском и Карачевском
районах это оседлая птица (Суслова, 1935 и 1937). В Козельском районе Калужской
БЕЛОСПИННЫЙ ДЯТЕЛ
593
обл. найден только раз зимой (Генглер и Кавелин, 1909). На территории
зимнего ареала, т. е. по Волге к югу до Сарепты и западнее Днепропетровска, а
также восточнее Волги появляется только зимой.
Биотоп. По преимуществу более или менее светлые лиственные леса,
предпочтительнее те, где много ветровальных старых березовых стволов. Во
многих местах особенно охотно селится (гнездится) в поймах рек. В Минской
обл. населяет почти исключительно лиственные леса, изредка смешанные,
совсем избегает хвойных. В Смоленской обл. охотнее держится в
смешанных сосново-лиственных и елово-лиственных насаждениях (чем больше
лиственных пород в лесу, тем лучше), встречается, однако, и в высокоствольных
чисто хвойных лесах (Огнёв, 1909), встречается и в редком лесу и даже на
отдельных деревьях на лугах (Федарака, 1928). В Псковской обл.—главным
образом лиственные леса. В Рязанской обл. — березовые леса, берёзово-
осиновые с примесью липы, местами смешанные леса. В Мордовской АССР
биотоп — смешанные насаждения и леса речных долин. В Воронежской обл.
смешанные леса и дубравы. Вблизи северной границы распространения
также предпочитает лиственные (урёма, гривы) и широколиственные леса
(Першаков, 1929). В горах Забайкалья заселяет поросшие лиственным лесом
поймы больших рек. По Амуру — преимущественно смешанные леса, чисто
хвойных лесов избегает. В Кентее кроме смешанных лесов также сосновые и
лиственничные. Уссурийский край — широколиственные леса долин и сопок, рощи
манчжурского ореха, со стороны рек довольно далеко заходит вглубь тайги.
Численность. Почти повсеместно уступает численно большому
пёстрому дятлу. В Минской обл. после большого пёстрого самый обыкновенный
из пёстрых дятлов. В Смоленской обл. значительно реже его, но в Вяземском
районе в связи с преобладанием там лиственных пород встречается чаще
большого пёстрого. Вообще говоря, в Белоруссии и в Смоленской обл. это
обыкновенный вид. В Ленинградской обл. редкая птица, не часто встречается в
Псковской обл. и редок в Калининской. Редок в Нижегородском крае (Воронцов, 1935).
Довольно редок в Костромской обл., в Рязанской — довольно обычен, в
Мордовской АССР значительно реже большого пёстрого дятла, а в Калужской обл.
редок. В Орловской и Тульской обл. очень редок. В юго-восточной части
Пензенской обл. нередкая птица, зимой встречается чаще. В Воронежской обл.
обыкновенен, также и Харьковской, однако реже большого пёстрого. В
Черниговской обл. это обыкновенная оседлая птица, обыкновенен он и в Полтавской
обл., однако численно заметно уступает другим пёстрым дятлам области. В
Подолии бывает многочисленен годами. По всему Уссурийскому краю обычная
птица, на Малом Хингане очень част, заменяя там большого пёстрого дятла;
по верхнему течению Амура редок, довольно редок в окрестностях Читы и
по верхнему течению Ингоды; в юго-западном Забайкалье весьма
обыкновенен, обычен в Кентее.
Экология. Размножение. Гнездится раньше других пёстрых дятлов
(особенно чем большой пёстрый), дупла долбит в гнилых берёзовых пнях (Благог
склонов, 1949). Дальность полётов от гнезда за кормом —около 200л*. Число
яиц в кладке 3—5. Размер: (18) 26,3—30,2 х 21— 21,4 мм, среднее 28 х 20 мм
(Нитхаммер, 1938). Длительность насиживания неизвестна. Птенцы выходят
из дупла в конце мая. В Харьковской обл. птенцы на взлёте найдены 11 июня
(Сомов, 1897). В Горьковском крае молодые появляются в начале июня (Сереб-
ровский, 1908). Осенние кочёвки приходятся главным образом на ноябрь
(Шнитников, 1913). В Уссурийском крае нелётные птенцы в гнезде встречаются
в середине июня, хорошо летающая молодая добыта 25 июня. Весенняя
трель его своеобразна: отдельные постукивания сливаются в один общий
протяжный звук, очень напоминающий скрипение от ветра большого сука дерева
(Огнёв, 1909).
38 Птицы СССР, т. Т
594
ОТРЯД ДЯТЛЫ
Линька. Линька крупного пера происходит в течение трёх летних
месяцев, от конца июня до начала сентября. В коллекциях Зоологического
музея Московского университета есть экземпляр от 14 июня, у которого шестое
маховое короче седьмого (Вологодская обл.) и в то же время экземпляры из
более южных мест (Переславль-Залесский) от 9 июля с подобным же состоянием
линьки. Восьмое маховое равно половине нормальной длины у самца от 28 июня
из Вяземского уезда. У самца от 30 июля
из Воронежской обл. шестое
первостепенное маховое равно одной трети своей
нормальной длины. В июле происходит
смена пятого и шестого первостепенных
маховых перьев. Экземпляр из
Ульяновска от 5 июля имеет пятое
первостепенное маховое равное половине нор
мальной длины. С подобным же
состоянием линьки и даже с несколько более
недоразвитым пятым маховым пером
(впрочем четвёртое уже выпало и
начинает расти вновь) есть экземпляр из
Калужской обл. от 2 августа. Недорос-
шее пятое маховое имеют также
экземпляры от 9 августа из Горьковского края.
С более продвинувшимся состоянием
линьки (четвёртое маховое несколько не
доросло) экземпляр взрослого самца из
Валдайского района от 21 июля. Из того
же места экземпляр от 2 августа, у
которого пятое маховое перо всё ещё короче
шестого. У остальных августовских птиц
линька в августе продвинулась дальше.
Самка от 5 августа из Мордовии имеет
четвёртое маховое короче третьего.
Меняется третье перо (короче второго) у
самца из Воронежской обл. от 17 августа,
то же наблюдается у самца от 22
августа из долины Суры. Одновременно в
той же области была добыта самка, у
которой меняется второе маховое перо.
Третье перо и одновременно второе
меняется у старой самки от 27 августа из
Боровического района Новгородской обл.
Наконец, у птиц, добытых в начале сентября, меняется второе маховое перо
(8 сентября, Горьковский край, самец). В Воронежской обл. самец с таким же
состоянием линьки добыт двумя неделями раньше — 24 августа. Таким
образом, есть птицы из Горьковскойобл., имеющие то же состояние линьки, как
воронежские, добытые двумя неделями раньше. С другой стороны есть птица из
Переславского района (Владимирской обл.), имеющая то же состояние линьки,
как и птица из Вологодской обл. добытая двадцатью пятью днями раньше.
Молодые птицы линяют несколько раньше старых. Молодой самец из
Воронежской обл. от 21 июня имеет пятое первостепенное перо равное по длине
второстепенным перьям. У молодого самца из-под Горок Бобруйской обл.
шестое перо имеет половину своей нормальной длины и всё крыло в общем не-
доросшее. Молодой самец от 9. VII из-под Переелавля-Залесского имеет
шестое перо недоросшее. А у молодой птицы из Слудского района от 7 августа
Рис. 146. Белоспинный дятел.
БЕЛОСПИННЫЙ ДЯТЕЛ
595>
меняется третье перо (короче второго). У самца от 30 июля (Сычевский район)
четвёртое перо короче второго. У самца от 9 августа из Переелавля четвертое
перо короче третьего. В том же состоянии линьки и молодая самка из
Мордовии, добытая того же числа. В августе встречаются взрослые птицы, у которых
линька закончилась. Линька рулевых перьев происходит в те же сроки, как
и линька первостепенных маховых перьев и в последовательности,
характерной для всех дятлов.
Питание. В течение всех сезонов — в основном насекомые-ксило-
фаги (личинки короедов, усачей, узкотелых златок и др.), в незначительном
проценте —- открыто живущие насекомые. Резких сезонных изменений состава
пищи нет, но во вторую половину лета ест также орехи, жолуди, ягоды
крушины, рябины и черемухи. Пищу добывает только в сильно ослабленных или
погибших деревьях, причем нередко полностью снимает с деревьев кору.
Кроме пользы, приносимой истреблением вредных насекомых, белоспинный
дятел полезен и тем, что «окориваемые» им деревья менее подвержены гниению
и лучше сохраняют все качества хорошего топлива.
Полевые признаки. В общем те же, что и у большого пёстрого дятла, от
которого довольно легко отличается большим распространением белого цвета на
спине. Довольно подвижная и доверчивая птица.
Описание. Размеры и строение. Формула крыла взрослой птицы чаще
4>5>3 и много реже 5>4>3. Длина крыла самцов (36 экз.) 141—153 мм, чаще всего
146—148 мм, самок (37 экз.) 139—154 мм; среднее 148 и 146 мм. Вес самцов 105—112 г,
самок 105 и 106 г. Длина тела самцов (23) 268—302 мм, самок (15) 266—290, размах крыльев
самцов (22) 447—490 мм, самок (14) 457—483 мм.
Окраска. Молодые в первом наряде очень сходны со старыми птицами,
но на верху головы красный цвет имеют только кончики перьев, розовый цвет на нижней
стороне тела несколько слабее выражен (как по интенсивности, так и по протяжению) и
чёрные полоски по бокам тела шире.
Окраска взрослой птицы в основе своей очень сходна с окраской всех пестрых дятлов,
У самца лоб грязновато-белый, иногда с буроватым налетом, особенно в передней части.
Реже лоб чистобелый. Весь остальной верх головы красный, причем основания перьев
темносерые, так что если кожа на голове растянута или птица линяет, этот цвет
выступает наружу, и красный цвет темени оказывается не сплошным. Бока головы, область
вокруг глаза и горло белые. Белая надглазная полоса отделяется от красного цвета
темени более или менее ясно выраженной черной полосой. Широкая чёрная полоса идет
от основания подклювья и, несколько расширяясь, очерчивает с обеих сторон белый
зоб. Грудь от белого с, розоватым налётом до розового цвета; по бокам тела чёрные
наствольные полосы, которые являются как бы продолжением указанной выше чёрной
полосы. В пограничной между областью груди и брюха области чёрные наствольные полосы
бывают расположены не только по бокам, но и по средней линии тела. Иногда наствольная
штриховка заходит также и в брюшную область, но более слабого тона и преимущественно
по бокам брюха. Брюхо и подхвостье красные, иногда розоватые. Зашеек и передняя часть
спины чёрные. Задняя часть спины белая, иногда на белых перьях задней части спины
замечаются чёрные кончики или предвершинные полосы. Верхние кроющие хвоста чёрные.
Маховые перья чёрные с характерными для всех пестрых дятлов белыми пятнами, имеющими
на второстепенных перьях вид поперечных, но не доходящих до стержня пера полос.
Верхние кроющие крыла чёрные, крупные из них с белым опятнением. Нижние кроющие, крыла
белые. Средние рулевые перья чёрные. Прочие все с большим количеством белого, причём-
крайние рулевые могут уже быть названы белыми, но с поперечными чёрными пятнами,
иногда редуцирующимися до недсходящих до стержня пера пятен. Клюв свинцово-серый,
ноги несколько темнее. Самка хорошо отличается от самца тем, что на голове у неё нет
красного цвета, верх головы у самки одноцветен с передней частью спины (т. е. черный). В
остальном самка окрашена так же как и самец.
Сибирский белоспинный дятел Dendrocopos leucotos uralensis M a 1 h.
Picus uralensis. Ma 1 h e г be. Monogr. Pic. I, 1861, стр. 792, Урал. Башкирия»
Распространение. Ареал. Северная граница распространения начинается
в области междуречья Вятки и Камы, охватывает Каму и пересекает Уральские
горы, так что Богословский Урал составляет на меридиане Урала самую се-
38*
596
ОТРЯД ДЯТЛЫ
верную часть гнездовой области. В Западной Сибири граница спускается
несколько к югу, к устью р. Парабель (Скалой, 1932), хотя во внегнездовое время
встречаются экземпляры и много севернее (нерегулярно осенью по рекам Таз
и Елагую; Скалой и Слудский, 1936). В бассейне Нижней Тунгуски не найден.
В верховьях Вилюя и Лены есть. Восточной границей следует считать Байкал.
В Забайкалье — номинальная форма. Южнее Байкала происходит
взаимопроникновение двух географических рас; как предполагал П. П. Сушкин,
возможно даже в области распространения западносибирской формы островное
нахождение колонии, не отличимой от европейских белоспинных дятлов.
Правильнее всего отнести к области распространения западносибирского бе-
лоспинного дятла весь Минусинский край (Минусинскую впадину), а Алтай
причислить к переходной области. В Зап. Сибири южная граница этого дятла
проходит по южной окраине лесостепья и выходит к Уралу примерно на
широте Орск —- Чкалов. На запад от Уральских гор встречается смешанная
популяция (которую всё же вернее отнести к D. I. uralensis). Граница выходит
на Волгу несколько южнее Куйбышева. Зимний ареал, вообще говоря, совпадает
с летним, расширяется на юг в сторону Алтая и, как уже было сказано, во
внегнездовое время (осенью) белоспинные дятлы распространяются также к
северу в область рек Таза и Елагуя.
Характер пребывания» В большинстве мест гнездового
ареала встречается круглый год, но должен быть отнесён к бродячим птицам.
Зимой он появляется и вне этого ареала: южнее, западнее и севернее Волги.
Биотоп. В Башкирской республике предпочитает чисто лиственные
леса, держится также в смешанных, в чисто хвойных отсутствует. В области рек
Самары и Бузулука встречается даже в самых безлесных местах, лишь бы было
несколько деревьев. На восточных склонах Южного Урала его местообитание
берёзовая лесостепь. По Эверсманну (1866), довольно обыкновенен и в хвойных,
но чаще все же в лиственных лесах, нередко в урёмах, а зимой и в садах (в урё-
мах степных рек его нет). По среднему течению реки Урала посещает густые
тальниковые заросли. В юго-восточной части ареала белоспинный дятел
связан с редкими лиственными лесами и зарослями широких речных долин.
Вообще говоря, это птица светлого леса, лиственного или смешанного;
в чисто хвойных участках и в тайге, как правило, не встречается, и по лесистым
горам распространён в вертикальном направлении незначительно. Охотно
селится в поречных рощах и островных лесах.
Экология. Размножение. Спаривание в западных частях ареала
происходит в начале апреля. В это время белоспинные дятлы усердно
перекликаются и барабанят. В кладке 4—6 яиц. 10-го июня в дупле почти совсем
гнилой ольхи на высоте примерно трёх метров от земли были найдены
пищащие птенцы.
Линька. Во второй половине июля попадаются особи, у которых
седьмое первостепенное маховое перо новое, а шестое недоросшее, есть экземпляры
от конца июля с недоросшим пятым маховым, тогда же встречаются птицы с
четвёртым маховым примерно в две трети его нормальной длины. Третье
первостепенное маховое меняется во второй половине июля и начале августа, а в
конце этого месяца и начале сентября меняется второе маховое. Майские
экземпляры не имеют ещё никаких признаков линьки; в конце сентября встречаются
уже полностью перелинявшие птицы. Что касается рулевых перьев, то средняя
пара их меняется во второй половине августа и даже в начале сентября.
Описание. Размеры и строение. Формула крыла взрослой птицы, как и у
номинальной фэрмы, преимущественно 4 > 5 > 3, много реже 5 > 4 > 3. Длина крыла самцов (30)
144—159, самок (21) 144—151 мм, среднее 151 и 148 мм. Вес самцов и самок (б) 10,35—113,8г.
Окраска сходна с окраской номинальной формы, но отличается значительно большим
развитием белого цвета и меньшей интенсивностью опятнения нижней стороны тела. Белые
БЕЛОСПИННЫЙ ДЯТЕЛ
597
пятна на больших кроющих крыла у описываемой формы занимают почти всю длину
открытой части этих перьев (у номинальной формы белый цвет занимает не больше половины
неприкрытой части пера). Большинство малых кроющих крыла в их неприкрытой
другими перьями части в заднем ряду белее с чёрным краем, вследствие чего образованное
ими белое пятно на крыле имеет большие размеры, чем у D. /. leucotos. Последние или
два последних третьестепенных маховых почти сплошь белые. Брюшная сторона тела
в основном более светлая, тёмная штриховка его более слабо выражена: штрихи имеют
меньшие размеры (уже), менее интенсивно окрашены и немногочисленны. В результате
указанных отличий птица с нижней стороны представляется более светлой, а с верхней
стороны с более сильно развитыми белыми полями. Описанные отличия формы uralensis
от leucotos наиболее ярко выражены главным образом у птиц из окрестностей Тюмени и
сев. Казахстана, также из сев. Приуралья. Большинство представителей описываемой
географической формы не обладает всей совокупностью указанных признаков и часто
бывает, что принадлежность их к названной форме определяется либо только по окраске
низа, тогда как верх очень сходен с окраской верха номинальной формы, либо низ
сразу выдаёт подвидовую принадлежность птицы; верхняя сторона тела неотличима
от окраски верха у европейских птиц. Наиболее светлые экземпляры, принадлежащие
к расе D. 1. uralensis, совершенно не отличимы от более тёмных D, L leucotos, особенно
много таких практически неопределимых птиц в пограничной области распространения
двух рас.
Камчатский белоспинный дятел Dendrocopos leucotos wznesenskti But.
Dendrocopus leuconotus woznesenskii. Бутурлин. Изв. Кавк. музея, 1907,
стр. 47, Камчатка.
Слабо изученная форма. Населяет (по Бутурлину), леса желтой берёзы
на Камчатке; редка.
Окраска. Наиболее светлая форма: вторая от середины пара рулевых
на концах помечена белым.
Курильский белоспинный дятел Dendrocopos leucotos subctrrls Stejn.
Dryobates subcirris. S t e j n e ge r. Proc. U. S. Nat. Mus. 1886, стр. 113, Хоккайдо.
Синоним. Dryobates leucotos yamashinae. Bergman n. Ark. f. Zool. 1931,,
стр. 4, Кунашири.
Распространение. Ареал: о-ва Кунашири и Хоккайдо.
Характер пребывания. Оседлый.
Биотоп. Пихтовые леса.
Численность. Довольно редок (Бергман, 1935).
Биология не изучена.
Окраска отличается от номинальной формы более или менее желтовато-бурым
налётом на нижней стороне тела. Подобный налёт заметен также на лбу, ушных партиях,
надхвостье и внешних рулях.
Буковинский белоспинный дятел Dendrocopos leucotos carpathlcus But.
Dendrocopus leuconotus carpathicus. Бутурлин. Изв. Кавк. музея, 1907, стр. 47,
Буковина.
Распространение. Ареал. Карпаты от южных отрогов Семиградских
гор, Буковина, восточные и западные Карпаты, Румыния, Венгрия и восточная
часть Чехословакии.
Характер пребывания. Оседлая птица.
Биотоп. Буковые леса. В чисто еловой тайге нет, но если среди
хвойных встречаются буки, встречается и этот дятел (Страутман, 1948).
Численность. Довольно обыкновенная птица.
Экология не изучена.
Описание. Размеры. Длина крыла (3) 149—152 мм. Окраска. Темнее
номинальной формы. Штриховка сильнее выражена.
598
ОТРЯД ДЯТЛЫ
Темный белоспинныйдятел Dendrocopos leucotos lilfordi Sharpe et Dresser.
Picas lilfordi. Sharpeet Dresser. Ann. et Mag. Nat. Hist. ser. 4, VIII, 1871,
стр. 436, Эпир.
Синонимы: Dryobates leucotos caacasicus. Tschchikwisch wili. Bull. Mus.
Georgie, 1929, стр. 281, Боржоми. — Dryobates lilfordi ponticus. Бутурлин. Птицы
Северного Кавказа, 1929, стр. 14, Хоста и сев.-зап. Закавказье, Чуруксу.
Распространение. Ареал: верховья р. Белой и Закавказье: Сочи, Ба-
тумский район, Манглиз, Боржом, Лагодехи, с. Привольное в Армении. Вне
нашей страны — Балканский п-ов до границы Австрии и устьев Дуная,
Малая Азия, известны залёты в Италию. Характер пребывания.
Рис. 147. Темный белоспинный дятел.
Оседлая птица. Биотоп. Широколиственные леса — дубовые, буковые и
другие. На склоне Главного Кавказского хребта до верхней границы леса (1700л/).
Численность. Редкая птица, но в Чуруксу обычен, чаще других дятлов.
Экология. Размножение не изучено. Линька. У экземпляра от
19 июля пятое перо недоросшее. Питание не изучено.
Описание. Размеры и строение. Длина крыла (9) 142—149 мм, среднее
145 мм. Окраска сходна с номинальной формой, но хорошо отличается тем, что задняя
часть спины вся з чёрных и белых поперечных полосах.
ВЕРТЛЯВЫЙ ИЛИ СРЕДНИЙ ДЯТЕЛ
599
пребывания. Оседлая и частично кочующая птица,
основном лиственные леса, реже хвойные. Сплошных
из-
10. Вертлявый или средний дятел Dendrocopos medias L.
Picus medius. Linnaeus. Syst. Nat., изд. X, 1758, стр. 114, Швеция.
Распространение. Ареал. Зап. и Средняя Европа, кроме Британских
островов, юга Франции и юга Испании; на восток до ю.-з. Швеции, Смоленской
и Воронежской обл.; Кавказ и Закавказье, Малая Азия, северная и
западная части Ирана.
Характер
Биотоп. В
высокоствольных лесов
бегает.
Численность.
Редкая птица, на Кавказе
довольно обычная.
Подвиды и варьирующие
признаки. Для характеристики
географических рас служит
главным образом степень
развития красного и жёлтого цвета в
оперении низа и интенсивность
развития тёмных полос на
груди. Шесть подвидов: D. т.
medias Linnaeus, 1758 —
Европа кроме Пиренейского
п-ова; D. т. lilianae W i t-
h e r b у, 1921 — Катабринские
горы (Пиренейский п-ов),
чёрный цвет верхней стороны чист,
без буроватого оттенка; D. т.
caucasicus В i a n с h i, 1904 —
Кавказ и Закавказье, красный
цвет брюха интенсивнее,
продольная штриховка боков тела
резко выражена; D. т. sane-
tijohannis В 1 a n f о г d, 1879—
юго-зап. Иран (Загрош),
штрихи на боках тела узкие, жёлтый
цвет на нижней стороне тела
образует узкую поперечную полосу; D. т. anatoliae Н а г t е г t, 1912 — Малая Азия, с
сильной примесью желтого цвета на брюшной стороне и развитием темного продольного
рисунка на боках.
Европейский средний дятел Dendrocopos medius medius L.
Распространение. Ареал. В СССР распространён только в самых
западных и юго-западных его частях. Есть в Калининградской обл. и в Литовской
ССР, в остальных странах Прибалтики отсутствует. Далее указан Заруд-
ным (1910) для окрестностей Торопца, Острова и Пскова, но южнее в
окрестностях Витебска и большей части Смоленской обл. его нет, в последней
известен только южнее Рославля. Старые данные о нахождении среднего
дятла в пределах б. Петербургской губ. не достоверны. Восточнее указан
Третьяковым для Калининской обл. (1914), хотя Давыдовым (1896) в Ржевском
уезде не найден. Южнее, в Московской обл., его нет. Есть в окрестностях
Козельска Калужской обл. и на юге Тульской обл. (бывш. Ефремовский уезд).
В Воронежской обл. к востоку до района Боброва включительно. От
юго-восточных частей Харьковской обл. граница распространения идёт в направлении
Азовского моря, до которого не доходит. В прилежащих к Крыму, Николаеву
и Одессе степях его нет, западнее Днепра есть в верховьях Ингула и Ингульца
(Чёрный лес), к западу от Днестра повсеместно. Вне СССР — Балканский п-ов
Карта 102. Распространение среднего дятла Dendrocopos
medius
1 —D. m. medius, 2 — D. m. lilianae, 3 — D. m. sanctijohannis*
4 — D. m. anatoliae; a — граница области распространения, д — не
вполне установленное нахождение птицы.
600
отряд дятлы
до Греции включительно, Швейцария, Франция, кроме юга; к северу —до
Бельгии и юго-зап. Швеции.
Характер пребывания. Для большинства мест гнездящаяся
и кочующая птица. В некоторых местах только зимой (Козельский р.
Калужской обл., Тульская обл. и др.). Время наиболее интенсивных осенних
кочёвок —конец сентября, октябрь и начало ноября (Минская обл., Шнит-
ников, 1913).
Биотоп. Предпочитает чисто лиственные леса: широколиственные
дубравы, широколиственные поймы, а также сосново-дубовые леса. Кроме того,
населяет смешанные сосново-лист-
венные и елово-лиственные
насаждения, чисто сосновых
насаждений избегает (Гомелыцина). В
Воронежской обл. преимущественно
лиственные леса из вяза, дуба и
чёрной ольхи в пойме речных
долин. На юге Белоруссии населяет
также грабовые леса, хотя и менее
охотно нежели дубовые; в Подолии
преимущественно в садах и на
окраинах лесов.
Численность. В
большинстве мест довольно редок,
значительно уступает в численности
большому пёстрому дятлу, только
в чисто лиственных лесах местами
его больше, нежели большого
пёстрого. Более или менее обыкнове-
нен — местами чаще большого
пёстрого и белоспинного дятлов — в
Воронежской обл. В Орловской,
наоборот, очень редок. Между
Днепром и Донцом на широте
Днепропетровска количество его
уменьшается (Вальх, 1911).
Экология. Размножение.
По наблюдениям в Харьковской
обл. средний дятел избирает для
гнездовья преимущественно дикие
плодовые деревья, разбросанные
по вырубкам и лесным полянам.
Дупла располагаются не выше Зг/2 м
и не ниже Р/4 м от земли. Кроме диких фруктовых деревьев для
гнездования используются также и дупла в бересте. Три слабо насиженных яйца
было найдено 24 мая, а кладка из шести ненасиженных яиц 25 мая. Яйца
имеют довольно тонкую скорлупу, обладают хорошим блеском. Размеры
(6) 25,5—26,1 х 19—19,3, в среднем 25,9 х 19,1 мм. Взрослые птенцы на взлете —
6 штук были замечены 22 июня (Сомов, 1897).
Линька. Смена крупного оперения происходит во второй половине лета,
начиная с июня. Птицы от 29 и 30 июня имеют 7-е первостепенное маховое
свежее и в ряде случаев недоросшее (примерно в одну треть нормальной длины).
У экземпляра от 15 июля 5-е и б-е маховые несколько короче нормальной длины,
от 25 августа не доросло 4-е маховое, а экземпляр от 24 августа имеет все
первостепенные маховые (за исключением первого) свежие, но второе маховое выросло
Рис.
Средний дятел.
ВЕРТЛЯВЫЙ ИЛИ СРЕДНИЙ ДЯТЕЛ 60Г
только наполовину. Птицы, добытые вне этого времени (от начала октября по
февраль включительно), полностью перелинявшие. Смена рулевых отмечена
на экземплярах от конца июня по 15 августа. Питание не изучено.
Добывает иногда крупных вредителей древесины (личинок дубового усача, ивового
древоточца).
Полевые признаки. Хорошо отличается от прочих пёстрых дятлов меньшей
величиной (кроме малого пёстрого). Чрезвычайно подвижная, можно сказать,,
непоседливая птица. Крик—очень часто повторяемое «кик-кик-кик-кик»;.
деревья долбит сравнительно редко и слабо.
Описание. Размеры и строение. Формула крыла взрослой птицы обычно-
4>5>3>6. Длина его у самцов (9) 126—132, самок (7) 126—135, среднее 129 и 130 мм.
Окраска. Молодые птицы отличаются от старых несколько более тусклой
раскраской оперения и более широким лбом. Характер раскраски взрослых в общем
тот же как у всех пёстрых дятлов. У самца верх головы красный, но передняя часть темени
и лоб грязнобелые. Бока головы (включая и^надбровную полосу, отделяющую красный цвет
темени от глаза) и низ её белые. Белый цвет боков головы отделяется от белого цвета нижней
её стороны буроватой полосой, идущей от основания клюва (точнее от основания подклювья)-
к чёрному пятну по бокам шеи. Зашеек и вся остальная верхняя сторона тела чёрные. По
бокам зашейка по большому белому пятну. Грудь с грязновато-жёлтым налётом. Брюхо и
подхвостье розовато-красные. Беловато-розовые бока тела с темными, иногда нерезко
выраженными наствольными штрихами. Первостепенные маховые перья чёрные с белыми пятнами
на наружных опахалах и часто с белыми концевыми пятнами. На внутренних опахалах тоже
есть белые пятна, но они едва доходят до середины ширины опахала. Сходно окрашены и
второстепенные маховые. Верхние кроющие крыла чёрные, плечевые перья белые, но с
тёмными основаниями, что впрочем остаётся незаметным. Нижние кроющие крыла белые.
Рулевые перья чёрные, но крайние пары в вершинной их части белые с чёрными пятнам»
и полосами. На третьей паре рулевых белый цвет имеется только на самом конце и немного
по краю внешнего опахала. Клюв и ноги тёмносерые. Основание подклювья несколько
светлее, с жёлтым оттенком. Самка отличается от самца менее интенсивным цветом верхней
стороны головы. Кроме того красная шапочка несколько меньше, чем у самца, и в задней части-
vмеет.оранжевый оттенок.
i
Кавказский средний дятел Dendrocopos me dins caucastcus Bianchi
Dendrocoptes medius caucosicus. Бианки. Ежег. Зоолог, музея А. Н., IX, 1904,
стр. 1, Сев. Кавказ.
Синонимы: Dendrocoptes medius colchicus. Б у т у р л и н. Орн. Вест., 1910, стр.
196, Кутаиси. — Dryobates medius transcaucasicus. В u t u r 1 i'n. Orn. Mon. 1912, стр. 64,
nom. emend. — Dryobates medius laubmanni. Gotz. Verhandl. Orn. Ges. Bayern, 1923,
стр. 321.
Распространение. Ареал. Кавказ и Закавказье. На севере ареал
включает в себя долину реки Кубани, восточнее — Северную Осетию и Ингушию. На
юге Грузию и Азербайджан, с запада на восток в общем от Батуми и Кутаиси
до Кубинского района Азербайджана. Известен для лесной зоны Армении и
частично заходит в Иран. Характер пребывания. Оседлая птица и
частично кочующая. Биотоп. Высокоствольные буковые леса и вообще
лиственные леса равнин и предгорий. В полосу хвойных лесов не заходит.
Численность. В Грузии обычная птица, не уступающая в количестве
большому пёстрому дятлу. В Северной Осетии и Ингушии немногочисленная птица,
в долине Куры и Алазани обычны.
Экология* Размножение. В месте слияния Куры и Алазани 11 мая
найдено гнездо. Молодые были вполне оперены и высовывали носы из дупла.
В Гурии гнездо с птенцами найдено 28 мая (Вильконский, 1893).
Самостоятельно державшаяся молодая птица добыта в дубняке 13 июня. На зимних кочёвках
появляется в больших количествах в Тбилиси.
Линька. Сведений о линьке у нас нет, так как находящиеся в
Зоологическом музее Московского университета экземпляры добыты вне времени
линьки (с октября по декабрь).
Описание. Размеры. Длина крыла салшов (8) 125—129 мм, среднее 128 мм. Длина-
крыла самки (1) 126 мм. По окраске сходен с предыдущим подвидом. Отличается от него
602
ОТРЯД ДЯТЛЫ
несколько более насыщенным цветом нижней стороны тела. А именно грудь более яркая
золотисто-жёлтая, брюшная область и подхвостье без розового оттенка, кирпично-красные,
у многих экземпляров на хвосте меньше белого. Продольная штриховка на боках тела резче
выражена, чем у номинальной формы.
11. Малый пестрый дятел Dendrocopos minor L.
Pirns minor. Linnaeus. Syst. Nat., Изд. X, 1758, стр. 114.
Распространение. Ареал. От Азорских о-вов, Алжира и Британии до
Камчатки, Сахалина, Кореи. На севере до 70° с. ш. в Скандинавии, границы
высокоствольного леса в Европейской части СССР, бассейна Таза и Ела-
гуя, верховьев Нижней Тунгуски и западной части бассейна Анадыря.
На юге — Манчжурия (частично), Монголия, Иран, Малая Азия,
Балканский п-ов.
Карта 103. Распространение малого пестрого дятла Dendrocopos minor
1 — D. m. minor, 2 — D. m. mongolicus, 3 — D. m. "kamtschatkensis, 4 — D. m. amurensis, 5 — D. m.
immaculatus, 6 — D. m. nojidoensis, 7 — D. m. hortorum, S — D. m. comminutus, 9 — D. m. buturlini,
10 ~D. m. danfordi, 11 —D. m. ledoucei, 12 — D. m. colchicus, 13 — D. m. quadrifasciatus, 14 — D. m.
morgani; a — граница области распространения, б — не вполне выясненная граница распространения,
г — переходные зоны между сибирскими подвидами.
Характер пребывай, и я. Гнездящаяся и кочующая птица.
Биотоп. Преимущественно лиственные и смешанные леса, на кочёвках
мелколесье. Численность. Обычная птица. На Анадыре редка.
Подвиды и варьирующие признаки. Подвиды различаются главным образом большей
или меньшей интенсивностью окраски тёмных частей оперения. Западные и южные
(Кавказ) подвиды темнее и в общей массе мельче, самые светлые й крупные — на северо-востоке
Сибири. Известно 14 слабо охарактеризованных подвидов: D. т. minor Linnaeus, 1758 —
номинальная форма, от Белоруссии и Скандинавии до северо-западной Сибири; D. т.
mongolicus В u t и г 1 i п, 1908 — от Омска до Зайсана и Хангая, светлее номинальной
формы; ZX т. kamtschatkensis Malherbe, 1861 — Восточная Сибирь, светлее
предыдущего подвида; D. т. amurensis В u t и г I i n, 1908 — советский Дальний Восток,
Хоккайдо, частично Манчжурия, темнее всех сибирских подвидов; D. т. immaculatus
S t e j n e g e r, 1884 — Камчатка, частично басе. Анадыря, самая светлая форма; D. ш.
nojidoensis Yamashina, 1930 —Сев. Корея, снизу тёмная, как кавказские дятлы;
D. т. hortorum Brehm, 1831 — Европа, к западу от номинальной формы без Балкан,
Италии и Пиренеев, темнее номинальной формы; D. т. comminutus H а г te r t, 1907 —
Англия и Ирландия, возможно Голландия, мельче номинальной формы; D. т. buturlini
Hartert, 1912 — Азорские о-ва, Испания, Италия, вероятно Балканский п-ов;
D. т. danfordi ,И а г g i 11, 1883 — возможно Балканский п-ов (без Румынии), Ма-
МАЛЫЙ ПЕСТРЫЙ ДЯТЕЛ 603
лая Азия, нижняя сторона тела буровата; D. т. ledouci Ma I he r b e, 1855 —
северный Алжир и северный Тунис, плохо охарактеризованная форма, близка к итальянским
дятлам; D. m. colchicus В u t и г 1 i n, 1908 — Кавказ и Закавказье без Талыша и
Ленкорани, темнее номинальной формы; D.m. quadrifasciatus R a d d e, 1913 — южное
побережье Каспийского моря, темнее кавказской формы; D, т. morgani Zarudny et
Loudon, 1904 — юго-зап. Иран (Загрош), отличается длинным клювом.
Восточноевропейский малый пестрый дятел Dendrocopos minor minor L.
Синонимы: Dryobates transitiuus. Loudon. Orn. Mon. 1914, стр. 77, Латвия.—
Dryobates minor lonnbergi. Domaniewski. Ann. Zool. Mus. Pol. 1927, стр. 77,
Лапландия. — Dryobates minor menzbieri. Domaniewski. Там же, 1927, стр. 71, Саратов. —
Dryobates minor obliotus. Гавриленко. Уч. Зап. Полт. гос. пед. ин-та, т. VI, 1948,
стр. 126, Украина.
Распространение. Ареал. Населяет Вост. Европу и часть Зап. Сибири.
Н а Скандинавском п-ове доходит до 70° с. ш. В СССР граница идет от
Варангер-фиорда к южному берегу Кольского залива и по границе лесной
растительности к Белому морю, охватывая Соловки, на меридиане п-ова Канин
почти до полярного круга (с. Несь). На Печоре — до северной границы
березовых лесов (67° с. ш.). В Зап. Сибири от Салехарда и Надыма; захватывая
частично бассейн Таза, граница выходит к Енисею несколько севернее Елагуя.
Южная граница южнее Тюмени и севернее Омска подходит к Уралу долиной
р. Илек. Встречается и у Актюбинска (Зарудный, 1888). Пойменными лесами
Волги проникает довольно далеко в дельту реки. Западнее через Воронежскую
обл. и Изюм область распространения выходит на Днепр недалеко от
Днепропетровска (Попов, 1937), включает в себя верховья Ингула и Ингульца
(Чёрный лес, Шевченко, 1940), проходит северной частью Винницкой обл. и
через Подольскую область и затем Польшу' выходит к Балтийскому морю.
Зимний ареал обширней гнездового. Встречается местами северней гнездовой
обл., на юге долетает до Гурьева, Ростова-на-Дону и до берегов Чёрного
моря. Как далеко продвигается в Западной Сибири — неизвестно ввиду
невозможности правильного подвидового определения кочующих зимних
экземпляров.
Характер пребывания. Гнездящийся подвид, во внегнездо-
вое время совершает более или менее значительные кочевки. Появляется в
местах, где летом его нет или мало (например Северный Урал, Портенко, 1937);
продвигается довольно далеко к югу. В окрестностях Чкалова появляется и
держится в большом количестве со второй половины сентября по середину
декабря, потом его становится меньше, а в марте и начале апреля опять много.
Биотоп. Широколиственные дубравы и урёма; парки, сады, не очень
большие старые смешанные леса, сырые и даже болотистые лиственные леса,
степные лесные островки (Башкирия); прибрежные таловые заросли, если
только в них есть пенёк, пригодный для устройства дупла; мелколесье.
Больших высокоствольных боров избегает. Зимой и на пролёте в городских садах,
бульварах и даже в бурьянах, на домах и заборах.
Численность. В Белоруссии довольно редок, в большинстве
других мест также уступает численно многим другим дятлам, в особенности
большому пёстрому, на севере чёрному. В Волынской обл. самый многочисленный
дятел. Много в окрестностях Тюмени и Тобольска.
Экология. Размножение. Для устройства дупел служат
лиственные, в той или иной степени повреждённые (гнилые) деревья: осина, ива,
берёза, рябина, ясень, вяз, плодовые деревья. Гнёзда устраивает на высоте от
полуметра до 10 м, но чаще всего 1—2 м. В дуплах кроме мелких щепочек
и гнилой древесины подстилки нет. Яйца, числом 5—9, фарфорово-белые с
значительным блеском скорлупы. Размеры: (5) 19,5—19,8 мм х 14,5 —14,7 мм
{Сомов, 1897); 17—21,6 х 13,5—15,6 (Хортлинг, Финляндия). Спаривание
604
ОТРЯД ДЯТЛЫ
на Южном Урале отмечено во второй трети апреля, яйца в гнездах в
середине апреля, последние свежие яйца найдены в конце первой половины июня
(Зарудный, 1897). 4 птенца, уже оперившихся и близких к вылету в
Ильменском заповеднике,—22 июня (Ушков). В Ленинградской обл.
насиживание начинается во второй половине мая, в середине июня уже оперившиеся
птенцы. 3 июля на севере Карелии найдено гнездо с яйцами, а 14 июля
в Хибинских горах хорошо летающий выводок. В окрестностях Пскова 5 совсем
свежих яиц найдено 11 апреля. В Белоруссии в середине мая ещё пустое
гнездо, 3 июня уже птенцы в нём. Кладка яиц
под Москвой в половине мая. Вылет молодых
в Костромской обл. 18 июня, тогда же
находили и не сильно насиженные яйца. В
Харьковской обл. вылет молодых в конце первой
декады июня. В низовьях Оби гнезда с
пищащими птенцами —17 июня. В течение 80
минут самец подлетает к гнезду с пищей 22
раза (Ильменский заповедник, Ушков).
Питание. Почти исключительно
насекомые, в большинстве вредители леса
(например личинки усачей, малый осиновый
скрипун и другие). Летом собирает главным
образом открыто живущих насекомых на
стволах и ветвях деревьев, зимой — скрытых
под корой. Часто долбит тонкие ветви и
сучья, не посещаемые другими дятлами.
В зимнее время приносит большую пользу
фруктовым садам.
Полевые признаки. Самый маленький из
наших дятлов (если не считать Дальнего
Востока), ростом немного больше воробья.
Подвижен, чаще встречается на боковых
сучьях и даже тонких ветвях, нежели на стволах.
В поисках корма не задерживается обычно на
одном дереве больше одной минуты. Долбит
сравнительно редко. В гнездовое время очень
скрытен и молчалив, в прочее время криклив.
Частый, не очень громкий, с несколько
жалобным оттенком крик его звучит как много раз
повторяемое «ки, ки, ки, ки, ки, ки ».
Описание. Размеры и строение. Длина тела 161—175 мм, размах крыльев
288—300 мм. Формула крыла 4>5>3>6 (иногда 5>4). Длина крыла самцов (27) 91—100,
самок (25) 93—99 мм; среднее 95,4 и 96 мм. Клюв самцов и самок в пределах 12,6—
15,5 мм. Вес 21—25 г.
Окраска. Молодые птицы обоего пола похожи на взрослых самцов, но имеют более
длинное 1-е маховое (около 2,1 ел), красный цвет на темени менее развит, чёрная окраска
буровата, нижняя сторона тела несколько грязнее и имеет больше отметин на боках.
Взрослый самец: лоб белый, но самый передний край его, точно так же как и густое опушение
надклювья, глинисто-буроватый. Верхняя сторона головы красная, слегка испещрённая белым,
вследствие того, что каждое перо имеет ближе к основанию белую поперечную полоску.
Зашеек чёрный. Бока головы.и надбровная полоска белые (отделена от красного цвета
темени узкой чёрной полосой). Щёки почти всегда буроваты. От подклювья к плечам идет
довольно широкая полоса. Передняя сторона спины и надхвостье чёрные, задняя сторона
спины белая с более или менее ясно выраженной поперечной полосатостью, иногда только
с чёрными пятнышками, иногда чисто белая. Маховые черные с белыми пятнами,
образующими на внутренних второстепенных поперечные белые полосы. Верхние кроющие крыла
чёрные, более крупные из них с белыми концевыми пятнами. Две средних пары рулевых
чёрные. Самые крайние имеют только основание внутреннего опахала (скрытое под нижними
Рис. 149. Малый пестрый дятел.
МАЛЫЙ ПЕСТРЫЙ ДЯТЕЛ 605
кроющими) чёрное, в остальном белые с одной или двумя чёрными поперечными полосами
(у наиболее светлых экземпляров полосы сильно редуцированы). На второй паре рулей
тёмное основание занимает почти половину внутреннего опахала, на белом опахале
поперечная тёмная полоса или едва заметное тёмное пятнышко. Третье рулевое чёрное, только
кончик внутреннего опахала и около трети длины внешнего опахала белые. Брюшная сторона
тела белая с очень слабо выраженным охристым или глинистым налётом. На боках тела
{главным образом на боках груди) слабо выраженная продольная исчерченность, иногда
более резкая, иногда её незаметно. Клюв серовато-чёрный, ноги зеленовато-серые, радужина
■красно-бурая. Самец в зимнем оперении несколько светлее, так'как белые тона оперения
чище и менее затронуты штриховкой. У самки верхняя сторона головы до темени белая,
задняя сторона головы чёрная, красного цвета нет. В остальном — как самец.
Западносибирский малый пестрый дятел Dendrocopos minor mongolicus But.
Leuconotopicus minor mongolicus. В u t u г 1 i п. Ежегодн. Зоолог, музея А. Н., 1908»
т. XIII, стр. 242, сев.-зап. Монголия.
Синоним. Xylocopus minor neglectus. Портенко. Фауна птиц внепол, части
Северного Урала, 1937, стр. 80, Омск.
Распространение. Ареал. От Омска к востоку до Байкала и на юго-
восток до Зайсана и Джунгарии. Южная граница идёт от Кокчетавских боров,
через Кулундинскую степь, охватывает Чёрный Иртыш и включает в себя Хан-
гай. Кентей и Забайкалье принадлежат уже следующей расе.
Характер пребывания. Гнездящаяся и бродячая птица.
Биотоп. Лиственные и смешанные леса, в хвойных лесах редок. Больших
тенистых лесов избегает. Гнездится в мелком лесу и даже среди кустарника с
отдельно стоящими высокими деревьями. В горы поднимается на Алтае до 1700 л*,
в Монголии до 1400 м.
Численность. В лесах близ Омска и восточнее многочисленная
птица. В горах Алтая распространён спорадично, местами отсутствует (в юго-
вост. Алтае, большей части таёжного северо-восточного и высоких частей
центрального Алтая, Сушкин, 1938). В Салаирском кряже редок.
Экология. Размножение. Почти не изучено. 28 июня у Онгудая
(Алтай), найдено гнездо с 2 птенцами и 3 яйцами. В то же приблизительно время
добыт вылетевший птенец у Катон-Карагая.
Линька с конца мая до конца сентября — начала октября.
Описание. Размеры и строение. По размерам едва ли отличим от
номинальной формы. Длина крыла самцов (19) 92—99, самок (17) 94—100 мм, среднее 96 и 96,1 мм.
Окраска. Несколько светлее номинальной формы. Нижняя сторона тела обычно
без тёмных пестрин, задняя сторона спины без чёрных полос и, как правило, без
чёрных пестрин; на боковых рулевых, как правило, одна чёрная полоса.
Восточносибирский малый пестрый дятел Dendrocopos minor kamtschatke nsls
Malh.
Picus kamtschatkensis. Malherbe. Mon. Pic. I, 1861, стр. 115, Охотск.
Распространение. Ареал. От верховьев Нижней Тунгуски северная
граница распространения пересекает Лену где-то севернее Олекминска и
идёт далее на восток к Охотскому морю. На юге граница проходит видимо по
Становому хребту, так что бассейн нижнего и среднего Амура принадлежит
уже другой расе, спускается на юг к Шилке и охватывает всё Забайкалье,
включая, видимо, и Кентей. Алтай и Саяны населены уже другой расой.
Характер пребывания. Гнездящаяся кочующая птица.
Биотоп. Лиственные, преимущественно разреженные, леса по берегам рек; горных
лесов избегает. Численность. Сравнительно редкая птица, на Витимском
плато отсутствует. В низовьях рек, впадающих в Байкал, и вблизи Читы
довольно обычна.
606
ОТРЯД ДЯТЛЫ
Экология. Размножение не изучено. Гнездо с громко кричащими
птенцами было найдено Гладковым на высоте 2,5 м в иве в пойме Ингоды (250 км
западнее Читы) 22 июня. Линька не изучена. Смена маховых кончается к
началу сентября (19 августа меняется 2-е и 3-е). Питание не изучено. В
желудке птицы, добытой 9 февраля в Тувинской обл., обнаружены личинки
усачей (Терновский, 1950).
Описание. Размеры и строение. Длина крыла самцов (22) 91—99 мм, самок
(11) 92—97 мм, среднее 96,3 и 95,6 мм.
Окраска. Отличия молодых от взрослых по окраске те же, что и у номинальной
формы. Взрослая птица в среднем светлее предыдущей расы. Штриховки на нижней стороне
тела нет, белые полосы на внутренних маховых шире, чёрных полос на двух крайних
рулевых нет, только пятна.
Амурский малый пестрый дятел Dendrocopos minor amurensis But.
Xylocopus minor amurensis. Buturlin. Ежег. Зоол. музея А, Н., 1908, т. XIII,
стр. 243, низовья Амура.
Синонимы: Xylocopus minor minutillus. Buturlin. Ежег. Зоол. музея А. Н.,
1908, т. XIII, стр. 246, Сидеми, п-ов Янковского. — Leuconotopicus minor yamashinai.
М о m i у a ma. Annot. Orn. Orient. 1928, 1, № 4, стр. 205, Южный Сахалин.
Распространение. Ареал. Бассейн среднего (от Шилки) и нижнего-
Амура, Сахалин, Уссурийский край, южная группа Курильских о-вон,
Иессо, Корея, Манчжурия, сев.-вост. Монголия (р. Тола). Характер
пребывания. Видимо бродячая птица. Биотоп и
численность. В области парковых лесов смешанного типа (Кумара, средний
Амур) весьма обыкновенен и в горах, и в долинах. Возле Джалиидм, где
господствуют лиственничные леса, придерживается долины Амура (Штсгмип),
На побережье чаще всего в пойменных лесах, в широколиственной тайго И
в чисто дубовых насаждениях, в чисто кедровом лесу не встречен. Встречается
также в кустарниковых зарослях леспидеции и орешника, но очень редко (13с-
лопольский).
Экология. Размножение. Кладка происходит в конце мая (20.V. 44
добыта самка, имевшая в яйцеводе яйцо в скорлупе). 13 июня найдены птенцы
в дупле, во второй половине июня птенцы уже летные.
Л и н ь к а. Не изучена. 5-е маховое меняется 5 июля;3-е и 2-е—26 августа.
Описание. Размеры и строение. Длинл крыла самцов (21) 88—99, самок
(11) 91—97, среднее 92 и 93,4 мм. Вес самцоп (4) 20,7—23,2, самок (1) 22 г.
Окраска. Значительно темнее всех сибирских подвидов. На задней стороне спины
темные пятна хорошо выражены, во многих случаях они сливаются в поперечные полосы.
Нижняя сторона тела имеет глинистый налет и продольную хорошо выраженную (особенно
на боках) темную штриховку. На крайней паре рулевых, как правило, 3 черных
поперечных полосы. Третья пара рулевых имеет на белом поле наружного опахала ясно выраженное
темное пятно. В общем по расцветке очень похож на номинальную форму, а по наличию
указанного темного пятна на D. т. hortorum и на D. т. colchicus.
Камчатский малый пестрый дятел Dendrocopos minor immaculatus Stejneger#
Dendrocopus immaculatus. S te j ne ge r. Pr. Biol. Soc. Wash., 1884, стр.98, Петропав"
ловск на Камчатке.
Распространение. Ареал. Западная часть Анадырского края: к северу
от Анадырской долины и в приморской полосе нет. Зимой на восток до р.
Великой. На Гижиге и на Пенжине в Коряцкой Земле и на Камчатке. Характер
пребывания. Гнездящийся и кочующий. Передвижения к югу в бассейн
Анадыря в сентябре и октябре, обратно в марте. Б и о т о п. На Анадыре
гнездится в тополевых лесках; во время кочевок — высокорослые древовидные
кустарники ивняка, встречается на оградах селений (Портенко, 1939). На
Камчатке — березовые леса водоразделов и лиственные леса речных долин.
МАЛЫЙ ПЕСТРЫЙ ДЯТЕЛ 007
Хвойных лесов избегает. Осенью залетает в стланцевые заросли.
Численность. В бассейне Анадыря редкая птица, на Камчатке обыкновенная,
зимой значительно чаще чем летом.
Экология. Размножение. В бассейне Анадыря барабанная дробь
зарегистрирована в мае и июне, птенцы 30 июня. На Камчатке откладывает
яйца в июне, вылет птенцов в середине — конце июля. 11 июня найдено в дупле
березы гнездо с 3 яйцами, 18 июля из гнезда был слышен писк птенцов (Аверин,
1948), 12 июля найдено гнездо с б подросшими птенцами. Помещалось в дупле
осины на высоте около 8 м; птенцы при постукивании уже вылезали из гнёзд
(Бергман, 1935).
Линька не изучена. 17 августа добыт экземпляр с 5-м недоросшим, а
1 сентября с недоросшими 2-м и 3-м маховыми.
Питание. Питается главным образом насекомыми. В одном желудке
от 7 апреля — 18 Chrysomelidae, около 50 личинок жуков и около 70 яиц
насекомых. Кормится в' одиночку, почти никогда не присоединяясь к стайкам
синиц (Портенко, 1939).
Описание. Размеры и строение. Длина крыла самцов (11) 92—99, самок
(5) 95—97 мм, среднее 95,6 и 96,2 мм. Вес 25,2 г (1 экз.).
Окраска очень светлая. Белый лоб шире, чем у других форм, на плечевых перьях
белого цвета больше, чем черного. Много белого на крыльях. Пестрин на нижней стороне
тела нет, крайние рулевые без или почти без черных отметин.
Кавказский малый пестрый дятел Dendrocopos minor colchicus But.
Xylocopusminor colchicus. В u t u r 1 i п. Ежег. Зоол. музея А. Н., 1908, т. XIII, стр. 249,.
Кубань.
Синонимы: Dryobates minor ernsti. Domaniewski. Acta Orn. Mus. Zool.
Pol. 1923, стр. 80, nom. emend. — Dryobates minor harterti. Doma niewski. Ann. Zool.
Mus. Pol. H. N. 1927, стр. 90, Тбилиси.
Распространение. Кавказ и Закавказье без Талыша и Ленкорани. Южна»
граница, где эта форма заменяется малоазиатским подвидом, точно не выяснена.
Характер пребывания. Частично оседлый, частично кочующий.
Биотоп и численность. Лиственные и смешанные леса и сады.
Всюду редок. В западном Предкавказье более част в грабовых и буковых
лесах. Поднимается в Предкавказье до верхней границы леса (2000 м). Зимой
только в поясе широколиственных лесов (Аверин и Насимович, 1937).
В Лагодехском заповеднике встречен до высоты 1400 м (Чхиквишвили, 1941).
Экология не изучена.
Описание. Размеры и строение. Мелкая форма. Длина крыла самцов (16)
83—89, самок (13) 83—91 мм, среднее 87 и 88,2 мм.
Окраска. В общем похож на номинальную форму, но несколько более темен, чем
сближается со среднеевропейским D. т. hortorum. Нижняя сторона тела с глинистым
налетом и хорошо выраженной темной продольной штриховкой. На спине либо поперечные
черные полосы, либо ясно выраженная пятнистость. Крайняя пара рулевых с тремя —
четырьмя черными поперечными полосами, часто полосатость имеется и на второй от края
паре рулевых, третья пара имеет ясно выраженное черное пятно на белом поле наружного
опахала.
алышский малый пестрый дятел Dendrocopos minor quadrifasciatus Radde
Picus minor var. quadrifasciatus, Радде. Орнитол. фауна Кавказа, 1884, cip 248,
Ленкорань.
Синоним. Leuconotopicus minor hynanus. Зарудный и Билькевич.
Орнитол. Вестник, 1913, стр. 21, Астрабад.
Распространение. Ареал. Талыш, Ленкорань, прилег ающие к
Каспийскому морю леса северных провинций Ирана, на восток до Астрабада.
Характер пребывания. Гнездящийся и вероятно кочующий подвид. Во
всяком случае экземпляры, добытые в Ленкорани, — зимние. Биотоп
Талышские леса. Зимой добыт в низменном буковом лесу. В Астрабаде —
в горных дубово-буковых лесах. Численность. Редок.
ОТРЯД ДЯТЛЫ
Экология не изучена. При вскрытии желудка зимнего экземпляра
обнаружены остатки насекомых (Тугаринов и Козлова-Пушкарева, 1935).
Описание. Размеры: крыло — самец (1) 81, самка (2) 83 мм; по Бутурлину (1936),
82—88 мм.
Окраска. Еще более темная раса, чем кавказская; довольно хорошо отличается
от нее и других форм этого вида отсутствием белого цвета на средних верхних кроющих
крыла. Низ бурый с хорошо выраженной продольной штриховкой.
РОД ОСТРОКРЫЛЫЕ ДЯТЛЫ JUNGIPICUS BONAPARTE 1864
Тип Picus hardwickii
12. Малый острокрылый дятел Jungipicus kizuki Temiu.
Picas kizuki. Temminck. Pi. Col. 1836, стр. 99, Киу-Сиу.
Распространение. Ареал. По восточным окраинам Азии от Курильских
ю-вов до о-вов Риу-Киу. На материке — Уссурийский край, Корея
Манчжурия, частично северо-вост. Китай. X а-
рактер пребывания. Строго оседлая
птица. Биотоп. Леса разнообразных типов
от жарких долин до субальпийских лесов.
Численность. Во многих местах
обычная птица.
Известны следующие подвиды: J. к. Лиг/-
lensis Bergman n, 1931 — Курильские о-ва; /.
к. seebohmi H а г g i 11, 1884 — Сахалин и
Хоккайдо; J. p. permutatus M e i s e, 1934 — Южно-
уссурийский край, сев.-вост. Манчжурия, сев.-вост.
Корея; J. к. acutirostris, Jamashina, 1931 —
Корея; J. к. nippon К и г о d a, 1922 — сев.
и вост. части Хондо, о-в Квельпарт, частично Корея;
J. Л. matsudairai К и го da, 1921 — о-ва Миаку-
шима и Якешима; — J. к. shikokuensis К и г о d a,
1922 —южная часть Хондо и Сикоку; /. к. ко-
tataki К и го da, 1922 — Цусима; J. к. amamii
К иго da, 1922 — Амами-Осима, Токуносима; /.
к. nigrescens Seebohm, 1887 — о-ва Риу-Киу
(Окинава и Ягацисима); J. Л. orii К uro da,
1923 —южн. Риу-Киу (Иромоту); J. к. witderi К
tiro d а, 1926 — северная часть Чжили (Китай).
Большинство перечисленных здесь подвидов
имеют очень узкое распространение, некоторые из
них очень плохо охарактеризованы.
Карта 104. Распространение малого
острокрылого дятла Jungipicus
kizuki
1 —J. k. kurilensis, 2 — J. k. seebohmi,
3 — J. k. permutatus, 4 — J. k. acutil-
rostris, 5 — J. k. nippon, 6 —J. k. mat-
«udairai, 7 — J. k. shikokuensis, 8 — J. k.
kotataki, 9 — J. k. amamii, 70 — J. k.
nigrescens, 77 — J. k. orii, 72— J. k.
wilderi; a — граница области
распространения.
Уссурийский малый острокрылый дятел
Jungipicus kizuki permutatus M e i s e,
Dryobates kizuki permutatus, Meise, Abh. u.
Mus., Dresd. XVIII, 1934, стр. 53
Ber.
Распространение. Ареал. Южные части
Уссурийского края. Частично Корея и
прилежащие части Китая и 'Манчжурии. Характер пребывании.
Оседлая птица. Б и о т7о п. Предпочитает насаждения мягких пород
деревьев. В гнездовое время преимущественно пойменные леса манчжурского
типа, где много мягких пород деревьев: бархат, липа, тополь и другие. В чисто
дубовых насаждениях и на отдельных твёрдых породах деревьев встречается
чрезвычайно редко. Во время кочевок обычен в смешанных, хвойно-лиственных
МАЛЫЙ ОСТРОКРЫЛЫЙ ДЯТЕЛ 60!)
лесах, посещает кустарниковые заросли даже открытой части поймы (Бело-
польский).
Численность. Довольно редкая птица.
Экология не изучена. Кладка во второй половине мая.
Питание не изучено. Вместе с синицами подбирает остатки еды
большого пёстрого, дятла. Выклёвывает еще не осыпавшиеся семена из толстых
травянистых растений.
Линька: маховые перья меняются в июле — августе. 7-е и 8-е в
первых двух третях июля, 1-е, 2-е и 3-е — в конце августа.
Полевые признаки. По величине сходен с малым дятлом. Подвижная и
общительная птица. Частый спутник стай синиц и поползней, может быть
встречен лазающим по толстым травянистым растениям (Велопольский).
Рис. 150. Малый острокрылый дятел.
Описание. Размеры и строение. Хорошо отличается от ранее
рассмотренных дятлов формулой крыла. Вершина его образована 3-м, 4-м и 5-м маховым, 2-е лишь
немного короче их и, как правило, длиннее 7-го. Длина крыла самцов (12) 80—90 мм, самок
(17) 80—93 мм, среднее 85,7 и 89,9 мм. Вес самцов 18,5—21,7, самок 22—25,9 г, среднее 20,1
и 23,4 г.
Окраска напоминает несколько окраску малого дятла, но спина, поясница и
крылья в черных и белых поперечных полосах. Голова сверху и с боков, а также средняя
часть зашейка буровато-серые. Бока зашейка белые, точно так же и полоса от клюва через
глаз. Бока зоба одноцветны с зашейком. Горло и средняя часть зоба белые. По бокам горла
по черной полоске, сливающейся с темным цветом боков шеи. Брюшная сторона тела
буроватая с густо посаженными темными продольными пестринами. Две средних пары рулевых
черные, остальные в черных и белых полосах. Радужина красная или коричневато-красная,
ноги оливково-серые, клюв темно серовато-роговой.
Сахалинский малый острокрылый дятел Yungipicus kizuki seebohmi H a r g.
Jungipicus seebohmi. H a r g i 11. The Ibis, 1884, стр. 100, Хоккайдо.
Синоним. Jungipicus kizuki ijimae. Так u-T s u к a s a. «Dobutsu Zashi», 1922,
стр. 292, Сахалин.
Распространение. Ареал. Южная половина Сахалина и Хоккайдо.
Характер пребывания. Оседлая птица. Биотоп не изучен.
Экология не изучена.
39 Птицы СССР, т. I
610
ОТРЯД ДЯТЛЫ
Описание. Мельче уссурийского (по Бутурлину, 1936, крыло
83—89,5 мм); окраской не отличается.
Курильский малый острокрылый дятел Yangipicas kizuki kurilensis Bergm.
Yungipicus kizuki kurilensis. Bergman n. Ark. f. Zool., 1931, стр.5,
Южно-Курильские о-ва.
Синонимы: Yungipicus kizuki 1 kurodae. Bergman, там же, 1931, стр. 5,
Иторофу, Курильские о-ва. — Dryobates kizuki nagamichii. Bergman, Zur Kenntnis
nordostasiat. Vogel, 1935, стр. 211, Иторофу.
Распространение. Ареал. О-ва Кунашири и Иторофу, Курильская
гряда. Характер пребывания. Оседлая птица. Биотоп.
Лиственные леса. В хвойных встречается только в случае, если к ним
примешаны лиственные деревья. Часто встречается в ольховых и тополевых
зарослях долин рек. Численность. Довольно многочисленен.
Экология не изучена.
Описание. Размеры и строение. Длинакрыла 85—93,5 мм (по Бергману, 1935).
Окраска отличается большим развитием белого цвета на верхней стороне тела. Серый
цвет на голове чистый, без примеси бурого.
13. Большой острокрылый
дятел Jangipicus nanus
Vigors
Picus nanus. Vigors, Proc. Zool. Soc.
London, 1831, стр. 172, зап. Гималаи.
Распространение. Д р е а л.
Уссурийский край, Корея, юго-восток
Манчжурии, Формоза, Китай к югу
до Юннаня и северного Тонкина,
Гималаи, Индо-Китай, Борнео, Суматра.
Характер пребывания.
Оседлая птица.
Биотоп и численность.
Обычная птица равнинных и горных
лесов.
Подвиды и варьирующие
признаки. J. n. doerriesi H a r g i 11,
1881 — Уссурийский край, Корея,
Манчжурия; J. п. scintilliceps Swinhoe,
1863 — северо-вост. Китай до нижней
Янцзы; J. п. kurodae La-Touch e,
1931 — юго-вост. Китай от вост. Юннаня;
J. n. nanus Vigors, 1832 — зап.
Гималаи; J. п. kaleensis Swinhoe, 1863 —
Формоза; J. n. omissus Rothschild,
1926 — Центральный и зап. Юннань;У« п.
swinhoei H а г t e r t, 1924 — южный
Юннань, Хайнань; J. n. canicapillus
В 1 у t h, 1845 — Бирма, Сиам, южн. Индо-
1845 — Малакка; J. п. aurantiiventris S а 1 v a d о г i,
Суматра; J. п. semicoronatus Ma 1 h e r b e, 1898 —
Карта 105. Распространение большого
острокрылого дятла Jungipicus nanus
7 — J. п. doerriesi, 2 — J. п. scintilliceps, 3—J. п.
kurodae, 4 — J. n. nanus, 5 — J. n. kaleensis, 6 — J.
n. omissus, 7 — J. n. swinhoei, 8 — J. n. canicapillus,
9 — J. n. auritus, 10 — J. n. aurantiventris, 77—J.
72 — J. n. semicoronatus; a — граница
области распространения.
n. volzi,
Китай; J. п. auritus Е у t о п,
Борнео; J. п. volzi Stresemann,
Сикким и Ассам.
Уссурийский большой острокрылый дятел Yungipicus nanus doerriesi Harg.
Yungipicus doerriesi. H a r g i 11. The Ibis, 1853, стр. 140, о-в Аскольд.
Распространение. Ареал. Южная часть Уссурийского края, о-в
Аскольд, восточная и юго-восточная Манчжурия, Корея. Характер пре-
ВЕРТИШЕЙКА 611
б ы в а н и я. Оседлая птица. Биотоп.
Лиственные леса долин и гор.
Численность. В низовьях Имана довольно
обычен. В других местах редок.
Экология. Размножение не
изучено. 4 мая добыт самец с сильно
развитыми семенниками, 6 мая самка с
большим наседным пятном (Спангенберг,
р. Иман).
Линька. Смена первостепенных
маховых заканчивается в сентябре.
Экземпляры от 8 и 10 сентября имеют 2-е
маховое перо (у некоторых и 3-е) недоросшее.
б-е перо меняется в середине июля.
Описание. Размеры и строение.
Формула крыла, как и у малого острокрылого
дятла. Длина крыла самцов и самок (11) 100—108 мм,
среднее 104 мм.
Окраска. Верх головы серый,
переходящий в черный на зашейке. От клюва через глаз к
зашейку тянется широкая белая полоса,
отделенная от серого цвета головы узкой черной линией.
Щеки буровато-серые. Передняя часть спины,
плечевые, мелкие и средние верхние кроющие крыла
и верхние кроющие хвоста черные. По сторонам
задней части головы несколько маленьких красных
перышек. Задняя часть спины бурая в черных
пятнышках. Маховые черные с белыми пятнами на
обоих опахалах, сливающимися на внутренних
второстепенных в сплошное белое поле. Средние
рулевые черные, смежные с ними с ржавчатыми р т Большой острокрылый
дятел.
отметинками на концах, остальные рулевые белые
с черными пятнами или черными полосками. Горло
беловатое, остальная брюшная сторона беловато
желтовато-бурая с многочисленными черными продольными полосами. Радужина красно-
бурая, ноги и клюв темносерые. Самка отличается от самца отсутствием красных
перышек на голове.
РОД ВЕРТИШЕЙКИ JYNX LINNAEUS, 1758
Тип Jynx tor quill а (ошибочно Junx и Yunx)
14. Вертишейка Jynx torquilla L.
Jynx torquilla Linnaeus. Syst. Nat. Изд. X, 1758, стр. 112, Швеция
Распространение. Ареал. Европа к северу до полярного круга в
Финляндии, на юг до Средиземного моря. Алжир. Азия — к северу до Берёзова и
Якутска (несколько севернее), к югу до Кашмира и Кореи. К востоку до
островов Японии. Зимой — Африка до Абиссинии и Сомали; Индия; Китай от
Фучжоу до Юннаня. На пролёте почти повсеместно между гнездовым и
зимним ареалом. Характер пребывания. Перелётная птица. Биотоп.
Леса разных типов, где придерживается опушек и полян. Глухих
однообразных лесов избегает. Численность. В большинстве мест более или
менее обыкновенная птица.
Подвиды и варьирующие признаки. Подвиды различаются по размерам и по общему
тону оперения. Известно 5 подвидов: J. t. torquilla Linnaeus, 1758 — номинальная
форма, вся Европа за исключением Италии, Далмации и Сардинии, Зап. Сибирь; J. t.
chinensis Hesse, 1911 — Азия к востоку от Байкала и от Кашмира, темнее и бурее номи--
39*
612
ОТРЯД ДЯТЛЫ
нальной формы; J. t. japonica Bonaparte, 1850 — Японские острова, мелкая
форма; J. t. tschusii Kleinschmidt, 1868 — Италия, Далмация, Сардиния, Сицилия,
темнее и мельче номинальной формы; J. t. mauretanica Rothschild, 1909 — сев.
Алжир.
Карта 106. Распространение вертишейки Jynx torquilla
* —J* t. torquilla, 2—J. t. chinensis, 3—J. t. japonica, 4 — J. t. tschusii, 5 — J. t. mauretanica.
a — граница гнездовий, г — область зимовок
Обыкновенная вертишейка Jynx torquilla toquilla L.
Синонимы: Jynx torquilla hyrcana. Зарудный. Орнитол. Вестн., 1913, стр.
140—141, Решт; — Jynx torquilla hartertu Поляков. Орнитол. Вестн., 1915, стр. 135—
136, Алтай, Енисейская г.: — Jynx torquilla sarudnii. Loudon. Orn. Mon. 1912, стр. 44,
Закаспийский край, пролет.
Распространение. Ареал. В Скандинавии до б5°^с. ш. и до полярного
круга в Финляндии. В СССР до южных берегов Белого моря, Холмогорского
района (ст. Тайга, озеро Б. Слободское) и несколько севернее Сыктывкара;
бассейна р. Камы до верховьев включительно (Воронцов, 1949). В Западной
Сибири до Берёзова на Оби и Верхне-Имбатского на Енисее, верхнее течение
Нижней Тунгуски, примерно до Ермбогачена и Вилюя, на востоке до Байкала
и Саян (экз. из окрестностей Иркутска переходны в сторону восточной формы).
Южная граница проходит, минуя Хангай, по северным склонам Танну-ола,
минует юго-восточный Алтай, включает в себя Зайсанскую котловину, идет к
Семипалатинску и, огибая Кулундинскую степь (и значительную часть Барайии-
ской степи) с востока и севера, направляется к Кокчетавским борам.* Отсюда
* Обычно принято считать вертишейку гнездящейся птицей всей Средней Азии, кроме
Закаспия. Однако точных доказательств гнездования ее здесь до настоящего орсмсии не
имеется (см. Иванов, 1940).
ВЕРТИШЕЙКА () 13
граница идет к низовьям Илека и к среднему течению Урала, далее к Еру-
слану, в пойме Волги проходит несколько южнее Красногвардейска, в
Донбассе около ст. Митякинской, на Днепре идет к югу до его порожистой части,
включая верховья Ингульца (Чёрный лес), и далее на северо-восток к Карпатам.
В СССР гнездится также в Крыму (горная и предгорная полоса) и на Кавказе.
В последнем редко и не везде, детали распространения её здесь требуют
дальнейшего выяснения. Вне СССР —Европа, без Италии и Далмации, Малая Азия,
сев. Иран. Зимний ареал — Африка к югу от Сахары, на юг примерно до
экватора; самые южные находки в южном Камеруне, Убанги, Уганде. На западе
отмечен на Золсугом берегу, в Сиерра-Леоне, Того, Биссао. На востоке обычна
в Кении и Уганде, в Судане, Сомали и Абиссинии (Гроте, 1930, 1937).
Кроме того, зимует в Индии. На пролёте всюду к югу от своей гнездовой
области в Европейской части СССР, на Кавказе и почти повсеместно в Средней
Азии.
Характер пребывания. Гнездящаяся перелётная, поздно
прилетающая птица. Появляется на территории Советского Союза в конце
апреля, начале мая. Под Киевом по многолетним наблюдениям самый ранний
прилёт 17 апреля, самый поздний 25 апреля (Данилович, 1933), около
Днепропетровска появляются 9 — 11 апреля, в Изюме 20—26 апреля, в окрестностях
Харькова в разные годы от 10 апреля по 2 мая, в окрестностях Орла с 20 по
25 апреля, Калуги 21 апреля, Москвы с 19 апреля по 22 мая, многолетнее
среднее 29 апреля (Паровщиков, 1941). В окрестностях Минска появляются 15—24
апреля, около Смоленска в самом конце апреля и начале мая, в Псковской обл.
первые появляются 27 апреля, обычно числа 3 мая, в Ленинградской обл.
в начале мая, в Горьковском крае — начало мая, как исключение в 1910 г. —
19 апреля (Серебровский, 1908). В Молотовской обл. пролёт 23 мая (Тепло-
ухов, 1891), в Самарской обл. 3^5 мая, в Башкирии 30 апреля. В Крыму
(Ялта) вертишейка добыта 9 апреля (Браунер, 1898). Сведений о сроках
весеннего пролёта на Кавказе нет (в Сев. Осетии —22 апреля). В Средней Азии на
Мургабе пролетает 20 апреля, а в Таджикистане во второй половине апреля,
самое раннее наблюдение 15 апреля (Ленинабад), самое позднее 28 апреля
(Гиссар, Иванов, 1940). У Джулека пролетают 19 апреля — 6 мая, под Гурь-
евом в конце апреля, около Чкалова первые появляются в первых числах мая,
валовый пролёт 10—15 мая. Летят широким фронтом, встречаются на
пролёте в пустыне Кызыл-Кум и Кара-Кум, в совершенно безлесных
местностях зап. Казахстана и в высоких горных хребтах на высотах около 2500 м
в альпийской зоне. Держатся во время пролета поодиночке, реже по нескольку
вместе. Осенний пролёт происходит в сжатые сроки в конце августа и в
октябре, но бывают и сильно запоздавшие с отлетом птицы. В Псковской обл. отлёт
в начале и середине августа, отсталые наблюдались 25 сентября, из Смоленской
обл. отлетают в начале сентября, из Куйбышевской 25 августа до конца
сентября. На Кавказе пролет отмечен в августе. В окрестностях Чкалова пролёт
начинается примерно с 2 августа, валовый —. в середине августа, отсталые
в середине октября. В низовьях Аму-Дарьи и на Теджене — в августе —•
сентябре, в Таджикистане самые ранние 14 и 18 августа (Искандер-Куль), самые
поздние 8 сентября (Куляб), главная масса летит с 31 августа по 7—8
сентября. Прилет на зимовки в Африке — главным образом в октябре, отлет оттуда
на север в марте — начале апреля.
Биотоп. Вертишейка — лесная птица, но глухих лесных массивов
избегает и нередко гнездится на одиночных деревьях, хотя поблизости есть
лес. Населяет не слишком густые лиственные и смешанные леса, также
фруктовые сады и парки, не стесняясь близости человека. На севере преимущественно
леса речных долин, в Башкирии главным образом в урёмах; где мало леса и по
вырубкам, заросшим кустами с отдельными старыми деревьями. В сосновых
614
ОТРЯД ДЯТЛЫ
лесах встречается много реже, чем в других типах насаждений, в ельниках
отсутствует. Иногда поселяется в глинистых оврагах вблизи леса
(Тульская обл., Сушкин, 1892). В Харьковской обл. биотоп — главным образом
опушки, поляны и просеки, встречается нередко и в самой глубине
однообразных лесов (Сомов, 1897). Заселяет островные леса, луга с редкой древесной
растительностью. В Днепропетровской обл. — степные сады и небольшие
рощи со старыми дуплистыми яблонями и грушами. На юге в Новороссийске
и Геленджике гнездится в приусадебных деревьях, на Сев. Кавказе в
поясе широколиственных лесов, по долинам рек заходит в пояс темнохвойных.
В Закавказье заселяет также дно долин с густыми зарослями и вьющимися
и колючими растениями, распространена однако до верхней границы леса
(Чхиквишвили, 1930). В Алтае распространена спорадично; избегает горной
тайги, предпочитает светлый лес; в чисто хвойных участках селится лишь
если они состоят из сосны или лиственницы (Сушкин, 1938). На пролете не
придерживаются определённых станций.
Численность. На Кавказе редка и распространена спорадично,
то же относится и к Алтаю. В большинстве других мест более или менее обыкно-
еенна. С установлением искусственных гнездилищ численность ее увеличивается
(Ушков).
Экология. Размножение. Первое время по прилёте, особенно если погода
неблагоприятна, держатся очень скрытно и не подают голоса. Дней через 7
(Харьковская обл., Сомов, 1897)—5 (Ильменский заповедник, Ушков)
начинается «пение», оно продолжается весь гнездовый период, постепенно
заканчиваясь к концу июля. Кричит самец обычно около дупла. Гнездовый
участок велик, ближе 200 м друг от друга птицы не селятся (Влагосклонов, 1949).
Для гнездования использует естественные пустоты и дупла, часто — покинутые
дупла дятлов и поползней. Поселяется и в искусственных дуплянках. Высота,
на которой расположено гнездо, колеблется от нескольких десятков см до
7 м. Для гнездования избирается ива и ольха, нередко осина, берёза и липа,
изредка дупла старых елей. Часто устраивает гнёзда не в вертикальных
стволах деревьев, а в пустотах толстых почти горизонтальных плодовых деревьев
в садах. Изредка устраивает гнёзда в норах глинистых оврагов, поблизости
однако от древесной растительности (Сушкин, 1938, Сомов, 1897). Гнездо может
быть устроено в дупле в стене деревянного здания (Карамзин, 1902) и даже за
тесовой обшивкой дома (Богачёв, 1927). Подстилки в гнезде не бывает, роль
последней играют кусочки гнилой древесины. В дощатых гнездовьях делает
настоящее гнездо (Влагосклонов, 1949). Если гнездо уже занято (например,
большой синицей), вертишейка выгоняет хозяина и выбрасывает яйца.
Известны случаи, когда яйца были отложены вертишейкой прямо на трупах птенцов
синицы. Откладывает от б до 12 яиц, сильно варьирующих по величине и форме
(правильно яйцевидная, удлинённая, элипсоидальная). В Поволжье
установлено в кладке только 8—»10 яиц (Карамзин, 1909, Эверсманн, 1866). Скорлупа
яиц нежная, матово-белого цвета, от просвечивающего белка кажется
розоватой, может быть блестяще-белого цвета (Аверин, 1910). Размер яиц (6) 22,9—
23,6 х 15—16,5 мм (Ушков), вес 3—3,2 г. Откладывание яиц начинается в
конце мая и, согласно Герхнеру (1928), за 6 дней птица может отложить 10 яиц.
8 начале мая начинается насиживание, которое длится И дней. Через 18—19
дней после вылупления, т. е. в конце июня, птенцы вылетают. Часть яиц (иногда
половина) остаётся болтунами.
Сидят птицы на гнезде крепко. Насиживает преимущественно самка,
самец заменяет её, когда она слетает кормиться. Более точные даты откладывания
яиц и насиживания следующие: в Минской обл. гнёзда с полными и ещё
вполне свежими кладками находили в середине июня, 23 июня находили
уже и сильно насиженные яйца. Близ Витебска начало кладки зарегистрировано
ВЕРТИШЕЙКА
615
23 мая, насиживание в том же гнезде с 1 июня, через 11 дней вывелись птенцы
и 30 июня вылетели (Федарака, 1926). Около Пропойска кладка из 8 яиц 10
июня, 2 июля крик птенцов из дупла. Близ Орши 25 июня оперившиеся птенцы
и 26 июня птенцы «в кисточках» (Бианки, 1914). В Псковской и Ленинградской
обл. полные кладки обычно находят в июне — начале июля. Точнее:
в Псковской обл. одно яйцо в гнезде 24 мая и 12 июня, 13 июня кладка в 11
совершенно свежих яиц и 1 июля сильно насиженные яйца. 25 июня птенцы
в гнёздах (Зарудный, 1910). В Новгородской обл. полная кладка в 9 яиц 12 июля
{Бианки, 1915). Под Москвой вылет из гнезда в разные годы от 2 до 12 июля.
В Подолии 3 мая найдено одно яйцо и 8 мая —> 10, в другом гнезде одно яйцо
26 мая и 6 июня — 10, в этом гнезде птенцы вывелись 18 июня (Герхнер, 1928).
Там же в начале июля отмечены лётные птенцы с невыросшими хвостиками
(Портенко, 1928). Под Киевом 22 мая найдено гнездо с 3 свежими яйцами (в этом
же гнезде 24 мая — 4 свежих яйца), 24 мая с 8 свежими яйцами, 24 июня —
гнездо с 5 яйцами и 4 вылупившимися птенцами; летающие молодые
появляются в середине июля (Шарлемань, 1915). В Харьковской обл. самая ранняя
кладка (из 4 ненасиженных яиц) за ряд лет наблюдений — 18 мая, самая
поздняя (из 9 ненасиженных яиц) — 19 июня (Сомов, 1897). В Орловской обл.
15 мая птенцы в пеньках, и 27 июня самостоятельные молодые (Ефимов, 1907).
В Тамбовской обл. молодые с недоросшими маховыми и рулями отмечены 1 июля
(Резцов, 1910), в Куйбышевской обл, вылет молодых по Карамзину (1909)—-
23 июля. На р. Еруслан старые кормят птенцов в 20-х числах июня; выводок,
летающий в кустах, 9 июля (Волчанецкий и Яльцев, 1934). На Урале
(Ильменский заповедник) кладка начинается 7—»8 июня, птенцы появляются 22—24
июня, через 12—13 дней птенцы выглядывают из домика, оставили гнездо 14 июля.
Следовательно, птенцы в гнезде были 20—22 дня (Ушков). В Крыму подлетки
и хорошо летающие молодые наблюдаются в середине июля (Браунер, 1898),
в Дагестане полные кладки в конце мая (Бёме, 1926). В Западной Сибири (На-
рымский край) недавно выведшиеся птенцы — 5 июля (Аникин, 1902); на юго-
восточной окраине Алтая 28 июня найдено гнездо с 7 совершенно свежими
яйцами, под Красноярском самостоятельные молодые отмечены в конце июля.
В сроках гнездования и откладки яиц в одной и той же местности наблюдаются
нередко значительные колебания (до 10—15 дней), связанные повидимому
с различным ходом метеорологических условий в разные годы. Бывают и
запоздалые (почти на месяц) кладки, связанные с тем, что первая кладка погибла.
Если взять из гнезда полную кладку, птица откладывает новую, иногда с тем же
количеством яиц, иногда меньше. В гнезде грязно, в период вывода птенцов
оно всё заполнено копошащимися птенцами, яйцами и скорлупой. Первое время
птенцы сидят в гнезде молча, по мере подрастания начинают издавать очень
слабые звуки, своеобразное «верещание». Крик подросших птенцов слышен
издалека, он напоминает крик птенцов большого пёстрого дятла, но громче и
хриплее. Притом кричат птенцы не равномерно и однотонно, а «залпом», одним
не очень продолжительным коленом, вскриком. За день или два до вылета
птенцы могут выходить на время из дупла, прячась обратно при первой
опасности. После вылета выводки не разбиваются в течение нескольких дней,
пока молодые не научатся самостоятельно добывать корм. Потом птицы
держатся в лесах и кустарниках в одиночку и не подают голоса. Брачный крик
прекращается обычно в конце мая.
Линька. Линяет вертишейка в конце июля и в августе, причем крупное
оперение сменяется очень быстро: выпадает, одновременно несколько
первостепенных маховых перьев. В конце июля и начале августа можно видеть птиц,
у которых второе, третье и четвертое первостепенные маховые сильно недорос-
шие. Многие птицы начинают отлёт ещё не закончив линьки. Линька молодых
птиц несколько более растянута.
616
ОТРЯД ДЯТЛЫ
Питание. Основная пища вертишейки — муравьи и их куколки, кроме
того в желудках бывают тли и вообще разные другие насекомые. На Алтае
добывали птиц, желудки которых были наполнены остатками кузнечиков
(Нестеров, 1909).
Полевые признаки. Небольшая, скромно окрашенная (серая) птичка,
размером примерно с малого дятла. В брачное время легко выдает себя
характерным криком, несколько напоминающим крик мелкого сокола (пустельги),
но слабее. Напуганная вертишейка характерно вертит головой и шеей
(откуда название) и издаёт своеобразное шипение.
Описание. Размеры и строение. Перья хвоста, в отличие от прочих
дятловых птиц, мягкие и не заострённые. Формула крыла часто 2>3>4, причём вершимы этих
перьев очень сближены, нередко бывает 3>4>2. Первое маховое перо очень коротко,
не выходит из-под верхних кроющих крыла; у молодых птиц оно больше,4 почти равно
одной трети длины второго пера. Клюв не приспособлен к долблению, тонко заострён
Рис. 152. Вертишейка.
на конце (на экземплярах коллекций самый кончик клюва часто бывает обломан). Длина
тела самцов и самок 183—197 мм, размах крыльев 289—307 мм. Длина крыла самцов
(бб) 77—97 мм, чаще всего 88—91 мм, самок (36) 83—93 мм, чаще всего 87—89 мм;
среднее 88,3 и 88,1 мм. Вес самца 37,2 г, самки 35 г.
t О к р а с к а. Молодые окрашены в общем так же, как взрослые птицы, но бледнее: опят-
нение нижней стороны тела менее интенсивно. Первое маховое длиннее верхних кроющих
крыла. Общий характер оперения взрослых сероватый с примесью бурого и
рыжевато-охристых тонов и мелкими тёмными и светлыми пестринами, создающими струйчатый характер
оперения. Верх головы светловато-серый с поперечной исчерченностью, образованной
наличием на перьях рыжеватых предвершинных и тёмнобурых вершинных полос. От
середины зашейка до середины спины идёт тёмное (продольное) пятно тёмнобурого и ржавчатого
цвета. Бока задней стороны шеи и передняя часть спины по обе стороны от тёмного пятна
светлосерые с мраморным рисунком. Задняя часть спины и надхвостье в общем того же цвета
с буроватой наствольной штриховкой на отдельных перьях. Плечевые перья, точно так же
как и третьестепенные маховые, сохраняя тот же «мраморный» тип разрисовки, имеют
общий тон рыжевато-охристый с тёмнобурым и светлоохристым опятнением. По бокам головы
буровато-ржавчатая полоса, идущая от глаза над ушными партиями. Подбородок и перья
по бокам основания клюва белые, горло и зоб охристые. И те и другие части оперения с
бурой поперечной штриховкой. Бурые штрихи на перьях зоба имеют в средней своЫЬ
части (у стержня пера) расширение (утолщение), переходящее в наствольную штрихонку,
Подобную же расцветку имеют и перья подхвостья. Остальной низ белый с бурыми пятил мм,
имеющими форму «птички», бока с охристым налётом. Подмышечные охристые с бурыми
поперечными полосами.
Первостепенные маховые бурые с рыжеватыми поперечными полосами на внешних
опахалах, второстепенные маховые окрашены сходно, но на наружных опахалах их рыжий
цвет преобладает над бурым. На внутренних опахалах (главным образом самых передних
первостепенных) могут быть белые пятна. Верхние кроющие крыла в общем одпоцпстпы
с третьестепенными маховыми (о которых речь была выше). Рулевые мягкие, не заострён-
ВЕРТИШЕЙКА
617
ные, цвет их серый с мраморным поперечным узором, образующим пять поперечных
полос. Клюв и ноги буровато-рогового цвета, иногда с зеленоватым, а на ногах с
желтоватым оттенком. Радужина красновато-бурая. Самка едва отличается по окраске от самца.
В общем она слегка серее сверху, черное опятнение верха выражено несколько слабее,
а поперечная полосатость низа несколько гуще.
Восточная вертишейка Jynx torquilla chinensis Hesse
Jynx iorguilla chinensis. H e s s~e. Orn. Mon. 1911, стр. 181, Китай, Циньдао.
Синонимы: Jynx torquilla intermedia. Stegmann, Bull. Br. Ornith. Club,
1927, XLVII, стр. 73, Сибирь. — Jynx torquilla incognita. Stachanow там же, 1933,
стр. 250, верх. Жёлтой Реки.
Распространение. А р е а л. От Забайкалья к востоку до Лены (с. Намское)
и до Охотского моря. Вероятно эта форма на Шантарских о-вах. Сахалин,
Приморье и Уссурийский край. Сев. Корея, Манчжурия, Алашань и
Кукунор, Кентей и возможно северные склоны Хангая. Зимой в Индии и в
Юннани.
Характер пребывания. Перелётная птица. Около Вилюя
первые появились 20 мая, много -^23 мая (Скалой), вблизи Читы 17 мая
(Павлов, 1948), в Кентее — первые отмечены 8 мая. Отлёт происходит в
августе — сентябре.
Биотоп. В бассейне Лены опушки лиственных лесов и в некоторых
случаях поселения (Скалой), на Амуре лиственные леса долины реки, в местах с
господством лиственничных лесов её нет совсем (Штегман, 1930). В Забайкалье
преимущественно речные долины, в Кентее держится в речных урёмах и по
южным склонам гор, где растут отдельные небольшие деревья и кусты.
Численность. На Лене довольно обыкновенна (в окр. с. Намского
радиусом в 2—3 км найдено 5 гнёзд). На Амуре сравнительно редка, под
Читой более обыкновенна, чем на Витимском плато. В лесах Кентея очень
обыкновенна.
Экология* Размножение. Гнёзда устраивает в дуплах лиственниц
и тополей, в ветлах и выгнивших пеньках на высоте от 1 до 7 м. Гнездится также
(Марха) в строениях, например, под самым коньком крыши сеновала, в
полом бревне надворной постройки и т. д. (Скалой). Гнездо неопрятное, издаёт
сильный, неприятный запах, подстилка состоит из сухой древесной трухи,
иногда с небольшой примесью мха. Под Якутском 21 июня ^гнёзда с 8 и с
10 яйцами, в первом гнезде птенцы вывелись 23 июня. 29 июня найдено гнездо
с 6 надклюнутыми яйцами и 3 только что выведшимися птенцами. Гнёзда с
птенцами на взлете найдены 14 и 15 июля, через пару дней покинуты. В
окрестностях Читы птенцы появляются также в конце июня. На р. Иман
(Уссурийский край) 25 мая обнаружена самка с яйцом в яйцеводе, 11 июня —
незаконченная кладка в 7 свежих яиц, 14 июня —кладка в 10 яиц в последней стадии
насиженности, 15 июня —кладка в 12 слабо насиженных яиц (Спангенберг).
Линька. Происходит энергично, в основном в августе у большинства
передвигающихся к югу птиц линька ещё не вполне закончена.
Питание составляют главным образом муравьи.
Описание. Размеры. Длина крыла самцов (7) 84—89, среднее 86 мм,? трех
самок 83—86 и 87 мм. Вес одного самца 37,4 г. Окраска сходна с номинальной формой,
но в свежем пере темнее и бурее сверху, черный рисунок верха более развит, горло и грудь
довольно резко отграничены от светлого подбородка.
Систематические замечания. Подвидовая принадлежность вертишеек, гнездящихся
в Якутии, остается неясной. Иванов (1929) считает, что их следует относить к
номинальной форме.
ОТРЯД ДЛИННОКРЫЛЫЕ
MACROCHIRES или MICROPODTFORMES
Характеристика отряда
Строение. Внешние признаки. Небольшие, частью
крошечные птицы с длинными крыльями, чрезвычайно быстрым полетом и окраской
то черно-бурой, то ярких металлически блестящих тонов.
Крыло диастатаксическое или эвтаксическое, его плечевой отдел сильно
укорочен, кистевой —• удлинен. Первостепенных маховых 10, второстепенных
сильно укороченных маховых 6—11. Рулевых — 10 (только у самцов колибри
Loddigesia mirabilis их 4). Контурные перья с побочным стержнем. Копчиковая
железа не оперена.
Анатомические признаки. Череп эгитогнатический или
схизогнатический, голоринальный, с несквозными ноздрями. Предсошник
имеется. На основной кости нет базиптеригоидных отростков. Шейных
позвонков 13—15. Грудина с очень короткими spina externa и interna, без вырезов,
но на заднем закругленном крае иногда с одной —двумя фонтанелями.
Вилочка U-образна и с коротким hypocleidium. Грудных ребер j4 пары.
Плечевая кость очень коротка, толста, с сильно развитыми fossa olecrani и condylus
radial is, а также с искривленным crista superior. В мускулатуре ноги
отсутствуют m. ambiens, ветвь В. мышцы caud — ilio — femoralis, также ветви
X и У мышцы caud — ilio — flexorius, однако мышцы пальцев развиты
хорошо. Для большинства характерна редукция правой сонной артерии, но у Сур-
seloides и некоторых видов рода Chaetura имеются обе сонные артерии. У
большинства видов слепых кишок нет, только у некоторых родов подотряда
колибри сохраняются их короткие рудименты.
Почти все виды длиннокрылых строят гнезда; некоторые из них
используют свои прошлогодние постройки, другие же занимают чужие. Птенцы иы-
клевываются слепыми и долгое время выкармливаются родителями.
В состав отряда входят две резко специализированные и экологически
сильно разнящиеся между собою группы: колибри Trochilin стрижи Apodcs,
В последнее время Лоу (1939) довольно веско подтвердил прежние
неоднократные предложения рассматривать колибри и стрижей в качестве двух пссьма
близких групп отряда Passeriformes. Однако в силу принятого плана издания мел
помещаем их в качестве подотрядов отдельного отряда длиннокрылых.
ХАРАКТЕРИСТИКА ОТРЯДА
619
Подотряд колибри Trochili
За немногим исключением птицы разнообразных, чрезвычайно ярких ирри-
зирующих окрасок, размерами от 210 до 30 мм. Вес мельчайших из них доходит
до 1,64—-1,8 г, вес их яиц около 0,2 г. Для всех представителей подотряда весьма
характерен чрезвычайно быстрый полет и остановки в воздухе, что достигается
частотой взмахов крыльев, доходящей до 30—50 в секунду; этим колибри
напоминают бражников. Крылья при этом кажутся туманными пятнами, а от их
ударов о воздух слышится особый жужжащий звук. У большинства выражен
половой диморфизм и более яркие самцы отличаются от самок еще и различными
украшениями: воротничками, хохолками, длинными перьями и пр.
Относительно узкое, длинное, уплощенное и слегка саблевидное крыло, как правило,
эвтаксического типа и, как исключение, диастатаксического. Наружные
первостепенные маховые чрезвычайно упругие. Второстепенных маховых только
6—7. Коготь на первом пальце крыла отсутствует. Побочные стволы перьев
коротки, дефинитивного пуха нет. Анизодактильного типа ноги малы и цевка
по своим размерам не превышает половины длины среднего пальца.
Чрезвычайно острые когти, наоборот, по своей длине почти равняются пальцам.
Варьирующий по форме и размерам клюв в большинстве случаев оказывается тесно
приспособленным к тем цветам, из которых разные виды колибри добывают себе
пищу. Щелевидные носовые отверстия находятся вблизи краев клюва.
У птенцов клюв короткий, похожий на клюв стрижей.
Анатомические признаки. Череп схизогнатический. Рожки
подъязычной кости весьма длинные. Шейных позвонков 14—15. Относительно
крупная грудина обладает значительной подвижностью и при акте дыхания
может подаваться вперед. Киль грудины длинный и высокий. Лопатка
достигает значительной длины. Укорочение плечевой и удлинение кисти доходит до
крайних известных у птиц пределов. Поднимающая крыло мышца —m. supraco-
racoideus крупна и по своим размерам почти равна мышце, опускающей крыло —
m. pectoralis major. Для нижней гортани характерны две пары трахео-брон-
хиальных мышц, но m. sterno-trachealis отсутствует. Конец узкого, могущего
далеко вытягиваться из клюва, языка глубоко расщеплен на две полоски, края
которых, заворачиваясь внутрь, образуют длинную трубку, при помощи
которой колибри сосут нектар цветов. Взрослые без зоба, но у птенцов таковой
имеется. Небольшой мускулистый желудок обладает гладкими стенками.
Желчного пузыря нет. Довольно грубо построенные легкие отличаются
небольшим количеством бронхов и парабронхов, а также относительно крупными
воздушными капиллярами. Но общая их дыхательная поверхность велика и
примерно в четыре раза превосходит таковую у голубей. Сердце значительных
размеров и по своему объему почти втрое больше желудка, почему оно и
занимает половину всей полости тела. Относительный вес сердца является
наибольшим среди класса птиц и достигает 2,85% веса тела. Относительно велики
яйцеводы, так как колибри по сравнению с незначительными размерами своего тела
откладывают довольно крупные яйца.
Образ жизни. Преимущественно дневные птицы, почти весь день
проводящие в воздухе. Однако имеются и сумеречные виды. Обитают в самых
различных биотопах — в лесах, рощах, садах, кустарниках, на лугах, в степях и
одинаково часто встречаются как на равнинах, так и в горах, но предпочитают
открытый ландшафт. Многие виды находятся в весьма тесной зависимости от
распределения тех растений, из цветов которых они добывают себе пищу.
Виды эти отличаются подчас очень узким распространением (Waldronia
milleri встречается только по склонам гор Айян-тепю и Дуида в Венецуэле,
в поясе 1200—1700 м; Amazilla hollandi известна лишь из верховьев
р. Гуйюни, на крайнем востоке той же Венецуэлы). Другие виды способны на-
620
ОТРЯД ДЛИННОКРЫЛЫЕ
ходить себе пищу на различных растениях и для них характерно широкое
распространение.
Имеются оседлые виды (преимущественно в тропических областях
Америки) и перелетные, приуроченные к умеренным широтам.
Гнезда строятся на ветвях деревьев, кустарников, иногда прикрепляются
к узким длинным листьям или на вайях высоких папоротников, зачастую па
различных травянистых растениях и даже на злаках. Материалом для
миниатюрных чашечковидных гнезд служат различные части растений, мхи,
лишайники и растительный пух. Кладка состоит из двух удлиненных матово-белых:
яиц, иногда сносимых через столь значительный промежуток времени, что нте
нец выклевывается только из одного яйца, в другом же зародыш замирает.
Насиживает яйца и выкармливает птенцов только самка. Сроки насиживании
14—19 дней. Птенцы появляются на свет слепыми, покрытыми лишь на спине
длинным, но редким ржавчатым эмбриональным пухом. Развиваются они
медленно и около месяца выкармливаются цветочным нектаром, который самка
вводит им в зоб.
Питание большинства взрослых, состоящее из нектара, мелких насекомых
и паучков, добывается своеобразно. Подлетев к цветку и остановившие!)
перед ним в воздухе, колибри вводит в цветок клюв и, не открывая его, лишь
слегка приподнимает вверх надклювье и высовывает вперед трубчато
сложенный конец языка. Затем сильными глотательными движениями через язык
и клюв нектар накачивается в полость рта, поступает в пищевод и далее,
минуя желудок, из proventriculus льется в двенадцатиперстную кишку. Мелкие
же насекомые и паучки при этом попадают в желудок. Другие виды кормятся
паучками и насекомыми, которых они, также останавливаясь в воздухе,
собирают с листьев и мелких веточек. Потребность в пище в связи с чрезвычайно!!
подвижностью у колибри огромная и в сутки примерно равна двойному весу
их тела. Ходить не могут и используют ноги лишь для сидения. Голос состоит
из свистящих звуков, но некоторым видам свойственно и довольно приятное
пение.
Систематика. Одно семейство Trochilidae, содержащее более
100родов из 319 видов, вместе с подвидами образующих около 600 форм.
Распространение. Огромное большинство видов свойственно тропическим
частям Америке: Мексике, где обитает до 20% всех форм, Центральной и
Южн. Америкам. К северу идут до 57°с. ш., к югу до Огненной Земли. В горы
поднимаются до 5000 м. Встречаются также на прилежащих к Америке
островах. Ископаемые остатки известны из плейстоцена Бразилии.
Подотряд стрижи Apodes
Морфология. Внешние" признаки. Небольшие, плотно сложенные
птицы с удлиненным телом, короткой шеей и уплощенной головой размерами от
330 до 120 мм. Длинные, узкие, саблевидно изогнутые крылья далеко заходят :ул
хвост. За исключением древесных стрижей (Hemlprocne), имеют оперении
жесткое, плотное, темнобурое, со слабым металлическим отблеском и, в
большинстве случаев, с наличием хотя бы небольших участков беловатого или
чистобелого цвета. Направленные в бок глаза спереди, сверху, отчасти исшпу
защищены короткими плотными перышками, у некоторых родов нависли»-
щими в виде киля. У большинства крыло эвтаксическое, но у родов Сур>
seloides, Hemiprocne и Streptoprocne оно диастатаксическое. Второстепенны к
маховых 7—11. Первый палец крыла вооружен когтем, отсутствующим у
Hemlprocne. Хвост разнообразен по форме и величине. В области лопаток спинная
птерилия раздваивается и между ее ветвями оказывается голое пространстио.
У некоторых представителей подотряда раздваивается и брюшная птерилия,
ХАРАКТЕРИСТИКА ОТРЯДА
621
не образующая однако внутренней горловой ветви. Перья с небольшим, но
хорошо развитым добавочным стержнем. Пух только на аптериях. Короткие,
но крепкие ноги с совершенно раздельными пальцами, положение которых
может быть трояким: 1) или все 4 пальца направлены вперед и слегка в
стороны, 2) или 3 пальца направлены вперед, а один назад, 3) или 2 пальца
направлены вперед, 2 назад. У представителей подсемейства Apodinae
наружный и средний палец с тремя фалангами. У остальных стрижей число фаланг
наружного и среднего пальцев нормальное. Сухожилия m. flexor perforans
profundus и т. flexor hallucis перед вхождением в пальцы полностью
сливаются друг с другом, затем дают четыре ветви (V тип). Пальцы заканчиваются
изогнутыми, сжатыми с боков, острыми когтями. Клюв в основании
треугольный, очень короткий,- несколько сжат в направлении сверху вниз. Ротовая
же щель чрезвычайно велика и доходит до уровня заднего края глаза, что дает
возможность широко раскрываться рту. Кожа толстая. Череп эгитогнатический,
с хорошо развитым предсошником. Шейных позвонков 13—14. Широкая,
суживающаяся кпереди грудина с короткими spina externa и interna, с сильным
гребнем и с задним краем или закругленными, или снабженными вырезками.
Самый короткий отдел передней конечности — плечевая кость; предплечье
длиннее, самый длинный отдел — кисть. По обеим сторонам уздечки языка
на пространстве почти от внутренней части вершины подклювья и до
отверстия верхней гортани расположены многочисленные подъязычные железы,
клейкий секрет которых в период гнездования выделяется чрезвычайно
обильно. Язык треугольный, короткий. Желчный пузырь имеется.
Образ жизни. Стрижи чрезвычайно активные дневные, птицы, быстрые
и верткие в движениях, неутомимые в полете. На земле же они довольно
беспомощны и лишь с трудом взлетают с горизонтальной поверхности. В своей
биологии стрижи весьма тесно связаны с воздушными насекомыми и в
умеренных широтах бывают только в периоды их максимального лёта.
Встречаются в различных биотопах: в равнинных и горных лесах, в
горах, предгорьях, на равнинах, в пустынях, в человеческих поселениях,
но, вообще говоря, предпочитают открытый ландшафт. Гнездятся
группами и колониями в самых различных условиях: в лесах — в дуплах и
выбоинах высоких деревьев; в горах — в трещинах, щелях и углублениях скал и
обрывов или в стенах пещер; в степях, пустынях и на побережьях морей —
в норах, щелях и углублениях отвесных обрывов и обнажений; в населенных
пунктах — под крышами, на карнизах, в щелях высоких и низких каменных и
деревянных строений.
Некоторые тропические виды прикрепляют свои гнезда к ветвям деревьев,
другие роют норки в песчаных склонах. Имеются, наконец, виды, занимающие
чужие гнезда. Материалом для постройки гнезда у многих видов служит
растительный пух, соломинки и перья, собираемые на лету и скрепляемые затем
клейким, твердеющим на воздухе секретом подъязычных желез. Гнезда же
саланган построены почти из одного только секрета последних. Этим же
секретом некоторые виды приклеивают и яйцо к гнезду.
Кладка состоит из 1—4, но по большей части из 2 белых яиц,
откладываемых с промежутками 48—72 часа (у некоторых видов только через 24 часа).
Насиживают оба родителя в течение 18—21 дня. В закрытых гнездах птенцы
выклевываются голыми, а в открытых гнездах, помещаемых на деревьях
(у Cipsiurus, у Hemiprocne), они покрыты густым белым эмбриональным пухом.
Постэмбриональное развитие чрезвычайно длительное и у некоторых наших
стрижей продолжается от б до 7 недель. Выкармливают птенцов оба родителя,
однако приносится корм не часто, но зато сразу помногу. Пища состоит из
насекомых. Непереваренные остатки пищи сбрасываются в виде погадок. Во время
отдыха стрижи прицепляются к откосам скал, карнизам зданий, стволам деревь-
622
ОТРЯД ДЛИННОКРЫЛЫЕ
ев или забираются в укрытия к гнезду, где садятся на брюхо. Линяют один
раз в году, иногда долго. Продолжительность жизни до 9 лет (по данным
кольцевания).
Перелетные виды на родине появляются только после окончательного
установления весны и улетают рано, что связано с наличием постоянного количества
насекомых в воздухе.
Хозяйственное значение стрижей окончательно еще не установлено и они,
повидимому, являются безразличными для нас птицами.
Систематика. Вместе с ископаемыми формами отряд стрижей
распадается на 3 семейства, в состав которых входят 20 родов и 86 видов, которые
с подвидами образуют около 220 форм. Появились стрижи, повидимому, с
начала кайнозоя, так как в эоцене уже представлено хорошо обособленное
семейство Aequialornithidae, из состава которого нам известны 3 рода и
несколько видов. Из верхнего эоцена —нижнего олигоценамы имеем виды —
Aequialornis leenhardti, Ае. gailicus и Cypselavus gailicus. Из верхнего оли-
гоцена — Limnatornis paludicola, Microtas ignotus и, возможно, Collocalia
incerta. Из верхнего миоцена — Cypselavus gailicus. В плейстоцене уже
появляются рецентные виды родов Microtas и Chaetura. Из известных в
настоящее время 17 современных родов упомянем Collocalia, Hirundapus, Strep-
toprocne, Aeronautes, Chaetura, Mearnsia, Cypseloides, Apus, Aerornis, Panyptila,
Cypsiurus и Hemiprocne. Из известных в настоящее время 79 рецентных видов и
212 форм в СССР встречаются один подвид вида Hirundapus caudUcutus, два
подвида Apus apus и по одному подвиду видов A. melba, A. pacificus и A affinis.
Все эти формы входят в состав семейства плотноперых стрижей Apodidae и
подсемейства черных стрижей Apodinae.
Распространение. За исключением холодных стран стрижи распространены
по всей суше земного шара и наиболее обильно представлены в субтропических
и тропических областях. Некоторые виды отличаются широким
распространением, как например, Cypsiurus parius, свойственный Эфиопской и Восточной
областям, для других же характерно весьма узкое распространение. Так,
Apus reichenowi найден только в небольшом районе Кении (Африка), a
Cypseloides cherrici обитает только в кратере вулкана Иразу в Коста-Рика (Ц.
Америка).
В СССР распространены всюду, кроме арктики и субарктики. Особенно
многочисленны в горах Средней Азии и Кавказа.
Таблица для определения видов стрижей.
1 (2) Спинная сторона без участков белого цвета. По своей окраске спина почти не
отличима от цвета головы и хвоста 3.
2 (1) На спинной стороне имеются участки белого цвета или же середина спины заметно
светлее головы и хвоста . . , . Я.
3 (4) Брюхо белое. Длина крыла не менее 215 мм ,
Белобрюхий с три ж Apus mcUm,
4 (3) Брюхо черно-бурое. Длина крыла не более 185 мм
Черный стриж Apus apus,
5 (6) На пояснице нет белой перевязи. Подхвостье белое. Стержни рулей выходит iki
опахал в виде твердых и острых игол. Колючехвостый стриж Hirund apus
caudaculus.
6 (5) На пояснице имеется белая перевязь. Подхвостье черное. Концы рулей не ныхи-
дят из опахал ♦ 7«
7 (8) Обрез хвоста почти ровный. Длина крыла менее 150 мм
Малый стриж Apus a/finis.
8 (7) Хвост с глубокой выемкой. Длина крыла более 170 мм
Белопоясный стриж Apus pacificus.
ЧЕРНЫЙ СТРИЖ
623
СЕМЕЙСТВО СТРИЖЕЙ APODIDAE
РОД СТРИЖИ APUS SCOPOLI, 1777
тип Hirundo apus
С и ноннмы: Micropus Meyer etWolf, 1810; Cypselus I 1 1 i g e г, 1811\
1. Черный стриж Apus apus L.
Hirundo apus Linnaeus. Syst. Nat., изд. X, 1758, стр. 192, Швеция.
Русское название. Стриж — широкораспространённое народное название
звукоподражательное.
Распространение. А р е а л. В гнездовое время лесная зона, зона открытых
сухих пространств и горная зона Евразии, а также горная зона с.-з. Африки; на
Карта 107. Распространение черного стрижа Apus apus.
1 — A. a. apus, 2 — A. a. pekinensis; а — граница оДпасти распространения, б— не вполне /выясненная
часть границы, г — зимовки.
юг до Марокко, о-вов Средиземного моря, с.-з. Гималаев, Ганьсу и
п-ова Ляодун. В зимнее время вся Африка от тропиков до ее южной оконеч*
ности и о-ва Мадагаскар.
624
ОТРЯД ДЛИННОКРЫЛЫЕ
Характер пребывания. Перелетная птица.
Биотоп. Леса, вертикальные обнажения скал, утесов и глинистых
береговых обрывов, в культурном ландшафте — строения; предпочитает
открытый ландшафт. На перелетах и зимовьях использует подобные же биотопы.
Подвиды и варьирующие признаки. Два подвида, слегка отличающиеся окраской: А.
a. apus Li n п., 1758, занимающий большую часть ареала на юге до с.-з. Африки,
Балканского п-ова, Сев. Кавказа и Забайкалья и A. a. pekinensis S w i nh о е, 1870,
распространенный в меньшей части ареала от Малой Азии до п-ова Ляодун. С трактовкой Питерса
(1945), объединяющего с видом A. apus ряд африканских форм, нельзя согласиться по
следующим соображениям: 1) окраска близких к A. apus африканских стрижей во
взрослом наряде отличается пестринами низа тела, 2) амплитуда размеров форм вида A. apus
значительно меньше таковой африканских форм. Поэтому африканских стрижей,
относимых Питерсом к виду A. apus, я рассматриваю как вид A. barbatus с подвидами А.
b. barbatus, A. b. nianzae, A. b. balstoni и A. b. mayottensis.
Европейский черный стриж Apus apus apus L.
Распространение. Ареал. Почти вся Европа, с.-з. Африка и Азия до
Байкала. В СССР насевере до бассейнаТуломы (67° 30' с. ш.), юго-западной части
Кольского п-ова и Кандалакшского залива, горла Белого моря и
Архангельска, Усть-Цыльмы на Печоре (где стриж еще довольно обыкновенен),
до 62° с. ш. на западном У рале, по всей Колве, до р. Ивдель на вост. Урале
(60°3(У с. ш.), до 61°50' с. ш., на М. Сосьве, до 59° с. ш. на р. Туртис Уватского
р-на Ханты-Мансийского нац. округа. Отмечен у устья р. Кеть под 58°30' с. ш.,
гнездится в Кетско-Сымском, Нижне-Тунгусском и Чуно-Ангарском зоогео-
графических округах (Тугаринов, 1927), но на самой Ангаре появляется только
залетом, нормально же гнездится лишь в Балаганской степи (Скалой и Слуд-
ский, 1933). Доходит до северной оконечности Байкала и м. Св. Нос на
восточном побережье Байкала. Отсюда граница резко понижается,
захватывает бассейн Чикоя и, не доходя до Онона, поворачивает на Танну-Ола, далее
идет по южным склонам Центрального Алтая, предгорьям и степям зап. Алтая,
Зайсанской котловины, Тарбагатая, Чингиз-Тау, Кызыл-Рая, северного берега
Аральского моря и Кара-Богаз-гола на Каспийском море, по ту сторону
которого занимает весь Кавказ, включая Закавказье, большую часть Малой Азии,
Крым, Балканский п-ов, все о-ва Средиземного моря, кроме Кипра, Тунис,
Алжир и Марокко. Западная граница идет от южной части последнего по
побережью Атлантического океана, о-вам Мадейра, Канарским, Британским и
Скандинавскому п-ову на север до 70° с. ш.
Залеты A. a. apus известны для Исландии, Азорских и Фарерских о-вов,
Шпицбергена (в августе 1891), пространства между о-вами Франца Иосифа и
Новой Землей, Мурманского побережья (25.VIII.1922 на Святой Нос), о-ва
Харлова (26.VI.1935), района на широте о-ва Колгуева (15.VI), северного о-ва
Новой Земли (на м. Желания в середине июля 1937 г.) южн. о-ва Новой
Земли (в губе Безымянной в первой половине августа 1949 г., Успенский). В
Сибири залеты указаны до 64° с. ш. на p.p. Елагуе и Верхнем Тазе, до 65° с/ ш.,
на Енисее, до 61° с. ш. на Лене. На юге залетом бывает на о-вах Черноуго моря
и в том числе на о-ве Джарылгач (в июне и июле). Зимний ареал A. a. apus
значительно меньше гнездового и захватывает экваториальную и южн. Африку
примерно от 10° с. ш. до ее южной оконечности и о-в Мадагаскар.
Характер пребывания. Перелетная птица. Перемещения с мест
зимовок начинаются в марте. Двигаясь широким фронтом, передовые в этом
же месяце достигают Пиренейского п-ова, Армянского нагорья и южн. Каспия.
Однако, пролет очень сильно растягивается и на зимовках часть стрижей
продолжает пребывать до конца мая и даже начала июня, отдельные же
неполовозрелые особи и вовсе не отлетают из Африки.
ЧЕРНЫЙ СТРИЖ
625
В пределах СССР передовые появляются в конце марта в Армении (28. III.
1930s окрестностях Еревана, Соснин и Ляйстер, 1942), в начале апреля в
Туркмении и вообще в Средней Азии, где пролет идет весь май и даже в первых
числах июня, массовое же появление приходится на вторую половину апреля —
конец апреля (на Сумбаре 2 апреля, у Кара-Богаз-гола 30. IV.39 — Исаков и
Воробьев, 1940; у Джаркента в конце апреля). В середине апреля передовые уже
на Черноморском побережье у Батуми и на северных склонах Главного
Кавказского хребта, но на местах гнездовий в разных районах Большого и Малого
Кавказа появляются только в конце апреля — начале мая. В начале апреля
передовые достигают южной Украины, Крыма, северного побережья Азовского
моря, около7-^8 мая линии Киев —Харьков —Хоперск с задержками в
отдельные годы до 20 мая, 13—21 мая — Белоруссии, Московской, юга Ярославской
(Плещеева озера) и Чкаловской областей, 16—22 мая Прибалтийских
республик, Ленинградской обл., Горьковской обл. и Забайкалья, 22 мая —
1 июня Карелии и 30 мая — б июня крайней северной границы своего ареала
на Кольском п-ове, на Печоре, на Среднем Урале, на р. Томи и на Енисее в
Минусинском крае. В более холодные таежные места этого последнего района
стрижи проникают только в первой трети июня, а массовый их прилет здесь
заканчивается к середине этого месяца, когда ими занимаются места гнездовий
(Сушкин, 1914). По одним данным весною стрижи появляются ночью, по
другим — утром или днем.
Осенний пролет протекает менее четко, чем весенний. В ряде
районов с мест гнездовья или из своих обычных кормовых участков стрижи
начинают исчезать уже с конца июля: 27. VII. 1928 из кормовых участков южнее
Киева (Шарлемань, 1930), 26. VII. 1906—с истоков Оки (Ефимов, 1907 и 1915),
30—31. VII с мест гнездовий в районе р. Малой Сосьвы (Самарин и Скалой,
1940). К концу августа —началу сентября передовые отмечаются уже в
Аравии (у Йемена), Египте и у северной окраины тропической зоны Африки, а в
сентябре у самой ее южной оконечности. Но в некоторые годы, с условиями
неблагоприятными для развития птенцов, стрижи задерживаются надолго даже у
северной границы своего ареала. Так, 24. IX. 1930 на р. Щугореза 64-й
параллелью при установившейся снежной погоде и температуре —8° С наблюдались
пролетные стайки, а 27. IX. 1930 при теплой погоде в 10° С отмечены
последние пролетные (Дмоховский, 1933). О такого же рода задержках в 1901 г
в Олонецком крае сообщает Исполатов. В нормальные же для вывода молодых
годы из разных мест своего ареала стрижи отлетают в общем в августе.
Северные гнездовые районы стрижи покидают в первой —второй декадах августа.
С Печоры исчезают 7—8 августа (Филатов, 1911, Самородов, 1949), с р. Ивдель
18 августа (Резцов, 1904), из Минусинского края в середине августа и в это
же время из Забайкалья. В более южных частях ареала ход осенних
передвижений таков: из Прибалтийских республик исчезают между 3 и 20 августа,
но могут задерживаться до 26 августа, из обл. в массе 2—23 августа, отдельные
особи остаются 24—26 августа, но дважды были встречены в октябре (17. X. 1893
и 23. X. 1899); из района Плещеева озера в 1931—34 гг. 10—46 августа, затем
дня через два начинается пролет, заканчивающийся к 20 августа; последние
у гнезда с птенцами — 1. IX. 1934 (Птушенко, 1937); из Московской обл. в
массе 10—12 августа, последние к 20 августа, но задерживаются до 14 сентября
(Лоренц, 1892); из Смоленской обл. между 10 августа и 7 сентября (Огнев,
1909); из Рязанской в конце августа, последние к 10 сентября, но Павлов (1879)
наблюдал стрижей еще 7. X. 1876; из Калужской к 1 сентября (Филатов, 1915);
из Тамбовской — к концу августа; из Орловской в среднем 14 августа с
крайними отклонениями от 8 до 18 августа; из Курской 10—24 августа по
наблюдениям Птушенко за 1926—1930 гг.; из Харьковской 12—17 августа,
последние 28—31, но встречаются еще до 25 сентября (Сомов, 1897); из сев. При-
40 Птицы СССР, т. I
626
ОТРЯД ДЛИННОКРЫЛЫЕ
азовья во второй трети сентября; с северных склонов Главного Кавказского
хребта в августе. Из Предкавказских равнин отлетают в начале августа, но
немногочисленные пролетные здесь, как и в южной и ю.-з. Украине,
встречаются еще в течение всего сентября. Из Армении улетают в конце августа.
Под Чкаловом на осеннем пролете — с конца июля до начала августа,
в северной части Киргизских степей 14—29 августа, но в области
Иргиза нормально встречаются еще до 13 сентября, а отдельные особи
даже до 21 сентября (Сушкин, 1908). В Арал о-Каспийских степях пролет с конца
августа и в сентябре, в Средней Азии — в сентябре, а именно: 4—10 сентября
в окрестностях Кара-Кала (Шестоперов, 1937), 2—13 сентября под Джаркентом
(Зарудный и Кореев, 1907), 9—12 сентября на Памире (Северцов, 1879). Пови-
димому, в удобных местах пролетные надолго задерживаются, так как известны
случаи находок в вост. Туркестане еще в ноябре. Самый отлет
характеризуется тем, что в ряде мест ему предшествует стаение. Покрутившись большой
стаей над местами гнездовья, стрижи затем внезапно исчезают. Из северных
частей ареала они отлетают сейчас же по вылете молодых, в южных они покидают
места гнездовий и кочуют по открытым местам, задерживаясь на 5—10 дней,
и затем окончательно скрываются. На смену им через несколько дней появляются
из более северных мест пролетные, занимающие те же кормовые места. Это
обстоятельство в ряде случаев создает впечатление позднего отлета стрижей.
Обычно пролетные движутся рассеянными стайками по 10—15, 30—40 особей
в зависимости от погоды и направления ветра на различной высоте, чаще всего
не выше ружейного выстрела. Летят молча, с умеренной скоростью, иногда
почти сплошным, но сильно рассеянным потоком, стайка за стайкой, иногда
поодиночке все в одном и том же направлении в течение одного-двух дней
(сообщение Спангенберга). В степных местах движутся прямо на юг, но иногда летят
вдоль ориентированных в южном направлении речных долин. Последние стайки
и одиночные молодые, летящие весьма поспешно, замечаются чаще всего
под вечер.
Биотоп. Селятся в равнинных и горных лесах, в открытых
равнинных местах с вертикальными обнажениями, среди лишенных
древесной растительности гор и в культурном ландшафте. В местах с
древесными насаждениями стрижами занимаются для гнездовья старые
высокоствольные или богатые дуплами разреженные леса, причем относительные
размеры последних» их высота над уровнем моря, характер входящих в них
ассоциаций и состав древесных пород для стрижа безразличны. Необходимым
условием гнездования стрижей в лесах является наличие в них отдельных
деревьев, подходящих для устройства гнезда. Близость открытых пространств
для стрижа необходима, и поэтому близ опушек, вырубок и гарей их гнездовья
встречаются чаще, чем в глубине леса. В открытых равнинных местах для
гнездовий выбираются береговые обрывы рек и морей, отвесные стены лёсса, в
полупустынях и пустынях — обрывы пересыхающих рек и чинки. В горах с этой
целью занимаются скалы, утесы, гребни и крутые склоны, причем на гнездовье
стрижи поднимаются на Байкале до 2000 м (Штегман, 1928), а на Кавказе
2250 м, долетая на кормежках до линии вечных снегов, на Алтае —до 1200 м,
в с.-з. Монголии до 1700 м (Сушкин, 1938). И, наконец, в культурном ландшафте
для гнездовья используются различного рода строения и сооружения, как в ни»
селенных пунктах, так и вне их.
Численность. Наличие и численность черных стрижей главным
образом обусловлены наличием подходящих гнездовых мест. Они обыкновении
поэтому в лесной и горной зонах, весьма богатых удобными для помещении
гнезд местами, и становятся более редкими и спорадично гнездящимися и ноне
открытых сухих пространств, где удобных для них мест меньше. Однако, именно
в этой зоне, за последние ЗОлет в связи с повсеместным сооружением высоких но-
черный стриж (У27
строек численность стрижей повсюду возрастает. На Алтае гнездятся только
в лесах (Щульпин, 1928). То же явление отмечалось для Урала. Однако в
настоящее время черные стрижи гнездятся и в высоких строениях уральских
городов. Но вообще говоря, численность черных стрижей в южной части
Украины, в Поволжье, в полупустынных и пустынных районах Казахстана, а
также в Забайкалье меньше, чем в других местах.
Экология. Размножение. Черные стрижи образуют постоянные пары.
К размножению приступают быть может только с третьего года своей жизни.
Гнездовые места занимаются вскоре после прилета, причем отдельные пары
ежегодно возвращаются в одно и то же гнездовое место. Селятся или отдельными
парами, что чаще наблюдается в лесах, а также в береговых обрывах, или
группами по 7—10—15 пар, или же сотенными колониями, что отмечается в гористых
районах. Устраиваются или отдельными колониями или вместе с другими
колониально гнездящимися птицами: белобрюхими стрижами, городскими
ласточками, голубями и пр. Иногда селятся в гнездах орлов, орлана белохвоста, скопы
и белых аистов (Зарудный, 1897). Местами гнезд в лесах являются дупла и
выбоины в самых различных высоких деревьях, при этом используют нередко
углубления и дупла, выдолбленные в стволе дерева дятлами. В обрывах
открытых равнин гнезда помещаются в различного рода щелях, углублениях или
в норах, сделанных другими птицами. В горах гнезда устраиваются в трещинах,
расщелинах скал, в пустотах или на выступах, прикрытых сверху
нависающими глыбами камней. И, наконец, в культурном ландшафте гнезда можно
находить в самых различных местах: в отверстиях, дырах, выбоинах и щелях стен,
под карнизами, под крышами высоких зданий, водонапорных и других башен,
колоколен, в щелях и трещинах железнодорожных и шоссейных мостов, под
крышами различных деревянных построек, чаще всего отдельно стоящих, где
их менее беспокоят, но иногда и под крышами веранд, домов, за наличниками
окон и проч. Гнездятся также в скворечниках и других дуплянках после
скворцов, иногда же гнезда отбиваются ими у воробьев, с которыми они вступают
в драки и изгоняют их даже в тех случаях, когда у последних имеются яйца или
птенцы (Зарудный, 1910).
Гнездо строится из различных материалов, поднимаемых ветром в воздух
и собираемых стрижами на лету: соломинок, мелких листочков, лоскутков,
ниток, волос, шерсти и проч., склеиваемых липким и быстро твердеющим в
воздухе секретом подъязычных желез. Все это складывается в небольшую
плоскую кучу, в которой лоточек обозначен мало заметным вдавлением. Величина
впервые сооруженного гнезда мала и доходит в диаметре до 9 см. Но старые,
неоднократно подправляемые и достраиваемые гнезда иногда достигают
15 см в поперечнике. Филатов (1915) в Калужской обл. в дуплах дубов
находил гнезда стрижей с птенцами без всякой подстилки.
Вместе с постройкой гнезда идет процесс спаривания. При этом самец более
или менее долго преследует самку, которая, наконец, замедляет свой полет.
В такой момент самец, перевернувшись в воздухе спиной вниз, подлетает к ней
снизу и на мгновение прикладывает свою клоаку к клоаке самки. В таком
положении птицы несколько секунд скользят в воздухе вниз, а затем
разлетаются (Огнев, 1948).
Полными данными о сроках гнездовых явлений мы пока не располагаем.
Есть основания считать, что у северной границы ареала стрижи приступают
к гнездовым делам сразу же по прилете, однако проходит около 15 дней прежде,
чем будет отложено ими первое яйцо. В такие же примерно сроки начинается
кладка в западных и центральных областях РСФСР, но на юге, в зоне открытых
степных пространств, между первым появлением стрижей и откладкой первого
яйца проходит около 25—30 дней и птицы не сразу по прилете берутся за
гнездовые дела.
40*
628
ОТРЯД ДЛИННОКРЫЛЫЕ
Первые яйца откладываются 15—21 июня на Печоре, 13—19 июня на
севере Ленинградской обл. и 9—15 июня в ее южных частях, 1—11 июня в
Московской и Харьковской обл., однако в Смоленской полная ненасиженная
кладка найдена 22. VI. 1902 (Огнев). Первые отложенные яйца в окрестностях
Вятки были находимы 9. VI. 1918 и 21. VI. 1921 (Плесский). Последующие яйца
в количестве 2—4 откладываются через день. Яйца продолговато-овальные, слабо
блестящие, белого цвета. Размеры: 22,8—27 х 16,0—17,24 лш, среднее
24,28 х 16,38 мм.* Насиживает около 18 дней одна самка, самец все это время
кормит ее.
Птенцов выкармливают оба родителя. В Ленинградской обл. общая
длительность кормлейия составляет около 19 часов в сутки и продолжается* около
35 дней. В начале родители кормят птенцов до 34 раза в день, а перед вылетом
только 4—6 раз (Промптов, 1940). Вес стрижа при вылуплении равен 2 г; его
рост заканчивается на 20—22-й день и вес в это время доходит до 43 г. Оперение
растет очень медленно. Зачатки маховых и рулевых появляются только на
8-й день и рост их, как и остального оперения, заканчивается лишь к самому
концу вылета на 38—40-й день.
Период размножения и явления роста птенцов приходится на время с
наибольшей продолжительностью дня (Вельский, 1947). Только после того, как
окончательно сформировалось перо, молодые покидают гнездо и сразу же
начинают летать. В северных и центральных частях ареала по вылете молодых
начинается их отлет с мест гнездовья, на юге же после вылета стрижи кочуют
в течение 10—15 дней вблизи мест гнездовья (Сомов, 1897), но держатся в этот
период в открытых местах и притом высоко в воздухе. Только ненастная
погода заставляет их спускаться вниз к земле.
Линька. Полная годовая, происходит вне пределов СССР зимой между
ноябрем и февралем.
Питание. Имеются лишь отрывочные данные, и питание стрижей у нас
по-настоящему еще не исследовано. Пищей стрижей Ленинградской обл.
являются главным образом двукрылые (Промптов, 1940). Для Череповецкого
края указываются комары и подденки, которых стрижи ловят над водоемами
(Богачев, 1927). Для средней полосы Европейской части СССР приводятся
жуки, бабочки, мухи, комары и оводы (Огнев, 1940). Для нагорных дубрав
Чувашской АССР —жуки, щелкуны и цикады (Першаков, 1932). Для
Харьковской обл. —летающие муравьи и мелкие насекомые (Сомов, 1897).
Имеются указания на нахождение в пищеварительном тракте стрижей пауков
и долгоножек.
Вообще говоря, стрижи в воздухе ловят самых разнообразных насекомых
и пауков, преимущественно мелких, или активно летающих или, что бывает
чаще', пассивно поднимаемых восходящими токами высоко над землею. Подолгу
собирая в воздухе различных насекомых, стрижи не глотают каждый отдельный
объект, а наполняют ими весь рот. Только склеив добычу секретом
подъязычных желез и сжав ее в комочек, птицы проглатывают все пойманное сраму,
Такими же комочками, покрытыми затвердевающей слизью указанных желе:»,
они выкармливают и своих птенцов, глубоко засовывая им в глотку шарики
из насекомых. В ясную и теплую погоду пища добывается высоко в воздухе1,
в ненастную и по вечерам — низко над полями, лугами и водоемами. В
холодную — в городах и вообще в населенных пунктах у строений и иногда иод
открытыми сводами железнодорожных построек.
В поисках пищи совершаются очень далекие вылеты, иногда до 4Г> км.
Такие вылеты на предкавказские равнины мне приходилось наблюдать дажи
* ПоданнымСпангенберга при2—3 яйцах в кладке их размеры относительно крушил,
при 4-х — размеры яиц мельче.
ЧЕРНЫЙ СТРИЖ
029
в 70 км от мест ближайших колоний. Временами в связи с наличием в
определенных местах насекомых эти вылеты имеют определенное направление, что
наблюдается в Волжско-Уральской степи и что иногда ошибочно принимается за
ранние перелеты.
Стриж нуждается в воде и пьет ее, летая у самой поверхности водоемов
с открытым ртом и зачерпывая ее подклювьем. Грудь и брюхо при этом
касаются воды, отчего на последней остается легкая борозда.
Полевые признаки. На полете от ласточек черные стрижи отличаются своей
несколько большей величиной, слегка серповидно изогнутыми крыльями и одно-
Рис. 153. Черный стриж.
тонной черной окраской. Вместе с тем они значительно подвижнее,
стремительнее и неутомимее ласточек. Движения их легки, быстры, разнообразны и
одинаково хорошо они мчатся в воздухе и машущим и планирующим полетом. За
18 летных часов при скорости только в 100 км в час стрижи могут за летний
день сделать до 1800/а*. Во всяком случае все светлое время дня от утренних до
вечерних сумерек они проводят в воздухе, добывая в нем корм или отдыхая
и играя на лету. Держатся черные стрижи и охотятся всегда небольшими
стайками.
Стриж — весьма крикливая птица и на полете постоянно издает резкое
пронзительное — «стри-и», от которого и происходит его русское название.
Беспомощные на земле, по которой ходить они не могут, хотя и в состоянии
взлетать с горизонтальной поверхности, стрижи всей своей организацией и
630
ОТРЯД ДЛИННОКРЫЛЫЕ
особенностями биологии приспособлены к воздуху больше, чем какие бы то ни
было другие наши птицы. В силу того, что стрижи отличаются великолепно
выраженными летательными способностями и в громадном большинстве случаев
устраивают свои гнезда в недоступных местах, у них нет сколько-нибудь
серьезных врагов. Однако не раз указывалось в литературе, что чеглок и сапсан могут
ловить стрижей. В культурном ландшафте их гнезда иногда разоряют кошки,
в лесах же белки. Помимо этого в их гнездах заводятся иногда
паразитические мухи.
Экономическое значение. Стриж полезен, хотя масштабы
этой пользы, повидимому, незначительны.
Описание. Размеры и строение. Клюв небольшой, 5—7 мм от начала
лобного оперения, разрез рта 13—17 мм. Крыло серпообразное. Первостепенных маховых 10,
формула крыла 2>1>3... Хвосте вырезкой. Когти загнутые, острые, цепкие. Длима
тела 180—210 мм; размах крыльев 385—420 мм. Длина крыла 165—180 мм, редко до 185 мм.
Хвост около 65—85 мм, вырез хвоста 30—35 мм, цевка 10—12 мм. Вес 40—45 г.
Окраска. Птенец в гнездовом наряде черновато-серый с белым горлом и
беловатыми каемками перьев. К моменту вылета каемки оббиваются, сохраняясь частично
на спине, надхвостье и нижней стороне тела. После первой весенней линьки птица
значительно темнеет, становится буровато-черной, горло белеет, каемки же перьев еще
хорошо выражены на брюхе, подхвостье и на нижних кроющих крыла. Взрослые самец
и самка буровато-черные со слабым зеленоватым отливом. Горло грязнобелое. Беловатые
каемки перьев сохраняются на сгибе крыла, иногда на нижних кроющих крыла. Летом перья
птицы значительно выгорают и общая окраска бледнеет. Клюв, ноги и радужина черные.
Азиатский черный стриж Apas a pus pekinensis S win hoe
Cypselus pekinensis. Swinhoe. Proc. Zool. London, 1870, стр. 435, Пекин.
Синонимы. Apus apas marwitzi. Reicheno'w. Orn. Mon. 1906, стр. 171, степь
Вембере в вост. Африке. — Apus apus turkestanensis. Зарудный, Орнитол. Вестник, 1911,
стр. 142, Туркестан.
Местные названия: узун-канат (у киргизов), ат-гарлывач (у туркмен).
Распространение. Ареал. От о-ва Кипра, сев. Ирана, равнин
Туркмении до Кара-Богаз-гола и берегов Аральского моря; далее на восток
северная граница не ясна; начинаясь от Кызыл-Рая и Зайсана, совпадает с
южной границей ареала. Южная граница от п-ова Ляодуна и центральных частей
провинции Чжили до Ланьчжоу и Бейка в провинции Гань-су, хребта Бурхан-
будда в верховьях Хуан-хе и Гималаев, где на гнездовье был найден в сев.
Качаре. Не установлен для южной части Белуджистана, Сеистана и побережья
Аравийского моря, но на юг обычен до Шираза, Керманшаха и отсюда через
Ирак до Палестины и Сирии.
На зимовках занимает, главным образом, вост. Африку от верховьев Белого
Нила до Калахари и Трансвааля. Рядом авторов указывается на зимовках в
зап. Индии, однако Бекером (1927) он здесь не найден.
Характер пребывания. Перелетная птица.
Даты. Из Африки отлёт начинается, повидимому, с конца февраля,
так как, с одной стороны, 5 марта найден в Камеруне, а, с другой стороны, в
начале марта появляется в пределах СССР. 17. III, 1935 уже был многочисленси
в Иране у Джизфуля. В Туркменской ССР в зап. Копет-Даге одиночный
отмечены б. III. 1942 у Дузлу-Олума и 8 марта у Кызыл-Арвата; 9. III. 1942 и
Ашхабаде, где 12 марта их уже было много (Дементьев, 1950), 25. III. 1886 — у
Артыка (Радде и Вальтер, 1889). Весеннее движение в Туркмении растягивается
до апреля: до 19. IV. 1892 у Казанджика и до 20—23. IV. 1892 у Ашхабада
(Зарудный, 1896). Близ Караганды стрижи появляются между 1 и 26. V (но
данным за 10-летие 1932—1942, Беме). Далее на восток у Джаркента одиночные
7. IV. 1900 и в массе 16. IV. 1900 (Зарудный и Кореев, 1907). В мае стрижи
появляются уже в Монголии: 9. V первые отмечены на Орок-норе, а 17. V
в Кентее (Козлова, 1930). В восточной части Монгольской Народной
республики прилет 9. V. 1941, 8. V. 1942, 17. V. 1943, 13. V. 1944 (Кучсрук).
ЧЕРНЫЙ СТРИЖ
631
Осеннее движение к местам зимовок в Туркмении начинается рано и
отсюда стрижи исчезают к началу июля. В конце июня 1942 встречались уже
редко и в небольшом количестве в предгорьях Копет-Дага, в Багире и Геок-
Тепе, а 1. VII. 1942 уже отлетели из Ашхабада (Птушенко). В конце июня —
начале июля исчезают из вост. Туркестана, в конце июля из Пекина; 30. VII
появляются на Андаманских о-вах, 17. VIII вюжн. Индии и в этом же месяце
идет пролет в Египте и Сомали. Более северные с мест гнездовья отлетают
позднее. 31 июля и 1 августа пролетные наблюдались в Китае — в с.-в. Нань-
Шане, в Монголии—-в степях по р. Чагрын-гол, а 10—20. VIII пролетные
стрижи почти ежедневно отмечались в долинах озер Цаган-нор, Далай-Дабасу
и Куку-нор; окончательно исчезает из Монголии к половине сентября
(Пржевальский, 1876). 21. VIII. 1902 пролетные во множестве держались в
Семиречье, в окрестностях У ял-Куль, наиболее же интенсивный пролет в
Семиречье наблюдается 23 августа—2 сентября; уже 10—20. IX. 1902 под
Джаркентом замечались лишь в небольшом количестве, а 21 сентября
отмечен одиночный (Зарудный и Кореев, 1906). Осенью летят иногда тысячными
стаями, что наблюдалось 13. IX. 1870 в 150 км южнее Урги. В густых
сумерках на высоте около 50 м над песками птицы летели несколько минут
огромной беспорядочной стаей в юго-восточном направлении (Пржевальский, 1876).
Биотоп. В зоне сухих открытых пространств занимает в большинстве
случаев вертикальные обнажения, иногда отрицательные по отношению к
общему рельефу местности, островные леса и поселения человека. В горной
зоне, — пояс лиственных лесов и горные луга до 2000—2500 м (Зарудный и
Кореев, 1906), но в горах Кара-Тау местами занимает населенные пункты
(Шапошников, 1931); а на кормежках доходит до 4000 м.
Численность. Везде более или менее обыкновенен, но колонии
разбросаны вдалеке одна от другой, что производит впечатление весьма
разреженного населения, хотя в некоторых городах с крупными строениями он много-
численен.
Экология. Размножение. Места гнездовий, как и у европейского
черного стрижа; в Средней Азии гнезда иногда строятся в могильниках. Близ
ст. Борзя в Забайкалье гнезда помещаются иногда за карнизами строений
разъездов; в Кентее обнаружены гнезда в дуплах лиственниц (Долгушин,
1941). В восточной части Монгольской Народной республики гнезда в
большинстве случаев встречаются под крышами строений и кумирен и только в
Далтыйн-хиде в скалах, вместе с гнездами Л. pacificus (Кучерук). Совместные
с другими стрижами колонии указаны также для Байкала, Хингана и Средней
Азии. Гнездятся также вместе с голубями, береговыми ласточками, даже
филином (Алфераки, 1891).
Первоначально сооруженное гнездо представляет собою довольно
миниатюрную постройку шириной в 9—10 см, высотой в 14—16 мм. Она плоска и,
как паутиной, смазана затвердевшим секретом подчелюстных желез, что
наблюдалось мною в Кара-Кумах в развалинах крепости Кызылча-кала. Но в случае,
если гнездо занимается несколько лет подряд, оно массивно и похоже на
беспорядочную кучу из соломинок и сухих листьев с выстилкой в середине из
перьев, шерсти и волос. В предгорьях Копет-Дага стройка гнезд идет во второй
половине апреля. 24—26 апреля в окрестностях Ашхабада Формозов
наблюдал спаривание стрижей в воздухе, а в конце апреля уже находили здесь
полные кладки (Шестоперов, 1937). В северных же Кара-Кумах, в стенах
крепости Кызылчакала 10. V. 1945 были еще только что отстроенные гнезда без
яиц (Птушенко). Наблюдений за другими явлениями гнездовой жизни почти
нет. Визг стрижей в гнездах в Монголии Кучерук слышал 9.VI 1.1941 в башне
Чингиз-хана и 12.VI 11.1942 в Мандалин-хиде. Хорошо летающие молодые
птицы на Ононе найдены в начале августа 1941 г. Долгушиньм.
632
ОТРЯД ДЛИННОКРЫЛЫЕ
Питание. Пища, как и у европейской формы, состоит из различных
насекомых и паучков, добываемых на лету в воздухе, но в связи с особенностями
распределения насекомых и воды на большей части ареала этого подвида, его
кормовые вылеты значительно длиннее, и зачастую его можно наблюдать в
нескольких десятках километров от гнездовой колонии. В широких долинах Тар-
багатая встречаются вдали от колоний, расположенных в горах (Хахлов, 1926).
В Кара-Кумах, в Репетеке 28 и 29 мая наблюдали охотящихся стрижей,
ближайшая колония которых имеется в Чарджоу, т. е. более чем в 60 км от мест
гнездовий. Стрижи прилетали с севера и, наловив добычи, улетали в северном же
направлении (Шестоперов, 1929). По встречам стрижей в Арал о-Каспийских степях
всегда безошибочно можно судить о близости водоемов, куда они прилетают
утолять жажду и ловить насекомых. Подобные кормовые вылеты в гористых
местах поража'ют своей стройностью. Во время насиживания кладок самками
все самцы соединяются в стаю, далеко отлетающую на охоту; возвращаясь,
все самцы одновременно влетают в гнезда, кормят самок и, как по сигналу,
стремительно и одновременно снова улетают (Зарудный, 1896).
Описание. Размеры и строение. Немного мельче номинальной формы. Длина
самцов (7) 180—190 мм, самок (13) 180—190 мм; размах самцов (7) 400—420 мм, самок (13)
395—415. Крыло самцов (16) 156—178, самок (19) 153,4—175,3, в среднем 170,9 и 168 мм.
Окраска. Несколько бледнее, чем у европейского стрижа. Лоб серовато-бурый,
отличающийся от цвета спины, горловое пятно больше и белее, чем у европейской формы, под-
хвостье светлее.
2. Белобрюхий стриж Apus melba L.
Hirundo melba. Linnaeus. Syst. Nat. изд. X, 1758, стр* 192, Гибралтар
Распространение. Ареал. Разорванный ареал вида отдельными своими
частями располагается в Евразии и Эфиопии. Большая, северная часть ареала
захватывает пространство от Пиренейского п-ова и Марокко до Кашмира,
Гималаев, Индии и Ассама. Меньшая приходится на Эфиопскую обл., где
белобрюхими стрижами занята вост. Африка от Абиссинии до Капской
Земли. Одна из форм заселяет о-в Мадагаскар.
Характер пребывания. Северная форма полностью или
частично перелетная, остальные оседлы.
Биотоп. Горная зона со всеми ее подзонами, а также скалистые и
обрывистые участки побережий и подгорных равнин.
Подвиды и варьирующие признаки. Семь подвидов: Apus melba melba L., 1758
(крупная форма)— от с.-з. Африки и Пиренеев до Ассама и Декканского нагорья; А. т. bakeri
Н а г t e r t, 1928 (более мелкая и темная форма) — южн. Индия и Цейлон; А. т. archeri
Н а г t e r t, 1928 — п-ов Сомали; А. т. striates Meinertzhagen, 1937 — Кения
в вост. Африке; A.m. maximus О g i 1 v i e-G rant, 1907 — горы Рувензори в тропической
Африке; А. т. africanus Temmink, 1815 — горы вост. Африки от Абиссинии до Капской
Земли (за исключением гор, указанных выше) и А. /п. willsi H а г t e r t, 1896 — о-в
Мадагаскар.
Евразийский белобрюхий стриж Apus melba melba L.
Синонимы. Apus melba tuneti. T s с h u s i. Orn. Jahrb. 15, 1904, стр. 123, Тунис. —
Apus melba petrensis. Bangs. Proc. Biol. Soc. Wash. 24, 1911, стр. 195, южн. Палестина
Распространение. Ареал. На севере Пиренейского п-ова, южн.
Франции, Альп, Балканского п-ова, Малой Азии, Крыма, Сев.Кавказа, Копет-
Дага, гор Средней Азии — на север до Кара-Тау, Гималаев, Ассама и
Декканского нагорья. На юг отсюда до сев. Белуджистана, Ирана (за исключением
Сеистана, а также побережий Аравийского моря и Персидского залинм),
Палестины, о-вов Средиземного моря, гор сев. Африки, Туниса, Алжира
и Марокко. Залеты известны для Канарских и Британских о-вов, сон.
Франции, Германии, о-ва Гельголанда, Дании, Чехословакии, а в СССР для
Днепропетровска, Донецкой и Харьковской обл., также для окрестностей
БЕЛОБРЮХИЙ СТРИЖ
633
г. Чкалова. Зимовки. Благодаря значительному сходству евразийского
белобрюхого стрижа с африканскими, его места зимовок до настоящего времени
с точностью еще не установлены и возможно будут выяснены только методом
кольцевания. На зимовках же указывается для южн. Индии, южн. Аравии,
Белого Нила в окрестностях Лаго, гор Рувензори, Дамараленда, в ю.-з. Африке.
Характер пребывания. В большей части ареала пролетная
птица, но стрижи, населяющие Гималаи, спускаются на зимовку в равнины
Индии и, в частности, из Восточных Гималаев в районы Качара и Силхета.
Данные по весеннему пролету недостаточны. С мест зимовок начинают двигаться
Карта 108. Распространение белобрюхого стрижа Apus melba
1 — A. m. melba, 2 — A. m. bakeri, 3— A.m. arched, 4 — A. m. striatus, 5 — A. m. maximus,
6 — A. m. africanus, 7 — A. m. willsi; a — граница области распространения, б — не вполне
выясненная часть границы, г — предполагаемая область зимовок.
уже с февраля, так как в конце этого месяца были отмечены в Сирии, но
основные сдвиги с зимовий происходят в марте, и в этот последний месяц белобрюхие
стрижи долетают до Алжира, южн. Аравии и пределов СССР, хотя еще и в
апреле идет пролет в сев. Африке и по о-вам Средиземного моря. Первые
в Туркмении появляются в конце марта, в Узбекистане — в начале апреля
(2 апреля в окрестностях г. Самарканда). Пролет в Средней Азии идет с апреля
и здесь они отмечены: 6.IV. 1917 на Больших Балханах (БилькевичиЗаруд-
ный, 1918), 14. IV. 1942 в окрестностях г. Ашхабада, 17. IV. 1946 в Бадхызе
(окрестности Навруз-абада, Дементьев, 1947), 19. IV. 1887 в окрестностях Тахта-
Базара (Радде и Вальтер, 1889), 22. IV. 1892 в Кара-Кумах у ст. Пески (Заруд-
ный, 1896). В начале мая первые появляются уже на северном склоне Главного
Кавказского хребта (5 мая в Осетии; Бёме, 1926), в Крыму первые наблюдаются
в первой половине мая. Сроки осеннего пролета известны мало.
634
ОТРЯД ДЛИННОКРЫЛЫЕ
С Сев. Кавказа (Осетия) исчезают в конце августа. В Рутульском округе
Дагестана огромная стая, перемещавшаяся на юго-запад, наблюдалась 22.VIII.
1922 (Красовский, 1932); из центрального же Дагестана местные отлетают II —
20 сентября (Россиков, 1884). Отлет из Туркмении совершается в сентябре —
начале октября (Дементьев, 1950), из Самарканда б сентября.
Рис. 154. Белобрюхий стриж.
Биотоп. Скалы, обрывы побережий морей, озер и рек, обрывы холмов,
утесы горных ущелий, населенные пункты (в Самарканде —. в щелях старых
мечетей — сообщение Бобринского). В Туркмении (Бадхыз, ущелье Кизыл-
Джар). Гептнер в июне 1942 г. нашел колонии белобрюхого стрижа в
вертикальных обрывах, располагающихся ниже окружающих обширных равнин. В
горной зоне до горных лугов, местами же обычен до 2000—2500 м.
Численное т'ь. Неравномерна. Колонии на далеких расстояниях
одна от другой, и местами белобрюхий стриж многочисленен (на северном склоне
Главного Кавказского хребта), местами вовсе отсутствует (в Аджарии
и Гурии). Даже в соседних районах численность неравномерна: на горе
Давида в окрестностях Тбилиси колония его невелика, в ущелье же р.
Храма имеются огромные колонии (Цветков, 1901). То же и в Средней Азии:
БЕЛОПОЯСНЫЙ СТРИЖ 635
в Кара-Тау он многочисленен, в хребте же Нура-Тау Мекленбурцев нашел
только одну гнездящуюся пару.
Экология. Размножение. Гнездится колониями в 20—30 или в
несколько сот пар (ущелье р. Храма) или, что реже, отдельными парами. Гнездо
строится из различных материалов, добываемых стрижем на лету. В нем
преобладают перья и шерсть на равнинах и части растений в высокогорных районах.
Гнездо или так мало, что не вмещает птицу, или, в случае если занимается по
нескольку лет подряд, представляет собою большую кучу, в диаметре почти
равную размерам стрижа. Кладка состоит в большинстве случаев из 2—3 яиц,
редко 4 (Бартельс, 1931). Яйца белые почти без блеска, более хрупкие, чем
у обыкновенного стрижа. Размеры (81) 30,1—34,3 мм х 17,8—-27,5, в среднем
31,1 х 19,2 мм (Джоурден).
Сроки откладки яиц мало известны. По сообщению Спангенберга 8. V. 1947
близ г. Самарканда в одном из гнезд добыто два ненасиженных яйца.
Откладываются яйца с промежутками не меньше, чем в 48 часов, и насиживание
начинается с первого яйца, что в ряде случаев влечет за собою
смертность младших птенцов. Роль самки и самца в насиживании недостаточно
выяснена. Насиживание от 18 до 21 дня. Выкармливание около 40 дней.
Кормят оба родителя до 24 раз в день. Принос корма птенцам и визг молодых
из гнезда отмечены мною 1. VII. 1914 на Кавказе в верхнем течении р. Кубани,
близ г. Клухори. 18 июля в Средней Азии, в Кугитанге из гнезда взят птенец
с еще недоросшими махами. Вылет птенцов из гнезда наблюдался в Дагестане
в конце первой трети июля 1892 г.
Питание. Пища состоит из различных, преимущественно мелких,
насекомых, добываемых в воздухе. Зачастую это комары и далее мошки,
собираемые в огромных количествах. Характерны весьма дальние кормовые вылеты на
равнины и речные долины. Хитии сбрасывается в виде погадок.
Полевые признаки. На лету ясно различимы светлое брюхо и более крупные,
чем у черного стрижа, размеры, что дает возможность сразу и безошибочно
различать оба этих вида. Полет быстрый, стремительный и более прямой, чем у
черного стрижа, но при ловле насекомых или при движениях над водою стриж
может летать медленнее и проявляет значительную поворотливость. Голос —
чистое, громкое и более мелодичное, чем у черного стрижа, «скри-и»... «скри-и».
В период размножения из гнезд часто слышен визг, который может
продолжаться почти всю ночь (в ночь с 1 на 2. VII. 1914, окрестности Клухори).
Описание. Размеры и строение. Клюв небольшой в 9—10 мм, но его разрез
доходит до 25 мм, что дает возможность птице очень широко раскрывать рот. Крылья
серповидные, формула 2 > 1 > 3... Хвост с вырезкой и разницей между средними и крайними
рулями от 15 до 26 мм. Пальцы очень цепкие с изогнутыми острыми когтями. Самцы
несколько крупнее самок. Крыло самцов (14) 210—225, самок (14) 213—227, в среднем
222 и 217,9 мм. Размах самца (1) 540 мм. Хвост 70—95 мм, цевка 15—18 1мм. Вес
95—110 г.
Окраска. У молодых темные перья низа и перья крыла с хорошо выраженными
беловатыми краями, с возрастом постепенно исчезающими, но всегда заметными в свежем
пере. Взрослые самец и самка сверху бурые, снизу белые. Поперек белого низа через
грудь идет бурая полоса. Испод крыла и подхвостье бурые. Клюв черный. Радужина
темная. Ноги розовато-желтые.
3. Белопоясный стриж Apas pacificus Latham
Hirundo pacijica. Latham. Ind. Orn. Suppl., 1801, Австралия
Распространение. Ареал. Разорванный ареал вида состоит из трех
участков. Первый, наибольший — от Зап. Сибири до Камчатки, Японии и
Ганьсу в Китае; второй захватывает почти целиком Гималаи; третий
наименьший —. центральные части Индостана.
636
ОТРЯД ДЛИННОКРЫЛЫЕ
Биотоп. Преимущественно горная зона, частью же—культурный
ландшафт.
Подвиды и варьирующие признаки. Три подвида, различающиеся между собою
размерами и окраской: Apas pacifictis pacificus Latham, 1811 — свойственный северной
части ареала; А. р. leuconyx В 1 у t h, 1845 — Гималаи и Декканское нагорье; A. a. cooki
Harrington, 1913 — Индостан.
Сибирский белопоясный стриж Apas pacificus pacificus Latham
Синонимы. Hirundo apus var. leucopyga. Pallas. Zoogr. Rosso-Asi;it. 1H11,
стр. 539, Байкал. — Micropus pacificus kurodae. Domaniewski. Acta Orn. Mns.
Zool. Polonici, 1, 1933, стр. 80; Япония. — Micropus pacificus kamtschaticus. Domani v-
wski. Acta Orn. Mits. Zool. Polonici, 1933, стр. 80, Петропавловск на Камчатке.
Русское название. Белопоясный стриж, белозадый стриж, белонадхвостпыи
стриж (книжные).
Распространение. Ареал. Северная граница распространения белопояс-
ного стрижа от гор Аир-Тау, озера Копчи, Зеренды и Щучьего в Акмолинской
обл. следует далее вдоль поднятия Казахской складчатой страны к зап.
Алтаю, откуда резко, почти к северу, поднимается к бассейну р. Томи,
среднему течению р. Чулышмана, к устью которого залетают только отдельные
стайки и особи, доходит до 58°3б' с. ш. на Енисее, до 64° с. ш. по среднему
течению Нижней Тунгуски (Ткаченко, 1937) и до этой же широты на Лене, где
гнездится в скалистых обрывах устья Вилюя и западнее по этой же реке в Сунтаре
(62°42' с. ш., Скалой). Однако Воробьевой (1928) при ее поездке в июне-
августе 1927 г. от Вилюйска до Сунтара и обратно белопоясный стриж ни для
одного пункта на этой реке не упоминается. К северу от устья Вилюя
распространен в системе Верхоянского хребта и по р. Батынтаю доходит
приблизительно до 67° с. ш. Встречается также на Саккырыре, в бассейне же р.
Колымы регулярно гнездится за полярным кругом в г. Средне-Колымске
(Скалой, 1927). Далее на восток найден в районе Пенжинской губы и по р. Пенжинс
(Дементьев, 1940), на всей Камчатке, но на Командорах, вопреки
утверждению Питерса, не гнездится (Бутурлин, 1936). На юг отсюда населяет острова
Японии, даже мелкие, например, «Семь островов Изу», Корею, Сев. Китай ол
Ян-цзы-цзяна, северную часть Сычуани, Ганьсу, Алашань, восточную,
северо-восточную и северную часть Монгольской Народной республики и
западный Алтай. Южная граница ареала в Казахской складчатой стране не ясна.
Зимою встречается в Ассаме (Манипур), Бирме и южной части Китая, п
Аннаме (в Фанранге),в Вьетнаме, в бухте Алонг, на о-ве Катба и Пуло-Кондоре,
и доходит до Австралии, где встречается в декабре — феврале.
Характер пребывания. Весеннее движение через азиатский
материк остается невыясненным, хотя, повидимому, часть птиц летит именно
таким образом.
Главная же их масса, как кажется, весною перемещается вдоль
побережья и островов воет, Азии и уже отсюда направляется в разные пункты
в глубь материка.
Данные по весеннему пролету ничтожны. Довольно рано первые прилетные
появляются в южн. Японии (окрестность г. Нагасаки, экз. добыт 18 марта).
Севернее, в центре о-ва Хондо (в районе вулкана Фуджияма) первые отмечены
3 апреля; массовый же прилет их здесь приходится на середину апрели,
(Ян, 1942). В конце этого месяца или в начале мая первые показываются и
Дальневосточном крае СССР. Массовое их появление в уроч. Славянке
Медведев наблюдал 23. V. 1912 и 19. V. 1913. Передовые же в это время, дшпдись
вдоль побережья, успевают добраться до Колымского хребта, перевалить его и
спуститься в бассейн р. Колымы, где в Средне-Колымске первые отмечаются
15 мая (Скалой, 1940). На материке местами также появляются рано. Весной
БЕЛОПОЯСНЫЙ СТРИЖ
637
1872 г. в юго-восточной Монголии Пржевальским первые в горах Муни-Ула
отмечены 24 апреля, но в Ганьсу они прилетают только в конце мая.
Сравнительно рано передовые оказываются в окрестностях г. Иркутска. По
многолетним наблюдениям Пуляевского (1937) сюда в среднем белопоясные
Карта 109. Распространение белопоясного стрижа Apus pacificus
7 — А. р. pacificus, 2 — А. р. leuconyx, 3 — А. р. cooki; a — граница гнездовой
области, б — не выясненная часть границы, г — зимовки.
стрижи прилетают 11 мая, с крайними отклонениями от 27 апреля (1930)
до 18 мая (1926). Массовое же появление здесь должно происходить
позднее. Валовой пролет в Чите идет в последней декаде мая. Повидимому,
массовое появление этих стрижей для с.-в. части Монгольской Народной
республики указывается Кучеруком 21. V. 1941, 29. V. 1942 и даже 4. VI. 1943.
В Якутию стрижи эти весной прилетают с востока, так как раньше всего они
показываются на восточных склонах горных хребтов, даже расположенных
далеко на севере, и уже затем на западных. В центральную Якутию первые
638
ОТРЯД ДЛИННОКРЫЛЫЕ
прилетают в середине или конце второй декады мая (15 мая в Новопокровск,
18 мая в Якутск). Только в конце мая или начале июня передовые достигают
системы Верхоянского хребта, причем на его восточных склонах они
наблюдаются уже в конце мая, а на западных — только в начале июня. Это
обстоятельство и затем то, что в Средне-Колымске первые стрижи отмечаются раньше,
чем в более южных местах Якутии, Скалой ставит в связь с движением бело-
поясного стрижа от морского побережья вост. Азии в глубь этой
страны.-Массовое появление этих
стрижей в Якутии
происходит на 5—8
дней позже прилета
передовых (Скалой,
1927). В Минусинском
крае появление
первых стрижей на Тубе
отмечено только 18
июня (Сушкин, 1914).
В Красноярске бело-
поясные стрижи
наблюдаются уже в
конце мая (30, V. 1924
и 31. V. 1921
—Шухов, 1925).
Еще меньшее
количество данных
имеется по осенним
передвижениям белопояс-
ных стрижей. Из
Якутии по данным Скало-
на они исчезают в
середине августа, 13.
VIII. 1937 в
центральной Якутии
количество их заметно
снизилось, 14. VIII. 1937
наблюдались
последние стайки, а 17. VIII,
1937 отмечен
одиночный. Из Урянхайской земли они исчезают к 21 августа (Сушкин, 1914); из
Прибайкалья в конце августа; в августе же из восточных частей Монгольской
Народной республики (14—22. VIII. 1941 и 27. VIII. 1942, из окрестностей Чойбал-
сана, Кучерук); с середины сентября с Шантарских о-вов (Дулькейт и Шуль-
пин, 1937) и Японии. Однако пролет их в Кобе (о-в Хондо), где они не гнездятся,
наблюдался 20 сентября, а 22 октября отмечена большая их стая у вулкана
Фуджияма (Ян, 1942). В пролетное время найден в Тарбагатае и в районе
Самарканда, последний очевидно залетный.
Биотоп. Культурный ландшафт с поселениями человека и горная зона,
в которой доходит до 4000 м над уровнем моря (Пржевальский).
Численность. Редок в Зап. Сибири, Алтае и Прибайкалье,
обычен в Забайкалье и снова уменьшается в численности в бассейне Амура.
Местами, как, например, в Ганьсу, численность одинакова и в верхнем и в
нижнем поясах гор.
Экология. Размножение. Колонии от нескольких до
нескольких сот пар. На равнинах занимает высокие постройки (старинные крепостные
Рис. 155. Белопоясный стриж.
МАЛЫЙ СТРИЖ
639
башни, церкви, заводские трубы), в щелях которых помещает гнезда, или же
устраивается под крышами высоких каменных строений. В гористых местах
колонии располагаются по неприступным скалистым берегам рек, или на
различного рода утесах, скалах и обрывах, иногда выше пояса тайги.
Необходимым условием устройства колонии является соседство значительного
водного пространства (Шухов, 1930). Гнездится часто вместе с городскими
ласточками. Интересно сообщение о колонии в гроте над р. Чарыш близ Усть-Канска,
где белопоясные стрижи гнездились совместно с черными (Кащенко, 1899),
но на Ононе всегда находили только отдельные колонии тех и других. По
сообщениям Гладкова, в том же Забайкалье городские постройки Читы
занимают A p. pacificus, в лесах же гнездится черный стриж. В Монгольской
Народной республике по наблюдениям Кучерука гнездится исключительно в
выходах скал. В Забайкалье же во всех высоких станционных сооружениях.
Гнездо строится из переносимых ветром сухих частей растений, стебельков
трав, соломинок, подхватываемых на лету, и представляет собою кучу,
склеенную секретом подчелюстных желез. Форма гнезда соответствует той щели или
трещине, в которой оно помещено. Кладка в Прибайкалье приходится на
середину июня и состоит из 2—3 яиц.
Размеры: 24—27,5; 16—17 мм (Хартерт, 1911). 15 июля близ Усть-
Канска шло выкармливание птенцов (Кащенко, 1899).
Питание. Пища состоит из различных насекомых, преимущественно
мелких. Птицы, зимующие в Бирме, ловят различных мелких пчел-
Линька происходит на зимовках. Детали неизвестны. Добытые у нас
в летнее время имеют мало обношенное оперение.
Полевые признаки. Узнается по белой перевязи в области надхвостья,
причем от городских ласточек, с которыми часто встречается вместе, отличается
большей величиной, стремительностью полета и голосом. Он в общем молчаливее
черного стрижа, голос его не так громок, пронзителен и резок, как у
последнего, и может быть передан, как двухсложное «сии-иррр». Но во время игр и
при преследовании самок этот стриж может быть так же шумен, как и черный.
В его колониях можно слышать почти беспрерывный крик, несущийся из
различных гнезд.
При дождях, туманах или при летних снегопадах в горах спускается в
низкие горные долины или даже вылетает на подгорные равнины, где и охотится.
При хорошей же погоде держится в горах и охотится часто у высочайших
вершин (Скалой, 1927). В равнинных местах у скал, где эти стрижи селятся
колониями, их можно видеть летающими в разных направлениях весь день, за
исключением утра и вечера, когда они вылетают на охоту за насекомыми в степи
или равнины.
В Сибири, в тех местах, где водится черный стриж, белопоясный
охотится вместе с последним, но летает вечером значительно дольше
черного и его можно еще встретить в 12 часов и даже в час ночи (Тачанов-
ский, 1891).
Описание. Размеры и строение. Длина крыла самцов (15) 165—182, самок
(11) 167—180, в среднем 176 и 173,5 мм. Длина самцов (7) в среднем 192 мм, самок (2)
197,5 мм. Размах крыльев самца (1)428, самки (1)420,5 мм. Вес самца (1) 42,35, самки
(1) 44,7 г. Хвост 70—86 мм с вырезом от 26 до 35, цевка 9—11 мм. Клюв 6,5—7,5 мм, его
разрез 16—17 мм.
Окраска. Молодая птица сходна по окраске со взрослой, но верхняя сторона
несколько пестрее. Взрослые самец и самка сверху почти чисто черные со слабым
зеленоватым отливом. Весною перья верха несут узенькие беловатые ободки. Поперек
поясницы широкая белая полоса с темными наствольями отдельных перьев. Горло и
подбородок белые с такими же наствольями. Остальной низ темнобурый, но каждое перо
заканчивается сначала темной предвершинной полосой, а затем белой изогнутой каймою.
Радужина темнобурая. Ноги и клюв черные.
640
ОТРЯД ДЛИННОКРЫЛЫЕ
4. Малый стриж Apus affinis Gray
Cypselus affinis. Gray. III. Ind. Zool., 1, ч. 2, табл. 35, фиг. 2, без текста и указания
местонахождения; предположительная terra typica — долина Ганга
Распространение. Ареал. Разорванный ареал вида слагается из двух
крупных площадей, к которым примыкает по небольшому участку. Северная
площадь охватывает пространство от южн. Средиземноморья через Иран,
Индию и Индо-Китай до о-ва Борнео. К этой площади примыкает южная око-
Карта 110. Распространение малого "стрижа Apus affinis
1 — A. a. galilejensis, 2 — A. a. affinis, 3 — A. a. nipalensis, 4 — A. a. singalensis, 5 — A. a. subfurcatus,
6 — A. a. abissinicus, 7 — A. a. bannermani;
а — граница области распространения, г — зимовки.
нечность Индии и о-в Цейлон. Южная площадь занимает Африку от Гамбии до
Сомали на севере и от Анголы до Ньяссы на юге. К ней примыкает ряд островов
Гвинейского залива.
Характер пребывания. Перелетная и оседлая птица.
Биотоп. Горная зона.
Подвиды и варьирующие признаки. Семь подвидов: A. a. galilejensis A n t i n о г i, 1855 —
от Марокко до Белуджистана; A. a. affinis Gray, 1830 — большая часть Индии; А, а.
nipalensis Н о d g s о n, 1863 — Неаполь, Бутан и северная часть Ассама; A. a. slnga-
lensis Madarasz, 1911 — оседлый вид Траванкора в южн. Индии и о-ва Цейлона; А. а.
subfurcatus В 1 у t h, 1849 — Суматра, Борнео и другие о-ва; A. a. abissinicus Strcubcl,
1930 — тропическая Африка от Гамбии до Эфиопии на севере и от северной части
Анголы до Ньяссы на юге; A. a. bannermani Hart., 1923 — о-в Фомы, Принсипе и
Фернандо По.
иглохвостый стриж
641
Западный малый стриж Apas affinis galilejensis Ant in.
Cypselus galilejensis. Antinori. Naumannia, 1855, стр. 307, Палестина
Синоним. Micropus koenigi. Reichenow. Orn. Monatsb., 2, 1894, стр. 191, Дже-
бель-Эль-Меда в Тунисе
Распространение. Ареал. Марокко, Алжир, Тунис, сев. Сахара,
Палестина, Сирия, Иран на север до самых южных частей СССР в Средней
Азии, на юг до северной границы Сеистана, в самом Сеистане отсутствует,
Белуджистан и Афганистан, в долине Инда заменяется индийским малым
стрижем, В пределах СССР достоверно
найден на гнездовье в Туркменской,
Таджикской и Узбекской ССР. В Туркмении найден
в Вами, Баба-Дурмазе и Дурянчи, однако,
гнездовье его здесь не установлено, а
указания Лоудона на встречи малого
стрижа значительно западнее у Казанджика и
в районе Куба-Даг требуют подтверждения.
В качестве обычной гнездящейся птицы
найден Радде и Вальтером (1889) в
следующих местах ю.-в. Туркмении: в Пуль-и-
Хатум, Зюльфагар*, Кешеф-Руд, в горах
Эльбир у Акар-Чешме. Кроме того стрижи
эти были многочисленны между Ак-Рабат и
Кунгруили. Гнездится в хребте Гязь-Гедык
в Бадхызе и появляется очевидно на
кормовых вылетах у колодца Адам-елен к северу
от указанного хребта (Дементьев, Рустамов,
Спангенберг, 1947). В Таджикистане
гнездование малого стрижа впервые установлено
наблюдениями Федченко в Пяндже и Пян-
джанкенте. Затем в 1910 г. он найден За-
рудным и Билькевичем в горах Шураб-
дара и Гами-дара и, наконец, обнаружен
Алексеевым (1911) в ущелье Ак-Капчигай
к юго-востоку от г. Байсуна и в ущелье
р. Кафирнигана. Гнездование малого
стрижа в Узбекистане в 1934 г.
установлено также на хребте Нура-Тау.
Указания Радде и Сатунина на нахождение
этого стрижа на Кавказе и, в частности,
в Армении сомнительны, т. к. позже здесь стриж этот не был обнаружен
(Ляйстер и Соснин, 1942). Зимует в Африке (сев. Нигерия, озеро Чад)
и в южн. Аравии. Биотоп. Горная зона.
Характер пребывания. Перелетная птица. Даты для СССР
еще не установлены. Численность. В ю.-в. Туркмении обычен, в
Таджикистане и Узбекистане малочисленен.
Экология. Размножение. Гнездятся отдельными парами или
колониями на скалах в ущельях. В хребте Нура-Тау занимают гнезда городских
ласточек и устраиваются в прохладных тенистых и сырых местах. Здесь по
наблюдениям Мекленбурцева (1937) с начала мая и по 24.V.1934 птицы
держались у колонии, поправляли гнезда, совершали брачные полеты, во время
которых самцы преследовали самок, но яиц не откладывали. В Туркмении, в
колонии около Пуль-и-Хатум 17.VI 1.1887 шло кормление молодых (Радде и
Вальтер, 1889). В некоторых случаях гнездятся в колониях вместе с горными
Рис. 156. Малый стриж.
41 Птицы СССР т. I
642
ОТРЯД ДЛИННОКРЫЛЫЕ
ласточками (Зарудный, 1896); в Тянь-Гедыке — вместе с черными и
белобрюхими стрижами (Дементьев, 1950). Линька. Не изучена.
Питание. На кормовых вылетах встречаются и охотятся вместе
с черным и белобрюхим стрижами. В Бадхызе на кормежку стрижи вылетали
в определенное место, в котором в 25 км от их гнездовий сохранилась
зеленая трава, изобиловавшая насекомыми. Последние беспрестанно поднимались
из травы в воздух и здесь становились жертвой малых, черных и белобрюхих
стрижей, постоянно сюда прилетавших.
Полевые признаки. Мельче остальных стрижей. От городских ласточек,,
с которыми имеет внешнее сходство, отличается серпообразным строением
крыльев и стремительностью своего полета. Летает значительно быстрее
черных стрижей, отличаясь от них вообще неровностью, выражающейся в
беспрестанных, часто совершенно неожиданных поворотах. Во время брачного
полета при преследовании самок самцами последние иногда поднимают крылья
над спиною и, не двигая ими, медленно по наклонной линии спускаются вниз,
чтобы затем снова стремительно подняться кверху. Крик малого стрижа
слышится так же часто, как и у черного, но его голос тише и не так резок
и пронзителен, как у черного.
Описание. Размеры и строение. Клюв и ноги коротки. Обрез хвоста почти
прямой. Длина крыла самцов (5) 130—147, самок (2) 125—137, в среднем 132,8 и 131,2 мм.
Длина хвоста 39—49 мм. Цевка 9—10 мм. Клюв от начала лобного оперения 6—7 мм
(Бутурлин, 1936). Окраска. Молодая птица несколько бурее взрослой, без отблеска.
Крылья и хвост грязнобурые. На перьях надхвостья, брюха и подхвостья как и на
вершинах маховых, беловатые ободки. Взрослый самец и самка. Лоб беловатый, голова,
шея, крылья и хвост бурые. Спина блестяще черная. Полоса поперек поясницы белая.
Горло белое, низ черно-бурый.
5. Иглохвостый стриж Hirundapus caudacutus Latham
Himndapus caudacuta. Latham. Ind. Orn. Suppl. стр. LVII, 1801, Австралия
Русское название. Иглохвостый, колючехвостый и колючий стриж, также
колючая касатка основаны на особенностях строения рулевых перьев
Распространение. Ареал. В состав разорванного ареала иглохвостого
стрижа входят: средняя и Вост. Сибирь, Монголия, Сев. Китай, Япония и
Курильские о-ва, о-в Формоза, пространство от зап. Гималаев до зап. Сычу-
ани на юг через Индо-Китай и Малакку до Суматры и Явы. Характер
пребывания. В северном участке ареала перелетная птица, зимующая
в Австралии, в южном — или совершает вертикальные миграции и кочевки,
или ведет оседлый образ жизни. Биотоп. Лесная и горная зоны с
горнолесной подзоной.
Подвиды и варьирующие признаки. Четыре подвида, отличающиеся размерами и
окраской оперения. Из них два более крупные Я. с. caudacutus Latham, 1801 и И. с. nudipes
Hodgson, 1836 имеют различную окраску лба (светлый у первого и темный у второго).
Два более мелкие подвида, встречающиеся в ю.-в. Азии, различаются окраской перьев
груди, темной у И. с. formosana Jamaschina, 1936 и серовато-белой у Н. с. cochin-
chinensis О u s t a 1 е t, 1878.
Северный иглохвостый стриж Hirundapus caudacutus caudacutus Latham
Распространение* Ареал. Самым западным пунктом северной границы
ареала является бассейн р. Васюгана, где почти под 58° 30' с. ш. и 78° и. д.
Башутин нашел иглохвостого стрижа в долине р. Черталы, левом притоке Иа-
сюгана; встречен также и на последнем. Далее на восток найден в долине р. Кеть
по Обь-Енисейскому каналу и южнее, в бассейнах рек Чулыма и Томи, причем
в долине последней- несомненно гнездится. Гнездится в Салаире, на Алтае в
иглохвостый стриж
643
сев.-восточных и северных предгорьях (Сушкин, 1938), в Саянах в пределах
Минусинского края. Яковлевым добыт в 80 км от г. Енисейска у дер.
Абалаково и наблюдался у дер. Додоновой. 22, VII. 1896 Кибортом добыт
в окрестностях г. Красноярска и затем неоднократно наблюдался и
добывался Шуховым (1924) в Красноярском районе (у деревень Торгашевой,
Кубековой, Березовки и др.). Шухов указывает, что в этом районе
иглохвостый стриж известен еще с
двадцатых годов XIX столетия. Сушкин
(1914) устанавливает его гнездованье
по окраине Мажарской тайги.
Встречается в окрестностях Иркутска.
Несомненно распространен по р.
Лене, так как неоднократно
отмечался и был добыт в окрестностях
г. Олекминска и однажды близ г.
Якутска. На восток отсюда
является обыкновенной птицей бассейна
Амура, найден на Сахалине и
Курильских о-вах, на Хоккайдо,
северной части о-ва Хондо, в Корее,
сев. Манчжурии, Сев. Китае,
юго-восточной части Монголии.
Характер пребывания.
Перелетная птица.
Даты. Австралийские места
зимовок начинает покидать в марте,
но далеко не одновременно, так как
весенние передвижения носят весьма
растянутый характер и в то время,
когда одни стаи в мае достигают
самых северных границ своего
гнездового ареала, другие пролетают
еще только по самой южной части
Китая. На Байкале появляются
во второй половине мая, на берегах
Шилки первые отмечены 23. V. 1857
(Радде), а на плато Тунка 10. V.
1859, на озере Ханка
Пржевальский наблюдал их 2. V. 1868 и
6. V. 1869. В районе этого озера
на весеннем перелете стрижи движутся разрозненными стайками или даже
в одиночку, летят низко, почти над самой землей, но время от времени
подымаются высоко вверх, делают круги в воздухе и затем снова движутся у самой
земли. Отлетают из Прибайкалья в конце августа, но в западной части ареала
у самой северной границы своего распространения наблюдаются еще в сентябре
(17. IX. 1921 на р. Кеть и 30. IX. 1927 на р. Васюган). Так же долго
задерживаются и на крайнем востоке ареала, и на о-ве Сахалине добывались
еще 3 и 4. X. 1906. В то же время в сентябре некоторые птицы вылетают
из пределов своего гнездового ареала (очевидно сбившиеся с направлений
движения, 4. IX. 1909 были встречены Тризна в Средней Азии на р. Карачик
в предгорьях Кара-Тау, а 19. IX. 1908 Зарудным также в Средней Азии
на болоте Джаман-Баткак). Пролет в общем идет через Китай, Японию и
продолжается до ноября. В декабре птицы появляются в Австралии и
Тасмании, где и зимуют в течение января и февраля. Осенний пролет-
Карта 111. Распространение иглохвостого
стрижа. Hirund apus caudacutus
1 — Н. с. caudacutus, 2 — Н. с. formosanus,
3 — Н. с. nudipes, 4 — Н. с. cochinchinensis; a —
граница области распространения, г — зимовки.
41*
644
ОТРЯД ДЛИННОКРЫЛЫЕ
вдоль побережья Уссурийского края иногда носит характер почти
беспрерывного разрозненного потока, причем птицы летят низко над землею и
временами останавливаются в устьях рек для охоты.
Биотоп. Менее, чем другие стрижи, связан с горным ландшафтом
и в горах, как и на равнинах, занимает лесистые места, разрезанные
открытыми пространствами. На охотах
за насекомыми посещает
одинаково и горные луга почти
вплоть до линии вечного снега
и низменные болотистые низины.
Численность.
Неравномерна; редок в западной части
ареала, в небольшом количестве
представлен в районе Байкала и
многочисленен по Амуру, Уссури
и на Сахалине, становясь снова
более редким в восточной части
Центральноазиатского нагорья.
Экология.
Размножение. Точных данных по
размножению нет. Местом для
гнезда в большинстве случаев
служат дупла высоких, чаще всего
хвойных деревьев, но в низовьях
Амура Золотарев в 1934 г. нашел
гнездо в старом тополевом пне
высотой в 6—7 м. В Японии
гнездится в дуплах вяза на высоте
в 5—6 м над землею
Пржевальский (1870) предполагает в р-не
озера. Ханка возможность
гнездования иглохвостого стрижа в
скалистых оврагах, что для
Прибайкалья Дыбовский и
Годлевский категорически отрицают
(Тачановский, 1891), однако для
р. Томи указываются
колониальные гнездования этой птицы
именно в высоких скалистых бе-
Рис. 157. Иглохвостый стри>^с. реговых обрывах (Иоганзен, 1915
и др.). То же отмечено и в
Японии. Брачный период в Уссурийском крае по Пржевальскому протекает
в конце мая — начале июня и знаменуется энергичным преследованием самок
самцами. Кладка происходит в конце первой — начале второй декады июня
и состоит чаще всего из двух белых яиц. Размеры: (19) 27,5 — 32,2 х 17,5—19,5,
среднее 22,74 х 18,18 мм (Хартерт, 1911).
Молодые покидают гнезда в конце июля—начале августа. С этого же
времени начинается период кочевок, длительность которого определяется
климатическими условиями районов гнездования.
Линька. Начинается на родине. Детали неизвестны. Добытые 23. VII.
1910 в Приморье взрослые птицы имели сильно выгоревшее оперение и
крайне обношенные концы первостепенных маховых. В конце июля у таких
птиц происходит смена второстепенных маховых. В августе же идет смена
перьев спины, новые из коих так же темны, как и у молодых (Бутурлин, 1913)»
ИГЛОХВОСТЫЙ СТРИЖ
645
Питание. Пища состоит из насекомых, добываемых в воздухе над
равнинами, речными долинами, водными пространствами, ущельями, горными
лугами и другими открытыми пространствами. В Уссурийском крае иглохво-
стыми стрижами ловятся главным образом мухи и жуки (Бутурлин, 1913),
также осы (Золотарев). Летние корма в Минусинском крае состоят
преимущественно из ос (Тугаринов, 1913), но фактов этих недостаточно и требуются
дальнейшие наблюдения. Воду эти стрижи пьют, как и другие, летая над
самой поверхностью водоемов и зачерпывая ее подклювьем.
Полевые признаки. Иглохвостые стрижи являются самыми быстрыми
птицами СССР и обладают изумительной скоростью полета, доходящей до 170 км
в час (Гладков, 1942). Вихрем проносясь у самой поверхности земли, в
мгновенье ока они оказываются в подоблачной выси, откуда с громадной скоростью
вновь возвращаются к земле. Когда они пролетают стайкой над головою,
слышен своеобразный звук. И вместе с тем их полет может быть легким, гибким
и грациозным, что особенно заметно, когда в погоне за насекомыми они
значительно медленнее движутся над водной поверхностью. На лету кажутся
черными. Чаще летают по нескольку особей, иногда до 20. В полете пребывают
весь день, стайки же их заметны по утрам и вечерам. Ночь проводят в дуплах
или трещинах скал, куда влетают, не уменьшая своей огромной скорости и
мгновенно исчезая в отверстии. В предгорных равнинах появляются в пасмурную
погоду и внизу обычны в то время, когда на вершинах выпадают осадки. Во
время своих внезапных появлений в Саянах летают и охотятся иногда вместе
с черными и белопоясными стрижами. Отличаются молчаливостью, но
временами издают особый негромкий крик.
Описание. Размеры и строение. Относительно крупный стриж с
непропорционально малым клювом и ровно обрезанным коротким хвостом, жесткие стволы рулевых
которого выходят в виде заострений длиною в 3—5 мм. Цевки коротки, не оперены. Первый
палец направлен назад, когти изогнутые, острые и цепкие. Длина крыла самцов (И) 194—
212, самок (4) 191—206, в среднем 206 и 197 мм. Размах крыльев самцов (1) 485 мм.
Хвост около 47,5—60 мм. Цевка около 15—18 мм. Клюв около 8—10 мм, его разрез 22—
25 мм. Окраска. Молодые. Однообразно темнодымчатые сверху, без белых пятен по
бокам лба. Концы белых нижних кроющих хвоста испещрены черно-бурыми отметинами.
Взрослые самец и самка бурые сверху и снизу. По бокам лба белые пятна,
сливающиеся у некоторых индивидов в одно. Середина спины светлее и внутренние опахала
.третьестепенных маховых белые. Крылья и хвост блестяще верные. Горло и подхвостье
белые.
ОГЛАВЛЕНИЕ
Стр.
Предисловие 3
Таблица для определения отрядов птиц 8
Отряд веслоногие (А. М. Судиловская)
Характеристика отряда • • • • 13
Таблица для определения видов веслоногих 19
Семейство баклановые 21
Род бакланы 21
1. Большой баклан 21
Материковый большой баклан 23
Атлантический большой баклан 33
2. Уссурийский баклан 34
3. Берингов баклан . . ' . 36
Западный берингов баклан 36
4. Краснолицый баклан 40
5. Длинноносый баклан 43
Атлантический длинноносый баклан 43
Средиземноморский длинноносый баклан 47
6. Малый баклан 48
Семейство пеликановые 52
Род пеликаны : 52
7. Кудрявый пеликан 52
8. Розовый пеликан . . 59
Семейство олуши . 64
Род олуша 64
9. Олуша 64
Атлантическая или северная олуша . 64
10. Красноногая олуша 66
Восточная красноногая олуша ............. ; 66
Семейство фрегатов 68
Род фрегаты 68
11. Большой фрегат . . . 68
Гавайский фрегат . . 68
Отряд хищные птицы (Г. П. Дементьев)
Характеристика отряда 70
Таблица для определения видов хищных птиц 81
Семейство соколиные 84
Род соколы 84
1. Сокол 84
Тундровый или белощекий сокол 86
Якутский сокол 90
Алеутский или черный сокол 92
Сахалинский или малый сокол 93
Русский сокол 94
ОГЛАВЛЕНИЕ 64 7
Сто.
Кавказский сокол . 97
Рыжеголовый или пустынный сокол, шахин 98
2. Балобан : . 100
Европейский балобан 102
Обыкновенный балобан 105
Сибирский балобан 108
Туркестанский балобан 110
Монгольский балобан 111
Тибетский балобан ИЗ
3. Лаггар 114
4. Средиземноморский сокол 116
Европейский средиземноморский сокол 117
5. Кречет П9
Лапландский кречет 119
Сибирский кречет 123
Восточносибирский или камчатский кречет 124
Алтайский кречет . 125
6. Чеглок , . . 127
Обыкновенный чеглок 128
Северный чеглок 133
7. Дербник 135
Обыкновенный дербник . ♦ . 136
Восточносибирский дербник 142
Тихоокеанский дербник 143
Центральноазиатский дербник 144
Степной дербник 145
8. Пустельга . . 147
Обыкновенная пустельга 147
Восточная пустельга 152
9. Степная пустельга 153
10. Кобчик 157
Западный или обыкновенный кобчик 157
Восточный или амурский кобчик 162
Семейство ястребиные 164
Род ястреба 164
11. Большой ястреб, ястреб тетеревятник 164
Среднеевропейский тетеревятник 166
Западносибирский тетеревятник 170
Камчатский большой ястреб 173
Южноуральский большой ястреб 176
Среднесибирский большой ястреб 178
Японский большой ястреб 179
Кавказский большой ястреб 180
12. Тювик 181
Туркестанский тювик 183
Европейский тювик 186
13. Малый перепелятник 188
Сибирский малый перепелятник 189
14. Перепелятник, малый ястреб 190
Обыкновенный малый ястреб или перепелятник i 191
Восточносибирский малый ястреб или перепелятник 196
Камчатский малый ястреб или перепелятник . . . .' 197
Гималайский малый ястреб или перепелятник 198
Род луни 199
15. Полевой лунь 199
Обыкновенный полевой лунь . 200
16. Луговой лунь ■ 203
17. Степной лунь 207
18. Пегий лунь . 211
19. Болотный или камышевый лунь 214
Обыкновенный болотный лунь •. 215
Восточный болотный лунь 219
Род коршун 221
20. Черный коршун 221
Европейский черный коршун 221
648
ОГЛАВЛЕНИЕ
Стр.
Сибирский черный коршун 227
21. Красный коршун ..." 229
Обыкновенный красный коршун 229
Род чернокрылые коршуны 231
22. Чернокрылый коршун 231
Индийский чернокрылый коршун 231
Род орланы 233
23. Орлан белохвост 233
Обыкновенный орлан белохвост 234
24. Белоголовый орлан 238
Северный белоголовый орлан 239
25. Орлан долгохвост 240
26. Тихоокеанский орлан 213
Род стервятники 240
27. Стервятник 240
Обыкновенный стервятник 240
Род сипы 250
28. Белоголовый сип 250
Обыкновенный белоголовый сип 251
Индийский белоголовый сип 254
Снежный гриф или кумай 254
Род Pseudogyps 250
29. Индийский гриф 250
Род серые грифы 258
30. Черный или серый гриф 258
Род бородачи . . . . 201
31. Бородач или ягнятник 201
Переднеазиатский бородач 202
Центральноазиатский бородач . '.- 20ГС
Род орлы 207
32. Беркут 207
Североевропейский беркут 209
Восточносибирский беркут 272
Южноевропейский беркут 273
Центральноазиатский беркут 274
33. Орел-могильник 275
Обыкновенный орел-могилышк 275
34. Степной орел 279
Европейский степной орел 280
Восточный степной орел 282
35. Большой подорлик или орел-крикун 284
36. Малый подорлик 288
Европейский малый подорлик 289
37. Ястребиный или длиннохвостый орел 291
Северный ястребиный или длиннохвостый орел 292
38. Орел-карлик 294
Европейский орел-карлик 295
Среднеазиатский орел-карлик 298
Род сарычи 299
39. Сарыч или канюк 299
Малый или рыжий сарыч 300
Кавказский сарыч 304
Восточноазиатский сарыч 305
40. Мохноногий канюк или зимняк 307
Западный или европейский мохноногий канюк ЗОН
Сибирский мохноногий канюк 312
Камчатский мохноногий канюк 314
41. Мохноногий курганник 315
42. Степной сарыч, курганник ! . ! ! 318
Восточный курганник 319
Род осоеды 322
43. Хохлатый осоед !!!!!! 322
Восточносибирский осоед ........... \ ] 323
44. Европейский осоед 325
ОГЛАВЛЕНИЕ 649
Стр.
Род ястребиные сарычи 330
45. Ястребиный сарыч 330
Род змееяды 333
46. Змееяд 333
Европейский змееяд 334
Туркестанский змееяд 336
Род скопы. . 338
47. Скопа 338
Обыкновенная скопа 339
Отряд совы (Г. П. Дементьев)
Характеристика отряда ." 342
Таблица для определения видов сов 346
Семейство настоящие совы '. 348
Род белые совы 348
1. Белая или полярная сова . . 348
Род филины 352
2. Филин 352
Среднеевропейский филин 354
Русский филин 357
Южный филин 358
Западносибирский филин 360
Енисейский филин 361
Якутский филин 362
Дальневосточный филин 362
Казахский филин 363
Туркменский филин , 364
Гималайский филин . 365
Род рыбные совы 366
3. Рыбная сова 366
Северная рыбная сова 366
Род совки 369
4. Ошейниковая совка 369
Уссурийская ошейниковая совка: 370
5. Пустынная совка 371
6. Обыкновенная сплюшка или зорька . ; 373
Европейская сплюшка 374
Сибирская сплюшка 376
7. Восточноазиатская совка . . . 377
Уссурийская совка 378
Род ушастые совы 380
8. Ушастая сова 380
Обыкновенная ушастая сова 381
9. Болотная сова 383
Обыкновенная болотная сова „■■ 385
Род иглоногие совы 390
10. Иглоногая сова 390
Уссурийская иглоногая сова , 390
Род мохноногие сычи 391
И. Мохноногий сыч 391
Европейский мохноногий сыч .. 392
Кавказский мохноногий сыч 394
Сибирский мохноногий сыч . 394
Большой мохноногий сыч 395
Туркестанский мохноногий сыч , 395
Род домовые сычи 396
12. Домовый сыч 396
Западный домовый сыч 397
Южноевропейский домовый сыч 399
Каспийский или пустынный домовый сыч 400
Горноазиатский домовый сыч 401
Монгольский домовый сыч 402
650 ОГЛАВЛЕНИЕ
Стр.
Род воробьиные сычи 402
13. Воробьиный сыч 402
Западный воробьиный сыч 403
Восточный воробьиный сыч 405
Род ястребиные совы ♦ . . 406
14. Ястребиная сова 406
Обыкновенная ястребиная сова 406
Тяньшанская ястребиная сова 409
Род неясыти 410
15. Бородатая неясыть 410
Лапландская бородатая неясыть 410'
16. Уральская или длиннохвостая неясыть 413
Западная длиннохвостая неясыть 414
Западносибирская длиннохвостая неясыть 416
Среднесибирская длиннохвостая неясыть 417
Якутская длиннохвостая неясыть 417'
Дальневосточная длиннохвостая неясыть 418
17. Обыкновенная или серая неясыть 41&
Восточноевропейская неясыть 420
Сибирская неясыть 423
Кавказская неясыть 423
Туркестанская обыкновенная неясыть 424
Семейство сипухи 425
Род сипухи . 425
18. Сипуха 425
Среднеевропейская сипуха . 427
Средиземноморская сипуха 429-
Отряд кукушки (А. М. Судиловская)
Характеристика отряда 430-
Таблица для определения видов кукушек • 435
Семейство кукушки . 436
Род хохлатые кукушки 436
1. Хохлатая или пятнистая кукушка 436
Род кукушки 438
2. Кукушка 438
Обыкновенная кукушка 440
Туркестанская кукушка 451
3. Глухая кукушка 453
Северная глухая кукушка 453
4. Малая кукушка 458
Китайская малая кукушка 459
5. Индийская кукушка 461
Обыкновенная индийская кукушка 462
6. Ширококрылая кукушка 463
Манчжурская ширококрылая кукушка 464
Отряд козодои (Е. П. Спангенберг)
Характеристика отряда 466
Таблица для определения видов козодоев . ' . . 469
Семейство настоящие козодои 409
Род козодой : . . 400
1. Обыкновенный козодой 4(И>
Северный обыкновенный козодой 471
Южный обыкновенный козодой 474
Пустынный обыкновенный козодой 470
Китайский обыкновенный козодой ■ 47К
2. Буланый козодой 478
Северный буланый козодой 479
3. Большой козодой 482
Северный большой козодой , 482
ОГЛАВЛЕНИЕ 651
Стр.
Отряд сизоворонки или ракши (А. М. Судиловская)
Характеристика отряда 486
Таблица для определения видов сизоворонок 490
Семейство сизоворонковые 491
Род сизоворонка 491
1. Сизоворонка 491
Обыкновенная сизоворонка 492
Туркестанская сизоворонка 498
Род широкороты 500
2. Восточный широкорот 500
Восточный азиатский широкорот . . '. 502
Семейство щуриковых 504
Род щурки 504
3. Золотистая щурка 504
4. Зеленая щурка 513
Обыкновенная зеленая щурка . 515
Семейство зимородковые 518
Род зимородок 518
5. Обыкновенный зимородок 518
Русский обыкновенный зимородок . 519
Западный обыкновенный зимородок . 525
Восточный обыкновенный зимородок 526
Род пегий зимородок 527
6. Малый пегий зимородок 527
Западный малый пегий зимородок 528
7. Большой пегий зимородок . 529
Японский большой пегий зимородок 530
Род красноносый зимородок < 531
8. Ошейниковый зимородок . . . ♦ 531
9. Красноносый зимородок 532
Белогрудый красноносый зимородок 533
Отряд удоды (А. М. Судиловская)
Характеристика отряда , 535
Семейство удоды 538
Род удоды 538
1. Удод 538
Отряд дятлы (Н. А. Гладков)
Характеристика отряда 548
Таблица для определения видов дятлов 550
Род желны 550
1. Желна или черный дятел 550
Обыкновенная желна 551
Западный черный дятел ' " \ \ , 555
Род зеленые дятлы 556
2. Зеленый дятел .....! 556
Северный зеленый дятел .....* 556
Среднеевропейский зеленый дятел 559
Южный зеленый дятел ] 560
3. Чешуйчатый дятел .....' 561
Мургабский чешуйчатый дятел. . . . ! . 561
4. Седой дятел \ 563
Обыкновенный седой дятел ' 564
Среднесибирский седой дятел . . . . . 567
Дальневосточный седой дятел ", 568
Род трехпалые дятлы ■ % 569
5. Трехпалый дятел '. . \ 569
Северный трехпалый дятел 570
Тянынанский трехпалый дятел . . . . 573
Камчатский трехпалый дятел , \ 573
Альпийский трехпалый дятел ' [ 574
652
ОГЛАВЛЕНИЕ
Стр.
Род пестрые дятлы . . .. . * 574
6. Большой пестрый дятел 574
Обыкновенный большой пестрый дятел >. • 575
Украинский большой пестрый дятел 581
Кавказский большой пестрый дятел 583
Талышский большой пестрый дятел . 584
Тянынанский большой пестрый дятел 584
Камчатский большой пестрый дятел 584
Уссурийский большой пестрый дятел 585
7. Белокрылый дятел 585
Горный белокрылый дятел "'.' '. . \\ \'.' . " 586
Равнинный белокрылый дятел 587
Темный белокрылый дятел 588
8. Сирийский дятел :....'. 589
Северный сирийский дятел ...... '. . .* . . . . ". . V . :: *. \' 590
Балканский сирийский дятел 590
9. Белоспинный дятел . . . . 591
Обыкновенный белоспинный дятел 592
Сибирский белоспинный дятел 595
Камчатский белоспинный дятел . 597
Курильский белоспинный дятел 597
Буковинский белоспинный дятел . 597
Темный белоспинный дятел 598
10. Вертлявый или средний дятел 599
Европейский средний дятел 599
Кавказский средний дятел 001
11. Малый пестрый дятел 002
Восточноевропейский малый пестрый дятел 003
Западносибирский малый пестрый дятел ....... v 005
Восточносибирский малый пестрый дятел 005
Амурский малый пестрый дятел 006
Камчатский малый пестрый дятел 006
Кавказский малый пестрый дятел 007
Талышский. малый пестрый дятел 007
Род острокрылые дятлы ' 008
12. Малый острокрылый дятел 008
Уссурийский малый острокрылый дятел 008
Сахалинский малый острокрылый дятел 009
Курильский малый острокрылый дятел 010
13. Большой острокрылый дятел (НО
Уссурийский большой, острокрылый дятел 010
Род вертишейки 011
14. Вертишейка . 611
Обыкновенная вертишейка . . : 612
Восточная вертишейка 617
Отряд длиннокрылые (Е. С. Птушенко)
Характеристика отряда 618
Таблица для определения видов стрижей 622
Семейство стрижей . . . 623
Род стрижи 623
1. Черный стриж . 623
Европейский черный стриж 624
Азиатский черный стриж 630
2. Белобрюхий стриж 632
Евразийский белобрюхий стриж 633
3. Белопоясный стриж 635
Сибирский белопоясный стриж 035
4. Малый стриж 039
Западный малый стриж 040
5. Иглохвостый стриж 042
Севеоный иглохвостый стоиж 042
ОПЕЧАТКИ
Страница
258
447
475
504
560
562
569
571
599
612
624
639
644
Строка
24 сверху
14 снизу
10 сверху
10 снизу
15 снизу
4 снизу
13 сверху
7 снизу
под рисунком
8 сверху
7 сверху
19 сверху
26—27 снизу
Напечатано
Тип Viltur. monachus
выбрасывающий из гнезда
других птенцов
(Сатунин, 1937)
Li nnac u 1758
в июле и позднее
состоит из 63*— яиц
Тип Picus tridactulus
7 декабря
3— D. m. sanctijohannis, 4 —
Jynxtorquilla toquilla
Сев. Кавказа
24—27, 5; 16—17 мм
над землею Пржевальский
(1870) предполагает в
районе озера. Ханка
Следует читать
Тип Vultur monachus
выбрасывающий из гнезда
яйца
(Сатунин, 1907)
Linnaeus, 1758
в июне и позднее
состоит из 3—6 яиц.
Picus tridactylus
— 7 июля
3 — D. m. caucasicus,
4—D.m. sanctijohannis, 5 —
Jynxtorquilla torquilla
Кавказа
24—27,5 X 16—17 мм
над землею. Пржевальский
(1870) предполагает в районе
озера Ханка.
Птицы Советского Союза, том I.