/

























































































Автор: Ересковский А.В. Вишняков А.Э.
Теги: porifera (sponges) систематика животных специальные зоологические науки биология
ISBN: 978-5-9906564-7-5
Год: 2015
Похожие




Текст
САНКТ-ПЕТЕРБУРГСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Кафедра зоологии беспозвоночных СПбГУ
«Конспекты по зоологии беспозвоночных»
Выпуск 3
А. В. Ересковский, А. Э. Вишняков
ГУБКИ (PORIFERA)
Учебное пособие
Товарищество научных изданий КМК
Санкт-Петербург - Москва ❖ 2015
Печатается по решению редколлегии серии
«Конспекты по зоологии беспозвоночных».
Рекомендовано к печати решением
Учебно-методической комиссии
биологического факультета СПбГУ
УДК 593.4(075.8)
ББК 28.691Д73-1
Е70
Ересковский А.В., Вишняков А.Э.
£70 Губки (Porifera)i Учебное пособие. — M.J Товарищество научных
изданий КМК, 2015. — 99 с.
Учебное пособие посвящено описанию одних из самых низкоорганизо-
ванных и в то же время процветающих в настоящее время животных
нашей планеты, губок (тип Porifera). Основной акцент делается на
описании морфо-функциональных особенностей этих организмов, их
многообразия, филогенетических отношений, цитологических особен-
ностей и пр. Особое место занимает глава, посвященная биологии раз-
вития губок, где дан подробный анализ всех этапов эмбриогенеза гу-
бок.
Библиогр. 95 назв. Ил. 36.
Рецензенты:
д.б.н. А.Н. Островский
д.б.н. Е.Н. Темерева
Ответственный редактор выпуска
А.И. Гранович
Редакционная коллегия серии:
А.И. Гранович, АЛ. Добровольский
Работа выполнена при поддержке
гранта СПбГУ Л? 1.38.209.2014
ISBN 978-5-9906564-7-5
© А.В. Ересковский, А.Э. Вишняков, 2015
Среди разнообразных групп многоклеточных животных губ-
ки занимают особое положение. Их план строения настолько
самобытен, что ему не найти аналогии в мире многоклеточных
животных. Уникальность строения связана с реализацией осо-
бого принципа взаимодействия со средой обитания. Подавляю-
щее большинство многоклеточных животных, взаимодействуя с
внешней средой, изолирует собственную внутреннюю среду ор-
ганизма. Губки реализуют обратный принцип — они обеспечи-
вают ток воды через собственное тело, используя характерную
только для них водо-канальную систему. Собственно, вокруг
этой водо-канальной системы, то есть проходящей сквозь тело
организма «внешней среде» и формируются пласты клеток, об-
разующие тело каждой губки. Уникальность губок среди много-
клеточных животных проявляется и во многих других призна-
ках; реализации «организменных» функций не специальными
системами многоклеточных органов, как у большинства мно-
гоклеточных животных, а отдельными клетками, то есть, на
«клеточном» уровне; отсутствии интеграционных систем, объ-
единяющих организм других многоклеточных в единое целое;
разнонаправленности и обратимости клеточных дифференци-
ровок. Все это признаки, указывающие на то, что целостность
губок как организмов, их «индивидуализация», находится на
более низкой ступени, чем у других многоклеточных животных.
Можно предположить, что в организации современных губок
проявляются особенности ранних этапов эволюции многокле-
точных животных. И это причина по которой поддерживается
интерес зоологов к этой группе животных. Точнее сказать одна
из причин.
Вероятно, не будет преувеличением сказать, что в послед-
ние годы интерес к губкам значительно вырос. Эта группа ока-
4
А. В. Ересковский, АЗ. Вишняков. Губки
зались в центре разработки целого ряда фундаментальных про-
блем. Я уже упомянул проблему ранней эволюции животных,
«корневой* группы многоклеточных, которая ныне решается
анализом молекулярно-генетических гомологий, в частности,
поиском у губок предполагаемых предшественников генов (ор-
тологов) известных для других многоклеточных. Помимо этого,
губки становятся модельными объектами в исследованиях про-
блемы целостности организма, распознавания «свое-чужое».
Можно упомянуть также проблемы клеточной дифференциров-
ки и межклеточных взаимодействий, проблемы становления
тканевой организации, проблемы симбиотических взаимодей-
ствий. Во всех этих случаях губки оказываются уникальной мо-
делью5 исследование которой может пролить свет на эволюцию
соответствующей важной функции.
Пристальное внимание к этой группе животных в последнее
время связано еще и с тем, что, помимо очевидных «фундамен-
тальных» загадок, губки оказались незаменимыми объектами
важнейших прикладных разработок. Исследования показыва-
ют, что различные виды губок и их симбионтов могут служить
источником целого спектра специфичных биологически актив-
ных веществ. Эти вещества могут влиять на рост клеток много-
клеточных животных, подавлять бактериальную активность,
выполнять другие важные функции. Они могут находить при-
менение в качестве основы медицинских препаратов, в частно-
сти, расширяя возможности эффективной противоопухолевой
терапии.
Наши представления о строении и функционировании губок
за последние десятилетия значительно расширились и углуби-
лись. Особенно это касается ультраструктуры, данных о видах
клеточных дифференцировках, о клеточных взаимодействиях
в теле губки. Значительно расширились представления об осо-
бенностях размножения губок, о разнообразии их личинок и о
разнообразных морфогенезах, приводящих к построению дефи-
нитивного тела. Накапливается все больше материала относи-
тельно функционирования организма губки в целом. Впервые
появляется возможность оперировать данными по геномам гу-
бок и структуре отдельных, важных в функциональном отноше-
нии генов.
5
Предлагаемое учебное пособие представляет современные
взгляды на уникальную группу многоклеточных животных. По-
явление этого пособия необходимо признать более чем своевре-
менным, поскольку имеющаяся учебная литература воспроиз-
водит устаревшие уже представления о морфо-функциональных
особенностях губок. Отрадно, что читатели получат информа-
цию, что называется, «из первых рук». Авторы — признанные
в России и за рубежом специалисты, изучающие губок в самых
разных аспектах на протяжении десятилетий.
Пособие адресовано студентам бакалавриата и магистрату-
ры биологических факультетов университетов по направле-
нию Биология и может быть использовано в качестве дополни-
тельной литературы. Можно рекомендовать пособие учителям
средних школ, а также преподавателям любых университетов,
в которых имеются курсы, посвященные биоразнообразию жи-
вотных. Книга может быть рекомендована всем интересующим-
ся эволюцией и разнообразием многоклеточных животных. До-
статочно простой и лаконичный стиль изложения позволяет
надеяться, что пособие будет интересно широкому кругу чита-
телей интересующихся биологией, зоологией, проблемами кле-
точной биологии и эволюции.
Объем пособия, к сожалению, не позволяет детально осве-
щать многие интереснейшие аспекты, связанные со строением,
функционированием и экологией губок. Именно с этим связано
то, что в ряде случаев авторы только обозначают отдельные на-
правления современных исследований губок. Однако уверен,
что и здесь пособие сыграет свою роль, стимулируя интерес чи-
тателя уже к самостоятельному и более детальному знакомству
с предметом.
А.И. Гранович
ВВЕДЕНИЕ
Что может быть проще и что может
быть целостнее, чем губки?!
Губки — загадочные организмы. Простой человек, не ис-
кушенный в зоологических знаниях, не сразу поймет, кто они
такие. Неподвижные (в человеческом восприятии времени), не
реагирующие внешне на какие-либо воздействия, часто не име-
ющие заметных структур, по которым обычно определяют при-
надлежность к той или иной категории существ. Если кому-ни-
будь показать губок и спросить, кто или что это такое, то можно
ожидать разные ответы — водоросли, грибы, растения и даже
выделения (экскреты) живых организмов. Но очень редко (и что
самое актуальное, правильно) можно услышать, что эти непо-
нятные образования — животные. Но именно так оно и есть.
Губки — это животные.
Какие они бывают, что является для них общим, и что позво-
ляет говорить об их многообразии организации, жизнедеятель-
ности, биологических особенностях и взаимосвязях с другими
представителями животных (Metazoa)? Ответы на эти вопросы
мы постараемся изложить в небольшом эссе.
Глава 1.
ОБЩАЯ ХАРАКТЕРИСТИКА ГУБОК
Если наблюдать за жизнью обитателей какого-либо морского
водоема, то среди прикрепленных к неподвижным субстратам
организмов можно увидеть разнообразные водоросли. Между
ними легко можно найти представителей животного мира: коло-
нии гидроидных полипов или кораллов, двустворчатых моллю-
сков, сидячих полихет или колонии мшанок. При ближайшем
рассмотрении окажется, что эти животные активно перемеща-
ются и заметно реагируют своим движением, если к ним при-
коснуться. Однако среди них можно часто обнаружить существ
разнообразной формы и расцветки, которые никак не будут себя
проявлять подвижностью, как бы мы ни старались их прости-
мулировать к этому. Скорее всего, ими окажутся животные, от-
носящиеся к типу Porifera (Spongia), или по-русски, губки. Эти
организмы — исключительно водные и живут в морях и пре-
сных водоемах.
Форма тела губок разнообразна. Они могут иметь вид плен-
ки, корки, быть комковидными или сферическими, трубчаты-
ми, ветвистыми, вееровидными и др. (рис. 1). При этом их осно-
вание достаточно прочно прикреплено к субстрату, а само тело
может характеризоваться специфичными для вида признаками.
Несмотря на большие различия в форме губок, можно опреде-
лить направленность расположения их тела, или полярность,
от нижней, или базальной прикрепленной части к верхней, или
апикальной. Такая полярность называется апикобазальной.
Размеры губок по этой оси сильно варьируют: от нескольких
миллиметров до 1,5-2 м.
Одна из их главных особенностей — отсутствие каких-либо
тканевых систем органов, которые традиционно рассматрива-
ются и сравниваются у других групп животных. Вся жизнеде-
ятельность губок определяется активностью отдельных клеток
и клеточных пластов, имеющих разное строение и функции.
Однако клеточные пласты губок, в большинстве случаев, не со-
ответствуют характеристикам, традиционно предъявляемым к
тканям. В связи с этим некоторые исследователи говорят о губ-
8
А,В, Ересковский, А.Э. Вишняков. Губки
Рис. 1. Разнообразие внешнего вида губок: а — Stelodoryx sp.;
б — Forcepia sp.; в — Poterion sp.; г — Polymastia hemisphaeri
cum; д — Guancha blanca; e — Sycon sp.; ж — Isodictya sp.
(из: Колтун, 1988).
ках как об организмах с «примитивными» тканями, имея в
виду, что тканевая организация эволюционно формировалась
постепенно и клеточные пласты губок, имея одни признаки
тканей (группы клеток, выполняющих сходную функцию), ли-
шены других (например, общности происхождения клеток пла-
ста). Иная точка зрения заключается в более строгой трактовке
понятия «ткань». В соответствии с этим взглядом говорить о
тканевом строении у животных можно только в том случае, если
Глава 1. Общая характеристика губок
9
все признаки ткани налицо. Следуя этой точке зрения, губки не
могут считаться животными обладающими тканями; их назы-
вают многоклеточными с «нетканевым» строением.
Отсутствие обособленного кишечника, нервной и мышечной
систем и гонад у губок не значит, что они не питаются, не раз-
множаются и не реагируют на изменения внешней среды или
внутренних процессов. Все эти функции им присущи, но они
реализуются качественно по-другому, не так, как мы привыкли
это видеть у других животных.
Обычно животные питаются, осуществляют газообмен, вы-
водят продукты обмена веществ, т.е. взаимодействуют с окру-
жающей средой за счет обособленных многоклеточных систем,
которые имеют характерные морфологические признаки и опре-
деленное положение в теле. Губки осуществляют эти процессы,
взаимодействуя одинаково с окружающей средой во всех ча-
стях своего тела. Они контактируют с водой своей поверхностью
и проводят воду через связанную систему полостей и каналов
(приводящих каналов и отводящих каналов), начинающихся
многочисленными крошечными входными отверстиями, ости-
ями, или порами, и заканчивающихся обычно одним или не-
сколькими крупными отверстиями, расположенными на конце
трубок, оскулюмами. Самый крупный канал, переходящий в
оскулюм называется атриумом или спонгоцелем (устаревшее
название, парагастральная полость) (рис. 2).
Вся совокупность каналов и полостей, проводящих воду че-
рез тело губок, называется водоносной системой. Поверхность
тела губки и каналы водоносной системы сформирована клет-
ками — пинакоцитами, обычно уплощенных, плотно прилега-
ющих друг к другу и, таким образом, формирующих клеточные
пласты, называемые пинакодерма. Клетки другого типа — хо-
аноциты, также располагаются группами и образуют слои (хо-
анодерма), выстилающие расширенные участки каналов водо-
носной системы. Эти расширения называют хоаноцитными, или
жгутиковыми камерами (рис. 2). Входное отверстие в хоано-
цитную камеру называется прозопиль, а выходное — апопиль.
Хоаноциты в свой апикальной части имеют тонкие цитоплазма-
тические выросты (микровилли), располагающиеся по кругу,
в центре которого находится жгутик. Именно согласованная ра-
10
А. В. Ересковский, А.Э. Вишняков. Губки
Рис. 2. Строение однооскулюмной губки (по: Weissenfels,
1989); ап — апопиль; ат — атриум; вк — выносящий канал;
м — мезохил; о — оскулярная трубка; п — пора; пк — при-
носящий канал; пп — прозопиль; сп — спикула; хк — хоано-
цитная камера. Стрелками показано направление движения
воды через водоносную систему.
бота этих жгутиков создает движущую силу, которая протяги-
вает воду от отверстий на поверхности губки — остий — до вы-
водного отверстия — оскулюма.
Слои пинакоцитов и хоаноцитов отграничивают внутреннее
пространство губки, которое непосредственно не сообщается
с водой. Оно называется мезохил и содержит клеточные и вне-
клеточные элементы (внеклеточный матрикс и скелет), выпол-
няющие разнообразные функции. Внеклеточные образования
имеют вид органических фибрилл коллагеновой природы, тол-
стых волокон белка спонгина и др. Неорганические структуры
мезохила — прямые или изогнутые, одно- и многоосные иголь-
чатые или другой формы тельца, называемые спикулами. Их
функция — опора и придание формы тела губки. Клетки мезо-
хила обычно не формируют скоплений. В мезохиле можно об-
наружить несколько типов клеток, что отражает различия в их
функциях. Кроме этого многие из них могут изменять форму и
передвигаться, используя псевдоподиальные выросты. Эта осо-
Глава 1. Общая характеристика губок
11
бенность порой подчеркивается в качестве определяющего при-
знака. Соответственно, всю совокупность клеток мезохила в
некоторых учебниках называют амебоциты (рис. 2). Однако, не-
смотря на это клетки мезохила очень многообразны, и для каж-
дой таксономической группы губок характерна определенная
совокупность типов клеток мезохила со своими функциями и
цитологическими свойствами.
Все участки тела губок, которые соприкасаются с водой и со-
стоят из хоаноцитов и пинакоцитов, потенциально способны
участвовать в газообмене, выводить продукты метаболизма и
поглощать вещества из внешней среды. Растворенные вещества,
имеющие питательную ценность, проходят через мембрану кле-
ток, а мелкие частицы (приблизительно до 1 мкм в своем боль-
шом сечении), поглощаются за счет фагоцитарной активности
клеток хоанодермы, а в некоторой степени и пинакодермы. Бла-
годаря такому способу питания губок рассматривают как орга-
низмы-фильтраторы, жизнь которых состоит в прокачивании
через тело при помощи водо-канальной системы окружающей
их воды, из которой они извлекают нужную им пищу, кисло-
род, минеральные вещества, сигнальные молекулы. Собственно,
с этим проходящим потоком воды так или иначе связаны все ос-
новные процессы жизнедеятельности губок.
Самый простой вариант организации губок заключается в
сочетании слоя пинакоцитов, окружающих губок снаружи, и
слоя хоаноцитов, образующих обширную выстилку внутри тела
губки. Наружный пинакоцитный слой смыкается со слоем хо-
аноцитов за счет узких приводящих каналов, стенки которых
также сформированы пинакоцитами, а хоаноцитная полость от-
крывается непосредственно оскулярным отверстием. Между на-
ружным слоем пинакоцитов и внутренним слоем хоаноцитов на-
ходится узкая прослойка мезохила со скелетными элементами и
отдельными клетками других типов.
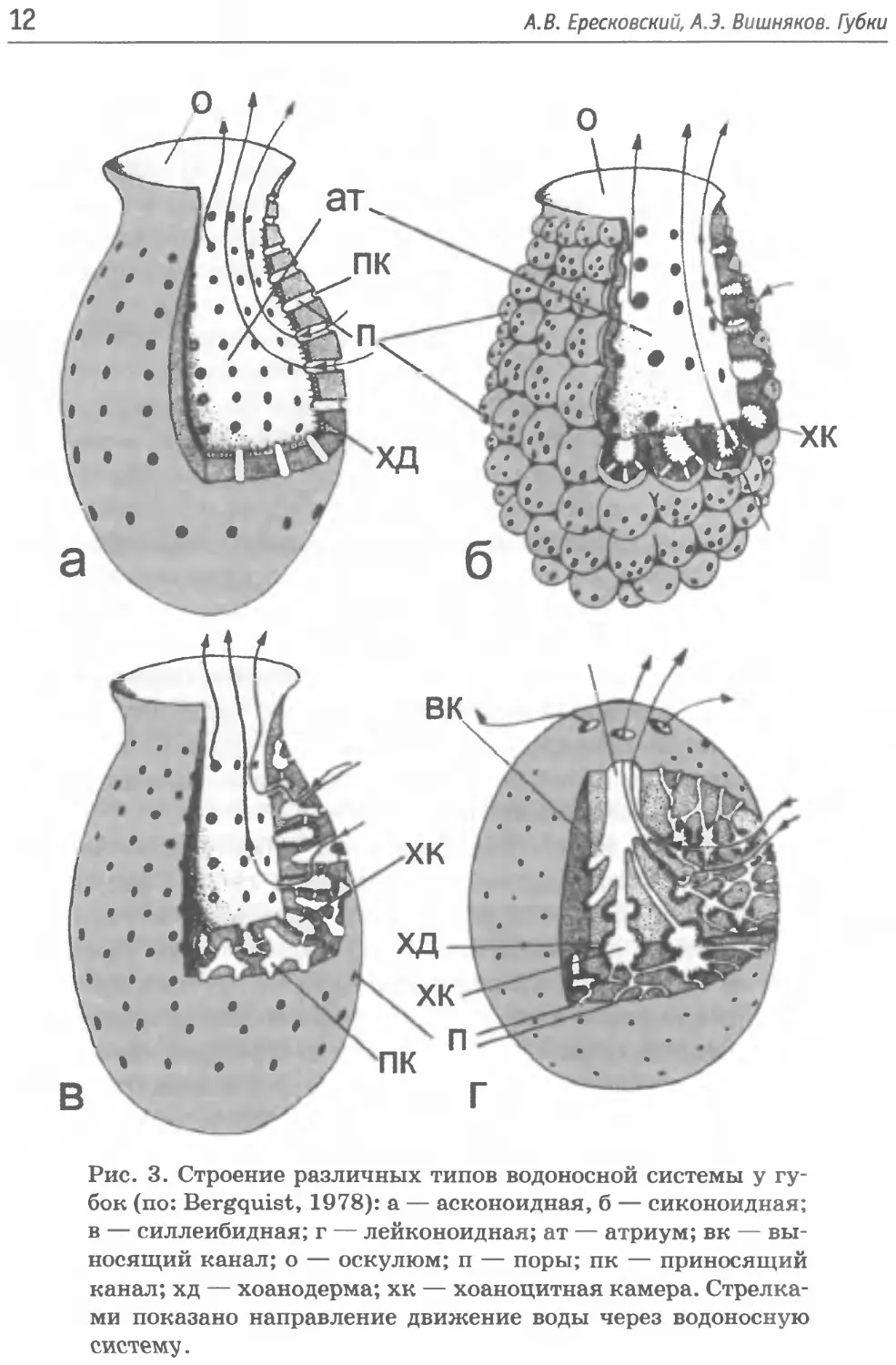
Такое строение водоносной системы принято называть аско-
ноидной (рис. За). Поскольку для создания такой конструкции
достаточно всего несколько сотен клеток каждого типа, то и раз-
меры губок с асконоидным строением достигают всего несколь-
ких миллиметров и обычно имеют вид анастомозирующих тру-
бочек.
12
А. В. Ересковский, А.Э. Вишняков. Губки
Рис. 3. Строение различных типов водоносной системы у гу-
бок (по: Bergquist, 1978): а — асконоидная, б — сиконоидная;
в — силлеибидная; г — лейконоидная; ат — атриум; вк — вы-
носящий канал; о — оскулюм; п — поры; пк — приносящий
канал; хд — хоанодерма; хк — хоаноцитная камера. Стрелка-
ми показано направление движение воды через водоносную
систему.
Глава 1. Общая характеристика губок
13
При сохранении асконоидной организации дальнейшее уве-
личение размеров возможно за счет увеличения атриальной по-
лости и/или за счет увеличения объема мезохила. Однако этого
в природе не происходит и связано это, по-видимому, с чисто
физическими причинами. Дело в том, что увеличение объема
атриума приводило бы к застою воды в центральной части этой
полости, поскольку жгутики выстилающих ее хоаноцитов спо-
собны перемещать только тонкий слой воды в непосредственной
близости от этих клеток. Увеличение объема мезохила при со-
хранении асконоидного варианта водоносной системы, очевид-
но, также не может происходить больше предела, который мы
наблюдаем у ныне живущих губок. Одно из объяснений — для
эффективного функционирования любой живой системы долж-
но соблюдаться оптимальное отношение площади поверхности
организма и его объема. При увеличении размеров любого тела
площадь поверхности увеличивается как квадратичная функ-
ция, тогда как объем, как кубическая. Когда площадь поверх-
ности губки превышает определенную величину, объем мезо-
хила должен увеличиться настолько, что поверхностный слой
клеток уже не сможет адекватно «обслуживать» протекающие
там процессы.
Однако большинство губок своими размерами значительно
превосходят асконоидных. При этом указанные выше конструк-
тивные проблемы решаются следующим образом. Сначала на-
ружный пинакоцитный слой и внутренний слой хоаноцитов
изгибаются навстречу друг другу. Хоаноциты образуют карма-
нообразные камеры, которые с одной стороны открываются в
атриум, выстилаемый уже не хоаноцитами, а пинакоцитами, а с
другой соединяются с внешней средой короткими приводящими
каналами. Такой вариант водоносной системы называется сико-
ноидным (рис. 36).
Дальнейшее усложнение водоносной системы и частичное
увеличение общих размеров тела губок происходит за счет того,
что поверхность наружного вогнутого пинакоцитного слоя сгла-
живается. При этом под пинакодермой появляются широкие по-
лости, сообщающиеся с внешней средой через узкие поры. Водо-
носная система такой конструкции называется силлеибидной
(рис. Зв).
14
А.В. Ересковский, А.Э. Вишняков. Губки
Последующее увеличение числа хоаноцитных камер, умень-
шение их диаметра, их погружение в пространство между
стенкой атриальной полости и слоем наружных пинакоцитов,
удлинение и создание сети приводящих и отводящих кана-
лов приводит к созданию лейконоидной водоносной системы
(рис. Зг), которая позволяет вырастать губкам до своего макси-
мального размера в 1-2 м.
В ходе развития и роста губок разных видов их водоносная
система может останавливаться на начальной асконоидной ста-
дии или продолжать развиваться до одной из трех последующих
форм — сиконоидной, силлеибидной или лейконоидной.
Указанные конструктивные варианты строения тела и во-
доносной системы встречаются в различных таксономических
группах Porifera. Таким образом, их значение для классифика-
ции губок невелико, хотя набор конструктивных вариантов и
используется как дополнительный признак при характеристике
таксонов.
Описанная выше последовательность строения водоносной
системы, представляет собой лишь ее конструктивное усложне-
ние и не является отражением эволюции этой системы у губок,
как это считалось ранее.
Глава 2.
ТАКСОНОМИЧЕСКАЯ СТРУКТУРА
ТИПА PORIFERA
То, что губки — это организмы, которые принципиально от-
личаются от всех других животных своим особым образом жиз-
ни и строением, конечно, позволяет выделять их в особую так-
сономическую группу, называемую тип Porifera, или несущие
поры. В нашей книге мы используем «ранговую» таксономиче-
скую систему, применяя названия традиционных таксонов —
тип, класс, отряд и т.д. На основе морфологических, эмбрио-
логических, молекулярных и цитологических данных можно
выделить четыре современных класса губок.
Класс Calcarea Bowerbank, 1864,
или Известковые губки
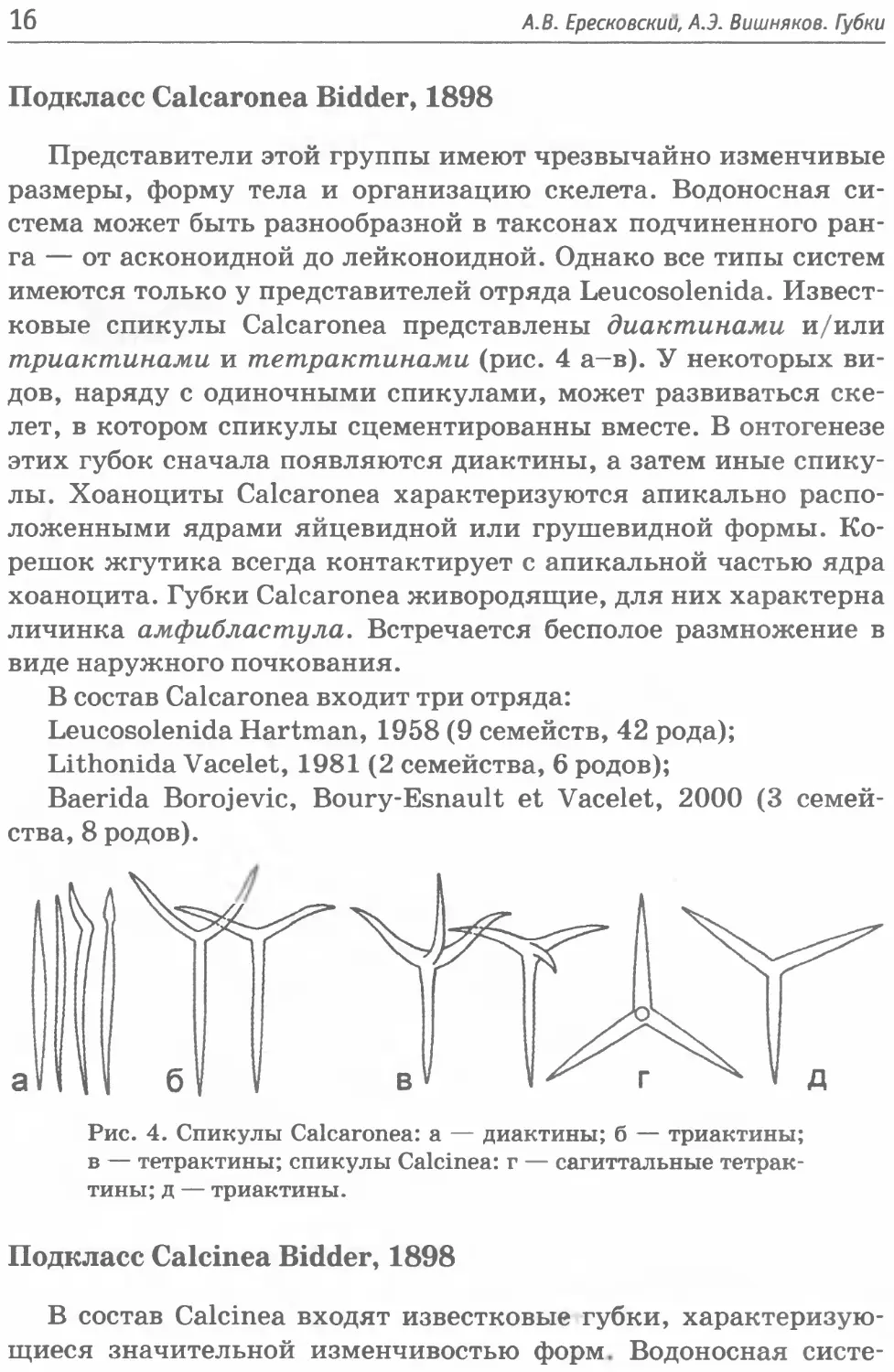
Представители Calcarea характеризуются тем, что их мине-
ральный скелет образован только карбонатом кальция. Элемен-
ты скелета располагаются в мезохиле свободно и имеют форму
двух-, трех- и четырехлучевых спикул (диактин, триактин, те-
трактин, соответственно) (рис. 4). Иногда имеется плотный ске-
лет из спикул, спаянных друг с другом. У разных видов известко-
вых губок присутствуют все варианты водоносной системы. Все
Calcarea исключительно морские и распространены во всех аква-
ториях от литорали до батиали. В ходе полового размножения у
них формируются однослойные личинки с полостью внутри.
Calcarea представляют собой монофилетическую группу
(Manuel et al., 2004; Voigt et al., 2012), в которую входит око-
ло 680 видов, что составляет 8,2% от всех описанных Porifera.
В настоящее время в пределах Calcarea выделяют две монофиле-
тичные группы — п/кл Calcinea и п/кл Calcaronea с пятью отря-
дами, 24 семействами и 77 родами (Manuel et al., 2002, 2003).
Известковые губки имеют два существенно различающихся
типа развития: «кальцибластульный тип», характерный для
Calcinea и «амфибластульный тип», характерный для Calcaro-
nea (см. гл. 7) (Ересковский, 2005).
16
А. В. Ересковский, А.Э. Вишняков. Губки
Подкласс Calcaronea Bidder, 1898
Представители этой группы имеют чрезвычайно изменчивые
размеры, форму тела и организацию скелета. Водоносная си-
стема может быть разнообразной в таксонах подчиненного ран-
га — от асконоидной до лейконоидной. Однако все типы систем
имеются только у представителей отряда Leucosolenida. Извест-
ковые спикулы Calcaronea представлены диактинами и/или
триактинами и тетрактинами (рис. 4 a-в). У некоторых ви-
дов, наряду с одиночными спикулами, может развиваться ске-
лет, в котором спикулы сцементированны вместе. В онтогенезе
этих губок сначала появляются диактины, а затем иные спику-
лы. Хоаноциты Calcaronea характеризуются апикально распо-
ложенными ядрами яйцевидной или грушевидной формы. Ко-
решок жгутика всегда контактирует с апикальной частью ядра
хоаноцита. Губки Calcaronea живородящие, для них характерна
личинка амфибластула. Встречается бесполое размножение в
виде наружного почкования.
В состав Calcaronea входит три отряда:
Leucosolenida Hartman, 1958 (9 семейств, 42 рода);
Lithonida Vacelet, 1981 (2 семейства, 6 родов);
Baerida Borojevic, Boury-Esnault et Vacelet, 2000 (3 семей-
ства, 8 родов).
Рис. 4. Спикулы Calcaronea: а — диактины; б — триактины;
в — тетрактины; спикулы Calcinea: г — сагиттальные тетрак-
тины; д — триактины.
Подкласс Calcinea Bidder, 1898
В состав Calcinea входят известковые губки, характеризую-
щиеся значительной изменчивостью форм. Водоносная систе-
Глава 2. Таксономическая структура типа Porifera
17
ма этих губок, как и у представителей Calcaronea может быть
асконоидной, сиконоидной, силлеибидной и лейконоидной.
Спикулы представлены три- и тетрактинами, лучи которых со-
измеримы по длине и расположены под равными углами друг
относительно друга (рис. 6г, д). У некоторых видов имеются
также спикулы, один луч которых длиннее других и распо-
ложен под иным углом, чем остальные лучи. У ряда предста-
вителей дополнительно может развиваться неспикульный ба-
зальный известковый скелет. В онтогенезе Calcinea сначала
появляются триактины, а затем иные варианты спикул. Ядра
хоаноцитов занимают в клетке базальное положение. Между
ядром и базальными структурами жгутика не имеется никаких
определенных топологических отношений. Calcinea являют-
ся живородящими губками, для которых характерна личинка
кальцибластула (целобластула). Для некоторых видов харак-
терно бесполое размножение в виде почкования.
Calcinea включает отряды:
Clathrinida Hartman, 1958 (7 семейств, 18 родов);
Murrayonida Vacelet, 1981 (3 семейства, 3 рода).
Класс Homoscleromorpha Bergquist, 1978
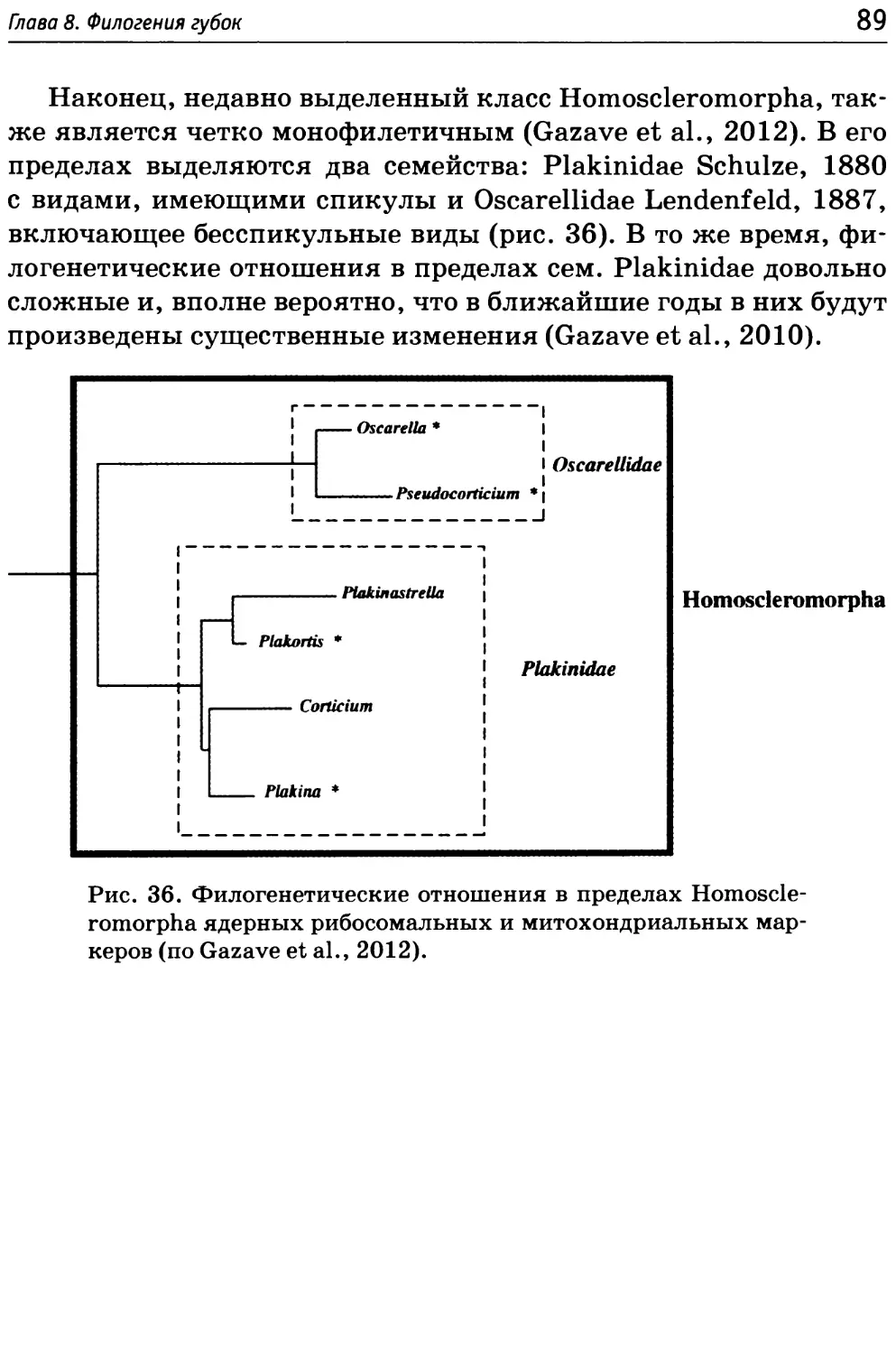
Homoscleromorpha — четко ограниченная группа губок, выде-
ляемая ранее как подкласс внутри Demospongiae. Современные
филогенетические исследования показали, что Homoscleromor-
pha представляет собой монофилетическую группу, независимую
от демоспонгий (Gazave et al., 2010, 2012). К Homoscleromorpha
относятся небольшие губки с гладкой поверхностью и формой
тела, варьирующей от корковой до комковидной. Простая орга-
низация этих губок позволила некоторым авторам рассматривать
их как «почти идеальный прототип организации губок». Однако
их водоносная система представлена двумя наиболее сложными
формами — силлеибидной или лейконоидной. Часто в базальной
части губок имеется обширная выносящая камера.
Неорганический кремнеземный скелет, если имеется, пред-
ставляет собой комбинацию мелких четырехостных спикул,
калтроп, и/или их дериватов (диоды и триоды) (рис. 5), распола-
гающихся в теле губки равномерно. Лучи спикул могут ветвить-
18
А. В. Ересковский, А.Э. Вишняков. Губки
Рис. 5. Спикулы Homoscleromorpha: а — калтроп; б, в — ло-
фокалтропы; г — диод.
ся и изгибаться. У некоторых видов спикулы отсутствуют, и их
форма тела поддерживается только фибриллярным коллагеном.
Ряд интересных особенностей Homoscleromorpha, связанных
с организацией их клеток, отличает их от всех других губок.
Так, базальная часть клеток хоанодермы и пинакодермы под-
стилается базальной пластинкой, сами эти клетки имеют между
собой специализированные клеточные контакты, а экзо- и эндо-
пинакоциты имеют жгутики.
Для них также характерно незначительное разнообразие
клеточных типов (не более 5), что существенно отличает пред-
ставителей Homoscleromorpha от Demospongiae на гистологиче-
ском уровне.
Если по анатомической организации Homoscleromorpha мо-
гут рассматриваться как наиболее просто устроенные предста-
вители Porifera, то наличие базальной пластинки, включающей
коллаген типа IV, тенасцин и ламинин, а также присутствие
межклеточных контактов в хоаноцитных и пинакоцитных сло-
ях принципиально выводит их на другой уровень организации и
сближает с животными, имеющими эпителиальные ткани.
Локализация базальной пластинки не ограничивается кле-
точными слоями хоанодермы и пинакодермы неразмножаю-
щихся губок. В период размножения она покрывает спермато-
цисту, а также присутствует у личинок (Ereskovsky et al., 2009).
Все Homoscleromorpha — живородящие губки, у которых
развивается личинка цинктобластула (см. гл. 7) (Ересковский,
Глава 2. Таксономическая структура типа Porifera
19
2005). Кроме этого у них отмечено бесполое размножение в виде
почкования и фрагментации. Homoscleromorpha обитают толь-
ко в морях, главным образом, на небольших глубинах.
Эта небольшая монофилетическая группа включает один от-
ряд Homosclerophorida, в котором выделяются два семейства и
6 родов.
Класс Demospongiae Sollas, 1885
В состав Demospongiae входят губки, скелет которых может
быть составлен только спонгиновыми волокнами или совокуп-
ностью спонгиновых волокон и кремневых спикул, которые, в
зависимости от размера, обычно разделяются на макросклеры
и микросклеры (рис. 6). Макросклеры — в основном одноосные
или четырехосные; микросклеры крайне разнообразны: много-
осные или одноосные, часто весьма причудливой формы. Осевая
нить спикулы находится в треугольной или шестиугольной по-
лости. Спонгин, также как и фибриллярный коллаген, имеется
у всех представителей класса, формируя отдельные волокна,
либо объединяя спикулы. У демоспонгий некоторых групп спи-
кульный скелет редуцирован. В этом случае на его месте раз-
вивается сложный органический спонгиновый скелет. Пред-
ставители нескольких групп Demospongiae не имеют никаких
скелетных элементов. Известно несколько мелких групп, особи
которых, в дополнение к другим скелетным элементам, имеют
базальный скелет, сильно обогащенный углекислым кальцием.
Форма тела губок Demospongiae может быть корковой, ло-
пастной, трубчатой, ветвистой, нитчатой, кубковидной. Ино-
гда губки обитают в толще известкового субстрата (сверля-
щие губки). Водоносная система подавляющего большинства
представителей Demospongiae — лейконоидного типа. Одна-
ко некоторые губки из таксона Poecilosclerida (Cladorhizidae,
Esperiopsidae) вовсе утратили водоносную систему, что связано
с необычным для губок способом питания — эти губки стали
плотоядными.
Личинки представлены главным образом паренхимулами, но
в некоторых группах имеются однослойные личинки. В преде-
лах группы представлены различные репродуктивные страте-
20
А. В. Ересковский, А.Э. Вишняков. Губки
Рис. 6. Спикулы Demospongiae: макросклеры (а-з) и мик-
росклеры (и-п): а — акантотилостиль; б — тилостиль; в —
тилота; г — стронгила; д-з — триены; и — сферастра; к —
сфероксиастра; л — метастра; м — сигмы; н — якорьки; о,
п — хелы.
гии, включая наружное развитие, яйцеживорождение и живо-
рождение.
Demospongiae включают 83,3% всех живущих губок (при-
близительно 6900 описанных видов). Большинство губок —
морские, но имеется несколько семейств, представители ко-
торых обитают в пресных водах всех континентов, исключая
Антарктиду.
В последние годы, благодаря углублению представлений
о филогении, связанном с применением молекулярных мето-
дов, таксономическая структура губок подверглась существен-
ной ревизии, при этом основные изменения затронули именно
класс Demospongiae (Redmond et al., 2013; Thacker et al., 2013).
В настоящее время в пределах Demospongiae выделяют четыре
Глава 2. Таксономическая структура типа Porifera
21
группы в ранге подклассов: Keratosa, включающий губок, не
имеющих кремневых спикул, Myxospongiae, Haploscleromorpha
(морские Haplosclerida) и Heteroscleromorpha.
Таксономическая структура Demospongiae представлена
ниже:
Подкласс Keratosa Grant, 1861:
Отряд Dendroceratida Minchin, 1900 (2 семейства),
Отряд Dictyoceratida Minchin, 1900 (5 семейств);
Подкласс Myxospongiae Haeckel, 1866, отряды:
Отряд Chondrillida Redmond et al., 2013 (2 семейства)
Отряд Verongida Bergquist, 1978 (4 семейства);
Подкласс Haploscleromorpha Cardenas, Perez et Boury-Es-
nault, 2012:
Отряд Haplosclerida Topsent, 1928 (морские) с подотрядами
Haplosclerina Topsent, 1928 (3 семейства) и Petrosina Boury-Es-
nault et van Beveren, 1982 (3 семейства);
Подкласс Heteroscleromorpha Cardenas, Perez et Boury-Es-
nault, 2012:
Отряд Agelasida Hartman, 1980 (5 семейств),
Отряд Lithistida (14 семейств),
Отряд Astrophorida Sollas, 1888 (8 семейств),
Отряд Spongillida — «пресноводные Haplosclerida» (8 се-
мейств),
Отряд Poecilosclerida Topsent, 1928 (25 семейств),
Отряд Hadromerida Topsent, 1894 (12 семейств),
Отряд Halichondrida Gray, 1867 (6 семейств),
Отряд Spirophorida Bergquist et Hogg, 1969 (3 семейства).
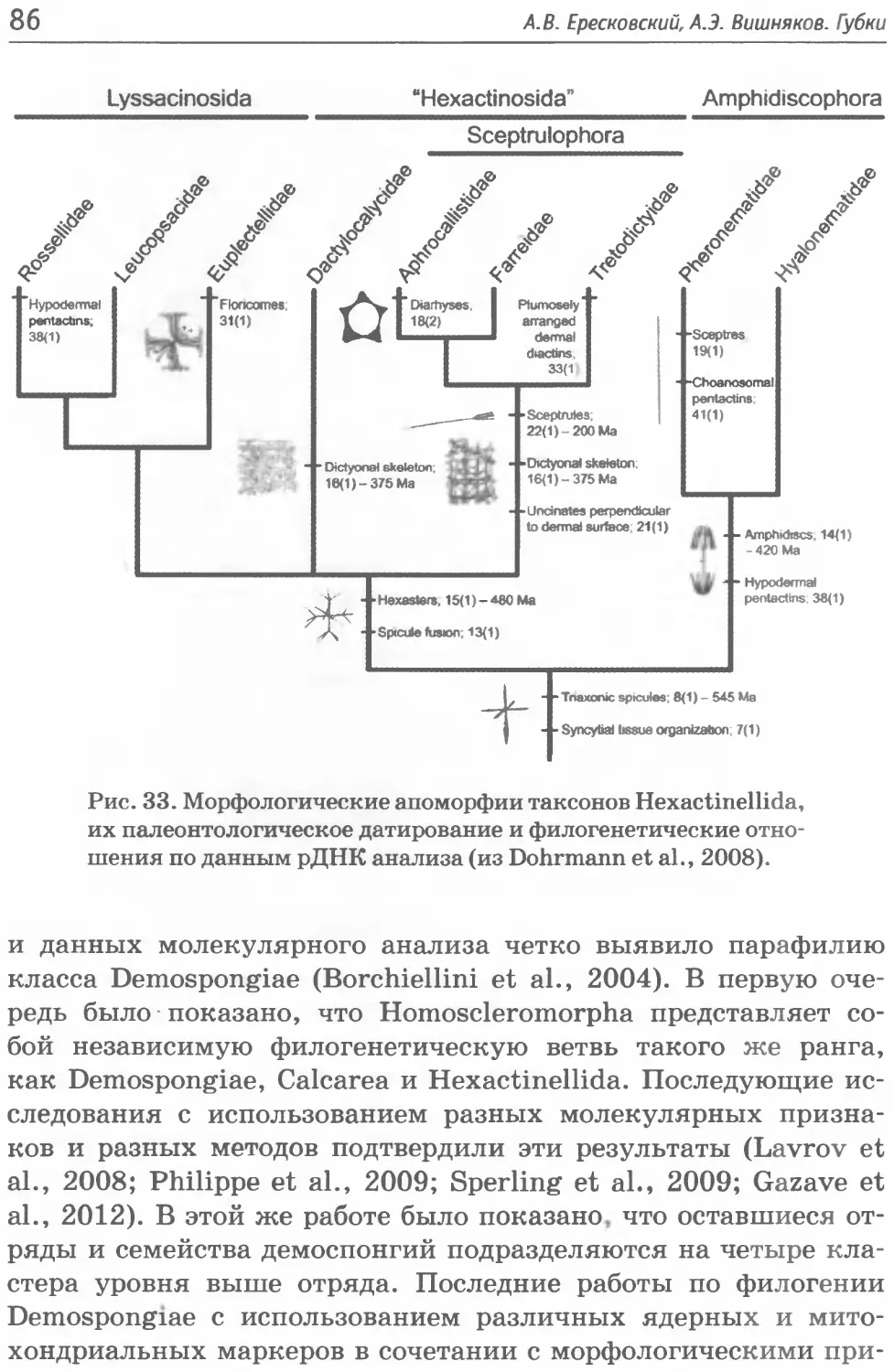
Класс Hexactinellida Schmidt, 1870
Губки этого таксона во многих книгах и учебниках имеют
еще одно название — стеклянные губки, или Hyalospongia. Это
связано с тем, что их скелетными элементами являются только
спикулы, состоящие из окиси кремния, как и у Demospongiae.
Однако для них типичны трехосные (или шестилучевые) спику-
лы и их производные, что, собственно, и дало другое название
группы Hexactinellida. Обычными спикулами являются гексак-
тины, представляющие собой три осевых элемента, пересека-
22
А. В. Ересковский, А.Э. Вишняков. Губки
Рис. 7. Спикулы и скелет Hexactinellida: а — дискоктастра;
б, в — триактинаны; г — пентактина; д — амфидиск; е — мо-
нактина; ж, з — фрагменты жестких скелетных структуры
(решеток) Hexactinellida, составленных спаянными макро-
склерами.
ющихся под правильными углами; потеря одного или больше-
го количества лучей приводит к формированию таких спикул
как пентактины, тетрактины (стаурактины), триактины
(тауактины) или диактины. Изредка встречаются монактины
(рис. 7). Осевая нить спикулы находится в четырехугольной по-
лости
Жесткие скелетные структуры (решетки) составлены макро-
склерами, часто спаянными друг с другом (рис. 7 ж, з). Губки со
спонгиновыми волокнами, отложениями кальция или бесске-
летные губки у гексактинеллид неизвестны.
Кремневый скелет дает опору для сложной живой синцити-
альной структуры — трабекулярного ретикулюма, — состав-
ные части которой, в зависимости от функций и расположения
в теле губок, имеют соответствующие названия: дермальная и
атриальная мембраны, внутренняя трабекулярная сеть первич-
ный ретикулюм и др. (см. гл. 3). Отдельные клетки, распола-
гающиеся в карманах или капсулах синцития, могут бьйь со-
единены особыми поровыми контактами (plagged junctions).
Воротничковые камеры большие, сиконоидного, силлеибидного
или лейконоидного типов.
Форма тела представителей Hexactinellida сильно варьирует.
Встречаются трубчатые, кубковидные, комковидные, отростча-
тые или лопастные губки с анастомозами либо без них, однако
корковые формы неизвестны.
Глава 2. Таксономическая структура типа Pohfera
23
Все представители класса живородящие. Характерная ли-
чинка — трихимелла имеет свойственный только ей тип разви-
тия (см. гл. 7).
Шестилучевые губки обитают исключительно в морях в диа-
пазоне глубин 5-6770 м. Класс включает два подкласса, 6 отря-
дов, 26 семейств, 166 родов и приблизительно 600 видов, что со-
ставляет 7,3% от всех описанных видов Porifera.
Подкласс Amphidiscophora Schulze, 1886
Отряд Amphidiscosida Schrammen, 1924 (3 семейства, 12 ро-
дов);
Отряд Hexactinosida Schrammen, 1903 (7 семейств, 41 род);
Подкласс Hexasterophora Schulze, 1886
Отряд Aulocalycoida Tabachnick et Reiswig, 2000 (2 семей-
ства, 9 родов);
Отряд Hexactinosida Schrammen, 1912(10 семейств, 43 рода);
Отряд Lychniscosida Schrammen, 1903 (2 семейства, 3 рода);
Отряд Lyssacinosida Zittel, 1877 (4 семейста, 58 рода).
Глава 3.
АНАТОМИЯ И КЛЕТОЧНЫЙ СОСТАВ ГУБОК
Являются ли губки организмами, имеющими тканевую ор-
ганизацию, — предмет дискуссий нескольких поколений био-
логов. История этого вопроса неоднократно обсуждалась в ли-
тературе (Bergquist, 1978; Короткова, 1981а, 1997; Simpson,
1984; Ересковский, 2005; Ereskovsky, Dondua, 2006; Leys,
2004).
Как известно, у многоклеточных выделяют четыре основных
типа тканей: (1) пограничные (эпителиальные), (2) ткани вну-
тренней среды, или соединительные (кровь, интерстициальные,
скелетные), (3) ткани нервной системы и (4) мышечные ткани.
У губок присутствуют совокупности клеток, которые по своим
функциям и некоторым структурным особенностями сходны
с первыми двумя типами тканей, что позволяет сравнивать их
между собой.
Эпителиальная организация представляет собой одну из
отличительных особенностей многоклеточных животных,
которые сохраняют покровный эпителий в течение всего
жизненного цикла. Для эпителиальных тканей Metazoa харак-
терны два основных первичных признака их системной органи-
зации: структурное объединение эпителиальных клеток в не-
прерывные пласты, функционирующие как целостные системы,
и полярность вследствие пограничного положения этих тканей.
Кроме этого есть еще две дополняющие особенности, такие как
наличие разнообразных оформленных межклеточных контак-
тов и базальная пластинка, подстилающая эпителий. Основная
функция эпителиев — защитная. Кроме этого некоторым*эпи-
телиям свойственна секреторная, всасывающая и другие функ-
ции. Под тканями внутренней среды подразумевается комплекс
тканей, образующих внутреннюю среду организма и поддержи-
вающих ее постоянство. Основными функциями этого комплек-
са тканей являются: (1) трофическая, (2) создание структурного
объединения составляющих организм клеток, (3) поддержание
постоянства химического и осмотического состава, (4) защитная
(иммунная) функция.
Глава 3. Анатомия и клеточный состав губок
25
Покровные клеточные слои — пинакодерма и хоанодерма —
и мезохил, как совокупность клеток и внеклеточных элементов
внутренней среды у губок более примитивны, чем их аналоги
у других Metazoa, как структурно, так и функционально, что
не позволяет рассматривать их как настоящие ткани. Клет-
кам губок свойственна способность к трансдифференцировкам
в другие типы клеток (Короткова, 1981а, 1997). Кроме того, у
Porifera нет единой категории тотипотентных или полипотент-
ных клеток. Так, у Demospongiae это археоциты и хоаноциты, у
Calcarea — хоаноциты, у Homoscleromorpha — хоаноциты и пи-
накоциты, у Hexactinellida, обладающих синцитиальной орга-
низацией, вероятно, археоциты.
Тем не менее, у отдельных таксонов более высокого порядка
можно обнаружить отдельные признаки, по которым прослежи-
ваются тенденции и направления эволюции становления насто-
ящих тканей.
Calcarea, Demospongiae и Homoscleromorpha — три класса
губок, имеющих клеточный уровень организации, на основа-
нии чего их объединяют в несистематическую группу Cellularia.
У класса Hexactinellida значительная часть тела представлена
объемной сетью синцитиальных образований, благодаря чему
их противопоставляют группе Cellularia и называют синцити-
альными губками, или Symplasta.
Организация губок группы Cellularia
Губки, представляющие клеточный тип организации име-
ют разные варианты строения, сводимые к четырем основ-
ным типам, описанным в главе 1. Значительные объемы воды
проводятся от остий до оскулюма по совокупности каналов и
хоаноцитных камер водоносной системы. Отграниченное хо-
анодермой и пинакодермой внутреннее пространство — мезо-
хил — иногда сопоставимо по объему со слоями клеток, но чаще
многократно его превышает. В мезохиле разных видов можно
обнаружить разные клетки, число типов которых у Cellularia по
разным источникам приближается к полутора десяткам. В слу-
чае асконоидной водоносной системы пространство атриальной
полости, выстланное хоаноцитами, можно условно назвать од-
26
А. В. Ересковский, А.Э. Вишняков. Губки
ной большой хоаноцитной камерой, у которой будет много «про-
зопилей», соответствующих внутренним отверстиям приводя-
щих каналов, и один «апопиль», совпадающий с оскулюмом.
В тех случаях, когда губки характеризуются иными варианта-
ми строения водоносной системы (сиконоидной, силлеибидной
и лейконоидной), то обычно в их теле можно выделить зону, в
которой располагаются хоаноцитные камеры — хоаносома.
У ряда видов губок между хоаносомоб и экзопинакоцитами на-
ходится слой мезохила, в котором хоаноцитные камеры отсут-
ствуй Эту часть губок принято называть эктосома. Часто экзо-
пинакоциты с прилегающим изнутри тонким слоем мезохила
выделяются на гистологических срезах большей плотностью
по сравнению с другими участками. Эту самую внешнюю часть
тела губок тогда называют кортикальным слоем, или кортек-
сом. Слой в базальной части губок, как и в эктосоме, хоаноцит-
ных камер обычно не имеет.
Клеточные слои Cellularia
Пинакодерма подразделяется на экзо-, базо- и эндопинако-
дерму. Экзопинакодерма покрывает тело губки снаружи. Базо-
пинакодерма развивается в основании губки и отвечает за при-
крепление ее к субстрату. Эндопинакодерма образует стенки
субдермальных полостей и каналов водоносной системы. Соот-
ветственно выделяют несколько разновидностей пинакоцитов.
Экзопинакоциты — покровные клетки губок, которые при
взгляде на губку снаружи выглядят как полигональные обра-
зования, а на продольных срезах имеют веретеновидную или
Т-образную форму (рис. 8).
Веретеновидные клетки описаны у многих Демоспонгий из
отрядов Spongillidae, Poecilosclerida, у всех Homoscleromorpha и
Calcarea. Т-образные экзопинакоциты описаны у многих Демо-
спонгий, а также у Calcarea.
Специализированные межклеточные контакты между эк-
зопинакоцитами у большинства видов не известны. Однако
между собой они связаны хорошо развитой адгезионной систе-
мой гликокаликса (Schiitze et al., 2001а). У Hippospongia com-
munis, Ephydatia fluviatilis, Sycon coactum, Oscarella lobularis
Глава 3. Анатомия и клеточный состав губок
27
Рис. 8. Пинакодерма и пинакоциты Cellularia: а — пинако-
дерма, состоящая из веретеновидных пинакоцитов (б); в —
пинакодерма, состоящая из Т-образных (г) пинакоцитов;
аг — аппарат Гольджи; ва — вакуоль; мт — митохондрия;
эпр — эндоплазматический ретикулюм; я — ядро (по: Корот-
кова, 1981).
и О. tuberculata в местах контактов экзопинакоцитов имеются
электронно-плотные утолщения мембран, напоминающие опо-
ясывающие десмосомы (Pavans de Ceccatty et al., 1970; Pottu-
Blumendil, 1975; Eerkes-Medrano, Leys, 2006; Ereskovsky,
Tokina, 2007; Ereskovsky et al., 2009). Экзопинакоциты способ-
ны к синтезу и выведению на свою поверхность компонентов
внеклеточного матрикса, в том числе и коллагена (Simpson,
1984). Экзопинакоциты Homoscleromorpha тесно ассоциирова-
ны с подстилающим их плотным фибриллярным слоем, вклю-
чающим коллаген IV, ламинин и тенасцин. Этот слой идентичен
базальной пластинке эпителиев позвоночных (Boute et al., 1996;
Ereskovsky et al., 2009). Уникальными особенностями экзопи-
накоцитов Homoscleromorpha являются наличие у них жгутика,
а также способность к синтезу спикул (Maldonado, Riesgo, 2007;
Ereskovsky et al., 2009).
В экзопинакодерме располагаются поры — многочислен-
ные микроскопические отверстия диаметром от 4 до 100 мкм,
через которые вода проникает в водоносную систему губки.
У большинства Demospongiae и у всех Homoscleromorpha поры
28
А.В. Ересковский, А.Э. Вишняков. Губки
представляют собой межклеточные отверстия. У Calcarea поры
формируются внутри отдельных клеток — пороцитов, цилинд-
рических трубчатых клеток (Eerkes-Medrano, Leys, 2006).
У Sycon coactum пороциты способны сокращаться в ответ на ме-
ханическое раздражение и на обработку химическими анестети-
ками (Eerkes-Medrano, Leys, 2006). У некоторых Demospongiae
также есть пороциты, которые имеют уплощенную форму с от-
верстием в центре или на периферии клетки. Отверстие способ-
но открываться и закрываться подобно сфинктеру.
Экзопинакодерма проявляет многие функции, свойственные
типичным эпителиям Eumetazoa, например, абсорбция, секре-
ция, транспорт, выделение, защита.
Базопинакоциты — уплощенные клетки, располагающиеся
на базальной поверхности губки. Основной их функцией явля-
ется прикрепление губки к субстрату за счет синтеза спонгина
и хитина. Базопинакоциты демоспонгий, имеющих массивный
арагонитовый скелет — Spirastrella wellsi (отр. Hadromerida),
Ceratoporella nicholsoni и Stromatospongia погас (отр. Agelasi-
da) — также участвуют в синтезе углекислого кальция (Reitner,
Gautret, 1996).
В базопинакоцитах пресноводных демоспонгий Ephydatia
muelleri и Spongilla lacustris был обнаружен упорядоченный ци-
тоскелет (Pavans de Сес catty, 1986; Wachtmann et al., 1990) и
между клетками этого типа у Е. muelleri были выявлены десмо-
сомоподобные контакты (Pavans de Ceccatty, 1986).
Эндопинакоциты — уплощенные, веретеновидные на срезах
клетки, выстилают приносящие и выносящие каналы губок.
Эндопинакоциты подразделяются на прозопинакоциты, обра-
зующие приносящие каналы, и апопинакоциты, образующие
выносящие каналы. У всех Homoscleromorpha и у некоторых
Demospongiae из отрядов Dictyoceratida и Dendroceratida они
имеют жгутики. Вероятно, наличие жгутиков у эндопинакоци-
тов связано с их участием в циркуляции воды по каналам водо-
носной системы. Базальная часть клеток часто формирует мно-
гочисленные псевдоподиальные выросты, с помощью которых
они закрепляются во внеклеточном матриксе.
Между собой эндопинакоциты контактируют по типу про-
стого прилегания. Однако у пресноводных губок в эндопинако-
Глава 3. Анатомия и клеточный состав губок
29
дерме трубки, ведущей к оскулюму (оскулярная трубка) клет-
ки объединены между собой десмосомоподобными контактами
(Masuda et al., 1998). Эндопинакоциты Oscarella (Homosclero-
morpha) объединены в пласты с помощью десмосомоподобных
контактов (Ereskovsky, Tokina, 2007).
В составе пинакодермы некоторых демоспонгий описаны осо-
бые сократимые клетки — миоциты. Они представляют собой
длинные веретеновидные клетки со скоплениями сократимых
цитоплазматических филаментов, встроенные в стенку оску-
лярной трубки Microciona prolifera и в сократимый край оску-
люма Tedania ignis (отр. Poecilosclerida) (Bagby, 1966). У пре-
сноводных губок пучки актиновых филаментов залегают в
эндопинакоцитах оскулярных трубок, а не в миоцитах (Masuda
etal., 1998).
Считается, что клетки с составе эпителиальных тканей зани-
мают относительно стабильное положение. Пинакоциты губок,
в отличие от эпителиальных клеток Eumetazoa, способны сокра-
щаться, а также принимать амебовидную форму и перемещать-
ся. Так, например, базопинакоциты Corvomeyenia carolinensis
(отр. Spongillida) перед распластыванием в новом месте ста-
новятся амебовидными, а отдельные экзо- и базопинакоци-
ты Ephydatia fluviatilis (отр. Spongillida) могут мигрировать
(Harrison, 1972; Weissenfels, 1978). Эта необычная для клеток
типичного эпителия способность, вероятно, связана с отсутстви-
ем настоящих десмосом между клетками пинакодермы у этих
губок, а также с наличием хорошо развитой сети цитоскелет-
ных актиновых микрофиламентов, ассоциированных с систе-
мой микротрубочек (Pavans de Ceccatty, 1986).
Все типы пинакоцитов способны фагоцитировать пищевые
частицы и выводить непереваренные остатки наружу.
Хоанодерма Cellularia состоит из клеток одного типа — хо-
аноцитов, обладающих жгутиком, который окружен ворот-
ничком из микроворсинок (рис. 9). В отличие от пинакодермы,
клетки хоанодермы могут иметь цилиндрическую, кубическую
и трапециевидную форму или быть слегка уплощенными.
Хоаноциты могут иметь различную форму как у разных гу-
бок, так и внутри одной губки. Их форма зависит от условий оби-
тания губки, физиологического состояния или от стадии онтоге-
30
А. В. Ересковский А.Э. Вишняков. Губки
Рис. 9. Хоанодерма (а) и хоаноциты Cellularia с апикально (б)
и базально (в) расположенным ядром (по: Короткова, 1981);
аг — аппарат Гольджи; мт — митохондрия; пв — пищевари-
тельная вакуоль; эпр — эндоплазматический ретикулюм; я —
ядро.
неза. Тем не менее, для этих клеток характерны общие признаки.
Жгутик отходит от небольшого бугорка или от кольцевидного
вдавливания на апикальной поверхности клетки. У некоторых
демоспонгий из отрядов Hadromerida и Halisarcida основание
жгутика окружено высоким цитоплазматическим манжетом.
Аксонема жгута имеет типичное для Eukaryota строение.
Базальный аппарат включает две центриоли, ориентированные
перпендикулярно, либо, как у Halisarcida, под косым углом
Глава 3. Анатомия и клеточный состав губок
31
друг к другу. Аппарат Гольджи находится между центриолями
и ядром, либо сбоку от ядра, когда оно располагается апикально
(рис. 96, в).
Воротничок хоаноцитов включает от 25 до 55 микровиллей.
Микровилли могут быть соединены между собой тонкими тяжа-
ми гликокаликса либо цитоплазматическими выростами, обра-
зующими тонкоячеистую сеть. Клетки этого типа обладают хо-
рошо развитым цитоскелетом, включающим F-актин и миозин.
Хоаноциты объединены в хоаноцитные камеры, которые у
Demospongiae подстилаются более или менее рыхлым слоем вне-
клеточного матрикса, включающего коллаген. Для хоанодермы
Homoscleromorpha, как и для их пинакодермы, характерно на-
личие настоящей базальной пластинки с коллагеном IV типа.
В базальной части хоаноцитов многих демоспонгий форми-
руются цитоплазматические выросты, которые не только за-
якоривают клетки во внеклеточном матриксе, но, переплетаясь
друг с другом, придают механическую стабильность хоанодер-
ме. Между собой хоаноциты имеют контакты по типу просто-
го прилегания в базальной, либо в средней части клетки. Хоа-
ноциты Homoscleromorpha: Oscarella tuberculata и О. lobularis
(Ereskovsky, Tokina, 2007), а также известковой губки Sycon
coactum (Eerkes-Medrano, Leys, 2006) образуют специализиро-
ванные межклеточные десмосомоподобные контакты.
Апопилярные клетки — особый тип клеток, переходных
между апопинакоцитами и хоаноцитами. Эти жгутиковые клет-
ки образуют границу между выносящим каналом и хоаноцит-
ной камерой. Апопилярные клетки имеют треугольную форму
на поперечном срезе. Их край, обращенный в камеру, несет гре-
бень микровиллей.
Апопилярные клетки описаны у ряда демоспонгий из отря-
дов Haplosclerida, Halisarcida, Dictyoceratida и Dendroceratida.
Они характерны для всех Homoscleromorpha (Ereskovsky, 2010).
Предполагается, что основной ролью апопилярных клеток явля-
ется регуляция потока в водоносной системе.
В состав хоаноцитных камер многих демоспонгий входят
также центральные клетки. У Suberites massa (отр. Hardo-
merida) центральная клетка, располагающаяся на выходе из
жгутиковой камеры, имеет неправильную разветвленную фор-
32
А. В. Ересковский, А.Э. Вишняков. Губки
Рис. 10. Центральная клетка (из: Simpson, 1984).
му с многочисленными выростами и сквозными отверстиями.
Внутри клетки проходит обширный канал, в который входят
жгутики хоаноцитов (рис. 10). Центральные клетки Pettina
fistulosa и Р. semitubulosa (отр. Haplosclerida) уплощены и име-
ют вид перфорированной пластинки, которая выстилает хоано-
цитную камеру поверх апикальной части хоаноцитов. При этом
жгутики хоаноцитов проходят в пространство камеры через от-
верстия в центральной клетке. Интересно, что эти клетки име-
ются не во всех хоаноцитных камерах одной губки. Считается,
что центральные клетки участвуют в регуляции биения жгути-
ков хоаноцитов в пределах одной камеры.
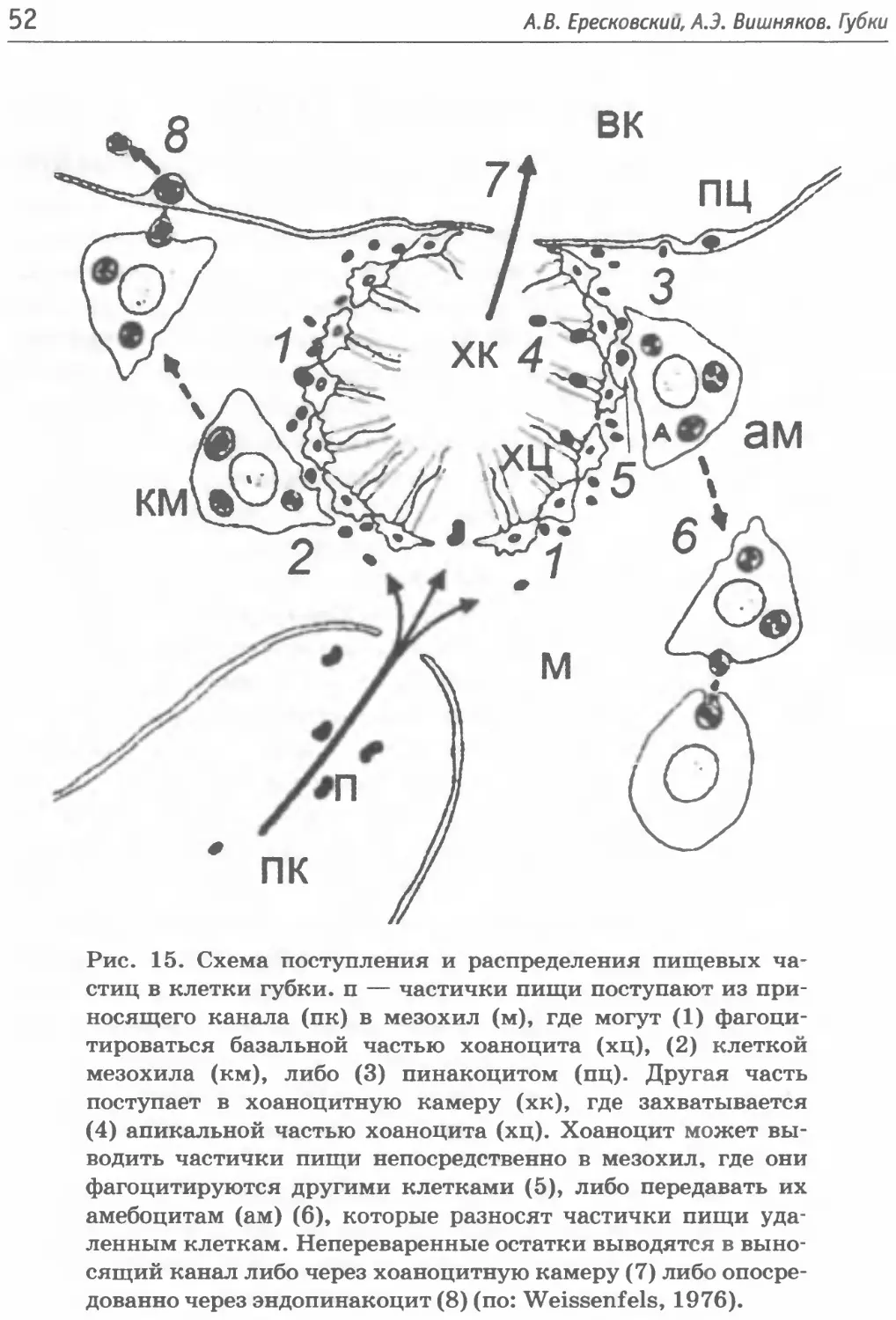
Клетки мезохила Cellularia
Мезохил — внутренняя среда губок, отграниченная от внеш-
ней слоями хоаноцитов и пинакоцитов и состоящая из скелет-
ных элементов, молекул внеклеточного матрикса (ВКМ) и кле-
ток, выполняющих разные функции, одна из которых, создание
ВКМ. По аналогии с другими животными можно выделить опор
но-соединительную и защитно-секреторную функции, которые
выполняются определенными группами специализированных
Глава 3. Анатомия и клеточный состав губок
33
клеток губок (Короткова, 1981а; Ересковский, 2005). Особенно-
стью всех клеток внутренней среды губок с клеточной организа-
цией можно считать то, что все они представляют собой популя-
ции свободных, мобильных, как неспециализированных, так и
способных к трансдифференцировкам клеток.
Клетки с опорно-соединительной функцией
Главная функция этой группы клеток состоит в формирова-
нии элементов органического и минерального скелета и ВКМ
мезохила.
Колленциты — подвижные клетки, секретирующие кол-
лаген у губок. Они представляют собой разновидность фи-
бробластов. Колленциты характеризуются звездчатой или
веретеновидной формой с ветвящимися псевдоподиями, безъ-
ядрышковым ядром и цитоплазмой, не содержащей специфич-
ные включения. Вполне возможно, что под колленцитами под-
разумевается гетерогенная популяция клеток (Simpson, 1984).
Эксперименты по пересадке фрагментов тела губок от одной осо-
би к другой показывают участие колленцитов в защитных реак-
циях (Custodio et al., 2004).
Лофоциты рассматриваются, в качестве разновидности кол-
ленцитов и, следовательно, являются разновидностью фибро-
бластов. Это амебовидные клетки, секретирующие элементы
внеклеточного матрикса и отвечающие за его организацию в
фибриллярные пучки (рис. 11а). Лофоциты имеют вид скольз-
ящих по поверхности субстрата клеток с четко выраженной
переднезадней полярностью. На переднем полюсе имеются ко-
роткие звездчатые псевдоподии. Задний конец оставляет за со-
бой шлейф в виде пучка коллагеновых фибрилл, объединенных
гликопротеиновым матриксом. Ядро содержит ядрышко, в ци-
топлазме имеется сильно развитый шЭПР, хорошо заметные
микротрубочки и микрофиламенты. Считается, что у Demospon-
giae колленциты и лофоциты являются производными архео-
цитной линии.
Склероциты секретируют минеральные элементы скелета —
спикулы (рис. 116). У кремневых губок эти клетки осуществля-
ют внутриклеточный синтез спикул. В зависимости от размера
34
А. В. Ересковский АЗ. Вишняков. Губки
Рис. 11. Клетки с опорно-соединительными функциями губок:
а — лофоцит; б — склероцит; в — спонгоцит; аг — аппарат
Гольджи; к — коллаген; сп — спикула; спо — спонгин; я — ядро.
спикул выделяют мегасклероциты и микросклероциты. Син-
тез спикулы осуществляется внутри вакуоли, сформированной
особой мембраной — силикалеммой (Simpson, Vaccaro, 1974).
Силикалемма не контактирует ни с клеточной мембраной, ни с
другими мембранными структурами клетки. На микрофотогра-
фиях она существенно отличается от плазматической мембраны
высокой электронной плотностью (Uriz et al., 2000). Возможно
это определяется транспортом молекул кремниевой кислоты и ее
димеров и тримеров в направлении к формирующейся спикуле.
На самых первых этапах спикулогенеза склероциты синтезиру-
ют аксиальный филамент (протеин силикатеин) с очень низкой
концентрацией кремния. В дальнейшем двуокись кремния от-
кладывается вокруг аксиального филамента, образуя концен-
трические слои, чередующиеся с нанофибриллами коллагена.
Единственный мегасклероцит не в состоянии полностью сфор-
мировать крупную спикулу, поэтому несколько склероцитов объ-
единяются для совместного синтеза (Uriz, 2006). У демоспонгий
склероциты представляют собой археоцитную линию клеток.
Склероциты известковых губок осуществляют внеклеточный
синтез спикул (Ledger, Jones, 1977) и имеют лишь одну морфо-
логическую категорию — мегасклероциты. В секреции спикулы
участвуют как минимум два склероцита, которые плотно кон-
тактируют друг с другом, формируя между собой пространство,
ограниченное собственной мембраной —внеклеточную вакуоль.
'*ава 3. Анатомия и клеточный состав губок
35
В месте контакта мембран склероцитов образуются септирован-
ные десмосомы (Ledger, Jones, 1977). По мере роста спикул и ее
лучей форма клеток изменяется, но изолированная вакуоль со-
храняется до окончания формирования спикулы.
Склероциты Calcarea дифференцируются из пинакоцитов и,
возможно, из хоаноцитов.
Спонгоциты отвечают за секрецию периспикулярного спон-
гина, а также формируют «роговой» скелет у Demospongiae, не
имеющих минерального скелета. Эти клетки мигрируют к скле-
роцитам, секретирующим спикулу, либо к свободным спикулам
и присоединяют к ним молекулы спонгина. Предполагается, что
эти клетки обладают положительным хемотаксисом к кремнию
(Garrone, 1978). В ходе развития органического скелета спонго-
циты группируются, формируя между собой пальцевидные кон-
такты (Garrone, 1978) (рис. Ив).
При образовании оболочки геммул у пресноводных губок
(отр. Spongillida) спонгоциты образуют плотный слой столбча-
тых клеток, между которыми располагаются микросклероциты
с геммосклерами (спикулами оболочки геммул) (Langenbruch,
1982). После формирования промежуточного спонгинового слоя
эти клетки дегенерируют, образуя пневматический слой оболоч-
ки геммулы (Langenbruch, 1982).
Считается, что у Demospongiae спонгоциты, как и склероци-
ты, представляют собой производные археоцитной линии.
Клетки с защитно-секреторной функцией
Все клетки этой группы можно условно подразделить на аме-
боидные клетки и клетки с включениями. Основными их функ-
циями являются защитная (фагоцитарная, бактерицидная), за-
пасающая, секреция основного вещества мезохила и функция
передачи частичек пищи и кислорода.
Амебоидные клетки
Археоциты — типичные для демоспонгий амебоидные клет-
ки с крупным ядром и ядрышком, способные к фагоцитозу
(рис. 12а). У Homoscleromorpha археоцитоподобные амебоидные
клетки встречаются не у всех видов.
36
А. В. Ересковский, А.Э. Вишняков. Губки
Рис. 12. Клетки губок, обладающие защитно-секреторными
функциями: а — археоцит; б — бактериоцит; в — вакуоляр-
ная клетка; г — глобулярная клетка; д — сферульная клетка;
е — гранулярная клетка; ж — серая клетка; з — микрогра-
нулярная клетка; б — бактерии; ва — вакуоль; г — глобула;
гр — гликогенные розетки; я — ядро.
В спонгиологической литературе нет четкого определения
археоцитов. В разное время различные авторы эту категорию
клеток называли тезоцитами, сферульными клетками, полибла-
стами или гиалиновыми амебоцитами, но чаще всего амебоцита-
ми или ядрышковыми амебоцитами (Simpson, 1984). Под архео-
цитами подразумевают как тотипотентные (стволовые) клетки,
морфологически сходные с подобными клетками у других жи-
вотных (такими, как i-клетки у книдарий или необласты у тур-
беллярий), так и любые амебоидные клетки, содержащие ядро с
ядрышком и многочисленные фагосомы. Это наиболее активно
делящаяся фракция свободных клеток.
Археоциты традиционно считаются тотипотентными клетка-
ми, активно участвующими как в половом и бесполом размно-
'газа 3. Анатомия и клеточный состав губок
37
жении, так и в восстановительных морфогенезах (Короткова,
1981а, 1997; Funayama, 2013).
Бактериоциты представляют собой подвижные амебоидные
клетки с вакуолями, включающими прокариотических микро-
симбионтов (рис. 126). Эти клетки пока описаны только для не-
которых видов Demospongiae. Условно бактериоциты можно
подразделить на две категории: (1) клетки, содержащие одну
обширную вакуоль, содержащую бактерий и (2) клетки, об-
ладающие многочисленными небольшими вакуолями, вклю-
чающими одну или несколько бактерий. Бактериоциты могут
достигать довольно крупных размеров, например, у Suberites
domuncula (отр. Hardomerida) их диаметр составляет около
20 мкм. Бактериоциты плотоядой губки Asbestopluma hypogea
(отр. Poecilosclerida) участвуют в переваривании пищи (Vacelet,
Duport, 2004). У Chondrosia reniformis (отр. Chondrosida) бакте-
риоциты с видоспецифическими бактериями проникают из тела
материнской губки в развивающийся зародыш, где они остают-
ся в интактном состоянии до конца метаморфоза, осуществляя
тем самым вертикальный перенос бактерий. Считается, что бак-
териоциты происходят от линии археоцитов (Simpson, 1984).
Клетки с включениями
Эта гетерогенная группа клеток характеризуется наличием в
цитоплазме специфических включений. От археоцитов они от-
личаются, в первую очередь, редуцированным шЭПР; у них не
выявлена активность кислой фосфатазы и нет фагосом. Ядрыш-
ко в ядрах отсутствует. Симпсон (Simpson, 1984) формально
подразделил эти клетки на две категории по размеру включе-
ний: на клетки с крупными включениями и клетки с мелкими
включениями.
Клетки, содержащие крупные включения
Вакуолярные клетки характеризуются тем, что в их цито-
плазме содержится одна или несколько крупных электрон-
но-прозрачных или светлых вакуолей (рис. 12в). У Oscarella
tuberculata (Homoscleromorpha) вакуолярные клетки формиру-
ются путем трансдифференцировки эндопинакоцитов. Они об-
38
А.В. Ересковский, А.Э. Вишняков. Губки
разуют своеобразный «гидроскелет» губки и участвуют в синте-
зе коллагена. У Tethya citrina и Т. aurantium (отр. Hadromerida)
вакуолярные клетки входят в состав формирующейся почки
при бесполом размножении. Вакуолярные клетки могут слу-
жить диагностическим признаком у близкородственных видов
Oscarella и Halisarca (Muricy et al., 1996; Ereskovsky, 2006,
2007; Ereskovsky et al., 2011).
Глобулярные клетки содержат небольшой объем свободной
цитоплазмы. Они имеют одну или две крупные глобулы неиз-
вестного химического состава и маленькое деформированное
ядро (рис. 12г). У Hymeniacidon heliophila (отр. Halichondrida)
глобулярные клетки активно участвуют в аллогенных рекон-
струкциях, однако их четких функций выявить не удалось
(Custodio et al., 2004).
Сферульные клетки, в отличие от глобулярных, заполнены
крупными округлыми включениями различной природы и хи-
мического состава (рис. 12д). Сферульные клетки характеризу-
ются тем, что вся их цитоплазма целиком заполнена крупными,
ограниченными мембранами включениями (сферулами). Не-
больших размеров ядро не имеет ядрышка и может быть дефор-
мировано сферулами. Содержимое сферул более или менее го-
могенное, хотя в их составе встречаются паракристаллиновые,
волокнистые, ламеллярные включения. Несмотря на некоторую
гетерогенность включений, сферулы четко отличаются от круп-
ных фагосом, т.е. не являются производными литических про-
цессов в клетке. Сферульные клетки могут быть диффузно рас-
пределены в мезохиле, но чаще они локализуются в эктосоме,
либо вдоль каналов водоносной системы.
Роль сферульных клеток довольно разнообразна. Показано,
что они содержат токсические метаболиты, участвуют в экскре-
ции, аккумулируют лектины, вовлечены в поддержание ВКМ.
Сферульные клетки часто проникают в развивающийся заро-
дыш на поздних стадиях развития либо в предличинку. Они
также являются важным компонентом развивающейся почкй
при бесполом размножении.
У ряда бесспикульных губок (Oscarellidae, Halisarcidae) эти
клетки имеют видоспецифичные морфологические особенности
и служат важным диагностическим признаком.
'*сва 3. Анатомия и клеточный состав губок
39
Гранулярные клетки, вероятно, являются разновидностью
сферульных клеток, поскольку никаких принципиальных от-
личий на ультраструктурном уровне они не имеют (рис. 12е).
В тех случаях, когда в мезохиле губки четко выделяются две
категории подобных клеток, различающихся лишь размерами
включений, клетки с меньшими включениями относят к грану-
лярным, а с крупными — к сферульным.
Цитоплазматические включения гранулярных клеток мезо-
хила Halisarca dujardini (отр. Chondrosida) содержат катион-
ные пептиды и протеины, которые представляют собой факто-
ры защиты от бактерий и низших грибов (Krylova et al., 2004).
Эти же клетки активно проникают в формирующуюся личинку
данного вида и сохраняются там до первых стадий метаморфоза.
Гранулярные клетки описаны и в составе паренхимул различ-
ных демоспонгий и амфибластул известковых губок (Ересков-
ский, 2005). Их цитологические особенности могут служить ди-
агностическим признаком у близкородственных видов.
Клетки, содержащие мелкие включения
Серые клетки иногда известны как гликоциты. Эти клетки
обнаружены у многих видов Demospongiae (Harrison, De Vos,
1991). Для серых клеток характерно наличие многочисленных
мелких, ограниченных мембранами овоидных ацидофильных
и осмиофильных включений, а также гликогеновых розеток
(рис. 12ж).
Основная роль серых клеток заключается в метаболизме гли-
когена и секреции коллагена. Вместе с археоцитами эти клетки
принимают активное участие в различных морфогенезах (поло-
вых, бесполых и регенерации).
Микрогранулярные клетки характеризуются тем, что их ци-
топлазма заполнена мелкими электронно-плотными гранулами
(рис. 12з). Роль и функции микрогранулярных клеток остают-
ся малоизученными. Одно можно сказать определенно, что эти
клетки не участвуют в биосинтезе коллагена (Simpson, 1984).
40
А.В. Ересковский, А.Э. Вишняков. Губки
Организация губок группы Symplasta
(класс Hexactinellida)
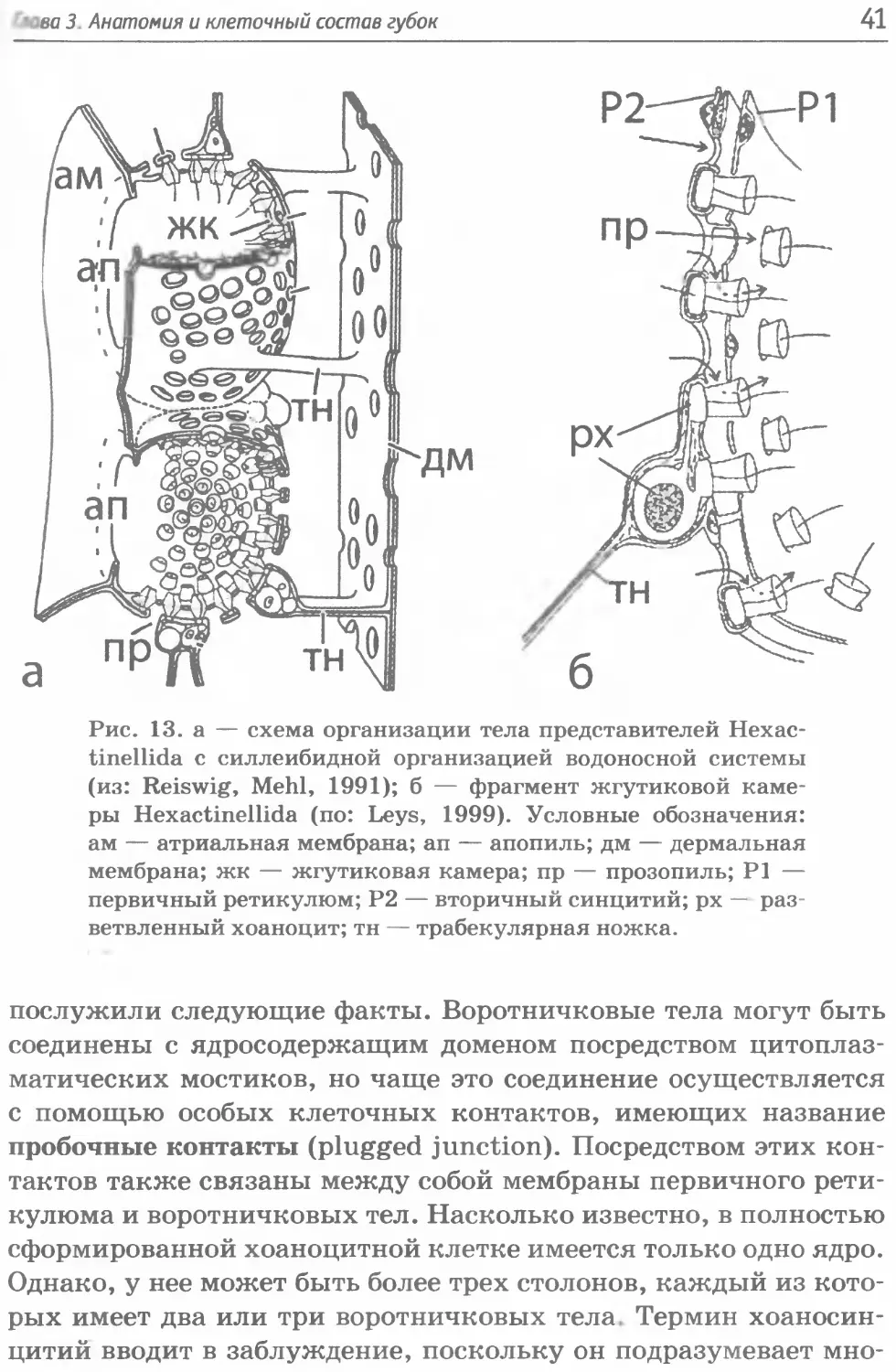
Hexactinellida — синцитиальные организмы. Это наклады-
вает отпечаток на многие их анатомические особенности, и в
значительной степени затрудняет терминологию при описании
этих губок. Их тело состоит из единого неразрывного синци-
тия, имеющего вид сети соединенных друг с другом плоскостей
и трубчатых тяжей и содержащего тысячи ядер, неспособных
к делению. Этот синцитий носит название трабекулярный ре-
тику люм (рис. 13а). Для облегчения понимания их устройства
можно вспомнить организацию эндоплазматического ретику-
люма эукариотических клеток, который имеет вид плоских ци-
стерн, соединенных между собой трубчатыми тяжами.
Снаружи тело губок ограничено плоскостным синцитием с
отверстиями (остиями), который называется дермальная мем-
брана. Аналогичное образование, выстилающее атриум — атри-
альная мембрана. Между собой эти мембраны соединены сетью
синцитиальных тяжей. Главный компонент, занимающий сре-
динное пространство между дермальной и атриальной мембра-
нами, — хоаносома, состоящая из жгутиковых камер. Трабеку-
лярные тяжи расходятся около жгутиковых камер (рис. 13а),
формируя поддерживающую сеть (первичный ретикулюм) для
воротничковых тел разветвленных хоаноцитов. Практически у
всех стеклянных губок также имеется ответвляющийся от пер-
вичного очень тонкий вторичный ретикулюм, располагающий-
ся в полости жгутиковых камер на уровне середины воротнич-
ков (рис. 136).
У Hexactinellida отсутствуют хоаноциты типичные для
Cellularia. Здесь они представлены ветвящимися клетками,
состоящими из базального ядросодержащего домена, посы-
лающего от себя выросты, заканчивающиеся воротничковы-
ми телами, или хоаномерами, сходными с апикальной ча-
стью типичных хоаноцитов (рис. 14) (Leys et al., 2007). Ранее
в научной литературе использовался термин хоаносинцитий
(choanosyncytium), — синцитиальное образование, выстилаю-
щую жгутиковые камеры. Однако позднее было предложено от-
казаться от этого термина (Leys et al., 2007). Основанием этому
юва 3 Анатомия и клеточный состав губок
41
Рис. 13. а — схема организации тела представителей Hexac-
tinellida с силлеибидной организацией водоносной системы
(из: Reiswig, Mehl, 1991); б — фрагмент жгутиковой каме-
ры Hexactinellida (по: Leys, 1999). Условные обозначения:
ам — атриальная мембрана; ап — апопиль; дм — дермальная
мембрана; жк — жгутиковая камера; пр — прозопиль; Р1 —
первичный ретикулюм; Р2 — вторичный синцитий; рх — раз-
ветвленный хоаноцит; тн — трабекулярная ножка.
послужили следующие факты. Воротничковые тела могут быть
соединены с ядросодержащим доменом посредством цитоплаз-
матических мостиков, но чаще это соединение осуществляется
с помощью особых клеточных контактов, имеющих название
пробочные контакты (plugged junction). Посредством этих кон-
тактов также связаны между собой мембраны первичного рети-
кулюма и воротничковых тел. Насколько известно, в полностью
сформированной хоаноцитной клетке имеется только одно ядро.
Однако, у нее может быть более трех столонов, каждый из кото-
рых имеет два или три воротничковых тела. Термин хоаносин-
цитий вводит в заблуждение, поскольку он подразумевает мно-
42
А. В. Ересковский, А.Э. Вишняков. Губки
Рис. 14. Разветвленные хоаноциты жгутиковой камеры
Hexantinellida (по Mackie, Singla, 1983). Условные обозначе-
ния: вт — воротничковое тело; ж — жгутик; я — ядро.
гоядерную структуру. Поэтому более уместно и точно говорить
об этих сложных клетках как о ветвящихся хоаноцитах. По
ультраструктуре хоаноциты стеклянных губок практически не
отличаются от подобных клеток других губок.
В том случае, когда губки имеют сиконоидный или силле-
бидный варианты водоносной системы, между хоаносомой и
дермальной мембраной находится небольшая сеть тяжей тра-
бекулярного синцития, называемая внешний периферический
трабекулярный ретикулюм (рис. 13а) (Leys et al., 2007). Вода
сразу после входных пор (остий) в дермальной мембране попа-
дает в объемные полости, отграниченные этим ретикулюмом, и
далее доходит до прозопилярных отверстий жгутиковых камер.
После этого, попав в пространство между первичным и вторич-
ным ретикулюмом, она проходит между микровил л ями ворот-
ничков и оказывается в жгутиковых камерах. Через крупные
апопили диаметром 20-35 мкм вода выходит в атриальную по-
лость и далее — во внешнюю среду. В случае лейконоидной
организации водоносной системы к развитому внешнему ре-
Глава 3. Анатомия и клеточный состав губок
43
тикулюму добавляется еще более сложный внутренний перифе-
рический ретикулюм, который располагается между атриаль-
ной мембраной и хоаносомой. Выносящие каналы водоносной
системы, окруженные этим ретикулюмом, по мере удаления от
хоаносомы постепенно увеличиваются в сечении и открываются
в атриальную полость отверстиями диаметром 100-200 мкм.
Синцитий трабекулярного ретикулюма функционально яв-
ляется аналогом клеточных пластов Cellularia, у которых пи-
накоциты, в сочетании с хоаноцитами, оконтуривают мезохил
со всех сторон. У стеклянных губок синцитий также окружает
мезохил, который описан только на электронно-микроскопиче-
ском уровне на срезах синцитиальных трабекул и пластов. Он
имеет вид тонкого фибриллярного пласта толщиной около 0,05-
ОД мкм между двумя слоями цитоплазмы (Leys et al., 2007).
Предполагается, что эта коллагеновая «мезоламелла» поддер-
живает трабекулы и, вероятно, секретируется самим трабеку-
лярным синцитием, а не свободными клетками. Среди волокон
мезохила находятся различные клетки и симбиотические бак-
терии.
Свободные клетки Symplasta
Свободные клеточные элементы стеклянных губок представ-
лены археоцитами, склероцитами и клетками с включениями.
Как и в случае с Cellularia, мы будем условно их делить на две
группы клеток — с опорно-соединительными и секреторно-за-
щитными функциями.
Клетки с опорно-соединительной функцией
Склероциты и склеросинцитий участвуют в формирова-
нии всех вариантов кремниевых спикул Hexactinellida. Однако
в большинстве случаев секреция скелетных элементов осущест-
вляется синцитиальными образованиями. Этот секретирующий
спикулы склеросинцитий получил название вторичного синци-
тия (Leys et al., 2007). Трабекулярный ретикулюм в этом слу-
чае называют первичным синцитием. Секреция кремниевых
спикул у Hexactinellida, как и у Demospongiae осуществляется
44
А. В. Ересковский, А.Э. Вишняков. Губки
в цитоплазме, то есть, внутри склеросинцития или склетоци-
тов. Формирующиеся спикулы, внутри которых располагается
аксиальный филамент, окружены снаружи силикалеммой. Ин-
тересно, что у зародышей Oopsacas minuta (отр. Lyssacinosida)
склероциты имеют псевдоподиальные выросты и соединяются
с окружающими клетками с помощью пробочных контактов
(Leys et al., 2007).
Клетки с защитно-секреторными функциями
Археоциты обычно собраны в кластеры или «скопления»
(congeries). Клетки имеют сферическую или округлую форму
диаметром от 3-5 до 8 мкм (Leys et al., 2007). Археоциты в кла-
стерах часто соединены друг с другом и с трабекулярным син-
цитием пробочными контактами. Округлая форма клеток и от-
сутствие псевдоподий свидетельствуют о том, что археоцитам
Hexactinellida, в отличие от соответствующих клеток других гу-
бок, не свойственна подвижность. Предполагается, что археоци-
ты стеклянных губок, как и у демоспонгий, представляют собой
пул полипотентных клеток.
К летки с включениями
Стеклянные губки имеют отдельные клетки разнообразных
типов с крупными и мелкими включениями, функция которых
неизвестна.
У нескольких видов Hexactinellida были обнаружены окру-
глые клетки, цитоплазма которых заполнена вакуолями, со-
держащими мелкодисперсный хлопьевидный материал. Эти
клетки были описаны как сферульные, или вакуолярные (Leys
et al., 2007). Кроме этого, описаны овальные или сферические
клетки размером от 5-7 мкм до 7-12 мкм, в цитоплазме кото-
рых имелась одна крупная гомогенная вакуоль, смещающая
ядро к одному из полюсов в узкую прослойку цитоплазмы. Ино-
гда внутри вакуоли находили кристалллоидные структуры, ве-
роятно, белкового происхождения. Эти клетки, получившие
название глобулярных, практически полностью окружены ци-
топлазматическими выростами трабекулярного синцития, обра-
зующими своеобразные карманы. Гранулярные клетки, около
'*ава 3. Анатомия и клеточный состав губок
8 мкм в сечении, имеют овальную форму. Они содержат много-
численные мелкие (1-2 мкм) электронно-плотные гранулы. Гра-
нулярные клетки не заключены в трабекулярный синцитий,
но способны частично проходить сквозь него и контактировать
с водой. В ряде случаев между мембранами синцития и клетка-
ми этого типа отмечены пробочные контакты (Leys et al., 2007)
В сходных с ними клетках, имеющих диаметр 10-25 мкм, отме-
чено высокое содержание ионов кальция. Предполагается так-
же, что эти клетки запасают лектины. Есть данные, показываю-
щие дифференцировку этих клеток из археоцитов.
Глава 4.
КОЛОНИАЛЬНОСТЬ, МОДУЛЬНОСТЬ
И ИНДИВИДУАЛЬНОСТЬ ГУБОК
Проблема колониальное™ и индивидуальности губок была
на протяжении всей истории спонгиологии одной из наиболее
дискуссионных. Многие исследователи относили губок, вне за-
висимости от уровня их организации, к колониальным орга-
низмам, рассматривая в качестве зооидов или особей в составе
колонии амебоидные или воротничковые клетки, хоаноцитные
камеры или оскулюм. Другие авторы, опираясь на общеприня-
тое представление о том, что колонии многоклеточных живот-
ных возникли в результате не доведенного до конца бесполого
размножения, в качестве зооидов у губок рассматривали оску-
люм с прилежащими участками водоносной системы. Противо-
положного мнения придерживались авторы, считавшие что
понятие «колониальность» не применимо к Porifera, поэтому
всякая морфологически обособленная губка является инди-
видом. По их мнению, образование оскулюмов происходит в
результате обычного роста, а не вследствие не доведенного до
конца бесполого размножения, свойственного образующейся
колонии. Третья группа авторов считала, что индивидуальность
у губок выражена крайне слабо, а признаки колониальное™ у
них еще не выработались, поэтому было предложено рассматри-
вать Porifera в качестве индивидуальных организмов (см.: Ере-
сковский, 2003, 2005).
Сложности, возникающие при интерпретации организации
губок с позиций колониальное™ или индивидуальности, в пер-
вую очередь связаны с уникальными особенностями морфофи-
зиологической организации этих организмов и спецификой их
индивидуального развития и жизненных циклов.
Долгое время основным критерием индивидуальности или
колониальное™ был сравнительно-анатомический план стро-
ения организма (Беклемишев, 1964). Однако с конца 70-х го-
дов XX столетия проблема колониальное™ вышла за пределы
сравнительной морфологии. Ее стали изучать с позиций особен-
ностей существования и физиологии организмов (Бигон и др.,
Глава 4. Колониальность, модульность и индивидуальность губок
47
1989; Марфенин, 1993). Было введено понятие биологического
модуля под которым подразумевается «любая из множествен-
ных частей организма, имеющая конструкционное значение».
Анализ характера размещения элементов водоносной си-
стемы губок (хоаноцитных камер, приносящих и выносящих
каналов, оскулюмов), их количества и объема в сочетании с
массой тканей губки и гидродинамическими условиями среды
обитания, привели У. Фрая (Fry, 1979) к формулировке поня-
тия «водоносный модуль» (aquiferrous module). Согласно авто-
ру, водоносным модулем является определенный объем губки,
обслуживаемый системой хоаноцитных камер и водоносных
каналов, ассоциированных с одним оскулюмом. Соответствен-
но, губка представляет собой не колонию с зооидами и не много-
оскулюмный индивид, а модульный организм.
В отличие от унитарных (одиночных) модульные организмы
состоят из набора основных повторяющихся конструктивных
элементов (модулей), количество которых варьирует в течение
жизненного цикла. Развитие модульных организмов не предо-
пределено какой-либо жесткой программой и в значительной
степени зависит от их связи со средой (Бигон и др., 1989; Мар-
фенин, 1993).
Морфологически обособленную губку, независимо от ее кон-
структивного уровня (аскон, сикон или лейкон) и количества
входящих в ее состав оскулюмов мы должны признать в каче-
стве индивида. Несомненно, существуют различные уровни ор-
ганизменной интеграции губок в зависимости от количества и
характера расположения водоносных модулей в составе их тела.
Это наиболее ярко проявляется в экспериментах по их восстано-
вительным морфогенезам (Короткова, 1997). Однако прямой за-
висимости уровня организменной интеграции от степени слож-
ности водоносной системы у губок нет.
Глава 5.
СИМБИОТИЧЕСКИЕ АССОЦИАЦИИ ГУБОК,
ПРОКАРИОТИЧЕСКИЕ СИМБИОНТЫ
Губки, как и любые другие существа, могут представлять со-
бой среду обитания для многих одноклеточных и многоклеточ-
ных организмов самых разных таксонов. Многоклеточные яв-
ляются комменсалами и обычно поселяются на поверхности, во
внутренних полостях губок, не препятствуя их жизнедеятель-
ности.
Многие губки в качестве одноклеточных симбионтов содер-
жат динофлагеллят, зоохлорелл, красных водорослей, фила-
ментозных зеленых водорослей и грибы.
Тем не менее, одной из самых примечательных черт губок
является наличие у них облигатных видоспецифичных, как
фотосинтезирующих так и неавтотрофных симбиотических
прокариот (эубактерий и архебактерий), поселяющихся в мезо-
хиле (Hentshel et al., 2006; Taylor et al., 2007; Webster, Taylor,
2012). Такие бактериальные симбионты, в большинстве случа-
ев свободно располагаются между элементами мезохила, хотя
описаны губки, у которых симбиотические бактерии обнаруже-
ны внутри специальных клеток — бактериоцитов. Количество
бактерий внутри своих хозяев может на два-три порядка превы-
шать количество бактерий в окружающей воде, а их биомасса
может составлять у некоторых видов морских губок до 60% био-
массы хозяев.
Разнообразие симбиотических прокариот в разных видах гу-
бок достаточно велико. Согласно современной систематике оно
включает в себя представителей около 20 бактериальных фи л,
одна из которых, Poribacteria, состоит исключительно из бакте-
рий, встречающихся только в губках.
Симбиотические бактерии могут принимать участие в раз-
нообразных физиологических процессах губок. Одной из важ-
нейших функций является их участие в рециклинге нераство-
римых протеинов и структурных перестройках органического
скелета и внеклеточного матрикса (Taylor et al., 2007). Многие
симбиотические бактерии синтезируют вещества, обладающие
~*ава 5. Симбиотические ассоциации губок, прокариотические симбионты
49
резким запахом и антибиотическими свойствами. Это использу-
ется губками для защиты от хищников и бактериальных патоге-
нов (Taylor et al., 2007). Симбиотические бактерии также могут
служить и дополнительным источником питания губок, как в
результате их прямого фагоцитоза, так и утилизации продуктов
их жизнедеятельности (Taylor et al., 2007).
Прокариотические симбионты могут проникать в тело губок
двумя способами. Первый связан с фильтрационной активно-
стью взрослых губок и захватом бактерий из внешней среды.
Во втором случае симбиотические бактерии переносятся от ма-
теринской губки к следующей генерации через яйца (у яйце-
кладущих видов) или личинок (у яйцеживородящих и живоро-
дящих) (Ereskovsky et al., 2005). Этот феномен, известный как
вертикальный перенос, говорит о физиологической значимости
симбиотических ассоциаций губок с прокариотическими эндо-
бионтами.
Глава 6.
ФИЗИОЛОГИЯ ГУБОК
Губки представляют собой «живые фильтры». Вся их жизне-
деятельность, все базовые физиологические процессы основаны
на этом фильтрационном способе жизнедеятельности.
Объем воды, проходящей за единицу времени, и скорость вы-
броса воды через отдельный оскулюм губки являются функцией
нескольких важных параметров: скорости внутреннего лами-
нарного потока, количества и объема хоаноцитных камер, вну-
треннего давления в хоаноцитных камерах, диаметра и длины
выводящих каналов, а также диаметра оскулярного отверстия
(Fry, 1979). Гидростатическое давление в пределах хоаноцит-
ных камер определяется особенностями функционирования хо-
аноцитов. Поэтому губки с небольшой массой тканей и тонкими
стенками тела могут оптимально функционировать, имея один,
как правило, широкий оскулюм и одну крупную хоаноцитную
камеру, имеющую низкое нагнетающее давление (Fry, 1979).
Примером могут служить асконоидные (Leucosolenia, Clathrina)
или сиконоидные (Sycon, Grantia) губки. Губки с большей мас-
сой тела требуют и более высокой пропускной способности по-
токов воды. В связи с этим у них повышается нагнетающее
давление, а увеличение диаметра выводящих каналов компен-
сируется увеличением их длины и диаметра оскулюма. У губок
с диффузно распределенными хоаноцитными камерами по «тка-
ням» одной толщины единственным фактором оптимизации
системы каналов может быть более или менее равномерное раз-
мещение приносящих и выносящих каналов. Объем и диаметр
каналов зависят от особенностей функционирования системы в
каждой особи и толщины стенки ее тела. В конечном итоге все
морфологическое разнообразие губок, количество, размер водо-
носных модулей определяется их объемом, который, в свою оче-
редь зависит от внешних условий, в первую очередь, гидродина-
мических.
мео 6. Физиология губок
51
Захват и переваривание пищи у губок
Porifera являются единственным, за исключением Placozoa,
типом Metazoa, у которых отсутствует фагоцитобласт в виде
кишечного эпителия. Нет ни пищеварительной паренхимы, ни
даже просто отдельной популяции клеток, специализирован-
ных для выполнения пищеварительных функций. В захвате
пищевых частиц принимают участие практически все покров-
ные клетки и многие клетки внутреннего пространства губок
(Ereskovsky, Dondua, 2006). Так, экспериментально была пока-
зана фагоцитарная и пищеварительная активность хоаноцитов,
археоцитов, экзопинакоцитов, эндопинакоцитов и базопина-
коцитов (Hahn-Kesel, Stockem, 1997). В одних экспериментах
использовались микроскопические латексные шарики и под-
крашенные бактерии, судьба которых от захвата и до экскреции
непереваренных остатков была прослежена на ультраструктур-
ном уровне. В других — использовалась флуоресцентная мет-
ка, а также иммуноцитохимические и биохимические методы.
Фагоцитарная активность также была описана у миоцитов и се-
креторных глобулярных клеток, у склероцитов и у лофоцитов.
Мелкие частички способны проникать между экзопинакоцита-
ми в мезохил. Однако механизм такого перемещения не изве-
стен. Некоторые клетки мезохила способны амебоидно выдви-
гать свои отростки между пинакоцитами в полость приводящих
каналов и фагоцитировать пищевые частицы, двигающиеся в
этих пространствах. Хоаноциты могут захватывать частицы
пищи как своей базальной, так и апикальной частью — в обла-
сти воротничка. После фагоцитоза хоаноциты и пинакоциты
способны передавать пищу археоцитам, либо выделять частич-
ки пищи путем экзоцитоза непосредственно в мезохил (рис. 15).
Для расщепления пищи клетки губок вырабатывают ряд фер-
ментов: амилазу, липазу, протеазу, эстеразу.
Однако губки могут быть и «плотоядными». Так, в 1995 г.
французскими исследователями Ж. Вас ле и Н. Бури-Эсно была
описана маленькая (около 15 мм) пещерная губка Asbestopluma
hypogea (отр. Poecilisclerida, сем. Cladorhizidae), у которой от-
сутствует водоносная система (Vacelet, Boury-Esnault, 1995).
Питание этих губок осуществляется исключительно за счет
52
А. В. Ересковский, А.Э. Вишняков. Губки
Рис. 15. Схема поступления и распределения пищевых ча-
стиц в клетки губки, п — частички пищи поступают из при-
носящего канала (пк) в мезохил (м), где могут (1) фагоци-
тироваться базальной частью хоаноцита (хц), (2) клеткой
мезохила (км), либо (3) пинакоцитом (пц). Другая часть
поступает в хоаноцитную камеру (хк), где захватывается
(4) апикальной частью хоаноцита (хц). Хоаноцит может вы-
водить частички пищи непосредственно в мезохил, где они
фагоцитируются другими клетками (5), либо передавать их
амебоцитам (ам) (6), которые разносят частички пищи уда-
ленным клеткам. Непереваренные остатки выводятся в выно-
сящий канал либо через хоаноцитную камеру (7) либо опосре-
дованно через эндопинакоцит (8) (по: Weissenfels, 1976).
‘*ж 6. Физиология губок
53
мелких (до 1 мм) ракообразных, которые запутываются в свое-
образной «ловчей сети», образованной длинными нитевидными
выростами пинакодермы с микросклерами в виде якорьков на
поверхности. При этом токсичные, паралитические или клей-
кие вещества не секретируются. В течение суток экзопинако-
лерма губки обрастает жертву. Пищеварение осуществляется
как внеклеточно, так и внутриклеточно, в течение нескольких
суток в мезохиле. После переваривания остается только ку-
тикула рачка, которая выводится во внешнюю среду (Vacelet,
Duport, 2004). Позднее подобный способ питания был опи-
сан и для других представителей (более 20 видов) семейств
Cladorhizidae и Esperiopsidae, обитающих на больших глубинах
(Vacelet, 2007).
Отсутствие обособленного фагоцитобласта составляет свое-
образную и неповторимую черту Porifera. Унаследованная от
протестов фагоцитарная функция, сохраняется у всех клеток
губок. Поэтому нет никаких оснований рассматривать хоа-
нодерму или систему каналов водоносной системы в качестве
производных фагоцитобласта (энтодермы). Закрепление фаго-
цитарной, а затем и более общей пищеварительной функции за
специализированными системами клеток явилось тем событи-
ем, которое открыло путь к возникновению всего многообразия
«настоящих многоклеточных» Eumetazoa.
Изменение формы тела и сенсорная активность
Традиционно принято считать, что у губок отсутствуют мы-
шечная и нервная системы и их элементы. Действительно, на
морфологическом уровне подобные структуры не обнаруже-
ны. Тем не менее, губки способны к изменению формы как все-
го тела, так и отдельных его участков, таких как оскулюм или
поры. Одновременно с этим меняется и объем тела губки. Недав-
но было показано, что эти изменения могут происходить в ответ
на внешние стимулы, либо быть эндогенными и иметь суточную
ритмичность (Renard et al., 2009; Nickel, 2010). Предполагает-
ся, что основную роль в таких трансформациях играет пинако-
дерма губки (Nickel, 2010). Что заставляет губок действовать
как единое целое и координировать морфологические измене-
54
А. В. Ересковский, А.Э. Вишняков. Губки
ния пока точно установить не удалось. В роли связующего звена
в передаче сигналов между клеткими при отсутствии нейронов,
могут выступать три известных в настоящее время механизма:
(1) клеточная проводимость, включающая межклеточную ком-
муникацию: (2) химическая диффузия и (3) электрическая про-
водимость (Renard et al., 2009; Nickel, 2010). Любой из этих
вариантов может иметь место, поскольку у этих просто устро-
енных животных выявлены биоэлектрические клеточные по-
тенциалы и нейротрансмиттеры. Анализ генома губок позволил
обнаружить гены, которые являются ортологичными генам,
вовлеченным в паттернизацию и функционирование нейро-сен-
сорной системы Eumetazoa. Интересно, что экспрессия этих ге-
нов (AmqbHLHl, AmqNotch и AmqDeltal) показана в специали-
зированных личиночных клетках паренхимулы Amphimedon
queeslandica (Demospongiae, отр. Haplosclerida) (см. Richards
et al., 2008). Таким образом, у губок есть очевидные признаки
зачатков нейро-мускульной активности, которые реализуются
фрагментарно и не оформлены в виде систем, которые можно
встретить у животных, имеющих ткани.
Глава 7.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ ГУБОК
Для губок характерно как бесполое, так и половое размноже-
ние. Половое размножение принципиально ничем не отличается
от аналогичных процессов у других многоклеточных животных.
Губки могут быть яйцеродящими (oviparous) и живородящими
(viviparous). В первом случае губки, как правило, раздельнопо-
лые, тогда как во втором, часто, гермафродиты.
Гаметогенез
Гонад у губок нет, и гаметогенез чаще всего не ограничен ка-
ким-либо определенным участком мезохила. Одной из особен-
ностей этого процесса у губок можно считать возникновение
половых клеток путем прямой трансформации из соматиче-
ских — ядрышковых амебоцитов или хоаноцитов. Основные же
стадии развития гамет сходны у губок с аналогичными процес-
сами у других животных (Ересковский, 2005).
Во время сперматогенеза у Hexactinellida, Demospongiae и
Homoscleromorpha формируются окруженные уплощенными со-
матическими клетками временные сферические образования —
сперматоцисты, в которых происходит сперматогенез. У Calca-
ronea сперматоцисты не формируются (Анакина, Короткова,
1989). Развитие мужских половых клеток в пределах цисты
обычно осуществляется синхронно. Однако у Homoscleromorpha
в пределах цисты наблюдается градиент созревания спермиев,
что, помимо других признаков, сближает эту группу губок с
Eumetazoa (Ересковский, 2005).
Сперматозоиды большинства губок относятся к примитив-
ному типу, для которого характерны большой объем цитоплаз-
мы и наличие нескольких митохондрий. Акросомы обычно нет
(рис. 15а). Однако на основании электронно-микроскопических
наблюдений, предпринятых за последние двадцать лет, установ-
лено, что среди Porifera имеются виды, представители которых
обладают сперматозоидами с акросомой, что ранее считалось
чертой, характерной для животных более высокоорганизован-
56
А. В. Ересковский, А.Э. Вишняков. Губки
Рис. 16. Сперматозоиды губок: а — примитивный спермий
Demospongiae; б — спермий с акросомой Homoscleromorpha; в
— атипичный спермий Calcaronea; ак — акросома; ж — жгу-
тик; мт — митохондрия; я — ядро.
ных групп. Так, акросомы присутствуют в сперматозоидах всех
Homoscleromorpha (рис. 166) и у представителей нескольких
видов отряда Poecilosclerida класса Demospongiae (Ereskovsky,
2010). Сперматозоиды Calcaronea атипичные, они имеют сфе-
рическую форму и лишены жгутика (рис. 16в).
Развивающиеся женские половые клетки располагаются в
хоаносоме диффузно или, иногда, мелкими группами. В ооге-
незе большинства живородящих губок не отмечено типично-
го образования желтка. У них преобладают различные формы
передачи питательного материала ооцитам от вспомогательных
клеток, из которого формируется вторичный желток. Нередко
происходит фагоцитоз целых клеток или их фрагментов, пре-
вращающихся в фаго л изосомы. У Homoscleromorpha и яйцекла-
дущих представителей Demospongiae вителлогенез осуществля-
ется за счет эндогенного синтеза.
Примитивными чертами организации ооцитов губок следу-
ет считать отсутствие сегрегации ооплазмы, а также отсутствие
яйцевых оболочек у яйцеживородящих губок. В то же время, у
яйцекладущих демоспонгий в ходе оогенеза формируется при-
митивная желточная оболочка.
•ава 7. Размножение и развитие губок
57
Эмбриогенез
Эмбриональное развитие живородящих губок происходит во
ременных выводковых камерах — особых капсулах, часто на-
зываемых фолликулами. Они формируются в конце вителло-
генеза из уплощенных клеток хоаноцитной или амебоцитной
природы и из эндопинакоцитов. По окончании развития стенки
выводковой камеры участвует в построении каналов водоносной
системы.
Дробление
У губок можно выделить четыре основных способа дробления
(рис. 17): (1) неупорядоченное (хаотическое) — Homoscleromor-
pha, большинство яйцеживородящих Demospongiae; (2) дробле-
Рис. 17. Основные типы дробления губок: а — неупорядоченное
(хаотическое); б — табличная палинтомия (инкурвационное
дробление); в — радиальное; г — полиаксиальное.
58
А. В. Ересковский, А.Э. Вишняков. Губки
ние с чертами радиальности — яйцекладущие Demospongiae,
Hexactinellida; (3) полиаксиальное — Halisarcidae (Demospon-
giae), Calcinea (Calcarea); (4) инкурвационное дробление —
Calcaronea (Calcarea). Примитивные черты дробления губок в
первую очередь связаны со слабой специализацией яйцеклетки.
У всех губок, за исключением Calcaronea, дробление обычно не
характеризуется устойчивостью пространственного расположе-
ния бластомеров. Синхронность делений либо вовсе отсутствует,
либо рано утрачивается.
Своеобразной особенностью раннего эмбриогенеза многих ви-
дов губок можно считать проникновение из мезохила в зародыш
эндосимбионтных бактерий. Особо следует отметить внедрение
в развивающийся зародыш губок соматических клеток мате-
ринского происхождения. Однако значениие этих клеток для
личинок и их метаморфоза до сих пор не исследованы.
У яйцекладущих демоспонгий рода Tetilla (отр. Spirophorida)
и Stylocordyla (отр. Hadromerida), для которых характерно пря-
мое развитие: вслед за дроблением сразу же наступает стадия
дифференцировки клеток и развитие молодой губки.
Эмбриональные морфогенезы у губок
Формирование многослойности зародышей Metazoa достига-
ется либо в результате перемещения отдельных клеток, и в этом
случае наблюдаются разные типы ингрессии и клеточной мигра-
ции, либо за счет движения пластов клеток, что является осно-
вой разного рода эпителиальных морфогенезов.
Морфогенетические движения клеток определяются слож-
ными и специфическими системами действия генов, которые
контролируют во времени и в пространстве синтез и деграда-
цию молекул адгезии, сборку и разборку элементов цитоскелета
и многие другие процессы, совокупность которых, в конечном
счете, обеспечивает движение отдельных клеток или клеточных
пластов (Дондуа, 2005). Эти механизмы универсальны.
Эволюционное появление систем «генов морфогенеза» от-
крыло путь для возникновения всего многообразия типов раз-
вития Metazoa. Очевидно, что эти системы используются и
для формирования многослойных зародышей Metazoa, в том
•ава 7, Размножение и развитие губок
59
Рис. 18. Различные типы морфогенезов Porifera, приводя-
щих к формированию личинки: а — морульная делямина-
ция (отряды Demospongiae: Dendroceratida, Dictyoceratida,
Halichondrida, Haplosclerida); 6 — клеточная деляминация
(Oopsacas minuta, Hexactinellida); в — инвагинация (Halt-
sarca dujardini, Demospongiae); г — мультиполярная им-
миграция (H. dujardini, Demospongiae); д — экскурвация
(Calcaronea); е — формирование бластулы за счет вселения
материнских клеток в морулу {Chondrosia reniformis, Demo-
spongiae); ж — поляризованная деляминация (Halichondrida,
Poecilosclerida); з — униполярная пролиферация (Vacele-
tia cripta, Demospongiae); и — мультиполярная эмиграция
(Homoscleromorpha).
60
А.В. Ересковский, А.Э. Вишняков. Губки
числе, многослойных зародышей Porifera. Так, при форми-
ровании личинок у губок описаны практически все типы
клеточных перемещений, характерные для Eumetazoa (Ере-
сковский, 2005). Действительно, здесь может иметь место мо-
рул ьная деламинация (кл. Demospongiae: отряды Dendrocera-
tida, Dictyoceratida, Halichondrida, Haplosclerida) (рис. 18a),
клеточная деламинация (кл. Hexactinellida) (рис. 186), инва-
гинация и мультиполярная иммиграция (кл. Demospongiae:
сем. Halisarcidae) (рис. 18в, г). В то же время, у губок описаны
и уникальные морфогенезы, которые не встречаются у других
многоклеточных животных. К ним относятся, например, экс-
курвация у Calcaronea (кл. Calcarea) (рис. 18д), формирование
псевдобластулы за счет вселения материнских клеток в дро-
бящийся зародыш у Chondrosia reniformis (кл. Demospongiae:
отр. Chondrosida) (рис. 18е), поляризованная деламинация, ра-
нее рассматривавшаяся как эпиболия (кл. Demospongiae: отр.
Poecilosclerida, Halichondrida) (рис. 18ж), униполярная проли-
ферация, которую некоторые авторы относили к униполярной
иммиграции (кл. Demospongiae: отр. Dictyoceratida) (рис. 18з),
и мультиполярная эмиграция (multipolar egression) у Homoscle-
romorpha (рис. 18и) (Ereskovsky, Dondua, 2006; Ересковский,
Дондуа, 2009).
Указанные морфогенетические движения отдельных клеток
или их пластов не связаны напрямую клеточной дифференци-
ровкой: они могут предшествовать дифференциации личиноч-
ных клеток, совпадать или следовать за ней (Ereskovsky, Don-
dua, 2006; Ereskovsky, 2010).
Сходство многих форм морфогенетических движений, в ходе
которых достигается достаточно сложная трехмерная организа-
ция тела животного, у губок и Eumetazoa очевидно. Важно под-
черкнуть, однако, что это сходство касается лишь особенностей
морфогенеза, но ни в коей мере не затрагивает сущности даль-
нейшей дифференциации, которая у Porifera и Eumetazoa имеет
глубокие и принципиальные различия.
В эмбриогенезе разных представителей Eumetazoa различа-
ют стадии дробления, гаструляции, гистогенеза и органогенеза.
Принципиальным отличием эмбрионального развития губок яв-
ляется отсутствие стадии формирования зародышевых листков,
Глава 7. Размножение и развитие губок
61
т.е. гаструляции. На поздних этапах дробления, которые обыч-
но называют бластулой или морулой, у губок начинаются про-
цессы дифференциации личиночных клеток (пограничных жгу-
тиковых клеток и склероцитов). У губок отсутствует гаструла,
которая у Eumetazoa представляет собой стадию становления
эктодермы и энтодермы (Иванова-Казас, 1995). Судьба специ-
ализированных личиночных клеток губок (покровных, скеле-
тогенных и др.) не гомологична судьбе зародышевых листков
Eumetazoa. Это, прежде всего, связано с тем, что у Porifera от-
сутствуют части тела, органы и ткани, гомологичные таковым
других Metazoa. У губок нет и не может быть эмбриональных
закладок типа эктодермы и энтодермы, или гомологичных
им групп клеток, детерминированных на развитие кишки и
кожных покровов, поскольку эти тканевые системы у них
отсутствуют (Короткова, 1981, 1997; Ересковский, 2005;
Ereskovsky, Dondua, 2006; Ereskovsky, 2010).
В литературе существует устойчивая мифологема. Говоря о
губках многие авторы, как бы закрывают глаза на то, что сущ-
ность гаструляции состоит в формировании первичной кишки
и образовании, по крайней мере, двух зародышевых листков —
эктодермы и энтодермы. Несмотря на давно доказанное и, в це-
лом, общепринятое представление о том, что первые двухслой-
ные животные, возникшие в ходе эволюции — это гребневики
и книдарии, тезис о наличии гаструляции у губок снова и снова
поднимается в литературе (Efremova, 1997; Leys, Degnan, 2002;
Maldonado, 2004; Leys 2004; Leys, Eerkes-Medrano, 2005). Одной
из причин этого служит, по-видимому, поразительное сходство
морфогенетических процессов, которые разворачиваются после
завершения дробления и у губок, и у книдарий.
Однако подобное сходство лишь подчеркивает универсаль-
ность механизмов формообразования, которые появились в
ходе эволюции независимо от процессов гаструляции. Можно с
полным основанием предполагать, что механизмы формообра-
зования возникали на самых ранних этапах эволюции Metazoa,
связанных с появлением Parazoa, многоклеточных животных,
потомков одноклеточных или колониальных Choanoflagellata.
Предполагается, что эти древнейшие Metazoa, ископаемые отпе-
чатки которых обнаружены в мезопротерозойских отложениях,
62
А.В. Ересковский, A.3. Вишняков. Губки
стали предшественниками губок и других более сложно органи-
зованных животных (Nielsen, 2008).
Таким образом, можно сделать вывод, что механизмы мор-
фогенетических движений возникли в ходе эволюции раньше
обособления покровного кинобласта и пищеварительного фаго-
цитобласта. Это свидетельствует о независимости процессов
пространственного распределения клеток и их спецификации
в формирующемся зародыше в виде эктодермальных и энтодер-
мальных тканей (Ereskovsky, Dondua, 2006; Ересковский, Дон-
дуа, 2009).
Личинки
К настоящему времени у губок описано восемь форм личи-
нок: целобластула, кальцибластула, цинктобластула, амфибла-
стула, дисферула, хоплитомелла, паренхимула и трихимелла
(рис. 19).
Наиболее просто устроены целобластульная и кальцибла-
стульная личинки, которые имеют однорядный жгутиковый
покров и обширную внутреннюю полость (бластоцель), запол-
ненную жидким бесструктурным веществом, отдельными сома-
тическими клетками материнского происхождения и симбио-
тическими бактериями (рис. 19а, б). Целобластульная личинка
описана у некоторых яйцекладущих демоспонгий. Разновид-
ностью целобластулы является кальцибластула известковых
губок подкласса Calcinea. В составе наружного слоя клеток
кальцибластул могут находиться отдельные безжгутиковые
клетки.
Амфибластульная личинка также однослойна: ее переднее
полушарие образовано жгутиковыми клетками небольшого раз-
мера, а заднее — безжгутиковыми крупными клетками с зерни-
стой цитоплазмой (рис. 19в). В области экватора располагаются
четыре клетки «креста». Небольшая внутренняя полость вклю-
чает различные материнские клетки и симбионтные бактерии.
Личинки амфибластульного типа описаны у известковых губок
подкласса Calcaronea.
Дисферула встречается в качестве одного из личиночных
морфотипов Halisarca (Demospongiae). Она характеризуется на-
Глава 7. Размножение и развитие губок
63
Рис. 19. Строение основных типов личинок губок: а — каль-
цибластульная личинка; б — целобластульная личинка; в —
амфибластула; г — дисферула; д — хоплитомелла; е — па-
ренхимула Poecilosclerida; ж — паренхимула пресноводных
Haplosclerida; з — трихимелла; и — цинктобластула; бм —
базальная мембрана; вн — внутренняя жгутиковая камера;
жк — жгутиковые клетки; кк — клетки креста; мк — мате-
ринские клетки; п — личиночная пинакодерма; пт — внутри-
ядерные паракристалические тела; рк — ресничные клетки;
с — личиночные спикулы; сб — симбиотические бактерии;
хк — личиночная хоаноцитная камера.
64
А.В. Ересковский, АЗ. Вишняков. Губки
личием замкнутого слоя (пузырька) жгутиковых клеток во вну-
тренней части личинки, образованного в результате инвагина-
ции поверхностного жгутикового покрова на последних этапах
личиночного морфогенеза (рис. 19г).
Хоплитомелла — уникальная планктонная личинка, описан-
ная у сверлящих губок родов Alectona и Thoosa (отр. Astrophor-
ida) (рис. 19д). Самая необычная особенность хоплитомеллы —
отсутствие покровных жгутиковых клеток. В то же время,
внутри личинки располагаются хоаноцитные камеры, разноо-
бразные клетки, коллагеновые волокна и спикулы — дискотри-
ены, которых нет у взрослых губок. Ранее этих личинок счита-
ли своеобразными плавающими почками и называли «armoured
gemmules» или «armouredbuds».
Паренхимульные личинки наиболее сложно организованы и
могут варьировать в своей организации даже в пределах одно-
го рода (рис. 18е, ж). Они описаны у подавляющего большин-
ства демоспонгий. Паренхимулы покрыты снаружи одно- или
многорядным слоем одножгутиковых клеток. Внутри имеют-
ся склероциты, археоциты, секреторные клетки, колленциты,
а также личиночный скелет, если он характерен для взрослой
губки. У всех пресноводных и некоторых мелководных морских
Haplosclerida паренхимула имеет в районе переднего полюса об-
ширную полость, выстланную эндопинакоцитами, а на проти-
воположном — хоаноцитные камеры и каналы (рис. 19ж). Па-
ренхимулы, выходящие во внешнюю среду, характеризуются
различной степенью дифференцировки как личиночных, так,
иногда, и дефинитивных клеток.
Трихимелла — личинка стеклянной губки Oopsacas minuta
(рис. 19з). Принципиальные отличия трихимеллы от парен-
химул следующие. Средняя часть личинки имеет пояс клеток,
несущих до 50 ресничек. Синцитиальный слой пинакодермы,
покрывающий всю личинку, покрывает и ресничные клетки.
Реснички выходят наружу через отверстия в синцитии. Кроме
этого, внутри личинки располагаются жгутиковые камеры, об-
разованные безъядерными воротничковыми телами, которые,
в отличие от таковых у дефинитивной губки, не связаны общей
цитоплазмой. Личинки имеют особые спикулы стауроактины,
отсутствующие у взрослых губок.
Глава 7. Размножение и развитие губок
65
Цинктобластула характерна для Homoscleromorpha. Внешне
она похожа на целобластулу, поскольку образована одним сло-
ем ресничных клеток и имеет обширную внутреннюю полость.
Однако клетки наружного слоя имеют десмосомы в своей апи-
кальной части, а со стороны полости подстилаются базальной
пластинкой. Эти признаки позволяют рассматривать их как
эпителиальные клетки. Кроме этого, в районе заднего полюса
личинки находится пояс клеток с внутриядерными паракри-
сталл ическими телами (рис. 19и). Внутренняя полость включа-
ет материнские клетки и симбиотических бактерий.
Личинкам всех типов, за исключением хоплитомеллы, свой-
ственна четкая переднезадняя полярность, которая проявляет-
ся в форме тела, в характере цилиатуры на разных полюсах, в
неравномерности окраски. На гистологическом уровне поляр-
ность личинки проявляется в характере распределения раз-
личных типов клеток, ориентации и положении спикул (у па-
ренхимул) и неравномерном распределении симбиотических
бактерий в полости личинки (цинктобластула). Ряд структур,
характерных для личинок, можно определить как провизорные
образования, то есть выполняющие важные функции только в
период личиночного развития, а впоследствии подвергающиеся
редукции. Прежде всего это относится к жгутиковому (реснич-
ному) покрову, выполняющему локомоторную (в период пла-
вания), прикрепительную (при оседании) и, возможно, трофи-
ческую функции. К этой же группе признаков можно отнести
личиночный скелет.
Все личинки губок, независимо от их морфологических осо-
бенностей, относятся к лецитотрофному типу, так как энерге-
тическим источником для них служат неизрасходованные в
процессе эмбриогенеза желточные гранулы и липидные кап-
ли. Обычно гранулы располагаются в базальных частях жгу-
тиковых клеток или во внутренних клетках личинки. Однако
у паренхимул некоторых демоспонгий (Halichondria panicea и
Spongilla lacustris) был описан фагоцитоз органических части-
чек из внешней среды, осуществляемый жгутиковыми клетка-
ми (Ivanova, Semyonov, 1997).
бб
А.В. Ересковский, А.Э. Вишняков. Губки
Метаморфоз
Под метаморфозом подразумевается кратковременная стадия
постэмбрионального развития, во время которой происходят ра-
дикальные морфологические и физиологические изменения ли-
чинки в связи с ее преобразованием в прикрепленную стадию.
Метаморфоз личинок губок включает в себя следующие про-
цессы: прикрепление личинки к субстрату, образование экзопи-
накодермы и базопинакодермы, развитие хоаноцитных камер
(или хоанодермы у асконоидных форм), развитие каналов и их
объединение с хоаноцитными камерами, образование пор и оску-
люма, синтез спонгина, спикул и основного вещества мезохила.
В полном объеме последовательность этих событий описана лишь
у немногих губок. Во всех известных случаях началом метамор-
фоза служит прикрепление личинки к субстрату и развитие по-
граничных слоев — экзопинакодермы и базопинакодермы.
Личинки губок прикрепляются к субстрату передним по-
люсом. Начальная адгезия личинки осуществляется благода-
ря секреции слизистого материала наружными жгутиковыми
клетками ее переднего полюса. Окончательное прикрепление
происходит после дифференцировки коллаген-секретирующих
клеток, ответственных за прикрепление.
Первый этап морфогенетических событий, происходящих
после прикрепления личинки, связан с развитием экзопинако-
дермы — наружного покрова постличинки. Формирование по-
кровных структур у губок в ходе метаморфоза может осущест-
вляться различными путями. У представителей подавляющего
большинства исследованных видов Porifera развитие покров-
ных структур происходит за счет объединения в клеточный
пласт отдельных амебоидных клеток, т.е. если проводить анало-
гию с Metazoa морфогенез осуществляется по типу мезенхимо-
эпителиального перехода. У Halisarcidae подобный морфогенез
вовлечен в развитие базопинакодермы, тогда как экзопинако-
дерма формируется по типу эпителиального морфогенеза. Все
покровные структуры Homoscleromorpha образуются в резуль-
тате трансдифференцировки пласта жгутиковых клеток цинк-
тобластулы без разрушения его эпителиальной структуры (Eres-
kovsky, 2010).
Глава 7. Размножение и развитие губок
67
В результате метаморфоза личинки обычно формируется од-
нооскулюмная особь, водоносная система которой отличается от
таковой у дефинитивной губки. В классе Calcarea губка на этой
стадии имеет асконоидный тип водоносной системы и называет-
ся олинтус, а. у Demospongiae и Homoscleromorpha — сиконоид-
ный или лейконоидный и называется рагон.
При изучении метаморфоза у губок авторы традиционно пы-
таются выяснить, есть ли преемственность между жгутиковыми
клетками личинки и хоаноцитами дефинитивных особей. Это
связано с попытками разрешения проблемы «инверсии зароды-
шевых листков» — «inversion of layers» у губок, поставленной
Деляжем (Delage, 1892). Данная гипотеза была положена авто-
ром в основу отделения Porifera от остальных Metazoa в подцар-
ство Enantiozoa.
Однозначного ответа нет и быть не может, поскольку сама
структура личинок губок крайне разнообразна. У губок одних
видов клетки, формирующие хоанобласты, появляются в ре-
зультате трансдифференцировки жгутиковых клеток личин-
ки. К этой группе относятся целобластульные личинки демо-
спонгий, кальцибластулы, цинктобластулы и амфибластулы.
У некоторых Demospongiae, например, Halisarca, хоанобласты
частично развиваются из жгутиковых клеток личинки, кото-
рые погружаются внутрь куколки (стадия метаморфоза у этих
губок), где и происходит их трансформация в хоанобласты.
Однако у ряда видов Demospongiae жгутиковые клетки личин-
ки (паренхимулы) не способны трансдифференцироваться и не
принимают участия в метаморфозе. Однако если на стадии ли-
чинки есть морфологически оформленные хоаноциты, то они
могут функционировать далее и у взрослых губок. Так, у пре-
сноводных и некоторых морских Haplosclerida личиночные хо-
аноцитные камеры во время метаморфоза непосредственно пере-
ходят в дефинитивные с последующей активной пролиферацией
хоаноцитов.
Судьба покровных жгутиковых клеток личинок губок во
время метаморфоза может существенно различаться. Соответ-
ственно, одни авторы считают, что существует преемственность
между жгутиковыми клетками личинки и хоаноцитами губки,
а другие полагают, что пласт жгутиковых клеток личинки пред-
68
А.В. Ересковский, А.Э. Вишняков. Губки
ставляет собой провизорный орган, а его клетки терминально
дифференцированы. В обобщающих работах по метаморфозу
губок игнорируется тот очевидный факт, что личинки предста-
вителей различных макрогрупп Porifera принципиально отли-
чаются по строению и клеточному составу. У губок имеются два
различающихся типа морфогенеза при метаморфозе.
Первый тип метаморфоза реализуется у паренхимульных
личинок, для которых характерно наличие различных клеток
во внутренней полости. Общая черта метаморфоза таких личи-
нок — деструкция их покровных клеток и формирование новых
покровов молодой губки внутренними клетками. Основная фор-
мообразующая роль при метаморфозе принадлежит внутренним
полипотентным клеткам личинки — археоцитам, которые так-
же являются источником хоаноцитов. Однако даже у паренхи-
мул в пределах одного таксона судьба жгутиковых клеток мо-
жет существенно различаться. Так, у всех изученных личинок
представителей отряда Poecilosclerida происходит полный или
частичный фагоцитоз жгутиковых клеток археоцитами. В то
же время источниками хоаноцитов могут быть также разные
клетки. У губок одних родов (Mycale, Hamigerd) хоанобласты
развиваются как из жгутиковых клеток, так и из археоцитов
(Borojevic, Levi, 1965). У губок из рода Microciona (Kaltenbach
et al., 1999) источником хоанобластов служат исключительно
археоциты. Различные источники хоаноцитов при метаморфозе
описаны и для пресноводных губок Spongillidae (Haplosclerida)
(Weissenfels, 1989).
Метаморфоз второго типа свойственен амфибластулам, каль-
цибластулам, целобластулам, цинктобластулам и личинкам
Halisarca. Этих личинок объединяет следующее: в ходе их ли-
чиночного морфогенеза дифференцируются только наружные
клетки, которым и принадлежит основная формообразующая
роль при метаморфозе. Формирование экзопинакодермы и вы-
стилки водоносной системы осуществляется наружными клет-
ками личинки, а их локализация относительно переднезадней
оси личинки соответствует базоапикальному положению в со-
ставе ювенильной губки.
Против экстраполяции гипотезы «извращения зародышевых
листков» на всех Porifera свидетельствуют также данные по
Глава 7. Размножение и развитие губок
69
развитию губок из рода Tetilla (отр. Spirophorida), Stylocordyla
(отр. Hadromerida) и семейства Thoosidae (отр. Astrophorida).
Губки Tetilla and Stylocordyla имеют прямой тип развития, и
жгутиковых клеток, аналогичных жгутиковым клеткам личи-
ночных покровов у них не образуется. При личиночном разви-
тии Thoosidae (Alectona и Thoosa) формируется личинка хопли-
томелла, у которой нет поверхностных жгутиковых клеток, а
внутри находятся личиночные хоаноцитные камеры. Соответ-
ственно и в данном случае «инверсия листков» исключена.
Типы развития
Хорошо известно, что губки характеризуются высокой пла-
стичностью и разнообразием морфологических структур. Тем не
менее, всегда можно выделить ряд морфологических характери-
стик, типичных для вида, рода, семейства, отряда и т.д. Каждая
структура формируется в результате предшествующего морфо-
генеза, поэтому возникает необходимость в сравнительном из-
учении всех стадий развития. Признаки, специфичные для так-
сонов разного ранга, можно обнаружить не только у взрослых
губок, но и на ранних этапах их онтогенеза.
Под типом развития понимается исторически сложившийся
комплекс коррелятивно связанных друг с другом морфогенети-
ческих процессов, как унаследованных, так и приобретенных
(Иванова-Казас, 1995).
Имеется восемь типов развития современных губок, назван-
ных по типу личинок, характерных для каждой группы (Ере-
сковский, 2005).
1. Амфибластульный тип развития (Calcarea, Calcaronea).
Яйцо изолецитальное, олиголецитальное, имеет овальную фор-
му, короткая ось яйца рассматривается как анимально-вегета-
тивная (рис. 20). Имеется комплекс специализированных пита-
ющих клеток. Дробление полное, асинхронное, неравномерное,
инкурвационное по типу табличной палинтомии. В течение
всего развития зародыш сохраняет бластульную организацию,
однако это «инвертированная» бластула — стомобластула.
Формирование амфибластульной личинки сопровождается вы-
70
А. В. Ересковский, А.Э. Вишняков. Губки
Рис. 20. Амфибластульный тип развития (Calcaronea); а —
яйцо; б — инкурвационое дробление; в — стомобл астула; г —
экскурвация; д — амфибластула; е — молодой олинтус.
ворачиванием (экскурвацией) сто моб л астулы. Переднезадняя
полярность зародыша проявляется рано — с первых циклов дро-
бления. Передний полюс личинки соответствует вегетативному
полюсу яйца и образован жгутиковыми клетками, на заднем по-
люсе находятся безжгутиковые зернистые клетки. Характерны
четыре клетки «креста». Корешки жгутиков поперечно исчер-
ченные. Специализированные контакты не выявлены. Метамор-
фоз осуществляется по мезенхимному типу: жгутиковые клет-
ки поодиночке погружаются внутрь, трансдифференцируясь в
хоаноциты, клетки мезохила, склероциты и эндопинакоциты,
а зернистые безжгутиковые клетки заднего полюса остаются на
поверхности, формируя экзо- и базопинакодерму. Переднезад-
няя ось личинки становится базоапикальной осью губки. Отме-
чено факультативное бесполое размножение в виде почкования.
2. Кальцибластульный тип развития (Calcarea, Calcinea).
Яйцо изолецитальное, олиголецитальное, без признаков поля-
ризации (рис. 21). Специализированные питающие клетки от-
сутствуют. Дробление полное, равномерное, полиаксиалъное,
имеется полость дробления. В течение всего развития зародыш
Глава 7, Размножение и развитие губок
71
Рис. 21. Кальцибластульный тип развития (Calcinea); а —
яйцо; б, в — полиаксиальное дробление; г — целобластула;
д — кальцибластула; е — начало метаморфоза; ж — олинтус.
сохраняет целобластульную организацию (рис. 16г). Передне-
задняя полярность проявляется поздно, в конце формирования
личинки — кальцибластулы. Жгутиковые клетки — основной
клеточный тип личинки. Кроме того, имеются бутылковидные
и вакуолярные клетки. Корешки жгутиков поперечно исчерче-
ны. Имеются специализированные межклеточные контакты.
Метаморфоз осуществляется по мезенхимному типу и сопрово-
ждается выселением части жгутиковых клеток внутрь полости
и трансдифференциацией их в хоаноциты и амебоциты. Остав-
шиеся на поверхности клетки трансформируются в пинакоци-
ты. Переднезадняя ось личинки становится базоапикальной
осью губки. Отмечено факультативное бесполое размножение по
типу почкования.
3. Трихимелльный тип развития (Hexactinellida). Яйцо изо-
лецитальное, олиголецитальное, без признаков поляризации
(рис. 22). Специализированные питающие клетки не выявле-
ны. Дробление полное, равномерное, асинхронное. В течение
72
А. В. Ересковский, А.Э. Вишняков. Губки
раннего развития зародыш сохраняет бластульную организа-
цию. Двухслойный морульный зародыш формируется путем
морульной деляминации. Переднезадняя полярность личинки
проявляется в ходе ее дифференцировки. В результате развития
формируется личинка трихимелла с поясом многожгутиковых
клеток и синцитиальными структурами. Корешки у жгутиков
отсутствуют. Имеются пробковые контакты. Метаморфоз не
описан. Встречается факультативное бесполое размножение.
Губки класса Demospongiae, характеризуются наибольшим
морфологическим разнообразием среди Porifera, и имеют крайне
широкий диапазон репродуктивных процессов. В результате срав-
нительного анализа у них можно выделить четыре типа развития.
Рис. 22. Трихимелльный тип развития (Hexactinellida); а —
яйцо; б, в —дробление; г — целобластула; д — начало кле-
точной деляминации; е — морфогенез; ж — предличинка; з,
и — трихимелла; fc — жгутиковые камеры личинки; та —
макромеры; тс — многожгутиковые клетки; mi — микроме-
ры; sc — спикулы (по: Leys et al., 2007).
Глава 7. Размножение и развитие губок
73
4. Дисферульный тип развития (Demospongiae, Halisarcida).
Яйцо изолецитальное, полилецитальное, полярность не выра-
жена (рис. 23). Специализированные питающие клетки не обна-
ружены. Дробление полное, равномерное, асинхронное, полиак-
сиальное, имеется полость дробления. В результате дробления
формируется целобластула либо морула. Последняя развива-
ется за счет мультиполярной иммиграции единичных клеток.
Переднезадняя полярность зародыша проявляется поздно — в
период цитодифференцировки поверхностного слоя клеток. Ха-
рактерен полиморфизм личинок: встречаются целобластулы,
паренхимулы и дисферулы. Внутренний клеточный слой дис-
ферулы образуется в результате инвагинации латеральной по-
верхности пред личинки. Личинки полностью покрыты жгути-
ками. У жгутика отмечается два корешка. Имеются адгезивные
контакты. Морфогенезы при метаморфозе носят смешанный ха-
рактер: экзопинакодерма развивается по эпителиальному типу
из жгутиковых клеток заднего полюса личинки, а базопинако-
дерма, эндопинакодерма и хоанодерма развиваются по мезен-
химному типу. Переднезадняя ось личинки становится базоапи-
кальной осью рагона. Бесполое размножение не известно.
Рис. 23. Дисферульный типа развития (Demospongiae:
Halisarcida); а — яйцо; б, в — полиаксиальное дробление; г —
мультиполярная иммиграция; д — инвагинация; е — цело-
бластула; ж — морулообразный зародыш; з — дисферульная
личинка; и — целобластульная личинка; к — паренхимуль-
ная личинка; л — молодой рагон.
74
А. В. Ересковский, А.Э. Вишняков. Губки
5. Паренхимульный тип развития (Demospongiae). Объ-
единяет яйцекладущих и яйцеживородящих Демоспонгий, для
которых характерно полное асинхронное, неупорядоченное
дробление, приводящее обычно к формированию морулы. В ре-
зультате последующего развития образуется личинка паренхи-
мула, имеющая диагностические особенности в ранге отряда.
Корешок жгутиков покровных клеток не исчерченный. Мета-
морфоз осуществляется по мезенхимному типу и сопровождает-
ся миграцией жгутиковых клеток внутрь, а внутренних — на-
ружу. В ходе метаморфоза происходит полный или частичный
фагоцитоз жгутиковых клеток личинки.
Можно выделить три подтипа паренхимульного развития.
Для первого подтипа (Halichondrida, Dendroceratida, мор-
ские Haplosclerida, Dictyoceratida и, частично, Hadromerida)
характерны следующие черты (рис. 24): а) изолецитальное, оли-
голецитальное или полилецитальное яйцо, без признаков по-
Рис. 24. Первый подтип паренхимульного развития (Demo-
spongiae: Halichondrida, Dendroceratida, морские Haploscle-
rida, Dictyoceratida и, частично, Hadromerida); а — яйцо;
б — хаотическое дробление; в — морула; г — паренхимула;
д — метаморфоз.
Глава 7. Размножение и развитие губок
75
ляризации; б) формирование равномерной аполярной морулы;
в) сегрегация морулы на поверхностный слой и внутреннюю
клеточную массу путем морульной деляминации; г) проявление
переднезадней полярности личинки во время ее цитодифферен-
цировки; д) бесполое размножение (если имеется) — почкова-
ние, фрагментация, геммулогенез.
Особенности второго подтипа (Poecilosclerida) (рис. 25)
определяются строением яйца упомянутой группы губок — те-
лолецитального, поляризованного и полилецитального. К дру-
гим специфическим чертам можно отнести: а) полное, асин-
хронное, неравномерное, дробление; б) формирование плотной,
поляризованной, неравномерной морулы; в) сегрегацию морулы
на поверхностный слой и внутреннюю клеточную массу путем
поляризованной деляминации; г) переход анимально-вегетатив-
ной полярности яйца в переднезаднюю ось личинки.
Третий подтип (пресноводные Haplosclerida — Spongillida)
отличается следующими признаками (рис. 26): а) наличием спе-
циализированных питающих клеток; б) сегрегации морулы пу-
Рис. 25. Второй подтип паренхимульного развития (Demo-
spongiae: Poecilosclerida); а — яйцо; б — хаотическое дробле-
ние; в — морула; г — паренхимула; д — метаморфоз.
76
А. В. Ересковский, А.Э. Вишняков. Губки
Рис. 26. Третий подтип паренхимульного развития (Demo-
spongiae: пресноводные Haplosclerida); а — яйцо пресновод-
ных представителей; б — дробление; в —морула; г — парен-
химула; д — метаморфоз; е — рагон.
тем поляризованной деляминации; в) паренхимула с высокой
степенью цитодифференцировки, а также с провизорными (жгу-
тиковый покровный слой, скелет, обширная полость, выстлан-
ная пинакоцитами) и дефинитивными (хоаноцитные камеры)
структурами; г) чередованием в жизненном цикле полового и
бесполого (геммулогенез) размножения.
6. Прямое развитие (Demospongiae) включает два подтипа.
Первый подтип — яйцекладущие раздельнополые губки отря-
да Spirophorida. Яйца мелкие изолецитальные, олиголециталь-
ные, без признаков поляризации (рис. 27). Специализирован-
ные питающие клетки не обнаружены. На поверхности яйца
имеются пучки коллагена. После проникновения спермия в
яйцо образуется оболочка оплодотворения. Дробление полное,
равномерное асинхронное, радиальное до четвертого цикла.
В результате дробления формируется рыхлая равномерная апо-
лярная морула. Морфогенез осуществляется путем морульной
деляминации. Морульный зародыш развивается непосредствен-
но в молодую губку минуя личиночную стадию. Бесполое раз-
множение по типу почкования.
Для второго подтипа (Hadromerida: Stylocordyla borealis, Spi-
rophorida: Craniella sp., Telilla polyura) также свойственно пря-
Глава 7. Размножение и развитие губок
77
Рис. 27. Прямой тип развития яйцекладущих демоспонгий
(Spirophorida); а — зигота в оболочке оплодотворения; б — ра-
диальное дробление; в — морула; г — развивающаяся губка.
Рис. 28. Прямой тип развития живородящих демоспонгий
(Hadromerida, Spirophorida); а — зигота; б — дробление; в —
морула; г — развивающаяся губка в мезохиле.
мое развитие. Представители характеризуются другими особен-
ностями по сравнению с иными подтипами паренхимульного
развития: а) наличием двух типов питающих клеток; б) формиро-
ванием неравномерной морулы; в) эмбриональным морфогенезом
по типу морульной деляминации; г) живорождением (рис. 28).
7. Целобластульный тип развития (Demospongiae: Chondro-
sida; Hadromerida — Polymastiidae) (рис. 29). Яйцекладущие
раздельнополые губки. Отсутствует фолликулярая оболочка,
яйца окружены слоем коллагена и слизи. Яйца изолецитальные
олиголецитальные, неполяризованные.
Дробление полное, равномерное может быть радиальным,
формирующим аполярную морулу, которая развивается в рав-
номерную целобластулу путем реарранжировки клеток. Перед-
незадняя полярность зародыша проявляется в период цитодиф-
ференцировки поверхностного слоя клеток. Целобластульная
78
А. В. Ересковский, А.Э. Вишняков. Губки
Рис. 29. Целобластульный тип развития (Demospongiae:
Chondrosida; Hadromerida — Polymastiidae); a — яйцо; б —
дробление; в — морула; г — целобластула; д — личинка цело-
бластула; е — метаморфоз.
личинка состоит из одного слоя жгутиковых клеток. Корешок
жгутика не исчерченный. Имеются адгезивные межклеточные
контакты. Метаморфоз сопровождается выселением части жгу-
тиковых клеток внутрь полости и трансдифференцировкой их
в хоаноциты и амебоциты. Оставшиеся на поверхности клетки
трансформируются в пинакоциты. Переднезадняя ось личинки
становится базоапикальной осью губки. Отмечено факультатив-
ное бесполое размножение по типу почкования.
8. Цинктобластульный тип развития (Homoscleromorpha)
(рис. 30). Яйцо изолецитальное, полилецитальное, поляр-
ность не выражена. Специализированные питающие клетки
не обнаружены. Дробление полное равномерное, асинхронное,
неупорядоченное, приводящее к формированию рыхлой, не-
поляризованной, равномерной морулы. Развитие целобласту-
лы осуществляется вследствие мулътиполярной эмиграции
(центробежного расхождения клеток морулы на периферию).
Переднезадняя полярность проявляется поздно — в конце ци-
тодифференцировки личинки. Личинка — цинктобластула,
образована однослойным жгутиковым эпителием. Среди жгу-
тиковых клеток встречаются редкие безжгутиковые. В районе
заднего полюса имеется пояс клеток с внутриядерным кристал-
лоидом. Корешок жгутика поперечноисчерченный. Характерна
базальная пластинка и опоясывающие десмосомы. Метаморфоз
осуществляется по эпителиальному типу: переднее полушарие
Глава 7. Размножение и развитие губок
79
Рис. 30. Цинктобластульный тип развития (Homoscleromor-
pha); а — яйцо; б — дробление; в — морула; г — целобласту-
ла; д — личинка цинктобластула; е — метаморфоз.
цинктобластулы инвагинирует, формируя карманообразные
впячивания жгутикового слоя с последующей их изоляцией в
хоаноцитные камеры. Жгутиковые клетки личинки служат ис-
точником всех клеток губки. Переднезадняя ось личинки стано-
вится базоапикальной осью рагона. Бесполое размножение осу-
ществляется по типу почкования и фрагментации.
Бесполое размножение (бластогенез)
У губок бесполое размножение встречается во всех классах и
реализуется путем фрагментации, геммулогенеза и почкования
(Ересковский, 2005)
Фрагментация рассматривается в качестве наиболее прими-
тивного способа бесполого размножения. Она может происхо-
дить в результате волновой активности при сильных штормах,
деятельности хищников (рыб, черепах), как следствие инфек-
ционных заболеваний, при которых губки распадаются на фраг-
менты, после разрыва или раскола субстрата, на котором обита-
ют губки, вследствие роста губок, главным образом, корковых,
80
А.В. Ересковский, А.Э. Вишняков. Губки
что приводит к распадению тела на фрагменты. Таким образом,
этот способ размножения не закономерен, а является следстви-
ем действия каких-либо случайных внешних факторов. Соот-
ветственно никаких цитологических изменений в составе губки
перед отделением фрагмента от тела родительской особи не про-
исходит. Фрагментация больше характерна для демоспонгий.
При этом чаще всего ей подвергаются ветвистые формы губок
или губки, имеющие выросты на теле.
Самые древние свидетельства наружного почкования у гу-
бок обнаружены в отложениях раннего венда, имеющих возраст
около 680 млн. лет. В южном Китае, в провинции Гуижоу в фос-
фатных отложениях были найдены мелкие (150-750 мкм) мо-
наксонные демоспонгии, на поверхности тела которых имелись
небольшие булавовидные выросты, которые авторы трактовали
как почки (Li et al., 1998).
Общей характеристикой почек, формирующихся у всех гу-
бок за исключением Radiospongilla cerebellata (отр. Haploscle-
rida) (Sailer, 1990) и Homoscleromorpha (Ereskovsky, Tokina,
2007), можно считать то, что на начальных этапах своего разви-
тия они представляют собой плотный конгломерат клеток, глав-
ным образом археоцитов, на поверхности материнского тела.
В основе формирования почек лежит миграция полипотентных
клеток (эпиморфоз) с последующей их дифференциацией в де-
финитивные клетки. Такая почка не имеет ни хоаноцитных ка-
мер, ни каналов, ни оскулюма. Отделившиеся почки оседают на
субстрат и прикрепляются к нему, после чего у них начинается
формирование водоносной системы и рост. Во время оседания
почки могут сливаться друг с другом, и в дальнейшем из них
формируются более крупные губки.
Почкование у Homoscleromorpha резко отличается от подоб-
ного бластогенеза у других губок. В основе развития почки ле-
жит эпителиальный морфогенез — эвагинация, т.е. формирую-
щаяся почка представляет собой не что иное, как вырост части
тела родительской губки. В ходе этого процесса не наблюдается
пролиферации, а клеточный состав почек качественно иденти-
чен таковому родительской губки. Подобный бластогенез уни-
кален среди губок и возможен, вероятно, благодаря эпителиаль-
ному строению тела Homoscleromorpha.
Глава 7. Размножение и развитие губок
81
Геммулогенез наиболее сложный тип бесполого размноже-
ния у губок. Для него характерно формирование особых струк-
тур — геммул, которые представляют собой покоящуюся во вре-
мя неблагоприятных условий стадию. Геммуллогенез является
также средством распространения губок. Геммулы состоят из
внутренней массы полипотентных клеток, тезоцитов (неактив-
ных археоцитов), заполненных резервным веществом, Снаружи
геммула окружена неклеточной защитной оболочкой.
Рис. 31. Геммулогенез пресноводных Haplosclerida (по: Lan-
genbruch, 1982): а — начало геммулообразования: концен-
трация археоцитов, трофоцитов и спонгобластов в мезохиле;
б — начало формирования геммульной оболочки; в — форми-
рование альвеолярного слоя, микропиле и дифференциация
тезоцитов; г — сформированная геммула. а — археоциты;
аж — археоциты с желточными пластинками; ам — амфи-
дискобласт с молодым амфидиском; ми — микропиле; ст —
столбчатый слой спонгоцитов; тр — трофоциты; хк — хоано-
цитная камера.
82
А. В. Ересковский, А.Э. Вишняков. Губки
У всех изученных губок геммулы развиваются из однотип-
ных клеточных источников — полипотентных ядрышковых
амебоцитов (археоцитов). Начинается геммулогенез с направ-
ленного перемещения этих клеток в определенные участки ме-
зохила. Здесь клетки образуют плотные агрегаты диаметром
250-500 мкм (рис. 31а). Часть клеток, называемых питающи-
ми, активно фагоцитируется своими собратьями-археоцитами,
в цитоплазме которых накапливаются характерные для этих
клеток желточные пластинки. У губок, содержащих в мезохи-
ле симбиотические зоохлореллы, археоциты захватывают и их;
в дальнейшем симбионты в интактном состоянии содержатся в
тезоцитах.
Вокруг клеточных агрегатов образуются два слоя клеток:
внутренний из уплощенных археоцитов и наружный из спон-
гиоцитов, формирующих столбчатый слой (рис. 316). Одно-
временно с этим микросклероциты мезохила со сформирован-
ными микросклерами мигрируют к развивающейся геммуле
и встраиваются в состав ее столбчатого слоя (рис. 31 в). Форми-
рование оболочки геммулы начинается на одном полюсе и по-
степенно распространяется к противоположному, где позже об-
разуется микропиле — участок оболочки, свободный от спикул
(Langenbruch, 1982). После этого клетки оболочки разрушают-
ся, оставляя микросклеры и волокна спонгина, которые уплот-
няются и образуют три слоя неклеточной оболочки. Под сфор-
мированной оболочкой находится однородная масса тезоцитов
(рис. 31г).
Геммулогенез сопровождается значительной, либо полной
дезорганизацией водоносной системы и мезохила материнской
губки.
Геммулы резистентны ко многим неблагоприятным факто-
рам. Они могут прорастать после двухмесячного содерживания
их при -80 °C или даже -100 °C (Barbeau et al., 1989). Показано,
что 25% геммул, выдержанных без воды в течение четырех ме-
сяцев при +5 °C, способны впоследствии развиваться в нормаль-
ную губку (Fell, Bazer, 1990).
Сигналом для прорастания обычно служат определенные
значения температуры, освещенности, влажности, ионного со-
става воды. В это время неклеточная оболочка в районе микро-
Глава 7. Размножение и развитие губок
83
пиле разрушается, тезоциты выходят наружу и, дифференци-
руясь, начинают формировать губку. Триггерным механизмом
прорастания геммул ряда видов служит фотосинтетическая ак-
тивность симбиотических водорослей (Kanayama, Kamishima,
1990).
Бесполое размножение в форме наружного почкования из-
редка встречается практически у всех групп губок, независимо
от их таксономического положения или местообитания. Однако
в этих случаях бластогенез можно рассматривать как факульта-
тивное явление.
Облигатное бесполое размножение отмечено лишь у предста-
вителей двух отрядов Demospongiae: (1) отр. Haplosclerida — у
пресноводных губок семейств Spongillidae, Potamolepidae, Meta-
niidae, Palaeospongillidae и у некоторых морских Chalinidae
формируются геммулы; (2) отр. Hadromerida — у представите-
лей сем. Suberitidae и Clionaidae осуществляется геммулогенез,
а у представителей сем. Polymastiidae и Tethyidae наружное
почкование.
Для видов, жизненный цикл которых включает бесполое
размножение, часто характерно закономерное чередование по-
лового и бесполого типов репродукции. Так, геммулогенез в
пределах одной особи начинается не раньше, чем пройдет ви-
теллогенез ооцитов. Это связано с конкуренцией за клеточный
материал и, в первую очередь, за полипотентные клетки (архе-
оциты). Прекращение или существенное снижение активности
бесполого размножения во время полового размножения свя-
зано с вовлечением археоцитов в оогенез и эмбриогенез особи.
Так, почкование представителей беломорской губки Polymastia
arctica наиболее интенсивно происходит в весенне-летние меся-
цы (в период начала гаметогенеза) и ослабевает в период вител-
логенеза ближе к осени (Plotkin, Ereskovsky, 1997).
Бывает, что усиление бесполого размножения следует за
половым и завершает активную фазу жизненного цикла по-
пуляций губок, обитающих в нестабильных условиях, под-
готавливая их к переживанию стрессовых или летальных для
дефинитивной губки периодов. Такой вариант чередования ха-
рактерен для многих пресноводных губок.
Глава 8.
ФИЛОГЕНИЯ ГУБОК
Филогенетические отношения губок вызывают в настоящее
время пристальный интерес в контексте построения общей фи-
логении многоклеточных животных. Ежегодно публикуются
данные о новых вариантах реконструкции филогенетических
отношений, выполненных на основе анализа ядерных, мито-
хондриальных генетических маркеров, или на их комбинациях.
Эти результаты сопоставляются с данными по филогении губок,
оцененной с помощью морфологических признаков. Многие
морфологические и цитологические признаки, и прежде всего
единый план организации, основный на уникальной водоносной
системе губок, указывают на то, что тип Porifera является моно-
филетической группой. Однако первые молекулярные исследо-
вания филогении губок с использованием 18S рибосомальной
ДНК показали, что Calcarea имеют больше общих черт сходства
с Eumetazoa, чем с Demospongiae (Borchiellini et al., 2001). Для
интерпретации этих данных была предложена гипотеза парафи-
лии губок.
Недавние масштабные исследования последовательностей
ДНК с использованием методологии expressed sequence tag
(EST) и анализа полного митохондриального генома губок убе-
дительно вернули гипотезу монофилии Porifera (Lavrov et al.,
2008; Philippe et al., 2009; Worheide et al., 2012) (рис. 32).
По-разному складывается современная ситуация с анализом
филогенетических отношений внутри Calcarea, Hexantinellida
и Demospongiae. Так, молекулярный анализ, основанный на
последовательностях 18S, 28S и митохондриальной 16S рибосо-
мальной ДНК и выполненный на 34 видах стеклянных губок,
позволил построить филогенетическое древо, хорошо согласую-
щееся с традиционной классификацией и ранее предложенной
филогенетической системой, основанной на морфологических
признаках.
Он четко подтвердил монофилию Hexactinellida и оказался
хорошо скореллированным с палеонтологическими данными
(рис. 33) (Dohrmann et al., 2008; 2013). Важно отметить также,
Глава 8. Филогения губок
85
Рис. 32. Схема современной гипотезы филогенетических от-
ношений Metazoa и Porifera (по Philippe et al., 2009).
что проведенный анализ показал близкие филогенетические от-
ношения Hexactinellida и Demospongiae.
Не изменились и представления о филогенетических отно-
шениях в пределах известковых губок: подтверждены монофи-
лия класса Calcarea, а также подклассов Calcinea и Calcaronea
(рис. 34) (Manuel et al., 2003; Voigt et al., 2012).
В то же время филогенетические отношения в пределах са-
мого крупного класса губок Demospongiae подверглись корен-
ному пересмотру. Сопоставление морфологических признаков
86
А. В. Ересковский, A.3. Вишняков. Губки
Lyssacinosida
“Hexactinosida”
Amphidiscophora
Sceptrulophora
Hypodermal
perrtacbns,
38(1)
Pfumosely
arranged
dermal
diactins
33(1
' -Sceptres
19(1)
- -Choanosomal
pentactins
41(1)
" "Floncomes
- -Hexasters, 15(1)-480Ma
- -Spicule fusion; 13(1)
' Dictyonal skeleton;
10(1) — 375 Ma
Dictyonal skeleton;
16(1)-375 Ma
-Sceptrutes,
22(1)-200 Ma
* -Undnates perpendicular
to dermal surface 21(1)
Tnaxoruc spicules; 8(1) - 545 Ma
• Syncybai tissue organization; 7(1)
Рис. 33. Морфологические апоморфии таксонов Hexactinellida,
их палеонтологическое датирование и филогенетические отно-
шения по данным рДНК анализа (из Dohrmann et al., 2008).
и данных молекулярного анализа четко выявило парафилию
класса Demospongiae (Borchiellini et al., 2004). В первую оче-
редь было показано, что Homoscleromorpha представляет со-
бой независимую филогенетическую ветвь такого же ранга,
как Demospongiae, Calcarea и Hexactinellida. Последующие ис-
следования с использованием разных молекулярных призна-
ков и разных методов подтвердили эти результаты (Lavrov et
al., 2008; Philippe et al., 2009; Sperling et al., 2009; Gazave et
al., 2012). В этой же работе было показано, что оставшиеся от-
ряды и семейства демоспонгий подразделяются на четыре кла-
стера уровня выше отряда. Последние работы по филогении
Demospongiae с использованием различных ядерных и мито-
хондриальных маркеров в сочетании с морфологическими при-
Глава 8. Филогения губок
87
Calcaronea
Leucosolenida II
Leucosolenida HL Leucosolenia
Llthonlda: Ptectroninia
Leucettida sensu lato
"Clathrinida"
sensu Hartman 1958
Рис. 34. Филогенетические отношения в пределах Calcarea
реконструированные на основании анализа морфологических
признаков и последовательностей 18S рРНК (Worheide et al.,
2012).
знаками подтвердили и уточнили гипотезу Борчиеллини с со-
авторами (Hille et al., 2012; Worheide et al., 2012; Redmond et
al., 2013; Thacker et al., 2013). В первую очередь, за вычетом
группы Homoscleromorpha, для остальных представителей была
подтверждена монофилия. Таким образом, крупным филогене-
88
А.В. Ересковский, А.Э. Вишняков. Губки
тическим кладам Demospongiae предложено придать статус под-
классов, как было показано в гл. 2. В частности, Keratosa = (от-
ряды Dendroceratida, Dictyoceratida), Myxospongiae = (отряды
Chondrosida, Verongida),
Haploscleromorpha = (морские Haplosclerida) и оставшиеся
демоспонгии = Heteroscleromorpha (отряды Agelasida, Astropho-
rida, Hadromerida, Halichondrida, Poecilosclerida, Spirophorida и
пресноводные Haploscerida) (рис. 35).
:□ Raspailiidae
Desmacellidae
Poecilosclerida sensu stricto
Suberitidae + Halichondriidae
Tethyidae + Hemiasterellidae
+ Timeidae + Trachycladidae
Clionaidae + Spirastrellidae
•Д Polymastiidae
। Д Axinellidae
। -J Dictyonellidae
Heteroscleromorpha
:3 Stelligeridae
“Д Agelasida
Spirophorida \ A v ...,
J Tetractinelhda
Astrophonda J
«=^3 Spongillina (freshwater sponges)
Haploscleromorpha (Marine haplosclerids)
-------------- Haplosclerina + Petrosina
Myxospongiae
Chondrosida (incl. Halisarca)
Verongida
Keratosa
Dendroceratida
Dysideidae *
Irciniidae I Dictyoceratida
spongiids I
thorectids J
Рис. 35. Филогения Demospongiae на основе 18S, 28S и 16S ми-
тохондриальных рибосомальных маркеров (по Hille et al., 2012;
Worheideet al., 2012; Redmond et al., 2013; Thacker et al., 2013).
Глава 8. Филогения губок
89
Наконец, недавно выделенный класс Homoscleromorpha, так-
же является четко монофилетичным (Gazave et al., 2012). В его
пределах выделяются два семейства: Plakinidae Schulze, 1880
с видами, имеющими спикулы и Oscarellidae Lendenfeld, 1887,
включающее бесспикульные виды (рис. 36). В то же время, фи-
логенетические отношения в пределах сем. Plakinidae довольно
сложные и, вполне вероятно, что в ближайшие годы в них будут
произведены существенные изменения (Gazave et al., 2010).
Рис. 36. Филогенетические отношения в пределах Homoscle-
romorpha ядерных рибосомальных и митохондриальных мар-
керов (по Gazave et al., 2012).
ЗАКЛЮЧЕНИЕ
Материал, приведенный в этой книге, это лишь малая толи-
ка современной информации о губках, — самых просто органи-
зованных и, в то же время, очень интересных и перспективных
для изучения животных.
Губки являются связующим звеном, между другими много-
клеточными животными и протистами. Благодаря этому по-
ложению, всестороннее изучение губок позволит восстановить
историю возникновения многоклеточных животных из проти-
стов на самых разных уровнях — от генетически-молекулярно-
го до структурно-функционального. С другой стороны именно
изучение этих просто организованных животных может помочь
понять какими путями шла эволюция, позволившая возникно-
вение более сложно организованных животных.
Разнообразные варианты строения тела губок, возникшие
на основе единого морфофункционального способа существо-
вания этих животных, иллюстрируют основные принципы ус-
ложнения и тенденции развития живых организмов вообще: от
мелких форм к крупным, от просто организованных — к более
сложным и дифференцированным.
Среди других групп животных губки занимают то место, где
природа могла пробовать, экспериментировать и предлагать то,
что впоследствии дало устойчивые комбинации, реализуемые
уже на принципиально ином уровне — тканевом и органном.
В экспериментальном плане губки представляют собой объек-
ты, удобные для изучения взаимодействия клеток на том уров-
не, когда клетки почти каждого типа способны, воспринимая
сигналы извне, менять свой жизненный путь и трансдифферен-
цироваться в другой, необходимый в настоящий момент онтоге-
неза.
Наличие у клеток губок больших потенций к трансдиффе-
ренцировкам усложняет переход самих животных к единоо-
бразным общим принципам, на которых можно было бы произ-
водить усложнения более высокого порядка — формирование
тканевых систем органов. Возможно, именно это и не позволило
губкам однозначно сформировать эпителии, как пограничные
Заключение
91
ткани, ограничившись лишь тем вариантом подобия, который
реализуется у губок класса Homoscleromorpha.
Обладая уникальной способностью к преобразованиям кле-
ток, губки могут возрождаться из мелких фрагментов и диссо-
циированных клеток. Разобранные на свои кирпичики-клетки,
они собираются в клеточные агрегаты — примморфы, которые
через некоторое время на основе все тех же сложных межкле-
точных взаимодействий создают организм губки, идентичный
тому, от которого эти клетки были взяты.
Сложная сеть межклеточных взаимодействий усложняется
еще и тем, что губки, по сути, не чисто эукариотические орга-
низмы. Значительная часть массы их тела составлена различ-
ными симбионтами — и, в первую очередь, прокариотическими.
Роль последних в жизнедеятельности губок очень значима, и
даже простое количество прокариотических клеток может гово-
рить само за себя, когда их общее число и биомасса могут быть
равными или даже превышать число и биомассу собственных
клеток губок. Благодаря длительной коэволюции, симбиоти-
ческие прокариоты обеспечивают многие ключевые моменты
жизни губок, такие, как рост, формообразование, обеспечение
питания и защиты. В сложном процессе взаимодействия клеток
губок и бактериальных симбионтов есть еще одна грань, кото-
рая интересна не только в фундаментальном плане, но имеет и
непосредственное практическое значение для человека. Губки,
при посредничестве симбиотических прокариот, синтезируют
огромное количество биологически активных веществ, которые
люди используют в медицине, научных экспериментах с други-
ми живыми организмами или при строительстве морских соору-
жений, которые должны быть защищены от морских обраста-
ний.
В данной работе мы обозначили лишь незначительную часть
научных проблем, которые ставят перед исследователями губки
и только наметили перспективы их исследования. Студентам и
молодым ученым мы предлагаем отнестись с еще большим вни-
манием к этой загадочной и перспективной группе животных, а
также попробовать оставить свой след в науке, изучая их.
СПИСОК ЛИТЕРАТУРЫ
Анакина Р.П., Короткова Г.П. 1989. Сперматогенез у баренцевоморской губ-
ки Leucosolenia complicata Mont. // Онтогенез. Т.20. С.77-86.
Беклемишев В.Н. 1964. Основы сравнительной морфологии беспозвоноч-
ных. М.: Наука. Т.1.
Бигон М., Харпер Д., Таусенд К. 1989. Экология. Особи, популяции и со-
общества. М.: Мир. Т.1.
Дондуа А.К. 2005. Биология развития. СПб.: Изд-во СПб. ун-та. Т.1, 2.
Ересковский А.В. 2003 Проблема колониальное™, модулярности и индиви-
дуальности губок и особенности их морфогенезов при росте и бесполом
размножении // Биология моря. Т.29. Вып.1. С.3-12.
Ересковский А.В. 2005. Сравнительная эмбриология губок (Porifera). СПб.:
Изд-во СПб. ун-та.
Ересковский А.В., Дондуа А.К. 2009. Проблема зародышевых листков у
губок и некоторые вопросы ранней эволюции Metazoa // Тр. СПб. о-ва
естествоисп. Cep.l. Т.97. 4.2. С.71-88.
Иванова-Казас О.М. 1995. Эволюционная эмбриология животных. СПб.:
Наука.
Колтун В.М. 1988. Развитие индивидуальности и становление индивида у
губок // Губки и книдарии: современное состояние и перспективы иссле-
дований. Л.: Зоол. ин-т АН СССР. С.24-34.
Короткова Г.П. 1981. Общая характеристика организации губок // Морфоге-
незы у губок. Л.: Изд-во Ленингр. ун-та. С.5-91.
Короткова Г.П. 1997. Регенерация животных. СПб.: Изд-во СПб. ун-та.
Марфенин Н.Н. 1993. Феномен колониальное™. М.: Изд-во Моск, ун-та.
Bagby R.M. 1966. The fine structure of myocytes in the sponges Microciona pro-
lifera (Ellis and Sollander) and Tedania ignis (Duchassaing and Michelotti) //
J. Morph. Vol. 118. P. 167-182.
Barbeau M.A., Reiswig H.M., Rath L.C. 1989. Hatching of freshwater sponge
gemmules after low temperature exposure: Ephydatia muelleri (Porifera:
Spongillidae) //J. Therm. Biol. Vol. 14. P.225-231.
Bergquist P.R. 1978. Sponges. Los Angeles: Univ California Press.
Borchiellini C., Manuel M., Alivon E., Boury-Esnault N., Vacelet J., Le Parco Y.
2001. Sponge paraphyly and the origin of Metazoa // J. Evol. Biol. Vol.244.
Pl 71-179.
Borchiellini C., Chombard C., Manuel M., Alivon E., Vacelet J., Boury-Esnault N.
2004. Molecular phylogeny of Demospongiae: implications for elassifeation
and scenarios of character evolution // Mol. Phylog. Evol. Vol.32. P.823-837.
Список литературы
93
Borojevic R., Levi С. 1965. Morphogenese experimentale d’une eponge a par-
tir de cellules de la larve negeante dissociee // Z. Zellforsch. Mikrosk. Anat.
Bd.68. P.57-69.
Boute N., Exposito J.Y., Boury-Esnault N., Vacelet J., Noro K., Miyazaki K.,
Yoshigato K., Garrone R. 1996. Type IV collagen in sponges, the missing link
in basement membrane ubiquity // Biol. Cell. Vol.88. P. 37^14.
Custodio M.R., Hajdu E., Muricu G. 2004. Cellular dynamics of in vitro alloge-
neic reactions of Hymeniacidon heliophila (Demospongiae: Halichondrida) //
Mar. Biol. Vol.144. P.999-1010.
Delage Y. 1892. Embryogenese des eponges silicieuses // Arch. Z. Exp. Gen.
Т.10. Р.345-Л98.
Dohrmann M., Janussen D., Reitner J., Collins A.G., Worheide G. 2008. Phylog-
eny and Evolution of Glass Sponges (Porifera, Hexactinellida) // Syst. Biol.
Vol.57. Р.388-Л05.
Dohrmann M., Vargas S., Janussen D., Collins A.G., Worheide G. 2013.
Molecular paleobiology of early-branching animals: integrating DNA
and fossils elucidates the evolutionary history of hexactinellid sponges //
Paleobiology. Vol.39. No.l. P.95-108.
Eerkes-Medrano D., Leys S.P. 2006. Ultrastructure and embryonic development
of a syconoid calcareous sponge // Invert. Biol. Vol. 125. P. 177-194.
Efremova S.M. 1997. Once more on the position among Metazoa — Gastrulation
and germinal layers of sponges // A.V. Ereskovsky, H. Keupp, R. Kohring
(eds.). Modem problems of Poriferan biology. Freie Univ. Berlin.
Ereskovsky A.V. 2006. A new species of Oscarella (Demospongiae: Plakinidae)
from the Western Sea of Japan // Zootaxa. Vol. 1376. P.37-51.
Ereskovsky A.V. 2007. A New Species of Halisarca (Demospongiae: Halisarcida)
from the Sea of Okhotsk, North Pacific // Zootaxa. Vol. 1432. P.57-66.
Ereskovsky A.V. 2010. The Comparative Embryology of Sponges. Dordrecht
Heidelberg London New York: Springer-Verlag.
Ereskovsky A.V, Borchiellini C., Gazave E., Ivanisevic J., Lapebie P., Perez
T., Renard E., Vacelet J. 2009. The Homoscleromorph sponge Oscarella
lobularis, a promising sponge model in evolutionary and developmental
biology // BioEssays. Vbl.31. P.89-97.
Ereskovsky A.V, Dondua A.K. 2006. The problem of germ layers in sponges
(Porifera) and some issues concerning early metazoan evolution // Zool. Anz.
Vol.245. P.65-76.
Ereskovsky A.V, Gonobobleva E.L., Vishnyakov A.E. 2005. Morphological
evidence for vertical transmission of symbiotic bacteria in the viviparous
sponge Halisarca dujardini Johnston (Porifera, Demospongiae, Halisarcida) //
Mar. Biol. Vol. 146. P.869-875.
94
А.В. Ересковский, А.Э. Вишняков. Губки
Ereskovsky A.V., Lavrov D.V., Boury-Esnault N., Vacelet J. 2011. Molecular and
morphological description of a new species of Halisarca (Demospongiae:
Halisarcida) from Mediterranean Sea with redescription of the type species
Halisarca dujardini // Zootaxa. Vol.2768. P.5-31.
Ereskovsky A.V., Tokina D.B. 2007. Asexual reproduction in homoscleromorph
sponges (Porifera; Homoscleromorpha) // Mar. Biol. Vol. 151. P.425-434.
Fell P.E., Bazer L.J. 1990. Survival of the gemmules of Anheteromeyenia ryderi
(Potts) following aerial exposure during winter in New England // Hydrobiol.
Vol. 190. P.241-246.
Fry W.G. 1979. Taxonomy, the individual and the Sponge // G. Larwood, B.R.
Rosen (eds.). Biology and systematics of colonial organisms. London, New
York: Acad. Press. P.39-47.
Funayama N. 2013. The stem cell system in demosponges: suggested involve-
ment of two types of cells: archeocytes (active stem cells) and choanocytes
(food-entrapping flagellated cells) // Dev. Genes Evol. Vol.223. P.23-38.
Garrone R. 1978. Phylogenesis of connective tissue. Morphological aspects and
biosynthesis of sponge intercellular matrix // Front Matrix Biol. Vol.5. P.l-
250.
Gazave E., Lapebie P, Renard E., Vacelet J., Rocher C., et al. 2010. Molecular
Phylogeny Restores the Supra-Generic Subdivision of Homoscleromorph
Sponges (Porifera, Homoscleromorpha) // PLoS ONE 5(12): el4290.
doi: 10.1371 /joumal.pone.0014290
Gazave E., Lapebie P., Ereskovsky A.V, Vacelet J., Renard E., Cardenas P,
Borchiellini C. 2012. No longer Demospongiae: Homoscleromorpha formal
nomination as a fourth class of Porifera // Hydrobiologia. Vol.687. P.3-10.
Hahn-Keser B., Stockem W. 1997. Detection of distinct endocytotic and phago-
cytotic activities in epithelial cells (pinacocytes) of freshwater sponges (Porif-
era, Spongillidae) //Zoomorphology. Vol. 117. P. 121-134.
Harrison F.W. 1972. The nature and role of the basal pinacoderm of Corvomey-
enia carolinensis Harrison (Porifera: Spongillidae) A histochemical and de-
velopmental study // Hydrobiologia. Vol.39. P.495-508.
Harrison F.W., De Vos L. 1991. Porifera // F.W. Harrison, J. A. Westfall (eds.).
Microscopic Anatomy of Invertebrates. Vol.2. Placozoa, Porifera, Cnidaria,
Ctenophora. New York: Wiley-Liss, Inc.
Hentshel U., Uscher K.M., Taylor M.W. 2006. Marine sponges as microbial fer-
menters // FEMS Microbiol. Ecol. VoL55. P. 167-177.
ill M.S., Hill A.L., Lopez J., Peterson K.J., Pomponi S., et al. 2013. Recons-
truction of Family-Level Phylogenetic Relationships within Demospongiae
(Porifera) Using Nuclear Encoded Housekeeping Genes // PLoS ONE 8(1):
e50437. doi:10.1371/joumal.pone.0050437
Список литературы
95
Ivanova L.V., Semyonov V.V. 1997. Feeding habits of the larvae of sponges //
A.V. Ereskovsky, H. Keupp, H.R. Kohring (eds.). Modem problems of Porif-
eran biology. Berliner Geowiss Abh., Freie Univ., Berlin.
Kaltenbach J.C., Kuhns W.J., Simpson T.L., Burger M.M. 1999. Intense concana-
valin A staining and apoptosis of periferal flagellated cells in larvae of the ma-
rine sponge Microciona prolifercr. significance in relation to morphogenesis //
Biol Bull. Vol. 197. P.271-273.
Kanayama Y., Kamishima N. 1990. Role of symbiotic algae in hatching of gem-
mules of the freshwater sponge Radiospongilla cerebellata И Zool. Sci. Vol.7.
P.649-655.
Krylova D.D., Aleshina G.M., Kokijakov V.N., Ereskovsky A.V. 2004. Antimi-
crobal properties of mesohylar granular cells of Halisarca dujardini John-
ston, 1842 (Demospongiae, Halisarcida) // Boll. Mus. 1st. Biol. Univ. Genova.
Vol.68. P.399—404.
Langenbruch P.F. 1982. Die Entstehung der Gemmula-Schalen bei Spongilla fra-
gilis Leidy (Porifera) // Zoomorphology. Vol.99. S.221-234.
Lavrov D.V, Wanga X., Kelly M. 2008. Reconstructing ordinal relationships
in the Demospongiae using mitochondrial genomic data // Mol. PhyL EvoL
Vol.49. P. 111-124.
Ledger P.W., Jones W.C. 1977. Spicule formation in the calcareous sponge Sycon
ciliatum // Cell Tissue Res. Vol. 181. P.553-567.
Leys S.P. 2004. Gastrulation in Sponges // C.D. Stem (ed.). Gastrulation. From
cell to embryon. New York: Cold Spring Harbor Lab Press. P.23-32.
Leys S.P. 1999. The choanosme of hexactinellid sponges // Invert. Biol. Vol. 118.
P.221-235
Leys S.P, Degnan B.M. 2002. Embryogenesis and metamorphosis in a haploscle-
rid demosponge: gastrulation and transdifferentiation of larval ciliated cells to
choanocytes//Invert. Biol. Vol. 121. P. 171-189.
Leys S.P., Eerkes-Megrano D. 2005. Gastrulation in Calcareous Sponges: In
Search of Haeckel’s Gastraea // Integr. Comp. Biol. Vol.45. P.342-351.
Leys S.P., Mackie G.O., Reiswig H.M. 2007. The Biology of Glass Sponges //
Advanc. Mar. Biol. VoL52. P. 1-145.
Li C.W., Chen J.Y, Hua ТЕ. 1998. Precambrian sponges with cellular struc-
tures // Science. Vol.279. P.879-882.
Mackie G.O., Singla C.L. 1983. Studies on hexactinellid sponges. I. Histology
of Rhabdocalyptus dawsoni (Lambe, 1873) // Phil. Trans. Roy. Soc. Lond.
Vol.301. P.365-400.
Maldonado M. 2004. Choanoflagellates, choanocytes, and animal multicellula-
rity// Invert. Biol. Vol. 123. P.1-22.
96
А.В. Ересковский, А.Э. Вишняков. Губки
Maldonado М., Riesgo А. 2007. Intra-epithelial spicules in a homosclerophorid
sponge // Cell Tissue Res. Vol.328. P.639-650.
Manuel M., Borojevic R., Boury-Esnault N., Vacelet J. 2002. Class Calcarea //
J.N.A. Hooper, R.W.M. van Soest (eds.). Systema Porifera: A Guide to the
Classification of Sponges. New York: Kluwer Academic/Plenum Publishers.
Manuel M., Borchiellini C., Alivon E., Le Parco Y, Vacelet J., Boury-Esnauly N.
2003. Phylogeny and evolution of Calcareous sponges: monophyly of Cal-
cinea and Calcaronea, high level of morphological homoplasy, and the primi-
tive nature of axis symmetry // Syst. Biol. Vol.52. P.311-333.
Manuel M., Borchiellini C., Alivon E., Boury-Esnauly N. 2004. Molecular phy-
logeny of calcareous sponges using 18S rRNA and 28S rRNA sequences //
M. Pansini, R. Pronzato, G. Bavestrello, R. Manconi (eds.). Sponge science in
the new millennium. Boll. Mus. 1st. Biol. Univ. Genova. Vol.68. P.449^161.
Masuda Y, Kuroda M., Matsuno A. 1998. An ultrastructural study of the contrac-
tile filament in the pinacocyte of a freshwater sponge // Y. Watanabe, N. Fuse-
tani (ed.). Sponge Sciences. Multidisciplinary perspectives. Tokyo: Springer-
Verlag. P.249-258.
Muricy G., Boury-Esnault N., Bezac C., Vacelet J. 1996. Cytological evidence for
cryptic speciation in Mediterranean Oscarella species (Porifera, Homosclero-
morpha) // Can. J. Zool. Vol.74. P.881-896.
Nickel M. 2010. Evolutionary emergence of synaptic nervous systems: what can
we learn from the non-synaptic, nerveless Porifera? П Invert. Biol. Vol. 129.
No.l.P.1-16.
Pavans de Ceccatty M., Thiney Y, Garrone R. 1970. Les bases ultrastructurales
des communications intercellulaires dans les oscules de quelques eponges //
W.G. Fry (ed.). The biology of Porifera. London: Academic Press. Vol.25.
P.449—466.
Pavans de Ceccatty M. 1986. Cytosceletal organisation and tissue patterns of epi-
thelia in the sponge Ephydatia muelleri // J. Morphol. Vol. 189. P.45-65
Philippe H., Derelle R., Lopez P., Pick K., Borchiellini K., et al. 2009.
Phylogenomics Revives Traditional Views on Deep Animal Relationships //
Curr. Biol. Vol. 19. P.706-712.
Plotkin A.S., Ereskovsky A.V. 1997. Ecological aspects of asexual reproduction
of the White Sea Sponge Polymastia mammillaris (Demospongiae, Tetracti-
nomorpha) in Kandalaksha Bay //A.V. Ereskovsky, H. Keupp, H.R. Kohring
(eds.). Modem problems of Poriferan biology. Berliner Geowiss Abh., Freie
Univ., Berlin. P. 127-132.
Pottu-Boumendil J. 1975. Ultrastructure, cytochimie, et comportements
morphogenetiques des vellules de I’eponge Ephydatia muelleri. These. Lyon:
Univ. Cl. Bernard.
Список литературы
97
Redmond N.E., Morrow С.С., Thacker R.W., Diaz M.C., et al. 2013. Phylogeny
and Systematics of Demospongiae in Light of New Small-Subunit Ribosomal
DNA(18S) Sequences // Integr. Comp. Biol. Vol.53. P.388^115.
Reiswig H.M., Mehl D. 1991. Tissue organization of Farrea occa (Porifera,
Hexactinellida) // Zoomorphology. Vol. 110. P.301-311.
Reitner J., Gautret P. 1996. Skeletal formation in the modem but ultraconserva-
tive chaetetid sponge Spirastrella (Acanthochaetetes) wellsi (Demospongiae,
Porifera) // Facies. Vol.34. P. 193-207.
Renard E., Vacelet J., Gazave E., Lapebie P., Borchiellini C., Ereskovsky A.V
2009. Origin of the neuro-sensory system: new and expected insights from
sponges // Integr. Zool. Vol.4. P.294-308.
Richards G.S., Simionato E., Perron M., Adamska M., Vervoort M., Degnan B.M.
2008. Sponge Genes Provide New Insight into the Evolutionary Origin of the
Neurogenic Circuit//Curr. Biol. Vol. 18. P.l 156-1161.
Sailer U. 1990. Formation and construction of asexual buds of the freshwater
sponge Radiospongilla cerebellata (Porifera, Spongillidae) // Zoomorphol-
ogy. Vol. 109. P.295-301.
Schiitze J., Krasko A., Diehl-Seifert B., Muller W.E.G. 2001. Cloning and expres-
sion of the putative aggregation factor from the marine sponge Geodia cydo-
nium // J. Cell Sci. Vol. 114. P.3189-3198.
Simpson T.L. 1984. The cell biology of Sponges. Springer-Verlag, New York.
Simpson T.L., Vaccaro C.A. 1974. An ultrastructural study of silica deposition in
the freshwater sponge Spongilla lacustris // J. Ultra Res. Vbl.47. P.296-309.
Sperling E.A., Peterson K.J., Pisani D. 2009. Phylogenetic-Signal Dissection of
Nuclear Housekeeping Genes Supports the Paraphyly of Sponges and the
Monophyly of Eumetazoa // Mol. Biol. Evol. Vol.26. No. 10. P.2261-2274.
Taylor M.W., Radax R., Steger D., Wagner M. 2007. Sponge-Associated Micro-
organisms: Evolution, Ecology, and Biotechnological Potential // Microbiol.
Mol. Biol. Rev. VoL7L P.295-347.
Thacker R.W., Hill A.L., Hill M.S., Redmond N.E., Collins A.G., et al. 2013.
Nearly Complete 28S rRNA Gene Sequences Confirm New Hypotheses of
Sponge Evolution // Integr. Comp. Biol. Vol.53. P.373-387.
Uriz MJ. 2006. Mineral skeletogenesis in sponges // Can. J. Zool. Vol.84. P.322-
356.
Uriz M.J., Turon X., Becerro M.A. 2000. Silica deposition in Demosponges: spic-
ulogenesis in Crambe crambe 11 Cell Tiss. Res. Vol.301. P.299-309.
Vacelet J. 2007. Diversity and evolution of deep-sea carnivorous sponges //
M.R. Custodio, G. Lobo-Hajdu, E. Hajdu, G. Muricy (eds.). Porifera Re-
search: Biodiversity, Innovation and Sustainability. Rio de Janeiro: Museu
Nacional. P.l07-115.
98
А.В. Ересковский, А.Э. Вишняков. Губки
Vacelet J., Boury-Esnault N. 1995. Carnivorous sponges // Nature. Vol.373.
P.333-335.
Vacelet J., Duport E. 2004. Prey capture and digestion in the carnivorous sponge
Asbestopluma hypogea (Porifera: Demospongiae) // Zoomorphology. Vol. 123.
P. 179-190.
Voigt O., Wulfing E., Worheide G. 2012. Molecular Phylogenetic Evaluation of
Classification and Scenarios of Character Evolution in Calcareous Sponges
(Porifera, Class Calcarea) // PLoS ONE 7(3): e33417. doi: 10.1371/journal.
pone.0033417.
Wachtmann D., Stockem W, Weissenfels N. 1990. Cytoskeletal organisation and
cell organelle transport in basal epithelial cells of the freshwater sponge Spon-
gilla lacustris // Cell Tiss. Res. Vol.261. P. 145-154.
Webster N.S., Taylor M.W. 2012. Marine sponges and their microbial symbionts:
love and other relationships // Environmental Microbiology. Vol. 14. No.2.
P.335-346.
Weissenfels N. 1978. Bau und Funktion des SuBawasserschwamms Ephyda-
tia fluvialis (Porifera). V. Das Nadelskelet und seine Entstchung // Zool. Jb.,
Anat. Bd.99. S.211-223.
Weissenfels N. 1989. Biologie und microscopishe Anayomie der SiiBswas-
sershwamme (Spongillidae). Stuttgart, New York: Fisher.
Worheide G., Dohrmann M., Erpenbeck D., Larroux C., et al. 2012. Deep Phy-
logeny and Evolution of Sponges (Phylum Porifera) // Advances in Marine
Biology. V0I.6I.P. 1-78.
ОГЛАВЛЕНИЕ
Введение ......................................................6
Глава 1. Общая характеристика губок............................7
Глава 2. Таксономическая структура типа Porifera..............15
Класс Calcarea Bowerbank, 1864, или Известковые губки.....15
Класс Homoscleromorpha Bergquist, 1978 ................... 17
Класс Demospongiae Sollas, 1885 .......................... 19
Класс Hexactinellida Schmidt, 1870 ........................21
Глава 3. Анатомия и клеточный состав губок ...................24
Глава 4. Колониальное^, модульность и индивидуальность губок .46
Глава 5. Симбиотические ассоциации губок, прокариотические
симбионты.................................................48
Глава 6. Физиология губок.....................................50
Глава 7. Размножение и развитие губок.........................55
Глава 8. Филогения губок......................................84
Заключение ...................................................90
Список литературы ............................................92