/



















































































































































































































Похожие




Текст
вш
HEINRICH WALTER
DIE VEGETATION
DERERDE
in oko-physiologischer
Betrachtung
*
Bd IL
Hie gemassigten
und
arktisehen Zonen
*
VEB Gustav Fischer Verlag
Jena 1968
Г. ВАЛЬТЕР
РАСТИТЕЛЬНОСТЬ
ЗЕМНОГО ШАРА
Эколого-физиологическая
характеристика
*
т. III
Тундры, луга, степи, внетропические пустыни
*
Перевод с немецкого
Ю. Я, Ретеюма
Под редакцией Т• А, Работнова
Издательство «Прогресс»
Москва 1975
Редакция литературы по географии
© Перевод на русский язык, издательство «Прогресс», 1975 г.
20901-702 Л
----------40—75
006(01)-75
Глава 1
АРКТИЧЕСКАЯ ЗОНА ТУНДРЫ И АЛЬПИЙСКИЙ ПОЯС ГОР
УМЕРЕННОЙ ГУМИДНОЙ ЗОНЫ
ОБЩИЕ СВЕДЕНИЯ О КЛИМАТЕ И ФЛОРЕ
Климатические условия в арктической тундровой области далеко
не однообразны. Значение имеют не только положение области
по широте, но и степень океаничности или континентальности ее кли-
мата, что наглядно иллюстрируют климадиаграммы (рис. 1—9). На
диаграммах указано число дней со средней суточной температурой выше
—10° (внизу) и выше +10° (у вершины температурной кривой). Если число
дней с температурой более 4-10° превышает 30, это означает, что метеоро-
логическая станция расположена еще в лесотундре или даже в северной
области хвойных лесов. Станции, на которых отмечены наиболее низкие
температуры (в Сибири и на Аляске), одновременно регистрируют резко
выраженный засушливый период. Тем не менее эти станции расположены
в пределах области хвойных лесов: температура летом здесь относительно
высока и корни растений по мере постепенного оттаивания почвы получа-
ют достаточно влаги.
Полярная граница леса отделяет арктическую зону тундры от полосы
лесов бореальной и умеренной зон точно так же, как альпийская граница
леса в горах отделяет альпийский пояс от субальпийского и горного поя-
сов. Арктическая зона почти сплошным кольцом охватывает высокие широ-
ты северного полушария; только между Гренландией, Исландией и Сканди-
навским полуостровом это кольцо прерывают сравнительно обширные мор-
ские пространства (рис. 10). Такое единство поверхности объясняет,
почему по флористическому составу и растительности тундра более одно-
родна, нежели бореальная зона хвойных лесов.
Значительно большее разнообразие присуще альпийским поясам
отдельных, расположенных далеко одна от другой горных систем умерен-
ной зоны (Альпы, Кавказ, горы южной Сибири и Скалистые горы в Север-
ной Америке). Вместе с тем во флористическом отношении у них можно
обнаружить много черт, общих с арктической зоной, ибо в период оледе-
нения альпийская флора гор в различных пунктах могла соприкасаться
с арктической флорой. Кроме того, все эти горные системы возникли
в третичное время, поэтому следует считать, что их флора произошла от
арктотретичной флоры северного полушария. В климатическом отношении
арктическая зона и альпийские пояса, особенно пояса гор, лежащих
южнее 50° широты, не вполне сходны х. Но у них есть одна важная общая
черта — кратковременность вегетационного периода. И дело здесь не
в длительности безморозного периода, а во времени, в течение которого
снежный покров отсутствует (так называемый «бесснежный» период).
1 А. Р i s е k, Pflanzen der Arktis und des Hochgebirges, Hb. d. Pflanzenphysiol.,
Bd. V, 1960, S. 376—414.
6
Глава 1
АНАДЫРЬ,ПОРТ (5м)
Рис. 1—6. Климадиаграммы субарктической и арктической областей Евразии.
Абиску — шведская часть Лапландии; Карские Ворота — пролив в Северном Ледовитом океане;
Анадырь — побережье Берингова моря; Хатанга — север Средней Сибири; Верхоянск — конти-
нентальный климат, полюс холода; Петропавловск - Камчатский — океанический климат. Вер -
хоянск и Петропавловск - Камчатский расположены еще в лесной зоне.
Цифры в середине диаграммы означают число дней со средней суточной температурой выше 4-10°
и выше —10°. Полярная граница древесной растительности приблизительно совпадает с изотермой
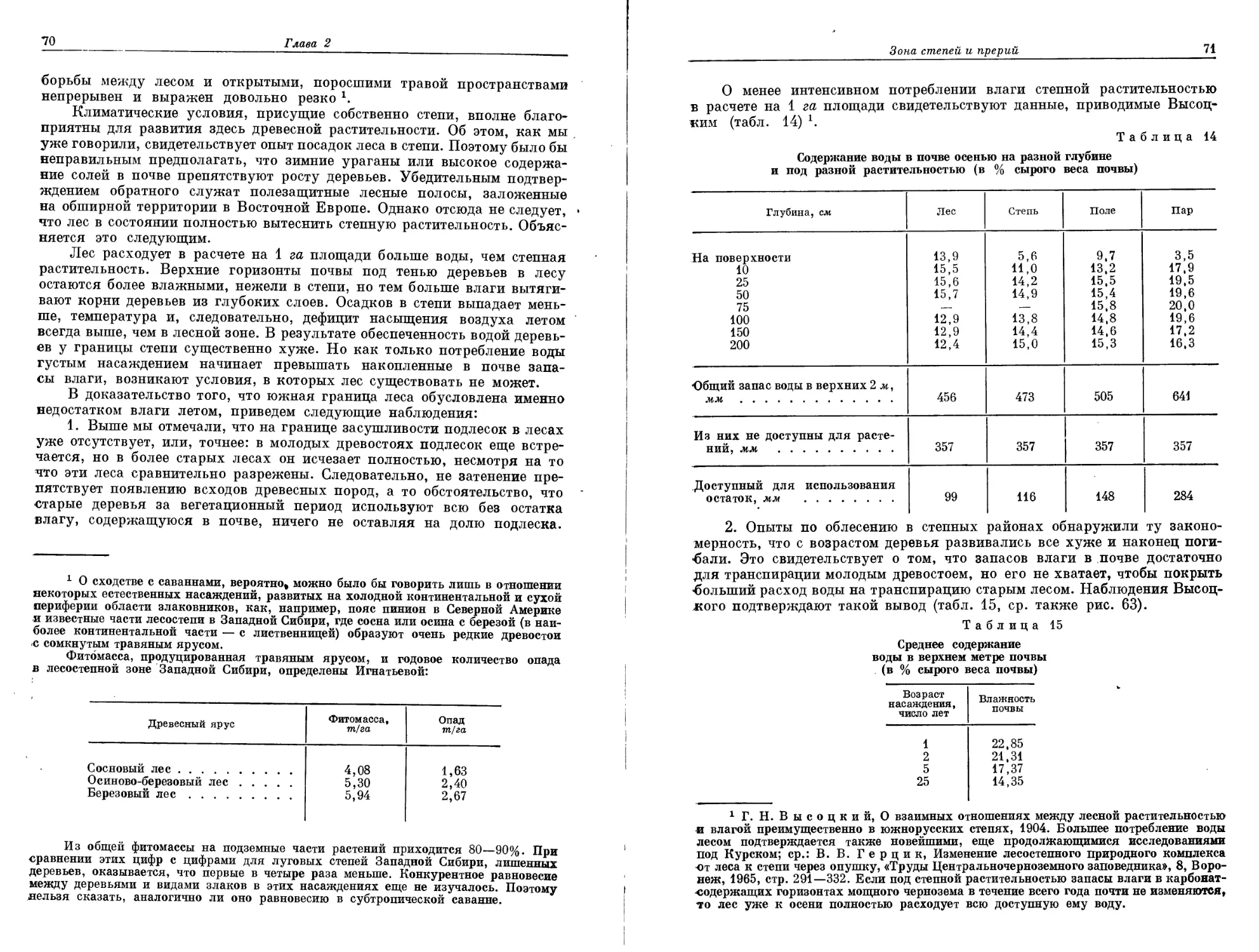
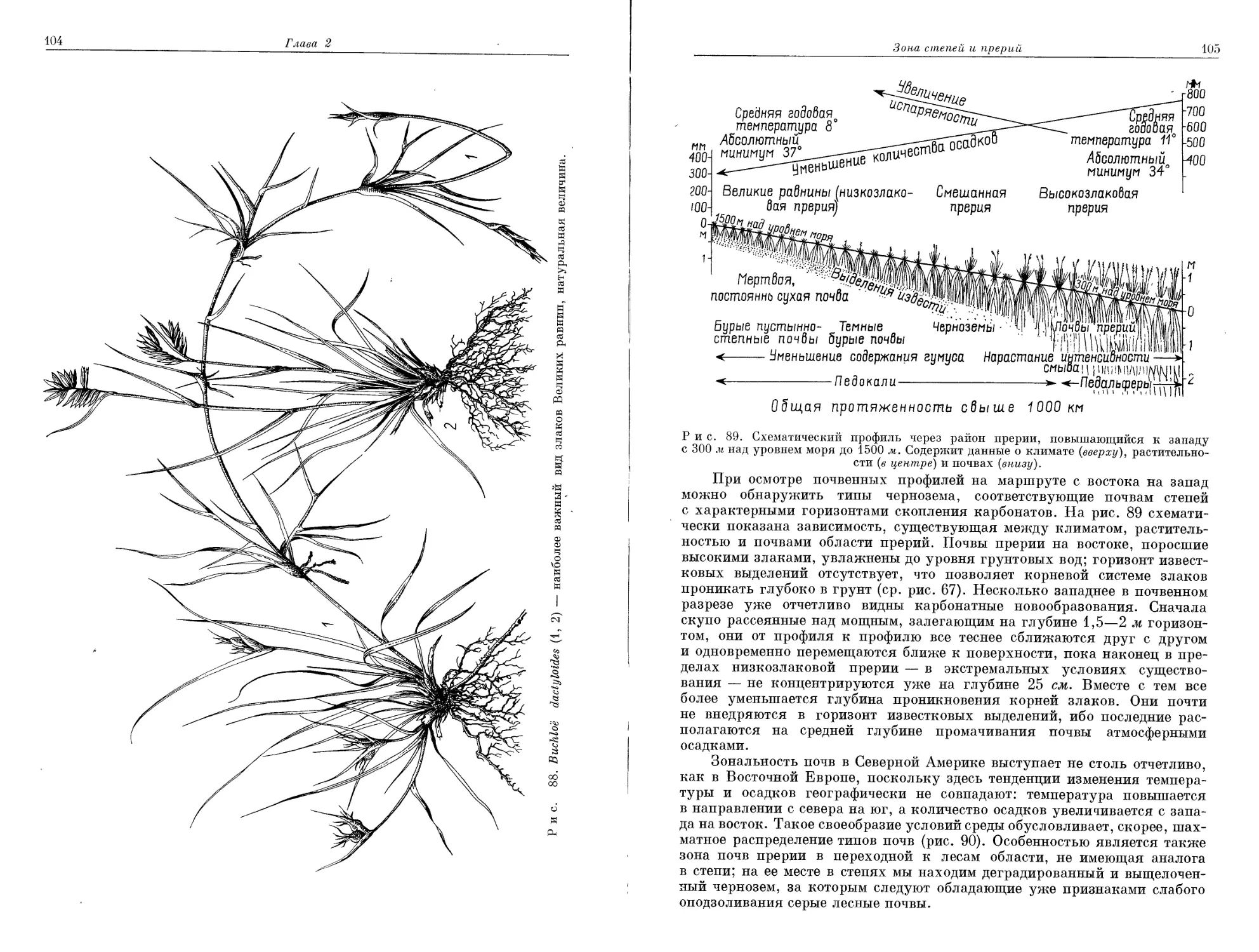
июля 4-10°.
Рис. 7—9. Климадиаграммы границы Субарктики и Арктики в Северной Америке.
Хопдейл — вблизи зоны тундры на востоке; Честерфилд — южная тундра, центральная часть
Сев. Америки; Форт-Юкон — Аляска, условия напоминают Верхоянск (рис. 5): в обоих районах
засушливый ^период ^вследствие наличия многолетней мерзлоты не препятствует росту деревьев.
Там, * где образуется сравнительно мощный снежный покров,
это время значительно короче безморозного периода. Каким
образом продолжительность «бесснежного» периода в Альпах при нормаль-
ном снежном покрове изменяется с высотой, количественно установил
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 7
Рис. 10. Полярная граница древесной растительности, ограничивающая арктическую
зону на юге (по Тихомирову, 1962).
Границу образуют: 1 — Betula tortuosa', 2 — В. егтапг', 3 — Ргсеа obovata', 4 — Р. татгапа', 5 —
Larix sibirica', 6 — L. dahurica.
еще Кернер (рис. 11). Напомним также, что процесс накопления снега,
следовательно, длительность «бесснежного» периода существенно зави-
сят от рельефа местности и направления господствующих ветров. Как
результат воздействия всех этих факторов формируется мозаичная струк-
тура распределения растительности 4. Рис. 12 иллюстрирует то положе-
ние, что конкурентная способность отдельных доминантных видов опре-
деляется в основном продолжительностью «бесснежного» периода.
Климат арктической зоны весьма разнообразен. Напри-
мер, на северо-западном побережье Европы господствует океаническо-
арктический климат с умеренно холодной зимой и прохладным летом.
1Н. Friedel, Schneedeckendauer und Vegetationsverteilung im Gelande,
«Mitt. Forstl. Bundes-Versuchsanst.». 59, Wien. 1961, S. 317—369.
8
Глава 1
ALectorietum
(наветренный склон)
Alectorietum
LoiseLeurletum
Vaccinieiurri ullginosi
Vaccinietum myrtilli
Rhododendretum ferruginel
Festucetum halleri
Рис. 11. Средняя продолжительность сохранения снежного пок-
рова и «бесснежного» периода в зависимости от высоты местности
в долине среднего Инна (по Кернеру).
Тонкая линия — на солнечных, жирная — на теневых склонах.
Рис. 12. Средняя продолжительность сохранения снежного пок-
рова (более 5 см) для некоторых растительных сообществ близ
Обергургля в Тироле (по Турнеру).
На севере же Сибири климат континентально-арктический, с весьма
суровой зимой и относительно теплым летом. Свойственные этому климату
очень низкие средние годовые температуры воздуха обусловливают
распространение многолетней мерзлоты, ареалы которой на территории
Сибири проникают глубоко в лесную зону.
Климат Арктики претерпевает изменения и в направлении с юга
на север: «бесснежный» период становится короче, дни летом длиннее,
но одновременно уменьшается высота стояния солнца в полдень. Поступле-
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 9
ние радиации на горизонтальную поверхность сокращается и почва прогре-
вается слабее, различия в температуре дня и ночи сглаживаются.
Именно в этом отношении горный климат особенно резко отличается
от климата Арктики. В горах прямая радиация с высотой увеличивается,
но увеличивается и собственное излучение земной поверхности, особенно
заметно ночью и на теневых склонах. Благодаря этому температурные
контрасты дня и ночи, а также солнечных и теневых склонов становятся
все более резкими. Высоко в горах зимой, прежде всего при часто возни-
кающих инверсиях температуры, обычно стоит солнечная погода, в Аркти-
ке же в это время года царит полярная ночь. Однако летом более длитель-
ный арктический день, по-видимому, компенсирует пониженную прямую
радиацию, что имеет значение для продуктивности растений. Но даже
в период, когда солнце не заходит, в Арктике отмечаются известные коле-
бания интенсивности освещения (58 000—4500 лк), температуры
(7,8—0,8°) и влажности (73—98%).
По количеству осадков Арктика также сильно отличается от альпий-
ского пояса. Годовая сумма атмосферных осадков в Арктике, особенно
в континентальных условиях, очень мала; в горах, напротив, она с высо-
той все более возрастает х. Этот контраст заметен главным образом зимой,
так как снежный покров в горах обычно намного мощнее, поэтому расте-
ния здесь защищены от мороза лучше, чем в Арктике. В вегетационный
период разница в общем количестве осадков такой большой роли не игра-
ет, поскольку из-за слабого испарения растения в Арктике не испытывают
недостатка влаги, в горах же избыточная вода стекает по склону.
Как бы ни было велико различие климатов Арктики и альпийского
пояса, все же отмечается известное сходство общих условий произраста-
ния растений. Оно обусловлено крайней непродолжительностью вегета-
ционного периода и проявляется в том, что в той и другой области одина-
ково хорошо растут многие общие виды, такие, как Dryas octopetala, Saxifra-
ga oppositifolia, Salix herbacea и др. Правда, речь, видимо, идет о различ-
ных экотипах или географических расах.
Почвы Арктики и альпийского пояса характеризуются рядом особен-
ностей. Сильное физическое и почти полное отсутствие следов химического
выветривания горных пород роднит их с почвами пустынь. Но в отличие
от последних почвы Арктики и альпийского пояса отличаются избытком
влаги и значительным накоплением органического вещества в форме
торфа или гумусовидных соединений. Конечно, это относится к районам,
где имеется хотя бы редкая растительность, отмершие остатки которой
в условиях низких температур и сильного увлажнения разлагаются край-
не медленно. В экстремальных условиях высокой Арктики и нивального
пояса гор растительный покров столь скуден, что сколько-нибудь значи-
тельного накопления органического вещества не происходит.
Другое типичное для той и другой области явление, связанное
с частым чередованием дней с морозом и оттепелью,— это солифлюкция
(рис. 13); в районах развития торфяных почв она наблюдается уже на
склонах с падением всего в несколько градусов. При этом возникают
своеобразные формы рельефа, сходные с тропинками, выбитыми скотом
на склонах, террасы, кочки или небольшие бугры 1 2. Чем реже
1 Общие климатические условия Альп и их предгорий характеризует карта
климадиаграмм, опубликованная Редером: Н. R eh d er, Flora, В, 156, 1966, S. 78—93.
2 Т. С. Е. F г i е s, Botanische Untersuchungen im nordlicben Schweden, Uppsala,
1913 (дисс.)
10
Глава 1
Сагастыр (устье Лены)
73°23' с.ш.
Цугшпитце (Альпы) 2964п
□< fflh ЕЗз
Рис. 13. Повторяемость переходов температуры через нуль в Арктике и
высоких горах (по Троллю).
1 — дни без мороза; 2 — дни с круглосуточным морозом з — дни с переходом температуры
через нуль.
растительность, тем интенсивнее протекают процессы солифлюкции.
В экстремально арктических районах на склонах образуются каменные
реки и полосчатые почвы, а на ровной местности — столь характерные
для Арктики каменные многоугольники или полигональные почвы. Анало-
гичные формы известны также в высоких горах. Но здесь склоны обычно
скалистые или обрывистые, поэтому большие ровные площади встречаются
очень редко.
ЛЕСОТУНДРА
Между бореальной зоной хвойных лесов и безлесной арктической
тундрой вклинивается зона лесотундры, в пределах которой лес и тундра
мозаикообразно заходят друг в друга. Сначала в лесу появляются мало-
приметные лишенные деревьев участки, которые к северу все более увели-
чиваются в размере, пока наконец в тундре не останутся лишь единичные
островки леса; последние все больше разреживаются, и вот уже на равнине
маячат лишь одинокие кустарникообразные уродливые деревца. Эта зона
борьбы между лесом и тундрой, в горах не превышающая нескольких
сотен метров в ширину, в равнинных районах Арктики простирается
на сотни километров.
Границу леса в областях с океаническим климатом образует береза
(в Фенноскандии Betula pubescens ssp. tortuosa, рис. 14; на Камчатке
В. ermani) \ на севере Европы и в Северной Америке — ель (Picea obovata
или Р. mariand), а в условиях континентального климата Сибири —
лиственница (Larix sibirica в Западной Сибири до Енисея и L. dahurica —
в Средней и Восточной Сибири, рис. 10).
В Лапландии мы наблюдали интересную особенность. На самых северных место-
обитаниях сосна (Pinus silvestris) посреди низкого кустарника Betula tortuosa представ-
ляла собой не угнетенные, уродливые, а прямые, хорошо развитые деревья. Рядом
с ними стояло много молодых деревец, почти лишенных хвои, хотя они и перезимовали
1 J. Bluthgen, Der skandinavische Fjallbirkenwald als Landschaftsformation,
«Peterm. Geogr. Mitt.», I960, S. 119—144; E. H u 11 e n, Kamtschatka, «Veg. Bilder»,
Reihe 23, H. I/II, 1932; В. H. Васильев, Растительность Анадырского края,
М. —Л., 1956.
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны И
Рис. 14. Betula tortuosa и единичные Pinus silvestris под Абиску, север ШвецииЦ
(фото Э. Вальтер).
под снегом. Здесь причиной ограничения ареала, по всей видимости, было уничтоже-
ние молодых деревьев снежным грибом (Lophodermium pinastri). Подрост получает воз-
можность успешно развиваться лишь тогда, когда деревьям удается достичь высоты,
при которой покрытые иголками конечные побеги выступают над снегом. Эти особен-
ности роста деревьев характерны для районов Арктики с океаническим климатом,
отличающихся относительным обилием снега и вместе с тем мягкой зимой. Такой кли-
мат благоприятен для развития снежного гриба, поражающего подрост хвойных;
этим, по-видимому, объясняется, почему границу произрастания деревьев^образуют
не хвойные деревья, а береза, для которой гриб не опасен. На развитии березы влияние
снега проявляется в том, что деревья принимают весьма причудливые формы роста
(рис. 15).
Тихомиров 1 объясняет образование полярной границы леса прежде
всего отсутствием его возобновления. Исследования показали, что деревья
у северной границы своего распространения плодоносят редко и семена
их обычно не прорастают. Большая часть не потерявших всхожести семян
поедается животными либо же штормовые ветры переносят их по снегу
далеко на север, где климатические условия уже не пригодны для роста
деревьев. Кроме того, острова тундры в зоне лесотундры обычно бывают
покрыты плотным слоем лишайников и мхов, затрудняющим прораста-
ние семян.
Неудивительно поэтому, что сеянцы деревьев встречаются только
на обнаженных минеральных почвах или на эродированных склонах.
Молодые деревца обычно группируются вокруг более старых древостоев,
где условия для возобновления оказываются благоприятнее (здесь больше
1 Б. А. Тихомиров, Безлесье тундры, ее причины и пути преодоления,
М.-Л., 1962.
12
Глава 1
Лишенная корковых
лишайников кустар-
никовая зона
Испытывающая дав-
ление снега базаль-
-кс=зона набуба снега
_]/Ь=термальная экстре-
мальная зона
а = Высота снега в конце
зимы
Рис. 15. Схема роста березы под снежным покровом (по Блютгену).
ная зона
снега, лучше защита от ветра, глубже оттаивает почва над многолетней
мерзлотой). Особенно важным фактором (имеющим значение и для поло-
жения альпийской границы леса) является деятельность человека; его
стада оленей, выпас которых особенно интенсивен в лесотундре, причиняют
растительности большой ущерб; животные наносят деревьям и механические
повреждения (обгладывают кору, скусывают побеги), человек вырубает их
на дрова. Поэтому граница леса смещается все дальше к югу г. Условия
роста для деревьев столь неблагоприятны, годичный прирост древесины
так мал, что потребление ее не восполняется естественным приростом.
Незначительное число способных к прорастанию семян и трудности
укоренения проростков, которое только и возможно, если в течение двух
лет подряд удерживаются оптимальные температуры, усугубляются чрез-
вычайно медленным ростом сеянцев: 20—25-летние деревца Larix sibirica
и Picea obovata едва возвышаются над ярусом травянистых растений. Их
ежегодный прирост в высоту обычно не превышает 1—2 см (максимум
3,5 см).
Сколь различными для развития деревьев могут быть условия разных
лет, иллюстрирует рис. 16. В благоприятные годы почки березы распуска-
ются в конце мая, в неблагоприятные — в июле. Решающее значение для
роста имеет температура лета, причем длина годичных побегов сосны
и интенсивность ее цветения отстают на год, так как почки закладываются
годом раньше (рис. 18).
В направлении к северу лесотундра сначала переходит в южную
кустарничковую тундру, где еще доминируют многие деревянистые кустар-
нички (рис. 17 и 19), затем в среднюю арктическую зону с лишь частично
сомкнутым растительным покровом (рис. 20) и, наконец, в высокую Аркти-
ку, где растут только некоторые виды мхов и лишайников (рис. 21).
1 По материалам Андреева, в районе Малоземельской тундры у границы леса
уничтожено 10—15% всех деревьев, из них 75% — стадами северных оленей, 25% —
вырубкой. Самый северный еще сохранившийся лиственничный лес находится в бас-
сейне Хатанги. Самая северная лиственница растет под 72°40' с. ш. Исландия до засе-
ления ее человеком была почти сплошь покрыта лесами, сегодня же здесь трудно найти
даже одиночное дерево. Однако успешно проводятся работы по искусственному обле-
сению территории, местами экзотическими породами.
5 .ИГ.
ЗО.И.
25.И.
20.И.
15.ЕГ.
1О.И
5.И.
31.У.
J--1--1-1--1-1--11---1-1--1-1--1-!__I.» I I___I I I 1 l I I i I
18853. 18903. 18953. 19003. 19053. ~ 1910г.
Рис. 16. Время распускания листьев березы в 1884—1909 гг.
в шведской части Лапландии (по Фризу).
Рис. 17. Кустарничковая тундра на галечниках; полуостров
Рыбачий, 69—70° с. ш. Важнейшие виды: Arctous alpina, Diapensia
lapponica, Empetrum hermaphroditum, Loiseleuria procumbens, Betula
папа, Vaccinium vitis-idaea и др. (фото Висниевского).
Рис. 18. Рост сосны в толщину (в мм)
в зависимости от температуры июля в
районе Инари (68°57' с. ш.), Финляндия
(по Хустиху).
1 — рост сосны; 2 — температура июля; з — дли-
на однолетних побегов в см и 4 — интенсивность
цветения в % (1938 г. = 100%), запаздываю-
щего на 1 год по сравнению с ходом темпера-
туры.
..2
—3
- -4
14
Глава 1
Рис. 19. Кустарничковая тундра (Betula папа} у Полярного Урала
(фото Поле)»
Рис. 20. Papaver radicatum в каменистой тундре, Западный Шпицберген
(фото Холмсена).
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 15
Рис. 21. Магдалене-фьорд на севере Западного Шпицбергена летом.
На переднем плане — моховая и лишайниковая тундра, на заднем — ледя-
ная пустыня (фото Гинцбергера).
ТУНДРА
Самая обширная область тундры простирается на севере Сибири.
Совершенно лишенная леса площадь составляет здесь 3 млн. км1 2 (рис. 22).
Кратким новейшим обобщением наших знаний об экологии растений
тундры мы обязаны Тихомирову х. Первые важные экологические наблю-
дения сделал Бэр в 1837 г. во время своей поездки на Новую Землю.
Собранные им сведения в 1860—1867 гг. были дополнены Мидцендорфом;
в последнее время начатую работу продолжают многочисленные иссле-
дователи, в частности в Гренландии 2.
В зоне тундры число дней со средней суточной температурой выше 0°
равно 55—118. Вегетационный период начинается в июне и заканчивается
в сентябре. Низкая температура летом частично объясняется большим
расходом тепла на таяние снега и оттаивание верхних слоев почвы. Роль
ветра в формировании снежного покрова и обусловленная воздействием же
ветра мозаичность размещения растительности напоминают обстановку
в горах. Все очевидцы непременно упоминают характерные для тех мест
зимние ураганы, при которых скорость ветра достигает 15—30 м/сек,
а отдельные его порывы — 60 м/сек. В океаническом климате Кольского
полуострова сумма осадков составляет 400 мм, в районах континенталь-
ной Сибири — обычно лишь 200—250 мм. Тем не менее климат Арктики
влажный, в частности очень высока повторяемость туманов на побережье.
На испарение расходуется только около 30% общей суммы осадков. Много-
1 Б. А. Тихомиров, Очерки по биологии растений Арктики, М.—Л., 1963.
2 Th. Sorensen, Temperature relations and phenology of the Northeast Green-
land flowering plants, «Meddel. om Gronland», 125, № 9, Kebenhavn, 1941; Summary
of the botanical investigations in N. E. Greenland, ibidem, 144, № 3, 1945; T. W. B o -
c h e r, Phytogeography of middle West Greenland, ibidem, 148, № 3, 1963.
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 17
летняя мерзлота препятствует просачиванию воды в грунт, поэтому почвы,
как правило, переувлажнены. Только каменистые почвы на возвышенно-
стях иногда успевают «просохнуть» в летние месяцы. Мощность снежного
покрова колеблется в пределах 10—50 см. Зимой сильные ветры сдувают
весь снег с высоких участков, в низинах же накапливаются огромные
массы снега. На Новой Земле из 41 вида растений только 5 способны
пережить зиму без повреждений, не будучи укрыты снегом. На таких
местообитаниях роль ветра, переносящего кристаллы льда и песок, особен-
но ощутима. Все возвышения гладко отшлифованы перевеваемым снегом
и песком. Солнце даже в полдень летом стоит невысоко, поэтому лучи
его нагревают только крутые южные склоны. Они получают тепла
в 4—5 раз больше, чем ровные поверхности, и поэтому часто представляют
собой настоящие «цветники». В летние месяцы благодаря длительному
освещению происходит непрерывный положительный фотосинтез и, следо-
вательно, быстрое накопление органического вещества. Мощность слоя
многолетней мерзлоты, местами превышающая 100-м, на развитие расте-
ний не влияет. Важна лишь глубина, на которую летом оттаивает почва.
Иногда это слой толщиной всего 10 см. Существенную роль играет плот-
ность растительного покрова, препятствующего нагреванию почвы солнеч-
ными лучами. В Арктике можно выделить несколько важнейших типов
местообитаний:
1. Каменистые и крутые горные склоны с обломочными осыпями,
предоставляющие их обитателям самые неблагоприятные условия для
жизни. В Сибири в расселинах скал ютятся единичные Роа abbreviata, Saxifra-
ga oppositifolia, Dryas punctata и др., а на скалистых поверхностях —
лишайники: виды Rhizocarpon, Gyrophora, Parmelia.
2. Ровные платообразные вершины гор с развитыми на них полиго-
нальными почвами, обычно свободные от снежного покрова. Они также
«лабо заселены. Растения появляются лишь там, где в трещинах скал
-сохраняется снег.
3. Крутые, покрытые слоем мелкозема склоны. Зимой склоны, обра-
щенные на юг, обычно укрыты толстым слоем снега, который весной быстро
•стаивает. Здесь раскинулись богатые цветковыми растениями луга с вида-
ми Cerastium, Draba, Erigeron, Taraxacum и др. На склонах северной
экспозиции снег часто лежит круглый год.
4. Ложбины, расположенные ниже снеговых пятен, постоянно увлаж-
няемые талой водой. В них создаются хорошие условия для произрастания
•Caltha arcttca, Ranunculus nivalis, R. sulphureus, Pleuropogon sabinii, Dupon-
iia jisheri и др.
5. Берега ручьев и рек, зимой покрытые снегом и при южной экспози-
ции рано оттаивающие. Это наиболее благоприятные местообитания. При
ледоходе растительный покров часто «сдирается», а на обнажившейся почве
поселяются разнообразные виды.
6. Заливаемые полыми водами озерные и речные террасы; здесь
растения начинают цвести до половодья и после спада воды продолжают
развиваться на покрытой илом почве. Такие местообитания служат для
многих растительных видов путями миграции.
7. Песчаные речные террасы. Представляют собой аналогичные место-
обитания, с той лишь разницей, что заселяют их растения с длинными
корневищами — Equisetum arvense, Rumex graminifolius и др.
8. Склоны с явлениями солифлюкции, покрытые растительностью,
постоянно нарушаемой «течением» почвы, поэтому здесь могут закрепиться
многие проростки.
2-0434
18
Глава 1
9. Заболоченные участки по берегам озер с относительно скудной^
растительностью (Arctophila fulva, Ranunculus pallasii, Pleuropogon sabiniiy.
Hippuris vulgaris, Comarum palustre и др.).
10. Морские побережья, где встречаются галофильные виды, на
песке — вместе с Elymus, на иле — с Atropis и Calamagrostis. Там, где-
влияние морской воды ослаблено, развиваются кустарничковые пустоши
с Empetrum hermaphroditum и Vactinium vitis-idaea (рис. 17).
Все это более или менее специфические местообитания. В основном же
бескрайние площади тундры покрыты Betula папа (рис. 19), низкорослыми
видами Salix или видами Eriophorum и Carex. Sphagnum в Арктике ника-
кой роли в составе растительности не играет. На коренных выходах
известняков встречаются кальциефильные виды (Lesquerella arctica, Thlaspi
cochleariforme и др.). Вокруг поселений распространена рудеральная-
растительность.
Биологический спектр арктической флоры Гренландии, по Раункиеру>.
характеризует табл. 1.
Таблица 1
Биологический спектр флоры Гренландии 1
С К н G НН Th Всего
51 27,7% 96 52,2% 29 15,8% 5 2,7% 3 1,6% 184 вида 100%
1 Ch — хамефиты; Н — гемикриптофиты; О —геофиты; НН — гело-или гидро-
фиты; Th—терофиты.
На более сухих местообитаниях в Арктике доминируют лишайники,,
а на влажных — мхи.
Приводимые метеорологами данные о температуре воздуха, измерен-
ной на высоте 2 ж от поверхности почвы, не дают правильного представле-
ния о жизненных условиях арктической растительности, так как темпера-
тура самих растений большей частью оказывается более высокой. К тому
времени, когда воздух нагреется до нуля, почва обычно уже оттаивает
на 0,5 м, поскольку температура приземного слоя воздуха (0—10 см над.
почвой) значительно выше, чем в 2 .и над ней. Поэтому наиболее оптималь-
ные условия для роста растений создаются у самой земли, в узком про-
странстве над и под поверхностью почвы х. В этой зоне жизненные процес-
сы в течение короткого вегетационного периода могут протекать достаточ-
но интенсивно, чтобы происходило продуцирование органического веще-
ства, а значит, и развитие у растений генеративных органов и созревание
семян (рис. 23).
Неверно представление, что арктические виды размножаются пре-
имущественно вегетативно. В вегетативном состоянии существуют только*
бореальные виды, проникающие в Арктику. Правда, в высокоширотной
Арктике семена созревают не каждый год, а лишь в наиболее благоприят-
ные годы. Краткость вегетационного периода определяет необходимость.
1 С. Н. Карандина, Распределение подземных частей растений в некоторых;
ассоциациях хибинской тундры, «Бот. ж.», 39, № 6, 1954, стр. 809—818.
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 19
Рис. 23. Saxifraga groenlandica, S. nivalis, рано зацветающие в теплом при-^'
земном слое воздуха, Западный Шпицберген (фото Холмсена).^
очень раннего цветения, поэтому цветочные почки часто закладываются
за год до цветения. В Гренландии это наблюдалось у половины из 184 ис-
следованных видов (Sorensen, 1941; см. примечание на стр. 15).
Цветочные почки переживают 8—10 зимних месяцев обычно во вполне
развитом состоянии, чтобы весной, как только растение освободится от
снега, тотчас распуститься. Обычно закладывается много почек на случай,
если часть их зимой или при поздних весенних заморозках погибнет.
Наблюдалось, что отдельные почки раскрывались уже осенью и в цвету-
щем состоянии уходили в зиму и что в неблагополучные годы цветение так
и не завершалось. Но часто цитируемое сообщение, что цветки Cochlearia
arctica невредимыми сохраняются под снегом и весной продолжают цвести,
основано на неправильном истолковании заметки Чельмана (ср. работу
Тихомирова). В действительности открывшиеся осенью цветки зимой поги-
бают, однако перезимовавшие почки раскрываются, как только появляют-
ся из-под снега. Наиболее чувствительны к морозу цветки х, затем листья
и травянистые стебли; напротив, очень устойчивы почки возобновления.
Температура под снегом в Арктике может понизиться до —30°, и все же
многие листья переживают зиму, оставаясь зелеными, и весной сразу начи-
нают фотосинтез. Правда, чем дальше к северу, тем число перезимовавших
зеленых листьев заметно уменьшается. У многих видов эти листья весной
отмирают, как только образуются молодые листочки. Иными словами,
они служат своего рода запасающими органами. Сокращается в северном
направлении и численность ветнозеленых видов. Их листья не живут
дольше 12—14 месяцев, но, даже отмершие, они еще некоторое время
1 Ночной заморозок они все же переносят; так, например, цветки Ranunculus
glacialis, в течение часа испытавшие температуру —10°, совершенно не пострадали
(см. работу Рассела, примечание на стр. 22).
2*
20
Глава 1
держатся на ветках. Вечнозеленые виды цветут поздно, плодоносят редко
и для севера Арктики нетипичны.
Весьма характерны для Арктики непериодические виды,
как, например, Bray a humilis (см. работу Сёренсена). Развитие их растяги-
вается на несколько лет и временно прерывается лишь зимой на любой
стадии развития. Тем самым эти виды не зависят от трудностей, созда-
ваемых коротким полярным летом, и цветут либо в начале, либо в конце
вегетационного сезона. Их цветочные почки могут быть заложены за 2 года
до цветения.
Большинство видов начинают цвести очень рано, и тем не менее
в Арктике также можно обнаружить типичную смену аспектов. На Таймы-
ре различают до 5 аспектов, правда, они нередко перекрываются, и тогда
образуется пестрая мозаика различно окрашенных участков. Цветки
преимущественно белые или желтые и не очень крупные: диаметр 30%
цветков не превышает 1 см и лишь 16% цветков имеют размер более 2 см.
Мелкие цветки обычно соединены в крупные соцветия, благодаря чему они
становятся более заметными для насекомых. Представлены энтомогамные
виды, но преобладают анемогамные. Распространение плодов или семян
происходит также преимущественно с помощью ветра (иногда до 84'%),
обычно путем переноса по поверхности снега, а также по воде (10%).
Ягоды встречаются главным образом в лесотундре. Из-за затрудненного
продуцирования органического вещества семена обычно мелкие, весом
менее 1 мг (приблизительно у 75% видов). У большинства видов семена
способны прорастать только после воздействия на них зимних морозов.
Прорастание весной позволяет всходам накопить к осени некоторое коли-
чество запасных веществ. Известна также вивипария (у 1,5% видов)—
как у злаков, так и у Polygonum, Stellaria, Cerastium. Жизнеспособность
семян достаточно высока. При проверке у 50% видов всхожесть составила
40% Ч Семена Salix reptans, перезимовав, обнаружили 100%-ную всхо-
жесть, хотя обычно семена Salix утрачивают ее в течение нескольких
дней; но летом и они за короткое время утрачивали ее.
Благодаря обильному образованию семян растения Арктики быстро
заселяют незанятые открытые пространства, как это наблюдается, напри-
мер, в нижнем течении Лены. Напротив, неблагоприятным местом для
прорастания семян являются моховой и лишайниковый покровы. Но если
растение уже укоренилось, тогда моховой покров служит ему хорошей
защитой от зимних холодов и ветра. В холодных пустынях с коротким
вегетационным периодом и низкими температурами в отличие от жарких
пустынь число однолетних видов (терофитов) очень невелико. Это Koenigia
islandica (крестоцветные), 3 вида Gentiana, Montia lamprosperma и 2 вида
Pedicularis. Большинство видов — гемикриптофиты (см. табл. 1) и хаме-
фиты, что и понятно, ибо развиваются они в нижних, более теплых слоях
воздуха, зимой же их защищает тонкий снежный покров. Большинство
видов достигает всего 1—20 см в высоту, у многих розеточных и полурозе-
точных растений листья плотно прилегают к теплой поверхности земли.
Растений-подушек здесь немного. Они развиваются, как и виды со
стелющимися побегами, преимущественно на местообитаниях, открытых
ветрам. Виды с листьями, опушенными белыми волосками, на Таймыре
также немногочисленны (не более 23%). Опушение улучшает температур-
1 N. Soyrinki, Studien uber die generative und vegetative Vermehrung der
Samenpflanzen in der alpinen Vegetation Petsamo-Lapplands, «Ann. Bot. Soc. Zool.-
Bot. Fenn. Vanamo», 11, № 1, 1938; 14, № 1, 1939.
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 21
ный режим листьев при облучении их солнечными лучами, так как
препятствует выдуванию теплого, прилегающего к листу слоя воздуха1.
Поскольку перепады температуры в самых нижних слоях воздуха и
верхних горизонтах почвы весьма резкие, разные части растений, к тому
же нередко затеняющие друг друга, на-
греваются по-разному (рис. 24) 1 2.
Росту растений весной могут способ-
ствовать так называемые «ледяные окна»:
при более раннем таянии снега вокруг
растения, нагреваемого проникающей
сквозь снежную толщу радиацией, в снегу
образуется полость, закрытая сверху тон-
кой, прозрачной корочкой льда; возникает
как бы парник в миниатюре, в котором
температура иногда бывает на 14° выше,
чем на поверхности. Когда в ледяном
окошке при таянии образуется отверстие,
температура в полости резко понижается
(рис. 25). Особенно быстро тает снег
вокруг растений-подушек.
На буграх морозного вспучивания,
например, в субарктических районах
Исландии, где эти бугры достигают 30—
—50 см в высоту и 75—200 см в диаметре,
уже проявляются различия в экспозиции,
влияющие на характер размещения отдель-
ных видов. Северную сторону предпочи- „ „
о?-? г т -л Рис. 24. Температура отдельных
тают Salix herbacea, Rhacomitrium, а также частей N ovosieversta giaciaiis и
лишайники Thamnolia и Stereocaulon", окружающего воздуха в 11 часов
южную занимают Calluna vulgaris, Vacci- 12 июля 1955 г. (по Тихомирову^
nium myrtillus, V. uliginosum, Arctosta-
phylos uva-ursi и Betula папа. Равномерно распределяются Empetrum
hermaphroditum, Festuca ovina, Thalictrum alpinum, Dryas octopetala. Но и
эти виды значительно лучше развиваются на южных склонах. Так,
100 верхушечных побегов Empetrum на вершине бугра весили 1,63 г
(сухой вес), на северном склоне — 1,80, а на южном — 2,54 г. Вес побегов
свободно стоящих экземпляров Empetrum на вновь заселяемых ею площа-
дях, то есть там, где конкуренция отсутствует, достиг 6,0 г 3.
Многие арктические виды обладают толстыми корнями, которые
служат «кладовыми» запасных веществ. Чем больше этот запас у растения,
тем быстрее оно развивается с весны. Среди арктических видов много
бобовых (Oxytropis, Astragalus, Hedysarum и др.). Они особенно многочис-
ленны в арктических районах Восточной Сибири. Корневые клубеньки
растений располагаются непосредственно под быстрее нагревающейся
1 Подобное опушение свойственно также видам горных растений тропиков, где
оно, по-видимому, имеет то же назначение (см. т. I, гл. 4). Аналогичную роль, вероят-
но, играет опушение у растений-подушек, характерных, правда, для районов с силь-
ными ветрами, например для субантарктических областей или Патагонии.
2 Е. Dahl. Heat exchange ecology of Koenigia, «Oikos», 14, 1963, p. 190—211.
3 H. L i e t h, Okologische Notizen von den Frostbuckeln der Mo-Vegetation im
nordostlichen Island, «Flora», 150, 1961, S. 166—169; В. Д. Александрова,
О влиянии экспозиции нанорельефа на почвенный микроклимат и на развитие расте-
ний в арктической тундре, «Труды Моск, о-ва испытателей природы, 3, 1960,
стр. 11—26.
22
Глава 1
Рис. 25. Небольшие, уже открытые «ледяные парники» в Арктике. Объясне-
ние см. в тексте (по Тихомирову).
поверхностью почвы. Способность фиксировать азот из воздуха имеет
очень большое значение для жизни растений Арктики, так как здешние
почвы бедны им и нитрификация почти отсутствует. Проблема азотного
питания более подробно исследована Расселом г. Ему удалось показать,
что содержание азота в почве является одним из важнейших факторов
формирования структуры растительного покрова на острове Ян-Майен
(71° с. ш.), лежащем в 320 км восточнее Гренландии. Здесь развиты бедные
азотом вулканические почвы на трахитовом базальте. Там, где азот в почве
вообще отсутствует, растут только мхи и лишайники. Удобряется почва
лишь экскрементами животных. Наблюдается отчетливая связь между
содержанием азота в почве и плотностью растительного покрова. Бактерий
в почве мало, но у большинства видов имеется микориза. Тихомиров отме-
чает распространение ностоковых, которые поселяются на голом субстрате
как растения-пионеры. Концентрация в них азота достигает почти 5%
сухого веса, так что можно считать, что они потребляют азот непо-
средственно из воздуха.
Арктические растения, даже травянистые виды, растут очень долго.
Так, возраст 100 специально исследованных экземпляров Novosieversia
glacialis колебался от 3 до 97 лет. Самому старому из найденных растений
с корневищем длиной 36 см и толщиной 0,7 см было 143 года, причем остат-
ки листьев сохранялись в течение 112 лет. О возрасте других видов опубли-
кованы данные, которые приведены в табл. 21 2.
Таблица 2
Предельный возраст различных видов растений Арктики
(число лет)
Betula папа ...................80
Salix myrsinites ..............89
Salix arctica.................200
Vaccinium uliginosum ... 93
Calluna ....................42
Loiseleuria.................64
Empetrum ...................97
Dry as .....................108
1 R. S. Russel (et al.), Physiological and ecological studies on an arctic vege-
tation I—III, «J. Ecol.», 28, 1940, p. 153—179, 269—309; H. G. W age r, Growth and
survival of plants in the Arctic, «J. Ecol.», 26, 1938, p. 390—410.
2 Б. А. Тихомиров, Г. И. Г а л а з и й, Определение возраста Sieversia
glacialis и некоторые вопросы продолжительности жизни растений, «Бот. ж.», № 3,
1952, стр. 332—348.
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 23
ЭЕ ОЛОГО-ФИЗ И О Л ОГИЧЕСК НЕ ИССЛ ЕДОВАНИЯ
В АРКТИЧЕСКОЙ ЗОНЕ
В условиях сурового климата Арктики произошел отбор совершенно
определенных морфолого-физиологических типов растений, которые, кон-
курируя друг с другом, заселили различные местообитания и оказались
жизнеспособными даже на самых северных выступах берега Северного
«Ледовитого океана. Как и растениям других зон, арктическим видам
овойственно продуцировать путем фотосинтеза необходимое количество
органического вещества. Здесь мы уже касаемся результатов эколого-
-физиологических исследований.
Обеспеченность водой не представляет для арктических растений
особой проблемы. Измерения интенсивности транспирации, произведенные
Штоккером 1 в Лапландии, показали, что сокращение транспирации не
происходит даже в ясную погоду. Еще ранее Столфельт установил, что
летом устьица растений остаются открытыми даже в полночь и что их
•«открытость» изменяется в зависимости от интенсивности дневного осве-
щения. Таким образом, газообмен осуществляется непрерывно. По мате-
риалам русских исследователей (цитируем по Тихомирову), осмотическое
.давление у большинства видов сравнительно низкое — не превышает
7—20 атм. И если все же некоторые виды отличаются известными «ксеро-
морфными» чертами, то это, как считает Дадыкин, связано с затрудненным
поглощением азота из холодной почвы. Стало быть, речь идет о явлениях
недостаточности, то есть о пейноморфозе (см. т. II, гл. 2). Представляется
особенно важным знать интенсивность фотосинтеза и величину нетто-
продуктивности.
Сначала Мюллер 1 2 установил в Годхавне (западная Гренландия,
€9° с. ш.), что интенсивность освещения в первой половине июля даже
в полночь (исключая дождливую погоду) составляет 450 лк и при темпера-
туре несколько выше нуля все еще превосходит компенсационную точку
у Salix glauca и Chamaenerium. Таким образом, листья растений продуци-
руют органическое вещество круглые сутки. Аналогичное мнение высказал
Уэйджер 3, исследовавший поведение Oxyria digyna, Ranunculus glacialis
и Saxifraga cernua в восточной Гренландии (68°30' с. ш.). Эти выводы под-
тв ерждаются и круглосуточными измерениями величины фотосинтеза
в естественных условиях (побережье у Мурманска), выполненными Косты-
чевым с сотрудниками 4. В качестве примера на рис. 26 приведена кривая
ассимиляции СО2 Rubus chamaemorus в ясный день. В пасмурную погоду
ночью кривая может временно опуститься ниже нулевой отметки. Макси-
мальная интенсивность фотосинтеза у различных растений не превышает
12 мг СО2/дм2-час, но, поскольку процесс фотосинтеза продолжается все
24 часа, растение за сутки ассимилирует около 100 мг СО2/дм2, что округ-
ленно соответствует 60 мг крахмала. Такая высокая продуктивность
1 О. Stocker, Transpiration und Wasserhaushalt in verschiedenen Klimazo-
nen I. Untersuchungen an der arktischen Baumgrenze in Schwedisch-Lappland, «Jb.
wiss. Bot.», 75, 1932, S. 494—549.
2 D. Muller, Die CO2-Assimilation bei arktischen Pflanzen und die Abhan-
gi gkeit der Assimilation von der Temperatur, «Planta», 6, 1928, S. 22—39.
3 H. G. W a g e r, On the respiration and carbon assimilation rates of some arctic
plants as related to temperature, «New Phyt.», 40, № 1, 1941.
4 S. Kostytschev, W. Tschesno ko v, К. В azyrina, Untersuchun-
,gen uber den Tagesverlauf der Photosynthese an der Kiiste des Eismeeres, «Planta»,
11, 1930, S. 160—168.
24
Глава 1
позволяет растениям в течение короткого вегетационного периода накопить
достаточно запасных веществ, чтобы пережить долгую зиму. Кроме того,
арктические виды обладают способностью в дневные часы использовать
сильное освещение как гелиофиты, а при низкой температуре ночью —
слабое освещение как теневые растения.
Другие советские исследователи в своих работах, посвященных обосно-
ванию границы леса на Кольском полуострове, отмечают аналогичную
интенсивность фотосинтеза
у Betula папа, Solidago
virgaurea и приблизитель-
но вдвое меньшую у ку-
старничков Vactinium uli-
ginosum, Empetrum и Phyl~
lodoce.
Рассел косвенным спо-
собом определил величину
нетто-ассимиляции на Ян-
24 6 12 18 Часы. -Майене, установив в 50 сре-
занных листьях каждого
Рис. 26. Ассимиляция СО2 Rubus chamaemorus растения содержание крах-
8-9 июля под Мурманском (по Костылеву и др.). мала и сахара и повторив
опыт через 6 дней. Листья,
воткнутые черешками во влажный песок, остались свежими. По увеличе-
нию концентрации углеводов была вычислена величина нетто-ассимиляции
Oxyria digyna, которая составила 0,3 г!дм1 2 за неделю. В листьях
и корнях Oxyria, Polygonum viviparum и Ranunculus glacialis содержалось
значительное количество ассимилятов. Устьица растений были постоянно
открытыми, причем в большей или меньшей степени в зависимости от
количества света. Результаты измерений позволяют сделать вывод о том,
что углеводов арктические растения получают достаточно; ограничено,
видимо, поступление азота. Интересно, что содержание углеводов у Oxyria
digyna в Арктике и Каракоруме (Гималаи) на высоте 3750 м оказалось
совершенно одинаковым Г
Годовая нетто-продуктивность растительного покрова в субарктической
зоне шведской части Лапландии под Абиску (вегетационный период
111 дней) определена в 2500 кг/га, или 250 г/ж2. Для сравнения в Англии,
где вегетационный период продолжается 205 дней, она составила
6900 кг/га 2, в северной Аляске (вегетационный период 70 дней) — 830 кг!га,
а в высокоширотной Арктике (вегетационный период 60 дней) — только
30 кг/га 3. Фитомасса арктического ивняка в Гренландии едва достигает
5,5 т/га сухого веса, а, например, 25-летнего букового леса — 63,7 т!га.
Климатические условия, коренным образом отличающиеся от климата
остальной части тундры и характеризующиеся резко выраженной конти-
нентальностью, мы находим в арктической степи восточной Гренландии
> (74° с. ш.), на крайних северных форпостах растительности в северной
Гренландии у залива Гекла (80° с. ш.) и во внутренних частях Ингольфс-
1 R. S. Russel, The effect of arctic and high mountain climates on the carbo-
hydrat content of Oxyria digyna, «J. Ecol.», 36, 1948, p. 91—95.
2 W. H. Pearsall, P. J. Newbould, Production ecology IV. Standing
crops of natural vegetation in the Sub-Arctic, «J. Ecol.», 45, 1957, p. 593—599.
3 L. C. Bliss, A comparison of plant development in microenvironments of
arctic and alpine tundra, «Ecol. Monogr.», 26, 1956, p. 303—337; W. J. Warren,
Arctic plant growth, «Advanc. Sci.», 13, 1956, p. 383—388.
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 25
фьорда (80°30' с. ш.) *. Районы ограничены с востока или юга материковым
ледником и расположены на высоте всего 200 м над уровнем моря либо-
на дне глубоких троговых долин. В период, совпадающий со временем
вегетации, эти районы находятся под влиянием нисходящих ветров (фён),.
поэтому в летние месяцы дождей здесь прак-
тически не бывает, но часты ураганы, пере-
вевающие огромные массы песка 1 2. Годовая
сумма осадков менее 100 мм, в отдельные
годы около 25 мм. Температура летом здесь
выше, чем на побережье с частыми тума-
нами, благодаря чему граница произраста-
ния растительности поднимается до абсолют-
ной отметки 1600 м.
ЗЕМЛЯ ПИРИ (9м) -15,0° 53
Р и с. 27. Климадиаграмма
Земли Пири, северная Грен-
ландия; крайне аридный аркти-
ческий климат.
Во внутренних районах Земли Пири
(северная Гренландия) летом 1949 г. про-
водились метеорологические наблюдения
(рис. 27, табл. 3).
Растительность Гренландии сравнитель-
но хорошо изучена. В сухих районах восточ-
ной Гренландии встречаются 129 видов сосудистых растений, а в анало-
гичных условиях северной Гренландии — 98. Но все это виды, обычные-
для Арктики, среди которых произошел естественный отбор засухо-
устойчивых форм. Доминируют Dryas integrifolia, близкая ей D. chamissonisr
Таблица 3
Метеорологические наблюдения на Земле Пири (северная Гренландия)
Май Июнь Июль Август Сентябрь
Средняя месячная тем- пература, °C .... Осадки, мм —8,6 0,2 2,5 6,8 6,0 3,7 3,7 Следы —5,7 0,1
а также Elyna myosuroidesvt образующие дерновины осоки — Carex nardinar
С. misandra и С. rupestris’, злаки почти полностью отсутствуют, лишь
изредка встречаются Роа glauca и Calamagrostis purpurascens. Широко-
распространены Saxifraga oppositifolia и виды Вгауа (крестоцветное),
реже — Salix arctica и как единичные экземпляры — Saxifraga aizoides,
Polygonum viviparum, Silene acaulis, Lesquerella arctica (крестоцветное)
и др. На влажных участках в восточной Гренландии местами преобла-
1 F. Н. Schwarzenbach, Die arktische Steppe in den Trockengebieten
Ost- und Nordgronlands, «Вег. Geobot. Inst. Rubel fur 1959», 1960, S. 42—64. Cp.:
T. W. В о c h e r, Climate, soil and lakes in continental West Greenland in relation to
plant life, «Medd. oin Gronland», 147, № 2, 1949, p. 1—63; Oceanic and continental
vegetational complexes in Southwest Greenland, ibidem, 148, № 1, 1954, p. 1—336;
G. Seidenfaden, Th. Sorensen, The vascular plants of Northeast Greenland,
ibidem, 101, № 4, 1937, p. 1—125.
2 Краткое, очень наглядное описание Гренландского ледника, его климата и вет-
ров читатель найдет в книге: F. Loewe, Das Inlandeis Gronlands, «Umschau», H. 16,
1966, S. 532—538.
26
Глава 1
дает Arctous alpina, тогда как Betula папа и Vaccinium uliginosum зани-
мают резко подчиненное положение.
В наиболее типичном развитии эта растительность представлена на
известняках и доломитах (освещаемые солнцем склоны), в котловинах,
наполненных песком и гравием, а также на речных террасах. Зимой здесь
скапливается навеваемый ветром снег, который весной быстро сходит —
и талая вода просачивается в почву. Поверхность почвы, однако, быстро
высыхает, но в глубине вода, вероятно, скапливается над слоем мерзлоты,
и растения используют ее в течение всего лета. В отличие от других райо-
нов высокой Арктики почва оттаивает здесь на глубину 70—100 см. Мерзлот-
ные явления почти не наблюдаются. У растений формируются мощные
стержневые корни. Например, у средних размеров экземпляра Вгауа
purpurascens корни достигают 1,1 м в длину, что в мерзлой почве было бы
невозможно. При резком недостатке влаги листья свертываются. В разгар
лета преобладает соломенно-желтый или бледно-бурый аспекты.
Таким образом, картина в целом напоминает условия, существующие
в теплых аридных областях, там, где скалистые и песчаные местообитания
обеспечивают растениям наиболее благоприятный водный режим. Сходство
подчеркивают образующиеся в засушливых районах Гренландии выцветы
соли на поверхности почвы (вследствие сильного испарения) и щелочная
реакция почв. На берегах высыхающих водоемов содержание соли в почве
может быть столь высоким, что здесь, в глубине суши, далеко от морского
побережья, успешно развиваются галофиты Carex maritima, С. lachenalii,
Armeria maritima ssp. labradorica. Источником соли, видимо, здесь, как
и во многих пустынях, являются подстилающие осадочные породы морского
происхождения. Из-за сухости климата даже в районах, сложенных кри-
сталлическими породами, почвы не обладают кислой реакцией, хотя
значения pH в них ниже, чем в почвах областей распространения известня-
ков. Поэтому мы находим здесь сходную растительность с той лишь разни-
цей, что добавляются виды, не связанные с определенным типом почвы
(Woodsia glabella, Potentilla nivea, Trisetum spicatum, Papaver radiatum
и др.), а также слабоацидофильные растения (Saxifraga nivalis, S. cernua,
S. caespitosa, Draba hirta, Campanula uniflora, Luzula confusa, Carex capil-
laris и др.). Лишь в восточной и северной Гренландии на известняках
растут Вгауа humilis, Carex giaciaiis, Saxifraga aizoides, Epilobiumarcticum.
Бессточные соленые озера в континентальной части западной Грен-
ландии (Bocher, 1949), характеризующиеся значением pH до 9,5, отно-
сятся к карбонатному или карбонатно-хлоридному типу засоления. Высо-
кая концентрация магния приближает эти водоемы к типу горько-соленых
озер; содержание калия в них выше, чем в большинстве других соленых
озер.
Если не считать различий, обусловленных разным широтным поло-
жением, то условия в континентальных районах Гренландии в некотором
•отношении напоминают обстановку засушливых нагорий Центральной
Азии.
В заключение приведем перечень некоторых фитоценологических
работ, посвященных арктическим и аркто-альпийским районам (эти работы
содержат также списки дополнительной литературы).
По Скандинавии назовем труды:
О. I. R onning, The vegetation and flora north of the arctic circle, «Tromso
Museums Skr.», 8, 1960, p. 50—72.
N. P о 1 u n i n, Plant succession in Norwegian Lapland, «I. Ecol.», 24, 1926,
ip. 372—391.
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 27
R. N ord h a gen, Die Flora und Vegetation des Sylenegebiets, «Norske Vidensk.
Akad. Skrift.», 1928, 1928.
0. Jjaerevoll, The plant communities of the Scandinavian alpine snow beds,
K. Norske Vidensk. Selskeb. Skrift., Trondheim, 1956.
Интересны также соответствующие разделы «The Plant cover of Sweden» в «Acta
Phytogeographica Suecica», 50, 1965, Uppsala.
Аркто-альпийская высокогорная растительность Северного Урала подробно
описана в книге: П. Л. Горчаковский, Высокогорная флора и растительность
Урала, Свердловск, 1966.
Растительность Исландии охарактеризована в работах:
Н. Hesmer, Waldvernichtung und Waldschaffung auf Island, «Forstarch.», 31,
1966, S. 109—119.
H. M. Hansen, Studies in the vegetation of Iceland, The botany of Iceland,
III Part, I, 10, Copenhagen, 1930.
F. W. A n d e г s e n, P. F a 1 c k, Observation on the ecology of the Central Desert
of Iceland, «I. Ecol.», 23, 1935, p. 406—421.
P. F a 1 c k, Further observation on. the ecology of Central Iceland, «I. Ecol.», 28,
1940, p. 1—41.
С растительностью Шпицбергена и Ян-Майена знакомят работы:
V. S. S u m m е г h а у е s, С. S. Elton, Further contributions to the ecology
of Spitzbergen, «I. Ecol.», 26, 1938, p. 193.
R. S. Russel (et al.), 1940. См. примечание на стр. 22.
Экология растений арктической и аркто-альпийской зон Северной Америки, вклю-
чая Гренландию, кратко описана в работе Кнаппа: R. К napp, Die Vegetation Nord-
und Mittelamerikas (см.: H. Walter, Vegetationsmonographien der einzelnen Gro6-
raume, Bd. I, Stuttgart und Jena, 1965). В работе приведена также соответствующая
литература.
ПРОДУКТИВНОСТЬ И КРУГОВОРОТ ВЕЩЕСТВ
В АРКТИЧЕСКОЙ ЗОНЕ
Плотность растительного покрова в арктической зоне в направлении
к северу все более уменьшается, пока наконец у границы сплошного
распространения снега и льда жизнь не прекращается совсем и продуци-
рование органического вещества не падает до нуля. Поэтому данные
табл. 4 характеризуют области Арктики с менее суровыми условиями
жизни, причем мы различаем истинную тундру, за пределами распростра-
нения кустарничков, кустарничковую тундру и лесотундру. Средние
величины получены Родиным с использованием всех имеющихся данных
Приведенные в таблице цифры позволяют сделать некоторые выводы.
Продуктивность и прирост органического вещества, в том числе в форме
подстилки, в Арктике невелики. Так же мало содержание золы. По этой
причине и круговорот зольных элементов и азота протекает несколько
замедленно. Тем не менее содержание азота по сравнению с количеством
зольных элементов весьма значительно. Среди последних преобладает
калий, а не кальций. Это и понятно, поскольку процесс химического вывет-
ривания в Арктике отличается заторможенностью, в то же время почвы
сильно выщелочены и бедны известью. Недостаток извести наряду с низ-
кими температурами затрудняет образование в почве мягкого гумуса, или
мулля. Гумификация, скорее, связана с образованием фульвокислот,
благодаря чему облегчается перемещение по почвенному профилю полу-
торных окислов. В подстилке накапливаются кремнекислота и алюми-
ний. Поглощающая способность почвы мала.
1 Л. Е. Родин, Н. И. Ба з и л е в и ч, Динамика органического вещества
и биологический кругооборот зольных элементов и азота в основных типах раститель-
ности земного шара, Л., 1965.
28
Глава 1
Таблица 4
Продуктивность и круговорот зольных элементов и азота
в арктической зоне (по Родину и Базилевич)
Показатель Тундра Лесотундра
арктическая кустарнич- ковая ельник- зеленомошник
Биомасса, т/га 5,0 28,0 137,26
в том числе:
зеленые части 1,0 3,2 8,86-
многолетние надземные части 0,5 1,7 99,23-
корни 3,5 23,1 29,17
Прирост (первичная продук-
т и в н о с т ь), т/га 1,0 2,38 5,62
в том числе:
зеленые части 0,28 0,95 4,04
многолетние надземные части 0,02 0,03 0,66
корни 0,70 1,40 0,92
Опад, т/га 0,95 2,27 5,31
в том числе:
зеленые части 0,26 0,90 3,84
многолетние надземные части 0,01 0,02 0,57
корни 0,68 1,35 0,90
Истинный прирост, т/га 0,05 0,11 0,31
Подстилка, ml га 0,35 83,0 Не о пр.
Химические элементы в био-
массе, кг!га 159,0 901,0 143 □,0
в том числе:
в зеленых частях 56,4 110,0 281,0
в многолетних надземных частях .... 8,3 26,0 576,0
Потребляется приростом химических элемен-
тов, кг/га 38,0 111,0 166,0
в том числе:
зелеными частями 18,9 47,0 138,0
Возвращается с опадом химических элемен-
тов, кг/га 37,0 108,0 157,0
Удерживается химических элементов в истин-
ном приросте, кг/га 1,0 3,0 9,0
Химические элементы в подстилке, кг/га . . 280,0 4200,0 Не опр.
Возвращается с опадом азота, кг!га 20,0 51,0 79,0
в том числе:
от зеленой части 8,0 20,0 63,0
Возвращается с опадом зольных элементов,
кг/га 17,0 57,0 78,0
Возвращается с опадом органогенов, кг/га . . 10,0 37,0 52,0
Возвращается с опадом био галогенов, кг/га 0,7 3,0 Не опр.
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 29
ВЗАИМОСВЯЗИ ЖИВОТНОГО МИРА
И РАСТИТЕЛЬНОГО НО ЕРОВ А ТУНДРЫ
Тундра — одно из немногих обширных пространств земного шара,
где население почти отсутствует и где воздействие человека проявляется
лишь ограниченно, отчего здесь еще сохранился первоначальный живот-
ный мир. Поэтому особый интерес представляет возможность изучить
в таких естественных условиях характер взаимосвязей, существующих
между животными и растениями тундры. В качестве объекта изучения
выбрана тундра Средней и Восточной Сибири г.
Выше уже отмечалось, что насекомые играют известную роль в опыле-
нии цветков в тундре. Тем не менее опыление многих энтомогамных расте-
ний тундры осуществляется главным образом ветром. Широко распростра-
нены также различные формы самоопыления.
Известное значение имеет в тундре, как и других местах, почвенная
фауна. Однако здесь мы остановимся только на роли позвоночных живот-
ных, которых в других зонах человеку редко удается свободно наблюдать.
Характерно, что большинство позвоночных животных зимой уходят
из тундры и мигрируют либо к югу, в лесотундру, либо, как почти все
птицы,— далеко на юг. В тундре перезимовывают лемминги (Lemnus
obensis и Dicrostonyx torquatus), в горной тундре Якутии и на Чукотском
полуострове также суслики и сурок (Citellus undulatus и Marmota сат-
tschatica), песец (Alopex lagopus) и снежная сова (Nyctea scandica) 1 2. Песцы
и совы обычно покидают самые северные, бедные добычей районы, так что
собственно постоянными обитателями тундры являются грызуны. Летом
в отличие от зимы тундра изобилует животными, здесь также гнездится
бесчисленное множество водоплавающей птицы.
Лемминги не впадают в зимнюю спячку, не делают они и запасов
пищи, а живут и даже размножаются под тонким, но твердым снежным
покровом. Особенно плотно заселены южные склоны с легкими и хорошо
дренируемыми почвами. Зимой они укрыты достаточно толстым слоем
снега. Распространены лемминги до побережья Северного Ледовитого
океана. Несмотря на небольшие размеры (вес около 50 г), потребность
-леммингов в пище очень велика: за сутки они съедают в 1,5—2 раза больше,
чем весят сами.
За год лемминг поглощает 40—50 кг растительной массы. Зимой
он устраивает гнездо из побегов осоковых вблизи своего пастбища, раз-
меры которого для каждой семьи колеблются от 100 до 200 ж2. В эту
пору года зверек питается зелеными частями Carex stans и Eriophorum,
а также злаками или листьями Dry as и Sieversia (Novosieversia), добывая
их из-под снега; не пренебрегает он и молодыми побегами Salix, карликовой
березы (Betula exilis) или Cassiope tetragona, верхушками Polytrichum
strictum. Однако основной пищей грызуну служат почки возобновления
осоковых.
Колонии леммингов на Таймырском полуострове располагаются
в канавоподобных углублениях или на склонах ручьев и занимают площадь
до 1—1,5 га, на которой стравливается около 90—94% растений. На таких
участках Eriophorum angustifolium зацветает лишь в те годы, когда числен-
1 Б. А. Тихомиров, Взаимосвязи животного мира и растительного покрова
тундры, М.—Л., 1959.
2 В тундре Гренландии и Канады перезимовывает также мускусный бык (Qvibos
.moschatus).
30
Глава 1
ность леммингов резко сокращается. Максимум размножения их происхо-
дит в среднем каждые 3 года.
Сухие части растений лемминги не трогают. Весной это «сено» (около
1—2 т!га) талые воды смывают, нагромождая из них большие валы;
со временем формируется микрорельеф из торфяных кочек. Между кустар-
ничками бросается в глаза лабиринт из ходов, маркирующих зимние
квартиры леммингов. Общая длина их на участке 100 м2 достигает 21 —
329 л?; из общей площади, занимаемой колонией, на ходы приходится
1—20%. Отсюда ясно, насколько значительные изменения структуры
растительного покрова могут быть вызваны жизнедеятельностью леммингов.
Покинув зимние гнезда, лемминги переходят на более высокие места
и выкапывают себе новые норы, выбрасывая при этом до 250 кг земли
на 1 га площади. Таким путем возникают нарушенные местообитания,
занимаемые характерным растительным сообществом, которое, однако,
вскоре вытесняется видами устойчивого сообщества, более соответствую-
щего данному местообитанию. Если представить себе, что это воздействие
леммингов продолжается тысячелетиями, то легко понять, каким важным
экологическим фактором оно является. То же самое можно сказать и о сус-
ликах, обитающих в тундре на северо-востоке Сибири. Их сложные под-
земные сооружения с многими входами и выходами можно найти повсюду
на склонах вдоль водотоков. Особенно густо заселены «окрестности»-
близ термальных источников на Чукотском полуострове. Эти сооружения
также несут на себе определенный растительный покров. Зиму суслики
проводят в спячке, в которую впадают, как только температура понижается
до 10—20° мороза, и просыпаются с первыми теплыми днями, но до начала
снеготаяния.
Не меньшее воздействие на растительность оказывают водоплавающие
птицы, особенно гуси (виды Anser и Branta). Прилетают они ранней весной,
когда еще лежит снег, и добывают из-под него зеленые части растений.
Предпочитают почки и молодые листья видов Oxytropis, но употребляют
и их подземные органы. Позже основной пищей гусям служат содержащие
крахмал корневища Eriophorum angustifolium. которые они выклевывают
из почвы. Растительный покров нарушается при этом на 50—80%. Обна-
жившаяся почва покрывается плотным слоем мха и на пологих склонах
особенно легко сползает, что снова ведет к нарушению растительного
покрова; возникшие в результате солифлюкции небольшие валообразные
возвыщения зарастают Dryas.
Прилетают гуси двумя волнами. Ранней весной появляются взрослые
птицы. Занятые выведением потомства, они лишь незначительно изменяют
растительный покров. Несколько позже, по окончании ледохода на реках,
прилетают более многочисленные стаи еще неполовозрелых птиц, срезаю-
щих растительность по берегам озер словно бритвой. Гуси, сбивающиеся
в стаи перед отлетом, на значительной площади общипывают все цветоч-
ные почки, так что цветения на следующий год не происходит.
Гораздо меньшие изменения претерпевает растительность там, где
гнездятся белые куропатки (Lagopus lagopus и L. mutus).
Места гнездовий и сбора птиц перед отлетом, естественно, хорошо-
удобряются. На таких участках можно найти многочисленные нитрофиль-
ные виды, например Rhodiola arctica, Stellaria humifusa, но также Pole-
monium, Erysimum, Myosotis, Draba, Papaver и др.
Неотъемлемый обитатель тундры — северный олень (Rangifer taran-
dus). Покидает он тундру^зимой лишь тогда, когда снежный покров ста-
новится слишком плотным и добыть из-под него пищу оленю трудно. Пере-
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 31*
зимовывают в тундре только отдельные небольшие группы животных,
но и они перекочевывают в гористые районы в поисках не покрытых
снегом участков.
Число диких северных оленей все больше сокращается, но увели-
чивается количество домашних; однако еще известны обширные терри-
тории, где дикие олени преобладают, как это иллюстрирует табл. 5.
Таблица 5
Число северных оленей на 100 н.н- в евразиатской тундре
Район Дикие Домашние Всего
Северный Таймыр 36 7,4 43,4
Северная Якутия 28,6 10,8 39,4
Северная Европа . . 0,4 72,2 72,6
Чукотский полу-
остров 0,1 67,7 67,8
Летом северные олени пасутся поодиночке, поэтому большого вреда
растительному покрову они не приносят. Осенью же, когда олени собира-
ются в стада, насчитывающие сотни и даже тысячи голов, вытаптывание
в местах передвижения стад становится заметным. Выпас ведет к умень-
шению участия лишайников и кустарников, к образованию [луговых
сообществ с Deschampsia arctica, D. caespitosa, Роа alpigena и P. pra-
tensis. Об отступании границы леса вследствие повреждения молодого-
подроста деревьев оленями, которые скусывают побеги и обгладывают
кору, мы уже говорили.
Хищники, такие, как песец, оказывают на растительность намного-
меньшее и, скорее, косвенное влияние. Они размножаются в те годы,
когда резко увеличивается количество леммингов, и число их сокращается,,
когда численность леммингов убывает. Почва вокруг песцовых нор особенна
сильно удобряется и поэтому обычно покрыта пышным разнотравьем.
Часто норы сплошь зарастают Artemisia tilesia и Cerastium maximum.
Остальные хищники более редки, и их воздействие на растительный мир-
едва заметно.
^АЛЬПИЙСКАЯ РАСТИТЕЛЬНОСТЬ
В Евразии высокие горы умеренной зоны расположены преимуще-
ственно между 40 и 50° с. ш. В Северной Америке они, напротив, бе&
перерыва простираются в северном направлении, вплоть до арктической
зоны. Горы Скандинавии, Урал и восточносибирские горы непосредственно-
граничат с арктической зоной, и их альпийский пояс существенно не от-
личается от тундры.
Наиболее подробно в геоботаническом отношении изучены Альпы х.
Поэтому мы рассмотрим их прежде других.
1 См. новейшую короткую сводку Элленберга в «Phytologie», Bd. IV, 2, 1963;.
фундаментальными работами или монографиями, посвященными отдельным районам,
являются: С. Schroter, Das Pflanzenleben der Alpen, Zurich, 1926; H. Brock-
mann-J erosch, Die Vegetation der Schweiz I, «Beitr. Geobot. Landesaufn.
Schweiz», 12, 1925—1929; J. Braun-Blanquet, H. J e n n y, Vegetationsgliede-
rung und Bodenbildung in der alpinen Stufe der Zentralalpen, «Denkschrift Schweiz.
32
Глава 1
Рис. 28. Pinus montana (темные участки на склонах) на горе Хейтер-
ванд в западном Тироле (фото Фабера).
Как мы уже видели, бореальная зона хвойных лесов весьма посте-
пенно через лесотундру переходит в арктическую зону. Альпийский пояс
от лесного также отделяет промежуточная зона — полоса «борьбы за суще-
ствование» у верхнего предела распространения древесных пород. С одной
стороны, это пояс криволесья и стлаников (Pinus montana, рис. 28; Alnus
viridis, Juniperus communis ssp. папа, на Кавказе также Betula, рис. 29),
который еще может быть отнесен к субальпийскому поясу, с другой —
пояс кустарничков (виды Rhododendron и Vaccinium), помещаемый уже
в нижний альпийский пояс. Отметим, что именно эта полоса «борьбы
за существование» особенно сильно изменена и расширена деятельностью
человека.
Выше по склонам гор альпийский пояс столь же постепенно переходит
в нивальный пояс. Последний простирается через снеговую линию до высо-
чайших горных вершин, где растут лишь единичные цветковые растения,
мхи, лишайники и водоросли (рис. 30) Ч
Для альпийского пояса в отличие от зоны тундры характерна скучен-
ность всего живого на небольших пространствах. Обширных ровных пло-
щадей здесь нет, поэтому мозаичность структуры распределения расти-
- Naturf. Ges.», 63, 1926, S. 183—349; Н. G a m s, Von den Follateres zur Dent de Moreles,
«Beitr. Geobot. Landesaufn. Schweiz», 15, 1927; E. Schmid, Erlauterungen zur Ve-
getationskarte der Schweiz, «Beitr. Geobot. Landesaufn. Schweiz», 39, 1961; R. S char-
fetter, Das Pflanzenleben der Ostalpen, Wien, 1938; E. A i c h i n g e r, Vegetations-
kunde der Karawanken, Pflanzensoziol. Bd. 2, Jena, 1933; H. G a m s, Die Pflanzenwelt.
Cm.: Landes- u. Volksk. Vorarlberg, Bd. I, Innsbruck, 1961, S. 135—172; P. О z e n d a,
Perspectives nouvelles pour I’etude phytogeographique des Alpes du Sud, Docum. Carte
Veget. Alp. IV, Grenoble, 1966.
1 H. Reisigl, H. Pitschman, Obere Grenzen von Flora und Vegetation
in der Nivalstufe der zentralen Otztaler Alpen (Tirol), «Vegetatio», 8, 1958, S. 93—128;
Zur Abgrenzung der Nivalstufe, «Phyton», 8, 1959, S. 219—224.
Рис. 29. Betula pubescens var. carpatica и В. raddeana c Rhododendron
caucasicum в подлеске выше границы древесной растительности; северный
склон центрального Кавказа, высота 2200 м (фото Щукина).
Верхняя граница
отЗельных узанерогамоб
Верхняя граница____—
ЗбуЗольных поЗушеч-
никоб
Мхи и лишайники
СНЕГОВАЯ ЛИНИЯ
Ж долин ые-
ушечники-
и ковры
„Пионерные“луга—>
Альпийский
луг с сомкну-
тым траво-
стоем
Отдельные
уродливые
деревья
Кустарнички
ГРАНИЦА ЛЕСА
Криволесье —
Разреженный^
лес
Сомкнутый выс
ствольный лес
Луга на путях
лавин--------
yMVВЫСОТНЫЕ ПОЯСА
ВЕРХНИЙ.
СРЕДНИЙ
НИВАЛЬНЫЙ
НИЖНИЙ
СУБНИВАЛЬНЫИ
ВЕРХНИЙ
СРЕДНИЙ
АЛЬПИЙСКИЙ
- НИЖНИЙ
ии-
СУБАЛЬПИИСКИИ
ВЕРХНИЙ,
ГОРНЫЙ
Рис. 30. Схема вертикальной поясности Альп (по Рейзиглю и Пичману,
из «Phytologie», Bd. IV, 2).
3-0434
34
Глава 1
СКАЛА
Раститель-
\ность
vkochineii
Лишайники
Растительность
В трещинах СНЕЖНЫЙ ПОКРОВ
скал — зимой
— РАННИМ ЛЕТОМ
АЛЬПИИСКИИ ПОЯС
ВЕТЕР
СнеговаяТ
_досы77йл0жбйнка
Кустарничка- Сильное
бая пустошь /бозбейстбие
' ветра „ „
Первичные луга
Паломощные почбы..
Мощные0 Низинное
почвы оолото
Расти-
тельность-
ключевин
грубый гумус
ГУ МУ СИ РОВ А ИНЫЕ
ПОЧВЫ
СМОРЕНА
ТОРФ
Рис. 31. Мозаичный характер распределения типов местообитаний и раститель-
ности в альпийском поясе на небольшой площади, схема (по Элленбергу, из
«Phytologie», Bd. IV, 2).
источников
тельности и взаимное проникновение отдельных сообществ друг в друга
выражены значительно более резко (рис. 31), чем в арктической зоне.
Что касается арктической границы леса, то легко заметить, что-
в областях с океаническим климатом она располагается значительна
южнее, а в районах с континентальным климатом — намного севернее.
Нечто похожее мы наблюдаем в горах. Граница распространения древес-
ной растительности в Центральных Альпах с их более континентальным
климатом лежит приблизительно на 500—600 м выше, чем на северной
окраине Альп, и выше, чем на их южном склоне; правда, следует учиты-
вать, что во всех трех случаях ее образуют разные виды деревьев. В соот-
ветствии с различными типами климата мы выделяем в Альпах три типа
вертикальной поясности (табл. 6).
Таблица 6
Типы климата и вертикальной поясности растительности
в Альпах (схема)
I Гельветский тип (северная окраина Альп) II Пеннинский тип (Центральные Альпы) III Инсубрийский тип (южная окраина Альп)
Поясность Альпийский пояс Ель Бук Дуб Альпийский пояс Сосна кедровая — лиственница Ель Сосна Отсутствует Альпийский пояс Бук Каштан — дуб пу- шистый (вечно- зеленый)
Климат Умеренный цен- тральноевро- пейский Континентальный (сухой) Субсредиземномор- ский
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 35
Более высокое положение границы древесной растительности в Цент
ральных Альпах нельзя, однако, объяснять только сменой древесных
пород, поскольку снеговая линия проходит здесь также выше; почти
повсеместно это превышение составляет 900 м. Большая масса горного
массива обусловливает более сильное нагревание поверхности почвы
и воздуха, особенно в летние месяцы, и тем самым создает благоприятные
условия для роста деревьев. Мы уже показали (рис. 18), что рост деревьев
у границы древесной растительности в основном определяется количеством
летнего тепла. В текущем столетии в период с 1910 по 1930 г. летние
месяцы в Фенноскандии близ арктической границы были особенно теплыми.
Это проявилось в усиленном росте деревьев в толщину, повышении урожая
зерновых в северной Финляндии, в распространении многих видов к северу
и в ряде других явлений биологии растений, описанных Эркамо Ч
В Альпах также доказано существование в прошлом более теплого периода,
вызвавшего отступание ледников. Континентальность климата у границы
древесной растительности повышает конкурентную мощность сосны кедро-
вой {Pinus cembra либо Р. sibirica) и лиственницы (Larix decidua, или
L. sibirica, или L. dahurica), поэтому в климатически наиболее экстре-
мальных лесных районах Центральных Альп и Сибири эти виды начинают
доминировать.
Климатические условия, существующие у альпийской границы дре-
весной растительности, подробно описаны выше. Их особенности с.увели-
чением высоты проявляются все более резко. Усиливается инсоляция;
рассеянная радиация, напротив, уменьшается, и температурные конт-
расты между солнечной и теневой сторонами, даже разделенными лишь
небольшим пространством, возрастают (табл. 7).
Таблица 7
Величина инсоляции в ясные дни в горах на разной высоте
(по Шрётеру) 1
Пункт измерений Температура на солнце Температура в тени Разность
2980 м, Дьяволецца 59,5 6,0 53 i 5
1800 м, Понтрезина 20 м, Уитби, Анг- 44,0 26,5 17,5
лия 37,8 32,7 5,1
1 Измерения произведены с помощью термометра с черным ртут-
ным шариком (в вакууме).
Сильное прогревание почвы, освещаемой солнцем, позволяет неко-
торым геофитам (как Crocus'), генетически представляющим собой среди-
земноморские элементы, цвести тотчас после стаивания снега (рис. 46).
С высотой сокращаются в размерах плакорные участки, возрастает кру-
тизна склонов и, следовательно, обостряются противоположности, обус-
ловленные экспозицией. Уменьшение давления воздуха для растений,
видимо, не имеет значения и вызывает лишь понижение парциального
1 V. Е г k a m о, Untersuchungen fiber die pflanzenbiologischen und einige andere
Folgeerscheinungen der neuzeitlichen Klimaschwankung in Finnland, «Ann. Bot. Soc.
«Vanamo», 28, № 3, Helsinki, 1956.
3*
36
Глава 1
давления С02 (на высоте 2000—2500 м оно составляет около 0,35—0,4 мг/л).
В отличие от условий среды в Арктике альпийский пояс Альп характе-
ризуется некоторыми особенностями:
1) регулярной сменой дня и ночи и связанными с этим резкими
колебаниями температуры в течение суток;
2) намного более интенсивной радиацией во все времена года;
3) отсутствием многолетней мерзлоты, поскольку почва под снежным
покровом даже зимой охлаждается ненамного ниже нуля, летом же в сол-
нечные дни она хорошо прогревается;
4) значительно большим количеством осадков, благодаря чему снеж-
ный покров зимой в Альпах во много раз толще и длительность «бесснеж-
ного» периода даже на весьма ограниченных участках колеблется в более
широких пределах;
5) преобладанием каменистых почв, частой сменой горных пород
и более резкими различиями флоры и растительности на карбонатных
и кристаллических породах.
В условиях значительной радиации различия в микроклимате при-
земного 'слоя воздуха и на высоте 2 м над поверхностью почвы еще более
разительны. Поэтому альпийские растения прилегают к земле, пожалуй,
еще плотнее, чем растения Арктики (рис. 32—34). Однако карликовый
рост альпийских растений отнюдь не является приспособительной модифи-
кацией, а зафиксирован генетически и, видимо, обусловлен отбором соот-
ветствующих мутантов. Температура самих растений на некоторых место-
обитаниях может более резко отличаться от температуры воздуха на высоте
2 м, чем от температуры воздуха на высоте листьев. Интересные сравнитель-
ные микроклиматические измерения и фенологические наблюдения Блисс
произвел в северной низкой части Аляски (69°22' с. ш., 100 м абс. высоты)
и в горах Сноуи-Рейндж (Вайоминг) (41°20' с. ш., высота 3300 м). Он при-
шел к выводу, что из двух местообитаний Арктика менее сурова, чем
высокогорье, но последнее гораздо суше и характеризуется очень боль-
шими колебаниями температуры; особенно велика разность между суточ-
ными максимумами и минимумами. Измерения на высотах 3800 и 4300 м
(горы Маунт-Эванс в Колорадо, США) проведены Солсбери и Спомме-
ром х. Граница леса проходит здесь на высоте 3500 м. Измеренная темпе-
ратура листьев, освещенных солнцем, была на 22° выше температуры
воздуха, а в тени скалы — на 3,5° ниже. Экстремальные измеренные темпе-
ратуры составляли +32,5 и 0°. Особенно сильно перегревался Geum turbi-
natum. Oxyria digyna была теплее окружающего воздуха лишь на 5—10°,
а растение-подушйа Arenaria obtusiloba — на 12°. В общем подушковидные
формы облучались интенсивнее, нежели пряморастущие растения. Непо-
средственные измерения температуры листьев показали, что в альпийском
поясе сведения о климате, получаемые на основе метеорологических данных,
еще менее, чем в Арктике, отражают истинный климат растений.
Места произрастания альпийских видов в Альпах в общем расположены
по высоте значительно ниже, чем в горах Колорадо, однако эту общую
закономерность нарушает Ranunculus giaciaiis, который поднимается
по склонам до отметки 4275 м (рис. 35). Суммарная радиация в Альпах
изменяется в зависимости от сезона. Если принять сумму общей радиации
1 L. С. Bliss, A comparison of plant development in microenvironmets of
arctic and alpine tundra, «Ecol. Monogr.», 26, 1956, p. 303—337; F. B. S alisbu r y,
G. G. S p о m m e r, Leaf temperature of alpine plants in the field, «Planta», 60, 1964,
p. 497—505; D. M. G a t e s, R. Janke. The energy environment of the alpine tundra,
«Oecologia Plant.», 1, 1966, p. 39—61.
Рис. 32. Шпалерный кустарник Salix retusa на известняковых скалах
в южном Тироле, высота около 2450 м над уровнем моря. В центре Carex firfita
(фото Шенка).
Рис. 33. Viola alpina на плато Раксальпе
(200Q м), Нижняя Австрия (фото Юраски).
38
Глава 1
Рис.
34. Hutchinsia alpina, там же, см. рис. 33.
Р и с. 35. Ranunculus glacialis в верхнеальпийском поясе
Альп (фото Геллера).
в Вене (202 м над уровнем моря) за 100%, то в Альпах на высоте 2000 м,
при средней облачности, она составит: зимой приблизительно 200—250%
(на высоте 2600—3000 м даже 250—300%), а в разгар лета всего лишь 110—
120 или 120—130%. При безветрии и в Альпах температура растений
в ясные дни может быть выше температуры воздуха на 10°. Каждый альпи-
нист знает такие теплые ниши на солнечных скалистых склонах, которые
можно обнаружить даже на больших высотах. Напротив, при сильном
ветре или в пасмурные дни разность температур невелика Ч Существенно
важны для жизни растений также самые низкие зимние температуры.
ХЕ. Winkler, Beitrage zur Klimatologie hochalpiner Lagen der Zentralalpen,
«Вег. Naturw.-Med. Ver. Innsbruck», 53, 1963, S. 209—223.
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 39
Морозы в Альпах не столь сильны, как в Арктике с ее длинной зимней
полярной ночью, и держатся недолго, так как днем воздух обычно теплеет.
Средняя минимальная температура самого холодного месяца в Альпах
на высоте 2000—3000 м не падает ниже —10° (—15°), абсолютный минимум
колеблется от —28 до —37°. Наинизшая температура в Центральной Европе
—51° измерена в Восточных Альпах у Лунца, на высоте всего 1270 м,
но в долине, в которой скапливается холодный воздух. Здесь при силь-
ном излучении даже в разгар лета бывают ночные заморозки.
ЭК ОЛОГО-ФИЗИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
В АЛЬПИЙСКОМ ПОЯСЕ
Морозоустойчивость альпийских кустарничков характеризуется
таким же годовым ходом и с ней связаны такие же явления закаливания
(рис. 36) и изнеживания, с которыми мы уже познакомились на примере
ели (ср. т. II, рис. 289). Хотя кустарнички поднимаются в горах намного
выше ели и кедровой сосны, однако они менее морозоустойчивы (рис. 37) х.
Это легко понять, если учесть, что кустарнички проводят зиму под снегом
и, следовательно, не испытывают влияния отрицательных температур.
Лишь Loiseleuria procumbens, растущая на открытых ветру участках
(рис. 38) и обычно не покрываемая снегом, почти столь же морозоустой-
чива, как Pinus cenibra. На наветренных склонах она, как правило, не под-
вержена воздействию даже самых низких температур, однако на такого
рода местообитаниях растение может испытать физиологическую сухость.
Но так как Loiseleuria плотно прилегает к почве, то ее листья, задержав-
шийся между ними снег и верхние сантиметры почвы зимой каждый раз,
когда на них падают прямые солнечные лучи, оттаивают, и листья и неглу-
бокая корневая система получают возможность поглотить некоторое коли-
чество влаги. То, что эта влага необходима растению, показали опыты:
срезанные ветки Loiseleuria, несмотря на ксероморфное строение, засы-
хают, если их вынести зимой из помещения, уже через 15 суток 1 2. Анало-
гично ведут себя Rhododendron, Vaccinium и Calluna" различие, однако,
в том, что эти виды перезимовывают под снегом в состоянии практически
полного насыщения влагой. Гораздо чаще они страдают от мороза на более
низких позициях, особенно если отсутствует снежный [покров, защи-
щающий растения от потерь воды.
Мы подошли к вопросу об особенностях водного режима альпийских
видов. Обильные и частые осадки в период после стаивания снега способ-
ствуют тому, что водный баланс альпийских растений практически всегда
уравновешен 3. Сухих местообитаний в альпийском поясе почти не встре-
1 W. Ulmer, Uber den Jahresgang der Frostharte einiger immergriiner Arten der
alpinen Stufe..., «Jb. wiss. Bot.», 84, 1937, S. 553—592; E. Gartellieri, Jahresgang
des osmotischen Wertes, der Transpiration und Assimilation einiger Ericacieen der
alpinen Zwergstrauchheide..., «Jb. Wiss. Bot.», 82, 1935, S. 460—506; A. P i s e k,
R. Schiessl, Die TemperaturbeeinfluBbarkeit der Frostharte von Nadelholzern und
Zwergstrauchern an der alpinen Waldgrenze, «Вег. Naturwiss. med. Ver. Innsbruck»,
47, 1946, S. 33—52. Краткую эколого-физиологическую характеристику растений
альпийского пояса см. в работе: W. Tranquillini, The physiology of plants at
high altitudes, «Ann. Rev. Plant Physiol.», 15, 1964, p. 345—362.
2 W. L archer, Frosttrocknis an der Waldgrenze und in der alpinen Zwergstrauch-
heide, «Veroff, Ferdinand. Innsbruck», 37, 1957, S. 49—81.
3 A. Pisek, E.Gartellieri, Zur Kenntnis des Wasserhaushaltes der Pflanze,
III. Alpine Zwergstraucher, «Jb. wiss. Bot.», 79, 1933, S. 131—190; A. P i s e k, E. В e г -
ger, Kutikulare Transpiration und Trockenresistenz isolierter Blatter und Sprosse,
-«Planta», 28, 1938, S. 124—155; U. Berger-Landefeldt, Der Wasserhaushalt
der Alpenpflanzen, «Bibl. Botanica», H. 115, 1936.
40
Глава 1
Рис.; 36. Интенсивное изменение величины осмотического давления у трех
альпийских кустарничков осенью в процессе закалки (штриховка); верхняя
кривая — повышение осмотического давления у тех же видов зимой вслед-
ствие физиологической сухости при низких температурах (по Пизеку, Зому
и Картельери, из работы Штейнера).
Рис. 37. Вверху: суточные минимальные температуры воздуха близ
Инсбрука (1960 м). Внизу: годовой ход морозоустойчивости некоторых
альпийских кустарничков и для сравнения Pinus cembra. Объяснение
см. т. II, рис. 289 (но Ульмеру, из «Phytologie», Bd. IV, 1).
тишь. Даже на кажущихся сухими скальных поверхностях или каменистых
осыпях, в трещинах скал и между камнями скапливается всегда влажный
мелкозем. Поэтому корни альпийских растений не проникают глубо-
ко \ Только у видов, растущих на щебнистых осыпях, формируется
1 М. Rochow, Wurzeluntersuchungen an subalpinen Grasnarben, «Вег. Geobot.
Inst. Rubel fur 1955», 1956, S. 50—64; A. E. H о 1 c k (et al.), Root habitat of certain
plants of the foothill and alpine belts of Rocky Mts-National Park, «Ecol. Monogr.»,
11, 1941, p. 327—345; A. M. Семенова-Тян-Шанская, Корневые системы
растений субальпийских лугов юга Осетии, «Труды Бот. ин-та им. Комарова, сер.
геоботаника», вып. 5, 1948, стр. 89—119.
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 41
Рис. 38. Размещение растительности в зависимости от высоты снеж-
ного покрова на открытом сильным ветрам бугре у перевала Бернина,
высота 2328 м (по Брокман-Ерошу).
1 — Carex curvula (отрастает поздно); 2 — Vaccinium uliginosum (растения надежно
укрыты снегом); 3 — Juniperus, слой снега тоньше, ощутимо воздействие ветра;
4 — заросль Agrostis alpina", 5 — растения (4) выдуты ветром; 6 — заросль Loiseleu-
ria, едва прикрыта снегом, обычно обнажена (из «Phytologie», Bd. Ill, 1).
широко распространенная корневая система, которая проникает между
свободнолежащими, иногда даже подвижными камнями, обеспечивает
прочное прикрепление растений к субстрату и одновременно закрепляет
осыпь. При этом втягивающие корни развиваются косо вверх по отно-
шению к поверхности осыпи. У растений, обитающих в трещинах скал
(например, Androsace helvetica), обычно образуется толстый стержневой
корень. В критические периоды растения снижают потерю воды, частично
закрывая устьица. Особенно велики расходы воды на транспирацию
у видов, растущих на наветренных участках (например, у Elyna). Однако
выполненные автором измерения величины испарения с помощью эва-
порометра Пише (зеленая бумага, диаметр 3 см) показали, что макси-
мальная интенсивность испарения на высоте приблизительно 2000 м
редко превышает 1 мл/час (рис. 39). Испарение днем (с 8 до 15 часов),
составляющее 7,5 мл/час, значительно превышает величину испарения
в Субарктике под Абиску, где в августе, несмотря на непрерывное дневное
освещение, за 24 часа оно достигло у почвы лишь 3,1—4,1 мл, а на высоте
1 м — 4,1—4,6 мл. Для июня в районе Абиску приводятся максимальные
значения в полдень 0,4—0,6 мл/час и в целом за сутки 5—6 мл. Величина
влаги, транспирируемой альпийским растительным покровом, в расчете
на 1 ж1 2 поверхности почвы также оказалась невысокой. Причина в том,
что сырая масса транспирирующих частей, приходящаяся на 1 м2 поверх-
ности, в альпийском поясе невелика, например на осыпи она составляет
50—60 г/м2, в местах скопления снега — 280 и лишь на кустарничковых
пустошах достигает 500—1000 г/м2. Но даже на последних суммарная
водоотдача за год не превышает приблизительно 200 мм, что составляет
только х/3 количества летних осадков и V4 годовой суммы осадков, выпа-
дающей в относительно сухих Центральных Альпах х. Соответственно
и осмотическое давление у альпийских видов всегда очень низкое 2. Обычно
оно колеблется в пределах 8—12 атм и никогда не достигает 20 атм,
даже у таких ксероморфных видов, как Dry as, Carex firma, Androsace
helvetica.
1 A. Pisek, E. C ar t elli eri, Der Wasserhaushalt einiger Pflanzenvereine,
«Jb. wiss. Bot.», 90, 1941, S. 255—291.
2 A. Pisek, H. S ohm, E.Cartellieri, Untersuchungen iiber osmotischen
Wert und Wassergehalt von Pflanzen und Pflanzengesellschaften der alpinen Stufe,
«Beih. Bot. Zbl.», 52, B, 1935, S. 634—675.
42
Глава 1
.......2 -------4
Рис. 39. Величина испарения в альпийском поясе (массив Дюр-
ренштейн, Нижняя Австрия) в безоблачный, почти безветренный
день (19 сентября).
1 — отвесная южная стена с Potentilla clusiana', 2 — там же, но в расселине
скалы Ranunculus alpestris', 3 — ковер из Dry as octopetala и Carex firma на
поверхности выровненной вершины; 4 — Dryadetum с многочисленными
лишайниками на северном склоне; 5 — Aconitum, Adenostyles и др. в карровой
расселине. Цифры означают температуру воздуха и влажность (Walter, 1927).
Интенсивность испарения на разных местообитаниях зависит от силы
ветра и, следовательно, от экспозиции склона, подобно тому как тесно
связаны друг с другом высота снежного покрова и характер рельефа.
Однако в нашем случае зависимость обратная х.
При изучении экологии альпийских и арктических видов возникает
вопрос, каким образом за столь короткий вегетационный период эти
растения могут продуцировать такое количество органического вещества,
которое оказывается достаточным, чтобы образовались плоды и семена
и были отложены запасные вещества на следующий год. Условия в альпий-
ском поясе несколько менее благоприятны, чем в тундре, ибо, хотя радиа-
ция здесь днем и интенсивнее, но ночи темные. Правда, температура
1 Е. Prutzer, Die Verdunstungsverhaltnisse einiger subalpiner Standorte,
«Mitt. Forstl. Bundes-Versuchsanst.», 59, Wien, 1981. S. 233—256.
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 43
Рис. 40. Интенсивность ассимиляции у Ranunculus giaciaiis (два листа: I и II)
в безоблачный день (7 марта) при максимальной температуре воздуха 17° на
высоте 2600 м под Инсбруком. Характерен большой разброс данных измере-
ний. Кривая средних значений проходила бы параллельно кривой освещен-
ности (по Картельери).
ночью в горах обычно сильно понижается, так что потери на дыхание
относительно невелики. Ассимилируют альпийские виды весьма энер-
гично (рис. 40). Они хорошо используют яркий свет в солнечные дни,
но и в ненастье их продуктивность достаточно высока. За час растения
ассимилируют 18—27, а за сутки 100—300 мг СО2М%1 2. Опыты произво-
дились с Ranunculus giaciaiis (рис. 35), Sieversia reptans и Doronicum
clusii на абсолютной высоте 2600 ж1. Продуцирование органической массы
в горах в общем протекает более интенсивно, чем на низких позициях.
Из вещества, образуемого одним листом за 15 дней, может развиться
новый лист такой же величины. Одного месяца благоприятной погоды
растению было бы достаточно, чтобы отложить запасные вещества на сле-
дующий сезон. Поскольку вегетационный период обычно длится 3 месяца,
то растения успевают накопить большое количество сухого вещества
и обильно плодоносят.
При сравнении поведения популяций растений одного и того же вида
в Арктике и в высоких горах на примере Oxyria digyna удалось доказать,
что в физиологическом отношении они различны 2. Фотосинтез растений
Арктики в фитотронах при низкой температуре характеризуется большей
интенсивностью по сравнению с высокогорными видами. Оптимум нетто-
продуктивности расположен при более низкой температуре, а ее максимум
достигается при меньшей интенсивности светового потока. С другой
стороны, интенсивность дыхания у арктических растений при любой
температуре выше, чем у высокогорных видов. Вместе с тем у популяций
растений из Арктики и из горных районов обнаруживаются определенные
морфологические различия. Их фотопериодизм соответствует продолжи-
тельности дня на естественном местообитании.
На северо-востоке США граница леса проходит намного ниже. Климат
здесь влажный, часты туманы. На горе Вашингтон (1917 м), самой высокой
вершине этого района, выделяют нижний альпийский пояс с Ledum groen-
landicum, Vaccinium uliginosum, V. vitis-idaea, Juncus trifidus и др. (про-
ективное покрытие 75%) и верхний альпийский пояс с Carex bigelowii,
Polytrichum juniperinum, Cetraria islandica и др. (проективное покрытие
1 Е. Cartellieri, Uber Transpiration und Kohlensaureassimilation an einem
hochalpinen Standort, «Sitzber. Akad. Wiss. Wien, math.-nat. KI.», Aft. I, 149, 1940,
S. 95—143.
2 H. A. Mooney, W. D. Billings, Comparative physiological ecology of
arctic and alpine populations of Oxyria digyna, «Ecol. Monogr.», 31, 1961, p. 1—29.
44
Глава 1
60—70%). По интенсивности фотосинтеза и дыхания местные растения
скорее сходны с арктическими видами, нежели с видами Скалистых гор.
Оптимальная температура нетто-ассимиляции для Carex bigelowii рав-
на 10—15°, для других видов 15—20° и только для Geum packii она составля-
ет 20-25° х.
В горах Медисин-Боу (Вайоминг) на высоте 3353 м было определена
также количество продуцированного сухого вещества 1 2. Альпийские луга
на одном из северных склонов с падением 11° состояли: выше снежника —
из Salix cascadensis, Geum turbinatum, Trifolium parryi, Artemisia scopulorumr
Carex drummondiana', ниже его — из Carex elynoides, Deschampsia caespitosa,
Sibbaldia procumbens и др. Зацвели растения через 3 недели после начала
роста, семена созрели через 7—8 недель. При двух укосах среднесуточное
накопление сухой массы составило 0,6 и 1,1 з/ж2 выше снежного пятна
и при более благоприятных условиях ниже снежника — 2,3 и 0,6 з/ж2,
что близко к значениям, наблюдаемым в арктических районах (ср. табл. 4).
Опытным путем была определена также фитомасса альпийских лугов,
(табл. 8).
Таблица 8
Фитомасса альпийских лугов на высоте
3353 м в Вайоминге, США (в г/л«2)
Общая Подземная
Влажное местообитание 1400 1100
Сухое местообитание . . 750 350
Надземная фитомасса большинства альпийских сообществ составляла
100—200 (14—348) з/ж2. Общий прирост за сутки в июне и июле на влаж-
ном местообитании был равен 11,1 г/ж2, а на сухом — 1,2 з/ж2. Опыты
с растениями в фитотронах в свою очередь показали, что интенсивность
фотосинтеза при температурах выше 25° резко понижалась.
В Альпах подробно изучались также условия роста растений в ниж-
нем альпийском поясе. Произрастающие здесь кустарнички отличаются
менее интенсивной ассимиляцией СО2 (ср. Gartellieri, 1935, примечание
на стр. 39). Но так как вегетационный период в нижнем поясе длительнее
Таблица 9
Предельный возраст кустарничков Альп и Пиренеев
(число лет)
Calluna vulgaris...........
Empetrum nigrum ...........
Rhododendron ferrugineum
Vaccinium myrtillus . . . .
Vaccinium uliginosum (пони-
жение) ....................
42 На Памире известны
30 почти 100-летние рас-
88 тения Artemisia
28
8
1 Е. В. Н’а d 1 е у, L. С. Bliss, Energy relationships of alpine plants on Mt.
Washington, «Ecol. Monogr.», 34, 1964, p. 331—357.
2 W. D. Billings, L. C. Bliss, An alpine snowbank environment and its
effect on vegetation, plant development and productivity, «Ecology», 40, 1959, p. 388—
397; D. S с о 11, W. D. В i 11 i n g s, Effects of environmental factors on standing crop
and productivity of an alpine tundra, «Ecol. Monogr.», 34, 1964, p. 243—270.
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 45
и листовая поверхность кустарничков больше, то и величина нетто-про-
дуктивности у них выше, чем у травянистых видов более высоких поясов.
Продолжительность жизни кустарничков относительно велика (табл. 9).
Для флористически различных альпийских лугов с сомкнутым тра-
вяным покровом (Альпы) определена величина годовой нетто-продуктив-
мости (см. «Phytologie», Bd. Ill, 1, S. 416 и приводимую ниже табл. 10).
Таблица 10
Годовая нетто-продуктивность альпийских растительных
сообществ (в г/м2)
Луга с сомкнутым покровом Луга с разреженным покровом
Caricetum ferrugineae . . 276 Dryadeto — Firmetum . . . 91
Seslerieto-semperviretum 180 Salicetum herbaceae 85
Луг с Festuca rubra var. com- 15
mutata 140 Oxyrietum 1
Nardetum 50 Осыпи известняковой гальки
.УСЛОВИЯ СУЩЕСТВОВАНИЯ АЛЬПИЙСКОЙ РАСТИТЕЛЬНОСТИ
В ЗАВИСИМОСТИ ОТ НЕКОТОРЫХ ФАКТОРОВ МЕСТООБИТАНИЯ
Здесь не место останавливаться на вопросах сложного подразделения
^альпийской растительности на отдельные сообщества в зависимости
от высоты и экспозиции, характера почвы и типа горной породы, а также
на особенностях обеспечения альпийских растений влагой, тем более что
все эти вопросы недавно были подробно рассмотрены Элленбергом
(см. «Phytologie», Bd. IV, 2). Познакомимся поближе только с сообществом
снеговых ложбинок. Это знакомство позволит понять, как формируется
растительность в условиях крайне короткого вегетационного периода,
иными словами в условиях неблагоприятного для развития растений
климата.
Особенно хорошо выражены снеговые ложбинки в верхнем альпий-
ском поясе горных цепей Альп, сложенных силикатными породами. Рас-
положены они обычно у подножия северных склонов, там, где зимой
накапливается много снега, медленно тающего летом и постепенно осво-
бождающего почву снизу вверх по склону. Здесь можно наблюдать ряд
зон, которые в направлении к остатку снежника освобождаются от снега
на все менее короткое время (рис. 41). Условия роста на этом местообитании
благоприятны, если не считать краткости «бесснежного» периода и обус-
ловленной талыми водами и северной экспозицией низкой температуры;
почва всегда хорошо увлажнена, относительно богата питательными
веществами, реакция среды слабокислая. Там, где снежный покров отсут-
ствует 3 месяца, формируется нормальный осоковый луг (Curvuletum)
с доминирующей Carex curvula. При сокращении «бесснежного» периода
сначала в большом количестве добавляется Ligusticum mutellina, затем
Gnaphalium supinum, причем одновременно появляется типичный для
снеговых ложбинок вид — Salix herbacea, который вскоре становится
господствующим. Salicetum herbaceae образует собственно сообще-
ство снеговых ложбинок и в крайних случаях довольствуется 8-недельным
периодом без снега. При столь коротком вегетационном сезоне возможности
для продуцирования органического вещества, естественно, весьма огра-
46
Глава?!
а — зимний снежный покров; б — остатки снега в конце июля; 1 — Polytri-
chetum sexangulare; 2 —Salicetum herbaceae; 3 — то же c Gnaphalium supinum',
4 — то же с Ligusticum mutellina', 5 — Garicetum curvulae.
ничены. Следовательно, все органы растения, не участвующие в накоп-
лении сухой массы, должны быть по возможности редуцированы. Преждо
всего это стебель, и особенно одревесневший стебель. У Salix herbacea
эти части столь сильно редуцированы, что почти не поднимаются над
поверхностью почвы, отчего листья растения лежат прямо на земле. Ее
подземные побеги чрезвычайно разветвлены, поэтому листья образуют
над почвой сомкнутый покров. На цветки и плоды расходуется наибольшая
доля органического вещества, поэтому ива начинает цвести только тогда,
когда период без снега после малоснежной зимы или особенно жаркого*
лета длится не менее 3 месяцев. Все другие травянистые виды Salicetum
herbaceae, как Gnaphalium supinum, Alchemilla pentaphylla, Arenaria
biflora, Soldanella pusilia, Sibbaldia procumbens, также отличаются низ-
корослостью, и у них хорошо выражена редукция осевых органов (рис. 42).
Корневая система развита слабо, поскольку почва всегда хорошо увлаж-
нена на всю глубину профиля. В типичные Salicetum herbaceae проникают
также мхи, прежде всего Polytrichum sexangulare-, при еще большем укора-
чивании «бесснежного» периода ни один из представителей покрытосемен-
ных не успевает образовать необходимое для своего существования орга-
ническое вещество в достаточном количестве. Мхи, напротив, ведут
более экономный образ жизни. Им не нужно формировать корней, доля
массы стебля у них тоже невелика, так что ассимилируют практически'
все части растения. Мало вещества расходуется и на образование органов,
размножения и коробочки (ассимилирующей некоторое количество СО2),
а также спор. Поэтому при дальнейшем сокращении времени вегетации
сообщество мхов Polytrichetum norvegicum=sexangularis сменяет Salicetum
herbaceae, причем к Polytrichum присоединяются некоторые другие мхи,
и прежде всего микотрофный печеночник Anthelia juratzkana, симбиоти-
чески связанный с грибом и сам похожий на налет плесени на почве.
Питается он, видимо, гетеротрофно органическим веществом почвы и встре-
чается еще там, где для нормальных мхов «бесснежный» период оказы-
вается слишком коротким. Было бы чрезвычайно интересно изучить,
физиологию Anthelia в культурах, содержащихся в холодных условиях.
Завершает экологический ряд снежных долинок снег, не тающий
в течение всего лета.
Эта комплексность дает представление о том, как слагалась бы вер-
тикальная поясность, если бы в горах с увеличением высоты могли
образовываться обширные плакорные пространства с равномерно рас-
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 47
Рис. 42. Снеговая ложбинка у пика Лагальб (2700 м), Западные
Альпы. Gentiana bavarica (справа), Arenaria btflora (вверху), Veronica
alpina (слева и между Gentiana), Gnaphalium supinum (маленькие листики
в центре) (фото Шенка).
пределенным снежным покровом и если бы продолжительность «бесснеж-
ного» периода постоянно сокращалась бы вплоть до границы вечных снегов.
Curvuletum в районах Альп, сложенных кристаллическими породами,
несомненно, представляет собой характерное и на всех участках домини-
рующее сообщество верхнего альпийского пояса. Выше следовало бы ожи-
дать обширные площади с Salix herbacea, и действительно, заросли Salix
можно обнаружить в местах скопления снега. Тем не менее большие ровные
пространства на этих высотах в Альпах почти не встречаются, однако'
их много в горах Скандинавии с их мягкими формами рельефа. Там сооб-
щества Salicetum herbaceae и Polytrichetum с господством Polytrichum
norvegicum, Webera commutata, Dicranum starkii, Anthelia juratzkana покры-
вают значительные территории, поздно освобождающиеся от снега. В бла-
гоприятные годы Polytrichetum бывает свободен от снега в течение месяца;
но случается, что сообщество остается под снегом 2 года и более. Непосред-
ственно у снеговой линии из толщи снега торчат обычно лишь скалы.
В местах скопления мелкозема развитие растительности нарушается соли-
флюкцией (рис. 43) или образованием полигональных почв (рис. 44;
Gjaerevoll, 1956, цитируется на стр. 27). Последние особенно широко
распространены в Скалистых горах Северной Америки на высоте 3000—
3300 м Последним представителем покрытосеменных является довольно
редко встречающийся Ranunculus pygmaeus высотой едва более 1 см.
Для арктического острова Акпаток (побережье Северной Америки) Полу-
нин намечает следующий ряд растительности снежников в направлении
сокращения «бесснежного» периода:
1 Р. L. J о h n s о n, W. D. Billings, The alpine vegetation of the Beartooth
Plateau in relation to cryopedonic processes and patterns, «Ecol. Monogr.», 32, 1962,
p. 105—135.
48
Глава 1
Рис. 43. Поперечный разрез солифлюкционной террасы
на высоте 3000 м. Погребенные горизонты гумуса —
свидетельство перемещения грунта. Скальные обломки
преимущественно скапливаются в лобовой части тер-
расы (по Джонсону и Биллингсу).
Рис. 44. Полигональные почвы с Anthelia jaratzkana на высоте 1100 м, швед-
ская часть Лапландии (фото Э. Вальтер).
1. Луга с Salix herbacea и Polygonum viviparum.
2. Сообщество Oxyria — Saxifraga.
3. Мхи c Cardamine spec.
4. Голая почва с чахлыми Saxifraga сегпиа.
Границу вечных снегов пересекают лишь водоросли (Haematococcus
nivalis), скопления которых часто образуют на снегу тонкий розовый
налет. Те же водоросли можно встретить на фирновых полях Альп.
В отличие от хионофильных сообществ, длительное время пребываю-
щих под снегом, на подверженных сильному воздействию ветра участках
склонов развиваются хионофобные сообщества, на которых снег никогда
подолгу не задерживается. Из цветковых растений на этих местообитаниях
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 49
особенно типичны Loiseleuria procumbens (в Арктике — также Diapensia
lapponica), а на известняках Dryas; выше в горах — это определенные виды
Carex и Juncus trifidus. На наиболее неблагоприятных местообитаниях
поселяются только лишайники. Но и они дают ряд по возрастанию числа
дней со снежным покровом:
Solo- Clado- (Cladonia rangiferina
rina nia -> s Cladonia silvatica —>
crocea alpestris {.Cetraria islandica
(Cetraria cuculata
Cetraria nivalis ->
* Thamnolia vermi-
.cularis
Alec-
toria
ochro-
leuca
Особое значение для распространения отдельных видов растений
в горах имеет химический состав горных пород. С ним связана реакция
почвы, в которой укореняются растения. Весьма резки флористиче-
ские различия Известняковых и Центральных Альп, сложенных силикат-
ными породами. Приведем в качестве примера растения, укореняющиеся
в расселинах скал, растения, приуроченные к обширным осыпям на скло-
нах, и виды разных травяных сообществ, описанные Гамсом в работе,
посвященной флоре^Форарльберга (Gams, 1961, см. примечание на стр. 32)
(табл. 11).
Отметим еще один фактор — привнос азота. Альпийские
почвы, особенно кислые гумусовые почвы, бедны усваиваемыми растениями
формами азота. При низкой температуре активность почвенных организмов
снижается и разложение гумусовых соединений протекает медленно.
Поэтому в нижнем альпийском поясе на грубогумусных почвах растут
те же кустарнички, какие характерны для омбротрофных болот. Обширные
площади занимает белоус (Nardus stricta); в верхнем альпийском поясе
его замещает Carex curvula. Всем этим видам свойственна «ксероморф-
ность», или, лучше сказать, пейноморфизм. Но если в почве повышается
«одержание азота, на ней тотчас развивается растительность иного состава.
Что это так, подтверждает пример со стойбищами. Это места,
где отдыхают дикие животные или в наше время обычно скот. Экскре-
менты животных обильно удобряют почву. Растительный покров стойбищ
состоит из быстро растущих высокорослых видов разнотравья с крупными
листьями, образующих так называемое высокотравье (рис. 45). Характер-
ными для Альп видами являются: виды Adenostyles и Aconitum, Peuceda-
пит ostruthium, Cicerbita alpina, Cirsium heterophyllum, Ranunculus plata-
nifolius, Epilobium alpestre, Saxifraga rotundifolia, Geranium silvaticum,
Veratrum album и др. На сильно «переудобренных» местах вокруг хижин
пастухов широко распространяется Rumex alpinus, крупные листья кото-
рого часто сплошь покрывают всю площадь. Выпас скота благоприятствует
также^распространению видов Crocus; как только сходит снег, их цветки
живописно украшают еще не зазеленевшие пастбища (рис. 46). Многие
из растений‘стойбищ на удобренных и более влажных местообитаниях
поднимаются высоко в альпийский пояс, подтверждая этим тот факт, что
«ксероморфный» облик большинства альпийских растений обусловлен
не столько неблагоприятными климатическими условиями, сколько недо-
статком азота в почве.
Ольха зеленая (Alnus viridis), на влажных склонах пояса криволесья
вытесняющая сосну горную (Pinus montana), имеет на корнях клубеньки,
образованные симбионтами-актиномицетами, ассимилирующими азот из воз-
духа. Благодаря этому почва на таких местообитаниях обогащена азотом,
и сопровождающими кустарник ольхи видами являются здесь виды высоко-
травья. Среди них многие представители внеальпийской флоры — обычно
-4-0434
50
Глава 1
Таблица 11
Базифильные и ацидофильные виды и типы растительных
сообществ Альп (по f Гансу)
Типы растений Преимущественно на карбонатных породах Безразличные к породе или
предпочитающие нейтральную предпочитающие силикатную
Л истостебе льные мхи Distichium capilla- сеит Encalypta streptocar- ра Tortella tortuosa Amphidium mougeotii Виды Rartramia Schistostega pennata
Папоротники Asplenium ruta-mu- raria, A. viride Cystopteris spp. Asplenium trichoma- nes Cystop teris fragilis Woodsia alpina Asplenium septentrio— nale Polypodium vulgare
Розеточные тра- вянистые мно- голетники Primula auricula Draba tomentosa Dianthus Silvester Draba dubia Primula hirsuta
Травянистые многолетники с удлиненными побегами Potentilla caulescens Hieracium humile Valeriana tripteris Viola biflora Bupleurum stellatum. Hieracium amplexi- caule
Подушечники Aretia (=Androsace) helvetica Petrocallis pyrenaica Saxifraga moschata и S. oppositifolia Saxifraga exarata в S. bryoides
Суккуленты Sedum album и S. atratum Sedum dasyphyllum Saxifraga aizoon Sempervivum arach- noideum Sedum annuum Saxifraga cotyledon S empervivum monta- num
Передвигающие- ся со щебнем Lastrea (==Dryopte- ris) robertiana Trisetum disticho- phyllum Thlaspi rotundifoli- um Valeriana montana, V. supina Petasites paradoxus Poa nemoralis Rumex scutatus Viola calcarata Hieracium staticifo- lium Calamagrostis villosa- и C. tenella Achillea moschata
Растения, пере- двигающиеся по щебню Arabis alpina Cerastium latifolium Galium helveticum Silene alpina Cerastium strictum Linaria alpina Arenaria biflora Cerastium uniflorum Trifolium pallescens
Растения, раз- двигающие ще- бень Dryopteris villarsii Rumex nivalis Doronicum grandi- florum Oxyria digyna Cryptogramma crispa. Doronicum clusii
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 51
Продолжение табл. 11
Типы растений Преимущественно на карбонатных породах Безразличные к породе или
предпочитающие нейтральную предпочитающие силикатную
Растения, покры- вающие щебень Gypsophila repens Saxifraga biflora Saxifraga oppositi- folia Salix retusa и S. serpyllifolia Salix herbacea
Закрепляющие осыпь растения Sesleria varia Carex firma Juncus monanthos Hutchinsia alpina Ranunculus parnas- sifolius Papaver sendtneri Athamanta cretensis Leontodon montanus Festuca violacea Carex sempervirens Juncus facquinii Hutchinsia brevicau- lis Ranunculus glacialis Carduus defloratus Poa laxa Luzula spadicea Juncus trifidus Peucedanum ostru- thium Hieracium intybaceum
Луга альпийско- го пояса (мат- ты) Agrostetum alpinae Seslerietum variae Festucetum violaceae Caricetum sempervi- rentis Festucetum halleri
Луга более высо- ких поясов Festucetum pumilae Caricetum firmae Elynetum myosuroi- dis Juncetum jacquinii Caricetum curvulae Juncetum trifidi
Вторичные тра- вянистые пу- стоши лесных поясов Caricetum ferrugi- neae и C. albae Seslerio-Sempervire- ta Brachypodietum pinnati Nardetum strictae Festucetum ovinae
мало разнящиеся между собой подвиды илиЪкотипы (Taraxacum officinale,
Роа annua, Milium effusum, Stellaria nemorum и др.); это свидетельствует
о том, что верхняя граница растительности часто бывает обусловлена
неблагоприятными условиями обеспечения растений минеральными веще-
ствами.
Напротив, на бедных, кислых, уплотненных и плохо аэрируемых
грубогумусных почвах распространяется устойчивый к вытаптыванию
скотом белоус (Nardus stricta). Борьбу с ним следует вести, применяя
загонный выпас после внесения удобрения (гюлле).
Аналогичные условия, описанные здесь преимущественно для Альп,
наблюдаются и в других горах умеренной зоны, в Карпатах \ на Кав-
казе, а также в Северной Америке и Азии; изменяется лишь флористиче-
ский состав отдельных сообществ (рис. 47).
В качестве примера поясности иного вида, но также зависящей
от длительности сохранения снежного покрова, приведем на рис. 48
схему для нижнего пояса гор Уайт-Маунтинс, Новая Англия (США).
Очень интересный анализ жизненного цикла и состава популяций
отдельных видов субальпийских лугов на относительно аридном Северном
1 N.fD о n it a, Vegetationsstufen in den Karpaten Rumanians, «Rev. Roumaine
de Biol.», Ser Bot., 10, 1965, S. 455—467.
4*
52
Глава 1
Рис. 45. Высокотравье на высоте 1800—2200 м в долине бокового притока
Балкарского Терека, северный склон Центрального Кавказа. На переднем
плане — цветущие Ligularia sibirica, справа и слева — соцветия Aconitum
orientals, на заднем плане справа — зонтики Eleutherospermum cicutarium (фото
Щукина).
Рис. 46. Только что освободившееся от снега пастбище с цветущими
Crocus albiflorus под Зефельдом (западный Тироль), на высоте 1200 м
(фото Рипенхаузена).
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 53
Рис. 47. Ковер из Phyllodoce aleutica в поясе кустарничков
(высота 2400 м) гор Сироума, среднее Хонсю. В центре снимка —
А петопе narcissi flor а, на заднем плане слева — крупные листья
Veratrum stamineum (фото Такенаки).
Кавказе произвел Работнов Пробные площадки были расположены
в 25—30 км южнее Кисловодска на высокогорном плато с отметками
1900—2130 м. Маломощный почвенный покров сформирован на известня-
ках. Холодная сырая весна сменяется прохладным дождливым летом,
осень сухая, зима умеренно холодная. Высота снежного покрова незначи-
тельна, заморозки случаются каждый месяц. Даже в августе была изме-
рена температура —6,7°. Ночи обычно холодные, лишь редко безросные.
Среди злаков преобладает Bromus variegatus, а также Festuca varia. Доминантным
разнотравьем являются Primula ruprechtii, Inula glandulosa, Betonica grandiflora,
Trollius patulus и Anemone fasciculata. На южных склонах распространена степная
растительность с господством Stipa pulcherrima, Festuca sulcata, Carex humilis (или
C. buschiorum) и угнетенным Brachypodium pinnatum. Луга на северных склонах богаты
разнотравьем (до 100 видов на площади 100 м1 2), много Anemone fasciculata. Впадины
занимает высокотравье с Aconitum, Veratrum. Heracleum и Dactylis glomerata. Можно
выделить 9 типов растительных сообществ. В военные годы, когда луга не выкашива-
лись, на них распространилась водяная крыса, обычно поселяющаяся у водотоков,
и своими норами существенно нарушила растительный покров.
При исследовании луговых сообществ недостаточно ограничиться
лишь составлением перечня видов и определением их относительного коли-
чества. Каждый вид в растительном сообществе представляет собой популя-
цию индивидуумов, весьма непохожих по возрасту и стадиям развития.
Жизненный цикл вида распадается на следующие стадии: 1) семена
в состоянии покоя; 2) проростки и вегетативные молодые растения;
3) генеративные, образующие семена особи; 4) стареющие, уже не цвету-
щие индивидуумы. Число способных к прорастанию семян, содержащихся
в верхних 5 см почвы, различно для разных типов лугов; их общее коли-
чество может достигать нескольких десятков тысяч на 1 м\
1 Т. А. Работнов, Жизненный цикл многолетних травянистых растений
в луговых ценозах, «Труды Бот. Ин-та АН СССР, сер. III (геобот.)», т. 6, Л., 1950.
54
Глава 1
г. Вашингтон, 1917 м
увеличение
Р и’с. 48. Распределение растительности в горах Уайт-Маунтинс
в зависимости от наветренности местоположения, воздействия
талых вод и высоты снежного покрова; Новая Англия, США.
Свободное поле слева: различные сообщества пустошей с видами
Vaccintum и Empetrum (по Харрису).
Прорастание семян одного вида обычно растягивается на многие
годы, причем всхожесть не утрачивается, а, напротив, нередко с течением
времени возрастает. Отношение числа проростков к числу образовавшихся
в том же году семян весьма мало: у Deschampsia caespitosa, например, оно
равно 1 : 27, у Carex goodenoughii 1 : 284 и у Juncus gerardi 1 : 8458.
Основным объектом исследования была размножающаяся семенами
Anemone fasciculata, возраст отдельных экземпляров которой легко устано-
вить. Количество семян этого вида, продуцированных одним растением,
составило (в скобках на 100 м2); в 1940 г.— 60, в 1945 г.— 68 (14 000),
в 1946 г._ 45 (8800), в 1947 г.— 13 (1170).
Причина старения растений неизвестна. Возможно, какую-то роль
играет почвоутомление, то есть утрата почвой определенной части питатель-
ных веществ или микроэлементов. Некоторые виды лугового сообщества
сохраняются лишь в результате постоянной смены места обитания в пре-
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 55
делах сообщества благодаря побегам размножения или обсеменениюJ1.
'Старея, растение теряет способность производить семена и либо сразу
погибает, либо продолжает еще некоторое время вегетативно существо-
вать, приобретая все более угнетенный вид. Избыток в почве азота уско-
ряет процесс старения: растение быстрее достигает стадии цветения,
но и скорее отмирает. На лугах происходит естественное удобрение почвы
язотом благодаря грибам, образующим ведьмины кольца. Средний возраст
Libanotis transcaucasica в пределах такого кольца составил 11—15, вне его
21—25 лет.
Опыты в культуре не позволяют судить о максимальном возрасте,
какого может достичь вид, объединенный с другими в растительное сооб-
щество. Вследствие конкуренции условия для развития растений в сооб-
ществе значительно менее благоприятны; часто им требуется очень много
времени, чтобы зацвести; цветут они далеко не каждый год и живут гораздо
дольше. Виды, возраст которых в культуре не превышает двух лет, в луго-
вом сообществе достигают возраста 10—20|или даже 40—50 лет. До 50—
'60 лет живут Gentiana lutea и Cyclamen europaeum.
Анализ популяции представлен в табл. 12.
Аналогичные анализы популяций выполнены и для других видов.
Так, Peucedanum pschavicum достигает 22 лет, цвести начинает спустя
7 лет после прорастания, однако цветущие экземпляры не составляют
Таблица 12
Возрастной состав популяции Anemone f asciculata на площадке
/размером 30 м2 в сообществе Bromus varlegatus 4- разнотравье
на влажном местообитании. Подсчет производился в период
с 24.YIII по;6.1Х.1945 г. (по Работнову)
Возрастные группы Число особей и % общего количества Из них взрослые вегетативные Из них цветущие Цветущие растения, % общего количества
Проростки 506; 32,0
1 год 158; 9,9 — —
‘2 года 74; 4,7 —• —
3 года 23; 1,4 — —
4 года 21; 1,2 — —
0—4 года 782; 49,2
5—9 лет 101; 6,4 2; 0,4 — —
10—14 100; 6,3 20; 4,1 — —
15—19 96; 6,0 67; 13,7 — —
20—24 139; 8,7 119; 24,7 5; 5,3 6,3
25—29 122; 7,6 102; 21,2 16; 17,1 13,1
30—34 115; 7,2 90; 18,5 24; 25,5 16,6
35—39 72; 4,5 51; 10,8 19; 19,2 25,0
40—44 44; 2,8 18; 3,7 24; 25,5 54,6
45 лет и более 21; 1,3 14; 2,9 7; 7,4 33,3
Всего 1592; 100 483; 100 95; 100
% общего количества 100 30,4 5,9
1 Н. L i е t h, Patterns of change within grassland communities, «The Biology of
'Weeds», Oxford, I960,
56
Глава 1
и 5% общего числа растений. Продолжительность жизни Pedicularis соп-
densata составляет 43 года, причем цветут уже 4-летние особи, 25% всех
растений образуют соцветия.
Структура популяции в разные годы в зависимости от погодных
условий различна. Ощутимые изменения даже в пределах отдельных сооб-
ществ вносит вмешательство человека (косьба, выпас скота); причина
этого в неодинаковой степени напряженности конкуренции между отдель-
ными видами.
Доля цветущих экземпляров”Л петопе в пяти типах сообществ приве-
дена в табл. 13.
Таблица 13
Доля цветущих растений Anemone в различных типах
сообществ (в %)
Годы Местообитания
I II III IV V
1945 . . 26,8 11,6 66,2 35,0 10,2
1946 . . 23,0 6,7 68,0 52,0 8,6
1947 . . 9,0 5,3 64,0 43,4 13,4
Местообитание III с большим числом цветущих особей оказалось
более влажным по сравнению с остальными. На количество цветущих
экземпляров повлияло также выжигание травы весной, кроме того, мно-
гие растения могли быть съедены грызунами.
Это небольшое число примеров мы привели с единственной целью
показать, сколь сложны условия взаимоотношения растений в луговых
сообществах. Поэтому агроному знать структуру популяций отдельных
видов так же необходимо, как лесничему знать состав древесных пород
охраняемого им леса. Правда, такого рода анализы луговых сообществ
отнимают очень много времени. Однако важно уже само представление
о чрезвычайной сложности состава растительного сообщества, образуе-
мого многими видами, а эта цель детальными исследованиями Работнова
достигнута.
АНТАРКТИДА И СУБ АНТАРКТИЧЕСЕ ПИАСТРОВ А
На покрытом льдом Антарктическом континенте растений практи-
чески нет. В этой части света обнаружены только два вида цветковых
растений: Colobanthus crassifolius (гвоздичные) и Deschampsia antarctica.
В последнее время на южной стороне залива Олений (64°10' ю. ш.,
60°57' з. д.) найден занесенный человеком Роа pratensis х. На лишенных
снега участках побережья (крутые скальные стены, осыпи на склонах)
встречаются только мхи, лишайники (всего 350 видов) и наземные водо-
росли. Особенно широко они распространены по берегам фьордов Земли
Грейама, благоприятных в климатическом отношении. Здесь выделяют
следующие типы сообществ: 1) Andreaea — Usnea', 2) Polytrichum — Dicra-
num; 3) Drepanocladus — Acrocladium — Brachythecium.
1 А. С о г t e, La primera fanerogama adventicia hallada'en el continente antartico,
«Institute Antdrtico Argentine. Contr.», № 62, 1961.
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 57
В настоящее время проводятся эколого-физиологические исследова-
ния у Мыса Халлетт (72°17' ю. ш., 170°18' в. д.) х. Ланге опубликовал
о них лишь предварительное сообщение. Мыс Халлетт — вулканическое
горное плато, круто обрывающееся к морю. У подножия северо-западного
склона сформировалась полоса пляжа, покрытая моренным обломочным
материалом и галечником. Выше его расположилась большая колония
пингвинов (рис. 49). В летнее время на пляже скапливается стекающая
со склона вода, удобренная экскрементами, что способствует развитию
пышного покрова из водорослей и мхов с преобладанием Prasiola crispa
и Bryum argenteum (рис. 49, внизу). На склоновых осыпях и скалистых
обнажениях фосфатно-нитратное поступление менее обильно. Мхи и водо-
росли здесь почти исчезают, уступая место различным лишайникам:
Caloplaca, Buellia, Rinodina или виды с мелкочешуйчатым слоевищем —
Xanthoria, Candelariella и др. (всего около 20 видов). Но распространение
и этих лишайников связано с талой водой, которая собирается преимуще-
ственно в ложбинах стока, углублениях или даже под свободно лежащими
камнями, в расщелинах и нишах, где остается лежать снег, тающий летом.
При опытах с антарктическим лишайником (Parmelia coreyi) Ланге уста-
новил 2, что максимальная продуктивность фотосинтеза за вычетом рас-
хода на дыхание наблюдается при 0—5°, что соответствует температуре
талой воды. Правда, по данным Рудольфа, лишайники в ясные летние
дни нагреваются до 32° (температура поверхности горных пород), но в этом
случае они, видимо, высыхают. Мхи редко образуют спорофиты; вместе
с тем у них известны разные формы вегетативного размножения3.
На нунатаках на высоте 1800 м и в 50 км от побережья обнаружена лишь
скромная, плохо различимая глазом растительность; Янечек предлагает
выделить этот «пояс микрофитов» из собственно пояса криптогамных
растений 4. Тем не менее во время экспедиции Бёрда (1934 г.) в горах
Королевы Мод на высоте 4700 м (86°03' ю. ш.) был собран ряд лишайников
(Alectoria, Buellia, Lecanora и др.).
В омывающие со всех сторон Антарктиду моря с их почти непрерыв-
ными штормами вкраплено всего несколько субантарктических островов,
образующих прерывистый пояс суши, лишь немного выступающий к северу
за 50° ю. ш. (рис. 50). На островах не растет ни единого дерева, хотя
по своему положению они соответствуют в северном полушарии Британ-
ским островам, Ньюфаундленду и Камчатке.
Температура летом недостаточна для произрастания здесь деревьев,
но зимы нехолодные. При постоянно ветреной и туманной погоде нередко
в течение всего года днем и ночью температура не поднимается более
чем на несколько градусов выше нуля (см. климадиаграммы на рис. 50).
Яркое описание острова Кергелен оставил Шимпер, посетивший остров
в качестве участника немецкой глубоководной экспедиции на судне
«Вальдивия».'
1 Е. D. R u d о 1 р h, Vegetation of Hallett Station area, Victoria Land, Antarctica,
«Ecology», 44, 1963, p. 585—586.
3 O. L. L an g e, DerCO2-Wechselder Flechtenbei tiefen Temperaturen, «Planta»,
04, 1965, S. 1—19.
_ тл 3nW'jC' 1S V e r e’ Antarctic bryophyta, «Bio. Science», 15, 1965, p. 283—285;
Д 1 k m ° 1 * * * R •’ Antarctic lichenes, «Bio. Science», 15, 1965, p. 285—287; E. D. R u -
n o Ipn, Terrestrial vegetation of Antarctica, «Antarc. Res.», Ser. 8, 1966, p. 109—
xr ттУ e, J. v a n Mieghem, Biogeography and ecology in Antarctica,
V and VI, The Hague, 1966.
. i Л,п e tschek, Die Mikrophytenstufe, «Anz. Osterr. Akad. Wiss., math.-
nat. KI.», 1964, s. 185—191.
58
Глава 1
Рис. 49. Ландшафт у мыса Халлетт (Антарктида). Верхний снимок', на переднем
плане — полоса плоского пляжа с мхами, выше колонии пингвинов — скалистый
крутой склон с осыпями и лишайниковой растительностью. Нижний снимок
(фото крупным планом): Bryum argenteum, растущий на галечнике пляжа. На са-
мом влажном участке (левее снежного пятна) — корковое слоевище Prasiola crispa
(фото Ланге).
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 59
Рис. 50. Климадиаграммы антарктической и субантарктической областей.
«С террасовидной возвышенности в окрестностях бухты Газелей вся сильно рас-
члененная поверхность Кергелена выглядит как пустыня, на серой, усеянной скаль-
ными глыбами почве которой, словно зеленые кочки, выступают то одиночные, высотой
до полуметра, то расположенные группами подушки Azorella Selago Hook, f., достигаю-
щие 1 м в поперечнике. Как и арктическая, антарктическая тундра имеет свои оазисы;
выделяются более или менее равномерной зеленой окраской склоны, обращенные к севе-
ру, зеленеют рытвины в почве, если только они не заполнены снегом и льдом; словно
изумруд, светятся на обычно серой поверхности моря маленькие островки в бухте...
Пустынный характер Кергелена обусловлен не физической сухостью почвы —
поверхность острова, хотя и каменистая, всегда увлажнена, на небольшой глубине
в почве также постоянно содержится определенный запас влаги; не имеет решающего
60
Глава 1
значения и низкая температура почвы, в противном случае северные склоны были бы
просто покрыты более густой растительностью, чем южные \ Ограничивает
развитие растений ветер, дующий преимущественно с
запада и часто достигающий ураганной силы. Кергелен
представляет собой ветровую пустыню, где основной неблаго-
приятный фон, создаваемый иссушающим воздействием ветра, усиливается низкой
температурой почвы. Сочетанию этих факторов и обязан своим своеобразием
здешний растительный мир. Пышное развитие растительности в углублениях почвы
можно объяснить только защищенностью этих мест произрастания от ветра, ибо недо-
статка влаги нигде нет, а повышенной температурой эти местообитания, конечно,
не отличаются. Внимательный осмотр подушек Azorella также подтверждает высказан-
ное предположение об отрицательной роли ветра: западная часть их менее развита,
чем другие, особенно восточная часть; лишь на последней растут Асаепа adscendens
Vahl, Agrostis antarctica Hook, f., Lycopodium saururus Link., на каждой подушке
Azorella. Разбросанные на открытых ветрам плоских возвышенностях обломки скал
затянуты покровом из бородатого лишайника (Neuropogon melaxanthus Ach.
и АГ. Taylori Hook, f.), плотность которого на разных сторонах глыб неодина-
кова. Примечательно, что особенно густо поросли лишайником не те стороны,
которые обращены в сторону наиболее частых дождей или штормовых сухих западных
и северо-западных ветров, напротив, на совершенно не защищенных от ветра место-
обитаниях эти поверхности безжизненны. Лишайники предпочитают восточные сторо-
ны; северные и южные бока глыб покрыты ими более редко.
Из приведенного описания явствует, что Асаепа adscendens Vahl. выбирает самые
благоприятные для жизни местообитания, предпочитая склоны восточной и северной
экспозиции. Как только условия существования улучшаются, подушки Azorella
сдвигаются плотнее и в конце концов образуют сомкнутый покров. По мере еще большей
оптимизации условий среды ковер из Azorella постепенно сменяется покровом из Асаепа.
Azorella господствует в пустыне, где Асаепа или не растет вовсе, или же развивается
угнетенно; Асаепа доминирует в оазисах и препятствует проникновению туда Azorella».
Наиболее широко распространена Azorella selago (рис. 51), наряду
с ней встречается Pringlea antiscorbutica, так называемая «кергеленская
капуста» (рис. 52), крупные листья которой моряки раньше употребляли
в пищу как противоцинготное средство; этот вид произрастает лишь
на защищенных от ветра позициях. Асаепа adscendens или другие виды
этого рода известны на всех субантарктических островах с аналогичным
типом растительности. Широко распространены туссоковые (кочкарные)
злаковники (виды Festuca и Роа). Встречаются также папоротники, как,
например, Lomaria alpina, Hymenophyllum peltatum, Polypodium australis,
P. vulgare, Cystopteris fragilis. Многочисленны мхи и лишайники; домини-
рует Neuropogon melaxanthus.
Растительность Антарктики обнаруживает весьма отчетливые связи
с флорой альпийского пояса южной оконечности Чили, острова Южный
Новой Зеландии и с горной растительностью Тасмании. Эколого-физиоло-
гические исследования в незаселенных районах не производились. Поэтому
ограничимся лишь ссылкой на некоторые работы 1 2.
1 Необходимо иметь в виду, что описываемый район расположен в южном полу-
шарии.— Прим. ред.
2 Н. Schenck, Verglei chende Darstellung der Pflanzengeographie der suban-
tarktischen Inseln, insbesondere fiber Flora und Vegetation von Kerguelen, Wiss. Ergebn.
'Deutsch. Tiefsee-Exp. «Valdivia», 1898/99, Jena, 1905; C. Skottsberg, Vegetations-
bilder aus Feuerland, von den Falklandinseln und von Siidgeorgien, «Veg. Bild.», R. 4,
H. 3/4, 1906; G. Skottsberg, I. The vegetation in South Georgia und IL Einige
Bemerkungen uber die Vegetationsverhaltnisse des Graham-Landes, «Wiss. Ergebn.
Schwed. Siidpolar-Exp. 1900/03», 4, Lief. 12, 13, Stockholm, 1912; E. Werth, Die
Vegetation der subantarktischen Inseln Kerguelen, Possession und Heard-Eiland. Cm.:
E. Drygalski, Deutsch. Siidpolar Exp. 1901/03, 8, Berlin—Leipzig, 1928; E. J. God-
ley, Notes on the vegetation of the Auckland Islands, «Proc. New Zeal. Ecol. Soc.», 12,
1965, p. 57—63; E. J. G о d 1 e y, The botany of southern Chile in relation to New Zea-
land and the Subantarctic, «Proc. Roy. Soc. B.», 152, 1960, p. 457—475; E. Aubert
de la Rue, Les lies Australes frangaises, «La Nature», № 3192, Paris, 1951.
Рис. 51. Плато южнее залива Газелей на острове Кергелен. На переднем плане —
подушки Azorella, selago’, на склоне слева виден плотный покров из Асаепа adscendens
и Azorella (фото Заксе).
Рис. 52. Группа плодоносящей Pringlea antiscorbutica (крестоцветные) с отмер-
шими прошлогодними соплодиями. На заднем плане — лужайка из Cotula plu-
mosa и Асаепа adscendens, вытесняющих подушки Azorella selago, справа —
отдельные дерновинки Festuca erecta’ скалы покрыты лишайниками (фото
Винтера).
62
Глава 1
Уже за пределами Антарктики лежит Гоф — маленький острой
в южной части Атлантического океана (40°19' ю. ш.), в’370 км юго-восточ-
нее архипелага Тристан-да-Кунья х. Длина острова всего 13 км при
ширине 5—6 км. Находится он в 2400 км от южной оконечности Африки
и в 3000 км от Южной Америки — это один из немногих островов, почти
не тронутых человеком и сохранивших поэтому первичную раститель-
ность. Климат здесь уже утрачивает черты, свойственные Субантарктике.
Его можно характеризовать как умеренный климат со средней годовой
температурой 11,7° (средние месячные максимальная и минимальная
температуры колеблются в пределах 2—24°). Постоянные западные ветры,,
порождаемые областями низкого давления, приносят более 3000 мм осад-
ков в год, которые на высотах свыше 450 м зимой выпадают в виде снега.
Влажность воздуха 81%, что позволяет говорить о здешнем климате как
сугубо океаническом.
Возник вулканический остров, вероятно, в третичном периоде.
За последние 2300 лет извержений не происходило. Пыльцевой анализ
показал, что в течение 5 тыс. лет растительность оставалась без изменений.
На острове лишь временно поселялись моряки, которые домашних живот-
ных не завозили. В наше время здесь создана метеорологическая станция,,
персонал’которой разводит овец и домашнюю птицу; однако влияние живот-
ных на растительность проявляется лишь в непосредственном окружении
станции. Случайно завезена мышь (Mus musculus).
Флора представлена 32 видами местных цветковых растений
и 12 видами случайно занесенных сорняков, из которых уже получили
распространение Rumex obtusifolius, Sonchus oleraceus, Роа annua, Holcus
lanatus, Agrostis stolonifera. На крутом побережье в местах отдыха пинг-
винов часто можно встретить Plantago major и Stellaria media.
Внутренняя часть острова представляет собой плато, возвышающееся
на 600 м над уровнем океана. Самая высокая вершина имеет отметку 910 м-
Растительный покров образуют туссоковые злаковники с Роа flabellata.
и Spartina arundinacea, нарушаемые выпасом овец. Зональной раститель-
ностью можно назвать кустарник из Phylica arborea (крушиновые) в окру-
жении низких древовидных папоротников (Blechnum palmiforme, Hystio-
pteris incisa). Обширные площади заняты влажными пустошами с господ-
ством Blechnum palmiforme, Empetrum nigrum, с злаками и осоковыми или
заболоченными участками, поросшими Rhacomitrium lanuginosum (30%}
и Empetrum rubrum (25%), а также Agrostis carmichaelii (20—40%). Эти
группировки постепенно сменяются растительностью скал — мхами,
лишайниками и Lycopodium saururus. Плоские долины вдоль водотоков
заняты настоящими торфяными болотами, их формированию способствуют
главным образом виды Sphagnum и Dicranoloma, а также Hepaticae.
На островах Тристан-да-Кунья растительность, по-видимому, имела
аналогичный характер. Однако в настоящее время она сильно нарушена
человеком и домашними животными. Туссоковые злаковники уничто-
жены; местами, как и в Новой Зеландии, территорию сплошь занимают
заросли Rumex acetosella.
Фолклендские (Мальвинские) острова расположены значительно
южнее, в Субантарктике, и отличаются поэтому более холодным клима-
том 1 2. Кустарник с Phylica здесь отсутствует, встречаются только кустар-
никовидные Hebe elliptica и Empetrum rubrum. На незаболоченных площа-
1 N. М. W а се* The vegetation of Gough Island, «Ecol. Monogr.», 31,1961, p. 337-367.
2H. Wilhelm y, Die Falkland-Inseln, «Geogr. Zschr.», 51, 1963, S. 53—62-
Арктическая зона тундры и альпийский пояс гор умеренной гумидной зоны 6?
дях первоначально ведущее положение, вероятно, занимали туссоковые
злаковники {Роа flabellate). Теперь они почти полностью уничтожены
'В результате выпаса овец. Неустойчива к выпасу!и Cortaderia pilosa
(рис. 53). Напротив, широко распространяется пустошь с Empetrum.
На сильно перетравленных участках остается только сообщество Blech-
num — Pernettia (вересковые), не представляющее кормовой ценности.
Рис. 53. Злаковник с господством Cortaderia pilosa на Фолклендских остро-
вах (фото Скотсберга).
Распространены также характерные для субантарктических районов
подушечники — Azorella, Donatia, Gaismardia, Astelia и др.
Годовое количество осадков невелико и составляет 600—700 мм,
но распределяются они в течение года довольно равномерно (240—245 дней
в году с осадками). Выпадают осадки обычно в виде моросящих дождей,
часто сопровождающихся туманами, которые могут стоять неделями.
Почва постоянно хорошо увлажнена. Средняя годовая температура
составляет 6,6°, причем средняя месячная температура самого холодного
месяца равна 2,8°, а самого теплого — ниже 10° (абсолютный максимум
Н-24,4°, минимум—11,1°); характерны почти непрерывно дующие сильные
ветры (в среднем в году бывает только 20 дней без ветра).
Группа островов Южная Георгия выделяется еще более прохладным
климатом, зимой здесь лежит снег. Преобладает Роа flabellate, на сырых
участках развивается злаковник с Festuca erecta, растут также Phleum
alpinum и Deschampsia antarctica. Местами распространена и Асаепа adscen-
dens.
Глава 2
ЗОНА СТЕПЕЙ И ПРЕРИЙ
ЛЕСОСТЕПЬ
Под степью (русский термин) мы понимаем тип растительности уме-
ренной зоны, в котором господствующее положение заняли
многолетние зимостойкие и способные переносить продолжительную
засуху злаки г. У растений отмечается длительный период покоя в холод-
ное зимнее время и более короткий — во время засухи поздним летом.
Особенно характерно полное отсутствие в зоне степей деревьев. Правда,
низкий кустарник в виде одиночных или собранных в небольшие группы
кустов встречается в некоторых степях, но обычно лишь на местообита-
ниях с каменистыми почвами. В Восточной Европе это виды Amygdalus
папа, Spiraea hyperictfolia, Caragana frutex и др.
Степная зона в евросибирской области протягивается широкой
полосой через всю Восточную Европу от устья Дуная до Урала и далее
в Западную Сибирь и Северный Казахстан; кроме того, степь распростра-
нена отдельными участками в котловинах южной, горной, части Средней
и Восточной Сибири до Амура 1 2. Вторая обширная область степей —
североамериканская прерия — простирается на среднем Западе Северной
Америки от Канады до Мексиканского залива, на юге она постепенно
переходит в субтропическую саванну. Наконец, к этому классу формации
можно причислить также южноамериканскую злаковую пампу с пата-
гонскими степями, ту ее часть, которая относится к умеренной зоне,
а также степи района Отаго во внутренних областях острова Южный
(Новая Зеландия); все эти районы мы рассмотрим ниже, в гл. 3.
Наше знакомство с растительностью степей и прерий мы начнем
с евросибирской степной зоны. На севере ее обрамляет лесная зона,
на югсГ-востоке она переходит в полупустыню. Граница с зоной лесов
нерезкая. Подобно тому как на севере лесной зоны в качестве переходной
полосы к тундре выделяют «лесотундру», на юге лесной зоны различают
«лесостепь» — переходную ступень к собственно степи. Лесостепь, просле-
живаемая через Восточную Европу и южную часть Сибири, представляет
собой, как и лесотундра, мозаичный макрокомплекс, в котором резко
отграничены друг от друга лиственные леса с типичными лесными видами
(рис. 54) и луговые степи со степными элементами (рис. 55). Следовательно,
неверно определять лесостепь как лес, в котором степные элементы рас-
пространены в нижнем ярусе. В последнем случае говорят о степных
1 В "субтропической и тропической климатических зонах также известны типы
растительности с преобладанием многолетних злаков. Мы называем их просто арид-
ными злаковниками, когда речь идет о засушливых районах, и гумидными злаков-
никами, если они распространены во влажных областях (см. т. I).
2 М. А. Рещчиков, Степи западного Забайкалья, «Сиб. отд. АН СССР, отде-
ление биологии», № 34, М., 1961»
Зона степей и прерий
65
Рис. 54. Лес из Quercus robur в лесостепной зоне^(весной), район
Воронежа (фото Калугина).
лесах или правильнее — об «остепненных лесах». Оба понятия не следует
путать. То, что Вендельбергер ошибочно назвал в Европе лесостепью,
в действительности является «остепненными лесами» Ч
Среди причин макромозаичного распределения степной и лесной
растительности известную роль играет рельеф, однако решающее значе-
ние принадлежит почве. Леса распространены на хорошо!дренируемых
серых лесных почвах, деградированных черноземах возвышенностей,
G. Wendelberger, Die Waidsteppen des pannonischen Raumes, «Veroff.
Geobot. Inst. Rubel», 35, Zurich, 1959, S. 77—113; E. M. L a v г e n k o, Die Felssteppen
der Umgebung von Wien und des uralten Mediterrangebietes, «Veroff. Geobot. Inst.
Rubel»,|35, Zurich, 1959, S. 114—127; R. S о 6, Das heutige Bild der Vegetation Ungarns,
«Ann. Bot. Fenn.», 3, 1966, S. 349—3.57; S. Pascovschi, N. siSonita, Vegetatia
lemnoasa din silvostepa romanici, Acad. Rep. Soc. Romania, Bucuresti, 1967.
5—0434
66
Глава 2
Рис. 55. Лесостепная зона в районе Воронежа. На переднем плане —
степь со Stipa lessingiana, на заднем — дубовый лес по оврагам (фото
Келлера).
получающих несколько большее количество осадков, на высоких право-
бережьях рек и вообще на склонах, глубоко расчлененных эрозионными
балками (рис. 56).
Напротив, мощные черноземные почвы являются местообитаниями
растительности луговой степи. Занимают они обширные плакорные про-
странства или низкие левобережья рек, на которых после таяния снега
весной еще длительное время застаивается вода — обстоятельство, небла-
гоприятное для развития деревьев. Существенную роль играет также
характер почвы: лес растет на более легких и грубых почвах, степи,
напротив, распространены на тонкозернистых лёссовых грунтах. Здесь,
в пограничной полосе, условия конкуренции между лесом и злаковником
столь неустойчивы, что при малейшем улучшении условий в сторону,
благоприятную для произрастания леса, он начинает вытеснять степь,
и наоборот. И дело не в том, что естественный рост деревьев в степях
якобы невозможен. Лесопосадки совершенно однозначно доказали обрат-
ное; но в условиях свободной конкуренции лес в степи утвердиться
не в состоянии, причем надо учесть, что распространению степей прежде
во многом способствовали степные пожары и пастьба крупных диких
животных. Этот вопрос мы еще рассмотрим подробнее. Наибольшие
пространства в пределах лесостепи лес занимает на высоких участках
Волыно-Подольской возвышенности правобережья Днепра, на Средне-
Русской возвышенности, на возвышенном правом берегу Волги и в высоком
Заволжье. Основной древесной породой является Quercus robur, в бассейна
Днестра — также Q. petraea, в долине Прута — Q. pubescens и на крайнем
северо-востоке лесостепной зоны — береза {Betula verrucosa).
Луговые степи в настоящее время почти полностью превращены
в пашни. Заповедные территории с первичным растительным покровом
сохранились только в районах Курска (Стрелецк), Воронежа (Хреновое),
Пензы. Небольшие площади луговой степи рассеяны по окраинам лесных
массивов. Но и сами леса большей частью уступили место пашне, поэтому
Зона степей и прерий
6?
Рис. 56. Взаимосвязь растительности, почвы и рельефа в лесостепной
зоне (по Танфильеву и Морозову, с большими изменениями).
1 — мощные слабо дренируемые черноземы на плакорных пространствах с расти-
тельностью луговой степи; 2 — хорошо дренируемые деградированные черноземы;
3 — темно-серые лесные почвы; 4 — водопроницаемые песчано-суглинистые лесные
почвы; 5 — светло-серые лесные почвы; в —солонцы на ровных террасах или вокруг
бессточных впадин, обогащенные содой; 7 — флювиогляциальные пески; 8 — морен-
ные отложения или лёссовидные суглинки; 9 — доледниковые отложения; 10 — аллю-
вий речных долин.
I — комплексы дубовых лесов на хорошо дренируемых возвышенностях или на
склонах; II — пойменные леса с дубом и др.; III — сосновые леса на бедных песча-
ных почвах со сфагновым болотом в переувлажненной низине; IV — сосново-дубовые
леса на суглинистых почвах; V — осиновые рощи в небольших низинах (подах),
где весной скапливаются и медленно просачиваются в грунт талые воды (почвы в
центральной части выщелочены); Уа — то же, но с ивовым кустарником; VI —овраж-
ный дубовый лес, по кромке степной кустарник.
сегодня лесостепь в целом менее облесена, чем прежде. По песчаным
почвам сосновые леса проникают глубоко в собственно степную зону.
Климадиаграммы (рис. 57—59, 60—62) позволяют четко различать
в климатическом отношении лесную зону, лесостепь и степную зону \
Общим является относительно благоприятное распределение осадков
в течение года. Максимум их приходится на лето. В зоне лесов засушливый
период отсутствует. На климадиаграммах лесостепной зоны всегда отчет-
ливо выделяется, правда обычно непродолжительный, сухой период
в конце лета, но засухи здесь не бывает. В степной зоне засуха наблю-
дается обычно весной и поздним летом, сухое время продолжается весь
летний период, прерывается оно только грозами, обычно в июне. Зима
во всех зонах в направлении к востоку становится все холоднее, лета
жарче, но в меньшей степени, поэтому средняя годовая температура
к востоку понижается.
Тот факт, что граница леса в результате вмешательства человека
местами значительно сместилась к северу и степь расширилась за счет
леса, дал некоторым авторам основание утверждать, что степь антропо-
генна по своему происхождению, иными словами — что вся степная
область первоначально была покрыта лесом. По этому вопросу известный
1 Н. Walter, Die Klimadiagramme der Waldsteppen und Steppengebiete in
Osteuropa, «Stuttg. Geogr. Stud.», 69, 1957, S. 253—263; A. В orhi d i, Klimadiagram-
me und klimazonale Karte Ungarns, «Ann. Univ. Sci. Budapest», Sect. Biol., 4, 1961,
S. 21-50.
5*
68
Глава 2
Рис. 57—59. Климадиаграммы лесостепной зоны; засушливый период в конце лета
в направлении к востоку становится все более резко выраженным.
Умань — на запад© зоны; Воронеж — центральная часть; Саратов — восточная часть (переходная
к степи).
Рис. 60—62. Климадиаграммы степной зоны, хорошо выражен период засухи.
Одесса — западная окраина зоны; Оренбург — Южный Урал; Семипалатинск — юг Сибири.
украинский почвовед Махов высказал (устное сообщение) следующее
мнение. В период максимального оледенения, когда ледник двумя язы-
ками, днепровским и донским, достиг Украины, растительность на месте
современной степной зоны представляла своего рода тундру, возникнове-
ние которой было обусловлено холодным и сухим климатом; зона тундры
простиралась до Крымских гор. В пещерах Крыма найдены следы обита-
ния там неандертальского человека, который спасался в них от арктиче-
ского холода; в пещерах обнаружены также древесный уголь из арктиче-
ской березы и кости арктических животных. После конца эпохи оледене-
ния тундру сменила степь. На территорию Украины с юга (с Балканского
полуострова) началась миграция видов растений нижнего горного пояса,
в том числе и ковылей (Stipa). Степная растительность распространилась
по поверхности лёссовой равнины. Присутствие в лёссе СаСО3, CaSO4,
а местами и NaCl также имеет свое объяснение. При выветривании горных
пород в Арктике происходило накопление растворимых солей, поскольку
многолетняя мерзлота препятствовала их выносу. Фён, дувший с ледника,
развевал обогащенные солями продукты выветривания, которые и способ-
ствовали образованию соленосного лёсса. Конечно, при этом следует
различать собственно процесс отложения лёсса и процесс его дальнейшего
преобразования и развития в разных климатических условиях.
Первая половина каждого межледникового периода, а следовательно,
и эпохи постгляциала в Восточной Европе была неблагоприятна для
произрастания леса как вследствие холодного сухого климата, так и из-за
развития процесса засоления почвы (типа сухих солончаков). Распростра-
нение березово-осиновых лесов в Полесье (лесная зона Украины) началось
лишь после того, как климат стал более теплым и влажным. Лиственные
Зона степей и прерий
69-
леса степной зоны Украины в период вюрмского оледенения сохранились
лишь в особо благоприятных районах. Такими рефугиумами служили:
теплая долина Днестра, правый берег Северного Донца, Донецкий кряж.
Продвижение лиственных лесов в пределы лесостепи началось только
в период климатического оптимума послеледникового времени (теплый,
влажный климат).
Существует возможность восстановить ареалы прежнего распростра-
нения леса в довольно точных границах, поскольку лес, занимая чернозем,
изменяет его свойства, причем физико-химические преобразования почвы
оказываются столь глубокими, что сохраняются тысячелетиями, даже
если лес исчезает намного раньше. Благодаря этому карты когда-то суще-
ствовавших лесов можно составить по состоянию до 5 тыс. лет назад.
Уместен вопрос, почему же лес не распространился на всю область лесостепи
и почему сегодня в окружении лесных почв встречаются довольно многочисленные
островки сохранившегося с древнейших времен чернозема. Объяснение’этому следую-
щее. Главное наступание леса произошло 5 тыс. лет назад, однако уже. в то время мест-
ное население занималось возделыванием культурных растений (трехпольное хозяй-
ство позднего неолита). Острова чернозема посреди лесных почв и есть остатки преж-
них пашен. Лес не мог занять эти площади, так как заселение в ту эпоху, как показали
раскопки, уже было сравнительно плотным и земля интенсивно распахивалась и воз-
делывалась. Изучение реликтовых почв позволяет проследить процесс сведения лесов
человеком и в более позднее время. Хозяйственная деятельность человека препятство-
вала продвижению леса дальше к югу, хотя условия среды способствовали тому: потеп-
ление в послеледниковый оптимум сменилось похолоданием и увеличением влажности.
Такой точки зрения придерживался Махов. То, что почвы, на которых
сегодня растет лес, в период послеледникового оптимума еще были степ-
ными образованиями, явствует, как мы увидим ниже, из профиля серых
лесных почв. В последних находят следы (кротовины) прежних нор сусли-
ка — грызуна, живущего только в степи. Кротовины обычно весьма много-
численны, так как суслики при наступании леса были вытеснены на неболь-
шие остатки степи. В конце концов лес захватил и эти остатки, чернозем
со временем деградировал и преобразовался в серую лесную землю.
ПОЧЕМУ СТЕПЬ БЕЗЛЕСНА?
В субтропической зоне между лесной зоной и злаковниками, развитие
которых обусловлено климатическими причинами, вклинивается широкая
полоса истинной саванны (см. т. I, стр. 275—301); в умеренной зоне такая
картина наблюдается редко. Даже в пределах лесостепи массивы леса
и луговые степи обычно резко отграничены друг от друга, порой их разде-
ляет лишь узкая полоска кустарника, растущего у опушки леса. Эта
характеристика может быть отнесена*и к американской прерии, распашка
которой началась лишь в последние сто лет.
Важную роль в североамериканской прерии играл такой фактор, как
широко практиковавшееся раньше индейцами выжигание сухого траво-
стоя. Но пожары часто случаются и в саванне, тем не менее деревья
образуют неотъемлемый элемент ее растительного покрова. В умерен-
ной зоне отсутствует тип столь характерного для саванны отдельно стоя-
щего устойчивого против огня дерева с зонтовидной кроной. Все дре-
весные породы «собраны» здесь в более или менее сомкнутые древостои
и весьма чувствительны к огню. Поэтому в умеренных широтах фронт
70
Глава 2
борьбы между лесом и открытыми, поросшими травой пространствами
непрерывен и выражен довольно резко
Климатические условия, присущие собственно степи, вполне благо-
приятны для развития здесь древесной растительности. Об этом, как мы
уже говорили, свидетельствует опыт посадок леса в степи. Поэтому было бы
неправильным предполагать, что зимние ураганы или высокое содержа-
ние солей в почве препятствуют росту деревьев. Убедительным подтвер-
ждением обратного служат полезащитные лесные полосы, заложенные
на обширной территории в Восточной Европе. Однако отсюда не следует,
что лес в состоянии полностью вытеснить степную растительность. Объяс-
няется это следующим.
Лес расходует в расчете на 1 га площади больше воды, чем степная
растительность. Верхние горизонты почвы под тенью деревьев в лесу
остаются более влажными, нежели в степи, но тем больше влаги вытяги-
вают корни деревьев из глубоких слоев. Осадков в степи выпадает мень-
ше, температура и, следовательно, дефицит насыщения воздуха летом
всегда выше, чем в лесной зоне. В результате обеспеченность водой деревь-
ев у границы степи существенно хуже. Но как только потребление воды
густым насаждением начинает превышать накопленные в почве запа-
сы влаги, возникают условия, в которых лес существовать не может.
В доказательство того, что южная граница леса обусловлена именно
недостатком влаги летом, приведем следующие наблюдения:
1. Выше мы отмечали, что на границе засушливости подлесок в лесах
уже отсутствует, или, точнее: в молодых древостоях подлесок еще встре-
чается, но в более старых лесах он исчезает полностью, несмотря на то
что эти леса сравнительно разрежены. Следовательно, не затенение пре-
пятствует появлению всходов древесных пород, а то обстоятельство, что
старые деревья за вегетационный период используют всю без остатка
влагу, содержащуюся в почве, ничего не оставляя на долю подлеска.
1 О сходстве с саваннами, вероятно, можно было бы говорить лишь в отношении
некоторых естественных насаждений, развитых на холодной континентальной и сухой
периферии области злаковников, как, например, пояс пинион в Северной Америке
и известные части лесостепи в Западной Сибири, где сосна или осина с березой (в наи-
более континентальной части — с лиственницей) образуют очень редкие древостои
с сомкнутым травяным ярусом.
Фитбмасса, продуцированная травяным ярусом, и годовое количество опада
в лесостепной зоне Западной Сибири, определены Игнатьевой:
Древесный ярус Фитомасса, т/га Опад т/га
Сосновый лес 4,08 1,63
Осиново-березовый лес 5,30 2,40
Березовый лес 5,94 2,67
Из общей фитомассы на подземные части растений приходится 80—90%. При
сравнении этих цифр с цифрами для луговых степей Западной Сибири, лишенных
деревьев, оказывается, что первые в четыре раза меньше. Конкурентное равновесие
между деревьями и видами злаков в этих насаждениях еще не изучалось. Поэтому
нельзя сказать, аналогично ли оно равновесию в субтропической саванне.
Зона степей и прерий
71
О менее интенсивном потреблении влаги степной растительностью
в расчете на 1 га площади свидетельствуют данные, приводимые Высоц-
ким (табл. 14) *.
Таблица 14
Содержание воды в почве осенью на разной глубине
и под разной растительностью (в % сырого веса почвы)
Глубина, см Лес Степь Поле Пар
На поверхности 13,9 5,6 9,7 3,5
10 15,5 11,0 13,2 17,9
25 15,6 14,2 15,5 19,5
50 15,7 14,9 15,4 19,6
75 — — 15,8 20,0
100 12,9 13,8 14,8 19,6
150 12,9 14,4 14,6 17,2
200 12,4 15,0 15,3 16,3
•Общий запас воды в верхних 2 м,
мм ............. 456 473 505 641
Из них не доступны для расте-
НИЙ, мм 357 357 357 357
Доступный для использования остаток, мм 99 116 148 284
2. Опыты по облесению в степных районах обнаружили ту законо-
мерность, что с возрастом деревья развивались все хуже и наконец поги-
бали. Это свидетельствует о том, что запасов влаги в почве достаточно
для транспирации молодым древостоем, но его не хватает, чтобы покрыть
•больший расход воды на транспирацию старым лесом. Наблюдения Высоц-
кого подтверждают такой вывод (табл. 15, ср. также рис. 63).
Таблица 15
Среднее содержание
воды в верхнем метре почвы
(в % сырого веса почвы)
Возраст насаждения, число лет Влажность почвы
1 22,85
2 21,31
5 17,37
25 14,35
1 Г. Н.Высоцкий, О взаимных отношениях между лесной растительностью
и влагой преимущественно в южнорусских степях, 1904. Большее потребление воды
лесом подтверждается также новейшими, еще продолжающимися исследованиями
под Курском; ср.: В. В. Г е р ц и к, Изменение лесостепного природного комплекса
ют леса к степи через опушку, «Труды Центральночерноземного заповедника», 8, Воро-
неж, 1965, стр. 291—332. Если под степной растительностью запасы влаги в карбонат-
содержащих горизонтах мощного чернозема в течение всего года почти не изменяются,
то лес уже к осени полностью расходует всю доступную ему воду.
72
Г лава
Рис. 63. Содержание воды в почве под посадками дуба раз-
ного возраста. Измерение произведено 24 августа в степи
под Великим Анадолом (по Храмову).
Рис. 64. Изменение содержания воды в почве
(в % сырого веса почвы) в течение года под зре-
лым насаждением в Шиповом лесу, лесостепная
зона (по Высоцкому).
То, чтоТприведенная в табл. 15 цифра 14,35% почти соответствует
влажности завядания|или содержанию воды в мертвом, то есть не прони-
занном корнями, слое почвы, есть доказательство того, что взрослые-
дер евьяТрасходуют практически всю доступную им воду (рис. 64). Отсюда
ясно, почему предельный возраст леса в степном районе не превышает
20—30 (иногда 40) лет. Это следует учитывать, прежде чем планировать
широкие посадки леса в степи.
Такого рода опытное облесение в открытом степном районе в Вели-
ком Анадоле еще в 1843 г. предпринял Графф; позднее опыт продолжил
Высоцкий. Высаживались древесные породы лесостепи (дуб, ясень, клеи
Зона степей и прерий
131
80м 107м
Рис. 65. Положение уровня грунтовых вод в Шиповом лесу (по Отоц-
кому).
Левая скважина — в степи, средняя — на вырубке, правая — под сомкнутым лесным
пологом.
остролистный, вяз), но также иноземные виды, такие, как белая акация
(Robinia pseudacacia'), шелковица (Morus alba), Gleditschia triacanthos,
желтая акация (Caragana arborescens) и др. Площадь посадок заняла почти
20 кл12. Первые 35—40 лет деревья росли прекрасно, и лесоводы были-
уверены в успехе. Требовалось лишь частое рыхление почвы в искусст-
венных лесных насаждениях и постоянное удаление травяного покрова.
Но вот наступил засушливый 1891 г. Деревья начали болеть и полностью'
уже не оправились; со временем весь лес погиб.
3. Если на границе между лесом и степью имеются не слишком глу-
боко залегающие грунтовые воды, то наблюдается постепенное пониже-
ние их уровня в направлении от опушки леса в глубь леса. Это означает,,
что лес расходует не только воду атмосферных осадков, но и энергично-
черпает влагу из более глубоких горизонтов почвы, понижая тем самым
зеркало грунтовых вод. В результате под лесом выпавшие на поверхность
почвы осадки не успевают просочиться до водоносного горизонта 1
(рис. 65).
Там же, где корни деревьев достигают уровня грунтовых вод, леса*
сохраняются даже в степных районах. Типичны для степной зоны также
галерейные леса и леса в оврагах.
4. В августе и сентябре степь выгорает, надземные части растений,-
за исключением почек возобновления, отмирают. К этому времени года
запасы воды в почве оказываются исчерпанными самими степными расте-
ниями, которые переносят летнюю засуху без ущерба для себя, однако
последствия ее для древесных растений оказываются катастрофическими.
На границе с лесостепью можно часто наблюдать, как уже в сентябре,
то есть до начала собственно листопада, засыхают листья на деревьях
и кустарнике. Нередко отмирают даже целые ветви, и деревья принимают
уродливый вид.
5. Все сказанное выше объясняет, почему в степных районах нет
лесов, хотя одиночные деревья не являются здесь редкостью. У отдельно
стоящего дерева корневая система, беспрепятственно развивающаяся во
1 Именно поэтому колодцы всегда закладывают не в лесу, а за его пределами..
74
Глава
Рис. 66. Древесная растительность (Pinus ponderosa) на скалистых грядах
в низкозлаковой прерии (фото Упхофа).
все стороны, пронизывает столь большой объем почвы, что дерево в тече-
ние года не испытывает недостатка воды. Если это так, то следовало бы
ожидать, что в естественных условиях степь примет саванноподобный
облик. Однако в умеренной зоне саванны не возникают по той причине,
что сеянцы лиственных древесных пород не выдерживают жесточайшей
конкуренции со злаками. Только там, где сомкнутый злаковый покров
•отсутствует, например на скалистых гребнях и скальных обрывах, в степ-
ных районах, а также в прерии, создаются предпосылки для роста деревьев
(рис. 66). Известно, что в аридных районах каменистые почвы характе-
ризуются сравнительно высокой влажностью (ср. т. I, стр. 315). Такие
почвы, сухие с поверхности, в глубине содержат запас влаги, вполне
достаточный для развития древесных растений. Напомним, что степные
-злаки обладают очень густой, разветвленной корневой системой, которая
охватывает, однако, лишь очень небольшой объем почвы, интенсивно его
используя (рис. 67) Ч Такая корневая система оказывается оптимальной
на тонкозернистой мощной почве, какой является чернозем, подстилаемый
лёссом. Каменистая же почва делает невозможным развитие интенсивной
корневой системы злаков. Здесь в более благоприятном положении ока-
зываются древесные растения с их экстенсивной корневой системой,
простирающейся далеко в горизонтальном и вертикальном направлениях.
Корни деревьев, используя расселины скал или трещины между камнями,
проникают глубоко в грунт и поглощают всегда имеющуюся на глубине
влагу (ср. т. I, стр. 284). На такого рода скалистых местообитаниях раз-
вивается редкий древостой, состоящий обычно из дуба или сосны, под
кронами которых вырастают лишь отдельные травянистые растения.
Следовательно, границу между лесом и степью наряду с климатом
определяет также и почва. На мощных тонкозернистых почвах степь
проникает через климатическую границу далеко к северу, лес, напротив,
используя каменистые почвы, выдвигается к югу.
1 R. Т. С о u р 1 a n d, R. Е. I о h n s о n, Rooting characteristics of native grass-
land species in Saskatchewan, «J. Ecol,», 53, 1965, p, 475—507.
Рис. 67. Корневая система у шести экземпляров Stipa spartea var. curtiseta, взятых с разных местообитаний, 4канадская прерия
(по Купленду и Джонсону).
1 южный склон, песчаная почва! 2 — ровная'поверхность на черноземе; 3—6 — более сухая зона развития, бурых почв. Максимальная мощность
почвенного слоя 150 см, верхний метр всегда густо пронизан корнями.
76
Глава 2
В степных районах Восточной Европы некоторое время назад весьма интенсивно»
закладывались лесные полосы, которые должны были служить защитой посевов от вет-
ра. Цель их — борьба с засухой, от которой каждые 3—5 лет страдало сельское хозяй-
ство южных? и’юго-восточных областей Восточной Европы. Задача состояла не в увели-
чении количества осадков, а в изменении климата приземного слоя воздуха в благо-
приятном для культурных растений направлении.
Ослаблением силы ветра предполагалось добиться:
1) более равномерного отложения снега зимой;
2) ослабления ветровой эрозии в сухие весенние месяцы, когда ветер беспрепят-»
ственно выдувает с полей еще голую почву, а вместе с ней и озимые (пыльные бури);
3) снижения иссушающего воздействия ветра в жаркие летние месяцы, то есть
сокращения потери влаги растениями в процессе транспирации.
При равномерном снежном покрове почва зимой глубоко не промерзает и весной
скорее оттаивает: талые воды проникают глубже, благодаря чему почва эродируется
менее интенсивно и в ней накапливается больше влаги.
Опыты, проведенные на небольших площадях, подтвердили ожидаемое благо-
приятное влияние ветрозащитных лесных полос, но при закладке последних в боль-
шом масштабе встретились определенные трудности. При посадке использовались,
не только местные древесные породы, как Quercus robur, Ulmus effusa, Acer platanoides,
A. tataricum, но и экзотические виды — Acer negundo, Fraxinus pennsylvanica, Robinia
pseudacacia, Gleditschia triacanthos, а также местами абрикос и шелковица. Из кустар-
ников в подлеске оказались пригодными Caragana arborescens, Amelanchier ovalis,
Elaeagnus angustifolia. Принятая вначале ширина полос 15—30 м позже в целях
уменьшения расходов была сокращена до 7,5 м и более. Но ветрозащитные лесные поло-
сы хорошо росли только при тщательном уходе, а на юге, несмотря на все усилия,
оставались низкими; их влияние было заметно лишь на расстоянии, в 20—30 раз
превышавшем высоту древостоя. Иными словами, сеть ветрозащитных полос должна
была быть очень частой, что на практике оказалось невыполнимым. Деревья в полосе
страдают от недостатка воды, о чем свидетельствует интенсивность транспирации,
оказавшаяся в 2—3 раза ниже, чем у древостоев в окрестностях Москвы, хотя испаряе-
мость в степи намного выше х. При организации лесополос оправдал себя посев желудей
гнездами по нескольку штук. В этом случае всходит сразу много сеянцев, причем
крайние из них в первые годы угнетаются степными злаками, но вместе с тем они защи-
щают срединный всход от конкуренции. Поэтому срединный сеянец развивается быст-
рее. Он йоднимается над злаками и со временем вырастает в дерево, тогда как краевые
сеянцы погибают 1 2. Было также установлено, что в посадках дуба корневые системы
отдельных деревьев срастаются одна с другой, образуется своеобразный «многостволь-
ный индивидуум».
Согласно устному сообщению Гродзинского (Киев), при полном
заповедании луговых степей[происходит деградация растительного покро-
ва, поскольку интенсивно накапливающаяся подстилка угнетает дерно-
винные злаки. Мертвый покров снижает испарение, благодаря этому
улучшается водный режим почвы, что в свою очередь способствует посе-
лению корневищных злаков — пырея и Calamagrostis epigeois. В услови-
ях отсутствия конкуренции со стороны степных злаков появляется воз-
можность прорастания семян сосны, которые заносятся ветром с соседних
скалистых возвышенностей, поросших сосной. Дернообразующие злаки
распространяются центробежно, однако самые старые части в центре
дерновины мало-помалу отмирают. Возникают лысые пятна, занимаемые
сорняками; правда, со временем их снова вытесняют дерновинные злаки:
Stipa, Festuca sulcata.
В естественных условиях накоплению подстилки препятствуют уме-
ренный выпас и случайные, вызванные ударом молнии, степные пожары.
До заселения степей человеком здесь паслись сайгаки и тарпаны (дикие
1 Л. А. И в н о в, А. А. С и л и н а, Ю. и Л. Ц е л ь н и к е р, О транспирации
полезащитных полос в условиях Деркульской степи, «Бот. ж.», 38, № 2, 1953,
стр. 167—184.
2 Е. Г. Орленке, О взаимоотношениях дуба в густых культурах* «Доклады
АН СССР», 102, № 4, 1955, стр. 841—844.
Зона степей и прерий
77
лошади), но преобладали главным образом бесчисленные крупные и мел-
кие грызуны; известную роль играли также саранчовые. Умеренную
пастьбу и периодические пожары следует рассматривать как естественные
факторы распространения злаковников.1 Мы не знаем ни одного злаков-
ника, где дикие животные не входили бы в состав естественного биогео-
ценоза (см. также гл. 3).
По данным Гродзинского, структура растительного покрова в степи в известной
мере зависит также от присутствия в почве тормозящих веществ, так называемых
Коллинов, выделяемых некоторыми видами растений. С. помощью опытных растений
(проростков редиса и степных злаков) удалось определить их концентрацию в почве,
По активности образования коллинов растения можно подразделить на три группы.
1. Наиболее активно выделяют коллины отдельные экземпляры видов, растущих
в степи, как, например, Salvia, Crambe tatarica, Verbascum lychnites, Serratula xeranthe-
moides, Adonis wolgensis и др. Стоят эти растения поодиночке, так как почва вокруг
них содержит много коллинов, которые препятствуют возникновению даже их соб-
ственных всходов. Поэтому в следующей генерации эти виды меняют местообитание
и, следовательно, не могут быть доминантами.
2. Приблизительно в 10 раз меньше коллинов выделяют корневищные злаки
(A gropyrum repens, Calamagrostis epigeois, Hierochloa odorata), а также подушечники,
например Thymus. Однако co временем под ними накапливается столько тормозящих
веществ, что растения сами себя отравляют, поэтому растущие в центре куста особи
постепенно отмирают. Эти виды часто доминируют, но лишь в течение ограниченного
времени, что, правда, не совпадает с наблюдениями над Calamagrostis, нередко
•сплошь занимающим лесные вырубки. В восточном Средиземноморье Thymus покрывает
обширные площади, тем не менее признаки вырождения не были замечены.
3. Преобладающие в степи виды злаков, такие, как Stipa, Festuca sulcata, Роа
angustifolia и др., вырабатывают очень небольшое количество коллинов, поэтому их
рост на одной и той же площади ничем не ограничен.
Для выявления присутствия коллинов в почве используют водные вытяжки,
которые содержат много соединений, оказывающих тормозящее влияние на развитие
растений. Это влияние в почвенных пробах, взятых в 10 см одна от другой, может быть
весьма различным. Для отдельных видов растений в большинстве случаев оно оказа-
лось неспецифическим, однако известны и строго индивидуальные зависимости. Так,
например, семена Salvia austriaca прорастают только на вытяжках из почвы под Festuca
и Stipa, но не всходят на вытяжке из почвы под A gropyrum', для S. tesquicola наблюдает-
ся обратная картина, тогда как семена S. nutans какой-либо закономерности не обна-
руживают. Соответственно S. austriaca растет только в сообществе Stipa — Festuca,
a S. tesquicola — только в травостое A gropyrum. Напротив, S. nutans встречается
в обеих группировках 1 2
ПОЧВЕНННЕ УСЛОВИЯ В СТЕПЯХ
Почвы восточноевропейской степи и почвы североамериканской
прерии во многом сходны между собой. Однако зависимость типа почвы
от климата в Восточной Европе проявляется более резко, так как здесь
сокращение количества осадков и повышение температуры происходят
в одном и том же направлении — с ССЗ на ЮЮВ. В Северной Америке,
напротив, осадки уменьшаются с востока на запад, а температура повы-
шается с севера на юг. Кроме того, местность к западу значительно под-
нимается; у подножия Скалистых гор низкозлаковая прерия расстилается
уже на высоте 1000—2000 м над уровнем моря. Познакомимся поближе
1 Значение степной фауны, подтверждаемое также палеонтологическими наход-
ками, в геоботанической литературе до настоящего времени мало учитывалось. На это
указывают Е.'М. Лавренко и А. А. Юнатов в статье «Залежный режим в степях как
результат воздействия полевки Брандта на степной травостой и почву», см.: «Бот. ж.»,
37, № 2, 1952, стр. 128—138.
2 А. М. Гродзинский, Аллелопатия в жизни растений и их сообществ,
основы химического взаимодействия растений, Киев, 1965.
78
Глава 2
Рис. 68. Карта типов почв степной области Восточной Европы и прилегаю-
щих|[лесных районов.
1 — подзолистые почвы; 2 — луга и болота; 3 — карбонатное засоление; 4 — серые лесные
почвы; черноземы; 5 — северные; 6 — мощные; 7 — обыкновенные, 8 — южные, 9 — каш-
тановые и бурые пустынно-степные^почвы; 10 — песчаные дюны; 11 — каменистые почвы.
п — поды (бессточные степные блюдца); с — засоленные почвы (солончаки) (из «Phytologle»,
Bd. Ill, 1).
с почвами восточноевропейской степи. Они изучены большим числом
русских почвоведов, и прежде всего Докучаевым (1883) и Глинкой (1913),
чьи труды заложили основы научного почвоведения. Из анализа почвен-
ной карты Восточной Европы (рис. 68) видно, что речь идет преимущест-
венно о разных подтипах чернозема. Только в лесостепной зоне под леса-
ми распространены серые лесные почвы. Материнская порода весьма
однородна и представлена мощным, занимающим всю область лесостепи'
и степи лёссовым покровом. Лёсс обогащен известью; более легкий на
севере области (только 5—15% глины, 60—80% пылевидных частиц),
к югу лёсс становится тяжелее (глины до 35—55%). Климат семиаридный,
характеризуется в районе лесостепи сухим периодом; в собственно степи
нередки даже засухи. Семиаридный климат, злаковая растительность-
и мощная, известьсодержащая подпочва — вот три определяющих усло-
вия формирования типичного чернозема. Климат не остается неизменным
Зона степей и прерий
79»
в пределах всей области. Аридность климата плавно возрастает по пере-
движения с ССЗ на ЮЮВ, вместе с тем и растительность приобретает) все-
более ксерофитный характер. Такая смена условий среды позволяет
подразделить черноземную область Восточной Европы на ряд подзон.
Мы различаем:
1) мощный чернозем;
2) обыкновенный чернозем и
3) южный чернозем.
Каждому типу почв свойствен характерный почвенный профиль-
(рис. 69). Темноокрашенный гумусовый горизонт мощного чернозема
прослеживается до глубины 170 см, содержание гумуса в горизонте Аг
составляет 5—6%; книзу горизонт постепенно переходит в более свет-
лый горизонт А2 х. Его сменяет горизонт А3 — слабоокрашенный гумусом
лёсс, а затем горизонт С — неизмененный лёсс с призматической струк-
турой. Выделения извести встречаются только в виде псевдомицелия.
Темный цвет почвы обусловлен хорошим перемешиванием гумусовых
соединений с минеральными частицами и насыщением поглощающего-
комплекса почвы известью. Два фактора способствуют созданию опти-
мальных условий для гумусообразования: густая корневая система зла-
ков, пронизывающая верхние слои почвы и после отмирания преобразую-
щаяся в гумус, и интенсивная жизнедеятельность населяющих почву
животных. Мелкие грызуны — и прежде всего суслики — проклады-
вают в почве глубокие ходы. Многочисленные дождевые черви и личин-
ки насекомых обеспечивают поступление надземных частей растений
в виде экскрементов в почву и превращение их в гумус. Количество
сусликов в украинских степях оценивается в 30 млн. Но надо пом-
нить, что каждый суслик, помимо кормовых злаков, съедает
в год 4—8 кг пшеницы. Тем самым он наносит урожаю большой урон.
Глубина норы суслика достигает 2 м. Необитаемые норки постепенно
заполняются сверху мелкоземом, содержащим частицы гумуса. В разрезе
лёссовой толщи такие норы, называемые «кротовинами», выделяются своим
черным цветом. Гумусовый горизонт черноземных почв обладает зернисто-
комковатой структурой и представляет собой идеальную пахотную землю.
Поэтому в настоящее время вся степная область, за исключением очень
небольших участков, занята пашней.
Во влажное время года свободная известь в почве растворяется
просачивающимися с поверхности дождевыми и талыми водами, погло-
щающими из почвы СО 2, и перемещается вместе с током воды вглубь.
Однако для того, чтобы смыть известь на всю глубину профиля, до уровня
грунтовых вод, осадков не хватает; более того, летом резко увеличивается
поглощение воды корнями растений. Раствор извести становится все
более насыщенным, и наконец известь кристаллизуется как карбонат.
В мощных черноземах карбонаты имеют форму белых тонких нитей,
налет которых в свежем почвенном профиле напоминает плесень. Поэтому
мы говорим о «плесневых карбонатах», или «псевдомицелии». Положение
их в разрезе маркирует среднюю глубину просачивания атмосферных
1 Запасы питательных веществ и ежегодно поступающее в почву в результате-
минерализации растений количество питательных веществ весьма велики. Удобрение-
фосфатом и калием результатов не дает, но внесение 40 кг нитрата на 1 га дает несом-
ненный эффект, причем стимулируется рост главным образом злаков, особенно корне-
вищных (Bromus и др.). Ср.: Н. И. Болотина, Е. А. В у л ь ф и у с, Питание-
растений и круговорот соединений азота, фосфора и калия в мощном целинном черно-
земе, «Труды Центральночерноземного заповедника», 8, Воронеж, 1965*,
«80
Глава 2
Р и с. 69. Профили почв лесостепной и степной зон Восточной Европы в направлении
с севера на юг (из «Phytologie», Bd. Ill, 1).
1 — горизонт вскипания; 2 — псевдомицелий; з — белоглазка; 4 — кротовины; 5 — пластинчатая
•структура лесных почв. Содержание гумуса в горизонте А, дано в %. Дальнейшие объяснения см.
в тексте.
осадков и проникновение корней растений. Такое переотложение угле-
кислых соединений ведет к тому, что в верхнем слое почвы они отсут-
ствуют; поэтому «горизонт вскипания» обычно залегает не у поверхности,
а в 20—30 см выше скопления карбонатов, у мощных черно-
13 е м о в — на глубине около 70 см. Чем меньше становится годовая
Зона степей и прерий
81
сумма осадков по направлению к югу, тем менее глубоко промачивают
они почву, тем выше располагаются горизонты вскипания и зона накоп-
ления карбонатов. Одновременно изменяется и их структура.
В обыкновенных черноземах под псевдомицелием
выделяется еще слой, содержащий твердые округлые конкреции СаСО3
(«белоглазка»). Поскольку продуктивность растительного покрова на обык-
новенных черноземах сокращается, гумусовый горизонт здесь менее
мощный (приблизительно до 75 см). К тому же сухой климат препятствует
разложению гумуса, из-за чего содержание его в почве в этих условиях
несколько возрастает (до 7—8%). Поэтому в континентальных районах
восточнее Днепра чернозем в тех же подзонах богаче гумусом, чем черно-
зем(западнее Днепра. Пересекая степную область с севера на юг, легко
заметить, как равномерно сокращается мощность гумусового горизонта,
а горизонты вскипания и накопления карбонатов поднимаются все
выше по профилю.
Южнее подзоны обыкновенного чернозема протягивается полоса
южного чернозема. Гумусовый горизонт здесь едва достигает
50 см] его окраска становится светлее, и содержание гумуса понижается
до 5%. Псевдомицелий уже отсутствует; встречается только белоглазка,
которая наблюдается с глубины 50 см (см. рис. 69, справа).
С увеличением сухости климата к югу продолжает изменяться
и характер почвы. Гумусовый слой становится тоньше и светлее, черная
окраска сменяется темно-бурой и, наконец, светло-бурой. В районах
аридного климата уже развиты каштановые почвы ив области
полупустынь и пустынь — очень бедные гумусом бурые почвы
исероземы.
Так как почвы аридных районов Восточной Европы повсеместно
подвержены засолению, к их описанию мы вернемся еще раз в разделе,
посвященном засоленным почвам.
Карта размещения зон чернозема в Восточной Европе (рис. 68)
в упрощенном виде отображает условия, выделенные Маховым на очень
детальной карте масштаба 1 : 750 000. Каждому типу почвы соответствует
определенная зона растительности (табл. 16).
Луговую степь принято относить к лесостепной зоне, в пределах кото-
рой на местообитаниях, хорошо обеспеченных водой, сначала развивается
дубовый кустарник с колючими кустарниками, а затем — дубово-гра-
бовые леса.
Такая растительность распространена на деградированных
черноземах. Мощность перегнойного горизонта не превышает
60 см, содержание гумуса снижается доЗ%. Карбонаты из почвы полностью
вымыты; поэтому реакция верхних слоев почвы кислая. Начинается
разрушение глинных минералов, образуются коллоидальные гумусные
кислоты, происходит миграция глины а также гидратов окиси железа
из верхних частей почвенного профиля в глубину, возникает иллю-
виальный горизонт В. Процесс оподзоливания проявляется отчетливее,
когда вырастает дубово-грабовый лес. Под таким лесом формируются
лесные почвы, темно- или светло-серые в зависимости от степени
оподзоливания. Гумусовый горизонт сокращается и светлеет (содержа-
ние гумуса часто падает до 1%), комковатая структура утрачивается,
реакция становится все более кислой, на поверхности почвы накапливается
подстилка (горизонт Ао); полуторные окислы (А12О3, Fe2O3) вместе с золя-
ми гумуса выносятся в глубь профиля и образуют четко выраженный
иллювиальный горизонт В; под перегнойным горизонтом располагается
6-0434
82
Глава 2
Таблица 16>
Зависимость между типом почвы и растительностью (по Клеопову)
I. Лесостепная зона
Тип почвы
Оподзоленные лесные
почвы на лёссах
а) светло-серые и серые лесные поч-
вы — влажные и более сухие
б) деградированный чернозем и его
южные варианты
Чернозем
а) северный чернозем
б) мощный чернозем
Торфяно-болотные почвы
а) сильнощелочной низинный торф
Засоленные почвы
а) содовый луговой солончак
б) содовый корковый солонец
Характер растительности
Лиственные леса
Carpineto-Nemoretum, на востоке Nemo re-
turn aegopodiosum и caricopilosum
Дубовые леса с кустарниковым ярусом
(Quercetum fruticosum) и дубово-терновый
кустарник
Луговая степь
Влажные разнотравные луговые степи
Типичная луговая степь
Сообщества болот
Крупноосоковые и тростниковые болота
Луга на засоленных почвах
Влажные гемигалофитные луга
Сухие гемигалофитные луга
II. Степная зона
Чернозем
а) обыкновенный чернозем (северный,
типичный и южный варианты)
б) каштановая почва, слабосолонцева-
тый чернозем
Степи со Stipa
Разнотравные степи со Stipa (гигро-мезо-
и ксерофитная бедная разнотравьем
степь со Stipa)
Сухая, бедная разнотравьем степь со Stipa.
III. Полынная степная зона
Засоленные почвы
а) сильносолонцеватая каштановая поч-
ва
б) хлоридно-сульфатный солончак и
комплекс корковых и корково-столб-
чатых солонцов
Сообщества полупустынь
Степи с A rtemisia — Stipa |
Сообщества галофитов на влажных и сухих
местообитаниях с мозаичной структурой
размещения
собственно подзолистый горизонт А2 — белесый, песчаный, с высоким
содержанием зерен кварца.
Островки леса на светло-серых лесных почвах особенно характерны
для Волыно-Подольской возвышенности, где количество осадков несколь-
ко увеличивается. Но то, что эти леса вторично обосновались на черноземе'
и оподзоливание произошло позднее, подтверждают многочисленные ста-
рые кротовины, которые легко заметить в нижней части почвенного про-
филя (рис. 69, слева) Ч
Такого рода морфологические изменения в свойствах почвы, обу-
словленные прогрессирующим оподзоливанием, подтверждаются данными
1 С другой стороны, степи заповедника в районе Курска — антропогенного про-
исхождения. На заповедной территории лес за 5 лет выдвинулся в степную раститель-
ность на 1—8 м. См.: Г. М. Зозулин, Н. М. Кузмарцова, Смещение границы
травяной растительности на постоянных пробных площадках Центральночернозем-
ного заповедника за 5 лет, «Бот. ж.», 36, 1951, стр. 240—248.
Зона степей и прерий
83
Черноземы Серые лесные почвы
мощные деградированные темные сильно оподзоленные
Рис. 70. Распределение гумуса (1), глинных минералов (2) и обмен-
ных Са и Mg (3) в почвенном профиле черноземов и серых лесных почв
(по Тюрину).
На оси абсцисс внизу — содержание гумуса в %, вверху — содержание глинных
минералов в % и Са + Mg в мжв.
химического анализа (рис. 70). В мощном черноземе содержание гумуса
с глубиной постепенно убывает, концентрация глинистых частиц не изме-
няется, количество обменных катионов (Са и Mg) весьма велико по всему
профилю, иными словами, гумус мощных черноземов полностью насыщен.
В профиле деградированного чернозема наблюдается быстрое уменьше-
ние содержания гумуса с глубиной; вместе с тем в верхних слоях почвы
несколько сокращается количество глины, накапливающейся на глубине
30—50 см', насыщение кальцием и магнием еще достаточное. Отчетливо
видны изменения в темно-серых лесных почвах, но наиболее резко они
выражены в светло-серых, сильно оподзоленных почвах, в которых боль-
шое содержание гумуса отмечается только в верхних 10 см. Глинистая
фракция из этих слоев частично вымыта и сконцентрирована на глубине
40—60 см. Количество обменных кальция и магния особенно мало в гори-
зонте А2 — признак, характерный уже для настоящих подзолистых
почв. Намного более сложные условия формирования почв в Северной
Америке иллюстрирует рис. 90. Почвы прерий соответствуют северному
чернозему.
ХАРАКТЕРИСТИКА СТЕПНОЙ РАСТИТЕЛЬНОСТИ
ВОС ТО ИНОЙ ЕВРОПЫ1
Каждой почвенной зоне отвечает определенный тип растительности.
Подробно изучая зависимости, существующие между почвой и раститель-
ностью на сохранившихся участках степи и в заповедниках, можно
1Г. Вальтер, В. Алехин, Основы ботанической географии, М. — Л.,
1936.
6*
84
Глава 2
с их помощью восстановить прежнюю растительность степей тех времен,
когда степь еще не была превращена в пашню. Такую реконструкцию
растительного^покрова Украины произвел, в частности, Клеонов.
Луговая степь на мощном черноземе
Южная граница зоны проходит от Кишинева на восток через Кре-
менчуг, Харьков, Балашов, Вольск, Куйбышев к Уфе.
Почва в полосе луговой степи после таяния снега хорошо увлажнена,
поэтому весной и ранним летом, когда температура воздуха день ото
дня быстро повышается, здесь развивается богатая растительность.
Можно установить такие характерные аспекты (рис. 71).
Рис. 71. Весенние аспекты в луговой степи; вертикальная
проекция, квадраты дециметровые (по Покровской).
Вверху. Начало апреля: бурый аспект с лиловыми пятнами Pulsatilla
patens; желтые — мужские колоски Carex humilis. Первые цветки Adonis.
В середине. Вторая половина апреля: желтый аспект от Adonis vernalis.
Кроме того, цветет нежно-голубой Hyacinthus leucophaeus. Степь еще не
зазеленела.
Внизу. Конец мая: голубой аспект от Myosotis silvatica, отдельные цвету-
щие экземпляры Anemone silvestris и желтые пятна Senecio campestris.
Зацветает ковыль.
Зона степей и прерий
85
I. Послезимний аспект. Снег в степи только что сошел и почва покрыта остатками
прошлогодних растений.
II. Предвесенний аспект. Достаточно нескольких теплых дней в апреле, чтобы
появились фиолетовые цветки прострела (Pulsatilla patens)’, одновременно начинает
«пылить» осока низкая (Carex humilis).
III. Ранневесенний аспект. Уже в первой половине мая повсюду появляются
крупные золотистые звезды Adonis vernalis, красиво выделяющиеся на фоне еще цвету-
щего прострела, между ними проглядывают нежные светло-голубые соцветия Hyacin-
thus leucophaeus.
IV. Средневесенний аспект. Степь уже зеленеет, раннецветущие виды исчезают.
Во второй половине мая их сменяют белые соцветия чины (Orobus albus = Lathyrus
pannonicus), лиловые цветки ириса (Iris aphylla) и крупные белые звезды Anemone
silvestris.
V. Поздневесенний аспект. В конце мая — начале июня степь снова преобра-
жается. Это ее самая красочная фаза: на зеленом фоне живописно разбросаны бесчис-
ленные нежно-голубые незабудки (Myosotis silvatica),. желтый крестовник весенний
(Senecio campester) и лютик (Ranunculus polyanthemus), появляются первые легкие
перья ковыля (Stipa Joannis) (рис. 71, внизу).
VI. Раннелетний аспект. Весенний период закончился, начинается фенологиче-
ское лето. Степь отливает фиолетово-голубым цветом от изобилия цветков лугового
шалфея (Salvia pratensis). Между ними колышутся на ветру длинные перистые ости
видов Stipa (рис. 72 и 73), поднимаются метелки высокого костра (Bromus riparius,
вид, близкий к В. erectus). Ранним утром повсюду светятся золотистые звезды козло-
бородника (Tragopogon pratensis), закрывающего свои цветки уже в полдень.
Рис. 72. Аспект луговой стеци в июне (по Вальтеру и Але-
хину, 1936).
Слева направо: Trifolium repens, Salvia pratensis, Festuca sulcata, Viola are-
naria, Trifolium montanum, Stipa Joannis, Carex vema, Ranunculus poly-
anthemus, Chrysanthemum leucanthemum, Hypochoeris maeulata, Anthoxan-
thum odoratum, Arenaria graminifolia, Filipendula hexapetala, Carex
montana, Agrostis canina, Viola canina, Scorzonera purpurea, Euphorbia
gracilis, Potentilla opaca, Festuca rubra, Astragalus danicus, Phlomis tuberosa,
Koeleria delavignei, Galium boreale, Echium rubrum, Carex montana. Ha
почве Thuidium abietinum. Квадраты дециметровые.
86
Глава 2
Рис. 73. Stipa Joannts в луговой степи, район Воронежа (фото Попова).
Рис. 74. Летний аспект луговой степи с многочисленными цветущими видами
разнотравья и Stipa. На заднем плане — осиновые кусты в небольших низинах
(фото Келлера).
Зона степей и прерий
87
VII. Среднелетний аспект. Во второй половине июня степь становится белой
от бесчисленных соцветий горного клевера (Trifolium montanum) и нивянника (Chry-
santhemum leucanthemum), степной таволжанки (F tlipendula hexapetala). В общий фон
вкраплены колокольчики (Carppanula sibirica и С. persicifolia), соцветия Knautia
arvensis, высокие свечки румянки (Echium rubrum) is др. (рис. 74).
VIII. Аспект разгара лета. В начале июля начинают доминировать розовые соцве-
тия эспарцета (Onobrychis arenarta). Однако пестрые краски уже блекнут, число цвету-
щих видов уменьшается, степь приобретает монотонную окраску. В это время цветет
желтый подмаренник (Galium verum).
IX. Позднелетний аспект. С середины июля степь уже не напоминает пестрый
ковер. Многие растения засыхают. Общий фон не сочный зеленый, а бурый. Цветущие
растения редки, например высокая с синими соцветиями живокость (Delphinium lit-
winowi).
X. Последствий аспект. Вспыхивающие в конце июля темно-красные свечки чеме-
рицы (Veratrum nigrum) возвещают о конце лета.
XI. Осенний аспект. В августе и сентябре цветущих растений в степи уже нет
совсем. Из-за большой сухости степь выгорает и кажется бурой и безжизненной. В таком
состоянии она уходит под снег.
Подробный флористический состав степи передает приводимое ниже
описание пробных площадок. Из этого списка видно, что многие растения
луговых степей востока Европы встречаются и в Центральной Европе,
где они предпочитают сухие теплые местообитания на карбонатных поч-
вах, выступая как компоненты так называемых «степных пустошей»
или остепненных лесов. Правда, иногда речь идет о других подвидах
(табл. 17).
Общее число видов, образующих степное сообщество Курского райо-
на, равно 220, из них 20 — злаки и 180 — собственно разнотравье х.
Следовательно, последнее резко преобладает. Некоторые виды разно-
травья к югу исчезают (Salvia pratensis, Chrysanthemum, Filipendula
hexapetala, Senecio campester, Myosotis silvatica, Pedicularis comosa, Vale-
riana rossica и др.). Напротив, типичные для южных степей виды разно-
травья {Salvia nutans, Phlomis pungens, Potentilla recta) здесь отсутствуют
или встречаются только на южных склонах. Но и среди злаков собственно
дерновинные злаки, прежде всего ковыли, также отступают на вто-
рой план.
Преобладают дернообразующие злаки {Bromus riparius, Agrostis tenui-
folia, Avena pubescens, A. schelliana). Типчак {Festuca sulcata), правда,
встречается часто, но лишь в виде подсева, цветет редко. Весьма типична
осока {Carex humilis). Поверхность почвы, не занятая злаками, покрыта
мхом Thuidium abietinum. Из луковичных геофитов встречаются только
Hyacinthus leucophaeus и Gagea erubescens. Растения, образующие форму
«<перекати-поле», почти полностью отсутствуют.
В соответствии с законом относительного постоянства местообита-
ний и смены биотопов (ср. т. I, стр. 317 и след.) растительность луговой
степи экстразонально проникает глубоко в гумидную область Централь-
ной Европы; но здесь она в качестве «степной пустоши» связана с наиболее
сухими местообитаниями, например с песчаными почвами Верхнерейнской
низменности под Майнцем, с маломощными [почвами на известняках
1 Ср. многочисленные описания участков степи, еще существовавших полстоле-
тия назад, в работах, посвященных охране природы: И. И. С п р ы г и н, Злаковые
степи Пензенской губернии, М., 1926; Б. А. Келлер, Степи центральной чернозем-
ной области. М. — Л., 1931. В последней работе приведены также многочисленные
данные об урожае сена (вес в воздушно-сухом состоянии), получаемом в разных степ-
ных сообществах. Урожаи в зависимости от интенсивности выпаса колеблются от
1000 до 5000 кг!га и достигают максимума, если преобладают виды Stipa, что вместе
с тем является признаком лишь слабого выпаса,
88
Глава 2
Таблица 17
Список видов и степень участия на пробной площадке (100 ле2)
в степном заповеднике южнее Курска
Злаки
3 Bromus erectus ssp. riparius
2 Festuca sulcata
2 Carex humilis
1 Agrostis canina ssp. tenuifolia
1 Avena pubescens
1 Poa pratensis ssp. tenuifolia
1 Stipa pennata ssp. Joannis
1 Carex praecox
+ Festuca pratensis
-j- Stipa stenophylla
+ Agropyrum glaucum ssp. intermedium
+ Agropyrum repens
+ Avena pratensis ssp. schelliana
+ Bromus inermis
+ Briza media
+ Koeleria gracilis
+ Phleum phleoides
+ Carex verna
-j- Luzula campestris ssp. pallescens
Бобовые
3 Trifolium montanum
2 Onobrychis arenaria
1 Lathyrus pannonicus
1 Trifolium alpestre
1 Vicia crace a
+ Anthyllis vulneraria ssp. polyphylla
+ Cytisus ruthenicus *
+ Lotus corniculatus
+ Medicago falcata
+ Trifolium campestre
+ Trifolium pratense
-J- Trifolium repens
Виды разнотравья
3 Filipendula hexapetala
2 Alectorolophus major
2 Вunias orientalis
2 Hypochoeris maculata
2 Tragopogon orientalis
2 Viola arenaria
1 Achillea millefolium ssp. setacea
1 Ajuga genevensis
1 Androsace septentrionalis
1 Arenaria graminifolia
1 Arenaria serpyllifolia
1 Asperula cynanchica
1 Asperula tinctoria
1 Campanula sibirica
1 Cerastium caespitosum
1 Erigeron acer
1 Galium verum
1 Inula hirta
1 Iris aphylla
1 Jurinea cyanoides
1 Knautia arvensis
1 Linum perenne
1 Myosotis silvatica
1 Pedicularis comosa
1 Plantago lanceolata
1 Plantago urvilleana *
1 Polygala comosa
1 Potentilla argentea
1 Primula officinalis
1 Salvia pratensis
1 Stachys recta
1 Thymus marschallianus
1 Valeriana rossica *
1 Vipla hirta
+ Adonis vernalis
4- Anthericum ramosum
+ Artemisia austriaca
- - Convolvulus arvensis
- - Delphinium litivinowi *
- - Dianthus capitatus *
- - Dracocephalum ruyschiana
- - Erysimum hieracifolium
- - Euphorbia gracilis *
- - Euphorbia virgata
- - Fragaria viridis
- - Galium boreale
- - Gentiana ligulata
- - Geranium sanguineum
- - Helichrysum arenarium
- - Hieracium pilosella
- - Hieracium spec.
- - Melampyrum cristatum
- - Nonnea pulla
- - Phlomis tuberosa
- - Plantago media
- - Polygonatum .officinale
- - Potentilla alba
- - Potentilla opaca
- - Prunella grandiflora
- - Prunella vulgaris
- - Pulsatilla patens
-- Rumex acetosa
+ Rumex acetosella
+ Salvia nutans *
+ Scabiosa ochroleuca
+ Scorzonera purpurea
+ Senecio campester
+ Senecio jacobaea
+ Serratula heterophylla *
+ Sisymbrium austriacum ssp. junceum
+ Stellaria graminea
+ Taraxacum vulgare
+ Trinia henningii *
,,Veratrum nigrum
Зона степей и прерий
89
Продолжение табл. 17
+ Berteroa incana
-j- Betonica officinalis
+ Calamintha acinos
-j- Campanula persicifolia
+ Carduus crispus ssp. hamulosus
+ Centaurea marschalliana *
+ Centaurea scabiosa
+ Veronica austriaca
-j- Veronica chamaedrys
+ Veronica prostata
+ Verbascum phoeniceum
+ Vincetoxicum officinale
* Виды, отсутствующие в Центральной Европе.
или лёссовыми склонами южной экспозиции х. В каждом случае на место-
обитании встречается только ограниченное число видов луговой степи.
К ним присоединяются субсредиземноморские виды (например, орхидеи),
которые в степи полностью отсутствуют; на юго-западе Центральной
Европы эти виды все более выдвигаются на передний план. Напротив,
ближе к восточной периферии региона, в окрестностях Вены, степные
элементы наблюдаются особенно часто 1 2. Луговая степь имеет свои вариан-
ты. На северном черноземе добавляются влаголюбивые виды, такие, как
Anthoxanthum odoratum, Calamagrostis arundinacea, Alopecurus pratensis,
Festuca rubra, Polygonum bistorta, Viola canina, виды Alchemilla и др.
В таких случаях часто говорят о «степных лугах». Перечисленные виды
отсутствуют на юге или появляются там лишь на северных склонах. Самая
южная полоса зоны луговой степи на менее мощном черноземе является уже
переходной к ковыльной степи. Здесь чаще встречается ковыль-тырса (Stipa
capillata). В широтном направлении также наблюдаются некоторые
различия, например в районе Тамбова в составе травостоя присутствуют
некоторые сибирские виды.
Ковыльная степь
Ковыльную степь можно подразделить на северную, богатую разно-
травьем зону, развитую на обыкновенном черноземе, и южную, бедную
разнотравьем зону, связанную с южным черноземом. Виды разнотравья
менее засухоустойчивы, чем степные злаки, и поэтому с увеличением
сухости климата они становятся все менее конкурентоспособными. Но все
они проникают в аридную область гораздо дальше, чем древесные расте-
ния, ибо, как и злаки, рано заканчивают вегетацию; во время засухи
их надземные части засыхают. Некоторые поздноцветущие виды разно-
травья,[например определенные зонтичные (как Eryngium или Peucedanum)
либо сложноцветные (как Artemisia, Centaurea или Linosyris), обладают
очень глубоко проникающей корневой системой или утолщенными стерж-
невыми корнями, в которых скапливаются запасы воды. Большая часть
их листьев, и прежде всего розеточные листья, к началу засухи засыхает,
соцветия же, расходующие мало влаги, сохраняются живыми. Опреде-
ленные виды, такие, как Artemisia и полукустарнички из маревых, даже
более засухоустойчивы, чем злаки, и сменяют их, как мы увидим ниже,
в полосе перехода степи в полупустыню.
1 Эти сухие луга обычно антропогенного происхождения; распространяются
они после - сведения леса на сухих местообитаниях; подробно описаны Элленбергом
(см. «Phytologie», Bd. IV, 2).
2Н. Nikfeld, Zur xerothermen Vegetation im Osten Niederosterreichs, «Verb.
Zool.-Bot. Ges.», Wien, 103/104, 1964, S. 152—181.
90
Глава 2
На границе между луговой и ковыльной степью доминантами ста-
новятся ковыли, ости которых в период цветения придают степи сереб-
ристый оттенок. Степь напоминает седое волнующееся море, когда траво-
стой колышется от легкого ветра (рис. 73). В это время года степь особенно
красива. Из ковылей важнейшими являются: Stipa lessingiana и на юге
S. ucrainica. В северных частях степи встречаются также Stipa stenophylla
и S. Joannis (из группы S. pennata), но сомкнутый покров эти виды обра-
зуют редко. Следует назвать и ковыль-тырсу {Stipa capillata). Большая
сухость климата в южной степной зоне обусловливает не только низкий
рост растений и ксероморфное строение ассимилирующих органов, но
и небольшую плотность растительного покрова. Поэтому на свободных
пространствах между многолетниками поселяются многочисленные одно-
летние и низшие растения. Терофиты особенно хорошо развиваются влаж-
ной весной и бесследно исчезают с началом сухого периода после созре-
вания семян.
В составе травостоя значительную роль играют также луковичные
геофиты (виды Tulipa, Ornithogalum, Gagea, Crocus и многие другие).
В разные времена года ковыльные степи, подобно луговым степям,
дают ряд аспектов. Приведем в качестве примера смену аспектов богато-
разнотравной ковыльной степи в районе Старобельска.
I. Предвесенний аспект. Тотчас после схода снега, во второй половине марта,
между остатками прошлогодних растений начинают зеленеть мхи (Syntrichia — Tortula
ruralis) и развивается густой покров сине-зеленой водоросли (Nostoc commune).
II. Ранневесенний аспект. Во второй половине апреля появляются цветущие
растения: разноцветные пятна низких Iris pumila, ярко-желтые крупные звезды Adonis
wolgensis и желтовато-зеленые звездочки Gagea pusilia и G. bulbifera. Весьма многочис-
ленны маленькие озимые однолетники (Draba verna и Holosteum umbellatum)-, на слабо-
засоленных почвах цветут тюльпаны (Tulipa schrenkii и Т. biebersteiniana).
III. Аспект разгара весны. Вскоре степь зеленеет, и на общем зеленом фоне «вспы-
хивают» многочисленные светлые соцветия Orobus albus и О. pallescens, Anemone Sil-
vestris, Valeriana tuberosa, желтые цветки Potentilla opaciformis н. P. patula. Но лучшим
украшением украинских степей служат пионы (Paeonia tenuifolia), предпочитающие
северные склоны. В это же время появляются первые ковыли (Stipa Joannis и S. ruben-
tiformis). Начинают цвести кустарники, образующие сплошные заросли на склонах
и каменистых почвах: терн (Prunus spinosa), миндаль степной (Amygdalus папа) и кара-
ганник кустарниковый (Caragana frutex).
IV. Раннелетний аспект. Во второй половине мая и до середины июня степь дости-
гает наибольшей красочности. Цветут ковыли (особенно Stipa lessingiana и S. rubenti-
formis), различные другие злаки (Festuca sulcata, Koeleria gracilis, Bromus riparius).
Между белыми метелками ковылей проступают синие соцветия шалфея (Salvia nutans),
белые цоцветия Filipendula hexapetala, Trifolium montanum и Arenaria graminifolia,
светло-голубые незабудки (Myosotis silvatica), голубые колокольчики (Campanula
steveni), красные головки юринеи (Jurinea arachnoidea) и высокие свечки румянки
(Echium rubrum).
V. Аспект разгара лета. Постепенно красочность степей тускнеет. Уже во второй
половине июня степь начинает приобретать бурый оттенок. Цветут лишь немногие
виды: желтые Galium verum и Centaurea ruthenica, беловатый подорожник (Plantago
media), фиолетово-красные Serratula radiata, Jurinea linearifolia и местами Salvia
nemerosa. В это время года цветет и большинство растений, образующих перекати-поле
(Statice tatarica, Serratula xeranthemotdes, Phlomis pungens, Statice latifolia и др.).
Преобладают желтый и голубой с красноватым оттенком цвета.
VI. Позднелетний аспект. Со второй половины июля до начала сентября в облике
степи доминируют жесткие, с металлическим блеском ости ковыля (Stipa capillata)
и многие зонтичные (Peucedanum ruthentcum, Ferula ferulago, Seseli tortuosum, Falcaria
vulgaris и др.). В это время цветет также грудница мохнатая (Linosyris villosa).
VII. Осенний аспект. С середины октября до снегопада степь безжизненна. Осен-
няя влага пробуждает к жизни только низшие растения (водоросли, мхи).
Еще южнее виды Filipendula hexapetala, Trifolium montanum, Anemone
silvestris, Echium rubrum и др., характерные для луговой степи, распростра-
нены только во влажных балках.
Зона степей и прерий
91
проекция, квадраты дециметровые (по Вальтеру и Алехину, 1936).
Слева направо: Stipa ucrainica, Geranium ucrainicum, Pyrethrum mil lefalia turn,
Stipa capillata (до цветения), Draba verna, Festuca sulcata (2 экземпляра),
S tip a I essingiana.
В качестве примера самой южной, бедной разнотравьем ковыльной
степи изберем степи обширного заповедника Аскания-Нова. Видовое
разнообразие этой степи в сравнении с северным вариантом значительно
беднее: около 12 видов на 1 м2 площади. Плотность травостоя показана
на вертикальной проекции пробной площадки (рис. 75). Важную роль
в травостое играет Festuca sulcata.
Ход развития по временам года здесь следующий.
I. Предвесенний аспект. В марте и начале апреля развиваются преимущественно
зазеленевшие уже осенью листья луковичного мятлика (Роа bulbosa var. vivipara)
и золотистые звездочки различных видов Gagea. Одновременно начинается цветение
весенних эфемеров (озимых однолетников): крупки весенней (Draba verna), вероники
(Veronica verna), незабудки (Myosotis arenaria) и ясколки (Cerastium ucrainicum).
II. Ранневесенний аспект. Приблизительно 15—20 апреля весенняя раститель-
ность достигает полного расцвета — появляются красные и желтые тюльпаны (Tulipa
schrenkii), желтые, лиловые, белые и коричневые низкие ирисы (Iris pumila).
III. Аспект разгара весны. Уже в мае степь пребывает в апогее своего развития:
цветут ковыли (Stipa lessingiana и У. ucrainica) и типчак (Festuca sulcata). Из видов
разнотравья выделяются крупные корзинки чертополоха (Carduus uncinatus) и темно-
лиловые гроздья коровяка (Verbascum phoeniceum). Это время фенологического лета.
IV. Раннелетний аспект. Уже вскоре злаки желтеют; однако число видов цвету-
щего разнотравья увеличивается (Phlomis pungens, Salvia nemorosa, Centaurea ruthenica,
Statice tatarica, Salvia aethiopis, Serratula xeranthemoides). В это время цветет также
ромашник тысячелистный (Chrysanthemum millefoltatum), часто растущий группами.
V. Аспект разгара лета. С середины июня до середины июля степь выгорает.
Только у немногих видов разнотравья еще развиваются соцветия.
VI. Позднелетний аспект. Иногда степь снова оживает, когда Stipa capillata
выбрасывает из колосьев длинные ости. Правда, полного развития она достигает лишь
при обильных дождях. В благоприятные годы Stipa вырастает до высоты груди челове-
ка, в сухие остается низкой. В это время могут зацвести также Centaurea diffusa, Allium
guttatum, A. paczoskianum, Peucedanum ruthenicum и др. Последними появляются
Linosyris villosa и Artemisia austriaca.
92
Глава 2
VII. Осенний аспект. Лишь с началом осенних дождей степь вновь зеленеет:
развертываются листья ковылей и овсяницы и всходят бесчисленные озимые однолет-
ки. В таком полузазеленевшем состоянии южные степи уходят под снег. Правда,
снежный покров сохраняется только в защищенных от ветра местах, со скошенных
участков снег сдувается
В доисторические времена степи Восточной Европы изобиловали
дикими животными. Напрашивается вопрос, какую роль играла пастьба
диких копытных в формировании растительного покрова степей. В наши
дни сохранившиеся в северных районах зоны степи обычно выкашиваются,
что оказывает, однако, не совсем такое же влияние, какое оказывала пасть-
ба диких животных. Косьба — одна из причин существования резкой
Рис. 76. Смена доминантов в ковыль-
ной степи по мере увеличения интен-
сивности выпаса А -> G (по Шалыту).
1 — виды Stipa', 2"— Festuca sulcata', 3 — Arte-
misia \austriaca.
границы между луговой степью и
лесом, так как [на сенокосах древе-
сные растения не возобновляются.
В первобытной степи низкорослые-
древесные виды Amygdalus папа, ви-
ды Spiraea и Caragana по опушкам
леса, вероятно, встречались более
часто.
Влияние выпаса изучалось в
южной степи Аскания-Нова х. Ин-
тенсивный выпас наносит степной
растительности тяжелый урон.
Первыми исчезают ковыли
(рис. 76). Из них наименее стоек
Stipa stenophylla и наиболее устойчив
Stipa capillata. Длительное время
сохраняется типчак (Festuca sulcata).
Совершенно не страдает луковичный
мятлик (Роа bulbosa var. vivipara),
который поэтому можно считать
типичным злаком пастбищ с чрез-
мерным выпасом. В больших коли-
чествах появляется однолетний сорняк Eragrostis minor.
На пастбищах исчезают также многие виды красиво цветущего степ-
ного разнотравья (например, Phlomis pungens, Ph. tuberosa, Iris pumila,
Jurinea linearifolia, Onosma tinctorium и др.). Напротив, широко распро-
страняются виды, не поедаемые скотом, например молочай (Euphorbia
segueriana) или Kochia sedoides. Таким образом, при интенсивном выпасе из
травостоя полностью выпадают многие степные растения, а на освободив-
шихся местах поселяются многочисленные сорняки, первоначально
чуждые степи (Artemisia austriaca, Centaurea solstitialis и др.). Однако
полный заповедный режим также неблагоприятен. Листья крупных
злаков в этом случае перед зимой не удаляются. Снег лишь придав-
ливает их к земле и тем самым затрудняет отрастание весной ранних
растений; в результате многие важные степные виды, утратив возмож-
ность возобновления, исчезают. Со временем все более начинает доми-
нировать ковыль-тырса (Stipa capillata). В конце концов образуется
столь плотная подстилка, что затрудняется возобновление семенами
и этого злака. В растительном покрове появляются лысые участки, где
1 М. С. Ш а л ы т, Степная растительность Аскании-Нова, «Сообщения Крым-
ского педагогия, ин-та», 7, Симферополь, 1938.
Зона степей и прерий
93
укореняются сорные растения; среди них первыми оказываются чертополо-
хи (Carduus hamulosus, Onopordon acanthium sl др.). Распространяется иног-
да полупустынный злак (Agropyron cristatum), образующий почти чистые
травостои. Растительный покров становится неоднородным, что всегда
служит одним из признаков нарушения равновесия степных сообществ.
Напрашивается вывод: для формирования типичной
степной растительности необходим умеренный
выпас, каким он был в доисторическое время. В результате такого
слабого выпаса: 1) в землю втаптываются семена степных растений, что
облегчает их прорастание; 2) уничтожаются пришлые сорные растения;
3) удаляется лишняя листовая масса злаков, в результате чего к зиме
сокращается слой отмерших растений. Все это, вместе взятое, способ-
ствует сохранению равновесия между различными компонентами степной
растительности и ее гармоничному развитию. Кроме того, выяснилось,
что степные растения очень чутко реагируют на различия в микрорельефе.
Уже в самом незначительном углублении, почва которого несколько боль-
ше увлажняется стекающей в него водой и одновременно обогащается
гумусом, поселяется иная растительность, носящая более северный харак-
тер по сравнению с растительностью окружающей степи на ровных место-
обитаниях. Эта особенность проступает еще более резко, когда мы знако-
мимся с более крупными блюдцеобразными, бессточными впадинами,
часто встречающимися именно в зоне южных степей, где их называют
«подами». В полосе перехода от плакорного пространства вниз, ко дну
впадины, легко обнаружить поясообразное расположение многих различ-
ных растительных ассоциаций, каждой из которых соответствует также
определенный тип почвообразования. Эта микрозональность напоминает
зональную смену растительности, наблюдаемую при пересечении Украи-
ны с юга на север (ср. стр. 131 и след.).
Микромозаичность, или микрокомплексность, растительности в сте-
пях^обусловлена жизнедеятельностью грызунов х. Норы крупных грызу-
нов — суслика (Citellus), байбака (Marnwta) и др. уходят очень глубоко
вниз, нередко достигая материнской породы. Выброшенная у выходного
отверстия земля лишена гумуса, богата известью, то есть обладает иными
свойствами, нежели верхние горизонты почвы. В районе распространения
солонцов таким образом часто выносятся на поверхность более засоленные
слои почвы. Поэтому вокруг нор отмечается иной состав растительности.
Если нор очень много и зверьки часто роют новые, то возникает отчетливо
выраженная микромозаичность растительного покрова. Требуется весьма
длительное время, чтобы почва и растительность на таких нарушенных
местообитаниях снова приняла свой обычный облик. Если степь распахи-
вается, то даже спустя много лет после начала распашки с самолета можно
заметить множество светлых пятен бедной гумусом почвы, выброшенной
из прежних нор.
Вероятно, даже большее значение имеют мелкие грызуны (виды Micro-
tus, Lagurus lagurus и др.). Правда, они перекапывают только верхние
слои почвы, так что почвенный профиль в целом нарушается лишь незна-
чительно. Опасность в другом: зверьки стравливают почти всю раститель-
ность вокруг своих нор, поедают даже подземные части дерновинных
1 Е. М. Л а в р еа. ко, Микрокомплексность и мозаичность растительного по-
крова степей как результат жизнедеятельности животных и растений, «Труды Бот. ин-та
АН СССР, сер. геоботаника», 8, 1952, стр. 40—70. См. также работу Е. М. Лавренко,
А. Юнатова, посвященную значению Microtus brandtii в формировании растительности
монгольских степей («Бот. ж.», 37, 1952, стр. 128—138).
94
Глава 2
злаков. Оголив все пространство вокруг нор, колония перекочевывает
на другое место. На площади, удобренной экскрементами и мочой, начи-
нается процесс, сходный с тем, какой происходит на заброшенной пашне;
заселение однолетними сорными растениями, двухлетними видами и сук-
цессией многолетних видов, которое продолжается до тех пор, пока не вос-
станавливается зональная растительность. Пастбищные угодья крупных
грызунов значительно обширнее, поэтому столь сильного локального
изменения растительности они не вызывают.
Мелкие грызуны каждые 4—5 лет размножаются особенно интенсивно,
а каждые 10 лет их численность достигает катастрофических размеров.
В степях Монголии на пространстве 500 тыс. га на 1 га площади насчитали
1—3 тыс. нор Microtus brandtii. Грызуны уничтожали 9/10 растительной
массы, так что для скота травы не оставалось. Мелкие грызуны интенсивно
изменяют растительность, но на непродолжительный срок; крупные гры-
зуны изменяют главным образом почву, но на очень длительное время.
С учетом высказанного становится понятным, почему растительный
покров степей отнюдь не столь однороден, как это может показаться на пер-
вый взгляд. В результате жизнедеятельности грызунов происходит извест-
ный круговорот растительных микрокомплексов, иными словами, отдель-
ные виды растений на определенных местопроизрастаниях постоянно
сменяют друг друга с течением времени. Такого рода чередование, по-види-
мому, является непременным условием длительного сохранения зональ-
ной растительности в неизменном составе на обширных площадях.
СЕВЕРОАМЕРИКАНСКИЕ ПРЕРИИ
Степная растительность, называемая в Северной Америке «прерией»,.
в средней части материка занимает огромную площадь, простирающуюся
от 54—56° с. ш. в северном Саскачеване и провинции Альберта до Мекси-
канского нагорья на юге. На востоке прерия проникает на территорию штата
Индиана, причем отдельные ее форпосты достигают штатов Огайо и Мичи-
ган. Западной границей области прерий принято считать восточный склон
Скалистых гор, хотя злаковники широко распространены также в обшир-
ных котловинах между горными хребтами; но здесь преобладают уже
зимние дожди, тогда как в собственно прерии и в степях Восточной Европы ‘
максимум дождей приходится на летние месяцы (рис. 77—82).
Основными растениями области прерий являются дерновинные злаки,
которые на востоке достигают большей высоты, а на сухом западе остаются
очень низкими. Соответственно в направлении с востока на запад разли-
чают три зоны:
1) высокозлаковую прерию, еще богатую разнотравьем; 2) смешан-
ную прерию с высокими и низкими злаками; 3) низкозлаковую прерию,
или Великие равнины.
На севере и северо-востоке прерия граничит с бореальной зоной хвой-
ных лесов, на востоке и юго-востоке — с зоной лиственных лесов, а на
западе — с горными лесами, образованными различными видами хвойных.
На юге злаковник постепенно переходит в субтропическую саванну, где
в качестве элементов древесной растительности примешиваются виды
Juniperus, Prosopis и Larrea.
Поскольку температура изменяется в направлении, перпендикуляр-
ном направлению уменьшения осадков, то есть с севера на юг, во всех
трех выделенных зонах, если пересечь их так же с севера на юг, можно
Зона степей и прерий
95-
Рис. 77—79. Климадиаграммы станций, расположенных у восточной границы обла-
сти прерий и лесной зоны. Характеризуют изменение климата в направлении с севера-
на юг.
Рис. 80—82. Климадиаграммы станций, расположенных у западной границы высоко-
злаковой прерии. Характеризуют изменение климата в направлении с севера на юг.
АБИЛИН (534м) 18,16 622
[61-45] а Техас
обнаружить различие в флористическом составе, связанное с изменением
теплового режима. Таким образом, в высокозлаковой, или настоящей,
прерии выделяют: на севере — прерию с Festuca scabrella или прерию со
Stipa — Agropyrum, в средней части— прерию с Andropogon scoparius —
А. gerardv, на юге — область с другими весьма многочисленными тепло-
любивыми видами злаков.
Сложный флористический состав растительности прерий обобщенно
описан в работе Кнаппа (R. Knapp, Die Vegetation Nord- und Mittelame-
rikas, 1964). Работа содержит карту разных типов прерий и обширный спи-
сок литературы. Поэтому ограничимся здесь лишь важнейшими данными х.
В североамериканской прерии в отличие от степей Восточной Европы
обращает на себя внимание резкое преобладание более тропических родов
злаков (Andropogon, Panicum и др.), цветущих поздним летом. Прерии
простираются значительно дальше к югу и связаны через Мексику с суб-
тропическими районами Центральной Америки. Восточноевропейские
степи, напротив, отделены от субтропических и тропических областей
обширными пустынями и горными сооружениями. На востоке Северной
Америки прерии в наши дни почти полностью распаханы. Возделываются
кукуруза (в последнее время также соя) и пшеница, на юге также сорго.
1 Ср. также новую карту растительности в работе: A. W. К u с h 1 е г, Potential
natural vegetation of the conterminous United States, «Amer. Geogr. Soc., spec, publ.»,
№ 36, 1964.
96
Глава 2
Западная, более сухая часть используется как экстенсивное пастбище,
причем на одну голову крупного рогатого скота здесь приходится
несколько гектаров пастбищных угодий.
Автор имел возможность в 1930 г. пересечь несколькими маршрутами
всю область прерий между 39° и 43° с. ш. и познакомиться с характером
растительности и почв.
На границе между зоной лиственных лесов и областью прерий, прохо-
дящей примерно по 40-й параллели, среднее годовое количество осадков
составляет 800 мм (на севере 500 мм, на юге 1000 мм). В пределах настоя-
щей прерии выпадает около 700 мм осадков, у границы с низкозлаковой
прерией — 470, на Великих равнинах — 420 мм. В сухие годы сумма
осадков может сократиться на 25%, а в годы катастрофической засухи
(1934—1941) она была еще меньше. К западу местность повышается
с 200—300 м над уровнем моря до 1000—2000 м у подножия Скалистых гор.
Температура, особенно зимних месяцев, резко варьирует в зависимости
от широты. Если на севере средняя температура января приближается
к —20° и абсолютный минимум почти достигает —50°, то на юге средняя
температура всех месяцев не опускается ниже 0°, а сильные морозы бывают
только при прорывах холода.
Высокозлаковая, или настоящая, прерия
Поскольку здесь выпадает большое количество осадков, почвы прерии
хорошо увлажнены. Поэтому облесение этой области представляется
делом вполне реальным. Однако при этом нельзя забывать, что некогда
в прерии паслись гигантские стада бизонов. Предполагают, что до засе-
ления прерий человеком численность бизонов достигала 50—60 млн.
голов. Учитывать нужно также пожары, частью специально проводимые
индейцами, частью вызванные ударом молнии. И тот, и другой факторы
уничтожают древесные растения и способствуют развитию злаковника.
Поэтому лес и прежде, по-видимому, не достигал климатической границы,
тем более что климат в течение послеледникового времени на Североаме-
риканском континенте, как и в Европе, неоднократно становился чрез-
мерно сухим.
Несмотря на столь большие различия в температуре между севером
и югом, определенные виды растений распространены повсеместно на всем
пространстве высокозлаковой прерии. Опыты разведения некоторых видов
злаков разного происхождения в культуре, проводившиеся под Линколь-
ном, показали, что обычно эти виды состоят из различных экотипов с раз-
ными потомственно закрепленными ритмами развития Ч
Решающее значение для развития растений прерии имеет не количе-
ство осадков, а влажность почвы. Поэтому холмистый рельеф высоко-
злаковой прерии обусловливает формирование нескольких ее вариантов:
а) настоящей прерии на возвышенностях (высокая прерия) с доминирую-
щими Andropogon scoparius (длина листьев 30—40см, а генеративных побе-
гов 50—100 см), Stipa spartea, Sporobolus heterolepis, Bouteloua curtipendula,
Agropyrum smithii, Koeleria cristata и др.; б) прерии на низменностях
(равнинная прерия) с более увлажненной почвой и преобладающими
1 С. McMillan, The role of ecotypic variation in the distribution of the central
glassland of North America, «Ecol. Monogr.», 29, 1959, p. 285—308; Texas grassland
communities under transplanted conditions, «Amerj J. Bot.», 48, 1961, p. 778—785.
Зона степей и прерий
97
высокорослыми злаками Andropogon gerardi (длина листьев 0,5—1 м, гене-
ративных побегов 1,5—2 ж), Sorghastrum nutans, Panicum virgatum, Elymus
canadensis и др.; в) прерии влажных впадин с Spartina pectinata, достигаю-
щей 2 м высоты (длина генеративных побегов до 3 ж). На востоке эти виды,
кроме того, занимают обширные низменные площади в долинах рек,
слишком влажные для земледелия. Такие высокозлаковые степи, в зарос-
лях трав которых мог, не спешиваясь, надежно укрыться всадник, якобы
были широко распространены в Восточной Европе во времена кочевников.
Их исчезновение сегодня объясняется, видимо, начавшейся вслед за раз-
витием земледелия сильной эрозией почв, которая повлекла за собой обра-
зование глубоких оврагов (балок), интенсивное врезание рек и, наконец,
осушение низменностей х. Исследователь прерии Уивер завершил труд
всей своей жизни обзорной работой, в которой подробно описал раститель-
ные сообщества прерии и последствия катастрофических засух 1 2.
Настоящая прерия, как и луговая степь, богата разнотравьем. Из-за
того, что растения цветут в разное время, здесь также наблюдается четкая
смена аспектов. В центральной части прерии жизнь просыпается в конце
марта; первыми зацветают Carexpensilvanica, Anemone, Erythronium, Ranun-
culus, Lomatium (зонтичные); затем следуют различные летние аспекты;
в начале сентября развитие большинства видов заканчивается. Виды
Andropogon цветут поздно, так же как ряд глубокоукореняющихся расте-
ний (Kuhnia, Liathris, Aster, Solidago, Vernonia). Позднелетний аспект
связан с глубоким увлажнением почвы летними дождями; горизонт карбо-
натных новообразований в почвенном профиле отсутствует, и осадки бес-
препятственно просачиваются вглубь. Однако апогей развития раститель-
ности прерии, как и степной растительности, приходится на начало лета
(июнь, рис. 83). Число цветущих видов в это время года равно:
В предвесеннее время . . 12 В начале лета . . 70
Весной..................40 В разгар лета . . 40
Разнообразие видов в прерии больше, чем в степи. Очень богато пред-
ставлены бобовые: виды Psoralea, Amorpha, Petalosternum, Astragalus,
Glycyrrhiza, Baptisia и др.
Причина отсутствия деревьев в восточной высокозлаковой прерии
часто становилась предметом острых дискуссий 3. Поскольку по климати-
ческим условиям и содержанию воды в почве рост деревьев здесь вполне
возможен, отсутствие их должно быть связано с иными причинами, напри-
мер с пожарами и выпасом. Известно, что ежегодное выжигание трав способ-
ствует образованию злакового покрова. Защита от огня, напротив, благо-
приятно сказывается на видах разнотравья: накапливающаяся с течением
времени большая масса органических остатков в виде подстилки затруд-
няет отрастание злаков, но не является препятствием для разнотравья.
Обжиг через год ведет к образованию смешанных травостоев из злаков
1 Такой же процесс наблюдался в Южной Америке, а в последние 50 лет в Юго-
Западной Африке, где вместо прежних обширных зарослей высоких злаков вдоль рек
сегодня тянутся безжизненные песчаные пространства (см.: Н. W а 1 t е г, О. Н. V о 1 к,
Die Grundlagen der Farmwirtschaft in Sudwestafrika, Stuttgart, 1954).
2 J. E. Weaver, North American prairie, Lincoln, 1954; J. E. Weaver,
W. J. Himmel, The environment of the prairie, «Cons, and Surv. Div. Univ. Nebras-
ka», Bull. 5, 1931, p. 1—50; J. E. Weaver, T. J. Fitzpatrick, Ecology and
importance of the dominants of tail-grass prairie, «Bot. Gaz.», 93, 1932, p. 113—150.
3 H. Walter, 1st die Prarie von Natur aus baumlos? «Geogr. Z.», 41, 1935,
S. 16—26.
7-0434
98
Глава 2
и разнотравья 1. При отсутствии пожаров могли бы укорениться сеянцы
деревьев, но этого не происходит.
С целью выяснения причины безлесья Уивер и Клементс провели ряд
экспериментальных исследований 1 2. Обширные пробные площади у гра-
ницы леса были обнесены изгородью, с тем чтобы защитить их от какого-
либо влияния деятельности человека. Многолетние наблюдения позволили
установить, что кустарники, узкой полосой расположившиеся перед лесом,
Рис. 83. Богатая разнотравьем настоящая прерия, июльский аспект; восточ-
ная часть штата Небраска. Видны светлые соцветия Achillea millefolium и
Erigeron ramosum, на заднем плане — кустистые Psoralea floribunda. Виды
Andropogon цветут поздно (фото Уивера).
спустя какое-то время после начала эксперимента начали медленно раз-
растаться и теснить растительность прерий. Так, например, Rhus glabra
образовал 7—9-метровые подземные побеги на глубине 10—30 см, которые
тянулись в злаковник и давали там надземные побеги. Эти побеги подни-
мались над злаками и ослабляли их рост, создавая дополнительное затене-
ние на почве. К R. glabra присоединялись другие кустарники, такие, как
Symphoricarpus orbiculatus, образующие плотные, дающие густую тень
группы, и Corylus americana, чьи побеги длиной 30—50 см простираются на
глубине 15 см. Под этим кустарниками злаки уже не растут. Как только
1 С. L. Kucera, M.Koelling, The influence of fire on composition of central
Missouri prairie, «Amer., Midland Natur.», 72, 1964, p. 142—147.
2 J. E. W e a v e r, A. F. T h i e 1, Ecological studies in the tension zone between
prairie and woodland, «Bot. Surv. Nebr.», N.S. 1, 1917, p. 1—60; F. E. Clements,
J. E.Weaver, H. C. Hanson, Plant competition, «Carneg. Inst, of Wash.», Publ.
398, 1929; J. E. Weaver, T.J. Fitzpatrick, The prairie, «Ecol. Monogr.», 4, 1934,
p. 109—295; F. W. Albertson, J. E. Weaver, Injury and death or recovery of
trees in prairie climate, «Ecol. Monogr.», 15, 1945, p. 393—433.
Зона степей и прерий
99
сомкнутость злакового покрова нарушается, семена лесных деревьев под
защитой кустарника получают возможность прорастать, а всходы разви-
ваться. Сеянцы деревьев, подрастая, в свою очередь начинают угнетать
кустарник. В результате по истечении 3—5 лет лес сомкнутым фронтом
надвинулся на прерию, отторгнув у нее полосу шириной 1 м. При благо-
приятных условиях возможно и более быстрое наступание.Однако следует
считаться и с периодами засухи. Так, в засушливое время 1934—1941 годов
лес был оттеснен несколько назад.
Семена и плоды лесных деревьев естественным путем попадают, конеч-
но, и за пределы леса, в злаковый покров прерии. Они прорастают здесь,
но лишь редко всходы древесных растений одерживают верх над злаками
в борьбе за свет и влагу. Уже в первые годы они, как правило, погибают.
Без предшествующей стадии кустарника иногда выживают лишь пророст-
ки дуба крупноплодного (Quercus macrocarpa). Они быстро образуют стерж-
невой корень, уходящий в почву через год на 1 ж, через 3 года — на
2,5 м, а у взрослого дерева достигающий длины более 5 м. Позднее разви-
ваются поверхностные боковые корни длиной 6—18 м, которые вступают
в борьбу за почвенную влагу и питание с корневой системой злаков и разно-
травья. Образуется редкий, главным образом вследствие широко раскинув-
шейся сети корней, древостой.
Клементс и Уивер не ограничились изучением поведения древесных
пород в пограничной полосе леса и прерии. Их интересовало также, могут
ли всходы древесных растений выдержать конкуренцию злаков в клима-
тических условиях настоящей прерии.
Для получения ответа на этот вопрос в злаковом покрове настоящей
прерии были проведены четыре борозды шириной 10 см каждая, в которые
посеяли семена и посадили молодые проростки деревьев и кустарни-
ков — обитателей полосы, разделяющей прерию и лес. В первой борозде
в соответствии с программой эксперимента древесные растения требова-
лось защитить от какой бы то ни было конкуренции. Для этого борозду
расширили до 40 см и взрыхлили почву; кроме того, удалили все нависаю-
щие над бороздой листья злаков. На второй борозде в полосе такой же
ширины близ поверхности земли были срезаны все надземные части расте-
ний прерии, но сама злаковая дернина осталась ненарушенной. Молодые
деревца получали весь свет, но их корни конкурировали с корнями расте-
ний прерии. У следующих двух борозд покров прерии сохранили целым,
но древесные растения в третьей опытной борозде поливались. Однако
полив шел на пользу и росшим по-соседству злакам. Они пышно раз-
растались, из-за чего древесные породы в третьей борозде развивались
хуже, чем в четвертой борозде, где они были полностью предоставлены
самим себе. Такие же опытные борозды были заложены в более низких
частях прерии (равнинная прерия) и непосредственно близ границы пре-
рии и леса. Почва в этих случаях была несколько более влажной. Резуль-
таты опыта однозначно показали, что перспектива развития была только
у древесных растений первой борозды каждой серии, где исключалась
всякая конкуренция. Во всех остальных бороздах корневая система
растений прерии перехватывала у древесных пород почвенную влагу и пита-
тельные вещества, обрекая их на гибель.
Эти опыты также подтвердили (ср. стр. 69), что, хотя безлесие пре-
рии'обусловлено прежде всего водным фактором, важную роль играет
и конкуренция злаков. Закрепиться постоянно древесные растения первой
борозды могут лишь там, где почва лучше и глубже увлажнена. Это наблю-
дается, например, в более низменных частях прерии (равнинная прерия)
7*
100
Глава 2
или в пограничной полосе леса и прерии. Отсюда понятно, почему лес
по эрозионным ложбинам проникает глубоко в область прерий (рис. 84).
Во-первых, водный режим здесь более благоприятен, и, во-вторых, зла-
ковый покров на склонах нередко бывает разрушен эрозией, вследствие
чего проростки древесных пород на оголенных поверхностях не испытывают
конкуренции со стороны злаков.
Итак, мы приходим к выводу, что прежние частые пожары в прерии —
независимо от того, были ли это искусственно пускавшиеся человеком
Р и с. 84. Прерия на востоке штата Небраска вблизи, границы с лесом.
Доминирует Stipa spartea. Древесные растения проникают по оврагам
далеко в пределы области прерий (фото Уивера).
палы или результат удара молнии,— несколько оттеснили лес к востоку
и что утраченную территорию лес сомкнутым фронтом может постепенно
при благоприятных условиях среды вернуть назад. Но совершенно неправ-
доподобно предположение, что лес может занять всю площадь настоящей
прерии до ее западной границы, где появляющийся в почве на определен-
ной глубине карбонатный горизонт указывает на увлажнение лишь ее
верхних слоев. Нельзя также забывать, что лесная растительность
на единицу поверхности почвы расходует намного больше воды по сравне-
нию с растительностью прерии. Даже если бы вся почва в пределах настоя-
щей прерии в какой-то момент оказалась насыщенной влагой, лесной
древостой вскоре израсходовал бы все запасы воды, содержавшиеся в почве.
Этот процесс можно наблюдать, например, при возделывании люцерны в пре-
рии. Первые годы она дает очень хороший урожай, но при повторных посевах он от
года к году быстро уменьшается. Это происходит отнюдь не из-за обеднения почвы
питательными веществами, а вследствие сокращения запасов воды в почве. Люцерна,
отличающаяся очень глубокой корневой системой, в культуре расходует за год
Зона степей и прерий
101
больше влаги, чем могут восполнить атмосферные осадки. От года к году люцерна
все более иссушает почву, в связи с чем сокращаются и урожаи. Аналогично дело
обстоит, вероятно, и с лесом.
Следовательно, искусственное облесение прерии на первых порах,
так же как и в восточноевропейской степи, может быть успешным, но испы-
тание временем оно не выдерживает. В восточной окраинной полосе пре-
рии местами распространены злаковые сообщества с многочисленными
лиственными деревьями. Предполагают, что до заселения прерии фер-
мерами в одном только Висконсине они занимали 3 млн. га площади.
Эта так называемая «дубовая саванна» («oak savanne» американских
исследователей) напоминает прежние облесенные пастбища Централь-
ной Европы; образовалась она, по-видимому, в результате частого выжи-
гания травы в прерии. Благодаря очень толстой коре доминирующая здесь
древесная порода Quercus macrocarpa оказывается весьма устойчивой
против огня. Сильно развитая корневая система позволяет дубу одержи-
вать победу в борьбе за существование со злаками. Даже длительную
засуху 1934—1941 гг. деревья перенесли без особого для себя вреда.
Такая открытая прерия с дубовыми «лесами» не соответствует тому,
что в Восточной Европе понимают под лесостепью. С последней, скорее,
можно было бы сравнить районы, где прерия в виде островов вкраплена
в массивы леса и где характер растительного покрова определяет тип
почвы и рельеф. В пограничной полосе можно наблюдать, как леса сначала
освобождают более сухие холмы и отступают в низины, чтобы затем по доли-
нам рек проникнуть глубоко в область прерии и занять эрозионные
ложбины. Гораздо дальше, чем Quercus macrocarpa и Tilia americana,
в глубь прерии проникают в штате Небраска Fraxinus pennsilvanica, Ulmus
americana, U. fulva, Platanus occidentalis и Acer saccharinum. В качестве
крайних форпостов леса в эрозионных руслах здесь выступают только
Acer negundo, Populus deltoides и некоторые виды Salix. Под пологом этих
деревьев поселяются многие теневыносливые растения лесных почв.
К факторам, способствующим сохранению высокозлаковой прерии в есте-
ственных условиях, относятся, как, впрочем, и во всех злаковниках уме-
ренной зоны, случайные, вызванные ударом молнии пожары и пастьба
крупных диких животных (здесь стада бизонов) и грызунов. Животный
мир является важной составной частью биогеоценоза области прерий.
Самые южные районы высокозлаковой прерии в Техасе имеют уже
совершенно иной облик. Здесь господствует почти субтропический климат
и появляется типичный для этих условий кустарник, который в пустыне
Сонора носит название «мескит» — виды Prosopis (см. т. I, стр. 338 и 341).
Мескитовая саванна в южном Техасе сменяет прерию. В результате интен-
сивного выпаса резко сокращается участие большинства многолетних
злаков, прежде всего Andropogon scoparius, местами слагающего до 72%
злакового покрова. Напротив, возрастает количество Prosopis. Иными сло-
вами, происходит известное зарастание территорий кустарниками (см.
т. I, стр. 289 и след.). С прекращением выпаса восстанавливается, через
стадию, в которой доминирует Stipa, естественная растительность. На
прежних пахотных землях уже через 20 лет в составе растительного
покрова можно обнаружить присутствие большинства типичных злаков
прерии Б
1 J. L. Launchbaugh, Vegetational changes in the San Antonio prairie as-
sociated with grazing, retirement from grazing and abandonment from cultivation, «Ecol.
Monogr.», 25, 1955, p. 39—57.
102
Глава 2
Рис. 85. Смешанная прерия на западе штата Небраска. Между
дерновинами высоких злаков Andropogon scopartus мелкие Bouteloua
gracilis, В. hirsuta (фото Уивера).
Рис. 86. Песчаные холмы в штате Небраска с растительностью смешан-
ной прерии. На переднем плане — Yucca glauca и A ndropogon scoparius,
за ними низкорослая Bouteloua gracilis (фото Уивера).
Зона степей и прерий
103
Смешанная прерия
Эта прерия представляет собой переходный тип между настоящей
и низкозлаковой прерией. В ней распространены и высокие и низкие злаки
(рис. 85). Как и в высокозлаковой прерии, здесь также заметны известные
различия во флористическом составе северной и южной частей. Число
разнотравья в западном направлении, по мере увеличения сухости кли-
мата, быстро уменьшается. При выпасе из травостоя сначала выпадают
высокие злаки. Со временем происходит деградация высокозлаковой пре-
рии и постепенное превращение ее в низкозлаковую. Поэтому в настоящее
время трудно провести четкую границу между данными двумя типами.
Высказывалось даже мнение, что низкозлаковая прерия собственно и сфор-
мировалась вследствие неумеренного выпаса, но справедливость этой
мысли сомнительна, поскольку здесь и раньше паслись стада крупных
диких животных. На увлажненных песчаных почвах смешанная прерия
проникает далеко на запад (рис. 86).
Низкозлаковая прерия, или Великие равнины
Низкозлаковая прерия развита в самой засушливой западной части
зоны прерии. Здесь абсолютно господствуют низкие злаки Bouteloua
gracilis и Buchloe dactyloides (рис. 87, 88); виды разнотравья редки; при
чрезмерном выпасе широко распространяются низкорослые виды Opuntia.
Эти кактусы хорошо переносят холодную зиму и поэтому прослеживаются
далеко на север, до Канады.
Рис. 87. Характерный облик сомкнутого травостоя ВисМоё dacty-
loides в низкозлаковой прерии восточного Колорадо (фото Уивера).
104
Глава 2
Зона степей и прерий
105
мн
400-
300-
200-
Ю0-
0
м
Средняя годоваяо
температура 8°
Абсолютный
минимум 37°^--~-z
Великие равнины (низкозлако- Смешанная
бая прерия) прерия
•^Ом Над
нм
^г800
Средняя''7й0п
годовая -600
температура 11°
Абсолютны^
минимум 34°
Высокозлаковая
прерия
-500
400
о
М
1
Мертвая, - °tyde
постоянно сухая почва '
Бурые пустынно- Темные
степные почвы бурые почбы
<------Уменьшение содержания гумуса Нарастание интенсивности
смыбац р|(и!м\/\|,|<
-<---------------П е 3 с кал и------------------>- Педалыреры
Общая протяженность свыше 1000 км
,-z
Рис. 89. Схематический профиль через район прерии, повышающийся к западу
с 300 м над уровнем моря до 1500 м. Содержит данные о климате (вверху}, растительно-
сти (в центре} и почвах (внизу}.
При осмотре почвенных профилей на маршруте с востока на запад
можно обнаружить типы чернозема, соответствующие почвам степей
с характерными горизонтами скопления карбонатов. На рис. 89 схемати-
чески показана зависимость, существующая между климатом, раститель-
ностью и почвами области прерий. Почвы прерии на востоке, поросшие
высокими злаками, увлажнены до уровня грунтовых вод; горизонт извест-
ковых выделений отсутствует, что позволяет корневой системе злаков
проникать глубоко в грунт (ср. рис. 67). Несколько западнее в почвенном
разрезе уже отчетливо видны карбонатные новообразования. Сначала
скупо рассеянные над мощным, залегающим на глубине 1,5—2 м горизон-
том, они от профиля к профилю все теснее сближаются друг с другом
и одновременно перемещаются ближе к поверхности, пока наконец в пре-
делах низкозлаковой прерии — в экстремальных условиях существо-
вания — не концентрируются уже на глубине 25 см. Вместе с тем все
более уменьшается глубина проникновения корней злаков. Они почти
не внедряются в горизонт известковых выделений, ибо последние рас-
полагаются на средней глубине промачивания почвы атмосферными
осадками.
Зональность почв в Северной Америке выступает не столь отчетливо,
как в Восточной Европе, поскольку здесь тенденции изменения темпера-
туры и осадков географически не совпадают: температура повышается
в направлении с севера на юг, а количество осадков увеличивается с запа-
да на восток. Такое своеобразие условий среды обусловливает, скорее, шах-
матное распределение типов почв (рис. 90). Особенностью является также
зона почв прерии в переходной к лесам области, не имеющая аналога
в степи; на ее месте в степях мы находим деградированный и выщелочен-
ный чернозем, за которым следуют обладающие уже признаками слабого
оподзоливания серые лесные почвы.
Р и’с. 90. Почвы США (по почвенной карте Министерства сельского хозяйства США),
Педалъферы: 1 — подзолистые почвы; 2 — серо-бурые оподзоленные лесные почвы; 3 — желтые и красные лесные почвы; 4 — горные почвы; 5 — почвы
прерий. Педокали: 6 — южные черноземы и темные бурые почвы; 7 — северные черноземы; 8 — каштановые и бурые пустынно-степные почвы; 9 —
северные и 10 — южные бурые почвы; 11 — сероземы; 12 — почвы в долинах рек Тихоокеанского побережья.
Зона степей и прерий
107
Р и с. 91. Распределение доступной для растений влаги в профиле почв прерий во время
жесточайшей засухи 1934 г.; район Линкольна, штат Небраска. Зимой пробы не отби-
рались (по Уиверу).
ПОСЛЕДСТВИЯ ЗАСУШЛИВЫХ ЛЕТ В ПРЕРИИ
В 1934—1941 гг. в области прерии природа осуществила эксперимент
крупнейшего масштаба. Засухоустойчивость растений прерии подверглась
при этом жесточайшему испытанию, последствия которого были тщательно
изучены Уивером (1954, см. примечание на стр. 97). Уже 1931—1933 гг.
были относительно сухими, но 1934 стал годом, самым бедным дождями со
времени организации здесь метеорологических наблюдений Ч Обычно содер-
жание воды в почве на глубине 30—120 см никогда не опускается ниже 5%
воды, доступной для растений. Ниже почва остается практически всегда
влажной. Весной 1934 г. впервые прошли пыльные бури. На полях запад-
ной периферии зоны прерии ветер взвихрил всю почву. Уже весен-
ние растения развивались плохо. В июне прерия пожелтела, растения
поднялись лишь до половины своей обычной высоты, виды с мелкой корне-
вой системой засохли. В июле прошли только слабые дожди, давшие всего
10 мм осадков. Почва даже на глубине 1,5 м не содержала и 2% влаги,
доступной для растений (рис. 91). Обильно цвели лишь Oenothera serru-
lata, Psoralea tenuifolia, Rosa, корни которых проникают вглубь до
6 м. В низинах растения еще сохраняли свежесть. Сухость сопровожда-
лась высокой температурой (до 44°), относительная влажность воздуха
временами уменьшалась до 3—5%. Но растения страдали главным обра-
1 L. A. S t о d d а г t, Osmotic pressure and water content of prairie plants, «Plant
Physiology», 10, 1935, p. 661—680.
108
Глава 2
зом от недостатка влаги в почве. В августе аспект стал бурым. Весной
1935 г. прерия снова зазеленела, но тщательная проверка показала, что
30—50% растений не образовали надземных побегов. Западнее урон
достиг еще больших размеров; Agropyrum smithii на некоторых местооби-
таниях почти полностью вытеснил все другие виды. Распространились
также некоторые глубокоукореняющиеся злаки и многочисленные припт-
лые сорные травы (Lepidium densiflorum, Conyza canadensis, Salsola kali).
Засуха с ее страшными пыльными бурями, на несколько дней совер-
шенно закрывавшими солнце, длилась 7 лет. Последствием ее было почти
полное исчезновение Andropogon scoparius. Напротив, Stipa spartea, бес-
препятственно размножаясь, занял обширные площади (см. рис. 84).
Bouteloua curtipendula, в восточной прерии составлявшая лишь 0,5%
растительного покрова, теперь нередко занимает 20—30% территории,
однако уступает в конкурентной борьбе Agropyrum. Последний оказался
чрезвычайно засухоустойчивым. Заметным стало и участие однолетних
злаков, которые обычно в прерии отсутствуют. Очень резко сократилось
число видов разнотравья, возросло только количество тех из них, которые
обладают подземными водозапасающими органами.
Таким образом многолетняя засуха полностью изменила состав расти-
тельного покрова прерии. После засухи настоящая прерия физиономиче-
ски соответствовала, скорее, смешанной прерии с ее пестрой мозаикой высо-
ких и низких злаков. В 1942 г. засуха наконец закончилась. После трех
хороших дождливых лет растительный покров снова ожил, но еще не стал
однородным. В последующие годы прежде всего были вытеснены одно-
летние виды, что позволило исконным растениям прерии начать распро-
страняться вширь. Распространение шло медленно и постепенно. В начале
растения образовали не более чем пятна вокруг переживших засуху экзем-
пляров, от которых иногда сохранялось только корневище, нередко лишь
часть его. Они давали новые растения, из-за слабой конкуренции хорошо
развивавшиеся. По-видимому, имело значение также высокое содержание
азота в почве, источником которого стала масса отмерших подземных
частей. Красивый, с голубыми соцветиями Sisyrinchium, которого не виде-
ли с 1934 г., в 1943 г. был в 7 раз многочисленнее, чем до засухи (видимо,
взошел из семян). Другие виды разнотравья восстанавливались медленнее.
Обычные сезонные аспекты первое время не проявлялись; однако продук-
ция сухой массы за период с 1940 по 1943 г. увеличилась в 2,7 раза. Весьма
устойчивым после засухи оказался Agropyrum smithii. Там, где он занимал
доминирующее положение, его трудно было вытеснить. Из видов Andro-
pogon быстро распространился A. gerardi, обладающий более глубоко опу-
скающимися корнями. Медленнее возрождался A. scoparius, отдельные
старые корневища которого иногда давали побеги. Вскоре он стал даже
доминантом и вытеснил Agropyrum. В 1945 г. численность разнотравья
снова возросла, но еще в 1953 г. следы засухи в районах западнее Миссури
были хорошо заметны; восточнее этой реки, где засуха не проявилась
столь сильно, по-видимому, восстановилось первоначальное состояние.
Результаты этого грандиозного природного эксперимента свидетель-
ствуют о том, что даже естественный растительный покров вследствие
колебаний климата никогда не находится в устойчивом состоянии, а стече-
нием времени несколько изменяется, причем равновесие смещается то в
одну, то в другую сторону. Периоды засухи, аналогичные описанному,
несомненно, были в зоне прерий и прежде. Их последствия нельзя видеть
сегодня, но, безусловно, они являются существенной причиной отсутствия
в прерии деревьев. Надо считаться с возможностью жестоких и длитель-
Зона степей и прерий
109
ных засух и в будущем. Чем суше климат, тем резче колеблются суммы
осадков от года к году и тем отчетливее проявляются последствия этих
колебаний.
В меньшем масштабе ущерб от засухи можно было наблюдать в Цен-
тральной Европе после засушливых 1947 и 1949 гг. В маловодном 1964 г.
луговая растительность характеризовалась изменениями, похожими на те,
которые преобразили облик североамериканской прерии в 1934 г. После
первого укоса злаки не отросли. Вся площадь имела бурый аспект, и толь-
ко у зонтичных с глубокоуходящими корнями {Heracleum, Daucusj, неко-
торых сложноцветных {Centaurea jacea, Cichoriuni), а также у чертополоха
и бобовых {Medicago, Melilotusj образовались соцветия; однако их розе-
точные листья засохли. Наибольшей засухоустойчивостью отличались
виды с глубокопроникающим стержневым корнем. Особенно свежими
выглядели цветущие розетки Plantago media и Р. major. Все это виды, кото-
рые на обычных влажных лугах отсутствуют. Растения умеренной зоны
также приспособлены к чередованию от года к году благоприятных и небла-
гоприятных условий влагообеспечения. В сухие годы концентрация клеточ-
ного сока увеличивается; повышение осмотического давления обусловли-
вает морфологическое изменение вновь образовавшихся побегов, сокраще-
ние транспирирующей поверхности и известную ксероморфность листьев.
Резко изменяется численное соотношение видов, так как засухоустойчи-
вые растения при недостатке влаги развиваются сравнительно лучше,
чем незасухоустойчивые формы. Последние заметно угнетаются. Но каче-
ственный состав растительного покрова первое время остается прежним,
изменение его происходит лишь в том случае, если необычные условия
стойко сохраняются на протяжении нескольких лет.
ЭКОЛОГО-ФИЗ ИОЛО Г И ЧЕС К НЕ И ССЛЕДОВАНИЯ
РАСТЕНИЙ СТЕНЕЙ И СТЕННЫХ ПУСТОШЕЙ
Степные растения нельзя ставить на одну ступень с субтропическими
растениями пустынь. В экологии это привело к длительным бесплодным
дискуссиям. Максимов, изучавший экологию только степных растений \
считал полученные им данные типичными для всех ксерофитов и думал,
что опровергает представления Шимпера. По Шимперу, сокращение интен-
сивности транспирации в засушливое время является важнейшим меха-
низмом приспособления ксерофитов к условиям затрудненного водоснаб-
жения; такое поведение характерно прежде всего для растений субтропи-
ческих пустынь с их намного более длительными периодами засухи по срав-
нению со степными районами.
Степные растения — преимущественно гемикриптофиты, почки возоб-
новления которых переживают холодную зиму у поверхности почвы,
чтобы дать побеги только весной. Следовательно, это травянистые виды,
4-месячный вегетационный период которых приходится на самое благо-
приятное время года — весну и раннее лето; испытываемые ими в этот
период моменты недостатка влаги кратковременны и преходящи. Сезон
вегетации растениям приходится использовать с максимальной интенсив-
ностью. Они быстро, с наименьшим возможным расходом материала раз-
1 N. A. Maximo w, Physiologisch-dkologishe Untersuchungen fiber die Diir-
reresistenz der Xerophyten, «Jb. wiss. Bot.», 62, 1923, S. 128—143; The plant in relation
to water, London, 1929.
но
Глава 2
вивают обширную листовую поверхность. Поэтому листья не склерофиль-
ные, а мягкие, без опорной ткани и толстой оболочки эпидермиса. Благо-
даря этому транспирация, даже при закрытых устьицах, полностью пре-
кратиться не может. По этой причине при временном недостатке влаги
водный режим растений нарушается, возникает острый водный дефицит
в тканях. Следовательно, в степных условиях должен был произойти отбор
таких видов, которые способны перенести нарастающий дефицит влаги
без отмирания при этом листьев. С улучшением условий обеспечения во-
дой листья оживают и вновь становятся способными к фотосинтезу. Та-
кое поведение Максимов считал типичным для всех ксерофитов. Однако
в условиях очень сильной засухи подобный механизм приспособления
оказывается несовершенным. В субтропиках, где холодный зимний пе-
риод отсутствует, для увеличения конкурентной мощности выгоднее
по возможности сохранить надземные органы, то есть образовать вечно-
зеленые ассимилирующие органы, устроенные таким образом, что при
недостатке влаги они почти полностью прекращают транспирировать.
Такие признаки приспособления мы находим у суккулентов и склеро-
филов, или они проявляются в развитии афильных форм у видов пустынь.
Мягколистные виды, подобные типу, описанному Максимовым, правда,
встречаются в Средиземноморской области, а единичными экземпля-
рами также в не слишком экстремальных условиях пустыни, но по
сравнению с другими жизненными формами они отходят здесь на вто-
рой план. Однолетние теплолюбивые поздноцветущие виды, как, напри-
мер, виды Aegilops, ведут себя очень характерно. Весной во время куще-
ния все листья имеют зеленую окраску. Они сильно транспирируют,
следовательно, способны и к интенсивному фотосинтезу. Однако в пе-
риод цветения транспирация сокращается, листья засыхают, так что ра-
стение в целом расходует очень незначительное количество воды.
Поэтому цветки, а также созревающие плоды, несмотря на увеличиваю-
щуюся сухость воздуха, получают достаточно влаги. Засыхание листьев
в период цветения происходит независимо от содержания воды в почве;
обычно это генетически закрепленный, приспособленный к летней засухе
ритм развития, свойственный вообще растениям засушливых областей
Как только начинают формироваться соцветия, к ним из листьев начинают
поступать питательные вещества, сами же листья в конце концов поги-
бают. У одних видов это происходит раньше, у других позже. У растений
с глубркопроникающей корневой системой листья сохраняются дольше,
чем у видов с поверхностными корнями.
Кроме того, для некоторых степных видов (Eryngium, Falcaria vulgaris,
видов Seseli, Phlomis pungens, Centaurea diffusa, Salsola kali) весьма харак-
терна жесткость стебля, поэтому отмершие соплодия, имеющие шаровид-
ные очертания, сохраняются как одно целое. Но у корневой шейки сте-
бель ослаблен, и в этом месте он легко ломается. Целые соплодия, под-
хватываемые ветром, перекатываются, сцепляются с другими такими
же соплодиями и образуют клубки диаметром около 1 м; эти перекати-
поле часто с большой скоростью катятся по степи. При сильных толчках
семена высыпаются и равномерно рассеиваются по всей поверхности.
Своеобразное впечатление производят эти перекати-поле, в большом коли-
честве вдруг появляющиеся издалека и так же быстро бесшумно исчезаю-
щие вдали.
1 Ю. С. Григорьев, Л. Е. Паузнер, Материалы по экологии видов
Aegilops L., «Бот. ж.», 48, № 5, 1963, стр. 640—660.
Зона степей и прерий
111
Рис. 92. Корневая система растений прерий (по Уиверу).
S — Sieversia ciliata’, W — Wyethia amplexicaule (сложноцветные); LI — Lupinus
leucophyllus, Lo — Lupinus ornatus, P — Poa sandbergii, Lm — Lep to taenia multifida,
A — Agropyrum spicatum.
Значение корневой системы для обеспечения водой растений северо-
американской прерии первым изучил Уивер1. Он установил, что из 43 ти-
пичных видов прерии 14% развивают свои корни только в верхних
60 см, у 21 % растений корни проникают на 1,5 м, у остальных 65% — еще
глубже (2,5—3,5 м и даже 6 м) (рис. 92).
Правда, эти данные относятся только к почвам прерии, увлажненным
до большой глубины. Корни разных видов располагаются тремя ярусами.
Благодаря этому конкуренция корней ослабляется, и становится понятным
большое богатство видами именно степных и прерийных сообществ. Глу-
бокой корневой системой обладают те виды, которые цветут поздним летом,
когда у видов с поверхностной корневой системой подземные побеги уже
засохли. Многие культурные растения этой области также способны уко-
реняться достаточно глубоко (например, у люцерны через 2 года корни
проникают до 7,5 м, через 6 лет — до 10 м). Однако если мы будем двигаться
в более сухие районы (смешанная и низкозлаковая прерия), то заметим, что
корневая система растений становится все более поверхностной, поскольку
корни опускаются лишь до верхней границы «мертвой почвы», не содержа-
щей доступной для растений влаги (табл. 18). Очень хорошо это можно
наблюдать и у пшеницы на полях.
Аналогичные исследования произведены в последнее время также
в канадской прерии Коплендом и Джонсоном (см. примечание на стр. 74).
1 J. Е. Weaver, The ecological relation of roots, «Carnegie Inst, of Wash.»,
Publ. 286, 1919; Root development in grassland formation, «Car. Inst, of Wash.», Publ.
292, 1920; J. E. Weaver, F. E. Clements, Plant ecology, New York, 1938.
112
Г лава 2
Таблица 18
Глубина проникновения корней и высота стеблей растений
в зависимости от осадков в североамериканской прерии (по Уиверу)
Зона растительности Количество осадков, мм Глубина про- никновения корней, см Высота стеблей, см
Высокозлаковая прерия 660—815 160 100
Смешанная прерия . . . 535—610 130 95
Низкозлаковая прерия 405—485 75 65
Сведениями о корневых системах растений восточноевропейских
степей мы располагаем благодаря весьма детальным исследованиям
Шалыта х.
Из этой работы мы заимствовали ряд рисунков, на которых показаны
распределение корневой массы в разных почвенных горизонтах, масса
надземных побегов растений и подстилки. Рис. 93 отражает картину разно-
травной ковыльной степи на обыкновенном черноземе. Остальные рисунки
характеризуют условия в южной степной зоне, а именно степи на кашта-
новых почвах, слабосолонцеватом черноземе, на столбчатом солонце,
в одном из подов с солодью (рис. 94). На отдельных разрезах показаны
строение корневых систем и их расположение на примере сильноразвет-
вленной корневой системы злаков (рис. 95—96) и слаборазветвленной,
часто со стержневым корнем, системы многолетних видов разнотравья
(рис. 97—101). Эфемеры и геофиты отличаются поверхностной корневой
системой, корни же злаков проникают на глубину до 1 м, поздноцвету-
щих видов разнотравья — на глубину 1,5—2 м и более. Однако основ-
ная масса корней злаков всегда расположена в верхних горизонтах поч-
вы, богатых гумусом, весной хорошо увлажняемых и вместе с тем особен-
но насыщенных питательными веществами. Глубина проникновения кор-
ней здесь меньше, чем в высокозлаковой прерии, почвы которой увлаж-
нены до уровня грунтовых вод; в черноземах же дождевая вода просачи-
вается только до определенной глубины.
Виды со стержневыми корнями укореняются более глубоко. В этом
случае вода, вероятно, просачивается вдоль корня ниже по профилю.
Так, стержневой корень Euphorbia seguieriana с отметки 240 см развивался
по ходу старого корня. Чрезвычайно интересную особенность наблюдал
Шалыт у растений со стержневыми корнями, разветвляющимися лишь
на значительной глубине. На ранней стадии от верхней части корня также
ответвляются боковые корешки, но по мере углубления корня верхние его
ответвления отмирают. Очень многие степные растения обладают способ-
ностью образовывать — при достаточном промачивании верхних горизон-
тов почвы — даже на старых частях корня эфемерные сосущие корешки,
в дальнейшем не утолщающиеся. Формируются они либо как настоящие
боковые корни, либо вырастают пучками из бородавкообразных бугорков
на старых корнях. Эти бугорки состоят из меристематической ткани, кото-
рая сохраняется у основания прежних боковых корней. После высыхания
1 М. С. Шалыт, Подземные части некоторых луговых, степных и пустынных
растений и фитоценозов, «Труды Бот. ин-та им. Комарова, сер. геоботаника», вып. 6,
1950; вып. 8, 1952.
Зона степей и прерий
ИЗ
Рис. 93. Профиль корневых систем растений Хому-
товской степи на обыкновенном черноземе (по Шалыту).
1 — Euphorbia segueriana", 2 — Festuca sulcata", 3 — Salvia nutans",
4 — Stipa lessingiana.
почвы эфемерные корни, наблюдавшиеся у трав, а также у кустарников
или деревьев аридных районов, отмирают. Такие корни живут от несколь-
ких дней до нескольких недель х. Чем ближе к поверхности распо-
1 М. С. Шалыт, Эфемерные корни растений, их распространение и дня пение,
«Вопросы ботаники», вып. 3, 1960, стр. 14—16: О. Stocker, Untersuchungen in der
ungarischen Alkalisteppe, «Jb. wiss. Bot.», 78, 1933, S. 751—856. Автор отмечает удиви-
тельную способность сосущих корней всех растений, растущих на быстровысыхающих
почвах, развиваться (в течение нескольких часов) после увлажнения.
-8 — 0434
114
Глава 2
Рис. 94. Распределение надземной массы и корней растительностиПв различных
степных сообществах (в г сухого веса) (по Шалыту).
1 — разнотравная Stipa lessingiana — Festucasulcata Хомутовская степь на южной разно-
видности обыкновенного чернозема; 2 — то же, с большим количеством Salvia nutans',
3—степь с Pyrethrum millefoliatum— Festuca sulcata на слабосолонцеватых черноземах
(Аскания-Нова)', 4 — степь с Agropyrum pectiniforme—Stipa lessingiana (остров Йуюктук,
Сиваш) на каштановых почвах; 5—полупустыня с Kochia prostrata —Festuca sulcata на столб-
чатых солонцах (Аскания-Нова); в—травостой Agropyrum pseudocaesium (Аскания-Нова) на
солодях.
а — живая надземная масса; б — мертвый покров; в — клубни, луковицы, корневища (у
Роа bulbosa надземные); г — мелкие корни; д — крупные корни.
лягается корневая система растений, тем раньше растения заканчивают
свое развитие, поскольку имеющиеся в их распоряжении запасы влаги
исчерпываются в этом случае быстрее. В результате скорость транспира-
ции, возрастающая в условиях хорошей обеспеченности растений водой,,
что свидетельствует об интенсивном газообмене и, следовательно, об интен-
сивном фотосинтезе, резко сокращается. По Бейдеман, у злаков Ногайской
степи уменьшение транспирации начинается уже в июне, но у Artemisia,.
имеющей более глубокие корни,— лишь в июле Напротив, у видов,
корни которых достигают горизонта грунтовых вод, максимум транспи-
1 И. Н. Бейдеман, К методике изучения водного режима растений,
«Бот. ж.», 41, № 2, 1956, стр. 212—219.
Зона степей и Прерий
115
Рис. 95. Корневая система Stipa lessingiana в сообществе с Agropyrum pecti-
niforme на слабосолонцеватой каштановой почве; остров Куюктук, Сиваш
(по Шалыту).
Рис. 96. Корневая система Festuca sulcata (там же).
рации отмечается именно во время засухи, в разгар лета. Ограничение
транспирации связано с закрытием устьиц, что обусловливает снижение
интенсивности фотосинтеза, прекращение продуцирования органического
вещества и задержку роста. В табл. 19 приведен расход воды на транспи-
рацию некоторыми растительными сообществами за 5 летних месяцев.
Таблица 19
Расход воды на транспирацию различными растительными сообществами
южной степи в течение вегетационного периода (по Бейдеман)
Растительное сообщество Транспирация, лии
Степь с Stipa — Festuca (раннее сокращение транспирации) Галофиты на слабоувлажненпой почве Болотная растительность и сообщества галофи- тов на сырой почве Древостои (Tamarix, Populus) на местообита- ниях с высоким уровнем грунтовых вод . . . 78—99 70-150 410—643 572—1143
8*
116
Глава 2
Рис. 97. Корневая система Artemisia austriaca. Пастбища на слабосолонцеватом чер-
ноземе в Аскании-Нова (по Шалыту).
Рис. 98. Корневая система Falcarta vulgaris в степи с господством Festuca и Stipa
на слабосолонцеватом черноземе, Аскания-Нова (по Шалыту).
Рис. 99# Корневая система Euphorbia seguieriana (= gerardiand)", условия, как на
рис. 98. Глубже 240 см корень идет по полости,
оставленной старым корнем.
В 1957—1962 гг. в Казахстане исследовались два местообитания в сте-
пи (Festuceto sulcatae — Stipetum) и одно в полупустыне с группировкой
Artemisia — Festuca. Особенно подробно была изучена экофизиология
Stipa lessingiana и S. kirghtsorum х. Зима в этой континентальной области
очень холодная (средняя температура января ниже —15°), поэтому расте-
ния в течение всего зимнего периода находятся в состоянии покоя. Еще
сильнее здесь летняя засуха, иногда начинающаяся уже в мае и продол-
жающаяся до конца августа или даже сентября (рис. 102). В это сухое
время года злаки снова пребывают в состоянии полупокоя.
1 В. М. Свешникова, О некоторых эколого-физиологических особенно-
стях ковылей — эдификаторов степных фитоценозов, «Проблемы ботаники», 6, 1962,
стр. 388—398; Е. М. Лавренко, В. М. Свешникова, О синтетическом изу-
чении’жизненных форм на примере степных дерновинных злаков, «Журнал общей
биологии», 26, № 3, 1965, стр. 261—275.
Зона степей и прерий
117
Рис. 100. Корневая си-
стема Kochia prostrata на
сильносолонцеватом чер-
ноземе в Аскании-Нова;
интенсивно выпасаемый
участок (по Шалыту).
Рис. 101. Стержневой корень
Statice (Limonium) latifolia
(верхняя часть); Хомутовская
степь с Festuca sulcata, Salvia
nutans на обыкновенном чер-
ноземе (по Шалыту).
Из двух видов Stipa вид Stipa kirghisorum является казахстано-сред-
неазиатским элементом с ограниченным распространением в горах Тянь-
Шаня, Памиро-Алая и Тарбагатая, тогда как S. lessingiana повсеместно
встречается в более сухих степях. В Казахстане он растет на солонцеватых
каштановых почвах, S. kirghisorum — на выщелоченных песчано-глини-
стых светлых каштановых почвах. Сезонное развитие Stipa lessingiana
показано на рис. 103. Вегетационный период в степи короче, чем’в более
жаркой полупустыне, но летняя засуха в последней длится дольше. В благо-
приятные годы, каким был, например, 1957 г., часть растений осенью цвела
второй раз, в засушливом же 1959 г. цветки вообще не образовались.
Отдельные дерновины S. lessingiana достигают 30—50-летнего возраста,
однако со временем центральная их часть отмирает и единый прежде куст
разделяется на несколько меньших. Базальные части растений занимают
118
Глава 2
лишь часть поверхности почвы (обычно менее 50%), но покрытие листьями
в дождливые годы составляет почти 100%, в засушливые — значительно
Рис. 102. Климадиаграмма пробной площадки в Казахстане
для времени производства опытов. Для сравнения приведена
климадиаграмма станции Алма-Ата, характеризующейся зна-
чительно большим количеством осадков.
меньше. Следовательно, злаки обладают способностью редуцировать
и вновь восстанавливать транспирирующую поверхность в зависимости
от условий влагообеспеченности, благодаря чему.растения выживают даже
в самые засушливые годы.
1957г.
1958г.
1957г.
1958г.
Рис. 103. Фенологический спектр Stipa lessingiana в Казахстане.
Вверху’, разнотравная степь; внизу, переходная зона к полупустыне; в каждом случае на
двух разных местообитаниях.
1 — вегетативная стадия; 2 — колошение; з — бутонизация; 4 — цветение 5 — плодоно-
шение; 6 — осыпание семян; 7 — период полупокоя (по Борисовой).
Зона степей и прерий
119
Водный режим того и другого вида изучался в течение всего вегета-
ционного периода. Условия местообитания S. kirghisorum более благо-
приятны. Содержание воды в листьях обоих видов в мае колебалось
в пределах 50—60%; у S', lessingiana в июне оно резко понизилось и в сен-
тябре составляло лишь 32%; у S. kirghisorum сокращение влагосодержа-
ния началось лишь в июле, но даже к сентябрю количество воды в расте-
нии упало только до 42% сырого веса. Возможно, эти различия объясняют-
ся неодинаковым строением механической ткани: у S. lessingiana листья
несколько более ксероморфны, к тому же развитие этого злака начинается
раньше. Увеличение же сухого веса обусловливает кажущееся уменьше-
ние содержания воды.
Показателем способности растений приспосабливаться к изменчивым
условиям влагообеспечения может служить величина осмотического дав-
ления (табл. 20).
Таблица 20
Осмотическое давление у видов Stipa (в атм)
Stipa Май Июнь Июль Август Сентябрь
S. lessingiana .... 26,5 37,1 38,8 51,3 52,6
S. kirghisorum .... 30,6 35,6 40,6 39,8 37,5
Водный режим S. kirghisorum более сбалансирован, что находит отра-
жение и в величине транспирации, в два раза превышающей транспира-
цию S. lessingiana. Потеря воды этим видом в июне за сутки составляла
около 10 г/г сырого веса, тогда как у S. lessingiana лишь немного более
5 г/г.
Растения регулируют интенсивность транспирации путем свертыва-
ния листьев, а еще раньше путем замыкания устьиц, расположенных
только на верхней поверхности листьев; при свертывании листа устьица
на их внутренней стороне оказываются закрытыми. У S. lessingiana
устьица расположены глубоко в ткани листа, сами листья обладают тол-
стой кутикулой и резко выраженным ксероморфным строением.
По сравнению с 5. kirghisorum S. lessingiana сокращает транспирацию
® природных условиях много раньше (табл. 21 и рис. 104). Поэтому этот
вид более засухоустойчив и проникает глубоко в пустыню, тогда как рас-
пространение S. kirghisorum ограничено горными степями. Festuca sulcata
ведет себя так же, как виды Stipa, но при наметившемся дефиците влаги
се листья засыхают быстрее. Поэтому в резко засушливых районах она
утрачивает свое фитоценотическое значение.
Таблица 21
Регулирование величины транспирации у видов Stipa
Stipa Устьица широко открыты днем Из всех листьев к 14 часам свернулись
весной летом весной летом
S. lessingiana . . . 6 часов 2 часа 60% 75%
S. kirghisorum . . . 9 часов 5 часов 25% 65%
120
Глава 2
°C
201 Stipa lessingiana _
’„та п стптгяш и
7 8 9 10 11 12 13 14
15 16
*4 Stipa kirghisorum
тапгта
17 18 19 20 21
18 19
7 8 9 10 11
Рис. 104. Дневной ритм свертывания листьев Stipa lessingiana и St. kirg-
hisorum в Казахстане в мае перед засухой и в сентябре к концу засушливого
периода (по Свешниковой).
Доля листьев: 1 — развернутых; 2 — полусвернувшихся; 3 — полностью свернувшихся.
Кривые — дневной ход температуры.
Динамика приспособления видов Stipa к осуществлению фотосинтеза
аналогична изменениям скорости транспирации. Весной до полудня фото-
синтез весьма интенсивен, после полудня может быть второй мак-
симум; в разгар же лета кривая фотосинтеза очень пологая. Цветки
Stipa lessingiana начинают открываться ночью в 3 часа и закрываться уже
в 7 часов утра. Известны также другие виды злаков, которые пылят
между '14 и 19 часами, как, например, S. kirghisorum и Festuca sulcata.
Многие виды Stipa факультативно клейстогамии, что связано с засухой
или с низкой температурой. Плоды созревают очень быстро. Чере»
8 дней после цветения зародыш S. lessingiana уже вполне развит; спустя
12—15 дней плоды поспевают и опадают. Все виды Stipa являются
анемохорами, за исключением S', capillata, относящегося к группе авто-
хорных растений. Энтомофауна степей относительно бедна. У степных
злаков, по-видимому, имеется эндотрофная микориза, но вопрос этот еще
недостаточно выяснен. Из ризосферы выделен Clostridium pasteurianum,
но Azotobacter, вероятно, отсутствует. Доказано постоянное присутствие
нитрифицирующих и денитрифицирующих бактерий.
В общем исследованиями установлено, что степные злаки хорошо
приспособились к существованию в климате степей и что они обладают
способностью соответствующим образом реагировать на резкие колебания
условий обеспечения водой от года к году. Именно эти свойства и позво-
лили злакам стать доминирующими видами в этой зоне.
Зона степей и прерий
12Т
Совершенно аналогичная картина наблюдается в североамерикан-
ской прерии с той только разницей, что видовой состав в прерии несколько-
иной. Виды Stipa распространены и здесь, но во влажной прерии они
играют по сравнению с видами Andropogon лишь второстепенную роль
и утрачивают всякое значение на Великих равнинах. Более широко рас-
пространены они вместе с видами Festuca только на крайнем севере обла-
сти прерий.
В прерии весна также очень благоприятное для растений время года.
Растительный покров напоминает цветущие свежие луга Центральной
Европы. Обеспеченность растений водой оптимальна, что находит свое
отражение в низком осмотическом давлении. В 1930 г. автором произведе-
ны в Небраске многочисленные измерения, но здесь мы приведем резуль-
таты лишь некоторых из них. По нашим данным, даже в разгар лета в обыч-
ные годы повышение концентрации клеточного сока относительно невели-
ко. Только в необычно сухом 1934 г. осмотическое давление у изучавших-
ся видов резко поднялось (табл. 22).
Таблица 22
Осмотическое давление у растений прерий (в атм)
Название растений До волны жары в мае — июне После волны жары в конце июля Максимальные значения в за- сушливом 1934 г. (по Стоддарту)
Белмонт 1 Вудлаун Белмонт Вудлаун
Stipa spartea . . Andropogon gerar- 12,1—15,3 11,6—13,5 20,2 19,8 33
di Psoralea floribun- 8,4 — 12,9 11,9 38
da 9,8-12,4 9,9—11,9 22,4 19,8 28 Amorpha canescens
Helianthus rigidus 7,3-7,7 8,7—9,1 15,1 11,9 37
1 Прерия Белмонт представляла собой несколько более сухое местообитание, чем прерия*-
Вудлаун.
В 1934 г. исключительно высокое давление (55 атм) отмечалось,
у Andropogon scoparius, корневая система которого проникает не столь
глубоко, как у A. gerardi. У Koeleria cristata измеренное осмотическое дав-
ление составило 35 атм, у Sporobolus heterolepis — 38, у Liatris scariosa —
37 атм. Дефицит влаги в листьях был также весьма значительным. Судя
по всему, растения прерий вели себя так же, как степные виды, исследо-
ванные Максимовым (об ущербе, нанесенном засухой, см. выше).
Поскольку недостаток влаги в низкозлаковой прерии (Великие рав-
нины) выражен гораздо острее, повышение осмотического давления летом
у здешних видов происходит значительно более резко. Весной основные
злаки, как Bulbitis dactyloides, Bouteloua gracilis, В. hirsuta и В. racemosa,
характеризуются очень низким, 8—12 атм, осмотическим давлением,
но летом оно поднимается до 40 атм и выше. Только у встречающихся
здесь суккулентов осмотическое давление оставалось по-прежнему низ-
ким (Opuntia polyacantha 6—8 атм, Yucca glauca 11—20 атм).
Результаты измерений осмотического давления, произведенных Бира-
ном в степях Анатолии, мы уже приводили (см. т. II, гл. 2).
Имеются материалы весьма многочисленных эколого-физиологиче-
ских исследований степных элементов, встречающихся в Центральной
122
Глава 2
Европе как реликты ксеротермического периода эпохи постгляциала.
В Паннонском бассейне Венгрии с его климатом лесостепи на вторичных
местообитаниях можно еще найти весьма обширные степные сообщества,
детально изученные Шоо и его учениками х. Встречаемость их под Веной,
а также облик степных пустошей Центральной Европы мы уже обсуждали
(ср. «Phytologie», Bd. IV, 2).<
Эти вклинивающиеся далеко в Центральную Европу степные элементы просле-
живаются: вверх по течению Днестра в междуречье Вислы и Варты, откуда, с одной
«тороны, через долину Одера в Тюрингенскую котловину, горы Кифхейзер и область
Магдебургская Бёрде, с другой — на север, к островам Эланд и Готланд в Балтийском
море.
Другой путь их миграции шел от долины нижнего течения Дуная и области
Добруджа, степи которых непосредственно примыкают к восточноевропейским, в Пан-
нонский и Венский бассейны, оттуда в южную Моравию и Чехию, а затем вверх по
течению Дуная к пустошам нижнего Изара, Франконской и Швабской Юре, в бассейн
Майна и Неккара, Верхнерейнскую низменность с Майнцской котловиной, где еще
встречаются Adonis vernalis, Onosma arenarium, Jurinea cyanoides, Stipa и др. На западе
к понтическим элементам все более примешиваются понтическо-средиземноморские
или субсредиземноморские элементы, проникшие в Центральную Европу с Балканско-
го полуострова, вдоль Роны и Соны, а также по Мозелю, и широко распространенные
еще в Тюрингенской котловине. К ним относятся не только субсредиземноморские
орхидеи, почти отсутствующие в восточноевропейских степях, так как не переносят
холодной зимы, но и Quercus pubescens, Coronilla emerus, Amelanchier ovalis, виды Teucri-
um£Anthericum, Globularia и Helianthemum, Dictamnus albus и многие другие.
В условиях гумидного климата Центральной Европы степные эле-
менты представляют собой экстразональную растительность, которая сохра-
няется на местообитаниях, сходных по условиям среды с восточноевропей-
скими луговыми степями 1 2.
Это прежде всего известняковые и лёссовые склоны южной экспози-
ции, сухие и лучше прогреваемые, либо же очень маломощные почвы,
формирующиеся, например, на гребнях известковых скал также южной
экспозиции, либо хорошо водопроницаемые известняковые осыпи на скло-
нах и песчаные почвы со щелочной реакцией в относительно бедных осад-
ками районах. Сухость этих местообитаний препятствует развитию на них
центральноевропейских лесных растений, поэтому степные виды не испы-
тывают здесь какой-либо конкуренции. Это естественные, лишенные леса
участки, где могут закрепиться субсредиземноморские элементы. Встре-
чаются даже альпийские элементы, как Sesleria coerulea, Saxifraga
aizoon, Daphne cneorum, Biscutella laevigata, но они предпочитают, скорее,
•северные склоны скальных гребней.
На таких узкоограниченных местообитаниях степные элементы сохра-
нились в Центральной Европе со времен сухого предбореального периода
или термического оптимума постгляциала, но уже очень давно человек
создал для них благоприятные вторичные безлесные местообитания бла-
годаря выжиганию травы, косьбе или выпасу овец. Таким образом эти
виды могли распространиться шире, и в настоящее время они являются
основным компонентом растительности сухих лугов, заброшенных вино-
градников, овечьих пастбищ или придорожных полос и межей в местах
с теплым микроклиматом.
1 R. S о 6, Systematische Ubersicht der pannonischen Pflanzengesellschaften,
«ActalBot. Acad. Sci. Hung.», 5, 1959, S. 473—500; Bibliographia phytosociologica:
Hungaria, «Excerpta bot.», Sect. В 2, 1960, p. 93—156.
2 H. D. Krause h, Mikroklimatische Untersuchungen an Steppenpflanzengesell-
schaften der Randhange des Oderbruches, «Arch. f. Natursch.», 1, 1962, S. 142—164;
A. Q u a n t i n, L’evolution de la vegetation a 1’etage de la chenaise dans le Jura meri-
dional, These, Paris, 1935.
Зона степей и прерий
123
Мы не можем подробно останавливаться здесь на разборе всех много-
численных флористических и фитоценологических работ, посвященных
описанию «степных пустошей» Центральной Европы (см. «Phytologie»,
Bd. IV, 2, S. 597—641 и список литературы). Отметим лишь, что эти
формации заинтересовали экологов мира главным образом в связи с пробле-
мой ксерофитов. Дело в том, что в их составе были обнаружены так назы-
ваемые «центральноевропейские ксерофиты», которые, правда, при более
тщательном изучении и сопоставлении с растениями пустынь оказались
всего лишь мезофильными светолюбивыми растениями. Этот курьезный
случай убеждает нас в том, что определение «ксерофит» часто следует
понимать лишь относительно; обычно «ксерофитами» называют виды,
растущие в том или ином районе на наиболее сухих местообитаниях.
Но именно изучение степных пустошей Центральной Европы, вызвав-
шее столь оживленные дискуссии, около 40 лет назад побудило начать
эколого-физиологические исследования во многих уголках Европы.
Назовем лишь несколько работ: Г. Вальтер и Э. Вальтер (район озера
Балатон), 1929 г.; Хейлиг (район Кайзерштуля), 1930—1931 гг.; Шандерль
(Вюрцбург), 1930 г.; Фольк (пески под Гейдельбергом), 1931 г.; Пизек
и Картельери (Инсбрук), 1931 г.; Штоккер (Хортобаги-Пушта), 1933 г.;
Бозиан (район Кайзерштуля), 1933—1934 гг.; Фрей (Вале), 1934 г.;
Мюллер-Штолль (Крайхгау), 1935 г.; Хертель (Вена), 1936 г.; Гофман
(Мёдлинг под Веной), 1936 г.; Брзоска (бассейн Одера), 1936 г.; Фольк
(известняковые гребни под Вюрцбургом), 1937 г.; Штубер (Валлис),
1938 г.; Прейс (Чехия), 1939 г.; Дёр (Мёдлинг, под Веной), 1941 г. Кроме
-того, следует отметить несколько более поздних публикаций: Арвидсон
(Эланд), 1951 г.; Ман (Галле), 1957 г.; Хорански (Венгрия), 1957 г.; Борн-
кам (Гёттинген), 1958 г.; Сметанкова (Прага), 1959 г.; Степанович-Весели-
цич (Белград), 1959 г.; Шоо и сотр. (Венгрия), 1961 г.; Рихновска (Чехия),
1961 и 1965 гг. 1
Особенности микроклимата степных пустошей привлекли к себе вни-
мание после классической работы Крауса (G. Kraus, Boden und Klima
auf kleinstem Raum, Jena, 1911). В дальнейшем исследования микрокли-
мата производились неоднократно. Они показали, что температура
на поверхности почвы пустошей уже весной при интенсивной инсоляции
может повышаться до 50°. Летом нередко отмечалась температура 60°.
На сухом замоховелом лугу на песчаных почвах под Гейдельбергом Фольк
в июле однажды измерил даже 70°. Высокая температура приземного слоя
воздуха обусловливает большой дефицит насыщения и соответственно
интенсивное испарение; стандартным эвапорометром Пише (зеленая бума-
га) на высоте растений в полдень была измерена величина 1,6—1,85 мл/члс.
В часы, когда дул горячий ветер, испарение превышало 2,5 мл!час. Эти
значения соответствуют условиям в пустыне. Разница состоит, однако,
в том, что в степных пустошах столь высокие показатели зафиксированы
лишь в полдень, в редкие жаркие безоблачные дни, тогда как в пустыне
экстремальные условия могут сохраняться в течение нескольких недель
и даже месяцев.
При рассмотрении климадиаграмм наиболее засушливых районов
Центральной Европы, составленных по средним многолетним данным,
можно заметить как крайний случай лишь незначительный по длитель-
ности сухой период; следовательно, в кчиматическом отношении эти райо-
1 Более'’подробные данные о перечисленных работах си. в «Phytologie», Bd. HI,
1; Bd. IV, 2.
124
Глава 2
Рис. 105. Экстразональная степная растительность на гипсо-
носных почвах, гора Кифхёйзер (по Мёйзелю, из «Phytologie»,
Bd. IV, 2).
Вверху: скалистая пустошь с Festuca glauca на освещенной солнцем
каменистой маломощной почве. Слева направо: Sedum boloniense, Alys-
sum montanum, Festuca glauca, Artemisia campestris, Sedum boloniense,
Potentilla arenaria, Festuca glauca.
Внизу: степная пустошь на мощных почвах. Слева направо: Scorzonera
purpurea, Stipa pennata, Scabiosa canescens, Carex humilis, Festuca glauca,
Fumana procumbens, Stipa sennata, Aster linosyris, Stipa capillata, Carex
humilis, Festuca glauca.
ны соответствуют пограничной полосе между лесом и лесостепью, приме-
ром чему является Магдебургская Бёрде или Майнцская котловина.
Правда, случаются и засушливые годы (например, 1964 г.), когда летом
долго не бывает дождя, тогда как обычно он идет в среднем через день.
В такие годы в течение некоторого периода создаются условия, которые
напоминают условия среды, существующие собственно в степной зоне.
Местообитания степной пустоши засушливое время преображает особенно
сильно, ибо запасы влаги в здешних почвах из-за их маломощности весьма
низкие. Почвенный покров степных пустошей чрезвычайно пестр. Здесь
можно встретить все типы почв от еще подвижных слаборазвитых поч-
венных образований на известняковых склоновых осыпях или на поверх-
ности известняковых скал, разбитых трещинами, через рендзины и перегной-
Зона степей и прерий
125
но-карбонатные почвы до мощных лёссовых почв х. Поэтому растительность
пустошей то очень разрежена, то образует сомкнутый покров. По этой
причине к степным пустошам относятся также сообщества-пионеры
на голых скальных площадках с суккулентами, например видами
Рис. 106. Скалистая пустошь в районе Фри-
дингена (долина Дуная, Швабская Юра)
с Saxifraga aizoon, Dianthus caesius, Vincetoxi-
cum officinale, Rumex scutatus, Festuca glauca,
Asplenium trichomanes (фото Фейхта).
Sedum и Sempervivum (рис. 105, вверху), либо с растениями, живущими
в трещинах скал (рис. 106) или закрепляющими осыпи {Dryopteris rober-
tiana, Rumex scutatus, Vincetoxicum officinale), а на мелкообломочных грун-
тах — также Sesleria coerulea и Teucrium chamaedrys.
Облик степной растительности приобетают только сообщества на мощ-
ных песчаных или лёссовых почвах (рис. 105, внизу). На таких местооби-
таниях наблюдается смена аспектов, вполне соответствующая их последо-
вательности в степях.
Весной первыми появляются терофиты и весенние геофиты {Pulsatilla,
Adonis vernalis), затем в рост трогаются злаки и разнотравье, обладающие
неглубокой корневой системой; в июне наступает апогей цветения; к раз-
гару лета часть весенних растений уже засыхает, у других видов созревают
1 На кислых породах преимущественно развиты верещатникоподобные сообще-
ства, не содержащие степных элементов; пример — сообщества на порфировых скалах
южного Шварцвальда.
126
Глава 2
плоды и только у видов с глубокопроникающими корнями начинается
цветение, которое у многих зонтичных и сложноцветных затягивается
до осени.
В благоприятных условиях весеннего увлажнения транспирация
растений степных пустошей протекает весьма интенсивно: кривая суточ-
ной транспирации параллельна кривой испарения. Но как только с
10 11 12 13 14 15 16 17 18 Часы
а-----&1 о-----оз а-----*5 •-----• 7
□-----о 2 ------ 4 а-----а 6
Рис. 107. Дневной ход интенсивности транспирации у раз-
личных видов степной пустоши в Крайхгау, земля Баден
; (по МюллерШтоллю, из «Phytologi е», Bd. Ill, 1).
Вверху: в начале сухого периода (10.VIII), внизу: во время сухого
периода (19.VIII). 1 — Prunella grandifora', 2 — Centaurea scabiosa;
3 — Heliantdemum chamaecistus: 4 — Genistatlinetoria", 5 — Geranium
sanguineum', 6 — Aster amellus', 7 — Teuctium chamaedrys.
наступлением засухи запасы влаги в почве иссякают и листья начинают
вянуть, растения сокращают транспирацию, закрывая устьица,— сначала
только в полуденное время, позже на весь день, исключая утренние часы
(рис. 107). Первыми снижают интенсивность транспирации виды, у кото-
рых корни пронизывают лишь верхние слои почвы. Если засуха затяги-
вается, листья вянут и засыхают, но у некоторых видов, например у
Euphorbia seguieriana, лишь желтеют. Степень обеспеченности водой нахо-
дит отражение в величине осмотического давления. При хорошем снабже-
нии водой его значения, так же как это наблюдается у степных и пре-
рийных растений, остаются низкими, но, как только начинает ощущаться
Зона степей и прерий
127
Рис. 108. Годовой ход осмотического давления у двух видов остепнен-
ной пустоши на песчаных почвах Верхнерейнской низменности (по опре-
делениям Фолька, заимствовано из Walter, 1931).
недостаток воды, осмотическое давление тотчас повышается (рис. 108),
причем мелкоукореняющийся Helichrysum реагирует на дефицит влаги
быстрее, чем глубокоукореняющаяся Artemisia. Слаботранспирирующие
лилейные гидростабильны, величина их осмотического давления изме-
няется мало, хотя они и относятся к видам, которые развивают повер-
хностную корневую систему (ср. поведение Anthericum, рис. 109).
Колебания осмотического давления отражают также различия в харак-
тере местообитаний. Например, у видов (при одновременном отборе проб)
на сухом известняковом гребне осмотическое давление всегда выше, чем
у тех же видов, произрастающих на более влажном лёссовом склоне
(рис. 109). В экстремальных условиях у растений степных пустошей
в редких случаях отмечалось очень1 высокое осмотическое давление,
превышавшее 80 атм.
При слабом, но сохраняющемся длительное время недостатке влаги
наблюдается активное регулирование растениями осмотического давле-
ния. Оно постоянно сохраняется на более высоком уровне, даже если
содержание воды в* почве увеличивается. Обычно это происходит летом
у растений на несколько более сухих местообитаниях. Например, у особей,
-128
Глава 2
поселившихся на известняковом гребне, осмотическое давление и после
.дождя всегда выше, чем у растений на лёссовых склонах. Наличие у расте-
ний механизма регуляций осмотического давления способствует тому, что
анатомо-морфологическая структура вновь развившихся листьев стано-
вится более ксероморфноц (рис. 110).
Засушливые годы, которые в Центральной Европе значительно менее
-экстремальны, чем даже средние по водности годы в восточной степи, тем
Рис. 109. Годовой ход осмогического давле-
ния у видов на сухом известняковом гребне (1)
и на лёссовом склоне в Крайхгау (2) ^(по
Мюллер-Штоллю).
Hipp. = Hippocrepis comosa', Hel. = Helianthemum
•chamaecistus’, Anther. = Anthericum ramosum. Значе-
ния на местообитании I выше, чем на местообитании 2.
la —> S. Joannis -> S. pulcherrima -> S.
не менее оказывают на расти-
тельный покров губительное воз-
действие — он практически весь
выгорает. Но этот урон имеет
временный характер. Подземные
части или почки возобновления
у поверхности почвы остаются
живыми, либо же запас семян
столь велик, что обеспечивает
семенное возобновление расте-
ний. Таким образом, происхо-
дят лишь количественные изме-
нения состава растительного по-
крова, в качественном же отно-
шении он весьма устойчив. Ины-
ми словами, видовой состав оста-
ется неизменным вне зависимо-
сти от количества осадков в
отдельные годы.
Имеются данные сравни-
тельного исследования не-
скольких видов Stipa, проведен-
ного Рихновска в Моравии Ч
Пробная площадка была вы-
брана на юго-западном склоне,
сложенном пермскими конгломе-
ратами. Наблюдениями установ-
лено, чтоксерофильные свойства
все более отчетливо проявлялись
у растений в следующей после-
довательности: S. stenophyl-
capillata. Микроклиматические
измерения показали аналогичное направление увеличения сухости место-
обитаний этих видов Stipa. Изучение величин транспирации и условий
обеспеченности растений водой во время засухи позволило установить,
что приспособленность злаков к недостатку влаги нарастает в той же
последовательности. Сильно пострадал только S. capillata. Правда, засуха
захватила этот поздноцветущий злак перед цветением, когда другие
травы уже плодоносили. Рихновска изучала также условия местообитания
Corynephorus canescens 1 2. В отличие от континентальных видов Stipa Согу-
1 М. Rychnovska, A contribution to the ecology of the genus Stipa, «Preslia»
(Praha), 37, 1965, p. 42—52; M. Rychnovska-Soudkova, Wasserhaushalt
einiger Sfipa-Arten am naturlichen Standort, «Ceskoslow. Akad. Ved.», 76, № 6, 1966.
2 M. Rychnovska, Corynephorus canescens (L.) P. Reauv. (Physiologisch-
•okologische Studie einer Pflanzenart), «Akad. d. Wiss. Prag.», 71, 1961.
Зона степей и прерий
обобщили для разных подзон степи Восточной Европы, Западной Сибири
и Алтая. Важнейшие показатели для 6 типов степи приведены в табл. 23.
Состав растительного покрова отдельных типов характеризуется преобла-
дающими видами.
I. ВОСТОЧНАЯ ЕВРОПА (РУССКАЯ РАВНИНА)
1. Bromus erectus ssp. riparius, В. inermis, Koeleria gracilis, Stipa Joannis, Calama-
grostis epigeios, Carex humilis, Medicago falcata, Filipendula hexapetala, Falcaria sioidfis,
Salvia pratensis, Centaurea marschalliana и др. на мощном, слабовыщелоченном чернот-
земе, Стрелецкая степь. ..}
2. Stipa ucrainica, Festuca sulcata, Galium verum, Falcaria sioides, Linosyris villosa
и др. на южном черноземе и темных каштановых почвах, Аскания-Нова. 1
II. ЗАПАДНАЯ СИБИРЬ И АЛТАЙСКИЙ КРАЙ
3. Stipa capillata, S. rubens, Festuca sulcata, Filipendula hexapetala, Thymus mar-
schallianus, Phlomis tuberosa, Libanotis intermedia и др. на выщелоченном черноземе
со средним содержанием гумуса, Приобское плато.
4. Stipa capillata, S. rubens, Festuca sulcata, Koeleria gracilis, Veronica spicata,
Medicago falcata и др. на южном черноземе, Алтайский край.
5. Stipa capillata, S. rubens, Festuca sulcata, Galium verum, Medicago romanica
и др. на южном черноземе, Алтайский край.
6. Festuca sulcata, Stipa capillata, S. lessingiana, Artemisia frigida, A. glauca и др.
на каштановых почвах, Алтайский край.
В степях большая часть надземной массы осенью отмирает, годовая
продукция сухого вещества почти равна годовому опаду, образующему
подстилку. Сохра чются только базальные части дерновин злаков
и части побегов, масса которых с возрастом растения увеличивается-
В итоге надземная фитомасса более чем вдвое превышает годовую продук-
цию. Наблюдения показывают, что величина этой ежегодно продуцируемой
надземной массы от года к году резко колеблется.
Так, например, для Аскании-Нова (бедная разнотравьем ковыльная
степь) приводятся следующие данные о надземном урожае (в кг/га)’.
Во влажный год . . 4530—6250
В сухой год .... 710—2700
Подземная фитомасса, напротив, остается неизменной. В этом прояв-
ляется способность растительного покрова приспосабливаться к иеремен-
чивым условиям влагообеспечения, о чем мы уже говорили: в сухое время
года транспирирующая и ассимилирующая листовая поверхность сокра-
щается, соответственно уменьшается и количество вновь образующегося
органического вещества.
Опытным путем для разных степных сообществ было установлено, что
доля подземной фитомассы возрастает с севера на юг, то есть в направле-
нии увеличения сухости климата. Поскольку основная часть надземной'
фитомассы ежегодно отмирает, доля ежегодного опада фитомассы, равно
как и сама фитомасса, уменьшается с юга на север. Мертвая подстилка,
покрывающая землю, так называемый степной войлок, достигает в луго-
вой степи 8—10 т!га, в сухой степи — только 3 тп!га, ибо в последнем слуг
чае продуцирование надземного сухого органического вещества происхо-
дит менее интенсивно. Однако объем подстилки не остается постоянным
в течение вегетационного периода и колеблется в больших пределах.
Весной и летом процессы разложения протекают наиболее энергично,
потому к началу засухи, в разгар лета, количество подстилки мини-
мально. Максимальное количество подстилки накапливается поздней
9*
132
Глава 2
осенью или в предвесеннее время. Концентрация азота в подстилке
относительно высокая. Зола злаков содержит кремнекислоту.
ТОПОГЕННЫЕ БОЛОТА И ЯВЛЕНИЯ ЗАСОЛЕНИЯ
В СТЕПНОЙ ЗОНЕ
Топогенные болота возникают повсеместно там, где грунтовые воды
располагаются очень близко к земной поверхности или где происходит
их излив на поверхность почвы. Разумеется, уровень грунтовых вод может
колебаться в известных пределах, но он не должен опускаться ниже гра-
ницы, доступной для корней болотных растений. Таким образом, расти-
тельный покров низинных, или плоских, болот не зависит от количества
атмосферных осадков и, следовательно, в известной мере также от клима-
та. Тем не менее его вряд ли следует относить к категории азональной
растительности, ибо климат оказывает определенное влияние на состав
грунтовых вод и косвенно на растительность болот.
В горных местностях, где грунтовые воды просачиваются на дневную
поверхность и на самых низких позициях рельефа заболачивают почву,
фактором, определяющим состав грунтовых вод, становятся различия
в исходных породах. В районах, сложенных известняками, грунтовые
воды содержат известь и имеют слабощелочную реакцию; в местах разви-
тия песчаников, а также там, где обнажаются граувакки, глинистые слан-
цы или граниты, грунтовые воды бедны известью и обычно слегка кислые.
В пределах же равнинных областей Восточной Европы, где грунтовые
воды почти не соприкасаются с подстилающими почву горными породами,
а их приток и отток затруднены, воздействие климата более значительно.
В условиях гумидного климата, например в бореальной зоне, количе-
ство осадков значительно превышает величину испаряемости. Поэтому раз-
витые здесь подзолистые почвы выщелочены. Просачивающаяся сквозь
почву и скапливающаяся в рытвинах вода почти не содержит растворен-
ных веществ; коллоиды гумуса окрашивают ее в буроватый цвет, реакция
воды кислая. В местах застаивания воды почва переувлажняется до преде-
лов полного насыщения водой, образуются болота с залежами торфа.
По мере поднятия болота вверх поверхностная его часть все более отда-
ляется от зеркала грунтовых вод и в этом случае увлажнение почвы проис-
ходит только за счет атмосферных осадков; начинается чисто омброгенное
развитие болота. Создаются условия для поселения видов Sphagnum.
Последнее же обеспечивает дальнейшее развитие болотного процесса
в направлении образования верховых болот (см. т. II, гл. 5). Чем ближе
между собой величины осадков и испаряемости и чем чаще поверхность
болота периодически несколько высыхает, тем меньше возможность обра-
зования верховых болот и тем меньше их доля в общей площади болот
(табл. 24).
Самые южные сфагновые болота распространены под Днепропетров-
ском и Кировоградом1. Южнее, количество осадков уменьшается, ис-
паряемость увеличивается, и хотя легкорастворимые соединения еще все
вымыты, в грунтовых водах происходит все большее накопление трудно-
растворимых солей кальция.
1 В северной части Венгерской низменности (Альфельд) также еще встречаются
сфагновые болота с Eriophorum vaginatum, Drosera rotundifolia, Vaccinium oxycoccus
и др. Ср.: T. Simon, Die Vegetation der Moore in den Naturschutzgebieten des nord-
lichen Alfold, «Acta Bot. Acad. Sci. Hungar», 6, 1960, S. 107—137.
Зона степей и прерий
133
Таблица 24
Доля верховых и плоских (низинных) болот
в общей площади болот Восточной Европы (в %)
Область Верховые болота Плоские болота
Северная бореальная 75 25
Междуречье Вятки и Ветлуги . . 66 34
Ленинградская 65 35
Центральная 50 50
Белоруссия, западная часть . . 45 55
Средняя Волга 40 60
Зона лесостепи 10 90
Степная зона 0 100
Реакция грунтовых вод становится все менее кислой, затем нейтраль-
ной и даже слабощелочной. Эта перемена находит отражение в составе
болотной растительности. Ацидофильные виды выпадают из растительного
покрова, но появляются базифильные. Если в гумидных районах в почве
всегда наблюдается нисходящее движение воды, то в семигумидцой области
летом из-за сильного испарения воды с поверхности почвы господствует
режим непромывного типа с восходящим током почвенной влаги. При
большом содержании в грунтовых водах растворенного Са(НСО3)2 воз-
никает необратимый процесс выпадения в осадок СаСО3. Торф в таких
болотах насыщен известью, что может стать причиной образования болот-
ного мергеля. Величина pH превышает 7, поэтому говорят о болотах
со щелочной реакцией или о засолении кар-
бонатом кальция. Щелочные болота покрыты преимущественно
высокорослыми, образующими крупные кочки видами Carex {Carex elata
ssp. omskiana и др.); в Венгрии их называют формацией «цзомбек». По-види-
мому, они типичны для пограничной полосы между лесной и лесостепной
зонами.
Об экологии видов низинных болот известно немного. Хотя развитие
этих видов связано с насыщенным водой субстратом, их листья могут
иметь несколько ксероморфное строение. Осмотическое давление у отно-
сительно высокорослых видов сравнительно большое, например у круп$
ных осоковых — свыше 15 атм, у Phalaris — более 20 и у Phragmites
на сухих местообитаниях почти 28 атм. Это явление, видимо, можно
объяснить низкой пропускной способностью водопроводящей системы
растений. Бросается в глаза большая ксероморфность образованных летом
верхних листьев в сравнении с нижними листьями А Затруднения в погло-
щении влаги, обусловленные плохой аэрацией почвы, по-видимому, зна-
чения не имеют, так как маленькие, с нежными листочками растения харак-
теризуются осмотическим давлением, колеблющимся в пределах 5—10 атм.
Снабжение корней кислородом в часто содержащей H2S и почти лишен-
ной О2 почве осуществляется через крупные межклетники, расположен-
ные в основании побегов. Чем влажнее местообитание, тем большее про-
странство в корнях занимают межклетники, тем меньше их удельный вес.
Низинные болота обычно используются как сенокос, дающий сено
1 R. Y а р р, Spiraea ulmaria L. and its bearing on the problem of xeromorphy iri
marsh plants, «Ann. Bot.», 26, 1912, p. 815—870.
134
Глава 2
для подстилки. Если выкашивание не производится, то уже в скором
времени болота зарастают лесом. Сначала укореняются различные ивы
(виды Salix), за ними следуют также древесные породы: на богатых пита-
тельными веществами болотах ольха (Alnus glutinosa), на обедненных
ими — береза (Betula). Низинные болота превращаются в заболочен-
ные леса.
С приближением к границе между гумидной и аридной областями,
то есть к полосе, где количество осадков соответствует испаряемости,
на равнинной местности становятся все более заметными явления засоле-
ния. Часто встречаются бессточные низины, вода из которых стекает
только в особенно полноводные годы. В таких условиях из почвы вымы-
вается лишь часть образующихся в процессе выветривания горных пород
солей натрия (Na2SO4). Поэтому в почвах с близкими грунтовыми водами,
в их гумусовом слое, происходит известное накопление ионов натрия.
В результате избытка натриевых солей кальций вытесняется из обменно-
способных частей почвы и замещается натрием. Так образуются гуматы
йатрия или же такие соединения, как Na2SiO3 и NaA102.
Во влажное время года почва глубоко и обильно промачивается водой.
Избыток влаги, особенно если поступающая с поверхности вода в гумусо-
вом горизонте обогащается СО2, обусловливает развитие реакции гидро-
лиза, при котором Na+ вытесняется Н+ и может произойти замещение
силиката^натрия Са(НСО3)2
Гумат натрия + Н2СО3 [ гумат Н + NaHCO3
Na2SiO3 + Сэ(НСО3)2 | CaSiO3 + 2NaHCO3
В том и другом случаях образуется легкорастворимая сода, которая
нисходящим током воды вымывается в горизонт грунтовых вод. Однако
в сухой сезон Na2GO3 накапливается на поверхности почвы и обусловли-
вает содовое засоление почв, характерное для лесостепной
зоны, в частности для райнов лесостепи Маньчжурии.
В Паннонском бассейне обширные площади прежде регулярно затап-
ливались водами крупных рек. Поэтому концентрации солей в почве не про-
исходило. После того как реки были зарегулированы и разливы прекра-
тились, на всех летом сильно высыхающих почвах равнин с высоким стоя,
нием грунтовых вод началось засоление содой, сформировались почвы «сик»-
в которых известную роль играют также сульфаты. Место прежних пой-
менных лесов заняли открытые сообщества пушты 1 2.
Такого рода «содовые» почвы с «содовыми озерами» известны в Цен-
тральной Европе в районе озера Нейзидлер-Зе, причем засолению в дан-
ном случае способствует вымывание солей из соленосных поверхностных
пластов горных пород третичного возраста 3. Вендельбергер подразделяет
1 В. А. К о в д а, Е. Н. Иванова: см. «Почвы СССР», т. I, 1939, стр. 299—
403; Р. J anitzky, Saiz- und Alkaliboden und Wege ihrer Verbesserung, Giessen, 1957.
2 0.Stocker, Untersuchungen in der ungarischen Alkalisteppe, «Jb. wiss. Bot.»,
78, 1933, S. 751—856. Особенно детальные исследования этих галофильных раститель-
ных сообществ произведены: Gy. Bodrogkozy, «Acta Bot. Acad. Sci. Hungar»,
6. 1960, S. 171—207; 8, 1962, S. 1—37; 11, 1965, S. 1—51; 12, 1966, S. 9—26. Почвен-
ную карту, на которой выделены почвы «сик», читатель найдет в работе: Н.-Р. В 1 u m е,
Bodenkundliche Studien im Pannon-Becken und seinen westlichen Randgebieten, «Schrift.
Landw. Fakult. Kiel», № 34, 1963.
3G. Wendelberger, Die Vegetation des Neusiedler See-Gebietes, «Szb.
Osterr. Akad. Wiss., math.-naturw. Kl.», Abt. I, 168, 1959, S. 305—314; O. S t о c k e r,
Einige Bemerkungen uber die Salzstandorte ostlich des Neusiedler Sees, «Verh. Zool.-Bot.
Ges. Wien», 100, 1960, S. 106—111; H. Franz, G. H и s z, Das Neusiedlerseebecken,
«Osterr. Bodenk. Ges.», H. 6, 1961, S. 52—75.
Зона степей и прерий
135
галофильные виды по их отношению к содержанию солей в почве на при-
мере видов, распространенных в окрестностях озера Нейзидлер-Зе Ч
Подразделим здесь (с некоторыми изменениями) растения по степени
галофильности на 5 групп — от Г 5 (на сильнозасоленных почвах) до Г 1
{растения, избегающие засоленных почв). Римские цифры за видовым
названием отражают степень потребности растений во влаге (от I —
на очень сухих местообитаниях до V — растущих в погруженном в воду
состоянии). Приводимая ниже классификация относится только к содо-
вому засолению.
Г 5 — Camphorosma annua I, Lepidium cartalagineum II, Suaeda maritima и S. salsa
II — IV.
Г 4 a) — Matricaria chamomilla-bayeri I, Salicornia europaea III, Puccinellia limosa
II — IV, Pholiurus pannonicus III — IV, Plantago tenuiflora III — IV, Suaeda
pannonica II, Cy perus pannonicus III — IV, Crypsis aculeata III — IV;
6) — (менее засоленные почвы) Puccinell'~ salinaria III — IV, Aster pannonicus
III — IV, Triglochin maritimum JV, Jcirpus maritimus IV, Scirpus tabernae-
montani IV, Plantago maritima III.
ГЗ — Artemisia maritima I, Limonium gmelini I, Taraxacum bessarabicum III, Scorzo-
nera parviflora III — IV, Juncus gerardi III — IV, Roripa kerneri III — IV,
Beckmannia erucaeformis III — IV, Bupleurum tenuissimum II, Trifolium
fragiferum III, Samolus valerandi III и др.
Г 2 — Potentilla anserina III, Glaux maritima III, Peucedanum officinale I, Odontites
rubra II —III, Lotus siliquosus II — III, Lotus corniculatus-tenuifolius. III,
Triglochin palustris III — IV, Cladium mariscus IV.
Г 1 — в эту группу отнесены все виды, избегающие засоленных почв.
ГО — (безразличны к засолению) Phragmites communis, Potamogeton pectinatus,
Lepidium ruderale встречаются и в сильнозасоленной воде, тогда как Myrio-
phyllum spicatum и Utricularia переносят лишь слабое засоление.
Систематизация галофильных видов при засолении почв хлоридами
иная. В этом случае Salicornia относится к наиболее солевыносливым
формам, Glaux также, видимо, должна быть причислена к более высокой
группе.
Однако подразделение, основанное лишь на приуроченности растений
к определенному типу местообитаний, еще не дает полного представления
об их экологии. Например, Artemisia maritima и Limonium gmelini попа-
дают в одну и ту же группу, поскольку в естественных условиях они обыч-
но встречаются вместе. Однако, как показал Штоккер, корни этих видов
•размещаются в совершенно разных почвенных горизонтах. У Artemisia
корни '^располагаются около поверхности, и поэтому растение страдает
ют летней засухи. Limonium, напротив, имеет стержневой корень, прони-
кающий до грунтовых вод и благодаря этому постоянно обеспечивающий
растение влагой.
Сообщества галофитов встречаются также в некоторых районах Цен-
тральной Европы с гумидным климатом. Однако развитие их здесь обуслов-
лено отнюдь не климатическими причинами. Галофиты поселяются в местах
выхода на поверхность естественных или искусственных соляных источ-
ников, и их распространение имеет лишь чисто локальное значение.
Более обширные площади засоленных почв располагаются на морских
побережьях. Здесь засоление почв вызвано воздействием соленых морских
вод или солоноватых вод суши (марши, кромки морских пляжей, лагуны;
ср. гл. 5).
В аридных условиях собственно лесостепной зоны содержание Na2SO4
и особенно NaGl в грунтов х водах все более повышается. В этом случае
1 G. Wendelberger, Zur Soziologie der kontinentalen Halophytenvegetation
Mitteleuropas, unter besonderer Berucksichtigung der Salzpflanzengesellschaften am
TJeusiedler See, «Osterr. Akad. Wiss., math.-naturw. KI.», Denkschr. 108, S. 5.
136
Глава 2
летом вследствие непрерывного испарения поднимающейся по капилля-
рам воды на поверхности почвы накапливается много солей, образующих
белую солевую корку. Развивается хлоридно-сульфатное
засоление и формируются настоящие солончаки, называемые в Аме-
рике «белыми щелочными почвами» в отличие от «черных щелочных почв»,
или содовых солончаков. Дело в том, что гуматы натрия легко пептизи-
руются и в воде образуются темные коллоидные гумусовые растворы
(черная корка). Напротив, в концентрированных растворах солей гуму-
совые вещества выпадают в осадок и поверхность почвы покрывается
скрепленной только солями коркой белого цвета1. Растения — индикаторы
содового и хлоридного засоления относятся к самым различным видам.
Для засоленных почв Восточной Европы можно назвать следующие имею-
щие индикаторное значение виды.
Содовое засоление (сильно увлажненные почвы): Trifolium fragiferum,
Triglochin maritimum, Juncus gerardi, Geranium collinum, Taraxacum bessarabicum, Aster
tripolium, несколько южнее также Scirpus maritimus, Puccinellia palustris, Peucedanum
latifolium, Senecio racemosus и др.
Засоление хлоридами: Salicornia europaea (= herbacea), Suaeda
prostrata, Limonium tomentellum, L. caspica, на солончаках в прибрежной полосе Черно-
го моря, кроме того, Halo спешит strobilaceum, Obione pedunculata, Bassia hirsuta, Suaeda
altissima, Salsola soda, Petrosimonia crassifolia, Frankenia hirsuta, Aeluropus litoralts,
Tamarix pallasii и др.
Приведенные данные указывают на особенно отчетливую связь рас-
тительности, свойственной местообитаниям с высоким уровнем грунтовых
вод, с определенным типом зональной растительности, развитие которой
обусловлено фактором «климат».
Резкогумидный климат: засоление отсутствует, преимущественно развиты
олиготрофные болота бореальной зоны.
Гумидный климат: засоление отсутствует, преимущественно распро-
странены эвтрофные болота зоны смешанных
лесов.
Субгумидный климат: засоление карбонатами кальция, «щелочные» боло-
та зоны, пограничной с зоной лесостепи.
Семиаридный климат: содовое засоление лесостепной зоны.
Аридный климат: засоление хлоридами собственно степной зоны
и зоны полупустыни.
В качестве примера приведем очень ровные речные террасы левобе-
режья Днепра с высоким уровнем грунтовых вод. Севернее Киева засоле-
ние не встречается; несколько южнее наблюдается накопление в почве
значительных количеств карбонатов кальция; на террасах выше Днепро-
петровска очень широко распространено содовое засоление; далее к югу
хорошо выражено засоление хлоридами, которое вблизи Черного моря
и особенно у Сиваша охватывает обширные площади.
Наряду с такого рода засолением сильно увлажненных почв известно-
засоление сухих почв. Оно сопровождается все большим участием
видов Artemisia (сначала A. austriaca, затем A. maritima), вследствие
чего степная растительность принимает облик полупустыни.
Слабое засоление почв, не испытывающих воздействия грунтовых
вод, в восточноевропейской степной области связано с приносом ветром
солевой пыли. Обширные площади, затопленные солоноватыми водами
Сиваша, летом высыхают и покрываются белой коркой соли. Ветер взвих-
1 К. К. G е d г о i z, Der absorbierende Bodenkomplex, Dresden, Leipzig, 1929.
Зона степей и прерий
137
ривает соль в воздух и переносит ее к северу, где она отлагается в пределах
зоны южного чернозема. Последний при дальнейшем увеличении аридно-
сти становится беднее гумусом, из-за чего горизонт А все более светлеет.
Такие почвы уже называют каштановыми. У этих почв горизонт
с пятнами карбонатных новообразований (белоглазка) по мере возраста-
ния сухости воздуха поднимается все выше по разрезу и, наконец, у свет-
лых типов почв залегает на глубине всего 30 см. Каштановые почвы в свою
очередь переходят i бурые почвы полупусты ни, а послед-
ние, уже в полосе па "тынь,— в лишенные гумуса сероземы.
Отложение солевой пыли на поверхности почвы в зоне южного черно-
зема и каштановых почв вызывает характерное изменение почвенного
профиля. Во влажное время года, после таяния снега, соль из верхних
горизонтов почвы вымывается, причем образовавшиеся соединения гуму-
са и натрия переходят в неустойчивую форму. Образуются золи гумуса,
которые вместе с полуторными окислами (Fe2O3, А12О3) перемещаются
в более глубокие горизонты почвы, где осаждаются и формируют гори-
зонт В. Процесс напоминает подзолистый период почвообразования с той
лишь разницей, что здесь пептизирующую функцию Н-иона выполняет
Na-ион; реакция почвы становится щелочной (pH более 9). Такие почвы
называются солонцеватыми (рис. Ill, 1 и 2).
Чем дальше к югу, тем большим становится количество солей, пере-
носимых пылью, горизонт А выщелачивается все более интенсивног
а горизонт В уплотняется. Летом при высыхании почвы образуются тре-
щины, которые рассекают ее на столбчатые отдельности. Такой тип струк-
турной почвы называют солонцом (рис. 111, 3 и 4).
Солонец имеет следующее строение:
Горизонт А полностью выщелочен, пепельно-серый, с листоватой структурой,
обогащен кремнеземом. Горизонт вмывания В очень темный, плотный, распадается
на хорошо выраженные столбчатые отдельности. Отсюда название «столбчатые солон-
цы». Если горизонт А имеет толщину всего несколько сантиметров, такую почву назы-
вают корковым солонцом.
Во влажном состоянии горизонт В, содержащий поглощенный натрий, сильно
набухает и становится водонепроницаемым, вследствие чего весной во время таяния
снега и после летних или осенних дождей почва совершенно размокает. Горизонт С,
в верхней части которого появляется белоглазка (Сх), содержит на глубине гипсовые
трубочки (С2) и, наконец, конкреции гипса (С3). Резкощелочная реакция горизонта В
свидетельствует о том, что произошло образование соды. Абсолютное содержание
солей в почве невелико.
На типичных солонцах злаки отступают на второй план, раститель-
ный покров становится более разреженным и принимает облик, харак-
терный для полупустыни.
Типичными растениями-индикаторами солонцов являются: Puccinellia convoluta,
Camphorosma annua, C. monspeliaca, Limonium alutacea, Artemisia maritima ssp. salina,
Kochia prostrata, Linosyris villosa, Lepturus pannonicus, Gypsophila stepposa, Petrosimo-
nium volvox и др.; на корковых солонцах преобладает Artemisia pauciflora.
На крайнем юге степной зоны, на солонцеватых почвах, нередко
встречаются бессточные впадины, сходные с долинами карстовых обла-
стей. Здесь под мощным лёссовым слоем залегают осадочные породы мор-
ского происхождения (третичные известняки). При вымывании содержа-
щихся в этих породах включений соли возникают полости, обусловливаю-
щие проседание грунта. Такие бессточные впадины называют «подами».
Это могут быть впадины небольших размеров, но могут быть и понижения
диаметром во много сотен метров и площадью 15 тыс. га.
Что касается образования подов, то Махов (устное сообщение) на осно-
вании данных бурения доказал, что современный рельеф в значительной
138
Глава 2
Рис. 111. Типичные профили слабо- и сильнозасоленных
почв в Восточной Европе.
1 — южный чернозем (слабосолонцеватый) со слабоуплотненным гори-
зонтом А2В; 2 — темно-каштановые почвы с отчетливо выраженным
горизонтом В; пятна белоглазки (СО, ниже аккумуляция гипса (труб-
ки С2 и друзы С3); в типах 1 и 2 горизонт At имеет пластинчатую струк-
туру; гв — горизонт вскипания; в % указано содержание гумуса;
S — светло-каштановые почвы (сильносолонцеватые): горизонт В тем-
ный, столбчатый и очень плотный, горизонт А беден гумусом, пластин-
чатый. в остальном такой же, как предыдущий; 4 — типичный столбча-
тый солонец: горизонт А пепельно-серый, пластинчатый, горизонт
В мощный; 5 — солонец, измененный поднимающимися грунтовыми
водами, белоглазка растворена, выделения гипса прослеживаются до
глубины 40 см, внизу горизонт оглеения(в); 6 —типичный солончак
при высоком стоянии грунтовых вод, с темным гумусовым горизонтом
At гипсовыми трубками в горизонте G, ниже — глеевый горизонт;
на поверхности часто образуется солевая корка.
мере соответствует рельефу третичного времени до начала процесса отло-
жения лёсса, когда западины уже существовали. Если во впадинах скап-
ливается вода, которая насыщает лёсс, то лёсс (вследствие переотложения
пылеватых частиц) «проседает», благодаря чему поды углубляются.
Весной талые воды заполняют понижения. Летом в центре пода
может еще сохраниться небольшое озерцо.
При длительном воздействии воды, медленно просачивающейся в со-
лонцеватую почву, ионы натрия вымываются и замещаются в гумусовом
комплексе ионами водорода. В результате формируются особенно сильно-
Зона степей и прерий
139
деградированные солоди, соответствующие подзолам; разница состоит
в том, что горизонт грубого умуса в них отсутствует.
Растительность подов обн* рущивает резко выраженную поясность.
<В подах южной степной зоны ковыльная степь равнинных местообитаний
по мере увеличения влажности почвы в направлении от края пода к его
середине образует следующие микрозоны:
1) луговая степь с отдельными ковылями (почва более богата гуму-
сом);
2) луговая растительность без ковылей, очень богатая разнотравьем
{наибольшее содержание гумуса в почве);
3) заливаемые луга с гигрофильными злаками, как Alopecurus praten-
sis (содержание гумуса в почве более низкое);
4) растительность низинных болот на выщелоченных солодях;
5) сообщества водных растений на самом глубоком месте впадины
с видами Potamogeton и Characeae.
Состав растительного покрова в самой глубокой части впадины непо-
стоянен, так как в засушливые годы поды обычно остаются сухими и степ-
ная растительность беспрепятственно проникает в их более низменные
.части. Растения низинных болот способны находиться в латентном состоя-
нии в почве или в виде семян до 10 лет. После дождливого года тотчас
снова появляются Butomus, Heleocharis, Elatine, Alisma, Limosella и раз-
личные виды Carex.
Особенно типичен пырей подовый, Agropyrum repens ssp. pseudocae-
sium, легко переносящий засуху, но и не страдающий от затопления.
Несколько севернее для подов уже характерны небольшие рощи осины
(Populus tremula), а северо-восточнее также березы (Betula). Поды имеют
важное хозяйственное значение, так как во время засухи служат хоро-
шими пастбищами для скота.
Солонцеватые степные почвы оказываются пригодными для богарного
земледелия, если удается нейтрализовать щелочность почв, указывающую
на присутствие соды в горизонте В. Осуществляют это внесением в почву
большого количества гипса, причем в процессе перестройки содержащихся
в почве Na2CO3 и CaSO4 образуются легкорастворимый и легковымывае-
мый Na2SO4 и нерастворимый СаСО3; последний улучшает структуру
почвы. .
Развитие солонцов может быть также следствием химического пре-
образования солончаков. Последние, как известно, связаны с высокостоя-
щими грунтовыми водами, причем к поверхности почвы поднимается толь-
ко слабоминерализованная грунтовая вода; при ее испарении соли накап-
ливаются в почве. Таким образом, концентрация солей в почвенном про-
филе увеличивается снизу вверх. Сначала выпадает слаборастворимый
гипс (CaSO4), в то время как NaCl и Na2SO4 образуют на поверхности
почвы солевую корку. Вследствие высокой концентрации солей гумусо-
вые вещества выпадают из раствора (см. рис. 111, 6).
Если в таком районе вследствие развития процессов эрозии уровень
грунтовых вод понижается, то весной всякий раз происходит вынос солей
из верхних гумусовых горизонтов, при этом гумусовые коллоиды, содер-
жащие натрий, переходят в раствор. Со временем на такого рода место-
обитаниях формируется солонец (рис. 111, 4). И наоборот, при подъеме
зеркала грунтовых вод солонец вновь преобразуется в солончак. Это
воздает чрезвычайную пестроту почвенного покрова. Очень часто он
представляет собой микромозаику различных типов почв с преимуществен-
ным развитием солонцов на возвышенных частях рельефа и с солончаками
140
Глава 2
в отрицательных формах, как это наблюдается, например, в Прикаспий-
ской полупустыне.
Эта область, бывшая некогда дном Каспийского моря, простирается
от устья Терека (Северный Кавказ) до излучины Волги у Волгограда,
отсюда до среднего течения Урала и далее проникает глубоко в пределы
Средней Азии.
Количество осадков здесь не превышает 250 мм, под Астраханью оно
колеблется в отдельные годы от 263-до 46 мм. Зимы еще холодные, снег
лежит до 5 месяцев. Прикаспийская низменность сложена мощными тол-
щами морских отложений. Соли, содержащиеся в донных осадках преж-
него моря, после выхода этих пород на сушу оказались выщелоченными.
Со временем происходило также перераспределение солей по земной
поверхности. Дождевые воды смывали их с возвышенных частей рельефа
и переносили на более низкие позиции. Неудивительно поэтому, что
микрорельеф при этом превращался в фактор, который подчас направлял:
развитие почвообразовательного процесса.
Бурые почвы повышений в верхнем метре почвенного профиля
содержат мало солей (0,2—0,4%), однако с глубиной их концентрация
увеличивается. Гумусосодержащими являются только верхние 30—40 см,
общее количество гумуса незначительно (2—3%). Горизонт вскипания
с СаСО3 часто расположен уже на отметке 25 см', следовательно, почва
увлажняется неглубоко. Покрытие почвы растениями не достигает 50%,.
но корни заполняют весь объем верхних горизонтов почвы (Шалыт, 1950,.
см. примечание на стр. 112). Влажной весной свободное пространство
между растениями временно занимают эфемеры. Сообщества, которые
были бы распространены на значительных площадях, почти отсутствуют,
так как каждое изменение микрорельефа влечет за собой изменение соста-
ва почв: развиваются либо солонцы, либо — во влажных впадинах —
солончаки. Соответственно варьирует и состав растительных группи-
ровок.
В типичной полупустыне среди злаков преобладает Festuca sulcata,
тогда как Stipa capillata, St. lessingiana и Koeleria gracilis наблюдаются
в виде отдельных экземпляров. Из мелких полукустарников особенно
многочисленны Pyrethrum achilleifolium (= Chrysanthemum millefoliatum
var. achilleifolium), Artemisia maritima var. incana и Kochia prostrata.
Разрозненно встречаются Agropyrum cristatum, Aer stvillosus, Achillea leptophylla,.
Artemisia austriaca, Erysimum versicolor, Stalice sareptana и др. Весенние эфемеры пред-
ставлены Alyssum desertorum, Carex stenophylla, Draba verna, Poa bulbosa var. vivipara,.
Ranunculus polyrrhizus, Tulipa biebersteiniana, Veronica verna и др. Некоторые одно-
летники развиваются только осенью, как Bassia sedoides (обильно), Ceratocarpus arenari-
us, Polygonum bellardi и Salsola tamariscina.
Кроме того, широко представлены низшие растения — сине-зеленые водоросли,
наземные лишайники, а также мхи.
В местах выпаса скота злаки исчезают, увеличивается только коли-
чество Роа bulbosa, а также однолетних и двухлетних видов. При болен
интенсивном выпасе исчезают и полукустарники, остаются только серо-
белые сорные растения Ceratocarpus, Bassia и, кроме того, Atriplex lacinia-
tum. На сильно деградированных участках сохраняются лишь низкорос-
лые, прилегающие к почве растения.
На солонцах растительность еще беднее: преобладают Artemisia
pauciflora, растущая совместно с Camphorosma monspeliaca и Kochia
prostrata (рис. 112). Эфемеры и осенние однолетники в основном те же.
Очень велико количество наземных лишайников; часто встречаются мхи,
Зона степей и прерий
141
Рис. 112. Растительность полупустыни на солонцах в конце августа под
Красноармейском: Artemisia pauciflora (голые кусты), Camphorosma monspeliaca,
Kochia prostrata и др. (фото Келлера).
«среди них виды Riccia. Лишайник Aspicilia местами распространен столь
широко, что в прежние времена кочевники собирали его и использовали
как заменитель хлеба.
Охарактеризуем еще вкратце растительность влажных засоленных
местообитаний на солончаках Ч
Местообитания вокруг соленых озер Баскунчак и Эльтон, характе-
ризующиеся экстремальными условиями среды, сложены твердой пова-
ренной солью и полностью лишены растительности. Соль выветривается
из горных пород пермского возраста и выносится в озера несколькими
соляными источниками. Растением-пионером на солончаках является
солерос, Salicornia herbacea, сопровождаемый обычно Halocnemum strobi-
laceum. Почва между растениями часто бывает покрыта солевой коркой,
но корни растений углубляются в слои почвы, содержащие меньше солей.
Тем не менее осмотическое давление у этих видов, как правило, очень
высокое (до 60 атм).
Другими наиболее типичными галофитами являются Suaeda maritima,
Obione pedunculatum, Limonium caspica и др. Несколько менее засоленные,
но еще влажные местообитания занимают Petrosimonia crassifolia и Atriplex
verructferum. Значительно суше и лишены на поверхности солей почвы
с Atriplex сапит и Anabasis salsa, часто простирающиеся на многие кило-
метры и только на 20% занятые растениями. Здесь уже развиты лишай-
ники и эфемеры (злаки Agropyrum prostratum и Colpodium humile), но
1 В. Keller, Die Vegetation auf den Salzboden der russischen Halbwiisten und
Wiisten, «Zschr. f. Bot.», 18, 1925/26, S. 113—137, а также многочисленные работы
этого автора н$ русском языке; см. также: «Veget. Bilder», R, 18, Н. 8 (Tafel 43—48.)
142
Глава 2
450
600
+2
60
^,4,0
Stf ^45“
10 0 ” ____-
-ь ин- Тайга НЬ|йлео ныи лес
нИ_^--1500
•85°-^^Г
щи^-гзо
____-190' , <
«“Г" KKan/№j£“90
+5O<3?Z
+11
Лесостепь
Степь Пустыня
<250
90-
30-
-9°-
Лесотундрс^.
Почвы Северные Подзолы и подзолис-
тунЭры подзолы тые долотные почвы
Серые лесные
почвы
и мощные
черноземы
Черноземы
и каштано-
вые почвы Сероземы
3^
Рис. 113. Схема изменения климата, растительности и почв на профиле Восточной
Европы в направлении с северо-запада на юго-восток (до Прикаспийской низменности)
(по Шенникову, с изменениями).
Мощные черноземы соответствуют лесостепи. Зачернен гумусовый горизонт, штриховкой выделен
иллювиальный горизонт В.
поселяются и многолетние виды, такие, как Artemisia maritima ssp. incana
или Limonium suffruticosum. Появление этих видов означает уже переход
к растительности солонцов.
Описанием растительности засоленных почв мы заканчиваем харак-
теристику степной зоны. Растительность полупустынь и пустынь уме-
ренной зоны будет описана подробнее в гл. 5.
КРАТКИЙ ОБЗОР КЛИМАТИЧЕСКИХ, РАСТИТЕЛЬНЫХ
И ПОЧВЕННЫХ ЗОН ВОСТОЧНОЙ ЕВРОПЫ
Нигде на Земле зональность растительности в условиях арктического
и умеренного климатов не выражена столь отчетливо, как на обширной
Восточно-Европейской равнине, которая монотонно, без каких-либо
особенно заметных возвышений, простирается от Северного Ледовитого
океана до Черного и Каспийского морей, пересекая около 25 градусов
по широте. Располагается она уже в области континентального климата
и достаточно далеко удалена от океана. Поэтому некоторые зоны, харак-
терные, скорее, для океанического климата, здесь выпадают полностью
или едва выражены, например зона жестколистной растительности обла-
стей с зимними дождями, лишь слабо намечающаяся в Южном Крыму,
или зона лиственных лесов, узкой полосой вклинивающаяся между зоной
смешанных лесов и степями. На остальном же пространстве без перерывов
прослеживается весь ряд природных зон — от зоны арктической тундры
до зоны пустынь. Однако, поскольку эти зоны мы уже рассмотрели в пре-
дыдущих разделах, а с пустынями подробнее познакомимся в гл. 5, здесь
Зона степей и прерий
. 14$
в кратком, схематическом обзоре отметим лишь важнейшие их черты
(рис. ИЗ).
Схема не претендует на большую точность и приведена лишь для
общей ориентировки.
В направлении от тундры к пустыне возрастает значение следующих
факторов:
1) годовой температуры,
2) солнечной радиации,
3) длительности вегетационного периода,
4) испаряемости.
Годовое количество осадков, сначала увеличивающееся] по мере
приближения к зоне смешанных лесов, уменьшается в зоне степей и пу-
стынь. Поэтому в левой части профиля (рис. 113) сумма осадков превышает
испаряемость (гумидный климат), тогда как в правой части испаряемость
больше количества осадков (аридный климат). Граница между гумидным
и аридным климатами совпадает с границей между лиственным лесом
и лесостепью. Зону лиственных лесов в Восточной Европе можно назвать-
семигумидной, зону же лесостепи уже семиаридной, ибо в последнейг
по мере движения с севера на юг впервые в бессточных впадинах отме-
чаются явления слабого засоления содового типа. С нарастанием аридно-
сти климата признаки засоления становятся все заметнее, площадь засо-
ленных почв в равнинных, плохо дренируемых местностях — все болен
обширной.
С этими изменениями климата теснейшим образом связана зональность
типов почв. Многолетняя мерзлота известна лишь там, где средняя годовая
температура значительно ниже нуля; в районах со снежным покровом, име-
ющим защитное значение, средняя годовая температура почвы превосходит
температуру воздуха. Уровень грунтовых вод, в общем, тем выше, чем
гумиднее климат. Гумусовый горизонт отличается наибольшей мощностью
в зоне лесостепи. Правда, в северном направлении его мощность не сокра-
щается столь равномерно, как это показано на схеме. В частности, в боре-
альной зоне, где низкие температуры обусловливают медленное разложе-
ние гумусных соединений, мощность перегнойного горизонта снова уве-
личивается, и только на крайнем севере Арктики или в пустыне он пол-
ностью отсутствует. На севере часто развиваются торфянистые почвы.
Процесс выщелачивания почв в гумидных областях обусловливает
формирование в профиле развитых здесь почв горизонта вмывания В
(показан косой штриховкой), в котором накапливаются глина и полу-
торные окислы. Напротив, в почвах аридного климата аккумулируется
СаСО3, что легко заметить по вскипанию почвы, если капнуть на нее
соляной кислотой. Горизонт вскипания залегает тем ближе к поверхности,
чем ариднее климат. В крайне аридных климатических условиях, кроме
того, происходит накопление гипса.
Судницын 1 определил сосущую силу почвы под древостоем в разных
растительных зонах. В бореальной зоне ее величина обычно была равна
0,5—2 атм, редко 5 атм. В зоне смешанного леса летом она составляла
2—5 атм, редко 10 атм. В лесостепной зоне эта величина достигала уже
5—10 атм и даже 20 атм. В степных районах почва под лесонасаждениями
летом сильно высыхает, что находит отражение в высоких значениях
сосущей силы: 5—20 атм, а в верхних горизонтах 50 атм; в южной степ-
1 И. И. Судницын, Новые методы оценки водно-физических свойств почв
и влагообеспеченности леса, М., 1966.
144
Глава 2
Таблица 25
Общая надземная фитомасса и годовая продукция надземного сухого вещества
в различных зонах растительности (в ц/га)
Растительные зоны (плакорные местообитания) Надземная фитомасса Годовая продукция
Зона тундры I. Арктическая полупустыня (Dryadetum) . . 6 4
II. Арктическая богатая разнотравьем тундра Caricetum 12 7
III. Северная тундра Salicetum с разнотравным и моховым покровом 30 12
IV. Южная тундра Betuletum с моховым по- кровом 32 12
V. Лесотундра (Picea и Betula) 73 14
Лесная зона VI. Северная тайга (Piceetum licheno-hyloco- miosum) 900 15
VII. Средняя тайга (Piceetum myrtillosum) . . . 1300 30
VIII. Южная тайга (Piceetum oxalidosum) .... 2200 50
IX. Зона широколиственного леса (Quercetum) 2600 56
Степная зона X. Богатые разнотравьем луговые степи . . . 15 15
XI. Типичная богатая разнотравьем степь с Fes- tuca sulcata и Stipa 12 12
XII. Типичная бедная разнотравьем степь с Fes- tuca sulcata и Stipa 10 10
XIII. Сухая степь с Artemisia, Festuca и Stipa 5 5
Полупустыни и пустыни XIV. Северная полынная полукустарничковая по- лупустыня со злаками 7 4
XV. Южная полынная полукустарничковая полу- пустыня с эфемерами 10 —
XVI. Древостои черного саксаула (Haloxylon aphyl- lum) 70 12
ной области соответствующие показатели увеличиваются до 5—50
и 100 атм. При исследовании почв под древесными насаждениями в зоне
полупустыни в верхнем метре почвенного профиля измерены значения
10—100 атм, а в верхних горизонтах даже свыше 200 атм', лишь на глу-
бине 1,5 м сосущая сила летом приблизительно постоянна и равна 15 атм',
таким образом, деревья могут всасывать воду лишь с большой глубины.
В табл. 25 сопоставлены величины надземной фитомассы и годовой
продукции надземного сухого вещества для отдельных растительных зон
и подзон х. Значения I—XIII относятся к Восточной Европе, XIV—XVI
ж пустыням Казахстана и Средней Азии.
Наибольшей продуктивностью характеризуются южная часть зоны
хвойных лесов и зона лиственных лесов.
1 Е. М. Лавренко, В. Н. Андреев, В. Л. Леонтьев, Профиль про-
дуктивности надземной части природного растительного покрова СССР от тундры
к пустыням, «Бот. ж.», 40, № 3, 1955. стр. 415—419.
Глава 3
ЗЛАКОВНИКИ УМЕРЕННОЙ КЛИМАТИЧЕСКОЙ ЗОНЫ
ЮЖНОГО ПОЛУШАРИЯ
ОБЩИЙ ОБЗОР УМЕРЕННОЙ ЗОНЫ ЮЖНОЙ АМЕРИКИ
К BOCTOF ’ ОТ АНД
Для того чтобы пондть характер растительности умеренной зоны
Южной Америки, познакомимся в общих чертах с ее климатическими
условиями. Высокие Кордильеры, сплошной стеной вздымающиеся
на севере Аргентины (Аконкагуа, около 7000 ж), образуют в этой части
страны резкий климатический раздел с Чили. Напротив, в южной ее
половине высота Анд резко уменьшается, так что дующие на этих широтах
(южнее 40 параллели) в течение всего года западные ветры свободно
проникают на аргентинскую сторону Анд, в Патагонию. Большую часть
несомой ими влаги ветры теряют на западной стороне Анд и на их восточ-
ном склоне, поэтому в Патагонию приходят уже холодные и сухие воздуш-
ные массы. В северной же половине Аргентины вследствие наблюдаемого
здесь эффекта отклонения пассатов преобладают северо-восточные ветры,
дующие с Атлантического океана. Они приносят теплые, влажные воздуш-
ные массы и летом проникают до восточных предгорьев Анд. При этом
максимальное количество дождей перемещается в течение года с востока
на запад и обратно х. На востоке, в районе Ла-Платы, наблюдаются весен-
ний и осенний максимумы на фоне относительно равномерного распределе-
ния осадков в течение года, тогда как на западе отмечается всегда более
четкий летний максимум, зимой же осадков выпадает мало. В целом
-среднее годовое количество дождей к западу убывает. В результате у во-
сточного подножия Анд складываются условия, напоминающие климат
пустыни. Аридность климата усиливается случайными фёнами, которые
приходят с запада и с большой высоты скатываются по восточному скло-
ну Анд.
Только на самом севере региона, в тропических провинциях Тукуман,
Жужуй и Сальта, восходящие ветры обусловливают в выдвинутых далеко
на восток Предкордильерах (Сьерра-де-Аконкия, более 5000 м) резкое
увеличение летних осадков, благодаря чему здесь сформировался свое-
образный «дождевой оазис» с субтропическими вечнозелеными дождевыми
лесами. На высоте 1400—2100 м распространение лесов со сбрасывающей
зимой листву Alnus jorullensis свидетельствует о том, что «оазис» приобрел
уже черты умеренного климата.
Эти напоминающие Голарктику ольховые леса подробно описаны Хюком 1 2. Сред-
няя годовая температура у нижней границы произрастания ольховых лесов еще равна
13,9°, однако регулярно с мая по сентябрь (редко до ноября), то есть в то время, когда
лес стоит без листвы, бывают заморозки. Иногда падает снег, но лежит недолго. Изред-
ка отмечается повреждение растений поздними заморозками. Наиболее низкая изме-
ренная температура равна —5°. Почва, окрашенная гумусом в темный цвет, местами
1 Устное сообщение X. Хофмана из Национальной метеорологической службы,
Буэнос-Айрес.
2 К. Hueck, Der Anden-Erlenwald (die Alnus jorullensis-Assoziation) in der
Provinz Tucuman (Argentinian), «Angew. Pflanzensoz. (Klagenfurt), Festschr. Aichinger»,
1, 1954, S. 512—572.
10-0434
Рис. 114. Карта растительности Аргентины (по Кабрере,
с небольшими изменениями).
1 — субтропические дождевые и пойменные леса; 2 — сухой лес «чако»;
3 — влажные колючие кустарники; 4 — сухие рощи из Prosopis calde-
nia и саванны; з — пампа с древесной растительностью (соответствует
лесостепи); в — злаковая пампа без деревьев; 7 — предпуна со столб-
чатыми кактусами; 8 — кустарниковая пустыня с господством Larrea',
9 — пуна — холодная пустыня; 10 — патагонская кустарничковая
полупустыня; 11 — патагонский злаковник (степь); 12 — альпийская
растительность Высоких Анд; 13 — вальдивийские и!субантарктиче-
ские дождевые леса.
Злаковники умеренной климатической зоны южного полушария
147
достигает мощности 30—40 см. Собственно гумусовый слой соответствует муллевым
почвам средних широт Центральной Европы. Значения pH обычно колеблются в пре-
делах 5,5—7 (4,5—7,5). В оптимальных условиях Alnus формирует чистые древостои;
во влажных ущельях к ней примешивается Sambucus peruviana. У Alnus jorullensis,
как и у всех других видов ольхи, на кбрнях имеются клубеньки величиной с кулак,
образованные фиксирующими атмосферный азот актиномицетами в качестве симбион-
тов. Кроме того, обнаружена микориза, образованная различными гименомицетами.
Листья начинают распускаться между 15 сентября и 15 октября, листопад приходится
на конец марта. Деревья достигают возраста 80 лет, но в одном случае на спиле насчи-
тали свыше 100 годовых колец.
Эти ольховые леса (леса «алисо») не представляют собой островоподобные есте-
ственные насаждения, а являются южной оконечностью ареала, простирающегося
от севера Мексики вдоль склонов Анд почти на 50 градусов по широте. Помимо упомя-
нутого вида Sambucus, здесь распрост± тены и другие голарктические роды (Ranuncu-
lus, Anemone, Thalictrum, Geranium). У нижней границы пояса можно встретить также
Juglans australis и Prunus tucumanensis*
Граница между климатическими областями, испытывающими влияние
теплых северо-восточных и холодных западных ветров, проходит в Арген-
тине, следуя преимущественно долине реки Рио-Негро (приблизительно
по 40° ю. ш.). Летом фронт нередко перемещается к северу, за Ла-Плату;
в это время года холодные массы воздуха в виде очень сильных порывистых
ветров, называемых здесь «памперо», проникают до Буэнос-Айреса,
вызывая на всем пути продвижения внезапное понижение температуры
(до 30° за сутки). Холодный ветер, сопровождающий непродолжительную
сильную грозу на большой высоте, приносит приятную прохладу после
изнуряющей духоты.
Из карты растительности Аргентины (рис. 114) видно, что к умерен-
ной зоне принадлежит зона пампы, южная часть области лесов с колючими
кустарниками («эспинор»), южная часть полупустыни с Larrea и вся Пата-
гония с Огненной Землей.^
ПРОБЛЕМА ПАМПЕР1
Аналогом обширных восточноевропейско-сибирских степей и северо-
американских прерий в южном полушарии, в пределах умеренной зоны
южнее 30° ю. ш., являются восточноаргентинская пампа 1 2 и небольшие
по площади туссоковые злаковники в провинции Отаго на острове Южный
(Новая Зеландия). Занимаемая пампой площадь равна приблизительно
0,5 млн. км2. В климатическом отношении пампа отличается от степи
и прерии более благоприятными температурными условиями, прежде
всего отсутствием холодного зимнего периода, хотя изредка в некоторых
местностях и бывают морозы (рис. 115—116).
Благодаря значительно более высокой температуре воздуха пампа
по сравнению с восточноевропейскими степями и прерией показательна
прежде всего тем, что здесь высаживают различные экзотические древес-
ные породы. Это не только хорошо растущие и в степи Robinia pseudacacia,
Gleditschia triacanthos, Acer negundo, а на влажных участках также Populus
alba, но и различные виды Eucalyptus и Melia azedarach, виды Casuarina
и Cupressus и даже разводимые в городских парках виды Citrus и Phoenix
1 Н. W а 11 е г, Das Pampaproblem und seine Losung, «Вег. Deut. Bot. Ges.», 79,
1966, S. 377—384; Das Pampaproblem in vergleichend bkologischer Betrachtung, «Erd-
kunde», 21, 1967, S. 181-203.
2 «Пампой» индейцы Южной Америки называют всякую лишенную деревьев рав-
нинную местность. Фитоценология использует это обозначение в более узком смысле’
под «пампой» понимают только восточноаргентинские злаковники.
10*
148
Глава 3
canariensis. В береговой зоне Ла-Платы с ее мягким климатом, в садах
вокруг Буэнос-Айреса растут такие тропические виды, как Ficus elastica,
Philodendron, Hibiscus sinensis, Bougainvillea и другие. Однако в холодные
зимы они вымерзают. Относительно благополучны для развития растений
и условия увлажнения атмосферными осадками: около 1000 мм в год
на северо-востоке пампы и приблизительно 500 мм у границы засушливо-
сти. Как уже упоминалось, осадки распределяются внутри года более
или менее благоприятно — максимум их приходится на осень, второй,
меньший,— на весну. Поэтому климат пампы в общем можно называть
Рис. 115—116. Климадиаграмма Буэнос-Айреса, северо-во-
сточная пампа, и для сравнения климадиаграмма Оклахома-Си-
ти, расположенного в юго-восточной прерии (США).
«гумидным». Но еще в 1872 г. Гризебах заинтересовался вопросом: почему
же при таком, казалось бы, выигрышном климате пампа уже в первона-
чальном состоянии была лишена деревьев? Этот вопрос неоднократно
ставили многие европейские ботаники и географы и отвечали ’на него
по-разному Г
Отсутствие деревьев представляется тем более странным, что пампа
у границы засушливости сменяется не полупустыней, как этого можно
было бы ожидать по аналогии со степью, а лесоподобными древостоями,
в которых доминирует Prosopis caldenia, колючий вид мимозовых. Иссле-
дователям казалось непонятным, почему эта древесная порода не рас-
пространилась по всей, более гумидной по сравнению с окраинными частя-
ми, области пампы. Так возникла широко обсуждавшаяся проблема
пампы.
Шмидер в 1927 г. высказал мысль, что пампа вообще является не есте-
ственным, а вторичным, антропогенно обусловленным злаковником,
образовавшимся на месте лесистого ландшафта вследствие частого выжи-
гания его индейцами 1 2. Произошло это якобы задолго до появления
на Южноамериканском континенте первых испанских поселенцев.
Его предложение не встретило поддержки, однако в последнее время
Элленберг опубликовал наблюдения, которые, по его мнению, подтвер-
1 A. G г i s е b а с h, Die Vegetation der Erde nach ihrer klimatischen Anordnung,
Bd. II, Leipzig, 1872; J. Frengue Hi, Rasgos principales de fitogeografia argentina,
«Rev. Museo de la Plata (N.S.) Bot.», 3, 1941, p. 65—181.
2 O. S c h m i e d e r, The pampa a natural or culturally induced grass-land? «Univ.
Calif. Publ. in Geography», 2, 1927, p. 255—270; Das Pampaproblem, «Peterm. Geogr.
Mitt.», 75, 1929, S. 246—247.
Злаковники умеренной климатической зоны южного полушария
14»
ждают гипотезу Шмидера *. Современная пампа отнюдь не лишена деревь-
ев. Многочисленные посадки упомянутых выше экзотических древесных
пород преобразили прежнюю картину ландшафта. Первичная раститель-
ность пампы пр ктически уже не существует. Почва повсеместно по
меньшей мере о, ин раз распахивалась. Поэтому в здешнем пейзаже
преобладают пашни или чаще сеяные пастбища с различными, преиму-
щественно европейскими, видами злаков и разнотравья, но нередко
и с многочисленными видами сорных растений, не поедаемых скотом,
например Cynara cardunculus, Carduus nutans, Raphanus sativus, видами
Rapistrum, Foeniculum vulgare и др. Таким образом, восстановить перво-
начально существовавшие в пампе условия конкуренции сегодня уже
не представляется возможным.
Напрашивается вопрос: действительно ли климат пампы; является
гумидным «лесным» климатом? Если сравнить климадиаграммы станций,
расположенных в прерии, с климадиаграммой Буэнос-Айреса, to обнару-
живается поразительное сходство с климадиаграммой южной части прерии.
Для Оклахомы (рис. 116) характерны та же средняя годовая температура
и то же количество осадков, правда на 100 мм меньше. Однако зима в пре-
рии холоднее, поэтому запасы влаги, накопившиеся в зимние месяцы
в виде снега, весной могут быть с пользой израсходованы растениями. Если
спроектировать территорию США на южное полушарие, повернув ее
вокруг экватора как оси (рис. 117), то легко обнаружить, что Буэнос-
Айрес по своему широтному положению точно соответствует Оклахоме.
Но Буэнос-Айрес лежит у моря и ход его температуры выровнен, тогда
как Оклахома расположена в глубине материка и характеризуется более
резкими колебаниями температуры. В Техасе прерия с уменьшением коли-
чества осадков переходит в саванну с Prosopis, то же наблюдается в пампе.
Однако одно это сходство не может рассматриваться как аргумент в пользу
предположения, что климат в пампе «лесной».
Для решения проблемы пампы не менее важно знание особенностей
геоморфологии и гидрографии области. Пампа распространена преиму-
щественно в провинции Буэнос-Айрес и лишь на севере и западе несколько
выступает за ее границы. Рельеф пампы в’основном равнинный (рис. 118);
только небольшая ее часть, вдоль Параны и Ла-Платы, волниста («рашра
undulada») и пересечена многочисленными, параллельными между собой
руслами рек. В остальном же территория представляет собой либо очень
слабо дренируемую, лежащую невысоко над уровнем моря равнину
(«pampa deprimida»), либо местность с отметками не более 200 м, заклю-
ченную между двумя горными системами, с редкими водотоками по окраи-
не («pampa alta») 1 2.
Для пампы, так же как для степи и прерии, типичны лёссовые почвы.
Лёсс пампы по своему гранулометрическому составу близок к лёссам
Европы или Северной Америки, но в отличие от последних беден СаСО3—
обычно менее 2% (редко 4, максимально 8, часто 0%). Лёсс имеет вулка-
ническое происхождение, состоит, особенно в тонких фракциях, преиму-
щественно из вулканического стекла. В большинстве случаев принесенный
ветром лёсс переотложен водой и обезглинен 3. Своеобразие пампы состоит
1Н. Ellenberg, Wald in der Pampa Argentiniens?, «Veroff. Geobot. Inst.
Rubel (Zurich)», H. 37, 1962, S. 39—52.
2 A. S iragusa, Geomorfologio de la Provincia de Buenos Aires, «GAEA (Buenos
Aires)», 12, 1964, p. 93—122. ?
3 J. Frenguelli, Loess у limos pampeanos, «Ser. Teen, у Didact.», № 7, La
Plata, 1955; M. E. T e r u g g i, The nature and origin of Argentin loess, «I. Sedim.
Petrol.», 27, 1957, p. 322—332. ;
150
Глава 3
Рис. 117. Сопоставление положения пампы и южной прерии. Территория США без
изменения широты спроектирована на южное полушарие.
Г— пампа или прерия; 2 — Великие равнины, или низкозлаковая прерия, которую нельзя сравни-
вать с пампой, так как она развита на высоте более 1000 м\ 3 — саванна с Prosopis на юге Техаса.
Злаковники у. -оенной климатической зоны южного полушария
151
Р[и с. 118. Геоморфология пампы в провинции Буэнос-Айрес (по Сирагузе, с изме-
нениями).
1 — «pampa undulada», хорошо дренируемая; 2а, б, в — «pampa deprimida», слабо дренируемая
рекой Рио-Саладо; большая часть территории не имеет стока; з — «pampa alta» на высоте около
200 м, дренируемая только по окраинам; 4 — «pampa arheica» с песчаными почвами и без систем стока.
и в том, что ее почвы на разной глубине подстилаются очень плотными,
твердыми слоями извести («тоска»), часто достигающими мощности в не-
сколько метров. Генезис и возраст этих слоев неодинаков, часть их обра-
зовалась в межледниковые периоды, часть в послеледниковое время
как известковые корь! Ч
1 A. Siragusa, Contribucion al conocimiento de las toscas de la Republica
Argentina, «GAQA (Buenos Aires)», 12, 1964, p. 123—148.
152
Глава 3
Но особенно важное значение имеет следующее обстоятельство.
При рассмотрении топографической карты, а тем более во время
поездки по стране в глаза бросаются многочисленные небольшие и более
крупные озера, все без исключения бессточные, которые местные жители
называют «лагунами» (рис/ 119). Еще больше здесь маленьких западин,
весной заполняющихся водой, но летом полностью высыхающих (рис. 120).
Ферворст (см. примечание на стр. 162) не без основания сравнивает их
со степными подами. Но если последние имеют все же некоторый подзем-
ный сток, то у западин пампы он отсутствует совершенно. Почвы и под-
стилающие их грунты — это тяжёлая водонепроницаемая глина. Высота
местности над уровнем моря незначительна, поэтому на большей части'
пампы нет достаточного уклона, необходимого для движения грунтовых,
вод. И то, что летом вода из западин исчезает, есть следствие не просачи-
вания ее в грунт, а испарения с поверхности.
Реакция воды, заполняющей лагуны и западины, несмотря на бед-
ность местного лёсса известью, резко щелочная (pH = 8—9 и более).
Все почвы с высоким стоянием грунтовых вод солонцеватые, часто харак-;
теризуются типичным профилем солонца, о чем свидетельствует также’]
состав растительного покрова (дернообразующий Distichlis и другие!
виды) х. ; J
Таким образом, условия среды в южноамериканской пампе во многом •
напоминают обстановку, с которой мы познакомились в лесостепной зоне-
Восточной Европы при рассмотрении почв с неглубоким залеганием;
грунтовых вод. Оба факта — бессточность котловин и содовое засоление]
почв пампы —'широко известны. Как ни странно, но никто не сделал:
отсюда вывода, что эти факты противоречат представлению о гумидном,
климате пампы. Ведь в условиях гумидного климата количество осадков I
должно превышать величину испарения с открытой водной поверхности.
Рис. 119. Участок карты, лист Долорес. «Pampa
deprimida» с многочисленными бессточными озе-
рами (штриховка) и высыхающими летом запа-
динами (точки).
1 Уивер также считает Distichlis spicata индикатором слабозасоленных, но сильно-
щелочных почв прерии с близким залеганием грунтовых вод и отсутствием стока.
Злаковники, умеренной климатической зоны южного полушария
153
Рис. 120. «Pampa alta» между Лап-
ридой и Хуаресом со множеством
пересыхающих летом мелких западин.
Широко распространены солонцы.
О гумидности климата пампы создают ложное представление влажная
духота в летние месяцы и климадиаграммы (см. рис. 115). Заштрихованная
часть диаграммы отражает лишь относительно более влажное время года,
но она отнюдь не означает, что в этот период осадков выпадает больше,
чем может испариться. С этим фактом мы столкнулись уже при знакомстве
с лесостепью. Поэтому на климадиаграмме для тех широт сухое время
года выделено с помощью вспомогательной кривой осадков, проведенной
при соотношении 1 : 3 (10° = 30 мм). Учитывая более высокую темпе-
ратуру в пампе, для лета следовало бы наносить кривую осадков при
соотношении 1 : 5 (10° — 50 мм), тогда летний сухой период нашел бы
свое графическое отражение.
Наш вывод, что в пампе речь идет не о гумидном, а уже о слабосемиа-
ридном климате, подтверждается измерениями испаряемости, произведен-
ными на метеорологических станциях Аргентины. Все полученные значе-
ния уменьшены путем умножения на коэффициент 0,7. Расчеты годового-
водного баланса (осадки минус испарение) для всех тех станций, которые
располагают данными о величине потенциального испарения, показали,
что только непосредственно в районе Ла-Платы водный баланс уравнове-
шен, на остальной части территории он отрицательный; в юго-западном
направлении, с уменьшением количества осадков и увеличением испаряе-
мости, превышение расходной части баланса над приходной возрастает
и в районе Буэнос-Айреса (рис. 121) составляет более 700 мм. Соответ-
ственно в западной пампе с ее уже отчетливой аридностью наряду с содо-
вым засолением наблюдается преимущественно хлоридно-сульфатный
тип химизма; значительную площадь здесь занимают соленые озера и засо-
ленные блюдца (ср. стр. 136).
154
Глава 3
Рис. 121. Водный баланс (количество осадков минус испаряемость) провинции
Буэнос-Айрес по показаниям метеорологических станций (в мм).
Близ Ла-Платы баланс уравновешен, на остальной территории отрицательный, при этом равно-
весие нарушается во все большей степени в направлении к юго-западу.
Использовавшаяся до настоящего времени формула Торнтвейта для
вычисления величины эвапотранспирации дает для пампы значения, зани-
женные по меньшей мере на 200 мм, неудивительно поэтому, что восточная
-925 I
555 X.
(-1334)
t ~244
-1501
i
-967
("1278)
40°
Ю.щ.
~228
-301
1210
-1108
-987
>+300 ।
+127
-424
-655
-572
(425)
-259
-416
-514
(-716
-424^
“554
/-440
-6261
(-600
-850 >
1-717
-394
Г789
-690
-107
-94
-175
-968
(-1500)
~555 \
(н*0бЗ) S
Рис. 122. Водный баланс территории Аргентины.
Положительный баланс — только на северо-востоке страны (провинции
Мисьонес, Корриентес), уравновешен в устье реки Уругвай и в бассейне
Ла-Платы; в остальных районах баланс отрицательный с возрастанием дефи-
цита в направлении к западу, однако в Патагонии у границы Чили (лес из
Nothofagus) осадки вновь превышают величину испаряемости. Дальнейшие
объяснения см. в тексте.
156
Глава 3
Рис. 123. Климадиаграммы Пергамино (влажная северная
пампа) и района Лопес —Хуарес («Pampa alta») с кривой
испаряемости Е и кривой температуры (t X 7) при соотно-
шении 10° = 70 мм или (t X 9) при соотношении 10° = 90 мм.
В районе Пергамино кривая осадков для апреля —июля располагает-
ся над кривой испаряемости, в районе г Лопес — Хуарес она всегда
ниже ее.
пампа представлялась исследователям как гумидная область. По Лауэру,
в Буэнос-Айресе все месяцы в году являются также гумидными 1.
В целом для всей территории Аргентины (рис. 122) годовой водный
баланс положителен (+600 мм) лишь на северо-востоке страны, в провин-
ции Мисьонес и некоторых частях провинции Корриентес. На остальной
территории дефициты обычно намного больше, чем собственно в пампе.
Поэтому во всех этих районах Аргентины распространены бессточные
площади с засоленными впадинами-блюдцами.
Если на климадиаграмму станции Пергамино, расположенной в север-
ной, влажной части пампы, нанести месячные значения испаряемости,
то обнаружится, что количество осадков несколько превышает величину
испарения только в зимние месяцы. Напротив, в летние месяцы, особенно
с ноября пр февраль, водный баланс резко отрицателен (рис. 123). Вместе
с тем оказывается, что даже при соотношении 10° = 50 мм кривая тем-
пературы будет располагаться ниже кривой испаряемости. Лишь при
соотношении 10° = 70 мм кривая температуры почти повторяет кривую
испарения, лишь с некоторым ее отставанием, поскольку динамика испа-
рения в течение года не остается неизменной: весной, при повышении
температуры и, следовательно, при больших дефицитах насыщения, испа-
рение быстро возрастает, осенью же, при понижении температуры и мень-
шем дефиците насыщения, оно интенсивно сокращается. В зимние месяцы
температурная кривая будет располагаться несколько выше, чем можно
было бы ожидать.
Таким образом, более глубокий климатический анализ позволяет
утверждать, что наиболее влажные районы пампы характеризуются
климатом, приблизительно соответствующим климату лесостепи, а более
1 W. Lauer, Humide und aride Jahreszeiten in Afrika und Siidamerika und ihre
Beziehung zu den Vegetationsgurteln, «Bonner Geogr. Abb.», H. 9, 1952.
Злаковники умеренной климатической зоны южного полушария
157
сухие — настоящим климатом степи. Количество осадков представляется
сравнительно большим, однако следует учитывать, что летом осадки
выпадают в виде сильных грозовых ливней вечером или ночью, поэтому
величина инсоляции, а стало быть и испарения, остается днем по-преж-
нему высокой. Эту особенность, к сожалению, нельзя отразить на кли-
мадиаграмме.
i
Лауэр для определения индекса аридности по Ма'ртонну пользовался формулой:
12 п
, где п — месячная сумма осадков, t — средняя температура отдельных
t + 10
месяцев; аридными считаются месяцы, для которых i < 20. Однако применительно
к пампе формула дает прямо противоположные результаты: в восточной пампе все
месяцы также оказываются влажными; несколько западнее хотя и отмечаются три
сухих месяца, но это июнь, июль и август, то есть время года (зима), когда все запади-
ны затоплены водой; летние же месяцы, когда водоемы высыхают, согласно формуле,
должны рассматриваться как «гумидные».
При всех этих рассуждениях нельзя также забывать, что многолетние средние
данные недостаточно точно характеризуют климат: образуемый ими ряд нивелирует
экстремальные случаи. Отсутствие же деревьев в этих районах, несомненно, связано
с вероятностью наступления крайне засушливых лет. Поэтому небезынтересно будет
ознакомиться с климадиаграммами, составленными по данным за 50-летний период
наблюдений. В этом случае на диаграммах выделяются не только периодически слу-
чающиеся в пампе засушливые годы, но и экстремально влажные годы (количество
осадков за месяц свыше 300 мм)
В каждом десятилетии практически случаются 2—3 года с сухим
летом (рис. 124). Часто в январе или феврале выпадает лишь несколько
миллиметров дождей, не имеющих никакого экологического значения.
В отдельные годы летом в течение двух месяцев осадков вообще не бывает.
Даже в лежащем на юге Энтре-Риос с октября 1949 г. по февраль 1950 г.,
то есть за 5 летних месяцев, выпало всего 135 мм осадков, что означает
катастрофическую засуху. В 1910—1911 гг. за 8 летних месяцев здесь
зарегистрировано 193 мм', в Асуле в 1943—1944 гг. в течение 4 месяцев
выпало только 80 мм.
Зимние месяцы тоже могут быть чрезвычайно сухими, например
в 1920 г. за 5 месяцев (с мая по сентябрь) выпало всего лишь 35 мм осадков.
Деревья, лишенные средств приспособления к недостатку влаги,
продолжительной засухи не переносят, они погибают, тогда как злаки
в состоянии покоя легче переживают сухое время года. Достаточно вспом-
нить последствия восьми засушливых лет в прерии (1934—1941 гг.),
которые нанесли урон даже травянистой растительности. Такие губи-
тельные засухи, видимо, служат важной причиной отсутствия деревьев
в южноамериканских злаковниках.
Еще один фактор, препятствующий росту деревьев,— это слабая
дренируемость прерийных почв. Как для лесостепи, так и для прерии
доказано, что древесная растительность может успешно конкурировать
со злаками только на хорошо дренируемых почвах. Относится этэ и к самой
восточной окраине пампы. Естественные древостои отсутствуют здесь
не'полпостью. Леса или, точнее, редколесье (в соответствии с английским
«woodland») покрывают хорошо дренируемые крутые склоны, обращенные
к< нижней террасе Параны, а также Ла-Платы; отдельные островки леса
встречаются на возвышенностях, прерывистой цепью протягивающихся
вдоль Атлантического побережья до Мар-дель-Платы. Эти гряды либо
сложены пластами ракушечника, маркирующими прежнюю береговую
линию, или водопроницаемыми известняками, либо представляют собой
неподвижные, закрепленные растительностью дюны; иными словами,
во всех случаях речь идет о почвах без явлений застоя воды.
ДОЛОРЕС (9м) 14,7е 912
Р ц с. 124. Клцмадиаграммы Долореса (восточная пампа): средняя за 50-летний период наблюдений и за отдельные
засушливые годы.
Злаковники умеренной климатической зоны южного полушария
159
Необходимо, однако, учитывать, что виды деревьев, с которыми
вступают в конкуренцию злаки пампы, нельзя сравнивать с древесными
породами зоны лиственных лесов северного полушария. В нашем случае
речь идет о летнезеленых или вечнозеленых видах субтродических деревь-
ев. У северной границы пампы, в провинции Энтре-Риос (между реками
1
I г из 4
1 2иЗ
5 6
Рис. 125. Схематический профиль через ландшафт в южной части провинции Энтре-
Риос; у северной границы участки пампы чередуются с древесными насаждениями.
1 — галерейный лес вдоль рукава Параны; 2 — затапливаемая площадь с зарослями Scirpus giganieus
и 3 — деревьями Erythrina crus-galli', 4 —туссоковый злаковник (Р asp alum prionitis) на местообита-
ниях с высоким стоянием грунтовых вод; 5 — заросли Acacia caven — Prosopis nigra (на более высо-
ких участках — Celtis spinosa) на склонах и в долинах; 6 — злаковая пампа без древесных растений
на почти равнинных, плохо дренируемых местообитаниях (плато).
Парана и Уругвай), они представлены колючими Prosopis nigra, Р. alga-
robillo или Acacia caven, с характерной зонтообразной кроной, деревом
«тала» {Celtis spinosa) и др. В переходной полосе от зоны древесных насаж-
дений к пампе можно наблюдать и типичное для лесостепи чередование
типов растительности: пампа занимает плоские поднятия х, а древостои —
склоны долин и берега водотоков (рис. 125, 126). Весьма своеобразным
растением в лесах вдоль Параны является «омбу» {Phytolacca dioica),
которое хотя и выглядит как мощное дерево с несообразно толстым ство-
лом, но настоящей древесины не образует. На эстансиях часто можно
встретить гигантские омбу, посаженные вблизи домов первыми испански-
ми поселенцами.
По мере удаления из области пампы и с переходом в районы редко-
лесья нач севере, где распространена пальма Trithrinax campestris, содовое
засоление наблюдается все реже и наконец исчезает. Хотя на равнинной
местности здесь еще встречается много болот, но все они имеют сток, расте-
ния же — индикаторы содового засоления — пропадают. Это уже при-
знаки гумидного климата.
Выше мы установили, что в области степей Восточной Европы суще-
ствуют очень тесные взаимосвязи между растительным покровом и типом
почвы. В пампе систематического картирования почв пока не произво-
дилось, но в ряде работ содержится описание типов почв, распростра-
ненных на территории пампы 1 2. Почвы более низко расположенных
1 Здесь был описан почвенный профиль с 154-сантиметровым слоем гумуса.
Почва напоминает мощный чернозем лесостепи, но вместе с тем обладает рядом черт,
которые роднят ее с вертизолями. то есть почвами с неблагоприятным водным режимом,
которые периодически то пересыхают, то переувлажняются (рис. 127). Горизонт В
с пятнами извести обладает полиэдрической структурой, обусловленной трещинами
усыхания. Эта почва является уже переходным типом к переменно увлажняемым поч-
вам субтропических и тропических злаковников, широко распространенным в Брази-
лии и связанным не с особыми условиями климата, а с определенными формами
рельефа.
2 J. Papadakis, Soils of Argentina, «Soil Science», 95, 1963, p. 356—366;
C. R. O.Miaczynski,M. Tschapek, Los suelos de estepa de la region pampeana,
'160
Глава 3
Рис. 126. Деградированная пампа с древесной растительностью в долинах
рек, Уругвай (фото Гаснера).
областей пампы хотя и отличаются характерным для почв злаковников
мощным темноцветным гумусовым горизонтом, однако не являются
типичными черноземами. Из-за того, что зима здесь мягкая, разложение
гумуса продолжается и в холодное время года, поэтому содержание его
в почве пампы ниже, чем в черноземе. Обогащение горизонта В глинными
минералами — обычное явление. Значения pH колеблются от 5,5 до 6,0.
Кубиена (письменное сообщение) по-разному называет здешние почвы:
влажные почвы, причем преимущественно заболоченные; «луговые почвы»,
если следовать терминологии североамериканских почвоведов; влажные
черноземы (почвы прерий) с обильным накоплением глины и аккумуляци-
ей гидроокиси железа. Почвам свойственны способность сильно набухать,
частые трещины усыхания и отчасти — благодаря развитию псевдооглее-
ния — признаки лессивирования.
/Характер гумусовых образований свидетельствует о формировании
в прошлом мощного горизонта гумуса, какой обычно возникает под тра-
вяной растительностью. Остатки лесного гумуса в области современных
злаковников не обнаружены. Развитые местами сухие почвы часто харак-
теризуются реликтовыми признаками, отражающими существование
в прежние эпохи влажных периодов. Нет принципиальной разницы между
почвами аргентинской пампы и почвами злаковников штата Риу-Гранди-
ду-Сул в Бразилии, однако последние, по-видимому, сохранились до на-
стоящего времени значительно более влажными.
На западе, у границы засушливости, пампа сменяется такими же
колючими древесными насаждениями, какие свойственны ее северной
окраине. Но доминантом здесь является Prosopis caldenia. Род Prosopis
в Аргентине представлен многими видами; распространение одной части
их связано с гумидным, другой — с аридным климатом. Пампа вклини-
вается между областью произрастания видов Prosopis в гумидных районах
«Rev. Invest. Agropecuar.», Ser. Ill, 2, № 3, Buenos Aires, 1965; C. G. В о n f i 1 s,
Rasgos principals de los suelos pampeanos, INTA, Buenos Aires, 1966 (с почвенной кар-
той в масштабе 1 : 4 000 000). 1
Злаковники умеренной климатической зоны южного полушария
161
Рис. 127. Почвенный разрез в «pampa undulada» севернее Гуале-
гуайчу (провинция Энтре-Риос). Почва, хотя напоминает мощный
чернозем или почву прерий (гумусовый слой мощностью 154 еж),
входит в особую почвенную группу «вертизоли» (Шлихтинг, устное
сообщение) (фото Э. Вальтер).
с годовыми осадками, превышающими 1000 мм (Р. nigra и Р. algarobillo),
и ареалом того же рода (Р. caldenia), но уже в аридных районах, получаю-
щих не более 500 мм осадков. С увеличением сухости климата появляется
Р. flexuosa, а в еще более сухих условиях — Р. alpataco, Р. striata (= glo-
bosa), Р. denudans и др.
Граница между пампой и кальденалем, то есть районом распростра-
нения естественных насаждений Prosopis caldenia, одновременно пред-
11-0434
162
Глава 3
ставляет собой фронт конкурентной борьбы. Наблюдалось, что в засуш-
ливые годы Р. caldenia наступает на пампу, тогда как во влажные годы
одерживают верх злаки пампы. Поэтому кальденаль не распространяется
за пределы аридных районов. Вместе с тем эта область характеризуется
иным распределением осадков по временам года: преобладают летние
осадки. Свою лепту вносят и почвы — они здесь песчаные, водопроницае-
мые и лучше дренируемые. Таким образом, пампа занимает территорию,
где климатические и эдафические условия благоприятствуют развитию
слабосемиаридного злаковника, который существует здесь уже со вре-
мени эпохи оледенения и отложения лёсса. Об этом свидетельствует факт
обогащения почв остатками окремнелых клеток эпидермиса злаков.
По данным Теругги, они составляют до 20% сухой массы почвы.
РАСТИТЕЛЬНОСТЬ ПАМПЫ
Реконструкция первоначального растительного покрова пампы сопря-
жена со значительными трудностями, тем более что заповедные терри-
тории здесь не были созданы. Устойчивые к выпасу европейские виды
сильно потеснили местную флору на площадях, где выпасают крупный
рогатый скот, лошадей и овец. В настоящее время пампа представляет
собой наиболее важный сельскохозяйственный район Аргентины: здесь
возделывают 95% пшеницы, сосредоточено 80% пахотных земель и 60%
всего поголовья скота. На территории пампы проживает 2/3 всего насе-
ления страны (включая несколько миллионов жителей столицы — Буэнос-
Айреса). Ничтожные остатки более или менее естественной растительности
можно найти лишь в полосе отчуждения железной дороги, по обе стороны
автомобильных дорог за пределами огражденных пашен или пастбищ
и по берегам водотоков.
Описанию растительности пампы посвящены монографии Пароди —
север провинции Буэнос-Айрес, Ферворста — низменность Саладо и Каб-
реры — западная часть пампы Ч
В северной части пампы зональная растительность была представ-
лена богатой видами группировкой Stipetum. Пароди насчитывает 26 видов
злаков, из которых 3 адвентивных и 46 широко распространенных видов
разнотравья. Из злаков чаще встречаются представители семейства
Paniceae, количество которых к югу сокращается. В то же время в этом
направлении возрастает число видов Festuceae и Agrostideceae, типичных
для умеренной зоны.
Важнейшие виды злаков: Stipa neesiana и S. papposa, Piptochaetium
(4 вида), Bothriochloa lagurioides, 2 вида Panicum и 2 вида Paspalum,.
Bromus unioloides, Briza triloba, Melica rigida, Poa lanigera, Eragrostis
lugens, Eleusine tristachys.
Разнотравье выделяется здесь менее резко, чем в северных луговых
степях или в высокотравной прерии, где оно определяет аспект сообще-
ства. Высота растений в пампе достигает 120 см. Различают 3 яруса траво-
стоя. Верхний образуют генеративные побеги злаков; средний, высотой
30 см, занимают листья злаков и разнотравья; нижний, высотой всего
5 см, состоит из ползучих или образующих розетки видов разнотравья.
1 L. R. Р а г о d i, Essayo fitogeografico sobre el Partido de Pergamino, «Revista
Facult. Agron. у Veter.», 7, 1930, p. 65—271; F. B. Vervoorst, Las comunidades
vegetales de la depresion del Salado (Prov. Buenos Aires); A. L. Cabrera, Apuntes
sobre la vegetacion del Partido de Pellegrini, «DAGI (Buenos Aires)», 3, 1945, p. 1—86.
Злаксвники умеренной климатической зоны южного полушария
163
Заморозки бывают редко, однако из-за многочисленных отмерших
листьев злаков пампа зимой приобретает монотонную желтую окраску г.
Лишь весной (конец сентября — начало октября), когда распускаются
молодые листья и появляются цветки весенних однолетников и геофитов
(Anemone decapetala var. foliosa, Nothoscordum montevidensis [лилейные]
и касатиковые Sisyrinchium platense, S. laxum, Cypella herbertii и Alophia
amoend), пампа совершенно преображается. Полного развития раститель-
ность пампы достигает в ноябре и декабре, когда определяющими аспект
становятся виды Stipa. Из них только Stipa papposa выбрасывает перистые
ости. В январе и феврале наступает период относительного покоя. Хотя
и в эти месяцы временами идут дожди, испарение при температуре, часто
достигающей 30°, протекает столь интенсивно, что растения вянут и рас-
тительный покров снова принимает желтоватый оттенок. В разгар лета
нередко случаются засухи, которые длятся несколько недель. На клима-
диаграммах, составленных по средним месячным данным, эти периоды
отражения не находят. И марте из-за множества серых метелок поздно-
цветущего Вothriochloa lagurioides пампа приобретает свежий серебристый
тон, а затем, ближе к зиме, вновь постепенно желтеет. Значение pH почвы
колеблется в пределах 5,5—6,0.
В районах с близко расположенными грунтовыми водами широко
распространен Paspalum quadrifarium в сообществе с Eryngium eburneum.
Этот злак образует большие плотные дерновины, которые в северном
полушарии известны только на низинных болотах, с образующими кочки
Carex. Такую форму роста трав в южном полушарии называют «туесок».
Если грунтовые воды стоят столь высоко, что вызывают содовое засоление
почв (pH = 8,0—9,0), то обширные площади на такого рода местообита-
ниях бывают заняты низким травостоем Distichlis. Переход к нему нередко
образует туссоковый злак Spartina montevidensis (pH = 7,5).
Вместе с Distichlis растут также некоторые другие виды растений — индикаторы
содового засоления: Hordeum stenostachys, Sporobulus poiretii, Puccinellia glaucescens
и др., а также виды разнотравья: Sida leprosa, Petunia parviflora, Lepidium parodi,
Sisyrinchium platense, Spergularia villosa и т. д. (см. работы Ферворста, Пароди, Каб-
реры) .
Вокруг небольших западин, время от времени заполняющихся водой, наблюдает-
ся весьма четко выраженная поясность растительности (Ферворст). Размер этих пони-
жений различен: от 50 м, в поперечнике и меньше. Наиболее глубокая часть их нахог-
дится на 1—1,5 м, ниже поверхности пампы.
1. Центральную часть впадины с открытой водой занимают Glyceria multiflora,
Alternanthera philoxeroides и осоковые; в более крупных впадинах, где вода стоит
дольше, распространены также Scirpus americanus, Jussieua repens, Lemna, Ricciocarpus
natans, Azolla filiculoides.
2. Второй пояс образуют болотные растения: Scirpus cernuus, Marsilia concinna,
Juncus microcephalus, Mentha pulegium, Cerastium humifusum, Ranunculus bonariensis
и часто Rumex crispus.
3. Во внешнем поясе преобладает Solanum malacoxylon (= glaucum), часто дости-
гающий в высоту 1—2 м, кроме того, встречаются Eryngium ebracteatum и несколько
видов Heleocharis.
Летом западины высыхают; почва приобретает серую окраску; видны
многочисленные следы копыт животных; растительность уничтожена,
торчат только стебли Solanum. Поскольку засоленные почвы очень широко
распространены, становится понятным, почему вода в большинстве рек,
пересекающих пампу, солоноватая на вкус. Об этом красноречиво говорят
даже названия рек — Рио-Саладо и Саладильо.
1 В это время цветут занесенные из Европы и менее требовательные к теплу
Stellaria media, Capsella bursa-pastoris, Роа annua, Coronopus didymus и др.
It*
164
Г лава 3
Из работы Рингеле мы заимствуем данные анализов воды рек Самборомббн и Рио-
Саладо х. Пробы взяты в нижнем течении рек.
Таблица 26
Содержание солей в воде рек пампы (в г/л)
Река pH Na+ Cl- sor Общая щелочность H2SO4 Постоянная щелочность СаСОз
Рио-Саладо 8,8 1,0112 1,2141 0,8420 0,3969 0,1600
Самборомбон 7,9 0,3638 0,3656 0,3867 0,2548 0,1550
Из таблицы явствует, что вода Рио-Саладо минерализована в большей степени
чем в Самборомбоне. Последняя течет по хорошо дренируемой и, следовательно, менее
засоленной волнистой пампе. Первая же берет начало в расположенной на сухом западе
«лагуне» Мар-Чикита и пересекает насыщенную солями низменность Саладо.
Автор определил значение pH в среднем течении Рио-Саладо. В воде росли
Zannichellia palustris и Enteromorpha, на влажной кромке берега — Salicornia ambigua,
Sesuvium portulacastrum, Heliotropium curassavicum, Cressa truchilense, Spergularia villo-
sa, Sida leprosa и Distichlis. Выцветы соли, усеивающие высокую часть склона, имели
pH = 9,5; слой светлого лёсса под ними — 8,5; проба, взятая в 1,5 м выше пронизанного
корнями слоя гумуса, дала величину pH, равную 7,0 и 7,5. Следовательно, влажная
почва содержит соду. В речной воде, напротив, много хлоридов и сульфатов. Легко-
растворимые соли частично вымываются из почвы и попадают в реки.
В «лагунах» с небольшой концентрацией соды формируются заросли
Scirpus caltfornicus с Jussieua repens и Alternanthera philoxeroides, среди
которых встречаются изредка Azolla и Hydrocotyle. В зоне повышенного
увлажнения, располагающейся перед береговыми дюнами, вода, если
сток отсутствует, солоноватая; часто наблюдаются заросли Juncus acutus,
которые занимают полосу шириной 3—4 км. При наличии дренажа через
песок вода пресная; в этом случае здесь поселяется типичное для дельты
Параны растение — Scirpus giganieus; к нему присоединяются Equiset ит
giganteum, Androtrichum trigynum с волосками в соплодиях, развившимися
из удлиненных тычиночных нитей, и лишь изредка — Phragmites commu-
nis. На участках, где грунтовые воды залегают близко к поверхности,
появляется Cortaderia selloana, высокая «трава пампы», которая в соб-
ственно злаковой пампе отсутствует. В какой мере Typha angustifolia
и Zizaniopsis bonariensis переносят содовое засоление, не исследовалось.
В ющной части провинции Санта-Фе, относящейся к пампе, известна
сильнозасоленная «лагуна» Ла-Пикаса 1 2. На песчаных солончаках бере-
говой зоны лагуны, покрытых белой коркой солей (хлоридно-сульфатный
тип засоления), растут солерос Salicornia ambigua, Suaeda maritima,
Sesuvium portulacastrum и др.; по мере удаления от берега концентрация
солей уменьшается, но проявляется содовое засоление, заметное по обра-
зованию на поверхности почвы черных корок; здесь доминирует Distichlis
spicata (pH = 9,6—10,0) либо Paspalum vaginatum (pH — 9,5—9,8); на
слабозасоленных щелочных почвах (pH = 8,2) поселяется Hordeum .
stenostachys совместно с видами, приуроченными обычно к содовым поч-
вам; на почвах с очень низким содержанием солей, где сульфаты уже
отсутствуют, обычно преобладает Spartina montevidensis (pH = 7,0).
1 Е. J. Ringuelet, Datos ecologicos sobre las aguas de los rios Samborombon
у Salado de Buenos Aires, «Notes Museo de la Plata», 1, 1935, p. 159—175.
2 A. Ragonese, La vegetacion de la Provincia de Santa Fe, «Darwiniana», 5,
1941, p. 369—416; A. R a g о n e s e, G. С о v a s, La flora halofila del sur de la Provincia
Santa Fe, «Darwiniana», 7, 1947, p. 401—496.
Злаковники умеренной климатической зоны южного полушария
165
Рис.
128. Южная часть туссоковой пампы
Буэнос-Айрес (фото Э.
со Stipa brachy chaeta,
Вальтер).
провинция
Мы столь подробно рассмотрели явления засоления по той причине,
что они служат показателем семиаридного климата. В провинции Энтре-
Риос засоление наблюдается даже на бессточных участках островов в дель-
те Параны при условии, что они длительное время не затапливаются вода-
ми реки х. Мы обнаружили солонцы западнее Консепсьон-дель-Уругвая,
но они отсутствуют несколько севернее, в лесах района Монтиэль, где
уже начинается гумидная субтропическая область.
В южном направлении видовой состав первичной пампы, вероятно,
становился беднее. Подробно эта область в литературе не описана, однако
автор видел здесь обширные площади, покрытые туссоковыми дерновин-
ными злаками Stipa brachychaeta и S. trichotoma (рис. 128). После распа-
хивания территории эти злаки спустя много лет появляются снова. Их
зависимость от высоты стояния грунтовых вод иллюстрирует рис. 129.
Туссоковые злаки — это форма роста, встречающаяся только в южном
полушарии и в альпийском поясе тропических высоких гор. Они обра-
зуют мощные дерновины высотой более 1 л, в которых старые жесткие
листья сохраняются и после отмирания, между ними пробиваются живые
вечнозеленые листья. Поэтому травостой из туссоковых злаков никогда
не имеет цвета свежей зелени; лишь редко после пожара, уничтожившего
ветошь, и появления новых листьев «туесок» приобретает зеленую окраску.
Такая форма роста связана с вполне морским характером климата, в кото-
ром холодное время года мягкое и многоснежное, а заморозки бывают
лишь очень редко. Среди туссоковых злаков встречаются роды, распро-
страненные также в северном полушарии, но не развившие эту форму
1 A. Burkart, Ojiada sinoptica sobre la vegetacion del delta del rio Parana,
«Darwiniana», 11, 1957, p. 457—561.
166
Г лава 3
роста в районах с холодной зимой (Stipa, Festuca, Calamagrostis, Роа
и др.). Для пастбищного использования такие злаки слишком жестки.
Поэтому их стремятся удалить с пастбища и заменить более мягкими
европейскими видами. Сделать это вполне удается, так как местные виды
не переносят постоянного интенсивного или частого выпаса. Легче всего
Scirpus
americanus
Рис. 129. Распределение растительности в зависимости от уровня грун-
товых вод. Paspalum. quadrifarium в отличие от Stipa trichotoma растет на
местообитаниях с более высоким зеркалом грунтовых вод; заросли Супага
cardunculus адвентивно замещают травостой Stipa brachychaeta.
распространяется Stipa brachychaeta, образующая у основания очень
много к лейстогамных цветков и обильно плодоносящая. На севере это
растение считают нежелательным вторженцем, однако не исключено, что
этот вид входил в естественный травостой первичной пампы.
Выше мы уже отмечали, что к факторам, способствующим формированию злаков-
ника, относятся умеренная пастьба диких животных и случайные пожары. Прежде
в пампе были шире распространены: маленький пампасский олень, гуанако (дикая
форма ламы), страус нанду, броненосцы, часто встречающиеся и в наше время, и раз-
личные грызуны, из них вискаша считается вредителем, с которым ведется борьба.
Несомненно, случались и пожары, вызванные ударом молнии. Позднее индейцы умыш-
ленно поджигали траву, стремясь изгнать белых пришельцев.
На поверхности бессточной «рашра alta», между Хуаресом и Лапри-
дой, развит едва заметный взгляду микрорельеф, обусловливающий
микромозаичное распределение растительности. Самые высокие позиции
занимает Stipa brachychaeta, на более пологих участках распространен
Stipa trichotoma, а в низинах, на солонцах,— Distichlis. После распашки
территории поверхность пашни выглядит пятнистой — все участки, на
которых рос Distichlis, выделяются светло-серым горизонтом А, свой-'
ственным солонцу, а места произрастания Stipa — темным гумусом.
На богатой гумусом пашне (pH — 5,5) кукуруза дала хорошие всходы,
на светлых же пятнах (pH = 8,5) проростков не было. Здесь лежали
плотные черные комки горизонта В, вывернутые с глубины плугом;
на пролысинах поселились индикаторы соды — Distichlis, Hordeum steno-
stachys, Puccinellia glaucescens.
Пересекая пампу на автомобиле, можно легко распознать уже издали
засоленные площади: на таких участках вдоль дорог обычно высаживают
Злаковники умеренной климатической зоны южного полушария
167
деревья Tamarix gallica. Другие древесные породы на почвах, содержа-
щих соду, не растут.
В «pampa undulada» засоление практически не выражено, так как
густая речная сеть препятствует образованию бессточных впадин. Застой
воды возможен только на речных террасах, если они окаймлены достаточ-
но высокими береговыми валами. Например, на террасах реки Ареко
близ Сан-Антонио-де-Ареко или реки Реконкиста у Кастелара (пригород
Буэнос-Айреса) можно видеть густой ковер из растений, которые хорошо
переносят содовое засоление.
В западном направлении, как уже отмечалось, климат становится
суше (Пеллегрини, 676 мм осадков), одновременно максимум дождей
смещается на лето. Почвы преобладают песчаные, местами песок собран
в невысокие, покрытые растительностью дюны. При чрезмерном выпасе
может начаться движение дюн. Изменяется состав растительности. Наи-
более широко распространен злак Роа ligularis, появляются новые виды
Stipa (S. latissimifolia, S. tenuissima, а на юге также S. dusenii) и Aristida
pollens. Для рыхлых песчаных почв дюн характерны Elionurus viridulus,
Panicum urvilleanum и Hyalis (Plazed) argentea (сложноцветные) с сереб-
ристо-серыми листьями.
АРИДНАЯ ДРЕВЕСНАЯ РАСТИТЕЛЬНОСТЬ
И ПОЛУПУСТЫНЯ С LARREA ЗАПАДНЕЕ ПАМПЫ1
Граница между провинциями Буэнос-Айрес и Ла-Пампа прежде рас-
сматривалась как рубеж между злаковниками и лесами, образованными
Prosopis caldenia. Ныне деревья вырублены, поэтому этот рубеж сместился
к западу. Только вокруг домов или на пастбищах сохранились отдельные
старые экземпляры, свидетели прежнего более широкого распространения
леса. В зоне отчуждения вдоль железнодорожных насыпей можно встре-
тить и молодую поросль. Сравнительно большие массивы леса еще удер-
живаются на пространстве между городами Санта-Роса и Викторика,
•а также севернее этой линии. В них наряду с Prosopis caldenia (кальден)
встречаютсяР. flexuosa, Geoffroea (=Gurliea) decorticans (чанар), Jodinarhom-
bifolia — древовидная Santalaceae (полупаразит, паразитирующий на Proso-
pis), Schinus longifolia, Ximenia americana и др. В низинах Prosopis caldenia
иногда образует старовозрастные насаждения, в которых деревья с мощ-
ными стволами достигают высоты 10 м и стоят приблизительно в 10 м
одно от другого. Под их пологом обычен кочкарник, состоящий из Stipa
tenuissima и S. gyneroides, которые покрывают около 50% площади. Эти
леса обманчиво сходны с насаждениями Acacia giraffae в Юго-Западной
Африке, в травяном покрове которых господствует Aristida. Этот факт
сходства не является исключением. Зоны растительности в области раз-
вития песчаных почв между Викторикой и Санта-Исабелем, в которой
количество осадков уменьшается с 500 до 300 мм в год, напоминают районы
Юго-Западной Африки с такой же годовой суммой осадков и такой же
песчаной почвой. К западу леса все более редеют и наконец переходят
в саванну с единичными на фоне травяного покрова деревьями; последнюю
сменяет кустарниковая саванна, причем число экземпляров Prosopis
flexuosa увеличивается, а виды Stipa постепенно выпадают из травостоя.
1 A. L. С a b г е г a, Fitogeografia. См.: La Argentina-Suma de Geografia, Bd. Ill, 2,
Buenos Aires, 1958.
168
Глава 3
В конце концов остаются лишь низкорослые Prosopis alpataco со стелю-
щимися по земле ветвями, у которых вверх торчат лишь верхушки побе-
гов, да прутьевидные кустарники Ephedra ochreata, Bredemeyera microphylla
(истодовые) и безлистные Neosparton aphyllum (вербеновые), Prosopis
striata (= globose), Cassia aphylla и др. На самых сухих местообитаниях,
на гребнях дюн, исчезают и эти древесные растения; здесь уже развит
чистый злаковник с господством Elionurus viridulus и с некоторыми травя-
нистыми псаммофитами в травостое. Дальше простирается территория
с обширными очагами засоленных почв, занимающая низменную долину
реки Рио-Атуэль.
Но уже в пределах зоны с Prosopis caldenia на местообитаниях, недо-
статочно увлажненных, распространены виды, характерные для зоны
полупустыни: Larrea divaricate, Condalia microphylla (крушиновые) и др.
Далее к западу, где количество осадков сокращается до 200 мм в год,
эта полупустыня доминирует в ландшафте, прежде всего на почвах, раз-
витых на базальтах. Мощность почвенного слоя весьма незначительна,
ибо уже на небольшой глубине залегает известковая корка, подобно
цементу связывающая скальные обломки в монолитный плотный слой.
Для полупустыни типичны виды Larrea (парнолистниковые), и прежде
всего L. divaricate, по-видимому идентичная Larrea tridendata, знакомой
нам по Аризонской пустыне. Распространена также Larrea cuneifolia,
облиствленные плоские побеги которой вытянуты строго в меридиональ-
ном направлении, и L. nitida. Все виды Larrea содержат смолы и издают
после каждого дождя характерный запах (креозотовый кустарник).
Растения отличаются чрезвычайной засухоустойчивостью.
Последовательность зон растительности приведена в табл. 27.
Полупустыня с Larrea в виде сплошного пояса длиной более 2000 км
и шириной 250—400 км пересекает самые засушливые части Аргентины
восточнее Анд приблизительно между 25° и 44,5° ю. ш. Таким образом,
на севере она простирается в аридную субтропическую зону и прослежи-
вается здесь в горах до высоты 3000 м над уровнем моря; на юге она гра-
ничит с патагонской полупустыней умеренной зоны, растительность
которой менее требовательна к теплу и потому образует в горах над полу-
пустыней с Larrea следующий, более высокий пояс, прослеживаемый
на север до Мендосы (см. рис. 114).
В зоне развития полупустыни с Larrea, вблизи больших полноводных
рек, стекающих с высокогорных районов Анд, раскинулись города-
оазисы: Неукен, Сан-Рафаэль, Мендоса, Сан-Хуан, Риоха и др. Города
окружены плодородными поливными площадями, на которых культиви-
руются главным образом семячковые и косточковые плоды и виноград.
Средняя годовая температура в этой обширной области колеблется от 19°
на севере до 14° на юге. Максимальная температура поднимается до 45°,
но с марта по октябрь бывают заморозки. Общей чертой всей территории
является засушливость, количество осадков колеблется в пределах
323—86 мм', дожди идут на севере чаще летом, в средней части зоны вес-
ной и осенью, а на юге зимой. Повсеместно растительность испытывает
длительный период засухи, к которой виды Larrea особенно устойчивы.
Поражает их исключительная приспособляемость к условиям среды:
в более влажных районах растения становятся выше, растут гуще и обра-
зуют менее ксероморфные листья; в сухих местностях растения низкие
и почти безлистные.
Флористический состав сопровождающей флоры также не везде
одинаков. На севере присоединяются элементы предпуны, такие, как
Злаковники умеренной климатической зоны южного полушария
169
Таблица 27
Схема последовательности зон растительности и изменения климата
в направлении: провинция Энтре-Риос — провинция Буэнос-Айрес —
провинция Ла-Пампа до Рио-Негро
Растительность Почвы Климат Годовое количество осадков, ММ
Леса с Prosopis nigra я Р. algorobillo Леса с Acacia caven и Celtis spinosa Гумидный >1000
Переходная зона от леса 1 макромозаика к злаковой пампе / Северо-восточная бога- тая видами злаковая пампа 1 собственно Сухая, менее богатая । пампа видами туссокозлако- вая пампа J Лёссовые Семиарид- ный 1000
Переходная зона, воз- современный можно саванноподоб- j культурный ная ландшафт Леса с Prosopis caldenia (с видами Stipa} Саванна с деревьями Prosopis caldenia (без видов Stipa) Кустарниковая саванна с Prosopis calde- nia — Р. flexuosa Prosopis alpataco л Neospartum aphyllum 1 кустарниковая Ephedra ochreata [ саванна Schinus polygamus л Злаковник c Elionurus viridulus, но без древесных растений, на сухих гребнях ДЮН Песчаные, ме- стами дюны; различия в рельефе не учтены Аридный 500 300
Кустарниковая полупустыня с Larrea Каменистые Аридный 200
кустарник из бобовых Zuccagnia punctata, Bougainvillea spinosa и др.;
на юге же патагонские элементы.
Многие сопровождающие виды являются конвергирующими^ кустар-
никами с прутьеобразными побегами и часто совсем лишены листьев.
Назовем здесь Cassia aphylla (Caesalpineae), Monttea aphylla (норичнико-
вые), Cyclolepis genistoides и Chuquiraga crinacea (оба сложноцветные),
Atamisquea emarginata (каперсовые) и др.
Особенно интересно появление Cercidium australe у подножия крутых
склонов, где известковая корка разламывается и, видимо, имеются более
благоприятные условия увлажнения. Этот вид близок Cercidium micro-
phyllum, распространенному в пустыне Сонора; на его стволе видна та же
типичная для этих растений зеленая кора.
На тот факт, что в этой области легко может произойти аккумуляция
солей в почве, указывает избирательно поселяющийся Atriplex lampa,.
а при сильном засолении почв — Atriplex undulata. Оба растения —1
170
Глава 3
ксерогалофиты. Во влажных низинах рядом с выцветами солей встречаются
ярко выраженные суккулентные гигрогалофиты — Allenrolfia, Salicornia,
Heterostachys, Suaeda и др. Вследствие попеременного высыхания и увлаж-
нения почвы здесь наблюдается образование полигональных корковых
почв. Из-за высокой концентрации солей в почвенном растворе сода
не образуется. Показатель pH солевой корки 7,0, влажной почвы под
ней — 6,5. Однако там, где почва накапливает меньше солей, например
у края бьющего из базальта источника, значение pH повышается до 9,0—
10,0. Здесь росли Atriplex lampa и Cyclolepis genistoides. Более подробных
определений содержания солей в почве и в клеточном соке Larrea и сопро-
вождающих видов не производилось. Южная граница полупустыни
с Larrea, по-видимому, обусловлена низкой температурой. В переходной
области четко намечается верхняя граница распространения Larrea
в горах, где она предпочитает хорошо прогреваемые склоны.
Из трех видов Larrea L. nitida наиболее требовательна к влаге. При-
ближаясь с влажного нагорья или со стороны патагонской полупустыни
к зоне с Larrea, встречаешь только этот вид. В северной части зоны расте-
ние приурочено к местообитаниям, где грунтовые воды залегают близко
к поверхности.
Весьма подробное эколого-физиологическое исследование видов Larrea
и кустарника из бобовых Zuccagnia осуществил Морелло х.
Листья этих видов имеют изолатеральное строение, но тем не менее
им не свойственна ни склерофильность, ни малакофильность. Клетки
многослойного эпидермиса крупные, стенки толстые, палисадная парен-
хима хорошо выражена и плотная. Устьица не погружены в ткань листа,
а даже несколько приподняты и поэтому лишены смолистого налета
(табл. 28).
Таблица 28
Число устьиц на 1 мм2 листовой поверхности кустарников;
северная полупустыня с Larrea (по Морелло)
Название растений Верхняя сторона Нижняя сторона
Larrea cuneifolia (гумидная область) . . 90 120
Larrera cuneifolia (аридная область) . . 120—170 140—190
Larrea divaricata 104 130
Larrea nitida 100 150
Zuccagnia punctata 110 140
Как явствует из таблицы, степень ксероморфности у всех четырех
видов одинакова. Большое число устьиц позволяет растениям интенсивно
транспирировать во влажное время года, кривые суточной транспирации
параллельны в этот период кривой испарения. В период засухи устьица
открываются только утром, вновь замыкаясь задолго до полудня. Кути-
кулярная транспирация днем, когда кутикула обезвожена, составляет
около 1% общей транспирации; ночью, когда кутикула менее обезвожена,
кутикулярная транспирация в два раза больше.
1 J. Morello, Estudios botanicos en las regiones aridas de la Argentina. I, II,
«Revista Agron. Noroeste Argent. Tucuman», 1, 1955, p. 301—370; 385—524.
Злаковники умеренной климатической зоны южного полушария
171
Наряду с уменьшением транспирации в период засухи наблюдается
'также сокращение общей листовой поверхности, которое проявляется
в том, что старые листочки опадают и остаются только молодые.
Для кустарников 18—20-летнего возраста приводятся данные, сумми-
рованные в табл. 29.
Таблица 29
Общая листовая поверхность одного куста (в ле2)
Название растений Дождливый период Засушливый период Поверх- ность после редукции, %
Larrea cuneifolia 1,6—2,0 0,4—0,8 25—40
Larrea divarlcata 2,0-2,5 0,5—1,0 25—40
Larrea nitida ....... 2,5 2,0 80
Zuccagnia punctata .... 2,5—3,0 1,5—2,0 60—67
Сокращение листовой поверхности особенно незначительно у L. nitida
(20%), связанной здесь с грунтовыми водами и потому даже в засуху
не испытывающей недостатка влаги. Zuccagnia также сравнительно слабо
сокращает площадь транспирирующей поверхности.
Такое уменьшение интенсивности транспирации и редукция листовой
поверхности обусловлены повышением осмотического давления при наступ-
лении засухи. Морелло не определил непосредственно величину осмоти-
ческого давления, но доказал увеличение дефицита насыщения. Слабая
транспирация и небольшая листовая поверхность позволяют растению
резко ограничить расход влаги на транспирацию во время затяжной
засухи (табл. 30).
Таблица 30
Потери влаги путем транспирации у Larrea cuneifolia
в дождливый период и в период летней засухи
Время года Транспирация, л/м^ Общая листовая поверхность Куста, Л4.2 Расход воды за сутки, л
Дождливое . . . 3,25—1,55 2,0—1,3 6,5—2,0
Засушливое . . 0,62—0,4 0,8—0,4 0,5—1,6
В табл. 31 сопоставлены суточные расходы влаги на транспирацию
четырьмя исследованными видами (по Морелло).
На основании приведенных цифр можно количественно оценить
потребление растениями влаги за год, приняв длительность дождливого
и засушливого периодов равной 6 месяцам каждый. В табл. 32 помещены
полученные результаты.
Larrea nitida характеризуется иными особенностями водного режима.
Редукция листовой поверхности и снижение интенсивности транспирации
в засушливое время года у этого вида незначительны. Но, принимая
во внимание, что потенциальная транспирация в сухой сезон больше,
Морелло приходит к заключчению, что в дождливый и засушливый периоды
172
Глава 3
Таблица 31
Дневное потребление воды одним кустом
Larrea (в л)
Название растений Дождливый период Засушливый период
Larrea cuneifolia . . Larrea divaricata . . Larrea nitida .... Zuccagnia punctata 6,5-2,0 8,0—7,5 6,75—6,5 8,5—7,5 0,5—0,16 1,75—0,5 Слабое 0,8—0,23
Таблица 32
Поглощение воды одним растением за год (в л)
Название растений Дождливый период Засушливый период Всего
Larrea cuneifolia Larrea divaricata Zuccagnia puncta- ta 360—1150 1350—1450 1350—1450 90—300 90—300 40—150 450—1450 1450—1750 1400—1600
растение расходует на транспирацию приблизительно одинаковое количе-
ство влаги, которое за год составляет около 2600 л.
Однако эти величины в расчете на один куст мало что говорят эко-
логу, поскольку размер кустов различен. Более важным представляется
сопоставление потерь влаги одним кустом с ее запасами в почве, также
в расчете’на один куст. Величина этих запасов зависит не только от коли-
чества атмосферных осадков, но и от характера и площади восприни-
мающей дождевую влагу поверхности почвы в радиусе распространения
боковых корней растения. Для Larrea cuneifolia эта площадь равна 70—
100, редко 150 ж2, для L. divaricata 60—110, для Zuccagnia 14—32 и для
Larrea nitida — 4 м2.
Столь незначительная площадь у названного последним вида объяс-
няется тем, что на севере он приурочен к местообитаниям с близким зале-
ганием грунтовых вод (фреатофит), где образует лишь один стрежневой
корень, без ответвляющихся от него боковых корней.
Расход влаги^на транспирацию одним кустом Larrea в расчете на 1 м2
площади почвы, пронизанной корнями, приведен в табл. 33.
Значения в литрах на 1 ^'означают одновременно и количество воды
в миллиметрах. Это позволяет непосредственно сопоставлять их с годовой
суммой осадков, которая в исследованном районе составляет в среднем
140 мм, то есть 140 л!м2. Разумеется, не вся атмосферная вода оказывается
доступной для растений.
Мы различаем следующие типы местообитаний:
1) горные склоны с интенсивным поверхностным стоком;
2) прилегающие к их подножию осыпи (бахадас) с почти ровной
поверхностью;
3) врезанные сухие долины, в которые после дождя с окружающих
возвышений скатываются дождевые потоки.
Часть атмосферных осадков стекает по склонам или испаряется
с поверхности почвы. Вода, просачивающаяся в грунт, частично исполь-
зуется эфемерами, которые в большом количестве появляются после1
Злаковники умеренной климатической зоны южного полушария
173
Таблица 33
Расход воды кустарниками в полупустыне с Larrea
Название растений Площадь, пронизанная корнями, л2 Транспирация, л!м^ Всего за год, л/лс2 = лип
дождливый период засушливый период
Larrea cuneifolia . . . 70 5,5-17 1-4 6,5-21
Larrea cuneifolia . . . 100 3,5—11,5 1-3 4,5—14,5
Larrea divaricata . . . 60 23,5—25 1,5-5 25—30
Larrea divaricata . . . ПО 12-13 1-3 13—16
Zuccagnia punctata . . 14 97—105 3—10 100—115
Zuccagnia punctata . . 32 43,5—45,5 1,5-4,5 45—50
Larrea nitida 4 325 325 650
каждого дождя. Промачивание почвы до уровня грунтовых вод возможно
лишь в долинах временных потоков.
Морелло следующим образом оценивает распределение атмосферной
влаги между отдельными компонентами водного баланса (табл. 34).
Таблица 34
Водный баланс полупустыни с Larrea
(в % количества осадков)
Элементы баланса Склоны Бахадас
Поверхностный сток Непосредственное испарение с 31—37 7—10
поверхности почвы 22—29 30—32
Инфильтрация в грунт 0—3,7 0-4
Транспирация эфемеров .... 8—13,7 10—18
Таким образом, доступное для кустарников количество воды состав-
ляет (в мм):
На склоновых местообитаниях . . 25—55
На бахадас..................... 50—80
В ложбинах стока................ 140 и более
Размещение четырех видов следующее: L. cuneifolia встречается
только на склонах; L. divaricata растет на благоприятных участках совме-
стно с L. cuneifolia, однако преимущественно заселяет бахадас, изредка
спускается в ложбины временного стока; Zuccagnia предпочитает послед-
ние, хотя иногда перебирается и на бахадас.
Если даже учесть весьма приблизительный характер вычислений,
то все же видно, что все названные виды вполне довольствуются тем коли-
чеством воды, которое содержится в почве их местообитаний. Максималь-
ному потреблению воды L. cuneifolia, равному 21 мм, на склонах соответ-
ствует минимальное количество воды в почве, составляющее 25 мм. Мак-
симальному расходованию влаги L. divaricata, не превышающему 30 мм,
соответствует минимальная влагоемкость почвы на бахадас, равная
50 мм', соответствующие величины для Zuccagnia — 115 мм и 140 мм
в ложбинах стока или 50 мм и 80 мм на бахадас.
174
Глава 3
У одной только L. nitida годовое потребление влаги, достигающее
650 мм, во много раз превышает общее количество осадков. Поэтому
данный вид растет лишь там, где ему доступны обильные грунтовые воды.
Разумеется, единственная причина, определяющая тот или иной
характер распределения растений по площади,— это разная конкурент-
ная мощность видов. Нужно предполагать, что конкурентоспособность
убывает в последовательности: L. nitida —Zuccagnia -> L. divaricata—^
-> L. cunetfolia. Каждый вид занимает оптимальное для него местообитание
и вытесняет более слабого конкурента на менее благоприятное. L. cunetfo-
lia в таком случае приходится довольствоваться особенно сухими место-
обитаниями.
ЗЛ А КОВНИГ И ПОЛУПУСТ БЕН Я В ПАТ АГОН ИИ 1
Растительность Патагонии часто называют «степью». Но мы будем
использовать это понятие в первоначальном его значении лишь по отно-
шению к восточноевропейско-сибирским злаковникам. Термин «степь»
нередко применялся для обозначения столь различных типов раститель-
ности во всех частях земного шара, что сегодня его употребление означает
лишь, что речь идет о тех аридных районах, где растительный покров,
более сомкнут, чем в пустыне.
Некоторые исследователи считают возможным называть «степью»
все злаковники, развитые в областях с холодной зимой, однако и это-
предложение не всегда приемлемо. Но как бы то ни было, в данном случае-
не следует говорить о «патагонской степи», ибо злаковники здесь хотя
и встречаются, но чаще они распространены в окраинных районах или
на влажных местообитаниях. Как правило, в растительном покрове
Патагонии доминируют деревянистые кустарнички, которые нередко
покрывают только 25% поверхности почвы. Поэтому обозначение «кустар-
ничковая полупустыня» намного точнее характеризует распространенный
здесь тип растительности («semidesierto patagonico», по Рагонезе).
Патагонская зона растительности охватывает всю южную часть терри-
тории Аргентины (см. рис. 114) и, постепенно все более сужаясь и вместе
с тем поднимаясь до абсолютной высоты 3000 м, вклинивается на севере
между Андами и полупустыней с Larrea. Поэтому в гористой и сильна
расчлененной предандской зоне возникают в зависимости от высоты,
экспозиции и типа горных пород чрезвычайно сложные растительные
макрокомплексы, в которых закономерно преобладают то патагонские,
то андские элементы, то виды, свойственные полупустыне с Larrea. Послед-
ние форпосты патагонской растительности достигают Мендосы, где повсе-
местно еще встречается характерный вид Mulinum spinosum (зонтичные),
образующий большие колючие подушки. Севернее патагонская зона пере-
ходит в физиономически очень похожую на нее растительность пуны.
Рассмотрим прежде всего климатические условия на поперечном
профиле северной Патагонии, проходящем от Сан-Карлос-де-Барилоче-
до Макинчао.
Между озерами Тодос-лос-Сантос, лежащем на чилийской стороне,
и Науэль-Уапи, на берегах которого расположен город Барилоче,—
на аргентинской, горная цепь Анд резко понижается. Высота перевала
1 A. Soriano, Los distritos floristicos de la Provincia Patagonica, «Rev. Inve-
stig. Agric.», 10, 1956, p. 323—347; О. В о e 1 c k e, Comunidades herbaceas del Norte de-
Patagonia у sus relaciones con la ganaderia, «Rev. Investig. Agric.», 11, 1957, p. 1 — 98..
Злаковники умеренной климатической зоны южного полушария
175
едва достигает 1000 м, отметка же Барилоче равна 853 м. Поэтому непре-
рывно дующий с океана западный ветер легко преодолевает горы и при-
носит обильные дожди также и на восточный склон Анд.
У западной оконечности сзера Науэль-Уапи, под Пуэрто-Блестом,
годовая сумма осадков еще составляет 4000 мм. В 45 км восточнее, у города
Барилоче, который лежит у самого подножия внешних низких отрогов
гор, количество осадков уже падает до 1081 мм в год. У реки Лимай,.
удаленной от Барилоче на 15 км к востоку, годовая сумма осадков едва
достигает 500 мм, а через 40 км, под Пильканиеу,— всего лишь 300 мм.
Таким образом, на расстоянии 100 км количество осадков умень-
шается с 4000 мм до 300 мм в год. Дальше к востоку убыль несколько
замедляется и вблизи Макинчао, еще через 150 км, за год выпадает только
173 мм. Всего 300 км отделяют Макинчао от берегов Атлантического
океана, но сумма осадков на этом расстоянии почти не изменяется. Места-
ми она, правда, сокращается до 160 мм, но на побережье, у Сан-Антонио,,
снова увеличивается до 200 мм. Изменение характера растительности
идет в соответствии с повышением степени аридности. В районе Пуэрто-
Бласта, где выпадает значительное количество осадков, развит вальдивий-
ский, богатый древесными породами дождевой лес из Nothofagus, типич-
ный для южного Чили (см. т. II, гл. 3). Однако в направлении к востоку
это видовое разнообразие быстро утрачивается. В древостое сохраняется
лишь Nothofagus dombeyi, а на больших высотах также N. ритШо, кото-
рый у верхней альпийской границы распространения образует криво-
лесье. На разных высотах встречается и N. antarctica, но обычно в пла-
корных условиях, с большой вероятностью заморозков или на заболочен-
ных участках. Еще перед Барилоче нотофагусовые леса исчезают. Далее
протягивается узкая полоса с доминирующим Austrocedrus (Libocedrus)
chilensis, который в зоне с Nothofagus растет только на сухих местообита-
ниях. Здесь он играет примерно такую же роль, как сосна в лесах Цен-
тральной Европы. В окрестностях Барилоче обнаруживаются только
редкостойные леса и заросли кустарников, образующие переходную зону
к собственно патагонской растительности. Доминируют Nothofagus antarc-
tica, Maytenus boaria, Aristotelia maqui, протейные Lomatia hirsuta и Emboth-
rium coccineum с яркими красными цветками (последнее растение повсе-
местно маркирует границу между лесом и аридной областью), кустарники
с прутьевидными побегами Diostea juncea (вербеновые), Ovidia pilo-pila
(волчниковые), Schinus patagonica, Buddleia globosa, низкие кустарники
Discaria serratifolia, Colletia spinosissima, Ribes cucullatum и R.magellani-
cum, Chacaya trinervis, Escallonia virgata и разные виды Berberis. Austocedrus
и Maytenus экстразонально растут и несколько восточнее на влажных
южных склонах, низкий же кустарник по долинам ручьев или по поймам
рек внедряется глубоко в патагонскую флористическую область Е
О весьма своеобразных климатических условия Патагонии дают
представление климадиаграммы городов Сан-Карлос-де-Барилоче, рас-
положенного у границы лесной растительности, и Макинчао, лежащего
у края полупустыни с Larrea. Для этих станций имеются также данные
об испаряемости (рис. 130—132).
Распределение осадков в Барилоче такое же, как в южном Чили:
резко выраженный максимум осадков зимой (южного полушария) и мини-
мум в январе. Экстремальная сухость слабо намечается только в январе,
но назвать климат гумидным все же нельзя. Средней сумме осадков —
1 Ср.: W. Eriksen, Landschaft, Nationalparks und Fremdenverkehr am
ostpatagonischen Andenrand, «Erdkunde», 21, 1967, S. 230—240.
176
Глава 3
1081 мм здесь противостоит значительная испаряемость, достигающая
1240 мм. За период с 1921 по 1950 г. среднее годовое количество осадков
составило 1169 мм. Следовательно, водный баланс здесь лишь слабо
отрицательный, что хорошо согласуется с положением района у границы
произрастания леса. Если нанести на Клима диаграмму значения средней
месячной испаряемости, то окажется, что кривая испаряемости будет
сравнительно точно соответствовать температурной кривой, построенной
в вертикальном масштабе 1:12. Заметим, что в области пампы требовалось
Рис. 130—132. Климадиаграммы Патагонии с кривой испаряемости Е, располагаю-
щейся вследствие постоянных сильных ветров высоко на графике, и кривыми темпера-
туры (t X 10) при соотношении 10° = 100 мм, (t X 12) при соотношении 10° = 120 мм
и (t X 15) при соотношении 10° = 150 мм.
Сан-Карпос-де-Барилоче — граница леса; Макинчао — очень сухая кустарничковая полупустыня;
Рио-Гранде, Огненная Земля, — вблизи границы между злаковником и сильно заболоченными суб-
антарктическими лесами.
лишь 7—9-кратное увеличение. Необыкновенно высокая испаряемость
в Патагонии объясняется воздействием на климат устойчивых сильных
западных ветров. Средняя скорость ветра в Барилоче в летние месяцы
колеблется в пределах 6,5—7 м/сек4. В Макинчао, расположенном на
250 км восточнее, скорость ветра значительно меньше (4—4,5 м/сек)-,
кривая испаряемости близко повторяет кривую температуры, вычерченную
с 10-кратным увеличением вертикального масштаба. Но количество
осадков здесь невелико, их слабый максимум отмечается в мае. На клима-
диаграмме отчетливо выделяется период сильной засухи, который длится
с октября до апреля; годовой водный дефицит равен почти 1000 мм. Столь
же высокий водный дефицит характерен для всей Патагонии, только
на крайнем юге он меньше: в районе Рио-Гранде, Огненная Земля, пре-
вышение расхода влаги над приходом составляет 385 мм (рис. 132), тогда
как в Ушуае, близ границы леса, водный баланс почти уравновешен,
насколько об этом можно судить по данным кратковременных наблюдений.
Таким образом, в Патагонии господствуют условия среды, аналогичные
1 В Эскеде, расположенном южнее Барилоче, средняя скорость 'ветра в те же
месяцы равна почти 8 .м/сек; кривая температуры вычерчивается при соотношении гори-
зонтального и вертикального масштабов 1 : 15, такое же соотношение использовано
при построении климадиаграммы для района Рио-Гранде на Огненной Земле (рис. 132).
Злаковники умеренной климатической зоны южного полушария
177
Рис. 133. Патагонский ландшафт; озеро Вьедма на восточном склоне Анд, зла-
ковый покров образован Stipa humilis (фото Дузена).
условиям в полупустыне, с той лишь разницей, что температура здесь
намного ниже. Средняя годовая температура в Барилоче равна 8,3°, на
Огненной Земле 5°. Колебания ее в течение года невелики. Казалось бы,
что можно говорить о почти морском климате, но с ним несовместима
большая сухость. Холодное время года длится 4—5 месяцев, однако сред-
няя суточная температура самого холодного месяца не опускается в Бари-
лоче ниже —1,3° (абсолютный минимум —15,4°), а в Макинчао — ниже
—3,6° (абсолютный минимум —24,5°). На Огненной Земле зима, пожалуй,
даже мягче: в Рио-Гранде соответственно —2,5° и —16,2°, в Ушуае —1,4°
и —11,6°. Количество осадков резко колеблется от года к году в Паль-
кание от 157 мм до 519 мм) и от сезона к сезону.
Почвы Патагонии маломощные, бедны известью и питательными веще-
ствами, обычно имеют слабокислую реакцию. В низинах, часто бессточных,
происходит засоление почвы, иногда весьма значительное.
В более влажных западных районах еще доминируют злаки: на воз-
вышенных участках — Festuca pallescens, обычно же Stipa speciosa л др.
Оба вида относятся к низкотуссоковым злакам, распространенным и в Но-
вой Зеландии, в Отаго (рис. 133). По мере движения в глубь засушливых
областей злаки отступают на второй план. Встречаются они прежде всего
на ровных песчаных территориях или по более сырым днищам долин, где
с увеличением увлажненности появляются осоковые и Juncus вместе
с занесенными европейскими видами {Taraxacum officinale, Trifolium repens,
Роа pratensis, Deschampsia flexuosa).
Сориано выделяет в Патагонии несколько флористических провинций:
субандскую, западную, центральную, провинцию в районе залива Сан-
Хорхе и огнеземельную. Нам придется ограничиться перечислением лишь
некоторых характерных для каждого подразделения видов.
В субандской и западной провинциях особенно приметны большие,
имеющие полусферическую форму подушки колючего и безлистного
12-0434
178
Глава 3
Рис. 134. Полусферические колючие подушки Mulinum spinosum (зонтичные}
в патагонской полупустыне. Белеют стволы погибшего в результате пожара Her-
berts heterophylla. Дерево справа — Nothofagus antarctica (фото Дузена).
Mulinum spinosum (зонтичные) (рис. 134). Распространению этого расте-
ния способствует интенсивный выпас овец, единственных домашних
животных в Патагонии. Таким же путем распространяется Senecio filagi-
noides. Чем суше становится климат по мере движения к востоку, тем
большую площадь начинает занимать Nassauvia glomerulosa (сложно-
цветные) совместно с другими видами. Но постепенно Nassauvia мельчает,,
степень покрытия почвы этим видом сокращается.
Часто почва обнажена на 60—70%. Преобладает невысокая (15—
20 см) Nassauvia (рис. 135), к ней^присоединяются виды Chuquiraga (слож-
ноцветные), Grindelia (сложноцветные) и Senecio, одиночные растения
Corinabutilon bicolor (мальвовые), Verbena ligustrina, Ephedra, Fabiana
(паслёновые) и др., достигающие 1 м высоты. Широко распространены
также Adesmia campestris (бобовые) и несколько видов Асаепа (розоцвет-
ные). На влажных участках выделяются большие плоские подушки
Azorella (зонтичные). Подушечные формы образуют также некоторые
сложноцветные, вербеновые или Cruckshanskia glacialis (мареновые). Такая
форма роста, по-видимому, образовалась как приспособление к существо-
ванию в климате с низкими температурами и постоянными сильными
ветрами (рис. 136).
Встречаются в Патагонии и миниатюрные шарообразные кактусы.
Из них наиболее распространены Maihuenia patagonica и Austrocactus*
последний сплошь покрывает освещаемые солнцем скалистые стены.
В северной Патагонии, где климат суше и теплее, снова появляются
высокие древесные растения. Прежде всего бросается в глаза Prosopis:
denudans, достигающий приблизительно высоты 2 м, затем поселяется
первый из видов Larrea — L. nitida, отмечающий начало собственна
полупустыни с Larrea, в растительном покрове которой доминируют
Злаковники умеренной климатической зоны южного полушария
179
Рис. 135. Патагонская полупустыня с Nassauvia glomerulosa, Verbena ligustrina,
Senecio, Stipa humilis. На заднем плане, справа,— кусты Prosopis. Район Ин-
хеньеро-Якобаччп, провинция Рио-Негро (фото Э. Вальтер).
Рис. 136. Патагонская полупустыня с растениями-подушками (Chuquiraga
aured) в окрестностях Рио-Чико, прогпнция Рио-Негро (фото Э. Вальтер).
12*
180
Глава 3
кустарники. Стало быть, древесные растения отнюдь не являются абсо-
лютным показателем гумидного климата. Отсутствие Larrea на юге области
объясняется неблагоприятными для ее роста слишком низкими темпера-
турами, здесь патагонская растительность покрывает все пространство
между цепью Анд и береговой линией Атлантического океана.
Растительность Огненной Земли обогащена злаками, доминирует
Festuca gracillima. Обширные болота (образованные преимущественно
осоковыми, но также Sphagnum, Drosera и Empetrum rubrum) распола-
гаются уже в области с холодным гумидным климатом. Этот тип расти-
тельности, как и тип растительности Фолклендских (Мальвинских) и дру-
гих островов Антарктики, уже можно рассматривать как особый вид
тундры (ср. гл. 1).
Из приведенного обзора явствует, что преобладающая часть терри-
тории Патагонии относится к полупустыне. Лишь небольшой район ее
можно было бы назвать злаковником и тем самым «патагонской степью».
Это узкая, вытянутая вдоль меридиана полоса у подножия Анд, которая
служит переходной зоной от альпийского пояса к нагорью Анд и непре-
рывность которой прерывается только между озерами Буэнос-Айрес и Лаго-
Архентино. Лишь на крайнем юге, южнее Ла-Эсперансы, злаковник зани-
мает всю территорию от Анд до Атлантического океана и простирается
до северных выступов лесов, образованных Nothofagus (см. рис. 114).
ТУССОКОВЫЙ ЗЛАКОВНИК НОВОЙ ЗЕЛАНДИИ
Климат Новой Зеландии на севере умеренно теплый, на юге — уме-
ренно холодный и влажный океанический. Поэтому первоначально лес
КРАЙСТЧЕРЧ (7м) 11,3°656
J30] . /1ГМ
Рис. 137. Климадиаграмма Александры (Отаго) и для сравне-
ния климадиаграмма Крайстчёрча.
Александра — аридная часть ландшафта котловин на острове Южный,
Новая Зеландия; Крайстчёрч — восточное побережье с едва намечающим-
ся сухим периодом.
покрывал оба острова. Лишь часть острова Южный, расположенная в дож-
девой тени Новозеландских Альп и обладающая семиаридным климатом,
и в прошлом представляла собой естественный злаковник. Эта область
лежит приблизительно между 43° и 45° ю. ш. (широта средней Патагонии)
и испытывает влияние постоянных западных ветров. На своем пути они
встречают горную преграду высотой более 3000 м и на восточной стороне
гор нисходят в виде сухого фёна. Сумма осадков почти во всей Новой
Зеландии превышает 1000 мм. На западной стороне гор она увеличи-
вается до 4000 мм, сокращаясь в ветровой тени гор до 500—300 мм (Алек-
сандра — 330 мм), однако лето здесь сухое (рис. 137). Злаковник зани-
Злаковники умеренной климатической зоны южного полушария
181
Рис. 138. Туссоковый злаковник на острове Южный, Новая Зеландия, высота 600 м
над уровнем моря. Основной травянистый вид здесь Festuca novae-zelandicae. В центре
(темные пятна) — кустарники Hebe, Coprosma, Discaria; в правой части снимка, ввер-
ху,— начало леса из Nothofagus (фото Симпсона и Скотт-Томпсона).
мает Кентерберийские равнины за Крайстчерчем, которыев настоящее время
большей частью превращены в пашни или сеяные луга, и предгорные кот-
ловинные ландшафты, как, например, котловины Маккензи, северного
и южного Отаго.
Речь идет о типичном туссоковом злаковнике, причем различают
низкий (30—50 см) злаковник на низменностях (рис. 138) и высокий (более
1,5 м) злаковник на возвышенных участках (в горах он носит название
«снежного» туссока) х.
Низкий туссоковый злаковник представляет собой зональную расти-
тельность семиаридного климата. Годовой сумме осадков 500 мм про-
тивостоит испаряемость, равная 1000 мм. Поэтому лес расти здесь не
может. Доминируют злаки Festuca novae-zelandiae и Роа caespitosa; прежде,
очевидно, был широко распространен дерновинный злак Agropyrum scab-
rum s.l. Верхушки листьев отдельных растений травостоя «туесок» почти
соприкасаются друг с другом, поэтому злаковник производит впечатление
сомкнутого покрова. Однако между базальными частями дерновин оста-
ются свободными значительные промежутки, которые частично заполняют
низкорослые, более нежные виды. В окраинной полосе области, особенно
на каменистых почвах, в злаковник внедряются такие кустарники, как
Discaria toumatoa (крушиновые), Carmichaelia (бобовые) и своеобразная,
напоминающая юкку Aciphylla (зонтичные).
1 L. Cockayne, The vegetation of New Zealand, Weinheim, 1958; V. D. Z о t о v,
Survey of the Tussock-grasslands of the South Island, New Zealand, «New Zeal. J. Sei.
and Technol.», 20, Wellington, 1947.
182
Глава 3
Первоначально злаковник, по-видимому, занимал более низкие пози-
ции (до 600 м над уровнем моря); над ним по мере увеличения количества
осадков и уменьшения испаряемости развивалась в качестве переходной
зоны кустарниковая стадия, а еще выше — лес, образованный Nothofagus.
В целях расширения пастбищных угодий лес неоднократно выжигали, что
способствовало распространению туссокового злаковника. Густо развет-
вленная, проникающая на глубину более 1 м корневая система придает
злакам большую конкурентную мощность. Ныне лишь небольшие рощи
леса во влажных ущельях или многочисленные остатки пней свидетель-
ствуют о существовании здесь в прошлом крупных массивов лесов. Пока-
зателем лесного климата является также орляк (Pteridium esculentum),
избегающий аридных районов. Туссоковые злаки — жесткие и хорошим
кормом служить не могут, особенно если на растениях сохраняются старые
листья. Поэтому фермеры каждый год выжигают весь травостой. Появ-
ляющиеся весной молодые листья злаков овцы охотно поедают. Однако
пожары и последующий выпас скота вредят туссоковым злакам, и они
со временем исчезают. В злаковнике образуются пролысины, которые
занимают европейские виды, главным образом Agrostis tenuis, Anthoxan-
thum odoratum, Holcus lanatus, но также Rumex acetosella, Hypochoeris
radicata и др. При постоянном выжигании и интенсивном выпасе происхо-
дит полная деградация пастбища и начинается сильная эрозия почвы х.
Таким путем наиболее засушливые районы Отаго превратились со вре-
менем в настоящую пустыню, почти лишенную растительности. Типичнйй
для сильно деградированных травостоев вид — Raoulia lutescens (scab-
weed) — образует широкие, до 1 л в диаметре, плоские плотные подушки,
напоминающие гигантские колонии плесневых грибов. Как пастбище
такие участки не представляют ценности, но распространение Raoulia
по крайней мере предохраняет почву от эрозии.
Естественная область распространения высокого туссокового злаков-
ника в горах начинается лишь выше границы леса на глинистых, слабо-
заболоченных почвах, развитых на граувакках и слюдяных сланцах.
Высота травостоя 1,5 ж (до 2 ж). Важнейшими видами являются Chionochloa
(= Danthonid) rigida, Ch. flavescens, Festuca matthewsii, а также Роа colensoi
и различные альпийские виды. В зависимости от экспозиции склонов
злаковник появляется на высотах от 750 до 2000 м. Зимой здесь уже
довольно холодно, поэтому снег обычно лежит 2—3 месяца. В наиболее
засушливых горах низкий туссоковый злаковник непосредственно пере-
ходит в высокозлаковый, причем даже там, где лес был уничтожен путем
раскорчевки, пожаров или пастьбы.
Высокозлаковые туссоковые злаковники как пастбища для овец
совершенно не пригодны. Поэтому их также выжигают, чтобы создать
летние пастбища для скота; в результате высокозлаковый злаковник
сменяется низкозлаковым. Таким образом, в настоящее время последний
может быть встречен не только на месте прежних лесов, но и там, где
раньше была распространена формация «туесок», образуемая высокими
злаками.
На острове Северный высокий туссоковый злаковник также занимает
обширные площади. Здесь он широко развит в альпийском поясе гор,
например на влажном вулканическом плоскогорье у подножия вулканов
1 Развитию эрозии способствовали также кролики, завезенные на Новую Зелан-
дию из Европы и быстро размножившиеся здесь. Ныне эта угроза ликвидирована
(см.: W. Е. Howard, The rabbit problem in New Zealand, «DSIR-Inform. Ser.»,
№ 16, 1958).
Злаковники умеренной климатической зоны южного полушария
183
Руапеху и Нгаурухоэ, на высоте 950 м над уровнем моря, где злаковник
покрывает мощные отложения пемзы и вулканического пепла. Развитие
-его здесь, видимо, обусловлено не климатом. Скорее всего, это переходная
стадия в процессе нового заселения растениями изверженного вулканиче-
ского материала. Лес очень медленно отвоевывает занятую им прежде
территорию. Высаженные лиственницы (Larix) быстро приспосабливаются
к новым для них условиям среды. Широко распространились в злаковнике
.адвентивные виды Calluna vulgaris и Erica lusitanica, на выжигаемых
площадях поселяется кустарник Leptospermum scoparium (миртовые),
а на влажных участках — Phormium tenax.
В противоположность низкому туссоковому злаковнику злаковник
•с высокорослыми видами относится не к умеренной степной зоне, а к впол-
не гумидным злаковникам альпийского пояса, которые южнее переходят
в субантарктические туссоковые кочкарники. Следует упомянуть, что
Роа caespitosa s.l. образует чистые травостои также в Австралии, в Снеж-
ных горах, где занимает ровные часто переувлажненные или подвержен-
ные заморозкам во все месяцы года местообитания и где условия для роста
видов эвкалиптов оказываются уже неблагоприятными.
Эколого-физиологические исследования этих злаковников не произ-
водились, но имеются фитоценологические работы, связанные с проблемой
рационального использования пастбищ А В естественных условиях в Но-
вой Зеландии паслись только бегающие птицы моа, которые довольство-
вались тем, что отщипывали листочки злаков у их основания, и которые
позже были полностью истреблены человеком.
ОГОНЬ КАК ЕСТЕСТВЕННЫЙ ЭКОЛОГИЧЕСКИЙ ФАКТОР
Почти во всех разделах этой книги было показано значение огня
как фактора формирования растительности. Лишь в гумидной области
лиственных лесов, в арктической зоне или альпийском поясе огонь как
природный фактор не играет какой-либо роли. Так, широкое распростра-
нение высокозлаковой прерии на восток, в пределы лесной зоны, обу-
словлено частыми пожарами в прерии, которую индейцы поджигали еще
до заселения ее белыми людьми. Проблема пампы и туссокового злаковни-
ка в Отаго также не может быть понята без того, чтобы не проанализиро-
вать воздействие огня на растительность, причем последний рассматри-
вался преимущественно как антропогенный фактор. Поэтому в заключение
небезынтересным будем обсудить, насколько такое представление является
правомерным, поскольку известно, что лесные и степные пожары могут
возникнуть и естественным путем в результате удара молнии. На первый
1 А. Р. Barker, An ecological study of tussock grassland, Hunters Hills, South
Canterbury, «DSIR Res. Bull.», 107, Wellington, 1953; Tussock Grassl. Res. Comm.:
The high-altitude snow-tussock grassland in South Island, New Zealand, «New Zeal. J.
Sci. a. Technol.», Sect. A, 36 (4), 1954, p. 335—364; L.B. Moore, Some Rumex acetosella
communities in New Zealand, «Vegetatio», 5/6, 1954, p. 268—278; A. F. M ar k, Grass-
land and shrubland on Maungatua, Otago, «New Zeal., J. Sci. a. Technol.», Sect. A, 37,
1955, p. 349—366; H. M. Drummond, E. H. L e a t h a m, Bibliography of New
Zealand tussock grasslands, «DSIR Res. Bull.», 132, Wellington, 1959; H. E. С о n n о r,
A tall-tussock grassland community in New Zealand, «New Zeal. J. Sci. a. Technol.»,
Sect. A, 43, 1961, p. 825—835; Tussock grassland communities in the Mackenzie Country,
South Canterbury, New Zealand, «New Zeal. J. Bot.», 2, 1964, p. 325—351; Tussock
grasslands in the Middle Rakaia Valley, Canterbury, New Zealand, «New Zeal. J. Bot.»,
3, 1965, p. 261—276.
184
Глава 3
взгляд, однако, кажется, что число таких пожаров в сравнении с количе-
ством палов, искусственно пускаемых человеком для выжигания сухой
травы, неизмеримо мало, чтобы приписывать им сколь-либо существенное
значение. Статистика в общем подтверждает такой взгляд. В одной из ста-
рых русских статистических сводок (см. т. II, табл. 62) упоминается,
например, что из 1535 лесных пожаров, которые возникли в южной части
зоны хвойных лесов, только 2% можно было связать с ударом молнии.
Однако при такого рода рассуждениях не учитывают, что регулярно
устраиваемые человеком палы, например в злаковниках, сильно умень-
шают возможность поджога травостоя молнией, так как там, где палы
пускаются ежегодно, накапливается слишком мало легковоспламеняю-
щегося сухого опада. В сельскохозяйственных же угодьях пожар, вызван-
ный молнией, обычно вообще не находит условий для широкого распро-
странения.
По-другому обстояло дело в естественном, не тронутом человеком
ландшафте. Вызванные молнией пожары, вероятно, и не были столь
частыми, как искусственные пожары в наше время, но они легко охваты-
вали обширные пространства и бушевали до тех пор, пока наконец не
угасали сами по себе. О роли естественных пожаров свидетельствуют
упомянутые выше (см. т. II, гл. 5) исследования сосны в заповеднике
Муддус (Швеция). Возможно, еще более убедительным является суще-
ствование иирофитов в Австралии, развитие которых в геологическом
прошлом без влияния пожара как фактора отбора трудно себе представить.
С другой стороны, отсутствие типичных пирофитов в других зонах расти-
тельности позволяет, скорее, отрицать значение огня как естественного
экологического фактора. Для решения этого вопроса особый интерес
имеет оценка статистических данных о пожарах в государственных лесах
США, произведенная Комареком г. Автор доказывает, что пожары, вызван-
ные ударом молнии, представляют весьма частое явление во всех зонах
растительности Северной Америки, за исключением чистых лиственных
лесов. Интерпретировались главным образом сведения, относящиеся
к лету 1965 г.
Летом в Северной Америке часто формируются резко выраженные
фронты между поступающими с севера полярными холодными воздушными
массами и движущимся с юга морским тропическим теплым воздухом.
Прохождение фронта сопровождается обычно сильными грозами. Один
такой фронт простирался 1—5 мая 1965 г. от Южной Дакоты до Филадель-
фии. В последующие дни он переместился к югу и 15 мая достиг Флориды.
За эти несколько дней в штатах Южная Дакота и Небраска в 9 лесных
округах были зарегистрированы пожары, возникшие вследствие удара
молнии; в Нью-Мехико зарегистрированы пожары в одном, в Аппалачах —
в 13, в районе Великих озер — в одном и во Флориде — в 13 лесных окру-
гах, всего 37 пожаров. При этом речь идет только о бесспорных случаях,
когда удар молнии и поднявшийся вслед за тем столб дыма наблюдались
непосредственно.
Особенно подвержены пожарам лесонасаждения в области песчаных
дюн штата Небраска, окруженной естественной высокозлаковой прерией.
Пожары в прерии столь часты, что это вызывает необходимость организа-
ции здесь сторожевых постов. Пожар в прерии, начавшийся 6 мая 1965 г.
1 Е. V. К от ar ek, The meteorological basis for fire ecology, Proceed. 5. Ann.
Tall Timber Fire Ecol. Confer., Tallahassee (Florida). 1966, p. 85—125; H. Walter,
Das Feuer als natiirlicher klimatischer Faktor, Soyrinki-Festschr., Aquilo. Ser. Botanica,
Oulu, Finnland, 1967.
Злаковники умеренной, климатической зоны южного полушария
185
из-за удара молнии, перекинулся на лесопосадки, быстро распростра-
нился на 20 км к западу и после того, как ветер переменился, 20-километ-
ровым фронтом двинулся на север, пока наконец не заглох у края сильно
стравленной пастбищной территории. По сообщению Службы леса, еже-
годно в прерии возникают в среднем 7 пожаров, связанных с прохождением
грозы; в одном году отмечено даже 28 случаев. Если сопоставить количе-
ство естественных пожаров с общей контролируемой площадью, то ока-
жется, что один пожар приходится на 5000 га. Таким образом, нет никакого
сомнения в том, что пожары в прерии еще до поселения здесь человека
представляли собой весьма распространенное явление. В настоящее время
в области прерий, преобразованной в сельскохозяйственные угодья, есте-
ственные пожары очень редки.
В лесах Аппалачей пожары, вызванные ударом молнии, также слу-
чаются достаточно часто. В Национальном парке Мононгахела (западная
Виргиния) с этой причиной связано 2 пожара из 30. Отсюда понятно
широкое распространение сосновых лесов в восточной части США.
Описанные выше погодные условия, сложившиеся летом 1965 г.,
следует считать для Северной Америки весьма типичными. Они могут
повторяться, ежегодно, несколько раз в течение года.
Особое значение имеет тот факт, что грозовые фронты формируются
и в аридных западных штатах Америки, когда теплые воздушные массы
устремляются из Мексиканского залива в горную местность и над хреб-
тами сталкиваются с холодным воздухом; при этом образуются более
локальные грозовые фронты. Летние дожди в Аризоне связаны именно
с ними. За время с 7 по 16 июля 1965 г. в Аризоне и Нью-Мехико зареги-
стрировано 536 пожаров, причиной которых был удар молнии.
Для растительности типа чаппараль важно то обстоятельство, что
такие грозы бывают и в Калифорнии, чаще в годы с очень сухим летом,
а также на пространстве севернее Калифорнии, вплоть до канадской
границы. В июле 1965 г. из многих лесных округов поступили сообщения
о более чем 5, а из некоторых даже о 28 пожарах, вызванных ударом
молнии.
14 августа в округе Элко (Невада) во время грозы возникло 30 пожа-
ров, которые, объединившись, образовали 5 обширных очагов пожара.
Причиненный ими ущерб оценивается в 1 млн. долларов.
В Национальном парке Титон (Вайоминг) в 1965 г. благодаря много-
снежной предшествовавшей зиме лесных пожаров не наблюдалось, но,
судя по средним многолетним данным, ежегодно здесь происходит 10 пожа-
ров, связанных с прохождением грозового фронта.
Аналогичные сведения приведены Комареком для всех районов
США, как для резко аридных, с годовой суммой осадков 200 мм, так и для
гумидных — с годовыми осадками 2500 мм. При этом нужно иметь в виду,
что национальные парки занимают лишь небольшую часть всей площади
страны.
Статистический учет, ведущийся Службой леса, весьма убедительно
показывает, что для хвойно-смешанных лесов, злаковников и жестко-
листной растительности, а также полупустынь Северной Америки огонь
следует рассматривать как вполне естественный климатический фактор.
В Калифорнии путем подсчета годовых колец у старых деревьев, повреж-
денных пожаром, удалось установить, что уже в XVI в. в лесах Сьерра-
Невады в среднем пожары возникали каждые 8 лет.
Следовательно, можно предположить, что и без участия человека
характер растительности наряду с прочими факторами среды определяется
186
Глава 3
также огнем. Поэтому искусственное предотвращение пожаров в запо-
ведниках нужно рассматривать, скорее, как нежелательное, противо-
естественное вмешательство *.
В заключение Комарек отмечает, что огонь играет известную роль
и в жизни диких животных. Зональную растительность (климаксное сооб-
щество) часто, и не без основания, считают «биологической пустыней».
Дикие животные находят достаточно корма только на участках с более
открытыми после пожара стадиями сукцессии.
Было бы весьма желательно провести аналогичные исследования
на заповедных территориях Южной Америки, Африки и Австралии.
Для 1955—1959 гг. лесной департамент (Претория) опубликовал
.достаточно точные данные, которые относятся к территории, охватываю-
щей 5% всей площади Южной Африки. Число пожаров, вызванных ударом
молнии (в скобках — процент от всех пожаров), равно: в 1955 — 10 (6,3),
1956 - 27 (11,7), 1957 - 35 (16,9), 1958 - 22 (13,2) и в 1959 - 36 (13,5) 1 2.
Особенно интересно было узнать, имеет ли значение молния как
причина пожара в области зимних дождей Капской провинции, где развита
растительность «финбош». Статистическими данными по этой провинции
мы не располагаем, но имеется письменное сообщение Боуза из Универси-
тета Вестерн-Кейп о том, что в горах Цедер (севернее Кейптауна) за послед-
ние 18 лет отмечено 14 пожаров, вызванных ударом молнии.
Таким образом, можно констатировать, что огонь представляет
собой естественный экологический фактор, в существенной мере опреде-
ливший и способствовавший распространению многих типов раститель-
ности, например жестколистной растительности в областях с преоблада-
нием зимних дождевых осадков, а также злаковников как в умеренной
и субтропической зонах, так и спорадически в тропиках. Регулярно
пускавшиеся человеком палы стали причиной дальнейшего распростра-
нения этих типов растительности за счет неустойчивых к пожарам лесных'
формаций. Однако было бы неправильно считать эти типы растительности
обусловленными лишь антропогенным фактором. Несомненно, естествен-
ные пожары и без вмешательства человека в известной степени всегда
благоприятствовали замене зональной растительности другими раститель-
ными группировками благодаря их «пирофильности», то есть способности
благоприятно реагировать на пожары. Пример тому — различные типы
лесов, образованных Plnus, или заросли жестколистных кустарников.
Поэтому прежде, чем решить вопрос о первичности ландшафта злаковников
или жестколистного кустарника, надо непременно изучить интенсивность
и длительность воздействия на растительность естественных пожаров.
1 W. Е. Wagner, Past fire incidents in Sierra Nevada forests, «J. Forestry»,
59 (10), 1961, p. 739—748.
2D.J. B. Killick. Anaccount of the plant ecology of the Cathedral Peak area
of Natal Drakensberg, «Bot. Surv. S-Afrika», № 34, Pretoria, 1963. Автор описывает
пожар, вызванный камнепадом. Драконовы горы сложены чрезвычайно твердыми
кварцитами, часто образующими отвесные стены. Обломок породы, упавший с крутого
склона, с большой силой ударяется о лежащие внизу кварцитовые глыбы, высекая
сноп искр, которые легко могут поджечь сухую траву.
Глава 4
АРИДНЫЕ ОБЛАСТИ ЦЕНТРАЛЬНОЙ АЗИИ
ОБЩИЙ ОБЗОР
Р и с. 139. Области пустынь умеренной клима-
тической зоны Азии (по Петрову).
Среднеазиатские пустыни: 1 — Ирано-Туранская (климат
региона субтропический, континентальный); II — Джун-
гаро-Казахстанская. Центральноазиатские пустыни:
III — собственно центральноазиатские (жаркие) и
IV — Тибетские высокогорные (холодные).
Афро-Азиатская аридная зона, простирающаяся от Атлантического
побережья на западе почти до Тихого океана на востоке (см. т. II,
рис. 5) и охватывающая большую часть территории аридных районов
земного шара, может быть подразделена на сладующие регионы 1 (ср.
рис. 139):
1)Судано-Синдский пояс, к которому относятся зона
Сахель южнее собственно Сахары, береговая полоса Аравийского моря
и пустыня Тар (Синд) на се-
веро-западе Индии;
2) Сахаро - Ара-
вийская область пу-
стынь, совершенно лишен-
ная осадков или лишь с не-
значительными осадками зи-
мой;
3) Ирано-Туран-
с к а я область .пу-
стынь, охватывающая на-
горную равнину северного
Ирана и южную часть Ара-
ло-Каспийской низменности;
4) Джунгаро-Ка-
захстанская область
полупустынь и пу-
стынь — переходная зона
к восточноевропейско-южносибирским степям, прослеживаемая от При-
каспийской низменности на западе до озера Балхаш на востоке;
5) Центральноазиатская область пустынь,
охватывающая территории Монголии и Северного Китая;
6) Высокогорные пустыни Тибета, западную грани-
цу которых образуют Гималаи и Памирское нагорье,— ярко выраженный
тип холодной пустыни, развитой на высоте около 4000 м.
В пределах Судано-Синдского пояса еще выпадают тропические лет-
ние дожди, хотя интенсивность их здесь весьма незначительна. Соответ-
ственно этому флора Тара (Синда), как и Судана, состоит из палеотропи-
ческих элементов, например из видов родов Acacia, Grewia, Salvadora,
1 М. П. Петров, Сравнительная характеристика ландшафтов пустынь Азии
и Северной Африки, «Вестн. ЛГУ, серия геол, и геог.» № 6, 1965, стр. 84—93; Types
de desert de 1’Asie centrale, «Ann. de Geogr.», 1963, p. 131—155; E. M. Л а врепко,
Основные черты ботанической географии пустынь Евразии и Северной Африки,
М,—Л., 1962.
188 Глава 4
Commiphora, Calotropis, Aerva, Crotalaria, Panicum, Eleusine, Pennisetum,
Aristida и др. Напротив, в Сахаро-Аравийской области пустынь уже пре-
обладают мезогенные элементы, которые возникли в пределах океана
Тетис еще до мощных складкообразований третичного периода и в настоя-
щее время распространены на всем пространстве от Атлантического океана
до Гималаев. Особенно много представителей этих элементов можно встре-
тить среди крестоцветных и гвоздичных. Мезогенные элементы служили
исходными формами, из которых развились следующие флоры: 1) сре-
диземноморская, заселившая область современного Средизем-
номорья от Атлантического океана до южной Анатолии и Ливана; 2) и р а -
но-туранская, представленная в Иране и Средней Азии; 3) с а -
харо-аравийская.
Во всей области господствует средиземноморский тип климата с зим-
ними дождями, обильными, однако, только в пределах собственно Сре-
диземноморья; к востоку количество атмосферных осадков убывает.
Судано-Синдская и Сахаро-Аравийская области в климатическом
отношении входят в зону субтропиков и рассмотрены в т. I. Остальные
чисто азиатские области пустынь уже отличаются холодной зимой, что
дает основание относить их к зоне умеренного климата.
Переход от субтропических пустынь к пустыням умеренной зоны
осуществляется постепенно, поэтому провести между ними резкую границу
не представляется возможным. Ближе всего, по мнению автора, к ней
подходит граница промышленной культуры финиковой пальмы (Phoenix
dactyliferd), плоды которой служат основой питания жителей субтропи-
ческих пустынь. Финиковая пальма без ущерба для себя переносит кратко-
временные понижения температуры ниже нуля, но страдает от более
длительных заморозков или резких, ежегодно повторяющихся прорывов
холода.
Граница возделывания финиковой пальмы пересекает под углом
Месопотамско-Иранской регион, причем большую, северную, часть Иран-
ского нагорья уже следует относить к Ирано-Туранской области. Послед-
няя характеризуется относительно мягкими зимами и служит переходной
зоной к области пустынь Казахстана, климат которой имеет четко выра-
женное холодное время года. Туранскую часть региона (юг Арало-Кас-
пийской низменности) вместе с территорией Казахстана в литературе
на русском языке принято называть Средней Азией в отличие от Централь-
ной Азии, расположенной восточнее. Казахстан простирается на восток
до предгорий Алтая, гор Тарбагатая и Джунгарского Алатау. Южнее
последнего узкая, но открытая полоса ведет в Джунгарию и далее в Цен-
тральную Азию. В Джунгарии еще отчетливо проявляется воздействие
циклонов североатлантического происхождения, что позволяет говорить
о наличии флористических связей ее со Средней Азией. Таким образом,
наблюдаемая часто тенденция объединять Джунгарию с Казахстаном имеет
под собой реальную почву (рис. 139). На остальной же части территории
Центральной Азии выпадение осадков связано с приходом восточноазиат-
ского муссона; выпадают осадки преимущественно летом, годовая сумма
их в восточном направлении постепенно возрастает. В свою очередь соб-
ственно Центральноазиатская область пустынь имеет более мелкие под-
разделения (рис. 140): 1) восточнее Джунгарии и южнее восточного отрезка
Монгольского Алтая начинается пустыня Гоби, занимающая большую
часть Монголии; 2) на юго-востоке, в громадной излучине Хуанхэ, лежит
область Ордос. Ее восточная, более богатая осадками часть имеет уже
степной характер, а западная представляет собой пустыню; 3) к западному
Аридные области Центральной Азии
189
Рис. 140. Обзорная карта Центральноазиатской пустынной области (по Петрову, с изменениями).
190
Глава 4
краю Ордоса прилегает песчаная пустыня Алашань, а дальше, до озера
Лобнор, простирается пустыня Бэйшань; 4) Таримскую котловину, или
Кашгарию, где выпадает очень мало осадков, занимает пустыня Такла-
Макан; 5) наконец, высоко приподнятый бассейн Цайдам ведет в обширную
область высокогорных пустынь Тибета.
Все физико-географические области пустынной зоны Азиатского
материка имеют значительные отличия в климате и растительности.
Некоторые данные о климате пустынь Азии приведены в табл. 35.
Таблица 35
Климат пустынь внутренней Азии (по Петрову)
Станция Высота над уровнем моря, м Температура воздуха, °C Сумма осадков за год, мм
январь июль средняя за год
Туранская низ- менность Иранское наго- рье .... Казахстан . . Джунгария 22—242 516—1773 —22—396 318—903 —6,9 до 4,8 1,0 до 6,0 —13,6 до —6,0 —22,0 до —15,2 27,1—32,3 27,9—33,0 25,1—28,6 23,9—25,4 11,0—17,1 15,4—20,0 7,5—12,1 4,3—6,8 82—244 76—250 103—184 96—247
Пустыня Гоби Алашань . . Бэйшань . . 991 1240—1563 1122—1776 — 18,1 —11,2 до —6,8 —15,6 до —9,8 23,3 22,6—24,5 21,1—26,3 3,8 7,0—9,6 4,3—9,6 107 39—219 42—74
Такла-Макан Цайдам .... 820—1459 2700—3000 —11,0 до —6,8 —13,1 до —11,8 24,4—27,3 16,7—17,8 9,8—11,6 2,3—3,5 9—63 13-26 -
Переходная область между субтропическими пустынями Африки
и пустынями умеренной климатической зоны Азии занимает большую
часть территории Передней Азии с Сирийской пустыней, Месопотамию,
Иран и крайний юг Афганистана. Растительность этого обширного региона
до настоящего времени еще мало изучена, поэтому нам придется ограни-
читься указаниями на некоторые последние публикации.
Средиземноморская растительность на восточном побережье Среди-
земного .моря в районе Дамаск — Хомс — Халеб (Алеппо) переходит
в степь со Stipa barbata, Роа sinaica, Artemisia sieberi, частично простираю-
щуюся до Евфрата. К полосе степей примыкает Сирийская пустыня, для
которой имеется новая карта ландшафтов А Вся описываемая область
в границах почти до 35° с. ш. относится к поясу субтропических пустынь.
У Пальмиры еще культивируется финиковая пальма, севернее более дли-
тельные заморозки уже препятствуют ее возделыванию, хотя средняя
температура января колеблется здесь в пределах 5—7° выше нуля. Осадки
(100—200 мм в год) выпадают зимой и весной (максимум в январе). В со-
ставе флоры преобладают ирано-туранские элементы: кустарнички (Arte-
misia sieberi, Hammada ramosissima, Noaea mucronata, Salsola rigidd) и эфе-
меры (Роа sinaica, Carex p achy sty lis). На поливных землях возможно зем-
леделие, например на равнинах с лёссовидными или песчаными почвами,.
Р Б. В. Виноградов, Л. Е. Родин, Ландшафты Сирийской пустыни,
«Труды Бот. ин-та АН СССР, серия III (геобот.)», вын. 16, 1964, стр. 301—312 (с картой);
Л. Е. Родин, Пастбища и геоботаническое районирование Сирийской Арабской
Республики, там же, стр. 253—300.
Аридные области Центральной Азии
191
причем на последних растительный покров несколько богаче. В низинах
появляются галофиты (Halocnemum, Frankonia, Aeluropus и др.), распро-
странены также Tamarix, Alhagi и Cynodon, связанные с менее минерализо-
ванными грунтовыми водами. К югу равнина постепенно поднимается
к складчатым горным возвышенностям, достигающим 1000 м над уровнем
моря. Здесь осадков выпадает несколько больше, поэтому уже встре-
чаются разрозненные древовидные кустарники (10—20 экземпляров на 1 га
площади), такие, как Pistacia mutica, Prunus tortuosa, Rhamnus palaestinae.
Расход воды в Евфрате, образующем восточную границу рассматри-
ваемой области, колеблется в пределах 250—4000 лР/сек. Очень широкая
пойма реки занята преимущественно пашней. Сохранились остатки лесов
из Populus euphratica, сомкнутость крон которых не превышает 0,6—0,7.
В старых руслах рек развиты засоленные почвы и барханные пески.
На заброшенных пашнях распространены Alhagi maurorum и A. pseudo-
alhagi, Lagonychium farctum, а также Peganum harmala. Характер расти-
тельности в Месопотамии аналогичен, разница состоит лишь в том, что-
естественная растительность нарушена здесь сильнее. Посадки финиковой
пальмы вдоль крупных рек в Иране прослеживаются до Аны на Евфрата
и севернее Самарры на Тигре, то есть также почти до 35° с. ш. х.
Восточнее начинаются сильно расчлененные горы Загрос высотой
более 4000 м. Количество осадков здесь возрастает. Лишь во внутренних
районах Ирана, представляющих собой ландшафты обширных котловин,
мы снова встречаем пустынные местности с суммой осадков, непревышающей
100 мм в год. Зона побережья на юге полностью входит в область суб-
тропических пустынь. Напротив, внутренние части страны подвержены
зимой прорывам холодных воздушных масс из Арктики, которые наблю-
даются в среднем 9—12 раз за зиму, причем 3—4 из них приносят особенна
сильные холода. Абсолютная минимальная температура колеблется от
-10 до -20°.
Обширная, идеально выровненная центральноиранская пустыня Деш-
те-Кевир лежит на высоте 800 м. Здесь, за исключением участков, совер-
шенно лишенных растительности или покрытых солевой коркой, распро-
странены галофиты Halocnemum strobilaceum, Salsola arbuscula или Halo-
xylon aphyllum, Salcola richteri, виды Tamarix и др. (рис. 141). Южнее
расстилается песчаная пустыня Деште-Лут. На рыхлых песках разви-
ваются виды Caligonum и Haloxylon persicum (рис. 142), на песчано-гравий-
ных грунтах — кустарнички из маревых, например Salsola rigida, Ephedra
strobilacea, Artemisia и др. (рис. 143). На глинистых почвах растительность
более разрежена, преобладают эфемеры, но распространены и Роа bulbosa
и Carex pachystylis', Cousinia представлена несколькими видами, полукустар-
ники растут только поодиночке. Не столь уж редки в Иране лишайни-
ково-водорослевые такыры (ср. стр. 208) 1 2. Эти типы растительности мы
подробнее опишем в разделе, посвященном Средней Азии.
Финиковая пальма, распространенная в Иране в зоне побережья
Персидского залива и Индийского океана, в пределах песчаной пустыни
Деште-Лут заходит далеко на север и достигает приблизительно 34° с. ш. 3
1 С. R е g е 1, Beitrage zur Pflanzengeographie des Iraq, I u. II, «Eeitr. Biol. d.
Pflanzen», 33, 1957, S. 313—324; 35, 1960, S. 1—21; К. К r e e b, Okologische Grundla-
gen der Bewasserungskulturen in den Subtropen mit besonderer Beriicksichtigung des
Vorderen Orients, Stuttgart, 1964.
2 M. П. П e т p о в, Иран, M., 1955; M. Z ohary, On the geobotanical structure
of Iran, «Bull. Res. Counc. of Israel, Sect. D. Bot.», Suppl. 11 D, Jerusalem, 1963.
3 H. В о b e k, Die natiirlichen Walder und Geholzfluren Irans, «Bonner Geogr.
Abh.», H. 8, 1951, S. 1—60.
192
Глава 4
Рис. 141. Окраинная часть солончаковой пустыни восточнее Кума (Иран)
с видами Salsola и тамариском (фото Габриэля).
Рис. 142. Область песков в Хорасане (Иран): кустарник из Haloxylon рег-
sicum', A rtemisia и два заметно выделяющиеся экземпляра высоких зонтич-
ных — Dorema ammoniacum (фото Габриэля).
В Афганистане смешиваются разные типы растительности. Северная
его часть непосредственно продолжает зону среднеазиатских полупустынь
и степей, в южную — вклиниваются иранские пустыни и кустарничковые
полупустыни; на востоке расположен ареал индо-гималайских лесов
и синдских сухих саванн; северо-восточный сектор принадлежит области
высокогорных пустынь Памира, а всю центральную часть страны занимают
степи с аридным горным климатом, напоминающим климат центральной
Анатолии Ч
1 О. Н. Volk, Klima und Pflanzenverbreitung in Afghanistan, «Vegetatio»,
5/6, 1954, S. 422—433; И. А. Линчевский, А. В. Прозоровский, К поз-
нанию растительности Афганистана, «Вот. ж.», 29, № 4, стр. 114—123.
Аридные области Центральной Азии
193
Рис. 143. Песчаная пустыня на высоте 2200 м с единичными деревцами Pista-
cia khinjuk, подушками Acantholimon и Artemisia; местность Серхад, Иран
(фото Габриэля).
Детальный обзор растительности засушливых территорий Азии мы
начнем с характеристики семиаридной степной области северного Казах-
стана, служащей непосредственным продолжением пояса степей Восточ-
ной Европы.
ОБЛАСТЬ СТЕПЕЙ И ПОЛУПУСТЫНЬ КАЗАХСТАНА
Интенсивное заселение территории северного Казахстана началось
лишь в последние десятилетия. Одновременно были начаты и ее экологи-
ческие исследования. Организован ряд стационаров, однако данными их
наблюдений мы еще не располагаем.
Обширную область, образующую переход от сибирских лесов на севе-
ре к среднеазиатским пустыням на юге, поделим для удобства описания
на северную и южную половины. Первая охватывает территорию между
55 и 50° с. ш. и простирается от Урала до предгорьев Алтая. Здесь выде-
ляются три подзоны * 1.
Лесостепная зона
. В узкой полосе, протягивающейся южнее 55° с. ш., условия среды
благоприятствуют росту деревьев. Годовая сумма осадков около 300—
325 мм; средняя температура июля +19°, января —19,5°. Почвы —
выщелоченный чернозем.
Леса занимают здесь 20—30 % (до 50 %) площади и приурочены к пло-
ским водоразделам или низинам. Они состоят из Betula verrucosa и Populus
Н. F. Neubauer, Versuch einer Kennzeichnung der Vegetationsverhaltnisse Afgha-
nistans, «Ann. Naturhist. Mus. Wien», 60, 1954—1955, S. 77—113.
1 Список основных растений северного Казахстана (под ред. Е. М. Лавренко),
«Труды Бот. ин-та им. Комарова, серия III (геобот.)», вып. 13, М.—Л., 1961; М.Г.По -
п о в, Растительный покров Казахстана, М., 1940.
13-0434
194
Глава 4
tremula; высота деревьев 12—15 ж, сомкнутость крон 0,6—0,7. В ярусе
кустарников встречаются Salix cinerea, S. livida, S. xerophila, Prunus
padus, Viburnum opulus, Rhamnus frangula. Кустарник у опушки леса
представлен Rosa cinnamomea, R. acicularis, Spiraea hypericijolia, S. crena-
ta и реже Prunus fruticosus. В ярусе травянистых растений распространены
Brachypodium pinnatum, Calamagrostis lanceolata, Rubus saxatilis. Степи
образуют Stipa pennata, S. rubens, Festuca sulcata, Koeleria gracilis. С ними
произрастают Роа angusttfolia, Helictotrichon schellianum, Agropyrum
repens, Bromus inermis, Calamagrostis epigeios, а из разнотравья Filipendula
hexapetala, Fragaria viridis, Phlomis tuberosa, Onobrychis sibirica и др.
2. Южная лесостепь простирается до 54° с. ш.; годовая сумма осадков
уменьшается здесь до 280—325 мм. Леса с древостоем высотой всего
6—8 м занимают только 10—20% территории и растут лишь в понижениях
рельефа.
В степях 90—95% всех видов составляют виды разнотравья. К пере-
численным выше видам присоединяются Phleum phleoides, Lathyrus tube-
rosus и виды Peucedanum.
Степная зона
Здесь мезофильные виды разнотравья отсутствуют; в качестве новых
элементов появляются Pulsatilla patens и Artemisia sericea. Почвы в север-
ной части — южный чернозем, в южной — каштановые. Количество'
осадков сокращается в пределах этой зоны с 250—300 мм до 200—250 мм
в год. На климадиаграммах, помимо сухого времени года, уже четко выде-
ляется период засухи. Доминируют: Stipa rubens, S. capillata-, кроме того,
распространены S. lessingiana, Festuca sulcata, Koeleria gracilis. Количеств»
разнотравья в южном направлении уменьшается. Встречаются виды
Tulipa и Ornithogalum, а также Salvia stepposa, виды Pyrethrum, Linosyris
и др.; на юге нередко можно заметить Caragana pumila. Обычны засолен-
ные почвы. На песчаных почвах распространены леса, образованные
Pinus silvestris, Equisetum hiemale, Galium boreale, Ramischia (Pirolo) secundar
Chimaphila (Pirolo) umbellata, Monotropa hypopitys и большим количеством
Cladonia.
На более высоких поднятиях, где сумма осадков увеличивается
до 300 мм, снова развита лесостепь с березово-осиновыми лесами.
Зона полупустынь
Это область с сухим климатом. Осадки не достигают и 200 мм’, засуха1
продолжается все лето; средняя температура июля 22 , января —17°.
В северном Казахстане полупустыня занимает небольшую территорию-
лишь на крайнем юге. Здесь распространены комплексы растительности
с видами Artemisia, а на засоленных почвах в низинах — с Lasiagrostis
1 Понятие «полупустыня» в русскую геоботаническую литературу введено
Б. Келлером. Этим термином обозначают область, переходную от злаковников зоны
степи к пустыне с ее обычно уже редкой растительностью. Типичных для полупустыни
элементов не существует, однако для нее характерно сокращение видов злаков и пре-
обладание кустарничков. Характерна микрокомплексность растительного и почвен-
ного покровов. См.: Ф. Я. Левина, К вопросу о зональности и подразделении евро-
пейских полупустынь, «Бот. ж.», 44, № 8, 1959, стр. 1051—1061.
Аридные области Центральной Азии
195
Рис. 144. Lasiagrostis {Stipa} splendens на глинистой почве в районе
озера Зайсан (фото Келлера).
splendens (рис. 144), Aneurolepidium, Psathyrostachys Halimodendron, Ni-
traria, Halocnemum, Kalidium. Эта растительность полупустыни особенно
характерна для южной половины Казахстана. Еще южнее она уступает
место среднеазиатским песчаным пустыням Кызылкум, Муюнкум и др.
Своеобразен характер расселения степного злака Stipa lessingiana.
В зоне сухих степей северного Казахстана этот вид часто встречается
на черноземе и бурых почвах, в более влажной северной части ареала он
распространен только на маломощных карбонатных почвах, развитых
на известняках. По мере увеличения сухости S. lessingiana начинает отда-
вать предпочтение песчаным или каменистым почвам, а в пустыне он
растет лишь во влажных низинах. Эти примеры еще раз подтверждают
правильность закона относительного постоянства местообитаний и смены
биотопов (ср. т. I, стр. 22 и 317).
В Казахстане ковыль обычно вырастает до 70 см в высоту, однако
в сухие годы поднимается над землей едва ли на 40 см. Зона кущения рас-
полагается в почве на глубине 3—4 см. Весной, в середине или в конце
апреля, в дерновинах возникают новые побеги; колошение происходит
в начале мая, а цветение начинается в первых числах июля. В конце
июня почва становится сухой, листья отмирают и растение впадает в состо-
ние покоя, которое длится до 1,5 месяца. В конце августа образуются
осенние листья. Таким образом, вегетационный период продолжается
около 4 месяцев (ср. стр. 117—119).
Выше (см. стр. 140) мы рассмотрели растительные комплексы полу-
пустынь, типичные для северной части Прикаспийской низменности. Позна-
комимся теперь с растительностью, расположенной несколько южнее обла-
сти песчаных пустынь, непосредственно граничащей с Казахстаном и слу-
жащей переходной зоной к пустыням Средней Азии. Этому району посвя-
щена капитальная монография Якубова х.
1 Т. Ф. Якубов, Песчаные пустыни и полупустыни Прикаспийской низмен-
ности. М., 1955; Б. А. Келлер (см. примечание на стр. 141).
13*
196
Глава 4
Прикаспийскую низменность ограничивают: на севере уступ Общего
Сырта, на западе — излучина Волги и возвышенность Ергени; на востоке
граница размыта. Территорию Прикаспия пересекают реки Волга и Урал;
реки Малый и Большой Узень моря не достигают, а теряются в песках
и засоленных блюдцах. Низменность понижается в сторону Каспийского
моря (отметка уровня Каспийского моря —26 м)1.
Вся эта область неоднократно подвергалась трансгрессии Каспийского
моря; воды его трижды заливали Прикаспийскую низменность. Периоди-
чески здесь возникали пресноводные озера. Позднее, когда море отсту-
пило, огромные площади низменности оказались покрыты песками аллю-
виального происхождения. Судя по минеральному составу песков, пески
были принесены речными потоками с Русской равнины и Уральских гор.
В настоящее время пески занимают площадь около 5 млн. га. Климат
на севере низменности полупустынный, годовое количество осадков не пре-
вышает 200—280 мм, причем в отдельные годы их сумма едва достигает
120—150 мм', испаряемость равна 1200 мм. На юге провинции уже рас-
полагается настоящая пустыня. Она полукругом охватывает Каспийское
море и получает в среднем только 140—215 мм осадков в год (минималь-
но 85 мм), тогда как испаряемость увеличивается до 1400 мм и лишь
непосредственно у побережья сокращается до 900—1000 мм (см. рис.
149-151).
Внутригодовое распределение осадков имеет два минимума (февраль—
март и август) и два максимума (июнь и сентябрь, иногда ноябрь — де-
кабрь). Около 60—80% осадков выпадает летом; тем не менее из-за высо-
кой температуры (в июле 25—26°, абсолютный максимум 40—41°) лето
здесь очень сухое. Относительная влажность воздуха в летние месяцы
часто равна лишь 30%, но возможно и 11%. Температура на поверхности
почвы достигает 78°. Зима холодная (средняя температура января от —8
до —9°, абсолютный минимум —40°). Из-за малой толщины снежного
покрова почва промерзает на глубину до 1 м. Наиболее сильно почва
бывает увлажнена весной, особенно если в предшествующую зиму выпало
много снега. Деятельность ветра выражена чрезвычайно сильно. Преобла-
дают восточные и юго-восточные ветры, но летом часто дуют также запад-
ные и северо-западные. Восточные ветры летом обусловливают развитие
пыльных бурь, при которых скорость ветра составляет более 15 м!сек',
взвихрёваемая ветром горячая пыль нагревает воздух до 40°. Энергичная
деятельность ветров вызывает движение песка, причем процессы развева-
ния продолжаются даже зимой.
По характеру эолового рельефа песчаные площади Северного При-
каспия можно подразделить на:
1) ровные песчаные поверхности (ашики);
2) полого бугристые пески с буграми высотой до 2—3 м, редко 5 м',
3) бугристые пески с буграми высотой 6—8 м, реже 10—15 м, между
которыми заключены котловины различной формы и размеров.
1 Уровень Каспийского моря с 1929 г. опустился на 2,5 м, береговая линия пере-
местилась на 25—60 км\ площадь акватории, в 1869 г. составлявшая 420 тыс. кл*2,
сократилась до 370 »л*2. Строительство гидросооружений на Волге и увеличение забора
воды на орошение, видимо, ускорили этот процесс. Обнажившееся между устьями
Волги и Сулака морское дно постепенно заселяется растениями, причем наблюдаются
следующие стадии сукцессии: Phragmites communis -> Puccinellia gigantea ->• Ephemere-
tum. Растительность второй террасы уже приняла степной характер; ср.: К). М. и
Н. В. Мирошниченко, Смены почвенно-растительного покрова в приморской
полосе северо-западного Прикаспия, «Бот. ж.», 51, 1966, стр. 520—541.
Аридные области Центральной Азии
197
Почти полностью лишены растительности грядово-барханные пески.
Барханы обычно располагаются рядами под углом к господствующим
ветрам, причем их наветренный склон пологий (13—15°), а подветрен-
ный — крутой (30—33°). Грядово-барханные пески образуются там, где
рельеф местности до накопления
песка был всхолмлен. Песок пере-
крывает эти неровности часто лишь
слоем толщиной несколько мет-
ров (рис. 145). Максимальная
Рис. 145. Схема образования песчаной
дюны у препятствия (возвышенность), ча-
сто перекрываемого лишь тонким слоем
песка (пунктир).
мощность эоловых песков,
по-видимому, не превышает
7-8 м.
Первоначально все пески, ве-
роятно, были закреплены расти-
тельностью, но со временем по-
стоянно возраставший выпас скота и попытки использовать песчаные
почвы под пашню привели к разрушению растительного покрова и спо*
собствовали развеванию песка.
На карте, составленной в 1840 г., отмечено лишь несколько районов
распространения сыпучих подвижных песков, но уже в 1864 г. они зани-
мали большую площадь. Между песками распространены обширные участ-
ки тяжелых, обычно засоленных почв; в ряде мест на них образовались
соленые озера. Соль в этих озерах имеет различное происхождение. Она
образовалась частью за счет выветривания морских осадочных породг
слагающих значительные площади в пределах Прикаспийской низменно-
сти, частью за счет эродирования соленосных пластов; некоторое коли-
чество солей привносят в озера также мелкие водотоки, питающиеся
источниками из пермских и юрских отложений, которые слагают возвы-
шенность Общий Сырт. Отсюда понятно, почему химический состав во-
ды в озерах весьма различен; в одних преобладают хлориды, в дру-
гих сульфаты. Некоторые озера представляют собой когда-то потеряв-
шие связь с морем лагуны (лиманы), в наши дни постепенно усыхаю-
щие.
Поскольку рельеф Прикаспийской низменности очень ровный, постоян-
ный горизонт грунтовых вод залегает на сравнительно небольшой глу-
бине. Эта вода в общем заметно минерализована. Кроме того, в пределах
распространения песков в различного рода депрессиях грунт нередко
содержит в себе слои пресной воды (рис. 146), см. след. стр.). Оголен-
ные песчаные дюны служат хорошими аккумуляторами влаги, так как
дождевая вода легко просачивается вглубь, за исключением лишь неболь-
шого количества, задерживаемого верхними слоями. Толща песка пред-
охраняет воду от быстрого испарения. В межбарханных понижениях
нередко образуются даже небольшие пресноводные озера, вокруг кото-
рых, подобно тому, как это наблюдается в ложбинах, пересекающих пески,,
растут Populus hybrida, Р. alba, Elaeagnus angustifolia, Salix caspica,
S. rosmarinifolia, реже Alnus glutinosa с подлеском, характерным для пой-
менных лесов. Правда, в большинстве случаев древесные виды представ-
лены только Calligonum aphyllum и видами Tamarix, в юго-восточной части
низменности — древовидным эндемиком Нalimodendron (Astragalus) атто-
dendron. Атмосферные осадки, просачиваясь в песок, часто накапливаются
на небольшой глубине в виде линз, плавающих на минерализованных
грунтовых водах (верховодка) (рис. 147). Все колодцы заложены обычно»
до глубины залегания пресной верховодки. Уровень грунтовых вод резко
198
Глава 4
колеблется в зависимости от количества осадков, выпавших в том или
ином году. Летом уровень снижается, в годы, особенно бедные осадками,
колодцы иногда полностью высыхают.
Рис. 146. Грунтовые воды в песчаном районе Прикаспийской низменности (по Яку-
бову).
Пресная вода накапливается под барханом в слое плотного песка, подстилаемом слоем суглинка,
ниже которого находится горизонт соленой воды. Растительность встречается в местах выхода прес-
ных вод на поверхность. Обычно это наблюдается в днищах часто высыхающих соленых озер.
Наиболее обводнены толщи песка, совершенно лишенные с поверхно-
сти растительности. По мере зарастания песков все большая часть филь-
трующихся в грунт осадков начинает расходоваться растениями на транс-
пирацию, что наглядно иллюстрирует рис. 148. Если в закрепленных
Рис. 147. Схема условий залегания грунтовых вод на, участке между Каспий-
ским морем и заливом Кара-Богаз-Гол (по Еловской).
В песках пресная вода залегает в виде линз непосредственно на поверхности минерали-
зованных грунтовых вод. Цифры означают удельный вес подземных вод, вскрытых скважи-
нами; самым высоким удельным весом обладает вода в заливе (1,160). Его уровень лежит
ниже зеркала Каспийского моря, вследствие чего наблюдается постоянный ток воды из
моря в залив.
растительностью песках содержание воды до глубины 4 м лишь несколько
превышает влажность завядания, то в песках, лишенных растительного
покрова, оно составляет уже 7—9%, а на глубине 4,8 м увеличивается
до 25%, что говорит о полном насыщении почвы влагой. Вот почему при
облесении песчаных пространств необходимо учитывать большой объем
влаги, потребляемой растениями. Насаждения Populus nigra или Salix
acuminata хорошо развиваются только в первые годы жизни, пока запасы
Аридные области Центральной Азии
199
влаги в почве достаточны; но со временем этот резерв иссякает, ибо транс-
пирация растений начинает превышать годовую сумму осадков. Табл. 36
иллюстрирует увеличивающееся иссушение почвы.
Рис. 148. Влажность голых и заросших песков
(по Якубову).
На оси ординат — глубина взятия проб. Вследствие большого
расхода воды на транспирацию запас влаги в заросших песках
на всех глубинах ниже.
Согласно данным таблицы, в течение 4 лет содержание воды в глубо-
ких горизонтах почвы осенью все более сокращалось: на глубине 1,5 м —
с 7,6 до 2,7%, на глубине 2 .и — с 8,9 до 2,9%. Одновременно понизился
уровень грунтовых вод с 2,9 м в 1914 г. до 3,4 м в 1918 г.; количество
Таблица 36
Содержание воды в песчаной почве под насаждением Populus nigra.
(в % сухого веса) (по Томашевскому)
Время отбора проб Глубина отбора проб, см
20 40 60 80 100 150 200
1914 г. 3.IV 9,6 10,5 8,3 8,6 9,0 9,4 9,6
17.IX 2,8 4,6 5,8 6,9 7,8 7,6 8,9
1915 12.IV 5,2 5,7 6,4 7,9 8,6 7,8 9,1
26.Х 7,2 4,3 3,6 3,9 3,8 4,3 7,8
1916 26.III 9,8 10,1 5,8 6,0 5,0 7,0 9,0
10.VIII 2,3 3,2 3,1 3,8 4,1 5,3 6,7
1917 7.IV 6,5 7,3 7,2 7,8 5,6 6,1 7,6
3.IX 1,6 1,8 2,2 2,5 2,6 3,8 4,0
1918 И.IX 5,7 6,5 6,4 5,2 4,0 2,6 3,5
23.Х 1,9 1,9 2,2 2,4 3,5 2,7 2,9
200
Глава 4
растворенных солей увеличилось с 2,6 г/л (1914 г.) до 5,3 г/л (1918 г.).
Ясно, что сплошное облесение песчаных почв в столь аридных условиях
невозможно; в лучшем случае допустима посадка узких ветрозащитных
полос.
Неудивительно поэтому, что лесов здесь нет; растительный покров-
имеет типичный полупустынный облик — господство видов Artemisia.
На песчаных почвах доминирует Artemisia maritima ssp. incana, к которой
в ряде мест примешиваются злаки Agropyrum sibiricum, A. ramosum,.
A. cristatum, Festuca sulcata (beckeri), Koeleria glauca и др. Весной появ-
ляются многочисленные виды эфемеров. Такое сообщество можно рассмат-
ривать как зональную растительность. Его водный баланс по отношению
к количеству влаги, поступившему в почву с осадками, уравновешен.
Там, где растительный покров разрушается, песок обретает подвижность.
Интенсивность развевания зависит от расчлененности рельефа. Особенна
энергично сдувается песок с крутых склонов, расположенных поперек
господствующих ветров. Переносимый ветром песок отлагается на защи-
щенных от ветра участках, где погребает под собой все живое. Места
погибших растений занимает Aristida pennata, которая обладает свойством
снова и снова отрастать из-под засыпавшего ее слоя песка. Только там,
где масса постоянно выдуваемого песка очень велика, образуются лишен-
ные растительности подвижные барханные пески, обычно отделенные
от развеваемого ветром участка склона полосой Aristida. Чем мощнее эро-
дируемый ветром слой песка, тем выше становятся барханы. Они могут
достичь высоты 20 ле, но в конце концов сливаются в барханные гряды
большой протяженности.
Различают несколько стадий формирования подвижных песков: рас-
смотрим их на примере бурых супесчаных почв. В разрезе под 15-санти-
метровым слоем рыхлого песка с тонкой поверхностной корочкой зале-
гает желтовато-бурый пласт более плотного песка толщиной 20—25 см?
его подстилает также песчаный слой, но прочно сцементированный СаСО3.
Стадия I. Пасущийся скот разрушает поверхностную корку,
и песок верхнего слоя развевается. При этом происходит сортировка
материала: пылеватые частицы почвы ветер относит на то или иное рас-
стояние от очагов развевания; более крупные фракции обычно остаются
на месте. Остальная часть выдутой массы сползает или скачкообразна
передвигается ветром; незначительные неровности рельефа или кусты
задерживают эту массу, так что позади них образуются небольшие вытя-
нутые по направлению ветра песчаные косы (первичные дюны). Наконец,
обнажается следующий слой песка, более устойчивый к выдуванию. Злаки
Agropyrum, Eragrostis minor, корни которых при этом обнажились, поги-
бают; высыхают также Artemisia maritima л Kochia prostrata. На невысоких
первичных дюнах поселяются Artemisia scoparia, Salsola ceratocarpus^
Corispermum, Alhagi. Если выпас скота прекратить, то через 3—5 лет расти-
тельный покров сомкнется и возникший благодаря навеванию и развева-
нию песка эоловый рельеф приобретет более мягкие очертания.
Стадия II. Если выпас скота продолжается, то поверхность дефля-
ции увеличивается. Ветер разрыхляет и выдувает второй слой песка, пока
наконец не обнаружится третий прочно сцементированный карбонатами
горизонт почвы. Развевание принимает крупные масштабы. В западинах
и понижениях усиливаются турбулентные движения. В этих условиях
разрушением захватывается и рыхлая песчаная коренная порода. Возни-
кают ландшафты выдутых котловин и навеянных дюн-бугров. В первых
сохраняются устойчивые к выдуванию псаммофиты, на вторых поселяются
Аридные области Центральной Азии
201
виды, которые без ущерба для себя переносят засыпание песком. Если
процессы развевания проявляются особенно резко, то образуются голые
подвижные барханные пески. Ветер переносит песчаные зерна вверх
по наветренному склону до гребня барханной гряды, откуда они скаты-
ваются по крутому подветренному склону. Так постепенно вовлекается
в движение вся гряда, погребающая под собой любое препятствие, встре-
чающееся на ее пути.
Поступление песка прекратится, как только будет достигнут базис
дефляции — песчаный горизонт, увлажненный благодаря капиллярному
подъему грунтовых вод к поверхности, —или будет вскрыт тяжелый глини-
стый слой, не развеваемый ветром. В этом случае на перевеянном песке
рано или поздно снова образуется сомкнутый растительный покров. Про-
цессы естественного зарастания песков протекают стадийно.
В качестве первого растения-пионера на подвижных песках Прикас-
пийской низменности поселяется Elymus giganteus в сочетании с Agrio-
phyllum arenarium (маревые). Затем появляются Salsola ruthenica, S. kali,
Corispermum aralo-caspicum. Все эти виды развивают мощную корневую
систему, простирающуюся в сторону на 5—7 ж, а в глубину — только
на 1 м. В понижениях между грядами закрепляются одиночные экзем-
пляры Heliotropium micranthum, Tournefortia sibirica (бурачниковые),
Chondrilla ambigua, Aristida pennata, Linaria odorata, Melilotus polonicus,
Artemisia scoparia и др. По окраинам барханных гряд можно встретить
отдельные кусты Calligonum и Tamarix.
Благодаря растительности пески уплотняются: в их верхнем гори-
зонте накапливаются органические вещества; содержание воды в почве
вследствие того, что растения интенсивно транспирируют, уменьшается.
Начинается разрастание Artemisia arenaria, пионерные же виды сохра-
няются лишь на гребнях барханов. Вскоре поселяются представители
зональной растительности: Artemisia maritima, Agropyrum sibiricum, Ephe-
dra, Bromus tectorum, Poa bulbosa. В ряде мест образуется моховой покров
из Tortula desertorum. Эта стадия зарастания уже служит переходной
к исходному состоянию. В травостое все более доминирует Artemisia та-
ritima, проникает Agropyrum desertorum, увеличивается количество эфе-
меров, присоединяется Eragrostis minor. Мхи {Tortula с Bryum caespito-
sum), цементируя песок, образуют корку. В процессе зарастания рельеф
песков во все большей степени выполаживается. В конце концов повы-
шения уже не превышают 0,6 м в высоту.
Так протекает сукцессия в зоне пустынь. В зоне полупустынь ход
аналогичен. В качестве растений-пионеров мы находим здесь наряду
с Elymus также Carex colchica или Calamagrostis epigeios и Glycyrrhiza
glabra. На периферии барханов, в котловинах с пресной верховодкой,
вырастают Populus и Elaeagnus. В другом же случае наблюдается переход-
ная стадия с Artemisia arenaria, а затем — несколько более богатое видами
сообщество Artemisia maritima с Euphorbia seguieriana, Jurinea polyclonos,
Achillea gerberi, Centaurea arenaria и др. На слаборазвитых дюнах вторич-
ное зарастание протекает соответственно быстрее.
Там, где навеянный песок покрывает влажную, засоленную глини-
стую почву, поселяются солеустойчивые галофиты. Песок засыпает их
снова и снова, но верхушки побегов спустя некоторое время вновь выхо-
дят на поверхность, так что в конечном итоге все кусты, которые прежде
росли на глинистой поверхности, оказываются на вершине «своей» дюны.
Это виды Tamarix, Nitraria schoberi и Halocnemum strobilaceum. Промежутки
между дюнами заселяют мелкие галофиты и эфемеры. При дальнейшем
202
Глава 4
навевании песка обеспеченность кустов водой ухудшается, и их место
занимают маревые, такие, как Kalidium, A triplex, Anabasis aphylla, Alhagi,
Corispermum и Peganum harmala (парнолистниковые).
ПУСТЫНИ СРЕДНЕЙ АЗИИ
Область пустынь Средней Азии, помимо части территории Казахста-
на, охватывает всю южную половину Арало-Каспийской низменности
и протягивается до подножия высоких гор на востоке. На юг она про-
стирается в Афганистан до Гиндукуша; от Ирана ее отделяет хребет Копет-
даг. Пустыни распространены лишь в наиболее низкорасположенных
частях Средней Азии; в направлении к горам растительность все более
принимает степной характер.
С севера область открыта воздействию полярных воздушных масс.
Поэтому зимой наблюдается в среднем 12—23 прорыва сильных холодов,
Рис. 149—151. Климадиаграммы станций в пустыне.
которые становятся причиной гибели пальм, таких, как Phoenix, Trachy-
carpus и даже Chamaerops, а также вечнозеленых садовых культур. Только
на крайнем юге, под Термезом, температура января в среднем держится
несколько выше 0°. Лето из-за сильной радиации жаркое. Поток радиации
в Ташкенте в декабре равен 5000 кал/см2, в июле — 24 500 кал/см2. Для
горизонтальной поверхности соответствующие значения равны 3860
и 20 190 кал/см2. Суммарная радиация за год достигает 130—140 ккал/см2,
число часов солнечного сияния 2500—3000 (летом около 90% максимально
возможного, зимой — менее 50%) Ч
Осадки в южной зоне среднеазиатских пустынь выпадают преимуще-
ственно зимой (средиземноморский тип), летом дождей совсем не бывает;
северные осадки распределяются по временам года более равномерно.
Годовая сумма осадков — менее 250 мм, однако в ряде мест она сокра-
щается до 100 мм и более (рис. 149—151). С низкой температурой
зимой связана относительно низкая испаряемость, равная 1000—1400
(до 2500 мм), благодаря чему в песчаных пустынях, даже там, где грун-
товые воды залегают глубоко, развивается заметная растительность.
Испарение с поверхности Каспийского моря, имеющего соленость 1,4%
(соленость Мирового океана 3,4%), составляет 986 мм. В заливе Кара-
Богаз-Гол испарение, несмотря на более высокую соленость воды в зали-
ве, повышается до 1100 мм.
В составе флоры Средней Азии преобладают ирано-туранские элемен-
ты. В Средней Азии и Египте насчитывается всего 10% общих видов. Фло-
ра сравнительно молодая, отмечается только плейстоценовый неоэндемизм.
1 См. раздел «Климат» в трудах симпозиума «Средняя Азия», М., 1948 (с картой
растительности).
Аридные области Центральной Азии
203
Рис. 152. Карта Средней Азии (по Коровину).
I — пустыни; сумма осадков менее 250 мм, летом дождей не бывает; 2 — полупустыня; сумма осад-
ков менее 250 мм, но осадки распределяются в течение года более или менее равномерно; 3 — степи;
сумма осадков более 250 .о, зима — самое сухое время года; 4 — то же, но осадки распределяются
по сезонам сравнительно равномерно; 5 — горные степи; сумма осадков более 250 мм, лето сухое;
6 — высокогорный климат.
Граница между континентальной пустынной среднеазиатской зоной
на юге и полупустынной на севере, где все сезоны года получают более
или менее одинаковое количество осадков, проходит приблизительно
по 44° с. ш. (рис. 152) х.
Области пустынь Средней Азии свойственно большое разнообразие
в характере почвообразующего субстрата, что, разумеется, накладывает
отпечаток и на облик растительности. Здесь распространены соленосные
морские образования третичного времени, лёссы ледниковых периодов
и аллювиальные пески, смытые плейстоценовыми реками с гор и отложен-
ные на низменности. Специфичны каменистые гипсофитные пустыни, в ко-
торых роль эдафического фактора особенно активна.
Соответственно могут быть выделены следующие формации расти-
тельности:
а) эфемеровая пустыня на лёссовидных, суглинистых почвах;
б) растительные сообщества каменистой гипсофитной пустыни;
в) формация галофитов на глинистых почвах;
г) псаммофитовая пустыня на песчаных почвах.
1 Е. П. Коровин, Растительность Средней Азии и южного Казахстана, Моск-
ва — Ташкент, 1934 (с картой растительности). См. также: Л. Е. Родин, 1963.
204
Глава 4
Эфемеровая пустыни
Пустыни этого типа распространены преимущественно на подгорных
лёссовых и лёссовых глинистых равнинах, например у подножия Копет-
дага. Развитые здесь суглинистые почвы не засолены и очень плодородны.
Весной они хорошо увлажнены на всю глубину профиля, но к концу мая
полностью высыхают и становятся твердыми, как камень. Поэтому веге-
тационный период весьма короток (с начала марта до середины мая).
В столь напряженных условиях среды развиваются лишь эфемерные виды:
геофиты или однолетники (терофиты).
Обширную площадь занимает эфемеровая пустыня западнее озера
Балхаш, где она известна под названием Голодная степь и подробно иссле-
дована.
Водный режим почвы в пустыне весной характеризует табл. 37.
Таблица 37
Содержание воды в почвах Голодной степи (в %)
Глубина, СЛ1 19.IV 24.IV 3.V 19.V
0—2 7,3 23,3 0,7 0,5
8-9 6,3 18,7 8,4 5,3
20—22 9,2 8,0 10,0 8,2
60 8,7 8,7 9,4 8,6
90 4,8 8,7 8,3 7,1
200 2,4 3,7 4,7 —
Глубина промачивания почвы, естественно, меняется от года к году
в зависимости от обилия осадков. Однако, поскольку в марте и апреле
каждые 4—5 дней идет дождь, верхние слои почвы долгое время остаются
влажными и лишь в конце мая быстро высыхают, когда дожди прекра-
щаются и почва до глубины 10 см нагревается до 30° и выше.
Весной условия для развития растительности весьма благоприятны.
Уже в начале марта, когда температура повышается, отрастают Carex
hostii (вид, близкий С. stenophylla) и Роа bulbosa — два важнейших вида
эфемеровой пустыни. Осока иногда занимает до 75—80% площади либо-
встречается в сообществе с мятликом, иногда же последний доминирует.
Carex hostii — маленькое растение, при оптимальных условиях достигаю-
щее высоты 20 см; листья его короткие и узкие. На глубине 5 см у неге
развивается корневище, годовой прирост которого не превышает 2—8 см,.
но общая длина может достичь 6 м. Отдельные побеги появляются в 3—
5 см один от другого. Многочисленные корни, отходящие от побегов,,
на глубине 5—15 см образуют густо разветвленную сеть, отдельные кор-
ни проникают на глубину 60 см. Роа bulbosa-vivipara также низкорослое-
растение, но образует дерновинки до 2—3 см в поперечнике; побеги с мно-
гочисленными луковичками у основания утолщены. В соцветии вместо-
семян также формируются луковички. Оба типа луковичек даже в гер-
барии через несколько лет после сбора остаются живыми и в сухом состоя-
нии безболезненно переносят высокую температуру.
К названным двум видам присоединяются виды, у которых наблю-
дается образование подземных многолетних органов (клубни, луковицы
или корневища). Это виды родов Bunias, Ranunculus, Haplophyllum (руто-^
Аридные области Центральной Азии
205
вые), Scorzonera, Eranthis, Tulipa, Gagea, Ixiolirion и Geranium tuberosum.
Реже встречаются Gentiana olivieri. В ряде мест можно заметить своеобраз-
ную Ferula foetida. Этот вид, как и другое зонтичное — Dorema aitchisonii,
достигает в высоту 2 м, цветет один раз спустя 6—7 лет после прорастания,
затем плодоносит и отмирает (см. рис. 142). Стержневой корень растений
диаметром 17—29 см проникает только на 20 см в глубину, где разветвляет-
ся; боковые корни вырастают до 2 м в длину и уходят вглубь на 80—
100 см. Корни одного растения пронизывают до 4 м3 почвы. Розетка листьев
покрывает приблизительно 1 м2 поверхности; соцветие шарообразное, дает
многочисленные, дурно пахнущие плоды (всего до 1 кг). Если отрезать
генеративный побег, из корня выступает смолистое вещество, применяемое
в медицине под названием Assa foetida.
Для всех многолетних эфемеров характерно вегетативное размноже-
ние, которое при коротком вегетационном периоде даже в сухие годы
обеспечивает сохранение вида в сообществе. Ranunculus severzovii с листья-
ми, покрытыми белыми волосками, цветет хотя и регулярно, но почти
не производит способных к прорастанию семян, а размножается подзем-
ными корневищами длиной 10 см, на концах которых образуются новые
растения. К этим геофитам присоединяются 40—50 однолетних эфемерных
видов, семена которых созревают за 30—45 дней, как, например, Holosteum
umbellatum, Erophila verna, Erodium cicutarium, виды родов Arenaria,
Papaver, Delphinium, Nigella, Malcolmia, Alyssum, Capsella, Veronica,
а также однолетние злаки из родов Bromus, Phleum, Agropyrum и т. д.
В годы с обильными дождями все эти виды достигают высоты 10—20 см,
в сухие же — либо их семена вообще не всходят, либо растения остаются
карликами высотой 1—2 см. В зависимости от количества и распределения
осадков, а также от температуры в соответствующий период создаются
благоприятные условия для прорастания тех или иных видов, так что
состав флоры однолетников резко меняется от года к году и от места
к месту.
Весной, когда растительный покров полностью сомкнут, пустыня
напоминает луг: пышно развиваются мезофильные растения, лишенные
каких-либо ксероморфных признаков. Но начиная с конца мая и на про-
тяжении 9 месяцев пустыня выглядит совершенно безжизненной. Своеоб-
разным исключением являются два мелких вида: Aphonopleura capillifolia
(зонтичные) и Diarthron vesiculosum (сладкоягодниковые). Цвести они начи-
нают тогда, когда солнце выжжет все живое и почва становится твердой,
как камень, причем Diarthron не высыхает до осени. Этот экологический
феномен пока не исследован; следовало бы установить, как глубоко про-
никают в почву корни этих растений.
Пространства эфемеровой пустыни используются преимущественно
в качестве весенних пастбищ (в течение 3 месяцев). Траву можно даже
выкашивать в конце апреля. С 1 за площади получают 0,5—2,5 т сухой
массы. Однако в годы с сухими веснами продуктивность пустыни как
пастбища весьма мала. Количество сена, получаемого при скашивании
травы в разное время года, отражено в табл. 38.
Как видно, наибольший урожай дает Роа bulbosa, но сено из осоки
содержит вдвое больше протеина. Carex hostii может полностью вытеснить
Роа bulbosa.
Подземная фитомасса также весьма велика, причем 95,5 % ее сосредото-
чено в верхних 10 см почвы, 4% — в следующем 10-сантиметровом слое
и 0,5% — на глубине до 60 см. Установлено, что на 1 м2 поверхности
почвы приходится более 2 кг корневищ, клубней и корней.
206
Глава 4
Таблица 38
Урожай сена (в ц/га) в Голодной степи (по Коровину)
Доминантный вид 19.IV 24.IV 29.IV 4.V 9.V 14.V 19.V
Carex hostii Carex hostii и 6,4 8,7 4,7 5,0 4,0 3,6 2,4
Роа bulbosa 12,3 13,9 15,4 12,9 11,6 9,5 7,2
Роа bulbosa 25,2 29,6 40,9 37,8 41,6 24,7 18,8
Благодаря равнинному рельефу и плодородным почвам эфемеровая
пустыня особенно пригодна для возделывания поливных культур, прежде-
всего хлопчатника. В настоящее время сохранились лишь небольшие
участки первичной эфемеровой пустыни.
Флористический состав эфемеровой пустыни изменяется, если в почве
присутствуют легкорастворимые соли. Одновременно пространственное
размещение растительности приобретает характерный микромозаичный
рисунок: при малейшей неровности рельефа соли смываются в небольшие
углубления почвы и там накапливаются, растительность же очень чутко
реагируетйна самые незначительные изменения содержания солей в почве.
Первым признаком засоления служит появление Salsola sclerantha*
S. carinata или Halocharis hispida. Семена этих видов прорастают поздно,
и растения начинают развиваться только тогда, когда другие эфе-
меры уже засохли. В дальнейшем даже очень слабое засоление почвы
ослабляет Carex hostii и на передний план выступает Роа bulbosa. Но кор-
невая система этого вида очень слабая, растение легко выдергивается
и потому чаще распространено пятнами. Одновременно появляются новые
однолетние, несколько более солеустойчивые виды, например виды с мяси-
стыми листьями, как Senecio coronopijolius, Lepidium perfoliatum, Spergu-
laria marginata, Sclerochloa dura, виды Statice и Plantago и др. Plantago
lachnantha в засохшем состоянии ведет себя так же, как иерихонская
роза {Anastatica hierochuntica'), ветви которой в период засухи при созре-
вании плодов загибаются вверх и внутрь, а после дождя резко распрям-
ляются, разбрасывая вокруг семена (см. т. I, стр. 485).
Широко распространены засоленные почвы в низменных местах Голод-
ной степи. Эта слабогалофитная эфемеровая пустыня служит переходным
звеном, с одной стороны, к полосе такыров, в исключительных условиях
среды совершенно лишенных растительности, а с другой — к типичной
галофитной пустыне, к рассмотрению которой мы еще вернемся.
Гипсовая пустыня
Гипсовая пустыня — это каменистая пустыня, иначе называемая тама-
дой (см. т. I, стр. 472). Занимает она чаще поверхности плато или плоско-
верхие возвышенности, сложенные соленосными породами третичного
и мелового времени — песчаниками, конгломератами или известняками
с прослоями гипса. Почвы каменистой пустыни покрыты с поверхности
тонкой коркой, под которой залегает рыхлый пылевидный горизонт,
на глубине 15—20 см уплотненный. Почвы содержат до 50% CaSO4, но
лишь следы легкорастворимых солей. Грунтовые воды залегают глубоко
Аридные области Центральной Азии
207
и для растений недоступны. Годовая сумма осадков составляет 120—
140 мм, в сухие годы — только 40—60 мм. Гипс поглощает, влагу и сохра-
няет ее, благодаря чему растения даже в засушливое время остаются актив-
ными. Однако на почвах, содержащих большое количество CaSO4, могут
произрастать лишь особо приспособленные растения. Степень покрытия
почвы чрезвычайно мала, часто 0,1%; нередко пустыня кажется совсем1
лишенной растительности. Только весной картину оживляют терофиты,
но уже в мае они исчезают. Несколько большее число растений можно-
найти в маленьких эрозионных ложбинках, где скапливается песок;
таким образом, условия напоминают северную Сахару. Растения имеют
ксероморфный облик, у них узкие листья или колючки, как, например,
у видов Gaillonia (мареновые), Sisymbrium subspinescens, Noaea spinosissi-
ma, Cousinia, Atraphaxis. Многие виды (зонтичных, рутовых, каперсовых,
губоцветных) содержат эфирные масла, у других, как у Zygophyllum,.
Otostegia (губоцветные), листья суккулентные, встречаются также типич-
ные суккулентные галофиты (Salsola) или галофиты, выделяющие соль.
(Statice, Reaumuria, Frankenia и др.). Общее число видов, включая ред-
кие, колеблется от 100 до 120. Некоторые из них распространены вплоть
до Африки, другие эндемичны.
Подробное описание отдельных типов растительных сообществ выхо-
дит за рамки данной книги, тем более что их экология и физиология
не исследовались. Упомянем лишь, что здесь нередки ассоциации, слу-
жащие переходным звеном к сообществам галофитов с господством Artemi-
sia maritima. Площади, на которых растительный покров несколько более-
сомкнут, используются для выпаса, причем преимущественно овец кара-
кульской породы. Важным кормовым растением является однолетний
Ceratocarpus arenarius (маревые), который не засыхает до осени и даетг
до 500 кг/га сухой массы. На гипсоносных почвах эта растительность
встречается также экстразонально, за пределами областей пустынного*
климата.
Пустыни с галофитами
Этот тип пустыни широко распространен в Средней Азии и Казах-
стане и на засоленных почвах азонально внедряется в область степей.
Галофитная растительность развита на почвах с неглубоким залеганием
грунтовых вод и поэтому сопровождает нижние течения многих рек, зани-
мает низины, куда с окрестных возвышений стекают воды атмосферных
осадков, и обрамляет соленые озера. Решающим экологическим фактором
служит здесь не вода, а соли (преимущественно хлориды). Лишь немногие-
виды приспособились переносить столь высокое содержание солей в поч-
ве. Особенно характерны широко распространенные маревые: Salicornia
herbacea, Halocnemum strobilaceum и Halostachys caspica, только Seidlitzia
rosmarinus является эндемиком (рис. 153). Кроме того, следует назвать
черный саксаул, Haloxylon aphyllum, а также другие виды, и в первую-
очередь представителей свинчатковых, гвоздичных, бурачниковых и гре-
бенщиковых, присутствие в растительном покрове которых, однако,
обычно малозаметно.
Большинство видов принадлежит к типу суккулентных галофитов.
Свойственное им высокое осмотическое давление обусловлено накоплением
в клеточном соке растений хлоридов. Часть этих видов представлена дре-
весными растениями, достигающими нередко «почтенного» возраста, как
Haloxylon aphyllum, Halostachys, Kalidium, Seidlitzia. Наряду с ними
208
Глава 4
Рис. 153. Солончаковая пустыня с Halocnemum strobilaceum в Туркмении (фото
Келлера).
встречаются и несуккулентные, выделяющие соль галофиты, к которым
относится, помимо Cresset cretica, также злак Aeluropus.
Галофиты могут иметь значение как кормовые растения лишь при
условии, что соли после отмирания тканей оказываются выщелоченными:
содержание протеина в сухой массе сравнительно высоко.
Крайним, обычно лишенным растительности типом солончаковой
пустыни являются шоры и такыры.
Шоры, или солончаковые пятна, образуются в понижениях рель-
ефа при очень высоком состоянии грунтовых вод. Солевые растворы, под-
нимающиеся по капиллярам до поверхности почвы, кристаллизуются,
и тогда на почве образуется толстая солевая корка, содержащая нередко
гипс. Высокая концентрация солей в почве препятствует произрастанию
каких бы то ни было растений. В качестве первого растения-пионера
на окраине шора появляется Halocnemum strobilaceum, образующий низ-
кие подушки, степень покрытия почвы которыми достигает 25%. Весной
талая вода застаивается между подушками, и тогда ландшафт напоминает
кочковатое болото. Распространены также Salicornia, Halostachys, Kali-
dium, Halopeplis pigmaea. В следующей зоне заметны виды Sucteda и Salsola,
местами Petrosimonia. По краю у впадины растут Anabctsis salsa и Artemisia
maritima.
Такыр — обширное плоское понижение рельефа с крайне разре-
женной растительностью или почти лишенное ее, которое периодически
затапливается талыми снеговыми, паводковыми или селевыми водами г.
После спада воды голая, очень плотная глинистая поверхность такыра
быстро высыхает и растрескивается на полигональные отдельности диа-
метром 15—25 см.
Такыры меньшей площади могут образоваться в песчаной пустыне
в результате развевания верхнего слоя песка и обнажения плотных глини-
стых, геологически более древних пластов. После каждого сильного дождя
на таких участках скапливается вода, образуя временные неглубокие
озера.
1 В. А. Ковда, Н. И. Базилевич, Л. Е. Родин и др., Такыры запад-
ной Туркмении и возможности их хозяйственного использования, М., 1956.
Аридные области Центральной Азии
209
Но самые обширные пространства такыры образуют в полосе равнин
у подножия среднеазиатских гор, например Копетдага, а также в древних
дельтах крупных рек. Здесь такыры приурочены к площадям распростра-
нения глинистых осадков делювиального или аллювиального происхож-
дения, принесенных во взвешенном состоянии реками, текущими с гор.
Воды содержат также известное количество растворимых солей -г- хло-
ридов и сульфатов (до 2 г/л), поэтому почвы такыров характеризуются
более или менее сильным засолением. Растительность такыров изучал
Родин х. Такыры в узком понимании представлены двумя основными раз-
новидностями: такырами водорослевыми и такырами лишайниковыми.
Высшие растения представлены видами эфемеров, которые возникают
из семян, в большом количестве приносимых водой. Чем сильнее выражено
засоление, тем меньше число прорастающих семян. Напротив, на почвах,
бедных солями, на 1 м1 2 насчитываются многие тысячи всходов.
Во влажное время года на такырах бурно разрастаются водоросли.
Установлено 92 вида сине-зеленых водорослей (преобладают Phormidium
и Microcoleus), 38 видов Chlorophyceae, кроме того, ряд жгутиковых и диа-
томей. Из гетеротрофных микроорганизмов чаще всего встречаются акти-
номицеты. Микрофауна имеет эфемерный характер и периодически разви-
вается в мелководных блюдцах, возникающих при затоплении такыров
атмосферными водами. Достаточно незначительной разности уровней,
чтобы вода распределилась по всей поверхности такыра неравномерно;
она переливается через края крупных водотоков, переходящих в малень-
кие промоины, и образует множество крайне мелких, глубиной всего
5—10 см, своеобразных «озер». Затапливаемая поверхность такыра часто
нагревается на 20° выше температуры воздуха, при этом создаются благо-
приятные условия для развития Cyanophyceae. Они образуют зеленую,
активно фотосинтезирующую оболочку, которая выделяете О2 в виде
пузырьков и вследствие поглощения СО2 придает воде днем щелочную реак-
цию. Известно, что эти водоросли способны связывать и элементарный
азот. Отсюда понятно, почему в сухой массе водорослей содержится 4,5%
азота. Общий вес водорослей, собранных с 1 га такырной площади, оцени-
вается в 500—600 кг. На возвышенных, не заливаемых полыми водами
участках поселяются лишайники Diploschistes albissimus, Squamaria len-
tigera и др. Растут они крайне медленно и поэтому сохраняются лишь
там, где не происходит отложения наилка. Увлажняются такие участки
только дождевыми осадками 2.
Так как поверхность такыра вследствие седиментации постоянно изме-
няется, затопление каждый раз захватывает все новые и новые участки;
поэтому сообщества водорослей и лишайников попеременно сменяют друг
друга.
В результате местных просадок грунта на такырах иногда образуют-
ся западины глубиной 30—60 см и диаметром 20—50 см, в которых время
1 Л. Е. Родин, Роль растительности в образовании такыров и их комплек-
сов. См. кн.: «Пустыни СССР и их освоение», т. II, М.—Л., 1954, стр. 392—413.
2 Низшие растения играют в открытых растительных сообществах аридных райо-
нов, как в степях, так и в полупустынях, значительно большую роль, чем предполага-
лось раньше. Чем экстремальнее условия, тем заметнее в растительном покрове преоб-
ладают Cyanophyceae и лишайники. Сообщества тайнобрачных отчетливо подразделяют-
ся в зависимости от свойств местообитания, особенно от степени увлажнения, засоле-
ния и’т. д. Ср. работы: В. С. Л ы с е н к о, Изучение низших растений в полупустынях
западного Прикаспия в связи с почвенно-геоботаническими исследованиями, «Бот.
ж.», 48, 1963, стр. 1610—1622; Э. А. Ш т и н а, Н. Н. Б о л ы ш е в, Сообщества водо-
рослей в почвах сухих и пустынных степей, «Бот. ж.», 48, № 5, 1963, стр. 670—680.
14-0434
210
Глава 4
от времени скапливается вода. Здесь почва увлажняется на глубину 3—
4 см, тогда как обычно воду поглощает лишь самая верхняя, набухающая
при этом глинистая корка; в таких увлажненных микроуглублениях
развивается более густой покров из цветковых растений. В зависимости
от степени засоления почв они представлены многолетними негалофитны-
ми либо галофитными видами. Аналогичная растительность формируется
и в эрозионных ложбинах, пересекающих такыры.
Глубокое промачивание почвы в западинах может привести к даль-
нейшему рассолению почвы: выщелачиваются нижние горизонты, просе-
дание грунта прогрессирует и размеры впадин увеличиваются.
В направлении от ландшафта такыров к подножию гор резко изме-
няется характер растительности и почв. Почвы такыров постепенно пере-
ходят в сероземы (серые пустынные почвы), засоление ослабевает, расти-
тельный покров становится все более сомкнутым, возрастает количество
видов. Как видно из табл. 39, можно выделить 3 зоны.
Таблица 39
Фитомасса растительности такыров и ее химический состав (по Родину)
Зоны Общий запас органического сухого веще- ства, кг/га Ежегодный опад органической массы, кг/га Содержание азота в опаде, кг/га Количество зольных эле- ментов в опа- де, кг/га
I Наиболее низкие
участки с глини-
отыми отложения-
ми; почвы засоле-
ны; эфемеров очень -
мало 100 100 1,5 6—7
II Средние участки.
Напочвенные во-
доросли или ли-
птайники; эфеме-
ров мало .... 300 300 9—10 20—22
III а) Эфемеровая расти-
тельность пятна-
ми (переходная
зона) 1200—1600 1200—1600 20—30 90—140
б) Полупустыня с Аг-
temisia и эфемера-
ми на сероземах 12 000 10 000 90 475
Пустыни с псаммофитами
Песчаные пустыни особенно характерны для Средней Азии. Здесь
расположены две крупнейшие в СССР пустыни: Каракумы («черные
пески»), между Каспийским морем и Амударьей, и Кызылкум («красные
пески») — в междуречье Амударьи и Сырдарьи.
Хотя эти песчаные территории получают не больше дождевой влаги,
чем соседние районы, они отнюдь не производят впечатления безрадост-
ной пустыни. В течение всего года на песках видна растительность: ран-
ней весной развиваются эфемеры, многие однолетние злаки, осока песча-
ная (Carex physodes), крестоцветные, бобовые, маревые и др. На неподвиж-
Аридные области Центральной Азии
211
ных песках эти растения покрывают около 50% площади, причем преоб-
ладает осока. Как только эфемеры исчезают, появляются виды летнего
разнотравья, начинают цвести кустарники. Многие из них сохраняют
активность до сентября.
Кустарники образуют интересную, чрезвычайно характерную для
Средней Азии экологическую группу.
Центром распространения рода Calligonum с 30 видами (семейство
гречишных) являются Средняя Азия и Казахстан. Это кустарники с без-
листными прутьевидными ветвями, по форме плодов подразделяющиеся
на 4 секции. Только один из видов встречается в Сахаре, немногие из них
известны на Кавказе, в Иране и Монголии. Оба вида эндемичного рода
Eremosparton (бобовые) также относятся к группе кустарников с прутье-
видными ветками. Кроме того, распространены 5 видов Ammodendron
(бобовые), близкие к ним 20 видов астрагала секции Ammodendron (кустар-
ник с серебристыми листьями, высотой до 1 м), Haloxylon persicum, 3 вида
Salsola, эндемик Smirnovia turkestana (бобовые) с маленькими сердцевидны-
ми листьями, древесный вид Artemisia, Ephedra и др.
Прутьевидная форма побегов, по-видимому, весьма характерна для
растений песчаных пустынь, иными словами для растений аридных райо-
нов на местообитаниях, относительно хорошо обеспеченных влагой. Рас-
тения с прутьевидными ассимилирующими органами интенсивно транспи-
рируют, но, поскольку их общая транспирирующая поверхность мала,
суммарная водоотдача оказывается низкой.
Псаммофит Haloxylon persicum (маревые) называют белым саксаулом
в отличие от черного саксаула (Я. aphyllum), являющегося галофитом.
Белый саксаул достигает в высоту 3—4 м при диаметре ствола до 35 см
(рис. 154).
Травянистые растения представлены приблизительно 100 видами.
Особо следует выделить многолетние злаки: 5 видов Aristida, Danthonia
и Cutandia, встречающиеся и в Сахаре, виды Convolvulus и род Acantho-
phyllum (гвоздичные). С экологией Carex physodes мы еще познакомимся.
Характерно большое участие однолетников.
Относительно большая плотность растительного покрова в песчаной
пустыне и своеобразие формирующих его видов свидетельствуют о том,
что в песчаной пустыне мы имеем дело с особыми экологическими усло-
виями. Обеспеченность растений водой здесь значительно лучше, чем
в других пустынях. Объясняется это хорошей водопроницаемостью пес-
чаных грунтов и наличием на небольшой глубине горизонта пресной воды,
которую предохраняют от испарения верхние, быстро высыхающие слои
песка; в этом случае восходящие токи влаги отсутствуют.
Чем меньше содержится в песке пыли и глины, тем ярче его водоудер-
живающие свойства. Вот почему дюнные сыпучие пески сохраняют воду
лучше, чем уплотненные. Количество атмосферных осадков в песчаной
пустыне достигает 172 мм!год, но в наиболее неблагоприятные годы оно
может сократиться до 24 мм.
Этих количеств атмосферной влаги, видимо, достаточно для обеспе-
чения водой крайне разреженного растительного покрова в пустыне,
однако в русской литературе, прежде всего Благовещенским х, указы-
вается еще на один источник влаги, а именно способность песков поглощать
(конденсировать и сорбировать) влагу из атмосферного и почвенного воз-
1 Е. Н. Благовещенский, О конденсации влаги в почвах пустынь. См.
кн.: «Пустыни СССР и их освоение», т. II, М.—Л., 1954, стр. 508—519.
14*
212
Глава 4
Рис. 154. Белый саксаул (Haloxylon persicum) в песчаной пустыне Ка-
ракумы (фото Петрова).
духа. Конденсация водяных паров в почве есть следствие значительных
колебаний температуры песчаной почвы. Опыты для доказательства
реальности процесса конденсации были проведены в Каракумах и в
Прибалхашье. На защищенной от осадков пробной площадке спустя 5 лет
определялось содержание влаги в почве. В верхних горизонтах оно состав-
ляло 0,2—0,7% сухого веса почвы, а на глубине 220 см — 1,5%. В Прибал-
хашье содержание воды в некоторой части профиля ненамного превыша-
ло максимальную гигроскопичность почвы; было предположено, что источ-
ником влаги является зеркало грунтовых вод. В другом опыте в грунт
была погружена изолированная снизу и с боков труба длиной 3 м и диа-
метром 1 м, заполненная мелкозернистым песком с содержанием воды
0,1—0,2%. Через 5 летних месяцев было произведено определение влаж-
ности по всей колонке. Влажность песка увеличилась до 0,3%, на глубине
50—60 см обнаружен максимум, равный 0,6%, а на глубине 200—230 см —
второй (1,1—1,9%). В данном случае влага является, вероятно, водой,
Аридные области Центральной Азии
213
поглощенной из атмосферы. Затем трубу' опорожнили и снова наполнили
сухим песком (влажность 0,2%). Через следующие 5 месяцев снова
было обнаружено повышение влажности до 0,5% с максимумом 0,7%
на глубине 50—70 см и вторым максимумом 0,7% на глубине 170—
180 см.
Из результатов опытов Благовещенский заключает, что такого рода
слабая конденсация почвенной влаги в пустынях имеет огромнейшее био-
логическое значение и что экологический эффект колебаний температуры
как причины конденсации трудно переоценить. Пары конденсируются там,
где колебания температуры затухают. При этом Благовещенский исходит
из Предположения, что вода атмосферных осадков в аридных областях
проникает в почву не глубже, чем на 2 м, хотя это и противоречит^ извест-
ному факту, подтвержденному непосредственными наблюдениями в Кара-
кумах, что в районах развития дюнных песков даже при количестве атмо-
сферных осадков 100 мм в год в толще песка возможно накопление прес-
ных грунтовых вод. Конечно, нельзя отрицать, что вследствие диффузии
и конденсации водяных паров в почвах, температурный режим которых
крайне изменчив, может происходить слабое увеличение содержания
влаги, но трудно согласиться с тем, что оно играет экологическую роль.
Во всех опытах использовался очень сухой песок, в результате конден-
сации не была достигнута даже точка завядания. Возникает вопрос, как
протекал бы опыт, если бы был применен песок с влажностью 1—1,5%..
Не произошла бы тогда потеря влаги?
Благовещенский замечает, что в лишенной осадков Ливийской пустыне
растительность (там, где она есть) встречается преимущественно на пес-
чаных и каменистых, а не на глинистых почвах и что это якобы возможно
лишь благодаря накоплению конденсационной влаги. Автор имел
возможность пересечь южную часть Ливийской пустыни, где вообще
не выпадает дождей, но не видел ни одного растения ни на песке, ни на ка-
менистых почвах. Стало быть, поступления влаги в почву в парообразной
форме и ее дальнейшего сжижения, несмотря на весьма резкие колебания
температуры, здесь не происходит. Поэтому нам представляется излитп-
ним предположение о существовании экологически важного процесса
конденсации влаги в почве пустынь (см. также стр. 271). Конечно, осадки
в пустынях не каждый год просачиваются глубоко в толщу песка; это
происходит лишь в особенно дождливые годы. Но, однажды проникнув
на большую глубину, вода сохраняется достаточно долгое время и исполь-
зуется растениями с глубокой корневой системой, причем запасы влаги
в отдельные дождливые годы периодически пополняются.
Против экологического значения процессов конденсации свидетель-
ствует и тот факт, что обширнейшая область распространения подвижных
дюн в пустыне Намиб южнее залива Уолфиш-Бей, где осадки вообще
не выпадают, совершенно лишена растительности, хотя условия для кон-
денсации в береговой полосе особенно благоприятны: почти каждую ночь
верхние 1—2 см песка увлажняются густым туманом и росой. Дюнная
фауна, питающаяся занесенными ветром органическими частицами, ночью
использует эту влагу. Но уже в утренние часы почва отдает ее в атмосфе-
ру; диффузии в более глубокие слои не происходит, хотя количество вла-
ги, оседающей из тумана и росы, за одну ночь обычно составляет 0,1 —
0,2 мм (см. т. I, стр. 377 и след.).
Итак, еще раз подчеркнем, что не следует переоценивать значение
конденсации как фактора, якобы создающего благоприятный водный
режим в песчаных районах. Если предположить, что скопившаяся в почве
214
Глава 4
доля осадков составляет 50%, то и тогда этого количества влаги будет
достаточно, чтобы покрыть расход на транспирацию лишь весьма скудной
растительности.
В большинстве случаев судить об условиях обеспечения растений
влагой лучше не на основе данных об интенсивности транспирации на еди-
ницу листовой поверхности, а зная величину водоотдачи в расчете на
на 1 га площади.
Верхние слои песка бывают влажными только весной. В них укоре-
няются эфемеры, которые исчезают, как только эти слои высыхают. Мно-
голетние растения поглощают воду из пластов, залегающих на глубине
1—1,5 м и сохраняющих влажность в течение всего года. От чрезмерного
испарения растения защищает незначительность транспирирующей поверх-
ности (редукция листьев, прутьевидная форма веток, превращение листьев
или побегов в колючки).
К суровым климатическим условиям среднеазиатских пустынь расте-
ния приспособились по-разному. Calligonum вовсе не образует листьев;
уже летом после созревания плодов этот вид сбрасывает свои зеленые
ассимилирующие короткие побеги («иголки»). То же наблюдается у сак-
саула. Лишен листьев и Eremosparton. Редуцирована листовая поверх-
ность у видов Salsola и Astragalus. Smirnovia сбрасывает свои мелкие
листочки перед созреванием плодов. Фотосинтез осуществляют во всех
этих случаях зеленые, обычно с толстой кутикулой побеги. У Acantho-
phyllum листья способны превращаться в колючки. У других пустынных
растений крупные весенние листья сменяются очень мелкими жесткими
летними.
Количество луковичных растений в пустынях Средней Азии невелико
(всего 7 видов), что объясняется подвижностью сыпучего субстрата. Перн-
отложение песка ветром неблагоприятно для этих растений. Псаммофиты
же — многолетние злаки и виды разнотравья (например, Ferula') — обла-
дают способностью все выше перемещать почки возобновления по мере
засыпания стебля песком. Побеги злаков легко прорастают сквозь слой
песка толщиной 15—20 см. Недостатка О2 в песчаных почвах никогда
не бывает.
Особенно хорошо приспособилась к засыпанию песком Aristida каге-
lini. Этот злак высотой до 1 м переносит засыпание до верхушек верхних
листьев; он упрямо прорастает сквозь слой песка и образует новые корни
на все более высоких уровнях. Так же ведут себя Agriophyllum (маревые)
и многие другие виды, растущие на подвижных песках. Часто псаммо-
фиты образуют почки возобновления на длинных горизонтальных корнях
или корневищах, которые дают побеги, если песок полностью засыпает
растение. Прутьевидная форма не способствует отложению песка около
растения; кроме того, эти виды энергично растут в длину: прирост иногда
превышает 1 м за вегетационный период.
Большинство видов растений песчаной пустыни — анемохоры, их
крылатые плоды, переносимые ветром с места на место, таким образом
избегают губительного засыпания песком.
На барханных песках юго-восточных Каракумов в верхней части
склона гряды встречается кустарник высотой 4—5 м, образованный вида-
ми Calligonum и Salsola, корни которых не достигают уровня грунтовых
вод. Покрытие почвы нередко достигает 60—65%. Однако древесные рас-
тения преимущественно растут поодиночке (рис. 155).
Флористический состав на подвижных и закрепленных песках совер-
шенно различен, часто встречаются викарирующие виды. Приведем назва-
Аридные области Центральной Азии
215
Рис. 155. Растительность барханных песков в Каракумах (по Коровину).
1 — пионерный травостой с Aristida и Tournefortia на постоянно перевеваемом гребне бархана', 2 —
верхняя часть склона бархана с кустарниками и низкими деревьями; 3 — богатое видами расти-
тельное сообщество нижней части склона; 4 — более бедный видами растительный покров с Carex
physodes и злаками в межгрядовых ложбинах.
ния соответствующих пар, причем
подвижных песков.
Aristida karelini — А.
Crucianella sabulosa — С.
Salsola paletzkana — S.
Convolvulus divaricatus — C.
первый вид всегда характерен для
minor (оба из таксона A. pennata)
filifolia
richteri
korolkovii
Для неподвижных песков особенно характерна осока песчаная (Carex
physodes), образующая очень длинные ползучие корневища (рис. 157).
Carex physodes близка к Carex hostii и связана с ней переходными формами.
Ее легко узнать по сильно вздутым мешочкам. Корневище находится
на глубине 10—15 см. Через каждые 5—10 см от него отходят надземные
побеги (до 400 на 1 м2, рис. 156), корни которых проникают в почву
на 25—30 см (до 60 см). Подземная масса достигает величины 1 кг!мг =
= 10 т!га, надземная фитомасса оценивается в 300 кг/га. Carex physodes
считалась одним из немногих пойкилогидрических покрытосеменных.
Предполагалось, что побеги летом не отмирают, а лишь полностью высы-
хают. но после дождя вновь зеленеют и возобновляют свой рост.
Эти приводимые Васильевым (1930, 1931) данные Янишевский (1937)
опровергает Ч Переживает засуху только корневище этого вида, причем
песок на его местообитании может нагреваться до 40—50°. В конце веге-
тационного периода клетки коры корневища отмирают и вместе с одре-
весневшей эндодермой образуют защитный чехол. Тонкостенные клетки
осевого цилиндра, окружающие проводящие пучки, содержат много крах-
мала, который летом переводится в жиры. В несколько более влажных
понижениях между дюнами травостой Carex остается зеленым дольше.
1 См.: В. К. Василевская, Формирование листа засухоустойчивых расте-
ний, Ашхабад, 1954.
216
Глава 4
Р и с. 156. Пески с плодоносящей Carex physodes в Каракумах близ Репетека
(фото Петрова).
Таким образом, с Carex physodes происходит то же, что с луковичками
Роа bulbosa var. vivipara, которые после 8 лет хранения в сухом виде
при увлажнении прорастают, или с клубнями Leontice eversmannii
(барбарисовые), жизнеспособность
которых в "воздушно-сухом состоя-
нии сохраняется в течение 4—5 лет.
В юго-восточных Каракумах
на сыпучих, барханных, песках
различают следующие типы ра-
стительных группировок, после-
довательность расположения кото-
рых схематически изображена на
рис. 155:
I — растения-пионеры на греб-
нях барханов;
II — кустарники в верхней ча-
сти склона барханной гря-
ды;
III — богатые видами разнотрав-
ные сообщества с крупными
Ferula oopoda и Rheum tur-
kestanicum;
IV — более однообразная расти-
тельность уплотненных пе-
сков в межбарханных пони-
жениях с Carex physodes
л многочисленными эфеме-
рами (терофиты и геофиты).
Рис.* 157. Carex physodes, V3 натураль-
ной величины (по Коровину).
Аридные области Центральной Азии
217
Рис. 158. Центральная часть Каракумов с редкими зарослями Haloxylon persi-
сит и Carex physodes в травяном ярусе (во время засухи свернута) (фото Петрова).
Монотонный’ландшафт бугристых и грядовых песков с подвижными
грядами и уплотненными ложбинами простирается в Каракумах на сотни
километров. Среди бугристых песков расположены плоские уплотненные
глинистые участки — такыры; подвижные, лишенные растительности
барханные пески занимают примерно 5% площади. Часто можно наблю-
дать взаимное проникновение различных типов пустыни, например в юго-
восточной части Каракумов, где рельеф более разнообразен. Здесь разли-
чают песчаные равнины, которые на юге переходят в обширные такыры,
а на севере — в ландшафт бугристых песков; кроме того, распространены
низины с сильно засоленными почвами (шоры) и участки с подвижными
барханами или барханными грядами. Встречаются и глубоко расчленен-
ные эрозией и дефляцией останцы третичного песчаникового* плато. Белый
саксаул {Haloxylon persicum) распространен только в бугристой песчаной
пустыне, где в мощной толще песка уже на небольшой глубине содержится
некоторый запас влаги (рис. 158) х.
Для естественных насаждений характерна отчетливая ярусность:
древесный ярус имеет высоту всего 2,5—3 м (до 4 м) и сомкнутость крон
0,2—0,3. Ярус кустарников (высота 1—1,5 м) более сомкнут (покрытие
40—60%). Под пологом древесного яруса развиваются кустарнички
(высотой 30—50 см), травы и эфемеры. При благоприятных условиях сре-
ды на 1 га площади насчитывается 300—600 экземпляров саксаула, запа-
сы древесины составляют 2 м?!га. Высоты саксаула достигают также Salso-
la richteri и виды Calligonum. Всего насчитывают 45—50 видов деревяни-
стых растений. Менее широко распространены травянистые многолетние
виды; напротив, здесь много (свыше 50 видов) эфемеров, причем особи
отдельных видов обычно растут группами. Общее число' видов на типич-
ных площадях колеблется от 30 до 80.
Фенологический спектр в Каракумах имеет свои особенности. Цвете-
ние деревьев и кустарников приходится на время с середины апреля
до середины мая (виды Salsola цветут в июне — начале июля), эфемеров —
1 С. А. Н и к и т и и, Древесная и кустарниковая растительность пустынь СССР,
М., 1966.
218
Глава 4
Рис. 159. Густые заросли черного саксаула {Haloxylon aphyllum) на слабозасолен-
ной влажной почве с богатой травянистой растительностью. Высокий кустовой
злак —Lasiagrostis splendens (фото Петрова).
с середины марта до мая, причем уже перед концом мая они исчезают.
Однолетние маревые живут до йюля, некоторые даже до поздней осени.
Различия в экспозиции местообитаний на барханных песках могут обусло-
вить смещение фенологических дат на неделю х.
Длительность вегетационного периода иллюстрирует табл. 40.
Таблица 40
Длительность безморозного периода
в песчаной пустыне Каракумы (Репетек)
Год Последний поздний заморозок Первый ранний заморозок Продолжи- тельность безморозного периода, ДНИ
1944 —2,9° 29.11 -1,8° 27. IX 210
1947 —0,7° З.Ш -2,6° 9.Х 219
1950 —0,2° 19. IV —2,6° 19.IX 169
1951 —3,0° 29.III —0,7е 15.Х 199
Значительно более широким экологическим распространением поль-
зуется Haloxylon aphyllum (черный саксаул). Прежде всего он менее чув-
1 Т. Д ы б с к а я, Рельеф и фенофазы развития травянистой растительности
песчаной пустыни, «Бюлл. Моск, о-ва испытателей природы, отд. биол.», 67, 1962,
стр. 133—136.
Аридные области Центральной Азии
219
ствйтелен к солям, хотя и не
растет на солончаках. Соот-
ветственно его спутниками
часто являются галофиты, на-
пример виды Tamarix на пло-
щадях с близко расположен-
ными грунтовыми водами. Не-
когда черный саксаул образо-
вывал густые заросли. Но так
как древесина саксаула может
быть использована для поде-
лок, насаждения его в настоя-
щее время большей частью
вырублены. На местообитани-
ях с неглубоким залеганием
грунтовых вод на 1 га пло-
щади насчитывается 2500—
— 5000 деревьев (выше 1 м).
В старых насаждениях число
экземпляров вследствие корне-
вой конкуренции уменьшается
(рис. 159). Возобновлению дре-
востоев препятствует выпас
скота, так как овцы скусы-
вают проростки деревьев.
В настоящее время черный
саксаул, достигающий более
крупных размеров, чем белый,
используют при создании
искусственных насаждений.
Некоторые авторы пред-
полагают, что конечной ста-
дией естественного развития
растительности в песчаной пу-
стыне должна быть также ра-
стительность галофитов. Они
исходят при этом из резуль-
татов наблюдений в юго-во-
сточной части Каракумов, где
было установлено, что в опаде,
образуемом Haloxylon persi-
сит, содержится известное
количество легкорастворимых
солей, которые со временем
накапливаются в почве. Таким
образом, чувствительный к
соли белый саксаул постепен-
но вытесняется более солеу-
стойчивым черным саксаулом
{Haloxylon aphyllum). Послед-
ний образует еще большую
массу опада, приблизительно
1550 кг!га, в результате мине-
Рис. 160. Схематический профиль через Каракумы в засушливый период (по Благовещенскому).
- подвижные барханные пески; 2 — древние уплотненные бугристые пески со слаборазвитым почвенным профилем; 3 — серые биотитсодержащие
ки; 4 — серые суглинистые пески; 5 — желтые суглинистые пески; 6 — суглинки; 7 — песчаный слой с содержанием воды более 1 % (установлено буре-
sm); 8 —пресные и 9 — соленые грунтовые воды; 10 — Haloxylon aphyllum’, 11 — Н. persicum. Отсутствующая часть профиля (4—7 км) соответствует
соседним участкам.
220
'лава 4
рализации которой в почву вносится 94 кг зольных веществ Это должно
привести к значительному увеличению концентрации солей в почве . и
как результат — к появлению галофитной растительности. Однако
такая сукцессия, видимо, имеет местный характер и ее не следует возво-
дить в ранг всеобщего правила. Дело в том, что в рассматриваемой
области на небольшой глубине залегает соленосный суглинистый слой,
из которого корни деревьев поглощают соли. Кроме того, засолению песка
способствуют термиты, которых здесь известно 8 видов; они переносят
соленосные частицы почвы из глубины на поверхность и нагромождают их
вокруг своих построек.
Размещение древостоев черного и белого саксаула в Каракумской
пустыне иллюстрирует рис. 160. Белый саксаул распространен на мощ-
ных толщах песка, черный — на песках, подстилаемых суглинками; изве-
стны, конечно, и исключения. Оба вида не связаны с горизонтом грун-
товых вод.
ИЗМЕНЕНИЯ РАСТИТЕЛЬНОСТИ ПЕСЧАНЫХ ПУСТЫНЬ
ПОД ВЛИЯНИЕМ ДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКА
Растительность песчаных пустынь непрерывно изменяется. Неподвиж-
ные пески могут вновь обрести подвижность, так как в естественных усло-
виях животные воздействуют на растительный покров и непосредственно,
и косвенно. Так, черепахи (Testudo horsfieldi) выкапывают себе норы дли-
ной до 1 м, в которых они проводят в спячке 10 месяцев. Увидеть их мож-
но часто лишь в течение 2—3 весенних месяцев. В это время года они
питаются многими эфемерами. Особенно большой ущерб растительному
покрову наносят многочисленные грызуны, разрушая и поедая многие
растения. Огромное влияние на состояние растительности оказывает
выпас животных. Еще 100—200 лет назад, когда огнестрельное оружие
не было столь широко распространено, в пустыне паслись большие стада
копытных; животных (Gazella subgutturosa, Saiga tatarica, Equus hemionusr
Ovis orientalis areal). В настоящее время эти животные стали редкими,
хотя в незаселенных районах иногда можно встретить стада джейранов
и сайгаков численностью 500—1000 голов. В Прибалхашье еще в начале
нашего столетия видели табуны диких лошадей численностью до 1000 голов.
В наши дни выпас крупных домашних животных также отрицательно
сказывается; на состоянии пастбищ. Чрезмерный выпас вызывает^ резкое
ухудшение экологических условий и даже разрушение сложившихся рас-
тительных сообществ. Поэтому в пустынях Средней Азии сегодня почти
невозможно найти следы первоначальной растительности. Даже в слабо-
заселенных районах растительный покров нигде не является естественным,
а повсюду преобразован и сформировался под длительным воздействием
человека. Вмешательство человека нарушает равновесие в природе, изме-
няет конкурентные взаимоотношения между видами, в результате чего
одни оказываются в более благоприятных условиях, а другие вытесняют-
ся 1 2. И всегда в конечном итоге формируется более аридный тип расти-
тельности; именно поэтому со стороны неспециалистов раздаются жалобы
на ухудшение климата в направлении увеличения его сухости и на насту-
1 По данным Родина, годовая масса опада всего сообщества Haloxylon aphyllum-
равна 4500 кг/га, а золы — 252 кг.
2 Л. Е. Родин, Динамика растительности пустынь. На примере Западной
Туркмении, М.—Л., 1961.
Аридные области Центральной Азии
221
лание пустыни. Археологические исследования показали, что современные
песчаные пустыни в неолите охотнее заселялись человеком, так как он
находил здесь источники воды, достаточное количество топлива и паст-
бища для скота. Потребность в топливе увеличилась, когда человек овла-
дел гончарным искусством и научился обжигать кирпич; расход древесины
в дальнейшем все возрастал с ростом населения, причем уничтожение дре-
весных растений, видов Artemisia и особенно обоих видов саксаула привело
к обнажению песков и образованию подвижных дюн.
Уже на ранней стадии охоты и собирательства человек нарушал рав-
новесие в природе. Он истреблял не только копытных животных, служив-
ших ему пищей, но и хищных зверей, мех которых он использовал, и тем
самым способствовал размножению грызунов, так как их численность под-
держивается в известных границах хищниками, для которых грызуны
составляют основу питания.
Из грызунов чрезвычайно сильно размножилась большая песчанка
(Rhombius opimus). На 1 га площади находят до 300 особей, а число выход-
ных отверстий местами превышает 2500 на 1 га. Вообще в песчаной пусты-
не водится множество видов грызунов. Они изменяют растительность
не только тем, что охотнее поедают определенные виды, но и, может быть
даже в большей мере, своей активной роющей деятельностью. В резуль-
тате в почве накапливаются органические вещества; глубоко залегающие
частицы почвы и подпочвы выносятся на поверхность; разрыхляется песок,
что ведет к его развеванию и образованию дюн даже в районах, где дея-
тельность человека непосредственно не проявляется.
Однако наибольший урон наносит пастбищное хозяйство. Песчаная
пустыня играет в наши дни большую роль как пастбище для каракуль-
ских овец, которые находят здесь корм в течение всего года, причем
в количестве до 1000 кг/га сухого веса, в среднем 400 кг!га. Кормовая
продуктивность пастбища сравнительно мало зависит от суммы осадков
в отдельные годы.
Пасущийся скот держится вблизи редких колодцев. Каждый колодец
окружен круговыми полосами различных растительных сообществ, нахо-
дящихся на той или иной стадии деградации. Разрушение растительного
покрова усугубляется тем, что вблизи ’колодцев стоят жилища пастухов,
которым нужны дрова; потребность в топливе удовлетворяется в первую
очередь за счет вырубок саксаула и вообще всего, что способно гореть.
Вокруг мест водопоя можно выделить следующую поясность:
1. Непосредственно у колодца, где отдыхают овцы (или козы,
верблюды), почва сплошь покрыта навозом и лишена растительного
покрова. Изменение всех условий среды здесь столь сильное, что даже
через несколько лет после прекращения пользования колодцем раститель-
ность никогда не восстанавливается в том виде, какой она имела прежде.
При небольшом количестве осадков выщелачивания унавоженной почвы
не происходит.
2. Далее идет кольцо барханов, по которым уже издалека можно
определить местоположение колодца. Растительность стравлена, голый
песок легко перевевается ветром. Сохраняются лишь немногие псам-
мофиты.
3. В следующем поясе степень покрытия увеличивается, но все же
песок закреплен слабо.
4. На некотором удалении от колодца уже появляются характерные
полукустарники, но полное покрытие песка Carex physodes еще не до-
стигнуто.
222
Глава 4
5. И лишь на очень значительном расстоянии от места водопоя влия-
ние выпаса на растительность почти исчезает.
Вблизи колодцев происходит не только нарушение естественной рас-
тительности. Под влиянием выпаса в пустынные сообщества внедряются
сорные виды, чуждые флоре пастбищных территорий. Так как скот их
не поедает, для них создаются особенно благоприятные условия произрас-
тания и они быстро распространяются. В колодезной зоне успешно раз-
растаются, например, Anabasis aphylla и Peganum harmala. Последний
является классическим примером. На всем Востоке даже спустя столетия
гармала отмечает местопребывания, испытавшие губительное антропо-
генное влияние. Человек при кочевках заносил семена этого нитрофиль-
ного растения на все новые места водопоя и отдыха скота.
При полном прекращении выпаса возникает своего рода «одряхление
настбищ», приводящее в конечном итоге к значительному снижению
кормовой продуктивности. Прекращается рыхление поверхности песка,
уменьшается его водопроницаемость и ухудшается аэрация верхних слоев
почвы. Почва уплотняется, и даже на песчаных субстратах возникает кор-
ка. Такое уплотнение угнетает рост Carex physodes, и ее замещает моховой
покров из Tortula desertorum. Активно поселяются лишайники (Placodium
tominii, Psora decipiens, Collema и др.), а также почвенные водоросли. Усло-
вия существования для травянистых растений становятся неблагоприят-
ными, так как осадки, перехватываемые низшими растениями, уже
не в состоянии просочиться в почву. Развитие моховых белосаксаульников
отмечается также на наиболее удаленных от колодца невыпасаемых терри-
ториях. Отсюда следует, что известное воздействие животных (прежде —
стад парнокопытных и диких лошадей) неотъемлемо от развития нормаль-
ного биогеоценоза.
Продолжавшееся столетиями хищническое использование земли (чрез-
мерный выпас) в Средней Азии и на Прикаспийской низменности привело
к образованию подвижных песков, ныне занимающих обширные про-
странства; вторичность этого явления не вызывает сомнений. Процессы
образования, перемещения и закрепления барханных песков весьма
детально исследовались на основанной в 1912 г. станции Репетек в Турк-
мении х. Наблюдения показали, что на процесс развевания песков суще-
ственное влияние оказывают условия рельефа. Несомый ветром песок
откладывается позади препятствия в виде невысокой дюны. При дальней-
шем поступлении песка в средней части дюны его накапливается больше,
чем по краям, которые перемещаются быстрее; таким образом, форма дюны
постепенно изменяется, она становится серповидной, то есть превращается
в бархан. Одиночные барханы встречаются относительно редко. Они дости-
гают 3—4 м в высоту и быстро передвигаются. Обычно же барханы собра-
ны в большие компактные группы, которые, перестраиваясь под ударами
воздушных струй, образуют извилистые барханные цепи, вытянутые
поперек направления господствующих ветров. Если же ветер временно
начинает дуть под углом к цепи, на гребне возникает местное воздушное
течение, направленное вдоль гребня. Воздушный поток захватывает
навеянные на вершину бархана песчаные зерна и переносит их вдоль
по гребню. Так начинается образование прямолинейных барханных гряд
высотой 5—15 м (до 40 м), протягивающихся параллельно одна другой
на многие километры.
1 «Труды Репетекской песчано-пустынной станции АН Туркм. ССР», т. 3, Ашха-
бад; 1955; Очерки природы Каракумов (под ред. В. Н. Кунина), М., 1955.
Аридные области Центральной Азии
223
Как показывают данные анализов, в механическом составе перевеян-
ных песков преобладает фракция мелкого песка (0,25—0,05 мм). Переме-
щение песчаных зерен начинается, как только скорость ветра достигает
5 м/сек (около 4 м/сек у поверхности земли). В дождливые дни движение
песка прекращается. Только при сильном ветре (свыше 10 м/сек), быстро
высушивающем верхний слой песка, снова начинается его перевевание.
В течение года гребень бархана, следуя направлению ветра, перемещается
то в одну, то в другую сторону; в сухие годы процесс развевания приобре-
тает более интенсивный характер. Труднее доказать, происходит ли пере-
движение дюны в целом на протяжении нескольких лет в определенном
направлении.
Как только процессы навевания прекращаются, на песке поселяются
единичные растения, которые, разрастаясь, все более закрепляют песок.
В Каракумах различают несколько стадий зарастания барханных песков,
отличающихся от схемы зарастания песков на Прикаспийской низменно-
сти. Здесь наблюдается такая последовательность:
1) стадия растения-пионера Arvstida karelini (из группы A. pennata)-,
2) стадия псаммофитных кустарников;
3) стадия травянисто-кустарникового растительного покрова;
4) конечная стадия растительности из эфемеров и осоки песчаной.
Смена этих стадий связана с ухудшением водного режима песков
и увеличением в их механическом составе доли пылеватых частиц (за счет
опада Calligonum и Haloxylon). Величина фракции пыли в песке под Ari-
stida pennata составляет всего 0,44%, но под Haloxylon persicum в верхних
10 см достигает почти 9% и даже на глубине 40 см еще превышает 4%.
Повышенное содержание пылеватых частиц, как известно, отрицательно
сказывается на аккумуляции влаги в почве.
Однако конечная стадия зарастания песков, в которой появляется
осока песчаная, обычно достигается только в межбарханных низинах.
В общем, сукцессия прекращается на третьей стадии — стадии бугристой
крупнокустарниковой пустыни. Объясняется это тем, что процесс смен
растительности постоянно нарушается активным вмешательством в этот
процесс целого ряда факторов окружающей среды, так что утверждается
стадия, соответствующая результирующей воздействий этих факторов
(климата, животных и растений).
Но особенно глубокие изменения естественная растительность испы-
тала в районах орошаемого земледелия, занимающих в Средней Азии
обширные площади. Мы оставляем в стороне основное отличие оазисов
от пустыни: преобладание здесь культурной растительности. Отметим
лишь, что на орошаемых землях происходит перераспределение поверх-
ностного стока. Вода, которая подводится на поля, теперь отсутствует
там, где прежде текла в естественных условиях. В результате растения,
которые прежде росли в обстановке достаточного увлажнения, оказы-
ваются сразу перенесенными в условия безводной пустыни, и наоборот.
Кроме того, в орошаемых районах быстро увеличивается плотность
населения, а следовательно, возрастает и потребность в топливе, которое
доставляют из окружающих оазис районов. Прежде дрова перевозили
на верблюдах в радиусе суточного перехода верблюда, то есть 30—50 км',
ныне, при использовании грузовых автомашин, этот радиус действия
увеличился до 120—150 км и даже 200 км.
При любом виде орошения необходим известный дренаж орошаемой
территории, если хотят избежать опасности засоления почв. Эту избыточ-
ную воду сбрасывают в водоприемники, что опять-таки вызывает изме-
224
Глава 4
нение растительности в самых различных направлениях. В очагах застоя
поверхностного стока вода энергично испаряется, очень быстро начинают-
ся процессы засоления почв и развивается резко выраженная раститель-
ность галофитов.
ПОЙМЫ РЕК СРЕДНЕЙ АЗИИ
Истоки Амударьи, крупнейшей водной артерии Средней Азии, рас-
положены на юге области, на северном склоне хребта Гиндукуш. Около
84% стока реки составляют талые воды ледников и снега нивального
поясаДПоэтому наблюдается два паводка: относительно небольшой весен-
ний паводок, обычно в конце апреля, когда в горных долинах начинает
таять снег и-проходят сильные дожди, и значительно более сильное летнее
половодье, наступающее, как правило, в конце июля, когда высокая тем- !
пература летом вызывает таяние ледников и снега в высокогорье. Во время |
половодья в среднем и^нижнем течении реки затапливаются обширные ]
территории, где отлагается слой ила толщиной до 20—30 см, способствую-
щий развитию пышной пойменной растительности х. Характер отложений I
зависит от скорости течения потока. В местах относительно сильного тече- I
ния осаждается только богатый слюдой песок. После спада воды здесь
остаются голые песчаные площади, заселяемые пионерными видами. Гли-
нистые осадки образуются там, где течение замедляется или вода длитель-
ное время застаивается. При быстром изменении скорости течения проис-
ходит отложение смеси песка и глины. Поскольку река очень часто меняет
свое русло, слагается необычайно пестрая картина различных типов почв.
Направление же процесса почвообразования имеет для пойменной рас-
тительности особенно важное значение. Большую роль играет также поло- i
жение уровня грунтовых вод; как правило, он расположен близко к днев-
ной поверхности, но под влиянием режима затопления залегание грунто-
вых вод может изменяться. В общем же растения в поймах рек, несмотря ।
на в целом пустынный климат, развиваются в течение всего года в опти-
мальных условиях обеспечения водой. Грунтовые воды пресные. Тем
не менее на возвышенных участках поймы, не заливаемых в половодье,
может произойти засоление почвы,’ если уровень грунтовых вод времена-
ми поднимается столь высоко, что поверхность почвы за счет восходящих
токов влаги постоянно увлажняется и поэтому интенсивно испаряет.
Обогащение солями при этом наблюдается только в самых верхних слоях
почвы. Аналогичный процесс возникает и в наиболее низких местах пой- |
мы, но соли здесь при следующем половодье снова растворяются, выно- |
сятся и отлагаются лишь в районе дельты, где нередко формируются
такырные “'почвы. Под очень густой пойменной растительностью засоле-
ние не происходит: во-первых, воздух вблизи земной поверхности почти
постоянно насыщен водяным паром, следовательно, испарение с поверх- '
ности почвы невелико; во-вторых, интенсивная транспирация растений 1
вызывает понижение зеркала грунтовых вод, в результате чего поверх-
ность почвы высыхает и капиллярное поднятие влаги прекращается.
Местное население Средней Азии различает три типа пойменной рас-
тительности: 1) пойменные леса, или тугаи; 2) высокозлаковую и крупно-
разнотравную растительность, или джангиль (ср. название «джунгли»
__________ >•
1 Б. Б. К е р б а’б а е в, “Растительность джангиль поймы Амударьи, «Труды
ин-та биол.», I, Ашхабад, 1964, стр. 22—74; С. А. Никитин, Древесная и кустар-
никовая растительность пустынь СССР, М., 1966. i
Аридные области Центральной Азии
225
в Индии), и 3) бедную растительность аллювиальных равнин, или кайре.
Пойменные леса в настоящее время занимают только около 8% площади
поймы Амударьи. Вследствие вырубки и пожаров большая часть их
нарушена. В состав растительности джангиль входят 96 видов, причем
некоторые из них эндемичны, например Marsilia aegyptiaca, Ophioglossum
bucharicum, два вида Typha, Saccharum spontaneum, Dichostylis homulosa
(осоковые), Bergia koghanii (Elatinaceae), Oxytropis riparia и сапрофитная
орхидея Eulophia turkestanica, принадлежащая тропическому роду и рас-
тущая в дерновинах Erianthus purpurascens.
Кайре заселяют многие виды растений-пионеров. Влажная поверх-
ность свежих наносов за короткое время покрывается бесчисленным мно-
жеством проростков (100—500 на 1 л*2). На песчаных почвах преимуще-
ственно встречается Typha pallida, на глинистых — Calamagrostis dubia.
Если растительность в последующие годы не погибает при половодье,
со временем складываются отчетливо выраженные растительные группи-
ровки с доминирующими Тypha, Calamagrostis или Phragmites. В процессе
естественной сукцессии позже начинает преобладать Saccharum spontaneum.
Но этот злак сохраняет доминирующее положение только в том случае,
если уровень грунтовых вод не опускается ниже 1,5—1,75 м от поверх-
ности земли и если занятая им площадь ежегодно затапливается, в иных
условиях его вытесняют другие виды — Equisetum ramosissimum, Gly-
cyrrhiza glabra, Erianthus purpurascens или Typha elephantina. Часто пре-
обладают солодка (Glycyrrhiza) или Erianthus. Образуются растительные
сообщества, где высокие злаки, такие, как Erianthus, Phragmites и Saccha-
rum, слагают верхний (высотой 2—4 м) ярус, нижний же — с Glycyrrhiza,
Typha или Alhagi — достигает лишь высоты 1—1,5 м. При этом подзем-
ная фитомасса, как правило, превышает надземную. Так, в густых траво-
стоях Glycyrrhiza под землей сосредоточено органического вещества 10—
15 т!га, а над землей только 3—4 т1га\ для Typha pallida соответственные
величины равны 25—30 и 8—10 т!га. Этот бурный рост и продуцирование
столь большой массы органического вещества возможны благодаря постоян-
ной влажности и высокой температуре воздуха.
Характерная особенность видов, произрастающих в зоне затопления
паводковыми водами, состоит в том, что после отложения толщи осадков
они обладают способностью образовывать новые корни на стебле на любой
высоте. Например, Saccharum переносит заиление высотой 30—40 см
(рис. 161). Аналогично приспособлены Clematis orientalis, Cynanchum sibi-
ricum, Tamarix. У видов более поздних стадий сукцессии эта особенность
выражена не столь резко. Так, на местообитаниях Erianthus, Glycyrrhiza
или Equisetum толщина ила не должна превышать 10—15 см. Затопление
не вредит растениям, если вода покрывает не более чем половину над-
земных частей и если паводок длится меньше 3 недель. Напротив, в засуш-
ливый период растения особенно чувствительны к резкому снижению
уровня грунтовых вод. Травостой тогда изреживается, и развивается
следующая стадия сукцессии — лес, или тугай. Тугайные заросли обра-
зуют Populus pruinosa, Р. diversifolia (рис. 162), Salix acmophylla и другие
виды, а также Elaeagnus angustifolia или Е. orientalis. Из лиан представ-
лены Clematis orientalis, Cynanchum sibiricum и Asparagus persicus. Древес-
ный ярус достигает высоты 13—15 м. Запас древесины составляет 90—
200 мЧга.
При засолении почвы появляются различные виды Tamarix и Hali-
modendron halodendron (бобовые); в подлеске виды стадии джангиль, к кото-
рым относятся также Cynodon dactylon и Imperata cylindrica, замещаются
15-„0434
104
20-
30-
40-
СМ
0
Рис. 161*. Подземная часть Saccharum sponta-
neum. Новообразование корневищ и корней на бо-
лее высоком уровне (на глубине 8—10 см) после-
того, как растение было погребено под отложе-
ниями мощностью 30 см (по Кербаеву).
Рис. 162. Одиночное дерево Populus diversifolia как остаток пойменного леса
вдоль старого русла Западного Узбоя (фото Петрова).
Аридные области Центральной Азии
227
галофитами Aeluropus litoralis, Karelinia caspica, Zygophyllum oxianum
el Halostachys caspica (маревые).
В заключение вкратце остановимся на фенологии пойменной расти-
тельности. Безморозный период в долине среднего течения Амударьи
длится с конца марта до начала ноября — в среднем 216 (190—246) дней,
В начале апреля пойма зеленеет. В конце апреля начинается бурный рост
растений. Glycyrrhiza цветет в первой половине мая, прежде чем злаки
не обгонят ее в росте. В начале июня преобладает розовый аспект, свя-
занный с цветением Tamarix и обильным плодоношением Glycyrrhiza.
Лишь в середине июля один за другим зацветают высокие злаки, что
придает пойме серебристо-белую окраску. Erianthus и Phragmites зацветают
только в первой половине августа. Последний аспект злаков с созре-
вающими плодами характеризуется темно-желтым оттенком.
В пойменных лесах цветение тополя дает первый заметный аспект.
В сравнении с песчаной пустыней развитие растений на влажных почвах
поймы происходит на месяц позднее.
ЭКОЛОГО-ФИЗИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ ВОДНОГО
РЕЖИМА РАСТЕНИЙ В СРЕДНЕАЗИАТСКИХ ПУСТЫНЯХ
Выше мы отметили, что большая часть фитомассы пустынных расте-
ний сосредоточена под землей. Это справедливо даже для более крупны^
деревьев. Поэтому исследованию корневой системы придается особое зна-
чение. Действительно, в приводимых ниже работах это должным образом
учитывается х. Для большинства видов составлены проекции корневых
систем. У эфемеров корневая система слабо развита, пронизывает только
верхние, влажные весной слои почвы. Другие виды песчаной пустыни
имеют как горизонтально распростертые корни, так и глубоко проникают
щие, которые достигают горизонтов почвы, содержащих даже летом
доступную для растений влагу. Таким образом известная обеспеченность
водой гарантирована многолетним видам в течение всего года.
Интенсивность транспирации пустынных растений определяется прежт
де всего объемом поглощаемой растением воды. При достатке влаги рас-
тения энергично транспирируют, что вместе с тем служит признаком бур-
ного газообмена и, следовательно, интенсивного фотосинтеза и продуци-
рования органического вещества. Известны результаты многочисленных
измерений величины транспирации. Первые несовершенные опыты про-
водились Арциховским, последующие, уже с помощью торзионных весов,—
Васильевым; данные весьма подробных исследований опубликовала Ко-
кина 1 2.
Она определяла интенсивность транспирации в конце июня, конце
июля и в середине сентября, то есть в самое сухое и жаркое время года.
Выявлена существенная разница между видами, растущими на подвижных
1 М. С. Шалыт, Подземные части некоторых луговых, степных и пустынных
растений и их сообществ, ч. II, «Труды Бот. ин-та АН СССР, серия III (геобот.)», 8,
Л., 1952, стр. 71—139.
2 V.M.Arcichowski, A. Osipov, Die Saugkraft der baumartigen Pflanzen
der zentralasiatischen Wiisten nebst Transpirationsmessungen am Saksaul, «Planta»,
14, 1931, S. 552—565; I. M. V as i 1 j e v, Uber den Wasserhaushalt von Pflanzen der
Sandwiiste im sudostlichen Kara-Kum, «Planta», 14, 1931, S. 225—309; С. И. К о к и -
н а, Водный режим и внутренние факторы устойчивости растений песчаной пустыни
Каракум, «Пробл. растениеводства и освоения пустынь», 4, 1935, стр. 99—196.
15*
228
Глава 4
песках, такими, как Smirnovia turkestanica и Ammodendron conollyi, транс-
пирация которых была весьма интенсивна, и видами, обитающими
на закрепленных песках,— Salsola subaphylla и Ephedra strobilacea. Послед-
ние росли более густо и транспирировали приблизительно в 10 раз сла-
бее. Другие виды, как видно из табл. 41, занимают промежуточное поло-
жение.
Таблица 41
Интенсивность транспирации у видов песчаной пустыни
(по Кокиной) (Smirnovia = 100)
Название растений Конец июня Конец июля Сентябрь
Smirnovia turkestanica . . . 100 100 100
Ammodendron conollyi .... 82,2 96,4 120,3
Calligonum caput-medusae . . 48,4 47,4 53,1
Haloxylon aphyllum 30,6 32,1 34,9
Haloxylon persicum 26,5 31,2 32,8
Salsola richteri 34,2 41,3 44,8
Salsola subaphylla 8,2 10,3 11,5
Ephedra strobilacea — 7,6 9,1
Smirnovia — монототипный эндемичный вид Средней Азии, широко
распространенный на дюнных песках. Этот кустарник из семейства бобо-
вых достигает] 1 м в’высоту, имеет простые, слабо опушенные листья,
которые появляются в начале апреля, вырастают до 2,5 см в длину, но
в начале лета опадают — остаются только позднее развившиеся более
мелкие листья; одновременно повышается осмотическое давление (ср.
Encelia, т. I, стр. 371). В августе сохраняются только мелкие, длиной 1 мм,
листья, которые растение позднее сбрасывает. Ammodendron, также являю-
щийся эндемиком, вырастает в высоту до 2,5 м‘, листья его густо опуше-
ны, и лишь часть их опадает. Это растение-пионер на голых песках \
Наибольшее количество влаги на транспирацию растение расходует
в июне, причем дневной максимум достигается уже к 10—11 часам. В тече-
ние лета, по мере ухудшения условий влагообеспечения и возрастания
водного дефицита, скорость испарения снижается; все виды растений
сокращают площадь транспирирующей поверхности, сбрасывая часть
листьев. Как показывает табл. 42, снижение интенсивности транспирации
особенно велико у Smirnovia.
Таблица 42
Интенсивность транспирации у видов песчаной пустыни
в разное время года
(июнь = 100)
Название растений Конец июля Сентябрь
Smirnovia turkestanica 76,6 42,2
Ammodendron conollyi 89,7 64,6
Calligonum caput-medusae . . . 87,5 56,2
Haloxylon aphyllum 85,2 53,3
Haloxylon persicum 90,0 54,5
Salsola richteri 92,6 57,5
Salsola subaphylla 95,8 66,6
1 Ю. С. Григорьев, Сравнительно-экологические исследования ксерофити-
зации высших растений, Л., 1955.
Аридные области Центральной Азии
229
Относительные значения интенсивности транспирации по дайным
Кокиной и Васильева совпадают, но абсолютные величины у Васильева
выше (табл. 43).
Таблица 43
Интенсивность транспирации у видов песчаной пустыни
в июне до полудня (в г/г сырого веса в час)
Название растений По Кокиной По Васильеву
Smirnovia turkestanica . . 1,075 2,93
Ammodendron conollyi . . 0,876 1,99
H aloxylon aphyllum .... 0,305 0,87
Haloxylon persicum .... 0,281 0,43
Salsola aphylla 0,073 0,20
Как видно из таблиц, все полученные Васильевым значения в 2—
3 раза превышают величины, определенные Кокиной, которая работала
с теми же видами и на том же местообитании несколькими годами позже.
Кокина считает данные Васильева завышенными и объясняет это тем, что
он измерял транспирацию очень маленьких веток (вес 0,5 г) и экспониро-
вал всего 1 минуту, вследствие чего ошибка резко возросла. Кокина
не подтвердила и вывода Васильева о том, что растения пустынь всегда
обеспечены влагой. Такому выводу противоречат следующие наблюдения:
в течение лета водный дефицит у пустынных видов нарастает, осмотиче-
ское давление повышается в полтора — два раза, транспирирующая
поверхность сокращается, вновь образующиеся листья обладают более
ксероморфным строением. Мы останавливаемся на этих определениях
столь подробно потому, что опубликованные на немецком языке данные
Васильева совершенно выпадают из рамок обычно получаемых значений
и поэтому часто цитируются, тогда как сомнения Кокиной в точности
определения абсолютных значений остаются неизвестными.
Тем не менее измерения Кокиной также показали, что листья Smir-
novia и Ammcdendron хорошо обеспечены влагой: листья на срезанных
ветках быстро засыхают. Оба вида растут на открытых местообитаниях,
где конкуренция между корнями отсутствует и где содержание воды в поч-
ве особенно высоко; поэтому поглощение влаги тем и другим видом про-
текает беспрепятственно.
В песчаных пустынях Средней Азии производилось также измерение
интенсивности фотосинтеза 1. Весьма сильная транспирация свидетель-
ствует об активном поглощении растениями СО2, как и об относительно
большом продуцировании органического вещества (см. табл. 25). Однако
приводимые Костычевым цифры трудно поддаются интерпретации. В тече-
ние дня он получил чрезвычайно высокие значения интенсивности асси-
миляции СО2, но вслед за тем установил выделение СО2. В настоящее вре-
мя известно, что этот процесс связан с перегревом листьев в испытатель- 1 * з
1 S. Kostytschev, Н. Kardo-Syssojeva, Untersuchungen uber den
Tagesverlauf der Photosynthese in Zentralasien, «Planta», 11,1930, S. 117—143; С. P. Фа-
з ы'л о в а. Фотосинтез некоторых полукустарников, кустарников и деревьев южных
Кызылкумов, Ташкент, 1962 (дисс.); Ю. С. Н асыр о в, М. А. Л о гино в, Потен-
циальный фотосинтез и коэффициент использования солнечной энергии растениями
Таджикистана, «Физиология и биофизика растений», 2, Душанбе, 1963.
230
Глава 4
ной камере и не соответствует истинным условиям в природе. Отметим, что
производились также эколого-анатомические исследования растений Сред-
ней Азии х.
Рассмотрим здесь материалы весьма подробного изучения водного
режима Мильской и Муганской степей, лежащих в Куро-Араксинской низ-
менности. Правда, эта область расположена не в Средней Азии, а в восточ-
ном Закавказье, но ее климат и растительность во многом напоминают
условия среды в Средней Азии. Милзскую степь ограничивает Кура
на севере и Араке на юго-востоке; Муганская степь простирается восточ-
нее долины Аракса до ^Каспийского моря. Еще в начале нашего летосчис-
ления эта низменность при более высоком стоянии уровня Каспийского
Рис. 163. Климадиаграмма
(неполная) опытной станции
в Мильской степи, восточный
Кавказ (но данным Бейдеман).
моря была морским заливом, который позд-
нее был заполнен преимущественно наноса-
ми Куры и Аракса и полностью осушился
при понижении уровня моря. В V —VIII вв.
область была густо заселена; разветвленная
сеть оросительных ’каналов питалась водами
Аракса. В XIII в. Чингисхан истребил почти
все местное население. Лишь в XV в. Тамер-
лан повелел восстановить оросительные ка-
налы, что позволило снова частично заселить
земли. В настоящее время площади под
поливными культурами резко расширяются. Это в свою очередь потре-
бовало проведения детальных экологических исследований2.
В апреле воды Аракса несут до 25 г!л взвешенных частиц, в сентяб-
ре — 12 г/л. Воды содержат 0,5 г!л растворенных солей (NaGl) и имеют
щелочную реакцию. Грунтовые воды, питаемые в основном Араксом,
в прибрежной части низменности залегают сравнительно высоко и содер-
жат хлориды. Климат весьма засушливый (см. климадиаграмму на
рис. 163). Время с июля до августа сухое, максимум дождей приходится
на октябрь — ноябрь. Годовое количество осадков колеблется от 227
до 369 мм.
Годовые потери воды на транспирацию у растений, корневые системы
которых достигают уровня грунтовых вод, составляют 1500 мм (заросли
Phragmites). Почвы представлены сероземами. В ряде мест активны про-
цессы засоления. Зональная растительность — полынная полупустыня;
на площадях с высоким уровнем грунтовых вод распространены поймен-
ные леса, образованные Populus hybrida, Morus alba и Salix australior,
болота {Phragmites, Bolboschoenus, Arundo) или луга {Cynodon, Glycyrrhiza)'
кроме того, наблюдаются сообщества галофитов: на влажных почвах —
Halocnemum, Halostachys, Kalidium, на более сухих — Artemisia meyeriana,
Salsola deniroides, а также сообщества псаммофитов. В полупустыне наря-
ду с Artemisia meyeriana растут Роа bulbosa var. vivipara и Capparis spino-
sa, много эфемерных и низших растений {Nostoc, Collema). Интерес пред-
ставляет произрастание здесь Nelumbium caspicum и Pistacia mutica. Эко-
логически своеобразна Salsola dendroides. Останавливаться здесь подроб-
нее на характеристике типов растительных сообществ и закономерностях
1 В. К. Василевская, Анатомические приспособления растений жарких
и холодных пустынь Средней Азии и Казахстана, «Пробл. современной ботаники»,
2, М., 1965, стр. 5—18.
2 И. Н. Бейдеман, С. Г. Беспалова, А. Т. Рахманин, Эколого-
геоботанические и агромелиоративные исследования в Куро-Араксинской низменно-
сти Закавказья, М.—Л., 1962.
Аридные области Центральной Азии
231
их размещения в зависимости от положения уровня грунтовых вод и сте-
пени засоления почв мы не имеем возможности.
Биология и фенология Salsola dendroides детально изучались; так же
подробно исследовались корневые системы этого вида и некоторых других
видов, корни которых всегда достигают водоносного горизонта: наряду
с Salsola dendroides это также галофиты Halocnemum strobilaceum, Halos-
tachys caspica, Tamarix ramosissima, кроме того, Glycyrrhiza glabra и Lago-
nychium forctum, распространенные лишь на слабозасоленных почвах.
Limonium scoparium, Cynodon dactylon, Camphorosma lessingii и Suaeda
microphylla столь обязательной связи с грунтовыми водами не обнару-
живают.
По степени и характеру влагообеспеченности различают 4 группы
растений:
1. Гидрофиты, например Phragmites, связанные с очень высоко
лежащими грунтовыми водами, нередко выступающими даже на поверх-
ность почвы.
2. Фреатофиты, развивающиеся на местообитаниях с несколько
более низким стоянием грунтовых вод, например Alhagi, Capparis, Nitra-
ria, Kalidium и выше названные виды.
3. Трихогидрофиты, корни которых не достигают уровня
грунтовых вод, но проникают до капиллярной каймы х.
4. Омброфиты, питающиеся исключительно накопленной^поч-
ве дождевой водой, например Artemisia meyeriana.
У фреатофитов максимальная транспирация в разгар лета в полдень
•составляет (в г на 1 г сырого веса в час):
Phragmites
Glycyrrhiza
Lagonychium
0,9 Alhagi
1,8 Capparis
1,9
1,06
1,15
корнями до зеркала грунтовых
У галофитов, также проникающих
вод, измерены следующие значения:
Tamarix 1,3 Halostachys 0,7
Halocnemum 0,6 Nitraria 0,5
Kalidium 0,7
Величина транспирации у других видов:
Cynodon 1,7 Camphorosma 0,6
Lycium 0,9 Salsola dendroides 0,5
Aeluropus 1,3 Salsola crassa 0,3
Limonium 0,9 Petrosimonia brachiata 0,3
Suaeda microphylla 0,7 Salicornia europaea 0,3
Atriplex tatarica 0,7 Suaeda confusa 0,3
Из омброфитов исследовалась только Artemisia meyeriana. Макси-
мальная интенсивность ее транспирации достигает 0,9 г!г -час, но в усло-
виях затрудненного водоснабжения составляет лишь 0,3 г/г-час. Сравне-
ние этих величин со значениями, полученными для других видов Arte-
misia, показало, что все полыни расходуют на транспирацию примерно
•одинаковое количество влаги (табл. 44).
Как следует из таблицы, у всех видов Artemisia интенсивность транс-
пирации летом и осенью резко сокращается, что связано с иссушением
1 Представляется спорным, можно ли действительно различить между собой
типы 2 и 3.
232
Глава 4
Таблица 44
Данные сравнительных измерений интенсивности
транспирации у видов Artemisia в период с апреля по сентябрь
(в г/г сырого веса в час)
Название растений и место произрастания IV V VI VII VIII IX
A. meyeriana, Муганская степь 0,62 0,46 0,61 0,30 0,17 0,20
A. pauciflora, центр. Ка- захстан 0,72 0,67 0,32 0,18 0,12 -—
A. pauciflora, зап. Казах- стан 0,48 0,27 0,24 0,17
A. semiarida, центр. Ка- захстан 0,60 0,58 0,31 0,17 0,11 —
почвы — в Муганской степи это наблюдается лишь в июле после прохож-
дения июньских дождей, а в Казахстане, где преобладают весенние дожди,
раньше. По Григорьеву (1955), осмотическое давление у A. semiarida
возрастает при этом с 25 до 40 атм и более.
Особенно интересны результаты вычисления суммарного потребления
влаги растительным покровом за год. Для этой цели вначале определяли
сырой вес транспирирующих частей растений в расчете на 1 га площади.
Полученные цифры умножили на соответствующие средние значения
за день и на количество дней в году.
В табл. 45 приведены величины суммарного потребления воды неко-
торыми растительными группировками в летние месяцы (проективное
покрытие 40—70%).
Таблица 45
Потребление воды растительным покровом в летние месяцы (в мм)
Не галофиты V VI VII , VIII IX Всего
Phragmites communis . . 484 348 251 345 81 1509 '
Glycyrrhiza glabra .... 25 203 257 223 29 737
Cynodon dactylon .... 7 61 78 69 4 219
Alhagi pseudoalhagi . . . 7 14 12 14 5 51
Artemisia meyeriana . . . 22 35 17 12 16 102
Хлопчатник, поливной 1 119 180 316 134 777
Галофиты
Tamarix ramosissima . . . 54 238 201 193 167 853
Salsola dendroides1 . . . 32 47 63 49 20 211
Halocnemum strobilaceum 16 88 136 116 52 408
Suaeda microphylla . . . 39 57 39 26 19 180
A eluropus repens 41 39 59 49 2 190
Salicornia europaea . . . 10 43 64 21 16 155
1 С марта по декабрь расход воды на транспирацию оценивается в 330 мм.
Аридные области Центральной Азии
233
Эти величины потребовались для того, чтобы составить карту расхода
влаги на транспирацию отдельными растительными сообществами Муган-
ской степи. Точность приведенных сведений не следует переоцениватьг
однако они все же дают представление о том, сколько грунтовой воды
потребляют растения. Сравнение этой карты с картой залегания подзем-
ных вод отчетливо показывает, что большое потребление влаги растения-
ми возможно лишь там, где их корни проникают до горизонта пресных
грунтовых вод. Над солеными грунтовыми водами растут только типич-
ные галофиты, у которых расход влаги на транспирацию сравнительно
мал. Интенсивно транспирирующий Tamarix переносит лишь очень сла-
боминерализованную воду. Там, где грунтовые воды недосягаемы для
корней растений, расход влаги на транспирацию растительным покровом
всегда должен быть меньше годовой суммы осадков.
В настоящее время большая часть площади Муганской степи занята
поливными культурами, причем избыточная вода сбрасывается по дре-
нажным канавам. В результате систематического полива содержание-
солей в обрабатываемых почвах сократилось, а на необрабатываемых
землях, наоборот, увеличилось. Растения, всасывая из почвы воду, погло-
щают с ней некоторое количество солей, которые накапливаются в транс-
пирирующих частях растения и после его отмирания снова, попадают
в почву. Если же естественный растительный покров потребляет больше-
влаги, чем поступает в почву с атмосферными осадками, то это свидетель-
ствует об активном подтоке грунтовых вод и, следовательно, о дополни-
тельном привносе солей. Эти соли, накапливаясь в почве, обусловливают
ее постепенное засоление. Действительно, за 20 лет на всех неорошаемых
площадях характер растительности изменился в сторону, свидетельствую-
щую об известном нарастании засоления,— сравнительно солевыносли-
вые растения сменились более резко выраженными галофитами. Посколь-
ку причиной такого засоления является транспирация растений, Бейде-
ман говорит о «биологическом» накоплении солей.
КРУГОВОРОТ ВЕЩЕСТВ И СОЛЕВОЙ РЕЖИМ
РАСТИТЕЛЬНОСТИ ПУСТЫНЬ 1
Для различных растительных сообществ среднеазиатских пустынь
были определены: 1) общий запас сухой органической фитомассы, вклю-
чая надземную и подземную (в большинстве случаев до глубины 50 см}
части; 2) количество ежегодного опада за счет отмирающих надземных
частей и корней; 3) общее количество ежегодно поступающих в почву
азота и зольных элементов.
Расчет величины 2 для однолетних растений трудности не представ-
ляет, в данном случае она равна величине 1. У многолетних же видов
нельзя непосредственно определить массу ежегодно отмирающих корней.
Эта доля исчислена исходя из средней продолжительности жизни много-
летних видов. Например, для Artemisia badghysi, достигающей в среднем'
20-летнего возраста, ежегодный опад принимался равным V20 части всего
количества многолетних корней. Для травянистых многолетних злаков
было принято, что ежегодно отмирает г/3 общей массы корней. Эти расчеты-
оказались сходными с данными непосредственного определения корневых
1 Л. Е. Родин, Динамика растительности пустынь. На примере Западной
Туркмении, М.—Л., 1961.
234
Г лава 4
Рис. 164. Фитомасса (общий запас сухой органической массы) и еже-
годный опад (надземный и подземный) в различных растительных сооб-
ществах среднеазиатской пустыни. Указаны общее количество опада (оп.)
в % фитомассы, содержание азота (N) и зольных элементов в % опада
£(по Родину).
1 — ассоциация Artemisia kemrudi—Anabasis salsa (on. 27%, N 1,5%, зольность 5,1 %);
2 — Artemisia badghysi с эфемерами (on. 86,%, N 0,9%, зольность 4,7%); 3 — чистая
заросль Anabasis salsa (on. 42%, N 2,3%, зольность 9,3%); 4 — однолетние галофиты
c Suaeda (on. 85%, N 1,3%, зольность 13,4%); 5 — чистая заросль Halocnemum
strobilaceum (on. 30%, N 1,8%, зольность 16,5%); в — Suaeda совместно с однолет-
ними галофитами (on. 40%, N 1,6%, зольность 11,2%); 7—чистая заросль Ana-
basis aphylla (on. 49%, N 2,8%, зольность 6,5%); 8 — чистый луг с Егеторугит
orientale на намытой лаве в предгорье (оп. 100%, N 1,2%, зольность 7,6%, много
SiO2); 9 —луг с Arfemisia и Роа bulbosa в западине (оп. 77%, N 1,3%, зольность
6,9%); 10 — ассоциация Aeluropus—Vida—Alopecurus в низине на луговом сероземе
(оп. 73%, N 1,7%, зольность 5,2%, много SiO2); 11 —заросли Haloxylon aphyllum
с участием Tamarix (1 %) над солеными грунтовыми водами (оп. 19%, N 1,9%, золь-
ность 6,3%); 12 — кустарниковые тугаи (Tamarix—Aeluropus) над солеными грунто-
выми водами (оп. 61%, N 1,1%, зольность 13%); 13 — тростниковое болото (Phragmi-
tes) в дельте Амударьи; надземная фитомасса может достигать максимально 100 т/га
(оп. 51%, N 1,5%, зольность И %; 58% золы составляет SiO2, 10% G1, 7,5% S, 14% К,
6% Ga, 2,5% Na); 14 — пойменный лес с Salix songarica—Calamagrostis pseudophrag-
mites (on. 9%, N 0,6%, зольность 5,4%); 15 — пойменный лес c Elaeagnus orientalis—
Alhagi persarum (on. 18%, N 1,4%, зольность 4,8%); 16 — пойменный лес c Populus
diversifolia—Halimodendron halodendron, поверхность почвы слабо засолена (on. 14%,
N 1,4%, зольность 5,6%); 17—пойменный лес с Populus diversifolia—Aeluropus,
засоление более заметно (оп. 17%, N 1,0%, зольность 6%); 18 — Alhagi persarum на
заброшенном орошаемом участке; когда запас влаги в почве израсходован, его заме-
щает Anabasis aphylla', корни исследованы лишь до глубины 25 см (оп. 30%, N 1,9%,
зольность 5%).
масс в степных сообществах. Для Роа bulbosa, ежегодно образующего мно-
жество отмирающих эфемерных корешков, для расчета было принято, что
•ежегодно в опад поступает половина общей массы корней. Всего Родиным
изучено 31 растительное сообщество. Это единственные в своем роде дан-
ные, имеющиеся для области пустынь. Поэтому мы остановимся на них
подробнее. На рис. 164 графически изображены общий запас сухой орга-
нитеской массы, надземная и подземная фитомассд в различных сообще-
Аридные области Центральной Азии
235
ствах пустынной растительности, а также количество ежегодно уходящего
в опад общего запаса органического вещества, слагаемого из массы отмер-
ших надземных частей и корней. Приведено также процентное содержание
азота и золы в опаде. В общем, из имеющихся данных можно сделать
следующие выводы:
1. У всех растительных сообществ пустыни доля подземной фитомассы
значительно превосходит надземную. Разница невелика только у сооб-
ществ эфемеров или у сообществ, обитающих на переувлажненных поч-
вах. В среднем на корни приходится 75% (67—91 %) от общего количества
органического вещества, в пойменных лесах — 50% (45—54%).
2. В структуре ежегодного опада наибольшую массу дают также
корни. Среди пустынных формаций саксаульники занимают первое место
по степени концентрации количества живого вещества в подземных орга-
нах растений.
3. Отмершая масса корней бедна золой. Основная доля зольных эле-
ментов содержится в опаде зеленых ассимилирующих органов листьев,
стеблей. На последние приходится 9—28% всего опада; лишь при хорошей
обеспеченности водой в пойменных лесах эта доля возрастает до 41—56%.
4. Чрезвычайно высокой зольностью отличаются злаки и галофиты.
У первых это обусловлено содержанием SiO2 (окремневший эпидермис),
у вторых связано с накоплением солей в клеточном соке растений.
5. Накопление главнейших элементов по-разному осуществляется
у трех групп галофитов: а) аккумулируют преимущественно NaGl,
б) накапливают относительно много сульфатов и в) содержат Na больше,
чем это соответствует эквивалентному количеству С1' и SO'j. Следова-
тельно, натрий присутствует в клеточном соке в виде солей органических
кислот.
6. Особую группу образуют галофиты, выделяющие соли. В образуе-
мый ими опад поступает относительно небольшое количество элементов,
но известно, что виды Tamarix и Aeluropus сбрасывают некоторое неучи-
тываемое количество соли на поверхность почвы в течение всего вегета-
ционного периода.
7. Содержание азота в опаде пустынных сообществ относительно высо-
кое. Доля азота в опаде в среднем составляет: в пустыне — 1,7% (в пой-
менных лесах — 0,9%), в степях — 1,2 и в бореальных лесах — 0,5%.
8. Содержание золы в опаде пустынных формаций (8%) превосходит
аналогичные показатели для опада лесных (3,3%) и степных (7,3%) сооб-
ществ.
9. Чем больше общий запас органической массы, тем меньшая ее
доля поступает в опад. В лесах органическое вещество как бы консерви-
руется в одревесневших стволе и корнях и снова вовлекается в круговорот
веществ лишь спустя длительное время. Напротив, в сообществах эфеме-
ров, создающих сравнительно небольшую фитомассу, опад ежегодно пол-
ностью минерализуется. Еще интенсивнее совершается биологический
круговорот в сообществах такырных водорослей. Косвенным путем опре-
делена их сухая масса в расчете на 1 га площади; она составила 0,5—0,6 т.
Можно предполагать, что в течение вегетационного периода кругооборот
веществ в этих своеобразных биоценозах водорослей совершается
неоднократно.
Доля высших растений на такырах чрезвычайно мала. На слабозарос-
ших такырах общая фитомасса высших растений достигает 0,15—0,3 т!га,
на сильнозаросших — увеличивается до 0,6—1,6 т!га. Опад в этом слу-
чае составляет 100%, так как живыми остаются только семена; зольность
236
Глава 4
опада равна 7,3—9,1%, содержание азота в нем достигает 2%. В заклю-
чение приведем составленную Родиным по различным источникам таблицу^
в которой сопоставлены между собой различные группы формаций по раз-
мерам фитомассы на единицу площади и по количеству ежегодного опада
(табл. 46).
Таблица 46
Биологический запас (общее количество органической массы)
и количество ежегодного опада в различных группах
формаций (в т/г)
Группы формаций Биологиче- ский запас Опад
Сосновые леса 200 7
Еловые леса 150—380 6-20
Липовые смешанные леса .... 160-220 5—10
Дубовые леса 160-500 6-27
Осинники 190-250 12-25
Остепненные луга 11-16 6-9
Луговые степи Разнотравно-типчаково-ковыль- 20—23 9-13
ные степи Пустынные сообщества Централь- ной Азии: 18—34 8—14
Древесные тугаи 70-80 7—13,5
Кустарниковые тугаи 6-11 3,5—6,5
Травяные тугаи 35 18
Пустынные луга 24-45 20—32
Саксауловые пустыни 55 10,5
Солянковые пустыни 2-12,5 1—5,5
Эфемерово-полынные пустыни 11—14 6-10,5
Полынные пустыни 4,5 1,2
Зарастающие такыры Водорослевые такыры с немно- 0,6—1,6 0,6-1,6
гими эфемерами 0,1—0,3 0,1—0,3
Для пустынь характерно повсеместное распространение засоленных
почв. Заметную роль в образовании засоленных почв играет отложении
пыли. В больших количествах соль содержится во всех морских осадоч-
ных отложениях, и прежде всего в часто обнажающихся на поверхности
морских третичных породах. Переотложение солей происходит обычно
с помощью воды; соли смываются на ниже расположенные участки рельефа
и интенсивно накапливаются в местах испарения капиллярной влаги.
Растения, произрастающие на засоленньгх местообитаниях, по лощают
вместе с водой соли, совершенно не нужные им для обмена веществ, такие,
как хлористый натрий и сернокислый натрий. Эти соли накапливаются
в транспирирующих органах растений, которые, как известно, одновре-
менно осуществляют процесс фотосинтеза. Поэтол у такие органы характе-
ризуются наивысшей зольностью, и прежде всего содержат много балласт-
ных солей (NaCl, Na2SO4). Для большинства растений такая аккумуля-
ция солей даже в небольших размерах оказывается вредной; поэтому
на засоленных почвах эти виды не выдерживают конкуренции. Галофиты,
напротив, легко переносят относительно большую концентрацию солей;
соли в известной мере даже стимулируют их рост. Эти виды успешно
развиваются на засоленных почвах и со временем начинают дажо
Аридные области Центральной Азии
237
Рис. 165. Схема распределения растительности в засоленной озерной де-
прессии в долине реки Чиж, западный Казахстан (по Ларину).
1 — вода; 2 — заросли тростника; 3 — Agropyrum repens или Calamagrostis (Gala-
tella glabra)', 4 —галофиты (Halocnemum, Suaeda, Atriplex, реже Aeluropus); 5 — Atro-
pis C Agropyrum cristatum или Galatella villosa', 6 — то же c Artemisia incana (г;ло-
фильное растение); 7 — Atriplex verrucifera’, 8 — комплексная полупустыня. Западина
обладает, по-видимому, некоторым подземным стоком, иначе засоление увеличива-
лось бы по направлению к центру, как это отражено на рис. 166.
доминировать, поскольку быстрее растущие, но менее солеустойчивые
виды по мере нарастания соленосности почвы один за другим выбывают
мз’конкурентной борьбы. Вокруг засоленных впадин наблюдается очень
резко выраженная поясность растительности, обусловленная активизацией
соленакопления в направлении к центру впадины (рис. 165 и 166). Там, где
содержание солей становится предельно высоким, растительность полно-
стью отсутствует. На этот факт мы неоднократно указывали в т. I и отме-
чали различия в количестве и качестве солей, поглощаемых растениями,
которые пронизывают своими корнями одну и ту же почву. Мы различаем
238
Глава 4
Artemisia terrae albae^:-
Salsola gemmascens й-
Anabasis salsa^
Haloxylon aphyllum^
Ё
Е
Е
'Haloxylon aphyllum
Artemisia terras albae
Nanophyton erinaceum
и
о
И
2
й
S
и
Limonium suffruticosum ~
Salsola arbuscula
Artemisia terrae albae
Salsola-gemmascens
Arte mis ш_ terrae_albae
Halocnemum strobilaceum
сев <----ююз
Halocnemum strobilaceum
Salsola gemmascens
Artemisia terrae albae
Halocnemum-strobilaceum
Вертикальный масштаб увеличен в 20 раз. В кружках показан химический состав (в %) водных вытяжек из почвы (места взятия проб
отмечены на профиле ямками). В центре впадины наблюдается присутствие только NaCl, к периферии увеличивается доля сульфатов и
в некоторой степени также растворимых карбонатов.
Аридные области Центральной Азии
23»
виды, накапливающие в своих транспирирующих органах преимуществен-
но NaCl и, следовательно, являющиеся суккулентами, и виды, которые
аккумулируют наряду с хлоридами сравнительно много сульфатов и поэто-
му отличаются меньшей суккулентностью или вовсе суккулентами
не являются (см. т. I, стр. 386, 405, 433). Эти рассуждения подтверждаются
данными Родина, осуществившего многочисленные определения состава
золы для растений среднеазиатских пустынь.
Из его весьма содержательных материалов мы приведем здесь только
часть анализов и ограничимся сравнением транспирирующих и ассимили-
рующих органов. Эту роль исполняют либо^листья, либо, при редукции
последних, фотосинтезирующие однолетние побеги; те и другие ежегодно
возобновляются (рис. 167, 168).
На графиках представлены только сумма зольных веществ (зольность)
в процентах от сухого веса и содержание тех элементов, которые в виде
легкорастворимых солей накапливаются в клеточном соке и обусловли-
вают повышение осмотического давления. Из анионов — это ионы хлора
и сульфата, из катионов — натрий; калий приведен лишь в тех случаях,
когда он составляет более 5% общего количества золы. Содержание Са
и Mg на графиках не отражено. Правда, эти элементы также могут суще-
ствовать в растворе, но большая их часть, видимо, присутствует в нерас-
творимой форме; так, например, кальций, на долю которого часто прихо-
дится более 10% и даже более 20% золы, преимущественно содержится
в растениях как щавелевокислый кальций, то есть в форме конечного
продукта обмена веществ.
У злаков общее количество зольных элементов вследствие окремнения
эпидермиса обычно резко повышено, что может создать впечатление
о высоком содержании солей в листьях злаков. Поэтому на графиках
всякий раз это окремнение отмечено. Для негалофитов показано также
содержание фосфора, имеющее в определении кормовой ценности расте-
ний особое значение. При выпасе скота на естественных пастбищах сле-
дует учитывать, что фосфор является элементом, который находится в рас-
тениях в минимальном количестве. У галофитов содержание фосфора
по сравнению с другими зольными компонентами всегда очень мало.
В работе Родина зольность выражена как процентное содержание Na2O, К2О,
Cl, SO3. Для удобства сравнения содержаний катионов и анионов, возможно, было бы
целесообразнее выразить эти значения в грамм-эквивалентах. Однако пробный расчет
показал, что общая картина графика изменилась бы мало. Столбцы для сульфата оказа-
лись бы несколько короче, чем для хлора. В меньшей мере это относится и к К по срав-
нению с Na. Относительная длина столбцов для С1 в сравнении с длиной столбцов Na
осталась бы почти такой же. Поэтому от пересчета мы отказались. Бросается в глаза,
что столбцы анионов у галофитов часто бывают длиннее столбцов катионов. Во всех
случаях, когда содержание калия для достижения равновесия недостаточно, следует
предполагать, что в клеточном соке имеются ионы кальция. -"I 77 7^
Для многих галофитов длина столбца натрия намного больше длины
столбцов хлора и сульфата. В этих случаях в клеточном соке должны
присутствовать значительные количества органических кислот. При озо-
лении образуется карбонат натрия, и зола будет отличаться резко щелоч-
ной реакцией. То же самое произойдет при разложении опада. Следова-
тельно, опад таких видов, минерализуясь, может вызвать щелочное содо-
вое засоление и солонцеватость почв. Поэтому мы называем эти виды ще-
лочными галофитами, хотя в живых органах клеточный сок,
несомненно, имеет кислую реакцию. На рис. 167 приведены данные анали-
зов золы галофитов. Мы подразделяем их на галофиты, накапливающие
соли, и галофиты, выделяющие соли. Последние через особые железки
240
Глава 4
Выделяющие Щелочные Сульфатные Хлоридные галофиты
соль галофиты галофиты галофиты
Рис. 167. Состав золы галофитов по данным Родина.
а — вычислено на золу без SiO2; содержание SiO2 в золе 65%; б,— то же, но содержа-
ние SiO2 в золе 36%. Объяснения см. в тексте.
Аридные области Центральной Азии
241
Разнотравье
Alhagi persarum
tlalcolmia africana
Lepidium perfoliaium
Artemisia kemrudica
Artemisia badghysi
Matricaria lamellata
Qlycyrrhiza glabra
Злаковидные растения
Phragmites communis
Paa bulbosa
Stipa szowitsiana
Carex physodes
Пойменные
dpefiecHbie растения
Salix songarica
Populus diversifolia
ELaeagnus orientalis
Рис. 168. Состав золы негал офитов по данным Родина.
Объяснения см. в тексте.
в листьях могут сбрасывать некоторое количество поглощенных солей
(прежде всего NaCl). Поэтому содержание в них золы оказывается суще-
ственно ниже. У Tamarix выделившаяся соль еще долго остается на поверх-
ности листьев. Неясно, учитывалось ли это обстоятельство при определе-
нии средней зольности. Возможно, что относительно высокое содержание
NaCl у Tamarix laxa, так же как и в последней пробе Aeluropus, объяс-
няется именно этой особенностью солевого режима.
В отношении галофитов, не выделяющих соль, обращает на себя вни-
мание крайне высокая зольность отдельных видов, иногда достигающая
х/3 всей массы сухого вещества; при этом соли калия занимают долю,
намного меньшую, чем соединения натрия. Лишь в Anabasis aphylla
и Salsola rigida они составляют около половины. Иначе обстоит дело
с анионами. На их основе можно выделить три типа галофитов, связанных
между собой переходными типами:
£•1) хлоридные галофиты, накапливающие много хлоридов
и мало сульфатов;
16-0434
242
Глава 4
2) сульфатные галофиты, аккумулирующие относительно
много сульфатов;
3) щелочные галофиты, в золе которых хлориды и сульфа-
ты присутствуют в сравнении с содержанием натрия в небольшом коли-
честве.
Поскольку растения в первую очередь освобождаются от NaCl, в золе
выделяющих соль видов Tamarix и Aeluropus наблюдается относительно
высокое содержание К2О и SO3.
Совершенно иную картину дает анализ золы негалофитов (рис. 168),
которые лишь в крайнем случае растут на слабозасоленных местообита-
ниях. Содержание золы только у солеустойчивых Matricaria и Phragmites
превышает 10%. Однако у последнего зола на 69% состоит из SiO2. Без
SiO2 зольность Phragmites окажется ниже 10% — величина, близкая
к содержанию золы у Stipa.
Некоторое количество NaCl поглощают и эти растения, обычно боль-
ше С1, чем Na. Содержание последнего в золе существенно меньше кон-
центрации калия; в большинстве случаев хлора в ней также меньше,
чем SO3. Однако следует учитывать, что часть SO3 связана с серой, кото-
рая содержится в белковых соединениях. Листья типичных видов древес-
ных тугаев, связанных с пресными грунтовыми водами, ведут себя как
листья настоящих негалофитов.
В заключение еще раз подчеркнем, что все изложенное выше отно-
сится лишь к транспирирующим органам растений. Содержание раствори-
мых неорганических солей в осевых и корневых органах даже у типичных
галофитов весьма и весьма мало.
ПУСТЫНИ ЦЕНТРАЛЬНОЙ АЗИИ^
Выше мы уже отмечали, что пустыни Центральной Азии отличаются
от среднеазиатских пустынь иными условиями атмосферного увлажнения:
сюда осадки приходят с востока, из муссонной области Китая, и выпадают
преимущественно летом. Зима и весна сухие. Поэтому здесь почти пол-
ностью отсутствуют весенние эфемеры, играющие столь большую роль
в растительном покрове Средней Азии. Количество осадков летом сокра-
щается в направлении с востока (Ордос — 250 мм) на запад (в западной
части пустыни Гоби оно не превышает 35 мм). В переходной области —
Джунгарии — атмосферные осадки более или менее равномерно распре-
делены внутри года, сумма их колеблется в пределах 97—193 мм. Один
из самых сухих районов Центральной Азии — Таримская впадина. Она
получает осадки как с западными (максимум в начале лета), так и восточ-
ными ветрами (максимум дождей в августе), тем не менее максимум осад-
ков не превышает 65 мм (Кашгария), а во впадине Лобнор составляет лишь
И мм. В общем, Центральная Азия засушливее Средней Азии, и отсут-
ствие влажного времени года весной отрицательно сказывается на разви-
тии растительности. Восточную периферию области центральноазиатских
пустынь дренирует река Хуанхэ, впадающая в Тихий океан; значительно
большая по размерам внутренняя часть бессточна. Продукты выветрива- -
ния накапливаются в понижениях рельефа и перевеваются ветром. Соот-
ветственно этому огромные площади здесь занимают песчаные пустыни.
1 М. П. П е т р о в, Пустыни Центральной Азии, т. I, Ордос, Алашань, Бэйшань,
М.—Л., 1966. Более раннюю литературу см.: Schimper-Faber, 1935. См.
также: Chi-Wu Wang, The forests of China, «Carbot. Found. Publ.», № 5, 1961.
Аридные области Центральной Азии
243
Своеобразна гидрография внутренних районов ЦентральнойтАзии. Много-
численные лишь периодически заполняющиеся водой сухие русла впадают,
образуя обширные дельты, в бессточные озера, часто меняющие свои
очертания и размеры. С окружающих высоких горных хребтов на пред-
горные равнины стекает сравнительно^много воды, поэтому запасы грун-
товых вод часто значительны. £ . 7- F'.-
Характер растительности в зависимости от сухости местообитания,
высоты залегания грунтовых вод и степени засоления почв* изменяется
от типичных аридных сообществ до сообществ, в^избытке обеспеченных
влагой. Там, где широко распространены пески, преобладают псаммо-
фиты. Флора бедна. На востоке господствуют восточнокитайско-монголь-
ские элементы, и среди них кустарниковидные псаммофиты родов Сага-
gana, Hedysarum, Artemsia и др.; кроме того, встречаются некоторые
эндемики, как, например, Tetraena mongolica (парнолистниковые), Атто-
piptanthus mongolicus и др. На западе^ флора^напоминает флору Средней
Азии.
Плато Ордос
Плато Ордос занимает северо-западную часть большой излучины реки
Хуанхэ (см. рис. 140); его южную границу образует Великая китайская
стена, проходящая вдоль края массивов барханных песков в зоне кон-
такта их со степями Лёссового плато (1600—1800 м абсолютной высоты).
Прежде степная растительность покрывала всю территорию Лёссового
плато, сегодня густо изрезанную системой; оврагов. Эрозия почв — след-
ствие земледелия — вызвала разрушение естественного растительного
покрова; сохранились лишь небольшие остатки его х. Климат резко кон-
тинентальный (рис. 169—170). Декабрь — февраль холодные (средняя
минимальная температура января —13,5°). Весна необычайно сухая,
с июня до сентября идут сильные дожди, приносимые теплыми и влажными
юго-восточными муссонными ветрами. Доминируют виды Stipa, но цен-
тральноазиатские элементы, как S. brevifolia, S. bungeana, виды Cleistoge-
ЮЙЛИНЬ (1121м) 8,7°405
Ге] Испаря-
L J i емость
j И689ММ-
Р и с. 169—170. Климадиаграммы Ордоса.
Юйлинь — близ границы степи; Дэнкоу — район пустыни.
’ F1 Е. М. Л а в р е и к о, Степная растительность лёссовых холмов в восточной
части провинции Ганьсу (Китай), «Изв. АН СССР, сер. биол.», 5, 1959, стр. 714—728.
16*
244
Глава 4
nes, кроме того, полукустарники, как Тапасе turn fruticulosum, реже Reau-
muria soongarica, Zygophyllum mucronatum, Convolvulus ammanii и др.
Из однолетников встречаются только цветущие осенью Artemisia pectindta,
виды Salsola и др. Покрытие почвы редко превышает 15% (до 30%). Речь
идет о крайних юго-восточных переселенцах Евроазиатской зоны пустынь
3
CD
2 3 4 3 4 3 754 5656 7568В
Рис. 171. Схематический геоботанический профиль через Ордос от гор Арбисо до
Лёссового плато (по Петрову).
1 — мелкокустарничковая пустыня в горах Арбисо; 2 — полупустыня с Reaumuria soongarica,
Caragana tragacanthoides, Stipa glareosa', 3 — ассоциация ордосской полыни (Artemisia ordosica);
4 — сообщество гидрогалофитов; 5 — остепненные полынники с Artemisia ordosica', 6 — остепненные
луга; 7 — кальцефитная группировка на выходах коренных меловых песчаников с древними карбо-
натными почвами (Gypsophila, Euphorbia, Haplophyllum); 8 — ксерофитная степь Лессового плато.
и степей, служащей переходным звеном к областям Восточной Азии с лет-
ними дождями. Почвы — не черноземы, бедны гумусом.
|$На плато Ордос широко распространены меловые и юрские песчаники,
отличающиеся очень слабой сопротивляемостью выветриванию. В условиях
очень засушливого климата третичного времени и плейстоцена корен-
ные породы подверглись энергичному физическому выветриванию. В ре-
зультате обширные площади оказались занятыми эоловыми песками. Засо-
ление почв слабое. Климат плато в направлении с юго-востока на северо-
запад становится все более аридным, растительность соответственно —
все более пустынной. Основной период засухи приходится на начало лета.
Центральная часть плато Ордос бессточна и изобилует небольшими озе-
рами. Вода озер содержит преимущественно Na2GO3 или Nad в зависимо-
сти от типа горных пород, слагающих верховья рек, питающих озера.
Грунтовые воды обычно пресные или слабосоленые. Смена растительных
группировок по широте от гор Арбисо до Лёссового плато показана
на рис. 171.
Цифрами на профиле обозначены следующие растительные группи-
ровки:
1. Сложенные древними породами горы Арбисо почти лишены расти-
тельности. Изредка заметны ассоциации кустарничков. Характерен энде-
мик Ammopiptanthus.
--*1 2. На грубообломочных осыпях подгорной полосы степень покрытия
почвы растительностью достигает 20—25%. Здесь встречаются ассоциации
Reaumuria soongorica, Caragana tragacanthoides, Zygophyllum xanthoxylon,
Salsola passerina, S. laricifolia и древнего реликта Центральной Азии
Tetraena mongolica.
3. На полузаросших песках развита широко распространенная
в Ордосе полынная полупустыня (покрытие 30—40%), представленная
Artemisia ordosica (рис. 172) и Pycnostelma lateriflorum (ластовнё-
вые).
4. Между участками полупустыни встречаются связанные с грунто-
выми водами засоленные луга, образованные Carex duriuscula, Anemone
rivularis, Potentilla anserina, Taraxacum chinensis и др., по окраинам луго-
вых понижений на опесчаненных грядах легко заметить высокий злак чий
(Lasiagrostis splendens, см. рис. 144).
Аридные области Центральной Азии
245
Рис. 172. Заросшие Artemisia ordosica пологогрядовые пески, Ордос (фото Петрова).
Рис. 173. Caragana korshinskii в барханных песках, Центральный Ордос (фото Попова).
5. В полынной полупустыне с увеличением количества осадков уже
появляются степные элементы, такие, как Stipa glareosa, S. bungeana,
Agropyrum mongolicum, Pennisetum flaccidum, Lespedeza dahurica, Thymus
serpyllum, Astragalus melilotus. Покрытие почвы достигает.^ 60—70%.
Artemisia ordosica на тяжелых почвах замещается A. frigida.
6. Во впадинах на влажных почвах развиваются луговые сообщества
с Agrostis alba, Polypogon lutosus, Arthraxon hispidum и др. с куртинами
кустарников (Salix, Hippophae rhamnoides, Rhamnus erythroxylori).
7. Кальцефильная растительность на выходах коренных меловых
пород с видами Gypsophila, Haplophyllum, Euphorbiajnlpp.
246
Глава 4
8. Степные сообщества на лёссах.
Вокруг соленых озер поселяются Nitraria schoberi (парнолистниковые)
и Kalidium gracile (маревые). Весьма скудна растительность незакреплён-
ных барханных песков (покрытие почвы 1—3%). Она представлена единич-
ными растениями Hedysarum mongolicum, Caragana korshinskii (рис. 173)
C. microphylla, Psammochloa villosa, Agriophyllum arenarlum.
Алашанъ
Эта значительно более обширная, чем плато Ордос, пустынная область
примыкает к нему с запада. На юге ее ограничивают горы Наныпань
с хребтом Рихтгофена; к северу область простирается примерно до широ-
кой депрессии, которая идет от озера Гашун-Нур на восток и по которой
пролегает древняя караванная дорога. Севернее этой впадины уже начи-
нается пустыня Гоби (см. рис. 140).
С самолета пустыня Алашань представляется сочетанием обширных
пространств с барханными песками, темных пятен зарослей саксаула,
перемежающихся со светлыми пятнами солончаков, обширных предгорных
равнин и довольно многочисленных островных низкогорий и холмистых
останцов размытого плато.
Климат Алашани обладает теми же чертами, что и климат Ордоса, но
западная часть пустыни более сухая и жаркая, чем восточная. Количество
осадков сокращается с 219 мм до 68 мм, испаряемость же возрастает
с 2400 мм до 3700 мм и более. Максимум дождей выпадает в августе. Сред-
няя годовая температура равна 8°, абсолютная минимальная температура
колеблется от —25 до —32°, максимальная — от 35 до 40°. Речная сеть
весьма^разветвленная, но русла рек наполняются водой только летом
в период муссонных дождей. Озера пустыни Алашань многочисленны.
Они либо бессточные (с NaCl), либо пресные. Последние располагаются
в глубоких межгрядовых понижениях с опресненными грунтовыми водами.
Стекающие с окраинных гор и просачивающиеся в песок атмосферные воды
создают дополнительный резерв питания грунтовых вод. В горах Алаша-
ни имеются даже многочисленные родники с пресной водой. Широкое рас-
пространение подвижных песков здесь, как и везде, обусловлено нера-
циональным выпасом и уничтожением древесных растений человеком
(рис. 174). В пески вкраплены цайдамы, влажные впадины, представляю-
щие собой остатки древней, засыпанной песком речной системы. Цайдамы
обычно окружены массивами барханных песков, ибо здесь особенно
ярко проявляется губительное для растительности воздействие выпаса.
На рис. 175 показаны распределение грунтовых вод цайдама и химиче-
ский состав солей в этих водах.
Растительный покров пустыни Алашань напоминает растительность
Ордоса. На кучевых песках с близким уровнем грунтовых вод широко
распространена Nitraria schoberi (см. рис. 180), сопровождаемая единич-
ными растениями видов Kalidium и Lycium, на более влажных участках
также Lasiagrostis splendens (см. рис. 144), а на заболоченных — Phragmi-
tes communis и Typha minima. Вдоль сухих русел рек можно заметить жал-
кие остатки пойменного леса из Populus diversifolia и Ulmus pumila
(рис. 176). Здесь на скоплениях речного аллювия встречаются также дре-
востои центральноазиатского вида Haloxylon ammodendron, которому, воз-
можно, идентичен черный саксаул (Н. aphyllum) Средней Азии, либо же
это викарирующие расы.
Аридные области Центральной Азии
247
Рис. 174. Барханные пески в пустыне Алашань (фото Петрова).
На каменистых почвах поселяются отдельные кустарники; помимо
свойственных плато Ордос, здесь распространены виды Ephedra, Calligo-
num mongolicum, Convolvulus fruticosus, Amygdalus mongolica, эндемики
Рис. 175. Распределение грунтовых вод, характер и степень мине-
рализации их в одном из цайдамов в пустыне Алашань
(по Петрову).
1 — подвижные барханные пески; 2 — полузаросшие и заросшие кучевые
пески; 3 — сульфатно-хлоридный солончак; 4 — хлоридный солончак; 5 — кар-
бонатно-хлоридный солончак; 6 — окраина подгорной равнины Холаныпаня.
Внизу — преобладающее засоление грунтовых вод: по периферии — карбо-
натное засоление, в центре — хлоридно-сульфатное.
Potaninia mongolica (розоцветные), Gymnocarpos przewalskii (гвоздичные),
Ammopiptanthus mongolicus и др. Резко отличный характер имеет расти-
тельность гор, окружающих пустыню Алашань с юга и востока. Так, высо-
кий хребет Холанывань имеет пять вертикальных поясов растительности:
248
Глава 4
Рис. 176. Ulmus pumila в центральной части пустыни Алашань. Это самая засухо-
устойчивая древесная порода Центральной Азии, наиболее глубоко проникающая
в аридные районы (фото Петрова).
Р и с. 177. Пояс хвойных лесов с Ptcea asperata в восточном Наныпане (фото Петрова).
Аридные области Центральной Азии
249
1500—1600 м пустыня с Reaumuria soongarica, Artemisia pectinata,
Stipa glareosa и др.
1600—1900 м степь с Stipa brevifolia, видами Caragana, Ephedra,
Athraphaxis, Allium mongolicum и др.
1900—2500 м мезофильный кустарник, представленный Lonicera, Rosa,
Rhamnus, Potentilla fruticosa, Cotoneaster, Spiraea trilo-
bata, Zizyphus jujuba и др.
2500—3000 м хвойный лес (сомкнутость крон 0,6—0,7); Picea asperata
(рис. 177) \ Pinus tabulae]ormis, Juniperus rigida (им
предшествуют Populus davidiana, Betula platyphylla)
с кустарником в подлеске и Syringa oblata в сухих ущель-
ях. На открытых лесных полянах произрастают— Polygo-
natum, Valeriana, Thalictrum, Aquilegia, Lilium tenuifolium.
выше 3000 м субальпийский кустарник c Potentilla fruticosa, Spiraea,
Salix, Caragana-, альпийские луга, образованные Poly-
gonum viviparum, Saussurea pygmaea, Dianthus, Ranunculus,
Delphinium и др.
Бэйшань
Область Бэйшань, примыкающая на западе к Алашани и простираю-
щаяся до обширной депрессии оз. Лобнор и Хамийской впадины на севе-
ро-западе, по строению своей поверхности резко отличается от районов,
Рис. 178. Комплексный ландшафтный профиль через пустыню Бэйшань с северо-
запада (Хами) на юго-восток (Аньси) (по Петрову).
1 — межгорные понижения, заполненные обломочным материалом, со смешаннокустарничковой
формацией; грунтовые воды местами поднимаются близко к поверхности (имеются колодцы и насе-
ленные пункты); 2 — скалистые древние низкогорья, ландшафт, напоминающий гамаду, сильно
разреженная растительность (преобладание трещинных грунтовых вод). У подножия гор — равни-
ны с сухими руслами и разреженной более богатой кустарниковой растительностью вдоль них (очень
глубокое залегание грунтовых вод).
описанных выше. Это обширная низкогорная страна, длиной около 500
и шириной 200 км, представляет собой древнюю поднятую глыбу (рис. 178).
Сложен массив метаморфическими докембрийскими породами, занимаю-
щими около половины всей его территории. Значительные площади в Бэй-
шане занимают также отложения каменноугольного и мелового возра-
ста. Бейшань является областью активной денудации, продолжающейся
уже длительное время. В рельефе местности преобладает беспорядочное
1 Этот восточноазиатский вид лишь в последнее время был выделен от близкой
ему Picea schrenkiana, встречающейся в восточном Тянь-Шане (см. рис. 188) и свиде-
тельствующей о миграции лесных деревьев не из Сибири, а из Восточной Азии.
250
Глава 4
сочетание низких гор и холмов с многочисленными котловинообразными
впадинами между ними, заполненными третичным и четвертичным обло-
мочным материалом. Предельная высота — 2800 м, высота низменньтх
окраинных районов не более 1000 м.
В климатическом отношении территория Бэйшаня представляет собой
пустыню, за исключением его центральной, наиболее высокой части, где
уже проявляется вертикальная поясность ландшафтов. Поскольку Бэй-
шань находится почти на границе раздела влияния атлантической и тихо-
океанской циркуляции, воздействие восточнокитайского муссона почти
не проявляется. Годовое количество осадков составляет 39—85 мм, выпа-
дают они летом в виде непродолжительных грозовых ливней. Испаряе-
мость превышает 3000 мм. Температурный режим несколько умереннее,
чем в низкорасположенных центральноазиатских депрессиях. Притока
вод с окружающих гор эта область не получает, более того, часть воды
стекает к ее окраинам. Значительная доля поверхностного стока просачи-
вается в рыхлую толщу наносов, заполняющих мульды, и скапливается
на водоупоре. Поэтому в таких местах иногда есть колодцы с питьевой
водой. Эти колодцы обусловили возникновение в еще древние времена
караванных дорог, в наши дни они снабжают водой местное редкое насе-
ление. Почвы Бэйшаня серо-бурые, нередко покрыты с поверхности панци-
рем из гальки, под которым залегает толща мелкозема, часто содержащая
прослои карбонатов и гипса.
Растительность Бэйшаня в основном представляет собой низкую
кустарниковую пустыню с обычными для Центральной Азии видами родов
Ephedra, Nitraria, Zygophyllum, Calligonum, Athraphaxis, Lycium, Cara-
gana, Haloxylon, Salsola, Anabasis, Tamarix, Reaumuria, Artemisia. Только
по обводняемым руслам рек встречается древесная и кустарниковая рас-
тительность (Populus, Elaeagnus) и другие фреатофиты (Alhagi, Lasiagros-
tis, Phragmites). На склонах наиболее высоких горных вершин растут уже
хвойные леса с Picea asperata, видами деревьев и кустарников, упомяну-
тыми нами при описании горной растительности Алашани.
Т арийская впадина и пустыня Т акла-Макан
(Кашгария)
Таримская впадина занимает площадь около 400 тыс. км2, прости-
рается по широте на 1300 км, а по долготе на 500 км. С трех сторон впади-
ну окружают мощные горные сооружения — на севере Тянь-Шань,
на западе Памир и Каракорум, на юге — Куньлунь (рис. 179). На востоке
Таримская впадина, сужаясь, заканчивается Лобнорской котловиной.
Абсолютная высота местности колеблется от 780 м на востоке до 1400 м
на западе. Всю центральную часть занимает труднопреодолимая, незасе-
ленная и наиболее засушливая в Азии песчаная пустыня Такла-Макан
(см. рис. 140).
Таримская впадина — экстрааридная часть Центральной Азии. Годо-
вая сумма осадков на западе у Кашгара равна 65 мм, на востоке — только
9—11 мм. Испаряемость достигает 2000—3000 мм. Воздух почти всегда
полон пыли, лето знойное, зима холодная (средняя температура января
колеблется от —6 до —10°, абсолютный минимум равен —27,6°). Тем
не менее область богата подземными водами. Питаемые горными ледника-
ми реки несут свои воды к южной части впадины, но вскоре теряются
в песках. Западную и северную окраины орошают водные артерии системы
Тарима, самой крупной и многоводной реки впадины; ее длина, включая
Аридные области Центральной Азии
251
Рис. 179. Северный склон Куньлуня (хр. Алтынтаг) на высоте около
2000 м. В Таримскую котловину впадают реки, текущие по глубоко вре-
занным долинам. Склоны гор крутые и голые, лишь местами с единичными
маленькими одревесневшими растениями (фото Хауде).
Яркенд, около 2000 км. В среднем река несет 1200—1300 мЧсек (в поло-
водье расход увеличивается вдвое); русло реки, обрамляемое широкой
поймой, постоянно смещается в сторону пустыни Такла-Макан, в этом же
направлении идет довольно активная фильтрация в пустыню речных вод.
Оканчивается река в бессточной Лобнорской котловине, образуя здесь
соленое озеро диаметром 100 км. Временами, когда Тарим отклоняется
в сторону, озеро полностью пересыхает. Блуждание русел — особенность
здешних рек. Это дало основание говорить о кочующих реках Центральной
Азии, постоянно меняющих свое русло и строящих новые и новые дельты.
Из-за небольшого количества осадков песчаная пустыня Такла-Макан
совершенно лишена растительности. Но эоловые пески пустыни как бы
«плавают» по насыщенной водой толще аллювиальных отложений, поэто-
му под пустыней предполагают существование на небольшой глубине
сплошного зеркала воды. С. Гедин при пересечении восточной части пусты-
ни в 1904 г. почти ежедневно, выкапывая колодец в самых глубоких лож-
бинах между барханами, находил на глубине 1,2—1,5 м пресную или соле-
ную воду. На таких местах иногда можно встретить кусты Tamarix или
тростник (Phragmites').
В целом же территория подвижных песков полностью лишена расти-
тельности, поскольку слишком малое количество атмосферных осадков,
по-видимому не превышающее 25 мм в год, не обеспечивает накопление
в песках достаточного запаса влаги. Своеобразные гидрографические усло-
вия Таримской впадины определяют и характер произрастающей здесь
растительности. Петров различает следующие типы растительности х:
1 М. П. П е т р о в, Пустыни Центральной Азии, т. II, Коридор Хэси, Цайдам,
Таримская впадина, Л., 1967.
252
Глава 4
Рис. 180. Мелкокучевые пески с Nitraria schoberi в долине реки Тарым. Слабоминера-
лизованные грунтовые воды лежат на небольшой глубине (фото Петрова).
Рис. 181. Haloxylon ammodendron, Таримская котловина (фото Петрова).
Аридные области Центральной Азии
253
Рис. 182. Пойменный лес, образованный Populus diverstfolia, в Таримской котловине
(фото Петрова).
1. Сильно разреженная мелкокустарничковая (Ephedra, Zygophyllum,
Gymnocarpus, Sympegna, Anabasis) пустынная растительность скалистых
склонов хребтов горного обрамления.
2. Весьма скудная псаммофитная растительность пустыни Такла-
Макан и пространств в полосе контакта речных долин с массивами
песков.
3. Галофильная растительность древних речных долин и дельт,
состоящая из однолетников (Salsola, Suaeda, Halogeton), кустарничков
(Kalidium, Halocnemum, Nitraria), кустарников, как Tamarix, Halostachys,
Nitraria schoberi (рис. 180) и редких древостоев Haloxylon ammodendron
(рис. 181).
4. Особенно характерна для Таримской впадины мезофильная древес-
ная и кустарниковая растительность в долинах современных рек и в оази-
сах окраинных районов (рис. 182). Мезофильная растительность Тарим-
ской котловины идентична тугайной растительности Средней Азии (см.
стр. 224). Но тугаи Тарима слабо освоены под земледелие, и поэтому их
растительность менее изменена деятельностью человека. Заросли Popu-
lus diverstfolia и Р. pruinosa, а у небольших рек — Ulmus pumila занимают
всю площадь поймы; открытые травостои или тростниковые заросли отсту-
пают на второй план. Очень часто лесообразующими породами являются
также Hippophae rhamnoides, Tamarix ramosissima, Elaeagnus.
В целом флористический состав растительности Таримской впадины
очень беден. Эндемичные виды, несмотря на резкую обособленность тер-
ритории, отсутствуют. Преобладают турано-джунгарские элементы, но
встречаются и монгольские.
254
Глава 4
Впадина Цайдам
Цайдамская котловина со всех сторон окружена высокими горными
поднятиями; от впадины Лобнор на севере она отделена массивом Алтын-
таг (5400 м). Цайдам лежит выше (2700—3000 м) Таримской впадины и об-
наруживает уже постепенный переход ландшафтов от равнин Центральной
Азии к высокогорным пустыням Тибета (Петров, 1967). Но несмотря
на значительную приподнятость и непосредственную близость к Тибету,
в составе флоры Цайдамской впадины обнаруживается большое участие
элементов, характерных для более низких районов Центральной Азии.
Собственно тибетская растительность начинается южнее 4000-метровых
горных цепей, отделяющих заполненную молодыми аллювиальными отло-
жениями впадину Цайдам от Тибета.
Количество осадков, выпадающих летом в Цайдаме, почти так же
мало, как и в Таримской впадине, а испаряемость столь же велика.
Но лето здесь из-за большей абсолютной высоты прохладнее и зима холод-
нее (средняя температура января колеблется от —10 до —15°, абсолютная
минимальная температура равна —31,7°). Средняя годовая температура
несколько превышает 0°.
Цайдамская высокогорная равнина обильно орошается реками, теку-
щими с пограничных гор, хотя крупных водных артерий здесь нет. Цен-
тральная часть впадины в плейстоцене была заполнена обширным Древне-
цайдамским озером, на месте которого в настоящее время распространены
обширные площади лишенных растительности засоленных почв с рядом
небольших соленых остаточных озер.
Рельеф вносит некоторое разнообразие в преимущественно пустынный
характер растительности Цайдама (покрытие почвы не превышает 5—7%).
Особенно скуден растительный покров каменистых или покрытых грубо
обломочным материалом местообитаний в краевых горах и их предгорьях,
а также в поясе барханных песков. Лишь в восточной, более обводненной
части впадины растительность на песчаных почвах значительно богаче,
основной растительной группировкой здесь являются полынные и полый,
Рис. 183. Полупустыня с Artemisia у подножия горных склонов, область
Цайдам (фото Петрова).
Аридные области Центральной Азии
255
Рис. 184. Панорама Алтынтага на высоте 3000 м. В долине реки на
почве обильные выделения соли. На террасе Tamarix и Astragalus (фото
Хауде).
но-солянковые ассоциации; проективное покрытие 70—80% (рис. 183).
На почвах с высоким уровнем пресных или слабо минерализованных грун-
товых вод развивается растительность пойм или засоленных лугов, очень
сходная с растительностью речных долин и солончаков Таримской впа-
дины (рис. 184).
стыня Гоби
Севернее плато Ордос, пустынь Алашань и Бэйшань, занимая всю
южную часть Монголии, простирается пустыня Гоби *. На востоке хребет
Хинган отделяет ее от более влажных степных и лесных областей2,
на западе, южнее Монгольского Алтая, она примыкает к Джунгарии, уже
характеризующейся несомненно среднеазиатскими чертами. На севере
четкая граница отсутствует, здесь пустыня Гоби постепенно переходит
в полупустыню, затем в монгольскую степь и, наконец, в горные степи
и лесные пояса северной Монголии и гор южной Сибири (рис. 185). Мон-
гольские степи образуют особую группу, которую можно сравнить с ланд-
шафтами восточноевропейско-казахстанской области.
Годовая сумма осадков в пустыне Гоби не превышаетг100—150 мм,
нередко она едва достигает 40 мм. В западной, наиболее сухой частиЪусты-
ни случаются годы, когда осадки вообще не выпадают.
Поскольку большая часть территории Гоби располагается на высоте
более 1000 м, неудивительно, что средняя годовая температура только
1 А. А. Ю н а т о в, Основные черты растительного покрова МНР, «Труды Мон-
гольской комиссии», вып. 39, М.— Л., 1950; И. П. Г е р а с и м о в, Е. М. Л а в р е н -
к о, Основные черты природы МНР, «Изв. АН СССР, серия геогр.», № 1, 1952.
2 Ср.: Chi- Wu Wang, The forests of China, «Cabot Found. Publ.» № 5,
1961; S. Danert, S. Geiler, P. Hanelt, Vegetationskundliche Studien in Nord-
ostchina (Mandschurei) und der Inneren Mongolei, Feddes Repert; BeihJ 139,1960.
256
Глава 4
Рис. 185. Карта растительности Монголии (по Юнатову).
альпийский пояс; 2 — горный пояс хвойных лесов; 3 — горные лесостепи и степи; 4 — злаковая степь; 5 — полупустыня; 6 — пустыня.
Аридные области Центральной Азии
257
на юге достигает нескольких градусов выше нуля. Зимы очень холодные,
сухие и безоблачные. Сплошной снежный покров не образуется; иногда
снег лежит пятнами или покрывает 40—80% поверхности почвы. Благо-
даря сильной инсоляции максимальная температура днем даже в холодное
время года может подняться выше 0°. Снег испаряется, не тая. Почва столь
сухая, что замерзания ее не происходит. Весна бездождная, поэтому весен-
ние эфемеры здесь отсутствуют. Развитие растительности начинается лишь
с первыми летними дождями. Появляются многочисленные поздноцвету-
щие однолетники: виды Salsola и другие маревые, из злаков Aristida
ascensionis, Pappophorum boreale, Eragrostis minor. Луковичные растения,
за исключением видов Allium (A. tenuissimum и A. polyrrhizum), не
представлены. Таким образом, собственно вегетационный период огра-
ничивается летом. Осенью рано наступают заморозки и растительность
отмирает; однако сухие побеги сохраняются всю зиму и служат пищей
скоту.
Вследствие сильной сухости в пустыне Гоби широко распространены
засоленные и гипсовые почвы. Тем не менее рельеф формируется под воз-
действием скорее воды, чем ветра; нижние части горных хребтов покрыты
мощным плащом обломочного материала. Только в северо-западной части
Гоби, в районе больших озер, известны древние озерные пески, перевеян-
ные ветром в подвижные дюны. Следовательно, пески в пустыне Гоби
занимают лишь ограниченные площади.
Растительный покров повсюду весьма скудный. Продуцирование орга-
нического вещества даже в северных степных районах достигает лишь
400—500 кг!га, в собственно пустыне — едва 100—200 кг!га. Центральная
часть Гоби — слабозасоленная, почти лишенная растительности щебни-
стая пустыня. Одиночные растения встречаются только в обычно сухих
долинах временных водотоков, известных нам по описаниям Сахары (ср.
т. I, стр. 475) х. По характеру растительности восточное Гоби отличается
от крайне засушливого запада.
Преобладающими видами на востоке являются кустарниковидные
Salsola passerina и Anabasis brevifolia. Вместе с ними встречаются Reaumu-
ria soongarica, Nitraria sphaerocarpa, Zygophyllum xanthoxylon, Convolvu-
lus gortschakowii, Amygdalus pedunculata, Tanacetum achilleoides, травяни-
стые виды Stipa gobica, Cleistogenes sinensis, Halogeton, Artemisia scopa-
ria и др.
Засоленные низины характеризуются Nitraria sibirica, Lasiagrostis
splendens, Peganum nigellastrum, Kalidium gracile. На участках с навеян-
ным песком растет Haloxylon ammodendron (см. рис. 181), достигающий
здесь северной границы своего распространения. Единственным деревом
на почвах с легкоподвижной влагой является Ulmus pumila (см. рис. 176).
На всей западной половине Гоби грунтовые воды нигде не подсту-
пают близко к поверхности, поэтому оазисов здесь нет. На каменистых
почвах растет преимущественно Haloxylon ammodendron — низкий, высо-
той 1,5—2 м, кустарник. Его сопровождают обычные для этих условий
древесные виды, которые можно найти и на нижних участках склонов гор.
Растительный покров местами столь редок, что даже на низменностях
на площади 100 ж2 трудно встретить 1—2 растения. Спорадически встре-
чается лишь Nitraria sphaerocarpa совместно с Ephedra przewalskii. Приме-
чательно появление «тора» (Nanophyton erinaceum), вида, очень часто
наблюдаемого западнее, в Джунгарии, и прослеживаемого до Казахста-
1 Е. М. Лавренко, Растительность Гобийских пустынь МНР и ее связь
с современными геологическими процессами, «Бот. ж.», № 9, 1957, стр. 1361—1382.
17-0434
258
Глава 4
на. Оазисы попадаются лишь в полосе,
переходной к Джунгарии. Там, где на
поверхность выходят грунтовые воды,
растут виды Carex, Phragmites communis,
Calamagrostis macilenta, Potentilla anse-
rina, Triglochin palustris, Epilobium hir-
sutum и развиты пойменные леса с Popu-
lus, Salix, Elaeagnus и рядом видов,
проникающих сюда с запада и восточнее
уже не встречающихся.
Переход монгольских пустынь! в
степи отмечен появлением перистых
видов ковылей, Carex duriuscula и луко-
вичных растений (Allium polyrrhizum);
реже можно заметить два ксероморфных
вида Artemisia и напоминающие их виды
Tanacetum, кроме того — Eurotia сега-
toides и Caragana pygmaea. На засолен-
ных почвах во впадинах развиваются
маревые.
Севернее, в собственно монгольских
степях, растительный покров стано-
вится более сомкнутым; в составе тра-
востоя начинают преобладать злаки,
такие, как виды Stipa и Aneurolepidium
(Agropyrum) pseudoagropyrum, к кото-
рым примешиваются виды Artemisia и
многие виды разнотравья. С северо-за-
пада в пустыню Гоби протягивается
хребет Монгольский Алтай, продолже-
нием которого служит Гобийский Алтай.
На склонах гор проявляется вертикаль-
ная поясность растительности, носящая,
правда, сильный отпечаток аридного
климата (рис. 186). Растительность
северных и южных склонов совершенно
различна г.
Над пустыней начинается пояс гор-
ной степи, над ним, на северных скло-
нах, следует полоса кустарников с ви-
дами родов Lonicera, Rhamnus, Spiraea,
Cotoneaster. Его сменяет пояс хвойных
лесов с различными видами Pinus, Picea
и Juniperus, присутствуют уже таежные
виды Picea obovata, Abies sibirica, Larix
sibirica. В лесах, образованных Pinus
sibirica (=cembra), в нижнем ярусе доми-
нирует Bergenia crassifolia, часто выса*
живаемая в парках Центральной Европы
как декоративное растение.
1 П. Б.Виппер, Леса юго-западного
Чентая, «Труды Монгольской комиссии», вып.
54, 1953.
Аридные области Центральной Азии
259
Рис. 187. Долина, ведущая к озеру Богдо-Ула (северо-восточный Тянь-
Шань). Начало елового леса на высоте 1600 м. Северный склон с богатой
злаковой и разнотравной растительностью, южный — с относительно
скудным растительным покровом (фото Хауде).
Во влажных ущельях встречаются даже насаждения из лиственных
пород {Populus, Betula)’, открытые участки между ними занимает высоко-
травье {Valeriana, Thalictrum, Aquilegia, Astragalus, Sanguisorba и др.).
Лишь на высоте 3000 м начинается альпийский пояс. На южных склонах
последовательность поясов почти аналогичная; но лесной пояс обычно
отсутствует и горные степи переходят непосредственно в субальпийские
степи, а последние — в альпийский пояс. В качестве примера вертикаль-
ной поясности в западных горах опишем поясность Тянь-Шаня, служаще-
го границей между Средней и Центральной Азией. Восточная часть этих
гор на значительной высоте еще относительно влажная, тогда как запад-
ная, распадающаяся на отдельные, веерообразно расположенные хребты
и резко обрывающаяся к жаркой Туранской низменности, отличается
весьма аридным климатом. Еще разительнее климатический контраст
между очень сухим, ступенеобразно спускающимся к Таримской впадине
южным склоном гор и северными горными цепями, вершины которых
овеваются влажными ветрами, что позволяет расти здесь сомкнутым лесам
(рис. 187, 188). Имеется весьма подробное описание растительности и фено-
логические наблюдения для Западного Тянь-Шаня х. Над поясом пустыни
выделяют степной пояс с доминирующими на темно-бурых степных поч-
вах Festuca sulcata и Helictotrichon {Avenastrum) desertorum. На высоте
2000—2500 м степь переходит в субальпийские луга на темноцветных
почвах, несколько увлажняемых при таянии расположенных выше снеж-
ных полей. На таких лугах уже преобладают влаголюбивые альпийские
элементы. Часто доминируют Polygonum hissaricum, Р. nitens, Geranium
1И. М. Культиасов, Экологические особенности высокогорных растений
Западного Тянь-Шаня, М., 1955.
17*
260
Глава 4
Рис. 188. Густые леса из Picea schrenkiana у озера Богдо-У ла на высоте
1800—2800 м, Восточный Тянь-Шань. На заднем плане — массив Богдо-
Ула (фото Хауде).
saxatile или Alopecurus songaricus. К ним примешиваются Trollius, Myo-
sotis, Potentilla, Rhodiola, Papaver, Allium, Oxytropis и др. или степные
элементы, как Festuca sulcata, Bromus turkestanicus, Tulipa dasystemon.
Вблизи снежных полей формируются альпийские снеговые ложбинки
с очень влажными, легко выщелачивающимися почвами, не похожие,
однако, на снеговые ложбинки Альп. Мхи и лишайники здесь отсутствуют.
Так как ложбинки расположены на значительном расстоянии одна от дру-
гой, состав их растительности весьма различен. Обычно доминируют А Шит
monadelphum или Potentilla tephroleuca; повсеместно встречаются Carex
melanantha, Oxytropis immersa, Cerastium cerastioides. Назовем, кроме того,
Primula algida, Anemone protracta, Lloydia serotina, Minuartia biflora,
Ranunculus rubrocalyx, Pedicularis amoena, Draba talassica, Poa alpina,
Taraxacum alpinum и др. Эти группировки весьма напоминают сходные
сообщества Альп или Кавказа. Для растений таких местообитаний харак-
терно, что рост их начинается еще под снегом, задолго до того, как он
стаивает. Из-под снега появляются бледные побеги, которые быстро зеле-
неют и в развитии догоняют растения, раньше освободившиеся от снеж-
ного покрова. На благоприятных местообитаниях число проростков необы-
чайно велико (свыше 200 на 0,5 ж2). Но уже в первые дни 80% молодых
растений погибают; сохранившиеся проростки подрастают и обладает
способностью переждать под снегом два-три года, чтобы после того, как
снег сойдет, снова развиться. Вероятно, это возможно благодаря свой-
ству растений уже очень рано закладывать запасающие органы.
Луговые сообщества поднимаются по склонам гор до абсолютной
высоты 2900—3100 м, в ряде мест языками вклиниваясь в ареалы снеж-
ных полей. На более сухих каменистых местообитаниях существует расти-
Аридные области Центральной Азии
261
тельность высокогорных ксерофитов. Наиболее типичным видом послед-
них является колючее подушечное растение Onobrychis echidna-, кроме
него, распространены Ziziphora clinopodioides, Richteria pyrethroides, Cousi-
nia caespitosa и др., имеющие крайне незначительную листовую поверх-
ность. Хотя растительный покров производит здесь впечатление очень
разреженного, все же и на высоте 2900 м на площадке размером 100 м1 2
можно насчитать до 27 видов. На южных склонах поверхности осыпи сте-
лется Juniperus turkestanica. Все перечисленные виды распространены
до гребней высоких гор. Даже на перевале высотой 3100 м встречены
Helictotrichon desertorum и Ziziphora clinopodioides высотой 1,5 м и более
20 сопровождающих видов. Отдельные стволы Juniperus достигают длины
20 м и 4—5 м в поперечнике. Высота ветвления зависит от мощности снеж-
ного покрова зимой, которая в свою очередь определяется направлением
ветра.
Поясность неодинакова в горах, получающих разное количество
осадков. В аридных условиях леса в схеме вертикальных поясов не пред-
ставлены. Формируются лишь редкие древостои Juniperus (на Копет-
даге J. turcomanica) или других хвойных. Там, где осадков выпадает
несколько больше, в обычную последовательность поясов иногда вкли-
нивается зона лиственных лесов; например, склоны южных гор Средней
Азии поросли лесами из диких яблонь или — на высоте 1000—2200 м при
сумме осадков 1000 мм в год — лесами, образованными Juglans regia.
Приведенные примеры характеризуют картину растительности лишь
в самых общих чертах. Каждый горный массив индивидуален как в кли-
матическом отношении, так и в последовательности вертикальных поясов
растительности; в них могут найти убежища эндемики даже из числа дре-
весных растений х. Экологические исследования в пустынных областях
Центральной Азии не производились, поэтому нам приходится ограни-
читься этим кратким обзором растительности.
ВЫСОКОГОРНЫЕ ПУСТЫНИ ЦЕНТРАЛЬНОЙ АЗИИ
К Центральной Азии относят также обширное Тибетское нагорье,
заключенное между горами Куньлунь на севере и главным Гималайским
хребтом на юге. Его отличительная особенность — значительная высота
над уровнем моря, в среднем около 4000 м', отдельные вершины поднимают-
ся до 6—7 тыс. м. В фитоценологическом и тем более в экологическом отно-
шении вся эта область практически является неизведанным белым пятном.
Исключение составляет лишь самая западная ее окраина, Памирское
нагорье, от которого в разных направлениях ответвляются многочислен-
ные хребты. Эта горная система экологически детально изучена, так как
здесь на абсолютной высоте 3864 м расположена Памирская биологиче-
ская станция, где русскими исследователями проводятся тщательные
многолетние экологические наблюдения (рис. 189). Такого рода станции
всегда являются непременным условием для детального экологического
изучения района. Лишь познакомившись с климатическими условиями
и характером растительности во все времена года, можно понять суть
целого ряда важных взаимосвязей, существующих в пределах отдельных:
биогеоценозов или целых экосистем. Об этом свидетельствует опыт Лабора-
1 И. В. Выходцев, Вертикальная поясность растительности Киргизии, М.,
1956; А. В. Куминова, Растительный покров Алтая, Новосибирск, 1960.
262
Глава 4
тории по изучению пустынь под Таксоном (Аризона) или опыт станции
Репетек в Каракумах. Кратковременные экспедиционные исследования
могут дать лишь общие исходные сведения, нередко создающие к тому же
одностороннюю картину увиденного.
Центральноазиатские высокогорные пустыни представляют собой
аналог пуны Южной Америки, хотя флора этих двух областей весьма
различна. Правда, пуна расположена между 15 и 28° ю. ш., то есть еще
Рис. 189. Холодная пустыня на высоте 4000 м, Памир. На галечнике
единичные кусты характерного памирского растения — терескена (Eurotia
ceratoides), ближе к реке — виды Carex и низкорослые сообщества Salix
охусагра (фото Шульца).
в пределах тропиков, тогда как Памир находится между 37 и 39° с. ш.,
а самая южная часть Тибета простирается лишь до 28° с. ш. Поэтому вре-
мена года в азиатских высокогорных пустынях значительно более резко
выражены и зимы холоднее. Тем не менее экологические условия в обеих
областях, видимо, более или менее одинаковы. Поскольку пуна в эколо-
гическом отношении совершенно не изучена, исследования, произведен-
ные на Памире, приобретают особое значение.
Памир
Памирское нагорье на востоке примыкает к высоким хребтам Кара-
корума и Куньлуня, на юго-западе граничит с Гиндукушем, а на севере
с горной системой Тянь-Шань. Памир представляет собой огромную высо-
когорную страну, пересеченную многочисленными хребтами, вытянутыми
преимущественно в широтном направлении и достигающими высоты
5000—6000 ж, хребты разделяются широкими долинами или озерными
котловинами, дно которых имеет отметки 3500—4000 м (рис. 190).
Аридные области Центральной Азии
263
Рис. 190. Памирский ландшафт. Вид на север через Алайскую долину шириной 20—
25 км (на высоте около 3000 м) на Заалайский хребет. На переднем плане — альпийская
степь с Festuca sulcata, видами Carex, Helictotrichum (Avenastrum) desertorum, Androsace
villosa, Ligularia и др. (фото Финстервальдера).
Расположенное в дождевой тени высочайших гор земного шара
Памирское нагорье получает очень мало осадков. В Мургабе в среднем
за 16 лет выпало только 66 мм осадков за год. На лежащей несколько
выше биологической станции за время с 1941 по 1951 г. определено, что
в среднем за год выпадает 102,6 мм (Максимум 144,8 мм, минимум 40,3 мм).
Распределение осадков на протяжении вегетационного периода иллюстри-
рует табл. 47. Наиболее дождливыми (10—20 мм) являются май, июнь,
июль и август, то есть летние месяцы.
Осадки выпадают ничтожными порциями (в 50% случаев от 0,2 до 0,5 мм)
и обычно в виде крупы или снега, редко в виде дождя. Лишь8.У1П. 1952 г.
выпало более 22 мм жидких осадков. Столь сильные дожди, вероятно,
случаются один раз за 15—20 лет, но для растительности их значение
трудно переоценить. Слабые осадки заметного влияния на водный режим
почв не оказывают. Зимой месячная их сумма редко превышает 2—3 мм.
Эти осадки увлажняют скелетные почвы лишь до глубины 1 м. Величина
осадков за отдельные дни года приведена в табл. 48.
Относительная влажность воздуха на Памире чрезвычайно низка —
летом в среднем за месяц около 50—70%, зимой — 20%. В условиях высо-
когорья и при сухом воздухе инсоляция днем и излучение ночью особенно
велики. Интенсивность солнечной радиации составляет 90% величины
солнечной постоянной. Облачность в условиях Памира составляет 39%,
264
Глава 4
Таблица 47
Распределение осадков весной и летом по данным
Памирской биологической станции (в мм)
Год Апрель Май Июнь Июль Август Сентябрь Всего за IV—IX
1939 - 0 0 18,0 36,6 0 65,4
1940 — 0 23,6 32,7 0 0 56,3
1941 7,5 0 42,0 28,1 0 5,9 83,5
1942 6,0 30,0 73,8 4,0 2,5 0 116,3
1943 15,3 36,4 10,6 12,0 22,8 8,7 105,3
1944 9,4 20,2 8,6 0 24,1 4,3 66,6
1945 18,1 15,0 19,9 3,4 23,5 6,0 85,9
1946 0,0 16,2 0,6 0 0 15,6 32,4
1947 7,0 3,6 10,5 35,2 4,5 0 60,8
1948 0,6 20,3 29,7 12,6 19,4 0,1 82,7
1949 5,0 6,6 30,6 2,0 26,7 0 70,9
1950 18,2 19,5 8,1 0 20,7 18,5 85,0
1951 —- 17,5 4,8 0,8 6,0 0,2 29,1
1952 0,1 13,7 2,8 24,5 59,5 1,7 102,3
1953 .— 40,7 36,6 19,4 11,0 24,0 131,7
1954 — 0,3 0,3 32,7 29,8 8,2 71,5
1955 — 37,8 6,9 6,2 0 0 51,0
Среднее (7,9) 15,3 18,9 13,9 16,6 5,8 70,5
Таблица 48
Осадки за период с 1.IX. 1951 г. по 31.VIII. 1952 г.
по данным Памирской биологической станции (в мм)
Дата Осадки | Дата Осадки J Дата Осадки | Дата Осадки
10.IX 2,0 10.11 0,8 28. V 5,7 20.VH 4,6
И.IX 3,1 18.11 0,9 29.V 3,2 26.VII 0,5
24. IX 0,9 4.III 2,0 31 .V 4,3 27.VII 0,5
ю.х 0,1 6.III 0,2 1.VI 2,8 30.VII 4,8
26.Х 0,2 13.III 0,3 4.VII 0,5 31.VII 0,9
26.XII 0,1 27.III 0,2 15.VII 1,0 8.VIII 22,2
13.1 0,3 8.IV 0,1 16.VII 2,8 23.VIII 2,5
4.II 0,4 27. IV 0,1 I 18.VII 1,6 24.VIII 5,7
8.II 9,5 18.V 0,2 19.VII 2,8 25.VIII 26.VIII 3,0 5,0
тогда как в Альпах на высоте 3100 м она достигает 65%. В году в сред-
нем наблюдается 45 пасмурных и 130 ясных дней.
Испарение с поверхности почвы на увлажненных площадях летом
иногда превышает 5 мм/сутки. Испарение с открытой водной поверхности,
вероятно, достигает 8 мм. Годовую испаряемость можно лишь оценить*
(свыше 1000 мм), поскольку соответствующие измерения не производи-
лись. Средняя годовая температура колеблется от —1,0 до —2,8°, но в лет-
ние месяцы температура воздуха на высоте 2 м от земли повышается до
20—22°, вблизи почвы до 28° и на поверхности почвы до 52°. Ночью тем-
пература падает ниже точки замерзания; в течение года отмечено только
Аридные области Центральной Азии
265
10—30 ночей без заморозков. Температурные условия у земной поверхно-
сти напоминают ход температуры в Высоких Альпах (ср., гл. 1). Экстре-
мальные температуры,
видимо, несколько смяг-
чаются очень сильными
ветрами, тем не менее
была зафиксирована
температура 68°. Сила
ветра в послеполуден-
ное время обычно равна
15 м/сек.
Годовая амплитуда
температур поверхности
почвы достигает 100°,
а средняя суточная 25—
Рис. 191. Хроноизоплеты температуры почвы на глу-
бинах от 0 до 3 м, Памирская биологическая станция,
лето 1940 г. (по Свешниковой, 1962).
— 30°. Даже на глубине
1 м температура может изменяться от 13,0 до
—8,5°. До глубины 4 м почва промерзает редко. Поэтому многолетняя
мерзлота встречается лишь фрагментарно, обычно в днищах долин или
озерных котловин. Максимальную и минимальную температуры в верхних
горизонтах почвы характеризует табл. 49 (см. также рис. 191).
Таблица 49
Экстремальные температуры в почве на территории Памирской
биологической станции (по Свешниковой)
Глубина, см Декабрь Январь Февраль Март
макс. мин. макс. мин. макс. МИН. макс. мин.
0 11,9 —30,5 —2,3 —42,7 7,8 —45,7 30,5 —39,4
20 —2,7 —17,3 —11,3 —29,4 -8,4 —28,3 2,1 —14,9
40 —3,0 —10,1 —7,9 —22,1 —9,9 —20,7 —1,1 —11,3
80 -0,9 -4,5 -3,7 —12,8 —8,1 —13,1 -3,8 -9,0
Глубина, см Апрель Май Июнь Июль
макс. мин. макс. мин. макс. мин. макс. мин.
0 37,1 —18,2 35,4 —11,0 50,0 —4,3 53,3 —0,6
20 11,4 —4,3 16,9 -2,3 22,4 7,6 24,8 10,1
40 9,1 — 13,0 2,4 19,0 9,4 21,3 10,4
80 3,4 —3,6 6,9 2,7 12,3 7,0 14,9 12,1
Глубина, см Август
макс. мин.
0 56,8 —4,2
20 24,3 10,1
40 20,2 14,1
80 16,3 13,1
266
Глава 4
Рис. 192. Высокогорная степь на ледниковой морене; южный склон
перевала Терсагар (3100 jw), Памир. Разреженный злаковый покров
(Stipa, Elijmus), местами Eurotia ceratoides и Ephedra. Правее центра
снимка высохшее соцветие Ferula asa-foetida (фото Финстервальдера).
Рис. 193. Конусы выноса у нижнего озера Ак, недалеко от Памир-
ского поста, 3700 м над уровнем моря. Холодная пустыня с поли-
гональными почвами, образовавшимися вследствие крайней засушли-
вости климата; видны редко стоящие кусты Eurotia ceratoides (маре-
вые) (фото Рейнига).
Аридные области Центральной Азии
267
В аридных условиях на обширных обломочных конусах выноса
формируются своеобразные серо-бурые, весьма бедные гумусом скелетные
почвы, близкие к сероземам пустынь и склонные к содовому засолению.
Растительность разных районов Памира характеризуется некоторым
своеобразием х. В южной его части она напоминает альпийские сухие луга
с Festuca sulcata и Роа laxa, а также некоторыми видами альпийского разно-
травья. В центральной части преобладают бедные разнотравьем ковыль-
ные степи (рис. 192), тогда как восточная область Памира носит пустын-
ный характер, причем доминируют здесь Eurotia ceratoides (рис. 193),
или Artemisia skorniakovii, либо же Tanacetum pamiricum. В долинах рек
распространены луга, а в районах же с более низкими абсолютными
отметками также кустарники (рис. 194). Из 150—200 встречающихся
здесь видов не менее 100 связаны с влажными местообитаниями. На пустын-
ных местообитаниях редко удается найти более 5—7 видов; обычно это
кустарнички высотой 10—15 см, покрытие почвы которыми редко дости-
гает 10%.
Приводимый в табл. 50 список видов дает представление о характере
растительности в окрестностях Памирской биологической станции. Боль-
шую часть области занимают субальпийские и альпийские пустыни.
Таблица 50
Некоторые важнейшие виды, произрастающие в окрестностях
Памирской биологической станции
Субальпийские пустыни (3 860 м) Альпийские пустыни (4700 м)
Eurotia ceratoides (рис. 193) Artemisia skorniakovii Artemisia pamirica Artemisia macrocephala Stipa glareosa Zygophyllum rosovii Christolea crassifolia Oxytropis chilliophylla Potentilla pamiroalaica Potentilla moorkroftii Astragalus borodini и др. (рис. 195) Tanacetum xylorrhizum Oxytropis poncinsii Oxytropis immersa Richteria pyrethroides Dracocephalum discolor Hedysarum cephalotes Smelowskia calycina Braya pamirica Parrya excapa Acantholimon hedinii Saussurea pamirica и др.
На несколько менее сухих местообитаниях развиваются степи с Роа litwinowiana,
Р. relaxa, Dracocephalum heterophyHum, Allium polyphy llum, Iris tianschanica, Solenan-
thus stylosus и Christolea pamirica.
В долинах ручьев, водоносность которых в августе резко уменьшается, значитель-
ную площадь занимают луга с Elymus dasystachys, Clynelymus nutans, Atropis hackeliana,
Hordeum turkestanicum, Poa pamirica, P. tibetica, Carex duriusculiformis, C. melanantha
и т. д.; Kobresia capillifolia, Polygonum viviparum, Gentiana prostrata, Swertia marginata,
Ranunculus spp., Clematis tangutica, Pedicularis abrotanifolia и многими другими видами.
Наконец, назовем еще растения снеговых пятен, произрастающие на постоянно
хорошо увлажненных местообитаниях: Primula nivalis, Androsace akbaitalense, Ranun-
culus rufosepalus, Thalictrum alpinum, Waldheimia tridactylites, Rheum denticulatum,
Rhodiola pamiroalaica, Oxygraphis glacialis, Rraya pamirica, Parry a excapa, Smelowskia
calycina, Neogaya simplex и др.
Этот перечень показывает, что речь идет не о собственно пустынной
флоре. К альпийским элементам здесь примешиваются степные компонен-
1 Г. М. Ладыгина, Н. П. Литвинова, Изучение растительных сооб-
ществ в высокогорье Памира, «Бот. ж.», 51, № 6, 1966, стр. 792—800; К. В. С т а н ю -
кович, Растительный покров Восточного Памира, М., 1949.
268
Глава 4
Рис. 194. Чистые заросли Hippophae rhamnoides высотой 5 ж на берегу
Танимаса; внутренний Памир, 3300 м над уровнем моря (фото Рикмерса).
Рис. 195. Astragalus sp. в холодной пустыне Памира, 3700 м (фото
Рейнига).
ты и только на самых сухих почвогрунтах поселяются представители полу-
пустыни, такие, как ксероморфная Artemisia skorniakovia и Eurotia ceratoi-
des (маревые), которая вместе с Nitraria schoberi и Sympegna regelis, видимо,
относится к галофитам, ибо нормальное осмотическое давление у этих
Аридные области Центральной Азии
269
видов колеблется в пределах 33—48 атм., а максимальное — от 42 до
58 атм. Хотя осадков выпадает крайне мало, сухость воздуха несколько
смягчается низкой температурой и, следовательно, слабым испарением
с поверхности увлажненной талыми водами почвы. Таяние снега начинает-
ся лишь в первой половине лета.
Вегетационный период начинается на Памире уже в мае; тем не менее
годовой прирост растений на сухих местообитаниях чрезвычайно мал,
у растений-подушек он составляет самое большее несколько миллиметров.
Столь низкая продуктивность определяет развитие особых жизненных
форм, преобладающих на Памире: кустарничков, растений-подушек, розе-
точных трав. Проходит очень много времени, пока растения начинают
цвести Ч Eurotia ceratoides зацветает только через 25 лет, Stipa glareosa
Рис. 196. Развитие Eurotia ceratoides (по Стешенко).
Возраст растений (слева направо): однолетнее, двулетнее, 3—5 лет, 8—12 и более чем 100 лет. Най-
дены экземпляры 250—300-летнего возраста, но и это, видимо, не предел.
и Acantholimon diapensioides — через 15 лет, Kobresia pamiroalaica и Carex
orbicularis — через 8 лет. Напротив, растущие на альпийских лугах
Trisetum spicatum и Atropis pamirica зацветают на первом году, a Blysmus
compressus, Astragalus heterodontus и другие — спустя 3—4 года. Как бы
компенсируя этот недостаток, главные виды растительности пустынь,
степей и формаций подушечников доживают до 100—300 лет, а некоторые
из них, например Acantholimon, вероятно, до 1000 лет. Виды же альпий-
ских лугов, такие, как Trisetum, Atropis, Primula algida, Swertia marginata,
достигают всего 10—15-летнего возраста.
Биологический запас органического вещества в ковыльных степях
составляет в среднем 200 (80—320) кг!га, в полынных полупустынях —
490 (260—880) и в пустынях с Eurotia — 720 (300—1350) кг!га. Большая
часть фитомассы Eurotia состоит из мертвой ткани. Этот вид отличается
весьма своеобразным ритмом развития (рис. 196).
Опубликованы результаты очень детального изучения корневых
систем растений Памира 1 2. Используя траншейный метод, было составле-
но 310 профилей корневых систем 33 видов. Наибольшая масса корней
сосредоточена на глубине 0—40 см. Температура этого слоя почвы летом
в течение приблизительно 3 месяцев превышает 10°. Правда, время от вре-
мени почва пересыхает. Общая же закономерность та, что колебания влаж-
ности в ее верхних горизонтах весьма значительны, тогда как водный
1 О. В. Заленский, А. П. Стешенко, О биологических особенностях
основных видов растительного покрова Памирского нагорья, «Тезисы докл. делегатов
съезда Всесоюзн. бот. о-ва», 7, 1957, стр. 9—12.
2 В. М. Свешникова, Корневые системы растений Памира, Сталинабад,
1952; Водный режим растений и почв высокогорных пустынь Памира, Душанбе, 1962.
270
Глава 4
режим более глубоких слоев до отметки 180 см отличается стабильностью.
В то же время эти слои холоднее. В общем, подземные части растений
в сравнении с надземными развиты сильнее, как это всегда наблюдается
у многолетних растений аридных районов (см. стр. 235). Глубина про-
никновения корневой системы иногда в 25—45 раз превышает высоту над-
земных побегов, сухая масса корней в 10—12 раз больше массы надземных
органов, а пронизанное корнями пространство может в 500—800 раз пре-
вышать объем, занятый побегами (табл. 51).
Таблица 51
Средняя (в скобках максимальная) протяженность
корневых систем у отдельных видов памирской пустыни (в см)
Название растений По гори- зонтали По вертикали
Eurotia ceratotdes Artemisia skorniakovii . . Zygophyllum rosovii . . . Christolea crasstfolia . . . 160—240 120—220 180—220 180—200 120—140 (180) 50—60 (120) 90—120 100—120
Корни Stipa glareosa опускаются в глубину всего на 50—80 см. Уко-
ренение луговых растений на влажной почве также неглубокое (20—60 см).
На содержании воды в почве зимние осадки практически не отражают-
ся. Снег ложится на уже замерзшую почву, легко сдувается или чаще —
испаряется. Поэтому сплошной снежный покров не образуется. Лишь там,
где происходит навевание снега, толщина снежного покрова иногда
составляет несколько дециметров. Но еще до наступления оттепели мощ-
ность таких сугробов в результате процессов сублимации быстро умень-
шается. Таяние снега начинается в марте; вода, соприкасаясь с холодной
почвой, снова замерзает, сковывая и делая непроницаемыми ее верхние
слои. Поэтому талые воды большей частью стекают без пользы по поверх-
ности мерзлой почвы или проникают вглубь лишь там, где почва рано
оттаивает. Увлажненная только с поверхности, почва весной быстро высы-
хает, чем и объясняется медленное развитие растений в начальной фазе
вегетационного периода. Обеспеченность водой растений улучшается лишь
после первых летних дождей. Некоторое количество влаги поступает
в почву снизу вследствие конденсации воды в сильно охлажденных зимой
горизонтах почвы. На рис. 197 отчетливо видно, как осенью и особенно
в период с ноября по декабрь содержание воды в каменистой почве на глу-
бине 1 м резко повышается, причем это увеличение влажности прослежи-
вается на некоторое расстояние вверх по разрезу, однако ближе к весне
влажность почвы снова уменьшается. Измерения температуры почвы
показали, что конденсация начинается тогда, когда температура почвы
до глубины 1 м падает ниже нуля, а глубже держится выше нуля. В рыхлой
каменистой почве в этом случае образуется своего рода иней. При промер-
зании грунта в январе или феврале до отметки 4 м содержание влаги
на глубине 1 м понижается. По нашему мнению, такая конденсация
не может иметь большого значения для растений, поскольку зимой актив-
ность корней прекращается. Следует также учесть, что на рис. 197 града-
ции влажности разнятся между собой на величину всего лишь в 1%,
но столь незначительная разница в условиях большой неоднородности
каменистых почв, несомненно, лежит в пределах возможного разброса
Аридные области Центральной Азии
271
Рис. 197. Динамика запаса влаги в почвах холодных пустынь Памира (по
Свешниковой). Объяснение см. в тексте.
значений. Летом, наоборот, верхние слои почвы увлажняются сильнее,
причем дождевыми осадками. Но вследствие развития процессов поверх-
ностного испарения и интенсивного поглощения влаги растениями эти
горизонты вскоре высыхают. Обычно содержание воды в почве не превы-
шает 2—3%, то есть оно никогда не достигает величины влажности завя-
дания.
Свешникова, ссылаясь на опыты Благовещенского в песчаной пустыне
(ср. стр. 213), придает процессам конденсации влаги в почве очень боль-
шое экологическое значение. Действительно, днем температура верхних
слоев почвы колеблется в значительных пределах, поэтому ночью песча-
ная почва которой свойственна высокая гигроскопичность, адсорби-
рует влагу. Но речь идет при этом о количестве воды, соответствующем
долям миллиметра атмосферных осадков. Однако Свешникова предпола-
гает, что растения тотчас поглощают эту сконденсировавшуюся влагу;
поэтому она умножает дневное количество воды на число дней вегетацион-
ного периода и получает величину 35 мм, что отвечало бы 30% годовой
суммы осадков. Более вероятно, однако, предположить, что то небольшое
впитанное почвой в ночные часы количество влаги днем снова испаряется.
Влажность воздуха над поверхностью почвы в полдень равна всего 5%,
в утренние часы она достигает 100% только после нескольких предшест-
вующих дождливых дней, а в засушливый период не превышает 50%, так
что выпадения росы не происходит. В водном балансе растений эти колеба-
ния влажности верхних горизонтов почвы, видимо, никакой роли не
играют. В равной мере это относится к конденсации влаги в более глу-
боких слоях почвы; при большой сухости почвы наличие такой конденса-
ции, правда, подтверждается (в среднем конденсируется 0,16 мм/сутки),
но далеко не установлено, используется ли эта влага корнями расте-
ний.
В общем динамика влажности щебнистых горных почв вследствие
их малой влагоемкости и неравномерности выпадения осадков носит далеко
не выровненный характер. Запасы влаги в верхнем метровом слое почвы
под пустынной растительностью в зависимости от количества осадков
колеблются в отдельные годы в пределах 24—104 мм. В 1940—1945
272
Глава 4
и 1950—1955 гг. максимальное содержание воды в почве за вегетационный
период не превышало 53—55 мм, а минимальное 28—30 мм’, иными сло-
вами, резерв доступной для растений влаги предположительно составлял
не более 26 мм и не менее 5 мм. Это очень немного, если учесть, что запасы
воды в верхнем метровом слое почвы в лесной зоне в среднем достигают
500 мм и даже в самых южных степях не сокращаются менее чем до 120—
170 мм.
Были определены размеры потребления почвенной влаги растениями
в пустыне с Eurotia и Artemisia. Пробные площадки представляли собой:
1) участки с неизмененной естественной растительностью {Eurotia, Artemisia,
Stipay, 2) участки, где все растения были удалены, почва не прикрыта;
3) участки, закрытые с целью предотвратить испарение. На одной части
пробных площадок растения имели угнетенный вид. Здесь эвапотранспи-
рация в 1950 г. за 45—50 летних дней составила около 4 мм. На других
пробных площадках растения хорошо развивались, что свидетельствовало
о лучших условиях увлажнения. Испарение равно 35 мм, транспирация—
16 мм. В 1951 г. за это же время величина транспирации в зависимости
от фазы развития растений составила 4—35 мм, а холодным летом 1952 г.—
только 3—6 мм. Основную долю влаги корни поглощали преимущественно
с глубины 20—50 см — горизонт, где сконцентрирована главная масса
корней. На глубине 1 м содержание воды в почве практически не изме-
нялось.
Данные наблюдений свидетельствуют о том, что, несмотря на общую
большую сухость, обеспеченность водой очень скудной памирской расти-
тельности вполне достаточна. Поэтому неудивительно, что осмотическое
давление, определенное Свешниковой у 113 видов, оказалось высоким
лишь у растений, занимавших крайне сухие местообитания. Пробы отби-
рались: 1) пустынные виды на высоте 3860 м, 2) пустынные виды на высоте
4300 м, 3) степные виды и 4) растения нивального пояса на высоте
4700 м. Как видно из табл. 52, наибольшая концентрация клеточного
сока отмечена у видов, произраставших на экстремально сухих место-
обитаниях.
Таблица 52
Величина осмотического давления у пустынных растений
Памира (в атм)
Название растений Осмотическое давление
среднее наивысшее наинизшее
Stipa glareosa 34,7 37,7 23,6
Artemisia skorniakovii 32,5 54,3 19,5
Richteria pyrethroides 24,6 30,8 19,0
Potentilla pamiroalaica 26,9 41,6 20,9
Christolea crassifolia 22,5 43,7 15,0
Oxytropis chiliophylla 24,2 34,2 16,6
Astragalus chadjanensis 22,6 32,4 19,2
Zygophyllum rosovii 25,5 38,4 19,4
Любопытно, что даже в самых неблагоприятных условиях существова-
ния получены лишь два значения осмотического давления, превысившие
30 атм.
Аридные области Центральной Азии
273
Возникает следующий ряд, составленный в направлении увеличения
осмотического давления (в атм):
Растения снежников....................... 10,5—18,8
Луговые растения ........................ 12,3—22,9
Степные растения......................... 16,2—22,2
Виды альпийских пустынь.................. 13,8—24,6
Виды субальпийских пустынь............... 17,1—34,7
Но и у видов, замыкающих ряд, величина осмотического давле-
ния в условиях хорошей влагообеспеченности значительно понижается
(9,4—23,6 атм). В общем, осмотическое давление у памирских растений
несколько более высокое, чем у растений Альп, где оно не достигает 20 атм
ни у одного из видов («Phytologie», Bd. Ill, 1, S. 301).
Бросаются в глаза резкие дневные изменения осмотического давления,
обусловленные действием прямых солнечных лучей на большой высоте.
У растений на сухих местообитаниях эти колебания достигают 12—14 атмг.
Отсюда следует, что все эти виды относятся к числу гидролабильных мягко-
листных растений и что собственно склерофильные виды здесь не встре-
чаются. Нет здесь и вечнозеленых видов. Так, например, совершенно
отсутствуют вересковые Альп и Арктики, а также Dryas, но распространен
Leontopodium alpinum, среднее осмотическое давление которого равно
18,4 атм (минимум 12,3 атм, максимум23,2 атм). Весьма типичны и много-
численны виды Artemisia, из которых A. macrocephala характеризуется
самым низким средним осмотическим давлением —15,8 атм (8,7—21,6 атм).
Однако дневной ход осмотического давления обусловлен отнюдь не накоп-
лением ассимилятов, как это считает Свешникова. Для этого количество
сахаров, накапливающихся в растениях за день, слишком мало.. Парал-
лельный ход кривых осмотического давления и содержания сахаров днем,
на который указывает Свешникова, возможен лишь при сгущении клеточ-
ного сока вследствие недостатка влаги. Увеличение концентрации сахаров
при накоплении продуктов ассимиляции, выраженное в атмосферах,
весьма мало. У видов с суккулентными листьями или запасающими воду
корнями дневные колебания осмотического давления невелики даже
в условиях Памира. Эти виды гидростабильны, осмотическое давление
у них более или менее постоянное и сравнительно низкое. У Zygophyllum
rosovii оно изменяется в пределах 19—24 атм, у Christolea crassifolia
(крестоцветные), а также у Gypsophila capitultfera — от 16 до 20 атм.
В еще большей степени уравновешен водный баланс у видов альпийских
лугов, растущих на постоянно влажных местообитаниях. Их осмотическое
давление в течение всего вегетационного периода почти не изменяется.
Летом осмотическое давление у всех видов несколько повышается,
особенно заметно на сухих местообитаниях. На протяжении года наиниз-
шие значения осмотического давления фиксируются в мае, а самые высо-
кие — в августе и сентябре. На первый взгляд это противоречит данным
определения содержания воды в растениях, ибо памирские виды больше
всего воды в листьях содержат летом, а наименьшее ее количество — засуш-
ливой весной.
При измерении получены следующие крайние значения запаса воды в листьях
растений, произрастающих в естественных условиях (в % сырого веса):
Пустынные растения (сухие местообитания)
Eurotia ceratoides............................... 54,4—75,2
Artemisia skorniakovii .......................... 49,9—72,7
Stipa glareosa................................... 46,5—67,0
1 При поливе растений в условиях естественного их произрастания колебания
осмотического давления в течение дня уменьшаются до 0,3—3,0 атм.
18-0434
274
Глава 4
Растения альпийских лугов (влажные местообитания)
Polygonum viviparum ............ 73,4—82,5
Potentilla multifida ............. 71,0—78,0
Hedysarum cephalotes........ 68,3—73,7
Суточные амплитуды не превышают в зависимости от характера местообитания
1,2-9,7о/о.
Наименьшее количество воды заключают в себе перезимовавшие части растений
(корни, почки возобновления) в зимние месяцы.
При вычислении содержания воды в процентах сырого веса необходи-
мо учитывать, что оно зависит от всей массы накопленного растением
сухого вещества и, таким образом, от возраста листьев. Поскольку расте-
ния Памира дают побеги лишь летом, определения запаса воды, сделанные
в летние^месяцы, относятся к совсем молодым листьям с тонкими стенками
клеток и с еще не вполне развитой механической тканью. Величина же
осмотического давления не зависит от толщины стенок клеток.
При сравнении величины осмотического давления у одного и того
же вида, но отобранных с разных мест произрастания, выявляется его
отчетливая зависимость от влажности почвы. Чем суше местообитание,
тем выше концентрация клеточного сока. Эта закономерность проявляется
и при сопоставлении видов одного рода. С увеличением высоты над уров-
нем моря осмотическое давление повышается, правда у особей с разных
местообитаний в разной степени. Однако трудно решить, связано ли это
с понижением температуры с высотой или с нарастанием сухости место-
обитаний.
По условиям гидратуры высокогорные растения Памира скорее сход-
ны со степными растениями, чем с видами жарких пустынь (ср. т. I). Хотя
на Памире очень сухо, но засушливые периоды относительно коротки;
периодом же абсолютного покоя является длительная суровая зима,
однако транспирация может прекращаться и в холодные летние ночи.
Проводились также измерения сосущей силы клеток методом Шарда-
кова. Тюриной получены чрезвычайно высокие отрицательные значения
тургора, что у малакофильных видов вряд ли возможно; однако полу-
ченные ею величины небезупречны.
При изучении интенсивности транспирации у растений Памира ника-
ких особенностей не обнаружено. Днем устьица всех растений открыты;
кривая дневного хода транспирации обычно характеризуется пиком
в полуденное время. Абсолютные значения сравнительно высоки, однако,
как и везде, существуют интенсивно и слабо транспирирующие виды.
Максимальные значения интенсивности транспирации соответствуют
величинам, определенным в умеренных полупустынях Средней Азии.
Представляют интерес определения расхода воды на транспирацию
в расчете на 1 а сырого веса. Наибольшие значения обнаружены у Poly-
gonum viviparum (22—34 г) и других растений альпийских лугов на постоян-
но влажных почвах (23—31 а). Наиболее вялый водообмен установлен
у суккулентов Christolea и Zygophyllum — лишь 3,4—4,2 а. У важнейших
видов пустыни, таких, как Eurotia ceratoides и Artemisia skorniakovii,
интенсивность водообмена достигает 6,8 а, но при искусственном поливе
увеличивается до 16—20 а. Таким образом, запас воды в листьях на про-
тяжении дня, иногда даже одного часа, обновляется несколько раз. Транс-
пирирующая масса Eurotia в течение вегетационного периода возрастает
в расчете на одно растение с 3 до 10 а (при поливе— до 60 а). У Artemisia
skorniakovii получены сходные значения (при поливе — до 33 а), но у. Stipa
Аридные области Центральной Азии
275
glareosa, напротив, они значительно меньше, а именно только с 0,8 до 2,5 г
(при поливе — почти до 4 г).
В среднем отдача воды одним растением за день на фоне постепенного
нарастания транспирирующей массы и роста испаряемости увеличивается
с мая по июль или август в следующих пределах (в г):
У растений пустынь
Eurotia ceratoides ........................ 2,5—50
Artemisia skorntakovii................... 12—80
Stipa glareosa .......................... 0,6—17
У суккулентных видов
Chrtstolea crassifolia..................... 16—75
Zygophyllum rosovii...................... 2,5—45
У растений альпийских лугов
A rtemisia pamirica........................ 23—320
Potentilla multifida .................... 12—140
Из приведенных данных следует, что суккулентные виды, обладая
большим сырым весом и незначительно транспирируя (в расчете на 1 г
сырого веса), потребляют тем не менее столько же воды, сколько и пустын-
ные виды; если же последние поливать, что ведет к их пышному развитию,
то потребление влаги одним растением за день увеличивается до 300 г
и более, что даже превышает расход воды растениями хорошо увлажнен-
ных альпийских лугов. Если учесть число экземпляров того или иного
вида на определенной площади, вес зеленой массы среднего экземпляра
и дневную величину транспирации отдельным экземпляром, то можно
вычислить общий примерный расход воды пустынными сообществами,
заселяющими конусы выноса за вегетационный период.
Этот расход в 1949—1952 гг. составил (в мм!год):
В верхней части конуса выноса............13—26
В сухой средней его части................ 7—14
Во влажной нижней части..................20—30
При поливе . ............................ 220 (макс. 325)
Из общей суммы расхода на долю Artemisia приходилось 45—70%,
Eurotia 20—30, Stipa 5—10, Christolea 5—8 и Zygophyllum 2—3%.
В такого рода расчетах не исключена вероятность больших ошибок.
Поэтому следует отметить, что названные цифры хорошо совпадают со зна-
чениями, полученными на основе определения уменьшения запасов воды
в почве (за 45—50 дней).
Тюрина приводит следующий годовой баланс водооборота (в мм):
Поступление воды: осадки (в среднем) 102
Потери воды: испарение с поверхности почвы 120
транспирация 21
Всего: 141
Дефицит (39 жм/год)|рассматривается авторами] расчета как подтвер-
ждение дополнительного поступления воды в почву в результате конден-
сации влаги в почве и на ее поверхности. Представляется, однако, что все
полученные значения слишком мало обоснованны, чтобы производить
с ними точный расчет баланса. С таким же успехом испарение с поверх-
ности почвы могло составить лишь 80 мм или осадки — 140 мм. В этом
случае баланс был бы уравновешен. Кроме того, конденсирующаяся
в почве влага в конечном счете также имеет атмосферное происхождение,
поскольку и грунтовые воды, и местный горизонт верховодки в свою
очередь питаются атмосферными осадками. Дефицит мог бы быть покрыт
и осадками необычно дождливых лет, которые глубже проникают в почву
18*
276
Глава 4
и там в течение длительного времени, исчисляемого годами, накапли-
ваются, как это установлено наблюдениями в египетско-аравийской пусты-
не (см. т. I, стр. 495 и след.).
Фотосинтез растений Памира изучал Благовещенский Б Виды Eurotia
ceratoides, Elymus dasystachys, Gentiana algida и Lepidium ассимилируют
весьма энергично, правда лишь до полудня. Интенсивность ассимиляции
днем составляет в среднем 25 мг/дм\ Из-за низкой температуры ночью
потери на дыхание минимальны. Растения приспособлены переносить
чередование заморозков и оттепели без повреждения фотосинтезирующего
аппарата.
Данные последующих исследований Глаголевой, которая использо-
вала высокие концентрации СО2, для вычисления нормальной интенсив-
ности ассимиляции, не пригодны. Однако они также свидетельствуют
о том, что все виды растений Памира способны к усиленному фотосин-
тезу. Из материалов других работ следует, что ассимиляция СО2 расте-
ниями альпийских лугов, листья которых имеют многослойный мезофилл,
чрезвычайно велика. Фотосинтез же маревых — их листья состоят литпь
из двух содержащих хлорофилл слоев — в два раза менее интенсивен 1 2.
В заключение подчеркнем, что проведенные исследования впервые
позволили познакомиться с эколого-физиологическими условиями высоко-
горных пустынь. Здесь выделяются два совершенно различных по проис-
хождению типа растительности:
1) растительность пустынь на сухих местообитаниях, сложенных
обломочным материалом (главные виды: Eurotia ceratoides и Artemisia
skorniakovii)’,
2) альпийская растительность, развитая на влажных местообитаниях.
Растительность пустынь в горах мигрировала из гипсометрически
ниже расположенных полупустынь. Некоторое сходство условий среды
облегчило процесс приспособления. Длительность вегетационного периода
в обоих случаях одинакова; разница состоит лишь в том, что период покоя
в высокогорье обусловлен долгой холодной зимой, а на более низких отмет-
ках — затяжной летней засухой и короткой холодной зимой. Поскольку
растения морозоустойчивы, смена местообитания для них существенного
значения не имела. Мощная корневая система позволила им заселить
также сухие местообитания. Короткий вегетационный период растения
пустынь используют полностью. Там, где влаги достаточно, растения
интенсивно транспирируют и ассимилируют; легко переносят они и боль-
шой водный дефицит и связанное с ним повышение осмотического давле-
ния. Осенью, с началом устойчивых холодов, растения сбрасывают листья
так же, как они делали это в полупустыне с наступлением засухи. К этому
времени стебли достаточно одревесневают, чтобы перенести долгую зиму.
Аналогично поведение Stipa glareosa, но в отличие от описанных выше
мягколистных видов 5. glareosa более склерофилен: растение регулирует
транспирацию также и в течение суток, рано закрывая устьица, поэтому
интенсивность его транспирации ниже, но вместе с тем сокращается
1 A. W. Blagowestschenski, Uber den Verlauf der Photosynthese im
Hochgebirge des Pamir, «Planta», 24, 1935, S. 276—287.
2T. А. Глаголева, Фотосинтез растений верхнего альпийского пояса
восточного Памира, «Бот. ж.», 47, № 11, 1962, стр. 22; А. А. Ф и л и п п о в, Суточные
и годовые колебания интенсивности фотосинтеза растений восточного Памира, «Труды
Бот. ин-та, серия IV», 13, 1959; О. В. Заленский, Максимальная потенциальная
интенсивность фотосинтеза растений Памира и других областей, «Труды Памирской
биол. станции», 1, Душанбе, 1963.
Аридные области Центральной Азии
277
и продуцирование сухого вещества. Ковыль на Памире уже близок к гра-
нице своего распространения. Часть листьев этого злака засыхает летом,
и растение плодоносит слабо или вообще не дает семян. Zygophyllum
и Christolea соответствуют больше типу суккулентных галофитов с урав-
новешенным водным балансом. Оба вида происходят из более низко рас-
положенных полупустынь, но в горах в растительном покрове на очень
слабозасоленных почвах большой роли не играют. В известной мере пере-
ходной группой ко второму типу растительности — альпийским видам,
характеризующимся более высокими требованиями к условиям влаго-
обеспечения,— являются полупустынные кустарники Artemisia pamirica,
Astragalus borodini и Dracocephalum heterophyllum, а также растения-
подушки Gypsophila capituliflora и Acantholimon diapensioides. Первые,
вероятно, относятся, скорее, к степным элементам, оба же последних
вида типичны для нагорий Передней Азии, менее засушливых, чем горы
Памира, и получающих больше осадков зимой. Растения собственно аль-
пийских лугов находят для себя на Памире подходящие температурные
условия и из-за отсутствия здесь сплошного снежного покрова распола-
гают достаточно длительным «бесснежным» периодом. Вообще же эти
виды встречаются чаще в горах с влажным климатом, а в условиях сухого
климата Памира — на почвах, постоянно содержащих необходимое количе-
ство влаги. Следовательно, и здесь речь, скорее, идет об экстразона льном
распространении растительности согласно закону относительного постоян-
ства местообитаний и смены биотопов. Каждая из рассмотренных групп
растений относится к иному, почти противоположному, типу зональной
растительности и потому занимает экстразонально на Памире также про-
тивоположные местообитания.
Отметим еще вкратце материалы сравнительного экологического изучения интерес-
ного рода, распространенного на Памиро-Алае,— Cousinia (сложноцветные) A Cousinia
близкородственна лопуху (Arctium) Центральной Европы; последний напоминает мезо-
морфные С. umbrosa и С. pulchella. Этот род характерен тем, что включает виды, обра-
зующие переход от мезоморфных растений до типичных растений-подушек. В Централь-
ной Азии мы различаем: 1) виды с крупными мягкими листьями, растущие в тени дере-
вьев в горах средней высоты; 2) виды с колючими листьями, образующие переходное
звено к группе 3) альпийских колючеподушечников, тдких, как С. splendida, С. fed-
tschenkoana, С. macilenta, распространенных в высокогорьях.
Виды первой группы отличаются высокой интенсивностью транспирации, зна-
чительным содержанием воды в листьях, низкой концентрацией клеточного сока и не-
большим водным дефицитом. С наступлением засухи их листья засыхают. Третья
группа, напротив, характеризуется слабой транспирацией, низким содержанием воды
в листьях и высоким осмотическим давлением. Водный дефицит может быть очень
большим. С началом засушливого периода водообмен резко сокращается, так что
растения переживают засуху в активном состоянии. У видов, принадлежащих к этой
группе, наблюдается либо превращение всех листьев в шипы, либо густое опушение
колючих листьев. Растения образуют глубоко проникающий в почву стержневой
корень.
Вторая группа растений в экологическом отношении занимает промежуточное
положение. Весной они весьма интенсивно транспирируют, но позже транспирация
сокращается, причем увеличиваются концентрация клеточного сока и водный дефицит.
Виды предпочитают солнечные местообитания и некоторое время без ущерба переносят
засуху, но затем засыхают. Следовательно, приспособление растений этой группы
к засухе носит прежде всего морфолого-анатомический характер. Проявляется оно
в сокращении листовой поверхности и во все более интенсивном развитии корневой
системы; экономно расходуя влагу, растения поддерживают водный баланс на одном
уровне даже в экстремальных условиях существования.
1 В. Ф. Купревич, Ю. С. Григорьев, Е. К. Висковска я, Водный
режим некоторых видов рода Cousinia Памиро-Алая, «Бот. ж.», № 1, 1949, стр. 16—33;
Ю. С. Григорьев, Сравнительно-экологические исследования ксерофитизации
высших растений, Л., 1955.
278
Глава 4
На высоте 2320 м, то есть на 1500 м ниже Памирской биологической
станции, на площади 93 га был заложен опытный сад с целью испытать
возможности возделывания полевых культур, плодовых и других деревьев
в суровых условиях Памира х. Оказалось, что неизвестные ранее в этом
районе культуры, такие, как картофель, различные виды капусты и корне-
плоды, а также кормовая свекла, при искусственном орошении дают очень
высокие урожаи; рассаду помидоров сначала выращивают в парниках;
хорошо растут клевер и эспарцет. Виноград плодоносит, если его укры-
вать на зиму. Обильный урожай дают абрикос, персик, слива, вишня, крас-
ная и черная смородина, малина. Большие сборы дают также черешня,
груша и яблоня. Но грецкий орех и шелковица не плодоносят. Доказана
возможность культивирования большинства лиственных деревьев умерен-
ной зоны, даже Robinia pseudacacia', однако некоторые из этих видов дают
побеги несколько раз в году, и поэтому их крона имеет неправильную
форму. Сеять ячмень и выращивать брюкву можно и значительно выше
в горах.
Экологическая обстановка в высокогорных пустынях Тибета, по-види-
мому, близка к только что описанным условиям среды на Памире.
Тибет
Это обширнейшее на земном] шаре горное поднятие ограничено
на севере хребтами КуньлушЛи Алтынтаг; на востоке нагорье прилегает
к провинциям Китая Ганьсу, Сычуань и Юньнань, на юге над ним стеной
возвышаются Гималаи, протягивающиеся от Памира до горных хребтов
северной Бирмы и северо-западного Юньнаня. К западу Тибетское нагорье
сужается и переходит в системы Гиндукуша и Памира. Его протяженность
по широте составляет 2000 км, с севера на юг — 1200 км; средняя высота
около 4200—4800 м. Ни в фитоценологическом, ни в экологическом отно-
шении нагорье не исследовано. В общих чертах известна только флора.
Альпийские виды обнаруживают тесные связи с гималайскими видами
(рис. 198), а виды высокогорных пустынь — с видами Центральной Азии.
Рельеф Тибета довольно разнообразен. Нагорье пересекают многочис-
ленные хребты, редко возвышающиеся, однако, более чем на 1000 м над
заполненными обломочным материалом котловинами, днища которых рас-
полагаются на высоте 4000—5000 м (рис. 199). Стекающие с гор талые
ледниковые и снеговые воды обусловливают широкое распространение
повышенно увлажненных площадей (болота на обломочном материале,
образовавшемся вследствие ^морозного выветривания). Так как в Тибете,
за исключением его крайних ^восточной и южной частей, сток к океану
отсутствует, реки обычно впадают в бессточные соленые озера; развиты
засоленные почвы. В то же время распространены ландшафты подвижных
песков. Западный Тибет отличается наиболее сухим и континентальным
климатом, но южная его окраина уже испытывает влияние муссонов
(рис. 200—201). Выделяют несколько областей растительности (рис. 202) 1 2.
1 А. В. Гурский, И. В. Каневска я, Л. Ф. Остапович, Основные
итоги интродукции растений в Памирском ботаническом саду, Сталинабад, 1953.
2 F. К. Ward, Asketch of the geography and botany of Tibet, «J. Linn. Soc.
(London)», 50,1935, p. 239—265; H. vonlW i s s m a n n, Stufen und Gurtel der Vegetation
und des Klimas in Hochasien und seinen Randgebieten, «Erdkunde, Arch. Wiss. Geogr.
(Bonn)», 14, 1960, S. 249—272; 15, 1961, S. 19—44; В. И. Г p у б о в, Опыт ботанико-
географического районирования Центральной Азии, Л., 1959.
Рис. 198. Виды Saussurea альпийской флоры Западного Тибета.^
280
Глава 4
I. Пустынная область
Джангтанг занимает всю се-
верную, западную и цент-
ральную части Тибетского
нагорья. Эта экстремально
суровая холодная пустыня
лежит на абсолютной высоте
5000 м. Над ней поднимаются
многочисленные, расходя-
щиеся к востоку горные
хребты, имеющие относитель-
ные отметки 300—400 м.
Средняя годовая температура
равна —5°. Только в июле
средняя температура поло-
жительная (+8°). Суточная
амплитуда колебаний темпе-
ратуры достигает 37°, средняя
скорость ветра равна м/сек,
годовая сумма осадков редко
превышает 100 мм. Расти-
тельность столь редкая, что
ее хватает лишь для прокорма
диких копытных животных.
Флора области очень мо-
лодая. В ледниковый период
весь прежний растительный
покров был уничтожен, при-
чем до настоящего времени
не известны какие-либо иско-
паемые остатки его. Все виды
растений мигрировали сюда
в послеледниковое время из
соседних районов. Домини-
руют центральноазиатские
виды: Eurotia ceratoides, Ко-
chia prostrata, Microula tibeti-
ca (бурачниковые), Reaumuria
trigyna, Rheum spiciforme,
Thylacospermum rupifragum
(гвоздичные), Chamaerhodos
sabulosa (розоцветные), Oxyt-
ropis aciphylla, Thermopsis
alpina (бобовые), виды родов
Astragalus, Acantholimon, Ar-
temisia, Ptilagrostis, Agropy->
rum, A venastrum, Festuca,
Elymus, Cleistogene и др. Из
древесных видов представлены
только Ephedra, Tanacetum и
Myricaria (рис. 203).
Во влажных низинах
преобладают широко распро-
Аридные области Центральной Азии
281
страненные голарктические роды (Carex moorcroftii, Juncos thomsoniiT
Triglochin palustre, Heleocharis и Eriophorum, а также Ranunculus tricuspis).
Рис. 200—201. Климадиаграмма центральноазиатских высо-
когорных пустынь.
Памирский пост — климат сходен с климатом Западного Тибета (пу-
стыня Джангтанг, где метеорологические станции отсутствуют); Лха-
са — юго-восток района, климат здесь уже испытывает влияние индий-
ского муссона (летние дожди).
На северных склонах заросли тибетской кобрезии (Kobresia tibetica)
образуют обширные бугристые болота («нака»); трава эта настолько жест-
Р и с. 202. Карта растительности Тибета (по Уорду).
I — пустыня Джангтанг; 1а — восточная часть с впадиной Цайдам и озером Кукунор;
II — внешнее плато; На — более влажный район со злаковой растительностью; III — район
глубоких речных долин: нижние ущелья и Ша — верхние, выше расположенные ущелья;
IV — китайский Тибет (переходная область).
кая, что прокалывает не только губы, но и толстую кожу на ступнях вер-
блюдов (рис. 204). Эндемичных видов здесь нет. Берега многочисленных
соленых озер поросли галофитами.
282
Глава 4
Рис. 203. Высокогорная пустыня в Западном Тибете на высоте
4600 м. Крупные кусты Myricaria elegans, перед ними Caragana pygmaea
(фото Цунмайера),
Более резко отличается от собственно Тибета его северо-восточная
часть, Цинхай — район озера Кукунор {1а на рис. 202, но без Цайдама).
Здесь флора необыкновенно богата и обнаруживает отчетливые связи
-с флорой Восточных Гималаев. В понижениях рельефа уже встречаются
многие виды кустарников {Rhododen-
dron, Acer, Philadelphus, Hydrangea,
Sorbus, Prunus, Sorbaria, Lonicera),
часть которых пережила период’оле-
денения в окраинных районах. Во
впадине Кукунор (3200 м) распро-
странены степи с господствомi Stipa,
Festuca, Роа bulbosa и Artemisia, а
также видами Astragalus и Oxytropis.
Южная, более высокая часть района
(Амдо) местами носит характер вы-
сокоальпийских разнотравных лугов.
II. Область внешнего нагорья
едва заметно отделяется от его части,
Рис. 204. Болота «нака» на северном
склоне как результат солифлюкции;
они представляют собой периодически
высыхающие или замерзающие моча-
жины, разделенные узкими низкими ва-
лами (по Тафелю).
описанной под пунктом I, но уже имеет сток в океан и содержит пресно-
водные озера. Здесь берут начало крупные речные системы Инда и Сатлед-
жа (рис. 206, карта-врезка), Салуина, Меконга, Янцзы и Хуанхэ. Осадков
выпадает несколько больше (250—500 мм), причем в основном летом.
Область заселена; в более низкорасположенных районах (3600—4200 м)
с мягким климатом по берегам рек раскинулись города. Западная часть
представляет собой полупустыню с галечными почвами, без древесной
Аридные области Центральной Азии
282
Рис. 205. Альпийский пояс в горах северо-западной Юньнани, высота
4225 м: Rhododendron adenophorum (крупные цветки), Rh. rupicola,
Anemone demissa, Cassiope selaginoides и др. (фото Хандель-Маццетти).
растительности; в лучшем случае в долинах можно встретить виды Juni-
perus, а непосредственно вблизи водотоков виды Salix и Populus, Ulmus
pumila или Hippophae. В долине Цангпо (верхнее течение Брахмапутры),
восточнее Лхасы, растут Berberis, Sophora, Moorcroftia, Buddleia, Poten-
tilla fruticosa; ниже по течению уже появляются леса. Флора здесь богаче,
известен 541 вид. В восточной части {На) начинается горная низкозлако-
вая степь, перемежаемая пятнами лугов альпийского облика с Spiraea
и Juniperus в долинах ручьев. На горных склонах и в долинах рек можно
заметить купы деревьев. На высоких горных хребтах альпийская флора
обогащена гималайскими видами родов Meconopsis (маковые), Primula,
Bentiana и многими другими.
III. Некогда эта область также представляла собой ландшафт нагорья.
В настоящее же время ее цоколь рассекают глубокие ущелья рек, разде-
ленные узкими, поднимающимися выше снеговой границы хребтами.
Восточнее Янцзы начинается китайская часть Тибета (IV), рельеф которой
в общих чертах аналогичен. Климат верхних, высокорасположенных
участков ущелий (II 1а) холодный и сухой, с летними дождями (500—
750 мм), тогда как в нижних ущельях климат теплее, здесь распростране-
ны пышно развитые леса; эти районы располагаются уже за пределами
собственно Тибета. В области II 1а распространены только хвойные
леса, но они избегают плоскогорий. Различают следующие пояса расти-
тельности:
2700—3600 м Пояс хвойных лесов из Picea и Juniperus, а также Populus,
многочисленные кустарники и травы (Eremurus, Primula;
Amphicome из семейства бигнониевых и др.).
284
Глава 4
3600—4200 м Субальпийский пояс с древовидным Rhododendron, Rerberis, Lonicera, Ribes, Clematis, Rosa и многими видами разно- травья (Incarvillea из семейства бигнониевых, лютиковые, Stellaria, Primula, Dracocephalum, Pedicularis и др.).
4200-4500 м Альпийский пояс с Meconopsis, Aster, низкорослыми видами Rhododendron (рис. 205) и др.
4500-5100 м Пояс карликовых растений-подушек с Primula, Gentiana, Lagotis и др.
5100-5400 м Нивальный пояс с видами Delphinium, Primula, Saussurea
(рис. 198) и др.
В нижних ущельях пояса растительности расположены на более
низких уровнях:
1500-2400 м Лесной пояс с Pinus khasia, Picea, Magnolia и т. д. Quercus, Acer, Ilex,
2400—3000 м Леса, образованные Picea, Taxus, Abies и древовидным Rhododendron.
3000-3600 м Леса из Abies webbiana (см. рис. 209) с примесью листвен-
ных деревьев, а также с древовидными Rhododendron.
Выше следуют субальпийский, альпийский и нивальный
пояса.
О сложном характере растительности горной системы Гималаев
мы попытаемся дать лишь общее представление на основании выполненных
Швейнфуртом описаний геоботанических профилей.
ВЕРТИКАЛЬНАЯ ПОЯСНОСТЬ ГИМАЛАЕВ
Центральноазиатские высокогорные пустыни обрамлены на юге высо-
чайшим горным сооружением земного шара — Гималаями, которые
на всем своем протяжении образуют резкий ландшафтный и климатиче-
ский рубеж. Это препятствие индийский муссон преодолеть не в силах;
только по отдельным долинам влажные ветры проникают до окраин сухого
Тибетского нагорья. На основании имевшихся далеко не полных сведений
Швейнфурт составил карту растительности этого огромного горного мас-
сива х. Ограничимся здесь рассмотрением двух профилей, которые про-
ведены через геоботанически лучше всего изученные районы и которые1
характеризуют вертикальную поясность растительности на южном и се-
верном склонах; один профиль проходит близ западной оконечности
Гималаев от Равалпинди до Нанга-Парбата (8125ж), а другой—на востоке
горной системы, от Дарджилинга через Канченджангу (8585 ж) к Джомо-
лунгме (8840 м) (рис. 206).
Познакомимся сначала с западным профилем, который проложен север-
нее Равалпинди. Если в Афганистане, западнее Кабула, еще проявляется
влияние области средиземноморских зимних дождей, то восточнее Кабула
уже заметен переход к муссонному климату с осадками, выпадающими
летом. В районе Равалпинди и реки Марри основной максимум осадков
приходится на лето, а второй, меныпий,— на весну. Лишь севернее глав-
ного хребта, у Гил гита, основной максимум наблюдается весной и вто-
рой — летом. В глубоких долинах осадков выпадает мало, но на большой
1 U. Schweinfurt h, Die horizontale und vertikale Verbreitung der Vege-
tation im Himalaya, «Bonner Geogr. Abh.», H. 20, 1957. В этой работе рассмотрены
также материалы К. Тролля по Нанга-Парбату.
s
о
£
5S ,± г е а х
й £ св d в
я 2 ? я я 5
Нй-ъ s
e в; о d
е* в
I и g г
। г- о « £-g
я
s
«
я
и
Ф
я
ф
s
я
S
Я
X I S' 2Q -
в I s00 з е
. и р ф
w н д Ф
9 s ।
s
о Э . эЯ w
gw S® 3 3
Sb ' ~°и«
я
’Я
ф
и
а
в
В
Я
л
Я
св
Я
Я
3
Я
V
в
a
и
и
&
co
я
св
К
К
Рн Я
ф
я
я
я
л
«
Я
£
В
я я
к д
CQ И
»wt
Я§« |
д«!1
Я 2 я ч
s Е®.
Э“§Ьсв
со Р "
Еч Ф Я
° §«
I ЯЯ
г»О
^5я
я S
я 5
B« fe
«Г I
3 I
я S’
я
чя8
S w®
SS§
Я.|§
»Я е §>
я
s
R
я
«
о
я
§
а
к S
П св
Я л
я
cu
Я
g
я
я
e
в
ЙЗ
И S
I §1 I §*я11
— « S § &S
»8
i§ я S о>
О Я 2 <7’ Я w
ф й ,. Я И в< з
R и и Я Т о g
§ я1^ EtS И!В
So
S R
Я н
я
g®
s' 3
я я
и®
д&н „ а
в ® g
я е § я
з В.« § 2
о я р,
а св о .г?
•S
® ® Ф и *'*
3 ЗуЧи..
Й Д (X *7? г—Ч
5gз
S Я .. 5 О ?
n
Е«
Д нч
D 3 •« - ч-
Я я я вь
r* Й
4 g
286
Г лава 4
Рис. 207. Лес, образованный Pinus roxburghii (— longifolia),
в Западных Гималаях (фото Марсдена). ,
высоте, где обширные площади заняты ледниками, их количество значи-
тельно увеличивается. Больше всего осадков всегда получают южные
склоны гор, испытывающие влияние^муссона/1$'
Рассмотрим характер растительности на участке профиля от Равал-
пинди до Кашмирской котловины, проложенном в направлении с юго-
запада на северо-восток.1
В окрестностях Равалпинди формация колючих кустарников практи-
чески уступила место культурному ландшафту, но непосредственно выше,
в горах можно обнаружить остатки субтропического вечнозеленого жест-
колистного леса, кустарник Dodonaea viscosa и Olea cuspidata, причем Acacia
modesta служит формой, переходной к колючим кустарникам. Заметно
некоторое сходство со средиземноморской растительностью, усиливаемое
появлением в сухих долинах временных ручьев Neriurrt oleander. На высоте
1000 м на северных склонах и на высоте 1500 м на южных, как это наблю-
Аридные области Центральной Азии
287
дается и в^континентальной части Средиземноморья, начинается сосновый
лес, представленный здесь Pinus roxburghii (рис. 207). Если по долине
реки Марри подняться еще выше, до уровня 2200 м, то попадаешь во фло-
ристически необыкновенно богатый,' более влажный смешанный дубово-
хвойный лес, образованный Quercus incana, Q. dilatata, Acer, Prunus,
Aesculus indica, Pieris ovalifolia (вересковые), Evonymus, а также Picea
morinda, Pinus excelsa, Abies webbiana, A. pindrow, Taxus baccata; ветви
и стволы деревьев покрыты Hedera helix. Видовое разнообразие напоми-
нает третичные реликтовые леса. В долине Джелама этот пояс особенно
хорошо развит^ даже на высоте более 3000 м. На самых нижних участках
склонов долины жестколистный лес, в котором встречается также древес-
ное растение Dalbergia sissoo (семейство бобовых), сопровождает реку
вплоть до Ури. Однако до отметки 2000 м он поднимается только по южным
склонам, на склонах же северной экспозиции Pinus roxburghii спускается
до уровня 800 м, а местами даже до дна долины. Выше Ури последователь-
ность вертикальных поясов резко нарушается. Вместо Pinus roxburghii
мы встречаем здесь Cedrus deodara (рис. 208). Леса, образованные гималай-
ским кедром, напоминают влажные пояса Средиземноморья в горах Атласа
и Тавра. Наконец, всю нижнюю часть склонов долины в пределах 1600—
3400 м занимает смешанный хвойный лес. Его сменяет субальпийский
березовый лес (Betula utilis), выше переходящий во влажные альпийские
кустарники и матты. Барамула расположена у входа в Кашмирскую кот-
ловину, ландшафты которой полностью окультурены.
Теперь совершим путешествие от северной окраины Трагбалской кот-
ловины в направлении на север мимо Нанга-Парбата до горных цепей Кара-
корума.
Самый нижний пояс начинается здесь кашмирским кустарникомг
обычно представленным колючими летнезелеными кустарниками Berberis,
Cotoneaster, Rubus, Ribes, Daphne, Jasminum, Fothergilla (гамамелидовые),
Indigofera, Plectranthus и др. Но уже ненамного выше 2000 м мы снова
встречаем влажный хвойный лес, образованный Pinus excelsa, Picea morinda
и Abies webbiana, который вверх по склону занимает все более обширную
площадь. Различия в растительности, обусловленные экспозицией место-
обитаний, повсюду выражены очень резко. На путях схода лавин растет
стелющийся кустарник Prunus. У перевала Трагбал (3608 м) можно заме-
тить низкий субальпийский березовый лес.
Границу леса образуют здесь на северных склонах Betula utilis
совместно с Abies webbiana (рис. 209), а на южных — Pinus excelsa, Picea
morinda и Juniperus.
В долине у Казалвана на высоте 2300 м состав растительности север-
ного и южного склонов совершенно различен. Северную сторону занимает
влажный хвойный лес. Южный же склон до отметок, значительно превы-
шающих 3000 м, покрывает полынная пустыня с редкими кустами Juni-
perus squamata. Привлекает внимание древовидный Juniperus macropoda',
в ряде мест полынные ассоциации сменяются пастбищем со скабиозой.
По лавинным ложбинам береза кое-где спускается далеко вниз к под-
ножию склона. Долины ручьев облюбовывают некоторые лиственные
деревья (Juglans regia, Corylus colurna, Salix oxycarpa, Populus balsamifera),
а на галечных речных отмелях растет знакомая нам по Центральной Азии
Hippophae rhamnoides. Такая последовательность вертикальных поясов
прослеживается без существенных изменений до Нанга-Парбата. На высоте
4000 м у перевала Камри раскинулись пышные альпийские разнотрав-
ные луга.
288
Глава 4
Рис. 208. Лес из Cedrus deodara в Западных Гималаях (фото Лейса).
Аридные области Центральной Азии
289
Рис. 209. Abies webbiana близ границы древесной растительности на
высоте 4000 м. На заднем плане — вершина Пандим (6711 м). (фото Селлы).
Особенно детальные геоботанические исследования в районе Нанга-
Парбата произвел Тролль (им составлена геоботаническая карта в мас-
штабе 1 :50 000). Этот мощный горный массив, расположенный севернее
главной цепи, возвышается над глубоко врезанной долиной реки Инд
более чем на 8000 м. Ниже 2000 м распространена пустыня центрально-
азиатского облика, характеризующаяся родами Capparis, Salsola, Stipa,
Echinops, Ephedra и другими. Местами пустыня поднимается высоко в горы
и переходит там в альпийские степи. На благоприятных скалистых место-
обитаниях еще встречаются Olea cuspidata, Daphne oleoides и Rosa webbiana:
Древесные растения, приуроченные к местам с грунтовым водным пита-
нием, вблизи русла Инда отсутствуют, так как колебания уровня воды
слишком велики. В полосе между 2000 и 3000 м снова преобладает полын-
ная полупустыня. Еще выше к Artemisia присоединяются деревья Juniperiis
19-0434
290
Глава 4
semiglobosa и Pinus gerardiana, так что картина в целом напоминает усло-
вия, существующие в области полынной полупустыни Северной Америки
(ср. т. II, рис. 97). Вместе с Artemisia встречаются известные нам по Пами-
ру Eurotia ceratoides и виды Kochia. Кроме того, распространены летнезеле-
ные кустарники {Rosa webbiana, Daphne oleoides, Ribtes orientalis, Colutea
arborescens, Fraxinus xanthoxyloides, Lonicera, Sophora, Berberis и т. д.),
а на песчаных почвах также Astragalus. На южных склонах эта полупусты-
ня проникает до высоты 4200 м и достигает альпийского пояса. В боль-
шинстве случаев между ними вклинивается пояс влажного хвойного леса,
у нижней границы которого Pinus gerardiana образует сравнительно
густые древостои. Здесь можно найти еще дуб каменный, Quercus ilex
s.l. (baloot), однако в районе Нанга-Парбата он далеко не столь широко рас-
пространен, как на крайнем северо-западе. К названным видам присоеди-
няются степные элементы, такие, как Thymus, Nepeta, Ranunculus, Pedicu-
laris и др. В субальпийском поясе полынная полупустыня напоминает,
скорее, луговую степь.
На галечных отмелях реки Астор (в по ясе Artemisia) растут Myricaria
germanica (как и в Альпах) и Hippophae, сопутствуемые Salix, Lonicera,
Ribes, Berberis, Spiraea, Rosa, Pirus, Cotoneaster, Evonymus, Viburnum и др.
В поясе влажного хвойного леса доминируют уже упомянутые хвой-
ные, а среди них живописные насаждения Cedrus deodara. На одном место-
обитании найден Taxus baccata. В качестве подлеска на крутых северных
склонах на скалистом грунте распространена Bergenia ligulata.
Границу леса на высоте 3800—3900 м образует Betula utilis, до этой
же высоты поднимаются Sorbus aucuparia и Salix wallichiana. Лес, обра-
зованный этими видами, переходит в кустарник из Salix hastata и Rhodo-
dendron hypenanthum, который на склонах южной экспозиции сменяется
насаждениями Juniperus semiglobosa, J. папа и J. squamata. На солнечных
склонах значительные площади занимает Ephedra gerardiana. Наибольшей
высоты из числа древесных растений достигают шпалерные кустарники
Polygonum affine или Salix flabellaris. Альпийские луга изобилуют цвету-
щими растениями. Тролль называет здесь следующие виды:
Iris hookeriana, Lloydia, Aquilegia jucunda, Trollius acaulis, Pulsatilla wallichiana,
Ranunculus hirtellus, Anemone rupicola, Thlaspi, Draba, Corydalis, Sibbaldia cuneata,
Potentilla argyrophylla, Polygonum viviparum, Nepeta, Thymus serpyllum, Pedicularis
bicornuta, Taraxacum, Chrysanthemum, Myosotis, Mertensia primuloides, Saussurea soro-
cephala, Aconitum rotundifolium, Saxifraga flagellaris, Geranium pratense, Trachydium
roylei, Androsace septentrionalis, Primula nivalis, Pirola rotundtfolia, Aster flaccidus,
Cousinia thomsoni, Leontopodium, а из видов разнотравья Phleum alpinum, Роа alpina,
Festuca^alatavica, F. ovina. Luzula sptcata, Kobresia.
Альпийский пояс простирается приблизительно до высоты 5000 м.
Здесь, в районе Нанга-Парбата, главная горная цепь Гималаев утрачивает
свое значение климатического рубежа. Осадки в верхних поясах проникают
далеко на север, до склонов Каракорума. Пояс пустынь прослеживается
там до отметки 2700 м, выше уже раскинулась полынная полупустыня.
В окрестностях Гил гита ее сопровождают обширные леса из Pinus gerar-
diana, более часто наблюдается здесь и Quercus ilex. Пояс влажного хвой-
ного леса сохраняется только на северных склонах; последними его отго-
лосками в долине Хунзы служат виды Juniperus. На северной оконечно-
сти профиля выделяют только три вертикальных пояса: пустыня, полын-
ная полупустыня и влажный альпийский пояс, у нижней границы которого
исчезает и береза, так что в горах древесная растительность уже отсут-
ствует полностью.
Аридные области Центральной Азии
291
В качестве второго примера рассмотрим составленный Швейнфуртом
так называемый Сиккимский профиль, который проходит в Восточных
Гималаях от Дарджилинга в направлении строго на север. После Гукера
(1848 г.) эту местность неоднократно посещали ботаники.
Строение второго профиля значительно более простое, чем первого.
Главная горная цепь здесь круто вздымается от предгорной равнины
к высочайшим вершинам; влажные воздушные массы, в течение всего года
перемещающиеся в основном с юга, из района Бенгальского залива,
на север, поднимаются по склонам гор, обильно увлажняя их. Горные цепи
обусловливают климатические контрасты на подветренной и наветренной
сторонах гор. Только через горный проход у Цумтана южные ветры еще
достигают нагорья Лхонак («Малый Тибет») и иногда приносят даже крат-
ковременные ливни в окраинные районы Тибетского нагорья. В Лхасе
с мая по сентябрь выпадает 500 мм дождевых осадков.
По мере увеличения континентальности климата снеговая линия
повышается с 4650 м на южном склоне Канченджанги до 5700—6000 м
на северной стороне вершины Канченгиао. Границы распространения
растений на севере также смещены приблизительно на 300 м по вертикали.
Пояса растительности на южном склоне гор прослеживаются от обла-
сти тропических лесов до снеговой линии.
Самый нижний пояс образует тропический влажный муссонный лес,
лес «сал» из Shorea robusta, древесной породы, к концу зимнего сухого
периода сбрасывающей листья. На пожарищах распространяется высо-
кий бамбук {Dendrocalamus hamiltonii). В долинах много пальм и Pandanus.
Обращают на себя внимание орхидеи Platy cerium, Lycopodium, Pothos
и Scitamineae. Приблизительно на высоте 900 м влажный муссонный лес
переходит в вечнозеленый тропический горный лес. Основные виды здесь:
Schima wallichii (чайные), Castanopsis tribuloides и С. indica (буковые),
а также древовидный папоротник A Isophila. Но уже в данном поясе заметны
роды и виды умеренной зоны: Quercus, Acer, Juglans regia, Vaccinium и др.
Выше начинается типичный высокогорный лес пояса облаков, о влажно-
сти которого свидетельствуют гименофилловые и мхи, покрывающие
сплошь стволы, ветви и скалы, а также множество пиявок.
Как и повсюду в тропиках, вертикальные пояса выделить здесь труд-
но, так как они постепенно переходят один в другой. В верхней части опи-
сываемого пояса уже резко преобладают роды умеренной зоны. В литера-
туре приводят следующие их них:
Magnolia campbelli, М. excelsa, Quercus pachyphylla, Qu. annulata, Qu. lanuginosa,
Qu. lamellosa, Machilus edulis (лавровые), Betula, Acer, Alnus, Ilex, Styrax, Berberis,
Michelia excelsa, M- cathcartii (магнолиевые), Rhododendron arboreum, Rh. dalhousiae,
Rh. barbatum, Rh. falconeri, Rh. campbelli, Rh. grande и т. д., Prunus, Pirus, Sorbus,
Pieris, Symplocos, Gleichenia, Hydrangea, Budaleia, Aralia, Skimmia и др.
Чрезвычайно велико видовое разнообразие древесного яруса и яруса
травянистых растений. Одних папоротников здесь насчитывается свыше
60 видов. Резкая верхняя граница распространения растений обусловлена
лишь морозным периодом; проходит она на высоте 1800—2000 м. Выше
начинают преобладать виды Rhododendron, в подлеске повсеместно пред-
ставлен низкий бамбук Arundinaria. Постепенно появляются первые хвой-
ные: Tsugadumosasi. Taxus baccata и, наконец, начинается царство хвойных.
На высоте 3000—3900 м склоны покрывает Abies densa. Rhododendron
и здесь широко распространен (рис. 210); встречаются также лиственные
деревья, например Retula, Pirus, Prunus, Acer, Alnus, Populus. Границу
леса образуют Abies densa и Juniperus pseudosabina. Стволы деревьев уве-
19*
292
Глава 4
tP и с. 210. Лес, образованный Rhododendron, в цолине Цему; Восточные Гималаи,
высота около 3500 м (фото Селлы).
шаны серым: бородатым лишайником. Betula встречается лишь единичными
экземплярами. Rhododendron постепенно становится все ниже и наконец
принимает почти кустообразную форму. На самых верхних позициях
раскинулись влажные альпийские луга с Senecio, Pedicularis, Aconitum,
Delphinium, Primula, Meconopsis (маковые), Saxifraga, Polygonum и т. д.
Узкий горный проход у Цумтана отделяет очень влажный нижний Сикким
от умеренно влажного верхнего. Пояса тропического леса здесь не раз-
виты. Их место занимает пояс умеренного хвойного леса, более континен-
тальный климат которого проявляется в распространении Larix griffithii.
Кроме того, представлены Tsuga, Picea morinda, Р. spinulosa, Р. morin-
doides', встречается Taxus, а также, правда реже, Juniperus. Перечислен-
ные выше лиственные деревья известны и в Восточных Гималаях, заметны
Fraxinus и Corylus, много кустарников: Rosa, Berberis, Sambucus, Rubus,
Rhododendron, Lonicera, Ilex, Viburnum, Evonymus, Spiraea, Ribes, Gaulthe-
ria (вересковые), Litsea (лавровые), Elaeagnus. Интересны американские
и восточноазиатские элементы, как Buddleia, Sassafras, Hydrangea, Mag-
nolia, Aralia, Panax, Camellia, Deutzia, Stauntonia (лардизабалевые^),
Skimmia (рутовые), Enkianthus (вересковые) и др. Флористический состав
и здесь еще необычайно богат. Род Rhododendron представлен в субальпий-
ском и альпийском поясах очень многими видами; субальпийский пояс
образуют виды древовидного Rhododendron, который с высотой все заметнее
отстает в росте, и, наконец, Rhododendron nivale на уровне 5400 м представ-
ляет собой уже крохотный кустарничек.
Аридные области Центральной Азии
293
Расположенные на широте Канченгиао перевалы создают значитель-
ные различия климатических условий даже в пределах альпийского поя-
са. Виды Rhododendron и луга, изобилующие цветами, исчезают, вместо
них развиты альпийские степи. Климатический рубеж разграничивает
между собой умеренно влажный верхний Сикким и сухой высокий Сик-
ким. На высоте 5100 м распространены полусферические подушки родов
Arenaria, Astragalus, Saussurea, Saxifraga, Alsine, Sedum, Rheum nobile
th. всего два древесных растения — Rhododendron nivale и Ephedra. Снеговая
линия поднимается здесь до 5700 м.
К сухому высокому Сиккиму на севере примыкает обширное одно-
образное Тибетское нагорье. Экологические условия соответствуют усло-
виям Памира. В окраинных районах растительность состоит из альпийских
и степных элементов.
Важнейшими родами альпийской степи являются: Caragana, Astra-
galus, Tanacetum, Artemisia, Thermopsis (бобовые), Draba, Lonicera, Ber-
beris, Rosa sericea, Potentilla fruticosa, Sophora moorcroftiana, Clematis
leptodermis, Wikstrbmia (волчниковые), Cotoneaster, Ceratostigma griffithii
(свинчатковые) и др.; Juniperus — единственный представитель в переле-
сках. Вдоль водных артерий встречаются небольшие древостои Salix
amygdalina, S. ораса, Populus alba, P. ciliata, Ulmus pumila, Hippophae
rhamnoides, Buddleia tsetangensis.
Эколого-физиологические исследования в этой труднодоступной
области не производились, поэтому мы ограничились тем, что лишь очень
кратко охарактеризовали сложное распределение растительности в Гима-
лаях, обусловленное сменой климатических условий по вертикали.
Глава 5
АЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ РАЗНЫХ КЛИМАТИЧЕСКИХ ЗОН
АЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ И КЛИМАТ
Выше мы рассматривали преимущественно зональную раститель-
ность, то есть тот естественный растительный покров, который наи-
более полно отражает климатические условия соответствующей при-
родной зоны. При этом нельзя было не учитывать влияние эдафического
фактора, но ведь известно, что тип почвы также в значительной мере опре-
деляется климатом. Правда, дифференцирующее воздействие материнских
пород заметно отражается и на характере растительности, однако это воздей-
ствие второстепенное, лимитирующим же фактором среды остается климат.
Под азональной растительностью мы понимаем совокупность расти-
тельных сообществ, в формировании которых влияние климата отступает
на второй план и главную роль играет почвенный фактор. Полностью
азональной растительности, которая при определенных почвенных усло-
виях встречалась бы в одинаковом развитии во всех климатических зонах,
не существует. Водные растения, правда, обнаруживают почти полную
независимость от климата, но для них имеет значение вероятность наступ-
ления в данной местности морозного периода. Именно по этой причине
тропические водные растения в умеренном климатическом поясе не встре-
чаются. Тем не менее и среди них известны виды, в большей или меньшей
мере требовательные к теплу. Так, Victoria regia и Eichhornia crassipes
придерживаются лишь районов с очень теплым климатом, где заморозков
вообще не бывает. С другой стороны, существуют водные растения умерен-
ной зоны, местонахождения которых обнаружены по всей земной поверх-
ности, например Ceratophyllum demersum, Lemna minor или Phragmites
communis. Широко распространены некоторые галофиты, например SalP
cornia, а также, если не понимать виды излишне ограниченно, Glaux mari-
tima и Triglochin maritimum. Другие галофиты, напротив, строго приуро-
чены к определенным климатам; пример — виды мангровой раститель-
ности (т. I, стр. 163 и след.).
Мы не будем придавать здесь слишком узкий смысл понятию «азо-
нальная растительность» и рассмотрим экологию тех растительных
сообществ, для которых влияние климата оказывается менее существен-
ным, чем воздействие почвенных условий, и которые никогда не выступают
как зональная растительность, чем и отличаются от экстразональных
группировок.
РАСТИ ТЕЛЬНОСТЬ РЕЧНЫХ НАНОСОВ1
Обширные низменности умеренных зон неотделимы от больших,
медленно текущих водных потоков, уровень которых с началом таяния
1 В. В. А ле хин, Наши поемные луга, М., 1925; В. Р. Вильямс, Почвове-
дение, М., 1939. [В учение В. Р. Вильямса о пойме в настоящее время внесены значи-
тельные коррективы.— Ред.]
Азональная . растительность разных климатических зон
295
снега быстро поднимается; в результате полыми водами периодически
затапливаются огромные пространства. Достаточно вспомнить речную
систему Миссури — Миссисипи в Северной Америке, или Днепр, Волгу
и крупные сибирские реки в СССР, или Хуанхэ в Китае. Половодье создает
по обе стороны русла реки особый рельеф и своеобразные почвы, которые
в свою очередь обусловливают развитие пойменных лесов или пойменных
лугов. Понять закономерность резкой смены одних растительных сооб-
ществ другими в поймах рек можно, лишь зная устройство поверхности
Рис. 211. Схематический профиль через долину реки с подмываемым (справа} и
отлогим (слева} берегами.
1 — русло реки в межень; 2 — область поймы (а — прирусловые дюны, б — центральная
пойма, в — притеррасная пойма); 3 — речная надлуговая терраса; 4 — коренной берег, под-
нимающийся к водоразделу. Пунктирная линия — уровень воды в половодье; точками обозна-
чены вертикальные вихревые токи в главных, и второстепенных течениях.
поймы, которое повторяется с небольшими вариациями в поймах боль-
шинства рек. Объясним ситуацию, складывающуюся в естественных усло-
виях, с помощью четырех схематических рисунков, заимствованных
с некоторыми изменениями у Вильямса.
Вследствие вращения Земли в северном полушарии правый берег
реки обычно крутой, а левый пологий; в южном полушарии наоборот.
В поперечном профиле типичной речной долины (рис. 211) выделяют:
1) собственно русло реки, 2) низменную зону затопления, или область
поймы (называемую также первой террасой), 3) собственно речную, над-
луговую, террасу, не заливаемую весенним половодьем (вторая терраса),
и 4) коренной берег, поднимающийся к водоразделу. Особый интерес
как местонахождение пойменных лесов и долинных лугов представляет
зона затопления. Более высоко приподнятая речная терраса, сложенная
песчано-галечным материалом, обычно покрыта сухолюбивыми расти-
тельными сообществами.
В зоне затопления происходит не только регулярный разлив воды
в половодье, но и процесс седиментации, то есть отложение взвешенных
в воде частиц песка или глины. Геоморфология зоны весьма сложная.
Различают три основные части: область, непосредственно прилегающую
к руслу реки, область центральной поймы и область притеррасной поймы,
расположенную у подножия высокой террасы или граничащую с корен-
ным берегом. Поверхность полосы затопления неровная и обычно при-
поднята у берега реки в виде прируслового вала. При подъеме воды весной
разлив покрывает прежде всего участки, примыкающие к руслу, и лишь
при дальнейшем повышении уровня вода затапливает всю пойму. Понятно,
что скорость течения воды не везде одинакова: чем ближе к руслу реки,
чем вода движется с большей скоростью. Такая разность скоростей обус-
ловливает известную сортировку осадков. Из воды, еще сравнительно
быстро текущей над прирусловым валом (рис. 211, 2а), осаждаются более
крупные частицы песка. После спада воды песок с поверхности быстро
296
Глава 5
высыхает, и ветер перевевает его, ссыпая в плоские бугры или в гряды
притеррасных дюн; ветер может разносить песок более или менее далеко-
в глубь поймы и даже навевать песок на высокую террасу или коренной
берег (рис. 212). При вступлении в область центральной поймы (рис. 211,.2)
вода замедляет свое течение и освобождается от фракций песка мельче
Рис. 212. Участок долины реки с поймой и песчаными дюнами. На
склоне террасы — овраги, в бортах которых выступает почвенная,
вода, питающая притеррасную речку.
Рис. 213. Пойма реки в период затопления ее паводковыми водами.
Видны горизонтальные вихревые токи в главном меандрирующем
русле и второстепенных течениях слева и справа от него.
0,25 мм и частиц глины. Масса осажденного материала здесь меньше, из-за
чего эта часть поймы всегда ниже и более выровнена, чем поверхность-
прирусловых дюн. Наиболее слабое отложение наилка наблюдается у под-
ножия террасы. В половодье, помимо главного течения, можно наблюдать,
боковое, направленное по средней, наиболее пониженной линии централь-
ной поймы (по ее тальвегу); по обе стороны от этой линии скорость воды
уменьшается и поэтому также происходит отложение значительного коли-
чества более грубозернистых элементов.
Азональная растительность разных климатических зон
297
Если полоса затопления располагается по обе стороны меандрирующе-
го русла реки, что обычно наблюдается в безлесных местностях, возни-
кают три течения: главное в русле и два вторичных (второстепенных) сле-
ва и справа от русла, которые направлены по тальвегам поймы (рис. 213,
пунктирные линии). Гряды, обрамляющие эти течения, сложены материа-
лом разной крупности; наиболее мелкие частицы осаждаются там, где
движение воды резко замедляется. Отложение наилка происходит ежегод-
но, и его отдельные слои накладываются один на другой — образуется
слоистая наносная почва. Предпосылкой ее формирования являются
быстрый подъем полых вод весной и почти одновременное затопление
Рис. 214. Обстановка в долине реки при медленном развитии половодья.
Течение реки слева направо. Стрелки показывают направление участков затопления пойм,
происходящее против общего уклона поверхности центральной поймы. Штриховка — подмы-
ваемые крутые склоны террасы; частый пунктир — области наибольшего скопления песков
поймы; редкий пунктир — песок, навеянный на левобережную террасу.
всей поймы. В настоящее время, когда в большинстве речных бассейнов
леса вырублены, образование слоистой поймы представляет собой обычное
явление.
В лесных районах уровень воды в реке весной поднимается очень
медленно; снег тает постепенно; вода не переливается через прирусловые-
валы, а проникает в область центральной поймы по ложам небольших
притеррасных речек (ручьев), обычно образующихся из источников, бью-
щих из подножия верхней террасы. Вода заливает пойму, поднимаясь
против общего уклона поверхности долины (рис. 214). Поэтому над пой-
менными лугами вода в половодье не течет, а стоит неподвижно. При таком
порядке поступления полой воды в осадок выпадают только мельчайшие
органические и глинистые частицы (крупность зерна около 0,005—0,001 мм).
После спада воды почва бывает покрыта 2—30-сантиметровым слоем полу-
жидкого ила, который после высыхания распадается в горизонт комков
в 2—3 см в диаметре. Таким путем возникает наносная почва зернистой
поймы х. Для развития растительного мира комковатая структура почвы
обладает рядом преимуществ: улучшается аэрация почвы, образующие
комки органические частицы содержат много азота и других питательных
солей. Следовательно, создаются весьма благоприятные условия для
роста растений.
1 Размер агрегатов почв «зернистой поймы» обычно значительно меньше.—
Прим. ред.
298
Глава 5
Иной режим складывается в почвах слоистой поймы. Слагающий
ее материал представляет собой тонкозернистый песок или пылевые
частицы, состоящие главным образом из кварца. Богатые питательными
веществами органические остатки оказываются погребенными под бес-
плодными осадками пыли, которые ложатся
Р ис. 215. Продолжительность затопления террито-
рии, выраженная в % от времени вегетации (1 мар-
та — 31 октября), и распространение растительных
•сообществ в пойме Дуная между Собом и Будапеш-
том (Karpati, Vargast, 1965).
1 — Fraxineto—Ulmetum; 2 — Lolio—Potentilletum anseri-
-nae; 3 — Alopecuretum pratensis; 4 — Festucetum pratensis;
5 — Agrostidetum. albae; 6 — Ranunculetum repentis; 7 — Sa-
licetum purpureae; 8 — Salicetum triandrae; 9 — Rorippo
Silvestri—Agrostidetum albae; 10 — Rumici—Alopecuretum
geniculati; 11 —растительные сообщества на илистых отло-
жениях; 12 — Сурего—Juncetum; 13 —Eliochari aciculari—
Schoenoplectetum supini; 14 — Caricetum > acutiformis-ripa-
riae; 15— Glycerio—Sparganietum neglecti; 10 —Scirpeto —
Phragmitetum.
столь плотно, что вместо
развития аэробного бакте-
риального процесса проис-
ходит гниение органиче-
ского вещества с образо-
ванием сероводорода. На
таких почвах условия су-
ществования для растений
достаточно суровые.
Водный фактор в зоне
затопления дифференци-
рован. Состоящие из пес-
чано-галечного грубозер-
нистого материала приру-
словые валы при спаде
воды быстро высыхают;
центральная пойма с ее
тонкозернистыми отложе-
ниями, выровненной по-
верхностью и, как прави-
ло, сравнительно высоким
залеганием грунтовых вод
представляет собой более
увлажненные местообита-
ния; наконец, притеррас-
ная пойма склонна даже
к заболачиванию, так как
здесь, грунтовые воды не-
редко выходят на поверх-
ность. Они дают начало
так называемым притер-
расным речкам, которые
ниже по течению вливаются в основное русло реки (рис. 214) х.
Таким образом, речные наносы могут представлять собой местообита-
ния как богатые питательными веществами, так и бедные ими; местооби-
тания как сухие, так и влажные либо даже сырые. Соответственно различ-
ными должны быть и растительные сообщества. Особенно важным факто-
ром, определяющим в конечном итоге характер размещения раститель-
ности в пределах поймы, является длительность ее затопления в поло-
водье (рис. 215).
Разумеется, разграничение зоны затопления на сравнительно сухую
область прирусловых валов, влажную центральную часть и переувлаж-
ненную притеррасную пойму весьма условное. В действительности нод-
1 Обширные болота в виде тростниковых зарослей сопровождают, например,
русло Рейна в пределах Верхнерейнской низменности. Переувлажнение вызвано здесь
водами, стекающими с прилегающих гор, и ровным рельефом, затрудняющим дренаж
почвы. В настоящее время большая часть этих болот осушена и превращена в пахотные
земли. Реки ограждены дамбами, зарегулированы и несут воду в Рейн кратчайЩим
путем.
Азональная растительность разных климатических зон
299
разделение поймы значительно сложнее. В каждой из названных трех
частей следует различать высокорасположенные, сухие местообитания,
местообитания со средними условиями увлажнения и низколежащие,
влажные местообитания. Общую картину осложняют многочисленные
находящиеся на разных стадиях заиления старицы, небольшие озера,
старые русла реки. Кроме того, повсюду проявляется различие в механи-
ческом составе наилка, который может быть представлен и крупной галь-
кой, и тончайшей глиной. В таких условиях растительный покров должен
выглядеть как сложная мозаика переплетения различнейших сообществ,
и мы зашли бы слишком далеко, пытаясь дать их детальный анализ.
Поэтому ограничимся рассмотрением растительности пойм лишь в общих
чертах, приняв за основу условия, наблюдаемые на обширной Восточно-
европейской равнине.
Среди злаков, распространенных на речном аллювии, выделяют три
типа: 1) корневищные злаки (обычно с длинными подземными корневи-
щами), например пырей {Agropyrum repens), костер безостный {Bromus
inermis) или вейник {Calamagrostis epigeios)’, 2) дернообразующие злаки
с короткими дуговидными корневищами и стеблями, слагающие редкий
луговой травостой, например овсяница красная {Festuca rubra), полевица
'(Agrostis canina) или мятлик {Роа trivialis), и 3) кустовые злаки с плотно-
стоящими прямыми стеблями, как у луговика {Deschampsia caespitosa),
типчака {Festuca sulcata) или у ковы лей (виды Stipa).
Эти три типа злаков заселяют разные части долинных лугов. Корне-
вищные злаки типичны для рыхлых песчаных отложений прирусловых
валов с подвижным субстратом; дернообразующие злаки занимают плотные
почвы центральной поймы; кустовые злаки либо как индикаторы плохой
аэрации и почвы появляются у подножия верхней террасы (например,
Deschampsia caespitosa), либо поселяются на самых высоких и сухих,
не заливаемых в половодье участках (пример — Festuca sulcata).
Можно установить следующий экологический ряд доминантов
в направлении возрастания влажности местообитания:
Festuca sulcata Bromus erectus бобовые —> Alopecurus pratensis-^-
то же совместно с Роа palustris Beckmannia erucaeformis Glyceria
fluitans -> Glyceria aquatica -+ Phragmites communis Scirpus lacustris ->
-> водная растительность открытой воды.
Два первых вида характерны для возвышенных участков, бобовые
и Alopecurus — для средних по высоте местообитаний, следующие три
вида — для пониженных мест; Glyceria aquatica, Phragmites и Scirpus
не образуют лугов, а относятся уже к болотной растительности.
Наибольшую площадь занимает центральная пойма, наиболее важна
она и в хозяйственном отношении. Заболоченная притеррасная часть поймы
как луг практически не используется, для нее очень характерны леса
из ольхи клейкой {Alnus glutionosa); из злаков здесь преимущественно
растут Deschampsia caespitosa или же Festuca rubra, Agrostis alba, осоки или
ситник {Carex vesicaria, С. rostrata, Heleocharis palustris и виды Juncus).
Наконец, растительность прирусловых валов обнаруживает признаки
крайнего непостоянства, многое зависит здесь от случая. Полые воды
и ветер постоянно «перемешивают» почву береговых валов, и растения
должны приспособиться к этой ненадежности субстрата. Непосредственно
у уреза воды на песчаных отмелях весьма типичен белокопытник {Petasites
spurius), корневища которого быстро захватывают все новые и новые уча-
стки. На некотором удалении от песчаной отмели протягивается полоса
кустарника, представленного разными видами ивы. Кроме того, здесь
300
Глава 5
охотно поселяются рудеральные растения и виды с мощными корневищами,
которые, размножаясь вегетативно, образуют чистые травостои, например
пырей. На самых сухих местообитаниях часто растут Medicago -falcate,,
Trifolium montanum, Galium verum, Achillea millefolium, Libanotis montana,
Rumex acetosa ssp. haplorrhizus, Potentilla argentea’, у многих из этих видов
развиты уходящие глубоко в почву стержневые корни. Растения южнцх
степей нередко проникают по береговым валам далеко в северные лесные
районы; северные же виды в свою очередь используют заболоченную при-
террасную пойму как путь, по которому они мигрируют к югу. Давно
известно, какую большую роль при расселении растений играли в после-
ледниковое время речные долины с их обширными аллювиальными рав-
нинами; в наши дни таким же способом в состав местной флоры внедряют-
ся чуждые элементы из соседних районов.
Важнейшей в геоботаническом и хозяйственном отношениях являет-
ся, как мы уже сказали, центральная часть поймы. Здесь развиты соб-
ственно пойменные (заливные) луга, ширина которых часто достигает
нескольких километров, а в долинах крупных рек нескольких десятков
километров. Эти часто кажущиеся однообразными обширные площади
занимает сплошной травянистый покров, состоящий то из одних злаков,
то из злаков со значительной примесью разнотравья. Растения нередко
достигают в высоту 1 м, на влажных участках с Phalaris arundinacea
они бывают в рост человека. Весьма велики, следовательно, и укосы. При
внимательном же рассмотрении оказывается, что растительный покров
пойменных лугов в соответствии с разнохарактерностью мест обитания
в пределах центральной поймы также неоднороден. Создается причудливая
и красочная картина: темно-красные пятна Agrostis перемежаются с серо-
вато-зелеными скоплениями Alopecurus*, темно-зеленый цвет придает лугу
Роа, а ярко-желтая окраска свидетельствует о резком доминировании
в травостое Ranunculus. Каждый участок ассоциации очерчен сравнитель-
но резко и своей границей передает изменение условий произрастания.
Для более сухих площадей пойменных лугов особенно характерны
злаки: Koeleria, Роа pratensis, Festuca rubra, Alopecurus pratensis, среди
которых видны многочисленные Filipendula hexapetala, Trifolium monta-
num, Galium verum и др. На местах среднего увлажнения наряду с Alopecu-
Таблица 53
Число видов на лугах реки Вороны (I) и реки Цны (II)
Доминирующие виды злаков на местообитаниях с увеличивающейся влажностью злаки осоки бобовые разнотравье всего
I II I II I II I II I II
Festuca sulcata 9 11 1 1 11 9 46 52 67 73
Bromuserectus . . 16 10 2 1 11 10 70 111 99 132
Agrostis canina 13 16 1 4 9 11 45 78 78 109
Alopecurus praten-
sis 18 16 3 2 7 10 38 76 66 104
Alopecurus praten-
sis и Роа palus-
tris 10 15 6 5 5 7 32 54 53 81
Poa palustris . . 12 11 8 8 5 2 34 39 59 60
Beckmannia eru-
caeformis . . . 10 13 7 5 1 1 31 30 49 49
Glyceria fluitans 3 6 3 2 0 0 10 15 16 23
Glyceria aquatica 3 4 3 2 0 0 10 10 16 16
Азональная растительность разных климатических зон
301
rus pratensis распространены обычные виды клевера {Trifolium pratense,
Т. repens, Т. hybridum), нивяник {Chrysanthemum leucanthemum), лютик
едкий {Ranunculus acer}, Sanguisorba officinalis, Rumex acetosa, Rhinanthus
major и др. Эти виды соответствуют растительности свежих тучных лугов
Центральной Европы.
В общем заливные луга представляют собой необыкновенно богатые
видами сообщества; бедные видами луговые ассоциации приурочены толь-
ко к самым влажным участкам и служат переходным звеном к низинным
болотам и топям. Это хорошо иллюстрирует составленная Алехиным
таблица, в которой сопоставлен состав пойменных лугов, окаймляющих
течение рек Вороны и Цны (табл. 53).
Из данных таблицы следует, что по числу видов злаки особенно
широко представлены на местообитаниях средней влажности, осоки —
на сырых, а бобовые, как и разнотравье,— на сухих местопроизрастаниях.
По количеству видов преобладает разнотравье, но по массе первое место
всегда занимают злаки. Совпадение числовых значений каждый раз хоро-
шее, хотя реки относятся к разным речным системам (Ворона входит
в систему Дона, а Цна — в систему Волги). При сравнении долинных
лугов разных ландшафтных областей легко заметить, что растительность
лугов испытывает на себе известное влияние той природной зоны, в кото-
рой протекает река. Это влияние тем больше, чем меньше река. И это не
должно казаться странным, ибо характер половодья зависит от климата
(количество атмосферных осадков) и коэффициента лесистости водо-
сборного бассейна. Кроме того, растительность любых пойм ощущает на
себе непосредственное воздействие макроклимата (именно он определя-
ет длительность вегетационного периода, температурный режим, степень
влажности и т. д.). Отсюда понятно, почему пойменные луга северных
областей по флористическому составу коренным образом отличаются от
пойменных лугов юга. Для последних, например, весьма характерно частое
засоление почвы и связанное с ним распространение галофитов. Первые
признаки засоления обнаруживаются уже при вступлении в зону лесосте-
пи (слабое садовое засоление). Бросается в глаза появление следующих
видов: Cirsium canum, Atropis distans, Trifolium fragiferum, Alopecurus
ventricosus. Южнее степень засоления увеличивается, о чем свидетельству-
ет, например, распространение Taraxacum bessarabicum, Centaurea glasti-
folia, Carex diluta, Juncus gerardi, Aster tripolium, Scorzonera parviflora,
Statice gmelini и др. Наконец, на юго-востоке Восточной Европы в долинах
уже можно видеть чистые травостои Salicornia herbacea, Suaeda maritima,
Salsola soda, Statice gmelini и др.
Луговая растительность изменяется в направлении с юга на север:
степные виды постепенно исчезают из ее состава и уступают место север-
ным видам, пока наконец в тундре уже не наблюдаются типичные аркти-
ческие элементы, как Phleum alpinum, Polygonum viviparum, Rubus arcticus
и др. Налицо влияние климата, которое, однако, проявляется в значитель-
но ослабленном виде.
В совершенно определенном направлении протекает развитие расти-
тельности на аллювиальных отложениях Нижней Волги. Половодье
наблюдается здесь не как обычно в начале вегетационного периода, а
делит его на две фазы. Наивысший уровень (+4 м) достигается 12—
14 июня. Весна начинается на юге в марте, задолго до наступления оттепе-
ли в верхнем течении Ч Поэтому здесь на затапливаемых участках распро-
1 Ледяной покров держится на Нижней Волге в среднем не более 110 дней,, выше
же Казани — около 140—150 дней.
302
Глава 5
странены весенние эфемеры, заканчивающие развитие до начала половодья
в верховье (конец апреля — начало мая); назовем лишь некоторые виды;
Ranunculus pedatus, Stenophragma thalianum, Androsace elongata, Draba
nemorosa, Cerastium anomalum; кроме того, в районе Волго-Ахтубинской
поймы повсеместно встречаются Alyssum desertorum, Chorispora tenella,
Agropyrum orientate, A. prostratum и др. После спада воды в конце июля
появляются и другие быстрорастущие виды, которые можно было бй
назвать летними эфемерами: Scirpus hamulosus, S. supinus, S. micheliana,
Marsilia aegyptica, Vandellia pyxidaria и др. Чтобы достичь стадии созрева-
ния семян, им нужно всего 10—15 дней. Некоторые многолетние растения,
застигнутые половодьем в средней фазе развития и затапливаемые водой
на три месяца, либо полностью утрачивают надземные побеги (Agropyrum
repens, Tournejortia sibirica'), либо же сбрасывают листья; живыми остаются
только стебли и верхушечные почки (Euphorbia uralensis, Cynanchum acu-
tum). Другие растения способны развивать водные формы (Butomus umbel-
latus, Lysimachia nummularia, Nasturtium brachycarpum). В дельте Волги
растет экзотический для этих мест индийский лотос (Nelumbium indicum),
высаженный, вероятно, в историческое время. Описанные условия на
Нижней Волге относятся к тому периоду, когда еще не были построены
гигантские волжские плотины и канал Волга — Дон. Сооруженные
в последнее время крупные водохранилища и оросительные системы совер-
шенно изменили режим половодий.
Подразделение речных пойм мы рассмотрели на примере рек Восточ-
ной Европы, так как здесь еще можно встретить условия, более или менее
близкие к природным. В остальной же части Европы нет ни одной крупной
реки, сток которой в разное время года не был бы сильно изменен различно-
го рода гидротехническими системами.
Но истинно природные, не потревоженные человеком условия нельзя
обнаружить даже в Восточной Европе. Вследствие вырубки лесов на водо-
сборной площади значительно повысились пики паводка и соответственно
резче обозначилась летняя межень. В корне изменился состав пойменной
растительности. Теперь повсюду преобладают луга, там где прежде были
пойменные леса, очень сильно отличающиеся по составу от лесов зональ-
ной растительности. Эти леса, первоначально сплошь покрывавшие пой-
мы, как и луга, заливаются полыми водами на более или менее длительное
время. Таким образом они получают дополнительную влагу и в качестве
галерейных лесов могут проникать глубоко в засушливые районы, где
осадков для произрастания зональной лесной растительности уже недоста-
точно. Такие пойменные леса мы отмечали даже в Средиземноморье и в
азиатских пустынях.
Важнейшим фактором среды, регулирующим развитие пойменных
лесов, является уровень грунтовых вод, амплитуда колебания которого
может достигать очень значительной величины. Мы различаем:
1) поймы, затапливаемые весенним разливом почти ежегодно и обле-
сенные мягкодревесинными породами, такими, как виды Salix и Populus'r
2) поймы, обычно заливаемые на более короткое время и облесенные-
твердодревесинными породами. Особенно типичны дуб, вяз и ясень
(рис. 216).
Нередко и ольха (Alnus glutinosa) образует в пойме чистые древостои,
но преимущественно на более влажных участках, где грунтовые воды
даже при низком уровне воды в реке стоят относительно высоко. Ольха
не является видом собственно пойменного леса, а может рассматриваться
как древесная порода заболоченного леса, не приуроченного к речным
Азональная растительность разных климатических зон
303
Рис. 216. Профили пойменной части долины главного рукава Дуная
в пределах Венгерской низменности (Karpati, Pecsi, 1959).
1 — сообщества карликовых сутей (Nanocyperion); 2 — тростниковые заросли
(Scirpeto — Phragmitetum); з — ивовый кустарник (Salicetum triandrae); 4 — пойма
с ивой и тополем (Salicetum albae-fragilis) (б — с Carex acutiformis, с — с Agrostis
alba); 5 — пойма с дубом и вязом (Querceto—Ulmetum) (а — обычные, б — с Aspe-
rula odorata, с — С Convallaria majalis).
долинам, а чаще распространенного вокруг озер или на низменностях
с беспрерывным восходящим током грунтовых вод. Колебания уровня
последних в заболоченных лесах обычно не достигают 1 м; разлив реки
наступает весной, когда тает снег; поэтому осаждение наилка здесь не
происходит; в результате же ежегодного поступления лесного опада
образуется слой торфа мощностью нередко 10—20 см.
Таким образом, в растительном покрове речных пойм наряду с лугами
даже в настоящее время важную роль часто играют леса. На недавно
отложенном аллювии рек, песчаных или галечных отмелях, ежегодно возо-
бновляются травостои однолетников. Вместе с однолетниками прорастают
семена многолетних луговых растений и древесных пород, преимущественно
видов ив. При условии, что летом эти местообитания не слишком высыхают
и в течение ближайших лет переотложения почвы не происходит, много-
летние виды быстро растут. Если особенно высокий паводок не уничтожает
древостоя полностью, начинается прогрессирующая сукцессия, ведущая
к длительно существующей стадии пойменного леса.
304
Глава 5
В естественных, не испытывающих влияния человека поймах рек
происходит непрерывная перегруппировка всех элементов поймы.
Постоянно возникают новые открытые участки и разрушаются старые
насаждения. Ландшафт поймы чрезвычайно динамичен.
В бореальной зоне, где вегетационный период для развития листвен-
ных пород слишком короток, пойменные леса слагают хвойные виды;
в поймах арктических рек древесная растительность отсутствует вообще.
В направлении же к югу лиственные пойменные леса простираются без
существенных изменений через несколько зон растительности, хотя места-
ми обнаруживают некоторые различия во флористическом составе.
Пойменные леса Центральной Европы подробно описаны в другой
работе («Phytologie», Bd. IV, 2, S. 322—381) 1. Останавливаться на
деталях мы здесь не имеем возможности. Отметим лишь общие для
пойм экологические условия. Каждая речная система характеризуется
присущими ей особенностями годового хода уровня воды. При этом не без-
различно, течет ли река с севера на юг, отчего снег в нижнем течении
стаивает раньше, или с юга на север, когда скатывающаяся с верховьев
волна паводка преждевременно взламывает лед в низовьях. Самый высо-
кий уровень у рек северного полушария, берущих начало в высоких
горах, обычно наблюдается в июле, когда начинают интенсивно таять
снега в горах.
Пойменные луга в наше время либо выкашиваются, либо служат
пастбищем — и в том и другом случае рост деревьев невозможен. Поэтому
возникает вопрос, покрылись бы эти луга лесами, если полностью исклю-
чить вмешательство человека; иными словами, существуют ли в лесной
зоне естественные пойменные луга.
Этот вопрос часто дискутировался. На неиспользуемых лугах обычно
очень скоро вырастают древесные растения, сначала ивовый кустарник,
а затем и другие древесные породы. Но сам этот факт еще не означает,
что луга со временем исчезли бы совсем. Указывалось на то, что при ледо-
ходе льдины снова и снова разрушали бы подрастающий лес. Однако
детальные исследования урона, наносимого древесной растительности
пойменных лесов половодьем и ледоходом, проведенные на реке Потомак
в штате Вашингтон, показали, что деревья лишь повреждаются, но не
погибают и отрастают вновь 1 2. Силы, вырывающие деревья с корнем
и ломающие стволы и ветви, необычайно велики. Максимальная скорость
1 Ср. также: R. Lauterborn, Die geographische und biologische Gliederung
des Rheinstroms, «Sitz.-Вег. Heidelb. Akad. Wiss., Math.-nat. Kl.», Abt. 3, (1916—1918),
S. 1—3; О. H. V о 1 k, Soziologische und okologische Untersuchungen an der Auenvege-
tation im Churer Rheintal, «Jhber. Naturw. Ges. Graubunden», 76, 1940, S. 1—51;
'E. Wendelberger-Zelinka, Die Vegetation der Donau-Auen bei Wallsee,
Weis, 1952; I. Karpati, V. Karpati, The periodic rhythm of the flood-plain
forests in the flood area of the Danube between Vac and Budapest in 1960, «Acta Bot.
Acad. Sci. Hung.», 8, 1962, S. 59—92; V. Karpati, Die zouologischen und okologi-
schen Verhaltnisse der Wasservegetation des D on au-Uberschwemmungsraum.es in Ungarn,
«Acta Bot. Acad. Sci. Hungar.», 9, 1963, S. 323—385; I. Karpati, V. Karpati,
G. Varga, Periodische Dynamik der zum Agropyro-Rumicion crispi gehorenden Ge-
.sellschaften des Donau-Uberschwemmungsgebietes zwischen Vac und Budapest im Jahre
1963, «Acta Bot. Acad. Sci. Hung.», 11, 1965, S. 165—196; H.-D. К г a u s c h, Vegeta-
tionskundliche Beobachtungen im Donaudelta, «Limnologica», 3, Berlin, 1965, S. 271 —
"313; P. Simon, Contribution a la connaissance de la vegetation du Delta du Danube,
«Ann. Univ. Sci. Budapest», Sect. Biol., 3, 1960, p. 307—333. Описание пойменной расти-
тельности больших североамериканских рек (см.: R. Knapp, Die Vegetation Nord-
und Mittelamerikas, Stuttgart u. Jena, 1965 (приведен список литературы)).
2 R. S. S i g a f о о s, Botanical evidence of floods and floodplain deposition,
-«Geolog. Surv.», Paper 485-A, Washington, 1964.
Азональная растительность разных климатических зон
305
Рис. 217. Судьба ясеня (Fraxinus pennsylvanica), растущего в пойме реки Потомак
под Вашингтоном, период наблюдений с 1928 по 1964 г. Крону дерева четыре раза
ломали половодье и ледоход (по Сигафусу).
течения Потомака в половодье достигает 6 м/сек (более 20 км/час)’, с такой
скоростью плывущий лес и льдины устремляются на каждое препятствие.
На рис. 217 отражены не только история бедствий ясеня (Fraxinus pennsyl-
vanica) в течение 27 лет, но и жизнестойкость дерева. Рис. 218 иллюстри-
рует образование побегов у дерева, которое было повалено ледоходом
и погребено под 70-сантиметровой толщей наносов. Время погребения под
осадком можно установить по срезам древесины, так как образовавшаяся
20-0434
306
Глава 5
после захоронения древесина ствола анатомически напоминает корневую
древесину.
Даже при длительном весеннем разливе древесная растительность
поймы не погибает. Опасным затопление становится тогда, когда про-
должительность нахождения деревьев под водой отнимает более 60%
времени вегетации. Наиболее чувствительны к затоплению старые и очень
молодые деревья. Fraxinus pennsylvanica и Ulmus americana перенесли
Рис. 218. Поваленный в половодье и погребенный под 70-сантимет-
ровым слоем наносов ствол Fraxinus pennsylvanica; пойма реки
Потомак. Из ствола взята проба для исследования анатомического
строения захороненной древесины (по Сигафусу).
затопление, длившееся целый год. Чем ближе стоят деревья к воде, особен-
но если они растут на подмываемом склоне, тем чаще они подвергаются
сокрушительным ударам плывущих льдин. Соответственно можно выде-
лить три зоны:
1) зону прибрежных кустарников,
2) зону низких, сильно ущербных деревьев и
3) зону высоких, с незначительными повреждениями деревьев на
большом удалении от реки.
Из приведенных фактов можно сделать вывод, что в зоне лиственных
лесов естественные луга как следствие половодья и ледохода образоваться
не могут. Хвойные породы более чувствительны к повреждениям, но,
как бы компенсируя этот недостаток, в бореальной зоне распространены
виды Salix и Betula, которые обрамляют русла рек и обладают всеми теми
же свойствами, какие присущи другим лиственным деревьям поймы.
Только там, где затопление при паводке ежегодно продолжается дли-
тельное время, леса в пойме должны бы отсутствовать, как, например,
в нижнем течении Днепра, где до сооружения больших плотин паводковыо
воды стояли так долго, что даже тростник не успевал зацвести. В нижнем
течении Волги до строительства плотин подъем воды на 1 м держался
до 92 дней, то есть около 3 месяцев (в среднем 77 дней). Отсутствие здесь
деревьев, возможно, также следует объяснять большой продолжитель-
ностью затопления.
Таким образом, в естественных условиях луга в поймах рек не были
бы столь обширными, какими они стали сегодня в результате их сельско-
хозяйственного использования. Тем не менее в пойме из-за регулярно-
Азональная растительность разных климатических зон
307
происходящего перемещения почвы постоянно возникают’местообитания,
где злаки и разнотравье современных лугов могут известное время удержи-
вать свои позиции, даже если отсутствует вмешательство человека. Кроме
того, в литературе не упоминается такой важный, способствующий форми-
рованию лугов фактор, каким является заселение рек бобром, первона-
чально широко распространенным как в Европе, так и в Северной Амери-
ке. Срезая необходимые им для постройки плотин деревья, растущие
в пойме реки, бобры освобождают тем самым площадь для луговой и высо-
котравной растительности. Тот, кто наблюдал в Йеллоустонском нацио-
нальном парке, как на реках, текущих по равнинной местности, одна
бобровая плотина сменяется тут же другой, и видел, сколь широка
лишенная деревьев полоса вдоль берега, тот не может недооценить этого
фактора.
К элементам пойменных ландшафтов относятся и многочисленные
старицы, которые, будучи отшнурованными от основного русла, превра-
щаются постепенно в озера. С их богатой водной растительностью мы
познакомимся в следующем разделе, причем сведем наш обзор к рассмотре-
нию общих, экологически важных проблем.
ПРЕСНОВОДНЫЕ РАСТЕНИЯ
И ИХ ЭКОЛОГО-ФИЗИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА
Изучение живого мира вод — задача двух отраслей биологии: гидро-
биологии и лимнологии, если на первом плане стоит исследование озер.
Веская причина позволила выделить учение о материковых водоемах
в самостоятельную область знания: внутренние водоемы представляют
собой четко ограниченные, несущие в себе большой запас информации,
замкнутые экосистемы. Поэтому они наиболее пригодны для изучения
закономерностей кругооборота веществ в природе и выявления различного
рода пищевых цепей. К тому же одно из звеньев пищевой цепи — рыбы —
особенно ценно в хозяйственном отношении.
Саму среду, в которой реализуется этот кругооборот,— воду легко
исследовать физико-химическими методами. Изучение же населяющих
водную среду живых существ требует более сложного подхода. Различные
водные организмы — растительные и животные — в процессе обмена веще-
ством оказываются тесно связанными друг с другом. Организмы, пассивно
переносимые водными течениями, называют планктоном. Центральной
группой растительного планктона являются одноклеточные и колониаль-
ные водоросли.
В нашу задачу не входит рассмотрение основ лимнологии и сложных
изменений факторов водных местообитаний (температуры, освещения,
химизма) в суточном и годовом ритмах, а также обсуждение жизнедея-
тельности планктонных организмов, поэтому отсылаем читателей к соот-
ветствующим обобщающим работам х. Ограничимся здесь эколого-физиоло-
1 F. Ruttner, GrundriB der Limnologie, Berlin, 1962; A. Thienemann,
Die Binnengewasser Mitteleuropas, «Die Binnengewasser», 1, 1925. См. также другие
тома этой серии. Ср.: F. Е. Round, Biologie der Algen, Stuttgart, 1968; F. Lenz,
Biologie der SuBwasserseen, Berlin, 1928; F. G e s s n e r, Die Binnengewasser, «Hdb.
d. Pflanzenphysiol.», 4, 1956, S. 179—232; G. E. Hutchinson, Treatise on Limnolo-
gy, v. I. 1957; «Жизнь пресных вод СССР», под ред. В. И. Жадина, М.— Л., 1949;
S. A. S е г п о w, Allgemeine Hydrobiologie, Berlin, 1958.
Характеристика растительности озер вблизи северной границы леса приведена
в работе: G. W a s s е n, Gardiken, Vegetation und Flora eines lapplandischen Seeufers,
«Kungl. Svenska Vetenskapakad. Avh.», № 22, Stockholm, 1966.
20*
308
Глава 5
гической характеристикой высших растений, которые произрастают в мел-
ководной прибрежной зоне крупных водоемов и которым лимнологи обычно
уделяют мало внимания. Подобающее цветковым растениям место отве-
дено только в большом сводном труде Геснера 1.
Низшие растения — первично водные организмы; обитающие *же
в воде цветковые растения произошли от наземных растений. Они лишь
вторично приноровились к необычной для них среде жизни и обладают
рядом приспособительных анатомо-морфологических признаков 1 2.
Недостаточное внимание лимнологов к высшим растениям, вероятно,
объясняется тем, что в живом состоянии эти растения практически лишь
частично вовлекаются в кругооборот веществ. Водные животные их не едят,
питаются ими разве только личинки некоторых насекомых, не относящихся
к собственно водной фауне. Только после отмирания высших растений их
остатки поступают в круговорот вещества. Цветковые растения занимают
некоторое особое положение также и в пространстве, ибо распространены
они только в мелководной прибрежной зоне — литорали. Тем не менее
продуцируемая ими здесь масса органического вещества достаточно
велика — до 2 т!га, а на густо поросших болотах она составляет даже
14—16 т/га.
По химическому составу воды различают несколько типов озер,
связанных между собой промежуточными типами:
1. Эвтрофные озера. Богатая питательными веществами вода;
содержание N свыше 1 мг/л и Р2О5 более 0,5 мг/л.
Значение pH около 7,0 или несколько больше. Вода обычно мутная,
дно илистое. Характерно большое поступление в озеро органических
соединений извне.
2. Олиготрофные озера. Вода бедна питательными веществами
и содержит лишь следы N и Р2О5. Выделяют два подтипа: а) вода
насыщена известью; pH = 7,5; цвет воды голубой до зеленого.
Такие озера встречаются в районах, сложенных известняками.
Примеры — озера в предгорьях Известняковых Альп или в областях
распространения молодых ледниковых морен; б) вода бедна из-
вестью; pH всегда ниже 7,0 (до 4,5); цвет воды зеленоватый до буро-
го. В водосборном бассейне таких озер развиты преимущественно
силикатные породы или пески (обычно в областях океанического
климата с выщелоченными почвами).
3. Дистрофине озера. Характеризуются большим количеством
принесенных извне гумусовых соединений; цвет воды желтый или
темно-бурый; pH ниже 5,0. К этому типу относятся также озерки
верховых болот. Дистрофные озера распространены преимуще-
ственно в бореальной зоне.
4. Галотрофные озера. Соленые озера бессточных областей
аридного климата. В зависимости от состава и содержания солей
различают: содовые озера с резко щелочной реакцией воды, хлорид-
ные и сульфатные озера. Соотношение химических элементов, как
правило, различное. Крупнейшим соленым озером является Кас-
пийское море; самыми большими озерами с особенно высокрй
соленостью — Мертвое море и Большое Соленое озеро в штате Юта
(США), а также озеро Туз в Анатолии.
1 F. Gessner, Hydrobotanik, Bd. I, Berlin, 1955; Bd. II, Berlin, 1959.
2 H. G 1 u c k, Wasserpflanzen, «Hdb. d. Naturwis.», 10, 1934, S. 575—590;
A. A r b e r, Water plants. A study of aquatic angiosperms, London, 1920; H. S c h e n c k,
Die Biologie der Wasserpflanzen, Bonn, 1886.
Азональная растительность разных климатических зон
309
Рис. 219. Схема последовательного расположения прибреж-
ных растительных сообществ на озере (Walter, 1927).
Справа налево: 1 — пояс свободно плавающих растений (не показаны);
2 — пояс погруженных растений; 3 — пояс растений с плавающими
листьями; 4 — заросли Scirpus lacustris и Phragmites', 5 — пояс
болотных растений, крупных кочкарных осок; 6 — низинное болото.
Наиболее широко распространены эвтрофные озера. Степень эвтроф-
ности всех озер увеличивается, если поблизости располагаются крупные
поселения, сточные воды которых сбрасываются в озеро. Типичный при-
мер — Цюрихское озеро. В последнее время этот процесс заметен и в отно-
шении Боденского озера.
Химизм озерных вод в известной мере зависит от глубины водоема.
Горные озера с крутыми берегами и, следовательно, с очень узкой лито-
ральной зоной, где нет условий для развития прибрежных макрофитов,
способных продуцировать много органического вещества, как правило,
олиготрофные (озера Альп); напротив, мелководные озера, окаймленные
широкой зоной гелофитов, всегда эвтрофные.
Имеют значение и колебания уровня воды. Как только они превыша-
ют известный предел (5 м), берега лишаются растительности. На илистом
грунте, осушенном в длительную межень, поселяются лишь эфемерные
однолетники.
Следует, однако, подчеркнуть, что биологическая типизация озер
не совпадает с их химической классификацией. Для водных и болотных
растений значение имеет прежде всего характер грунта. Приспособйвшиеся
к жизни в воде высшие растения, гидрофиты, укореняются на разной глу-
бине 1. В связи с этим у берегов озер наблюдается весьма отчетливая пояс-
ность растительности.
Выделяют следующие основные пояса (рис. 219).
1. Свободно плавающие растения, называемые также плеустон:
а) растения, плавающие ниже поверхности воды, например Сега-
tophyllum, Utricularia или Lemna trisulca;
б) растения, плавающие на поверхности воды, например Lemna
minor, Hydrocharis, Salvinia, Azolla или также Stratiotes, кончики
листьев которых поднимаются над водой.
1 S. Н е j п у, Okologische Gharakteristik der Wasser- und Sumpfpflanzen in den
slowakischen Tiefebenen, Bratislava, 1960.
310
Глава 5
Рис. 220. Пояс растений с плавающими листьями (Nuphar lutea) на
озере Федер. Справа — болотные растения (A Its та и др.), за ними заро-
сли камыша (Scirpus lacustris) (фото Штаудахера).
2. Погруженные в воду, или подводные, растения. Лишь немногие
из них цветут под водой (Najas), у большинства же цветки возвышаются
над поверхностью ^воды и опыляются ветром или насекомыми (виды Pota-
mogeton, Hottonia palustris, виды Ranunculus).
Рис. 221. Trapa natans, однолетний водяной орех, в одном из рука-
вов реки Моравы, Нижняя Австрия (фото Мортона).
Азональная растительность разных климатических зон
311
Рис. 222. Заросли Phragmites совместно с Scirpus lacustris у берегов
олиготрофного, богатого известью нижнего Лунцского озера в Нижней
Австрии; далее зона с Potamogeton natans (фото Кравани).
3. Растения с плавающими листьями, которые, однако, могут иметь
и[подводные листья (рис. 220). К ним относятся виды Nymphaea и Nuphar,
Nymphaeoides (=Limnanthemum), Potamogeton natans (богатые известью
олиготрофные озера), Ranunculus aquatilis, однолетний Trapa natans
(рис. 221), произрастающий в более теплой воде, и многие другие.
4. Выступающие из воды и образующие «плавни» растения (рис. 222).
Максимальная глубина, на которой они еще встречаются, различна; для
Scirpus lacustris — это 5 м, для Phragmites — 1,2—2 м. Typha связана
с резко эвтрофной средой, Cladium mariscus—с богатыми известью и олиго-
трофными водами.
5. Болотные растения, или гелофиты, не являются собственно водны-
ми растениями, они растут только на постоянно повышенно влажной
почве. Это уже пояс, переходный к осоковому кочкарнику (Magnocarice-
tum, рис. 223).
6. Низинные болота или заболоченные леса, почва которых время
от времени выступает из-под воды; к этому типу лесов в Центральной
Европе принадлежат болотистые леса-ольшаники (рис. 224). На юго-
востоке США широко распространены заболоченные леса, образованные
Taxodium (болотный кипарис) и Nyssa. Так как семена этих древесных
пород в воде не прорастают, образованные ими леса встречаются лишь
там, где почва высыхает по меньшей мере каждые 10—20 лет. Продолжи-
тельность жизни Taxodium составляет 1000 (до 3000) лет; высота дерева
может превысить 40 м. Своеобразны причудливо изогнутые дыхательные
корни болотного кипариса, выступающие из болотной почвы.
Особенно четко выражена поясность в мелководных эвтрофных озерах.
Такое распределение растительности поясами часто рассматривают как
ряд автогенных первичных сукцессий (т. I, стр. 20), ведущих к зарастанию
озер. Предполагают, что в результате отложения на дне продуцируемого
312
Г лава 5
Рис. 223. Пояс крупных кочкарных осок на озере Федер (Верхняя,
Швабия) при высоком уровне воды. Вдали открытая водная поверхность
озера (фото Штаудахера).
Рис. 224. Заболоченный лес-ольшаник (A Inus glutinosa) в южной Швеции
в межень. Деревья (пневая поросль) растут группами на кочках, между
которыми видны болотные растения (фото Лундегорда).
растениями органического вещества глубина водоема в направлении от
берега постепенно уменьшается, благодаря чему пояса растительности
непрерывно наступают от края водоема к открытой воде. Стадии зараста-
ния (или заиления) водоемов могут быть выделены путем изучения ненару-
шенных озерных отложений.
Однако заиление озера происходит относительно быстро лишь тогда,
когда впадающие в озеро реки приносят большое количество взвешенных
Азональная растительность разных климатических зон
313
Рис. 225. Заиление
(заштриховано) озера
Санджерв в восточной
Прибалтике, идущее
с северо-запада ;на
юго-восток (стрелки
указывают направле-
ние господствующих
ветров). Частичное
заиление, наблюдае-
мое на юго-западной
окраине водоема,
связано с впадением
в озеро реки (по
Клинге).
недостатка в воде,
частиц, которые осаждаются в местах, не подверженных влиянию ветра
и не испытывающих поэтому удара волн (рис. 225). Роль самих растений
в этом процессе относительно невелика. Когда озеро обмелеет настолько,
что может развиться тростниковое болото или топь, образуется торф,
состоящий главным образом из корневищ тростника (Phragmites) или
корней крупных видов осок. В заболоченных лесах также возможно раз-
витие процесса заторфовывания.
Гидрофиты продуцируют в неэвтрофных типах озер столь мало орга-
нического вещества, что оно практически никакой роли в общем круго-
вороте веществ не играет. Свободно плавающие растения, черпающие
питательные соли непосредственно из толщи воды,
в «малокормных» водоемах отсутствуют. В эвтроф-
ных озерах они могут сохраняться лишь там, где
ветер и волны не выбрасывают их на берег, обрекая
на засыхание. Гибель части растений большого урона
сообществу не наносит, так как размножаются они
очень быстро — особенно Lemna minor, которая
в небольших, богатых питательными веществами
водоемах за короткое время может образовать столь
плотный покров, что погруженные растения здесь
из-за недостатка света не развиваются.
Глубина, до которой опускаются гидрофиты
в озерах, зависит исключительно от прозрачности
воды в озере (рис. 226). Нижняя граница их рас-
пространения проходит там, где световое доволь-
ствие сокращается до 2% полного дневного света.
В эвтрофных озерах эта граница пролегает на глу-
бине 8 м или менее. В олиготрофном Боденском
озере на глубине 30 м погруженные цветковые
растения, правда, уже не встречаются, но домини-
руют Chara и Nitella, образующие нередко сплош-
ные заросли. На рис. 227 показано поясное разме-
щение растительности в различных частях Боден-
ского озера.
Поскольку гидрофиты никогда не испытывают
лимитирующим в их развитии становится фактор питания. Чтобы осу-
ществлять фотосинтез, водным растениям нужен не только свет, но и СО2,
который погруженные растения способны усваивать только в растворен-
ном виде. Имеющийся в воде СО2 находится в равновесии с атмосферным
СО2, концентрация которого в воздухе относительно мала. Иная картина
наблюдается в водах, богатых известью,— содержащийся в них бикарбонат
кальция диссоциирует: Са(НСО3)2 ** Са++ + 2НСО3.
У растений, способных усваивать бикарбонатный ион СО2, запас
последнего оказывается вполне достаточен для осуществления фотосинте-
за. Правда, при этом образуются ионы ОН1(НСОд СО2 + ОН")
и реакция воды становится резко щелочной.
Рутнер доказал, что водные мхи, например Fontinalis, ассимилируют
только несвязанный СО2. Если же они будут ассимилировать СО2 в воде,
содержащей двууглекислую известь, то pH повысится лишь до 8,8, ибо
свободного СО2 в воде практически уже не остается. Поставив такой же
опыт с погруженными цветковыми растениями, мы обнаружим, что они
способны поглощать и СО2, образующийся при расщеплении бикарбоната
кальция; в результате возникает избыток ионов ОН и величина pH
314
Глава 5
Рис. 226. Максимальная прозрачность воды (черные колонки)
различных озер Центральной Европы и нижняя граница распро-
странения литоральной растительности (белые колонки) (по Тине-
ману).
Максимальная прозрачность воды Боденского озера по сравнению с положе-
нием нижней границы появления растительности прибрежной зоны сравни-
тельно^ мала, однако средняя прозрачность (величина более важная) очень
значительна.
повышается до 10,2—10,4. Наземные растения, ассимилирующие при
опыте в воде, ведут себя, как мхи; среди водных растений аналогичное
поведение отмечено у Callitriche, которая по строению листьев не соответ-
ствует погруженным растениям г.
В олиготрофных озерах с кислой реакцией бикарбонат кальция
отсутствует, поэтому растения могут использовать только свободно раство-
ренный в воде СО 2, из-за чего условия их роста оказываются крайне
неблагоприятными. По этой причине в таких водах развивается лишь
1 F. R u 11 п е г, Zur Frage der Karbonatassimilation der Wasserpflanzen I und II,
«Osterr. Bot. Zschr.», 94, 1947; 95, 1948.
Азональная растительность разных климатических зон
315
Северный 5ерег боЗенскоео озера
и Е
I Е4 3: Е <5
S а ° EStqjo ~'г$95? S?
-10 °
-20 1
о
-30 g
-40 f
Й
со
-50 £
Е с§
И II °
Обрыв у Ибер-
лингского
Высокий уровень ОЗбрД
Низкий уровень Schizothrix-
vhulla Rhizoclonium
Aegagropila
profunda
Зона
Gongrosira
codiolifera
и др.
Нормальная кривая уровня озера
I ДШЫ1ИИ1МД1Д2]1
50м-
30м-
40м-
0-
10м-
20м-
os Ей
eg Й2<з
Йс сй<и
С сз
S Js>s
1
Е
Е
оз.Унтер-Зе у Рейхенау
Chara aspera и
Nitella syncarpa
t. *** г» г* jurmtw _ -
Высокий
уровень
г Низкий уро-
LSS
Рис. 227. Профиль через Боденское озеро и пояса растительности на его берегах
(по Гамсу),
неболыпое число малоспособных к конкуренции видов, которые хотя
и не избегают богатой известью воды, однако в ней они угнетаются
другими энергично размножающимися видами. Иными словами, такие
растения не являются собственно ацидофильными. Правда, Lobelia и
Isoetes, два характерных вида растительности озер с кислой реакцией
воды, по-видимому, не обладают способностью потреблять ионы бикарбо-
ната вТпроцессе фотосинтеза.
Резкое возрастание величины pH, как результат фотосинтезирующей
деятельности погруженных растений в богатых известью водах, приводит
к*распаду бикарбоната кальция (HGO3 + ОН~-> СО" + Н2О) и, следова-
тельно, способствует осаждению СаСО3 (Са++ + СО3 -> СаСО3). В олиго-
трофных, богатых известью озерах углекислый кальций выпадает из
раствора в виде тонкой взвеси, которая оседает в тихих участках водоема
и ведет к образованию толщ озерного мела. Если же СаСО3 кристаллизует-
ся на зеленых частях растений, образуется известковый туф. В карстовых
районах отложения туфа или травертина в текучих водах бывают доста-
точно мощными (рис. 228). Правда, участие растений в этом процессе лишь
пассивное г.
В грунтовых водах содержание свободной углекислоты обычно очень
высокое, благодаря чему в районах, сложенных известняками, они могут
1 I. Matoni ckin, Z. Pavlet ic, Entwicklung der Lebensgemeinschaften
und ihre Bedeutung fur die Bildung und Erhaltung der Kalktuff-Wasserfalle, «Arch.
Hydrobiol.», 58, 1962, S. 467—473.
316
Глава 5
вызвать явления карста, то есть процесс химического растворения горных
пород- При выходе грунтовых вод на поверхность в виде известковистого
источника большая часть СО2 улетучивается; таким образом бикарбонат
кальция теряет СО2 согласно уравнению: Са(НСО3)2 — СО2 -> СаСО3 +
+ Н2О, и СаСОч кристаллизуется в форме кальцита. Следовательно, рець
идет о процессе чисто физического свойства. Отложение же туфа на расту-
щих в воде растениях, особенно на мхах, объясняется тем, что водные
Рис. 228. Мощные отложения известкового туфа, обусловленные разрастанием
Cratoneuron commutatum, в большом водопаде между озерами Галовац и Градинско-
Езеро, Плитвицские озера, Хорватия (фото Э. Вальтер).
растения, разрастаясь, чисто механически увеличивают поверхность отда-
чи углекислого газа в атмосферу. Вместе с тем растения при фотосинтезе
извлекают из воды СО2, благоприятствуя тем самым образованию туфа;
однако такой процесс приобретает некоторое значение, по-видимому, лишь
в стоячих водах и там, где распространены погруженные растения (харо-
вый туф), так как в проточной воде туф осаждается даже на неживых
предметах.
В карбонатных водах погруженные растения снабжаются СО2 лучше,
чем наземные, ибо концентрация СО2 в воде выше и находится он в ней
уже в растворенном виде. При этом львиную долю СО2 поглощает,слой
клеток, непосредственно соприкасающихся с водой, то есть эпидермис,
который в данном случае содержит больше всего хлоропластов. Внутренние
клетки получают СО2 в значительно меньшем количестве. Тонкие или
сильно рассеченные листья, создающие большую общую поверхность
и характерные для видов погруженных растений, возможно, являются
приспособлением к этим условиям.
Азональная растительность разных климатических зон
317
Кислород, освобождающийся при фотосинтезе, накапливается в меж-
клетниках в газообразном виде; им снабжаются незеленые дышащие части
растения, прежде всего корни, к которым он диффундирует по межклетни-
кам. Напомним, что в воде кислорода содержится гораздо меньше, чем
в воздухе.
Икусима 1 изучал количество органического вещества, продуцируемо-
го густой зарослью Vallisneria denseserrulata в оросительной канаве под
Токио. Сначала он измерил площадь листовой поверхности в каждом слое
воды толщиной 20 см (подобно тому, как это делают в луговом травостое),
затем определил индекс освещенности при четырех типах погоды на раз-
ной глубине, интенсивность фотосинтеза в зависимости от величины осве-
щения и интенсивность дыхания. Верхние части листьев Vallisneria ведут
себя как солнечные листья; максимум интенсивности фотосинтеза дости-
гается при силе света 60 тыс. лк, нижние — как теневые листья, с макси-
мумом фотосинтеза уже при 20 тыс. лк.
Наибольшая нетто-продуктивность, то есть количество сухого веще-
ства за вычетом затрат на дыхание, приходится на долю верхних,
богатых хлорофиллом частей листьев. У нижних частей листьев баланс
вещества положителен только в ясные или малооблачные дни. Фитомасса
травостоя составляла 212 а/ж2.
В июле и августе суточное количество продуцируемого сухого веще-
ства составляло (в г/ж2):
Июль Август
Средняя нетто-продуктивность 1,7 3,9
Максимальная брутто-продуктивность 4,8 11,0
Общая продуктивность в ясные, малооблачные, облачные и дождли-
вые дни в июле выражалась соотношением 100 : 87 : 60 : 40, а в августе —
100 : 92 : 77 : 60. С учетом погодных условий было высчитано количество
органической массы, накопленной зарослью V. denseserrulata в отдельные
месяцы (табл. 54).
Таблица 54
Средняя за месяц продуктивность травостоя V alllsnerla (в г/л<‘2)
Показатель Июнь Июль Август Сентябрь Октябрь
Общая продуктивность 63 104 291 160 91
Расход на дыхание . . . 49 53 171 102 64
Нетто-продуктивность 14 51 120 58 27
Отношение чистой про- дукции к общей, % 22 49 41 36 30
Оптимальный индекс листовой поверхности при четырех типах погоды
в июле был равен 8—10, 6—8, 3—5 и 2—3; в августе же 6—6,5, 5,5—6,
5,5 и 4.
Кислород в ассимилирующих частях растений находится в межклетни-
ках под известным давлением, благодаря чему объем растения увеличи-
вается, а объемный вес уменьшается. Этим объясняется, почему стебли
1 I. Ikusima, Ecological studies on the productivity of aquatic plant commu-
nities II, «The Bot. Mag.,», 19, № 931, Tokyo, 1966, p. 7—19.
318
Глава 5
Ceratophyllum demersum, плавающей в толще воды и лишенной корней
травы, днем поднимаются над водной поверхностью, а вечером, когда
кислород израсходован, снова погружаются вглубь. Осенью этот вид дает
побеги с большим объемным весом, которые опускаются на дно водоема
и в таком состоянии переживают зиму.
Корни для погруженных растений большого значения не имеют, они
служат лишь средством прикрепления к субстрату. Так, Ceratophyllum
вообще не образует корневой системы, питательные вещества листья
всасывают непосредственно из воды. Однако известное количество пита-
тельных веществ, которыми дно водоема богаче, чем толща воды, растение
все же добывает с помощью корней. Об этом свидетельствует опыт, постав-
ленный в аквариуме (табл. 55).
Таблица 55
Общая длина стеблей и боковых побегов растений (в еле),
укорененных в естественном грунте и в бедном питательными
веществами песке, а также растений, не имеющих корней в почве
(по Снеллу)
Название растений С корнями в грунте Закрепив- шиеся над грунтом С корнями в песке Закрепив- шиеся над песком
Elodea canadensis . . 308 177 114 114
Potamogeton densus 3671 819 1032 718
Возникает вопрос, поглощают ли корни также воду. Потери воды
на транспирацию у погруженных видов не происходит. Тем не менее у
у этих растений удалось обнаружить небольшое корневое давление.
Выделение воды листьями осуществляется через апикальные отверстия,
или водные устьица (гидатоды), но такие приспособления могут и отсут-
ствовать, например у Elodea. Ток воды через гидатоды, вероятно, служит
для переноса питательных веществ, но он очень слаб, и поэтому прово-
дящая ткань сильно редуцирована. Отсутствует также механическая ткань;
так как удельный вес водных растений с большими, наполненными возду-
хом межклетниками меньше 1, побеги в воде всплывают. У погруженных
растений нет камбия, хорошо различимых проводящих пучков и одревес-
невших клеточных оболочек. На сосудах образуются только маленькие
кольчатые трахеиды. Определенное плазмолитическим методом осмоти-
ческое давление колеблется в пределах 9—11 атм. Клетки, естественно,
полностью тургесцентны.
Прежде считалось, что у погруженных частей растений кутикула
не развита. Однако в последнее время с помощью электронного микроско-
па удалось обнаружить чрезвычайно тонкую кутикулу со множеством
мелких пор. Обмену веществом с окружающей средой она, по всей види-
мости, не препятствует.
Не столь резко выражено приспособление к водной среде у растений
с плавающими листьями, верхняя поверхность которых соприкасается
с атмосферой. Поэтому на наружной стороне у них можно видеть обычную
несмачиваемую наружную кожицу с многочисленными устьицами
(у Nuphar насчитывается 410 устьиц на 1 мм2 листовой поверхности).
Но устьица, как правило, не закрываются и ночью. Сокращать транспи-
рацию растению нет необходимости, так как оно может поглощать воду
непосредственно нижней стороной листа. На последней устьица отсутству-
ют, но имеются особые группы клеток — гидропоты, которые, видимо,
Азональная растительность разных климатических зон
319
служат как для водообмена, так и обмена веществом. Ряска, например,
засыхает, если поверхность воды покрыть тонкой пленкой парафинового1
масла, хотя корешки растения остаются полностью погруженными в воду.
У кувшинковых также доказано наличие корневого давления. Вели-
чина вполне достаточна, чтобы снабжать водой полностью выступающие
над поверхностью воды плавающие листья. Однако, если черешки листьев
срезать, то растение засыхает, так как при этом, по-видимому, разрывают-
ся водяные нити в сосудах. Интересно, что длина черешков листьев
соответствует глубине данного водоема. Представляется, что фотосинтез!
листовых пластинок каким-то образом регулирует рост черешков в длину.
До тех пор пока плавающий лист находится в воде и, следовательно,,
в отличие от погруженных растений не в состоянии ассимилировать СО2,
черешки продолжают расти в длину. Рост прекращается, как только лист
развертывается на поверхности воды и начинает активно фотосинтезиро-
вать. Аналогичный механизм действует, вероятно, и у тростника и сход-
ных с ним по экологии видов. Это предположение' обосновывается тем
фактом, что выступающая над водой часть побегов'независимо от глубины
воды на местообитании всегда имеет одну и ту же длину.
То, что корневища и корни, живущие в анаэробных условиях, в иле
снабжаются кислородом через листья, можно доказать на примере Nuphar,
исследуя воздух, заключенный в межклетном пространстве. Выяснено,
что в листьях в течение суток происходит изменение содержания О2 и СО2.
Наблюдается это изменение и в воздухе межклетников корневищ (табл. 56).
Таблица 56
Содержание кислорода и углекислого газа в воздухе
межклетников Nuphar (по Геснеру)
Части растений Содержание Ог, % Содержание СОг, %
до полудня после полудня до полудня после полудня
Черешки ли- стьев .... 0,9 16,1 7,5 0,6
Корневища . . 0,2 13,7 18,1 7,7
Зимой, когда поступление'О2 в корневища прекращается, начинается
интрамолекулярное дыхание. Если в это время выкопать корневище,
почувствуется резкий запах спирта, количество которого обратно про-
порционально поступившему в корневище количеству. О2.
Каков же механизм, подачи кислорода из листьев в, корневища и кор-
ни? Для чистой диффузии путь в 1—2 м слишком длинен. Если срезанный
с частью черешка лист Nymphaea положить на воду, можно наблюдать
странное явление. При нагревании воды до 25—30° и ярком освещении
из среза черешка начинают выступать пузырьки газа, причем часто такой
процесс длится не один и не два дня, так что в итоге удается собрать
несколько литров воздуха.
Геснер дает этому явлению такое объяснение: упругость водяного па-
ра, содержавшегося в межклетниках листа, при нагревании последнего
возрастает, вследствие чего парциальное давление кислорода оказывается
ниже атмосферного. Кислород диффундирует через устьица в лист; возни-
кает небольшое избыточное давление. Но вызванное им истечение смеси
320
Глава 5
водяного пара и воздуха будет осуществляться легче по крупным ходам
межклетников, ведущим к черешку листа, чем по мелким отверстиям
устьиц, которые способствуют только диффузии. Следовательно, истечение
будет продолжаться до тех пор, пока упругость водяных паров сохра-
няется повышенной. Энергию на скрытую теплоту испарения дает радиа-
ция. Такой «диффузионный насос», видимо, мог бы проталкивать более
богатый О2 воздух в корневище и неповрежденного растения.
Укореняющиеся в воде растения, ассимилирующие органы которых
полностью выступают из воды, от наземных видов отличаются лишь тем,
что постоянно хорошо обеспечены водой и растут на плохо аэрируемой
почве.
И в данном случае на примере детально исследованных экземпляров
Cladium mariscus удалось показать, что у этих растений развита непре-
рывная система межклетников, прослеживаемая от листьев до корней х.
Доля наполненных воздухом полостей (в %) оказалась равной в общем
объеме
оснований быстро растущих листьев .............. 6
оснований зеленых, медленно растущих листьев... 26
оснований увядших листьев.......................56
корней (отрезок длиной 2 см) ................... 60
корневищ (отрезок длиной 1 см)..................38
В данном случае кислород в подземные органы растений поступает
из оснований нижних отмерших листьев, причем содержание О2 в корнях
равно 16—18%, даже если в окружающей среде его концентрация состав-
ляет только 0—1%. Если увядшие листья отрезать, содержание О2 в кор-
нях понизится до 10—15%, а при удалении и живых листьев — до 3—4%,
даже при условии, что корни располагаются в обогащенной кислородом
воде. В этом отношении они отличаются от корней риса, которые хотя
и получают О2 от выступающих из воды частей растения, но, кроме того,
сами способны поглощать кислород непосредственно из воды. Все корни
болотных растений остаются живыми и в анаэробных условиях, но процесс
активного роста проявляется лишь тогда, когда становится возможным
аэробное дыхание.
Процесс транспирации у растений, высоко возвышающихся над водой,
также начинается при открытых устьицах — необходимом условии фото-
синтеза. Сравнительные измерения показали, что растения транспирируют
весьма активно; в расчете на единицу лисп-овой поверхности интенсивность
транспирации составляет 50—60% величины испарения с открытой водной
поверхности. Поскольку дефицит насыщения воздуха даже над зеркалом
озера вследствие охлаждающего воздействия ветра может быть значитель-
ным, то и абсолютные значения транспирации также могут быть доста-
точно большими. Лишь внутри густых зарослей тростника и сходных с ним
растений влажность воздуха резко увеличивается, так как сила ветра
здесь ослабевает. У надводных листьев хотя и имеется нормально развитая
кутикула, однако кутикулярная транспирация у них обычно протекает
все же интенсивнее, чем у наземных растений.
Если определить размер водоотдачи не отдельных листьев, а всего
травостоя, то окажется, что его транспирация в несколько раз превышает
испарение с открытой водной поверхности. Расход воды на транспирацию
зарослями тростника под Берлином в августе, то есть в фазу полного раз-
1 V. М. С on wa у, Studies in autecology of Cladium mariscus Rs Br. Ill, «New
Phytologist», 36, 1937, p. 64—96.
Азональная растительность разных климатических зон
321
вития растений, в 7 раз превышал величину испарения с водной поверх-
ности такой же площади
На основе данных'непосредственного измерения испарения и отноше-
ния испарение/транспирация для тех дней, когда производилось измере-
ние, получены приблизительные значения, характеризующие изменение
транспирации в течение года (табл. 57).
Таблица 57
Расход воды на транспирацию (в кг!м- = мм) сообществом
Phragmitetum communis — Glycerietum’ aquaticae под Берлином (по Киндлю)
I II Ш IV V VI VII VIII IX X XI XII
Glycerietum (1951 r.) ... 0 0 0 20 144 362 334 321 218 120 42 27
Phragmitetum (1950 г») ... 0 0 0 19 86 188 256 384 240 120 12 0
Эти цифры позволяют определить примерный расход воды раститель-
ным покровом в год. Эта величина оказалась равной 1500 мм. Если учесть,
что среднее годовое количество осадков в районе Берлина составляет
576 мм, то становится ясно, что заросли тростника расходуют на транспи-
рацию приблизительно в два с половиной раза больше воды. Конечно,
принимая во внимание ориентировочный характер расчетов, речь может
идти только о порядке величин.
Взводном балансе гелофитов и болотных растений известную роль
играет уже водный режим местообитания. В общем, их обеспеченность
водой в сырых местах хорошая, соответственно низко и осмотическое
давление. Но при очень интенсивной транспирации в солнечные дни все же
проявляется некоторая напряженность подачи воды к листьям; осмоти-
ческое давление обнаруживает отчетливый дневной ход 1 (табл. 58).
« Таблица 58
Колебания осмотического давления (в атм) у болотных
растений. Озеро Балатон, 6 сентября (по Г. и Э. Вальтер) 1 2
Название растений 8—9 час. 17—18 час. Разность
Iris pseudacorus .... 9,3 9,3 0,0
Polygonum amphibium 9,5 9,9 0,4
Epiloblum hirsutum . . 10,1 10,4 0,3
Lysimachia vulgaris . . 10,6 11,0 0,4
Carex acutiformis . . . 12,4 14,7 2,3
Phragmites communis 17,0 19,2 2,2
Cladium mariscus . . . — 14,6 —.
1 J. Kiendl, Zum Wasserhaushalt des Phragmitetum communis und des Glyce-
rietum aquaticae, «Вег. Deutsch. Bot. Ges.», 66, 1953, S. 246—-362; Zur Transpirations-
messung an Sumpf-und Wasserpflanzen, «Вег. Deutsch. Bot. Ges.», 67, 1954, S. 243—248.
2H. Walter, E. Walter, Okologische Untersuchungen des osmotischen
Wertes bei Pflanzen aus der Umgebung des Balatons (Plattensees) in Ungarn, «Planta»,
3, 1928, S. 571—624; W. R. Miiller-Stoll, Wasserhaushaltsfragen bei Sumpf- und
emersen Wasserpflanzen, «Вег. Deutsch. Bot. Ges.», 56, 1938, S. 355—367.
21-0434
322
Глава 5
Аналогичную картину получил Мюллер-Штолль на Рейнской равнин©
под Карлсруэ. В умеренно теплые летние дни дневные значения обычна
превосходят утренние не более чем на 2—5%; однако в очень жаркие дни
амплитуда была весьма значительной (табл. 59).
Таблица 59
Колебания осмотического давления (в атм)
в крайне жаркий день в августе. Рейнская низменность (по Мюллеру-ШтоллюУ
Название растений 7 час. 14 час. Разность
Glyceria aquatica . . 10,0 11,1 1,1
Typha angustifolia 8,8 10,4 1,6
Phragmites communis 19,6 21,3 1,7
Scirpus lacustris . . 9,4 11,6 2,2
Такое непостоянство осмотического давления свидетельствует о томг
что поступление воды по стеблям болотных растений в моменты интенсив-
ного ее расходования оказывается уже недостаточным. Этим, вероятно,
объясняется, почему осмотическое давление высоких гелофитов, например
тростника (до 3 м), достигает столь относительно высоких значе-
ний — 20 атм. Соответственно и листья растений имеют ксероморф-
ное строение.
В течение вегетационного периода осмотическое давление гелофитов
постепенно возрастает, например у Phragmites до 28 атм, что, по-видимо-
му, можно отнести на счет известной концентрации солей в клеточном соке,,
которая должна возникнуть как результат интенсивной транспирации
этих видов.
Особенность гелофитов состоит в том, что их осмотическое давление
на сухих местообитаниях не повышается, а понижается. Впервые мы обна-
ружили это в альпийском поясе на горе Пайкс-Пик (США) 1 (табл. 60).
Таблица 60
Размер листьев и величина осмотического давления у гелофитов
на сырых и сухих местообитаниях (Пайкс-Пик, Колорадо, июнь 1930 г.)
Название растений В воде На галечнике
размер листьев, СЛ1 осмотиче- ское давление, атм размер листьев, СЛ4 осмотиче- ское давление, атм
Caltha leptosepala 6 13,7 3—4 11,7
Caltha leptosepala 6 12,6 3 12,4
Polygonum bistorta 15 11,4 6 9,9
Это на первый взгляд странное положение вещей подтверждено Мюл-
лером-Штоллем в отношении всех исследованных им гелофитов. Даже
растения, растущие на борту осушительных канав, отличались более
низким осмотическим давлением, чем растения, находившиеся в воде.
1 Н. W а 11 е г, Die Hydratur der Pflanze, Jena, 1931.
Азональная растительность разных климатических зон
323
Более детальная проверка показала, что лишь на немногим более
сухих местообитаниях осмотическое давление, как обычно, повышалось,
однако при еще большей сухости наблюдалось уменьшение его величи-
ны (табл. 61).
Таблица 61
Осмотическое давление (в атм) у гелофитов
при возрастающей сухости местообитания (по Мюллеру-Штоллю)
Название растений В воде У уреза воды На сухом месте
Polygonum amphibium . . 9,5 13,5 7,1
Alisma plantago 11,2 12,0 9,6
Tris pseudacorus 13,5 14,2 12,7
Phragmites communis . . . 19,6 21,9 17,8
Вместе с тем растения сухих местообитаний отличаются угнетенным
ростом. Можно предложить следующее объяснение. На местообитаниях
с лишь едва заметными признаками сухости устьица у гелофитов еще
открыты, поэтому фотосинтез осуществляется беспрепятственно; поскольку
возникает лишь слабый водный дефицит, осмотическое давление возрастает
незначительно. Однако, как только потеря влаги увеличивается, устьица
закрываются, о чем свидетельствует резкое сокращение интенсивности
транспирации. Благодаря этому растение не засыхает, но фотосинтез
зрекращается и растение начинает голодать, расходуя составные части
клеточного сока, регулирующие осмотическое давление. Для некоторых
видов удалось доказать, что содержание сахаров в клеточном соке на сухих
местообитаниях понижается. Правда, у Glyceria и Phragmites оно не изме-
нилось. В данном случае, по-видимому, расходовались другие вещества,
возможно органические кислоты. Поведение гелофитов соответствует здесь
поведению стеногтщрических ксерофитов.
Многие гидрофиты образуют наземные формы, у которых изменяется
структура листьев, а осмотическое давление возрастает (табл. 62).
Таблица 62
Осмотическое давление (в атм)
у наземных и водных форм одного и того же вида
(по Вальтеру и Геснеру)
Название растений Подводные листья Надводные листья
Batrachium divari- catum 10,0
Potamogeton grami- neus 11,0
Myriophyllum spica- tum 11,4.
Hippuris vulgaris . . — 9,2
Alisma plantago . . — 11,8
Nuphar luteum . . . — 9,3
Trapa natans .... 6,8 15,9
Veronica anagallis 5,3 6,6
Hippuris vulgaris . . 9,6 15,0
Ranunculus baudotii 8,5 19,5
21*
324
Глава 5
Напротив, по данным Мюллера-Штолля, осмотическое давление ниже
у надводных листьев. Вполне возможно, что и у наземных форм сначала
происходит’рост осмотического давления, а затем, при увеличении сухости
среды,— его уменьшение. Но также вероятно, что в данном случае полу-
ченные значения надо рассматривать всего лишь как заниженные.
На берегах больших озер с сильным прибоем растительность обычно
отсутствует. Прибой формирует здесь более или менее широкий отлогий
берег — пляж. Под действием волнения песок и галька постоянно пере-
отлагаются, на месте остаются только крупные камни. Характерно обра-
зование на таких мелководных участках у берегов богатых известью олиго-
трофных озер «бороздчатых» камней (рис. 229) Речь идет о корках изве-
сти, ^выделяемой обитающей на камнях Scytonema\ в этих корках животные
организмы — по одним авторам, а по
Рис. 229. Бороздчатый камень на
берегу Боденского озера у Лангенар-
гена (по Кирхнеру).
другим — растворяющие известь виды
Nostoc «проедают» извилистые бороздки.
Встречаются па пляжах также рако-
вины улиток или обломки их либо же
фрагменты раковин гастропод, сцемен-
тированные слоистой известковой мас-
сой, образованной Cyanophyceae, и ока-
танные в зоне прибоя. Вообще Cyano-
phyceae играют большую роль'в пляжной
полосе озер. Различные виды растений,
укореняющиеся между камнями, кон-
центрируются в той или иной зоне в
зависимости от высоты последней' над
средним уровнем воды, то есть в зави-
симости от того, как часто эта полоса
берега смачивается водой при волнении.
Здесь мы уже подходим к проблеме зна-
чения движения воды для развития рас-
тительности озер. При описании верхо-
вых болот (т. II, стр. 400) мы указы-
вали, что проточность воды способствует большему потреблению расте-
ниями питательных веществ, благодаря чему даже в воде, бедной ими,
могут расти требовательные к фактору питания виды.
Геснер также доказал, что некоторая циркуляция воды в водоеме
в какой-то мере стимулирует фотосинтез. В стоячих водах интенсивность
ассимиляции со временем падает. Как известно, водные растения извлека-
ют СО2, необходимый для осуществления фотосинтеза, из бикарбоната
кальция, причем освобождению углекислоты предшествует образование
гидроокиси кальция: Са(НСО3)2 — СО2 = Са(ОН)2. Последний накапли-
вается на поверхности листьев и вызывает резко щелочную реакцию, так
что при добавлении в воду фенолфталеина она окрашивается у поверхно-
сти листьев в красный цвет. Движение воды способствует удалению Са(ОН)2
с листьев и привносу новой порции Са(НСО3)2. У нерасчлененных листьев
этот процесс может вызвать увеличение интенсивности фотосинтеза на
35%, а у сильно рассеченных — на 15%, в среднем, следовательно,
на 25% по сравнению с интенсивностью фотосинтеза в стоячей воде.
:С. Schroter, О. Kirchner, Die Vegetation des Bodensees, Bd. I, II,
Lindau, 1896—1902; E. Baumann, Die Vegetation des Untersees (Bodensee), «Arch,
f. Hydrobiol. und Planktonk.», Suppl. Bd. I, V, 1911, S. 1—554.
Азональная растительность разных климатических зон
325
В реках с быстрым течением чисто механический эффект от перемеще-
ния частиц воды затрудняет развитие в них водной растительности.
Скорость потока определяет характер грунта г:
Скорость течения, см/сек 120 35 20 12
Грунт галька гравий песок ил
Мы различаем ленитические биотопы, со слабым течением воды
в заливах озер и на отмелых берегах рек, которые легко заселяются расте-
ниями, и лотические биотопы, с сильным течением и подвижным субстра-
том (галька и гравий), обычно не занятые растительностью. Нагроможде-
ния камней в руслах рек обусловливают, с одной стороны, образование
порогов, а с другой — тихих, с медленным движением воды мест, где
1М
Рис. 230. Водная растительность в каменистом русле реки
Урт, Арденны (по Ван ден Бергену из «Phytologie», Bd. IV, 2).
R —'Ranunculus aquatilis', M — Myriophyllum spec, (растет в местах с за-
медленным течением); L —Lemanea spec, (красные водоросли); F — Fonti-
nalis antipyre tica‘, В — Brachythecium rivulare', C —Cinclidotus fontinaloides
(F, В и G —:мхи).
осаждаются песок и ил, дающие растениям возможность укорениться.
Поэтому по течению реки перемежаются участки, поросшие раститель-
ностью и лишенные ее. Такое чередование наблюдается не только в про-
странстве, но и во времени, поскольку река, отложив в одном месте нано-
сы, может спустя какое-то время размыть их и переместить ниже по тече-
нию (рис. 230). В свою очередь растения, закрепившись на дне, в известной
мере корректируют сортировку и отложение речного материала. Таким
образом, условия существования в быстротекущих водах характеризуются
большой сложностью и непостоянством.
Это обстоятельство обусловливает интенсивный отбор: в проточных
водах обитает лишь сравнительно небольшое число видов растений. Для
такой территории, как Англия, называют только 46 видов, для' областей
меньшей площади, например Верхняя Бавария или Шлезвиг-Гольштейн,—
всего 28
1 R. W. В u t с ha г, A preliminary account of the vegetation of the River Itchen,
«J. Ecol.», 15, 1927, p. 55—65; Studies on the ecology of rivers I. On the distribution of
macrophytic vegetation in the rivers of Britain, «J. Ecol.», 21,1953, p. 58—91; H. R о 11,
Die Pflanzengesellschaften ostholsteinischer Fliehgewasser, «Arch. Hydrobiol.», 34,
1938, S. 159—305; G. Van den Bergen, Contribution a I’etude des groupements
vegetaux notes dans la vallee de 1’Ourthe en amont de Laroche-en-Ardenne, «Bull. Soc.
Roy. Bot. Belgique», 85, 1953, p. 195—277.
326
Глава 5
£Р и с. 231. Podostemonaceae (Apinagia warmingiana) в водопаде Сальто-Гранде
на реке Паранапанема; южная Бразилия, высота 360 м над уровнем моря’.
Виды этого семейства растут только в реках с очень быстрым течением (фото
Ветштейна).
Виды, способные противостоять сильному течению: Potamogeton flui-
tans, Ranunculus trichophyllus, а также погруженные формы Butomus
umbellatus, Glyceria fluitans и Stum erectum.
В водотоках с более медленным движением воды встречаются: Elodea
canadensis, Potamogeton crispus, P. lucens, P. nitens, P. perfoliatus и погру-
женные формы Glyceria maxima (=aquaticd), Lysimachia nummularia,
Sparganium ramosum и Phalaris arundinacea.
Особенность текучих вод состоит в том, что заселение их раститель-
ностью происходит не в результате укоренения проростков, а принесенны-
ми течением из водоемов со стоячей водой вегетативными побегами, кото-
рые укореняются в реках и вегетативно размножаются. Поэтому в умерен-
ной зоне нет видов, которые являлись бы собственно видами проточных
вод, в отличие от тропиков с характерными для них растениями семейства
подостемоновых (рис. 231). Не существует также видов, связанных с пото-
ком определенной скорости, тем более что речь идет не о средней, а о пре-
дельной скорости течения. Многие виды в реках всю зиму остаются зеле-
ными, другие отмирают и весной снова дают побеги из корневищ и сто-
лонов.
Все погруженные формы имеют очень похожие лентовидные листья,
что крайне затрудняет определение вида. Эти листья соответствуют первич-
ным листьям. Растение как бы остается на ранней стадии развития и часто
не достигает фазы цветения. По-видимому, движение воды сдерживает
рост растения в длину — чем течение сильнее, тем короче листья или
их черешки.
Азональная растительность разных климатических зон
327
СОЛОНОВАТЫЕ ВОДЫ И СОЛЕНЫЕ ОЗЕРА
Важным, подчас определяющим развитие водной растительности
фактором является количество растворенных в воде солей. Выше мы рас-
смотрели условия среды, существующие в водоемах, воды которых содер-
жат бикарбонат кальция.
Теперь объектом нашего внимания станут солоноватые воды, основ-
ным компонентом которых является хлорид натрия. Зона солоноватых
вод чаще формируется в устьевых участках рек, впадающих в море, где
к пресной воде может примешиваться соленая морская вода.
Солоноватой называют смесь пресной и соленой воды, содержание
солей в которой превышает О,5°/оо. Солоноватые воды подразделяют на:
1) олигогалинные 0,5—5°/оо» 2) мезогалинные 5—18, 3) полигалинные
18—30, 4) эугалинные 30°/оО и выше (до солености морской воды).
Согласно такому определению, Балтийские внутреннее море является
солоноватым морем Поскольку испарение с водной поверхности Балтий-
ского моря приблизительно соответствует количеству выпавших на нее
атмосферных осадков, привнос воды в море многочисленными реками,
составляющий 472 км?! год, должен вызвать отток избыточной воды
в Северное море. Если при этом не произошло полного опреснения Бал-
тийского моря, то это объясняется одной причиной: более тяжелая вода
Северного моря в виде противотечения устремляется по дну пролива
Большой Бельт в Балтийское море.
У южного края Швеции содержание солей в воде еще равно 17—24°/00,
у острова Хиддензе оно понижается до 8,6°/Оо, У западной оконечности
Финского залива становится еще ниже, а у восточной — соленость почти
нулевая. Именно эта особенность делает Балтийское море неповторимым
объектом для изучения проблем, связанных с солоноватыми водами.
Растительность последних отличается бедностью видового состава.
Из числа морских, равно как и пресноводных, организмов развилось
очень немного видов, способных жить в солоноватой воде. Поэтому суще-
ствует своего рода «барьер солености», разделяющий между собой пресно-
водную и морскую флоры. Бедность видами солоноватых вод связана со
спецификой условий обитания в них: для зоны солоноватых вод обычно
характерны резкие кратковременные изменения содержания солей выводе.
При увеличении концентрации солей типичные пресноводные растения
начинают быстро поглощать соль, что для них оказывается небезвредным;
в то же время, как показал Геснер, у Ranunculus haudotii, для которого
солоноватые воды являются естественной средой обитания и протоплазма
которого слабопроницаема для солей (особенность, по-видимому, харак-
терная вообще для всех галофитов 1 2), выравнивание содержания солей
во внешней среде и в клеточном соке вакуолей происходит достаточно
медленно для того, чтобы протоплазма сумела приспособиться к избыточ-
ному поступлению солей в клетки растения.
В отдельных частях современного Балтийского моря соленость воды
колеблется лишь в незначительных пределах. Однако море в целом пре-
терпело определенную эволюцию: постледниковое соленое Иольдиевое
море (до 8800 г. до н. э.) сменилось пресноводным Анциловым озером
(до 5000 г. до н.э.) и затем снова относительно соленым Литориновым
1 Балтийское море как среда для организмов очень подробно описано в работе
Геспера: F. G е ssner, Meer und Strand, 2. Aufl., Berlin, 1957.
2 G. Repp, Die Salztoleranz der Pflanzen, «Osterr. Bot. Zschr.», 104, 1958.
S. 454—490.
328
Глава 5
морем (до 2000 г. до н. э.). При каждом изменении солености в бассейн
мигрировал новый «набор» видов, прежние же виды вымирали. Сохранить-
ся могли лишь эвригалинные виды, то есть виды, относительно нейтраль-
ные к величине солености. Ими могут быть как животные, так и расти-
тельные организмы. Особенно характерна для солоноватых вод зеленоцвет-
ная Enteromorpha intestinalis (Chlorophyceae). В Атлантическом океане
у побережья Франции насчитывается 136 видов бурых водорослей, в Бал-
тийском море у острова Хиддензе — только 27; для багрянок приводятся
соответственно цифры 299 и 18. Charales встречаются в Балтийском море
(так же как в Каспийском), но полностью отсутствуют в других морях.
Перечень видов Phaeophyceae и Rhodophyceae приведен в работе Геспера
(т. II). Мы ограничимся здесь перечислением видов покрытосеменных.
Только в заметно соленых частях Балтийского моря можно найти:
Zostera marina, Zostera папа, Ruppia maritima, Ruppia spiralis, Zannichellia
palustris var. major, Scirpus parvulus.
Менее требовательны к концентрации солей Potamogeton vaginatus,
Potamogeton pectinatus, Najas marina, Ranunculus baudotii, Zannichellia
palustris, Scirpus maritimus, Scirpus tabernaemontani.
Все другие встречающиеся в Балтийском море цветковые растения
являются лишь факультативными галофитами, то есть они могут расти
в слабосоленой воде, но развиваются наиболее пышно в пресной воде.
Приведем несколько примеров такой толерантности по данным Лютера,
который подробно изучал этот вопрос на побережье южной Финляндии
в районе Экенеса (рис. 232) х.
Пресные воды
Появляются
Исчезают
Chara aspera
Potamogeton pectinatus
Scirpus maritimus
Najas marina
Chara tomentosa
Potamogeton fillformis
Fucus vesiculosus
Scirpus parvulus'
Chara baltica
Ruppia spiralis
Zostera marina
Fisstdens julianus
Fontinalis hypnoides
Potamogeton obtusifolius
Sagittaria sagittifolia
Lemna minor
Nuphar luteum
Elodea canadensis
Lemna trisulca
Butomus umbellatus
Fontinalis antipyretica
Nitella flexilis
Potamogeton nitens
Callitriche autumnalis
Ranunculus circinatus
Соленость 6°/oq (Балтийское море, Финский залив^
Рис. 232. На схеме показана приблизительная градация солености, переносимой
разными видами водных растений (в том числе некоторыми мхами и водорослями,
живущими в прибрежных водах района Экенеса) (по Лютеру).
Слева — нижняЯ граница распространения солоноватоводных растений; справа — верхний предел
распространения пресноводных видов при постепенном увеличении солености моря до2"6%о-
1 Н. Luth е r,*Verbreitung und Okologie der hoherenWasserpflanzen' im Brack-
Wasser der Ekenasgegend in Siidfinnland I-II, «Acta. Bot. Fenn.», 49, 50, 1951.
Азональная растительность разных климатических зон
329
Из приведенной схемы следует,, что граница раздела проходит на уров-
не 5°/оо: факультативные галофиты не встречаются выше ее, а облигат-
ные — ниже ее.
Черное море также представляет собой бассейн солоноватых вод
глубиной свыше 2000 м. Но соленость поверхностных слоев воды в нем
равна уже 17—18°/00» а на глубине более 100 м повышается до 22,5°/00.
Флора Черного моря полностью соответствует морской флоре. Лишь
в мелководном Азовском море соленость падает до 11,2°/00; водорослевая
флора здесь исключительно бедна видами (табл. 63).
Таблица 63
Число видов некоторых семейств морских водорослей
(по Касперсу)
Семейство Средиземное море Черное море Азовское море
соленость, o/oo
около 38 около 18 около 11
Chlorophyceae . . . 88 54 19
Phaeophyceae . . . 94 64 4
Rhodophyceae . . . 289 103 10
Морским солоноватым водам противостоят водоемы с концентрирован-
ными солевыми растворами, например содержащие NaCl Большое Соленое
озеро (штат Юта), соленость которого колеблется в'.зависимости от напол-
нения озерной чаши от 150 до ЗЗО°/оо; озеро Резайе (Урмия) в Иране (За-
падный Азербайджан) с соленостью более ЗОО°/Оо и Мертвое море в грабене
Иордана с общей соленостью 270—ЗЗО°/оо и, значительной концентрацией
магния (4—6°/оо в виде MgBr2). В такого рода водоемах обитает лишь
несколько видов низших организмов.
Меньшей соленостью наряду с высоким содержанием карбонатов
и хлоридов отличается озеро Ван в Анатолии. К цему можно применить
термин «содовое озеро» с величиной pH == 9,3—9,6. Высшие водные расте-
ния в озере также, отсутствуют. Напротив, озеро Нейзидлер-Зе у австрий-
ско-венгерской границы с pH = 8,4 и соленостью 1,5°/Оо (максимальная
соленость 13,5°/оо) обнаруживает столь слабую концентрацию бикарбоната
натрия, что в нем прекрасно себя чувствуют даже цветковые растения,
например Ч
Potamogeton pectinatus^ Zannichellia palustris ssp. pedicellata, Myriophyllum sptca-
tum, Utricularia vulgaris, виды Ranunculus и в прибрежной зоне — полоса зарослей
Phragmites шириной до 1 км, кроме того, Scirpus lacustris, S. tabernaemontani, Juncus
maritimus, на влажной почве также Triglochin maritimum, Heliocharis palustris, H.
uniglumis, Puccinellia salinaria, Aster iripolium ssp. pannonicus, Trifolium fragiferum,
Lepidium cartilagineum.
В умеренной зоне Евразии и Северной Америки, а также Южной
Америки и Австралии имеется большое число соленых озер с различной
степенью концентрации солей в них. Состав озерной флоры резке арьиру-
1 G. Wendelberger, Die vegetation auf den Salzboden des Neus.edler Sees,
«Denkschr. Osterr. Akad. Wisse, math.-nat. KL», 108, 5, 1950, S. 1— 180; G. R epp,
Okologische Untersuchungen im Halophytengebiet am Neusiedler See, «Jbc wiss,, Bot.»,
88, 1939, S, 554-632.
330
Глава 5
ет в зависимости от химического состава воды и величины солености.
Самое крупное на земном шаре озеро —Каспийское море (площадь
370 тыс. кл12), как и Аральское море (67 769 «л2), является остатком
древнего океана Тетис. Средняя соленость!Каспийского моря — 12,85°/оо,
то есть она ниже солености Мирового океана. По своему химизму вода
Каспийского моря отличается от воды Мирового океана повышенным содер-
жанием сульфатов, по всей вероятности поступающих из гипсовых отло-
жений, развитых на берегах Волги. Цветковые растения представлены
только Zostera папа; вместе с тем в Каспийском море распространены
7 видов бурых водорослей (в Аральском морелих{нет) и 25^видов багрянок
(в Аральском море — 11).
МА ЕРОФИ TH АЯГ'Р АСТИТ ЕЛЬНОСТЬ^МОРЕЙ
Крупнейшими солеными водными бассейнами на Земле являются
моря, образующие единый Мировой океан. Но подобно тому, как это
наблюдается в лимнологии, океанология, которая в еще большей степени,
чем озероведение, представляет собой самостоятельную отрасль науки,
уделяет лишь небольшое внимание относительно узкой береговой зоне.
Мы же ограничимся рассмотрением только этой зоны и закрепившихся
в ней макрофитов, которые объединяются понятием «бентос». Поскольку
планктон остается вне поля нашего зрения, отсылаем интересующихся
им читателей к работам Геснера х.
Соленость Мирового океана относительно велика; она колеблется
в пределах 32—38%0 и равна в среднем 350/00. Химический состав морской
воды весьма постоянен; основными компонентами морской соли являются
11 элементов (табл. 64); данные приведены в расчете не на литр, а на кило-
грамм, с тем чтобы исключить воздействие колебаний температуры.
Таблица 64
Химический состав морской воды с соленостью 35°/00 (по Калле)
Катионы г/кг мгкв/кг Анионы г/кг мгкв/кг
Натрий . . . Калий .... Магний . . . Кальций . . Стронций . . 10,752 0,390 . 1,295 0,416 0,013 467,56 9,98 106,50 20,76 0,30 Хлор .... Бром .... Фтор .... Сульфат . . . Бикарбонат Борная кисло- та 19,345 0,066 0,0013 2,701 0,145 0,027 545,59 0,83 0,07 56,23
Сумма катионов 605,10 Сумма сильных анионов 602,72
Таким образом, имеющийся избыток катионов по сравнению с содер-
жанием сильных анионов составляет 2,38 мэкв!кг. Угольная и борная
кислоты столь слабые, что не образуют с катионами нейтральных солей.
Поэтому реакция морской воды щелочная; pH = 8,1—8,3. Из-за избытка
оснований морская вода легко насыщается углекислым газом воздуха.
1 F. Gessner, Hydrobotanik, Bd. I, Berlin, 1955; Bd. II, Berlin, 1959; Meer
und Strand, Berlin, 1957; К. К a 11 e, Der Stoffhaushalt des Meeres, Leipzig, 1943.
Азональная растительность разных климатических зон
331
В результате содержание в ней СО2 может в 100—200 раз превышать
концентрацию углекислоты в чисто физическом растворе. Морская вода
соответствует раствору бикарбоната натрия и потому обладает такими же
буферными свойствами, как последний; значения pH в ней почти не изме-
няются. Мировой океан «регулирует» содержание СО2 в атмосфере: воды
океана поглощают СО2, когда его количество в воздухе увеличивается,
и отдают, когда концентрация его в атмосфере уменьшается. Приводим
солевой состав морской воды при температуре 20° (в г/л):
NaCl .............28,014 СаСО3..........0,122
MgCl2............. 3,812 КВг ...........0,101
MgSO4 ............. 1,752 SrSO4..........0,028
CaSO4 ............. 1,283 Н3ВО3..........0,028
K2SO4............. 0,816 Следы других солей
Помимо перечисленных основных элементов, обнаружены следы
34 других, в том числе (в жг/ж3): N — 1000, Р — 60, J — 50, Си — 5
и даже золота — 0,004.
Некоторые морские организмы обладают способностью накапливать
в своем теле отдельные микроэлементы: например, крупные водоросли
ассимилируют в тканях значительное количество йода.
Обращает на себя внимание совершенно иной химический состав
морской воды по сравнению с составом воды рек, которые, собственно,
и выносят в море соли. Причину этого различая нужно искать в свойстве
глинистых частиц адсорбировать катионы тем сильнее, чем больше их
атомный вес. Поэтому калий и кальций осаждаются с глиной морских
отложений, тогда как натрий преимущественно остается в растворе.
Морская соль и морские осадки должны, следовательно, содержать все
те элементы, которые освобождаются при выветривании первичных гор-
ных пород или выбрасываются при извержении вулканов. Мировой
океан — своего рода бессточное озеро в аридной области, в которое смы-
ваются все соли из его окружения и из которого в виде пара удаляется
только вода.
Но если морские соли не представляют собой выпавшую в осадок часть
принесенных реками продуктов^выветривания, то отсюда следует, что в пе-
риод, предшествовавший началу процессов выветривания на Земле, вода
в морях была пресная и что концентрация в ней солей в течение истории
Земли непрерывно возрастала. Как известно, жизнь зародилась в воде,
и эта вода, по-видимому, была пресной; этим, вероятно, можно было бы
объяснить, почему основные компонентьГморской соли — натрий и хлор —
не играют никакой роли в жизни растений. Таким образом, приспособле-
ние морских водорослей Jk обитанию в соленосной среде, наверное, пред-
ставляет собой вторичное явление, аналогичное явлению приспособления
наземных галофитов к существованию на засоленном субстрате.
К прикрепленным морским макрофитам относятся преимущественно
водоросли, причем большая часть видов Chlorophyceae и практически все
Phaeophyceae и Rhodophyceae, из которых лишь несколько мелкорослых
видов можно найти в пресных водах. Немногие виды высших растений,
которые обитают в море, растут в погруженном состоянии. Это исключи-
тельно покрытосеменные, принадлежащие к семействам Potamogetonaceae
и Hydrocharitaceae. Из видов первого семейства следует назвать широко
распространенную Zostera marina, цветки которой опыляются в воде длин-
ными нитевидными зернами пыльцы; встречающуюся в Средиземном море
Posidonia oceanica (рис. 233); Phyllospadix (западное побережье Северной
Америки, Япония), а также тропические роды Xymodocea, Halodule и Al-
332
Г/ава 5
.Рис. 233. «Морские травы».
1"— Zostera marina (J/2 нат. величины); 2 — ее соцветие (нат. величина);
3 — Posidonia oceanica из'Средиземного моря ('/2 нат. величины). Во-
локнистые остатки ее листьев образуют своеобразные «шарики», в
изобилии встречающиеся на песчаных пляжах.
thenia. Виды семейства Hydrocharitaceae: Halophila, Enhalus и Thalassia
распространены только в теплых водах тропических морей.
Присутствие в море вблизи побережья Posidonia легко заметить по вы-
брошенным на пляж волокнистым клубкам размером с теннисный мяч.
Образуются клубки из отломленных корневищ и остатков листьев, пере-
катываемых течением по песчаному дну и принимающих форму шара.
Морские покрытосеменные растут изолированно от морских крупных
бурых водорослей. Они пронизывают своими корневищами песчаный
грунт и часто образуют обширные подводные луга. У бурых же водорослей
имеются маленькие когтевидные отростки, с помощью которых они могут
прикрепляться только к твердому субстрату; иногда водоросли образуют
Азональная растительность разных климатических зон
333
на камнях корковидные налеты (рис. 234). Поэтому бурые водоросли весь-
ма характерны для растительности скалистых морских побережий. Лишь
зеленые водоросли Caulerpa дают побеги с ризоидами и способны расти
на песчаной фации.
Видовой состав водорослевой растительности отдельных морей харак-
теризуется известным различием х. Поскольку важным экологическим
Рис. 234. Водоросли на скалах острова Гельголанд при отливе. Вни-
зу — крупные слоевища Fucus platy carpus, в середине — корки Ralf-
sia verrucosa, вверху — белые известковые домики морских желудей
(Balanus).
^фактором является температура воды, в тропических морях распростране-
ны иные водоросли, нежели в холодных северных водах.
Однако более существенное значение для процесса заселения морских
побережий растениями имеет характер субстрата. Во всех морях богатые
водорослями местообитания (скалистые побережья) чередуются с местооби-
таниями, почти лишенными водорослевой растительности (песчаные и или-
стые берега). На западногерманском побережье Северного моря скалистый
остров Гельголанд представляет, пожалуй, единственное место, где можно
изучать жизнь водорослей.
Наконец, не следует упускать такой немаловажный и специфический
фактор среды, как приливы и отливы, ибо в приливной зоне растения
попеременно и по-разному подвергаются воздействию моря: то покрывают-
ся водой, то выступают из нее и обсыхают.
Приливные явления — подъем (прилив) и падение (отлив) уровня
моря — происходят дважды в течение лунного дня (24 часа 50 мин.). Осо-
бенно велика разность уровней в полнолуние и новолуние (сизигийные
приливы); намного меньше эта амплитуда в первую и последнюю четверти
Луны (квадратурный прилив). Два раза в год приливы (так называемые
1 F. Е. Round, Biologie der Algen, Stuttgart, 1968.
334
Глава 5
равноденственные приливы) достигают наибольшей высоты. Кроме того,,
амплитуда приливов колеблется в зависимости от очертаний береговой
линии и направления подхода приливной волны к побережью (ср. т. I,
стр. 164, и рис. 112).
На скалистых побережьях наблюдается избирательное вертикальное
распределение растительности, особенно резко бросающееся в глаза
у крутых, обрывистых берегов. Обычно различают три основные зоны:
1. Супра литораль, которую море никогда не заливает, а лишь оро-
шает брызгами в зоне прибоя; этот пояс отсутствует в тихих заливах, где
нет волнения.
2. Литораль, периодически затопляемую приливами. Пребывание
водорослей над водой в этом поясе тем длительнее, чем выше над уровнем
малой воды сизигийных приливов они растут. Ширина литорали зависит
от амплитуды приливов. Максимальная высота сизигийного прилива равна
в Патагонии 10 л«, в Бретани — 12 и на полуострове Новая Шотландия —
16 Л1. Обычно же подъем воды’во время прилива не превосходит несколько-
метров. В Средиземном море приливы почти не выражены и обычно пере-
крываются колебаниями уровня воды, вызванными ветром. Литораль здесь
практически отсутствует (подъем уровня воды у Сплита равен 25 cjk,
у Триеста — 98 см.) i
3. Суб литораль, простирающуюся ниже уровня малой воды равноден-
ственных приливов; иными словами, эта зона постоянно пребывает под водой.
Границы между этими тремя зонами не резкие, так как высота подъ-
ма уровня воды при сизигийном приливе значительно больше, чем во вре-
мя квадратурного прилива.
Сообщества водорослей в этих трех зонах совершенно различны.
Трудно назвать вид, который бы встречался во всех трех поясах. Мы не бу-
дем приводить здесь всего перечня видов водорослей, характерных для
различных типов морских побережий, а ограничимся рассмотрением двух
профилей. На рис. 235 показана последовательность расположения видов
растительности у западной стены острова Гельголанд, сложенной из блоков
твердого гранита и потому дающей водорослям устойчивую опору. Цифры
на профиле указывают высоту уровня^моря над уровнем малых квадратур-
ных вод; средняя высота квадратурного прилива равна + 2,20 м, сизигий-
ного прилива + 2,50 м. В зоне брызг (супралитораль) (до отметки
+ 4,70 м) растет черная* Calo thriz scopulorum, под ней примитивная Bangia
fuscopurpurea (Rhodophyceae), еще глубже — Enteromorpha Ungulata, кото-
рая вместе с Porphyra atropurpurea и Enteromorpha compressa обитает уже
на границе с литоралью. К сообществу этой зоны принадлежат также
корковидные Ralfsia verrucosa и некоторые прикрепляющиеся к скале
животные, например б а ляну сы х.
1 Самую верхнюю границу супралиторали провести трудно. В сильный шторм
брызги залетают далеко вверх на берег. В Шотландии наряду с Calothrix scopulorum
встречаются другие Cyanophyceae, прежде всего лишайники (виды Verrucaria) и неко-
торые мелкие зеленые водоросли (Blindia minima, Prasiola stipitata). Пока не ясно,
можно ли относить к супралиторали зону лишайников с Xanthoria parietina и видами
Lecanora, к которым иногда присоединяется и Rhacomitrium lanuginosum, спускающую-
ся на защищенных побережьях на 0,5—1 м над средним уровнем прилива, посколь-
ку названные виды уже не связаны с зоной брызг. Разговор касается также тех видов
цветковых растений, которые переносят увлажнение мельчайшими капельками соленой
морской воды и поэтому часто поселяются на скалистых береговых местообитаниях,
где ослаблена конкуренция с более чувствительными к соленой воде видами. Некото-
рые из этих видов распространены исключительно на прибрежных скалах, как, напри-
мер, Mertensia maritima (бурачниковые) на севере или Asplenium marinum (до северной
Испании) и Crithmum maritimum на побережье Средиземного моря.
Азональная растительность разных климатических зон
335
Рис. *235. Схема размещения водорослей на западной
стене острова Гельголанд (по Ниенбургу).
В литорали достаточно дифференцированные горизонты занимают
прежде всего виды Fucus'. F. platycarpus (2,0—1,0 м над уровнем малой
воды сизигийного прилива), F. vesiculosus (1,4—1,1 ж) и F. serratus, которая
опускается до уровня сизигийных вод (± 0,5 м). Скорее уже к сублитора-
ли относятся крупные ламинарии и многие виды мелких водорослей.
Необыкновенно четко выступают зоны литорали на очень слабо наклон-
ной, шириной почти 200 м абразионной террасе у острова Гельголанд
(рис. 236), где во время’квадратурных отливов на короткое время обнажа-
ются из-под воды даже заросли ламинарий (рис. 237) х.
На рис. 238 приведен профиль побережья Ла-Манша. Поясность
растительности здесь очень напоминает описанную выше, причем в самой
верхней части зоны*брызг местами развиты корковые лишайники (Verru-
caria). На побережье Адриатического моря 18 различных видов семейства
Cyanophyceae образуют в супралиторали сходную зональность 1 2: непо-
средственно над поверхностью воды растут известковые водоросли, под
1 Подробное описание водорослевой растительности и животного мира гельго-
ландской литорали и сублиторали читатель найдет в работе Геснера: F. G е s sn е г,
Meer und Strand, Berlin, 1957.
2 A. Ginzberger, Der EinfluB des Meerwassers auf die Gliederung der sud-
dalmatinischen Kiistenvegetation, «Osterr. Bot. Zschr.», 1925, S. 1—14.
336
Глава 5
Рис. 236. Fucus serratus на абразионной террасе острова Гельголанд во время отливая
Рис. 237. Часть абразионной террасы Гельголанда с Laminaria digitata, высту-
пающая из воды только во время очень сильного отлива (фото Шмидта).
Азональная растительность разных климатических зон
337
+14-
+13-'
+12-
+11-
+10-
+9-'
+8-
+7-
+6-'
+5-
+4-
+3-
+2-
+1-
0--
-1-
-2-
-3-.
-4-
-5-
-6-
-7-
-8-
-9-
-10-
-11-
-12-
-13-
-14-
-16-
Ч^Корковые водоросли и лишайники
% (Prasiola-Verrucaria)
Ш Нитчатые водоросли (Bangia-Urospora)
Верхние пластинчатые приливах
водоросли (Porphyга-Enteromorpha) (сизигия)
(Peivetia) „
Ж, LF«™ . лПпК£,
platycarpus) иООО TWU-------
(F.vesiculosus) , Приливах
\ J (коаоратура)
Литоральные^ (ДзсорЬу Пит)
ламинарии \ пластинчатые водоросли
\ (Rnodymenia)
(F.serrahis)
Малая \
--вода при-------
приливах (квавратураД
Литоральные кустистые
боЗоросли Малая ffetoW
приливах
'х^^~х^'воворосли (сизигия)
X Сублиторальные \ ’%«
' \ ламинарии \
\ Кожистые '
ламинарии
(L. hyperboreajr-
filaria)
(Laminaria
digitata)
Известковые
конкреции
Сублиторальные
кустистые ''
водоросли
Рис. 238. Горизонты водорослей на скалистом побережье Па-де-Ка-
ле, характеризующемся особенно большой амплитудой приливов.
Справа — открытое морским течениям побережье, слева — более
защищенный участок берега. Вертикальный масштаб сильно уве-
личен (по Ниенбургу).
ними — бурые водоросли Cystosira (рис. 239) 4. Аналогичные условия
обнаруживаются на побережьях всех других материков с той лишь раз-
1 В сублиторали острова Мальта до глубины 15 м встречаются преимущественно
бурые водоросли, затем, до глубины 75 м,— зеленые водоросли, а в условиях очень
слабой освещенности, на глубине 89—90 м,— багрянки; спектральный состав света
на размещение видов растительности якобы не влияет. Уничтожение водорослей игло-
кожими имеет существенное значение для жизни биоценозов в верхней сублиторали,
до отметки 15 м (определена также фитомасса на разной глубине); ср.: A. W. Lak-
rum, Е. A. Drew, R. N. Crosse t, The vertical distribution of attached marine
algae in Malta, «J. Ecol.», 55, 1967, p. 361—371.
22-0434
338
Г лава 5
Рис. 239. Известковые водоросли Lithophyllum tortuosum, образующие широ-
кую светлую полосу [выше уровня воды на побережье Адриатического моря
Непосредственно под ними едва заметны Cystosira (фокусовые). Над светлой
полосой выделяется темная или совершенно черная зона с Entophy salts granu-
losa, Gleocapsa deusta, Calothrix scopulorum (сине-зеленые водоросли). Еще
выше — лишенный растительности пояс с естественным цветом горной поро-
ды (фото Шиллера).
ницей, что состав флоры не остается неизменным. Например, в южной
половине Тихого океана в верхних горизонтах сублиторали широко рас-
пространена крупнейшая из водорослей — Macrocystis pyrifera (Phaeo-
phyceae), достигающая 70 в длину и весящая несколько сот килограммов.
Благодаря свойству выделять’известь водоросли могут иметь значение для
образования горных пород. Так, тропические виды Lithothamnion (Rhodo-
phyceae) в сообществе с рифообразующими животными играют существен-
ную роль в построении и укреплении рифов, в частности коралловых
(рис. 240).
Поясность водорослевой растительности на морских побережьях пред-
определяется конкурентными взаимоотношениями слагающих ее видов
и теснейшим образом связана со способностью растений переносить крат-
ковременное высыхание.
Наиболее суровые условия складываются для водорослей зоны брызг,
обладающих малой конкурентной мощностью. При устойчивой безветрен-
ной солнечной погоде Bangia обсыхают, и поверхность почвы вокруг них
в такие дни бывает покрыта кристаллами соли, которая, однако, в после-
дующие дождливые дни полностью смывается. В сухой период содержимое
клеток нитчатых водорослей сильно уменьшается в объеме, при этом вну-
тренние слои стенок клеток заметно набухают; при смачивании же содер-
жимой расширяется и сдавливает стенки клеток, отчего они кажутся
неестественно тонкими. При слишком частой и быстрой смене высыхания
и увлажнения отдельные клетки растения отмирают, но сохранившиеся
Азональная растительность разных климатических зон
339
Рис. 240. Банка, образованная красной известковой водорослью с Lithothamnium
во время отлива, побережье острова Тимор (фото Вебер-ван-Боссе).
обеспечивают регенерацию ткани, причем образуются даже новые резоиды,
с помощью которых водоросли прикрепляются к субстрату Ч С другой
стороны, Bangia не выдерживают длительного пребывания в морской воде
и погибают через 21 день полного затопления, причем нити уже через 6 су-
ток заметно бледнеют. По-видимому, водоросли зоны брызг не могут жить
в литорали, и тем более в сублиторали.|
Но и для водорослей зоны литорали длительное пребывание вне воды
при отливе может повлечь за собой частичное обсыхание, сильный нагрев
солнечными лучами, а в холодные зимы — также замерзание. Установлено
что Fucus vesiculosus в арктических водахВне' замерзает при температуре
до — 40° в течение 6 месяцев, тогда как Ulva pertusa погибает уже при
температуре — 15°. На побережьях Швеции крайняя граница морозо-
стойкости водорослей колеблется от —16 до —20°. Напротив, глубинные
водоросли, никогда не испытывающие воздействия очень низких темпера-
тур, при замерзании отмирают. Водоросли, растущие в приливной
зоне, повреждаются от повышения температуры морской воды до 30°,
но при высыхании они более устойчивы к перегреванию 1 2.
Мы не располагаем данными о том, что водорослям, испытывающим
воздействие температур, свойственных их естественным местообитаниям,
наносится необратимый вред. Обсыхание толстых слоев Fucus на воз-
духе представляет собой медленный процесс обезвоживания; напротив,
набухание, то есть поглощение воды, протекает очень быстро.
1 Н. W а 11 е г, Zur Biologie der Bangia fusco-purpurea Lyngb., «Flora», 116, 1923,
S. 316—322; Protoplasma- und Membranquellung bei Plasmolyse, «Jb. wiss. Bot.», 62,
1923, S. 145—322.
2 R. В i e b 1, Protoplasmatische Okologie der Pflanzen, «Protoplasmatologia»,
Bd. XIIi, Wien, 1962.
22*
340
Глава 5
Вопрос о том, влияет ли пребывание водорослей вне воды на процесс
фотосинтеза, изучался разными исследователями *.
Обобщая полученные ими данные, можно констатировать, что водо-
росли приливной зоны тем легче переносят обсыхание, чем длительнее их
обычное времянахождение вне воды. Например, усыхание на 10% не при-
носит вреда Fucus vesiculosus, тогда как F. serratus, растущий в более
глубокой зоне, потеряв такое же количество воды, заметно повреждается.
Водоросли, оказавшиеся вне воды, при особенно хорошем освещении
некоторое время еще продолжают ассимилировать СО2. Нетто-ассимиля-
ция у Fucus platycarpus мало изменяется при нагревании до приблизительно
32°. Вместе с тем водоросли приливной зоны приспособились к интен-
сивному освещению. Более того, ассимиляция видов Fucus при кратковре-
менном их обсыхании даже возрастает, однако она уменьшается при
сильном обезвоживании. Поэтому в солнечные дни можно наблюдать, что
фотосинтез уже через два часа пребывания растения на воздухе прекра-
щается. Следовательно, облачные дни для продуцирования органического
вещества благоприятнее, тем более что оптимальная ассимиляция дости-
гается уже при освещенности 21—33 тыс. лк. Этим, вероятно, можно объ-
яснить, почему виды Fucaceae на побережье Британских островов в север-
ном направлении поселяются все выше над уровнем малой воды;'' дело
в том, что к северу дни становятся более облачными и повторяемость
туманов увеличивается.
Максимальная интенсивность ассимиляции СО2 у Fucus platycarpus
равна 14,9 л«г/<?л&2 -час. Количество СО2, ассимилированного различными
видами во время отлива, по данным Штоккера и Хольдхейде, приведено
в табл. 65.
Таблица 65
Ассимиляция СО2 во время отлива (в мг)
Название растений Ha 1 Q«2 листовой поверхности На 1 г сухого вещества
Porphyra atropurpurea ...*.. 4,5 23
Enteromorpha linza 6 25
Fucus platycarpus 37 16
Fucus vesiculosus 33 16
Fucus serratus 51 27
Водоросли зоны брызг при отливе обычно быстро высыхают, и процесс
фотосинтеза у них полностью прекращается. Поэтому количество ассими-
лированного ими во время отлива СО2 в расчете на 1 дм2 листовой поверх-
ности весьма незначительно. G другой стороны, слоевища этих водорослей
очень тонки. Следовательно, для построения ткани стенок клеток им тре-
буется "мало ассимилятов, поэтому их продуктивность в расчете на 1 г
сухого вещества оказывается равной продуктивности ламинарий литорали.
1 О.Stocker, W. Holdheide, Die Assimilation Helgolander Gezeitenalgen
wahrend der Ebbezeit, «Zschr. f. Bot.», 32, 1938, S. 1—59; J. Kaltwasser, Assimi-
lation und Atmung von Submersen als Ausdruck ihrer Entquellungsresistenz, «Protoplas-
ma», 29, 1938, S. 498—535; C. Montfort, Die Trockenresistenz der Gezeitenpflanzen
und die Frage der Ubereinstimmung von Standort und Vegetation, «Вег. Deutsch. Bot.
Ges.», 55, 1937, S. 85, 95; E. С о n w a y, The modification and raising of intertidal algal
zones on exposed jocky coasts of British Isles, 8. Gongr. Int. Botan., 1954.
Азональная растительность разных климатических зон
341
Если Fucus vesiculosus с содержанием воды, равным 10% его максималь-
ного значения, оставить на воздухе больше чем на сутки, а затем дать сно-
ва набухнуть, то после двухсуточного высушивания интенсивность его
ассимиляции еще достигнет 70% первоначальной величины, но уже спустя
трое суток она составит только 20% (F. platycarpus — 40%); после четы-
рехсуточного пребывания вне воды баланс вещества становится отрицатель-
ным и растение погибает. Отсюда следует, что водоросли этого вида при-
способились лишь к определенной длительности отлива, свойственной
зоне литорали, и поэтому за ее пределами не встречаются. Fucus serratus,
обитающий в нижней части литорали, не переносит уже трехсуточного
высыхания, а для ламинарий, растущих в сублиторали, пребывание на воз-
духе даже в течение 1—2 часов оказывается гибельным.
Интенсивность дыхания растений после того, как в них восстановится
обычное содержание воды, остается сначала несколько повышенной, но со
временем, если, конечно, водоросли не были повреждены летально, при-
ходит в норму.
Монтфорт исследовал для сравнения поведение Fucus vesiculosus,
горизонт обитания которого в Балтийском море находится между средним
уровнем полных сизигийных вод и глубиной 8—10 м уже в зоне сублитора-
ли. Оказалось, что глубинные формы менее устойчивы к высыханию. После
обретения вновь тургесцентности интенсивность дыхания у них сохраняется
на более высоком уровне, и проходит гораздо больше времени, прежде чем
восстановится нормальная ассимиляция. Таким образом, у растений осуще-
ствляется и функциональное приспособление к условиям внешней среды,
хотя преобладают наследственно закрепленные приспособительные свойства.
Следует еще упомянуть, что виды Fucus, оказавшись'на воздухе, изли-
вают наружу содержимое половых органов; у Dictyota приливы и отливы
обусловливают определенную ритмичность этого /процесса (14 дней или
1 месяц), которая сохраняется и у растений в культуре, причем даже
у вновь образовавшихся ответвлений таллома, никогда не испытывавших
воздействия отливов. Большая часть сперматозоидов мужских особей
и яйцеклеток женских извергается в течение одного часа.
Но приливы и отливы не являются единственным фактором среды,
определяющим законы распространения водорослей. Значение имеют
также субстрат и его сопротивляемость выветриванию. На легко разру-
шающихся породах можно обнаружить только быстрорастущие виды.
Ascophyllum nodosum встречается на побережье острова Гельголанд только
на немногих твердых гранитных глыбах или искусственных каменных
стенах и избегает местообитаний на песчанике. На последнем Ralfsia ver-
rucosa встречается лишь там, где горная порода менее интенсивно осве-
щается солнцем и не столь быстро поддается разрушению. Важны также
сила прибоя и экспозиция побережья. Porphyra растет в зоне брызг на се-
верной стороне, так как не переносит обсыхания на солнце. Вообще надо
отметить, что водоросли, по-видимому, очень чувствительны к прямому
воздействию солнечных лучей; прежде всего это относится к Rhodophyceae,
поселяющимся на наибольшей глубине в сублиторали.
В Средиземноморской области главными вегетационными периодами
для водорослей, обитающих вблизи поверхности моря, являются весна
и зима, для глубинных же видов — лето и осень, когда улучшаются усло-
вия освещения х. Таким образом, и в морях различия в характере место-
1 G. Berthold, Uber die Verteilung der Algen im Golf von Neapel..., «Mitt.
Zool. Stat. Neapel», 3, 1882; G. Funk, Die Algenvegetation des Golfs von Neapel
«Veroff. Zool. Stat. Neapel», Suppl. 7, 1927,
342
Глава 5
обитаний имеют для растений большое значение. Весьма многочисленны
водоросли-эпифиты, то есть мелкие водоросли, и прежде всего из семей-
ства Rhodophyceae, использующие в качестве опоры талломы крупных
водорослей (например, ламинарий).
В заключение — несколько слов о Саргассовом море. Оно находится
в Атлантическом океане и заключено между Северным экваториальным
и Антильским течениями и Гольфстримом. Образует море скопление водо-
рослей Sargassum (тропические, сильно рассеченные виды Fucaceae),
оторванных прибоем с побережий Вестиндских и Багамских островов.
Нам удалось наблюдать с корабля это «море» в виде массы свободно пла-
вающих буро-желтых водорослей. Видимо, это единственное место на зем-
ном шаре, где макрофитная растительность в больших количествах встре-
чается в пелагиали, то есть на открытой поверхности океана (Роунд, см.
примечание на стр. 333).
ЗАСОЛЕННЫЕ МАРШИ НА МОРСКИХ ПОБЕРЕЖЬЯХ
В отличие от скалистых морских берегов, где растительность вплоть
до литорали представлена водорослями, глинистые и песчаные морские
побережья почти исключитзльно покрыты цветковыми растениями.
В сублиторали здесь раскинулись луга «морской травы» (Potamogeto-
пасеае), о которых мы уже упоминали (см. рис. 233). Нижняя литораль
обычно лишена растительности; напротив, в верхней литорали различные
галофильные виды покрытосеменных образуют весьма своеобразные расти-
тельные сообщества, которые мы рассмотрим в этом разделе. Водоросли
обычно представлены свободноплавающими в воде формами. Fucus vesi-
culosus лишь иногда прикрепляется к раковинам мидий. Крупные водоро-
сли никакой роли на этих побережьях не играют. В изобилии появляются
микроскопических размеров водоросли (Gyanophyceae и диатомеи), кото-
рые густо покрывают дно литорали, но тем не менее едва различимы глазом.
Засоленные марши — явление, свойственное только умеренной зоне.
В тропиках на местообитаниях, периодически затопляемых морем, повсе-
местно распространена мангровая растительность (т. I, гл. 3). Марши
связаны с глинистыми побережьями, едва заметно наклоненными в сто-
рону открытого моря. Они образуются там, где прибой наблюдается только
во время очень сильных штормов, то есть в широких устьях рек или под
защитой расположенных перед ними островов, на участках, где илистые
частицы осаждаются на дно. Такие покрытые преимущественно илом пло-
щади литорали называют «ваттами». Отлогие побережья ватты окайм-
ляют поясом шириной до’нескольких километров. О маршах говорят лишь
тогда, когда растительный покров ватта становится столь густым, что
приобретает облик галотрофного луга. Различают внешний марш,
периодически заливаемый морем, и внутренний марш, прости-
рающийся от крайней границы приливов. Почвы внутреннего марша опрес-
нены дождевой водой, и поэтому галофильные растения здесь отсутствуют.
На побережьях Европы внутренние марши в настоящее время ограждены
дамбами, которые'защищают их от штормовых нагонов. Осадки, отложен-
ные на ваттах, либо поступают сюда с участков побережья, размываемых
морем (рис. 241), либо представляют собой материал, принесенный во взве-
шенном состоянии реками. Обычно взвеси отлагаются в устьях рек, спо-
собствуя^формированию аллювиальной дельты. Мельчайшие же частицы
выносятся в море, где под действием электролитов коагулируют, образуют
Азональная растительность разных климатических зон
343
хлопья и затем приливной волной выносятся на ваттовую полосу. Здесь
на дно сначала опускаются более крупные частицы, последними — самые
мелкие. Поэтому различают песчаные и илистые ватты, причем последние
располагаются несколько выше по профилю берега. Состав ила: 50—65%
тонкозернистого и пылеватого песка, 10—25% глины, 7—10% извести
(раковины моллюсков) и значительное количество органического вещества
(5—10%), содержащего азот и фосфорную кислоту. Органическое вещество
Р£и с. 241. ^Ваттовое море у острова Нордштранд (острова Халли-
ген). Марши крутым уступом обрываются к морю; размываемый
здесь грунт уносится течением и в виде ила осаждается на закры-
тых от ветра участках побережья. На заднем плане — судострои-
тельная верфь с волнозащитными сооружениями (фото Мёллера).
поступает с раскинувшихся перед ваттами пространств, покрытых «мор-
ской травой». Годовая продуктивность Zostera marina, по некоторым
данным, составляет 5 т!га (в сухом весе), то есть она близка к продуктив-
ности наиболее урожайных лугов Центральной Европы.
Поверхность ваттов и маршей не абсолютно ровная. Во время прилива
вода поднимается очень медленно, почти незаметно, но при отливе она
стекает по извилистым каналам-руслам, бороздящим поверхность марше-
вой низменности (рис. 242). Эти русла в верховьях представляют собой
мелкие углубления, которые затем сливаются, углубляются и наконец
превращаются каждая в широкую промоину; по ней, столь глубокой, что
она никогда не осушается, вода скатывается с ваттовой полосы в море.
По этой своеобразной, сильно разветвленной «речной системе» в глубь
берега проникают также воды прилива, заполняют ее и переливаются через
ее берега. Принесенный ими материал осаждается, нагромождая вдоль
водных артерий береговые валы.
На маршах встречаются также углубления, которые заполняются
водой только при сизигийных приливах или штормовых нагонах и которые
негосушаются во время отлива, етом вода испаряется, и на обнажившем-
ся дне впадины появляются выцветы солей. Мы подошли к вопросу о соле-
носности маршевых почв. Море, затапливая почву маршей, вносит в нее
344
Глава 5
соли. В сырую погоду при отливе дожди вымывают содержащиеся в почве
соли, причем тем интенсивнее, чем дольше почва остается незатопленной
морем. Следовательно, можно предполагать, что концентрация солей в поч-
ве уменьшается в сторону суши. Иная картина наблюдается в солнечную,
жаркую погоду: на осушенной во время отлива площади остатки воды
вследствие испарения и транспирации растений уходят в атмосферу, и поч-
венный раствор становится более концентрированным. В прилив морская
вода перемещает в более глубокие горизонты почвы накопившиеся на ее
поверхности соли. Концентрация почвенного раствора может превысить
Рис. 242. Маршевые луга с протокой на острове Нордш-
транд. Цветущие Armeria maritima ближе к воде сменяются Рис-
cinellia maritima (фото ^Мёллера).
даже соленость морской воды, особенно значительно во впадинах, длитель-
ное время остававшихся сухими. Если зима дождливая, а лето сухое,
можно заметить годовой ход содержания солей в почве. Таким образом,
в зависимости от климата на маршах складываются весьма сложные
условия солевого режима. В гумидной умеренной области количество солей
в почве, как правило, уменьшается по направлению в глубь суши, что
находит отражение в поясном размещении растительности.
>ТО растительности маршей написано много работ. Назовем здесь лишь несколько
обобщающих трудов, содержащих описания отдельных районов. Чэпмен дает общий
обзор растительности маршей по всем материкам, особенно подробно останавливаясь
на растительности Британских островов; краткое резюмирование условий в районе
Северного моря, основанное на использовании литературы на немецком языке, чита-
тель найдет в «Phytologie», Bd. IV, 2. Гильнер изучал засоленные луга западного побе-
режья Швеции. Засоленные марши побережий Средиземного моря исследованы Браун-
Бланке и его сотрудниками. Список литературы, посвященной маршам восточного
побережья Северной Америки, приведен в работе Штейнера. В дальнейшем изложении
мы использовали материалы перечисленных выше авторов и некоторые другие работы 1.
1 V. J. Chapman, Salt marshes and salt deserts of the world, London, 1960;
Vegetation and salinity, Salm, and Arid., The Hague, 1966, p. 23—42; V. Gillner
Азональная растительность разных климатических зон
345
Описания растительности засоленных^ маршей имеются почти для
всех областей земного шара, но работ, посвященных эколого-физиоло-
гическим исследованиям в этой зоне, на которые мы могли бы сослаться,
опубликовано немного.
Поясность маршевой растительности в каждом отдельном районе имеет
свои особенности. Они могут быть связаны с климатом и флорой, почвой
и скоростью накопления ила, с процессами поднятия или опускания побе-
режья и со многими другими, порой трудно устанавливаемыми причинами.
Приведем лишь несколько примеров (табл. 66).
Таблица 66
Основные ассоциации засоленных маршей, начиная с внешней зоны
Северо-Западная Европа Южная Англия (влажная серия) Восточная Северная Америка
Salicornietum Puccinellietum Juncetum gerardi Artemisietum maritimae Spartinetum townsendii Влажный засоленный марш Juncetum gerardi или maritimi Scirpetum maritimae Граница явно засоленной почвы Spartinetum glabrae Distichletum Juncetum gerardi Ivetum
Festucetum rubrae Phragmitetum Panicetum virgati
i Весьма проста последовательность ассоциаций в Арктике. За внешним
поясом с Puccinellia phryganodes следует внутренний пояс, образованный
различными видами Carex (С. salina, С. subspathacea, С. glareosa}, за кото-
рым начинается внутренний марш с преобладанием Agrostis и Festuca.
Совершенно иная зональность наблюдается на средиземноморском
побережье Франции. Здесь важное место занимают кустарникообразные
виды Salicornia (S. fruticosa, S. radicans}, Inula crithmoides, Atriplex por-
tulacoides, виды Limoniumiz Juncus acutus (рис. 243). Приливы и отливы
почти не выражены.
Южная Америка, Африка и Новая Зеландия принадлежат к разным
флористическим царствам, однако засоленные марши умеренной климати-
ческой зоны в этих районах обнаруживают много общего с засоленными
маршами голарктического флористического царства, так как значитель-
ное число галофитов являются космополитами.
Vegetations- und Standortsuntersuchungen in den Strandwiesen der schwedischen West-
kiiste, «Acta Phytogeogr. Suec.», 43, 1960; J.Iversen, Biologische Pflanzentypen als
Hilfsmittel in der Vegetationsforschung, Kopenhagen, 1936; H. W a 11 e r, Einfiihrung
in die allgemeine Pflanzengeographie Deutschlands, Jena, 1927; W. G. В e e f t i n k,
De zoutvegetatie van ZW-Nederland beschouwd in Europees verband, «Meded. Landbouwh.
Wageningen», 65, 1965, S. 1—167; Vegetation and habitat of the salt marshes and beach
plains in the South-Western part of the Netherlands, «Wentia», 15, 1966, p. 83—108;
J. Braun-Blanquet, Prodrome des groupements v eget aux, Fasc. 1, Ammophile-
talia.et Salicomietalia Medit., Montpellier, 1933; M. J. Adrian i, Recherches sur la
synecologie de quelques associations halophiles mediterraneennes, «SIGMA», № 32,
1934; R.Molinier, La vegetation des rives de I’Etang de Berre, «SIGMA», № 103,
Montpellier, 1947; M. Steiner, Die Pflanzengesellschaften der Salzmarschen in den
nord5stlichen Vereinigten Staaten von Nordamerika, «Rep. spec. nov. rrgni vegetab.»,
Beih. 81/G. 1935.
346
Глава 5
Pji с. 243. Лагуна у Фабрека (южная Франция). Засоленные марши покрыты
кустарникообразными видами Salicornia (фото Э. Вальтер).
Что расчленение маршевой растительности зависит от высоты место-
обитания над уровнем малой воды, показал Гильнер, проведя точную ниве-
лировку нижней границы распространения Puccinellietum (рис. 244).
Рис. 244. Нижняя граница нахождения ассоциации Puccinellietum mariti-
mae (7) и распространение Salicornietum (2) и Ruppia maritima (3) в зависи-
мости от высоты местообитания у западного побережья Швеции. На оси орди-
нат — высота над нулем мареографа в Варберге (по Гильнеру).
Азональная растительность разных климатических зон
347
Рис. 245. Salicornia herbacea как растение-пионер на илистой отмели
в устье Эльбы (фото Колюмбе).
За одним исключением, это сообщество повсюду западной Швеции'рас-
полагается примерно на одной и той же высоте; само сообщество занимает
пояс шириной 15—20 см по вертикали. Верхняя граница [засоленного
марша, фиксируемая Festuca rubra, проходит в 30—40 см над нижней гра-
ницей распространения Puccinellia. *
Пионером на ваттах Северо-Западной Европы является однолетнее
растение солерос {Salicornia herbacea s. 1.)ЛВ настоящее время выделяют
несколько элементарных видов этого растения (рис. 245, 246). На
сухих засоленных местообитаниях растение называют S. еигораеа,
на ваттах же встречается полиплоидная S. strictissima. Последняя способ-
ствует осаждению ила только тогда, когда образует достаточно густые
заросли х. Для того чтобы закрепиться в почве, проросшим семенам соле-
роса требуется 2—3-дневное пребывание вне воды. За это время корешочки
проникают в ил столь глубоко, что отливная волна оказывается’не в силах
унести их. Поэтому самым благоприятным периодом для прорастания
семян являются особенно сильные сизигийные отливы.
Наиболее эффективно способствует накоплению ила жизнедеятель-
ность мелких рачков {Corophium volutar) и различных моллюсков, а на
песчаной почве червей пескожилов {Arenicola marina), которые оставляют
в почве плотные комочки экскрементов (Gessner, 1957). На побережье
Северного моря насчитывали до 12 тыс. жилых трубок рачков на 1 м*
площади. Сцеплению частиц почвы помогает также слизь, выделяемая
Cyanaphyceae, Chlorophyceae и диатомеями, которые служат пищей
улиткам.
Однако действительное связывание ила начинается лишь после посе-
ления на нем многолетнего растения Puccinellia maritima (= Festuca thalas-
sica = Glyceria maritima = Atropis maritima).
1 E. Wohlenberg, «Wiss. Meeresunters, Abt. Helgoland», 19, 3, 1933; 19,
4, 1933; «Helgoland, wiss. Meeresunters.», 1, 1937, S. 1—92.
348
Глава 5
Как видно на рис. 247, к нему вскоре присоединяются другие солевы-
носливые виды. Складывается богатое видами сообщество, и солерос
вытесняется. Среди новых видов следует назвать солончаковую астру
(Aster tripolium), имеющую корни с множеством межклетников и потому
растущую даже на плохо аэрируемой почве, Armeria maritima, Plantago
maritima, Triglochin maritimum, Spergula-
P и c. 246. Отдельное растение
Salicornia herbacea в V4 нат. ве-
; личины (фото Штоккера).'
ria, ОЫопе и др. (рис. 248, 249). В следую-
щей зоне вместе с Juncus gerardi и J. mariti-
mus местами распространены те же виды,
а также Glaux maritima и др. На возвышениях
(особенно охотно — на берегах русел, пере-
секающих марши) поселяются Artemisia
maritima, Centaurium pulchellum, Trifolium
fragiferum, Potentilla anserina и др. С некото-
рыми из этих видов мы знакомы как с расте-
ниями — индикаторами содового засоления.
Сведения о том, происходит ли в слабо засо-
ленных и богатых гумусом маршевых почвах
образование соды, в литературе отсутствуют.
Против этого свидетельствует величина pH:
она несколько выше 7, а в ряде мест даже
равна 6.
Очень сходные сообщества засоленных маршей
известны на восточном побережье Японского острова
Хоккайдо Ч В глубокой суб литорали растет Zostera
marina, в верхней — Z. папа', на илистом грунте
поселяется Salicornia brachystachya, далее следует
пояс Puccinellia kurilensis совместно со Spergularia
marina, а несколько выше — пояс Glaux maritima
var. obtusifolia. Переходную область к внутреннему
маршу образуют Juncus gracillimus в ассоциации
с Potentilla egedei var. grandis и Eleocharis kamtscha-
tica; к ним присоединяется Phragmites communis,
обычно угнетенный; нормально он начинает разви-
ваться лишь тогда, когда прекращается влияние
солей, признаком этого служит появление Calamag-
rostis epigeios и Inula japonica.
Приведенная в табл. 66 серия сообществ из Англии представляет осо-
бый случай. За последние десятилетия в этом районе распространился
Spartina townsendii — амфидиплоидный гибрид европейской «S'. stricta
и американской S. alterniflora. Гибрид образовал густые заросли, вытес-
нил из верхней сублиторали и нижней литорали все другие виды и уже
в 1906 г. поселился на побережье Франции. Вщелях увеличения толщи ила
на побережье Западной Германии высадили S. townsendii, но растет она
плохо, возможно потому, что’ил здесь более песчаный. Для наращивания
суши эти виды оказались малопригодными, тац как образуемые ими густые
кусты сужают водный поток при отливе, тем самым повышая его скорость
и усиливая эрозионную силу. Вторая особенность избранного для Англии
примера состоит в том, что ряд не ведет к обычному лугу с Festuca и Agro-
stis, а заканчивается зоной пресноводного переувлажнения.
Третий пример в табл. 66 (побережье штата Коннектикут, США) сви-
детельствует о том, что здесь виды галофильных злаков играют значитель-
1 А. М i у a w a k i, Т. O h b a, Studien fiber Strand-Salzwiesengesellschaften auf
Ost-Hokkaido (Japan), «Sci. Rep. Yokohama Nat. Univ.», Sect. II, 12, 1965, S. 1—25.
Азональная растительность разных климатических зон
349
Профиль Меиры5 ю 15 2д 25 30 35 40
Galium verum , . .
Carex parviflora (= nigra)
Viela canina
Paa pratensis
Euptwrbia palustris
Lptus .corniculatus
Linana vulgaris
carex recta , ,
Qphioglossum vulgatum _
Angelica archangelica var. litoralis
Lemtoaon autumnalis
Agropyron repens
Armeria maritima
Potentilla anserina
Viola cracca
Atriplex latifolia
Agrostis stolonifera
Festuca rubra
Plantago maritima
Juncus gerardi
Glaux maritima
Spergularia .marginata
Scirpus mantlmus
Aster tripolium
Triglochin marlttmum
Puccinellia maritima
Salicornia strictissima .......
Ruppia maritima-------- .
100
80
60
40
20
0
Рис. 247. Поясность ^'маршевой растительности на открытом западном побе-
режье Швеции (по Гильнеру, из «Phytologie», Bd. IV, 2).
Профиль имеет длину 45 л»; пунктирная линия — данные нивелировки (в см) в соответствии
с масштабом справа; вертикальные линии — суточные колебания уровня воды в период с апреля
по сентябрь; толщина горизонтальных линий отражает обилие отдельных видов.
Рис. 248. Растения маршей на побережье Северного моря в
апреле. Перед цветущей Cochlearta anglica виден Triglochin
maritimum, справа — листья Plantago jnaritima (фото Штоккера).
350
Глава 5
Рис. 249. Марши в бухте Яде-Бузен на Север-
ном море. Вдали — открытая водная поверх-
ность; на повышенных участках заметны серые
листья Artemisia maritima', на переднем пла-
не — соцветия Armeria maritima (фото Ницшке).
но большую роль и формируют густые, местами высокие травостои. В ка-
честве растения-пионера выступает Spartina glabra. Переход от этой зоны
к низкозлаковому лугу с господством Distichlis spicata образует Spartina
patensх. Видов разнотравья на этих маршах меньше. Появляющаяся
на*границе с внутренним маршем Iva oraris представляет собой кустовид-
ное' растение из семейства сложноцветных. Солерос развивается только
в чрезмерно засоленных впадинах.
Чрезвычайно детальные эколого-физиологические исследования севе-
роамериканских маршей провел Штейнер.
Тесные взаимосвязи отдельных поясов маршевой растительности
с уровнями приливов не обнаружены, но установлена их зависимость
от содержания солей в почвенном растворе. Ведь экологическая особен-
ность? всех маршей состоит именно в высокой концентрации соленосного
почвенного раствора. Осмотическое давление морской воды приблизитель-
но соответствует 20 атм. На сырых засоленных маршах определены зна-
1 С этим видом мы познакомились, рассматривая почвы пампы с содовым засо-
лением (см. стр. 152).
Азональная растительность разных климатических зон
351
чения до 31 атм, на экстремальных местообитаниях — еще более высо-
кие. Шимпер, исходя из предположения, что растениям трудно извлекать
воду из солевых растворов, называл засоленные почвы «физиологически
сухими» и ставил галофиты на одну ступень с /ксерофитами физически
сухих почв, считая, что галофиты, как и ксерофиты, в условиях затруднен-
ного водообмена должны сокращать интенсивность транспирации.
Поэтому эколого-физиологические исследования растений засоленных
маршей были начаты с измерения величины транспирации. Предполага-
лось выяснить, транспирируют ли галофиты менее интенсивно, чем виды
растений, заселяющих незасоленные почвы г. Как и можно было ожидать,
при вычислении интенсивности транспирации на единицу листовой поверх-
ности разница оказалась практически неустановимой, тогда как при рас-
чете на единицу сырого веса или на содержание воды в теле растения потери
воды суккулентными галофитами были меньше.
Важность того, что принять за «точку отсчета», выступает особенно
отчетливо, когда сопоставляются виды одного рода, выросшие в одинако-
вых условиях среды, например галофиты Plantago maritima и Р. coronopus
и негалофильные Р. lanceolata и Р. major (табл. 67).
Таблица 67
Данные сравнительных измерений интенсивности транспирации
у четырех видов Plantago, выросших в отсутствие NaCl (по Шратцу)
Виды Plantago coronopus maritima lanceolata major
Транспирация в расчете на пло- щадь, mzIcm2 8.45—9.20 час. 9.20—10.05 час. 10.25—11.05 час. 11.35—12.20 час. 6,24 8,89 10,11 13,41 6,17 5,68 10,01 9,44 4,47 5,62 6,56 7,51 2,49 3,42 5,47 5,60
Транспирация в расчете на сы- рой вес, мг/г 8.45—9.20 час. 9.20—10.05 час. 10.25—11.05 час. 11.35—12.20 час. 1,56 2,22 2,76 3,38 1,54 1,42 2,25 2,36 2,45 3,08 3,95 4,51 2,35 2,26 4,98 5,29
Отношение транспирации в рас- чете на вес к транспирации в расчете на площадь 0,252 0,250 0,548 0,944
Дальнейшие исследования показали, что при повышении содержания
солей в почвенном растворе все растения сокращают транспирацию^
причем увеличение содержания NaCl оказывает гораздо большее влияние,
чем это соответствует его осмотической концентрации. Однако транспира-
ция для этих растений на сырых засоленных почвах в экологическом отно-
шении отнюдь не является решающим фактором. *
1 М. J. Adrian i, Der Wasserhaushalt der Halophyten, «Hdb. d. Pflanzenphy-
siol.», Bd. 3, 1956, S. 902—914; Die Halophyten, Bd. 4, 1958, S. 709—736; 0. Sto-
cker, Das Halophytenproblem, «Ergeb. d. Biol.», 3, 1928, S. 265—354; E. Schratz,
Beitrage zur Biologie der Halophyten I-IV, «Jb. wiss. Bot.», 80, 1934, S. 112—142; 81,
1935, S. 50—93; 83, 1936, S. 133—149; 84, 1937, S. 593—638. .
352
Глава 5
Для всех без исключения галофитов было установлено, что они погло-
щают хлориды и накапливают их в клеточном соке транспирирующих
органов, поэтому осмотическое давление клеточного сока вместе с раство-
ренными неэлектролитами всегда оказывается выше концентрации почвен-
ного раствора *; тем самым осмотические силы последнегожомпенсируют-
ся (табл. 68).
Таблица 68
Содержание NaCl в почвенном растворе корневой зоны,
осмотическое давление и доля хлоридов (NaCl) в отжатом соке
растений засоленных маршей под Неаполем (по Еналю)
Название растений Сок растений Содержание NaCl в поч- венном рас- творе, атм
осмотическое давление, атм доля хлоридов, атм
4 triplex portulacoides . . . 41,1 36,4 27,7
Salicornia fruticosa .... 41,3 31,7 20,6
Inula crithmoiaes 24,7 17,6 17,0
Statice limonium 23,5 18,5 10,5
Juncus acutus 19,4 11,9 9,3
Plantago coronopus .... 11,7 7,7 4,0
Еще одна важная проблема при изучении взаимоотношений галофи-
тов с внешней средой — это определение солеустойчивости их протоплаз-
мы и роль корней в ругулировании процесса поступления солей в растение.
Растения всасывают из почвы не чистую воду, иначе в них не происхо-
дило бы накопления солей. Однако не может быть речи и о поглощении
неизмененного почвенного раствора, так как в противном случае это при-
вело бы спустя всего несколько дней к пересыщению клеточного сока
хлоридами. Следовательно, растения?берут из почвы разбавленный раствор.
Но каким образом происходит регуляция его концентрации, пока не уста-
новлено.
Во всяком случае, при эколого-физиологических исследованиях гало-
фитов одного определения соленосности почвы недостаточно, необходимо
также знать содержание солей в клеточном соке, с которым соприкасается
протоплазма. Кроме того, растения поглощают соли не в том соотношении,
в каком они находятся в почвенном растворе, а предпочитают хлориды
(в меньшей степени — сульфаты), [причем натрий частично заменяется
калием.
Наиболее подробными исследованиями в этом направлении мы обяза-
ны Штейнеру1 2. По его данным, концентрация солей в почвах маршей
1 Это справедливо только в отношении транспирирующих органов растений.
Корни и стебли, выполняющие^ функцию проводящих систем, часто содержат более
низкое количество соли (ср. стр. 242).
На побережье Средиземного моря у Монпелье между 21.XI и 25.VI получены
следующие значения осмотического давления (в атм)'. ОЫопе portulacoid.es 25,8—62,4;
Salicornia fruticosa 34,1—46,5; Statice limonium 20,9—39,1; Juncus marttimus 15,0—
29,0; Spartina versicolor 16,1—40,7.
Cp.: J. Braun-Blanqjiet, Lagunenverlandung und Vegetationsentwicklung
an der i'ranzosischen Mittelmeerkiiste bei Palavas, ein Sukzessionsexperiment, «SIGMA»,
Commun. № 141, Montpellier, 1958.
2M. Steiner, Zur Okologie der Salzmarschen der norddstlichen Vereinigten
Staaten von Nordamerika, «.Tb. wiss. Bot.», 81, 1935, S. 94—202; Die Zusammensetzung
des Zellsaftes bei hoheren Pflanzen in ihrer okologischen Bedeutung, «Erg. d. Biol.»,
16, 1939, S. 189—234.
Азональная растительность разных климатических зон
353
восточного побережья Северной Америки не одинакова в разных гори-
зонтах профиля. Поэтому, чтобы разобраться в тонкостях солевого режи-
ма растений, важно знать глубину проникновения корней в грунт; напри-
мер, корни Salicornia опускаются только на 10 см от поверхности почвы,
большинство других видов — до 30 см, корни Juncus gerardi — на глубину
100 см.
Если в растительном покрове наблюдается известная поясность или
если даже на самой маленькой площадке обнаруживается мозаичность
растительности, то в каждом случае всегда удается доказать различие
в степени засоленности почвы. Дифференциацию растительности опреде-
ляют в первую очередь колебания солености, обусловленные погодными
условиями, и максимальные значения концентрации солей в почве. Экстре-
мальные местообитания в исследованном районе, приурбченные к высы-
хающим летом западинам, занимает Salicornia. Максимальные концентра-
ции в корневой зоне солероса достигают 50—60 атм (у поверхности поч-
вы — 198 атм); амплитуды в зависимости от погоды могут быть весьма
значительными (30—40 атм). При еще большем засолении растительность
не развивается.
Гильнер проследил на западном побережье Швеции динамику содер-
жания хлоридов в почве в течение всего вегетационного периода и также
установил отчетливую зависимость отдельных видов от степени и качества
засоления субстрата (рис. 250).
При определении осмотического давления у растений засоленных
маршей оказалось, что оно колеблется в пределах 20—40 атм, а у видов,
растущих за пределами засоленных маршей, никогда не достигает 20 атм,
а местами даже 10 атм. Но высокие значения осмотического давления у гало-
фитов связаны с накоплением солей в клеточном соке, и поэтому они не мо-
гут быть сопоставлены с показателями осмотического давления у ксеро-
фитов. Высокое осмотическое давление галофитов отнюдь не означает, что
произошло обезвоживание протоплазмы, оно обусловливает лишь усиле-
ние суккулентности. Некоторое дополнительное поглощение солей даже
способствует росту галофитов; только когда концентрация солей в кле-
точном соке превысит определенное, пороговое, значение, растения повре-
ждаются. Это сверхпредельное количество солей зависит от солеустойчи-
вости протоплазмы и для отдельных видов различно.
Очень высокое пороговое значение обнаружено у Salicornia. Шратц
нашел, что сырой вес солеросов в расчете на 1 м? площади возрастает
параллельно увеличению содержания солей в грунтовых водах до 6%
(табл. 69).
Таблица 69
Вес растений Salicornia в расчете на 1 ле2 почвы
в зависимости от солености грунтовых вод
Содер- жание солей, % 1,6—2,0 2,1—2,5 2,6—3,0 3,1—3,5 3,6—4,0 4,1—5,0 5,1—6,0 6,0
Вес, г 58 85 223 232 242 250 250 293
Однако замечено, что с ростом минерализации грунтовых вод число
экземпляров растений уменьшалось, отдельные же особи развивались осо-
бенно хорошо. Тем не менее продукция сухого вещества при высоком
23-0434
354
Глава 5
Рис. 250. Содержание солей и воды в почве засоленных маршей под
различными типами растительных сообществ; западное побережье Шве-
ции (по Гильнеру).
Вверху — количество осадков; внизу — суточные колебания уровня воды относи-
тельно нуля мареографа в Варберге в период с апреля по сентябрь.
1 — засоленные почвы без растительности; 2 — Salicornietum; 3 — Puccinellietum;
4 — Juncetum gerardi c Centaurium (= Erythraea). В сентябре марш был затоплен
морем.
содержании солей, видимо, должна сократиться, ибо растения становятся
суккулентнее, сочнее и больше накапливают солей. То и другое отражается
на сыром весе.
Доля осмотического давления, которая обусловлена присутствующи-
ми в клеточном соке хлоридами, составляет у видов засоленных маршей
более 67%, а у Salicornia может достигать даже 88%. Доля нехлоридов
заставляет вспомнить состав клеточного сока растений, обитающих на не-
засоленных почвах (рис. 251).
В среднем осмотическое давление у растений засоленных маршей при-
близительно на 20 атм выше, чем у растений низинных болот. Эта разница
Азональная растительность разных климатических зон
355
примерно соответствует среднему содержанию хлоридов в клеточном соке
галофитов, а также величине осмотического давления морской воды, обу-
словливающей засоление почв маршей.
Весьма отчетливую зависимость Шратц обнаружил также между кон-
центрацией солей в растениях пляжей и дюн (в процентах NaCl) и содер-
жанием солей в почвах, однако эти
величины непосредственно не сопо-
ставимы, если они не выражены
в атмосферах (Шратц, см. примеча-
ние на стр. 351).
Определения осмотического дав-
ления и содержания хлоридов в кле-
точном соке галофитов, растущих на
почвах разной степени засоления,
позволили установить, что на почвах,
бедных хлоридами, галофиты погло-
щают непропорционально много хло-
ридов, вследствие чего осмотическое
давление у них повышается быстрее,
чем концентрация почвенного раство-
ра. Благодаря этому облегчается про-
цесс поглощения влаги растениями,
и, возможно, этим же можно объя-
снить наблюдаемое у галофитов сти-
мулирование роста небольшим коли-
чеством солей. Но если концентра-
ция почвенного раствора становится
очень высокой, что наблюдается, на-
пример, на местообитаниях солероса
в замкнутых понижениях, то рост
осмотического давления замедляется
и обеспеченность растений водой
ухудшается. В конце концов дости-
гается состояние, при котором поступ-
ление солей прекращается. При
дальнейшем увеличении концентра-
ции почвенного раствора растения
вянут, то есть происходит сгущение
клеточного сока как результат умень-
шения влагосодержания. Это повы-
шение осмотического давления ска-
Р и с. 251. Величина осмотического дав-
ления (вся колонка) и концентрация
солей в клеточном соке (в атм) у расте-
ний засоленных маршей Северной Аме-
рики (по Штейнеру).
1 — Spartina glabra', 2 — Sp. patens; 3 — Dis-
tichlis spicata; 4 — Juncus gerardi; 5 — Sali-
cornia mucronata; 6 — S. europaea; 7 — Plantago
decipiens; 8 — Atriplex hastata; 9 — Aster subu-
latus (Na не определялся); 10 — Limonium са-
го linianum; 11 —Suaeda linearis; 12 — Iva
oraria. Только у 8 видов растений содержа-
ние Na превышает долю С1.
зывается на растении отрицательно;
оно принимает угнетенный вид, обычно имеет красную окраску и нако-
нец погибает х. Таким образом, на этой последней стадии галофиты
ведут себя так же, как и виды, обитающие на незасоленных почвах.
На маршах Северной Америки в условиях сильного засоления встре-
чаются Salicornia europaea и более солеустойчивая S. mucronata; S. еиго-
1 Окрашивание в красный цвет обусловлено бетацианом и является признаком
азотного голодания. Необходимость препятствовать процессу поглощения солей корня-
ми (при высокой концентрации солей в почвенном растворе), видимо, затрудняет также
потребление азота. Если растения опрыскать 1%-ным раствором мочевины, они сохра-
няют зеленый цвет. Ср.: Р. Hoffmann, Н. Sacher t, Der EinfluB von Harnstoff
auf die Entwicklung von Salicornia..., «Вег. Deutsch. Bot. Ges.», 80, 1967, S. 437—446.
23*
356
Глава 5
раеа повреждается, если осмотическое давление превышает 35 атм;
S. mucronata даже при осмотическом давлении 49 атм развивается нор-
мально.
Поскольку галофиты вместе с водой всегда поглощают некоторое, хотя
и небольшое, количество солей, следовало бы ожидать, что осмотическое
давление у них в течение вегетационного периода будет непрерывно повы-
шаться. Однако этот рост отмечен лишь у Juncus gerardi. Напрашивается
вопрос, каким образом регулируют содержание солей другие виды рас-
тений.
Штейнер среди исследованных им видов растений засоленных маршей
различал три типа.
1. Суккулентные галофиты препятствуют повышению содержания
хлоридов в клеточном соке, постоянно увеличивая суккулентность. Сле-
довательно, одновременно с накоплением солей возрастает степень
оводненности’ тканей растения. Это так называемый регулирую-
щий тип.
2. "У несуккулентных галофитов, к которым относятся виды Spartina
и Distichlis, а также Limonium, имеются железы, активно выделяющие
избыточную соль. Это солевыделяющий тип.
3. Третий, кумулятивный, тип представлен Juncus gerardi,
возможно, к нему принадлежит и Triglochin maritimum. Эти виды накапли-
вают соли, причем у Juncus gerardi осмотическое давление непрерывно
повышается с 23 атм в мае до 40 атм в августе, хотя вид растет на почве,
относительно бедной солями. При этом содержание воды в растении летом
несколько снижается. Достигнув максимального, порогового, обезвожи-
вания, побеги отмирают.
Приобретение галофитами признаков суккулентности наитеснейшим
образом связано с поглощением растениями хлоридов. У всех видов,
даже негалофильных, обитающих на засоленных почвах, в той или иной
степени проявляется суккулентность, поскольку эти виды по своей гене-
тической конституции предрасположены к образованию такой анатоми-
ческой структуры. Однако Juncus, как и Phragmites, накапливая хлориды,
не становятся собственно суккулентами, это относится и ко всем злаковид-
ным растениям. Единственным известным нам суккулентным злаком
является Spinifex hirsutus, встречающийся на западном побережье Австра-
лии. На свойство хлоридов вызывать суккулентность обратил внимание
еще Лесаж х. Как показали опыты в культуре, произведенные Б. Келле-
ром и в большем масштабе Ван Эйком, коэффициент суккулентности воз-
растает тем больше, чем выше концентрация хлоридов в питательном рас-
творе; при этом катионы Na, К, Mg, Са существенной роли не играют,
но важное значение имеют анионы: например, Salicornia в культуре
на сульфатных растворах такой же концентрации не обнаруживает при-
знаков суккулентности 1 2. Однако известно, что ионы хлора обусловлива-
ют набухание белка, ионы же сульфата вызывают его обезвоживание.
Соответственно и галофиты, накапливающие в клеточном соке, помимо
хлоридов, много сульфатов, несуккулентны или лишь слабо суккулентны.
1 Р. М. L е s a g е, Recherches experimentales sur les modifications des feuilles chez
les plantes maritimes, «Rev. Gen. Rot.», 2, 1890, p. 55, 106, 163; Extension du charactere
aquis et faits d’heredite dans le Lepidium sativum arrose a 1’eau sallee, «Gpt. rend. hebd.
Seances Acad. Sci.», 180, 1925, p. 154.
2 M. van E i j k, Analyse der Wirkung des NaCl auf die Entwicklung, Sukkulenz
und Transpiration bei Salicornia herbacea..., «Extr. Rec. Trav. Rot. Neerl.», 36, 1939,
S. 559—567.
Азональная растительность разных климатических зон
357
Ван Эйк, напротив, считает, что Salicornia в культуре на нитратном рас-
творе не является суккулентным растением, хотя ионы нитрата в большей
мере способствуют набуханию, чем ионы хлора; он упускает при этом
из виду, что все дело в степени концентрации ионов в среде, с которой
протоплазма соприкасается, то есть в размерах концентрации ионов
в клеточном соке; непосредственно воздействовать питательный раствор
может только на корневые волоски. Нитраты же, поглощенные растением,
используются для построения белка и лишь редко и в небольшом количе-
стве накапливаются в клеточном соке.
При эколого-физиологических исследованиях нельзя органичиваться
регистрацией обстановки вне растения, всегда необходимо выяснять
условия, в которых находится протоплазма. Что касается температуры,
то ее различия между вцюшней средой и растением несущественны; напро-
тив, водный режим и солевой баланс внешней и внутренней сред могут
быть резко неодинаковыми.
Строгонов 1 в последнее время снова подчеркнул, что большое влия-
ние на закономерности роста и формирование анатомо-морфологической
структуры растений оказывает не только общее содержание солей в почве,
но и их химический состав. По его мнению, качество засоления играет
главную роль в явлении солеустойчивости растений. Строгонов отмечает
избирательное воздействие ионов S0'4' и С1' на растения, но ограничивается
в своих исследованиях анализом солей в почве, не определяя содержания
сульфатов или хлоридов в листьях растений. В пустыне Намиб мы наблю-
дали, что хлоридные и сульфатные галофиты могут расти вместе на одной
почве, ибо одни виды накапливают в своих органах только хлориды, а дру-
гие, помимо хлоридов, также большее или меньшее количество сульфатов.
Первые из них — суккулентные растения, вторые, скорее, ксероморфные
(«Phytologie», Bd. III. 1, S. 487). Строгонов"поставил опыты с культурны-
ми растениями и галофитами, высаженными в вегетационных сосудах.
Использовалось одностороннее засоление субстрата: в почву сосудов вно-
сились либо сульфаты, либо хлориды. Контролем служили растения на’не-
засоленной почве. Отмеченные Строгоновым анатомо-физиологические^ раз-
личия у растений, выращенных на разном субстрате, приведены в табл. 70.
Анализ таблицы позволяет сделать некоторые выводы. Так, увеличе-
ние сухого веса 100 ел*2 листовой поверхности при засолении почвы, несом-
ненно, объясняется главным образом накоплением солей в листьях.
В условиях хлоридного засоления интенсивность водообмена снижается
гораздо сильнее, чем при сульфатном засолении; вместе с тем более резко
проявляется суккулентность (содержание воды на единицу листовой поверх-
ности более чем вдвое превышает содержание на контроле, при сульфат-
ном засолении — только на 24%). Кроме того, оказалось, что при смесях
солей, приближающихся к наблюдаемому в природе, осмотическое давле-
ние у растений с преимущественно сульфатного засоления не отличалось
от давления на контроле (9,6 атм), тогда как на хлоридном местообитании
оно было значительно выше (13,4 атм)', вместе с тем вязкость протоплазмы
под воздействием хлористых солей в девять раз превысила вязкость про-
топлазмы контрольного растения и в два с половиной раза вязкость при
засолении почвы сернокислыми соединениями, что служит явным призна-
ком изменения свойств протоплазмы ионами хлора. Гидратация прото-
плазмы при этом увеличивается. Факты свидетельствуют о том, что при
1 Б. П. Строгонов, Физиологические основы солеустойчивости растений,
М., 1962.
358
Глава 5
Таблица 70
Изменение развития и интенсивности транспирации хлопчатника
в зависимости от типа солевого состава почвы.
Влажность почвы = 70% ее полной влагоемкости;
концентрация солей = 1 % на абсолютно сухую почву.
Время наблюдения — август
Контроль Сульфатное засоление Хлоридное засоление
Общее содержание воды, % веса сырой массы 78,16 76,48 83,34
Сырой вес 100 см2 листовой поверхности, г Количество воды в 100 см2 листовой поверх- 2,08 2,53 4,00
ности, г 1,60 1,98 3,40
Количество сухого вещества в 100 см2 листо-
вой поверхности, г 0,48 0,55 0,60
Интенсивность транспирации, г!м2‘час . . . Интенсивность возмещения транспирацион- 188,4 243,6 109,8
ной воды, % общего содержания воды
в листьях за час 120,6 135,0 35,4
Площадь листьев одного растения, см2 . . . 3628 2505 685
Сухой вес корней одного растения, г ... . 4,34 3,22 1,46
хлоридном засолении анатомо-структурные изменения идут в направлении
галосуккулентности, а при сульфатном — в сторону галоксероморфности.
Засоление почв создает неблагоприятные условия для роста, развития
и формирования урожая культурных растений, причем засоление хлори-
дами — в значительно большей степени, чем сульфатное засоление; напро-
тив, рост Salicornia herbacea стимулируется ионами хлора (при содержа-
нии солей, равном 1% на абсолютно сухую почву) (табл. 71).
Таблица 71
Рост и развитие Salicornia herbacea на засоленной почве
(по Строгонову)
Показатели роста и развития Контроль Сульфатное засоление Хлоридное засоление
Число члеников на главном стебле . . 18,7 17,0 21,8
Сырой вес растения, г 1,33 3,19 6,36
Сухой вес растения, г1 0,33 0,60 0,82
1 Видимо, включая содержащиеся в нем соли.
Иная реакция на качество засоления наблюдается у Suaeda glauca'.
в условиях хлоридного засоления ее рост и развитие угнетаются (табл. 72).
У растений хлопчатника, томата, подсолнечника и конских бобов
присутствие ионов хлора обусловливает суккулентность листьев и более
слабое развитие ксилемы, тогда как ионы SO4 способствуют формированию
ксероморфной структуры. Напротив, листья ячменя при сульфатном засо-
лении по сравнению с контрольными растениями почти не изменяются, под
влиянием же засоления хлоридами они становятся уже и тоньше.
Азональная растительность разных климатических зон
359
• Таблица 72
Рост и развитие 'Suaeda glauca на засоленной почве
(по Строгонову)
Показатели роста и развития Конт- роль Сульфатное засоление Хлоридное засоление
Высота растения, см . . 39,8 51,2 30,6
Число боковых ветвей 20,6 33,3 15,6
Сырой вес растения, г 8,5 22,6 10,7
Воздействие ионов хлора специфичное и отличается от влияния
сульфатных ионов.
Данные детальных исследований фотосинтеза галофитов, растущих
на засоленных маршах, нам неизвестны. При повышенной концентрации
солей в почве галофиты часто приобретают бледно-зеленую окраску, но
световая компенсационная точка у таких растений не повышена. Миера
(дисс., Бомбей, 1967) показал, что бледно-зеленые суккулентные листья
Clerodendron inerme (в условиях засоления), содержавшие меньше хлоро-
филла по сравнению с контрольными растениями на «пресных» почвах,
на свету тем не менее ассимилировали большее количество 14СО2. Это дает
основание говорить о предполагаемом стимулировании ионами хлора
процесса фотосинтеза. При определении состава ассимилятов оказалось,
что хотя растения на засоленных почвах содержат меньше сахаров и орга-
нических кислот, но количество вновь образовавшихся аминокислот
у них выше. Таким образом, процесс фотосинтеза у галофитов может в из-
вестной мере отличаться от нормального. На изолированные хлоропласты
свеклы и шпината NaCl оказывает характерное вредное влияние, вызывая,
в частности, их обезвоживание х. Кроме того, Ван Эйк отмечает, что интен-
сивность дыхания у галофитов при поглощении ими солей увеличивается
и что, следовательно, речь идет об активном процессе. Было бы интересно
изучить механизм поглощения солей галофитами более детально.
Очень важной проблемой является проблема прорастания семян
и развитие проростков в условиях засоления. Следовало бы подробнее
изучить вопрос о возможном содержании солей в семенах галофитов.
Шратц в семенах Aster tripolium и Salsola kali хлоридов не обнаружил.
В общем, повышенные концентрации солей в окружающей среде тор-
мозят прорастание семян. Помимо того, на негалофиты NaCl оказывает
особое, дополнительно тормозящее влияние, которое не проявляется у га-
лофитов, выросших на засоленных почвах1 2.
Опыты показали, что семена растений засоленных маршей лучше всего
прорастают в чистой воде либо в воде, содержащей х/8 часть морской воды;
лишь для Salicornia установлен резко выраженный оптимум прорастания
при содержании х/2—3/4 частей морской воды. Artemisia maritima и Sper-
gularia marginata, обитающие на высоком засоленном марше, в 1 %-ном
1 К. A. Santarius, R. Ernst, Das Verhalten von Hill-Reaktion und Photo-
phosphorylierung isolierter Chloroplasten in Abhangigkeit vom Wassergehalt, «Planta»,
73, 1967, S. 91—108.
2 C. Montfort, W. Brandrup, Physiologische und pflanzengeographische
Seesalzwirkungen II, «Jb. wiss. Rot.», 66, 1927, S. 902—946; III, «Jb.», 67, 1928, S. 105—
173; E. Schratz, Beitrage zur Biologie der Halophyten I. Zur Keimungsphysiologie,
4<Jb. wiss. Bot.», 80, 1934, S. 112—142; I. A. U n g a r, Influence of salinity on seed germi-
nation in succulent halophytes, «Ecol.», 43, 1962, p. 763—764.
360
Глава 5
Рис. 252. Рост корней галофитов и
негалофитов в растворах морской соли
разной концентрации (по Монтфорту и
Брандрупу, из «Phytologie», Bd. IV, 2).
растворе NaCl имеют всхожесть, равную соответственно 8 и 4%. Семена
большинства других видов еще прорастают в 2 %-ном растворе NaCl.
В нормальной морской воде прорастают*только семена Salicornia (они не
утрачивают всхожесть и в 10 %-ном растворе NaCl) и Spartina townsendii,
то есть семена растений-пионеров на почвах ваттов, где концентрация поч-
венного раствора никогда не падает ниже солености морской воды. У дру-
гих видов семена трогаются в рост обычно в дождливое время года (осень,
весна), когда на незатопляемых морем участках происходит опреснение
почвы дождевыми водами. В Среди-
земноморье зимний влажный сезон
также особенно благоприятен для
ростовых процессов. Коль скоро кор-
ни проростка углубились в почву,,
накопление соли на ее поверхности
летом на развитии растения уже не-
отражается.
Благоприятствует прорастанию,,
по-видимому, и низкая температура
зимой; так, например, семена Spergu-
laria, выдержанные при температур©
—10°, дали всходы даже в 3%-ном
растворе NaCl.
Семена, не прораставшие в со-
левых растворах, будучи перенесены
в воду, дружно прорастали. Можно
предположить, что NaCl препятствует
набуханию семян. Следовательно,
вполне вероятно, что семенная ко-
жура под влиянием низких темпе-
ратур становится более проницае-
мой, из-за чего тормозящее воздействие NaCl не проявляется.
Подрастающие проростки галофитов очень скоро обретают определен-
ную солеустойчивость, что, очевидно, связано с началом накопления солей
в клеточном соке. Неодинаковый рост корней негалофильных видов и га-
лофитов в солевых растворах разной концентрации Монтфорт схематиче-
ски отразил на рис. 252. Сравнение Aster tripolium и Salicornia позволяет
утверждать, что существуют разные типы галофитов. Относительную соле-
устойчйвость корней Aster tripolium удается установить лишь в том случае,
когда для опыта используются семена растений, произраставших на засо-
ленной почве. Корни Aster из семян растений, выросших в отсутствие засо-
ления, ведут себя так же, как негалофильные виды. Следовательно, пре-
дыстория развития играет у галофитов большую роль. Она проявляется
уже при прорастании семян. Интенсивность всхожести семян растений,
выращенных на субстрате, не содержавшем солей, оказалась наилучшей
в водной среде. Семена растений того же вида, но с засоленной почвы, име-
ли большую всхожесть, если их помещали в слабый раствор NaCl.
РАСТИТЕЛЬНОСТЬ ПЕСЧАНЫХ ПЛЯЖЕЙ
И БЕРЕГОВЫХ ДЮН
Познакомимся теперь с растительностью песчаных побережий, для
которых характерны сильный прибой и подвижность грунта. В более глу-
бокой части сублиторали, где волновые течения уже не проявляются^
Азональная растительность разных климатических зон
361
могут поселиться «морские травы». Оторванные части растений (Zostera f
Posidonia и др.), перепутанные движением воды в клубки, нередко выбра-
сываются морем на берег.
Литораль, постоянно перемываемая волнами, как и супралитораль,
совершенно лишена растительности. Пески, которые скапливаются на пля-
жах, являются продуктом разрушения морем берегов, сложенных из песка
либо из материалов, богатых песком (например, ледниковые отложения).
Песчаная фракция легко транспортируется морскими течениями, но, как
только скорость течения ослабевает, песок быстро осаждается. Поэтому
песчаные пляжи образуются прежде всего у отмел ого берега. Отсюда
высохший песок иногда переносится ветром в глубь суши.
Контур береговой линии песчаного побережья зависит от того, направ-
лены волны и ветер перпендикулярно берегу или под углом к нему.
В первом случае происходит формирование прибрежного песчаного рифа,
располагающегося на некотором расстоянии от материка. Причем благо-
даря интерференции прямых и отраженных волн образуются многочислен-
ные песчаные банки (мелководья), цепочкой протягивающиеся параллель-
но береговой линии.
Если же ветер и волны подходят к берегу под углом, происходит
транспортировка песка вдоль размываемого побережья; наблюдается сме-
щение береговой линии с образованием кос и отмелей. Песчаные зерна
переносятся волнами на берег под углом, но скатываются назад перпен-
дикулярно берегу. При многократном повторении этого процесса каждая
песчинка совершает ломаный путь, поступательно перемещаясь при этом
вдоль берега. Там, где береговая линия отступает в глубь суши, песок
отлагается на продолжении простирания береговой линии. Возникает
песчаный мыс, а затем коса, которая может нарастать со скоростью до 6 м
в год. Когда коса своим концом снова причленяется к берегу, говорят об об-
разовании пересыпи, отшнуровывающей участок моря. Такой обособлен-
ный участок, который в условиях гумидного климата со временем опрес-
няется, называют «б о д д е н о м», или «г а ф ф о м», либо же «л а -
г у н о й», «э т а н о м» или «лиманом» в аридных районах, где
вследствие интенсивного испарения и воздействия морской воды, филь-
трующейся сквозь песок, его соленость увеличивается.
Описанные процессы на всех материках протекают одинаково. Анало-
гичны также процессы формирования береговых песчаных дюн, в которых
принимает участие и растительность. На песчаном побережье высшие рас-
тения поселяются выше намывной полосы, граничащей непосредственно
с морем и отмеченной выброшенными на берег водорослями и пучками
«морской травы», то есть на участке пляжа, затапливаемом лишь в самые
сильные зимние штормы. Первыми укореняются однолетние виды. Почва
здесь из-за разложения погребенных под песком водорослей, вынесенных
на сушу штормами, хорошо удобрена х. Если к песчаному пляжу примы-
кает пояс дюн, то, как известно, даже в аридных районах под ними скап-
ливается пресная грунтовая вода (ср. стр. 198 и т. I, стр. 525). Последняя
1 Если подводные луга из «морской травы» остаются затопленными водой, разло-
жение органического вещества происходит путем анаэробного и аэробного гниения,
обусловливающего появление серобактерий. Если же органический материал лежит
на открытом воздухе, процессы распада протекают различно, в зависимости от воды
и солей в массе водорослей и «морской травы». Сначала в процессе разложения участву-
ют солевыносливые бактерии, позднее актиномицеты и грибы. Ср.: R. S u с к о w,
W. Schwartz, Die Mikrobenassoziationen der Seegrasbanke, «Z. f. Allg. Mikrobio-
logie», 1, 1960, S. 71—77.
362
Глава 5
Рис. 253. Первичная дюна на песчаном пляже острова Бор-
кум (Северное море), образованная Agropyrum junceum (фото
Бенекке).
просачивается в море непосредственно под перемываемой морем песчаной
полосой, так что вблизи берега уровень грунтовых вод располагается
на глубине всего одного или нескольких дециметров от поверхности,
вода более или менее солоноватая. Поэтому первые пионерные растения
в большинстве своем нитрофильные и вместе с тем солеустойчивые; в Цен-
тральной Европе это, например, Cakile maritima (космополит), Salsola kali,
а также пляжные экотипы Atriplex hastata, Matricariamaritima, Polygonum
aviculare и др. На побережье Средиземного моря наряду с Cakile и Polygo-
num maritimum первыми поселяются Matthiola sinuata, Sporobolus arenari-
us, Суperus capitatus, Euphorbia peplis, Inula viscosa, Diotis maritima,
Crithmum maritimum, виды Glaucium и др. Аналогичные сообщества можно
встретить и на других морских побережьях. Однако важно отметить, что
вслед за пионерными видами вскоре может появиться и многолетний злак—
Agropyrum junceum, который на плоских побережьях дает начало образо-
ванию дюн (рис. 253).
Зерна дюнного песка имеют совершенно определенную величину;
на участках, характеризующихся сильными ветрами, размер зерен колеб-
лется в пределах 1—2 мм, в местах затишья — от 1 до 0,5 мм и мельче.
Все пески состоят почти исключительно из зерен кварца с примесью
обломков раковин. Вследствие непрерывного перемещения песка морски-
ми волнами все более мягкие минералы перетираются, остаются только
твердые зерна кварца. Этим береговые дюны отличаются от дюн в пусты-
нях, в которых в зависимости от исходной выветривающейся породы пред-
ставлены различные минералы силикатных пород. Только в том случае,
если источником песка на морском побережье является какой-либо один
строго локализованный участок, состав песка может быть иным, например
Азональная растительность разных климатических зон
363
черный песок вулканических островов. При скорости ветра у поверхности
земли 4 м/сек зерна песка перекатываются либо же перемещаются «прыж-
ком». Перед непроницаемым для ветра высоким препятствием, например
на опушке густого леса, ветровой поток завихривается, энергия ветра
здесь расходуется полностью и поэтому в самом лесу воздух уже обретает
неподвижность. Песок прекращает движение и слагает дюну, растущую
вместе с лесом в высоту; если лес вырубают, процесс перевевания песка
возобновляется с новой силой (рис. 254).
Если же препятствие преодолимое, например дерновина злаков, то
сила ветра в нем и за ним ослабевает. Песок откладывается тогда в самом
Рис. 254. Образование дюны перед лесом (по Самуэльсону).
1 — песок; 2 — лес; 3 — находящаяся под воздействием ветра зона лесной опушки
и вершин деревьев (густое переплетение ветвей). Стрелка указывает направление
ветра.
кусте, а в ветровой тени образуется песчаный «хвост», вытянутый в направ-
лении ветрового потока (см. рис. 253). Таким способом развивается невы-
сокая первичная дюна Б Засыпанная песком дерновина дает боковые побе-
ги, пробивающиеся сквозь песок и задерживающие вновь поступающие
порции песка, и дюна увеличивается в размерах. Из-за роста дюн в высо-
ту песок увлажняется только дождевыми водами, поэтому почвенный рас-
твор почти не содержит солей. Лишь некоторое их количество поступает
с брызгами морской воды во время прибоя. Присутствие извести обуслов-
ливает значение pH около 7,5. В этих условиях к Agropyrum junceum
может присоединиться менее солеустойчивый Elymus arenarius (рис. 255).
На слабозасоленных почвах побережья Балтийского моря Agropyrum
обычно отсутствует, и первичные дюны, или «преддюны», здесь образует
Elymus. На песке, содержащем еще меньшее количество солей, поселяется
более чувствительный к ним злак Ammophila arenaria 1 2. Этот вид имеет
сравнительно крупные размеры, образует более густые дерновины и растет
быстрее; он выстесняет все другие виды и ускоряет развитие дюн. Ветер
1 Образованию дюн на побережьях Балтийского моря посвящены работы:
К. Н. Р a u 1, Morphologie und Vegetation der Kurischen Nehrung I u. II, «Nova Acta
Leopoldina», N.F. 13, 1944, S. 215—378; 16, 1953, S. 261—378.
2 W. В e n e c k e, Zur Biologie der Strand- und Diinenflora I. Vergleichende
Versuche uber die Salztoleranz von Ammophila arenaria Link., Elymus arenarius L. und
Agropyrum junceum L., «Deutsch. Bot. Ges.», 48, 1930, S. 127—139; W. В e n e c k e,
A. Arnold, Zur Biologie der Strand- und Diinenflora II. Der Salzgehalt der natiirli-
chen Standorte von Agropyrum junceum P.B. und Ammophila arenaria Link, auf dem
Sandstrande von Norderney, «Вег. Deutsch. Bot. Ges.», 49, 1931, S. 363—381; A. A r-
n о 1 d, Beitrage zur okologischen und chemischen Analyse des Halophytenproblems,
«Jb. wiss. Bot.», 83, 1936, S. 105—132.
364
Глава 5
Рис. 255. Распространение двух важнейших видов дюнных злаков в Европе. Дюны с Ammophila являются хорошим
примером ареала азональной растительности (по Майеру и Вильгельму).
Азональная растительность разных климатических зон
365
Рис. 256. Вторичная, или «белая», дюна на побережье Север-
ного моря. Вид со стороны суши. Ammophila arenaria покры-
вает только переднюю часть дюны. На заднем плане — пес-
чаный пляж с первичной дюной и открытое море с волнами
прибоя (фото Банекке).
сдувает с пляжа весь сухой песок, а море выбрасывает все новые и новые
порции песка. Формируются многочисленные высотой до 10л€ «белые дюны»,
сливающиеся в конце концов в один непрерывный береговой вал, подмы-
ваемый морем при штормовых нагонах. Песок белых дюн, поросших рас-
тительностью только со стороны суши, еще не закреплен (рис. 256),
а медленно перемещается внутрь страны. Тем не менее, здесь уже может
поселиться ряд других растений, не переносящих сильного засыпания
песком, — Honckenia peploides, Eryngium maritimum, Hieracium umbella-
tum, Lathyrus maritimus, Oenothera и др., обычно представленных песчаны-
ми экотипами. Вал дюн во время шторма может быть на отдельных участ-
ках прорван. Тогда песок выдувается и Ammophila погибает (см. рис. 259).
Не сдерживаемый больше растениями, песок в середине прорыва переве-
вается в глубь суши с большей скоростью, чем по краям. Таким образом
образуются дюны, имеющие в плане форму параболы. Но рано или поздно
песок снова закрепляется, чаще всего благодаря поселению Carex arenaria
(рис. 257) или Salix repens (рис. 258), которые без ущерба для себя перено-
сят как легкое засыпание песком, так и развевание его (рис. 259). Здесь
может пустить корни и облепиха (Hippophae rhamnoides), знакомая нам
по Центральной Азии. Ее кусты обычно развиваются в углублениях
поверхности, где вследствие выдувания песка почва становится несколько
влажнее.
Из-за медленного перемещения белых дюн в глубь суши или, что
наблюдается чаще, в результате нового отложения песка со стороны моря
песчаный пляж расширяется, происходит нарастание суши за счет моря х.
1 R.M.M. Crawford, D. Wishart, A multivariate analysis of the develop-
ment of dune slack vegetative in relation to coastal accreation at Tentsmuir. Fife, «J.
Ecol.», 54, 1966, p. 729—743.
366
Глава 5
Рис. 257. Низкорослая Carex arenaria более крупными
экземплярами Ammophila (фото Бенекке).
Рис. 258. Кустарник Salix repens, слева — обнажившиеся кор-
невища растения. На заднем плане — третичная дюна с лишенным
растительности (ветровая эрозия) участком, на котором высажены
дюнные злаки; в середине участка — вторично эродированное пят-
но; остров Боркум, Северное море (фото Бенекке),
Азональная растительность разных климатических зон
367
Salix repens
Рис, 259, Условия роста различных видов дюнных расте-
ний на подвижных песках (по Массару, с изменением).
Salix repens легко выносит засыпание песком и выдувание; при
развевании образует корневые отпрыски; поведение Carex arenaria
аналогично, новое корневище располагается выше или ниже перво-
начальной поверхности; Ammophila arenaria переносит очень силь-
ное навевание на нее песка, но погибает при развевании его; Hip-
pophae rhamnoides отмирает, будучи погребенной песком, при выду-
вании же дает корневые отпрыски.
Как только передний край достигнет определенной ширины, закладывается
новый дюнный вал, через некоторое время третий и т. д, Возникает широ-
кий пояс дюн. Дюнные валы, располагающиеся вдали от моря, испытыва-
ют уже ослабленное влияние ветра; кроме того, внешние валы перехва-
тывают весь песок, несомый с пляжа. Поэтому внутренние дюны становят-
ся все более неподвижными и легко заселяются растениями. Однако сла-
гающий их песок по своему составу не похож на песок белых дюн, так как
атмосферные осадки, особенно в условиях дождливого гумидного климата,
быстро выщелачивают его. Известь интенсивно вымывается; сорбция же
песчаных зерен столь слабая, что песок обедняется питательными веще-
368
Глава 5
отвами; кислотность почвы увеличивается. Ammophila начинает чахнуть,
лак как поступление свежего песка прекращается. Его место занимает
Festuca rubra, причем особая песчаная форма вида. Если в песке еще содер-
жится некоторое количество извести, появляется Koeleria glauca, вместе
с ней Helichrysum arenarium, Anthyllis vulneraria, Thymus и др., а при сла-
бокислой реакции почвы также булавоносец (Corynephorus canescens),
Hieracium pilosella, Jasione montana и др. Почва во все большей степени
покрывается мхами, такими, как Tortula (Syntrichia) ruralis, Rhacomitrium
canescens, и лишайниками (Cornicularia aculeata и др.), скрывающими под
собой белый песок. Так формируются «серые», или третичные, дюны
(рис. 258). Развитие «белой» дюны в «серую» длится на побережье Северно-
го моря приблизительно 10—20 лет. В течение этого времени значение pH
уменьшается до 6,0—5,0. Вместе с тем благодаря растениям верхние слои
песка постепенно обогащаются органическим веществом. Эта стадия харак-
теризуется появлением сначала Empetrum nigrum, а затем и вереска (Callu-
па vulgaris). Возникает кустарничковая пустошь со множеством лишайни-
ков (виды Cladonia, Cetraria), которую называют «бурой» дюной; создают-
ся благоприятные условия для проростков семян сосны (Pinus silvestris),
так что конечным членом ряда развития является сосновый лес. Искус-
ственное насаждение сосны позволяет ускорить этот процесс. В гумидной
атлантической области смена сукцессий может привести к формированию
дубового леса на кислых почвах.
Благодаря свойству песчаных дюн аккумулировать дождевые осадки
уровень грунтовых вод под ними поднимается, что часто вызывает забола-
чивание междюнных понижений. В первой междюнной котловине вода еще
обычно солоноватая, что способствует заселению ее Scirpus maritimus
и S. tabernaemontani. В следующей ложбине, где песок еще содержит
известь, образуется низинное болото или заболоченный лес. Если же раз-
витие идет в направлении стадии кустарничков, что свидетельствует о бед-
ности грунтовых вод питательными веществами, начинается формирование
омброгенного болота с видами сфагнума, переходного или верхового
болота (рис. 260). Таким образом, ненарушенный дюнный ландшафт бла-
годаря многочисленности и разнообразию слагающих его растительных
сообществ фитоценологически очень интересен г.
Приведенное здесь в качестве примера развитие дюнного ландшафта
типично для условий центральноевропейских побережий Северного и Бал-
тийского морей (ср. «Phytologie», Bd. IV, 2). Но в принципе оно одинаково
на всех широтах. Меняются только виды растений, да в аридных районах
менее заметен процесс выщелачивания песка.
В Средиземноморье на белых дюнах распространены Ammophila
arenaria ssp. arundinacea совместно c Medicago marina, Echinophora spinosa,
Eryngium maritimum, Scleropoa maritima, Convolvulus soldanella, Malcolmia
litorea и др.; на невыщелоченных серых дюнах, помимо названных видов,
встречается характерный вид Crucianella maritima (мареновые) в сочетании
с красиво цветущим Pancratium maritimum (амариллисовые), Koeleria villo-
sa, Artemisia campestris, Scabiosa maritima, Medicago litoralis, Euphorbia
paralias, Bromus villosus и экотипами растений гариги, таких, как Helichry-
sum stoechas, Teucrium polium и др. Поскольку выщелачивания не происхо-
дит, а лишь уменьшается с удалением от моря подвижность песка, расти-
1 Ср. карту в работе: Е. van der Maarel, V. Westhoff, The vegetation of
the dunes near Oostvoorne (The Netherlands), Wentia, H. 12, Amsterdam, 1964; V. W e s t-
h о f f, C. G. van Leeuwen, M. J. Adrian i, Enkele aspecten van vegetatie en
bodem der duinen van Goersee..., Jaarb. Wetensch. Genootsch., Goersee~O verfl., 1961.
Азональная растительность разных климатических зон
369
тельность отдельных зон дюнного ланд-
шафта не обнаруживает сдоль резких
различий. Поселяются здесь также дере-
вянистые виды, как Ephedra distachya,
или виды маквиса Pistacia lentiscus,
образующая полусферические кусты.
Стадия леса наблюдается редко, так как
растительный покров в целом сильно
нарушен. Вполне возможно, что утра-
тившие подвижность дюны служили
естественным местообитанием для пинии
(Pinus pined). В южной Анатолии мы
видели такой лес. Сейчас трудно уста-
новить, могла ли зональная раститель-
ность с Quercus ilex стать доминирующей
и на дюнных песках Средиземноморья,
ибо эти пески, даже если отбросить
такой положительный факт, как их сла-
бую выщелоченность, бедны питатель-
ными веществами и обладают малой
влагоемкостью. Большую роль, возмож-
но, играли бы Erica arborea и виды
Cistus.
Для песчаных пляжей восточного побе-
режья Северной Америки указываются сле-
дующие виды растений (ср.: R. Knapp, Die
Vegetation Nord- und Mittelamerikas, Stuttgart
u. Jena, 1965):
Cakile edentula, Atriplex arenaria, Eu-
phorbia polygonifolia, Arenaria peploides ssp.
robusta, Salsola kali, Xanthium canadense, Cen-
chrus tribuloides, Bradburya virginiana (сложно-
цветные), то есть сообщество, аналогичное
описанному выше для Центральной Европы.
На перевеваемых ветром дюнах растут:
Ammophila breviligulata, Artemisia caudata,
Elymus canadensis, Lathyrus maritimus, Oeno-
thera oakesiana.
Лес на песчаных дюнах образуют раз-
ные виды Pinus.
На Тихоокеанском побережье Аме-
рики дюнные злаки, по всей видимости,
не играют столь большой роли. Назы-
вают здесь лишь Elymus vancouveriensis
и Agrostis pollens. В образовании дюн
участвует Ambrosia (Franserid) chamisso-
nis (сложноцветные); такое же значение
она имеет на берегах Чили (ср.: A. Koh-
ler, Ambrosia chamissonis* Less Greene,
ein Neophy t der chilenischen Pazifik-
kiiste, «Вег. Deutsch. Bot. Ges.», 79,
1966). В Калифорнии и Чили сукцессия
ведет к формированию вечнозеленого
жестколистного кустарника.
24-0434
370
Глава 5
На Атлантическом побережье Аргентины, напротив, снова в изоби-
лии появляются типичные дюнные злаки. Это Spartina ciliata и Panicum
racemosum, при слабом навевании песка также Роа lanuginosa, Senecio
crasstfolius и Calycera crassifolia. Уже на белых дюнах растут Tamarix
и искусственно высаживаемая австралийская Acacia longifolia (с филло-
диями). При облесении дюн используют Pinus maritima — характерный
вид для песчаных местностей на юго-западе Франции.
Дюнная растительность Южной Африки описана Марлотом (см.:
R. М а г 1 о t h, Das Kapland, «Wiss. Ergeb. d. Deutsch. Tiefsee-Exp.»,.
Bd. II, Teil III, Jena, 1908).
На песчаных пляжах распространены солеустойчивые виды Sporobolus pungens,
образующего маленькие дюны, и Sebaca ambigua var. crassa (горечавковые). В некото-
ром отдалении от кромки берега образуются метровые дюны, поросшие Agropyrum
distichum и Eragrostis cyperoides. Свернутые, жесткие, острые, как иголка, листья этих-
злаков похожи на листья Eragrostis spinosa, занимающего песчаные площади на боль-
шом расстоянии от моря. На влажных участках встречается Scirpus nodosus, выбрасы-
вающий жесткие метелкообразные листья. Из однолетних видов сравнительно широко
распространены Senecio elegans с мясистыми курчавыми листьями и лиловыми цветка-
ми, а также Senecio maritimus, украшенный желтыми цветками.
Собственно дюны высотой 3—6 м, часто занимающие обширные пространства,
образуются с помощью кустарника Chymococca empetroides (волчанковые), имеющего
маленькие листья и красные ягоды. Аналогичную роль в формировании дюн играют
Myrica cordifolia или Rhus crenata и др. Естественные древостои на дюнах образует
ветроустойчивый Sideroxylon inerme (сапотовые). Число различных мелких видов расте-
ний дюн чрезвычайно велико; назовем здесь только Pelargonium capitatum.
Для засоленных лугов типичен Eragrostis glabrata, к нему присоединяются Falkia
repens (вьюнковые), Frankenia capitata, Plantago carnosa, Statice scabra. На особенно
сильно засоленных участках растут Salicornia fruticosa и Chenolea diffusa (маревые),
на несколько более высоких и менее соленосных местообитаниях встречается злак
Stenophragmum glabrum, а также Samolus valerandi, S. campanuloides Polypogon mon-
speliensis.
Дюны Юго-западной Австралии описаны в т. II, гл. 3«
Почти повсюду на морских песчаных побережьях в настоящее время
можно встретить подвижные, лишенные растительности дюны. Подобно
пустынным формам, они имеют вторичное происхождение. Иными словами,
фактором, обусловившим их возникновение, является деятельность чело-
века, и прежде всего выпас скота. Животные разрушают растительный
покров, отчего песок серых дюн снова приходит в движение. К еще более
тяжелым последствиям приводит вырубка леса на высоких дюнах, в резуль-
тате чего подвижными становятся такие огромные массы песка, что дюнные
растения оказываются уже не в силах задержать их. Такая катастрофа
произошла, например, в свое время на Курской косе, в юго-восточной
части побережья Балтийского моря, где подвижные дюны засыпали нес-
колько деревень и угрожали еще ряду селений, пока особыми, дорогостоя-
щими мерами не удалось снова закрепить песок. Этот показательный при-
мер подробно, в его историческом развитии, исследован Паулем (см. при-
мечание на стр. 363).
Экологию дюнных растений в первую очередь определяет такой важ-
нейший, способствующий жесткому естественному отбору фактор среды,
с которым мы уже познакомились,, как движение песка. Именно с ним свя-
зано засыпание растений песком или развевание песка и обнажение их
корней. Следующий, особенно существенный в гумидных областях, фак-
тор — это процесс вымывания растворимых составных частей песчаной
почвы. Сначала происходит удаление морских солей, вследствие чего гало-
фильные виды замещаются быстрее растущими, но более чувствительными
Азональная растительность разных климатических зон
371
к засолению почвы растениями. В табл. 73 приведены значения осмотиче-
ского давления и содержание хлоридов (в пересчете на NaCl) у дюнных
растений, определенные Еналем под Неаполем х.
Таблица 73
Осмотическое давление и содержание хлоридов у дюнных растений
Название растений Дата Осмотическое давление, атм Содержание NaCl
атм %
Песчаный пляж
Sporobolus arenarius 7.VI 25,0 16,0 64
Anthemis maritima 18.Ill 11,3 6,9 61
Echinohpora spinosa 7.VI 14,2 4,3 30
Частично закреплен- ные дюны
Ammophila arenaria 7.VI 13,8 5,3 38
Ammophila arenaria 23.VIII 17,9 7,3 41
Medicago marina 7.VI 14,9 3,8 25
Eryngium maritimum 7.VI 11,1 2,6 23
Маквис на песке
Cistus salviaefolius 18.Ill 15,6 4,1 26
Cistus salviaefolius 7.VI 14,4 3,9 27
Lonicera implexa 7.VI 16,9 3,6 21
Rosmarinus officinalis 18.Ill 18,2 2,7 15
Pistacia lentiscus 18.Ill 24,6 4,5 18
Phillyrea latlfolia 18.Ill 23,1 3,4 15
Pinus pinaster 18.Ill 21,9 6,9 31 1
Quercus ilex 18.Ill 26,8 2,2 8
Засоленный марш (для сравнения)
Atrlplex portulacoides 2.IV 28,9 22,7 79
A triplex portulacoides 30.IV 37,0 25,7 70
Atriplex portulacoides 24.V 41,0 25,6 63
A triplex portulacoides 17.VI 46,6 34,3 73
A triplex portulacoides 4.VII 55,2 39,8 72
Plantago coronopus 30.IV 11,2 5,4 48
Plantago coronopus 24 .V 11,4 5,4 48
Plantago coronopus 17.VI 11,8 7,6 64
Plantago coronopus 15.VII 17,2 11,4 66
Inula viscosa (у границы марша) 24. V 10,3 3,7 40
Phillyrea latifolia (вне марша) . . 27.11 21,7 2,1 10
Phillyrea latifolia 30.IV 23,7 0,9 4
Phillyrea latifolia 24.V 26,3 2,4 9
1 Согласно графику — только 20%.
Из табл. 73 следует, что в клеточном соке растений дюн также содер-
жатся хлориды, но на них приходится только 23—41 % веществ, влияющих
1 М. Ona 1, Zusammensetzung des Zellsaftes einiger Salzmarschen- und Dunen-
pflanzen in der Umgebung von Neapel, Beitr. z. Phytologie (Waiter-Festschrift), Stutt-
gart, 1964, S. 89—100; Vergleichende okologische Untersuchungen bei Halophyten und
Glykophyten in der Nahe von Neapel, «Rev. Faculted. Sci. Univ. Istanbul», Ser. B., 31,
1966, S. 209—248.
24*
372
Глава 5
на величину осмотического давления. Почва этих местообитаний сравни-
тельно бедна солями, однако при сильном шторме распыленная морская
вода смачивает листья растений. Следовательно, чтобы выжить, они долж-
ны обладать известной солеустойчивостыо.
Нередко на морских побережьях можно видеть деревья с сильно дефор-
мированными стволами, ветки которых сохранились только на подветрен-
ной стороне. Чаще всего повреждение растения не есть результат механи-
ческого действия ветра, а вызвано накоплением соли, которая осаждается
на дереве из брызг морской воды. На стороне, обращенной к ветру, губи-
тельное влияние соли, естественно, проявляется сильнее. Phillyrea lati-
folia, растущая на дюнах за пределами засоленных почв маршей, при оди-
наковом осмотическом давлении содержит на наветренных местообитаниях
15% хлоридов, а на более защищенных от ветра участках вблизи марша —
в среднем только 4—9% (см. табл. 73).
Назовем еще один важный экологический фактор. Им являются усло-
вия обеспечения дюнных растений водой. Песок дюн на определенной,
доступной для растений глубине всегда влажный, даже в областях с зим-
ними дождями и очень длительным засушливым периодом летом. Одна-
ко полевая влагоемкость бедного гумусом дюнного песка невелика, около
10%. Кроме того, чрезвычайно интенсивно протекает процесс испарения,
усугубляемый иссушающим воздействием ветра, а в безветренные ясные
дни — нагреванием поверхности песка до 60° х. Неудивительно поэтому,
что все дюнные растения обладают ксероморфными признаками; жесткие
свернутые листья (Ammophila), голубой восковой налет (Eryngium, Cram-
be, Elymus), мясистые листья (Нопскепуа, Crambe), волосяной покров
(Hippophae, Salix repens). Такого рода приспособления свидетельствуют
о том, что растения вынуждены в определенное время, обычно в полдень,
резко сокращать расход воды на транспирацию, с тем чтобы компенсиро-
вать недостаток поступления влаги в эти часы. Однако развитию сукку-
лентности или ксероморфности могут способствовать также поглощение
растением некоторого количества солей или недостаток азота в песке дюн.
Водный режим дюнных растений детально еще не исследован. Срав-
нительные измерения интенсивности транспирации в последнее время про-
изведены Шратцем (табл. 74) 2. Следует учесть, что условия постановки
опыта не всегда оставались неизменными.
Таблица 74
Средняя интенсивность транспирации (в мг!мип на 1 г сырого
веса) у центральноевропейских растений дюн и маршей
(по Шратцу)
Phragmites .... 17,3 Aster 5,0
Salix repens .... 17,0 Cakile 4,1
Erica tetralix . . . 11,0 Puccinellia .... 3,4
Hippophae 10,3 Spergularia .... 3,1
Agropyrum .... 10,5 Atriplex 3,0
Ammophila . . . . 7,2 Plantago maritima 2,9
Elymus 6,9 Honckenya 2,7
Limonium 7,4 Suaeda 2,6
A grostis 7,3 Salsola 2,1
Glaux 6,4 S alicornia 1,5
1 D. Radulescu-Ivan, Beitrage zur okologischen Charakteristik der
Sanddiinenvegetation der Schwarzmeerkuste, «Rev. Roum. Biol. Botan.», 10, Bucarest,
1965, S. 301—309.
2 См. примечание на стр. 351.
Азональная растительность разных климатических зон 373
Обобщенные данные Шратца позволяют сделать лишь вывод о том, что
транспирация в расчете на сырой вес растений с увеличением степени
суккулентности сокращается.
НЕСКОЛЬКО СЛОВ О НЕОБХОДИМОСТИ
СОЗДАНИИ ЗАПОВЕДНИКОВ
В этой книге мы рассматривали преимущественно естественную расти-
тельность, ибо лишь она находится в равновесии с окружающей ее природ-
ной средой. Культурные же и полукультурные формации, относящиеся
к азональной растительности, так же как и растительность, пребывающая
на разных стадиях деградации, возникли прежде всего при прямом или
косвенном участии человека.
Сегодня, путешествуя по странам и материкам, с горечью замечаешь,
что естественная растительность повсюду быстро исчезает с лица Земли.
Она уже полностью уничтожена во многих районах земного шара. Поэтому
сегодня очень важно отчетливо представлять себе необходимость приня-
тия немедленных мер к охране природной растительности, в противном
случае она исчезнет бесследно и там, где еще сохранилась.
Взрывной характер роста народонаселения неизбежно влечет за собой
все большее расширение ареала культурных земель. Технические средства
позволяют сегодня осуществлять проекты, проведение которых в жизнь
прежде казалось невозможным,— это и распашка или осушение обшир-
ных площадей, и сооружение плотин на крупнейших реках, и использо-
вание воды для производства энергии или для орошения пустынь. И как
результат этих преобразований — коренное изменение гидрологических
условий территории.
Применение минеральных удобрений позволяет получать урожаи
на землях, некогда представлявших собой пустоши с естественным расти-
тельным покровом. Однако в большинстве случаев было бы рациональнее
повышать урожаи на уже используемых культурных землях в сильно пере-
населенных районах, чем уничтожать последние остатки девственной рас-
тительности.
Для того чтобы осуществлять планирование производства продуктов
питания в будущем, требуется знать величину возможного оптимального
продуцирования органического вещества в отдельных климатических
зонах. Мы вправе предположить, что в природных условиях в разных кли-
матах зональную растительность образуют такие естественные сообщества,
которые при оптимальных условиях, создаваемых для них данным кли-
матом, продуцируют максимум сухого вещества. Достаточно, например,
вспомнить рассмотренные в трех томах ряды формаций, которые, с одной
стороны, идут от тропического дождевого леса гумидной климатиче-
ской области через субтропические и умеренные дождевые леса к бореаль-
ным лесам и тундре, а с другой — в пределах каждой определяемой тем-
пературой климатической зоны дают ряд, развивающийся в направлении
от зональной гумидной растительности через субгумидную и семигумидную
к редкой растительности аридного климата. Лимитирующими факторами
в гумидных условиях являются температура и солнечная радиация или
поступление СО2, а в аридных — прежде всего вода.
Главной задачей осуществляемой в настоящее время Международной
биологической программы должна быть разработка надежной основы для
оценки максимально возможного продуцирования сухого вещества, что
374
Глава 5
удастся осуществить только^путем детальных исследований экосистем
отдельных зональных растительных сообществ. Не менее важно решить
вопрос о том, будет ли достигнут абсолютный предел производства орга-
нической массы, либо же человек сможет преодолеть естественную гра-
ницу, например совершенствуя ’методы удобрения или создавая улучшен-
ные селекционные сорта.
Лимитирующим фактором часто является доступный растениям свя-
занный минеральный азот, необходимый для построения живого вещества
растений. Почти повсеместно отмечается также недостаток фосфора, запа-
сы же фосфатов на Земле ограниченны. Когда имеющиеся залежи их будут
выработаны, положение с фосфорным удобрением станет критическим. Его
вряд ли хватит в достаточном количестве для подкормки им всех бедных
фосфором почв, к которым относятся почти все почвы Австралии и боль-
шинство почв Африки. Иначе дело обстоит с азотом. Запасы элементарного
азота, бесполезного для организмов, за исключением немногих азотофик-
саторов, практически неисчерпаемы. Если энергии достаточно, можно
выработать неограниченное количество водорода, чтобы, соединив его
с азотом, получить аммиак, а из последнего нитрат. Следовательно, азот-
ное питание растений можно было бы довестиТдоТоптимального и таким
путем значительно увеличить ассимилирующую листовую поверхность.
Однако у зональной растительности такая оптимальная листовая поверх-
ность, по-видимому, уже сформировалась. Ведь известно, что беспредель-
ное увеличение индекса листовой поверхности неосуществимо. Как только
листовая поверхность достигнет таких размеров, что освещенность ниж-
них листьев будет соответствовать световой компенсационной точке, азот-
ный баланс этих листьев окажется отрицательным, так как потери на ды-
хание ночью уже не компенсируются ассимиляцией в дневное время.
Но Международная биологическая программа при решении вопросов,
связанных с первичной продукцией, сталкивается с большой трудностью,
которая выражается в том, что во многих климатических областях зональ-
ная растительность полностью уничтожена. Это произошло, например,
на большей части Европы, почти во всех густонаселенных районах с зим-
ними дождями; во всех злаковниках, развитых на почвах, оказавшихся
особенно пригодными для земледелия, а также в тех обширных тропиче-
ских областях, где первичные леса вследствие пожаров и выпаса скота
сегодня повсеместно преобразованы в саванну, и т. д. На всех этих терри-
ториях в качестве объектов изучения приходится довольствоваться в луч-
шем случае естественными вторичными сообществами, которые, по всей
видимости, более или менее соответствуют первоначальной зональной рас-
тительности — но именно только более или менее.
Отсюда видно, сколь важно создать во всех климатических областях,
где это еще возможно, обширные заповедники зональной растительности.
Существенного сокращения полезной площади это не вызовет, а для науч-
ных исследований и практического решения первоочередного вопроса
о повышении продуктивности ландшафта такие заповедные территории
имеют неоценимое значение.
Движение в защиту природы в последние десятилетия повсюду приоб-
рело широкий размах, но преимущественно в районах, где уже не осталось
нетронутых земель. Жизнь снова и снова убеждает в том, что человек
начинает ценить значение природы для физического здоровья и поддержа-
ния душевного равновесия лишь тогда, когда он прежде разрушил ее.
Но и поняв это, охрану природы часто путают с реконструкцией ланд-
шафта, считая, что природу нужно переделывать в определенном направ-
Азональная растительность разнщх климатических зон
375
лении. Нередко национальные парки создают в местностях с красивым
ландшафтом с единственной целью — доставить людям эстетическое удо-
вольствие. Сколь ни похвальны такие начинания, это еще не охрана при-
роды в подлинном смысле этого слова. Естественный ландшафт нередко
однообразен и поэтому не привлекателен для горожанина. Природу мож-
но сохранить неизмененной лишь в том случае, если запретить массовые
посещения заповедников.
Такие заповедники, находящиеся на разной ступени заповедания,
необходимо создавать повсюду там, где еще сохранились остатки естествен-
ной растительности, даже если местным властям необходимость в них еще
не кажется первоочередной. И это не должны быть только районы, из-за
плохих почв или сложного рельефа непригодные’для сельского хозяйства
или лесонасаждений; непременно следует охранять также местности с оп-
тимальными условиями для роста растений. Организация таких предста-
вительных резерватов зональной растительности во всех частях земного
шара также является одним из важных пунктов Международной^ биоло-
гической программы. Будущие поколения также должны располагать
возможностью проводить биологические исследования в естественных
условиях.
Если же эти требования не удастся узаконить с помощью соответствую-
щих правительственных решений, разрушение последних остатков есте-
ственной растительности ускорится и ближайшие поколения будут зна-
комиться с зональной растительностью только теоретически, как^это про-
исходит сегодня в густонаселенной Центральной Европе. Здесь различают
естественную растительность, которая, вероятно, сформировалась бы после
прекращения какого-либо вмешательства человека, и первичную, суще-
ствовавшую до этого вмешательства, о которой имеются лишь приблизи-
тельные представления.
Мы стремились в этих трех книгах обобщить главное в наших знаниях
относительно эколого-физиологической характеристики зональной рас-
тительности основных климатических зон. Мы опирались преимуществен-
но на собственный опыт, почерпнутый из наблюдений на разных^матери-
ках; собран обширный материал, но, чтобы не выйти за рамки объема,
пришлось, к сожалению, отказаться от описания многих интересных под-
робностей.
УКАЗАТЕЛЬ РУССКИХ ТЕРМИНОВ
Аапа II, 402, 403, 405, 408—411
Австралия I, 437—466
Центральная
влажные местообитания I, 463,
466
засушливые области
климат I, 438—442
растительность I, 443—450
экологические исследования
I, 450—466
Южная
климат II, 188
растительность II, 195—206
эколого-физиологические ис-
следования II, 188—195
Автохорные растения III, 120
Акклиматизация древесных пород в тро-
пиках I, 123
Аланг-аланг I, 163
Алашань, пустыня III, 190, 246—249
Аллелопатические взаимовлияния рас-
тений I, 24
Альпийская растительность
Австралии II, 217
Альп III, 45—51
Анд I, 207
Арктики III, 31
Новой Зеландии II, 220—222
Субантарктических островов III,
62
Тасмании II, 218—220
Явы I, 205—207
Альпийские матты II, 218—221; III, 287
Альпийский пояс I, 222, 223, 225, 226,
229; II, 80, 134, 210, 216, 217, 223;
III, 9, 34, 36, 42, 43, 44, 47, 49
Анатолия II, 97
Анды I, 207—219
климат 1, 209
растительность I, 209—215, 422
экологические исследования I,
209__________211
Анемохоры I, 485; II, 277; III, 120, 214
«Антарктическая тундра» II, 23
Антарктические подушечники II, 218,
220
Антропогенный фактор I, 17; II, 25,
145, 336, 337; III, 183, 186, 222
Аргентина III, 146, 153, 155, 156, 160
Арег, пустыня I, 474
Аридные области, I, 302
круговорот воды I, 314, 506, 507,
513
субтропиков
климат I, 305
экологические группы расте-
ний I. 320—331
Центральной Азии
климат III, 190, 191
общий обзор III, 187
растительность III, 190—193
Аризона
вертикальная поясность II, 129
экологические исследования II,
121—123
энсинальная растительность II,
116—120
Арктическая тундровая зона
климат III, 5—9
почвы III, 9, 10
продуктивность III, 27, 28
растительность III, 19—22
эколого-физиологические иссле-
дования III, 23—26
«Аррайан» II, 150
Аскания-Нова III, 92
Асимметрия зон растительности I, 72, 73
Ассоциации
засоленных маршей III, 345
полынно-солянковые III, 255
полынные III, 255
Атакама, пустыня I, 420, 421
Бата II, 42
«Беллото» II, 136
«Бесснежный» период III, 7, 8, 36, 45 —
47, 277
Биогеоценоз II, 259, 260, 276, 280, 281
Биогеоценозы хвойных лесов II, 358
Биологический спектр арктической фло-
ры Гренландии III, 18
Биомасса
животных II, 276
лесов II, 269
почвы II, 278
Биотип I, 123
Биотоп I, 316—318, 325
Бодден III, 361
Болота
распространение II, 403—405
растительность II, 405, 407
378
Указатель русских терминов
типы II, 400—402, 408, 409
экология II, 412—422
Болота, типы
бугристое II, 403, 409—411; III,
281
верещатниковое II, 252
верховое II, 406
влажных тропиков I, 156
грядово-мочажинное II, 408
злаковое I, 162
мезотрофное II, 402,'408
низинное III, 311
олиготрофное I, 98; II, 401, 405
омброгенное Ш, 368
островное II, 402, 403
подушечниковое II, 151
покровное II, 245, 248, 252, 253,
403
солигенное II, 401—403, 405
топогенное II, 400; III, 132
тростниковое III, 313
эвтрофное I, 98; II, 401
«Больдо» II, 136
Бореальная зона II, 23, 25, 233, 320,
329, 334, 347, 348, 356, 358, 359,
364, 382, 399, 400, 405; III, 5, 32
Бореальная зона Европы
хвойные леса
биогеоценозы II, 358—373
климат II, 341, 344
типы II, 349, 352—355
Бореально-неморальная зона
смешанные леса
климат II, 334, 336
типы II, 336, 337
Брутто-ассимиляция II, 296
Брутто-прирост II, 271
Брутто-продуктивность II, 272, 287,
363
Брутто-фотосинтез II, 283
«Буферность» растительности I, 311 •
Бэйшань, пустыня III, 190, 249, 250
Вади I, 475, 476, 477, 500, 502, 532
Ватты III, 342, 343, 347
Вельд II, 170
Верещатники II, 96, 242, 246—259
Вертикальная поясность растительно-
сти II, 49, 123, 128
Австралии II, 206, 216
Альп II, 378; III, 34
: Анд II, 149
Аризоны II, 129
Гималаев III, 284
Кипра II, 81
Монголии III, 258
Новой Зеландии II, 206, 221
Северной Анатолии II, 241
Средиземноморья II, 77—79, 86
Тасмании II, 200
Тенерифе II, 90—93
Тропиков I, 193, 194
Чили II, 140, 152, 154
Вечнозеленые субтропические дождевые
леса
общие условия I, 195
растительность I, 196—198
Виды
викарирующие I, 252, 333; II, 20,
109; III, 214, 246
гигроморфные I, 131, 155, 312,
420, 484
гидростабильные III, 273
двулетние I, 39, 40
мезофильные III, 194
многолетние I, 40, 41, 352; III,
217, 232, 270, 302, 303
непериодические Ш, 20
однолетние I, 39, 40, 501, 506,
509, 510, 530, 537; II, 114,
115, 134, 138; III, 110, 218,
244, 253, 257, 303, 361
пирофильные III, 186
стеногидрические I, 327; II, 9, 10
теневыносливые II, 296, 301
эвригалинные III, 328
эвригидрические I, 327, 373
энсинальные II, 121
Внетропическая растительная зона II,
22, 23
Водный баланс II, 71
верхового болота II, 412
елового леса II, 248
Водный режим растений I, 45
Альп III, 39—42
аридных областей I, 306—313
пустынь III, 227—233
Средиземноморья II, 49, 50, 52, 54
степей III, 230
Возраст арктических растений III, 22
Вудленд II, 185
Выпас, влияние III, 92
Высокогорные формы растений II, 19
Галоксероморфность III, 358
Галосуккулентность III, 358
Галофиты I, 165, 171, 185, 331, 341,
389, 391, 395, 400, 404, 405, 422,425,
433, 447, 452, 455, 457, 479, 4’83, 486,
488, 491, 500, 515, 516, 522, 532,
537, 538, 546; II, 130, 187; III, 18,
26, 115, 134, 135, 141, 191, 203, 207,
210, 211, 219, 227, 231, 233, 235, 236,
239, 241, 242, 281, 294, 301, 342, 351,
357, 370
несуккулентные III, 356
облигатные III, 329
суккулентные I, 482; II, 207;
III, 277, 351, 356
сульфатные III, 357
факультативные III, 328, 329
хлоридные III, 357
Гамада I, 472, 473, 498, 526, 527, 529
Гарига II, 32, 42, 83, 110
Гаруа I, 411, 415, 417
Гафф III, 361
Гелиофиты III, 24
Гелофиты III, 311, 322, 323
Гемикриптофиты I, 537; II, 44, 187;
III, 18, 20, 109
Указатель русских терминов
379
Гемиэпифиты I, 133, 137
Генотип II, 10
Геофиты I, 271, 312, 325, 326, 419,
424, 474, 483, 541; II, 9, 44, 47, 62,
63, 93, 101, 102, 134, 162, 165, 303—
307, 313; III, 18, 35, 87, 90, 112, 125,
163, 204, 205, 216
Гетеротрофные организмы II, 280; III,
209
Гигрогалофиты I, 331, 463, 484; II, 131
Гигрофиты III, 301, 354
Гидатоды III, 318
Гидратура плазмы I, 48—50, 53, 111,
115, 131, 232, 234, 311, 318, 320—
324, 327, 349, 373, 417, 435, 483, 486,
488, 537, 538; II, 10, 50, 60, 65, 146,
291
Гидроноты III, 318
Гидростабильность I, 327; II, 9, 52, 58,
60
Гидрофиты III, 18, 231, 309, 313, 323,
Гималаи III, 284—293
Гиперпаразиты I, 155
Гоби, пустыня III, 188, 255—257
Голарктическое флористическое цар-
ство II, 133
Гоф, остров III, 62
Граница древесной растительности I,
75, 203, 216, 219; II, 19, 130, 148,
152, 216, 220; III, 34—36, 289
Австралии II, 217
Средиземноморья II, 78
Тасмании II, 211
Граница засушливости II, 93, 110; III,
160, 280
Граница леса I, 219; II, 93, 154, 389,
394; III, 10, 33, 34, 36, 43, 67, 290,
291
альпийская II, 378, 387, 388,
390, 398; III, 12
арктическая III, 25, 34
климатическая II, 388, 394; III,
74
полярная III, 5, 7, 11
потенциальная II, 388
фактическая II, 388
Гумидная область I, 287, 299, 302, 304;
II, 11, 13
Гумидная умеренная зона II, 77
Дамбо I, 270, 271
Деревья
бутылочные I, 188, 257, 258, 260
вечнозеленые I, 40, 42, 232; II,
116, 119, 149, 240
гвоздичные I, 480
летнезеленые II, 152
листопадные I, 123, 127
путешественников I, 130
«Тала» III, 159
удушите л и II, 144
хлебные I, 480
Джангиль III, 224, 225
«Джаррах» II, 183—185
Динамика растительного покрова
в Египте I, 522, 523
Долина туманов II, 153
Доминанты I, 13, 18
Дюны III, 221, 222, 223, 228, 360—373
Европейская зона лиственных лесов II,
233, 234
Египетско-аравийская пустыня
климадиаграмма I, 508
растительность I, 498—502
условия среды I, 495
экологические исследования I,
502—515
Жестколистная растительность II, 94,
170
Австралии II, 176—187
Калифорнии II, 106—116, 123
Капской провинции II, 156—168
Средиземноморья II, 25, 27, 34—
38, 50, 56, 58
Среднего Чили II, 136—139
Жизненные формы I, 39—43, 106, 287;
II, 13; III, 269
Закалка растений II, 320, 322—324
Закон относительного постоянства ме-
стообитаний и смены биотопов I,
316; II, 120; III, 87, 195
Закономерности вертикальной поясно-
сти в тропиках I, 193, 194
Засоленные марши III, 342—360, 371
372
ассоциации III, 345
поясность растительности .III,
345, 349
эколого-физиологические. иссле-
дования растений III, 351—
360
Зимний покой хвойных бореальной зо-
ны II, 382
Злаки
дерновинные III, 94, 181, 299
дюнные III, 364, 366, 369, 370,
372
колючие I, 458
корневищные III, 299
кустовые III, 299
туссоковые I, 214; III, 165, 182
Злаковники I, 158, 163, 242, 243, 245,
246, 266, 267, 269, 270, 276, 277,
285, 427, 436, 442, 446, 500; II, 110,
130, 245; III, 66, 69, 77, 94, 96, 184
альпийские II, 217, 220, 221,
389; III, 183
Новой Зеландии III, 180—183
Средиземноморья II, 99
тропические I, 270—275, 277, 285
туссоковые III, 60, 62, 63, 147,
180—183
умеренной зоны III, 145—186
380
Указатель русских терминов
Зона
вечнозеленых дубов II, 47
вечнозеленых умеренных дожде-
вых лесов южного Чили II,
132, 149
влажных муссонных и лавровых
лесов 1, 73
жестколистной растительности
II, 35, 71, 140, 148, 155
зимних дождей II, 23, 73
лесостепная III, 64, 66, 67, 69
193, 194
летнезеленых лесов II, 145
лиственных лесов II, 71, 94, 95,
320
полупустынь
общий обзор III, 194
растительность III, 195
сукцессии III, 201
травяных степей I, 73
хвойных лесов II, 123
Зональность растительности
Восточной Азии II, 232
субтропиков I, 296—301
тропиков I, 193, 194
циркумполярная II, 22
Зоомасса леса II, 275
Идеальный континент I, 73, 74; II, 23
Индекс листовой поверхности I, 307;
II, 188, 271—273, 284, 296, 297;
III, 317, 374
Индо-тихоокеанская область I, 182
Ирландия II, 245
Каатинга I, 252, 258, 259
Казахстан III, 193—202
Кайре III, 225
Кактусы I, 210, 336, 337, 339, 340,
352—364, 420; II, 141; III, 178
Калифорния II, 106—116
Кальдена ль III, 162
Канарские острова II, 89, 92
Капская провинция II, 156—175
климадиаграммы II, 157
почвы II, 158
растительность II, 160—168
экологотфизиологические иссле-
дования II, 173—175
Каракумы III, 210, 214—220
Карру, плато II, 166—169, 171
Карру, пустыня I, 423—435
климадиаграмма 1, 424
растительность I, 424—429
экология растений I, 429—430
Карты распространения
лесов Центральной Европы II,
237
пуны Центральных Анд I, 217
чапарраля Калифорнии II, 117
энсина ля Аризоны II, 117
Карты растительности
Австралии I, 446
Аргентины III, 146
Восточной Африки I, 252
Палестины I, 543
Сев. Америки II, 229
Тибета III, 281
Тенерифе II, 91
Каулифлория I, 115, 118, 119, 239?
Кейп, п-ов II, 157
«Кеньоа» II, 134
Кергелен, о-в III, 57—60
«Керри» II, 180—183 х
«Киллай» II, 136
Кладодии I, 360, 361
Климадиаграммы I, 60—70
Австралии I, 439; II, 172, 216
Азии II, 228
Америки II, 228, 334
Анд I, 209
Анкары II, 99
Антарктики III, 59
Арктики III, 6
Африки I, 172; II, 157
Европы II, 234, 235
Калифорнии II, 106, 107
Новой Зеландии II, 215
пустынных областей I, 305, 334,
378, 412, 470, 497, 508
Сибири II, 344
Средиземноморья II, 27, 30, 78
Тасмании II, 211
тропических лесов I, 235, 260
Чили II, 132
Климакс I, 19, 22, 23, 275
Климат
аридный III, 136, 143, 161, 162,
244, 259
арктический II, 23
горный III, 192
гумидный II, 77; III, 122, 132,
135, 136, 143, 148, 149, 152,
159, 361
континентальный III, 34, 35, 278,
292
морской II, 247
океанический II, 35, 244, 245,
323; III, 34
пустынный III, 224
семиаридный III, 152, 162, 165,
180, 181
средиземноморский II, 96, 106,
226; III, 188
субсредиземноморский II, 106
субтропический III, 101
Климатические зоны земного шара I,
53—60
карта годовых осадков I, 59
Климатограммы I, 64—67
Клинотипы II, 326
«Конуэ» II, 150
Коллины III, 77
Колхида II, 240, 241
Колючеподушечники I, 300; II, 80, 81,
85
Компенсационная точка II, 312, 313
Конкурентная мощность I, 36, 286;
II, 56, 111, 127, 154, 301, 315
Конкурентная способность вида I, 14,
30, 34, 35, 39, 41, 43, 165, 233, 234;
Указатель русских терминов
381
II, 95, 96, 293, 301, 323, 325, 334,
343, 344; III, 7, 89, 174, 182
Конкурентные взаимоотношения I, 15
23, 25, 26, 37—39, 104, 285—288,
509; II, 20, 71—76, 154, 296, 297,
315, 316, 352, 365—370
Консументы I, 43, 44
Континуум I, 12, 13
Копрофаги II, 276
Корневые выделения I, 24
Корневая система растений
еловых лесов II, 360
Памира III, 269—270
прерий III, 111, 112
пустынь I, 354, 431, 483—485,
503
саванн I, 295
степей III, 75, 113—117
Корни
воздушные I, 162
досковидные I, 99, 103, 106, 107,
162, 239
дыхательные 1,176,178—180; III,
311
коленчато-изогнутые I, 177
метловидные I, 162
ходульные I, 107, 108, 162, 168
179, 181
эфемерные III, 112, 113
Криптофиты I, 537
Критическая фаза развития растений I
373
Круговорот веществ в лесной экосисте-
ме II, 284, 287
Ксерогалофиты I, 331, 433, 459, 531;
II, 131; III, 170
Ксерофиты I, 325—327, 329, 331, 373,
389, 483, 537, 540; II, 9, 10; III,
123, 323, 351, 353
Ксилоподии I, 254
Культурный ландшафт II, 25, 136, 148
Кустарники II, 56; III, 211, 217, 223
267, 287, 290, 292
Кустарнички I, 424, 434—435, 537;
III, 39, 217, 253, 269
Кызылкумы III, 210
Лагг II, 408
Лагуна III, 361
Ледниковые реликты II, 421
«Ледяные окна» III, 21, 22
«Ленга» II, 152
Леса
болотные I, 162
буковые II, 96, 235, 236
вальдивийские II, 149, 150
вечнозеленые дубовые II, 34
вечнозеленые эвкалиптовые II,
188
галерейные I, 401; II, 134; III, 73,
302
горные I, 79, 193; III, 291
девственные I, 220; II, 149, 338—
340, 345, 357
диптерокарповые I, 101, 244
дождевые I, 193, 195—203; II,
148—151, 212
дубовые II, 27, 32—38, 125, 224,
231, 261—269; III, 287
жестколистные II, 110, 112, 126,
136, 138; III, 287
западноевропейские II, 96
индо-гималайские III, 192
кактусовые I, 340
колхидские II, 240
ксерофитные II, 136
лавровые II, 90, 92
листопадные I, 411; II, 133
мескитовые I, 342
муссонные I, 243; III, 291
нотофагусовые III, 175, 182
остепненные III, 64
первичные I, 118, 189—190, 203
пойменные III, 224, 225, 230, 235,
246, 253, 258, 302—304
пихтово-буковые Центральной
Европы II, 337
саванновые I, 246
средиземноморские II, 35
«туманов» II, 140, 144
хвойные II, 123, 343; III, 287
Центральной Европы II, 237
Лес как экосистема II, 259
Лесостепная зона III, 64, 66—67, 69,
193, 194
Лесотундра III, 10—12, 27, 32
условия произрастания древес-
ных пород III, 11, 12
Лианы I, 100, 133—137, 140, 242, 243,
245, 250, 251, 260; II, 135, 172, 193,
196, 197, 204, 221; III, 225
Листопад II, 324
Листья
световые I, 33, 323; II, 10, 283,
291—293, 295—297, 380
скобки I, 142, 145
теневые I, 33, 323; II, 10, 146,
291—293, 296—298, 313, 380
Литораль III, 334, 339—342, 361
«Литре» II, 136
Литофиты I, 544
Лишайники I, 381, 391, 418, 470, 482;
II, 134, 140, 145, 150, 410; III, 11,
12, 18, 21, 22, 31, 32, 34, 49, 56, 57,
60, 62, 141, 292
Лом, растительность I, 418—422
Луга
альпийские III, 269, 290, 292
альпийские разнотравные III, 287
галотрофные III, 342
маршевые III, 344
морской травы III, 342, 343
пойменные III, 300, 301, 304, 306
субальпийские III, 259
туссоковые I, 214, 217, 222, 223;
II, 221, 222
Луканга I, 156
Маквис II, 26, 27, 31—33, 37—39, 41
46-48, 55, 60, 71, 86, 107; III
369—370
382
Указатель русских терминов
Макрофиты III, 330
«Мамора» II, 37
Мангровая растительность
климат I, 167, 172
поясность I, 165—167, 171, 176,
177
распространение I, 163, 164
экология I, 166—180
Матты III, 51
«Мбуга» I, 270
Мезо-гигрофиты II, 354
Мезофиты II, 12, 354
«Мескит» III, 101
Микориза II, 112, 364, 365, 413; III, 22
Микромозаичность растительности
пампы III, 166
степи III, 93
Микрофиты III, 331
Микроэндемы I, 470
«Миомбо» I, 249
Мирмекохоры II, 277
Мозаичность растительности I, 187, 231,
260; II, 212, 238, 402; III, 352
Альп III, 32
Арктики III, 7, 15
лесов III, 65
степей III, 65
«Мопане» I, 247
Морозное высыхание II, 320, 325—328,
389, 391, 395, 399; III, 39
Морозоустойчивость растений II, 67—
- 69, 320—323, 326, 384, 385, 389, 390;
III, 39, 40, 276
Моховые белосаксаульники III, 222
Мульга I, 300, 432, 458—460, 462
Мхи II, 150, 207; III, 11, 12, 18, 22, 32,
46, 56, 57, 60, 62, 141, 291, 325
Нака III, 281
Намиб, пустыня I, 374—410
карта I, 375
климадиаграмма I, 378
режим осадков I, 374—381
экология растений I, 382—410
Нангапарбата III, 289
Нанофанерофиты I, 42
Негалофиты II, 206; III, 351, 359,
366
Неджеб, пустыня I, 529—539
карта растительности I, 529
типы растений 1, 537
экологические исследования I,
529—536
Некрофаги II, 276
Неморальная зона II, 30, 227, 334
климадиаграммы II, 228, 234, 235
продуктивность лиственных лесов
II, 269—290
характеристика лиственных лесов
II, 227—242
эколого-физиологические иссле-
дования II, 291—319
Нетто-ассимиляция II, 253, 281, 296,
306, 309, 380; III, 24
Нетто-диссимиляция III, 24
Нетто-продуктивность I, 44, 245; II, 73,
74, 286, 287, 296, 363; III, 24, 317
Нитрофилы I, 405; III, 30
Новая Зеландия III, 180—183
«Ньире» II, 152
Оазисы I, 419, 420, 468, 469. 479—481;
III, 145, 223, 253, 258
Область степей и полупустынь Казах-
стана III, 193—202
«Облачное покрывало» гор II, 159
Облесение пустынь I, 316
Огненная Земля III, 147, 177
Огонь как экологический фактор I, 274,
289; III, 183
Озера
галотрофные III, 308
дистрофные III, 308
олиготрофные III, 308, 311, 313,.
314, 324
эвтрофные III, 308, 309, 311, 313
Оконные водоросли I, 381
Оконные растения I, 381, 393
Омброфиты III, 231
Ордос, плато III, 243—246
Осмотическое давление растений
Австралии I, 461, 462; II, 190, 191
Альп III, 301
аридных областей I, 322—323
болот III, 321
дюн III, 371
засоленных маршей III, 353
Памира III, 272, 273
пустынь I, 487, 522, 531, 535
тропического дождевого леса 1г
110—112, 132, 137, 138, 150,.
151
Осмотическое давление
галофитов I, 433, 434
лиан I, 137
мангров I, 172, 175
опунций I, 362, 363
склерофилов I, 436; II, 51, 52,.
174, 175
суккулентов I, 394, 432
эфемеров I, 350
Основные показатели дубовых лесов
лесостепи II, 285
«Пальмиет», формация II, 165
Памир III, 262—278
климат III, 263—265
растительность III, 267
Пампа III, 147, 160, 169
климадиаграмма III, 148, 156
общая характеристика III, 149,.
152
почвы III, 160
проблема III, 147—149
растительность III, 162—167
Памперо III, 147
Папоротники III, 291
Парамос I, 75, 207, 213—217
Парковый ландшафт I, 266—268, 278
Указателъ русских терминов
385
Патагония III, 174—178
климат III, 175, 176
почвы III, 177
растительность III, 174—180
«Пеймо» II, 136
Пейноморфоз II, 12, 193, 224, 225, 422
«Перекати-поле» III, 87
Периодичность развития II, 19, 44
Песчаные области Прикаспия
грунтовые воды III, 198
общий обзор III, 196
растительность III, 197
Пино II, 154
Пирофиты I, 289, 292
Плавающие газоны I, 157
Плавающие острова I, 160
Плавни III, 311
Плейотропия II, 19
Плеустон III, 309
Пневматофоры I, 176, 178—180
Подушечники, формация III, 269
Поды III, 137—139
Пожары I, 200, 246, 254, 269, 274, 277,
289, 297; II, 26, 47, 49, 114, 115,
126, 128, 161, 196, 198, 206, 208,
334, 337, 343, 355—358
Показатель облиственности (ПО) I, 32—
34, 44, 245
Показатель успешного роста (УР) I,
34, 35
Полукустарники II, 112, 115
Полупустыня II, 128; III, 290
Казахстана III, 193
кустарниковая I, 346, 347; II, 134
с Larrea III, 168
Патагонская III, 179—180
полынная III, 230, 244, 245, 289,
290
солянковая I, 443, 445, 450
Популяция II, 13, 19
Почвенные зоны земного шара I, 70, 71
Пояс
альпийский III, 34, 47, 49, 284,
290, 292, 322
бамбуковых зарослей I, 225
нивальный III, 224, 289
подушковидных растений I, 209,
210;II, 222;III, 280
пуны I, 75; 209—211
субальпийский III, 284
тумана II, 219
хвойных горных лесов II, 374—
379
энсинальный II, 119, 121, 128
Прерия
злаковая III, 96, 103—106
североамериканская III, 64, 69,
77, 94—96, 121
смешанная III, 103
Пресноводная растительность III, 307—
327
Прикаспийская низменность III, 196—
202
Приспособление растений к факторам
среды II, 223—226
Продолжительность
дня в тропиках I, 84
сумерек в тропиках I, 84
Продуктивность I, 16, 31—36, 39—40,
234, 245; II, 396—398; III, 9, 18,
20, 23, 24, 129—131
альпийского пояса III, 42, 43
Арктики III, 27, 28
древесного яруса II, 281, 282,
298, 364
злаковников I, 308
ковыльных степей III, 269
лесных трав I, 311
лесов I, 118, 245; II, 73—75, 283,
284, 286, 361
лиственных деревьев II, 73, 75—
76
лиственных трав I, 311
луговых злаков I, 28—29
полупустынь III, 269
различных жизненных форм I, 40'
растительного покрова II, 269
степных сообществ III, 129—132.
сфагновых мхов II, 415
Продуценты I, 43, 44; II, 275
Профильные диаграммы леса
вечнозеленого I, 103
дождевого I, 101, 102
смешанного диптерокарпового I,
101
смешанного умеренного дождево-
го II, 208
умеренного дождевого леса I,
201; II, 208
Псаммофиты II, 354; III, 168, 211, 214,
223, 243, 246
Пузыревидные волоски I, 452, 491, 492
Пузырчатый эпидермис I, 384
«Пуна» I, 75, 209—212, 214, 217, 421;
II, 134; III, 262
Пустынные области Центральной Азии
III, 187, 242, 261, 262
Пустыни
аллохтонные I, 473
альпийские III, 267
бугристые III, 217, 223
галофитные III, 207—210
гипсовые III, 206, 207
горные холодные II, 91
кактусовые I, 353
каменистые I, 345, 472, 473, 498,
527, 529; II, 27, 86
каменистые гипсофитные III,
203
кустарниковые II, 128, 132; III,
250
песчаные I, 474—475; III, 193,
210, 211, 214, 221, 227, 229,
242, 250
полынные III, 287
псаммофитовые III, 203, 210
солончаковые III, 192
солянковые I, 432
субальпийские III, 287
щебнистые I, 473; III, 257
эфемеровые I, 418; III, 204—206
384
Указатель русских терминов
Радиационный режим I, 83—85; II,
263, 302, 303, 391, 392; III, 9
Распространение болот в Финляндии
II, 411
Растения
альпийские III, 274, 277
анемогамные III, 20
ацидофильные II, 39; III, 132
базифильные I, 99; III, 132
вечнозеленые II, 93, 110, 114,
119, 145, 146, 383; III, 20
гигроморфные I, 131, 155, 312,
420, 484
гидрофильные I, 479
гомойогидрические I, 48, 323,
324, 326
длинного дня I, 85, 124
дюн III, 221—228, 360—371
жестколистные II, 52, 56, 58, 59,
64, 93, 108, 112, 137, 138;
III, 112
кальцефильные III, 18
кальцефобные II, 258
ксероморфные I, 131, 155, 312;
II, 11—13; III, 258
листопадные I, 40, 41, 232—233,
254, 371; II, 93, 110, 119, 120,
130
малакофильные I, 231, 326, 327,
371, 435; II, 54, 61, 224, 333
мезоморфные I, 420
мезофильные III, 123, 194, 205
нитрофильные I, 405
пионеры III, 201, 223, 225, 228,
350, 362
подушковидные I, 74, 174, 178,
209, 210, 222, 422, 446, 470,
482, 485; II, 88, 134, 210—212,
220—222; III, 36, 182, 269, 277
пойкилогидрические I, 47, 53,
147, 246, 260, 323, 324, 351,
352, 357, 395, 425, 428, 434,
441,.470, 482; II, 64, 326; III,
90 91 91е»
пустынные III, 272, 273, 275
стеногидрические I, 327
степные III, 272
удушители I, 137, 138, 198; II,
213
энсинальные II, 121
энтомогамные III, 20, 29
Растительное сообщество I, 11—19, 27
климаксовое I, 225
пионерное I, 20; II, 260
Р астительность
азональная I, 22, 285; II, 237;
III, 294
аридная III, 167—174, 220
галофитная III, 220, 224, 253
зональная I, 19, 22, 274, 277,
285, 292—293; II, 31—37, 128,
235, 238, 296, 300, 341; III,
200, 230, 294, 369, 373—375
кальциефитная III, 245
лома I, 418—422; II, 140
луговая III, 301
макрофитная III, 330
мазофитная III, 253'
первичная I, 188; III, 62
песчаных пляжей III, 360
пойменная III, 299
псаммофитная III, 253
речных наносов III, 294—307
склерофильная II, 27, 107
степная III, 68, 70
субсредиземноморская II, 96
сухих местообитаний тропиков I,
187—189
туманов II, 134
экстразональная I, 22, 242, 283,
285; II, 237
экстремная II, 139
энсинальная II, 116
эфемерная III, 223
Растительные зоны земли, карта II, 24
«Раули» II, 145, 146
Per I, 473
Реликты I, 466, 493, 494, 528; II, 77,
89, 125, 145, 154, 155, 242; III, 244
третичные II, 76, 88, 150, 240
Реностер II, 166, 167
Рестинги I, 187
Рефугиум II, 327, 374, 375; III, 69
«Робле» II, 145, 146
Роса I, 89, 518—520
Ротвальд II, 338—340
Рюллен II, 408
Саванна
понятие I, 276—278
растительность I, 279, 293—301
экологические исследования I,
27g_____283
Саванна I, 190, 256, 264, 269, 275—285,
288, 296—301, 345, 442, 467, 493;
II, 116, 170; III, 69, 70, 74
древесная I, 280
дубовая III, 101
зональная I, 277, 279
колючекустарниковая I, 277
кустарниковая I, 283; III, 169
мескитовая III, 101
пальмовая I, 157, 278
производная I, 190, 276—278
синдская III, 192
сухая I, 276
термитная I, 269
эвкалиптовая I, 263, 264, 442
Саванноподобные древостои I, 200
Саксаульники, формация III, 235
Санта-Каталина, горы II, 116—121
Сапрофаги II, 276—278
Саргассово море III, 342
Сахара, пустыня I, 467—494
биология растений I, 482—485
климадиаграмма I, 470
почва и растительность I, 471 —
479, 493, 494
экологические исследования I,
485—492
Световое довольствие II, 298, 302, 307—
313; III, 313
Указатель русских терминов
385
Световой минимум II, 296, 297
Себхи I, 479
Сезонный листопад I, 232, 234
Серир, пустыня I, 473
Симпсон, пустыня I, 447
осадки I, 448
положение I, 447
почва и растительность I, 449
Синай, пустыня I, 525—528
растительность I, 527—528
экологические условия I, 525—
526
эндемы I, 528
Синзоохоры II, 277
Синузии I, 106
Сирийская пустыня III, 190
Склерофилы I, 283, 326, 364—369, 420,
436, 446, 447, 459, 460, 546; II, 27,
52, 54, 56—58, 60—62, 65, 67—69,
71—73, 76, 224, 333; III, 110, 128
Смена аспектов II, 34, 319
биотопов I, 316—318
злаковников саваннами I, 288
растительности после пожара II,
162
растительного покрова I, 296—
301
Снежные горы II, 216
«Снежный туесок» III, 181
Солоноватые воды и соленые озера III,
327—330
Сонора, пустыня I, 332—374
климадиаграмма 1, 334
растительность I, 338—347
условия среды I, 332—338
экологические группы растений
I, 347
Сообщества растений
альпийские II, 139
аридные III, 243
галофитов III, 230
жестколистных древесных расте-
ний II, 23
злаковые I, 190
псаммофитов III, 230, 235
снеговых ложбинок III, 45, 47
степные II, 99; III, 246
суккулентов II, 91, 138
хионофильные III, 48
хионофобные III, 48
эфемеров III, 235
Соотношение физиологических и эко-
логических оптимумом произраста-
ния растений I, 30
Спинифекс I, 232—234, 458—460
Средней Азии пустыни
климат III, 202
общий обзор III, 202
растительность III, 203
солевой режим III, 241
Средиземноморская область степей Ана-
толии II, 97
Средиземноморье
вертикальная поясность II, 77—
93
климат II, 27—31, 73, 78
25—0434
растительность II, 25—27, 31—
49, 71—77
схема растительных зон I, 542
эколого-физиологические иссле-
дования II, 49—69
Стадии деградации растительности II,
24, 38, 39, 42
Степная зона Восточной Европы III, 64
климат III, 68
почвы III, 76—83, 139
продуктивность III, 129—131
растительность III, 83—89
эколого-физиологические иссле-
дования III, 109
явление засоления III, 132—142
Степные пустоши
микроклимат III, 123
почвы III, 124, 125
растительность III, 125
эколого-физиологические иссле-
дования III, 126
Степи
Афганистана III, 192
альпийские III, 289, 293
восточноевропейские III, 77, 80
горные III, 258, 283
ковыльные III, 89—94, 267
луговые III, 64, 66, 69, 76, 82,
84—87, 89, 90, 92, 97
Монголии III, 258
Степь
безлесие III, 69
климат III, 68
общий обзор III, 66—69
почвы III, 77—83
распространение III, 64
Структура растений
гигроморфная I, 233
ксероморфная I, 214, 231, 233,
405, 409, 459; II, 9—11, 56,
62, 193, 291, 422; III, 39, 49,
127, 128, 133, 170, 229, 322,
372
склерофильная II, 56—59
Субантарктические острова III, 56
Субантарктические туссоковые кочкар-
ники III, 183
Сублитораль III, 334—335, 338, 341, 342
Суборь II, 336
Субсредиземноморская зона
климат II, 93, 94
растительность II, 96
Сугрудки II, 336
Суккуленты I, 373, 379, 386, 391, 393—
395, 401, 419, 424—427, 429, 431,
433—434, 470, 482, 509; II, 10, 11,
62, 63, 138, 168, 224; III, 50, 110,
121, 125, 239, 274, 275
Сукцессии I, 17, 19—21, 189, 190; II,
42, 114, 115, 161, 164, 357; III, 201,
225, 311
Суммарное испарение в еловых лесах
различных зон II, 362
Супралитораль III, 334, 335, 361
Схема обмена энергией и веществом
биогеоценоза II, 260
386
Указатель русских терминов
Схема пищевых цепей и круговорот
углерода II, 278
Схема растительных зон земли, карта
II, 24
Сьерра-Невада III, 126, 127
Сэдд, болота I, 157, 160
Такла-Макан, пустыня III, 190, 250—
253
Такыры III, 208—210, 217, 235
Тасмания II, 206
Тенерифе, остров
карта растительности II, 91
климадиаграмма II, 90
профиль II, 91
Тепловой баланс средиземноморских
видов II, 65
Термитники I, 105, 248, 277
Терофиты I, 43, 325, 476; II, 44, 62,
101, 138, 187; III, 18, 20, 90, 125,
204, 216
Тибести, нагорье I, 493—494
Тибет III, 278—284
климат III, 280
общая характеристика III, 278
растительность III, 280
Тип растений
береговой II, 17
горный II, 17
долинный II, 17
предгорный II, 17
«Тола», формация II, 134
Томилляры II, 42
Транспирация
верещатников II, 257
древесных пород II, 275
дубовых насаждений II, 274
кактусов I, 355, 357
растений Анд I, 210
Памира III, 274, 275
пустынь I, 484, 490, 507, 508,
530, 531, 539; III, 228
саванн I, 282, 283
тропиков I, 85, 113, 114, 117,
129, 131
склерофилов I, 436; II, 55
суккулентов I, 429
Третичный период II, 133
Трихогидрофиты III, 231
Тропическая зона I, 56
Тропический дождевой лес
климат I, 79—91
общий обзор I, 76, 78
почвы I, 91
структура I, 99—106
экология растений I, 107—155
Тугаи III, 224, 225
Туман как экологический фактор I,
378, 411; II, 139
«Туесок» I, 213; III, 183
«Ульмо» II, 149, 150
Умеренные дождевые леса
Австралии II, 206
Новой Зеландии II, 206, 211
Тасмании II, 206
Фанерофиты I, 42, 106 \
Фенологическая интерцепция II, 166-
Фенологический спектр
видов дубово-грабовых лесов II,
319
геофитов II, 304
Каракумов III, 217
пойменной растительности III,
227
Физиологический оптимум произраста-
ния I, 28—30
Физиологическая сухость, см. Мороз-
ное высыхание
Филлодии II, 194
«Финбос» II, 160
Финбош III, 186
Фитомасса III, 24, 129—131
альпийских лугов III, 44
древесного яруса II, 271, 362
леса II, 269—271, 275, 276, 370,
371
подлеска II, 277
пустынь III, 205, 225, 227
различных зон II, 270
растительных такыров III, 210
степных сообществ III, 129—131
травяных сообществ II, 269, 271
Фитоценоз II, 280
Фитофаги II, 276, 278
Флора
альпийская III, 5, 283
арктотретичная II, 240
степная III, 85—87
Флористическое подразделение Ближ-
него Востока I, 540
Фолклендские острова III, 62—63
Фреатофиты III, 172, 231, 250
Фригана II, 42
Хамефиты I, 42; III, 18, 20
Хасмофиты I, 498, 500, 544
Хвойные леса бореальной зоны II, 334
Хироптерогамия I, 119
Хула, болота I, 541
Цайдам, пустыня III, 254, 255
«Ципрес» II, 154
Циркумполярная безлесная тундра II,.
23
Чапарраль I, 256; II, 27, 107, 109—117,.
119—120, 124, 129, 130, 133, 136;.
III, 185
Чили
деление лесной зоны II, 149
леса II, 148
профиль растительности II, 149'
Среднее
климадиаграммы II, 132
растительность II, 136—139
условия среды II, 133, 139—144
эколого-физиологические ис-
следования II, 146, 147
Южное
климат II, 148
растительность II, 149—156-
Указатель русских терминов
387
Чилийско-Перуанская пустыня I, 411—
422
растительность лома I, 418—422
туман, влияние I, 411—417
Шибляк II, 94
Эвапотранспирация II, 274, 286, 361;
III, 272
Эвкалипты I, 232, 233, 261, 445
Экваториальная зона I, 56, 79, 163,
194 195 294
Экоклины II, 13, 19—21, 326, 374
Экологическая зависимость I, 14
Экологический оптимум I, 28—30
Экологическое равновесие I, 14, 17
Экологический ареал кустарников Цен-
тральной Европы II, 300
Эколого-физиологические исследования
травяного покрова лиственных
лесов II, 301—319
хвойных пород II, 379—399
широколиственных пород II,
291-____301
Экосистема I, 43, 71; II, 259—260, 269,
275 290
Экотипы II, 13, 14, 17, 19, 20; III, 9, 51,
96, 362, 365
Экстремальные местообитания III, 272,
353
Экстремальные температуры II, 320
Элементы флоры
альпийские II, 77; III, 182, 267,
293
американские III, 292
андские III, 174
антарктические I, 197; II, 133,
149, 206, 211, 218, 219
арктические II, 242, 246; III, 301
аркто-альпийские II, 85
атлантические II, 245
атлантическо-европейские II, 244
атлантическо-средиземноморские
II, 245
бореально-атлантические II, 242
бореальные II, 77, 96, 97; III, 18
восточноазиатские III, 292
восточномонгольские III, 243
восточносубсредиземноморские
II, 96
голарктические I, 349, 371
евразиатские II, 245
европейские III, 178, 182
западносредиземноморские 11,
243, 245
иберийские II, 242, 243, 245
иберийско-атлантические II, 242
индо-малайские I, 233, 260—261
ирано-туранские I, 528, 534, 539;
III, 190, 202
иранские II, 99
казахстано-среднеазиатские III,
117
ксеротермические II, 94
мавритано-иранские I, 542
монгольские III, 253
неотропические I, 349; II, 133
палеотропические III, 187
патагонские III, 169, 174
понтические III, 122
понтическо-средиземноморские
III, 122
сахаро-аварийские I, 470, 492,
534, 539; II, 90
сахарские I, 470
сахельские I, 493
средиземноморские I, 471, 493,
494, 527, 528; II, 89, 96, 240,
242—246; III, 35
средиземномо рско-атлантиче окне
II, 243, 245
степные III, 89, 245, 277, 290, 293
субантарктические II, 146
субатлантические II, 245; III, 128
субсредиземноморские II, 71; III,
89, 122
судано-синдские I, 493, 541
суданские I, 470; II, 240, 329
тропические II, 89, 133, 156
турано-джунгарские III, 253
центральноазиатские III, 28, 246
центральноевропейские II, 77,
245, 329, 375
эуатлантические II, 242, 245
эусредиземноморские II, 243
Эндемизм
Сахары I, 470
Синайского полуострова I, 528
Чили II, 133, 144, 145, 147
Эндемы II, 89, 92, 93, НО, 115, 138;
III, 225, 228, 243, 244, 247, 253
Эндогенная ритмика развития I, 125;
II, 306
Эндозоохоры I, 485; II, 277
Энсинальная растительность в Аризоне
II, 117
Энтомофауна III, 120
Эпизодический дождь I, 469, 475, 482
Эпизоохоры I, 485; II, 272
Эпифиллы I, 152—154
Эпифиты I, 100, 125, 133, 134, 138—145,
147—149, 152, 153, 181, 187, 193,
196, 197, 202, 204, 221, 222, 242, 243,
245, 254, 260; II, 144, 150, 221
Эрг I, 474, 475
Эспинор III, 147
Этан III, 361
Эфемеры I, 304, 311—313, 316, 325,
327, 386, 432, 447, 500, 506—509,
513, 515, 516, 520, 521, 530, 531, 537;
II, 102, 134, 136; III, 112, 139, 141,
173, 190, 191, 200, 201, 204—206,
210, 214, 216, 217, 220, 223, 227,
230, 242, 302
Эффект «Соссюра» I, 149, 150, 362
Южное побережье Крыма II, 96, 97
Ява I, 203—207
25*
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ РАСТЕНИЙ
И РАСТИТЕЛЬНЫХ СООБЩЕСТВ
Abies I, 347; II, 77, 78, 80, 123, 232, 246;
III, 284, 285
alba ( = pectinata) II, 78, 311,
320, 346, 347, 374, 376, 383
balsamea II, 347, 348
borisii-regis II, 374
bornmuelleriana II, 316, 374
cephalonica II, 374
cilicica II, 80, 81, 374
concolor I, 346; II, 120, 123, 126,
127
densa III, 291
grandis II, 125
magnifica II, 123, 127
maroccana II, 88
nordmanniana II, 374
numidica II, 88, 374
pectinata (cm. A. alba)
pindrow III, 287
pinsapo II, 374
sachalinensis I, 245
sibirica II, 239, 325, 343, 344, 346,
374; III, 258,
webbiana III, 284, 285, 287, 289
Abronia I, 351; II, 115
Abrotanella II, 218, 220
forsterioides II, 212
Abutilon darwini I, 117, 327
Acacia I, 85, 157, 200, 211, 213, 233,
247, 278, 283, 285, 300, 344, 419,
462, 470, 476, 479, 493, 510, 533,
537, 541; II, 86, 177, 180, 193, 194,
203—207; III, 187, 285
abyssinica I, 319
olofo IT 4 QZ
albida I,’ 296, 297, 319, 401, 484,
541
aneura I, 300, 432, 445, 446, 447,
450, 454, 456, 458, 459-462,
464, 466
arabica I, 484
armata II, 194
bakeri II, 205
benthamii I, 286
burkittii I, 450, 454, 456
cambagei I, 449
campylacantha I, 319
catechu I, 481
caven II, 136, 138; III, 159
constricta I, 338, 340
cyanophylla (= saligna) I, 113,
114, 522, 526; II, 166
cyclopis II, 166, 202
decipiens II, 194
detinens I, 292
drepanolobium I, 272, 319
ehrenbergiana (= flava) I, 296
319
estrophiolata I, 466
fistula I, 319
flava (cm. A. ehrenbergiana)
floribunda II, 192
giraffae I, 280, 287, 291, 320, 390
401; III, 167
greggii I, 341
haematoxylon 1, 320
harpophylla I, 463
hebecladoides I, 319
horrida I, 390
kempeana I, 462
laeta I, 493
linearis II, 192
longifolia I, 464; III, 370
malacocephala I, 272
marginata II, 194
melanoxylon I, 133; II, 138, 147,
172
mellifera I, 296, 297, 319
modesta III, 286
nilotica I, 273
orfota I, 319
pentadenia II, 180
pulchella II, 180, 181
pygnantha II, 191
raddiana I, 296, 319, 484, 501,
533, 535, 539
saligna (cm. A. cyanophylla)
Senegal I, 296, 318, 319
seyal I, 272, 296, 297, 319, 484
sieberiana I, 319
spirocarpa I, 253, 533, 539
stenocarpa I, 493
sundra I, 481
tetragonophylla I, 449
tortilis I, 296, 318, 319, 501, 502,
538, 541
urophylla II, 180
xanthoploea I, 158
Acaena II, 218; III, 178
adscendens III, 60, 61, 63
Acalypha maginata I, 481
tricolor I, 117
390
Указателъ латинских названий растений
Acanthocarpus preissiii II, 202
Acanthococcos I, 254
Acantholimon II, 88, 89, 101, 104; III,
269, 280
diapensioides III, 269, 277
hedinii III, 267
Acanthophyllum III, 211, 214
Acanthosicyos horrida I, 401, 402
Acanthus ilicifolius I, 164
Acer II, 12, 72, 110, 148, 277; III, 282,
284, 287, 291
campestre II, 240, 261, 300, 319,
325
grandidendata II, 130
laetuna II, 240
macrophyllum II, 110, 111
monspessulanuna II, 83, 93, 330,
331
negundo III, 101, 147
obtusatum II, 93
opalus II, 93, 331
platanoides II, 233, 321, 322, 325,
336
pseudoplatanus II, 233, 239, 299,
317, 319, 321, 322, 325
saccharinum II, 101
tataricuna II, 325
Achillea borealis II, 19
gerberi III, 201
millefolium II, 17, 18, 19, 21;
III, 98, 300
millefolium ssp. setacea III, 88
santolina II, 80, 102
Aciphylla II, 218, 222; III, 181
Aconitum III, 42, 49, 53, 292
excelsum II, 353
orientale III, 52
Acrostemon II, 158
Acrostichum aureum I, 166
Acrotriche affinis II, 196
Actaea spicata II, 331, 353
Actiniopteris radiata I, 246, 395
Actinostrobus II, 200
arenarius II, 187
Adansonia dagitata I, 247, 249, 251, 266
Adenia globosa I, 251, 253
pechuelli (Echinithamnus) I, 391,
394
schweinfurthii I, 137
Adenium obesum I, 254
soco tranum 1, 255
Adenathera pavonina I, 481
Adenocarpus II, 93, 242
mannii I, 222
Adenostoma II, 112, 114, 115
fasciculatum II, 108, 109, 112, 113
Adenostyses III, 42, 49
Adesmia bedwellii II, 144, 145
campestris III, 178
horridiuscula I, 421
Adianthum I, 131
capillus-veneris I, 132, 352, 494,
498, 528; II, 246
hispidulum I, 466
Adonis flammeus II, 102
vernalis III, 84, 85, 88, 122, 125
wolgensis III, 76, 90
Adoxa moschatelliana II, 319
Aechmea pectinata I, 149
Aegialitis I, 168
Aegiceras I, 168
corniculatum I, 164, 180
floridum I, 164
Aegilops III, 110
Aegopodium podagraria II, 239, 261, 315,
319, 338, 349, 367
Aeluropus III, 191, 231, 235, 241
lagopoides I, 521
litoralis I, 480, 490; III, 136, 227,
240
repens III, 232
Aeonium (Sempervivum) II, 92
Aerva I, 476; III, 188
Afframomum (Kaempferia) I, 132
Aesculus II, 12, 110, 233
californica II, 110
indica III, 287
Aethionema II, 80
Aextoxicum punctatum II, 144, 146, 149
Agathis I, 197; II, 211
Agathosma capitatum II, 163
Agauria salicifolia I, 221, 223
Agave I, 346
palmeri II, 119
schottii II, 119
sisalana I, 131, 269
tabulate I, 188
Ageratum conyzoides I, 191
Agonis flexuosa II, 182, 190
juniperina II, 182
Agriophyllum III, 214
arenarium III, 201, 246
Agropyrum II, 43; III, 95, 205, 280, 372
cristatum II, 354; III, 93, 200
desertorum III, 201
glaucum ssp. intermedium III, 88
junceum I, 521; III, 362, 363
mongolicum III, 245
orientale III, 302
pectiniforme III, 114, 115
prostratum III, 141, 302
pseudocaesium III, 114
pseudoagropyrum (cm. Aneurole-
pidium)
ramosum III, 200
repens III, 76, 77, 88, 194, 299,
302, 349
repens ssp. pseudocaesium III, 139
scabrum III, 181
sibiricum III, 200, 201
smithii III, 96, 108
spicatum III, 111
Agrostis II, 31, 49; III, 300, 345, 348
alba II, 349; III, 245, 299
alpina III, 4
canina III, 85, 299, 300
canina ssp. tenuifolia III, 88
carmichaelii III, 62
muscosa II, 221
pallens III, 369
stolonifera III, 62, 349
tenuifolia III, 87
Указатель латинских названий растений
391
tenuis III, 182
volkenisii I, 222, 245
vulgaris II, 370
Aizoon dinteri I, 382, 386
Ajuga chia II, 102
genevensis III, 88
reptans II, 353
Albizzia I, 91, 247, 297
adianthifolia I, 242
anthelminthica I, 320
ayemeri I, 318
distachya II, 180
fastigiata I, 110, 113
julibrissin II, 240
lebbek I, 479
lophanta II, 166
zygia I, 242
Alchemilla I, 230; III, 89
argyrophylla I, 222
johnstonii I, 225
novaehollandiae II, 218
pentaphylla III, 46
Alectoria III, 57
ochroleuca III, 49
Alectrolophus major III, 88
Aleurites moluccensis I, 481
Alhagi III, 191, 202, 225, 231, 250
camelorum I, 469; II, 104
maurorum I, 479, 532; III, 191
persarum III, 241
pseudoalhagi III, 191, 232
Alaria III, 337
Alisma III, 139, 310
plantago III, 323
Allanblackia stuhlmannii I, 110, 113
Allenrolfia III, 170
occidentalis II, 130
Alliaria officinalis II, 290, 307
Allionia incarnata I, 349, 350
Allium I, 516; II, 62
сера II, 305
guttatum III, 91
mongolicum III, 249
paczoskianum III, 91
polyphyHum III, 267
polyrrhizum III, 257, 258
roseum I, 521
tenuissimum III, 257
ursinum II, 250, 304, 305, 312, 313,
319
Alnus I, 199; II, 76, 258; III, 291
acuminata II, 120
glutinosa II. 247, 299, 317, 319,
325; III, 134, 197, 299, 302, 312
incana II, 233, 325
jorullensis III, 145
viridis I, 43; II, 388; III, 32, 49
Aloe I, 252, 295, 425, 428, 429, 432;
II, 158, 168
asperifolia I, 391, 393—395
dichotoma I, 396
dinteri I, 394
ferox II, 167
plicatilis II, 168
rubrolutea I, 393, 394
Alopecurus III, 300
pratensis III, 89, 139, 299, 300
ventricosus III, 301
Alophia amoena III, 163
Alphitonia zizyphoides I, 121
Alpinia nutans I, 481
Alsine III, 293
Alsophila III, 285, 291
australis I, 133
Alstonia constricta I, 260
scholaris I, 481
Alstroemeria II, 134, 139
Althenia III, 332
Alternanthera philoxeroides III, 164
Altingia excelsa I, 203, 204, 205
Alyssum III, 205
desertorum III, 302
montanum III, 124
(Ptilotrichum) spinosum II, 85
Amaranthus I, 443
palmeri I, 349, 350
retroflexus II, 102
Ambrosia chamissonis (Franseria) II,
115; III, 369
Amelanchier II, 110, 130
ovalis II, 93, 300; III, 122
Ammodendron III, 211, 229
conollyi II, 12; III, 228, 229
Ammophila I, 459; III, 372
arenaria I, 521, 522; II, 202, 203,
204; III, 363—368, 371
arenaria ssp. arundinacea III, 368
breviligulata III, 369
Ammopiptanthus mongolicum III, 243,
247
Amoora rohituca I, 481
Amorpha III, 97
Amor pho phallus I, 128
Ampelocissus volkensii I, 136, 137
Amphicome III, 283
Amygdalus I, 542
communis I, 124; II, 12, 99
mongolica III, 247
nana III, 64, 90, 92
pedunculata III, 257
vulgaris I, 522
Anabaena I, 450
Anabasis I, 526, 527, 533, 535, 539; III,
250, 253
aphylla III, 202, 222, 240, 241
articulata I, 326, 521
brevifolia III, 257
salsa III, 141, 238, 240
set if era I, 500
Anacampseros I, 425, 433
albissima I, 391
Anacardium occidentale I, 481
Anagyris foetida I, 537
Ananas sativa I, 131, 133
Anaphalis javanica I, 206, 208
Anaptychia ciliaris II, 33
Anarrhinum pubescens I, 528
Anarthrophyllum II, 139
Anastatica hierochuntica I, 485, 498;
III, 206
Anchusa italica II, 102
Andromeda polifolia II, 405, 420
392
Указатель латинских названий растений
Andropogon I, 214, 245, 447; III, 95, 97,
98, 121
annulatus I, 490
gerardi III, 95, 97, 108, 121
hirtus I, 494; II, 162
laniger I, 490
scoparius III, 95, 96, 101, 102,
108, 121
Androsace ( = Aretia)
Androsace I, 205; II, 80
akbaitalense III, 267
elongata III, 302
helvetica III, 41
septentrionalis III, 88
villosa III, 263
Androtrichum trigynum III, 164
Anemone II, 247, 304, 306, 308; III, 56,
97
alpina II, 218
decapetala var. foliosa III, 163
demissa III, 283
fasciculata III, 53, 54
hepatica II, 310, 311, 319
narcissiflora III, 53
nemorosa I, 15; II, 250, 304, 306,
307, 310, 311, 319
ranunculoides II, 319, 331
rivularis III, 244
silvestris III, 84, 85, 90
sphenophylla I, 350, 351
thomsonii I, 222
Aneurolepidium III, 195
(Agropyrum) pseud о agro pyrumllll,
258
Angelica silvestris II, 349, 353
Angianthus cunniaqhamii II, 202
Angiopteris thysmanniana I, 133
Angraecum I, 148 ц
Anisospenna passiflora I, 136
Anodopetalum biglandulosum II, 207
Anogeissus schimperi I, 297, 298
Antennaria II, 259
dioica II, 349, 354
Anthelia juratzkana III, 46
Anthemis maritima III, 371
melampodina I, 501
ovata I, 501
Anthericum III, 122, 127
divaricatum II, 202
ramosum III, 88
Antbohryum triandrum I, 422
Anthocleista orientalis I, 109, 110, 113
Anthospermum II, 162, 173
Anthoxanthum odoratum III, 85, 89, 182
Anthriscus silvestris I, 16
Anthurinum venosum I, 188
Anthyllis vulneraria III, 368
vulneraria ssp. polyphylla III, 88
Antidesma bunius I, 481
Aphidium mougeotii III, 50
Aphyllanthes III, 45
mospeliensis II, 49, 62
Apicra I, 425
Apinagia warmingiana III, 326
Apodytes II, 158
Apollonias canariensis II, 92
Aponogeton distachyon II, 165
Aquilegia III, 249, 259
Arabidopsis kneuckeri I, 528
Arabis alpina III, 50
Arachnitis uniflora II, 156
Aralia III, 292
papyrifera I, 481
Araucaria II, 154, 206
araucana (= imbricata) II, 146,
149, 154, 155
bidwillii I, 196
cunninghamii I, 196; II, 205
Arbutus II, 57, 119, 121
andrachne I, 547; II, 60, 96, 240
arizonica I, 346; II, 120
menziesii II, 110, 126
unedo II, 41, 50, 52, 56, 64, 68,
240, 242, 332
Archontophoenix cunninghamii I, 261
Arctophila fulva III, 18
Arctostaphylos II, 109, 115, 119—121,
254, 354
grandulosa II, 112, 114
glauca II, 109, 114
manzanita II, 109, 113
pungens I, 346; II, 119
tomentosa II, 108, 109
uva-ursi II, 251, 254, 349; III, 21
Arctotheca II, 203—205
nivea II, 202
Arctous alpina (— Arctostaphylos) II,
254, 354; III, 13, 26
Areca catechu I, 130
lutescens I, 481
Arenaria III, 205, 293
biflora III, 46, 47, 50
graminifolia III, 85, 88, 90
marina III, 347
obtusiloba III, 36
peploides ssp. robusta III, 369
pungens II, 85
serpyllifolia III, 88
Aretia (== Androsace) helvetica III, 50
Argania II, 35
Argemone mexicana I, 401, 428
Argyroderma I, 425
Argyrolobium abyssinicum I, 494
linnaeanum (= Cytisus argenteus)
II, 245
Aricfno TT 4 R4
Aristida I,’ 280, 389, 474, 541; III, 167,
188, 211, 215
arenaria I, 459, 464
ascensionis III, 257
brevifolia I, 447, 390
ciliata I, 282
hochstetteriana I, 389
karelini III, 214, 215, 223
minor III, 215
namaquensis I, 447
nitidula I, 466
pennata III, 200, 201, 223
plumosa I, 515, 527
pungens I, 469, 475, 490, 515
scoparia I, 527
uniplumis I, 287, 292, 294
Указателъ латинских названий растений
393
Aristolochia I, 134; II, 34
Aristotelia magui II, 137; III, 175
Armeria maritima III, 344, 348
maritima ssp. labradorica II, 26
Artemisia I, 506, 538, 541; II, 114; III,
89, 114, 127, 191, 200, 211, 221,
243, 250, 258, 272, 273, 280, 282, 289,
290, 293
afra I, 222
arenaria III, 201
austriaca III, 88, 91, 92, 136
badghysi III, 233, 241
californica II, 110
campestris III, 124, 127, 368
caudata III, 369
caucasica II, 80
fragrans I, 542; II, 80, 101—103
fragrantissima I, 513
frigida III, 131, 245
glauca III, 131
herba-alba I, 326, 480, 521, 523,
528, 537, 539, 541, 542; II, 110
judaica I, 494, 501, 528
kemrudica III, 241
macrocephala III, 267, 273
maritima III, 135, 136, 201, 207,
344, 348, 350, 359
maritima ssp. incana III, 140, 142,
200
meyeriana III, 230, 231 232
monosperma I, 326, 501, 515, 516,
526-528, 537—539
nevadensis II, 87
ordosica III, 244, 245
pamirica III, 267, 275, 278
pauciflora III, 137, 140, 232
pectinata III, 244, 249
scoparia III, 201, 257
scopulorum III, 44
semiarida III, 232
sericea III, 194
skorniakovii III, 267, 268, 270,
272—276
sieberi III, 190
terrae-alba III, 238
tilesia III, 31
tridentata I, 347; II, 128—131
Arthraerua leubnitziae I, 386—389, 402,
403, 408, 410
Arthraxon hispidum III, 245
Arthrocnemum I, 369; II, 187
arbusculum II, 205
glaucum I, 402, 405, 406, 454,
463-465, 492, 521, 526, 539
halocnemoides II, 205
Artocarpus integrifolia I, III, 113
Arum II, 62
italicum II, 34
maculatum II, 239, 306, 319
Arundinaria I, 245; III, 291
Arundo III, 230
donax I, 245; II, 92
Asarum II, 319
europaeum II, 239, 310—312, 315,
338
Ascophyllum III, 337
nodosum III, 341
Aspalathus II, 161, 173
Asparagus I, 425, 432
aculeatus II, 33
acutifolius II, 54, 332
capensis II, 161
persicus III, 225
Asperula II, 11, 310, 311
cynanchica III, 88
odorata II, 239, 308, 311, 316»
331, 353
tinctoria III, 88
Asphodelus II, 47, 62, 63, 162
cerasifer II, 41, 44, 45, 63
microcarpus I, 516, 518, 521
Aspicilia III, 141
Aspidistra I, 133
elatior I, 133
Aspidium filix-mas II, 309
Asplenium II, 144
adianthum-nigrum II, 33
cicutarium I, 132
flabellifolium II, 218
macrophyllum I, 151
marinum III, 334
nidus I, 131, 132, 140, 144, 151»
187
protensum I, 151
ruta-muraria III, 50
septentrionale III, 50
sinuatum I, 151
theciferum I, 151
theciforme I, 146
trichomanes II, 308; III, 50, 125-
unitum I, 132
viride III, 50
Astelia II, 151; III, 63
alpina II, 212, 218, 220, 221
Astre I, 174; III, 97, 284, 372
linosyris III, 124
pannonicus III, 135
tripolium III, 136, 301, 348, 349»
359, 360
Astericus graveolens I, 498, 500, 514
pygmaeus I, 487
Astragalus (Tragacantha) I, 470, 494,
527; II, 88, 89, 101
Astragalus III, 21, 97, 259, 268, 280,
282, 290, 293
ammodendron (= Halimodendron
ammodendron)
atropurpureus II, 101
borodini III, 277
chadjanensis III, 272
danicus III, 85
forskalei I, 501
heterodontus III, 269
melilotus III, 245
Astrebla I, 442, 443, 446, 449
Astroloma conostephioides II, 197
Atalaya I, 466
Atamisquela emarginata III, 169
Athiamanta cretensis III, 51
Athanasia parviflora II, 165
Atherosperma II, 207
moschata I, 200, 201; II, 207, 208
394
Указатель латинских названий растений
Athrotaxis II, 210
cupressoides II, 220
selaginoides II, 220
Athyrium filix femina II, 319, 353
Atractylis serratuloides I, 529
Atragene sibirica II, 353
Atraphaxis III, 207, 249, 250
Atriplex I, 422, 425, 432, 434, 450, 451,
456, 486, 521; II, 130, 131, 205; III,
202, 372
arenaria III, 369
canescens I, 346, 347, 456—458
canum III, 141
confertifolia I, 456, 457; II, 131
halimus I, 478, 487, 500, 538, 539
hastata III, 362
inf lata I, 464
isatidea II, 203
laciniatum III, 140
lampa III, 169
latifolia III, 349
mauretanica I, 546
mollis I, 491, 492
mural is I, 405
nummularia I, 444, 449, 466, 451
nutallii II, 131
paludosa I, 464; II, 206
portulacoides I, 521; III, 345,
352, 371
ragodioides I, 444
spongiosa I, 452
tatarica III, 231
undulata III, 169
verruciferum III, 141
vesicaria I, 443, 445, 449—454,
465
Atropis III, 18, 269
distans III, 301
maritima (— Puccinellia mariti-
ma)
nackeliana III, 267
pamirica III, 269
Aucuba japonica II, 327
Aulax pinifolia II, 173
Austrocactus III, 178
Austrocedrus ( = Libocedrus) chilensis II,
139, 154, 155, 156; III, 175
Avena barbata II, 110
fatua II, 110
pratensis ssp. schelliana III, 88
pubescens III, 87, 88
schelliana III, 87
Avicennia I, 167—169, 171—174, 178,
180; II, 206
marina var. resinifera II, 205
A venastrum III, 280
desertorum (= Helictotrichon de-
sertorum)
-A.Za.rSL II 136
Azolla I,’ 158, 162, 209; III, 164, 309
diapensioides II, 210
Azorella I, 209, 210; II, 134, 139, 218;
III, 63, 178
selago III, 60, 61
Azotobacter I, 153, 450; II, 280; III,
120
Babbagia I, 456
Baccharis I, 211, 343
petiolata II, 133
rosmarinifolia II, 137
tola II, 134
Baeckia II, 218
ericaea II, 199
polystoma I, 466
Baijerinckia I, 153
Balanites I, 157, 476
aegyptiaca I, 272, 296, 297, 484,
541
Balanophora elongata I, 155
Ballota hirsuta I, 494
Bambusa I, 193, 297
arundinacea I, 126, 133
vulgaris I, 114
Bangia fuscopurpurea II, 334, 335
urospora III, 337, 338
Banksia I, 200,.289; II, 185, 189, 199, 207
attenuate II, 190, 203
ericifolia II, 178
marginata II, 178, 200
menziesii II, 190, 191, 203
ornata II, 196—201
prostrata II, 187
repens II, 187
serrata II, 178
spinulosa II, 178
Baptisia III, 97
Barbella javanica I, 152
Barringtonia I, 183, 185
Bartramia III, 50
Bassia I, 449—451, 453, 454, 464, 465;
II, 206; III, 136, 140
Batrachium divaricatum III, 323
Bauera II, 220
Bauhinia I, 233, 401, 408
heterophylla I, 481
Beaumontia grandiflora I, 481
Beckmannia erucaeformis III, 135,299,300
Begonia I, 128, 129, 133
hybrida I, 133
meyeri-johannis I, 222.
zimmermannii I, 129
Beilschmiedia I, 197, 198
miersii II, 136
Bellis perennis I, 16
silvestris II, 63
Berberis I, 199; II, 139, 156; III, 175,
283, 284, 287, 290, 292, 293
cretica II, 81
heterophylla III, 178
vulgaris II, 300
Bergenia crassifolia III, 258
Bergenia ligulata III, 290
Bergeranthus glenensis I, 429, 433
Bergia koghanii III, 225
Berlinia schefferi I, 110
Berteroa incana III, 89
Berzelia abrotanoides II, 163
lanuginosa II, 162, 165
Beta II, 147
Betonica grandiflora III, 53
officinalis III, 89
Betula II, 233, 247, 317; III, 32, 134,
Указатель латинских названий растений
395
139, 259, 285, 291, 306
ermani II, 7, 10
exilis III, 29
humilis II, 325
папа II, 405, 410; III, 13, 14, 18,
21, 22, 24, 26
papyrifera II, 347
pendula II, 299
platyphylla III, 249
pubescens II, 246, 250, 299, 325;
III, 10, 33
raddeana III, 33
resinifera II, 347
tortuosa III, 7, 11
utilis III, 287, 290
verrucosa II, 246, 325; II, 66, 193
Bignonia magnifera I, 481
purpurea I, 481
Bilbergia amoena I, 150
Biscutella laevigata III, 122
Blaeria I, 222
Blechnum australe II, 161
capense II, 172
palmiforme III, 62
pennamarina II, 218
spicant II, 252
Blennodia pterosperma I, 449
Blindia minima III, 334
Blysmus compressus III, 269
Bobartia spathacea II, 167
Boerhavia I, 343, 349
Boquila trifoliata II, 150
Bolbophyllum I, 148, 260
Bolboschoenus III, 230
Bombax I, 188, 242, 481
Borassus I, 266, 267, 273
aethiopica I, 157
flabelliformis I, 481
Borbonia II, 161
Boronia coerulescens II, 199
Borya nitida II, 190
Boscia I, 266, 296
albitrunca I, 320
foetida I, 283, 295
salicifolia I, 493
senegalensis I, 296
Bossiaea II, 191, 218
amplexicaulis II, 180
JBoswellia, I, 251, 297
Bothriochloa lagurioides III, 162, 163
Botryceras laurina II, 171
Bougainvillea I, 481; II, 136; III, 148
spinosa III, 169
Bouteloa I, 349
curtipendula III, 96, 108
gracilis III, 102, 103, 121
hirsuta III, 102, 121
racemosa III, 121
Brabejum II, 158
stellatifolium II, 157, 165
Brachychiton I, 233, 258, 260
Brachyome II, 218
Brachypodium II, 45, 63
distachyon II, 44
pinnatum III, 194
ramosum II, 43, 62, 63
silvaticum II, 239, 308, 319
Brachysiphon II, 165
Brachystegia I, 249, 271
Brachythecium rivulare III, 325
rutabulum II, 33
Bradburya virginiana III, 369
Brassaia actinophylla I, 481
Brassica nigra II, 114
Braya III, 25
humilis III, 20, 26
purpurascens III, 26
pamirica III, 267
Bryum argenteum III, 57, 58
caespitosum III, 201
Breutelia elongata II, 221
pendula II, 218
Brillantaisia I, 131
Briza triloba III, 162
Brodiaea I, 350; II, 114
Bromeliaceen I, 103, 121, 134, 138, 141,
146, 254, 258
Bromus III, 205
erectus III, 299, 300
erectus ssp. riparius III, 88,
131
inermis III, 88, 131, 194, 299
mollis II, 110
rigidus II, 110
riparius III, 85, 87, 90
rubens I, 494; II, 114
sterilis II, 290
tectorum II, 114; III, 201
tomentellus II, 101
unioloides III, 162
variegatus III, 53
villosus III, 368
Bruguiera I, 166, 171, 179, 180
caryophylloides I, 164
eriopetala I, 164
gymnorrhiza I, 164, 179, 180
parviflora I, 164, 175, 180, 181
Bubunapus III, 20
Buchea I, 494
Buchloe dactyloides III, 103, 104
Buddleia I, 481; II, 158; III, 283, 292
globosa III, 175
tsetangensis III, 293
Buellia III, 57
Bufonia murbechii II, 88
Bilbilis bactyloides III, 121
Bumelia 1, 482
Bunias III, 204
orientalis III, 88
Bupleurum aureum II, 239
rigidum II, 62
spinosum II, 85
stellatum III, 50
tenuissimun III, 135
Burkea I, 246, 298
Butea I, 481
Butomus III, 139
umbellatus III, 302, 326, 328
Buxus II, 58
sempervirens II, 12, 33, 50, 53,
71, 72, 93, 240, 329, 383
Byblis II, 157
396
Указатель латинских названий растений
Cacalia hastata (= Senecio') II, 239
Cadaba rotundifolia I, 297
Caesalpinia crista I, 184
tinctoria I, 419
Cakile III, 372
edentula III, 369
maritima II, 202; III, 362
Calamagrostis I, 210, 211, 213, 214; II,
349; III, 18, 166
arundinacea III, 89
dubia III, 225
epigeios II, 355, 368; III, 76, 131,
194, 201, 299
lanceolata III, 194
langsdorffii II, 367
macilenta III, 258
purpurascens III, 25
tenella III, 50
villosa III, 50
Calamagrostis — Vaccinium myrtillus II,
370
Calamintha acinos III, 89
clinopodium II, 315
Calamus indicus I, 85, 133, 161
Calathea zebrina I, 132
Caldcluvia paniculata II, 150
Calicotome villosa II, 244
Calligonum III, 191, 201, 211, 214, 217,
223, 250
aphyllum III, 197
azel I, 475
caput-medusae III, 228
comosum I, 475, 498, 534—537,
539
mongolicum III, 247
Callistemon II, 218
lanceolatum I, 289, 481
sieberi II, 218
Callitriche III, 314
Callitris I, 471; II, 171
glauca I, 466
quadrivalvis (== Tetraclinis arti-
culate) II, 82, 83
Calluna II, 248, 254, 256, 257—259, 349;
III, 39
vulgaris II, 53, 244, 245, 250—
253, 316, 328, 354, 405, 419,
420; III, 21, 22, 183, 368
Calocephalus II, 203, 204
brownii II, 202
Calochortus II, 114
Calodendrum II, 171
Calophyllum inophyllum I, J117, 183
Calophaca III, 57 । f *
Calothamnus quadrifid us II, 202
Calothrix I, 420
scopulorum III, 334, 335, 338
Calotropis I, 476; III, 188
procera I, 479, 484, 541
Caltha II, 217
arctica III, 17
introloba II, 218
leptosepala III, 322
Calvaria inermis (= Sideroxylon) II,
165
Calycera crassifolia III, 370
Calycotome spinosa II, 64
Calypso bulbosa II, 353
Camellia III, 292
Campanula latifolia II, 319
persicifolia II, 353; III, 87, 89
sibirica III, 87, 88
stevenii III, 90
trachelium II, 319
uniflora III, 26
Camphorosma II, 101; III, 135, 137 231
lessingii III, 231
monspeliaca III, 137, 140
Campylopus II, 165
Canavalia I, 182
Cannelariella III, 57
Cannistrum cyathiforme I, 150
Canthium lucidum I, 260
Capparis I, 420, 479; III, 231, 289
decidus I, 290
mitchellii I, 460
spinosa I, 487, 488, 498; III, 230*
zeylanica I, 481
Capsella III, 205
Caragana III, 92, 243, 249, 250, 293
arborescens III, 73
frutex III, 64, 90
korshinskii III, 245, 246
micro phylla III, 246
pygmaea III, 258
pumila III, 194
tragacanthoides III, 244
Caralluma I, 251, 253, 424, 470
lateritia I, 393, 395
sinaica I, 527
Cardamine hirsute II, 218
pratensis I, 16
Carduus crispus ssp. hamulosus III, 89>
defloratus III, 51
hamulosus III, 93
keniensis I, 231
nutans III, 149
uncinatus III, 91,
Carex II, 34, 217, 414, 417; III, 18, 49,.
139, 258, 263
acutiformis III, 321 ,
arenaria III, 365—367
binervis II, 251, 252
buschiorum (= C. humilis) III,' 53
capillaris III, 26
cinerea II, 252
curvula III, 41, 45, 49
digitate II, 353
diluta III, 301
distachya II, 33
drummondiana III, 44
duriuscula III, 244, 258
duriuseuliformis III, 267
echinata II, 253
elata ssp. omskiana III, 133
elynoides III, 44
finna III, 37, 42
fusca II, 417
gaudichaudiana II, 218
glareosa III, 345
goodenoughii III, 54
gracilis III, 26
Указателъ латинских названий растений
397
hostii III, 204—206, 215
humilis (= buschiorum) III, 53,
84, 85, 87, 88, 124, 131
lachenalii III, 20
limosa II, 253, 405, 407
maritima III, 26
melanantha III, 267
misandra III, 25
monostachya I, 222, 224, 225
montana III, 85
mooreroftii III, 281
muricata II, 315
nardina III, 25
nigra (= parviflora)
orbicularis III, 269
pachystylis III, 190, 191
parviflora (= nigra) III, 349
pensilvanica III, 97
physodes III, 210, 211, 215, 216,
221, 222, 241
pilosa II, 261
pilulifera II, 251
praecox III, 88
recta III, 349
remota II, 319
runssorensis I, 228
rostrata III, 299
rupestris III, 25
salina III, 3, 45
sempervirens III, 51
silvatica II, 319
stans III, 29
subspathacea III, 3, 45
verna III, 85, 88
vesicaria III, 299
Carica I, 130, 133
candicans I, 419
papaya I, 130, 481
Carissa edulis I, 493
ferox I, 425
grandiflora I, 481
ovata I, 260
Carlina corymbosa II, 44
Carludovica palmata I, 131
Camegiea I, 337—340, 353—357, 359,
363, 366
Carmichaelia III, 181
Carpino — Nemoreto — Pinetwn II, 336
355
myrtillosum II, 355
Carpino — Querceta II, 336, 337
Pinetum II, 337
Carpinus II, 72, 240, 241, 277, 310, 311
betulus I, 14; II, 233, 238, 239,
299, 300, 317, 319, 325, 336,
337
orientalis II, 78, 93
Carpobrotus (= Mesembryanthemum) I,
369; II, 187, 203—205
aequilaterus II, 202
Carya II, 231
Caryota arens I, 481
Cassandra II, 405
Cassia I, 459, 494
aphylla III, 169
artemisioides I, 481
coquimbensis II, 144
desolata I, 449, 450
fistula I, 117, 123, 481
lanceolata I, 484
nodosa I, 481
odovata I, 484
siamea I, 481
timorensis I, 117
Cassinia II, 221
Casysiope II, 243
selaginoides III, 283
tetragona III, 29
Casstha capensis I, 155, 183, 186; II, 161
glabella II, 202
Castanea I, 42; II, 72, 78, 148, 233, 241
javanica I, 205
sativa (— vesca) II, 317
Castanopsis I, 150, 244, 245
chryso phylla II, 110, 126
indica III, 291
tribuloides III, 291
Casuarina I, 419, 445, 464, 465, 480; II,
11, 193, 200, 201; HI, 147
decussata II, 180
equisetifolia I, 114, 183, 456
freiseriana II, 190, 203
humilis II, 187
lepidophloia I, 450, 455, 456
preisii II, 185
pusilia II, 196, 200
stricta II, 206
Catalpa II, 12
Catesbaea spinosa I, 481
Catophractes I, 283, 294, 295, 436
Caulerpa III, 333
Cavanillesia arborea I, 257
Ceanothus II, 109, 110, 112, 126
cuneatus II, 108, 109, 113
fendleri II, 120
leucodermis II, 112, 114
papillosus II, 108
velutinus II, 109, 130
Cecropia I, 103
Cedrela serrulata I, 117
toona I, 481
Cedrus II, 77, 123, 326; III, 285
atlantica II, 68, 83, 84
deodara II, 68; III, 287, 288, 290
libani I, 547
libanotica II, 80, 81
Ceiba pentandra I, 299
Celastrus balocularis I, 260
Celmisia II, 218
coriacea II, 222
hectori II, 223
longifolia II, 220, 221
Celtis II, 158, 172
australis II, 93
pallida I, 338, 341, 344
rhamnifolia II, 171
Cenchrus I, 447
biflorus I, 494
tribuloides III, 369
Cenociphium lanceolatum II, 173
Centaurea II, 101; III, 89
aegyptiaca I, 501, 510, 514
398
Указатель латинских названий растений
arenaria III, 201
diffusa III, 91, НО
glastifolia III, 301
jacea III, 109
marschalliana III, 89, 131
ruthenica III, 90, 91
scabiosa III, 89
solistialis III, 92
Centaurium malzacianum I, 528
pulchellum III, 348
Cephalaria lucantha II, 63
Cephalotaxus drupacea I, 480
Cephalotus follicularis II, 182
Ceramium rubrum III, 335
Cerastium III, 17, 20
anomalum III, 302
caespitosum III, 88
glomeratum II, 218
latifolium III, 50
maximum III, 31
strictum III, 50
ucrainicum III, 91
uniflorum III, 50
Cerasus microcarpa II, 12
Ceratonia II, 37, 68, 80, 86
siliqua I, 119, 522, 547; II, 35, 36,
41, 60
Ceratocarpus III, 140
arenarius III, 207
Ceratophyllum I, 158, 479; III, 309
demersum III, 294, 318
Ceratopteris pteridoides I, 162
Ceratostigma griffithii III, 293
Cerbera I, 186
Cercidium (= Parkinsonia) I, 338, 340,
343, 344
australe III, 169
microphyllum I, 340, 353, 364,
367, 368; III, 170
Cercis II, 110
siliquastrum I, 119; II, 47, 93
Cercocarpus betulaefolius II, 109, 130
ledifolius II, 113
Cereus I, 419
leucostele I, 259
megalanthus I, 256
Cerinthe II, 102
Ceriops I, 164, 166
candolleana I, 164
roxburghiana I, 164
Cestrum parqui II, 136
Ceterach cordata I, 428
of ficinarum II, 64, 242
Cetraria II, 354, 366, 407; III, 368
cuculata III, 49
islandica III, 43
nivalis III, 49
Chacaya trinervis III, 175
Chaerophyllum silvestre II, 349
temulum II, 319
Chamaecyparis nutkaensis II, 383
Chamaedaphne calyculata II, 405
Chamaegigas interpidus I, 399, 400
Chamaenerium III, 23
angustifolium II, 248, 349, 368—
370
Ghamaerhodos sabulosa III, 280
Chamaerops III, 202
humilis II, 36, 68
Chara III, 313
aspera III, 328
baltica III, 328
tomentosa III, 328
Cheilanthes I, 260, 428, 434
lindheimeri I, 351, 352
multifida II, 161
pteridioides I, 494
tenuifolia I, 464
wrightii I, 352
Cheiranthus scoparius II, 93
Cheiridopsis I, 426
Chelidonium ma jus II, 290
Chenopodiaceae I, 300, 442, 449, 456,
463, 470, 482, 485, 492
Chenopodium I, 419, 425, 457, 530; II,
11
album I, 518
auricomum I, 421, 449, 452
murale I, 518
Chilanthus I, 427, 436
Chimaphila (= Pirola) umbellata II, 355
III, 194
Chionochloa (= Danthonia) flavescens II,
221; III, 182
rigida III, 182
Chlorophora excelsa I, 243, 299
Chlorophyceae III, 329, 331, 347
Chondrilla ambigua III, 201
Chordaria III, 335
Chorisia speciosa I, 481
Christolea III, 275
crassifolia III, 267, 270, 272-274,
277
pamirica III, 267
Chrysanthemum II, 218
coronarium I, 516
leucanthemum I, 16; III, 85, 87,
301
millefoliatum III, 91
Chrysobalanus icaco I, 185
Chrysocoma II, 167
Chrysophyllum cainito I, 482
Chrysothamnus II, 130
Chuquiraga II, 139; III, 178
aurea III, 179
crinacea III, 169
Chusquea coleu II, 150
quila II, 150
Cicerbita alpina III, 49
Cichorium III, 109
Cinclidotus fontinaloides III, 325
Cinnamomum camphora I, 481
ceylanicum I, 111
Circaea alpina II, 353
lutetiana II, 319
Cirsium canum III, 301
ellenbergii II, 80
heterophyllum II, 353; III, 49
Cissus I, 252, 260
cramerianus I, 395
juttae I, 397
subaphylla I, 255
Указатель латинских названий растений
399'
Cistus II, 32, 41, 44, 53, 54, 61, 64, 91,
93, 333; III, 369
albidus II, 32, 41, 52, 53, 67, 69
aurantium I, 114
crispus II, 242
incanus var. villosus II, 330
ladaniferus II, 61, 242
laurifolius II, 61, 69, 99
monspeliensis II, 61, 63, 64
salviaefolius I, 545; II, 41, 48, 64,
67; III, 37
tauricus II, 96
trifoliatus II, 68
villosus II, 61
Citharexylum quadrangulare I, 482
Citrullus I, 403
ecirrhosus I, 401, 466
colocynthis I, 479, 484, 487, 488,
500, 501
vulgaris II, 67
Citrus III, 147
aurantium I, 114, 133
Cladium I, 160; II, 206
mariscus II, 165; III, 135,311, 320,
321
Cladonia I, 418, 420; II, 93, 242, 354,
366, 407; III, 194 368
alpestris III, 49
islandica III, 49
rangiferina II, 420; III, 49
silvatica II, 254; III, 49
Cladosporium macrocarpum II, 413
Cladothrix lanuginosa I, 349, 350
Cleistogenes sinensis III, 257, 280
Clematis I, 341; II, 34, 180; III, 284
flammula II, 33, 71
leptodermis III, 293
orientalis III, 225
tangutica III, 267
vitalba II, 300
Clenelymus nutans III, 267
Cleome arabica I, 501
Clerodendron 184, 482
Cliffortia II, 157, 161
falcata II, 173
ruscifolia II, 161, 167
Clinogyne ugandensis I, 132
Clistoyucca brevifolia II, 129
Clostridium I, 450
pasteurianum III, 120
Cluytia polyfolia II, 171
pubescens II, 171
rubricaulis II, 171
Coccoloba unifera I, 182
Cochlearia anglica III, 349
arctica III, 19
Cochlospermum I, 233
Cocos nucifera I, 117, 481
romanzoffiana I, 481
Codonanthe gracilis I, 149
Coffea I, 112, 116; II, 13
arabica I, 111—113, 481
liberica I, 111—113
robusta I, 111—113, 124, 299
Coix lacrimans I, 132
Colchicum II, 101
Coleogyne I, 346, 347
ramosissima II, 129
Coleonema album II, 163
Collema III, 222, 230
Colletia II, 138
spinosissima III, 175
Colliguaja II, 136
integerrima II, 137
odorifera II, 137
Collospermum II, 222
Colobanthus crassifolius III, 56
Colocasia I, 129, 131, 135
cochlearia I, 133
odorata I, 133
Colophospermum mopane I, 247
Colpodium humile III, 141
Colubrina asiatica I, 184
Colutea arborescens III, 290
cilicica II, 99
Comarum palustre III, 18
Combretum primigenium I, 268
Commiphora I, 247, 260; III, 188
berberidifolia I, 397
dulcis I, 401
glaucens I, 395
oblanceolata I, 395
Condelia lycioides I, 341
Conopodium denudatum II, 304
Conospermum stoechadis II, 187
Convallaria majalis II, 311, 353
Convolvulus III, 211
althaeoides II, 67
ammanii III, 244
arvensis I, 421; III, 88
divaricatus III, 215
fruticosus III, 247
gortschakowii III, 257
lanatus I, 501
korolkovii III, 215
soldanella II, 115; III, 368
spicatus I, 528
Conyza canadensis III, 108 U
Copaifera mopane I, 297
Coprosma II, 221, 222; III, 181
Cordia alba I, 481
gharaf I, 541
heterophylla I, 481
myxa I, 481
Cordyline indivisa II, 222
Coriaria myrtifolia II, 64
plumosa II, 221
pteridioides II, 221
ruscifolia II, 150
Corinabutilon bicolor III, 178
Corispermum III, 202
aralo-caspicum III, 201
Cornicularia aculeata III, 368
Cornulaca I, 469, 474
Cornus mas II, 93, 300, 330
sanguinea II, 300, 319
sibirica II, 239
suecica II, 352, 353
Coronilla emerus II, 332, 333; III, 122.
varia II, 102
Corophium volutar III, 347
Cortaderia pilosa III, 63
400
Указатель латинских названий растений
selloana 111, 164
•Corydalis 1, 15; 11, 80, 277, 304
cava И, 307, 308, 319, 331
Corylus 11, 110; 111, 292
americana 111, 98
avellana 11, 239, 247, 250, 261,
297, 300, 319, 355
columa 111, 287
Corynephorus canescens 111, 128, 129,
368
Corypha umbraculifera 1, 126
Cosotropis 111, 285
Costus afer 1, 133
subbiflorus 1, 129, 132
Cotinus coggygria 11, 240, 330
Cotoneaster 11, 300; 111, 249, 285, 287,
290, 293
nummularia 11, 81
orbicularis 1, 528
Cotula plumosa 111, 61
Cotyledon 1, 424—426, 432
cristata 1, 379
orbiculata 1, 391, 394; 11, 168
paniculata 11, 167, 168
Couroupita quianensis 1, 119
Cousinia 111, 191, 207
Covillea glutinosa (= Larrea tridentata)
Crambe 111, 372
tatarica 111, 76
Crassula 1, 425, 432
arborea 11, 168
decipiens 1, 379
natans 11, 165
platyphylla 1, 429, 433
Crataegus 1, 542; 11, 99, 300, 319
monogyna 11, 250
sibirica 11, 239
sinaica 1, 528
Cratoneuron commutatum 111, 316
Cratystylis 1, 464
Crepis 11, 80
biennis 1, 16
paludosa 11, 319, 353
Crithmum maritimum 111, 334, 362
Crocus 11, 101; 111, 35, 49, 90
albiflorus 111, 52
Crotalaria 1, 222, 449, 494; 111, 188
aegyptiaca 1, 501
Croton 1, 255, 420
Crucianella filifolia 111, 215
maritima 1, 523; 111, 368
sabulosa 111, 215
Cruckshanskia giaciaiis 111, 178
Cryophy turn (= Mesembryanthemum)
cristallinum 1, 385, 402, 405, 406; 11,
92
Cryptandra spathulata 1, 466
Cryptocarya 11, 146
rubra 11, 136
Cryptogramma crispa 111, 50
Cryptomeria japonica 11, 346
Cryptostegia madagascariensis 1, 481
Cucumis melo 11, 67
Cucurbita digitata 1, 350
Culcasia 1, 137
Cunonia II, 158, 160, 171
capensis 11, 172
Cupaniopsis 11, 206
anacardioides 11, 205
Cupressus 1, 471; 11, 68, 77, 86; 111, 147
arizonica 11, 119
dipreziana 1, 484
goveniana 11, 116
horizontalis 1, 547
macrocarpa 11, 116, 117
sempervirens 1, 528; 11, 68, 81, 82
Curtisia 11, 172
faginea 11, 172
Curvuletum 111, 47
Cuscuta 1, 155
epithymum 11, 252
planiflora 1, 494
Cussonia 1, 273, 427, 436
thyrsiflora 11, 171
Cutandia 111, 211
Cyanidium 1, 420
Cyathea usambariensis 1, 131, 132
Cycas 1, 480
circinalis 1, 131, 133
Cyclamen europaeum 111, 55
neapolitanum 11, 332
Cyclolepis genistoides 111, 169, 170
Cyclosorus 1, 466
Cylicomorpha parviflora 1, 107, 110, 132
Cynanchum 1, 260
acutum 111, 302
sibiricum 111, 225
vincetoxicum 11, 307, 315
Cynara cardunculus 111, 149, 166
Cynodon 111, 191, 230
dactylon 1, 222, 490; 111, 225,
231 232
Cypella horbertii 111, 163
Cyperus 1, 296, 470; 111, 135
capitatus 111, 362
papyrus 1, 159, 160, 541
rotundus 1, 158
Cyrtandra 1, 128, 205
Cystopteris 111, 50
fragilis 11, 218; 111, 50, 60
Cystosira 111, 337, 338
Cytinus hypocistis 11, 64
Cytisus argenteus (= Argyrolobium lin-
naeanum)
balansae 11, 85
candicans 11, 242
linifolius 11, 242
ruthenicus 11, 338, 354;}111, 88
sessiliflorus 11, 245, 330
triflorus 11, 242
Cyttaria 11, 154
Daboecia cantabrica 11, 244
Dacrydium 1, 196, 198; 11, 213
beforme 11, 221
cupressinum 11, 154, 212, 221
fonkii 11, 154
Daeryodes 1, 102
Dactylis glomerata 11, 319, 369; 111, 53
hispanica 1, 520
Dactyloctenium I, 443
Указатель латинских названий растений
401
Dactylorchis maculata ssp. ericetorum II,
252
Daemia cordata 1, 501
Dalbergia 1, 135, 298, 480
sissoo 111, 287
Danae racemosa 11, 24
Daniella 1, 242
Danthonia (= Chionochloa)
fragilis 1, 475
micrantha 11, 162
Daphne 111, 285, 287
cneorum 111, 122
gnidinm 11, 40, 64
mezereum 11, 239, 300
oleoides 111, 289, 290
pontica 11, 240
Dasylirion 1, 344, 346
wheeleri 11, 119
Datura 1, 349, 401, 405
Daucus 111, 109
Davallia canaricum 11, 242
Daviesia 11, 186
divaricata 11, 190
Delphinium 111, 205, 249, 284, 292
litwinowi 111, 87, 88
orientale 11, 102
scaposum 1, 350, 351
Dendrobium 1, 260; 11, 222
crumenatum 1, 125, 126
Dendrocalamus hamiltonii 111, 291
Dendromecon rigidum 11, 109
Dendrosonchus 11, 92
Dentaria bulbifera 11, 331
Deschampsia 1, 214, 228
arctica 111, 31
antarctica 111, 56, 63
caespitosa 1, 222; 11, 319, 349,
353, 369, 370; 111, 31, 44, 54,
299
flexuosa 11, 250 , 251, 253 , 254,
316, 329, 349, 353; 111, 177
Desmodium 1, 184
Deutzia 111, 292
Dianthus 111, 249
brachyanthus 11, 87
caesius 111, 125
capitataus 111, 88
Silvester 111, 50
sinaicus 1, 528
Diapensia lapponica 111, 13, 49
Diarthron vesiculosum 111, 205
Dichanthium 1, 443
Dichostylis hamulosa 111, 225
Dichrostachys nutans 1, 290, 291
Dicksonia antarctica 1, 196, 200; 11, 201,
206—209
Dicranoloma 111, 62
Dicranum scoparium 11, 352
starkii 111, 47
undulatum 11, 352
Dictamnus albus 111, 122
Dictyota 111, 341
Didymaotus 1, 425
Dierama pendulum 1, 222
Digitalis purpurea 11, 316, 328
Digitaria 1, 443, 466
26-0434
milanjana 1, 272
Diosma 11, 173
succulentum 11, 163
Diospuros 1, 243
ebenus 1, 481
lotus 11, 240
melanoxylon 1, 243
Diostea juncea 11, 156; 111, 175
Diotis maritima 111, 362
Diplachne fusca 1, 158
paucinervis 1, 403, 405
Diplacus glutinosus 11, 109
Diplopappus 1, 424
Diploschistes albissimus 111, 209
Diplotaxis acris 1, 494, 498, 515
harra 1, 498, 513, 514
Dipterocarpus incanus 1, 122
tuberculatus 1, 238
Disa uniflora 11, 165
stairsii 1, 222
Discaria serratifolia 11, 156; 111, 175
toumatoa 111, 181
Dischidia 1, 144
imbricata 1, 148
Diselma archeri 11, 154, 220
Disylium racemosum 1, 245
Distichum capillaceum 111, 50
Distichlis 11, 130, 133; 111, 152, 163,
166, 356
humilis I, 422
spicata 11, 206; 111, 164, 350, 355
Dodonaea 1, 425, 449, 459, 466; 11, 167;
111, 285
thunbergii 11, 171
viscosa 1, 482; 111, 286
Donatia 11, 151, 220; 111, 63
Dorema aitschisonii 111, 205
ammoniacum 111, 192
Doronicum clusii 111, 43, 50
grandiflorum 111, 50
Dorycnium suffruticosum 11, 62, 94
Draba 11, 80; 111, 17, 293
dubia 111, 50
hirta 111, 26
nemarosa 111, 302
tomentosa 111, 50
verna 111, 90, 91
Dracaena draco 11, 92
Dracocephalum 111, 284
discolor 111,
heterophyHum 111, 277
ruyschianum 11, 354; 111, 88
Dracontomelum mangiferum 1, 107
Dracophyllum 11, 221, 222
traversii 11, 219
Drepanium cupressiforme 11, 33
Drimys winteri 11, 144, 149, 150
Drosanthemum 1, 382
Drosera 11, 185, 187, 196, 207, 416, 422;
111, 180
anglica 11, 253, 405
cuneata 11, 164
intermedia 11, 405
pallida 11, 182
rotundifolia 11, 253, 405
stolonifera II, 182
402
Указатель латинских названий растений.
trinervia II, 165
uniflora II, 149
Drosophyllum lusitanicum II, 242, 244
Dryandra mucronulata II, 178
nivea II, 187
prostrata II, 187
Dryadetum III, 42, 144
Dryas III, 29, 30
chamissonis III, 25
integrifolia III, 25
octopetala II, 246; III, 9, 21, 22,
42, 49
punctata III, 17
Drynaria I, 142, 145, 149, 151
Dryopteris II, 247
austriaca ssp. dilatata II, 319
filix-mas II, 311, 353
linnaeana II, 319, 353
phegopteris II, 353
robertianum III, 125
spinulosa II, 353
villarsii III, 50
Dupontia fischeri III, 17
Dysoxylum I, 115, 118
Earina II, 222
Echinocactus I, 419
Echinocereus I, 354, 364
Echinochloa pyramidalis I, 157
stagnina I, 157
Echinophora II, 104
spinosa III, 368, 371
Echinops III, 289
banaticus III, 129
bovei I, 494
spinosus I, 498, 500, 502, 520,
521, 523
Echiochilon fruticosum I, 523
Echium bourgaeanum II, 93
pustulatum II, 45
rubrum III, 85, 87, 90
sericeum I, 521, 523
vulgare II, 147, 148
Ehretia hottentotica I, 427
membranifolia I, 260, 424
obtusifolia I, 493
Ehrharta villosa II, 165
Eichhornia I, 157, 158
azurea I, 162
crassipes I, 157; III, 294
Ekebergia II, 171
Elaeagnus III, 201, 250, 253, 258, 292
angustifolia III, 197, 225
orientalis III, 225, 241
Elaeis (= Olpalme) I, 96, 242, 251, 273
guinensis I, 481
Elaeodendron II, 172
Elatine III, 139
Elatostema acuminatum I, 128
Elegia II, 165, 175, 176
stipularis II, 174, 175
Elettaria cardamomum I, 130, 133
Eleusine III, 188
tristachys III, 162
Eleutherospermum cicutarium III, 52
Elodea canadensis III, 318, 326, 328
Elymus III, 18, 201, 266, 280, 372
arenarius III, 363, 364
canadensis III, 97, 369
dasystachys III, 267, 276
giganteus III, 201
vancouveriensis III, 369
Elyna III, 41
myosuroides III, 25
Elytropappus rhinocerotis II, 166
Embothrium coccineum II, 156, 175
Empetrum II, 243, 259, 410; III, 24, 54,
63
hermaphroditum II, 254; III, 13,.
18, 21, 22
nigrum II, 251—253, 405; III, 44,
62, 180, 368
rubrum II, 149; III, 62
serrulatum II, 171
Encalypta streptocarpa III, 50
Encelia II, 61
farinosa I, 339, 368
Encephalartos I, 131, 133, 187, 425
hildenbrandtii I, 131
lehmannii I, 133
Endocarpon I, 498
Endymion (= Scilla) non-scriptus II, 250
Engelhardtia spicata I, 205
Enhalus III, 332
Enkianthus III, 292
Enteromorpha compressa III, 334
intestinalis III, 328
Ungulata III, 334, 335
linza III, 340
Entodon schreberi II, 419
Entophysalis granulosa III, 338
Epacris II, 218
paludosa II, 218
petrophila II, 218
serpyllifolia II, 218
Eperua I, 98, 102
Ephedra I, 476, 498, 537; III, 178,’201,
211, 247, 249, 250, 253, 266, 280, 289,
293
alata I, 475, 527, 528
altissima I, 494
distachya III, 369
gerardiana III, 290
przewalskii III, 257
strobilacea III, 191, 228
trifurca I, 338
viridis I, 346, 347
Ephemeropsis tjibodensis I, 154
Epidandrum ellipticum I, 150
Epilobium II, 221
alpestre III, 49
arcticum III, 26
hirsutum III, 258, 321
montanum II, 369
Equisetum arvense III, 17
fluviatile II, 409
giganteum III, 164
hiemale III, 194
pratense II, 353
ramosissimum I, 528; III, 225
silvaticum II, 353
Указатель латинских, названий растений
403
Eragrostis I, 245, 280, 282, 428, 447,
462, 470
bipinnata I, 479, 541
lehmanniana I, 282
lugens III, 162
minor III, 92, 201
paspaloides I, 282
spinosa I, 402, 447
Eranthis III, 205
Eremia II, 158
Eremophila I, 447, 449, 450, 454, 456,
459, 462, 464—466
Eremosparton III, 211, 214
Eriachne aristidea I, 449
Erianthus I, 479; III, 227
purpurascens III, 225
Erica II, 41, 83, 91, 157, 161, 173, 242,
255
arborea I, 222, 223; II, 41, 53, 55,
64, 92, 157, 240, 243; III, 369
australis II, 243
ciliaris II, 244
cinerea II, 53, 242, 243, 251, 253,
254, 258, 259
lusitanica II, 243; III, 183
mackiana II, 243
mediterranea II, 244
multiflora II, 40, 41, 49, 50, 53,
243
scoparia II, 41, 53, 55, 64, 243, 259
terminalis II, 243
tetralix II, 244, 251, 253, 256, 258,
259, 405; III, 372
umbellata II, 243
vagans II, 244, 257, 259
verticillata II, 46
Ericameria arborescens II, 109
ericoides II, 115
Ericinella I, 224
Erigeron III, 17
acer III, 88
ramosum III, 98
Eriocaulon I, 160
Eriocephalus I, 424, 429; II, 158, 167
Eriodendron I, 481
Eriogonum II, III
parvifolium II, 115
Eriophorum II, 251, 409, 417; III, 18, 29,
281
angustifolium II, 253, 405, 411;
III, 29, 30
scheuchzeri II, 411
vaginatum II, 252, 253, 405, 407,
420
Erodium I, 349, 350, 419, 420, 516
arborescens I, 498, 499
cicutarium II, 110; III, 205
glaucophyllum I, 498, 500
pulverulentum I, 487, 494, 501,
505, 507
Erophila verna III, 205
Eryngium II, 44, 62,101; III, 89, 110, 372
campestre II, 44, 62, 104
eburneum III, 163
maritimum III, 365, 386, 371
Eryobothria japonica I, 481; II, 136
Eryophyllum stoechadifolium II, 115
Erysimum III, 30 ;
hieracifolium III, 88 ।
Erythrina I, 246, 273
indica I, 121
Erythronium III, 97
Erythroxylon I, 260
Escallonia II, 136
arguta II, 137
virgata HI, 175
Espeletia grandiflora I, 213
Eucalyptus I, 199, 200, 233, 263, 419,
456, 462, 526; II, 177, 182, 206; III,
147
alba I, 233
astringens II, 180
baxteri II, 188, 195
calophylla II, 180, 183, 191
camaldulensis (= rostrata) I, 445,
460, 480; II, 49, 166, 177, 179
citriodora I, 114; II, 49
cladocalyx II, 166
clavigera I, 233
coccifera II, 179, 210—212, 220
confertifolia I, 233
coolabah I, 449
dichromophloia I, 466
diversicolor I, 445; II, 166, 172,
177, 180, 183, 185, 190
gamophylla I, 460, 462
gigantea I, 199, 200; II, 206, 207,
219
globulus II, 147
gomphocephala II, 49, 184, 203
grandis II, 177, 179
lesouefii I, 464, 465
longicornus II, 186
loxophleba II, 186
marginata II, 183, 184, 190, 191
melanophloia I, 463
microcorys I, 262
niphophila II, 216, 217
obliqua I, 200; II, 188, 206—208,
219
odora II, 191
patens II, 184
pauciflora II, 179
populnea I, 463
regnans I, 42, 199, 200, 445; IE,
177, 203, 206, 207, 219
rostrata (= camaldulensis)
rudis II, 49, 184
saligna II, 179
salmonophloia II, 180, 186
salubris II, 180
sideroxylon II, 49
simondsii I, 200; II, 207
subcrenulata II, 220
tereticornis II, 49
viminalis II, 49, 179
Euchylaena tomentosa I, 449
Euclea I, 297, 424, 425, 427, 436
acutifolia II, 171
polyandra II, 171
racemosa II, 171
pseudebenus I,, 401
26*
404
Указатель латинских названий растений
undulata II, 167
Eucryphia cordifolia II, 146, 149, 150
Eugenia gambolana I, 481
javanica I, 481
rosea I, 481
uniflora I, 481
Eulophia turkestanica III, 225
Eupatorium pallescens I, 191
Euphorbia I, 393, 425; 433, 470; II, 9, 44,
63, 158; III, 244, 245
aleppica II, 104
biglandulosa II, 96
brachiata I, 391, 394
canariensis II, 90
cervicornis I, 338
characias II, 33, 34, 62, 63
comuta I, 501
cuneata I, 253
dendroides II, 32
dinteri I, 393—395
heterophylla I, 349, 350
gracilis III, 85, 88
lactiflua I, 419
lignosa I, 393
mauretanica I, 426; II, 167, 168
myrsinites II, 104
nicaeensis II, 63
paralias I, 521; III, 368
peplis III, 362
polygonifolia III, 369
querichii I, 395
qunnnifera I, 396
segueriana II, 354; III, 92, 112,
113, 126, 201
tinctoria II, 104
tirucalli I, 251
uralensis III, 302
virgata III, 88
wheeleri I, 449
Euphrasia II, 217
alsa II, 218
Eupteria (Pteridium) aquilinum II, 308
Eurotia III, 272
ceratoides III, 258, 266—270,
273—276, 280, 290
lanata II, 131
Euryale I, 159
Eurynchium circinnatum II, 33
Euryops I, 224, 225, 424
Eustachys I, 282
Evonymus III, 287, 290, 292
europaea II, 239, 300, 319
japonica II, 12, 136
verrucosa II, 239
Exocarpus spartea II, 202
Exotheca I, 222
Fabiana densa I, 421
imbricata II, 156
Fagelia II, 161
Fagetum nudum II, 315
Fagonia arabica I, 501
bruguieri I, 469
glutinosa I, 487
kahirina I, 500, 514
mollis I, 498, 500
Fagraea fragrans I, 125
Fagus I, 42; II, 78, 241, 277, 278, 294,
309—311, 323—325, 388, 408
moesica II, 21
orientalis II, 21, 80, 240
silvatica I, 14, 23, 124; II, 21, 78,
233, 235—237, 239, 247, 296,
300, 317, 319, 321, 322, 325,
331, 336
taurica II, 21
Falcaria sioides III, 131
vulgaris III, 90, 110
Farsetia aegyptiaca I, 498, 501, 502,
514
Fascicularia bicolor II, 150
Faurea II, 157
Felicia II, 161
Ferocactus wislizenii I, 338, 339, 354,
356—360
Ferula I, 483; III, 214
asa-foetida III, 266
ferulago III, 90
foetida III, 205
oopoda III, 216
Festuca I, 210, 211, 214, 231, 422; III,
60, 115, 116, 121, 129, 166, 280, 282,
345, 348
abyssinica I, 225
erecta III, 61, 63
gigantea II, 319
glauca III, 124, 125
gracillima III, 180
indigesta II, 88
kilimanjarica I, 225
matthewsii III, 182
novazeelandiae II, 181, 221
nubigena I, 206
orthophylla I, 212
ovina II, 250, 254, 256; III, 21
ovina var. vallesiaca II, 101
pallesccens III, 177
pilheri ssp. supina I, 231
pratensis II, 370; III, 88
rubra III, 85, 89, 299, 300, 347,
349, 368
scabrella III, 95
sulcata II, 102; III, 53, 76, 77,
85, 87, 90—92, 113, 114, 117,
119, 120, 131, 140, 194, 200,
259, 263, 267, 299, 300
thalassica (-Puccinellia mariti-
ma)
vaginata III, 129
varia III, 53
violacea III, 51
Ficaria vema II, 304
Ficus I, 138, 256, 297, 399; II, 13, 78
bengalensis I, 138
carica var. rupestris I, 528
capensis II, 171
elastica III, 133, 148
hircana II, 240
myrmecophila I, 141
nitida (= retusa) I, 111, 138, 139
platypoda I, 466
Указатель латинских названий растений
405
pseudosycomorus I, 498, 528
rhynchospora I, 138
salicifolia I, 484, 494
subcalcarata I, 138
sycomorus I, 479
ulugurensis I, 138
vesiculosus III, 328, 335
Filago I, 537
Filipendula hexapetala III, 85, 87, 88,
90, 131, 194, 300
ulmaria II, 319, 338, 349
Fissidens julianis III, 328
Fitzroya II, 154
cupressioides II, 149, 154
Flagellaria I, 85
Fleury a lanceolata I, 132
Flindersia I, 104
Flotovia dacanthoides II, 149
Flourencia turifera II, 144
Fockea damarana I, 250, 251, 425
Foeniculum vulgare III, 149
Fontinalis III, 313
antipiretica III, 325, 328
hypnoides III, 328
Forstera II, 221
Fothergilla III, 287
Fouquieria I, 232, 326, 337, 338, 341,
353, 364—366
Fourcroya gigantea I, 131
Fragaria vesca II, 310, 311, 315, 353
viridis III, 88
Frangula II, 247
viridis III, 194
Frankenia I, 464, 478; III, 191, 207
hirsuta III, 136
pauciflora II, 205
pulverulenta I, 480, 487, 491
revoluta I, 521
thymifolia I, 487
thymoides I, 488
Franseria I, 341
bipinnatifida (= Ambrosia cha-
missonis)
dumosa 1, 342
Fraxinus I, 343; II, 277, 278; III, 292
excelsior II, 233, 239, 247, 250,
261, 298, 299, 309, 317, 319,
325, 336
omus II, 56, 71, 78, 93, 330, 331
pennsilvanica III, 101, 305, 306
toumeyi II, 119
xanthoxyloides III, 290
Fredolia aretioides I, 470, 482, 485, 488
Freycinetia I, 134, 197
Freylinia undulata II, 117
Frullania nervosa II, 93
Fuchsia II, 138
excorticata II, 221
lycioides II, 144
Fucus platycarpus III, 333, 335, 337,
340, 341
serratus III, 335, 337, 340, 341
vesiculosus III, 335, 337, 339—342
Fumana procumbens III, 124
Gagea II, 101; III, 90, 91, 205
bulbifera III, 90
erubescens III, 87
lutea II, 319
pusilia III, 90
Gaillonia III, 207
Gaimardia II, 151
Gaismardia III, 63
Galanthus nivalis II, 304—306
Galenia africana I, 426
Galeopsis II, 290
Galinsoga I, 419
Galium II, 11, 101, 349
aparine II, 290, 319
boreale III, 85, 88, 194
helveticum III, 50
pyrenaicum II, 87
saxatile II, 251
schultesii II, 353
silvaticum II, 319
sinaicum I, 528
verum III, 87, 88, 131, 300, 349
Garry a elliptica II, 109
wrightii II, 119, 120
Gasteria I, 425
Gastrochilus panduratum I, 240
Gaultheria II, 220, 221; III, 292
fragrantissima I, 208
nummularioides I, 208
Gaussia princeps I, 188,
Gazania varians I, 403
Geigeria I, 494
Geijera salicifolia I, 260
Gelocapsa deusta III, 338
Genista II, 242
anglica II, 245, 252, 258, 259
cinerea II, 245
germanica II, 245
pflosa II, 245, 257
(Cytisus) purgans II, 245
sagitalis II, 245
saharae I, 475
scorpius II, 41, 45, 245
tinctoria II, 245, 354
Gentiana I, 205; II, 218; III, 283, 284
algida III, 276
bavarica III, 47
ligulata III, 88
lutea III, 55
olivieri III, 205
Geoffroea (— Gourliea) decorticans IIt
134; III, 167
Geranium III, 136, 259
libanoticum II, 81
robertianum II, 307, 309, 319
sanguineum III, 88, 129
silvaticum II, 353; III, 49
tuberosum III, 205
ucrainicum III, 91
Geum packii III, 44
rivale II, 349
turbinatum III, 36, 44
urbanum II, 315, 319
Glaucium II, 102; III, 362
Glaux III, 135; 372
maritima III, 135, 294, 348, 349
Glechoma hederacea II, 319
406
Указатель латинских названий растений
Gleditschia triacanthos III, 73, 147
Gleichenia I, 190; II, 222
alpina II, 212
Globularia III, 122
alypum I, 494; III, 40
arabica I, 520, 527
Glyceria aquatica (= maxima) III, 299,
300, 322, 323, 326
fluitans III, 299, 300, 320
maritima (= Puccinellia mariti-
ma)
Glycyrrhiza III, 97, 227, 230
glabra III, 201, 225, 231, 232, 241
Gnaphalium supinum III, 45—47
Gomphocarpus fruticosus I, 428
Gompholobium II, 182
Gonioma camassi II, 171, 172
Goodenia cycloptera I, 449
Goodyera repens II, 355
Gourliea decorticans (= Geoffroea decor-
ticans) II, 134; III, 167
Grammatophyllum speciosum I, 142
Grevillea I, 91; II, 177, 186
juncea I, 462
juncifolia I, 449, 460
stenobotrya I, 449
Grewia III, 187
Grindelia III, 178
Griselinia'?II, 221
litoralis I, 137; II, 221
lucida I, 137
scandens II, 144
Guettardia speciosa I, 184
Guevina avellana II, 149
Gunnera I, 75; II, 221
Gymnocarpus III, 253
decander I, 516, 520
przewalskii III, 247
Gymnopteris I, 352
Gymnoschoenus sphaerocephala (= Me-
somelaena sphaerocephala)
Gymnosporia (= Maytenus) II, 160
acuminata II, 172
buxifolia II, 161
laurina II, 160
Gypsophila III, 244, 245
capillaris I, 501
capitulifera III, 273, 277
repens III, 51
stepposa III, 137
Gyrocarpus I, 232
americana I, 250, 258
Gyrophora III, 17
Haberlea II, 64
Haemanthus coccineus II, 161
Hagenia abyssinica I, 222, 223
Hakea I, 289, 460, 466; II, 57, 177, 190
acicularis II, 166
amplexicaulis II, 180
leucoptera I, 449
multilineata I, 369
pectinata II, 166
platysperma II, 199
prostrata II, 190
saligna II, 192
suaveolehs II, 193
Halimium II, 242
Halimocnemis mollissima III, 240
Halimodendron III, 195
(= Astragalus) ammodendron III,
197
halodendron III, 225
Halleria lucida II, 172
Halocharis hispida III, 206
Halocnemum III, 191, 195, 253
strobilaceum I, 478, 521, 522, 526;
III, 136, 141, 191, 201, 231,
232, 238, 240
Halodule III, 331
Halogeton III, 253, 257
alopecuroides I, 501
Halophila III, 332
Halostachos III, 253
Halostachys III, 230
belangeriana III, 240
caspica III, 227, 231
Haloxylon I, 473, 488, 510, 536; II, 86,
269; III, 223, 250
ammodendron III, 246, 252, 253,
257
aphyllum III, 144, 191, 211, 218,
219, 228, 229, 238, 240, 246
articulatum I, 326, 488, 529, 537,
539
persicum I, 534, 535, 537, 538;
III, 144, 191, 192, 211, 217,
219, 223, 228, 229
salicornicum I, 501, 502, 507,
526, 527, 533, 535, 539
Hammoda ramosissima III, 190
Haplopappus II, 136
Haplophyllum III, 204, 244, 245
Hapoptelia grandis I, 243
Hardenbergia II, 180
comptoniana II, 190, 203
Hartogia capensis II, 171
Hastia II, 221
Haworthia I, 425, 432
Hebe II, 221, 222, 389; III, 181
elliptica III, 62
Hedera helix I, 233; II, 33, 53, 56, 83,
310, 311, 319, 329, 383; III, 287
Hedysarum III, 21, 243
cephalotes III, 267, 274
mongolicum III, 246
Heleocharis III, 139, 166, 281
palustris III, 299
Helianthemum III, 122
chamaecistus (= nummularium)
II, 16; III, 129
elipticum I, 494, 523
kahiricum I, 498, 520
lippii I, 494
nummularium (chamaecistus)
Helianthocereus pasacana I, 336
Helianthus annuus I, 117; II, 272
rigidus III, 121
Helichrysum I, 222, 225, 230, 447; II,
165, 221; III, 127
arenarium II, 354; III, 88,127, 368
Указатель латинских названий растений
407
conglobatum I, 520
cordatum II, 190
newii I, 222, 225
stoechas II, 64; III, 368
stuhimannii I, 228
Helictotrichon (= A venastrum) deserto-
rum III, 259, 263
schellianum III, 194
Heliotropium arabianense I, 514
curassavicum I, 405, 406
europaeum II, 67
luteum I, 500, 501
micranthum III, 201
stenophyllum II, 135, 144
Helipterum I, 443, 447; II, 165
Helleborus foetidus II, 329, 330
Hemitelia II, 172
capensis II, 173
Hepatica nobilis II, 315
Heracleum III, 53, 109
Hereroa (= Mesembryanthemum) bossii
I, 391-394
Hermannia candidissima I, 434
Herpotrichia nigra II, 396
Heterodendron I, 450, 456
Heteropogon contortus I, 338
Heteromeles arbutifolia II, 109
Heteromorpha arborescens II, 171
Heterostacnys III, 170
Hevea brasiliensis I, 116
Hibbertia II, 200
hypericoides II, 191
glaberrina I, 466
sericea II, 198
stricta II, 191, 197
Hibiscus I, 466, 470
coulteri I, 338
sinensis III, 148
tiliaceus I, 183
Hieracium III, 88
amplexicaule III, 50
humile III, 50
intybaceum III, 51
pilosella III, 88, 368
staticifolium III, 50
umbellatum III, 365
Hierochloa odorata III, 77
Hillia ulei I, 147
Hippeastrum II, 134
uniflorum II, 139
Hippomane mancinella I, 183
ШррорЬаё rhamnoides III, 245, 268,
287, 293, 365, 367
Hippuris vulgaris III, 18, 323
Hohenbergia catingae I, 259
Hoodia I, 380, 424
currori I, 391, 393—395
Holcus mollis II, 308
lanatus III, 62, 182
Holoschoenus romanus II, 65
Holosteum umbellatum III, 90, 205
Honckenya II, 372
peploides III, 365
Hoppea I, 125
ferrea I, 99
Hordeum I, 479
murinum I, 350
stenostachys III, 164, 166
turkestanicum III, 267
vulgare I, 518
Hortensia scandens II, 150
Hottonia palustris III, 310
Hovea II, 218
Humiria I, 98
Humulis lupulus II, 300, 319
Hutchinsia alpina III, 38, 51
brevicaulis III, 51
Hyacinthus leucophaeus III, 84, 87
Hydnophytum I, 141, 181
Hydrangea III, 282, 292
Hydrylla verticillata I, 158
Hydrocharis III, 309
Hydrocotyle III, 164
calyculata II, 187
Hydrodoa bossiana I, 382, 383, 386
Hylocomium splendens II, 251, 252,
254, 259, 352
Hymenoclea I, 343
Hymenophyllaceen I, 75, 86, 147, 193,
200, 204, 220, 221
Hymenophyllum II, 144
capense II, 173
marlothii II, 173
peltatum III, 60
polyanthos I, 147
tunbridgense II, 173, 246
Hyoscyamusmuticus I, 474, 501, 505, 506
Hyparrhenia rufa I, 157
Hypecoum II, 102
Hypericum I, 222; II, 101
perforatum II, 310
pulchrum II, 252
Hyphaene I, 266, 273, 297
thebaica I, 157,; 296, 318, 479,
484, 493
Hypnum cupressiforme II, 251
Hypochoeris glabra II, 110
maculata III, 85, 88
radicata II, 2,18; III, 182
Hypocyrta radicans I, 149
Hypolaena fastigiata II, 198
Hypolitis sparsisora II, 173
Hystiopteris incisa III, 62
Idria I, 372, 373
columnaris I, 366, 367
Ifloga I, 516
fontanesii I, 501
Ilex I, 233; II, 158; III, 284, 292
aquifolium II, 50, 56, 71, 83,
240, 246, 247, 300, 319, 329
canariensis II, 92
mitis I, 220
paraguajensis I, 111
platyphylla II, 92
Impatiens I, 87, 221
noli tangere II, 308, 319
parviflora I, 132
valleriana I, 129
Imperata I, 95
arundinacea I, 163
408
Указатель латинских названий растений
cylindrica I, 192, 273; III, 225
Incarvillea III, 284
Indigofera III, 287
cytisoides II, 165
Inula crithmoides I, 521; III, 352
glandulosa III, 53
hirta III, 88
viscosa III, 362, 371
Iphiona I, 513
mucronata I, 514
Ipomoea pes carpae I, 182, 183
reptans I, 160
Iriartea orbignyana I, 108
Iris II, 62
aphylla III, 88
cnamaeiris II, 44, 62
pseudacorus III, 321, 323
pumila III, 90—92
sisyrinchium I, 527
tianschanica III, 267
Iseilema I, 443
Isoberlinia (= Julbemardia) I, 246, 249
doka I, 298
Isoetes II, 64; III, 315
Iva oraris III, 350
Ixiolirion III, 205
excelsa I, 542; II, 80, 96, 97, 99
foetidissima I, 542; macropoda III, 287 monosperma II, 128 II, 80, 99
nana III, 290 oxycedrus II, 34, 42, 44, 45, 52,
53, 81, 89, 96
pachyphloea II, 119 phoenicea I, 527; II, 49, 53, 83
procera I, 220, 347
pseudosabina III, 291
rigida III, 249
scopulorum II, 128
semiglobosa III, 290
squamata III, 287, 290
thurifera II, 83
utahensis II, 128, 129
virginiana II, 334
Jurinea arachnoidea III, 90
cyanoides II, 354; III, 88, 122
linearifolia III, 90, 92
polyclonos III, 201
Jussiena I, 162
repens I, 159, 160; III, 164
Justicia arenicola I, 403
Japandariba spruceana I, 147
Jasione montana III, 368
Jasminium III, 287
fruticans II, 96
Jatropha I, 338, 366
Jodina rhombifolia III, 167
Jubaea II, 146
spectabilis II, 138
Jucca I, 344, 346
glauca III, 102, 121
macrocarpa II, 119
whipplei II, 112, 113
Juglans I, 199; II, 12, 233
major II, 119
regia II, 240; III, 287, 291
Julbemardia (— Isoberlinia)
Juncel III, 177
Juncellus I, 158
laevigatus I, 403, 405
Juncus III, 299
acutus I, 479, 526; III, 164, 345,
352
arabicus I, 528
gerardi III, 54, 135, 136, 301, 348,
349, 353, 355, 356
jacquindii III, 51
maritimus I, 174; II, 206; III, 348
monanthos III, 51
squarrosus II, 252, 253
stygius II, 409
thomsonii III, 281
trifidus III, 43, 49, 51
Juniperus I, 471; II, 32, 77, 80, 86, 96,
97, 123, 249; III, 41, 94, 258, 283,
285, 287, 290, 292, 293
communis II, 250, 251, 255
communis ssp. папа II, 254; III, 32
drupacea II, 80
Kaempferia (= Afframomum) I, 132
Kageneckia II, 136
angustifolia II, 137
oblonga II, 137
Kalidium III, 195, 202, 230, 231, 253
caspicum III, 240
gracile III, 246, 257
Kallstroemia I, 349, 350, 354, 355
Karelinia caspica III, 227, 240
Kennedya prostrata II, 191
Keteleeria I, 244
Kiggelaria II, 171
Kingia australis II, 184, 185
Kleinia (= Senecio) I, 429, 433
neriifolia II, 90
Kniphofia I, 222
Knoltonia capensis II, 173
Kobresia capillifolia III, 267
pamiroalaica III, 269
tibetica III, 281
Kochia I, 432, 445, 450, 451, 453—455,
457, 458, 464, 465; III, 290
georgeii I, 455
glomerata I, 464
indica I, 516, 517
lanosa I, 455
oppositifolia II, 205
planifolia I, 454, 464
prostrata III, 114, 137, 140, 280
pyramidata I, 454, 464
sedifolia I, 432, 445, 453—455
sedoides III, 92
tomentosa I, 454, 464
triptera I, 454, 464
vestita I, 458
Koeleria III, 300
cristata III, 96, 121
delavignei III, 85
glauca II, 354; III, 129, 200, 368
Указатель латинских- названий растений
400-
gracilis I, 222, 225; III, 88, 90,
131, 140, 194
villosa III, 368
Koenigia islandica III, 20
Knautia arvensis III, 87, 88
Kuhnia III, 97
Kunzea II, 218
Lachnaea II, 173
Lachnostylis II, 172
Lagonychium faretum III, 191, 231
Lagotis III, 284
Laminaria digitata III, 336, 337
Laminaria hyperborea III, 337
Laminaria saccharina III, 335
Lamium II, 80
Lamium galeobdolon II, 312, 315, 319,
353
Lamium maculatum II, 307, 315, 319
Lantana camara I, 191
Laretia II, 139
compacta II, 134
Larix II, 232, 294, 380; III, 183, 285
dahurica II, 343, 345, 346; III, 7,
10, 35
decidua I, 43; II, 298, 325, 375,
377, 378, 391, 398; III, 35
europaea (= decidua) II, 317, 346
griffithii III, 292
kampferi II, 345
laricina II, 347
leptolepis ( L. kaempferi ) II, 345
sibirica II, 239, 343, 346; III, 7,
10, 12, 35, 258
Larrea I, 338, 339; III, 94, 147, 170, 178
cuneifolia III, 170—174
divaricata II, 116; III, 170—174
nitida III, 171—174, 178
tridentata (Covillea glutinosa) I,
337, 341, 342; II, 116, 128;
352, 364, 365, 368
Lasiagrostis III, 195, 250
bromoides II, 96
splendens III, 218, 244, 246, 257
Lasiurus hirsutus I, 501
Lastrea (= Dryopteris) robertiana III,
50
Lathraea sguamaria I, 15
Lathyrus maritimus III, 365, 369
montanus II, 310
pannonicus III, 88
tuberosus III, 194
vemus II, 239, 307, 310, 316
Launaea I, 537
(Zollikoferia) spinosa I, 483, 502,
521; II, 90, 91
Laurelia aromatica II, 146, 149
serrata II, 149, 150
Laurus II, 60
canariensis II, 92
nobilis I, 100, 547; II, 41, 53,
56, 60, 68, 72, 92, 240
Lavandula I, 494; II, 42, 53
coronopifolia I, 513, 514
latifolia II, 41, 45, 49, 53, 63
pubescens I, 527
stoechas II, 40, 41, 64, 67, 24?
Lecanora I, 498; III, 57
Lecidea I, 498
Ledum groenlandicum III, 43
palustre II, 353, 405, 410
Lemaireocereus I, 353, 363
Lemna I, 158, 162, 479
minor III, 294, 309, 313, 328
trisulca III, 309, 328
Lemanea spec. Ill, 325
Leontice enersmannii III, 216
Leontodon montanus III, 51
Leontopodium alpinum III, 273
Lepidium I, 443; III, 135, 276
deusiflorum III, 108
draba II, 102
perfoliatum III, 206, 241
Lepidosperma carphoides II, 196
gladiatum II, 202
laterale II, 198
Leptadenia spartium I, 296
Leptocereus I, 189
Leptospermum I, 200; II, 207, 218-
myrsinoides II, 191, 200
scoparium II, 210, III, 183
Leptotaenia muttifida III, 111
Lesguerella arctica III, 18, 25
Lespedeza dahurica III, 245
Leucadendron II, 162, 173
argenteum II, 159, 160
levisancus II, 163
Leucopogon II, 218
Leucospermum caniculatum II, 173
conocarpum II, 160, 161, 174
resiniferum II, 192
Liatris scariosa III, 97, 121
Libanotis intermedia III, 131
montana III, 300
transcaucasica III, 55
Libocedrus bidwillii II, 121
chilensis (Austrocedrus) II, 149r
154
decurrens II, 123, 126
uniferum (Pilgerodendron) II, 153
Ligularia III, 263
sibirica III, 52
Ligustrum japonicum II, 136
mutellina III, 45
vulgare II, 300, 330
Lilium tenuifolium III, 249
Limnanthemum I, 159; II, 165
Limnophyton I, 160
Limodorum abortivum II, 64
Limoniastrum I, 486
feei I, 482, 488
guyonianum I, 487
monopetalum I, 523
Limonium I, 526; III, 137, 345, 356, 372
caspica III, 136, 141
gmelini III, 135
(Statice) pruinosa I, 500
scoparium III, 231
suffruticosum III, 149, 238
Limosella III, 139
Linaria aegyptiaca I, 501
410
Указатель латинских названий растений
alpina III, 50
corifolia II, 102
cymbalaria II, 328
genistifolia II, 354
glacialis II, 87
nevadensis II, 87
odorata III, 201
vulgaris III, 349
Lindsaea ensifolia I, 466
Linnaea borealis II, 353
Linosyris III, 89, 194
villosa III, 90, 91, 131, 137
Linam perenne II, 354; III, 88
Liperia litoralis I, 403
Lippia wrightii I, 338
Liquidambar II, 233
orientalis II, 75, 76
Liriodendron tulipifera I, 124
Lissochilus I, 132
Listera cordata II, 252, 353
Lithocarpus I, 219, 244; II, 126
Lithops I, 305, 381, 392, 394, 425, 428,
430, 431, 433
Lithophyllam tortuosam III, 338
Lithothamnion III, 338, 339
Lithraea caustica II, 136, 137
Litsea III, 292
Livistona-Palmen I, 466
Loasa urens I, 419
Lobelia III, 315
deckenii I, 224, 225
keniensis I, 231
kamuriae I, 132
telekii I, 212
wollastonii I, 228
Loiseleuria procambens I, 23; II, 254;
III, 13, 22, 39, 41, 49
Loliam rigidam II, 185
Lomaria alpina III, 60
Lomatia ferruginea II, 149
hirsuta II, 149; III, 175
Lomatiam III, 97
hirsutam II, 156
Lonchitis hirsuta I, 132
Lonchocarpus I, 243, 297
Lonicera II, 34; III, 249, 284, 290, 292,
293
etrusca II, 33, 330, 331
implexa II, 33, 52, 64, 67; III, 371
periclymenam II, 300
tatarica II, 239
xylosteam II, 239, 300
Lophira alata I, 107
procera I, 107
Lophocereus I, 353, 363
Lophodermiam pinastri II, 396
Loranthus I, 155, 167, 181
Lotus comiculatus II, 88, 349; III, 135
scoparius II, 111
Loudetia simplex I, 271
Lucama valparadisea II, 136
Lamnitzera I, 164
litorea I, 164
racemosa I, 164
Lupinus alopecuroides I, 214
chamissonis II, 115
omatus III, 111
teucophyllus III, 111
Luzula II, 217, 221, 277
campestris II, 218
campestris ssp. pallescens III, 88
confusa III, 26
luzuloides II, 316
pilosa II, 319, 353
silvatica II, 329
spadicea III, 51
Lychnis flos-cuculi I, 16
Lyciarn I, 341, 344, 422, 424, 428, 464;
II, 167; III, 231, 246, 250
arabicam I, 500, 502, 514, 533,
535, 537, 539
australe I, 454
europaeam I, 521
tetrandram I, 401, 405
Lycopodiam II, 22, 222; III, 291
annotinam II, 250, 353
cernam I, 151
clavatam I, 151, 202, 222; II, 353
complanatam II, 355
saururus III, 62
selago II, 354
Lygeam spartam I, 490, 545, 546; II, 84
Lysimachia nummularia III, 302, 326
vulgaris II, 353; III, 321
Maba hamilis I, 260
Macaranga I, 103, 110, 113
usambarica II, 113
usambariensis II, 110
Macrocystis pyrifera II, 151; III, 338
Macrozamia fraseri II, 185, 190
macdonnellii I, 466
Maerua I, 476, 479
crassifolia I, 296, 484, 541
Magnolia III, 284, 292
grandiflora II, 12
Mahonia aquifoliam II, 383
Maihuenia patagonica III, 178
Majanthemam bifoliam II, 349, 353
Malcolmia III, 205
Malcolmia africana III, 241
litorea III, 368
Mallephora mollis I, 433
Malus II, 233
silvestris II, 247, 250
Malva I, 516
parviflora I, 494, 501, 505
rotundifolia I, 494
silvestris I, 349, 350
Mammillaria I, 354, 364
Mangifera indica I, 121
Manihot utilissima I, 130
Maniltoa gemmipara I, 109
Maranta arundinacea I, 130
Marattia fraxinea I, 131, 132
Marrubiam vulgare I, 350
Marsdenia australis 466
Marsilia 11, 64
aegyptiaca III, 225, 302
Martynia I, 349
Massonia I, 429
Указатель латинских названий растений
411
Matricaria III, 242
chamomilla-bayeri III, 135
lamellata III, 241
maritima III, 362
Matthiola I, 516
himilis I, 521
livida I, 501
sinuata III, 362
Maurocenia frangularia II, 172
Maxillaria picta I, 149, 150
Maytenus II, 136; III, 175
boaria II, 149, 156; III, 175
Meconopsis III, 283, 284, 292
Medicago III, 109
falcata III, 88, 131, 300
hispida II, 110
litoralis III, 368
marina III, 368, 371
romanica III, 131
Medinilla I, 204
Melaleuca I, 200, 464, 466; II, 11, 207
cuticularis I, 187
leucadendron, I, 190
Melampyram I, 15
cristatam III, 88
pratense II, 316
Melandriam dioecam II, 319
Melastoma malabaricam I, 191
Melianthus comosus I, 428
Meliea II, 101
nutans II, 353
regida III, 162
uniflora II, 358
Melilotus II, 101; III, 109
polonicus III, 201
Mentha microphylla I, 528
Menyanthes II, 409
trifoliata II, 253
Mercurialis perennis I, 15; II, 250, 304,
315, 316, 319 .
Mertensia maritima III, 334
Mesembryanthemam I, 294, 424—426,
428, 431, 432, 483; II, 158, 165, 168,
205
chilense II, 115
crystallinam I, 521
ferox I, 433
forskalei I, 501, 505, 507, 509,
515, 516
hamatam I, 429, 433
nodifloram I, 521
salicornioides I, 382—386
saxicolam I, 433
c.f. spinosam I, 429, 433
unidens I, 429, 433
Mesomelaena II, 249
(Gymnoschoenus) Sphaerocephala
I, 200; II, 207, 210
Metalasia II, 157, 167
Metrosideros II, 158, 213, 221
diffusa II, 222
lucida II, 221
robusta I, 137, 198; II, 221
viminalis II, 192
Microcachrys tetragona II, 154, 220
Microcoleus III, 209
Microcycas calocoma I, 188
Microlepis speluncae I, 132
Micromeria I, 211; II, 102
Microula tibetica III, 280
Mielichhoferia II, 144
Miliam effusam I, 15; II, 239, 308, 315,
319, 349; III, 51
spinosam II, 139; III, 178
Mimetes II, 160
Mimusops I, 298
Miscanthidiam I, 156
Mitraria coccinea II, 150
Moehringia trinervia II, 290
Molinia II, 308
coerulea II, 65, 251—254, 256,
259, 355
Molinam spinosam III, 174
Mollugo I, 349
Moltkia II, 102
ciliata I, 475
Monotrapa hypopitys II, 365; III, 194
Monstera II, 13
deliciosa I, 131; 133; 137
Montia II, 221
aphylla III, 169
lamprospenna III, 20
Mora excelsa I, 102
Moringa aptera I, 541
ovaliifolia I, 395
peregrina I, 528
Moorcroftia III, 282
Morns alba III, 73, 230
Muehlenbeckia II, 136
cunninghatii 1, 449
Mulgediam macrophyllam II, 239
Meraltia II, 157
Musa I, 117, 204, 130, 133
paradisiaca I, 130
sapientiam I, 133
textilis I, 117
Musanda I, 103
smithii I, 96
Muscari I, 516; II, 101
Mycetanthe I, 155
Myoporum I, 450; II, 203, 204
insulate II, 202
platycarpam I, 454, 450
Myosotis III, 30
arenaria III, 91
silvatica III, 84, 85, 87, 88, 90
Myrceugenia II, 136
correifolia II, 144
exsuca II, 156
Myrceuginella apiculata II, 150, 155
Myrianthus arboreus I, 109, 113
Myrica I, 273
burmannii II, 171
califomica II, 110
fa ya II, 92
gale II, 253, 257, 258, 300
Myricaria III, 280
elegans III, 282
germania III, 290
Myriophyllam I, 158; III, 325
spicatam III, 135, 323
Myrmecodia tuberosa I, 141, 142, 143
412
Указатель латинских названий растений
Myrothamnus I, 323
flabellifolia I, 395, 398, 399
Myrsine gilliana II, 171 '
Myrtus I, 547; II, 240 г **
communis II, 41, 46, 64, 67
nivellei I, 494
Myzodendron II, 154
punctulatum II, 152
Najas III, 310
major I, 466
marina III, 328
Nananthus vittatus I, 428, 430, 432, 433
Nanophyton erinaceum III, 238, 257
Nardus stricta II, 250—254, 256; III, 49,
51
Narthecium ossifragum II, 253, 405
Nassauvia glomerulosa III, 178, 179
Nasturtium brachycarpum III, 302
Nelumbium I, 159, 160
caspicum III, 230
indicum III, 302
Nemoreto — Pinetum alnosum II, 355
Neogaya simplex III, 267
Neottia nidus-avis I, 15; II, 239
Nepenthes I, 144, 160, 187
Nepeta III, 290
septemcrenata I, 528
tibestica I, 494
Nephrolepis cordifolia I, 466
Neptunia I, 162
Nerium II, 112; III, 285
oleander I, 487; II, 68, 76; III, 286
Nesaea fleckii I, 405
Neuropogon melaxanthus III, 60
Nicotiana glauca I, 401, 405, 419, 428
rustica I, 117
velutina I, 466
Nidorella I, 312
Nigella III, 205
Nipa fraticans I, 164, 180, 181
Nitella III, 313
flexilis III, 328
Nitraria III, 195, 201, 231, 250
retusa ( = tridentata) I, 491, 492,
500, 515, 521, 526, 528, 539
schoberi I, 449; III, 246 , 252, 253,
268
sibirica III, 257
sphaerocarpa III, 257
tridentata (N. retusa)
Nivenia II, 173
Noaea I, 537, 538; II, 101
macrantha I, 326
mucronata I, 521, 527; III, 190
spinosissima III, 207
Nolina microcarpa I, 344, 346; II, 119
Nonnea pulla III, 88
Nostoc I, 450; III, 230, 324
Notenia amaniensis I, 132
Nothofagus I, 75, 197—200, 443; II, 140,
154, 206, 207, 212, 213, 214, 221,
222, 247, 270, 388, 389; III, 180, 181
alpina ( — procera) II, 149
antarctica II, 146, 149, 151, 152,
153, 156; III, 175, 178
betuloides II, 146, 151—153
cliffortioides II, 220
cunninghamii I, 199, 201; II, 206?
208, 209, 219, 220
dombeyi II, 145, 146, 149, 150,
151, 152, 155, 156; III, 175
fusca II, 215, 220
gunnii II, 207
menziesii II, 220
nervosa II, 145, 149
nitida II, 146, 151
obliqua II, 133, 145, 149
pimilio III, 175
procera ( = nervosa) II, 145, 149
pumilio II, 146,149,151—153,155.
pumilio-antarctica II, 149
Notholaena I, 352, 395
maranthae II, 64
vellea I, 527
Nothopanax II, 221
colensoi I, 137; II, 221, 222
simplex I, 137; II, 221
Nothoscordium montevidensis III, 163
Nuphar III, 311, 318, 319
lutea III, 310, 323, 328
Numphaea I, 159; II, 165; III, 311, 319
coerulea I, 159
lotus I, 159
Nyssa III, 311
Obione III, 348
pedunculatum III, 136, 1415
Ochradenus baccatus I, 514, 533, 535, 539"
Ocotea bullata II, 172
foetens II, 92
Oenothera I, 351; III, 365
oakesiana III, 369
Oidiodendrum griseum II, 413
Olea II, 37, 57, 60, 68, 158, 160, 172;
III, 285
capensis ( — O. laurifolia)
chrysophylla I, 220
cuspidata III, 286, 289
europaea I, 518, 522; II, 52, 55r
56, 59, 68, 69, 76
exasperate II, 171
laperrini I, 484
laurifolia ( = capensis) II, 172
oleaster II, 35
verrucosa I, 424, 436; II, 160
Oleandra africana I, 151
Olearia I, 200, 459; II, 203, 204, 207,
218, 221, 222, 389
axillaris II, 202
Olinia II, 160, 172
cymosa II, 172
Olneya tesota I, 339, 368
Onobrychis arenarius III, 87, 88
cornuta II, 81
sibirica III, 194
Ononis minutissima II, 63
vaginalis I, 523; II, 45
Onopordon acanthinun III, 93
Onosma arenarium III, 122
aucheriana II, 102
Указатель латинских названий растений
413
tinctorium Ill, 92
Cphioglossum bucharicum III, 225
vulgatum III, 349
Ophrys II, 94
Cpuntia I, 212, 422; III, 103
acanthocarpa I, 354
arborescens I, 336
arbuscula I, 354, 363
bigelovii I, 354
blakeana I, 361, 363
discata I, 360
engelmanii I, 362
ficus-indica II, 92
fulgida I, 339, 340, 354
phaeacantha I, 361, 362
polyacantha I, 346, 347; III, 121
subulata I, 211
toumeyi I, 363
versicolor I, 338, 362, 363
Crchideen I, 103, 155, 244
Oreobolus II, 151, 218
Oreocereus I, 421
Origanum syriacum I, 528
vulgare III, 129
Orites II, 218
acicularis II, 220
Omithogalum III, 90, 194
altissimum II, 167
Orobus albus ( — Lathyrus pannonicus)
III, 85, 90
pallescens III, 90
vernus II, 353
Oroya borchersii I, 211
Oryza perennis I, 162
Osmitopsis II, 165
Osmunda regalis II, 246
Ostrya II, 240
carpinifolia II, 93, 330, 331
Osyris abyssinica (= O. compressa)
compressa (abyssinica) I, 427, 436
Othonna dentata II, 165
protecta I, 391, 394
Otostegia III, 207
Oxalis I, 419, 424; II, 139, 161
acetosella II, 250, 257, 308, 309
313, 319, 349, 353, 362, 367
corniculata I, 494
natans II, 165
Oxylobium II, 218
Oxyria digyna III, 23, 24, 36, 43, 50
Oxytenanthera I, 297
Oxytropis III, 21, 30, 282
aciphylla III, 280
chilliophylla III, 267, 272
immersa III, 267
poncincii III, 267
riparia III, 225
Pachycereus I, 333, 335, 359, 363
Pachystima II, 130
Pachycormus discolor I, 367
Pachypodium I, 432
giganteum (P. lealii)
namaquanum I, 395
lealii (P. giganteum) I, 397
Paconia tenuifolia III, 90
Palmen I, 190, 266
Panax III, 292
Pancratium maritimum III, 368
sickenbergeri I, 501
trianthum I, 483
Pandanus I, 100, 161, 193; III, 291
tectorius I, 183, 186
Panicum I, 256, 296, 299, 447, 493, 509;
III, 95, 162, 188
racemosum III, 370
turgidum I, 476, 500—502 , 507,
515, 516, 526, 527
virgatum III, 97
Papa ver III, 30, 205
radicatum III, 14, 26
sendtneri III, 51
Pappea II, 158
Pappophorum III, 257
Parietaria obtusa I, 350
ramiflora II, 328
Parinarium holstii I, 110
Paris quadrifolia II, 319, 353
Parkinsonia (Cercidium) I, 401, 408
aculeata I, 368
Parmelia II, 33; III, 17
conspersa II, 164
coreyi III, 57
Parrotia persica II, 240
Parrya excapa III, 267
Parsonsia I, 270
Pasania densiflora II, 110
Paspalum III, 162
quadrifarium III, 163, 16>6
repens I, 162
vaginatum III, 164
Passerina II, 161
Paxiodendron usambarense I, 110, 221
Pedicularis III, 20, 284, 290, 292
abrotanifolia III, 267
comosa III, 87, 88
condensata III, 56
Pedilanthus macrocarpus I, 353
Peganum harmala I, 487; II, 102; III, 191,
202, 222
nigellastrum III, 257
Pegolettia II, 167
Pelargonium I, 424; II, 161, 162, 203, 204
cucullatum II, 165
drummondii II, 202
Pellaea hastata I, 246, 398, 428
mucronata I, 352
viridis I, 132
Pelvetia III, 337
Pemphis I, 184
Penaea II, 165, 173
Penicereus I, 354
Pennisetum III, 188
cladestinum I, 158, 272, 273, 422
dichotomum I, 476, 490, 500, 501,
£4 q_____Kx к
flaccidum III, 245
Pentacme I, 241, 243, 244
Pentaschistis II, 165
borussica I, 225
Pentstemon wrightii I, 350
414
Указатель латинских названий растений
Pentzia I, 424, 429; II, 158, 167
Peperomia II, 172
coquimbensis II, 144
crystalline I, 141, 419
retusa II, 173
Perezia diversifolia II, 139
Pergularia tomentosa I, 514
Periploca laevigata I, 484
Persea indica II, 92
lingue II, 146
Pertusaria lactea II, 164
Pesania I, 244
Petalidium variabile I, 403
Petalostemum III, 97
Petasites paradoxus III, 50
spurius III, 299
Petrocallis pyrenaica III, 50
Petrosimonia eranchiata III, 231
crassifolia III, 136, 141
Peucedanum III, 89, 135, 194
arenarium II, 354
ostruthium III, 49, 51
pschvaicmn III, 55
ruthenicum III, 90, 91
Phacelia brachyloba II, 114
tanacetifolia I, 350
Phaeophyceae III, 329, 331
Phalaris III, 133
arundinacea III, 300, 326
Phelodendron III, 148
Pherosphaera ( = Dacrydium) hookeri II,
220
Philadelphus III, 282
Phillyrea II, 34, 57
angustifolia II, 33, 45, 50, 55, 64
latifolia II, 332; III, 371, 372
media II, 33, 40, 41, 52, 54, 55,
60, 240
Philippia I, 98, 228
excelsa 1, 222
Phleum II, 101; III, 205
alpinum III, 63, 301
phleoides III, 88, 194
pratense II, 14
Phlomis II, 101
aurea I, 528
herba-venti II, 102
lychnitis II, 41, 44, 52 , 53
pungens III, 87, 90, 91, 92, 110
tuberosa III, 85, 88, 92, 131, 194
viscosa I, 326
Phoenix II, 13; III, 147, 202
canariensis II, 92, 136
dactylifera I, 469, 479, 484, 522;
III, 188
reclinata I, 131, 133, 156, 161
Pholiurus pannonicus III, 135
Phormidium III, 209
Phormium colensoi II, 222
tenax II, 214; III, 183
Photinia arbutifolia II, 112
Phragmites I, 160; III, 133, 225, 227,
230, 231, 242, 250, 251, 311, 313,
356, 372
communis I, 157, 403, 405, 479,
488, 521; III, 135, 164, 232,
246, 258, 294, 299, 322, 323
Phycopeltis I, 153
Phylica arborea III, 62
buxifolia II, 165
paniculate II, 163
Phyllachne colensoi II, 223
Phyllanthus fluitans I, 162
Phyllocactus I, 141
Phyllocladus I, 196, 200; II, 207, 219 —
222
alpinus II, 221
Phyllodoce II, 243; III, 24
aleutica III, 53
Phyllospadix III, 331
Phyllota II, 201
pleurandroides II, 197
remota II, 200
Physica I, 498
Physocarpus malvaceus II, 130
Phyteuma spicatum II, 319
Picea I, 347; II, 78, 232, 294, 380, 388„
418, 419; III, 258, 283-285
abies ( = excelsa) II, 20, 78, 238„
298, 325, 328, 335, 343, 346,
347, 367, 374, 376, 383, 384
aristata II, 122
asperate III, 248—250
engelmannii II, 120, 122, 130, 38S>
excelsa (P. abies) II, 20, 78, 317,.
385, 386
fennica II, 21, 343
glauca II, 347
mariana II, 347; III, 7, 10
medioxima II, 21, 344
morinda III, 287, 292
morindoides III, 292
obovata II, 21, 325, 343; III, 7,
10, 12, 258
omorica II, 88, 376
rubens II, 347, 348
sitchensis II, 123, 125, 254
spinulosa II, 292
uralensis II, 21, 344
Piceetum herbosum II, 352, 400
hylocomiosum II, 373
myrtillosum II, 352; III, 144
nemorosum II, 352
oxalidosum II, 352, 353; III, 144
polytrichosum II, 352
sphagnosum II, 352, 400
typicum II, 352
vacciniosum II, 352
Picemmorinda III, 287
Pickeringia montana II, 108
Pieris ovalifolia III, 287
Pilgerodendron ( = Libocedrus) II, 153
uviferum II, 149, 153, 154
Pilularia II, 64
Pimelea II, 218
Pinguicula chilensis II, 149
Pinetum cladinosum II, 354, 355
herbosum II, 354
hylocomiosum II, 354, 355
myrtillosum II, 354, 355, 364
nemorosum II, 354, 355
oxalidosum II, 354, 355
Указатель латинских названий растений
415.
polytrichosum II, 354
sphagnosmn II, 354, 400
steposum II, 354
substepposum II, 354
uligino-ledosmn II, 355
vacciniosmn II, 354, 355
Pinus I, 83, 205; II, 11, 77, 80, 123, 299;
III, 258, 285
aristata II, 122, 388
banksiana II, 347
brutia II, 47, 48, 80, 81
canariensis II, 91
caribaea II, 179
cembra (== sibirica) I, 43; II, 325,
375, 377, 383—385, 391, 394,
395, 397, 398; III, 35, 39
cembra ssp. sibirica II, 343, 344,
346
cembroides II, 119, 120
contorta II, 123, 124
edulis II, 128
excelsa III, 287
flexilis II, 388
gerardiana III, 290
halepensis II, 46, 47, 49, 53, 60,
83, 86, 166, 316
hartwegii II, 388; I, 219
jeffreyi II, 127
junnanensis II, 232
khasia III, 284
lambertiana II, 123, 126
longifolia (P. roxburghii)
maritima ( = pinaster) II, 45, 47,
53, 166, 172, 183; III, 370
massoniana II, 232
merkusii I, 190, 244, 245
monophylla II, 128
montana I, 43; II, 300, 346, 375,
378, 388, 391, 402; III, 32, 49
nigra II, 241, 329
nigra ssp. pallasiana II, 80, 99
nigra ssp. poiretiana II, 78, 95,
96
pallasiana II, 96
pence II, 88, 375, 377
pinaster ( = maritima) II, 78, 95,
183, 246; III, 371
pinea II, 48, 49, 64, 68, 166, III,
369
pithyusa var. stankevici II, 96
ponderosa II, 123,126, 129; III, 74
pumila II, 346
pungens II, 334
radiata II, 115, 116, 147, 148, 166,
172, 183
resinosa II, 334
rigida II, 334
roxburghii ( = longifolia) 111,285,
287
salzmanni II, 53
sibirica ( = cembra) II, 239, 346,
405; III, 35, 258
sibirica v. humistrata II, 346
silvestris I, 15; II, 13, 20, 78, 95,
96, 241, 246, 250, 257, 298,
317, 325, 328, 335, 336, 346,
355, 374, 375, 402, 405; III.
11, 154, 368
strobus II, 329, 334, 335
tabulaeformis III, 249
uncinata II, 78
virginiana II, 834
Piper capensis I, 132
umbellatus I, 132
Piptadenia buchanani I, 110, 1
Piptochaetium III, 162
Pirola chlorantha II, 355
media II, 353
minor II, 353
( = Moneses) uniflora II, 353
rotundifolia II, 353
( = Ramischia) secunda II, 338.
353
Pirns II, 233; III, 290, 291
amygdaliformis II, 93
elaeagrifolia II, 93, 99, 292, 293-
mains II, 330
rotundifolia II, 310
Pistacia I, 476; II, 86; III, 285
atlantica I, 484, 542
lentiscus I, 547; II, 35, 37, 41,
46, 50, 51, 55, 64, 67, 83, 332
mutica I, 542; II, 96
palaestina II, 60, 71
lentiscus III, 369, 371
mutica III, 191, 230
terebinthus I, 482; II, 33, 41, 51,
60, 71, 240, 332
vera II, 12
Pistia I, 157, 160, 162
Pittosporum I, 443; II, 158
conifolium II, 222
phillyraeoides II, 192
Pituranthos I, 476, 483
tortuosus I, 501, 506, 513, 514,
516, 520, 521, 522
Placodium tominii III, 222
Plagianthus II, 187
holmsii I, 464, 465
Plagiosetum refractum I, 449
Plagiothecium undulatum II, 252
Plantago I, 209, 210, 350, 351, 470; III,
135, 206
ablicans I, 516, 521
coronopus II, 206; III, 351
nivalis II, 87
ovata I, 498, 501, 515
rigida I, 210
coronopus III, 351, 352, 371
lanceolata III, 88, 206, 351
major III, 62, 109, 351
maritima III, 135, 348, 349, 351,
372
media III, 88, 90, 109
urvilleana III, 88
Platan us I, 344; II, 120
orientalis II, 76; III, 101
wrightii II, 119
Platycerium I, 142, 181; III, 291
willinckii I, 145
Platylophus trifoliata II, 171, 172
Plectrachne I, 291, 300, 446, 458, 459
416
Указатель латинских названий растений
Plectranthus 111, 287
Plectronia 11, 171
Pleiogynium 1, 258
Pleomele (=Dracaena) papahu 1, 129,131
Pleuropogon sabinii 111, 17, 18
Pleurozium schreberi 11, 251, 254, 352,
410
Pluchea chingoyo 11, 133
Poa 11, 217, 218; 111, 60, 166, 300
abbreviate 111, 17
alpigena 111, 31
angustifolia 111, 77, 194
annua 11, 218; 111, 51, 62
bulbosa 11, 101; 111, 191, 201,
204—206, 241, 282
caespitosa 111, 181, 183
colensoi 11, 221; 111, 182
flabellata 111, 62, 63
glauca 111, 25
gymantha 1, 422
lanigera 111, 162
lanuginosa 111, 370
laxa 111, 51, 267
litwinowiana 111, 267
nemoralis 11, 319, 353; 111, 50
palustris 111, 299, 300
pamirica 111, 267
pratensis 11, 218; 111, 31, 56, 177,
300, 301, 349
relaxa 111, 267
sandbergi 111, 111
sinaica 1, 228; 111, 190
trivialis 11, 315
tibetica 111, 267
trivialis 111, 299
Podalyria calyptrata 11, 165
Podocarpus 1, 75, 196, 222, 223, 273, 274;
11, 158, 171, 212
alpinus 11, 218
andinus 11, 149, 154
chilina ( = salignus) 11, 154
dacrydioides 11, 214, 221
falcatus 11, 172
hallii 11, 221
latifolius 11, 172
nivalis 11, 221
nubigenus 11, 149, 154
salignus ( = chilina) 11, 149, 154
Podonosma syriacum 1, 544
Pohlia natans 11, 251
Polemonium 111, 30
Polygala comosa 111, 88
myrtifolia 11, 165
Polygonatum 111, 249
latifolium 11, 239
multiflorum 11, 319, 331
officinale 111, 88
Polygonum 111, 20, 292
affine 11, 290
amphibium 11, 14; 111, 321, 323
aviculare 111, 362
barbatum 1, 160
bistorta 111, 89
hispidum 1, 162
hissaricum 111, 259
maritimum 111, 362
nitens 111, 259
viviparum 111, 24, 25, 48, 249,
267, 274, 301
Polylepis 1, 75, 210, 216
tomentella 1, 421
Polypodium 1, 181; 11, 247
australis 111, 60
isoetifolia ( = P. phymatodes)
lanceolatum 1, 146; 11, 173
phymatodes ( = isoetifolia) 1, 151
verrucosum 1, 139
vulgare 111, 50, 60
Polypogon lusosus 111, 245
Polyporus robustus 11, 69
Polyscias elegans 1, 258
polybotrya 1, 110
Polystichum 1, 199
vestitum 11, 215
Polytrichum commune 11, 218, 248, 249
juniperinum 11, 242; 111, 43
norvegicum 111, 47
piliferum 11, 242
sexangulare 111, 46
strictum 11, 407; 111, 29
Pomaderris 1, 200; 11, 207
Populus 11, 76, 119, 277; 111, 115, 201,
250, 258, 259, 283, 291, 302
alba 11, 166; 111, 147, 197, 293
balsamifera 11, 347; 111, 287
canescens 11, 166
ciliata 111, 293
davidiana 111, 249
deltoides 111, 101
diversifolia 111, 225, 226, 241, 246,
253
euphratica 1, 484; 111, 191
hybrida 111, 197, 230
nigra 11, 293; 111, 198
pruinosa 111, 225, 253
tremula 11, 233, 250, 293, 300, 317;
111, 139, 194
tremuloides 11, 120, 347
Porlieria 11, 138, 144
Porphyra 111, 341
atropurpurea 111, 334, 340
Portulacaria 1, 425; 11, 158
afra 11, 168
Posidonia 111, 361
oceanica 111, 331, 332
Potanimia mongolica 111, 247
Potamogeton 1, 158, 479; 11, 165; 111,
139, 310
crispus 111, 326
densus 111, 318
filiformis 111, 328
fluitans 111, 326
gramineus 111, 323
lucens 111, 326
natans 111, 311, 326, 328
obtusifolius 111, 135, 328
pectinatus 111, 135, 328
perfoliatus 111, 326
vaginatus 111, 328
Potentilla 11, 101
alba 111, 88
anserina 111, 135,244, 258, 348, 349
Указатель латинских названий растений
417
arenaria ill, 124
argentea 111, 88, 300
caulescens 111, 50
clusiana 111, 42
erecta 11, 251
fruticosa 111, 249, 283, 293
moorkroftii 111, 267
multifida 111, 274, 275
nivea 111, 26
opaca 111, 85, 88
opacifonnis 111, 90
pamiroalaica 111, 267, 272
patula 111, 90
recta 111, 87
reptans 11, 65
sterilis 11, 316, 319
Poterium spinosum 1, 326, 545; 11,
42
Pozoa 11, 139
Pothos 111, 291
Prasiola crispa 111, 57, 58
Premna 1, 184
Prenanthes purpurea 11, 307
Primula 1, 205; 111, 283, 284, 292
algida 111, 269
auricula 111, 50
boveana 1, 528
elatior 11, 319
hirsuta 111, 50
nivalis 111, 267
officinalis 111, 88
ruprechtii 111, 53
veris 1, 124
vulgaris 11, 25
Pringlea antiscorbutica 1, 74; 111, 60, 61
Prionium serratum 11, 165
Prosopis 1, 337—339, 341, 342, 344, 345,
421, 480; 11,116; 111, 94,101,149,179
algarobillo 111, 159, 161
alpataco 111, 161
caldenia 111, 148, 160—162, 167
denudans 111, 161, 178
flexuosa 111, 161, 167
globosa ( = Prosopis striata)
juliflora 11, 133
nigra 111, 159, 161
striata ( = globosa) 111, 161
tamarugo 11, 133
Prostranthera 11, 218
Proteaceae 1, 75, 200, 255, 289, 453;
11, 157, 162, 164, 165, 177, 185, 187,
193, 195, 199, 225
Protea 11, 57, 157, 173
arborea II, 160, 174, 175, 176
cynaroides 11, 162
grandiflora 11, 160, 161, 162, 164
incompta 11, 161
kilimandscharica 1, 222, 273, 274
mellifera ( = P. repens) 11, 610
nerifolia 11, 160, 174
repens 11, 160, 174
Proustia pungens 11, 144
pyrifolia 11, 136
Prunella grandiflora 111, 88
vulgaris 11, 353; 111, 88
Prunus 1, 199; 11, 110; 111, 282, 287, 291
27—0434
avium 11, 233, 239, 250, 319
fruticosus 111, 194
ilicifolia 11, 109, 112, 113, 114
laurocerasus ( = lusitaniea) 1, 99;
11, 72, 240, 242, 327, 382, 383
mahaleb 11, 93, 330 >
padus 11, 239, 250, 300, 317, 319;
111, 194
spinosa 11, 250, 300; 111, 90
tortuosa 111, 191
virens 11, 120
Psammochloa villosa 111, 246
Psathyrostachys 111, 195
Pseudotsuga 11, 120, 329, 380
douglasii 11, 317
macrocarpa 11, 126
menziesii ( = taxifolia) 11, 110,
123, 125—127, 130, 148
taxifolia 11, 298
Pseudowintera 11, 221
Psidium quajava 1, 191, 526
Psila boliviensis 1, 421
Psilocaulon 1, 428, 433
Psilotum 1, 202
Psoralea 1, 443; 111, 97
aphylla 11, 173
bituminosa 11, 45, 64, 67, 96
decipiens 111, 222
floribunda 111, 98, 121
pinnata 11, 165, 173
Pteridium 1, 190; 11, 180, 182
aquilinum 1, 95, 131, 132, 222;
11, 243, 245, 247, 252, 308,
338, 349, 355
esculentum 111, 182
Pteridopappus 11, 220
Pteris biaurita 1, 132
Pterocarya 11, 240
Pterocarpus 1, 247, 298
Pteronia 1, 424; 11, 158, 167
Pterospartum 11, 242
tridentatum 11, 242
Ptilagrostis 111, 280
Ptilium crista-castrensis 11, 352
Ptilotrichum ( = Alyssum) spinosum 11,
85
Ptilotus latifolius 1, 449
Puccinellia 111, 372
limosa 111, 135
glaucescens 111, 166
maritima ( = Festuca thalassi-
ca = Glyceria maritima —
= Atropis maritima) 111, 344,
347, 349, 372
phryganoides 111, 345
Pulmonaria mollissima 11, 239
obscura 11, 239
officinalis 11, 315, 319, 353
Pulsatilla 111, 125
patens 111, 84, 85, 88, 194
Purshia 11, 130
Pycnophyllum, 1, 209, 210, 422
Pycnostelma lateriflorum 111, 244
Pyrenacantha 1, 251
Pyrethrum till, 194
santolinoides I, 124, 528
418
Указатель латинских названий растений
Pyrus I, 137
Pyrrosia confluens II, 205, 206
Puya chilensis II, 135
Querceto-Aceretum campestris II, 263,
267
Querceto—Betuletum II, 272
Betuletum molinietosum II, 308
carpinetum II, 272, 308
Flaxinetum II, 263, 267
Pinetum ledosum II, 355
Pinetum myrtillosum II, 355
Pinetum pteridiosum II, 355
Quercetum ilicis II, 33, 43
ilicis gallopovinciales II, 33
Quercus I, 42, 83, 219, 244, 245; II, 72,
86, 96, 126, 240, 241, 277, 278, 311,
317, 334; III, 284, 291
aegilops I, 542; II, 64
agrifolia II, 110, 115
alba II, 231
alnifolia II, 81
arizonica II, 119
borealis II, 298
calliprinos II, 60, 81
castanopsis III, 285
cerris II, 93
chrysolepis II, 110, 112, 126
coccifera I, 547; II, 32, 37, 41,
44, 45, 51, 55, 60, 64, 83, 119
coccifera var. calliprinos I, 544,
547
dilatata III, 287
dumosa II, 108, 109, 112, 114
emoryi II, 118, 119, 120
engelmannii II, 110
fendleri II, 129
gamblei II, 120, 130
griffithii II, 232
hypoleuca II, 119, 120
ilex I, 471; II, 26, 31—36, 41—46,
48, 50—52, 55, 56, 59, 60, 64,
66, 68, 69, 72—76, 78, 82, 83,
89,136, 225, 240, 332, 333, 398;
III, 285, 290
incana III, 287
induta I, 205
infectoria I, 542; II, 81
ithaburensis I, 542, 544; II,’ 37
keloggii II, 110
libani II, 80
lobata II, 110
lusitanica (Q. mirbeckii)
lusitanica ssp. canariensis II, 242
lusitanica ssp. faginea II, 83
macrocarpa III, 99, 101
macrolepis I, 542
mirbeckii ( = Q. lusitanica) II,
242
oblongifolia II, 118, 119, 121
pedunculate I, 124; II, 325
petraea ( = Q. sessilis) I, 14; II,
95, 233, 246, 250, 299, 300,
311, 321, 322, 336; III, 66
pseudomoluccana I, 205
pubescens II, 56, 59, 64, 71—76,
78, 93, 95, 96, 99, 240, 246,
299, 330, 331, 398; III, 66, 122
pyrenaica ( = Q. toza) II, 78, 93,
246
reticulata II, 119, 120
robur I, 14; II, 73, 95, 166, 233,
238, 239, 246, 250, 261, 299,
300, 317, 319, 321, 322, 335;
III, 65, 66
sessilis ( = Q. petraea)
suber II, 35, 37, 50, 55, 56, 64,
66, 78, 83, 225, 243
subznoHis II, 120
toza ( = Q. pyrenaica)
turbinella II, 129
wislizenii II, 110, 126
Quesnelia testudo I, 150
Quillaja saponaria II, 136, 137
Quincnamalium II, 139
Rafflesia I, 106, 155
arnoldi I, 155
patma I, 105
Ralfsia verrucosa III, 333, 334 341,
Ramalina I, 498; II, 33
Ramischia (Pirola) secunda III, 194
Ramondia II, 64
pyrenaica II, 64
Randonia I, 474
Ranunculus II, 80, 217, 218, 221; III, 97,
204, 249, 290, 300, 310
acer I, 16; III, 301
acetosellaefolius II, 87
alpestris III, 42
aquatilis III, 311, 325
auricomus II, 319
baudotii III, 323, 327, 328
ficaria I, 15; II, 304, 306, 319
gracialis III, 23 , 24, 36, 38, 43 , 51
illyricus II, 102
muelleri II, 218
nivalis III, 17
pallasii III, 18
parnassifolius III, 51
pedatus III, 302
platanifolius III, 49
pygmaeus III, 47
rufosepalus III, 267
severzovii III, 205
sulphures III, 17
trichophyllus III, 320
tricuspis III, 281
Raoulia II, 221
lucescens III, 182
Raphanus sativus III, 149
Raphia I, 156, 161
Raphithamnus carpus II, 149
spinosus II, 144
Rapistrum III, 149
Ravenala I, 129
madagascariensis I, 130
Reaumuria I, 207, 250
hirtella I, 498, 500, 527
muricata I, 480
palaestina I, 529, 537, 539
soongarica III, 244, 249, 257
Указатель латинских названий растений
419
trigyna III, 280
Reseda I, 470
lutea II, 102
Restionaceen I, 200; II, 157, 162, 164,
165, 174, 177, 225
Restio II, 157, 165
australis II, 218
Retama I, 536
duriaei I, 539
retam I, 475, 483, 488, 501, 502,
506, 514, 527, 529, 534—537,
539
Rhacomitrium III, 21
lanuginosum II, 252—254; III, 62,
259, 334
lanuginosum var. pruinosum II,
221
Rhagodia baccata II, 202
spinescens I, 449
Rhamnus I, 542; II, 34, 66; III, 249, 258
alaternus II, 33, 52, 55, 67, 96,
332
cathartica II, 300
crocea II, 109, 112, 114
erythroxylon III, 245
frangula II, 300; III, 194
palaestinae III, 191
Rheum denticulatum III, 267
nobile III, 293
spiciforme III, 280
turkestanicum III, 216
Rhigozum II, 167; I, 295, 426, 436
Rhinanthus major III, 301
Rhipogonum II, 221
Rhipsalis I, 138, 144, 150, 189
cassytha I, 149, 151
Rhizocarpon III, 17
Rhizophora I, 166, 171, 174, 176, 179
conjugata I, 177
mangle I, 170, 174, 175, 178
mucronata I, 164, 176, 180, 181
Rhodiola aretica III, 30
pamiroalaica III, 267
Rhododendron I, 187, 205, 219; II, 227,
388; III, 32, 39, 282, 284, 285, 291—
293
adenophorum III, 283, 284
caucasicum III, 33
ferrugineum III, 44
hypenanthum III, 290
javanicum I, 204
nivale III, 292, 293
ponticum II, 72, 240, 241
ponticum ssp. baeticum II, 242,
243
retusum I, 208
rupicola III, 283
Rhodomela III, 335
Rhoiocissus II, 172
capensis II, 172
Rhus II, 109, 130, 158, 161, 162
burchellii I, 436
ciliata I, 436
coriaria II, 96, 240
cotinus II, 93
erosa I, 427, 436
glabra III, 98
laevigata II, 171
lancea I, 401, 424, 428
villosa I, 493
Rhynchospora I, 160
alba II, 253, 405
jusca II, 253
Rhynia II, 13
Ribes II, 156; III, 284, 287, 290, 292
alpinum II, 300
cucullatum III, 175
grossularia II, 319
magellanicum III, 175
nigrum II, 300
orientalis III, 290
uva crispa II, 300
Riccia I, 428; III, 141
Ricciocarpus I, 162
Richea continentis II, 218
scoparia II, 220
Richteria pyrethroides III, 267, 272
Ricinus I, 401
communis I, 512
Rinodina III, 57
Robinia II, 148
pseudacacia II, 166, 289, 290, 317;
III, 73, 147, 278
Rochea coccinea II, 165
Roridula II, 157
Roripa kerneri III, 135
Rosa II, 34; III, 249, 284, 290, 292
acicularis III, 194
arabica I, 528
canina II, 300
cinnamomea III, 194
fendleri II, 120
glutinosa II, 81
pomifera II, 300
sempervirens II, 33
sericea III, 293
webbiana III, 289, 290
Rosmarinus II, 32, 41, 60, 61, 83
It officinalis II, 41, 42, 49, 53, 64,
67; III, 371
Royena ( = diospyros) I, 424, 427 , 428
glabra II, 171
lucida II, 173
rugosa II, 173
Rubia aucheri II, 81
peregrina II, 33, 52, 64, 242
Rubus I, 85, 134; II, 34, 329; III, 287,
292
arcticus III, 301
caesius II, 300, 319
chamaemorus II, 246, 405; III, 23
cissoides II, 221
idaeus II, 239, 300, 319, 368, 369
saxatilis II, 349, 353; III, 154
serpens II, 300
Rulingia I, 466
Rumex I, 350, 421, 494
acetosa III, 88, 301
acetosa ssp. haplorrhizus III, 300
acetosella II, 218, 329; III, 62,
88, 182
alpinus III, 49
27*
420
Указатель латинских названий растений
graminifolius III, 17
nivalis III, 50
obtusifolius III, 62
scutatus III, 50, 125
vesicara I, 498
Ruppia maritima III, 328, 349
spiralis III, 328
Ruscus aculeatus II, 33, 52, 54, 67, 69,
240, 332
hypoglossum II, 240
pontica II, 96
Ruta angustifolia II, 63
Saccharum spontaneum I, 163; III, 225,
226
officinarum I, 130
Sagittaria I, 160; III, 328
Saintpaulia usambariensis I, 133
Salaxis II, 158
Salicetum j herbaceae III, 45, 46, 47
Salicornia !, 369, 405, 464, 478, 526; II,
130; III, 135, 170, 294, 346, 353, 356,
357, 359, 360, 372
ambigua III, 164
australis I, 174; II, 205
europaea III, 135, 136, 231, 232,
240, 347, 355, 356
fruticosa I, 480, 492, 522; III,
345, 352
herbacea I, 522; III, 141, 301,
347, 348, 358
mucronata III, 355, 356
pulvinata I, 422
radicans III, 345
strictissima III, 347
Salix I, 343; II, 166, 233, 277; III, 18,
20, 29, 101, 134, 245, 249, 258, 283—
290, 302, 306
acmophylla III,'225
acuminata III, 198
alba II, 325
amygdalina III, 293
arctica III, 22, 25
aurita II, 300
australis III, 230
capensis I, 424, 428
caprea II, 239, 325
cascadensis III, 44
caspica III, 197
cinerea II, 300; III, 194
flabellaris III, 290
glauca III, 23
hastata III, 290
herbacea II, 246; III, 9, 21, 45—
48, 51
livida III, 194
myrsinites III, 22
opaca III, 293
oxycarpa III, 287
purpurea II, 325
reptans III, 20, 365—367, 372
retusa III, 37, 51
rosmarinifolia III, 197
serpyllifolia III, 51
songarica III, 21
taxifolia II, 120
wallichiana III, 290
xerophila III, 194
Salsola I, 294, 402, 405, 406, 425, 428,
433, 434, 443, 449, 453, 470, 476,
478, 537; III, 192, 207, 211, 214, 217,
289, 372
aphylla I, 281
arbuscula III, 191, 238
autrani I, 539
carinata III, 206
cracca III, 231
dendroides III, 230—232, 240
gemmascens III, 238
inermis I, 529, 530, 539
kali III, 108, 110, 201, 359, 362,
369
laricifolia III, 244
paletzkina III, 215
passerina III, 244, 257
richteri III, 191, 215, 217, 218
rigida III, 190, 191, 240, 241
ruthenica III, 201
sclerantha III, 206
soda III, 136, 301
subaphylla III, 228
tetrandra I, 521, 532, 533, 538
turcomanica III, 240
villosa I, 537, 538
zeyheri II, 167
Salvadora I, 408; III, 187
persica I, 401, 484, 541
Salvia I, 494; II, 102, 111, 114; III, 76
aegyptiaca I, 510
aethiopis III, 91
austriaca III, 77
cleistogama I, 435
lanigera I, 516, 521
nemorosa III, 90, 91
nutans III, 87, 88, 90, 113, 114,
117
pratensis III, 85, 87, 88, 131
stepposa III, 194
Salvinia I, 158, 162; III, 309
Sambucus I, 199, 343; III, 292
nigra II, 300, 319
racemosa II, 300
Samolus I, 174; III, 135
Sanguisorba II, 101; III, 259, 301
minor II, 315
Sanicula europaea I, 15; II, 319 $ 331
Sansevieria I, 249, 253, 281, 393
Santalum II, 202
Sapindus drummondii II, 119
Sarcinanthus utilis I, 134
Sarcobatus vermiculatus II, 130, 131
Sarcocaulon I, 391, 424
Sarcostemma australis I, 466
Sargassum III, 342
Sarmienta repens II, 144
Sarothamnus II, 242, 249, 258, 327
balticum II, 242
catalanicus II, 242
scoparius II, 64, 245, 300, 328, 329
welwitschiae II, 242
Satureja montana II, 61
virgata II, 137
Указатель латинских названий растений
421
Saussurea III, 284, 293
dridactyla III, 279
kuntdiana III, 279
pamirica II, 267
pygmaea III, 249
thomsoni III, 279
wernerioides III, 279
Saxegothea conspicua II, 149, 154
Saxifraga III, 292, 293
aizoides III, 25, 26
aizoon III, 50, 122, 125
biflora III, 51
bryoides III, 50
caespitosa III, 26
ceraua III, 23, 26, 48
cotyledon III, 50
exarata III, 50
moschata III, 50 r
nivalis III, 26
oppositifolia II, 246; III, 9, 17,
25, 50, 51
rotundifolia III, 49
Scabiosa II, 101
canescens III, 124
maritima III, 368
ochroleuca III, 88
Scaevola II, 202—205
crassifolia II, 202
spinosa I, 464
Schefflera volkensii I, 221
Scheuchzeria painstris II, 405, 407, 409
Schima wallichii I, 205; III, 291
Schinus II, 136
longifolia III, 167
molle II, 133
patagonica II, 156; III, 175
terebinthifolius I, 522
Schismus calycinus I, 501, 507
Schistostega pennata III, 50
Schizaea fistulosa II, 149
pectinata II, 165
Schizanthus II, 139
Schizolobium excelsum I, 127
Schizothrix I, 420
Schmidtia quinqueseta I, 280
Schoenia I, 447
rosea I, 449
Schoenus nigricans II, 253, 405
tepperi II, 196
Schorea robusta III, 291
Schotia II, 158
Schilla II, 308
bifolia (Endymion) II, 239, 304
nutans ( = Endymion nonscriptus)
II, 304
Scirpus I, 158, 479; II, 165
americanus III, 166
califomicus III, 164
darvlus III, 328
giganteus III, 164
hamulosus III, 302
holoschoenus I, 528
lacustris III, 299, 310, 311, 322
maritimus III, 135, 136, 328, 349.
368
micheliana III, 302
nodosus II, 202
parvulus III, 328
supinus III, 302
tabernaemontani III, 135, 328,
368
Sclerocarya I, 250, 268, 298
Sclerochloa dura III, 206
Scleropoa maritima III, 368
Scleropodium purum II, 33
Scolymus hispanicus II, 104
Scorobolus heterolepis III, 96
Scrophularia nodosa II, 319
Scorzonera III, 205
parviflora III, 135, 301
purpurea III, 85, 88, 124
Scutia myrtina II, 172
Scyphiphora hydrophyllacea I, 164
Scytonema II, 93; III, 324
Secamone alpini II, 172
Sedum I, 150; II, 44, 45, 63; III, 125, 293
album III, 50
altissimum II, 62
annuum III, 50
atratum III, 50
boloniense III, 124
dasyphyllum III, 50
nicaeense II, 44, 62, 63
reflexum II, 328
Selaginella I, 86, 101, 128, 132, 193,
220, 221, 244, 323, 338, 351, 352
denticulate II, 64
rupicola II, 120
selaginoides II, 246
Selinicereus I, 189
Sempervivum I, 150; III, 125
arachnoideum III, 50
montanum III, 50
Senecio I, 74, 230, 231, 350, 470; II,
221, 222, 389; III, 136, 178, 179, 292
calcarius I, 219
campester III, 84, 85, 87, 88
coronopifolius I, 494; III, 206
cottoni I, 225
crassifolius III, 370
filaginoides III, 178
fuchsii II, 309
jacobaea III, 88
keniodendron I, 212
kilimanjari I, 224
kirkii II, 222
longiflorus ( = Kleinia) I, 391,
394, 395
majeri-johannis I, 231
neriifolia ( = Kleinia) II, 90
vernalis II, 102
Sequoia II, 109, 123, 125, 154
gigantea ( = seguoiadendron) II,
109, 123, 125, 154, 383
sempervirens I, 42; II, 109, 123—
125, 270
Serratula heterophylla III, 88
radiata III, 90
xerahthemoides III, 76, 90, 91
Seseli III, 110
tortuosum III, 90
Sesleria coerulea III, 122, 125
422
Указатель латинских названий растений
varia 111, 51
Seslerietum anatolicae 11, 80
Sesuvimn j digynum 1, 389
portulacastrum 1, 182; 111, 164
Setaria 1, 245
incrassata 1, 157
Shorea 1, 125, 243; 111, 291
Sibbaldia procumbens 111, 44, 46
Sicyos 1, 350
Sida 1, 443, 449
Sieglingia decumbens 11, 250
Sieversia ( = Novosieversa) 111, 29
ciliata 111, 111
Silene 1, 494
acaulis 111, 25
alpina 111, 50
leucophylla 11, 528
nocteolens 11, 93
schimperiana 11, 528
Siler trilobium 11, 239
Simmondsia chinensis ( = califomica) 1,
370
Sisymbrium austriacum 111, 88
bourgaeanum 11, 93
reboudianum 1, 494
subspinescens 111, 207
Sisyrinchium 111, 108
laxum 111, 163
pratense 111, 163
Sium erectum 111, 326
Skimmia 111, 292
Smelowskia calycina 111, 267
Smilax 11, 92, 240
aspera 11, 33, 41, 50, 52, 54, 67,
332
kraussiana 1, 137
Smirnovia turkestana 111, 211, 214, 228,
229
Solanum 1, 419; 111, 163
andigenum 1, 421
elaeagnifolium 1, 349
incanum 1, 541
melongea 11, 67
Soldanella pusilia 111, 46
Solenanthus stylosus 111, 267
Solidago virgaurea 11, 353; 111, 24, 97
Solorina crocea 111, 49
Sonchus arvensis 1, 420
asper 1, 349, 350
oleraceus 1, 419; 111, 62
Sonneratia 1, 171, 174, 176, 181
acida 1, 164, 176
alba 1, 164, 175
Sophora 111, 283, 290
moorcroftiana 111, 293
tetraptera 11, 146
Sorbaria lonicera 111, 282
Sorbus 11, 72, 299; 111, 282
aria 11, 300
aucuparia 11, 233, 246, 250, 251;
111, 290
domestica 11, 71, 72, 93
intermedia 11, 336
torminalis 11, 56, 71, 93, 239, 300
Sparmannia africana 1, 117; II, 171
Sorghastrum nutaus 111, 97
Sparganium ramosum 111, 326
Spartina 11, 130; 111, 356
alterniflora 111, 348
arundinacea 111, 62
ciliata 111, 370
glabra 111, 350, 355
montevidensis 111, 163, 164
patens 111, 350
pectinata 111, 97
sownsendii 111, 348
stricta 111, 348
townsendii 111, 348, 360
Spartium 11, 92
iunceum 11, 53, 56, 64, 244
Spathelia brittonii, 1, 184
Spartocytisus 11, 91, 111
supranubium 11, 93
Spathodea campanulata 1, 120, 121
Spatholobus 1, 238
Spergularia 111, 348, 372
marginata 111, 206, 349, 359, 360
Sphaeralcea 1, 350
Sphagnum 1, 98, 156, 160, 225, 228;
11, 248, 249, 252, 253, 256, 338, 362,
405—408, 413, 415, 417-421; 111,
18, 62, 132, 180
acutifolium 11, 149, 406, 407
apiculatum 11, 406
balticum 11, 406
cristatum 11, 218
cuspidatum 11, 406, 415
dusenii 11, 415
fuscum 11, 406, 407, 410, 415
magellanicum ( = medium) 11,
406, 407, 415—417, 420
medium (Sph. magellanicum)
papillosum 11, 406, 415
parvifolium 11, 415, 420
pulchrum 11, 406
recurvum 11, 416, 417
rubellum 11, 406, 407, 415, 417
tenellum 11, 406
Sphenopus divaricatus 1, 521
Spinifex 1, 182, 291
hirsutus 11, 202—205; 111, 350
litoreus 1, 184
longifolius 11, 202—204
Spiraea 111, 92, 249, 258, 283, 290, 292
crenata 111, 194
hypericifolia 111, 64, 194
trilobata 111, 249
Sporobolus 1, 428; 11, 130
arenarius 111, 362, 371
heterolepis 111, 96, 121
pungens 1, 403
rigens 1, 429
robustus 1, 158
spicatus 1, 158
virginicus 11, 206
Spyridium globulosum 11, 202
Squamaria leutigera 111, 209
Stachys aegyptiaca 1, 500, 513, 514
arvensis 1, 419, 420
officinalis 11, 33
recta 111, 88
palaestina 1, 545
Указатель латинских названий растений
423
silvatica 11, 319
Staechelina dubia II, 49
Stapelia 1, 424; -11, 158
Staphyllea pinnata 11, 93
Statice 1, 486; 111, 206, 207
delicatula 1, 487
gmelini 111, 301
latifolia 111, 90
limonium 111, 352
pruinosa ( = Limonium) 1, 521
tataria 111, 90, 91
Stauntonia 111, 292
Stelechocarpus burahol 1, 117
Stellaria 111, 20, 284
graminea 111, 88
holostea 11, 239, 308, 310, 319, 353
humifusa 111, 30
media 1, 419; 111, 62
nemorum 11, 319, 349; 111, 51
Stenomesson coccineum 1, 419
Stenophragma thalianum 111, 302
Sterculia 1, 297, 318, 399
Stereocaulon 111, 21
vesuvianum 11, 221
Stereospermum 1, 273, 297
Steversia reptans 111, 43
Stible 11, 173
Stipa 1, 210, 211, 214; 11, 110; 111, 68,
76, 77, 86, 91, 95, 101, 115, 116,
119—122, 166, 243, 258, 266, 272,
282, 289, 299
barbata 111, 190
brachychaeta 111, 165, 166
brevifolia 111, 243, 249
bungeana 111, 245
capillata 111, 89—92, 124, 128,
131, 140, 194
glareosa 111, 244, 245, 249, 267,
269, 270, 272, 273, 275, 276
goliba 111, 257
gyneroides 111, 167
humilis 111, 179
ichu 1, 210
Joannis 111, 85, 86, 90, 128, 129
lagascal 1, 521, 541; 11, 101
lessingiana 111, 66, 90, 91, 113,
114, 131, 140, 194, 195
neesiana 111, 162
nitida 1, 450
papposa 111, 162, 163
parviflora 1, 490
pennata 11, 354; 111, 124, 194
pulcherrima 11, 101, 102; 111, 53,
128
rubens 111, 131, 194
rubentiformis 111, 90
semibarbata 11, 195
spartea 111, 96, 100, 108, 121
speciosa 111, 177
stenophylla 111, 88, 90, 92, 128
szovitziana 1, 541; 111, 241
tenuissima 1, 541, 545, 546; 11,
84; 111, 167
tibestica 1, 494
trichotoma 111, 165, 166
ucrainica 111, 90, 91, 131
Stirlingia 11, 189, 190, 191
Stoebe 11, 157, 161
Stomatium erminium 1, 433
Stratiotes 111, 309
Streptanthus arizonicus 1, 350
Streptocarpus ovata 1, 132
Strobilanthes cemuus 1, 12, 125, 205
Strychnos 1, 260, 298
Styphelia cunninghami 11, 192
Styrax officinalis 11, 37 ,71
Suaeda 1, 478, 522, 526; 11, 130; 111, 135,
170, 253, 372
asphaltica 1, 537
atrumentifera 1, 428, 433, 434
australis 11, 205
confusa 111, 231
fruticosa 1, 402, 405, 406, 521,
522, 546
glauca 111, 358, 359
maritima 111, 135, 141, 164, 301
microphylla 111, 231, 232, 240
monoica 1, 173, 281
moquini 1, 341
palaestina 1, 537
pruinosa 1, 521
salsa 111, 135
vermiculata 1, 487, 528; 11, 90
Subularia 1, 228
Succisa pratensis 11, 14
Sweetia dasycarpa 1, 250
Sympegua 111, 253
regelis 111, 268
Symphoricarpus 11, 130; 111, 98
Sympieza 11, 158
Symplocos 1, 245
Synadenium carinatum ( = Euphorbia)
1, 132
Syndesmanthus 11, 158
Syntrichia (Tortula) ruralis 111, 90
Syringa 11, 139; 111, 249
Syzygium cordatum 1, 273
Tabemaemontana holstii 1, 113
Taeniophyllum zollingeri 1, 153
Tagetes minuta 1, 428
Tumarindus 1, 298, 318
Tamarix 1, 297, 316, 401, 408, 425, 479,
500, 510, 526, 537, 539; 111, 115, 191,
197, 201, 219, 225, 227, 241, 250,
253, 285, 370
aphylla ( — T. articulata)
articulata ( = T. aphylla) 1, 465,
475, 476, 478, 500, 523, 526,
528
gallica 1, 479; 111, 167
lonxa 111, 240
mannifera 1, 527
nilotica 1, 479
pallasii 111, 136
ramosissima 111, 231—233, 235,
240, 253
tetragyna 1, 528
Tamus communis 11, 240
Tanacetum 111, 258, 280, 293
adulleoides 111, 257
I
424
Указатель латинских названий растений
fruticulosum III, 244
pamiricum III, 267
xylorrhizum III, 267
Taraxacum II, 80; III, 17
bessarabicum III, 135, 136, 301
officinale I, 16; II, 15, 218; III,
51, 177
vulgare III, 88
Tarchonanthus I, 436
Tavaresia grandiflora I, 393
Taxodium III, 311
Taxus II, 154; III, 284, 292
baccata II, 247, 329, 383; III,
287 290 291
Tectonia grandis I, 122, 190, 240, 243,
244
Templetonia retusa II, 202
Tephrocactus floccosus I, 210, 211
Tephrosia I, 470
Terminalia catappa I, 183, 185, 186,
243—245, 320, 391
Tessaria absinthoides II, 133
Testudinaria elephantipes I, 425
Tetraclinis II, 171, 202
articulata ( = callitris quadriva-
lis) II, 83
Tetraena mongolica III, 243, 244
Tetragonia I, 281; II, 203, 204
schenckii I, 281
zeyheri II, 202
Tetrameles I, 239
Teucrium I, 326, 466; II, 102;'III, 122
chamaedrys II, 33, 61; III, 125,
129
flavum II, 332, 333
polium I, 494, 527; II, 61; III, 368
scorodonia II, 257, 314, 316, 328,
329
Thalassia III, 332
Thalictrum III, 249, 259
alpinum III, 21, 267
Thamnolia III, 21
vermicularis III, 49
Thea sinensis I, 113
Themeda I, 245, 272—274, 428, 466
Thermopsis III, 293
alpina III, 280
Theobroma I, 116
Thesium hystrix I, 429
Thespesia populnea I, 185
Thlaspi cochleariforme III, 18
rotundifolium III, 50
Threlkeldia diffusa II, 202
Thrinax microcarpa I, 188
Thrixspermum I, 146
Thuidium abietinum III, 85, 87
Thuja occidentalis II, 347
plicata II, 123, 125, 126
Thylacospermum rupifragum III, 280
Thymelaea hirsuta I, 516, 520, 522, 526
passerina II, 40
Thymus I, 326; II, 43, 61, 101, 259; III,
77, 290, 368
bovei I, 527
decussatus P, 528
glabrescens III, 129
marschalliamus III, 88, 129
serpyllum II, 329; III, 131, 245
vulgaris II, 41, 42, 44, 45, 51, 52,
61
Tilia II, 72, 148, 241, 277, 278, 317
americana III, 101
cordata II, 233, 238, 261, 298,
299, 319, 321, 336
platyphyllos II, 233, 321, 325
tomentosa II, 240
Tilieto — Querceta II, 336
Tillandsia I, 212, 213, 413, 414; II, 139»
pallacea I, 211
usneoides I, 146, 152
werdermannii II, 141
Titanopsis I, 425
Todea barbara II, 165
Tortella tortuosa III, 50
Tortula III, 201
desertotum III, 201, 222
ruralis ( = Syntrichium) II, 354;
III, 368
Tournefortia III, 215
argentea I, 183
sibirica III, 201, 302
Trachycarpus III, 202
fortunei II, 68
Tradescantia zebrina I, 133
Traganum nudatum I, 469, 478, 487
Tragopogon orientalis III, 88
pratensis I, 16; III, 85
Trapa I, 159
natans III, 310, 311, 323
Trema I, 260
Trentepohlia I, 46, 153; II, 134
occulta II, 164
Trevoa II, 138
trinerva II, 136
Trianthema I, 349, 350
Tribulus I, 401, 349, 350
alatus I, 501
Trichocaulon I, 424
dinteri I, 391, 393, 394
pedicellatum I, 391, 394
Trichocereus chilensis II, 135, 144
Trichocladus crinitus II, 172
Trichodesma 1, 449
Trichophorum caespitosum II, 251, 253,
405
Trientalis europaea II, 252, 349, 353
Trifolium alpestre III, 88
campestre III, 88
fragiferum III, 135, 136, 301, 348
hybridum III, 301
montanum III, 85, 87, 88, 90, 300
pallescens III, 50
parryi III, 44
pratense III, 88, 301
repens II, 248; III, 85, 88, 177, 301
subterraneum II, 185
Triglochin I, 174
maritimum III, 135, 294, 348, 349,
356
palustris III, 135, 258, 281
Trigonella stellata I, 498, 501, 505, 515,
516
Указатель латинских названий растений
425
Trinia henningii III, 88
Triodia I, 291, 300, 369, 442, 445, 449,
458, 459, 462, 466
Tripogon exiguus I, 206
Tripteris I, 424
arborescens I*, 403
pachypteris I, 429
Triset um III, 269
distichophyllum III, 50
spicatum III, 26, 269
Trithrinax campestris III, 159
Triticum camnum II, 349
Triuridaceen I, 155
Trollius patulus III, 53
Tropaeolum II, 139
Trymalium spathulatum II, 180, 190
Tsuga III, 285, 292
canadensis II, 334, 383
dumosa III, 291
heterophylla II, 123, 125—127
mertensiana II, 127
Tulipa III, 90, 194, 205
biebersteiniana III, 90
shrenkii III, 90, 91
Turraca I, 260
Tussilago farfara I, 231
Typha I, 160, 466, 479; II, 165; III, 166,
225 311
angustifolia III, 164, 322
elephantina III, 225
[W -«
UapacaJI, 242
Ugni molinae II, 150
Ulex II, 242, 249, 258, 259
europaeus II, 245, 251, 257
gallii II, 245
minor II, 252
nanus II, 245
parviflorus II, 64, 245
Ulmus americana III, 101, 306
campestris II, 76
effusa I, 107
fulva III, 101
glabra II, 247, 250, 336
laevis II, 233, 261
montana II, 325, 339
pumila III, 246, 248, 253, 257
293
scabra II, 233, 282, 293, 299
Ulva pertusa III, 339
Umbilicaria rubiginosa II, 164
Umbilicus pendulinus II, 62, 242
Umbellularia califomica II, 110, 126
Uncinia II, 218
Urospermum dalechampii II, 63
Urtica I, 160, 419
dioica II, 290, 308, 319, 329, 349
Usnea I, 222, 223, 244, 498; II, 144
Utricularia I, 160, 479; II, 207; III, 135,
309
capensis II, 164
lusitanica II, 242
menziesii II, 187
minor II, 253
schimperi I, 150
Vaccinium I, 205, 219; II, 243, 250, 388;.
Ill, 32, 39, 291
arctostaphylos II, 240
myrtillus II, 248, 251—254, 259,
308, 316, 328, 329, 347, 349,
353, 361, 362, 399, 418, 420;.
Ill, 21, 44, 54
oxycoccus II, 405, 417, 420
uliginosum II, 405, 419, 420; III,
94 99 94 9R
varingiifolium I, 188, 206
vitis idaea II, 251, 252, 254, 349,.
353, 362, 405, 420; III, 13, 18
43
Valenzuellia trinervia II, 139
Valeriana III, 249, 259
montana III, 50
rossica III, 87, 88
tripteris III, 50
Vallisneria denseserrulata III, 317
Vandellia pyxidaria III, 302
Vanilla I, 134
Varthemia iphionoiles I, 544, 545
Vaucheria I, 47
Vauguelinia californica II, 119
Ventilago viminalis I, 466
Veratrum III, 53
album III, 49
nigrum III, 87, 88
stamineum III, 53
Verbascum lychnitis III, 76
phoeniceum III, 89, 91
Verbena ciliata I, 350
ligustrina III, 178
Veronica II, 80, 217; III, 97, 205
alpina III, 47
anagallis I, 528; III, 323
austriaca III, 89
chamaedrys II, 353; III, 89
hederifolia II, 290, 319
montana II, 319
officinalis II, 349, 353
prostrata III, 89
spicata III, 131
verna III, 91
Verrucaria I, 498; III, 334, 335
Veteveria nigritana I, 157
Viburnum III, 290, 292
lantana II, 300
opulus II, 250, 300; III, 194
tinus II, 33, 35, 50—53, 62, 68,
332, 333
Vicia cracca III, 88, 348
dumetorum II, 64
faba I, 479
sepium II, 319, 353
Victoria regia I, 159, 162; III, 294
Vinca herbacea II, 101
minor II, 328, 329
Vincetoxicum officinale III, 89, 125
Viola II, 34, 134, 277, 309, 310
abyssinica I, 221
alpina III, 37
arenaria III, 85, 88
biflora III, 50
calcarata III, 50
426
Указатель латинских названий растений
canina III, 85, 89, 349
cheiranthifolia II, 91, 93
hirta III, 88
mirabilis II, 239
nevadensis II, 87
riviniana II, 315
scotophylla II, 33
silvatica II, 311
Virgilia capensis II, 171
Viscaria vulgaris II, 315
Viscum album II, 329
capense I, 436
rotundifolium I, 436
Visnea mocanera II, 92
Vitex agnuscastus II, 96
Vitis arizonica II, 119
silvestris II, 300
Vittaria lineata I, 151
isoetifolia II, 172
Viviania rosea II, 139
Vossia cuspidate I, 157
Vriesea I, 146
altodaserrae I, 149
inflate I, 149, 150
Waitzia I, 447
Waldheimia tridactyloides III, 267
Watsonia rosea II, 162, 164
Webera commutata III, 47
Weinmannia II, 221
trichosperma II, 150, 151
Weissia verticillata II, 93
Welwitschia mirabilis I, 406—408
Werneria I, 209
Whitfieldia longifolia I, 110
Widdringtonia II, 158, 171
cupressoides II, 160
Wikstoromia III, 293
Wistaria II, 136
sinensis I, 134
Woodsia alpina III, 50
glabella III, 26
Wyethia amplexicaule III, 111
Xanthium canadense III, 369
Xanthoria III, 57, 334
Xanthorrhoea I, 264; II, 189, 195, 198,
201
australis II, 195—197; 200
preisii II, 184, 185, 190
semiplana II, 191
Xerotes dura I, 466
Ximenia americana III, 167
Xylocarpus I, 164, 179
granatum I, 164, 180, 181
moluccensis I, 164, 180
Xylomelum I, 289
puriforme II, 200
Xymodocea III, 331
Xyris I, 160
Yucca I, 344, 346
macrocarpa II, 119
whipplei II, 112, 113
Zannichellia palustris III, 328
Zelkowa II, 240
ZiHa I, 476, 483
macroptera I, 488
spinosa I, 477, 500, 501, 502, 506,
513-515, 527, 529, 534-536,
539
Zizaniopsis bonariensis III, 164
Zizyphus I, 296, 476; II, 86
lotus I, 488, 541
jujuba I, 522; II, 12; III, 249
spina-christi I, 296, 541
Zollikoferia arborescens I, 488
( = Launea) spinosa II, 90, 91
Zostera III, 361
marina III, 328, 331, 332, 343
nana III, 328, 330
Zuccagnio punctata III, 169—174
Zygochloa paradoxa I, 449
Zygophyllum I, 424, 454, 456, 464, 465,
478, 479, 480, 526, 528; III, 207, 250,
253
album I, 469, 474, 478
coccineum I, 500, 502, 513, 515
dumosum I, 326, 526, 527, 529,
530, 531, 537, 538
fontanesii II, 90, 91
mucronatum III, 244
oxianum III, 227
rosovii III, 267, 270, 272
simplex I, 382, 385, 386, 469, 479,
498, 501, 505, 513, 515
stapffii I, 386, 389, 402, 403
xanthoxylon III, 244, 257
ОГЛАВЛЕНИЕ
Глава 1
АРКТ ЧЕСКАЯ ЗОНА ТУНДРЫ И АЛЬПИЙСКИЙ ПОЯС ГОР
УМЕРЕННОЙ ГУМИДНОЙ ЗОНЫ
ОБЩИЕ СВЕДЕНИЯ О КЛИМАТЕ И ФЛОРЕ (5). ЛЕСОТУНДРА (10). ТУНДРА
<15). ЭКОЛОГО-ФИЗИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ В АРКТИЧЕСКОЙ
ЗОНЕ (23). ПРОДУКТИВНОСТЬ И КРУГОВОРОТ ВЕЩЕСТВ В АРКТИЧЕСКОЙ
ЗОНЕ' (27).ВЗАИМОСВЯЗИ ЖИВОТНОГО МИРА И РАСТИТЕЛЬНОГО ПОКРОВА
ТУНДРЫ (29). АЛЬПИЙСКАЯ РАСТИТЕЛЬНОСТЬ (31). ЭКОЛОГО-ФИЗИОЛО-
ГИЧЕСКИЕ ИССЛЕДОВАНИЯ В АЛЬПИЙСКОМ ПОЯСЕ (39). УСЛОВИЯ СУЩЕ-
СТВОВАНИЯ АЛЬПИЙСКОЙ РАСТИТЕЛЬНОСТИ В ЗАВИСИМОСТИ ОТ^НЕКО*
Т(5РЫХ ФАКТОРОВ’МЕСТООБИТАНИЯ (45). АНТАРКТИДА ЩСУБАНТАРКТИ-
ЧЕСКИЕ ОСТРОВА (56).
Глава 2
ЗОНА СТЕПЕЙ И ПРЕРИЙ
ЛЕСОСТЕПЬ (64). ПОЧЕМУ СТЕПЬ БЕЗЛЕСНА? (69). ПОЧВЕННЫЕ УСЛО-
ВИЯ В СТЕПЯХ (77). ХАРАКТЕРИСТИКА СТЕПНОЙ РАСТИТЕЛЬНОСТИ
ВОСТОЧНОЙ ЕВРОПЫ (83). СЕВЕРОАМЕРИКАНСКИЕЗ ПРЕРИИ (94). ПО-
СЛЕДСТВИЯ ЗАСУШЛИВЫХ ЛЕТ В ПРЕРИИ (107). ЭКОЛОГО-ФИЗИОЛОГИ.
ЧЕСКИЕ ИССЛЕДОВАНИЯ РАСТЕНИЙ СТЕПЕЙ И СТЕПНЫХД1УСТО111ЕЙ (109).
ПРОДУКТИВНОСТЬ и КРУГОВОРОТ ВЕЩЕСТВ В СТЕПНОЙ ЗОНЕ (129).
ТОПОГЕННЫЕ БОЛОТА И ЯВЛЕНИЯ ЗАСОЛЕНИЯ В СТЕПНОЙ ЗОНЕ (132).
КР АТКИЙ 8ОБЗОР КЛИМАТИЧЕСКИХ, ^РАСТИТЕЛЬНЫХ И ПОЧВЕННЫХ
ЗОН ВОСТОЧНОЙ ЕВРОПЫ (142).;
Глава 3
ЗЛАКОВНИКИ УМЕРЕННОЙ КЛИМАТИЧЕСКОЙ ЗОНЫ
ЮЖНОГО ПОЛУШАРИЯ
ОБЩИЙ ОБЗОР УМЕРЕННОЙ ЗОНЫ ЮЖНОЙ АМЕРИКИ К ВОСТОКУ ОТ ^АНД
(145). ПРОБЛЕМА ПАМПЫ (147). РАСТИТЕЛЬНОСТЬ ПАМПЫ (162). АРИДНАЯ
ДРЕВЕСНАЯ* РАСТИТЕЛЬНОСТЬ И ПОЛУПУСТЫНЯ С .LARREA 53АПАДНЕЕ
ПАМПЫ (167). ЗЛАКОВНИК И ПОЛУПУСТЫНЯ В ПАТАГОНИИ (174).ТУССО-
КОВЫЙ ЗЛАКОВНИК НОВОЙ ЗЕЛАНДИИ (180). ОГОНЬ КАК ЕСТЕСТВЕННЫЙ
ЭКОЛОГИЧЕСКИЙ ФАКТОР (183).
Глава 4
АРИДНЫЕ ОБЛАСТИ ЦЕНТРАЛЬНОЙ АЗИИ
ОБЩИЙ ОБЗОР (187). ОБЛАСТЬ СТЕПЕЙ И ПОЛУПУСТЫНЬJKA3AXCTAHA
(193). ПУСТЫНИ СРЕДНЕЙ АЗИИ (202). ИЗМЕНЕНИЯ РАСТИТЕЛЬНОСТИ
ПЕСЧАНЫХ ПУСТЫНЬ ПОД ВЛИЯНИЕМ ДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКА (220).
ПОЙМЫ РЕК СРЕДНЕЙ АЗИИ (224). ЭКОЛОГО-ФИЗИОЛОГИЧЕСКИЕГ ИССЛЕ-
ДОВАНИЯ; ВОДНОГО РЕЖИМА РАСТЕНИЙ В СРЕДНЕАЗИАТСКИХ ПУСТЫ-
НЯХ (227). КРУГОВОРОТ ВЕЩЕСТВ И СОЛЕВОЙ<РЕЖИМ РАСТИТЕЛЬНОСТИ
ПУСТЫНЬ (233). ПУСТЫНИ ЦЕНТРАЛЬНОЙ^АЗИИ (242). ВЫСОКОГОРНЫЕ
ПУСТЫНИ ЦЕНТРАЛЬНОЙ АЗИИ (261). ВЕРТИКАЛЬНАЯ ПОЯСНОСТЬ ГИМА-
ЛАЕВ (284).
Глава, 5
АЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ РАЗНЫХ КЛИМАТИЧЕСКИХ ЗОН
АЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ И КЛИМАТ (294). РАСТИТЕЛЬНОСТЬ
РЕЧНЫХ НАНОСОВ (294). ПРЕСНОВОДНЫЕ РАСТЕНИЯ И ИХ ЭКОЛОГО-
ФИЗИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА (307). СОЛОНОВАТЫЕ ВОДЫ
И СОЛЕНЫЕ ОЗЕРА (327). МАКРОФИТНАЯ РАСТИТЕЛЬНОСТЬ МОРЕЙ (330).
ЗАСОЛЕННЫЕ МАРШИ НА МОРСКИХ ПОБЕРЕЖЬЯХ (342). РАСТИТЕЛЬНОСТЬ
ПЕСЧАНЫХ ПЛЯЖЕЙ И БЕРЕГОВЫХ ДЮН (360). НЕСКОЛЬКО СЛОВ О НЕОБ-
ХОДИМОСТИ СОЗДАНИЯ ЗАПОВЕДНИКОВ (373).
УКАЗАТЕЛЬ РУССКИХ ТЕРМИНОВ (37 6).
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ РАСТЕНИЙ И РАСТИТЕЛЬНЫХ СООБ-
ЩЕСТВ (387).
Г. Вальтер
РАСТИТЕЛЬНОСТЬ
ЗЕМНОГО ШАРА
т. III
Редактор Л. Бабинцева
Младший редактор М. Бродовская
Художник В. Кузянов
Художественный редактор В. Пузанков
Технические редакторы Р. Медведева и Л. Жарова
Корректор В. Пестова
Сдано в производство 14.VII 1974 г.
Подписано к печати 2.VI 1975 г.
Бумага 70X1081/16 тип. № 1 Бум. л. 131/2-
Печ. л. 37,84. Уч.-изд. л. 34,21. Изд. № 17565
Цена 3 р. 26 к. Заказ 0434. Тираж 11000
Издательство «Прогресс»
‘Государственного комитета
Совета Министров СССР по делам издательств,
полиграфии и книжной торговли
Москва, Г-21, Зубовский бульвар, 21
Ордена Трудового Красного знамени Московская типография № 7 «Искра революции»
Союзполиграфпрома при Государственном комитете Совета Министров СССР
по делам издательств, полиграфии и книжной торговли
Москва, К-1, Трехпрудный пер., 9